
Cátedra de Biología - Facultad de Agronomía y...
22
Cátedra de Biología - Facultad de Agronomía y Agroindustrias, UNSE de 1 1 Producción e identificación de cultivos transgénicos. Introducción . Desde hace muchos años, el hombre manipula genéticamente los cultivos, mediante técnicas de mejoramiento tradicionales. De este modo se transfiere a la progenie, mediante cruzamientos, las características genéticas de interés. Lamentablemente, estos métodos poseen algunos inconvenientes, como ser la reducción del “pool de genes”, la incompatibilidad sexual, y el tiempo necesario para la transferencia de las características deseadas, que en especies perennes o altamente heterocigotas, puede demandar décadas. La combinación de técnicas de biología molecular, cultivo de tejidos e ingeniería genética, representa una herramienta poderosa para introducir nuevas características en una determinada planta. De este modo, los genes oriundos de diferentes especies vegetales, animales, o microorganismos, pueden ser introducidos de forma controlada en un genoma vegetal receptor. Para tal fin, es necesario localizar y aislar el gen responsable de la característica de interés. Una vez conseguido esto, el gen deberá ser incorporado en un vector, para luego ser introducido al genoma de la planta. Las células vegetales son “totipotentes”, o sea que poseen la potencialidad de desarrollarse, produciendo nuevas plantas. Los adelantos producidos en las investigaciones de los reguladores de crecimiento vegetales, y las técnicas de cultivo de tejidos, han posibilitados la regeneración de muchas especies a partir de una única célula. En la actualidad se conocen varias técnicas que permiten la obtención de plantas transgénicas. Entre ellas se destaca la que utiliza como vector a Agrobacterium tumefaciens, con la cual se han obtenido resultados positivos, especialmente en dicotiledóneas. Estas bacterias del suelo, transfieren naturalmente parte de su genoma a las células vegetales en el momento de la infección, y su ADN puede ser manipulado mediante técnicas de ingeniería genética para introducir en él los genes de interés. De este modo, en el proceso de infección de células vegetales, la bacteria se encargará de introducirlo en el genoma de la planta. Posteriormente, la regeneración de las células que contienen ese ADN foráneo, dará origen a plantas transgénicas, transfiriendo el gen integrado a su progenie de forma mendeliana. Otros métodos, permiten introducir a la célula genes foráneos, mediante mecanismos físicos o químicos. En la electroporación de protoplastos, por ejemplo, se aplica un pulso de alto voltaje a una solución conteniendo ADN y protoplastos en suspensión. La descarga eléctrica hace que se abran poros en la membrana plasmática, permitiendo la entrada del ADN. Este método exige el desarrollo de técnicas de cultivo de tejidos para cada especie y cultivar en particular. Otro método de transformación directa, es la biobalística, que utiliza microproyectiles impulsados a alta velocidad, para introducir el ADN en el genoma celular. Esta técnica presenta la ventaja de poder ser utilizada en tejidos intactos, prescindiendo de las técnicas de cultivo de tejidos, cuyo desarrollo en algunas especies presenta serias dificultades. La transformación genética ha permitido por ejemplo, obtener plantas de papa resistentes a virosis, algodón resistente a insectos, tomates de madurez tardía, flores ornamentales con nuevos patrones de pigmentación, etc. Estas técnicas serán desarrolladas con mayor detalle, en las próximas secciones. Obtención de vectores para transformación directa .
Transcript of Cátedra de Biología - Facultad de Agronomía y...

Cátedra de Biología - Facultad de Agronomía y Agroindustrias, UNSE
de 1 1
Producción e identificación de cultivos transgénicos. Introducción.
Desde hace muchos años, el hombre manipula genéticamente los cultivos, mediante técnicas de mejoramiento tradicionales. De este modo se transfiere a la progenie, mediante cruzamientos, las características genéticas de interés.
Lamentablemente, estos métodos poseen algunos inconvenientes, como ser la reducción del “pool de genes”, la incompatibilidad sexual, y el tiempo necesario para la transferencia de las características deseadas, que en especies perennes o altamente heterocigotas, puede demandar décadas.
La combinación de técnicas de biología molecular, cultivo de tejidos e ingeniería genética, representa una herramienta poderosa para introducir nuevas características en una determinada planta. De este modo, los genes oriundos de diferentes especies vegetales, animales, o microorganismos, pueden ser introducidos de forma controlada en un genoma vegetal receptor. Para tal fin, es necesario localizar y aislar el gen responsable de la característica de interés. Una vez conseguido esto, el gen deberá ser incorporado en un vector, para luego ser introducido al genoma de la planta.
Las células vegetales son “totipotentes”, o sea que poseen la potencialidad de desarrollarse, produciendo nuevas plantas. Los adelantos producidos en las investigaciones de los reguladores de crecimiento vegetales, y las técnicas de cultivo de tejidos, han posibilitados la regeneración de muchas especies a partir de una única célula.
En la actualidad se conocen varias técnicas que permiten la obtención de plantas transgénicas. Entre ellas se destaca la que utiliza como vector a Agrobacterium tumefaciens, con la cual se han obtenido resultados positivos, especialmente en dicotiledóneas. Estas bacterias del suelo, transfieren naturalmente parte de su genoma a las células vegetales en el momento de la infección, y su ADN puede ser manipulado mediante técnicas de ingeniería genética para introducir en él los genes de interés. De este modo, en el proceso de infección de células vegetales, la bacteria se encargará de introducirlo en el genoma de la planta. Posteriormente, la regeneración de las células que contienen ese ADN foráneo, dará origen a plantas transgénicas, transfiriendo el gen integrado a su progenie de forma mendeliana.
Otros métodos, permiten introducir a la célula genes foráneos, mediante mecanismos físicos o químicos. En la electroporación de protoplastos, por ejemplo, se aplica un pulso de alto voltaje a una solución conteniendo ADN y protoplastos en suspensión. La descarga eléctrica hace que se abran poros en la membrana plasmática, permitiendo la entrada del ADN. Este método exige el desarrollo de técnicas de cultivo de tejidos para cada especie y cultivar en particular. Otro método de transformación directa, es la biobalística, que utiliza microproyectiles impulsados a alta velocidad, para introducir el ADN en el genoma celular. Esta técnica presenta la ventaja de poder ser utilizada en tejidos intactos, prescindiendo de las técnicas de cultivo de tejidos, cuyo desarrollo en algunas especies presenta serias dificultades.
La transformación genética ha permitido por ejemplo, obtener plantas de papa resistentes a virosis, algodón resistente a insectos, tomates de madurez tardía, flores ornamentales con nuevos patrones de pigmentación, etc.
Estas técnicas serán desarrolladas con mayor detalle, en las próximas secciones. Obtención de vectores para transformación directa.

