
8 fisiologia esfuerzo-deporte
80
te la consecuencia de un menor ritmo de sudora- ción. Las glándulas sudoríparas individuales de los niños forman sudor más lentamente que las de los adultos y son menos sensibles a los incremen- tos de la temperatura del centro del cuerpo que los adultos. Aunque los chicos jóvenes pueden acli- matarse al ejercicio en ambientes calurosos, su rit- mo de aclimatación es menor que el de los adultos. No se dispone de datos sobre la aclimatación de las niñas. Sólo unos pocos estudios se han centrado en el ejercicio de los niños en ambientes fríos. De la poca información disponible, los niños parecen tener una mayor pérdida conductiva de calor que los adultos, debido a una mayor relación superfi- cie/masa. Esto debe imponerles un mayor riesgo de sufrir hipotermia. Por lo que se refiere a la tensión por el calor y por el frío, se han llevado a cabo pocos estudios sobre niños, y las conclusiones extraídas de di- chos estudios han sido contradictorias algunas veces. Es preciso investigar más en esta área para determinar los riesgos con los que se enfrentan los niños que hacen ejercicio en ambientes calu- rosos y en ambientes fríos. Mientras tanto, es aconsejable adoptar un enfoque conservador. Se FISIOLOGÍA DEL ESFUERZO Y DEL DEPORTE 572 Figura 16.12 Rendimiento máximo a nivel nacional en Estados Unidos de chicos y chicas de 10 años de edad hasta 17-18 años, en las siguientes pruebas de natación y atletismo: (a) 100 m de natación; (b) 400 m de natación; (c) carrera de 100 m, y (d) carrera de 1.500 m. Los récords se obtuvieron de USA Track & Field (diciembre de 2001; http://www.usatf.org) y United States Swimming (noviem- bre de 2001; http://www.usaswimming.org). Chicos Chicas Récord en tiempo (s) Récord en tiempo (min) Récord en tiempo (s) Récord en tiempo (min) 10 y menos 10 y menos 10 y menos 11-12 13-14 15-16 17-18 11-12 13-14 15-16 17-18 10 y menos 11-12 13-14 15-16 17-18 11-12 13-14 15-16 17-18 65 63 61 59 57 55 53 51 49 47 0 13,0 12,6 12,2 11,8 11,4 11,0 10,6 10,2 0 5,0 4,8 4,6 4,4 4,2 4,0 3,8 0 5,1 4,9 4,7 4,5 4,3 4,1 3,9 3,7 0 a c d b Grupos por la edad Grupos por la edad Grupos por la edad Grupos por la edad
-
Upload
victor-soto -
Category
Documents
-
view
1.808 -
download
0
Transcript of 8 fisiologia esfuerzo-deporte

te la consecuencia de un menor ritmo de sudora-ción. Las glándulas sudoríparas individuales de losniños forman sudor más lentamente que las delos adultos y son menos sensibles a los incremen-tos de la temperatura del centro del cuerpo que losadultos. Aunque los chicos jóvenes pueden acli-matarse al ejercicio en ambientes calurosos, su rit-mo de aclimatación es menor que el de los adultos.No se dispone de datos sobre la aclimatación de lasniñas.
Sólo unos pocos estudios se han centrado enel ejercicio de los niños en ambientes fríos. De lapoca información disponible, los niños parecen
tener una mayor pérdida conductiva de calor quelos adultos, debido a una mayor relación superfi-cie/masa. Esto debe imponerles un mayor riesgode sufrir hipotermia.
Por lo que se refiere a la tensión por el calory por el frío, se han llevado a cabo pocos estudiossobre niños, y las conclusiones extraídas de di-chos estudios han sido contradictorias algunasveces. Es preciso investigar más en esta área paradeterminar los riesgos con los que se enfrentanlos niños que hacen ejercicio en ambientes calu-rosos y en ambientes fríos. Mientras tanto, esaconsejable adoptar un enfoque conservador. Se
FISIOLOGÍA DEL ESFUERZO Y DEL DEPORTE
572
Figura 16.12 Rendimiento máximo a nivel nacional en Estados Unidos de chicos y chicas de 10 años de edad hasta 17-18 años, en lassiguientes pruebas de natación y atletismo: (a) 100 m de natación; (b) 400 m de natación; (c) carrera de 100 m, y (d) carrera de 1.500m. Los récords se obtuvieron de USA Track & Field (diciembre de 2001; http://www.usatf.org) y United States Swimming (noviem-bre de 2001; http://www.usaswimming.org).
ChicosChicas
Réc
ord
en
tiem
po
(s)
Réc
ord
en
tiem
po
(min
)
Réc
ord
en
tiem
po
(s)
Réc
ord
en
tiem
po
(min
)
10 ymenos
10 ymenos
10 ymenos
11-12 13-14 15-16 17-18 11-12 13-14 15-16 17-18
10 ymenos
11-12 13-14 15-16 17-18 11-12 13-14 15-16 17-18
65
63
61
59
57
55
53
51
49
47
0
13,0
12,6
12,2
11,8
11,4
11,0
10,6
10,20
5,0
4,8
4,6
4,4
4,2
4,0
3,8
0
5,1
4,9
4,7
4,5
4,3
4,1
3,9
3,7
0
a
c d
bGrupos por la edad Grupos por la edad
Grupos por la edad Grupos por la edad
03 Cap 16-21 • 549-776 17/1/07 16:38 Página 572

no se establecen adecuadamente, los deportistaspueden ser inducidos a reducir su peso muy pordebajo del nivel óptimo. Por lo tanto, es de im-portancia crítica establecer estándares de pesoadecuados.
Los estándares de peso corporal deben ba-sarse en la composición corporal del deportista.Una vez determinada la composición corporal, lacantidad de masa magra se usa para calcular loque debe pesar el deportista para un nivel especí-fico de porcentaje de grasa corporal. Considere-mos un ejemplo en que el objetivo es hacer queuna mujer nadadora de 72,5 kg con un 25% degrasa corporal descienda hasta el 18%, tal comose muestra en la tabla 14.3. Sabemos que su pesoobjetivo estará compuesto por un 18% de grasa ypor un 82% de masa magra. Por lo tanto, paracalcular su peso objetivo con un 18% de grasacorporal, dividiremos su masa magra por el 82%,que es la fracción de su peso objetivo que tiene queestar representado por su masa magra. Este cálculo(54,4 kg dividido por 0,82) nos da un peso objeti-vo de 66,3 kg, por lo que esta mujer necesita per-der 6,2 kg.
Por lo tanto, el establecimiento de estándaresde peso debe sustituirse por el de estándares deporcentaje de grasa corporal para cada deporte.Teniendo esto presente, ¿cuál es el porcentaje degrasa corporal recomendado para un deportistade elite en un deporte dado? Para cada deportedebe establecerse un valor o un intervalo de valo-res óptimos de grasa relativa fuera de los cualeses probable que el rendimiento del deportista sedeteriore. Y puesto que la distribución de la grasa
PESO CORPORAL, COMPOSICIÓN CORPORAL Y DEPORTE
497
Peso 72,5 kg
Grasa relativa 25%
Peso de grasa 18,2 (72,5 kg × 0,25%)
Peso magro 54,4 kg (72,5 kg - 18 kg de peso
de grasa)
% objetivo de grasa 18% (= 82% de masa magra)
Peso objetivo 66,3 kg (= 54,4 kg/0,82)
Objetivo de pérdida de peso 6,35 kg
�TABLA 14.3 Cálculo del peso objetivo para la obten-ción del rendimiento óptimo en una nadadora
Béisbol/softball 8-14 12-18
Baloncesto 6-12 10-16
Culturismo 5-8 6-12
Piragüismo/kayak 6-12 10-16
Ciclismo 5-11 8-15
Esgrima 8-12 10-16
Fútbol americano 6-18 ––
Golf 10-16 12-20
Gimnasia 5-12 8-16
Hípica 6-12 10-16
Hockey sobre hielo/hierba 8-16 12-18
Orientación 5-12 8-16
Pentatlón ––– 8-15
Frontón con raqueta 6-14 10-18
Remo 6-14 8-16
Rugbi 6-16 ––
Patinaje sobre hielo 5-12 8-16
Esquí 7-15 10-18
Saltos de esquí 7-15 10-18
Fútbol 6-14 10-18
Natación 6-12 10-18
Natación sincronizada –– 10-18
Tenis 6-14 10-20
Pista y campo
Pruebas de carrera 5-12 8-15
Pruebas de campo 8-18 12-20
Triatlón 5-12 8-15
Balonvolea 7-15 10-18
Levantamiento de pesos 5-12 10-18
Lucha libre 5-16 ––
% de grasa
Deporte Hombres Mujeres
�TABLA 14.4 Intervalo de valores para el porcentaje degrasa corporal de deportistas masculinos y femeninos envarios deportes
02 Cap 07-15 • 221-548 17/1/07 16:31 Página 497

Adaptaciones respiratoriasal entrenamientoIndependientemente de lo eficaz que sea el siste-ma cardiovascular suministrando cantidadesadecuadas de sangre a los tejidos, la capacidadde resistencia se verá perjudicada si el sistemarespiratorio no aporta suficiente oxígeno para sa-tisfacer las demandas. El funcionamiento del sis-tema respiratorio no suele limitar el rendimientoporque la ventilación puede incrementarse enmayor medida que la función cardiovascular. Pe-ro, al igual que con el sistema cardiovascular, elsistema respiratorio experimenta adaptacionesespecíficas al entrenamiento de fondo para ma-ximizar su eficacia. Consideremos algunas deellas.
Ventilación pulmonarDespués del entrenamiento, la ventilación pul-monar permanece esencialmente invariable o sereduce levemente en reposo, y disminuye ligera-mente a ritmos de esfuerzo submáximos estanda-rizados. Pero la ventilación pulmonar máximaaumenta sustancialmente. Normalmente, los in-crementos en sujetos no entrenados son desde unritmo inicial de 120 l/min hasta un ritmo de apro-ximadamente 150 l/min después del entrena-miento. Los ritmos de ventilación pulmonar nor-malmente aumentan hasta unos 180 l/min endeportistas muy entrenados. Dos son los factoresque pueden ser responsables del incremento en laventilación pulmonar máxima después del entre-namiento: el mayor volumen oscilante y la mayorfrecuencia respiratoria en el ejercicio máximo.Los estudios recientes revelan que el entrena-miento de los músculos inspiratorios puede me-jorar significativamente el rendimiento. En un es-tudio con 14 remeras competitivas, 11 semanasde entrenamiento de los músculos inspiratoriosmejoraron la distancia recorrida en un esfuerzomáximo durante 9,6 kilómetros remando (3,5%)y en la marca para completar una contrarreloj de5.000 m (3,1%).46
FISIOLOGÍA DEL ESFUERZO Y DEL DEPORTE
312
i El volumen sanguíneo aumenta comoconsecuencia del entrenamiento de fon-do.
i El incremento está ocasionado princi-palmente por un aumento en el volumende plasma sanguíneo.
i El número de glóbulos rojos puede au-mentar, pero la ganancia de plasma esnormalmente mucho más elevada, loque da lugar a una porción fluida de lasangre relativamente mayor.
i El aumento del volumen plasmático incre-menta la viscosidad de la sangre, lo cualmejora la circulación y la disponibilidadde oxígeno.
i Determinadas investigaciones han de-mostrado que los cambios en el volumende plasma guardan una estrecha relacióncon los producidos en el volumen sistó-lico y en el nVO2 máx, lo que provocaque el incremento del volumen de plas-ma inducido por el ejercicio sea uno delos efectos más significativos del entre-namiento.
Los grandes deportistas con un alto nivelde entrenamiento de fondo, tales comolos remeros, pueden tener índices de ven-tilación pulmonar máxima de 240 l/min,más del doble del ritmo normal de indivi-duos no entrenados.
Generalmente, la ventilación no se consideraun factor limitante del rendimiento en los ejer-cicios de resistencia. No obstante, existen prue-bas que indican que, en un determinado momentode la adaptación de una persona muy entrenada,la capacidad del sistema pulmonar para transpor-tar oxígeno no podrá satisfacer las demandas delas extremidades y del sistema cardiovascular.9
Esto causa lo que se ha denominado hipoxemiaarterial inducida por el ejercicio, en la que la sa-turación arterial de oxígeno disminuye por deba-jo del 96%. La hipoxemia arterial leve inducidapor el ejercicio se define como una saturación ar-terial del 93% al 95%, moderada del 88% al 93%y grave de menos del 88%.10
02 Cap 07-15 • 221-548 17/1/07 16:29 Página 312

de, nuestra resistencia aeróbica y, por lo tanto,nuestro rendimiento, puede incrementarse sus-tancialmente. Ésta es la premisa que subyace ba-jo el dopaje sanguíneo.
Efectos demostrados
Ekblom y colaboradores27 provocaron una con-moción en el mundo deportivo a comienzos de ladécada de 1970. En uno de sus estudios, retiraronentre 800 y 1.200 ml de sangre de sus sujetos yluego volvieron a inyectarles los glóbulos rojosunas 4 semanas más tarde. Los resultados mos-traron una mejora considerable en el nVO2 máx(9%) y en el tiempo en la cinta ergométrica(23%). Inmediatamente después, durante unospocos años, varios estudios no lograron confir-mar estos originales descubrimientos, pero otrosvarios mostraron un efecto ergogénico.
Por lo tanto, la literatura de las investigacio-nes estuvo dividida sobre la eficacia del dopajesanguíneo hasta que en 1980 se produjo un im-portante adelanto como consecuencia de un estu-dio realizado por Buick y colaboradores.13 Seexaminó a 11 corredores de fondo muy entrena-dos en diferentes momentos durante el estudio:
(1) antes de la extracción de sangre; (2) des-pués de dar al cuerpo un tiempo apropiado pararestablecer los niveles normales de glóbulos ro-jos, pero antes de la reinfusión de la sangre extraí-da; (3) después de una simulada reinfusión de 50ml de una solución salina (un placebo); (4) des-pués de la reinfusión de 900 ml de sangre que ori-ginalmente se habían extraído y preservado conge-lándolos; (5) después de que los niveles elevadosde glóbulos rojos hubiesen vuelto a la normali-dad.
Tal como se muestra en la figura 15.5, los in-vestigadores hallaron un incremento sustancialen el nVO2 máx y en el tiempo en la cinta ergomé-trica después de la reinfusión de los glóbulos ro-jos (grupo A) y ningún cambio después de la fal-sa reinfusión (grupo B). Este incremento en elnVO2 máx persistió durante un máximo de 16 se-manas, pero los incrementos del tiempo en la cin-ta ergométrica se redujeron antes de transcurridoslos primeros 7 días.
Maximización de los beneficios
¿Por qué fue un gran adelanto el estudio de Buicky colaboradores? Gledhill30, 31 ayudó a explicar
las controversias que originaban los primeros es-tudios. Muchos de los estudios iniciales que nohallaron ninguna mejora con el dopaje sanguíneohabían reinyectado sólo pequeños volúmenes deglóbulos rojos y la reinfusión se efectuó antes depasadas 3 ó 4 semanas de la extracción de sangre.Primero, parece ser que hay que extraer y rein-yectar 900 ml de sangre, o incluso más cantidad.Los incrementos en el nVO2 máx y en el rendi-miento no son tan grandes cuando se usan volú-menes más pequeños. De hecho, algunos estudiosque usan volúmenes pequeños no han logrado ha-llar ninguna diferencia.
Segundo, parece que hay que esperar por lomenos de 5 a 6 semanas y posiblemente hasta 10semanas antes de la reinfusión. Esto se basa en eltiempo requerido por el cuerpo para restablecer suhematocrito previo a la extracción de sangre.
Por último, estudios iniciales refrigeraron lasangre retirada. El tiempo máximo de almacenajebajo refrigeración es de, aproximadamente, 5 se-manas. Además, cuando la sangre es refrigerada,aproximadamente el 40% de los glóbulos rojos sedestruyen o se pierden. Estudios posteriores hanusado la congelación para almacenar la sangre. Lacongelación permite un tiempo de almacenaje ca-si ilimitado y solamente se pierden aproximada-mente el 15% de los glóbulos rojos.
Gledhill30, 31 concluyó que el dopaje sanguí-neo produce mejoras significativas en el nVO2
máx y en la capacidad de resistencia cuando sehace bajo condiciones óptimas:
• Un mínimo de 900 ml de reinfusión de san-gre.
• Un intervalo mínimo de 5 a 6 semanas en-tre la extracción y la reinfusión.
• Almacenaje de la sangre por congelación.
Gledhill ha demostrado también que estasmejoras son la consecuencia directa del mayorcontenido de hemoglobina de la sangre, no delmayor gasto cardíaco debido a un incremento delvolumen del plasma.
Dopaje sanguíneo y capacidad de resistencia
¿Produce el incremento del nVO2 máx y del tiempoen la cinta ergométrica, como consecuencia deldopaje sanguíneo, una mejora en la capacidad deresistencia? Diversos estudios han analizado estacuestión. En un estudio se observaron tiempos decarrera de 8 km sobre cinta ergométrica en un gru-po de 12 corredores de fondo experimentados.63
AYUDAS ERGOGÉNICAS Y RENDIMIENTO
533
02 Cap 07-15 • 221-548 17/1/07 16:32 Página 533
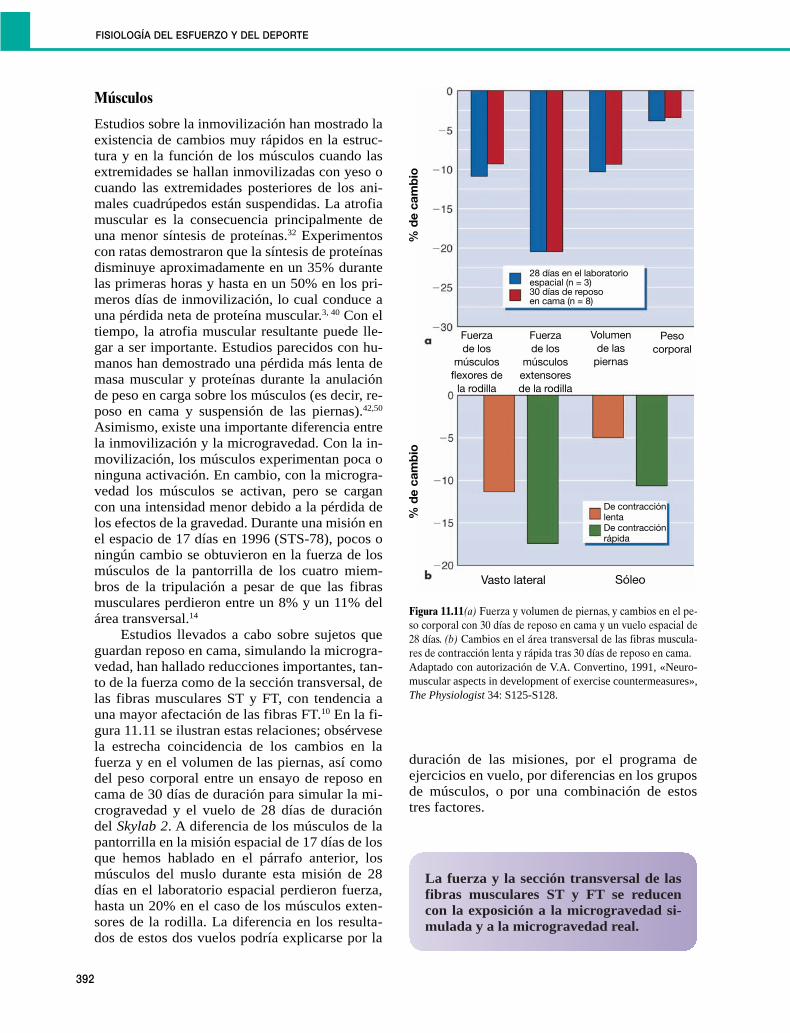
Músculos
Estudios sobre la inmovilización han mostrado laexistencia de cambios muy rápidos en la estruc-tura y en la función de los músculos cuando lasextremidades se hallan inmovilizadas con yeso ocuando las extremidades posteriores de los ani-males cuadrúpedos están suspendidas. La atrofiamuscular es la consecuencia principalmente deuna menor síntesis de proteínas.32 Experimentoscon ratas demostraron que la síntesis de proteínasdisminuye aproximadamente en un 35% durantelas primeras horas y hasta en un 50% en los pri-meros días de inmovilización, lo cual conduce auna pérdida neta de proteína muscular.3, 40 Con eltiempo, la atrofia muscular resultante puede lle-gar a ser importante. Estudios parecidos con hu-manos han demostrado una pérdida más lenta demasa muscular y proteínas durante la anulaciónde peso en carga sobre los músculos (es decir, re-poso en cama y suspensión de las piernas).42,50
Asimismo, existe una importante diferencia entrela inmovilización y la microgravedad. Con la in-movilización, los músculos experimentan poca oninguna activación. En cambio, con la microgra-vedad los músculos se activan, pero se cargancon una intensidad menor debido a la pérdida delos efectos de la gravedad. Durante una misión enel espacio de 17 días en 1996 (STS-78), pocos oningún cambio se obtuvieron en la fuerza de losmúsculos de la pantorrilla de los cuatro miem-bros de la tripulación a pesar de que las fibrasmusculares perdieron entre un 8% y un 11% delárea transversal.14
Estudios llevados a cabo sobre sujetos queguardan reposo en cama, simulando la microgra-vedad, han hallado reducciones importantes, tan-to de la fuerza como de la sección transversal, delas fibras musculares ST y FT, con tendencia auna mayor afectación de las fibras FT.10 En la fi-gura 11.11 se ilustran estas relaciones; obsérvesela estrecha coincidencia de los cambios en lafuerza y en el volumen de las piernas, así comodel peso corporal entre un ensayo de reposo encama de 30 días de duración para simular la mi-crogravedad y el vuelo de 28 días de duracióndel Skylab 2. A diferencia de los músculos de lapantorrilla en la misión espacial de 17 días de losque hemos hablado en el párrafo anterior, losmúsculos del muslo durante esta misión de 28días en el laboratorio espacial perdieron fuerza,hasta un 20% en el caso de los músculos exten-sores de la rodilla. La diferencia en los resulta-dos de estos dos vuelos podría explicarse por la
duración de las misiones, por el programa deejercicios en vuelo, por diferencias en los gruposde músculos, o por una combinación de estostres factores.
FISIOLOGÍA DEL ESFUERZO Y DEL DEPORTE
392
Figura 11.11(a) Fuerza y volumen de piernas, y cambios en el pe-so corporal con 30 días de reposo en cama y un vuelo espacial de28 días. (b) Cambios en el área transversal de las fibras muscula-res de contracción lenta y rápida tras 30 días de reposo en cama.Adaptado con autorización de V.A. Convertino, 1991, «Neuro-muscular aspects in development of exercise countermeasures»,The Physiologist 34: S125-S128.
28 días en el laboratorio espacial (n = 3)30 días de reposo en cama (n = 8)
% d
e ca
mb
io
Fuerzade los
músculosflexores de
la rodilla
Fuerzade los
músculosextensoresde la rodilla
Volumende las
piernas
Pesocorporal
De contracción lentaDe contracción rápida
% d
e ca
mb
io
Vasto lateral Sóleo
La fuerza y la sección transversal de lasfibras musculares ST y FT se reducencon la exposición a la microgravedad si-mulada y a la microgravedad real.
02 Cap 07-15 • 221-548 17/1/07 16:30 Página 392

hematíes a lo largo de años de entrenamiento de-vuelve la relación entre la masa celular y el volu-men total a niveles más normales.
Este cambio en la proporción entre plasma ycélulas, resultante de un incremento en la porciónfluida, reduce la viscosidad de la sangre. Ellopuede facilitar el movimiento de la sangre por losvasos sanguíneos, particularmente a través de losvasos más pequeños, como son los capilares. De-terminadas investigaciones han demostrado quela baja viscosidad de la sangre aumenta el aportede oxígeno a las masas musculares activas.
Tanto la cantidad total de hemoglobina (valo-res absolutos) como el número total de glóbulosrojos en deportistas altamente entrenados general-mente son superiores a lo normal, aun cuando losvalores relativos son inferiores a los normales.Ello asegura que la sangre tenga una capacidad detransporte de oxígeno más que suficiente para sa-tisfacer las necesidades del cuerpo en todo mo-mento. El índice del recambio metabólico de he-matíes también podría ser mayor con unentrenamiento intenso. Esto tal vez sea una venta-ja, siempre que la producción sea igual o mayorque la destrucción, dado que las células jóvenesson más eficaces en el transporte de oxígeno.42
Volumen de plasma, volumen sistólico y nVO2 máx
Un incremento del volumen de plasma es uno delos cambios más significativos que se producencon el entrenamiento de fondo. Recordemos denuestra anterior discusión que el incremento delvolumen de plasma es un factor importante en elaumento del volumen sistólico resultante del en-trenamiento. El volumen sistólico, a su vez, afec-ta al consumo de oxígeno.
Cuando el volumen de plasma aumenta, tam-bién lo hace el volumen sanguíneo. En conse-cuencia, entra más sangre en el corazón. Cuandoentra más sangre en el corazón, el volumen sistó-lico aumenta. A intensidades máximas de esfuer-zo, el nVO2 máx suele permanecer relativamenteestable, por lo que un mayor volumen sistólicopermite que el gasto cardíaco máximo se incre-mente. El aumento del gasto cardíaco máximohace que haya más oxígeno disponible para losmúsculos activos, permitiendo así que aumente elnVO2 máx.
ADAPTACIONES CARDIOVASCULARES Y RESPIRATORIAS AL ENTRENAMIENTO
311
Figura 9.10 Aumento en la volemia total y el volumen plasmá-tico con un entrenamiento de fondo. Hay que reparar en que,aunque el hematocrito (porcentaje de hematíes) disminuye del44% al 42%, el volumen total de hematíes aumenta un 10%.
Volumensanguíneo = 5 l
2,8Volumen
plasmático
2,2Hematíes
Preentrenamiento Posentrenamiento
2,4Hematíes
3,3Volumen
plasmático
Hematocrito = 44%
Volu
men
(l)
Hematocrito = 42%
Volumensanguíneo = 5,7 l
El incremento del volumen sanguíneocon el entrenamiento aeróbico de fondose debe al gran aumento del volumenplasmático y a un pequeño incrementodel número de hematíes; ambos cam-bios facilitan la llegada de oxígeno a losmúsculos activos.
Esta secuencia de acontecimientos –un au-mento del volumen de plasma que conlleva unaumento del volumen sistólico, que, a su vez,conduce a un aumento en el nVO2 máx– se ha de-mostrado que también tiene lugar en individuosno entrenados cuyos volúmenes de plasma sehan incrementado usando una infusión de dex-trano al 6%.24 Esta secuencia también funciona ala inversa: una reducción del volumen de plasmapor el abandono de los entrenamientos por partede deportistas muy entrenados, o situando ahombres sanos en una posición de tendido con lacabeza hacia abajo (–5°) durante 20 h se obtie-nen reducciones significativas en el volumen sis-tólico y disminuciones equivalentes en el nVO2
máx.8, 18
02 Cap 07-15 • 221-548 17/1/07 16:29 Página 311

01 Crédits+Cap 01-06 • 001-220 17/1/07 16:20 Página 126

Evitar el ayuno y las dietas de choqueNuestro jugador de fútbol americano debe perderunos 2 kg por semana durante las 4 semanas si-guientes, por lo que decide embarcarse en unadieta de choque, y elige cualquier dieta de las queestán de moda, sabiendo que una persona puedeperder unos 3 kg por semana con tales dietas. Noes un caso aislado. Muchos deportistas descubrenque tienen un peso excesivo y que no están enforma porque han comido en exceso y reducidosu actividad al finalizar la temporada y suelen es-perar hasta las últimas semanas antes de la fechaen que deben presentarse para atajar el problema.En nuestro ejemplo, el jugador de fútbol america-no puede ser capaz de perder 9 kg en 4 semanascon la dieta de choque. Pero una gran parte de es-ta pérdida de peso lo será de agua corporal y muypoca de grasa acumulada. Varios estudios han in-formado de que se producen sustanciales pérdidasde peso con dietas muy bajas en calorías (500kcal por día o menos); pero de la pérdida de peso,más del 60% procede de tejidos magros y menosdel 40% de depósitos de grasa.
Aunque una gran parte del peso de los juga-dores de fútbol americano se pierde disminuyen-do las reservas de agua, también se pierde unasustancial cantidad de proteínas. Asimismo, lamayoría de las dietas de choque se basan en unaimportante reducción de la ingestión de hidratosde carbono. Esta menor ingestión es insuficientepara abastecer las necesidades del cuerpo encuanto a hidratos de carbono, y en consecuencialas reservas de estos se agotan. Puesto que la acu-mulación de agua acompaña el almacenaje de hi-dratos de carbono, las reservas de agua tambiénse reducen cuando disminuyen las reservas deaquellos. Por cada gramo de hidratos de carbonoque emplea el cuerpo, se pierden aproximada-mente 2,6 g de agua. El contenido total de glucó-geno del cuerpo es de 800 g a 1.000 g, por lo quela depleción del glucógeno almacenado provoca-ría una pérdida aproximada de 2.080-2.600 g deagua, en torno a 2-2,6 kg.
Además, el cuerpo depende en mayor medi-da de los ácidos grasos libres para obtener ener-gía cuando se agotan las reservas de hidratos decarbono. En consecuencia, los cuerpos cetónicos,un subproducto del metabolismo de los ácidosgrasos, se acumulan en la sangre y producen unacondición conocida como cetosis. Esta condiciónincrementa todavía más la pérdida de agua. Unagran parte de esta pérdida de agua se produce du-rante la primera semana de dieta.
La figura 14.8 ilustra estos cambios en elagua, la grasa, las proteínas y los hidratos de car-bono del cuerpo durante un período de 30 días deayuno. El agotamiento de los hidratos de carbonose produce antes de transcurridos 3 días. El con-tenido de proteínas cae significativamente en elmismo período de tiempo. El contenido de aguadel cuerpo disminuye abruptamente durante losprimeros 3 días. El ritmo de pérdida de grasa,aunque disminuye algo en el segundo y tercerdías, permanece relativamente constante duranteel resto del período de ayuno.
Para la mayoría de la gente es imposibleperder más de 2 kg de grasa por semana, inclusoen condiciones de ayuno completo. Esto se pue-de demostrar fácilmente. Si 0,5 kg de grasa (teji-do adiposo) tienen un equivalente calórico de3.500 kcal, se necesita un déficit de 3.500 kcalpara perder 0,5 kg de grasa. El ritmo metabólicoen reposo del jugador profesional de fútbol ame-
PESO CORPORAL, COMPOSICIÓN CORPORAL Y DEPORTE
499
Figura 14.8 Composición de la pérdida de peso con 30 días deayuno.
Pér
did
a d
e p
eso
(g/2
4 h)
Días de ayuno
Hidratos de carbono
Proteínas
Grasas
Agua
0 3 5 10 15 20 25 300
100
250
500
1.000
1.500
2.000
02 Cap 07-15 • 221-548 17/1/07 16:31 Página 499

mizar la fuerza generada por un músculo agonis-ta, es necesario reducir al mínimo el grado de co-activación.14 La reducción en la coactivación po-dría explicar una porción de aumento de la fuerzaatribuida a factores neuronales, pero su contribu-ción sería pequeña.
La codificación del índice es otro factorpotencial que puede aumentar la producciónde fuerza de un músculo después del entrena-miento. La codificación del índice es un térmi-no empleado para describir la frecuencia deactivación o la frecuencia de descarga de uni-dades motoras. Aunque no se haya estudiadobien, hay ciertas pruebas que lo respaldan co-mo un factor posible.17 También se han apre-ciado cambios en la morfología de la uniónneuromuscular con el aumento y reducción delos niveles activos que tal vez estén directa-mente relacionados con la capacidad de pro-ducción de fuerza del músculo.11
Ahora que hemos examinado los factoresneurológicos potenciales que contribuyen a au-mentar la fuerza con entrenamiento resistido, pres-temos atención a la hipertrofia muscular. Estu-diaremos la forma en que los músculos aumentande tamaño y cómo ello contribuye al incremento dela fuerza, o a la capacidad de producción de fuerzadel músculo.
Hipertrofia muscular¿Cómo aumenta el tamaño de un músculo? Dostipos de hipertrofia pueden tener lugar: temporaly crónica. La hipertrofia temporal es este abul-tamiento del músculo que tiene lugar durante unasola serie de ejercicios. Es el resultado principal-mente de la acumulación de fluidos (edema) enlos espacios intersticiales e intracelulares delmúsculo. Este fluido lo pierde el plasma sanguí-neo.42 La hipertrofia temporal, como su nombreindica, dura tan sólo un corto período de tiempo.El fluido vuelve a la sangre al cabo de unas horasde haber finalizado el ejercicio.
La hipertrofia crónica se refiere al incre-mento en el tamaño muscular que se produce me-diante el entrenamiento resistido a largo plazo.Esto refleja verdaderos cambios estructurales enel músculo como resultado de un aumento delnúmero de fibras musculares (hiperplasia) o co-mo consecuencia de un aumento del tamaño delas fibras musculares individuales existentes (hi-pertrofia). Las teorías que intentan explicar lacausa subyacente de este fenómeno son muy con-
trovertidas. Sin embargo, es de importancia el re-ciente descubrimiento que indica que el compo-nente excéntrico del entrenamiento es importantepara los incrementos máximos en la seccióntransversal de la fibra muscular.14, 30 En estudiosllevados a cabo con sujetos que realizaban sóloacciones concéntricas o excéntricas, se observó,después de 36 sesiones de entrenamiento, que elentrenamiento excéntrico producía un aumentoen la fibra FT y de la fuerza aproximadamente 10veces mayor que el alcanzado con el entrena-miento concéntrico. Entonces, el entrenamientosólo con acciones concéntricas podría limitar almúsculo hipertrofiado. Echemos un vistazo ados mecanismos de los que se ha postulado queaumentan el tamaño muscular con el entrenamien-to resistido: la hipertrofia y la hiperplasia.
Hipertrofia de las fibras
Las primeras investigaciones indicaban que elnúmero de fibras musculares de cada uno denuestros músculos queda establecido en el naci-miento o poco después, y que este número per-manece invariable a lo largo de la vida. Si esto escierto, entonces la hipertrofia crónica sólo puedeser la consecuencia de la hipertrofia de fibrasmusculares individuales. La explicación de ellopuede radicar en:
• un mayor número de miofibrillas;
• más filamentos de actina y de miosina;
• más sarcoplasma;
• más tejido conectivo, o
• cualquier combinación de lo anterior.
Tal como se observa en las micrografías dela figura 3.4, el entrenamiento intenso contra re-sistencia puede incrementar notablemente la sec-ción transversal de las fibras musculares. En esteejemplo, la hipertrofia de las fibras es producidaprobablemente por un mayor número de miofibri-llas y de filamentos de actina y de miosina, queproporcionarán más puentes cruzados para laproducción de fuerza durante la contracción má-xima. Estos espectaculares agrandamientos de lasfibras musculares no ocurren, sin embargo, en to-dos los casos de hipertrofia muscular.
La hipertrofia de las fibras individuales me-diante entrenamiento resistido parece ser pro-
ADAPTACIONES NEUROMUSCULARES AL ENTRENAMIENTO RESISTIDO
99
01 Crédits+Cap 01-06 • 001-220 17/1/07 16:20 Página 99

