
Fisiologia I. Bloque 1-Introduccion a La Fisiologia
62
Fisiología I Alberto Gómez Esteban Fisiología I Alberto Gómez Esteban 2º Medicina
-
Upload
victoria1999 -
Category
Documents
-
view
60 -
download
1
Transcript of Fisiologia I. Bloque 1-Introduccion a La Fisiologia

Fisiología I
Alberto Gómez Esteban
Fisiología I
Alberto Gómez Esteban 2º Medicina
Fisiología I
Alberto Gómez Esteban 2
Bloque I
Introducción a la fisiología
− Tema 1. Homeostasis, medio interno y sistemas de control biológico
− Tema 2. Membrana plasmática
− Tema 3. Mecanismos generales de transporte
− Tema 4. Potenciales de membrana
− Tema 5. Células excitables
− Tema 6. Fibras nerviosas y conducción del impulso
− Tema 7. Fisiología de la comunicación celular
− Tema 8. Sinapsis electroquímicas
− Tema 9. Fisiología general del músculo esquelético
− Tema 10. Fisiología general del miocardio
− Tema 11. Fisiología general del músculo liso
− Tema 12. Mecanismos de secreción celular
− Tema 13. Introducción a la fisiología de los receptores sensoriales

Fisiología I
Alberto Gómez Esteban 3
Tema 1. Homeostasis. Medio interno. Sistemas de control biológico
Medio interno
El medio interno esta principalmente formado por el agua corporal, que es el elemento mayoritario del organismo a cualquier edad y sexo. A pesar de ser el mayoritario en el organismo, existen diferencias en la proporción de agua corporal dependiendo de dichos factores, disminuyendo la proporción de agua corporal (en general) con la mayor edad; también hay mayor proporción de agua en los hombres que en las mujeres.
Es el medio en el que se llevan a cabo todas las reacciones químicas fisiológicas y como se ha mencionado antes varía con la edad y sexo a pesar de ser el componente mayoritario del organismo.
Se reparte en líquido intracelular (LIC - 2/3 del total de agua) y liquido extracelular (LEC - 1/3 del total de agua).
El LIC o líquido intracelular es el agua que se encuentra en el interior de todas las células del organismo. El LEC o líquido extracelular se encuentra repartido entre el plasma que circula por los vasos sanguíneos (5% del LEC) y el líquido intersticial o intersticio (15% del LEC).
Ambos líquidos establecen medios de distinta composición debido a que se hallan separados por barreras de permeabilidad selectiva. Todos los compartimentos tienen casi todos los elementos en común, pero en distinta concentración, lo que favorece los gradientes electroquímicos.
Existen dos barreras de permeabilidad selectiva:
− Membrana plasmática. Separa el liquido intracelular del intersticio
− Paredes capilares. Separan el intersticio del plasma
El medio interno se utiliza en fisiología como sinónimo exacto de líquido extracelular. Este término se aplica debido a que el líquido extracelular presenta una gran constancia a pesar de hallarse en equilibrio dinámico, manteniéndose la composición química. Lógicamente el medio interno excluye el líquido intracelular.
Las células al estar vivas continuamente captan elementos del medio interno para garantizar su supervivencia; en caso de isquemia en el tejido (se suspende la renovación del medio interno), se produce masiva muerte celular, lo que demuestra claramente la dependencia de las células del medio interno.
En el medio interno las células eliminan sus deshechos metabólicos así como captan nutrientes y oxígeno para continuar con este metabolismo. El funcionamiento celular, por tanto, va a quedar reflejado en la composición del medio interno, siendo esta un útil instrumento diagnóstico.

Fisiología I
Alberto Gómez Esteban 4
El medio interno es constante en cuanto a composición y similar en todos los tejidos corporales. Además es un medio dinámico que se mantiene en continua renovación, precisamente para asegurar su constancia.
Nuestras células requieren unas condiciones ambientes determinadas que son proporcionadas por el medio interno. El conjunto del organismo trabaja encaminado a mantener la constancia del medio interno en un proceso que se denomina homeostasis.
Uno organismo se encuentra en homeostasis cuando su medio interno tiene:
1. Concentraciones óptimas de nutrientes, agua, electrolitos, etc…
2. Temperatura óptima, que en estado de salud es una constante cercana a los 37º C
3. Presiones óptimas de O2 (pO2), CO2 (pCO2) y las presiones hidrostáticas, osmóticas, etc...
Un organismo será mas independiente del medio externo cuanto mayor sea su capacidad de mantener estable su medio interno.
Los sistemas corporales trabajaran por tanto a favor de mantener el medio interno, pero beneficiándose al mismo tiempo de él.
Mecanismos homeostáticos
Toda persona esta sometida a continuos cambios y perturbaciones del medio externo, lo que obliga a una respuesta para el mantenimiento correcto del medio interno.
Puede haber desajustes a nivel de los distintos niveles de organización fisiológicos, como los que afectan a:
− Células. Obliga a la actuación de orgánulos
− Tejidos enteros. Obliga a la actuación de aparatos y sistemas enteros.
Para regular estos mecanismos son precisos sensores para conocer las condiciones del medio. Estos sensores se pueden identificar con receptores sensoriales, que se localizan tanto a nivel interno, que aportan información acerca del medio interno como a nivel externo que hacen lo propio en el medio externo.
El estrés orgánico corresponde a la perturbación que modifica el estado homeostático a distinto nivel según la intensidad de la perturbación.
La homeostasis puede ser definida como el mantenimiento de las constantes del organismo por la acción coordinada de los procesos biológicos.

Fisiología I
Alberto Gómez Esteban 5
En un organismo sano hay muchas situaciones de estrés fácilmente superables. Si ese organismo se ve desbordado por esa situación y no es capaz de reajustar su actuación, sobreviene la enfermedad, que puede llegar a requerir de ayuda externa, como la actuación clínica o farmacológica para restaurar la salud.
Sistemas de control biológico
Las estructuras corporales emplean un sistema de “lazo cerrado” para mantener la constancia del medio interno. Este sistema garantiza que no exista ni exceso ni defecto de respuesta.
En caso de estrés orgánico se produce una entrada de información mediante los sensores del organismo; esta información llega a un sistema corporal que provoca una respuesta adecuada.
Los sistemas de control biológico pueden basarse en la retroalimentación negativa o la retroalimentación positiva.
Retroalimentación negativa
La retroalimentación negativa o por termostato es el más frecuente de los dos sistemas, y es la única forma de mantener un valor constante en el organismo.
La retroalimentación negativa es contraria al estimulo que la originó, por lo que tiene que descender si el estimulo era algo alto, lo que permite mantener las constantes vitales en el rango de normalidad.
*Ejemplo*
Una ascensión a una montaña con poco oxigeno supone una situación de estrés orgánico que requiere una respuesta adaptativa.
Fisiología I
Alberto Gómez Esteban 6
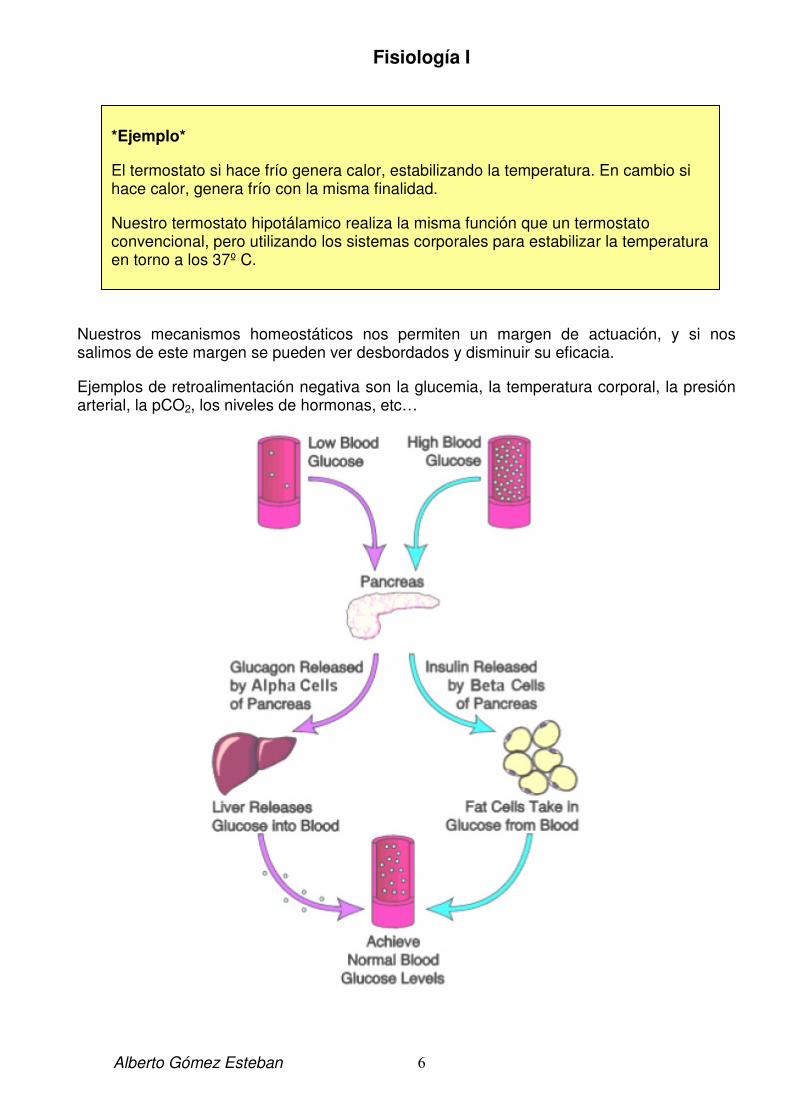
Nuestros mecanismos homeostáticos nos permiten un margen de actuación, y si nos salimos de este margen se pueden ver desbordados y disminuir su eficacia.
Ejemplos de retroalimentación negativa son la glucemia, la temperatura corporal, la presión arterial, la pCO2, los niveles de hormonas, etc…
*Ejemplo*
El termostato si hace frío genera calor, estabilizando la temperatura. En cambio si hace calor, genera frío con la misma finalidad.
Nuestro termostato hipotálamico realiza la misma función que un termostato convencional, pero utilizando los sistemas corporales para estabilizar la temperatura en torno a los 37º C.

Fisiología I
Alberto Gómez Esteban 7
Existen intervalos de normalidad en los cuales la constante es estable. Normalmente si el sistema actúa por defecto el límite se aproxima a la zona mas baja del intervalo y viceversa. Dentro de esos intervalos, la constante se considera estable.
Estos intervalos de normalidad también se basan en edad, peso, sexo, etc… Ya que pueden variar dependiendo de estos factores.
Retroalimentación positiva
Es menos frecuente que la retroalimentación negativa, pero no obstante también sirve para regular determinados momentos en los que necesitamos más de lo mismo. Es fundamental que este limitada en el tiempo para que no se exageren sus efectos.
Un feedback positivo correría el peligro de llegar a un círculo vicioso, que seria una situación patológica de mayor o menor gravedad según las características del mismo.
Algunos ejemplos de círculos viciosos son:
− Choque hipovolémico. Si perdemos un volumen amplio de sangre (∼ 2L) considerando el sistema circulatorio como el circuito cerrado que es, el corazón recibirá menos sangre, y por lo tanto, bombeará menos sangre en cada latido. Esto causa que la perfusión sea cada vez menor, con el consiguiente debilitamiento del corazón y su perdida de eficacia. Esto puede finalizar con la muerte si no es solucionado a tiempo.
− Aumento excesivo de la temperatura. El aumento de temperatura causa una mayor velocidad del ritmo metabólico, lo cual a su vez hace subir la temperatura aun más, pudiendo llegar a causar la desnaturalización de las enzimas y fallo metabólico
Algunos ejemplos de procesos de retroalimentación positiva fisiológicos son:
− Potencial de acción. Se encuentran regulados en las neuronas mediante canales de voltaje que permiten el paso de iones Na+. Estos canales responden a aumentos de potencial en el interior celular, es decir, cada vez que entra un Na+ aumenta el potencial, lo que causa a su vez que entre más sodio debido a los canales.
− Coagulación sanguínea. La coagulación normal es un proceso que sirve para mantener una perfusión sanguínea constante y es una cascada de acciones. Cuando se rompe un vaso se libera un factor de coagulación lo cual activa a su vez otros, lo cual causa una reacción en cadena que causa que el fibrinógeno se transforme en fibrina y la sangre coagule para reparar el vaso.
− Oxigenación de la hemoglobina. La hemoglobina es una molécula con un efecto similar al alostérico, es decir, cuando permite la entrada de una molécula de oxigeno, la facilidad para captar más oxigeno aumenta, y viceversa a la hora de expulsarlo debido a cambios de conformación en el interior de la molécula de hemoglobina.

Fisiología I
Alberto Gómez Esteban 8
Participación en la homeostasis de los sistemas corporales
El sistema cardiovascular tiene como función fundamental mantener el medio interno en continua renovación en todos los órganos mediante la llegada de sangre nueva a todos los tejidos.
El sistema linfático colabora con el anterior en la recuperación de líquido que tiende a quedarse en el intersticio, lo que previene el edema (acumulación excesiva de líquido intersticial que provoca hinchazón). El sistema linfático también es el portador de células implicadas en la línea defensiva
El aparato respiratorio sirve para la incorporación de oxigeno del exterior a nuestro organismo así como la eliminación del principal metabolito de deshecho: el CO2
El aparato digestivo sirve para incorporar nutrientes de los alimentos y se encarga de la incorporación de agua y electrolitos. Es un sistema de procesamiento compuesto por una serie de órganos que procesan el alimento para convertirlo en nutrientes utilizables por todos los tejidos.
El sistema urinario es también fundamental para mantener la homeostasis debido a que ayuda a eliminar los deshechos metabólicos disueltos en agua en forma de orina.
El esqueleto junto con los músculos (sistema musculoesquelético) sirve para mantener la temperatura y sobre todo posibilita el movimiento para alimentarnos y escapar de un ambiente desfavorable.
El sistema nervioso junto con el endocrino son los principales sistemas reguladores:
− El sistema nervioso nos proporciona información de los cambios y da las órdenes motoras al resto de los sistemas, actuando sobre los músculos esquelético, cardiaco y liso. Colabora estrechamente con el endocrino
− El sistema endocrino es un sistema de regulación metabólico. También colabora con el sistema nervioso en tareas como la regulación en la contracción del músculo liso.
El aparato reproductor es difícil de englobar homeostáticamente pero podemos afirmar que su misión es ayudar a la perpetuación de la especie.

