
6.1 FENOTIPO DE LAS MUTANTES - Universidad de las...
23
6. RESULTADOS 6.1 FENOTIPO DE LAS MUTANTES CAX 6.1.1 Germinación de las semillas Las semillas de Arabidopsis thaliana, tanto las tipo silvestre como las mutantes, iniciaron su germinación alrededor de las 24 horas después de haber sido sembradas. Sin embargo, no fue sino hasta las 96 horas cuando se observó que más del 90% de las semillas habían germinado. Las semillas de las plantas WT y CAX1- OE mostraron mayor porcentaje de germinación (superior al 70%) desde las 24 horas después de haberse sembrado. Las plantas con menor porcentaje de germinación inicial fueron las mutantes cax1 (alrededor del 16%), mientras que las mutantes cax3 presentaron un porcentaje de germinación inicial aproximado del 50% (Tabla 6.1). Tabla 6.1 Porcentaje de germinación de las semillas de las plantas en estudio a las 24 y 96 horas después de haber sido sembradas. Tipo de planta % germinación a las 24 hrs % germinación a las 96 hrs WT 74.1 93.9 cax1 17.0 96.8 cax3 53.3 100.0 CAX1-OE 70.7 91.7 35
Transcript of 6.1 FENOTIPO DE LAS MUTANTES - Universidad de las...

6. RESULTADOS
6.1 FENOTIPO DE LAS MUTANTES CAX
6.1.1 Germinación de las semillas
Las semillas de Arabidopsis thaliana, tanto las tipo silvestre como las
mutantes, iniciaron su germinación alrededor de las 24 horas después de haber sido
sembradas. Sin embargo, no fue sino hasta las 96 horas cuando se observó que más
del 90% de las semillas habían germinado. Las semillas de las plantas WT y CAX1-
OE mostraron mayor porcentaje de germinación (superior al 70%) desde las 24
horas después de haberse sembrado. Las plantas con menor porcentaje de
germinación inicial fueron las mutantes cax1 (alrededor del 16%), mientras que las
mutantes cax3 presentaron un porcentaje de germinación inicial aproximado del
50% (Tabla 6.1).
Tabla 6.1 Porcentaje de germinación de las semillas de las plantas en estudio a las 24 y 96 horas después de haber sido sembradas.
Tipo de planta % germinación a las 24 hrs % germinación a las 96 hrsWT 74.1 93.9 cax1 17.0 96.8 cax3 53.3 100.0
CAX1-OE 70.7 91.7
35

6.1.2 Fenotipos observados
Respecto a los fenotipos de cada una de las plántulas de A. thaliana, fue
difícil diferenciar a las tipo silvestre (WT) de las sobreexpresoras CAX1-OE, pues
tuvieron tamaños similares, tanto en el ancho de sus cotiledones como en el largo de
las raíces (datos no mostrados). Ambos tipos de plántulas fueron las de mayor
tamaño de los cuatro tipos de plántulas estudiadas (Figura 6.1.b y 6.1.d). La mutante
cax1 fue la más fácil de distinguir, pues sus plántulas fueron las de menor tamaño,
con cotiledones angostos y doblados en comparación con los cotiledones grandes y
extendidos de las plántulas WT (Figura 6.1.e); además la longitud de su hipocótilo
fue aproximadamente dos milímetros menor que el de WT (Figura 6.1.a).
Finalmente, las mutantes cax3 mostraron un tamaño intermedio entre las plántulas
cax1 y WT; con cotiledones extendidos pero de menor tamaño que las WT y un
hipocótilo largo como las tipo silvestre (Figura 6.1.c).
Después de una semana de crecimiento, las plántulas de A. thaliana
alcanzaron una longitud de aproximadamente un centímetro y medio, y presentaron
sus primeras hojas; sus raíces midieron alrededor de un centímetro con una buena
verticalidad respecto al vector de gravedad. Algunas de estas plántulas comenzaron
a generar sus primeras raíces secundarias, justo por debajo del sitio en el que se
inició la raíz principal (datos no mostrados). No se encontraron diferencias claras
entre el fenotipo de las raíces de las mutantes cax3 y CAX1-OE respecto a la WT,
pero las raíces de las plántulas cax1 fueron más pequeñas que las de la WT. De
manera interesante, todas las raíces presentaron mayor densidad de pelos radicales
36

en el extremo superior y las plántulas cuya raíz se ‘enterró’ en el agar desarrollaron
un menor número de pelos radicales en esta zona.
(a) (b) (c)
(d)
(e)
Figura 6.1 Fenotipos de las plántulas de Arabidopsis thaliana a la edad de una semana de crecimiento: (a) cax1, (b) WT, (c) cax3, (d) CAX1-OE. (e) Las mutantes cax1 muestran menor tamaño que las plántulas WT y diferencias claras entre sus cotiledones.
37