Cátedra de Biología - Facultad de Agronomía y Agroindustrias, UNSE
de 2 2
Los vectores utilizados en la transformación de plantas mediante métodos directos, son plásmidos bacterianos en los que han sido clonados los genes a ser introducidos en el genoma vegetal. Estos vectores son de tamaño variable (2 a 50 kb).
Una vez identificados los genes y las respectivas secuencias reguladoras, ellos son agrupados, formando una construcción que consta de un promotor, una secuencia codificadora, y una señal de terminación. Esas construcciones son luego introducidas en un vector adecuado, o sea en un plásmido bacteriano.
Un vector contiene básicamente, los genes de interés a ser introducidos, los genes marcadores para transformación y selección, un origen de replicación, y un gen que confiere a las bacterias resistencia a un antibiótico. Uno de los genes marcadores más utilizados es el gus (uidA), debido a que su expresión puede ser fácilmente detectada mediante métodos enzimáticos, colorimétricos y de fluorescencia.
Como se mencionó, para la selección de las células transformadas se utilizan genes que confieren resistencia a antibióticos. Los más utilizados son el gen neo (npt II), que confiere resistencia a la canamicina, geneticina o paromomicina y el gen hpt, que confiere resistencia a la higromicina. También se puede recurrir a genes que confieren resistencia a herbicidas, como el bar, que codifica a la enzima fosfinotricina acetiltransferasa (PAT), confiriendo resistencia a la fosfinotricina.
Existen diferentes métodos de aislamiento de plásmidos bacterianos. Todos ellos siguen tres etapas básicas: crecimiento de la bacteria y amplificación del plásmido, concentración y lisis de las células bacterianas, y purificación del plásmido. El método de aislamiento del plásmido por lisis alcalina ha sido empleado exitosamente en diferentes linajes de bacterias. Se basa en la naturaleza circular de los plásmidos y en el alto peso molecular del ADN cromosómico. Cuando un extracto celular se expone a un pH alcalino (cercano a 12), el ADN linear (cromosómico) se desnaturaliza mientras que el ADN circular (plásmido) permanece intacto. Luego el extracto es neutralizado con una solución de acetato de sodio, en presencia de una alta concentración salina, lo que hace que el ADN cromosómico precipite, junto con las proteínas.

Cátedra de Biología - Facultad de Agronomía y Agroindustrias, UNSE
de 3 3
Métodos de transformación:
Los biólogos pueden recurrir a tres métodos, para introducir los genes de interés en la célula vegetal. Ellos son: electroporación de protoplastos, biobalística e infección con Agrobacterium. - Electroporación de protoplastos:
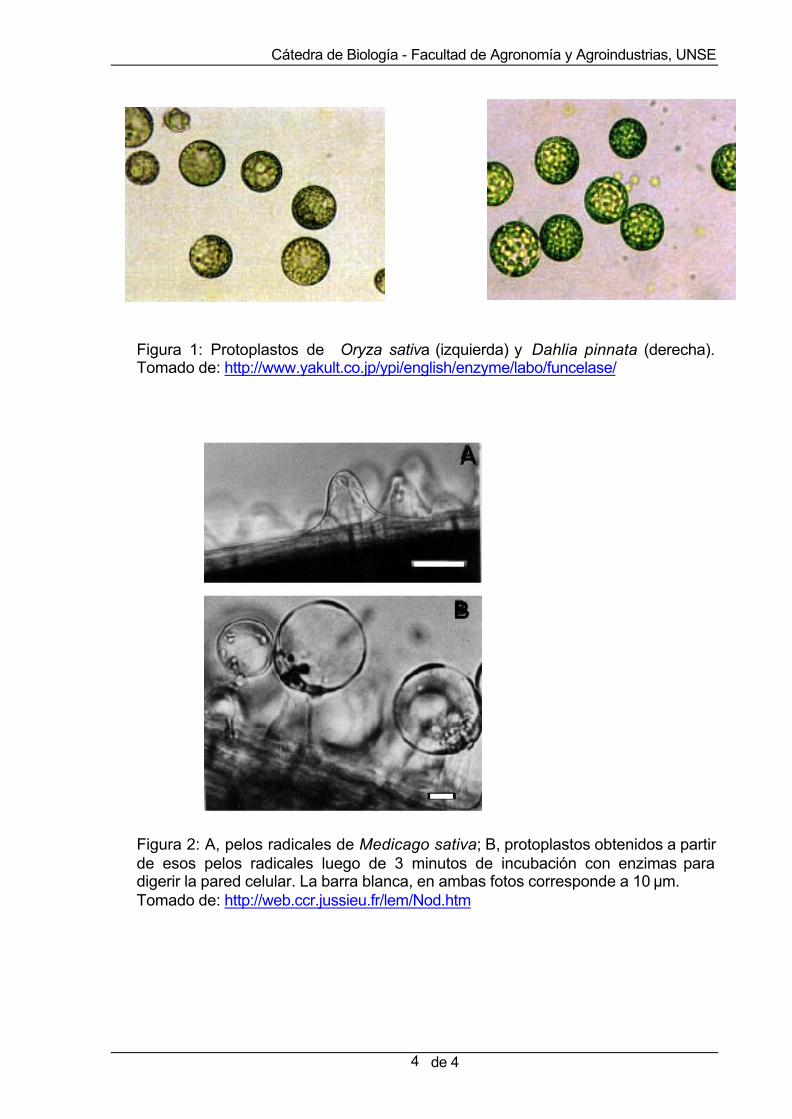
Se trata de un método utilizado para introducir macromoléculas en células vegetales. Llamamos protoplastos a las células vegetales desprovistas de su pared celular (Figuras 1 y 2). Cuando estos protoplastos son cultivados “in vitro”, pueden reconstituir su pared, dividirse por mitosis y regenerar una planta entera.
La obtención de protoplastos requiere de la incubación del tejido vegetal en un medio de digestión compuesto de enzimas pectocelulolíticas, como las que digieren la celulosa, las hemicelulosas, y las pectinas, que son los principales constituyentes de las paredes celulares. Es importante que el pH de las soluciones empleadas favorezca la actividad de las enzimas empleadas, sin comprometer la viabilidad de las células.
Luego de la digestión de la pared celular, se procede a la purificación y determinación del número de protoplastos intactos.
La electroporación consiste en la inducción de poros reversibles en las membranas celulares, que permiten el pasaje de iones y moléculas.
La electroporación de protoplastos se realiza inmediatamente después de su purificación. Para ello, una suspensión de protoplastos se incuba con los plásmidos en los que están clonados los genes de interés, y los genes marcadores, que posibilitan la identificación de las células transformadas.
La mayoría de los aparatos de electroporación (electroporadores) utilizan descargas de capacitores, para producir descargas de alto voltaje. La intensidad del pulso está determinada por el voltaje aplicado y la conductividad del medio. El grado de permeabilidad de la membrana, por su parte, depende del campo eléctrico aplicado y del tipo celular. Altos niveles de permeabilidad facilitan la entrada del ADN, pero disminuye la viabilidad de las células.
La eficiencia de la transformación puede estimarse mediante dos parámetros: * Frecuencia absoluta de transformación: nº de colonias transformadas
nº inicial de protoplastos
* Frecuencia relativa de transformación: nº de colonias transformadas . 100 nº total de colonias obtenidas
La eficiencia de la transformación varía considerablemente entre especies, y entre cultivares de la misma especie. Se han reportado frecuencias absolutas de transformación en el rango de 10-6 a 10-3.
Cátedra de Biología - Facultad de Agronomía y Agroindustrias, UNSE
de 4 4
Figura 1: Protoplastos de Oryza sativa (izquierda) y Dahlia pinnata (derecha). Tomado de: http://www.yakult.co.jp/ypi/english/enzyme/labo/funcelase/ Figura 2: A, pelos radicales de Medicago sativa; B, protoplastos obtenidos a partir de esos pelos radicales luego de 3 minutos de incubación con enzimas para digerir la pared celular. La barra blanca, en ambas fotos corresponde a 10 µm. Tomado de: http://web.ccr.jussieu.fr/lem/Nod.htm