Fisiología I
Alberto Gómez Esteban 9
Tema 2. La membrana plasmática
Introducción
La membrana plasmática o celular es la barrera que delimita la célula y separa el LIC del LEC.
En resumen se trata de una bicapa lipídica que sirve para establecer una frontera entre dos medios acuosos. La estructura de la membrana sigue el modelo del mosaico fluido en el cual existe doble capa de fosfolípidos y otras estructuras lipídicas como el colesterol, además de proteínas que se encuentran inmersas en esa bicapa.
Lípidos
El carácter anfipático de los fosfolípidos junto con el hecho de que se trate de una doble capa permite la separación ya que ambas monocapas se disponen enfrentadas.
Cuando una molécula ha de pasar por esta frontera es común que se encuentre con un lípido ya que es el componente mayoritario de esta membrana.
Proteínas
Las proteínas son el componente mayoritario en peso de la membrana (55% del peso). Las proteínas de membrana le proporcionan numerosas características funcionales a la célula, como formar parte de canales de paso selectivo de sustancias. También serán receptores de mensajeros químicos. Muchas proteínas integrales tienen por si mismas función enzimática o están adosadas a proteínas con función enzimática.
En la membrana celular dispondremos de distintos tipos de proteínas con distintas funciones. Estas proteínas hacen que la membrana no sea universal ya que cambian de un tipo celular a otro. Definen las características de la célula como su permeabilidad o el hecho de ser células diana.
Glúcidos
El tercer componente son los glúcidos que están asociados a lípidos y a proteínas.
Es frecuente que los glúcidos estén expuestos hacia la superficie exterior fundamentalmente, y también tienen su papel aunque sean minoritarios los que forman parte de las proteínas. Sirven de etiquetaje y reconocimiento celular.
Función de la membrana
Resumida la función de esta membrana es el de mediar controladamente el paso de sustancias del medio interno hacia el interior celular. Esto mismo esta en concordancia con que se encuentra separando dos medios de distinta composición (en el sentido de concentración).

Fisiología I
Alberto Gómez Esteban 10
Los principales cationes son el sodio y el potasio:
� El sodio es mayoritario en el medio interno
� El potasio es mayoritario en el citosol.
Dado que la membrana tiene una composición definida con función especifica, presenta una permeabilidad selectiva. Es una barrera que tiene que servir de intercambio y paso. El organismo renueva el medio interno, el cual dota de nutrientes a la célula. La permeabilidad selectiva por tanto deja pasar más fácilmente unas sustancias que otras.
Factores que influyen en la permeabilidad
A priori los factores que influyen en la permeabilidad son principalmente 4:
Tamaño de la sustancia que va a atravesar la membrana, el cual va en relación a su peso molecular (PM). Las sustancias pequeñas y de menor peso molecular atraviesan con mayor facilidad la membrana. También la forma influye en este factor (moléculas globulares atraviesan mas fácilmente que las fibrosas).
Los gases atraviesan muy fácilmente la membrana plasmática por su tamaño y peso molecular.
Liposolubilidad, debido a que la mayor parte de la superficie expuesta esta formada por lípidos, por lo que si es capaz de trasladarse entre éstos, atravesará la membrana con más facilidad. Esto hace que moléculas orgánicas de un PM relativamente elevado puedan atravesar la membrana a pesar de ser grandes.
Carga. A pH y temperaturas fisiológicas hay numerosas partículas cargadas, sales inorgánicas disociadas, etc… Por tanto la carga es condicionante ya que estas sustancias solo pueden pasar por regiones de la membrana sin carga o con carga inversa.
Presencia de transportadores, que diferencian unas células de otras y permiten el paso de sustancias que en relación a los tres puntos anteriores no podían pasar (por ejemplo moléculas grandes e hidrosolubles). Estos transportadores son proteínas que permiten atravesar la membrana a metabolitos esenciales como la glucosa.
El LIC es más rico en potasio, fosfatos y proteínas.
El LEC es más rico en sodio, cloruro y muy pocas proteínas.

Fisiología I
Alberto Gómez Esteban 11
Tema 3. Mecanismos generales de transporte
Introducción
La membrana plasmática media el transporte de sustancias para entrar en la célula desde el medio interno, así como para salir de la célula al intersticio. Este movimiento bidireccional es imprescindible para la supervivencia de la célula.
En términos generales los transportes transmembrana pueden ser pasivos o activos.
En los procesos pasivos la célula no tiene que aportar energía porque la sustancia que se va a desplazar lo va a hacer a favor de gradiente, debido a su energía cinética.
Esta sustancia se desplazara mediante un gradiente electroquímico (de carga o concentración) o de presión y se dirige de + a – intentando llegar a deshacer el gradiente.
En los procesos activos se requiere energía ya que el movimiento es contra gradiente electroquímico o de presión. Muchos procesos metabólicos generan energía que luego se va a utilizar en transportes activos en sustancias que la célula desea introducir.
Un ejemplo de proceso activo se da en las células del intestino delgado en las cuales es preciso que los nutrientes entren contra gradiente para que no prosiga el transito intestinal sin producirse digestión y los nutrientes lleguen al medio interno y puedan ser aprovechados por los tejidos.
Procesos pasivos
Los procesos pasivos o de difusión no requieren de energía. El proceso de difusión es el proceso por el cual se expande un gas o una molécula en solución ya que tiende a ocupar el máximo espacio posible. Existe por tanto un flujo neto de donde hay más sustancia a donde hay menos sin mediar en éste aporte energético.
En los procesos pasivos se aplica la Primera ley de Fick que indica hacia donde se produce ese flujo neto. La formula del flujo neto enuncia que:
Tx
CADJN s ···
∆
∆=
− D. Difusibilidad. Capacidad de esa sustancia concreta a moverse en esa
membrana concreta = PM
1
− A. Área. El flujo neto es proporcional al área. Los tejidos con mayor capacidad de intercambio amplían mediante orgánulos el área de intercambio para mejorar la difusión.

Fisiología I
Alberto Gómez Esteban 12
− ∆∆∆∆C. Gradiente de concentración. Se refiere a la diferencia de concentración entre los dos medios. Esta variable nos indica la dirección y el sentido del flujo neto. Indica asimismo que cuando C1 = C2 no existe flujo neto = C1 – C2
− ∆∆∆∆x. Distancia. Se trata del grosor/distancia que debe atravesar el soluto. Lógicamente tejidos que realicen muchas difusiones, deben tener membranas finas y cercanía entre las estructuras de difusión
− T. Temperatura. Influye en todos los factores anteriores, alterando la fluidez de la membrana. Es constante en estado de salud.
Esta ley es muy importante en fisiología.
Para diferenciar estos procesos de difusión distinguimos entre la difusión simple y la difusión facilitada por un transportador de membrana. Ambos son procesos pasivos en los cuales la sustancia se mueve a través de la membrana favorecida por su gradiente.
Difusión simple
Por difusión simple pasaran todas las sustancias capaces de pasar por los espacios intermoleculares como moléculas liposolubles (alcoholes, ácidos grasos, vitaminas liposolubles, esteroides…) y/o moléculas pequeñas (O2, CO2, urea…).
Si la molécula que pasa por difusión simple es el agua, hablamos de ósmosis.
Ósmosis
El agua no es una molécula grande por lo tanto pasa fácilmente por todas las regiones proteicas (poros acuosos, AQP). En la mayoría de los casos como el agua se mueve por difusión, aplicamos la idea de que el agua se mueva por ósmosis en función de su gradiente de concentración.
Si disponemos de un recipiente de dos medios de distinta concentración de soluto ambos separados por una membrana de permeabilidad selectiva.
El agua es el disolvente que se encuentra en ambos compartimentos y puede atravesar la barrera (no así el soluto). El agua se puede mover, igualando las concentraciones, lo que se conoce como ósmosis.
La única manera de oponerse al movimiento de ósmosis sería aplicar en el compartimento más diluido una fuerza con un émbolo la cual seria igual a la presión osmótica y de signo opuesto.
En esta situación aparece el concepto de presión osmótica (π) que es la fuerza atractiva que se ejerce sobre las moléculas de agua (disolvente) desde la solución más concentrada, y que promueve el movimiento del agua desde la solución más concentrada a la más diluida para romper el gradiente.
V
TRn ··=π

Fisiología I
Alberto Gómez Esteban 13
n. Número de partículas
R. Constante de los gases perfectos
T. Temperatura. Como decíamos en un individuo sano es constante
V. Volumen
Unidades empleadas.
� Osmolalidad. Número de osmoles / Kilogramo de disolución
� Osmolaridad. Número de osmoles / Litro de disolución
*Osmol. Partícula osmóticamente activa. No es preciso que coincida con mol. La coincidencia o no depende de si es un ión que se disocia en disolución*
La osmolaridad intracelular debe ser igual a la extracelular. El sodio es el catión mas abundante y forma la gran mayoría de las sales extracelulares.
Al comparar osmolaridades resulta el término de medida que es la tonicidad. Podemos hablar de los siguientes líquidos comparados con el plasma:
− Medio hipertónico. Presenta una osmolaridad mayor que el plasma sanguíneo. En estos medios la membrana semipermeable hace que el agua salga de la célula, y causa que las células (eritrocitos) queden con forma de ruedas dentadas (proceso de crenación).
− Medio hipotónico. Presenta una osmolaridad menor que el plasma sanguíneo. Causaría la entrada de agua a la célula (eritrocito) que ganaría turgencia.
Esto se utiliza en pruebas de resistencia celular, en el que se colocan muestras de eritrocitos en disoluciones con concentraciones salinas progresivamente diluidas y observaríamos cuando sedimentan (mantienen la forma) o bien se produce hemólisis. Esto permite estudiar el envejecimiento eritrocitario (eritrocatéresis) en el cual la hemólisis es prematura.
− Medio isotónico. Presenta la misma osmolaridad que el plasma sanguíneo. Nos permite mantener eritrocitos en un medio isotónico en caso de realizar una analítica.
Filtración
En el proceso de filtración el líquido se mueve por gradiente de presión, es decir, sale o entra a presión a través de la membrana.
Como todo proceso de difusión se aplica la 1ª ley de Fick, previamente vista, pero hablando de un proceso en el que el movimiento es por gradiente de presión:
Tx
PADJ N ···
∆
∆=

Fisiología I
Alberto Gómez Esteban 14
En estos procesos se produce arrastre por solvente que salen por la presión del líquido. El incremento de presión marca el sentido del movimiento, ya que siempre hemos de tener una presión mayor en la zona de salida que en la zona de entrada.
La aplicación fisiológica de este proceso de filtración es la salida de líquido de los capilares sanguíneos que están en estrecha proximidad de los tejidos, y se encargan del intercambio circulatorio.
Los capilares sanguíneos en la región próxima a las arterias todavía tienen una presión hidrostática relativamente alta, lo suficientemente grande como para que se favorezca la salida de agua de ellos. Hablamos normalmente de esto como ultrafiltración. En la región venosa se produce lo contrario, es decir, la presión es menor que en los capilares del lado arterioso, y se produce la entrada de fluido.
Esta ley se denomina Ley de Starling que se basa en las presiones hidrostáticas para enunciar la salida de sangre en la porción arterial, y la reentrada en la región venosa.
Además de la presión hidrostática actúa la presión osmótica que actúa en sentido inverso a la hidrostática, ésta es debida a las proteínas disueltas en el plasma, las cuales por no encontrar presentes en el intersticio generan una presión osmótica que promueve la salida de fluido del capilar. Esto ocurre también en la filtración renal, en los glomérulos.

Fisiología I
Alberto Gómez Esteban 15
En los capilares arteriosos disponemos de una presión hidrostática mayor que en el intersticio, pero también disponemos de una presión osmótica u oncótica, debida al efecto osmótico de las proteínas, presentes en un solo compartimento (el vascular). Si la presión hidrostática del vaso es mayor que la oncótica, entonces puede salir líquido hacia el intersticio.
Si la concentración plasmática de proteínas es baja, sale agua excesiva al intersticio en el lado arterioso y se reabsorbe menos agua de la necesaria en el capilar venoso. En estos casos se da el edema o acumulación excesiva de líquidos en el intersticio. También se puede dar por un aumento de presión venosa que impida la reabsorción de fluidos.
Canales iónicos
La difusión mediante canales iónicos se considera un transporte pasivo debido a que el ión que atraviesa no interacciona con la proteína canal de manera específica, además de moverse a favor de gradiente.
El concepto de gradiente debe ser ampliado ya que al hablar de un ión con carga, no solo hablamos de afinidad en cuanto a concentración, sino en cuanto a carga eléctrica (hablando ya de gradiente electroquímico), es decir, una molécula cargada positivamente tiende a verse atraída por un campo eléctrico negativo y viceversa.
Este transporte se trata de un trasiego bidireccional que sigue la ley de Fick. La estructura de la proteína de la membrana permite el movimiento a un lado o a otro. Estos mecanismos de transporte son muy eficaces (108 moléculas/segundo)
Existen algunos tipos de estos canales que son regulables, lo que quiere decir que el canal puede estar abierto o cerrado. En estado normal el poro se encuentra abierto, pero en otros momentos, como estado de reposo celular la proteína se cierra a modo de compuerta. La proteína tiene una estructura en forma de ala, que puede plegarse o desplegarse.
*Clínica*
La filtración tiene una aplicación en las membranas de diálisis. Si disponemos de una solución separada por una membrana de diálisis, que permite separar moléculas grandes de moléculas pequeñas que difunden.
La aplicación clínica se da en la hemodiálisis que se basa en crear un riñón artificial mediante este proceso de diálisis. Se da en un cartucho de diálisis en el que le llega la sangre “contaminada” de un paciente, es depurada, y vuelve a ser prefundida al paciente.