ye nuestro cuerpo, y proporcionan generalmentepoca energía para la función celular. Al pasar dela realización de un esfuerzo muscular suave aotro más duro, se emplean progresivamente máshidratos de carbono, dependiendo menos de lasgrasas. En los ejercicios máximos de corta dura-ción, el ATP se genera casi exclusivamente a par-tir de los hidratos de carbono.
Hidratos de carbonoLa dependencia de nuestros músculos respecto alos hidratos de carbono durante el ejercicio estárelacionada con la disponibilidad de hidratos decarbono y con que el sistema muscular esté biendesarrollado para su metabolismo. Los hidratosde carbono se convierten en última instancia englucosa, un monosacárido (azúcar de una solaunidad) que es transportado por la sangre a los te-jidos activos, donde se metaboliza. En reposo, laingesta de hidratos de carbono es absorbida porlos músculos y el hígado, y luego se convierte enuna molécula de azúcar mucho más compleja:glucógeno. Éste se almacena en el citoplasmahasta que las células lo utilizan para formar ATP.El glucógeno, depositado en el hígado, cuando senecesita, se convierte nuevamente en glucosa quees transportada por la sangre a los tejidos activosdonde es metabolizada.
Las reservas de glucógeno en el hígado y enlos músculos son limitadas y pueden agotarse amenos que la dieta contenga una razonable canti-dad de hidratos de carbono. Por lo tanto, depen-demos fuertemente de nuestras fuentes dietéticasde almidones y azúcares para reponer nuestrasreservas de hidratos de carbono. Sin una inges-tión adecuada de ellos, los músculos y el hígadopueden quedar desprovistos de su principal fuen-te de energía.
GrasasLas grasas y las proteínas también se usan comofuentes energéticas. Nuestro cuerpo acumula mu-cha más grasa que hidratos de carbono. Tal comose ve en la tabla 4.1, las reservas energéticas delcuerpo en grasas es mucho mayor que las de hi-dratos de carbono. Pero las grasas son menos ac-cesibles para el metabolismo celular, porque pri-mero deben ser reducidas desde su formacompleja (triglicéridos) a sus componentes bási-cos: glicerol y ácidos grasos libres. Sólo estos seusan para formar ATP.
Tal como se ve en la figura 4.1, se obtienesustancialmente más energía de una cantidad de-terminada de grasa (9 kcal/g) que de la mismacantidad de hidratos de carbono (4 kcal/g). Noobstante, el ritmo de liberación de energía de es-tos compuestos es demasiado lento para satisfa-cer todas las demandas de energía de la actividadmuscular intensa.
ProteínasEl proceso por el que las proteínas o las grasas seconvierten en glucosa recibe el nombre de gluco-neogénesis. Alternativamente, las proteínas pue-den convertirse, a través de una serie de reaccio-nes, en ácidos grasos. Esto recibe el nombre delipogénesis.
Las proteínas pueden aportar entre el 5% y el10% de la energía necesaria para mantener unejercicio prolongado. Sólo las unidades más bási-cas de las proteínas (los aminoácidos) puedenusarse para obtener energía. Un gramo de proteí-na genera alrededor de 4,1 kcal.
Ritmo de liberación de energíaPara ser útil, la energía libre debe liberarse a partirde compuestos químicos a un ritmo controlado.Este ritmo viene parcialmente determinado por laelección de la fuente primaria de combustible.Grandes cantidades de un combustible determina-do pueden hacer que las células dependan más deesta fuente que de otras alternativas. Esta influen-cia de la disponibilidad de energía recibe la deno-minación de efecto de acción de masa.
Enzimas específicas proporcionan un mayorcontrol estructurado del ritmo de liberación deenergía libre. Muchas de estas proteínas especia-les facilitan la descomposición (catabolismo) delos compuestos químicos. El camino por el que
METABOLISMO, ENERGÍA Y SISTEMAS BÁSICOS DE ENERGÍA
131
Las reservas de hidratos de carbono en elhígado y en los músculos esqueléticos estánlimitadas a menos de 2.000 kcal de energía,o el equivalente de la energía necesariapara correr aproximadamente 32 km.Las reservas de grasa, no obstante, sue-len superar las 70.000 kcal de energíaacumulada.
01 Crédits+Cap 01-06 • 001-220 17/1/07 16:21 Página 131

temperaturas corporales se desvíen fuera de suintervalo normal de variación de entre 36,1 y37,8 oC.
La temperatura corporal refleja la existenciade un cuidadoso equilibrio entre la producción yla pérdida de calor. Siempre que este equilibrio sealtera, la temperatura de nuestro cuerpo cambia.Recordemos del capítulo 4 que una gran parte dela energía que genera nuestro cuerpo se degrada acalor, la forma más baja de energía. Todos los teji-dos metabólicamente activos producen calor quepuede usarse para mantener la temperatura internacorporal. Pero si la producción de calor corporalsupera la pérdida de calor del cuerpo, nuestratemperatura interna se eleva. Nuestra capacidadpara mantener una temperatura interna constantedepende de nuestra capacidad para equilibrar elcalor que obtenemos del metabolismo y del am-biente con el calor que pierde nuestro cuerpo. Enla figura 10.1 se muestra este equilibrio. Exami-
nemos ahora los mecanismos mediante los cualesse transfiere el calor corporal al ambiente.
Transferencia del calor corporalPara que el cuerpo pueda transferir calor al am-biente, el calor corporal debe poder acceder alexterior. El calor de las partes profundas del cuer-po (el núcleo) es transportado por la sangre haciala piel (la cáscara). Una vez que el calor se acercaa la piel, puede ser transferido al ambiente porcualquiera de los cuatro mecanismos siguientes:conducción, convección, radiación y evapora-ción. Estos se ilustran en las figuras 10.1 y 10.2.
Conducción y convección
La conducción del calor supone la transferenciade éste desde un material a otro a través del con-
REGULACIÓN TÉRMICA Y EJERCICIO
335
Figura 10.2 Eliminación de calor por la piel. El calor se dirige a la superficie del cuerpo a través de la sangre arterial y también por con-ducción a través del tejido subcutáneo. Cuando la temperatura de la piel es mayor que la del ambiente, el calor se elimina por conducción,convección, radiación y evaporación del sudor.
Radiación Evaporación delsudor
Convección
Conducción
Calor producido enlos músculosCalor en la
sangre
Glándulasudorípara
02 Cap 07-15 • 221-548 17/1/07 16:29 Página 335

Un estudio indicó que las áreas medias de fi-bras musculares de los músculos vasto externo ydeltoides eran menores en un grupo de culturistasde alto nivel que en un grupo de referencia dehalterófilos de competición, y eran casi idénticasa las de estudiantes de educación física y a las depersonas que no seguían programas de entrena-miento de fuerza. Esto indica que la hipertrofiade fibras musculares individuales no tenía unaimportancia crítica para las ganancias en masamuscular de los culturistas.57
Se obtuvieron resultados similares en un es-tudio posterior que comparaba culturistas muyentrenados y controles activos, pero no entrena-dos. Las secciones de fibra muscular de los suje-tos entrenados eran similares a las de los sujetosde control, a pesar del hecho de que los primerostenían circunferencias mucho mayores en las ex-tremidades.38 Los investigadores también encon-traron más fibras musculares por unidad motoraen los culturistas entrenados que en el grupo decontrol no deportista. Dado que estos hombres te-nían una circunferencia muscular sustancialmen-te mayor, pero una sección transversal normal defibras musculares, estos descubrimientos indicanque el número de fibras musculares aumentó.Una explicación alternativa es que estos deportis-tas tenían un mayor número de fibras al nacer.
• Estudiantes femeninas de educación física:4.400 µm2.
Las diferencias entre estos estudios puedenexplicarse por la naturaleza de la carga del entre-namiento o del estímulo. Se cree que el entrena-miento con intensidades elevadas o con grandesresistencias produce una mayor hipertrofia de lasfibras, especialmente de las fibras FT, que el en-trenamiento con intensidades o resistencias másbajas.35
Sólo ha habido un estudio longitudinal quehaya demostrado la posibilidad de hiperplasia enhombres sometidos previamente a entrenamientoresistido a nivel recreativo.39 Después de 12 se-manas de entrenamiento intensificado contra re-sistencia, pareció haber un aumento significativodel número de fibras musculares en el bíceps bra-quial de varias de las 12 personas. De este estu-dio se deduce que la hiperplasia puede producirseen seres humanos, pero sólo en algunas personasy en ciertas condiciones de entrenamiento.
De la información precedente, parece serque la hiperplasia puede darse en animales y po-siblemente en humanos. ¿Cómo se forman estascélulas nuevas? Como se ve en la figura 3.6, sepostula que las fibras musculares individualestienen capacidad para dividirse en dos células,cada una de las cuales se desarrolla en una fibramuscular funcional. Más recientemente se ha es-tablecido que es probable que las células satéli-tes, que son células madre miógenas implicadasen la regeneración del músculo esquelético, inter-vengan en la generación de nuevas fibras muscu-lares. Estas células suelen activarse después delesiones musculares y, como veremos más ade-lante en este capítulo, las lesiones musculares sonproducto de un entrenamiento intenso, sobre todoel entrenamiento de acciones excéntricas.3 Las le-siones musculares pueden causar una cascada derespuestas, en la que las células satélites se acti-van y proliferan, migran a la región dañada, y sefusionan con las miofibrillas existentes, o se com-binan y fusionan para producir nuevas fibras.31
Esto se muestra en la figura 3.7.
Integración de la activación neuronal y la hipertrofia fibrilarLos estudios realizados hasta la fecha sobre lasadaptaciones del entrenamiento resistido mues-tran que los incrementos iniciales en la fuerza vo-
FISIOLOGÍA DEL ESFUERZO Y DEL DEPORTE
102
Ha quedado ampliamente demostradoque la hiperplasia de las fibras muscula-res se produce en modelos animalesusando distintos medios para inducir lahipertrofia muscular. Por otra parte, sólounos pocos estudios aportan evidenciasde hiperplasia en seres humanos.
Por otro lado, al menos un estudio ha encon-trado grandes diferencias en la sección de las fi-bras musculares al comparar a culturistas con estu-diantes de educación física masculinos yfemeninos.44 Las secciones medias de fibra delvasto externo para cada uno de los tres grupos erancomo sigue:
• Culturistas: 8.400 µm2.
• Estudiantes masculinos de educación físi-ca: 6.200 µm2.
01 Crédits+Cap 01-06 • 001-220 17/1/07 16:20 Página 102

reducción del glucógeno muscular y hepático ge-neralmente produce un cambio en el peso corpo-ral de 0,45 a 1,36 kg. Algunos científicos hanpropuesto que las reservas de glucógeno muscu-lar y del hígado pueden controlarse registrando elpeso del deportista a primeras horas de la mañanainmediatamente después de levantarse, despuésde haber vaciado la vejiga y antes de tomar el de-sayuno. Una súbita reducción de peso puede re-flejar una incapacidad para reemplazar el glucó-geno, un déficit en el agua corporal, o ambascosas.
Los deportistas que deben entrenarse o com-petir en pruebas agotadoras durante días sucesi-vos deben reemplazar las reservas de glucógenomuscular y hepático con la mayor rapidez posi-ble. Aunque el glucógeno hepático puede agotar-se totalmente al cabo de 2 horas de ejercicio al70% del nVO2 máx, se recupera antes de transcu-rridas unas pocas horas cuando se toma una dietarica en hidratos de carbono. Por otro lado, la re-síntesis del glucógeno muscular es un procesolento, que precisa varios días para volver a la nor-malidad después de una sesión de ejercicio ago-tador como puede ser el maratón (figura 13.10).7,
50 Estudios llevados a cabo a finales de los años1980 revelaron que la resíntesis del glucógenomuscular alcanzaba su mayor velocidad cuandolos individuos tomaban al menos 50 g (aproxima-damente, 0,7 g por kg de peso corporal) de gluco-sa cada 2 horas después del ejercicio.8, 34 Tomarcantidades superiores a éstas no parece que ace-lere el reemplazo del glucógeno muscular. Du-rante las primeras 2 horas después del ejercicio,el ritmo de resíntesis del glucógeno muscular os-cila entre el 7% y el 8% por hora (de 7 a 8 mmolpor kg de músculo por hora), lo cual es algo másrápido que el ritmo normal de entre el 5% y el6% por hora.33 Por lo tanto, un deportista que serecupera de una prueba agotadora que requierecapacidad de resistencia debe ingerir suficienteshidratos de carbono tan pronto como sea posibledespués del ejercicio. También se postuló queañadir proteínas y aminoácidos a los hidratos decarbono ingeridos durante el período de recupera-ción mejoraría la síntesis de glucógeno muscularpor encima del nivel conseguido sólo con hidratosde carbono. Varios estudios posteriores no fueroncapaces de confirmarlo,36, 53 pero un estudio más re-ciente ha demostrado claramente que un suple-mento de hidratos de carbono y proteínas es máseficaz para recuperar el glucógeno muscular des-pués del ejercicio que sólo un suplemento de hi-dratos de carbono.32
Diseño de bebidas deportivasAnteriormente, hemos mencionado que ingerirsoluciones de hidratos de carbono durante elejercicio puede beneficiar el rendimiento al ase-gurar una cantidad adecuada de combustible pa-ra la producción de energía y de fluidos para la
FISIOLOGÍA DEL ESFUERZO Y DEL DEPORTE
470
i De un estudio sobre corredores de am-bos sexos, se dedujo que la mayoría se-guía una alimentación sana, que cubríala RDR de la mayoría de los nutrientes,pero su composición era distinta de laesperada, con un 50% del total de las ca-lorías procedente de hidratos de carbo-no, un 36% de grasas, y un 14% de pro-teínas.
i Muchos deportistas han adoptado dietasvegetarianas y parecen rendir bien. Sinembargo, debe tenerse mucho cuidadocon las fuentes proteicas y el consumode niveles adecuados de hierro, zinc,calcio y vitaminas.
i La comida de precompetición deberíaconsumirse no antes de las 2 horas pre-vias a la competición, y debería ser bajaen grasas y fácilmente digerible. Unacomida líquida de precompetición bajaen grasas y rica en hidratos de carbonopresenta ventajas.
i La carga de hidratos de carbono aumen-ta en gran medida el contenido de glucó-geno de los músculos, lo cual, a su vez,potencia el rendimiento de fondo.
i Después de una prueba o entrenamientode fondo, es importante consumir unacantidad sustancial de hidratos de carbo-no que reemplace el glucógeno consu-mido durante la actividad. El cuerpo pa-rece estar preparado para reemplazar elglucógeno durante las primeras horasdespués del entrenamiento o competi-ción, porque los niveles de glucogeno-sintasa están al máximo.
02 Cap 07-15 • 221-548 17/1/07 16:31 Página 470

EE.UU. no tenían un buen papel en la competi-ción internacional debido en gran parte a queeran más débiles que sus competidores. Lenta-mente, las investigaciones demostraron que lasmujeres pueden beneficiarse considerablementede los programas de entrenamiento de fuerza yque las ganancias de fuerza no van acompañadasgeneralmente de grandes aumentos en el volu-men muscular.
Un estudio comparó las respuestas al entrena-miento de 47 mujeres y 26 hombres que volunta-riamente participaron en programas idénticos deentrenamiento de pesos con resistencia progresiva.El programa se llevó a cabo dos veces por semana,40 min al día, durante un total de 10 semanas. Lasganancias de fuerza fueron las siguientes:
• Fuerza en press de banca: 29% en las muje-res y 17% en los hombres.
• Fuerza en press de piernas: 30% en las mu-jeres y 26% en los hombres.
La circunferencia muscular aumentó sólo li-geramente en las mujeres, pero los hombres exhi-bieron una clásica hipertrofia muscular.65 Por lotanto, la hipertrofia no es una consecuencia nece-saria ni un prerrequisito para las ganancias defuerza muscular. Diversos estudios han confirma-do estos resultados.
Las mujeres tienen el potencial para desarro-llar sustancialmente más fuerza de la que normal-mente se identifica en la mujer media, general-mente sedentaria. Pero, ¿serán capaces algunavez las mujeres de conseguir tener la misma fuer-za que los hombres para todas las regiones im-portantes del cuerpo? Tal como hemos tratadopreviamente en este capítulo, las proporcionesentre fuerza y peso de las piernas entre los sexosson similares, lo que sugiere que las cualidadeshistológicas y bioquímicas de los músculos sonlas mismas, con independencia del sexo. Esto seconfirmó mediante tomografías computarizadasde la parte superior de los brazos y de los muslosde estudiantes de educación física de ambos se-xos y de culturistas masculinos.52 Aunque ambosgrupos de hombres tenían niveles absolutos defuerza mucho mayores que las mujeres, no se ha-llaron diferencias entre los grupos cuando lafuerza se expresaba por unidad de área muscularcruzada (figura 18.6).
Debido a sus menores niveles de testostero-na, las mujeres tienen menos masa muscular to-tal. Si la masa muscular es el principal determi-nante de la fuerza, entonces las mujeres están en
clara desventaja. Pero si los factores neurales sontan o incluso más importantes que el tamaño, en-tonces el potencial de ganancia de fuerza absolu-ta es considerable. Asimismo, las mujeres puedenconseguir una hipertrofia muscular significativa.Esto se ha demostrado en mujeres culturistas queno han usado esteroides anabólicos. Además, enun estudio, hombres y mujeres mostraron aumen-tos de un porcentaje similar tanto en la fuerza co-mo en la hipertrofia muscular después de 16 sema-nas de entrenamiento resistido.15 Los mecanismosbásicos que permiten mayores niveles de fuerzano han sido definidos con claridad, por lo que nopodemos sacar conclusiones en este momento.En general, aunque las mujeres suelen crear sus-tancialmente menos masa muscular que los hom-bres como respuesta a un estímulo dado, algunasmujeres adquieren más masa muscular que algu-nos hombres. Al igual que la capacidad aeróbica,de la que hablamos previamente, se produce algúnsolapamiento entre mujeres y hombres.
Teniendo todo esto presente, resulta intere-sante reparar en los récords mundiales masculi-nos y femeninos de halterofilia por categorías depeso. La figura 18.7 muestra estas plusmarcasmundiales para el peso total levantado (la sumade la arrancada y los dos tiempos) de 2002. Seaprecia en esta figura que los hombres son bas-tante más fuertes en todas las categorías por elpeso. En la parte más baja de la clasificación, loshombres levantan casi 100 kg más que las muje-res, aunque también levantan 120 kg más en lascategorías más altas por el peso. Parte de la dife-rencia se explica porque, para un peso corporaldado, es muy probable que los hombres tenganmás masa magra. Además, pocas mujeres partici-pan en la halterofilia de competición. Cuanto ma-yor sea el número de participantes, mayor es laposibilidad de conseguir plusmarcas más altas.No obstante, las diferencias son tan grandes quedeben intervenir otros factores.
FISIOLOGÍA DEL ESFUERZO Y DEL DEPORTE
622
Las mujeres pueden experimentar im-portantes aumentos de fuerza (del 20%al 40%) como consecuencia del entrena-miento resistido, y la magnitud de estoscambios es similar a la observada en loshombres. Probablemente, estas ganan-cias se deben en mayor medida a factoresneurales, puesto que el incremento de lamasa muscular suele ser pequeño.
03 Cap 16-21 • 549-776 17/1/07 16:38 Página 622

Figuras 0.0, 0.6a, 0.7a Por cortesía de AmericanCollege of Sports Medicine Archives. Reserva-dos todos los derechos.
Figura 0.1, 0.2, 0,3a-b, 0.4a-b, 0.5a-c, 0.6b,0.7b-c, 0.8a-c, 0.9a-b, 0.12, 1.1a-c, 1.10a-b,1.11, 1.12a-c, 1.13a-b, 3.4a-b, 4.20a-b, 6.1, 6.3a-b, 6.6, 10.3a-b, 17.6, 19.1, 19.2, 19.4a-b, 20.7,21.9a-c, y fotografías de la contraportada delos autores: Por cortesía de los autores.
Figura 0.10, 0.11, 3.2, 4.12, 9.4a-b, 12.11, 12.13,y 19.3 © Tom Roberts.
Figura 1.4, 2.2a, 7.4a, y 20.3a-b © Custom Me-dical Stock Photo.
Figura 3.1, 3.15, 3.16 a-c, 14.05, 14.06, 18.12,19.5 y 19.8 © Human Kinetics.
Figura 3.10 y 3.11a-b De F.C. Hagerman, R.S.Hikida, R.S. Staron, W.M. Sherman y D.L. Cos-till, 1984. “Muscle damage in marathon runners,”Physician and Sportsmedicine 12:39-48.
Figuras 0.8 Fotógrafo/Bruce Coleman, Inc.
Figuras 0.9 y 19.9 © Empics.
Figura 11.9 Fotografía por cortesía de DennisGraver.
Figura 11.14. Por cortesía de NASA.
Figura 14.2a Tom Pantages.
Figura 14.3a-b Fotografía por cortesía de Holo-gic, Inc.
Figura 14.4. Fotografía por cortesía de Life Me-asurement, Inc.
Figura 18.10a-b Reproducido de J. Bone MinerRes 1986, 1:15-21 con autorización de la Ameri-can Society for Bone and Mineral Research. Fo-tografías por cortesía de D.W. Dempster.
Figura 19.10 © 2002 iPhotoNews.com
Introducción: Fotografía por cortesía de Cos-med, Srl; © Empics; Fotografía por cortesía delos autores.
Capítulo 1: © Custom Medical Stock Photo;© Human Kinetics; Fotógrafo/ Bruce Coleman,Inc.
Capítulo 2: © Custom Medical Stock; © HumanKinetics; © Empics.
Capítulo 3: © Human Kinetics; © Empics;© Human Kinetics.
Capítulo 4: © Human Kinetics; © Tom Roberts;© Human Kinetics.
Capítulo 5: © Empics; © Human Kinetics;© Empics.
Capítulo 6: © Human Kinetics; © Human Kine-tics; Fotografía por cortesía de los autores.
Capítulo 7: Custom Medical Stock Photo;© Human Kinetics; © Human Kinetics.
Capítulo 8: Fotógrafo/Bruce Coleman, Inc.;© Human Kinetics; © Human Kinetics.
Capítulo 9: © Empics; © Human Kinetics;© Human Kinetics.
Capítulo 10: Fotografía por cortesía de los auto-res; Fotógrafo/Bruce Coleman, Inc.; © HumanKinetics.
Capítulo 11: © Empics; © Digital Vision; Corte-sía de NASA.
Capítulo 12: Fotógrafo/Bruce Coleman, Inc.;© Empics; © Human Kinetics.
Capítulo 13: © Human Kinetics; © Human Ki-netics; © Human Kinetics.
Capítulo 14: Tom Pantages; © Human Kinetics;© Human Kinetics.
XIV
C R É D I T O S F O T O G R Á F I C O S
01 Crédits+Cap 01-06 • 001-220 17/1/07 16:17 Página XIV

Bibliografía
1. Bar-Or, O. (1983). Pediatric sports medi-cine for the practitioner: From physiologic prin-ciples to clinical applications. Nueva York:Springer-Verlag.
2. Bar-Or, O. (1989). Temperature regulationduring exercise in children and adolescents. EnC.V. Gisolfi & D.R. Lamb (eds.), Perspectives inexercise science and sports medicine: Youth,exercise and sport (págs. 335-362). Carmel, IN:Benchmark Press.
3. Bass, S.L. (2000). The prepubertal years:A uniquely opportune stage of growth when theskeleton is most responsive to exercise? SportsMedicine, 30, 73-78.
4. Bjorntorp, P. (1986). Fat cells and obesity.En K.D. Brownell & J.P. Foreyt (eds.), Handbo-ok of eating disorders: Physiology, psychology,and treatment of obesity, anorexia, and bulimia(págs. 88-98). Nueva York: Basic Books.
5. Boisseau, N., & Delamarche, P. (2000).Metabolic and hormonal responses to exercise inchildren and adolescents. Sports Medicine, 30,405-422.
6. Brooks, G.A., Fahey, T.D., White, T.P. yBaldwin, K.M. (2000). Exercise physiology: Hu-man bioenergetics and its applications, 3ª ed.Mountain View, CA: Mayfield.
7. Clarke, H.H. (1971). Physical and motortests in the Medford Boys’ Growth Study. Engle-wood Cliffs, NJ: Prentice-Hall.
8. Daniels, J., Oldridge, N., Nagle, E, &White, B. (1978). Differences and changes innVO2 among young runners 10 to 18 years of age.Medicine and Science in Sports and Exercise, 10,200-203.
9. Eriksson, B.O. (1972). Physical training,oxygen supply and muscle metabolism in 11-13-year old boys. Acta Physiologica Scandinavica,(Supl. 384) 1-48.
10. Fleck, S.J., & Kraemer, W.J. (1987). De-signing resistance training programs 2ª ed.Champaign, IL: Human Kinetics.
11. Froberg, K., & Lammert, O. (1996). De-velopment of muscle strength during childhood.En O. Bar-Or (ed.), The child and adolescent ath-lete (pág. 28). Londres: Blackwell Science.
12. Inbar, O., & Bar-Or, O. (1986). Anaero-bic characteristics in male children and adoles-cents. Medicine and Science in Sports and Exer-cise, 18, 264-269.
13. Kraemer, W.J., & Fleck, S.J. (1993).Strength training for young athletes. Champaign,IL: Human Kinetics.
14. Krahenbuhl, G.S., Morgan, D.W., &Pangrazi, R.P. (1989). Longitudinal changes indistance-running performance of young males.International Journal of Sports Medicine, 10, 92-96.
15. Mahon, A.D., & Vaccaro, P. (1989). Ven-tilatory threshold and nVO2max changes in chil-dren following endurance training. Medicine andScience in Sports and Exercise, 21, 425-431.
16. Malina, R.M. (1989). Growth and matu-ration: Normal variation and effect of training.En C.V. Gisolfi & D.R. Lamb (eds.), Perspecti-ves in exercise science and sports medicine:Youth, exercise and sport (págs. 223-265). Car-mel, IN: Benchmark Press.
17. Malina, R.M., & Bouchard, C. (1991).Growth, maturation, and physical activity.Champaign, IL: Human Kinetics.
18. Pfeiffer, R.D., & Francis, R.S. (1986).Effects of strength training on muscle develop-ment in prepubescent, pubescent, and postpubes-cent males. Physician and Sportsmedicine, 14(9),134-143.
19. Ramsay, J.A., Blimkie, C.J.R., Smith,K., Garner, S., MacDougall, J.D., & Sale, D.G.(1990). Strength training effects in prepubescentboys. Medicine and Science in Sports and Exer-cise, 22, 605-614.
20. Robinson, S. (1938). Experimental stu-dies of physical fitness in relation to age. Ar-beitsphysiologie, 10, 251-323.
21. Rogers, D.M., Olson, B.L., & Wilmore,J.H. (1995). Scaling for the nVO2-to-body size re-lationship among children and adults. Journal ofApplied Physiology, 79, 958-967.
22. Rowland, T.W. (1985). Aerobic responseto endurance training in prepubescent children: Acritical analysis. Medicine and Science in Sportsand Exercise, 17, 493-497.
23. Rowland, T.W. (1989). Oxygen uptakeand endurance fitness in children: A developmen-
NIÑOS Y ADOLESCENTES EN EL DEPORTE Y EN EL EJERCICIO
575
03 Cap 16-21 • 549-776 17/1/07 16:38 Página 575

3. Marieb, E.N. (1995). Human anatomyand physiology (3.ª ed.). Nueva York: Benja-min/Cummings.
4. Petajan, J.H., Gappmaier, E., White, A.T.,Spencer, M.K., Mino, L., & Hicks, R.W. (1996).Impact of aerobic training on fitness and qualityof life in multiple sclerosis. Annals of Neurology,39, 432-441.
5. Pette, D., & Vrbova, G. (1985). Neuralcontrol of phenotypic expression in mammalianmuscle fibers. Muscle and Nerve, 8, 676-689.
Lecturas seleccionadas
Bawa, P. (2002). Neural control of motor out-put: Can training change it? Exercise and SportSciences Reviews, 30, 59-63.
Binder, M.D., Heckman, C.J., & Powers,R.K. (1996). The physiological control of moto-neuron activity. En L.B. Rowell & J.T. Shepherd(Eds.), Handbook of physiology: Section 12.Exercise: Regulation and integration of multiplesystems (págs. 3-53). Nueva York: Oxford Uni-versity Press.
Christensen, N.J., & Galbo, H. (1983). Sym-pathetic nervous activity during exercise. AnnualReview of Physiology, 45, 139-153.
Cope, T.C., & Pinter, M.J. (1995). The sizeprinciple: Still working after all these years.News in Physiological Sciences; 10, 280-286.
Edgerton, V.R., Bodine-Fowler, S., Roy, R.R.,Ishihara, A., & Hodgson, J.A. (1996). Neuromus-cular adaptation. En L.B. Rowell & J.T.Shepherd (Eds.), Handbook of physiology: Sec-tion 12. Exercise: Regulation and integration ofmultiple systems (págs. 54-88). Nueva York: Ox-ford University Press.
Emonet-Denand, F., Hunt, C.C., & Laporte, Y.(1988). How muscle spindles signal changes ofmuscle length. News in Physiological Sciences,3, 105-109.
Enoka, R.M. (2001). Neuromechanics of hu-man movement (3ª ed.). Champaign, IL: HumanKinetics.
Enoka, R.M., & Stuart, D.G. (1984). Henne-man’s “size principle”: Current issues. Trends inNeurosciences, 7, 226-227.
Gandevia, S.C. (1996). Kinesthesia: Roles forafferent signals and motor commands. En L.B.Rowell & J.T. Shepherd (Eds.), Handbook ofphysiology: Section 12. Exercise: Regulation andintegration of multiple systems (págs. 128-172).Nueva York: Oxford University Press.
Gielen, C.C.A.M., & Denier van der Gon, J.J.(1990). The activation of motor units in coordi-nated arm movements in humans. News in Phy-siological Sciences, 5, 159-163.
Hasan, Z., Enoka, R.M., & Stuart, D.G.(1985). The interface between biomechanics andneurophysiology in the study of movement: So-me recent approaches. Exercise and Sport Scien-ces Reviews, 13, 169-234.
Henneman, E., & Mendell, L.M. (1981).Functional organization of motoneuron pool andits inputs. En V.B. Brooks (ed.), Handbook ofphysiology. Section I, Volume II, The nervous sys-tem: Motor control, Part I (págs. 423-507). Bet-hesda, MD: American Physiological Society.
McArdle, W.D., Katch, F.I., & Katch, V.L.(2001). Exercise physiology: Energy, nutrition,and human performance (5ª ed.). Baltimore: Wi-lliams & Wilkins.
O’Donovan, M.J. (1985). Developmental regu-lation of motor function: An uncharted sea. Medici-ne and Science in Sports and Exercise, 17, 35-43.
Powers, S.K., & Howley, E.T. (2001). Exerci-se physiology: Theory and application to fitnessand performance (4ª ed.). Madison, WI: Mc-Graw-Hill.
Prochazka, A. (1996). Proprioceptive feed-back and movement regulation. En L.B. Rowell& J.T. Shepherd (Eds.), Handbook of physiology:Section 12. Exercise: Regulation and integrationof multiple systems (págs. 89-127). Nueva York:Oxford University Press.
Proske, U. (1997). The mammalian musclespindle. News in Physiological Sciences; 12, 37-42.
Roy, R.R., Baldwin, K.M., & Edgerton, V.R.(1991). The plasticity of skeletal muscle: Effectsof neuromuscular activity. Exercise and SportSciences Reviews, 19, 269-312.
FISIOLOGÍA DEL ESFUERZO Y DEL DEPORTE
88
01 Crédits+Cap 01-06 • 001-220 17/1/07 16:19 Página 88

Los filetes contienen un alto porcentaje de grasa,que requiere varias horas para ser plenamente di-gerida; durante la competición, el sistema diges-tivo entrará en competición con los músculos porel aporte sanguíneo disponible. Asimismo, la tensiónnerviosa suele ser alta antes de una gran competi-ción, por lo que incluso el filete de mejor calidadno se puede disfrutar en momentos así. El fileteresultaría más gratificante y tendría menos pro-babilidades de trastornar el rendimiento si el de-portista lo comiese la noche anterior o después dela competición. Pero si excluimos el filete, ¿quédebe comer el deportista antes de competir?
Aunque la comida tomada unas pocas horasantes de la competición puede contribuir poco alas reservas de glucógeno muscular, puede ase-gurar un nivel normal de glucosa en sangre y pre-venir el hambre. Esta comida debe contener sóloentre 200 y 500 kcal y estar compuesta principal-mente por hidratos de carbono que se digierencon facilidad. Los alimentos tales como los cerea-les, los zumos de frutas y las tostadas se digierencon bastante rapidez y no harán que el deportistase sienta repleto durante la competición. En ge-neral, esta comida debe tomarse al menos con 2horas de antelación a la competición. Los ritmoscon los que se digieren y absorben los alimentosen el cuerpo son bastante individualizados, por loque la programación de la comida previa a lacompetición puede depender de experiencias an-teriores. En un estudio con ciclistas de fondo, sepracticó una prueba larga hasta el agotamiento al70% del nVO2 máx y en dos condiciones distintas,con 14 días entre una y otra prueba: un desayunode 100 g de hidratos de carbono 3 horas antes delejercicio y ayuno antes del ejercicio. Los ciclistasque desayunaron se ejercitaron 136 minutos antesde agotarse, en comparación con los 109 minutosde los que ayunaron, lo cual manifiesta la impor-tancia de la comida previa a la competición.47
Una comida líquida previa a la competiciónpuede tener menos probabilidades de producir in-digestión nerviosa, náuseas, vomitos y calambresabdominales. Este tipo de alimentos se distribu-yen comercialmente, y en general se ha descu-bierto que son útiles tanto antes como entre la ce-lebración de las distintas pruebas. No obstante, aligual que con cualquier comida previa a unacompetición, se deben evitar en la hora final dela competición. Con frecuencia, es difícil encon-trar tiempo para alimentar a los deportistas cuan-do deben participar en múltiples pruebas prelimi-nares y finales. En estas circunstancias, unacomida líquida con poco contenido de grasa y
una elevada proporción de hidratos de carbonopuede ser la única solución.
Carga y sustitución del glucógeno muscularEn el análisis precedente hemos establecido quedietas diferentes pueden influir notablemente enlas reservas de glucógeno muscular y que la ca-pacidad de resistencia depende en gran medidade estas reservas. La teoría es que cuanto mayores la cantidad de glucógeno almacenado, mejores la capacidad de resistencia potencial, porque lafatiga se retrasará. Por lo tanto, el objetivo de undeportista es comenzar una sesión de ejercicio ode competición con tanto glucógeno muscularcomo sea posible.
Basándose en estudios de biopsias muscula-res llevados a cabo a mediados de la década de1960, Åstrand3 propuso un plan para ayudar a loscorredores a almacenar la mayor cantidad posiblede glucógeno. Este proceso se conoce como car-ga del glucógeno. Según el régimen de Åstrand,los deportistas deben prepararse para la competi-ción completando una serie de entrenamientoagotador 7 días antes de la prueba. Durante los 3días siguientes, deberían comer grasas y proteí-nas casi exclusivamente para privar a los múscu-los de hidratos de carbono, lo que, a su vez, in-crementa la actividad de la glucogenosintasa, unaenzima responsable de la síntesis del glucógeno.Los deportistas deben tomar entonces una dietarica en hidratos de carbono durante los días res-tantes. Dado que la actividad de la glucogenosin-tasa aumenta, la mayor ingestión de hidratos decarbono produce un mayor depósito de glucóge-no muscular. La intensidad y el volumen del en-trenamiento durante este período de 6 días debereducirse considerablemente para prevenir unagotamiento adicional del glucógeno muscular, ymaximizar, por lo tanto, las reservas de glucóge-no hepático y muscular.
Este régimen ha demostrado que eleva lasreservas de glucógeno muscular hasta el doble desu nivel normal, pero es más bien poco prácticopara la mayoría de los competidores altamente en-trenados. Durante los 3 días de baja ingestión dehidratos de carbono, los deportistas suelen en-contrar difícil el entrenamiento. Con frecuenciase sienten irritables e incapaces de llevar a cabotrabajos mentales y generalmente muestran sig-nos de bajo contenido de azúcar en sangre, talescomo debilidad muscular y desorientación. Ade-
FISIOLOGÍA DEL ESFUERZO Y DEL DEPORTE
468
02 Cap 07-15 • 221-548 17/1/07 16:31 Página 468