Cátedra de Biología - Facultad de Agronomía y Agroindustrias, UNSE
de 5 5
- Biobalística
Esta técnica fue propuesta por Sanford en 1987, para introducir material genético en el genoma nuclear de plantas superiores. En los últimos años se ha usado para transformar bacterias, protozoos, hongos, algas, insectos y tejidos animales.
La biobalística, utiliza microproyectiles a alta velocidad, para introducir ácidos nucleicos y otras moléculas en células y tejidos. Este proceso también se conoce con los nombres de “bombardeo con microproyectiles”, “gen gun” (pistola génica), “aceleración de partículas”, etc.
Se utilizan micropartículas de 0,2 a 4 µm de diámetro, cubiertas con secuencias de ácidos nucleicos, aceleradas a velocidades superiores a los 1500 km/h. Estas partículas (de oro ó tungsteno) penetran la pared y la membrana plasmática, alojándose aleatoriamente en los organoides celulares. Posteriormente, el ADN se libera de las partículas y se integra en el genoma nuclear del organismo receptor.
Se han desarrollado varios mecanismos para lograr la aceleración de las micropartículas, entre los que se destaca uno que funciona mediante la descarga de helio de alta presión. En la Figura 3 se aprecia un esquema de una pistola génica.
Figura 3: Fotografía mostrando una pistola génica. Tomado de: http://www.mdc-berlin.de/~gfactor/Methods.htm

Cátedra de Biología - Facultad de Agronomía y Agroindustrias, UNSE
de 6 6
- Transformación mediante infección con Agrobacterium. Este método se basa en el uso de una bacteria del género Agrobacterium como herramienta para introducir el gen de interés en el genoma de la planta. Antes de describir la técnica propiamente dicha, veamos algunas características de la especie bacteriana. i) Características del género Agrobacterium
Se trata de bacterias aeróbicas y Gram positivas, comúnmente encontradas en el suelo. No forman esporas, poseen forma de bacilo (Figura 4 A) y se mueven por medio de 1 a 6 flagelos. En particular, Agrobacterium tumefaciens, es el agente etiológico de la “agalla de la corona” y A. rhizogenes el de la “raíz en cabellera”, ambas diseminadas en casi todos los tipos de suelos, cultivados o no.
La infección de una planta con Agrobacterium, comienza con la penetración de la bacteria en el tejido vegetal, a través de una lesión producida por insectos, tratamientos culturales, heladas, etc. Las bacterias son atraídas por sustancias liberadas por las células lesionadas, tales como aminoácidos, azúcares y fenoles. Una vez en contacto con las células vegetales, las bacterias sintetizan microfibrillas de celulosa, propiciando una mejor fijación.
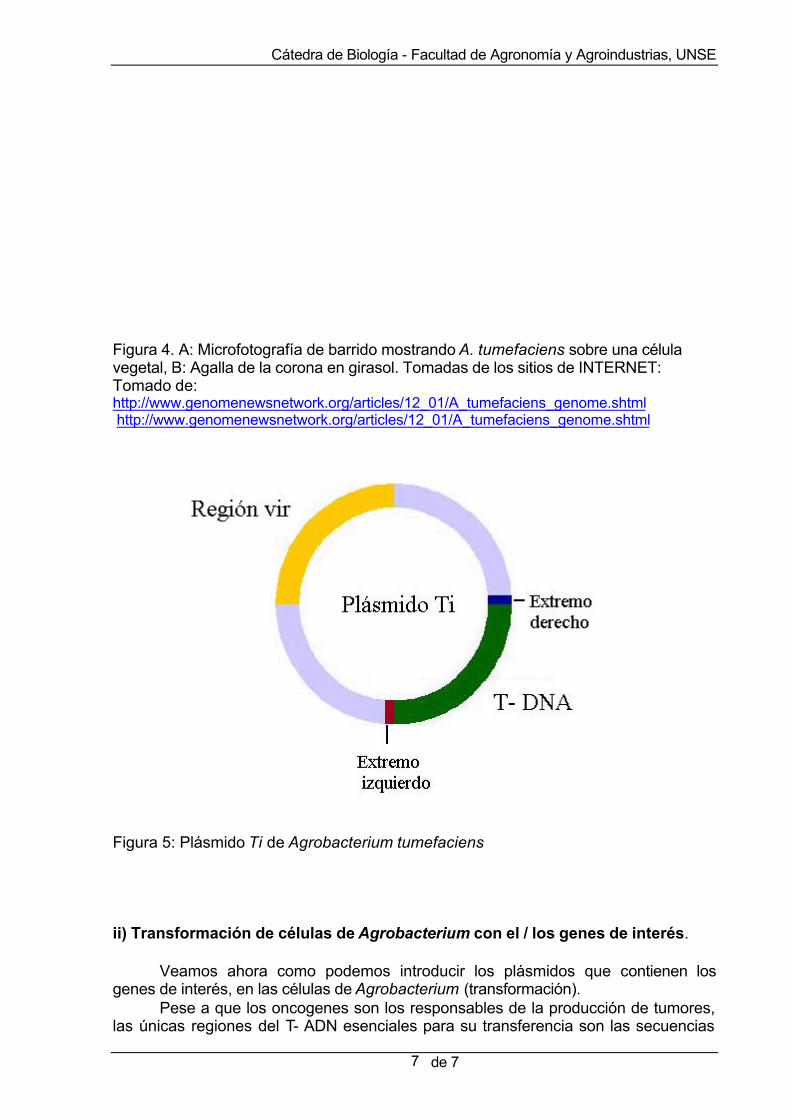
Las moléculas liberadas a través de las heridas también son responsables por la activación de genes localizados en la región de virulencia (región vir) del plásmido Ti de la bacteria (Ti: inductor de tumores) que es un plásmido de alto peso molecular, presente en todos los linajes patogénicos de Agrobacterium (Figura 5). La región vir contiene aproximadamente 25 genes, que codifican proteínas que promueven la transferencia de otra región del plásmido Ti de la bacteria, hacia la célula infectada. Esa región se denomina T-ADN, y se integra en forma estable, al genoma vegetal.
Una vez en el genoma vegetal, los genes de T-ADN (también conocidos como oncogenes) son transcriptos, codificando enzimas involucradas en la síntesis de hormonas vegetales, tales como las auxinas y citocininas. Como consecuencia de ese desbalance hormonal, las células transformadas proliferan desordenadamente, produciendo un tumor que se conoce vulgarmente con el nombre de “agalla de la corona” (Figura 4 B). En el caso de A. rhizogenes, la expresión de los oncogenes conduce a la producción de raíces en la región donde se produjo la herida, síntoma denominado “raíces en cabellera”.
Nótese que en la naturaleza se producen plantas transgénicas, sin la intervención del hombre, cuando el T-ADN bacteriano es “inyectado” a la célula vegetal, para luego integrarse y expresarse en el genoma hospedante.
El T-ADN también posee genes que codifican enzimas responsables por la síntesis de opinas, que son aminoácidos y carbohidratos modificados. Las opinas producidas por las células transformadas son utilizadas exclusivamente por la bacteria como fuente de energía.
Los linajes salvajes, en general no son utilizados para la obtención de plantas transgénicas, debido a las alteraciones fisiológicas que ellos producen.
Cátedra de Biología - Facultad de Agronomía y Agroindustrias, UNSE
de 7 7
Figura 4. A: Microfotografía de barrido mostrando A. tumefaciens sobre una célula vegetal, B: Agalla de la corona en girasol. Tomadas de los sitios de INTERNET: Tomado de: http://www.genomenewsnetwork.org/articles/12_01/A_tumefaciens_genome.shtml http://www.genomenewsnetwork.org/articles/12_01/A_tumefaciens_genome.shtml
Figura 5: Plásmido Ti de Agrobacterium tumefaciens ii) Transformación de células de Agrobacterium con el / los genes de interés.
Veamos ahora como podemos introducir los plásmidos que contienen los genes de interés, en las células de Agrobacterium (transformación).
Pese a que los oncogenes son los responsables de la producción de tumores, las únicas regiones del T- ADN esenciales para su transferencia son las secuencias

