
biofertilizantes
134
“LA BIOFERTILIZACION COMO TECNOLOGIA SOSTENIBLE” 25-NOVIEMBRE-2004 RIO BRAVO, TAM., MEXICO
-
Upload
latinplast -
Category
Documents
-
view
246 -
download
6
description
cultivos
Transcript of biofertilizantes

“LA BIOFERTILIZACION COMO TECNOLOGIA SOSTENIBLE”
25-NOVIEMBRE-2004
RIO BRAVO, TAM., MEXICO


SIMPOSIO DE BIOFERTILIZACIÓN
“LA BIOFERTILIZACIÓN COMO TECNOLOGÍA SOSTENIBLE”
COMITÉ ORGANIZADOR
ARTURO DÍAZ FRANCO, FELIPE SERRANO MEDINA Y NICOLÁS MALDONADO MORENO
COORDINACIÓN
JAVIER GONZÁLEZ QUINTERO, IDALIA GARZA CANO, VÍCTOR PECINA
QUINTERO, LEOPOLDO GARZA GUAJARDO, MANUEL ALVARADO CARRILLO, JAIME SALINAS GARCÍA E HIPÓLITO CASTILLO TOVAR
MEMORIA
EDICIÓN
ARTURO DÍAZ FRANCO, NETZAHUALCÓYOTL MAYEK PÉREZ, ALBERTO MENDOZA HERRERA Y NICOLÁS MALDONADO MORENO
DISEÑO DE PORTADA
LEOPOLDO GARZA GUAJARDO Y ARTURO DÍAZ FRANCO
Primera edición: 2004 Para la presente edición, Campo Experimental Río Bravo, INIFAP. Carretera Matamoros-Reynosa km 61, Río Bravo, Tam., México. Foto de portada: Colonización de Glomus intraradices; con autorización de P. Tiwari.

ii
CONTENIDO
Página TRABAJOS COMPLETOS
Biotecnología de los hongos micorrízicos arbusculares…………………………. Ronald Ferrera-Cerrato y Alejandro Alarcón
1
Impacto de la micorriza arbuscular en la productividad del sorgo en Tamaulipas. Arturo Díaz Franco, Idalia Garza Cano, Víctor Pecina Quintero y Agustín Magallanes Estala
10
Biofertilización bacteriana del pasto buffel……………………………………… C. Loredo-Osti, D. Espinosa V., R. Ferrera-Cerrato, J. Castellanos R. y J. Pérez
22
Biofertilizantes; mcorrizas y bacterias promotoras de crecimiento…………….. Víctor Olalde Portugal y Rosalinda Serratos
31
Efectos de labranza y biofertilización en propiedades del suelo que afectan la sostenibilidad de producción de frijol……………………………………………. Jaime Roel Salinas García, Arturo Díaz Franco e Idalia Garza Cano
35
Respuesta de la biofertilización en el crecimiento y rendimiento de sorgo de grano en Linares, Nuevo León…………………………………………………… Juan Martínez Medina
42
Biotecnología de los hongos etomicorrízicos…………………………………… Jesús Pérez-Moreno
53
Evaluación de la inoculación de simbiontes en soya (Glycine max) bajo condiciones de campo……………………………………………………………. Ponciano Pérez García, Arturo Díaz Franco y Nicolás Maldonado Moreno
66
Biofertilizantes microbianos: Antecedentes del programa y resultados de validación en México……………………………………………………………..
Juan Francisco Aguirre Medina
71
Aislamiento, selección, producción y evaluación de un inoculante basado en cepas nativas de Azospirillum en el Norte de Tamaulipas……………………….. Alberto Mendoza H., Antonia Cruz H. y Cuauhtemoc Jacques H.
87

iii
RESÚMENES Respuesta de variedades de cacahuate (Arachis hypogea L.) a la fertilización química y biológica en un suelo regosol…………………………………………. Arturo Durán Prado, Víctor López Galván y Oscar Hugo Tosquy Valle
102
Respuesta de germoplasma de frijol a la fertilización química y biológica…………………………………………………………………………… Arturo Durán Prado, Víctor López Galvan y Javier Cumpián Gutiérrez
103
Respuesta de variedades de frijol a la fertilización química y biológica en un suelo fluvisol de Veracruz………………………………………………………... Arturo Durán Prado, Víctor López Galván y Oscar Hugo Tosquy Valle
104
Respuesta de variedades de soya a la inoculación con micorriza Glomus intraradix en Veracruz…………………………………………………………… Arturo Durán Prado, Oscar Hugo Tosquy Valle y Víctor López Galván
105
Efectividad de micorriza arbuscular en genotipos de pasto buffel (Cenchrus ciliaris)…………………………………………………………………………… Arturo Díaz Franco, Idalia Garza Cano y Asunción Méndez Rodríguez
106
Influencia de micorriza arbuscular en el crecimiento y rendimiento de cártamo... Arturo Díaz Franco, Alfredo Ortegón Morales e Idalia Garza Cano
107
Biofertirrigación: tecnología sustentable del Siglo XXI…………………………. Lina Hernández Flores, Juan M. Covarrubias Ramírez, Rodrigo Aveldaño Salazar y Juan José Peña Cabriales
108
Respuesta del maíz y sorgo a la fertilización biológica………………………….. Víctor Pecina Quintero, Arturo Díaz Franco e Idalia Garza Cano
109
Efecto de la micorriza arbuscular en sorgo bajo dos condiciones de humedad….. Víctor Pecina Quintero, Arturo Díaz Franco e Idalia Garza Cano
110
Efecto de una composta y ácidos fúlvicos en la producción de lilies (Lilium sp) en el sureste de Coahuila…………………………………………………………. Ma. del Rosario Zúñiga Estrada, Juan M. Covarrubias Ramírez y Rubén López Cervantes
111
Transferencia tecnológica del maíz QPM y el biofertilizante en la Huasteca Hidalguense………………………………………………………………………. Juan Pablo Pérez Camarillo, Guadalupe Zacatenco González, René Galván Parra y Rodrigo Aveldaño Salazar
112

iv
Producción y evaluación de un biofertilizante (Azospirillum spp) para el noreste de México IPN-CBG…………………………………………………………….. Jesús G. García, Alberto Mendoza, Cuauhtemoc Jacques, Antonia Cruz y Felipe Serrano
113
Respuesta del sorgo a la inoculación de Glomus intraradices en campo……….. Agustín Magallanes Estala
114
Rentabilidad del sorgo mediante la inoculación de simbiontes en suelo con y sin fertilización química……………………………………………………………... Agustín Magallanes Estala, Arturo Díaz Franco y Víctor Olalde Portugal
115
Evaluación de biofertilizantes en cártamo……………………………………….. Agustín Magallanes Estala, Alfredo S. Ortegón Morales y Arturo Díaz Franco
116
Interacción de Azospirillum brasilense, nitrógeno y azúcar en canola de riego bajo labranza convencional y de conservación…………………………………... Mario Cepeda Villegas, Eulalio Venegas González y Blanca Gómez Lucatero
117
Respuesta del maíz al tratamiento con Azospirillum brasilense y nitrógeno bajo labranza de conservación………………………………………………………… Mario Cepeda Villegas y Eulalio Venegas González
118
Efecto estimulante de bacterias esporuladas sobre crecimiento y desarrollo del chile jalapeño (Capsicum annuum) en invernadero y campo……………………. Nicolasa García Licona, Gabriel Gallegos Morales, Melchor Cepeda Siller y Fco. Daniel Hernández Castillo
119
Resultados preliminares de la evaluación de biofertilizantes en maíz QPM……. José Ernesto Cervantes Martínez
120
Efecto de biofertilizantes sobre el rendimiento del maíz………………………… César A. Reyes Méndez y Miguel Ángel Cantú Almaguer
121
Evaluación combinada de inoculantes microbiológicos y fertilizantes químicos en el cultivo de sorgo…………………………………………………………….. Othón Martín Carrillo Rendón, María Esther Salazar Durán, Ismael Machuca Orta y Cuauhtemoc Jacques Hernández
122
Biofertilización en sorgo de temporal en la zona media de San Luis Potosí…….. Andrés Ramiro Córdova y Cesario Jasso Chaverría
123
Simbiosis Rhizobium-micorriza arbuscular y uso de brassinoesteroides en frijol. Cesario Jasso-Chaverría y Miguel Ángel martínez-Gamiño
124
Efecto del biofertilizante y la preparación del suelo en la producción de maíz, sorgo y sorgo x Sudán en la zona media Potosina……………………………….. Miguel Ángel Martínez Gamiño y Cesario Jasso Chaverría
125

v
Biofertirrigación por goteo a base de guano en cultivos diversos bajo un sistema hidropónico con producción de tilapia en Jalisco, Nayarit………………………. Alberto Betancourt Vallejo, Pedro D. Flores Peña, Víctor M. González Velásquez, Rafael Quezada Morales, Víctor Jiménez García y Roberto Gómez Aguilar
126

1
BIOTECNOLOGIA DE LOS HONGOS MICORRIZICOS ARBUSCULARES
Ronald Ferrera-Cerrato1, Alejandro Alarcón1, 2
1Microbiología de Suelos. Especialidad de Edafología. Instituto de Recursos Naturales. Colegio de Postgraduados. Carretera México-Texcoco, km 36.5. Montecillo, Estado de México 56230.
México. 2Department of Horticultural Sciences. Texas A&M University. College Station, Texas. 77843-2133 USA
[email protected], [email protected], [email protected]
LA SIMBIOSIS MICORRIZICA La simbiosis micorrízica1 se refiere a la asociación mutualista que se establece
entre plantas y específicos grupos de hongos que habitan en el suelo y en la rizosfera. De este modo, se tienen identificados siete diferentes tipos de simbiosis micorrízicas, las cuales tienen repercusión en lo que respecta a la evolución, fisiología y adaptación ecológica de las plantas que habitan los ecosistemas terrestres (Smith y Read, 1997): 1) simbiosis ectomicorrízica, la cual se forma específicamente entre miembros de familias botánicas como Cupresaceae, Pinaceae, Betulaceae, Fagaceae, entre otras, con cierto grupo de hongos que pertenece a las clases Basidiomycetes, y Ascomyceks, principalmente; 2) la micorriza orquideoide, que forma entre orquídeas y hongos del género Rhizoctonia; 3) micorriza monotropoide; 4) micorriza arbutoide; 5) micorriza ericoide; 6) ectoendomicorriza. No obstante, una de las simbiosis micorrízicas que tiene vasto avance científico y biotecnológico es aquella que se forma entre aproximadamente 150 especies de hongos de Glomeromycota con más del 80% del total de las plantas terrestres que se conocen hoy en día, la cual se denomina 7) micorriza (vesículo-)arbuscular.
En la actualidad, la simbiosis micorrízica arbuscular tiene enorme trascendencia ya que en diversos estudios se ha demostrado el efecto benéfico de los hongos micorrízicos arbusculares (HMA) en el mejoramiento de la nutrición, aprovechamiento de agua, crecimiento y adaptación de las plantas ante diversas condiciones de estrés intuido tanto por factores bióticos como por factores abióticos (Augé, 2001; Jeffries et al., 2003).
ECOLOGÍA DE LA MICORRIZA ARBUSCULAR Los HMA son microorganismos rizosféricos cosmopolitas por lo que se pueden
encontrar en la mayoría de los biomas terrestres (Smith y Read, 1997). No obstante, la distribución y presencia de estos simbiontes es afectada en aquellos suelos con diferente grado de fertilidad, diferente manejo agrotecnológico (ejemplo, aplicación de insumos fertilizantes, biocidas o plaguicidas), o por diversos agentes de perturbación tales como el pastoreo el cual produce compactación del suelo, o por impacto de procesos industriales como la minería, petroquímica, o deposición de desechos industriales (Abbott y Robson, 1991; Sieverding, 1991; Sylvia et al., 1993; Nadian et al., 1998; Trejo et al. , 1998; Jeffries et al., 2003). No obstante, estas condiciones desfavorables inducen una selección de HMA 1El término micorrízico se utiliza con base en el diccionario de Micología del Dr. Miguel Ulloa (1991)a y el diccionario de Font Quer (1982)b: a) micorrízico, ca, micorrizógeno, na: mycorrhizal, mycorrhizogenous (del gr. mykes, hongo; rhíza, raíz, y génos origen, de gennáo, engendrar, producir): hongo que es capaz de formar una micorriza, que interviene en su constitución. b) micorrízico, ca: adj. Relativo a la micorriza, perteneciente a ella.

2
más aptos para tolerar condiciones ambientales adversas. En este particular caso, estos HMA ecológicamente adaptados pueden ser sujetos de manipulación para la producción de inoculante con fines de restauración o remediación de áreas altamente perturbadas.
El manejo sustentable de agro ecosistemas además, contribuye en la regulación de la diversidad, composición e interacciones de la microflora y de la macro fauna que se establecen en el suelo y en la rizosfera de las plantas (Linderman, 1993; Harinikumar y Bajyaraj, 1994; Thimm y Larink, 1994; Ferrera-Cerrato, 1995; González y Ferrera-Cerrato, 1996; Ferrera-Cerrato y Alarcón, 2001; Rilling y Steinberg, 2002; Jefrries et al., 2003). Otros factores que determinan no solo la distribución pero también la funcionalidad o efectividad de los HMA se relacionan con la vegetación predominante, la cual participa como reservorio de estos hongos, y la variación estacional (Smith y Read, 1997; Sanders y Fitter, 1992). El genotipo tanto de los hospedantes como el de los HMA involucrados en la simbiosis, ha recibido especial atención ya que esta interacción determina la respuesta de la inoculación con los HMA. En este sentido, se han identificado variaciones en la respuesta de las plantas a la inoculación de HMA ya sea nativos o exóticos y se ha llegado a establecer que no es apropiado generalizar que los HMA siempre estimulan el crecimiento de las plantas, tanto en ambientes naturales como controlados. Por esta razón, se ha propuesto definir a la simbiosis micorrízica arbuscular como un continuo del parasitismo al mutualismo (Johnson et al., 1997; Klironomos, 2003). Ante tal situación, la selección de HMA debe enfocarse a aspectos que se relacionen con el objetivo de mejorar el crecimiento y desarrollo de las plantas (por ejemplo, mejorar la tolerancia y adaptación a condiciones adversas, mejoramiento del aprovechamiento de fertilizantes, etc.). Para tal fin, se debe tener mayor estudio y entendimiento de los aspectos ecológicos que podrían influir en el establecimiento y efectividad de los HMA (Sanders y Fitter, 1992).
FISIOLOGÍA DE LA SIMBIOSIS MICORRIZICA ARBUSCULAR
Los HMA son considerados como simbiontes obligados, de tal forma que la simbiosis con plantas le permite captar fotosintatos que favorecen el desarrollo y propagación de los HMA. El mutualismo, que en la mayoría de los casos caracteriza a la simbiosis HMA-planta (Dakessian et al., 1986; González y Ferrera-Cerrato, 1990; Rapparini et al., 1994; Ferrera-Cerrato y Alarcón, 1998; Alarcón y Ferrera-Cerrato, 2003), es determinado por la integración morfológica y estructural de ambos simbiontes, lo cual define el intercambio bidireccional de nutrimentos entre ambos simbiontes. La colonización de las células corticales permite al HMA la posibilidad de obtener fuentes de carbono, el cual es considerado la “divisa de cambio” que la planta aporta al hongo con el fin de tener mayor capacidad de captación de nutrimentos minerales necesarios para sus diversas etapas de desarrollo (Bago et al., 2000).
La mayor respuesta de las plantas a la inoculación de HMA se presenta cuando se utilizan sustratos o suelos con limitada disponibilidad nutrimental. El mejoramiento del aprovechamiento y captación nutrimental por efecto de HMA depende de factores inherentes a la planta así como factores del suelo (Gianinazzi-Pearson y Azcón-Aguilar, 1991). En el caso de las plantas, el sistema radical tiene un papel trascendental en la absorción y aprovechamiento de nutrimentos desde la solución de suelo hasta los tejidos internos de ellas (Gregory, 1992). No obstante, los aspectos fisiológicos de la absorción de iones por la raíz dependen de la movilidad de los iones, y de la selectividad por algunos iones, lo cual depende de la especie vegetal. De esta forma, aspectos como la morfología de la raíz (longitud, diámetro, distribución, cantidad de pelos absorbentes, etc.), determina la capacidad de explorar mayor volumen de suelo y con ello, captar nutrimentos minerales.

3
La simbiosis micorrízica representa una alternativa biológica para las plantas para satisfacer sus requerimientos por nutrimentos y agua a partir del suelo, ya que la extensiva red de hifas que se desarrollan en el suelo, es capaz de explorar mayor volumen de suelo y llegar a sitios ricos en nutrimentos donde la raíz por si misma seria incapaz de penetrar (Smith y Read, 1997; Schachtman et al., 1998). El entendimiento de los factores ambientales que favorecen o afectan a las cepas de HMA es fundamental para manipular y controlar ciertas condiciones de cultivo que los hongos requieren para establecerse y producir mayor cantidad de propágulos. De esta forma, se mantiene la actividad fisiológica del hongo que a corto y largo plazo contribuye en beneficios para las plantas inoculadas.
APLICACIONES DE LOS HONGOS MICORRÍZICOS ARBUSCULARES
El beneficio que aporta la simbiosis micorrízica arbuscular en plantas ha sido bien
documentado, dando especial énfasis en los que respecta a la promoción del crecimiento y nutrición de plantas, especialmente aquellas de interés hortícola, frutícola y forestal (Alarcón y Ferrera-Cerrato; 1999; Davies et al., 2000; Jeffries et al., 2003). La aplicación de los HMA es factible de realizarse en los diferentes sistemas de propagación de plantas: semillas, cultivo de tejidos o estacas (Alarcón y Ferrera-Cerrato, 1999; Davies et al., 2000).
En la mayoría de los casos, los HMA incrementan el crecimiento y estado nutricional de las plántulas, y mejoran la etapa de aclimatación y adaptación de vitro-plantas a condiciones ambientales y su productividad (Alarcón et al., 2000, 2001). La inoculación de HMA ha contribuido a la adaptación y crecimiento de plantas en condiciones ambientales extremas como lo son sitios erosionados, sitios con baja fertilidad, sitios con problemas de salinidad, y en sitios de zonas áridas o con problemas de contaminación por diversos agentes orgánicos e inorgánicos (Ferrera-Cerrato y Alarcón, 2004). Otra aplicación de los HMA con repercusión ecológica se dirige a la rehabilitación de áreas afectadas por deposiciones volcánicas, como se ha demostrado en Japón (Saito y Marumoto, 2002).
Uno de los aspectos que ha recibido poca atención se refiere al efecto de los HMA en plantas utilizadas en interiores. En este sentido, existe un reporte en el que se evaluaron diferentes cepas de HMA sobre la adaptación y crecimiento de siete géneros de plantas. En dicho estudio se reportan variaciones de la respuesta de las plantas al establecimiento de los HMA que fueron desde la promoción hasta la inhibición del crecimiento de algunas de ellas (Busch y Lelley, 1997).
A pesar de las beneficios de los HMA no solo en la producción sostenible de los cultivos sino también en lo que respecta a la conservación ambiental, uno de los problemas de la producción de inóculo de estos simbiontes mutualistas, estriba en su condición natural de biotrofismo obligado, es decir, que estos micosimbiontes son dependientes de su establecimiento en un sistema radical vivo para satisfacer sus requerimientos nutricionales y completar su ciclo biológico. Con base en lo anterior, a continuación se describen brevemente, diferentes sistemas de cultivo y propagación de HMA con fines de producción de inoculo a diferente escala.
PROCESO BIOTECNOLÓGICO DE LA PRODUCCIÓN DE INÓCULO DE HMA
En la mayoría de los sistemas de producción de plantas se ha acudido a la
utilización de inoculante a base de esporas en suelo y raíz, suelo-inóculo, o raíces colonizadas por los HMA. Uno de los aspectos que deben ser considerados en los diferentes métodos de producción de inoculante micorrízico, es la selección de la planta

4
hospedante. Generalmente para obtener mayor cantidad de propágulos de HMA se ha acudido a utilizar gramíneas como plantas trampa, ya que producen mayor cantidad de raíces las cuales son susceptibles de ser colonizadas y utilizar estos propágulos como fuente de inóculo de HMA. La combinación HMA-planta trampa es un factor determinante en la propagación de una cepa fúngica en especial. En este sentido, se ha mencionado que existe cierto grado de especificidad de algunos hongos por un determinado genotipo de un mismo género de planta hospedante (Boyetchko y Tewari, 1995).
La producción de inóculo mediante el uso de arcillas expandibles permite obtener mayor cantidad de propágulos de HMA (micelio, esporas y raíces colonizadas), así como mantener el inoculante viable durante largos periodos de almacenamiento de hasta cinco años, incluso a temperatura ambiente (Gruntwald-Stocker y Dehne, 1989; Aboul-Nasr, 1997).
El uso de soportes orgánicos e inorgánicos para la producción de inóculo micorrízico arbuscular, representa en la actualidad una excelente alternativa de bajo costo (Jarsfter y Sylvia, 1992; Saito, 1990; Saito y Marumoto, 2002) ya que se puede lograr el escalamiento masivo de la producción de inoculante para ser utilizado en los diferentes sistemas agrícolas y hortícolas (Alarcón et al., 2004). Para este caso, es necesario tener un buen cuidado de las condiciones de cultivo de las plantas trampa, ya que el inóculo debe estar libre de microorganismos fitopatógenos. Por tal razón, se requiere de la aplicación de plaguicidas que además de erradicar los organismos plaga, deben permitir el desarrollo de la simbiosis micorrízica. Algunos fungicidas comúnmente utilizados para erradicar enfermedades de tipo fúngico y que han mostrado tener poco efecto en la producción de esporas de HMA, son el benomilo (sistémico) y mancozeb (contacto) aplicados en dosis recomendadas (Cabello, 1994).
En la actualidad existen diferentes métodos para la propagación de cepas, así como para la producción de inoculantes a base de HMA. Básicamente, los sistemas de producción de inoculante micorrízico arbuscular consisten en la utilización de una planta hospedante que permita el establecimiento y reproducción de los hongos tanto en raíces como en la rizosfera. Estos sistemas pueden considerar el uso de suelo o cualquier substrato o material inerte, así como el uso de sistemas más sofisticados en invernadero que permitan la propagación de los HMA en un sistema radical el cual se asperja o impregna con específicas soluciones nutritivas (Cooper, 1975; Zobel et al., 1976; Mosse y Thompson, 1984; Sylvia y Hubbel, 1986; Heinzeman y Weritz, 1990; Sylvia, 1999; Alarcón y Ferrera-Cerrato, 2001).
La aplicación de fertilizantes en los cultivos trampa o incluso en sistemas aeropónicos e hidropónicos, tiene especial repercusión en la colonización de los HMA así como en la producción de esporas y micelio externo. Uno de los elementos críticos que influyen en establecimiento de la colonización por HMA en el sistema radical, es el fósforo (Jasfter y Sylvia, 1992). Sin embargo, algunas cepas o complejos micorrízicos pueden ser tolerantes a altas concentraciones de este elemento (Alarcón y Ferrera-Cerrato, 2003), por lo que pueden mantener su capacidad de promover el crecimiento y producir incluso mayor cantidad de propágulos. Por otra parte, la aplicación de soluciones nutritivas es muy común para la producción de inóculo en cultivos con plantas trampa. En este sentido, el uso de la solución nutritiva de Hoagland a 20 y 50%, ha permitido la producción de mayor cantidad de raíz colonizada, así como producción de esporas (Chen et al., 2003).
El cultivo aeropónico de plantas (Zobel et al., 1976) ha permitido la obtención de inoculantes de HMA. Este sistema consiste de la aspersión de solución nutritiva a diversos intervalos y duraciones, al sistema radical que esta colonizado por el o los HMA. No obstante, las plantas hospedantes deben ser precolonizadas con HMA y ser mantenidas en invernadero o cámaras de crecimiento, de tal forma que después de cierto periodo las

5
plantas puedan ser transferidas al sistema de cultivo aeropónico. De este modo, se ha logrado producir inoculantes de hongos como Glomus intraradices Schenck et Smith, Entrophospora kentinensis Wu et Liu, G. clarum Nicol. et Schenck. G. etunicatum Becker et Gerd. (Sylvia y Hubbel, 1986; Wu et al., 1995; Paiva et al., 2003).
La producción de inoculantes de HMA no esta sujeta a la propagación de una sola especie fúngica, sino también es posible crear cultivos mixtos de especies que han sido previamente seleccionadas como eficientes en al promoción del crecimiento de diversas plantas. Por otra parte, es posible la propagación de consorcios naturales de HMA no seleccionados por el hombre, pero aislados de la rizosfera de plantas de sitios específicos y cuyas especies se expresan naturalmente (Chamizo et al., 1998). En algunos reportes se ha mencionado que la efectividad de los HMA es mayor cuando se inoculan cultivos mixtos o consorcios naturales, comparado con los efectos obtenidos por la inoculación individual de cada especie fúngica que integra al cultivo mixto o al consorcio (Mehrotra y Baijal, 1995; Ferrera-Cerrato y Alarcón, 2003).
En la actualidad se cuenta ya con un sistema de propagación de HMA in vitro, en el que es posible obtener micelio y esporas libres de otros microorganismos. Como se ha venido mencionando, la propagación de estos hongos requiere de la presencia de una raíz que suministre fuentes de carbono indispensables para el desarrollo del hongo. Bajo este sistema de cultivo, se han utilizado raíces transformadas de Daucus carota (en las que se inserta el plásmido inductor de la proliferación de raíces que se encuentra en Agrobacterium rhizogenes), o bien raíces sin transformar de Lycopersicum esculentum (Becard y Fortín, 1988; Berbara, 2000). Este sistema fue implementado desde 1975 y a la fecha se ha venido perfeccionando de tal modo que ya se cuenta con bancos de germoplasma de algunos HMA propagados en cultivo in vitro o cultivo monoxénico, como también se le conoce. Este tipo de cultivo de HMA ofrece enormes ventajas para el estudio fisiológico, bioquímico y taxonómico de estos hongos, por lo que se tiene información fundamental de la biología de estos micosimbiontes (Lammers et al., 2001; Declerck et al., 2001; Bago et al., 2002).
Además de las ventajas mencionadas del cultivo monoxénico, es posible también producir inoculante para fines de investigación o incluso para su aplicación en menor escala. Este tipo de sistemas, dado las condiciones de esterilidad, reactivos para la preparación de medio de cultivo, y de equipo de laboratorio, representa un alto costo. Lo anterior representa una desventaja para producir inoculante a gran escala, aunque con base a la obtención de propágulos como esporas (Declerck et al., 2001), es posible aplicar menor cantidad de ellos a las plantas. Otra desventaja es que solo algunas especies de HMA, principalmente del género Glomus, han sido exitosamente cultivadas in vitro, por lo que se requiere de mayor conocimiento respecto a la composición de medios de cultivo, raíz hospedante y condiciones de cultivo para propagar el mayor número de especies de estos hongos simbióticos.
La aplicación comercial de inoculantes de HMA es factible realizarla al inocular sustratos o el sistema radical de las plantas con el suelo inoculo procedente de cultivos trampa. No obstante, para el caso de inoculante producido por sistemas aeropónicos o vía cultivo monoxénico, se ha ideado la suspensión de los propágulos micorrízicos en soluciones de alginato de sodio al 2% y posteriormente, procesadas para su polimerización en una solución de CaCl2 0.1 M (Declerck et al., 1996), y cuyas perlas resultantes contendrán los propágulos micorrízicos para ser utilizados en la inoculación de plantas.
El uso de inoculantes de HMA contenidos en alginato ha resultado apropiado para inducir crecimiento y mejorar la nutrición de plantas de banano, así como proliferar en el sustrato comercial del vivero (Jaizme-Vega et al., 2003). El uso de alginatos, ya sea películas o perlas, como una técnica de inoculación representa ventajas que se relacionan

6
con la protección y retención de los propágulos micorrízicos en suelos arenosos o en sustratos artificiales inorgánicos en los que el inóculo puede perderse fácilmente por lixiviación, al momento de aplicar los riegos (Calvet et al., 1996). En México, este tipo de inoculante así como su presentación en tabletas, ha sido producido a nivel experimental por el grupo de investigación de la Facultad de Agronomía, Universidad Veracruzana, Xalapa, Veracruz (Dra. Dora Trejo, comunicación personal).
PERSPECTIVAS Y CONCLUSIONES
El conocimiento de los procesos involucrados en el establecimiento de la simbiosis
micorrízica, permite que el éxito de la colonización de las raíces por los hongos benéficos pueda realizarse al tiempo o edad apropiada en que las plantas hospedantes la necesitan. Este conocimiento será la base de los lineamientos a seguir en el manejo y producción de inoculante micorrízico arbuscular, con la finalidad de maximizar los beneficios que esta simbiosis aporta a sus macrosimbiontes. Así también, la relativa tolerancia de algunos endófitos a perturbaciones específicas en sus nichos proveerá la identificación, a través de la selección, de aquellos hongos con características para persistir en ambientes desfavorables para el crecimiento de plantas.
Asimismo, se requiere de mayor vinculación entre la industria y expertos de la simbiosis micorrízica con el fin de coadyuvar en el mejoramiento de sistemas de producción de inoculante, así como en la calidad de los mismos. De esta forma, se podrá ofrecer productos biofertilizantes que beneficien a las plantas inoculadas, ya sea en condiciones de invernadero, vivero o en condiciones de campo, donde sea factible realizar la inoculación de HMA, especialmente cuando se intenta fortalecer los programas de restauración y recuperación de suelos perturbados, degradados o contaminados. De igual manera, se requiere de mayor estudio respecto a la aplicabilidad de HMA en el crecimiento y desarrollo de plantas destinadas a decorar interiores en edificios públicos, cuyas condiciones potencialmente representan estrés para ellas, y la simbiosis micorrízica arbuscular podría contribuir en aminorar sus efectos adversos.
Por otra parte, se hace especial énfasis en la utilización de cepas de HMA de origen mexicano que puedan ser reintroducidas a través de su inoculación en plantas destinadas a sitios con climas específicos. Con ello, se puede reducir el impacto sobre la diversidad de HMA nativos lo cual aun es desconocido, por efecto de la introducción de cepas de HMA alóctonos o y de las que procedan del extranjero. Para tal fin, se requiere estudiar detalladamente las interacciones de HMA nativos versus alóctonos en microcosmos, con el objeto de predecir la competencia y el posible desplazamiento de especies de HMA, sobre todo por aquellos géneros fúngicos que se caracterizan por tener mayor “agresividad” para colonizar hospedantes. Agradecimientos. Este trabajo forma parte del proyecto correspondiente al programa de colaboración en investigación aplicada: SISIERRA-CONACYT 2002-0301 “Escalamiento del inóculo micorrízico arbuscular para la sostenibilidad de viveros de cítricos tolerantes al VTC, en el Estado de Campeche”.
LITERATURA CITADA
Abbott, L.K. y A.D. Robson. 1991. Factors influencing the occurrence of vesicular arbuscular mycorrhizas. Agric. Ecosystems Environ. 35:121-150.
Aboul-Nasr, A. 1997. Inoculum production for vesicular arbuscular mycorrhizal fungi on expanded clay. Alex. J. Agric. Res. 42:169-176.

7
Alarcón, A. y R. Ferrera-Cerrato. 1999. Manejo de la micorriza arbuscular en sistemas de propagación de plantas frutícolas. Terra 17:171-191.
Alarcón, A. y R. Ferrera-Cerrato. 2001. Biofertilizantes: importancia y manejo en la agricultura. Rev. Agricultura Técnica de México. 26:63-75.
Alarcón, A. y R. Ferrera-Cerrato. 2003. Aplicación de fósforo e inoculación de hongos micorrízicos arbusculares en el crecimiento y estado nutricional de Citrus volkameriana Tan & Pasq. Terra 21:91-99.
Alarcón, A., R. Ferrera-Cerrato, M.C. González-Chávez y A. Villegas-Monter. 2000. Hongos micorrízicos arbusculares en la dinámica de aparición de estolones y nutrición de plantas de fresa cv. Fern obtenidas por cultivo in vitro. Terra. 18:211-218.
Alarcón , A.; M.C. González-Chávez, R. Ferrera-Cerrato, y A.Villegas-Monter. 2001. Efectividad de Glomus fasciculatum y Glomus etunicatum en el crecimiento de plántulas de Vitis vinifera L. obtenidas por micropropagación. Terra. 19:29-35.
Alarcón, A.; J.J. Almaraz S., R. Ferrera-Cerrato, M.C.A. González-Chávez, M.E. Lara-Hernández, M.J. Manjarrez M., R. Quintero L. y S. Santamaría R. 2004. Manual: Tecnología hongos micorrízicos en la producción de especies forestales en vivero. R. Ferrera-Cerrato, A. Alarcón, y M.E. Lara H. (Eds.). Colegio de Postgraduados, Montecillo. SEMARNAT-PRONARE, México. 98 p. Se puede consultar en: http://www.conafor.gob.mx/programas_nacionales_forestales/pronare/sire/publicaciones/PARTE1%20MICORRIZAS.pdf
Augé, R.M. 2001. Water relations, drought and vesicular-arbuscular mycorrhizal symbiosis. Mycorrhiza 11:3-42.
Bago, B.; C. Azcón-Aguilar, Y. Shachar-Hill, y P.E. Pfeffer. 2000. El micelio externo de la micorriza arbuscular como puente simbiótico entre la raíz y su entorno. In. A. Alarcón y R. Ferrera-Cerrato (Eds.).Ecología, fisiología y biotecnología de la micorriza arbuscular. Ed. MundiPrensa. Mexico. pp. 78-92.
Bago, B.; W. Zipfel, R.M. Williams, J. Jun, R. Arreola, P.J. Lammers, P.E. Pfeffer, y Y. Shachar-Hill. 2002. Translocation and utilization of fungal storage lipid in the arbuscular mycorrhizal symbiosis. Plant Physiol. 128:108-124.
Becard, G. y J.A. Fortín. 1988. Early events of vesicular-arbuscular mycorrhiza formation on Ri T-DNA transformed roots. New Phytol. 108:211-218.
Barbara, R.L.L. 2000. Potencialidades no estudo com fungos micorrizicos arbusculares em sistemas radiculares transgénicos. In. A. Alarcón y R. Ferrera-Cerrato (Eds.). Ecología, fisiología y biotecnología de la micorriza arbuscular. Mundi Prensa. México. pp.118-123.
Boyetchko, S.M. y J.P. Tewari. 1995. Susceptibility of barley cultivars to vesicular-arbuscular mycorrhizal fungi. Can. J. Plant Sci. 75:269-275.
Busch, E, y J.I. Lelley. 1997. Use of endomycorrhizal fungi for plant cultivation on buildings. Angew. Bot. 71:50-53.
Cabello, M.N. 1994. Efecto de la aplicación secuencial de fungicidas sobre hongos formadores de micorrizas vesículo-arbusculares en condiciones de campo. Bol. Micol. 9:1-7.
Calvet, C.; A. Camprubi, y R. Rodríguez-Kábana. 1996. Inclusion of arbuscular mycorrizal fungi in alginate films for experimental studies and plant inoculation. HortScience 31:285.
Chamizo, C.A., R. Ferrera-Cerrato y L. Varela. 1998. Identificación de especies de un consorcio del género Glomus. Rev. Mex. Micol. 14:37-40.
Chen, N; Y.S. Wang, X.L. Li, M.Q. Zhang, L.J. Xing, G. Feng, y X.H. Ni. 2003. Effects of nutrient solution strength on development of arbuscular mycorrhizal fungi. Mycosistema 22:394-401.
Cooper, A.J. 1975. Crop production in re-circulation nutrient solutions. Sci. Hortic. 3:251-258. Dakessian, S.; M.S. Brown y G.J. Bethlenfalvay. 1986. Relationship of mycorrhizal growth
enhancement and plant growth with soil water and texture. Plant Soil 94:439-443. Davies, Jr. F.T.; A.A. Estrada-Luna, T.L. Finnerty, y V. Olalde-Portugal. 2000. Applications of
mycorrhizal fungi in plant propagation systems. In. A. Alarcón y R. Ferrera-Cerrato (Eds.).Ecologia, fisiologia y biotecnología de la micorriza arbuscular. Ed. MundiPrensa. Mexico. pp. 123-140.

8
Declerck, S.; D.G. Strullu, C. Plenchette, y T. Guillemette. 1996. Entrapment of in vitro produced spores of Glomus versiforme, associated with Ri T-DNA transformed carrot roots. J. Biotechnol. 48:51-57.
Declerck, S.; D. D’or, S. Cranenbrouck, y E. Le Boulengé. 2001. Modelling the sporulation dynamics of arbuscular mycorrhizal fungi in monoxenic culture. Mycorrhiza 11:225-230.
Ferrera-Cerrato, R. 1995. Efecto de Rizosfera. In. R. Ferrera-Cerrato y J. Pérez-Moreno (Eds.). Agromicrobiología. Elemento útil en la agricultura sustentable. Colegio de Postgraduados en Ciencias Agrícolas. Montecillo, Estado de México. pp. 36-53.
Ferrera-Cerrato, R y A. Alarcón. 1998. Importancia de la simbiosis en la agricultura.. Programa y Resúmenes. II Symposium Nacional de la Simbiosis Micorrízica. Colima, Colima. México. 4-6 de noviembre de 1998. pp. 1-2
Ferrera-Cerrato, R. y A. Alarcón. 2001. La microbiología del suelo en la agricultura sostenible. Rev. Ciencia ErgoSum 8:175-183.
Ferrera-Cerrato, R. y A. Alarcón. 2003. The effectiveness of a Mexican endogenous arbuscular mycorrhizal fungi consortium on two cultivars of Carica papaya L. depends on the interaction of its own three fungal components. In. Proceedings of the Fourth International Conference on Mycorrhizae. ICOM4 August 10-15, 2003. P. 167.
Font Quer, P. 1982. Diccionario de Botánica. Editorial Labor. Barcelona. pp. 713-714. Gianinazzi-Pearson, V. y C. Azcón-Aguilar. 1991. Fisiología de las micorrizas vesículo-
arbusculares. In. J. Olivares y J.M. Barea (Coordinadores) Fijación y Movilización biológica de nutrientes. Vol. 11 Fijación de N y Micorrizas. Consejo Superior de Investigaciones Científicas. Madrid, España. pp. 175-202.
González, C.M.C y R. Ferrera-Cerrato. 1990. Effect of vesicular-arbuscular mycorrhizae on tissue culture derived plantlets of strawberry. HortScience 25:903-905.
González-Chávez, M.C. y R. Ferrera-Cerrato. 1996. Ecología de la endomicorriza vesículo-arbuscular en un agroecosistema sostenible de maíz en el trópico húmedo de México. Micol. Neotrop. Apl. 9:53-66.
Gregory, J.P. 1992. Crecimiento y funcionamiento de las raíces. In. A. Wild (Coordinador) Condiciones del suelo y desarrollo de las plantas según Russeli. Versión Española de P. Urbano T y C. Rojo H. Ediciones Mundi-Prensa. Madrid. pp. 121-175.
Grundwaldt-Stocker, G. y H.W. Dehne. 1989. The use of vesicular-arbuscular mycorrhizal fungi on plant production. II. Characterization of inocula on inorganic carrier material. J. Plant Disease Protection. 96:615-626.
Harinikumar, K.M. y D.J. Bagyaraj. 1994. Potential of earthworms, ants, millipedes, and termites for dissemination of vesicular-arbuscular mycorrhizal fungi in soil. Biol. Fertil. Soils 18:115-118.
Heinzeman, J. y J. Weritz. 1990. Rockwool: a new carrier system for mass multiplication of vesicular-arbuscular mycorrhizal fungi. Angew. Botanik 64:271-274.
Jaizme-Vega, M.C.; A.S. Rodríguez-Romero, C. Marín-Hermoso y S. Declerck. 2003. Growth of micropropagated bananas colonized by root-organ culture produced arbuscular mycorrhizal fungi entrapped in Ca-alginate beads. Plant and Soil 254:329-335.
Jasfter, A.G. y D.M. Sylvia. 1992. Inoculum production and inoculation strategies for vesicular arbuscular mycorrhizal fungi. In. B. Metting (Ed.). Soil microbial ecology, application in agricultural and environmental management. Marcel Dekker, Inc. NY. pp. 349-377.
Jeffries, P., S. Gianinazzi, S. Perotto, K. Turnau, y J.M. Barea. 2003. The contribution of arbuscular mycorrhizal fungi in sustainable maintenance of plant health and soil fertility. Biol. Fertil. Soils 37:1-16.
Johnson, N.C., J.H. Graham, y F.A. Smith, 1997. Functioning of mycorrhizal associations along the mutualism-parasitism continuum. New Phytol. 135:575-586.
Klironomos, J.N. 2003. Variation in plant response to native and exotic arbuscular mycorhizal fungi. Ecology 84:2292-2301.
Lammes, J.P.; J. Jun, J. Abubaker, R. Arreola, A. Gopalan, B. Bago, C. Hernandez-Sebastia, J.W. Allen, D.D. Douds, P.E. Pfeffer, y Y. Sachar-Hill. 2001. The glyoxylate cycle in an arbuscular mycorrhizal fungus. Carbon flux and gene expression. Plant Physiol. 127:1287-1298.

9
Linderman, R.G. 1993. Effects of rnicrobial interactions in the mycorrhizosphere of plant growth and health. In. R. Ferrera-Cerrato y R. Quintero L. (Eds.). Agroecología. Sostenibilidad y Educación. Centro de Edafología, Colegio de Postgraduados. Montecillo, Estado de México. pp. 138-152.
Mehrotra, V.S. y U. Baijal. 1995. Effects of single and mixed inocula of VAM fungi on the growth and yield of sunflower (Helianthus annuus L.). Philippine J. Sci. 124:183-201.
Mosse, B. y J.P. Thompson. 1984. Vesicular-arbuscular endomycorrhizal inoculum production. I. Exploratory experiments with beans (Phaseouls vulgaris) in nutrient flow culture. Can. J. Bot. 62:1523-1530.
Nadian, H.; S.E. Smith, A.M. Alston, R.S. Murray y D. Siebert. 1998. Effects of soil compaction on phosphorus uptake and growth of Trífolíum subterraneum colonized by four species of vesicular- arbuscular mycorrhizal fungi. New Phytol. 139:155-165.
Paiva, L.M., M.A. Silva, P.C. Silva, y L.C. Maia. 2003. Glomus clarum e Glomus etunicatum: cultivo em solo e aeroponia. Revista Brasil. Bot. 26:257-262.
Rapparini, F.; R. Baraldi, G. Bertazza, B. Branzanti y S. Predieri. 1994. Vesicular-arbuscular mycorrhizal inoculation of micropropagated fruit trees. J. Hortic. Sci. 69:1101-1109.
Rilling, M.C. y P.D. Steinberg. 2002. Glomalin production by an arbuscular mycorrhizal fungus: a mechanism of habitat modification. Soil Biol. Biochem. 34:1371-1374.
Saito, M. 1990. Charcoal as a micro-habitat for VA mycorrhizal fungi, and its implication. Agric. Ecosyst. Environ. 29:341-344.
Saito, M. y T. Marumoto. 2002. Inoculation with arbuscular mycorrhizal fungi: the status quo in Japan and the future prospects. Plant Soil 244:273-279.
Sanders, I.R. y A.H. Fitter. 1992. The ecology and functioning of vesicular-arbuscular mycorrhizas in co-existing grassland species. New Phytol. 120:517-524.
Schachtman, D.P.; R.J. Reid y S.M. Ayling. 1998. Phosphorus uptake by plants: from soil to cell. Plant Physiol. 116:447-453.
Sieverding, E. 1991. Vesicular-arbuscular mycorrhiza management in tropical agrosystems. Technical Cooperation, GTZ. Eschborn, Germany. 371 p.
Smith, S.E. y D.J. Read. 1997. Mycorrhizal symbiosis. 2nd Edition. Academic Press. London. Sylvia, D.M. 1999. Mycorrhizal symbiosis. In. Principles and applications of soil microbiology.
D.M. Sylvia, J.J. Fuhrmann, P.G. Hartel, y D.A. Zuberer (Eds.) Prentice Hall. New Jersey, USA. pp. 409-426.
Sylvia, D.M. y D.H. Hubbel. 1986. Growth and sporulation of vesicular-arbuscular mycorrhizal fungi in aeroponic and membrane systems. Symbiosis 1:259-267.
Sylvia, D.M.; D.O. Wilson, J.H. Graham, J.J. Maddox, P. Millner, J.B. Morton, H.D. Skipper, S.F. Wright y A.G. Jarstfer. 1993. Evaluation of vesicular-arbuscular mycorrhizal fungi in diverse plants and soils. Soil Biol. Biochem. 25:705-713.
Thimm. T. y O. Larink. 1994. Grazing preference of some collembola for endomycorrhizal fungi. Biol. Fertil. Soils 18:1-3.
Trejo, A.D.; E. Hernández A. y R. Ferrera-Cerrato 1998. Ecología y comportamiento de la endomicorriza arbuscular en el cultivo del café (Coffea arabica L.). In. R. Zulueta R., M.A. Escalona A. y D. Trejo A. (Eds.). Avances de la Investigación Micorrízica en México. Universidad Veracruzana. México. pp. 41-52.
Ulloa, M. 1991. Diccionario ilustrado de Micología. UNAM. México p. 185. Wu, C.G., Y.S. Liu, y L.L. Hung. 1995. Spore development of Entrophospora kentinensis in an
aeroponic system. Mycologia 87:582-587. Zobel, R.W.; P.D. Tredice y J.G. Torrey. 1976. Methods for growing plants aeroponically. Plant
Physiol. 57:344-346.

10
IMPACTO DE LA MICORRIZA ARBUSCULAR EN LA PRODUCTIVIDAD DEL SORGO EN TAMAULIPAS
Arturo Díaz Franco, Idalia Garza Cano, Víctor Pecina Quintero
y Agustín Magallanes Estala*
Campo Experimental Río Bravo, INIFAP. Apdo. Postal 172, Río Bravo, Tam. 88900.
En la región semiárida del norte de Tamaulipas se siembra la mayor superficie de sorgo (Sorghum bicolor) en el país, es un monocultivo que ocupa alrededor de 750 mil hectáreas de temporal. Esta región cuenta con suelos degradados, que en general son carentes de N, P y materia orgánica (Durán, 1992). Sin embargo, la fertilización química no es una práctica generalizada, y cuando se aplica, las dosis son inferiores a las requeridas, debido a que representa una inversión de alto costo y riesgo en la eficacia, por la errática humedad en el suelo (Díaz M., 2002). La alteración del agroecosistema ha traído como consecuencia, que la productividad y rentabilidad del sorgo, haya decrecido de forma considerable (Salinas, 2002).
La biofertilización incluye como agentes promotores del crecimiento vegetal, a hongos y bacterias benéficas del suelo (micorrizas y rizobacterias), así como desechos orgánicos a través de compostas y vermicompostas (Alarcón y Ferrara, 2000). Dentro de este contexto, las micorrizas vesículo arbusculares (MVA), han tenido tal repercusión que ya existen en el mercado diferentes productos comerciales (Sylvia, 1999, 2004), aunque muchos de ellos tienen un alto costo. González (2002) señaló que, como una aplicación biotecnológica, las plantas necesitan la reintroducción de especies micorrízicas eficientes en situaciones como ausencia o baja población de MVA nativas, en suelos erosionados, degradados, contaminados o en suelos con especies micorrízicas ineficientes. Por lo anterior, el propósito del presente documento es dar a conocer la influencia de la inoculación de semilla de sorgo, con la MVA Glomus intraradices, su impacto en la productividad y experiencias en la validación de esa tecnología como una práctica para hacer una producción sostenible.
Características de las MVA
La actividad de las MVA, a través de la extensa red de hifas que los hongos generan, incrementa la superficie de exploración radical y beneficia a la planta con una mayor absorción de P y de microelementos; aunque de manera indirecta y en menor proporción, también existe absorción de N. En la simbiosis se establecen diversos procesos fisiológicos y bioquímicos, de modo que la planta puede presentar cambios en la morfología de la raíz e incrementos de la actividad fotosintética; otros aspectos relacionados es la capacidad de las MVA de producir hormonas como ácido abscísico, giberelinas, auxinas y citoquininas (Alarcón y Ferrara, 1999; Smith y Gianinazzi, 1988; Sylvia, 2004). También la colonización micorrízica contribuye a que la planta tolere condiciones adversas de humedad, salinidad, metales pesados, así como a la acción
* A partir de julio de 2003 labora en el INIFAP-Campo Experimental Pachuca.

11
protectora en contra de algunos fitopatógenos del suelo. Las respuestas pueden variar en función al grado de dependencia entre los endófitos y la planta hospedante, por lo que la efectividad micorrízica depende de la respuesta de la planta a la colonización (Alarcón y Ferrara, 1999; Sylvia, 2004).
Los estudios locales con inoculación de semilla de sorgo, han determinado que G. intraradices (cepa INIFAP), destacó como un simbionte eficiente, no así cuando se combinó con otros simbiontes o con fitohormona (Díaz, 2003; Díaz M., 2002; Garza et al., 2003; Magallanes y Díaz, 2002). También se ha demostrado su efectividad por el incremento en la producción de otros cultivos regionales como maíz (Zea mays) y okra (Abelmoschus esculentus) (Alvarado et al., 2003; Díaz et al., 2002). De igual forma, en otras regiones del país el mismo endófito fue promotor del crecimiento y de rendimiento, inclusive igual o superior a la fertilización química, en maíz, trigo (Triticum aestivum), tomate (Lycopersicon esculentum), soya (Glycine max), frijol (Phaseolus vulgaris) y naranjo (Citrus sinensis) (Durán et al., 2001; González et al., 2002; Irizar et al., 2003).
El hongo MVA G. intraradices, pertenece a la clase Zygomicetos, orden Glomales, suborden Glomineae y a la familia Glomaceae. Posee esporas unicelulares que nacen de una sola hifa, son globosas o subglobosas (93 µm), de color blanco, crema a amarillo-café, cuentan con tres paredes; además de tener hifas cilíndricas (Morton, 2002).
Actualmente la multiplicación masiva de G. intraradices, así como el proceso para su comercialización, la ha realizado la Dra. Ma. Peña del Río en el Campo Experimental General Terán, INIFAP, General Terán, N. L., mediante camas reproductoras donde se utiliza sorgo como planta hospedera. Esta metodología es descrita por Durán et al. (2001), y con ella se obtiene el sustrato infectado con una concentración de ≥200 esporas g-1.
Inoculación de la semilla de sorgo
La dosis de sustrato micorrízico utilizado para la inoculación de semilla de sorgo ha sido la indicada por Irizar et al. (2003), quienes recomiendan 1 kg de sustrato para la semilla necesaria en una hectárea (aproximadamente 8 kg); y como adherente durante la mezcla, utilizan 60 ml de carboxi metil celulosa. La mezcla de la semilla puede ser en un tanque tratador de 200 Lt u otro tipo de mezcladora mecánica. El procedimiento sugerido para la inoculación de la semilla de sorgo es: 1) se mezcla el adherente en 0.7 a 0.9 Lt de agua; 2) se vacía la semilla en la mezcladora y se adiciona la solución del punto 1, posteriormente se mezcla; y 3) se agrega el sustrato micorrízico y se mezcla hasta que la semilla se cubra uniformemente. Aunque la inoculación se puede realizar al momento de la siembra, es conveniente que ésta se programe desde que el productor adquiere la semilla (uno o más meses antes de la siembra); cuando ese sea el caso, se debe de secar la semilla bajo sombra antes de almacenarla.
Influencia de Glomus intraradices en sorgo
A) Resultados de invernadero.
En un estudio de invernadero se evaluó la inoculación de la G. intraradices en nueve híbridos comerciales de sorgo: Asgrow Coral, RB-Patrón, RB-3030, Pioneer 83G66,

12
Pioneer 82G63, Pioneer 8443, DK-55, DK-52 y Wac 690. Los resultados mostraron que las variables altura de planta, la biomasa seca y el peso de raíz de los nueve híbridos de sorgo, manifestaron un incremento significativo con la inoculación micorrízica, comparado con el testigo, sin inoculación. En promedio, el endófito incrementó 8.4% la altura de planta, 12.3% la biomasa seca, y 10.8% el peso de la raíz (Cuadro 1).
Cuadro 1. Influencia de la inoculación con G. intraradices en algunas características de planta de nueve híbridos comerciales de sorgo en invernadero.
Inoculación Altura de planta
(cm)
Biomasa seca
(g)
Peso de raíz
(g)
Con 46.29 16.44 10.86
Sin 42.70 14.63 9.80
DMS (p<0.05) 1.78 0.72 0.69
Por otro lado, el parámetro índice de clorofila registró una respuesta variable entre el híbrido de sorgo y la inoculación micorrízica (Cuadro 2); en todos los casos se acrecentó el contenido de clorofila, aunque solamente en los híbridos RB-Patrón y Pioneer 83G66, el valor de la diferencia con y sin la inoculación no fue significativa. Lo anterior demuestra que la manifestación de la micorriza, en función a la clorofila, no presenta consistencia entre los híbridos de sorgo. Se registró la mayor colonización micorrízica radical en los híbridos Pioneer 8442 y Wac 690; ésta colonización solo se correlacionó significativamente (r=0.67*) con la diferencia obtenida en el índice de clorofila con y sin la inoculación (Cuadro 2).
Otros estudios de invernadero han constatado que la inoculación de MVA en sorgo promueve el crecimiento y desarrollo de la planta. Abdel y Mohamedin (2000) informaron que la inoculación de sorgo con G. intraradices (cepa LAP8), incrementó significativamente el crecimiento, los pigmentos fotosintéticos y la proteína, comparado con el sorgo no inoculado.
B) Evaluaciones en campo El tipo de suelo y la micorrización. Para conocer la respuesta del sorgo a la
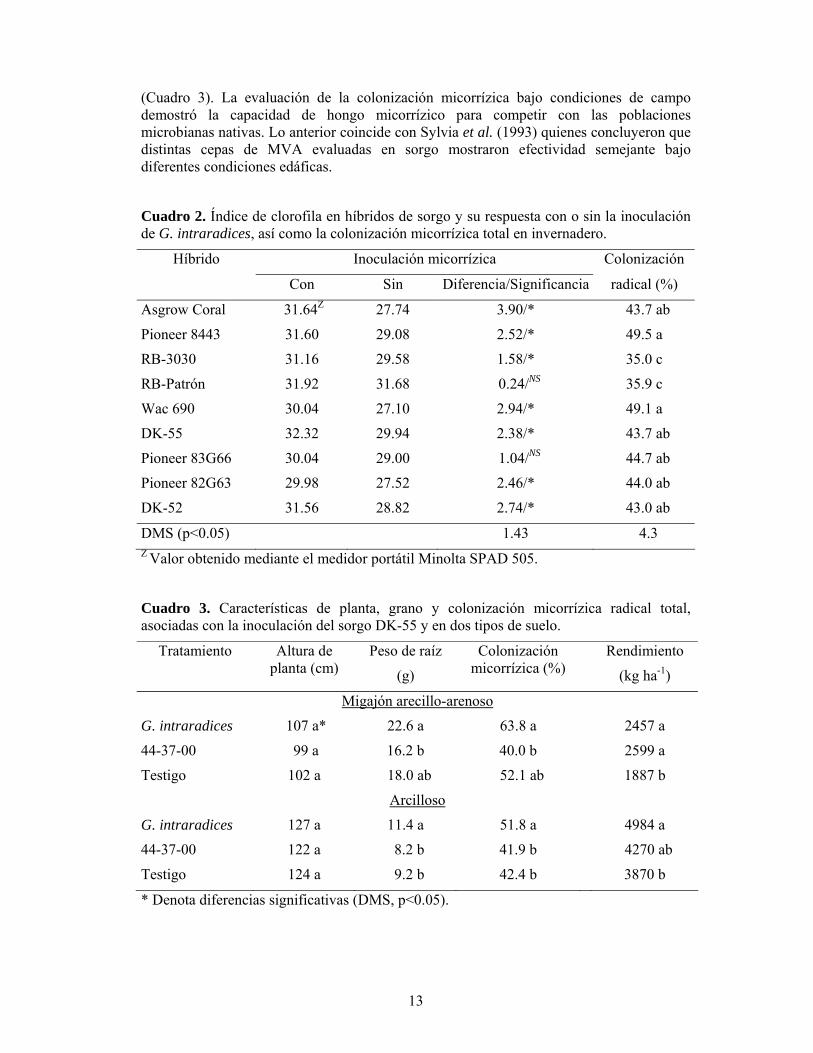
inoculación de G. intraradices en sorgo asociada con el tipo de suelo, se realizó un trabajo de campo, en condiciones de temporal, donde se seleccionaron dos suelos, uno de textura arcillosa y el otro migajón arcillo arenoso (Díaz M., 2002). Sin embargo, ambos suelos fueron alcalinos (pH 8), con bajo contenido en materia orgánica, N, P y alto en K. En ese estudio se utilizaron como tratamientos, la dosis de fertilización química empleada por el productor, que consistió de la adición en presiembra de 44 y 33 kg ha-1 de N y P, respectivamente, la semilla inoculada con G. intraradices y el testigo absoluto. Los resultados indicaron que el sorgo respondió de forma similar a los tratamientos independientemente de la textura del suelo; el promedio de las variables de ambos sitios se encuentran en el Cuadro 3. Con excepción de la altura de planta, la inoculación con el simbionte incrementó el peso de raíz, la colonización micorrízica y el rendimiento de grano. Con relación al rendimiento, la MVA igualó o superó a la fertilización química
13
(Cuadro 3). La evaluación de la colonización micorrízica bajo condiciones de campo demostró la capacidad de hongo micorrízico para competir con las poblaciones microbianas nativas. Lo anterior coincide con Sylvia et al. (1993) quienes concluyeron que distintas cepas de MVA evaluadas en sorgo mostraron efectividad semejante bajo diferentes condiciones edáficas.
Cuadro 2. Índice de clorofila en híbridos de sorgo y su respuesta con o sin la inoculación de G. intraradices, así como la colonización micorrízica total en invernadero.
Híbrido Inoculación micorrízica Colonización
Con Sin Diferencia/Significancia radical (%)
Asgrow Coral 31.64Z 27.74 3.90/* 43.7 ab
Pioneer 8443 31.60 29.08 2.52/* 49.5 a
RB-3030 31.16 29.58 1.58/* 35.0 c
RB-Patrón 31.92 31.68 0.24/NS 35.9 c
Wac 690 30.04 27.10 2.94/* 49.1 a
DK-55 32.32 29.94 2.38/* 43.7 ab
Pioneer 83G66 30.04 29.00 1.04/NS 44.7 ab
Pioneer 82G63 29.98 27.52 2.46/* 44.0 ab
DK-52 31.56 28.82 2.74/* 43.0 ab
DMS (p<0.05) 1.43 4.3 Z Valor obtenido mediante el medidor portátil Minolta SPAD 505.
Cuadro 3. Características de planta, grano y colonización micorrízica radical total, asociadas con la inoculación del sorgo DK-55 y en dos tipos de suelo.
Tratamiento Altura de planta (cm)
Peso de raíz
(g)
Colonización micorrízica (%)
Rendimiento
(kg ha-1)
Migajón arecillo-arenoso
G. intraradices 107 a* 22.6 a 63.8 a 2457 a
44-37-00 99 a 16.2 b 40.0 b 2599 a
Testigo 102 a 18.0 ab 52.1 ab 1887 b
Arcilloso
G. intraradices 127 a 11.4 a 51.8 a 4984 a
44-37-00 122 a 8.2 b 41.9 b 4270 ab
Testigo 124 a 9.2 b 42.4 b 3870 b
* Denota diferencias significativas (DMS, p<0.05).

