
CARACTERIZACIÓN Y VALORACIÓN DEL POTENCIAL … · dado, dañado o destruido (s e r 2004) ......
26
45 Recepción: 10 de septiembre de 2008 / Aprobación: 31 de octubre de 2008 Revista Colombia Forestal Vol. 11:45-70 / Diciembre 2008 CARACTERIZACIÓN Y VALORACIÓN DEL POTENCIAL DE REGENERACIÓN DEL BANCO DE SEMILLAS GERMINABLE DE LA RESERVA FORESTAL CÁRPATOS (GUASCA, CUNDINAMARCA) 1 Edgard Ernesto Cantillo Higuera 2 Viviana Castiblanco Gutiérrez 3 David Fernando Pinilla Mondragón 4 Claudia Liliana Alvarado 5 1 Resumen de los resultados del Proyecto de Investigación financiado por el Centro de Investigaciones de la Universidad Distrital Francisco José de Caldas. 2 Universidad Distrital Francisco José de Caldas, Bogotá, Colombia, [email protected]. 3 Universidad Distrital Francisco José de Caldas, Bogotá, Colombia, [email protected]. 4 Universidad Distrital Francisco José de Caldas, Bogotá, Colombia, [email protected]. 5 Universidad Distrital Francisco José de Caldas, Bogotá, Colombia, [email protected]. Palabras clave: bosque andino, banco de semillas, restauración ecológica, fenología, dispersión. Key Words: Andean forest, seed bank, ecological restoration, phenology, scattering. RESUMEN En la Reserva Forestal Cárpatos se caracterizó y va- loró el banco de semillas germinable (BSG) y se de- finieron los mecanismos y estrategias de dispersión de la reserva, que posteriormente determinaron la estructura espacial, temporal y dinamogenética de los estados serales, así como de sus poblaciones. La vegetación en pie registró 61 especies, repre- sentadas por 53 géneros y 36 familias; paralela- mente en el BSG se identificaron 55 especies, con de 50 géneros y 27 familias. El volumen mínimo de la unidad de muestreo fue de 1.500 cm 3 , con un total de 13.150 semillas germinadas, mientras que la densidad total para el BSG fue de 1.553 semillas por m 2 . Las especies emergidas en el BSG estuvie- ron constituidas en su mayoría de porte herbáceo y rasante –aproximadamente 70% del total de in- dividuos y de especies–; Phytolacca bogotensis, Digitalis purpurea y Ageratina sp. fueron las es- pecies dominantes en los tres ensayos del banco. Otras especies de importancia fueron Carex sp., Poa annua, Rubus floribundum, Bulbostylis sp. y Borreria sp. Las especies de porte leñoso variaron en cada ensayo de acuerdo a la fenofase de cada una de ellas; las más representativas fueron Ces- trum mutisi, Clusia ducu, Myrsine coriacea, Axi- naea macrophylla y Miconia denticulata. En cuanto a fenología y dispersión, se encontró que la vegetación en pie presentó a la zoocoria como la estrategia de dispersión dominante en todas las parcelas, los máximos de floración se presentaron durante la estación seca, entre diciembre y marzo, mientras que los máximos registros de fructifica- ción correspondieron al periodo de la estación de lluvias más prolongada, entre junio y agosto. ABSTRACT In the Cárpatos Forest Reserve the bank seed germinate (BSG) was marked and valued and the dispersion mechanisms and strategies of the reserve defined. They subsequently determined the spatial, dinamogenetic and temporary structures of the seral states and their populations. For the vegetation on foot 61 species were recorded, represented by 53 genera and 36 families, alongside the BSG were 55 species, represented by 50 genera and 27 families. The minimum volume was 1.500 cm 3 , for a total of 13.150 germinating seeds; the total density for BSG was 1.553 seeds per m 2 . The species emerged in the BSG were mostly herbaceous and flush –approximately 70% of all individuals and species–; Phytolacca bogotensis, Digitalis purpurea and Ageratina sp. were the dominant species in the three
Transcript of CARACTERIZACIÓN Y VALORACIÓN DEL POTENCIAL … · dado, dañado o destruido (s e r 2004) ......

45Recepción: 10 de septiembre de 2008 / Aprobación: 31 de octubre de 2008 Revista Colombia Forestal Vol. 11:45-70 / Diciembre 2008
CARACTERIZACIÓN Y VALORACIÓN DEL POTENCIAL DE REGENERACIÓN DEL BANCO DE
SEMILLAS GERMINABLE DE LA RESERVA FORESTAL CÁRPATOS (GUASCA, CUNDINAMARCA)1
Edgard Ernesto Cantillo Higuera2
Viviana Castiblanco Gutiérrez3
David Fernando Pinilla Mondragón4
Claudia Liliana Alvarado5
1 ResumendelosresultadosdelProyectodeInvestigaciónfinanciadoporelCentrodeInvestigacionesdelaUniversidadDistritalFranciscoJosédeCaldas.
2 UniversidadDistritalFranciscoJosédeCaldas,Bogotá,Colombia,[email protected] UniversidadDistritalFranciscoJosédeCaldas,Bogotá,Colombia,[email protected] UniversidadDistritalFranciscoJosédeCaldas,Bogotá,Colombia,[email protected] UniversidadDistritalFranciscoJosédeCaldas,Bogotá,Colombia,[email protected].
Palabras clave: bosqueandino,bancodesemillas,restauraciónecológica,fenología,dispersión.
Key Words: Andeanforest,seedbank,ecological restoration,phenology,scattering.
RESUMEN
EnlaReservaForestalCárpatossecaracterizóyva-loróelbancodesemillasgerminable(bsg) y se de-finieronlosmecanismosyestrategiasdedispersióndelareserva,queposteriormentedeterminaronlaestructuraespacial,temporalydinamogenéticadelos estados serales, así comode suspoblaciones.La vegetación en pie registró 61 especies, repre-sentadas por 53 géneros y 36 familias; paralela-mente en el bsg se identificaron55especies,conde50génerosy27familias.Elvolumenmínimodelaunidaddemuestreofuede1.500cm3,conuntotalde13.150semillasgerminadas,mientrasqueladensidadtotalparaelbsgfuede1.553semillasporm2.Lasespeciesemergidasenelbsg estuvie-ronconstituidasensumayoríadeporteherbáceoyrasante–aproximadamente70%deltotaldein-dividuos y de especies–; Phytolacca bogotensis, Digitalis purpurea y Ageratina sp. fueron las es-peciesdominantesen los tresensayosdelbanco.Otras especies de importancia fueron Carex sp., Poa annua, Rubus floribundum, Bulbostylis sp. y Borreria sp.Lasespeciesdeporteleñosovariaronen cada ensayo de acuerdo a la fenofase de cada unadeellas; lasmás representativas fueron Ces-trum mutisi, Clusia ducu, Myrsine coriacea, Axi-naea macrophylla y Miconia denticulata.
Encuantoafenologíaydispersión,seencontróquelavegetaciónenpiepresentóa lazoocoriacomolaestrategiadedispersióndominanteentodaslasparcelas,losmáximosdefloraciónsepresentarondurantelaestaciónseca,entrediciembreymarzo,mientrasque losmáximos registrosde fructifica-cióncorrespondieronalperiododelaestacióndelluviasmásprolongada,entrejunioyagosto.
ABSTRACT
In the Cárpatos Forest Reserve the bank seedgerminate (bsg) wasmarked and valued and thedispersionmechanismsandstrategiesofthereservedefined.Theysubsequentlydeterminedthespatial,dinamogeneticandtemporarystructuresoftheseralstatesandtheirpopulations.Forthevegetationonfoot61specieswererecorded, representedby53generaand36families,alongsidethebsg were 55 species,representedby50generaand27families.Theminimumvolumewas1.500cm3, fora totalof13.150germinatingseeds;thetotaldensityforbsgwas1.553seedsperm2.Thespeciesemergedin the bsg were mostly herbaceous and flush–approximately70%ofallindividualsandspecies–;Phytolacca bogotensis, Digitalis purpurea and Ageratina sp.werethedominantspeciesinthethree

46 Revista Colombia Forestal Vol. 11 - Diciembre 2008
CaracterizaciónyvaloracióndelpotencialderegeneracióndelbancodesemillasgerminabledelaReservaForestal...
trialsBank,otherimportantspecieswereCarex sp., Poa annua, Rubus floribundum, Bulbostylis sp.and Borreria sp. The species ofwoody habit usuallyvariedineachtrialaccordingtothephenophaseofeachspecies,themostrepresentativespecieswereCestrum Mutisia, Clusia ducu, Myrsine coriacea, Axinaea macrophylla and Miconia denticulata.
As for phenology and dispersal, it was foundthat vegetation stands submitted to the zoocoriaas the dominant scattering strategy in all fields;themaximum flowering occurred during the dryseason, from December to March, though thegreatestfruitingrecordscorrespondtothelongestrainyseason,fromJunetoAugust.
INTRODUCCIÓN
Los ecosistemas altoandinos y de páramo se en-cuentran actualmente sometidos a una gran trans-formacióngeneradaporlaexpansióndelafronteraagrícolayganadera.Antelaurgentenecesidadderestablecerlosecosistemasandinos,larestauraciónecológica, definida como el proceso asistido derecuperacióndeunecosistemaquehasidodegra-dado,dañadoodestruido(ser2004),seconvierteenelmecanismoquepermiteobtenerelrestableci-mientodelosatributosestructuralesyfuncionalesde losecosistemasdeteriorados.Así, larecupera-cióndeestosecosistemasdependerádel“potencialde regeneración in situ”,esdecir,delosremanen-tesbióticossupervivientesa laperturbación(Uhl&Saldarriaga1986,citadosporSaldarriaga1994).Comopartedeestepotencialbiótico,elbancodesemillasgerminableylalluviadesemillasrepre-sentanlasprincipalesestrategiasquepermitenqueunindividuosereemplaceporotroenlassiguientesgeneraciones(Sorzano2001).Elbancodesemillasgerminable (bsg) tiene la tarea de mantener una reserva natural de diversidad genética o memoria genéticadelosefectosselectivosdeambientespa-sados(McGraw&Vavrek1989);ésteserelacionade manera directa con las fases del ciclo de vida de lasplantasofenofases,queincluyenelcrecimien-todeyemasflorales,laaperturafloral,laproduc-ción de frutos inmaduros y la maduración de los mismos (vanSchaik1993), convirtiéndoseenunrequisitoindispensableparalarestauracióndelos
ecosistemas(Newstrowet al. 1994).Ladispersióndesemillas,consideradacomoelúltimopasodelcicloreproductivodelasplantas,eselprocesome-dianteelcual lassemillassontransportadas lejosdelparentalaunsitiofavorableparasucrecimien-toydesarrollo(Herreraet al.1997).Elprocesodedispersiónpermitelacolonizacióndenuevoshábi-tats,reducelacompetenciaintraespecíficayfaci-lita losprocesosdegerminación (Morales2002).Enestacontribuciónselogrócaracterizaryvalorarelpotencialderegeneracióndelbsg en la Reserva Forestal Cárpatos; nuestros objetivos fueron: (1)Evaluarladiversidaddelasespeciespresentesenel bsgparalosdiferentesestadosseralesdelare-serva.(2)Establecersiexisteestacionabilidadparalasfenofasesdeproducciónylostiposdedisper-sión,ydeterminarlaincidenciadeéstosenelbsg.(3)Determinarlasprincipalesformasdedispersióndelassemillaseidentificarcuálestiposdeespeciessonlasmayoresaportantesparalapreservacióndela dinámica del bosque. (4)Cuantificar el aportedelaavifaunaalosprocesosdedispersióndese-millasenclarosdebosquesecundario.(5)Evaluarlasdiferenciasencuantoaespecie,númeroyger-minabilidaddelasplántulasproducidasporbsg,endiferentesperiodosdelaño.
METODOLOGÍA
DESCRIPCIÓN DE LA ZONA DE ESTUDIO
LaReservaForestalCárpatoshacepartedelbos-quealtoandinoyestálocalizadaenelcostadono-roccidental de la jurisdicción de la CorporaciónAutónomaRegionaldelGuavioenelmunicipiodeGuasca,Cundinamarca,a97kmdeBogotá(Figura1),enlaveredaConcepción,cercadelsitiodeno-minadoPotreritos,ubicadoenelextremoorientaldelmunicipiomencionado,enellímitesurdelmu-nicipioGuatavitay aloccidentedemunicipiodeJunín.Altitudinalmentesehallaentre los2.600y3.000msnm. Se localiza geográficamente en lascoordenadas:latitudNorte4º47minutos,longitudOeste73º40minutos;sutopografíaesmontañosa,destacándosecomoaccidentesorográficoslosce-rrosdeLosRobles.Porsuubicación, laestaciónPotreritoseslaquemejortipificalaprecipitaciónenlareserva.Laprecipitaciónpromedioanuales

