
› docs › ... · Hiểu Biết Về Các Tác Động Điều Hòa Miễn Dịch Của Các...
16
Makrides M, Ochoa JB, Szajewska H (eds): The Importance of Immunonutrition. Nestlé Nutr Inst Workshop Ser, vol 77, pp 75–90, (DOI: 10.1159/000351388) Nestec Ltd., Vevey/S. Karger AG., Basel, © 2013 Tóm Tắt Bruno Pot • Benoît Foligné • Catherine Daniel • Corinne Grangette Hệ Vi Khuẩn và Pro-/Prebiotics Hiểu Biết Về Các Tác Động Điều Hòa Miễn Dịch Của Các Probiotics (Lợi Khuẩn) Phòng thí nghiệm về vi khuẩn acid lactic và tính miễn dịch của niêm mạc, Trung tâm nghiên cứu về nhiễm khuẩn và miễn dịch Lille, Viện Pasteur de Lille, INSERM U1019 - CNRS UMR 8204 - Đại học Lille Nord de France, Lille, Pháp Hệ vi sinh vật ở đường ruột được biết đến như một động lực cho sự phát triển và duy trì hệ miễn dịch. Trong khi các thay đổi quan trọng trong thành phần hệ vi sinh có thể ảnh hưởng đến tính năng miễn dịch trong một thời gian dài, các thay đổi không thường xuyên trong ngắn hạn cũng có thể được nhận thấy. Việc nhận ra các thay đổi ngắn hạn mở ra triển vọng lớn cho việc sử dụng các can thiệp dinh dưỡng, hướng đến việc điều chỉnh tính năng miễn dịch theo hướng mong muốn. Probiotics được thảo luận ở đây là một cách khả thi để đạt được mục tiêu này. Có vẻ như các tác động không chỉ đặc trưng cho chủng chuyên biệt, mà sẽ phụ thuộc vào nhiều yếu tố môi trường làm cho hệ thống miễn dịch dễ tiếp nhận hoặc không bị ảnh hưởng bởi việc đưa vào một chủng probiotic. Các tương tác giữa các probiotic, và mặt khác là các tương tác giữa probiotic và các tế bào ruột hoặc các tế bào miễn dịch, là tác động qua lại phức tạp, hiếm khi qua trung gian của một cơ chế đơn lẻ. Vì thế, điều hòa miễn dịch thông qua dinh dưỡng là một hiện tượng phức tạp cần có sự xem xét cẩn thận, cần tìm hiểu tính năng miễn dịch, cũng như biết rõ các cơ chế hoạt động của probiotic. Chỉ có thể đạt được điều này sau khi thực hiện các thử nghiệm lâm sàng thích hợp, với các dữ liệu được thiết lập thích hợp, để chứng minh tính hiệu quả của từng chủng/hỗn hợp riêng biệt. Bản quyền thuộc Nestec Ltd. 2013, Vevey/S. Karger AG, Basel Mở Đầu Các tác động của probiotic có thể được cho là do hoạt tính của hệ vi sinh, sự trao đổi chất hoặc miễn dịch. Mặc dù hầu hết các tác động này có thể được nghiên cứu trong phòng thí nghiệm, chúng sẽ bị ảnh hưởng nặng nề bởi các điều kiện môi
Transcript of › docs › ... · Hiểu Biết Về Các Tác Động Điều Hòa Miễn Dịch Của Các...

M akrides M, Ochoa JB, Szajewska H (eds): The Importance of Immunonutrition.Nestlé Nutr Inst Workshop Ser, vol 77, pp 75–90, (DOI: 10.1159/000351388) Nestec Ltd., Vevey/S. Karger AG., Basel, © 2013
Tóm Tắt
Bruno Pot
•
Benoît Foligné
•
Catherine Daniel
•
Corinne Grangette
Hệ Vi Khuẩn và Pro-/Prebiotics
Hiểu Biết Về Các Tác Động Điều Hòa Miễn Dịch Của Các Probiotics (Lợi Khuẩn)
Phòng thí nghiệm về vi khuẩn acid lactic và tính miễn dịch của niêm mạc, Trung tâm nghiên cứu về nhiễm khuẩn và miễn dịch Lille, Viện Pasteur de Lille, INSERM U1019 - CNRS UMR 8204 - Đại học Lille Nord de France, Lille, Pháp
Hệ vi sinh vật ở đường ruột được biết đến như một động lực cho sự phát triển và duy trì hệ miễn dịch. Trong khi các thay đổi quan trọng trong thành phần hệ vi sinh có thể ảnh hưởng đến tính năng miễn dịch trong một thời gian dài, các thay đổi không thường xuyên trong ngắn hạn cũng có thể được nhận thấy. Việc nhận ra các thay đổi ngắn hạn mở ra triển vọng lớn cho việc sử dụng các can thiệp dinh dưỡng, hướng đến việc điều chỉnh tính năng miễn dịch theo hướng mong muốn. Probiotics được thảo luận ở đây là một cách khả thi để đạt được mục tiêu này. Có vẻ như các tác động không chỉ đặc trưng cho chủng chuyên biệt, mà sẽ phụ thuộc vào nhiều yếu tố môi trường làm cho hệ thống miễn dịch dễ tiếp nhận hoặc không bị ảnh hưởng bởi việc đưa vào một chủng probiotic. Các tương tác giữa các probiotic, và mặt khác là các tương tác giữa probiotic và các tế bào ruột hoặc các tế bào miễn dịch, là tác động qua lại phức tạp, hiếm khi qua trung gian của một cơ chế đơn lẻ. Vì thế, điều hòa miễn dịch thông qua dinh dưỡng là một hiện tượng phức tạp cần có sự xem xét cẩn thận, cần tìm hiểu tính năng miễn dịch, cũng như biết rõ các cơ chế hoạt động của probiotic. Chỉ có thể đạt được điều này sau khi thực hiện các thử nghiệm lâm sàng thích hợp, với các dữ liệu được thiết lập thích hợp, để chứng minh tính hiệu quả của từng chủng/hỗn hợp riêng biệt.
Bản quyền thuộc Nestec Ltd. 2013, Vevey/S. Karger AG, Basel
Mở Đầu
Các tác động của probiotic có thể được cho là do hoạt tính của hệ vi sinh, sự trao
đổi chất hoặc miễn dịch. Mặc dù hầu hết các tác động này có thể được nghiên cứu
trong phòng thí nghiệm, chúng sẽ bị ảnh hưởng nặng nề bởi các điều kiện môi

76 P ot · Foligné · Daniel · Grangette
trường (ở miệng, ruột, âm đạo), lối sống (thuốc, chế độ ăn uống, stress, vệ sinh)
và các tình trạng của vật chủ (cơ sở di truyền, trẻ sơ sinh so với người trưởng
thành hoặc cao tuổi; khỏe mạnh hay bệnh tật). Do đó, việc sàng lọc trong phòng
thí nghiệm (in vitro) các chủng có các đặc tính probiotic mong muốn cần được
kiểm định trên cơ thể (in vivo) và các phân tích cuối cùng trong các thử nghiệm
lâm sàng nhằm hướng tới một dân số được xác định rõ ràng. Vì việc kiểm định
trên lâm sàng này rất tốn kém và mất thời gian, việc lựa chọn các chủng thích hợp
nhất vào giai đoạn bắt đầu từ nguồn chọn chủng là rất quan trọng.
Viêm, trong nhiều trường hợp, là một phản ứng tự phát tức thời đối với các tổn
thương hay nhiễm trùng cấp tính của cơ thể. Tuy nhiên, nhiều dạng khác nhau của
tình trạng viêm mạn tính đã được mô tả là gây ra hoặc bị gây ra bởi các bệnh lý
khác nhau. Kể từ lúc các triệu chứng ban đầu thường gặp chưa xuất hiện trên lâm
sàng, giai đoạn đầu của bệnh hầu hết vẫn không được chú ý; nhưng trong một thời
gian dài như vậy, tình trạng viêm ở mức thấp có thể dẫn đến các rối loạn chuyển
hóa nghiêm trọng, các bệnh qua trung gian miễn dịch hoặc thậm chí là các vấn đề
trên tim hay tâm thần. Trong hơn 50 năm qua, chúng ta đã chứng kiến một sự gia
tăng kinh khủng của dạng này ở các bệnh lý mạn tính,1 một quan sát có liên quan
đến sự thiếu hụt các tiếp xúc đầy đủ của hệ miễn dịch ở giai đoạn đầu đời (giả
thuyết về vệ sinh). [2]
Song song với đó, các dự án nghiên cứu gần đây như MetaHit [3,4] hoặc Dự
án Quần thể vi khuẩn Người (http://commonfund.nih.gov/hmp/) đã cho thấy tầm
quan trọng của hệ vi sinh ở nhiều khía cạnh của sức khỏe và bệnh tật của con
người. Nghiên cứu tập trung vào số lượng (nhiều hơn gấp 10 lần số tế bào và gấp
150 lần số gien hiện có trong cơ thể người), [5] chất lượng và sự đa dạng hay khả
năng phục hồi của hệ vi sinh6 trong IBD, đái tháo đường tuýp 1 hoặc béo phì. Kết
quả là, thành phần của hệ vi sinh có liên kết rộng rãi với các rối loạn qua trung
gian miễn dịch, tương tự như các suy luận trước đó của các giả thuyết về vệ sinh.
