
Ecología Aplicada. Vol. 2
146
ISSN 1726-2216 UNIVERSIDAD NACIONAL AGRARIA LA MOLINA FACULTAD DE CIENCIAS Maestría Ecología Aplicada LIMA PERÚ ECOLOGÍA APLICADA Revista del Departamento Académico de Biología Volumen 2, Nº 1 / Diciembre 2003 ea
Transcript of Ecología Aplicada. Vol. 2

ISSN 1726-2216
UNIVERSIDAD NACIONAL AGRARIA LA MOLINA
FACULTAD DE CIENCIAS Maestría
Ecología Aplicada
LIMA PERÚ
ECOLOGÍA APLICADA
Revista del Departamento Académico de Biología Volumen 2, Nº 1 / Diciembre 2003
ea

ECOLOGIA APLICADA Revista del Departamento Académico de Biología
UNIVERSIDAD NACIONAL AGRARIA LA MOLINA Av. La Molina s/n - La Molina - LIMA – PERU
Apartado postal 456 Lima 100 http://www.lamolina.edu.pe/ecolapl/
EDITOR
Germán Arellano Cruz, Blgo., Mg. Sc. Universidad Nacional Agraria La Molina
CONSEJO EDITOR Ph.D. Lucrecia Aguirre Universidad Nacional Agraria La Molina (Perú) Ph.D. Consuelo Arellano Universidad Nacional Agraria La Molina (Perú) Dr. Antonio Brack Programa de las Naciones Unidas Para el Desarrollo (Perú) Mg. Sc. Aldo Ceroni Universidad Nacional Agraria La Molina (Perú) Mg. Sc. Mercedes Flores Universidad Nacional Agraria La Molina (Perú) Dra. Maura Kufner Universidad Nacional de Córdoba (Argentina) Dr. Jaime Mendo Universidad Nacional Agraria La Molina (Perú) Dra. Pia Parolin Universität Hamburg (Alemania)
Ph.D. Cesar Rodríguez University of Toronto (Canadá) Dra. Lily Rodríguez Centro de Conservación, Investigación y Manejo de Áreas Naturales Cordillera Azul (Perú) Dra. Inés Redolfi Universidad Nacional Agraria La Molina (Perú) Dr. Edgar Sánchez Universidad Nacional Agraria La Molina (Perú) Mg. Sc. Juan Torres Universidad Nacional Agraria La Molina (Perú) Dra. Graciela Vilcapoma Universidad Nacional Agraria La Molina (Perú) Mg. Sc. Martha Williams Universidad Nacional Agraria La Molina (Perú) Dra. Doris Zúñiga Universidad Nacional Agraria La Molina (Perú)
La Revista Ecología Aplicada, es un medio de expresión legítimo de la ciencia de la Ecología para investigadores peruanos y extranjeros, el concepto de esta revista reúne todas las características principales para cumplir la función de la comunicación científica: la calidad, el registro para la protección de los derechos de autor, al conocimiento revelado con los resultados de la investigación y la función de archivo relacionada con el almacenamiento, accesibilidad que aseguran la estabilidad de la información. Constituye un espacio para la discusión y la publicación de los avances de los trabajos científicos o técnicos en Ecología y la problemática ambiental que pueden ser divulgados como artículos originales, notas científicas y artículos de revisión, que son sometidos al arbitraje y revisión por pares. ISSN 1726-2216 Título clave: Ecología aplicada Título clave abreviado: Ecol. apl. Hecho el depósito legal 2002-5474 Revisión del texto en Inglés: Martha Williams Diagramación: Germán Arellano Portada: Bosque tropical estacionalmente seco. Piura – Perú. Fotografía: Zulema Quinteros © Departamento Académico de Biología Universidad Nacional Agraria La Molina Apartado Postal 456. Lima 100 Telf. 3495647 - 3495669 Anexos 271 – 273
Financiamiento Vol. 2: Vicerrectorado Académico. Universidad Nacional Agraria La Molina (UNALM) (26%). Escuela de Post Grado (UNALM) (14%). Maestría Ecología Aplicada (UNALM) – Cursos Modulares (47%). Autofinanciamiento (13%). Información para la preparación de manuscritos y suscripciones: Correo electrónico: [email protected] Apartado Postal 456. Lima 100 Departamento Académico de Biología Universidad Nacional Agraria La Molina. Av. La Molina s/n La Molina Lima – Perú.

ECOLOGÍA APLICADA
Revista del Departamento Académico de Biología de la Universidad Nacional Agraria La Molina
__________________________________________________________________________________________ Volumen 2, número 1 Diciembre 2003 __________________________________________________________________________________
Contenido
Artículos originales
1 Aldo Ceroni Stuva. Composición florística y vegetación de la Cuenca la Gallega. Morropón. Piura………………………………………………………………………..
Pag. 1 – 8
2 Aldo Ceroni Stuva. Distribución de las leguminosas de la parte alta de la Cuenca la Gallega. Morropón. Piura………………………………………………………………
Pag. 9 - 13
3 Tina Lerner Martínez, Aldo Ceroni Stuva y Claudia E. González Romo. Etnobotánica de la Comunidad Campesina “Santa Catalina de Chongoyape” en el bosque seco del área de conservación privada Chaparrí – Lambayeque……………………………….
Pag. 14 -20 4 Cruz Márquez, Hernán Vargas, Howard Snell, André Mauchamp, James Gibbs y
Washington Tapia ¿Por qué tan pocas Opuntia en la Isla Española-Galápagos?..........
Pag. 21 -29 5 M. T. F. Piedade, P. Parolin y W. J. Junk. Estratégias de dispersão, produção de frutos
e extrativismo da palmeira Astrocaryum jauari Mart. Nos Igapós do Rio Negro: implicações para a ictiofauna………………………………………………………….
Pag. 31 -40 6 Percy A. Zevallos Pollito e Ymber Flores Bendezú. Caracterización morfológica de
plántulas de “uña de gato” Uncaria tomentosa (Willdernow ex Roemer & Schultes) D.C. y U. guianensis (Aublet) Gmelin del Bosque Nacional Alexander von Humboldt……………………………………………………………………………….
Pag. 41-46 7 Carlos Tavares Corrêa y Ana Sabogal de Alegría. Estabilización de dunas litorales
utilizando Sesuvium portulacastrum L. en el Departamento de La Libertad, costa norte del Perú…………………………………………………………………………...........
Pag. 47-50 8 Jaime Mendo y Matthias Wolff. El impacto de El Niño sobre la producción de concha
de abanico (Argopecten purpuratus) en Bahía Independencia, Pisco, Perú…………...
Pag. 51-57 9 Alfredo Giraldo Mendoza y Germán Arellano Cruz. Resiliencia de la comunidad
epígea de Coleoptera en las Lomas de Lachay después del evento El Niño 1997-98….
Pag. 59-68 10 Sidney Novoa S., Viviana Castro C., Aldo Ceroni S. e Inés Redolfi P. Relación entre
la hormiga Camponotus sp. (Hymenoptera: Formicidae) y una comunidad de cactus (Cactaceae) en el Valle del Río Chillón………………………………………………..
Pag. 69-73 11 Francisco Fontúrbel Rada. Algunos criterios biológicos sobre el proceso de
eutrofización a orillas de seis localidades del Lago Titikaka…………………………..
Pag. 75-79 12 Gisella Matos Cuzcano y Doris Zúñiga Dávila. Viabilidad de cepas de Rizobios en
inoculantes basados en soportes no estériles……………………………………………
Pag. 81-85 13 Pedro O. Ruiz y Charles B. Davey. Efectos del manejo de suelos de laderas en hongos
formadores de micorrizas arbusculares y en bacterias fijadoras de nitrógeno en ultisoles sujetos a erosión pluvial en la Amazonía Peruana……………………………
Pag. 87-92 14 Luis A. Arévalo, Julio C. Alegre y Raúl Fasabi. Efecto del fósforo sobre el
establecimiento del Centrosema macrocarpum Benth dentro de una plantación de pijuayo (Bactris gasipaes H.B.K.) en un ultisol del trópico húmedo………………….
Pag. 93-97 15 Abelardo Calderón Rodríguez y Román Felipe Serpa Ibáñez. Efectos del Paraquat
sobre el crecimiento y la morfología de la microalga Dunaliella tertiolecta…………..
Pag. 99-102 16 José Alberto Iannacone Oliver, Neil Salazar Capcha y Lorena Alvariño Flores.
Variabilidad del ensayo ecotoxicológico con Chironomus calligraphus Goeldi (Diptera: Chironomidae) para evaluar cadmio, mercurio y plomo…………………….
Pag. 103-110 17 José Alberto Iannacone Oliver y Lorena Alvariño Flores. Efecto ecotoxicológico
agudo del mercurio sobre larvas del “muy muy” Emerita analoga (Stimpson) (Decapoda: Hippidae) procedentes de cuatro localidades de lima……………………..
Pag. 111-115

18 José Iannacone, Jaime Mansilla y Karen Ventura. Macroinvertebrados en las lagunas de Puerto Viejo, Lima – Perú…………………………………………………………..
Pag. 116-124
19 Augusto Fachín Terán y Eduardo Matheus von Mülhen. Reproducción de la taricaya Podocnemis unifilis Troschel 1848 (Testudines: Podocnemididae) en la Várzea del Medio Solimões, Amazonas, Brasil. ……………………………………………….
Pag. 125-132 20 Armando H. Escobedo Galván y Fernando Mejía Vargas. El “cocodrilo de tumbes”
(Crocodylus acutus Cuvier 1807): estudio preliminar de su estado actual en el Norte de Perú……………………………………………………………………………….....
Pag. 133-135 21 Armando H. Escobedo Galván. Períodos de actividad y efecto de las variables
ambientales en cocodrilos (Crocodylus acutus Cuvier 1807): evaluando los métodos de determinación de la fracción visible………………………………………………...
Pag. 136-140 Notas científicas
22 Shaleyla Kelez, Ximena Vélez-Zuazo y Camelia Manrique. New evidence on the loggerhead sea turtle Caretta caretta (Linnaeus 1758) in Peru……………………….
Pag. 141-142
23 Martha Williams de Castro, Alexandra Castillo y Constanza Rosas. Observaciones preliminares sobre comportamiento prenatal y postnatal de Rattus rattus var. norvegicus (Linneo 1758) en el laboratorio…………………………………………...
Pag. 143-148

Ecología Aplicada, 2(1), 2003 ISSN 1726-2216 Depósito legal 2002-5474
COMPOSICIÓN FLORÍSTICA Y VEGETACIÓN DE LA CUENCA LA GALLEGA. MORROPÓN. PIURA
Aldo Ceroni Stuva1
Resumen
Se estudió la composición florística y la vegetación de la cuenca La Gallega (Morropón, Piura). Las plantas fueron recolectadas entre los 165 y 3,100 msnm. Se registraron los rangos altitudinales de los diferentes pisos de vegetación. El total de especies colectadas pertenecen a 56 familias, siendo Asteraceae, Fabaceae, Lamiaceae y Solanaceae, las mejor representadas. Algunas especies como Acacia macracantha, Cordia lutea, Eriotheca discolor y Erythrina edulis, fueron las de mayor distribución altitudinal. La cuenca La Gallega también presenta una gran diversidad de ecosistemas encontrándose 6 pisos de vegetación: piso de cactáceas columnares y bosque seco, bosque caducifolio, bosque siempre verde, vegetación arbustiva y herbácea, monte ribereño y vegetación de arbustos bajos. Palabras clave: Composición florística, vegetación, rangos altitudinales, pisos de vegetación.
Abstract The floristic composition and the vegetation of the La Gallega basin (Morropón, Piura) were studied. The plants were collected between 165 and 3,100 masl. The altitudinal ranges of the different vegetation floors were registered. The total amount of collected species belong to 56 families, being Asteraceae, Fabaceae, Lamiaceae and Solanaceae, the best represented. Species such as Acacia macracantha, Cordia lutea, Eriotheca discolor and Erythrina edulis, were those of greater altitudinal distribution. La Gallega basin also presents a great diversity of ecosystems composing 6 vegetation floors: columnar cactus and dry forest, deciduous forest, evergreen forest, shrubby and herbaceous vegetation, riverside mountain vegetation and low shrub vegetation. Key words: Floristic composition, vegetation, altitudinal ranges, vegetation floors.
Introducción Los estudios de composición florística y vegetación son muy importantes por cuanto permiten conocer las especies que conforman un área geográfica, así como su distribución y fisonomía. La composición florística del norte peruano es conocida parcialmente gracias a investigaciones interesadas en conocer algunos tipos de hábitats, como el caso de los desiertos y formaciones de lomas de Ferreyra (1977), Dillon & Rundel (1990) y Rundel et al. (1991) y de los bosques montanos de Dillon (1994), Dillon et al. (1995), Gentry (1992), Sagástegui (1994) y Young & Valencia (1992), citados por Sagástegui et al. (1999). La falta de reportes sobre estudios florísticos y distribución de especies vegetales en áreas como la cuenca La Gallega, ubicada en la sierra norte del Perú, impulsaron a realizar el presente estudio. Además, los Andes del norte presentan una pronunciada discontinuidad como si las montañas se curvaran de NW a NE y se fragmentan haciendo que las vertientes del Amazonas y del Pacífico estén separadas por uno de los pasos más bajos de toda la cordillera, llamado el Abra de Porculla o Depresión de Huarmaca o Deflexión de Huancabamba. La región que circunda a esta deflexión ha sido ampliamente reconocida como una fuente de diversidad primaria en muchos grupos
de plantas. En la región de la deflexión (Piura, Cajamarca y Amazonas) contamos con alrededor de 715 especies endémicas, que representan alrededor del 20% del endemismo para todo el país (Sagástegui et al., 1999). Asimismo, en el Perú la complejidad de cadenas andinas genera una sucesión de pisos ecológicos diversos desde el mar tropical, desierto, bosque seco, bosques templados, jalca, valles cálidos, bosques de neblina y bosques tropicales amazónicos depositarios de una composición florística muy rica representada por cerca de 17,144 especies, distribuidas en 2,458 géneros y 224 familias (Brako & Zarucchi, 1993; Sagástegui et al., 1999). La cuenca La Gallega no ha sido exactamente visitada por expediciones botánicas, sin embargo, hubo algunas que pasaron muy cerca de ella y que han dado a conocer la vegetación en general con su respectiva composición florística. Entre ellas tenemos por ejemplo la Ex-pedición Geográfico-Científica para sudamérica, en 1802, a cargo del insigne naturalista alemán ALEXANDER VON HUMBOLDT y del botánico francés AIMEE BONPLAND y en 1912, la del insigne botánico alemán AUGUSTO WEBERBAUER, al estudiar los Andes peruanos al norte del grado 6 (Weberbauer, 1945). Estudios taxonómicos y etnobotánicos han sido centrados en la
____________________ 1 Departamento Académico de Biología. Universidad Nacional Agraria La Molina. Av. La Molina s/n. Apartado postal 456. Lima 100.Perú. Fax: 3496015. Correo electrónico: [email protected]

COMPOSICIÓN FLORÍSTICA Y VEGETACIÓN DE LA CUENCA LA GALLEGA. MORROPÓN. PIURA
Diciembre 2003 __________________________________________________________________________________________
2
parte alta de la cuenca por haberse observado que esta zona presenta una vegetación rica en especies muchas de las cuales son utilizadas por los pobladores, representando así una fuente de recursos poten-cialmente importantes (Ceroni, 1998, 2002). El presente trabajo tuvo como objetivo aportar al conocimiento de la composición florística y la vegetación de la cuenca La Gallega, para lo cual se registró la distribución altitudinal de las especies, así como de los pisos de vegetación. Materiales y métodos Ubicación La cuenca La Gallega está ubicada al suroeste de la microregión andina central, en el Departamento de Piura, dentro de la gran Cuenca del Piura. La microregión andina central se ubica entre los 4°90' y 5°10' LS y entre los 79°30' y 80°10' LO (Figura 1).
Figura 1. Cuenca la Gallega Santo Domingo Morropón , Piura - Perú.
Caracterización de la zona de estudio Área La cuenca La Gallega está formada por el río La Gallega que nace en el lado sur de los Altos, en el cerro Peña de Huaycas. Su longitud es de 37 Km. Tiene una superficie de 17,897 Ha. y un área de 185. 60 Km2 (CIGA 1990). La cuenca puede ser dividida en tres zonas: baja (entre Morropón y El Puente), media (entre Paltashaco y Chungayo); y alta (entre Chungayo y Los Altos, en la divisoria de aguas). Clima Las partes bajas son áreas con escasas lluvias, de menos de 450 mm/año. Las partes medias con pre-cipitaciones moderadas de 450 a 650 mm/año. Mien-tras que las partes más altas son húmedas con preci-pitaciones de 800 a 1,000 mm/año. La humedad ambiental también hace notar sus diferencias entre las partes bajas, medias y altas. Las primeras son más secas, mientras que la última es muy húmeda y con nieblas casi estables durante el verano. Suelos En las partes bajas de la cuenca los suelos son básicamente arenosos-cascajosos y superficiales en la margen derecha. En el lado izquierdo los suelos son más profundos. En las partes medias van apareciendo suelos arcillosos-arenosos y arenosos. En las partes altas, como Quinchayo, los suelos también son arcillosos y pedregosos. A la altura de Santo Domingo, los suelos son arcillosos-limosos y de color pardo. Fisiografía La cuenca está formada por numerosas quebradas que han excavado profundamente sus lechos. Las prin-cipales quebradas son por la margen derecha: El Palto, de La Cruz, Santo Domingo, Chungayo, Tuñalí, Jacanacas y Caracucho. Por la margen izquierda: El Guabo, Huancas, Sural, Paltashaco, La Ensillada y del Guineo. El ancho máximo del valle es de 11 Km. en su sector alto y el mínimo de 6 Km. en su sector bajo. En el fondo del valle hay terrazas escalonadas con un ancho medio de 1 Km. Entre este lugar y Paltashaco, el valle desaparece en una encañada de laderas con pendientes superiores al 45%. Pasando los 600 m, las paredes que acompañan el fondo del valle se van abriendo dando lugar a un valle en "V". Luego las gra-dientes del terreno se hacen más suaves de 20% a 30%. A la altura de Santo Domingo se abre una repisa y el relieve aquí tiene una pendiente de 10% al 15%. Ecología De acuerdo al Mapa Ecológico del Perú (INRENA, 1995) la cuenca La Gallega se encuentra comprendida entre 8 zonas de vida: la parte baja entre Morropón y El Puente: matorral desértico-Tropical (md-T), matorral desértico-Premontano Tropical (md-

ALDO CERONI STUVA
Ecología Aplicada Vol. 2 Nº 1, pp. 1-8 __________________________________________________________________________________________
3
PT), monte espinoso-Tropical (mte-T) y monte espinoso-Premontano Tropical (mte-PT); la parte media entre Paltashaco y Chungayo: bosque seco-Premontano Tropical (bs-PT); y la parte alta entre Chungayo y Los Altos, en la divisoria de aguas: bosque seco-Montano Bajo Tropical (bs-MBT), bosque húmedo-Montano Bajo Tropical (bh-MBT) y bosque muy húmedo-Montano Tropical (bmh-MT). Metodología Se realizaron viajes de recolección de las especies botánicas, registrándose las altitudes, así como la especie o especies dominantes en cada piso altitudinal. Los viajes de recolección se realizaron durante 1991, 1994 y 1995 en los meses de abril, agosto, octubre y noviembre, a fin de tener una mayor información acerca de los cambios de vegetación entre las épocas seca y húmeda. Las plantas fueron recolectadas entre los 165 y 3,100 msnm. en las tres zonas correspondientes a las partes baja, media y alta de la cuenca. Se tomó además la altitud de una serie de puntos referenciales de la cuenca (Tabla 1). El material botánico fue posteriormente determinado en el Herbario Weberbauer del Departamento Académico de Biología de la Universidad Nacional Agraria La Molina (MOL) y en el Herbario del Museo de Historia Natural de la Universidad Nacional Mayor de San Marcos (USM). Se levantó un perfil del recorrido rea-lizado en la cuenca mostrando los pisos de vegetación. Resultados y discusión Composición Florística Los resultados de la composición florística nos muestra la gran diversidad biológica de esta cuenca. Se registraron un total de 191 especies pertenecientes a 54 familias de angiospermas y 2 familias de plantas vasculares sin semilla (Tabla 2). Entre las familias de angiospermas 50 fueron de dicotiledóneas y 4 de monocotiledóneas. Las familias mejor representadas fueron: Asteraceae, Fabaceae, Lamiaceae y Solanaceae con 19, 29, 13 y 18 especies, respectivamente (Figura 2).
Asteraceae19
Solanaceae18
Otras109
Fabaceae29Lamiaceae
13 Figura 2. Número de Especies de las familias de angiospermas mejor representadas en la cuenca La Gallega. Morropón. Piura.
Tabla 1. Relación de puntos referenciales de la cuenca la gallega. Santo Domingo. Morropón. Piura.
LUGAR ALTITUD (msnm)
Baja Morropón Piedra del Toro Caracucho Faycal El Puente
165 260 330 475 565
Media Paltashaco Pambarumbe El Bronce y Tuñalí *
885 1,110 1,230
Alta Chungayo Santiago Santo Domingo Yumbe San Miguel Ñoma Cabuyal Tiñarumbe Quinchayo Chico Bosque de Tiñarumbe Quinchayo Grande Quinchayo Alto Chicope La Cordillera Los Altos
1,350 1,350 1,475 1,500 1,550 1,640 1,750 1,770 1,800 1,960 1,970 2,325 2,900 3,000 3,100
* Medida tomada al frente. Pisos de Vegetación De acuerdo a Weberbauer (1945) se puede encontrar 6 pisos de vegetación (Figura 3), con las siguientes especies dominantes:
Figura 3. Pisos de vegetación en el perfil del recorrido en la cuenca La Gallega. Morropón. Piura. (a) Piso de Cactáceas Columnares y Bosque Seco; (b) Bosque Caducifolio; (c) Bosque Siempre Verde; (d) Vegetación Arbustiva y Herbácea; (e) Monte Ribereño y (f) Vegetación de Arbustos Bajos.

COMPOSICIÓN FLORÍSTICA Y VEGETACIÓN DE LA CUENCA LA GALLEGA. MORROPÓN. PIURA
Diciembre 2003 __________________________________________________________________________________________
4
a. Piso de Cactáceas Columnares y Bosque Seco (165 - 350 msnm). En Morropón, a 177 m aproximadamente, se ob-serva en ladera cactáceas columnares, entre ellas Neoraimondia arequipensis “gigantón” así como también Cordia lutea “overo". En Piedra del Toro, a 260 m se presenta un Piso de Cactáceas Columnares mezclado con "overo". b. Bosque Caducifolio (350 - 1,000 msnm). En Caracucho a 350 m aparecen especies caduci-folias como Loxopterigium huasango "hualtaco"; Bursera graveolens "palo santo"; Erythrina edulis "pajul" y otras como Bougainvillea peruviana "papelillo" y Senna atomaria "mataperro". A 495 m aparece un bosque de Ceiba sp. "ceibo" y a 533 m continúan las bombacáceas mezcladas con Erythrina edulis "pajul"; Acacia macracantha "faique"; Cordia lutea "overo"; así como cactáceas y bromeliáceas. En El Puente a 565 m persisten las especies caducifolias, pero esta vez comparten el hábitat con Prosopis pallida "algarrobo"; Eriotheca discolor "pasallo blanco" y Cochlospermum vitifolium "pasallo negro". A 885 m en Paltashaco o Santa Catalina de Mossa, es muy frecuente Tillandsia usneoides "barbas de viejo" sobre los árboles de Ceiba sp. "ceibo", ya más dispersos. El Bosque Seco sigue un poco más arriba mezclándose finalmente con cactáceas y "tilansias", hasta los 1,000 m. c. Bosque Siempre Verde (1,200 - 1,300 msnm). A partir de los 1,000 m. se va dejando atrás el Bosque Caducifolio. Entre El Bronce y Tuñalí, a 1,230 m, hay un notable cambio de vegetación, de un Bosque Caducifolio dominado por Eriotheca discolor "pasallo" a un Bosque Siempre Verde dominado por Acacia macracantha "faique". d. Vegetación Arbustiva y Herbácea (1,400 - 1,700 msnm) Por encima de los 1,300 m el Bosque Siempre verde es reemplazado por una vegetación arbustiva mezclada con herbáceas, en donde destacan especies como Lycopersicon peruvianum "hierba del huisco", Fulcaldea laurifolia "flor de agua", Styrax cordatus "piñán", Cestrum aff. auriculatum "sauco", Buddleja americana "Santa María" y Passiflora manicata "granadilla de ratón", entre otras. e. Monte Ribereño (1,700 - 2,500 msnm). Por Quinchayo Grande hacia el Alto, desde los 1,780 hasta los 2,130 m aproximadamente, se puede encontrar vegetación de Monte Ribereño, con especies como Ocotea cernua "puchuguero"; Peperomia blanda "jergo"; Myrcianthes discolor "lanche"; Baccharis latifolia "chilca"; Mauria sp. "shimir" y Erythrina edulis "pajul", entre otras. Por Ñoma, a los 2,000 m aproximadamente, se observa vegetación natural con especies como Peperomia blanda "jergo"; Ocotea cernua "puchuguero" y Erythrina edulis "pajul". También hay vegetación secundaria con plantas espinosas como Duranta mutisii "cashaquiro".
f. Vegetación de Arbustos Bajos (2,700 - 3,100 msnm). Llegando a las partes más altas hay un notable cambio de vegetación quedando atrás las especies arbóreas y arbustivas o haciéndose más dispersas y apareciendo una vegetación más baja. Es característica la presencia de arbustos bajos como Arcytophyllum ericoides "pichana", Satureja cf. panicera "llantén serrano", Passiflora cf. mixta "granadilla de campo", Achyrocline alata "vira-vira", Salvia corrugata "salvia real" y Baccharis grandicapitulata "huacum", entre otras. Tal como se aprecia en la descripción anterior, la cuenca La Gallega presenta una importante diversidad vegetal tanto por el número de especies encontradas, así como, por el número de pisos de vegetación que va desde piso de cactáceas columnares y bosque seco, de zonas cálidas y desérticas, pasando por un bosque transicional y bosque siempre verde correspondiente a zonas más húmedas, hasta llegar a una vegetación de arbustos bajos propia de zonas altoandinas. Es importante resaltar que las cuatro familias mejor representadas corresponden al 42% de las especies registradas, aproximadamente la mitad de la composición florística de la cuenca. Estas familias son las que comprenden el mayor número de especies en el norte de Perú (Sagástegui et al., 1999). También entre las especies algunas de ellas alcanzan la mayor distribución altitudinal, encontrándose en más de un piso de vegetación, como Acacia macracantha, Cordia lutea, Eriotheca discolor y Erythrina edulis, siendo esta última la de mayor rango de distribución. Desde el punto de vista fitogeográfico algunas leguminosas registradas aquí son importantes y características de pisos altitudinales de las vertientes occidentales del extremo norte. En los pisos medios entre los 1,000 y 2,200 m, en los montes de arbustos solos o con algunos árboles que alternan con estepas de gramíneas o con herbazales mezclados, destacan entre otras leñosas Amicia glandulosa, Caesalpinia spinosa y especies de Dalea e Inga. En los pisos superiores entre los 2,700 y 3,300 m, en las estepas de gramíneas con arbustos dispersos destacan Fiebrigiella gracilis, Vicia andicola y especies de Lupinus, entre otras (Weberbauer, 1945). Todo esto especialmente en la cordillera situada entre Lagunas, Frías y Ayabaca, lugares cercanos al área del presente estudio. Otro aspecto importante de la composición florística de la cuenca es la presencia de especies endémicas para el Perú, como Caesalpinia spinosa, Lupinus albert-smithianus y Lupinus mutabilis, en la parte alta (Ceroni, 1998). La cuenca La Gallega también representa una valiosa fuente de recursos fitogenéticos de importancia económica. En un estudio etnobotánico realizado en la parte alta se registraron 86 especies útiles, de las cuales 46 tienen uso medicinal para 32 dolencias diferentes. Estas especies corresponden a 8 categorías de uso:

ALDO CERONI STUVA
Ecología Aplicada Vol. 2 Nº 1, pp. 1-8 __________________________________________________________________________________________
5
Alimenticias-Bebidas-Frutales, De carpintería y Construcción, De higiene, Forrajeras, Medicinales, Para la buena suerte, Para leña y Para reforestar (Ceroni, 2002). Conclusiones 1. Se registraron un total de 191 especies correspon-dientes a 56 familias botánicas, indicando esto una importante diversidad específica. 2. Las familias mejor representadas fueron: Asteraceae, Fabaceae, Lamiaceae y Solanaceae con 19, 29, 13 y 18 especies, respectivamente. 3. La cuenca La Gallega presenta una gran diversidad de ecosistemas encontrándose 6 pisos de vegetación entre los 165 y 3,100 msnm. 4. Existen especies de amplia distribución altitudinal como Acacia macracantha, Cordia lutea, Eriotheca discolor y Erythrina edulis, siendo esta última la de mayor rango de distribución. 5. La cuenca La Gallega representa un importante ecosistema por albergar especies endémicas para el Perú y ser una fuente de recursos fitogenéticos por la importancia económica de las especies presentes. Agradecimientos El autor expresa su agradecimiento al M.Sc. Juan Torres por sugerir el área de estudio y apoyar en las primeras visitas de reconocimiento, a la Bach. Alina Pace, la Blga. Susana Gushiken y el Blgo. Omar Carrión por su apoyo en las salidas de campo, a la Ing. Luz María Jiménez, por los contactos necesarios para contar con guías durante los recorridos en el área de estudio, a la Dra. Elida Carrillo, Dra. Magda Chango, Dra. Graciela Vilcapoma, M.Sc. Mercedes Flores, Bach. Arturo Granda y al Dr. Carlos Reynel por las facilidades dadas en los Herbarios MOL y UNMSM, y a la Dra. Elsa Fun por el apoyo financiero y logístico de la Central Peruana de Servicios (CEPESER) de Piura. Sin todos ellos hubiera sido difícil poder realizar el presente estudio. Litertura citada Brako L. & Zarucchi J. 1993. Catalogue of the
Flowering Plants and Gymnosperms of Perú. Monograhs in Systematic Botany from the Missouri Botanical Garden. Vol 45.
Ceroni A. 1998. Estudio Taxonómico de la Familia Leguminosae de la Cuenca La Gallega. Santo Domingo. Parte Alta (Morropón, Piura). Tesis para optar el Grado de Magíster en Botánica Tropical. Universidad Nacional Mayor de San Marcos.
. 2002. Datos Etnobotánicos del Poblado de Huaylingas. Cuenca La Gallega. Morropón. Piura. Revista Ecología Aplicada. Vol. 1. (1): 65 -70.
CIGA. 1990. Cuaderno de Geografía Aplicada. Proyecto de Desarrollo Rural Integral de la Sierra Central del Departamento de Piura. PUC-ORSTOM. Primera parte: El Medio Natural.
Ferreyra R. 1979. Sinopsis de la Flora Peruana. Gymnospermas y Monocotiledóneas. Universidad Nacional Mayor de San Marcos. Lima.: 2-6
INSTITUTO NACIONAL DE RECURSOS NATURALES (INRENA). 1995. Mapa Ecológico del Perú. Guía explicativa. Lima-Perú.
MacBride J.F. 1936 - 1971. Flora of Perú. Botanical Series. Field Museum of Natural History. U.S.A.
Sagástegui A. 1995. Diversidad Florística de Contumazá. Editorial Libertad E.I.R.L. Universidad Antenor Orrego de Trujillo. Perú.
, Dillon M., Sánchez I., Leiva S. & Lezama P. 1999. Diversidad Florística del Norte de Perú. Tomo I. World Wildlife Fund Inc. UPAO.
Soukup J. 1987. Vocabulario de los Nombres Vulgares de la Flora Peruana y Catálogo de los Géneros. Editorial Salesiana. Lima. Perú.
Weberbauer A. 1945. El Mundo Vegetal de los Andes Peruanos. Estudio Fitogeográfico. Estación Experimental Agrícola de La Molina. Ministerio de Agricultura. Lima.: 6, 30, 31.
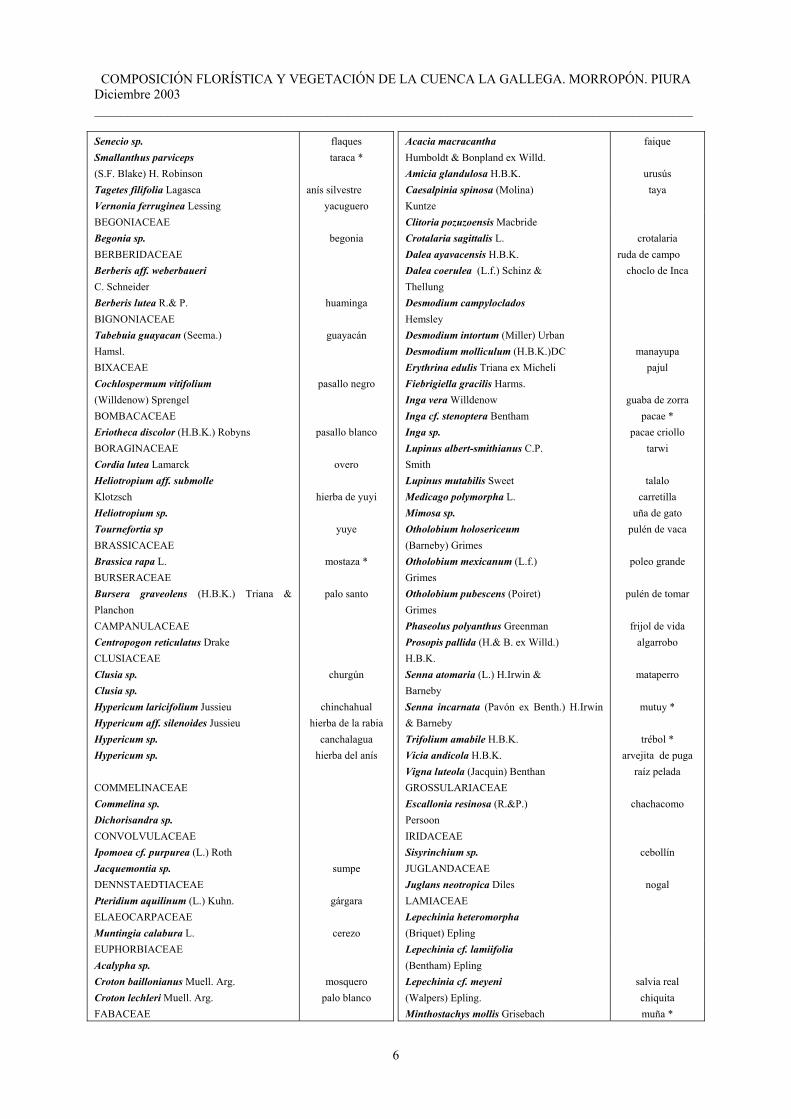
Tabla 2. Plantas colectadas en la cuenca La Gallega Santo Domingo. Morropón.
FAMILIA / GÉNERO / ESPECIE NOMBRE COMÚN
ACANTHACEAE Dyschoriste sp. AMARANTHACEAE Alternanthera dominii Schinz Alternanthera villosa H.B.K. Alternanthera sp. Amaranthus sp Iresine sp. ANACARDIACEAE Loxopterygium huasango Spruce Mauria sp. ASTERACEAE Achyrocline alata (H.B.K.) DC Baccharis latifolia (R.& P.)Persoon Baccharis grandicapitulata Hieron Bidens pilosa L. Diplostephium sp. Eupatorium sp. Fulcaldea cf. laurifolia (Humb.& Bonp.) Poir.ex Less. Fulcaldea laurifolia (Humb.& Bonp.) Poir.ex Less. Gnaphalium cf. dombeyanum DC. Gnaphalium cf. elegans H.B.K. Hymenostephium sp. Hypochoeris sp. Liabum floribundum Lessing Matricaria recutita L. Senecio aff. usgorensis Cuatrecasas
abrojo quínua negra
hualtaco shimir
vira-vira
chilca huacum
amor seco huancuncillo
flor de agua
flor de agua
vira-vira hierba Julia achicoria
yutuz manzanilla
COMPOSICIÓN FLORÍSTICA Y VEGETACIÓN DE LA CUENCA LA GALLEGA. MORROPÓN. PIURA
Diciembre 2003 __________________________________________________________________________________________
6
Senecio sp. Smallanthus parviceps (S.F. Blake) H. Robinson Tagetes filifolia Lagasca Vernonia ferruginea Lessing BEGONIACEAE Begonia sp. BERBERIDACEAE Berberis aff. weberbaueri C. Schneider Berberis lutea R.& P. BIGNONIACEAE Tabebuia guayacan (Seema.) Hamsl.
flaques taraca *
anís silvestre
yacuguero
begonia
huaminga
guayacán
BIXACEAE Cochlospermum vitifolium (Willdenow) Sprengel BOMBACACEAE Eriotheca discolor (H.B.K.) Robyns BORAGINACEAE Cordia lutea Lamarck Heliotropium aff. submolle Klotzsch Heliotropium sp. Tournefortia sp BRASSICACEAE Brassica rapa L. BURSERACEAE Bursera graveolens (H.B.K.) Triana & Planchon CAMPANULACEAE Centropogon reticulatus Drake CLUSIACEAE Clusia sp. Clusia sp. Hypericum laricifolium Jussieu Hypericum aff. silenoides Jussieu Hypericum sp. Hypericum sp. COMMELINACEAE Commelina sp. Dichorisandra sp. CONVOLVULACEAE Ipomoea cf. purpurea (L.) Roth Jacquemontia sp. DENNSTAEDTIACEAE Pteridium aquilinum (L.) Kuhn. ELAEOCARPACEAE Muntingia calabura L. EUPHORBIACEAE Acalypha sp. Croton baillonianus Muell. Arg. Croton lechleri Muell. Arg. FABACEAE
pasallo negro
pasallo blanco
overo
hierba de yuyi
yuye
mostaza *
palo santo
churgún
chinchahual hierba de la rabia
canchalagua hierba del anís
sumpe
gárgara
cerezo
mosquero palo blanco
Acacia macracantha Humboldt & Bonpland ex Willd. Amicia glandulosa H.B.K. Caesalpinia spinosa (Molina) Kuntze
faique
urusús taya
Clitoria pozuzoensis Macbride Crotalaria sagittalis L. Dalea ayavacensis H.B.K. Dalea coerulea (L.f.) Schinz & Thellung Desmodium campyloclados Hemsley Desmodium intortum (Miller) Urban Desmodium molliculum (H.B.K.)DC Erythrina edulis Triana ex Micheli Fiebrigiella gracilis Harms. Inga vera Willdenow Inga cf. stenoptera Bentham Inga sp. Lupinus albert-smithianus C.P. Smith Lupinus mutabilis Sweet Medicago polymorpha L. Mimosa sp. Otholobium holosericeum (Barneby) Grimes Otholobium mexicanum (L.f.) Grimes Otholobium pubescens (Poiret) Grimes Phaseolus polyanthus Greenman Prosopis pallida (H.& B. ex Willd.) H.B.K. Senna atomaria (L.) H.Irwin & Barneby Senna incarnata (Pavón ex Benth.) H.Irwin & Barneby Trifolium amabile H.B.K. Vicia andicola H.B.K. Vigna luteola (Jacquin) Benthan GROSSULARIACEAE Escallonia resinosa (R.&P.) Persoon IRIDACEAE Sisyrinchium sp. JUGLANDACEAE Juglans neotropica Diles LAMIACEAE Lepechinia heteromorpha (Briquet) Epling
crotalaria
ruda de campo choclo de Inca
manayupa pajul
guaba de zorra
pacae * pacae criollo
tarwi
talalo carretilla
uña de gato pulén de vaca
poleo grande
pulén de tomar
frijol de vida
algarrobo
mataperro
mutuy *
trébol * arvejita de puga
raíz pelada
chachacomo
cebollín
nogal
Lepechinia cf. lamiifolia (Bentham) Epling Lepechinia cf. meyeni (Walpers) Epling. Minthostachys mollis Grisebach
salvia real chiquita muña *

ALDO CERONI STUVA
Ecología Aplicada Vol. 2 Nº 1, pp. 1-8 __________________________________________________________________________________________
7
Minthostachys tomentosa (Bentham) Epling Minthostachys sp. Salvia corrugata M.Vahl. Salvia pauciserrata Bentham Salvia cf. oppositiflora R.& P. Salvia cf. rhombifolia R.& P. Salvia sp. Salvia sp. Satureja pulchella (H.B.K.) Briquet LAURACEAE Ocotea cernua (Nees) Mez Persea caerulea (R.& P.) Mez LILIACEAE Bomarea distichifolia (R.& P.) Baker Bomarea sp. Nothoscordum cf. andicola Kunth LOASACEAE Caiophora sp. LOGANIACEAE Buddleja americana L. LORANTHACEAE Psittacanthus cucullaris (Lamarck) Blume LYCOPODIACEAE Lycopodium clavatum L. Lycopodium thyoides Willd. LYTHRACEAE Cuphea strigulosa H.B.K. Cuphea aff. ciliata R.& P. MALVACEAE Sida sp. MELASTOMATACEAE Axinaea cf. oblongifolia (Cogniaux) Wurdack Brachyotum sp.
poleo madural lúcuma
polen de vaca salvia real
hierba de la sarna hierba del quinde
llantén serrano
puchuguero paltón
hierba del chucaque
Santa María
suelda con suelda
purga trencilla
hierba del toro hierba del toro
windum
illirque
arcillesca
Brachyotum sp. Miconia aff. alpina Cogniaux Miconia sp. Tibouchina laxa (Desrousseaux) Cogniaux MORACEAE Ficus sp. MYRSINACEAE Myrsine oligophylla Zalbruckner Myrsine sp. MYRTACEAE Eucalyptus globulus Labillardiere Eugenia myrobalana DC. Myrcia sp. Myrcianthes discolor (H.B.K.) Mc Vaugh Myrcianthes cf. discolor (H.B.K.)
zarcilleja chiquita chuchilla
higuerón
yuto
alcanfor arrayán lanche lanche
lanche de
Mc Vaugh Myrcianthes sp Psidium guajava L. NYCTAGINACEAE Boerhavia sp. Bougainvillea peruviana H.& B. ONAGRACEAE Fuchsia sp. Oenothera rosea Aiton OXALIDACEAE Oxalis sp. Oxalis sp. PAPAVERACEAE Argemone subfusiformis Ownbey PASSIFLORACEAE Passiflora ligularis A.L.Jussieu Passiflora manicata (A.L.Jussieu) Persoon Passiflora cf. mixta L.f. PHYTOLACCACEAE Phytolacca weberbaueri H.Walter PIPERACEAE Peperomia blanda (Jacquin) H.B.K. Peperomia trinervis R.& P.
pava lanche de Dios
guayabo
papelillo
fucsia * hierba de la rabia
cardo santo
granadilla silvestre
granadilla de ratón
granadilla de campo
yumbe
jergo" jergo"
Piper aff. hispidum Swartz Piper sp. POACEAE Cortaderia sp. Chusquea sp Rhipidocladum sp. POLYGALACEAE Monnina salicifolia R.& P. POLYGONACEAE Muehlenbeckia hastulata (J.E.Smith.) I.M.Johnston Muehlenbeckia volcanica (Bentham) Endlicher Rumex conglomeratus Murray PROTEACEAE Oreocallis grandiflora (Lamarck) R.Brown ROSACEAE Rubus robustus C.Presl. Rubus sp. RUBIACEAE Arcytophyllum ericoides (Willd.ex R.& S.) Standley Cinchona sp. Galium hypocarpium (L.) Endlicher ex Grisebach
cordoncillo colorado
cordoncillo blanco
cortadera
suro
clarín
bejuco colorado
hierba dulce
putaya
cucharillo
zarzamora zarzamora
pichana
cascarilla perlilla

COMPOSICIÓN FLORÍSTICA Y VEGETACIÓN DE LA CUENCA LA GALLEGA. MORROPÓN. PIURA
Diciembre 2003 __________________________________________________________________________________________
8
Palicourea latifolia Krause Spermacoce tenuior L. RUTACEAE Ruta graveolens L. SAPOTACEAE Pouteria lucuma (R.& P.) Kuntze SCROPHULARIACEAE Alonsoa linearis (Jacquin) R.& P. Alonsoa meridionalis (L.f.) Kuntze Calceolaria nivalis H.B.K. Calceolaria cf. ballotifolia Kraenzlin Calceolaria cf. Chelidonioides H.B.K.
hierba de araña
ruda
lúcumo silvestre
hierba del oso hierba de la quemazón zapatito * zapatito *
zapatito *
Calceolaria cf. engleriana Kraenzlin SOLANACEAE Acnistus arborescen (L.) Schlechtendal Brugmansia candida Persoon Brugmansia sp. Cestrum affine H.B.K. Cestrum aff. auriculatum L'Héritier Cestrum sp. Iochroma grandiflorum Bentham Lycopersicon peruvianum (L.) Miller Nicandra physalodes (L.) Gaertner Solanum americanum Miller Solanum piurae Bitter Solanum cf. capirense Dunal Solanum cf. chrysotrichum Schlechtendal Solanum sp. Solanum sp. Solanum sp. Solanum sp. Streptosolen jamesonii (Bentham) Miers STYRACACEAE Styrax cordatus (R.& P.) A.DC. URTICACEAE Boehmeria aff. caudata Swartz Phenax hirtus (Swartz) Weddell VERBENACEAE Duranta mutisii L.f. Lippia cf. alba (Miller) N.E.Brown Verbena sp.
zapatito *
chin-chin
guarguar guarguar
sauco
sauco chin-chin serrano hierba del huisco
chilalo
hierba mora *
palito negro ambuyuco
espino
mermelada *
piñán
chogolo
cashaquiro mastrante Verbena
(*) Nombres comunes de fuentes bibliográficas

Ecología Aplicada, 2(1), 2003 ISSN 1726-2216 Depósito legal 2002-5474
DISTRIBUCIÓN DE LAS LEGUMINOSAS DE LA PARTE ALTA DE LA CUENCA LA GALLEGA. MORROPÓN. PIURA.
Aldo Ceroni Stuva1.
Resumen Se estudió la distribución de las leguminosas de la parte alta de la cuenca La Gallega (Morropón, Piura). Las plantas fueron recolectadas entre los 1,350 y 3,100 msnm. Se registraron los rangos altitudinales de las especies. Las 21 especies colectadas pertenecen a 3 subfamilias y 9 tribus. La subfamilia Papilionoideae y la tribu Phaseoleae presentaron una mayor distribución. La especie con menor distribución fue Caesalpinia spinosa, encontrada sólo en los alrededores de Santo Domingo, mientras que las especies más ampliamente distribuidas fueron Erythrina edulis y Vigna luteola. También se encontraron 10 nuevos registros de distribución de leguminosas para el Perú. Palabras clave: Distribución, rangos altitudinales, familia, subfamilia, tribu.
Abstract The leguminous plants distribution in the highest part of the La Gallega basin (Morropón, Piura) was studied. The plants were collected between 1,350 and 3,100 masl. Altitudinal ranges of the species were registered. The 21 collected species belong to 3 subfamilies and 9 tribes. The greatest distribution corresponded to the Papilionoideae subfamily and to the Phaseoleae tribe. Caesalpinia spinosa was the species with the narrowest distribution and was found only near Santo Domingo, while species such as Erythrina edulis and Vigna luteola were those of the widest distribution. We also found 10 new records of leguminous plants distribution in Perú. Key words: Distribution, altitudinal ranges, family, subfamily, tribe.
Introducción Una de las familias botánicas más importantes en la flora tropical es el de las Leguminosae o Fabaceae. Esta familia incluye numerosas especies con una gran variedad de portes y formas de vida. Muchas de sus especies tienen una enorme importancia económica para el hombre (Heywood, 1985). En muchos países en desarrollo, las leguminosas alimenticias constituyen la principal fuente de proteína en su alimentación (Bruno, 1990). Un estudio de distribución de las especies de esta familia es muy importante por cuanto permite por un lado reforzar la situación taxonómica de las especies y por otro lado, conocer la ubicación de recursos vegetales tanto para el uso directo, industrial, así como, fuente de recursos fitogenéticos. Estudios sobre leguminosas en el Perú se hallan en forma fragmentaria en diversas obras a nivel nacional e internacional, tales como los de MacBride (1943), Gómez (1960), Arce (1970), Carrillo (1974), Cerrate (1979), Ferreyra (1987), Barneby (1988), Tovar (1990), Brako & Zarucchi (1993), Flores (1997) y Reynel & Pennington (1997), todos ellos en estudios florísticos y revisión de géneros (Flores, 1997; Ceroni, 1998). Las leguminosas son también una de las familias con mayor número de especies para el norte peruano. Además, por ser los Andes del norte más bajos en altitud, debido al Abra de Porculla o Deflexión de Huancabamba, es una región ampliamente reconocida
como una fuente de diversidad primaria y endemismo en muchos grupos de plantas (Sagástegui et al., 1999). El presente trabajo tuvo como objetivo aportar al conocimiento de la distribución de las leguminosas de la parte alta de la cuenca La Gallega, para lo cual se registró el rango altitudinal de las especies, en cada una de las localidades. El estudio ha sido centrado en la parte alta por haberse observado que esta zona presenta una vegetación más rica en especies, muchas de las cuales son utilizadas por los pobladores. Materiales y métodos Ubicación La cuenca La Gallega está ubicada al suroeste de la microregión andina central, en el Departamento de Piura, dentro de la gran Cuenca del Piura. La microregión andina central se ubica entre los 4°90' y 5°10' LS y entre los 79°30' y 80°10' LO (Figura 1). Caracterización de la zona de estudio Área La cuenca La Gallega está formada por el río La Gallega que nace en el lado sur de los Altos, en el cerro Peña de Huaycas. Su longitud es de 37 Km. Tiene una superficie de 17,897 Ha. y un área de 185. 60 Km2 (CIGA, 1990). La parte alta es aquella comprendida entre Chungayo y Los Altos, en la divisoria de aguas, es decir entre 1,350 y 3,100 msnm.
_____________________________ 1 Departamento Académico de Biología. Universidad Nacional Agraria La Molina. Av. La Molina s/n Lima Perú. Apartado postal 456. Lima 100 Perú. Fax: 3496015. Correo electrónico: [email protected].