14
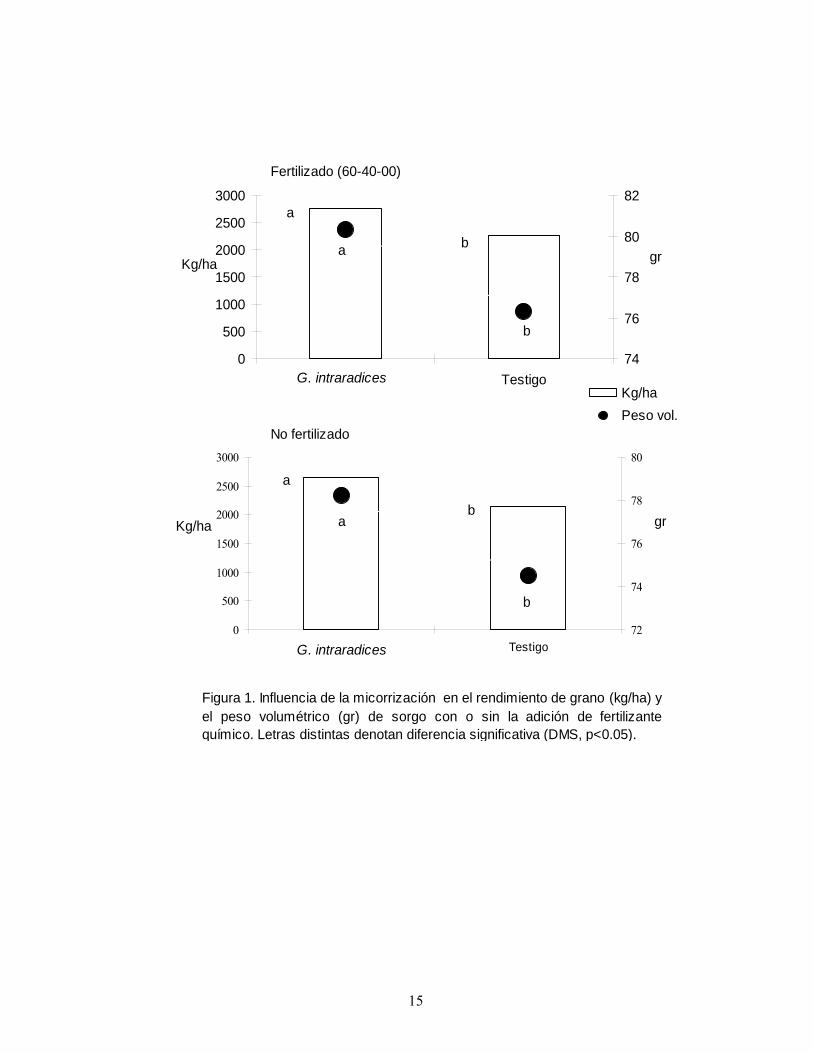
La fertilización química y la micorrización. Se realizó un estudio en condiciones de temporal, para determinar la influencia de la fertilización química en la efectividad de la inoculación micorrízica del sorgo. Los tratamientos fueron con o sin la inoculación de la semilla sorgo con G. intraradices, los cuales se compararon en un suelo fertilizado en presiembra con la adición de 60 y 40 kg ha-1 de N y P, respectivamente (Montes y Aguirre, 1992) y otro sin fertilizar. Los resultados indicaron que la efectividad de la MVA tuvo una respuesta significativa, favorable y similar en las dos condiciones de evaluación. El hongo micorrízico fue capaz de incrementar el peso volumétrico y el rendimiento de grano de sorgo en el rango de 4.9-5.2% y 22-23%, respectivamente (Figura 1). Estos resultados son coincidentes con los de Montaño et al. (2001), quienes indicaron que la actividad de MVA en trigo no estuvo influenciada por el nivel de N adicionado. Estudios de Bressan et al. (2001) informaron que la respuesta del sorgo varía según el nivel de P en el suelo, así como la cepa de MVA inoculada. Por el contrario, Johnson y Pfleger (1992) y Garza (2002), concluyeron que G. intraradices es insensible a la fertilización química; en particular, Garza (2002) determinó que el endófito aumentó significativamente el contenido de P foliar en sorgo, aún en suelos deficientes de P.
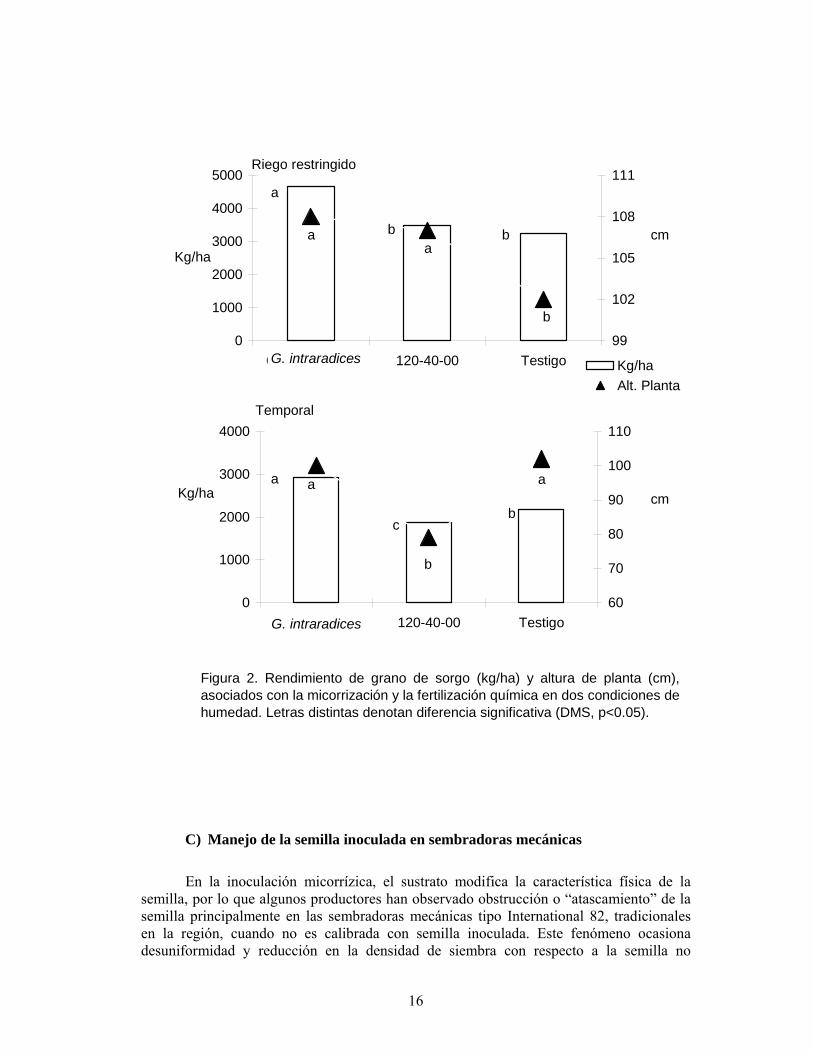
El riego restringido y la micorrización. La escasez de agua para uso agropecuario en la región ha traído consigo restricciones en las áreas irrigadas, dentro de estas medidas esta la aplicación de un riego de auxilio o riego restringido para los cultivos. Por lo que se realizó un estudio para conocer la influencia del riego restringido y la inoculación micorrízica de la semilla de sorgo (Díaz, 2003). La inoculación de semilla con G. intraradices; fertilización química a razón de 120 y 40 kg ha-1 de N y P, respectivamente (Aguirre et al., 1992); y el testigo absoluto, se evaluaron en condición de riego restringido (riego en floración) y en temporal. La mayor altura de planta en riego restringido la registraron la inoculación de MVA y la fertilización química, mientras que en temporal, la mayor altura fue semejante entre el simbionte y el testigo; al parecer la disminución de la altura en las plantas con fertilización química en temporal, se debió a la competitividad de la humedad entre la planta y el fertilizante, ya que éste presenta características higroscópicas. Este mismo efecto impactó negativamente en el rendimiento de grano en temporal. Por el contrario, la micorrización incrementó significativamente en rendimiento de grano, comparado con la fertilización química y el testigo, en las dos condiciones de humedad evaluadas (Figura 2). En maíz, Sylvia et al. (1993) informaron que la respuesta a la inoculación con G. etunicatum, fue proporcional en los tres gradientes de humedad en el suelo estudiados.
15
0
500
1000
1500
2000
2500
3000
G. intraradices Testigo74
76
78
80
82
Kg/haPeso vol.
0
500
1000
1500
2000
2500
3000
G. intraradices Testigo72
74
76
78
80
G. intraradices
G. intraradices
Fertilizado (60-40-00)
No fertilizado
Figura 1. Influencia de la micorrización en el rendimiento de grano (kg/ha) yel peso volumétrico (gr) de sorgo con o sin la adición de fertilizantequímico. Letras distintas denotan diferencia significativa (DMS, p<0.05).
a
ba
b
a
ba
b
Kg/ha gr
Kg/ha gr
16
0
1000
2000
3000
4000
5000
G. intraradices 120-40-00 Testigo99
102
105
108
111
Kg/haAlt. Planta
0
1000
2000
3000
4000
G. intraradices 120-40-00 Testigo60
70
80
90
100
110
Riego restringido
Temporal
G. intraradices
G. intraradices
a
b baa
b
a
cb
a
b
a
Figura 2. Rendimiento de grano de sorgo (kg/ha) y altura de planta (cm),asociados con la micorrización y la fertilización química en dos condiciones dehumedad. Letras distintas denotan diferencia significativa (DMS, p<0.05).
Kg/hacm
Kg/ha cm
C) Manejo de la semilla inoculada en sembradoras mecánicas
En la inoculación micorrízica, el sustrato modifica la característica física de la semilla, por lo que algunos productores han observado obstrucción o “atascamiento” de la semilla principalmente en las sembradoras mecánicas tipo International 82, tradicionales en la región, cuando no es calibrada con semilla inoculada. Este fenómeno ocasiona desuniformidad y reducción en la densidad de siembra con respecto a la semilla no

17
inoculada y un consecuente decremento en el rendimiento. Por lo anterior, se realizó un estudio (Díaz y Pecina, 2003) donde se utilizó la sembradora International 82. En ésta se evaluaron tres factores: 1) dos tamaños de semilla; 2) el tiempo de la inoculación (0, 12 y 24 h); y 3) platos de siembra con 24, 30 y 40 orificios. Se consideró como densidad testigo u óptima el tirar un rango de 25-27 semillas m–1 lineal (Montes y Aguirre, 1992).
Los resultados revelaron que los tres factores están involucrados en la obstrucción de la semilla. Con la semilla más grande de sorgo se registró mayor obstrucción. La inoculación implica el humedecimiento de la semilla, por lo que al paso del tiempo esta se hidrata y tiende a hincharse, lo cual provoca la obstrucción de la semilla. Cuando se siembra semilla recién inoculada o sin inocular, no hay obstrucción al paso de la semilla, mientras que en semilla de 12 y 24 h después de inoculada, la densidad de siembra se reduce a 20.8 y 13.9 semillas m-1, respectivamente. También se demostró que un mayor número de orificios en los platos de siembra facilita que la semilla fluya sin obstrucción en la sembradora. Se observó que platos con un mínimo de 30 orificios resultaron óptimos para obtener la densidad de siembra esperada. Por lo anterior, se concluyó que para obtener el número deseado de semillas de sorgo por metro lineal, la calibración de la sembradora debe hacerse con semilla inoculada.
Adicionalmente y en las mismas sembradoras mecánicas, dada la fricción con el sustrato, el productor ha experimentado que los platos de plástico se desgastan prematuramente después de sembrar 120-140 hectáreas; aunque el reemplazo de los platos tiene un bajo costo (aprox. $5.00 ha-1 para sembradora de seis botes), representa una desventaja. Para evitar el desgaste, el productor puede adicionar una taza de talco agrícola (aprox. 80 g) en cada bote de la sembradora.
D) Resultados de validación
Durante el periodo de 2000 al 2003, se establecieron parcelas de validación en el norte de Tamaulipas, bajo diferente condición de humedad, además de que el híbrido y el manejo del cultivo fue a criterio del productor. En cada parcela se comparó la inoculación micorrízica de la semilla de sorgo con un testigo absoluto y en la mayoría de los casos con la fertilización química utilizada por el productor. Los resultados indicaron que la micorrización del sorgo aumentó de manera consistente la producción de grano en el rango de 18 a 36%, con relación al testigo, y en los casos donde se comparó la fertilización química, el simbionte también igualó o superó el rendimiento con esa práctica (Cuadro 4). La información generada de las parcelas de validación, constatan los resultados de campo experimentales señalados anteriormente. Además, representan un mayor incremento en el rendimiento de grano de sorgo comparado con el 15% registrado por Acosta et al. (2001) en la región, mediante la inoculación mixta de Azospirillum brasilense y G. intraradices. Con relación a la fertilización química, Durán et al. (2001), coinciden al informar que el híbrido de maíz H-512 inoculado con G. intraradices, produjo 12% de incremento sobre el tratamiento con fertilización química, el cual llevó 138 y 46 kg ha-1 de N y P, respectivamente.
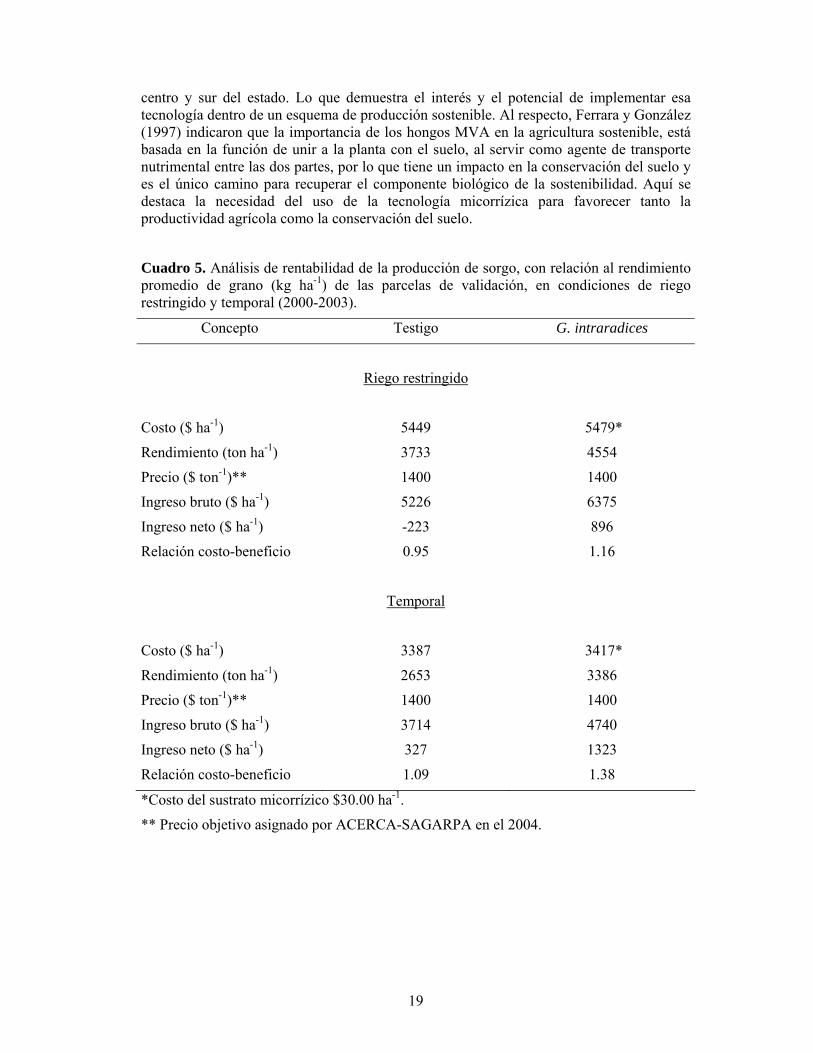
El análisis económico de la producción en función al rendimiento promedio de grano de sorgo, obtenido de las parcelas de validación, tanto en condiciones de riego restringido como de temporal, determinó que la inoculación de la semilla con G. intraradices fue una práctica redituable en la producción de sorgo. En condiciones de riego restringido, la micorriza aumentó la rentabilidad de la producción en 22% (relación
18
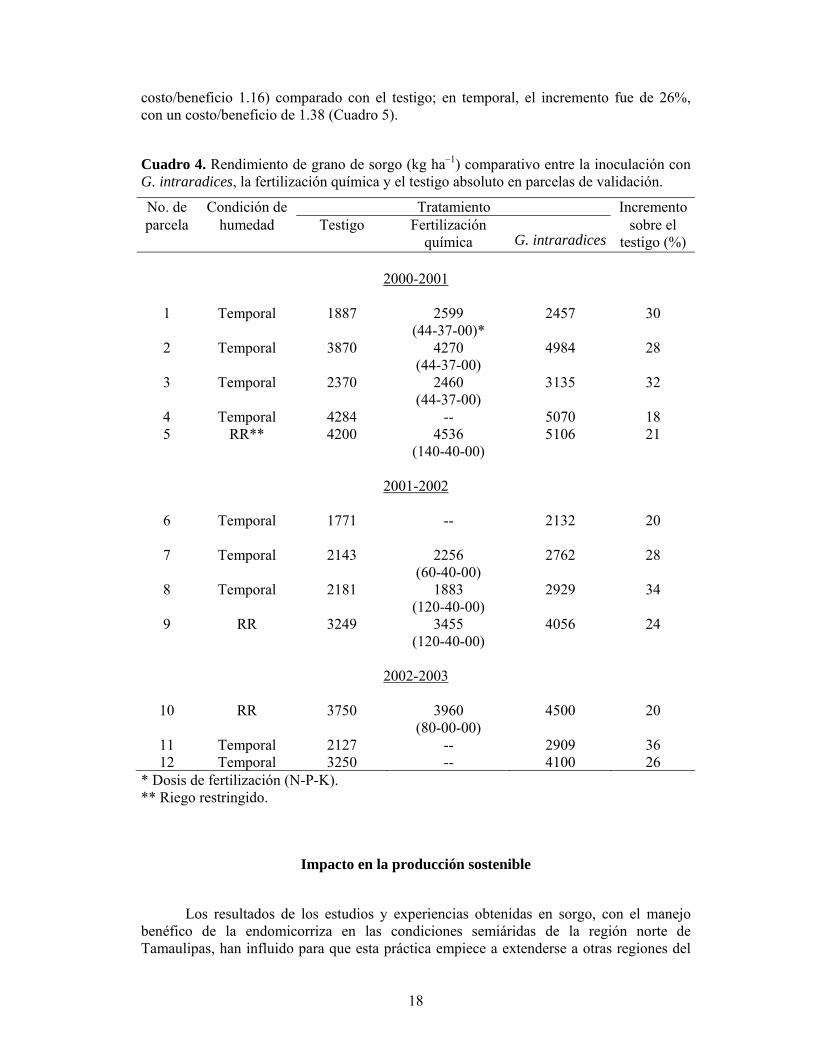
costo/beneficio 1.16) comparado con el testigo; en temporal, el incremento fue de 26%, con un costo/beneficio de 1.38 (Cuadro 5).
Cuadro 4. Rendimiento de grano de sorgo (kg ha–1) comparativo entre la inoculación con G. intraradices, la fertilización química y el testigo absoluto en parcelas de validación.
No. de Condición de Tratamiento Incremento parcela humedad Testigo Fertilización
química G. intraradices sobre el
testigo (%)
2000-2001
1 Temporal 1887 2599 (44-37-00)*
2457 30
2 Temporal 3870 4270 (44-37-00)
4984 28
3 Temporal 2370 2460 (44-37-00)
3135 32
4 Temporal 4284 -- 5070 18 5 RR** 4200 4536
(140-40-00) 5106 21
2001-2002
6 Temporal 1771 --
2132 20
7 Temporal 2143 2256 (60-40-00)
2762 28
8 Temporal 2181 1883 (120-40-00)
2929 34
9 RR 3249 3455 (120-40-00)
4056 24
2002-2003
10 RR 3750 3960
(80-00-00) 4500 20
11 Temporal 2127 -- 2909 36 12 Temporal 3250 -- 4100 26
* Dosis de fertilización (N-P-K). ** Riego restringido.
Impacto en la producción sostenible
Los resultados de los estudios y experiencias obtenidas en sorgo, con el manejo benéfico de la endomicorriza en las condiciones semiáridas de la región norte de Tamaulipas, han influido para que esta práctica empiece a extenderse a otras regiones del
19
centro y sur del estado. Lo que demuestra el interés y el potencial de implementar esa tecnología dentro de un esquema de producción sostenible. Al respecto, Ferrara y González (1997) indicaron que la importancia de los hongos MVA en la agricultura sostenible, está basada en la función de unir a la planta con el suelo, al servir como agente de transporte nutrimental entre las dos partes, por lo que tiene un impacto en la conservación del suelo y es el único camino para recuperar el componente biológico de la sostenibilidad. Aquí se destaca la necesidad del uso de la tecnología micorrízica para favorecer tanto la productividad agrícola como la conservación del suelo.
Cuadro 5. Análisis de rentabilidad de la producción de sorgo, con relación al rendimiento promedio de grano (kg ha-1) de las parcelas de validación, en condiciones de riego restringido y temporal (2000-2003).
Concepto Testigo G. intraradices
Riego restringido
Costo ($ ha-1) 5449 5479*
Rendimiento (ton ha-1) 3733 4554
Precio ($ ton-1)** 1400 1400
Ingreso bruto ($ ha-1) 5226 6375
Ingreso neto ($ ha-1) -223 896
Relación costo-beneficio 0.95 1.16
Temporal
Costo ($ ha-1) 3387 3417*
Rendimiento (ton ha-1) 2653 3386
Precio ($ ton-1)** 1400 1400
Ingreso bruto ($ ha-1) 3714 4740
Ingreso neto ($ ha-1) 327 1323
Relación costo-beneficio 1.09 1.38
*Costo del sustrato micorrízico $30.00 ha-1.
** Precio objetivo asignado por ACERCA-SAGARPA en el 2004.

20
Literatura citada
Abdel, F.G. and Mohamedin, A.H. 2000. Interactions between a vesicular-arbuscular mycorrhizal
fungus (Glomus intraradices) and Streptomyces and their effects on sorghum plants. Biol. & Fertility Soils 32(5): 401-409.
Acosta N., S., A. Díaz F., A. Magallanes E. y M. Silva S. 2001. La inoculación de Azospirillum brasilense y Glomus intraradices en la producción comercial de sorgo. Memorias XXVIII Congreso Nacional de Fitopatología. Resumen F-15.
Aguirre R., J., H. Williams A. y R. Rodríguez H. 1992. La producción de sorgo de riego para el norte de Tamaulipas. En: Manual de Cultivos del Norte de Tamaulipas. SARH, INIFAP, PIFSV. Pp: 45-53.
Alarcón A., y R. Ferrara C. 2000. Biofertilizantes: Importancia y utilización en la agricultura. Agr. Téc. Méx. 26(2): 191-203.
Alarcón A., y R. Ferrara C. 1999. Manejo de la micorriza arbuscular en sistemas de propagación de plantas frutícolas. Terra. 17(3): 179-191.
Alvarado C., M., A. Díaz F. y J. Morales B. 2002. Efecto de la aplicación de simbiontes en la producción de okra y maíz para elote. Memoria XXXI Congreso Nacional de la Ciencia del Suelo.
Bressan, W., Siquiera, J.O., Vasconcellos, C.A., Purcino, A.A. 2001. Mycorrhizal fungi and phosphorus on growth, yield and nutrition of intercropped grain sorghum and soybean. Pesquisa Agrop. Brasileira 36(2): 315-323.
Díaz F., A. 2003. Inoculación de sorgo con Glomus intraradices y Azospirillum brasilense en riego restringido y temporal. Memoria de Resúmenes VII Simposio Internacional y II Congreso Nacional de Agricultura Sostenible. P. 35.
Díaz F., A. y V. Pecina Q. 2004. Manejo óptimo de semilla de sorgo inoculada con micorriza durante la siembra. INIFAP- Campo Experimental Río Bravo. Publicación Especial No. 28. Pp: 20-21.
Díaz F., A., E. Tavárez L. y J. Ramírez. 2002. Validación del efecto de simbiontes en la producción comercial de elote. Biotam 13(2): (en impresión).
Díaz M., 2002. Influencia de la biofertilización en las características de planta, de grano e infección micorrízica del sorgo (Sorghum bicolor), en dos condiciones edáficas. Tesis Lic. UAMRA, UAT.
Durán A., M. 1992. Suelos. En: Hess M., L. y D. Pérez D. (eds.). Manual de Cultivos del Norte de Tamaulipas. SARH-Patronato para la Investigación, Fomento y Sanidad Vegetal. Pp: 11-16.
Durán P., A., J. Aguirre M., G. González Cu., M. Peña R., y E. Schonhoven. 2001. Producción in vivo de micorriza arbuscular Glomus intraradix con Brachiaria bryzantha como hospedero en camas reproductoras. INIFAP-Campo Experimental Cotaxtla. Folleto Técnico 29. 28 p.
Ferrara C., R. y M. C. González C. 1997. La biotecnología micorrízica en la producción agrícola, frutícola y hortícola. En: Ruiz H., J., D. Guzmán y J. Peña C (eds.). Perspectivas de la Microbiología en México. IPN. P. 325-343.
Garza C., I. 2002. Evaluación de biofertilizantes en el cultivo de sorgo, como un recurso natural sostenible. Tesis MC. UAMRA, UAT.
Garza C., I, A. Díaz F., A. Ramírez L. e I. Machuca O. 2003. Validación de Glomus intrardices y brassinoesteroides en la productividad de sorgo. Memoria de Resúmenes VII Simposio Internacional y II Congreso Nacional de Agricultura Sostenible. P. 120.

21
González Ch., M. 2002. Producción y control de calidad de inoculantes de hongos micorrízicos arbusculares. En: Pérez M., J., J. Alvarado L. y R. Ferrara C. Producción y Control de Calidad de Inoculantes Agrícolas y Forestales. Comité Mexicano de Inoculantes Agrícolas y Forestales. Pp. 36-46.
González S., D., G. Tsuzuki R. y R. Ramírez G. 2002. Efecto de la inoculación de Azospirillum y hongos micorrízico arbusculares en el crecimiento de Lycopersicon esculentum. En: Pérez M., J., J. Alvarado L. y R. Ferrara C. Producción y Control de Calidad de Inoculantes Agrícolas y Forestales. Comité Mexicano de Inoculantes Agrícolas y Forestales. Pp. 100-101.
Irizar G., M., Vargas P., Garza D., Tut C., Rojas M., Trujillo A., García R., Aguirre D., Martínez J., Alvarado S., Grageda O., Valero J. y Aguirre J. 2003. Respuesta de cultivos agrícolas a los biofertilizantes en la región central de México. Agr. Téc. Méx. 29(2): 213-225.
Johnson, C.N. and Pfleger, F.L. 1992. Vesicular-arbuscular micorrhizae and cultural stresses. In: Bethlenfalvay, G.J. and Linderman, R. G. Micorrhizae in Sustainable Agriculture. ASA Special Publication No. 54. Pp: 71-99.
Magallanes E., A. y A. Díaz F. 2002. Eficiencia de micorriza arbuscular y hormona esteroidal en la producción de sorgo. Memoria XXXI Congreso Nacional de la Ciencia del Suelo.
Montaño A., N., Quiroz G. y Cruz, F., G. 2001. Colonización micorrízica arbuscular y fertilización mineral de genotipos de maíz y trigo cultivados en un andisol. Terra 19(4): 337-344.
Montes G., N. y J. Aguirre R. 1992. La producción de sorgo de temporal en el norte de Tamaulipas. En: Manual de Cultivos del Norte de Tamaulipas. SARH-Patronato para la Investigación, Fomento y Sanidad Vegetal. Pp: 54-63.
Morton, B.J. 2002. Glomus intraradices (reference accession UT126). http://invam.caf.wvu.edu.
Salinas G., J. 2002. Labranza de conservación de suelo y agua en el norte de Tamaulipas. INIFAP-Campo Experimental Río Bravo. Publicación Especial 25. Pp: 9-10.
Smith, S.E. and Gianinazzi V. 1988. Physiological interactions between symbionts in vesicular-arbuscular mycorrhizal plants. Ann. Rev. Plant Physiol. Plant Mol. Biol. 39: 221-224.
Sylvia, D.M. 1999. Fundamentals and applications of arbuscular mycorrhizae: A “biofertilizer” perspective. In: Siqueira, J.O. and Moreira F.M. Soil Fertility, Soil Biology, and Plant Nutrition Interrelationships. Sociedade Brasileira de Ciencia do Solo. Pp: 705-723.
Sylvia, D.M., Hammond, L.C., Bennett, J.M., Hass, J.H., Linda, S.B. 1993. Field response of maize to a VAM fungus and water management. Agron. Jour. 85(2): 193-198.
Sylvia, D.M. 2004. Overview of mycorrhizal symbioses. http://ifas.ufl.edu/mycorrhiza.htm.
Sylvia, D.M., Wilson D.O., Graham, J.H., Madox, J.J., Millner, P., Morton J.B., Skipper, H.D., Wright, S.F., Jarstfer, A.G. 1993. Evaluation of vesicular-arbuscular mycorrhizal fungi in diverse plants and soils. Soil Biology & Biochemistry 25(6): 705-713

22
BIOFERTILIZACION BACTERIANA DEL PASTO BUFFEL
Loredo Osti C.1, Espinosa V. D.2, Ferrera-Cerrato R.2, Castellanos R. J.3 y Pérez J. J.4
1 C. Exp. San Luis del INIFAP ([email protected]); 2 Programa en Edafología del Colegio de Postgraduados; 3 C. Exp. Bajío-INIFAP; 4 Programa en Ganadería del Colegio de
Postgraduados
INTRODUCCIÓN
El pasto Buffel (Cenchrus ciliaris L.) es la gramínea más difundida en las resiembras de pastizales de zonas áridas y semiáridas en México (Alcalá, 1995). Este pasto representa una opción para incrementar la producción de forraje en los agostaderos y en las tierras agrícolas marginales, de acuerdo a estudios realizados por el Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP) en San Luis Potosí; su establecimiento ha sido validado con éxito en el Proyecto Estatal de Reconversión Productiva (Beltrán y Loredo, 2002).
Los principales problemas que enfrenta la siembra del pasto buffel en condiciones de temporal, son la escasa precipitación y la baja fertilidad del suelo. A este respecto, el pasto demanda altas cantidades de nitrógeno durante su ciclo y existen evidencias favorables acerca de la respuesta que presenta a la fertilización nitrogenada (González et al., 1990). No obstante, solamente se han reportado resultados exitosos con esta práctica, cuando la precipitación anual es superior a 350 milímetros, lo que limita el uso de fertilizantes químicos en zonas de escasa precipitación. Además, en esas zonas los productores enfrentan fuertes restricciones de capital y no pueden invertir en la aplicación de fertilizantes nitrogenados orgánicos o inorgánicos.
En las últimas décadas se han realizado diversas investigaciones que destacan la importancia de las bacterias de la rizosfera. Muchas de ellas estimulan el crecimiento de gramíneas como caña de azúcar, trigo, cebada, sorgo, maíz y pastos tropicales. Los mecanismos de acción de las bacterias promotoras del crecimiento vegetal (BPCV) son variados e incluyen la fijación biológica de nitrógeno atmosférico (Döbereiner et al., 1995), producción de sustancias reguladoras del crecimiento (Arshad y Frankenberger, 1998), solubilización de minerales y nutrimentos (Crowley et al., 1991), incremento en el volumen de la raíz (Bowen y Rovira, 1999), inducción de resistencia sistémica a patógenos (van Peer et al., 1991), inhibición del crecimiento de organismos antagónicos (Utkhede et al., 1999) e interacción sinérgica con otros microorganismos (Bashan et al., 1996).
La información sobre el uso de BPCV en el establecimiento del pasto buffel en zonas áridas y semiáridas es escasa. Tampoco hay estudios que indiquen cuales rizobacterias se asocian en forma natural a esta gramínea en condiciones de aridez. Sin embargo, la inoculación del pasto buffel con BPCV, podría tener un impacto significativo en su establecimiento y producción, dado el efecto favorable que se ha obtenido en otras gramíneas. Además, puede ser una opción accesible a los productores de esas zonas, debido a su bajo costo, comparado con la fertilización química. Por lo anterior, se considera importante investigar el efecto de las BPCV en el establecimiento y producción del pasto buffel en condiciones de temporal deficiente. Los objetivos de esta investigación fueron:

23
• Estimar el efecto rizosférico del pasto buffel sobre las poblaciones de bacterias, hongos y actinomicetos.
• Evaluar y caracterizar bacterias fijadoras de nitrógeno atmosférico (BFN) de la raíz del pasto buffel, creciendo en condiciones de temporal deficiente.
• Determinar el efecto de la biofertilización con bacterias promotoras del crecimiento vegetal, sobre el desarrollo del pasto buffel y sobre su nutrición nitrogenada.
MATERIALES Y MÉTODOS
Efecto rizosférico del pasto buffel
En el Campo Experimental San Luis del Instituto Nacional de Investigaciones Forestales Agrícolas y Pecuarias (INIFAP) (precipitación media anual: 240 mm, msnm: 1870, suelo franco-arenoso con densidad aparente: 1.2 g cm-3, C.E: 0.8 mmhos cm-1 y pH: 7.5), se obtuvieron veinte muestras de suelo y raíz de pasto buffel. Se estimaron las poblaciones de microorganismos en el suelo libre de raíces (SLR), suelo rizosférico (SR) y rizoplano (R), mediante la siembra en placa, de acuerdo a lo descrito por Zuberer (1994). Los medios de cultivo utilizados fueron agar nutritivo, Rennie, Papa Dextrosa Agar + Rosa de Bengala y Czapek, para bacterias totales, bacterias diazotróficas, hongos y actinomicetos respectivamente.
Los cultivos se incubaron a 28oC. A los tres días se cuantificaron las unidades formadoras colonias (ufc) de bacterias y hongos y a los 10 días las ufc de actinomicetos. Para estimar el efecto rizosférico, se obtuvo la relación R/S (Barea y Azcón-Aguilar, 1982), considerando las ufc del R sobre SLR, así como las ufc del SR sobre SLR. Se realizaron análisis de varianza bajo un diseño Completamente al Azar y pruebas de medias Tukey (α=0.05).
Evaluación y caracterización de las poblaciones de bacterias de la raíz del pasto buffel en el Altiplano Potosino
En tres sitios del Altiplano Potosino localizados en los municipios de Villa Hidalgo, Villa de Reyes y Soledad de Graciano Sánchez, S.L.P., se colectaron veinte plantas de pasto buffel con raíz completa. En el laboratorio, se separó la raíz de la parte aérea, se lavó con agua corriente y se enjuagó con agua destilada estéril. Se pesaron por duplicado 10 g de raíz. Los primeros 10 g fueron empleados para determinar materia seca (secado en horno a 65ºC, durante 48 h). En condiciones de asepsia, con los 10 g restantes, se prepararon diluciones (10-1, 10-2, 10-3, 10-4, 10-5 y 10-6), de acuerdo al procedimiento descrito por Zuberer (1994). De las diluciones 10-4, 10-5 y 10-6, se tomó una alícuota de 0.1 ml y se vertió sobre placas con medio de cultivo. Por cada dilución se prepararon tres repeticiones. Los medios de cultivo utilizados fueron: medio de carbono combinado, libre de nitrógeno o medio Rennie (Rennie, 1981) medio para Azotobacter y Derxia (Kole et al., 1988), y medio para Pseudomonas (Holguín et al., 1996). Los cultivos se incubaron a 28oC. A los tres días se estimaron las poblaciones de bacterias (ufc g-1 de materia seca de raíz). Se realizaron análisis de varianza bajo un diseño factorial 3x4 completamente al azar y pruebas de medias (Tukey; α=0.05). Con este procedimiento se obtuvieron 72 cepas.
Para describir la morfología colonial, cada cepa se resembró por triplicado en agar nutritivo, se incubó a 28oC durante siete días; desde el primer día se llevaron registros para

24
conocer la velocidad de crecimiento. Una vez que el cultivo se desarrolló, se describió considerando diámetro, color, consistencia, forma, elevación y borde (Seeley y vanDemark, 1972; Carpenter, 1977). Para caracterizar morfología microscópica, se realizó la tinción de Gram y frotis de cultivo fresco, observando las preparaciones en el microscopio (100x). Se evaluó la producción de indoles, mediante la prueba SIM (sulfuro-indol-movilidad) (UASLP, 1995).
Se estimó la capacidad para fijar N2 con la prueba de reducción de acetileno (C2H2) (Weaver y Danso, 1994). Para ello, una muestra de cada cultivo se sembró por triplicado en tubos que contenían medio semisólido libre de N. Se incubó a 28oC durante 24 h; enseguida se reemplazó el 10% del volumen de aire interno de cada tubo con C2H2 y nuevamente se incubó por 24 h. Se extrajeron 5 ml de gas, que fueron depositados en tubos vacutainer al vacío de 5 ml; de ahí se tomó una muestra de 0.5 ml de gas la cual fue inyectada a un cromatógrafo de gases, equipado con un detector de conductividad térmica y un detector de ionización de flama. Los resultados se analizaron bajo un diseño completamente al azar con tres repeticiones, utilizando el procedimiento ANOVA del Statical Analysis System (SAS, 1989); cuando se encontraron diferencias estadísticas entre tratamientos se realizaron pruebas de comparación de medias (Tukey, α≤ 0,05).
Efecto de la inoculación sobre el desarrollo y nutrición nitrogenada del pasto buffel
El experimento se realizó en un invernadero del C. E. San Luis del INIFAP con temperatura promedio de 25oC. Los factores de prueba fueron dos: la inoculación con cepas diazotróficas seleccionadas en la etapa anterior (1R2, 2CE3, 3R1 y testigo sin inocular) y dos niveles de fertilización (solución nutritiva completa y solución nutritiva sin nitrógeno). El inóculo con concentración de 1x107 células ml-1 de acuerdo a la escala de McFarland, se aplicó a la base de la planta, el día del trasplante del pasto a macetas de polietileno negro de 22x12 cm, con 1.0 kg de arena de río estéril, cribada en tamiz de 2.0 mm de abertura. La solución nutritiva se preparó con el procedimiento descrito por Sánchez et al., (2001) y se aplicó manteniendo la humedad por encima del 40% de humedad aprovechable.
Se evaluó altura de planta, número y diámetro de tallos, producción de biomasa aérea, volumen de raíz y biomasa radical, en muestreos destructivos realizados a los 15, 25, 32, 40, 68 y 82 días después del trasplante. Para el análisis estadístico se utilizó el procedimiento ANOVA del Sistema SAS, en un diseño completamente al azar, con arreglo factorial 4x2x6 (cepa x nitrógeno x fecha de evaluación ó CxNxF).
Para estimar el contenido de N, se utilizó un medidor portátil SPAD-502, cuyas lecturas tuvieron una correlación alta con la concentración de N (Loredo, 2004). La medición de las unidades SPAD se hizo en la parte media de la hoja recientemente madura, localizada dos hojas abajo de la hoja bandera, durante seis evaluaciones (a los 15, 25, 32, 40, 68 y 81 días después del trasplante). A las lecturas de unidades SPAD obtenidas de esta forma, se les aplicó un modelo obtenido previamente (Loredo, 2004). Se utilizó un diseño factorial 4x2x6: (CxNxF; C = tres cepas y un testigo, N = con o sin nitrógeno y F = seis fechas de muestreo); se tuvieron cinco repeticiones y se realizaron pruebas de medias (Tukey; α=0.05).

25
RESULTADOS Y DISCUSIÓN
Efecto rizosférico del pasto buffel
Se encontraron diferencias altamente significativas en todos los grupos microbianos para el nivel de poblaciones estimado. El rizoplano presentó el mayor número de poblaciones de microorganismos (con excepción de las poblaciones de hongos), seguido del suelo rizosférico (Cuadro 1). El de menor población de microorganismos fue el suelo libre de raíces. Estos resultados indican que la población de microorganismos si varía en función de la zona rizosférica, lo cual concuerda con lo expresado por Ferrera-Cerrato (1995), quien señala que en la rizósfera existe un flujo de compuestos orgánicos producto de la fotosíntesis, que son exudados por la raíz, lo cual hace de este sitio una zona ideal para el crecimiento de microorganismos.
Cuadro 1. Poblaciones microbianas en la rizosfera de pasto buffel. Unidades formadoras de colonias por gramo de suelo
Zona rizosférica Hongos
x102 Actinomicetos
x105 Bacterias
x105 BFN x105
R 0.00 b 6.18 a 29.14 a 113.05 a SR 28.28 a 3.85 b 16.97 a 19.23 b S 6.20 b 2.96 b 2.41 b 1.38 b
R: Rizoplano; SR: Suelo rizosférico; S: Suelo libre de raíces Valores con la misma literal entre filas no difieren estadísticamente; Tukey (α=0.05).
Las diferencias entre las poblaciones de actinomicetos no fueron tan acentuadas; no se observó diferencia en el nivel de poblaciones entre el suelo rizosférico y suelo sin raíces. El grupo de microorganismos más numeroso fue el de las bacterias fijadoras de nitrógeno, incluso mayor que el de bacterias totales. Esto puede deberse a que el medio utilizado para evaluar bacterias totales no sea el más conveniente, ya que de acuerdo a Buckman y Brady (1977), no todos los microorganismos crecen de igual forma en los medios de cultivo utilizados en el laboratorio, debido a que ciertas especies pueden vivir muy bien en el suelo y crecer precariamente en cualquiera de los medios utilizados. Cualquier modificación de la humedad, la materia orgánica o el pH del suelo, puede influir para cambiar los cálculos de 10 a 100 veces (Alexander, 1994) y uno de los factores de mayor influencia es el pH (Forster, 1995; Gregory y Hinsinger, 1999).
Evaluación de las poblaciones de bacterias de la raíz del pasto buffel en el Altiplano Potosino
Las poblaciones bacterianas en la raíz del pasto buffel variaron de 6.3 x 106 a 1.26 x 108 ufc g-1 raíz. Resultados similares fueron reportados por Malik et al., (1997) en pasto Kallar. Se encontraron diferencias estadísticas (α=0.01), debidas al efecto del sitio de muestreo, medio de cultivo y a la interacción de ambos factores (Cuadro 2).
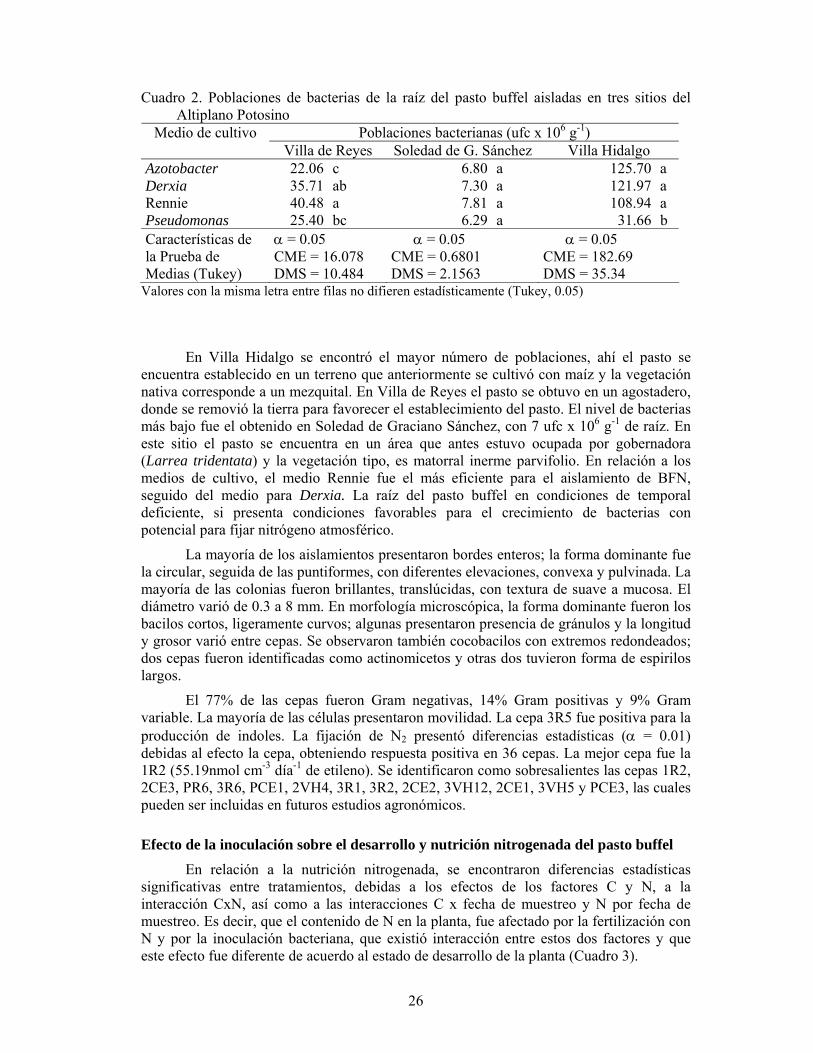
26
Cuadro 2. Poblaciones de bacterias de la raíz del pasto buffel aisladas en tres sitios del Altiplano Potosino
Poblaciones bacterianas (ufc x 106 g-1) Medio de cultivo Villa de Reyes Soledad de G. Sánchez Villa Hidalgo
Azotobacter 22.06 c 6.80 a 125.70 a Derxia 35.71 ab 7.30 a 121.97 a Rennie 40.48 a 7.81 a 108.94 a Pseudomonas 25.40 bc 6.29 a 31.66 b Características de la Prueba de Medias (Tukey)
α = 0.05 CME = 16.078 DMS = 10.484
α = 0.05 CME = 0.6801 DMS = 2.1563
α = 0.05 CME = 182.69 DMS = 35.34
Valores con la misma letra entre filas no difieren estadísticamente (Tukey, 0.05)
En Villa Hidalgo se encontró el mayor número de poblaciones, ahí el pasto se encuentra establecido en un terreno que anteriormente se cultivó con maíz y la vegetación nativa corresponde a un mezquital. En Villa de Reyes el pasto se obtuvo en un agostadero, donde se removió la tierra para favorecer el establecimiento del pasto. El nivel de bacterias más bajo fue el obtenido en Soledad de Graciano Sánchez, con 7 ufc x 106 g-1 de raíz. En este sitio el pasto se encuentra en un área que antes estuvo ocupada por gobernadora (Larrea tridentata) y la vegetación tipo, es matorral inerme parvifolio. En relación a los medios de cultivo, el medio Rennie fue el más eficiente para el aislamiento de BFN, seguido del medio para Derxia. La raíz del pasto buffel en condiciones de temporal deficiente, si presenta condiciones favorables para el crecimiento de bacterias con potencial para fijar nitrógeno atmosférico.
La mayoría de los aislamientos presentaron bordes enteros; la forma dominante fue la circular, seguida de las puntiformes, con diferentes elevaciones, convexa y pulvinada. La mayoría de las colonias fueron brillantes, translúcidas, con textura de suave a mucosa. El diámetro varió de 0.3 a 8 mm. En morfología microscópica, la forma dominante fueron los bacilos cortos, ligeramente curvos; algunas presentaron presencia de gránulos y la longitud y grosor varió entre cepas. Se observaron también cocobacilos con extremos redondeados; dos cepas fueron identificadas como actinomicetos y otras dos tuvieron forma de espirilos largos.
El 77% de las cepas fueron Gram negativas, 14% Gram positivas y 9% Gram variable. La mayoría de las células presentaron movilidad. La cepa 3R5 fue positiva para la producción de indoles. La fijación de N2 presentó diferencias estadísticas (α = 0.01) debidas al efecto la cepa, obteniendo respuesta positiva en 36 cepas. La mejor cepa fue la 1R2 (55.19nmol cm-3 día-1 de etileno). Se identificaron como sobresalientes las cepas 1R2, 2CE3, PR6, 3R6, PCE1, 2VH4, 3R1, 3R2, 2CE2, 3VH12, 2CE1, 3VH5 y PCE3, las cuales pueden ser incluidas en futuros estudios agronómicos.
Efecto de la inoculación sobre el desarrollo y nutrición nitrogenada del pasto buffel
En relación a la nutrición nitrogenada, se encontraron diferencias estadísticas significativas entre tratamientos, debidas a los efectos de los factores C y N, a la interacción CxN, así como a las interacciones C x fecha de muestreo y N por fecha de muestreo. Es decir, que el contenido de N en la planta, fue afectado por la fertilización con N y por la inoculación bacteriana, que existió interacción entre estos dos factores y que este efecto fue diferente de acuerdo al estado de desarrollo de la planta (Cuadro 3).
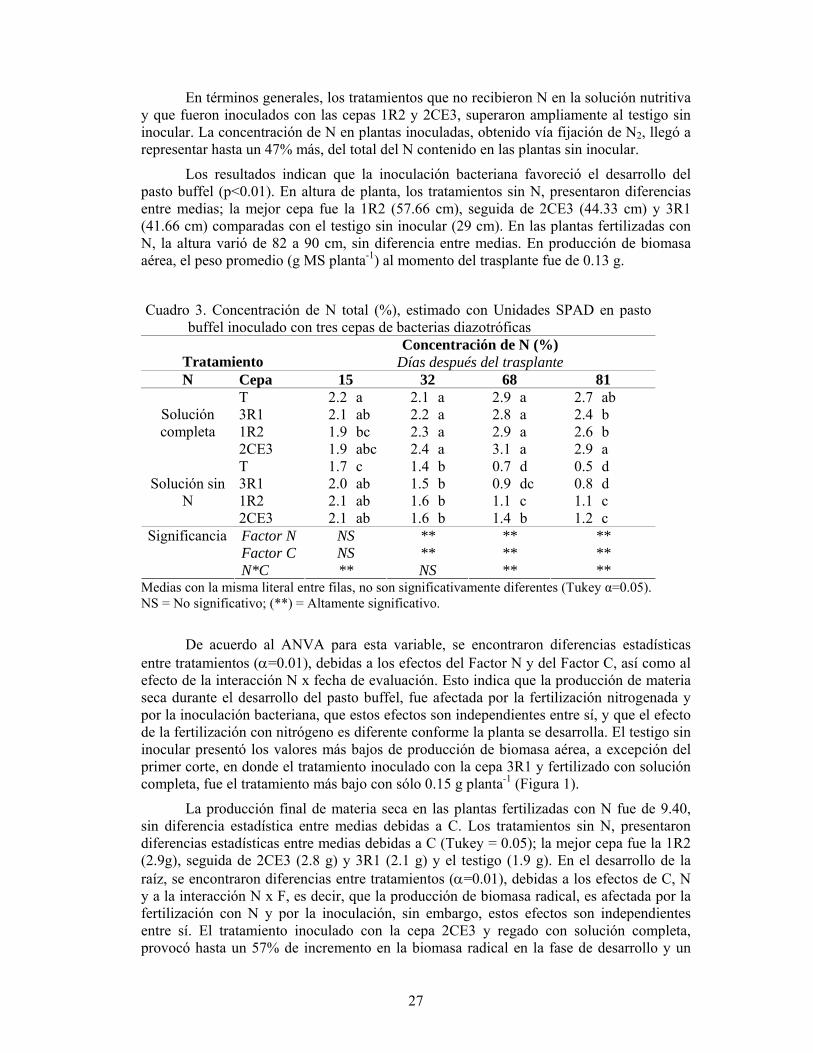
27
En términos generales, los tratamientos que no recibieron N en la solución nutritiva y que fueron inoculados con las cepas 1R2 y 2CE3, superaron ampliamente al testigo sin inocular. La concentración de N en plantas inoculadas, obtenido vía fijación de N2, llegó a representar hasta un 47% más, del total del N contenido en las plantas sin inocular.
Los resultados indican que la inoculación bacteriana favoreció el desarrollo del pasto buffel (p<0.01). En altura de planta, los tratamientos sin N, presentaron diferencias entre medias; la mejor cepa fue la 1R2 (57.66 cm), seguida de 2CE3 (44.33 cm) y 3R1 (41.66 cm) comparadas con el testigo sin inocular (29 cm). En las plantas fertilizadas con N, la altura varió de 82 a 90 cm, sin diferencia entre medias. En producción de biomasa aérea, el peso promedio (g MS planta-1) al momento del trasplante fue de 0.13 g.
Cuadro 3. Concentración de N total (%), estimado con Unidades SPAD en pasto buffel inoculado con tres cepas de bacterias diazotróficas
Concentración de N (%) Tratamiento Días después del trasplante N Cepa 15 32 68 81
T 2.2 a 2.1 a 2.9 a 2.7 ab 3R1 2.1 ab 2.2 a 2.8 a 2.4 b 1R2 1.9 bc 2.3 a 2.9 a 2.6 b
Solución completa
2CE3 1.9 abc 2.4 a 3.1 a 2.9 a T 1.7 c 1.4 b 0.7 d 0.5 d 3R1 2.0 ab 1.5 b 0.9 dc 0.8 d 1R2 2.1 ab 1.6 b 1.1 c 1.1 c
Solución sin N
2CE3 2.1 ab 1.6 b 1.4 b 1.2 c Factor N NS ** ** ** Factor C NS ** ** **
Significancia
N*C ** NS ** ** Medias con la misma literal entre filas, no son significativamente diferentes (Tukey α=0.05). NS = No significativo; (**) = Altamente significativo.
De acuerdo al ANVA para esta variable, se encontraron diferencias estadísticas entre tratamientos (α=0.01), debidas a los efectos del Factor N y del Factor C, así como al efecto de la interacción N x fecha de evaluación. Esto indica que la producción de materia seca durante el desarrollo del pasto buffel, fue afectada por la fertilización nitrogenada y por la inoculación bacteriana, que estos efectos son independientes entre sí, y que el efecto de la fertilización con nitrógeno es diferente conforme la planta se desarrolla. El testigo sin inocular presentó los valores más bajos de producción de biomasa aérea, a excepción del primer corte, en donde el tratamiento inoculado con la cepa 3R1 y fertilizado con solución completa, fue el tratamiento más bajo con sólo 0.15 g planta-1 (Figura 1).
La producción final de materia seca en las plantas fertilizadas con N fue de 9.40, sin diferencia estadística entre medias debidas a C. Los tratamientos sin N, presentaron diferencias estadísticas entre medias debidas a C (Tukey = 0.05); la mejor cepa fue la 1R2 (2.9g), seguida de 2CE3 (2.8 g) y 3R1 (2.1 g) y el testigo (1.9 g). En el desarrollo de la raíz, se encontraron diferencias entre tratamientos (α=0.01), debidas a los efectos de C, N y a la interacción N x F, es decir, que la producción de biomasa radical, es afectada por la fertilización con N y por la inoculación, sin embargo, estos efectos son independientes entre sí. El tratamiento inoculado con la cepa 2CE3 y regado con solución completa, provocó hasta un 57% de incremento en la biomasa radical en la fase de desarrollo y un
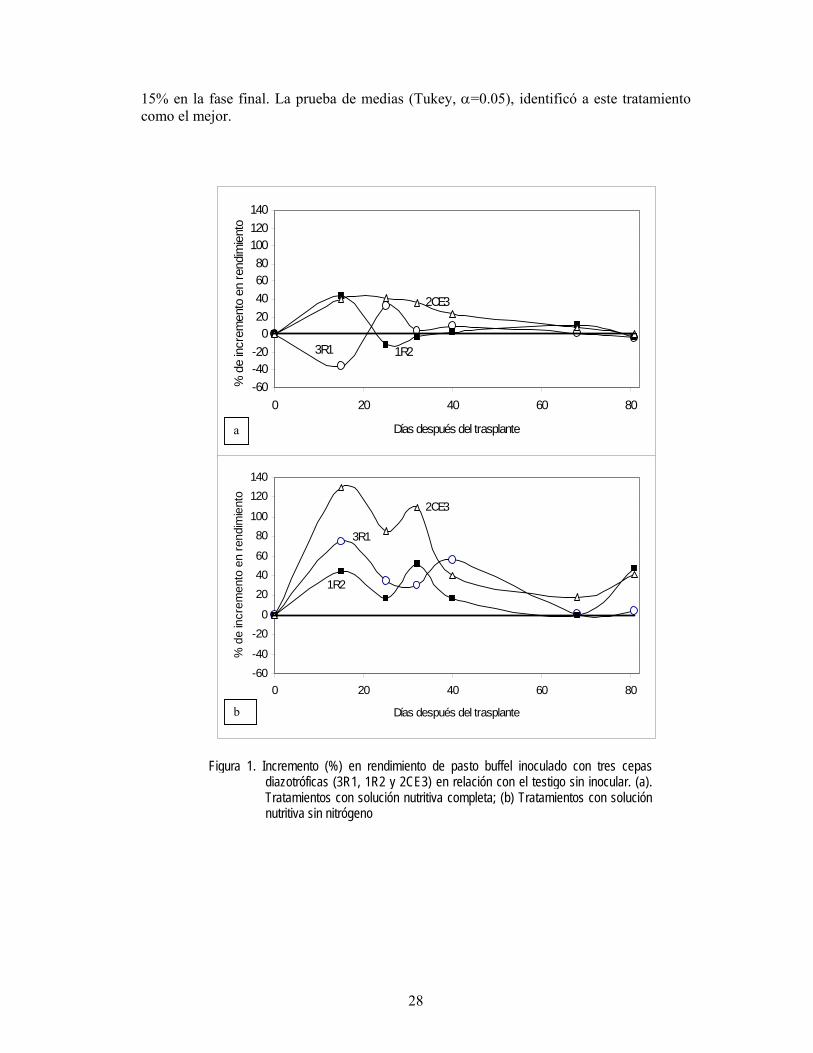
28
15% en la fase final. La prueba de medias (Tukey, α=0.05), identificó a este tratamiento como el mejor.
3R1 1R2
2CE3
-60-40-20
020406080
100120140
0 20 40 60 80
Días después del trasplante
% d
e in
crem
ento
en
rend
imie
nto
3R1
1R2
2CE3
-60-40
-200
204060
80100
120140
0 20 40 60 80
Días después del trasplante
% d
e in
crem
ento
en
rend
imie
nto
a
b
Figura 1. Incremento (%) en rendimiento de pasto buffel inoculado con tres cepas diazotróficas (3R1, 1R2 y 2CE3) en relación con el testigo sin inocular. (a). Tratamientos con solución nutritiva completa; (b) Tratamientos con solución nutritiva sin nitrógeno

29
CONCLUSIONES
• La rizosfera del pasto buffel tiene un efecto favorable para el crecimiento de los microorganismos. El efecto rizosférico es más evidente en las poblaciones de bacterias fijadoras de N2 y menos notorio en las poblaciones de actinomicetos y hongos.
• La inoculación con bacterias diazotróficas incrementa la concentración de N en la planta del pasto buffel, cuando las condiciones de N mineral son muy deficientes. Si la planta tiene cubiertos los requerimientos de N, el efecto de la inoculación no es evidente sobre la concentración de N.
• La inoculación bacteriana favorece el desarrollo aéreo del pasto buffel en condiciones de deficiencia de N, sin embargo en la raíz el crecimiento es favorecido independientemente de la condición de N en el sustrato.
LITERATURA CITADA
Alcalá, G. C. H. 1995. Origen geográfico y características biológicas del pasto buffel. pp 9-14. In: Guía para el establecimiento, manejo y utilización del zacate buffel. Patronato del Centro de Investigaciones Pecuarias del Estado de Sonora A. C. Hermosillo, Son. México.
Arshad, M. and W. T. Frankenberger Jr. 1998. Plant growth-regulating substances in the rhizosphere: Microbial production and functions. Advances in Agronomy 62: 45-151.
Barea J. M. y Azcón-Aguilar C. 1982. La Rizosfera: Interacciones microbio-planta. Anales de Edafología y Agrobiología. XI.I (7-8): 1517-1532.
Bashan, Y., Holguín G. y Ferrera-Cerrato R. 1996. Interacciones entre plantas y microorganismos benéficos. II. Bacterias asociativas de la rizosfera. Terra 14 (2) 195-210.
Beltrán, L. S. y Loredo, O. C. 2002. Reconversión de áreas agrícolas marginales a praderas de pasto buffel. Pub. No. 36. SAGAR-INIFAP-Campo Experimental Palma de la Cruz. 16 p.
Bowen G. D. y Rovira A. D. 1999. The Rhizosphere and Its Management to Improve Plant Growth. Advances in Agronomy V 66: 1-102.
Carpenter, P. L. 1977. Microbiology. W.B. Sounders Company. Philadelphia. E.U.A.
Crowley, D.E., Wang Y. C., Reid C.P.P. and Szaniszlo P.J. 1991. Mechanisms of iron acquisition from siderophores by microorganisms and plants. Plant and Soil 130: 179-198.
Döbereiner, J., Urquiaga S., Boddey R.M. and Ahmad N. 1995. Alternatives for nitrogen of crops in tropical agriculture. Nitrogen economy in tropical soil. Fertilizer Research 42:339-346.
Ferrera-Cerrato R. 1995. Efecto de Rizosfera. En: Ferrera-Cerrato R. y Pérez M.,J. Editores. Agromicrobiología, elemento útil en la agricultura sustentable. Colegio de Postgraduados en Ciencias Agrícolas, Montecillo, Estado de México. p 36-53.
González, A., Eguiarte, S. A. y Rodríguez R. P. 1990. Respuesta productiva durante cinco años de fertilización al zacate buffel. En: Resúmenes de las Memorias del VI Congreso Nacional sobre Manejo de Pastizales. Monterrey, N.L. p 20.
Holguín, G., Bashan Y., y Ferrera-Cerrato R. 1996. Interacciones entre plantas y microorganismos benéficos. III. Procedimientos para el aislamiento y caracterización de hongos micorrízicos y rizobacterias promotoras del crecimiento en plantas. Terra 14 (2) 211-224.
Kole, M. M., Page, W. J. and Altosaar. 1988. Distribution of Azotobacter in Eastern Canadian soils and in association with plant rhizospheres. Canadian Journal of Microbiology 34:815-817

30
Loredo O. C. 2004. Biofertilización bacteriana del pasto buffel en el Altiplano Potosino. Tesis de Doctorado. Colegio de Postgraduados.
Malik, K. A., R, Bilal, S. Mehnaz, G. Rasul, M.S. Mirza and S. Ali. 1997. Association of nitrogen-fixing, plant-growth-promoting rhizobacteria (PGPR) with kallar grass and rice. Plant and Soil 194:37-44.
Rennie, R.J. 1981. A single medium for isolation of acetylene reduction (dinitrogenfixing) bacteria from soil. Can. J. Microbiol. 27:8-14.Rennie, 1981
Sánchez, G.P., L. I. Trejo T., y L. Fernández P. 2001. Manual de prácticas de nutrición vegetal. EDA-651. Colegio de Postgraduados. Especialidad de Edafología. Área de nutrición vegetal. 32 p.
SAS, 1989. SAS/STAT Users Guide. Release 6.12 ed. Statistical Analysis System Institute, Inc., Cary, N.C.
Seeley, H. W. and P.J. VanDemark. 1972. Microbes in Action. A laboratory manual of microbiology. Ed. Freeman and Company. p 11-20
Utkhede, R. S., Koch C.A. and Menzies J.G. 1999. Rhizobacterial growth and yield promotion of cucumber plants inoculated with Pythium aphanidermatum. Can. J. Plant Pathol. 21: 265-271.
Van Peer, R., Niemann, G. J. and Schippers, B. 1991. Induced resistance and phytoalexin accumulation in biological control of Fusarium wilt of carnation by Pseudomonas sp. strain WCS417r. Phytopathology 81:728-734.
Weaver, R. W. and S. K.A. Danso. 1994. Dinitrogen fixation. In: Methods of Soil Analysis. Part 2. Microbiological and Biochemical Properties. Soil Science Society of America, Inc. p 1019-1043.
Zuberer D. A. 1994. Recovery and Enumeration of Viable Bacteria. In: Methods of Soil Analysis. Part 2. Microbiological and Biochemical Properties. P 119-144

31
BIOFERTILIZANTES: MICORRIZAS Y BACTERIAS PROMOTORAS DE CRECIMIENTO
Víctor Olalde Portugal y Rosalinda Serratos
CINVESTAV, Unidad Irapuato. Irapuato, México.
La producción agrícola requiere de una gran cantidad de agroquímicos
generalmente derivados del petroleo. En muchas de las ocasiones son contaminantes del ambiente y/o pueden generar daños a la salud de los animales incluido el hombre. Ejemplo de ello, es la fertilización con nitrógeno, el cual, no es aprovechado por la planta, algunos autores indican que en el mejor de los casos se aprovecha el 44%. El resto se puede lixiviar, volatilizar o realizar transformaciones que pueden dar origen a compuestos contaminantes, como las nitrosaminas. Estas nitrosaminas pueden originar cancer. Por otra parte, para el control de enfermedades se requiere de la aplicación de pesticidas, dentro de estos los funguicidas son de los más utilizados. Muchos de ellos, generan alteraciones muy graves al hombre.
Existen alternativas para reducir los riesgos sin menos cabo de la productividad,
éstas en parte pueden ser utilizar microorganismos. Considerando que los microorganismos son los grandes actores del suelo y que pueden sustentar el desarrollo saludable de las plantas.
Los microorganismos realizan diferentes reacciones químicas, mineralización,
inmovilización, oxidaciones, reducciones, fijaciones de algunos gases como nitrógeno, volatilización, solubilización etc. Estas reacciones finalmente se reflejan en las siguientes acciones; formación de humus, degradación de materia orgánica, control microbiano, fijar nitrógeno, etc.
En esta ocasión me referiré a las micorrizas y al control biológico.
De entre las diversas asociaciones mutualistas planta-microorganismo, la micorrícica es, con diferencia, la que se encuentra más ampliamente extendida sobre la superficie terrestre (alrededor del 90% de las plantas terrestres descritas la forman, Smith y Read, 1997) y sin duda la más antigua (400 millones de años, Simon et al. 1993, Taylor et al., 1995). Existen diferentes tipos de micorrizas, sin embargo en esta ocasión describiremos a la arbuscular debido a su importancia en la agricultura. Las micorrizas arbusculares (MA), ocurren de manera natural en la gran mayoría de las angiospermas, entre las que se incluyen plantas de alto interés agrícola, hortícola y de zonas templadas y tropicales (Smith y Read, 1997). En contraste con la enorme diversidad de las plantas huésped potenciales, los hongos formadores de MA (HMF) se reducen a pocas familias y no mas de 130 especies (Walker, 1992). El ciclo de vida de los HMA se inicia con la germinación en el suelo de sus propágulos o esporas, que desarrollan las conocidas como hifas o tubos de germinación. Estas hifas germinativas crecen al azar en busca de una raíz susceptible de ser colonizada.
Las plantas durante su desarrollo están expuestas a diferentes factores que influyen sobre su fisiología. La planta ante estos factores realiza una serie de ajustes para tratar de mantener su salud y funcionalidad. Entre ellas la apertura de los estomas es de las mas

32
activas y rápidas de las respuestas. Estas respuestas dependen del tipo de planta, de las condiciones en que se desarrollan etc.
Por otra parte, se sabe que la mayoría de las plantas se asocian con algunos hongos benéficos. Estos hongos se ha mencionado le ayudaron a las plantas a invadir la tierra, lo que les permitió adaptarse y sobrevivir a la adversidad de las condiciones de esa época. Se ha demostrado que las plantas que se encuentran en condiciones de estrés se logran desarrollar mejor cuando tienen micorriza.
Las plantas que se encuentran creciendo en suelos deficientes en fósforo soluble,
su gran mayoría lo hace debido a su capacidad de asociarse a estos hongos.
Se ha demostrado que plantas de jitomate y chile creciendo en 22 ppm de fósforo pero que presentan micorriza son del mismo vigor y contenido de fósforo que las plantas que crecen en 44 ppm de fósforo y que no tienen micorriza (Salas-Galván, 2000.). Sin embargo, la fisiología de las plantas de jitomate parece ser diferente, ya que las plantas con micorriza presentaron más altas tasas fotosintéticas. También ajustan el metabolismo fotosintético de tal suerte que las plantas con micorriza toleran mas los incrementos de temperatura, y por lo tanto reducen la fotorespiración ( Luna y col, 2003). Como se mencionó anteriormente, las plantas con micorriza pueden alterar la apertura de los estomas. Por lo que, otro beneficio de las plantas con micorriza es su capacidad para mejorar sus relaciones hídricas, por lo que hemos podido reducir un riego en trigo reduciendo adicionalmente el fósforo.
Esta mejora, se debe a que las plantas con micorriza, ajustan la apertura de los estomas de tal suerte que la perdida de agua se reduce, también se incrementa que la concentración de fitohormonas y los osmoreguladores. El resultado se refleja en el uso eficiente del agua, esto es un incremento de fotosíntesis con un menor gasto de agua.
Adicional a esto, también los hongos micorrícicos, mejoran la calidad de las plantas producidas in vitro, al momento del proceso de aclimatación. En este sentido, el mejoramiento es debido a dos efectos al menos, incremento de la absorción de nutrimentos y a la mas rápida actividad de los estomas. Cabe mencionar, que las plantas se desarrollan con mayor velocidad. Todos estos efectos no resultarían tan importantes, si esto no se reflejará el rendimiento y/o la calidad de los productos. Ejemplo de esto, es el jitomate cuyos frutos que provienen de plantas con micorriza tiene mas vida de anaquel, y mejor color.
En cuanto a vida de anaquel estos frutos sufren menos contaminación y por lo tanto deterioro y reducen menos su peso por pérdida de agua. Cuando se evaluó la expresión de genes de maduración se observa que los frutos provenientes de plantas con micorrizas, la expresión de la poligalacturonasa (enzima involucrada en la degradación de la pared celular) y de la ACC oxidasa (enzima involucrada en la producción de etileno) se reducen. Esta reducción en la expresión de estos genes se refleja en la mejor calidad de los frutos. Efectos parecidos se observaron en chile. Aunque estos ensayos se realizaron en condiciones adecuadas de humedad se realizaron otros ensayos en condiciones de estrés hídrico. Resultando que los frutos de chile pueden mantener una buena calidad, en plantas con algún tipo de hongo micorrícico. Tec-Myc puede mantener esta calidad, mientras que G.fasciculatum no la mantiene.