Cátedra de Biología - Facultad de Agronomía y Agroindustrias, UNSE
de 8 8
de aproximadamente 25 pb localizadas en sus extremidades. De este modo, los genes presentes en el T- ADN pueden ser eliminados y sustituidos, sin alterar el proceso de transferencia. Además de las extremidades del T- ADN, la región vir también es esencial para la transferencia.
La preparación de un linaje de Agrobacterium para ser utilizado como vector para la transformación de plantas transgénicas, incluye dos etapas:
En la primera etapa, se debe obtener un “linaje desarmado”, en el cual el T-ADN original, con sus oncogenes fue eliminado por un proceso de doble recombinación. Actualmente disponemos de linajes desarmados, obtenidos a partir de linajes salvajes de diferentes orígenes.
La segunda etapa, implica la preparación de un vector conteniendo el T-ADN con los genes de interés. Debido a su gran tamaño (aproximadamente 200 kb), el plásmido Ti no puede ser manipulado directamente, por lo que se emplean plásmidos más pequeños (vectores), que resultan fáciles de manipular. Estos vectores contienen las extremidades del T-ADN, entre las cuales se clonan los genes de interés.
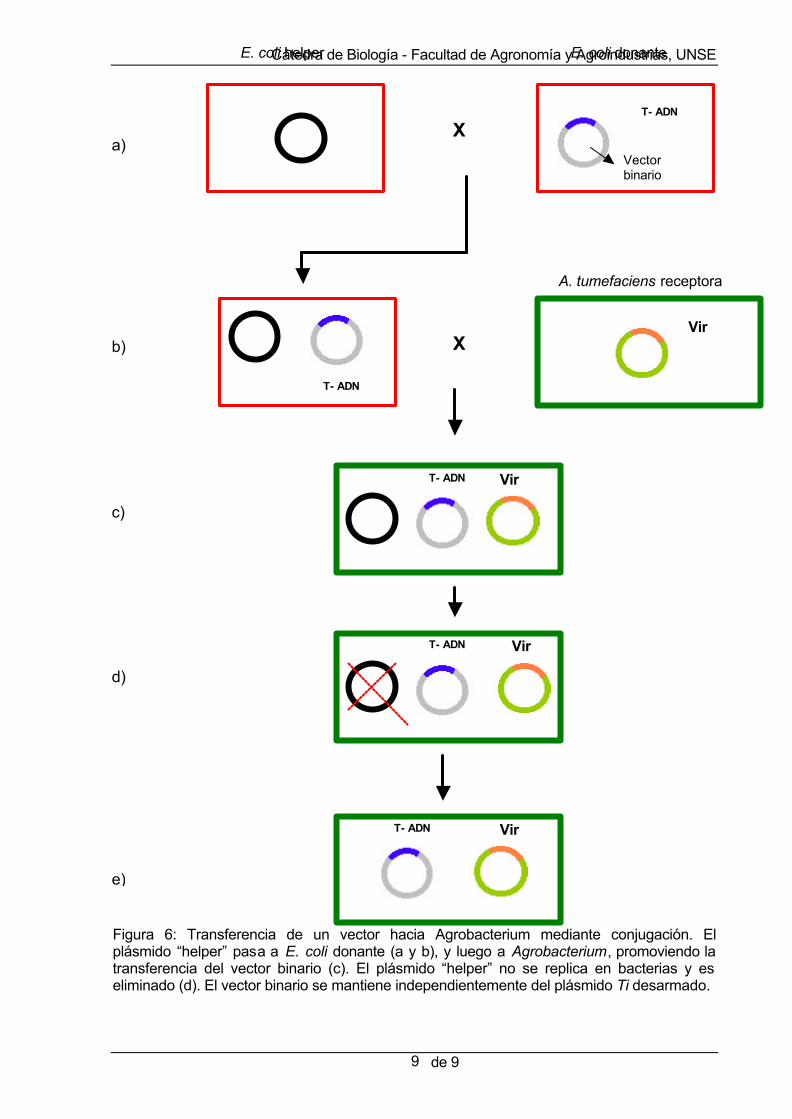
Una vez obtenido el vector, este debe ser transferido al Agrobacterium mediante un proceso que se conoce con el nombre de transformación. Se han desarrollado tres métodos de transformación: conjugación, electroporación y shock térmico: a) Conjugación: se trata de un método simple y eficiente que no requiere de equipamiento específico y prescinde de la preparación de ADN plasmidial de E. coli. En el protocolo más común, se realiza un cocultivo de dos linajes de E. coli (“helper” y “donante”) y un linaje de Agrobacterium (receptora), según puede apreciarse en la Figura 6. El linaje “helper” suministra las funciones de movilización (mob) y transferencia (tra) de plásmidos, entre bacterias compatibles, mientras que el linaje “donante”, contiene el vector que será transferido. Durante la conjugación, el plásmido ‘helper” es transferido al linaje donante, promoviendo posteriormente su movilización hacia Agrobacterium, junto con el vector. El plásmido “helper” no se replica en Agrobacterium, y por lo tanto es eliminado. Luego de la transferencia, se seleccionan los linajes recombinates de Agrobacterium mediante el uso de antibióticos apropiados. La principal desventaja de la conjugación es la posibilidad de que ocurran alteraciones en el plásmido introducido, debido a la recombinación con el plásmido ‘helper”.
Cátedra de Biología - Facultad de Agronomía y Agroindustrias, UNSE
de 9 9
Figura 6: Transferencia de un vector hacia Agrobacterium mediante conjugación. El plásmido “helper” pasa a E. coli donante (a y b), y luego a Agrobacterium, promoviendo la transferencia del vector binario (c). El plásmido “helper” no se replica en bacterias y es eliminado (d). El vector binario se mantiene independientemente del plásmido Ti desarmado.
T- ADN
E. coli helper E. coli donante
X
T- ADN
X Vir
A. tumefaciens receptora
Vir T- ADN
T- ADN Vir
T- ADN Vir
a) b) c) d) e)
Vector binario