DISTRIBUCIÓN DE LAS LEGUMINOSAS CUENCA LA GALLEGA. MORROPÓN. PIURA.
Diciembre 2003 __________________________________________________________________________________________
10
Figura 1. Cuenca La Gallega. Santo Domingo. Morropón. Piura. Clima La parte alta es húmeda con precipitaciones de 800 a 1,000 mm/año. La humedad ambiental también hace notar sus diferencias entre la parte baja, media y alta de la cuenca. Esta última es muy húmeda y con nieblas casi estables durante el verano. Suelos La parte alta presenta suelos arcillosos y pedregosos. A la altura de Santo Domingo, los suelos son arcillosos-limosos y de color pardo. Fisiografía La cuenca está formada por numerosas quebradas que han excavado profundamente sus lechos. El ancho máximo del valle es de 11 Km. en su sector alto y el mínimo de 6 Km. en su sector bajo. En el fondo del valle hay terrazas escalonadas con un ancho medio de 1 Km. A la altura de Santo Domingo se abre una repisa y el relieve aquí tiene una pendiente de 10% al 15%.
Ecología De acuerdo al Mapa Ecológico del Perú (INRENA, 1995) la parte alta de la cuenca La Gallega se encuentra comprendida entre 3 zonas de vida: bosque seco-Montano Bajo Tropical (bs-MBT), bosque húmedo-Montano Bajo Tropical (bh-MBT) y bosque muy húmedo-Montano Tropical (bmh-MT). Metodología Se realizaron viajes de recolección de las especies botánicas, registrándose las altitudes, así como la especie o especies dominantes en cada lugar. Los viajes de recolección se realizaron durante 1991, 1994 y 1995 en los meses de abril, agosto, octubre y noviembre. Las plantas fueron recolectadas entre los 1,350 y 3,100 msnm., en los siguientes lugares: Santo Domingo, Ñoma, Yumbe, Portachuelo, Chungayo, Cabuyal, Tiñarumbe, Chicope, Quinchado Chico, Quinchado Grande, Quinchado Alto y Las Pircas. El material botánico fue posteriormente determinado en el Herbario Weberbauer del Departamento Académico de Biología de la Universidad Nacional Agraria La Molina (MOL) y en el Herbario del Museo de Historia Natural de la Universidad Nacional Mayor de San Marcos (USM). Resultados y discusión Especies colectadas Las leguminosas colectadas en la parte alta de la cuenca La Gallega fueron un total de 21, pertenecientes a las tres subfamilias: 1 Mimosoideae; 1 Caesalpinioideae y 19 Papilionoideae (Tabla 1). Asimismo, las especies de la última subfamilia pertenecen a 9 tribus (Tabla 2). Distribución de las especies Las especies de leguminosas encontradas en el área de estudio se distribuyen en su mayoría de manera amplia. Sin embargo, a fin de conocer mejor esta distribución, se destaca la ubicación de los diferentes lugares de colección para cada una de ellas. Así la Mimosoideae Inga vera Willdenow, se encontró en las partes bajas y medias del área de estudio, en Santo Domingo y Ñoma, entre los 1,500 y 1,800 msnm. La Caesalpinioideae Caesalpinia spinosa (Molina) Kuntze, se encontró sólo en los alrededores de Santo Domingo a 1,500 msnm, la cual aparentemente tendría una distribución más restringida (Figura 2). Las Papilionoideae presentaron una mayor distribución en toda el área de estudio desde Chungayo hasta las Pircas, cubriendo un rango altitudinal entre 1,350 y 3,100 msnm. (Figuras 2, 3 y 4). Entre ellas, la tribu Phaseoleae fue la de mayor distribución ya que sus representantes fueron encon-trados en la mayoría de los puntos de colección: Chungayo, Yumbe, Santo Domingo, Tiñarumbe, Ca-buyal, Ñoma, Quinchayo Chico y Quinchayo Grande,

ALDO CERONI STUVA
Ecología Aplicada Vol. 2 Nº 1, pp. 9-13 __________________________________________________________________________________________
11
cubriendo un rango altitudinal entre 1,350 y 1,900 msnm (Figura 4). En cuanto a las especies mismas, las que presentaron mayor distribución, considerando el número puntos de colección y la distancia entre los puntos, fueron: Amicia glandulosa H.B.K., Dalea coerulea (L.f.) Schinz & Thellung, Desmodium campyloclados Hemsley, Erythrina edulis Triana ex Micheli, Fiebrigiella gracilis Harás., Inga vera Willdenow, Lupinus mutabilis Sweet, Otholobium holosericeum (Barneby) Grimes, Phaseolus polyan- thus Greenman y Vigna luteola (Jacquin) Bentham; siendo las más ampliamente distribuidas Erythrina edulis Triana ex Micheli y Vigna luteola (Jacquin) Bentham.
Figura 2. Lugares de colección de la Mimosoideae, Caesalpinoideae y las Papilionoideae de las Tribus Psoraleeae y Amorpheae en la parte alta de la cuenca La Gallega. Morropón. Piura. Al hacer una revisión de la distribución en el Perú de las especies encontradas aquí, basada principalmente en la información dada por Brako & Zarucchi (1993) y la obtenida de los herbarios USM y MOL, se encuentra que varias especies no están reportadas para Piura o para la zona específica de Santo Domingo en Morropón. Por lo tanto especies como Crotalaria sagittalis L., Clitoria pozuzoensis Macbride, Dalea coerulea (L.f.) Schinz & Thellung, Desmodium molliculum (H.B.K.) DC., Inga vera Willdenow, Medicago polymorpha L., Otholobium
holosericeum (Barneby) Grimes, Phaseolus polyanthus Greenman, Vicia andicola H.B.K. y Vigna luteola (Jacquin) Bentham, serían casos de nuevos reportes de distribución para el Perú (Ceroni, 1998). Entre ellas, por ejemplo, al consultar con uno de los autores del estudio del Género Inga en el Perú (Reynel & Pennington, 1997) para la respectiva determinación, fue interesante conocer que la especie Inga vera Willdenow, también llega hasta Piura.
Figura 3. Lugares de colección de las Papilionoideae de las Tribus Desmodieae, Aeschynomeneae, Crotalarieae y Genisteae en la parte alta de la cuenca La Gallega. Morropón. Piura. Conclusiones 1. Se registraron un total de 21 especies de leguminosas, pertenecientes a 3 subfamilias y 9 tribus. 2. La subfamilia Papilionoideae presenta una mayor distribución desde Chungayo hasta las Pircas, cubriendo un rango altitudinal entre 1,350 y 3,100 msnm. 3. La tribu Phaseoleae presenta una mayor distribución, con representantes encontrados en la mayoría de puntos de colección, en un rango altitu-dinal entre 1,350 y 1,900 msnm. 4. La especie con menor distribución fue Caesalpinia spinosa, encontrada sólo en los alrededores de Santo Domingo, mientras que las especies más ampliamente distribuidas fueron Erythrina edulis y Vigna luteola.

DISTRIBUCIÓN DE LAS LEGUMINOSAS CUENCA LA GALLEGA. MORROPÓN. PIURA.
Diciembre 2003 __________________________________________________________________________________________
12
5. Las especies: Crotalaria sagittalis, Clitoria pozuzoensis, Dalea coerulea, Desmodium molliculum, Inga vera, Medicago polymorpha, Otholobium holose-riceum, Phaseolus polyanthus, Vicia andicola y Vigna luteola, constituyen nuevos registros de distribución para el Perú. 6. Las localidades con mayor concentración de leguminosas fueron Santo Domingo, Ñoma y Quinchayo Grande, a 1,475, 1,640 y 1,970 msnm, respectivamente.
Figura 4. Lugares de colección de las Papilionoideae de las Tribus Vicieae, Trifolieae y Phaseoleae en la parte alta de la cuenca La Gallega. Morropón. Piura. Agradecimientos El autor expresa su agradecimiento al M.Sc. Juan Torres por sugerir el área de estudio y apoyar en las primeras visitas de reconocimiento, a la Bach. Alina Pace, la Blga. Susana Gushiken y el Blgo. Omar Carrión por su apoyo en las salidas de campo, a la Ing. Luz María Jiménez, por los contactos necesarios para contar con guías durante los recorridos en el área de estudio, a la Dra. Elida Carrillo, Dra. Magda Chango, Dra. Graciela Vilcapoma, M.Sc. Mercedes Flores y al Dr. Carlos Reynel por las facilidades dadas en los Herbarios MOL y UNMSM, y a la Dra. Elsa Fun por el apoyo financiero y logístico de la Central Peruana de Servicios (CEPESER) de Piura. Sin todos ellos hubiera sido difícil poder realizar el presente estudio.
Tabla 1. Lista de leguminosas colectadas en la parte alta de la cuenca La Gallega. Santo Domingo. Morropón. Piura.
S U B F A M I L I A / N O M B R E C I E N T Í F I C O
N O M B R E C O M Ú N
M I M O S O I D E A E I n g a v e r a W i l l d e n o w
g u a b a d e z o r r a
C A E S A L P I N I O I D E A E C a e s a l p i n i a s p i n o s a ( M o l i n a ) K u n t z e
t a y a
P A P I L I O N O I D E A E A m i c i a g l a n d u l o s a H . B . K . C l i t o r i a p o z u z o e n s i s M a c b r i d e C r o t a l a r i a s a g i t t a l i s L . D a l e a a y a v a c e n s i s H . B . K . D a l e a c o e r u l e a ( L . f . ) S c h i n z & T h e l l u n g D e s m o d i u m c a m p y l o c l a d o s H e m s l e y D e s m o d i u m i n t o r t u m ( M i l l e r . ) U r b a n D e s m o d i u m m o l l i c u l u m ( H . B . K . ) D C . E r y t h r i n a e d u l i s T r i a n a e x M i c h e l i F i e b r i g i e l l a g r a c i l i s H a r m s . L u p i n u s a l b e r t - s m i t h i a n u s C . P . S m i t h L u p i n u s m u t a b i l i s S w e e t M e d i c a g o p o l y m o r p h a L . O t h o l o b i u m h o l o s e r i c e u m ( B a r n e b y ) G r i m e s O t h o l o b i u m m e x i c a n u m ( L . f . ) G r i m e s O t h o l o b i u m p u b e s c e n s ( P o i r e t ) G r i m e s P h a s e o l u s p o l y a n t h u s G r e e n m a n V i c i a a n d i c o l a H . B . K . V i g n a l u t e o l a ( J a c q u i n ) B e n t h a m
u r u s ú s
c r o t a l a r i a
r u d a d e c a m p o c h o c l o d e l
I n c a
m a n a y u p a
p a j u l
t a r w i t a l a l o
c a r r e t i l l a p u l é n d e v a c a
p o l e o g r a n d e
p u l é n d e t o m a r
f r i j o l d e v i d a a r v e j i t a d e
p u g a r a í z p e l a d a
Tabla 2. Tribus de las Papilionoideae colectadas en la parte alta de la cuenca La Gallega. Morropón. Piura.
T R I B U E S P E C I E D E S M O D I E A E A E S C H Y N O M E N E A E P S O R A L E E A E A M O R P H E A E C R O T A L A R I E A E G E N I S T E A E V I C I E A E T R I F O L I E A E P H A S E O L E A E
D e s m o d i u m c a m p y l o c l a d o s H e m s l e y D e s m o d i u m i n t o r t u m ( M i l l e r . ) U r b a n D e s m o d i u m m o l l i c u l u m ( H . B . K . ) D C . A m i c i a g l a n d u l o s a H . B . K . F i e b r i g i e l l a g r a c i l i s H a r m s . O t h o l o b i u m h o l o s e r i c e u m ( B a r n e b y ) G r i m e s O t h o l o b i u m m e x i c a n u m ( L . f . ) G r i m e s O t h o l o b i u m p u b e s c e n s ( P o i r e t ) G r i m e s D a l e a a y a v a c e n s i s H . B . K . D a l e a c o e r u l e a ( L . f . ) S c h i n z & T h e l l u n g C r o t a l a r i a s a g i t t a l i s L . L u p i n u s a l b e r t - s m i t h i a n u s C . P . S m i t h L u p i n u s m u t a b i l i s S w e e t V i c i a a n d i c o l a H . B . K . M e d i c a g o p o l y m o r p h a L . C l i t o r i a p o z u z o e n s i s M a c b r i d e E r y t h r i n a e d u l i s T r i a n a e x M i c h e l i P h a s e o l u s p o l y a n t h u s G r e e n m a n V i g n a l u t e o l a ( J a c q u i n ) B e n t h a m

ALDO CERONI STUVA
Ecología Aplicada Vol. 2 Nº 1, pp. 9-13 __________________________________________________________________________________________
13
Literatura citada Brako L. & Zarucchi J. 1993. Catalogue of the Flowe-
ring Plants and Gymnosperms of Perú. Monograhs in Systematic Botany from the Missouri Botanical Garden. Vol 45.
Bruno H.J. 1990. Leguminosas Alimenticias. Distribuidora “Faede”. S.A. Lima. Perú.
Ceroni A. 1998. Estudio Taxonómico de la Familia Leguminosae de la Cuenca La Gallega. Santo Domingo. Parte Alta (Morropón, Piura). Tesis para optar el Grado de Magíster en Botánica Tropical. Universidad Nacional Mayor de San Marcos.
CIGA. 1990. Cuaderno de Geografía Aplicada. Proyecto de Desarrollo Rural Integral de la Sierra Central del Departamento de Piura. PUC-ORSTOM. Primera parte: El Medio Natural.
Flores M. 1997. La Familia Leguminosae en el Valle del Chillón – parte media y alta. Tesis para optar el Grado de Magíster en Botánica Tropical.
Universidad Nacional Mayor de San Marcos. Heywood V.H. 1985. Las Plantas con Flores.
Editorial Reverté S.A. España. : 145-148. INSTITUTO NACIONAL DE RECURSOS
NATURALES (INRENA). 1995. Mapa Ecológico del Perú. Guía explicativa. Lima-Perú.
MacBride J.F. 1936 - 1971. Flora of Perú. Botanical Series. Field Museum of Natural History. U.S.A.
Sagástegui A., Dillon M., Sánchez I., Leiva S. & Lezama P. 1999. Diversidad Florística del Norte de Perú. Tomo I. World Wildlife Fund Inc. UPAO.
Soukup J. 1987. Vocabulario de los Nombres Vulgares de la Flora Peruana y Catálogo de los Géneros. Editorial Salesiana. Lima. Perú.
Weberbauer A. 1945. El Mundo Vegetal de los Andes Peruanos. Estudio Fitogeográfico. Estación Expe-rimental Agrícola de La Molina. Ministerio de Agricultura. Lima.: 6, 30, 31.

Ecología Aplicada, 2(1), 2003 ISSN 1726-2216 Depósito legal 2002-5474
ETNOBOTÁNICA DE LA COMUNIDAD CAMPESINA “SANTA CATALINA DE CHONGOYAPE” EN EL BOSQUE SECO DEL ÁREA DE CONSERVACIÓN PRIVADA CHAPARRÍ – LAMBAYEQUE
Tina Lerner Martínez1, Aldo Ceroni Stuva2 y Claudia E. González Romo3.
Resumen Resultan particularmente escasos los estudios etnobotánicos en los bosques secos del noroeste peruano como también los referidos a comunidades campesinas o mestizas sujetas a una mayor influencia modificadora del mercado. Asimismo, son pocos los ejemplos sobre comunidades campesinas que logran proteger sus terrenos y establecerlos como unidades de conservación reconocidas por el estado. En este sentido, como punto de partida para la formulación de proyectos de desarrollo dentro del Área de Conservación Privada Chaparrí, se llevó a cabo un estudio durante enero y setiembre del 2002, con la participación de los pobladores de la comunidad de Santa Catalina de Chongoyape, para identificar los recursos vegetales que se utilizan localmente. Se registraron 122 taxa de plantas útiles (103 géneros y 45 familias botánicas), además 8 géneros nuevos de plantas alimento de la “pava aliblanca” (Penelope albipennis). Por otro lado, se describen los vocablos y las expresiones locales referidas a las plantas. Además, Se recopiló información histórica y arqueológica para evidenciar que algunas plantas se siguen utilizando desde épocas precolombinas en el bosque seco del noroeste peruano. Palabras claves: Bosque seco, Chaparrí, etnobotánica, Santa Catalina de Chongoyape
Abstract There is a lack of ethnobotanic studies conducted in the dry forests of northern Perú and in peasant communities subjected to man¨s major influencing action. Likewise, examples that show the influence of market economy on peasant communities are scarce, particularly those that show how a peasant community can establish a national conservation area in its territory as a means of protecting their land. In order to build the foundations of a future development project, an ethnobotany study, from January to September 2002 was conducted in the Chaparrí Private Area of Conservation which included the participation of the Santa Catalina de Chongoyape community. The goals were to identify local plants used by the population. 122 taxa of useful plants were registered (103 genera and 45 botanical families were identified). 8 new genera of plants used as food by the white-winged guan (Penelope albipennis) were found. A description of the local vocabulary and expressions was made as well as a compilation of historical and archaeological data records to establish the traditional uses of the plants of the zone. Key words: Chaparrí, dry forest, ethnobotany, Santa Catalina de Chongoyape
Introducción Los estudios de etnobotánica perse desarrollados en la zona norte del Perú son escasos (La Torre, 1998; Schjellerup et al., 2000; Ceroni, 2002; Fernández & Rodríguez, 2003) y para los bosques secos semidensos y de sabana de Lambayeque no existen antecedentes. Por otro lado, uno de los aspectos más descuidados de la Etnobotánica es el estudio de los usos de las plantas por los campesinos o mestizos. A diferencia de las sociedades indígenas, que generalmente viven en núcleos geográfica y económicamente aislados de las sociedades nacionales, los grupos campesinos viven en comunidades conectadas con mercados regionales, nacionales e, incluso, internacionales; y representan la mayor parte de la población, de las así llamadas, culturas tradicionales (Toledo, 1993). Santa Catalina de Chongoyape es una comunidad campesina agrícola
y ganadera, asentada en el bosque seco de Lambayeque a 60 kilómetros de la ciudad de Chiclayo, en un lugar clave de tránsito de costa a sierra y viceversa. Se diferencia por haber creado en sus territorios la primera Área de Conservación Privada en el Perú. El 86% del territorio comunal (34,413 hectáreas) ha sido asignado para su conservación desde diciembre de 1999. En ella se vienen realizando programas de reintroducción de animales endémicos en peligro de extinción, como es el caso de la “pava aliblanca” (Penelope albipennis Taczanowski). Dentro de sus objetivos, además de preservar la biodiversidad y prestar distintos “servicios ambientales” (mantenimiento del ciclo hidrológico, conservación de suelos, mitigación del calentamiento global) está el desarrollo de la población local contemplando una política de
_____________________________ 1 Herbario MOL. Universidad Nacional Agraria La Molina. Av. La Molina s/n Lima - Perú. Apartado postal 456. Lima 100. Correo electrónico: [email protected]. 2 Departamento Académico de Biología Facultad de Ciencias. Universidad Nacional Agraria La Molina. Apartado 456. Lima 100. Correo electrónico: [email protected] 3 Universidad Autónoma de Tamaulipas México. 13 Blvd. López Mateos 928, Cd. Victoria, Tamaulipas México. Correo electrónico: [email protected]

TINA LERNER, ALDO CERONI Y CLAUDIA E. GONZÁLEZ
Ecología Aplicada Vol. 2 Nº 1, pp. 14-20 __________________________________________________________________________________________
15
aprovechamiento sostenible de su flora (Angulo, 2000). En este sentido, un estudio etnobotánico caracterizado por la participación comunal, que explora la diversidad vegetal y los conocimientos sobre las plantas, es necesario como punto de partida en el proceso de desarrollo. La investigación debe propiciar que los campesinos contemplen las alternativas para el aprovechamiento de su diversidad vegetal (recursos genéticos y bioquímicos) y las ventajas comparativas que representan la conservación de estos ecosistemas neotropicales. Además, es importante revalorar los conocimientos sobre el bosque seco que han persistido ante el proceso de aculturación, deforestación y avance de la frontera agrícola, y en cierta medida buscar su protección para el beneficio de la comunidad. Por lo expuesto, el presente estudio se refiere al registro de la utilización de los recursos vegetales por parte de la Comunidad Campesina de Santa Catalina de Chongoyape en el año 2002. Los datos están complementados con información histórica y cultural que brindan un contexto más amplio para el análisis sobre el conocimiento de las plantas y su estado de conservación, en el bosque seco del noroeste peruano. Los objetivos principales fueron: 1) identificar las
plantas útiles de la zona y documentar sus usos; 2) aportar al conocimiento de la “etnoclasificación local”; y 3) documentar el uso tradicional de las plantas en el bosque seco del noroeste peruano. Materiales y métodos Ubicación La Comunidad Campesina Santa Catalina de Chongoyape se localiza dentro del Área de Conservación Privada Chaparrí, entre los 6º28’39’’ – 6º 06’53’’ LS y 79º16’58’’ y 79 30’50’’ LO, en el Distrito de Chongoyape, Provincia de Chiclayo, Departamento de Lambayeque (Figura 1). Descripción de la zona de estudio Área de Conservación Privada Chaparrí Fue creada en diciembre del 2001 constituyéndose como la primera Área Natural Protegida de Conservación Privada del Perú con una extensión de 34,413 hectáreas y ocupando 86% del territorio de la comunidad. Inicia sus programas de conservación con el proyecto de reintroducción de la “pava aliblanca” Penelope albipennis a su ambiente natural (Angulo, 2002).
Figura 1. Mapa de ubicación de la zona de estudio.

ETNOBOTÁNICA DE LA COMUNIDAD CAMPESINA “SANTA CATALINA DE CHONGOYAPE”
Diciembre 2003 __________________________________________________________________________________________
16
Fisiografía e hidrología La zona comprende planicies desérticas (ca. 200 m de altitud) y bosque seco semi-denso (ca. 600 m) en las laderas del contrafuerte andino occidental. Ocupa la cuenca media del Río Chancay-Lambayeque cuyas nacientes están en la laguna de Yanahuanca, en el departamento de Cajamarca. Clima Presenta un rango medio de temperatura entre 28.2 ºC y 14.7 ºC, una precipitación media anual de 100 mm (registrándose más del 80 % de este valor durante la estación húmeda). El fenómeno de “El Niño”, asociado al aumento de la temperatura (+2 ºC) y precipitaciones (10 veces) en el departamento de Lambayeque, no ocurrió ese año (Angulo, 2002). Suelos La región posee tierras aptas para protección, que reúnen las condiciones mínimas requeridas para cultivo, pastoreo y producción forestal. Sin embargo, hay terrenos que permiten el uso continuo o temporal para sostener una ganadería económicamente productiva (ONREN, 1982). Ecología La zona de estudio corresponde a la ecorregión del bosque seco ecuatorial (Brack & Mendiola, 2000) y presenta las siguientes zonas de vida: matorral desértico – Tropical, matorral desértico - Premontano Tropical, monte espinoso - Premontano Tropical (ONER, 1976). Además se consideran dos comunidades vegetales, bosque ralo semicaducifolio y bosque semidenso caducifolio (CDC, 1992). Metodología Se desarrollaron entrevistas semi-estructuradas contemplando dos grupos de entrevistados. Uno constituido por tres colaboradores locales claves (personas adultas con reconocida experiencia en el campo) para las caminatas etnobotánicas. Éstas se llevaron a cabo con cada colaborador en lugares representativos del bosque seco semi-denso caracterizados por una mayor diversidad florística y menor impacto antrópico. El segundo grupo, compuesto por 25 personas (niños, jóvenes y ancianos), para las entrevistas etnobotánicas en la zona de las viviendas y chacras. Se utilizaron listas de plantas de la zona, descripciones orales de las características más conspicuas de las mismas, fotografías y material botánico montado. Se tomó en cuenta para el registro de información el permiso de los colaboradores para colectar plantas, escribir, tomar fotografías y grabar 15 testimonios orales. El proyecto obtuvo la aprobación del presidente de la comunidad y se estableció un acuerdo de cooperación, verbal y por escrito, con colaboración de la gente local y el compromiso de retorno de los resultados obtenidos en el estudio a la comunidad. Para la documentación del uso tradicional de algunas plantas útiles del
presente estudio se utilizaron registros arqueológicos de las culturas pre-colombinas que habitaban los bosques secos del noroeste peruano: material orgánico encontrado en tumbas (alimento, ofrendas funerarias u artefactos antiguos) y representaciones de plantas en cerámica (Yacovleff & Herrera, 1934; Cabieses, 1974; Lumbreras, 1974; Hocquenghem, 1987; Polia, 1989; Ostolaza, 1998; Elera, 1990; Cárdenas et al., 1995; Correa, 2000). Además, los hallazgos científicos del Proyecto Arqueológico de Sicán (Universidad de Princeton New Yersey - Pontificia Universidad Católica del Perú – Museo Nacional Sicán, Lambayeque) en el valle del Río La Leche con respecto al uso de la vegetación combustible y la reconstrucción experimental de hornos alfareros u orfebres (Shimada & Merkel, 1991; Shimada et al., 1994; Shimada & Wagner, 2001). Se consideró además material histórico recopilado a partir de la conquista española (Valdizán & Maldonado, 1923; Mateos, 1954; Kauffman, 1973; Rostworowski, 1981,1988). Resultados y discusión Registro Etnobotánico En este estudio se reporta 122 taxa de plantas útiles (103 géneros y 45 familias botánicas) de las cuales una es un helecho y el resto angiospermas. La mayor cantidad de especies fueron silvestres principalmente distribuidas en el bosque ribereño restringido a las faldas de los cerros. En segundo lugar, las domesticadas en los llanos cercanos a los caseríos y luego las malezas que crecen en hábitats perturbados sin depender de los humanos para su sobrevivencia. En menor proporción se encontraron plantas asilvestradas y semicultivadas (Figura 2). Las familias botánicas más representativas en cuanto a número de especies útiles fueron Fabaceae con 17,2%, Poaceae con 9%, Asteraceae, con 6,5%, Cactaceae con 5,7%, Solanaceae con 5,7%, Convolvulaceae con 3,3% y Anacardiaceae con 2,5% (Tabla 1). Las tres familias mejores representadas son las que presentan mayor número de géneros y especies neotropicales (Gentry, 1993; Maas et al., 1998; La Torre, 1997; Schjellerup et al., 2000). Asimismo, se registraron 10 categorías de uso: alimenticia con 38 especies útiles, medicinal con 37, combustible con 16, construcción de viviendas, cercos y corrales con 21, tecnología local con 22, comerciales con 16 y agroforestería con 9. También se reconocieron 8 especies melíferas, 40 que sirven de forraje a los animales domésticos y 21 como alimento de la “pava aliblanca” (Penelope albipennis) (Figura 3). La mayor proporción de especies comestibles y medicinales refleja los satisfactores básicos de la población (Hernández et al., 1991; Schjellerup et al., 2000; Ceroni, 2002), y las especies forrajeras, el potencial ganadero de la zona (ONERN, 1982).

TINA LERNER, ALDO CERONI Y CLAUDIA E. GONZÁLEZ
Ecología Aplicada Vol. 2 Nº 1, pp. 14-20 __________________________________________________________________________________________
17
Cultivadas27%
Silvestres63%
Malezas7%
Asilvestradas
2%
Semicultivada1%
Figura 2. Porcentajes de especies cultivadas y asilvestradas en Santa Catalina de Chongoyape. En cuanto al alimento de la “pava aliblanca” (P. albipennis) en el Área de Conservación Privada Chaparrí se reportan 8 nuevos géneros de plantas: Bastardia, Corchorus, Loxopterygium, Melocactus, Momordica, Prunus, Solanum y Verbesina, (Ortiz, 1980; Ortiz & Purisaca, 1981; Ortiz & Díaz, 1997; Zevallos & Ríos, 1998). Se tomó en cuenta a los individuos en estado de semicautiverio o en proceso de reintroducción que son monitoreados periódicamente en el área de conservación. Estos crácidos están iniciándose en el forrajeo de plantas silvestres, principalmente las de quebrada (como son los árboles Pithecellobium excelsum (Kunth) C.Martius y Muntingia calabura L., y las arbustivas Acnistus arborescens (L.) Schlecht. y Carica parviflora (A. DC.) Solms, entre otras); por otro lado dos especies cultivadas (Coffea arabica L. y Prunus avium L.) forman parte de la alimentación en semicautiverio. Si bien existen conocimientos sobre el uso de los vegetales, en muchos casos éstos no tienen un carácter práctico. Algunos testimonios locales refieren la pérdida de la utilización del vegetal, que observaron o aprendieron de las generaciones pasadas, por la escasez del recurso, un mal manejo o su reemplazo por productos sintéticos, entre otras causas. El uso de las plantas y la aplicación del conocimiento están cada vez más limitados por la lejanía y escasez del recurso vegetal. En efecto, el bosque seco de Lambayeque está en proceso de desertificación dada la ampliación de la frontera agrícola y tala indiscriminada (Proyecto Algarrobo, 1997). En el caso de las plantas medicinales, existe una tendencia a reemplazar las prácticas etnomedicinales por la utilización de medicamentos sintéticos. Sin embargo, cuando la medicina moderna falla y la enfermedad es grave, los pobladores acuden donde los “curanderos”
campesinos que preservan una gran farmacopea basada en plantas nativas (Prance, 1991).
construcción9%
forrajeras17%
alimenticias17%
medicinales 16%
comerciales7%
alimento de la pava aliblanca
9%tecnología local10%
agroforestería 4%
combustibles7%
melíferas4%
Figura 3. Porcentajes de especies vegetales según la categoría de uso
Tabla 1. Número de especies de plantas útiles por familiaNº Familia Nº de especies
1 Acanthaceae 1
2 Amaranthaceae 1
3 Anacardiaceae 3
4 Annonaceae 2
5 Apocynaceae 3
6 Arecaceae 1
7 Asteraceae 8
8 Asclepiadaceae 1
9 Bixaceae 1
10 Bombacaceae 1
11 Boraginaceae 2
12 Burseraceae 1
13 Cactaceae 7
14 Capparaceae 3
15 Caricaceae 2
16 Convolvulaceae 5
17 Cucurbitaceae 3
18 Cyperaceae 1
19 Elaeocarpaceae 1
20 Euphorbiaceae 2
21 Fabaceae 21
22 Lamiaceae 2
23 Lauraceae 1
24 Liliaceae 1
25 Loranthaceae 1

ETNOBOTÁNICA DE LA COMUNIDAD CAMPESINA “SANTA CATALINA DE CHONGOYAPE”
Diciembre 2003 __________________________________________________________________________________________
18
26 Malvaceae 2
27 Moraceae 1
28 Musaceae 2
29 Myrtaceae 2
30 Nyctaginaceae 3
31 Passifloraceae 3
32 Piperaceae 2
33 Poaceae 11
34 Pteridophyta 1
35 Rosaceae 2
36 Rubiaceae 1
37 Rutaceae 2
38 Sapindaceae 1
39 Solanaceae 7
40 Scrophulariaceae 3
41 Sterculiaceae 1
42 Tiliaceae 1
43 Ulmaceae 1
44 Verbenaceae 2
45 Vitaceae 1
Etnoclasificación local Se registraron 24 vocablos del habla local en 7 categorías etnobotánicas (Lerner, 2003). Se encontraron voces de origen español y quechua que denotan el carácter mestizo de los pobladores de la comunidad de Chongoyape dada la migración desde la sierra de Cajamarca (Tocmoche y Santa Cruz) a Chongoyape. De origen español tenemos “tabardillo” que significa insolación; americanismos como “chacla” que significa en el Perú, caña muy fuerte; del quechua “quincha”, kincha, que significa pared de ramas o cañas; empalizada; pared; cerca; muro; corral; y “siso”, sisa que significa polen, entre otras. Además, el léxico está influenciado por el entorno predominantemente agrícola y ganadero del valle costero y muchos de los nombres asignados localmente a las plantas, son de carácter descriptivo y resultan de una conjunción de información diversa (formas, hábitos, usos, etc.) sobre cada especie. Documentación del uso tradicional de las plantas en el bosque seco del noroeste peruano Las fuentes arqueológicas e históricas utilizadas logran establecer parcialmente enlaces tradicionales en el uso de las plantas del bosque seco dado que no es posible la verificación botánica de la especie. En este sentido, se sugiere que, desde tiempos precolombinos hubo continuidad en la utilización de Prosopis pallida (H & B ex Willd.) H.B.K. como combustible y en la construcción, como también en las prácticas curanderas el uso de Echinopsis peruviana (Britton & Rose) Friedrich & G.D. Rowley desde el periodo
Cupisnique (1500-100 a.e.c) hasta la Colonia (1535-1824). Asimismo, el cultivo y la alimentación con Zea mays L. desde el periodo Moche (0-800 e.c) hasta la Colonia. Actualmente, encontramos que la especie preferida como leña para cocinar y como postes en la construcción es Prosopis pallida (H & B ex Willd.) H.B.K. “algarrobo”, además de Loxopterigium huasango Spruce ex Engles “hualtaco”, debido a su gran rendimiento como combustible y alta durabilidad como elemento estructural de la vivienda. Asimismo, Zea mays L. “maíz” constituye la base alimentaria, junto a Oryza sativa L. “arroz”, plantas a las cuales el campesino le destina la mayor fuerza de trabajo por su valor comercial. Por otro lado, el cactus alucinógeno Echinopsis peruviana (Britton & Rose) Friedrich & G.D. Rowley “San Pedro” es utilizado en la zona por seis curanderos, quienes guardan la sabiduría acerca del proceso de colecta y preparación de la planta y la sanación de los pacientes enfermos. Sin embargo, la dimensión sacramental y la modalidad de su uso han cambiado según las tendencias y exigencias del medio; actualmente, el aspecto económico y monetario juega un papel determinante en las sesiones de curación. Por otro lado, el manejo sostenible de las plantas combustibles ha ocurrido desde el periodo Sicán (900-1100 e.c) hasta el Chimú (1100 e.c– 1480) dada la producción alfarera y orfebre a gran escala. Los antiguos consideraban una mayor diversidad de especies combustibles (arbóreas, arbustivas y herbáceas), una forma racional de cosecha de la leña (principalmente ramas) y la eficiencia de los hornos. También, la abundancia de los árboles, sobre todo del “algarrobo” por su alto poder calorífico, fue probablemente el factor clave para la presencia de centros cerámicos y metalúrgicos (Shimada & Wagner, 2001). Además, se debe considerar que el manejo racional fue posible dada la organización social-política-religiosa pre-colombina y la menor densidad poblacional con respecto a nuestra era. Conclusiones 1. En el Área de Conservación Privada Chaparrí se encontraron 124 taxa de plantas útiles pertenecientes a 104 géneros y 45 familias botánicas, todas angiospermas y una sola pteridofita. Y en cuanto a las familias botánicas, Fabaceae y Poaceae registraron mayor número de especies útiles. 2. Se reportan 40 taxa de plantas forrajeras, 38 alimenticias, 37 medicinales, 22 utilizadas en la tecnología local, 21 alimento de la “pava aliblanca” (Penelope albipennis), 20 usadas en la construcción, 16 combustibles, 9 agroforestales y 8 melíferas, lo que permite identificar qué especies son importantes para la gente de Santa Catalina de Chongoyape para su manejo, conservación y administración dentro del Área Natural Protegida de la comunidad.

TINA LERNER, ALDO CERONI Y CLAUDIA E. GONZÁLEZ
Ecología Aplicada Vol. 2 Nº 1, pp. 14-20 __________________________________________________________________________________________
19
3. Las categorías de uso mejor representadas con especies vegetales fueron las forrajeras, alimenticias y medicinales. Esto se relaciona con las principales actividades económicas del poblador, la agricultura y la ganadería, sin embargo las prácticas etnomedicinales, entre otros conocimientos, están siendo afectadas por la ampliación de la frontera agrícola y los recursos silvestres están cada vez más lejanos y escasos. 4. Bastardia, Corchorus, Loxopterygium, Melocactus, Momordica, Prunus, Solanum y Verbesina, son los 8 nuevos géneros de plantas que se reportan como alimento de la “pava aliblanca” (Penelope albipennis) encontrados en la zona de semicautiverio y reintroducción de esta ave en el Área de Conservación Privada Chaparrí. 5. Los 24 vocablos del habla local reportados en 7 categorías etnobotánicas son voces de origen español y quechua, denotando el carácter mestizo de los pobladores de la comunidad de Santa Catalina de Chongoyape. 6. Las fuentes arqueológicas e históricas utilizadas logran establecer parcialmente enlaces tradicionales en el uso de las plantas del bosque seco dado que no es posible la verificación botánica de la especie. En este sentido, se sugiere principalmente, desde tiempos precolombinos, la continuidad en la utilización de Prosopis pallida como combustible y en la construcción, de Zea mays en la alimentación y cultivo, y de Echinopsis peruviana en las prácticas curanderas. Agradecimientos Los autores expresan su agradecimiento a todos los amigos de Santa Catalina de Chongoyape que colaboraron en el estudio de campo, especialmente a Pedro Cáceres, José Cruz, Felipe Diaz e Iván Vallejos. A la Asociación Cracidae Perú por el apoyo financiero, especialmente a Fernando Angulo por la logística. A los Herbarios de la Universidad Nacional Agraria La Molina (MOL), Universidad Nacional Mayor de San Marcos (USM), Universidad Privada Antenor Orrego y Universidad Nacional de Trujillo por su acogida. A nuestro amigo David Goldstein por su hospitalidad en el valle de La Leche y apoyo con la información del Proyecto Arqueológico de Sicán. Literatura citada Angulo F. (editor). 2002. Área de Conservación
Privada Chaparrí. Plan Maestro. Brack A. & Mendiola C. 2000. Ecología del Perú.
PNUD. Ed. Bruño. Lima, Perú. Cabieses F. 1974. Dioses y enfermedades: la medicina
en el antiguo Perú. Tomo I. Ed. Arte gráfico. Lima.
Cárdenas J., Rodríguez J. & Aguirre L. 1995. El material orgánico en Huaca de la Luna. Páginas 129-149. en: Investigaciones en la Huaca de la
Luna. Ed. Facultad de Ciencias Sociales de la Universidad de la Libertad, Trujillo. Perú.
Ceroni A. 2002. Datos etnobotánicos del poblado de Huaylingas, cuenca la Gallega, Morropón, Piura. Revista Ecología Aplicada Departamento Académico de Biología.UNALM. 1(1): 65-70.
CENTRO DE DATOS PARA LA CONSERVACIÓN (CDC). 1992. Estudio de Conservación de la Diversidad Natural de la Región Noroeste del Perú. UNALM.
Correa R. 2000. Arquitectura rural en la costa norte Túcume: continuidad y mestizaje. Revista de Turismo y patrimonio. Universidad San Martín de Porres. Lima. (1)1: 69-81.
Elera C. 1990. El complejo cultural cupisnique: antecedentes y desarrollo de su ideología. Páginas 229-257. en: Senri Ethnological studies Nº37 `El mundo ceremonial andino´. National Museum of Ethnology. Japón.
Fernández A. & Rodríguez E. 2003. Etnobotánica del Perú pre-hispano. Public. Del Orthus. Bot. Truxillense (HBT) UNT. Perú (en prensa).
Gentry A.H. 1993. A field guide to the families of woody plants of Northwest South America (Colombia, Ecuador, Peru) with supplementary notes of herbaceous taxa. Washington, D.C.: Conservation International.
Hernández L., González C. & González F. 1991. Plantas útiles de Tamaulipas, México. Anales del Instituto de Biología de la UNAM. Ser. Bot. 62(1): 1-38.
Hocquenghem A. 1987. Iconografías mochica. PUCP Fondo editorial. Lima.
Kauffman D.F. 1973. Manual de Arqueología peruana. Ed. Peisa. Lima.
La Torre M. 1997. Etnobotánica de los Recursos Vegetales Silvestres del Caserío de Yana cancha, Distrito de Cumucho, Provincia de Celendín, Departamento de Cajamarca. Tesis para optar al título de Bióloga. UNALM.
Lerner T. 2003. Etnobotánica de los Recursos Vegetales de la Comunidad ‘Santa Catalina de Chongoyape’, Microcuenca del Río Chanchay, Distrito de Chongoyape, Provincia de Chiclayo, Departamento de Lambayeque. Tesis para optar el grado de Biólogo. Universidad Nacional Agraria La Molina.
Lumbreras L.G. 1974. Los orígenes de la civilización en el Perú. Ed. Milla Batres. Lima-Perú.
Maas J.M., Westra L.Y.Th. & Farjon A. 1998. Familias de Plantas Neotropicales, una guía concisa de las familias de plantas vasculares neotropicales. (Germany): Koeltz Scientific Books.
Mateos P.F. (Editor). 1954. Historia del nuevo mundo. Obras del Padre Bernabé Cobo. Compañía de Jesús. Madrid.

ETNOBOTÁNICA DE LA COMUNIDAD CAMPESINA “SANTA CATALINA DE CHONGOYAPE”
Diciembre 2003 __________________________________________________________________________________________
20
OFICINA NACIONAL DE EVALUACIÓN DE RECURSOS NATURALES (ONERN). 1976. Mapa Ecológico del Perú. Guía explicativa. Lima. Mapa escala 1:1’000,000.
_______. 1982. Clasificación de las tierras del Perú. Lima. Mapa a escala 1: 1’000,000.
Ortiz E. 1980. Estudio preliminar sobre la pava aliblanca (Penelope albipennis). Lima. No publicado.
_______ & Purisaca J. 1981. Estudio preliminar sobre la Pava aliblanca (Penelope albipennis) Taczanowski 1877. Primer Simposio de la familia Cracidae. UNAM. México.
_______ & Diaz V.R. 1997. Estudio de Campo y Reevaluación de la Población de Pava Aliblanca (Penelope albipennis). Pp. 218-232 En: The Cracidae: their Biology and Conservation (S.D. Strahl, S. Beaujon, D.M. Brooks, A.J. Begazo, G. Sedaghatkish and F. Olmos). Hancock House Publ.
Ostolaza C. 1998. Etnobotánica V: La cultura Moche. Quepo, Sociedad Peruana de Cactus y Suculentas. (Lima). 12: 62-68.
Polia M. 1989. Las lagunas de los encantos: Medicina tradicional andina del Perú septentrional. CEPESER. Piura, Perú.
Prance G. 1991. What is Ethnobotany today? Journal of Ethnopharmacology. 32: 206-209.
PROYECTO ALGARROBO. 1997. Políticas para la gestión sostenible de los bosques secos de la costa norte del Perú (en prensa).
Schjellerup I., Espinoza C., Quipuscoa V. & Fjeldsa J. (editores). 2000. Yanchalá – La gente y la biodiversidad. DIVA, Technical Report 10. Ecología y Desarrollo 9.
Shimada I. & Merkel J.F. 1991 Copper Alloy Metallurgy in Ancient Perú. Scientific American. 265(1): 80-6
_________, Elera C., Chang V., Neff H., Glascock M., Wagner U. & Gebhard R. 1994. Hornos y producción de cerámica durante el periodo formativo en Batán Grande, costa norte del Perú. Páginas 67-372. en: Tecnología y organización de la producción de cerámica prehispánica en los Andes. PUCP. Lima.
__________ & Wagner U. 2001. Peruvian Black Pottery Production and Metalworking: A Middle Sicán Craft Workshop at Huaca Sialupe. MASCA Journal. 26(1): 25-30.
Rostworowski De Diez Canseco M. 1981. Recursos Naturales Renovables y Pesca Siglos XVI y XVII. Instituto de Estudios Peruanos, Lima.
________________. 1988. Historia del Tahuantinsuyu. Instituto de Estudios Peruanos. Lima.
Toledo V. 1993. La Racionalidad Ecológica de la Producción Campesina. Páginas 197-218. en: Ecología, Campesinado e Historia. Sevilla, E. y González de Molina, M. (editores). La Piqueta, Madrid.
Valdizán H. & Maldonado A. 1923. La medicina popular peruana. Tomo II. Editorial Torres – Aguirre. Lima.
Yacovleff E. & Herrera F. 1934. Revista del Museo Nacional (Lima). 3: 241-323.
Zevallos P.A. & Ríos J. 1998. Determinación botánica de 24 especies arbóreas del Departamento de Lambayeque. Páginas 319-342. en: Bosques secos y desertificación. Memorias del seminario internacional. Proyecto Algarrobo – INRENA. Lambayeque, Perú.

Ecología Aplicada, 2(1), 2003 ISSN 1726-2216 Depósito legal 2002-5474
¿POR QUÉ TAN POCAS Opuntia EN LA ISLA ESPAÑOLA-GALÁPAGOS? Cruz Márquez1, Hernán Vargas1, Howard Snell2, André Mauchamp3, James Gibbs4 y Washington Tapia5
Resumen
El presente trabajo intenta explicar por qué Opuntia megasperma var. orientalis (Howell 1933) es actualmente rara en la isla Española, Galápagos. Se hace una descripción detallada de cómo los principales componentes del ecosistema de la isla pudieron haber interactuado, desde el pasado hasta el presente, y haber moldeado las etapas de sucesión ecológica dando como resultado la rareza actual del cactos Opuntia. Desconociendo si esta especie de cactos fue abundante o rara en el pasado, nuestra investigación parte del estudio de la situación actual de aquellos elementos del ecosistema que interactuan, ya sea como dispersadores, consumidores o competidores de Opuntia y que pudieron haber influido, como tales, en el pasado. Proporcionamos datos actuales de densidades de cactos y tortugas gigantes de Española. Se analiza el impacto que las cabras pudieron haber tenido en la Opuntia y el ecosistema en general y cómo otros elementos, tales como las tortugas, las aves y especies de plantas pueden actualmente limitar la recuperación natural de los cactos. Concluimos que el efecto de las cabras, por sí sólo, no explica totalmente la rareza actual de O. megasperma var. orientalis en Española, sino que adicionalmente hay otros factores involucrados. Palabras Claves: Pocas, Opuntia, Española, islas Galápagos.
Abstract This article attempts to explain why Opuntia megasperma var. orientalis is now rare in Española Island, Galapagos. We give a detailed description of the principal components of the island ecosystem that could have interacted, from the past to the present, and could have molded the stages of ecological succession, resulting in the present rarity of Opuntia cactus. Since we do not know whether this species of cactus was abundant or rare in the past, our investigation begins with the study of the current situation of those elements in the ecosystem that interact, whether as dispersal agents, consumers, or competitors of Opuntia, and could have had an influence as such in the past. We provide current data on the densities of cactus and Española giant tortoises. We analyze the impact that goats could have had on Opuntia and on the ecosystem in general and how other elements, such as tortoises, birds, and plant species, currently limit the natural recuperation of the cactus. We conclude that the effect of goats, by itself, does not completely explain the present rarity of O. megasperma var. orientalis on Española, but that there are additional factors involved. key Words: Española- Opuntia few, Galapagos Island.
Introducción
1 Estación Científica Charles Darwin, Casilla 17013891 Quito, Isla Santa Cruz, Galápagos- Ecuador. Correo electrónico: [email protected] y [email protected] Fax: 593-4-564-636 2 Department of Biology, University of New Mexico, Albuquerque, New Mexico, 87131 - EE. UU. Correo electrónico: [email protected] / [email protected] 3 Le Grand Mas d'Avignon, Le Sambuc, 13200 Arles Francia. Correo electrónico: [email protected] 4 Department of Environmental and Forest Biology, State University of New York College of Environmental Science É Forestry, Syracuce, New York 132CO USA. Correo electrónico: [email protected] 5 Parque Nacional Galápagos, Puerto Ayora, Isla Santa Cruz, Galápagos - Ecuador, Fax: 593-5-526, Correo electrónico: [email protected]
Existen pocos estudios sobre el papel que desempeñan los dispersadores y consumidores nativos en el mantenimiento del equilibrio de los ecosistemas en las islas Galápagos. Paralelamente, con la introducción de especies exóticas a las islas, se sospecha, principalmente que los herbívoros como las cabras, pueden producir cambios substanciales en la estructura y funcionamiento de los ecosistemas
insulares. Se ha culpado a las cabras, por ejemplo, de limitar el reclutamiento de varias especies de plantas y particularmente de los cactos del género Opuntia (Calvopiña, 1979; Hoeck, 1984). Sin embargo no se ha comparado o evaluado los impactos diferenciales causados por los dispersadores y consumidores nativos versus los similares introducidos.

POCAS Opuntia EN LA ISLA ESPAÑOLA
Diciembre 2003 __________________________________________________________________________________________
22
En el presente trabajo se intenta responder a la pregunta “¿porqué actualmente el cacto Opuntia megasperma var. orientalis (Howell 1933) es raro en la isla Española?”. Para lograr responder a esta pregunta se realizó una revisión bibliográfica de la situación presente y pasada de los cactos en la isla. En el análisis partimos de la situación actual que muestra que los cactos son muy escasos en la isla. Para explicar la escasez de los cactos en la isla Española se trabajó con diferentes escenarios y actores en el pasado y presente: Pasado: 1. Los cactos nunca fueron abundantes debido a
factores naturales. Esta posibilidad no se puede reconfirmar ni rechazar debido a la ausencia de datos. Solamente existe un registro en 1901, (Beck, 1903; Slevin, 1931) que sugiere la presencia local de un parche de cactos al Este de la isla.
2. Las tortugas gigantes y pinzones redujeron la pequeña población natural de cactos. Esta reducción fue acelerada por la presencia de las cabras.
Presente: 1. Los pinzones y tortugas terrestres gigantes limitan
la germinación, dispersión y reclutamiento de los cactos.
2. El incremento de la cobertura de la planta nativa Prosopis juliflora (Styward, 1825) DC, limita el reclutamiento de los cactos por competencia interespecífica.
3. Los cactos han llegado a ser escasos o raros en la población de Opuntia en la isla Española, lo cual ha limitado el intercambio de polen y por tanto la descendencia.
Se evaluó los efectos positivos que tienen los dispersadores como los efectos negativos que pueden tener los consumidores de los cactos. Los consumidores y dispersadores de las semillas de Opuntia, en las Islas Galápagos, desde hace miles de años fueron los herbívoros nativos tales como las tortugas gigantes Geochelone elephantopus (Harland 1827) y las iguanas terrestres Conolophus subcristatus (Gray 1831) y C. Pallidus (Heller 1903) (Dawson, 1964, 1966; Christian et al., 1984; Snell & Snell, 1988; Hicks & Mauchamp, 1995, 1996; Estupiñan & Mauchamp, 1995) y especies de aves como los cucuves Nesomimus sp, pinzón de cactos Geospiza conirostris y G. scandens, (Ridguay 1890) y palomas Zenaida galapagoensis (Gould 1837) (Bowman, 1961; Grant & Grant, 1979, 1982, 1989; Márquez com. pers., 1994). Los cactos del género Opuntia son componentes claves de los ecosistemas de las zonas costeras, áridas y de transición (10-200 msnm) de las Islas Galápagos. Las 6 especies y 14 subespecies son endémicas del Archipiélago (Hicks & Mauchamp, 1996). Además
son parte importante en la dieta de las tortugas terrestres gigantes Geochelone elephantopus (Harlan 1827), iguanas terrestres Conolophus subcristatus y C. pallidus, cucuves Nesomimus macdonaldi (Ridgway 1890), pinzones Geospiza conirostris, G. scandens, y G. fortis (Ridgway 1890). Estos últimos, especialmente, el gran pinzón de cactos, remueve excrementos de tortugas y el substrato buscando semillas, las trituran y las comen, de lo cual escapan pocas (Darwin, 1845; Bowman, 1960; Black, 1973). Las cabras fueron introducidas en la isla Española probablemente entre 1885-1895; ya que, en 1897 la expedición de Webster Harris encontró y reportó presencia de cabras (Rothschild & Hartert, 1899). Las cabras estuvieron presentes en la isla hasta la década de los 70 cuando fueron exterminadas por el Parque Nacional Galápagos (Calvopiña & de Vries, 1975; Smith, 1976, 1978; Calvopiña, 1979; Hoeck, 1984). Fueron especialmente comunes en la década de los 60 cuando se registraron grandes poblaciones y seguramente causaron cambios significativos en las comunidades vegetales (Eliasson, 1968; Van der Werff, 1982,). Las cabras fueron erradicadas de la isla Española en 1978 (Smith, 1978). La recuperación de la flora en la isla Española fue progresiva después de la eliminación de las cabras cimarronas (Hamann, 1979, 1982; Schofield, 1989), a excepción de los cactos, que aún se mantienen limitados a pequeños parches en la isla Española (Fig. 1). En los siglos XVI y XVII, Española tuvo una gran población de tortugas gigantes, de la cual sólo quedaron 14 individuos, por la cacería de los balleneros (Townsend, 1925). Puesto que la especie estuvo amenazada, entre 1964 y 1974, el tamaño poblacional de las tortugas en Española, era muy reducido, fue imperativo remover a cautiverio, los 14 últimos ejemplares adultos (12 hembras y 2 machos), para reproducirlos y restaurar la población con jóvenes desarrollados en cautividad. El Centro de Crianza esta dirigido por la Estación Científica Charles Darwin (ECCD) y Parque Nacional Galápagos (PNG) en Santa Cruz (Bacon, 1978; MacFarland et al., 1974; Márquez & Rea, 1991; Márquez et al., 1999). La reproducción y crianza bajo condiciones de cautiverio, permitió en 1975, iniciar la repoblación de la isla con tortugas juveniles y hasta la actualidad se han repatriado 1213 individuos. Los objetivos de este trabajo, fueron censar con marcación recaptura, los individuos de la población de tortugas gigantes repatriadas (tortugas crecidas en cautiverio y trasladadas a la isla), cuantificar los cactos juveniles, menores de ocho cladodios en el hábitat de las tortugas, en la isla Española. Se decidió contar los cactos menores de 8 cladodios, por ser los de mayor presencia y abundancia en la isla, mientras los mayores de 8, fueron raros o inexistentes, en el área donde se marcaron y recapturaron las tortugas.