47Revista Colombia Forestal Vol. 11 - Diciembre 2008
Edgard Ernesto Cantillo Higuera / Viviana Castiblanco Gutiérrez / David Fernando Pinilla Mondragón / Claudia Liliana Alvarado
de 1.714.3mm y lamediamensual es de 142.9mm. El régimen pluviométrico es monomodal-biestacional, concentrándose las mayores lluviasentreabrilyagosto,periodoenelcuallosvientosalisiosprovienendelEste,pudiéndoseinferirqueéstosvienencargadosdehumedady,por efectosorográficos, descargan parte de ésta en la zonadeestudio.Elperiodosecosepresentaentresep-tiembreymarzoyelmesmássecocorrespondeaenero,con47mmdeprecipitación.Sustierrassoninfluenciadas por los ríosGuavio y Sueva, entreotros (Cantilloet al. 2004).Engeneral, son sue-losjóvenes,coninfluenciadecenizavolcánicaensusprimeroshorizontes.Elrelievehainfluidoenladispersiónyacumulacióndelascenizasycons-tituyeunfactorimportanteparalasusceptibilidadalaerosión.Seencontraronbásicamentedostiposdesuelos:Typic Humitropept y Typic Dystropept,quesonmoderadamenteprofundosasuperficiales,bien drenados, de alta acidez, friables, con bajoscontenidosdecarbono,deunafertilidadmedianaabaja(Corpoguavio2000).
FASE DE CAMPO
Composición de la vegetación
Seestablecierontresparcelasde50x50m,cadauna en un estado de desarrollo sucesional de la re-servaasí:bosquesecundarioclímax,bordedebos-que y borde de bosque empatado con claros. Encadaparcelaserealizóuninventariodelosindivi-duosmásrepresentativosencuantoamayortalla,áreabasalycoberturadecadaunadelasespeciespresentes,identificadasporCantilloet al. (2004).
Banco de semillas germinable
Premuestreo(determinacióndelvolumenmínimo).Eltamañoóptimodelaunidaddemuestreofuedeter-minadoatravésdeunacurvadeespecies(Simpsonet al.1989);enlosbosquesaltoandinos,deacuerdoalopropuestoporJaimesyRivera(1990),launidadmuestralsecalculóapartirdeunacurvaespeciesvs.volumen;losvolúmenesseextrajeronalolargodelareservaconunidadesmuestralesde1.000,1.500,
Figura 1. Localizacióndeláreadeestudio.

48 Revista Colombia Forestal Vol. 11 - Diciembre 2008
CaracterizaciónyvaloracióndelpotencialderegeneracióndelbancodesemillasgerminabledelaReservaForestal...
2.000,2.500y3.000cm3.Cadavolumencontócontresrepeticionesyaquellosquepresentaronlosma-yoresvaloresdeespeciesporvolumensedefinieroncomoelvolumenmínimoamuestrear.
Muestreo.Cadaparcelade50x50m fue subdi-vididaencuadriculasde10x10m.Setomóunamuestradesueloencadaunodelos36puntosdeintercepcióndelaparcela,aunaprofundidadde15cm.Almomentodetomarlasmuestrasseretirólacoberturavegetal,asícomolahojarasca,briofitosyhepáticas.Cadamuestrasedepositóenbolsasplás-ticasqueseetiquetaron,sesellaronysetrasporta-ronalinvernadero.Paracadamuestrasetomóunareplica a una distancia aproximada de unmetro.Entotalsetomaron72muestrasdesueloporcadaparcelay216paracadaensayo.Serealizarontresmuestreos con intervalo de cuatro meses entre cada uno,parauntotalde648muestras,conuntiempototalempleadodeunaño.
Fenología y dispersión de semillas
Este estudio se realizó en dos tipos de coberturavegetal:elprimerodentrodelbosque,conelregis-trodelasfenofasesapartirdelregistroquincenaldetrampasdevueloyladeterminacióndelosprin-cipalessíndromesdedispersióndesemillasde lavegetaciónsuperiorapartirdeobservacióndirectayreferenciasbibliográficas,paralavegetaciónenpieyelbsg;elsegundoenclarosdebosquesecun-dariopormediodelainstalacióndeperchasartifi-cialesparadeterminarlaincidenciadelaavifaunaenladispersiónlocaldesemillas;lainstalacióndelas perchas se realizó a diferentes distancias delbordedelbosque(cercanía,medianíaylejanía);elnúmerodeperchasporensayofuedediezycadaunotuvounaréplica,parauntotalde60perchas.Elmonitoreoenamboscasos, tantoen fenologíacomoenladispersióndesemillas,serealizócadadossemanasduranteunperiododeunaño.
FASE DE ANÁLISIS
Vegetación en pie
Seconstruyerongráficasdetodaslasvariablesto-madas para determinar el comportamiento de la
vegetación en los diferentes estados serales y la di-námicadeloshábitosdentrodecadaunodeellos.Paralelamente,seelaborarongráficas,porestrato,delcomportamientodelacoberturadelasespeciesparalastresparcelas.
Banco de semillas germinable
SerealizóunanálisisdevarianzamultifactorialpormediodelprogramaStatgraphicsPlus5.1,conelfinde determinar diferencias espacio-temporales delbsg.Adicionalmente, se efectuó el análisis geoes-tadístico por medio de la generación y posteriorrelación de variogramas de correlación espacial atravésde losprogramasGeo-eas 1.2.1ySurfer32v8.Estosvariogramasayudanadeterminarsilasva-riablesevaluadaspresentanalgúntipodeestructuraespacial.Delamismamanera,secrearonmapasdedistribuciónpara cadaparcela, según los datos deabundanciaobtenidosenelbsg,pormediodelpro-gramaSurfer32v8;estosprogramasfueronutiliza-dosporMontenegro(2000)yAcosta(2004)ensusrespectivostrabajossobrebsg.Larealizacióntantodelosvariogramascomodelosmapasdesuperfi-cie se efectuó con la suma de los datos de los tres muestreosparacadapuntomuestral;deestamane-ra, aquellos puntos donde se presentaronmayoresabundanciassonresaltadospormediodepicosquedefinentendenciasagrupadas;análogamente,seeli-minaron datos gregarios poco representativos delensayoengeneral (Rossiet al. 1995,Montenegro2000).Paraanalizarladominanciayladiversidaddelasespeciesdelbancodesemillasseconstruye-ronmodelosdeacuerdoalosugeridoporMagurran(1989).Estosmodeloshansidoproducidospordi-versosestudiosdebancosdesemillasenelpaís,en-tre losquesedestacan losdePosada&Cárdenas(1999),Montenegro(2000)yAcosta(2004).Selle-vóacabolapruebadediferenciasignificativaparaestablecercuálesparcelaspresentabanmayorsimi-litud(Montenegro,2000).
Fenologia y dispersión
EnlastrampasyperchasseutilizaronpruebasdeconcordanciadeKendall para este tipodedistri-bución; los análisis se realizaron por medio delprograma spss 7.5 paraWindows y Statistica, de

49Revista Colombia Forestal Vol. 11 - Diciembre 2008
Edgard Ernesto Cantillo Higuera / Viviana Castiblanco Gutiérrez / David Fernando Pinilla Mondragón / Claudia Liliana Alvarado
Zoocoria Anemocoria Barocoria
PORCENTAJE DE ESPECIES POR TIPO DE DISPERSIÓN
Anemocoria30%Barocoria
2%
Zoocoria68%
Figura 2. Porcentajedeespeciesenpieportipodedispersión.
acuerdo a lo sugerido y realizado por Morales(2002)ySánchez(2003).
RESULTADOS
COMPOSICIÓN DE LA VEGETACIÓN
Entotal,paralavegetaciónenpiedelastrespar-celasseencontraron61especies,representadaspor53génerosy36familias.Lasfamiliasmásrepre-sentativasfueron:Lauraceae(11%),Melastomata-ceae(10%),Compositae(7%)yMyrsinaceae(5%)(Anexo1).Lazoocoriasepresentócomoelsíndro-mededispersióndominantecon68%,seguidadelaanemocoriaconel30%ylabarocoriaconel2%derepresentatividad(Figura2).
Elnúmerodefamiliasenlasparcelasaumentaenformaproporcionalconrespectoaladisminucióneneldesarrollodelasetapasséricas(Figura3),yaquemientraslaparceladebosqueclímaxcontieneel 54% de los registros, la parcela de borde conclarocontieneel70%delasfamilias.
Laparceladebosqueclímaxregistró35especiesde28génerosy20familias;porsuparte,enlapar-celadebordeseencontraron34especiesde30gé-nerosy23familias,mientrasqueenlaparcelade
bordedebosqueconclarosehallaron41especiesde37génerosy26familias.
Larelacióndelastresparcelasportipodedisper-sión(Figura4)mostrólazoocoriacomoelsíndro-medominanteparalastresparcelas,variandoentreel60%(bordeconclaro)y80%(bosqueclímax);comosegundaestrategiasepresentalaanemocoriaconunafluctuaciónentreel35%(bordeconclaro)y15%(bosqueclímax),seguidaporlabarocoria,la cual no esmuy frecuente y presenta un valorbajo(<3%)endondeparticipa.
Entre el dappromedioporparcelaylaalturapro-mediopor parcela (Figura 5) se presenta una re-lacióndirectamenteproporcional, al presentar unincrementoensusregistrosamedidaquelacomu-nidadsetornamásmadura.
BANCO DE SEMILLAS GERMINABLE (BSG)
Premuestreo
Losmayores registros de número de especies sepresentaronen lasmuestrasde1.500cm3,por loqueéstesedefiniócomoelvolumenmínimoparalareserva(Figura6).

50 Revista Colombia Forestal Vol. 11 - Diciembre 2008
CaracterizaciónyvaloracióndelpotencialderegeneracióndelbancodesemillasgerminabledelaReservaForestal...
Muestreos
En total a partir de los tresmuestreos realizadosa lo largodeunañogerminaron13.150 semillaspertenecientes a 55 especies de 50 géneros y 27familias;deltotaldeespeciessóloocho,queequi-valenal2.34%deltotaldelosindividuos,seiden-tificaronhastafamilia;larelacióndetalladadelasespeciesseencuentraenelAnexo2. La densidad total del bsgfuede1.529semillas/m2.
Lasespeciesmásabundantesparaelbancodese-millasdeacuerdoconlaFigura7 fueron en su or-den Phytolacca bogotensis, Carex albolutenscens, Digitalis purpurea, Borreria sp. y Ageratina sp.,convaloresentreel15%yel6%delaabundanciarelativatotal,esdecir,entre2.024y789semillasgerminadas.
0
1 0
2 0
3 0
4 0
5 0
6 0
7 0
% E
SPEC
IES
BOSQUE CLIMAX BORDE DE BOSQUE BORDE/CLARO
PARCELAS
% ESPECIES POR PARCELA
Figura 3. Porcentajedeespeciespresentesporparcela.
0102030405060708090
%
Zoocoria Anemocoria Barocoria
SÍNDROME DE DISPERSIÓN POR PARCELA
BOSQUE CLIMAX BORDE DE BOSQUE BORDE/CLAROPARCELAS
Figura 4. Síndromededispersiónporparcela.