33
Efectos adicionales y no por ello menos importantes es la resistencia a enfermedades que se genera por la colonización de las raíces por parte de estos hongos. Hemos reducido la incidencia de marchitez en chile en un 60%, pudrición blanca en ajo y cebolla en un 55%. Se tienen diferentes explicaciones a esta protección, un incremento de las defensas de la planta, como enzimas hidrolíticas, un incremento de las fitoalexinas, competencia de por los sitios de infección, aumento de la raíz y por lo tanto se tiene menor efecto de los patógenos sobre la planta (Rojas Andrade, y col 2003),
Otro grupo microbiano, que incrementa el desarrollo y productividad de las plantas es el de las bacterias promotoras del crecimiento. Este grupo de bacterias puede estimular a las plantas de dos maneras; directa a través de la producción de metabolitos como la producción de fitohormonas, fijación de nitrógeno u otro, o uno indirecto en el que la bacteria controla patógenos de la planta. Este último puede ser debido a la producción de compuestos antifúngicos, sideróforos, enzimas líticas, incrementa los mecanismos de defensa de la planta, etc (Glick y col. 1999).
En cuanto a la producción de fitohormonas las más estudiadas son las auxinas., las cuales estimulan el desarrollo de las raíces de las plantas. Otro mecanismos para estimular el desarrollo de la raíz, es a través de la disminución del ACC que es precursor del etileno, hormona que detiene el crecimiento de la raíz. Algunas bacterias promotoras del crecimiento pueden utilizar este substrato (ACC) como fuente de carbono y nitrógeno, por lo que se disminuye la producción de etileno y por lo tanto la raíz sigue creciendo. Otro mecanismo, que puede incrementar la estimulación de la raíz es a través de pequeños péptidos que regulan en la planta la producción de fitohormonas. En cuanto, a fijadores de nitrógeno existen algunas bacterias como Azospirillum que pueden fijar todo o casi todo el nitrógeno requerido por las plantas. Este proceso es debido a que bacterias tienen el equipo enzimático necesario para tomar el N atmosférico y lo transforman en nitrógeno asimilable por la planta.
Por otra parte, pueden combatir enfermedades producidas por Fusarium,
Rhizoctonia, Verticillium, Sclerotium etc. Esto se puede deber como se mencionó en el párrafo anterior, estas bacterias pueden producir enzimas como las quitinasas o glucanasas que degradan la pared celular de estos hongos. Otro manera de impedir el ataque de los patógenos es a través de la producción de sideróforos, compuestos que quelan el Fe, elemento necesario para que los patógenos ataquen las plantas. Así mismo, algunas bacterias como Bacillus subtilis producen algunos péptidos con actividad antifúngica. Finalmente, algunas otras bacterias pueden producir elicitores que estimulan los mecanismos de defensa de la planta, lo que se conoce como resistencia sistémica inducida. Estos efectos se pueden reflejar en incrementos en los rendimientos y acortar los tiempos de producción. En el caso de papa, cuando se inocula un litro por hectárea se aumentan los rendimientos entre 8-10 ton por ha. Un 30 % en la producción de jitomate y chile. Adicionalmente, hemos observado que se incrementa la calidad de los frutos.
Finalmente, cuando se inoculan los dos tipos de microorganismos (hongos micorrícicos y bacterias promotoras de crecimiento) y los resultados son más espectaculares. En chile se puede incrementar 40%, en un tiempo de cosecha menor. En pimiento morrón se incremento la calidad de estos en un 30% mas de los de calificación de

34
primera. Es reconocido que las bacterias promotoras de crecimiento se desarrollan de manera significativa en la denominada micorrizósfera.
Cabe de mencionar que la productividad de manera sustentable puede ser una realidad con el tiempo, si dedicamos más esfuerzos al estudio de la flora microbiana y otros productos de origen natural.
LITERATURA CITADA
1. Glick B.R., Patten C.L, Holguin G. Penrose D.m: 1999. Biochemical and Genetic Mechanisms
Used by Plant Growth Promoting Bacteria. Imperial College Press. 2. Rojas-Andrade, C.M. Cerda-García-Rojas, J.T. Frías-Hernández, L. Dendooven, V. Olalde-
Portugal and A. C. Ramos-Valdivia. 2003. Arbuscular mycorrhizal fungus induced changes in the concentration of trigonelline in a semi-arid leguminous plant (Prosopis laevigata) during the pre-symbiotic phase. Mycorrhiza:13:49-52.
3. Salas Galván Ma.E. 2000. Crecimiento, intercambio de gases y carbohidratos en plantas de
tomate con micorriza y diferentes niveles de fósforo. Tesis doctorado, CINVESTAV. 4. Simon L, Bousquet J, Lévesque R, Lalonde M. 1993. Origin and diversification of
endomycorrhizal fungi and coincidence with vascular land plants. Nature 363: 67-69 5. Smith SE, Read DJ. 1997. Mycorrhizal symbiosis. Academic Press, San Diego. 6. Taylor TN, Remy W, Hass H, Kerp H .1995. Fossil arbuscular mycorrhizae from the early
devonian. Mycologia 87: 560-573. 7. Vargas Luna, L. Madueño G. Gutierrez, JJ. Bernal Alvarado, M. Sosa, JL: Gonzalez, Solis., S.
Sánchez Rocha. S., Olalde Portugal V., JJ Alvardo Gil. P.Campos. 2003. Photorespiration and temperature dependence of oxigen evolution in tomato plants monitored by open photoacoustic technique. Review Scientific Instrument. 74:707-708.
8. Walker C. 1992. Systematics and taxonomy of the arbuscular endomycorrhizal fungi
(Glomales) a possible way forward. Agronomie 12:.

35
EFECTOS DE LABRANZA Y BIOFERTILIZACION EN PROPIEDADES DEL SUELO QUE AFECTAN LA SOSTENIBILIDAD
DE PRODUCCIÓN DE FRIJOL
Jaime Roel Salinas García, Arturo Díaz Franco, Idalia Garza Cano
Campo Experimental Río Bravo, INIFAP. Río Bravo, México.
INTRODUCCIÓN
El frijol (Phaseolus vulgaris L.) es una leguminosa que ha estado presente en la dieta de muchos pueblos del mundo y forma, junto con otros productos, la base de su alimentación desde siglos atrás. El consumo de frijol en el mundo no se ha generalizado como sucede con otros granos como el trigo o el maíz, sino más bien se trata de un grano cuyo consumo se encuentra localizado en algunos países de América y África (ASERCA, 1997). El frijol es una de las fuentes de alimentación mas importante por su gran valor alimenticio; el grano de frijol contiene un 20-25% de proteínas, 1.6% de lípidos, cerca de 60% de hidratos de carbono y además, minerales como calcio, fósforo, hierro, entre otros (SEP, 1991). En México, el frijol ocupa el segundo lugar en superficie de siembra, después del maíz (Zea mays L.) (Williams et al., 1995). El frijol adquiere gran importancia en el norte de Tamaulipas, según las demandas y precio en el mercado, por lo que se considera una alternativa de producción dentro de una rotación de cultivos (gramínea-leguminosa) ya que reporta utilidades económicas y aporta nitrógeno al suelo (Díaz y Reyes, 1993).
En años recientes se ha incrementado el interés por la sostenibilidad de la agricultura de la región norte de Tamaulipas. Los sistemas de producción de cultivos con manejo sostenible mantienen el suelo en una buena condición física, biológica y química. De acuerdo con el censo agropecuario de 1991, se resume que en México, las estrategias de manejo de los cultivos varían dependiendo de las condiciones ambientales y socioeconómicas, ya que esto influye para la selección y rotación de cultivos, métodos de labranza y uso de fertilizantes (INEGI, 1991).
El uso de sistemas de labranza de conservación en México se inició en la década de los setentas con el objetivo de promover la sostenibilidad de los agro ecosistemas agrícolas. En México se siembran aproximadamente 650 mil hectáreas con los sistemas de labranza de conservación, lo que representa el 3.25% de la superficie cosechada (Claveran-Alonso et al., 2001). Labranza de conservación se refiere a las diferentes prácticas de labranza que utilizan y manejan los residuos de cosecha para cubrir la superficie del suelo, alterando al mínimo posible la composición, la estructura y la biodiversidad natural del suelo y lo protege de la degradación por erosión.
El empleo tradicional de fertilizantes químicos, trae consecuencias directas en la contaminación del suelo, subsuelo, agua y atmósfera (Peña y Grageda, 1997), efectos que no se le ha dado la importancia que se merece. La fertilización biológica o “biofertilización” constituye una alternativa viable (Alarcón y Ferrera, 2000) que puede complementar o sustituir la fertilización química. Dentro de este concepto, las micorrizas vesículo-arbusculares son hongos que han tenido un impacto como biofertilizantes en la agricultura (Sylvia, 1999; Alarcón y Ferrera, 2000). El objetivo de esta investigación fue

36
evaluar el efecto de los sistemas de labranza y la biofertilización como prácticas de sostenibilidad en la producción de fríjol.
MATERIALES Y MÉTODOS
Descripción del sitio
Esta investigación se condujo en el Campo Experimental Río Bravo, INIFAP, en el Municipio de Río Bravo, Tamaulipas (25o 57´ N 98o 01´ O). El suelo es arcilloso (28% arena, 41% arcilla y 31% limo), con un contenido de materia orgánica de 1.2%, nitrógeno inorgánico (NO3) 25 p.p.m, fósforo disponible 12.3 p.p.m., potasio extractable 470 p.p.m. y un pH moderadamente alcalino 7.8 (1:2 suelo/agua). El clima es cálido subtropical, con veranos húmedos y calurosos e inviernos secos, la temperatura media anual es de 23 oC, y la precipitación media anual es de 635 mm.
Diseño experimental
En este sitio y durante lo últimos tres años se han evaluado cuatro tratamientos de labranza y en el presente año, dos tratamientos de fertilización, en un diseño experimental de bloques al azar con arreglo de parcelas divididas. Los tratamientos de labranza constituyeron la parcela grande mientras que los tratamientos de fertilización constituyeron la parcela chica. Tratamientos de labranza: labranza tradicional (barbecho), labranza mínima (subsuelo-bordeo y destronque-bordeo) y labranza cero (herbicida-siembra). La preparación del suelo en los tratamientos de labranza se realizó durante el mes de julio y agosto antes de que se iniciara el periodo de lluvias. En el tratamiento de labranza cero la maleza se controló con dos aplicaciones de glifosato (600 g/ha i.a.) y 2-4 D Amina (600 g/ha i.a.) durante el periodo de descanso (julio-enero). Tratamientos de fertilización: fertilización química comprendió la aplicación de la formula 40-20-00 en presiembra (Pérez y Cortinas, 1994) y como fertilización biológica, se inoculó la semilla con el hongo micorrízico Glumus intraradices a razón de 1 kg del sustrato en 60 kg de semilla (ha), mezclada con 60 ml de adherente y 700 ml de agua. La siembra se realizó con humedad residual el 17 de febrero de 2003 con la variedad de fríjol Pinto Anzalduas 91, a una densidad de siembra de 20 plantas por metro.
Muestreo de suelo
Se colectó una muestra de suelo por unidad experimental en la etapa de floración del fríjol (abril 2003). Las muestras se tomaron en la parte central del surco utilizando una pala angosta y se dividieron en segmentos de 0 a 5, 5 a 10 y 10 a 15 cm de profundidad, se colocaron en bolsas de plástico y se trasladaron al laboratorio para su preparación. Las muestras de suelo se secaron al aire y se molieron para ser tamizadas a través de una malla de 2 mm para su análisis químico.
Análisis fisicoquímico del suelo
El contenido de materia orgánica se determinó por el método de oxidación húmeda con dicromato de potasio y el exceso de dicromato añadido es valorado con sulfato ferroso amoniacal (Nelson y Sommers, 1982). El contenido de nitrógeno inorgánico (NO3) se determinó colorimetricamente con el método fenol disulfonico, utilizando una solución extractora de sulfato de plata y sulfato de cobre para extraer los nitratos, esta solución se evapora a sequedad, posteriormente se disuelve y se trata con hidróxido de amonio, formando una solución amarilla usada para la determinación colorimétrica de los nitratos. El contenido de fósforo disponible se determinó con el método Olsen (Olsen el al., 1954).

37
El potasio extractable (con acetato de amonio) se determino por el método Peech (Pratt, 1965). La textura del suelo se determinó con el método del hidrómetro (Gee y Bauder, 1986).
RESULTADOS Y DISCUSIÓN Materia orgánica
Los tratamientos de labranza afectaron significativamente el contenido de materia orgánica (M.O.) en las tres profundidades de suelo (Cuadro 1). Los cambios se observaron principalmente en la primera profundidad de 0-5 cm. Donde el tratamiento de labranza cero fue estadísticamente superior al resto de los tratamientos, seguido por los tratamientos de labranza mínima (destronque-bordeo y subsuelo-bordeo) y el tratamiento de labranza barbecho que mostró los valores más bajos. En las siguientes profundidades 5-10 y 10-15 cm el tratamiento de labranza-cero superó en 22% en el contenido de M.O. al resto de los tratamientos. Debido a que no hubo inmersión de suelo que pudiera incorporar los residuos y con ello provocar una descomposición más acelerada. De acuerdo con Doran (1980), Maskina et al. (1993) y Salinas et al. (1997), los residuos de cosecha incorporados al suelo ayudan a mantener e incrementar la M.O., la cual es importante para la estructura del suelo, conservación de humedad y la actividad microbiana; sin embargo el microambiente formado por la cobertura del suelo con residuos de cosecha es menos oxidativo (tratamiento de labranza cero) que donde los residuos son incorporados al suelo (labranza mínima y labranza convencional).
Los tratamientos de fertilizantes no afectaron significativamente el contenido de M.O. en las tres profundidades de suelo (Cuadro 2). Probablemente debido a que las plantas tratadas con los fertilizantes produjeron similar cantidad de biomasa o área foliar que las plantas fertilizadas. Nitratos
Los tratamientos de labranza afectaron significativamente el contenido de nitratos en las tres profundidades de suelo (Cuadro 1). El tratamiento de labranza barbecho superó a los demás tratamientos por 21.92 %, mientras que el tratamiento de labranza cero consistentemente mostró los valores mas bajos. La baja cantidad de nitratos en el tratamiento de labranza cero, indicó que el proceso de incorporación de los residuos de cosecha acelera la mineralización de la M.O. propiciando que los nitratos puedan perderse por precolación o ser tomados, por la maleza, provocando una disminución de estos, comparado con los otros tratamientos de labranza. Los niveles de nitratos son incrementados al sumarse tanto los nitratos mineralizados como el aplicado en forma de fertilizante. A pesar de que la tasa constante de mineralización orgánica es mayor en los suelos barbechados a mediano plazo, la acumulación gradual de una mayor fuente de nitratos en los sistemas de labranza de conservación puede compensar este efecto (Rice et al., 1986; Doran, 1987; Franzluebbers et al., 1995).
Los tratamientos de fertilizantes no afectaron significativamente el contenido de nitratos en las tres profundidades de suelo (Cuadro 2). Aunque el tratamiento de fertilizante químico fue ligeramente superior al fertilizante biológico, esto es probable por la cantidad de nitratos aplicados en el fertilizante.
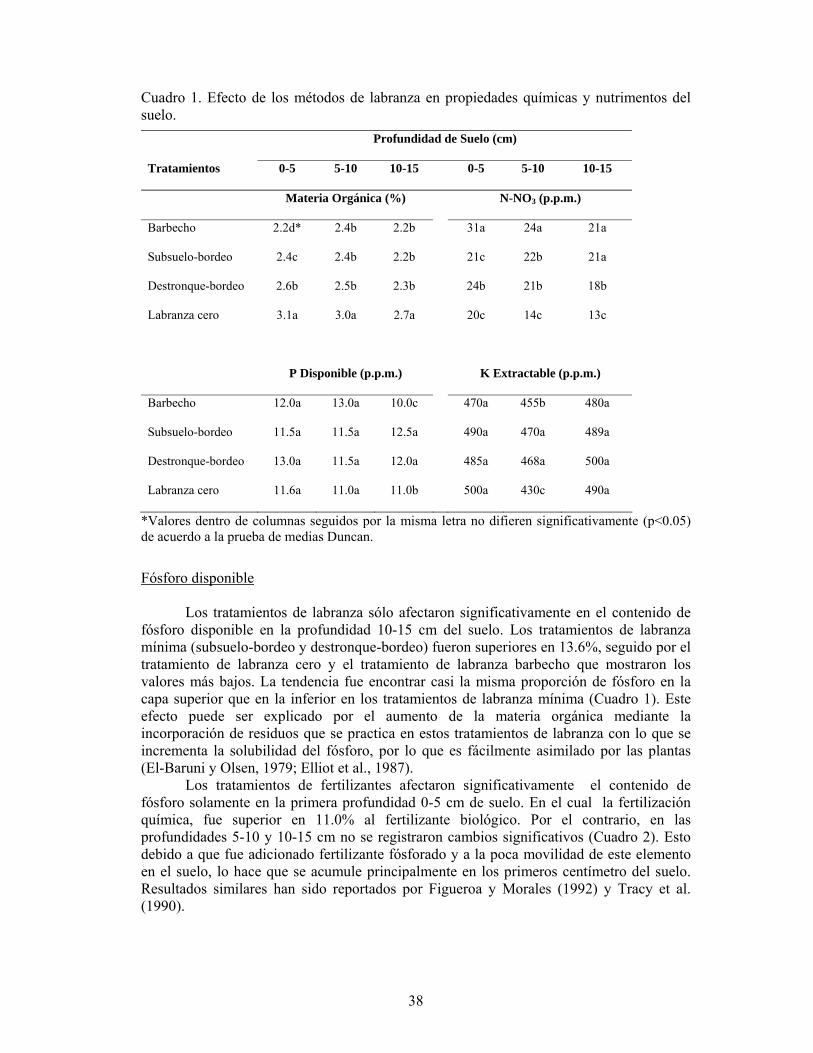
38
Cuadro 1. Efecto de los métodos de labranza en propiedades químicas y nutrimentos del suelo.
Profundidad de Suelo (cm)
Tratamientos 0-5 5-10 10-15 0-5 5-10 10-15
Materia Orgánica (%) N-NO3 (p.p.m.)
Barbecho 2.2d* 2.4b 2.2b 31a 24a 21a
Subsuelo-bordeo 2.4c 2.4b 2.2b 21c 22b 21a
Destronque-bordeo 2.6b 2.5b 2.3b 24b 21b 18b
Labranza cero 3.1a 3.0a 2.7a 20c 14c 13c
P Disponible (p.p.m.) K Extractable (p.p.m.)
Barbecho 12.0a 13.0a 10.0c 470a 455b 480a
Subsuelo-bordeo 11.5a 11.5a 12.5a 490a 470a 489a
Destronque-bordeo 13.0a 11.5a 12.0a 485a 468a 500a
Labranza cero 11.6a 11.0a 11.0b 500a 430c 490a
*Valores dentro de columnas seguidos por la misma letra no difieren significativamente (p<0.05) de acuerdo a la prueba de medias Duncan.
Fósforo disponible
Los tratamientos de labranza sólo afectaron significativamente en el contenido de fósforo disponible en la profundidad 10-15 cm del suelo. Los tratamientos de labranza mínima (subsuelo-bordeo y destronque-bordeo) fueron superiores en 13.6%, seguido por el tratamiento de labranza cero y el tratamiento de labranza barbecho que mostraron los valores más bajos. La tendencia fue encontrar casi la misma proporción de fósforo en la capa superior que en la inferior en los tratamientos de labranza mínima (Cuadro 1). Este efecto puede ser explicado por el aumento de la materia orgánica mediante la incorporación de residuos que se practica en estos tratamientos de labranza con lo que se incrementa la solubilidad del fósforo, por lo que es fácilmente asimilado por las plantas (El-Baruni y Olsen, 1979; Elliot et al., 1987).
Los tratamientos de fertilizantes afectaron significativamente el contenido de fósforo solamente en la primera profundidad 0-5 cm de suelo. En el cual la fertilización química, fue superior en 11.0% al fertilizante biológico. Por el contrario, en las profundidades 5-10 y 10-15 cm no se registraron cambios significativos (Cuadro 2). Esto debido a que fue adicionado fertilizante fósforado y a la poca movilidad de este elemento en el suelo, lo hace que se acumule principalmente en los primeros centímetro del suelo. Resultados similares han sido reportados por Figueroa y Morales (1992) y Tracy et al. (1990).

39
Cuadro 2. Efecto del tipo de fertilización en propiedades químicas y nutrimentos del suelo. Profundidad de Suelo (cm)
Tratamientos 0-5 5-10 10-15 0-5 5-10 10-15
Materia Orgánica (%) N-NO3 (p.p.m.)
Fertilización química 2.7a* 2.5a 2.3a 25a 23a 20a
Fertilización biológica 2.6a 2.4a 2.2a 23a 21a 18a
P Disponible (p.p.m.) K Extractable (p.p.m.)
Fertilización química 12.5a 12.0a 12.0a 495a 475a 480a
Fertilización biológica 11.0b 11.0a 11.0a 490a 460a 475a
*Valores dentro de columnas seguidos por la misma letra no difieren significativamente (p<0.05) de acuerdo a la prueba de medias Duncan.
Potasio extractable
Los tratamientos de labranza solo afectaron significativamente en el contenido de
potasio en la profundidad 5-10 cm del suelo. En el que fueron superiores los tratamientos de labranza mínima (destronque-bordeo y subsuelo-bordeo) por 4.8%, seguido por el tratamiento de labranza barbecho y el tratamiento de labranza cero que mostró los valores mas bajos. En la primera y última profundidad no registraron diferencia significativa entre los tratamientos de labranza (Cuadro 1). Resultados similares han sido reportados por Muzilli (1983), quien mencionó que la disponibilidad del potasio es menos afectada por los sistemas de labranza.
Los tratamientos de fertilizantes no afectaron significativamente el contenido de potasio en las tres profundidades de suelo (Cuadro 2). Esto debido probablemente a que las micorrízas exudan sustancias químicas al suelo, lo que ayuda a disolver algunos nutrientes como el fósforo y potasio (Robson et al. 1994).
CONCLUSIONES
Los residuos de cosecha sobre la superficie del suelo, promovidos por los tratamientos de labranza de conservación, mejoraron los indicadores de la sostenibilidad del suelo, respecto al tratamiento de labranza tradicional de barbecho. Los tratamientos de labranza de cero y mínima (subsuelo-bardeo y destronque-bordeo) incrementaron la concentración de materia orgánica, nitratatos, fósforo y potasio, en la capa superficial del suelo (0-5 min). Los tratamientos de fertilización no afectaron estadísticamente, los indicadores de sostenibilidad del suelo (materia orgánica, nitratos, fósforo y potasio). Los tratamientos de labranza mínima subsuelo-bordeo y destronque-bordeo y el uso de fertilizantes biológicos promueven la sostenibilidad del suelo.

40
REFERENCIAS Alarcón, A. y Ferrara-Cerrato, R. 2000. Biofertilizantes: Importancia y utilización en la agricultura.
Agr. Téc. Méx. 26 (2): 191-203.
ASERCA. 1997. Producción mundial de frijol. Claridades Agropecuarias 44: 24-32.
Claveran-Alonso, R., Fregoso-Tirado, L.E. and Sánchez-Brito, C., 2001. La labranza conservacionista en México. I World Congress on Conservation Agriculture. Madrid, Spain.
Díaz, F., A. y Reyes M.C.A. 1993. Situación de la Agricultura en el Norte de Tamaulipas. IV Encuentro Internacional de Investigadores en Economía Agrícola. UAT. p. 280-290.
Doran, J. W. 1980. Microbial changes associated whit residue management whit reduced tillage.
Soil Sci. Am. J. 44:518-523 . Doran, J.W. 1987. Microbial biomass and mineralizable nitrogen distribution in no tillage and
plowed sofis. Biol. Fertil. Soils. 5:68-75. El-Baruni, B., and S.R. Olsen 1979. Effects of manure on solubility of phosphorus in calcarcous
sofis. Soil Sci. 12: 219-225. Elliot, L.F., R.I. Papendick and D.F. Bezdicek. 1987. Cropping practices using legumes with
conservation tillage and soil benefits. P. 812-889. In J.F. Power (Ed.), The role of legumes in conservation tillage systems. Soil Conserv. Soc. Am., Ankeny, IA.
Franzluebbers A. J., F. M. Hons and D. A. Zuberer. 1995. Tillage and crop effects on seasonal soil carbon and nitrogen dynamics. Soil Sci. Soc. Am. J. 59:1618-1624.
Gee, G.W., Bander, J.W., 1986. Particle Size Analysis. In! Klute, A., Campbell, G.S., Jackson, R.D., Mortland, M.M., Nielson, D.R. (Eds) Methods of Soil Analysis, Part 1. Physical and Mineralogical Methods, 2nd Edition. Soil Sel. Soc. Am. Book Series, Number 5, Part 1, Madison, W.I. pp. 383-411.
INEGI, 1991. VII Censo Agropecuario. Base de datos en disco compacto. México. SSP.
Maskina, M.S., J.F. Power., J.W. Doran and W.W. Wilhelm. 1993. Residual effects of notillage crop residue on com yield and nitrogen uptake. Soil Sci. Soc. Am. J. 57:1555-1560.
Muzilli, O. 1983. Influencia do Sistema de Plantio Directo, Comparado ao Convencional, sobre a Fertilidade da Camada a Rável do solo. R. Bras. Cl. Solo, Campinas. 7:95-102.
Nelson, A.W., And L.E. Sommers. 1982. Total carbon, Organic Carbon and Organic Matter. P. 539-580. In A.L. Page, R.H. Miller, and A.R. Keeny (ed.) Methods of Soil Analysis. Partz, 2nd Ed. Asvon. Monogr. 9, ASA and SSSA, Madison, WI.
Olsen, S.R., C.V. Cole, F.S. Watanable and L.A. Dean. 1954. Estimation of Available Phosphorus in Soils by Extraction sith Sodium Bicarbonate. USDA circular. 939:1-9.
Peña C., J. y 0. Grageda C. 1997. Dinámica del nitrógeno en el ecosistema agrícola. En: Ruiz H., J., D. Guzmán P. y J. Peña C. (Ed). Perspectivas de la Microbiología en México. IPN. pp. 345-366.
Pérez G., P. y H. Cortinas E. 1994. Guía para cultivar frijol en el norte de Tamaulipas. Campo Experimental Río Bravo, INIFAP. Folleto 6. 18 p.
Pratt P.F. 1965. Total Potassium, Exchangeable, Potassium and Availability Indexes. P.1022-1034. In C.A. Block, A.D. Evans, J.L. Write, L.E. Ensminger, and E. Clork (Ed) Methods of Soil Analysis. Part I. Agron. Monogr. 9. ASS and Ssa, Madison, WI.

41
Rice, C.W., M.S. Smith and R.L. Blevins. 1986. soil nitrogen availability after long term continuos no-tillege and conventional tillage corn production. Soil Sci. Soc. Am. J. 50:1206-1210.
Robson, A.D., L.K. Abbott, And N. Malajezuk. 1994. Management of Micorrizas in Agriculture, Horticulture and Forestry. Kluwer Academic Publishers, Boston.
Salinas-Garcia, J. R., J. E. Matocha and F. M. Hons. 1997. Long-term tillage and nitrogen fertilization effects on soil properties of an Alfisol under dryland corn/cotton producction. Soil Tillage Res. 42:79-93.
SEP. 1991. Frijol y chícharo. David B. Parsons. 2 Ed. México: Trillas.
Sylvia, D.M. 1999. Fundamentals and applications of arbuscular mycorrhizae: A “biofertilizer” perspective. In: Siqueira, J.O. and Moreira F.M. (eds.). Soil Fertility, Soil Biology, and Plant Nutrition Interrelationships. Sociedade Brasileira de Ciencia do Solo, pp: 705-723.
Tracy, P. W., D. G. Westfall, E. T. Elliot, G. A. Peterson and C. V. Cole. 1990. Carbon, nitrogen, phosfhorus and sulfur mineralization in plow and no.till cultivation. Soil Sci. Soc. Am. J. 54:457-461.
Williams, A., H., Rodríguez, R., H. y Montes G. N. 1995. 20 años de investigación en sorgo en el campo experimental Río Bravo, Tamaulipas México. Germen. 8:35.

42
RESPUESTA DE LA BIOFERTILIZACIÓN EN EL CRECIMIENTO Y RENDIMIENTO DE SORGO DE GRANO EN LINARES, NUEVO
LEON
Juan Martínez Medina
Campo Experimental General Terán. General Terán, Nuevo León, México
INTRODUCCION
En México millones de hectáreas sembradas por campesinos de subsistencia se fertilizan muy poco o incluso nada, lo cual reduce de manera importante los rendimientos potenciales de los cultivos; sin embargo, la solución a este problema no es fácil ya que los fertilizantes industriales, a pesar de las ventajas, su excesivo uso ha causado efectos negativos de contaminación del ambiente y a la vez el encarecimiento de los productos agropecuarios. Uno de los retos tecnológicos que enfrenta la agricultura es conseguir alternativas que permitan reducir las aplicaciones de fertilizantes químicos, sin afectar los niveles de productividad y rentabilidad de los cultivos. Actualmente existe un concepto de agricultura basada en sistemas biológicos, en el cual se considera la producción y conservación de los recursos de una manera sostenible. En este contexto se encuentran los biofertilizantes los cuales son microorganismos que viven en el suelo asociados directa o indirectamente a las raíces de las plantas, capaces de fijar nitrógeno ya sea en forma simbiótica o libre, solubilizar el fósforo, producir estimuladores de crecimiento y reducir enfermedades fungosas causadas por nemátodos. Los principales microorganismos utilizados como biofertilizantes son bacterias (Azospirillum, Rhizobium y Bradyrhizobium), y hongos micorrízicos del género Glomus (Quilambo, 2003; Bashan y Olguin, 1997; Bashan, 1998; Bashan y Levanony; 1990). En las gramíneas el Azospirillum es una bacteria que vive en asociación con las plantas en la zona radical que es capaz de infectar diversos cultivos de importancia como trigo, maíz, arroz, sorgo y mijo; uno de los principales mecanismos a través de los cuales esta bacteria promueve el crecimiento de las plantas es su capacidad para producir hormonas de crecimiento vegetal, además de que fija nitrógeno en bajas cantidades. Por su parte, el término micorriza significa literalmente hongo-raíz y se refiere a una estructura radicular resultado de una asociación entre los pelos radiculares de una planta y hongos específicos del suelo; casi todas las plantas forman micorrizas y el principal beneficio es incrementar la longevidad de los pelos radicales e incrementar el área de absorción de nutrientes mayores y menores, especialmente P, K, Cu y Zn, confiriendo además tolerancia a condiciones adversas. En general, los biofertilizantes permiten poner al alcance de los agricultores, productos con alta efectividad, con los que se sustituye hasta 50% del fertilizante nitrogenado industrial en el caso de los fijadores asociativos, hasta el 80% en el de los simbióticos, mientras que los microorganismo solubilizadores de fósforo permiten sustituir hasta el 70% del fertilizante fosfórico; en términos de producción, los rendimientos se incrementan hasta en un 30%. (INIFAP s/f)

43
Así, la biofertilización es una forma viable, sostenible y económica de mejorar la fertilidad del suelo y elevar la producción, por lo que la Secretaría de Agricultura, Ganadería y Desarrollo Rural a través del Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias, puso en marcha un Programa de Biofertilización, iniciando con 750,000 ha de maíz, trigo, cebada, sorgo, soya y frijol en 1999. En Nuevo León, en el 2000 dicho programa tuvo una cobertura de 1,000 hectáreas en la zona sur, a base de Azospirillum aplicado principalmente en tierras de temporal de productores de maíz de escasos recursos. En el resto de la entidad, el INIFAP estableció parcelas de validación principalmente en sorgo y maíz tanto en condiciones de riego como de temporal, usando como biofertilizante Azospirillum y Micorriza (Glomus intraradices); en menor escala se ha usado Rhizobium en el cultivo de frijol. Aunque los resultados han sido variables en términos de rendimiento, las respuestas en el crecimiento y desarrollo de plantas y raíces han sido más evidentes a favor de los biofertilizantes. En este contexto, el propósito del trabajo fue evaluar el efecto de la aplicación de Azospirillum brasilense y Micorriza (Glomus intraradices) en el crecimiento y rendimiento de sorgo para grano, en parcelas comerciales de productores en la zona centro de Nuevo León.
METODOLOGÍA Localización. El trabajo se desarrolló en el Ejido La Petaca, Linares, N.L.; se encuentra ubicado geográficamente entre las coordenadas 24o 51’, 24o 52’ de latitud norte y 99o 31’, 99o 32’ de longitud oeste, a 300 msnm. Los suelos son vertisoles, profundos, y de buen drenaje; la precipitación anual es de 586 mm (1985-1998), temperatura máxima de 20.5oC a 36.3oC y mínima de 9.2oC a 23.9oC, con una evaporación de 2,035 mm anuales. Selección de las parcelas y establecimiento del cultivo. En la localidad existen productores ejidales con un patrón de cultivos de sorgo, maíz y trigo con superficies promedio de 3 ha, en condiciones de riego restringido (generalmente con un riego de auxilio). Fueron seleccionados tres productores que mostraron interés por la biofertilización en el cultivo de sorgo grano; las fechas de siembra fueron el 12 de febrero, 16 de febrero y 2 de marzo de 2000, las cuales se identificaron como Parcela 1, 2 y 3, respectivamente. En todos los casos se utilizó la mezcla de Micorriza (Glomus intraradices) y Azospirillum brasilense aplicada directamente a la semilla para sembrar una hectárea (con biofertilizante). Esta superficie fue cubierta con una bolsa de cada biofertilizante con un peso aproximado de 1 kg y 380 g para Micorriza y Azospirillum, respectivamente; la superficie restante (aproximadamente 2 ha) fue sembrada sin biofertilizante (tratamiento testigo). En el caso de la Parcela 1, además de la mezcla, se incluyeron tratamientos correspondiendo a semilla inoculada con Micorriza y Azospirillum en forma separada. La variedad de sorgo fue GS-66-Y.
44
Manejo del cultivo y variables de respuesta. En la zona, las siembras se realizan en forma tradicional consistente en dos pasos de rastra, surcado, riego, rastreo, siembra y cultivo (escarda); con está secuencia se manejaron dos lotes, mientras que en la Parcela 1 se usó labranza de conservación. En todos los casos se aplicó solo el riego para la siembra, manejándose luego bajo temporal.
Se realizaron muestreos para estimar la altura de planta, peso seco total y peso seco de raíz (en la Parcela 2 en etapa de grano ‘lechoso-masoso,’ en la Parcela 3 la planta estaba en la fase de ‘embuche’); esta determinación se realizó en cinco puntos tomando tres plantas en cada uno. Al final del ciclo, en las tres Parcelas, para estimar rendimiento de grano se cosecharon las panojas existentes en 5 metros de surco, repetido 4 veces en cada tratamiento; de cada muestra fueron seleccionadas 10 plantas a las que les registró la altura, longitud de panoja, peso seco total, peso seco de la parte aérea, peso seco de panoja, peso seco de hojas y tallo, peso seco de raíz. La información fue analizada con un modelo estadístico completamente al azar, utilizando el Paquete de Diseños Experimentales FAUANL, Versión 2.5 (Olivares, 1995). Este análisis no se realizó en la Parcela 1 para los pesos secos total, de panoja y raíz debido a que los datos se tomaron solo en una muestra por tratamiento.
RESULTADOS Y DISCUSIÓN
Efecto del biofertilizante en etapa vegetativa del sorgo. De acuerdo con los análisis de varianza, se encontraron diferencias estadísticas en las variables medidas, lo anterior se observa en el Cuadro 1 en el que se consignan los valores de los cuadrados medios y los coeficientes de variación. Cuadro 1. Cuadrados medios y coeficiente de variación para variables medidas. INIFAP-CEGET, 2000.
Parcela 2 Parcela 3
Variabe C.M. Trat. C.M. Error C.V. (%)
C.M. Trat. C.M. Error C.V. (%)
Altura
P.seco total
P. seco raíz
Area foliar
327.195313 *
483024414 *
18.496017 *
214154.2500*
17.260742
7.335510
0.253500
7310.9375
4.8
12.4
14.6
14.6
39.606445 *
18.911255 *
0.319988 *
5.424967
1.776225
0.046667
7.1
11.7
8.4
* Significativo al 0.05
Los resultados de los análisis de varianza son indicativos de un comportamiento diferencial de la planta de sorgo al ser sometida a los tratamientos considerados. Los efectos expresados favorecieron a las plantas que estuvieron con biofertilización; en el Cuadro 2 se presentan los valores numéricos mostrados por las plantas en cada tratamiento en las Parcelas 2 y 3.

45
Cuadro 2. Efecto de la biofertilización en algunas características de la planta de sorgo en etapa vegetiva. INIFAP-CEGET, 2000.
Variable Parcela 2 Parcela 3
Sin Biofert. Con Biofert. % Sin Biofert. Con Biofert. %
Altura de planta (cm)
Peso seco total (g)
Peso seco raíz (g)
Area foliar (cm2)
80.4 b
14.7 b
2.0 b
439.3 b
91.8 a
28.6 a
4.8 a
732.0 a
14.1
94.5
140.0
66.6
30.9 b
9.8 b
2.3 b
-
34.9 a
12.9 a
2.7 a
-
12.9
31.6
17.3
-
De acuerdo con los datos, los efectos combinados de los biofertilizantes posibilitaron que las plantas de ambas parcelas acumularon una mayor cantidad de materia seca total y que fueran capaces de desarrollar mayor masa radical, los incrementos fueron del orden de 94.5% y de 140% para esas características respectivamente, en una de las parcelas; las plantas de la Parcela 3, mostraron incrementos menos espectaculares. El efecto de los biofertilizantes en el incremento de la altura de la planta fue más estable en ambas parcelas. En la Figura 1 se muestra gráficamente lo anterior.
Fig. 1 Etapa vegetativa en la parcela 2 y 3. Efecto del biofertilizante en el comportamiento final del sorgo. No se encontraron diferencias estadísticas en los atributos considerados en las plantas de las Parcelas 1 y 2; sin embargo, las diferencias si se detectaron cuando se analizaron las plantas de la Parcela 3, excepto en las variables de rendimiento de grano/parcela y panojas cosechadas. Como se recordará, en la Parcela 1 se consideraron 3 tratamientos de biofertilización mas el testigo, en el Cuadro 3 se presentan los valores promedio para cada una de las características consideradas.
46
Cuadro 3. Efecto del biofertilizante en el comportamiento final de la planta de sorgo en la parcela 1. INIFAP-CEGET, 2000.
Trat. Altura (cm)
Peso Seco/planta (g)
Panoja Rend/parc (g)
Total Raíz Long. (cm) Peso (g) Cosech.
Azospirillum
Micorriza
Azosp.+Micorr.
Testigo
96.9
101.9
102.0
101.0
63.0
56.1
44.4
46.1
10.2
8.0
8.2
6.2
18.9
18.6
20.0
18.4
26.3
27.3
28.7
20.8
43.0 b
55.0 ab
47.0 b
70.0 a
816.7
956.0
933.5
812.0
DMS 16.6
En términos generales, para el caso de la Parcela 1, existió una respuesta favorable en el comportamiento final de la planta cuando se aplicó biofertilizantes (Cuadro 3); los datos sugieren que las plantas utilizan dichos productos para incrementar la materia seca tanto total como de raíz y de la panoja, parece ser que esta última característica es determinante en el rendimiento de grano. Así, la planta incrementó su peso seco total entre 21.6 y 36.6% cuando se usó Micorriza y Azospirillum, respectivamente, aumentando su masa radical en 29 y 64%; cuando se usó la mezcla de esos productos se obtuvo un 37.9% de incremento en el peso de la panoja y un 14.9% en el peso de grano por parcela; este rendimiento de grano fue de 17.7% con el uso de biofertilizante a base de Micorriza y apenas de 0.5% cuando se aplicó Azospirillum. Existen muchos reportes que indican la capacidad del género Azospirillum para promover el crecimiento en las gramíneas (Döbereiner, 1983); de la misma manera son múltiples los trabajos realizados sobre el papel de las micorrizas en la nutrición (mejorando la disponibilidad del P) y en la eficiencia en el uso del agua en un amplio rango de cultivos (Daft and Nicolson 1969, Bethenfalvay et al. 1982, Abbott et al. 1983, Diederichs 1990). Sin embargo, relativamente poca investigación se ha realizada para evaluar el efecto combinado de la inoculación con micorrizas y Azospirillum. El hecho de que el tratamiento Azospirillum + micorriza haya tenido menor efecto en el peso seco total por planta en relación a la inoculación individual, es consistente con lo señalado por Diederichs y Manske (1990) para trigo, sin embargo, en sorgo y mijo para el mismo carácter, observaron un efecto sinérgico cuando se aplicaron ambos microorganismos al momento de la siembra. En relación al rendimiento de grano en las últimas especies señaladas, fue menor, sugiriendo una competencia por la misma fuente de carbohidratos entre la bacteria y el hongo. En la Figura 2 se muestra el efecto a nivel de panoja de cada tratamiento.
47
Fig. 2. Efecto de la aplicación de biofertilizantes en la panoja de sorgo. A, Azospirillum; B, Micorriza; C, Azospirillum+Micorriza; D, Testigo.
Lo encontrado en la Parcela 1 es consistente con el comportamiento mostrado por el sorgo en la Parcela 3, tal como se aprecia en el Cuadro 4.
Cuadro 4. Efecto del biofertilizante en el comportamiento final de la planta de sorgo en la parcela 3. INIFAP-CEGET, 2000.
Trat. Altura (cm)
Peso seco/planta (g) Panoja Rend. parcela
(g)
Total Aérea T. hojas Panoja Raíz Long. (cm)
Cosech.
Con Biofert.
Sin Biofert
63.7a
55.6b
41.7a
24.6b
35.9a
21.6b
19.0a
12.5b
16.9a
9.12b
5.7a
2.9b
17.0a
13.6b
28.0
25.0
213.0
134.2
DMS 7.8 10.2 8.7 6.0 3.6 1.7 1.6
T. hojas =Tallo y hojas
A B
C D

48
El incremento en el peso seco total, de la raíz y de la panoja fueron los mas favorecidos por la aplicación de biofertilizante: 69.5%, 85.5% y 96%, respectivamente, en comparación al testigo; el peso de grano por parcela fue superior en un 58.7%. En la Figura 3 se observa el efecto a nivel de raíz y de planta completa.
Fig. 3. Efecto de la biofertilización en el desarrollo de raíz y de planta de sorgo, parcela 3.
No obstante lo anterior, aunque el patrón general de la respuesta en el comportamiento final de la planta de sorgo en la 2 fue igual en términos estadísticos en ambos tratamientos, se observó un ligero incremento en las características relacionadas con el peso seco de la planta y en el tamaño de la panoja donde no se aplicó biofertilizante (Cuadro 5). En la Figura 4 se aprecian los efectos señalados. Cuadro 5. Efecto del biofertilizante en el comportamiento final de la planta de sorgo en la parcela 2. INIFAP-CEGET, 2000.
Trat. Peso seco/planta (g) Panoja Rend. parcela (g)
Total Aérea T. hojas Panoja Raíz Long. (cm)
Cosech.
Con Biofert.
Sin Biofert
34.5
40.0
29.3
34.4
18.5
21.9
10.7
12.4
5.2
5.6
16.2
16.9
41.0
43.0
240.2
229.5
T. hojas =Tallo y hojas

49
Fig. 4. Efecto de la biofertilización en el desarrollo de raíz y planta de sorgo, parcela 2. Sin embargo, el efecto de la aplicación del biofertilizante fue positivo en el rendimiento de grano en un 4.6%, la explicación posible para este comportamiento pudiera relacionarse con un tamaño mas grande del grano considerando que se cosecharon una menor cantidad de panojas.
El impacto en el rendimiento de grano de sorgo por efecto del biofertilizante.
En relación al rendimiento de grano, aun y la ausencia de evidencia estadística, fue favorable con la biofertilización (Azospirillum + Micorriza); aunque el incremento varió en cada una de las parcela (5.5, 14.9 y 58.7%) el promedió fue de 27%. En el Cuadro 6 y en la Figura 5 muestra el rendimiento de grano por hectárea en cada parcela.
Cuadro 6. Rendimiento de grano (kg ha-1) de sorgo por efecto de la biofertilización con Azospirillum y micorriza. INIFAP-CEGET, 2000.
Tratamiento Parcelas
1 2 3
Azospirillum 2,040
Micorriza 2,390
Azospirillum+Micorriza 2,332 677 532
Testigo 2,030 572 335
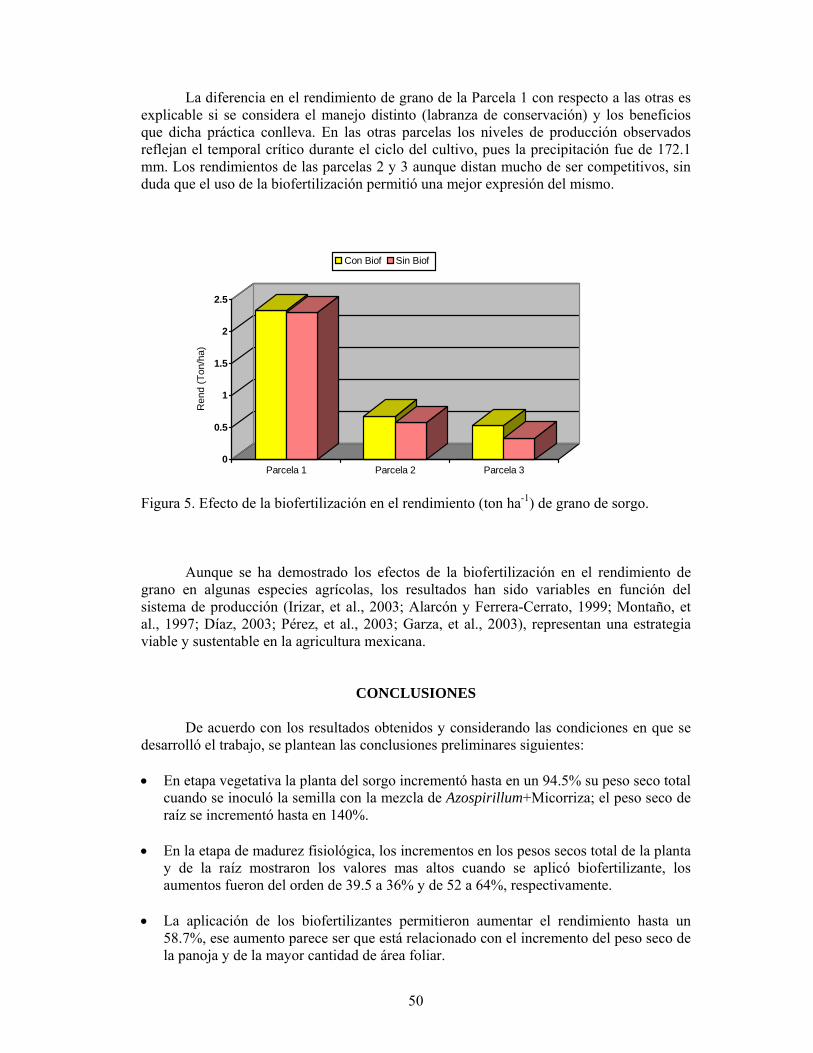
50
La diferencia en el rendimiento de grano de la Parcela 1 con respecto a las otras es explicable si se considera el manejo distinto (labranza de conservación) y los beneficios que dicha práctica conlleva. En las otras parcelas los niveles de producción observados reflejan el temporal crítico durante el ciclo del cultivo, pues la precipitación fue de 172.1 mm. Los rendimientos de las parcelas 2 y 3 aunque distan mucho de ser competitivos, sin duda que el uso de la biofertilización permitió una mejor expresión del mismo.
0
0.5
1
1.5
2
2.5
Ren
d (T
on/h
a)
Parcela 1 Parcela 2 Parcela 3
Con Biof Sin Biof
Figura 5. Efecto de la biofertilización en el rendimiento (ton ha-1) de grano de sorgo. Aunque se ha demostrado los efectos de la biofertilización en el rendimiento de grano en algunas especies agrícolas, los resultados han sido variables en función del sistema de producción (Irizar, et al., 2003; Alarcón y Ferrera-Cerrato, 1999; Montaño, et al., 1997; Díaz, 2003; Pérez, et al., 2003; Garza, et al., 2003), representan una estrategia viable y sustentable en la agricultura mexicana.
CONCLUSIONES
De acuerdo con los resultados obtenidos y considerando las condiciones en que se desarrolló el trabajo, se plantean las conclusiones preliminares siguientes: • En etapa vegetativa la planta del sorgo incrementó hasta en un 94.5% su peso seco total
cuando se inoculó la semilla con la mezcla de Azospirillum+Micorriza; el peso seco de raíz se incrementó hasta en 140%.
• En la etapa de madurez fisiológica, los incrementos en los pesos secos total de la planta
y de la raíz mostraron los valores mas altos cuando se aplicó biofertilizante, los aumentos fueron del orden de 39.5 a 36% y de 52 a 64%, respectivamente.
• La aplicación de los biofertilizantes permitieron aumentar el rendimiento hasta un
58.7%, ese aumento parece ser que está relacionado con el incremento del peso seco de la panoja y de la mayor cantidad de área foliar.

51
• El uso de los biofertilizantes potencializa la producción de grano bajo el sistema de
labranza de conservación, donde es posible obtener hasta 2,390 kg ha-1 contra 667 kg ha-1 en el sistema tradicional, con un riego de presiembra y 172 mm de precipitación
BIBLIOGRAFIA
Abbot, L.K., A.D. Robson and I.R. Hall. 1983. Introduction of Vesicular-arbuscular Micorrhizal Fungi into Agricultural Soils. Australian J. Agric. Res. 34:741. Alarcón, A. y R. Ferrera-Cerrato. 1999. Manejo de la Micorriza Arbuscular en Sistemas de Propagación de Plantas Frutícolas. Terra 17: 179: 191 Bashan, Y. 1998. Inoculants of Plant Growth-Promoting bacteria for Use in Agricultura. Biotechnology Advences, 16 (4): 729-770. Bashan, Y y Holguin G. 1997. Azospirillum-Plant Relationships: Enviromental and Physiological Advances (1990-1996). Can. J. Microbiol. 43: 103-121. Bethlenfalvay, G.S. and R.N. Ames. 1987. Comparison of two Methods for Quantifying Extraradical Mycelium of Versicular-arbuscular Micorrhizal Fungi. Soil Sci. Soc. Am. J. 51:834-837. Daft, M.J. and T.H. Nicolson. 1969. Effect of Endogone Mycorrhizae on Plant Growth. II. Influence of Soluble Phosphate on Endophyte and Host in Maize. New Phytol. 68:945-952. Díaz, F. A. 2003. Inoculación de Sorgo con Glomus intraradices y Azospirillum brasilense en Riego Restringido y Temporal. Memoria de Resúmenes, VII Simposio Internacional y II Congreso Nacional de Agricultura Sostenible. Diederichs, C. 1990. Improved Growth of Cajanus cajan L. In an Unsterile Tropical Soil by three Mycorrhizal Fungi. Plant and Soil. 123:261-266. Diederichs and G.G. B. Manske. 1991. The Role of Micorrhizal Fungi in Crop Nutrition in the Warmer Regions. In: Wheat for Nontraditional, Warm Areas. A Proceedings of the International Conference, july 29-august 3, 1990, Foz do Iguacu, Brazil. Dobereiner, J. 1983. Ten Years of Azospirillum. Experentia Suplementum 48:9-23. Garza, C. I., A. Díaz F. A. Ramírez de L e I. Machuca O. 2003. Memoria de Resúmenes, VII Simposio Internacional y II Congreso Nacional de Agricultura Sostenible. INIFAP. S/f. Biofertilizante. Irizar, G. M.B., P. Vargas V., D. Garza G., C. Tut., I. Rojas M., A. Trujillo C., R. García S., D. Aguirre M., J.C. Martínez G., S. Alvarado M., O. Grajeda C., J. valero G y J.F. Aguirre M. 2003. Respuesta de cultivos agrícolas a los biofertilizantes en la región central de México. Agric. Tec. Méx. 29: 213-225. Montaño, A. N.M. 2001. Colonización micorrízica arbuscular y fertilización mineral de genotipos de maíz y trigo cultivados en un andisol. Terra 19: 337-344.

52
Olivares S, E. 1995. Notas de Diseños Experimentales con Aplicación a la Experimentación Agrícola y Pecuaria. Facultad de Agronomía, UANL. Marín, N.L. Pérez, G. P., A. Díaz F., A. Ramírez de L e I. Garza C. 2003. Efecto de Biofertilizantes sobre el Rendimiento y Proteína de Soya. Memoria de Resúmenes, VII Simposio Internacional y II Congreso Nacional de Agricultura Sostenible Quilambo, O.A. 2003. The Vesicular-arbuscular Mycorrhizal Simbiosis. Afr. J. Biotechnol. Vol 2 (12): 539-546.

53
BIOTECNOLOGÍA DE LOS HONGOS ECTOMICORRÍZICOS
Jesús Pérez-Moreno
Microbiología, Edafología-IRENAT, Colegio de Postgraduados, Montecillo, Texcoco, Estado de México. CP 56230. MEXICO. [email protected]
Introducción
La producción de inoculantes basados en hongos ectomicorrízicos (ECM), ha cobrado una enorme importancia a nivel mundial por diversas razones, dentro de las que se incluyen de manera importante las siguientes: i) existe una búsqueda de tecnologías que originen un menor deterioro ambiental, y se ha intentado entonces buscar alternativas biológicas, al utilizar microorganismos benéficos importantes para la nutrición vegetal y evitar así la utilización excesiva de fertilizantes químicos (Pérez-Moreno y Ferrera-Cerrato, 1996); ii) se ha demostrado que los hongos ECM juegan un importante papel tanto en la nutrición de las plantas (Read y Pérez-Moreno, 2003) como en su protección contra patógenos (Whipps, 2004); y iii) estudios básicos desarrollados en el último siglo han permitido cultivar algunos hongos ECM en medios sintéticos, lo que ha facilitado su manipulación (Pérez-Moreno et al., 2002). Así, a la fecha la producción de inoculantes basados en hongos ECM ha cobrado una enorme importancia en los países con tradición forestal, como Suecia, Finlandia, Canadá y Estados Unidos de América. En países como México, este tipo de acercamiento se encuentra en desarrollo y posee un enorme potencial, como será discutido en el presente escrito.
En términos generales, la micorriza es una de las asociaciones simbióticas de mayor importancia ecológica y fisiológica establecida por las raíces de las plantas. Las asociaciones micorrízicas se encuentran distribuidas actualmente, en todos los ecosistemas terrestres. En ellos, estas simbiosis juegan un importante papel en su funcionamiento y mantenimiento, tanto en las regiones templadas (Allen, 1993; Heijden et al., 1998; Read y Pérez-Moreno, 2003) como en las tropicales (Newberry et al., 1997; Moyersoen et al., 1998 a y b; Pérez Moreno y Ferrera-Cerrato, 1997). Una de las asociaciones micorrízicas más importantes es la ectomicorriza, término propuesto por Peyronel et al. (1969). En esta simbiosis, el hongo asociado cubre las raíces cortas, formando un manto (o vaina). Las hifas crecen, de este manto, hacia afuera en el sustrato y hacia dentro, entre los espacios intersticiales de las células corticales de la raíz, así forman, un complejo sistema intercelular denominado“red de Hartig”. Generalmente, no existe penetración intracelular a las plantas asociadas (Smith y Read, 1997). Por lo tanto, las estructuras diagnósticas, de este tipo de micorriza son: i) el manto fúngico, ii) la red de Hartig y iii) el micelio externo vegetativo que emerge a partir de las raíces.
El presente escrito presenta información relacionada con la distribución biogeográfica y relevancia ecológica de la simbiosis ECM. Posteriormente, se presentan aspectos generales de la biotecnología de los hongos ECM, se describen los principales tipos de inóculo ECM, se discuten las ventajas y limitaciones del uso de cada uno de ellos y finalmente se analizan sus perspectivas en México.