6.2 RESPUESTA GRAVITRÓPICA
Para estudiar posibles alteraciones en la respuesta gravitrópica de las raíces
de las mutantes cax, se indujo la respuesta gravitrópica en plántulas de una semana
de edad crecidas verticalmente. Tanto las raíces de las plántulas tipo silvestre como
las de las mutantes respondieron al estímulo gravitrópico aplicado, es decir, todas
las raíces desarrollaron una curvatura en su raíz como consecuencia del giro de 90°
respecto al vector de gravedad (Figura 6.2). Sólo algunas de las raíces que quedaron
hundidas en el agar mostraron una curvatura menos pronunciada o casi
imperceptible en comparación con las que lograron crecer por encima del medio
sólido. Además, el mayor cambio de ángulo de las raíces se observó durante las
primeras cuatro horas después del giro y fue más o menos constante a partir de las
seis horas (Figura 6.3)
Las plántulas WT generaron una mayor curvatura en su raíz, alcanzando un
ángulo mayor a los 70° a las 24 horas después del giro; mostraron además una
respuesta más rápida durante las primeras ocho horas de estimulación (Figura 6.4).
Entre las mutantes, las plantas cax1 mostraron la curvatura más grande a las 24
horas después del giro, quedando por arriba de las mutantes cax3 y de las
sobreexpresoras CAX1-OE, pero por debajo de la WT; sin embargo, las plántulas
cax1 desarrollaron la curvatura inicial más pequeña en las primeras ocho horas
después del estímulo gravitrópico (Figura 6.4). Las plántulas CAX1-OE fueron las
mutantes que mostraron una respuesta gravitrópica menor. Finalmente, las plántulas
cax3 fueron las únicas en presentar raíces con ángulos de giro mayores que los de la
WT en dos de los cuatro experimentos de estimulación gravitrópica (28-07-04 y 05-
38

08-04) y dentro de las primeras seis horas de giro; sin embargo, a las 24 horas
después del giro desarrollaron curvaturas menores a la WT.
Figura 6.2 Respuesta de las plántulas de A. thaliana al estímulo gravitrópico. Las puntas de las raíces desarrollan una curvatura de acuerdo al nuevo vector de gravedad.
39
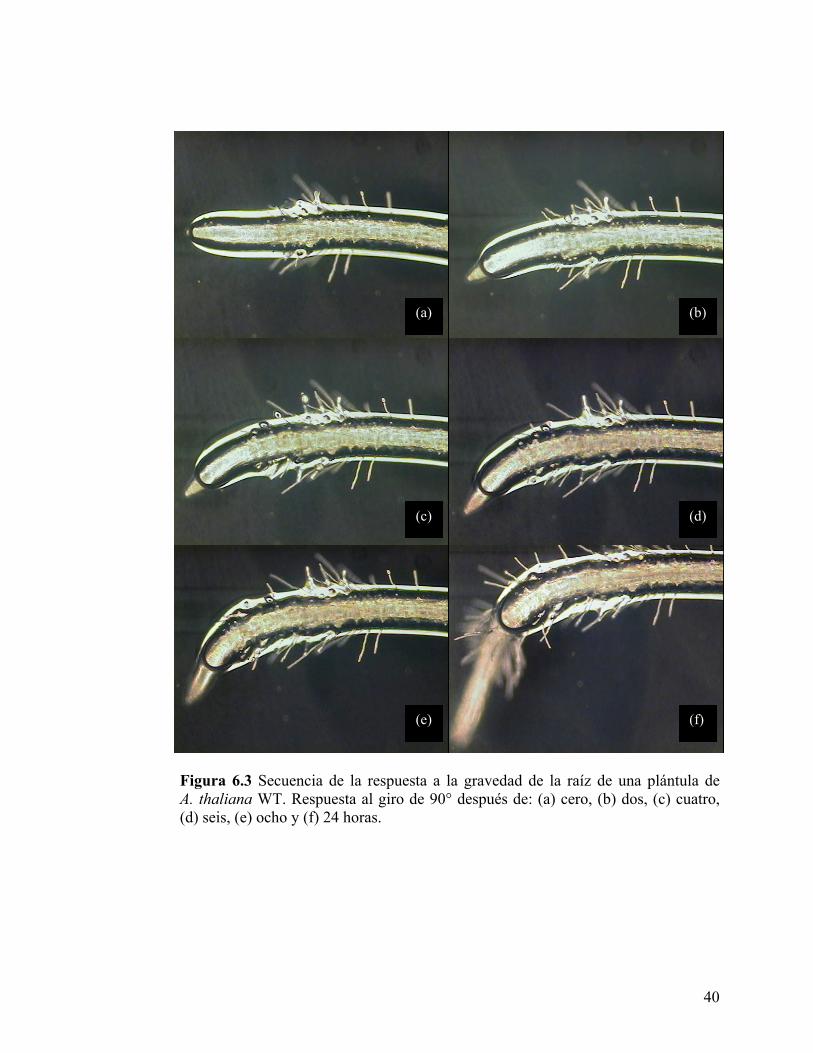
(a) (b)
(c) (d)
(e) (f)
Figura 6.3 Secuencia de la respuesta a la gravedad de la raíz de una plántula de A. thaliana WT. Respuesta al giro de 90° después de: (a) cero, (b) dos, (c) cuatro, (d) seis, (e) ocho y (f) 24 horas.
40