15. ¿Qué medidas podrían ser adecuadas paraayudar al astronauta en el momento del regre-so a la Tierra?
Bibliografía
1. Adams W.C., Bernauer, E.M., Dill, D.B.,& Bomar, J.B. (1975). Effects of equivalent sealevel and altitude training on nVO2 and runningperformance. Journal of Applied Physiology, 39,262-266.
2. Bert, P. (1943). La pression barometrique(MA. Hitchcock & F.A. Hitchcock, Trans.). Co-lumhus, OH: College Book Co.
3. Booth, F.W. (1982). Effect of limb immo-bilization on skeletal muscle. Journal of AppliedPhysiology, 52, 1.113-1.118.
4. Brosnan, M.J., Martin, D.T., Hahn, A.G.,Gore, C.J., & Hawley, J.A. (2000). Impaired in-terval exercise responses in elite female cyclistsat moderate simulated altitude. Journal of Ap-plied Physiology, 89, 1.819-1.824.
5. Bungo, M.W., Goldwater, D.J., Popp,R.L., & Sandler, H. (1987). Echocardiographicevaluation of space shuttle crewmembers. Jour-nal of Applied Physiology. 62, 278-283.
6. Buskirk, E.R., Kollias, J., Piconreatigue,E., Akers, R., Prokop, E., & Baker, P. (1967).Physiology and performance of track athletes atvarious altitudes in the United States and Peru.En R.F. Goddard (ed.), The effects of altitude onphysical performauce (págs. 65-71). Chicago:Athletic Institute.
7. Clark, J.M., & Lambertsen, C.J. (1971).Pulmonary oxygen toxicity: A review. Pharma-cology Review, 23, 37-133.
8. Convertino, V.A. (1987). Potential bene-fits of maximal exercise just prior to return fromweightlessness. Aviation, Space, and Environ-mental Medicine, 58, 568-572.
9. Convertino, V.A. (1990). Physiologicaladaptations to weightlessness: Effects on exerci-se and work performance. Exercise and SportSciences Reviews, 18, 119-166.
10. Convertino, V.A. (1991). Neuromuscularaspects in development of exercise countermea-sures. Physiologist, 34, S125-S125.
11. Convertino, V.A., Doerr, D.F., & Stein,S.L. (1989). Changes in size and compliance ofthe calf after 30 days of simulated microgravity.Journal of Applied Physiology, 66, 1.509-4.512.
12. Cotes, J.E. (1968). Lung function: As-sessment and application in medicine (2.ª ed.).Filadelfia: Davis.
13. Daniels, J., & Oldridge, N. (1970). Ef-fects of alternate exposure to altitude and sea le-vel on world-class middle-distance runners. Me-dicine and Science in Sports, 2, 107-112.
14. Fitts, R.H., Widrick, J.J., Knuth, S.T.,Blaser, C.A., Karhanek, M., Trappe, S.W., Trap-pe, T.A., & Costill, D.L. (1997). Force-velocityand force-power properties of human muscle fi-ber after spaceflight [Abstract]. Medicine andScience in Sports and Exercise, 29, S190.
15. Forster, P.J.G. (1985). Effect of differentascent profiles on performance at 4200 m eleva-tion. Aviation, Space, and Environmental Medici-ne, 56, 785-794.
16. Green, H.J., Sutton, J., Young, P., Cy-merman, A., & Houston, C.S. (1989). OperationEverest II: Muscle energetics during maximal ex-haustive exercise. Journal of Applied Physiology,66, 142-150.
17. Grover, R., Reeves, J., Grover, E., & Le-athers, J. (1967). Muscular exercise in youngmen native to 3,100 m altitude. Journal of Ap-plied Physiology, 22, 555-564.
18. Groves, B.M., Reeves, J.T., Sutton, J.R.,Wagner, P.D., Cymerman, A., Malconian, M.K.,Rock, P.B., Yound, P.M., & Houston, C.S. (1987).Operation Everest II: Elevated high-altitude pul-monary resistance unresponsive to oxygen. Jour-nal of Applied Physiology, 63, 521-530.
19. Hackett, P.H., Rennie, D., & Levine,H.D. (1976). The incidence, importance, andprophylaxis of acute mountain sickness. Lancet,2, 1.149-1.154.
20. Hackett, P.H., Roach, R.C., Schoene,R.B., Harrison, G.L., & Mills, W.J., Jr. (1988).Abnormal control of ventilation in high-altitude
FISIOLOGÍA DEL ESFUERZO Y DEL DEPORTE
400
02 Cap 07-15 • 221-548 17/1/07 16:30 Página 400

Bust Developer, el Astro-Trimmer Exercise Belty el Slim-Skins Vacuum Pants. El primer instru-mento afirmaba añadir de 5 a 8 cm al busto antesde transcurridos entre 3 y 7 días, y los dos últi-mos instrumentos afirmaban eliminar centíme-tros del abdomen, caderas, nalgas y muslos encuestión de minutos. Los tres instrumentos no lo-graron producir ninguno de los cambios al serevaluados en estudios científicos seriamente con-trolados.37, 38
Con frecuencia, quienes están considerandoreducir su peso se estremecen al pensar en incre-
mentar sus actividades físicas, y ¿quién no prefe-riría obtener resultados inmediatos en lugar de te-ner que esperar el momento oportuno? Pero hayque hacer frente a la realidad. Para obtener losbeneficios del ejercicio es necesario trabajar deverdad.
Actividad física y reducción de losriesgos para la saludDurante la década de 1990 se descubrió una rela-ción importante que sugiere otro beneficio sus-tancial de la forma física y la actividad. En el ca-so de personas obesas o con sobrepeso, su riesgogeneral de muerte por enfermedad se reduce mu-cho si son físicamente activas y están en forma.2,
34 Es una buena noticia para los que parecen con-denados a mantenerse obesos o con sobrepeso:un estilo de vida activo y niveles de forma físicamoderados o altos reducen mucho el riesgo demuerte por enfermedades degenerativas crónicas,como enfermedad coronaria y diabetes.
OBESIDAD, DIABETES Y ACTIVIDAD FÍSICA
733
Baja: 50% 1,50 l/min 0,85 50 50 110 110 220
Alta: 75% 2,25 l/min 0,90 67 33 222 110 332
% kcal kcal para 30 min
Intensidad del ejercicio V·O2 medio R media Hidratos de carbono Grasa Hidratos de carbono Grasa Total
�TABLA 21.4 Estimación de las calorías utilizadas procedentes de las grasas y de los hidratos de carbono para una sesión de en-trenamiento aeróbico de baja y de alta intensidad
Figura 21.11. Ritmo de oxidación de grasas a distintas intensi-dades de ejercicio, expresado como porcentaje del nVO2 máx.Reproducido con autorización de J. Acten, M. Gleeson, y A.E.Jeukendrup, 2002, «Determination of the exercise intensity thatelicits maximal fat oxidation», Medicine and Science in Sportsand Exercise 34: 92-97.
Ritm
o d
e ox
idac
ión
de
las
gras
as (g
/min
)
nVO2 máx
45 55 65 75 85 95
0,7
0,6
0,5
0,4
0,3
0,2
0,1
0
i La inactividad es una causa importan-te de obesidad en EE.UU., quizá inclusomás importante que comer en exceso.
i La energía consumida por la actividadincluye el ritmo constante de consumoenergético durante la actividad y tam-bién la energía consumida después delejercicio, ya que el ritmo metabólicopermanece elevado durante algún tiem-po después del cese de la actividad.
03 Cap 16-21 • 549-776 17/1/07 16:39 Página 733

tance exercise in humans. American Journalof Physiology, 273, E99-E107.
Richardson, R.S. (1998). Oxygen transport: Airto muscle cell. Medicine and Science inSports and Exercise, 30, 53-59.
Romijn, J.A., Coyle, E.F, Sidossis, L.S., Zhang,X.-J., & Wolfe, R.R. (1995). Relationshipbetween fatty acid delivery and fatty acidoxidation during strenuous exercise. Journalof Applied Physiology, 79, 1.939-1.945.
Scott, C.B., Roby, F.B., Lohman, T.B., & Bunt,J.C. (1991). The maximally accumulatedoxygen deficit as an indicator of anaerobiccapacity. Medicine and Science in Sportsand Exercise, 23(5), 618-624.
Sjøgaard, G. (1991). Role of exercise-inducedpotassium fluxes underlying muscle fatigue:A brief review. Canadian Journal of Physio-logy and Pharmacology, 69, 238-245.
St. Clair Gibson, A., Lambert, M.I., & Noakes,T.D. (2001). Neural control of force outputduring maximal and submaximal exercise.Sports Medicine, 31, 637-650.
Tipton, K.D., Ferrando, A.A., Williams, B.D., &Wolfe, R.R. (1996). Muscle protein metabo-lism in female swimmers after a combina-tion of resistance and endurance exercise.Journal of Applied Physiology, 81, 2.034-2.038.
Vandewalle, G.P., & Monod, H. (1987). Standardanaerobic exercise tests. Sports Medicine, 4,268-289.
METABOLISMO, ENERGÍA Y SISTEMAS BÁSICOS DE ENERGÍA
169
01 Crédits+Cap 01-06 • 001-220 17/1/07 16:21 Página 169

levantamiento. No obstante, en el momento enque llegue a un ángulo de 100º en su amplitud to-tal de movimiento, estará levantando sólo el 70%de lo que podría levantar en ese ángulo. Por tan-to, con pesas libres sólo se cargan al máximo lospuntos más débiles de la amplitud del movimien-to y se aporta una resistencia relativamente ligeraen la amplitud media (90º a 140º). Por eso laspersonas que practican flexiones con los dos bra-zos reducen mucho su amplitud de movimientocuando empiezan a cansarse. Simplemente tratande alejarse de la porción más débil de su ampli-tud de movimiento. La razón es que con pesas li-bres se levanta el peso máximo para la porciónmás débil de la amplitud del movimiento, lo cualsupone que la porción más fuerte nunca se pone aprueba. Sin embargo, como veremos más adelan-te, esto no significa que las pesas libres no seanla mejor forma de entrenar.
Con un dispositivo de resistencia variable, sedisminuye la resistencia en los puntos más débi-les de la amplitud del movimiento y se aumentaen los puntos más fuertes. Ésta es la base de va-rios populares instrumentos de entrenamiento re-sistido. La teoría subyacente es que el músculopuede entrenarse más plenamente si se le fuerza aactuar a porcentajes constantes más elevados desu capacidad a lo largo de cada punto de su am-plitud de movimiento. La figura 3.15 ilustra uninstrumento de resistencia variable en el que unapolea altera la resistencia a lo largo de la ampli-tud del movimiento.
ADAPTACIONES NEUROMUSCULARES AL ENTRENAMIENTO RESISTIDO
113
Figura 3.14 Variación de la fuerza respecto al ángulo de losmúsculos flexores del codo durante una flexión con los dos bra-zos. La fuerza es óptima en un ángulo de 100º. La capacidad dedesarrollo de fuerza máxima del grupo de músculos en un ángulodado se ofrece como un porcentaje de la capacidad en el ángulo óp-timo de 100º.
Figura 3.15 Un dispositivo de entrenamiento contra resistencia va-riable que usa una leva para alterar la resistencia a lo largo de laamplitud del movimiento.
La capacidad de un músculo o grupo demúsculos para generar fuerza varía a lolargo de toda la amplitud del movimiento.
01 Crédits+Cap 01-06 • 001-220 17/1/07 16:20 Página 113

Actividad hormonal y equilibriode fluidos posejercicioLas influencias hormonales de la aldosterona yde la ADH persisten entre 12 y 48 horas despuésdel ejercicio, reduciendo la producción de orina yprotegiendo el cuerpo de una mayor deshidrata-ción.3,4 De hecho, el estímulo prolongado porparte de la aldosterona de la reabsorción de Na+
hace que la concentración de Na+ en el cuerpollegue a un nivel superior al normal después deuna sesión de ejercicios. En un esfuerzo por com-pensar esta elevación en los niveles de Na+, hayuna mayor cantidad del agua que ingerimos quese traslada al compartimiento extracelular.
Tal como se muestra en la figura 5.14, los in-dividuos que se someten a días repetidos de ejer-cicio y a deshidratación muestran un significati-vo incremento en el volumen del plasma quecontinúa elevándose durante el período de activi-dad. Cuando las series diarias de actividad se in-terrumpen, el exceso de Na+ y de agua es excreta-do por la orina.
La mayoría de los deportistas que se some-ten a un entrenamiento pesado tienen un volumenaumentado del plasma, que diluye varios consti-tuyentes de la sangre. La verdadera cantidad desustancias dentro de la sangre sigue invariable,pero están dispersas en un mayor volumen deagua (plasma), por lo que se diluyen. A este fenó-meno se le llama hemodilución.
FISIOLOGÍA DEL ESFUERZO Y DEL DEPORTE
194
Figura 5.14 Cambios en el volumen plasmático durante 3 días deejercicio repetido y deshidratación. Se ejercitaron con calor losdías 3-6. Apréciese el repentino declive del volumen de plasmacuando dejaban de entrenar (sexto día). Los cambios en la hemo-globina y el hematocrito reflejan la expansión y contracción delvolumen plasmático durante y después del período de 3 días deentrenamiento.
Volumende plasma
Descarga delexceso de H2O
Volumen deglóbulos rojos
Hemoglobina +hematocrito
% d
e ca
mb
io
Días
La hemoglobina es una de las sustanciasdiluidas por la expansión del plasma.Por esta razón, algunos deportistas queverdaderamente tienen niveles normalesde hemoglobina puede parecer que es-tán anémicos como consecuencia de estahemodilución inducida del Na+. Estacondición, que no debe confundirse conla verdadera anemia, puede remediarsecon unos pocos días de reposo, dandotiempo para que los niveles de aldoste-rona vuelvan a su estado normal y paraque los riñones descarguen el Na+ y elagua extras.
ConclusiónEn este capítulo, nos hemos centrado en la fun-ción que desempeña el sistema endocrino en laregulación de los procesos fisiológicos queacompañan al ejercicio. Nos hemos concentradoen la función de las hormonas en el metabolismode la glucosa y de las grasas, y también en el man-tenimiento del equilibrio de los fluidos. Nuestrointerés principal en este capítulo ha sido lo quesucede durante el ejercicio agudo. En el capítulosiguiente, examinaremos las adaptaciones en losprocesos metabólicos del cuerpo que tienen lugarcomo consecuencia del entrenamiento.
Expresiones clave
activación génica directaadenosinmonofosfato cíclico (AMPc)
01 Crédits+Cap 01-06 • 001-220 17/1/07 16:22 Página 194

ocasionados por un menor tiempo de llenado ven-tricular. Un estudio observó que el tiempo de lle-nado ventricular se redujo desde 500 ó 700 ms enreposo hasta alrededor de 150 ms con ritmos car-díacos más altos (aproximadamente entre 150 y200 latidos/min).24 Por lo tanto, con ritmos cre-cientes de esfuerzo que van llevando a frecuenciascardíacas máximas, el tiempo diastólico de llenadopuede reducirse lo bastante como para limitar elllenado. En consecuencia, el volumen diastólicofinal puede nivelarse o comenzar a disminuir.
Para que el mecanismo de Frank-Starlingfuncione, la cantidad de sangre que entra en losventrículos debe aumentar. Para que esto ocurra,el retorno de sangre venosa al corazón debe in-crementarse. Esto puede suceder rápidamentecon la redistribución de la sangre por la activa-ción simpática de las arterias y de las arteriolasen áreas inactivas del cuerpo y por la activaciónsimpática general del sistema venoso. Asimismo,los músculos son más activos durante el ejer-cicio, por lo que su acción de bombeo se incre-menta. Además, la respiración aumenta, por loque los cambios en las presiones intratorácica eintraabdominal también lo hacen. Todos estoscambios incrementan el retorno venoso.
Hemos expuesto dos factores que contribuyena aumentar el volumen sistólico al incrementar laintensidad del ejercicio: el aumento del retorno ve-noso y el aumento de la contractilidad ventricular.Un tercer factor que también contribuye al incre-
mento del volumen sistólico durante el ejercicio esla reducción de la resistencia periférica total debi-da al aumento de la vasodilatación de los vasossanguíneos que conducen a los músculos esquelé-ticos activos. Esta reducción de la resistencia peri-férica total permite al ventrículo izquierdo con-traerse con menor resistencia, lo cual facilita elvaciamiento de la sangre de esta cavidad.
Gasto cardíacoDespués de haber tratado los dos componentesdel gasto cardíaco, podemos reunir esta informa-ción para entender lo que le sucede al gasto car-díaco durante el ejercicio. Puesto que los cambiosen el gasto cardíaco son el producto de la frecuen-cia cardíaca y del volumen sistólico, aquellos sonpredecibles con el incremento de los niveles deesfuerzo, tal como se ve en la figura 7.14. El valoren reposo para el gasto cardíaco es aproximada-mente de 5,0 l/min. El gasto cardíaco aumenta enproporción directa con el incremento de la inten-sidad del ejercicio hasta al menos 20 ó 40 l/min.El valor absoluto varía con el tamaño del cuerpo yel acondicionamiento de resistencia. No obstante,la relación lineal entre el gasto cardíaco y el ritmode esfuerzo no debe sorprendernos puesto que elpropósito principal del aumento del gasto cardía-co es satisfacer la incrementada demanda de oxí-geno de los músculos.
CONTROL CARDIOVASCULAR DURANTE EL EJERCICIO
245
Figura 7.12 Cambios en el volumen ventricular izquierdo diastólico final, en el volumen sistólico final y en el volumen sistólico en reposo ydurante la realización de un esfuerzo de baja intensidad, de intensidad intermedia y de intensidad máxima cuando el sujeto se halla (a) enposición supina y (b) en posición erguida.Adaptado con autorización de L.R. Poliner y col., 1980, «left ventricular performance in normal subjects: A comparison of the responses to exercise in theupright and supine position», Circulation 62: 528-534.
a b
Volu
men
(ml)
Reposo Bajo Intermedio Máximo
Intensidad del esfuerzo (supino)
Reposo Bajo Intermedio Máximo
Intensidad del esfuerzo (erguido)
VDF
VS
VSF
Volu
men
(ml)
VDF
VS
VSF
02 Cap 07-15 • 221-548 17/1/07 16:28 Página 245

DensitometríaLa densitometría supone medir la densidad delcuerpo del deportista. La densidad (D) se definecomo la masa dividida por el volumen:
Dcorporal = masacorporal ÷ volumencorporal
La masa corporal es la escala de peso del de-portista. El volumen corporal puede obtenersemediante varias técnicas, pero la más común es elpesaje hidrostático, en el cual el deportista es
pesado estando totalmente sumergido en el agua.La diferencia entre el peso del deportista dadopor una báscula y el peso debajo del agua, unavez corregido por la densidad del agua, igualael volumen del cuerpo. Este volumen tiene quevolverse a corregir teniendo en cuenta el volu-men de aire atrapado en el cuerpo. La cantidad deaire atrapado en el tracto gastrointestinal es difí-cil de medir, pero afortunadamente es un volu-men pequeño (unos 100 ml) y se suele pasar poralto. No obstante, el gas atrapado en los pulmonesdebe medirse porque su volumen suele ser gran-de, dando un promedio de 1.500 ml en los hom-bres adultos jóvenes y de 1.200 ml para las muje-res adultas jóvenes, dependiendo del tamaño.
La figura 14.2 ilustra la técnica de pesajehidrostático usada para dos jugadores profesio-nales de fútbol americano de estatura y pesoidénticos, pero composiciones corporales sustan-cialmente diferentes. La tabla de la figura mues-tra los cálculos de su composición corporal, y de-muestra que Dave es casi dos veces más grasoque Jack. Para que estos dos deportistas alcancenun valor de porcentaje de grasa corporal del 10%,
PESO CORPORAL, COMPOSICIÓN CORPORAL Y DEPORTE
485
Figura 14.2 Uso de la técnica de pesaje debajo del agua en dos jugadores profesionales de fútbol americano del mismo peso pero de distin-ta composición corporal.
Jack
6,50 kg 5,0 kg
Dave
Variable Jack Dave
Estatura 188 cm 188 cm Peso 93 kg 93 kg Peso debajo del agua 6,5 kg 5,0 kgVolumen 86,5 l 88,0 lDensidad 1,075 g/ml 1,057 g/mlGrasa relativa 10,5% 18,4%Peso de grasa 9,7 kg 17,1 kg Peso magro 83,3 kg 75,9 kg Peso objetivo con 92,6 kg 84,3 kg
un 10% de grasaPérdida de peso para 0,4 kg 8,7 kg
alcanzar el pesoobjetivo
Volumen = peso - peso debajo del agua.Densidad = peso ÷ volumen.
02 Cap 07-15 • 221-548 17/1/07 16:31 Página 485

Acción estática
Los músculos pueden actuar también sin mover-se. Cuando esto ocurre, el músculo genera fuer-za, pero su longitud permanece estática (invaria-ble). Esto recibe la denominación de acciónestática (figura 1.16), ya que el ángulo de la arti-culación no cambia. Ha recibido también la de-nominación de acción isométrica. Sucede, porejemplo, cuando intentamos levantar un objetoque es más pesado que la fuerza generada pornuestro músculo, o cuando sostenemos el pesode un objeto manteniéndolo fijo con el codo fle-xionado. En ambos casos, percibimos que losmúsculos están tensos, pero que no pueden mo-ver el peso por lo que no se acortan. En esta ac-ción, los puentes cruzados de miosina se formany son reciclados, y producen fuerza, pero la fuer-za externa es demasiado grande para que los fila-mentos de actina se muevan. Permanecen en suposición normal, de modo que el acortamientono puede tener lugar. Si se pueden reclutar sufi-cientes unidades motoras como para producir lanecesaria fuerza para superar la resistencia, una
acción estática puede convertirse en una accióndinámica.
Acción excéntrica
Los músculos pueden producir fuerza inclusocuando se alargan. Este movimiento es una ac-ción excéntrica. Puesto que el movimiento arti-cular se produce, ésta es también una acción di-námica. Un ejemplo de esto es la acción delbíceps braquial cuando el codo se extiende parabajar un gran peso. En este caso, los filamentosde actina son arrastrados en dirección contrariaal centro de la sarcómera, esencialmente esti-rándola.
Generación de fuerza
La fortaleza de nuestros músculos refleja su capa-cidad para producir fuerza. Si se tiene fuerza paralevantar un peso de 150 kg en press de banca, esque los músculos son capaces de producir sufi-ciente fuerza para superar una carga de 150 kg.Incluso cuando están descargados (no intentando
FISIOLOGÍA DEL ESFUERZO Y DEL DEPORTE
56
Figura 1.16 Acciones de los músculos agonistas, antagonistas y sinergistas durante la flexión del codo.
Acción concéntrica:el músculo se acorta
Acción estática:la longitud del músculono cambia
Acción excéntrica:el músculo seelonga
Músculo braquiorradial(sinergista)
Músculo trícepsbraquial (antagonista)
Músculo braquial(agonista)
Músculo bícepsbraquial (agonista)
01 Crédits+Cap 01-06 • 001-220 17/1/07 16:19 Página 56

Actividadfísica para
la salud y laforma física
P A R T E
VII�
�
�
En anteriores partes de este libro, nos hemos concentradoen las bases fisiológicas de la actividad física y en cómopodemos mejorar nuestro rendimiento. Ahora, en la parteVII, apartaremos nuestra atención del rendimiento depor-tivo y la dirigiremos hacia un área especial de la fisiologíadel esfuerzo y del deporte: la actividad física para la saludy el fitness. En el capítulo 19, Programación de ejer-cicios para la salud y el fitness, trataremos de cómo dise-ñar un programa de ejercicios que pueda mejorar la saludy el fitness. Consideraremos los componentes esenciales,cómo adaptar el programa a las necesidades específicas decada individuo y la función única de la actividad física pa-ra la rehabilitación de pacientes enfermos. En el capítuloXX, Enfermedades cardiovasculares y actividad física,examinaremos los tipos más importantes de enfermedadescardiovasculares, sus bases fisiológicas y cómo la activi-dad física puede beneficiar a quienes tienen estas enfer-medades. En el capítulo 21, Obesidad, diabetes y activi-dad física, examinaremos las causas de la obesidad y dela diabetes, los riesgos para la salud asociados con cadauno de estos aspectos y cómo con la actividad física sepueden controlar ambos trastornos.
647
03 Cap 16-21 • 549-776 17/1/07 16:38 Página 647

elevada de H+ inhiba los procesos contráctiles.Como se expuso en el capítulo 4, una vez que elpH intracelular se sitúa por debajo de 6,9, las enzi-mas glucolíticas críticas empiezan a inhibirse.Cuando el pH alcanza 6,4, ya no es posible la de-gradación de glucógeno.
Curiosamente, bajo circunstancias similares(tales como en la realización de esprines hasta elagotamiento), los sujetos que siguen entrena-mientos de resistencia no acumulan tanto lactatomuscular ni experimentan los inusualmente bajosvalores de pH observados en las personas que si-guen entrenamientos de esprín. Esta diferencia esdifícil de explicar, pero el pH muscular no parecelimitar el rendimiento en esprín de las personasque siguen entrenamientos de resistencia.
Especificidad de la respuesta al entrenamiento
La tabla 6.2 muestra las actividades de enzimasmusculares seleccionadas a partir de los tres sis-temas de energía para hombres no entrenados,hombres entrenados anaeróbicamente y hombresentrenados aeróbicamente. Esta tabla explica quelos músculos entrenados de forma aeróbica tie-nen una actividad enzimática glucolítica signifi-cativamente menor. Por tanto, podrían tener me-nos capacidad para el metabolismo anaeróbico odepender menos de la energía obtenida de la glu-
cólisis. Se necesitan más estudios para explicarlas implicaciones de los cambios musculares queacompañan al entrenamiento anaeróbico y aeró-bico, si bien la tabla muestra con claridad el altogrado de especificidad de los estímulos dados delentrenamiento.
FISIOLOGÍA DEL ESFUERZO Y DEL DEPORTE
216
Enzimas aeróbicasSistema oxidativo
Succinatodeshidrogenasa 8,1 8,0 20,8a
Malatodeshidrogenasa 45,5 46,0 65,5a
Carnitina palmitiltransferasa 1,5 1,5 2,3a
Enzimas anaeróbicasSistema ATP-PC
Creatincinasa 609,0 702,0a 589,0
Miocinasa 309,0 350,0a 297,0
Sistema glucolítico
Fosforilasa 5,3 5,8 3,7a
Fosfofructocinasa 19,9 29,2a 18,9
Lactatodeshidrogenasa 766,0 811,0 621,0
a Presenta una significativa diferencia con los valores no entrenados.
Entrenados EntrenadosNo entrenados anaeróbicamente aeróbicamente
�TABLA 6.2 Actividades enzimáticas musculares seleccionadas (mmol·g–1·min–1) para hombres no entrenados, entrenados anae-róbicamente y entrenados aeróbicamente
i Las series de ejercicio anaeróbico mejo-ran el rendimiento anaeróbico, pero estebeneficio parece que se debe más a ga-nancias en fuerza que a mejoras en elfuncionamiento de los sistemas anaeró-bicos de energía.
i El entrenamiento anaeróbico tambiénaumenta la eficacia del movimiento, yun movimiento más eficaz requiere me-nos consumo energético.
i Aunque el esprín es anaeróbico por na-turaleza, parte de la energía usada du-rante la realización de series de esprinesmás prolongados proviene de la oxida-ción, por lo que la capacidad aeróbicamuscular puede incrementarse tambiéncon este tipo de entrenamiento.
01 Crédits+Cap 01-06 • 001-220 17/1/07 16:22 Página 216

Esto se muestra en la figura 12.7a. La figura12.7b muestra las interacciones del encéfalo y elsistema inmunitario con estos dos ejes.
Recientemente, se ha propuesto un papelprincipal de las citocinas en el síndrome por so-breentrenamiento, respaldando así el modelo deArmstrong y VanHeest de la figura 12.7.31 La ele-vación del número de citocinas circulantes es
producto de los traumatismos en el músculo es-quelético, el hueso y las articulaciones asociadoscon el sobreentrenamiento.
Inmunidad y sobreentrenamiento
Nuestro sistema inmunológico nos proporcionauna línea de defensa contra las bacterias, los pa-
ENTRENAMIENTO PARA EL DEPORTE
417
Figura 12.7 (a) Papel del hipotálamo y los ejes simpático-adrenal (SAM) e hipotalámico-hipofisario-adrenal (HPA) como posiblesmediadores del síndrome por sobreentrenamiento. (b) Las interacciones del sistema inmunológico con este modelo, en el que las cito-cinas desempeñan un importante papel potencial en la modelación del sobreentrenamiento.Símbolos: ACTH = adrenotropina; ADR = adrenalina (epinefrina); CRH = hormona liberadora de corticotropina; GH = hormona delcrecimiento; IL-1 = interleucina-1; IL-6 = interleucina 6; NA = noradrenalina; P = sustancia P; PRL = prolactina; FNT = factor de ne-crosis tumoral; Y = neuropéptido Y; βE = endorfina β.Adaptado con autorización de L.E. Armstrong y J.L. VanHeest, 2002, «The unknown mechanism of the overtraining syndrome»,Sports Medicine 32: 185-209.1
Factores estresantes
Eje SAM Hipotálamo Eje HPA
CRH
ACTH
ACTH
CRH
GH, PRL, βENA, ADR, Y, P
NA, ADR
NA
ADR
Adenohipófisis
Cortezasuprarrenal
Inhibe larespuesta
inmunológica y lainflamación
Médulasuprarrenal
Órganos dedestino
Aumenta laactividad
neurológica
Estimula lamovilización/redistribución de
la energía y aumenta larespuesta cardiovascular
Cortisol
Hipotálamo
Glándulassuprarrenales
Cortisol
Adenohipófisis
Inmunocitos
Citocinas (IL-1, IL-6, FNT)
Bazo, timo y otros órganosdel sistema inmunológico
Nerviosespinales
simpáticos
Factoresestresantes
Nervios espinalessimpáticos
02 Cap 07-15 • 221-548 17/1/07 16:30 Página 417