51Revista Colombia Forestal Vol. 11 - Diciembre 2008
Edgard Ernesto Cantillo Higuera / Viviana Castiblanco Gutiérrez / David Fernando Pinilla Mondragón / Claudia Liliana Alvarado
Paraelcasode lasespeciesarbóreas, lasmásre-presentativas fueron Cestrum mutisi, Buddleja bullata, Solanum inopinum, Axinaea macrophylla y Miconia denticulata, entreotras;estasespeciesindividualmentepresentanunporcentajepocosig-nificativo,puestodasestánpordebajodel2.2%deltotaldelaabundanciarelativa.
Lostotalesdesemillasgerminadasporparcelapre-sentanvaloresdisímiles,queaumentanconformea
unamayorheterogeneidadpresenteenlacobertu-ravegetal (Figura8).Este comportamientopare-ceindicarqueentremásdesarrollosucesionaldelecosistema,mayoreslaposibilidaddeaportetantodelnúmerocomodelavariabilidaddeespeciesdesemillasyaseaporlluviaoporestrategiadedis-persióndesdelugaresaledaños.
El comportamiento de muestreos varió, se re-gistraron losmenores valores para elmuestreo I
0
2
4
6
8
10
12
14
16
18
PARCELAS
ALTURA PROMDAP PROM
ALTURA PROM Vs DAP PROM
BOSQUE CLIMAX BORDE DE BOSQUE BORDE/CLARO
Figura 5. Relacióndealturapromediovs.dappromedioporparcela.
0
2
4
6
8
10
12
14 REPETICIÓN 1REPETICIÓN 2 REPETICIÓN 3
DETERMINACIÓN DEL VOLUMEN MíNIMO
VOLUMEN DE SUELO (cm3)
No. S
P GE
RMIN
ADAS
1000 1500 2000 2500 3000
Figura 6. DeterminacióndelvolumenmínimodemuestreoparalaReservaForestalCárpatos.

52 Revista Colombia Forestal Vol. 11 - Diciembre 2008
CaracterizaciónyvaloracióndelpotencialderegeneracióndelbancodesemillasgerminabledelaReservaForestal...
(noviembre-diciembre), con 3.214 semillas ger-minadas, hasta 5.378 semillas en el muestreo iii (junio-julio,Figura9).
Noobstantelavariabilidaddelaportedesemillasentrelosmuestreos,elincrementoenelnúmerodesemillas germinadas en las parcelas fue en igualproporción.Alsituarcronológicamentelosmues-treosycompararlosconelrégimendeprecipitación
delareserva,seencontróunarelacióndirectamen-teproporcionalentrelaprecipitaciónyelnúmerodesemillasinventariadas,teniendoencuentaqueelprimermuestreo fue realizadoafinalesdeno-viembre,elsegundoenmarzoyelterceroenjulio(Figuras10y11).
EnelAnexo3 se indican de manera resumida los valores de los índices de Margalef & Shannon
0
2
4
6
8
10
12
14
16
%
Ageratina sp. Carex albolutenscens
Digitalis purpurea
Phytolaccabogotensis
Poa annua
ESPECIES
ABUNDANCIA RELATIVA DEL BSG POR ESPECIE
Figura 7. Abundanciarelativadelbsgporespecie.
0
1000
2000
3000
4000
5000
6000
7000
8000
No. S
EMILL
AS G
ERMI
NADA
S
BOSQUE CLIMAX BORDE DE BOSQUE BORDE/ CLARO
No. SEMILLAS GERMINADAS POR PARCELA
Figura 8. Númerodesemillasgerminadasporparcelaparabsg.

53Revista Colombia Forestal Vol. 11 - Diciembre 2008
Edgard Ernesto Cantillo Higuera / Viviana Castiblanco Gutiérrez / David Fernando Pinilla Mondragón / Claudia Liliana Alvarado
hallados por muestreo y por parcela; a partir deellossepuedendeducirlosmodelosderiquezayabundanciadelbsg de acuerdo a lo sugeridoporMagurran(1989).Singularmente,losmayoresre-gistrostantoenriquezaydiversidadloobtuvieronlasparcelasdeborde,debidoposiblementealahe-terogeneidadde lacomposiciónde lascoberturasboscosasenestoslugares.
Latipificacióndelbsgpresentóalazoocoriacomoestrategiadominantededispersiónconun55%del
totaldelasespecies,mientrasqueelrestante45%loconstituyólaanemocoria(Figura12).Nosepre-sentóningunaespecieconestrategiabarocoria.
Ladistribucióndeltipodesemillasparaelbsg re-gistróel84%porpartedeespeciesquepresentansemillasdicotiledóneas,frenteaun16%quecons-tituyenlasespeciesconsemillamonocotiledóneas.Conrespectoaltipodesemillalarelaciónfuede3:1(9.500semillasdicotiledóneasfrenteasólo3.500semillas de unmonocotiledón).El tipo de banco
0
1000
2000
3000
4000
5000
6000
7000
8000
No. S
EMILL
AS G
ERMI
NADA
S
BOSQUE CLIMAX BORDE DE BOSQUE BORDE/ CLARO
No. SEMILLAS GERMINADAS POR PARCELA
Figura 9. Númerodesemillasgerminadaspormuestreoparaelbsg.
RÉGIMEN DE PRECIPITACIÓN
0
50
100
150
200
250
300
Enero
Febrero
Marzo
Abril Mayo Junio
Julio
Agosto
Septiem
bre
Octubre
Noviem
bre
Diciembre
mm
Figura 10. RégimendeprecipitacióndelaReservaForestalCárpatos.

54 Revista Colombia Forestal Vol. 11 - Diciembre 2008
CaracterizaciónyvaloracióndelpotencialderegeneracióndelbancodesemillasgerminabledelaReservaForestal...
paralareservapresentaunatendenciapermanentealcontenermásdelamitaddeltotaldelasespeciespresentesdelensayo;lostiposdebancotransitorioypseudopersistentepresentaronvaloresmenores,aunquecontaronconunaparticipaciónsignificati-vaaltenerel31%yel13%,respectivamente,deltotaldelasespeciesregistradas(Figura13).
Lazoocoriasepresentacomolaestrategiadomi-nanteenelaporteylluviadesemillas(Figura14),tantodemaneraespacial (localy regional)comotemporal(bancopersistenteytransitorio).
El análisismultifactorial del comportamiento espa-cio-temporaldelbsgmostróquenohubodiferencias
significativasentreparcelasymuestreos,perosíexis-tiódiferenciaentrelacomposiciónmuestraldecadaunadelasparcelas(f=0.0005;g.l.=2;P=0.05).
Aunquelavariaciónparacadaparcelafuediferen-teenlacomposiciónynúmerodeespecies,todasmostraron lamisma tendenciade incrementopormuestreo; además todas presentaron losmayoresregistros de emergencia de plántulas en los pun-tosdondelacoberturaeramásheterogénea(Figura15).Pormediodelapruebadediferenciasignifica-tivaseestablecióquelasparcelasqueserelacionancon el bordepresentanmayor asociaciónque lasdelbosquemaduro.
0
500
1000
1500
2000
2500
3000
3500No
. DE
SEMI
LLAS
GER
MINA
DAS
1 2 3MUESTREOS
No. DE SEMILLAS GERMINADAS POR MUESTREO EN CADA PARCELA
BOSQUE CLIMAXBORDE DE BOSQUEBORDE / CLARO
Figura 11. Distribuciónespacio-temporaldelaabundanciadesemillasgerminadasparabsg.
TIPOS DE DISPERSIÓN DEL BSG
Zoocoría55%
Anemocoria45%
Figura 12. Distribucióndelossíndromesdedispersiónparaelbsg.

55Revista Colombia Forestal Vol. 11 - Diciembre 2008
Edgard Ernesto Cantillo Higuera / Viviana Castiblanco Gutiérrez / David Fernando Pinilla Mondragón / Claudia Liliana Alvarado
% DE ESPECIES POR TIPO DE BANCO
56% 13%
31%
Peristente Pseudopersistente Transitorio
Figura 13. Distribucióndelnúmerodeespeciesportipodebanco.
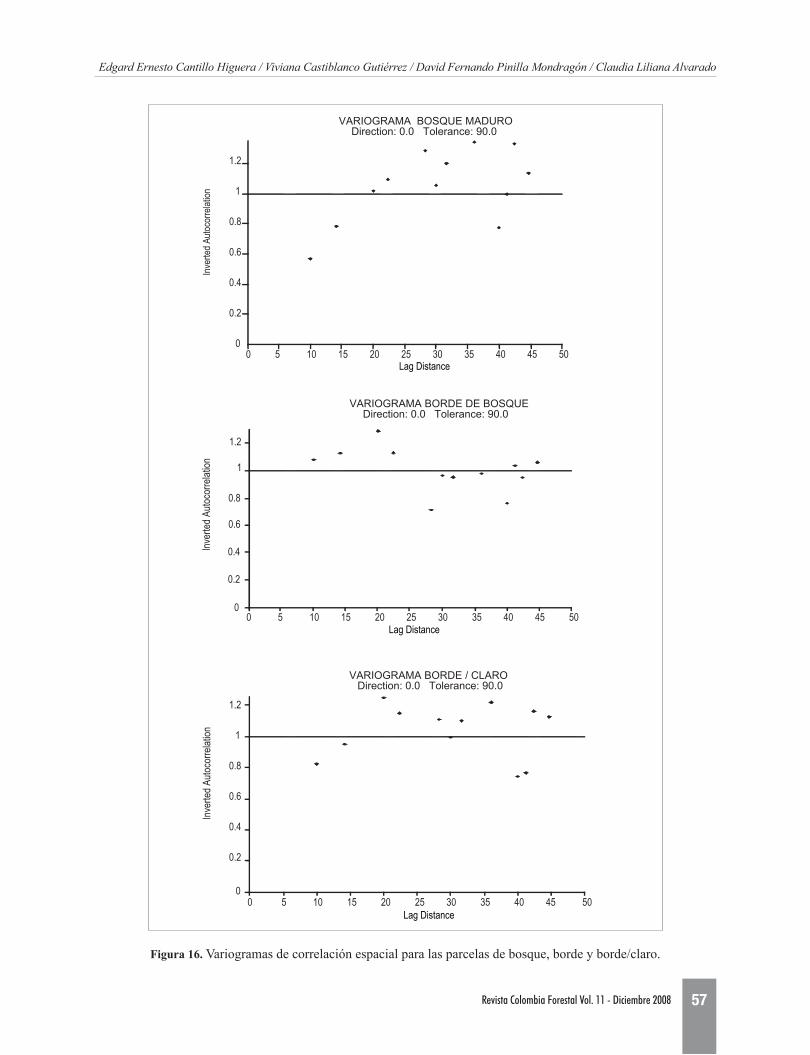
El comportamiento entre lasunidadesmuestralesdelasparcelasfuediferente,tantodemaneraespa-cialcomotemporal;así,algenerarlosvariogramasdecorrelaciónespacialsepresentandiferenciasenlaestructuraespacialdelospuntosmuestralesparacadaparcela(Figura16).
Igualmente, losmapasde superficie efectuadosapartirde losdatosdeabundanciaobtenidosenelbsg presentaronun comportamientodisímil entrelasparcelas,variandoasíelnúmero,ladisponibi-lidad y la dispersión de las plántulas emergentesparacadaunadeellas(Figura17).
0
10
20
30
40
50
60
70
%
PERSISTENTE PSEUDOPERSISTENTE TRANSITORIO
TIPO DE BANCO VS DISPERSIÓN
Zoocoría Anemocoría
Figura 14. Distribucióndeltipodedispersiónportipodebanco.
La variación en la composición de los registrosfuedisímilconrespectoalosmuestreosporloqueestructuralmenteexisteunadinámicaespacio-tem-poral en la germinación de semillas (Figura 18),apesardequelasmuestrasfuerontomadasenelmismolugar(parcela)ytiempo(muestreo).
FENOLOGÍA Y DISPERSIÓN
Losperiodosde fenofasespara la reservaparecentener una relación estrecha con el régimen local de precipitación(Anexo4);losmáximosdefloraciónsepresentarondurantelaestaciónseca(diciembrea