0 5 10 15 20 250
10
20
30
40
50
60
70
80
90
CAX1 CAX3 OE WT
Ángu
lo d
e cu
rvat
ura
de la
raíz
(Gra
dos)
Tiempo después del giro (Horas)
(a)
0 5 10 15 20 250
10
20
30
40
50
60
70
80
90
CAX1 CAX3 OE WT
Ángu
lo d
e cu
rvat
ura
de la
raíz
(Gra
dos)
Tiempo después del giro (Horas)
(b)
Figura 6.4 Respuesta gravitrópica general de las raíces de A. thaliana. Experimentos: (a) 28-07-04, (b) 05-08-04
41

0 5 10 15 20 250
10
20
30
40
50
60
70
80
90
CAX1 CAX3 OE WT
Ángu
lo d
e cu
rvat
ura
de la
raíz
(Gra
dos)
Tiempo después del giro (Horas)
(c)
0 5 10 15 20 250
10
20
30
40
50
60
70
80
90
CAX1 CAX3 OE WT
Ángu
lo d
e cu
rvat
ura
de la
raíz
(Gra
dos)
Tiempo después del giro (Horas)
(d)
Figura 6.4 Respuesta gravitrópica general de las raíces de A. thaliana. Experimentos: (c) 25-09-04, (d) 02-10-04.
42

6.2.1 Resultado del análisis estadístico de la respuesta gravitrópica
De los cuatro experimentos seleccionados para el análisis estadístico, en los
dos primeros, 28-07-04 y 05-08-04, y después de la aplicación de las pruebas t-
Student correspondientes (IC 95%), no se encontró ninguna diferencia en las
respuestas gravitrópicas entre las plántulas WT y las mutantes cax. Para el
experimento 28-07-04, sólo en una de las 15 pruebas de t-Student se encontró una
diferencia en el ángulo de giro entre la mutante cax1 y la WT a las ocho horas
(p<0.011) después de la aplicación del estímulo gravitacional.
En el experimento 25-09-04 se encontraron diferencias significativas entre
las tres mutantes y la WT. Para la mutante cax3 se encontró que la respuesta al
graviestímulo fue distinta de la WT a las dos (p<0.045), cuatro (p<0.018), seis
(p<0.005) y ocho (p=0.000) horas después del giro. La mutante cax1 sólo resultó
diferente a la WT en las respuestas de las seis (p<0.049) y ocho (p<0.013) horas
después del giro. Finalmente, las plantas CAX1-OE mostraron un giro en su raíz
distinto a la WT a las seis (p<0.032), ocho (p<0.001) y 24 (p<0.006) horas después
de la estimulación gravitrópica. Asimismo, para el experimento 02-10-04 se
observaron respuestas distintas entre las tres mutantes y la tipo silvestre. La mutante
cax3 respondió de forma diferente a la WT a las cuatro (p<0.008), seis (p<0.029) y
24 (p<0.024) horas después de giro. La mutante cax1 giró de forma distinta a la WT
a las dos (p<0.015), cuatro (p=0.000), seis (p<0.009) y ocho (p<0.034) horas
después del giro. Y la mutante CAX1-OE respondió diferente a la WT a las cuatro
(p<0.018), seis (p<0.021), ocho (p<0.025) y 24 (p<0.003) horas después del giro.
43