Estudios experimentales sobre animales hanrelacionado la obesidad con factores hereditarios(genéticos). Estudios recientes en humanos lleva-dos a cabo por el Dr. Albert Stunkard y colabora-dores en la Universidad de Pennsylvania hanmostrado una influencia genética directa sobre laestatura, el peso y el IMC.28, 29, 30 Un estudio de laUniversidad Laval, en Quebec, ha facilitado po-siblemente la prueba más sólida hasta la fecha deun componente genético significativo para laobesidad.5 Los investigadores tomaron 12 parejasde gemelos hombres adultos jóvenes monocigóti-cos (idénticos) y los alojaron en una sección ce-rrada de un dormitorio bajo observación las 24 hdel día, durante 120 días consecutivos. Las dietasde los sujetos fueron monitorizadas durante los pri-meros 14 días para determinar sus valores inicia-les de ingestión calórica. A lo largo de los 100días siguientes, los sujetos fueron alimentados con1.000 kcal por encima de las iniciales durante 6días de cada 7. En el séptimo día, a los sujetosse les alimentaba solamente con sus kcal dietéti-cas de inicio. Por lo tanto, fueron sobrealimentadoscon 1.000 kcal durante 84 de los 100 días. Los ni-veles de actividad también fueron controlados es-trechamente. Al final del período de estudio, talcomo se muestra en la figura 21.5, el verdaderopeso ganado variaba ampliamente, desde 4,3 has-ta 13,3 kg, una variación que significaba el triplede ganancia de peso para una sobrealimentacióncon las mismas calorías. Sin embargo, la respues-ta de los dos gemelos en cualquiera de las parejasde gemelos fue muy similar; las variaciones másimportantes se produjeron entre parejas distintasde gemelos. Resultados similares se hallaron paralas ganancias de masa grasa, porcentaje de grasacorporal y grasa subcutánea.
La obesidad se ha relacionado también expe-rimental y clínicamente tanto con traumas fisio-lógicos como psicológicos. Los desequilibrioshormonales, los traumas emocionales y las alte-raciones en los mecanismos homeostáticos bási-cos han demostrado estar directa o indirectamen-te relacionados con el inicio de la obesidad.Factores ambientales, tales como los hábitos cul-turales, una actividad física inadecuada y dietasinapropiadas también contribuyen a la obesidad.
Por lo tanto, la obesidad es de origen com-plejo, y sus causas específicas indudablementedifieren de una persona a otra. Reconocer esto esimportante para el tratamiento de la obesidadexistente y para la prevención de su aparición.Atribuir la obesidad únicamente a la gula es in-justo y psicológicamente perjudicial para las per-sonas preocupadas por su problema y que estánintentando corregirlo. De hecho, varios estudioshan demostrado que las personas con obesidad enrealidad comen menos, aunque también realizanuna actividad física mucho menor, que las perso-nas de su mismo sexo y edad con contenidos degrasa corporal medios.
OBESIDAD, DIABETES Y ACTIVIDAD FÍSICA
721
Investigaciones recientes confirman queexiste un significativo componente gené-tico en la etiología de la obesidad. Sinembargo, es posible ser obeso debido bá-sicamente al estilo de vida elegido, en au-sencia de una historia familiar (genética)de obesidad. También es posible estar re-lativamente delgado, incluso si se tieneuna predisposición genética para la obe-sidad, mediante niveles adecuados de ac-tividad y con la dieta.
Figura 21.5 Similar ganancia de peso entre gemelos en respuesta aun incremento en la dieta diaria de 1.000 kcal durante 84 días. Lospuntos representan el peso ganado por cada uno de los gemelos;los valores del gemelo A se arrojan en el eje y mientras que losdel B están en el x.Adaptado con autorización de C. Bouchard y cols., 1990, «Theresponse to long-term overfeeding in identical twins», New En-gland of Medicine 322: 1477-1482. Copyright © 1990 Massa-chusetts Medical Society. Reservados todos los derechos.
Cam
bio
en
el p
eso
co
rpo
ral
del
gem
elo
A (k
g)
Cambio en el peso corporaldel gemelo B (kg)
03 Cap 16-21 • 549-776 17/1/07 16:39 Página 721

limitante del rendimiento y la causa más impor-tante de la fatiga durante la realización de ejer-cicios máximos de corta duración.
Tal como se ve en la figura 4.22, el restable-cimiento del pH anterior al ejercicio después deun esprín agotador requiere alrededor de 30 ó 35min de recuperación. Incluso cuando se ha res-taurado un pH normal, los niveles de lactato ensangre y músculo pueden seguir siendo elevados.No obstante, la experiencia ha demostrado queun deportista puede continuar ejercitándose a in-tensidades elevadas incluso con un pH muscularinferior a 7,0 y un nivel de lactato superior a 6 ó 7mmol/l, cuatro o cinco veces el valor de reposo.
Actualmente, algunos entrenadores y fisiólo-gos del deporte están intentando usar medicionesdel lactato sanguíneo para cuantificar la intensi-dad y el volumen del entrenamiento necesariopara producir un estímulo de entrenamiento ópti-mo. Estas mediciones proporcionan un índice deintensidad del entrenamiento, pero pueden no te-ner relación con los procesos anaeróbicos o conel estado de acidosis en los músculos. Cuando segeneran H+ y lactato en los músculos, ambos di-funden fuera de las células. Entonces se diluyenen los fluidos corporales y son transportados aotras áreas del cuerpo para ser metabolizados. Enconsecuencia, los valores del lactato en sangre
dependen de los ritmos de producción, difusión yoxidación. Existe una diversidad de factores quepueden influir en estos procesos, por lo que losvalores del lactato tienen un uso cuestionable pa-ra programar el entrenamiento.
Fatiga neuromuscularHasta ahora sólo hemos considerado factoresdentro de los músculos que pueden ser responsa-bles de la fatiga. También hay pruebas que indi-can que bajo determinadas circunstancias la fati-ga puede ser el resultado de una incapacidad paraactivar las fibras musculares, una función del sis-tema nervioso. Tal como se ha indicado en el ca-pítulo 2, el impulso nervioso se transmite a travésde la placa motora para activar la membrana delas fibras y hacer que el retículo sarcoplasmáticode las fibras libere calcio. Éste, a su vez, se enla-za con la troponina para iniciar la contracciónmuscular. Examinemos dos posibles mecanismosnerviosos que pueden alterar este proceso y con-tribuir posiblemente a la fatiga.
Transmisión nerviosa
La fatiga puede ocurrir en la placa motora, queimpide la transmisión del impulso nervioso a lamembrana de las fibras musculares. Estudiosefectuados a principios de este siglo determina-ron con claridad tales insuficiencias en la trans-misión del impulso nervioso en los músculos fa-tigados. Esta insuficiencia puede suponer laintervención de uno o más de los procesos si-guientes:
• La liberación o síntesis de la acetilcolina,el neurotransmisor que transmite el impul-so nervioso desde el nervio motor hasta lamembrana muscular, puede reducirse.
• La colinesterasa, la enzima que descom-pone la acetilcolina una vez que se hatransmitido el impulso, puede volverse hi-peractiva, impidiendo la concentración desuficiente acetilcolina para iniciar un po-tencial de acción.
• La actividad de la colinesterasa puede vol-verse hipoactiva (inhibidora), permitiendoque la acetilcolina se acumule excesiva-mente y paralice la fibra.
FISIOLOGÍA DEL ESFUERZO Y DEL DEPORTE
164
Figura 4.22 Cambios en el pH muscular durante un ejercicio deesprín y recuperación. Préstese atención a la caída espectaculardel pH muscular durante el esprín y la recuperación gradual hastael nivel normal después del esfuerzo. Nótese que pasan casi 30min hasta que el pH vuelve al nivel previo al ejercicio.
pH
mus
cula
r
Esprín
7,1
7,0
6,9
6,8
6,7
6,6
6,5
00 5 10 15 20 25 30 35
Recuperación (min)
01 Crédits+Cap 01-06 • 001-220 17/1/07 16:21 Página 164

ronas, conducen impulsos eléctricos por la médu-la espinal hasta el brazo. Al llegar al músculo bí-ceps, estas neuronas liberan mensajeros químicosque salvan el espacio entre el nervio y el múscu-lo, excitando cada neurona un número de mioci-tos o fibras musculares. Los impulsos nerviososuna vez que salvan este espacio, se extienden to-do a lo largo de las fibras musculares y entran enellas por pequeños poros. Una vez dentro de lasfibras musculares, el impulso activa los procesosde contracción de estas fibras, lo cual afecta amoléculas de proteínas específicas –actina y mio-sina– y a un elaborado sistema de energía queaporta la fuente energética necesaria para realizaruna contracción y las siguientes. Es a este niveldonde otras moléculas, como el adenosintrifosfa-to (ATP) y la fosfocreatina (PC) son críticas paraaportar la energía necesaria para la contracciónenergética.
La actividad física es un proceso complica-do. Los científicos deben examinar cada ajusteque hace el cuerpo observando estos hechos tantoindividual como colectivamente. En este capítu-lo, analizaremos cómo enfocar esta tarea.
Enfoque de la fisiologíadel esfuerzo y del deporteLa fisiología del esfuerzo y del deporte ha evo-lucionado a partir de la anatomía y de la fisiolo-gía. La anatomía es el estudio de la estructurade un organismo, o morfología. Con la anato-mía aprendemos la estructura básica de diver-sas partes del cuerpo y sus relaciones. La fisio-logía es el estudio de la función del cuerpo. Enfisiología estudiamos cómo funcionan nuestrossistemas orgánicos, tejidos y células, y cómo seintegran sus funciones para regular nuestrosambientes internos. Puesto que la fisiología secentra en las funciones de las estructuras, nopodemos discutir fácilmente sobre fisiologíasin entender la anatomía.
La fisiología del esfuerzo es el estudio decómo las estructuras y funciones de nuestroscuerpos se ven alteradas cuando estamos expues-tos a series agudas y crónicas de ejercicio. La fi-siología del deporte aplica además los conceptosde la fisiología del ejercicio al entrenamiento deldeportista y a mejorar su rendimiento deportivo.Por lo tanto, la fisiología del deporte deriva de lafisiología del esfuerzo.
Consideremos un ejemplo para ayudarnos adistinguir entre dos ramas estrechamente relacio-nadas de la fisiología. En la fisiología del esfuerzo,mediante una investigación considerable, ahoraconocemos mejor el modo en que nuestros cuer-pos extraen energía de los alimentos que tomamospara permitir que se inicien las acciones muscula-res y se mantenga el movimiento. Hemos aprendi-do que las grasas son nuestra principal fuente deenergía cuando estamos en reposo y durante larealización de ejercicios de baja intensidad, peroque nuestros cuerpos usan proporcionalmente máshidratos de carbono cuando la intensidad del ejer-cicio se incrementa, hasta que los hidratos de car-bono se convierten en nuestra principal fuente deenergía. Los ejercicios prolongados de intensidadelevada pueden reducir sustancialmente las reser-vas de hidratos de carbono de nuestros cuerpos, locual puede contribuir al agotamiento.
La fisiología del deporte, entonces, toma es-ta información y, comprendiendo que el cuerpotiene unas reservas energéticas limitadas de hi-dratos de carbono, intenta encontrar modos de:
• aumentar los depósitos de hidratos de car-bono del cuerpo (carga de hidratos de car-bono);
• reducir el ritmo con el que el cuerpo utilizalos hidratos de carbono durante el rendi-miento físico (ahorro de hidratos de carbo-no), y
INTRODUCCIÓN
3
La fisiología del esfuerzo ha evoluciona-do a partir de su disciplina madre, la fi-siología. Se ocupa del estudio de cómo seadapta fisiológicamente el cuerpo al es-trés agudo del ejercicio, o a la actividadfísica, y al estrés crónico del entrena-miento físico. La fisiología del deporteha evolucionado a partir de la fisiologíadel esfuerzo. Aunque se realizaron va-rios estudios de fisiología a mediados dela década de 1980, podría argüirse que lafisiología del ejercicio como la conoce-mos hoy era un apéndice de la fisiologíadel deporte, ya que muchos de los prime-ros estudios fisiológicos se centraron enlos deportistas y en el rendimiento de-portivo.
01 Crédits+Cap 01-06 • 001-220 17/1/07 16:18 Página 3
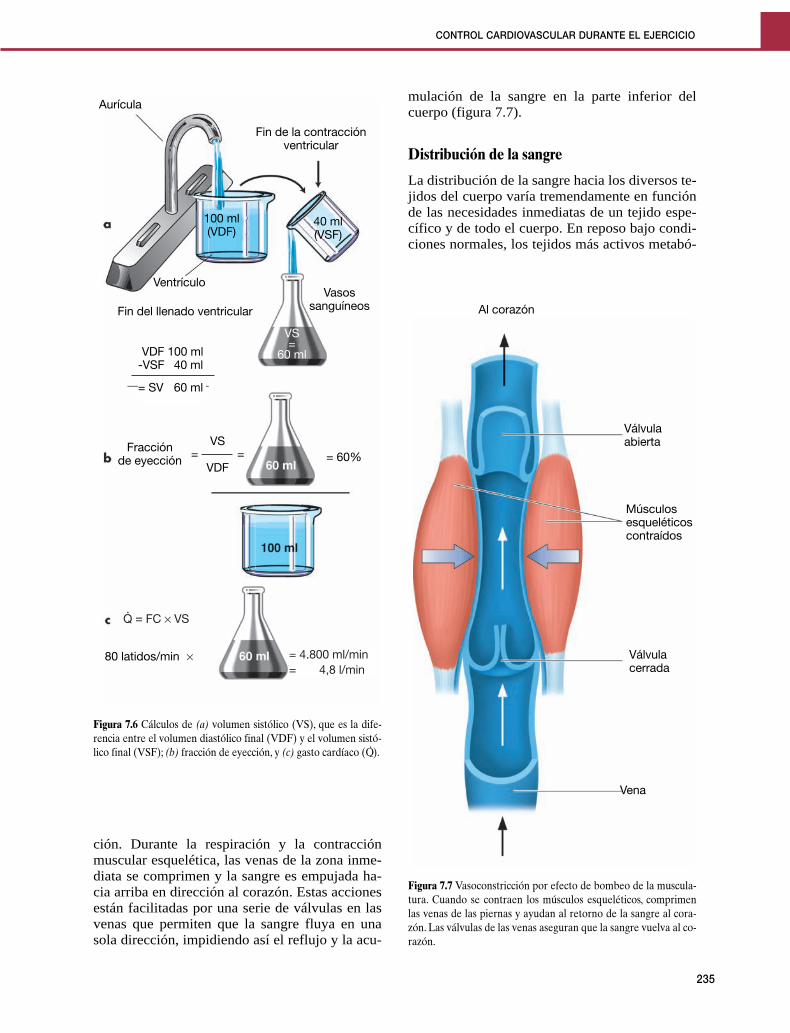
ción. Durante la respiración y la contracciónmuscular esquelética, las venas de la zona inme-diata se comprimen y la sangre es empujada ha-cia arriba en dirección al corazón. Estas accionesestán facilitadas por una serie de válvulas en lasvenas que permiten que la sangre fluya en unasola dirección, impidiendo así el reflujo y la acu-
mulación de la sangre en la parte inferior delcuerpo (figura 7.7).
Distribución de la sangre
La distribución de la sangre hacia los diversos te-jidos del cuerpo varía tremendamente en funciónde las necesidades inmediatas de un tejido espe-cífico y de todo el cuerpo. En reposo bajo condi-ciones normales, los tejidos más activos metabó-
CONTROL CARDIOVASCULAR DURANTE EL EJERCICIO
235
Figura 7.6 Cálculos de (a) volumen sistólico (VS), que es la dife-rencia entre el volumen diastólico final (VDF) y el volumen sistó-lico final (VSF); (b) fracción de eyección, y (c) gasto cardíaco (Q·).
Aurícula
Fin de la contracciónventricular
Ventrículo
100 ml(VDF)
40 ml(VSF)
VS=
60 ml
Fin del llenado ventricular
VDF 100 ml-VSF 40 ml
= SV 60 ml
Fracciónde eyección
Vasossanguíneos
VS
VDF= = = 60%
80 latidos/min × = 4.800 ml/min= 4,8 l/min
Q· = FC × VS
Figura 7.7 Vasoconstricción por efecto de bombeo de la muscula-tura. Cuando se contraen los músculos esqueléticos, comprimenlas venas de las piernas y ayudan al retorno de la sangre al cora-zón. Las válvulas de las venas aseguran que la sangre vuelva al co-razón.
Al corazón
Válvulaabierta
Músculosesqueléticoscontraídos
Válvulacerrada
Vena
02 Cap 07-15 • 221-548 17/1/07 16:28 Página 235

2. Linealidad.
3. Adiposidad.
La complexión de cada deportista es unacombinación única de estos tres componentes.Los deportistas de ciertos deportes exhiben gene-ralmente una predominancia de un componentesobre los otros dos. El culturista muestra princi-palmente muscularidad, el pívot de baloncesto de218 cm que pesa tan sólo 82 kg muestra lineali-dad y el luchador de sumo muestra adiposidad.La mayoría de los deportistas están equilibradosentre muscularidad y linealidad, pero la muscula-ridad tiende a dominar en los deportistas mascu-linos.5
El tamaño corporal hace referencia a la al-tura y a la masa (peso) de un individuo. La formacorporal se clasifica frecuentemente como baja oalta, grande o pequeña, pesada o ligera. Las dis-tinciones entre estas categorías pueden variardependiendo de las necesidades específicas de ren-dimiento, por lo que el tamaño corporal debe con-siderarse en relación con el deporte específico, laposición del deportista o el tipo de prueba. Porejemplo, entre los hombres una estatura de 190,5cm sería escasa para un jugador profesional de ba-loncesto, pero alta para un corredor de fondo. Del
mismo modo, en el fútbol americano profesional,un peso de 104 kg sería excesivo para un defensacentral, correcto para un defensa lateral, pero es-caso para un defensa extremo.
La composición corporal hace referencia ala composición química del cuerpo. La figura14.1 ilustra cuatro modelos de composición cor-poral. Los dos primeros dividen el cuerpo en susvarios componentes químicos y anatómicos; losdos últimos simplifican la composición corporalen dos componentes. La principal diferencia en-tre estos dos últimos modelos es la terminologíade masa magra y masa grasa. Behnke propusooriginalmente el concepto de masa magra, defini-da para incluir la masa magra y la grasa esencial(la cantidad de grasa necesaria para la supervi-vencia).10Aunque este modelo es conceptualmen-te sólido, presenta problemas de medición: no esposible diferenciar entre grasa esencial y noesencial. En consecuencia, la mayoría de loscientíficos han adoptado el modelo de dos com-ponentes que incluye la masa grasa y la masa ma-gra, que es el modelo usado en este libro. La ma-sa grasa se discute con frecuencia en términos deporcentaje de grasa corporal, que es el porcen-taje de la masa corporal total que se compone degrasa. La masa magra simplemente se refiere atodo el tejido corporal que no es grasa.
PESO CORPORAL, COMPOSICIÓN CORPORAL Y DEPORTE
483
Figura 14.1 Cuatro modelos de composición corporal. Hay una ligera diferencia en el componente de lípidos en los cuatro modelos. En elmodelo anatómico, el tejido adiposo comprende grasa y la matriz celular y el agua presente en el tejido adiposo. En el modelo de Behnke,la masa corporal magra incluye algo de grasa, que Behnke denominó grasa esencial. Por tanto, la grasa en este modelo es la grasa total me-nos la grasa esencial.Adaptado con autorización de J.H. Wilmore, 1992, Body weight and body composition. En Brownell, Rodin y Wilmore (eds.) Eating,body weight, and performance in athletes: Disorders of modern society (Filadelfia, PA: Lippincott, Williams, y Wilkins), 77-93.
Grasa
Proteínas
CHO
Agua
Minerales
Modelo químico
Tejido adiposo
Músculo
Órganos
Hueso
Otros
Modelo anatómico
GrasaMasa grasa
Masa magra
Grasa esencial
Masa corporal magra
Modelo de 2componentes de Behnke
Modelo de 2componentes
02 Cap 07-15 • 221-548 17/1/07 16:31 Página 483

Adaptaciones fisiológicas a losejercicios de entrenamientoTal como hemos visto en capítulos anteriores, lafunción fisiológica básica en reposo y durante elejercicio cambia sustancialmente con el entrena-miento físico. En esta sección, investigaremos có-mo se adaptan las mujeres al ejercicio crónico, ydestacaremos aquellas áreas en las que las res-puestas pueden diferir de las de los hombres.
Composición corporalCon el entrenamiento de fondo cardiorrespirato-rio, tanto los hombres como las mujeres experi-mentan:
• pérdidas de la masa corporal total;
• pérdidas de masa grasa;
• pérdidas de grasa relativa, y
• ganancias de masa magra (MM).
Las mujeres generalmente ganan mucha me-nos masa magra que los hombres. Con la excep-
ción de la MM, la magnitud del cambio en lacomposición corporal parece estar más relaciona-da con el consumo total de energía asociado conlas actividades de entrenamiento que con el sexode los participantes. Por lo que se refiere a laMM, se gana significativamente más en respues-ta al entrenamiento de fuerza que con el entrena-miento de fondo, y la magnitud de estas respues-tas es mucho menor en las mujeres, debidoprincipalmente a diferencias hormonales.
El tejido óseo y conectivo experimentan al-teraciones con el entrenamiento, pero no se cono-cen bien. En general, los estudios con animales ylos limitados estudios con seres humanos hallanun incremento en la densidad de los huesos lar-gos que sostienen el peso corporal. Esta adapta-ción parece ser independiente del sexo, al menosen poblaciones jóvenes y de mediana edad. Estu-diaremos algunas excepciones a esto más adelan-te en este capítulo.
El tejido conectivo parece fortalecerse con elentrenamiento de fondo, y en esta respuesta no sehan hallado diferencias entre los sexos. La posibi-lidad de que las mujeres sean más susceptibles alas lesiones que los hombres mientras participanen actividades físicas y deportivas ha producidopreocupación sobre las diferencias específicas decada sexo en cuanto a la integridad de las articula-ciones y la fuerza de los ligamentos, tendones yhuesos. Desgraciadamente, la literatura de inves-tigación contribuye poco a confirmar o desmentirla validez de tales preocupaciones. Donde se hanobservado diferencias en el índice de lesiones, esposible que dichas lesiones estén más relaciona-das con el nivel de acondicionamiento que con elsexo del participante. Quienes están menos prepa-rados físicamente son más propensos a lesionarse.Ésta es un área extremadamente difícil para obte-ner datos objetivos, pero es un tema importanteque es preciso definir mejor.
FuerzaHasta los años 1970, la programación de entrena-miento de fuerza para niñas y mujeres no se con-sideraba apropiada. A las mujeres no se las creíacapaces de ganar fuerza debido a sus niveles ex-tremadamente bajos de hormonas anabólicasmasculinas. Paradójicamente, muchas personastemían también en general que el entrenamientode fuerza masculinizaría a las mujeres. Sin em-bargo, durante los años 1960 y 1970 se hizo evi-dente que muchas de las mejores deportistas de
DIFERENCIAS SEXUALES EN EL DEPORTE Y EL EJERCICIO
621
probablemente debido a su menor conte-nido de hemoglobina, por lo que seaporta menos oxígeno a los músculosactivos por unidad de sangre.
i Las diferencias en las respuestas respira-torias entre los hombres y las mujeres sedeben principalmente a diferencias en eltamaño corporal.
i Pasada la pubertad, el nVO2 máx de lamujer media es de sólo entre el 70% y el75% del que presenta el hombre medio.No obstante, una gran parte de esta dife-rencia puede deberse a los estilos de vi-da menos activos de las mujeres. Lasinvestigaciones con deportistas muy en-trenados revelan que una gran parte dela diferencia se debe a la mayor masagrasa de las mujeres.
i En el umbral anaeróbico se halla poca oninguna diferencia entre los sexos.
03 Cap 16-21 • 549-776 17/1/07 16:38 Página 621

FISIOLOGÍA DEL ESFUERZO Y DEL DEPORTE
XII
Al final del capítulo, se relacionan las expre-siones clave para verificar la comprensión denuestro vocabulario. Las cuestiones para estudiarpermiten examinar nuestros conocimientos delcontenido del capítulo. También aparecen re-ferencias numeradas a lo largo de todo el texto, aligual que varias lecturas seleccionadas, que pro-porcionan información adicional sobre cualquiertema de especial interés dentro del capítulo. Porúltimo, al final del libro hallaremos un glosariocompleto que incluye definiciones de todas lasexpresiones clave, un índice minucioso y una ta-bla de conversiones y de equivalentes métricospara una fácil referencia.
Muchos lectores leerán este libro únicamen-te porque es un texto obligatorio para un cursoque se les exige. Pero tenemos la esperanza deque la información les seduzca tanto como paracontinuar estudiando esta área relativamente nue-va y emocionante. Pretendemos al menos incre-mentar el interés y la comprensión del lector so-bre las maravillosas capacidades de su cuerpopara llevar a cabo esfuerzos físicos, para adaptar-se a situaciones de estrés y para mejorar sus ca-pacidades fisiológicas. Lo que el lector aprendaaquí será práctico no sólo para todo aquel que si-ga una carrera en la ciencia del ejercicio o del de-porte, sino también para cualquiera que quiera seractivo, sano y que desee estar en buena forma.
Expresiones clave
conducción saltatoriadespolarizaciónengramahiperpolarizaciónhuso muscularimpulso nerviosointegración sensomotoraneurotransmisorórgano tendinoso de Golgipotencial de acciónpotencial de membrana en reposopotencial graduadopotencial postsináptico excitatoriopotencial postsináptico inhibitorioprincipio de la movilización ordinariareflejo motorsinapsissumaciónterminal del axónumbralunión neuromuscularvaina de mielina
Cuestiones para estudiar
1. Nombrar las diferentes regiones de una neurona.
2. Explicar el potencial de membrana en reposo. ¿Qué loproduce? ¿Cómo se mantiene?
3. Describir un potencial de acción. ¿Qué se precisa an-tes de que se dispare un potencial de acción? Unavez se ha disparado, ¿cuál es la secuencia de aconte-cimientos?
4. Explicar cómo se transmite un impulso eléctrico des-de una neurona presináptica hasta una neurona post-sináptica. Describir una sinapsis y una unión neuro-muscular.
5. ¿Cómo se genera un potencial de acción en una neu-rona postsináptica?
6. ¿Cuáles son las divisiones principales del sistemanervioso? ¿Cuáles son sus funciones más importan-tes?
7. ¿Qué centros del cerebro tienen funciones importan-
tes en el control del movimiento, y cuáles son estasfunciones?
8. ¿En qué se diferencian los sistemas simpático y para-simpático? ¿Cuál es su trascendencia para llevar acabo actividades físicas?
9. Explicar cómo se producen los movimientos en res-puesta al contacto con un objeto caliente.
10. Describir la función de los husos musculares en elcontrol de la acción muscular.
11. Describir la función del órgano tendinoso de Golgien el control de la acción muscular.
12. ¿Qué es una unidad motora, y cómo se movilizan lasunidades motoras?
Bibliografía
1. Edstrom, L., & Grimby, L. (1986). Effect of exer-cise on the motor unit. Muscle and Nerve, 9, 104-126.
2. Guyton, A.C., & Hall, J.E. (1996). Textbook ofmedical physiology (9.ª ed.). Filadelfia Saunders.
3. Marieb, E.N. (1995). Human anatomy and phy-siology (3.ª ed., p. 387) Nueva York: Benjamin/Cum-mings.
4. Petajan, J.H., Gappmaier, E., White, A.T., Spen-cer, M.K., Mino, L., & Hicks, R.W. (1996). Impact of ae-robic training on fitness and quality of life in multiplesclerosis. Annals of Neurology, 39, 432-441.
5. Pette, D., & Vrbova, G. (1985). Neural control ofphenotypic expression in mammalian muscle fibers.Muscle and Nerve, 8, 676-689.
Lecturas seleccionadas
Binder, M.D., Heckman, C.J., & Powers, R.K. (1996).Thephysiological control of motoneuron activity. EnL.B. Rowell & J..T. Shepherd (Fds.), Handbook ofphysiology: Section 12. Exercise: Regulation and in-tegration of multiple systems (págs. 3-53). NuevaYork: Oxford University Press.
Christensen, N.J., & Galbo, H. (1983). Sympathetic ner-vous activity during exercise. Annual Review ofPhysiology, 45, 139-153.
FISIOLOGÍA DEL ESFUERZO Y DEL DEPORTE
Bibliografíasobre el materialusado en elcapítulo
Lecturasadicionalesseleccionadaspara ampliarconocimientos
Preguntas paraprobar tus
conocimientos
Lista detérminos clave
empleados en elcapítulo
01 Crédits+Cap 01-06 • 001-220 17/1/07 16:17 Página XII

bilidad pueden limitar el rendimiento. Tambiénhemos aprendido que nuestras necesidades meta-bólicas varían considerablemente. En el capítulosiguiente, volveremos nuestra atención hacia laregulación del metabolismo al concentrarnos en elsistema endocrino y en la regulación hormonal.
Expresiones clave
acetilcoenzima A (acetil-CoA)ácidos grasos libresadenosindifosfato (ADP)adenosintrifosfatasa (ATP-asa)adenosintrifosfato (ATP)agotamientoanaeróbicobetaoxidación (oxidación ß)cadena de transporte de electronescaloría (cal)calorimetría directacalorimetría indirectacalorímetrocapacidad oxidativa (QO2)ciclo de Krebsconsumo máximo de oxígeno ( nVO2 máx)exceso de consumo de oxígeno posejerciciofatigafosfocreatina (PC)glucógenoglucólisisglucólisis anaeróbicahidratos de carbonoíndice metabólico en reposo (IMR)inicio de la acumulación de lactato en sangrekilocaloría (kcal)lactatometabolismo aeróbicometabolismo anaeróbicorelación de intercambio respiratorio (R)ritmo metabólico basal (RMB)sistema ATP-PCsistema glucolíticosistema oxidativo
transformación de Haldaneumbral del lactato (UL)
Cuestiones para estudiar
1. ¿Cuál es la función de la PC?
2. Describir la relación existente entre el ATP yla PC durante la realización de un esprín.
3. ¿Por qué los sistemas energéticos ATP-PC yglucolítico se consideran anaeróbicos?
4. ¿Qué función desempeña el oxígeno en elproceso del metabolismo aeróbico?
5. Describir los productos de desecho de la pro-ducción de energía a partir del ATP-PC, de laglucólisis y de la oxidación.
6. ¿Qué es la relación de intercambio respirato-rio (R)? Explicar cómo se usa para determi-nar la oxidación de los hidratos de carbono yde las grasas.
7. ¿Cuál es la relación entre el consumo de oxí-geno y la producción de energía?
8. ¿Qué es el umbral del lactato?
9. ¿Cómo podemos usar las mediciones de con-sumo de oxígeno para estimar nuestra efica-cia al hacer ejercicio?
10. ¿Por qué los deportistas con valores de nVO2
max elevados rinden mejor en las pruebas deresistencia que quienes tienen valores másbajos?
11. ¿Por qué se expresa frecuentemente el consu-mo de oxígeno como milímetros de oxígenopor kilogramo de peso corporal por minuto(ml·kg-1·min-1)?
12. Describir las posibles causas de la fatiga du-rante la realización de series de ejercicio queduran entre 15 y 30 s , y entre 2 y 4 h.
Bibliografía
1. Åstrand, P.-O., & Rodahl, K. (1986). Text-book of work physiology: Physiological bases ofexercise (3.ª ed.). Nueva York: McGraw-Hill.
FISIOLOGÍA DEL ESFUERZO Y DEL DEPORTE
166
01 Crédits+Cap 01-06 • 001-220 17/1/07 16:21 Página 166

en el último cuarto de juego han ignorado elcomponente de resistencia en sus programas deentrenamiento. Puede decirse lo mismo de lospracticantes de la mayoría de deportes. Pero an-tes de ver específicamente cómo la resistenciapuede mejorar el rendimiento, debemos entenderqué es dicha capacidad de resistencia.
Resistencia: muscular y cardiorrespiratoriaResistencia es un término que describe dos con-ceptos separados pero relacionados: la resistenciamuscular y la resistencia cardiorrespiratoria. Cadauna de ellas contribuye de una manera especial alrendimiento deportivo, por lo que cada una difiereen importancia para los diferentes deportistas.
Para los corredores de distancias cortas, laresistencia es la cualidad que les permite sosteneruna elevada velocidad a lo largo de toda la dis-tancia de, por ejemplo, las carreras de 100 o de200 m. Esta cualidad es la resistencia muscular:la capacidad de un músculo o de un grupo mus-cular para sostener ejercicios de alta intensidad,repetitivos o estáticos. Este tipo de resistencia es-tá representado también por el halterófilo, el bo-xeador y el luchador. El ejercicio o la actividadpueden ser de naturaleza rítmica o repetitiva, comoel press de banca para el levantador de pesos ygolpear rápidamente para el boxeador. O la acti-vidad puede ser más estática, como, por ejemplo,una acción muscular sostenida cuando un lucha-dor intenta inmovilizar a un oponente contra eltapiz. En cualquier caso, la fatiga resultante estáreferida a un grupo muscular específico y la du-ración de la actividad no suele ser superior a 1 ó2 min. Nuestra resistencia tiene una fuerte rela-ción con nuestra fuerza muscular y nuestro desa-rrollo anaeróbico.
Mientras que la resistencia muscular hace re-ferencia a la capacidad de músculos individuales,la resistencia cardiorrespiratoria guarda rela-ción con el cuerpo como un todo. Específicamen-te se trata de la capacidad del cuerpo para soste-ner ejercicios prolongados rítmicos. Este tipo deresistencia es típico del ciclista, el corredor defondo o el nadador de fondo que pueden comple-tar grandes distancias a un ritmo muy rápido.Nuestra resistencia cardiorrespiratoria está muyrelacionada con el desarrollo de nuestros siste-mas cardiovascular y respiratorio, y, por lo tanto,con nuestro desarrollo aeróbico.
En el capítulo 6 hemos analizado las adapta-ciones al entrenamiento de la resistencia muscular.En este capítulo nos concentraremos en la resis-tencia cardiorrespiratoria. Ahora que entendemoseste concepto, estamos preparados para examinarsus bases fisiológicas. Centraremos nuestra aten-ción principalmente en el entrenamiento de la re-sistencia, pero ocasionalmente haremos referenciaal entrenamiento explosivo y al de resistencia es-pecíficamente por su nombre.
Evaluación de la resistenciacardiorrespiratoriaPara estudiar los efectos del entrenamiento sobrela resistencia, necesitamos un medio que poda-mos usar fácilmente para controlar su mejora du-rante el programa de entrenamiento.
nVO2 máx: potencia aeróbicaLa mayoría de los científicos del deporte consi-deran el nVO2 máx como representante de la po-tencia aeróbica, como la mejor manera de mediren laboratorio la resistencia cardiorrespiratoria.Recordemos del capítulo 4 que el nVO2 máx sedefine como el ritmo más alto de consumo deoxígeno alcanzable durante la realización de ejer-cicios máximos o agotadores. Si incrementamosla intensidad de nuestro ejercicio más allá delpunto en que se alcanza el nVO2 máx, nuestro con-sumo de oxígeno se estabilizará o se reducirá li-geramente, incluso con mayores aumentos de laintensidad, lo cual manifiesta que se ha alcanzadola nVO2 máx.
Con el entrenamiento de la resistencia, pue-de suministrarse y consumirse más oxígeno queen un estado no entrenado, como se expuso en elcapítulo 6. Sujetos previamente no entrenadosmuestran incrementos medios del nVO2 máx del15%-20% después de un programa de entrena-miento de 6 meses. Estas mejoras nos permitenejecutar actividades de resistencia con un nivel
ADAPTACIONES CARDIOVASCULARES Y RESPIRATORIAS AL ENTRENAMIENTO
297
La resistencia cardiorrespiratoria es lacapacidad del cuerpo para sostener ejer-cicios prolongados.
02 Cap 07-15 • 221-548 17/1/07 16:29 Página 297

minales del axón o fibrillas terminales. Laspuntas de estos terminales se dilatan en diminu-tos bulbos conocidos como protuberancias sinápti-cas. Estas protuberancias albergan numerosasvesículas (sacos) llenas de sustancias químicas,conocidas como neurotransmisores, que se usanpara la comunicación entre una neurona y otracélula. (Esto se tratará más tarde con mayor de-talle.) La estructura de la neurona permite a losimpulsos nerviosos entrar en la neurona por lasdendritas y, en menor medida, a través del so-ma, y luego viajar por el soma y la eminencia
FISIOLOGÍA DEL ESFUERZO Y DEL DEPORTE
66
Figura 2.2 (a) Microfotografía de una neurona y (b) su estructura.
Núcleo
Nucléolo
Cuerposde Nissl
Eminencia axónica
Nódulo deRanvier
Neurilema
Impulso
Vaina demielina
AxónNódulos
de RanvierVaina demielina
Terminaciónaxónica
Corpúsculo terminal
Soma
Dendritas
b
a
01 Crédits+Cap 01-06 • 001-220 17/1/07 16:19 Página 66

Visión general del capítulo
No podemos vivir sin oxígeno. Nuestras célulasdependen de él para sobrevivir, y, tal como hemosaprendido en el capítulo 4, el oxígeno es esencial pa-ra la producción de la energía que alimenta todaslas actividades de nuestro cuerpo. La capacidadde resistencia depende del aporte de cantidadessuficientes de oxígeno a nuestros músculos y deun consumo celular adecuado de este gas una vezque llega allí. Pero al mismo tiempo, los procesosmetabólicos que tienen lugar en nuestros múscu-los activos generan otro gas, el dióxido de carbo-no, que, a diferencia del oxígeno, es tóxico. Laactividad celular normal requiere oxígeno, perose ve dificultada cuando los niveles de dióxido decarbono aumentan.
Las necesidades cruciales de nuestros músculospara un aporte adecuado de oxígeno y respecto a unaadecuada eliminación del dióxido de carbono sonsatisfechas por el aparato respiratorio. Tal como he-mos visto en el capítulo anterior, el sistema cardio-vascular transporta estos gases. Pero el aparato res-piratorio lleva oxígeno a nuestros cuerpos y noslibera del exceso de dióxido de carbono. En este ca-pítulo nos centraremos en este sistema. Comenzare-mos con una visión general de las fases que intervie-nen en la respiración y en el intercambio de gases, yluego examinaremos cómo se regulan estos proce-sos. Consideraremos cómo funciona el aparato res-piratorio cuando estamos haciendo ejercicio y dequé manera puede limitar el rendimiento. Por últi-mo, analizaremos la especial función del aparatorespiratorio en el mantenimiento del equilibrio aci-dobásico en nuestro cuerpo y la trascendencia de es-te equilibrio durante la actividad física.
Esquema del capítulo
Ventilación pulmonar 264Inspiración 265Espiración 266
Difusión pulmonar 267Membrana respiratoria 267Presiones parciales de los gases 267Intercambio de gases en los alvéolos 268
Transporte de oxígeno y de dióxido de carbono 271Transporte de oxígeno 271Transporte de dióxido de carbono 273
Intercambio de gases en los músculos 274Diferencia arteriovenosa de oxígeno 274Factores que influyen en el transportey consumo de oxígeno 275Eliminación del dióxido de carbono 276
Regulación de la ventilación pulmonar 277Mecanismos de regulación 277Ventilación pulmonar duranteel ejercicio 279Irregularidades respiratorias durante el ejercicio 280
Ventilación y metabolismo energético 282Equivalente ventilatorio para el oxígeno 282Punto de máxima tensión ventilatoria tolerable 283Umbral anaeróbico 283
Limitaciones respiratorias al rendimiento 285Regulación respiratoria del equilibrio
acidobásico 286Conclusión 290
REGULACIÓN RESPIRATORIA DURANTE EL EJERCICIO
263
02 Cap 07-15 • 221-548 17/1/07 16:29 Página 263