CRUZ MÁRQUEZ, HERNÁN VARGAS, HOWARD SNELL, ANDRÉ MAUCHAMP, JAMES GIBBS Y WASHINGTON TAPIA
Ecología Aplicada Vol. 2 Nº 1, pp. 21-29 __________________________________________________________________________________________
23
Según Hicks & Mauchamp (1996), los parches de cactos adultos en Española, se agrupan en parches puntuales, donde son raros o inexistentes los cactos juveniles.
Figura 1. Una pareja de cabras sombreando bajo cuatro troncos de cactos Opuntia echios var. barringtonensis en la zona costera de la isla Santa Fe (Las cabras fueron eliminadas exitosamente en esta pequeña isla árida en 1971. Foto: O. Hamann, 1975). Materiales y métodos Área de Estudio Se estudiaron las Opuntia y las tortugas terrestres en la isla Española (-1º 38’ N; -89º 67’ O) con una superficie de 60.48Km2, una altitud de 206 msnm, contiene el 0.77 % con relación a la superficie total del archipiélago de Galápagos, es de levantamiento geológico (no de origen volcánico), con una precipitación anual de 10 mm en años secos y de 600 mm en años lluviosos. La temperatura promedio anual es de 23.8oC. La cobertura herbácea está compuesta por Galactea striata (Jacq 1900), Galactea tenuiflora (Wight & Arn 1834), Phaseolus mollis (Hook 1847), Rhynchosia minima (Medik 1787) y otras especies herbáceas, no incluye la cobertura arbórea de Cordia lutea (Lam 1791), Prosopis juliflora (Styward 1825)
vegetal dominante en la isla, misma que por su estructura y follaje denso disminuye el ingreso de los rayos solares a los estratos inferiores, favoreciendo de esta manera la germinación y el crecimiento de los cactos. Los pocos parches de cactos adultos son de pocos individuos, y poco comunes en la isla. Especies de Aves: cuatro de pinzón de cacto, una de cucuve (Nesomimus macdonaldi Ridgway 1890), una de paloma silvestre (Zenaida galapagoensis Gould 1841), una de Gavilán (Buteo galapagoensis Gould 1837). Especies de Reptiles: una de culebra (Phillodryas biserialis Steindachner 1876), una de tortuga gigante (G. hoodensis VanDenburgh 1914) y una de lagartija (Microlophus delanoni Van Denburgh 1912, Figura 2). Se cuantificaron los cactos juveniles menores de ocho cladodios entre el 18 al 26 de noviembre de 1994. Los cactos totales de cada nivel de cladodio, y sector fueron comparados. Los datos fueron obtenidos, mediante marcación y recaptura de las tortugas, al mismo tiempo, siete personas registraron en forma detallada la regeneración de los cactos menores de ocho cladodios, usando ocho transectas generalmente utilizadas para marcar y recapturar las tortugas: Se ubicó, cuatro transectas en la zona Las Tunas y cuatro en El Caco. La longitud de las transectas fue medida con carretes de hilo de 500 m c/u y varió de 1.200 - 1.500 m y el ancho con una cinta métrica de 50 m y varió desde 170-200 m. A cada cacto encontrado, se le contó el número de cladodios desde uno hasta ocho; no se contaron los juveniles mayores de ocho cladodios, por ser muy raros o inexistentes. Se observó también, el comportamiento de alimentación de aves, cactos juveniles comidos por tortugas, la abundancia de Prosopis en las transectas y una revisión bibliográfica. El muestreo de los cactos, fue realizado en los sitios donde actualmente se repatrían y distribuyen las tortugas (Figura 2). Se anota que, con base a las observaciones realizadas en los varios viajes al campo, que los dos sitios usados para la repatriación de las tortugas, representan posiblemente las áreas de mayor abundancia de cactos en la isla. La revisión bibliográfica abarca el período comprendido entre 1897 y el presente. Se entrevistaron a varias personas lo que incluye antiguos pescadores colonos de las islas, guardaparques y científicos, quienes por una u otra razón visitaron estas dos islas en el pasado.

POCAS Opuntia EN LA ISLA ESPAÑOLA
Diciembre 2003 __________________________________________________________________________________________
24
Los datos de abundancia y densidad de Opuntia por sector, fueron analizados con el test de Fisher, mientras las estimaciones de la población y densidad de las tortugas se realizaron mediante el método de Jolly Seber (Krebs, 1989). Resultados Cuantificación y Densidad de los Cactos Juveniles: Las Tunas fue diferente de El Caco en la abundancia de los cactos en los ocho niveles de cladodios (Tabla 1, F = 8.004, P < 0.01). Las Tunas fue diferente de El Caco en las densidades de los cactos en los ocho niveles de cladodios (Tabla 2, F = 11.353, P < 0.0025). Los promedios en los distintos niveles de los juveniles menores de ocho cladodios fueron: Las Tunas 8 ± 3.7 (rangos = 2 - 15), y en El Caco fue de 4 ± 1.1 (rangos = 3 - 7);. Las densidades, tanto para Las Tunas como para El Caco, los valores mostraron diferencias estadísticamente significativas (F = 1.47, P < 0.05). Por otro lado, las densidades de los cactos de uno a dos y de siete a ocho cladodios, fueron mucho más bajas, que en las demás categorías. Las densidades generales fueron de 61 cactos/Km2 (Las Tunas) y 35 cactos/Km2 (El Caco), pero la densidad general incluida las dos zonas, fue 48 cactos/Km2. Población y Densidad de las Tortugas En Las Tunas la población de tortugas estimada fue 91 individuos. En El Caco se estimó una población de 103 animales. La población general fue
407 individuos, incluidos machos, hembras y juveniles. La diferencia del tamaño de la población estimada en cada área fue relativamente mínima. La densidad general estimada fue 127 tortugas/Km2. Mientras la densidad de tortugas en Las Tunas fue de 65 tortugas/Km2, en El Caco fue de 74 tortugas/Km2, o sea no presentó diferencias estadísticamente significativas. Observaciones de Fauna Reptiles Las tortugas siempre estuvieron a la sombra o junto a los troncos de los cactos adultos donde caen flores, frutos o cladodios de Opuntia que pueden ser consumidos (Figura 3a-c). Las tortugas se alimentaron de cladodios y frutos caídos como también de los cactos derribados, por sí solos o por las tortugas. En ninguna de las transectas de las dos zonas de muestreo, se encontró cactos juveniles mordidos y comidos por las tortugas. Las tortugas adultas, además de comer las yemas tiernas de regeneración reciente en cactos caídos y accesible en los troncos de los cactos; mordieron los troncos de los cactos adultos hasta derribarlos y luego los comieron (Figura 4 a-b y Figura 5a-d). También es importante anotar, que las tortugas podrían fácilmente ejercer presión en la población de Opuntia al consumir los cactos juveniles. Por otro lado, la hipótesis acerca de la variedad de cactos, la distancia entre los árboles de cactos adultos y la limitación del polen, está por estudiarse.
Figura 2. Área de estudio y distribución de los cactos (Opuntia megasperma var. orientalis) juveniles menores de 8 cladodios, mismos lugares donde se marcaron y recapturaron las tortugas gigantes (Geochelone hoodensis) en la isla Española.
Figura 2. Área de estudio y distribución de los cactos (Opuntia megasperma var. orientalis) juveniles menores de 8 cladodios, mismos lugares donde se marcaron y recapturaron las tortugas gigantes (Geochelone hoodensis) en la isla Española.

CRUZ MÁRQUEZ, HERNÁN VARGAS, HOWARD SNELL, ANDRÉ MAUCHAMP, JAMES GIBBS Y WASHINGTON TAPIA
Ecología Aplicada Vol. 2 Nº 1, pp. 21-29 __________________________________________________________________________________________
25
Figura 3a-c. Tres diferentes tortugas (Geochelone hoodensis), bajo la sombra de tres diferentes cactos (Opuntia megasperma var. orientalis), esperando que caigan cladodios, flores o frutos, donde obtienen agua y nutrientes para sobrevivir los años ecos en la isla Española (Fotos: Cruz Márquez 2000).
Figura 4a-b. Tortuga terrestre gigante macho (Geochelone microphyes) rompiendo y comiendo cladodios de Opuntia insularis en la ladera Oeste del volcán Darwin (Foto: Hernán Vargas 1999).
Figura 5a-d. Cuatro cactos Opuntia megasperma var. orientalis (a) tres derribados por descomposición de sus raíces por efecto de El Niño 1997-98, y uno por mordeduras de tortugas (Geochelone hoodensis), en la base del tronco (Foto: Cruz Márquez, 2000).

POCAS Opuntia EN LA ISLA ESPAÑOLA
Diciembre 2003 __________________________________________________________________________________________
26
Aves Los pinzones de cactos G. conirostris (Ridgway 1890) removieron en su totalidad los excrementos secos y semi-secos de las tortugas y buscaron, trituraron y comieron las semillas de los cactos que estos contenían. El pinzón terrestre Geospiza fuliginosa (Ridgway 1890) fue observado buscando alimento en los mismos sitios usados por G. conirostris. Sin embargo, fue desplazado de las fuentes de alimentación por G. conirostris. Se observó a los cucuves (N. macdonaldi), perforar con el pico y comer el contenido de los frutos maduros de O. megasperma y luego regurgitar las semillas. Las palomas comieron las semillas secas localizadas en los sectores circundantes de los troncos de los cactos adultos. Tabla 1. Abundancia de las Opuntia megasperma var. orientalis, Cactácea juveniles en los habitats de las tortugas en la isla Española.
Opuntia/Número de cladodios Zona 1 2 3 4 5 6 7 8 Total Las Tunas 12 9 22 16 12 12 3 6 92
El Caco 5 7 12 8 7 7 8 6 60
Total 17 16 34 24 19 19 11 12 152 F = 8.004, P < 0.01 Tabla 2. Densidad de Opuntia megasperma var. orientalis, Cactácea juveniles (Opuntia/Km2), clasificados por números de cladodios en Las Tunas y El Caco.
Opuntia/Número de cladodios Zona 1 2 3 4 5 6 7 8 Las Tunas 8 6 15 11 8 8 2 4 El Caco 3 4 7 5 4 4 5 4 F = 11.353, P < 0.0025. La variación de las densidades alrededor del promedio, aplicando el coeficiente de variación (C.V.), fue 47.6%, para Las Tunas y 25.9%, para El Caco. Observaciones de Cambio Florístico En la isla Española ningún cacto juvenil estuvo junto a un adulto (Fig. 6). En 1984, los claros observados sin Prosopis, eran más grandes y permitían facilidad para caminar, mientras que, diez años después (1994), los claros son cada vez más pequeños y dificultan el tránsito del hombre (Márquez, datos no publicados). Los pinzones no comieron las semillas de P. juliflora que se encontraban en los excrementos de las tortugas. Los cactos juveniles rara vez fueron encontrados donde P. juliflora fue dominante, no se encontró ningún cacto juvenil bajo los matorrales de P. juliflora. Las pocas y pequeñas colonias de Opuntia adultas que actualmente existen en la isla Española, están ubicadas donde hay poca dominancia de P. juliflora. Entre 1984-1985, se derribaron 8 a 10 cactos viejos que murieron en toda la isla Española, por
efecto tardío del fenómeno El Niño 1982-1983. En 1999-2000 se derribaron entre 10-20 cactos adultos por efecto tardío del fenómeno de El Niño 1997-1998. Los cactos son plantas suculentas de áreas desérticas, por tanto las lluvias abundantes por largo tiempo, pudren las raíces y se derriban en años posteriores.
Figura 6. Una pequeña colonia de cactos juveniles (Opuntia galapageia var. galapageia en la zona árida de vegetación en la isla Pinta en 1980 (Hamann 1984). Discusión Aunque El Caco tiene mayor área, presenta menos cactos juveniles por km2 que Las Tunas. La mínima diferencia en los números y densidades de Opuntia encontrada entre los dos sitios pudiera ser explicada por diferencias en la cobertura vegetal. 1) En el Caco y Las Tunas, no se encontró cactos jóvenes consumidos por tortugas, pero no descartamos dicha posibilidad. Las hierbas permanentes son muy escasas y las plántulas de cactos estarían más expuestas a la depredación de las tortugas. 2) En Las Tunas se observan más hierbas permanentes y lianas, que se asume protegen los cactos juveniles de los depredadores y por ende sirven como refugios. Esta asunción, de que la cobertura herbácea (tipo liana) favorece el reclutamiento de los cactos, apoya los resultados obtenidos en las estimaciones de población y densidad de tortugas, que son aproximadamente similares en los dos sitios. Sin embargo la abundancia y densidad de los cactos son diferentes, entre las dos áreas. La baja abundancia y densidad de O. megasperma var. orientalis encontrada en el Caco (Tablas 1 y 2), sugiere, que en ausencia de la cobertura herbácea, las tortugas ejercen un efecto limitante más fuerte en el reclutamiento de los cactos juveniles. Los extremos, en los niveles (1-2) y (7-8) cladodios, muestran cantidades bajas en número de individuos, mientras los niveles intermedios (3-6), presentan cantidades superiores en número de individuos, esto podría estar influenciado por factores como, 1) Las plántulas de 1-2 cladodios, son más

CRUZ MÁRQUEZ, HERNÁN VARGAS, HOWARD SNELL, ANDRÉ MAUCHAMP, JAMES GIBBS Y WASHINGTON TAPIA
Ecología Aplicada Vol. 2 Nº 1, pp. 21-29 __________________________________________________________________________________________
27
difíciles de ver, se protegen en poca vegetación herbecía, y se asume que están en mayor número. 2) Los niveles 7-8, son más visibles, asumiéndose que son más escasos, porque recibieron mayor acción depredadora por parte de las tortugas y los pocos sobrevivientes observados probablemente, escaparon protegiéndose entre lianas y otras hierbas. 3) Los niveles intermedios, son más abundantes; pero presumiblemente se encuentran vía presión depredadora, hasta llegar a tallas mayores de 8 cladodios, cuando hayan desarrollado prolongadas y gruesas espinas que los protegerá de la agresión depredadora de las tortugas. Similar explicación tiene los datos de la tabla de las densidades. A pesar de que las tortugas pueden limitar el reclutamiento de los cactos juveniles, experimentos recientes han demostrado que la tasa de germinación en semillas de cactos es más rápida y se incrementa entre 60-80%, luego de pasar por el tracto digestivo de tortugas e iguanas terrestres (Wiggins & Focht, 1967; Arp, 1973; Christian et al., 1984; Estupiñan & Mauchamp, 1995). Mientras la tasa de germinación de las semillas que no pasaron por el tracto digestivo fue significativamente inferior, alrededor del 20% en promedio (Hicks & Mauchamp, 1996). De acuerdo a los resultados de estos experimentos se puede deducir: 1) La ausencia de tortugas, podría retardar la germinación de las semillas, pero eventualmente una determinada cantidad podría germinar cuando las condiciones son favorables (años lluviosos). Sin embargo ahora la población de cactos es baja, presumiblemente todas las semillas podrían estar comidas por las tortugas. Luego esto podría servir como un banco de semillas viables. Las tortugas comen las semillas, ellas germinan después de pasar por el tracto digestivo y luego las tortugas comen las plántulas. 2) Que la disminución al mínimo de la población de tortugas en la isla Española, y posteriormente la ausencia de ellas in situ, puede haber causado también una disminución de los cactos. En Española, el efecto del pinzón G. conirostris y G. fuliginosa, sobre los cactos es más intenso puesto que las afectan: 1) destruyendo las semillas cuando se alimentan, 2) cortando los estigmas de las flores y de esa manera reducen la producción de semillas (Grant & Grant, 1981; Grant, 1996). Este comportamiento incrementaría las probabilidades de extinción de la especie de Opuntia y asociado a ello, los pinzones de cactos. Tampoco se conoce detalladamente el papel como dispersadores que desempeñan otras especies de aves tales como las palomas, cucuves y pinzones (e.g. Geospiza fuliginosa Gould 1837). La presencia de herbívoros tiene un efecto medible en Opuntia; si los herbívoros estuvieron presentes en grandes densidades, podrían haberse comido los juveniles y retoños que producen plantas nuevas en los cactos derribados (Snell et al., 1994). Probablemente esto es
lo que pudo haber ocurrido en el pasado (XVI - XVIII) en la isla Española, cuando hubo una alta densidad de tortugas gigantes. En el pasado, el tamaño de la población de cactos en Española, se cree que fue reducida al mínimo por el pastoreo de las cabras cimarronas (D. Anderson comn. Pers. 1995). Sin embargo, la regeneración fue lenta a pesar de la erradicación total de los herbívoros introducidos. Hamann (1979, 1984), indica que en la isla Pinta, O. galapageia (Hensl 1837) virtualmente fue una de las pocas especies de plantas sobrevivientes, después de la devastación de la vegetación por las cabras introducidas. En la vegetación de Santa Fe, tan sólo los cactos (Opuntia) y los palo santo (Bursera graveolens Trian & Planch 1872) quedaron en pie después de la acción destructora de las cabras (Hamann, 1982; Black 1973). Algunos autores como Harris (1973) afirman que las cabras fueron las causantes de la disminución al mínimo de las Opuntia en la isla Española. Si las cabras fueron los causantes de la disminución de los cactos de Española, uno se puede preguntar por qué no causaron el mismo efecto en la isla Santa Fe. Estas dos islas hace dos o tres décadas mantuvieron grandes poblaciones de cabras cimarronas. Sin embargo, Española aún mantiene mínimos parches de cactos con muy pocos adultos y juveniles; mientras que en Santa Fe, la dominancia de los cactos adultos y juveniles de todos los tamaños es alta. Las afirmaciones de Hamann y nuestras observaciones actuales en Española y Santa Fe, indican que, las cabras no fueron las únicas causantes de la reducción al mínimo de los cactos en Española. En Española, las cabras mordieron los troncos de unas pocas Opuntia adultas hasta derribarlas (Llerena, Sanmiguel, Chapi, comn. Pers. 1996). Los cactos juveniles, también fueron comidos por las cabras y no sobrevivía ninguna (Chapi comn. Pers. 1996). La erradicación de las cabras cimarronas en la isla Española fue hace aproximadamente dos décadas; desde entonces y hasta la actualidad debería existir gran cantidad de parches de bosques de cactos juveniles con por lo menos 7 a 8 cladodios, sobre todo las que germinaron en la época de El Niño de 1983. Sin embargo, no es común encontrar cactos de estas tallas. Esto sugiere que además de las cabras, existieron y existen otros factores que consumieron y limitaron el reclutamiento de los cactos en la isla Española. Conclusión Las cabras no fueron el único factor causante de la disminución de los cactos Opuntia de Española. Las tortugas y el pinzón de cactos también contribuyeron en la disminución y están actualmente limitando la recuperación de las poblaciones de cactos. La

POCAS Opuntia EN LA ISLA ESPAÑOLA
Diciembre 2003 __________________________________________________________________________________________
28
cobertura herbácea favorece el reclutamiento de los cactos juveniles. Se sugiere que Prosopis juliflora, por competencia interespecífica, limita la recuperación natural de O. megasperma var. orientalis. Se sugiere también hacer un estudio ecológico futuro de los principales elementos del ecosistema de la isla Española. Una vez realizado el estudio ecológico determinar la viabilidad de realizar experimentos ex situ. Sería muy importante, estudiar zonas excluidas de tortugas o excluir un parche de Opuntia de otro cercano a uno libre, y observar el efecto sobre los juveniles y el resultado será apreciado a largo plazo entre 10 - 20 años. Agradecimientos Nuestros agradecimientos a Telmo Zúñiga, Sinecio Olaya, Jorge Serrano y Edwin Jiménez (PNG) por su colaboración en el censo de las tortugas y en el contaje de los cactos juveniles. A Córley McMullen y Ole Hamann, por la revisión y comentario del manuscrito. A los directivos del PNG y de la ECChD, por la logística y financiamiento del viaje a Española. Literatura citada Arp G.K. 1973. The Galápagos Opuntia: Another
interpretation. Noticias de Galápagos. 21: 33-37. Bacon J.P. 1978. A tortoise goes Home. ZooNooz.
51(2): 4-7. Beck R.H. 1903. In the home of the giant tortoise.
Annual Report of the New York. Zoological Society. 7: 160-174.
Black J. 1973. Galápagos: Archipiélago del Ecuador. Quito: Imprenta Europa.
Bowman R.I. 1960. Report on a Biological reconnaissance of the Galapagos Islands during 1957 UNESCO, Paris.
--------. 1961. Morfological differentiation and adaptation in the Galapagos finches. Univ. Calif. Publ. Zool.
Calvopiña L.H. 1979. Estudios y protección de la vegetación nativa de la Isla San Salvador y erradicación de los chivos salvajes en la Isla Española. Inf. Annual ECChD.
Calvopiña L.H. & de Vries T.J. 1975. Estructura de la población de Cabras Salvajes (Capra hircus L.) y los daños causados en la vegetación de la Isla San Salvador Galápagos. Rev. PUCE. 8: 219-241.
Krebs Ch.J. 1989. Ecological Mathodology. Harper & Row, Publishers, New York.
Christian K.A., Tracy C.R. & Porter W.P. 1984. Diet, digestion and food preferences of galapagos land iguanas. Herpetologica. 40: 205 -212.
Darwin C. 1845. Journal of Research into the Nature History and Geology of the coutries visited during el voyage of the HMS Beagle round the World under the command of Capt. Fitz Roy, R. N. from 1832-1836. 2nd ed. Murray, London.
Dawson E.I. 1964. Cacti in the Galápagos Island. Noticias de Galápagos. 4: 12-13.
-------. 1966. Cacti in the Galápagos. In the Galápagos edited by R.I. Bowman. University of Calif. Press., Berkeley and Los Angeles.
Eliasson U. 1968. On the influence of introduced animals on the natural vegetation of the Galapagos Islands. Not. Galápagos. 11: 19-21.
Estupiñan S.B. & Mauchamp A. 1995. Interacción planta-animal en la dispersión de Opuntia de Galápagos. II Congreso Nacional de Botánica Quito, Ecuador.
Grant B.R. & Grant P.R. 1979. The Feeding Ecology of Darwin’s Ground finches. Noticias the Galapagos. 30: 14-18.
--------. 1981. Exploitation of Opuntia cactos bay Birds on the Galápagos. Oecologia (Berl). 49: 179-187.
--------. 1982. Niche shifts and competition in Darwin finches. Geospiza conirostris and its congeners. Evolution. 36: 637-657.
Grant P.R. & Grant B.R. 1989. The slow recovery of Opuntia megasperma on Española. Noticias de Galápagos. 48: 13-15.
--------. 1996. Pollen digestion by Darwin,s Finches andits importance for early breeding. Ecology. 77(2): 489-499.
Hamann O. 1979. Regeneration of vegetation on Santa Fe and Pinta Island, Galápagos after the elimination of goats. Biological Conservation. 15: 215-236.
--------. 1982. Cambios en la vegetación de las Islas Galápagos durante el período de 1966 a 1973. Compendio de Ciencias en Galápagos, edic. Ulrike Eberhardt.: 179-199.
-------. 1984. Changes and Threats to the vegetation. In R. Perry, ed., Key Environments: Galápagos, Pergamon Press, Oxford.: 115-131.
Harris M.P. 1973. The Galapagos Avifauna. Condor. 75: 265-278.
Hicks D.J. & Mauchamp A. 1995. Size dependent predation by feral mammal on Galápagos Opuntia. Noticias de Galápagos. 55: 15-17.
Hicks D.J. & Mauchamp A. 1996. Evolution and Conservation Biology of the Galápagos Opuntias (Cactaceae). Haseltonia. 4: 89-101.
Hoeck H.N. 1984. Introduced Fauna. In R. Perry, ed., Key Environments: Galapagos, Pergamon Press Oxford.: 233-245.
MacFarland C.G., Villa J. & Toro B. 1974. The Galápagos giant tortoises (Geochelone elephantopus) Part. I: Status of the surviving populations. Biological Conservation. 5: 118-133.
Márquez C. & Rea S. 1991. Gemelos en las tortugas terrestres gigantes de las Islas Galápagos. Rev. Geograf. 29: 109-115.

CRUZ MÁRQUEZ, HERNÁN VARGAS, HOWARD SNELL, ANDRÉ MAUCHAMP, JAMES GIBBS Y WASHINGTON TAPIA
Ecología Aplicada Vol. 2 Nº 1, pp. 21-29 __________________________________________________________________________________________
29
Márquez C., Cayot L. & Rea S. 1999. La crianza de tortugas gigantes en cautiverio: Un manual operativo. Imprenta A & B Editores, Quito.
Rothschild W. & Hartert E. 1899. A review of the ornithology of the Galápagos Islands. With notes on the Webster-Harris expedition. Novit. Zool. VI.: 85-205.
Slevin J.R. 1931. Log of the Schooner “Academy” on a voyage of Scientific research to the Galápagos Islands 1905-1906. Occasional papers of the California Academy of Sciences. 17: 1-162.
Smith G.T.C. 1976. News from Academy Bay. Noticias de Galápagos. 25: 1-4.
--------. 1978. New from Academy Bay. Noticias de Galápagos. 27: 1-6.
Snell H. & Snell H. 1988. Seleción Natural de la morfología de Opuntia en Isla Plaza Sur. Inf. Anual de la ECCHD.: 27-29.
Snell H., Snell H. & Stone P. 1994. Acelerated mortality of Opuntia on Isla Plaza Sur: Another threat from an introduced vertebrate? Noticias de Galápagos. 53: 19-20.
Schofield E. 1989. Effects of introduced plant and animals on island vegetation: examples from the Galápagos Archipelago. Conservation Biology. 3: 227-238.
Townsend C.H. 1925. The Galápagos Tortoises in their relation to the whaling industry: a study of old logbooks. Zoologica. 4: 55-135.
Van der Werff H. 1982. Effects of feral pigs and donkeys on the distribution of selected food plants. Noticias de Galápagos. 36: 17-18.
Wiggins Y.L. & Focht D.W. 1967. Seeds and seedlings of Opuntia echios J.T. Howell var. gigantea Dawson. Cactus and Succulent Journal 34(3 & 4).: 67-74; 99-105.

Ecología Aplicada, 2(1), 2003 ISSN 1726-2216 Depósito legal 2002-5474
ESTRATÉGIAS DE DISPERSÃO, PRODUÇÃO DE FRUTOS E EXTRATIVISMO DA PALMEIRA Astrocaryum jauari MART. NOS IGAPÓS DO RIO NEGRO: IMPLICAÇÕES PARA A ICTIOFAUNA
M. T. F. Piedade1, P. Parolin2 y W. J. Junk2
Resumo
Astrocaryum jauari é a palmeira mais freqüente nos igapós do Rio Negro, na Amazônia brasileira, medindo em torno de 20 m de altura, encontrada raramente isolada e comumente formando touceiras com 4 a 6 indivíduos de tamanhos distintos, oriundos a partir de um único rizoma. A abscisão dos frutos ocorre durante o pico das cheias e inicio da descida das águas, sendo estes consumidos por peixes. A palmeira tem valor comercial sendo dela extraído palmito que, por cerca de 20 anos, até 1998, constituiu a base da produção industrial de palmito na Amazônia Central. Este estudo visou estabelecer a dinâmica de produção sazonal e biomassa de frutos de A. jauari em relação aos níveis de inundação de sua área de ocorrência e suas implicações em termos da dispersão e distribuição local da palmeira. Observações sistemáticas foram conduzidas na Estação Ecológica doArquipélago das Anavilhanas, perfazendo uma área total de levantamento de 5.000 m2. Determinou-se a densidade relativa de A. jauari em relação ao nível e duração da inundação anual. A produção de frutos foi acompanhada semanalmente em plantas individuai. Dados sobre peixes foram obtidos por meio de coletas, entrevistas e literatura. A. jauari ocorre das porções mais elevadas às mais baixas da planície alagável, inundadas de 270 a 340 dias/ano. 125 indivíduos de A. jauari ocorreram nos 5.000 m2 amostrados, equivalendo a uma densidade de 250 ind/ha nas Anavilhanas. Apenas plantas adultas diretamente expostas à luz produziram frutos, cada estipe 4 cachos ao ano, com cerca de 90 cm de comprimento, e 106 frutos por cacho. Cada fruto pesou em média 17 g, equivalendo à produção de 6,60 kg/palmeira/ano (peso fresco). 16 espécies de peixes exploram os frutos da palmeira, 10 possíveis dispersoras, dentre elas, 3 também predadoras. Pela ausência de estruturas para flutuação, quando na água os propágulos afundam e, se as condições forem hipóxicas, ocorrerá a morte do fruto e semente. Todas as sementes coletadas boiando estavam podres, descartando a hidrocoria como estratégia de dispersão e, apontando a ictiocoria como mecanismo para a colonização de novos habitats por A. jauari. O elevado conteúdo calórico de vitamina A pode ser atrativo para a busca desse recurso pelos peixes. Em porções alagadas por curtos períodos os frutos podem ficar acumulados próximos a planta adulta (barocoria), ou deslizar pelo relevo em virtude do seu peso, possivelmente com auxílio da ação da água (barocoria + deslizamento), aumentando a densidade local da planta. As espécies ocorrendo nas mesmas faixas de distribuição de A. jauari apresentam a hidrocória e ictiocoria como estratégias de dispersão predominantes, e grande tolerância a prolongados períodos de inundação. Na principal área de extrativismo da espécie no passado, no Município de Barcelos, a densidade de plantas também variou muito entre ilhas, porém, com densidades superiores àquelas encontradas neste estudo, variando de 250 a 2.000 ind/ha. Nesse local, um único trabalhador pode derrubar e pré-limpar até 90 palmeiras na cheia (meses de maio, junho) e cerca de 25 na vazante (dezembro, janeiro). O rendimento por palmeira é de 0.5 a 1 lata de palmito processada. Supondo-se um número médio de 10 trabalhadores na área, de 900 a 250 palmeiras são exploradas ao dia, o que pode corresponder de 1a 0.5 ha derrubados. Considerando as densidades da palmeira e sua produção de frutos calculada para as ilhas Anavilhanas, que foi de 165 kg, o extrativismo sem plantio de reposição, implicaria na exclusão deste recurso para a ictiofauna daquele Arquipélago, com conseqüências negativas para as populações regionais e para o comércio íctico. Palavras chave: Astrocaryum jauari, ictiocoria, extrativismo, igapós, produção de frutos, Amazônia, áreas alagáveis
Abstract Astrocaryum jauari is the most frequent palmin the floodplains (igapó) of the Rio Negro, in Brazilian Amazonia. It grows to 20 m of height and is rarely found isolated. More commonly, 4-6 individuals of different sizes stand together which originate from the same rhizome. Fruit abscission occurs at highest water levels and dispersal occurs by fish. The palm has commercial
1 INPA/Max Planck, C.P. 478, 69011 Manaus, Am, Brasil. 2 Max-Planck-Institute for Limnology, Tropical Ecology, P.O. Box 165, 24302 Plön, Germany.

ESTRATÉGIAS DE DISPERSÃO, PRODUÇÃO DE FRUTOS E EXTRATIVISMO DA PALMEIRA Astrocaryum jauari MART. NOS IGAPÓS DO RIO NEGRO: IMPLICAÇÕES PARA A ICTIOFAUNA
Diciembre 2003 _________________________________________________________________________________________
32
value: until 1998, for 20 years ‘palmito’ was the basis of the industrial production in Central Amazonia. In the present study, seasonal dynamics and biomass of fruit production in A. jauari are described in relation to water level and annual flood duration. Systematic studies were performed at the Ecological Station of the Anavilhanas Archipelago, with an inventory area covering 5.000 m2. Fruit production was monitored at weekly intervals and biomass was determined. Data on the dispersing fishes was obtained by collection in the field and by interviews of local people. A. jauari occurs from the higher levels to the lowest in the flooding gradient, being waterlogged up to 270-340 days/year. 125 individuals of A. jauari occorred in the sampled 5.000 m2, which is equivalent to a density of 250 ind/ha in the Anavilhanas. Only adult trees which were directly exposed to light produced fruits. 16 species of fish feed on the fruits of A. jauari. Ten of these act as possible dispersers, 3 as predators. The high contents of vitamine A may be attractive for the fishes. Since the fruits do not have structures enhancing floatation, hydrochoric dispersal does not occur and the seeds succumb to the hypoxic conditions under water. In habitats with short periods of flooding, the fruits may be accumulated near the adult plant (barochory). Other trees occurring in the habitats colonized by A. jauari have hydrochoric and ichthyochoric dispersal syndromes and show high flooding tolerance. In the main area of extractivism of the past, in the Município Barcelos, the density of the species varied between the islands, and had densities between 250 and 2.000 ind/ha. Here, one worker could fell up to 90 palms at high water (May, June) and up to 25 in the low water period (december, january). For each tree, 0.5 to 1 bin of processed bin of “palmito” is produced. Assuming a mean number of ten workers in the area, 900 to 250 palms are exploited per day, corresponding to 1-0.5 ha destroyed forest. If we consid the densities of the palm and its productivity as calculated for the Anavilhanas islands (whcih was 165 kg), extractivism without replacement of new palm trees implies a fast extinction and exclusion of this ressource for the ichthyofauna of the archipelago with all consequences for the local people. Key words: Astrocaryum jauari, ichthyochory, extractivism, igapó, fruit production, Amazonia, floodplains.
Introdução Dentre as poucas palmeiras de ocorrência nas áreas de inundação dos igapós do Rio Negro (Prance, 1980), a mais freqüente é Astrocaryum jauari (Figura 1A y B), que mede em torno de 20 metros de altura, e apresenta o caule ornamentado por banda de acúleos, na extremidade do qual há uma pequena porém densa cabeça de folhagem. O número de folhas contemporâneas é de 6 a 14 (Boer, 1965), tendo cada uma delas o comprimento médio de três metros. A. jauari pode ser encontrada raramente isolada e mais freqüentemente formando grandes grupamentos nas porções marginais dos rios e lagos, ou mais interiorizada na floresta alagada (Correa, 1969; Goulding, 1980). Quando agrupada, o número de indivíduos varia em geral de 4 a 6, existindo na mesma touceira indivíduos de tamanhos distintos, sendo os clones formados a partir de um único rizoma (Boer, 1965). Quando as águas no igapó atingem sua altura máxima e durante o início de sua descida, A. jauari elimina seus frutos. Estes são ovóides, medindo de 3 a 4 cm, apresentando coloração entre verde e amarelada (Figura 1B). Assim como se verifica com outras plantas dessas comunidades, os frutos, ao caírem na água podem ser comidos por peixes, sendo então predados, caso sejam triturados, ou possivelmente
transportados até locais adequados à sua germinação (Gottsberger, 1978; Goulding, 1980; Junk, 1980). Comercialmente também pode ser extraído de A. jauari um palmito bastante apreciado por ser tenro e de sabor agradável, tendo sido a produção industrial de palmito da Amazônia Central até 1998, derivada exclusivamente dessa espécie. A partir desse ano, devido a facilidades de extração, transporte e constância na produção, o extrativismo passou a ser feito em plantações de Bactris gasipaes, popularmente chamada de pupunha (com. pess. Sr. Alírio Rocha, SHARP S/A). Para os peixes da família Characidae, comercialmente importantes na região, em particular a pirapitinga (Colossoma bidens) e o tambaqui (Colossoma macropomum), os frutos de A. jauari constituem, durante a fase de alagação, respectivamente o primeiro e o segundo item alimentarpreferenciais (Goulding, 1980), contribuindo de maneira singular a alimentação regional, fortemente sustentada por proteína animal oriunda de peixe. Este estudo objetivou determinar a dinâmica de produção e a biomassa de frutos de A. jauari em relação aos níveis de inundação de sua área de ocorrência, as implicações desses aspectos em temos da dispersão e distribuição local da palmeira, bem

M. T. F. PIEDADE, P. PAROLIN Y W. J. JUNK
Ecología Aplicada Vol. 2 Nº 1, pp. 31-40 ______________________________________________________________________________________
33
como o impacto de seu extrativismo nas cadeias alimentares desses ambientes. Área de estudo A base principal de dados de fenologia, produção de frutos e densidade da palmeira foi obtida de agosto de 1980 a novembro de 1982. Observações confirmatórias e informações sobre o extrativismo continuaram a ser acumuladas até 1998. A coleta de dados sobre fenologia e produção de frutos e densidade da espécie foi feita no Arquipélago de Anavilhanas, Rio Negro, Amazônia Central, na Estação Ecológica Flutuante do Instituto Brasileiro de Meio Ambiente (IBAMA), distante aproximadamente 100 km da cidade de Manaus (Figura 2). Tomando como base dados climáticos de Manaus, segundo Ribeiro (1976), a região é caracterizada por um clima tropical quente e úmido (Afi de Köppen), com temperatura média inferior nunca abaixo de 18ºC e o mês mais árido com uma umidade relativa superior a 60%. A precipitação média anual para a região varia de 1000 a 2500 mm, com a existência de uma estação seca no verão (Irmler, 1975). O nível do rio na área de estudo oscila ao longo do ano, com padrão e amplitude similares aqueles obtidos para o Rio Negro na altura da cidade de Manaus, apresentando um desnível médio de 10m entre os picos das cheias e secas (Schmidt, 1973), que determina a presença de uma fase terrestre e uma fase aquática ao longo do ano, de grande importância biológica (Junk et al., 1989). A vegetação da área apresenta em média três estratos e um sub-bosque pouco denso. As árvores
mais altas apresentamentre 15 e 20 m de altura, e as emergentes ao redor de 25 m. Durante as águas altas, dependendo do relevo das ilhas, a vegetação dessas áreas permanece em submersão total ou parcial, indicando a adaptação dos indivíduos a esse regime (Gessner, 1955; Fittkau, 1967; Irmler, 1973). Uma outra adaptação típica da vegetação desses ambientes é a eliminação dos frutos em sincronia com os níveis de águas altas, sendo por isso freqüentes a hidro e ictiocoria como mecanismos de dispersão (Gottsberger, 1978). Metodologia As observações foram centralizadas em dois sítios amostrais, próximos a sede flutuante do IBAMA (Figura 2). Para verificar as áreas de ocorrência e densidade relativa de A. jauari em relação às demais espécies presentes nos sítios amostrais, foram feitos dois transectos, um no sítio amostral 2 e outro no sítio amostral 3 (Figura 3). Os transectos estenderam-se do litoral da ilha até a porção do barranco, percorrendo toda a superfície transversal em cada um deles. Assim, foi realizado no sítio 2 um transecto de 20 x 115 m (2300 m2), e no sítio 3 um transecto de 20 x 135 m (2.700 m2). Essas faixas foram divididas em quadrados de 20 m, em cada um dos quais procedeu-se ao levantamento de todos os indivíduos com DAP superior a 10 cm, sendo feito um mapeamento para verificar a posição relativa de A. jauari e demais espécies presentes. Com os valores encontrados determinou-se a densidade relativa (DR) de cada espécie presente por faixa de 20 m, segundo Cottam &
Figura 1. Astrocaryum jauari no ambiente natural. A Detalhe da árvore, B cacho de frutos (Fotos: F. Wittmann).

ESTRATÉGIAS DE DISPERSÃO, PRODUÇÃO DE FRUTOS E EXTRATIVISMO DA PALMEIRA Astrocaryum jauari MART. NOS IGAPÓS DO RIO NEGRO: IMPLICAÇÕES PARA A ICTIOFAUNA
Diciembre 2003 _________________________________________________________________________________________
34
Curtis (1956), onde DR = n x 100/ nt (n = número de indivíduos da espécie; nt = número de indivíduos de todas as espécies presente na área amostrada).
Destacou-se a presença de A. jauari em relação ao nível de inundação. O cálculo do número de dias de alagação ao qual as comunidades vegetais estão submetidas por faixa de relevo, foi realizado tomando como base a curva do valor médio de dez anos de dias de inundação confeccionada por Junk et al. (1983), com as cotas do Rio Negro, na altura de Manaus, fornecidas pela PORTOBRÁS. As demais espécies presentes foram agrupadas por família, e ordenadas de forma decrescente, conforme o número de espécies componentes. Para a obtenção de dados sobre a dinâmica de produção de frutos e aspectos de dispersão, foram realizadas observações gerais em todo o Arquipélago, bem como em todos os indivíduos dos 3 sítios amostrais (Figura 3), e, particularmente em 3 indivíduos marcados nos sítios 1, 2 e 3, em um total de 9 indivíduos acompanhados. Os registros foram feitos pelo menos uma vez ao mês e pelo menos uma vez por semana, durante a fase de queda de frutos. Com o auxilio de rapixé, procedeu-sea coleta de sementes de A. jauari flutuando na água, num total
superior a 250, além de coletas casuais. Todas as sementes foram quebradas para verificar as condições da amêndoa. Além das observações gerais, foram coletados dez cachos de frutos de palmeiras diferentes, sempre marginalmente dispostas. Os cachos coletados foram medidos, tendo sido contado o número total de frutos presentes. A seguir, de cada um dos cachos retirou-se uma sub-amostra de 20 frutos, dos quais foi tomado o peso fresco utilizando-se uma pesola. Para verificar as formas de exploração dos frutos pela fauna do igapó, observou-se mais de 3000 frutos no solo (ataque pós-dispersão) e mais de 1000 frutos em cachos (ataque pré-dispersão). Como parte de outros estudos em andamento, a coleta de peixes realizada sistematicamente durante as cheias na área da Estação Ecológica, forneceu-se dados a respeito da ingestão de frutos por peixes. A esses dados foram acrescentados os da bibliografia e de entrevistas com moradores da região. Dados sobre o extrativismo da palmeira até de 1998, foram fornecidos por técnicos da Fábrica de Palmito Jauari, SHARP do Brasil S.A., Manaus/AM. Resultados A distribuição de indivíduos de A. jauari, variou bastante entre os dois sítios amostrais. No sítio amostral 2, apenas 4 indivíduos adultos de A. jauari foram encontrados. Estes mediam acima de 14 m de altura e frutificaram durante o período de estudo. Desse total, 3 distribuíram-se nos 40 primeiros metros da porção litoral em direção a parte mais elevada da ilha. O quarto indivíduo foi encontrado na porção do barranco, em uma altura equivalente a faixa de ocorrência da porção litoral. O número de plântulas apresentou-semaior ao redor dos adultos, em particular nas porções litorais, as menos íngremes do relevo. As faixas de ocorrência de A. jauari, foram as que apresentaram maior diversidade de espécies nessa estação, tendo sido encontradas 29 espécies associadas (Tabela 1 y 2). Contudo, ao longo da ilha, observou-se a substituição de dominantes conforme o tempo de alagação e a altura do relevo. Neste sítio amostral, observou-seque a palmeira se distribuiu em uma faixa que permanece inundada, pelo menos na base do caule e sistema radicular, de 270 a 340 dias, o mesmo ocorrendo com as espécies a ela associadas. O sítio amostral 3 apresentou uma densidade muito mais elevada de indivíduos de A. jauari. O número total de indivíduos com mais de 14 m de altura foi de 121. Neste sítio, onde a porção litoral apresentou-se muito plana, os adultos de A. jauari situaram-se apenas a partir dos primeiros 60 m em relação ao litoral da ilha. Na porção mais isolada do relevo predominaram indivíduos clonais, enquanto que nos 20 m anteriores e no barranco, predominaram adultos isolados com plântulas e jovens ao seu redor. Considerando toda a faixa de ocorrência da palmeira
Figura 2. A Arquipélago de Anavilhanas, Rio Negro, Amazônia Central, distante aproximadamente 100 km da cidade de Manaus; B sítios amostrais próximos a Estação Ecológica Flutuante do Instituto Brasileiro de Meio Ambiente (IBAMA).