54
Distribución biogeográfica y relevancia ecológica de los hongos ECM La simbiosis ECM se establece principalmente entre alrededor de 3000 especies de
plantas angiospermas y gimnospermas y 5000 especies de hongos, principalmente de los grupos de los Basidiomycetes y Ascomycetes (Malloch et al., 1980; Molina et al., 1992). Esta simbiosis es característica de familias de árboles y arbustos, los cuales incluyen a especies pertenecientes a las familias Pinaceae, Cupressaceae, Fagaceae, Betulaceae y Salicaceae (Malloch et al., 1980). La mayoría de las plantas que forman ectomicorriza, son especies dominantes en los bosques templados boreales y australes del globo (Cuadro 1, Figura 1). Así, grandes zonas del Hemisferio Norte se encuentran cubiertas por pináceas que forman simbiosis ECM, principalmente adscritas a los géneros Abies, Larix, Pinus y Picea. En dichas áreas la ECM es la simbiosis dominante en biomas donde dominan suelos tipo “moder” o “mull”, con altos contenidos orgánicos, lo que ha promovido cierta capacidad saprofítica en ellos (Pérez-Moreno y Read, 2000, 2001a y b). En zonas más cercanas al Ecuador se encuentran Cedrus, Juniperus y Quercus. Algunas angiospermas de estas zonas, como Alnus, Betula, Carpinus, Carya, Castanea, Fagus, Ostrya, Populus, Quercus, Tylia y Salix, también son ECM (Harley y Smith, 1983; Smith y Read, 1997).
Las plantas que establecen la simbiosis ECM forman una franja que va desde Japón, a través de Eurasia, hasta Norteamérica (Read et al., 2004). En el continente asiático, esta distribución incluye zonas del Himalaya, China, Indonesia y Australia (Figura 1). En el continente americano, se ha reconocido que todas las especies de Nothofagus de Sudamérica son ECM (Meyer, 1973; Palfner, 2001). Además, algunos árboles tropicales y subtropicales en América, Africa, Asia y Australia también establecen simbiosis ECM. En los trópicos, por lo menos, se han reportado 46 géneros, distribuidos en 13 familias (Cuadro 2), capaces de formar ectomicorriza (Alexander y Högberg, 1986; Lee et al., 1997; Moyersoen et al., 1998a y b; Pérez-Moreno, 1998; Newberry et al., 2000).
Dentro de las especies de hongos ECM, se incluyen todas las especies pertenecientes a importantes familias de Basidiomycetes dentro de las que se incluyen Amanitaceae, Russulaceae, Paxillaceae, Boletaceae y Strobilomycetaceae, las cuales parecen haber evolucionado como micobiontes. Algunas especies, como Pisolithus tinctorius (Pers.) Cooker, tienen la habilidad de formar simbiosis ECM con una gran cantidad de plantas, incluyendo especies vegetales de diferentes familias. En contraste, otras especies fúngicas se asocian con un restringido número de especies vegetales que integran a un género o una familia; por ejemplo, Suillus coloniza y forma simbiosis restringida en la familia Pinaceae (Molina et al., 1992).
Existen estudios de campo en donde se estima que de 10 a 15% de la producción total de los fotosintatos de los fitobiontes de ecosistemas templados se transfiere a los simbiontes fúngicos (Vogt et al., 1991). A pesar de este elevado costo energético, la presencia de los micobiontes es obligada en los bosques naturales templados, lo cual denota la relevancia de la simbiosis para el mantenimiento de estos sistemas (Brundett, 1991). Una de las principales razones de esta presencia obligada, se fundamenta en la actividad de la ectomicorriza en la absorción nutrimental en los árboles de zonas templadas (Vogt et al., 1982). Además, dicha simbiosis proporciona a los fitobiontes mayor captación de agua, fitohormonas (Harley y Smith, 1983; Marks y Kozlowski, 1973; Smith y Read, 1997), incremento en la longevidad de las raíces (Marshall y Perry, 1987) y protección contra patógenos (Whipps, 2004). Esta estrecha relación entre simbiontes origina, frecuentemente, una dependencia de las plantas asociadas, lo que origina que la ECM usualmente es una relación obligada. Géneros completos de árboles como Abies, Betula, Pinus, Picea, Fagus y Quercus, que cubren extensas áreas del planeta, no podrían

55
sobrevivir en condiciones naturales en ausencia de la simbiosis ECM (Meyer, 1973; Smith y Read, 1997; Pérez-Moreno, 1998). Figura 1. Distribución de los principales biomas terrestres y tipos de micorrriza dominantes en cada uno de ellos. ECM = ectomicorriza, MER = micorriza ericoide, NM = no micorrízicas, MA= micorriza arbuscular. Apréciese que los dos tipos de micorriza más importantes desde el punto de vista biogeográfico, a nivel mundial, son la ectomicorriza y la micorriza arbuscular (Fuente: Read, Leake y Pérez-Moreno, 2004). Aspectos biotecnológicos
Desde el siglo XVIII se efectuaron intentos para utilizar hongos específicos para formar ectomicorrizas en plántulas de encinos en Europa. El objetivo inicial de dichas inoculaciones fue incrementar la producción natural de trufas, que son hongos que tradicionalmente han tenido un alto valor económico por su enorme importancia gastronómica en países mediterráneos como Italia, España y Francia y que se asocian con dichos árboles. Las trufas eran colocadas en los “cajetes” donde las plántulas de los encinos eran sembradas en nuevas plantaciones (Malencon, 1938; Trappe, 1977). Interesantemente esto ocurrió mucho antes de que el mismo vocablo “micorriza” fuera acuñado, hecho que se dio en 1885 (Frank, 1885). Desde esta fecha, hasta finales de 1970 el interés de la aplicación práctica de los hongos micorrízicos, en general, se concentró principalmente en la utilización de los hongos ECM en la producción forestal (Mikola, 1980). Las fuentes principales de inóculo ECM que han sido utilizadas,

56
estuvieron constituidas en un inicio, por las esporas o los carpóforos ECM molidos, frescos o secos, y después por cultivos miceliales (Vozzo and Hackslayo, 1971; Marx, 1980; Mikola, 1980; Brundrett et al,. 1996). En el caso de los inoculantes basados en esporas, se han utilizado principalmente Gasteromycetes, debido al gran número de esporas que pueden llegar a producir, el cual pueden ser del orden de 106 a 109 esporas por gramo de cuerpos fructíferos molidos (Marx y Bryan, 1975). Cuadro 1. Géneros típicos de plantas y hongos de zonas templadas y boreales que establecen simbiosis ectomicorrízicas. * Hongos asociados Plantas asociadas
Basidiomycotina Alpova, Amanita, Astraeus, Boletus Cantharellus, Cortinarius, Entoloma Gastroboletus, Gauteria, Gomphidius, Hebeloma, Hygrophorus, Hymenogaster, Hysterangium, Inocybe, Laccaria, Lactarius, Leccinum, Lycoperdon, Martellia, Paxillus, Pisolithus, Rhizopogon, Rozites, Russula, Scleroderman, Suillus, Tylopilus, Tricholoma, Xerocomus
Abies, Alnus, Betula Corylus, Eucalyptus, Fagus, Larix, Picea, Pinus, Populus, Pseudotsuga, Quercus, Salix, Tsuga
Ascomycotina Balsamia, Elaphomyces, Genea, Geopora, Helvella, Hydnotria, Sphaerosporella, Tuber
Abies, Betula, Corylus, Eucalyptus, Fagus, Larix, Pinus, Populus, Pseudotsuga, Quercus, Salix, Tsuga,
Zygomycotina Endogone
Eucalyptus, Pinus, Pseudotsuga
Deuteromycotina Cenococcum
Abies, Larix, Pinus, Polygonum, Pseudotsuga
* Basado principalmente en Harley y Smith (1983), Miller (1984), Castellano y Molina (1989) y Smith y Read (1997).

57
Cuadro 2. Familias y géneros de plantas tropicales que establecen simbiosis ectomicorrízicas y hongos asociados.* Plantas asociadas
Hongos asociados
Leguminosae Acacia, Afzelia Aldinia, Anthonota, Aphanocalyx, Berlinia, Brachystegia, Didolotia, Eperua, Gilbertiodendron, Inga, Instia, Isoberlinia, Julbernardia, Macrolobium, Microberlinia, Monopetalanthus, Ormosia , Paraberlinia, Paramacrolobium, Pericopsis, Tetraberlinia Casuarinaceae Casuarina, Allocasuarina
Amanita, Cantharellus, Gyrodon,Gyroporus, Inocybe,Lactarius, Porphyrellus, Russula, Scleroderma, Sclerogaster, Strobilomyces, Xerocomus Amanita , Elaphomyces, Hysterangium, Laccaria, Paxillus, Pisolithus, Scleroderma, Thelephora
Dipterocarpaceae Anisoptera, Dipterocarpus, Dryobalanopus, Hopea, Marquesia, Monotes, Shorea, Vateria
Agaricus, Amanita, Boletus, Cantharellus, Geastrum, Lactarius, Russula, Sclerodema
Euphorbiaceae Uapaca
Amanita, Austrogautieria, Boletellus, Cantharellus, Coltricia, Elasmomyces, Lactarius, Pulveroboletus, Russula, Scleroderma, Tubosaeta, Xerocomus
Fagaceae Castaneopsis, Quercus, Lithocarpus, Pasania Gnetaceae Gnetum Myrtaceae Campomanesia, Eucalyptus, Eugenia, Melaleuca, Tristania Nyctaginaceae Neea, Pisonia, Torrubia Polygonaceae Coccoloba Pinaceae Pinus
Boletus, Boletellus, Gyroporus, Leccinum, Tylopilus nr Cenococcum, Hebeloma, Laccaria, Pisolithus, Scleroderma, Setchelliogaster, nr Cantharellus Amanita, Boletus, Coltricia, Gyroporus, Hebeloma, Lycoperdon, Pisolithus, Rhizopogon, Scleroderma,Suillus, Thelephora, Tylopilus
Proteaceae Faurea Sapotaceae Glycoxylon Sapindaceae Allophyllus, Nephelium
Nr nr nr
* Recopilado principalmente de Alexander (1985; 1990), Alexander y Högberg (1986); Amakiri y Ojobo (1985), Ba et al.(1990), Cabrera et al. (1990), Ekwebelam y Odeyinde (1985), Founoune et al. (2002); Green y Newbery (2001a y b); Högberg (1982), Högberg y Piearce (1986), Ivory (1980), Janos (1983; 1985), Lee et al., (1990; 1997); Moyersoen et al. (1998a y b), Muchovej et. al. (1990), Newberry et al. (2000), Onguene y Kuyper (2002), Pérez-Moreno (1998), Rives et al. (1990); Theodorou y Redell (1991), Thoen y Ducousso (1990). nr= no reportados.

58
Actualmente, en la preparación de inoculantes comerciales basados en hongos ECM, se continúan utilizando diversas especies de hongos Gasteromycetes, dentro de los géneros Pisolithus, Scleroderma y Rhizopogon, y adicionalmente algunos Agaricales, dentro de los géneros Laccaria, Suillus y Paxillus (Cuadro 3). Tanto las esporas como los cultivos miceliales de dichos hongos han sido utilizados para inocular semillas o plantas de importancia forestal tanto en regiones templadas como tropicales (Mikola, 1980; Castellano y Molina, 1989; Brundrett et al., 1996). A la fecha existen diversas compañías a nivel mundial que efectúan la comercialización de diversos productos que contienen propágulos de diversos hongos ECM. Sin embargo, el hongo ECM que ha sido utilizado con mayor frecuencia en la preparación de dichos inoculantes comerciales, es Pisolithus tinctorius (Pers.) Coker et Couch (sinónimo P. arhizus (Scop.: Pers.) Rauschert). Este tiene una amplia distribución a nivel mundial, y forma simbiosis ECM con una gran cantidad de géneros de angiospermas y gimnospermas (Cairney y Chambers, 1997), dentro de los cuales se incluyen, principalmente, Abies (Vaario et al., 1999), Acacia (Duponnois y Plenchette, 2003), Afzelia (Ba et al., 1994), Castanea (Martins et al., 1996; 1997), Castanopsis (Tam y Griffiths, 1994), Dipterocarp (Kanchanaprayudh et al., 2003), Eucalyptus (Dell et al., 1994; Xianheng et al., 1998), Picea (Vodnik y Gogala, 1994), Pinus (Tam y Griffiths, 1994; Rincón et al., 2001), Pseudotsuga (Parladé et al., 1995) y Shorea (Kanchanaprayudh et al., 2003). Como podría esperarse, este hongo ha sido localizado asociado con dichas plantas, en un amplio espectro de habitats, los cuales incluyen bosques, áreas urbanas, huertos y suelos erosionados, mineros o con altas concentraciones salinas (Malloch y Kuja, 1979). La amplia plasticidad ecológica de esta especie, ha sido uno de los factores de éxito de su utilización biotecnológica en productos comerciales. Esta especie, que según la evidencia experimental más reciente es al parecer en realidad un complejo de especies (Anderson et al., 1998), ha sido utilizada ampliamente en programas de inoculación forestal durante más de 30 años y ha demostrado ser particularmente efectiva en mejorar el crecimiento vegetal en sitios con baja precipitación o que presentan altas temperaturas (Mikola, 1980; Marx et al., 1985). En México, en el Colegio de Postgraduados, se han efectuado diversas investigaciones tendientes a la utilización de cepas nativas de hongos ECM tendientes a su utilización en plantaciones comerciales, principalmente de pinos y eucaliptos, y en programas de reforestación. Los hongos que han sido estudiados han incluido principalmente: Amanita caesarea, Amanita rubescens, Boletus edulis, Laccaria laccata, Lactarius deliciosus, Lactarius salmonicolor, Hebeloma fastibile, Suillus
Tipos de inoculantes ECM
Debido al carácter obligatorio de la simbiosis ECM con la mayoría de la plantas
con las que se establece, desde principios de siglo se ha intentado su inoculación controlada (Norkrans, 1950, 1953; Melin y Nilsson, 1950, 1953; Mikola, 1970). Para inducir la micorrización en especies de importancia forestal, se han utilizado principalmente tres tipos de inóculo: i) tierra de monte, ii) esporas de hongos ECM y iii) micelio ECM. Enseguida se describen cada uno de ellos y las posibles ventajas o desventajas de los mismos.
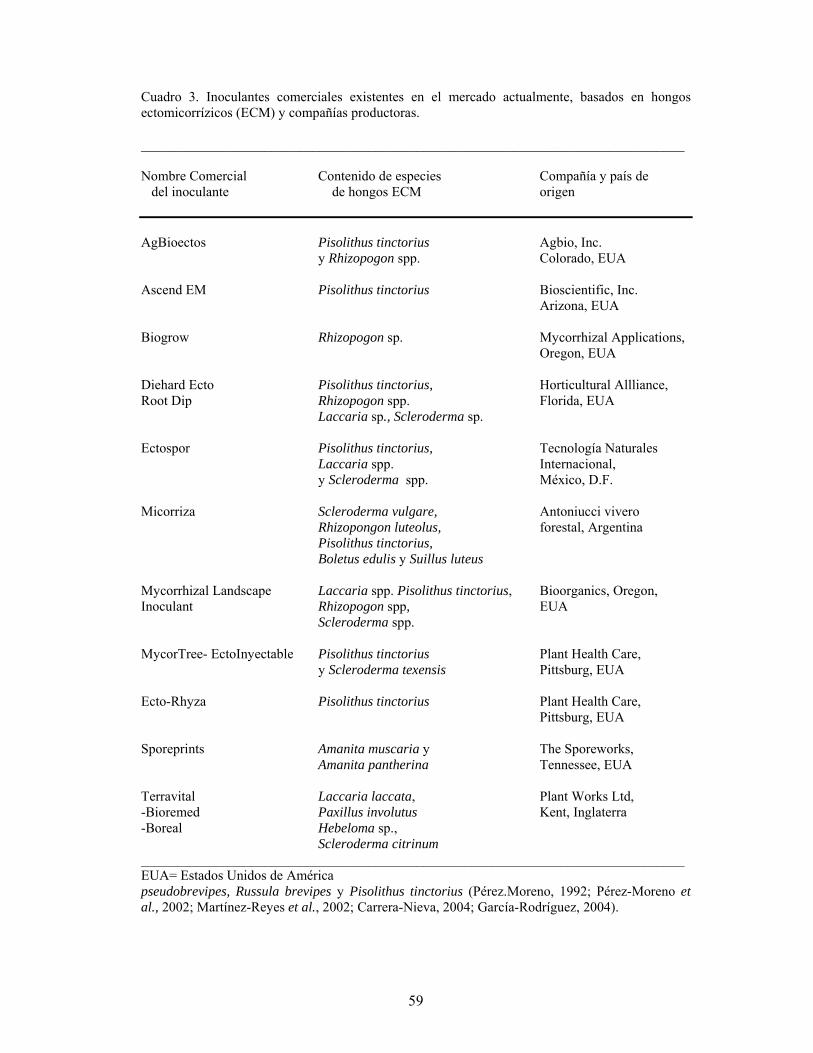
59
Cuadro 3. Inoculantes comerciales existentes en el mercado actualmente, basados en hongos ectomicorrízicos (ECM) y compañías productoras. _______________________________________________________________________________ Nombre Comercial Contenido de especies Compañía y país de del inoculante de hongos ECM origen AgBioectos Pisolithus tinctorius Agbio, Inc. y Rhizopogon spp. Colorado, EUA Ascend EM Pisolithus tinctorius Bioscientific, Inc. Arizona, EUA Biogrow Rhizopogon sp. Mycorrhizal Applications,
Oregon, EUA Diehard Ecto Pisolithus tinctorius, Horticultural Allliance, Root Dip Rhizopogon spp. Florida, EUA
Laccaria sp., Scleroderma sp. Ectospor Pisolithus tinctorius, Tecnología Naturales
Laccaria spp. Internacional, y Scleroderma spp. México, D.F.
Micorriza Scleroderma vulgare, Antoniucci vivero
Rhizopongon luteolus, forestal, Argentina Pisolithus tinctorius, Boletus edulis y Suillus luteus
Mycorrhizal Landscape Laccaria spp. Pisolithus tinctorius, Bioorganics, Oregon, Inoculant Rhizopogon spp, EUA Scleroderma spp. MycorTree- EctoInyectable Pisolithus tinctorius Plant Health Care,
y Scleroderma texensis Pittsburg, EUA Ecto-Rhyza Pisolithus tinctorius Plant Health Care, Pittsburg, EUA Sporeprints Amanita muscaria y The Sporeworks, Amanita pantherina Tennessee, EUA Terravital Laccaria laccata, Plant Works Ltd, -Bioremed Paxillus involutus Kent, Inglaterra -Boreal Hebeloma sp.,
Scleroderma citrinum _______________________________________________________________________________ EUA= Estados Unidos de América pseudobrevipes, Russula brevipes y Pisolithus tinctorius (Pérez.Moreno, 1992; Pérez-Moreno et al., 2002; Martínez-Reyes et al., 2002; Carrera-Nieva, 2004; García-Rodríguez, 2004).

60
i) Tierra de monte. Las capas superficiales del suelo forestal (por lo común, el material orgánico que abarca los primeros 10 cm) se han utilizado por mucho tiempo como fuente de inóculo ECM, en los países denominados del tercer mundo. Esta es la fuente que tradicionalmente ha sido usada en México con mayor frecuencia. Las principales desventajas de esta fuente son las siguientes: 1) necesidad de grandes cantidades de suelo, con la subsecuente erosión y deterioro ambiental en las áreas de extracción; 2) inconsistencia en la calidad del inóculo, debido principalmente a la distribución heterogénea de los propágulos micorrízicos en condiciones naturales; y 3) probabilidad de introducción de microorganismos patógenos y malezas en las áreas de producción de plántulas (Pérez-Moreno et al., 2002). Por estas razones, diversos autores (e. gr. Castellano y Molina, 1989) consideran que este método no resulta recomendable, a menos que no se cuente con otras formas de inóculo.
ii) Esporas de hongos ECM. Las esporas de hongos ECM se han usado ampliamente en la producción de plantas en los denominados países desarrollados (Castellano, 1994). Con mayor frecuencia, se han utilizado cuerpos fructíferos de Gasteromycetes, como Pisolithus, Scleroderma o Rhizopogon, los cuales se producen en abundancia y contienen grandes cantidades de esporas. En el caso de hongos que producen esporas en menor cantidad, como Russula, Suillus, Laccaria, Hebeloma, Boletus y Amanita, existe el potencial de utilizar, de manera adicional a las esporas, los píleos de los cuerpos fructíferos como fuente de inóculo. En este sentido, en el Colegio de Postgraduados, el autor del presente escrito desarrolla una línea de investigación que tiene como objetivo generar información básica, relacionada con el uso de esporas para especies de hongos y plantas ECM mexicanas, tendiente al desarrollo de esta biotecnología en México.
Por lo general, se recomienda el uso de esporas frescas siempre que sea posible; sin embargo, se ha demostrado que esporas refrigeradas a 5°C pueden mantener una viabilidad aceptable hasta por un periodo de un año. Las esporas de los hongos ECM se han aplicado en soluciones con agua destilada estéril, mezcladas con arena (usualmente en proporciones 1/100, peso/peso), como recubrimientos en semillas o comprimidas con arcillas que actúan como “acarreadores” (Brundett et al., 1996). En todos los casos, resulta importante hacer conteos del número de esporas aplicadas, mediante el uso de un contador de glóbulos rojos o hematocitómetro.
Un criterio, recomendado por Trappe (1977), al seleccionar las especies a emplearse como fuente de inóculo ECM, es la comestibilidad de las mismas. En México, se conocen más de 220 especies de hongos comestibles silvestres (Villarreal y Pérez Moreno, 1989). Más de 100 de estas especies son objeto de autoconsumo, principalmente en la población rural del país; y más de diez de ellas, tienen un alto valor en los mercados internacionales en Estados Unidos de América, Canadá y diversos países europeos y del sureste asiático (Pérez Moreno y Ferrera-Cerrato, 1996). Si se desarrollara un programa de inoculación con especies de hongos comestibles en las especies de árboles utilizadas en plantaciones o reforestaciones, se podría tener, previo a los aprovechamientos forestales, un producto forestal no maderable de alto valor como alimento.
Las principales desventajas de la utilización de esporas como fuente de inóculo son la corta disponibilidad de cuerpos fructíferos durante el año, alrededor de dos a tres meses, y las posibles variaciones genéticas de las esporas al variar el sitio o temporada de recolección. A pesar de ello esta fuente de inóculo tiene amplias perspectivas de uso en México, debido principalmente a su bajo costo y a su relativa facilidad de manejo.
iii) Micelio de hongos ECM. Esta es una de las fuentes de inóculo que ha recibido mayor atención en las últimas décadas. El micelio fúngico producido en condiciones de cultivo estériles puede utilizarse directamente como inóculo o puede mezclarse con

61
diversos “acarreadores” inertes como granos de cereales, mezclas de turba o vermiculita (Brundett et al., 1996) o arcillas. La principal limitación de esta técnica, junto con su alto costo, es que el aislamiento de los hongos ECM en medios sintéticos se encuentra, a la fecha, muy limitado. De alrededor de 5000 especies de hongos ECM conocidos ( Molina et al., 1992), menos de 2% se han aislado exitosamente. Dentro de géneros ECM importantes, como Cortinarius, Russula, Lactarius e Inocybe, se han aislado sólo un número reducido de especies. A pesar de ello, esta fuente de inóculo es la que más se emplea actualmente en los países con amplia tradición forestal.
La principal desventaja de esta fuente de inóculo ECM es su alto costo. Sin embargo, tiene la ventaja de que puede disponerse de inóculo en cualquier época del año, lo cual facilita su manejo y aplicación práctica. Actualmente, existen diversas compañías internacionales que comercializan micelio de hongos ECM, esporas o una mezcla de ambos (Cuadro 3). Independientemente de las limitaciones que pudiera tener la producción de inoculantes basados en micelio o esporas de hongos ECM, las ventajas que presentan, originan que esta biotecnología tiene un gran potencial de aplicación en México.
Literatura Citada Alexander, I.J. 1985. Mycorrhizas of West African forest trees. In Molina R. (Ed.) Proceedings of
the 6th North American Conference on Mycorrhizae. Oregon State University and United States Department of Agriculture, Corvallis, Oregon, EEUU. pp. 447.
Alexander, I.J. 1990. The ecology of ectomycorrhizal legumes in Africa. In: 8th North American Conference on Mycorrhizae. Jackson, Wyoming, EEUU. pp. 1.
Alexander, I.J. and P. Högberg. 1986. Ectomycorrhizas of tropical angiospermous trees. New Phytol. 102: 541-549.
Allen, M. 1993. The ecology of mycorrhizae. Cambridge University Press. Cambridge, England. Amakiri, M.A. and L.I. Ojobo. 1985. Nitrogen, fertilization and ectomycorrhizal formation of
Pinus caribea (Morelet) seedlings. In Molina R. (Ed.) Proceedings of the 6th North American Conference on Mycorrhizae. Oregon State University and United States Department of Agriculture, Corvallis, Oregon, EEUU. pp. 33.
Anderson, I.C., S.M. Chambers and J.W.G. Cairney. 1998. Molecular determination of genetic variation in Pisolithus isolates from a defined region in New South Wales, Australia. New Phytol. 138: 151-162.
Ba, A.M., J. Garbaye and J. Dexheimer. 1990. Influence of soil propagules on the sequence of ectomycorrhizal infection on Afzelia africana Sm. seedlings. In 8th North American Conference on Mycorrhizae. Jackson, Wyoming. EEUU. pp. 11.
Ba, A.M., J. Garbaye and J. Dexheimer. 1994. The influence of culture conditions on mycorrhiza formation between the ectomycorrhizal fungus Pisolithus sp. and Afzelia africana Sm. Seedlings. Mycorrhiza 4: 121-129.
Brundett, M. 1991. Mycorrhizas in natural ecosystems. Adv. Ecol. Res. 21: 171- 313. Brundett, M., N. Bougher, B. Dell, T. Grove and N. Malajczuk. 1996. Working with mycorrhizas
in forestry and agriculture. Australian Center for International Agricultural Research. Canberra, Australia.
Cabrera, T., H. Iglesias, M. Ruiz, S. Herrera y A.M. Sotolongo. 1990. Micorrizas de Coccoloba uvifera Jacq. En Resúmenes del V Congreso Latinoamericano de Botánica, La Habana, Cuba. pp. 26.
Cairney, J.W.G. and S.M. Chambers. 1997. Interactions between Pisolithus tinctorius and its hosts: a review of current knowledge. Mycorrhiza 7: 117-131.
Carrera-Nieva, A. 2004. Inoculación con hongos ectomicorrízicos en Pinus patula Schl. et Cham. y Pinus greggii Engelm. en invernadero. Universidad Autónoma Chapingo, Tesis de Maestría en Ciencias Forestales, Chapingo, México. 195 pp.

62
Castellano M. A. 1994. Current status of outplanting studies using ectomycorrhiza-inoculated forest trees. pp. 261-281. In: Pfleger F., Linderman B. (eds.) The American Phytopathological Society, St. Paul, Minnessota.
Castellano M.A. and R. Molina. 1989. The biological component: nursery pests and mycorrhizae. pp. 101-167. In: Landis T.D., R.W. Tinus, S.E. McDonald and J.P. Barnett (eds.). The container tree nursery manual, Vol 5. USDA, Washington, D.C.
Dell, B., N. Malajczuk , N. L. Bougher and G. Thomson. 1994. Development and function of Pisolithus and Scleroderma ectomycorrhizas formed in vivo with Allocasuarina, Casuarina and Eucalyptus. Mycorrhiza 5: 129-138.
Duponnois, R. and C. Plenchette. 2003. A mycorrhiza helper bacterium enhances ectomycorrhizal and endomycorrhizal symbiosis of Australian Acacia species. Mycorrhiza 13: 85 – 91.
Ekwebelam, S.A. and M.A. Odeyinde. 1985. Field response of Pinus species inocu1ated with ectomycorrhizal fungi in Nigeria. En Molina R. (Ed.) Proceedings of the 6th North American Conference on Mycorrhizae. Oregon State University and United States Department of Agriculture, Corvallis, Oregon, EEUU. pp. 220.
Founoune, H., R. Duponnois, A.M. Ba and F. El Bouami F. 2002. Influence of the dual arbuscular endomycorrhizal/ectomycorrhizal symbiosis on the growth of Acacia holosericea (A. Cunn. ex G. Don) in glasshouse conditions. Annals of Forest Science 59 : 93-98.
Frank, A.B. 1885. Ueber die auf Wurzelsymbiose beruhende Ernährung gewisser Baume durch unterirdische Pilze (On the root-symbiosis-depending nutrition through hypogeous fungi of certain trees). Ber. Deut. Bot. Gesell. 3: 128-145 (traducido por by J.M.Trappe, Proc. 6th N. Amer. Conf. Myc.:18-25).
Garcia-Rodríguez, L. 2004. Propagación micelial del hongo ectomicorrízico Pisolithus tinctorius, e inoculación de brinzales de Eucalyptus urophylla S.T. Blake. Tesis de Maestría, Colegio de Postgraduados, Montecillo, México (por concluirse).
Green, J.J. and D.M. Newbery. 2001a. Light and seed size affect establishment of grove-forming ectomycorrhizal rain forest tree species. New Phytologist 151 : 271-289.
Green, J.J. and D.M. Newbery. 2001b. Shade and leaf loss establishment of grove-forming ectomycorrhizal rain forest tree species. New Phytologist 151 : 291-309.
Harley, J. L. and S. E. Smith. 1983. Mycorrhizal symbiosis. Academic Press. London. Heijden, M. G., A. van der, J.N. Klironomos, M. Ursic, P. Moutoglis, R. Streitwolf-Engell, T.
Boller, A. Wiemken and I.R. Sanders. 1998. Mycorrhizal fungal diversity determines plant biodiversity, ecosystem variability and productivity. Nature 396: 69-72.
Högberg, P. 1982. Mycorrhizal associations in some woodland and forest trees and shrubs in Tanzania. New Phytol. 92: 407-415.
Högberg, P. and O.D. Piearce. 1986. Mycorrhizas in Zambian trees in relation to host taxonomy, vegetation type and successionaI patterns. J. Ecol. 74: 775-785.
Ivory, M.H. 1980. Ectomycorrhizal fungi of lowland tropical pines in natural forests and exotic plantations. In Mikola P. (Ed.) Tropical Mycorrhizal Research. Clarendon Press, Oxford, Reino Unido. pp. 107-117.
Janos, D.P. 1983. Tropical mycorrhizas, nutrient cycles and plant growth. In Sutton, S.L., T.C. Whitmore and A.C. Chadwick (Eds.) Tropical rain forest: ecology and management. Blackwell Scientific Publications. Oxford, Reino Unido. pp. 327-345.
Janos, D.P. 1985. Mycorrhizal fungi: agents or symptoms of tropical community composition. In Molina, R. (Ed.) Proceedings of the 6th North American Conference on Mycorrhizae. Oregon State University and United States Department of Agriculture, Corvallis, Oregon, EEUU. pp.98-103.
Kanchanaprayudh, J., Z. Zhou, S. Yomyart, P. Sihanonth and T. Hogetsu. 2003. Molecular phylogeny of ectomycorrhizal Pisolithus fungi associated with pine, dipterocarp, and eucalyptus trees in Thailand. Mycoscience 44: 287 –294.
Lee, S.S. 1990. The mycorrhizal association of the Dipterocarpaceae in the tropical rain forests of Malaysia. Ambio 19: 383-385.
Lee, L.S., I.J. Alexander and R. Watling. 1997. Ectomycorrhizas and putative ectomycorrhizal fungi of Shorea leprosula Miq (Dipterocarpaceae). Mycorrhiza 7: 63-81.

63
Malencon, G. 1938. Les truffes européennes. Revue Mycologie 3 : 1-92. Malloch, D. and A.L. Kuja. 1979. Occurrence of the ectomycorrhizal fungus Pisolithus tinctorius
in Ontario. Canadian Journal of Botany 57: 1848- 1849. Malloch, D.W., K.A. Pirozynski and P.H. Raven. 1980. Ecological and evolutionary significance of
mycorrhizal symbioses in vascular plants (A review). Proc. Natl. Acad. Sci. USA 77: 2113-2118.
Marks, G.C. and T.T. Kozlowski. (eds.) 1973. Ectomycorrhizae: their ecology and physiology. Academic Press, New York.
Martinez-Reyes, M., J. Pérez-Moreno, A. Aldrete y V.M. Cetina-Alcala. 2002. Potencial biotecnológico de especies mexicanas de hongos etomicorrízicos comestibles para inoculantes forestales a base de esporas. In: Pérez-Moreno J., J. Alvarado-López R. Ferrera-Cerrato (Eds.) Producción y control de calidad de inoculantes agrícolas y forestales. Colegio de Postgraduados y Comité Mexicano de Inoculantes Agrícolas y Forestales. Montecillo, Texcoco, Estado de México. pp. 103-104.
Martins, A., A. Casimiro and M.S. Pais. 1997. Influence of mycorrhization on physiological parameters of micropropagated Castanea sativa Mill. Plants. Mycorrhiza 7: 161- 165.
Martins, A., J. Barroso and M.S. Pais. 1996. Effect of ectomycorrhizal fungi on survival and growth of micropropagated plants and seedlings of Castanea sativa Mill. Mycorrhiza 6: 265-270.
Marx, D.H. 1980. Ectomycorrhizal fungus inoculations: a tool for improving forestation practices. In: Mikola, P. (ed.) Tropical mycorrhizal research. Oxford University Press, Oxford, United Kingdom. pp. 13-71.
Marx, D.H. and W.C. Bryan. 1975. Growth and ectomycorrhizal development of loblolly pine seedlings in fumigated soil infested with the fungal symbiont Pisolithus tinctorius. Forest Science 21: 245-254.
Marx, D.H., A. Hedin and S.F.P. Toe. 1985. Field performance of Pinus caribaea var. hondurensis seedlings with specific ectomycorrhizae and fertilizer after three yr on a savanna site in Liberia. Forest Ecology and Management 13: 1-25.
Melin, E. and H. Nilssson. 1950. Transfer of radioactive phosphorus to pine seedlings by means of mycorrhizal hyphae. Physiologia Plantarum 3: 88-92.
Melin, E. and H. Nilssson. 1953. Transfer of labeled nitrogen from glutamic acid to pine seedlings through the mycelium of Boletinus variegatus (S.W.) Fr. Nature 171: 134.
Meyer, F.H. 1973. Distribution of ectomycorrhizae in native and man-made forests. pp. 79-105. In: Marks, G.C. and T.T. Kozlowski. (eds.). Ectomycorrhizae: their ecology and physiology. Academic Press. New York.
Mikola, P. 1970. Mycorrhizal inoculation in afforestation. International Review of Forest Research 3: 123-196.
Mikola, P. (Ed.). 1980. Tropical mycorrhizal research. Oxford University Press, Oxford, United Kingdom.
Miller, O.K. Jr. 1984. Taxonomy of ecto and ectendomycorrhizal fungi. In: N.C. Shenck (Ed.) Methods and principles of mycorrhizal research. 2nd edition. The American Phytopathological Society, Saint Paul, Minnesota. pp. 91-102.
Molina, R., H. Massicote and J.M. Trappe. 1992. Specificity phenomena in mycorrhizal symbioses: community-ecological consequences and practical implications. pp. 357-423. In: Allen, M.F. (ed.). Mycorrhizal functioning an integrative plant-fungal process. Chapman and Hall, New York.
Moyersoen, B., A.H. Fitter and I.J. Alexander. 1998a. Spatial distribution of ectomycorrhizas and arbuscular mycorrhizas in Korup National Park rain forest, Cameroon, in relation to edaphic parameters. New Phytol. 139: 311-320.
Moyersoen, B., I.J. Alexander and A.H. Fitter. 1998b. Phosphorus nutrition of ectomycorrhizal and arbuscular mycorrhizal tree seedlings from a lowland tropical rainforest in Korup National Park, Cameroon. J. Trop. Ecol. 14: 47-61.

64
Muchovej, R.M.C., M.C.M. Kasaya and J.J. Muchovej. 1990. Effect of time of inoculation on ectomycorrhizal colonization of Eucalyptus grandis by Pisolithus tinctorius or Paxillus involutus. In 8th North American Conference on Mycorrhizae. Jackson, Wyoming, EEUU. pp. 219.
Newberry, D.M., I.J. Alexander and J.A. Rother. 1997. Phosphorus dynamics in a lowland African rain forest. The influence of ectomycorrhizal trees. Ecol. Monogr. 67: 367-409.
Newberry, D.M., I.J. Alexander and J.A. Rother. 2000. Does proximity to conspecific adults influence the establishment of ectomycorrhizal trees in rain forest? New Phytol. 147: 401-409.
Norkrans, B. 1950. Studies in growth and cellulolytic enzymes of Tricholoma. Symbolae Botanicae Upsaliensis 11.pp. 126.
Norkrans, B. 1953. The effect of glutamic acid, aspartic acid, and related compounds on the growth of certain Tricholoma species. Physiol. Plant. 6: 584-593.
Onguene, N.A. and T.W. Kuyper. 2002. Importance of the ectomycorrhizal network for seedling survival and ectomycorrhiza formation in rain forests of south Cameroon. Mycorrhiza 12: 13-17.
Palfner, G. 2001. Taxonomische studien an ektomykorrhizen aus den Nothofagus-Wäldern Mittelsüdchiles. Bibliotheca Mycologica Band 190. J. Cramer: Berlin, Stuttgart.
Parladé, J., I. F. Álvarez and J. Pera. 1995. Ability of native ectomycorrhizal fungi from northern Spain to colonize Douglas-fir and other introduced conifers. Mycorrhiza 6: 51-55.
Pérez-Moreno, J. 1992. Aspectos ecológicos, aislamiento de cepas, síntesis de micorriza y pruebas “in vitro” contra patógenos de diversos hongos ectomicorrízicos del Parque Nacional Zoquiapan, Edo. de México. Tesis de maestría en Ciencias, Colegio de Postgraduados, Montecillo, Texcoco, Estado de México.
Perez-Moreno, J. 1998. La ectomicorriza una simbiosis mutualista en el sostenimiento de Gaia, el planeta viviente. pp. 93-120. In: Ferrera-Cerrato, R. y J. Perez-Moreno. (eds.). Manejo de agroecosistemas sostenibles. Universidad Veracruzana. Xalapa, Ver.
Pérez-Moreno, J. and D. J. Read. 2000. Mobilization and transfer of nutrients from litter to tree seedlings via the vegetative mycelium of ectomycorrhizal plants. New Phytol. 145: 301-309.
Pérez-Moreno, J. and D. J. Read. 2001b. Nutrient transfer from soil nematodes to plants: a direct pathway provided by the mycorrhizal mycelial network. Plant Cell Environ. 24: 1219-1226.
Pérez-Moreno, J. and D. J. Read. 2001a. Exploitation of pollen by mycorrhizal mycelial systems with special reference to nutrient recycling in boreal forests. Proc. R. Soc. London B 268: 1329-1335.
Pérez-Moreno, J. and R. Ferrera-Cerrato. 1996. (eds.). Nuevos horizontes en agricultura: agroecología y desarrollo sostenible. Colegio de Postgraduados. Montecillo, estado de México.
Pérez-Moreno, J. and R. Ferrera-Cerrato. 1996. Sustainability and economic importance of the edible ectomycorrhizal mushrooms. pp. 172-183. In: J. Pérez-Moreno y R. Ferrera-Cerrato. (eds.). Nuevos horizontes en agricultura: agroecología y desarrollo sostenible. Colegio de Postgraduados. Montecillo, estado de México.
Pérez-Moreno, J. and R. Ferrera-Cerrato. 1997. Mycorrhizal interactions with plants and soil organisms in sustainable agroecosystems. pp. 91-112. In: Brussaard, L. y R. Ferrera Cerrato. (eds.). Soil ecology in sustainable agricultural systems. CRC Lewis Publishers, Boca Raton, Florida.
Pérez-Moreno, J., J. Alvarado-Lopez y R. Ferrera-Cerrato (Eds.). 2002. Producción y control de calidad de inoculantes agrícolas y forestales. Colegio de Postgraduados y Comité Mexicano de Inoculantes Agrícolas y Forestales. Montecillo, Texcoco, Estado de México.107 pp.
Peyronel, B., B. Fassi, A. Fontana and J.M. Trappe. 1969. Terminology of mycorrhizae. Mycologia 61: 410-411.
Read, D.J. and J. Pérez-Moreno., 2003. Mycorrhizas and nutrient cycling in ecosystems a journey toward relevance? New Phytologist 157: 475-492.

65
Read, D.J., J.R. Leake and J. Pérez-Moreno. 2004. Mycorrhizal fungi as drivers of ecosystem processes in heathland and boreal forest biomes. Can. J. Bot. 82: 1243-1263.
Rincón, A., I.F. Alvarez and J. Pera. 2001. Inoculation of containerized Pinus pinea L. seedlings with seven ectomycorrhizal fungi. Mycorrhiza 11: 265 – 271.
Rives, C., F. Herrera, F. Ruiz, J.L. Ortíz, C. González y O. Alvarez. 1990. Producción del hongo comestible Cantharellus cinnabarinus (Schw.) Schw. en uverales de la isla de la Juventud, Cuba. En Resúmenes del V Congreso Latinoamericano de Botánica. La Habana, Cuba. pp. 196.
Smith, S.E. and D.J. Read. 1997. Mycorrhizal symbiosis. 2nd edition. Academic Press. London. Tam, P.C.F. and D. A. Griffiths. 1994. Mycorrhizal associations in Hong Kong Fagaceae VI.
Growth and nutrient uptake by Castanopsis fissaseedlings inoculated with ectomycorrhizal fungi. Mycorrhiza 4: 169 – 172.
Theodorou, C. and P. Redell. 1991. In vitro synthesis of ectomycorrhizas on Casuarinaceae with a range of mycorrhizal fungi. New Phytol. 118: 279-288.
Thoen, D. and M. Ducousso. 1989. Mycorrhizal habit and sc1erogenesis of Phlebopus sudanicus (Gyrodontaceae) in Senegal. Agric. Ecosystems Environ. 28: 519-523.
Trappe J.M.1977. Selection of fungi for ectomycorrhizal inoculation in nurseries. Ann. Rev. Phytopathol. 15: 203-222.
Trappe, J. M. 1977. Selection of fungi for ectomycorrhizal inoculation in nurseries. Annual Review of Phytopathology 15: 203-222.
Vaario, L.M., M. Tanaka, Y. Ide, W.M. Gill and K. Suzuki. 1999. In vitro ectomycorrhiza formation between Abies firma and Pisolithus tinctorius. Mycorrhiza 9: 177 – 183.
Villarreal, L. and J. Pérez Moreno. 1989. Los hongos comestibles silvestres de México, un enfoque integral. Micol. Neotrop. Apl. 2: 77-114.
Vodnik, D. and N. Gogala. 1994. Seasonal fluctuations of photosynthesis and its pigments in 1-year mycorrhized spruce seedlings. Mycorrhiza 4: 277-281.
Vogt K.A:, D.A: Publicover, D.J. Vogt. 1991. A critique of the role of ectomycorrhizas in forest ecology. Agric. Ecosystems Environ. 35: 171-190.
Vogt, K.A., C.E. Grier, C.E. Meier and R.L. Edmons. 1982. Mycorrhizal role in net primary production and nutrient cycling in Abies amabilis ecosystems in Western Washington. Ecology 63: 370-380.
Vozzo, J. A. and E. Hacskaylo. 1971. Inoculation of Pinus caribaea with ectomycorrhizal fungi in Puerto Rico. Forest Science 17: 239-245.
Whipps, J.M. 2004. Prospects and limitations for mycorrhizas in biocontrol of root pathogens. Can. J. Bot. 82: 1198-1227.
Xianheng L., N. Malajczuk and B. Dell. 1998. Mycorrhiza formation and growth of Eucalyptus globulus seedlings inoculated with spores of various ectomycorrhizal fungi. Mycorrhiza 8: 81 – 86.

66
EVALUACIÓN DE LA INOCULACIÓN DE SIMBIONTES EN SOYA (Glycine max) BAJO CONDICIONES DE CAMPO
Ponciano Pérez García, Arturo Díaz Franco y Nicolás Maldonado Moreno.
INIFAP-Campo Experimental Río Bravo. Apdo. 172, Río Bravo, Tam., México, 88900. [email protected]
INTRODUCCION
Una de las estrategias para diversificar la agricultura en la zona norte de Tamaulipas, lo representa la soya (Glycine max L. Merrill), un cultivo viable y con antecedentes de buena productividad; aunado a esto, se dispone con la tecnología de producción para las áreas potenciales de la zona. Actualmente tiene un mercado asegurado, así como diversos apoyos para su comercialización; esto representa un incentivo para el productor. Además, un punto importante de este cultivo es que, por su fecha de siembra y cosecha (julio-noviembre), bajo condiciones de riego y buen temporal, no afecta la siembra de maíz o sorgo en el ciclo de otoño-invierno, lográndose la rotación ideal, gramínea–leguminosa.
De las prácticas agronómicas de producción de soya, la inoculación de la semilla con la rizobacteria fijadora de nitrógeno Bradyrhizobium japonium (Elizondo, 1999 y Jiménez et al., 1991) y el tratamiento de la semilla con fungicidas para la protección de la plántula contra el complejo de patógenos del suelo (Maldonado, 2002, Sine, 1993 y Wendell, 1988), son procedimientos comunes en las regiones productoras de soya. La efectividad protectora de los fungicidas está bien documentada, en especial cuando se mezclan dos o más de ellos (Sine, 1993 y Wendell, 1988). No obstante, la efectividad de la simbiosis con B. japonium, puede tener una respuesta variable y depende de diversos factores, como el ambiente, el genotipo y la cepa utilizada (Jiménez et al., 1991 y Tarafdar y Rao, 1997). Por otro lado, el beneficio de la inoculación de la semilla asociado con el tratamiento con fungicidas no está claro (Revellin et al., 1993). La inoculación de micorriza vesículo arbuscular (MVA), representa una práctica poco común en soya. Durán et al. (2001), señalaron que la MVA Glomus intraradices en la variedad Huastecas-200, produjo 21.6% más rendimiento que el tratamiento con fertilización química (60-40-00). Además, existen evidencias de que la combinación de MVA y B. japonium inducen una sinergia (Crop Watch, 2002; Dubey, 1999 y Young et al., 1988 ). Considerando la escasa información de estos simbiontes en soya y en esta zona, así como aprovechando siembras comerciales de este cultivo durante el ciclo primavera-verano, se realizó la presente validación para determinar el efecto de la inoculación de los simbiontes y el tratamiento a la semilla, sobre la productividad de la soya en campo.
MATERIALES Y MÉTODOS
El estudio se estableció en dos localidades bajo condiciones de temporal, en el “Pozo 4”, Valle Hermoso y en “La Jarita”, Matamoros, Tam., el 19 y 24 de julio de 2003, respectivamente. La siembra fue mecanizada en suelos sin fertilización química y en

67
ambos sitios se usó la variedad Huasteca-200. Los tratamientos evaluados fueron: 1) inoculación a la semilla con B. japonium (Nitragin, Liphatech Inc.) a razón de 400 g/ha (B); 2) inoculación a la semilla con MVA Glomus intraradices (INIFAP) a dosis de 1 kg/ha (M); 3) Vitavax 200® (20% carboxin y 20% thiram), 250 ml en 100 kg de semilla (V); 4) combinación de B + M; 5) combinación de V + B; 6) combinación de B + M + V; y 7) el testigo absoluto.
Los tratamientos se distribuyeron en franjas en una superficie de 16 surcos (0.86 m)
de 150 m de longitud en la “La Jarita” y de seis surcos (0.86 m) por 250 m de longitud en el “Pozo 4”. La densidad de población en la primera localidad fue de 18 plantas/m lineal (208,800 plantas/ha), mientras que en la segunda fue de 20 plantas/m (232,000 plantas/ha).
Dentro de cada tratamiento se marcaron ocho y cinco subparcelas (repeticiones) de
cuatro surcos de 5.0 m de longitud, en “La Jarita” y “Pozo 4”, respectivamente. En floración, se realizó un muestreo destructivo de cinco plantas en los surcos laterales de cada subparcela, en ellas se cuantificó la biomasa fresca, el número y peso de nódulos y biomasa seca. En madurez, se determinó la altura de planta, el número de vainas y el rendimiento de grano. Los valores del número y peso de los nódulos se transformaron mediante √x+1. La productividad (kg/ha) se determinó cosechando los dos surcos centrales (0.86 m) por 5.00 m de longitud (8.6 m 2). El análisis estadístico se realizó mediante comparaciones de parcelas apareadas (prueba de t, p < 0.05).
RESULTADOS Y DISCUSIÓN
En el Cuadro 1 se anotan las características de planta y rendimiento de soya con la variedad Huasteca-200, asociadas con la inoculación de simbiontes en las dos localidades del norte de Tamaulipas. Se observa que en las dos localidades, la biomasa fresca y seca se incrementaron significativamente con los simbiontes, cuando fueron inoculados solos o en combinación. La diferencia en promedio de biomasa fresca y seca entre la localidad “La Jarita” (104.33 y 28.87) y la de “Pozo 4” (74.56 y 18.70) se atribuye a la diferencia en el vigor o grosor del tallo entre una y otra localidad, esto debido principalmente a que en “La Jarita” la población promedio de plantas por metro lineal fue de 18 y en el “Pozo 4” fue de 20. Además se refleja en la altura de planta promedio registrada en una y otra localidad, que para “La Jarita” fue de 64.03 y para el “Pozo 4” de 90.16 cm.
El mayor número y peso de nódulos en ambas localidades se registraron en los
tratamientos con simbiontes solos y asociados; y los menores valores se observan para el tratamiento con el fungicida Vitavax 200 y el testigo absoluto, esto se debe a la pobre población de cepas afines al cultivo, debido a la ausencia de siembras de soya en esta región. Este fenómeno también se ha observado en soya en otras regiones donde tradicionalmente no se siembra este cultivo (Jiménez et al., 1991). Respecto al tratamiento de Vitavax 200, puede anotarse que es evidente que cuando se combinó con los simbiontes, se redujo significativamente el número y peso de los nódulos, y en la mayoría de los casos, el fungicida influyó negativamente en las demás variables observadas.
El Vitavax 200 es de uso común para el control de dampig-off en algunas regiones productoras de soya (Revellin et al., 1993; Wendell, 1988 y Maldonado, 2002). Sin embargo, existen evidencias de que algunos fungicidas tienen un alto nivel de toxicidad, como el thiram y captan, que afectan la multiplicación de B. japonicum, por el contrario,

68
fungicidas como el metalaxyl no tienen efecto adverso sobre la bacteria fijadora de nitrógeno (Crop Watch, 2002).
Respecto al número de vainas en ambas localidades, los menores valores se
observaron para el tratamiento con Vitavax 200 y el testigo absoluto; los mayores valores se registraron en los tratamientos con simbiontes solos o asociados. En la localidad “La Jarita”, la combinación de simbiontes (B + M) incrementaron el rendimiento de grano en un 271 % en relación al testigo absoluto. Lo anterior coincide con lo informado por Young et al., (1988) quienes consignaron un incremento del 126 % con la mezcla de los dos simbiontes. Otros autores han informado un efecto sinérgico con la combinación de rizobacteria-micorriza MVA en soya y otras leguminosas. En la localidad del “Pozo 4”, el incremento de la misma combinación, fue de 398 % en relación al testigo absoluto.
CONCLUSIONES
1) De acuerdo con el rendimiento reportado con algunos tratamientos (1954 kg/ha), el
cual es similar al de otras regiones productoras de soya, se reafirma que este cultivo representa un alternativa potencial para el norte de Tamaulipas.
2) La literatura lo anota y los datos de este estudio lo confirman, que el efecto antagónico del fungicida Vitavax 200, principalmente con base de thiram, cuestiona el uso de este fungicida para el tratamiento a la semilla en esta y otras zonas productoras de soya.
3) Por la importancia adquirida y su efecto sinérgico de la MVA, se sugiere la evaluación de estos productos bajo diferentes condiciones ambientales para generar una recomendación específica para la zona norte de Tamaulipas.
REFERENCIAS
Crop Watch. 2002. Focus on soybean production, plus army cutworm. http://cropwatch.unl.edu/archives/2002/crop02-5.htm
Danielshylton, K.D.M. and Ahmad, M.H. 1994. Inoculation response in kidney beans (Phaseolus vulgarus) to vesicular-arbuscular mycorrhizal and rhizobia in non-sterilized soil. Biol & Fert. Soils. 18(2): 95-98.
Dubey, S.K. 1999. Response of soybean (Glycine max) to biofertilizers with and without nitrogen, phosphorus and potassium on swell-shrink soil. Indian J. Agron. 43 (3): 546-549.
Elizondo B., J. 1999. Uso de biofertilizantes a base de Bradyrhizobium japonicum para mejorar la producción de soya. Campo Experimental Sur de Tamaulipas. INIFAP. Desplegable Nº 3.
Jiménez J. A., Olalde P. V. y Peña C. 1991. Evaluación en campo de Bradyrhizobium japonicum en dos variedades de soya (Glycine max). Terra 9 (1): 79-86.
Maldonado M. N. 2002. El cultivo de soya de invierno en el sur de Tamaulipas. Campo Experimental Sur de Tamaulipas, INIFAP. Desplegable para productores Nº 2.
Revellin C., Leterme, P. and Catroux, G. 1993. Effect of some fungicides treatments on the survival of Bradyrhizobium japonicum and on the nodulation and yield of soybean. Biol. & Fert. Soils. 16: 211-214.
Sine, C . 1993. Plant Health Guide. Meister Publishing Co. 178 p.
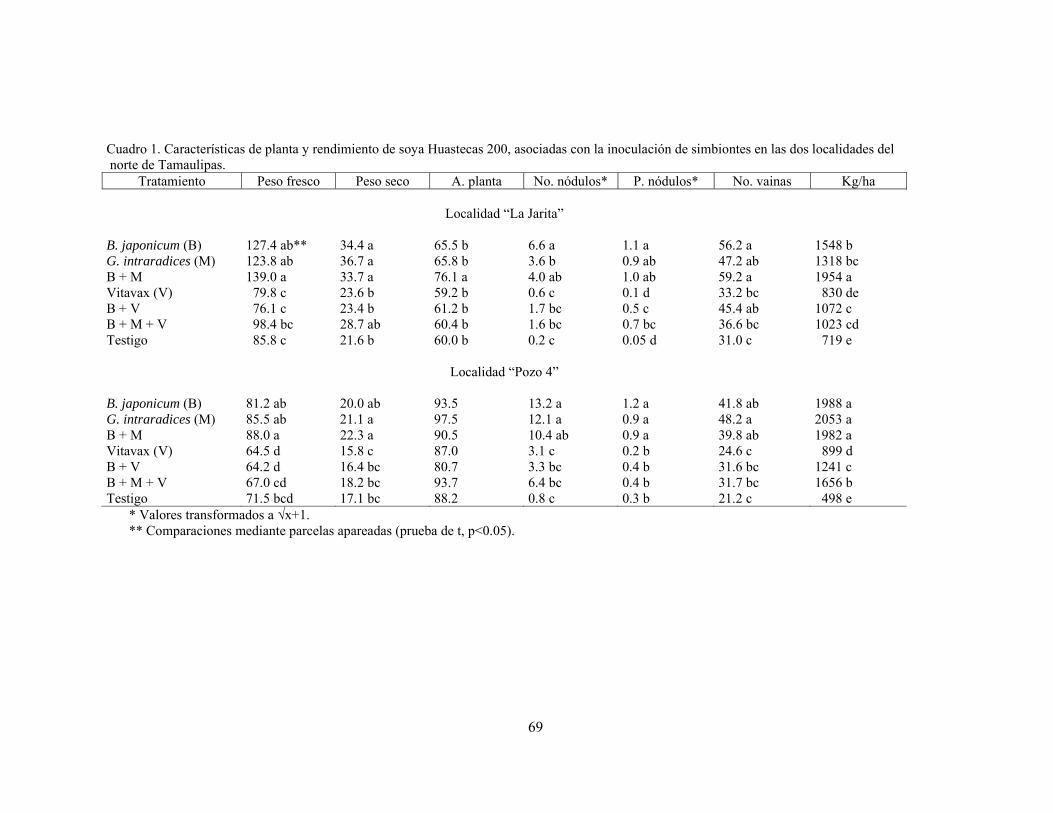
69
Cuadro 1. Características de planta y rendimiento de soya Huastecas 200, asociadas con la inoculación de simbiontes en las dos localidades del norte de Tamaulipas.
Tratamiento Peso fresco Peso seco A. planta No. nódulos* P. nódulos* No. vainas Kg/ha
Localidad “La Jarita”
B. japonicum (B) 127.4 ab** 34.4 a 65.5 b 6.6 a 1.1 a 56.2 a 1548 b G. intraradices (M) 123.8 ab 36.7 a 65.8 b 3.6 b 0.9 ab 47.2 ab 1318 bc B + M 139.0 a 33.7 a 76.1 a 4.0 ab 1.0 ab 59.2 a 1954 a Vitavax (V) 79.8 c 23.6 b 59.2 b 0.6 c 0.1 d 33.2 bc 830 de B + V 76.1 c 23.4 b 61.2 b 1.7 bc 0.5 c 45.4 ab 1072 c B + M + V 98.4 bc 28.7 ab 60.4 b 1.6 bc 0.7 bc 36.6 bc 1023 cd Testigo 85.8 c 21.6 b 60.0 b 0.2 c 0.05 d 31.0 c 719 e
Localidad “Pozo 4”
B. japonicum (B) 81.2 ab 20.0 ab 93.5 13.2 a 1.2 a 41.8 ab 1988 a G. intraradices (M) 85.5 ab 21.1 a 97.5 12.1 a 0.9 a 48.2 a 2053 a B + M 88.0 a 22.3 a 90.5 10.4 ab 0.9 a 39.8 ab 1982 a Vitavax (V) 64.5 d 15.8 c 87.0 3.1 c 0.2 b 24.6 c 899 d B + V 64.2 d 16.4 bc 80.7 3.3 bc 0.4 b 31.6 bc 1241 c B + M + V 67.0 cd 18.2 bc 93.7 6.4 bc 0.4 b 31.7 bc 1656 b Testigo 71.5 bcd 17.1 bc 88.2 0.8 c 0.3 b 21.2 c 498 e
* Valores transformados a √x+1. ** Comparaciones mediante parcelas apareadas (prueba de t, p<0.05).

70
Tarafdar, J. C. and Rao, A. V. 1997. Response of arid legumes to VAM fungal inoculation. Simbiosis 22 (3): 265-274.
Wendell, H. C. 1988. Texas Plant Disease Control Handbook Chemical Control Supplement. Texas Agr. Ext. Serv. and Texas A & M University System. Publ. B-1140 A.
Young, C. C., T. C. Juang, and C. C. Chao. 1988. Effects of Rhizobium and vesicular-arbuscular mycorrhiza inoculations on nodulation, symbiotic nitrogen fixation, and soybean yield in subtropical-tropical field. Biol. & Fertil. Soils 6: 165-169.

71
BIOFERTILIZANTES MICROBIANOS: ANTECEDENTES DEL PROGRAMA Y RESULTADOS DE VALIDACIÓN EN MÉXICO
Juan Francisco Aguirre Medina
Campo Experimental Rosario Izapa, INIFAP
Introducción
La agricultura siempre ha estado relacionada con la sociedad para satisfacer sus necesidades alimenticias (Hernández, 1988) y en la actualidad, la búsqueda de una mayor productividad por unidad de área, como consecuencia del incremento de nuestra población, ha originado la alteración de las comunidades vegetales. Esta actividad antropocéntrica ha acelerado la degradación de los sistemas agrícolas y como consecuencia, los sistemas de producción enfrentan el reto de mantener una producción sostenida.
Describir y entender los sistemas biológicos de cada cultivo para lograr un impacto en la producción y conservar el medio, consolida la importancia de estudiar el sistema planta-microsimbionte. La unidad e interdependencia de esta relación es poco conocida y poco apreciada. El sistema radical ha sido llamado, el componente olvidado (Davidson, 1978), la mitad escondida (Waisel et al., 2002), aunque para muchas plantas representa mucho más que la parte aérea. El sistema radical, además de las funciones de absorción y transporte de agua y nutrientes al sistema vascular y el anclaje, pone a la planta en contacto con la rizosfera. Esta región es el único ambiente bajo la influencia de las raíces de las plantas (Alexander, 1977) que favorecen la proliferación de microorganismos (Balandreau y Knowles, 1978). Existen evidencias que la relación se ha desarrollado desde que inició la colonización de la superficie terrestre por las plantas y, algunas de las primeras algas en ocupar la tierra, estaban asociadas a microorganismos, entre los cuales se encontraban los que serían las micorrizas (Azcón y Barea, 1980; Selosse y Le Tacon, 1998). La relación simbiótica planta-microorganismo ha contribuido al mantenimiento, funcionamiento y la estabilidad de los ecosistemas mediante su influencia en la diversidad de las especies en las comunidades vegetales (Read, 1998), sin embargo la actividad agrícola, ha sido una prueba crucial para la recirculación de los componentes de la naturaleza (Altieri et al, 1984), como es el caso de los nutrimentos.
La nutrición de las plantas se ha estudiado y desarrollado con mayor entusiasmo en dos grandes vertientes, una de ellas, la más tradicional en los últimos 50 años, mediante productos químicos sintéticos y la otra, explorando la capacidad que tienen algunos microorganismos para fijar nitrógeno y transportar fósforo y otros nutrimentos. Últimamente además, se ha puesto de manifiesto el interés por la fermentación de los residuos orgánicos para abonar los cultivos mediante el desarrollo de compostas o bien con la utilización de lombrices para producir la vermicomposta.
De los nutrimentos esenciales para el desarrollo de las plantas, el nitrógeno es el más importante y el más utilizado en la agricultura. Aún cuando, abunda en la atmósfera terrestre, es inerte y solamente puede ser aprovechado cuando se combina industrialmente

72
con ciertos elementos, como el hidrógeno, mediante la utilización de altas cantidades de energía proveniente del petróleo ó el carbón, o bien, mediante su incorporación a los sistemas biológicos cuando es fijado por algunos microorganismos que realizan una simbiosis mutualística con las plantas superiores. En este caso, la energía utilizada proviene del sol, vía la fotosíntesis de las plantas.
Los fertilizantes químicos sintéticos utilizados para la producción de alimentos, han estado cumpliendo su función hasta cierto límite. Han sido fundamentales en el abastecimiento de la demanda alimenticia de nuestra creciente población nacional, sin embargo, en la última década nuestra sociedad reclama mayor producción de alimentos de alta calidad proteínica, pero, producidos sin agroquímicos. Esta nueva aptitud ha favorecido el desarrollo de tecnologías de producción menos contaminantes y ecológicamente más racionales.
Con estos antecedentes y después de la crisis energética mundial de los años setenta, el estudio de las bacterias asociadas a las plantas avanzó rápidamente en algunos países europeos y asiáticos y en menor grado en México y otros países latinoamericanos (Okon y Labandera, 1994). Sin embargo en los últimos años, se han obtenido resultados satisfactorios en México con la aplicación masiva de diversos microorganismos en campo en cultivos anuales (Aguirre, 2001) y algunos perennes tropicales en vivero con Azospirillum, y hongos micorrizógenos (Mendoza y Aguirre, 2002; Moroyoqui y Aguirre, 2002).
El uso de los recursos microbiológicos del suelo en los sistemas agrícolas son una alternativas para mejorar el balance biológico en el suelo y reducir el uso de fertilizantes químicos y de otros agroquímicos en los sistemas de producción, principalmente los fertilizantes nitrogenados y fosfatados. En este sentido, el uso de microorganismos en la agricultura han sido denominado inoculantes (Sylvester-Bradley et al., 1985), fertilizantes bacterianos (Dommergues, 1978) y últimamente biofertilizantes. En general constituyen un componente vital de los sistemas sostenibles al crear un medio económicamente atractivo y ecológicamente aceptable de reducir las aplicaciones de algunos agroquímicos y de mejorar, y hacer más eficientes, los nutrientes disponibles en cada hábitat. Realizan un amplio rango de actividades, como mejorar la disponibilidad y transporte de nutrimentos, favorecer el crecimiento de las raíces, la fijación del nitrógeno, la solubilización de minerales, la producción de estimuladores del crecimiento vegetal y el biocontrol de patógenos. (Barea y Azcón, 1983; Rovira et al., 1983).
Los “biofertilizantes microbianos” pueden utilizarse en los cultivos anuales, las praderas de gramíneas y leguminosas, hortalizas y frutales. Son una alternativa de gran validez para los campesinos que no fertilizan o lo hacen con pequeñas cantidades. Estos productos son recomendados en la Agenda 21 como resultado de la llamada Cumbre de la Tierra, firmada en Río de Janeiro en 1992. Son considerados como biotecnologías “apropiables”, que es un término creado para las herramientas biotecnológicas que contribuyen al desarrollo sostenible por ser técnicamente factibles dentro del nivel científico-técnico de un país y que proveen beneficios tangibles a los destinatarios y además por ser ambientalmente seguras y socioeconómica y culturalmente aceptables.