Fisiología I
Alberto Gómez Esteban 16
Los canales iónicos tienen cierta especificidad:
� Carga. El polo proteico por el que pasa un catión debe ser aniónico y viceversa. Esto significa que son necesarios distintos canales para aniones que para cationes.
� Tamaño del ión. Permite cierta discriminación. Por ejemplo algunos canales permiten el paso más facilitado de K+ que de Na+ debido a que el Na+ pasa hidratado, y de mayor tamaño, dificultando por tanto el paso.
Existen sustancias que bloquean algún tipo de canal (el bloqueo de los canales de Na+ se pueden bloquear por tetrodotoxina) las cuales se pueden utilizar a modo de drogas.
Se nombran de acuerdo al ión que transportan y si están regulados. También se denominan “ionóforos” y “permeasas”.
Los canales iónicos como decíamos pueden estar regulados en ciertos casos, cambiando su conformación espacial. Los métodos de regulación son:
� Ligando. Son moléculas que no son el ión a transportar que tienen un sitio de unión en la proteína de membrana y su unión a la misma produce el cambio conformacional.
Ejemplo. Canal de sodio regulado por acetilcolina, es frecuente en la unión neuromuscular.
� Voltaje. Es un tipo de canal que se encuentra en el axón neuronal, de importancia en la transmisión del impulso nervioso, que se basa en cambios de potencial en la célula.
Partimos de una situación de reposo (canal de sodio cerrado) que pasa a una situación de excitación con el canal abierto.
Las canalopatías tienen una interesante aplicación clínica. Afectan a tejidos excitables (fundamentalmente nervioso y muscular). Se han encontrado en afectaciones como migrañas, mialgias… Y fundamentalmente en patologías paroxísticas (intermitentes) que cursan con crisis convulsivas.
Difusión facilitada
Es un tipo de transporte mediado, que a diferencia de los transportes citados hasta ahora, la difusión facilitada junto con los transportes activos utilizan una proteína transportadora con la que tiene que interaccionar la sustancia que se va a transportar. Se trata de un transporte mediado.
Cualquiera de los transportes mediados por el mero hecho de la interacción sustancia-transportador, cumple una serie de características comunes:
1. El transportador facilita el transporte (de ahí el nombre). La glucosa al ser grande e hidrosoluble tiene dificultad para atravesar la bicapa, lo que requiere de un carrier de membrana para su entrada en la célula.

Fisiología I
Alberto Gómez Esteban 17
2. Es un proceso saturable, debido a que cuando todos los canales están ocupados por sustancia a transportar, la velocidad se estabiliza en una meseta.
3. Es inhibible, normalmente con una molécula semejante que se une al mismo sitio que la original, de modo que si tenemos una sustancia compitiendo con la que es preciso transportar, el transporte de la molécula se dificulta.
4. Es muy especifico debido a que lo que hace que la proteína permita el paso de la sustancia, es la unión de esa molécula a un “sitio receptor”, lo que causa el cambio conformacional del carrier.
Existen varios transportadores para la glucosa (proteínas GLUT):
− Glut 4. Se encuentra en el músculo esquelético, cardiaco y tejido adiposo. Es activado por insulina. A este tipo de transporte se le denomina uniporte.
La insulina actuando sobre sus receptores aumenta el transporte de glucosa, debido a que favorece la translocación, es decir, la presencia de transportadores en la membrana plasmática, retrasando la saturación.

Fisiología I
Alberto Gómez Esteban 18
Procesos activos
Primario
Se produce contra gradiente, el aporte de energía viene en forma de ATP del cual depende directamente.
Se subdivide en dos tipos de transportadores dependientes de ATP:
− Transportadores iónicos ATPasa. Normalmente ellos mismos presentan la actividad ATPasa, son los más frecuentes. Se subdividen en dos tipos:
� Tipo P. El fósforo del ATP fosforila la proteína. Esto ocurre en numerosos transportadores como la bomba de sodio
� Tipo V. Se encuentran en orgánulos. Un ejemplo es la ATPasa de H+ en endosomas y lisosomas.
− Transportadores con casete de unión al ATP. Están peor caracterizados y la idea de este tipo de transportadores es que el ATP esta unido a un dominio aminoacílico de esta proteína transportadora.
Se han descrito transportadores de este tipo para colesterol, hierro, iones cloruro. Un ejemplo es el CFTR (regulador transmembrana de la fibrosis quística) que es un transportador de cloruro y sodio que en caso de mutación, provoca la patología de la fibrosis quística.
La bomba de sodio potasio (Na+-K+-ATPasa) es una ATPasa tipo P. Tiene actividad enzimática y transporta sodio y potasio. Se conoce muy bien su estructura proteica, siendo multimérica, en la que se compone de una subunidad α, una β y una γ.
La subunidad αααα de esta proteína es la que realiza casi todas las actividades, ya que tiene actividad ATPasa y unión a reguladores, mientras que el resto de subunidades se encargan del mantenimiento estructural.
Su mecanismo de acción se basa en los siguientes pasos:
1. Se unen tres sodios intracelulares (3Na+)
2. Se produce la hidrólisis del ATP, que fosforila a la proteína
3. Se unen dos potasios extracelulares (2K+)
4. Al defosforilarse la proteína, los dos potasios entran al citosol y los tres sodios salen al intersticio
Este transportador es muy importante por su universalidad ya que se encuentra en todas las células y genera gradientes iónicos causando que haya mucho sodio en el exterior, y mucho potasio en el interior. Es una bomba electrógena ya que intercambia tres cargas positivas por dos, lo que causa que la célula quede polarizada, dejando el interior celular más negativo.

Fisiología I
Alberto Gómez Esteban 19
Es importante en el mantenimiento del volumen celular ya que ayuda a regular la ósmosis. Si hay alteraciones en la bomba de sodio, los niveles de sodio intracelulares aumentan, este nivel actúa atrayendo el agua al interior de la célula que aumenta de tamaño pudiendo llegar a explotar.
Secundario
Se trata de un transporte que se realiza dependiente de un transporte activo primario. En cualquiera de los casos trata de mover sustancias contra gradiente, pero en el secundario, la energía para el movimiento contra gradiente se debe a otra sustancia que se mueve a favor de su gradiente. Es la energía potencial de esa sustancia que se mueve a favor de gradiente la que permite el transporte activo secundario.
Muy frecuentemente la energía la proporciona la entrada de sodio en la célula a favor de gradiente electroquímico, arrastrando consigo una molécula, o permitiendo que salga de la célula. Todas las células que utilicen al sodio como molécula primaria, requieren de la Na+-K+-ATPasa.
Hay dos posibilidades te transporte activo secundario:
− Cotransporte. Las sustancias (contra gradiente, y a favor de gradiente) se mueven en la misma dirección.
Se da por ejemplo para la entrada de aminoácidos de la luz intestinal al enterocito mediante la entrada de sodio. También se da en los transportadores SGLT que transportan glucosa al interior del enterocito en contra de gradiente, y mediando de nuevo la entrada de sodio.
En los túbulos renales (Asa de Henle) disponemos de una bomba Na+-K+-2Cl-.
− Contratransporte. Ambas sustancias se mueven en dirección contraria, es decir si entra sodio a la célula, la sustancia contra gradiente sale de la célula.
Se utiliza por ejemplo para evitar la acidosis intracelular extrayendo protones a costa de introducir sodio en la célula.
En el miocito se utiliza para extraer calcio en contra de gradiente.
Otro ejemplo frecuente es el intercambio anión-anión, en este caso el cloruro con el bicarbonato. El cloruro se introduce en la célula a costa de extraer bicarbonato. Este tipo de transporte se da en el estómago en el que se exporta bicarbonato a la sangre, a costa de extraer cloruro y protones (HCl) al lumen gástrico.

Fisiología I
Alberto Gómez Esteban 20
Endocitosis y exocitosis
En la endocitosis mediante la formación de una vesícula se introduce material desde el exterior de la célula
En cambio en los procesos de exocitosis algún producto celular se libera al medio interno, al exterior de la célula, por ejemplo como medio de secreción.
En ambos procesos hay un gran número de orgánulos membranosos implicados.
Endocitosis
Hay varios tipos de endocitosis dependiendo del tamaño de lo que se introduzca.
1. Fagocitosis. Se introducen partículas grandes. Es importante en el sistema defensivo y mediante el actúan los leucocitos. Tras la entrada de la vesícula, ésta se funde con un lisosoma primario (lo que da como resultado un lisosoma secundario), lo que causa la destrucción de la partícula endocitada.
2. Pinocitosis. Se introducen partículas en disolución que pueden ser incluso macromoléculas como proteínas.
3. Mediada por receptor. Es específica ya que se produce en regiones determinadas de membrana (indentaciones) que reconocen lo que se va a introducir. Las indentaciones suelen tener una proteína llamada clatrina para revestir las vesículas formadas. En la vesícula también encontramos una GTPasa llamada dinamina.
Ejemplos de este sistema se dan en los receptores de LDL-colesterol, y asociados a sinapsis nerviosas.
Exocitosis
Es el proceso principal por el que la célula exporta sustancias sintetizadas en la propia célula.
Algunas de estas sustancias se encuentran en vesículas formadas en el Aparato de Golgi. Esta secreción es dependiente de calcio, y para el movimiento y traslado de la vesícula, así como para la expulsión de la sustancia, se requiere de un entramado proteico formado por proteínas dependientes de GTP o bien proteínas v-SNARE (se encargan de la vesícula) y t-SNARE (se encuentran en la membrana).
Otra aplicación de la exocitosis no es verter sustancias, sino para traslocar transportadores sintetizados en el interior de la célula, a la membrana. Disponemos de transportadores en el interior de la célula. De recibir la señal adecuada se desplazan a la membrana fusionándose con esta y sumando sus transportadores.
Transportes en epitelio
Son necesarios para la digestión y para recuperar nutrientes en la nefrona tras la filtración.

Fisiología I
Alberto Gómez Esteban 21
Los epitelios especializados tienen membranas orientadas, es decir, son células polarizadas (no en el sentido de cargas sino de orientación). Las células epiteliales tienen membranas luminales con distinta estructura y composición que la del resto de la membrana (basolateral). Esto permite que haya distintos transportadores en ambas membranas.
Las células epiteliales suelen estar unidas entre ellas por uniones estrechas (tight junctions) en las que hay varios tipos de proteínas implicadas, sobre todo de la familia de las claudinas y ocludinas, que se encuentran en estas regiones de unión y evitan el paso de sustancias entre células adyacentes.
Cuando una sustancia se debe mover a través del epitelio lo puede hacer por dos posibles vías:
− Vía transcelular. Atraviesa la célula, lo que implica transporte luminal y transporte basolateral.
− Vía paracelular. Sustancias que son capaces de atravesar las uniones estrechas. No todas las sustancias pueden ni las mismas sustancias en todas las uniones estrechas de un mismo tracto.

Fisiología I
Alberto Gómez Esteban 22
Tema 4. Potenciales de membrana
Generalidades
Se trata de una forma de comunicación entre células, que parte de las bases de permeabilidad de membrana, canales iónicos, etc…
La difusión de iones se ve afectada por su carga eléctrica y si existe un campo, se verán afectados por el. Esto se conoce como efecto Donnan el cual afirma que si existe un campo eléctrico los cationes se mueven hacia las cargas negativas y viceversa.
Modelo de Donnan
El efecto Donnan se explica si observamos dos disoluciones separadas por una membrana semipermeable. Llamamos a los medios A y B en los que hay el mismo número de cationes proteicos y cloruro que de aniones potasio. Si la membrana permite pasar potasio y cloruro, pero no proteínas cargadas existe igualdad de cargas pero si existe un gradiente de cloruro que fuerza a difundir de B (donde está mas concentrado) hacia A y se genera una diferencia de potencial.
El movimiento de unión cloruro que antes estaba en el otro compartimento con cargas neutralizadas, conlleva un cambio de cargas, ahora A es negativo, y B es positivo. Esto detiene el paso de cloruro lo que causa que no se iguale la concentración. El potasio se ve atraído por el campo de A- para igualar las cargas, moviéndose y desequilibrando su gradiente químico para igualar el eléctrico.
El principio de Donnan enuncia que en presencia de iones no difusibles se regula el movimiento de aquellos que si pueden difundir.
A
B
B
A
Cl
Cl
K
K
][
][
][
][−
−
+
+
=
Ecuación de Gibbs-Donnan
La solución al efecto Donnan, causa que exista una desigualdad a pesar de haberse detenido el flujo neto. Esto causa que se genere una pequeña diferencia de cargas a ambos lados de la membrana debido a las partículas cargadas no difusibles, lo que también se suma a los efectos de la bomba de Na+/K+

Fisiología I
Alberto Gómez Esteban 23
Potenciales
El VM, potencial de membrana se trata de la diferencia de potencial que presenta la membrana en cada momento:
VM= Vcito – VLEC
En los tejidos excitables hablamos del potencial de reposo VR, que se trata del potencial existente en una célula excitable cuando esta en reposo. Relacionado con este concepto nace el del potencial de acción VA, que es el potencial de membrana cuando la célula excitable es estimulada.
El interior de la membrana es siempre más negativo que el exterior. Esto se puede comprobar midiendo con un voltímetro, usando dos electrodos que situaríamos en proximidad de la membrana, uno fuera y otro dentro. Esta diferencia de potencial a ambos lados de la membrana en reposo en algunas neuronas humanas es de -90 mV.
El potencial de reposo VR significa que no hay excitación, pero si existe diferencia de potencial (VR ≠ 0) debido a:
1. Los gradientes existentes en el interior de la célula, debido a los mecanismos de transporte activo, pasivo y a la existencia de un campo eléctrico.
2. La membrana en reposo presenta distinta permeabilidad para iones difusibles: aunque exista un campo eléctrico la permeabilidad para algunas partículas es muy baja aunque vayan a favor de gradiente electroquímico.
El Ex ó potencial de equilibrio siendo X la especie iónica, es el potencial de equilibrio de un ión X. Se trata del potencial que tiene la membrana celular cuando el flujo neto para ese ión es nulo, lo que responde tanto al gradiente químico como al eléctrico. Cuando la presión osmótica iguala a la atracción eléctrica de ese ión, se da esta circunstancia.
)][
][·log(
3,2
e
i
QK
K
z
RTW
+
+−=
Ecuación de Nernst. Va respecto al K+
WE= - z·F·EQ
Si JN=0 → WQ = - WE
)][
][·log(
··3,2
)][
][·log(
··3,2··
e
i
K
e
i
K
K
K
F
TRE
K
K
z
TREFz
+
+
+
+
+
+
−=
−=
z. Carga iónica

