Células y Tejidos Inmunitario
38
CELULAS Y TEJIDOS DEL SISTEMA INMUNITARIO El Sistema inmune está conformado por una serie de órganos, tejidos y células esparcidas de manera amplia por todo el cuerpo. Desde el punto de vista de sus características estructurales podemos encontrar órganos macizos como el timo, el bazo y los ganglios linfáticos y estructuras tubulares como los vasos linfáticos que se encuentra intercomunicando algunos de los órganos mencionados anteriormente. Si se toma en cuenta las funciones que realizan, entonces se pueden clasificar dichos órganos en primarios y secundarios. En los primeros tienen lugar la generación de las células que conforman al sistema inmune (linfopoyesis) y además existe un microambiente idóneo de modo que los linfocitos adquieren su repertorio de receptores específicos para cada tipo de antígeno. Mientras que los segundos se encargan de hospedar las células capacitadas funcionalmente para interactuar con microorganismo o antígeno, atrapados por estos órganos, en un entorno adecuado para que las mismas interactúen con dichos agentes extraños al organismo y los eliminen. Como ya habíamos dicho anteriormente, estos órganos están interconectados por vasos sanguíneos y vasos linfáticos, de forma tal que se constituye un sistema unitario, entrelazado y bien comunicado. Estos vasos transportan las células del sistema inmune, de las cuales el actor principal es el linfocito 1.Fagocitos: - Origen: Su origen se encuentra en su antecesor ontogenético que es la célula pruripotencial mielo-monocitica (CFU- GM) que se diferencia en dos líneas
-
Upload
jade-rosangel-sb -
Category
Documents
-
view
34 -
download
0
Transcript of Células y Tejidos Inmunitario

CELULAS Y TEJIDOS DEL SISTEMA INMUNITARIO
El Sistema inmune está conformado por una serie de órganos, tejidos y células esparcidas de manera amplia por todo el cuerpo. Desde el punto de vista de sus características estructurales podemos encontrar órganos macizos como el timo, el bazo y los ganglios linfáticos y estructuras tubulares como los vasos linfáticos que se encuentra intercomunicando algunos de los órganos mencionados anteriormente. Si se toma en cuenta las funciones que realizan, entonces se pueden clasificar dichos órganos en primarios y secundarios. En los primeros tienen lugar la generación de las células que conforman al sistema inmune (linfopoyesis) y además existe un microambiente idóneo de modo que los linfocitos adquieren su repertorio de receptores específicos para cada tipo de antígeno. Mientras que los segundos se encargan de hospedar las células capacitadas funcionalmente para interactuar con microorganismo o antígeno, atrapados por estos órganos, en un entorno adecuado para que las mismas interactúen con dichos agentes extraños al organismo y los eliminen. Como ya habíamos dicho anteriormente, estos órganos están interconectados por vasos sanguíneos y vasos linfáticos, de forma tal que se constituye un sistema unitario, entrelazado y bien comunicado. Estos vasos transportan las células del sistema inmune, de las cuales el actor principal es el linfocito
1.Fagocitos:
- Origen: Su origen se encuentra en su antecesor ontogenético que es la célula pruripotencial mielo-monocitica (CFU-GM) que se diferencia en dos líneas
- Localización: Son células presentes en la sangre y otros tejidos animales.
- Estructura :
- Función: La función de los fagocitos es de captar microorganismos y restos celulares (incluye particular inútiles y dañinas para el organismo) e introducirlos en su interior con el fin de eliminarlos.
- Otras: Existen muchos tipos de células capaces de efectuar la fagocitosis; las células del sistema inmune que la realizan son de vital importancia en la defensa del organismo contra las infecciones

2.Neutrofilos:
- Origen:
- Localización:
Los neutrifilos normalmente se encuentran en el torrente sanguíneo
- Estructura: Su estructura consta de un núcleo lobulado y gran cantidad de gránulos y lisosomas en su citoplasma con diferentes contenido.
- Función: Su función principal es fagocitar y destruir bacterias, hongos y participar en el inicio del proceso inflamatorio.
- Otros: Se llaman neutrófilos porque no se tiñen con colorantes ácidos ni básicos, por lo que su citoplasma se observa rosa suave.
3.FAGOCITOS MONONUCLEARES
EN INUNIDAD INNATA:
Tienen como función fagocitar los microbios y producir citosinas que atraviesan otras células inflamatorias y las activen.
Tienen su origen en la medula ósea. Circundan por la sangre, maduran en los tejidos para así quedar activadas.
El primer tipo de célula que se forma es el mastocito (no está diferenciado por completo). Tiene un diámetro de 10-15 um.
Tiene núcleos reniformes. Tiene un citoplasma granular abundante, en el que se encuentran lisosomas, vacuolas fagocíticas y un citoesqueleto de filamentos.
Después de entrar a los tejidos esos se van a diferenciar en macrófagos.estos últimos tiene un citoplasma abundante (células epiteliales)

Los macrófagos se reúnen entre si y forman células gigantes mononucleadas, como pro ejemplo:
SNC MICROGLIOCITOS.HIGADO CELULAS DE KUPFFERVIAS RESPIRATORIAS PULMONARES,MACROFAGOS ALVEOLRES, FAGOCITOS MULTINUCLEARES OSTEOBLASTOS
Desde el punto de vista filogénico son los mediadores más antiguos.
Responden a microbios casi con tanta velocidad como los neutrófilos pero su supervivencia es mayor.
Los macrófagos no están diferenciados por completo, y pueden sufrir división en el foco inflamatorio.
Son células efectoras dominantes durante las últimas etapas de respuesta inmunitaria innata, 1 ó 2 días después de la infección.
4.Mastocitos
-Participan en las respuestas inmunitarias innatas y adaptativas
-Presentan gránulos citoplasmáticos con mediadores inflamatorios y antimicrobianos
-Protegen contra helmintos y causantes de enfermedades alérgicas
• Origen:
-Derivan de la médula ósea
-Residen en la mayoría de tejidos adyacentes a los vasos sanguíneos y nervios, y bajo epitelios de órganos linfáticos
-Expresan un receptor del Fc con una alta afinidad der la IgE
• Estructura:
-Forma variable
-Núcleos redondeados
-Citoplasma rico en gránulos rodeados de membrana, que contienen proteoglicanos y cuerpos lipídicos
-Los proteoglicanos contienen ácidos