73
Antecedentes del programa
La agricultura nacional se desarrolla en poco mas de 17 millones de hectáreas en cultivos anuales, de las cuales 11 millones (65%) se cultivan bajo temporal (SAGAR, 1997). El 79 % de la superficie se cultiva con gramíneas como maíz, sorgo, avena, cebada, trigo y arroz que en su conjunto cubren cerca del 79% de la superficie nacional de temporal. Si consideramos al frijol en este análisis, la superficie supera el 95%, con excepción de las áreas de temporal con alto potencial productivo. Bajo estas condiciones de agricultura de temporal que la mayoría es de subsistencia con suelos marginales, los productores no aplican fertilizantes químicos a sus cultivos o lo hacen en pequeñas cantidades y, es en estos sistemas de producción donde se presenta la alternativa de utilizar algunos recursos microbiológicos del suelo para favorecer la nutrición mineral de las plantas y reducir o hacer más eficiente, cuando sea el caso, los fertilizantes químicos que se utilizan en diferentes sistemas agroecológicos.
Los biofertilizantes en México han sido poco utilizados por los agricultores, su uso en la producción agrícola nacional ha tenido su mayor relevancia en las leguminosas, como el caso de la soya y en algunos casos el frijol. Las gramíneas no habían sido sometidas en grandes extensiones con alguna simbiosis vía comercial en su sistema radical. La aplicación de estos productos, los biofertilizantes, que integran los microorganismos, Azospirillum brasilense, Rhizobium etli, Bradyrhizobium japonicum y el hongo micorrízico Glomus intraradices, en las áreas menos favorecidas, representan una oportunidad para incrementar la producción y productividad de los cultivos a bajo costo sin la contaminación de los mantos freáticos y a la vez favorecer la conservación de los recursos naturales.
La amplia exploración en ambientes y sistemas de manejo son una excelente herramienta para la promoción de esta componente ante los diferentes agentes de cambio en el país, autoridades del sector y por supuesto ante los productores de los diferentes cultivos. En todos los casos se pretende establecer un vínculo más directo entre el sector productivo y la nueva opción tecnológica que se les presenta. Este proceso, pretendió además, mostrar las ventajas y eficiencias en el desarrollo vegetal (aéreo y radical) y reproductivo de los cultivos biofertilizados con alguno de los microsimbiontes o con ambos, en comparación con los diferentes testigos seleccionados por los investigadores, que pueden ser la reducción de la fertilización química ó la suspensión total de la misma. Las validaciones se desarrollaron de acuerdo a la pertinencia con algún productor junto o por separados los microsimbiontes, en varios cultivos, y en diferentes ambientes agroecológicos y sistemas de producción.
Desarrollo del programa
La validación de una componente tecnológica tiene la característica de realizarse en superficies más grandes que las desarrolladas en los procesos experimentales y además, de ser manejada agronómicamente por el productor. En este caso, la componente biofertilizantes microbianos se implementó como seguimiento al Programa Nacional de Biofertilizantes desarrollado por le Gobierno Federal a partir de 1999. Este programa se originó como una alternativa para favorecer la nutrición de las plantas por medios biológicos mediante el planteamiento de diferentes estrategias, entre las cuales se consideró la exploración de diversas regiones agroecológicas de México. El proceso de producción de

74
biofertilizantes se realizó a través de la Fundación Mexicana para la Investigación Agropecuaria y Forestal A.C., y su distribución a los productores, mediante el programa del Gobierno Federal Alianza para el Campo. Los microorganismos utilizados masivamente como biofertilizante fueron las bacterias Azospirillum brasilense2, Rhizobium etli1, Bradyrhizobium japonicum y el hongo micorrizógeno Glomus intraradices. Se evaluaron durante los ciclos agrícolas Primavera-Verano de 1999, Otoño-Invierno 1999-2000 y Primavera-Verano 2000. El número de parcelas por estado y ciclo agrícola se presenta en el Cuadro 1.
Metodología de evaluación
Las inoculaciones de la semilla se hicieron al momento de la siembra utilizándose carboxi metil celulosa como adherente. La cantidad de biofertilizante fue de 280 g por 20-25 kg de semilla, para el caso de las bacterias y de 1-2 kg de micorriza-arbuscular para la misma cantidad de semilla. El número de bacterias contenido en el producto fue de al menos 100 x 106 y la micorriza con 1000 esporas/gramo de suelo y 90-95 % de infección en raíces.
Los tratamientos evaluados fueron el biofertilizante con un microorganismo o bien la combinación de dos de ellos. En algunos casos, se aplicó una dosis de fertilidad recomendada por el INIFAP en la región, una parte de la misma o bien sin aplicación de fertilizante químico. Las parcelas fueron de al menos una hectárea y se establecieron bajo condiciones de temporal. En todos los casos el manejo del cultivo estuvo a cargo del productor cooperante y los científicos del INIFAP solo dieron seguimiento técnico a las parcelas. Las variedades utilizadas fueron diferentes entre localidades de acuerdo a la decisión del productor.
Resultados
Se presentan los resultados de rendimiento por cultivo en las diferentes regiones donde se ejecutó el programa. Frijol
El rendimiento de las diferentes variedades de frijol fue superior cuando se biofertilizó con un microorganismo o la combinación de dos de ellos, Rhizobium etli y/o Glomus intraradices en comparación, con el testigo sin aplicación de biofertilizante. Las diferencias encontradas en porcentaje van del 16 al 70% con algún microsimbionte solo. Resultados importantes sin la aplicación de fertilizante han sido obtenidas en diversas regiones de México, como en Chiapas (Camas, 2000), en Guerrero (Cruzaley, 2000), Veracruz (Durán et al., 2001), Coahuila (Covarrubias, et al., 2000) y Yucatán (Uribe,
2 Proporcionadas por el Centro Nacional de Fijación de Nitrógeno-UNAM, mediante convenio.
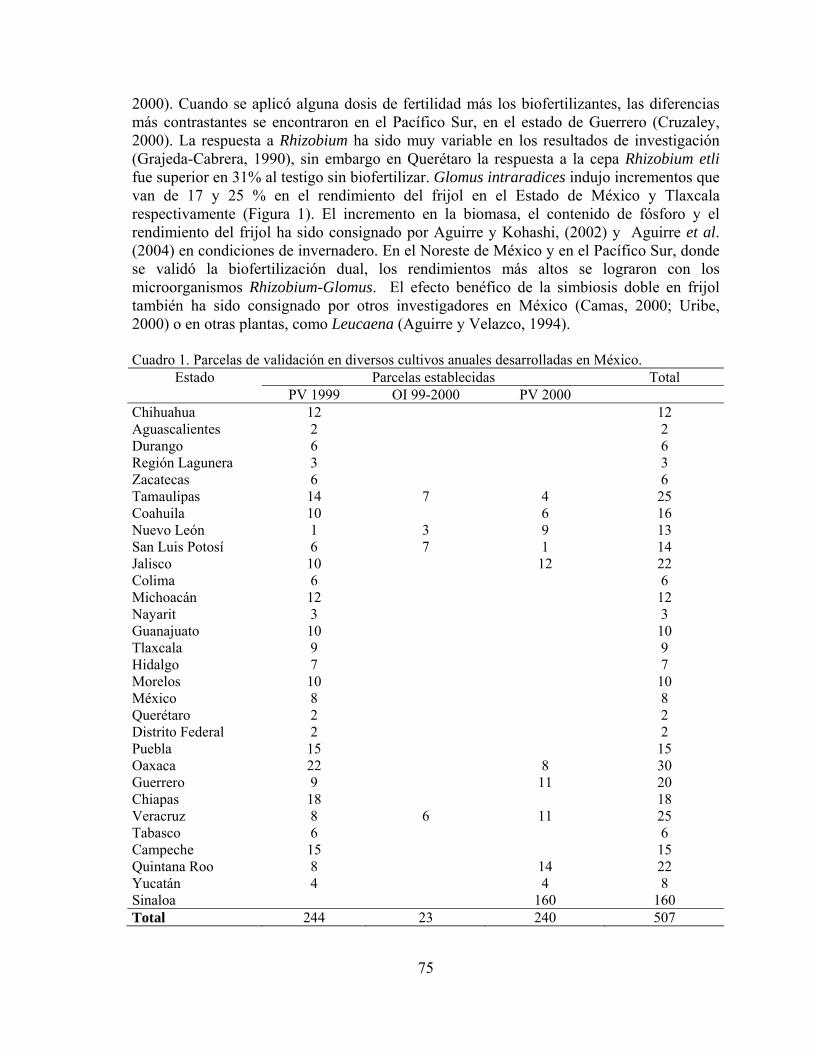
75
2000). Cuando se aplicó alguna dosis de fertilidad más los biofertilizantes, las diferencias más contrastantes se encontraron en el Pacífico Sur, en el estado de Guerrero (Cruzaley, 2000). La respuesta a Rhizobium ha sido muy variable en los resultados de investigación (Grajeda-Cabrera, 1990), sin embargo en Querétaro la respuesta a la cepa Rhizobium etli fue superior en 31% al testigo sin biofertilizar. Glomus intraradices indujo incrementos que van de 17 y 25 % en el rendimiento del frijol en el Estado de México y Tlaxcala respectivamente (Figura 1). El incremento en la biomasa, el contenido de fósforo y el rendimiento del frijol ha sido consignado por Aguirre y Kohashi, (2002) y Aguirre et al. (2004) en condiciones de invernadero. En el Noreste de México y en el Pacífico Sur, donde se validó la biofertilización dual, los rendimientos más altos se lograron con los microorganismos Rhizobium-Glomus. El efecto benéfico de la simbiosis doble en frijol también ha sido consignado por otros investigadores en México (Camas, 2000; Uribe, 2000) o en otras plantas, como Leucaena (Aguirre y Velazco, 1994).
Cuadro 1. Parcelas de validación en diversos cultivos anuales desarrolladas en México.
Parcelas establecidas Total Estado PV 1999 OI 99-2000 PV 2000
Chihuahua 12 12 Aguascalientes 2 2 Durango 6 6 Región Lagunera 3 3 Zacatecas 6 6 Tamaulipas 14 7 4 25 Coahuila 10 6 16 Nuevo León 1 3 9 13 San Luis Potosí 6 7 1 14 Jalisco 10 12 22 Colima 6 6 Michoacán 12 12 Nayarit 3 3 Guanajuato 10 10 Tlaxcala 9 9 Hidalgo 7 7 Morelos 10 10 México 8 8 Querétaro 2 2 Distrito Federal 2 2 Puebla 15 15 Oaxaca 22 8 30 Guerrero 9 11 20 Chiapas 18 18 Veracruz 8 6 11 25 Tabasco 6 6 Campeche 15 15 Quintana Roo 8 14 22 Yucatán 4 4 8 Sinaloa 160 160 Total 244 23 240 507
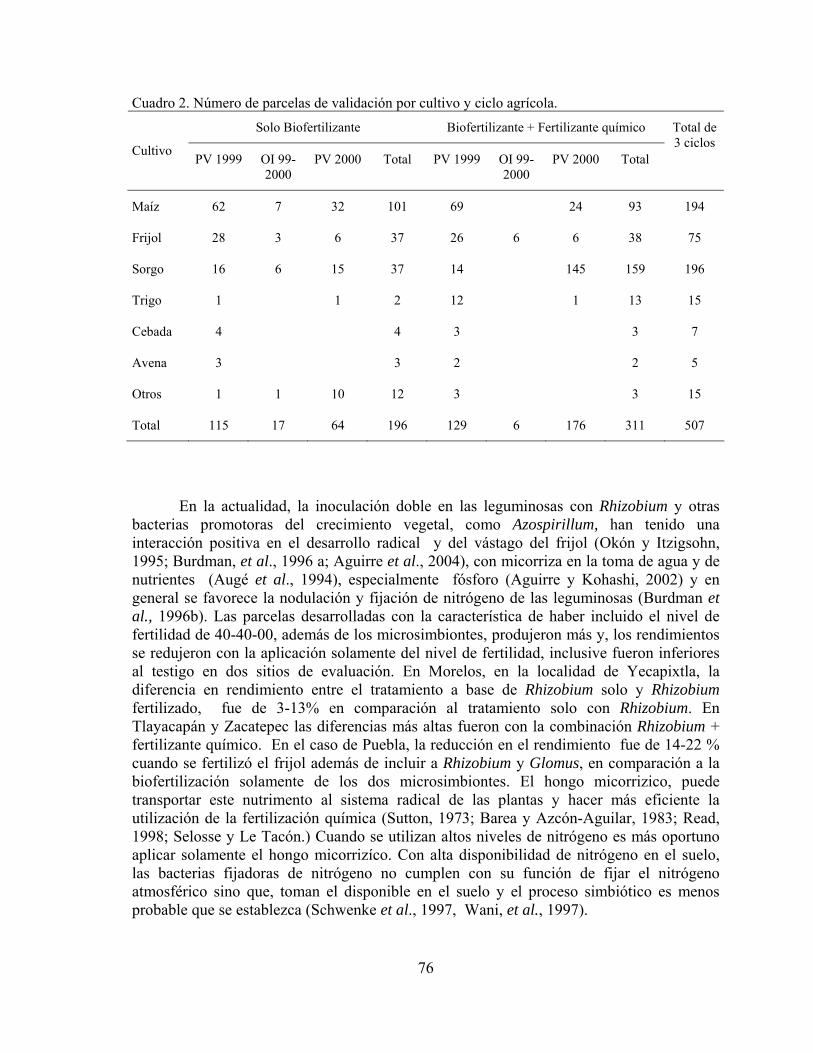
76
Cuadro 2. Número de parcelas de validación por cultivo y ciclo agrícola.
Solo Biofertilizante Biofertilizante + Fertilizante químico
Cultivo PV 1999 OI 99-2000
PV 2000 Total PV 1999 OI 99-2000
PV 2000 Total
Total de 3 ciclos
Maíz 62 7 32 101 69 24 93 194
Frijol 28 3 6 37 26 6 6 38 75
Sorgo 16 6 15 37 14 145 159 196
Trigo 1 1 2 12 1 13 15
Cebada 4 4 3 3 7
Avena 3 3 2 2 5
Otros 1 1 10 12 3 3 15
Total 115 17 64 196 129 6 176 311 507
En la actualidad, la inoculación doble en las leguminosas con Rhizobium y otras bacterias promotoras del crecimiento vegetal, como Azospirillum, han tenido una interacción positiva en el desarrollo radical y del vástago del frijol (Okón y Itzigsohn, 1995; Burdman, et al., 1996 a; Aguirre et al., 2004), con micorriza en la toma de agua y de nutrientes (Augé et al., 1994), especialmente fósforo (Aguirre y Kohashi, 2002) y en general se favorece la nodulación y fijación de nitrógeno de las leguminosas (Burdman et al., 1996b). Las parcelas desarrolladas con la característica de haber incluido el nivel de fertilidad de 40-40-00, además de los microsimbiontes, produjeron más y, los rendimientos se redujeron con la aplicación solamente del nivel de fertilidad, inclusive fueron inferiores al testigo en dos sitios de evaluación. En Morelos, en la localidad de Yecapixtla, la diferencia en rendimiento entre el tratamiento a base de Rhizobium solo y Rhizobium fertilizado, fue de 3-13% en comparación al tratamiento solo con Rhizobium. En Tlayacapán y Zacatepec las diferencias más altas fueron con la combinación Rhizobium + fertilizante químico. En el caso de Puebla, la reducción en el rendimiento fue de 14-22 % cuando se fertilizó el frijol además de incluir a Rhizobium y Glomus, en comparación a la biofertilización solamente de los dos microsimbiontes. El hongo micorrizico, puede transportar este nutrimento al sistema radical de las plantas y hacer más eficiente la utilización de la fertilización química (Sutton, 1973; Barea y Azcón-Aguilar, 1983; Read, 1998; Selosse y Le Tacón.) Cuando se utilizan altos niveles de nitrógeno es más oportuno aplicar solamente el hongo micorrizíco. Con alta disponibilidad de nitrógeno en el suelo, las bacterias fijadoras de nitrógeno no cumplen con su función de fijar el nitrógeno atmosférico sino que, toman el disponible en el suelo y el proceso simbiótico es menos probable que se establezca (Schwenke et al., 1997, Wani, et al., 1997).
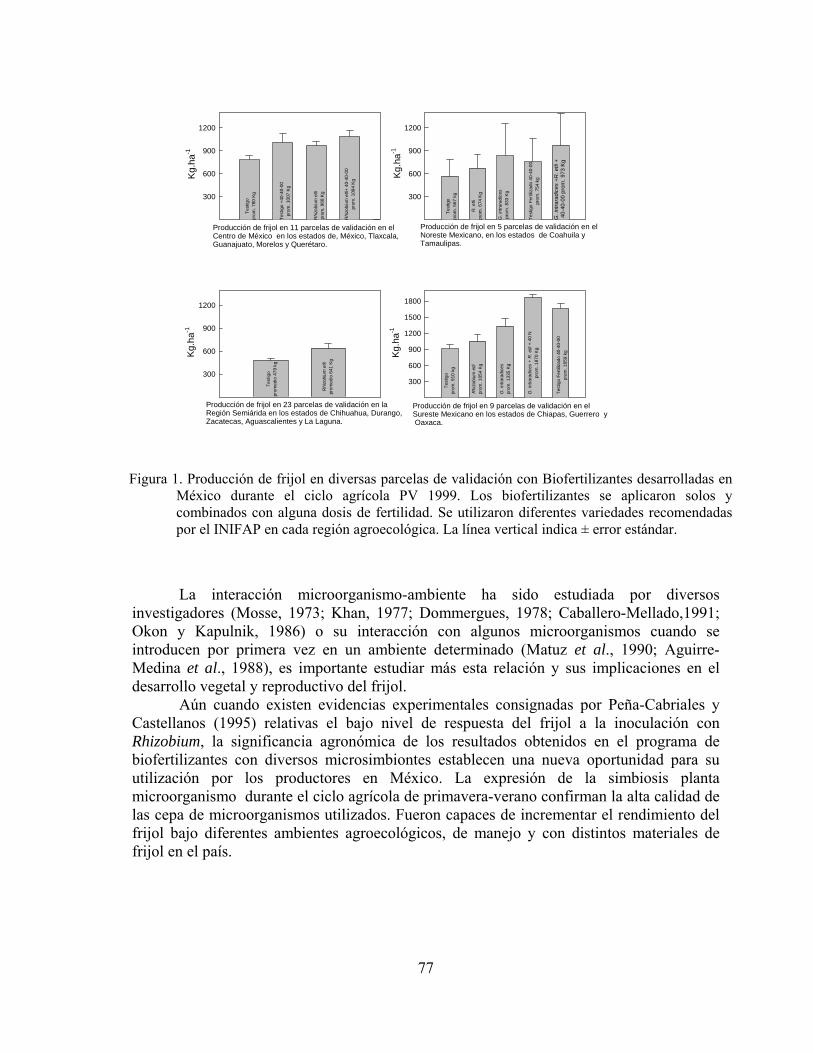
77
La interacción microorganismo-ambiente ha sido estudiada por diversos investigadores (Mosse, 1973; Khan, 1977; Dommergues, 1978; Caballero-Mellado,1991; Okon y Kapulnik, 1986) o su interacción con algunos microorganismos cuando se introducen por primera vez en un ambiente determinado (Matuz et al., 1990; Aguirre-Medina et al., 1988), es importante estudiar más esta relación y sus implicaciones en el desarrollo vegetal y reproductivo del frijol.
Aún cuando existen evidencias experimentales consignadas por Peña-Cabriales y Castellanos (1995) relativas el bajo nivel de respuesta del frijol a la inoculación con Rhizobium, la significancia agronómica de los resultados obtenidos en el programa de biofertilizantes con diversos microsimbiontes establecen una nueva oportunidad para su utilización por los productores en México. La expresión de la simbiosis planta microorganismo durante el ciclo agrícola de primavera-verano confirman la alta calidad de las cepa de microorganismos utilizados. Fueron capaces de incrementar el rendimiento del frijol bajo diferentes ambientes agroecológicos, de manejo y con distintos materiales de frijol en el país.
Kg.h
a-1300
600
900
1200
Producción de frijol en 11 parcelas de validación en el Centro de México en los estados de, México, Tlaxcala, Guanajuato, Morelos y Querétaro.
Kg.h
a-1
300
600
900
1200
Producción de frijol en 5 parcelas de validación en elNoreste Mexicano, en los estados de Coahuila y Tamaulipas.
Kg.h
a-1
300
600
900
1200
Test
igo
prom
edio
479
kg
Rhi
zobi
um e
tlipr
omed
io 6
41 K
g
Producción de frijol en 23 parcelas de validación en la Región Semiárida en los estados de Chihuahua, Durango, Zacatecas, Aguascalientes y La Laguna.
Kg.h
a-1300
600
900
1200
1500
1800
Test
igo
prom
. 910
kg
Rhi
zobi
um e
tlipr
om. 1
054
Kg
Producción de frijol en 9 parcelas de validación en elSureste Mexicano en los estados de Chiapas, Guerrero y Oaxaca.
G. i
ntra
radi
ces
prom
. 133
5 Kg
G. i
ntra
radi
ces
+ R
. etli
+ 4
0 N
prom
. 187
0 Kg
Test
igo
Ferti
lizad
o 40
-40-
00pr
om. 1
659
kg
Test
igo
prom
. 780
Kg
Rhi
zobi
um e
tli+
40-4
0-00
prom
. 108
4 Kg
Test
igo
+40-
40-0
0pr
om. 1
007
Kg
Rhi
zobi
um e
tlipr
om. 9
68 K
g
Test
igo
prom
. 567
kg
R. e
tlipr
om. 6
74 K
g
G. i
ntra
radi
ces
prom
. 833
Kg
G. i
ntra
radi
ces
+R. e
tli +
40
-40-
00 p
rom
. 973
Kg
Test
igo
Ferti
lizad
o 40
-40-
00pr
om. 7
54 k
g
Figura 1. Producción de frijol en diversas parcelas de validación con Biofertilizantes desarrolladas en México durante el ciclo agrícola PV 1999. Los biofertilizantes se aplicaron solos y combinados con alguna dosis de fertilidad. Se utilizaron diferentes variedades recomendadas por el INIFAP en cada región agroecológica. La línea vertical indica ± error estándar.
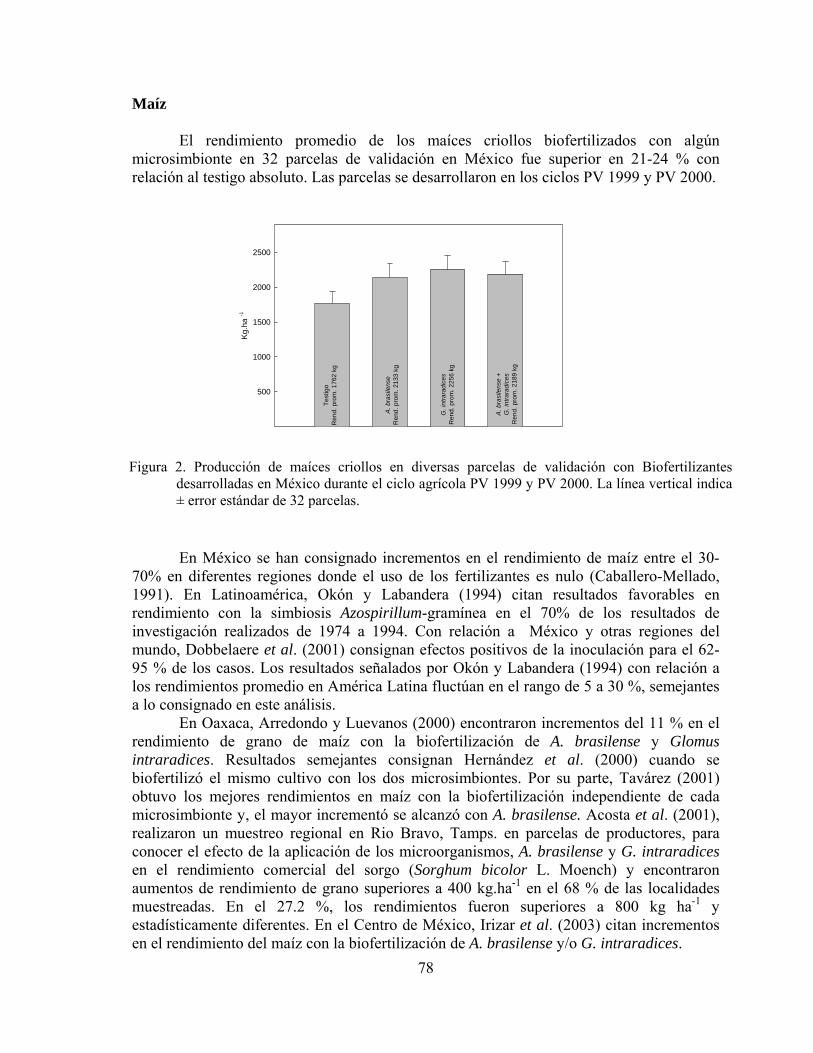
78
Maíz El rendimiento promedio de los maíces criollos biofertilizados con algún
microsimbionte en 32 parcelas de validación en México fue superior en 21-24 % con relación al testigo absoluto. Las parcelas se desarrollaron en los ciclos PV 1999 y PV 2000.
En México se han consignado incrementos en el rendimiento de maíz entre el 30-70% en diferentes regiones donde el uso de los fertilizantes es nulo (Caballero-Mellado, 1991). En Latinoamérica, Okón y Labandera (1994) citan resultados favorables en rendimiento con la simbiosis Azospirillum-gramínea en el 70% de los resultados de investigación realizados de 1974 a 1994. Con relación a México y otras regiones del mundo, Dobbelaere et al. (2001) consignan efectos positivos de la inoculación para el 62-95 % de los casos. Los resultados señalados por Okón y Labandera (1994) con relación a los rendimientos promedio en América Latina fluctúan en el rango de 5 a 30 %, semejantes a lo consignado en este análisis.
En Oaxaca, Arredondo y Luevanos (2000) encontraron incrementos del 11 % en el rendimiento de grano de maíz con la biofertilización de A. brasilense y Glomus intraradices. Resultados semejantes consignan Hernández et al. (2000) cuando se biofertilizó el mismo cultivo con los dos microsimbiontes. Por su parte, Tavárez (2001) obtuvo los mejores rendimientos en maíz con la biofertilización independiente de cada microsimbionte y, el mayor incrementó se alcanzó con A. brasilense. Acosta et al. (2001), realizaron un muestreo regional en Rio Bravo, Tamps. en parcelas de productores, para conocer el efecto de la aplicación de los microorganismos, A. brasilense y G. intraradices en el rendimiento comercial del sorgo (Sorghum bicolor L. Moench) y encontraron aumentos de rendimiento de grano superiores a 400 kg.ha-1 en el 68 % de las localidades muestreadas. En el 27.2 %, los rendimientos fueron superiores a 800 kg ha-1 y estadísticamente diferentes. En el Centro de México, Irizar et al. (2003) citan incrementos en el rendimiento del maíz con la biofertilización de A. brasilense y/o G. intraradices.
Kg.h
a -1
500
1000
1500
2000
2500
Test
igo
Ren
d. p
rom
. 176
2 kg
A. b
rasi
lens
eR
end.
pro
m. 2
133
kg
G. i
ntra
radi
ces
Ren
d. p
rom
. 225
6 kg
A. b
rasi
lens
e +
G. i
ntra
radi
ces
Ren
d. p
rom
. 218
9 kg
Figura 2. Producción de maíces criollos en diversas parcelas de validación con Biofertilizantes desarrolladas en México durante el ciclo agrícola PV 1999 y PV 2000. La línea vertical indica ± error estándar de 32 parcelas.
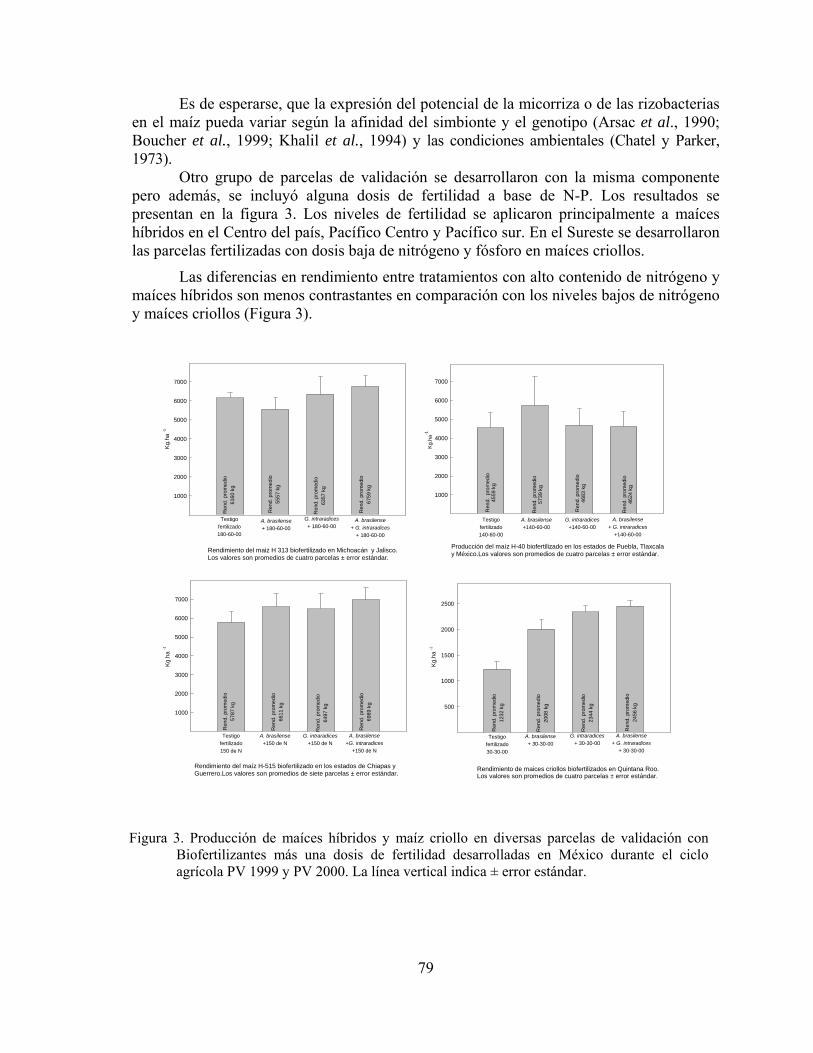
79
Es de esperarse, que la expresión del potencial de la micorriza o de las rizobacterias en el maíz pueda variar según la afinidad del simbionte y el genotipo (Arsac et al., 1990; Boucher et al., 1999; Khalil et al., 1994) y las condiciones ambientales (Chatel y Parker, 1973).
Otro grupo de parcelas de validación se desarrollaron con la misma componente pero además, se incluyó alguna dosis de fertilidad a base de N-P. Los resultados se presentan en la figura 3. Los niveles de fertilidad se aplicaron principalmente a maíces híbridos en el Centro del país, Pacífico Centro y Pacífico sur. En el Sureste se desarrollaron las parcelas fertilizadas con dosis baja de nitrógeno y fósforo en maíces criollos.
Las diferencias en rendimiento entre tratamientos con alto contenido de nitrógeno y maíces híbridos son menos contrastantes en comparación con los niveles bajos de nitrógeno y maíces criollos (Figura 3).
Kg.h
a-1
1000
2000
3000
4000
5000
6000
7000
Testigofertilizado140-60-00
A. brasilense+140-60-00
G. intraradices+140-60-00
A. brasilense+ G. intraradices
+140-60-00
Producción del maíz H-40 biofertilizado en los estados de Puebla, Tlaxcalay México.Los valores son promedios de cuatro parcelas ± error estándar.
Ren
d. p
rom
edio
4559
kg
Ren
d. p
rom
edio
5739
kg
Ren
d. p
rom
edio
4683
kg
Ren
d. p
rom
edio
4624
kg
Kg.h
a -1
1000
2000
3000
4000
5000
6000
7000
Testigofertilizado180-60-00
A. brasilense+ 180-60-00
G. intraradices+ 180-60-00
A. brasilense+ G. intraradices
+ 180-60-00
Ren
d. p
rom
edio
6160
kg
Ren
d. p
rom
edio
5557
kg
Ren
d. p
rom
edio
6357
kg
Ren
d. p
rom
edio
6759
kg
Rendimiento del maiz H 313 biofertilizado en Michoacán y Jalisco.Los valores son promedios de cuatro parcelas ± error estándar.
Kg.h
a -1
1000
2000
3000
4000
5000
6000
7000
Testigofertilizado150 de N
A. brasilense+150 de N
G. intraradices+150 de N
A. brasilense+G. intraradices
+150 de N
Rendimiento del maíz H-515 biofertilizado en los estados de Chiapas y Guerrero.Los valores son promedios de siete parcelas ± error estándar.
Ren
d. p
rom
edio
5787
kg
Ren
d. p
rom
edio
6611
kg
Ren
d. p
rom
edio
6497
kg
Ren
d. p
rom
edio
6989
kg
Kg.h
a -1
500
1000
1500
2000
2500
Testigofertilizado30-30-00
A. brasilense+ 30-30-00
G. intraradices+ 30-30-00
A. brasilense+ G. intraradices
+ 30-30-00
Rendimiento de maices criollos biofertilizados en Quintana Roo.Los valores son promedios de cuatro parcelas ± error estándar.
Ren
d. p
rom
edio
1232
kg
Ren
d. p
rom
edio
2008
kg
Ren
d. p
rom
edio
2344
kg
Ren
d. p
rom
edio
2456
kg
Figura 3. Producción de maíces híbridos y maíz criollo en diversas parcelas de validación con Biofertilizantes más una dosis de fertilidad desarrolladas en México durante el ciclo agrícola PV 1999 y PV 2000. La línea vertical indica ± error estándar.
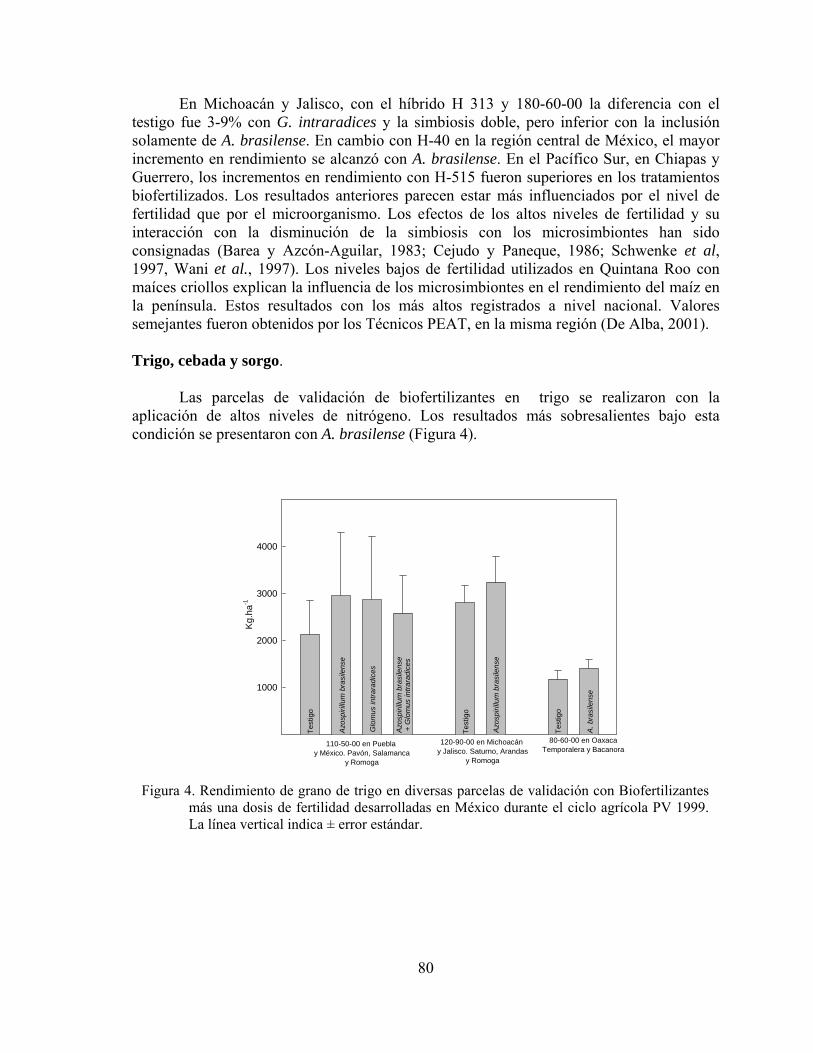
80
En Michoacán y Jalisco, con el híbrido H 313 y 180-60-00 la diferencia con el testigo fue 3-9% con G. intraradices y la simbiosis doble, pero inferior con la inclusión solamente de A. brasilense. En cambio con H-40 en la región central de México, el mayor incremento en rendimiento se alcanzó con A. brasilense. En el Pacífico Sur, en Chiapas y Guerrero, los incrementos en rendimiento con H-515 fueron superiores en los tratamientos biofertilizados. Los resultados anteriores parecen estar más influenciados por el nivel de fertilidad que por el microorganismo. Los efectos de los altos niveles de fertilidad y su interacción con la disminución de la simbiosis con los microsimbiontes han sido consignadas (Barea y Azcón-Aguilar, 1983; Cejudo y Paneque, 1986; Schwenke et al, 1997, Wani et al., 1997). Los niveles bajos de fertilidad utilizados en Quintana Roo con maíces criollos explican la influencia de los microsimbiontes en el rendimiento del maíz en la península. Estos resultados con los más altos registrados a nivel nacional. Valores semejantes fueron obtenidos por los Técnicos PEAT, en la misma región (De Alba, 2001). Trigo, cebada y sorgo.
Las parcelas de validación de biofertilizantes en trigo se realizaron con la aplicación de altos niveles de nitrógeno. Los resultados más sobresalientes bajo esta condición se presentaron con A. brasilense (Figura 4).
Kg.h
a-1
1000
2000
3000
4000
110-50-00 en Puebla y México. Pavón, Salamanca
y Romoga
120-90-00 en Michoacán y Jalisco. Saturno, Arandas
y Romoga
80-60-00 en OaxacaTemporalera y Bacanora
Test
igo
Azos
piril
lum
bra
sile
nse
Glo
mus
intra
radi
ces
Azos
piril
lum
bra
sile
nse
+ G
lom
us in
trara
dice
s
Test
igo
Test
igo
Azo
spiri
llum
bra
sile
nse
A. b
rasi
lens
e
Figura 4. Rendimiento de grano de trigo en diversas parcelas de validación con Biofertilizantes más una dosis de fertilidad desarrolladas en México durante el ciclo agrícola PV 1999. La línea vertical indica ± error estándar.
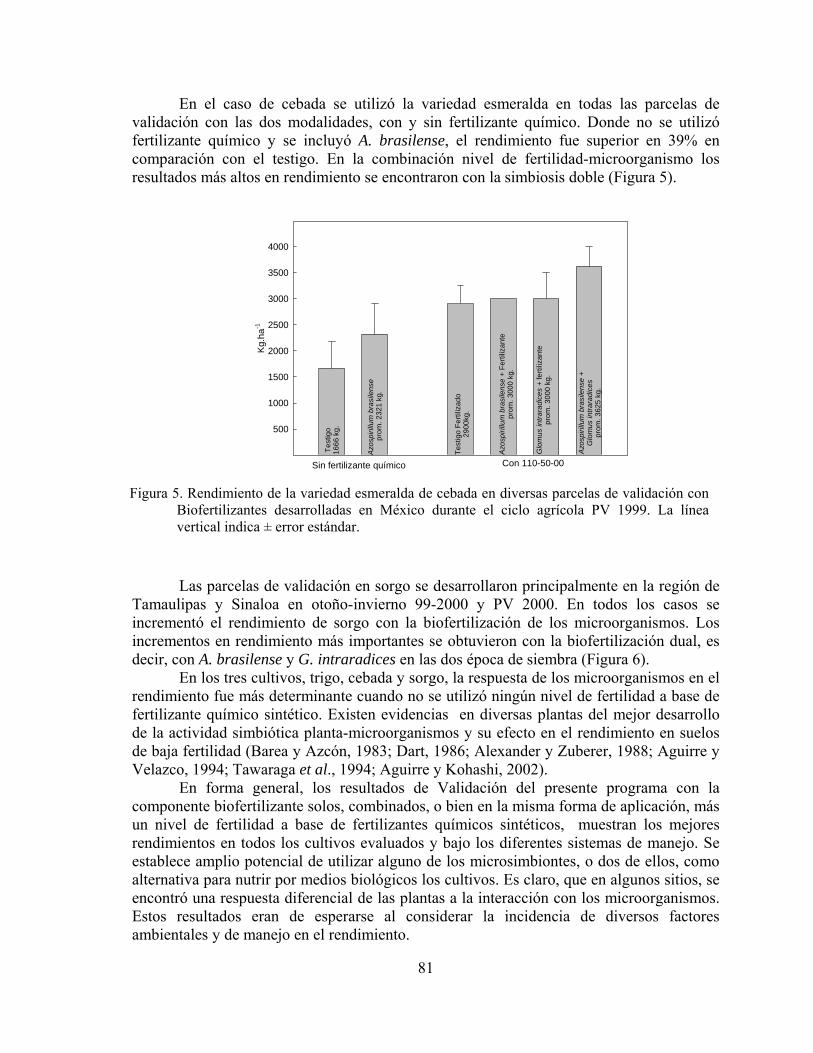
81
En el caso de cebada se utilizó la variedad esmeralda en todas las parcelas de validación con las dos modalidades, con y sin fertilizante químico. Donde no se utilizó fertilizante químico y se incluyó A. brasilense, el rendimiento fue superior en 39% en comparación con el testigo. En la combinación nivel de fertilidad-microorganismo los resultados más altos en rendimiento se encontraron con la simbiosis doble (Figura 5).
Las parcelas de validación en sorgo se desarrollaron principalmente en la región de
Tamaulipas y Sinaloa en otoño-invierno 99-2000 y PV 2000. En todos los casos se incrementó el rendimiento de sorgo con la biofertilización de los microorganismos. Los incrementos en rendimiento más importantes se obtuvieron con la biofertilización dual, es decir, con A. brasilense y G. intraradices en las dos época de siembra (Figura 6).
En los tres cultivos, trigo, cebada y sorgo, la respuesta de los microorganismos en el rendimiento fue más determinante cuando no se utilizó ningún nivel de fertilidad a base de fertilizante químico sintético. Existen evidencias en diversas plantas del mejor desarrollo de la actividad simbiótica planta-microorganismos y su efecto en el rendimiento en suelos de baja fertilidad (Barea y Azcón, 1983; Dart, 1986; Alexander y Zuberer, 1988; Aguirre y Velazco, 1994; Tawaraga et al., 1994; Aguirre y Kohashi, 2002).
En forma general, los resultados de Validación del presente programa con la componente biofertilizante solos, combinados, o bien en la misma forma de aplicación, más un nivel de fertilidad a base de fertilizantes químicos sintéticos, muestran los mejores rendimientos en todos los cultivos evaluados y bajo los diferentes sistemas de manejo. Se establece amplio potencial de utilizar alguno de los microsimbiontes, o dos de ellos, como alternativa para nutrir por medios biológicos los cultivos. Es claro, que en algunos sitios, se encontró una respuesta diferencial de las plantas a la interacción con los microorganismos. Estos resultados eran de esperarse al considerar la incidencia de diversos factores ambientales y de manejo en el rendimiento.
Kg.h
a-1
500
1000
1500
2000
2500
3000
3500
4000
Test
igo
1666
kg.
Azos
piril
lum
bra
sile
nse
+G
lom
us in
trara
dice
spr
om. 3
625
kg.
Sin fertilizante químico Con 110-50-00
Azos
piril
lum
bra
sile
nse
prom
. 232
1 kg
.
Test
igo
Ferti
lizad
o29
00kg
.
Azos
piril
lum
bra
sile
nse
+ Fe
rtiliz
ante
prom
. 300
0 kg
.
Glo
mus
intra
radi
ces
+ fe
rtiliz
ante
prom
. 300
0 kg
.
Figura 5. Rendimiento de la variedad esmeralda de cebada en diversas parcelas de validación con Biofertilizantes desarrolladas en México durante el ciclo agrícola PV 1999. La línea vertical indica ± error estándar.
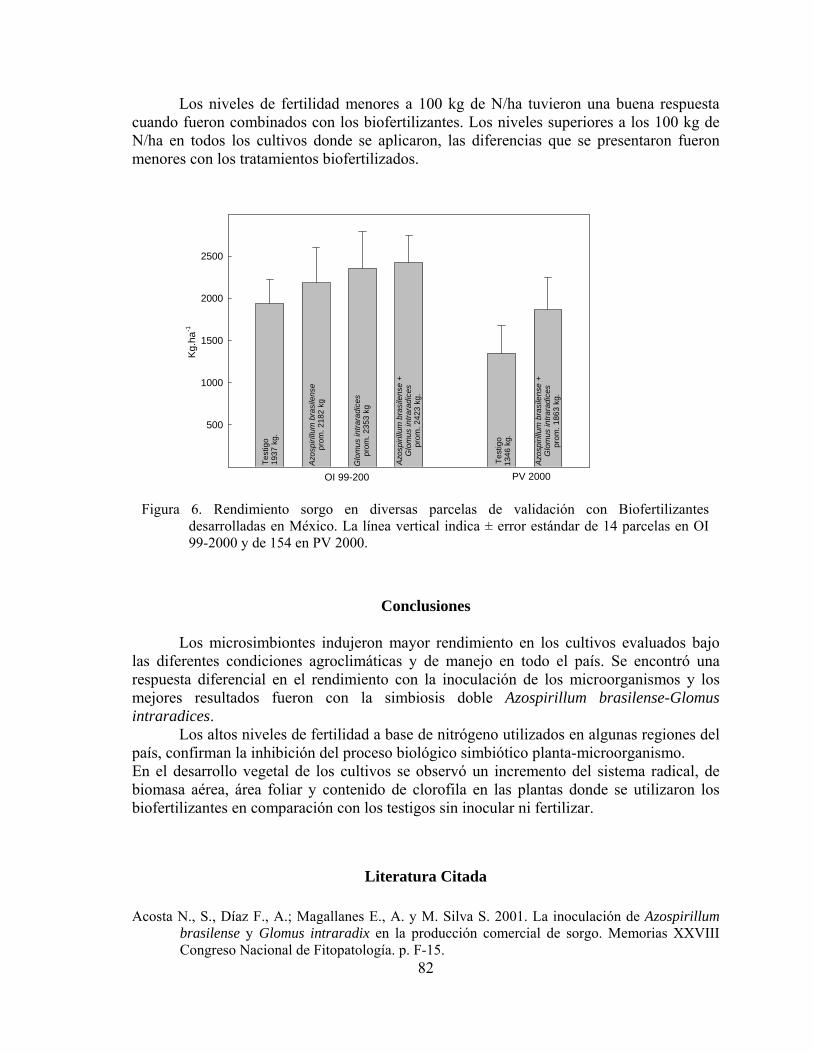
82
Los niveles de fertilidad menores a 100 kg de N/ha tuvieron una buena respuesta cuando fueron combinados con los biofertilizantes. Los niveles superiores a los 100 kg de N/ha en todos los cultivos donde se aplicaron, las diferencias que se presentaron fueron menores con los tratamientos biofertilizados.
Conclusiones
Los microsimbiontes indujeron mayor rendimiento en los cultivos evaluados bajo las diferentes condiciones agroclimáticas y de manejo en todo el país. Se encontró una respuesta diferencial en el rendimiento con la inoculación de los microorganismos y los mejores resultados fueron con la simbiosis doble Azospirillum brasilense-Glomus intraradices.
Los altos niveles de fertilidad a base de nitrógeno utilizados en algunas regiones del país, confirman la inhibición del proceso biológico simbiótico planta-microorganismo. En el desarrollo vegetal de los cultivos se observó un incremento del sistema radical, de biomasa aérea, área foliar y contenido de clorofila en las plantas donde se utilizaron los biofertilizantes en comparación con los testigos sin inocular ni fertilizar.
Literatura Citada Acosta N., S., Díaz F., A.; Magallanes E., A. y M. Silva S. 2001. La inoculación de Azospirillum
brasilense y Glomus intraradix en la producción comercial de sorgo. Memorias XXVIII Congreso Nacional de Fitopatología. p. F-15.
Kg.h
a-1
500
1000
1500
2000
2500Te
stig
o19
37 k
g.
Azos
piril
lum
bra
sile
nse
prom
. 218
2 kg
Glo
mus
intra
radi
ces
prom
. 235
3 kg
Azos
piril
lum
bra
sile
nse
+G
lom
us in
trara
dice
spr
om. 2
423
kg.
OI 99-200 PV 2000
Test
igo
1346
kg.
Azos
piril
lum
bra
sile
nse
+G
lom
us in
trara
dice
spr
om. 1
863
kg.
Figura 6. Rendimiento sorgo en diversas parcelas de validación con Biofertilizantes desarrolladas en México. La línea vertical indica ± error estándar de 14 parcelas en OI 99-2000 y de 154 en PV 2000.

83
Aguirre, M.J.F., M. Valdés y R.M. Silvester-Bradley. 1988. Simbiosis entre rizobios y cuatro leguminosas tropicales adaptadas en Chiapas, México. Pasturas Tropicales. Vol. 10 (3): 18-21.
Aguirre-Medina, J. F. y E. Velazco-Zebadua. 1994. Componentes morfológicos y fisiológicos del rendimiento en Leucaena leucocephala al inocularse con micorriza VA y/o Rhizobium loti J . Agricultura Técnica en México. Vol. 20 (1) : 43-45.
Aguirre, M. J. F. 2001. Programa Nacional de Biofertilizantes del INIFAP. Informe de labores. Dirección General de la División Agrícola. México, D. F. (Impreso).
Aguirre, M.J.F. y J. Kohashi-Shibata. 2002. Dinámica de la colonización micorrízica y su efecto sobre los componentes del rendimiento y el contenido de fósforo en frijol común. Agricultura Técnica en México. Vol 28 (1): 23-33.
Aguirre, M.J.F., J. Kohashi-Shibata, Trejo-López, C., Acosta-Gallegos, J.A. y Cadena-Iñiguez, J. 2004. La inoculación de Phaseolus vulgaris L. con tres microorganismos y su efecto en la tolerancia a la sequía (en prensa).
Alexander, M. 1977. Introduction to soil microbiology. Second edition. John Wiley and Sons. U.S.A. p. 423-437.
Altieri, M.A., K.D. Letourneau and J.R. Davies. 1984. The requirements of sustainable agro ecosystems. In: Agricultural sustainability in changing world order. G.R. Douglass (ed). Westview Press. Boulder Colorado. USA. p. 175-189.
Alexander, D. B. and D. A. Zuberer. 1988. 15N2 Fixation by bacteria associated with maize roots at a low partial O2 pressure. Applied and Environmental Microbiology. p. 1748-1753.
Arredondo V., C. y Luevanos A., A. 2000. Inoculación con Azospirillum brasilense y Glomus intraradices en maíz de temporal en los valles centrales de Oaxaca. Memoria XVIII Congreso Nacional de Fitogenetica. p. 151.
Arsac, J. F., Lamothe, C., Mulard, D. and Fages, J. 1990. Growth enhancement of maize (Zea mays L.) through Azospirillum lipoferum inoculations: effect of plant genotype and bacterial concentration. Agronomie 10: 640-654.
Azcón-G. de Aguilar, C. y Barea, J.M. 1980. Micorrizas. Investigación y Ciencia. Barcelona, España. 47: 8-16 p.
Balandreau, J. and R. Knowles. 1978. The rhizosphere. In: Interactions between non-pathogenic soil microorganisms and plants. Y.R. Dommergues and S.V. Krupa (eds). Elsevier. The Netherlands. p. 243-268.
Barea, J.M., and C. Azcón. 1983. Mycorrhiza and their significance on nodulating nitrogen fixing plants. Advances in agronomy. 36: 1-54.
Boucher, A.; Dalpe, Y. and Charest, C. 1999. Effect of arbuscular mycorrhizal colonization of four species of Glomus on physiological responses of maize. Jornal of Plant Nutrition. 22(4):783-797.
Burdman, S., S. Sarig, J. Kigel and Y. Okon. 1996a. Field inoculation of common bean(Phaseolus vulgaris L.) and chick pea (Cicer arietinum L.) with Azospirillum brasilense strain Cd. Symbiosis. 21, 41-48.
Burdman, S., H. Volpin, J. Kigel, Y. Kapulnik and Y. Okon. 1996b. Promotion of nodgene inducers and nodulation in common bean (Phaseolus vulgaris) roots inoculated with Azospirillum brasilense Cd. Applied and Environmental Microbiology 62, 3030-3033.
Caballero-Mellado, J. 1991. Experiencias de inoculación de maíz y trigo con Azospirillum a nivel intensivo y de investigación. En: Memorias del III congreso Nacional de la Fijación Biológica del Nitrógeno, efectuada del 2 al 5 de diciembre de 1991. Centro de Fijación Biológica del Nitrógeno. UNAM. Cuernavaca Morelos, Méx. p. 20-24.

84
Camas, G. R. 2000. Programa de Validación de Biofertilizantes en Chiapas. En; Informe anual de labores PV 1999, OI 1999-2000. Campo Experimental Centro de Chiapas. Centro de Investigación Regional del Pacífico Sur. (Impreso).
Cejudo, J. F. and A. Paneque. 1986. Short-term nitrate (nitrite) inhibiting of nitrogen fixation in Azotobacter chroococcum. Journal of Bacteriology 165: 240-243.
Chatel, D.L. and Parker, C.A. 1973. Survival of field-grown rhizobia over the dry summer period in Western Australia. Soil Biology and Biochemistry 5, 415-423.
Cruzaley, S. R. 2000. Validación de Biofertilizantes en cultivos básicos en el Estado de Guerrero. En: Informe anual de labores de la Dirección de Vinculación del INIFAP en el Estado de Guerrero. Centro de Investigación Regional del Pacífico Sur. (Impreso).
Covarrubias, R.J. M., I. Sánchez, V. y E. Rodríguez, C. 2000. Biofertilizantes: Simbiosis doble Azospirillum-Micorriza-arbuscular y Rhizobium-Micorriza en trigo, maíz y frijol en Coahuila. In; Informe anual de Labores del Campo Experimental Zaragoza, Coah. Centro de Investigación Regional del Norte Centro. (Impreso).
Davidson, R.L. 1978. Root systems. The forgotten component of pastures. In: Plant relations in pastures. J.R. Wilson (ed) CSIRO. Australia. p. 86-94.
Dart, P. J. 1986. Nitrogen fixation associated with non – leguumes in agriculture. Plant and Soil. 90:303 – 334.
De Alba, R. 2000. Programa de Validación de Biofertilizantes en el CIRSE. En; Informe anual de labores PV 1999, OI 1999-2000. Centro de Investigación Regional del sureste. (Impreso).
Dobbelaere, S., Croonenborghs, A., Thys, A., Ptacek, D., Vanderleyden, J., Dutto, P., Labandera-Gonzalez, C., Caballero-Mellado, J., Aguirre Juan Francisco, Kapulnik, Y., Brener, S., Burdman, s., Kadouri, D., Sang, S. and Okón J. 2001. Responses of agronomically important crops to inoculation with Azospirillum. Aust. J. Plant Physiol.. 28 (9): 871-879.
Dommergues Y.R. 1978. Impact on soil management and plant growth. In: Interactions between non-pathogenic soil microorganisms and plants. Y.R. Dommergues and S.V. Krupa (eds). Elsevier. The Netherlands. p. 443-458.
Durán, P. A., J.F. Aguirre-Medina y V. López, G. 2001. Modulo de Investigación en frijol ciclo otoño-invierno 2000-2001. En; Informe anual de labores del Campo Experimental Cotaxtla. Centro de Investigación Regional del Golfo. (Impreso).
Grajeda-Cabrera.O.A., 1990. Cinética de la fijación de nitrógeno en frijol común Phaseolus vulgaris L. Tesis de Maestría en Ciencias. Centro de Investigación y de Estudios Avanzados del Instituto Politécnico Nacional. Unidad Irapuato. 115 p.
Hernández, X. E. 1988. Biología Agrícola. Los conocimientos biológicos y su aplicación a la Agricultura. Consejo Nacional para la Enseñanza de la Biología A.C. CECSA. México, DF. 62 p.
Hernández M., L.; Trejo A., D.; Quinto T., Y. y Escalona A., M. 2000. Interacción entre Azospirillum brasilense y hongos micorrízicos arbusculares en el rendimiento de maíz bajo condiciones de campo. Resúmenes Reunión Iberoamericana y III Simposio Nacional Sobre Simbiosis Micorrízica. p. 33.
Irizar-Garza, M.B.G., P. Vargas-Vázquez, D. Garza-García, C. Tut y Couoh, I. Rojas-Marínez, A. Trujillo-Campos, R. García-Silva, D. Aguirre-Montoya, J.C. Martínez-González, S. Alvarado-Mendoza, O. Grajeda-Cabrera, J. Valero-Garza, y J.F. Aguirre-Medina. 2003. Respuesta de cultivos agrícolas a los biofertilizantes en la región central de México. Agricultura Técnica en México. Vol 29(2): 213-225.
Khan, A.G. 1977. Growth effects of VA mycorrhiza on crops in the field. In: Endomycorrhizas. F.S. Sanders, B. Mosse and P.B. Tinker, P.B. (eds.), Academic Press. London. p. 419-435.
Khalil, S., Loynachan, T. and Tabatabai, M. 1994. Mycorrhizal dependency and nutrient-uptake by improved and unimproved corn and soybean cultivars. Agr. J. 86: 949-958.

85
Matus, J.M., M. Valdés y J.F. Aguirre-Medina. 1990. Capacidad competitiva de cepas de rizobio en la formación de nódulos en Leucaena leucocephala. Pasturas Tropicales. Vol. 12 (3): 22-24.
Mendoza, L.A. y Aguirre M.J.F. La biofertilizacoión del cacao Theobroma cacao L. con Azospirillum brasilense y Glomus intraradices en etapa de vivero. Avances de resultados. Primer Congreso Internacional de Investigación y Transferencia de Tecnología Agropecuaria Chiapas 2002. Realizado del 19 al 22 de febrero del 2002 en Tuxtla Gutiérrez, Chiapas (Memorias).
Moroyoqui-Ovilla, D.M. y Aguirre, M.J.F. 2002. Avances en el desarrollo vegetativo del café var. Oro azteca con diferentes sustratos y dos microsimbiontes en vivero. Primer Congreso Internacional de Investigación y Transferencia de Tecnología Agropecuaria Chiapas 2002. Realizado del 19 al 22 de febrero del 2002 en Tuxtla Gutiérrez, Chiapas (Memorias).
Mosse, B. 1973. Advances in the study of vesicular-arbuscular mycorrhiza. Annual Review of Phytophatology. II . 171-196.
Okon, Y. and Y. Kapulnik. 1986. Development and function of Azospirillum-inoculated roots. Plant and Soil. 90, 3-16.
Okon, Y. and C. Labandera. 1994. Agronomic Applications of Azospirillum evaluation of 20 years world wide field inoculation. Soil Biology. 26 (12): 1591-1601.
Okon, Y. and R. Itzigsohn. 1995. The development of Azospirillum as a commercial inoculant for improving crop yields. Biotechnology Advances 13, 415-424.
Peña-Cabriales J. J. and J. Z. Castellanos. 1993. Effects of water stress on N2 fixation and grain yield of Phaseolus vulgaris L. Plant and Soil 152(1): 151-155.
Read, D. 1998. Plants on the web. Nature. 396: 22-23. Rovira, A. D.; G.D. Bowen and R.C. Foster. 1983. The significance of rhizosphere microflora and
mycorrhizas in plant nutrition. In: Encyclopaedia of plants. Spring Verlag. p. 61-93. Schwenke, G.D., D.F. Herridge., R.D. Murison, W. Felton and H. Marcellos. 1997. Management of
crop residues and nitrogen fertility for improved dryland cereal production can benefit soil organic fertility. In: Extending nitrogen fixation research to farmer s fields. Procc. Of a International Workshop on Managing Legume Nitrogen Fixation in the Cropping systems of Asia, 20-24 Aug 1996. ICRISAT, Asia Center. Rupela, O., Johansen, C and Herridge, D.F. (eds) Andhra Pradesh. India. p. 154-165.
Selosse, M.A. and Le Tacon, F. 1998. The land flora: a phototroph-fungus partnership? Tree: 15-20. Sylvester-Bradley, R., J.A. Kipe-Nolt, y D.J. Harris. 1985. Evaluación, selección y manejo de
sistemas leguminosa-rizobio para aumentar la fijación de nitrógeno. Guía de Estudio del Curso Intensivo realizado del 18 de noviembre al 13 de diciembre de 1985 en la Universidad Federal de Río Grande do Sul. Departamento do solos. Faculdade do agronomía. Porto Alegre, Brasil. 79 p.
Sutton, C.J. 1973. Development of Vesicular-arbuscular mycorrhizae in crop plant. Canadian Journal of Botany . 51 (12): 2487-2493.
Tavárez L., E. 2001. Inoculación de los simbiontes Azospirillum brasilense y Glomus intraradices y su efecto en la productividad del maíz en Río Bravo, Tamaulipas. Tesis Profesional. UAMRA, Universidad Autónoma de Tamaulipas. 59 p.
Uribe, V.G. 2000. Parcelas de Validación establecidas por Técnicos PEAT, ZINDER e Investigadores del INIFAP en Yucatán. En: Informe anual de Labores del Campo Experimental Mocochá. Centro de Investigación Regional del Sureste. (Impreso).
Waisel, Y., A. Eshell and U. Kafkafi. 2002. Plant roots. The hidden half. Third edition revised and expanded. Marcel Dekker New York. 1120 p.
Wani, S. P., O.P. Rupela and K.K. Lee. 1997. soil mineral nitrogen concentration and its influence on biological nitrogen fixation of grain legumes. In: Extending nitrogen fixation research to farmer s fields. Procc. Of a International Workshop on Managing Legume Nitrogen

86
Fixation in the Cropping systems of Asia, 20-24 Aug 1996. ICRISAT, Asia Center. Rupela, O., Johansen, C. and Herridge, D.F. (eds) Andhra Pradesh. India. p. 183-198.
Tawaraga, K., K. Sasai and T. Wagatsuma. 1994. Effect of phosphorus application on the contents of aminoacids and reducing surged in the rhizosphere and vesicular – arbuscular mycorrhizal infection of white clover. Soil Science and Plant Nutrition 4: 539-543.