Fisiología I
Alberto Gómez Esteban 24
F. Constante de Faraday
El VM entonces dependería de tres factores
� Permeabilidad de la membrana a ese determinado ión
� Concentraciones intra y extracelulares del ión
� Carga iónica
La membrana neuronal en reposo tenemos canales de fuga de K+ en los que el sodio entra con dificultad elevadísima en comparación al potasio. En estos casos cuando la bomba de Na+/K+ detecta un aumento de concentración de sodio, lo bombea al exterior para mantener el gradiente.
Para hallar el potencial de membrana de células permeables a múltiples iones aplicaremos la ecuación de Goldmann-Hopkin que tiene en cuenta a las tres especies iónicas principales que contribuyen al potencial de membrana:
−=
−++
−++
iCleNaeK
eCliNaiK
MClPNaPKP
ClPNaPKPV
][·][·][
][·][·][·log61
La permeabilidad de un ión se debe a la presencia de canales para ese ión. Existen canales con distinto grado de especificidad, regulables, etc… En reposo el canal abierto es un canal catiónico que permite fácilmente el paso de potasio (canal de fuga de potasio) el cual tiende a salir por gradiente químico.
El cloruro por su carga tiene muy poca atracción hacia el citosol debido a que en éste existen numerosos aniones permanentemente.
El potencial de membrana VM que tiene una célula en cada momento es muy próximo al potencial de equilibrio del ión más permeable que es el potasio (EK+).
Según parece otra variación de la ecuación de Nernst es la siguiente:
−=
=→
−=
−+
−+
−+
−+
e
i
X
e
i
X
X
XE
mVe
kT
X
X
e
kTE
][
][·log4,61
7,26][
][·log·3,2
/
/
/
/
k. Constante de Boltzmann

Fisiología I
Alberto Gómez Esteban 25
En una neurona grande tiene un potencial estándar de -86mV que es un valor muy próximo al EK (-94mV). Este valor si se combina al ENa (+61mV) da el valor que conocemos, lo que indica que el potencial de membrana de una neurona se debe fundamentalmente al potasio, y en menor medida al sodio.
La baja contribución del sodio se debe a que la membrana en reposo es muy poco permeable al sodio, lo que dificulta su entrada a pesar de que pasaría a favor de gradiente.
El potencial en reposo se debe en su mayor parte a la Na+-K+-ATPasa ya que intercambia tres cargas positivas por dos, es decir, es electrógena.
*Cuadro aclaratorio*
En una neurona en reposo existen tres principales potenciales en reposo:
� Potencial del potasio. Se trata de una diferencia de potencial de -94 mV y se trata del potencial existente cuando la membrana es permeable únicamente al potasio. Fisiológicamente se da en la fase de hiperpolarización de los potenciales de acción (estudiada a fondo en la unidad siguiente).
� Potencial de sodio + potasio. Se trata de una diferencia de potencial de -86 mV cuando difunde únicamente sodio y potasio. La membrana es muy poco permeable al sodio en condiciones normales y por ello la contribución del sodio es muy inferior a la del potasio.
� Potencial de sodio + potasio (con actividad de Na+/K+/ATPasa). Se trata de la diferencia de potencial presente en las neuronas en reposo y es de -90 mV. Se da cuando además de difundir iones sodio y potasio, actúa la Na+/K+/ATPasa.

Fisiología I
Alberto Gómez Esteban 26
Tema 5. Células excitables
Introducción
En todas las células existe una diferencia de potencial a ambos lados de la membrana. En células no excitables existe igualmente un potencial negativo en el interior, pero suele suponer menos diferencia, y es bastante estable.
Las células excitables son aquellas cuya función se basa en el cambio de potencial. Estas células son capaces de generar potenciales de acción, es decir, cambios de potencial en su membrana. Hablamos en el caso contrario de potencial de reposo. Éstas son las células nerviosas, musculares y ciertos tipos de células secretoras.
En este tema hablaremos sobre todo de células nerviosas, y la sinapsis neuromuscular.
Potencial de acción
Un tejido excitable para que abandone el potencial de reposo y su potencial pase a ser de acción necesita recibir un estímulo. Para mayor regulación, el potencial de acción se produce si el estimulo tiene cierta intensidad (valor umbral). Si no tenemos esa intensidad, hay un ligero cambio de potencial, pero no un potencial de acción
Los tipos de estímulos que pueden recibir los tejidos excitables son:
� Químicos. Se trata de moléculas ligando que regulan el canal iónico neuronal, modificando la permeabilidad de la membrana a algunos iones. La mayoría de estas sustancias son neurotransmisores, aunque en el músculo liso también hay hormonas capaces de modificar el potencial
� Eléctricos. Requieren un flujo iónico entre células mediante canales. También se puede dar de forma exógena mediante electrodos, desfibriladores, etc…
� Mecánicos. Los receptores se denominan mecanoreceptores. Detectan presiones, cambios espaciales, fricciones, etc…
Los potenciales de acción son requeridos para la transmisión del impulso nervioso independientemente del tipo de entrada.

Fisiología I
Alberto Gómez Esteban 27
Potencial de acción neuronal
Un cambio de potencial se debe a un cambio de permeabilidad iónica, es decir, en la región del axón en la que el potencial es inverso al habitual, se ha debido producir una corriente iónica de cargas positivas. Suele ser un cambio brusco y transitorio (la duración es de milisegundos, o incluso inferior). Implica una retroalimentación positiva muy breve.
El sodio es el ión encargado de la electrogénesis del potencial. El sodio se encuentra en el intersticio en contra de gradiente y cuando se abren los canales iónicos se produce la entrada masiva de sodio, cambiando la proporción de cargas entre el citosol y el intersticio.
El potencial de acción es un potencial de membrana de una célula excitable que podemos dividir en cuatro etapas:
1. Fase de reposo. Existe un potencial debido a gradientes, etc… En reposo el interior es más negativo que el exterior.
2. Fase de despolarización. Comienza a entrar sodio por canales regulados por neurotransmisor hasta llegar a un potencial más positivo en el interior que en el exterior. El sodio entra por gradiente electroquímico.
3. Fase de repolarización. Los canales de sodio se cierran con muchísima rapidez hasta llegar a tener el interior negativo frente al exterior.
4. Fase de hiperpolarización. Se trata de un estado en el que el interior es más negativo de lo que debería hasta llegar a la repolarización definitiva.

Fisiología I
Alberto Gómez Esteban 28
Dentro de estas 4 etapas distinguimos los siguientes puntos:
1. Estimulo umbral. Se trata de un estimulo que alcanza un valor umbral. Si el estimulo alcanza un potencial umbral, se desencadena el potencial de acción.
Hasta que llega al valor umbral, el estimulo crea un ligero potencial, pero cuando llega a cierto valor de electropositividad, se desencadena el potencial de acción de manera muy rápida, dándose el siguiente paso.
2. Potencial invertido. La célula llega a invertir sus cargas. En algunos casos la célula se despolariza a 0, pero lo normal es que llegue a ser un electropositiva. El sodio es el ión responsable de ese cambio de potencial.
El potencial invertido suele estar por debajo de +61 mV (ENa) pero suele ser positivo.
3. Postpotenciales. El potencial de la membrana se acerca al potencial de equilibrio del potasio.
Cuando termina el potencial de membrana se regresa al estado de VR de -90 mV.
Primero el estimulo hace que se alcance un potencial umbral, que es un valor concreto distinto en cada neurona, y se trata del potencial que requieran los canales iónicos para su apertura.
Los canales iónicos necesitan un valor de voltaje concreto para su apertura, lo que se conoce como valor umbral, si no se alcanza, se da el llamado potencial electrotónico ó sinérgico, que se extingue antes de generar un potencial de acción.
La entrada del propio sodio al llegar al valor umbral, produce que vaya entrando cada vez más sodio mediante un sistema de retroalimentación positiva hasta llegar a la fase de despolarización. Una vez llegado un momento se inactiva el poro de entrada de sodio impidiendo completamente la entrada de sodio.
Los canales de sodio tienen dos compuertas: una de activación y una de inactivación.
� El poro activo tiene las dos compuertas abiertas.
� En estado de reposo la compuerta exterior esta cerrada
� En estado inactivación la compuerta interior se cierra, tras dejar entrar sodio.

Fisiología I
Alberto Gómez Esteban 29
Para que se produzca la despolarización es preciso que salgan cargas positivas. Esto se consigue gracias a canales de potasio regulados por voltaje que no están abiertos en reposo. En la fase de repolarización comienza a salir sodio debido a la Na+-K+-ATPasa y a salir potasio por gradiente debido a los canales de potasio, es decir, se produce la salida masiva de cargas positivas.
Cuando se cierran los canales de potasio regulados por voltaje se reestablecen las condiciones iniciales de potencial, en torno a los -90 mV.
Cuando comienza el potencial de acción en el canal de sodio se abre la compuerta de activación de forma muy rápida y causa que el sodio difunda a muchísima velocidad, la compuerta de inactivación se cierra al llegar a cierto potencial (+35 mV), y se cierra de manera más lenta de forma que el valor de potencial asciende aun más en torno al ENa.
Los canales de potasio se activan lentamente al llegar a un potencial de +35 mV (el mismo al que se cierran los canales de sodio) y se cierran al llegar a los -90 mV. El cierre lento provoca la fase de hiperpolarización, con potenciales más negativos de los habituales.
Esto se ha estudiado por dos métodos:
− Pinzamiento de parche de membrana. Permite apreciar las corrientes iónicas y la conductancia al ión sodio y la conductancia al ión potasio.
− Bloqueantes de canales. Los canales iónicos se pueden bloquear por determinadas sustancias. Si bloqueamos el canal de potasio con tetraetilamonio nos muestra una gráfica de la conductancia de sodio.
Si bloqueamos el canal de sodio con tetrodotoxina (localizada en órganos de algunos peces) o bien anestésicos locales como la provocaína podemos medir la conductancia del potasio. .
La recuperación de la neurona tras el potencial de acción supone la reactivación de los canales de voltaje, y el restablecimiento iónico mediante la Na+-K+-ATPasa.
En el potencial de acción hablamos de periodos refractarios como aquellos momentos en los que en cierta porción de la membrana no puede darse una nueva despolarización. Si una neurona esta despolarizada no puede despolarizarse más, y hablamos que es refractaria a otro estimulo, es decir, no responde a el.
La tetrodotoxina es un poderoso agente neurotóxico localizado en los órganos del fugu, un pez muy apreciado en la gastronomía japonesa que contiene esta neurotoxina que paraliza los músculos hasta que el afectado muere por asfixia.
No existe antídoto para la tetrodotoxina, solo se brinda apoyo respiratorio hasta que cesen sus efectos.

Fisiología I
Alberto Gómez Esteban 30
Hablamos de periodo refractario absoluto y relativo:
� Absoluto. Ningún estimulo puede llegar a desencadenar el potencial de acción.
� Relativo. Es mas difícil llegar al potencial de acción, y se requiere un estimulo con mayor valor umbral que el que normalmente se necesitaría.
El periodo refractario absoluto se debe a todo el tiempo que los canales de sodio están abiertos y algunos empiezan a inactivarse, pero es absoluto, debido a que todos los canales de sodio tienen que inactivarse y luego cerrarse para que vuelva a producirse un nuevo potencial de acción. Así garantizamos la intensidad del potencial de acción de una membrana, pero si llega un nuevo estimulo cuando los canales de sodio están cerrados para restablecer la normalidad, no se desencadenará un nuevo potencial.
El periodo refractario relativo ofrece dificultad a la aparición de un potencial de acción debido a que pueden quedar canales de sodio activos sin que se haya establecido del todo el periodo de reposo. Aunque existe poca diferencia, es más difícil establecer un potencial de acción debido a que quedan canales de sodio que aun permanecen en ciclo. El periodo refractario relativo finaliza al llegar a la fase de hiperpolarización.

Fisiología I
Alberto Gómez Esteban 31
Tema 6. Fibras nerviosas y conducción del impulso
Generalidades
El potencial de acción es la medida en una zona de la membrana del potencial de membrana, que se da normalmente en el axón neuronal.
Las membranas despolarizadas forman parte de una misma célula, y los cambios iónicos por tanto afectaran a las zonas adyacentes. Si se produce un aumento de concentración de sodio, éste tendera a difundir al resto de la célula.
En una neurona el potencial de acción se produce en la colina o cono axónico, en la que hay muchos canales iónicos y menor valor umbral, por lo que es más probable que se produzca el potencial de acción, y en caso de que se produzca, que éste se desencadene con mayor velocidad. La corriente iónica se propaga hasta el terminal axónico por conducción del potencial de acción.
Mientras se conduce el potencial a lo largo del axón, la señal no sufre decremento suponiendo que exista la misma densidad de canales iónicos en la fibra, el potencial se autoregenera en cada segmento.
El potencial de acción es ¿unidireccional?, es decir, esta diseñado para que si se integra en el cono axónico, se dirija únicamente hacia el terminal axónico. Esto es posible debido a que el axón se repolariza en la dirección inversa a la de propagación. La unidireccionalidad se debe al periodo refractario. Esto se denomina conducción ortodrómica.
El potencial de acción es un potencial “todo o nada” es decir, si el estimulo llega a un valor umbral, se desencadena hasta el final el potencial de acción, pero si no llega a cierto valor, el potencial no se desencadena.
No hay potenciales grandes, pequeños o medianos, sino un único potencial.