Nhiều khả năng làm ảnh hưởng đến chất lượng của hệ vi sinh thông qua dinh
dưỡng, trị liệu hoặc thậm chí cấy phân đã mở ra nhiều triển vọng cho nghiên cứu
sâu hơn, không chỉ tập trung vào thành phần và sự đa dạng của hệ vi sinh, mà còn
vào tiềm năng điều hòa miễn dịch của can thiệp được xem xét. Vì vậy, việc sử
dụng các phương pháp chống viêm khả thi, như ở khía cạnh can thiệp vào chế độ
ăn uống, đã gây được nhiều sự chú ý. Một trong các can thiệp này bao gồm các
chủng probiotic thích hợp. Trong chương này, chúng tôi sẽ tập trung vào một số
nguồn sàng lọc sẵn có có ích để phân tách các chủng có tiềm năng điều hòa miễn
dịch khác nhau, cũng như để hỗ trợ trong việc đưa ra cơ chế giải thích cho các
khác biệt đã được quan sát thấy.
Tầm Quan Trọng của Việc Điều Hòa Miễn Dịch

Như đã đề cập ở trên, hệ thống miễn dịch của động vật có vú bao gồm một bộ hoàn
chỉnh các tế bào miễn dịch và các phân tử truyền tín hiệu tương tác với các vi sinh
vật và các kháng nguyên xung quanh chúng ta. Vì có đến 1014 vi khuẩn, ruột đã
phát triển thành cơ quan miễn dịch chính của chúng ta. Do đó, không còn nghi
ngờ gì khi cho rằng ruột là một cơ quan dễ tiếp nhận, thích hợp cho các can thiệp
dinh dưỡng nhằm thay đổi hoặc điều chỉnh các đáp ứng miễn dịch của vật chủ.
Khả năng miễn dịch bẩm sinh cho phép phản ứng nhanh chóng và triệt để với
các kích thích gây ra bởi các tác nhân lây nhiễm. Các đáp ứng là phản ứng
viêm tự nhiên và chủ yếu gồm các tế bào thực bào (các đại thực bào, các bạch
cầu trung tính, các tế bào giết tự nhiên).
Hệ thống đáp ứng miễn dịch, sau khi được hoạt hóa bởi các tế bào trình diện
kháng nguyên, sẽ huy động các tế bào T và B đặc hiệu, thông qua việc sản xuất
các phân tử truyền tín hiệu đặc hiệu (các cytokine, chemokine), sẽ hỗ trợ điều
chỉnh các phản ứng miễn dịch, cả tự nhiên và thích nghi. Các tế bào B tiết ra
các kháng thể (tạo ra miễn dịch thể dịch), trong khi các tế bào T được chia
thành các tế bào T giúp đỡ (CD4+, còn được gọi là Th) và các tế bào T gây độc
(CD8+). Vì các tế bào trình diện kháng nguyên trong niêm mạc ruột tiếp tục
lấy mẫu các kháng nguyên ở trong lòng ống ruột, thách thức chính của hệ
thống miễn dịch ở ruột là phải liên tục cân bằng qua sự phòng vệ với sự dung
nạp: lựa chọn để hoạt hóa các tế bào T đáp ứng để tăng cường sự phòng vệ
chống lại các tác nhân gây bệnh, hoặc thúc đẩy sự biệt hóa thành các loại tế
bào T điều hòa khác nhau để tạo ra khả năng dung nạp. Các cơ chế cho phép
dung nạp phải bảo vệ vật chủ khỏi phản ứng viêm quá mức trong quá trình
nhiễm trùng, hoặc giúp tái cân bằng lại hệ thống miễn dịch khi bị rối loạn. Vì
thế, các tế bào miễn dịch bẩm sinh sẽ cần phải liên tục xác định “bản thân” so
với “không bản thân” bằng cách sử dụng các chỉ dấu bề mặt đặc hiệu. Bảng 1
liệt kê các loại phân tử liên quan đến vi sinh vật hoặc tác nhân gây bệnh của vi
khuẩn phổ biến nhất, cùng với các thụ thể nhận dạng tương ứng của chúng
(PRRs) được tìm thấy trong và trên các tế bào miễn dịch. Được biết đến nhiều
nhất là các thụ thể giống Toll có trên bề mặt, và các thụ thể giống NOD
(protein chứa vùng oligome hóa gắn kết với nucleotide) có ở trong nhân của
các tế bào miễn dịch. Trong số các kháng nguyên được tìm thấy,
lipopolysaccharides (LPS) của thành tế bào vi khuẩn gram âm thuộc nhóm
làm tăng các phản ứng viêm mạnh mẽ nhất. LPS được cho là tham gia vào tình
Dưới đây, chúng tôi mô tả rất ngắn gọn một số cơ chế đã được biết hiện nay là
có liên quan đến sự tương tác giữa vật chủ và hệ vi sinh vật. Nhiều đánh giá rộng
hơn của các quá trình này có thể được tìm thấy ở nghiên cứu của Delcenserie và
cộng sự [7] và của Lebeer và cộng sự. [8]
77
Các Cơ Chế Tham Gia
–
–
Các Tác Động Điều Hòa Miễn Dịch Của Các Probiotics

trạng viêm sinh lý mạn tính, gặp điển hình ở các bệnh nhân béo phì.9 Ngược
lại, peptidoglycans (PGNs) từ vi khuẩn Gram dương (như lactobacilli) có thể
tham gia vào hoạt tính chống viêm của probiotic. [10]
Các tác động của probiotic có thể được truyền thông qua sự tương tác của
chúng với PRRs trong các tế bào tua (DCs) hoặc trong các tế bào biểu mô của
ruột. DCs, được hoạt hóa bởi probiotic, tạo ra các cytokine và biểu hiện các
phân tử được kích thích; đồng thời sẽ cho phép phân cực các tế bào T thuần
khiết (naïve) theo hướng biệt hóa thành các tế bào T điều hòa (Tregs), đáng
chú ý là Tregs CD4+CD25+FoxP3, [10, 11] hoặc các tế bào kích thích Th112
sẽ sản xuất ra các cytokine tiền viêm như interferon-γ (IFN-γ), yếu tố gây hoại
tử khối u α (TNF-α) và interleukin-2 (IL-2) và IL-12. Do đó, các tế bào Th1 sẽ
kích thích sự thực bào và giúp chống lại các vi khuẩn gây bệnh, nhưng chúng
cũng có thể có liên quan đến các bệnh lý qua trung gian miễn dịch, như viêm
khớp hoặc đa xơ cứng. Các kháng nguyên không phải của probiotic có thể
phân tế bào T theo hướng biệt hóa thành các tế bào Th2, tạo ra các cytokine
như IL-4, IL-5, IL-6 và IL-13, giúp chống lại các ký sinh ngoại bào, nhưng
cũng được phát hiện trong các phản ứng dị ứng. Vì cả hai loại tế bào Th1 và
Th2 có tác động trái ngược nhau, sự cân bằng giữa việc sản xuất các cytokine
78 P ot · Foligné · Daniel · Grangette
–
TLR/PRR
PAMPs hoặc MAMPs ở vi khuẩnLipopolysaccharide TLR4 Vi Khuẩn Gram âmLipoteichoic acid TLR2 Vi khuẩn Gram dươngPeptidoglycan TLR2-NOD1
or NOD2Hầu hết các vi khuẩn
Triacyl lipopeptides TLR1 or TLR2 Hầu hết các vi khuẩnDiacyl lipopeptides TLR2 or TLR6 Mycoplasma spp.Glycan ligands (surface layers) DC-SIGN many bacteriaPorins TLR2 NeisseriaFlagellin TLR5 Salmonella TyphimuriumFimbriae (mannose glycoproteins) TLR4 Escherichia coliCpGDNA TLR Tất cả các vi khuẩnUnknown TLR11 Vi khuẩn gây bệnh Đ.tiểu
PAMPs or MAMPs in fungiZymosan TLR2 or TLR6 Saccharomyces cerevisiaePhospholipomannan TLR2 Candida albicansMannan TLR4 C. albicansO-linked mannosyl residues TLR4 C. albicansβ-Glucans TLR2 C. albicans
PAMPs = Các loại phân tử liên quan đến tác nhân gây bệnh; MAMPs = Các loại phân tử liên quan đến vi sinh vật; TLR = thụ thể giống Toll.
Bảng 1. Các kháng nguyên của vi khuẩn và nấm với các thụ thể miễn dịch tương ứng của chúng
Các loại

của Th1 và Th2 sẽ xác định chiều hướng của một phản ứng miễn dịch. Trong quá trình này, các tế bào Treg và các tế bào T giúp đỡ tuýp 3 (các tế bào Th3) có thể can thiệp. Các tế bào Treg có các đặc tính điều hòa miễn dịch qua trung gian của IL-10 và TGF-β, [13] trong khi các tế bào Th3 chủ yếu tiết ra TGF-β.14 Các cytokine IL-10 và TGF-β, với sự hỗ trợ của các tế bào T CD4+ từ GALT sẽ thúc đẩy sự biệt hóa các tế bào B thành các tế bào tiết kháng thể IgA trong huyết thanh. Các tế bào niêm mạc ruột sẽ tiết ra các kháng thể IgA vào lòng ống ruột non, nơi chúng bất hoạt các kháng nguyên và các tác nhân gây bệnh (các virus, vi khuẩn, độc tố). [15]
Một nhóm khác gồm các tế bào Th Cd4+, tế bào Th17, được đặc trưng bằng việc sản xuất chủ yếu IL-17 (ngoài IL-21 và IL-22) và bằng sự biệt hóa các cytokine TGF-β và IL-6,16 có liên quan vào sự phục hồi các bạch cầu trung tính và các đại thực bào ở mô bị nhiễm.Các tế bào biểu mô chuyên biệt, chẳng hạn như các tế bào Paneth ở dưới các nếp nhăn của ruột non, có thể sản xuất các defensin, trong khi các tế bào hình ly (globlet) sản xuất chất nhầy để bảo vệ biểu mô. Các defensin là một họ lớn gồm các peptide kháng khuẩn góp phần bảo vệ các bề mặt niêm mạc bằng cách gắn vào màng tế bào vi sinh vật, nơi chúng tạo thành các các cấu trúc giống như lỗ rỗng, làm khiếm khuyết màng tế bào vi khuẩn. IL-22 được tiết ra bởi các tế bào Th17 không chỉ có thể làm tăng tiết β-defensin 2 và β-defensin [3,17] hỗ trợ trực tiếp trong việc bảo vệ chống lại nhiễm trùng do vi khuẩn, mà còn thông qua các đường truyền tín hiệu của các cytokine này sẽ liên kết (và phối hợp?) với các nhánh khác của hệ thống phòng vệ của vật chủ.