(Ver Apéndice C). En general, se encontró que la respuesta de las raíces de las
mutantes cax al estímulo de la gravedad fue menor a la respuesta de la línea
silvestre.
6.3 RESPUESTA ESTOMATAL
6.3.1 Tratamiento de la epidermis de A. thaliana con ABA
La incubación de la epidermis recién aislada de las hojas de A. thaliana en
solución de MES/KOH 10mM a pH 5.5 con KCl 50mM, bombeada con aire libre de
CO2 y colocada bajo la luz de una lámpara, ocasionó la apertura de los estomas a
partir de los 50 minutos de incubación, aunque la mayor apertura se alcanzó hasta
después de las dos horas. Sin embargo, es importante mencionar que fue difícil
medir los cambios en la apertura de los estomas.
Para determinar los efectos del ABA sobre la apertura de los estomas de las
diferentes plantas bajo estudio, inicialmente se expusieron epidermis aisladas a 5
µM de ABA –concentración que causa el cierre de los estomas en Commelina
communis. Esta concentración de ABA no causó una clara respuesta estomatal de
cierre en ninguna de las plantas, por lo que se decidió realizar un tratamiento
simultáneo de diferentes segmentos de epidermis, expuestos a diferentes
concentraciones de ABA (1 µM, 10 µM y 100 µM). Sorprendentemente, ninguna de
estas concentraciones de ABA causó cambio alguno en la apertura estomatal de
dichas epidermis; es decir, no se obtuvo el esperado cierre de los estomas inducido
44

por el ácido abscísico (Figura 6.5). Es importante recordar que los estomas de
dichas epidermis fueron previamente estimulados para abrirse, mediante su
incubación por cerca de dos horas en la solución amortiguadora.
6.3.2 Dificultad para analizar la respuesta estomatal en epidermis
La superficie de la epidermis de A. thaliana es irregular, por lo que los
estomas quedan en distintos planos, algunos inclinados y otros paralelos a la
superficie de soporte sobre la que la epidermis fue montada, lo cual hizo difícil la
medición del ancho del poro estomatal (Figura 6.6). Además, los estomas no se
distribuyen uniformemente en la epidermis, por lo que se encuentran unas zonas con
escasos estomas y otras donde son más abundantes, a lo largo de toda la superficie
de la hoja. Por otro lado, debido al pequeño tamaño de los estomas de A. thaliana
fue necesario utilizar el objetivo 40X para poder distinguir la apertura de los
mismos.
Como consecuencia de la irregularidad de la superficie de la epidermis, el
análisis de la respuesta estomatal al ABA, que consistía en medir el ancho del poro
estomatal, no fue posible en la mayoría de los casos, como se muestra en el ejemplo
de la Figura 6.7. Para poder resolver este problema, se decidió medir el ancho de las
dos células guarda (como se refiere en la sección 5.2.2 de Material y Métodos)
esperando que este parámetro fuera indicador del ancho del poro; pues se sabe que
antes de que el estoma se cierre, las células guarda pierden su turgencia y por tanto
se encogen (dejan de estar hinchadas).
45