56 Revista Colombia Forestal Vol. 11 - Diciembre 2008
CaracterizaciónyvaloracióndelpotencialderegeneracióndelbancodesemillasgerminabledelaReservaForestal...
A A
B
0 5 10 15 20 25 30 35 40 45 500
5
10
15
20
25
30
35
40
45
50
A A
A
A
A
B
C CD
0
5
10
15
20
25
30
35
40
45
50
AB
C
D
E
0
5
10
15
20
25
30
35
40
45
50
0 5 10 15 20 25 30 35 40 45 50
0 5 10 15 20 25 30 35 40 45 50
Figura 15. Mapasdecoberturaparalasparcelasdebosque,bordeyborde/claro.
57Revista Colombia Forestal Vol. 11 - Diciembre 2008
Edgard Ernesto Cantillo Higuera / Viviana Castiblanco Gutiérrez / David Fernando Pinilla Mondragón / Claudia Liliana Alvarado
0 5 10 15 20 25 30 35 40 45 50Lag Distance
0
0.2
0.4
0.6
0.8
1
1.2
Inver
ted A
utoco
rrelat
ion
Direction: 0.0 Tolerance: 90.0VARIOGRAMA BOSQUE MADURO
Direction: 0.0 Tolerance: 90.0VARIOGRAMA BORDE DE BOSQUE
Direction: 0.0 Tolerance: 90.0VARIOGRAMA BORDE / CLARO
0 5 10 15 20 25 30 35 40 45 50Lag Distance
0
0.2
0.4
0.6
0.8
1
1.2
Inver
ted A
utoco
rrelat
ion
0
0.2
0.4
0.6
0.8
1
1.2
Inver
ted A
utoco
rrelat
ion
0 5 10 15 20 25 30 35 40 45 50Lag Distance
Figura 16. Variogramasdecorrelaciónespacialparalasparcelasdebosque,bordeyborde/claro.

58 Revista Colombia Forestal Vol. 11 - Diciembre 2008
CaracterizaciónyvaloracióndelpotencialderegeneracióndelbancodesemillasgerminabledelaReservaForestal...
51015202530354045505560657075
Parcela Bosque Maduro
20406080100120140160180200220240260280300320340
Parcela Borde de Bosque
50100150200250300350400450500550600650700750800850900950100010501100
Parcela Borde / Claro
Figura 17. Mapasdesuperficieporparcela.
marzo),mientrasquelosregistrosdefructificacióncorrespondieronalperiododelaestacióndelluviasmásprolongada (entre losmesesde junio a agos-to), independientementeentremeseslafloraciónyfructificaciónguardaronunarelacióninversamenteproporcionalencuantoalasincroníadesusproduc-ciones(W=0.028;X2 =0.333;n=12).
Laduracióndelasfenofasesentreespeciestambiénvarió,puesmientrasespeciesdecomunidadesma-duras(Hedyosmon bomplandianum, Clusia ducu y Ocotea callophylla,entreotras)presentaroncicloscortosyespecíficosdereproducción,lasespeciesquecorrespondenaestadossucesionalesinicialescomo Phytolacca bogotensis correspondieron a periodos de floración y fructificación constantesduranteelcicloanual.
Las perchas artificiales fueron exitosas ya quedurante todo el tiempo del ensayo contaron convisitas en mayor o menor proporción. El mayorregistro de número de perchas visitadas ocurriódurantelaépocaseca,dondelosrecursossonmáslimitados;elnúmerodesemillas,númerodeespe-ciesdispersadasyvisitasalasperchaspresentaronsusmáximosregistrosdurantelaépocadelluvias.Se identificarondoceespeciesque fuerondisper-sadasdirectamentedesdelasperchas,entrelasquese destacan Rubus floribundum, Arcytophyllum sp.y Phytolacca bogotensis;delamismamanera,seidentificarontreceespeciesdeaves(anexo5).
DISCUSIÓN DE RESULTADOS
COMPOSICIÓN DE LA VEGETACIÓN
Especies comoMyrsine coriacea, Miconia squa-mulosa, Drymis granadensis, Clusia multiflora, Weinmannia pinnata y Myrsine guianesis, perte-necientesacomunidadesmaduras,fuerontambiénregistradasporCantilloet al.(2004)eneláreadeestudioyporJaimes&Rivera(1990)yMontene-gro(2000)enestudiosrealizadosalamismaalti-tud;de lamismamanera, especiesasociadasconlasprimerasetapasdedesarrollo,talescomoAge-ratina sp., Carex sp., Poa sp.,Ruagea sp.,Arcyto-phyllum sp. yChusquea scandensformanpartede

59Revista Colombia Forestal Vol. 11 - Diciembre 2008
Edgard Ernesto Cantillo Higuera / Viviana Castiblanco Gutiérrez / David Fernando Pinilla Mondragón / Claudia Liliana Alvarado
123456789101112131415161718192021222324
246810121416182022242628303234
024681012141618202224262830323436
Muestreo 1 Bosque Maduro Muestreo 2 Bosque Maduro Muestreo 3 Bosque Maduro
51015202530354045505560657075
5101520253035404550556065707580859095100105110
10
20
30
40
50
60
70
80
90
100
110
120
130
140
150
160
Muestreo 1 Borde de Bosque Muestreo 2 Borde de Bosque Muestreo 3 Borde de Bosque
102030405060708090100110120130140150160170180190200210220230
20406080100120140160180200220240260280300320340360380400420
20406080100120140160180200220240260280300320340360380400420440460480500520540
Muestreo 1 Borde Claro Muestreo 2 Borde Claro Muestreo 3 Borde Claro
Figura 18. Dinámicaespacio-temporaldelbsg.

60 Revista Colombia Forestal Vol. 11 - Diciembre 2008
CaracterizaciónyvaloracióndelpotencialderegeneracióndelbancodesemillasgerminabledelaReservaForestal...
la vegetación de los ecosistemas altoandinos regis-trados por Posada&Cárdenas (1999),Montene-gro(2000),Olaya(2000),Pedraza(2000),Acosta(2004)yMahecha(2004),entreotrosautores.
Lamenorvariabilidaddehábitosyestratossepre-sentóenlaparceladebosquemaduro,dondecercadel95%deltotaldelasespeciescorrespondióalosestratossuperiores,mientrasqueunapequeñami-noríaserefirióahierbas(Ageratinasp.) y helechos (Cyathea sp.). Por su parte, la parcela que tuvomayornúmerodehábitoscorrespondióalbordedebosque,alcontarconochode losnueve tiposdevidaregistrados: arbóreo(68%)yarbustivo(18%),quefueronlosmásrepresentativos.Ladistribucióndeabundancia,tantoparalavegetacióncomoparael bsg,fuemayorennúmerodeespeciesysemillasque la registrada por Montenegro (2000), Olaya(2000) yAcosta (2004). Esta distribución pareceajustarsealmodelonormal,yaque,deacuerdoaMagurran(1989),enningunacomunidadtodaslasespeciespuedenserigualmenteabundantes.Lave-getaciónenpietuvoalazoocoriacomoelsíndro-mededispersióndominante,propiadevegetacióndehábitatshúmedos,concordandoconlopublica-doporHowe&Smallwood (1982), que sitúan aestaestrategiaenhábitatshúmedos,mientrasquelasestrategiasdedispersiónabióticacomolaane-mocoriasepresentanenagrupamientosmarcadosgeográficaytemporalmentedecarácterseco.
BANCO DE SEMILLAS GERMINABLE
Volumen mínimo y densidad del banco
Ladeterminacióndelvolumenmínimopresentóunvalormenor(1500cm3)aldeterminadoporJaimes&Rivera(1990),quecorrespondíaa2000cm3 y que se constituía como la unidad muestral idealpara estudios de bsg de los bosques altoandinos.Pormediodelhallazgodeunnuevovolumenmí-nimosepuedeafirmarquelosecosistemasaltoan-dinosnopresentanunpatrónespecifico,existiendoun alto grado de heterogeneidad en sus compo-nentes (unidad muestral, densidad, abundanciay diversidad), como lo han demostrado estudiosanterioresentrelosquesedestacanlosdePosada
&Cárdenas(1999),Montenegro(2000)yAcosta(2004). La densidad total del banco de semillasgerminablefuede1529semillas/m2,valormenoralos determinados en varios estudios en ecosistemas altoandinos,comoeselcasodeJaimes&Rivera(1990)con1.813semillas/m2yeldeMontenegro(2000)con3.954semillas/m2;sinembargo,elva-lorresultósermayorqueelencontradoporAcosta(2004)enlareservaforestaldeCogua,elcualfuede 1.397 semillas/m2.Este valor está en el inter-valode1.000a6.000semillas/m2 queGuevarayGómez-Pompa(1972)atribuyenabosquessecun-dariosparaeltrópico.
Dinámica de la emergencia de plántulas en el bsg
Phytolacca bogotensis, Digitalis purpurea, Age-ratina sp., Carex sp., Cestrum mutisi, Buddleja bullata y Solanum inopinum fueron las especiesprimordialesqueconformanelbsg en la reserva,especiesquesecuentandentrodelosestudiosdePosada y Cárdenas (1999), Montenegro (2000),Olaya(2000)yAcosta(2004).Lasfamiliasdema-yorrelevanciaencuantoaabundanciafueron:Cy-peraceae,Phytolaccaceae y Compositae,mientrasquelasfamiliasqueregistraronmayornúmerodeespecies fueronCompositae,Rubiaceae y Melas-tomataceae.Deigualmaneraqueenlavegetaciónen pie, la participación de los estratos arbóreo yarbustivofueimportantealcontenercercadel40%deltotal;sinembargoelhábitohierbafueelmásdominante para el banco al contener el 43% delasespecies.EstecomportamientoconcuerdaconlopublicadoporConell(1978),queafirmaqueeldisturbiopromueveeldominioyladispersióndeespecies pertenecientes a las etapas sucesionalestempranas.Losregistrosmásaltosdelaabundan-cia de semillas emergentes coincidieron con las fluctuaciones temporalesde laprecipitación; estacaracterísticapuedeestarrelacionadaconeleven-todequedespuésdelperiodolluvioso,lamayoríadesemillasyahansidodispersadasyseencuen-tran en el suelo prestas a germinar, coincidiendoconloafirmadoporBonilla&Zuloaga(1994),queregistranlosmáximosdefructificaciónenépocassecasylosdeemergenciaduranteelperiododellu-vias.Ladiversidadyriquezafueronmayoresenlas