5. BASOFILOS
Los basófilos se originan en la médula ósea, en los leucoblastos que al diferenciarse dan origen a los leucocitos entre los que se encuentran los basófilos .Conforman el tipo de leucocito menos abundante en sangre. Tiene núcleo irregular, difícil de ver por la granulación basófila que lo cubre casi siempre. Tamaño semejante al de los segmentados.
Los basófilos se encuentran generalmente en áreas tales como los pulmones y el hígado, donde hay un gran volumen de sangre, y es posible que la heparina liberada ayude a prevenir la formación de coágulos sanguíneos diminutos de la formación.
Los basófilos son los responsables del inicio de la respuesta alérgica.
Los gránulos de los basófilos son gruesos pero escasos. Son células de
unos 10 μm de diámetro y su núcleo tiene una forma que recuerda a una
S, su núcleo es globulado y se divide de 1 a 3 células.
Tienen una activa participación en la respuesta inmunitaria, a través de la
liberación de histamina, serotonina en bajas concentraciones, y otras
sustancias químicas.
Tiene gránulos de dos clases:
Gránulos azurófilos: Contienen lisosomas, que a su vez estos
contienen hidrolasas ácidas.
Gránulosespecíficos o secundarios: contienen histamina , heparin
ayleuco-trienos
Los basófilos además de poseer gránulos en su interior, poseen
receptores de IgE (inmunoglobulina E),
aquella inmunoglobulina relacionada con las alergias. Es por eso que el
basófilo participa en la respuesta inflamatoria

6. EOSINOFILO
Un eosinófilo es un leucocito de tipo granulocito pequeño derivado de la médula ósea, que tiene una vida media en la circulación sanguínea de 3 a 4 días antes de migrar a los tejidos en donde permanecen durante varios días. Su desarrollo en la médula ósea es estimulado por diversasinterleucinas, como la IL-5, la IL-3 y el factor estimulante de colonias granulocito-macrófago. Es característico su núcleo bilobulado, al igual que sus distintivos gránulos citoplásmicos. Estas proteínas granulares son responsables de muchas funciones proinflamatorias, principalmente en lapatogénesis de las enfermedades alérgicas,como célula efectora de hipersensibilidad inmediata, así como en la muerte de parásitos.
Una de las enzimas más importantes que contienen sus gránulos es la histaminasa, que se encarga de hidrolizar la histamina, regulando así la respuesta alérgica.
Los eosinófilos poseen movimiento ameboide y tienen débil actividad fagocítica. Su tamaño es semejante al del neutrófilo (10-12 μm). Constan de un núcleo bilobulado y su citoplasma está casi completamente ocupado por unos 20 gránulos acidófilos, pues se tiñen de color naranja o marrón anaranjado con las coloraciones panópticas. Los gránulos están formados por un núcleo electrodenso rodeado por una matriz electrotransparente, y contienen cuatro clases principales de proteínas: proteína básica mayor (MBP), proteína catiónica del eosinófilo (ECP), peroxidasa del eosinófilo (EPO) y neurotoxina derivada del eosinófilo (EDN). Los eosinófilos son capaces de sintetizar de nuevo otros productos, como mediadores lipídicos (PAF, LTC4), citocinas (IL-3, IL-5, GM-CSF), quimiocinas (eotaxina) y óxido nítrico (NO).
Los eosinófilos interaccionan con otras células por la expresión de múltiples receptores en su superficie. Además, son células fagocitarias que demuestran especial afinidad por los complejo santígeno-anticuerpo, por lo que la mayoría de los eosinófilos son atraídos por quimiotaxis. También los eosinófilos pueden ser atraídos por sustancias liberadas de los basófilos, como la histamina.

Los eosinófilos pueden regular la respuesta alérgica y las reacciones de hipersensibilidad mediante la neutralización de la histamina por la histaminasa, y a su vez producir un factor inhibidor derivado de los eosinófilos para inhibir la desgranulación de las células cebadas o de los basófilos, que contienen sustancias vasoactivas.
Los eosinófilos juegan un papel de defensa del huésped frente a microorganismos no fagocitables, poseen una función citotóxica (por sus proteínas granulares), inmunoreguladora(por las citocinas que libera) y son capaces de participar en la reparación y remodelación tisular (liberando TGF-β)
7. Células presentadoras de Ag
Células dendríticas
Origen: Las células dendríticas se originan a partir de células de la médula ósea. Existen evidencias experimentales que avalan varios orígenes posibles de estas células:
a. Un progenitor común con las células mieloides CD34+13, que bajo el estímulo de determinadas citoquinas puede generar dos tipos de poblaciones precursoras, CD1a y CD14, las cuales maduran a células dendríticas de distintas características. La primera dará origen a células con gránulos de Birbeck, y los antígenos Lag y E-catherina, marcadores típicos de las células de Langerhans y las segundas podrán diferenciarse a macrófagos o a células dendríticas carentes de gránulos y de los mencionados antígenos y que, por el contrario expresan CD2, CD9 y CD68.
b. El monocito sanguíneo bajo el estímulo de GM-CSF e IL-4 puede dar origen a células dendríticas inmaduras.
Localización: Se encuentran en órganos linfoideos y no linfoideos y también circulantes en linfa aferente y sangre periférica, con diferentes nombres según su ubicación, pero guardando características y funciones similares entre sí.
En órganos linfoideos: Se las encuentra en las regiones T dependientes de los ganglios linfáticos y bazo donde se las conoce como células interdigitantes (CID). En el bazo son más numerosas, ya que hay nidos de ellas en la periferia del área de linfocitos T, donde están posicionadas como puentes a través de los cuales deben pasar los linfocitos para entrar en el torrente sanguíneo. Las células foliculares dendríticas (CFD), se encuentran en los centros germinales de los folículos secundarios de las áreas de linfocitos B, de ganglios linfáticos y bazo, siendo parte integral del microambiente del folículo. También están presentes en el timo, sobre todo en la región medular.