Fisiología I
Alberto Gómez Esteban 32
El potencial de acción se puede propagar mediante dos formas, la conducción continua y la conducción saltatoria.
Conducción continua
Se produce en las fibras amielínicas. En estas fibras la conducción se produce punto a punto, despolarizándose regiones adyacentes debido a canales iónicos regularmente repartidos por todo el axón.
En una fibra amielínica aumenta la velocidad del impulso si se aumenta el diámetro de la fibra nerviosa (aumenta la conductividad).
Conducción saltatoria
Se produce en fibras mielínicas. En esta propagación se ahorra energía y el impulso viaja a mayor velocidad, esto es debido a que el impulso se regenera solo en los nódulos de Ranvier que son los que tienen los canales iónicos. En la región internodal apenas hay canales iónicos.
La difusión de sodio se da del axoplasma al nodo de Ranvier. Ahorra energía debido a que entra menor cantidad de sodio (más concentrado en los nodos) y se requiere menos actividad de la bomba de sodio/potasio.
Si los nodos de Ranvier están lo bastante separados, el sodio avanza mucho tramo solo por difusión y la inactivación llega solo a zonas muy concretas, lo que significa mayor velocidad del impulso.
La distancia internodal no puede ser ilimitada, ya que la concentración de sodio requiere un mínimo (es decir un potencial de acción suficiente) para que se desencadene la regeneración del impulso, es decir, para que se abran los canales de sodio del nodo siguiente.
En las fibras con mielina no suelen haber canales de potasio (se prescinde de la 4ª etapa).

Fisiología I
Alberto Gómez Esteban 33
Tema 7. Fisiología general de la comunicación celular
Generalidades
En términos generales, la comunicación celular se trata de pasar un mensaje de una célula a otra.
Por ejemplo si a una neurona llega un potencial de acción al terminal, se utilizara para causar un efecto en otra célula que se encuentre en esa vía.
En términos generales en las células corporales la comunicación se podría agrupar en una clasificación en tres dimensiones:
� Comunicación paracrina. Las células vierten un mensajero químico dirigido a células adyacentes (muchas veces en el mismo órgano). Ese vertido se realiza a la matriz extracelular y se dirige a células próximas.
Así funcionan los factores de crecimiento tisular, que se encuentran en los propios tejidos.
� Transmisión sináptica. Tras propagarse la señal en el axón, ésta llega al terminal, que vierte por exocitosis al neurotransmisor. Es similar a la comunicación paracrina, pero exclusivo de neuronas.
� Comunicación endocrina. Se realiza un vertido al medio interno. Los intermediarios son glándulas y el mensajero se denomina hormona y normalmente circula por la sangre.
Sinapsis
Se trata de la interfase especializada entre una neurona y otra célula nerviosa, muscular o glandular. Este término se utiliza cuando la señal parte de una neurona.
La sinapsis es el punto de actuación farmacológica y el punto vulnerable donde se producen alteraciones patógenas.
En una sinapsis destacamos de entrada tres componentes principales:
1. Elemento presináptico. Es de donde sale la sinapsis.
2. Elemento postsináptico. Es la célula o parte de la célula que recibe la señal.
3. Hendidura sináptica. Es la oquedad que queda entre ambos elementos pre y postsinápticos por donde viaja la sinapsis.

Fisiología I
Alberto Gómez Esteban 34
Clasificación de la sinapsis
− Neuroneurales. Entre dos células nerviosas
� Axodendrítica. Es la más frecuente. El elemento presináptico es el axón de otra neurona que manda un mensaje a la dendrita del elemento postsináptico.
� Axosomáticas. En ocasiones el axón presináptico hace sinapsis sobre el soma de la neurona.
� Axoaxónicas. El presináptico media la liberación de sustancias del postsináptico
� Dendrodendríticas. Se produce en algunas células sensoriales entre dos dendritas.
− Neuromusculares. El elemento postsináptico es una célula muscular de cualquier tipo.
− Neuroglandulares. El elemento postsináptico es una glándula.

Fisiología I
Alberto Gómez Esteban 35
Tema 8. Tipos de sinapsis
Generalidades
Podemos clasificar la sinapsis en función del tipo de señal de comunicación entre una célula y la otra. En estos casos hablamos de sinapsis eléctricas frente a sinapsis químicas.
La sinapsis química es la más frecuente y esta mediada por un neurotransmisor.
La sinapsis eléctrica es mas primitiva y frecuente en el estadio embrionario, aunque también existe en el adulto. Se trata de una prolongación del efecto de los canales iónicos, y por tanto requiere una corriente iónica mediada por GAP junctions (uniones en hendidura) de ambas células.
Sinapsis eléctricas
Cuando la comunicación se realiza mediante corrientes iónicas hablamos de sinapsis eléctricas y requieren uniones GAP en hendidura que se encuentran en ambas células, de forma que hay una continuidad de esas proteínas que operan de manera similar a un canal iónico
Esas proteínas multiméricas forman un hemicanal en cada célula que denominamos conexón, y esta compuesto por 6 subunidades del monómero conexina. Son uniones de baja resistencia (muy permeables a los iones) que permiten que prácticamente se propague el potencial de acción. El ión que más frecuentemente pasa por estas uniones es el ión sodio.
En muchas de estas uniones GAP tenemos posibilidad de estado abierto o estado cerrado. Que el estado cerrado pase a estado abierto depende de un cambio de potencial. La despolarización de una de las células permitirá la despolarización de la siguiente.
En principio la estructura es muy simétrica. Este tipo de unión se da en cardiomiocitos y miocitos lisos (en este caso no sería sinapsis ya que no interviene un elemento neuronal), es decir, se da entre dos estructuras muy semejantes.
Sólo en algunos casos (sinapsis eléctricas) se produce una utilización de las uniones GAP unidireccional, y este proceso se denomina rectificación.

Fisiología I
Alberto Gómez Esteban 36
Sinapsis químicas
Una sinapsis se considera química, cuando la señal que atraviesa la hendidura sináptica es química y se denomina neurotransmisor que será vertido por exocitosis, y se almacena en el terminal en vesículas.
En el elemento postsináptico disponemos de receptores para el neurotransmisor que se vierte desde el elemento presináptico. Para que se produzca el efecto el neurotransmisor se debe unir al receptor postsináptico. Esto produce un cambio biológico que causa un cambio de potencial en la célula postsináptica.
Las diferencias con la sinapsis eléctrica es que:
− Las sinapsis químicas son más lentas que las eléctricas debido a que requieren más actividad celular que las eléctricas que solo requieren la apertura del canal GAP.
− Las sinopsis químicas son unidireccionales. El elemento presináptico es el que tiene las vesículas y el elemento postsináptico es el que tiene los receptores, lo que garantiza que no se pueda dar en dirección contraria.
− Las sinapsis químicas son más versátiles ya que permite regular la cantidad de neurotransmisor, receptores. Asimismo el receptor puede causar distintos efectos, permeabilizando la membrana a iones positivos y causando una despolarización, o bien a aniones y causando una hiperpolarización.
La sinapsis elegida para los primeros estudios fue la unión neuromuscular. Son sinapsis muy amplias cuyo neurotransmisor es la acetilcolina (ACh) es decir, se trataba de una sinapsis colinérgica.
Las neuronas tienen una zona de recepción de señales que son las dendritas y el soma. En esas zonas existen canales regulados por ligandos.
En el axón se da la propagación y los canales están regulados por voltaje, siempre de sodio y en ocasiones también de potasio por voltaje. En los botones del terminal existen canales de voltaje también para el ión calcio.
En la neurotransmisión en primer lugar llega un potencial de acción al elemento presináptico conducido de acuerdo con la naturaleza de la fibra. En el botón disponemos de canales de calcio regulados por voltaje, los cuales se abren al llegar el potencial de acción, y se difunde calcio al interior del terminal presináptico.
El calcio es un ión implicado en el proceso de exocitosis que, además de producir la despolarización interacciona con las vesículas y proteínas dependientes de calcio (calmodulina).
La interacción con calcio produce la secreción de vesículas con acetilcolina desde unas vesículas secretorias que denominamos vesículas sinápticas.

Fisiología I
Alberto Gómez Esteban 37
Una vez disponemos de ACh en la hendidura sináptica esta se une a su receptor específico en el elemento postsináptico, que se trata de una proteína integral de membrana la cual reconoce la ACh.
En el músculo esquelético el receptor de ACh se trata de un ionóforo (un ionóforo es aquel canal que además de ser proteína receptora, también es canal iónico) de sodio regulado por ligando (precisamente el ligando es la acetilcolina). Esto causa un potencial postsináptico que denominamos PPM (potencial de placa motora).
En el músculo una vez alcanzamos un potencial umbral se da un efecto mecánico que es la contracción muscular.
Este proceso es un sistema muy especializado de comunicación celular de forma que el elemento presináptico, liberando un neurotransmisor, provoca un cambio en el elemento postsináptico.
Vesículas sinápticas
Las vesículas se forman en las estructuras del cuerpo celular fluyen por transporte axonal, hasta llegar al terminal axónico. Esa vesícula que viaja no contiene neurotransmisor. En general el neurotransmisor se sintetiza en el citoplasma del terminal, y de ahí se introduce a la vesícula.
El paso de exocitosis es un proceso que intervienen multitud de proteínas, entre ellas proteínas del complejo SNARE (target-SNARE y vesicle-SNARE) y hay una serie de pasos consecutivos que transportan las vesículas hasta un sitio activo.
Las vesículas suelen estar almacenadas en grandes cantidades en una zona de reserva. Hay otras vesículas mas próximas a la hendidura que se sitúan ya en el sitio activo, y luego hay un punto concreto que es donde las vesículas son liberadas.
Normalmente las vesículas se funden con la membrana del terminal, aunque a veces esta unión es muy leve (sistema de beso-y-escape).
En este entramado hay inhibidores como la toxina botulínica que se une al elemento presináptico y no permite la exocitosis.
El funcionamiento del sistema nervioso es pulsátil, de forma que la actuación sobre el receptor debe ser puntual. Lo fisiológico es que existan sistemas de terminación del neurotransmisor. Una forma drástica es hidrolizar la molécula por una enzima específica.
En este tipo de canales receptores de acetilcolina además de sodio, puede difundir el potasio.

Fisiología I
Alberto Gómez Esteban 38
En el caso de la ACh tenemos la acetilcolinesterasa que hidroliza la ACh en acetato y colina. Es frecuente que la acetilcolinesterasa se encuentre en la matriz extracelular y compite con el receptor de ACh.
Las formas posibles de terminación de un neurotransmisor son:
1. Difusión lejos de la membrana postsináptica. Al ir bajando el gradiente con el tiempo, la concentración de neurotransmisor deja de ser significativa.
2. Degradación enzimática. La acción de una enzima que rompe o modifica el neurotransmisor para inutilizarlo.
3. Recaptación en la terminación presináptica. Tiene lugar por transporte activo secundario. Esto ocurre en numerosas aminas biógenas y catecolaminas
En el caso de la unión neuromuscular mediada por acetilcolina no hay recaptura de ACh, pero el terminal presináptico si que tiene un sistema de recaptura de la colina una vez la ACh ha sido hidrolizada, para sintetizar nuevo neurotransmisor.
La síntesis de acetilcolina se da tras unir AcetilCoA del metabolismo celular a la colina recapturada o sintetizada, dando lugar a acetilcolina mas CoA libre. La enzima encargada es la colina-acetiltransferasa.
Existe un sistema de endocitosis llevado a cabo por el terminal presináptico, por el que se reciclan las vesículas. En el terminal disponemos de endosomas Este es el sistema de vesículas revestidas de clatrina mediado por receptor.
Al llegar un potencial de membrana la vesícula sináptica que se encuentra almacenada se va a desplazar liberándose. Una vez liberado el neurotransmisor y vacía la vesícula, ésta se reviste de clatrina. Cuando se ha alejado de la membrana pierde el revestimiento de clatrina y vuelve a rellenarse de neurotransmisores, situándose de nuevo en el sitio de reserva y aumentando la disponibilidad de neurotransmisor, evitando la fatiga neuronal.
La liberación del neurotransmisor es cuántica, debido a que las vesículas suelen tener una cantidad exacta de neurotransmisor, es decir, liberan múltiplos de una cantidad exacta de neurotransmisor.

Fisiología I
Alberto Gómez Esteban 39
Receptores de neurotransmisor
En una sinapsis química es clave el receptor del neurotransmisor. Existen dos clases de receptores:
� Ionotrópico. Se trata de una proteína integral de la membrana que sirve ella misma como canal iónico.
� Metabotrópico. Existe también en las células para receptores hormonales. Se trata de receptores asociados a proteínas G. Se produce una cascada de mensajeros intracelulares, lo que hace que este proceso sea más lento, con mayor tendencia a la amplificación, pero más perdurable. Esta relacionado con procesos de aprendizaje y expresión génica.
El objetivo final es el mismo: un cambio de permeabilidad para producir una corriente iónica.
Potenciales postsinápticos
Por medio de cualquier tipo de receptor mediante la unión del neurotransmisor se produce un potencial postsináptico, lo que significa un cambio de potencial en esa sinapsis activa por medio del neurotransmisor.
En la unión neuromuscular se produce la entrada de sodio lo que provoca una despolarización cuyo efecto es la contracción muscular.
*Ejemplo*
Receptor nicotínico de ACh (Ionotrópico)
La nicotina es agonista de este receptor. Se trata de una proteína multimérica que es al mismo tiempo un poro. También es receptor de la familia de los muscarínicos (su análogo estructural es la muscarina) de la acetilcolina.
Este receptor va a tener un efecto inhibitorio (hiperpolarizante) dificultando el potencial de acción dado que favorece la activación de un canal de potasio.
Esto causa que la ACh tenga efectos tanto excitatorios como inhibitorios dependiendo del receptor con el que interaccione.
La ACh en el miocardio es inhibitoria en el cardiomiocito, ralentizando la frecuencia cardiaca y la fuerza de la contracción, pero en el músculo liso es estimulatoria, favoreciendo el peristaltismo.