Cátedra de Biología - Facultad de Agronomía y Agroindustrias, UNSE
de 10 10
b) Electroporación: Consiste en someter a las células del linaje receptor de Agrobacterium a un pulso de alto voltaje, generado por un capacitor, en presencia del vector. Como consecuencia del campo eléctrico generado, la membrana plasmática se desestabiliza, produciendo poros por los que se produce el pasaje de macromoléculas. Se trata de un método muy eficiente y relativamente simple, que se utiliza con mucha frecuencia. Su principal limitante es la necesidad de contar con el equipamiento de laboratorio: electroporador y una fuente de alta tensión. c) Shock térmico (“freeze – thaw”): este método se basa en la permeabilización de la membrana en condiciones extremas de temperatura (de –186 a 37ºC), permitiendo el pasaje del vector hacia el Agrobacterium receptor.
En los métodos directos de transformación (electroporación y shock térmico),
en los cuales el vector es transferido directamente hacia la célula receptora, la probabilidad de que el ADN se altere es mínima, en comparación con el método de conjugación. iii) Obtención de plantas transgénicas mediante Agrobacterium.
El conocimiento de las bases moleculares del mecanismo de infección por Agrobacterium, y el desarrollo de técnicas de biología molecular, permitieron desarrollar protocolos para introducir en las plantas genes exógenos, utilizando aquella bacteria como vector. Como se ha mencionado en los ítems anteriores, esto fue posible gracias a la obtención de vectores derivados del plásmido Ti ó Ri, y su introducción en linajes de Agrobacterium “desarmados”, en los cuales los oncogenes fueron eliminados. Desde entonces, este sistema de transformación fue ampliamente utilizado, en un gran número de especies. Algunas de las razones que justifican su universalidad, son la alta eficiencia de transformación, el bajo costo operacional y la simplicidad de las técnicas.
La mayoría de los protocolos desarrollados, son una modificación del propuesto por Horsch en 1985 para transformar discos foliares de tabaco (Nicotina tabacum) y consta de las siguientes etapas (Figura 7): - Infección: Consiste en el cultivo del explanto vegetal en un medio líquido ó sólido, junto con el linaje desarmado de Agrobacterium que contiene el ó los genes a ser introducido/s en la planta. El tiempo de cultivo puede variar desde algunas horas, hasta varios días. En la elección del explanto debe tenerse en cuenta la facilidad de reproducción “in vitro”. De este modo, dependiendo de las especie y del cultivar, se han utilizado como explantos discos foliares, segmentos de tallos o raíces, tubérculos, cotiledones, protoplastos, embriones somáticos, etc.
Generalmente se realiza una herida en el explanto, antes o durante el contacto con la bacteria, ya que a través de ella se liberan las sustancias que inducen a los genes de virulencia del Agrobacterium. Durante esta etapa, se produce la unión de la bacteria a la región donde se encuentra la herida, la inducción de los genes de la región vir, y la transferencia del T-ADN hacia el genoma vegetal. - Selección: En una segunda etapa el explanto es transferido a un medio de regeneración apropiado conteniendo uno ó más antibióticos descontaminantes, que permiten eliminar a las células de Agrobacterium, que a partir de ese momento ya no son requeridas.

Cátedra de Biología - Facultad de Agronomía y Agroindustrias, UNSE
de 11 11
Normalmente se adiciona al medio de cultivo un agente de selección, que es responsable de la inhibición del crecimiento de las células no transformadas. El efecto nocivo de este agente, en las células transformadas, es anulado por el producto de expresión del gen marcador de selección (generalmente una enzima). De este modo, sólo las células transformadas serán capaces de desarrollarse en este medio de cultivo selectivo.
Debemos tener en cuenta que ningún sistema de selección es totalmente eficiente, de modo que pueden producirse algunos “escapes” (plantas no transformadas, que a pesar de no expresar el gen marcador, consiguen regenerarse en presencia del agente de selección). - Enraizamiento: Durante las semanas siguientes, los brotes resistentes al agente de selección son aislados a medida que regeneran, y transferidos a un medio de enraizamiento, rico en auxinas (hormonas vegetales, como el ácido 3-indol acético, 3- indol – butírico, etc, que se caracterizan por inducir la diferenciación de raíces). - Rustificación: Una vez enraizados, los brotes potencialmente transformados son aclimatados y llevados a invernáculo. - Análisis molecular: Para comprobar la integración y expresión del gen foráneo en el genoma de la planta, es necesario un exhaustivo análisis molecular, mediante técnicas como Northern Blot, Southern Blot, Western Blot, etc, que serán descriptas más adelante.
Posteriormente, también deben realizarse estudios genéticos de segregación, para demostrar la estabilidad de la integración.
Figura 7: Etapas de la transformación con Agrobacterium. Adaptado de: http://www.cipotato.org/Market/Belgtech/breakthr.htm