(a) (b)
Figura 6.5 Respuesta nula de los estomas de A. thaliana WT a la estimulación del ácido abscísico (ABA). (a) Control, 0 min. (b) Después de 60 min de incubación en ABA 10 µM.
Figura 6.6 Topografía de la epidermis de las hojas de A. thaliana WT obtenida por microscopía de barrido (2000X). Adquirida en: http://remf.dartmouth.edu/images/ ArabidopsisSEM/
46
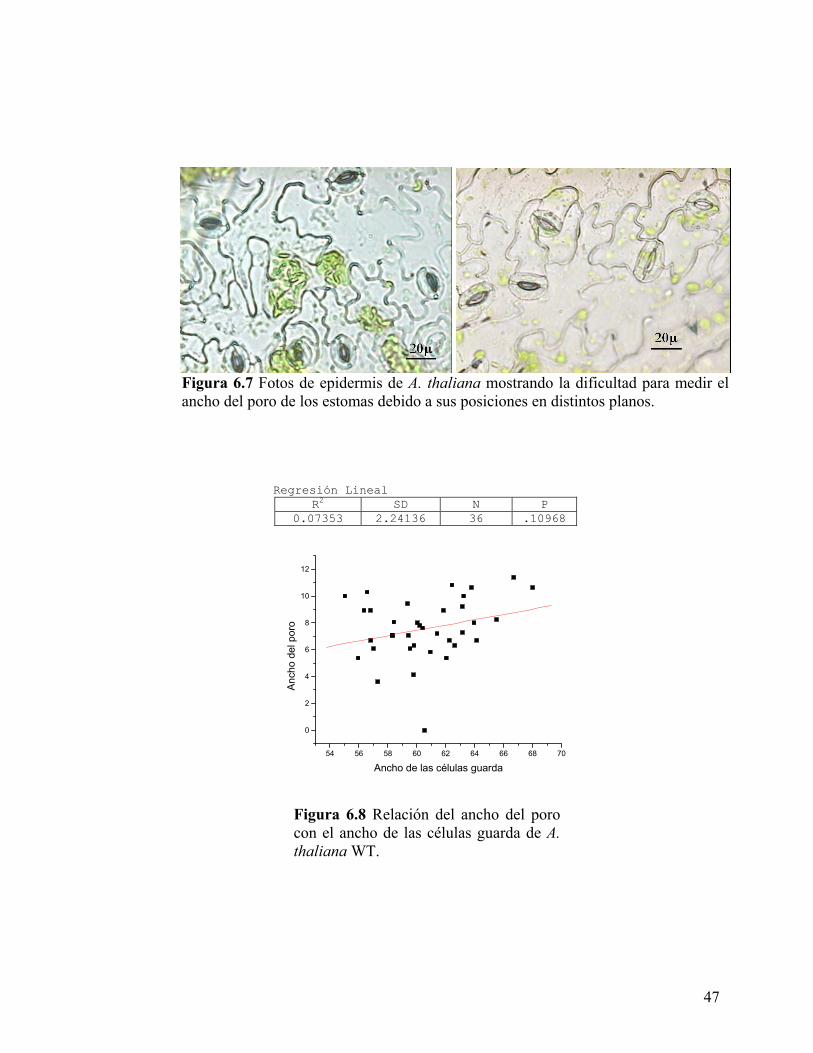
Figura 6.7 Fotos de epidermis de A. thaliana mostrando la dificultad para medir el ancho del poro de los estomas debido a sus posiciones en distintos planos.
Regresión Lineal R2 SD N P
0.07353 2.24136 36 .10968
54 56 58 60 62 64 66 68 70
0
2
4
6
8
10
12
Anch
o de
l por
o
Ancho de las células guarda
Figura 6.8 Relación del ancho del poro con el ancho de las células guarda de A. thaliana WT.
47