87
AISLAMIENTO, SELECCIÓN, PRODUCCIÓN Y EVALUACIÓN DE UN INOCULANTE BASADO EN CEPAS NATIVAS DE Azospirillum
EN EL NORTE DE TAMAULIPAS
Mendoza H. Alberto1, Cruz M. Antonia1, Jacques-Hernández Cuauhtémoc2.
(1) Lab. Biotecnología Vegetal I, (2) Transferencia de Tecnología, Instituto Politécnico Nacional, Centro de Biotecnología Genómica, Reynosa, Tamps. México.
RESUMEN La generación de desarrollos tecnológicos basados en oportunidades o necesidades reales de los diferentes sectores productivos es uno de los objetivos torales del Centro de Biotecnología Genómica (CBG). Desde la creación del CBG, el laboratorio de Biotecnología Vegetal I cuyo responsable es el Dr. Alberto Mendoza, definió como una línea de investigación el aprovechamiento de microorganismos benéficos del suelo y uno de los resultados tangibles fue el aislamiento de diversas cepas de bacterias Gram (-) del género Azospirillum. Está ampliamente documentado que estas bacterias colonizan la rizósfera de las gramíneas y estimulan el crecimiento de los cultivos debido a su capacidad de producción de sustancias promotoras de crecimiento y al incremento en la fijación de nitrógeno atmosférico (Iglesias et. al. 2001). En el presente trabajo se resumen los resultados más relevantes obtenidos en el CBG. De ellos se subraya, el aislamiento de un grupo de bacterias de la especie Azospirillum brasilense con características superiores en condiciones in vitro (BV-112, BV-191, BV-180, BV-181 y CBG-497), la producción de biomasa bacteriana de la cepa CBG-497, la formulación de inoculantes semisólidos con estas cepas, la evaluación en campo del mismo grupo de cepas, así como la evaluación extensiva de la cepa CBG-497 en una superficie superior a las 2000 ha de sorgo, para determinar su efecto sobre el rendimiento de grano. Concluyendo que con los resultados obtenidos se tienen suficientes elementos para definir una tecnología de fabricación de inoculantes bacterianos rentable, con posibilidad de ser aprovechada en beneficio del agricultor.
INTRODUCCIÓN
La fijación biológica de N2 por leguminosas en simbiosis con Rhizobium spp. ha sido estudiada por mas de un siglo, desde su descubrimiento por Hellriegel y Wilfarth (Bhote et. al., 1998). Reportes esporádicos de fijación de N2 (no simbiótico) asociado con pastos tropicales aparecieron hace más de dos décadas, pero su descubrimiento quedó en

88
duda hasta el desarrollo de la técnica de reducción de acetileno que permitió la estimación precisa in situ de la actividad de la nitrogenasa (Graham et al. 1998). Numerosas bacterias diazótrofas fijadoras de N2 han sido identificadas y clasificadas en diversos géneros incluyendo Klebsiella, Azobacter, Azospirillum, Clostridium, Pseudomonas entre otras. Estas bacterias han sido aisladas de rizosfera, suelo, superficie de raíz y del interior de la raíz donde ocasionalmente se encuentran en gran cantidad (Bashan y Holguin, 1997). Bacterias del género Azospirillum son organismos fijadores de nitrógeno que viven en asociación cercana con la rizósfera de las plantas (Bashan, et al. 1993). En muchas especies de plantas estudiadas la colonización bacteriana se localiza en la zona de elongación de la raíz. Miembros de Azospirillum estimulan la densidad y longitud de los pelos radiculares, la taza de aparición de raíces laterales y la superficie del área de la raíz. La intensidad de sus efectos en la morfología de la raíz es dependiente de la especie de la planta y del cultivar pero sobre todo de la concentración del inóculo de Azospirillum. En muchos casos, la concentración mínima es de 1 x 107 colonias formando unidades (UFC) por semilla (Bashan, 1998). Los efectos de la inoculación por Azospirillum parecen estar determinados principalmente por los estados tempranos del desarrollo de la planta durante las primeras semanas después de la colonización óptima de las raíces (Burdman, et al. 2000). Sin embargo, la inoculación con Azospirillum puede producir aumentos significativos en la producción bajo ciertos ambientes y condiciones de suelo aun en la ausencia de una eficiente fijación de N2. Consecuentemente, los estudios en busca de efectos benéficos adicionales gracias a las inoculaciones con Azospirillum para crecimiento de plantas han aumentado. Actualmente, el énfasis se ha puesto en la asociación de la bacteria con las raíces de las plantas y en elucidar los mecanismos de la mutua interacción (Bashan y Holguin, 1997). Azospirillum ha sido un organismo modelo para el estudio de las interacciones planta-bacteria y es probablemente la bacteria de la rizósfera no-simbiótica más estudiada. Azospirillum ha sido aislada de una amplia variedad de plantas incluyendo muchos pastos y cereales en todo el mundo, de climas tropicales, templados y fríos; plantas desérticas, cultivos de maíz inundados y suelos afectados por sales (Neyra, 1981). Las asociaciones pueden ser descritas como una colonización de rizósfera, rizoplano y el interior de la raíz. Esta colonización es el resultado de un enriquecimiento selectivo del microorganismo mejor adaptado al nicho ecológico formado por la raíz ambiental (Bashan y Holguin, 1997). Azospirilla es quimiotácticamente atraído hacia la rizósfera por un exudado radicular y su alto mecanismo versátil le permite sobrevivir bajo condiciones ampliamente variantes que ocurren en el suelo. Azospirillum spp; puede adherirse a la superficie celular de la raíz, penetrar y colonizar espacios intercelulares dentro del interior de la raíz (Bashan y Holguin, 1994). Las cepas de Azospirillum tienen potencial para producir bacteriocinas las cuales inhiben componentes de la microflora. También, ellas producen sideróforos los cuales son

89
muy eficientes para la adquisición de Fe++. Algunas cepas de Azospirillum penetran las raíces de su hospedero y enseguida se establecen en gran número en los espacios intercelulares entre la epidermis y el cortex o en las capas corticales (Okon et al., 1997) y aun en el sistema vascular (Patriquin et al.; 1983). En adición, varios Azospirilla son dotados con una muy eficiente asimilación de Fe en la rizósfera, por medio de eso toman este elemento no disponible para otros microorganismos sin capacidad siderófora (Kloepper, et al., 1980). El fenómeno de promoción de crecimiento de la planta debido a la asociación de la bacteria Azospirillum con las raíces de las plantas es de considerable interés económico y científico. Hasta ahora beneficios de Azospirillum en Poaceae tales como maíz, sorgo y trigo han sido bien establecidos y los resultados de muchos experimentos con al respecto han sido publicados. Sin embargo, la mayoría de los estudios de campo han sido desarrollados en zonas templadas y suelos con alto contenido orgánico, por lo tanto en suelos ácidos, esos suelos son los más comunes en los cuales Azospirillum spp ha sido reportado en la naturaleza (Glick, 1995). Además, los mejores resultados han sido obtenidos con cepas aisladas del mismo hospedero en este ambiente adaptado (cepas homólogas) y experimentos no comparativos se han realizado usando diferentes cepas bajo diferentes condiciones ambientales (Bashan y Holguin, 1997). El mayor problema que ha impedido la aplicación a gran escala de Azospirillum como un Biofertilizante es que los efectos benéficos en las semillas dependen de múltiples factores, muchos de los cuales no son conocidos o sujetos de controlar bajo condiciones de campo (ejemplo: temperatura, lluvia, difusión de O2, pH del suelo, suelos pobres y la presencia de microorganismos competentes, entre otros). Como lo mencionado por Okon y Labandera, (1994), esto es una falta de información importante que por otro lado puede indicar un suceso de fracaso de Biofertilizantes concerniente a su promoción o capacidad de producción. Esto también ha sugerido que muchas más cepas necesitan ser estudiadas para cada cereal (Caballero et al., 1992).
OBJETIVO El objetivo de este trabajo fue aislar, seleccionar y evaluar cepas de Azospirillum brasilense nativas de suelos alcalinos típicos del noreste del país, que sean la base de productos formulados que a manera de fertilizantes biológicos, bioestimulantes o inoculantes demuestren en evaluaciones en campo, una clara capacidad para favorecer el desarrollo vegetativo e incrementar la productividad en gramíneas, principalmente del sorgo y maíz que se cultivan en las diferentes condiciones agro ecológicas típicas (región semiárida y suelos alcalinos) del norte del Estado de Tamaulipas. Es necesario destacar que se pretende que el conjunto de metodologías empleadas este proyecto sean reproducibles en diversas zonas de siembra de los cultivos de interés.

90
MATERIALES Y MÉTODOS
Muestras de suelo y tejido para aislamiento de cepas de Azospirillum El aislamiento de la bacteria se realizó a partir de muestras suelos, raíces y tejido de maíz y sorgo, en ambos ciclos de siembra (Primavera-Verano y Otoño-Invierno), de parcelas de temporal y riego, donde se sembraron los híbridos Asgrow-Jaguar (maíz) y Pioneer-3025 (sorgo), localizadas en los municipios Tamaulipecos de Río Bravo, Díaz Ordaz y Miguel Alemán. Se colectaron muestras de suelo (arcillo-arenoso, materia orgánica baja <1%, pH 7.8) y de raíz de cada planta, se guardaron en bolsas limpias de polietileno y se transportaron inmediatamente al laboratorio donde fueron procesadas. Se colectaron plantas adultas de los cultivos dentro de un área de 30,000 m2, con una distancia aproximada de 75 a 100 m. entre plantas.
Aislamiento de cepas de Azospirillum El aislamiento de Azospirillum de suelo se realizó haciendo diluciones en una solución estéril de NaCl2 al 0.85%. Las diluciones de 10-3 y 10-4 fueron plaqueadas por triplicado en medio Rojo Congo. Para el aislamiento de rizósfera se tomó un gramo de raíz; el suelo intacto adherido a la raíz fue resuspendido en 1 ml de solución estéril de NaCl2 0.85% (Baldini y Dobereiner, 1980). El aislamiento de Azospirillum de tejido de plantas fue hecho de 5 g de tejido molido con 5 ml de NaCl2 al 0.85% estéril y plaqueado en cantidades iguales en Rojo Congo (Caballero et al 1992). Estos cultivos fueron incubados a 30ºC por 48-72 horas. Se hicieron dos diferentes pruebas después para verificar las colonias típicas de Azospirilla. Después de 72 horas, se observaron pequeñas colonias rojas representativas, indicando la presencia de Azospirillum spp. (Rodríguez Cáceres, 1982) y las colonias del primer aislamiento fueron inoculadas en condiciones de microaerofílicas, en el medio NFB (Caballero et al., 1992).
Identificación de cepas de Azospirillum
La identificación fenotípica de las cepas obtenidas fue realizada de acuerdo a los criterios aceptados para el género (Tarrand et al. 1978) y fue confirmado por el análisis genético descrito por Giovannetti en 1990, y Han & New en, 1998, el cual consiste en hacer un análisis tipo “Fingerprint” del ADN del 16S rDNA mediante la digestión del gen con la enzima AluI y los perfiles genéticos se observaron en geles de agarosa al 1%, teñidos con bromuro de etidio.
Determinación de la actividad nitrogenasa
Para determinar la actividad de nitrogenasa, se utilizó el método de la reducción de acetileno en medio NFb semigelificado (Dobereiner y Marriel, 1976) mediante cromatografía de gases de acuerdo a las condiciones descritas por Mascarúa-Esparza et al (1988).

91
Producción in vitro de Ácido Indol Acético (AIA)
A la fecha existe poca información disponible sobre variaciones en la producción de la fitohormona responsable del beneficio en promoción del crecimiento en gramíneas como lo es el ácido-indol acético (AIA), por lo cual se evaluó la producción de ésta fitohormona en las cepas de Azospirillum aisladas. Cada cepa de A. brasilense se creció durante 18 h a 32 ºC en medio de malato adicionado con 0.2% de NH4Cl. Posteriormente cada aislamiento de A. brasilense se inoculó en matraces de 125 ml conteniendo 50 ml de caldo succinato-fructuosa de sales adicionado con 100 mg de triptofano y crecido durante 48 h a 30 ºC (Jain y Patriquin, 1985). Las condiciones de cultivo de A. brasilense, extracción y cuantificación del AIA se hizo de acuerdo al método descrito por Mascarúa-Esparza, et al., en 1988 y Fuentes-Ramírez et al., 1993.
Producción de la biomasa bacteriana
La cepa de Azospirillum CBG-497, seleccionada por su capacidad de producción de ácido-indol acético in vitro y la capacidad de incrementar la producción de sorgo en bioensayos, fue elegida para aplicación extensiva con motivos de evaluación. Esta cepa fue multiplicada en medio líquido Luria Bertani (LB) suplementado con succinato de sodio en matraces Erlenmeyer y en fermentadores de 3 y 20 L. Los matraces inoculados con la bacteria se agitaron a 250 rpm a 30 ºC hasta que el crecimiento alcanzó la fase estacionaria. En el fermentador se empleó un inóculo de 2% v/v preparado en matraz Erlenmeyer y transferido al fermentador en fase logarítmica, cosechando la bacteria en el estado estacionario, cuando la fuente de carbono se agotó. El oxígeno disuelto se mantuvo a un mínimo de 15%, manipulando la agitación y la aireación de acuerdo a lo requerido. El pH al inicio de la fermentación se ajustó a 7.0 y no se controló en las corridas batch, solo en una corrida donde se controló con hidróxido de amonio. En las corridas Fed-Batch fueron pH-Stat, ya que el pH se empleó como parámetro de control para adición de la fuente de carbono que consistió en una solución de ácido succínico / succinato de sodio a un pH de 2.5. La temperatura se controló a 29 ºC. El crecimiento de la bacteria durante el curso de la fermentación se monitoreó determinando la densidad óptica de caldo de fermentación a una longitud de onda de 660 nm. Cada punto de densidad óptica equivale a 8x109 UFC/ml.
Producción del Biofertilizante
Se elaboró el volumen de inoculante para un total de 2150 Ha. La presentación del inoculante fue en turba (Redi earth Marca Scott) como soporte previamente esterilizado y posteriormente inoculado con la bacteria de manera tal que al momento de la preparación se tenía 1x106 UFC/semilla para sorgo y 6x107 UFC/semilla para maíz (UFC = Unidades Formadoras de Colonias o Bacterias). La presentación final fue en cajas con 10 dosis de 420 gr. y cada dosis fue suficiente para tratar un saco de 50 lb. (22.7 Kg.) de semilla de sorgo o dos sacos de 50 lb. de maíz (de 60000 semillas) de maíz.
Distribución y aplicación de Biofertilizante La evaluación del inoculante o Biofertilizante en el ciclo Otoño-Invierno 2002-2003, se entregaron un total de 860 dosis para su aplicación de una dosis por saco de

92
semilla que equivale a 2150 hectáreas de acuerdo con una densidad de siembra promedio de 2.5 Ha/saco. La distribución se realizó a través de Patronato para la Investigación Fomento y Sanidad Vegetal (PIFSV) y de los Centros de Apoyo de Desarrollo Rural (CADERs) de manera tal que se establecieron parcelas con el inoculante en los municipios de Reynosa, Matamoros, Río Bravo, Valle Hermoso, San Fernando y Abasolo.
Determinación del rendimiento de grano
Se seleccionaron parcelas donde se realizaron muestreos para estimar el rendimiento. Se establecieron las coordenadas geográficas de cada parcela con un geoposicionador global marca Garmin modelo e-trex. Se colectaron las plantas de 5 metros lineales seleccionados al azar de la superficie sin y con el inoculante. Se contaron las plantas, se pesó la planta y la panoja, se midió el tallo y la altura de la planta. Con estos datos se realizó una estimación de la producción escalada a toneladas de grano/ hectárea convertido a porcentaje de incremento con respecto a su testigo y estos datos son los reportados.
Evaluación de cepas de Azospirillum en CBTa 98
Alternativamente se evaluaron diferentes cepas de Azospirillum que mostraron alto potencial in vitro y en bioensayos en invernadero. Esta evaluación se realizó en condiciones más controladas en la parcela del CBTa 98 en el vaso en Villa Cárdenas, municipio de Matamoros (97º 48.945' latitud norte, 25º 54.735' longitud oeste y altitud promedio 11 metros sobre el nivel del mar). Las cepas evaluadas fueron; BV-112, BV-191, BV-180, BV-181 y CBG-497, empleando el híbrido de sorgo Dekalb DK-52.
RESULTADOS
Aislamiento de Azospirillum de suelo y rizósfera de maíz
En la figura 1, se muestra la forma típica de las colonias de Azospirillum brasilense obtenidas empleando en el medio de selectivo Rojo Congo. La determinación de poblaciones de Azospirillum en esos suelos orgánicos pobres y alcalinos (pH 7.8) arrojó un promedio de 3 x 105 UFC por gramo de suelo. Esta concentración fue más baja que la reportada en suelos ricos, predominantemente ácidos, de zonas templadas. En muchos casos, la concentración mínima de la bacteria requiere de establecer una efectiva interacción planta-microorganismo reportándose que sea de 1 x 107 UFC/planta.
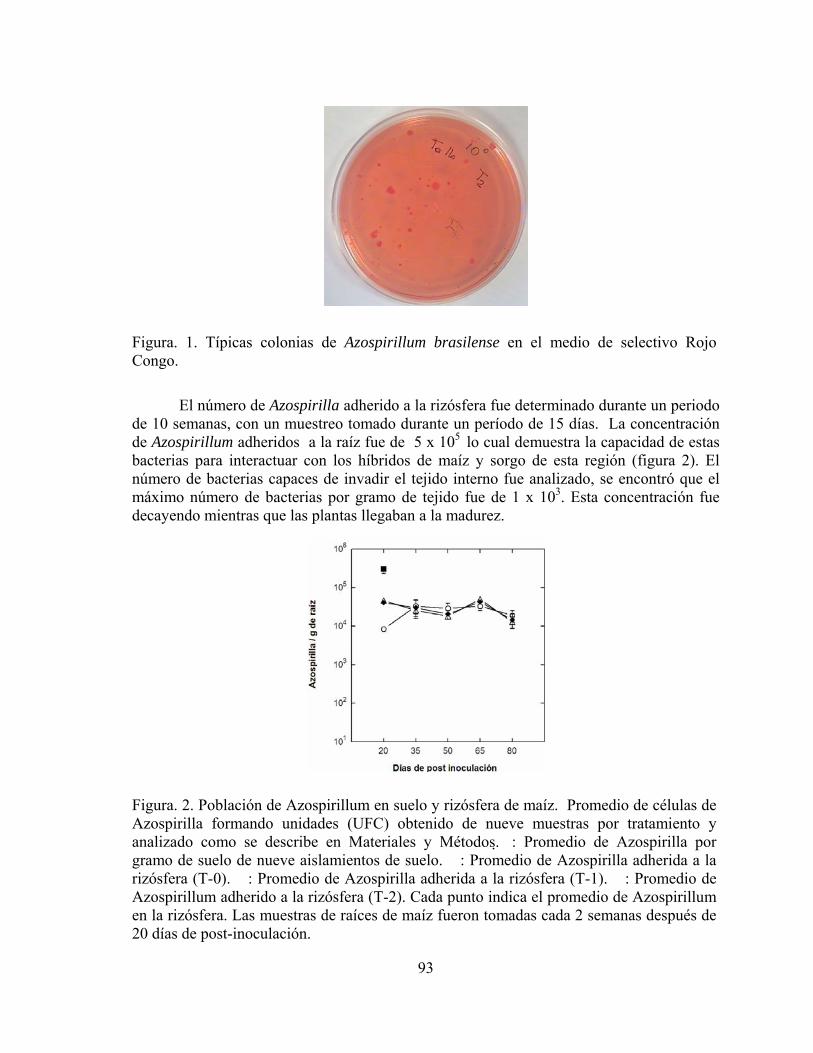
93
Figura. 1. Típicas colonias de Azospirillum brasilense en el medio de selectivo Rojo Congo.
El número de Azospirilla adherido a la rizósfera fue determinado durante un periodo de 10 semanas, con un muestreo tomado durante un período de 15 días. La concentración de Azospirillum adheridos a la raíz fue de 5 x 105 lo cual demuestra la capacidad de estas bacterias para interactuar con los híbridos de maíz y sorgo de esta región (figura 2). El número de bacterias capaces de invadir el tejido interno fue analizado, se encontró que el máximo número de bacterias por gramo de tejido fue de 1 x 103. Esta concentración fue decayendo mientras que las plantas llegaban a la madurez.
Figura. 2. Población de Azospirillum en suelo y rizósfera de maíz. Promedio de células de Azospirilla formando unidades (UFC) obtenido de nueve muestras por tratamiento y analizado como se describe en Materiales y Métodos.�: Promedio de Azospirilla por gramo de suelo de nueve aislamientos de suelo. �: Promedio de Azospirilla adherida a la rizósfera (T-0). �: Promedio de Azospirilla adherida a la rizósfera (T-1). �: Promedio de Azospirillum adherido a la rizósfera (T-2). Cada punto indica el promedio de Azospirillum en la rizósfera. Las muestras de raíces de maíz fueron tomadas cada 2 semanas después de 20 días de post-inoculación.

94
Producción in vitro de Ácido Indol Acético (AIA)
Se obtuvieron en el orden de 200 aislados, mismos a los que se determinó su producción de ácido-indol acético, de acuerdo a la metodología arriba descrita. Se aislaron cepas que incluso no producían esta fitohormona. Las cepas o aislados más destacados se muestran en la figura 3.
Figura. 3. Producción de ácido indol acético (AIA) de los aislados o cepas destacadas. La cepa CBG 497 se le seleccionó tomando entre otros parámetros, esta característica destacada.
Producción de la biomasa bacteriana
Cinética de crecimiento de Azospirillum en bioreactores de 3 y 20 litros se muestra en la figura 4. Empleando el sistema Fed-batch se logró la concentración máxima de biomasa bacteriana, de 9 unidades de densidad óptica, equivalente a 7.2x1010 bacterias/ml, y el mínimo fue 7.5 de densidad óptica equivalente a 6x1010 bacterias/ml logrado en sistema batch. Aunque estas concentraciones bacterianas son suficientes para hacer rentable la producción de inoculantes basados en esta bacteria, la etapa de fermentación debe ser estudiada a profundidad ya que aun existen muchos elementos que pueden ser sujetos a optimización, por ejemplo, el régimen Fed-batch, que hemos iniciado a estudiar como un sistema idóneo para incrementar sustancialmente la biomasa bacteriana.
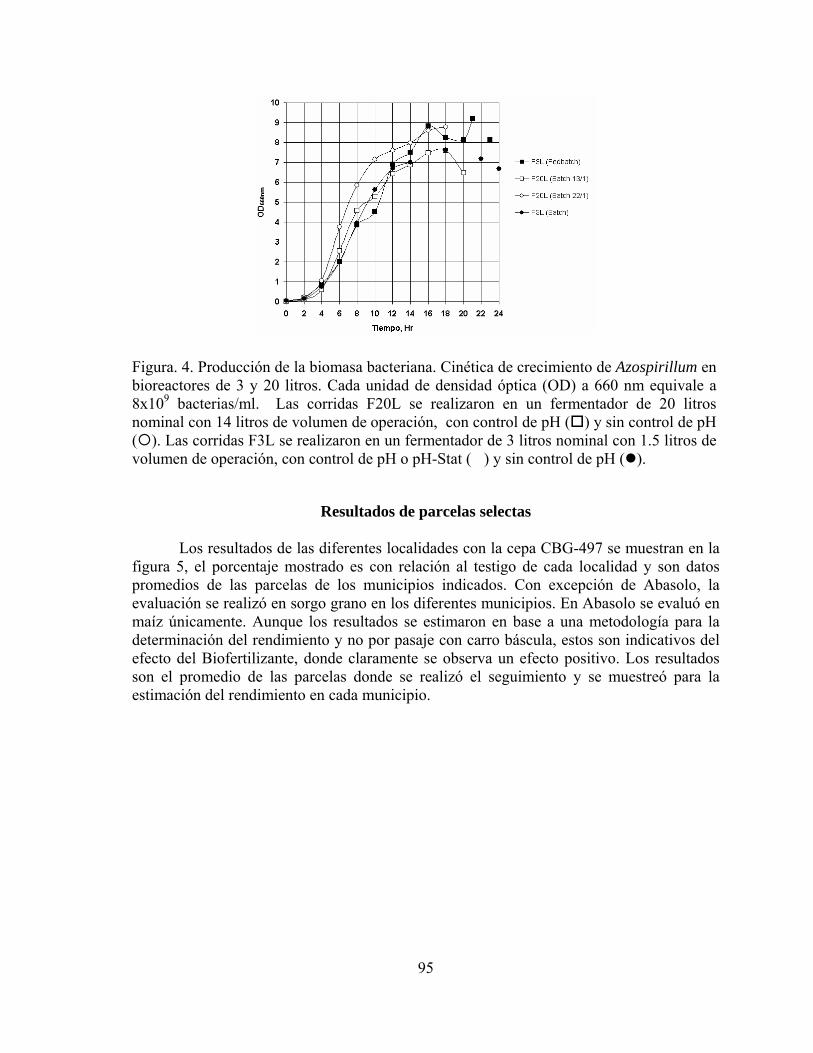
95
Figura. 4. Producción de la biomasa bacteriana. Cinética de crecimiento de Azospirillum en bioreactores de 3 y 20 litros. Cada unidad de densidad óptica (OD) a 660 nm equivale a 8x109 bacterias/ml. Las corridas F20L se realizaron en un fermentador de 20 litros nominal con 14 litros de volumen de operación, con control de pH ( ) y sin control de pH ( ). Las corridas F3L se realizaron en un fermentador de 3 litros nominal con 1.5 litros de volumen de operación, con control de pH o pH-Stat (�) y sin control de pH ( ).
Resultados de parcelas selectas
Los resultados de las diferentes localidades con la cepa CBG-497 se muestran en la figura 5, el porcentaje mostrado es con relación al testigo de cada localidad y son datos promedios de las parcelas de los municipios indicados. Con excepción de Abasolo, la evaluación se realizó en sorgo grano en los diferentes municipios. En Abasolo se evaluó en maíz únicamente. Aunque los resultados se estimaron en base a una metodología para la determinación del rendimiento y no por pasaje con carro báscula, estos son indicativos del efecto del Biofertilizante, donde claramente se observa un efecto positivo. Los resultados son el promedio de las parcelas donde se realizó el seguimiento y se muestreó para la estimación del rendimiento en cada municipio.

96
Figura. 5. Incremento en el rendimiento de producción de grano (datos estimados) cepa CBG-497. Porcentaje con respecto al Testigo.
El análisis estadístico por la prueba de medias de Tukey se muestra en la Tabla 1. El incremento observado en el rendimiento de grano fue estadísticamente significativo en los diferentes municipios donde se realizó la evaluación. Es necesario subrayar que en ninguna parcela se observaron resultados negativos.
Tabla 1. Prueba de medias por prueba de Tukey para la evaluación en municipios Tratamiento Kg. grano Ha-1 %Incre.*Río Bravo 4687 a 4.6 Matamoros 4687 a 11.6 Valle Hermoso 4581 b 7.2 Reynosa 4240 c 15.0 Promedio 4296 c 12.5 Testigo (promedio) 3773 d - San Fernando 3284** e 23.5 *Incremento en el rendimiento de grano con respecto al testigo. ** En San Fernando en rendimiento en el testigo fue de 2235 Kg./Ha Valores con las mismas letras son estadísticamente iguales (Tukey, α=0.05)

97
Resultados de parcela del CBTa 98
Los resultados obtenidos en la parcela del CBTa 98 se muestran en la siguiente Figura 6. Se incluyó para fines comparativos el Biofertilizante del INIFAP también distribuido por el PIFSV que está formulado con el hongo endomicorrízico arbuscular Glomus intraradix.
Figura. 6. Incremento en el rendimiento de producción de sorgo. Porcentaje con respecto al Testigo. Evaluación realizada en parcela del CBTa 98, localizada en el vaso en Villa Cárdenas, municipio de Matamoros (97º 48.945' latitud norte, 25º 54.735' longitud oeste y altitud promedio 11 metros sobre el nivel del mar). El análisis estadístico por la prueba de medias de Tukey se muestra en la Tabla 2. El incremento observado en el rendimiento de grano fue estadísticamente significativo en los formulados elaborados con las cepas CBG-497, BV-180, BV-181, Micorriza y Micorriza + CBG-497, por otro lado, a pesar de que el incremento en el rendimiento observado por el formulado multicepa fue de 6.7% con respecto al testigo, este resultado fue estadísticamente no significativo o dicho de otra manera, fue estadísticamente igual que el testigo. Aquí es claro que no todas las cepas que en experimentos in vitro pueden ser potencialmente elegibles, muestran resultados positivos en pruebas de campo. En esta evaluación se demuestra que el Laboratorio Vegetal I del CBG cuenta con un mayor número de cepas biológicamente eficientes en la estimulación del desarrollo y producción de grano de sorgo. También se confirma que el rendimiento de grano fue menor en la aplicación conjunta de Micorriza Glomus intraradix y Azospirillum cepa CBG 497 (al menos bajo las condiciones de esta evaluación) que en las aplicaciones individuales de ambos microorganismos.
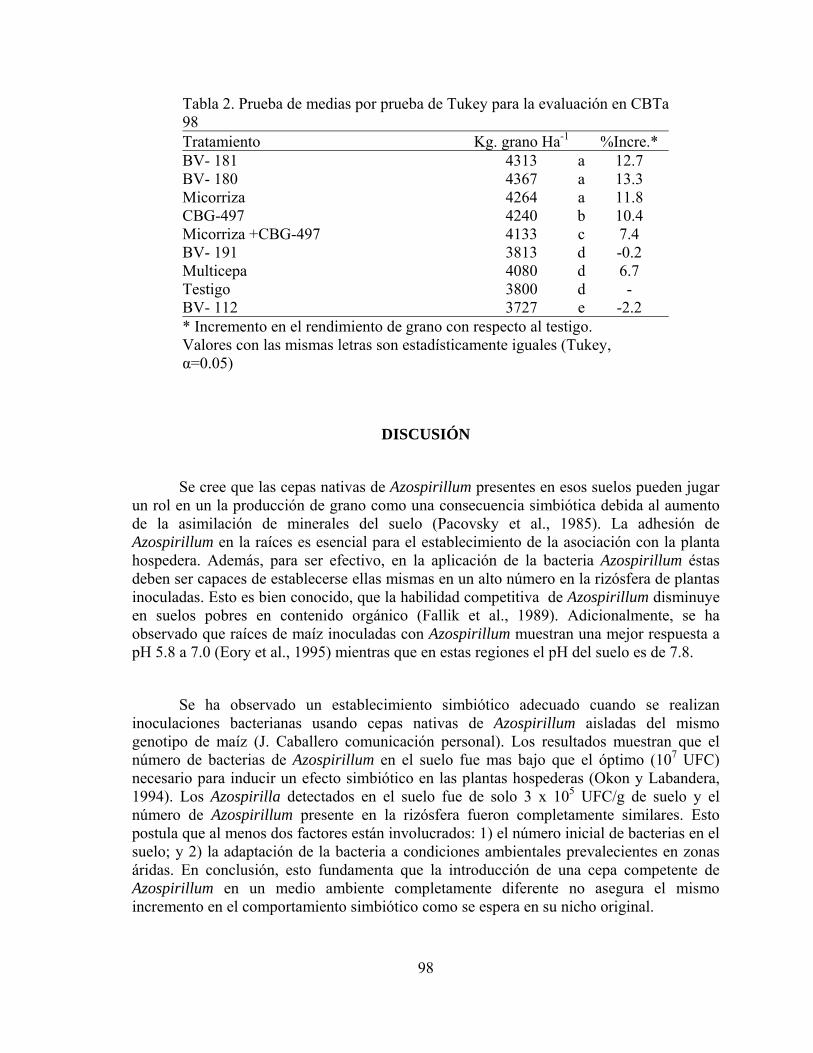
98
Tabla 2. Prueba de medias por prueba de Tukey para la evaluación en CBTa 98 Tratamiento Kg. grano Ha-1 %Incre.* BV- 181 4313 a 12.7 BV- 180 4367 a 13.3 Micorriza 4264 a 11.8 CBG-497 4240 b 10.4 Micorriza +CBG-497 4133 c 7.4 BV- 191 3813 d -0.2 Multicepa 4080 d 6.7 Testigo 3800 d - BV- 112 3727 e -2.2 * Incremento en el rendimiento de grano con respecto al testigo. Valores con las mismas letras son estadísticamente iguales (Tukey, α=0.05)
DISCUSIÓN
Se cree que las cepas nativas de Azospirillum presentes en esos suelos pueden jugar un rol en un la producción de grano como una consecuencia simbiótica debida al aumento de la asimilación de minerales del suelo (Pacovsky et al., 1985). La adhesión de Azospirillum en la raíces es esencial para el establecimiento de la asociación con la planta hospedera. Además, para ser efectivo, en la aplicación de la bacteria Azospirillum éstas deben ser capaces de establecerse ellas mismas en un alto número en la rizósfera de plantas inoculadas. Esto es bien conocido, que la habilidad competitiva de Azospirillum disminuye en suelos pobres en contenido orgánico (Fallik et al., 1989). Adicionalmente, se ha observado que raíces de maíz inoculadas con Azospirillum muestran una mejor respuesta a pH 5.8 a 7.0 (Eory et al., 1995) mientras que en estas regiones el pH del suelo es de 7.8.
Se ha observado un establecimiento simbiótico adecuado cuando se realizan inoculaciones bacterianas usando cepas nativas de Azospirillum aisladas del mismo genotipo de maíz (J. Caballero comunicación personal). Los resultados muestran que el número de bacterias de Azospirillum en el suelo fue mas bajo que el óptimo (107 UFC) necesario para inducir un efecto simbiótico en las plantas hospederas (Okon y Labandera, 1994). Los Azospirilla detectados en el suelo fue de solo 3 x 105 UFC/g de suelo y el número de Azospirillum presente en la rizósfera fueron completamente similares. Esto postula que al menos dos factores están involucrados: 1) el número inicial de bacterias en el suelo; y 2) la adaptación de la bacteria a condiciones ambientales prevalecientes en zonas áridas. En conclusión, esto fundamenta que la introducción de una cepa competente de Azospirillum en un medio ambiente completamente diferente no asegura el mismo incremento en el comportamiento simbiótico como se espera en su nicho original.

99
El porcentaje de incremento en el rendimiento de grano de sorgo con respecto al testigo en la evaluación con la cepa CBG-497 en las parcelas seleccionadas de los diferentes municipios fue del orden de 10%, similar al resultado en la evaluación realizada bajo condiciones mas controladas en la parcela del CBTa 98, donde el peso de grano fue obtenido por pesaje con carro báscula. Por lo que se asume que el procedimiento de estimación por muestreo al azar de cinco metros lineales tiene un aceptable porcentaje de error y es representativo. En maíz donde se contó con riego en el desarrollo del cultivo, el efecto de la cepa CBG-497 sobre el rendimiento fue muy significativo.
Con respecto a los resultados obtenidos de rendimiento de grano y tipo de suelo, los resultados más bajos en porcentaje de incremento en producción de grano con respecto a su testigo, fueron en Río Bravo y Valle Hermoso, municipios donde las parcelas evaluadas se realizaron en suelo arenoso y no franco limoso como en el resto de las parcelas. De aquí surge la pregunta; ¿la eficacia biológica de la cepa CBG-497 es menor en suelos arenosos con respecto a su eficacia biológica en suelos francos limosos?. Esta pregunta puede ser contestada definitivamente si se continúa la evaluación en campo de la cepa CBG-497 y se realiza igual manera la evaluación de las cepas que mostraron buenos resultados (BV-180 y 181).
En la evaluación realizada del CBTa 98, se encontró que las cepas BV-181 y BV-180 tienen comportamiento igual o mejor que la cepa CBG-497 y la Micorriza INIFAP (Glomus intraradix). De igual manera a la información proporcionada por personal del CERIB-INIFAP, se confirma el rendimiento de grano fue menor en la aplicación conjunta de Micorriza Glomus intraradix y Azospirillum cepa CBG 497. En caso de la formulación multicepa, el rendimiento de grano fue mayor que el testigo, pero inferior a los mejores resultados, probablemente sea el reflejo del empleo de las cepas BV-112 y BV-191 que no tuvieron un efecto positivo sobre el cultivo, ya que el incremento en el rendimiento porcentual promedio de efecto individual de la cuatro cepas que constituyeron la formulación multicepa, es similar al incremento en rendimiento porcentual observado de este tratamiento con la formulación multicepa (5.9% vs. 6.7% respectivamente).
Los resultados indican que el empleo de un inoculante basado en Azospirillum es una adecuada alternativa para obtener un incremento mínimo del 10% en rendimiento de grano por hectárea sin emplear fertilización nitrogenada. Por lo anterior es necesario continuar con la evaluación en campo de inoculantes o Biofertilizantes formulados para confirmar los resultados obtenidos. Esto nos dará la oportunidad de poder profundizar en el conocimiento de la producción microbiana en bioreactores de mayor escala, así como en las diferentes alternativas de formulación, por ejemplo; el desarrollo de inoculantes líquidos que resuelvan los problemas de aplicación o formulados multicepa y multiespecie. También se podrán evaluar en mayor superficie las cepas que mostraron buenos resultados de productividad además de la influencia de los factores agrobiológicos con los híbridos de sorgo y maíz más comúnmente empleados. Finalmente el continuar con actividades en campo empleando Biofertilizantes, promoverá el empleo de estos productos por parte de los agricultores en beneficio directo a la producción agrícola.

100
REFERENCIAS
Baldini VLD Döbereiner J. (1980). Hots-plant specificity in the infection of cereals with
Azospirillum spp. Soil Biol Biochem. 12:433-439.
Bashan Y, Holguin G, Lifshitz R .(1993). Isolation and characterization of plant growth-promoting rhizobacteria. In: Glick BR, Thompsom JE (eds) Methods in plant molecular biology and biotechnology. CRC, Boca Raton, Fla. Pp 331-345.
Bashan Y, Holguin G. (1994). Root-to-root travel of the beneficial bacterium Azospirillum brasilense. Appl Environ Microbiol 60:2120-2131.
Bashan Y, Holguin G. (1997). Azospirillum-plant relationships: environmental and physiological advances (1990-1996). Can J Microbiol 43:103-121.
Bashan Y. (1998). Inoculants of plant growth-promoting bacteria for use in agriculture. Biotechnol Adv 16:729-770.
Bothe H., Körsgen, H., Lehmacher, T., and Hundeshagen, B. (1992). Differential effects of Azospirillum, auxins and combined nitrogen on the growth of the roots.
Burdman S,Okon Y, Jurkevitch E, (2000). Surface characteristics of Azospirillum brasilense in relation to cell aggregation and attachment to plant roots. Crit. Rev. Microbiol 26:91-110
Caballero-Mellado J, Carcaño Montiel M. G, Mascarua-Esparza M. A. (1992). Field Inoculation of Wheat (Triticum aestivum) with Azospirillum brasilense under temperate climate. Symbiosis 13:243-253.
Dobereiner J, IE Marriel (1976). Ecological distribution of Spirillum lipoferum Beijerinck, Can J Microbiol 22: 1464-1473
Eory VJ, Momo FR, Alvarez M. (1995). Growth and survival of Azospirillum in roots and maize rhizospheres with different leaves of acidity. Rev. Argent. Microbiol 27: 99-105
Fallik E, Okon Y, Epstein E, Goldman A, Fischer M. (1989). Identification and quantification of IAA and IBA in Azospirillum brasilense inoculated maize roots. Soil Biol. Biochem 21:147-153
Fuentes-Ramírez LE, T.Jiménez-Salgado, I.R Abarca-Ocampo, J Caballero-Mellado. (1993). Acetobacter diazotrophicus, an indoleacetic acid producing bacterium isolated from sugarcane cultivars of Mexico. Plant and Soil. 154: 145-150
Glick R. (1995). The enhancement of plant growth by free-living bacteria. Can J. Microbiol 41:109-117
Graham PH, J Bale, D Baker, M Fried, J Roskoski, KT Mackay , E. Craswell (1998) The contribution of biological nitrogen fixation to plant production: An overview of the symposium and its implications. Plant and Soil 108: 1-6.
Iglesias, María C. Cracogna, Mariano F. Fogar, Mariela N. Romero, Edgar. (2001). Inoculación con Azospirillum ssp. en cultivos comerciales de girasol en una media loma baja. Comunicaciones Científicas y Tecnológicas. Univ. Nac. Del Nordeste. Republica de La Argentina. Boletín. 75.
Jain, D.K. D.G. Patrinquin. (1985). Characterization of a substance produced by Azospirillum which causes branching of wheat root hair. Can J. Microbiol. 31:206-210.
Kloepper JW, Leong J, Teintze M, Schroth MN. (1980). Enhanced plant growth by siderophores produced by plant growth-promoting rhizobacteria. Nature 286:885-886.

101
Mascarúa-Esparza, R. Villa-González and J. Caballero-Mellado. (1988). Acetylene reduction and indoleacetic acid production by Azospirillum isolates from Cactaceous plants. Plant and Soil 106: 91-95.
Neyra, C.A., L. De Vasconcelos, and L. Miller, (1980). Related Articles, Links Free-living and symbiotic characteristics of chlorate resistant mutants of Rhizobium. Can J Microbiol. 26(3):338-42.
Okon Y, Albrecht SL, Burris RH. (1977). Methods for growing Spirillum lipoferum and for counting it in pure culture and in association with plants. Appl. Environ Microbiol 33:85-88.
Okon Y, Labandera-Gonzales CA. (1994). Agronomic applications of Azospirillum: an evaluation of 20 years worldwide field inoculations. Soil Biol Biochem 26:1591-1601.
Pacovsky R S, Paul E A , Bethlenfalvay G J. (1985). Nutrition of sorghum plants fertilized with nitrogen or inoculated with Azospirillum brasilense. Plant Soil 85:145-148.
Patriquin, DG, Döbereiner, J Jain, D K. (1983). Sites and processes of association between diazotrophs and grasses. Can J Microbiol 29:900-915.
Rodriguez Cáceres EA. (1982). Improved medium for isolation of Azospirillum spp. Appl. Environ. Microbiol 44, 990-991.
Tarrand, J. J., N.R. Krieg and J. A Döbereiner. (1978). Taxonomic study of the Spirillum lipoferum. group, with description of a new genus Azospirillum gen. nov. and two species. Azospirillum lipoferum (Beijerinck) comb. nov. and Azospirillum brasilense sp. nov. Can. J. Microbiol. 24: 967-980.

102
Respuesta de Variedades de Cacahuate (Arachis hypogaea L.) a la Fertilización Química y
Biológica en un Suelo Regosol
Arturo Durán Prado1*, Victor López Galván1 y Oscar Hugo Tosquy Valle1
INTRODUCCION. El cultivo de cacahuate se siembra en México en 70,858 ha con una producción de 91,453 toneladas y un rendimiento promedio de 1.7 ton/ha. Para cubrir la demanda nacional se importan anualmente más de 7,000 toneladas. Veracruz cuenta con 213,000 ha de suelos regosoles apropiados para la producción de esta oleaginosa, sin embargo, en este tipo de suelo la concentración de los nutrimentos es sub-óptima, por lo que la inoculación con micorriza representa una alternativa para mejorar el desarrollo de las plantas, además de que se pueden reducir las dosis de fertilizante químico. La estrategia para lograr una eficiente manipulación de la asociación Rhizobium-raíz-micorriza consta de los siguientes pasos: 1) Inoculación con cepas de micorriza seleccionada, 2) Selección de genotipos que respondan favorablemente a la colonización con cepas seleccionadas, y 3) Establecer los cultivos en suelos apropiados para la proliferación de micorriza en la rhizosfera y con alto potencial para la función simbiótica (Lee y Wani, 1991). El objetivo de este trabajo fue evaluar la respuesta de cinco variedades de cacahuate a la biofertilización con micorriza Glomus intraradix.
MATERIALES Y MÉTODOS. En el ciclo primavera-verano 2001 en el ejido Las Lomas municipio de Úrsulo Galván, Ver., se estableció un experimento para evaluar cinco variedades de cacahuate y dos tipos de fertilización. Se empleo el diseño experimental de bloques al azar con cuatro repeticiones y arreglo de tratamientos en factorial 5X2. Las variedades evaluadas fueron. 1) Río Balsas (erecta), 2) A-18 (erecta), 3) Ranferi Díaz (erecta), 4) RF-214 (semi-rastrera) y 5) Huitzuco-93 (rastrera). Los tratamientos de fertilización consistieron en: 1) Fertilización química (40-40-00 kg/ha de N, P2O5 y K2O, respectivamente), empleando como fuentes sulfato de amonio (20.5% de N) y superfosfato de calcio simple (19.5% de P2O5) antes de la siembra y 2) Biofertilización con micorriza-arbuscular (Glomus intraradix) en dosis de 1.0 kg/ha aplicada a la semilla antes de la siembra + 1/3 de fertilización química. Como parámetro de respuesta se consideró el rendimiento de grano ajustado al 8% de humedad. Se realizó análisis de varianza y para determinar la rentabilidad de los tratamientos de fertilización 1 Campo Experimental Cotaxtla. INIFAP. Apdo. Postal # 429, Veracruz, Ver. [email protected]
se utilizó el análisis de beneficios brutos y netos propuesto por Perrin y Anderson (1988).
RESULTADOS Y DISCUSION. Se detectó significancia (α=0.01) en el factor Variedad y no hubo efecto significativo para el factor Tipo de Fertilización, ni para la interacción Variedad x Tipo de Fertilización. El máximo rendimiento de cacahuate con cáscara, se obtuvo con la variedad erecta Río Balsas con 2,281 kg/ha, la cual fue estadísticamente similar a la variedad semi-rastrera RF-214 con 1,936 kg/ha y superior en 100% a Huitzuco-93 en 117% a la variedad A-18 y en 126% a Ranferi Díaz. El costo del tratamiento de fertilización química 40-40-00, cubierto con las fuentes: sulfato de amonio ($400.00/ha) y superfosfato de calcio triple ($200.00/ha) y dos jornales para su aplicación ($70.00/jornal) fue de $740.00. Por su parte, el tratamiento de G. intraradix + 1/3 de fertilización química tuvo un costo total de $281.99/ha, que considera $133.00/ha de fertilizante nitrogenado + $66.66/ha de fertilizante fosfatado + $12.00/ha del costo de la micorriza-arbuscular. El costo total de producción del cultivo de cacahuate con fertilización química fue de $4,708.00/ha y de $4,250.00 empleando G. intraradix inoculado a la semilla + 1/3 de la fertilización química. Con una utilidad neta de $ 13, 040.00/ha y una relación beneficio/costo de 1: 4.06, el mejor tratamiento fue sembrar la variedad erecta Río Balsas con semilla inoculada con Glomus intraradix + 1/3 de la fertilización química.
CONCLUSIONES. Las mejores variedades en cuanto al rendimiento fueron Río Balsas y la línea RF-214. Los rendimiento de cacahuate no se afectan si en lugar de la dosis normal de fertilización NPK (40-40-00) se utiliza biofertilizante Glomus intraradix + 13-13-00 de N, P2O5 y K2O, lo que permite reducir el costo de esta practica en 62%. La mejor opción económica fue la combinación de Rio Balsas biofertilizada con Glomus intraradix + 1/3 de la dosis de fertilización química.
LITERATURA CITADA
Lee, K. K. and P. Wani, S. 1991. Possibilities for Manipulating Mycorrhizal Assocciations in Crops. In: ICRISAT (International Crops Research Institute for the Semi-Arid Tropics), Phosphorus nutrition of grain legumes en the semi-arid tropics. (Johansen, C, Lee, K. K., and Sahrawat, K. L. eds.). Patancheru, India. p. 107-116. Perrin, R. y J. Anderson, 1988. CIMMYT. Manual Metodológico de Evaluación Económica. La formulación de recomendaciones a partir de datos agronómicos. 78 p.

103
Respuesta de Germoplasma de Frijol a la Fertilización Química y Biológica
Arturo Durán Prado1* Víctor López Galván1 y Javier
Cumpián Gutiérrez2
INTRODUCCION. El frijol (Phaseolus vulgaris L.) se cultiva en suelos marginales de baja fertilidad y con bajo fósforo y pH (4.5), además se fertiliza sólo el 25% de la superficie sembrada anualmente en el estado de Veracruz, el cual produce un promedio de 25,506 ton/año con un déficit anual de más de las 100 mil toneladas de grano. En condiciones de temporal, en promedio, para el ciclo de primavera-verano se siembran 11,825 ha con un rendimiento de 591 kg/ha y en el ciclo otoño-invierno 35,151ha con un rendimiento de 620 kg/ha. El nitrógeno atmosférico puede ser fijado por el frijol para maximizar los rendimientos, Acuña et. al, (1995) reportan a las cepas de Rhizobium leguminosarum biovar phaseoli 4050, CR 477 y BAT 761 como responsables en el incremento de la producción de frijol de 30% a 60% a través de ambientes y países centroamericanos. Existe evidencia de la variación genética del frijol en cuanto a su eficiencia en el uso del fósforo del suelo y tolerancia a la toxicidad de aluminio y manganeso como lo citan Thung et. al, (1990). El objetivo de este trabajo fue identificar la respuesta de 46 genotipos de frijol a la biofertilización con bacterias Rhizobium leguminosarum biovar phaseoli y fertilización química.
MATERIALES Y MÉTODOS. En un suelo acrisol (pH= 4.5) del Campo Experimental Papaloapan se estableció el 11 de octubre, en el ciclo O-I 1996-1997, un experimento parcelas divididas con 3 repeticiones. En la parcela grande se utilizaron tres tratamientos: 1) Biofertilización con Rhizobium leguminosarum biovar phaseoli (151+ CR 477+613) en dosis de 1.0 kg/ha inoculada a la semilla antes de la siembra, recomendadas por Acuña (1995) + (00-120-00 kg/ha de N, P2O5 y K2O respectivamente); 2) Fertilización química (F. Q.) (90-120-00 kg/ha de N, P2O5 y K2O respectivamente) y 3) Fertilización química (120-120-00 kg/ha de N, P2O5 y K2O respectivamente). En las parcelas chicas se probaron 46 genotipos tolerantes a bajo fósforo.
RESULTADOS. Los resultados del análisis de varianza reportó diferencias significativas (α=0.05) en rendimiento de grano para el tratamiento 2) Fertilización química (90-120-00) con 1365 kg/ha, y 3) Fertilización química (120-120-00) con 1299 kg/ha mientras que el tratamiento biofertilización + (00-120-00) fue de 963 kg/ha. Los resultados indican que hay tres genotipos eficientes en la fijación de N2 con aumentos 1 Campo Experimental Cotaxtla. INIFAP. Apdo. Postal # 429, Veracruz, Ver. [email protected] 2 CE Papaloapan. INIFAP.
de rendimiento de 15.45% para BAT-1467 sobre el mismo genotipo en el tratamiento 3 (120-120-00) y de 14.25% para TLP-22 en el mismo genotipo sobre el tratamiento 2 (90-120-00); DOR-446 igualó al tratamiento 2 de F. Q. (90-120-00), los genotipos fueron BAT-1467 con 1596 kg/ha; TLP-22 con 1411 kg/ha y DOR-446 con 1278 kg/ha. El genotipo menos eficiente con estas cepas fue 632 con 779 kg/ha el cual fue superado en 105%; 81.12% y 64.05% respectivamente. Los genotipos que reportaron el mayor rendimiento con el tratamiento 3) F. Q. (120-120-00) fueron NAG-161 y 632 ambos con 1953 kg/ha; DOR-446 con 1799 kg/ha y A-774 con 1783 kg/ha superaron al genotipo G-3513 que menor respuesta produjo 1227 kg/ha en 59.16%; 59.16% y 46.61% y 45.31% respectivamente (Cuadro 1).
Cuadro 1. Respuesta en rendimiento (kg/ha) de genotipos de frijol a la biofertilización y fertilización química.
Genotipo T1 T2 T3 Contraste*
A-774
NAG-161
Sequía-12
BAT-1467
TLP-18
N. Tacaná
DOR-446
Sesentano
DOR-60
CHIS 3A-1
632
TLP-22
G-3513
1345
1076
1407
1596
1142
1122
1278
1027
1107
1021
779
1411
1113
1818
1781
1748
1768
2140
1945
1279
1393
1813
1619
1314
1235
1615
1783
1953
1597
1386
1368
1502
1799
1596
1243
1458
1953
1382
1227
a,b
a,b,c
a,b,c
a,b,c
a,b,c
a,b,c
b,c
a,b,c
a,b,c
a,b,c
a,b,c
a,c
a,c
*)= Significativo al 5%. a= T1 vs. T2; b= T1 vs. T3 y c= T2 vs. T3.
CONCLUSIONES. Se identificaron tres genotipos eficientes en la fijación biológica de N2: BAT-1467, TLP-22 y DOR-446. La fertilización química logra en general mayor rendimiento que la biofertilización en el germoplasma evaluado. Con los genotipos seleccionados se puede sustituir del 50% al 100% la F. Q. nitrogenada al suelo, mediante el uso del biofertilizante Rhizobium.
LITERATURA CITADA
Acuña, et al. 1995. Evaluación y validación de cepas de Rhizobium bajo condiciones limitantes. En: Proyecto R4. Informe técnico. PROFRIJOL. 9 p. Thung, M.; J. Ortega y O. Erazo. 1990. Metodología de selección por eficiencia en el uso de fósforo y por tolerancia a la toxicidad por aluminio y del manganese en el frijol (Phaseolus vulgaris L.). En: Sorgo para sueloa ácidos. Salinas et al. (eds.) INTSORMIL. ICRISAT. CIAT. p. 210.

104
Respuesta de Variedades de Frijol a la Fertilización Química y Biológica en un Suelo
Fluvisol de Veracruz
Arturo Durán Prado1*, Víctor López Galván1 y Oscar Hugo Tosquy Valle1
INTRODUCCION. El frijol (Phaseolus vulgaris L.) es la leguminosa de más baja capacidad de nodulación y fijación de N2 atmosférico (Burdman et al., 2000). Además las variedades varían en su habilidad para fijar el nitrógeno atmosférico y para sustraer el fósforo y otros nutrimentos del suelo (Graham, 1981), lo que aunado a deficientes prácticas de fertilización, ya que la mayoría de los productores no fertilizan o lo hacen de forma inadecuada, provocan que los genotipos no puedan maximizar sus rendimientos (Piha y Munns, 1987). Por lo anterior, el objetivo de este trabajo fue determinar la respuesta de seis variedades de frijol a la biofertilización con bacterias Rhizobium leguminosarum biovar phaseoli y micorriza-arbuscular Glomus intraradix.
MATERIALES Y MÉTODOS. Durante el ciclo O-I 2000-2001, se estableció un experimento en un suelo fluvisol del Campo Experimental Cotaxtla. El diseño estadístico utilizado fue bloques al azar con cuatro repeticiones y arreglo de tratamientos en parcelas divididas. En la parcela grande se utilizaron seis genotipos de frijol: 1) Negro INIFAP, 2) Negro Medellín, 3) Negro Tropical, 4) Negro Tacaná, 5) Línea UCR-55 y 6) Línea DOR-448. En la parcela chica se probaron siete tratamientos de fertilización: 1) Rhizobium + Glomus intraradix + 2/3 de la fertilización química recomendada (26-26-00 kg/ha de N, P2O5 y K2O, respectivamente); 2) Sin biofertilizantes + 40-40-00 kg/ha de N, P2O5 y K2O; 3) Testigo absoluto no fertilizado; 4) Rhizobium + G. intraradix sin Fertilización química; 5) Rhizobium + G. intraradix + 1/3 de fertilización química (13-13-00 kg/ha de N, P2O5 y K2O), 6) Rhizobium (Sin fertilización química) y 7) G. intraradix (Sin fertilización química). Para Rhizobium la dosis fue de 300 g/ha y para G. intraradix fue de 1.0 kg/ha, inoculando la semilla antes de la siembra. Se midió y analizó el rendimiento de grano y como prueba de separación de medias se utilizó Tukey al 5%.
RESULTADOS. Se detectaron diferencias altamente significativas (α= 0.05) para Genotipos, Fertilización y su interacción. Con rendimientos superiores a 1000 kg/ha, los mejores genotipos fueron: Negro Tacaná, Negro INIFAP y DOR-448. A su vez, el tratamiento de fertilización que proporcionó el mayor rendimiento de grano fue Rhizobium + G. intraradix + 26-26-00 con 1226 kg/ha, el cual fue estadísticamente similar a 40-40-00 de N, P2O5 con 1103 1 Campo Experimental Cotaxtla. INIFAP. Apdo. Postal # 429, Veracruz, Ver. [email protected]
kg/ha y superior al resto de los tratamientos. Estos dos tratamientos superaron en 16.9 y 14.2%, respectivamente, al testigo no fertilizado (Cuadro 1).
Cuadro 1. Rendimiento promedio de grano (kg/ha) de tratamientos de fertilización. Ciclo O-I 2000-2001.
Tratamiento Tukey (0.05)1
Rhizobium + G. intraradix + 2/3 de fertilización química
1226 a
Fertilización química (40-40-00) 1103 a b
Rhizobium + G. intraradix + 1/3 de fertilización química
977 b c
Rhizobium + G. intraradix 968 b c
Testigo absoluto (no fertilizado) 966 b c
Rhizobium 809 c
G. intraradix 801 c 1 Los tratamientos con letras similares son estadísticamente iguales.
La acción conjunta de ambos factores tuvo como mejores combinaciones a Negro Medellín biofertilizado con Rhizobium + G. intraradix + 26-26-00 con 1389 kg/ha, Negro Tacaná no biofertilizado + 40-40-00 con 1310 kg/ha y esta misma variedad biofertilizada con los dos microorganismos + 26-26-00, al igual que los genotipos Negro INIFAP y DOR-448 que obtuvieron ambos un rendimiento de 1250 kg/ha. La línea UCR-55, fue el genotipo que menos respondió a la biofertilización.
CONCLUSIONES. La respuesta a la fertilización biológica y química fue diferente para algunos genotipos. Los mayores rendimientos de grano se obtuvieron con Negro Medellín y Negro Tacaná, inoculadas con ambos biofertilizantes y fertilizadas con 2/3 de la dosis de fertilización química recomendada.
LITERATURA CITADA
Graham, P. H. 1981. Some problems of nodulation and symbiotic nitrogen fixation in Phaseolus vulgaris. A review Field Crops Res. 4: 93-112. Piha, M. I. and D. N. Munns. 1987. Nitrogen fixation capacity of field-grown bean compared to other grain legumes. Agronomy Journal. 79: 690-696. Burdman, S. et al. 2000. Improvement of legume crop yields by co-inoculation with Azospirillium and Rhizobium. The Otto Warburg Center for Agricultural Biotechnology. The Hebrew University of Jerusalem, Israel.