Trong nhiều thập kỷ qua, hàng trăm nghiên cứu đã tìm kiếm trên diện rộng tiềm
năng của các chủng probiotic (tiềm năng) để tương tác với một loạt các tế bào
miễn dịch bằng cách dùng vô số các mô hình in vitro, in vivo hoặc ex vivo (ngoài
cơ thể sống). Bảng 2 liệt kê các bài báo đã được chọn là có liên quan, được xuất
bản từ giữa tháng 1 đến tháng 10 năm 2012, đã nghiên cứu hoặc đánh giá các cơ
chế khả thi của việc điều hòa miễn dịch ở probiotic. Sự đa dạng của các dữ liệu,
các mô hình và các sinh vật phản ánh rõ nét sự phức tạp của loại nghiên cứu này.
Tính năng miễn dịch của bất kỳ chủng nào được dùng ở người sẽ phụ thuộc vào
môi trường, cũng như vào hệ vi sinh vật xung quanh (các vi sinh vật hội sinh, các
tác nhân gây bệnh, các thực khuẩn, các độc tố). Do đó, việc kiểm định cuối cùng
cho một chủng được chọn sẽ được xuyên suốt trong các thử nghiệm lâm sàng trên
người, khẳng định hiệu quả trong các dân số mục tiêu nhất định. Vì việc kiểm
định trên lâm sàng tốn kém, sự hiểu biết về cơ chế khả thi là một yếu tố có giá trị
trong việc xác định bối cảnh của thử nghiệm lâm sàng và để cân nhắc các điểm
kết thúc. Dưới đây là một ví dụ của một cách tiếp cận khả thi, bắt đầu bằng việc
sàng lọc một lượng (lớn) các chủng và kết thúc bằng một phân tử tinh khiết chịu
trách nhiệm cho hiệu quả chống viêm được quan sát thấy trên chủng được chọn.
79
–
–
Ảnh Hưởng của Probiotic Trên Hệ Miễn Dịch
Các Tác Động Điều Hòa Miễn Dịch Của Các Probiotics
80 P ot · Foligné · Daniel · Grangette
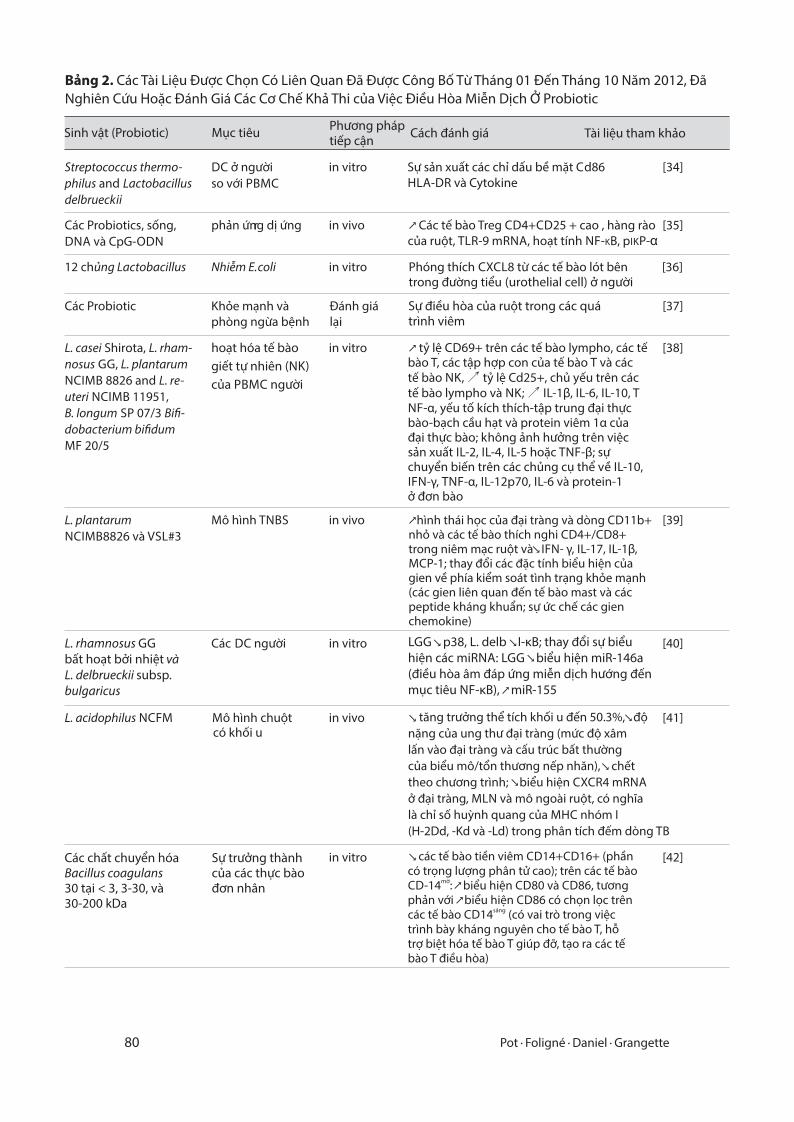
Streptococcus thermo-philus and Lactobacillus delbrueckii
DC ở ngườiso với PBMC
in vitro Sự sản xuất các chỉ dấu bề mặt Cd86HLA-DR và Cytokine
[34]
Các Probiotics, sống,DNA và CpG-ODN
phản ứng dị ứngr in vivo ↗ Các tế bào Treg CD4+CD25 + cao , hàng ràocủa ruột, TLR-9 mRNA, hoạt tính NF-KB, pIKP-α
[35]
12 chủng Lactobacillus Nhiễm E. coli in vitro Phóng thích CXCL8 từ các tế bào lót bêntrong đường tiểu (urothelial cell) ở người
[36]
Các Probiotic Khỏe mạnh vàphòng ngừa bệnh
Đánh giálại
Sự điều hòa của ruột trong các quá trình viêm
[37]
L. casei Shirota, L. rham-nosus GG, L. plantarum
NCIMB 8826 and L. re-uteri NCIMB 11951, B. longum SP 07/3 Bi�-dobacterium bi�dum
MF 20/5
hoạt hóa tế bào giết tự nhiên (NK) của PBMC người
in vitro ↗ tỷ lệ CD69+ trên các tế bào lympho, các tế bào T, các tập hợp con của tế bào T và các tế bào NK, ↗ tỷ lệ Cd25+, chủ yếu trên các tế bào lympho và NK; ↗ IL-1β, IL-6, IL-10, TNF-α, yếu tố kích thích-tập trung đại thực bào-bạch cầu hạt và protein viêm 1α của đại thực bào; không ảnh hưởng trên việc sản xuất IL-2, IL-4, IL-5 hoặc TNF-β; sự chuyển biến trên các chủng cụ thể về IL-10, IFN-γ, TNF-α, IL-12p70, IL-6 và protein-1 ở đơn bào
[38]
L. plantarum
NCIMB8826 và VSL#3Mô hình TNBS in vivo ↗ hình thái học của đại tràng và dòng CD11b+
nhỏ và các tế bào thích nghi CD4+/CD8+ trong niêm mạc ruột và IFN- γ, IL-17, IL-1β, MCP-1; thay đổi các đặc tính biểu hiện của gien về phía kiểm soát tình trạng khỏe mạnh (các gien liên quan đến tế bào mast và các peptide kháng khuẩn; sự ức chế các gien chemokine)
[39]
L. rhamnosus GG bất hoạt bởi nhiệt và L. delbrueckii subsp.bulgaricus
Các DC người in vitro
↘
↘ ↘↘
↘
[40]
L. acidophilus NCFM Mô hình chuộtcó khối u
in vivo ↘ ↘
↘↘
[41]
in vitro ↘
↗↗
[42]
Bảng 2. Các Tài Liệu Được Chọn Có Liên Quan Đã Được Công Bố Từ Tháng 01 Đến Tháng 10 Năm 2012, Đã Nghiên Cứu Hoặc Đánh Giá Các Cơ Chế Khả Thi của Việc Điều Hòa Miễn Dịch Ở Probiotic
Sinh vật (Probiotic) Mục tiêu Phương pháp tiếp cận Cách đánh giá Tài liệu tham khảo
LGG p38, L. delb I-κB; thay đổi sự biểu hiện các miRNA: LGG biểu hiện miR-146a (điều hòa âm đáp ứng miễn dịch hướng đến mục tiêu NF-κB), miR-155
tăng trưởng thể tích khối u đến 50.3%, độ nặng của ung thư đại tràng (mức độ xâm lấn vào đại tràng và cấu trúc bất thường của biểu mô/tổn thương nếp nhăn), chết theo chương trình; biểu hiện CXCR4 mRNA ở đại tràng, MLN và mô ngoài ruột, có nghĩa là chỉ số huỳnh quang của MHC nhóm I (H-2Dd, -Kd và -Ld) trong phân tích đếm dòng TB
Các chất chuyển hóa Bacillus coagulans 30 tại < 3, 3-30, và 30-200 kDa
Sự trưởng thành của các thực bàođơn nhân
các tế bào tiền viêm CD14+CD16+ (phần có trọng lượng phân tử cao); trên các tế bào
mờCD-14 : biểu hiện CD80 và CD86, tương phản với biểu hiện CD86 có chọn lọc trên
sángcác tế bào CD14 (có vai trò trong việc trình bày kháng nguyên cho tế bào T, hỗ trợ biệt hóa tế bào T giúp đỡ, tạo ra các tế bào T điều hòa)
81
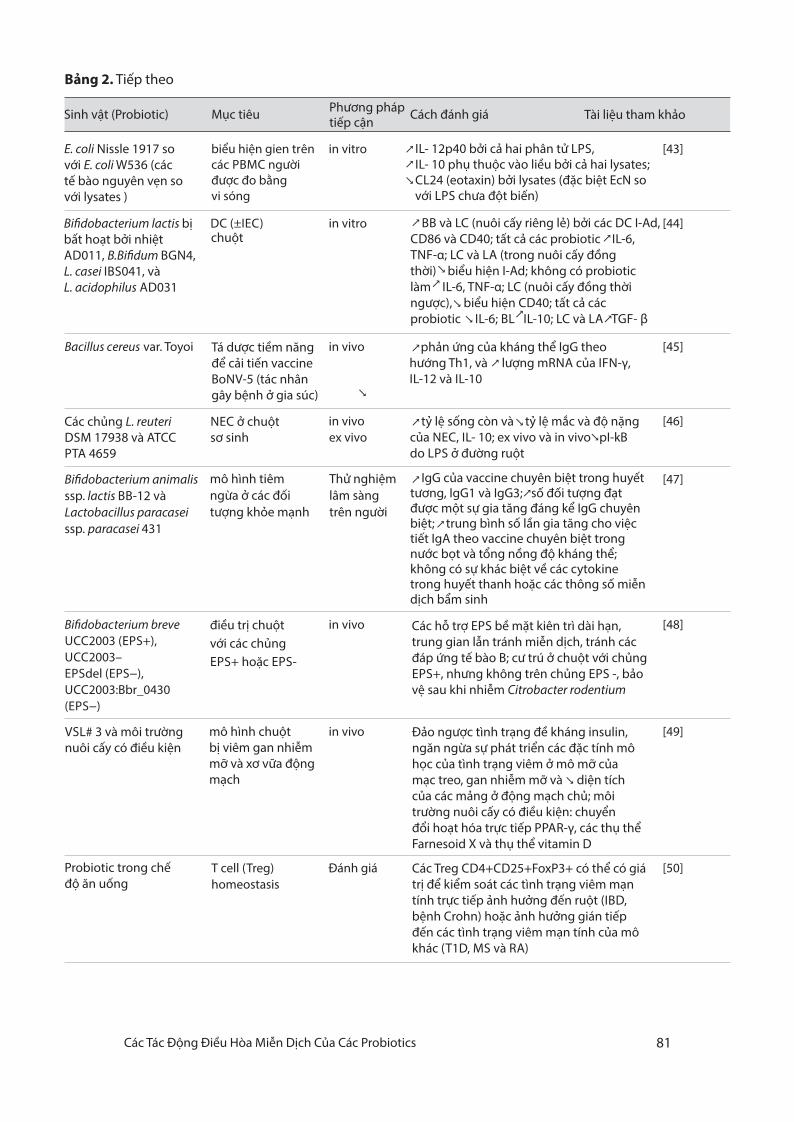
Bảng 2. T iếp theo
biểu hiện gien trên các PBMC người được đo bằng vi sóng
in vitro ↗↗↘