Optimizacióndel
rendimientodeportivo
P A R T E
V�
�
�
�
Ahora sabemos la forma en que el cuerpo responde a una se-sión de ejercicio intenso, sabemos cómo se adapta al en-trenamiento crónico y cómo se ajusta a las condicionesmedioambientales extremas. Ahora podemos aplicar esosconocimientos para mejorar el rendimiento deportivo. En laquinta parte, nos centramos en la preparación óptima de losdeportistas para la competición. En el capítulo 12, Entrena-miento para el deporte, exponemos los efectos del cambiodel volumen del entrenamiento y cómo demasiado o pocoentrenamiento empeoran el rendimiento. En el capítulo 13,Nutrición y deporte, evaluamos las necesidades dietéticasde los deportistas y analizamos las propuestas que se han he-cho sobre la suplementación nutricional y la manipulaciónde la dieta para mejorar el rendimiento. En el capítulo 14,Peso corporal, composición corporal y deporte, aborda-mos los temas de la evaluación de la composición corporal,la relación entre la composición corporal y el rendimientodeportivo, y el empleo de baremos de peso. También deter-minamos los medios más eficaces para perder con éxito elexceso de grasa corporal pero manteniendo la masa magra,lo cual permite al deportista obtener un peso competitivoóptimo y, probablemente, mejorar su rendimiento. En el ca-pítulo 15, Ayudas ergogénicas y rendimiento, estudiamosdistintos agentes farmacológicos, hormonales y fisiológicosque se han propuesto para mejorar el rendimiento. Examina-mos los beneficios potenciales, los efectos demostrados y losriesgos para la salud asociados con su consumo.
405
02 Cap 07-15 • 221-548 17/1/07 16:30 Página 405

Visión general del capítulo
El intenso esfuerzo y el consumo energético delentrenamiento y de la competición deportivosimponen exigencias inusuales sobre la dieta delos deportistas. De hecho, algunos de ellos, talescomo nadadores y fondistas, pueden encontrarsecon problemas para equilibrar su consumo ener-gético con las exigencias calóricas del entrena-miento. Por estas razones, la principal preocupa-ción dietética de muchos deportistas es la cantidadde comida que consumen, más que lo que comen.Pero, en su búsqueda del éxito, la mayoría de losdeportistas han buscado en algún momento unalimento mágico que les diese un rendimientotriunfal. Tal como vimos en el capítulo preceden-te, todas las sustancias que tienen estas propie-dades de mejora del rendimiento se llaman ergo-génicas. Desgraciadamente, las manipulacionesdietéticas se basan en el testimonio facilitado pordeportistas que tienen más éxito, en estudios deinvestigación mal diseñados, en afirmaciones noválidas de la propaganda comercial y en erroresde interpretación de la investigación nutricional.Pocas áreas de la ciencia del ejercicio están máscargadas de manías pasajeras que el campo de lanutrición deportiva. Con demasiada frecuencia,esto conlleva el empleo de prácticas nutricionalessin fundamento.
En este capítulo, examinaremos las sustan-cias que ingerimos y su importancia más allá desu función bioenergética. Vamos a centrarnos enla composición óptima de la dieta y en las espe-ciales necesidades dietéticas del deportista. Yexaminaremos cuidadosamente cómo la nutri-ción puede afectar al rendimiento, centrándonosen las potenciales propiedades ergogénicas de losdiversos nutrientes y disipando muchos mitos.
Esquema del capítulo
Las seis clases de nutrientes 440Hidratos de carbono 441Grasas 446Proteínas 449Vitaminas 451Minerales 454Agua 457
Equilibrio del agua y de los electrólitos 459Equilibrio del agua en reposo 459Equilibrio del agua durante el ejercicio 459Deshidratación y rendimiento durante el ejercicio 460Equilibrio de electrólitos durante el ejercicio 462Reemplazo de las pérdidas de fluidos corporales 464
Dieta del deportista 466Dieta vegetariana 467Comida previa a la competición 467Carga y sustitución del glucógeno muscular 468
Diseño de bebidas deportivas 470Tipos de hidratos de carbono 471Concentración de hidratos de carbono 472Rehidratación con bebidas deportivas 473Lo más adecuado 473
Conclusión 474
NUTRICIÓN Y DEPORTE
439
02 Cap 07-15 • 221-548 17/1/07 16:31 Página 439

Bibliografía
1. Bar-Or, O. (1987). The Wingate anae-robic test: An update on methodology, reliabilityand validity. Sports Medicine, 4, 381-394.
2. Bergman, B.C., Wolfel, E.E., Butter-field, G.E., Lopaschuk, G.D., Casazza, G.A.,Horning, M.A., & Brooks, G.A. (1999). Activemuscle and whole body lactate kinetics after en-durance training in men. Journal of Applied Phy-siology, 87, 1.684-1.696.
3. Bouchard, C., An, P., Rice, T., Skin-ner, J.S., Wilmore, J.H., Gagnon, J., Pérusse,L., Leon, A.S., & Rao, D.C. (1999). Familialaggregation of nVO2max response to exercisetraining: Results from the HERITAGE FamilyStudy. Journal of Applied Physiology, 87,1.003-1.008.
4. Brooks, G.A., & Mercier, J. (1994). Ba-lance of carbohydrate and lipid utilization duringexercise: The “crossover” concept. Journal ofApplied Physiology, 76, 2.253-2.261.
5. Costill, D.L. (1985). Practical problems inexercise physiology research. Research Quar-terly, 56, 379-384.
6. Costill, D.L. (1986). Inside running: Ba-sics of sports physiology. Indianápolis: Bench-mark Press.
7. Costill, D.L., Coyle, E., Dalsky, G.,Evans, W., Fink, W.W., & Hoopes, D. (1977). Ef-fects of elevated plasma FFA and insulin on mus-cle glycogen usage during exercise. Journal ofApplied Physiology, 43, 695-699.
8. Costill, D.L., Coyle, E.F., Fink, W.F., Les-mes, G.R., & Witzmann, F.A. (1979). Adapta-tions in skeletal muscle following strength trai-ning. Journal of Applied Physiology: RespiratoryEnvironmental Exercise Physiology, 46, 96-99.
9. Costill, D.L., Fink, W.J., Ivy, J.L., Get-chell, L.H., & Witzmann, F.A. (1979). Lipid me-tabolism in skeletal muscle of endurance-trainedmales and females. Journal of Applied Physio-logy, 28, 251-255.
10. Costill, D.L., Thomas, R., Robergs,R.A., Pascoe, D.D., Lambert, C.P., Barr, S.I., &Fink, W.J. (1991). Adaptations to swimming trai-ning: Influence of training volume. Medicine andScience in Sports and Exercise, 23, 371-377.
11. Essen, B., Hagenfeldt, L., & Kaijser, L.(1977). Utilization of blood-borne and intramus-cular substrates during continuous and intermit-tent exercise in man. Journal of Physiology, 265,480-506.
12. Farrell, P.A., Wilmore, J.H., Coyle, E.F.,Billing, J.E., & Costill, D.L. (1979). Plasma lac-tate accumulation and distance running perfor-mance. Medicine and Science in Sports andExercise, 11, 338-344.
13. Gollnick, P.D., Armstrong, R.B., Sau-bert, C.W., IV, Piehl, K., & Saltin, B. (1972).Enzyme activity and fiber composition in skele-tal muscle of untrained and trained men. Journalof Applied Physiology, 33, 312-319.
14. Green, H.J., Jones, S., Ball-Burnett, M.,Farrance, B., & Ranney, D. (1995). Adaptationsin muscle metabolism to prolonged voluntaryexercise and training. Journal of Applied Physio-logy, 78, 138-145.
15. Greiwe, J.S., Hickner, R.C., Hansen, P.A.,Racette, S.B., Chen, M.M., & Holloszy, J.O.(1999). Effects of endurance exercise training onmuscle glycogen accumulation in humans. Journalof Applied Physiology, 87, 222-226.
16. Harmer, A.R., McKenna, M.J., Sutton,J.R., Snow, R.J., Ruell, P.A., Booth, J., Thompson,M.W., MacKay, N.A., Stathis, C.G., Crameri,R.M., Carey, M.F., & Eager, D.M. (2000). Skeletalmuscle metabolic and ionic adaptations during in-tense exercise following sprint training in humans.Journal of Applied Physiology, 89, 1.793-1.803.
17. Hermansen, L., & Wachtlova, M. (1971).Capillary density of skeletal muscle in well-trai-ned and untrained men. Journal of Applied Phy-siology, 30, 860-863.
18. Holloszy, J.O., & Coyle, E.F. (1984).Adaptations of skeletal muscle to enduranceexercise and their metabolic consequences. Jour-nal of Applied Physiology, 56, 831-838.
19. Holloszy, J.O., Oscai, L.B., Mole, P.A.,& Don, I.J. (1971). Biochemical adaptations toendurance exercise in skeletal muscle. En B.Pernow & B. Saltin (eds.), Muscle metabolismduring exercise (51-56). Nueva York: PlenumPress.
20. Ingjer, F. (1979). Capillary supply andmitochondrial content of different skeletal mus-cle fiber types in untrained and endurance trained
FISIOLOGÍA DEL ESFUERZO Y DEL DEPORTE
218
01 Crédits+Cap 01-06 • 001-220 17/1/07 16:22 Página 218

Estándares de pesoTanto los entrenadores, como los deportistas,siempre están buscando aquello que les propor-cione la ventaja, por pequeña que sea, que lespuede dar la victoria. Una vez que un entrenadoro un deportista encuentra algo que funciona yque mejora el rendimiento, la noticia se difundecon rapidez. El amplio uso de los esteroides ana-bólicos es un ejemplo clásico. Lo que se inició afinales de la década de 1940 y principios de la de1950 como experimentación entre un pequeñonúmero de culturistas y de levantadores de pesose ha extendido en todo el mundo del deporte, enel que la mayoría de los deportistas de elite deciertos deportes son consumidores habituales deesteroides. Un fenómeno similar ha acompañadoel interés de los deportistas en el empeño por es-tar delgados.
El deportista de elite ha intentado desde hacetiempo representar las características físicas y fi-siológicas más deseables para el rendimiento enun deporte o actividad. Teóricamente, la base ge-nética del deportista de elite y los años de intensoentrenamiento se han combinado para proporcio-nar el perfil deportivo final para este deporte. Es-tos deportistas de elite crean las normas o mode-los que otros aspiran a alcanzar.
En la década de 1970, nosotros (JHW) tuvi-mos la oportunidad de someter a pruebas a mu-chas de las deportistas de elite de campo y pista
de EE.UU.35 Además de las pruebas en cinta er-gométrica, a todos los deportistas se les evaluó lacomposición corporal. En la figura 14.7 se ofre-cen los resultados de estas evaluaciones. Obser-vando solamente a las corredoras de fondo, mu-chas de ellas estaban por debajo del 12% de grasacorporal. Las dos mejores corredoras de fondo te-nían solamente un 6% de grasa corporal. Una deellas había ganado seis campeonatos interna-cionales consecutivos de cros y la otra poseía loque entonces era el mejor tiempo del mundo enmaratón. Partiendo de estos resultados, podíamostener la tentación de sugerir que cualquier corre-dora de fondo, niña o mujer, debería tener entreun 6% y un 12% de porcentaje de grasa corporalsi aspiraba a llegar a ser una corredora de catego-ría mundial. No obstante, una de las mejores co-rredoras de fondo de EE.UU. de aquel entonces,que había llegado a la cima hacía menos de 2años, tenía un porcentaje de grasa corporal del17%. Además, una de las mujeres de este estudiotenía un porcentaje de grasa corporal del 37% yestableció el mejor tiempo del mundo para la ca-rrera de las 50 millas (74,5 km) antes de transcu-rridos 6 meses de su evaluación. Es más que pro-bable que ninguna de estas mujeres hubieseobtenido alguna ventaja si se le hubiese forzado areducir su peso para tener una grasa corporal del12% o inferior.
PESO CORPORAL, COMPOSICIÓN CORPORAL Y DEPORTE
493
quieren fuerza, potencia y resistenciamusculares, pero puede ser un inconve-niente para los que practican deportesque requieren capacidad de resistencia,porque estos deportistas han de podermover su masa corporal total duranteextensos períodos de tiempo.
i El grado de adiposidad tiene más in-fluencia en el rendimiento que el pesocorporal total. En general, cuanto mayores la grasa corporal total, peor es el ren-dimiento. Entre las posibles excepcionesa esta premisa figuran los levantadoresde grandes pesos, los luchadores de su-mo y los nadadores.
Figura 14.7 Porcentaje de grasa corporal en mujeres deportistas depista y campo. Datos de Wilmore y col. (1977)35.
Gra
sa r
elat
iva
(%)
Edad (años)
6% de grasalas mejoresparticipantes
37% de grasa
DistanciaDistancia media en esprínDisco-jabalinaLanzamiento de peso
02 Cap 07-15 • 221-548 17/1/07 16:31 Página 493

constriñan este vaso de modo que pueda pasarmenos sangre a través de él.
En condiciones normales, los nervios simpáti-cos transmiten impulsos continuamente a los vasossanguíneos, lo que mantiene los vasos en un estadode constricción moderada para mantener una ten-sión arterial adecuada. Este estado de constricciónparcial recibe la denominación de tono vasomotor.Cuando la estimulación simpática aumenta, unamayor constricción de los vasos sanguíneos en unárea específica reduce el flujo de sangre hacia es-ta área y permite que vaya más sangre hacia otraspartes. Pero si la estimulación simpática disminuyepor debajo de lo necesario para mantener el tono, laconstricción de los vasos en esta área se reduce, porlo que los vasos se dilatan e incrementan el flujo desangre hacia esta área. En consecuencia, la estimu-lación simpática ocasionará vasoconstricción en lamayoría de los vasos, pero el flujo de sangre se vealterado por el incremento o la reducción de la in-tensidad de la vasoconstricción relativa al tono va-somotor normal.
El sistema simpático también puede producirdirectamente vasodilatación mediante algunas desus fibras. Un tipo diferente de fibra simpáticaabastece algunos vasos sanguíneos en los múscu-los esqueléticos y en el corazón. La estimulaciónde estas fibras produce vasodilatación e incre-menta el flujo de la sangre hacia los músculos yel corazón. Este sistema funciona durante la clá-sica respuesta de «lucha o huida» e incrementa elflujo de sangre hacia los músculos esqueléticos yhacia el corazón en momentos de crisis. Esta res-puesta también es activa durante el ejercicio,cuando los músculos esqueléticos y el corazónestán trabajando más duro, necesitando muchamás sangre que cuando están en reposo.
del cuerpo a otra. Pero la distribución de la san-gre por el cuerpo varía no solamente por los teji-dos que están siendo abastecidos, sino tambiénpor la localización de la sangre en el sistema vas-cular. En reposo, el volumen sanguíneo se distri-buye por el sistema vascular tal como se ve en lafigura 7.8. La mayor parte de la sangre se locali-za en los canales venosos de retorno (venas, vé-nulas, senos venosos). Por lo tanto, el sistema ve-noso proporciona una gran reserva de sangrefácilmente disponible para satisfacer las necesi-dades aumentadas. Cuando esta necesidad se pre-senta, la estimulación simpática de las vénulas yde las venas constriñe estos vasos. Esto ocasionauna rápida redistribución de la sangre desde lacirculación venosa periférica devolviéndola alcorazón, y luego hacia aquellas áreas que tienenmayores necesidades. No solamente se desvía san-gre desde otros tejidos, sino que se envía mássangre a la circulación arterial desde el sistemavenoso, asegurando con ello un incremento sus-tancial del flujo de sangre a un área necesitada.
CONTROL CARDIOVASCULAR DURANTE EL EJERCICIO
237
El riego sanguíneo se puede controlar anivel local del tejido mediante autorre-gulación intrínseca y control neural ex-trínseco. El sistema nervioso simpáticodesempeña un papel importante en la re-conducción del riego sanguíneo de áreaspoco necesitadas a otras muy necesita-das.
Redistribución de la sangre venosa
Ahora hemos discutido los mecanismos que con-trolan la redistribución de la sangre desde un área
Figura 7.8 Distribución de la sangre por el sistema vascular cuandoel cuerpo se halla en reposo.
9% en lospulmones
13%en las
arterias
PulmónPulmón
Cerebro
7% en elcorazón64%
en las venas
7% en las arteriolas y en los capilares
02 Cap 07-15 • 221-548 17/1/07 16:28 Página 237

Problemas sanitarios relacionadoscon el exceso de peso y la obesidad El sobrepeso y la obesidad se asocian con un au-mento de la mortalidad.6 Esta relación es curvilí-nea, como se aprecia en la figura 21.7. Se produ-ce un gran incremento del riesgo cuando el IMCsupera los 30 kg/m2, aunque valores del IMC en-tre 25 y 30 se asocian con un aumento del riesgode muchas enfermedades. La mortalidad excesi-va asociada con la obesidad y el sobrepeso se re-laciona con las siguientes enfermedades:
• Cardiopatías.
• Hipertensión.
• Diabetes.
• Ciertos tipos de cáncer.
Con el gran aumento de la prevalencia de laobesidad en EE.UU. durante los últimos 40 años,no sorprende observar también una prevalenciamuy elevada del síndrome metabólico (ver capítulo20) en los adultos estadounidenses. Los datos de laencuesta del tercer National Health and NutritionExamination Survey (NHANES-3), realizado entre1988 y 1994, demostraron que el 23,7% de losadultos en EE.UU. cumplen los criterios del sín-drome metabólico. En adultos de más de 60 años,la prevalencia fue superior al 40%. De acuerdocon las tendencias en la obesidad, la prevalencia
FISIOLOGÍA DEL ESFUERZO Y DEL DEPORTE
722
Una predisposición genética a la obesidad: los indios Pima
Ha quedado establecido con claridad que la genética es un factor importante en el desarrollo de laobesidad. El doctor Claude Bouchard, de la Laval University de Quebec, ha realizado muchos estu-dios sobre la herencia de la obesidad, y ha llegado a la conclusión de que la herencia de la masa gra-sa o la grasa corporal relativa (porcentaje de grasa) es en torno al 25% de la varianza ajustada a laedad y el sexo.4 ¿Tener predisposición genética a la obesidad significa que estés destinado a ser obe-so? ¡La respuesta es no! Hemos aprendido mucho del estudio de los indios Pima, que han vivido du-rante al menos 2.000 años cerca del río Gila en el desierto de Sonora en lo que hoy es el sur de Ari-zona. Hasta el paso al siglo XX, los indios Pima eran aparentemente un pueblo esbelto y sanofísicamente activo que comía una dieta sana. Al pasar a vivir en las reservas, dejaron de cultivar, yempezaron a seguir una dieta occidental rica en grasas y a consumir alcohol, se volvieron obesos,con una prevalencia de obesidad de aproximadamente el 70%.19 Asociada con su obesidad tienenuna prevalencia alta de diabetes. Es una población tan poco usual respecto a su prevalencia extrema-damente alta de obesidad y diabetes que los National Institutes of Health han establecido un centroespecial de investigación en Phoenix, Arizona, para estudiar sus problemas de salud.
Es interesante que haya otro grupo de indios Pima que viven en el norte de México. Se hanmantenido activos trabajando en granjas sin usar maquinaria motorizada. Además, consumen una
dieta rica en hidratos de carbono y baja en gra-sas. Han conseguido mantenerse relativamentedelgados. El IMC de estos dos grupos de indiosse ve en la figura 21.6. La base de la cuestión esque se puede tener una predisposición a la obesi-dad, pero, con una dieta y ejercicio correctos, sepuede mantener un peso corporal relativamentenormal. Los indios Pima nos han enseñado unalección importante.
Figura 21.6 IMC de los indios Pima que viven en Arizona y nor-te de México.
Hombres
Mujeres
Índ
ice
de
mas
a co
rpo
ral
´
03 Cap 16-21 • 549-776 17/1/07 16:39 Página 722

una meseta o comienza a declinar. Veamos variosejemplos.
La mayoría de los estudios sobre el entrena-miento excesivo se han realizado con nadadores.Por este motivo, el material de las próximas dossecciones deriva de investigaciones practicadascon nadadores, si bien dicha información es apli-cable a casi todas las formas de entrenamiento.
Volumen de entrenamientoEl volumen del entrenamiento puede aumentarseincrementando su duración o la frecuencia de lassesiones de entrenamiento. Las investigacionesmuestran que entrenarse nadando 3 ó 4 horas aldía, 5 ó 6 días por semana, no proporciona mayo-res beneficios que cuando el entrenamiento se li-mita solamente a 1 hora ó 1 hora y media al día.7
De hecho, un entrenamiento excesivo como ésteha demostrado que reduce significativamente lafuerza muscular y los resultados en los esprinesde natación.
Pocos estudios han comparado la prepara-ción física y los beneficios para el rendimientode una sesión diaria de entrenamiento frente asesiones múltiples.6, 22, 33 Los estudios realizadoshasta la fecha no revelan evidencias científicasde que las sesiones diarias múltiples mejorenmás la forma física ni el rendimiento que unaúnica sesión diaria. Esto se comprueba con losdatos de la figura 12.3, donde aparecen las res-puestas de dos grupos de nadadores que entrena-ron una vez al día (grupo 1) o dos veces (grupo2) durante un período de 6 semanas dentro de unprograma de entrenamiento de 25 semanas.6 To-dos los nadadores comenzaron el programa si-guiendo el mismo régimen de entrenamiento:una vez al día. Pero durante las semanas 5 a 11,el grupo 2 aumentó su entrenamiento a dos vecesdiarias. Pasadas 6 semanas de distintos regíme-nes, ambos grupos reanudaron el programa deuna sesión diaria. La frecuencia cardíaca y losvalores de lactacidemia de todos los nadadoresdisminuyeron bruscamente con el inicio del en-trenamiento, pero no hubo diferencias significa-tivas en las respuestas de ambos grupos debidoal cambio del volumen de entrenamiento. Losnadadores que entrenaron dos veces al día nomostraron mejoras adicionales sobre los que en-trenaron sólo una vez diaria. De hecho, sus nive-les de lactato en sangre (figura 12.3a) y sus fre-cuencias cardíacas (figura 12.3b) fueron un pocomayores con el mismo ritmo fijo de natación,
aunque las diferencias no fueron estadísticamen-te significativas.
Para determinar la influencia del entrena-miento excesivo a largo plazo, se compararon lasmejoras en el rendimiento de los nadadores quese entrenaron dos veces al día para una distanciatotal de más de 10.000 m al día con las mejorasde quienes nadaron aproximadamente la mitad deesta distancia en una sola sesión diaria. En la fi-
ENTRENAMIENTO PARA EL DEPORTE
411
Figura 12.3 Cambios en (a) los niveles de lactato en sangre de na-dadores y (b) de la frecuencia cardíaca durante una prueba están-dar de 366 m durante 25 semanas de entrenamiento. Entre la se-manas quinta y decimoprimera, un grupo entrenó dos veces diarias(grupo 2) y el otro entrenó una vez al día (grupo 1). Aprécieseque las diferencias en los niveles de lactato en sangre y en la fre-cuencia cardíaca de los dos grupos no fueron significativas, lo cualsugiere que entrenar una vez diaria es igual de efectivo, si no máseficaz, que hacerlo dos veces al día.
Grupo 1Grupo 2
Lact
ato
en
sang
re (m
mo
l/l)
Semanas de entrenamiento
Semanas de entrenamiento
Frec
uenc
ia c
ardí
aca
dura
nte
el e
jerc
icio
(lat
idos
/min
)
02 Cap 07-15 • 221-548 17/1/07 16:30 Página 411
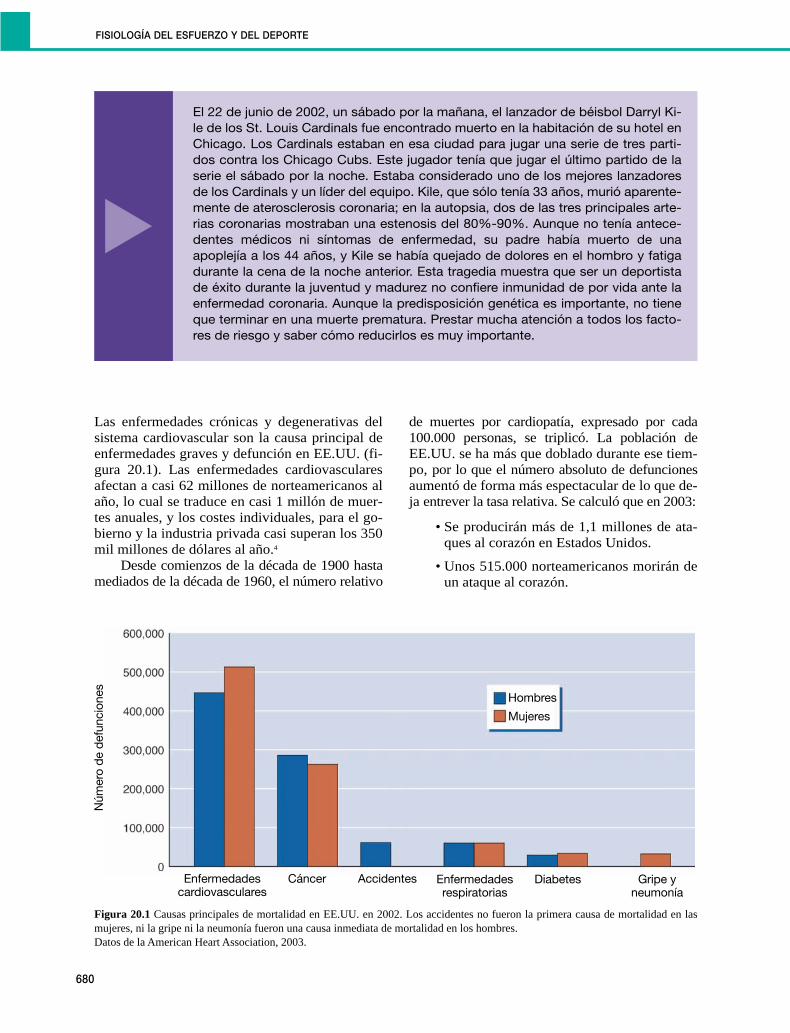
Las enfermedades crónicas y degenerativas delsistema cardiovascular son la causa principal deenfermedades graves y defunción en EE.UU. (fi-gura 20.1). Las enfermedades cardiovascularesafectan a casi 62 millones de norteamericanos alaño, lo cual se traduce en casi 1 millón de muer-tes anuales, y los costes individuales, para el go-bierno y la industria privada casi superan los 350mil millones de dólares al año.4
Desde comienzos de la década de 1900 hastamediados de la década de 1960, el número relativo
de muertes por cardiopatía, expresado por cada100.000 personas, se triplicó. La población deEE.UU. se ha más que doblado durante ese tiem-po, por lo que el número absoluto de defuncionesaumentó de forma más espectacular de lo que de-ja entrever la tasa relativa. Se calculó que en 2003:
• Se producirán más de 1,1 millones de ata-ques al corazón en Estados Unidos.
• Unos 515.000 norteamericanos morirán deun ataque al corazón.
FISIOLOGÍA DEL ESFUERZO Y DEL DEPORTE
680
El 22 de junio de 2002, un sábado por la mañana, el lanzador de béisbol Darryl Ki-le de los St. Louis Cardinals fue encontrado muerto en la habitación de su hotel enChicago. Los Cardinals estaban en esa ciudad para jugar una serie de tres parti-dos contra los Chicago Cubs. Este jugador tenía que jugar el último partido de laserie el sábado por la noche. Estaba considerado uno de los mejores lanzadoresde los Cardinals y un líder del equipo. Kile, que sólo tenía 33 años, murió aparente-mente de aterosclerosis coronaria; en la autopsia, dos de las tres principales arte-rias coronarias mostraban una estenosis del 80%-90%. Aunque no tenía antece-dentes médicos ni síntomas de enfermedad, su padre había muerto de unaapoplejía a los 44 años, y Kile se había quejado de dolores en el hombro y fatigadurante la cena de la noche anterior. Esta tragedia muestra que ser un deportistade éxito durante la juventud y madurez no confiere inmunidad de por vida ante laenfermedad coronaria. Aunque la predisposición genética es importante, no tieneque terminar en una muerte prematura. Prestar mucha atención a todos los facto-res de riesgo y saber cómo reducirlos es muy importante.
�
Figura 20.1 Causas principales de mortalidad en EE.UU. en 2002. Los accidentes no fueron la primera causa de mortalidad en lasmujeres, ni la gripe ni la neumonía fueron una causa inmediata de mortalidad en los hombres.Datos de la American Heart Association, 2003.
Núm
ero
de
def
unci
ones Hombres
Mujeres
Enfermedadescardiovasculares
Cáncer Accidentes Enfermedadesrespiratorias
Diabetes Gripe yneumonía
03 Cap 16-21 • 549-776 17/1/07 16:39 Página 680

término fatiga para describir las sensaciones ge-nerales de cansancio y las reducciones acompa-ñantes del rendimiento muscular.
La mayoría de los esfuerzos para describirlas causas y los puntos subyacentes de la fatiga secentran en:
• los sistemas energéticos (ATP-PC, glucóli-sis y oxidación),
• la acumulación de desechos metabólicos,
• el sistema nervioso y
• la insuficiencia del mecanismo contráctilde las fibras.
Ninguno de estos puntos puede explicar porsí solo todos los aspectos de la fatiga. Por ejem-plo, aunque la falta de energía disponible puedereducir la capacidad de los músculos para gene-rar fuerza, los sistemas energéticos no son total-mente responsables de todas las formas de fatiga.La sensación de cansancio que experimentamoscon frecuencia al final de una jornada laboral tie-ne poco que ver con la disponibilidad de ATP. Lafatiga puede ser también el resultado de la altera-ción de la homeostasis debida a la tensión am-biental. Quedan muchas preguntas por respondersobre la fatiga.
Sistemas energéticos y fatigaLos sistemas energéticos son un área que obvia-mente hay que explorar cuando se consideran lasposibles causas de la fatiga. Cuando nos sentimosfatigados, con frecuencia lo expresamos dicien-do: «No tengo energía». Pero esta utilización deltérmino energía se aleja mucho de su significadofisiológico. ¿Qué función desempeña la energía,en este sentido más estrecho, en la fatiga duranteel ejercicio?
Agotamiento de la fosfocreatina
Recordemos que la fosfocreatina (PC) se usa ba-jo condiciones anaeróbicas para reconstruir elATP altamente energético conforme se va usan-do, y de este modo mantener las reservas de ATPdel cuerpo. El estudio de biopsias musculares demuslos humanos ha demostrado que, durante laejecución de contracciones máximas repetidas,la fatiga coincide con el agotamiento de la PC.
METABOLISMO, ENERGÍA Y SISTEMAS BÁSICOS DE ENERGÍA
159
i Nuestro metabolismo aumenta con la in-tensidad incrementada del ejercicio, pe-ro nuestro consumo de oxígeno es limi-tado. Su valor máximo es el nVO2 máx.El éxito en el rendimiento aeróbico estárelacionado con un elevado nVO2 máx ycon la capacidad para rendir durante lar-gos períodos con un porcentaje mayordel nVO2 máx.
i El exceso en el consumo de oxígenodespués del ejercicio supone la eleva-ción del índice metabólico por encimade los niveles en reposo y que ocurredespués del ejercicio durante el períodode recuperación.
i El umbral del lactato es ese punto en quela producción de lactato en sangre co-mienza a superar la capacidad del cuer-po para eliminarlo, lo cual se traduce enun rápido incremento de las concentra-ciones de lactacidemia por encima delos niveles en reposo durante el ejerciciode intensidad creciente.
i Por lo general, las personas con umbra-les de lactato más elevados, expresadoscomo un porcentaje de su nVO2 máx, soncapaces de un mejor rendimiento de fon-do.
i La capacidad de ejercitarse en activida-des aeróbicas de fondo también se aso-cia con una elevada economía de esfuer-zos, o con un nVO2 bajo para el mismoíndice de trabajo.
La fatiga y sus causas¿Cuál es exactamente el significado del términofatiga durante el ejercicio? Las sensaciones defatiga son notablemente diferentes al hacer ejer-cicio hasta el agotamiento en pruebas que duranentre 45 y 60 s, tales como las carreras de 400 m,que las experimentadas durante la realización deesfuerzos musculares agotadores (tales como lascarreras de maratón). Normalmente, usamos el
01 Crédits+Cap 01-06 • 001-220 17/1/07 16:21 Página 159

Resumiendo, se ha demostrado que beber líqui-dos antes y durante el ejercicio reduce en granmedida los efectos negativos de ejercitarse conmucho calor. La ingesta adecuada de líquidosatenúa el aumento de la temperatura central delcuerpo y la frecuencia cardíaca que normalmentese aprecia cuando una persona se ejercita en am-bientes calurosos. Esto se ejemplifica en la figura10.10.
El American College of Sports Medicine(ACSM) ha facilitado unas orientaciones de ca-rácter general para ayudar a los corredores a pre-venir estas lesiones relacionadas con el calor.1 Enla tabla 10.4 aparece una lista modificada de es-tas recomendaciones. Aclimatación al ejercicio
en ambientes calurosos¿Cómo podemos prepararnos para actividadesprolongadas en ambientes calurosos? ¿Nos hacemás tolerantes al estrés por calor el entrenamien-to en ambiente caluroso? Muchos estudios haninvestigado estas cuestiones y han llegado a la
FISIOLOGÍA DEL ESFUERZO Y DEL DEPORTE
350
Figura 10.10 Efectos de la ingesta de líquidos en la temperatu-ra central del cuerpo (rectal) durante una carrera de 2 horas. Aquienes participaron se les dieron líquidos en una prueba, ycompletaron una segunda prueba sin líquidos en un día distinto.Hay que reparar en que la ingesta de líquidos no tiene mucha in-fluencia hasta pasados unos 45 minutos, después de lo cual laretención de calor corporal se reduce en comparación con laprueba en que no se bebieron líquidos.Adaptado con autorización de D.L. Costill y col., 1970, «Fluidingestion during distance running», Archives of EnvironmentalHealth 21: 520-525.
Ingesta de líquidosSin líquidos
Tiempo de la carrera (min)
Tem
per
atur
a re
ctal
(º C
)
i El estrés por el calor implica algo másque la mera temperatura del aire. Quizásel modo más preciso de medirlo es con
la temperatura del globo húmedo, quemide la temperatura del aire e informa so-bre el potencial de intercambio de calorpor conducción, convección, evapora-ción y radiación en un ambiente especí-fico.
i Los calambres por calor están produci-dos probablemente por pérdidas de flui-dos y minerales resultantes de una exce-siva sudoración.
i El síncope por calor es la consecuenciade la incapacidad del sistema cardiovas-cular para satisfacer adecuadamente lasnecesidades de los músculos activos yde la piel. Se produce por un menor vo-lumen de sangre, causado generalmentepor una excesiva pérdida de fluidos y deminerales debida a una fuerte y prolon-gada sudoración. Aunque en sí mismono constituye una amenaza para la vida,el síncope por calor puede degenerar engolpe de calor si no se trata.
i El golpe de calor se produce por la insu-ficiencia de los mecanismos termorregu-ladores del cuerpo. Si se deja sin tratar,irá progresando y será fatal.
i Cuando se planea hacer ejercicio en am-bientes calurosos, deben tomarse variasprecauciones. Entre ellas citaremos lacancelación de las pruebas si el estrésambiental por el calor es demasiado ele-vado (THG superior a 28 oC), el vestirropa apropiada, el estar alerta a las seña-les de hipertermia y el procurar una ade-cuada ingesta de fluidos.
02 Cap 07-15 • 221-548 17/1/07 16:29 Página 350