Cátedra de Biología - Facultad de Agronomía y Agroindustrias, UNSE
de 12 12
Genes Marcadores
El gen gus, que codifica para la enzima ß- glucuronidasa (GUS) es muy utilizado como gen marcador, en plantas transgénicas. Su amplia difusión se debe a la rapidez, simplicidad y versatilidad de los métodos de detección de la actividad enzimática, y al hecho de que la mayoría de las plantas no presenta una actividad endógena significativa. Además, GUS no requiere de una cofactor, es bastante estable y resistente a diversos solventes y detergentes, y es activa en un amplio rango de pH (5 a 9). También puede ligarse a otras secuencias tanto en la región N, como en la C terminal, lo que le permite ingresar a cloroplastos y otros organoides.
Otros genes utilizados como marcadores son el lucA de la luciferasa, el npt II ó neo, de la neomicina fosfotransferasa II, el gen cat del cloranfenicol acetiltransferasa. A diferencia de los ensayos para la detección de NPTII y CAT, que requieren del uso de radioisótopos, y del utilizado por la luciferasa, que utiliza un luminómetro, el ensayo de GUS no requiere equipamientos específicos.
GUS puede ser determinado mediante tres metodologías: - Ensayo histoquímico: es un método cualitativo que se basa en la disociación del sustrato 5-bromuro-4-cloro-3-indolil.ß-D-glucoronideo (X-gluc) por la ß-glucuronidasa. El producto de esta reacción, en presencia de oxígeno, forma dímeros que se manifiestan como un precipitado color azul (Figura 8).
El ensayo histoquímico no permite cuantificar la actividad enzimática, aunque la eficiencia de la transformación puede ser cuantificada mediante el porcentaje del área en la que el gen se expresa. Figura 8: Brotes de Eucalyptus occidentalis expresando el gen gus (zonas azuladas). Fuente: http://www.ffp.csiro.au/tigr/molecular/molbiology.html

Cátedra de Biología - Facultad de Agronomía y Agroindustrias, UNSE
de 13 13
- Ensayo flourimétrico: es un método muy sensible, utilizado para identificar y cuantificar la actividad de la enzima ß-glucuronidasa en extractos de tejidos vegetales. Se emplea un sustrato que es hidrolizado por la enzima, cuyo producto de reacción presenta fluorescencia a pH alcalino. - Ensayo espectrofotométrico: la enzima transforma al sustrato en un producto que a pH alcalino presenta un color amarillo, cuya intensidad es proporcional a la concentración. Este ensayo no es muy empleado, debido a que su sensibilidad es mucho menor a la del método flourimétrico. No obstante es un método simple, eficiente y representa una buena alternativa cuando no se cuenta con un flourímetro. Identificación de plantas transgénicas.
En los párrafos anteriores, mencionamos que la última etapa del proceso de obtención de plantas transgénicas consiste en detectar si el gen foráneo se ha integrado al genoma vegetal, y si se expresa. Esto exige la extracción del material genético, y el empleo de técnicas de biología molecular, para su identificación. Extracción del ADN en tejidos vegetales.
Actualmente contamos con numerosos protocolos que permiten extraer el ADN de diferentes especies y tejidos vegetales. La diversidad de técnicas refleja la variabilidad en la composición química de los vegetales, que imposibilita el desarrollo de un protocolo universal. Pese a eso, los fundamentos de cada etapa son comunes a todos los métodos.
Primero, el material vegetal es congelado mediante la adición de nitrógeno líquido, y macerado para producir la rotura de las membranas y paredes celulares. El polvo así obtenido es resuspendido en un buffer de extracción, a pH 8 - 9, que no es óptimo para la actividad de las enzimas hidrolíticas (el pH óptimo para las enzimas lipolíticas y lipoxigenasas es de 5 - 6, y el de las ADNasas es de 7). También se adiciona EDTA (ácido etilen diamino tetracético), un quelante de iones divalentes, como el Mg+2 y el Ca+2, y por lo tanto un poderoso inhibidor de las ADNasas que usan estos iones como cofactores. Para evitar los efectos no deseados de la oxidación de diferentes productos se suele incluir en los tampones de extracción, compuestos como el PVP (polivinilpirrolidona) y albúmina sérica bovina (BSA) en concentraciones de 1 a 2%. El primero, es un antioxidante que inhibe la acción de compuestos fenólicos, y la BSA actúa absorbiendo los polifenoles, evitando la acción de estos compuestos que oxidan al ADN e impiden la acción de las enzimas de restricción. Para evitar la oxidación de los compuestos fenólicos, también se suele adicionar ß-mercaptoetanol, un agente reductor que desnaturaliza peroxidasas y polifenoloxidasas, impidiendo la acción de estas enzimas sobre el ADN.
Una vez que se ha resuspendido el contenido celular, se realiza una desproteinización, mediante una ó más extracciones con solventes orgánicos (fenol ó cloranfenicol isoamílico), seguido por centrifugación, para separar la fase orgánica de la acuosa. Estos solventes desnaturalizan a las proteínas, que quedan en la interfase, y actúan sobre contaminantes como los ARNs y carbohidratos, que se mantienen en la fase acuosa. Algunos protocolos incorporan proteasas antes de la extracción con solventes orgánicos, lo que facilita la separación del ADN de las proteínas de la cromatina.