[43]
DC (±IEC) in vitro↗
↗
↗↗
↗
↗↗
↘
↘
↘
↘
↘
↘
[44]
Bacillus cereus var. Toyoi in vivo [45]
in vivoex vivo
↘
[46]
↗↗
↗
in vivo
[47]
[48]
[49]
T cell (Treg) homeostasis
Đánh giá [50]
Sinh vật (Probiotic) Mục tiêu Phương pháp tiếp cận
Cách đánh giá Tài liệu tham khảo
E. coli Nissle 1917 so với E. coli W536 (các tế bào nguyên vẹn so với lysates )
IL- 12p40 bởi cả hai phân tử LPS,IL- 10 phụ thuộc vào liều bởi cả hai lysates; CL24 (eotaxin) bởi lysates (đặc biệt EcN so với LPS chưa đột biến)
BB và LC (nuôi cấy riêng lẻ) bởi các DC I-Ad, CD86 và CD40; tất cả các probiotic IL-6, TNF-α; LC và LA (trong nuôi cấy đồng thời) biểu hiện I-Ad; không có probiotic làm IL-6, TNF-α; LC (nuôi cấy đồng thời ngược), biểu hiện CD40; tất cả các probiotic IL-6; BL IL-10; LC và LA TGF- β
Bi�dobacterium lactis bị bất hoạt bởi nhiệt AD011, B.Bi�dum BGN4, L. casei IBS041, và L. acidophilus AD031
chuột
Tá dược tiềm năng để cải tiến vaccine BoNV-5 (tác nhân gây bệnh ở gia súc)
phản ứng của kháng thể IgG theo hướng Th1, và lượng mRNA của IFN-γ, IL-12 và IL-10
Các chủng L. reuteri DSM 17938 và ATCC PTA 4659
NEC ở chuột sơ sinh
↗ tỷ lệ sống còn và tỷ lệ mắc và độ nặng của NEC, IL- 10; ex vivo và in vivo pI-kB do LPS ở đường ruột
Bi�dobacterium animalis ssp. lactis BB-12 vàLactobacillus paracasei ssp. paracasei 431
mô hình tiêm ngừa ở các đối tượng khỏe mạnh
Thử nghiệm lâm sàng trên người
IgG của vaccine chuyên biệt trong huyết tương, IgG1 và IgG3; số đối tượng đạt được một sự gia tăng đáng kể IgG chuyênbiệt; trung bình số lần gia tăng cho việc tiết IgA theo vaccine chuyên biệt trong nước bọt và tổng nồng độ kháng thể; không có sự khác biệt về các cytokine trong huyết thanh hoặc các thông số miễn dịch bẩm sinh
Bi�dobacterium breveUCC2003 (EPS+), UCC2003–EPSdel (EPS−), UCC2003:Bbr_0430 (EPS−)
in vivođiều trị chuộtvới các chủngEPS+ hoặc EPS-
Các hỗ trợ EPS bề mặt kiên trì dài hạn, trung gian lẫn tránh miễn dịch, tránh các đáp ứng tế bào B; cư trú ở chuột với chủng EPS+, nhưng không trên chủng EPS -, bảo vệ sau khi nhiễm Citrobacter rodentium
VSL# 3 và môi trường nuôi cấy có điều kiện
mô hình chuột bị viêm gan nhiễm mỡ và xơ vữa động mạch
Đảo ngược tình trạng đề kháng insulin, ngăn ngừa sự phát triển các đặc tính mô học của tình trạng viêm ở mô mỡ của mạc treo, gan nhiễm mỡ và diện tích của các mảng ở động mạch chủ; môi trường nuôi cấy có điều kiện: chuyển đổi hoạt hóa trực tiếp PPAR-γ, các thụ thể Farnesoid X và thụ thể vitamin D
Các Treg CD4+CD25+FoxP3+ có thể có giátrị để kiểm soát các tình trạng viêm mạn tính trực tiếp ảnh hưởng đến ruột (IBD, bệnh Crohn) hoặc ảnh hưởng gián tiếp đến các tình trạng viêm mạn tính của mô khác (T1D, MS và RA)
Probiotic trong chế độ ăn uống
Các Tác Động Điều Hòa Miễn Dịch Của Các Probiotics

82 P ot · Foligné · Daniel · Grangette
Bảng 2. Tiếp theo
in vitro
Enterococcus faecalis
CECT7121dị ứng bởi lòng trắng trứng
in vitro
↘↘
↗ ↗
L. acidophilus Lafti-L10, L. casei CRL-431, L. plantarum WCFS1 L. rhamnosus GG
đánh giálại
[32]
[51]
[52]
Sinh vật (Probiotic) Mục tiêu Phương pháp tiếp cận
Cách đánh giá Tài liệu tham khảo
L. reuteri ATCC PTA 6475 và các đột biến
nhận dạng hợp chất bằng khối phổ NMR HI-LIC-HPLC
Histamine đã kích thích tăng nồng độ cAMP, điều này ức chế truyền tín hiệu MEK/ERK MAPK xuôi dòng qua protein kinase A cùng với sự ức chế sản xuất TNF bằng việc điều hòa phiên mã
nồng độ IgE chuyên biệt, nồng độ IgG2a kháng OVA, không thay đổi nồng độ IgG kháng OVA và IgG1, tỷ lệ gia tăng của các tế bào trí nhớ, các cytokine Th2 IL-4, IL-5 và IL-13, không thay đổi sự tiết IL-10, IL-12 và IFN-γ
sự biểu hiện gien của vật chủ
Các tình trạng của ống tiêu hóa và chế độ ăn của vật chủ có thể ảnh hưởng đến sự biểu hiện gien của vi khuẩn có liên quan đến sự tổng hợp hoặc sửa đổi các phân tử của vi sinh vật với sự liên quan rõ ràng với việc truyền tín hiệu của vật chủ, bao gồm các gien có chức năng trong việc tổng hợp CPS, d-alanyl hóa LTA và sản xuất các protein biểu hiện trên bề mặt tế bào
Cách Sàng Lọc Để Có Probiotic Tốt Hơn
Trong năm 2007, Foligné và cộng sự18 đã mô tả nét tương đồng giữa một xét
nghiệm sàng lọc in vitro đơn giản sử dụng các tế bào đơn nhân ở máu ngoại vi
(PBMCs) của người và một phương pháp in vivo sử dụng mô hình chuột bị viêm
đại tràng do acid trinitrobenzen sulfonic (TNBS). [18] Trong khi hầu hết các
chủng đều cho thấy có sự trùng hợp lớn ở cả hai phương pháp, sự thay đổi đáng kể
đã được tìm thấy ở hầu hết các chi LAB và nấm men ở tiềm năng tạo ra IL-10 (chỉ
dấu kháng viêm) và IL-12 (chỉ dấu tiền viêm) của chúng. [18-22]