En órganos no linfoideos: Las células de Langerhans (CL) son las CD mejor caracterizadas. Están situadas en los epitelios planos estratificados de piel y mucosas, hallándoselas en varias fases del ciclo celular, por lo que se deduce que proliferan in situ6, 7. Migran por vía linfática y esta migración sería utilizada para llevar antígenos de la piel a los linfocitos T helper situados en los ganglios linfáticos.En linfa aferente: Se las conoce como células veladas, representando CL migrantes en tránsito desde la piel, al ganglio linfático donde se transformarán en CID8.En sangre periférica: Las CD son muy escasas ya que representan menos del 2% de las células mononucleares.
Estructura: Las células dendríticas se caracterizan por la gran densidad de moléculas del MHC de clase II que presentan en la membrana plasmática. Las células de Langerhans, un tipo especializado de células dendríticas, también presentan una elevada concentración de moléculas del MHC de clase II. De ese modo parece que la inducción de los gránulos de Birbeck sería una consecuencia de la función de captura de antígenos de la langerina, que permitiría conducirlo a los gránulos y activar de ese modo al procesamiento del antígeno por la ruta no clásica. Según el microambiente en que se encuentren, las células dendríticas expresan diversos marcadores de superficie,
como por ejemplo las citocinas IL-12, IL-1 y TNF-α.
Sólo las células dendríticas inmaduras que patrullan las regiones periféricas tienen la
típica forma de estrella, que les viene dada por unos procesos citoplasmáticos llamados dendritas, que miden más de 10 μm y que sobresalen del cuerpo de la célula. En las células vivas, estas proyecciones están en constante movimiento, retorciéndose, contrayéndose y volviéndose a extender por el lado contrario. Este fenotipo inmaduro se caracteriza por la escasez de proteínas del MHC y la ausencia completa de moléculas B7 coestimuladora. Durante su migración hacia los órganos linfáticos secundarios, después de haber capturado un antígeno durante una infección, la morfología de las células dendríticas cambia. Las dendríticas son remplazadas por pliegues y proyecciones membranales semejantes a velos, por lo que se las puede llamar células veliformes.
Función: Su función principal es tramitar el material del antígeno y presente lo en la superficie a otras células del sistema inmune, así funcionando como las células de antígeno-presentación.Las células Dendríticas están presentes en pequeñas cantidades en los tejidos que están en contacto con el ambiente externo, principal la piel (donde hay un tipo dendrítico especializado de la célula llamado las células de Langerhans) y el forro interior de la nariz, de los pulmones, del estómago y de los intestinos. Pueden también ser encontradas en un estado no maduro en la sangre.

Otros:
Células de Langerhans: Las células de Langerhans son un tipo de células dendríticas que residen en la epidermis
Células dendríticas intersticiales: Las células dendríticas intersticiales (IDC) son un tipo DC cercano a las DC linfoides. Se distinguen de los macrófagos tisulares por su fenotipo inmunológico, así como sus características citoquímicas y funcionales.
Células dendríticas plasmacitoides: Las células dendríticas plasmacitoides (PDC) son un subtipo raro de células dendríticas en circulación que se encuentran en la sangre y los órganos linfoides periféricos.
Células veliformes: Las células veliformes deben su nombre a los numerosos procesos en forma de velo que presentan en la superficie. Se encuentran en los senos linfoides y linfáticos aferentes.
Células dendríticas interdigitales: Las células dendríticas interdigitales derivan de la médula ósea. Se las encuentra en todo el cuerpo, pero sobre todo en las regiones de linfocitos T de los ganglios linfáticos, en la capa linfocítica periarteriolar del bazo, el timo, las amígdalas y las placas de Peyer.
Células dendríticas foliculares: Las células dendríticas foliculares (FDC) se encuentran en los folículos linfáticos de los órganos linfoides secundarios y órganos linfoides terciarios.
.

Células dendríticas foliculares.
Origen:
La mayoría de las CDF no derivan de la médula ósea, se desconoce su origen, pero se hallan en los centros germinales de los folículos linfocíticos y se las considera células accesorias de la respuesta inmune. Las CDF captan antígenos (Que son proteínas provenientes de sustancias extrañas para el organismo) que forman complejos con anticuerpos o productos del complemento y luego muestran estos antígenos sobre su superficie para que sean reconocidos por los linfocitos B, de esta manera maduran diferenciándose en linfocitos B de memoria o formadores de anticuerpos.
Localización:
Las células foliculares dendríticas (CFD), se encuentran en los centros germinales de los folículos secundarios de las áreas de linfocitos B, de ganglios linfáticos y bazo, siendo parte integral del microambiente del folículo. También están presentes en el timo, sobre todo en la región medular. Las células foliculares dendríticas están ubicadas en los centros germinales de los folículos secundarios38.
Estructura:
No tienen moléculas MHC-II en su superdicie, pero presentan gran cantidad de receptores para el complemento (CR1 y CR2) y para las IgG (el Fcg R). Los inmunocomplejos (complejos Ag-Ac) llegan a las áreas de células B de estos órganos linfoides secundarios, y allí quedan retenidos un cierto tiempo: se unen a los receptores para Fc de estas células, que son muy abundantes en sus "perlas" (engrosamientos esféricos espaciados regularmente a lo largo de sus prolongaciones). Parece que estas células desempeñan un papel esencial en el desarrollo de las células B de memoria.
Función:
Las células presentadoras de antígeno más potentes que existen y con la capacidad única de activar linfocitos T colaboradores que no han tenido contacto antigénico previo. No sólo son importantes en la regulación de respuestas inmunógenas efectivas, sino también en la inducción de fenómenos de tolerancia inmunológica, necesarios para evitar la aparición de procesos autoinmunes.
Tras la captación y procesamiento de los antígenos, las CDs se dirigen principalmente a los órganos linfoides, donde se produce la presentación de antígenos a los linfocitos T, proceso en el cual no sólo es necesaria la interacción del CMH-II con el receptor de linfocitos T sino también la interacción de otras moléculas accesorias como las moléculas coestimuladoras y las moléculas de adhesión.

Otros:
CPA para linfocitos T:
Origen: Son un subgrupo de linfocitos (a su vez un tipo de leucocito) que tienen un papel muy importante en establecer y maximizar las capacidades de defensa del sistema inmunitario.
Localizacion: Se sitúan agregados de tejido linfoide, difusos en la lámina propia y o en nódulos como las amígdalas y adenoides (en la nasofaringe) o las placas de Peyer (en el intestino).
Estructura: pertenecen al grupo de leucocitos que son conocidos como linfocitos. Estas células tienen núcleos de forma ovoide que ocupan la mayoría del espacio intracelular. Compuestos por una infiltración de linfocitos y nódulos linfoides localizadas, no encapsuladas, en la mucosa de los diferentes sistemas de nuestro cuerpo.
Funcion: Las células presentadoras de antígeno (CPAs) presentan el fragmento de antígeno sobre sus moléculas MHC de clase II (MHC II). Es entonces cuando los linfocitos T colaboradores reconocen ese complejo molecular, con la ayuda de la expresión de sus co-receptores CD4 (CD4+). La activación de un linfocito T inactivo, ocasiona que libere citoquinas y otras señales de estimulación las cuales provocan un aumento en la actividad de macrófagos, linfocitos citotóxicos (CD8+) y linfocitos B, siendo éstos últimos los que producen anticuerpos.