3. Costill, D.L., Daniels, J., Evans, W., Fink,W., Krahenbuhl, G., & Saltin, B. (1976). Skeletalmuscle enzymes and fiber composition in maleand female track athletes. Journal of AppliedPhysiology, 40, 149-154.
4. Costill, D.L., Fink, W.J., Flynn, M., &Kirwan, J. (1987). Muscle fiber composition andenzyme activities in elite female distance run-ners. International Journal of Sports Medicine, 8,103-106.
5. Costill, D.L., Fink, W.J., & Pollock, M.L.(1976). Muscle fiber composition and enzymeactivities of elite distance runners. Medicine andScience in Sports, 8, 96-100.
6. Gollnick, P.D., & Hodgson, D.R. (1986).The identification of fiber types in skeletal mus-cle: A continual dilemma. Exercise and SportSciences Reviews, 14, 81-104.
7. Gollnick, P.D., Piehl, K., & Saltin, B.(1974). Selective glycogen depletion pattern inhuman muscle fibers after exercise of varying in-tensity and at varying pedal rates. Journal ofPhysiology, 241, 45-47.
8. McComas, A.J. (1996). Skeletal muscleform and function. Champaign, IL: Human Kine-tics.
Lecturas seleccionadas
Bouchard, C., Simoneau, J.A., Lortie, G., Bou-lay, M.R., Marcotte, M., & Thibault, M.C.(1986). Genetic effects in human skeletalmuscle fiber type distribution and enzymeactivities. Canadian Journal of Applied Phy-siology and Pharmacology, 64, 1245-1251.
Buchthal, F., & Schmalbruch, H. (1970). Con-traction times and fiber types in intact mus-cle. Acta Physiologica Scandinavica, 79,435-452.
Burke, R.E., & Edgerton, V.R. (1975). Motor unitproperties and selective involvement in mo-vement. Exercise and Sport Sciences Re-views, 3, 31-81.
Coggan, A.R., Spina, R.J., Rogers, M.A., King,D.S., Brown, M., Nemeth, P.M., & Holloszy,
J.O. (1992). Characteristics of skeletal mus-cle in master athletes. Journal of AppliedPhysiology, 68, 1896-1901.
Essen-Gustavsson, B., & Borges, O. (1986). His-tochemical and metabolic characteristics ofhuman skeletal muscle in relation to age. Ac-ta Physiologica Scandinavica, 126, 107-114.
Gordon, T., & Pattullo, M.C. (1993). Plasticity ofmuscle fiber and motor unit types. Exerciseand Sport Sciences Reviews, 21, 331-362.
Heckman, C.J., & Sandercock, T.G. (1996).From motor unit to whole muscle propertiesduring locomotor movements. Exercise andSport Sciences Reviews, 24, 109.
Hultman, E. (1995). Fuel selection, muscle fibre.Proceedings of the Nutrition Society, 54,107-121.
Karlsson, J. (1977). Skeletal muscle fibres andmuscle enzyme activities in monozygousand dizygous twins of both sexes. Acta Phy-siologica Scandinavica, 100, 385-392.
Komi, P.V., & Karlsson, J. (1979). Physical per-formance, skeletal muscle enzyme activities,and fibre types in monozygous and dizygoustwins of both sexes. Acta Physiologica Scan-dinavica, 462(Supl.), 1-28.
Roy, R.R., Baldwin, K.M., & Edgerton, V.R.(1991). The plasticity of skeletal muscle: Ef-fects of neuromuscular activity. Exercise andSport Sciences Reviews, 19, 269-312.
Staron, R.S. (1997). Human skeletal muscle fibertypes: Delineation, development and distri-bution. Canadian Journal of Applied Phy-siology, 22, 307-327.
Staron, R.S., Hagerman, F.C., Hikida, R.S., Mu-rray, T.F., Hostler, D.P., Crill, M.T., Ragg,K.E., & Toma,K. (2000). Journal of Histo-chemistry and Cytochemistry, 48, 623-629.
Trappe, S.W., Williamson, D.L., Godard, M.P.,Porter, D.A., Rowden, G. & Costill, D.L.(2000). Effect of resistance training on sin-gle muscle fiber contractile function in oldermen. Journal of Applied Physiology, 89,143-152.
Trappe, S.W. (2001). Master athletes. Internatio-nal Journal of Sport Nutrition and ExerciseMetabolism, 11, S194-S205.
FISIOLOGÍA DEL ESFUERZO Y DEL DEPORTE
60
01 Crédits+Cap 01-06 • 001-220 17/1/07 16:19 Página 60

tejercicio (CEOP), 154-155, 728consumo máximo de oxígeno
(nVO2máx); adaptaciones al entre-namiento, 209, 315, 319f, 412,590-594; efectos del envejeci-miento, 153, 320, 558-560, 590;contaminación atmosférica y,289; en altitud, 370; en deportis-tas, 320t, 618f; dopaje en sangre,534f; en niños, 558-561, 567; yel nivel de preparación física,318; descripción 151-152; con eldesentrenamiento, 427, 430; du-ración del ejercicio, 661; heren-cia y, 318-319; mejora, 200, 201,204; masa del ventrículo izquier-do y, 299; con la exposición a lamicrogravedad, 393, 397; paramonitorizar los cambios en el en-trenamiento, 213; en personasnormales, 320t, en personas in-sensibles, 320-321; interaccionescon el volumen plasmático, 311;en personas sensibles, 320-321;escala para diferencias de tama-ño, 320, 320t, 618-620, 623; in-teracciones con el volumen sistó-lico, 311; durante el período deafinamiento, 423
consumo submáximo de oxígeno(nVO2), 314, 619
contaminación del aire, 289contenido venoso de oxígeno, 253contracción ventricular prematura
(CVP), 229control glucémico, 734control motor, 81control neural extrínseco, 236-237control neuroendocrino, regulación
del, 75controles de investigación, 27-28convección, 335-336, 337tConvertino, V.A., 395Cooper, Kenneth, 10, 651, 651fcorazón. Ver también miocardio:
anatomía, 225f; corazón de de-portista, 299-300; aumento de ta-maño, 723; control extrínseco dela actividad, 229-230; sistema deconducción intrínseca, 227f; re-torno de la sangre, 234; tamaño,299-300, 302, 557, 559, 620; es-tructura y función, 224-234
corazón de deportista, 299-300corpulencia, 354, 482, 611-612corteza cerebral, 74, 80, 81, 98,
340corteza motora primaria, 75
corteza promotora, 80cortisol, 178t, 182, 186t, 185, 188f,
190f, 416Costill, David, L., 212, 343, 516,
539Cousteau, Jacques, 385Coyle, E.F., 251, 255, 325, 428CR. Ver fibras de contracción rápida
(CR)creatina, 509t, 542-543creatincinasa (CK), 133, 212, 419crecimiento, 552-553, 556f, 573crecimiento del cartílago, 553Crick, Francis, 18cromo, 456t, 509tcuadrado lumbar, 266culturismo, 483Cureton, T.K., 11curva de disociación de oxígeno-he-
moglobina, 272, 272fCVP (contracción ventricular pre-
matura), 230
DDaniels, J., 380daños musculares, 108f-109fdarbepoetina, 472defectos septales, 686deficiencia de estrógenos, 634deficiencia de oxígeno, 154definición y período de afinamiento,
423-424dehidroepiandrosterona (DHEA),
527DeMar, Clarence, 580densidad corporal, 484-487, 611densidad mineral ósea, 634densitometría, 485-487depleción de sustratos, 495deportes, costes energéticos de dis-
tintas actividades, 158deportistas femeninas, 611-612,
614-620, 621-624, 639depósitos lipídicos (estrías grasas),
682desarrollo, 552. Ver también creci-
mientodesarrollo fetal, 553desensibilización, 176desentrenamiento cardiovascular,
589desentrenamiento, 425-432Desert Research laboratory, 2deshidratación, 180, 368, 371, 459-
462, 462t, 465f, 494. Ver tambiénequilibrio hídrico
desmosomas, 227desoxihemoglobina, 272
desplazamiento del aire, y compo-sición corporal, 585
despolarización, 67desuso, principio de, 22. Ver tam-
bién atrofia; desentrenamientodesviación cardiovascular, 250-
251, 342desviación nVO2, 151deuda de oxígeno, 154. Ver tam-
bién consumo excesivo de oxíge-no postejercicio
deuterio (2H), 149dexedrina, 593DHEA (dehidroepiandrosterona),
527diabetes del adulto, 734diabetes gestacional, 734diabetes juvenil, 734diabetes mellitus, 654t, 689, 693,
723, 734-738diabetes mellitus insulinodepen-
diente, 734diabetes mellitus no insulinodepen-
diente, 734diafragma, 265dianocitos, 172diástole, 231diencéfalo, 75dieta: de los deportistas, 466-470;
equilibrio en la, 501, 726; creci-miento óseo y, 553-554; dieta dechoque, 499-500; para la diabe-tes, 736; ejercicio combinadocon, 727-729; masa magra y,729f; regulación de la ingesta dealimentos, 74-75; peso de re-ferencia y, 719-720
dieta de choque, 499-500dieta vegetariana, 467dieta zona, 471dietas especiales, 726dietas muy hipocalóricas, 726diferencia arteriovenosa de oxíge-
no (diferencia a-vO2), 253, 274,313, 558, 616
diferencias específicas del sexo.Ver también sexo: medicionesantropométricas y, 612, 613t;composición corporal y, 611-612, 621, 723-724; adaptacionescardiovasculares y respiratoriasy, 623; respuestas cardiovascula-res y, 616-617; por factores am-bientales, 639-640; lesiones,621; adaptaciones metabólicas y,623-624; respuestas metabólicasy, 618-620; adaptaciones neuro-musculares y, 621-622; respues-
FISIOLOGÍA DEL ESFUERZO Y DEL DEPORTE
762
03 Cap 16-21 • 549-776 17/1/07 16:39 Página 762

este necesario incremento en el suministro deglucosa al plasma.
Regulación del metabolismode las grasas durante el ejercicioAunque las grasas generalmente contribuyen enmenor proporción que los hidratos de carbono alas necesidades de energía de los músculos du-rante el ejercicio, la movilización y la oxidaciónde los ácidos grasos libres son críticas para elrendimiento en las series de ejercicio de resisten-cia. Durante tales actividades, las reservas de hi-dratos de carbono se agotan y nuestro cuerpodebe depender con mayor intensidad de la oxida-ción de las grasas para la producción de energía.Cuando las reservas de hidratos de carbono sonbajas (poca glucosa en sangre y poco glucógenomuscular), el sistema endocrino puede acelerar laoxidación de las grasas (lipólisis), asegurando asíque se puedan satisfacer las necesidades muscula-res de energía. La lipólisis se estimula también porla elevación de los niveles de adrenalina y nora-drenalina.
Recordemos que los ácidos grasos libres sealmacenan como triglicéridos en las células gra-sas y dentro de las fibras musculares. Los trigli-
céridos deben descomponerse para liberar losácidos grasos libres, que entonces son transporta-dos a las fibras musculares. El ritmo de consumode ácidos grasos libres por los músculos activosguarda una fuerte relación con la concentraciónde ácidos grasos libres en sangre. El incrementode esta concentración aumenta el consumo celu-lar de ácidos grasos libres. Podemos suponer quela concentración incrementada de estos ácidos ensangre hace aumentar la oxidación de ácidos gra-sos libres, ya que el mayor consumo celular deestos proporciona más ácidos grasos libres parala oxidación.1 Por lo tanto, el ritmo de descompo-sición de los triglicéridos puede, en parte, deter-minar el ritmo al que los músculos usan las gra-sas como fuente de combustible durante elejercicio.
Los triglicéridos son reducidos a ácidos gra-sos libres y glicerol por una enzima especial lla-mada lipasa, que es activada por al menos cuatrohormonas:
1. Cortisol.
2. Adrenalina.
3. Noradrenalina.
4. Hormona del crecimiento.
Además de la función del cortisol en la glu-coneogénesis, esta hormona acelera también lamovilización y el uso de ácidos grasos libres pa-ra la obtención de energía durante el ejercicio.La figura 5.11a ilustra los cambios en las con-centraciones de ácidos grasos libres y cortisol ensangre durante la realización de ejercicios pro-longados. Los niveles de cortisol en sangre al-canzan su nivel más elevado al cabo de 30-45min de ejercicio, y decrecen luego hasta nivelescasi normales. Pero la concentración de ácidosgrasos libres en sangre continúa elevándose du-rante la actividad, lo cual quiere decir que la li-pasa debe continuar siendo activada por otrashormonas. Las hormonas que continúan esteproceso son las catecolaminas y la hormona delcrecimiento. Tal como se ve en la figura 5.11b,los niveles en sangre de estas hormonas siguenelevándose durante el ejercicio, incrementandoprogresivamente la liberación de ácidos grasoslibres y la oxidación de las grasas. Las hormonastiroideas también contribuyen a la movilizacióny metabolismo de los ácidos grasos libres, peroen menor grado que las hormonas previamentemencionadas.
REGULACIÓN HORMONAL DEL EJERCICIO
189
Figura 5.10 Cambios en los niveles en plasma de glucosa e insuli-na durante ciclismo prolongado al 65%-70% del V·O2 máx. Re-párese en el declive gradual de la insulina durante el ejercicio, locual sugiere un aumento de la sensibilidad a la insulina duranteesfuerzos prolongados.
GlucosaInsulina
Glu
cosa
(mm
ol/l)
Tiempo (min)
0 30 60 90 120 150 180
6,0
5,8
5,6
5,4
5,2
5,0
4,8
4,6
0
18
16
14
12
10
8
0
Insu
lina
(µun
idad
es/m
l)
01 Crédits+Cap 01-06 • 001-220 17/1/07 16:22 Página 189

• pulso y respiración rápidos;
• generalmente, hipertensión (tensión arterialelevada);
• confusión, e
• inconsciencia.
Si se deja sin tratar, el golpe de calor progre-sa hasta el coma, y la muerte le sigue con rapi-dez. El tratamiento supone enfriar rápidamente elcuerpo de la persona en un baño de agua fría ohielo, o envolver el cuerpo en sábanas húmedas yabanicar a la víctima.
Este trastorno se produce por la insuficien-cia de los mecanismos termorreguladores delcuerpo. La producción de calor por el cuerpo du-rante el ejercicio depende de la intensidad delejercicio y del peso corporal, por lo que los de-portistas más pesados están sometidos a un ma-yor riesgo de sobrecalentamiento que los depor-tistas de menor peso cuando hacen ejercicio conla misma intensidad, suponiendo que ambos ten-gan aproximadamente el mismo nivel de aclima-tación al calor.
Para el deportista, el golpe de calor no es unproblema asociado solamente con condicionesextremas. Diversos estudios han revelado tem-peraturas rectales superiores a 40,5 oC en corre-dores de maratón que completaron con éxito ca-rreras llevadas a cabo bajo condiciones térmicasrelativamente moderadas (p. ej., 21 oC y 30% dehumedad relativa).3, 22 Incluso en competicionesmás cortas, la temperatura corporal central puedealcanzar niveles peligrosos para la vida. En fechatan temprana como es el año 1949, Robinson18
observó temperaturas rectales de 41 oC en corre-dores que competían en pruebas que duraban so-lamente unos 14 min, tales como las carreras de5.000 m. Después de una carrera de 10.000 mejecutada con una temperatura del aire de 29,5 oC,una humedad relativa del 80% y un sol brillante,un corredor que sufrió un síncope tenía una tem-peratura rectal de 43 oC.2 Sin atención médicaadecuada, tales fiebres pueden provocar dañospermanentes en el sistema nervioso central o lamuerte. Afortunadamente, este corredor fue re-frescado rápidamente con hielo y se recuperó sincomplicaciones.
FISIOLOGÍA DEL ESFUERZO Y DEL DEPORTE
348
Figura 10.9 Signos premonitorios de mialgia térmica, insolación y golpe de calor. ©PepsiCo 1995. Reproducido con autorización.
Cefalea y náuseas
Escalofríos o carnede gallina
Cese de la sudación
Desvanecimientoso mareos
Pulso rápido y fuerte
Piel seca y caliente
Confusión
Piel pálida y fría
Debilidad
Sed
Sudación profusa
Fatiga
Calambres musculares
Term
opat
íaS
igno
sp
rem
onito
rios
Mialgia térmica
Aumento de la gravedad
Insolación Golpe de calor
02 Cap 07-15 • 221-548 17/1/07 16:29 Página 348

son generalmente capaces de realizar actividadesque requieren sólo intensidades moderadas de fuer-za muscular. Por ejemplo, abrir el tapón de una ja-rra que tiene una determinada resistencia es unatarea que pueden realizar fácilmente el 92% de loshombres y las mujeres de edades comprendidasentre los 40 y los 60 años. Pero después de los 60años el número de personas que no pueden hacerloaumenta espectacularmente hasta el 68%. Cuandoestos individuos llegan a la franja de edad com-prendida entre los 71 y los 80 años, sólo el 32%será capaz de abrir la jarra.48
Datos similares, mostrados en la figura 17.5,describen cambios en la fuerza de las piernas enadultos que envejecen. La fuerza de extensiónde la rodilla en hombres y mujeres de un nivel deactividad normal disminuye rápidamente una vezpasados los 45 o los 50 años. Pero el entrena-miento de fuerza de los músculos extensorescapacita a los hombres ancianos para obtener me-jores resultados a los 60 años de edad que lamayoría de hombres de 30 años con un nivel de ac-tividad normal. En un estudio transversal dehombres y mujeres japoneses con edades entre 20y 84 años, se registraron declives parecidos en lafuerza de los músculos flexores y extensores dela rodilla. El torque máximo, una medida de lafuerza, se determinó a distintas velocidades deacción muscular usando un aparato isocinético (ver
capítulo 3, figura 3.2). La disminución de la fuerzacon la edad mantuvo una estrecha correlación conla reducción del área transversal de los músculosimplicados.1
Las pérdidas de fuerza muscular relaciona-das con la edad son la consecuencia principal-mente de una pérdida sustancial de masa muscu-lar que acompaña al envejecimiento o de unamenor actividad física, como expusimos antes eneste capítulo. La figura 17.6 muestra una ima-gen tomográfica computarizada (TC) de la partesuperior del brazo en 3 hombres de 57 años depeso corporal similar (de entre 78 y 80 kg). Ob-sérvese que el sujeto sedentario tenía una fuerzamuscular sustancialmente menor y más grasaque los otros. El sujeto entrenado en natacióntenía menos grasa y músculos tríceps notable-mente mayores que el sujeto no entrenado, perosus músculos bíceps, que casi nunca se usan alnadar, no eran muy distintos. Sin embargo, am-bos músculos aumentaron de tamaño en el casodel hombre que practicaba entrenamiento de lafuerza. Las diferencias entre estos tres hombresprobablemente se puedan atribuir a una combina-ción de genética y del volumen y tipo de entre-namiento.
Ahora sabemos que el envejecimiento tieneun acusado efecto sobre la masa total de músculoy sobre la fuerza, pero ¿qué ocurre con los tipos de
FISIOLOGÍA DEL ESFUERZO Y DEL DEPORTE
584
Figura 17.5 (a) La capacidad de levantarse de una posición sedente empeora a los 50 años, y a los 80 resulta imposible para algunas perso-nas. (b) Cambios en la fuerza máxima de extensión de la rodilla en hombres entrenados y desentrenados a distintas edades. Apréciese quelos hombres mayores (p. ej., 60 a 70 años) con entrenamiento de la fuerza pueden tener la fuerza de extensión de la rodilla igual o mayorque personas que sólo tienen 30 años. CVM = contracción voluntaria máxima.Datos de 17.5b procedentes de Human Performance Laboratory, Ball State University.
Fuerza máximade la pierna
Hombres con entrena-miento de la fuerzaHombres no entrenados
Rápido
Lento
a b
100
90
80
70
60
50
40
30
20
10
0
240
220
200
180
160
140
120
100
80
60
40
020 30 40 50 60 70 80 90 0 20 30 40 50 60 70 80
Edad Edad
% c
ontr
acci
ón v
olun
taria
máx
ima
com
par
ada
con
la d
e lo
s 20
año
s
Fuer
za m
áxim
a d
e ex
tens
ión
de
la ro
dill
a (N
m)
Levantarsedesde
posiciónsedente
03 Cap 16-21 • 549-776 17/1/07 16:38 Página 584

mo de la tensión de las fibras musculares. Cuan-do los músculos no se usan, la frecuencia de suestimulación neurológica se reduce y la movili-zación normal de las fibras se altera. Por lo tanto,parte de la pérdida de fuerza asociada con elabandono de los entrenamientos puede ser laconsecuencia de una incapacidad para activar al-gunas fibras musculares.
Las investigaciones indican que, después delabandono de los entrenamientos, un deportistapuede retener la fuerza y la potencia muscularesganadas durante períodos de hasta 6 semanas. Si,en lugar de esto, se sigue entrenando una vez ca-da 10 ó 14 días, los deportistas pueden mantenergeneralmente la fuerza ganada durante muchomás tiempo. Evidentemente, los músculos re-quieren una estimulación mínima para retener lafuerza, la potencia y el tamaño ganados duranteel entrenamiento.
Esto tiene implicaciones extremadamenteimportantes para el deportista lesionado. Éste pue-de ahorrarse mucho tiempo y esfuerzo durante larehabilitación llevando a cabo incluso un nivelbajo de ejercicio con la extremidad lesionada, ycomenzando a los pocos días de iniciada la re-cuperación. Las acciones isométricas sencillasson muy efectivas para la rehabilitación, porquesu intensidad puede graduarse y no es precisomover la articulación. No obstante, cualquierprograma de rehabilitación debe diseñarse en co-operación con el médico supervisor y el fisiotera-peuta.
Superficialmente, todos estos descubrimien-tos parecen contradecir las observaciones ante-riores de que períodos de total inactividad, talescomo los de inmovilización de una extremidad,producían notables pérdidas en la fuerza, la po-tencia y la masa musculares. Pero, aparentemen-te, la mayoría de los individuos que abandonarono redujeron sus regímenes de entrenamiento hi-cieron suficiente ejercicio al caminar, subir esca-leras, arrastrar, empujar y levantar como parapermitirles retener la mayor parte de la fuerza ga-nada con el entrenamiento de fuerza. No obstan-te, con la inmovilización, virtualmente no hayninguna activación para estimular los procesoscontráctiles. La fuerza y la movilidad se pierdenrápidamente bajo estas condiciones.
Cambios en la resistencia muscularLa resistencia muscular disminuye después detan sólo 2 semanas de inactividad. En este mo-mento, no se dispone de suficientes pruebas para
determinar si esta disminución del rendimientoes la consecuencia de cambios en los músculoso en la capacidad cardiovascular. En esta sección,examinaremos los cambios musculares que se sa-be que acompañan el abandono de los entrena-mientos y que pueden producir una disminuciónen la resistencia muscular.
Las adaptaciones musculares localizadas quetienen lugar durante períodos de inactividad estánbien documentadas, pero el conocimiento de lafunción exacta que desempeñan estos cambios enla pérdida de resistencia muscular todavía se nosescapa. Conocemos casos de poscirugía en losque después de una semana o dos de inmoviliza-ción con yeso, las actividades de las enzimas oxi-dativas tales como la succinatodeshidrogenasa yla citocromooxidasa disminuyen entre un 40% yun 60%. Datos recogidos de nadadores, que semuestran en la figura 12.15, indican que el poten-cial oxidativo de los músculos se reduce muchomás rápidamente que el consumo máximo de oxí-geno de los sujetos con el abandono de los entre-namientos. Debería esperarse que la reducción dela actividad de las enzimas oxidativas empeore elrendimiento muscular, y esto está mucho más rela-cionado con la capacidad submáxima de resisten-cia que con la capacidad aeróbica máxima o con elnVO2 máx.
ENTRENAMIENTO PARA EL DEPORTE
427
Figura 12.15 Reducción en los porcentajes del V·O2 máx, la activi-dad de la succinatodeshidrogenasa (SDH) en los músculos y la ac-tividad de la citocromooxidasa durante 6 semanas de desentrena-miento. Estos interesantes resultados sugieren que el potencialmetabólico de los músculos experimenta un declive, a pesar deque las pruebas del V·O2 máx muestren pocos cambios durante es-te período de desentrenamiento.
nVO2 máxSDHCitocromooxidasa
% d
e ca
mb
io c
on
el d
esen
tren
amie
nto
Semanas de desentrenamiento
02 Cap 07-15 • 221-548 17/1/07 16:30 Página 427

• histología y ultraestructura anormales delos músculos.
Armstrong desarrolló un modelo de inicioretrasado de la inflamación muscular que propo-nía la siguiente secuencia de acontecimientos:
1. Una tensión elevada en el sistema contrác-til-elástico de los músculos da como resul-tado lesiones estructurales en ellos y ensus membranas celulares.
2. Las lesiones en las membranas celularestrastornan la homeostasis del calcio en lasfibras lesionadas, produciendo necrosis(muerte celular), que llega a su punto álgidoaproximadamente a las 48 horas de reali-zado el ejercicio.
3. Los productos de la actividad macrófaga ylos contenidos intracelulares (tales comola histamina, las quininas y los K+) se acu-
mulan fuera de las células. Entonces, estassustancias estimulan las terminacionesnerviosas libres de los músculos. Este pro-ceso parece acentuarse con el ejercicio ex-céntrico, en que se distribuyen grandesfuerzas en secciones transversales relati-vamente pequeñas de los músculos.
Análisis globales recientes han proporciona-do una comprensión mucho mejor de las causasde la inflamación muscular. Ahora estamos segu-ros de que ésta es el resultado de lesiones o dañosen el propio músculo, generalmente en las fibrasmusculares y posiblemente en el sarcolema.6 Estedaño pone en marcha una cadena de aconteci-mientos entre los que se halla la liberación deproteínas intracelulares y un incremento en elmovimiento de las proteínas musculares. En el da-ño y en el proceso de reparación intervieneniones calcio, lisosomas, el tejido conectivo, radi-cales libres, fuentes energéticas, reacciones infla-
ADAPTACIONES NEUROMUSCULARES AL ENTRENAMIENTO RESISTIDO
109
Figura 3.11 (a) Micrografía electrónica que muestra la distribución normal de los filamentos de actina y de miosina y la configuración de laslíneas Z en los músculos de un corredor antes de una carrera de maratón. (b) Una muestra muscular tomada inmediatamente después de unacarrera de maratón presenta el estiramiento de las líneas Z causado por las acciones excéntricas de la carrera.Reimpreso de Hagerman y col. (1984).
a b
01 Crédits+Cap 01-06 • 001-220 17/1/07 16:20 Página 109

• artritis, y
• fibrosis quística.
Más recientemente, ha aumentado el énfasissobre el uso del ejercicio en la rehabilitación depacientes trasplantados, incluidos los que han su-frido trasplantes de corazón, de hígado y renales,porque ayuda a aliviar algunos efectos secunda-rios de los medicamentos y a mejorar la salud engeneral.
PROGRAMACIÓN DE EJERCICIOS PARA LA SALUD Y LA FORMA FÍSICA
673
Los ejercicios de entrenamiento se hanconvertido en una parte extremadamenteimportante de los programas de rehabili-tación para un cierto número de enferme-dades. Aunque los mecanismos fisiológi-cos específicos que explican los beneficiosde los ejercicios de entrenamiento paracada una de estas enfermedades todavíano se han definido claramente, hay mu-chos beneficios generales para la saludque parecen mejorar el pronóstico delpaciente.
i El ejercicio es una parte vital de la reha-bilitación cardiopulmonar y asimismo esesencial para la rehabilitación de pacien-tes con enfermedades tales como el cán-cer, la obesidad, la diabetes, las enfer-medades renales, la artritis y la fibrosisquística.
i El tipo y detalles del programa de reha-bilitación dependen del paciente, de laenfermedad específica de que se trate yde su alcance.
i Cada sesión de actividad relacionadacon la capacidad de resistencia debe irseguida por enfriamientos y estiramien-tos para prevenir la acumulación de san-gre en las extremidades y la inflamaciónmuscular.
i Los ejercicios de flexibilidad deben reali-zarse lentamente, y el mejor momento paraincluir esta fase del programa es inmedia-tamente después del componente de capa-cidad de resistencia.
i El entrenamiento resistido debe iniciarsecon un peso de la mitad de nuestra 1-RM.Éste es el peso apropiado si podemos le-vantarlo 10 veces. Si lo levantamos másveces, es que necesitamos más peso, y si lolevantamos menos de ocho veces, necesita-mos menos peso.
i En nuestro programa de ejercicios debenincluirse actividades recreativas para dis-frutar y relajarse.
Ejercicio y rehabilitaciónde personas con enfermedadesEl ejercicio se ha convertido en un componenteimportante de los programas de rehabilitaciónpara un cierto número de enfermedades. Los pro-gramas de rehabilitación cardiopulmonar, quecomenzaron en los años cincuenta, se han con-vertido en los más visibles. Tremendos avancesen la rehabilitación cardiopulmonar han llevado ala formación de una asociación profesional. LaAmerican Association of Cardiovascular andPulmonary Rehabilitation, y una revista de inves-tigación profesional, el Journal of Cardiopulmo-nary Rehabilitation.
El ejercicio es también una parte importantede la rehabilitación de los pacientes con:
• cáncer;
• obesidad;
• diabetes;
• enfermedades renales;
La manera en que se utiliza el ejercicio parala rehabilitación de personas enfermas es alta-mente específica de la naturaleza y el alcance dela enfermedad. Por esto, queda fuera del alcancede este capítulo entrar en detalles específicos pa-ra cada enfermedad, pero muchas de las referen-cias y lecturas seleccionadas relacionadas al finalde este capítulo proporcionan más detalles sobreel establecimiento de programas de ejercicio paraestados de enfermedades específicas y los valo-res clínicos de estos programas.5, 15, 25, 30, 32, 33
03 Cap 16-21 • 549-776 17/1/07 16:39 Página 673

del movimiento que en las carreras. Parte de laenergía gastada al nadar, por ejemplo, se usa parasostener el cuerpo sobre la superficie del agua ypara generar suficiente fuerza para superar la re-sistencia del agua al movimiento. Aunque la
energía necesaria para la natación depende del ta-maño y de la flotabilidad del cuerpo, la aplica-ción eficaz de la fuerza contra el agua es el prin-cipal determinante de la economía en la natación.
METABOLISMO, ENERGÍA Y SISTEMAS BÁSICOS DE ENERGÍA
157
Determinar la capacidad anaeróbica
No existe un método aceptable para determinar la capacidad anaeróbica. Se han descrito varios mé-todos, pero su validez se ha puesto en entredicho y, en el mejor de los casos, sólo ofrecen una esti-mación aproximada de la capacidad anaeróbica. Los primeros intentos por determinar la capacidadanaeróbica midieron el nivel de lactato en sangre después de un ejercicio agotador.14,16 Aunque sue-le aceptarse que el nivel de lactato en sangre es una indicación de glucólisis, no da un cálculo cuan-titativo de la producción anaeróbica de energía. El déficit máximo de oxígeno o EPOC fue pro-puesto también como un índice de la capacidad anaeróbica pero posteriores investigaciones nocorroboraron esto. En 1988, Medbø y col.17 propusieron el uso de las mediciones del consumo deoxígeno durante y después de un ejercicio submáximo y máximo para calcular la capacidad anaeró-bica. Estudios posteriores confirmaron que esta prueba da una estimación válida de la capacidadanaeróbica. Otras pruebas que parecen prometedoras son el Wingate anaerobic test2 y la prueba dela potencia crítica12. A pesar de las limitaciones inherentes a estos métodos, nos proporcionan indi-cadores indirectos del potencial metabólico de la capacidad anaeróbica.
Características de los deportistas de éxito en las pruebas aeróbicas de fondo
De nuestra exposición sobre las características metabólicas de los deportistas de fondo en este ca-pítulo, y de las características de sus tipos de fibras musculares dadas en el capítulo 1, está claroque para tener éxito en actividades aeróbicas de fondo se necesita lo siguiente:
• Un valor elevado del nVO2 máx.
• Un umbral de lactato elevado cuando se expresa como porcentaje del nVO2 máx.
• Una gran economía de esfuerzo, o un valor bajo de nVO2 para el mismo índice de trabajo.
• Un elevado porcentaje de fibras musculares de ST.
De los limitados datos de que disponemos, estas cuatro características parecen estar en el or-den apropiado de importancia. Como ejemplo, el umbral de lactato es el mejor elemento predicti-vo que tenemos o el ritmo de carrera cuando pensamos en un grupo de corredores de fondo de eli-te. Sin embargo, estos corredores ya tienen un elevado nVO2 máx. Aunque la economía de esfuerzoes importante, no varía tanto entre corredores de elite. Finalmente, tener un elevado porcentaje defibras musculares de ST ayuda pero no es esencial. El ganador de la medalla de bronce en un ma-ratón olímpico tenía sólo un 50% de fibras de ST en su músculo gastrocnemio, uno de los princi-pales músculos usados para correr.
01 Crédits+Cap 01-06 • 001-220 17/1/07 16:21 Página 157