105
1500
2000
2500
3000
3500
4000
sin conG. intraradix
RG (k
g ha
-1)
Huasteca 100Huasteca 200
Respuesta de Variedades de Soya a la Inoculación con Micorriza Glomus intraradix en Veracruz
Arturo Durán Prado1*, Oscar Hugo Tosquy Valle1 y
Víctor López Galván1
INTRODUCCIÓN. La micorriza arbuscular debe tener la capacidad para sobrevivir bajo condiciones ambientales cambiantes, incluso durante períodos de tiempo cuando las plantas hospederas estén ausentes (Abbott y Robson, 1985). Para las regiones tropicales húmedas, se ha establecido la hipótesis, de que debido a la existencia de una diversidad de plantas y cultivos que pueden servir como hospederos de la micorriza, la capacidad de la micorriza para permanecer en estado latente a largo plazo, no se ha expresado completamente y que los cultivos afectan de manera diferenciada las poblaciones de micorriza. Por lo tanto se necesita entender mejor todos los factores que afectan la asociación con micorriza para optimizar su eficiencia en la promoción del crecimiento de los cultivos (Troeh y Loynachan, 2003). El objetivo de este trabajo fue conocer la respuesta de dos variedades de soya a la aplicación del biofertilizante Glomus intraradix, en un suelo utilizado alternadamente con soya y maíz.
MATERIALES Y MÉTODOS. El experimento se estableció el 24 de julio de 2003, en el municipio de Medellín de Bravo, Ver., en un suelo de textura migajón arcillosa, ligeramente ácido y con niveles medio y bajo de nitrógeno y fósforo, respectivamente. Se evaluaron dos variedades comerciales de soya: Huasteca 100 y Huasteca 200, generadas por el Campo Experimental Sur de Tamaulipas del INIFAP; y dos tratamientos de fertilización: 60-40-00 de N, P y K, respectivamente y 60-00-00 + Glomus intraradix inoculado a la semilla previo a la siembra en dosis de 1 kg ha-1. Se midió el rendimiento de grano (RG), el cual se transformó en kilogramos por hectárea ajustado al 14 % de humedad. Este parámetro se analizó utilizando el diseño de bloques al azar con cuatro repeticiones en factorial completo regular 2 x 2 y en cada uno de los factores y su acción conjunta en que se detectó significancia se aplicó la prueba de Tukey al 0.05.
RESULTADOS Y DISCUSIÓN. Se detectó efecto significativo en los factor Variedad y en la interacción Variedad x Fertilización. El Cuadro 1 muestra que la variedad más productiva fue Huasteca 200 que superó en 9.5% el rendimiento obtenido por Huasteca 100. Ambas fuentes de fertilización fosfatada proporcionaron un rendimiento estadísticamente similar. Sin embargo, la mejor alternativa de aplicación es la micorriza, ya que los fertilizantes químicos además de ser costosos, contaminan el suelo y el ambiente (Castellanos, 1995). 1 Campo Experimental Cotaxtla. INIFAP. Apdo. Postal # 429, Veracruz, Ver. [email protected]
Cuadro 1. Efecto de variedad y fertilización en el rendimiento de grano de soya.
Factor Rendimiento (kg ha-1) Variedad
Huasteca 200 3399.3 a Huasteca 100 3077.2 b
Fertilización 60-00-00 + G. intraradix 3324.3 a
60-40-00 3152.2 a Comparador Tukey 0.05 309.2
La respuesta a la inoculación con G. intraradix en sustitución de la fertilización fosfatada sólo fue significativa en la variedad Huasteca 200 que incrementó su rendimiento de grano en 557 kg ha-1 (Figura 1).
Figura 1. Efecto de la biofertilización con G. intraradix en el rendimiento de dos variedades de soya.
CONCLUSIONES. Las variedades varían en su respuesta a la inoculación con G. intraradix. Huasteca 200 biofertilizada produjo rendimientos superiores que con la fertilización fosfatada, mientras que Huasteca 100 obtuvo rendimientos similares con ambas fuentes de fertilización.
LITERATURA CITADA Abbott, L. K. and A. D. Robson. 1985. Assessing the potential for widescale VA mycorrhizal inoculation. In: R. Molina (ed.) Proc. North Am. Conf. on Mycorrizae, 6th, Bend, OR. 25-29 June 1984. Forest Res. Lab.; Oregon State Univ., Corvallis. p. 76-79. Castellanos, J. Z. 1995. Potencial de la fijación biológica de nitrógeno en las leguminosas de grano y su influencia en los sistemas de cultivo. In: Memoria del IV Congreso Nacional de la Fijación Biológica de Nitrógeno. Cuernavaca, Mor. p. 25. Troeh, Z. I. and T. E. Loynachan. 2003. Endomycorrhizal fungal survival in continuos corn, soybean and fallow. Agronomy Journal 95: 224-230.
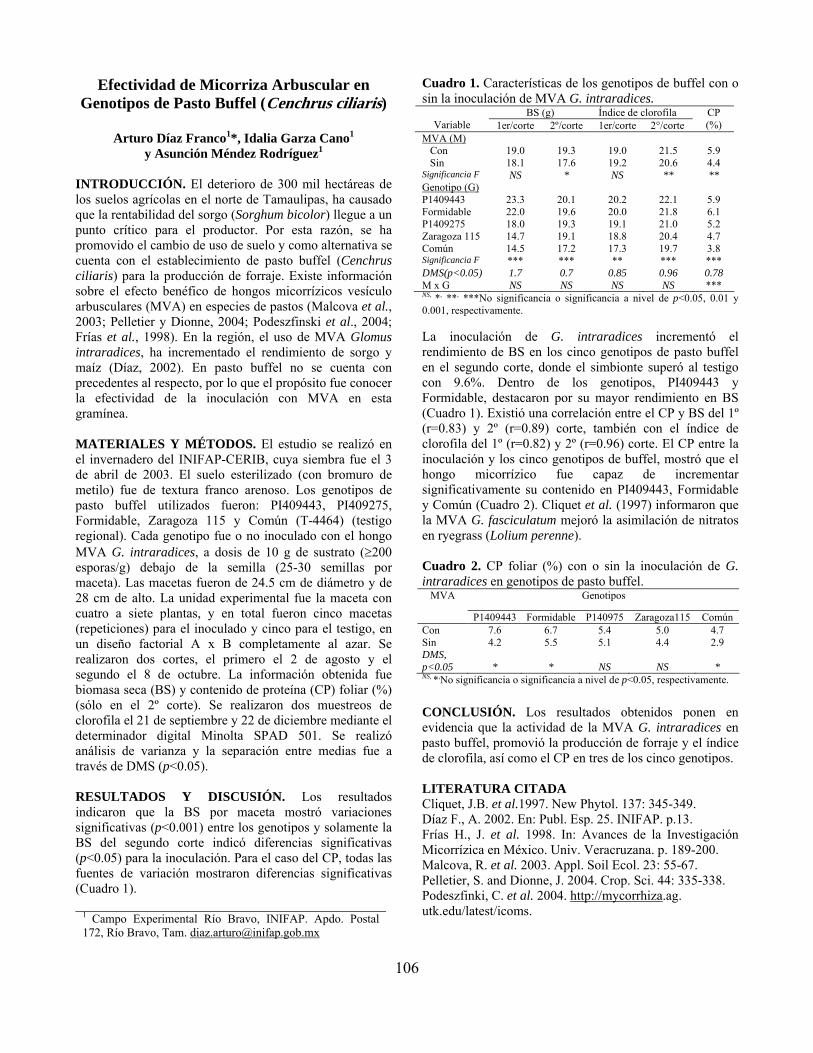
106
Efectividad de Micorriza Arbuscular en Genotipos de Pasto Buffel (Cenchrus ciliaris)
Arturo Díaz Franco1*, Idalia Garza Cano1
y Asunción Méndez Rodríguez1
INTRODUCCIÓN. El deterioro de 300 mil hectáreas de los suelos agrícolas en el norte de Tamaulipas, ha causado que la rentabilidad del sorgo (Sorghum bicolor) llegue a un punto crítico para el productor. Por esta razón, se ha promovido el cambio de uso de suelo y como alternativa se cuenta con el establecimiento de pasto buffel (Cenchrus ciliaris) para la producción de forraje. Existe información sobre el efecto benéfico de hongos micorrízicos vesículo arbusculares (MVA) en especies de pastos (Malcova et al., 2003; Pelletier y Dionne, 2004; Podeszfinski et al., 2004; Frías et al., 1998). En la región, el uso de MVA Glomus intraradices, ha incrementado el rendimiento de sorgo y maíz (Díaz, 2002). En pasto buffel no se cuenta con precedentes al respecto, por lo que el propósito fue conocer la efectividad de la inoculación con MVA en esta gramínea. MATERIALES Y MÉTODOS. El estudio se realizó en el invernadero del INIFAP-CERIB, cuya siembra fue el 3 de abril de 2003. El suelo esterilizado (con bromuro de metilo) fue de textura franco arenoso. Los genotipos de pasto buffel utilizados fueron: PI409443, PI409275, Formidable, Zaragoza 115 y Común (T-4464) (testigo regional). Cada genotipo fue o no inoculado con el hongo MVA G. intraradices, a dosis de 10 g de sustrato (≥200 esporas/g) debajo de la semilla (25-30 semillas por maceta). Las macetas fueron de 24.5 cm de diámetro y de 28 cm de alto. La unidad experimental fue la maceta con cuatro a siete plantas, y en total fueron cinco macetas (repeticiones) para el inoculado y cinco para el testigo, en un diseño factorial A x B completamente al azar. Se realizaron dos cortes, el primero el 2 de agosto y el segundo el 8 de octubre. La información obtenida fue biomasa seca (BS) y contenido de proteína (CP) foliar (%) (sólo en el 2º corte). Se realizaron dos muestreos de clorofila el 21 de septiembre y 22 de diciembre mediante el determinador digital Minolta SPAD 501. Se realizó análisis de varianza y la separación entre medias fue a través de DMS (p<0.05). RESULTADOS Y DISCUSIÓN. Los resultados indicaron que la BS por maceta mostró variaciones significativas (p<0.001) entre los genotipos y solamente la BS del segundo corte indicó diferencias significativas (p<0.05) para la inoculación. Para el caso del CP, todas las fuentes de variación mostraron diferencias significativas (Cuadro 1).
1 Campo Experimental Río Bravo, INIFAP. Apdo. Postal 172, Río Bravo, Tam. [email protected]
Cuadro 1. Características de los genotipos de buffel con o sin la inoculación de MVA G. intraradices.
BS (g) Índice de clorofila Variable 1er/corte 2º/corte 1er/corte 2°/corte
CP (%)
MVA (M) Con 19.0 19.3 19.0 21.5 5.9 Sin 18.1 17.6 19.2 20.6 4.4 Significancia F NS * NS ** ** Genotipo (G) P1409443 23.3 20.1 20.2 22.1 5.9 Formidable 22.0 19.6 20.0 21.8 6.1 P1409275 18.0 19.3 19.1 21.0 5.2 Zaragoza 115 14.7 19.1 18.8 20.4 4.7 Común 14.5 17.2 17.3 19.7 3.8 Significancia F *** *** ** *** *** DMS(p<0.05) 1.7 0.7 0.85 0.96 0.78 M x G NS NS NS NS *** NS, *, **, ***No significancia o significancia a nivel de p<0.05, 0.01 y 0.001, respectivamente. La inoculación de G. intraradices incrementó el rendimiento de BS en los cinco genotipos de pasto buffel en el segundo corte, donde el simbionte superó al testigo con 9.6%. Dentro de los genotipos, PI409443 y Formidable, destacaron por su mayor rendimiento en BS (Cuadro 1). Existió una correlación entre el CP y BS del 1º (r=0.83) y 2º (r=0.89) corte, también con el índice de clorofila del 1º (r=0.82) y 2º (r=0.96) corte. El CP entre la inoculación y los cinco genotipos de buffel, mostró que el hongo micorrízico fue capaz de incrementar significativamente su contenido en PI409443, Formidable y Común (Cuadro 2). Cliquet et al. (1997) informaron que la MVA G. fasciculatum mejoró la asimilación de nitratos en ryegrass (Lolium perenne). Cuadro 2. CP foliar (%) con o sin la inoculación de G. intraradices en genotipos de pasto buffel.
MVA Genotipos
P1409443 Formidable P140975 Zaragoza115 Común Con 7.6 6.7 5.4 5.0 4.7 Sin 4.2 5.5 5.1 4.4 2.9 DMS, p<0.05
*
*
NS
NS
*
NS, *,No significancia o significancia a nivel de p<0.05, respectivamente. CONCLUSIÓN. Los resultados obtenidos ponen en evidencia que la actividad de la MVA G. intraradices en pasto buffel, promovió la producción de forraje y el índice de clorofila, así como el CP en tres de los cinco genotipos. LITERATURA CITADA Cliquet, J.B. et al.1997. New Phytol. 137: 345-349. Díaz F., A. 2002. En: Publ. Esp. 25. INIFAP. p.13. Frías H., J. et al. 1998. In: Avances de la Investigación Micorrízica en México. Univ. Veracruzana. p. 189-200. Malcova, R. et al. 2003. Appl. Soil Ecol. 23: 55-67. Pelletier, S. and Dionne, J. 2004. Crop. Sci. 44: 335-338. Podeszfinki, C. et al. 2004. http://mycorrhiza.ag. utk.edu/latest/icoms.

107
Influencia de Micorriza Arbuscular en el Crecimiento y
Rendimiento de Cártamo
Arturo Díaz Franco1*, Alfredo Ortegón Morales1 e Idalia Garza Cano1
INTRODUCCIÓN. El cártamo (Carthamus tinctorius) es un cultivo de invierno con potencial para entrar en el sistema de producción agrícola del norte de Tamaulipas; con ello se tendría una alternativa de cultivo diferente al monocultivo de sorgo (Sorghum bicolor). Aunque se cuenta con información sobre la producción de cártamo (Ortegón, 2003), se desconoce el efecto de la micorrización sobre la productividad del cultivo. En esta región, la micorriza vesículo arbuscular (MVA) Glomus intraradices ha tenido un impacto favorable en la producción de sorgo y maíz, al incrementar sus rendimientos (Díaz F., 2002; Díaz M., 2002). Por lo anterior, el propósito del estudio fue el de conocer, en condiciones de campo, el efecto de la inoculación de MVA en dos variedades de cártamo. MATERIALES Y MÉTODOS. Dos experimentos se establecieron el 29 de noviembre y 6 de diciembre de 2001 en Matamoros (“El Vaso”, temporal) y Río Bravo, Tam. (CERIB, riego restringido), respectivamente. El diseño fue parcelas divididas en bloques al azar con cuatro repeticiones. En cada localidad se evaluaron las variedades de cártamo Guayalejo y Tantoan 91, como parcelas grandes; en las parcelas chicas se incluyeron los tratamientos: a) MVA G. intraradices, 1 kg/ha, inoculación a la semilla; b) fertilización química, 100-40-00 (urea y superfosfato, respectivamente); y c) testigo absoluto. La parcela experimental fue de tres surcos de 6 m y 0.82 m de ancho. La aplicación de la fertilización química se realizó durante la siembra. En la localidad Río Bravo un riego de auxilio se aplicó en floración. Los datos obtenidos por planta fueron: altura, número de capullos, peso de grano y colonización micorrízica radical (%). Además, el rendimiento de grano por parcela. Se realizó análisis conjunto de las dos localidades. RESULTADOS Y DISCUSIÓN. Los resultados del análisis de varianza combinado, indicaron que para las variables de planta, no existieron diferencias significativas entre tratamientos, variedades ni en las interacciones, y sólo se observaron diferencias significativas entre las localidades (p<0.001), excepto en el porcentaje de colonización micorrízica (Cuadro 1). Esta fluctuó de 14-22% y es considerada baja comparativamente con la registrada por Díaz M. (2002) en sorgo (52%), en la misma región, donde la MVA promovió el crecimiento y el rendimiento de ese cultivo. Para el rendimiento de grano se
1 Campo Experimental Río Bravo, INIFAP. Apdo. Postal 172, Río Bravo, Tam. [email protected]
observaron diferencias significativas en las localidades y entre las variedades; no obstante, la separación de medias (DMS, p<0.05) no diferenció el rendimiento de las variedades (Cuadro 2). Cuadro 1. Características de cártamo en las localidades del norte de Tamaulipas. Promedio de las vars. Guayalejo y Tantoan 91. Planta Localidad Altura
(cm) Col.
micor. (%)
No. capullos
Peso de grano
(g) Matamoros 77.8 15 14.1 15.7 Río Bravo 113.1 18 19.3 20.1 Diferencia F *** NS *** *** ***Denota diferencias a nivel de p<0.001. Cuadro 2. Rendimiento de grano en las variedades de cártamo obtenido de las dos localidades. Variable Rendimiento (kg/ha) Localidad Matamoros 678 bZ
Río Bravo 1094 a Variedad Guayalejo 779 a Tantoan 91 873 a Z Valores con la misma letra no denotan diferencias (DMS, p<0.05). Las escasas precipitaciones registradas (<85 mm) en las localidades durante el ciclo fueron determinantes para que en la de Río Bravo, donde se aplicó un riego de auxilio, el cártamo manifestara el mayor crecimiento y rendimiento. En esa condición se registraron los valores más altos en las variables estudiadas (Cuadro 1 y 2). CONCLUSIONES. Los resultados indicaron que la inoculación de G. intraradices o la fertilización química en cártamo, no tuvieron influencia en el crecimiento y rendimiento. Por el contrario, el cártamo mostró una respuesta esperada en función a la humedad donde fue cultivado. LITERATURA CITADA Díaz F., A. 2002. Incremento de la productividad del sorgo y maíz mediante la biofertilización. En: Publ. Esp. No. 25. INIFAP-CERIB. p. 13-14 Díaz M., R. 2002. La biofertilización en las características de planta, de grano e infección micorrízica del sorgo, en dos condiciones edáficas. Tesis Profesional. UAMRA, UAT. 62 p. Ortegón M., A. 2003. Cártamo: Cultivo de alternativa para el norte de Tamaulipas. Desplegable No. 25. INIFAP-CERIB.

108
Biofertirrigación: Tecnología Sustentable del Siglo XXI
Lina Hernández Flores 1, Juan M. Covarrubias
Ramírez2, Rodrigo Aveldaño Salazar1 y Juan José Peña Cabriales3.
INTRODUCCION. Para el año 2040 la población mundial será de 7.7 billones de personas y se necesitaran 2,500 millones de toneladas de alimentos para ella; la producción de alimentos requiere de 108 millones de toneladas de nitrógeno (N), pero para producir industrialmente este volumen se necesita de un volumen considerable de combustible. Como consecuencia, se incrementan los costos de producción de N y se aumenta la contaminación ambiental cuando algunos fertilizantes industriales son usados inadecuadamente ya que generan contaminación por volatilización y lixiviación (Covarrubias et al., 2002). En fertirrigación, los fertilizantes líquidos o solubles tienen un costo superior de cuatro a seis veces por kilogramo de nutrimento respecto a los fertilizantes sólidos tradicionales. El uso de microorganismos promotores del crecimiento (BPC) aplicados en solución a través del sistema de riego, pero sin fertilización, se define como biofertirriego (González, 1999), pero estos microorganismos no proporcionan el total de la demanda nutrimental del cultivo, por lo que implica la necesidad de aplicar fertilizantes en forma alterna a los microorganismos y/o aplicarlos durante la siembra o al inicio del desarrollo vegetativo del cultivo, etapas donde la colonización en la raíz es más exitosa por la exudación de ácidos orgánicos como malato y citrato (Vessey, 2003). El objetivo es dar a conocer esta nueva tecnología utilizada en varios cultivos, con sus logros y problemas encontrados en la agricultura de México. RESULTADOS Y DISCUSION. En maíz, con fertilizaciones de 140-80-00 en Villadiego, Gto, de 120-80-00 en El CEPAB, Ags, y de 80-00-00 en El CESAJ, Mich. se han aislado organismos diazótrofos como Azospirillum brasilense con una capacidad de fijar N2 de 9.5; 26.9 y 30.3 nMol h-1 mL-1, respectivamente (Hernández, y Covarrubias, 2004). Bajo biofertilización, fertirrigación y biofertirrigación, la fertirrigación tuvo 4.8 % mayor rendimiento de grano que la biofertilización y la biofertirrigación es mayor en 1.2 % que la biofertilización, pero los costos de producción son menores en 25 % que la fertirrigación. Se utilizó 1x109 mL-1 de A. brasilense aplicando 1 L cada 7 días en 6 ocasiones (Covarrubias et al., 2002). En papa, con fertilizaciones de 150-400-200 en El Tunal, y de 120-400-200 en el CESAL, ambos en Arteaga, Coah., se aisló A. brasilense con 21 y 26 nMol h-1 mL-1, respectivamente (Hernández y Covarrubias, 2004). La biofertirrigación mostró un rendimiento de 32.9 t ha-1 [email protected] 2 [email protected] [email protected]
versus 27.9 t ha-1 (15.2 % menos) que la fertirrigación, se aplico 1x105 UFC mL-1 de A. brasilense; 1x108 UFC mL-1 de Bacillus licheniformes y 1x109 UFC mL-1 de Pseudomonas aureofaciens, P. fluorescens y P. putida a una dosis de 2 L por aplicación (Covarrubias et al., 2002). Inoculando papa con Bacillus subtilis en un suelo rico en fósforo (865 kg P2O5 ha-1) con 1x103 UFC mL-1, a una dosis de 4 mL por planta, se incrementó la biomasa en 31.7 % y la absorción de P en 27.5 %, respecto al suelo sin BPC (Covarrubias et al., 2004). En manzano, bajo fertirrigación con 80-80-80-14S en los Lirios, Arteaga, Coah., se aisló A. brasilense con 18.9 nMol h-1 mL-1 (Hernández y Covarrubias, 2004). En esta huerta la fertirrigación (FER) se comparó con la biofertirrigación (BIO) aplicando A. brasilense y Glomus intraradices, a razón de 1.5x105 UFC g-1de turba para la bacteria en bolsa de 0.38 kg y el hongo con 20 % de inóculo en suelo en bolsa de 1 kg, ambas bolsas disueltas en 20 L de agua y aplicando 0.6 L por árbol a 0.3 m de profundidad (Covarrubias et al., 2003); en rendimiento se presentó diferencia significativa a favor de BIO respecto a FER, en 2001 con 8.4 % (86.3 vs.79.6 kg árbol-1) y en 2002 con 11.8 % (97.7 vs. 87.4 kg árbol-1). CONCLUSIONES. La biofertirrigación es una opción tecnológica que favorece la agricultura sustentable. LITERATURA CITADA Covarrubias R., J.M.; R. Núñez E. y L. Hernández F. 2002. Biofertigation: A new sustainable technology. In: IV International symposium on cleaner bioprocesses and sustainable development. Veracruz, Ver. CD-ROM. Covarrubias R., J. M.; L. Hernández F. y R. Aveldaño S. 2003. Biofertirrigacion del manzano en la Sierra de Arteaga, Coahuila. En: Memoria X Congreso Nal. Ciencias Hortícolas, Chapingo, Mex. 10: 297. Covarrubias R., J. M.; S. Castillo A.; J. A. Vera N.; R. Núñez E.; P. Sánchez G.; R. Aveldaño S. y J.J. Peña C. 2004. Medición de la absorción y eficiencia en el uso de fósforo en papa cv. Alpha con 32P. Agrociencia. En prensa. González M., A. 1999. Biofertirriego, un nuevo concepto en la agricultura. En: Memoria 4º Simposium internacional de fertirrigación. ICID, A.C. ed. Guadalajara, Jal. p 195. Hernández F., L. y J. M. Covarrubias R. 2004. Biodiversidad genética de Azospirillum brasilense en maíz, manzano y papa. Agricultura Técnica de México. En prensa. Vessey, J. K. 2003. Plant growth promoting rhizobacteria as biofertilizers. Plant and Soil 255:571-586.

109
Respuesta del Maíz y Sorgo a la Fertilización Biológica
Víctor Pecina Quintero1*, Arturo Díaz Franco1 e Idalia
Garza Cano1 Agustín Magallanes Estala
INTRODUCCIÓN. En el norte de Tamaulipas a pesar de que se ha observado un impacto favorable de los biofertilizantes en la producción de sorgo y maíz, este efecto ha sido inconsistente debido probablemente al tipo de suelo y cepa de microorganismo utilizado en la inoculación, ya que los incrementos en los rendimientos han variado de un 13 al 30 % (Rodelas et al., 1999; Díaz, 2002). Por lo anterior y debido a que existen en el mercado diferentes tipos de biofertilizantes y hormonas vegetales, fue de nuestro interés realizar el presente estudio con el objetivo de conocer la respuesta del sorgo y maíz a diferentes fertilizantes biológicos MATERIALES Y MÉTODOS. El presente estudio se realizó en la localidad Pedro Ochoa del Municipio de Río Bravo, Tam., el 27 de enero del 2002, para lo cual se establecieron dos ensayos. En el primero se utilizó el híbrido de maíz Pioneer 3025 y se evaluaron siete tratamientos: 1) micorriza vesículo arbuscular (MVA) G. intraradices ( ≥200 esporas g-1 de suelo e infecciones del 95%) en dosis de 1000 g para 20 kg de semilla ha-1; 2) dos aspersiones de brassinoesteroides CIDEF-4 (B) antes de floración; 3) mezcla de MVA+B; 4) Flower Saver a razón de 1 kg ha-1; 5) Burize, 40 g de esporas ha-1; 6) fertilización química a la siembra (120 kg ha-1 de N y 40 kg ha-1 de P; y 7) testigo absoluto. En el caso de sorgo se utilizó el híbrido Asgrow Coral y se evaluaron los tratamientos 1,2,3,4,6, y 7, anteriormente descritos. El diseño fue de bloques al azar con cinco repeticiones. La parcela experimental de maíz fue de seis surcos (0.84 m) de 8 m de longitud; mientras que la de sorgo fue de cuatro surcos de 5 m. En maíz los cuatro surcos centrales se dividieron entre dos para cuantificar peso de elote y rendimiento de grano. En sorgo se midió el rendimiento de grano y se muestrearon cinco plantas por parcela para estimar el porcentaje de colonización micorrízica en ambos cultivos. Se realizó un análisis de varianza y la diferenciación entre medias fue mediante DMS (p<0.05). RESULTADOS Y DISCUSIÓN. En maíz, el análisis de varianza para rendimiento de elote indicó diferencias estadísticas entre tratamientos de biofertilizantes como ha sido reportado (Rodelas et al., 1999), resultando la MVA y la fertilización química estadísticamente iguales, superando a los demás tratamientos incluyendo al testigo (Cuadro 1). Mientras que en rendimiento de grano los tratamientos de MVA, brassinoesteroides, fertilización química y Burize superaron a la mezcla MVA+B, Flower Saver y testigo. Con excepción del Flower Saver todos los
1 Campo Experimental Río Bravo, INIFAP. Apdo. Postal 172, Río Bravo, Tam. [email protected]
tratamientos de biofertilizantes incrementaron la colonización micorrízica. En sorgo, el rendimiento de grano se incrementó significativamente con la inoculación de la MVA presentando el mayor rendimiento (29%) sobre el testigo aunque fue estadísticamente igual con los demás tratamientos de fertilizantes biológicos. Así mismo el mayor porcentaje de colonización micorrízica la presentó la MVA (Cuadro 2).
Cuadro 1. Productividad de maíz y colonización micorrízica asociados a la aplicación de diferentes fertilizantes biológicos.
Tratamiento Rendimiento (kg ha-1) elote grano
Infección radical (%)
MVA (M) 13,450 a 3896 a 49.0 a Brassino. (B) 12,175 b 3370 ab 47.9 a
M + B 12,425 b 2972 b 47.8 a Fert. Química 12,748 ab 3481 ab 45.4 a Flower Saver 12,075 b 3314 b 38.8 b
Burize 12,500 b 3526 ab 43.5 a Testigo 11,225 c 3277 b 37.7 b
DMS (p<0.05) 728 558 5.7 * Valores con la misma letra no denotan diferencias (DMS, p<0.05). Cuadro 2. Productividad de sorgo y colonización micorrízica asociados a la aplicación de diferentes fertilizantes biológicos.
Tratamiento Rendimiento grano (kg ha-1)
Infección radical (%)
MVA (M) 4018 a 64.3 a Brassino. (B) 3647 a 60.9 b
M + B 3990 a 62.4 b Fert. Química 3669 a 56.4 c Flower Saver 3523 a 55.1 c
Testigo 3082 b 50.2 d DMS (p<0.05) 540 1.8
* Valores con la misma letra no denotan diferencias (DMS, p<0.05). CONCLUSIONES. Los resultados indicaron diferencias significativas entre los diferentes fertilizantes biológicos comerciales, observándose una mejor respuesta de los cultivos de maíz y sorgo a G. intraradices. Además, la MVA siempre fue igual estadísticamente a la fertilización química, lo que corrobora que la práctica de inoculación a la semilla con la MVA, puede ser un manejo rutinario que ayude a ser sostenible la producción de maíz y sorgo en la zona norte de Tamaulipas.
LITERATURA CITADA Díaz F., A. 2002. Incremento de la productividad de sorgo y maíz mediante la biofertilización. En: Publ. Esp. No. 25. INIFAP-CERIB. p. 13-14. Rodelas, M. B.; González. J.; Martínez, M. V.; Pozo, C. y Salmeron, V. 1999. Influence of Rhizobium-Azospirillum and Rhizobium-Azotobacter combined inoculation on mineral composition of faba bean (Vicia faba L.). http://link.springerny.com/link/service/journals/00374/bibs/9029002/90290165.htm.

110
Efecto de la Micorriza Arbuscular en Sorgo Bajo Dos Condiciones de Humedad
Víctor Pecina Quintero1*, Arturo Díaz Franco1 e Idalia
Garza Cano1
INTRODUCCIÓN. Se ha observado que la micorriza vesículo arbuscular (MVA) promueve el rendimiento, el área foliar, la longitud y peso de raíz, en diferentes cultivos como maíz, cilandro, betabel, haba, etc., no detectándose diferencias estadísticas en comparación con la fertilización química (Sánchez, 2004). En esta región, la MVA Glomus intraradices ha tenido un impacto favorable en la producción de sorgo y maíz, al incrementar los rendimientos de un 13 a 30 % (Díaz, 2002). No obstante que los estudios se han realizado bajo condiciones de temporal no existe información del efecto que puede tener la MVA bajo diferentes condiciones de humedad. Por lo anterior, el objetivo del presente estudio fue el de conocer el efecto de la inoculación de MVA en sorgo bajo dos condiciones de humedad. MATERIALES Y MÉTODOS. El presente estudio se realizó en terrenos del Campo Experimental Río Bravo, el 6 de febrero del 2003, para lo cual se utilizó el híbrido de sorgo RB-Patrón. El diseño experimental fue parcelas divididas en bloques al azar con cuatro repeticiones. La parcela mayor representó la condición de humedad: a) un riego de auxilio en hoja bandera y b) temporal. La parcela menor los tratamientos de fertilización: a) MVA G. intraradices ( ≥200 esporas g-1 de suelo e infecciones del 95%) en dosis de 1000 g para 8 kg de semilla ha-1; b) fertilización química (80 kg ha-1 de N) y c) testigo absoluto. La parcela experimental fue de cuatro surcos de 6 m y 0.80 m de ancho. La fertilización química se realizó durante el último cultivo con amoniaco anhídro previo al riego de auxilio, práctica común en la región. Los datos obtenidos fueron altura de planta, diámetro del tallo, el rendimiento y porcentaje de proteína del grano. Se realizó análisis de varianza y la diferenciación entre medias fue mediante DMS (p<0.05). RESULTADOS Y DISCUSIÓN. El análisis de varianza indicó diferencias significativas en altura de planta, rendimiento de grano y porcentaje de proteína para la condición de humedad (H), mientras que entre tratamientos de fertilización (F), solo hubo diferencias estadísticas en rendimiento y porcentaje de proteína, no así en la interacción (HxF) (Cuadro 1). La altura del sorgo se incrementó cuando fue aplicado el riego de auxilio independientemente del tratamiento de fertilización, lo que demuestra que la humedad es un factor limitante en la asimilación de nutrientes, que se refleja en el desarrollo de las plantas (Cuadro 2). Por el contrario el diámetro de tallo permaneció constante con relación a la condición de
1 Campo Experimental Río Bravo, INIFAP. Apdo. Postal 172, Río Bravo, Tam. [email protected]
humedad y tratamiento de fertilización (Cuadro 1). Así mismo como era de esperarse, la aplicación de un riego de auxilio durante la etapa de hoja bandera incrementó el rendimiento de grano comparativamente con el testigo bajo temporal (Cuadro 2). Resultó evidente la efectividad de la inoculación a la semilla con la MVA G intraradices en ambas condiciones de humedad, ya que incrementó significativamente el rendimiento de grano en 21% en función al testigo. La adición de N durante el último cultivo con amoniaco anhídro antes del riego de auxilio no tuvo impacto en el rendimiento y fue semejante su respuesta a la del testigo, lo que demuestra lo ineficiente de la fertilización química en esta etapa. De igual forma la disponibilidad de humedad incrementó el contenido de proteína en el grano hasta un 28% en comparación con el testigo (Cuadro 2), y no hubo diferencias estadísticas entre la inoculación con la MVA y la fertilización química, lo que coincide con lo reportado por Sánchez (2004). Cuadro 1. Cuadrados medios del análisis de varianza para cuatro características agronómicas de sorgo evaluadas en el norte de Tamaulipas.
Fuente de variación
Altura planta
Diámetro de tallo
Proteína (%)
Rendto kg ha-1
Humedad (H) 48.5** 0.003 10.7* 37.5** Fertilización (F) 0.6 0.12 6.8** 7.9**
H x F 0.006 0.14 0.4 1.2 *, ** Significativo a nivel de p<0.05 y p<0.01, respectivamente. Cuadro 2. Principales parámetros evaluados en sorgo en función de los tratamientos y condición de humedad. Condición de Humedad
Altura (cm)
Proteína (%)
Rendimiento Kg ha-1
Temporal 115 b 7.3 b 3227 b Riego restringido
129 a
9.4 a
4966 a
DMS p<0.05 10.8 2 887 CONCLUSIONES. Los resultados indicaron que la inoculación de la semilla de sorgo con la MVA G. intraradices tiende a incrementar el rendimiento y porcentaje de proteína en el grano y es efectiva tanto en riego restringido como temporal. LITERATURA CITADA Díaz F., A. 2002. Incremento de la productividad de sorgo y maíz mediante la biofertilización. En: Publ. Esp. No. 25. INIFAP-CERIB. p. 13-14. Sánchez, C. Ma. de Jesús. Efecto de las micorrizas arbusculares en el desarrollo de los cultivos agrícolas. XV Congreso Mexicano de Botánica y Micología. http://www.socbot.org.mx/resumenes/resumen529.html.

111
Efecto de una Composta y Acidos fúlvicos en la Producción de lilies (Lilium sp) en el sureste de
Coahuila
Ma. del Rosario Zúñiga Estrada1*, Juan M. Covarrubias Ramírez2 y Rubén López Cervantes3
INTRODUCCION. El lilies (Lilium sp) es un cultivo ornamental que representa una alternativa de producción para los agricultores de las comunidades rurales del sureste de Coahuila, por su alta demanda en el mercado regional, debido a su gran variedad de colores y su durabilidad en florero. Sin embargo, se ha detectado que uno de los factores que influye de manera determinante en la calidad de la flor es el manejo adecuado de la nutrición y las características del suelo en que se desarrolla el cultivo (1). Es conocido que la adición de materia orgánica al suelo, aumenta el contenido de este compuesto, mejora la estabilidad estructural e incrementa el contenido de poros, por donde circula el agua y por consiguiente provoca una mayor disponibilidad de elementos nutritivos para las plantas (2). Por lo anterior, el propósito del estudio fue evaluar el efecto de una composta, elaborada a base de gallinaza y los ácidos fúlvicos extraída de ella, en la calidad de la flor de corte de lilies MATERIALES Y METODOS. El trabajo se estableció en el ejido Guadalupe Victoria, mpio. de Saltillo. Coah. en febrero de 2004 bajo condiciones de invernadero rústico. Se utilizaron bulbos del tipo asiático cv. “Elite” a una densidad de 50 bulbos/m2. Se evaluaron tres dosis de composta (0.5; 1.0 y 1.5 t ha-1) y tres de ácidos fúlvicos (0.2; 0.4 y 0.6 mL L-1 de agua) combinados con fertilización química: 100-120-80 (N-P2O5-K2O). El diseño fue completamente al azar con arreglo factorial completo. El análisis estadístico fue a través de un análisis de varianza (ANVA) y la prueba de medias por Tukey (p ≤ 0.05). Se usó el paquete para computador MINITAB, versión 13 para WINDOWS. Las variables evaluadas fueron: días a brotación, longitud de tallo, de inflorescencia y de primer botón, diámetro de primera flor, número de flores abiertas y número de botones por planta. RESULTADOS Y DISCUSION. El análisis de varianza indicó que las variables días a brotación, diámetro de primera flor y número de flores abiertas, no presentaron diferencia significativa entre tratamientos ni en las interacciones. Sin embargo, las variables longitud de tallo, de inflorescencia, de primer botón y número total de botones por planta si presentan diferencia significativa ____________________________ 1 MC. Investigador del Campo Exp. Saltillo-CIRNE-INIFAP [email protected] 2 MC. Estudiante de Doctorado. CIRNE-INIFAP. 3 Dr. Profesor-Investigador del Departamento de Suelos. UAAAN. [email protected]
(Cuadro 1); donde se observa que el tratamiento con la mayor dosis de composta (1.5 t ha-1) y ácidos fúlvicos (0.6 mL L-1) tuvo el mayor efecto en las características de calidad de la flor de lilies demandadas por el mercado: longitud de tallo y número total de flores por planta; esto puede deberse a que al mezclar composta en la capa superior del suelo, en donde se desarrollan las raíces de los bulbos (20 cm), se logra tener una capa superficial con adecuada capacidad de retención de humedad y mayor porosidad, lo que permitió mayor disponibilidad de nutrimentos a la planta. El análisis económico, mostró que la cosecha realizada a las 10 semanas después de la siembra, coincidió con una de las fechas en donde la flor de corte alcanza el mayor precio de venta (10 de mayo). Cuadro 1. Resultados de variables vegetativas y productivas en lilies asiático cv. Elite INIFAP - CIRNE-CESAL. 2004.
Longitud (cm)
Factor
Dosis
Tallo
Inflorescencia
Botón
Número Botones/
planta
Composta 1.5* 91.0 a 10.9 a 8.8 a 9 a t ha-1 1 88.4 b 9.3 b 7.7 b 7 b
0.5 87.2 b 8.0 c 6.2 c 7 b
Acidos Fúlvicos 0.6 93.0 a 9.9 a 8.1a 8 .3 a mL L-1 0.4 88.6 b 9.4 b 7.4 b 7 .3 b
0.2 85.1 c 8.9 b 7.1 b 7 .3 b * Diferente literal entre columnas indica diferencia significativa CONCLUSIONES. 1). Con la adición de 1.5 t ha-1 y 0.6 mL L-1 se obtienen las mejores características de calidad en la flor de lilies como son: longitud de tallo y número de botones por planta. 2). La producción de lilies demostró ser una alternativa viable para los productores rurales del sureste de Coahuila. BIBLIOGRAFIA 1. Collicut, M. L. and Ronald W. G. 1995. Northern Beauty Lily. HortScience 30 (3): 652-653 2. Kikuchi, K. 2003. Evaluation of organic materials and their effective use in a era sustanaible agriculture. Hohhaído International Centre, Obihiro (HICO) p. 1-15.
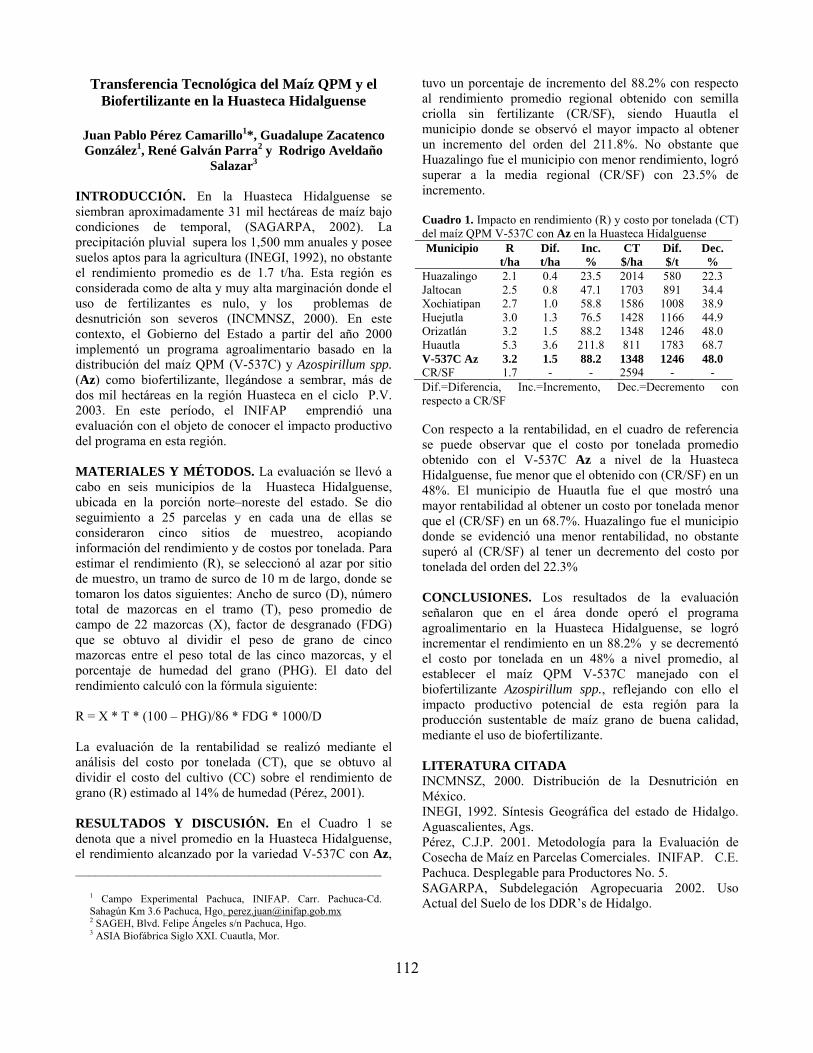
112
Transferencia Tecnológica del Maíz QPM y el Biofertilizante en la Huasteca Hidalguense
Juan Pablo Pérez Camarillo1*, Guadalupe Zacatenco González1, René Galván Parra2 y Rodrigo Aveldaño
Salazar3
INTRODUCCIÓN. En la Huasteca Hidalguense se siembran aproximadamente 31 mil hectáreas de maíz bajo condiciones de temporal, (SAGARPA, 2002). La precipitación pluvial supera los 1,500 mm anuales y posee suelos aptos para la agricultura (INEGI, 1992), no obstante el rendimiento promedio es de 1.7 t/ha. Esta región es considerada como de alta y muy alta marginación donde el uso de fertilizantes es nulo, y los problemas de desnutrición son severos (INCMNSZ, 2000). En este contexto, el Gobierno del Estado a partir del año 2000 implementó un programa agroalimentario basado en la distribución del maíz QPM (V-537C) y Azospirillum spp. (Az) como biofertilizante, llegándose a sembrar, más de dos mil hectáreas en la región Huasteca en el ciclo P.V. 2003. En este período, el INIFAP emprendió una evaluación con el objeto de conocer el impacto productivo del programa en esta región. MATERIALES Y MÉTODOS. La evaluación se llevó a cabo en seis municipios de la Huasteca Hidalguense, ubicada en la porción norte–noreste del estado. Se dio seguimiento a 25 parcelas y en cada una de ellas se consideraron cinco sitios de muestreo, acopiando información del rendimiento y de costos por tonelada. Para estimar el rendimiento (R), se seleccionó al azar por sitio de muestro, un tramo de surco de 10 m de largo, donde se tomaron los datos siguientes: Ancho de surco (D), número total de mazorcas en el tramo (T), peso promedio de campo de 22 mazorcas (X), factor de desgranado (FDG) que se obtuvo al dividir el peso de grano de cinco mazorcas entre el peso total de las cinco mazorcas, y el porcentaje de humedad del grano (PHG). El dato del rendimiento calculó con la fórmula siguiente: R = X * T * (100 – PHG)/86 * FDG * 1000/D La evaluación de la rentabilidad se realizó mediante el análisis del costo por tonelada (CT), que se obtuvo al dividir el costo del cultivo (CC) sobre el rendimiento de grano (R) estimado al 14% de humedad (Pérez, 2001). RESULTADOS Y DISCUSIÓN. En el Cuadro 1 se denota que a nivel promedio en la Huasteca Hidalguense, el rendimiento alcanzado por la variedad V-537C con Az, ______________________________________________
tuvo un porcentaje de incremento del 88.2% con respecto al rendimiento promedio regional obtenido con semilla criolla sin fertilizante (CR/SF), siendo Huautla el municipio donde se observó el mayor impacto al obtener un incremento del orden del 211.8%. No obstante que Huazalingo fue el municipio con menor rendimiento, logró superar a la media regional (CR/SF) con 23.5% de incremento. Cuadro 1. Impacto en rendimiento (R) y costo por tonelada (CT) del maíz QPM V-537C con Az en la Huasteca Hidalguense Municipio R
t/ha Dif. t/ha
Inc. %
CT $/ha
Dif. $/t
Dec. %
Huazalingo 2.1 0.4 23.5 2014 580 22.3 Jaltocan 2.5 0.8 47.1 1703 891 34.4 Xochiatipan 2.7 1.0 58.8 1586 1008 38.9 Huejutla 3.0 1.3 76.5 1428 1166 44.9 Orizatlán 3.2 1.5 88.2 1348 1246 48.0 Huautla 5.3 3.6 211.8 811 1783 68.7 V-537C Az 3.2 1.5 88.2 1348 1246 48.0 CR/SF 1.7 - - 2594 - - Dif.=Diferencia, Inc.=Incremento, Dec.=Decremento con respecto a CR/SF Con respecto a la rentabilidad, en el cuadro de referencia se puede observar que el costo por tonelada promedio obtenido con el V-537C Az a nivel de la Huasteca Hidalguense, fue menor que el obtenido con (CR/SF) en un 48%. El municipio de Huautla fue el que mostró una mayor rentabilidad al obtener un costo por tonelada menor que el (CR/SF) en un 68.7%. Huazalingo fue el municipio donde se evidenció una menor rentabilidad, no obstante superó al (CR/SF) al tener un decremento del costo por tonelada del orden del 22.3% CONCLUSIONES. Los resultados de la evaluación señalaron que en el área donde operó el programa agroalimentario en la Huasteca Hidalguense, se logró incrementar el rendimiento en un 88.2% y se decrementó el costo por tonelada en un 48% a nivel promedio, al establecer el maíz QPM V-537C manejado con el biofertilizante Azospirillum spp., reflejando con ello el impacto productivo potencial de esta región para la producción sustentable de maíz grano de buena calidad, mediante el uso de biofertilizante. LITERATURA CITADA INCMNSZ, 2000. Distribución de la Desnutrición en México. INEGI, 1992. Síntesis Geográfica del estado de Hidalgo. Aguascalientes, Ags. Pérez, C.J.P. 2001. Metodología para la Evaluación de Cosecha de Maíz en Parcelas Comerciales. INIFAP. C.E. Pachuca. Desplegable para Productores No. 5. SAGARPA, Subdelegación Agropecuaria 2002. Uso Actual del Suelo de los DDR’s de Hidalgo.
1 Campo Experimental Pachuca, INIFAP. Carr. Pachuca-Cd. Sahagún Km 3.6 Pachuca, Hgo. [email protected] 2 SAGEH, Blvd. Felipe Ángeles s/n Pachuca, Hgo. 3 ASIA Biofábrica Siglo XXI. Cuautla, Mor.

113
Producción y Evaluación de un Biofertilizante (Azospirillum spp.) Para el Noreste de México
IPN-CBG
Jesús G. García1*, Alberto Mendoza1, Cuauhtemoc Jacques1, Antonia Cruz1, Felipe Serrano1
INTRODUCCIÓN. Estudios diversos con microorganismos del suelo han demostrado que bacterias del genero Azospirillum spp, pueden aportar del 30 al 50% de los requerimientos de nitrógeno para algunas plantas de importancia económica como son el maíz, sorgo y caña de azúcar (1). Estas bacterias también producen fitohormonas naturales que provocan un mayor crecimiento de la raíz y permite una mayor área de asimilación de nutrientes (2 y 3). El empleo de las diversas especies de Azospirillum, ha sido reportado para varios cultivos, donde se han obtenido una amplia gama de resultados positivos, y donde se ha llegado a contundentes conclusiones, entre las que se destaca el mínimo número de bacterias por gramo o mililitro de formulado situado en 109 cfu/g (4).
MATERIALES Y MÉTODOS. En el presente trabajo, se realizó la evaluación en campo de un biofertilizante elaborado con la cepa CBG-497 de Azospirillum brasilense, que fue aislada en el laboratorio de Biotecnología Vegetal CBG, a partir de suelo de rizosfera y tallo de cereales de la región, en los ciclos O.I. 2002-2004 en diferentes municipios, en lotes representativos de las diferentes condiciones agro ecológicas típicas de zona Norte del Estado de Tamaulipas. La cepa CBG-497, fue seleccionada por su capacidad de producción in vitro de Ácido Indol Acético se inoculó para su evaluación en campo en una superficie del orden de 2000 hectáreas, en el ciclo O.I. 2002-2003. El biofertilizante fue formulado en turba a una concentración de 1x106 bacterias por semilla. En el ciclo O.I. 2003-2004 la superficie sembrada con el biofertilizante fue con la Tecnología aplicada por la empresa Biotec Internacional, la cual se realizo un proceso de liofilización, obteniendo concentraciones de 1 x 108 colonias formando unidades (cfu/g). El producto se aplico en 3000 hectáreas. Conjuntamente con la evaluación de campo se establecieron experimentos con diferentes cepas de Azospirillum brasilense en sorgo. Simultáneamente con las evaluaciones agronómicas de la cepa CBG-497, se realizaron diferentes trabajos sobre el escalamiento para la producción y elaboración del Biofertilizante, encontrándose los parámetros óptimos para producción de esta bacteria, optimizándose medios de cultivo y demás 1Centro de Biotecnología Genómica, Instituto Politécnico Nacional. Blvd. del Maestro. Col. Narciso Mendoza. Reynosa, Tam. 77810. E-mail [email protected]
factores que actúan en el proceso de producción en fermentador. En cuanto al proceso de divulgación de aplicación de esta técnica, preparó material documental del uso, manejo y aplicación del biofertilizante, el cual fue promovido reuniones técnicas donde asistieron productores y personal técnico del PIFSV y de SAGARPA. RESULTADOS. En el ciclo O.I 2002-2003 se presento un incremento en el rendimiento de grano/ha en los diferentes municipios con un promedio de 12.6% para el cultivo de sorgo, y 30% para el cultivo de maíz, al que se le aplico fertilización nitrogenada. En el ciclo O.I. 2003-2004 el comportamiento de los materiales de sorgo no presentó aumentos en el rendimiento, únicamente se encontró un aumento del 5% de rendimiento para el híbrido 83G15. En cuanto a los lotes de maíz evaluados en los diferentes municipios, presentan un 10% promedio de incremento. El cuanto al comportamiento de la cepas evaluadas en los dos ciclos, presentaran incrementos del 18% en el ciclo 2002-2003 y de 12% para el ciclo 2003-2004 con respecto al testigo.
CONCLUSIONES.
El uso de Azospirillum como biofertilizante, puede ser benéfico para tener incrementos en los rendimientos de grano en los cultivos de sorgo y maíz, mayormente si para la elaboración de los biofertilizantes se utilizan cepas nativas. Finalmente, es necesario continuar con evaluaciones en campo lo que, promoverá el empleo de estos productos por parte de los agricultores en beneficio directo a la producción agrícola. BIBLIOGRAFÍA. 1. Bashan, Y. and Holguin, G. (1994). Root-to Root travel of the beneficial bacterium. Applied and Environmental Microbiology. Vol. 60. No. 6: 2120-2131. 2. Eory, V.J., Momo, F.R. & Álvarez, M. (1995). Growth and survival of Azospirillum in roots and maize rhizospheres with different leaves of acidity. Rev. Argent. Microbial. 27: 99-105. 3. Vande Broek, A., Lambertch, M. & Vanderleyden, J. (1999) Auxins upregulate expression of the indole-3-pyruvate decarboxylase gene in Azospirillum brasilense. J. Bacteriol. 181: 1338-1342. 4. Okon, Y and Itzigshon, R. (1995). The development of Azospirillum as a commercial inoculant for improving crops yields. Biotechnology Advances. Vol. 13. No. 4: 415-424.