M. T. F. PIEDADE, P. PAROLIN Y W. J. JUNK
Ecología Aplicada Vol. 2 Nº 1, pp. 31-40 ______________________________________________________________________________________
35
em este sítio amostral, encontrou-se 27 espécies associadas a ela, distribuídas na porção litoral e do barranco. Da mesma forma que no sítio amostral 2,observou-se a substituição de espécies dominantes, conforme o tempo de inundação. Nesta área A. jauari ocorreu numa faixa de relevo submetida de 30 a 200 dias de inundação ao ano, dependendo da posição mais ou menos elevada da ilha. Considerando as duas áreas de estudo, num total de 5.000 m2 amostrados, foram encontrados 125 indivíduos, o que fornece uma densidade para A. jauarinas Ilhas Anavilhanas, de 250 indivíduos por hectare. A freqüência de produção de flores e frutos de A. jauari é anual. A grande maioria das palmeiras adultas inicia a produção de flores quando as águas do rio estão descendo, especialmente durante os meses de agosto e setembro. A fase de floração é rápida, sendo logo substituída pela presença de pequenos frutos que vão desenvolver-se lentamente, amadurecendo apenas quando as águas do rio sobem novamente, sofrendoabcisão principalmente quando elas atingem seus níveis mais altos, nos meses de maio, junho e julho. Tanto as fases de floração quanto os primeiros estágios de frutos jovens transcorrem durante os meses nos quais as águas estão descendo e a precipitação ainda não é muito intensa. O tempo gasto para a produção de frutos é em média de 9 meses. Verificou-se também que as palmeiras, ainda que possivelmente velhas pela presença de caule glabro e repleto de liquens (Boer, 1965), não produziram cachos caso estivessem abaixo do dossel ou em qualquer posição na qual não recebessem luz. Em áreas isoladas, mesmo aquelas pequenas e aparentemente jovens, já frutificavam. Cada palmeira individual de A. jauari, produz anualmente ao redor de 4 cachos, que medem cerca de 90 cm de comprimento. O número médio de frutos por cacho é de 106 e o peso médio por fruto é de 17 g. Devido a esse peso e a ausência de estruturas bem desenvolvidas para a flutuação, como é comum em outras palmeiras como Cocos nucifera, todos os frutos afundam ao cair na água. Por sua vez, todas as sementes coletadas boiando apresentavam a amêndoa podre. Como conseqüência das condições topográficas, após a queda, os frutos podem ficar acumulados
próximo a planta adulta (Barocoria), ou deslizar pelo relevo em virtude do seu peso, possivelmente com auxílio da ação da água (Barocoria + Deslizamento). Os frutos encontrados nessa circunstâncias são em menor número e isolados em pontos esparsos do relevo insular. Um total de 16 espécies de peixes exploram os frutos da palmeira, pertencentes a cinco famílias, das quais Characidae é a mais representada. Desse total de espécies, 10 foram categorizadas como possíveis dispersoras (Ictiocoria), embora 3 delas (Colossoma macropomum, Colossoma bidens e Prachtocephalus hemiliopterus), possam também atuar como predadoras, já que além de engolirem frutos inteiros, podem fragmentá-los com suas possantes mandíbulas. Durante este estudo, não observamos indícios de utilização dos frutos de A. jauari por nenhum outro vertebrado. Quanto a exploração econômica da espécie, as informações fornecidas pela Fábrica de Palmito Jauari – SHARP do Brasil S.A., dão conta que na região do Município de Barcelos, onde o extrativismo ocorre, a densidade de plantas também varia muito entre ilhas daquela localidade, porém, as densidades são muito superiores àquelas encontradas no Arquipélago de Anavilhanas (250 indivíduos por hectare), sendo da ordem de 250 até 2.000 indivíduos por hectare. Nesse local, um único trabalhador pode derrubar e pré-limpar até 90 palmeiras na cheia (meses de maio, junho) e cerca de 25 na vazante (dezembro, janeiro). Supondo-se um número médio de 10 trabalhadores na área, de 900 a 250 palmeiras são exploradas ao dia, o que pode corresponder de 1a 0.5 ha. O rendimento por palmeira é de 0.5 a 1 lata de palmito processada. Atualmente, desde outubro de 1998, devido a dificuldades em se manter constância na produção a produção de palmito da SHARP S.A., foi totalmente substituída por pupunha (Bactris gasipaes), plantadas em 45 ha manejados em um total de 225.000 palmeiras. Discussão Nos sítios amostrais verificou-se uma nítida substituição de comunidades ao longo da topossequência das ilhas, como resposta as variações do número de dias de alagação, e em função da compactação do solo, que aumenta em direção as
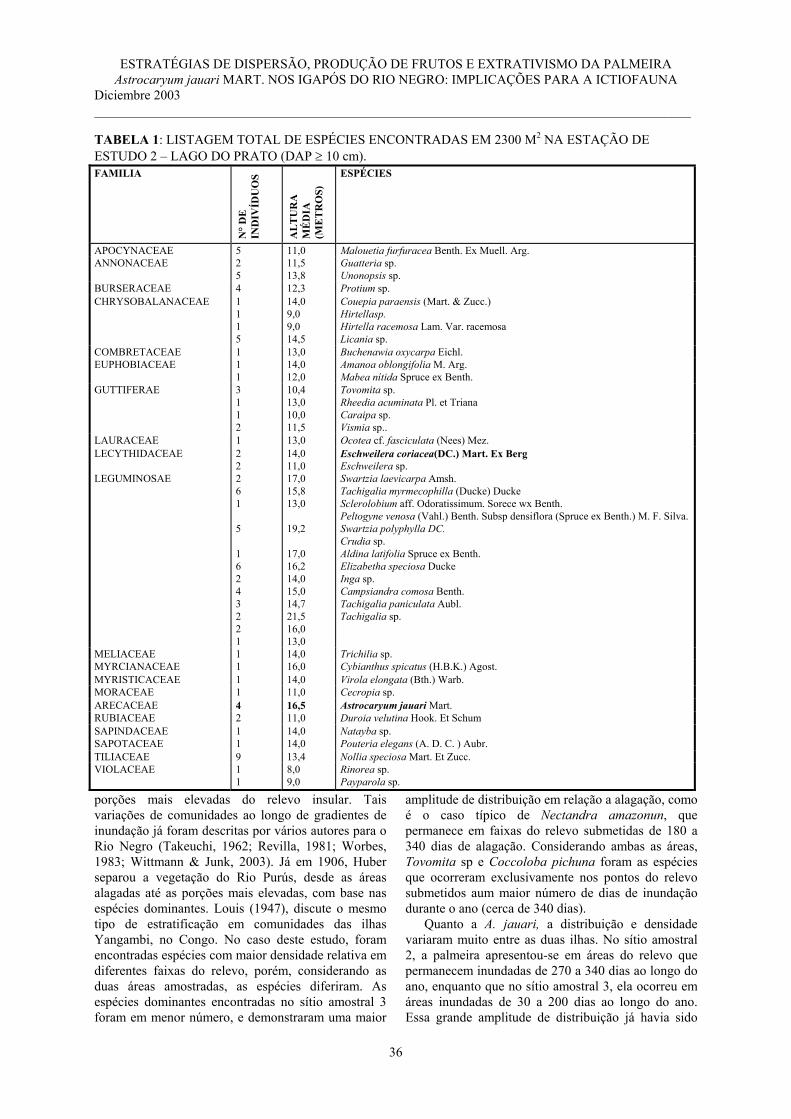
ESTRATÉGIAS DE DISPERSÃO, PRODUÇÃO DE FRUTOS E EXTRATIVISMO DA PALMEIRA Astrocaryum jauari MART. NOS IGAPÓS DO RIO NEGRO: IMPLICAÇÕES PARA A ICTIOFAUNA
Diciembre 2003 _________________________________________________________________________________________
36
porções mais elevadas do relevo insular. Tais variações de comunidades ao longo de gradientes de inundação já foram descritas por vários autores para o Rio Negro (Takeuchi, 1962; Revilla, 1981; Worbes, 1983; Wittmann & Junk, 2003). Já em 1906, Huber separou a vegetação do Rio Purús, desde as áreas alagadas até as porções mais elevadas, com base nas espécies dominantes. Louis (1947), discute o mesmo tipo de estratificação em comunidades das ilhas Yangambi, no Congo. No caso deste estudo, foram encontradas espécies com maior densidade relativa em diferentes faixas do relevo, porém, considerando as duas áreas amostradas, as espécies diferiram. As espécies dominantes encontradas no sítio amostral 3 foram em menor número, e demonstraram uma maior
amplitude de distribuição em relação a alagação, como é o caso típico de Nectandra amazonun, que permanece em faixas do relevo submetidas de 180 a 340 dias de alagação. Considerando ambas as áreas, Tovomita sp e Coccoloba pichuna foram as espécies que ocorreram exclusivamente nos pontos do relevo submetidos aum maior número de dias de inundação durante o ano (cerca de 340 dias). Quanto a A. jauari, a distribuição e densidade variaram muito entre as duas ilhas. No sítio amostral 2, a palmeira apresentou-se em áreas do relevo que permanecem inundadas de 270 a 340 dias ao longo do ano, enquanto que no sítio amostral 3, ela ocorreu em áreas inundadas de 30 a 200 dias ao longo do ano. Essa grande amplitude de distribuição já havia sido
TABELA 1: LISTAGEM TOTAL DE ESPÉCIES ENCONTRADAS EM 2300 M2 NA ESTAÇÃO DE ESTUDO 2 – LAGO DO PRATO (DAP ≥ 10 cm). FAMILIA
N° D
E
IND
IVÍD
UO
S
AL
TU
RA
M
ÉD
IA
(ME
TR
OS)
ESPÉCIES
APOCYNACEAE 5 11,0 Malouetia furfuracea Benth. Ex Muell. Arg. ANNONACEAE
2 5
11,5 13,8
Guatteria sp. Unonopsis sp.
BURSERACEAE 4 12,3 Protium sp. CHRYSOBALANACEAE 1
1 1 5
14,0 9,0 9,0 14,5
Couepia paraensis (Mart. & Zucc.) Hirtellasp. Hirtella racemosa Lam. Var. racemosa Licania sp.
COMBRETACEAE 1 13,0 Buchenawia oxycarpa Eichl. EUPHOBIACEAE 1
1 14,0 12,0
Amanoa oblongifolia M. Arg. Mabea nítida Spruce ex Benth.
GUTTIFERAE 3 1 1 2
10,4 13,0 10,0 11,5
Tovomita sp. Rheedia acuminata Pl. et Triana Caraipa sp. Vismia sp..
LAURACEAE 1 13,0 Ocotea cf. fasciculata (Nees) Mez. LECYTHIDACEAE 2
2 14,0 11,0
Eschweilera coriacea(DC.) Mart. Ex Berg Eschweilera sp.
LEGUMINOSAE 2 6 1 5 1 6 2 4 3 2 2 1
17,0 15,8 13,0 19,2 17,0 16,2 14,0 15,0 14,7 21,5 16,0 13,0
Swartzia laevicarpa Amsh. Tachigalia myrmecophilla (Ducke) Ducke Sclerolobium aff. Odoratissimum. Sorece wx Benth. Peltogyne venosa (Vahl.) Benth. Subsp densiflora (Spruce ex Benth.) M. F. Silva. Swartzia polyphylla DC. Crudia sp. Aldina latifolia Spruce ex Benth. Elizabetha speciosa Ducke Inga sp. Campsiandra comosa Benth. Tachigalia paniculata Aubl. Tachigalia sp.
MELIACEAE 1 14,0 Trichilia sp. MYRCIANACEAE 1 16,0 Cybianthus spicatus (H.B.K.) Agost. MYRISTICACEAE 1 14,0 Virola elongata (Bth.) Warb. MORACEAE 1 11,0 Cecropia sp. ARECACEAE 4 16,5 Astrocaryum jauari Mart. RUBIACEAE 2 11,0 Duroia velutina Hook. Et Schum SAPINDACEAE 1 14,0 Natayba sp. SAPOTACEAE 1 14,0 Pouteria elegans (A. D. C. ) Aubr. TILIACEAE 9 13,4 Nollia speciosa Mart. Et Zucc. VIOLACEAE
1 1
8,0 9,0
Rinorea sp. Payparola sp.

M. T. F. PIEDADE, P. PAROLIN Y W. J. JUNK
Ecología Aplicada Vol. 2 Nº 1, pp. 31-40 ______________________________________________________________________________________
37
apontada por Huber (1906), para populações de A. jauari do rio Purus. Uma vez que a espécie apresenta grande tolerância a inundação, a diferença no número de indivíduos e na faixa de distribuição entre as duas ilhas amostradas, possivelmente deve-se a diferenças no tempo de colonização entre elas.
A. jauari sincroniza sua floração com o período no qual a precipitação é pouco intensa, e, quando os índices pluviométricos maiores são obtidos, os frutos já estão em processo de crescimento, um processo longo que dura em torno nove meses, até o pico das águas altas, período mais intenso de abcisão dos mesmos. Possivelmente esse longo período é necessário em virtude do alto conteúdo energético e vitamínico dos frutos exigir um longo processo de bombeamento dos escassos nutrientes desses solos pobres (Sioli & Klinge, 1962; Furch, 1997). Cada
palmeira produz em torno de 6,60 kg de frutos ao ano (peso fresco). Entretanto, a pressão para uma sincronia na produção de frutos nas fases de águas mais altas pode ser mais importante, por ser compatível com os mecanismos de dispersão da espécie. Também o alto valor energético e nutritivo
dos frutos pode explicar a grande procura desse recurso pelos peixes. O elevado conteúdo calórico de vitamina A pode ser especialmente vantajoso se consumido em grande quantidades durante as cheias, para estocar reservas na forma de gordura para as fases de deslocamento, de desova ou de águas baixas, quando os recursos alimentares são escassos. Confirmando a preferencia dos peixes por esse recurso, Goulding (1980) encontrou em apenas um indivíduo de Colossoma macropomum 52 frutos inteiros, e em um indivíduo de Colossoma bidens 1 kg de frutos de A. jauari. A constatação de que todos os frutos encontrados boiando estavam podres, indica que a hidrocoria não se verifica com A. jauari. Ao cairem na água, os pesados frutos vão para o fundo, podendo eventualmente ser transportados por correstes submersas, mas não por flutuação. Nesses locais, eles poderão germinar, em função da profundidade e do nível da água durante a fase terrestre quando o local da queda é muito profundo, condições hipóxicas levam ao apodrecimento do fruto e semente que passa então a boiar. Corner (1966) salienta ser freqüente a “aparente” dispersão por flutuação em alguns gêneros de palmeiras, citando inclusive Astrocaryum entre eles. Assim, o aumento local na densidade de espécie é decorrente da barocoria e da barocoria + deslizamento, que associadas à propagação vegetativa propiciam o aparecimento de grandes massas de indivíduos da espécie nas ilhas Anavilhanas e densidades ainda maiores na região de seu extrativismo em Barcelos.
Uma vez que a hidrocoria não se verifica, a distribuição da espécie a longas distancias para a colonização de novos habitats, é decorrente da ictiocoria. Segundo Van der Pijl (1969,1972), a ictiocoria é um mecanismo de dispersão primitivo que em áreas inundáveis da Amazônia e Borneo se mantêm nos grupos de plantas mais antigos. O ruido da queda dos frutos na água, atraindo os peixes, contribui para o sucesso desse mecanismo.
TABELA 2: LISTAGEM TOTAL DE ESPÉCIES ECONTRADAS EM 2700 M2 NA ESTAÇÃO DE ESTUDO 3 LAGO MIUÁ (DAP ≥ 10 cm).
FAMILIA
N° D
E
IND
IVÍD
UO
S
AL
TU
RA
M
ÉD
IA
(ME
TR
OS)
ESPÉCIES
ANACARDIACEAE 1 18,0 Spondias sp. ANNONACEAE 1 13,0 Guatteria cf amazonica R. E. Fr. BURSERACEAE 2 12,0 Protium sp. CARYOCARACEAE 4 17,3 Caryocar microcarpum Ducke COMBRETACEAE 4 12,0 Buchenavia Fanshawei Exell &
Maguire CRYSOBALANACEAE 2 22,5 Couepia paraensis (Mart. &
Zucc.) Benth. EUPHORBIACEAE 63
1 1 5 14 1
16,2 14,0 14,0 17,0 18,6 10,0
Alchornea schomburgkiana Klotz. Amanoa cf oblongifolia M. Arg. Piranhea trifoliata Baill. Hevea spruceana Mull.Arg. Mabea nítida Spruce ex. Benth. Croton cuneatus Kl.
FLACOURTIACEAE 1 17,0 Banara guianensis Aubl. CLUSIACEAE 3 16,0 Calophylum brasiliense Camb. LAURACEAE 50 17,3 Nectandra amazonum Nees LECYTHIDACEAE 7 14,8 Eschweilera sp. LEGUMINOSAE 25
1 2 3 3 8 1 1
15,4 13,0 15,0 17,0 17,5 17,7 12,0 13,0 15,0
Macrolobium acaciifolium Benth. Sclerolobium sp. Ormosia excelsa Spr. Ex Benth. Tachigalia sp. Tachigalia myrmecophilla (Ducke) Ducke Inga sp. Pterocarpus santalinoides L. Hérit. Campsiandra augustifolia Spr. Ex Bth. Macrolobium multyngum (DC.) Benth.
MELASTOMATACEAE 1 13,3 Tococa subciliata (DC.) Triana MYRISTICACEAE 2
7 12,0 20,3
Virola elongata (Benth.) Warb. Virola surinamensis (Rol.) Warb.
ARECACEAE POLYGONACEAE
121 27
15,6 12,6
Astrocaryum jauari Mart. Coccoloba pichuna Huber
STYRACACEAE 1 13,0 Styrax guianensis A. D. C.

ESTRATÉGIAS DE DISPERSÃO, PRODUÇÃO DE FRUTOS E EXTRATIVISMO DA PALMEIRA Astrocaryum jauari MART. NOS IGAPÓS DO RIO NEGRO: IMPLICAÇÕES PARA A ICTIOFAUNA
Diciembre 2003 _________________________________________________________________________________________
38
Bibliografia Ab'Saber A.N. 1967. Problemas geomorfológicos da
Amazônia brasileira. Atas do Simpósio sobre a Biota Amazônica 1 (Geociências).: 35-67.
Adis J. 1977. Programa mínimo para análises de ecossistemas: artrópodos terrestres em florestas inundáveis da Amazônia Central. Acta Amazônica. 7(2): 223-229.
A.O.A.C. 1975. Handbook of chemical analysis. 10ª ed. Washington , Association of Official Analytical Chemist.
Boer J.E.W. 1965. Palmae. In: Ianjouw, J. (ed.) Flora of Suriname . vol. 5. pate 1. Leiden E. J. Brill. : 125-141.
Braga P.I.S. 1979. Subdivisão fitogeográfica, tipos de vegetação conservação e inventário florístico da floresta amazônica. Acta Amazônica. 9(4): 53-80 .
Corner E.J.H. 1966. The natural history of palms. London, Weidenfeld and Nicholson .
Corréa M.P. 1969. Dicionário de plantas üteis do Brasil. vol. 4. IBDF. Rio de Janeiro.
Cottam G. & Curtis J.T. 1956. The use of distance measures in phytosociological sampling. Ecology. 7(3): 451-460.
Coatinho L.M. & Lamberti A. 1971. Respiração edáfica e produtividade primária numa comunidade amazônica de terra firme. Ci. e Cult. 23(3): 411-419.
Cronquist A. 1974. The evolution and classification of flowering plants Boston, Houghton Mifflin Co.
Ducke A. & Black G.A. 1953. Notas sobre a fitogeografia da Amazônia brasileira. Bol.Têc.Inst. Agron. Norte. 29: 1-62.
Duvigneaud F. 1946. La variabilité des associations vêgitales. Bull. Soc. Dot. Belg. 78: 107-134.
Erwin T.L. & Adis J. 1982. Amazonian inundation forests their role as short-term refuges and generators of species richness and taxon pulsesIn: Prance , G.T. ed.-Biologicaldiversification in the tropics . New York ,Columbia.University Press. : 358-371.
Fittkau E.J. 1964. Remarks on limnology of Central Amazon rain-forest stream. Verh.Internat. Verein.Limnol. 15: 1092-1096.
Fittkau E.J. 1967. On the ecology of amazonian rain-forest streams. Atas do Simpósio sobre a Biota Amazônica 3(Limnologia).: 97-108.
Fittkau E.J. 1971. Esboço de uma divisão ecológica da região amazônica . In: Idrobo, J.M. (ed.). II Simpósio y Foro de Biologia Tropical Amazônica. Bogotá, Editorial PAX.: 365-372.
Fittkau E.J., Junk W.J., Klinge H. & Sioli H. 1975a. Substrate and vegetation in the Amazon region.In: Dierschke, Ii. (ed.). Vegetation und substrat. Vaduz, J. Cramer.: 73-90.
Fittkau E.J., Irmler U., Junk W.J., Reiss F. & Schmidt G.W. 1975b. Productivity, biomass, and
population dynamics in amazonian water bodies . In: Golley, F. B.& Medina, E. (eds.). Tropical ecological systems: trends in terrestrial and aquatic research. New York, Springer Verlag.: 289-311.
Fleming T.H. & Iieithaus E.R. 1981. Frugivorous bats, seed shadows and the struture of tropical forests. Biotropica. 13(2): 45-53.
Furch K. 1997. Chemistry of várzea and igapó soils and nutrient inventory in their floodplain forests. In: The Central Amazon floodplain: Ecology of a pulsing system. Junk W.J. (ed.). Ecological Studies 126, Springer Verlag, Heidelberg. : 47-68.
Gessner F. 1955. Hydrobotanik. Band I: Energiehaushalt. Berlin, VEB Deutscher Verlag der Wissenschaften.
Gibbs R.J. 1967. The geochemistry of the Amazon river system. Part I. The factors that control the salinity and the composition and concentration of the suspended solids. Geol. Soc. Am. Bull. 78: 1203-1232.
Gottsberger G. 1978. Seed dispersal by fish in the inundated regions of Humaitá, Amazônia.Biotropica. 10 (3): 170-183.
Goulding M. 1979. Ecologia da pesca no rio Madeira. Manaus, INPA.
Goulding M. 1980. The fishes and the forest. Los Angeles, Univ. of California Press.
Harper J.L. 1977. Population biology of plants. New York, Academic Press.
Huber J. 1906. La végétation de la vallée du rio Purus ( Amazone ). Bulletin de L'Herbier Boissier, 2me série. 4(4): 249-276.
Illies J. 1967. Versuch einer allgemeinen biozönotischen Gliederung der Fliessgewässer Int. Revue Hydrobiol. 46(2): 205-213.
Irio G. & Adis J. 1979. Evolução de florestas amazô-nicas inundadas, de igapó - um exemplo do rio Tarumã-mirim. Acta Amazonica. 9(2): 299-303.
Irmler U. 1973. Population-dynamic and physiological adaptation of Pentacomia egregia CHAUD. (Col. Cicindelidae) to the amazonian inundation forest. Amazoniana. 4(2): 219-227.
Irmler U. 1975. Ecological studies of the aquatic soil invertebrates in three inundation forests of Central Amazônia. Amazoniana. 3(3): 337-409.
Irmler U. 1977. Inundation-forest tipes in the vicinity of Manaus. Biogeographica. 8: 17-29.
Irmler U. 1978. Matas de inundação da Amazônia Central em comparação entre águas brancas e pretas. Ci. e Cult. 30(7): 813-821.
Janzen D.H. 1967. Synchronization of sexual reprodution of trees within the dry season in Central América. Evolution. 21(3): 620-637.
Janzen D.H. 1969. Seed-eaters versus seed size, number, toxicity and dispersal. Evolution. 23(1): 1-27.

M. T. F. PIEDADE, P. PAROLIN Y W. J. JUNK
Ecología Aplicada Vol. 2 Nº 1, pp. 31-40 ______________________________________________________________________________________
39
Janzen D.H. 1970. Herbivores and the number of tree species in tropical forests. Am. Nat. 104 (940): 501-528.
Janzen D.H. 1971. Seed predation by animals . A. Rev. Ecol. Syst. 2: 465-492.
Janzen D.H. 1972. Association of a rain-forest palm and seed-eating beetles in Puerto Rico. Ecology. 53(2): 258-261.
Janzen D.H. 1974. Tropical blackwater rivers, animals, and mast fruiting by the Dipterocarpaceae. Biotropica. 6 (2): 69-103.
Junk W.J. 1970. Investigations on the ecology and production-biology of the "floating meadows" (Paspalo Echinocloetum) on the Middle Amazon . I . The floating vegetation and its ecology . Amazoniana. 2(4): 449-495.
Junk W.J. 1980 . Áreas inundáveis - Um desafio para limnologia. Acta Amazonica. 10: 775-795.
Junk W.J. 1983. Ecology of swamps of the Middle Amazon. _In: Goodall, D.W. ed. - Ecosystems of the world . Amsterdam, Elsevier Scientific Publishing Co. : 269-294.
Junk W.J. & Furch K. 1980. Química da água e macrófitas aquáticas de rios e igarapés da bacia amazônica e nas áreas adjacentes . Parte I. Trecho Cuiabá - Porto Velho - Manaus. Acta Amazonica. 10(3): 611-633.
Junk W.J. , Klinge H. , Revilla J.C., Worbes M. & Furch B. 1983. A floresta inundável . In : INPA - Relatório anual.: 194-198.
Junk W.J., Bayley P.B. & Sparks R.E. 1989. The flood pulse concept in river-floodplain systems. In: Proceedings of the International Large River Symposium. Dodge D.P. (ed.). Can. Publ. Fish. Aquat. Sci. 106: 110-127.
Keel S.H.K. & Prance G.T. 1979. Studies of the vegetation of a white and black-water igapó ( Rio Negro Brazil ) . Acta Amazônica. 9 (4): 645-655.
Klinge H. 1965. Podzol soils in the Amazon basin. J. Soil Sci. 16(1): 95-103.
Leenheer J.A. & Santos H. 1980. Considerações sobre os processos de sedimentação na água preta ácida do rio Negro (Amazônia Central). Acta Amazônica. 10(2): 343-355.
Leite A.M.C. & Rankin J.M. 1981. Ecologia de sementes de Pithecolobium racemosumDucke. Acta Amazônica. 11 (2): 309-318 .
Louis L. 1947. L’origine et la vegétation des iles du fleuve de la région de Yangambi . C.R. Sem. Ar. Yangambi. 12: 924-933.
Luizão F.J. 1982. Produção e decomposição da liteira em floresta de terra-firme da Amazónia Central . Aspectos químicos e biológicos da lixiviação e remoção dos nutientes da liteira. Tese de Mestrado Mánaus , INPA/ FUA.
Machado-Allison A.J. 1982. Studies on the systematics of the subfamily Serrasalminae (
Pisces-Characidae). Ph. D. Thesis. Washington, The George Washington University.
Martius K.F.P. von. 1823-1850. Historia naturalis palmarum. vol 1-3. Lipsiae, T.O. Weigel.
Moore H.E.Jr. 1973a . Palms in the tropical forest ecosystems of Africa and South Amarica . In: Meggers , D. J. ; Ayensu , E. S. ; Duckworth , W. D. eds. - Tropical forest ecosystems in Africa and South America : a comparative review `. Washington , Smithsonian Institution Press. : 63-88.
Moore H.E. Jr. 1973b. The major groups of palms and their distribution. Ithaca , Cornell University.
Odum E.P. 1971. Ecologia. México , Nueva Editorial Interamericana.
Pianka E.R. 1970. On r-and K- selection . Am. Nat. 104: 592-597.
Pearson D. 1975. Laboratory techniques in food analysis. London, Butterworth & Co.
Pires J.M. 1974. Tipos de vegetação da Amazônia. Brasil Florestal. 5(17): 49-58.
Prance G.T. 1978. The origin and evolution of the Amazon flora. Interciência. 3(4): 207-222.
Prance G.T. 1980. A terminologia dos tipos de florestas amazônicas sujeitas à inundação. Acta Amazônica. 10(3): 495-504.
RADAMBRASIL. 1978. Levantamento de recursos naturais. Folha SA. 20. Manaus. Rio de Janeiro, Departamento Nacional de Produção Mineral.
Rankin J.M. 1978. The influence of predation on tree species abundance in two adjacent rain forest communities in Trinidad, West Indies. Ph.D. Thesis. Ann Arbor, University of Michigan.
Revilla J.D.C. 1981. Aspectos florísticos e fitossocio-lógicos da floresta inundável (igapó). Praia Grande, rio Negro, Amazonas, Brasil. Dissertação de Mestrado. Manaus, INPA/FUA.
Ribeiro M.C.L.B. 1983. As migrações dos jaraquis (Pisces, Prochilodontidae) no rio Negro, Amazonas, Brasil. Dissertação de Mestrado. Manaus, INPA/FUA.
Ribeiro M.N.G. 1976. Aspectos climatológicos de Manaus. Acta Amazonica. 6(2): 229-233.
Rodrigues W.A. 1961. Estudo preliminar de mata de várzea alta de uma ilha do baixo rio Negro de solo argiloso e úmido. Publicação do INPA - série Botânica. 10.
Sarukhán J. 1974. Studies on plant demography: Ranunculushepens L., R. bulbosus L. e R. acris L. II. Reproductive strategies and seed population dynamics. J. Ecol. 62(1): 151-177.
Schmidt G.W. 1973. Primary production of phytoplancton in the three types of amazonian waters. II. The limnology of a tropical flood-plain lake in Central Amazonia (Lago do Castanho). Amazoniana. 4(2): 139-203.

ESTRATÉGIAS DE DISPERSÃO, PRODUÇÃO DE FRUTOS E EXTRATIVISMO DA PALMEIRA Astrocaryum jauari MART. NOS IGAPÓS DO RIO NEGRO: IMPLICAÇÕES PARA A ICTIOFAUNA
Diciembre 2003 _________________________________________________________________________________________
40
Schmidt G.W. 1976. Primary production of phytoplancton in the three types of amazonian waters. IV. On the primary productivity of phytoplancton in a bay of the lower rio Negro (Amazonas, Brazil). Amazoniana. 5(4): 517-528.
SEMA. 1982. Programa de estações ecológicas. Brasília, Ministério do Interior.
Sioli H. 1951. Alguns resultados e problemas da limnologia amazônica. Bolm tëc. Inst. agron. N. 24: 2-44.
Sioli H. 1964. General features of the limnology of Amazonia. Verh. Internat. Verein. Limnol. W5(2): 1053-1058.
Sioli H. 1965. A limnologia e sua importância em pesquisas na Amazônia. Amazoniana. 1(1): 11-35.
Sioli H. 1968. Hydrochemistry and geology in the brazilian Amazon region. Amazoniana. 1(3): 267-277.
Sioli H. 1973. Principais biótopos de produção primária nas águas da Amazônia. B. geogr. (R. Janeiro). 32(236): 131-142.
Sioli H. 1975. Tropical river: the Amazon. In: Whitton, B. A. ed. - River ecology. Berkeley, Univ. of California Press. : 461-488.
Sioli H. 1976. A limnologia na região amazônica brasileira. Anais do I Encontro Nacional sobre Limnologia, Piscicultura e Pesca Continental. Belo Horizonte. : 153-169.
Sioli H. & Klinge H. 1962. Solos, tipos de vegetação e águas na Amazônia Brasileira. B. Mus. Paraen. Emilio Goeldi, nova sér., Avulsa. 1: 27-41.
Takeuchi M. 1962. The structure of the amazonian vegetation. VI. Igapó. Journ. Fac. Sci. Univ. Tokyo, III. 8(7): 297-304.
Yeaton R.I. 1979. Intraespecific competition in a population of the stilt palm, Socratea duríssima (Oerst.) Wendl. on Barro Colorado island, Panamâ. Biotropica. 11(2): 115-158.
van der Pijl L. 1969. Evolutionary action of tropical animals on the reproduction of plants. Biol. J. Linn. Soc. 1: 85-96.
van der Pijl L. 1972. Principes of dispersal plants. Berlin, Springer-Verlag. 161 p. in higher planta. Berlin, Springer-Verlag.
Whitmore T.C. 1975. Tropical rain forest of the far east. Oxford, Clarendon Press.
Wilson D.E. & Janzen D.H. 1972. Predation of Scheelea palm by bruchid beetles: seed density and distance from the parent palm. Ecology. 53: 954-959.
Wittmann F. & Junk W.J. 2003. Sapling communities in Amazonian white-water forests. Journal of Biogeography. 30(10): 1533-1544.
Worbes M. 1983. Vegetationskundliche Untersuchungen zweier Überschwemmungswälder in Zentralamazonien - vorläufige Ergebnisse. Amazoniana. 8(l): 47-65.
Worbes M. 1984. Periodische Zuwachszonen an Bäumen zentralamazonischer Überschwemmungswälder. Naturwissenschaften. 71:157.

Ecología Aplicada, 2(1), 2003 ISSN 1726-2216 Depósito legal 2002-5474
CARACTERIZACIÓN MORFÓLOGICA DE PLÁNTULAS DE “UÑA DE GATO” Uncaria tomentosa (Willdernow ex Roemer & Schultes) D.C. y U. guianensis (Aublet) Gmelin del BOSQUE
NACIONAL ALEXANDER VON HUMBOLDT. Percy A. Zevallos Pollito1 e Ymber Flores Bendezú2
Resumen
Con el propósito de identificar en el medio natural, los primeros estadíos de crecimiento de las plántulas de “Uña de Gato”, bejucos trepadores con valor medicinal representativos del Bosque Nacional Alexander Von Humboldt se describen las plántulas de Uncaria tomentosa (Willdernow ex Roemer & Schultes) D.C. y U. guianensis (Aublet) Gmelin obtenidas mediante germinación en vivero semidescubierto. Entre otros caracteres observados, útiles para el reconocimiento taxonómico, se tienen las modalidades de germinación, estructura y organización de los cotiledones, del hipocótilo y el epicótilo, complementados en la descripción de los protófilos y metáfilos. Palabras claves: “Uña de Gato”, plántulas
Abstract In order to identify, in the natural environment, the first growth stages of “Uña de Gato”
(representative shrubs of medicinal importance from the Alexander Von Humboldt National Forest), seedling growth of Uncaria tomentosa (Willdenow ex Roemer & Schultes) DC. and U. guianensis, (Aublet) Gmelin is described. The seeds were germinated under semigreenhouse conditions. Useful characters for taxonomic identification are among others: type of germination, cotyledon, hypocotyl and epycotyl structure and organization, coupled with the description of the protophylls and metaphylls. Key Words: “Uña de Gato”, seedling. _______________________________________________________________________________
1 Profesor de la Facultad de Ciencias Forestales de la Universidad Nacional Agraria La Molina. Apartado Postal 456. Lima 100 Perú. 2 Investigador del Instituto Nacional de Investigaciones Agropecuarias (INIA)- Pucallpa.
Introducción Uno de los principales problemas al que se
enfrenta en los programas de reforestación de las zonas tropicales de América Latina, es el de no contar con semillas suficientes en calidad y en cantidad que satisfagan la demanda actual creciente. Una alternativa de solución a este problema se encuentra en el empleo de regeneración natural procedente de árboles semilleros.
La “Uña de Gato” debido a sus principales propiedades medicinales se ha convertido en un “Boom económico” (Zavala et al., 1993), trayendo consigo una enorme y creciente demanda de la corteza en mercados nacionales e internacionales; sin embargo la actividad extractiva, ha puesto a las poblaciones naturales en una situación vulnerable, de peligro, emprendiéndose una fuerte campaña de reposición. Lamentablemente poco es lo que se conoce de su propagación y el manejo de sus semillas, por lo que es necesario trabajar con regeneración natural.
El empleo de regeneración natural requiere de información detallada sobre las características morfológicas determinantes de las especies seleccionadas; sin embargo, existe escasa información al respecto. En América Latina se conocen estudios aislados y dispersos (Ricardi et al., 1987; Duke, 1965; De Castro Oliveira et al., 1987) y en el caso específico del Perú encontramos el trabajo de Díaz & Ríos
(1993) quienes describieron las plántulas de 11 especies forestales de la zona de Pucallpa.
La mayor parte de especies forestales del bosque tropical húmedo peruano no cuentan con estudio de la germinación y del desarrollo de la plántula, conocimiento necesario para su manejo y utilización en programas de recuperación de la foresta. Al respecto, diversos ensayos llevados a cabo demuestran que el empleo de regeneración natural es una alternativa factible y promisoria (Zevallos et al., 1991; Flinta, 1960; Pacho et al., 1984). Los objetivos del presente estudio son:
Describir el proceso de germinación de las semillas y la morfología de las plántulas de Uncaria tomentosa y U. guianensis, en su desarrollo.
Proporcionar información adicional para contribuir a la identificación in situ de dichas especies.
Revisión de literatura. Taxonomía
De acuerdo a Cronquist (1981) las especies de “Uña de Gato” la podemos clasificar:
División : Magnoliophyta Clase : Magnoliopsida Sub-Clase : Asteridae Orden : Rubiales Familia : Rubiaceae Género : Uncaria Schreb

CARACTERIZACION MORFOLOGICA DE PLANTULAS DE UÑA DE GATO
Diciembre 2003 __________________________________________________________________________________________
42
Especies : Uncaria guianensis (Aublet) Gmelin. Uncaria tomentosa (Willd. ex Roemer & Schultes ) DC.
Importancia taxonómica y silvicultural de la caracterización de plántula
Es consenso entre los autores que han abordado la caracterización de plántulas que, esta permite analizar caracteres morfológicos de la regeneración natural en sus primeros estadíos y ofrecen la posibilidad que en esta etapa juvenil se presenten estructuras u órganos primitivos que desaparecen al continuar el desarrollo, pero que pueden tener extraordinario valor para establecer relaciones de parentesco o conexiones filogenéticas con grupos cuyos órganos adultos ya no lo presentan (Díaz & Ríos, 1993; Burger, 1972, Ricardi et al., 1987).
La importancia del reconocimiento in situ de las plántulas de especies forestales es bien conocida en la Silvicultura Tropical. Una técnica exitosa de manejo de bosques basado en regeneración natural depende de la disponibilidad de semillas y plántulas de las especies deseables. El reconocimiento de tales plántulas brinda una herramienta para incrementar la densidad de especies deseables así como predecir la composición del bosque en los sistemas silviculturales (Lampretch, 1990; Barrera, 1992).
En nuestro país el empleo de regeneración natural para la obtención de plantones no es algo nuevo. Ha sido empleado muchas veces cuando no había disponibilidad de semillas. Al respecto del uso de regeneración natural para reforestar áreas degradada Schwyzer (1982) determinó que es más barato reforestar con regeneración natural que con plantones producidos en vivero. Germinación
La germinación de las plantas con semilla puede ser definida como la secuencia de eventos morfogenéticos que resultan en la transformación de un embrión en una plántula (Barner, 1975). ISTA (1991) define la germinación como la emergencia y desarrollo a partir del embrión de la semilla, de aquellas estructuras esenciales que, para la clase de semilla que se está analizando, indican la capacidad de desarrollo bajo condiciones favorables del suelo para producir una planta normal.
Para algunos autores como Duke (1965), Burger (1972) y De Castro Oliveira et al. (1987) hay dos tipos de germinación: fanerocotilar en la cual los cotiledones emergen de la semilla y criptocotilar en la cual los cotiledones no emergen de la semilla. Sin embargo esta clasificación no indica si los cotiledones llegan a emerger o sobresalir por encima de la superficie del suelo.
En tal sentidos otros autores (Ricardi et al., 1987; Barrera, 1992; Díaz & Ríos, 1993) reconocen la germinación de tipo epígea cuando los cotiledones salen de la semilla y se exponen al aire y germinación
hipogea cuando los cotiledones permanecen en la envoltura de la semilla y no emergen a la luz. Otros autores como Flores et al. (1990) emplean ambos tipos de clasificación en forma simultánea.
Algunos autores distinguen un tercer tipo de germinación denominado semicriptocotiledonar (Duke, 1969) o semihipogea (Díaz et al., 1993). Según estos últimos autores la germinación semihipógea se da cuando ocurre primero la emergencia del eje de la plántula, que desarrolla los protófilos sobre el epícotilo, luego desarrolla el hipocótilo que eleva consigo ligeramente a los cotiledones al ras de suelo. Eje de la plántula
En la plántula ya desarrollada se puede distinguir que el eje principal se divide en dos partes: un segmento que está por debajo del punto de inserción de los cotiledones y otro que está por encima de dicho punto. El segmento que esta debajo se denomina hipocótilo (De “hypo”: debajo y “cotilo”: cotiledón) y el segmento que está por encima, que prácticamente va a dar origen a toda la parte aérea de la planta, se denomina epicótilo (Del prefijo “epi”. Encima y “cotilo”: cotiledón). (Ricardi et al., 1987; Barrera, 1992; De Castro Oliveira et al., 1987; Díaz et al., 1993). Duke (1969) indica además que el hipocótilo es la porción del eje en donde ocurre la transición de raíz a tallo probablemente debido a que en algunos casos hipocótilo es totalmente subterráneo. En realidad es posible distinguir ambos segmentos del eje de la planta a nivel del embrión pero como afirma Duke (1969) después de la germinación la distinción entre epicótilo e hipocótilo se hace notoria; así mismo, afirma que en algunas especies leñosas el epicótilo y el hipocótilo tienen diferente indumento y textura. Estadíos de desarrollo
Según Díaz et al. (1993) de acuerdo a los elementos foliares que van apareciendo en forma sucesiva es posible definir los siguientes estadíos de desarrollo: cotiledones, protófilos y matafilos. El estadío de cotiledones ocurre desde la germinación de la semilla hasta el desarrollo pleno de los cotiledones. El estadío de protófilos ocurre desde los cotiledones desarrollados hasta las primeras hojas. El estadío de metáfilos ocurre después de los protófilos, cuando la plántula muestra hojas similares a las de forma adulta. Los protofilo también denominados eófilos (Burger, 1972; Flores et al, 1990; Duke, 1969) son las primeras hojas que aparecen después de los cotiledones. Pueden ser similares o diferentes a los metáfilos.
Metodología Ubicación y descripción del área de estudio
Ubicación: El estudio se realizó en el Campo Experimental Alexander Von Humboldt ubicado a 86 km. De la Ciudad de Pucallpa en el distrito de Irazola, provincia de Padre Abad, departamento de Ucayali, Geográficamente se encuentra a 8º 22” de Latitud sur y 74º 28”de Longitud oeste (Figura 1).

PERCY ZEVALLOS E YMBER FLORES
Ecología Aplicada Vol. 2 No 1, pp. 41-46 __________________________________________________________________________________________
43
Clima: El área representa una temperatura media máxima de 31°C y una temperatura media mínima de 21°C, mientras que la temperatura media total es de 26°C. La precipitación anual es superior a 4,000 mm. Y la humedad relativa media es 75%.
Ecología: El área se ubica en las zonas ecológicas de bosque húmedo tropical (bh-T) y bosque húmedo premontano tropical (bh-PT).
Fisiografía: La zona se halla entre los 200 a 340
msnm de altitud y presenta un relieve plano a colina. Método
Selección de especies. Para la selección de especies se ha tenido en cuenta los siguientes criterios: a) Importancia y uso actual o potencial de la especie, b) Existencia de bejucos semilleros en los rodales semilleros de la Subestación Alexander Von Humboldt y c) Disponibilidad de semillas en cantidades suficientes.
Sobre la base de criterios mencionados se han seleccionado 02 especies denominadas la zona: “Uña de gato espina curvada” y “Uña de gato espina recta”.
Colección e identificación de muestras botánicas. La colección en la Subestación Alexander Von Humboldt. Los especimenes se identificaron en el Herbario de la Subestación y en el Herbario Dendrológico de la Facultad de Ciencias Forestales de la Universidad Nacional Agraria La Molina. Las especies determinadas son:
Uncaria guianensis (Aubl.) Gmel. “uña de gato espina curvada" Uncaria tomentosa (Willdenow ex Roemer & Schultes) DC. “uña de gato espina recta, ambas de la familia Rubiaceae.
Colección de procesamiento de semillas. Los frutos fueron coleccionados de dos formas: mediante el escalamiento de árboles o recogidos del suelo. Las semillas extraídas fueron secadas al aire libre y seleccionadas, luego se procedió a la siembra inmediata.
Siembra de las semillas, Los almácigos se prepararon utilizando tierra procedente del bosque, la cual fue posteriormente tamizada y desinfectada. En general no se empleó ningún tratamiento progerminativo ya que se deseaba que la germinación sea lo más natural posible. Sin embargo en caso de Uncaria tomentosa las semillas fueron puestas a remojar en agua hasta lograr que germinen. Este proceso duró 15 días. Se prepararon camas de almácigo (1 m2 por especie) para la realización de los ensayos de germinación (100 semillas por especie, a 10 cm. X 10cm. de separación para evitar que se dañaran las raíces cuando se extrajeran las plántulas para su posterior tratamiento).
Formulario descriptor. Para la elaboración del formulario se revisaron los trabajos de Ricardi et al. (1987); Diaz et al. (1993) y De Castro Oliveira (1987). El formulario fue utilizado para la descripción morfológica de las plántulas en sus distintos estadíos. Tipo de germinación
Hipógea, epígea o semihipógea. Eje de la plántula
Correspondiente al hipocótilo y epicótilo. 01 Sección transversal: redonda, cuadrada 02 Longitud 03 Diámetro 04 Superficie: lenticelada, lisa, etc. 05 Color 06 presencia de exudaciones o látex.
Elementos foliares Correspondiente a los cotiledones, protófilos y
metáfilos. 01 Número 02 Disposición: alternos, opuestos, etc. 03 Duración: persistente, deciduos, etc. 04 Consistencia: carnosos, coriáceos, papiráceos,
etc. 05 Inserción: sésiles, peciolados, etc. 06 Divisiones: simples, pinnadas, etc. 07 Forma: elípticos, aovados, etc. 08 Apice: obtuso, acuminado, etc. 09 Base: truncada, asimétrica, redonda, etc. 10 Margen: entero, aserrado, ciliado, etc. 11 Tamaño 12 Color 13 Dirección: rectos, inclinados, etc. 14 Superficie: glabra, tomentosa, puberulenta, etc. 15 Marcas o Huellas
Figura 1. - Ubicación de la Estación Experimental Alexander Von Humbolt.

CARACTERIZACION MORFOLOGICA DE PLANTULAS DE UÑA DE GATO
Diciembre 2003 __________________________________________________________________________________________
44
16 Venación : broquidódroma, eucamptódroma, etc.
17 Presencia de estípulas: aciculares, redondas, etc.
18 Exudaciones: látex, resinas, etc. Observaciones realizadas. Según la velocidad de
desarrollo de las plántulas de cada especie, se realizaron observaciones semanales, quincenales y mensuales.
Asimismo se tomaron muestras para prensarlas, secarlas y montarlas convenientemente. Se hizo ilustraciones considerando las características más representativas de las plántulas en sus diversos estadíos.
Resultados
Se logró estudiar plántulas de dos especies pertenecientes al género Uncaria Schreb de la familia Rubiaceae: 4.1 Uncaria guianensis (Aublet) Gmelin (Uña de gato espina curvada) 1.Tipo germinación Epígea 2. Eje de la plántula (a los 60 días)
Radícula: raíz primaria axonomorfa, larga, blanca; raíces secundarias numerosas, muy largas y ramificadas.
Hipocótilo: cilíndrico, de 3-4 mm de largo y 1 mm de diámetro, superficie glabra, blanco rojizo.
Epicótilo: ligeramente cuadrangular; de 3-4 mm de largo, superficie glabra, rojizo. 3. Estadíos
Cotiledones (a los 30 días): cotiledones 2, epígeos, opuestos, foliáceos. Peciolo con 1-2 mm de largo, blanco verdusco; lámina entera, ovada, 2-2.5 mm de largo por 1.5-1.8 mm de ancho, ápice obtuso o redondeado, base redondeada o truncada con estrechamiento en el peciolo, nervaduras rudimentarias, escasamente visible la nervadura central, ambas caras verdes claro y glabras.
Protófilos (a los 45 días): opuestos, simples; peciolos glabros de 2-4 mm de largo; láminas enteras, ovadas o elípticas, ápice agudo u obtuso, base atenuada. El tamaño de los protófilos es creciente, los primeros poseen una dimensión de 6-7 mm de largo por 4-5 de ancho, ambas caras verdes claro y glabras, el haz posee una superficie brillante con nervadura central ligeramente rojiza. Venación eucamptódroma.
Estípulas connatas, redondas. Metáfilos (a los 90 días): opuestos, simples.
Peciolos ligeramente aplanados por arriba y convexos por abajo, 0.4- a más mm, rojizos; lámina entera, ovadas o elípticas, ápice agudo o muy ligeramente acuminado, base atenuada, haz verde oscuro con una superficie brillante y las nervaduras de color rojo, principalmente la nervadura central, el envés es de color verde más claro, ambas caras son glabras, nervaduras prominentes en ambas superficies,
venación eucamptódroma. Estípulas connadas, interpeciolares (Figura 2). 4.2 Uncaria tomentosa (Willdenow ex Roemer & Schultes) DC. (Uña de gato espina recta). 1. Tipo de germinación Epígea 2. Eje de la plántula (a los 60 días)
Radícula: raíz primaria axonomorfa, profunda, blanca; raíces secundarias numerosas, muy largas y ramificadas.
Hipocótilo: cilíndrico, 2-3 mm de largo, 1 mm de diámetro, superficie glabra, blanco verdusco.
Epicótilo: ligeramente Cuadrangular; 1-2 mm de largo, superficie glabra, blanco verdusco. 3. Estadíos
Cotiledones (a los 30 días): cotiledones 2, epígeos, opuestos, foliáceos. Peciolo 1-2 mm de largo, blanco verdusco; lámina entera, ovada o ligeramente elíptica, 1.5-2.0 mm de largo por 0.8-1.0 mm de ancho, ápice obtuso o redondeado, base redondeada o truncada con estrechamiento en el peciolo, nervaduras rudimentarias, escasamente visible la nervadura central, ambas caras verdes claro y glabras.
Protófilos (a los 45 días): opuestos, simples; peciolos glabros de 2-4 mm de largo; láminas enteras, ovadas, ápice agudo u obtuso, base atenuada. El tamaño de los protófilos es creciente con el tiempo, los primeros poseen una dimensión de 6-7 mm de largo por 4-5 mm de ancho, ambas caras verdes claro
Figura 2. - Uncaria guianensis (Aublet) Gmelin RUBIACE. A) Inicio de la germinación (15 días). B) Plántula de 5 meses mostrando los metáfilos. C) Plántula de 2 meses.

PERCY ZEVALLOS E YMBER FLORES
Ecología Aplicada Vol. 2 No 1, pp. 41-46 __________________________________________________________________________________________
45
y glabras. Venación eucamptódroma. Estípulas connatas, interpeciolares, triangulares.
Metáfilos (a los 90 días): opuestos, simples. Peciolos ligeramente aplanados por arriba y convexos por abajo, 0.4- más mm; lámina entera, oblongas u ovadas, ápice agudo o muy ligeramente acuminado, base atenuada, ambas caras verdes claro y glabras, nervaduras prominentes en ambas superficies, venación eucamptódroma.
Estípulas connata, interpeciolares (Figura 3).
Discusión De la plántula
Una plántula es una planta nacida a partir de una semilla y no por reproducción vegetativa. En la mayoría de los casos el término “plántula” se refiere a individuos muy jóvenes. En este documento se describen 3 estadíos de desarrollo. Dicha clasificación es arbitraria, estuvo basada principalmente en los trabajos de Diaz & Rios (1993), Ricardi et al. (1987) y De Castro Oliveira et al. (1987). El primer estadío se definió para plántulas que muestren los cotiledones expuestos y plenamente desarrollados por lo que fue denominado como estadío de COTILEDONES. En este estadío puede o no haber presencia de la primera o primeras hojas verdaderas. El segundo estadío fue definido para las plántulas que poseían las primeras hojas (PROTOFILOS) plenamente desarrolladas. El tercer estadío fue definido para las plántulas que poseían ya hojas similares a las de la planta adulta
(METAFILOS). En la descripción, la secuencia y número de hojas son mencionados.
Se encontraron similitudes y diferencias entre las plántulas de las especies estudiadas. Uncaria tomentosa y Uncaria guianensis poseen similar tipo de germinación, disposición foliar, sistema radicular, forma de las estípulas, pero difieren principalmente en la forma, color y brillo de las hojas.
El tiempo que se requiere para que las plántulas alcancen los estadíos mencionados varía según la especie. Se requiere un minucioso cuidado para aquellas especies en que las plántulas alcancen los diversos estadíos en un tiempo relativamente corto. En el presente trabajo se ha incluido la edad de la plántula al momento de hacerse las descripciones y los dibujos, pero debe acotarse que la uniformidad en el desarrollo de las plántulas también depende de la especie. De las dimensiones
La altura y tamaño de las plántulas son frecuentemente inservibles para identificarlos, pero estas medidas son importantes para la impresión general que se haga de la planta, observación también realizada por Burger (1972). Las dimensiones que se dan fueron determinadas en los especímenes observados en todos los estadíos:
a) Longitud y diámetro del hipocótilo y del epicótilo excluyendo la raíz. b) Dimensiones de los elementos foliares: cotiledones, protófilos y metáfilos. Debe anotarse que haciendo una comparación entre las plántulas del presente ensayo y las que se encontraron dentro del bosque como parte de la regeneración natural, se observó que las primeras tenían por lo general mayor desarrollo y vigorosidad de sus diferentes partes que las segundas, debido probablemente a la mayor intensidad de luz que reciben. Asimismo, en algunos casos el tamaño de un órgano presente en la plántula es mayor que el que se halla en las plantas adultas de la misma especie. Color
El color de las partes y órganos de la planta puede ser característico, pero puede también ser cambiante, dependiendo principalmente de la edad de la plántula y de su contenido de humedad. Por ejemplo: el cambio de rojo a marrón en las nervaduras de Uncaria guianensis (Aublet) Gmelin, cuando se van secando. En este documento los colores descritos fueron observados en plántulas vivas o en material fresco recién colectado. Tipo de germinación
Las especies muestran una germinación de tipo epígea. Ellas presentan los cotiledones de color verde, lo cual evidencia su carácter fotosintético, además son muy persistentes y caen tardíamente debido al aumento del grosor del tallo (hipócotilo).
Conclusiones
Las plántulas de las dos especies poseen similar tipo de germinación, disposición foliar, sistema
Figura 3. - Uncaria tomentosa (Willdenow ex Roemer & Schultes) DC. RUBIACE.A, B y C) Secuencia de la germinación de las semillas hasta el estadío de cotiledones. D) Prótofilos (60 días). E) Plántula de 5 meses mostrando los metáfilos.

