Dig y Abrosción de Carbohidratos
12
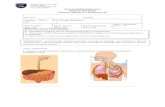
DIGESTIÓN Y ABROSCIÓN DE CARBOHIDRATOS El platillo, el cual consiste en pollo asado y puré de papa cuenta con los siguientes tipos de carbohidratos: almidón, lactosa y glucosa. Este último pasará directamente, esto es, sin modificación alguna al intestino delgado para ser absorbido. La digestión, concretamente del almidón, comienza en la cavidad bucal. La masticación es un proceso mecánico que hace disminuir el tamaño de los gránulos de almidón, aumentando la exposición para las enzimas hidrolíticas. La primera de ellas en actuar es la α-amilasa salival (ptialina), que actúa sobre los enlaces α (1→4). Su actuación permanece en el bolo alimenticio, incluso en el estómago por un tiempo, deteniendo su actividad cuando el pH ácido gástrico alcanza el grumo alimenticio, ya que el pH óptimo de esta enzima es vecino a la neutralidad. La ptialina puede hidrolizarse hasta 30-40% de almidón ingerido, obteniéndose maltosa, maltotriosa y dextrosas límites (compuestos con 3-5 unidades de glucosa en uniones α (1→4) y 1 ramificación en α (1→6)). Posteriormente, la hidrólisis del almidón restante se reinicia gracias a la amilasa pancreática tras el vaciamiento gástrico, fundamentalmente en duodeno y yeyuno. El resultado de la actividad amilásica es la producción de
-
Upload
jaqueline-odair -
Category
Documents
-
view
222 -
download
1
description
Digestión y absorción de chos
Transcript of Dig y Abrosción de Carbohidratos

DIGESTIÓN Y ABROSCIÓN DE CARBOHIDRATOS
El platillo, el cual consiste en pollo asado y puré de papa cuenta con los siguientes tipos de
carbohidratos: almidón, lactosa y glucosa. Este último pasará directamente, esto es, sin
modificación alguna al intestino delgado para ser absorbido.
La digestión, concretamente del almidón, comienza en la cavidad bucal. La masticación es
un proceso mecánico que hace disminuir el tamaño de los gránulos de almidón,
aumentando la exposición para las enzimas hidrolíticas. La primera de ellas en actuar es la
α-amilasa salival (ptialina), que actúa sobre los enlaces α (1→4). Su actuación permanece
en el bolo alimenticio, incluso en el estómago por un tiempo, deteniendo su actividad
cuando el pH ácido gástrico alcanza el grumo alimenticio, ya que el pH óptimo de esta
enzima es vecino a la neutralidad. La ptialina puede hidrolizarse hasta 30-40% de almidón
ingerido, obteniéndose maltosa, maltotriosa y dextrosas límites (compuestos con 3-5
unidades de glucosa en uniones α (1→4) y 1 ramificación en α (1→6)).
Posteriormente, la hidrólisis del almidón restante se reinicia gracias a la amilasa pancreática
tras el vaciamiento gástrico, fundamentalmente en duodeno y yeyuno. El resultado de la
actividad amilásica es la producción de más dextrinas límite, maltotriosa, maltosa y algunas
moléculas de glucosa.
Finalmente, la hidrólisis total de los productos de la digestión pancreática se completa en el
intestino, gracias a la actividad de las enzimas del borde en cepillo de la mucosa. La
glucoamilasa hidroliza las uniones α (1→4) de los oligómeros de glucosa, mientras que las
uniones α (1→6) de los puntos de ramificación son hidrolizados por la isomaltasa.
En el intestino se lleva a cabo también la hidrólisis de los disacáridos. La maltasa actúa
sobre la maltosa para generar dos moléculas de glucosa, mientras que la lactosa es
hidrolizada por la lactasa a glucosa y galactosa.