DIFERENCIAS SEXUALES EN EL DEPORTE Y EL EJERCICIO
613
Pliegues cutáneos (mm)
Escápula 13,2 15,3 17,3 14,1 13,9 20,2
Tríceps 12,8 18,8 22,2 7,9 13,6 18,5
Axilar medio 10,7 13,3 16,9 11,7 15,5 24,8
Pecho 14,0 14,0 11,4 20,6
Suprailíaco 17,2 15,3 17,3 19,3 15,2 22,0
Abdominal 15,1 22,8 29,6 16,0 20,6 30,0
Muslo 31,8 28,8 33,1 14,9 17,4 22,2
Rodilla 7,0 17,4 17,3 5,3
Circunferencias (cm)
Cabeza 55,0 57,5
Cuello 31,8 38,5
Hombros 101,9 99,7 100,9 117,0 112,5 114,8
Pecho 85,2 84,6 87,1 97,4 91,4 96,3
Busto 87,8 87,7 90,8
Abdomen 75,3 75,0 82,7 84,0 81,0 91,1
Caderas 95,9 93,1 97,5 96,9 94,4 98,4
Muslo 57,0 56,5 57,6 58,0 57,1 59,0
Rodilla 36,1 37,7
Pantorrilla 35,1 33,9 34,4 37,6 36,5 36,9
Tobillo 21,1 20,8 20,8 22,7 22,1 22,1
Deltoides 30,7 36,3
Bíceps, flexionado 27,2 27,0 28,6 33,2 32,6 34,0
Bíceps, extendido 25,0 29,1
Antebrazo 23,5 23,8 24,4 27,6 28,3 29,2
Muñeca 14,9 14,8 15,1 17,0 16,7 17,4
Diámetros (cm)
Longitud de la cabeza 19,0 19,9
Anchura de la cabeza 14,9 15,5
Biacromial 36,5 36,8 36,7 40,4 41,1 41,5
Bideltoides 42,1 41,4 41,8 47,6 46,9 47,4
Pecho 25,8 27,8 28,6 29,3 31,8 33,0
Biilíaco 28,4 29,9 31,2 28,4 29,6 31,4
Bitrocantéreo 32,1 34,0 35,3 32,9 33,6 35,1
Rodilla 8,9 9,3 9,6 9,5 9,8 10,1
Tobillo 6,3 7,1
Codo 6,0 7,0
Muñeca 4,9 5,1 5,2 5,6 5,9 6,0
Envergadura del brazo 165,8 181,7
Longitud del pie 24,1 26,7
Longitud de la mano 17,3 19,1
a Datos de Wilmore y Behnke (1969 y 1970).67, 68
b Datos de Pollock y col (1975 y 1976).48, 49
Mujeres Hombres
Jóvenes De mediana edad Jóvenes De mediana edad
Wilmore y Behnkea Pollock y colb Pollock y colb Wilmore y Behnkea Pollock y colb Pollock y colb
(n = 128) (n = 83) (n = 60) (n = 133) (n = 95) (n = 84)
�TABLA 18.2 Mediciones antropométricas para hombres y mujeres jóvenes y de mediana edad
03 Cap 16-21 • 549-776 17/1/07 16:38 Página 613

relativo de la inactividad física es similar al riesgoasociado con los otros tres factores de riesgo im-portantes de enfermedades de las arterias corona-rias. Además, el porcentaje de la población total deEE.UU. que es físicamente inactiva supera en mu-cho el porcentaje de los otros tres grupos de riesgoimportantes: quienes fuman, los hipertensos o losque tienen altos niveles de colesterol.9 Esto se ilus-tra en la figura 20.8. Los resultados de estos estu-dios epidemiológicos desempeñaron un impor-tante papel en la American Heart Associationen 1992 para declarar la inactividad física comoun factor primario de riesgo para las enfermedadesde las arterias coronarias.
A mediados de los años 1980 surgió otra im-portante preocupación: ¿qué nivel de actividadfísica o de preparación es necesario para reducirel propio riesgo de sufrir enfermedades de las ar-terias coronarias?32 Por los estudios epidemioló-gicos no estaba totalmente claro qué nivel de pre-paración o de actividad era efectivo. De hecho, amediados de los años 1980 apenas se había co-menzado a diferenciar entre el nivel de actividady la preparación física, y se definía la preparaciónfísica personal mediante el nVO2 máx de una per-sona. Retrospectivamente, la distinción entre es-tos dos términos fue crucial ya que una personapuede ser activa pero no estar en buena forma(bajo nVO2 máx), o estar en buena forma ( nVO2
máx elevado) pero ser inactiva. El Dr. RonaldLaPorte y colaboradores en la Universidad dePittsburgh contribuyeron de forma eficaz a rediri-gir el pensamiento y las investigaciones posterio-res en esta área.32 El Dr. LaPorte señaló que, ba-sándose en diversos estudios epidemiológicos,los niveles de actividad asociados con un bajoriesgo de enfermedades de las arterias coronariaseran generalmente bajos y ciertamente no al nivelque hubiese incrementado la capacidad aeróbica.Estudios posteriores lo han confirmado.8, 35, 36 Ni-veles bajos de actividad, tales como andar o lajardinería, pueden proporcionar considerables be-neficios reduciendo el riesgo de enfermedades delas arterias coronarias. Es probable que el ejer-cicio más vigoroso aporte beneficios incluso ma-yores.34
FISIOLOGÍA DEL ESFUERZO Y DEL DEPORTE
696
A partir de estudios epidemiológicos seha establecido que la inactividad físicadobla el riesgo de sufrir enfermedad delas arterias coronarias. Sin embargo,ahora es igualmente claro que la activi-dad de baja intensidad es suficiente parareducir el riesgo de sufrir esta enferme-dad. Los beneficios para la salud no re-quieren ejercicios de alta intensidad.
Figura 20.8 Porcentajes de la población de EE.UU. con un mayor nivel de enfermedades de las arterias coronarias basándose en los princi-pales factores de riesgo.Reproducido con autorización de Caspersen CJ: «Physical activity and coronary heart disease». Physicians Sportsmedicine 1987;15(11): 43-44 © The McGraw-Hill Companies.
% d
e la
po
bla
ció
n d
e E
E.U
U.
que
co
rre
ries
go
Factor de riesgo:Proporción deriesgo:Niveles de riesgo:
Tensión arterial Colesterol sérico Fumar cigarrillos Inactividad física
2,1 2,4 2,5 1,9≥ 150 mmHg frente a ≥ 268 mg/dl frente a 1 paquete al día frente a Sin actividad frente a
< 130 mmHg < 218 mg/dl no fumar 20 min, 3 veces por semana
10% 10%18%
59%
03 Cap 16-21 • 549-776 17/1/07 16:39 Página 696

cicios en carga pueden ser continuos en al-gunas circunstancias, si bien las actividadessin carga del peso corporal como el ciclismoo la natación reducen el riesgo de lesión.
• Hay que tener cuidado de no practicar ejer-cicios en los que pueda producirse la pérdidadel equilibrio o un traumatismo abdominal.
• Como el embarazo requiere 300 kcal adicio-nales (1.255 kJ) de energía diarias, las muje-res que hagan ejercicio deben prestar aten-ción especial a la dieta para asegurar unaingesta calórica adecuada.
• La disipación del calor es de particular im-portancia durante el primer trimestre, por loque las mujeres que hagan ejercicio debenllevar la ropa adecuada, asegurarse de que laingesta de líquido sea suficiente y seleccio-nar las condiciones medioambientales ópti-mas.
• Las actividades regulares de ejercicio du-rante el embarazo deben reanudarse de for-ma gradual en el puerperio, ya que los cam-bios asociados con el embarazo puedenpersistir de 4 a 6 semanas.
En 2002, ACOG publicó una corta «Opinióndel Comité» sobre el ejercicio durante el embara-zo y el puerperio, que en esencia respalda suspautas previas.2 Además, apoyaron la recomenda-ción actual de los Centers for Disease Controland Prevention y el American College of SportsMedicine para no embarazadas, que estableceque las personas deberían llegar a 30 minutos omás al día de ejercicio moderado casi todos, si notodos, los días de la semana (ver capítulo 19).Además, afirman que el submarinismo con esca-fandra autónoma debería evitarse durante el em-barazo, porque el feto corre un riesgo mayor dedisbarismo. Asimismo, existe un mayor riesgocuando las embarazadas se ejercitan a alturas porencima de 1.828 metros.
OsteoporosisMantener un estilo de vida saludable puede retra-sar el principal proceso de envejecimiento queafecta a las mujeres: la osteoporosis. La osteopo-rosis se caracteriza por una disminución del con-tenido de minerales en los huesos, lo cual causaporosidad ósea (figura 18.10). La osteopenia, co-mo descubrimos en el capítulo 17, describe la pér-
dida de masa ósea con el envejecimiento. La os-teoporosis es una pérdida más grave de masa óseapor el deterioro de la microarquitectura del hueso,y causa fragilidad esquelética y aumento del ries-go de fracturas.44 Estos cambios aumentan el ries-go de fractura que, generalmente, comienza al fi-nal de la década de los 30 años de edad. Laposibilidad de que se produzcan estas fracturas seincrementa entre 2 y 5 veces con el inicio de lamenopausia. Los hombres también sufren osteo-porosis, pero en un grado menor debido a que lapérdida de minerales de los huesos se produce aun ritmo más lento. Queda mucho por aprendersobre la etiología de la osteoporosis; sin embar-go, en la mujer posmenopáusica tres factores sonlos que principalmente contribuyen a ella:
• insuficiencia de estrógeno;
• ingestión inadecuada de calcio, y
• actividad física inadecuada.
Aunque el primero de estos riesgos es unaconsecuencia directa de la menopausia, los dos úl-timos reflejan modelos dietéticos y de ejercicio alo largo de la vida.
Además de las mujeres posmenopáusicas,las mujeres con amenorrea y las que tienen ano-rexia nerviosa también sufren osteoporosis debi-do a una ingestión insuficiente de calcio o a bajosniveles de estrógeno, o posiblemente a ambas co-sas. En un estudio sobre mujeres con anorexia,los investigadores hallaron que las densidades desus huesos eran significativamente menores encomparación con las de los controles.52 Cann ycolaboradores10 fueron los primeros en declararla existencia de contenidos minerales óseos sus-tancialmente menores en las mujeres físicamenteactivas diagnosticadas de amenorrea hipotalámi-ca.
En un segundo estudio, las densidades de loshuesos radiales y vertebrales de 14 mujeres depor-tistas con amenorrea fueron comparadas con las de14 mujeres deportistas con una menstruación nor-mal (eumenorrea).19 Los investigadores descu-brieron que la actividad física no protegía al grupocon amenorrea de pérdidas significativas de ladensidad ósea. Los valores de la densidad ósea deeste grupo a una edad media de 24,9 años eranequivalentes a los de mujeres con una edad mediade 51,2 años. En un estudio de seguimiento, se ha-llaron incrementos de la densidad mineral de loshuesos vertebrales en las mujeres que habían sido
FISIOLOGÍA DEL ESFUERZO Y DEL DEPORTE
634
03 Cap 16-21 • 549-776 17/1/07 16:38 Página 634

dejará menos sangre en el ventrículo izquierdodespués de la sístole. Esto se multiplica por ladisminución de la tensión arterial causada por elentrenamiento, con lo cual se reduce la resisten-cia periférica general.
Esta mayor contractilidad, junto con el ma-yor retroceso elástico resultante del mayor llena-do diastólico, incrementa la fracción de eyecciónen el corazón entrenado. Entra más sangre en elventrículo izquierdo, y un mayor porcentaje de loque entra es expulsado con cada contracción, porlo que el volumen sistólico se incrementa.
Estos cambios en el volumen sistólico estánbien ilustrados por un estudio en el que hombresancianos fueron sometidos al entrenamiento de suresistencia durante un período de 1 año.12 Su fun-ción cardiovascular fue evaluada antes y despuésdel entrenamiento. Durante una hora cada día, 4días a la semana, corrieron sobre cintas ergométri-
cas y pedalearon en bicicletas ergométricas. Elejercicio se ejecutó con intensidades de entre el60% y el 80% del nVO2 máx, con breves series deejercicio que superaban el 90% del nVO2 máx.
En la figura 9.5 se muestran los resultados. Elvolumen diastólico final aumentó en reposo y du-rante la realización de ejercicios submáximos. Lafracción de eyección aumentó, y esto se asociócon un menor volumen sistólico final, sugiriendoambos una mayor contractilidad del ventrículo iz-quierdo. El nVO2 máx aumentó un 23%, indicandouna mejora sustancial de la resistencia.El volumen sistólico en reposo y durante el ejer-cicio no es meramente una función del estado deentrenamiento de una persona. También refleja eltamaño del cuerpo. Las personas de mayor tama-ño tienen volúmenes sistólicos mayores. Recor-dar esto es muy importante cuando se comparanlos volúmenes sistólicos de distintas personas.
FISIOLOGÍA DEL ESFUERZO Y DEL DEPORTE
302
Medición del tamaño del corazónLa medición del tamaño del corazón ha despertado durante años el interés de los cardiólogos porque los cora-zones hipertrofiados suelen constituir un estado patológico indicador de la presencia de una enfermedad cardio-vascular. Más recientemente, los científicos del ejercicio se han interesado por el tamaño del corazón en relacióncon la forma física y el rendimiento de los deportistas. ¿Cómo se mide el tamaño del corazón? Los estudios ini-ciales emplearon radiografías para calcular el tamaño del corazón, y la información obtenida se limitaba a unaspocas mediciones básicas. Desde la década de 1970, los estudios sobre deportistas y personas que participabanen actividades deportivas de fondo han recurrido a las ecocardiografías para medir con mayor precisión el tama-ño del corazón y sus cavidades. La ecocardiografía se basa en la técnica de los ultrasonidos, en la que las ondassonoras de alta frecuencia se disparan al corazón a través de la pared torácica. Estas ondas sonoras se emitencon un transductor colocado en el pecho, y una vez que entran en contacto con las distintas estructuras del cora-zón, rebotan y vuelven a un sensor que captura una imagen en movimiento del corazón. Se visualiza el tamañode las cavidades del corazón, el espesor de sus paredes y la acción de las válvulas cardíacas. Hay varias formas deecocardiografía: ecocardiografía modo M, que proporciona una imagen monodimensional del corazón; ecocar-diografía bidimensional, que ofrece una imagen en dos dimensiones de las estructuras del corazón, y la ecocar-diografía Doppler, que se emplea con más frecuencia para la medición del riego sanguíneo. Las figuras 9.4a y bmuestran la obtención de una ecocardiografía bidimensional y el ecocardiograma resultante.
a b
Figura 9.4 Ecocardiografía bidimensional, (a) procedimiento y (b) el ecocardiograma resultante.
02 Cap 07-15 • 221-548 17/1/07 16:29 Página 302

do sangre en el espacio comprendido entre el ce-rebro y el cráneo. En ambos casos, el riego san-guíneo más allá del punto en que se ha producidola rotura disminuye porque la sangre abandona elvaso en el lugar de la lesión. Asimismo, cuandola sangre se acumula fuera del vaso, impone pre-sión al frágil tejido cerebral, lo cual puede alterarla función de éste. Las hemorragias cerebralescon frecuencia son la consecuencia de aneurismas,que aparecen en puntos débiles de la pared delvaso que se hincha hacia fuera, y los aneurismascon frecuencia se producen por causa de la hiper-tensión o por lesiones ateroscleróticas en la pareddel vaso.
Al igual que con los ataques al corazón, laapoplejía produce la muerte del tejido afectado.Las consecuencias dependen en gran medida dela localización y el alcance de la apoplejía. El da-ño cerebral producido por una apoplejía puedeafectar a los sentidos, el habla, el movimientocorporal, los modelos de pensamiento y la memo-ria. La parálisis de un lado del cuerpo es frecuen-te, al igual que la incapacidad para verbalizar lospensamientos. La mayor parte de los efectos deuna apoplejía son indicativos del lado del cerebroque ha sido dañado. Éstos se relacionan en la ta-bla 20.2.
Insuficiencia congestiva del corazónLa insuficiencia congestiva del corazón es untrastorno clínico en el que el músculo cardíacose debilita demasiado para mantener un gastocardíaco adecuado para satisfacer las demandasde oxígeno del cuerpo. Esto suele ser la conse-cuencia de que el corazón ha sido lesionado oha sido sometido a un esfuerzo excesivo. La hi-pertensión, la aterosclerosis y el ataque cardía-co figuran entre las posibles causas de este tras-torno.
Cuando el gasto cardíaco es inadecuado, lasangre comienza a acumularse en las venas. Estohace que se acumule un exceso de fluidos en elcuerpo, particularmente en las piernas y en los to-billos. Esta acumulación de fluidos (edema) pue-de afectar también a los pulmones (edema pul-monar), alterando la respiración y produciendofalta de aliento. La insuficiencia congestiva delcorazón puede progresar hasta el punto de lesio-nar el corazón de modo irreversible, y el pacientese convierte en un candidato para un trasplantecardíaco.
Otras enfermedades cardiovascularesOtras enfermedades cardiovasculares son las en-fermedades vasculares periféricas; las enferme-dades de las válvulas del corazón; las enfermeda-des reumáticas del corazón, y las enfermedadescardíacas congénitas.
Las enfermedades vasculares periféricasimplican a las venas y a las arterias sistémicas, enlugar de a los vasos coronarios. La arteriosclero-sis se refiere a numerosos procesos en los que lasparedes de las arterias se espesan, endurecen ypierden elasticidad. La aterosclerosis es una for-ma de arteriosclerosis. La arteriosclerosis obs-tructiva, en la que la arteria queda completamen-te ocluida, es otra forma. Entre las enfermedadesvenosas periféricas figuran las venas varicosas yla flebitis. Las venas varicosas son la consecuen-cia de la incapacidad de las válvulas de las venas,que dejan que la sangre retroceda y se acumuleen las venas haciendo que éstas se agranden, seretuerzan y se vuelvan dolorosas. La flebitis es lainflamación de una vena y también es muy dolo-rosa.
Las enfermedades de las válvulas del cora-zón afectan a una o más de las cuatro válvulas quecontrolan la dirección del flujo de la sangre haciadentro y hacia fuera de las cuatro cámaras del co-razón. Las enfermedades cardíacas reumáticasson una forma de enfermedad cardíaca valvular enque una infección por estreptococos ha producidofiebre reumática aguda, generalmente en niños deedades comprendidas entre los 5 y los 15 años. Lafiebre reumática es una enfermedad inflamatoriadel tejido conectivo que afecta comúnmente al co-razón, específicamente a las válvulas cardíacas. Lalesión en las válvulas suele producir dificultades
ENFERMEDADES CARDIOVASCULARES Y ACTIVIDAD FÍSICA
685
Lado izquierdo paralizado Lado derecho paralizado
Déficit espaciales y perceptuales Déficit del habla ydel lenguaje
Estilo de comportamiento: Estilo de comportamiento:rápido, impulsivo lento, precavido
Déficit de la memoria: lenguaje Déficit de la memoria:representación
Lesiones en el lado Lesiones en el ladoderecho del cerebro izquierdo del cerebro
�TABLA 20.2 Los efectos de las lesiones cerebrales encada lado del cerebro
03 Cap 16-21 • 549-776 17/1/07 16:39 Página 685

diante la oxidación en CO2 y H2O, proporcionan-do así la energía necesaria para restablecer las re-servas de glucógeno.
Con esta teoría, se creía que los componentesrápidos y lentos de la curva reflejaban la actividadanaeróbica que había tenido lugar durante el ejer-cicio. Se pensaba que, examinando el consumo deoxígeno posejercicio, se podía estimar la intensi-dad de la actividad anaeróbica producida.
No obstante, estudios más recientes han llega-do a la conclusión de que la explicación clásica delexceso de consumo de oxígeno posejercicio es de-masiado simplista. Por ejemplo, durante la faseinicial del ejercicio se toma algo de oxígeno de lasreservas del oxígeno (hemoglobina y mioglobina).Este oxígeno debe reponerse durante la recupera-ción. Asimismo, la respiración sigue siendo eleva-da temporalmente después del ejercicio, en partecomo un esfuerzo para eliminar el CO2 que se haacumulado en los tejidos como subproducto delmetabolismo. La temperatura del cuerpo tambiénse eleva, lo cual mantiene elevados los ritmos me-tabólico y respiratorio, requiriendo así más oxíge-no, y los niveles elevados de noradrenalina y deadrenalina durante el ejercicio tienen efectos simi-lares.
Por lo tanto, intervienen más factores de losque permite la teoría clásica. La deuda de oxíge-no depende de muchos factores, aparte de mera-mente reconstruir el ATP y el PC y eliminar ellactato producido por el metabolismo anaeróbico.Los mecanismos fisiológicos responsables delexceso de consumo de oxígeno posejercicio ne-cesitan ser definidos con claridad.
Umbral del lactato
Muchos investigadores consideran que el umbraldel lactato es un buen indicador del potencial deun deportista para el ejercicio de resistencia. Elumbral del lactato (UL) se define como el pun-to en que el lactato sanguíneo comienza a acumu-larse por encima de los niveles de reposo duranteel ejercicio de intensidad creciente. Durante laactividad entre leve y moderada, el lactato san-guíneo permanece sólo ligeramente por encimadel nivel de reposo. Con esfuerzos más intensos,el lactato se acumula más rápidamente. Fijémo-nos en la figura 4.17. A velocidades bajas al co-rrer, los niveles de lactato en sangre permanecenen, o cerca, de los niveles de reposo. Pero cuandola velocidad al correr aumenta por encima deaproximadamente 1,4 m/s, los niveles de lactatoen sangre aumentan con rapidez. Este punto deinflexión en la curva representa el umbral del lac-tato.
Se cree que el umbral del lactato refleja lainteracción de los sistemas anaeróbicos y aeróbi-cos de energía. Algunos investigadores sugierenque el umbral del lactato representa un desvíosignificativo hacia la glucólisis anaerobia, queforma lactato. Por consiguiente, el incremento re-pentino del nivel de lactato en sangre con el au-mento del esfuerzo también se denomina umbralanaerobio. No obstante, la concentración de lacta-to en la sangre está determinada no sólo por la pro-ducción de lactato en el músculo esquelético uotros tejidos, sino también por la eliminación de lalactacidemia en el hígado, el músculo esquelético,
METABOLISMO, ENERGÍA Y SISTEMAS BÁSICOS DE ENERGÍA
155
Figura 4.17 Relación entre la intensidad del ejercicio (velocidad en carrera) y la concentración de lactato en sangre. Las muestras desangre se tomaron de la vena del brazo de un corredor y se analizó el contenido de lactato después de haber corrido a distintas veloci-dades durante 5 minutos. UL = Umbral del lactato.
Velocidad del tapiz rodante (km/h)
Lact
acid
emia
UL
01 Crédits+Cap 01-06 • 001-220 17/1/07 16:21 Página 155

31. Kramsch, D.M., Aspen, A.J., Abramo-witz, B.M., Kreimendahl, T., & Hood, W.B.(1981). Reduction of coronary atherosclerosis bymoderate conditioning exercise in monkeys onan atherogenic diet. New England Journal of Me-dicine, 305, 1.483-1.489.
32. Laporte, R.E., Adams, L.L., Savage,D.D., Brenes, G., Dearwater, S., & Cook, T.(1984). The spectrum of physical activity, cardio-vascular disease and health: An epidemiologicperspective. American Journal of Epidemiology,120, 507-517.
33. Lee, C.D., & Blair, S.N. (2002). Cardio-respiratory fitness and stroke mortality in men.Medicine and Science in Sports and Exercise, 34,592-595.
34. Lee, I.-M., & Paffenbarger, R.S., Jr.(1996). Do physical activity and physical fitnessavert premature mortality? Exercise and SportSciences Reviews, 24, 135-171.
35. Leon, A.S., & Connett, J. (1991). Physi-cal activity and 10.5 year mortality in the Multi-ple Risk Factor Intervention Trial (MRFIT). In-ternational Journal of Epidemiology, 20,690-697.
36. Leon, A.S., Connett, J., Jacobs, D.R., &Rauramaa, R. (1987). Leisure-time physical acti-vity levels and risk of coronary heart disease anddeath. Journal of the American Medical Associa-tion, 258, 2.388-2.395.
37. Marieb, E.N. (1995). Human ana-tomy and physiology (3ª ed.). Redwood City,CA: Benjamin / Cummings.
38. McNamara, J.J., Molot, M.A., Stremple,J.F., & Cutting, R.T. (1971). Coronary arterydisease in combat casualties in Vietnam. Journalof the American Medical Association, 216,1.185-1.187.
39. Montoye, H.J., Metzner, H.L., Keller,J.B., Johnson, B.C., & Epstein, F.H. (1972). Ha-bitual physical activity and blood pressure. Medi-cine and Science in Sports, 4, 175-181.
40. Morgan, W.P. (1994). Physical activity,fitness and depression. En C. Bouchard, R.J.Shephard, & T. Stephens (eds.), Physical activity,fitness, and health (págs. 851-867). Champaign,IL: Human Kinetics.
41. Morris, J.N., Adam, C., Chave, S.P.W.,Sirey, C., Epstein, L., & Sheehan, D.J. (1973).
Vigorous exercise in leisure-time and the inci-dence of coronary heart-disease. Lancet, 1, 333-339.
42. Morris, J.N., Heady, J.A., Raffle, P.A.B.,Roberts, C.G., & Parks, J.W. (1953). Coronaryheart-disease and physical activity of work. Lan-cet, 265, 1.053-1.057, 1.111-1.120.
43. Morris, J.N., Pollard, R., Everitt, M.G.,Chave, S.P.W., & Semmence, A.M. (1980). Vigo-rous exercise in leisure-time: Protection againstcoronary heart disease. Lancet, 2, 1.207-1.210.
44. O’Connor, G.T., Buring, J.E., Yusuf, S.,Goldhaber, S.Z., Olmstead, E.M., Paffenbarger,R.S., & Hennekens, C.H. (1989). An overview ofrandomized trials of rehabilitation with exerciseafter myocardial infarction. Circulation, 80, 234-244.
45. Oldridge, N.B., Guyatt, G.H., Fischer,M.E., & Rimm, A.A. (1988). Cardiac rehabilita-tion after myocardial infarction: Combined expe-rience of randomized clinical trials. Journal ofthe American Medical Association, 260, 945-950.
46. Paffenbarger, R.S., Hyde, R.T., Wing,A.L., & Hsieh, C.-C. (1986). Physical activity,all-cause mortality, and longevity of collegealumni. New England Journal of Medicine, 314,605-613.
47. Petruzzello, S.J., Landers, D.M., Hat-field, B.D., Kubitz, K.A., & Salazar, W. (1991).A meta-analysis on the anxiety-reducing effectsof acute and chronic exercise: Outcomes and me-chanisms. Sports Medicine, 11, 143-182.
48. Pollock, M.L., & Wilmore, J.H. (1990).Exercise in health and disease: Evaluation andprescription for prevention and rehabilitation (2.ªed.). Filadelfia: Saunders.
49. Powell, K.E., Thompson, P.D., Casper-sen, C.J., & Kendrick, J.S. (1987). Physical acti-vity and the incidence of coronary heart disease.Annual Reviews in Public Health, 8, 253-287.
50. Ross, R. (1986). The pathogenesis of at-herosclerosis-An update. New England Journalof Medicine, 314, 488-500.
51. Seals, D.R., & Hagberg, J.M. (1984).The effect of exercise training on human hyper-tension: A review. Medicine and Science inSports and Exercise, 16, 207-215.
ENFERMEDADES CARDIOVASCULARES Y ACTIVIDAD FÍSICA
707
03 Cap 16-21 • 549-776 17/1/07 16:39 Página 707

combinado con la estatura o de los pliegues cutá-neos para estimar la prevalencia del sobrepesoy de la obesidad. Peso relativo es un términoempleado para expresar el porcentaje por el queun individuo tiene exceso o insuficiencia de peso.Este valor se determina generalmente dividiendoel peso de la persona por el peso medio (de lastablas de peso estándar) para la categoría de com-plexión media, basándose en la estatura. El resul-tado se multiplica entonces por 100 para expre-sarlo en porcentaje. Por ejemplo, usando la tabla21.1, un hombre que pesa 104 kg y que tiene unaestatura de 183 cm tendrá un peso relativo del
142%C [104 kg / 73 kg] × 100 = 142%). La cifrade 73 kg es el valor medio para la categoría decomplexión media de un hombre de 183 cm (en-tre 70 y 77 kg).
El índice de masa corporal (IMC) es otro va-lor frecuentemente usado para estimar la obesidad.El IMC de una persona se determina dividiendo elpeso corporal en kilogramos por el cuadrado de laestatura corporal en metros. Por ejemplo, el hombreque pesa 104 kg y mide 183 cm, tendrá un IMCde 32 kg por m2. Generalmente, el IMC está rela-cionado con la composición corporal. Está alta-mente correlacionado con el porcentaje de grasa
OBESIDAD, DIABETES Y ACTIVIDAD FÍSICA
713
Hombres a partir de los 25 años de edad
Estatura con calzado Peso ideal (lb y kg) talones 1 pulg. Complexión Complexión Complexión
(2,54 cm) pequeña media grande
Pies cm Pulg. cm lb kg lb kg lb kg
5 152 2 5,08 112-120 50-54 118-129 54-59 126-141 57-64
5 152 3 7,62 115-123 52-56 121-133 55-60 129-144 59-65
5 152 4 10,16 118-126 54-57 124-136 56-62 132-148 60-67
5 152 5 12,7 121-129 55-59 127-139 58-63 135-152 61-69
5 152 6 15,24 124-133 56-60 130-143 59-65 138-156 63-71
5 152 7 17,78 128-137 58-62 134-147 61-67 142-161 64-73
5 152 8 20,32 132-141 60-64 138-152 63-69 147-166 67-75
5 152 9 22,86 136-145 62-66 142-156 64-71 151-170 68-77
5 152 10 25,4 140-150 64-68 146-160 66-73 155-174 70-79
5 152 11 27,94 144-154 65-70 150-165 68-75 159-179 72-81
6 183 0 0 148-158 67-72 154-170 70-77 164-184 74-83
6 183 1 2,54 152-162 69-73 158-175 72-79 168-189 76-86
6 183 2 5,08 156-167 71-76 162-180 73-82 173-194 78-88
6 183 3 7,62 160-171 73-78 167-185 76-84 178-199 81-90
6 183 4 10,16 164-175 74-79 172-190 78-86 182-204 83-93
Mujeres a partir de los 25 años de edad
Estatura con calzado Peso ideal (lb y kg) talones 1 pulg. Complexión Complexión Complexión
(2,54 cm) pequeña media grande
Pies cm Pulg. cm lb kg lb kg lb kg
4 122 10 25,4 92-98 42-44 96-107 44-49 104-119 47-54
4 122 11 27,94 94-101 43-46 98-110 44-50 106-122 48-55
5 152 0 0 96-104 44-47 101-113 46-51 109-125 49-57
5 152 1 2,54 99-107 45-49 104-116 47-53 112-128 51-58
5 152 2 5,08 102-110 46-50 107-119 49-54 115-131 52-59
5 152 3 7,62 105-113 48-51 110-122 50-55 118-134 54-61
5 152 4 10,16 108-116 49-53 113-126 51-57 121-138 55-63
5 152 5 12,7 111-119 50-54 116-130 53-59 125-142 57-64
5 152 6 15,24 114-123 52-56 120-135 54-61 129-146 59-66
5 152 7 17,78 118-127 54-58 124-139 56-63 133-150 60-68
5 152 8 20,32 122-131 55-59 128-143 58-65 137-154 62-70
5 152 9 22,86 126-135 57-61 132-147 60-67 141-158 64-72
5 152 10 25,4 130-140 59-64 136-151 62-68 145-163 66-74
5 152 11 27,94 134-144 61-65 140-155 64-70 149-168 68-76
6 183 0 0 138-148 63-67 144-159 65-72 153-173 69-78
Nota. En los pesos relacionados se incluye el peso de la ropa que se lleva en el interior de un local. Para saber el peso sin ropa hay que deducir entre 5 y 7 li-bras (entre 2,26 y 3,18 kg) para los hombres y entre 2 y 4 libras (entre 0,9 y 1,81 kg) para las mujeres.
Preparado por la Metropolitan Life Insurance Company, deducidos principalmente de datos del Build and Blood Pressure Study, Society of Actuaries, 1959.
�TABLA 21.1 Tablas estándar (1959) para el peso de hombres y mujeres según su estatura y complexión corporal.
03 Cap 16-21 • 549-776 17/1/07 16:39 Página 713

Difusión pulmonarLa difusión pulmonar, que es el intercambio degases que tiene lugar en los alvéolos, no varía enreposo ni durante la realización de ejercicios sub-máximos estandarizados después del entrena-miento. No obstante, aumenta durante la realiza-ción de ejercicios máximos. El flujo de sangrepulmonar (la sangre que viene del corazón en di-rección a los pulmones) parece aumentar despuésdel entrenamiento, especialmente el flujo hacia lasregiones superiores de los pulmones, cuando unapersona está sentada o de pie. Ello incrementa laperfusión pulmonar. Más sangre es llevada a lospulmones para el intercambio de gases y, al mis-mo tiempo, la ventilación aumenta por lo que selleva más aire a los pulmones. Ello significa quehabrá más alvéolos interviniendo activamente enla difusión pulmonar. El resultado neto es que ladifusión pulmonar aumenta.
Diferencia arteriovenosa de oxígenoEl contenido de oxígeno en la sangre arterialcambia muy poco con el entrenamiento. Auncuando la cantidad total de hemoglobina aumen-ta, la cantidad de ésta por unidad de sangre es lamisma o incluso ligeramente menor. La diferen-cia arteriovenosa de oxígeno (dif. a--vO2), no obs-tante, aumenta con el entrenamiento, especial-mente con niveles máximos de ejercicio. Esteincremento es la consecuencia de un menor con-tenido de oxígeno venoso mezclado. Ello signifi-ca que la sangre que vuelve al corazón, que esuna mezcla de sangre venosa de todas las partesdel cuerpo, no sólo de los músculos activos, con-tiene menos oxígeno del que habría en una perso-na no entrenada. Ello refleja una mayor extrac-ción de oxígeno al nivel de los tejidos y unadistribución más efectiva del volumen sanguíneototal (una mayor cantidad de sangre va a los teji-dos activos).
En resumen, el aparato respiratorio tiene unagran habilidad para llevar cantidades adecuadasde oxígeno al interior del cuerpo. Por esta razón,el sistema respiratorio casi nunca es un limitadorde los resultados relativos a la capacidad de resis-tencia. No es extraño que las adaptaciones al en-trenamiento más importantes observadas en elaparato respiratorio se manifiesten con claridaddurante la realización de ejercicios máximoscuando todos los sistemas están siendo forzadosal máximo.
Adaptaciones metabólicasal entrenamientoAhora que hemos visto los cambios producidospor el entrenamiento en el sistema cardiovasculary el aparato respiratorio, estamos preparados paraver cómo estos sistemas se integran con el meta-bolismo en los tejidos activos. Puesto que lasadaptaciones metabólicas al entrenamiento setrataron en el capítulo 6, nuestra discusión secentrará brevemente en:
• el umbral del lactato;• el índice de intercambio respiratorio, y• el consumo de oxígeno.
ADAPTACIONES CARDIOVASCULARES Y RESPIRATORIAS AL ENTRENAMIENTO
313
Aunque la mayor parte del incrementodel nVO2 máx es producto del aumento delgasto cardíaco y el riego sanguíneo de losmúsculos, el aumento de la diferenciaa--vO2 también desempeña un papel cla-ve. Este aumento en la diferencia a--vO2
es atribuible a una distribución más efi-caz de sangre arterial de los tejidos inac-tivos a los activos, para que más cantidadde la sangre que retorna a la aurícula de-recha pase por los músculos activos.
i El efecto combinado del mayor volumenoscilante y del mayor ritmo de la respi-ración es el incremento de la ventilaciónpulmonar en los esfuerzos máximos des-pués del entrenamiento.
i La difusión pulmonar a ritmos de es-fuerzo máximos aumenta, probablemen-te debido a la mayor ventilación y a lamayor perfusión pulmonar.
i La diferencia a--vO2 aumenta con el en-trenamiento, especialmente a nivelesmáximos de esfuerzo, lo cual refleja unamayor extracción de oxígeno por los te-jidos y una más efectiva distribución dela sangre.
02 Cap 07-15 • 221-548 17/1/07 16:29 Página 313

entrenamiento, se puede aumentar el número derepeticiones por serie.
Para entrenar los músculos y aumentar lafuerza, el énfasis debe ponerse en ejercitarse conmayores resistencias (mayor intensidad) y menosrepeticiones. Para desarrollar el aguante físico delos músculos, el énfasis se pone en usar menosresistencia y más repeticiones. Para aumentar lapotencia, parte de la sesión debe potenciar la ve-locidad del movimiento. El número de seriesprescrito en el entrenamiento resistido ha sidotradicionalmente tres series. Sin embargo, estu-dios más recientes muestran que se obtienen me-joras similares en la fuerza con una sola serie. Es-to, además de la forma de entrenar los músculos,se aborda con más detalle en este capítulo másadelante, en la sección «Diseño de programas deentrenamiento resistido».
Ahora que hemos tratado los principios bási-cos del entrenamiento resistido, echemos un vis-tazo a la respuesta de los músculos a este tipo deentrenamiento.
Ganancias de fuerza mediante el entrenamiento resistidoEl sistema neuromuscular es uno de los sistemasdel cuerpo que más responde al entrenamiento.Los programas de entrenamiento resistido pue-den generar aumentos sustanciales en la fuerza.En 3 a 6 meses, se pueden apreciar mejoras entreel 25% y el 100%, a veces incluso más. Estas me-joras parecen ser parecidas cuando comparamoshombres con mujeres, niños con adultos, y ancia-nos con adultos jóvenes y de mediana edad. Elmúsculo es muy plástico, aumenta rápidamentede tamaño y fuerza con el ejercicio, y disminu-ye de tamaño y fuerza cuando se inmoviliza. Elresto de este capítulo se dedica a la forma en quese producen estos cambios. ¿Cómo llegar a sermás fuertes? ¿Qué adaptaciones fisiológicas seproducen que permiten desarrollar más nivelesde fuerza?
Mecanismos de las gananciasde la fuerza muscularDurante muchos años se supuso que las ganan-cias de fuerza eran el resultado directo de incre-mentos en el tamaño muscular (hipertrofia). Estasuposición era lógica porque la mayoría de los
que se sometían regularmente a entrenamientosde fuerza eran hombres, que con frecuencia de-sarrollaban músculos grandes y abultados. Asi-mismo, una extremidad inmovilizada con yesodurante semanas o meses comienza a perder tama-ño (atrofia) y fuerza casi inmediatamente. Lasganancias de tamaño muscular son generalmenteparalelas a ganancias de fuerza, y las pérdidas enel tamaño muscular tienen una alta correlacióncon pérdidas de fuerza. Por lo tanto, estamos ten-tados de concluir que existe una relación causa-efecto entre el tamaño y la fuerza muscular. Noobstante, esta última implica mucho más que elmero tamaño muscular.
Numerosos informes de los medios de co-municación indican que la gente puede realizarhazañas sobrehumanas de fuerza al estar someti-dos a una gran tensión psicológica. Las camisasde fuerza se diseñaron específicamente para con-trolar a pacientes en los hospitales mentales querepentinamente perdían los estribos y que era im-posible frenar. Incluso el mundo de los deportesse enorgullece de haber presentado ejemplos decompeticiones deportivas sobrehumanas, como lade Bob Beamon con un salto de 8,9 m en los Jue-gos Olímpicos de 1968, ¡un salto que excedió elanterior récord del mundo en casi 60 cm! Nor-malmente, los récords del mundo se superan porunos centímetros, o más frecuentemente por me-ras fracciones de centímetro. El récord de Beamonestuvo imbatido hasta 1991.
Las mujeres experimentan similares ganan-cias de fuerza a las de los hombres que participanen el mismo programa de entrenamiento, pero lasmujeres no sufren tanta hipertrofia (ver capítulo18). De hecho, algunas mujeres han doblado sufuerza sin ningún cambio observable en su tama-ño muscular. Por lo tanto, las ganancias de fuerzano requieren hipertrofia.
Esto no significa que el tamaño muscularcarezca de importancia en el potencial de fuerzafinal del músculo. El tamaño es extremadamen-te importante, tal como revelan los récordsmundiales en levantamiento de peso de compe-tición masculinos y femeninos existentes, quese muestran en la figura 3.3. A medida que lacategoría del peso aumenta (implicando un ma-yor tamaño muscular), también lo hace el regis-tro del peso total levantado. No obstante, ejem-plos de fuerza sobrehumana y estudios sobremujeres indican que los mecanismos asociadosa las ganancias de fuerza son muy complejos ypor ahora no se entienden del todo. ¿Cómo, en-tonces, pueden explicarse las ganancias de fuer-
FISIOLOGÍA DEL ESFUERZO Y DEL DEPORTE
96
01 Crédits+Cap 01-06 • 001-220 17/1/07 16:20 Página 96