Fisiología I
Alberto Gómez Esteban 40
Si pensamos en una sinapsis neuroneural el potencial postsináptico se recibe en el cuerpo celular, se integra en el cono y se propaga por el axón, de forma que el cuerpo celular está recibiendo señales de entrada directamente o bien a través de las dendritas. Este potencial postsináptico se diferencia del potencial de acción:
� Su amplitud puede variar
� Decrece según nos alejamos de la sinapsis debido a que la difusión de sodio es local y la concentración del mismo disminuye cuando nos alejamos de dicha sinapsis.
� Pueden sumarse, es decir, no es obligatorio que la descarga de un potencial postsináptico produzca un potencial de acción pero pueden sumarse varios receptores postsinápticos para provocarlo
La sumación espacial requeriría la descarga simultanea de dos botones sinápticos que se encuentren próximos, de forma que generen un gradiente de iones (y despolarización) mayor.
La sumación temporal requiere solo una entrada, que descargue repetidamente por lo que puede llegar a alcanzar el potencial umbral.
Una de las posibilidades que tenia la sinapsis química era la versatilidad, de forma que el cambio de acción que provoca puede ser:
� Despolarización. Conduce al potencial de acción y decimos que es excitatorio. Se producen debido a la entrada de cationes (Na+ y Ca2+)
� Hiperpolarización. Dificulta el potencial de acción y decimos que es inhibitorio. Se producen debido a la entrada de cationes (Cl-) o bien salida de K+.
En este caso cuando realizamos sumación de dos terminales, no es indiferente que el cambio de acción de uno o más terminales sea excitatorio a que sea inhibitorio.
Existen otras posibilidades como modificar el metabolismo interno de la neurona postsináptica destinados a aumentar el número de potenciales inhibitorios o bien disminuir el número de excitatorios.

Fisiología I
Alberto Gómez Esteban 41
Tipos de inhibición
Existe la posibilidad de realizar una inhibición presináptica y otra postsináptica. Un potencial postsináptico también puede ser inhibitorio, alejando la carga del potencial umbral.
− Postsináptica. El neurotransmisor liberado por el terminal presináptico realizará una inhibición postsináptica
− Inhibición presináptica. El elemento postsináptico de la sinapsis axoaxónica es otro axón que va a ser el elemento presináptico del elemento presináptico de la segunda sinapsis.
Si liberara un neurotransmisor excitatorio desde el segundo elemento, la sinapsis del primer elemento reduce el efecto de dicho neurotransmisor. Esto se ha observado como receptores postsinápticos en terminales que consideramos presinápticos de otra sinapsis posterior.
Si una neurona va a tener sobre ella multitud de elementos presinápticos, va a ser muy posible que más de uno de ellos descargue a la vez, por lo que la neurona postsináptica enviará la porción resultante. El impulso se integra en el cono o colina axónica, donde se produce o no el potencial de acción.
La integración del impulso no se da como suma algebraica, sino que en un determinado milisegundo puede ser que se esté recibiendo una cantidad suficientemente elevada de potenciales excitatorios tanto que la suma de ellos es mayor que el umbral, con lo que el resultado es que se produce un potencial de acción.
Si hay una elevada cantidad inhibitoria, el efecto es modular una excitación y por tanto no se produce el potencial de acción, cortándose la comunicación sináptica en dicho punto.
Si la excitatoria es superior a la inhibitoria pero inferior al valor umbral, no se da el potencial de acción (todo o nada), sino que se da el estado de facilitación es decir, el descenso del valor umbral para que un segundo estímulo muy próximo en el espacio o en el tiempo pueda llegar a conseguir el potencial de acción. Esta es la base que permite la sumación espacial o temporal antes explicada.

Fisiología I
Alberto Gómez Esteban 42
Neurotransmisores
El neurotransmisor (NT) es el detonante de todo el complejo que incidiendo sobre sus receptores es el que causa el potencial postsináptico.
Son mensajeros químicos presentes en las neuronas y que parten de ellas. Los criterios que se aplican para afirmar que una sustancia química es un neurotransmisor son los siguientes:
1. Se encuentra en el terminal presináptico en cantidades apropiadas, es decir, una cantidad que pueda ser responsable de un cambio en el elemento postsináptico.
2. Existencia de mecanismos capaces de explicar la síntesis y terminación del neurotransmisor
3. Debe liberarse como respuesta a la despolarización del terminal, es decir, que se podría liberar el neurotransmisor artificialmente, aplicando una descarga eléctrica exógena sobre el terminal presináptico.
Hay asimismo varios tipos de neurotransmisores agrupados:
Neurotransmisores de bajo peso molecular
A este grupo pertenecen los NT clásicos. Son moléculas de tamaño discreto.
Se sintetizan en el citoplasma y en general lo hacen mediante enzimas específicas de síntesis. Suelen ser asimismo de respuesta rápida y se encuentran en vesículas sinápticas más pequeñas (como resultado de su liberación cuántica).
Pueden coexistir con un neuropéptido, es decir, en la vesícula no solo encontraremos un solo tipo de neurotransmisor. El terminal para saber que NT liberar cambia la frecuencia de los potenciales, en estas frecuencias los neuropéptidos responderían a un tren o ráfaga de impulsos.
Ésteres
Acetilcolina
Las sinapsis se denominan colinérgicas y los receptores pueden ser:
− Muscarínicos. En general son metabotrópicos
− Nicotínicos. En general son ionotrópicos
Se localiza en las sinapsis sobre la unión neuromuscular sobre el músculo esquelético y la fracción del sistema nervioso que actúa sobre el mismo es el sistema nervioso somático (contiene la ACh)
Lo encontraremos asimismo en las neuronas preganglionares en el SN autónomo y en neuronas postganglionares del SNPS y escasas SNS así como en el encéfalo (corteza motora) y en los ganglios basales, aunque en el encéfalo es poco importante.

Fisiología I
Alberto Gómez Esteban 43
Aminas
Dopamina
Se sintetiza a partir de la tirosina por la enzima DOPA-descarboxilasa y se degrada por recaptación específica como todas las aminas biogénicas en este caso por el transportador de dopamina (DAT).
Es un neurotransmisor del sistema nervioso central que se encuentra en vías responsables de funciones motoras (sistema nigroestriado) y respuestas emocionales (sistema mesolímbico).
Las neuronas dopaminérgicas son afectadas por el alcohol, la marihuana, la morfina, la cocaína y las anfetaminas, que actúan inhibiendo la recaptura, prolongando los efectos de la dopamina.
Norepinefrina / Noradrenalina / Epinefrina / Adrenalina
Las sinapsis se denominan adrenérgicos y son importantes reguladores periféricos del SN autónomo sobre los efectores.
La noradrenalina es más típica como neurotransmisor que puede verse reforzada por los efectos de la adrenalina. Ambos neurotransmisores (adrenalina y noradrenalina) utilizan los mismos receptores de la familia α y de la familia β, que tienen efectos opuestos.
Hay vías adrenérgicas en el SNC como neurotransmisor excitatorio y es el neurotransmisor mas frecuente en la neurona simpática postganglionar.
La liberación de estos neurotransmisores ayuda a superar el estrés orgánico.
Estas aminas biógenas a partir de la tirosina forman un grupo que siempre finalizan su acción con la recaptura especifica del neurotransmisor y tras su recaptura pueden ser degradados por enzimas como la monoaminooxidasa (MAO) o bien la catecol-o-metiltransferasa (COMT) o bien pueden ser reutilizados.
Serotonina (5OH-Triptamina)
Es una molécula derivada del triptófano. Sus receptores se denominan como 5-HT, y hay alrededor de 15 tipos, de efecto mayoritariamente metabotrópico frecuente en las aminas biogénicas.
Es un transmisor del SNC que regula funciones vegetativas como la termorregulación, la regulación del ánimo, apetito, inducción al sueño, etc…
En el tratamiento para la depresión, el tratamiento va encaminado a los receptores de serotonina

Fisiología I
Alberto Gómez Esteban 44
Histamina
Es sobre todo una sustancia paracrina que participa en el estimulo inflamatorio, pero se ha visto que hay terminaciones que la utilizan como neurotransmisor. Se nombra como H1, H2 o H3.
Se localiza en el SNC, sobre todo en el hipotálamo.
Esta implicado en la termorregulación, en el ciclo sueño-vigilia, y en las regulaciones endocrinas.
Aminoácidos
GABA y Glicina
Son normalmente inhibitorios. Son abundantes en encéfalo como todos los aminoácidos con función neurotransmisora.
− GABA. Es muy conocido e importante. Sus receptores se denominan con un subíndice. Es el principal NT inhibitorio del SNC. Hay 3 receptores caracterizados:
� GABAA. Es un receptor postsináptico típico inhibitorio (canal de cloruro) cuya característica es tener el sitio de unión del GABA y un sitio que conoce sustancias ansiolíticas antiinsomnes como las benzodiacepinas y para hipnóticos como los barbitúricos.
� GABAB. Es metabotrópico presináptico, reduciendo la liberación de neurotransmisores excitatorios como el glutamato.
− Glicina. Es un neurotransmisor encontrado frecuentemente en interneuronas de la medula espinal, y se utiliza normalmente en la inhibición de músculos antagonistas.
La estricnina se une al receptor de la glicina y produce asfixia.
Glutamato y Aspartato
Son aminoácidos muy semejantes dicarboxílicos que actúan sobre el mismo tipo de receptores y se denominan como los análogos estructurales. Ambos aminoácidos son normalmente excitatorios.
Gases
NO
El óxido nítrico (NO) ha sido descrito recientemente. Se sintetiza en reacciones celulares, no se almacena en vesículas y carece de receptores en el elemento postsináptico debido a que difunde a través de la membrana de las células post y presinápticas.

Fisiología I
Alberto Gómez Esteban 45
Se sabe que tiene un efecto sobre la musculatura, sobre todo la de los vasos sanguíneos (vasodilatador), aunque también en el aparato digestivo y es secretado por el SNP. El efecto vasodilatador del NO es potenciado por el sildenafil (viagra).
En el SNC se relaciona con la conducta a largo plazo y la memoria
CO
Purinas y nucleótidos púricos
Adenosina
Tiene un efecto vasodilatador
Uridina y ATP
Se trata de un antagonista de la cafeína.
La uridina y el ATP no suelen tener un papel autónomo, sino que actúan como cotransmisrores. Normalmente coexisten en el terminal con otro transmisor con el que se vierten a la vez.
Lípidos
Endocannabinoides
Se trata de “marihuana endógena” cuyo efecto en el cerebro es la analgesia y reducción de sentimientos adversos (ansiedad fundamentalmente). El hecho de que sean análogos del cannabis causa el efecto estupefaciente de esta sustancia.
Otros
Prostaglandinas y neuroesteroides
Neurotransmisores de mayor peso molecular (neuropéptidos)
El grupo de neuropéptidos son neurotransmisores de alto peso molecular. No suelen ser proteínas muy grandes pero si son péptidos formados por cierta cantidad de aminoácidos lo que causa que tengan un cierto peso molecular, es decir:
� Tienen mayor peso molecular que los neurotransmisores pequeños.
� Su acción es más lenta pero más prolongada. Frecuentemente presentan receptores metabotrópicos
� Tienen una potencia mayor, lo que causa que su actuación se de a concentraciones más bajas.

Fisiología I
Alberto Gómez Esteban 46
Se sintetizan en los ribosomas del retículo endoplásmico rugoso (RER). A pesar de ser moléculas grandes se sintetizan en forma de péptidos precursores de mayor tamaño.
Las vesículas que contienen estos neuropéptidos se forman en el soma, donde se dirigen por transporte axonal hacia el terminal, actuando de la forma que ya hemos descrito. Durante el tiempo que dura el transporte axonal, se suele producir el procesamiento, reduciendo su tamaño y convirtiéndose en el péptidos de tamaño definitivo.
En el terminal se almacena en vesículas presinápticas que no se reciclan, y se liberan por exocitosis dependiente de calcio.
Los neuropéptidos terminan su acción por proteolisis (rompiendo sus enlaces peptídicos) y también muchas veces por difusión.
Tienen su propio receptor especifico en el elemento postsináptico.
Su descubrimiento cambio los principios de neurotransmisor. Pueden coexistir con un neurotransmisor clásico, y se liberan por trenes de potencial, es decir, cuando la frecuencia del potencial de acción que entrante es mayor.
Actúan a veces como neurotransmisor y muchas otras veces como neuromoduladores, despolarizando o hiperpolarizando para apoyar otro neurotransmisor, es decir, si no son capaces de dar una respuesta aislada, muy frecuentemente dan respuestas integradas con otros neurotransmisores.
Opiáceos endógenos
Encefalinas y endorfinas
Son agonistas del LSD junto con otros fármacos y drogas alucinógenas. Su papel general es el de ser analgésicos, por ejemplo aumentan durante el parto reduciendo el dolor. Otra función es la de actuar en las vías de recompensa (p.e. después del ejercicio).
Como analgésicos regulan la liberación del neuropéptido conocido como sustancia P.
Otros
Sustancia P
Se trata del neurotransmisor típico del dolor.
Neuropéptido Y
Se trata del neuropéptido más abundante encuentra en mayor cantidad en el encéfalo. Participa en la respuesta a la agresión y es un potente estimulador del apetito. Es inhibido por la hormona leptina.