CARACTERIZACION MORFOLOGICA DE PLANTULAS DE UÑA DE GATO
Diciembre 2003 __________________________________________________________________________________________
46
radicular y forma de estípulas; pero difieren en la forma, color y brillo de las hojas.
Las especies presentan una germinación de tipo epígea
3. En ambas especies los cotiledones tienen formas ovadas a ligeramente elípticas, los protófilos de ovadas a elípticas y los metáfilos en U. guianensis (Aublet) Gmelin ovadas o elípticas y en U. Tomentosa (Willdenow ex Roemer & Schultes) DC. oblongas u ovadas.
4. Las dos especies presentan raíz primaria axonomorfa, largas y raíces secundarias numerosas, largas y ramificadas.
5. El hipocótilo es cilíndrico para las dos especies; asimismo el epicótilo es ligeramente cuadrangular, sin embargo el color en U. Guianensis (Aublet) Gmelin es rojizo y el U. tomentosa (Willdenow ex Roemer & Schultes) DC es blanco verdusco.
6. Las dimensiones de las diferentes partes de las plántulas son frecuentemente inservibles para la identificación de las especies estudiadas; sin embargo, las características morfológicas si tienen valor taxonómico.
Recomendaciones
Se debe continuar con los estudios de caracterización de plántulas, sobre todo de aquellas especies que poseen un uso actual o potencial o de aquellas endémicas del Perú. Cuando ya se tenga un número considerable de especies descritas se puede elaborar claves de identificación como aquellas que se emplean para plantas adultas.
Se puede usar plántulas a partir de regeneración natural para disminuir los costos de plantación.
Bibliografía Barrera T.E. 1992. Plántulas de algunas especies
leñosas y connaturalizadas del Bosque Subandino Silvania-Cundinamarca, Colombia. En : Agronomía colombiana. 9: 131-160.
Barner H. 1975. Tree seed development and germination. En: FAO/DANINA training course of forest seed collection and handling Vol.2 Roma, Italia.: 24-41.
Burger-H. D. 1972. Seddling of some tropical trees and shrubs mainly of South East Asia Centre for Agricultural Publishing and Documentation. Wageningen, Bélgica.
Cronquist A. 1981. An Integrated System of Classification of Flowering Plant. New York Botanical Garden. New York.
De Castro Oliveira E. & Pereira T.S. 1987. Euphorbiaceae: Morfología da germinacão de algumas espécies I. Revista Brasileira da Semestes Ano 9 – n 1 Brasilia, Brasil.: 9-29.
Diaz-G. J. & Rios-T. J. 1993. Identificación de la regeneración natural de árboles tropicales por la morfología de sus estadíos iniciales. Revista
Forestal del Forestal del Perú Lima, Perú. 20(1): 35-61.
Duke-A. J. 1969. On tropical tree seedlings I. Seeds, seeling and sitematics. Ann. Missouri Botanical Garden 56 (2) Missouri, USA. : 125-165.
Flinta C. 1960. Prácticas de Plantaciones Forestales en América Latina. FAO Roma, Italia.
Flores E.M. & Benavides C.E. 1990. Germinación y morfología de la plántula de Hymenaea courbaril L. (Caesalpinaceae). Revista de Biología Tropical (Costa Rica). 38(1): 91-98.
Font Quer P. 1985. Diccionario de bótanica. Ed. Labor S.A. Barcelona, España.
INTERNACIONAL SEED TESTING ASSOCIATION (ISTA). 1991. Tree and Shrub Seed Handbook. Zuruch, Suiza p.v.
Lamprecht H. 1986. Silvicultura en los trópicos: los ecosistemas forestales en los bosques tropicales y sus especies arbóreas; posibilidades y métodos para un aprovechamiento sostenido. Deustsche Gesellschaft Fürtechnische Zugammenarbeit. Eschborn, República Federal de Alemania.
Pacho M.A. & Tipto R.H. 1984. Evaluación de prendimiento de plantas de regeneración natural. Instituto Superior Técnico Suiza. Pucallpa, Perú.
Ricardi M., Hernandez C. & Torres F. 1987. Morfología de plántulas de árboles de los bosques del estado Mérida. Talleres Gráficos de la Universidad de Los Andes. Mérida, Venezuela.
Rios-T. J. 1982. Prácticas de Dendrología Tropical. UNALM-COTESU Lima, Perú.
Schwyzer A. 1982. La regeneración natural puede ser un agente para la reforestación. PROFORESTAL. Lima, Perú.
Zavala-C. C. & Cevallos-P. P. 1996. Taxonomía distribución y status del Género Uncaria en el Perú – Uña de Gato. Universidad Nacional Agraria La Molina. Lima.
Zevallos-P. P. & De La Cruz-S. H. 1991. Tratamientos pregerminativos y repique de regeneración natural en vivero con cinco especies forestales de Cajamarca. Revista Forestal del Perú Lima, Perú. 18(1): 39-46.

Ecología Aplicada, 2(1), 2003 ISSN 1726-2216 Depósito legal 2002-5474
ESTABILIZACIÓN DE DUNAS LITORALES UTILIZANDO Sesuvium portulacastrum L. EN EL DEPARTAMENTO DE LA LIBERTAD, COSTA NORTE DEL PERÚ.
Carlos Tavares Corrêa1 y Ana Sabogal de Alegría2
Resumen
Se presentan resultados preliminares sobre la utilización de Sesuvium portulacastrum L. en la estabilización de dunas litorales. El estudio se llevó a cabo en el balneario de Santa Elena (7°28’52”S y 79°33’22”W), distrito de San Pedro de Lloc, Provincia de Pacasmayo, Departamento de la Libertad. Inicialmente, se realizaron ensayos de propagación en vivero con estacas terminales e intermedia – basales, en los siguientes tratamientos de suelo: arena pura y mezclas proporcionales de arena/humus y arena/humus/tierra de chacra. Posteriormente, 115 plantas enraizadas propagadas en viverofueron trasplantadas a una parcela de 20 x 5 metros, dispuestas en 3 filas alternadas por filas con 160 estacas terminales sin raíz. En ambos casos, el 75% de las plantas fueron tratadas con fertilizantes de liberación lenta y 25% no recibieron tratamiento. Se construyó una barrera porosa para protección de las plantas jóvenes de la erosión eólica. Cuarenta días después de la siembra, 138 plantas sobrevivieron y 142 se murieron. Entre las supervivientes, 118 fueron tratadas y 20 no recibieron cualquier tratamiento. Además, 107 eran estacas terminales sin raíz y solamente 31 eran plantas con raíz propagadas en vivero. Al cabo de cuatro meses del inicio del experimento, se logró una cobertura vegetal promedio de 60% y la formación inicial de dunas. Los resultados iniciales señalan que el mejor tratamiento es el de estacas terminales tratadas con fertilizante de liberación lenta y el empleo de medidas preventivas contra la acción del oleaje. Palabras clave: Sesuvium portulacastrum L., “verdolaga”, estabilización de dunas litorales, estacas terminales, fertilizante de liberación lenta, Perú.
Abstract Preliminary results on the utilisation of verdolaga (Sesuvium portulacastrum L.) on the foredune stabilisation are presented. The study was carried out at Santa Elena (7°28'52''S and 79°33'22''W), northern Peru. Initially, propagation tests were made with terminal, medium and basal stem cuttings in plant pots using the following soil treatments: pure sand, sand/humus and sand/humus/loam. Then, 115 plants with well developed roots were transplanted to a 20 x 5 meters plot near the beach, in three rows separated by rows of 160 rootless terminal cutting stems. In both cases, 75% of the plants were treated with a slow release fertiliser and 25% did not receive any treatment. A porous fence was built for the protection of the young plants from the abrasive action of the sand grains. Forty days after sowing, 138 plants survived and 142 died. Among the survivors, 118 were treated with a slow release fertiliser and 20 were not. In addition, 107 were rootless terminal stakes and only 31 were nursery propagated plants (plant pots). Four months after the beginning of the experiment, a vegetal cover of 60% and an initial dune formation were obtained. Initial conclusions suggest the utilisation of terminal cutting stems treated by slow release fertilisers and the use of porous fence for the prevention of deflation and against wave erosion. Key words: Sesuvium portulacastrum L., verdolaga, Foredune stabilization, cutting stems, slow release fertilizer, Perú.
1 Centro de Investigación en Geografía Aplicada, Pontificia Universidad Católica del Perú, Apartado Postal 1761, Lima 100, Perú. Correo electrónico: [email protected] 2 Sección Química, Pontificia Universidad Católica del Perú, Apartado Postal 1761, Lima 100, Perú. Correo electrónico: [email protected]
Introducción La costa peruana está situada en una región favorable a la formación de dunas debido a su clima desértico, a la disponibilidad de sedimentos, al régimen de vientos y a condiciones topográficas adecuadas. Según el INRENA (1996), 46,703.7 km2 de los terrenos desérticos están cubiertos por depósitos de sedimentos eólicos en forma de mantos de arena y dunas, concentrados principalmente en el desierto de
Sechura y en las Pampas de Villacurí, pero se encuentran diseminadas por toda la costa peruana, donde se concentra, aproximadamente, el 54% de la población del País. La gran movilidad de las arenas por el viento le otorga a las dunas un carácter extremadamente dinámico. Históricamente, el avance de las dunas se constituye en una constante amenaza a centros poblados, tierras agrícolas, infraestructura vial, entre
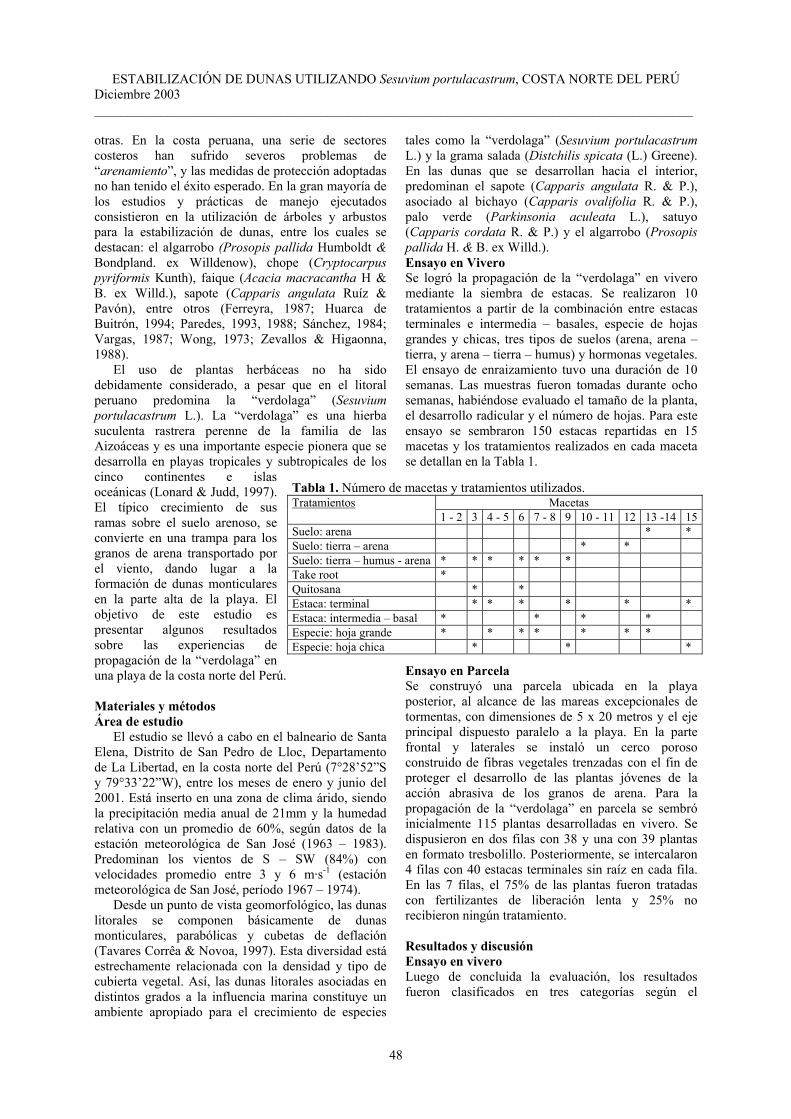
ESTABILIZACIÓN DE DUNAS UTILIZANDO Sesuvium portulacastrum, COSTA NORTE DEL PERÚ
Diciembre 2003 __________________________________________________________________________________________
48
otras. En la costa peruana, una serie de sectores costeros han sufrido severos problemas de “arenamiento”, y las medidas de protección adoptadas no han tenido el éxito esperado. En la gran mayoría de los estudios y prácticas de manejo ejecutados consistieron en la utilización de árboles y arbustos para la estabilización de dunas, entre los cuales se destacan: el algarrobo (Prosopis pallida Humboldt & Bondpland. ex Willdenow), chope (Cryptocarpus pyriformis Kunth), faique (Acacia macracantha H & B. ex Willd.), sapote (Capparis angulata Ruíz & Pavón), entre otros (Ferreyra, 1987; Huarca de Buitrón, 1994; Paredes, 1993, 1988; Sánchez, 1984; Vargas, 1987; Wong, 1973; Zevallos & Higaonna, 1988). El uso de plantas herbáceas no ha sido debidamente considerado, a pesar que en el litoral peruano predomina la “verdolaga” (Sesuvium portulacastrum L.). La “verdolaga” es una hierba suculenta rastrera perenne de la familia de las Aizoáceas y es una importante especie pionera que se desarrolla en playas tropicales y subtropicales de los cinco continentes e islas oceánicas (Lonard & Judd, 1997). El típico crecimiento de sus ramas sobre el suelo arenoso, se convierte en una trampa para los granos de arena transportado por el viento, dando lugar a la formación de dunas monticulares en la parte alta de la playa. El objetivo de este estudio es presentar algunos resultados sobre las experiencias de propagación de la “verdolaga” en una playa de la costa norte del Perú. Materiales y métodos Área de estudio El estudio se llevó a cabo en el balneario de Santa Elena, Distrito de San Pedro de Lloc, Departamento de La Libertad, en la costa norte del Perú (7°28’52”S y 79°33’22”W), entre los meses de enero y junio del 2001. Está inserto en una zona de clima árido, siendo la precipitación media anual de 21mm y la humedad relativa con un promedio de 60%, según datos de la estación meteorológica de San José (1963 – 1983). Predominan los vientos de S – SW (84%) con velocidades promedio entre 3 y 6 m·s-1 (estación meteorológica de San José, período 1967 – 1974). Desde un punto de vista geomorfológico, las dunas litorales se componen básicamente de dunas monticulares, parabólicas y cubetas de deflación (Tavares Corrêa & Novoa, 1997). Esta diversidad está estrechamente relacionada con la densidad y tipo de cubierta vegetal. Así, las dunas litorales asociadas en distintos grados a la influencia marina constituye un ambiente apropiado para el crecimiento de especies
tales como la “verdolaga” (Sesuvium portulacastrum L.) y la grama salada (Distchilis spicata (L.) Greene). En las dunas que se desarrollan hacia el interior, predominan el sapote (Capparis angulata R. & P.), asociado al bichayo (Capparis ovalifolia R. & P.), palo verde (Parkinsonia aculeata L.), satuyo (Capparis cordata R. & P.) y el algarrobo (Prosopis pallida H. & B. ex Willd.). Ensayo en Vivero Se logró la propagación de la “verdolaga” en vivero mediante la siembra de estacas. Se realizaron 10 tratamientos a partir de la combinación entre estacas terminales e intermedia – basales, especie de hojas grandes y chicas, tres tipos de suelos (arena, arena – tierra, y arena – tierra – humus) y hormonas vegetales. El ensayo de enraizamiento tuvo una duración de 10 semanas. Las muestras fueron tomadas durante ocho semanas, habiéndose evaluado el tamaño de la planta, el desarrollo radicular y el número de hojas. Para este ensayo se sembraron 150 estacas repartidas en 15 macetas y los tratamientos realizados en cada maceta se detallan en la Tabla 1.
Ensayo en Parcela Se construyó una parcela ubicada en la playa posterior, al alcance de las mareas excepcionales de tormentas, con dimensiones de 5 x 20 metros y el eje principal dispuesto paralelo a la playa. En la parte frontal y laterales se instaló un cerco poroso construido de fibras vegetales trenzadas con el fin de proteger el desarrollo de las plantas jóvenes de la acción abrasiva de los granos de arena. Para la propagación de la “verdolaga” en parcela se sembró inicialmente 115 plantas desarrolladas en vivero. Se dispusieron en dos filas con 38 y una con 39 plantas en formato tresbolillo. Posteriormente, se intercalaron 4 filas con 40 estacas terminales sin raíz en cada fila. En las 7 filas, el 75% de las plantas fueron tratadas con fertilizantes de liberación lenta y 25% no recibieron ningún tratamiento. Resultados y discusión Ensayo en vivero Luego de concluida la evaluación, los resultados fueron clasificados en tres categorías según el
Tabla 1. Número de macetas y tratamientos utilizados. Macetas Tratamientos
1 - 2 3 4 - 5 6 7 - 8 9 10 - 11 12 13 -14 15Suelo: arena * * Suelo: tierra – arena * * Suelo: tierra – humus - arena * * * * * * Take root * Quitosana * * Estaca: terminal * * * * * * Estaca: intermedia – basal * * * * Especie: hoja grande * * * * * * * Especie: hoja chica * * *

TAVARES CORRÊA Y ANA SABOGAL DE ALEGRÍA
Ecología Aplicada Vol. 2 Nº 1, pp. 47-50 __________________________________________________________________________________________
49
desarrollo de la planta como: bueno, regular y malo, tal como se presenta en las tablas 2a y 2b.
Se observa con evidencia que los mejores tratamientos tanto en velocidad de crecimiento como en número de hojas corresponden a estacas terminales y suelo con arena – tierra – humus. El 86% del total de estacas terminales mostraron un buen crecimiento, mientras que un 56% de los tratamientos con arena – tierra – humus dieron buenos resultados. Las estacas intermedia – basales mostraron un claro retraso en el crecimiento, tanto de la planta y las raíces, cuanto el número de hojas. No se notó una clara diferencia en los diversos tratamientos de suelo. Del total de tratamientos aplicados, el 60% de los que correspondían a arena – tierra, y a 100% de arena, resultaron deficientes en el crecimiento, mientras que un 33% correspondieron a los tratamientos de suelo con arena – tierra – humus.
Ensayo en parcela En la tabla 3 se presenta el estado de las plantas al cabo de cuarenta días de la siembra (del 21 de febrero al 1 de abril del 2001). Pasado este período, 138 plantas sobrevivieron y 137 se murieron. Entre las supervivientes, 118 fueron tratadas con fertilizantes de liberación lenta y 20 no recibieron tratamiento. Además, 107 eran estacas terminales sin raíz (96 con fertilizante de liberación lenta) y solamente 31 eran plantas con raíz propagadas en vivero (22 con fertilizante de liberación lenta). Posteriormente, se realizó una siembra de reposición con 60 estacas terminales sin raíz tratadas con fertilizante de liberación lenta, contabilizándose un total de 298 plantas en 7 filas. En la evaluación del 23 de junio del 2001 no fue posible contar individualmente el número de plantas existentes dado el grado de desarrollo que se encontraban. En este sentido, se realizó una evaluación de la cobertura vegetal, cuyo promedio en la parcela fue de 60 %. La acción del oleaje en la parte SE de la parcela destruyó parte de la vegetación y la cobertura fue solamente de 15%. En las zonas no afectadas, particularmente las de NW, la cobertura alcanzó el 85%. Además, se observó la formación de 3 pequeñas dunas incipientes en las partes SW, central y NE de la parcela, con cobertura superior a 60%. Hay una influencia directa entre la duna incipiente y la protección de las plantas. La mayor cobertura vegetal (85 %) coincide con la formación de dunas; mientras que la menor cobertura vegetal coincide con la falta de desarrollo de dunas y con la entrada del oleaje. Finalmente, se realizó nueva siembra de reposición en un total de 56 hoyos con dos esquejes terminales en cada uno tratados con fertilizante de liberación lenta. Dunas estabilizadas naturalmente Luego del ensayo de enraizamiento se realizó una evaluación de la cobertura vegetal que permitió comparar e interpretar los resultados finales alcanzados en la parcela. En la tabla 4 se muestra el estado de la cobertura vegetal de las dunas con S.
portulacastrum en medio natural, obtenido en 21 de febrero del 2001. Se nota una clara influencia del mar sobre la cobertura vegetal en las 5 dunas medidas. La menor densidad de plantas ocurrió en la cara oeste de la duna, que está directamente expuesta a la acción eólica y marina. En la cara oeste se acumula la arena fresca transportada por el viento y es afectada cíclicamente por la marea y oleaje, lo que limita el desarrollo de la “verdolaga”. Estos resultados permitieron evaluar comparativamente el desarrollo de S. portulacastrum en la parcela.
Tabla 2a. Clasificación del desarrollo de la planta por macetas según tipo de suelo. Clasificación Arena -
tierra – humus
Arena – tierra
arena total Macetas
Bueno 5 1 1 7 3, 4, 5, 6, 8, 12, 15
Regular 1 0 1 2 2, 13 Malo 3 2 1 6 1, 7, 9, 10,
11, 14 Total 9 3 3 15
Tabla 2b. Clasificación del desarrollo de la planta por maceta según tipo de estaca. Clasificación terminal intermedia-
basal total Macetas
Bueno 6 1 7 3, 4, 5, 6, 8, 12, 15
Regular 0 2 2 2, 13 Malo 1 5 6 1, 7, 9, 10,
11, 14 total 7 8 15 15
Tabla 3. Evaluación del estado de las plantas entre 22/02 y 31/03/01 Con tratamiento Sin tratamiento
Fila Vivas Muertas Total Vivas Muertas Total TOTAL Tipo de
tratamiento/estaca1 22 8 30 3 7 10 40 Hidrosorb/terminal
sin raíz 2 5 24 29 3 6 9 38 Hidrosorb/con raíz 3 23 7 30 3 7 10 40 Hidrosorb/terminal
sin raíz 4 10 19 29 5 5 10 39 Hidrosorb/con raíz 5 26 4 30 4 6 10 40 Hidrosorb/ terminal
sin raíz 6 7 22 29 1 8 9 38 Hidrosorb/con raíz 7 25 5 30 1 9 10 40 Hidrosorb/ terminal
sin raíz Totales 118 89 207 20 48 68 275

ESTABILIZACIÓN DE DUNAS UTILIZANDO Sesuvium portulacastrum, COSTA NORTE DEL PERÚ
Diciembre 2003 __________________________________________________________________________________________
50
Según la evaluación de 23 de junio del 2001, la cobertura promedio de “verdolaga” en la parcela (60%) fue superior al promedio de la cobertura en las dunas naturales (56.6 %). Aunque los resultados obtenidos son aceptables, se puede obtener mayor cobertura vegetal con medidas preventivas contra la acción del oleaje, como por ejemplo, sembrar a partir del límite superior de las mareas excepcionales. Tabla 4. Medición de la cobertura vegetal de dunas naturales realizado el 21 de febrero del 2001. Medición Cara Oeste Cima Cara Este 1 25 % 40 % 20 % 2 55 % 90 % 80 % 3 30 % 100 % 70 % 4 40 % 100 % 80 % 5 30 % 40 % 50 % Promedio 36 % 74 % 60 % Promedio de cobertura vegetal general: 56.6 % Conclusión Tanto en vivero como en parcela se recomienda la propagación de la “verdolaga” (Sesuvium portulacastrum L.) a partir de estacas terminales. Los diversos tratamientos de suelo en vivero no mostraron diferencias entre si. Cuando trasplantadas al medio natural, solamente el 30% de las plantas lograron sobrevivir aun siendo tratadas con fertilizante de liberación lenta. Después de 4 meses de siembra en parcela se logró una cobertura vegetal promedio con “verdolaga” del 60% lo cual supera ligeramente el promedio en las dunas naturales que alcanzan el 56%. Mejores resultados se pueden lograr con medidas preventivas contra la acción del oleaje. Agradecimientos Proyecto financiado con apoyo de la Dirección Académica de Investigación de la Pontificia Universidad Católica del Perú (Proyecto DAI n° 113.0121). Se agradece al Centro de investigación en Geografía Aplicada (CIGA) e Instituto de Estudios Ambientales (IDEA) por el apoyo logístico. Los autores agradecen al Ingeniero Zaniel Novoa, Hildebrando Palacios, Eduardo Voysest, María Teresa Román, Fabiola Luján y Alonso Velazco por el apoyo durante los trabajos de campo.
Literatura citada Ferreyra R. 1987. Estudio sistemático de los
algarrobos de la Costa Norte del Perú. Ministerio de Agricultura, INFOR - CUD - CANADA - CONCYTEC. Lima, Perú.
Huarca de Buitrón T. 1994. Métodos de control de la erosión eólica y estabilización de dunas en la pampa de Villacurí, Ica. Tesis para optar el grado de Magister Scientiae. Universidad Nacional Agraria La Molina, Escuela de Post-grado, especialidad en suelos.
INRENA. 1996. Mapa de erosión de suelos en el Perú. Guía explicativa. Instituto Nacional de Recursos Naturales, INRENA, Lima, Perú.
Lonard R.I. & Judd F.W. 1997. The biological flora of coastal dunes and wetlands. Sesuvium portulacastrum (L) L. Journal of Coastal Research. 13(1): 96-104.
Paredes C. 1988. Informe técnico sobre plantaciones de algarrobos en Lambayeque, Ministerio de Agricultura - Unidad Agraria III, Lambayeque - DFF. Chiclayo, Perú.
Paredes C. 1993. Estudio de diecisiete especies forestales nativas para la fijación de dunas. CONCYTEC, Lima.
Sánchez L. 1984. Revisión taxonómica y análisis cuantitativa del género Prosopis en el Departamento de Lambayeque. Tesis en Ingeniería Agronómica, Universidad Nacional Pedro Luis Gallo, Lambayeque, Perú.
Tavares Corrêa C.H. & Novoa Z. 1997. Dinámica de dunas en la costa norte del Perú: problemas y alternativas de solución. Anales VI Congreso Internacional de Geógrafos Latinoamericanistas. Arequipa, Perú.: 125-126.
Vargas C. 1987. Revisión de las leguminosas naturales del Departamento de Lambayeque y zonas adyacentes. Tesis Biología, Universidad Nacional Pedro Luis Gallo, Lambayeque, Perú.
Wong J. 1973. Algunas leguminosas como malezas en el Departamento de Lambayeque. Tesis en Ingeniería Agronómica, Universidad Nacional Pedro Luis Gallo, Lambayeque, Perú.
Zevallos P. & Higaonna R. 1988. Valor pecuario y apícola de 10 especies forestales en las zonas secas y semisecas de Lambayeque. Revista Zonas Áridas, CIZA – UNALM. 5: 31-43.

Ecología Aplicada, 2(1), 2003 ISSN 1726-2216 Depósito legal 2002-5474
EL IMPACTO DE EL NIÑO SOBRE LA PRODUCCIÓN DE CONCHA DE ABANICO (Argopecten purpuratus) EN BAHÍA INDEPENDENCIA, PISCO, PERÚ
Jaime Mendo1 y Matthias Wolff2
Resumen La Bahía Independencia puede ser considerada a nivel mundial como una de las zonas mas productivas en invertebrados bentónicos, cuyas capturas están sujetas a fluctuaciones fuertes originadas por la ocurrencia del fenómeno El Niño y La Niña y por la falta de medidas de manejo adecuadas. Durante períodos con calentamientos fuertes los desembarques anuales alcanzan las 50000 t en un área de aproximadamente 150 km² y durante años fríos estas se mantienen alrededor de 500 a 1000 t. El presente estudio analiza los cambios de los desembarques en la Bahía Independencia observados durante las dos últimas décadas y discute los principales factores que afectan los “booms” de concha de abanico. En este sentido se usa información sobre capturas, temperatura y aquella relacionada con el crecimiento, reproducción, prelación, densidad media y condiciones de oxígeno procedente de trabajos publicados y no publicados. La relación entre la captura anual y la temperatura promedio durante el periodo reproductivo de la concha de abanico durante los últimos 20 años, muestra que la producción es afectada positivamente solo con la ocurrencia de El Niño fuertes como en 1983 y 1998. Los resultados muestran que los “booms” de concha de abanico durante estos El Niño se debe al efecto combinado de (1) incremento en la actividad reproductiva a través de una aceleración de la maduración y un incremento de la frecuencia de desove; (2) acortamiento del periodo larval e incremento en la supervivencia larval; (3) incremento en el rendimiento en crecimiento individual; (4) incremento en la supervivencia de juveniles y adultos debido a la reducción de la biomasa de predadores; (5) incremento en la capacidad de carga de los bancos de conchas debido a elevados niveles de oxígeno. En el marco de discusión sobre los factores que contribuyen al desarrollo de la población de concha de abanico durante el evento El Niño, no podemos dejar de considerar en detalle el impacto de la pesquería. Considerando que el volumen de captura no solo depende de la abundancia relativa de la especie, la estrategia de pesca aplicada fue analizada con el fin de explicar los diferentes niveles de producción de concha de abanico durante El Niño 1983 y 1998. Palabras clave: El Niño, desembarques, manejo, Argopecten purpuratus, Perú
Abstract Independencia Bay can be considered as one of the most productive invertebrate fishing grounds worldwide. One of the most important exploited species is the scallop (Argopecten purpuratus) with strong catching fluctuations related to El Niño and La Niña events and to inadequate management strategies. During strong warming periods annual landings reach up to 50 000t in an area of about 150 km2 and during cold years they remain around 500 to 1000 t. This study analyses the changes in scallop landings at Independencia Bay observed during the last two decades and discusses the main factors affecting the scallop proliferations during the El Niño events. In this way data on landings, sea surface temperature and those related to growth, reproduction, predation, mean density and oxygen concentration from published and unpublished papers are used. The relationship between annual catches and average water temperature over the preceding reproductive period of the scallop over the past 20 year’s period, showed that scallop production is affected positively only with strong El Niño such as those of 1983 and 1998. Our review showed that the scallop stock proliferation can be traced to the combined effect of (1) an increase in reproductive output through an acceleration of gonad maturation and a higher spawning frequency; (2) a shortening of the larval period and an increase in larval survival; (3) an increase in the individual growth performance; (4) an increase in the juvenile and adult survival through reduction of predator biomass; (5) an increase in carrying capacity of the scallop banks due to elevated oxygen levels.
1 Facultad de Pesquería, Universidad Nacional Agraria la Molina, Apto. 456, Lima 100, Perú Correo electrónicol: [email protected] 2 Center for Tropical marine Ecology, ZMT, Fahrenheitstraße 1, 28359 Bremen, Alemania e-mail:[email protected]

EL NIÑO SOBRE LA PRODUCCIÓN DE CONCHA DE ABANICO (Argopecten purpuratus), PISCO, PERÚ
Diciembre 2003 __________________________________________________________________________________________
52
When discussing the factors that contribute to the development of the scallop population during an El Niño event, the fishery must be considered in detail. Since the volume of the catch not only depends on the relative abundance of the species, fishing efforts and strategies were analyzed in order to explain different levels of scallop production during El Niño 1983 and 1998. Key words: El Niño, landings, management, Argopecten purpuratus, Peru
Introducción Las aguas de la costa peruana están consideradas como las más productivas del mundo por sus altos niveles de productividad primaria y secundaria originado por los afloramientos de aguas ricas en nutrientes hacia la superficie (Rhyther, 1969). La pesquería artesanal costera, con una producción anual de cerca de 200 000 t, aparece como insignificante comparada con la bien conocida pesquería de pequeños pelágicos, la cual en algunos años supera los 10 millones de toneladas anuales, (Espino & Wosnitza-Mendo, 1988). Sin embargo, esta pesquería que explota alrededor de 170 especies de peces y 30 especies de invertebrados de alto valor comercial (Estrella & Guevara, 1998) da empleo a miles de pescadores y a sus familias. La mayoría de los invertebrados bentónicos son capturados por buzos que colectan estos manualmente del fondo, algunos son desprendidos del sustrato (Concholepas, Fisurela sp.) y otros como el pulpo son extraídos con un gancho de sus refugios. El centro de la pesquería de buceo es la Bahía Independencia
situada al sur de Pisco (Figura 1) y una de las especies comerciales más importantes es la concha de abanico (Argopecten purpuratus) cuyos volúmenes de desembarque se incrementan explosivamente durante eventos El Niño fuertes como el de los años 1983/84 o 1997/98. En 1983/84, bajo el impacto del fenómeno El Niño, la temperatura superficial del mar en los bancos de concha de abanico de 16°C durante un verano normal se incrementaron hasta casi los 25°C; con ello muchas áreas fueron recolonizadas y la población de concha de abanico se incrementó enormemente de unos cientos de toneladas a cerca de 30 000 t en pocos meses (Wolf, 1985; 1987). Estudios realizados sobre la dinámica poblacional de esta especie en años normales y con El Niño (Wolff & Wolff 1983; Wolff 1985, 1987, Mendo et al., 1988; Mendo & Yamashiro 1988; Mendo & Jurado, 1993) han sugerido que el crecimiento poblacional de concha de abanico se debe principalmente a: 1) incremento en la tasa de crecimiento; 2) la reducción del periodo larval y así la
mortalidad larval; 3) incremento de la mortalidad natural de predadores; 4) incremento de los productos sexuales y 5) reducción de los competidores filtradores. Sin embargo, solo algunos de estos factores han sido evaluados con información relevante. Así mismo aun cuando se presumía que durante El Niño 1997/98 en términos de producción sería similar al de El Niño 1983/84, los desembarques fueron mucho menores probablemente debido a inapropiadas estrategias de explotación aplicadas por pescadores y empresarios con la consecuente pérdida de biomasa y económica (Wolff & Mendo, 2000; Mendo & Wolff, 2002; Skrabonja & Mendo, 2002; Proleón & Mendo, 2002) El presente estudio analiza los cambios de los desembarques de la concha de abanico con relación a los cambios de temperatura en la Bahía Independencia durante las ultimas dos décadas y muestra evidencias de los cambios de algunos factores bióticos y abióticos como crecimiento, reproducción, predación, densidad media y condiciones de oxígeno considerados como causales del incremento de la producción durante los eventos fuertes de El Niño en 1983 y 1998.
Isla Independencia(La Vieja)
IslaSta. Rosa
BAHIA INDEPENDENCIA
ElAncla
PlayaCanastones
PtaCarhuaz
Tunga
MorroQuemado
76° 15' W 76° 10' W
14° 10' S
14° 15' S
14° 20' S
LAGUNA GRANDE
Figura 1. Mapa de la Bahía Independencia

JAIME MENDO Y MATTHIAS WOLFF
Ecología Aplicada Vol. 2 Nº 1, pp. 51-57 __________________________________________________________________________________________
53
Materiales y métodos Algunas características del área de estudio y la pesquería La Bahía Independencia (Figura 1) esta localizada a 250 Km al sur de Lima. Tiene una superficie de alrededor de 150 km2, una profundidad promedio de cerca de 25 m y esta fuertemente influenciada por el núcleo de afloramiento del sur (14°S-16°S) (Zuta et.al., 1978). La mayor parte del sustrato de la bahía es arenoso o pedregoso, pero existen también áreas con sustrato rocoso en la parte sur de la bahía. Los vientos que soplan de sur a norte, son fuertes (hasta 7 nudos) en las tardes y causan una mezcla constante en la columna de agua. Las temperaturas durante el año fluctúan normalmente entre 12°C (fondo) y 16°C (superficie) y los niveles de oxígeno se encuentran cerca del nivel de saturación en aguas someras (< 15m) y bajan drásticamente a valores críticos para invertebrados (<1ml l-1) a 30m de profundidad (Wolff 1985). Durante eventos fuertes de El Niño como el de 1982/83 y 1997/98 las temperaturas alcanzan valores > 20°C, la transparencia se incrementa a >10m al igual que el oxígeno cerca del fondo (>6ml/l) (Wolff 1985, Wolff & Mendo, 2000). La pesquería de concha de abanico se concentra a una profundidad que oscila entre 5 y 30 m, la cual varía con la zona. La producción diaria de los buzos depende mucho de la visibilidad por lo que solo operan durante las horas con luz (usualmente entre 9:00 y 14:00). La mayoría de los buzos permanecen sumergidos entre 3 y 5 horas al día. Base de datos usados en el presente estudio Los datos de desembarque anual y biomasa de concha de abanico fueron tomados de informes publicados por el Instituto del Mar del Perú (Flores et al., 1994; Estrella & Guevara, 1998; Mendo et al., 1987; 1989) y de informes no publicados por Mendo & Wolff). La serie de tiempo de temperatura del mar superficial registrado en base diaria en la Isla La Vieja (Figura 2) fue usada para calcular las anomalías de temperatura mensuales, las cuales fueron relacionadas con los desembarques de concha de abanico. Se asumió que la variación de las capturas podría ser explicada por las anomalías de temperatura tal como lo sugiere Arntz & Fahrbach (1991) y Wolff (1985). Ya que no se observó una clara relación entre la temperatura y las capturas incluso cuando se trabajo con desfases en las series de tiempo o cuando los datos de temperatura y las capturas fueron promediados a diferentes intervalos de tiempo, se trato de correlacionar la temperatura promedio de los meses de mayor actividad reproductiva de la concha de abanico (Noviembre-Abril) y las capturas acumuladas de los subsiguientes doce meses (Mayo-Abril) bajo el supuesto que la temperatura afecta significativamente
la reproducción y el desarrollo y supervivencia larval de la concha de abanico (Wolff 1987, 1988).
10,0
15,0
20,0
25,0
79 81 83 85 87 89 91 93 95 97 99 01
Año
Tem
pera
tura
(°C
) Finalmente, datos publicados sobre crecimiento de la concha de abanico (Wolf, 1987; Yamashiro & Mendo, 1988; Mendo & Jurado, 1993: Mendo, 1999; Porst, 2002; Skrabonja & Mendo, 2002; Mendo et al,. 2002) fueron usados para calcular y relacionar el índice de rendimiento en crecimiento (φ’) con la temperatura. El índice de rendimiento en crecimiento fue calculado usando los parámetros de la ecuación de von Bertalanffy K y Loo (φ’ =logK+2logLoo) (Pauly & Munro 1984). Así mismo datos de biomasa de predadores, índice gonadosomático y concentración de oxígeno registrados durante las evaluaciones de la población de concha de abanico (Mendo et al., 1987, 1988, 1989; Yamashiro et al., 1990; Rubio et al., 1998; Samame et al., 1985), fueron revisados y reanalizados con la finalidad de explicar las causales de los cambios en la producción de concha de abanico durante los eventos El Niño. Resultados y discusión Relación entre la temperatura superficial del mar y las capturas de concha de abanico La Figura 3 muestra las capturas mensuales de concha de abanico y las anomalías mensuales de temperatura superficial del mar durante el periodo 1983 a 2002. Las temperaturas máximas en el periodo se presentan en los años 1983 y 1998 y los mayores volúmenes de captura se presentan aproximadamente un año después de estos máximos de las anomalías de temperatura. También se puede observar un pico en el período 1985/86, sin embargo este no correlaciona con altas temperaturas del año anterior. Asimismo, en el año 1992 las temperaturas fueron 5°C por arriba de lo normal, y aun así no se registró un efecto sobre los desembarques. Ello nos permite concluir que en
Figura 2. Serie de tiempo mensual de la temperatura superficial del mar en la Bahía Independencia durante 1979 a 2001. Las flechas indican los eventos del El Niño 1983 y 1998.

EL NIÑO SOBRE LA PRODUCCIÓN DE CONCHA DE ABANICO (Argopecten purpuratus), PISCO, PERÚ
Diciembre 2003 __________________________________________________________________________________________
54
muchos casos los niveles altos o bajos de captura de concha de abanico no son explicados por los cambios de temperatura y solo existe una correlación muy alta cuando se trata de periodos con eventos fuertes de El Niño. La Figura 4 muestra esta correlación significativa entre los desembarques de concha de abanico y la temperatura promedio durante el periodo reproductivo de este recurso previo a la temporada de pesca.
Figura 3. Relación entre las anomalías mensuales de la temperatura superficial del mar (ºC, barras) y las capturas de concha de abanico (t, línea) en la Bahía Independencia. La predicción de las capturas de concha de abanico después del periodo reproductivo basado en la temperatura durante ese periodo tiene limitaciones fuertes, y solo podría ser usado para niveles de impacto fuertes originados durante eventos fuertes de El Niño. Es preciso mencionar que los intentos de correlacionar la temperatura (valores absolutos y anomalías) con las capturas (usando desfases de 3 y 6 meses entre ambos) a nivel mensual, estacional y semestral, no mejoraron los índices de correlación. Esto nos lleva a la siguiente interpretación: 1) se requiere de varios meses de elevada temperatura en el verano para incrementar significativamente la actividad reproductiva y el reclutamiento de la concha de abanico (determinado por el promedio de temperatura durante el periodo reproductivo) y 2) se requiere mas tiempo para que el cohorte incremente su biomasa y pueda ser convertido en capturas. En algunos años esto podría tomar entre 3 y 4 meses y en otros (de bajo reclutamiento y/o crecimiento más lento) podría tomar por lo menos 12 meses. El modelo de regresión entre ambas variables solo explica el 75% de la variabilidad inherente a los datos. El 25% de la variabilidad no explicado por el modelo posiblemente se deba a que no se usaron datos de biomasa en lugar de capturas, los cuales no estuvieron disponibles. Así mismo es importante mencionar que los volúmenes de captura no solo dependen de la abundancia de la especie, sino (por lo menos parcialmente) también de su valor en el mercado no tomado en cuenta en este estudio. Ocasionalmente los buzos colectan especies con poca abundancia si los precios son altos.
y = 4633,1x - 70958R2 = 0,7468
0
10000
20000
30000
40000
50000
14 16 18 20 22 24
Temp erat ura p ro medio ( °C )
92
83
98
Figura 4. Relación entre la temperatura promedio del mar durante el periodo reproductivo y los subsiguientes desembarques de concha de abanico en la Bahía Independencia. La naturaleza multifuncional del fenómeno El Niño puede ser una razón mas. Nosotros consideramos solo la temperatura como variable predictiva aun cuando sabemos que otros factores
bióticos y abióticos podrían influenciar el desarrollo poblacional de concha de abanico y de otras especies del ecosistema. Las altas concentraciones de oxígeno en los bancos de concha de abanico registradas durante los años El Niño, podrían ser el resultado de vientos fuertes poco usuales que mezclan las aguas de la bahía y de la intrusión de Aguas Subtropicales Superficiales en la Bahía con alta concentración de oxígeno (Com. Pers. O. Morón). Una conclusión importante es que el incremento de la población de concha de abanico necesita condiciones favorables de temperatura por varios meses tal como en los años 1983 y 1998. Aun cuando en 1992 se registró un periodo de calentamiento este no originó una explosión de la población de concha de abanico en la bahía. Factores que afectan la producción de concha de abanico durante El Niño La relación entre el índice de rendimiento en crecimiento (φ’) y la temperatura muestran claramente que la concha de abanico crece mejor bajo condiciones de calentamientos del mar (Figura 5a). Igualmente el índice gonadosomático durante El Niño se incrementa considerablemente (Figura 5b), lo cual sugiere que la actividad reproductiva en general se incrementa en condiciones de mar calientes. Wolff (1988) encontró que el tiempo de recuperación entre dos sucesivos desoves fue significativamente mas corto durante El Niño 1983 indicando que la maduración fue acelerada por las altas temperaturas del mar. También Disalvo et al. (1984) demostró en experimentos de laboratorio que las larvas desarrollaron dos veces mas rápido a 25°C que a 19°C y con una alta tasa de supervivencia. Por otro lado la Figura 5c muestra la relación directa entre la concentración de oxígeno y las densidades de
0,02,04,06,08,0
82 84 86 88 90 92 94 96 98 00Año
Ano
mal
ia
posi
tiva
(°C
)
0
5000
10000
Cap
tura
(t)
TSM-Anomalias positivasCaptura

JAIME MENDO Y MATTHIAS WOLFF
Ecología Aplicada Vol. 2 Nº 1, pp. 51-57 __________________________________________________________________________________________
55
concha de abanico, lo cual sugiere que las elevadas densidades de concha de abanico durante El Niño se deben por lo menos en parte a los elevados niveles de oxígeno. Finalmente la biomasa de los principales predadores de concha de abanico en un año con el evento El Niño (1998) es mucho menor que durante un año normal (1988) (Figura 5d), hecho que fue reportado también para El Niño 1983/84 por Wolff (1987). Nuestra revisión sugiere que el “boom” del stock de concha de abanico es debido al efecto combinado de: (1) incremento en la actividad reproductiva a través de una aceleración de la maduración y un incremento de la frecuencia de desove; (2) acortamiento del período larval e incremento en la supervivencia larval; (3) incremento en el rendimiento en crecimiento individual; (4) incremento en la supervivencia de juveniles y adultos debido a la reducción de la biomasa de predadores; (5) incremento en la capacidad de carga de los bancos de conchas debido a elevados niveles de oxígeno. Es conocido que el calentamiento de El Niño conduce a acelerar el metabolismo de concha de abanico (crecimiento y reproducción) y por lo tanto una fuente rica en alimento debe estar disponible para sostener el crecimiento y el desarrollo de la población. Sin embargo, durante el evento El Niño 97/98 la producción primaria decrece significativamente, tal como se puede apreciar en la Fig. 6. Dos explicaciones (no necesariamente exclusivas) al respecto pueden ser planteadas: 1) en años normales, el alimento para especies filtradoras se encuentra en exceso y lejos del nivel limitante y permanece así aun si los niveles bajan durante El Niño, 2) la concha de abanico usa una fuente de alimento diferente bajo condiciones de El Niño, posiblemente Bacterias enriquecidas con detritus en suspensión, las cuales periódicamente son suspendidas en la columna de agua por las corrientes de marea. De esta manera Bacterias y Protozoos podrían ser la principal fuente de alimento durante El Niño, una hipótesis que todavía no ha sido probada. En este contexto es interesante notar que durante el evento fuerte de El Niño 1983/84 grandes stocks de langostinos fueron introducidos en la zona de pesca de Pisco por una lengua de agua tropical caliente proveniente del norte de Perú y Ecuador. Tanto los langostinos y la concha de abanico no mostraron signos de escasez de alimento. El alimento definitivamente no fue un factor limitante a pesar de la reducción de la producción primaria. Al discutir los factores que afectan el desarrollo de la población de concha de abanico durante un evento fuerte de El Niño, es importante considerar a la pesquería en detalle. Tal como se mencionó antes, los volúmenes de captura no solo dependen de la abundancia relativa de la especie sino también del
valor del mercado y obviamente del esfuerzo pesquero y la estrategia de pesca que se le aplica. Si la pesquería permite crecer a la concha de abanico hasta un tamaño adulto (>65mm) respetando así la talla mínima legal de captura, el cohorte anual entonces estaría en la capacidad de formar su biomasa y de desovar varias veces. De esta manera los procesos reproductivos y los reclutamientos serian importantes y más intensos que en años normales y el incremento del stock se podría prolongar por más tiempo. Este fue el caso durante el evento El Niño 1983/84, cuando la concha de abanico de Bahía Independencia no fue explotada hasta Agosto de 1983, cuando la mayoría de las conchas ya habían crecido a tamaños >70mm (Wolff, 1987). A partir de entonces, aun cuando la biomasa fue reducida fuertemente por la pesquería, continuaron los pulsos fuertes de la población en los años 1984 y 1985 (Figura 3) debido a la existencia de una biomasa desovante alta en el cohorte original de 1983. Sin embargo, esta situación no duro mucho y luego mayormente debido a la implementación de una línea de exportación a fines de 1983, la presión pesquera se incremento en muchos casos sin restricciones ni cuotas de captura. Durante el ultimo evento fuerte de El Niño en 1998 la figura fue diferente a 1983: la concha de abanico fue colectada tan pronto aparecían los nuevos reclutas en los bancos y el esfuerzo de pesca aplicado fue enorme (Wolff & Mendo, 2000; Mendo & Wolff, 2002). Como un resultado de esta sobreexplotación por crecimiento, la biomasa del stock no se incremento como durante 1983 a pesar de las altas densidades de reclutas existentes (>300/m²) en Junio de1998 (Mendo & Wolff, observaciones de buceo). La experiencia acumulada y el análisis realizado nos permite concluir que el incremento de la producción anual de la población de concha de abanico en Bahía Independencia depende de la ocurrencia de eventos fuertes de El Niño y que el rendimiento total obtenido depende fuertemente de la estrategia de pesca, es decir, de la talla de primera captura, la cual debería ser mayor de 65mm de altura. Mendo & Wolff (2002) han planteado algunas medidas de manejo adicionales que deberían tomarse en cuenta en el caso de una nueva aparición de un evento fuerte de El Niño. Agradecimientos Esta publicación fue concluida durante una pasantía del primer autor en el Centro de Ecología Marina Tropical (ZMT) de la Universidad de Bremen. Nuestro agradecimiento a aquellas personas e instituciones que proporcionaron alguna información relevante para este estudio.