6.3.3 Relación entre el ancho del poro y el ancho de las dos células guarda.
Para poder analizar la respuesta estomatal, se decidió medir el ancho de las
dos células guarda que forma cada estoma y saber si existía una correspondencia
con el ancho del poro. Para ello se midieron el ancho del poro y el ancho de las
células guarda de los estomas que se encontraban en el mismo plano del
portaobjetos donde se montó la epidermis. Graficando la distancia entre los
extremos externos de dos células guarda contra el ancho del poro se obtuvo la
gráfica de la Figura 6.8. El análisis de los datos obtenidos mostró que no hubo una
relación lineal entre el ancho del poro y el ancho de las células guarda de los
estomas (r2 = 0.074). Por lo tanto, la distancia entre los extremos de dos células
guarda de un mismo estoma no fue un parámetro que nos permitiera inferir en
forma indirecta el grado de apertura del poro estomatal.
6.3.4 Respuesta a la Fusicoccina
Debido a la falta de respuesta de los estomas expuestos al ABA, se procedió
a determinar si esto era debido a que las células guarda pudieran estar dañadas,
mediante el estudio del efecto de la Fusicoccina sobre la apertura de los estomas.
Las epidermis recién aisladas de las plantas de A. thaliana WT se incubaron en
fusicoccina (5 µM) esperando que el poro del estoma se abriera, como se ha
reportado ampliamente en la literatura (Eun y Lee, 2000), pero no se observó
ninguna modificación en la apertura estomatal (Figura 6.9). Como se mencionó
anteriormente, se encontraron las mismas dificultades para hacer la medición del
ancho del poro debido a la irregularidad topográfica de la superficie epidermal.
48

6.3.5 Viabilidad de las células de las epidermis aisladas de A. thaliana.
Como consecuencia de la falta de respuesta de las células guarda a los
tratamientos de ABA y fusicoccina, se procedió a verificar la viabilidad de las
células de las epidermis aisladas mediante la ayuda de la tinción con el colorante
vital Rojo Neutro, el cual puede permear a través de las membranas celulares.
Después de incubar las epidermis recién aisladas de A. thaliana por quince minutos
en una solución al 0.01% de Rojo Neutro, se observó que al menos un 50% de las
células epidérmicas estaban vivas; lo cual indicaba que durante el proceso de
aislamiento de las epidermis, un gran número de células se dañaron (Figura 6.10.a).
Cabe señalar que las células que se tiñeron mayormente fueron las células guarda de
los estomas (Figura 6.10.a). También se tiñeron con Rojo Neutro epidermis que
habían sido incubadas en la solución amortiguadora de MES/KOH 10mM y KCl
50mM, bombeada con aire libre de CO2 y bajo luz. Estas epidermis se tiñeron
menos que aquellas recién aisladas de las hojas, lo que indicaba que la viabilidad de
las células epidérmicas disminuyó con el tiempo (datos no mostrados). Finalmente
se tiñeron epidermis tratadas con ABA por cerca de dos horas, después de haber
estado incubadas en la solución amortiguadora libre de CO2 y bajo luz, por al menos
una hora; es decir, se usaron epidermis que habían sido aisladas aproximadamente
tres horas antes. En estas muestras se observó que las células guarda y sólo unas
pocas células epidérmicas, que curiosamente estaban cercanas o junto a las células
guarda, estaban aún viables (teñidas de rojo; Figura 6.10.b).
Estos resultados indican que la viabilidad de las células epidérmicas en
epidermis aisladas disminuyó conforme transcurrió el tiempo. Es importante
49

recalcar que inicialmente sólo alrededor del 50% de las células epidérmicas
presentes en la epidermis aislada estaban vivas.
6.3.6 Respuesta estomatal en hojas completas de A. thaliana
Dado que no se observó ninguna respuesta en los estomas de las epidermis
aisladas ante los tratamientos con ABA y fusicoccina como consecuencia de un
posible daño a las células epidérmicas, se decidió realizar el ensayo con ABA pero
ahora sobre las hojas completas de A. thaliana. El objetivo de estos estudios fue
comprobar si el efecto mecánico del desprendimiento de la epidermis tenía un
efecto negativo sobre la actividad de las células guarda.
Las hojas en las que los estomas no se encontraban abiertos se incubaron en
la solución amortiguadora de MES/KOH 10mM con KCl 50mM, bombeada con
aire libre de CO2 y bajo iluminación. Después de aproximadamente una hora de
incubación, se observó un ligero aumento en la apertura de los estomas.
Posteriormente estas hojas se incubaron en solución de ABA 10 µM por al menos
una hora pero no se observó un cierre claro de los estomas (Figura 6.11).
El análisis de la respuesta estomatal en las epidermis unidas al mesófilo, fue
aún más complicado que el de las epidermis aisladas. En general, fue difícil
observar claramente la apertura de los poros, ya que un mismo estoma parecía estar
abierto o cerrado, según el plano de enfoque que se empleara. (Ver Figura 6.12.
Comparar con Figura 6.7). Por estas razones ya no se realizaron más ensayos con
hojas completas de A. thaliana, a pesar de que aparentemente se evitaba la afección
50