61Revista Colombia Forestal Vol. 11 - Diciembre 2008
Edgard Ernesto Cantillo Higuera / Viviana Castiblanco Gutiérrez / David Fernando Pinilla Mondragón / Claudia Liliana Alvarado
parcelasdondehubomayorheterogeneidaddees-pecies,delamismamaneraocurrióconlosmues-treos;estarelacióndevariaciónespacio-temporalfuetambiénidentificadaenlasparcelasrealizadasporMontenegro(2000).El tipodesemilladomi-nantefueladicotiledónea,coincidiendoconloqueafirmanMcGraw&Varek (1989): que las semi-llas monocotiledóneas son en proporción menosabundantesquelasdicotiledóneas.EstatendenciaconcuerdaconloexpuestoporRice(1989),quienafirmaquecualquiergrupodehierbasdicotiledó-neas sonmásdadas a formar bancosde semillasquelospastos.
Persistencia y tipo de dispersión del bsg
Elbancodesemillaspresentóunaporteconstan-tedurantetodoelaño,porloque,deacuerdoconlo publicado porGarwood (1989), se puede afir-marquelatasaderenovacióndelassemillasenelbancoesigualomenorquelatasademortalidad,tendenciaquepermitelapersistenciatemporaldelbancodesemillas.Losresultadosarrojadosenelpresente trabajo concuerdan con lo determinadopor Montenegro (2000), al encontrar que el bsg estácompuestoprincipalmenteporsemillaspersis-tentesypseudopersistentes, ymuypocasde tipotransitorio, ya que, de acuerdo conThompson et al.(1998),loslugaresconpresenciadedisturbiospresentan bancos con tendencia a la persistenciatemporal.Lareservamuestraunatendenciaapre-sentar un bsgconstanteconlamayoríadelasespe-ciesyel89%deltotaldelaabundanciadelbanco;los tiposdebancotransitorioypseudopersistentetuvieron una participación significativa en espe-ciesmasnoennúmerodeemergencias.Unapo-siblecausadelapersistenciapuededeberseaqueelbancopresentaenlamayoríadesucomposiciónaespeciesherbáceas,concordandoconlohalladoconMontenegro(2000)yAcosta(2004).Estetipodehábitotiendealapersistencia,yaquegerminanporpulsos,característicaquelesayudaaperdurartrasdisturbios(Bertiller1991,Grime1989).Otrasvariables que permiten relacionar la persistenciadelbancoserelacionanconelgradodedisturbioyeltipodedispersión;teniendoencuentalocitadoporThompsonet al. (1989)y lo identificadoporMontenegro(2000),sepuedeconfirmarqueáreas
quehansufridoprocesosdeperturbación,comolareserva,revelanunatendenciaapresentaraltaca-pacidaddepersistenciaenlassemillasyporendeensubanco.Elbancodesemillaspersistentepre-sentóabundanciassimilarespara lazoocoriay laanemocoria, el banco pseudopersistente presentóun marcado predominio de la anemocoria sobrela zoocoria y, finalmente, en el banco transitoriolazoocoriasepresentócomolaestrategiamásre-levante frente a la anemocoria. La capacidad depersistencia y el tipo de dispersión pueden dar aentenderporquéparaelensayolosmayoresregis-troscorrespondenaemergentesanemócoros,sisetieneencuentaqueelmáximodelasabundanciassepresentóenlasparcelasasociadasconelborde;estorelacionaloafirmadoporSchuppet al. (1989)ycorroboradoporMontenegro(2000),queindicanquelassemillasanemócorassevenfavorecidasenetapastempranasdelasucesiónopormediodelallegadade éstas, productodeunflujode aire in-tensificadoenunclaroreciéncreado,mientraslaszoócorasarribanydominanenetapasmásevolu-cionadasdelasucesión(Brown1992).
En el bancode semillas predominó levemente elnúmero de especies de dispersión zoócora sobrela anemócora; estos resultados concuerdan conestudios realizadosen los trópicosporSwaing&Hall(1983)enbosquesdeGhanayporYounget al.(1987,citadoenGarwood1989),enunbosquedeCostaRica,dondeparaamboscasoselbancofuedominadoporlazoocoria,debidoposiblemen-teaqueestosbosquesnohabíansufridodisturbiosdesdeañosatrásal estudio.Estacaracterística sepresentadeigualmaneraparalaReservaForestalCárpatos,lacualvienefluctuandodesdeunprocesodesucesiónsinintervencionesadisturbiosperiódi-cos.Este,pues,esotroindicadordequelareservaseencuentraenunaetapadedesarrollosucesionalasociado a lamadurez de la comunidad, ya que,deacuerdoaloreferidoporHowe&Smallwood(1986), Jaimes & Rivera (1990), Brown (1992),Vargas(1998)yMontenegro(2000),enlasprime-rasetapasdelasucesiónexisteunpredominioporpartedelasespeciesanemócorasqueaprovechanel disturbio, mientras que las especies zoócorasempiezana llegarposteriormente, sobre todopormediodelasaves.

62 Revista Colombia Forestal Vol. 11 - Diciembre 2008
CaracterizaciónyvaloracióndelpotencialderegeneracióndelbancodesemillasgerminabledelaReservaForestal...
Vegetación y bsg
Seregistraronparaeltotaldelestudio83especies;61especiesparalavegetaciónenpiey55especiesregistradas en el bsg.Estosvaloressuperansignifica-tivamentelosregistrosdeJaimes&Rivera(1990),Posada & Cárdenas (1999), Montenegro (2000),Olaya(2000),Acosta(2004)yCardona(2004),quesonlosestudiosmásrelevantesrealizadosdebancosparalosecosistemasaltoandinos.Lavegetaciónenpieyelbancodesemillascomparten34especies;lavegetaciónpresentóbancoenel44%desusespeciesquerepresentael75%deltotaldelaabundanciadelbanco,contrarioalohalladoporMontenegro(2000),queafirmaque lacomposicióndesubanco revelala conformación de la vegetación en estados suce-sionalesanterioresalestudio.Dichocomportamien-todelbancopuededeberse,deacuerdoaPickett&McDonnell (1989) y Rico-Gray & García-Franco(1992),aqueelnúmerodeespeciesencomúnentreelbancoylavegetaciónesbajoenáreasmuyper-turbadas(Gibson&Brown1991),mientrasqueenbosquesperturbadosconpresenciadecomunidadesmadurasestacifraaumenta(Fenner1995).Enotraspalabras, los bancos de semillas pueden presentarunaimagendual,comoloafirmanGrandinyRydin(1998)yFalinska(1999),quecalificanalbancodesemillas como la memoria ecológica sucesional del lugar,oafirmanqueéstepuedeconsiderarsecomounespectrodelavegetaciónactualendiferentesestadossucesionales.Paraelpresenteestudioenparticularaplican ambos postulados, pues el banco tambiénpresentaespeciesexclusivas;sinembargo, lacom-posicióndelbancopresentaensumayoríaunreflejodeldinamismoyevolucióndelavegetaciónactual.
Dinámica espacio-temporal del bsg
Lasparcelasquepresentaron lasmayoresoscila-ciones con respecto a la normalidad en cuanto alaabundancia,sontambiénlasqueposeenmayordiversidadyriqueza;paraestecasoestatendenciaserelacionahacialasparcelasdebordeydisminu-ye de manera gradual hacia la comunidad vegetal madura,concordandoconloconcluidoporAcos-ta (2004).La diversidad y riqueza del banco fuemayorenlasparcelasdondeexistiómásheteroge-neidaddeespecies,delamismamaneraqueocu-rrióconlosmuestreos.Estarelacióndevariación
espacio-temporal fue también identificada en lasparcelasrealizadasporMontenegro(2000)yAcos-ta(2004);elbsgtuvomayordinámicaenregistrosdeabundanciaanivelespacialenaquellospuntosdelasparcelasdondelascoberturasvegetalessonmás heterogéneas, dominadas en sumayoría porvegetacióncorrespondientealosestadosprimariosdeldesarrollosucesional.
FENOLOGÍA Y TIPO DE DISPERSIÓN
Fenofases de producción
Los patrones fenológicos de la floración y fructifi-caciónparecenestarinfluenciadosdirectamenteporlosfactoresbióticosyabióticos(Hilty1980),porloquesepuedeafirmarqueexisteunacorrelaciónen-trelaestacionalidaddelasfenofasesdeproducciónyelrégimendeprecipitación;asílocorroboranestu-diosrealizadoseneltrópicocomolosefectuadosporSmythe(1970)yFleminget al. (1987)yotrosreali-zadosenecosistemasaltoandinoscomolosdeMo-rales(2002)ySánchez(2003).Lafloración,aunqueesconstantedurantelosdocemesesdelaño,parecepresentarunpatrónrespectoasumáximaproduccióncon la época de menos precipitación (Carabias &Guevara,1985,Morales2002); así lo citanautoresendiversos trabajospara el trópico tanto en tierrasbajas(Croat1975,Mori&Kallunki1976yMarquis1988)comoenaltas(Bonilla&Zuloaga1994).Lafenofasedeproduccióndeflorespuedeestarasociadaalaépocasecaparaaumentarelporcentajedeéxitodepolinización(Fagua2002,Sánchez2003),deestamanera,aumentanlasprobabilidadesdegeneraciónyproduccióndefrutosdurantelatemporadadelluvias(Williamset al.2002).
Fenofases de fructificación y síndromes de dispersión
De acuerdo a Cuatrecasas (1934) y Howe & Smallwood (1982) el comportamiento esperado enlosbosqueshúmedosesquelazoocoriapredominecomoestrategiasobrelaanemocoria.Estedictamenconcuerdaconlodeterminadoenesteestudio,dadoquelazoocoriasepresentócomolaestrategiadomi-nanteparalavegetaciónenpieyelbancodesemillas,

63Revista Colombia Forestal Vol. 11 - Diciembre 2008
Edgard Ernesto Cantillo Higuera / Viviana Castiblanco Gutiérrez / David Fernando Pinilla Mondragón / Claudia Liliana Alvarado
presentandofructificaciónconregistrossuperioresal65%deltotaldelosindividuosdurantetodoelaño.Esteporcentajeesdeesperarseparalazoocoriasisetieneencuentaquelosvertebradosyenespeciallasavesse reconocencomodispersoresdesemillasdemásdel50%de las especiesdeárbolesyarbustostropicales(Fleminget al.1987,Fonseca2001).
Existió estacionalidadde laproducciónde frutosconrespectoalrégimendeprecipitación,compor-tamientocontrarioalobservadoporBonilla&Zu-loaga(1994)enunbosquealtoandino;sinembargo,otrosestudiosllevadosacaboeneltrópico(Howe&Smallwood1982yWikander1984)yotrosrea-lizadosenbosquesaltoandinos(Montenegro2000,Morales 2002 y Sánchez 2003) concuerdan conla tendencia de estacionalidad de las fenofases.Laperiodicidaddefenofasesentreespeciesvaría;así,especiespertenecientesacomunidadesmadu-raspresentancicloscortosyespecíficosderepro-ducción.Lasespeciesquecorrespondenaestadossucesionales iniciales presentaron fenofases deproducciónmáslargas;estavariabilidadperiódicaesapoyadaporMarquis(1988),quienafirmaquegeneralmentelasespeciespertenecientesaestadossucesionales tempranos presentan ciclos fenofá-sicosmás extensos que las que se encuentran encomunidadesmaduras.
Incidencia de la avifauna en la dinámica de dispersión
Durantelosdocemesesdelañosecontóconregis-trosdehecesyvisitasentodoslosensayos(cercanía,medianía, lejanía).Este patrón permite identificarquelasavessonlasresponsablesdirectasdeléxitodel establecimiento de algunas especies de plan-tasemergentesenlosclaros,comolodescribenensusestudiosMartínez&Álvarez(1986)yMurray(1987).Paradeterminarlaefectividaddeladisper-siónadiferentesescalasydistanciasdelbosqueesprecisoevaluaraspectoscuantitativosrelacionadoscon las visitas y el número de semillas encontra-dos(Jordano&Schupp2000).Paraesteestudiosepresentaronlosmayoresregistrosentodoslosas-pectos (especiesde semillas, númerode semillas,númerodevisitasyespeciesdeaves)paralasper-chasqueseencontrabanenlacercaníadelbosque
y disminuyó gradualmente su dinámica amedidaqueaumentabaladistanciadelborde.Elcompor-tamientodeladinámicadelasperchasfrentealadistanciaconcuerdaconlopublicadoporHowe&Primack (1975), que sugiereque ladistancia a lacual son trasportadas las semillas depende de lospatronesdemovimientoyforrajerodelasaves,asícomoladistanciaydireccióndelosvuelosdependedeladistribucióndefrutos,semillasysusparentales(Howe&Primack1975,Herrera&Jordano1981);enotraspalabras,lasprobabilidadesdedispersiónporpartedelasavesaumentaproporcionalmentealadisponibilidadyproximidaddelrecursoqueporlogeneralseasociaconelbosque.Latotalidaddelasvariablespresentaronunaestrecharelaciónconelrégimendeprecipitación,mostrandounaestacio-nalidaddefinidaenlacuallospatronesfenológicosdelasplantastienenlugarcuandolascondicionesclimáticas son favorables para la polinización delasfloresydispersióndelosfrutosporpartedelosfrugívoros (van Schaik et al. 1993), razón por lacual la temporadademayorproducciónde frutos(julio),quecoincideconelpicodeprecipitación,seconviertetambiénenlaépocademayordispersióndesemillasyvariabilidaddeespeciesdispersorasydispersadas, reafirmándose losugeridoporPoulinet al.(1999)cuandoestablecequelasincroníaenlaproduccióndefrutosentreindividuosyespeciespermiteatraerenconjuntoaunmayornúmerodedispersorespotenciales.
Lamayordispersiónregionalocurrióenlaépocaseca,contrarioaloesperado,dadoqueesteperiodoeseldemenorofertadefrutosmaduros;larespues-ta ante esta tendencia es que durante la estaciónsecayante laescasezdeotrosrecursosalimenti-cioscomoinsectos,lasavestienenquecambiarlaconstituciónde sudietapasandoa ser frugívorostemporales(Janzen1983,Levey1988,Poulinet al. 1999,Sánchez2003).Asímismo,durantelaescasezdel recurso las aves tienenque recorrer distanciasmáslargasparaconseguirsusustento.Lamayoríade las especies y familias registradas en las per-chas pertenecen a estados de desarrollo inicial ointermedio,comoeselcasodeArcytophyllum sp., Phytolacca bogotensis, Rubus floribundum, Mico-nia squamulosa y Ocotea sp., entreotras;entrelasfamilias se registró a Melastomataceae,Rubiaceae,