Sự thay đổi này được minh họa trong hình 1a, 8 chủng LAB khác nhau được so
sánh về tỷ lệ IL-10:IL-12. Các chủng Lactobacillus salivarius, Lactobacillus
rhamnosus và Lactobacillus casei có các đặc tính kháng viêm tiềm ẩn (tỷ lệ
cytokine > 15), trong khi hai chủng Lactobacillus plantarum rõ ràng là không có
cùng tiềm năng (tỷ lệ cytokine khoảng 5). Các chủng được nghiên cứu
Lactobacillus acidophilus, Lactococcus lactis và Streptococcus gordonii có thể
hoàn toàn không có tác dụng kháng viêm, vì đặc tính cytokine của chúng trên
PBMCs của người có xu hướng là bình thường hơn là tiền viêm (tỷ lệ cytokine <
5). Nhìn chung, các chủng có tỷ lệ IL-10:IL-12 cao có khả năng kháng viêm, và
thích hợp hơn cho các ứng dụng trên các cơ thể sống có nhu cầu làm giảm phản
ứng viêm; trong khi các chủng làm tăng cytokine tiền viêm nhiều hơn có thể được

Hình 1. a. Tỷ lệ cytokine IL-10:IL-12 thu được trên PBMCs ở người. Dữ liệu là kết quả trung bình của 4 người hiến máu khỏe mạnh khác nhau. b, c. Tác động bảo vệ của các chủng vi khuẩn acid lactic chống lại viêm đại tràng do TNBS ở chuột BALB/c. Các cột đại diện cho tỷ lệ phần trăm của khả năng bảo vệ (giảm về điểm trung bình trong thang điểm viêm Wallace có thể quan sát được ở chuột được điều trị bằng vi khuẩn so với điểm trung bình của nhóm chuột TNBS chứng, n = 10). Chỉ số viêm đại tràng được đánh giá 48 giờ sau khi chỉ định TNBS. Mỗi cột đại diện cho một thí nghiệm độc lập. Các so sánh giữa các nhóm TNBS chứng và các nhóm BMDCs không điều trị tương ứng (DC) đã được tính toán bằng cách sử dụng kiểm định Mann-Whitney U (*p < 0.05, **p < 0.01, ***p < 0.001). b. Các tác động của các điều trị bằng cách uống vi khuẩn acid lactic sống trong viêm đại tràng cấp tính do TNBS ở chuột BALB/c. c. Các tác động của điều trị bằng IP. Các thử nghiệm IP thiếu các chủng L. casei và L. plantarum. Để biết chi tiết hơn về thử nghiệm hãy tham khảo nghiên cứu của Macho-Fernandez và cộng sự. [10]
83
0
–25
25
50
75
**
***
0
–20
40
20
60
80
*****
***
*******
****
**
*
**
Kh
ả n
ăn
g b
ảo
vệ (
%)
Kh
ả n
ăn
g b
ảo
vệ (
%)
0
10
5
15
20
IL-1
0:IL-
12
L. sa
livar
ius
L. rh
amno
sus
L. ca
sei
L. pl
anta
rum
1
L. pl
anta
rum
2
L. ac
idop
hilu
s
L. la
ctis
S. g
ordo
nii
c
b
a
Các Tác Động Điều Hòa Miễn Dịch Của Các Probiotics

84 P ot · Foligné · Daniel · Grangette
quan tâm để hoạt hóa hệ thống miễn dịch, như để chống một số loại nhiễm trùng nhất định. [12]
Để kiểm định bản chất kháng viêm của các chủng, 8 chủng này đã được nghiên cứu sâu hơn trong một mô hình in vivo động vật bị viêm đại tràng. Mô hình viêm đại tràng do TNBS18 đã được sử dụng. Việc sử dụng 5 ngày liên tiếp có thể tạo ra một mức bảo vệ nhất định chống lại phản ứng viêm gây ra bởi TNBS. Sự bảo vệ được đo bằng một điểm số có thể quan sát được, [23] điểm số mô học [24] hoặc các thông số miễn dịch khác (myeloperoxidase ở ruột, nồng độ murine IL-6 hoặc protein amyloid A trong huyết thanh). Myeloperoxidase là thành phần của các hạt trong tế bào chất của bạch cầu trung tính; và do đó, đo hoạt tính của nó là một cách để đo trực tiếp sự hiện diện của bạch cầu trung tính và là một chỉ số gián tiếp của tình trạng viêm ở mô.
Trong hình 1b, các kết quả thu được trong mô hình viêm đại tràng do TNBS được biểu thị cho 8 chủng đã được nghiên cứu trước đó với mô hình PBMC. Sự bảo vệ có thể quan sát thấy đã được ghi nhận, thể hiện ở phần trăm của điểm số Wallace trung bình ở 10 con chuột được điều trị bằng TNBS không được bảo vệ, khẳng định ba nhóm chủng khác nhau đã được xác định bằng cách sử dụng mô hình PBMC ở người. [18] Các cột khác nhau trong hình 1b trình bày các thí nghiệm được lặp lại.
Mặc dù các mô hình này đang được quan tâm như một công cụ sàng lọc ban đầu, chúng không cho thấy bất kỳ thông tin nào về các cơ chế khả thi có tham gia. Câu hỏi đầu tiên mà chúng tôi cố gắng trả lời có liên quan tới bản chất của các tế bào miễn dịch tham gia trong việc truyền tín hiệu. Do đó, bốn trong số các vi khuẩn từ các mô hình trước đây đã được chỉ định tiêm vào màng bụng (IP) các con chuột đã được tiêm TNBS, thay vì sử dụng bằng ống thông dạ dày. Kết quả của thử nghiệm này, được thể hiện trong hình 1c, cho thấy việc sử dụng đường IP tạo ra các mức độ bảo vệ hầu như giống hệt nhau ở chuột, và xảy ra trong một khoảng thời gian không quá 2 giờ. Điều này đưa ra khả năng là DCs có thể đã tham gia vào sự bảo vệ này. [25]
Để kiểm tra giả thuyết này, các chủng L. salivarius (LS33) và L.acidophilus NCFM (được in đậm trong hình 1) đã được chọn làm các chủng có các đặc tính điều hòa miễn dịch trái ngược nhau được so sánh kỹ hơn trong các nghiên cứu về cơ chế.