Otros: puede observarse durante una infección por VIH, virus que infecta las células que son tipo CD4+ (incluidos los linfocitos T colaboradores). Hacia el final de la infección por VIH, el número de células T CD4+ funcionales cae, lo que lleva al estado sintomático de la infección conocido como síndrome de inmunodeficiencia adquirida (SIDA)
EN INMUNIDAD ADAPTATIVA:
Funcionan como CPA en respuestas de la inmunidad adaptativa, a cabo de los linfocitos T
Los macrófagos que contiene microorganismo en su interior después de haberlos ingerido exhiben los antígenos microbianos a los linfocitos T efectores diferenciados, estos van a activar a los macrófagos para destruir microrganismos.
Los linfocitos T estimulados por un antígeno activan a los macrófagos en la inmunidad celular para acabar con los microorganismos fagocitados.
En la humoral, los anticuerpos recubren a los microbios favorecen en su fagocitosis a través de receptores de superficie en los macrófagos.
8.Linfocitos T
Origen :Los Linfocitos T se originan en médula ósea a partir de una célula precursora pluripotencial hematopoyética (StemCell) de la cual se originan la UFC (Unidad Formadora de Colonias), que va a dar luego origen a las líneas eritroidea y mieloidea, y a un precursor linfoideo, a partir del cual se originarán células destinadas a ser Ly B, que continúan su desarrollo en médula ósea, y células destinadas a ser Ly T, que migran al timo, donde serán procesados y sufrirán diferenciación.
Sólo el 5% de los linfocitos presentes en el timo permanecen viables para una adecuada respuesta inmune y pasan a la circulación (el 95% restante muere por mecanismos de apoptosis celular). Los Ly T que dejan el timo y alcanzan el torrente sanguíneo, van a diseminarse por todo el organismo para alojarse en el tejido linfoideo corporal, más extensamente en los ganglios linfáticos, vigilante ante la entrada de algún agente agresor.
Los linfocitos T se desarrollan de células madre en la médula ósea. Temprano en la vida del feto, células inmaduras migran al timo, un órgano especializado del Sistema Inmunológico en el pecho.

En el timo, los linfocitos inmaduros se desarrollan a linfocitos T maduros ("T" por el Timo). El Timo es esencial para este proceso, y los linfocitos T no se pueden desarrollar en el feto si no tiene Timo. Linfocitos T maduros dejan el Timo y se van a otros organos del Sistema Inmunológico, como el baso, nodos linfáticos, médula ósea y la sangre.
Funciones
Los linfocitos T (algunas veces llamadas células T) son otro tipo de células inmunológicas. Los linfocitos T no producen anticuerpos moleculares. Las funciones especializadas de los linfocitos T son :
1) atacar directamente antígenos extraños como virus, hongos, tejidos transplantados
2) para actuar como reguladores del Sistema Inmunológico.
Cada linfocito T reacciona con un antígeno específico, así como cada anticuerpo reacciona con un antígeno específico. De hecho, los linfocitos T tienen moléculas en la superficie que son como anticuerpos que reconocen antígenos.
Tipos
La variedad de linfocitos T es tan grande que el cuerpo tiene linfocitos T que pueden reaccionar contra virtualmente cualquier antígeno. Los linfocitos T también varían con respecto a su función. Hay :
1) linfocitos T destructores ("killer" o "effector")
2) linfocitos T de ayuda ("helper")
3) linfocitos T supresores ("suppressor").
Cada uno juega distintas partes en el Sistema Inmunológico.
Los linfocitos T destructores son los linfocitos que destruyen al micro-organismo invasor. Estos linfocitos T protegen al cuerpo de bacterias especificas y virus que tienen la habilidad de sobrevivir y reproducirse en las células del cuerpo. Los linfocitos T destructores también responden a tejidos extraños en el cuerpo, como por ejemplo un hígado transplantado. Los linfocitos T destructores migran al sitio de la infección o al tejido transplantado. Cuando llegan, los linfocitos T destructores se fijan a su blanco y lo destruyen.

Los linfocitos T de ayuda, ayudan a los linfocitos B a producir anticuerpos y ayudan a los linfocitos T destructores en el ataque a sustancias extrañas. Los linfocitos T de ayuda hacen más efectiva la función de los linfocitos B, provocando una mejor y más rápida producción de anticuerpos. Los linfocitos T de ayuda también hacen más efectiva la función de destrucción de los linfocitos T destructores.
Por otra parte los linfocitos T supresores, suprimen o apagan a los linfocitos T de ayuda. Sin esta supresión, el Sistema Inmunológico seguiría trabajando después de la infección. Juntos los linfocitos T de ayuda y supresores actúan como el termostato de todo el sistema de linfocitos y los dejan prendidos el tiempo suficiente - no mucho tiempo y no muy poco tiempo.
Maduración y selección de las células T
Como pasa con todas las células linfocíticas, las células T provienen de una célula progenitora hematopoyética. Las células progenitoras de los linfocitos T migran desde la médula ósea hacia el timo, donde tiene lugar todo el proceso de maduración, generando células que expresan TCR y CD4 o CD8. Varias etapas definen esa diferenciación
Reorganización de los genes TCR
Pro-T. La célula pro-T es el primer precursor de la línea de linfocitos T. Estas células proliferan por efecto de la interleuquina IL-7 producida por las células del timo. En el núcleo de algunas de estas células, se reordenan los genes que codifican la cadena β del TCR, por recombinación mediada por la V(D)J recombinasa. El TCR no se expresa aún en la superficie del pro-T. Tampoco se expresan en la membrana celular las moléculas accesorias CD4 ni CD8. Por esa razón las células pro-T se denominan CD4-/CD8- o dobles negativas.
- El reordenamiento de la cadena β ocurre al activarse uno de los dos alelos que codifican la cadena β. El otro alelo es inhibido durante el reordenamiento del alelo activado (esto se denomina "exclusión alélica"). Si no se produce una cadena β completa en una célula pro-T, esa célula muere.