Cátedra de Biología - Facultad de Agronomía y Agroindustrias, UNSE
de 14 14
Una vez terminada la etapa de desproteinización, el ADN puede ser precipitado o ser sometido a diversos tratamientos que lo separan de otros contaminantes. Identificación de plantas transgénicas mediante la PCR (reacción de polimerización en cadena).
La reacción de polimerización en cadena, o reacción en cadena de la polimerasa (PCR) es una técnica que presenta numerosas aplicaciones en el campo de las ciencias biológicas. Ella permite generar cien billones de copias de una única molécula de ADN, en sólo una tarde.
Puede ser considerado como un método “in vitro” para producir grandes cantidades de un fragmento específico de ADN, de tamaño y secuencia definido, a partir de una pequeña cantidad de ácido nucleico. El número de fragmentos amplificados se duplica en cada ciclo de reacción, siendo por lo tanto un proceso exponencial (Figuras 9 y 10).
La reacción de amplificación es catalizada por la enzima ADN polimerasa, que extiende un pequeño fragmento de ADN de cadena simple llamado “primer” o cebador que se encuentra ligado a una cadena molde de ADN. Esta enzima va adicionando nucleótidos, al extremo 3’ del primer, complementarios a la secuencia de la cadena molde.
Los primers o cebadores, son sintetizados artificialmente. La técnica comienza con la separación de las dos cadenas del ADN molde,
como consecuencia de la elevación de la temperatura del medio de reacción (desnaturalización del ADN). Luego se disminuye la temperatura, para que los cebadores se apareen al ADN molde, y la polimerasa pueda actuar, extendiendo un nuevo fragmento. El fragmento producido en la reacción anterior, sirve también como molde para la próxima reacción de PCR.
Por lo tanto, la PCR implica ciclos repetidos de desnaturalización del ADN, unión de los primers al ADN molde, y polimerización de la cadena complementaria al molde.
Inicialmente, la enzima utilizada en la PCR era ADN polimerasa de E. coli, que por ser termolábil, debía ser adicionada en cada ciclo de reacción, pues era destruida durante la etapa de desnaturalización. Actualmente se utiliza la ADN polimerasa aislada de la bacteria termófila Thermus aquaticus, llamada comúnmente Taq polimerasa. Esta enzima permite la realización de múltiples ciclos de reacción, lo que posibilitó la automatización de la reacción. La Taq polimerasa soporta la temperatura de 95ºC, y es más activa entre los 70 y 75ºC, temperatura a la cual el apareamiento entre los primers y el ADN es más específico.
Por lo tanto, para realizar una PCR, se necesita una muestra de ácido nucleico con el fragmento a ser amplificado (molde), la Taq polimerasa, los primers o cebadores, los nucleótidos que constituirán el nuevo fragmento, y los cofactores de la reacción.
La PCR posee la ventaja de requerir pequeñas cantidades del ADN a ser amplificado. Es tan sensible que el ADN aislado de una única célula (por ejemplo protoplastos vegetales) es suficiente para la detección de secuencias específicas de genes. La PCR también puede ser empleada para la clonación del ADN, secuenciación, cuantificación de secuencias específicas, análisis de la expresión génica por la amplificación a partir de ARNm, análisis de la estructura del genoma, análisis de la interacción del ADN – proteína, evolución molecular, identificación de

Cátedra de Biología - Facultad de Agronomía y Agroindustrias, UNSE
de 15 15
mutaciones, diagnóstico de patógenos y enfermedades hereditarias, identificación de anormalidades cromosómicas y mutaciones somáticas específicas.
El resultado de la amplificación del ácido nucleico por PCR, puede ser fácilmente visualizado por electroforesis en geles de agarosa. El éxito de la amplificación depende de las condiciones de la reacción, de la pureza de las drogas utilizadas, y de los diferentes parámetros de la reacción (cantidad de enzima, concentración de nucleótidos, concentración de cebadores, temperatura de desnaturalización, tiempo de desnaturalización, número de ciclos, tamaño de los fragmentos del ADN a ser amplificado, etc). Figura 9: Fundamentos de la PCR. Adaptado de: http://www.aeu.es/Ponencias/sole/Capitulo2.htm
Fragmento de cromosoma que posee el gen que queremos amplificar
Desnaturalización, se adicionan los cebadores en un baño de nucleótidos
Se adiciona la polimerasa, y se forma la doble cadena de ADN
Se repite el ciclo

Cátedra de Biología - Facultad de Agronomía y Agroindustrias, UNSE
de 16 16
Figura 10: Esquema que representa la reacción en cadena de la polimerasa (PCR). En el primer ciclo, la elevación de la temperatura a 94 ºC, produce la desnaturalización del ADN (separación de las cadenas). Al descender la temperatura a 55 ºC, se produce la hibridación de los iniciadores en cada una de las cadenas. Al llegar a los 74 ºC, comienza la acción de la polimerasa, que extiende los iniciadores con los nucleótidos libres que se han agregado a la reacción. Los ciclos adicionales incrementan exponencialmente el número de moléculas de ADN Adaptado de: http://www.aeu.es/Ponencias/sole/Capitulo2.htm Análisis de la integración del ADN mediante la técnica de “Southern Blot”:
La técnica de Southern Blot permite detectar fragmentos específicos de ADN, en muestras de composición compleja, como los son los ADNs genómicos. El método fue propuesto por Southern en 1975, y actualmente se utiliza con muy pocas modificaciones.
Entre otras aplicaciones, esta técnica se utiliza para analizar secuencias de ADNs foráneos, introducidos en el genoma vegetal. El método consta básicamente de las siguientes etapas (Figura 11): - El ADN vegetal es extraído de las células y digerido con una ó más enzimas de restricción. - Los productos de la digestión del ADN son separados de acuerdo a su tamaño, mediante electroforesis en geles de agarosa. - El ADN, que ahora se encuentra en el gel, es desnaturalizado, y transferido a una membrana de nitrocelulosa. Cabe señalar que las posiciones relativas de los
Desnaturalización (94 ºC)
Hibridación (55 ºC)
Síntesis (74 ºC)
Desnaturalización (94 ºC)
Hibridación (55 ºC)
Síntesis (74 ºC)

Cátedra de Biología - Facultad de Agronomía y Agroindustrias, UNSE
de 17 17
fragmentos de ADN en el gel se mantienen cuando el ADN es transferido a la membrana. - El ADN es fijado a la membrana mediante la exposición a la luz ultravioleta, o por medio de altas temperaturas (80ºC). - El ADN ligado a la membrana es hibridado con una sonda de ADN ó ARN, que posee homología con la secuencia de interés. La posición de los fragmentos homólogos a la sonda puede ser detectada mediante autorradiografía o colorimetría, dependiendo del tipo de sonda utilizada. Figura 11: Representación esquemática de la técnica de Southern blotting: adaptado de: http://www.science2discover.com/Southernblot.htm