Trong thử nghiệm tiếp theo, DCs tủy xương nguyên sơ (BMDCs) đã được phân lập từ tủy xương của chuột BALB/c khỏe mạnh và được kích thích trong 16 giờ trong in vitro ở LAB tương ứng. Sau đó, DCs trong môi trường có probiotic được rửa sạch và được tiêm lại vào chuột qua đường IP. Sau đó, thực hiện trị liệu với TNBS, mức độ bảo vệ đã được đo bằng cách sử dụng các dữ liệu truyền thống đã được đề cập ở trên. Các kết quả đạt được đã khẳng định những phát hiện trước đó: BMDCs được kích thích với chủng LS33 có thể bảo vệ chuột khỏi viêm đại tràng (bảo vệ 58%), trong khi BMDCs được kích thích với chủng NCFM thì không. [25] Tiếp theo, bằng cách sử dụng chuột loại bỏ gien Nod2-/-, chúng ta có thể thấy rằng việc bảo vệ cần có sự truyền tín hiệu của Nod2, vì chủng LS33 đã không bảo vệ những con chuột này, trái ngược với chuột WT. [10]
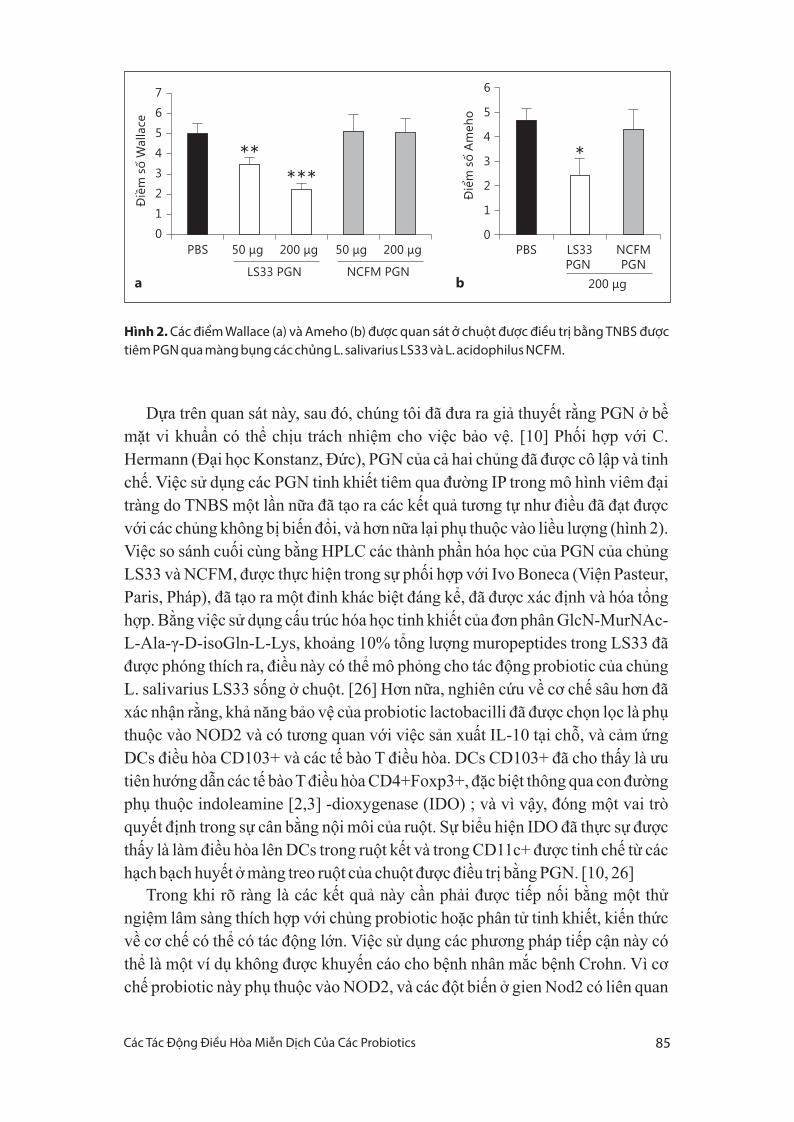
Dựa trên quan sát này, sau đó, chúng tôi đã đưa ra giả thuyết rằng PGN ở bề
mặt vi khuẩn có thể chịu trách nhiệm cho việc bảo vệ. [10] Phối hợp với C.
Hermann (Đại học Konstanz, Đức), PGN của cả hai chủng đã được cô lập và tinh
chế. Việc sử dụng các PGN tinh khiết tiêm qua đường IP trong mô hình viêm đại
tràng do TNBS một lần nữa đã tạo ra các kết quả tương tự như điều đã đạt được
với các chủng không bị biến đổi, và hơn nữa lại phụ thuộc vào liều lượng (hình 2).
Việc so sánh cuối cùng bằng HPLC các thành phần hóa học của PGN của chủng
LS33 và NCFM, được thực hiện trong sự phối hợp với Ivo Boneca (Viện Pasteur,
Paris, Pháp), đã tạo ra một đỉnh khác biệt đáng kể, đã được xác định và hóa tổng
hợp. Bằng việc sử dụng cấu trúc hóa học tinh khiết của đơn phân GlcN-MurNAc-
L-Ala-γ-D-isoGln-L-Lys, khoảng 10% tổng lượng muropeptides trong LS33 đã
được phóng thích ra, điều này có thể mô phỏng cho tác động probiotic của chủng
L. salivarius LS33 sống ở chuột. [26] Hơn nữa, nghiên cứu về cơ chế sâu hơn đã
xác nhận rằng, khả năng bảo vệ của probiotic lactobacilli đã được chọn lọc là phụ
thuộc vào NOD2 và có tương quan với việc sản xuất IL-10 tại chỗ, và cảm ứng
DCs điều hòa CD103+ và các tế bào T điều hòa. DCs CD103+ đã cho thấy là ưu
tiên hướng dẫn các tế bào T điều hòa CD4+Foxp3+, đặc biệt thông qua con đường
phụ thuộc indoleamine [2,3] -dioxygenase (IDO) ; và vì vậy, đóng một vai trò
quyết định trong sự cân bằng nội môi của ruột. Sự biểu hiện IDO đã thực sự được
thấy là làm điều hòa lên DCs trong ruột kết và trong CD11c+ được tinh chế từ các
hạch bạch huyết ở màng treo ruột của chuột được điều trị bằng PGN. [10, 26]
Trong khi rõ ràng là các kết quả này cần phải được tiếp nối bằng một thử
ngiệm lâm sàng thích hợp với chủng probiotic hoặc phân tử tinh khiết, kiến thức
về cơ chế có thể có tác động lớn. Việc sử dụng các phương pháp tiếp cận này có
thể là một ví dụ không được khuyến cáo cho bệnh nhân mắc bệnh Crohn. Vì cơ
chế probiotic này phụ thuộc vào NOD2, và các đột biến ở gien Nod2 có liên quan
85
PBS
Điề
m s
ố W
allace
Điể
m s
ố A
meh
o
50 μg 200 μg 50 μg 200 μg
LS33 PGN NCFM PGN
0
1
2
3
4
5
6
7
0
1
2
3
4
5
6
***
** *
200 μg
PBS LS33PGN
NCFMPGN
a b
Hình 2. Các điểm Wallace (a) và Ameho (b) được quan sát ở chuột được điều trị bằng TNBS được tiêm PGN qua màng bụng các chủng L. salivarius LS33 và L. acidophilus NCFM.
Các Tác Động Điều Hòa Miễn Dịch Của Các Probiotics

tính nhạy cảm ở bệnh Crohn, nên một kết cục tích cực trong các nghiên cứu lâm
sàng sẽ không được đảm bảo đầy đủ. Phát hiện này dường như xác nhận cho các
quan sát trước đó rằng một số chủng probiotic có đặc tính tốt không thể hiện kết
cục lâm sàng như mong đợi ở những bệnh nhân mắc bệnh Crohn.
Các cơ chế khác có liên quan đến hoạt tính kháng viêm. Sokol và cộng sự27 cho
thấy Faecalibacterium prausnitzii có liên quan cao hơn tới nguy cơ tái phát bệnh
Crohn ở hồi tràng sau phẫu thuật. Trong các tế bào Caco-2 được chuyển nạp một
gien chỉ thị hoạt tính NF-κB, F. prausnitzii không có tác dụng, trong khi đó các
chất nổi trên bề mặt đã loại bỏ hoạt tính NF- κB. Sự kích thích PBMCs bởi F.
prausnitzii làm giảm đáng kể nồng độ IL-12 và IFN-γ và làm tăng IL-10. Sử dụng
bằng đường uống chủng hoặc chất nổi trên bề mặt đã bảo vệ chuột khỏi bệnh viêm
đại tràng do TNBS gây ra. Hiện nay, không rõ những hợp chất nào được phóng
thích có ở bề mặt chịu trách nhiệm cho tác động này.
Bifidobacteria cũng đã được chứng minh là có các tác động kháng viêm.
Ivanov và cộng sự28 đã mô tả một serpin từ một chủng Bifidobacterium longum,
một loại vi khuẩn Gram dương yếm khí cư trú tự nhiên trong ống tiêu hóa ở
người. Serpin của B. longum đã được thấy là ức chế hiệu quả các protease giống
elastase ở sinh vật có nhân thật. Khi ức chế elastase của bạch cầu trung tính, bạch
cầu tham gia vào quá trình viêm cấp ở ruột, chủng này sẽ là một tác nhân kháng
viêm mạnh mẽ.29 Một chủng L. lactis tái tổ hợp biểu hiện quá mức serpin của
bifidobacteria này đã cho thấy làm giảm đáng kể tình trạng viêm trong mô hình
chuột TNBS [Arigoni và cộng sự trao đổi tại Hội nghị lần thứ 8 về vi khuẩn acid
lactic, từ 28/08 đến 01/09 năm 2005, tại Egmond aan Zee, Hà Lan].
Một chủng Bifidobacterium breve đã phóng thích ra các yếu tố hòa tan làm
giảm bài tiết các cytokine tiền viêm của các tế bào miễn dịch.30 Sự bài tiết
chemokine do TNF-α (CXCL8), sự thay đổi các đường truyền tín hiệu của NF-
κB và AP-1 đều đã khẳng định rằng chủng này và môi trường tự do không nằm
trong tế bào (CFM) của nó gây ức chế sự bài tiết CXCL8 ở các tế bào biểu mô,
phụ thuộc vào thời gian và liều lượng, bao gồm các con đường phiên mã của AP-1
và NF-κB. Hơn nữa, chủng này và CFM của nó đã thể hiện khả năng bảo vệ trong
một mô hình chuột bị viêm đại tràng do TNBS. Tác động tương tự đã đạt được ở
DCs được điều trị với CFM. Những kết quả này cho thấy rằng chủng B. breve có
thể điều hòa xuống các tiến trình gây viêm ở cấp độ biểu mô bằng cách ức chế các
quá trình phosphoryl hóa tham gia vào các quá trình viêm, và bằng việc điều hòa
khả năng bảo vệ của DCs. Một lần nữa dường như không phải các tế bào, mà
chính các yếu tố hòa tan được tiết ra, chịu trách nhiệm cho tác động này. Các yếu
86 P ot · Foligné · Daniel · Grangette
Các Cơ Chế Khác Đang Được Nghiên Cứu

tố này rõ ràng góp phần tích cực trong việc cân bằng nội môi ở ruột bằng cách làm
giảm sản xuất chemokine.