64 Revista Colombia Forestal Vol. 11 - Diciembre 2008
CaracterizaciónyvaloracióndelpotencialderegeneracióndelbancodesemillasgerminabledelaReservaForestal...
Lauraceae y Rosaceae. Esta tendencia concuerdaconlaestrategiadecontinuidadenlafructificaciónquepresentanalgunasespeciesporlacompetenciaparaatraerelmayornúmerodedispersorespoten-ciales (McKey 1975), en la que existe una granofertadefrutospequeñosconsemillaspequeñasymaduraciónsincrónica(Morales2002).Estepatrónatraeadiversosfrugívorosgeneralistasquesecon-centranenaquellasespeciesdemayorescosechas(Murray1988),queresultanseraquellasasociadasalasetapasinícialesdelasucesión(Foster&Jan-son1985),comoocurreparaesteestudio.
CONCLUSIONES
Puedeafirmarsequelasparcelasrelacionadasconelborde,sobretodolaquecontieneunclaro,pre-sentanuncomportamientomásdinámico,definidopor la heterogeneidad en sus coberturas, la cualpermitemayordistribuciónyparticipacióndedife-rentesespecies,hábitosyestrategiasdedispersión,indicandoqueenlugaresconvegetaciónséricain-termedialaprobabilidaddeencontrarmásdiversi-dadynúmerodesemillasesmayor.
La relación entre la zoocoria y la anemocoria esinversamenteproporcionalencuantoasupartici-paciónpresencialenlaReservaForestalCárpatos;deestamanera,encomunidadesqueseencuentranenetapasinícialesdedesarrollosucesionallaane-mocoriajuegaunpapelimportanteyamedidaqueseavanzahacialascomunidadesmadurassupar-ticipaciónvadisminuyendogradualmente,permi-tiendoa lazoocoriaprimarcomolaestrategiadedispersiónmásimportanteparalareserva.
Ladeterminacióndelvolumenmínimoindicóquenoexisteunpatrónespecíficoparaloscomponen-tesque integranelbancodesemillasde losbos-quesaltoandinos.
El banco de semillas de la reserva presenta unatendenciaa serconstanteespacio-temporalmente,alposeerunabuenacantidaddesemillas latentesdistribuidasentodoslosestadosseralesyencual-quier época del año, independientemente de lasfenofasesenqueseencuentrenlasespeciesdelavegetaciónenpie.Deigualmanera,esimportante
resaltar la alta tasade renovaciónde semillas, alpresentarseelbancotransitorioenelsegundolugarderelevancia.
El bsgtuvomayordinámicaenregistrosdeabun-danciaanivelespacialenlospuntosdondelasco-berturasvegetalessepresentanmásheterogéneasyaniveltemporalenlosperiodosquecoincidenconlasépocasdemayorprecipitación,debidoaquelamayoríadesemillasyahansidodispersadasyseencuentranenelsuelodispuestasagerminar.
Los patrones de fructificación, frente al tipo dedispersión,presentaronestacionalidad respectoalrégimen de precipitación. Las especies zoócoraspresentaronunaestrecharelaciónconlastempora-dasdemayorhumedad,mientrasquelasespeciesanemócorasparecenasociarsealperiodoanterioralasépocadelluvias,cuandoelambientesetornamás seco y la brisa, aunque constante, es suave,casocontrarioalabarocoriaqueparecerespondermejoralaintensidaddelluviayviento.
Elensayode lasperchaspuedeconsiderarseexi-toso,dadoquedurantelosdocemesesdelañosecontó en mayor o menor medida con registros de hecesyvisitasentodoslosensayos(cercanía,me-dianíaylejaníadelbosque).
Contrario a lo esperado, elmayor alcance en tér-minos de dispersión (número de visitas y área dedispersión)sepresentódurante laépocademayorescasezderecursos;estatendenciaserelacionaconelhechodequealgunasespeciesdeavespresentanestrategiasalimenticiasestacionalesyporqueanteladisminucióndelaofertadefrutosparasusustentolasavesaumentanelradiodeinteracción,variandoladispersióndeunfenómenolocalaunoregional.
AGRADECIMIENTOS
Al centro de investigaciones de la UniversidadDistritalFranciscoJosédeCaldas,porsu respal-do económico. A Corpoguavio por aprobar estainvestigación en sus predios.A la familiaRodrí-guezOtálorapor su incondicional apoyodurantelafasedecampo.AJavierBurgosporsusvaliosasapreciaciones.

65Revista Colombia Forestal Vol. 11 - Diciembre 2008
Edgard Ernesto Cantillo Higuera / Viviana Castiblanco Gutiérrez / David Fernando Pinilla Mondragón / Claudia Liliana Alvarado
REFERENCIAS BIBLIOGRÁFICASAcosta, M. 2004.Efectosobreelbancodesemillas
germinableenunfragmentodebosquealtoan-dino(páramodeGuerrero).Trabajodegrado.UniversidadNacionaldeColombia.Pg.89.
Bertiller, B. 1991.SeasonalVariationintheSeedBankofaPatagonianGrasslandinRelationtoGrazing andTography. Journal ofVegetationScience 2:47-54.
Bonilla, A. & G. Zuloaga. 1994.Fenologíadeal-gunasespeciesdeplantasdelpáramo“ElGra-nizo”,Monserrate(Cundinamarca,Colombia).Academia Colombiana de Ciencias Exactas,Físicas yNaturales. Bogotá. Colección JorgeÁlvarezLleras6:485-501.
Brown, D. 1992.EstimatingtheCompositionofaForestSeedBank:aComparisonof theSeedExtractionandSeedlinkEmergenceMethods.CanadianJournalofBotany 70:1603-1612.
Cantillo, E., K. Rodríguez & A. Avella. 2004.Caracterizaciónflorística, estructural, diversi-dad y ordenación de la vegetación en la Reser-vaForestalCárpatos,Guasca,Cundinamarca.UniversidadDistrital Francisco JoséDeCal-das.Bogotá.Pg.150.
Carabias, J. & S. Guevara. 1985.Fenologíadeunaselvatropicalhúmedayenunacomunidadderivada:LosTuxtlas,Veracruz.En:Gómez-Pompa,A.&S.delAmo(eds.).Investigacio-nes sobre la regeneración de selvas altas enVeracruz,México.VolumeniiPágs.27-78.
Cardona, A. 2004.Potencialderegeneracióndelbancodesemillasgerminableendostiposdebosque subandino: Implicaciones para la res-tauración (Reserva Biológica Cachalú, Enci-no,Santander).Trabajodegrado.UniversidadDistritalfranciscoJosédeCaldas.Bogotá.
Conell, H. 1978.DiversityinTropicalRainForestsandCoralReefs.Science1999:1302-1310.
corpoguavio. 2000.PlandeRecuperaciónyMa-nejoForestal,predioCárpatos.Bogotá.
Croat, T. 1975. Phenological Behavior of HabitandHabitatClassesonBarroColoradoIsland. Biotropical 7:270-277.
Cuatrecasas, J. 1934.Observaciones geobotáni-casenColombia.TrabajosdelMuseoNacionaldeCienciasNaturales.Madrid.Seriebotánica27. Pg.143.
Fagua, C.2002.EstrategiadereproducciónsexualdeunapoblacióndeE. grandiflora en el Par-queNacionalNatural deChingaza.Universi-dadNacional deColombia.DepartamentodeBiología.Bogotá.
Falinska, K.1999.SeedBankDynamicsinAban-donedMeadowsduringaTwentyYearPeriodin the Bialowieza National Park. Journal ofEcology87:461-475.
Fenner, M. 1995.EcologyofSeedBanks.En:Ki-gel,J..SeedDevelomentandGermination.Inc.NewYork.Págs.507-527.
Fleming, T., R. Bridtwisch & G. Whitesides. 1987.PatternsofTropicalVertebrateFrugivoreDiversity.AnnualReviewofEcologyandSys-tematics18:91-108.
Fonseca, T. 2001. Dinámica de la dispersión desemillas por aves en un pastizal con perchasartificiales en comunidades de vegetación al-toandina.Tesisdepregrado.PontificiaUniver-sidadJaveriana.Bogotá.
Foster, S. & C. Janson. 1985.TheRelationshipsbet-weenSeedSizeandEstablishmentConditionsinTropicalWoodyPlants.Ecology66:773-780.
Garwood, N. 1989. Tropical Soil Seed Banks.EcologyofSoilSeedBank.AcademicPress.SanDiego,California.Págs.149-209.
Gibson, D. & K. Brown. 1991. The Effects ofGrazingonLocalColonizationandExtinctionduringEarlySuccession.JournalofVegetationScience2:291-300.
Grandin, U. & H. Rydin. 1998.AtributesoftheSeedBankafteraCenturyofPrimarySucces-sion on Islands in Lake Hjälmaren, Sweden.JournalofEcology86:293-333.
Grime, J. 1989.SeedBankinEcologicalPerspecti-ve.En:Leck,M.A.,V.T.ParkeryR.L.Simpson(eds.). Ecology of Soil Seed Bank.AcademicPress.SanDiego,California.Págs.15-27.