Pre-T. Si la recombinación VDJ tiene lugar con éxito y se sintetiza una cadena β, ésta se expresa en la superficie celular, unida a una proteína invariante denominada pre-Tα, para formar el complejo pre-TCR. Por tanto, la célula pre-T es la misma célula pro-T a partir del momento en que la cadena β del TCR se expresa en la superficie celular.
La expresión de la cadena β marca el estadio pre-T y al mismo tiempo comienza el reordenamiento de la cadena α. En las pre-T, aún siguen sin expresarse en la membrana celular las moléculas accesorias CD4 ni CD8, así que las células pre-T son CD4-/CD8- o dobles negativas.
- La expresión en microdominios de la membrana celular de la cadena β unida a la proteína pre-Tα se conoce como pre-TCR y estimula el reordenamiento de uno de los alelos que codifican la cadena α. Si una célula no es capaz de sintetizar una cadena α y el TCR completo, esa célula muere.
Timocito doble-positivo. Estas son las células supervivientes, que expresan tanto la cadena α como β, es decir un TCR completo, CD3 y tanto CD4 como CD8. Por ello se denominan dobles positivas. Ello da paso a los mecanismos de selección positiva y negativa.
Selección positiva de los timocitos
En esta fase tiene lugar la puesta en marcha del mecanismo de restricción CMH: los linfocitos T de un individuo concreto sólo pueden detectar un antígeno si éste viene presentado por una molécula del complejo mayor de histocompatibilidad (CMH) del mismo individuo. Los diferentes clones de timocitos dobles positivos expresan diferentes tipos de TCR αβ. Si el TCR de una célula T reconoce una molécula CMH en el timo (que por definición es una molécula CMH presentando un péptido propio del individuo), esa célula T es seleccionada para sobrevivir: por eso se habla de selección positiva. Para asegurarse de que el linfocito T durante su maduración será expuesto a todo tipo de péptido propio, las células epiteliales medulares del timo expresan numerosos genes, que codifican la mayor parte de las proteínas presentes en los tejidos periféricos.
Las células que no son capaces de reconocer un complejo "péptido propio-CMH" en el timo mueren por apoptosis. Estas células no serían útiles al individuo, porque serían incapaces de ver los péptidos presentados por las moléculas de CMH en los tejidos periféricos.

Elección entre las líneas T CD4+ y CD8+
Durante el proceso de selección positiva, las células T que reconocen complejos péptido-CMH clase-I preservan la expresión de CD8, el coreceptor que se une a la molécula CMH-I, y pierden la expresión de las moléculas de CD4. A la inversa, las células que reconocen complejos péptido-CMH clase-II preservan la expresión de CD4 y pierden la de CD8.
Así, lo que se obtiene al final del proceso de selección positiva son timocitos simples positivos, que son o bien CD8+, restringidos para ver CMH-I, o bien CD4+, restringidos para CMH-II.
Además, durante este proceso, las células T también devienen segregadas funcionalmente: las células T CD8+ pueden convertirse en linfocitos T citotóxicos cuando se activan, mientras que las células T CD4+ serán linfocitos T cooperadores.
Selección negativa de los linfocitos
Los linfocitos dobles positivos inmaduros cuyos receptores reconocen fuertemente los complejos "péptido:CMH" en el timo también sufren apoptosis. Este es el proceso de selección negativa, que sirve para eliminar linfocitos que podrían reaccionar de forma dañina contra proteínas propias que se expresan en el timo. Por ello, se dice que este mecanismo permite el establecimiento de la tolerancia central, al asegurar que las proteínas propias no serán atacadas por los linfocitos T.
Las células T que han pasado los procesos de selección positiva y negativa son linfocitos maduros, que presentan las siguientes características:
son simples positivos CD4+ o CD8+ están restringidos para las moléculas CMH propias (CD4+:CMH-II,
CD8+:CMH-I) son tolerantes para las proteínas propias son vírgenes (naïves): no han encontrado nunca un antígeno extraño.
Los linfocitos maduros salen del timo y se distribuyen por la periferia a través del sistema circulatorio, donde pueden encontrar una célula que presente un complejo "péptido extraño:CMH", capaz de activar el linfocito y desecadenar una respuesta inmune.
Es interesante destacar que la capacidad de reconocer los antígenos extraños por parte de los linfocitos T no está sometida a selección, sino que es el producto del azar: las células T que reconocen los complejos "péptido propio:CMH propio" de forma débil puede que reconozcan fuertemente antígenos extraños, procedentes de microorganismos, en la periferia del organismo.

Activación de linfocitos T
La activación de linfocitos T tiene dos consecuencias generales:
En el ganglio linfático, la activación de los linfocitos T conduce a la activación de células efectoras inmunes.
En los téjidos periféricos, la activación de linfocitos T conduce a la erradicación del microorganismo del foco infeccioso
Una de las primeras respuestas detectables de los linfocitos T a la presentación y reconocimiento antigénico por células presentadoras de antígeno (CPA) es la secreción de citocinas, en especial la Interleucina-2 (IL-2), la cual actúa como factor de crecimiento sobre el mismo linfocito T que lo secreta, por tener ésta receptores para la IL-2. Bajo el efecto de la IL-2, la célula T sufre una proliferación numérica exponencial, denominada expansión clonal, la cual es el fundamento de la memoria inmunitaria. La expansión clonal es seguida por una diferenciación celular, produciendo linfocitos CD4 -encargados de la activación de macrófagos, linfocitos B y otras células- y linfocitos CD8 -las cuales eliminan ciertas «células diana» infectadas y también activan macrófagos en los tejidos afectados.
Receptor de la IL-2
La Interleucina-2 es producida por el cuerpo durante una respuesta inmune,3 4 cuando un antígeno (sea una molécula o un microorganismo) es reconocido por receptores antigénicos sobre CPA (usualmente una célula dendrítica). La presentación y unión de un pequeño segmento del antígeno, por el HMC, a una célula T por intermedio de su receptor (TCR) estimula la secreción de IL-2 y al mismo tiempo, la expresión de receptores de IL-2 (IL-2R). La subsecuente interacción de IL-2 con IL-2R, estimula el crecimiento, diferenciación y supervivencia de las seleccionadas —por el tipo de antígeno— células citotóxicas, por medio de la activación de genes específicos.5 6 7 Esto asegura que solo la célula T con el receptor específico al péptido antigénico sea la activada.
Una vez que IL-2 se une a los dominios externos de su receptor, IL-2R, y los dominios internos son activados, la señal de activación continúa hasta que el complejo IL-2/IL-2R es internalizada y degradada. Sin embargo, cada célula tomará el irrevocable cometido de replicar su ADN y pasar por la mitosis y citocinesis solamente cuando un número crítico de IL-2R han sido expresados y activados.8