Hệ vi sinh vật đường ruột tham gia phần lớn vào sự phát triển và duy trì hệ miễn
dịch của chúng ta. Một hệ thống “cân bằng miễn dịch" có thể không tồn tại, vì hệ
vi sinh, với tất cả thành phần đều có thể biến đổi trong ruột, tương tác liên tục với
hệ miễn dịch. Trong một thời gian dài, mỗi hệ thống miễn dịch sẽ biểu hiện các
phản ứng khác nhau với một kháng nguyên đặc biệt hoặc các kết hợp nhiều kháng
nguyên. Ví dụ, người hiến máu khỏe mạnh sẽ phản ứng khác nhau theo thời gian
khi PBMCs của họ được tiếp xúc với một tập hợp các vi khuẩn (dữ liệu không
được trình bày). Do đó, việc duy trì giám sát khả năng kháng viêm và điều hòa của
hệ thống miễn dịch đường ruột là rất quan trọng. Các yếu tố bên ngoài, bao gồm
cả các yếu tố dinh dưỡng, có thể ảnh hưởng đến các cơ chế điều hòa miễn dịch
này. Trong chương này, chúng tôi chỉ tập trung vào một vài cơ chế khả thi, bao
gồm các tế bào điều hòa, các cytokine, chemokine, defensin, và một số các tác
nhân khác kiểm soát các phản ứng miễn dịch, chống lại sự nhiễm trùng và độc tố,
hoặc gây ra sự bất thường qua trung gian miễn dịch. Khi các cơ chế kiểm soát này
thất bại, can thiệp từ bên ngoài là cần thiết. Các probiotic có tiềm năng hỗ trợ quá
trình này. Trong khi một vài cơ chế đang được làm sáng tỏ, vẫn còn có nhiều thứ
cần được làm rõ hơn.
Một ví dụ cuối cùng về yếu tố hòa tan không xác định với tiềm năng kháng
viêm đã được mô tả bởi Jones và Versalovic [31] cho các chủng Lactobacillus
reuteri. Những phần nổi trên bề mặt lấy từ các màng sinh học của L. reuteri đã
được thêm vào các tế bào bạch cầu đơn nhân THP-1 ở người trong điều kiện có
hoặc không có LPS. LPS kích thích sản xuất TNF-α tiền viêm bởi các tế bào THP-
1. Các chủng L. reuteri ATCC PTA 6475 và ATCC PTA 5289, không phải là hai
chủng khác nhau của cùng một loài, đã ức chế khả năng sản xuất TNF-α khi nuôi
cấy như các màng sinh học. Các tác giả đã lưu ý rằng các khả năng tương đối để ức
chế TNF-α của người ở các tế bào bạch cầu đơn nhân có tương quan trực tiếp với
các khả năng tương đối để tổng hợp và hình thành các màng sinh học trên các bề
mặt polystyrene. Gần đây, có ý kiến đề nghị rằng [32] histamine được sản xuất
bởi chủng ATCC PTA 6475 đã kích thích gia tăng nồng độ cAMP, ức chế tín hiệu
xuôi dòng MEK/ERK MAPK thông qua protein kinase A, và ức chế sản xuất
TNF sau khi điều hòa phiên mã .
Các khảo sát này lại một lần nữa cho thấy sự phức tạp của các điều kiện khả thi
sẽ thúc đẩy hoặc ức chế tác động kháng viêm, và nhấn mạnh tầm quan trọng của
các thử nghiệm lâm sàng được thiết kế tốt trong một dân số mục tiêu được kiểm
soát tốt, với một sản phẩm được kiểm soát tốt.
87
Kết Luận
Các Tác Động Điều Hòa Miễn Dịch Của Các Probiotics

Mặc dù việc hiểu biết đầy đủ các cơ chế này là hết sức cần thiết, không có vẻ gì
là các probiotic, vì sự phức tạp của tất cả các tác nhân của môi trường, sẽ mang lại
giải pháp cho tất cả các bệnh lý qua trung gian miễn dịch. Hơn nữa, không có vẻ gì
là chỉ một mình probiotic sẽ có thể phục hồi các hệ miễn dịch bị “tổn hại nghiêm
trọng”. Các kết quả tốt nhất đã đạt được ở các trẻ nhỏ, có thể là vì sự phát triển của
hệ miễn dịch vẫn cho phép các thay đổi trong thời gian dung nạp qua đường uống
và sự miễn dịch vẫn đang phát triển. Điều này có thể sẽ trở nên khó khăn hơn ở độ
tuổi lớn hơn.
Dù vậy, không còn nghi ngờ gì nữa, hiểu biết về cơ chế sẽ cho phép chọn ra
các chủng tốt hơn, thiết kế các thí nghiệm lâm sàng hiệu quả hơn, với các dữ liệu
phù hợp với các tác động mong muốn trên dân số mục tiêu. Điều này có nghĩa là
các vi khuẩn khác, ngoài lactobacilli và bifidobacteria truyền thống, có khả năng
trở nên quan trọng, đặc biệt là trong bối cảnh điều trị. Thuật ngữ dược vi sinh
(pharmabiotics) đã được đặt ra [33] và có thể sẽ được tiếp tục xây dựng trong
tương lai để bao gồm cả các chủng bị giết bằng nhiệt hoặc chiếu xạ, cũng như các
vi khuẩn bị biến đổi di truyền hay trao đổi chất. Các chủng bị biến đổi di truyền
hay trao đổi chất sẽ làm tối ưu việc sử dụng các nghiên cứu cơ chế, để cung cấp
các hiểu biết trên cơ sở phân tử cho các tác động mong muốn. Không còn nghi
ngờ gì nữa, tương lai của các *- vi sinh (*-biotics) là hết sức khả quan.
88 P ot · Foligné · Daniel · Grangette
Tài Liệu T ham Khảo
1 Bach JF: The effect of infections on suscepti-
bility to autoimmune and allergic diseases.
N Engl J Med 2002; 3 47:
9 11–920.
2 Guarner F, Bourdet-Sicard R, Brandtzaeg P,
et al: Mechanisms of disease: the hygiene hy-
pothesis revisited. Nat Clin Pract Gastroen-
terol Hepatol 2006; 3 :
2 75–284.
3 Qin J, Li R, Raes J, et al: A human gut micro-
bial gene catalogue established by metage-
nomic sequencing. Nature 2010; 4 64:
5 9–65.
4 Arumugam M, Raes J, Pelletier E, et al: En-
terotypes of the human gut microbiome.
N ature 2011;
4 73:
1 74–180.
5 Zhu B, Wang X, Li L: Human gut microbi-
ome: the second genome of human body.
Protein Cell 2010; 8 :
7 18–725.
6 Lozupone CA, Stombaugh JI, Gordon JI, et
al: Diversity, stability and resilience of the
human gut microbiota. Nature 2012; 4 89:
2 20–230.
7 Delcenserie V, Martel D, Lamoureux M, et al:
Immunomodulatory effects of probiotics in
the intestinal tract. Curr Issues Mol Biol
2008; 1 0:
3 7–54.
8 Lebeer S, Vanderleyden J, De Keersmaecker
SCJ: Host interactions of probiotic bacterial
surface molecules: comparison with com-
mensals and pathogens. Nature Rev Micro-
biol 2010; 8 :
1 71–184.
9 Delzenne NM, Cani PD: Interaction between
obesity and the gut microbiota: relevance in
nutrition. Ann Rev Nutr 2011; 3 1:
1 5–31.
Tuyên Bố Công Khai
Các tác già tuyên bố là không có bất kỳ xung đột lợi ích nào cho bài báo này.

89
10 Macho-Fernandez E, Valenti V, Rockel C, et al: Anti-inflammatory capacity of selected
lactobacilli in experimental colitis is driven
by NOD2-mediated recognition of a specific
peptidoglycan-derived muropeptide. Gut
2011; 6 0:
1 050–1059.
11 Kwon H-K, Lee C-G, So J-S, et al: Generation
of regulatory dendritic cells and
CD4+Foxp3+ T cells by probiotics adminis-
tration suppresses immune disorders. Proc
Natl Acad Sci USA 2010; 1 07:
2 159–2164.
12 Mohamadzadeh M, Olson S, Kalina WV, et al:
Lactobacilli activate human dendritic cells that
skew T cells toward T helper 1 polarization.
Proc Natl Acad Sci USA 2005;
102:
2880–2885.
13 Lavasani S, Dzhambazov B, Nouri M, et al: A
novel probiotic mixture exerts a therapeutic
effect on experimental autoimmune encepha-
lomyelitis mediated by IL-10 producing regu-
latory T cells. PLoS One 2010; 5 :e9009.
14 Weiner HL: Oral tolerance: immune mecha-
nisms and the generation of Th3-type TGF-
beta-secreting regulatory cells. Microbes In-
fect 2001; 3 :
9 47–954.
15 Lefrancois L, Puddington L: Intestinal and
pulmonary mucosal T cells: local heroes fight
to maintain the status quo. Ann Rev Immu-
nol 2006; 2 4:
6 81–704.
16 Korn T, Bettelli E, Oukka M, Kuchroo VK:
IL-17 and Th17 cells. Ann Rev Immunol
2009; 2 7: 4 85–517.