Fisiología I
Alberto Gómez Esteban 47
Vasopresina y oxitocina
Son dos hormonas típicas, pero también se liberan en algunas sinapsis actuando sobre la conducta sexual y el aprendizaje. Se encuentran el tallo encefálico y la médula espinal.
Otros
La lista sigue creciendo con hormonas hipofisarias, factores de liberación hipotalámicos, etc…

Fisiología I
Alberto Gómez Esteban 48
Tema 9. Fisiología general del músculo estriado esquelético. Unidad motora
Generalidades
Hay tres grandes tipos de músculos con características a grandes rasgos diferenciales, sencillos.
� Músculo esquelético. Se encarga en general de levantar peso, y movilizar el esqueleto. Se trata de un músculo estriado y voluntario. Es estriado gracias a sus bandas claras (I) y oscuras (A). Puede contraerse y relajarse voluntariamente gracias a su inervación por las fibras motoras del sistema nervioso somático
� Músculo cardiaco (miocardio). Se trata de un músculo involuntario y estriado. Tiene una estructura especial con un tejido autoexcitable regulado por un sistema marcapasos, y va a tener un movimiento automático. Se regula por el sistema nervioso autónomo.
� Músculo liso. Tiene muchas menos estrías que los dos anteriores. Es músculo involuntario y se encuentra en las paredes de las estructuras internas huecas, manteniendo su tono. Se encuentra en vasos sanguíneos, aparato digestivo, órganos abdominales, folículos pilosos, etc…
Las funciones del músculo son producir movimiento. El músculo esquelético forma parte del aparato locomotor, pero el resto de los músculos también producen movimiento de bombeo, peristaltismo, etc… El músculo liso se encarga de mantener el tamaño de los órganos huecos.
También se encargan de la termogénesis, sobre todo el músculo esquelético (responsable en un 75%). En caso de frío, se producen contracciones involuntarias del músculo esquelético.
Propiedades
El músculo es excitable debido a cambios de potencial que serán responsables de su contracción.
La contractilidad es la propiedad del músculo consistente en acortarse y engrosarse para levantar peso. A su vez el músculo también es extensible (observable en los músculos antagonistas) estos al contraerse los agonistas no solo se relajan sino que se distienden.
El músculo también es un tejido elástico, es decir, en condiciones fisiológicas recupera su estructura inicial tras contraerse o distenderse. Esto es de vital importancia en el diafragma
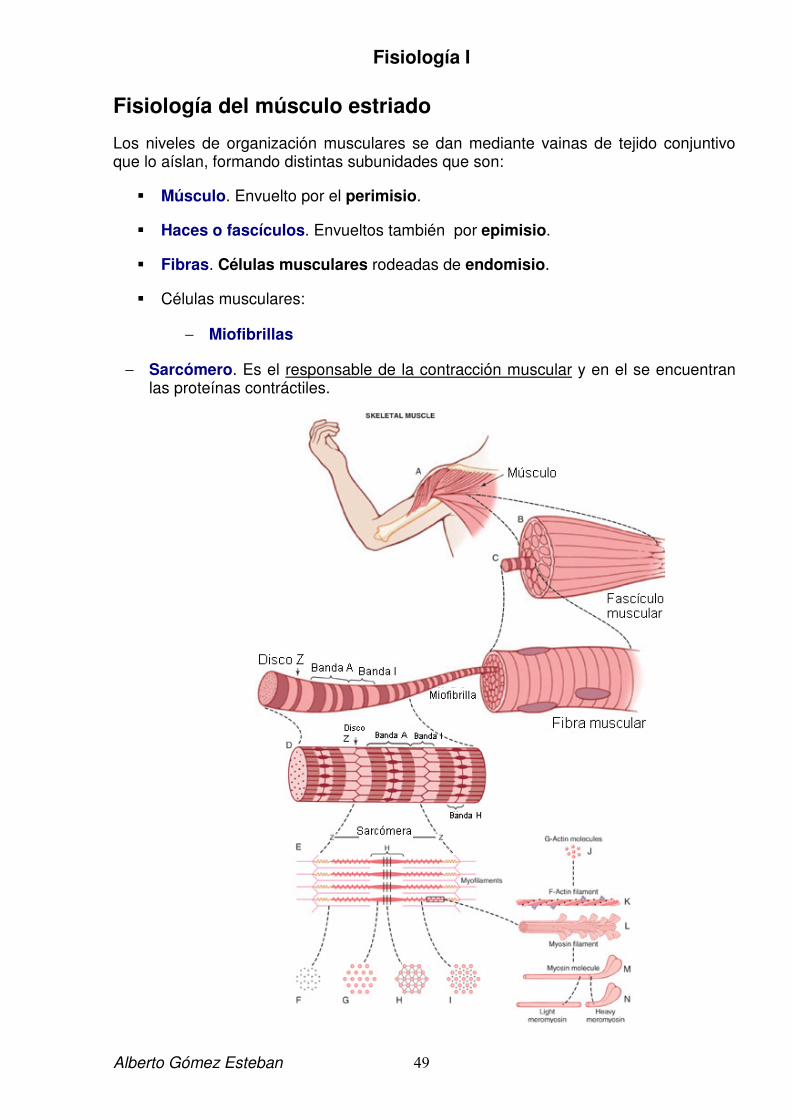
Fisiología I
Alberto Gómez Esteban 49
Fisiología del músculo estriado
Los niveles de organización musculares se dan mediante vainas de tejido conjuntivo que lo aíslan, formando distintas subunidades que son:
� Músculo. Envuelto por el perimisio.
� Haces o fascículos. Envueltos también por epimisio.
� Fibras. Células musculares rodeadas de endomisio.
� Células musculares:
− Miofibrillas
− Sarcómero. Es el responsable de la contracción muscular y en el se encuentran las proteínas contráctiles.

Fisiología I
Alberto Gómez Esteban 50
En el sarcómero encontramos las proteínas de la acción muscular. Encontramos filamentos gruesos de miosina, y filamentos finos de actina.
La teoría en la que se basa la contracción muscular (acortamiento y engrosamiento) es la teoría del deslizamiento de los filamentos finos de actina sobre los filamentos gruesos de miosina. El aspecto del sarcómero es de dimensiones más reducidas debido a éste deslizamiento.
El deslizamiento implica la formación de puentes cruzados de enganche entre la miosina y la actina. Ese deslizamiento es porque la miosina engancha los filamentos finos, atrayéndolos hacia el centro del sarcómero. Estos enganches generan puentes cruzados, llevando los filamentos finos de ambos lados hacia el centro del sarcómero.
En la miosina encontramos estructuras globulares (cabezas de la miosina) que se dirigen hacia los filamentos de actina, y se encarga de formar los puentes cruzados. La miosina dispone de muchas cabezas dirigidas en todas direcciones.
La contracción muscular requiere ATP que tiene un papel directo. La cabeza de la miosina tiene actividad ATPasa. Su conformación activa depende de ir unida a ADP, lo que causa la formación del puente cruzado causado por su afinidad por el filamento de actina. Al unirse el puente cruzado puede desplazar el filamento fino hacia el centro del sarcómero debido a que la cabeza es capaz de cambiar de conformación una vez se ha unido al filamento fino. El movimiento del filamento de actina unido a la cabeza de miosina se denomina golpe de fuerza.
El golpe de fuerza va a ser unidireccional. La cabeza desplaza el filamento, y se suelta por la unión de una molécula de ATP, lo que restablece el estadio inicial, y posibilita el inicio de un nuevo golpe de fuerza. Una contracción muscular requiere varios golpes de fuerza (múltiples tirones) que ejercen su fuerza en la misma dirección. Las cabezas que han realizado ya un golpe de fuerza tienen un breve periodo de inactividad mientras hidrolizan el ATP, que aprovechan otras cabezas para realizar sucesivos golpes de fuerza.
El ATP cuando se hidroliza en forma de ADP causa la afinidad de la miosina por el filamento fino, lo que provoca un cambio conformacional en bisagra sobre el cuello de la miosina lo que causa el golpe de fuerza. La unión de nuevo ATP causa el fin de la afinidad por la actina.
*Rigor mortis*
La relajación requiere ATP, si existe ausencia de ATP debido al fin del metabolismo basal, no se produce el suelte del filamento de actina por parte de la cabeza de miosina y se produce la rigidez muscular debido a que no existe relajación.

Fisiología I
Alberto Gómez Esteban 51
Cuando se habla de factores indispensables para la contracción muscular, se habla del ATP (que produce el puente cruzado) así como otro factor imprescindible que es el ión Ca2+.
El calcio interviene en la unión de los filamentos igual que lo hace el ATP. Los filamentos finos como ya hemos mencionado, están formados fundamentalmente por actina. En este filamento fino tenemos una estructura filamentosa que es la actina F, así como la nebulina que se encarga de localizar la actina en el sarcómero. La actina aparte de ir acompañada por nebulina va acompañada por una serie de proteínas que también intervienen en la contracción como es la tropomiosina, otra proteína filamentosa que sigue la estructura helicoidal de la actina, y también por otra proteína globular que se llama troponina, en la cual encontramos tres subunidades:
− Subunidad T. De anclaje a la tropomiosina.
− Subunidad I. Inhibe los sitios de unión a la miosina a la actina
− Subunidad C. Tiene 4 sitios de unión para calcio. La unión de estos cuatro cationes permite la contracción muscular, reduciendo el efecto inhibitorio y dándose el anclaje a la miosina.
El calcio por tanto modifica la troponina para que ésta produzca un desplazamiento de la tropomiosina y así que se puedan unir las cabezas de la miosina a los lugares de anclaje de la actina.
Se llega a la contracción muscular debido a la excitación mediante despolarización del músculo, que es un tejido excitable. Los fenómenos eléctricos por tanto anteceden la acción mecánica. Esto obedece a una sinapsis neuromuscular, ya que necesita la acción excitatoria de la neurona del sistema nervioso somático.
La fibra muscular es una célula grande, y la despolarización de la membrana afecta fundamentalmente a las regiones adyacentes, por lo que tiene poco efecto sobre el sarcoplasma en su conjunto, por lo tanto es preciso que existan unas invaginaciones de la membrana que son los túbulos T los cuales se internan en el interior del miocito, acercándose a las miofibrillas.
En este proceso interviene el retículo sarcoplásmico (RS), que se encarga de almacenar calcio en grandísimas cantidades. Dicho esto la contracción se sucede en una serie de pasos:
1. Se libera acetilcolina del elemento presináptico que se une a los canales catiónicos del elemento postsináptico muscular, el cual causa la entrada de sodio, y causa la despolarización del miocito (proceso estudiado en detalle en el tema anterior).
2. Los potenciales de acción recorren los túbulos T, lo que provoca un cambio de voltaje que activa canales de estos túbulos T.
3. En los túbulos T existen canales de calcio que se van a abrir en estas circunstancias. La entrada de calcio extracelular provoca a su vez la apertura de canales lentos de calcio presentes en el retículo sarcoplásmico. Estas dos acciones se traducen en el aumento de la concentración intracelular de calcio

Fisiología I
Alberto Gómez Esteban 52
(sobre todo el proveniente del RS, ya que se ha demostrado que la contracción muscular se puede iniciar con bajísimas concentraciones de calcio extracelular).
En el espacio intracelular y unida a canales del retículo sarcoplásmico encontramos el receptor para una sustancia denominada rianodina que es directamente responsable de la apertura de los canales de calcio del RS. También observamos otro receptor asociado a los canales de voltaje de los túbulos T que es el DHPR que es el receptor de dihidropiridina que se activa siempre que hay una contracción muscular, el cual a su vez activa el receptor de la rianodina provocando la apertura de canales de calcio del RS lo que causa a su vez la liberación masiva de calcio por difusión que es el que principalmente se utiliza en la contracción muscular.
4. El calcio se une a la troponina y permite la contracción muscular.
La contracción muscular se da por tanto como resultado de un potencial de acción en la fibra muscular. En el miocito estriado en reposo presenta un potencial más electronegativo que en las neuronas. El potencial de acción del músculo es ligeramente más prolongado que el neuronal, pero entraría dentro de los potenciales rápidos (en comparación con los del ventrículo cardiaco por ejemplo), y se debe a la apertura y cierre de canales que se regulaban con cierta rapidez.
En la relación nervio somático-músculo esquelético, veremos el concepto de unidad motora. La última neurona que forma la unidad motora será la neurona espinal. El concepto de unidad motora es la idea del conjunto que forma la motoneurona con todas las células musculares que inerva, y que responderán a sus potenciales de acción. En la unidad motora:
− La sinapsis se sitúa en el centro de la fibra muscular.
− Las células musculares reciben una única terminación axónica.
− Se trata de una sinapsis colinérgica, es decir, se produce liberación de acetilcolina.
− La neurona se trata de una motoneurona medular que parte de la corteza motora y es el último eslabón de transmisión nerviosa neuromuscular. Esta motoneurona medular α cuyo soma surge de las raíces ventrales de la medula espinal.
− El axón que es único se ramifica en varios botones sinápticos que son componentes de la misma unidad motora.
− En un músculo cualquiera tenemos distintos grados de tensión y estos distintos grados de tensión dependen de la cantidad de unidades motoras activadas. Normalmente los efectos son mayores con una contracción gradual asincrónica.
En una unidad motora existe una relación que se denomina coeficiente de inervación que es la relación entre la neurona motora y la cantidad de fibras musculares sobre las que hace sinapsis.

Fisiología I
Alberto Gómez Esteban 53
*Ejemplos fisiológicos*
En la laringe se da un control 3
1. Este es un control fino, debido a que una neurona
inerva tres fibras musculares, lo que permite un gran control del movimiento
En músculos potentes como el bíceps braquial se da 150
1.
En músculos grandes y potentes (gastrocnemio) se puede llegar a dar 1000
1. Esto
da lugar a contracciones muy intensas con una única unidad motora activada, y a poco control sobre el grado de intensidad de la contracción.

Fisiología I
Alberto Gómez Esteban 54
Tema 11. Fisiología general del músculo cardiaco
Generalidades
El músculo cardiaco es el músculo estriado involuntario que se encuentra en las paredes del corazón. Su función es realizar el bombeo sanguíneo para propulsar la sangre varios metros.
Se trata de un músculo con sarcómeros estriados semejantes a los del músculo esquelético. Las células son cortas y ramificadas, lo que permite tener un entramado de células que estarán interconectadas entre ellas por uniones GAP en hendidura, que forman estructuras denominadas discos intercalares. Esas uniones permiten el paso de iones con total facilidad, generando un sincitio funcional. En este sentido tenemos un sincitio auricular y un sincitio ventricular.
El músculo cardiaco tiene un mecanismo de acción para la contracción relativamente parecido al músculo estriado con algunas diferencias. En este caso si que es importante la cantidad de calcio que entra del medio extracelular, debido a que los retículos sarcoplásmicos del miocito cardiaco tienen menor desarrollo.
Algunos medicamentos cardiacos van encaminados a regular el trasiego de calcio extracelular para regular por tanto la contracción del miocardio.