sobre la viabilidad de las células epidérmicas ante el desprendimiento de la
epidermis.
Figura 6.9 Ausencia de respuesta de los estoma de A. thaliana WT a la estimulación por fusicoccina. (a) Control, 0 min. (b) Después de 60 min de incubación en fusicoccina 5µM.
(a)
(b)
Figura 6.10 Viabilidad de las células epidérmicas en segmentos de epidermis aislada de A. thaliana tipo silvestre. La coloración roja de las células indica que están vivas. (a) Tinción sobre epidermis recién aislada; células guarda viables. (b) Tinción sobre epidermis tratada por dos horas con ABA 10 µM.
51

(a) (b)
Figura 6.11 Respuesta al ABA de los estomas de A. thaliana en hojas intactas (epidermis unida al mesófilo). (a) Control, 0 min. (b) Después de 60 min de incubación en ABA 10 µM.
Figura 6.12 Hoja de A. thaliana mostrando la dificultad para distinguir los estomas y por lo tanto, para medir el ancho del poro de los mismos.
52

6.3.7 Tratamientos en Commelina communis
Debido a lo difícil que fue obtener observaciones claras con los estomas de
A. thaliana bajo los diferentes tratamientos, fue necesario comprobar que el
procedimiento experimental era el adecuado. Por lo tanto se procedió a realizar
estudios similares con otra especie que ha sido empleada ampliamente para el
estudio de la fisiología de los estomas. Esta planta fue Commelina communis la cual
permite el fácil aislamiento de la epidermis, mantiene la viabilidad de casi todas las
células epidérmicas, con células guarda relativamente grandes y que facilita la
medición de los cambios en la apertura de los poros de los estomas.
La incubación de las epidermis aisladas de C. communis que no mostraban
los estomas abiertos en la solución amortiguadora de MES/KOH 10mM con KCl
50mM, bombeada con aire libre de CO2 y bajo la luz, permitió que la apertura del
poro de los estomas aumentara. Una vez abiertos los estomas, se llevó a cabo el
tratamiento con ABA, incubando las epidermis por alrededor de una hora en una
solución de ABA 10 µM. Este tratamiento causó el cierre en los poros estomatales;
el cambio en la apertura del poro fue claro hasta las dos horas de tratamiento
(Figura 6.13).
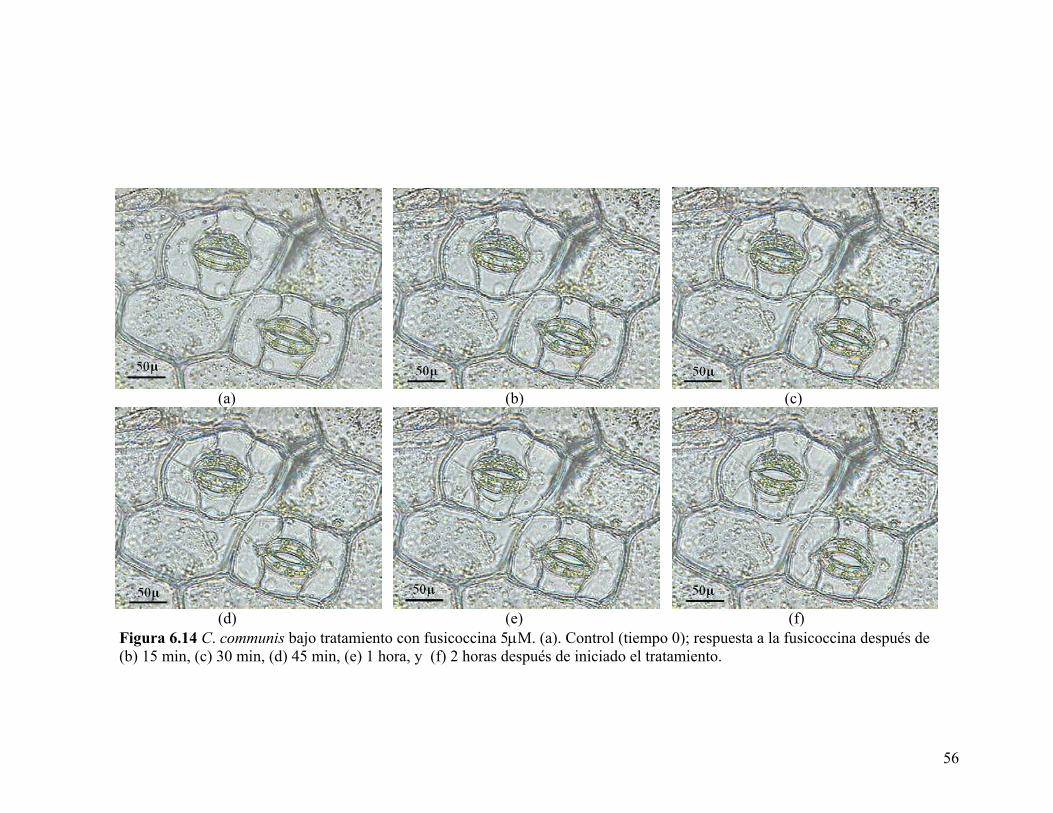
En experimentos adicionales, se estudió el efecto de la fusicoccina sobre la
apertura del poro de los estomas de C. communis. La incubación en una solución de
fusicoccina 5 µM se realizó inmediatamente después del aislamiento de la
epidermis, donde se pudo observar que la apertura de los estomas aumentó después
de media hora de incubación, aunque la apertura fue mayor a las dos horas de
tratamiento (Figura 6.14). Estos resultados contrastan con los observados en A.
53