66 Revista Colombia Forestal Vol. 11 - Diciembre 2008
CaracterizaciónyvaloracióndelpotencialderegeneracióndelbancodesemillasgerminabledelaReservaForestal...
Guevara, S. & A. Gómez-Pompa. 1972. SeedfromSurfaceSoilsinaTropicalRegionofVe-racruz,Mexico.JournaloftheArnoldArbore-tum53(3):312-335.
Herrera, C. & P. Jordano. 1981.Prunus mahaleb andBirds:TheHighEfficiency in theSystemof a Temperate Fruiting Tree. EcologicalMonographs51:203-218.
Herrera, R., D. Ulloa, O. Valdés–Lafont,A. Prie-go, A., & Valdés, A. 1997. Ecotechnologies for the SustainableManagement of Tropical Di-versity.Naturey Resources33:2-7.
Hilty, S. 1980.FloweringandFruitingPeriodicityinaPremontaneRainforestinPacificColom-bia.Biotropical 12:292-306.
Howe, H. & R. Primack.1975. DifferentialSeedDispersalbyBirdsoftheTreeCasearia nitida (Flacurtaceae).Biotropical7(4):278-283.
Howe, H. & J. Smallwood. 1982.EcologyofSeedDispersal.AnnualReviewofEcologyandSys-tematics13:201-208.
Jaimes, V. & D. Rivera. 1990.Bancodesemillasy tendencias en la regeneración natural de un bosque altoandino en la región deMonserrate(Cundinamarca, Colombia).Trabajo de grado.FacultaddeBiología.UniversidadNacionaldeColombia.Bogotá.Pg.56.
Janzen, D. 1983.Birds.En:Janzen,D.(ed).Cos-taRicaNaturalHistory.UniversityofChicagoPress.Págs.502-549.
Jordano, P. & E. Schupp. 2000.SeedDispersalEffectiveness: The Quantity Component andPatterns of Seed Rain for Prunus mahaleb.EcologicalMonographs70:591-615.
Levey, D. 1988.SpatialandTemporalVariationinCostaRicanFruitandFruit-EatingBirdAbun-dance.EcologicalMonographs58:251-269.
Magurran, A. E. 1989.Diversidadecológicaysumedición.EdicionesVedra.Barcelona.Pg.200.
Mahecha. G. 2004. La vegetación del territoriocar.car(Cundinamarca).Bogotá.Pg.620.
Marquis, R. 1988. PhenologicalVariation in theNeotropicalUnderstoryShrubPiper arienum:
CausesandConsequences.Ecology69:1552-1565.
Martínez-Ramos, N. & E. Álvarez-Buylla. 1986.SeedDispersal,GapDynamics andTreeRe-cruitment: the Case of Cecropia obtusifoliaatLosTuxtlas,Mexico.En:Estrada,A.&H.Fleming.FrugivoryandSeedDispersal. JunkPublishers.LaHaya.Págs.333-346.
Mc Graw, J. & M. Varek.1989.TheRoleofBu-riedViableSeedsinArticandAlpinCommu-nities.En:Leck,M.,V.ParkeryL.Simpson(eds.).EcologyofSoilSeedBank.AcademicPress,SanDiego,California.Págs.91-105.
Mckey, D. 1975.TheEcologyofCoevolvedSeedDispersalSystems.En:Gilbert,L.&P.Raven(eds.). Coevolution of Animals and Plants. UniversityofTexasPress.Págs.159-191.
Montenegro, A. 2000.Estrategiasdedispersiónyregeneraciónporbancodesemillasendosco-munidadesdebosquealtoandinoenColombia.Trabajodegrado.FacultaddeBiología.Uni-versidadNacional.Bogotá.Pg.132.
Morales, N. 2002. Ecología de la dispersión desemillasporavesdeBrunellia colombiana en unbosquealtoandino(ParqueNacionalNatu-ralChingaza).Tesis.UniversidadNacionaldeColombia.Pg.85.
Mori, S. & J. Kallunki. 1976.PhenologyandFloralBiologyofGustavia superba(Lecythidaceae) in CentralPanama.Biotropical 8:184-192.
Murray, G. 1987. Selection for Optimal FruitCropSizeinBird-DispersedPlants.AmericanNaturalist129:18-31.
Newstrom, L., G. Frankie, H. Baker y R. Collwell. 1994. Diversity ofLong-termFloweringPat-terns.En:McDade,K.,H.Bawa,H.Hespen-heide & G. Hartshorn (eds.). Ecology andNatural History of aNeotropical Rainforest..Págs.142-160.
Olaya, A. 2000. Efecto de borde sobre el bancodesemillasgerminabledelbosquealtoandino(EmbalsedeSanRafael,LaCalera,Cundina-marca).Tesisdegrado.PontificiaUniversidadJaveriana.Bogotá.

67Revista Colombia Forestal Vol. 11 - Diciembre 2008
Edgard Ernesto Cantillo Higuera / Viviana Castiblanco Gutiérrez / David Fernando Pinilla Mondragón / Claudia Liliana Alvarado
Pedraza, L., 2000.Las plantas conflores de losalrededoresdelalagunadeChisacá(Sumapaz,Colombia).TrabajodegradoenBiología.Uni-versidad Nacional de Colombia. Facultad deCiencias.Bogotá.Pg.237.
Piquett, S. & M. McDonnell. 1989. SeedBankDynamics in Temperate Deciduous Forests.En:Allessio,M.,V. ParkerV&R. Simpson(eds.).EcologyofSoilSeedBanks.AcademicPress.SanDiego,California.Págs.123-146.
Posada, C. & C. Cárdenas. 1999.Ecologíadelosbancos de semillas en una comunidad vegetaldepáramo sometida adisturbiosporquemaypastoreo (ParqueNacionalNatural Chingaza).Trabajodegrado.UniversidadNacionaldeCo-lombia.FacultaddeBiología.Bogotá.Pg.190.
Poullin, B., J. Wright, G. Lefebvre & O. Cal-derón. 1999. Interspecific Synchrony andAsynchrony in the Fruiting Phenologies ofCongerericBird-DispersedPlants inPanama.JournalofTropicalEcology15:213-227.
Rice, J. 1989.ImpactsofSeedBanksonGrasslandCommunity Structure and Population Dyna-mics. En: Leck,M.,V. Parker yR. Simpson(eds.).EcologyofSoilSeedBank. Academic Press.SanDiego,California.Pg.211-230.
Rico-Gray, V. & J. García-Franco. 1992.Vegeta-tionandSoilSeedBankofSuccessionalStagesinTropicalLowlandDeciduousForests.Jour-nalofVegetationScience3:617-624.
Rossi, R, P. Lavelle & E. Tondof. 1995.StatisticSoil Biology. GeostaticalAnalysis. EuropeanJournalofSoilBiology31(4):173-181.
Saldarriaga, J. G. 1994.RecuperacióndelaselvadetierrafirmeenelaltoríoNegro,AmazoniaColombiana-Venezolana.Tropenbos..Pg.201
Sánchez, M., 2003.Fenologíareproductivaydis-persióndesemillasdelarbustoaltoandinoMo-nina salicifolia. Tesis de grado. UniversidadNacionaldeColombia.Bogotá.Pg.65.
Schupp, E., H. Howe, C. Augspurger & D. Le-vey. 1989. Arrival and Survival in TropicalTreefallGaps.Ecology70(3):562-564.
Society for Ecological Restoration (ser). 2004.ser International Primer on Ecological Res-toration. Society for Ecological Restoration[disponible en: http://www.ser.org/content/ecological_restoration_primer.asp].
Simpson, R., M. Allessio & V. Parker. 1989.SeedBank: General Concepts andMethodologicalIssues.En:Leck,M.,V.ParkeryR.Simpson(eds.).EcologyofSoilSeedBank. Academic Press.SanDiego,California.Págs.3-8.
Smythe, N. 1970. Relationships between FruitingSeasonsandSeedDispersalMethodsinaNeo-tropicalForest.AmericanNaturalist104:25-35.
Sorzano, C. 2001. RegeneraciónreproductivaenunsitioabandonadoeinvadidoporPteridium aquilinumKhun.Implicacionesparalarestau-ración. Reserva Biológica Cachalú. Encino,Santander,Colombia.Trabajodegrado.Ponti-ficiaUniversidadJaveriana.FacultaddeCien-cias.DepartamentodeBiología.Bogotá.
Swaing, D. & J. Hall. 1983.EarlySuccessiononClearedForestLandinGhana.JournalofEco-logy 71:601-627.
Thompson, K., J. Bakker & R. Bekker. 1998.Ecological Correlates of Seed Persistence inSoilintheNorth-WestEuropeanFlora.JournalofEcology86:163-169.
van Schaik, C. 1993.The Phenology ofTropicalForest: Adaptative, Significance and Conse-quences for Primary Consumers. Annual Re-viewofEcologyandSystematics24:353-377.
Vargas, O.1998.Unmodelodesucesión/regene-racióndelospáramosdespuésdelasquemas.Caldasia19(1;2):331-345.
Wikander, T. 1984.MecanismosdedispersióndediásporasdeunaselvadeciduaenVenezuela.Biotropical 16(4):276-283.
Williams-Linera, G. & y J. Meave. 2002. Pa-trones fenológicos. En: Guariguata, M. y G.Kattan(eds.).Ecologíayconservacióndebos-ques neotropicales. Libro universitario regio-naldeCostaRica.Págs.407-431.

68 Revista Colombia Forestal Vol. 11 - Diciembre 2008
CaracterizaciónyvaloracióndelpotencialderegeneracióndelbancodesemillasgerminabledelaReservaForestal...
Ane
xo 1
. Registrogeneraldelasespeciesp
aralavegetaciónenpie.
ZO
OC
OR
IA
AN
EM
OC
OR
IA
BA
RO
CO
RIA
BO
SQ
UE
CL
IMA
XB
OR
DE
DE
BO
SQ
UE
B
OR
DE
/ C
LA
RO
Agera
tina
Agera
tina s
p.
CO
MP
OS
ITA
E
Aguaca
tillo
Pers
ea m
utis
iLA
UR
AC
EA
EA
jicill
oD
rym
is g
ranadensi
s W
INT
ER
AC
EA
EA
marg
oso
Buddle
ja b
ulla
taB
UD
DLE
JAC
EA
EA
marillo
Nect
andra
molli
sLA
UR
AC
EA
EA
rraya
nM
yrci
ante
s le
oco
xyla
MY
RT
AC
EA
EB
eju
coA
iouea d
ubia
LA
UR
AC
EA
EC
aca
oC
leth
ra la
nnata
CLE
TH
RA
CE
AE
Cansa
bra
zoA
rcyt
ophyl
lum
sp.
RU
BIA
CE
AE
Cedrillo
Bru
nelli
a in
tegrifo
liaB
RU
NE
LLIA
CE
AE
Chilc
oB
ach
aris
bogote
nsi
sC
OM
PO
SIT
AE
Chite
Hyp
ericu
m ju
nip
ericu
mH
YP
ER
ICA
CE
AE
Chucu
aV
iburn
um
triphyl
lum
CA
PR
IFO
LIA
CE
AE
Chusq
ue
Chusq
uea s
candens
PO
AC
EA
EC
ilantr
on
Dilo
dendru
m s
p.
SA
PIN
DA
CE
AE
Co
jinP
aepala
nth
us
sp.
ER
IOC
AU
LA
CE
AE
Cord
onci
lloP
iper
bogote
nsi
sP
IPE
RA
CE
AE
Cort
adera
Care
x alb
olu
tensc
ens
CY
PE
RA
CE
AE
Cuch
aro
Myr
sine g
uia
nensi
sM
YR
SIN
AC
EA
EC
ucu
bo
Cest
rum
mutis
iS
OLA
NA
CE
AE
Cura
pill
oE
ndlic
heria s
p.
LA
UR
AC
EA
ED
igita
lD
igita
lis p
urp
ure
aS
CR
OF
ULA
RIA
CE
AE
Dora
dill
aT
ibuch
ina g
ross
aM
ELA
ST
OM
AT
AC
EA
EE
nce
nill
aW
ein
mannia
balb
issi
ana
CU
NO
NIA
CE
AE
Ence
nill
o a
caci
oW
ein
mannia
pin
nata
CU
NO
NIA
CE
AE
Est
ora
que
Beils
chm
iedia
pendula
LA
UR
AC
EA
EG
alli
nazo
Nect
andra
sp.
LA
UR
AC
EA
EG
am
uzo
Oco
tea c
allo
phyl
laLA
UR
AC
EA
EG
aque
Clu
sia m
ulti
flora
CLU
SIA
CE
AE
Gra
niz
oH
edyo
smum
bom
pla
ndia
num
CH
LO
RA
NT
HA
CE
AE
Guaba m
ora
da
Phyt
ola
cca b
ogote
nsi
sP
HY
TO
LA
CC
AC
EA
EG
uasg
uin
Penta
calia
pulc
hella
CO
MP
OS
ITA
EH
ele
cha
Cya
thea s
pC
YA
TH
EA
CE
AE
Hele
cho
Pte
ridiu
m s
p.
HIP
OLE
PID
AC
EA
EH
ele
cho m
ach
oD
ryppte
ris
sp.
AS
PID
IAC
EA
EH
ueso
Geis
santh
us
andin
us
MY
RS
INA
CE
AE
Junco
Juncu
s bru
neus
JUN
CA
CE
AE
Mano d
e o
soO
reopanax
floribundum
AR
ALIA
CE
AE
Moquill
oS
aura
uia
urs
ina
AC
TIN
IDIA
CE
AE
Mora
Rubus
floribundum
R
OS
AC
EA
EO
rquín
Clu
sia d
ucu
CLU
SIA
CE
AE
Palm
iche
Geonom
a s
p.
AR
EC
AC
EA
EP
ast
o
Cam
ala
gro
stis
efu
saP
OA
CE
AE
Past
o p
aja
Poa s
p.
PO
AC
EA
EP
ate
galli
na
Ruagea g
labra
ME
LIA
CE
AE
Raya
do b
lanco
Ilex
danie
lisA
QU
IFO
LLIA
CE
AE
Raya
do c
olo
rado
Stir
ax
tom
ento
saS
TY
RA
CE
AE
Sanalo
todo
Bacc
haris
tric
uneata
CO
MP
OS
ITA
ET
ach
uelo
Sola
num
inopin
um
SO
LA
NA
CE
AE
Tin
tillo
Palic
oure
a o
blo
nga
RU
BIA
CE
AE
Tin
toP
alic
oure
a li
neariflo
raR
UB
IAC
EA
ET
ote
Kyl
lingia
sp.
CY
PE
RA
CE
AE
Tre
mentin
o
Myr
sine c
oriace
ae
MY
RS
INA
CE
AE
Tunill
aM
iconia
dentic
ula
taM
ELA
ST
OM
AT
AC
EA
ET
uno c
hill
on
Axi
naea m
acr
ophyl
laM
ELA
ST
OM
AT
AC
EA
ET
uno e
smera
ldo
Mic
onia
squam
ulo
saM
ELA
ST
OM
AT
AC
EA
ET
uno r
ojo
Mic
onia
ferr
ugín
ea
ME
LA
ST
OM
AT
AC
EA
ET
uno r
oso
Centr
onia
colo
mbia
na
ME
LA
ST
OM
AT
AC
EA
EU
va c
am
aro
na
Macl
eania
rupest
ris
ER
ICA
CE
AE
Uva
de a
nis
Cave
ndis
hia
cord
ifolia
ER
ICA
CE
AE
Vara
bla
nca
Aegifi
la b
ogote
nsi
sV
ER
BE
NA
CE
AE
42
18
135
34
41
68,8
5245902
29,5
0819672
1,6
39344262
57,3
7704918
55,7
3770492
67,2
1311475
PO
RC
EN
TA
JE P
AR
A C
AD
A U
NA
DE
LA
S V
AR
IAB
LE
S E
VA
LU
AD
AS
PA
RC
EL
A E
N Q
UE
SE
EN
CO
NT
RO
TO
TA
L E
SP
EC
IES
61
ES
TR
AT
EG
IA D
E D
ISP
ER
SIÓ
N
NO
MB
RE
CIE
NT
IFIC
OF
AM
ILIA
N
OM
BR
E C
OM
UN