EL NIÑO SOBRE LA PRODUCCIÓN DE CONCHA DE ABANICO (Argopecten purpuratus), PISCO, PERÚ
Diciembre 2003 __________________________________________________________________________________________
56
Literatura citada Arntz W. & Fahrbach E. 1991. El Niño-
Klimaexperiment der Natur. Berlin, Birkhäuser Verlag.
DiSalvo L.H., Alarcón E., Martinez E. & Uribe E. 1984. Progress in mass culture of Argopecten purpuratus (Lamarck 1819) with notes on its natural history. Rev.Chilena de Hist.Nat. 57: 33-45
Espino M. & Wosnitza-Mendo C. 1988. La pesquería artesanal y la variabilidad de los recursos en el Perú. p. 121-144. En C. Wosnitza-Mendo, M. Espino y M. Veliz (eds.) La Pesquería Artesanal en el Perú durante Junio de 1986 a Junio de 1988. Inf. Inst. Mar del Perú. 93.
Estrella C. & Guevara R. 1998. Informe estadístico anual de los recursos hidrobiológicos de la pesca artesanal por especies, artes, caletas y meses durante 1997. Inf. Inst. Mar del Perú. 132.
Flores M.,Vera S., Marcelo R. & Chirinos E. 1994. Estadísticas de los desembarques de la pesquería marina Peruana 1983-1992. Inf. Inst. Mar del Perú. 105.
Mendo J. 1999. Effect of some environmental variables on age and growth of the Peruvian scallop (Argopecten purpuratus) in suspended and bottom cultures, Independencia Bay, Pisco, Perú. Scientific Report. International Foundation for Science.
y = 0,1227x + 1,9608R2 = 0,8986
3,6
3,8
4
4,2
4,4
4,6
14 15 16 17 18 19 20 21 22Temperatura (°C)
Indi
ce d
e cr
ecim
ient
o
(a)10
15
20
25
j j a s o n d e f m a m j
Mes
Indi
ce g
ondo
som
atic
o 83/84
94/95
(b)
0
20
40
60
80
0 1 2 3 4 5 6
Concentracion de oxigeno (ml/l)
Indi
vidu
os/m
²
8386/8795/96
(c)0
0,4
0,8
1,2
1,6
Jun-98 Mar-99 May-99 Mar-00Fecha de evaluacion
Indi
vidu
os /m
²
Thais chocolataCancer polyodon
El Niño
(d)
Figura 5. (a) Relación entre el índice de rendimiento en crecimiento (φ’) de la concha de abanico y la temperatura del mar (°C); (b) Índice gonadosomático durante El Niño y condiciones normales; (c) Concentración de oxigeno (ml/l) versus densidad (n/m²) de concha de abanico; (d) Densidad (n/m²) de predadores de concha de abanico durante El Niño y condiciones normales en la Bahía Independencia.
0
2000
4000
6000
8000
10000
12000
14000
16000
sep-91
ene-92
may-92
sep-92
ene-93
may-93
sep-93
ene-94
may-94
sep-94
ene-95
may-95
ago-95
dic-95 abr-96
ago-96
dic-96 abr-97
ago-97
dic-97 abr-98
ago-98
Mes
celu
las/
ml
Figura 6. Número de células de fitoplancton (n/ml)
registradas en Bahía Independencia durante 1995 a 1998 (Datos proporcionados por Noemí Ochoa)

JAIME MENDO Y MATTHIAS WOLFF
Ecología Aplicada Vol. 2 Nº 1, pp. 51-57 __________________________________________________________________________________________
57
Mendo J. & Jurado E. 1993. Length-based growth parameter estimates of the Peruvian scallop (Argopecten purpuratus). Fisheries Research. 15: 357-367.
Mendo J., Yamashiro C., Rubio J., Kameya A., Jurado E., Maldonado M. & Guzmán S. 1989. Evaluación de la población de concha de abanico (Argopecten purpuratus) en la Bahía Independencia, Pisco, Perú (23 de Setiembre-9 de Octubre de 1987). Inf. Inst. Mar Perú-Callao. (94).
Mendo J., Valdivieso V., Yamashiro C., Jurado E., Morón O. & Rubio J. 1987. Evaluación de la población de concha de abanico (Argopecten purpuratus) en la Bahía Independencia, Pisco, Perú (17 de Enero-4 de Febrero de 1987). Inf. Inst. Mar Perú-Callao. (91).
Mendo J., Valdivieso V. & Yamashiro C. 1988. Cambios en densidad, numero y biomasa de la concha de abanico (Argopecten purpuratus) en la Bahía Independencia (Pisco, Perú) durante 1984-1987. pp. 163-168. En: Salzwedel, H. y A. Landa. (eds.). Recursos y dinámica del ecosistema de afloramiento Peruano. Bol. Inst. Mar Perú-Callao, Vol. extr.
Mendo J. & Wolf M. 2002. Pesquería y manejo de la concha de abanico (Argopecten purpuratus) en la Bahía Independencia. En: J. Mendo y M. Wolf (eds) Memorias de la I Jornada Científica de la Reserva Nacional de Paracas, 28-31 Marzo Pisco. del 2001. Univ. Nac. Agraria La Molina.
Pauly D. & Munro J.L. 1984. Once more on the comparison of growth in fish and invertebrates. Fishbyte. 2(1): 21.
Proleon J. & Mendo J. 2002. Estrategia adoptada por los pescadores artesanales ante el reclutamiento masivo de concha de abanico (Argopecten purpuratus) en la Bahía Independencia, 1997-2000 Pisco. En: J. Mendo y M. Wolf (eds) Memorias de la I Jornada Científica de la Reserva Nacional de Paracas, 28-31 Marzo Pisco. del 2001. Univ. Nac. Agraria La Molina.
Rubio J., Taipe A. & Aguilar S. 1998. Evaluación de la concha de abanico (Argopecten purpuratus) en Bahía Independencia, 15-27 de Julio 1998. Resumen Ejecutivo, Inst. Mar Perú – Callao.
Ryther J.H. 1969. Photosynthesis and fish production in the sea. Science. 166: 72-76.
Samame M., Benites C., Valdivieso V., Mendez M., Yamashiro C. & Morón O. 1985. Evaluación del recurso concha de abanico (Argopecten
purpuratus) en la Bahía Independencia y otros bancos naturales de la provincia de Pisco, en mayo 1985. Informe interno Inst. Mar Perú.
Skrabonja A. & Mendo J. 2002. Impacto de la actividad de repoblamiento, sobre el rendimiento biológico de la concha de abanico (Argopecten purpuratus) en Bahía Independencia En: J. Mendo y M. Wolf (eds) Memorias de la I Jornada Científica de la Reserva Nacional de Paracas, 28-31 Marzo Pisco. del 2001. Univ. Nac. Agraria La Molina.
Wolff M. & Wolff R. 1983. Observations on the utilization and growth of the pectinid Argopecten purpuratus in the fishing area of Pisco, Perú. Bol. Inst. Mar Peru. 7(6): 197-235.
Wolff M. 1985. Abundancia masiva y crecimiento de pre-adultos de la concha de abanico Peruana (Argopecten purpuratus) en la zona de Pisco bajo condiciones de "El Niño" 1983. pp. 87-90. En: Arntz, W., A. Landa y J. Tarazona (eds.). "El Niño" y su impacto en la fauna marina. Bol. Inst. Mar Perú-Callao, Vol. extr.
Wolff M. 1987. Population Dynamics of the Peruvian scallop Argopecten purpuratus during the El Niño Phenomenon of 1983. Can. J. Fish. Aquat. SCI. 44: 1684-1691.
Wolff M. 1988. Spawning and recruitment in the Peruvian scallop Argopecten purpuratus. Mar. Ecol. Prog. Ser. 42: 213-217.
Wolff M. & Mendo J. 2000. Management of the Peruvian Bay Scallop (Argopecten purpuratus) metapopulation with regard to environmental change. Aquatic Conserv: Mar. Freshw. Ecosyst. 10: 117-126.
Yamashiro C., Rubio J., Jurado E., Maldonado M., Auza E., Ayon P. & Antonieti E. 1990. Evaluación de la población de concha de abanico, Argopecten purpuratus, en la Bahía Independencia, Pisco, Perú. Inst. Mar. Perú. Inf. Nro. 48.
Yamashiro C. & Mendo J. 1988. Crecimiento de la concha de abanico (Argopecten purpuratus) en la Bahía Independencia, Pisco, Perú. pp. 163-168. En: Salzwedel H. y A. Landa (eds.) Recursos y dinámica del ecosistema de afloramiento peruano. Bol. Inst. Mar Perú-Callao, Vol. extr.
Zuta S., Rivera T. & Bustamante A. 1978. Hydrologic aspects of the main upwelling areas off Perú. P.235-257. In R. Boje and M. Tomczak. Upwelling Ecosystems. Springer-Verlag, Berlin.

Ecología Aplicada, 2(1), 2003 ISSN 1726-2216 Depósito legal 2002-5474
RESILIENCIA DE LA COMUNIDAD EPÍGEA DE COLEOPTERA EN LAS LOMAS DE LACHAY DESPUÉS DEL EVENTO EL NIÑO 1997-98
Alfredo Giraldo Mendoza1 y Germán Arellano Cruz1
Resumen
El presente estudio describe la resiliencia de una comunidad epígea de Coleoptera residente en formaciones de lomas ante los cambios provocados por la ocurrencia del evento El Niño 1997-98. La comunidad de Coleoptera fue evaluada 17 veces en el transcurso de cinco años (1998 – 2002) usando trampas pitfall que fueron sistemáticamente distribuidas en un área mixta de loma de herbáceas y loma tipo parque en la Reserva Nacional de Lachay. Para detectar las manifestaciones de la resiliencia a través del periodo de estudio fueron empleadas las siguientes medidas: abundancia, riqueza, diversidad, composición y abundancias relativas de las morfoespecies, abundancias relativas de los gremios, proporción carabidae:tenebrionidae (C:T) y un índice porcentual para medir estacionalidad (I.P.). Las manifestaciones más importantes de la resiliencia fueron detectadas al comparar las evaluaciones correspondientes al primer y segundo muestreo de cada año. Para las evaluaciones del 1er muestreo, fueron observadas tendencias progresivas de disminución de la diversidad, aumento de la abundancia relativa de saprófagos y aumento de la abundancia relativa de tenebriónidos. Para las evaluaciones del 2do muestreo, se observaron diferencias marcadas entre la evaluación de 1998 y las evaluaciones de años posteriores, consistentes en una disminución en la diversidad, un aumento en la abundancia relativa de omnívoros y un aumento en la abundancia relativa de carábidos. La abundancia y la riqueza carecieron de patrones claros de resiliencia. Asimismo, la resiliencia significó un aumento progresivo de la estacionalidad, como fue indicado por los valores de I.P. calculados con la diversidad y la proporción C:T. La medida de resiliencia más efectiva fue la proporción C:T, por tal motivo se recomienda el uso de ésta y de otras proporciones de este tipo para la evaluación de perturbaciones. Palabras clave: Comunidad epígea de coleópteros, resiliencia, El Niño 1997-98, formaciones de lomas, Reserva Nacional de Lachay.
Abstract
The present study describes the resilience of a epigeic beetle assemblage inhabiting in a lomas formations in the face of the changes caused by the occurrence of the El Niño 1997-98. The beetle assemblage was evaluated 17 times since 1998 until 2002 using pitfall traps that were systematically distributed in a mixed area of herbaceous lomas and type park lomas in the Lachay National Reserve. For detecting the manifestations of the resilience across the time, the following measures were employed: abundance, richness, diversity, composition and relative abundance of the morphospecies, relative abundance of the guilds, carabidae:tenebrionidae proportion (C:T) and a percentage index to measure seasonality (I.P.). The most important manifestations of the resilience were detected comparing the evaluations carried out in the 1st and 2nd sampling of every year. For the 1st sampling evaluations, progressive tendencies of decrease of the diversity, increase of the relative abundance of detritivores and increase of the relative abundance of darkling beetles were observed. For the 2nd sampling, marked differences were observed between the 1998´s evaluation and the evaluations of later years, these were a marked decrease in the diversity, an increase in the relative abundance of omnivores and an increase in the relative abundance of ground beetles. The abundance and the richness lacked clear patterns of resilience. Also, the resilience meant a progressive increase of the seasonality, as it was indicated by the I.P. values calculated with the diversity and the C:T proportion. The most effective measure of resilience was the C:T proportion, for that reason the use of this kind of proportions is recommended for the evaluation of perturbations. Key words: Beetle epigeic assemblage, resilience, El Niño 1997-98, lomas formations, Lachay National Reserve.
Introducción Se denomina perturbación al desplazamiento de alguna propiedad de una comunidad o ecosistema
lejos de su valor típico, el cual se considera generalmente que representa un valor de equilibrio o al menos un valor de estado estable para dicha
_____________________________ 1 Laboratorio de Control Biológico y Ecología de Artrópodos del Departamento Académico de Biología. Universidad Nacional Agraria La Molina. Av. La Molina s/n Apartado 456. Lima 100. Perú. Direcciones electrónicas: Giraldo [email protected] / Arellano [email protected]

RESILIENCIA DE COLEOPTERA EN LACHAY DESPUÉS DEL EVENTO EL NIÑO 1997-98
Diciembre 2003 ___________________________________________________________________________
60
propiedad (Huston, 1994). El término disturbancia, que suele considerarse equivalente al de perturbación, es sólo un caso particular que consiste en cualquier proceso o condición externa a la fisiología natural de los organismos vivientes que produce una repentina pérdida de biomasa en una escala de tiempo significativamente más corta que la de su acumulación (Margalef, 1991; Huston, 1994). La respuesta de los sistemas afectados por tales cambios es la resiliencia, término que cuenta con un gran número de definiciones, las cuales pueden sintetizarse como “el retorno al estado o dinámica de referencia, después de una disturbancia temporal” (Grimm & Wissel, 1997). Definiciones previas incluyen la planteada por Pimm (1984): “la rapidez con la cual las variables retornan hacia su equilibrio después de una perturbación” y aquella acuñada en el contexto del manejo adaptativo (Holling, 1996): “la habilidad de un sistema para absorber cambio y variación sin saltar a un estado diferente donde las variables y procesos que controlan su estructura y comportamiento cambien súbitamente.” Las lomas son formaciones vegetales que se desarrollan a modo de parches a lo largo del desierto hiperárido que se extiende en la costa oeste de Sudamérica (5°00´- 29°55´ L.S.), debido a la confluencia de un complejo conjunto de condiciones físicas y meteorológicas que favorecen la condensación de neblinas en las elevaciones andinas cercanas al mar (Dillon & Rundel, 1990). Tales condiciones persisten desde hace aproximadamente 5000 años, interrumpidas periódicamente por los eventos El Niño, los cuales producen un incremento significativo en las precipitaciones cada 2 o hasta 8.5 años (Rodbell et al., 1999). Cada uno de estos eventos constituye una oportunidad para la expansión de las lomas fuera de sus límites normalmente restringidos, lo cual hace de El Niño un elemento crucial en la dinámica y persistencia de estas formaciones vegetales (Dillon & Rundel, 1990). El área protegida de las lomas de Lachay ha sido el escenario de un numeroso conjunto de estudios acerca de los efectos de El Niño 1997-98 sobre las diferentes comunidades bióticas que habitan en dicho espacio (Alfaro et al., 2000). Más recientemente, la resiliencia de las comunidades de aves (Véliz, 2002) y de herbáceas (Tovar, 2003) han sido objeto de análisis detallado a partir de muestreos realizados por el Laboratorio de Ecología de Procesos – Dpto. de Biología (Universidad Nacional Agraria La Molina, Lima - Perú) entre los años 1998 y 2002. En tales muestreos, se colectó además abundante material de Coleoptera que sirvió para realizar una descripción de los patrones de variación espacio-temporal de dicha comunidad (Giraldo, 2002), en la cual el proceso de resiliencia sólo fue mencionado de modo implícito y en la que asimismo, no se incluyó las evaluaciones
realizadas por el mismo equipo durante los meses de Junio y Octubre del 2002. El presente trabajo se propone determinar las manifestaciones de la resiliencia en los patrones de variación temporal de la comunidad epígea de Coleoptera, frente a las perturbaciones provocadas por el evento El Niño 1997-98, a partir de datos obtenidos entre los años 1998 y 2002 en las lomas de Lachay. Materiales y métodos Área de estudio: Los muestreos fueron realizados en la Reserva Nacional de Lachay ubicada a 105 Kms. al norte de la ciudad de Lima (11° 21´ 00´´-11° 21´ 58´´ L.S., 77° 22´ 25´´-77° 22´ 28´´ L.O.). De su superficie total de 5,070 Ha, el área de estudio abarcó aproximadamente unas 430 ha comprendidas entre el Cerro Redondo, la Quebrada Yerbabuena y el Puquial, con un rango altitudinal de 200–600 m.s.n.m. Este espacio constituye una muestra representativa de las biocenosis denominadas loma de herbáceas y loma tipo parque (Brack, 1976). Las condiciones meteorológicas típicas de la zona incluyen una temperatura invernal promedio de 13°C, temperaturas veraniegas que superan los 25°C y precipitaciones que oscilan entre los 40 y 100 mm (Brack & Mendiola, 2000). Los diferentes niveles de precipitación que se presentan a lo largo del año permiten distinguir cuatro épocas (Saito, 1976; Torres & López Ocaña, 1981): seca (Enero – Marzo), inicios de época húmeda (Abril – Junio), época húmeda (Julio – Septiembre) e intermedia (Octubre – Diciembre). Muestreo: Se realizaron cuatro evaluaciones por cada año para representar las épocas señaladas anteriormente, aunque durante los años 2001 y 2002 sólo pudieron realizarse tres y dos evaluaciones respectivamente. Se emplearon trampas pitfall para evaluar la comunidad de coleópteros presente en las 31 estaciones de muestreo que fueron distribuidas sistemáticamente dentro del área de estudio. Las trampas fueron envases de plástico transparente con un diámetro de 9 cms y una capacidad aproximada de 572.5 cm3, enterrados al ras del suelo y llenos a la mitad de su capacidad con una mezcla de 7 partes de agua por 1 de formol al 40%, además de un poco de detergente. El tamaño de la unidad muestral fue de dos trampas por estación con un esfuerzo temporal de siete días, el cual resultó satisfactorio para medir la diversidad de la entomofauna (Ramírez et al., 2002). Los especimenes fueron contabilizados y registrados en su mayoría como morfoespecies, las cuales se asignaron a sus respectivas familias, empleando las claves de Britton (1973). No obstante, algunos de ellos pudieron determinarse a nivel de tribu, género o incluso especie, sea mediante comparación directa con ejemplares de colecciones ó mediante el uso de claves (Giraldo, 2002). Dichos especímenes se encuentran depositados en el Laboratorio de Control Biológico y Ecología de

ALFREDO GIRALDO Y GERMÁN ARELLANO
Ecología Aplicada Vol. 2 Nº 1, pp. 59-68 __________________________________________________________________________________________
61
Artrópodos – Departamento de Biología de la Universidad Nacional Agraria La Molina Lima Perú (UNALM) y en el Museo de Entomología Klaus Raven de la misma institución. Como registro de los efectos meteorológicos de El Niño se contó con los datos de precipitación total mensual, temperatura promedio mensual y humedad promedio mensual proporcionados por la estación meteorológica de la Reserva. Estos registros indican un aumento extraordinario de la precipitación durante 1998 en tanto, la temperatura y la humedad atmosférica no superaron el rango de años normales. Análisis de los datos: Para evaluar el modo de acción de la resiliencia en la comunidad de Coleoptera se consideraron variables comunitarias, medidas de variación en las abundancias relativas de las morfoespecies y el índice porcentual propuesto por Tovar (2003). Las variables comunitarias consideradas fueron la abundancia, la riqueza y la diversidad. La abundancia es el número de individuos capturados, la riqueza es el número de morfoespecies y la diversidad se calculó a partir de las abundancias de las morfoespecies usando el índice de Shannon (Krebs, 1989). Cada variable es presentada a través del promedio de los valores obtenidos en las 31 unidades muestrales por cada fecha de evaluación, los cuales cuentan con su respectivo intervalo de confianza calculado a partir de la distribución t (Steel & Torrie, 1988). Los cambios en las abundancias relativas se evaluaron a través de tres medidas diferentes: un análisis cluster, la variación en las abundancias relativas de gremios y la variación en la proporción carábidos: tenebriónidos. El análisis cluster permitió agrupar las evaluaciones realizadas en base a la composición y abundancia relativa de las morfoespecies presentes en cada una de ellas. Los datos fueron transformados a la escala logarítmica, antes de ser procesados usando la técnica de unión promedio de clusters (average linkage clustering) y el coeficiente de correlación de Pearson (Krebs, 1989). Las familias de coleópteros se agruparon en gremios, denominados categorías tróficas en Giraldo (2002), de acuerdo a las preferencias alimenticias predominantes de cada una de ellas (Britton, 1973; Crowson, 1981). Los cuatro gremios resultantes son: fitófagos, omnívoros, predatores y saprófagos. El gremio de fitófagos incluye consumidores de los diferentes órganos vegetales, el de predatores a especialistas y generalistas que actúan sobre diversos tipos de presas y, el de saprófagos a consumidores de cualquier tipo de materia orgánica en descomposición o de los microorganismos que habitan en ella. El gremio de omnívoros incluye sólo a los carábidos Notiobia peruviana Dejean y Pterostichus sp., puesto
que incluyen materia vegetal en su dieta (Crowson, 1981; Noonan, 1981). La proporción carábidos: tenebriónidos (C:T) se calculó a partir de las abundancias registradas para estas familias en cada fecha de evaluación. Una vez calculados, los valores fueron transformados a la escala logarítmica, de modo que valores positivos indicaran un predominio de carábidos, mientras que el caso contrario sea señalado por los valores negativos. Paoletti & Cantarino (2000) han resaltado el uso de la proporción Carabidae: Tenebrionidae como un adecuado bioindicador de condiciones ambientales áridas, según el cual un predominio en la abundancia relativa de Tenebrionidae parece estar directamente relacionada a la prevalencia de condiciones áridas con pobre cobertura de vegetación. El índice porcentual (I.P.) propuesto por Tovar (2003) es la diferencia porcentual existente entre los valores de una variable cualesquiera calculada para el 3er muestreo y el 1er muestreo de cada año. Este índice permite medir la estacionalidad comparando los dos periodos del año caracterizados por las condiciones meteorológicas más extremas en el curso de un año normal. Se expresa mediante la siguiente fórmula: I.P. = (valor 3er muestreo– valor 1er muestreo) x 100/valor 3er muestreo. Las medidas consideradas en el cálculo de este índice son la abundancia, la riqueza, la diversidad y la proporción C:T correspondientes a los cuatro primeros años, pues durante el año 2002 no se realizó la evaluación de la época seca. El análisis cluster y las correlaciones que se requirieron fueron realizados con el programa Statistica versión 6.0. Resultados y discusión La comunidad epigea de Coleoptera que fue muestreada incluyó 22,953 especímenes, correspondientes a 66 morfoespecies y 21 familias. Las familias dominantes, es decir las que incluyeron más del 85 % de la abundancia total colectada fueron en orden descendente Tenebrionidae, Carabidae, Curculionidae, Staphylinidae y Elateridae. Las abundancias que presentaron las familias en cada evaluación se presentan en el Anexo 1. Al comparar las abundancias promedio correspondientes a las evaluaciones realizadas en una misma época del año se observan las siguientes diferencias significativas P ≤ 0.05 (Figura 1): El mayor valor de Febrero-2001 con respecto a los demás del 1er muestreo, el mayor valor de Octubre-2002 con respecto a los demás del 3er muestreo, el menor valor de Agosto-1999 con respecto a los registrados en Septiembre-2001 y Octubre-2002 entre las del 3er muestreo, así como el menor valor de Diciembre-1999 con respecto a los demás del 4to muestreo. En ninguno de los casos se observan tendencias

RESILIENCIA DE COLEOPTERA EN LACHAY DESPUÉS DEL EVENTO EL NIÑO 1997-98
Diciembre 2003 ___________________________________________________________________________
62
(b) 2do muestreo
indi
vidu
os/u
nida
d m
uest
ral
8
12
16
20
24
28
32
36
MAY_98 MAY_99 JUN_00 JUN_02
(d) 4to muestreo
indi
vidu
os/u
nida
d m
uest
ral
0
10
20
30
40
50
60
70
80
DIC_98 DIC_99 DIC_00 DIC_01
(c) 3er muestreo
indi
viduo
s/un
idad
mue
stra
l
0
40
80
120
160
200
AGO_98 AGO_99 SEP_00 SEP_01 OCT_02
(a) 1er muestreo
indi
vidu
os/u
nida
d m
uest
ral
-20
20
60
100
140
180
FEB_98 FEB_99 FEB_00 FEB_01
Figura n° 1. Cambios en la abundancia promedio de la comunidad de Coleoptera de cada muestreo entre los años 1998 y 2002. Intervalos de confianza a partir de la distribución t (α = 0.05).
(a) 1er muestreo
n° e
spec
ies/
unid
ad m
uest
ral
1.5
2.5
3.5
4.5
5.5
6.5
7.5
FEB_98 FEB_99 FEB_00 FEB_01
(b) 2do muestreo
n° e
spec
ies/
unid
ad m
uest
ral
1.5
2.5
3.5
4.5
5.5
6.5
7.5
MAY_98 MAY_99 JUN_00 JUN_02
(d) 4to muestreo
n° e
spec
ies/
unid
ad m
uest
ral
3
4
5
6
7
8
9
DIC_98 DIC_99 DIC_00 DIC_01
(c) 3er muestreo
n° e
spec
ies/
unid
ad m
uest
ral
4.5
5.5
6.5
7.5
8.5
9.5
10.5
11.5
AGO_98 AGO_99 SEP_00 SEP_01 OCT_02
Figura n° 2. Cambios en la riqueza promedio de la comunidad de Coleoptera de cada muestreo entre los años 1998 y 2002. Intervalos de confianza a partir de la distribución t (α = 0.05).

ALFREDO GIRALDO Y GERMÁN ARELLANO
Ecología Aplicada Vol. 2 Nº 1, pp. 59-68 __________________________________________________________________________________________
63
ascendentes o descendentes que cubran todo el periodo de evaluación. Al comparar las riquezas promedio correspondientes a las evaluaciones realizadas en una misma época del año se observan las siguientes diferencias significativas P ≤ 0.05 (Figura 2): el menor valor de Febrero-2000 con respecto al de Febrero-2001 entre los del 1er muestreo, el menor valor de Mayo-1998 con respecto a los demás del 2do muestreo, el menor valor de Agosto-1998 con respecto a los registrados en Septiembre-2001 y Octubre-2002 entre los del 3er muestreo, además de el menor valor de Diciembre-1999 con respecto a los registrados en Diciembre-2000 y Diciembre-2001 entre los del 4to muestreo. En ninguno de los casos se observan tendencias ascendentes o descendentes que cubran todo el periodo de evaluación. Al comparar las diversidades promedio correspondientes a las evaluaciones realizadas en una misma época del año se observan las siguientes diferencias significativas P ≤ 0.05 (Figura 3): El mayor valor de Febrero-1998 con respecto a los registrados en Febrero-2000 y Febrero-2001 entre los del 1er muestreo, el menor valor de Mayo-1998 con respecto a los demás del 2do muestreo y asimismo, los menores valores de Agosto-1998 y Septiembre-2000 con respecto a los registrados en Septiembre-2001 y Octubre-2002. En el caso del primer muestreo se observa una tendencia descendente que cubre todo el periodo de evaluación. En el dendrograma elaborado con los datos de abundancia de las
morfoespecies (Figura 4), a una distancia de unión de 0.4 se distinguen cinco asociaciones: La 1ra asociación incluye a todas las evaluaciones realizadas durante 1998, la 2da asociación incluye las evaluaciones de Mayo, Agosto y Diciembre de 1999, la 3ra asociación incluye las evaluaciones correspondientes al 3er y 4to muestreo de los últimos tres años, la 4ta asociación incluye las evaluaciones correspondientes al 2do muestreo de los años 2000 y 2002. y finalmente, la 5ta asociación incluye las evaluaciones correspondientes al 1er muestreo de 1999, 2000 y 2001. Al interior de la 3ra asociación se distinguen claramente dos grupos, uno correspondiente al 3er muestreo y el otro al 4to muestreo. Asimismo, en la 5ta asociación existe una distinción muy marcada entre la evaluación de
(a) 1er muestreo
bits
/ind.
/uni
dad
mue
stra
l
0.4
0.8
1.2
1.6
2.0
2.4
FEB_98 FEB_99 FEB_00 FEB_01
(b) 2do muestreo
bits
/ind.
/uni
dad
mue
stra
l
0.4
0.8
1.2
1.6
2.0
2.4
MAY_98 MAY_99 JUN_00 JUN_02
(c) 3er muestreo
bits
/ind.
/uni
dad
mue
stra
l
1.5
1.7
1.9
2.1
2.3
2.5
AGO_98 AGO_99 SEP_00 SEP_01 OCT_02
(d) 4to muestreo
bits
/ind.
/uni
dad
mue
stra
l1.2
1.4
1.6
1.8
2.0
2.2
2.4
DIC_98 DIC_99 DIC_00 DIC_01
Figura n° 3. Cambios en la diversidad promedio de la comunidad de Coleoptera de cada muestreo entre los años 1998 y 2002. Intervalos de confianza a partir de la distribución t (α = 0.05).
Figura n° 4. Dendrograma de las 17 fechas de evaluación (matriz de abundancia de morfoespecies) Coeficiente de correlación de Pearson
Distancia de unión
FEB_01 FEB_00 FEB_99 JUN_02 JUN_00 DIC_01 DIC_00
OCT_02 SEP_01 SEP_00 DIC_99
AGO_99 MAY_99 MAY_98
DIC_98 AGO_98 FEB_98
0.0 0.1 0.2 0.3 0.4 0.5 0.6 0.7

RESILIENCIA DE COLEOPTERA EN LACHAY DESPUÉS DEL EVENTO EL NIÑO 1997-98
Diciembre 2003 ___________________________________________________________________________
64
Febrero-1999 y las otros dos. Resulta importante señalar que las distancias de unión al interior de la 3ra, 4ta y 5ta asociación, presentan diferentes magnitudes. Dichas magnitudes son mayores entre las evaluaciones correspondientes al 1er y 2do muestreo, que entre las evaluaciones del 3er muestreo y más aún con respecto a la existente entre las evaluaciones del 4to muestreo.
1er muestreo
0
30
60
90
1998 1999 2000 2001
Sp
Pr
Om
Ft
a)
2do muestreo
0
30
60
90
1998 1999 2000 2002
b)
3er muestreo
0
30
60
90
1998 1999 2000 2001 2002
c)
4to muestreo
0
30
60
90
1998 1999 2000 2001
d)
Al comparar las abundancias relativas de los gremios (Figura 5) se observan los siguientes cambios entre las evaluaciones correspondientes a cada muestreo: Entre las evaluaciones del 1er muestreo, la
de 1998 se distinguió por el predominio compartido de fitófagos y predatores, en contraste con el predominio de saprófagos registrado durante los años 1999, 2000 y 2001. Entre las evaluaciones del 2do muestreo, la de 1998 presentó un predominio de omnívoros, la de 1999 un predominio compartido de fitófagos y predatores, mientras que las de los años 2000 y 2001 se caracterizaron por un predominio compartido de omnívoros y predatores. Entre las evaluaciones del 3er muestreo, las de 1998 y 2001 presentaron un predominio compartido de fitófagos y omnívoros, las de 1999 y 2002 un predominio compartido de fitófagos y predatores, mientras que la del año 2000 un predominio compartido de fitófagos y saprófagos. Entre las evaluaciones del 4to muestreo, la de 1998 presentó un predominio de fitófagos, las de 1999 y 2001 un predominio compartido de fitófagos y saprófagos, en tanto la del año 2000 presentó un predominio compartido de fitófagos y predatores. Al comparar las proporciones carábidos: tenebriónidos (Figura 6) se observan los siguientes cambios entre las evaluaciones correspondientes a cada muestreo: Entre las evaluaciones del 1er muestreo, la de 1998 se distinguió por un alto valor positivo, en contraste con los valores cada vez más negativos registrados para los años 1999, 2000 y 2001. Todas las evaluaciones del 2do muestreo presentaron valores positivos con una clara tendencia descendente desde 1998 hasta el 2002. Entre las evaluaciones del 3er muestreo, la de 1998 se distinguió por su altor valor positivo, en tanto el resto de evaluaciones mostraron valores positivos ó negativos muy cercanos o incluso iguales a cero (año 2000). Todas las evaluaciones del 4to muestreo presentaron valores positivos o negativos cercanos a cero.
Figura n°6. Cambios en la proporción C:T de cada muestreo entre 1998 y 2002
-2.00
-1.00
0.00
1.00
2.00
1ro 2do 3ro 4to
1998 1999 2000 2001 2002
El índice porcentual (Figura 7) muestra una tendencia ascendente y continua durante el periodo 1998 – 2001, cuando es calculado a partir de los valores de diversidad promedio y de la proporción C:T. En cambio, cuando éste es calculado a partir de los valores de riqueza promedio, dicha tendencia se interrumpe en el año 2000 por el menor valor del índice obtenido para el año 2001. En el caso de la
entre los años 1998 y 2002. Ft=fitófagos, Om= omnívoros,Pr=predatores, Sp= saprófagos.
Figura n° 5. Cambios en la abundancias relativas de los gremios de la comunidad de Coleoptera de cada muestreo

ALFREDO GIRALDO Y GERMÁN ARELLANO
Ecología Aplicada Vol. 2 Nº 1, pp. 59-68 __________________________________________________________________________________________
65
abundancia promedio, se observa la misma tendencia pero fuertemente alterada por el valor negativo que adquirió el índice el año 2001. En todos los casos, obviando el signo del índice, la diferencia entre los valores de las distintas medidas empleadas aumentó a través del tiempo.
Figura n°7. Variación temporal del índice porcentual durante el periodo 1998-2001
-1.00
-0.50
0.00
0.50
1.00
1998 1999 2000 2001
Abund. prom Riq. prom.proporción C:T Div. prom.
En función de los resultados expuestos anteriormente, las manifestaciones de la resiliencia en la comunidad examinada pueden sintetizarse en: - Para la diversidad se distinguieron tres manifestaciones distintas, dependientes del porcentaje de abundancia alcanzado por las especies dominantes, las cuales incluyen no menos del 30 % de la abundancia por muestreo. Entre las evaluaciones del 1er muestreo, se produjo una pérdida gradual de equidad acompañada por una serie de cambios en las especies dominantes: dominancia del carábido Calosoma rufipenne Dejean (Febrero-1998), dominancia compartida del tenebriónido Ammophorus rubripes Solier y el clámbido Clambus sp. (Febrero-1999) y dominancia del tenebriónido Evaniosomus orbygnianus Guerin (Febrero-2000 y Febrero-2001). En el caso de las evaluaciones del 2do muestreo, las condiciones cambiaron desde una con baja equidad provocada por la dominancia del carábido Pterostichus sp. (Mayo-1998) hasta alcanzar niveles de mayor equidad, en los cuales se presentaron dominancias compartidas de Pterostichus sp. con una especie indeterminada de Elateridae (1999) y una de Coccinellidae (2000 y 2002). Para las evaluaciones del 3er muestreo, se produjeron dos descensos debido a las dominancias de Pterostichus sp. (1998) y de una especie indeterminada de Curculionidae - Naupactini (2000) respectivamente. - Para la composición y abundancias relativas de las morfoespecies se observó un retorno a las condiciones que corresponden normalmente a cada estación del año, tal como queda ilustrado por la 3ra (época húmeda e intermedia), 4ta (inicios de época húmeda) y 5ta (época seca) asociaciones. Todo ello luego de atravesar por dos etapas anómalas, la 1ra asociación (todo el año 1998) y la 2da asociación (el año 1999 excepto por Febrero). Dichas asociaciones responden
en última instancia a los ciclos vitales y niveles de actividad anual que caracterizan a las poblaciones durante un año con estacionalidad normal, los cuales deben reorganizarse en función de los cambios provocados por el EN. - Para las abundancias relativas de los gremios, se observó también un retorno a las condiciones que corresponden normalmente a cada estación del año. Para las evaluaciones correspondientes al 2do, 3er y 4to muestreo significó un descenso en el predominio de gremios cuya abundancia relativa es importante en años normales, pero claramente menor a la alcanzada durante 1998 ó 1999. Los casos destacables al respecto, están dados por el predominio de omnívoros (Mayo-1998) y el de fitófagos (Diciembre-1998). Para las evaluaciones correspondientes al 1er muestreo significó la recuperación del predominio del gremio de saprófagos, tras haber sido desplazados por los tres gremios restantes durante 1998. En consecuencia, las condiciones ambientales con un balance hídrico favorable y consiguiente incremento en las estructuras vegetales (por incidencia estacional de neblinas o precipitaciones provocadas por El Niño) son requisitos para la proliferación de los gremios de fitófagos, omnívoros y predatores. En cambio, el gremio de saprófagos encuentra condiciones para su proliferación en la época seca de años normales, cuando queda disponible gran cantidad de materia vegetal en estado de descomposición. - Para la proporción C:T, la manifestación de la resiliencia fue bastante clara, el retorno de una condición en la cual los tenebriónidos predominan sobre los carábidos durante la época seca y con las abundancias de ambas familias casi equiparadas durante el resto del año, en lugar del notorio predominio de carábidos observado en las tres primeras evaluaciones de 1998. La clara manifestación de la resiliencia a través de esta proporción, no es sorprendente puesto que los cambios en la diversidad señalados anteriormente tuvieron como causa principal cambios en las abundancias de algunas especies de carábidos y tenebriónidos. Asimismo, con respecto a las abundancias registradas para los gremios, los carábidos tuvieron una contribución importante en el gremio de predatores y fueron integrantes exclusivos del gremio de omnívoros, en tanto los tenebriónidos constituyeron el componente mayoritario del gremio de saprófagos. - Para el índice porcentual (I.P.), la resiliencia se manifestó en un aumento progresivo de valores, el cual señala que la estacionalidad fue alterada y que la recuperación de ésta, al menos para la diversidad y la proporción C:T, no había concluido transcurridos tres años desde la ocurrencia de El Niño. Para la abundancia y la riqueza, resulta difícil distinguir un patrón de resiliencia claro debido a la frecuencia con la cual los intervalos de confianza de sus valores se traslapan y, a la ausencia de tendencias

RESILIENCIA DE COLEOPTERA EN LACHAY DESPUÉS DEL EVENTO EL NIÑO 1997-98
Diciembre 2003 ___________________________________________________________________________
66
que cubran todo el periodo de estudio. No obstante, es posible sugerir dos patrones para estas variables comunitarias: una tendencia al incremento de valores hacia el final del periodo de evaluación y, la ocurrencia de los menores valores para el año 1999 e inicios del 2000. El primer patrón se ve ilustrado por el 1er y 3er muestreo para la abundancia y, por el 2do, 3er y 4to muestreo para la riqueza. El segundo patrón es válido para la abundancia entre Mayo-1999 y Febrero-2000, mientras que para la riqueza es válido sólo para Diciembre-1999 y Febrero-2000. De los patrones propuestos, el primero de ellos resulta contradictorio para la abundancia puesto que se esperaba una respuesta positiva acorde con el aumento de cobertura vegetal propiciado por El Niño (Tovar, 2003). La relación directa entre abundancia de artrópodos y diferentes medidas de desarrollo vegetal ha sido observada previamente, así por ejemplo con la estructura vertical de la vegetación (Dennis et al., 1998) ó con la productividad primaria (Ayal & Merkl, 1994). Para la riqueza, el primer patrón implicaría que la mayor variabilidad meteorológica de un año normal estaría ofreciendo un escenario adecuado para la coexistencia de más especies que un lapso de tiempo meteorológicamente homogéneo provocado por El Niño. El segundo patrón parece indicar que estas variables fueron negativamente afectadas por los bajos niveles de precipitación que caracterizaron a 1999, aunque como se indica más adelante sus correlaciones con la precipitación carecen de significación estadística. Pueden distinguirse tres características importantes en la resiliencia de la comunidad de coleópteros. Primero, los efectos son discernibles sobretodo en los muestreos correspondientes a la primera mitad del año. Esto guarda correspondencia con la duración de las precipitaciones provocadas por El Niño, las cuales sobrepasaron los niveles normales hasta Agosto de 1998. Segundo, la duración del proceso de resiliencia puede fijarse en dos años, es decir el año 2000 puede considerarse como un año normal, aunque no obstante las diferentes medidas empleadas indican que la estacionalidad se acentuó durante los años 2001 y 2002. Tercero, las variables comunitarias se mostraron menos sensibles a la perturbación de El Niño que las medidas de cambio en las abundancias relativas. Así lo demuestran las tendencias obtenidas con el índice porcentual (I.P.), así como la correlación de éstas con la precipitación total mensual, la cual es significativa para la proporción C:T (r = 0.69, p = 0.005) y no significativa para las variables comunitarias (abundancia r = -0.04, p = 0.89; riqueza r = 0.28, p = 0.32; diversidad r = 0.35, p = 0.19). Es preciso indicar, que si bien la precipitación es indicada frecuentemente como el agente causal de los patrones observados, el establecimiento formal de una relación causa-efecto exigiría en este caso establecer la demora de respuesta (time-lag) de la comunidad de
Coleoptera frente a esta variable meteorológica. Para tal efecto, se hubiera requerido un muestreo continuo cuyos datos servirían para efectuar un análisis de series de tiempo. Longcore (1999) reporta que la abundancia media mensual de 50 morfoespecies de artrópodos epigeos, tuvo en promedio una demora de respuesta de 2.8+ 2.1 meses, con respecto a la precipitación total mensual en un ambiente costero del sur de California. La diferente sensibilidad de variables comunitarias y medidas de cambio en las abundancias relativas frente a la ocurrencia de perturbaciones, ha sido previamente reconocida en otras comunidades de coleópteros epígeos. Es el caso de Werner & Raffa (2000) y Jankielsohn et al. (2001), quienes afirman que la composición y/o los cambios en las abundancias relativas fueron más sensitivas para caracterizar los tipos de hábitat y los regímenes de manejo evaluados por ellos, que la riqueza específica, la equidad o la diversidad medida con los índices de Simpson y Shannon. Comparando de modo preliminar la resiliencia de la comunidad epigea de Coleoptera con las descritas para las otras comunidades evaluadas simultáneamente en las lomas de Lachay, se observa como principal diferencia que las variables comunitarias fueron más sensibles, así en el caso de la vegetación herbácea (Tovar, 2003) la densidad, la cobertura y la diversidad estuvieron correlacionadas significativamente con la precipitación y, en el caso de la avifauna (Véliz, 2002) la abundancia, la riqueza y la diversidad aumentaron de modo notable durante 1998. A semejanza de la comunidad de Coleoptera, en la comunidad de herbáceas se produjeron incrementos en la abundancia relativa de algunas especies, en este caso las solanáceas Nicotiana paniculata, Nolana humifusa y Lycopersicon peruvianum (Cano et al., 1999). En tanto, la comunidad de aves fue similar a la de Coleoptera en vista de los incrementos registrados en las abundancias relativas de algunos gremios durante 1998, en este caso fueron los gremios de insectívoros y predatores (Véliz, 2002). Por otra parte, para la comunidad de arañas epigeas se cuenta sólo con un primer avance acerca de su resiliencia, en base al análisis de los datos correspondientes a 1998 y 1999 (en este volumen). La proporción Carabidae: Tenebrionidae se destaca como una eficiente medida del grado de aridez ambiental, no sólo porque en el presente caso tuvo resultados más claros que las variables comunitarias, sino además porque se fundamenta en las distinta relación que cada una de estas familias ha establecido con respecto a los recursos hídricos disponibles en los ambientes terrestres. En efecto, los carábidos han tenido su mayor radiación adaptativa en ecosistemas húmedos, siendo su diversidad menor en ambientes templados áridos (Erwin, 1985). Esto guarda relación con el linaje ancestral de carábidos, cuyo hábitat

ALFREDO GIRALDO Y GERMÁN ARELLANO
Ecología Aplicada Vol. 2 Nº 1, pp. 59-68 __________________________________________________________________________________________
67
habría sido las áreas ribereñas de ecosistemas tropicales húmedos durante el Pérmico (Erwin, 1979). Por su parte, los tenebriónidos han llegado a ser el grupo dominante de coleópteros en todas las regiones desérticas del mundo (Britton, 1973; Crowson, 1981), paralelamente al desarrollo de un conjunto de adaptaciones de tipo estructural, etológico, fisiológico y morfométrico, cuyas expresiones más notables se encuentran entre las especies del desierto de Namib (Hamilton & Seely, 1976; Crowson, 1981; Nicolson et al., 1984; Nicolson, 1990). Finalmente, cabe señalar que proporciones tales como la C:T presentan ventajas metodológicas en relación a las otras medidas de variación en las abundancias relativas. Dichas ventajas son la capacidad de ser expresadas mediante una sola cifra (fácil de graficar y susceptible de contar con un intervalo de confianza) y además, la reducción considerable del esfuerzo que demanda la determinación de numerosas familias (con sus respectivas especies) ó la formación de gremios (que implica muchas veces atribuir hábitos generales a especies cuya historia natural es desconocida). Por ello, para la evaluación de perturbaciones resultaría conveniente el uso de alguna proporción que confronte la abundancia de grupos taxonómicos o funcionales de requerimientos opuestos frente a una variable ambiental impactada. Conclusiones La resiliencia de la comunidad epígea de Coleoptera frente a El Niño 1997-98 estuvo definida por la recuperación de los patrones de variación temporal típicos de las diferentes especies a lo largo del año, es decir la estacionalidad que caracteriza normalmente sus ciclos vitales y niveles de actividad anual. Dichos patrones fueron alterados notoriamente entre Febrero y Agosto de 1998, produciéndose con respecto a las condiciones de un año normal, cambios ascendentes y descendentes en los valores de diversidad, desplazamiento del gremio de saprófagos en favor del resto de gremios y aumentos en la proporción C:T. Sin embargo, los cambios abarcaron los dos primeros años cuando se examinan a través de las abundancias relativas de las morfoespecies y los gremios. Mas aún, al menos para la diversidad y la proporción C:T, la recuperación de la estacionalidad parece ser un proceso inconcluso luego de transcurridos tres años a partir de la ocurrencia de dicho evento. Agradecimientos Al Consejo Nacional de Ciencia y Tecnología (CONCYTEC) por el financiamiento otorgado al proyecto en el marco de la Red de Impacto Biológico “El Niño” RIBEN. Al personal de la Reserva Nacional de Lachay, por las facilidades brindadas para la
realización de las evaluaciones durante las gestiones de David Velarde e Iris Zárate. Literatura citada Alfaro L., Barrera R. & Velarde D. 2000. Reporte
Lachay. INRENA-Dirección General de Áreas Naturales Protegidas y Fauna Silvestre. Lima – Perú.
Ayal Y. & Merkl O. 1994. Spatial and temporal distribution of tenebrionid species (Coleoptera) in the Negev Highlands, Israel. Journal of Arid Environments. 27: 347-361.
Brack A.J. 1976. Ecología Animal. Primera Parte: Sinecología. Editor Pedro Aguilar F.
________ . & Mendiola C. 2000. Ecología del Perú. Programa de las naciones unidas para el desarrollo (PNUD). Editorial Bruño. Lima-Perú.
Britton E.B. 1973. Coleoptera (Beetles). In: The Insects of Australia. Melbourne University Press.
Cano A., Roque J., Arakaki M., Arana C., La Torre M., Llerena N. & Refulio N. 1999. Diversidad florística de las Lomas de Lachay (Lima) durante el evento “El Niño 1997-98”. Revista Peruana de Biología, Vol.Extraordinario.: 125-132.
Crowson R.A. 1981. The Biology of the Coleoptera. Academic Press Inc. (London).
Dennis P., Young M.R & Gordon I.J. 1998. Distribution and abundance of small insects and arachnids in relation to structural heterogeneity of grazed, indigenous grasslands. Ecological Entomology. 23(3): 253-264.
Dillon M.O. & Rundel P.W. 1990. The botanical response of the Atacama and peruvian desert floras to the 1982-83 El Niño Event. En: Global Ecological Consequences of the 1982-83 El Niño-Southern Oscillation. Glynn, P.W. (ed.) Elsevier, New York.
Erwin T.L. 1979. Thoughts on the evolutionary history of ground beetles: hypoteses generated from comparative faunal analyses of lowland forest sites in temperate and tropical regions. En: Erwin TL, GE Ball & DR Whitehead (eds) Carabid beetles, their evolution, natural history, and classification: 539-592. Dr. W. Junk Publishers, The Hague, The Netherlands.
__________. 1985. The taxon pulse: a general pattern of lineage radiation and extinction among carabid beetles. En: Ball GE (ed) Taxonomy, phylogeny and biogeography of beetles and ants: 437-488. Dr. W. Junk Publisher, The Hague, The Netherlands.
Giraldo A. 2002. Análisis de los patrones de variación espacio-temporal de las poblaciones de coleópteros en la Reserva Nacional de Lachay durante el periodo 1998 - 2001. Tesis para optar el Título de Biólogo – UNALM.
Grimm V. & Wissel C. 1997. Babel, or the ecological stability discussions: an inventory and analysis of

RESILIENCIA DE COLEOPTERA EN LACHAY DESPUÉS DEL EVENTO EL NIÑO 1997-98
Diciembre 2003 ___________________________________________________________________________
68
terminology and a guide for avoiding confusion. Oecologia. 109: 323-334.
Hamilton W.J. & Seely M. 1976. Fog basking by the Namib Desert beetle, Onymacris unguicularis. Nature. 262: 284-285.
Holling C.S. 1996. Surprise for science, resilience for ecosystems, and incentives for people. Ecological Applications. 6(3): 733-735.
Huston H. 1994. Biological diversity, the coexistance of species in changing landscapes. Cambridge University Press.
Jankielshon A., Scholtz C.H. & Louw S. 2001. Effect of habitat transformation on Dung beetle assemblages: A comparison between a South African Nature Reserve and neighboring farms. Environmental Entomology. 30(3): 474-483.
Krebs J. 1989. Ecological Methodology. University of British Columbia. Harper Collins Publishers.
Longcore T.R. 1999. Terrestrial Arthropods as Indicators of Restoration Success in Coastal Sage Scrub. A dissertation for the degree Doctor of Philosophy in Geography. University of California- Los Angeles.
Margalef R. 1991. Reflexiones sobre la Diversidad y significado de su expresión cuantitativa. Diversidad Biológica: 105-114. Fundación Araces, Madrid.
Nicolson S. 1990. Water relationships of the Namib tenebrionid beetles. En Seely, M. K. ed., Namib ecology: 25 years of Namib research. Transvaal Museum Monograph. 7: 173-178.
__________ , Bartholomew G.A. & Seely M.K. 1984. Ecological correlates of locomotion speed, morphometrics and body temperature in three Namib Desert tenebrionid beetles. S. Afr. Tydskr. Dierk. 19(3): 131-134.
Noonan G. 1981. South American species of the subgenus Anisotarsus Chaudoir (genus Notiobia Perty: Carabidae: Coleoptera). Part I: Taxonomy and Natural History. Contributions in Biology and Geology, 44: 1-84. Milwaukee Public Museum.
Paoletti M.G. & Cantarino C.M. 2000. The use of invertebrates in evaluating rural sustainability.
Chapter 3 in: Interchanges of Insects. M. Irwin and Y. Robert (eds.). : 33-52.
Pimm S.L. 1984. The complexity and stability of ecosystems. Nature. 307: 321 – 326.
Ramírez D., Pérez D., Sánchez E. & Arellano G. 2002. Esfuerzo de muestreo para la evaluación de la diversidad colectada en pitfall en la Reserva Nacional de Lachay-Perú. Ecología Aplicada. 1(1): 37-42.
Rodbell D.T., Seltzer G.O., Anderson D. M., Abott M. B., Enfield D.B. & Newman J.H. 1999. An 15 000 year record of El Niño – driven alluviation in southwestern Ecuador. Science. 283: 516 – 520.
Saito C. 1976. Bases para el establecimiento y manejo de una unidad de conservación en las Lomas de Lachay, Perú. Tesis para optar el título de Ingeniero Forestal- Universidad Nacional Agraria La Molina, Lima Perú.
Steel R. & Torrie J. 1988. Bioestadística: Principios y procedimientos. Mc Graw-Hill Interamericana de México S.A. de C.V.
Torres J. & López Ocaña C. 1981. Productividad primaria en las Lomas de la Costa Central del Perú. Separata del Boletín de Lima. (14): 1 – 11.
Tovar C. 2003. Análisis de la Resiliencia de las comunidades herbáceas de las Lomas de Lachay, provincia de Huaura, Departamento de Lima después de su perturbación por el evento El Niño 1997-1998. Tesis para optar el Título de Bióloga –UNALM.
Véliz C. 2002. Resiliencia de comunidades de aves en la Reserva Nacional de Lachay luego del evento El Niño 1997 – 98. Tesis para optar el Título de Bióloga , Universidad Nacional Agraria La Molina, Lima – Perú.
Werner S.M. & Raffa K.F. 2000. Effects of forest management practices on the diversity of ground-ocurring beetles in mixed northern hardwood forest of the Great Lakes Region. Forest Ecology and Management. 139: 135-155.
TotalEspecies 1er 2do 3er 4to 1er 2do 3er 4to 1er 2do 3er 4to 1er 3er 4to 2do 3er
Tenebrionidae (16) 7 8 7 96 172 17 91 53 170 121 313 198 3788 312 441 91 428 6313Carabidae (5) 437 485 575 66 49 140 43 86 16 239 314 259 47 655 173 184 854 4622Curculionidae (10) 225 60 320 431 7 90 262 26 2 21 599 633 19 400 733 35 698 4561Staphylinidae (7) 6 9 108 351 31 75 136 37 5 54 99 161 19 195 245 59 546 2136Elateridae (1) 16 1 95 30 4 10 89 2 0 0 21 23 1 211 23 45 1349 1920Coccinellidae (5) 72 1 108 18 23 18 139 43 9 223 66 13 0 185 59 103 273 1353Phalacridae (1) 97 1 17 9 1 1 1 1 0 0 19 4 0 30 1 2 456 640Mycetophagidae (2) 21 24 196 24 46 24 70 13 1 6 12 6 3 18 1 22 4 491Clambidae (1) 0 0 4 1 117 14 36 41 28 6 9 13 3 3 5 1 0 281Mordellidae (2) 1 0 0 11 5 4 21 73 6 0 0 7 17 1 17 5 0 168Chrysomelidae (1) 31 10 32 33 4 1 2 2 0 0 4 3 0 18 0 1 4 145
Lathridiidae (2) 4 0 2 3 2 12 24 13 0 12 2 16 0 3 4 6 0 103Anthicidae (2) 0 0 0 5 5 8 8 3 1 7 1 2 4 3 7 3 6 63Bostrichidae (1) 0 0 0 0 1 1 1 5 0 4 0 0 0 1 0 23 2 38Pselaphidae (1) 1 5 0 2 0 0 0 0 0 1 0 0 0 2 0 4 14 29Nitidulidae (3) 9 0 3 0 0 1 0 0 0 2 1 0 0 0 0 7 0 23Trogidae (1) 0 0 0 0 0 0 0 0 0 0 0 2 1 1 0 5 7 16
Histeridae (2) 3 0 1 1 2 0 1 0 1 0 0 0 2 0 0 0 0 11
Colydiidae (1) 0 0 0 0 1 0 0 0 1 0 0 1 0 0 2 0 0 5Cleridae (1) 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 1Scarabaeidae (1) 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 1
Anexo n° 1. Abundancias de las familias de Coleoptera colectadas en la Reserva Nacional de Lachay entre los años 1998 y 2002Los números entre paréntesis indican el número de morfoespecies diferenciadas en cada familia.
1998 1999 2000 2001 2002