CD28 y B7
La activación de las células T ocurre a través de la interacción, tanto del receptor de la célula T (TCR) y de la molécula CD28 con el complejo mayor de histocompatibilidad (MHC) y la familia de receptores B7 sobre la CPA, respectivamente. Se requieren ambas interacciones para la producción de una respuesta inmune eficaz. En la ausencia de la co-estimulación del CD28, las señalizaciones dirigidas por el TCR reultará en anergia. Desde este punto de activación, las vías de señalización del linfocito T, ocupan muchas proteínas.
IL-12, IL-4
Las moléculas Interleucina-12 e Interleucina-4 juegan un papel importante en la diferenciación de linfocitos T. IL-12 está involucrada en la diferenciación de células CD4+ en células TH1, lo cual es importante en la activación de macrófagos (por medio del interferón gamma) y la destrucción de patógenos. A su vez, IL-4 participa en la diferenciación de linfocitos T en subpoblaciones células TH2.
CD40 y CD40L
La molécula del CD40 es una proteína co-estimuladora presente en CPA (un linfocito B, por ejemplo), el cual se une a su ligando, CD40L (CD154) sobre los linfocitos T, activando a la célula, en especial CD4+. Esta interacción CD40:CD40L, a su vez potencia la capacidad de moléculas co-estimuladoras sobre las CPA para actuar en la diferenciación de células T.
9. LINFOCITOS B
Los linfocitos B son los leucocitos de los cuales depende la inmunidad mediada por anticuerpos con actividad específica de fijación de antígenos. Las células B, que constituyen un 5 a 15% del total de linfocitos, dan origen a las células plasmáticas que producen anticuerpos.
Los linfocitos son de dos tipos principales, atendiendo a su origen y función: células T, que se diferencian inicialmente en el timo, y las células B, que se diferencian en el hígado y bazo fetal, y en la médula ósea del adulto (la 'B' proviene del latín Bursa fabricii, el órgano en el cual se desarrollan los linfocitos B en las aves). Durante su desarrollo, los linfocitos T y B adquieren receptores específicos para antígenos, el de las células B se le conoce como receptor de linfocito B (BCR).

Origen:
Los Linfocitos B son un tipo celular que cumple múltiples funciones en el mantenimiento de la inmunidad y ante la reexposición de noxas . Los linfocitos B se originan y maduran en medula ósea pero una vez que hayan completado estos cambios se ubican en los ganglios linfáticos, donde se activan en presencia de un agente extraño, con la ayuda de otro tipo celular, los Linfocitos T CD4 + o Linfocitos T helper; aunque bajo ciertas circunstancias pueden hacerlo en ausencia de estos. Las células B durante su maduración expresan diferentes moléculas de superficie que son útiles para su identificación y conocimiento de su capacidad funcional. Este trabajo busca ser una guía útil para el conocimiento del origen, maduración y activación de los Linfocitos B.
Función:
Los linfocitos B son los protagonistas principales de la respuesta inmune humoral e intervienen en la formación de anticuerpos , proteínas globulares complejas -conocidas también como inmunoglobulinas - que presentan en su estructura combinaciones tridimensionales precisas capaces de interactuar con moléculas que el cuerpo reconoce como extrañas o no propias.
Algunos linfocitos B "patrullan" el cuerpo humano y otros son sésiles; se aglomeran en los ganglios linfáticos, el bazo y otros tejidos linfoides, donde están expuestos a la sangre y la linfa circulantes. Los linfocitos B son células pequeñas, redondas, que no se dividen. Insertos en su membrana y, sobresaliendo de su superficie, se encuentran los anticuerpos con especificidad para reconocer a un determinado antígeno.
Cuando un linfocito B particular se encuentra en un órgano linfoide con el antígeno para el cual es específico, por complementariedad, los anticuerpos de su superficie interactúan con el antígeno. Esto activa al linfocito B, lo que provoca que la célula se agrande, se divida y que las células hijas -o plasmocitos- adquieran la capacidad de realizar una producción activa de anticuerpos. La proliferación de linfocitos B activados ocurre frecuentemente en los ganglios linfáticos, razón por la cual éstos se agrandan durante una infección. Las células hijas que resultan de la activación de linfocitos B se diferencian en dos tipos, uno de los cuales es la célula plasmática.

Maduración:
La maduración linfocitaria , son los procesos y cambios irreversibles con el fin de obtener linfocitos funcionalmente activos a partir de una célula madre pluripontencial.
Génesis de los Linfocitos
En el hígado fetal y en la médula ósea, después del nacimiento existe una célula llamada “célula madre hematopoyética pluripotencial” (CMHP), de las cuales derivan todas las células de la sangre.
Un grupo de estas CMHP se encuentran en estado quiciente (G0) , para mantener así un número constante, aunque los mismos descienden con la edad. Sin embargo la mayoría se diferencia a “células madres comprometidas”, estas son células que ya no pueden generar cualquier tipo celular, sino un grupo en especial, por eso se la denomina comprometidas. De esta manera tenemos células comprometidas que originan Eritrocitos, otras granulocitos y las “células madres linfoides” (CML) que solamente pueden originar linfocitos T y B
Activación:
La activación de linfocitos B es una combinación de su proliferación y diferenciación terminal en células plasmáticas. El reconocimiento de linfocitos B no es el único elemento requerido para la activación de células B. Aquellas que aún no han sido expuestas a antígeno, pueden ser activadas de manera dependiente o independiente de las células T.