Frecuencia cardíacaAhora que hemos examinado un aspecto del gas-to cardíaco, podemos volver nuestra atención a laotra mitad de la ecuación: la frecuencia cardíaca.Determinados estudios en los que el consumo deoxígeno del corazón ha sido controlado directa-mente han demostrado que la frecuencia cardía-ca, en reposo y durante el ejercicio, es un bueníndice de la intensidad con la que está trabajandoel corazón. Puesto que el músculo activo exigemás oxígeno que el músculo en reposo, no es sor-prendente que el consumo de oxígeno del cora-zón, y, por lo tanto, la intensidad del esfuerzo querealiza, estén directamente relacionados con elritmo de la contracción del corazón. Examine-mos cómo afecta el entrenamiento al ritmo delcorazón.
Frecuencia cardíaca en reposo
La frecuencia cardíaca en reposo se reduce nota-blemente como consecuencia del entrenamientode fondo. Si somos una persona sedentaria conuna frecuencia cardíaca inicial en reposo de 80latidos/min, esta frecuencia cardíaca se reduciráaproximadamente en una pulsación por minutocada semana durante las primeras semanas de en-trenamiento. Por lo que, después de 10 semanasde entrenamiento moderado de resistencia, nuestrafrecuencia cardíaca en reposo deberá pasar de 80 a70 latidos/min. Los mecanismos verdaderamenteresponsables de esta reducción no se conocen deltodo, pero el entrenamiento parece incrementar laactividad parasimpática en el corazón, reduciendoal mismo tiempo la actividad simpática.
ADAPTACIONES CARDIOVASCULARES Y RESPIRATORIAS AL ENTRENAMIENTO
303
Un corazón más fuerte y la disponibili-dad de un mayor volumen de sangre pa-recen ser los factores responsables de losincrementos del volumen sistólico en re-poso y durante la realización de ejer-cicios submáximos y máximos después dela ejecución de un programa de entrena-miento de fondo.
i Después del entrenamiento de fondo, elvolumen sistólico aumenta en reposo,durante la realización de ejercicios a unnivel submáximo y durante la ejecuciónde esfuerzos máximos.
i Un factor importante que lleva a aumen-tar el volumen sistólico es un incremen-to del volumen diastólico final, por cau-sa probablemente de un aumento delplasma sanguíneo.
i Otro factor importante es la mayor con-tractilidad ventricular izquierda. Ésta esproducida por la hipertrofia del músculocardíaco y por un mayor retroceso elás-tico, que son el resultado de un mayorestiramiento de la cámara con más llena-do diastólico. La reducción de la tensiónarterial general disminuye la resistenciaal flujo de la sangre bombeada fuera delventrículo izquierdo.
Figura 9.5 Diferencias en (a) volumen diastólico final (VDF), (b) volumen sistólico final (VSF) y (c) fracción de eyección con intensidadescrecientes de esfuerzo en individuos antes y después del entrenamiento de resistencia.Adaptado con autorización de A.A. Ehsani y col., 1991, «Exercise training improves left ventricular systolic function in older men, Circulation 83: 96-103.
a b c
VD
F (m
l)
Esfuerzo (vatios)
PreentrenamientoPosentrenamiento
VS
F (m
l)
Esfuerzo (vatios)
% f
racc
ión
de
eyec
ció
n
Esfuerzo (vatios)
02 Cap 07-15 • 221-548 17/1/07 16:29 Página 303

FISIOLOGÍA DEL ESFUERZO Y DEL DEPORTE
36
Cuando nuestros corazones laten, cuando una co-mida que hemos tomado se desplaza a lo largo denuestro intestino y cuando movemos cualquierparte de nuestro cuerpo, los músculos intervie-nen. La miríada de funciones del sistema muscu-lar las ejecutan solamente tres tipos de músculos(figura 1.1): liso, cardíaco, esquelético.
Los músculos lisos reciben la denominaciónde músculos involuntarios, porque no están di-rectamente bajo nuestro control consciente. Sehallan en las paredes de la mayoría de vasos san-guíneos, capacitándolos para contraerse o dilatar-se a fin de regular el riego sanguíneo. Se hallantambién en las paredes de la mayoría de órganosinternos, permitiéndoles contraerse y relajarse,quizá para mover la comida a través del tracto di-gestivo, para expulsar la orina o para dar a luz aun niño.
El músculo cardíaco se halla solamente en elcorazón, abarcando la mayor parte de la estructurade éste. Comparte algunas características con los
músculos esqueléticos, pero,igual que los músculos lisos,
no se halla bajo nuestrocontrol consciente. El
músculo cardíaco secontrola a sí mismo,con un mero afina-miento por partede los sistemasnervioso y endo-crino. El músculo
cardíaco se analizaplenamente en el ca-
pítulo 7. Generalmente,
prestamos sólo atención aaquellos músculos que pode-
mos controlar conscientemente.Estos son los músculos esqueléti-cos, o voluntarios, denominadosasí porque se unen a y mueven elesqueleto. Conocemos muchos deestos músculos por sus nombres–deltoides, pectoral, bíceps–, peroel cuerpo humano contiene más de
Jeremy Schill, de 9 años de edad, pesaba solamente 29,5 kg, pero esto no le impi-dió levantar la parte posterior del coche familiar de 1.860 kg del pecho de su pa-dre. El coche se había deslizado del gato mecánico mientras Rique Schill estabatrabajando debajo de él, atrapándolo debajo del eje posterior. Cuando Jeremycomprendió que su padre se estaba asfixiando lentamente, el muchacho levantólo bastante el coche como para permitir respirar a su padre y para que su madrepudiese poner otro gato debajo del parachoques posterior.
�
Figura 1.1 Microfotografíasde músculo (a) esquelético,(b) cardíaco y (c) liso.
a
b
c
01 Crédits+Cap 01-06 • 001-220 17/1/07 16:19 Página 36

Aunque la herencia es un factor de riesgo dehipertensión, probablemente desempeña un papelmucho menos importante que muchos de losotros factores propuestos. Debemos recordar quelos factores de estilo de vida son frecuentemen-te muy similares dentro de una misma familia.
Recientemente, los científicos han demostra-do un gran interés por la existencia de una posi-ble relación entre hipertensión, obesidad, diabe-tes del tipo II y las enfermedades cardíacascoronarias a través del camino común de la resis-tencia a la insulina o de la deteriorada acción dela insulina. Pero la obesidad también ha sido es-tablecida como un factor independiente de riesgode hipertensión. Numerosos estudios han mostra-do reducciones sustanciales en la tensión arterialcon la pérdida de peso en los pacientes hiperten-sos. Asimismo, aunque la ingestión de sodio seha relacionado tradicionalmente con la hiperten-sión, esta relación se limita probablemente aquienes son sensibles a la sal.
ENFERMEDADES CARDIOVASCULARES Y ACTIVIDAD FÍSICA
693
El síndrome metabólico
El síndrome metabólico es un término usado para relacionar la enfermedad coronaria, la hiperten-sión, la diabetes tipo II y la obesidad del tren superior del cuerpo con la resistencia a la insulina yla hiperinsulinemia. Este síndrome también se ha denominado síndrome x y síndrome de la civili-zación. No está del todo claro dónde se inicia el síndrome, pero se ha observado que la obesidadde la parte superior del cuerpo se asocia con la resistencia a la insulina, y que ésta está muy rela-cionada con el aumento del riesgo de enfermedad coronaria, hipertensión y diabetes tipo II. Seconvirtió en un tema principal de investigación en la década de 1990, y los resultados de esa in-vestigación deberían ayudar a entender mejor la fisiopatología de estas enfermedades y sus inte-rrelaciones.
i Los factores de riesgo de enfermedadesde las arterias coronarias que no pode-mos controlar son la herencia (e historiafamiliar), el sexo masculino y el enveje-cimiento. Los factores de riesgo quepueden controlarse son los niveles altosde lípidos en sangre, la hipertensión, elfumar cigarrillos, la inactividad física, laobesidad, la diabetes y el estrés. Los fac-tores primarios de riesgo son los que handemostrado estar fuertemente relaciona-
dos con las enfermedades. Para las en-fermedades de las arterias coronariasson el fumar, la hipertensión, los niveleselevados de lípidos en sangre, la inacti-vidad física y la obesidad.
i Se cree que el LDL-C es el responsablede la acumulación de colesterol en lasparedes arteriales. El VLDL-C está cadavez más implicado en la generación deenfermedades de las arterias coronarias.Sin embargo, el HDL-C actúa como unbasurero, llevándose el colesterol acu-mulado en las paredes de los vasos. Portanto, los niveles elevados de HDL-Cproporcionan cierto grado de proteccióncontra la enfermedad coronaria.
i La razón entre colesterol total y HDL-Cpuede ser el mejor indicador de riesgopersonal para las enfermedades de lasarterias coronarias. Los valores inferio-res a 3,0 reflejan un bajo riesgo, y losvalores superiores a 5,0 reflejan un ries-go elevado.
i Entre los factores de riesgo de hiperten-sión que no se pueden controlar secuentan la herencia, el envejecimientoy la raza. Los que podemos controlarson la resistencia a la insulina, la obesi-dad, la dieta (exceso de sodio), el uso deanticonceptivos orales y la inactividadfísica.
03 Cap 16-21 • 549-776 17/1/07 16:39 Página 693

biantes dentro del abdomen y del tórax compri-men la sangre en las venas, lo que estimula su re-torno mediante una acción de ordeño. Ésta es unaparte esencial del retorno venoso. Del mismomodo, las contracciones musculares durante elejercicio producen también este tipo de acción deordeño para facilitar el retorno venoso.
La difusión pulmonar requiere dos cosas:aire que lleve oxígeno hacia los pulmones y san-gre que reciba el oxígeno y elimine el dióxidode carbono. Durante la ventilación pulmonar sellevó aire a los pulmones; ahora debe producirseel intercambio de gases entre este aire y la san-gre.
La sangre de la mayor parte del cuerpo re-gresa a través de la vena cava al lado pulmonar(derecho) del corazón. Desde el ventrículo dere-cho, esta sangre es bombeada a través de la arte-ria pulmonar hacia los pulmones, abriéndose ca-mino al final hacia los capilares pulmonares.Estos capilares forman una densa red alrededorde los sacos alveolares. Estos vasos son lo bas-tante pequeños para que los glóbulos rojos debanpasar por ellos de uno en uno, exponiendo cadacélula al tejido pulmonar circundante. Aquí esdonde tiene lugar la difusión pulmonar.
Membrana respiratoriaEl intercambio de gases entre el aire en los alvéo-los y la sangre en los capilares pulmonares tienelugar a través de la membrana respiratoria (tam-bién llamada membrana alveolocapilar). Estamembrana, representada en la figura 8.3, se com-pone de:
• la pared alveolar;
• la pared capilar, y
• sus membranas subyacentes.
La membrana respiratoria es muy delgada,midiendo tan sólo entre 0,5 y 4,0 µm. En conse-cuencia, los gases en los casi 300 millones de al-véolos están muy próximos a la sangre circulantea través de los capilares. No obstante, esta mem-brana ofrece una barrera potencial para el inter-cambio de gases. Examinemos cómo se produceel repetido intercambio de gases.
Presiones parciales de los gasesEl aire que respiramos es una mezcla de gases.Cada uno ejerce una determinada presión propor-cional a su concentración en la mezcla de gases.Las presiones individuales de cada gas en unamezcla reciben el nombre de presiones parcia-les. Según la ley de Dalton, la presión total deuna mezcla de gases es igual a la suma de las pre-
REGULACIÓN RESPIRATORIA DURANTE EL EJERCICIO
267
i La ventilación pulmonar (respiración) esel proceso por el que se introduce y seextrae aire de los pulmones. Tiene dosfases: inspiración y espiración.
i La inspiración es un proceso activo me-diante el cual el diafragma y los múscu-los intercostales externos incrementansus dimensiones, y, por lo tanto, el volu-men de la caja torácica. Esto reduce lapresión en los pulmones y lleva aire ha-cia ellos.
i La espiración normal es un proceso pa-sivo. Los músculos inspiratorios se rela-jan y el tejido elástico de los pulmonesse encoge, devolviendo la caja torácica asus dimensiones normales, más peque-ñas. Ello incrementa la presión en lospulmones y fuerza al aire a salir de es-tos.
i La inspiración y la espiración forzadas olaboriosas son procesos activos, depen-dientes de acciones musculares.
Difusión pulmonarEl intercambio de gases en los pulmones, deno-minado difusión pulmonar, sirve para dos finali-dades importantes:
1. Reemplaza el aporte de oxígeno de la san-gre que se ha agotado al nivel de los teji-dos donde se utiliza para la producción deenergía oxidativa.
2. Elimina el dióxido de carbono de la san-gre venosa que regresa.
02 Cap 07-15 • 221-548 17/1/07 16:29 Página 267

Equilibrio de electrólitosdurante el ejercicioTal como se ha mencionado con anterioridad, lafunción corporal normal depende del equilibrioentre el agua y los electrólitos. Hemos discutidolos efectos de la pérdida de agua sobre el rendi-miento.
Ahora podemos volver nuestra atención hacialos efectos del otro componente de este delicadoequilibrio: los electrólitos. Cuando nuestro cuer-po pierde grandes cantidades de agua, como porejemplo durante el ejercicio, el equilibrio entre elagua y los electrólitos puede verse alterado conrapidez. En las secciones siguientes, examinare-mos los efectos del ejercicio sobre el equilibriode los electrólitos. Nuestro punto de atención se-rán las dos rutas principales de pérdida de elec-trólitos: la sudoración y la producción de orina.
Pérdida de electrólitos en el sudor
El sudor humano es un filtrado del plasma de lasangre, por lo que contiene muchas sustanciasque se hallan en ella, tales como el sodio (Na+), elcloro (Cl–), el potasio (K+), el magnesio (Mg2+) yel calcio (Ca2+). Aunque el sudor tiene un gustosalado, contiene muchos menos minerales que elplasma y otros fluidos corporales. De hecho, esagua en un 99%.
El sodio y el cloro son los iones predominan-tes en el sudor y en la sangre. Tal como se ve enla tabla 13.8, las concentraciones de sodio y decloro en el sudor son aproximadamente un terciode las encontradas en el plasma y cinco veces lasencontradas en los músculos. También se mues-tra la osmolaridad de cada uno de estos tres flui-dos, que es la proporción de solutos (tales comolos electrólitos) en relación con el fluido. La con-
FISIOLOGÍA DEL ESFUERZO Y DEL DEPORTE
462
excreción en un esfuerzo por prevenir ladeshidratación.
i Cuando la deshidratación es superior al 2%del peso corporal, el rendimiento físicoprolongado se ve notablemente perjudi-cado. Asimismo, en respuesta a la des-hidratación, la frecuencia cardíaca y latemperatura corporal aumentan.
Función fisiológicaCardiovascularVolumen sanguíneo
/volumen plasmático ↓ ↓ªGasto cardíaco ↓ ?Volumen sistólico ↓ ?Frecuencia cardíaca ↑ ?
MetabólicaCapacidad aeróbica: V·O2 máx ↔, ↓ ↔ªPotencia anaeróbica:
test de Wingate ↔, ↓ ↔, ↓Capacidad anaeróbica:
test de Wingate ↔, ↓ ↔, ↓Lactato en sangre, valor máximo ↓ ↓ªCapacidad de amortiguación
de la sangre ↓ ?Umbral del lactato, velocidad a LT ↓ ?Glucógeno en músculos e hígado ↓ ↓Glucemia durante el ejercicio posible ↓ ?Degradación de proteínas
con el ejercicio posible ↑ ?
Termorregulación y equilibrio hídricoElectrólitos, músculo y sangre ↓ ↔Temperatura central
durante el ejercicio ↑ ?Tasa de sudoración ↓, inicio diferido ?Riego sanguíneo cutáneo ↓ ?
RendimientoFuerza muscular ↔, ↓ ↔, ↓Resistencia muscular ↔, ↓ ↔, ↓Potencia muscular ? ↓b
Velocidad de movimiento ? ?Duración de la carrera hasta
agotamiento ↓ ?Trabajo total realizado ↓ ↓ªPruebas de simulación de lucha libreª ↓ ↔, ↓b
Nota. Los datos de esta tabla se obtuvieron de las siguientesrevisiones:Fogelholm (1994)25, Horswill (1994)29, Keller, Tolly & Freedson(1994)38, y Opplinger y col. (1996)44.
↓ = reducción ; ↑ = incremento; ↔ = ningún cambio conocido o vueltaa los valores normales; ? = desconocido.
ª De Burge, Carey y Payne (1993).10
b De Oopik y col. (1996)43.
Variables Deshidratación Rehidratación
�TABLA 13.7 Alteraciones de la función fisiológica y elrendimiento por deshidratación
02 Cap 07-15 • 221-548 17/1/07 16:31 Página 462

Cuando sobreviene la muerte durante el ejer-cicio a personas de 35 años o mayores, ésta sueleser la consecuencia de una arritmia cardíaca cau-sada por aterosclerosis de las arterias coronarias.Por otro lado, los que tienen menos de 35 añostienen más probabilidades de morir de una cardio-miopatía hipertrófica (corazón enfermo agranda-do, por lo general transmitido genéticamente), deun aneurisma aórtico o de miocarditis (inflama-ción del miocardio).
Expresiones clave
apoplejía
arteriosclerosis
aterosclerosis
enfermedad cardíaca congénita
enfermedad cardíaca reumática
enfermedad de las arterias coronarias
enfermedad vascular periférica
enfermedades cardíacas valvulares
estrías de grasa
factor de crecimiento derivado de las plaquetas
factores de riesgo primarios
fisiopatología
hipertensión
infarto cerebral
infarto de miocardio
insuficiencia cardíaca congestiva
isquemia
lípidos en sangre
lipoproteínas
lipoproteínas de alta densidad (HDL)
lipoproteínas de alta densidad colesterol (HDL-C)
lipoproteínas de baja densidad colesterol (LDL-C)
lipoproteínas de muy baja densidad (VLDL)
lipoproteínas de muy baja densidad colesterol(VLDL-C)
placa
síndrome metabólico
tensión arterial diastólica
tensión arterial sistólica
triglicéridos
FISIOLOGÍA DEL ESFUERZO Y DEL DEPORTE
704
i Las muertes durante el ejercicio sonraras, aunque suelen recibir una granatención informativa.
i Las muertes durante el ejercicio en per-sonas de más de 35 años suelen debersea una arritmia cardíaca resultante de unaaterosclerosis.
i Las muertes durante el ejercicio en per-sonas de menos de 35 años suelen de-berse a una cardiopatía hipertrófica, unaneurisma aórtico o una miocarditis.
ConclusiónEn este capítulo, hemos visto lo importante quees la actividad física en la prevención de las en-fermedades cardiovasculares, especialmente delas enfermedades de las arterias coronarias y lahipertensión. Hemos discutido la prevalencia deestos trastornos, los factores de riesgo asociadoscon cada uno de ellos y cómo la actividad físicapuede ayudar a reducir nuestros riesgos persona-les. En el capítulo siguiente, continuaremos exa-minando los efectos del ejercicio sobre nuestrasalud, dirigiendo nuestra atención hacia la obesi-dad y la diabetes.
03 Cap 16-21 • 549-776 17/1/07 16:39 Página 704

52. Siscovick, D.S., Weiss, N.S., Fletcher,R.H., & Lasky, T. (1984). The incidence of pri-mary cardiac arrest during vigorous exercise.New England Journal of Medicine, 311, 874-877.
53. Steinberg, D., Parthasarathy, S., Carew,T.E., Khoo, J.C., & Witztum, J.L. (1989). Be-yond cholesterol: Modifications of low-densitylipoprotein that increase its atherogenicity. NewEngland Journal of Medicine, 320, 915-924.
54. Tanasescu, M., Leitzmann, M.F., Rimm,E.B., Willett, W.C., Stampfer, M.J., & Hu, F.B.(2002). Exercise type and intensity in relation tocoronary heart disease in men. Journal of theAmerican Medical Association, 288, 1.994-2.000.
55. Thompson, P.D. (1982). Cardiovascularhazards of physical activity. Exercise and SportSciences Reviews, 10, 208-235.
56. Tipton, C.M. (1984). Exercise, training,and hypertension. Exercise and Sport SciencesReviews, 12, 245-306.
57. Tipton, C.M. (1991). Exercise trainingand hypertension: An update. Exercise and SportSciences Reviews, 19, 447-505.
58. Tran, Z.V., & Weltman, A. (1985). Diffe-rential effects of exercise on serum lipid and li-poprotein levels seen with changes in bodyweight. Journal of the American Medical Asso-ciation, 254, 919-924.
59. Williams, P.T. (2001). Physical fitnessand activity as separate heart disease risk factors:A meta-analysis. Medicine and Science in Sportsand Exercise, 33, 754-761.
60. Wilmore, J.H., Constable, S.H., Stan-forth, P.R., Tsao, W.Y., Rotkis, T.C., Paicius,R.M., Mattern, C.M., & Ewy, G.A. (1982). Pre-valence of coronary heart disease risk factors in13- to 15-year-old boys. Journal of Cardiac Re-habilitation, 2, 223-233.
61. Wilmore, J.H., & McNamara, J.J.(1974). Prevalence of coronary heart disease riskfactors in boys 8 to 12 years of age. Journal ofPediatrics, 84, 527-533.
Lecturas seleccionadas
American Heart Association. (1996). Cardiovas-cular preparticipation screening of compe-titive athletes: Exercise, sudden death. Medi-cine and Science in Sports and Exercise, 28,1.445-1.452.
Andersen, L.B., & Hippe, M. (1996). Coronaryheart disease risk factors in the physicallyactive. Sports Medicine, 22, 213-218.
Caspersen, CJ. (1989). Physical activity epide-miology: Concepts, methods, and applica-tions to exercise science. Exercise and SportSciences Reviews, 17, 423-473.
Franklin, B.A., & Shephard, R.J. (2000). Avoi-ding repeat cardiac events. Physician andSportsmedicine, 28(9), 31-58.
Harris, S.S., Caspersen, C.J., DeFriese, G.H., &Estes, E.H., Jr. (1989). Physical activitycounseling for healthy adults as a primarypreventive intervention in the clinical set-ting. Journal of the American Medical Asso-ciation, 261, 3.590-3.598.
Laughlin, M.H., & McAllister, R.M. (1992).Exercise training-induced coronary vascularadaptation. Journal of Applied Physiology,73, 2.209-2.225.
Leon, A.S. (ed.). (1997). Physical activity andcardiovascular health: A national consen-sus. Champaign, IL: Human Kinetics.
Morris, C.K., & Froelicher, V.F. (1993). Cardio-vascular benefits of improved exercise capa-city. Sports Medicine, 16, 225-236.
Morris, J.N., & Hardman, A.E. (1997). Walkingto health. Sports Medicine, 23, 306-332.
Nieman, D.C. (1998). The exercise-health con-nection. Champaign, IL: Human Kinetics.
Pollock, M.L., & Schmidt, D.H. (eds.). (1995).Heart disease and rehabilitation (3.ª ed.).Champaign, IL: Human Kinetics.
Schaible, T.F., & Scheuer, J. (1985). Cardiacadaptations to chronic exercise. Progress inCardiovascular Disease, 27, 297-324.
FISIOLOGÍA DEL ESFUERZO Y DEL DEPORTE
708
03 Cap 16-21 • 549-776 17/1/07 16:39 Página 708

Ejercicio en ambientes fríosLa creciente participación en actividades deporti-vas como el triatlón, la inmersión con escafandra,las carreras, el ciclismo y la natación de fondo hamotivado un nuevo interés y preocupación por elejercicio en ambientes fríos. Además, algunasocupaciones requieren que los empleados traba-jen en condiciones de frío que pueden limitar surendimiento. Por estos motivos, la comprensiónde las respuestas fisiológicas y de los riesgos pa-ra la salud asociados con el estrés por el frío sontemas importantes en la ciencia del ejercicio.Aquí definimos el estrés por el frío como cual-quier condición ambiental que produce una pér-dida de calor corporal que amenaza la homeosta-sis. En el análisis siguiente nos centraremos enlos dos principales estresores del frío: el aire y elagua.
El hipotálamo tiene una temperatura fija deaproximadamente 37 oC, pero las fluctuacionesdiarias en la temperatura corporal pueden ser dehasta 1 oC. Una reducción en la temperatura de lapiel o de la sangre facilita un impulso al centrotermorregulador (el hipotálamo) para activar losmecanismos que conservarán la temperatura cor-poral y que aumentarán la producción de calor.Los principales medios con los que nuestro cuer-po evita un excesivo enfriamiento son: los esca-lofríos, la termogénesis de origen distinto a losescalofríos y la vasoconstricción periférica.
Puesto que estos mecanismos de produccióny conservación de calor con frecuencia son inade-cuados, debemos valernos de la ropa y de la grasasubcutánea para facilitar el aislamiento del am-biente de nuestros tejidos corporales profundos.
Los escalofríos –las contracciones muscula-res no controladas tratadas antes– pueden haceraumentar 4 ó 5 veces el ritmo de producción decalor corporal en reposo. La termogénesis deorigen distinto a los escalofríos supone la esti-mulación del metabolismo por el sistema nervio-so simpático. El creciente ritmo metabólico au-menta la intensidad de la producción de calorinterno.
La vasoconstricción periférica se producecomo consecuencia de la estimulación simpáticade los músculos lisos que rodean las arteriolas dela piel. Esta estimulación hace que los músculos secontraigan, lo cual contrae las arteriolas y reduceel riego sanguíneo hacia la periferia del cuerpo,previniendo una pérdida innecesaria de calor. Elritmo metabólico de las células epiteliales también
REGULACIÓN TÉRMICA Y EJERCICIO
353
Los cambios cardiovasculares general-mente tienen lugar en los primeros 3 ó 5días, pero los relativos a los mecanismosde sudoración requieren mucho mástiempo, hasta 10 días.
i La exposición repetida al estrés por ca-lor produce una mejora gradual en nues-tra capacidad para perder el exceso decalor. Este proceso de adaptación se lla-ma aclimatación al calor.
i El ritmo de sudoración aumenta en áreasque están bien expuestas y que son las máseficaces para facilitar la pérdida de calor.Esto reduce la temperatura de la piel, e in-crementa el gradiente térmico desde la par-te interna a la externa del cuerpo, lo que fa-cilita la pérdida de calor.
i La temperatura central y la frecuenciacardíaca durante el ejercicio se reducen,mientras que el volumen sistólico au-menta con la aclimatación al calor. Elvolumen plasmático se eleva, lo cualcontribuye a un aumento del volumensistólico que ayuda a enviar más sangrea los músculos activos y a la piel cuandoes necesario.
i La aclimatación al calor reduce el ritmode uso del glucógeno muscular y retrasala aparición de la fatiga.
i La aclimatación al calor requiere hacerejercicio en ambientes calurosos, y nomeramente exponerse al calor.
i El grado de aclimatación al calor conse-guido depende de las condiciones a lasque hemos estado expuestos durante ca-da sesión, la duración de la exposición yel ritmo de producción de calor interno.
02 Cap 07-15 • 221-548 17/1/07 16:30 Página 353

Los pulmones no están directamente adheri-dos a las costillas. Más bien, están suspendidospor los sacos pleurales. Estos sacos envuelven lospulmones y contienen una fina capa de fluidopleural que reduce la fricción durante los movi-mientos respiratorios. Además, estos sacos estánconectados a los pulmones y a la superficie inte-rior de la caja torácica, haciendo que los pulmo-nes adopten la forma y el tamaño de la caja cuan-do el pecho se expande y se contrae.
Estas relaciones entre los pulmones, los sa-cos pleurales y la caja torácica determinan el flu-jo del aire hacia dentro y fuera de los pulmones.Examinemos las dos fases implicadas: inspira-ción y espiración.
InspiraciónLa inspiración es un proceso activo que implicaal diafragma y a los músculos intercostales exter-
nos. En la figura 8.2 se muestra la dinámica de lainspiración. La figura 8.2a muestra las dimensio-nes en reposo de los pulmones y de la caja toráci-ca. La figura 8.2b indica los movimientos que seproducen durante la inspiración. Los músculosintercostales externos mueven las costillas y elesternón. Las costillas oscilan hacia arriba y aba-jo, de modo muy similar al movimiento del asade un cubo. Al mismo tiempo, el diafragma secontrae y se aplasta hacia el abdomen. Estas ac-ciones se muestran en la figura 8.2b.
Estas acciones aumentan las tres dimensio-nes de la caja torácica, que expanden a su vez lospulmones. Cuando ocurre esto, el aire del interiortiene más espacio que llenar, por lo que la pre-sión dentro de los pulmones se reduce, tal comose muestra en la figura 8.2c. En consecuencia, lapresión en los pulmones (presión intrapulmonar)es inferior a la presión del aire fuera del cuerpo.Puesto que el tracto respiratorio está abierto ha-
REGULACIÓN RESPIRATORIA DURANTE EL EJERCICIO
265
Figura 8.1 (a) La anatomía del sistema respiratorio, que muestra las vías respiratorias (es decir, fosas nasales, faringe, tráquea y bron-quios). (b) La imagen aumentada del alvéolo muestra las regiones de intercambio de gases entre el alvéolo y la sangre pulmonar en los ca-pilares.
Alvéolo
Capilares pulmonares
Red capilar en la superficiedel alvéolo
Pulmones
Bronquiolo
Faringe
Cavidad nasal
Bronquiosprimarios
LaringeTráquea
02 Cap 07-15 • 221-548 17/1/07 16:29 Página 265

Tratamiento del síndromede sobreentrenamientoAunque las causas del deterioro del rendimientocon el síndrome de sobreentrenamiento no estánclaras, aparentemente, la intensidad o la velocidaddel entrenamiento es un factor estresante máspotente que el volumen de entrenamiento. La re-cuperación después de sufrir el síndrome de so-breentrenamiento es posible mediante una nota-ble reducción en la intensidad del entrenamientoo mediante un reposo absoluto. Aunque la mayo-ría de los entrenadores recomiendan unos pocosdías de entrenamiento fácil, los deportistas sobre-entrenados se recuperan con mayor rapidez sidescansan completamente entre 3 y 5 días o sihacen ejercicios de baja intensidad. En algunoscasos, puede ser preciso aconsejar a los deportis-tas para ayudarles a hacer frente a otros motivosde tensión en sus vidas que pueden estar contri-buyendo a esta condición.
El mejor modo de minimizar el riesgo de apa-rición del síndrome de sobreentrenamiento es se-guir procedimientos de entrenamiento cíclico, y al-ternar períodos de entrenamiento fácil, moderado yduro. Aunque la tolerancia individual varía tremen-damente, incluso los deportistas más fuertes tienenperíodos en los que son susceptibles de padecer elsíndrome de sobreentrenamiento. En general, 1 ó 2días de entrenamiento intenso deben ir seguidos porun número igual de días de entrenamiento aeróbicosuave. Asimismo, una semana o dos de entrena-miento duro deben ir seguidas por una semana deesfuerzo reducido con poco o ningún énfasis en elejercicio anaeróbico.
Los deportistas que practican deportes querequieren tener capacidad de resistencia (como,por ejemplo, los nadadores, los ciclistas y los co-rredores) deben prestar una atención especial a suconsumo de hidratos de carbono. Días repetidosde entrenamiento duro producen una reduccióngradual del glucógeno muscular. A menos que es-tos deportistas consuman una cantidad extra dehidratos de carbono durante estos períodos, susreservas de glucógeno muscular y hepático pue-den agotarse. En consecuencia, las fibras muscu-lares más intensamente movilizadas no serían ca-paces de generar la energía necesaria para elejercicio.
FISIOLOGÍA DEL ESFUERZO Y DEL DEPORTE
422
Figura 12.11 Corredor equipado con un controlador de la frecuen-cia cardíaca. La cinta en el pecho recoge y transmite impulsos eléc-tricos desde el corazón hasta el dispositivo de memoria que llevaen la muñeca. Después de la carrera, se muestran los registros parainterpretarlos.
i El sobreentrenamiento es intentar hacerun mayor esfuerzo del que físicamentesomos capaces de hacer. El sobreentre-namiento lleva a una reducción de la ca-pacidad de rendimiento.
i Los síntomas del síndrome de sobreen-trenamiento son subjetivos, y muchos deellos también se presentan en el entrena-miento regular, lo cual dificulta la pre-vención y el diagnóstico del sobreentre-namiento.
i Entre las posibles explicaciones del sín-drome de sobreentrenamiento estáncambios en el funcionamiento de las di-visiones del sistema nervioso autónomo,respuestas endocrinas alteradas y supre-sión de la función inmunológica.
i Muchos signos y síntomas potencialesdel sobreentrenamiento podrían usarse
02 Cap 07-15 • 221-548 17/1/07 16:30 Página 422

Capítulo 15: © Human Kinetics; © Empics;© Human Kinetics.
Capítulo 16: © Fotógrafo/Bruce Coleman, Inc.;© Human Kinetics; © Human Kinetics.
Capítulo 17: © Human Kinetics; © Human Ki-netics; © Human Kinetics.
Capítulo 18: © Human Kinetics; © Empics;© Human Kinetics.
Capítulo 19: © Human Kinetics; © 2002 iPhoto-News.com; © Human Kinetics.
Capítulo 20: © Human Kinetics; © Custom Me-dical Stock; Jim Whitmer.
Capítulo 21: © Human Kinetics; © Human Ki-netics; Por cortesía de Lifescan de Johnson yJohnson.
CRÉDITOS FOTOGRÁFICOS
XV
01 Crédits+Cap 01-06 • 001-220 17/1/07 16:17 Página XV


