Ecología, Aplicada 2(1), 2003 ISSN 1726-2216 Depósito legal 2002-5474
RELACIÓN ENTRE LA HORMIGA Camponotus sp. (Hymenoptera: Formicidae) Y UNA COMUNIDAD DE CACTUS (CACTACEAE) EN EL VALLE DEL RÍO CHILLÓN
Sidney Novoa S.1 , Viviana Castro C.2, Aldo Ceroni S.2 e Inés Redolfi P.1
Resumen
Las observaciones se realizaron en un ecosistema de zona árida en el Valle del Río Chillón, Lima-Perú, el mes de Julio de 2003. En 4 cuadrantes de 15 x 15 m cada uno, se determinó la relación de la hormiga Camponotus sp. con la comunidad de cactáceas. La riqueza de especies de cactus fue S = 8 y la hormiga forrajea únicamente en el exterior de los botones florales de la especie Neoraimondia arequipensis ssp. roseiflora. Los resultados sugieren que la hormiga es un bioindicador de nectarios externos en flor al demostrar el éxito de la colecta de alimento líquido en la prueba de presión del gáster. Así mismo, se obtuvo una correlación altamente significativa entre el número de botones florales de N. arequipensis y el número de hormigas forrajeando en los botones florales, utilizando el coeficiente de correlación de Pearson. Estas observaciones preliminares del subproyecto dentro del Proyecto Cactáceas de Lima son de gran importancia en el entendimiento de ecosistemas con escasos elementos bióticos y variables abióticas extremas, lo cual sería un interesante aporte para el conocimiento y comprensión de ecosistemas más complejos. Palabras clave: Cactus, hormiga, interacción, desierto.
Abstract Observations were made in an arid ecosystem in the Chillón River Valley, in Lima – Perú, on July 2003.The relation of the cacti community with the Camponotus sp. ant was determined in four 15 x 15 meters quadrants. The richness of the cacti species was S = 8 and it was found out that the ant forages only in the outside of the Neoraimondia arequipensis ssp. roseiflora bottom flowers. The results suggest that the ant acts as a bioindicator of the presence of external flower nectarines as the successful gaster pressure test conducted in the collected liquid content indicated. A highly significant correlation using the Pearson coefficient, between the number of bottom flowers in N. arequipensis and the foraging ants was obtained. These results on preliminary observations of the Lima’s Cactaceae Sub Proyect are very important in this ecosystem with limited biotic elements and extreme abiotic variables. It is an interesting tool for gaining knowledge and for the comprehension of complex ecosystems. Key words: Cacti, ant, interaction, desert.
1 Laboratorio de Control Biológico y Ecología de Artrópodos. Departamento Académico de Biología. Universidad Nacional Agraria La Molina. Av. La Molina s/n Apartado Postal 456. Lima 100. Perú. Dirección electrónica: [email protected] / [email protected] 2 Jardín Botánico “Octavio Velarde Núñez”. Departamento Académico de Biología. Universidad Nacional Agraria La Molina. Av. La Molina s/n Apartado Postal 456. Lima 100. Perú. Correo electrónico: [email protected] / [email protected]
Introducción El Valle del Río Chillón (Departamento de Lima, en las Provincias de Lima y Canta), presenta fondos profundos de zonas llanas cortadas por abruptos costados en quebradas de fuerte pendientes y estrechas gargantas. El clima en las partes media y baja es suave y templado, mientras que en las partes más altas es mas bien frío (ONERN, 1975). En la parte mas baja del piso, la vegetación natural está constituida de cactáceas columnares y otra vegetación reducida. En este sistema, la comunidad de cactus conforma el nivel trófico dominante y es el eje de todos los procesos. En estudios realizados en una de estas especies, Neoraimondia arequipensis ssp. roseiflora (Werd. & Backbg.) Rauh. Ostolaza et al. (1985), destaca la presencia de negras y grandes hormigas invadiendo
los brotes florales, flores abiertas y frutos. La presencia de hormigas en ecosistemas de zonas áridas debe ser importante objetivo de estudio por las variables puntuales extremas, en un ambiente tan agreste de escasos recursos, en donde la especificidad del nicho ecológico se incrementa. Las hormigas intervienen en el ciclo de nutrientes en la naturaleza, enriquecimiento de los suelos y en una gran diversidad de interacciones tróficas, tanto que se considera que el flujo de energía que pasa a través de ellas es superior al que pasa a través de los vertebrados homotermos que viven en el mismo hábitat (Adlung, 1966; Detrain, 1990; Weseloh, 1996). Esto se debe a que están entre los insectos más altamente sociales por el funcionamiento con sistemas de castas, división de labor, cooperación entre individuos en la cría de los

LA HORMIGA Camponotus sp. Y UNA COMUNIDAD DE CACTUS EN EL RÍO CHILLÓN
Diciembre 2003 __________________________________________________________________________________________
70
jóvenes, empleo de sustancias químicas (feromonas) para comunicarse y a que la reproducción se realiza mediante una casta separada (North, 1996). Las hormigas tienen regímenes alimenticios muy variados, así se las menciona como insectívoras, fitófagas, nectarívoras, granívoras y omnívoras (Retana et al., 1987; Alsina et al., 1988; Dejean et al., 1997) por lo que son consideradas importantes indicadores de los procesos en la dinámica de los ecosistemas. La relación hormiga-cactácea ha sido ya estudiada en otras zonas desérticas del mundo, como la relación entre la hormiga Crematogaster opuntiae Buren en la cactácea Opuntia acanthocarpa Engelmann & Bigelow en el noreste de Phoenix, Arizona (Pickett & Clark, 1979), dos especies de hormigas Crematogaster depilis Wheeler e Iridomyrmex pruninosus Royer en Ferocactus cylindraceus (Engelmann) Orcutt (Ruffener & Clark, 1986; Hunt, 1999). En el Perú, los únicos antecedentes que citan esta interacción son las observaciones de Ostolaza et al. (1985) ya mencionada y de Pizarro (1997) quien indica la visita de hormigas al “ayrampo” Opuntia soehrensii Britton & Rose, en especial en época de verano, cuando sus frutos maduran. El presente trabajo, parte inicial de un subproyecto dentro del Proyecto Cactáceas de Lima, tiene por objetivo determinar la relación entre la “hormiga negra” Camponotus sp. con la comunidad de cactáceas en el Valle del Río Chillón, Perú, considerando que este sistema de zona árida, con escasos elementos, podría ser un interesante medio para el conocimiento y comprensión de ecosistemas más complejos. Materiales y métodos Lugar de investigación El área de la investigación fue en el Cerro Umarcata, el cual se encuentra a una distancia de 6 kilómetros de Santa Rosa de Quives, a la altura del kilómetro 69 de la carretera Lima – Canta, a una altitud de 1260 m.s.n.m. a 11°37’39’’ L.S. y 76°46’9’’ L.O. en el Valle del Río Chillón, Departamento de Lima, en las Provincias de Lima y Canta (Figura 1). La vegetación natural está dominada por comunidades de cactáceas. La temperatura media anual está entre los 5 ºC a los 19ºC. La precipitación es prácticamente nula en la parte baja y se hace evidente a partir de los 2000 m.s.n.m., con rango de precipitación de 20 a 850 mm en promedio anual. La estación de lluvias comprende los meses de Diciembre a Marzo y la estación seca, los meses de Mayo a Setiembre (ONERN, 1975).
Metodología Se realizaron dos visitas al lugar: una previa para la colecta de especimenes de la hormiga y observar su comportamiento y la segunda para la toma de datos. En la visita previa se determinó el lugar de forrajeo de la hormiga y se realizó la prueba para comprobar el éxito de la colecta de alimento líquido por la hormiga, para lo cual se presionó suavemente el gáster de 30 individuos según la técnica utilizada por diferentes autores (Cherix, 1981; Retana et al., 1986, 1987; Alsina et al., 1988; Cerdá et al., 1989; Cavia, 1989). Se colectó 20 individuos de la “hormiga negra” en alcohol 70%, para su posterior determinación en el Museo de Entomología de la UNALM como
Camponotus sp. En la segunda visita, se trazaron 4 cuadrantes de 15 x 15 m cada uno seleccionados aleatoriamente. En el interior de cada cuadrante se determinó la riqueza de especies (S) de cactáceas. En cada individuo de cactus se contabilizó el número de botones florales y el número de individuos de la hormiga Camponotus sp. forrajeando sobre los botones. El muestreo se efectuó en el mes de Julio de 2003 y las observaciones de la presencia de la hormiga entre las 11:00 y 13:00 hs. El trazado de cuadrantes y toma de datos fue realizado por 10 observadores. Los datos se procesaron en el Laboratorio de Control Biológico y Ecología de Artrópodos y en el Jardín Botánico ¨Octavio Velarde Nuñez¨ de la Universidad Nacional Agraria La Molina. Para el análisis de los datos se calculó el coeficiente de correlación lineal de Pearson entre el número de botones florales y el número de hormigas forrajeando por planta, y se realizó la prueba estadística t-Student (9)gl, en el paquete estadístico Minitab 12.1 para probar la hipótesis de no correlación entre ambas variables (Hp: ρ = 0).
Figura 1. Ubicación del área de estudio en el Valle del Río Chillón. 1:100,000 (IGN, 1992).

SIDNEY NOVOA, VIVIANA CASTRO, ALDO CERONI E INÉS REDOLFI.
Ecología Aplicada Vol. 2 Nº 1, pp. 69-73 __________________________________________________________________________________________
71
Resultados y discusión Se determinó una riqueza S = 8 de cactus en la zona en estudio, con el mayor número de individuos (n = 201) pertenecientes a la especie Melocactus peruvianus Vaupel. En esta especie se destaca una estructura diferente llamada “cefalio” en lugar de botones florales. Otras especies también abundantes son Mila nealeana Backeberg y Haageocereus acranthus (Vaupel) Backeberg. Las especies con menor número de individuos fueron: Cleistocactus acanthurus ssp. faustianus (Backeberg) Ostolaza, H. pseudomelanostele ssp. aureispinus (Rauh & Backberg) Ostolaza, Neoraimondia arequipensis ssp. roseiflora (Backeberg) Ritter & Erort, Opuntia pachypus Schumann y Espostoa melanostele (Vaupel) Borg. (Tabla 1). En esta comunidad, únicamente la especie N. arequipensis tuvo presencia de la hormiga, mientras que las otras siete especies no presentaron obreras de Camponotus sp. forrajeando o visitando la planta. Cuatro especies de cactáceas presentaron botones florales: H. acranthus (n = 10), H. pseudomelanostele (n = 29), Cleistocactus acanthurus (n = 5) y N. arequipensis con el mayor número de botones florales (n = 83). Por otro lado, Camponotus sp. (n = 131) se presentó en 67 botones florales de N. arequipensis (Tabla 1). La presencia de la hormiga únicamente en N. arequipensis en relación a las ocho especies de la comunidad, el alto porcentaje de hormigas visitando a estos cactus cuando forman botones florales, así como su distribución sobre los botones florales (Tablas 2 y 3), puede deberse al hecho que esta especie presenta espacios más amplios y sin espinas entre las costillas (Figura 3) por los
cuales pueden desplazarse rápidamente las obreras menores de Camponotus sp., con una longitud del cuerpo de 0.66 cm., que no les permitirían visitar las otras siete especies de cactus, por la gran densidad de espinas que poseen cubriendo inclusive las costillas.
Camponotus sp. forrajeaba únicamente en la zona exterior de los botones florales de N. arequipensis en donde permanecía largo tiempo (hasta 20 minutos) y no formaba pistas al desplazarse por las ramas. Por otro lado, la prueba de presión del gáster demostró gran abundancia de alimento líquido transportado por las hormigas, lo que unido a la observación del lugar de forrajeo en la zona externa en los botones florales confirman la relación hormiga-nectarios externos, también observada en México (Pickett & Clark, 1979; Ruffener & Clarck, 1986).
Figura 3. Relación de la hormiga Camponotus sp. (Hymenoptera: Formicidae) en Neoraimondia arequipensis ssp. rosiflora en la comunidad de cactus (Cactaceae) en el Valle del Río Chillón. Fotos: Diego De la Barra (2003).
0
5
10
15
20
25
30
35
40
0 10 20 30 40
No. de hormigas
No.
de
boto
nes f
lora
les
Figura 2. Correlación entre botones florales de Neoraimondia arequipensis ssp. roseiflora y Camponotus sp., en el Valle del Río Chillón: Cerro Umarcata.

LA HORMIGA Camponotus sp. Y UNA COMUNIDAD DE CACTUS EN EL RÍO CHILLÓN
Diciembre 2003 __________________________________________________________________________________________
72
Con este resultado se planteó la posibilidad de la existencia de un grado de correspondencia entre la presencia de hormiga y la presencia de botones florales en esta especie de cactácea por lo que se realizó un análisis de correlación entre el número de botones florales por planta y el número de hormigas forrajeando por planta, en los individuos (11 cactus) que alcanzaron la edad adulta, es decir que son capaces de producir botones florales. Dicho análisis arrojó un valor de r = 0.828 (tcal = 4.43, 9 g.l., P < 0.001). Entonces, se concluye que para un α = 0.05, las evidencias muestrales indican que existe una correlación significativa entre el número de botones por planta y el número de hormigas forrajeando por planta en N. arequipensis ssp. roseiflora (Figura 2). Esta relación significativa (P < 0.001) entre Camponotus sp. y N. arequipensis es de gran importancia en este ecosistema de zona árida en donde las variables abióticas son extremas y el éxito de la biocenosis depende de finas interacciones entre las variables bióticas. Estas observaciones preliminares permiten elaborar una serie de preguntas que deberán ser absueltas en la continuación del subproyecto iniciado. Así, se esperaría que los hormigueros de Camponotus sp. estén ubicados al pie de los individuos de N. arequipensis, lo que contribuiría a un mayor aporte de nutrientes, necesarios para esta cactácea de mayor tamaño y biomasa en relación a las otras siete especies. Igualmente, las flores de N. arequipensis serían visitadas por otros artrópodos que conforman una delicada red trófica, posiblemente la única o más importante de este ecosistema. Tabla 1. Lista de especies, número de individuos, número de botones florales y número de hormigas en las cactáceas del Valle del Río Chillón: Cerro Umarcata.
Especie de cactáceas (Anderson, 2001)
N° i
ndiv
iduo
s
Pres
enci
a
horm
igas
N
° bot
ones
flo
rale
s N
° bo
tone
s co
n ho
rmig
as
N° t
otal
ho
rmig
as
Opuntia pachypus 17 no 0 0 0 Mila nealeana 62 no 0 0 0 Haageocereus acranthus 32 no 10 0 0 H. pseudomelanostele spp. aureispinus 57 no 29 0 0 Cleistocactus acanthurus spp. faustianus 40 no 5 0 0 Melocactus peruvianus 201 no 0 0 0 Neoraimondia arequipensis spp. roseiflora 28 si 83 67
131
Espostoa melanostele 9 no 0 0 0
Tabla 2. Porcentajes de plantas de Neoraimondia arequipensis spp. roseiflora con botones florales, cactus con hormigas y botones florales con hormigas, en el Valle del Río Chillón: Cerro Umarcata.
Porcentaje de Plantas con
botones florales
Porcentaje de plantas con
botones y con hormigas
Porcentaje de botones florales con hormigas
32.28% 100% 80.72%
Tabla 3. Distribución de Camponotus sp. sobre los botones florales de Neoraimondia arequipensis spp. roseiflora, en el Valle del Río Chillón: Cerro Umarcata.
Media Mediana Desviación estándar Máximo Mínimo
1.916 2.000 1.669 8.000 .0.000
Conclusiones 1. La hormiga Camponotus sp. interacciona únicamente con la especie, Neoraimondia arequipensis ssp. roseiflora, de las ocho que conforman la comunidad de cactáceas en el Valle del Río Chillón. 2. Las observaciones de campo y el análisis de datos muestran que existe una relación altamente significativa entre la hormiga Camponotus sp. y la presencia de botones florales de Neoraimondia arequipensis ssp. roseiflora. 3. Camponotus sp. podría ser denominada como un bioindicador de nectarios externos en Neoraimondia arequipensis ssp. roseiflora, por lo que se continuarán los estudios en la anatomía de la flor. 4. Estudios sobre la fenología de esta planta podrían descartar la mayor época de floración y por lo tanto el aumento de los visitantes florales como Camponotus sp. Agradecimientos Los autores expresan su agradecimiento a la Dra. Consuelo Arellano Ugarte, por la revisión del manuscrito y sugerencias en la presentación del análisis estadístico. A la Dra. Cecilia Díaz-Castelazo, del Departamento de Ecología Vegetal, Instituto de Ecología de Veracruz, México y al Dr. Carlos Ostolaza, Presidente de la Sociedad Peruana de Cactus y Suculentas ¨SPECS¨ por facilitar bibliografía sobre hormigas-cactáceas. A la Bachiller Silvia Castro por la determinación del género Camponotus. A los estudiantes de Biología de la UNALM que colaboraron en la toma de datos. Esta investigación

SIDNEY NOVOA, VIVIANA CASTRO, ALDO CERONI E INÉS REDOLFI.
Ecología Aplicada Vol. 2 Nº 1, pp. 69-73 __________________________________________________________________________________________
73
fue financiada por el FEDU (Fondo Investigación Universitaria) de la Universidad Nacional Agraria La Molina. Literatura citada Anderson E.F. 2001. The Cactus Family. Timber
press. Adlung K.G. 1966. A critical evaluation of the
european research on use of red wood ants (Formica rufa group) for the protection of forests against harmful insects. Dpto. Agricultural Research Service. París. 57: 167-189.
Alsina A., Cerda X., Retana J. & Bosch J. 1988. Foraging ecology of the aphid-tending ant Camponotus cruentatus (Hymenoptera, Formicidae) in a savanna-like grassland. Misc. Zool. 12: 195-204.
Cavia V. 1989. Régimen alimenticio de la hormiga Formica subrufa (Hymenoptera: Formicidae). Ses. Entomol. ICHN-SCL. VI: 97-107.
Cerda X., Retana J., Bosch J. & Alsina A. 1989. Exploitation of food resources by the ant Tapinoma nigerrimum (Hym. Formicidae). Acta Oecologica. 10(4): 419-429.
Cherix D. 1981. Contribution á la biologie et á l’écologie de Formica lugubris. Le probléme des super-colonies. Thesis, Lausanne.
Dejean A., Ngnegueu P.R., Durand J.L. & Bourgoin T. 1997. The influence of ants (Hymenoptera: Formicidae), particularly tramp species, on the proliferation of a maize pest. Sociobiology. 30(1): 85-93.
Detrain C. 1990. Field study on foraging by the polymorphic ant species, Pheidole pallidula. Ins. Soc. 37 (4): 315-332.
Hunt C. 1999. CITES. Cactaceae Checklist. 2nd. Ed. Instituto Geográfico Nacional. 1992. Carta Nacional
1:100000. Hoja 24-j. Lima, Perú. North R. 1996. Ants. World Wildlife Series. London. Oficina Nacional de Evaluación de Recursos
Naturales. 1975. Informe Técnico del Proyecto de Irrigación Marcapomacocha, Lima. Perú.
Ostolaza C., Mitich W.L. & King J.M. 1985. Neoraimondia arequipensis var. roseiflora (Werd & Backbg) Rauh. Cactus and Succulent. Journal (U.S). 57: 60-64.
Pickett C.H. & Clark W.D. 1979. The function of extrafloral nectarines in Opuntia acanthocarpa (Cactaceae). American Journal of Botany. 66: 618-625.
Pizarro J.O. 1997. Cactus y animales: Simbiosis maravillosa. Quepo. 11: 21-28.
Retana J., Bosch A., Alsina A. & Cerda X. 1987. Foraging ecology of the nectarivorous ant Camponotus foreli (Hymenoptera, Formicidae) in a savanna-like grassland. Misc. Zool. : 187-193.
Retana J., Cerda X., Alsina A. & Bosch J. 1986. Importancia del alimento sólido y del alimento líquido en el régimen trófico de la hormiga Cataglyphis cursor (Formicidae). Ses. Entom. ICHN-SCL.4:136-146.
Ruffener G.A. & Clark W.D. 1986. Extrafloral nectar in Ferocactus acanthodes (Cactaceae): composition and its importance to ants. American Journal of Botany. 73: 185-189.
Weseloh R.M. 1996. Effect of suplemental foods on foraging behavior of forest ants in Connecticut. Env. Ent. 25 (4): 848-853.

Ecología Aplicada, 2(1), 2003 ISSN 1726-2216 Depósito legal 2002-5474
ALGUNOS CRITERIOS BIOLÓGICOS SOBRE EL PROCESO DE EUTROFIZACIÓN A ORILLAS DE SEIS LOCALIDADES DEL LAGO TITIKAKA
Francisco Fontúrbel Rada1
Resumen
Se evaluaron las condiciones ambientales de 6 sitios a orillas del lago Titikaka (La Paz, Bolivia), encontrándose diferentes grados de eutrofización. Se emplearon 4 indicadores de evaluación rápida: macrófitas flotantes y sumergidas, fitoplancton y propiedades organolépticas, complementándolos con análisis de pH y micro-meso fauna. Se encontraron diferentes grados de afectación por el proceso eutrófico en parte debido a la influencia humana y la contaminación de los sitios de estudio. Se analizó la relevancia de cada indicador en este caso particular. Palabras clave: Lago Titikaka, eutrofización, indicadores biológicos, fitoplancton.
Abstract Environmental conditions were evaluated for 6 study sites at Titikaka’s lake margins (La Paz, Bolivia), where I founded 4 different degrees of eutrophic processes. 4 fast–evaluation indicator parameters were used: floating and submerged macrophytes, phytoplankton and organoleptic properties, which were complemented with pH and micro-middle fauna analysis. Different affectations degrees of eutrophic process were founded in associated in part to human influence and contamination at the study sites. Indicator relevancy was analyzed for each particular case. Key words: Titikaka Lake, eutrophication, biologic indicators, phytoplankton.
Introducción En la actualidad, la eutrofización cultural (asociada a la hipertrofia) es uno de los problemas con mayor repercusión sobre los cuerpos de agua y el medio ambiente (Dolbeth et al., 2003). La constante descarga de desechos sólidos y líquidos en lagos y lagunas ha ocasionado el desarrollo de este complejo problema en cuerpos de agua de todo el mundo, y el Lago Titikaka no es la excepción. La eutrofización es un fenómeno complejo que involucra factores climáticos, físicos y biológicos (Dolbeth et al., 2003; Howarth et al., 2000) , ocasionando cambios en la diversidad y la abundancia de las especies (Agatz et al., 1999; Barrón et al., 2003; Dolbeth et al., 2003; Lang , 1997; Weithoff et al., 2000). Si bien las implicancias de este proceso son muy complejas (Kim et al., 2001) –incluso a nivel de metabolismo (Macek et al., 2000)– algunos factores indicadores simples (Schroll, 2002) pueden ser usados para una detección y evaluación rápida del proceso de eutrofización. Esta trabajo hace referencia a un hecho no esperado durante un proyecto de investigación realizado en el Lago Titikaka, donde se documentó la presencia de indicios de eutrofización en los seis sitios estudiados (Fontúrbel et al., 2003), y por lo tanto pretende dar un informe preliminar de la situación observada.
Materiales y métodos Sitios de muestreo Se tomaron 6 sitios de muestreo (Tabla 1 y Figura 1) a orillas de la parte boliviana del Lago Titikaka, escogidos de acuerdo a su relevancia y aspectos paisajísticos.
Tabla 1: Identificación y localización de los sitios de estudio
Sitio Localidad Coordenadas
1 Ajaría Grande 16º1'58,71''S 68º45'43,87''W
2 Toke-Pucuru 16º1'16,81''S 68º49'25,84''W
3 Uricachi 16º3'40,18''S 68º50'16,84''W
4 Huatajata 16º12'35.58''S 68º41'11,54''W
5 Sorejapa 16º12'29,14''S 68º38'28,12''W
6 Huarina 16º13'26,58''S 68º35'9,66''W Análisis de parámetros En base a referencias de Pérez (1998, 2002) se analizaron los siguientes parámetros: pH del agua, abundancia y diversidad de fitoplancton, vegetación sumergida, vegetación flotante, micro y meso fauna, y adicionalmente se registró el grado de contaminación. Para todos los casos se tomaron muestras por triplicado, las que fueron analizadas en los
___________________________ 1 Unidad de Post–Grado Universidad Loyola. Maestría en Ingeniería de Medio Ambiente y Recursos Naturales. Av. Busch # 1191, La Paz, Bolivia. Autor: Casilla postal # 180, La Paz, Bolivia. Av. 20 de Octubre # 2005, Dep. 301, La Paz, Bolivia. Correo electrónico: [email protected]

ALGUNOS BIOCRITERIOS DE EUTROFIZACIÓN A ORILLAS DEL LAGO TITIKAKA.
Diciembre 2003 _________________________________________________________________________________________
76
laboratorios de Calidad Ambiental, Suelos y Limnología del Instituto de Ecología de la Universidad Mayor de San Andrés. Indicadores de eutrofización Se tomaron como indicadores de eutrofización: (1) la presencia y cobertura de las macrófitas flotantes Lemna spp. y Azolla spp., (2) la diversidad y cobertura de la vegetación sumergida, (3) la cantidad y diversidad de fitoplancton, y (4) las propiedades organolépticas del agua (turbidez y olor). Resultados Los 6 sitios estudiados presentaban algún tipo de contaminación (Tabla 2), los diferentes parámetros indicadores empleados mostraron diferentes grados de eutrofización (Tabla 3). El grado estimado de eutrofización para cada sitio también se muestra en la Tabla 3.
Discusión Si bien los 6 sitios de estudio registrados presentan algún grado de contaminación, la manifestación de los
procesos eutróficos no está directamente relacionada con la contaminación ni con la cantidad de habitantes de las poblaciones cercanas ya que, por la extensión del lago, los procesos eutróficos todavía se va dando de manera localizada, variando de un sitio a otro de acuerdo con la capacidad de resilencia, asociada la presencia de algunas macrófitas, principalmente Schoenoplectus californicus ssp. totora. Para esta evaluación preliminar se emplearon indicadores sencillos de evaluación situacional rápida como sugiere Schroll (2002), en base a ciertos criterios biológicos, que para este caso son: (1) presencia y cobertura de macrófitas flotantes, especialmente Lemna spp. y Azolla spp., características de los procesos eutróficos (Rodríguez, 2002), normalmente invasoras (RAMSAR, 1999) que se desarrollan por el incremento de carbono orgánico, nitrógeno y fósforo provenientes de los desechos orgánicos (Ferreira, 2001); (2) presencia de macrófitas sumergidas (Barrón et al., 2003; Dolbeth et al., 2003; McCook, 1999) que se reducen en abundancia y diversidad (García, 2003) por la falta de luz y oxígeno (Weisner et al., 1997), constituyéndose en un indicador relativamente bueno del avance del proceso eutrófico a mediano plazo (RAMSAR, 1999; Rodríguez, 2002); (3) abundancia y diversidad de fitoplancton, ya que los procesos eutróficos suelen producir un aumento de los productores primarios (Agatz et al., 1999; Dolbeth et al., 2003; Weisner et al., 1997; Weithoff et al., 2000), pero con una considerable reducción de la diversidad por la desaparición de los organismos estenotolerantes a ciertos nutrientes (Agatz et al., 1999, Kim et al., 2001; Weithoff et al., 2000), ocasionando una reducción de
Tabla 2: Descripción breve de la contaminación encontrada en cada uno de los sitios de estudio. Sitio Contaminación 1 Heces animales y residuos sólidos 2 Presencia de algunos restos plásticos en las orillas
3 Fuerte contaminación por desechos humanos, residuos sólidos y líquidos
4 Contaminación extrema por todo tipo de residuos y desechos
5 Escasa contaminación por residuos sólidos
6 Contaminación intermedia localizada, presencia de residuos sólidos
Fig. 1: Mapa de ubicación de los sitios de estudio. Los sitios más poblados están marcados con el ícono en forma de casa. Fig. 1: Mapa de ubicación de los sitios de estudio. Los sitios más poblados están marcados con el ícono en forma de casa.
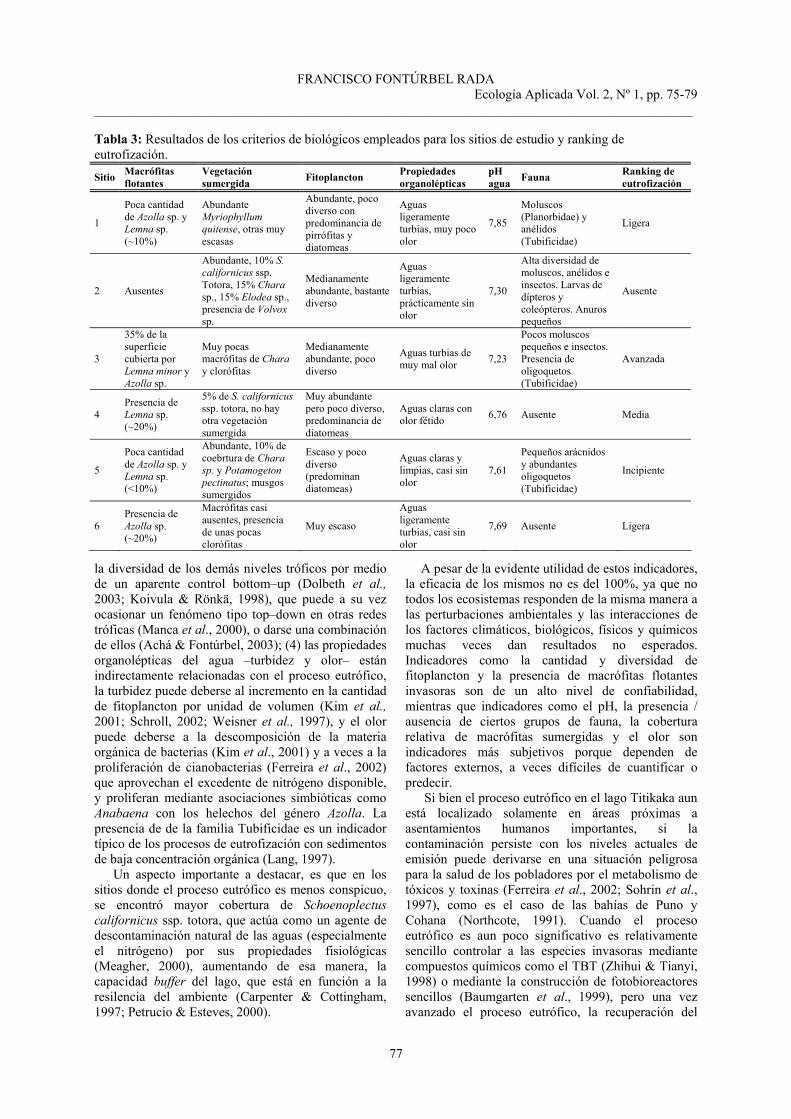
FRANCISCO FONTÚRBEL RADA
Ecología Aplicada Vol. 2, Nº 1, pp. 75-79 __________________________________________________________________________________________
77
la diversidad de los demás niveles tróficos por medio de un aparente control bottom–up (Dolbeth et al., 2003; Koivula & Rönkä, 1998), que puede a su vez ocasionar un fenómeno tipo top–down en otras redes tróficas (Manca et al., 2000), o darse una combinación de ellos (Achá & Fontúrbel, 2003); (4) las propiedades organolépticas del agua –turbidez y olor– están indirectamente relacionadas con el proceso eutrófico, la turbidez puede deberse al incremento en la cantidad de fitoplancton por unidad de volumen (Kim et al., 2001; Schroll, 2002; Weisner et al., 1997), y el olor puede deberse a la descomposición de la materia orgánica de bacterias (Kim et al., 2001) y a veces a la proliferación de cianobacterias (Ferreira et al., 2002) que aprovechan el excedente de nitrógeno disponible, y proliferan mediante asociaciones simbióticas como Anabaena con los helechos del género Azolla. La presencia de de la familia Tubificidae es un indicador típico de los procesos de eutrofización con sedimentos de baja concentración orgánica (Lang, 1997). Un aspecto importante a destacar, es que en los sitios donde el proceso eutrófico es menos conspicuo, se encontró mayor cobertura de Schoenoplectus californicus ssp. totora, que actúa como un agente de descontaminación natural de las aguas (especialmente el nitrógeno) por sus propiedades fisiológicas (Meagher, 2000), aumentando de esa manera, la capacidad buffer del lago, que está en función a la resilencia del ambiente (Carpenter & Cottingham, 1997; Petrucio & Esteves, 2000).
A pesar de la evidente utilidad de estos indicadores, la eficacia de los mismos no es del 100%, ya que no todos los ecosistemas responden de la misma manera a las perturbaciones ambientales y las interacciones de los factores climáticos, biológicos, físicos y químicos muchas veces dan resultados no esperados. Indicadores como la cantidad y diversidad de fitoplancton y la presencia de macrófitas flotantes invasoras son de un alto nivel de confiabilidad, mientras que indicadores como el pH, la presencia / ausencia de ciertos grupos de fauna, la cobertura relativa de macrófitas sumergidas y el olor son indicadores más subjetivos porque dependen de factores externos, a veces difíciles de cuantificar o predecir. Si bien el proceso eutrófico en el lago Titikaka aun está localizado solamente en áreas próximas a asentamientos humanos importantes, si la contaminación persiste con los niveles actuales de emisión puede derivarse en una situación peligrosa para la salud de los pobladores por el metabolismo de tóxicos y toxinas (Ferreira et al., 2002; Sohrin et al., 1997), como es el caso de las bahías de Puno y Cohana (Northcote, 1991). Cuando el proceso eutrófico es aun poco significativo es relativamente sencillo controlar a las especies invasoras mediante compuestos químicos como el TBT (Zhihui & Tianyi, 1998) o mediante la construcción de fotobioreactores sencillos (Baumgarten et al., 1999), pero una vez avanzado el proceso eutrófico, la recuperación del
Tabla 3: Resultados de los criterios de biológicos empleados para los sitios de estudio y ranking de eutrofización.
Sitio Macrófitas flotantes
Vegetación sumergida Fitoplancton Propiedades
organolépticas pH agua Fauna Ranking de
eutrofización
1
Poca cantidad de Azolla sp. y Lemna sp. (~10%)
Abundante Myriophyllum quitense, otras muy escasas
Abundante, poco diverso con predominancia de pirrófitas y diatomeas
Aguas ligeramente turbias, muy poco olor
7,85
Moluscos (Planorbidae) y anélidos (Tubificidae)
Ligera
2 Ausentes
Abundante, 10% S. californicus ssp. Totora, 15% Chara sp., 15% Elodea sp., presencia de Volvox sp.
Medianamente abundante, bastante diverso
Aguas ligeramente turbias, prácticamente sin olor
7,30
Alta diversidad de moluscos, anélidos e insectos. Larvas de dípteros y coleópteros. Anuros pequeños
Ausente
3
35% de la superficie cubierta por Lemna minor y Azolla sp.
Muy pocas macrófitas de Chara y clorófitas
Medianamente abundante, poco diverso
Aguas turbias de muy mal olor 7,23
Pocos moluscos pequeños e insectos. Presencia de oligoquetos (Tubificidae)
Avanzada
4 Presencia de Lemna sp. (~20%)
5% de S. californicus ssp. totora, no hay otra vegetación sumergida
Muy abundante pero poco diverso, predominancia de diatomeas
Aguas claras con olor fétido 6,76 Ausente Media
5
Poca cantidad de Azolla sp. y Lemna sp. (<10%)
Abundante, 10% de coebrtura de Chara sp. y Potamogeton pectinatus; musgos sumergidos
Escaso y poco diverso (predominan diatomeas)
Aguas claras y limpias, casi sin olor
7,61
Pequeños arácnidos y abundantes oligoquetos (Tubificidae)
Incipiente
6 Presencia de Azolla sp. (~20%)
Macrófitas casi ausentes, presencia de unas pocas clorófitas
Muy escaso
Aguas ligeramente turbias, casi sin olor
7,69 Ausente Ligera

ALGUNOS BIOCRITERIOS DE EUTROFIZACIÓN A ORILLAS DEL LAGO TITIKAKA.
Diciembre 2003 _________________________________________________________________________________________
78
lago sería un proceso largo, difícil y costoso (Myrbo & Ito, 2003). Sería recomendable realizar un estudio más profundo del avance del proceso eutrófico en el lago Titikaka, tomando en cuenta los parámetros indicadores mencionados y realizando mediciones de parámetros fisicoquímicos y de diversidad que permitan construir un modelo de simulación dinámica (Heiskary, 2000) que sería de mucha utilidad para el monitoreo del proceso, y también debería plantearse un modelo social–ambiental para gestión del problema como sugieren Janssen & Carpenter (1999). Conclusiones Se percibió que los sitios cercanos a asentamientos humanos están más expuestos a la contaminación y a la descarga de contaminantes ricos en nutrientes, y están desarrollando, todavía de manera localizada y temprana, procesos de eutrofización. El uso de los indicadores planteados se muestra como una herramienta útil para la evaluación rápida de la eutrofización localizada a orillas del Lago Titikaka. Sin embargo, para un estudio integral estos indicadores deberían complementarse con otro tipo de estudios fisicoquímicos y de diversidad más profundos, puesto que los indicadores biológicos a veces son contradictorios y no son 100% confiables por la influencia de externalidades ambientales. Es recomendable el proseguir con estudios más profundos y detallados sobre esta problemática, que permitan construir un modelo socio–ambiental adecuado para la gestión de esta problemática. Agradecimientos A Gabriela Mirones por su apoyo y ayuda incondicional. Al equipo de trabajo del proyecto Telmatobius: Dr. E. Richard, G. García, R. Rocha, M. Ferrufino y N. Brun. Literatura citada Achá D. & Fontúrbel F. 2003. La diversidad de una
Comunidad, ¿Está controlada por Top-Down, Bottom-Up o una combinación de estos?. Revista de Biología.Org. 13: 1-16.
Agatz M., Asmus R.M. & Deventer B. 1999. Structural changes in the benthic diatom community along a eutrophication gradient on a tidal flat. Helgol Mar Res. 53: 92-101.
Barrón C., Marbà N., Duarte C.M., Pedersen M.F., Lindblat C, Kersting K., Moy F. & Bokn T. 2003. High organic carbon export precludes eutrophication responses in experimental rocky shore communities. Ecosystems. 6: 144-153.
Baumgarten E., Nagel M. & Tischner R. 1999. Reduction of the nitrogen and carbon content in swine waste with algae and bacteria. Appl Microbiol Biotechnol. 52: 281-284.
Carpenter S. & Cottingham K. 1997. Resilience and Restoration of Lakes. Conservation Ecology. 1, art 2.
Dolbeth M., Pardal M.A., Lilleblo A.I., Azeiteiro U. & Marques J.C. 2003. Short- and long-term effects of eutrophication on the secondary production of an intertidal macrobenthic community. Marine Biology. 10 (1007): 1133-1135.
Ferreira A. 2001. Disponibilidade de nitrogênio, alterações nas características químicas do solo e do milho pela aplicação de efluente de esgoto tratado. MSc., Universidade de São Paulo, São Paulo.: 126.
Ferreira F., Góis M.H., Marques C., Rosas A., Kingwell P. & Simões I. 2002.Ocorrência de cianobactérias em três praias fluviais, localizadas na rede hidrográfica da bacia do Rio Tâmega- Praia Fluvial Aurora, itetos e Pontinha. 6º Congresso da água, Porto.: 2.
Fontúrbel F., García G., Brun N., Ferrufino M. & Rocha R. 2003. Propuesta de producción sostenible de Telmatobius culeus (Anura: Leptodactylidae) en el lago Titikaka, mediante la implementación conjunta de un sistema de control de calidad ambiental. En: Fontúrbel, F. & G. García (eds.), Propuestas Para Un Desarrollo Sostenible: Lago Titikaka Por Estrategas K. Publicaciones Integrales, La Paz.: 19-52.
García P. 2003.Macrófitos acuáticos en los humedales andaluces. Revista Medio Ambiente. Junta de Andalucía, Sevilla.
Heiskary S. 2000. Simplified Lake Eutrophication Modeling: Using the Ecoregionbased model MINLEAP. Minnesota Pollution Control Agency Minnesota Pollution Control Agency, Minnesota.: 19.
Howarth R., Swaney D.P., Butler T.J. & Marino R. 2000. Climatic control on eutrophication of the Hudson River Estuary. Ecosystems. 3: 210-215.
Janssen M. & Carpenter S. 1999. Managing the Resilience of Lakes: A Multiagent Modeling Approach. Conservation Ecology. 3, art 15.
Kim B., Park J.H., Hwang G., Jun M.S. & Choi K. 2001. Eutrophication in reservoirs of South Korea. Limnology. 2: 223-229.
Koivula K. & Rönkä A. 1998. Habitat deterioration and efficiency of antipredator strategy in a meadow-breeding wader, Temminck's stint (Calidris temminckii). Oecologia. 116: 348-355.
Lang C. 1997. Oligochaetes, organic sedimentation, and trophic state: how to assess the biological recovery of sediments in lakes? Aquat.sci.. 59: 26-33.
Macek T., Macková M. & Kás J. 2000. Exploitation of plants for the removal of organics in environmental remediation. Biotechnology Advances. 18: 23-34.
Manca M., Ramoni C. & Comoli P. 2000. The decline of Daphnia hyalina galeata in Lago Maggiore: a

FRANCISCO FONTÚRBEL RADA
Ecología Aplicada Vol. 2, Nº 1, pp. 75-79 __________________________________________________________________________________________
79
comparison of the population dynamics before and after oligotrophication. Aquat.sci. 62: 142-153.
McCook L.J. 1999. Macroalgae, nutrients and phase shifts on coral reefs: scientific issues and management consequences for the Great Barrier Reef. Coral Reefs. 18: 357-367.
Meagher R. 2000. Phytoremediation of toxic elemental and organic pollutants. Current Opinion in Plant Biology. 3: 153-162.
Myrbo A. & Ito E. 2003.Eutrophication and remediation in context: High-resolution study of the past 200 years in the sedimentary record of Lake McCarrons (Roseville, Minnesota). USGS-WRRI 104B National Grants Competition and the Center for Agricultural Impacts on Water Quality, Minnesota.: 5.
Northcote T. 1991. Eutrofización y problemas de polución. En: Dejoux, C. & A. Iltis (eds.), El Lago Titicaca: Síntesis Del Conocimiento Limnológico Actual. Hisbol - ORSTOM, La Paz.: 563-572.
Pérez E. 1998. Dieta y ciclo gametogénico anual de Telmatobius culeus (Anura: Leptodactylidae) en el lago Titicaca (Huiñaimarca). Licenciatura, Universidad Mayor de San Andrés, La Paz. : 140.
Pérez E. 2002.Evaluación de la población de la rana gigante en Bolivia. Subcontrato 21.22 del Proyecto de Conservación de la Biodiversidad en la Cuenca del sistema TDPS, ALT-IE-FUNDECO, La Paz.: 114.
Petrucio M. & Esteves F.A. 2000. Uptake rates of nitrogen and phosphorus in the water by Eichhornia crassipes and Salvinia auriculata. Rev. Brasil. Biol. 60: 229-236.
RAMSAR. 1999.Ramsar COP7 DOC. 24: Especies invasoras y humedales. RAMSAR COP, San José. : 9.
Rodríguez J.A. 2002.¿Cuál es la flora característica de un proceso eutrófico? Sierra de Baza. Vol. 2003.
Schroll H. 2002. Indicators of the Long-term Eutrophication of a Danish lake (Karlsø), and Water Pollution Management. The Journal of Transdisciplinary Environmental Studies. 1: 1-10.
Sohrin Y., Matsui M., Kawashima M., Hojo M. & Hasegawa H. 1997. Arsenic Biogeochemistry Affected by Eutrophication in Lake Biwa, Japan. ICR Annual Report. 4: 14-15.
Weisner S.E.B., Strand J.A. & Sandsten H. 1997. Mechanisms regulating abundance of submerged vegetation in shallow eutrophic lakes. Oecologia. 109: 592-599.
Weithoff G., Lorke A. & Walz N. 2000. Effects of water-column mixing on bacteria, phytoplankton, and rotifers under different levels of herbivory in a shallow eutrophic lake. Oecologia. 125: 91-100.
Zhihui S. & Tianyi C. 1998. Toxicity of Tributyltin to Lemna minor L. and Azolla filiculoides Lamk. Bull. Environ. Contam. Toxicol. 60: 318-322.

Ecología Aplicada, 2(1), 2003 ISSN 1726-2216 Depósito legal 2002-5474
VIABILIDAD DE CEPAS DE RIZOBIOS EN INOCULANTES BASADOS EN SOPORTES NO ESTÉRILES
Gisella Matos Cuzcano1 y Doris Zúñiga Dávila1
Resumen
Se evaluó la supervivencia de las cepas PLL113 de Bradyrhizobium sp. y PLC213 de Rhizobium sp. en dos inoculantes elaborados en base a turba y una mezcla de suelo-compost no estériles almacenados a 4°C durante seis meses. Las poblaciones de ambas cepas disminuyeron durante el almacenamiento en ambos tipos de inoculantes, sin embargo la turba permitió una mejor supervivencia. Se registraron evidencias de antagonismos o competencia entre las bacterias mesófilas totales (BMT) y mohos contaminantes. Así mismo se observaron correlaciones negativas entre las poblaciones de rizobios y el resto de microorganismos, lo que indica su pobre competitividad frente a los contaminantes. La calidad microbiológica comprendida como la concentración mínima de rizobios por gramo de los inoculantes fue satisfactoria durante las siete primeras semanas de almacenamiento. Palabras claves: Rhizobium, compost, turba, nódulos.
Abstract The survival of Bradyrhizobium sp. PLL113 and Rhizobium sp. PLC213 in soil-compost and peat based unsterile inoculants stored at 4 °C during 6 months was evaluated. The populations of both strains decreased during storage in both type of inoculants, although better survival was observed in peat. Evidence of antagonisms or competition between total contaminant bacteria and moulds was registered. In addition, negative correlations were observed between rhizobial populations and the rest of microorganisms, which indicated its poor competitiveness against contaminants. The inoculants microbiological quality was satisfactory during the first seven weeks of storage. Key words: Rhizobium, compost, peat, nodules.
1 Laboratorio de Ecología Microbiana y Biotecnología “Marino Tabusso”, Universidad Nacional Agraria La Molina. Apartado Postal 456, Lima 100, Perú. Correo electrónico : [email protected] / [email protected]
Introducción Las leguminosas tienen la capacidad de establecer una asociación simbiótica con bacterias de la familia Rhizobiaceae, comúnmente conocidas como rizobios. Estas bacterias inducen en la planta la formación de estructuras especializadas, denominadas nódulos, en donde se alojan como simbiontes intracelulares y fijan nitrógeno atmosférico que es utilizado por la planta. Para obtener un buen rendimiento en el cultivo de leguminosas es necesaria la utilización de fertilizantes nitrogenados, los cuales elevan los costos de producción afectando la economía de los productores y consumidores. Una alternativa a este problema es la utilización de inoculantes bacterianos que porten rizobios viables y seleccionados por su infectividad y efectividad simbióticas (Hamdi, 1985). Actualmente, los inoculantes en base a soportes sólidos son los más ampliamente difundidos. Se utilizan varios tipos de soportes como turba (Smith, 1992), carbón mineral (Paczkowski & Berryhill, 1979), mezclas suelo-vermicompost (Benedetti et al., 1991), suelo-compost (Ormeño-Orrillo & Zúñiga-Dávila, 1999a), suelo-carbon vegetal (Beck et al., 1993) y suelo-paja de arroz (Pramakik & Iswaran,
1973). En muchos casos se recurre a la esterilización de estos soportes en autoclave (Ormeño-Orrillo & Zúñiga-Dávila, 1999b) o por irradiación gamma (Strijdom & Jansen van Rensburg, 1981). Sin embargo esta práctica eleva los costos de producción de los inoculantes y por otro lado la esterilización por calor, puede conllevar a la liberación o producción de compuestos tóxicos para los rizobios. La fabricación del inoculante debe estar respaldado por un estricto control que asegure la calidad del producto, el control del inoculante se realiza en distintas fases del proceso parra comprobar las características cualitativas y cuantitativas de la población rizobiana del inoculante. Según la legislación brazileña el fabricante de inoculantes debe cumplir con los siguientes requisitos: empleo de cepas recomendadas por organismos de investigación y garantía mínima de que el producto contiene 10 milones de rizobios por gramo. El objetivo del presente trabajo fue evaluar la viabilidad de cepas de rizobios durante un almacenamiento prolongado en dos inoculantes no estériles elaborados en base a turba y una mezcla de suelo-compost.