Activación dependiente de células T
Cuando una célula B ingiere un patógeno, adhiere parte de las proteínas del organismo a la proteína del complejo mayor de histocompatibilidad tipo II. Este complejo es llevado a la superficie de la membrana celular, donde puede llegar a ser reconocido por los linfocitos T, los cuales son compatibles con estructuras similares sobre la membrana de los linfocitos B. Si las estructuras sobre la célula B y T son compatibles, el linfocito T activará al linfocito B, el cual producirá anticuerpos en contra de segmentos que el patógeno lleva sobre su superficie.
La mayoría de los antígenos son T-dependientes, es decir, requieren cooperadores para la producción máxima de anticuerpos. Con un antígeno T-dependiente, la primera señal proviene del entrecruzamiento del antígeno y el receptor de la célula B (BCR), y la segunda señal viene de una co-estimulación que provee una célula T. Los antígenos T-dependientes contienen proteínas sobre las MHC-II de las células B que son presentadas a células T llamadas TH2. Cuando una célula B procesa y presenta el mismo antígeno a la célula TH, ésta secreta citocinas que activan a la célula B. Estas citocinas producen la proliferación y diferenciación en células plasmáticas. El cambio al isotipoIgG, IgA, e IgE y la generación de células de memoria ocurren en respuesta a antígenos T-dependientes.
Activación T-independiente
Muchos antígenos son T-independientes, es decir, puede emitir ambas señales a la célula B. Los ratones sin timo (ratones atímicos que no producen células T), pueden responder en contra de antígenos, llamados T-independientes. Muchas de las bacterias tienen epítopos repetitivos de carbohidratos que estimulan a las células B, por medio de los llamados receptores reconocedores de patrones, para que respondan sintetizando IgM en ausencia de cooperación de un linfocito T. Hay dos tipos de activación T-independientes:
Tipo 1, o activación policlonal T-independiente;Tipo 2, en la que los macrófagos presentan antígenos a la célula B de tal modo que causa el entrecruzamiento de BCR requerido.

10.Órganos linfoides primarios
A) Medula ósea
• Origen:
se encuentra en el interior del hueso esponjoso. Dentro de la red tridimensional que forma el hueso esponjoso, existe otra red que forman las fibras reticulares y reticulocitos que constituyen el estroma de la médula ósea.
• Localización:
la médula ósea es una estructura en el interior de los huesos largos, vértebras, costillas, esternón, huesos del cráneo, cintura escapular y pelvis. Todas las células sanguíneas derivan de una sola célula madre hematopoyética pluripotencial ubicada en la médula ósea.• Estructura:
• Función:
la función principal de la médula ósea es la hematopoyesis, o sea, la producción sostenida de los tres tipos de células de la sangre: células rojas o hematíes, las células blancas o leucocitos y las plaquetas. Todas éstas proceden de la división y maduración de una célula precursora común denominada célula madre que reside en el interior del hueso. Conocida también como progenitor hematopoyético tiene capacidad de autorrenovación y diferenciación, al ser capaz de generar una o más subseries de células maduras.

B) TIMO
Origen: se origina de la tercera bolsa branquial en elfeto, que aparece completamente desarrollada en el tercer mes de gestación (de 12 a 15 g), y continúa creciendo hasta la pubertad donde alcanza su máximo crecimiento (entre 30 y 40 g). Luego involuciona atrofiándose de forma progresiva y constante, produciéndose el reemplazo del tejido tímico con tejido adiposo y conectivo areolar y alcanzando, en la edad adulta, unos 10 ó 15 g, siendo sustituido buena parte de él por tejido adiposo.
• Localización:
Entre los pulmones, por detrás del esternón, por delante de la tráquea y recubriendo los grandes vasos del corazón.
• Estructura:
Está formado por dos lóbulos rodeados por cápsula de tejido conjuntivo. A su vez, los lóbulos están divididos en lobulillos separados entre sí por trabéculas de tejido conjuntivo. Cada lobulillo tímico está relleno de células linfoides denominadas timocitos, dispuestas en una corteza de gran densidad celular y una médula (interior) de menor densidad celular. Desde la corteza hasta la médula existe un gradiente de diferenciación, de modo que en la corteza se encuentran los timocitos más inmaduros, mientras que en la médula se localizan los timocitos en fases madurativas más avanzadas. Tanto la corteza como la médula están rellenas de una red de células no linfoides que constituyen el estroma tímico.
• Función:
El timo ejerce una clara influencia sobre el desarrollo y maduración del sistema linfático y en la respuesta inmunitaria defensiva de nuestro organismo. También puede influir en el desarrollo de las glándulas sexuales. El timo es un órgano linfoide primario en el cual tiene lugar la diferenciación de los linfocitos indiferenciados (linfoblastos T) que salieron de la médula ósea; ingresan en el timo y van colonizando diferentes zonas del mismo, al tiempo que maduran y se diferencian. La primera área colonizada es el córtex superficial. De ésta pasan al córtex profundo y finalmente a la médula del timo. A lo largo de este recorrido, los linfoblastos T adquieren los receptores antigénicos específicos y aprenden a no atacar a los antígenos propios del individuo (autoantígenos), convirtiéndose en linfocitos T maduros.

11.Órganos linfoides secundarios:
A. proporcionan el entorno para que los linfocitos interaccionen entre sí, otras células accesorias, para que entren en contacto con el antígeno.
B. diseminan la respuesta inmune al resto del cuerpo.
Los órganos linfoides secundarios son:
1) Sistemas linfático
2) Ganglio linfático
3) Bazo
12.-SISTEMA LINFÁTICO:
Origen: El sistema linfático tiene su origen en los capilares situados en la trama de casi todos los órganos, donde nacen con fondo ciego
Localización:
Estructura:
El sistema linfático está formado por:- Los capilares linfáticos.- Los ganglios.- El bazo.- Las amígdalas y las adenoides.- El timo.- El quilo o sistema linfático intestinal
Función:
Entre sus funciones están:- El filtrar los microbios que causan enfermedades o sea que son patógenos.- Producir glóbulos blancos y anticuerpos.- Participar en la distribución de los líquidos y los nutrimentos para que lleguen a todo el cuerpo.- Recoger el exceso de líquido y de proteínas de los diferentes tejidos para evitar que se congestionen.