Los monosacáridos resultantes de la digestión del pollo y la papa serán entonces solamente
moléculas de glucosa y galactosa, las cuales van a ser absorbidas principalmente en
duodeno y yeyuno proximales.
La absorción de la glucosa y galactosa se lleva a cabo por un mecanismo de transporte
activo de membrana mediado por un transportador (proteína transportadora) y es
dependiente de Na+ habiendo una estimulación recíproca del transportador de los azúcares
por Na+ y de este ion por los azúcares. La proteína transportadora tiene dos lugares de
unión para el Na+, y uno para el azúcar.
El Na+ se mueve a favor de un gradiente electroquímico al interior de la célula. La energía
liberada por este transporte a favor de un gradiente químico y eléctrico de Na+ es utilizado
para transportar la glucosa en contra de un gradiente de concentración.
En el enterocito existe una ATPasa Na+-K+ dependiente que crea un gradiente
electroquímico para el Na+, al sacar éste fuera de la célula, que propicia el que el catión se
mueva desde la luz intestinal hacia el enterocito. Tras su entrada en el enterocito, la glucosa
y la galactosa pasan a los capilares de la mucosa por transporte facilitado o difusión simple.
Como resultado de la absorción intestinal, la glucosa llega al hígado por la vena porta. Parte
de la glucosa absorbida también llega a la circulación sanguínea periférica para ser
reconocida por receptores pancreáticos y estimular a las células β.

DIGESTIÓN Y ABSORCIÓN DE LÍPIDOS
En la digestión y absorción se pueden distinguir varias etapas: emulsión de la grasa
alimentaria, digestión intraluminal, solubilización micelar, absorción, formación de
quilomicrones en el enterocito y transporte desde estos a la circulación.
El alimento contiene los siguientes lípidos: ácidos grasos libres, triglicéridos, fosfolípidos y
colesterol.
Después de la ingesta del alimento el quimo se mezcla con la lipasa lingual. La enzima
permanece activa cuando el pH es bajo y degrada ácidos grasos de cadena corta sobre todo
procedentes de triglicéridos. Posteriormente actúa una lipasa gástrica, que con la motilidad
gástrica asegura que los nutrientes se mezclen con las enzimas en forma apropiada y que

los lípidos se transformen en partículas más pequeñas. En consecuencia, los lípidos que
entraron por el tubo digestivo quedan emulsificados (mezcla de lípidos con sustancias a
base de agua), luego de atravesar el estómago. Los ácidos grasos de cadena corta liberados
en el estómago debido a su hidrólisis en el C1 o C3 pueden absorberse en forma directa en
la corriente sanguínea venosa a través de la pared gástrica.
Controlados por el píloro del estómago los lípidos emulsificados se vierten en el duodeno,
donde se agregan jugos pancreáticos y bilis. Los ácidos biliares se unen con las partículas
lipídicas y forman cargas negativas en sus superficies, lo que permite que la colipasa se
adhiera a los triglicéridos. A partir de allí la lipasa pancreática (triacilglicerol esterasa),
inhibida por los ácidos biliares se une con la colipasa y cataliza la hidrólisis de los
triacilgliceridos en las posiciones 1 y 3. Hay muchas otras enzimas pancreáticas que
funcionan de acuerdo con el mismo principio:
Colesterol esterasa. Hidroliza ésteres de colesterol.
Fosfolipasas A1 y A2. Hidrolizan fosfolípidos en las posiciones 1 y 2,
respectivamente. La fosfolipasa A2 es la más importante.
El tamaño de las partículas lipídicas disminuye a medida que avanza la hidrólisis. Junto con
los ácidos biliares los productos resultantes de la lipólisis se ensamblan en forma
espontánea para constituir partículas denominadas micelas mixtas.
Los productos de la hidrólisis llegan a la membrana del borde en cepillo como micelas
mixtas, donde las células de la mucosa los absorben por difusión pasiva. Una vez en el
interior de la célula epitelial, se lleva a cabo una resíntesis en el retículo endoplásmico liso
de los compuestos que fueron ingeridos.
Dentro de la luz del intestino, los 1-monoacilgliceroles se hidrolizan hacia ácidos grasos y
glicerol y los 2-monoacilgliceroles se reacilan hacia triacilgliceroles por medio de la vía del
monoacilglicerol. El glicerol liberado en la luz intestinal no se reutiliza, sino que pasa hacia
la vena porta; el glicerol liberado dentro del epitelio se reutiliza para la síntesis de
triacilglicerol por medio de la vía del acido fosfatidico.