114
Respuesta del Sorgo a la Inoculación de Glomus intraradices en Campo
Agustín Magallanes Estala1*
INTRODUCCIÓN. Los estudios realizados con la inoculación micorrízica del hongo vesícilo arbuscular Glomus intraradices en semilla de sorgo, han mostrado efectividad de la práctica con manifestaciones variables en la promoción del crecimiento de raíz, aumento de clorofila, de P foliar, de biomasa, de infección micorrízica radical y además, incremento del rendimiento de grano. Con el propósito de darle seguimiento a los resultados obtenidos en sorgo, se realizó un estudio de validación comercial de la inoculación micorrízica, comparativamente con la aplicación de Biozyme a la semilla, práctica común en la región.
MATERIALES Y MÉTODOS. Con el apoyo del Sr. Daniel Bone, productor cooperante, se estableció una parcela de validación en un suelo de textura migajón arenoso, pH ligeramente alcalino, alto en nitrógeno y mediano en fósforo. en el poblado de Anahuac, municipio de Valle Hermoso, Tam. La siembra fue mecanizada, con sembradora International 82 y ésta se estableció el 3 y 4 de febrero de 2003 en un terreno sin fertilización química, en la cual se utilizó el híbrido de sorgo 83G63. Los tratamientos fueron a) inoculación a la semilla de G. intraradices, a razón de 1 kg de sustrato, 60 ml de adherente y 700 ml de agua, en 8 kg de semilla utilizada por hectárea; b) Biozyme® a dosis de 200 ml/100 kg de semilla; y c) testigo absoluto. Cada tratamiento se sembró en franjas y ocupó una superficie de 24 surcos de 0.86 m y 1000 m de largo (1020.64 m2). En madurez se realizaron cinco submuestreos en parcelas de dos surcos de 3 m de longitud donde se midió la altura en 10 plantas y se estimó el rendimiento de grano, ajustado al 12% de humedad. Los datos se analizaron estadísticamente.
RESULTADOS Y DISCUSIÓN. La mayor promoción en la altura de planta se observó con el tratamiento consistente en la inoculación de G. intraradices en la semilla de sorgo. Sin embargo, ese tratamiento y la aplicación de Biozyme en la semilla, incrementaron significativamente y de manera similar el rendimiento de grano con relación al testigo absoluto (Cuadro 1). Los resultados son coincidentes con los obtenidos con anterioridad y demuestran la efectividad de la inoculación micorrízica en sorgo. Existen antecedentes de que el Biozyme tiene impacto en la producción, lo cual fue confirmado en el presente estudio. No obstante, el costo del producto es superior al del sustrato micorrízico utilizado. 1 Campo Experimental Pachuca-INIFAP.
Cuadro 1. Altura de planta y rendimiento de grano de sorgo (83G63) influenciados por los tratamientos en la localidad Anáhuac, Valle Hermoso, Tam. Ciclo O-I 2003.
Tratamiento Altura de planta (cm)
Rendimiento (kg/ha)
G. intraradices 123 a 2909 a Biozyme 114 b 2813 a Testigo 118 b 2127 c Prueba de t * ** *,** Significancia a nivel de p<0.05 y 0.01, respectivamente
CONCLUSONES.
De acuerdo a los resultados obtenidos del presente estudio se observó la efectividad de la aplicación micorrízica en sorgo. Para el rendimiento de grano no existieron diferencias significativas con respecto a la inoculación micorrízica y la aplicación de Biozyme, con la desventaja de este último de presentar un costo mayor a la del sustrato micorrízico utilizado, lo cual impacta considerablemente en los costos de producción del cultivo.
LITERATURA CITADA.
1) Acosta N., S., Díaz F., A., Magallanes E., A., y Silva S., M. 2000. La biofertilización en el norte de Tamaulipas. Campo Experimental Río Bravo, INIFAP. Informe 1999/2000. 9 p.
2) Garza C., I., Díaz F., A., Pecina Q., V., Ramírez L., A., y Olalde P., V. 2001. Evaluación de simbiontes y hormona esteroidal en sorgo. Memorias XXVIII Congreso Nacional Fitopatología. P. F-14.
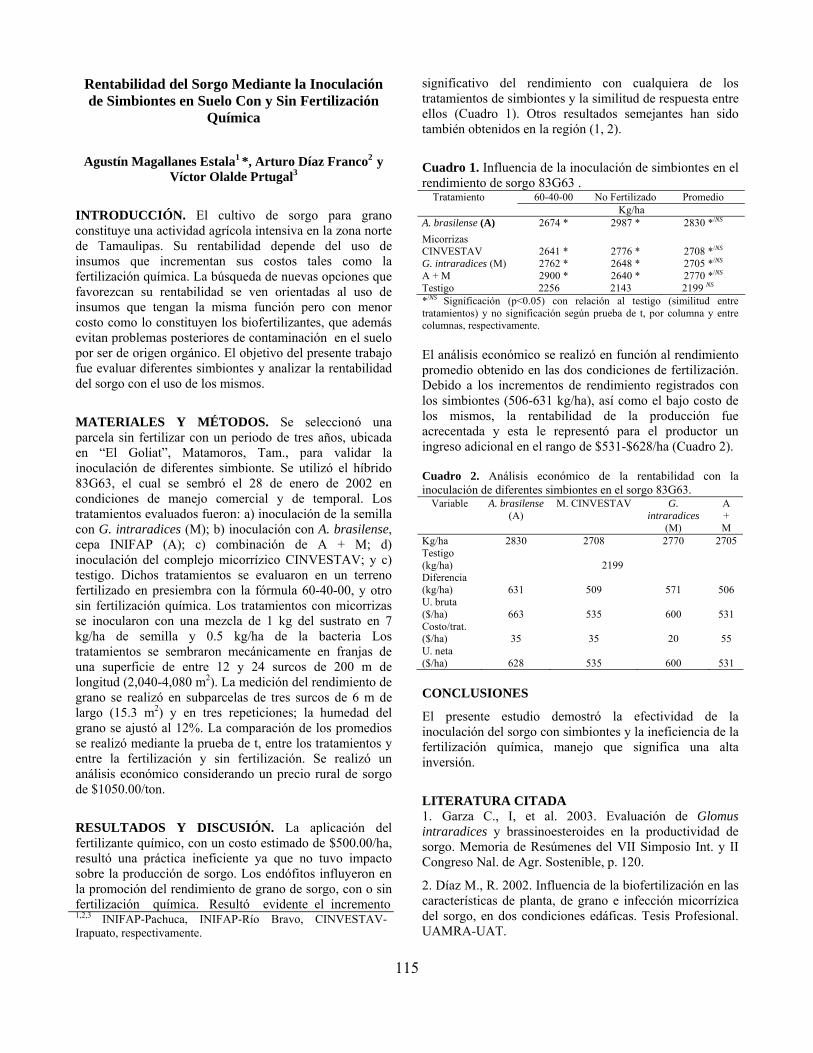
115
Rentabilidad del Sorgo Mediante la Inoculación de Simbiontes en Suelo Con y Sin Fertilización
Química
Agustín Magallanes Estala1 *, Arturo Díaz Franco2 y Víctor Olalde Prtugal3
INTRODUCCIÓN. El cultivo de sorgo para grano constituye una actividad agrícola intensiva en la zona norte de Tamaulipas. Su rentabilidad depende del uso de insumos que incrementan sus costos tales como la fertilización química. La búsqueda de nuevas opciones que favorezcan su rentabilidad se ven orientadas al uso de insumos que tengan la misma función pero con menor costo como lo constituyen los biofertilizantes, que además evitan problemas posteriores de contaminación en el suelo por ser de origen orgánico. El objetivo del presente trabajo fue evaluar diferentes simbiontes y analizar la rentabilidad del sorgo con el uso de los mismos.
MATERIALES Y MÉTODOS. Se seleccionó una parcela sin fertilizar con un periodo de tres años, ubicada en “El Goliat”, Matamoros, Tam., para validar la inoculación de diferentes simbionte. Se utilizó el híbrido 83G63, el cual se sembró el 28 de enero de 2002 en condiciones de manejo comercial y de temporal. Los tratamientos evaluados fueron: a) inoculación de la semilla con G. intraradices (M); b) inoculación con A. brasilense, cepa INIFAP (A); c) combinación de A + M; d) inoculación del complejo micorrízico CINVESTAV; y c) testigo. Dichos tratamientos se evaluaron en un terreno fertilizado en presiembra con la fórmula 60-40-00, y otro sin fertilización química. Los tratamientos con micorrizas se inocularon con una mezcla de 1 kg del sustrato en 7 kg/ha de semilla y 0.5 kg/ha de la bacteria Los tratamientos se sembraron mecánicamente en franjas de una superficie de entre 12 y 24 surcos de 200 m de longitud (2,040-4,080 m2). La medición del rendimiento de grano se realizó en subparcelas de tres surcos de 6 m de largo (15.3 m2) y en tres repeticiones; la humedad del grano se ajustó al 12%. La comparación de los promedios se realizó mediante la prueba de t, entre los tratamientos y entre la fertilización y sin fertilización. Se realizó un análisis económico considerando un precio rural de sorgo de $1050.00/ton.
RESULTADOS Y DISCUSIÓN. La aplicación del fertilizante químico, con un costo estimado de $500.00/ha, resultó una práctica ineficiente ya que no tuvo impacto sobre la producción de sorgo. Los endófitos influyeron en la promoción del rendimiento de grano de sorgo, con o sin fertilización química. Resultó evidente el incremento 1,2,3 INIFAP-Pachuca, INIFAP-Río Bravo, CINVESTAV-Irapuato, respectivamente.
significativo del rendimiento con cualquiera de los tratamientos de simbiontes y la similitud de respuesta entre ellos (Cuadro 1). Otros resultados semejantes han sido también obtenidos en la región (1, 2).
Cuadro 1. Influencia de la inoculación de simbiontes en el rendimiento de sorgo 83G63 . Tratamiento 60-40-00 No Fertilizado Promedio Kg/ha A. brasilense (A) 2674 * 2987 * 2830 */NS
Micorrizas CINVESTAV
2641 *
2776 *
2708 */NS
G. intraradices (M) 2762 * 2648 * 2705 */NS
A + M 2900 * 2640 * 2770 */NS
Testigo 2256 2143 2199 NS
*/NS Significación (p<0.05) con relación al testigo (similitud entre tratamientos) y no significación según prueba de t, por columna y entre columnas, respectivamente. El análisis económico se realizó en función al rendimiento promedio obtenido en las dos condiciones de fertilización. Debido a los incrementos de rendimiento registrados con los simbiontes (506-631 kg/ha), así como el bajo costo de los mismos, la rentabilidad de la producción fue acrecentada y esta le representó para el productor un ingreso adicional en el rango de $531-$628/ha (Cuadro 2). Cuadro 2. Análisis económico de la rentabilidad con la inoculación de diferentes simbiontes en el sorgo 83G63.
Variable A. brasilense (A)
M. CINVESTAV G. intraradices
(M)
A + M
Kg/ha 2830 2708 2770 2705 Testigo (kg/ha)
2199
Diferencia (kg/ha)
631
509
571
506
U. bruta ($/ha)
663
535
600
531
Costo/trat. ($/ha)
35
35
20
55
U. neta ($/ha)
628
535
600
531
CONCLUSIONES
El presente estudio demostró la efectividad de la inoculación del sorgo con simbiontes y la ineficiencia de la fertilización química, manejo que significa una alta inversión.
LITERATURA CITADA 1. Garza C., I, et al. 2003. Evaluación de Glomus intraradices y brassinoesteroides en la productividad de sorgo. Memoria de Resúmenes del VII Simposio Int. y II Congreso Nal. de Agr. Sostenible, p. 120.
2. Díaz M., R. 2002. Influencia de la biofertilización en las características de planta, de grano e infección micorrízica del sorgo, en dos condiciones edáficas. Tesis Profesional. UAMRA-UAT.
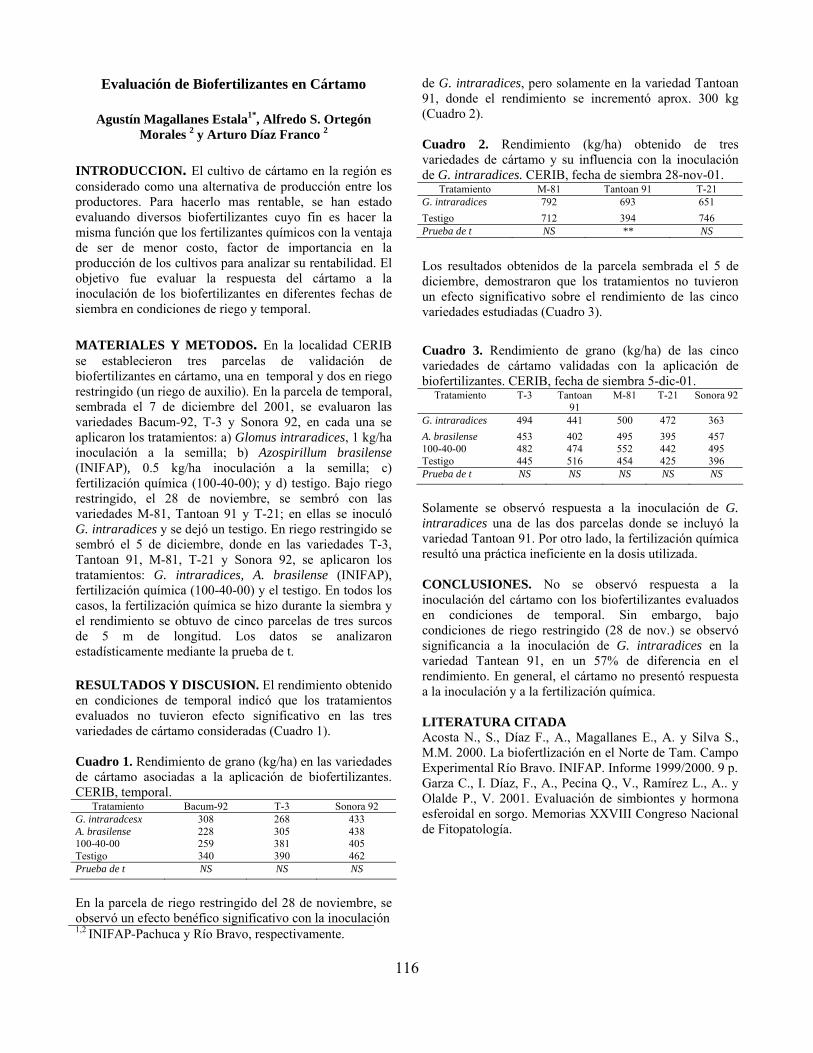
116
Evaluación de Biofertilizantes en Cártamo
Agustín Magallanes Estala1*, Alfredo S. Ortegón Morales 2 y Arturo Díaz Franco 2
INTRODUCCION. El cultivo de cártamo en la región es considerado como una alternativa de producción entre los productores. Para hacerlo mas rentable, se han estado evaluando diversos biofertilizantes cuyo fin es hacer la misma función que los fertilizantes químicos con la ventaja de ser de menor costo, factor de importancia en la producción de los cultivos para analizar su rentabilidad. El objetivo fue evaluar la respuesta del cártamo a la inoculación de los biofertilizantes en diferentes fechas de siembra en condiciones de riego y temporal. MATERIALES Y METODOS. En la localidad CERIB se establecieron tres parcelas de validación de biofertilizantes en cártamo, una en temporal y dos en riego restringido (un riego de auxilio). En la parcela de temporal, sembrada el 7 de diciembre del 2001, se evaluaron las variedades Bacum-92, T-3 y Sonora 92, en cada una se aplicaron los tratamientos: a) Glomus intraradices, 1 kg/ha inoculación a la semilla; b) Azospirillum brasilense (INIFAP), 0.5 kg/ha inoculación a la semilla; c) fertilización química (100-40-00); y d) testigo. Bajo riego restringido, el 28 de noviembre, se sembró con las variedades M-81, Tantoan 91 y T-21; en ellas se inoculó G. intraradices y se dejó un testigo. En riego restringido se sembró el 5 de diciembre, donde en las variedades T-3, Tantoan 91, M-81, T-21 y Sonora 92, se aplicaron los tratamientos: G. intraradices, A. brasilense (INIFAP), fertilización química (100-40-00) y el testigo. En todos los casos, la fertilización química se hizo durante la siembra y el rendimiento se obtuvo de cinco parcelas de tres surcos de 5 m de longitud. Los datos se analizaron estadísticamente mediante la prueba de t. RESULTADOS Y DISCUSION. El rendimiento obtenido en condiciones de temporal indicó que los tratamientos evaluados no tuvieron efecto significativo en las tres variedades de cártamo consideradas (Cuadro 1). Cuadro 1. Rendimiento de grano (kg/ha) en las variedades de cártamo asociadas a la aplicación de biofertilizantes. CERIB, temporal.
Tratamiento Bacum-92 T-3 Sonora 92 G. intraradcesx 308 268 433 A. brasilense 228 305 438 100-40-00 259 381 405 Testigo 340 390 462 Prueba de t NS NS NS
En la parcela de riego restringido del 28 de noviembre, se observó un efecto benéfico significativo con la inoculación 1,2 INIFAP-Pachuca y Río Bravo, respectivamente.
de G. intraradices, pero solamente en la variedad Tantoan 91, donde el rendimiento se incrementó aprox. 300 kg (Cuadro 2). Cuadro 2. Rendimiento (kg/ha) obtenido de tres variedades de cártamo y su influencia con la inoculación de G. intraradices. CERIB, fecha de siembra 28-nov-01.
Tratamiento M-81 Tantoan 91 T-21 G. intraradices 792 693 651 Testigo 712 394 746 Prueba de t NS ** NS
Los resultados obtenidos de la parcela sembrada el 5 de diciembre, demostraron que los tratamientos no tuvieron un efecto significativo sobre el rendimiento de las cinco variedades estudiadas (Cuadro 3).
Cuadro 3. Rendimiento de grano (kg/ha) de las cinco variedades de cártamo validadas con la aplicación de biofertilizantes. CERIB, fecha de siembra 5-dic-01.
Tratamiento T-3 Tantoan 91
M-81 T-21 Sonora 92
G. intraradices 494 441 500 472 363 A. brasilense 453 402 495 395 457 100-40-00 482 474 552 442 495 Testigo 445 516 454 425 396 Prueba de t NS NS NS NS NS
Solamente se observó respuesta a la inoculación de G. intraradices una de las dos parcelas donde se incluyó la variedad Tantoan 91. Por otro lado, la fertilización química resultó una práctica ineficiente en la dosis utilizada. CONCLUSIONES. No se observó respuesta a la inoculación del cártamo con los biofertilizantes evaluados en condiciones de temporal. Sin embargo, bajo condiciones de riego restringido (28 de nov.) se observó significancia a la inoculación de G. intraradices en la variedad Tantean 91, en un 57% de diferencia en el rendimiento. En general, el cártamo no presentó respuesta a la inoculación y a la fertilización química. LITERATURA CITADA Acosta N., S., Díaz F., A., Magallanes E., A. y Silva S., M.M. 2000. La biofertlización en el Norte de Tam. Campo Experimental Río Bravo. INIFAP. Informe 1999/2000. 9 p. Garza C., I. Díaz, F., A., Pecina Q., V., Ramírez L., A.. y Olalde P., V. 2001. Evaluación de simbiontes y hormona esferoidal en sorgo. Memorias XXVIII Congreso Nacional de Fitopatología.
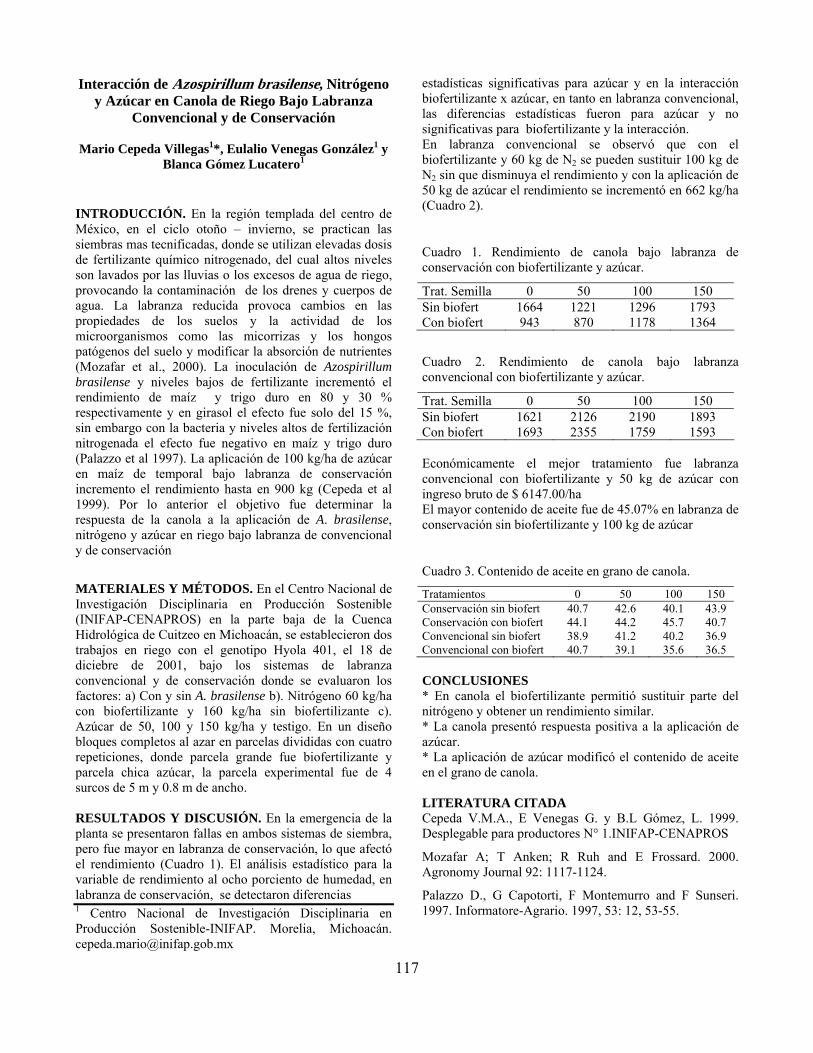
117
Interacción de Azospirillum brasilense, Nitrógeno y Azúcar en Canola de Riego Bajo Labranza
Convencional y de Conservación Mario Cepeda Villegas1*, Eulalio Venegas González1 y
Blanca Gómez Lucatero1
INTRODUCCIÓN. En la región templada del centro de México, en el ciclo otoño – invierno, se practican las siembras mas tecnificadas, donde se utilizan elevadas dosis de fertilizante químico nitrogenado, del cual altos niveles son lavados por las lluvias o los excesos de agua de riego, provocando la contaminación de los drenes y cuerpos de agua. La labranza reducida provoca cambios en las propiedades de los suelos y la actividad de los microorganismos como las micorrizas y los hongos patógenos del suelo y modificar la absorción de nutrientes (Mozafar et al., 2000). La inoculación de Azospirillum brasilense y niveles bajos de fertilizante incrementó el rendimiento de maíz y trigo duro en 80 y 30 % respectivamente y en girasol el efecto fue solo del 15 %, sin embargo con la bacteria y niveles altos de fertilización nitrogenada el efecto fue negativo en maíz y trigo duro (Palazzo et al 1997). La aplicación de 100 kg/ha de azúcar en maíz de temporal bajo labranza de conservación incremento el rendimiento hasta en 900 kg (Cepeda et al 1999). Por lo anterior el objetivo fue determinar la respuesta de la canola a la aplicación de A. brasilense, nitrógeno y azúcar en riego bajo labranza de convencional y de conservación
MATERIALES Y MÉTODOS. En el Centro Nacional de Investigación Disciplinaria en Producción Sostenible (INIFAP-CENAPROS) en la parte baja de la Cuenca Hidrológica de Cuitzeo en Michoacán, se establecieron dos trabajos en riego con el genotipo Hyola 401, el 18 de diciebre de 2001, bajo los sistemas de labranza convencional y de conservación donde se evaluaron los factores: a) Con y sin A. brasilense b). Nitrógeno 60 kg/ha con biofertilizante y 160 kg/ha sin biofertilizante c). Azúcar de 50, 100 y 150 kg/ha y testigo. En un diseño bloques completos al azar en parcelas divididas con cuatro repeticiones, donde parcela grande fue biofertilizante y parcela chica azúcar, la parcela experimental fue de 4 surcos de 5 m y 0.8 m de ancho. RESULTADOS Y DISCUSIÓN. En la emergencia de la planta se presentaron fallas en ambos sistemas de siembra, pero fue mayor en labranza de conservación, lo que afectó el rendimiento (Cuadro 1). El análisis estadístico para la variable de rendimiento al ocho porciento de humedad, en labranza de conservación, se detectaron diferencias 1 Centro Nacional de Investigación Disciplinaria en Producción Sostenible-INIFAP. Morelia, Michoacán. [email protected]
estadísticas significativas para azúcar y en la interacción biofertilizante x azúcar, en tanto en labranza convencional, las diferencias estadísticas fueron para azúcar y no significativas para biofertilizante y la interacción. En labranza convencional se observó que con el biofertilizante y 60 kg de N2 se pueden sustituir 100 kg de N2 sin que disminuya el rendimiento y con la aplicación de 50 kg de azúcar el rendimiento se incrementó en 662 kg/ha (Cuadro 2).
Cuadro 1. Rendimiento de canola bajo labranza de conservación con biofertilizante y azúcar.
Trat. Semilla 0 50 100 150 Sin biofert 1664 1221 1296 1793 Con biofert 943 870 1178 1364
Cuadro 2. Rendimiento de canola bajo labranza convencional con biofertilizante y azúcar.
Trat. Semilla 0 50 100 150 Sin biofert 1621 2126 2190 1893 Con biofert 1693 2355 1759 1593 Económicamente el mejor tratamiento fue labranza convencional con biofertilizante y 50 kg de azúcar con ingreso bruto de $ 6147.00/ha El mayor contenido de aceite fue de 45.07% en labranza de conservación sin biofertilizante y 100 kg de azúcar
Cuadro 3. Contenido de aceite en grano de canola.
Tratamientos 0 50 100 150 Conservación sin biofert 40.7 42.6 40.1 43.9 Conservación con biofert 44.1 44.2 45.7 40.7 Convencional sin biofert 38.9 41.2 40.2 36.9 Convencional con biofert 40.7 39.1 35.6 36.5 CONCLUSIONES * En canola el biofertilizante permitió sustituir parte del nitrógeno y obtener un rendimiento similar. * La canola presentó respuesta positiva a la aplicación de azúcar. * La aplicación de azúcar modificó el contenido de aceite en el grano de canola.
LITERATURA CITADA Cepeda V.M.A., E Venegas G. y B.L Gómez, L. 1999. Desplegable para productores N° 1.INIFAP-CENAPROS
Mozafar A; T Anken; R Ruh and E Frossard. 2000. Agronomy Journal 92: 1117-1124.
Palazzo D., G Capotorti, F Montemurro and F Sunseri. 1997. Informatore-Agrario. 1997, 53: 12, 53-55.
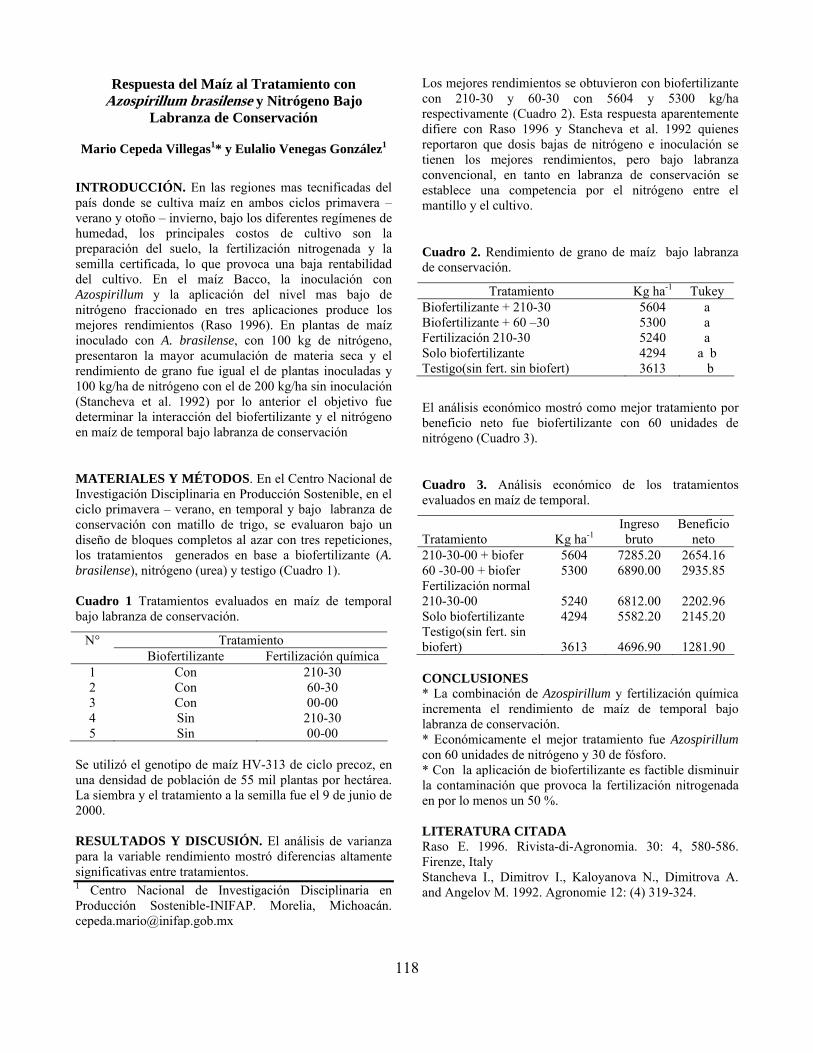
118
Respuesta del Maíz al Tratamiento con Azospirillum brasilense y Nitrógeno Bajo
Labranza de Conservación Mario Cepeda Villegas1* y Eulalio Venegas González1
INTRODUCCIÓN. En las regiones mas tecnificadas del país donde se cultiva maíz en ambos ciclos primavera – verano y otoño – invierno, bajo los diferentes regímenes de humedad, los principales costos de cultivo son la preparación del suelo, la fertilización nitrogenada y la semilla certificada, lo que provoca una baja rentabilidad del cultivo. En el maíz Bacco, la inoculación con Azospirillum y la aplicación del nivel mas bajo de nitrógeno fraccionado en tres aplicaciones produce los mejores rendimientos (Raso 1996). En plantas de maíz inoculado con A. brasilense, con 100 kg de nitrógeno, presentaron la mayor acumulación de materia seca y el rendimiento de grano fue igual el de plantas inoculadas y 100 kg/ha de nitrógeno con el de 200 kg/ha sin inoculación (Stancheva et al. 1992) por lo anterior el objetivo fue determinar la interacción del biofertilizante y el nitrógeno en maíz de temporal bajo labranza de conservación
MATERIALES Y MÉTODOS. En el Centro Nacional de Investigación Disciplinaria en Producción Sostenible, en el ciclo primavera – verano, en temporal y bajo labranza de conservación con matillo de trigo, se evaluaron bajo un diseño de bloques completos al azar con tres repeticiones, los tratamientos generados en base a biofertilizante (A. brasilense), nitrógeno (urea) y testigo (Cuadro 1). Cuadro 1 Tratamientos evaluados en maíz de temporal bajo labranza de conservación.
N° Tratamiento Biofertilizante Fertilización química
1 Con 210-30 2 Con 60-30 3 Con 00-00 4 Sin 210-30 5 Sin 00-00
Se utilizó el genotipo de maíz HV-313 de ciclo precoz, en una densidad de población de 55 mil plantas por hectárea. La siembra y el tratamiento a la semilla fue el 9 de junio de 2000. RESULTADOS Y DISCUSIÓN. El análisis de varianza para la variable rendimiento mostró diferencias altamente significativas entre tratamientos. 1 Centro Nacional de Investigación Disciplinaria en Producción Sostenible-INIFAP. Morelia, Michoacán. [email protected]
Los mejores rendimientos se obtuvieron con biofertilizante con 210-30 y 60-30 con 5604 y 5300 kg/ha respectivamente (Cuadro 2). Esta respuesta aparentemente difiere con Raso 1996 y Stancheva et al. 1992 quienes reportaron que dosis bajas de nitrógeno e inoculación se tienen los mejores rendimientos, pero bajo labranza convencional, en tanto en labranza de conservación se establece una competencia por el nitrógeno entre el mantillo y el cultivo.
Cuadro 2. Rendimiento de grano de maíz bajo labranza de conservación.
Tratamiento Kg ha-1 Tukey Biofertilizante + 210-30 5604 a Biofertilizante + 60 –30 5300 a Fertilización 210-30 5240 a Solo biofertilizante 4294 a b Testigo(sin fert. sin biofert) 3613 b
El análisis económico mostró como mejor tratamiento por beneficio neto fue biofertilizante con 60 unidades de nitrógeno (Cuadro 3).
Cuadro 3. Análisis económico de los tratamientos evaluados en maíz de temporal.
Tratamiento Kg ha-1 Ingreso bruto
Beneficio neto
210-30-00 + biofer 5604 7285.20 2654.16 60 -30-00 + biofer 5300 6890.00 2935.85 Fertilización normal 210-30-00 5240 6812.00 2202.96 Solo biofertilizante 4294 5582.20 2145.20 Testigo(sin fert. sin biofert) 3613 4696.90 1281.90 CONCLUSIONES * La combinación de Azospirillum y fertilización química incrementa el rendimiento de maíz de temporal bajo labranza de conservación. * Económicamente el mejor tratamiento fue Azospirillum con 60 unidades de nitrógeno y 30 de fósforo. * Con la aplicación de biofertilizante es factible disminuir la contaminación que provoca la fertilización nitrogenada en por lo menos un 50 %.
LITERATURA CITADA Raso E. 1996. Rivista-di-Agronomia. 30: 4, 580-586. Firenze, Italy Stancheva I., Dimitrov I., Kaloyanova N., Dimitrova A. and Angelov M. 1992. Agronomie 12: (4) 319-324.
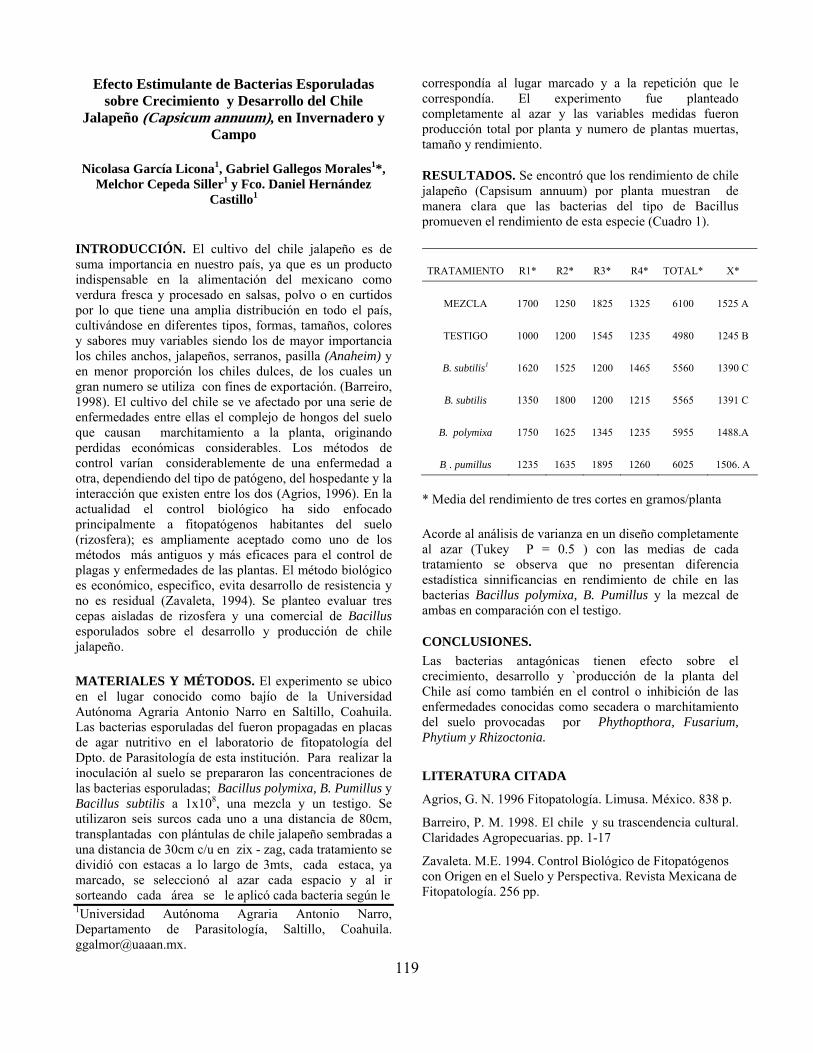
119
Efecto Estimulante de Bacterias Esporuladas sobre Crecimiento y Desarrollo del Chile
Jalapeño (Capsicum annuum), en Invernadero y Campo
Nicolasa García Licona1, Gabriel Gallegos Morales1*,
Melchor Cepeda Siller1 y Fco. Daniel Hernández Castillo1
INTRODUCCIÓN. El cultivo del chile jalapeño es de suma importancia en nuestro país, ya que es un producto indispensable en la alimentación del mexicano como verdura fresca y procesado en salsas, polvo o en curtidos por lo que tiene una amplia distribución en todo el país, cultivándose en diferentes tipos, formas, tamaños, colores y sabores muy variables siendo los de mayor importancia los chiles anchos, jalapeños, serranos, pasilla (Anaheim) y en menor proporción los chiles dulces, de los cuales un gran numero se utiliza con fines de exportación. (Barreiro, 1998). El cultivo del chile se ve afectado por una serie de enfermedades entre ellas el complejo de hongos del suelo que causan marchitamiento a la planta, originando perdidas económicas considerables. Los métodos de control varían considerablemente de una enfermedad a otra, dependiendo del tipo de patógeno, del hospedante y la interacción que existen entre los dos (Agrios, 1996). En la actualidad el control biológico ha sido enfocado principalmente a fitopatógenos habitantes del suelo (rizosfera); es ampliamente aceptado como uno de los métodos más antiguos y más eficaces para el control de plagas y enfermedades de las plantas. El método biológico es económico, especifico, evita desarrollo de resistencia y no es residual (Zavaleta, 1994). Se planteo evaluar tres cepas aisladas de rizosfera y una comercial de Bacillus esporulados sobre el desarrollo y producción de chile jalapeño. MATERIALES Y MÉTODOS. El experimento se ubico en el lugar conocido como bajío de la Universidad Autónoma Agraria Antonio Narro en Saltillo, Coahuila. Las bacterias esporuladas del fueron propagadas en placas de agar nutritivo en el laboratorio de fitopatología del Dpto. de Parasitología de esta institución. Para realizar la inoculación al suelo se prepararon las concentraciones de las bacterias esporuladas; Bacillus polymixa, B. Pumillus y Bacillus subtilis a 1x108, una mezcla y un testigo. Se utilizaron seis surcos cada uno a una distancia de 80cm, transplantadas con plántulas de chile jalapeño sembradas a una distancia de 30cm c/u en zix - zag, cada tratamiento se dividió con estacas a lo largo de 3mts, cada estaca, ya marcado, se seleccionó al azar cada espacio y al ir sorteando cada área se le aplicó cada bacteria según le 1Universidad Autónoma Agraria Antonio Narro, Departamento de Parasitología, Saltillo, Coahuila. [email protected].
correspondía al lugar marcado y a la repetición que le correspondía. El experimento fue planteado completamente al azar y las variables medidas fueron producción total por planta y numero de plantas muertas, tamaño y rendimiento.
RESULTADOS. Se encontró que los rendimiento de chile jalapeño (Capsisum annuum) por planta muestran de manera clara que las bacterias del tipo de Bacillus promueven el rendimiento de esta especie (Cuadro 1).
TRATAMIENTO R1* R2* R3* R4* TOTAL* X*
MEZCLA 1700 1250 1825 1325 6100 1525 A
TESTIGO 1000 1200 1545 1235 4980 1245 B
B. subtilis1 1620 1525 1200 1465 5560 1390 C
B. subtilis 1350 1800 1200 1215 5565 1391 C
B. polymixa 1750 1625 1345 1235 5955 1488.A
B. . pumillus 1235 1635 1895 1260 6025 1506. A
* Media del rendimiento de tres cortes en gramos/planta Acorde al análisis de varianza en un diseño completamente al azar (Tukey P = 0.5 ) con las medias de cada tratamiento se observa que no presentan diferencia estadística sinnificancias en rendimiento de chile en las bacterias Bacillus polymixa, B. Pumillus y la mezcal de ambas en comparación con el testigo.
CONCLUSIONES. Las bacterias antagónicas tienen efecto sobre el crecimiento, desarrollo y `producción de la planta del Chile así como también en el control o inhibición de las enfermedades conocidas como secadera o marchitamiento del suelo provocadas por Phythopthora, Fusarium, Phytium y Rhizoctonia.
LITERATURA CITADA
Agrios, G. N. 1996 Fitopatología. Limusa. México. 838 p.
Barreiro, P. M. 1998. El chile y su trascendencia cultural. Claridades Agropecuarias. pp. 1-17
Zavaleta. M.E. 1994. Control Biológico de Fitopatógenos con Origen en el Suelo y Perspectiva. Revista Mexicana de Fitopatología. 256 pp.

120
Resultados Preliminares de la Evaluación de Biofertilizantes en Maíz QPM
José Ernesto Cervantes Martínez1*
INTRODUCCION. El maíz como la mayoría de gramíneas, demanda alta fertilización N-P para proporcionar altos rendimientos. Dado el alto costo de dicho insumo, éste es mas usado en aquellas áreas dotadas de riego y donde el clima es más favorable para obtener rendimientos altamente rentables; sin embargo, en otras áreas donde el clima no es tan favorable para alta productividad de maíz y el agua es restringida o usada en cultivos más rentables, la fertilización es mínima y frecuentemente nula. Por lo tanto, es importante buscar otras opciones de fertilización baratas y efectivas que complementen las necesidades del maíz para que su cultivo sea rentable aún en condiciones de estrés por clima y suelo que caracterizan a la mayor parte de los sistemas de producción de maíz mexicanas, una de las cuales es el sur de Tamaulipas. Con base en lo anterior y tomando en cuenta ciertos antecedentes del efecto positivo de la aplicación de biofertilizantes sobre la producción de maíz y sorgo (Díaz, 2002), algodón (Salgado, 2000) y soya (Elizondo, 2000), se consideró prudente inocular semilla de maíz con algunos de tales biofertilizantes y evaluar su respuesta en el rendimiento de elote y grano de maíz QPM en condiciones de campo bajo riego.
MATERIALES Y METODOS. El estudio se realizó en terrenos del Campo Experimental Sur de Tamaulipas ubicado en el km. 55 a 2 km. de Est. Cuauhtémoc, Tam. La fecha de siembra fue el 14 de octubre de 1999, la cual fue mecanizada. Se utilizó un diseño de bloques completos al azar con cinco repeticiones en parcelas de ocho surcos de 16.0 m de longitud a 0.90 m de ancho. Los tratamientos (800 g/ha inoculación a la semilla) fueron: 1. Azospirillum sp., 2. micorriza arbuscular, 3. Azospirillum + micorriza, 4. Brassinoesteroides (fitohormona), y 5. testigo sin biofertilizante. Se utilizó un híbrido trilineal de maíz QPM (CML176 x CML149 x CML186) y se fertilizó con 120-40-0, aplicando la mitad a la siembra y la otra distribuida en los dos riegos de auxilio proporcionados. Se obtuvo información sobre rendimiento de elote (dos surcos de parcela útil, 9.0 m2) y de grano seco (14% de humedad).
RESULTADOS Y DISCUSION. Los resultados del análisis de varianza (datos no presentados) mostraron que para rendimiento de elote y grano hubo diferencias significativas entre tratamientos (P≤0.05). Los valores medios de elote y grano aparecen en el Cuadro 1, donde se puede apreciar que el mejor tratamiento correspondió a 1 Campo Experimental Sur de Tamaulipas, INIFAP. Apdo. Postal 31, Altamira, Tam. 89610. [email protected]
Brassinoesteroides con rendimiento de 17.1 y 7.2 t/ha de elote y grano, respectivamente. Estos rendimientos corresponden a 15.3 y 14.8% de incremento con relación al testigo sin biofertilizante. Azospirillum fue el tratamiento que le siguió en rendimientos al promediar 17.0 y 6.9 t/ha de elote y grano, respectivamente. Cuadro 1. Rendimiento de elote y grano (t/ha) de maíz de calidad proteica inoculado con varios biofertilizantes.
Rendimiento Biofertilizante Elote % Grano %
Azospirillum (Azos)
Micorriza (Mico)
Azos + Mico
Brassinoesteroides
Testigo
17.0 a*
14.3 c
16.4 b
17.1 a
14.8 c
14.7
3.7
10.4
15.3
-
6.9 a
5.5 c
5.9 bc
7.2 a
6.2 b
10.2
12.0
6.3
14.8
-
* Rendimientos con la misma letra no son diferentes significativamente según la prueba de Tukey (P<0.05).
El tratamiento con micorriza tuvo los promedios de rendimiento más bajos junto con la combinación de Azospirillum más micorriza. De acuerdo a lo observado (datos no presentados), la semilla tratada con micorriza tuvo problemas para germinar, razón por la cual la población de plantas fue inferior a la emergida con los otros tratamientos. Esto se debió a que los platos de la sembradora depositaron menos semilla por metro lineal, dado que el biefertilizante adherido aumento su tamaño. Por otro lado, los tratamientos con Brassionesteroides y Azospirillum (solo) no tuvieron ese problema porque tales productos son más solubles y no aumenta el volumen de la semilla como en el caso de micorriza. Por el contrario, en ambos tratamientos el maíz mostró buena población de plantas y en general buen vigor y porte agronómico.
CONCLUSIONES. Brassinoesteroides y Azospirillum provocaron incrementos en rendimiento de elote y grano de aproximadamente 15%. Dichos incrementos pueden considerarse sustanciales si se corroboran los resultados en más evaluaciones; razón por la cual es necesario reforzar la investigación en biofertilizantes y determinar la factibilidad de su uso como fertilización complementaria en maíz.
LITERATURA CITADA
Díaz F., A. 2002 Incremento en la productividad del sorgo y maíz mediante biofertilización. Publ. Esp. No. 25 INIFAP-CERIB. P 13-14. Elizondo B., J. 2000. Evaluación de biofertilizantes Bradirrizobium japonicum, Glomus intradices y una hormona esteroidal en soya. En: 12º. Encuentro de Investigación Científica y Tecnológica de Golfo de México. P 6. Salgado S., E. 2000. Evaluación de biofertilizantes en Algodonero. En: 12º. Encuentro de Investigación Científica y Tecnológica de Golfo de México. P 8.

121
Efecto de Biofertilizantes Sobre el Rendimiento del Maíz
César A Reyes Méndez1* y Miguel Angel Cantú
Almaguer1
INTRODUCCIÓN. El maíz ocupa el 43% de la superficie cultivada en México (7.9 millones de hectáreas), donde el principal problema es el temporal irregular que provoca rendimientos de grano bajos. En el norte de Tamailipas con el propósito de hacer más eficiente la nutrición del maíz bajo condiciones de temporal, se están aplicando biofertilizantes como un recurso para incrementar los rendimientos de este cereal y bajar los costos de producción. Estudios iniciales en esta región, indican que la aplicación de micorriza vesículo arbuscular (MVA) y Glomus intraradices ha tenido un impacto favorable en la producción de sorgo y maíz, al incrementar sus rendimientos (Díaz F., 2002; Díaz M., 2002). Por lo anterior, el objetivo del presente trabajo fue el de conocer, en condiciones de campo, el efecto de los biofertilizantes sobre el rendimiento del grano de maíz. MATERIALES Y MÉTODOS. En la localidad de Río Bravo, Tam. bajo condiciones de temporal, se evaluó el efecto de los biofertilizantes. El diseño fue de bloques al azar con cuatro repeticiones. Se utilizó el híbrido de maíz H-437, donde se estudiaron los siguientes tratamientos: a) fertilización química (140-40-00); b) fertilización química más MVA G. intraradices, 1 kg/ha, inoculación a la semilla; c) fertilización química más Azospirillum brasilense; d) MVA G. intraradices; e) Azospirillum brasilense; y f) testigo absoluto. La parcela experimental fue de cuatro surcos de 5 m y 0.82 m de ancho. La fertilización química fue a base de urea y superfosfato de calcio triple, aplicándose esta durante la siembra. Los datos obtenidos por planta fueron: floración masculina y femenina, altura de planta y mazorca, calificación de sanidad de mazorca, porciento de acame de tallo y rendimiento de grano por parcela. El análisis estadístico se realizó mediante el sistema SAS (SAS, 1996). RESULTADOS Y DISCUSIÓN. Los resultados del análisis de varianza indicaron, que para las variables estudiadas se encontraron diferencias significativas entre tratamientos, (Cuadro 1), excepto la altura de planta y el porcentaje de mazorcas sanas. Los bajos coeficientes de variación nos indican la confiabilidad de los resultados obtenidos. Los mejores rendimientos de grano se obtuvieron al aplicar la fertilización química, sola o combinada con alguno de los biofertilizantes mientras que al aplicar solamente Glomus o Azospirillum el rendimiento se abatió en 9.8% y 34.2% respectivamente,
1 Investigadores del Campo Experimental Río Bravo, INIFAP. Apdo. Postal 172, Río Bravo, Tam. [email protected]
(Cuadro 2). La aplicación de Azospirillum como única fuente de nutrimento presentó los más bajos rendimientos de grano, inclusive que el testigo absoluto, sin embargo no mostró síntomas de deficiencias nutricionales en el follaje, mientras que el testigo sí las presentó. Cuadro 1. Nivel de significancia y coeficiente de variación de diferentes variables agronómicas.
Variables Significancia C.V % Rendimiento ** 9.2 Floración ** 0.8 Altura de planta * 2.4 Altura de mazorca NS 9.9 Acame de tallo ** 38.8 Mazorcas sanas NS 21.8
**Diferencia altamente significativa, *Diferencia significativa, NS sin diferencias significativas. Cuadro 2. Respuesta de los biofertilizantes sobre el rendimiento del maíz. Río Bravo. OI.02-03 Tratamiento Rendimiento (t ha-1 ) 140-40-00 5.26 140-40-00 + Azospirillum 5.06
140-40-00 + Glomus 4.86 Azospirillum 4.79 0.05 Testigo absoluto 4.52 Glomus 3.92 * Valores unidos por la línea no difieren estadísticamente (DMS, p<0.05). La asociación de fertilizante químico mas biofertilizante ya sea Azospirillum o Glomus no favorece la producción de maíz ya que su rendimiento de grano se ve afectado en 3.84% y 7.67%, respectivamente, sin afectarse sus componentes del rendimiento, observando además que la aplicación de biofertilizante si tiene efecto sobre algunos de los componentes del rendimiento en comparación al testigo absoluto. CONCLUSIONES. Los mejores rendimientos de grano se debieron a la aplicación de la fertilización química. El uso de G. intraradices o de A .brasilense no presentan efecto sobre el incremento del rendimiento en maíz, ni en sus componentes. LITERATURA CITADA Díaz F., A. 2002. Incremento de la productividad del sorgo y maíz mediante la biofertilización. En: Publ. Esp. No. 25. INIFAP-CERIB. p. 13-14 Díaz M., R. 2002. La biofertilización en las características de planta, de grano e infección micorrízica del sorgo, en dos condiciones edáficas. Tesis Profesional. UAMRA, UAT. 62 p. Statistical Analysis Systems Institute (SAS Institute). 1996. SAS/STAT User’s Guide. Release 6.12. SAS, Cary, N.C., USA.
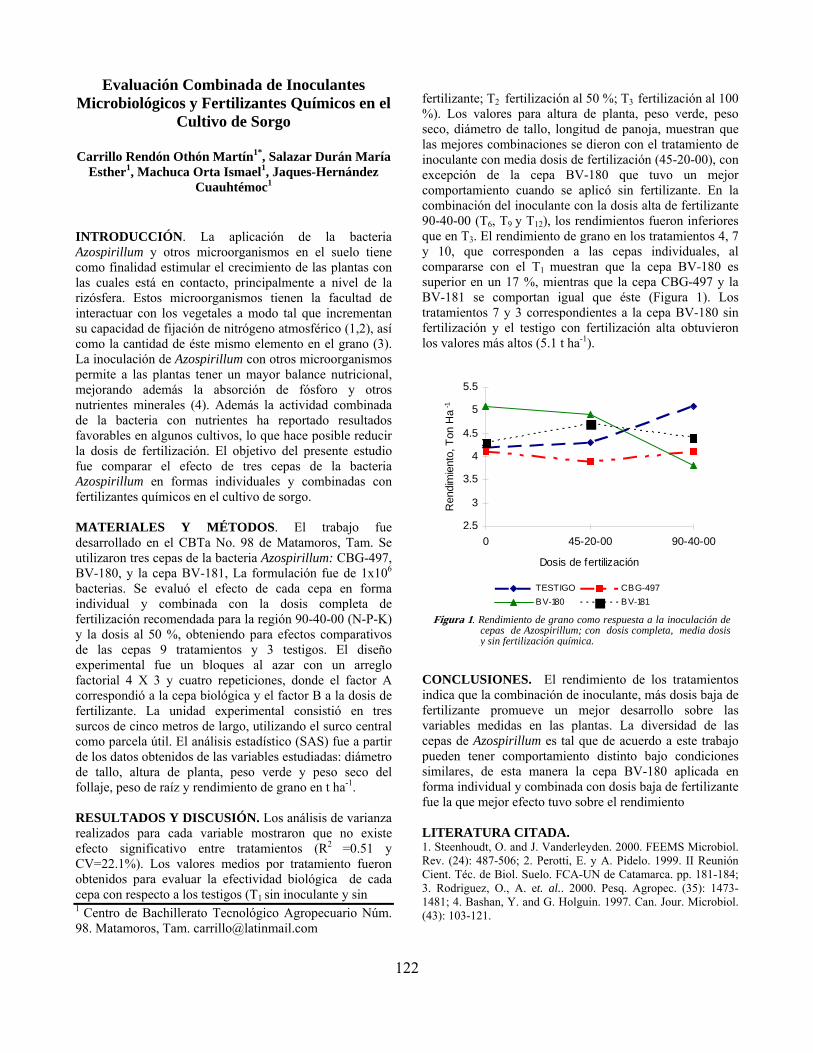
122
Evaluación Combinada de Inoculantes Microbiológicos y Fertilizantes Químicos en el
Cultivo de Sorgo
Carrillo Rendón Othón Martín1*, Salazar Durán María Esther1, Machuca Orta Ismael1, Jaques-Hernández
Cuauhtémoc1
INTRODUCCIÓN. La aplicación de la bacteria Azospirillum y otros microorganismos en el suelo tiene como finalidad estimular el crecimiento de las plantas con las cuales está en contacto, principalmente a nivel de la rizósfera. Estos microorganismos tienen la facultad de interactuar con los vegetales a modo tal que incrementan su capacidad de fijación de nitrógeno atmosférico (1,2), así como la cantidad de éste mismo elemento en el grano (3). La inoculación de Azospirillum con otros microorganismos permite a las plantas tener un mayor balance nutricional, mejorando además la absorción de fósforo y otros nutrientes minerales (4). Además la actividad combinada de la bacteria con nutrientes ha reportado resultados favorables en algunos cultivos, lo que hace posible reducir la dosis de fertilización. El objetivo del presente estudio fue comparar el efecto de tres cepas de la bacteria Azospirillum en formas individuales y combinadas con fertilizantes químicos en el cultivo de sorgo. MATERIALES Y MÉTODOS. El trabajo fue desarrollado en el CBTa No. 98 de Matamoros, Tam. Se utilizaron tres cepas de la bacteria Azospirillum: CBG-497, BV-180, y la cepa BV-181, La formulación fue de 1x106 bacterias. Se evaluó el efecto de cada cepa en forma individual y combinada con la dosis completa de fertilización recomendada para la región 90-40-00 (N-P-K) y la dosis al 50 %, obteniendo para efectos comparativos de las cepas 9 tratamientos y 3 testigos. El diseño experimental fue un bloques al azar con un arreglo factorial 4 X 3 y cuatro repeticiones, donde el factor A correspondió a la cepa biológica y el factor B a la dosis de fertilizante. La unidad experimental consistió en tres surcos de cinco metros de largo, utilizando el surco central como parcela útil. El análisis estadístico (SAS) fue a partir de los datos obtenidos de las variables estudiadas: diámetro de tallo, altura de planta, peso verde y peso seco del follaje, peso de raíz y rendimiento de grano en t ha-1. RESULTADOS Y DISCUSIÓN. Los análisis de varianza realizados para cada variable mostraron que no existe efecto significativo entre tratamientos (R2 =0.51 y CV=22.1%). Los valores medios por tratamiento fueron obtenidos para evaluar la efectividad biológica de cada cepa con respecto a los testigos (T1 sin inoculante y sin 1 Centro de Bachillerato Tecnológico Agropecuario Núm. 98. Matamoros, Tam. [email protected]
fertilizante; T2 fertilización al 50 %; T3 fertilización al 100 %). Los valores para altura de planta, peso verde, peso seco, diámetro de tallo, longitud de panoja, muestran que las mejores combinaciones se dieron con el tratamiento de inoculante con media dosis de fertilización (45-20-00), con excepción de la cepa BV-180 que tuvo un mejor comportamiento cuando se aplicó sin fertilizante. En la combinación del inoculante con la dosis alta de fertilizante 90-40-00 (T6, T9 y T12), los rendimientos fueron inferiores que en T3. El rendimiento de grano en los tratamientos 4, 7 y 10, que corresponden a las cepas individuales, al compararse con el T1 muestran que la cepa BV-180 es superior en un 17 %, mientras que la cepa CBG-497 y la BV-181 se comportan igual que éste (Figura 1). Los tratamientos 7 y 3 correspondientes a la cepa BV-180 sin fertilización y el testigo con fertilización alta obtuvieron los valores más altos (5.1 t ha-1). CONCLUSIONES. El rendimiento de los tratamientos indica que la combinación de inoculante, más dosis baja de fertilizante promueve un mejor desarrollo sobre las variables medidas en las plantas. La diversidad de las cepas de Azospirillum es tal que de acuerdo a este trabajo pueden tener comportamiento distinto bajo condiciones similares, de esta manera la cepa BV-180 aplicada en forma individual y combinada con dosis baja de fertilizante fue la que mejor efecto tuvo sobre el rendimiento LITERATURA CITADA. 1. Steenhoudt, O. and J. Vanderleyden. 2000. FEEMS Microbiol. Rev. (24): 487-506; 2. Perotti, E. y A. Pidelo. 1999. II Reunión Cient. Téc. de Biol. Suelo. FCA-UN de Catamarca. pp. 181-184; 3. Rodriguez, O., A. et. al.. 2000. Pesq. Agropec. (35): 1473-1481; 4. Bashan, Y. and G. Holguin. 1997. Can. Jour. Microbiol. (43): 103-121.
2.5
3
3.5
4
4.5
5
5.5
0 45-20-00 90-40-00
Dosis de fertilización
Ren
dim
ient
o, T
on H
a -1
TESTIGO CBG-497BV-180 BV-181
Figura 1. Rendimiento de grano como respuesta a la inoculación de cepas de Azospirillum; con dosis completa, media dosis y sin fertilización química.

123
Biofertilizacion en Sorgo de Temporal en la Zona Media de San Luis Potosí
Andrés Ramiro Córdova1* Cesario Jasso Chaverria1
INTRODUCCION. En el estado de San Luis Potosí el sorgo ocupa una superficie que varia de 8,500 a 9,000 ha anualmente, bajo condiciones de temporal con un rendimiento de 950 a 1,000 kg/ha. Para la siembra se utiliza semilla híbrida y una mecanización de la siembra a la cosecha; se puede encontrar como monocultivo, o bien en la rotación sorgo-maíz. Dentro de los factores que afectan la producción, se encuentra el poco o nulo uso de fertilizantes durante el desarrollo del cultivo, afectando la producción de grano, esto es debido a los costos del fertilizante y a lo incierto del temporal en la región. Una buena opción para incrementar la producción de este cultivo es la utilización de biofertilizantes o fertilizantes biológicos aplicados al momento de la siembra. Bashan y Ferrera en 1996 indican que la inoculación con la bacteria Azospirillum tiene un efecto favorable al utilizarse como inoculante en muchas especies vegetales. Así también las bacterias Azotobacter y Rhizobium contribuyen en la capacidad de sintetizar hormonas vegetales reguladoras de crecimiento. Los hongos micorrízicos viven en simbiosis con la raíz y su función consiste en contribuir eficazmente a solubilizar el fósforo existente en el suelo (Gianinazzi, 1986 citado por Aguirre 1999). Este estudio tuvo como propósito validar la aplicación de biofertilizantes en el cultivo de sorgo. MATERIALES Y METODOS. La parcela de validación (una ha) se estableció en el municipio de Cerritos, S.L.P. La siembra se realizó el 21 de julio de 1999; se utilizó el Híbrido de se sorgo Esmeralda. La distancia entre surcos fue de 0.84 m y la densidad de población de 160 000 plantas por ha. Los tratamientos de biofertilizantes utilizados fueron: 1) Inoculación con Azospirillum, 2) Inoculación con hongo micorrízico, 3) Inoculación con Azospirillum+ hongo micorrízico, 4) Inoculación con Brassinoesteroide y 5) Testigo sin biofertilizantes y sin fertilización químico. La aplicación del biofertilizante Azospirillum y hongo micorrízico, así como la mezcla de ambos se realizó al momento de la siembra, mientras que el Brassionoesteroide se aplicó con aspersora manual en la etapa de floración es decir a los 72 días después de la siembra. No se aplicaron fertilizantes convencionales en la siembra ni durante el desarrollo del cultivo. La precipitación pluvial ocurrida de enero a octubre fue de 306.5 mm, de los cuales 90.5 mm ocurrieron en mayo y junio, 192.5 mm en Julio y 114 mm se presentaron en el periodo de agosto-octubre. El tipo de suelo fue un vertisol de textura arcillosa, pH alcalino, materia orgánica de 5.5%, fósforo aprovechable de 14 ppm y 0.26% de nitrógeno total. Se utilizó como parcela útil 2 surcos de 5 m de
1 Campo Experimental San Luis, INIFAP.
total. Se utilizó como parcela útil 2 surcos de 5 m de longitud con 5 repeticiones seleccionadas al azar y el análisis se efectuó bajo un diseño completamente al azar. Los parámetros evaluados fueron altura de planta, longitud y diámetro de panoja, días a floración, rendimiento de grano y rendimiento de forraje verde.
RESULTADOS Y DISCUSION. Los resultados obtenidos nos indican que para las variables rendimiento de grano y forraje (Cuadro 1) los tratamientos con biofertilizantes fueron estadísticamente superiores al testigo (P<0.05).
Cuadro 1. Producción de grano y forraje en sorgo de temporal con el uso de biofertilizantes en San Luis Potosí.
* Valores con la misma letra no denotan diferencias (DMS. P<0.05)
Para la variable longitud de panoja los tratamientos con biofertilizantes alcanzaron de 23.4 a 24.3 cm de largo mientras que el testigo obtuvo 20.7 cm. El ancho de panoja también manifestó una tendencia positiva en los tratamientos con biofertilizantes siendo Azospirillum y el hongo micorrízico los más sobresalientes con 5.4 y 5.0 cm de ancho de panoja mientras que el testigo alcanzo un promedio de 4.3 cm. En el rendimiento de forraje verde se presentó una tendencia similar a las demás variables medidas.
LITERATURA CITADA Aguirre M. J. F. 1999. Micorriza-Arbuscular. Revisión bibliográfica. INIFAP 18 pp.
Bashan, Y., Holguín. G y Ferrera-Cerrato. R 1996 (b) Interacciones entre plantas y microorganismos benéficos. II. Bacterias asociativas de la rizosfera. Terra 14: 195-210.
Gianinazzi-Pearson 1986. The physiology of improved phosphorus nutrition in micorrizal plants. In: Physiological and Genetics Aspect of Mycorrhizae. INRA. París. pp 101-109.
Tratamiento Rendimiento grano (t ha-1)
Rendimiento forraje verde (t ha-1)
Azospirillum 4.18 a 11.6 a
Hongo micorrízico 4.22 a 11.3 a
Azospirillun + hongo micorrízico 4.30 a 11.7 a
Brassinoesteroide 4.14 a 13.9 a
Testigo 3.07 b 9.2 b

124
Simbiosis Rhizobium-Micorriza Arbuscular y Uso de Brassinoesteroides en Frijol
Cesario Jasso Chaverría1 y Miguel A. Martínez
Gamiño1
INTRODUCCION. El frijol (Phaseolus vulgaris) es el cultivo de mayor importancia económica y social en el estado de San Luis Potosí. La superficie sembrada anualmente en riego es de 10,500 ha; con rendimiento medio de 1.15 ton ha-1. Esta producción es considerada baja, dado el potencial de rendimiento de las variedades que actualmente existen el mercado y a las condiciones agro-climáticas de las zonas productoras de fríjol en el estado. En su producción destaca el escaso uso de fertilizantes químicos y aún más limitado, es el uso de fertilizantes de tipo biológico como las bacterias fijadoras de nitrógeno y hongos que solubilizan algunos nutrientes del suelo. Bashan et al, en 1996 indican que la inoculación mixta de Azotobacter con Rhizobium o Azospirillum incrementa el desarrollo de planta, así como también incrementa la fijación de nitrógeno. Las hormonas más conocidas como las auxinas, citocininas, giberelinas, etileno y ácido abscísico han sido utilizadas de manera extensiva en varios cultivos, los Brassinoesteroides son el primer miembro de un nuevo grupo de substancias reguladoras del crecimiento que se encuentran en las plantas y promueven su crecimiento (Gregory, 1981; Clouse y Sasse, 1998), son un grupo de productos naturales biológicamente activos basados en una estructura esteroidal. El objetivo de este estudio fue evaluar el efecto de la aplicación de dos microorganismos y una hormona esteroidal en el rendimiento de frijol. MATERIALES Y METODOS. El trabajo se realizó durante la primavera de 1999, en la localidad El Refugio, Ciudad Fernández, S.L.P. Los tratamientos evaluados fueron: 1. Rhizobium, 2. Micorriza arbuscular, 3. Brassinoesteroide, 4. Rhizobium + micorriza arbuscular, 5. Rhizobium + Brassinoesteroide, 6. Micorriza arbuscular + Brassinoesteroide, 7. Micorriza arbuscular + Rhizobium + Brassinoesteroide y 8. Testigo. El diseño experimental fue bloques al azar con cuatro repeticiones. La aplicación de los tratamientos de inoculación se realizó a la semilla al momento de la siembra y el brassinoesteroide en la antesis del cultivo. Para la siembra se utilizó la variedad Negro Huasteco 81 con densidad de 180 mil plantas ha-1. Las principales variables registradas fueron: días a floración, días a madures fisiológica, altura de planta, cobertura, vainas por planta, granos por vaina, peso de 100 granos y rendimiento de grano. 1 Campo Experimental San Luis, INIFAP. Apdo. Postal 1538-B, San Luis Potosí, SLP. [email protected]
RESULTADOS Y DISCUSION. Los resultados mostraron diferencias estadísticas significativas (p≤0.05) para altura de planta, número de vainas por planta y para el rendimiento de grano. Al realizar la prueba de Tukey (p≤0.05) se encontró que la altura de planta se incrementó significativamente al inocular la semilla con Rhizobium, una situación similar ocurrió al aplicar Brassinoesteroide o bien con la combinación Micorriza arbuscular + brassinoesteroide.
Cuadro 1. Altura de planta y rendimiento de grano en frijol de riego con la aplicación de dos microorganismos y una hormona esteroidal.
* Valores con la misma literal no son estadísticamente diferentes (Tukey P<0.05)
Para la variable rendimiento de grano, los tratamiento con microorganismos y brassinoesteroide fueron estadísticamente superiores al testigo, presentándose el mayor rendimiento en el tratamiento de micorriza arbuscular, cuyo valor superó en 68.38% al rendimiento del tratamiento testigo.
CONCLUSIONES. La inoculación de microorganismos y la aplicación de brassinoesteroides en frijol de riego, contribuyeron significativamente a incrementar el rendimiento de grano.
LITERATURA CITADA
Bashan, Y., G. Holguin. y R. Ferrera-Cerrato. 1996 (b) Interacciones entre plantas y microorganismos benéficos. II. Bacterias asociativas de la rizosfera. Terra 14 (2) 195-210; Clouse, S.D. and J. Sause. M. 1998. Brassinoesteroids: Essential regulators of plant growth and development. Annual Review of Plant Physiology and Plan Molecular Biology. 49: 427-451; Gregory, L.E. 1981. Scceleration of plant growth throught sedd treatment with brassinoesteroids. American Journal of Botany. 68: 586-588.
Tratamiento
Altura de planta
(cm)
Grano
(t ha-1)
Rhizobium 52.3a 2.701a
Micorriza arbuscular 51.7ab 2.728a
Brassinoesteroide 52.9a 2.560 a
Rhizobium + Micorriza arbuscular 51.0ab 2.472 a
Rhizobium + brassinoesteroide 50.4ab 2.428a
Micorriza arbuscular + brassinoesteroide 52.9a 2.466a
Micorriza arbuscular + Rhizobium+ brassinoesteroide 48.9ab 2.623a
Testigo 46.9b 1.680b

125
Efecto del Biofertilizante y la Preparación del Suelo en la Producción de Maíz, Sorgo y Sorgo x
Sudán en la Zona Media Potosina
Miguel Angel Martínez Gamiño1∗ y Cesario Jasso Chaverría1
INTRODUCCION. El uso de fertilizantes químicos en cultivos de temporal incrementa el riesgo financiero dada la incertidumbre de obtener cosechas debido a lo aleatorio de las lluvias. En la Zona Media de San Luis Potosí, la mayoría de los productores no fertiliza por esta razón, aún cuando se tiene información científica de la bondad del fertilizante en el rendimiento de los cultivos. Los biofertilizantes son una alternativa para incrementar el rendimiento sin necesidad de arriesgar por el alto costo del producto. Por otra parte, la preparación del suelo debe ser reorientada hacia prácticas que no destruyan su estructura y que permitan mantener los actuales rendimientos pero con menor costo (Abelson, 1995; Hamblin, 1995). El objetivo de este estudio fue evaluar el efecto en el uso de biofertilizantes con métodos de preparación del suelo bajo condiciones de temporal en la producción de maíz, sorgo y sorgo x sudán en la Zona Media de San Luis Potosí. MATERIALES Y METODOS. El trabajo se estableció en el Municipio de Cerritos, S.L.P., en un suelo tipo Vertisol. La precipitación media anual es de 550 mm y la temperatura media es de 20oC. Se evaluó un biofertilizante, producido por INIFAP, para gramíneas en base a Azospirillum, en dosis de un kilo de producto por la cantidad de semilla a emplear de maíz, sorgo y sorgo x sudán por hectárea. Para maíz se sembró un genotipo criollo regional 12 kg ha-1, en sorgo se empleó el híbrido Esmeralda 10 kg ha-1 y en sorgo x sudán Sweet Siux 12 kg ha-1. .Se combinaron los tratamientos con y sin biofertilizantes con los siguientes métodos de preparación del suelo: 1). labranza tradicional (LT) consistente en un barbecho + rastra, 2). labranza reducida (LR) con un solo paso de rastra y 3). labranza de conservación (LC) con el uso del multiarado. Cada tratamiento de labranza se estableció en parcelas de 3,333 m2, las cuales se subdividieron con los dos tratamientos de biofertilizantes. En todos los tratamientos no se aplicó fertilizante químico. El rendimiento de grano y rastrojo se evaluó de cinco puntos de muestreo seleccionados al azar en cada tratamiento y los datos se analizaron estadísticamente de acuerdo a un diseño completamente al azar. RESULTADOS Y DISCUSION. No se detectaron diferencias estadísticas (p≤0.05) entre los tratamientos (Cuadro 1). En sorgo x sudán, el rendimiento de forraje en los tratamientos con biofertilizante superó en promedio con 738 kg ha-1 al obtenido en los tratamientos sin
1 Campo Experimental San Luis, INIFAP. [email protected]
biofertilizante. Esta diferencia es atractiva para los productores dado que el costo del producto fue de 40 pesos por hectárea. En maíz, la producción de forraje tuvo una tendencia favorable en un 35% con el uso del biofertilizante, mientras que este efecto fue de solo 5% en el cultivo de sorgo. En los métodos de labranza rastra fue la opción más rentable al ahorrarse el costo del barbecho. Este tratamiento presenta la desventaja de generar una capa compacta a 20 cm de profundidad, mientras que el multiarado tiene la ventaja de no invertir el suelo, favorecer la infiltración del agua de lluvia y evitar la compactación que se genera al usar únicamente la rastra. Cuadro 1. Rendimiento de sorgo, maíz y sorgo x sudán con biofertilizante y diferentes métodos de labranza.
Tratamientos Sorgo grano
Maíz forraje
Sorgo x sudán
t ha-1 Multiarado con biofertilizante
1.364a
1.227a
2.981a
Multiarado sin biofertilizante
1.451a
1.028a
2.425a
Rastra con Biofertilizante
1.767a
1.350a
3.131a
Rastra sin biofertilizante 1.484a 0.787a 1.896a B+R con biofertilizante 1.506a 1.083a 3.318a B+R sin biofertilizante 1.456a 0.890a 2.892a
Medias con la misma literal son estadísticamente iguales, (p≤0.05).
CONCLUSIONES. No se detectaron diferencias en el uso del biofertilizante con diferentes métodos de preparación del suelo, aún cuando se observó una respuesta favorable. LITERATURA CITADA Abelson, P.H. 1995. Sustainable agriculture and the 1995 Farm Bill. Science 267:943. Hamblin, A. 1995. The Concept of Agricultural Sustainability. In. Andews, J.H. and I. Tommerup (eds). Advances in Plant Pathology, Vol. 11. Academic Press, New York. P 1-19.

126
Biofertirrigación por Goteo a Base de Guano en Cultivos Diversos Bajo un Sistema Hidropónico con Producción de Tilapia en Xalisco, Nayarit
Alberto Betancourt Vallejo1*; Pedro D. Flores Peña1,
Víctor M. González Velásquez1, Rafael Quezada Morales1, Víctor Jiménez García1 y Roberto Gómez
Aguilar1
INTRODUCCION. En función directa del crecimiento demográfico del país, se necesita hacer mas eficiente la utilización de los recursos suelo y agua. Una de las alternativas viables es la biofertirrigación en sistema hidropónico (Resh, 2001), su principal ventaja es la simplicidad, dado que no se utilizan formulas químicas en el substrato sino guano como base del sistema en riego por goteo y complementado con fertilización foliar a base de urea. El sistema hidropónico tradicional se aplica usando los elementos mayores y micro elementos en substratos inertes (Resh, 2001), estos ocasionan acidez de la solución, desbalance de los elementos y dificultades de manejo. Por la razón anterior se propone utilizar una solución orgánica que permita además de la producción de cultivos diversos la producción de peces de valor comercial como la Tilapia que es de fácil manejo y reproducción con los objetivos siguientes: a) establecer un diseño en riego por goteo de bajo costo y con estructura de una tecnología moderna; b) utilizar guano en la solución nutritiva, adicionado con un catalizador orgánico y complementado con fertilización foliar; c) capacitar a estudiantes y profesionales en las etapas de un diseño en riego por goteo de alta eficacia, sencillo en su operación y con la carga hidrostática adecuada para el control de riego contra reloj; y d) producir en pequeña escala la Tilapia.
MATERIALES Y METODOS. El sistema se estableció en cuatro módulos tipo invernadero en el año 2004 localizadas en la Unidad Académica de Agricultura de la Universidad Autónoma de Nayarit. El agua de riego proviene de un pozo profundo de buena calidad física, química y bacteriológica. Se utilizo tubería de color negro resistentes a los rayos UV, la línea principal de poliducto negro hidráulico de 50.8 mm de diámetro, el tanque fertilizador con válvulas de control y de paso para agua de entrada y de salida, con filtro de mallas, la cintilla es del tipo T tape 5000 de 5/8” de diámetro con emisiones cada 30 cm. En cada uno de los cuatro módulos se dispone de 80 líneas de riego de 6 m de longitud, esta 80 líneas se fraccionan para la operación equilibrada y estática de la carga hidráulica por lo que se divide este módulo en cuatro secciones de riego donde cada sección de riego se controle 1 Facultad de Agricultura, Universidad Autónoma de Nayarit. [email protected]
con un Piezometro, una válvula de paso y 20 líneas de riego (Losada, 2000). Los cultivos establecidos en octubre 1 del 2004 fueron: jitomate variedad ‘Tequila’ de hábito indeterminado, un jitomate de habito determinado, chile serrano, y pepino tipo criollo, en botes circulares de 20 litros y aserrín como sustrato. Otros cultivos se irán adicionando bajo el sistema de cultivos acompañantes para evitar daños de plagas y enfermedades, las Tilapias por requerir temperaturas del agua de 28ºC se establecerán el mes de marzo del 2005.
RESULTADOS Y DISCUSION. Los cuatro cultivos establecidos mostraron hasta la fecha muy buen aspecto, crecimiento y sanidad, con la salvedad de que en el sustrato de aserrín se mostraron pequeñas deficiencias de N que fueron corregidas con aplicaciones foliares de la fórmula líquida 20-10-2 en proporción 1:20, estos resultados preliminares concuerdan con (Resh, 2001), las mejores respuestas se observaron en aserrín con guano dejado a la intemperie 15-30 días previo al transplante. CONCLUSIONES. De acuerdo a los objetivos planteados, el sistema es promisorio para productores que requieran de un sistema simplificado de producción de cultivos orgánicos bajo riego por goteo, dado que se puede producir eficientemente con costos bajos de operación y manejo. LITERATURA CITADA Resh, H. M. 2001 Cultivos hidropónicos. Nuevas técnicas de producción. Ediciones Mundi-Prensa. 5a. Ed. p 285-314. Losada Villasante A. 2000. El riego. Fundamentos hidráulicos. Ediciones Mundi-Prensa 3a. Ed. 1-18

127


