17 Kreymborg K, Becher B: IL-22 vs IL-22: the
tissue matters. Open Autoimmun J 2010;
2:
181–186.
18 Foligné B, Nutten S, Grangette C, et al: Cor-
relation between in vitro and in vivo immu-
nomodulatory properties of lactic acid bacte-
ria. World J Gastroenterol 2007; 1 3:
2 36–243.
19 Cousin JF, Deutsch S-M, Perez Chaia A, et al:
Interactions between probiotic dairy propi-
onibacteria and the intestinal epithelium.
Curr Immunol Rev 2012; 8 :
2 16–226.
20 Foligné B, Dewulf J, Vandekerckove P, et al:
Probiotic yeasts: anti-inflammatory potential
of various nonpathogenic strains in experi-
mental colitis in mice. World J Gastroenterol
2010; 1 6:
2 134–2145.
21 Foligné B, Dewulf J, Breton J, et al: Probiotic
properties of non-conventional lactic acid
bacteria: immunomodulation by Oenococcus
oeni . Int J Food Microbiol 2010; 1 40:
1 36–145.
22 Li CY, Lin HC, Lai CH, et al: Immunomodu-
latory effects of Lactobacillus and Bifidobacte-
rium on both murine and human mitogen-
activated T cells. Int Arch Allergy Immunol
2011; 1 56:
1 28–136.
23 Wallace JL, MacNaughton WK, Morris GP,
Beck PL: Inhibition of leukotriene synthesis
markedly accelerates healing in a rat model of
inflammatory bowel disease. Gastroenterol-
ogy 1989; 9 6:
2 9–36.
24 Ameho CK, Adjei AA, Harrison EK, et al:
Prophylactic effect of dietary glutamine sup-
plementation on interleukin 8 and tumour
necrosis factor alpha production in trinitro-
benzene sulphonic acid induced colitis. Gut
1997; 4 1:
4 87–493.
25 Foligné B, Zoumpopoulou G, Dewulf J, et al:
A key role of dendritic cells in probiotic func-
tionality. PLoS One 2007; 2 :e313.
26 Macho Fernandez E, Pot B, Grangette C:
Beneficial effect of probiotics in IBD. Are
peptidogycan and NOD2 the molecular key
effectors? Gut Microbes 2011; 2 :
1 –7.
27 Sokol H, Pigneur B, Watterlot L, et al: Faecali-
bacterium prausnitzii is an anti-inflammatory
commensal bacterium identified by gut micro-
biota analysis of Crohn disease patients. Proc
Natl Acad Sci USA 2008;
105:
16731–16736.
28 Ivanov D, Emonet C, Foata F, et al: Serpin
from the gut bacterium Bifidobacterium long-
um inhibits eukaryotic elastase-like serine
proteases. J Biol Chem 2006; 2 81:
1 7246–
17252.
29 Sanchez B, Urdaci MC, Margolles A: Extra-
cellular proteins secreted by probiotic bacte-
ria as mediators of effects that promote mu-
cosa-bacteria interactions. Microbiology
2010; 1 56:
3 232–3242.
30 Heuvelin E, Lebreton C, Grangette C, et al:
Mechanisms involved in alleviation of intesti-
nal inflammation by Bifidobacterium breve
soluble factors. PLoS One 2009; 4 :e5184.
31 Jones SE, Versalovic J: Probiotic Lactobacillus
reuteri biofilms produce antimicrobial and
anti-inflammatory factors. BMC Microbiol
2009; 9 :
3 5.
32 Thomas CM, Hong T, van Pijkeren JP, et al:
Histamine derived from probiotic Lactobacil-
lus reuteri suppresses TNF via modulation of
PKA and ERK signaling. PLoS One 2012;
7 :e31951.
33 Colin H: Probiotics and pharmabiotics. Bio-
eng Bugs 2010; 1 :
7 9–84.
Các Tác Động Điều Hòa Miễn Dịch Của Các Probiotics

90 P ot · Foligné · Daniel · Grangette
34 Tiscornia I, Sanchez-Martins V, Hernandez
A, Bollati-Fogolin M: Human monocyte-de-
rived dendritic cells from leukoreduction sys-
tem chambers after platelet pheresis are func-
tional in an in vitro co-culture assay with
intestinal epithelial cells. J Immunol Methods
2012; 3 84:
1 64–170.
35 Zhong Y, Huang J, Tang W, et al: Effects of
probiotics, probiotic DNA and the CpG oli-
godeoxynucleotides on ovalbumin-sensitized
Brown-Norway rats via TLR9/NF-κB path-
way. FEMS Immunol Med Microbiol 2012;
6 6:
7 1–82.
36 Karlsson M, Jass J: Lactobacilli differently
regulate expression and secretion of CXCL8
in urothelial cells. Benef Microbes 2012; 3 :
1 95–203.
37 Rupa P, Mine Y: Recent advances in the role
of probiotics in human inflammation and gut
health. J Agric Food Chem 2012; 6 0:
8 249–
8256.
38 Dong H, Rowland I, Yaqoob P: Comparative
effects of six probiotic strains on immune
function in vitro. Br J Nutr 2012; 1 08:
4 59–
470.
39 Mariman R, Kremer B, van Erk M, et al: Gene
expression profiling identifies mechanisms of
protection to recurrent trinitrobenzene sul-
fonic acid colitis mediated by probiotics. In-
flamm Bowel Dis 2012; 1 8: 1 424–1433.
40 Giahi L, Aumueller E, Elmadfa I, Haslberger
AG: Regulation of TLR4, p38 MAPkinase,
IκB and miRNAs by inactivated strains of lac-
tobacilli in human dendritic cells. Benef Mi-
crobes 2012; 3 :
9 1–98.
41 Chen CC, Lin WC, Kong MS, et al: Oral in-
oculation of probiotics Lactobacillus acidoph-
ilus NCFM suppresses tumour growth both
in segmental orthotopic colon cancer and
extra-intestinal tissue. Br J Nutr 2012; 1 07:
1 623–1634.
42 Benson KF, Redman KA, Carter SG, et al:
Probiotic metabolites from Bacillus coagulans
TMGanedenBC30 support maturation of anti-
gen-presenting cells in vitro. World J Gastro-
enterol 2012; 1 8:
1 875–1883.
43 Güttsches AK, Löseke S, Zähringer U, et al:
Anti-inflammatory modulation of immune
response by probiotic Escherichia coli Nissle
1917 in human blood mononuclear cells. In-
nate Immun 2012; 1 8:
2 04–216.
44 Kim JY, Park MS, Ji GE: Probiotic modula-
tion of dendritic cells co-cultured with intes-
tinal epithelial cells. World J Gastroenterol
2012; 1 8:
1 308–1318.
45 Roos TB, de Lara AP, Dummer LA, et al: The
immune modulation of Bacillus cereus var.
Toyoi in mice immunized with experimental
inactivated bovine herpesvirus type 5 vaccine.
Vaccine 2012; 3 0:
2 173–2177.
46 Liu Y, Fatheree NY, Mangalat N, Rhoads JM:
Lactobacillus reuteri strains reduce incidence
and severity of experimental necrotizing en-
terocolitis via modulation of TLR4 and NF-
κB signaling in the intestine. Am J Physiol
Gastrointest Liver Physiol 2012; 3 02:G608–
G617.
47 Rizzardini G, Eskesen D, Calder PC, et al:
Evaluation of the immune benefits of two
probiotic strains Bifidobacterium animalis
®ssp. lactis, BB-12 and Lactobacillus paraca-
sei ssp. paracasei, L. casei 431 ® in an influen-
za vaccination model: a randomised, double-
blind, placebo-controlled study. Br J Nutr
2012; 1 07:
8 76–884.
48 Fanning S, Hall LJ, Cronin M, et al: Bifido-
bacterial surface-exopolysaccharide facilitates
commensal-host interaction through im-
mune modulation and pathogen protection.
Proc Natl Acad Sci USA 2012;
1 09:
2 108–2113.
49 Mencarelli A, Cipriani S, Renga B, et al:
VSL#3 resets insulin signaling and protects
against NASH and atherosclerosis in a model
of genetic dyslipidemia and intestinal inflam-
mation. PLoS One 2012; 7 :e45425.
50 Issazadeh-Navikas S, Teimer R, Bockermann
R: Influence of dietary components on regu-
latory T cells. Mol Med 2012; 1 8:
9 5–110.
51 Castro MS, Azpiroz MB, Molina MA, et al:
Preliminary studies on the prevention of the
ovalbumin-induced allergic response by En-
terococcus faecalis CECT7121 in mice. Int
Arch Allergy Immunol. 2012; 1 57:
1 1–20.
52 Bron PA, van Baarlen P, Kleerebezem M:
Emerging molecular insights into the interac-
tion between probiotics and the host intesti-
nal mucosa. Nat Rev Microbiol 2011; 1 0:
6 6–
78.










![PHIẾU ĐĂNG KÝ - FTMSGlobal Academy€¦ · THÔNG TIN HỌC VIÊN ƯU ĐÃI KHÁC [MIỄN PHÍ]dịch hồ sơ đk ban đầu [GIẢM 10%]tiền sách khi đăng ký học ≥](https://static.fdocuments.es/doc/165x107/5f5a2a3b0a9f6c78e164c4f8/phiu-ng-k-ftmsglobal-academy-thng-tin-hoec-vin-u-fi-khc-min.jpg)