Los ácidos grasos de cadena larga se esterifican para dar triacilglicerol en las células de la
mucosa, y junto con los otros productos de la digestión de lípidos, se secretan como
quilomicrones hacia los linfáticos y entran en el torrente sanguíneo por medio del conducto
torácico. Los ácidos grasos de cadena corta y media se absorben principalmente hacia la
vena porta hepática como ácidos grasos libres.
El colesterol se esterifica principalmente en la mucosa intestinal antes de ser incorporado
hacia quilomicrones. El colesterol no esterificado y otros esteroles se transportan de manera
activa hacia afuera de las células de la mucosa, hacia la luz del intestino.

DIGESTIÓN Y ABSORCIÓN DE PROTEÍNAS
La digestión de las proteínas comienza en el estómago gracias a la acción de las pepsinas.
Estas enzimas degradan los enlaces peptídicos dentro de grupos R presentes en fenilalanina
o tirosina. La mucosa intestinal secreta estas endopeptidadas como precursores inactivos
denominados pepsinógenos y estos se activan en medios con pH ácido. Además los ácidos

facilitan el ataque catalítico de las enzimas a través de la desnaturalización de proteínas.
Así pues, las enzimas gástricas producen polipéptidos u oligopéptidos.
En el duodeno las pepsinas se desactivan debido al aumento del pH. Las endopeptidasas
(tripsina, quimotripsina, y elastasa) y exopeptidasas (carboxipeptidasas) pancreáticas
hidrolizan péptidos de cadena larga para transformarlos en fragmentos más cortos o
aminoácidos individuales. Estas enzimas también se secretan como precursores inactivos:
el tripsinógeno se activa para formar tripsina gracias a la acción de la enzima
enteropeptidasa. La tripsina tiene como función hidrolizar enlaces peptídicos presentes en
arginina y lisina, la quimotripsina actúa sobre aminoácidos aromáticos, y la elastasa actúa
de manera preferencial sobre aminoácidos alifáticos neutros.
El paso digestivo final depende de enzimas que se encuentran en la membrana del borde en
cepillo. Estas enzimas son aminopeptidasas (degradan péptidos desde el extremo amino),
tripeptidasas y dipeptidasas (hidrolizan tripéptidos y dipéptidos respectivamente). Tras la
digestión combinada de las proteasas, los productos que entran en el enterocito son
aminoácidos, dipéptidos, tripéptidos y tetrapéptidos.
La absorción intestinal de aminoácidos se realiza por diversos mecanismos de transporte.
Para la entrada en el enterocito desde la luz intestinal, a través de la membrana apical
existen sistemas de transporte de dos tipos, unos dependientes de Na+ y otros
independientes de él. Además, estos sistemas son específicos de grupos de aminoácidos que
tienen ciertas características químicas.
En todos los casos se trata de un transporte activo con gasto de energía metabólica.
En la membrana basolateral del enterocito existen otros sistemas de transporte de
aminoácidos, diferentes a los descritos en el borde en cepillo, que permiten el paso de éstos
hacia la sangre de los capilares mucosales.

En el borde en cepillo existe un sistema de transporte que tiene afinidad por dipéptidos y
tripéptidos, haciendo que estos sean incorporados en el enterocito. Se trata de un transporte
activo secundario que establece un gradiente de potencial electroquímico para el Na+.