13.-Ganglio linfático:
Origen: El sistema linfático comienza como tubos con extremos ciegos o bulbos terminales linfáticos que se conocen como vasos linfáticos que se comunican con masas de tejidos linfáticos que son los que dan origen a los ganglios linfáticos y se continúa en un entramado de vasos linfáticos interconectados. El líquido de los tejidos que penetra en un vaso linfático se denomina LINFA.
Estructura:
Los nodos linfáticos tienen un tamaño menor a 1 cm de diámetro, con forma de riñón y están formados por una corteza externa con una cápsula de tejido conjuntivoque rodea al nodo y una zona medular. De la cápsula emergen trabéculas medias, que dividen al nodo internamente.
Función:
su función esta dada por el carácter protector que ofrece a todos los órganos corporales, para ayudar a eliminar productos de desecho, exceso de líquidos y productos que se encuentren almacenados en los tejidos que en ocasiones pueden ser nocivos para los mismos.
Bazo:
Origen:
El desarrollo embrionario del bazo se inicia alrededor de la quinta semana, por diferenciación mesenquimatosa del meso-gastrio dorsal entre el estómago y páncreas, migran y se fusionan estos remanentes mesenquimatosos dando lugar al bazo
Localización:
En el ser humano, el bazo es el mayor de los órganos linfáticos, está peritonizado, se sitúa habitualmente en el hipocondrio izquierdo de lacavidad abdominal, detrás del estómago y debajo del diafragma, unido a él por ligamento frenoesplénico.
Estructura:
El bazo es un órgano de forma ovalada de unos 12 cm de largo, situado a la izquierda y ligeramente detrás del estómago Presenta una depresión, el hilio, que marca el punto de entrada y de salida de los vasos sanguíneos y de los linfáticos eferentes del bazo.

Este órgano está rodeado por una cápsula elástica que contiene músculo liso y de la cual nacen las trabéculas que penetran en él
Función:
Defensa: elimina gérmenes de la sangre, destruyéndolos por fagocitosis.
Hematopoyesis: monocitos y linfocitos completan su desarrollo en el bazo.
Destrucción de hematíes y plaquetas: lo hacen por fagocitosis; a su vez separan las moléculas de hemoglobina de los gl. Rojos destruidos, recuperando hierro y globina.
Depósito de sangre: almacena una considerable cantidad de sangre.
14.SISTEMA INMUNITARIO REGIONAL
Son los componentes del sistema inmunitario con funciones especializadas en una localización anatómica particular, entre estos tenemos al Sistema inmunitario cutáneo y al sistema inmunitario de las mucosas.
Sistemas inmunitarios regionales
Malt
• Origen:
Compuesto por linfocitos y nódulos linfoides. El nódulo linfático y los centros germinativos de producción de linfocitos no aparecen en los ganglios hasta justo antes o después del nacimiento. Estos se producen como resultado de proliferación de células mesenquimatosas relacionadas con los plexos capilares linfáticos.
• Localización:
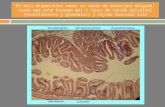
Son agrupaciones de tejido linfoide no encapsulado que forma parte de una serie de localizaciones linfoides repartidas por el organismo, situadas en la lámina propia (tejido conectivo) y las áreas mucosa y submucosas de aparatos digestivo (GALT), respiratorio (nariz: NALT, bronquios: BALT) y genitourinario.Comprende las amígdalas lingual, palatina y faríngea, las placas de peyer del intestino delgado y el apéndice

• Estructura:
La estructura de los MALT es variable dependiendo del lugar donde se encuentre. Puede estar formado por un conjunto de células linfoides (linfocitos entre otros) más o menos mezcladas, o formando estructuras inmunológicas más complejas como las Placas de Peyer del recubrimiento intestinal.Este tejido linfático es de tipo difuso. La estructura más conocida de la MALT son las Placas de Peyer, que se localizan en el intestino.
• Función:
- capaces de responder localmente frente a cualquier agresión del medio externo.
- Produce anticuerpos para las mucosas
- establecer una óptima vigilancia y protección específica a través de todo el organismo.
• Otras:
Ejemplos de tejidos MALT son las amígdalas faríngeas (en la faringe), las amígdalas linguales (en la parte más posterior de la lengua), las amígdalas palatinas (cuya inflamación se conoce como anginas), el apéndice, etc. Las tres primeras se conocen como anillo de Waldeyer.
La importancia de estos tejidos esta en ser la primera línea de defensa de esta gran superficie, para lo cual el organismo tiene una reserva mayor de células plasmáticas en los tejidos MALT que las suma de las existentes en bazo, ganglios y médula ósea
Galt
• Origen:
Comprende linfocitos y nódulos linfoides. El nódulo linfático y los centros germinativos de producción de linfocitos no aparecen en los ganglios hasta justo antes o después del nacimiento. Estos se producen como resultado de proliferación de células mesenquimatosas relacionadas con los plexos capilares linfáticos y luego migran a las diferentes partes del cuerpo donde madurara y cumplirán sus función a lo largo de la vida.
• Localización:
situada en la mucosa gastrointestinal y submucosa. Se compone de folículos linfoides a todo lo largo del tubo gastrointestinal casi todos están aislados entre si. situadas en la lámina propia de la mucosa del intestino delgado

• Estructura:
El mayor componente de la barrera mucosa intestinal compromete a la capa de células epiteliales, impermeabilizadas estas por medio de las uniones apretadas y por el revestimiento mucoso protector. se divide en dos compartimentos:
a) GALT organizado, inductor de la respuesta inmunitaria intestinal constituido por folículos linfoides aislados, folículos linfoides asociados o placas de Peyer y ganglios linfáticos mesentéricos
b) GALT difuso, efector de la respuesta inmunitaria -integrado por poblaciones de linfocitos dispersas en el entramado epitelial o en la lámina propia intestinal
• Función:
- promueve la tolerancia hacia los antígenos dietéticos inofensivos. actúa protegiendo contra los efectos potencialmente dañinos de los constituyentes intraluminales.
- permite la absorción de los nutrimentos dietéticos.
- detección y eliminación de bacterias patógenas, virus y parásito
- previene la respuesta potencialmente dañina a macromoléculas antigénicas persistentes
- contribuye a la defensa en contra de la fijación y penetración de antígenos
- constituyen una importante primera línea de defensa
- se encarga de procesar los antígenos que interaccionan con la mucosa intestinal y de diseminar la respuesta inmunitaria
• Otras:
- El GALT es capaz de discriminar de forma eficaz entre patógenos invasivos y antígenos inocuos.
- Los folículos linfoides de las placas de peyer están compuestos por células B rodeadas de una región mas laxa de células T
- El recubrimiento del íleon lo constituye un epitelio cilíndrico simple, las regiones adyacentes inmediatas a los folículos linfoides están recubiertas de células escamosas que se conocen como células M.