thaliana donde los estomas permanecieron con la misma apertura a lo largo del
tratamiento (Figura 6.8).
La medición del ancho del poro de los estomas fue mucho más sencilla en C.
communis, pues la superficie de la epidermis de esta especie es lisa, los estomas son
de mayor tamaño que los de A. thaliana (pudiéndose observar definidamente con el
objetivo de 40X) y se encuentran distribuidos uniformemente en toda la hoja.
Por otro lado, se analizó la relación entre el ancho del poro y el ancho de las
células guarda de C. communis como en el caso de A. thaliana (Ver sección 6.3.3),
para confirmar que nuestra propuesta de análisis era válida. La gráfica que se
muestra en la Figura 6.15, resume los datos obtenidos de varias epidermis de C.
communis, donde se puede observar que la relación ancho de dos células guarda
contra ancho del poro fue lineal. El análisis de regresión lineal de estos datos mostró
que la relación entre dichos anchos es confiablemente lineal (r2 = 0.87); es decir, el
ancho de las células guarda se puede emplear para estimar el ancho del poro
formado entre ellas.
Finalmente, se comprobó la viabilidad de las células epidérmicas de C.
communis mediante la tinción con Rojo Neutro sobre las epidermis recién aisladas y
se pudo observar que casi el 100% de las células presentes se tiñeron de rojo, lo cual
indica la viabilidad de todas las células, tanto epidérmicas como estomatales (Figura
6.16). Después de dos hora de haberse mantenido las epidermis en solución
MES/KOH 10mM a pH 5.5 con KCl 50mM, burbujeada con aire libre de CO2 y
bajo iluminación, la viabilidad de las células epidérmicas de C. communis no
mostró un cambio significativo.
54

Figura 6.13 C. communis, respuesta estomatal al ABA 10 µM (a) 30 min y (b) Dos horas en incubación.
(b)
(a)
55
56
(a) (b) (c)
(d) (e) (f)
Figura 6.14 C. communis bajo tratamiento con fusicoccina 5µM. (a). Control (tiempo 0); respuesta a la fusicoccina después de (b) 15 min, (c) 30 min, (d) 45 min, (e) 1 hora, y (f) 2 horas después de iniciado el tratamiento.

Regresión lineal
R2 SD N P .87277 1.58335 22 <0.0001
80 85 90 95 1004
6
8
10
12
14
16
18
20
22
24
Anc
ho d
el p
oro
Ancho de las células guarda
Figura 6.15 Relación entre el ancho del poro y el ancho de las células guarda de C. communis
Figura 6.16 Viabilidad de las células epidérmicas en trozos de epidermis recién aislada de C. communis La coloración roja de las células indica que están vivas; las distintas intensidades de rojo se presentan por diferencias en el pH.
57


