69Revista Colombia Forestal Vol. 11 - Diciembre 2008
Edgard Ernesto Cantillo Higuera / Viviana Castiblanco Gutiérrez / David Fernando Pinilla Mondragón / Claudia Liliana Alvarado
Ane
xo 2
. Registrogeneraldelasespeciesp
araelb
sg.
ZO
OC
OR
IA
AN
EM
OC
OR
IA
BO
SQ
UE
CL
IMA
XB
OR
DE
DE
BO
SQ
UE
B
OR
DE
/ C
LA
RO
Age
ratin
aA
ge
ratin
a s
p.
CO
MP
OS
ITA
E
Am
arg
oso
Bu
dd
leja
bu
llata
BU
DD
LE
JA
CE
AE
Ca
nsa
bra
zo
Arc
yto
ph
yllu
m s
p.
RU
BIA
CE
AE
Ca
rriz
oB
ulb
osty
lis s
p.
CY
PE
RA
CE
AE
Ch
ilco
Ba
cch
aris b
og
ote
nsis
CO
MP
OS
ITA
EC
hite
Hyp
ericu
m ju
nip
ericu
mH
YP
ER
ICA
CE
AE
Ch
ucu
aV
ibu
rnu
m t
rip
hyllu
mC
AP
RIF
OL
IAC
EA
EC
iro
Ba
cch
aris s
p.
CO
MP
OS
ITA
EC
ojin
Pa
ep
ala
nth
us s
p.
ER
IOC
AU
LA
CE
AE
Co
jinP
ae
pa
lan
thu
s k
ars
ten
iiE
RIO
CA
UL
AC
EA
EC
ort
ad
era
Ca
rex a
lbo
lute
nsce
ns
CY
PE
RA
CE
AE
Cu
cu
bo
Ce
str
um
mu
tisi
SO
LA
NA
CE
AE
Cu
rap
illo
En
dlic
he
ria
sp
.L
AU
RA
CE
AE
De
dil
La
esta
dia
sp
.C
OM
PO
SIT
AE
Dig
ita
lD
igita
lis p
urp
ure
aE
SC
RO
FU
LA
RIA
CE
AE
Do
rad
illa
Tib
uch
ina
gro
ssa
ME
LA
ST
OM
AT
AC
EA
EE
ncenill
aW
ein
mannia
balb
issia
na
CU
NO
NIA
CE
AE
Ga
lium
G
aliu
m s
pR
UB
IAC
EA
EG
am
uzo
Oco
tea
ca
llop
hylla
LA
UR
AC
EA
EG
era
nio
Ge
ran
ium
sp
.G
ER
AN
IAC
EA
EG
ua
ba
mo
rad
aP
hyto
lacca
bo
go
ten
sis
PH
YT
OL
AC
CA
CE
AE
Gu
asgu
inP
en
taca
lia p
ulc
he
llaC
OM
PO
SIT
AE
Ind
ete
rmin
ad
a 1
Sp
1C
OM
PO
SIT
AE
Ind
ete
rmin
ad
a 2
Sp
2S
OL
AN
AC
EA
EIn
de
term
ina
da
3S
p 3
RU
BIA
CE
AE
Ind
ete
rmin
ad
a 4
Sp
4R
OS
AC
EA
EIn
de
term
ina
da
5S
p 5
RU
BIA
CE
AE
Ind
ete
rmin
ad
a 6
Sp
6C
OM
PO
SIT
AE
Ind
ete
rmin
ad
a 7
Sp
7C
OM
PO
SIT
AE
Ind
ete
rmin
ad
a 8
Sp
8C
YP
ER
AC
EA
EJu
nco
Ju
ncu
s b
run
eu
sJU
NC
AC
EA
EM
ad
re d
e a
gu
ah
yd
roco
tyle
bo
np
lan
dii
UM
BE
LIF
ER
AE
Mo
raR
ub
us f
lorib
un
du
m
RO
SA
CE
AE
Orq
uín
Clu
sia
du
cu
CL
US
IAC
EA
EP
asto
p
oa
an
nu
aP
OA
CE
AE
Pa
sto
C
am
ala
gro
stis e
fusa
PO
AC
EA
EP
asto
ga
llin
a
Ag
rostis s
p.
PO
AC
EA
EP
asto
pa
jaP
oa
sp
.P
OA
CE
AE
Pa
tega
llin
aR
ua
ge
a g
lab
raM
EL
IAC
EA
ER
an
un
cu
loR
an
un
cu
lus s
p.
RA
NU
NC
UL
AC
EA
ER
aya
do
co
lora
do
Stira
x t
om
en
tosa
ST
YR
AC
EA
ER
um
ex
Ru
me
x a
ce
tose
llaP
OL
YG
ON
AC
EA
ES
an
alo
tod
oB
acch
aris t
ricu
ne
ata
CO
MP
OS
ITA
ET
ach
ue
loS
ola
nu
m in
op
inu
mS
OL
AN
AC
EA
ET
intillo
Pa
lico
ure
a o
blo
ng
aR
UB
IAC
EA
ET
ote
Kylli
ng
ia s
p.
CY
PE
RA
CE
AE
Tre
me
ntin
o
Myrs
ine
co
ria
ce
ae
MY
RS
INA
CE
AE
Tu
nill
aM
ico
nia
de
nticu
lata
ME
LA
ST
OM
AT
AC
EA
ET
un
o c
hill
on
Axin
ae
a m
acro
ph
ylla
ME
LA
ST
OM
AT
AC
EA
ET
un
o e
sm
era
ldo
Mic
on
ia s
qu
am
ulo
sa
ME
LA
ST
OM
AT
AC
EA
ET
un
o r
oso
Ce
ntr
on
ia c
olo
mb
ian
aM
EL
AS
TO
MA
TA
CE
AE
Uvo
Ca
ve
nd
ish
ia s
p.
ER
ICA
CE
AE
Va
leria
na
Va
leria
na
sp
.V
AL
ER
AN
IAC
EA
EV
ara
bla
nca
Ae
gifila
bo
go
ten
sis
VE
RB
EN
AC
EA
EV
ira
Bo
rre
ria
sp
.R
UB
IAC
EA
E30
25
28
43
47
54,5
4545455
45,4
5454545
50,9
0909091
78,1
8181818
85,4
5454545
PA
RC
EL
A E
N Q
UE
SE
EN
CO
NT
RO
TO
TA
LE
S5
5P
RO
ME
DIO
NO
MB
RE
CO
MU
N
NO
MB
RE
CIE
NT
IFIC
OF
AM
ILIA
E
ST
RA
TE
GIA
DE
DIS
PE
RS
IÓN

70 Revista Colombia Forestal Vol. 11 - Diciembre 2008
CaracterizaciónyvaloracióndelpotencialderegeneracióndelbancodesemillasgerminabledelaReservaForestal...
Anexo 3. CuadroresumendelosíndicesdeMargalefyShannonparabsg.
Índices de Margalef y Shanon por parcela
Índice Bosque clímax Borde de bosque Borde/Claro
Margalef 0,8679 1,2425 1,2493
Shannon 0,4618 1,2707 1,47
Índices de Margalef y Shanon por muestreo
Índice Muestreo I Muestreo II Muestreo III
Margalef 4,8295 5,4602 5,8207
Shannon 1,7986 1,7362 2,0548
Anexo 4. Relaciónmensualdelafloraciónyfructificaciónrespectoalrégimendeprecipitaciónyvientos.
Mes No. de flores No. de frutos Precipitación Viento
Enero 362 119 50 2,1
Febrero 358 131 70 2,1
Marzo 264 127 105 2,3
Abril 276 146 180 2
Mayo 223 192 240 2,2
Junio 156 244 225 2,3
Julio 112 251 270 2,6
Agosto 176 226 190 2,1
Septiembre 201 215 105 2,2
Octubre 169 238 115 2
Noviembre 208 178 105 2
Diciembre 258 103 60 2,1
Promedio 230,25 180,8333333 1715 2,1
Anexo 5. Especiesobservadasenlasperchasartificiales.
Nombre común Orden Familia Nombre científico
Vencejo Cypseloides rutilos APODIDAE APODIFORMES
Gorrión Zonotrichia capensis FRINGILLIDAE PASSERIFORMES
Gorrión del bosque Atlapetes pallidinucha FRINGILLIDAE PASSERIFORMES
Chamicero, fringilino Atlapetes brunneinucha FRINGILLIDAE PASSERIFORMES
Gorrión chamicero Atlapetes schistaceus FRINGILLIDAE PASSERIFORMES
Hornero, funarido Margarornis squamiger FURNARIIDAE PASSERIFORMES
Tordo, toche Icterus chrysater giraudii ICTERIDAE PASSERIFORMES
Tangará Hemispingus melanotis THRAUPIDAE PASSERIFORMES
Tangará Iridosornis rufivertex THRAUPIDAE PASSERIFORMES
Atrapamoscas Ochthoeca fumicolor TYRANNIDAE PASSERIFORMES
Atrapamoscas Mecocerculus leucophrys TYRANNIDAE PASSERIFORMES
Mirla negra Turdus fuscater TYRANNIDAE PASSERIFORMES


