Cátedra de Biología - Facultad de Agronomía y Agroindustrias, UNSE
de 18 18
Esta técnica suministra la prueba molecular de la integración del gen exógeno en el genoma de la planta. La PCR también puede ser empleada para tal fin, pero su confiabilidad es menor, ya que por causa de su alta sensibilidad, puede detectar “falsos positivos”, mediante la amplificación de pequeñas cantidades de ADN contaminante. Análisis del ARN mediante la técnica de “Northern Blot”.
La técnica de Northern Blot fue desarrollada por Alwine y colaboradores, en 1977, y es análoga al Southern Blot. Permite determinar la cantidad y tamaño de ARNs específicos presentes en preparaciones de ARN total, mensajero, o ribosómico. El método se basa en la hibridación de secuencias complementarias de ácidos nucleicos de cadena simple. Permite estudiar la expresión de secuencias específicas de ADN, y posteriormente correlacionarla con las características morfológicas y fisiológicas de la planta. De este modo, se emplea frecuentemente en estudios de expresión génica, en los cuales se pueden detectar los genes transcriptos en los diferentes tejidos (expresión espacial) y/o en las diferentes etapas del desarrollo (expresión temporal). También es empleada para monitorear la expresión génica del ADN exógeno en plantas transgénicas.
En esta técnica el ARN es transferido desde el gel, hacia un soporte sólido (membrana), y luego hibridado con una sonda específica de ADN ó ARN, marcada mediante radiactividad ó por métodos no radiactivos Detección de proteínas mediante la técnica de ELISA
Al final de la década de 1950, surgió una nueva perspectiva para la detección de proteínas mediante inmunoensayos. Un trabajo pionero fue el conducido por Yalow y Berson (1959), quienes describieron un método para la detección de insulina mediante un radioinmunoensayo. Posteriormente se introdujeron varias modificaciones, que mejoraron considerablemente la eficiencia de esta técnica. Entre ellas cabe mencionar la introducción de métodos no radiactivos. Estos ensayos denominados ELISA (enzyme-linked immunosorbent assay) además de usar reactivos enzimáticos, presentan alta sensibilidad, son de fácil desarrollo, rápidos y producen resultados reproducibles. Los sistemas ELISA facilitaron la detección de varias proteínas y virus, y fueron ampliamente utilizados en el diagnóstico clínico. Posteriormente, fueron implementadas por todas las áreas de análisis biológicos.
La técnica permite identificar una proteína particular, presente en una población, utilizando extractos crudos o semipurificados. De este modo, mediante ELISA, es posible detectar una proteína codificada por un gen exógeno de una planta transgénica. Además se trata de un método rápido, que permite el análisis de un gran número de muestras.
Se han utilizado anticuerpos conjugados de forma covalente a diferentes enzimas, como la fosfatasa alcalina, peroxidasa, ß-galactosidasa, glucoamilasa, ureasa, penicilinasa, etc (Tabla 1). Entre ellas la fosfatasa alcalina ha sido la enzima más utilizada, pues presenta una rápida actividad catalítica, alta especificidad y es fácil de obtener y conjugar al anticuerpo. Además, su sustrato es inocuo y relativamente estable.
Cátedra de Biología - Facultad de Agronomía y Agroindustrias, UNSE
de 19 19
Tabla 1: Enzimas y sus respectivos sustratos cromogénicos, comúnmente usados en ensayos de ELISA. Enzima sustrato Color inicial/final Pico de
absorbancia (nm) Peroxidasa ß-galactosidasa Fosfatasa alcalina
ABST OPD TMB ONPG PNPP
Incoloro/verde Incoloro/marrón Incoloro/amarillo Incoloro/amarillo Incoloro/amarillo
410 492 450 410 405
Existen varios tipos de ELISA, pero para la detección de proteínas en plantas transgénicas los más empleados son los métodos directos e indirectos. Método indirecto (Figura 12): Los extractos proteicos de las plantas que se desean analizar, son colocados en una placa especial para ELISA. En esta etapa, se ligan a la placa varias proteínas presentes en el extracto, entre ellas la proteína “blanco” (aquella que queremos detectar). Luego se adiciona el anticuerpo primario y se incuba, para después eliminar mediante lavados sucesivos las moléculas de anticuerpo que no se hayan ligado a las proteínas. Posteriormente se adiciona un anticuerpo secundario conjugado a una enzima (por ejemplo, fosfatasa alcalina o peroxidasa). Después de un período de incubación, cuando el anticuerpo secundario se liga al primario, se lava nuevamente para eliminar los anticuerpos que no se hayan ligado. Por último, se adiciona el sustrato, para que la enzima actúe sobre él, modificándolo y alterando su color (ensayo calorimétrico), o permitiendo que se produzca fluorescencia en determinadas longitudes de onda (ensayo fluorimétrico). Método directo: las etapas son semejantes a la del método indirecto. El extracto proteico es colocado en la placa, seguido de un lavado. En este caso se utiliza un solo anticuerpo específico para la proteína de interés (proteína blanco) conjugado a una enzima. Las etapas posteriores, de lavado y revelado son iguales a las mencionadas en el método indirecto.
Adicionar extracto proteico
Lavar – bloquear - lavar

Cátedra de Biología - Facultad de Agronomía y Agroindustrias, UNSE
de 20 20
Figura 12: Esquema representativo de las etapas del método de ELISA indirecto. Adaptado de Brasileiro & Carneiro, 1998. Detección de proteínas mediante la técnica de “Western Blot”.
Western Blot, ó immunoblot es un método que permite la detección de proteínas, luego de su separación a través de electroforesis en gel de poliacrilamida y transferencia a una membrana de nitrocelulosa. La detección se hace mediante anticuerpos, que reaccionan específicamente con la proteína de interés, seguido por reacciones colorimétricas ó radiográficas. De este modo, la técnica combina la resolución de la electroforesis con la especificidad de la detección inmunológica.

Cátedra de Biología - Facultad de Agronomía y Agroindustrias, UNSE
de 21 21
Ésta técnica es una poderosa herramienta para el análisis de plantas transgénicas, ya que permite evaluar la expresión de un “transgen”. Permite, por ejemplo, determinar la presencia y cantidad de una determinada proteína, y si ella mantiene sus características de peso molecular, o identificar el lugar donde se expresa en la planta (por ejemplo, en la raíz, tallo, semilla, etc).
La técnica se usa también para identificar y caracterizar proteínas de interés agronómico. Bibliografía - Alberts, B; Jonson, A; Lewis, J; Raff, M; Roberts, K; Walter, P. 2002. Molecular Biology of the Cell. Garland Science. - Brasileiro, A.C.; CamposCarneiro, V.T. 1998. Manual de transformação genética de plantas. Empresa Brasileira de Pesquisa Agrompecuária. - Hartl, D; Jones, E.2001. Genetics. Analysis of genes and Genomes. 5th. edition. Jones and Bartlett Publishers.

Cátedra de Biología - Facultad de Agronomía y Agroindustrias, UNSE
de 22 22
Universidad Nacional de Santiago del Estero Facultad de Agronomía y Agroindustrias
Cátedra de Biología
Producción e identificación de cultivos transgénicos
Diego Ariel Meloni (Prof. Adjunto) Julia Andrea Lescano (Pasante Cátedra de Biología)


