Fisiología I
Alberto Gómez Esteban 55
Tema 12. Fisiología general del músculo liso
Generalidades
El músculo liso esta formado por células fusiformes con un único núcleo. No contiene sarcómeros por lo que al microscopio carece de aspecto estriado. También tiene mayor proporción de actina. Los filamentos finos de actina son muy largos con una estructura que alcanza la membrana plasmática, uniéndose a ella por los llamados cuerpos densos, donde se encuentra envuelta por proteínas estructurales.
El miocito liso tendrá un aspecto muy característico al contraerse debido al anclaje de la actina a su membrana (cuando se contrae, tendrá un aspecto globuloso), además se acorta de forma mucho mayor que el músculo esquelético.
En el músculo liso también es preciso el calcio, cuyos canales se activan dando como consecuencia de la despolarización del miocito liso dando lugar también a la contracción en un proceso más lento. Las contracciones son graduales, lentas y sostenidas, pero parte de la lentitud se debe a la actividad ATPasa más lenta.
En el músculo liso hay un estado especial conocido como estado de cierre/cerrojo que significa que el músculo se puede mantener contraído mediante tiempos prolongados sin requerir incremento de ATP (esto tendrá importancia por ejemplo en esfínteres). Debido al estado de cierre el proceso digestivo se dará como un proceso en el que no se requiere una cantidad de ATP proporcional a la que se necesitaría si la pared digestiva estuviera constituida por músculo esquelético.
Existen dos tipos de músculo liso:
� Músculos unitarios. Recuerdan al sincitio cardiaco
� Músculos multiunitarios. Son similares al esquelético

Fisiología I
Alberto Gómez Esteban 56
Músculo liso unitario
Es aquel en el cual multitud de fibras de miocitos lisos se comportan como una única unidad. Se encuentra en la mayoría del tubo digestivo, así como del genitourinario.
El conjunto se despolariza siempre que se despolariza una única célula, que decimos que tiene actividad marcapasos. La célula marcapasos se inerva por sinapsis neuromuscular, y el resto lo hacen por uniones GAP con ésta célula marcapasos o con otras sucesivas que ya se hayan despolarizado por la propagación del potencial. Esto causa que no todos los miocitos lisos deban estar inervados.
La innervación de este tipo de músculo proviene de varicosidades axónicas, es decir, estructuras bulbosas que recuerdan a los botones sinápticos en los que existen vesículas de neurotransmisores, pero que se encuentran como ensanchamientos a lo largo de todo el axón. Estas sinapsis se denominan sinapsis “de paso”.
El músculo liso también se despolariza por acción de hormonas o de factores locales (acción paracrina).
Estas células de músculo liso suelen presentar actividad miogena. Esto ocurre en el músculo digestivo, pero es de mayor importancia en las paredes vasculares, es decir, el estiramiento (distensión) causa que el músculo liso responda con una contracción.
El músculo liso unitario está inervado por el sistema nervioso autónomo en vez del somático. También factores paracrinos y hormonas son capaces de generar una contracción muscular, en ocasiones sin producir despolarización de membrana, sino mediante cascadas de transducción con segundos mensajeros (p.e. inositol trifosfato).
El músculo liso unitario es el músculo liso visceral, y en las vísceras se recibe doble innervación autónoma (simpática y parasimpática) y a nivel de potencial de membrana, ambas inervaciones hacen lo contrario, es decir, si una de ellas es excitatoria, la otra tendrá acción inhibitoria hiperpolarizando la membrana. No necesariamente la simpática es siempre excitatoria y la parasimpática inhibitoria (o viceversa) sino que depende del efecto que causen.
Músculo liso multiunitario
Es aquel en el que cada una de las células recibe innervación independiente, y también suele recibir innervación de varicosidades axónicas. Al tener inervación directa las fibras de músculo liso multiunitario dispone de pocas o ninguna uniones comunicantes.
La excitación nerviosa se da sobre cada una de las células, y ejemplos de este tipo de músculo liso están:
� Músculos ciliares del cristalino
� Músculos erectores del pelo
� Músculos del iris

Fisiología I
Alberto Gómez Esteban
Se parecen a una unidad motora 1:1 (una porción de la neurona sinapta con una única fibra)
Contracción del músculo liso
La proporción de filamentos finos/gruesos en el músculo liso destaca por tener mayor número de filamentos finos de actina, de los que hay varios tamaños. La contracción es dependiente de calcio, pero el músculo liso carece de troponina, aunque si existe la tropomiosina.
Los iones calcio que entran para la contracción muscular se unen a otra proteína calciodependiente que es la calmodulina. Este complejo activa una enzima que fosforila la cabeza de la miosina, ésta enzima se trata de la miosina cinasa (MLCK). Este paso es imprescindible para la formación de los puentes cruzados.
Una vez fosforilada se da el ciclo de golpe de fuerza dependiente de ATP ya estudiado en el músculo esquelético. La contracción del miocito liso es más potente que la del músculo esquelético.
También existe otra enzima, la miosina fosfatasa que se encarga de defosforilar e inactivar la cabeza ligera. La proporción entre ambas enzimas (MLCK y miosina fosfatasa) es de vital importancia para determinar la velocidad de la contracción. La miosina fosfatasa se activa cuando bajan los niveles de calcio.
Contracción fásica
En el músculo liso hablamos de contracción fásica en estos ciclos de unión/desunión de puentes cruzados. El músculo se acorta por ejemplo para vaciar una víscera hueca.
La contracción fásica depende de la actividad de la MLCK y es muy potente y efectiva.
Contracción tónica
La contracción tónica puede ser casi permanente que se debe al estadío de cierre en el que hay cierto grado de contracción sin apenas gasto de ATP. La contracción tónica mantiene las dimensiones del órgano y evita que éste se colapse.
La contracción tónica es regulable desde fuera, aumentando o reduciendo dicho tono.
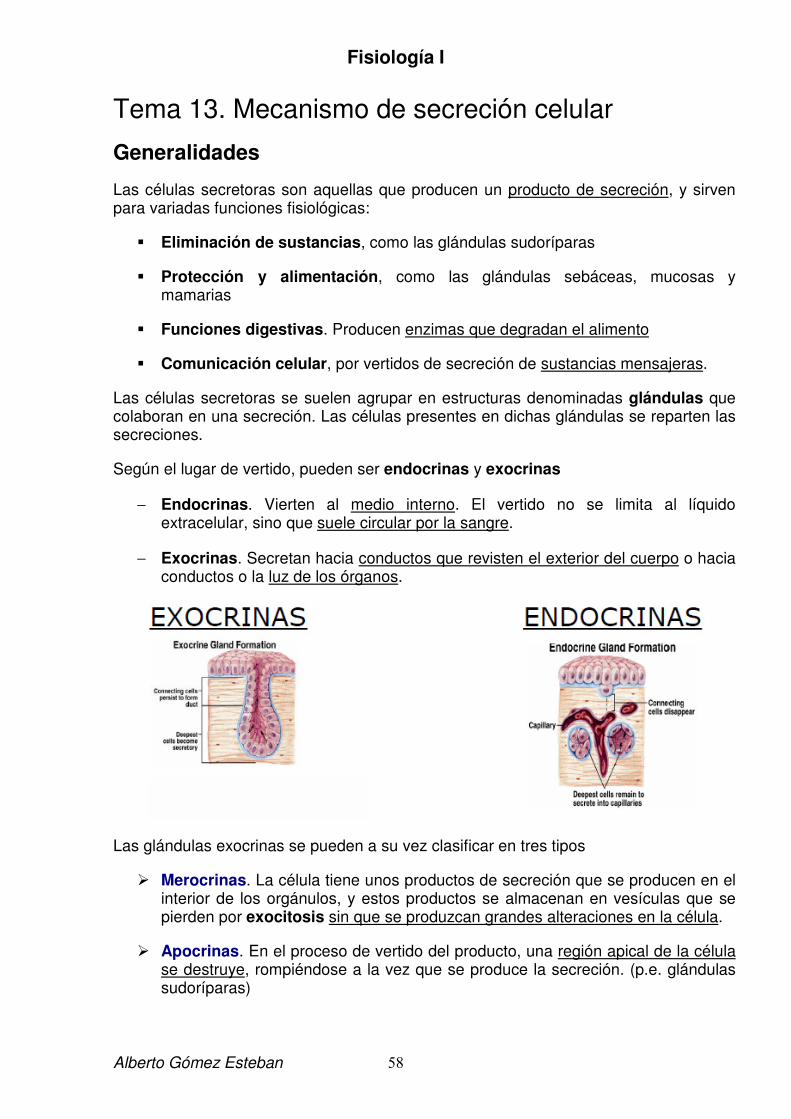
Fisiología I
Alberto Gómez Esteban 58
Tema 13. Mecanismo de secreción celular
Generalidades
Las células secretoras son aquellas que producen un producto de secreción, y sirven para variadas funciones fisiológicas:
� Eliminación de sustancias, como las glándulas sudoríparas
� Protección y alimentación, como las glándulas sebáceas, mucosas y mamarias
� Funciones digestivas. Producen enzimas que degradan el alimento
� Comunicación celular, por vertidos de secreción de sustancias mensajeras.
Las células secretoras se suelen agrupar en estructuras denominadas glándulas que colaboran en una secreción. Las células presentes en dichas glándulas se reparten las secreciones.
Según el lugar de vertido, pueden ser endocrinas y exocrinas
− Endocrinas. Vierten al medio interno. El vertido no se limita al líquido extracelular, sino que suele circular por la sangre.
− Exocrinas. Secretan hacia conductos que revisten el exterior del cuerpo o hacia conductos o la luz de los órganos.
Las glándulas exocrinas se pueden a su vez clasificar en tres tipos
� Merocrinas. La célula tiene unos productos de secreción que se producen en el interior de los orgánulos, y estos productos se almacenan en vesículas que se pierden por exocitosis sin que se produzcan grandes alteraciones en la célula.
� Apocrinas. En el proceso de vertido del producto, una región apical de la célula se destruye, rompiéndose a la vez que se produce la secreción. (p.e. glándulas sudoríparas)

Fisiología I
Alberto Gómez Esteban 59
� Holocrinas. La célula se degrada completamente en la secreción y debe reponerse en la división celular desde el estrato basal del tejido. (p.e. endometrio, glándulas sebáceas).
En conjunto, las secreciones de ambos tipos de glándulas en general suelen ser almacenadas en el aparato de Golgi, y que dicha liberación vaya precedida de un cambio de potencial de membrana muchas veces calciodependiente, de forma que el detonante para la secreción suele ser el abandono del potencial de reposo.
Implica un proceso de exocitosis mediado por calcio, de forma semejante a como se producía en el proceso de neurotransmisión. Cuando en este proceso intervienen las proteínas del complejo SNARE, el calcio interviene en muchos de los pasos.
Las moléculas secretadas varían mucho en tamaño, desde pequeñas moléculas como el CO (30 daltons) a otras muy grandes como la tirotropina (28.000 daltons). El lugar de síntesis intracelular (RER, REL…) depende de la naturaleza química de la sustancia a secretar.

Fisiología I
Alberto Gómez Esteban 60
Tema 13. Introducción a la fisiología de los receptores sensoriales.
Generalidades
En términos generales los receptores sensoriales son células nerviosas especializadas en la captación de estímulos, tan especializadas que su morfología en ocasiones suele no recordar a una neurona, aunque también puede servir de receptor sensorial el terminal dendrítico de una neurona.
Los receptores sensoriales registran continuamente los cambios del ambiente para permitir la actuación de los mecanismos homeostáticos. Captan variaciones del medio externo e interno.
Los receptores sensoriales son la primera unidad de una vía sensorial que es la vía que recoge ese estimulo y lo lleva al SNC. El hecho de que el estimulo sea consciente o no depende de la región del sistema nervioso que alcance (si alcanza el nivel cortical, el estimulo se convierte en una percepción consciente).
La vía sensorial requiere:
1. Estimulación. Un cambio del ambiente que estimule una vía sensorial.
2. Traducción. Es la función del receptor sensorial. Se trata de la captación de un tipo de estimulo y lo transforma en un potencial de acción para comenzar la comunicación. Corre a cargo de un receptor sensorial, especialmente sensible a una modalidad de estimulo.
3. Generación y conducción. Tras disponer de un potencial de acción, este debe ser conducido a través de la vía sensorial hasta alcanzar el SNC.
4. Integración. Se trata del procesamiento de la información recibida y queda a cargo del SNC.
El receptor sensorial recibe un estimulo y lo transforma en un impulso nervioso mediante una función de traducción del estimulo.
El receptor sensorial es la terminación dendrítica. A veces se agrupan en órganos terminales de neuronas sensoriales encargados siempre de una modalidad de estimulo. El receptor siempre responde al estimulo con un potencial de receptor. Cada receptor sensorial tiene un valor umbral para generar este potencial de receptor, que es menor para el estimulo para el que esta especializado.
El potencial de receptor se diferencia del potencial de acción a que éste primero recuerda a un potencial potsináptico sobre el que funciona la sumación espacial y temporal. El potencial de receptor puede ser insuficiente por si solo para alcanzar el umbral.

Fisiología I
Alberto Gómez Esteban 61
Tipos de receptores sensoriales
Los receptores sensoriales fueron clasificados por Sherrington en tres principales grupos en función de la localización en el cuerpo:
� Exteroceptores. Reciben información del exterior del organismo
� Interoceptores. Reciben información del medio interno
� Propioceptores. Se encuentran en músculos, tendones y órganos del equilibrio e informan acerca de la tensión que tienen dichos músculos.
La clasificación de Mountcastle se basa en los estímulos que excitan el receptor:
� Mecanorreceptores. Captan estimulos mecánicos
� Quimiorreceptores. Detectan concentraciones químicas
� Termorreceptores. Aportan información acerca de la temperatura del organismo
� Fotorreceptores. Detectan la luz y los encontramos principalmente en el ojo
� Nociceptores. Son las terminaciones nerviosas del dolor
Arco reflejo
El arco reflejo es la compenetración entre la unidad motora y el receptor sensorial, cuya finalidad es el mantenimiento de la homeostasis.
La idea del arco reflejo se trata de una vía de comunicación desde donde se da el estimulo hasta el órgano responsable de ejecutarlo (desde el receptor sensorial hasta el órgano efector, que puede ser un músculo u otro). Tendrá un total de 5 componentes

Fisiología I
Alberto Gómez Esteban 62
1. Receptor sensorial. Recibe el estimulo
2. Vía aferente. Conduce la información hasta un centro integrador del SNC
3. Centro integrador. La capacidad integradora es exclusiva del SNC, pero no es preciso que dicho estímulo o integración sean conscientes; sólo lo serán si llegan a nivel de la corteza cerebral.
4. Vía eferente. Conduce la información integradora desde el SNC que actúa sobre un efector.
5. Efector. Lleva a cabo la acción de mantenimiento de homeostasis.


