VIABILIDAD DE RIZOBIOS EN INOCULANTES BASADOS EN SOPORTES NO ESTÉRILES
Diciembre 2003 _________________________________________________________________________________________
82
Materiales y métodos Microorganismos y condiciones de cultivo Se utilizaron las cepas PLL113 de Bradyrhizobium sp. y PLC213 de Rhizobium sp. aisladas de nódulos de pallar (Phaseolus lunatus L.) (Matos-Cuzcano et al., 1998) y pertenecientes a la colección del Laboratorio de Ecología Microbiana y Biotecnología “Marino Tabusso” de la Universidad Nacional Agraria La Molina (UNALM). Los cultivos stock fueron mantenidos a 4°C en tubos con agar extracto de levadura manitol (LMA) (Beck et al., 1993) en plano inclinado. Las cepas fueron sembradas independientemente en 4 frascos Roux que contenían 25 mL de LMA en plano inclinado. Los cultivos se incubaron a 28°C por tres y siete días para la cepa de Rhizobium y Bradyrhizobium, respectivamente. Los inóculos se obtuvieron agregando 20 mL de solución salina (0,85 %) estéril a los frascos y realizando una suspensión homogénea de las bacterias. Elaboración de los soportes Los soportes se elaboraron utilizando suelo recolectado en el campus de la UNALM, compost adquirido en el Programa de Leguminosas – UNALM y turba donada por el Intituto Nacional de Investigación Agraria (INIA). El suelo y el compost se tamizaron a través de una malla de 0,16 mm. Los tres materiales se colocaron independientemente en bolsas de tela y fueron pasteurizados a vapor fluente por una hora en tres días consecutivos incubándolos a 37 °C entre cada tratamiento para permitir el crecimiento de la microflora sobreviviente (Alder & Simpson, 1982). Luego del tercer tratamiento se incubaron nuevamente a 37 °C para disminuir su humedad. El compost y el suelo se mezclaron en la proporción de 2:3 (p:p) y la mezcla resultante se empaquetó en bolsas de polietileno (200 g/bolsa). La turba también fue dispensada en bolsas en igual cantidad. Elaboración de los inoculantes Las bolsas con los soportes pasteurizados fueron impregnadas en condiciones asépticas con 40 mL de las suspensiones bacterianas, dejando como controles una bolsa de cada soporte sin inocular. Cada inoculante fue posteriormente separado en bolsas de 100 g cada una, obteniéndose 2 bolsas de inoculante de cada tipo para ambas cepas. Luego de un periodo de maduración de 24 h (Materon & Weaver, 1985), los inoculantes se almacenaron en refrigeración a 4°C. Evaluaciones Luego de la maduración y a los 47, 76, 96 y 176 días de almacenamiento se evaluaron las poblaciones de rizobios, bacterias mesófilas totales (BMT), mohos y levaduras determinando el número de unidades formadoras de colonia (UFC) por el método de
dilución decimal y recuento en placa (Speidel & Wollum, 1980). Los medios de cultivo utilizados fueron LMA suplementado con Rojo Congo e incubado a 28°C, agar Plate Count incubado a 28°C y agar Sabouraud incubado a temperatura ambiente para rizobios, BMT, y mohos y levaduras, respectivamente. Para el análisis, los datos se expresaron en base a peso seco y se transformaron a log10 UFC/g. Inoculación en plantas Se agregó 1 g de los inoculantes almacenados por 176 días a plantas de pallar cultivadas en tubos de ensayo (25 × 150 mm) con agar Sandman (CIAT, 1988) en plano inclinado. Las plantas se mantuvieron en una cámara de crecimiento con fotoperíodo de 12 h y se regaron con solución Sandman diluida 1:4 (v:v) cada dos o tres días. Al control con nitrógeno se le agregó 103,9 mg de urea por litro de solución Sandman. A los 30 d se finalizó el ensayo y se determinaron el número de nódulos y peso seco de la parte aérea. Resultados y discusión La población inicial de rizobios presente en los inoculantes fue de 1010 a 1011 UFC/g para la cepa PLC213 y de 109 a 1010 UFC/g para la cepa PLL113. Al evaluar la población a través del tiempo se observó un descenso paulatino de ambas cepas de ambos inoculantes (Tabla 1, Figuras 1 y 2). La viabilidad de las dos cepas utilizadas fue mayor en el soporte turba en comparación al soporte suelo-compost hasta los 176 días. Así también, se observó que la cepa PLC213 sobrevivió mejor que la cepa PLL113 en el soporte suelo-compost, mientras que lo contrario sucedió en el soporte turba. La pasteurización y secado aplicado inicialmente a los soportes redujo la población inicial de BMT hasta 105 UFC/g y a 102 UFC/g para mohos. La mayor proporción de microorganismos detectados en el medio Plate Count fueron bacilos esporulados Gram positivos lo que cabria esperarse luego del tratamiento térmico (Alder & Simpson, 1982), mientras que en agar Sabouraud sólo se obtuvo desarrollo de mohos filamentosos mas no así de levaduras. Las poblaciones de ambos grupos microbianos aumentaron durante los primeros 47 días de almacenamiento en ambos soportes. A los 76 días se observó una tendencia hacia el aumento de las poblaciones de BMT y a una disminución de los mohos presentes. Por el contrario, a los 96 días las BMT disminuyeron o se mantuvieron estables mientras que las poblaciones de mohos aumentaron. Al final del experimento (176 días) todas las poblaciones disminuyeron (Fig. 3 y 4). Vistos en conjunto, estos resultados indicarían una competencia o antagonismo entre ambos grupos microbianos. El incremento inicial de ambas poblaciones se explicaría por la abundante presencia de nutrientes en ambos

GISELLA MATOS Y DORIS ZÚÑIGA
Ecología Aplicada, Vol. 2, pp 81-85 __________________________________________________________________________________________
83
soportes que permitirían su desarrollo. Al disminuir la concentración de estos nutrientes (76 días), las bacterias tendrían una ventaja sobre los mohos por su mayor velocidad de crecimiento. Sin embargo al agotarse los nutrientes iniciales (96 días) los mohos podrían competir mejor por su capacidad de degradar residuos celulósicos y sustancias orgánicas complejas.
El agotamiento de los nutrientes y/o la acumulación de catabolitos tóxicos explicarían la disminución de ambas poblaciones a los 176 días.
Tabla 1. Viabilidad de las cepas PLC213 de Rhizobium sp. y PLL113 de Bradyrhizobium sp. en inoculantes no estériles a base de suelo-compost (S-C) y turba (T) almacenados a 4 ºC.
Viabilidad (%)a Tiempo PLC213 PLL113 (días) S-C T S-C T
2 100 100 100 100 47 84,1 75,6 63,6 93,2 76 49,0 66,1 47,7 74,9 96 53,6 61,9 41,6 63,5 176 52,4 59,1 35,1 63,8
a Porcentaje de sobrevivencia en base a Log10 de UFC/g
Las interacciones descritas anteriormente también se manifestaron entre los rizobios y las poblaciones de BMT y mohos. Sin tomar en cuenta la ultima evaluación en la cual todas poblaciones disminuyeron, se obtuvieron correlaciones negativas entre las poblaciones de rizobios, BMT y mohos (Tabla 2). El caso extremo lo presentó la interacción entre cepa PLL113 de Bradyrhizobium sp. y las BMT en el soporte suelo-compost (r = -0,93). Esta cepa fue la que presentó la menor supervivencia en ese soporte, en el cual también se cuantificaron las mayores poblaciones de BMT y estas se mantuvieron elevadas por más tiempo. En comparación, Ormeño-Orrillo &
Zúñiga-Dávila (1999a) observaron una mejor supervivencia de la cepa PLL113 en un soporte similar pero esterilizado. Los resultados obtenidos indican que los rizobios son pobres competidores frente al resto de la microflora presente en los soportes utilizados para elaborar inoculantes. Si bien el uso de soportes estériles conlleva a mayores costos de producción de los inoculantes, su uso se justifica ya que permite eliminar la competencia y/o antagonismo de hongos y otras bacterias, y la predación por protozoarios (Van Schreven, 1970).
A los 176 días de almacenamiento se evaluaron la infectividad y efectividad de los inoculantes en plántulas de pallar. A pesar de mantener una población mayor a 106 UFC/g, la cepa PLC213 de Rhizobium sp. no indujo la formación de nódulos. Las plantas tratadas con los inoculantes con la cepa PLL113 de Bradyrhizobium sp. a base de suelo-compost y turba produjeron un promedio de 51,3 y 130,4 nódulos/planta, respectivamente. Así mismo, el peso seco de estas plantas fue mayor que el de los controles no inoculados (con y sin nitrógeno). Estos resultados pueden explicarse por la naturaleza de los determinantes genéticos que contienen la información para el proceso simbiótico en cada una de las cepas. En el género Rhizobium los genes para la nodulación y fijación de nitrógeno se encuentran en sus plásmidos, mientras que en el género Bradyrhizobium, dichos genes están integrados en su
Tabla 2. Coeficientes de correlación entre las poblaciones de rizobios y de BMT y mohos en dos inoculantes a base de suelo-compost (S-C) y turba (T). BMT Mohos Rizobio S-C T S-C T PLL113 - 0,93 - 0,62 - 0,28 - 0,52 PLC213 - 0,22 - 0,20 - 0,37 - 0,70
PLC213
3
4
5
6
7
8
9
10
11
12
0 30 60 90 120 150 180
Tiempo (días)
Log
UFC
/g
Suelo-CompostTurba
Figura 1. Viabilidad de la cepa PLC213 de Rhizobium sp. en inoculantes no estériles a base de suelo-compost y turba almacenados a 4°C.
PLL113
3
4
5
6
7
8
9
10
11
12
0 30 60 90 120 150 180
Tiempo (días)
Log
UFC
/g
Suelo-Compost
Turba
Figura 2. Viabilidad de la cepa PLL113 de Bradyrhizobium sp. en inoculantes no estériles a base de suelo-compost y turba almacenados a 4°C.

VIABILIDAD DE RIZOBIOS EN INOCULANTES BASADOS EN SOPORTES NO ESTÉRILES
Diciembre 2003 _________________________________________________________________________________________
84
cromosoma (Jordan, 1984). Al ser el plásmido un elemento genético accesorio, es probable que la cepa de Rhizobium PLC213 lo haya perdido durante el periodo de almacenamiento. Por el contrario, la cepa de Bradyrhizobium PLL113 mantuvo su capacidad infectiva y efectiva debido a la estabilidad inherente de poseer esta información en el cromosoma.
SUELO-COMPOST
1
2
3
4
5
6
7
0 30 60 90 120 150 180
Tiempo (días)
Log
UFC
/g
Inoculante PLC213Inoculante PLL113
Control
Figura 3. Variación de las poblaciones de BMT (líneas contiguas) y mohos (lineas punteadas) en los inoculantes a base de suelo-compost. Los estándares de calidad para el número de rizobios que debe tener un inoculante sólido varían de acuerdo a las legislaciones vigentes en cada país entre 107 y 109 UFC/g (Boonkerd, 1991; Rodríguez-Navarro et al., 1991; Thompson, 1996). Los inoculantes analizados en este estudio se adecuaron a estos estándares durante los primeros 47 días de almacenamiento. Esto representa un periodo de vida útil corto en comparación con otros tipos de materiales como la turba (Somasegaran, 1985) o el carbón mineral (Paczkowski & Berryhill, 1979). Fue interesante observar que la turba utilizada no resultó ser un buen soporte teniendo en cuenta que se han reportado periodos de vida útil de hasta 6 meses para inoculantes basados en este material. Sin embargo, turbas de mala calidad pueden acortar bastante este periodo útil (Pramakik & Iswaran, 1973; Sparrow & Ham, 1983).
T U R B A
1
2
3
4
5
6
0 30 60 90 120 150 180
Tiempo (días)
Log
UFC
/g
Inoculante PLC213noculante PLL113
Contr ol
Figura 4. Variación en las poblaciones de BMT (líneas continuas) y mohos (lineas punteadas) en los inoculantes a base de turba. Conclusiones 1. La turba permitió una mejor supervivencia de los rhizobios en comparación a la mezcla suelo-compost. 2. En relación a estándares internacionales, la calidad microbiológica (según la legislación brasileña la concentración mínima de rizobios debe ser 100 millones por gramo ) de los inoculantes fue satisfactoria durante las primeras 7 semanas de almacenamiento. 3. Se recomienda la utilización de soporte estériles pues los contaminantes pueden inhibir la población rhizobiana presente en el inoculante. Literatura citada Alder V.G. & Simpson R.A. 1982. Sterilization and
disinfection by heat methods. pp. 433-453. En: Principles and Practice of Disinfection, Preservation and Sterilization. A.D. Russell, W.B. Hugo y G.A.J. Ayliffe (Eds.). Blackwell Scientific Publications. Oxford.
Beck D.P., Materon L.A. & Afandi F. 1993. Practical Rhizobium-Legume Technology Manual. Technical Manual N° 19. International Center for Agricultural Research in the Dry Areas (ICARDA). Alepo.
Benedetti E., Zúñiga D. & Carbajal N. 1991. Viabilidad y efectividad de diez inoculantes de Rhizobium almacenados durante seis meses a 4°C. p. 113. En: Segunda Reunión de Expertos en Leguminosas de Grano de la Zona Andina. Cali.
Boonkerd N. 1991. Inoculant quality control and standards in Thailand. pp. 121-130. En: Report on the Expert consultation on legume inoculant production and quality control. J.A. Thompson (ed.). Roma.
CIAT (Centro Internacional de Agricultura Tropical). 1988. Simbiosis Leguminosa-Rizobio. Manual de Métodos de Evaluación, Selección y Manejo Agronómico. Cali.

GISELLA MATOS Y DORIS ZÚÑIGA
Ecología Aplicada, Vol. 2, pp 81-85 __________________________________________________________________________________________
85
Hamdi Y.A. 1985. La fijación del nitrógeno en la explotación de los suelos. Boletín de suelos de la FAO N° 49. FAO. Roma.
Jordan D.C. 1984. Family III. Rhizobiaceae Conn 1938. pp. 234-256. En: Bergey's Manual of Systematic Bacteriology, vol. I. N.R. Krieg and J.G. Holt (Eds.). The Williams & Wilkins Co. Baltimore.
Materon L.A. & Weaver R.W. 1985. Inoculant maturity influences survival of rhizobia on seed. Appl. Environ. Microbiol. 49: 465-467.
Matos-Cuzcano G., Ormeño-Orrillo E. & Zúñiga-Dávila D. 1998. Diversidad de los rizobios que nodulan el cultivo de pallar (Phaseolus lunatus L.) en la costa central del Perú. Ecología. 1: 42-46.
Ormeño-Orrillo E. & Zúñiga-Dávila D. 1999a. Supervivencia y características simbióticas de Bradyrhizobium sp. inoculado en una mezcla de suelo y compost. Biota. 17(99): 41-49.
Ormeño-Orrillo E. & Zúñiga-Dávila D. 1999b. Optimización del tiempo de esterilización de soportes basados en suelo y compost en la producción de inoculantes para leguminosas. Revista Peruana de Biología. 6(2): 181-184.
Paczkowski M.W. & Berryhill D.L. 1979. Survival of Rhizobium phaseoli in coal-based legume inoculants. Appl. Environ. Microbiol. 38: 612-615.
Pramanik M. & Iswaran V. 1973. Survival of Rhizobium japonicum in various carriers. Zbl. Bakt. Abt. II. 128: 232-239.
Rodríguez-Navarro D.N., Temprano F. &. Orive R. 1991. Survival of Rhizobium sp. (Hedysarum
coronarium L.) on peat-based inoculants and inoculated seeds. Soil Biol. Biochem. 23: 375-379.
Smith R.S. 1992. Legume inoculant formulation and application. Can. J. Microbiol. 38: 485-492.
Somasegaran P. 1985. Inoculant production with diluted liquid cultures of Rhizobium sp. and autoclaved peat: evaluation of diluents, Rhizobium sp., peats, sterility requirements, storage, and plant effectiveness. Appl. Environ. Microbiol. 50: 398-405.
Sparrow S.D. & Ham G.E. 1983. Survival or Rhizobium phaseoli in six carriers materials. Agron. J. 75: 181-184.
Speidel K.L. & Wollum A.G. 1980. Evaluation of leguminous inoculant quality. Technical Bulletin N° 266. North Caroline Agricultural Research Service. Raleigh.
Strijdom B.W. & Jansen Van Rensburg H. 1981. Effect of steam sterilization and gamma irradiation of peat on quality of Rhizobium inoculants. Appl. Environ. Microbiol. 41: 1344-1347.
Thompson J.A. 1996. ¿Estamos haciendo el mejor uso de la tecnología de inoculantes?. pp. 417-436. En: XVIII Reunión Latinoamericana de Rhizobiología. J. Pijnenborg, D. Ruíz y W. Siles (Eds.). Santa Cruz de la Sierra.
Van Schreven D.A. 1970. Some factors affecting growth and survival of Rhizobium sp. in soil-peat cultures. Plant Soil. 32: 113-130.

Ecología Aplicada, 2(1), 2003 ISSN 1726-2216 Depósito legal 2002-5474
EFECTOS DEL MANEJO DE SUELOS DE LADERAS EN HONGOS FORMADORES DE MICORRIZAS ARBUSCULARES Y EN BACTERIAS FIJADORAS DE NITRÓGENO EN
ULTISOLES SUJETOS A EROSIÓN PLUVIAL EN LA AMAZONIA PERUANA Pedro O. Ruiz1 y Charles B. Davey2
Resumen
En un ensayo de vivero se evaluó la colonización con micorrizas arbusculares y la nodulación, en plantas de frejol caupí (Vigna unguiculata L. Walp) en suelos de ladera bajo cultivos en callejones (alley-cropping), cultivos continuos con bajos insumos, bosque secundario y suelo sin cobertura, a los 5, 15, 30 y 45 días después de la germinación de la semilla (DDGS). La colonización micorrícica y la nodulación presentaron una gran variación en los estados iniciales del crecimiento de las plantas de caupí a pesar de que ninguno de los sistemas estudiados recibieron fertilización fosfatada. A niveles similares y bajos de disponibilidad de fósforo en el suelo, en suelos diferentes, altos niveles de colonización micorrícica (sistema de cultivos continuos con bajos insumos) pueden ser ventajosos para el crecimiento de la planta y la nodulación. La nodulación estuvo ausente en las plantas de caupí que crecieron en el suelo de bosque secundario probablemente debido al efecto represivo de altos niveles de nitrógeno en el suelo o a la falta de rizobios para esta especie. En el suelo sin cobertura, las plantas de caupí tuvieron niveles muy bajos de colonización micorrícica. La formación de nódulos fue esencialmente nula, probablemente debido a los efectos de la pérdida de suelo por las lluvias. Los resultados obtenidos sugieren la necesidad de reintroducir los microorganismos del suelo como parte de programas de recuperación de suelos y/o revegetación de áreas severamente alteradas como los taludes de carreteras, derechos de vía de gasoductos y oleoductos, áreas expuestas por actividad minera y otras. Palabras clave: Laderas, erosión, micorrizas arbusculares, nodulación, revegetación, Amazonía
Abstract Arbuscular mycorrhizal colonization and nodulation in cowpea (Vigna unguiculata L.Walp) plants growing in soils under alley-cropping, low-input continuous cropping, secondary forest, and bare soil on a slope were evaluated at 5, 15, 30 and 45 days after seed germination (DASG) under greenhouse conditions. Mycorrhizal colonization and nodulation in cowpeas differed greatly at their early growth stages in spite that any of the systems studied received phosphorus fertilizers. At similar low levels of soil available phosphorus in different soils, high levels of mycorrhizal colonization (low-input continuous cropping system) may be advantageous for plant growth and nodulation. Nodulation was absent in cowpeas growing in the secondary forest soil probably due to repressing effects of high levels of nitrogen in soil or to the lack of rhizobia for cowpeas. In the bare soil plot, cowpeas had a very low level of mycorrhizal colonization. Nodule formation was essentially zero, probably as effects of soil loss due to rainfall. Results obtained suggest the need for re-introduction of soil microorganisms for revegetation and land reclamation programs in severely disturbed soils like road banks, gas and oil pipelines, mining exposed areas, and others. Key words: Slopes, erosion, arbuscular mycorrhizae, nodulation, revegetation, Amazon
1 Consultor. AndeStudio S.A. Manejo de Suelos, revegetación, micorrizas. Correo electrónico: [email protected] 2 Professor Emeritus. Soil Science and Forestry Departments. North Carolina State University. Raleigh, N.C., U.S.A.
Introducción En los trópicos húmedos bajos, la tasas de deforestación debido a la agricultura migratoria ha tenido un gran aumento en la última década debido principalmente a la gran presión demográfica y a la necesidad de producir cultivos alimenticios, afectando seriamente la productividad de los suelos por el acortamiento de los períodos de barbecho y la resultante reducción en los rendimientos de los cultivos (Sánchez et al., 1990). En la Amazonía del
Perú, esta situación se agrava por las grandes áreas dedicadas a la producción de coca (Dourojeanni, 1990). Los suelos de la Amazonía peruana, predominantemente Ultisoles, son ácidos y deficientes en nutrientes (Sánchez, 1976) y se encuentran mayormente en áreas onduladas con laderas que tienen del 2 al 50 % de pendiente (Benites, 1983). Estas áreas son particularmente propensas a una severa erosión pluvial, un problema muy serio y diseminado

MICORRIZAS ARBUSCULARES Y BACTERIAS FIJADORAS DE NITROGENO EN AMAZONIA PERUANA
Diciembre 2003 __________________________________________________________________________________________
88
ampliamente en muchos países tropicales en desarrollo (Stocking, 1988). Estudios conducidos en los trópicos húmedos bajos indican que la remoción del suelo superficial con la consecuente pérdida de la materia orgánica, nutrientes y la degradación de sus propiedades físicas y químicas, afectan notablemente la producción de cultivos (Alegre & Fernández, 1991) y en muchos casos, la aplicación de fertilizantes no compensa la pérdida del suelo superficial (Lal, 1988). El impacto de las pérdidas por erosión en las propiedades biológicas del suelo, sin embargo, no se conoce claramente. Entre estas, un importante aspecto constituye la capacidad del suelo para soportar una interacción simbiótica efectiva entre microorganismos benéficos del suelo y las raíces de las plantas. Entre las interacciones más importantes se involucra la relación simbiótica de las plantas con los hongos micorrícicos y con los rizobios (Habte, 1989; Rivera et al., 2000). Desde que el crecimiento de las plantas es afectado en forma significativa por los microorganismos del suelo, un claro entendimiento del impacto de la erosión en la productividad del suelo no se puede conseguir a menos que su impacto sobre las poblaciones microbianas sea también claramente entendido (Amaranthus & Trappe, 1993; Franco et al. 1997; Fernandes et al., 2001; Wu et al., 2002). En años recientes, el interés por los cultivos en callejones (alley-cropping) para la producción sostenible de alimentos y para la prevención de la pérdida del suelo en terrenos en pendientes en suelos ácidos ha aumentado en forma significativa (Palm, 1988; Szott et al., 1991a, b), donde las leguminosas constituyen un componente importante del sistema. La mayoría de las leguminosas, por otro lado, representan un caso especial entre la comunidad de plantas ya que ellas están asociadas íntimamente con micorrizas arbusculares y con rizobios. Se conoce muy poco acerca de los efectos de la pérdida del suelo superficial en esta asociación tripartita de leguminosa-micorriza-rizobio en sistemas con bajos insumos en suelos ácidos del trópico. Investigación pertinente en Ultisoles en la Amazonía peruana no existe considerando la gran proporción de áreas agrícolas en tierra en pendientes propensas a erosión severa, especialmente cuando son manejadas de manera inapropiada. El principal objetivo del presente estudio fue el de evaluar la colonización micorrícica y la nodulación en plantas de frejol caupí (Vigna unguiculata (L.) Walp) en Ultisoles bajo sistemas de cultivos en callejones (alley-cropping), cultivos continuos, bosque secundario y suelo sin cobertura en una pendiente en la Amazonía del Perú. Materiales y métodos Se condujo un experimento de vivero en la Estación Experimental de Yurimaguas, localizada en
la cuenca Amazónica del Perú (76º 05´ O, 05º 45´ S, a 180 m.s.n.m.). El área recibe una precipitación anual promedio de 2200 mm y tiene una temperatura anual promedio de 25 ºC. Los suelos predominantes pertenecen a la serie Yurimaguas, clasificados como franco-finos, silíceos, isohipertérmicos Paleudults Típicos. Los suelos se colectaron del experimento de campo “Pérdidas por erosión y escorrentía en bosques, cultivos con bajos insumos y cultivos en callejones, en laderas” iniciado en el lugar mencionado en 1988. El experimento se estableció en una ladera con aspecto norte con una pendiente entre 15-20%. Los tratamientos fueron instalados en parcelas de escorrentía de 22 m de largo por 2 m de ancho y fueron los siguientes: 1) cultivos continuos con bajos insumos con una rotación de arroz de secano con frejol caupí sembrados en líneas en sentido contra la pendiente (los residuos fueron dejados en la superficie del suelo); 2) líneas de guaba (Inga edulis) sembradas en sentido contra la pendiente con una separación de 4 m. Entre las líneas se sembró una rotación de arroz de secano-frejol caupí; las podas de los árboles y los residuos se colocaron en la superficie del suelo. Se incluyeron adicionalmente dos parcelas testigo (una de bosque secundario y un suelo sin cobertura). Se hizo una evaluación de la pérdida de suelo en el referido experimento durante el período de Diciembre de 1988 a Agosto de 1989. El área recibió 1317 mm de lluvia total (nueve meses) y la pérdida de suelo fue de 2.32, 0.60, 0.07 y 13.00 Mg ha-1 para los cultivos continuos, cultivos en callejones, bosque secundario y suelo sin cobertura, respectivamente. Para el presente experimento, se colectó suelo superficial a una profundidad de 20 cm, de los bordes de las parcelas de escorrentía de cuatro posiciones de la ladera y luego mezclado. El suelo fue secado al aire, tamizado y mezclado. Porciones de 1.7 kg de suelo (peso seco) se colocaron en bolsas de plástico de 2 kg. Las características químicas de los suelos al momento de la colecta se muestran en la Tabla 1. El experimento tuvo un diseño de parcelas divididas con los tratamientos de manejo de suelo como parcelas principales y épocas de cosecha como sub-parcelas. Los tratamientos de manejo de suelo incluyen: 1) cultivos continuos con bajo insumos (BI); 2) cultivos en callejones (CC); 3) barbecho de bosque secundario (BS); y 4) suelo sin cobertura (SC). Las plantas se cosecharon a los 7, 15, 30 y 45 días después de la germinación de la semilla (DDGS). Se sembraron 4 semillas de frejol caupí en cada bolsa y luego se dejó una después de la germinación. Los tratamientos de manejo de suelo tuvieron tres repeticiones por cada época de cosecha y fueron distribuidos en mesas de vivero en un diseño completo randomizado. Las plantas se regaron cuidadosamente para mantener la humedad del suelo en un 80% de su capacidad de retención de agua (por peso) y fueron re-

PEDRO O. RUIZ Y CHARLES B. DAVEY
Ecología Aplicada Vol. 2 Nº 1, pp. 87-92 __________________________________________________________________________________________
89
distribuidas cada dos semanas para minimizar efectos posicionales. Tabla 1. Características químicas de los suelos de los tratamientos en estudio (0-15 cm de profundidad).
Aci
dez
cam
biab
le
Ca Mg K CEIC
Tra
tam
ient
o
Aci
dez
(pH
)
P (m
g/L
)
cmol (+) / L Satu
raci
ón
de A
l (%
)
Cultivos en callejones
4.4 18.7 4.37 0.4 0.10 0.10 5.02 86.3
Cult. continuos bajos insumos
4.4 15.1 4.58 0.5 0.12 0.07 5.30 85.5
Bosque secundario 4.4 19.4 3.55 0.34 0.14 0.08 4.05 87.8
Suelo sin cobertura 4.2 14.1 5.88 0.2 0.09 0.08 6.28 93.4
En cada época de cosecha, se evaluaron 3 plantas por tratamiento. La parte área fue cortada con tijeras a ras del suelo, secada a 70ºC por 48 h y pesada. El suelo remanente y la raíz fueron colocados en bandejas de plástico donde se separó manualmente la raíz y luego se lavó. Se contaron los nódulos activos de la raíz principal. Las raíces fueron pesadas y una sub-muestra fue secada a 70ºC durante 48 h para determinar el peso seco. Se colectó el 10% del peso fresco de las raíces para estimar la longitud total de la raíz y el porcentaje de colonización micorrícica, el que se determinó mediante el método de Giovanetti & Mosse (1980) luego que las raíces fueran blanqueadas en KOH caliente y teñidas con azul de tripano en lacto-glicerina (Phillips & Hayman, 1970). Los promedios de peso seco total, y la longitud de raíces colonizadas por micorrizas, fueron comparados como tranformaciones log; los promedios del porcentaje de colonización micorrícica como tranformaciones arcsin; los promedios de nodulación como transformaciones de raíz cuadrada x+1. Los datos transformados fueron luego analizados mediante el procedimiento ANOVA del Sistema de Análisis Estadístico (SAS) (1988). A menos que se indique lo contrario, se usó un nivel de significación de p < 0.01 para comparar los promedios debido a una variabilidad anticipada de los suelos. El efecto lineal del fósforo en la biomasa total, nodulación, porcentaje de colonización micorrícica y longitud de raíces con micorriza, se probaron usando el procedimiento GLM (SAS, 1988). Resultados y discusión La diferencia en el porcentaje de colonización micorrícica entre los tratamientos de manejo de suelo fue significativa a través del tiempo (p = 0.0027). El porcentaje de colonización micorrícica en el sistema de bajos insumos (BI) fue significativamente mayor que en el de cultivos en callejones (CC), bosque
secundario (BS) y suelo sin cobertura (SC) (p = 0.0002), respectivamente (Figura 3). La longitud total de raíces con micorrizas fue también significativamente más grande que en el trt. BI que en los otros trts. (p = 0.002)., pero la interacción de este parámetro con la época de cosecha no fue significativa (Figura 2). Los niveles más altos de colonización micorrícica encontrados en el tratamiento BI, con relación a los otros tratamientos, se deben probablemente a la presencia de especies más efectivas o compatibles de hongos MA en el suelo. En el campo, es probable que durante aproximadamente 4 años de cultivos continuos de caupí en rotación con arroz de secano, haya ocurrido una selección natural de hongos micorrícicos adecuados para el caupí (Abbott & Robson, 1991). En el tratamiento CC, en condiciones de campo, el caupí también fue incluido como un cultivo rotacional, sin embargo, es posible que haya ocurrido una asociación preferencial hongo MA / planta hospedera a favor de Inga edulis la que se sembró en líneas como parte del sistema. Esta asociación preferencial es posible que ocurra probablemente debido a la mayor permanencia del árbol en el campo. En el trt. BS, los niveles bajos de colonización encontrados a lo largo del experimento, están probablemente relacionados a la falta de un simbionte adecuado para el caupí, teniendo en cuenta que la mayoría de especies en los barbechos de bosque secundario en la región de Yurimaguas están altamente colonizados por micorrizas. Los niveles muy bajos de colonización micorrícica en el trt. SC es posible que estén relacionados a otros factores diferentes a los del mismo bosque. Estos factores se relacionan a los efectos de la pérdida de suelo por su exposición permanente, sin una cobertura vegetal, a la ocurrencia de lluvias durante aproximadamente 4 años (Carpenter et al., 2001). La pérdida de suelo fue de 13 Mg ha-1 en un período de 9 meses (Diciembre de 1988 a Agosto de 1989) (Alegre & Fernández, 1991). En los trópicos húmedos bajos, los propágulos de los hongos MA se encuentran mayormente en la capa hojarasca (Rose & Paranka, 1987; Rivera, 2003) y en el suelo superficial (20 cm) donde proliferan las raíces absorbentes (Redhead, 1977). Entonces es probable que las poblaciones de hongos se hayan reducido como consecuencia de la pérdida de suelo. Las plantas de caupí en este suelo, sin embargo, tuvieron colonización micorrícica leve, debido probablemente a la presencia de raíces colonizadas de otras plantas en áreas adyacentes o a la ocurrencia de propágulos de micorrizas a profundidades mayores por debajo del suelo superficial (Zajicek et al., 1986). El análisis de variancia de la biomasa total no mostró diferencias significativas para todos los tratamientos con el tiempo. El peso total de las plantas de caupí sin embargo, fue más alto en el trt. BI en los

MICORRIZAS ARBUSCULARES Y BACTERIAS FIJADORAS DE NITROGENO EN AMAZONIA PERUANA
Diciembre 2003 __________________________________________________________________________________________
90
otros tratamientos a los 45 DDGS (Figura 2). El crecimiento de las plantas en los tratamientos BS y SC fueron los más bajos entre los tratamientos estudiados. Se encontraron diferencias significativas en la nodulación entre los tratamientos (p = 0.0007) y también con el tiempo (p = 0.0001). No se observó nodulación en ninguno de los tratamientos No se observó nodulación en ninguno de los tratamientos a los 7 y 15 DDGS (Figura 3). A los 30 DDGS se observaron nódulos solamente en los tratamientos de cultivos en callejones y sistema de bajos insumos (1 y 4, respectivamente) y similarmente a los 45 DDGS (2 y 7, respectivamente). Se debe señalar que en general, la nodulación en estos tratamientos fue más baja de lo que se esperaba. La presencia de rizobios compatibles en estos dos tratamientos probablemente dieron cuenta por la nodulación observada. Esto es posible debido a que el caupí fue sembrado durante 4 años antes que los suelos se colectaran para el presente experimento. La nodulación estuvo ausente por completo en los tratamientos de bosque secundario y suelo sin cobertura. En el tratamiento de bosque secundario la ausencia de nódulos se debió probablemente a la falta de rizobios adecuados para el caupí en el suelo, resultando en la inhibición de la actividad de los rizobios por los altos contenidos de nitrógeno en el suelo, de acuerdo con Vitousek & Sanford (1986) y Palm & Meléndez (1991). En el caso el suelo sin cobertura, la ausencia de nódulos, se debe posiblemente a los efectos de la erosión al reducir las poblaciones de rizobios o a la baja disponibilidad de nutrientes, principalmente fósforo y calcio y a la acidez y contenidos altos de aluminio (Aziz & Habte, 1990). El efecto lineal del fósforo no fue significativo para la biomasa total de la planta, el porcentaje de colonización micorrícica y la longitud de raíces con micorrizas. Más bien fue significativo solamente para la nodulación (p = 0.0812). Esta prueba puede indicar que a niveles similares de P en el suelo, la producción más alta de biomasa en trt. BI con relación a los otros tratamientos, se debió a los efectos de los niveles más altos de colonización micorrícica. Es posible también que los altos niveles de colonización en el trt. BI influyó en la nodulación del caupí. Se debe señalar que los valores de producción de biomasa y de nodulación en las plantas de caupí en el trt. BI presente experimento fueron comparables con los obtenidos en un sistema de bajos insumos de un experimento previo en fechas de cosecha similares. Asimismo, a los 45 DDGS, no se observó floración en ninguno de los tratamientos, tal como se observó en un tratamiento con fertilización. Estas observaciones sugieren que los niveles de P en el suelo probablemente no son los más adecuados para el crecimiento del caupí.
Conclusiones El presente bioensayo mostró los efectos del manejo del suelo en una ladera en el crecimiento, colonización micorrícica y nodulación en caupí. A pesar de que ninguno de los tratamientos estudiados recibió aplicaciones de fertilizantes, los niveles de colonización micorrícica y de nodulación tuvieron grandes diferencias en los estados iniciales del crecimiento del caupí. A niveles similares de P disponible en el suelo en suelos diferentes, niveles altos de colonización micorrícica pueden ser ventajosos para el crecimiento de la planta y para la nodulación. La formación de nódulos en el bosque secundario estuvo ausente debido probablemente a los altos niveles de nitrógeno en este suelo o a la falta del rizobio adecuado para el caupí. La pérdida de suelo en las áreas en laderas por efectos de la erosión pluvial parece afectar no sólo las propiedades físicas y químicas del suelo sino también las biológicas, tales como las poblaciones de hongos micorrícicos y bacteria rizobiales. La conducción de este tipo de bioensayos puede ser útil para evaluar la ocurrencia de la colonización micorrícica y la nodulación en especies leguminosas seleccionadas para sistemas con bajos insumos en suelos ácidos y deficientes en P en los trópicos húmedos bajos. Asimismo, los resultados obtenidos, indican la necesidad de introducir las poblaciones microbianas al suelo en programas de recuperación de tierras y/o revegetación de áreas severamente alteradas tales como los taludes de las carreteras, derechos de vía de oleoductos y gasoductos, áreas expuestas por la actividad minera y otras (Rosales et al., 1997; Raja et al., 2001; Dodd et al., 2002; Cuenca et al., 2002). Agradecimientos El presente estudio constituye un capítulo de la Tesis de Grado para optar el Doctorado en Ciencias del Suelo en la Universidad Estatal de Carolina del Norte. El autor agradece al Dr. Charles B. Davey, a la Misión Carolina del Norte y al personal profesional de la Estación Experimental de Yurimaguas, Loreto. Literatura citada Abbott L.K. & Robson A.D. 1991. Factors influencing
the occurrence of vesicular-arbuscular mycorrhizas. Agric. Ecosyst. and Environ. 35: 121-150.
Alegre J.C. & Fernandes E.C.M. 1991. Runoff and erosion losses under forest, low-input and alley-cropping in slopes. pp. 227-228. In: T. McBride (Ed.). TropSoils Technical Report 1988-1989. North Carolina State University. Raleigh, North Carolina.
Amaranthus M.P. & Trappe J.M. 1993. Effects of erosion on ecto- and VA-mycorrhizal inoculum potential of soil following forest fire in southwest Oregon. Pl. Soil. 150: 41-49.
Aziz T. & Habte M. 1990. Stimulation of mycorrhizal activity in Vigna unguiculata through low level

PEDRO O. RUIZ Y CHARLES B. DAVEY
Ecología Aplicada Vol. 2 Nº 1, pp. 87-92 __________________________________________________________________________________________
91
fertilization of an Oxisol subjected to imposed erosion. Commun. in Soil Sci. Planta Anal. 21: 493-505.
Benites J.R. 1983. Alternativas para los terrenos abandonados del cultivo de la coca en el Alto Huallaga. CIPA XVI-Estación Experimental de Yurimaguas. Programa de Suelos Tropicales. Serie de Separatas No 4. Yurimaguas, Perú.
Carpenter F.L., Mayorga S.P., Quintero E.G. & Schroeder M. 2001. Land use and erosion of a Costa Rican Ultisol affect soil chemistry, mycorrhizal fungi and early regeneration. Forest Ecology and Management. 144(1-3): 1-17.
Cuenca G., De Andrade Z., Lovera M., Meneses E., Márquez M. & Machuca R. 2002. The use of mycorrhizal native shrubs in the rehabilitation of degraded areas from La Gran Sabana State. Venezuela. Interciencia. 27(4): 165.
Dodd J.C., Dougall T.A., Clapp J.C. & Jeffries P. 2002. The role of arbuscular mycorrhizal fungi in plant community establishment at Samphire Hoe, Kent, UK - the reclamation plattform created during the building of the Channel tunnel between France and UK. Biodiversity and Conservation. 11(1): 39.58.
Dourojeanni M.J. 1990. Amazonía: Qué hacer? Centro de Estudios Teológicos de la Amazonía (CETA). Iquitos, Perú.
Fernández E.C.M., Styger E. & Rakotondramasy H. 2001. Mycorrhiza fungal diversity as impacted by the loss of endemic, rainforest plants in Madagascar. ICOM3. Adelaide, Australia.
Franco A.A. & DeFaria S.M. 1997. The contribution of N-2 fixing tree legumes to land reclamation and sustainability in the tropics. Soil Biol. Biochem. 29(5-6): 897-903.
Giovanetti M. & Mosse B. 1980. An evaluation of techniques for measuring vesicular-arbuscular mycorrhizal infection on roots. New Phytol. 84: 489-500.
Habte M. 1989. Impact of simulated erosion on the abundance and activity of indigenous vesicular-arbuscular mcorrhizal endophytes in an Oxisol. Biol. Fert. Soils. 7: 164-167.
Lal R. 1988. Soil erosion research on steep lands. In: pp. 45-53. W.C. Moldenhauer and N.W. Hudson (Eds.). Conservation Farming on Steep Lands. SWCS, Ankeny, IA.
Palm C.A. 1988. Mulch quality and nitrogen dynamics in an alley cropping system in the Peruvian Amazon. Ph. D. Dissertation. Soil Science Dept. North Carolina State University. Raleigh, NC. U.S.A.
Palm C.A. & Meléndez G. 1991. Sustainable agriculture: soil carbon and nitrogen dynamics. In: T. McBride (Ed.). TropSoils Annual Report 1988-1989. North Carolina State University. Raleigh, North Carolina. : 205-206.
Phillips J.M. & Hayman D.S. 1970. Improved procedures for clearing roots and staining parasitic
and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. Trans. Brit. Mycol. Soc. 55:158-161.
Raja P., Herdhawati J., Nazam C.A. & Zubaidah I. 2001. Rehabilitation of Malasyan highway landscape plants using commercial arbuscular mycorrhizal fungi. ICOM3. Adelaide, Australia.
Redhead J.F. 1977. Endotrophic mycorrhizas in Nigeria: species of the Endogonaceae and their distribution. Trans. Brit. Mycol. Soc. 69: 275-280.
Rivera E.L. 2003. Presencia de estructuras características de hongos micorrícicos arbusculares en hojas en procesos de descomposición ¿Se puede considerar un hecho común en la naturaleza?. Unidad de Biotecnología Vegetal. Universidad Javeriana. Bogotá, Colombia.
Rivera E.L., Campos R. & De Martínez A.L. 2000. Mycotrophic status of wax laurel useful for the recuperation of eroded soils in Tomine Reservoir in Colombia. Suelos Ecuatoriales. 30 (2): 188-193.
Rosales J.G., Cuenca G., Ramírez N. & Andrade Z. 1997. Native colonizing species and degrades land restoration in La Gran Sabana, Venezuela. Restoration Ecology. 5(2): 147.155.
Rose S.L. & Paranka J.E. 1987. The location of roots and mycorrhizae in tropical forest litter. In: D.M. Sylvia, L.L. Hung, and H.J. Graham (Eds.). Mycorrhizas in the Next Decade. Proc. 7th North American Conference on Mycorrhizae (NACOM). Gainesville, Florida.
Sanchez P.A. 1976. Properties and management of soils in the tropics. Wiley, New York.
Sanchez P.A., Palm C.A. & Smyth T.J. 1990. Approaches to mitigate tropical deforestation by sustainable soil management practices. Developments in Soil Science. 20: 211-220.
Statistical Analysis System. 1988. SAS/STAT User's Guide. SAS Institute, Cary, NC, USA.
Stocking M.A. 1988. Socio-economics of soil conservation in developing countries. J. Soil Water Conserv. 43: 381-385.
Szott L.T., Palm C.A. & Sanchez P.A. 1991a. Agroforestry in acid soils in the humid tropics. Adv. Agron. 45: 275-301.
Szott L.T., Fernandes E.C.M. & Sanchez P.A. 1991b. Soil-plant interactions in agroforestry systems. For. Ecol. Mgmt. 45: 127-152.
Vitousek P.M. & Sanford R.L. 1986. Nutrient cycling in moist tropical forest. Ann. Rev. Ecol. 17: 136-167.
Zajicek J.M., Daniels Hetrick B.A. & Owensby C.E. 1986. The influence of soil depth on mycorrhizae colonization of forbs in the tallgrass prairie. Mycologia. 76: 316-320.
Wu T.H., Hao W.Y., Lin X.G. & Shi Y.Q. 2002. Screening of arbuscular mycorrhizal fungi for the revegetation of eroded red soils in subtropical China. Plant and Soil. 239 (2): 25-235.

MICORRIZAS ARBUSCULARES Y BACTERIAS FIJADORAS DE NITROGENO EN AMAZONIA PERUANA
Diciembre 2003 __________________________________________________________________________________________
92
.Anexos
SE1 = Error estándar para una diferencia de promedios entre dos fechas para un tratamiento (raíz cuadrada 2MSEb / 3)
SE2 = Error estándar para comparar dos de estas diferencias entre tratamientos (raíz cuadrada 4MSEb / 3)
Figura 1. SE1 = 0.18 SE2 = 0.25 Figura 2. SE1 = 0.12 SE2 = 0.17 Figura 3. SE1 = 4.51 SE2= 6.38 Figura 4. SE1 = 0.08 SE2= 0.11
Figura 1. Peso seco total de plantas de caupí. Días después de la siembra (DDGS)
0
200
400
600
800
1000
1200
CC BI BS SC
mg
/ pla
nta
7 DDGS 15 DDGS 30 DDGS 45 DDGS
Figura 2. Longitud total de raíces de caupí colonizadas por hongos micorrícicos. Días
después de la siembra (DDGS)
0
200
400
600
800
1000
CC BI BS SCcm
/ pl
anta
5 DDGS 15 DDGS 30 DDGS 45 DDGS
Figura 3. Porcentaje de colonización micorrícica en plantas de caupí. Días
después de la germinación de la semilla (DDGS)
0
20
40
60
80
100
CC BI BS SC
Porc
enta
je d
e co
loni
zaci
ón
7 DDGS 15 DDGS 30 DDGS 45 DDGS
Figura 4. Nodulación en plantas de caupí. Días después de la germInación de la
semilla (DDGS)
012345678
CC BI BS SC
Núm
ero
de n
ódul
os /
plan
ta
5 DDGS 15 DDGS 30 DDGS 45 DDGS

Ecología Aplicada, 2(1), 2003 ISSN 1726-2216 Depósito legal 2002-5474
EFECTO DEL FÓSFORO SOBRE EL ESTABLECIMIENTO DEL Centrosema macrocarpum Benth DENTRO DE UNA PLANTACIÓN DE PIJUAYO (Bactris gasipaes H.B.K.) EN UN ULTISOL DEL
TRÓPICO HÚMEDO. Luis A. Arévalo1, Julio C. Alegre1 y Raúl Fasabi2.
Resumen
En una plantación de Pijuayo (Bactris gasipaes H.B.K.) de cuatro años de edad se sembró una leguminosa forrajera centrosema (Centrosema macrocarpum Benth.) como cobertura del suelo, con niveles crecientes de Fósforo (P) con el fin de encontrar la dosis adecuada que permita lograr un rápido establecimiento de la cobertura, para su posterior utilización como un sistema silvopastoril. Cuatro niveles de P fueron probados: 0, 20, 40 y 80 Kg/ha usando roca fosfórica como fuente de P. Estas dosis se aplicaron al voleo, antes de la siembra de la cobertura, al inicio del experimento. La respuesta a las dosis de P se midió con el diseño de Bloques Completamente Randomizados (BCR). Los niveles iniciales de P en el suelo fueron bajos, incrementándose significativamente en función a las dosis crecientes de P. Similar tendencia se observó en la concentración de P tanto en los tejidos del centrosema como en los tejidos de las plantas de pijuayo. Hubo una buena acumulación de P en la biomasa de centrosema y reciclaje en el suelo. El establecimiento y producción de biomasa del centrosema, se incrementó a los 150 días de sembrado para todos los tratamientos pero el desarrollo del centrosema que recibió 20 Kg de P/ha cubrió el suelo más rápido y presentó la mayor producción de biomasa sobre la base de materia seca que los demás tratamientos. Como no hubo diferencias entre las dosis de P adicionadas al suelo, se recomienda la aplicación de 20 Kg de P/ha para la fase de establecimiento del centrosema. Palabras Claves: Silvopasturas, pijuayo, centrosema, roca fosfórica, suelos ácidos, voleo.
Abstract A leguminous forage centrosema (Centrosema macrocarpum) was planted as a cover crop in a 4 year peach palm plantation (Bactris gasipaes K.) at different increasing rates of phosphorous (P) in order to determine the adequate doses needed to get rapid cover establishment for future use as a silvopastoral system. Four rates of P as rock phosphate were tested: 0, 20 40 and 80 kg/ha. This fertilizer was uniformly sprayed before planting the cover crop at the beginning of the experiment. A randomized complete block design was used to find the response to the different fertilizer rates. The initial levels of soil P were low and increased significantly as a function of the increasing levels of applied P. Similar trends were observed in the tissue of centrosema and peach palm. There was a good P accumulation in the centrosema biomass and nutrient cycling of the soil. The establishment and production of centrosema biomass increased after 150 days of planting in all treatments but the development of centrosema with the rates of 20 kg/ha covered the soil faster and presented greater biomass production than the others. Because there were non significant differences between the establishment rates of P, it is recommended to apply only 20 kg/ha during centrosema establishment. Key words: Silvopastoral, peach palm, centrosema, rock phosphate, acid soils.
1 World Agroforestry Centre. Carretera Federico Basadre Km 4.200 Pucallpa, Correo electrónicol: [email protected] 1 World Agroforestry Centre. Avda. La Molina No. 1895 Lima, Perú. Correo electrónico: [email protected] 2 Universidad Nacional de la Amazonía Peruana, Yurimaguas, Loreto, Perú.
Introducción El Fósforo (P) es uno de los principales elementos nutritivos para las plantas y animales que restringe en mayor grado la producción agropecuaria en la región de la Amazonía Peruana (Toledo & Morales, 1979). Este problema puede corregirse mediante la aplicación de abonos químicos fosfatados de alta solubilidad como el super fosfato simple (SFS) o el super fosfato
triple (SFT) o con la aplicación de fuentes menos solubles (Fenster & Leon, 1979), pero al mismo tiempo más económicas como es el caso de las rocas fosfóricas (Alegre & Chumbimune, 1992). Solucionado la deficiencia de P en el suelo, la producción de biomasa tanto de las gramíneas como de las leguminosas se incrementará y por ende la productividad del sistema.

EFECTO DEL FÓSFORO SOBRE Centrosema macrocarpum Benth DENTRO DE UNA PLANTACIÓN DE PIJUAYO (Bactris gasipaes H.B.K.)
Diciembre 2003 __________________________________________________________________________________________
94
El uso directo de la roca fosfórica como fertilizante ha sido una práctica ampliamente probada bajo diferentes condiciones de suelos y cultivos. Los diferentes trabajos realizados, tanto en la amazonía peruana como en otros países latinoamericanos (Ensminger et al., 1967; Alegre & Chumbimune, 1992; Gighuru & Sánchez, 1988; Toledo & Morales, 1979; Fenster & Leon, 1979; Vela, 1991) demuestran las excelentes propiedades de la roca fosfórica; cuya utilización en la fertilización de los suelos pobres en P ofrece magnificas perspectivas. Una alternativa viable para incrementar la producción de carne y leche en los trópicos húmedos parecen ser los sistemas agrosilvopastoriles donde se asocian cultivos de corto ciclo, árboles, pasturas y animales (Nair, 1993). La incorporación de los árboles dentro de las pasturas o viceversa incrementará la productividad de la tierra (Arévalo et al., 1998) por los productos adicionales que proveerá, tales como frutas, madera, carbón, leña, en adición al forraje y ganado. En la Región Amazónica existen muchas plantaciones de pijuayo que son manejados por el agricultor sin una cobertura o pastura adecuada, pero con exceso de carga animal. Esto determina que el suelo se degrade, causando la muerte de las raíces por el pisoteo de los animales, así como la reducción significativa en la producción de frutos. En investigaciones preliminares se encontró que la asociación de pijuayo con el centrosema (Centrosema macrocarpum Benth) y manejo de los animales con rotación de potreros ofrecía una buena alternativa en los trópicos húmedos del Perú (Arévalo et al., 1998). Sin embargo la productividad del sistema estuvo afectada probablemente por el déficit de nutrientes en el suelo, sobre todo del P. Una de las ventajas que presentan las leguminosas como centrosoma, es su establecimiento como cobertura en áreas degradadas (Sánchez, 1982). Una de las formas de acelerar su establecimiento y mejorar su calidad nutritiva para su uso como forraje es mediante la aplicación de P al suelo. El objetivo del ensayo fue determinar la dosis adecuada de P para el establecimiento del Centrosema macrocarpum Benth dentro de una plantación de pijuayo, para su posterior utilización al pastoreo. Materiales y métodos Descripción del sitio El ensayo se inició en Mayo de 1996 en el Centro de Investigación Yurimaguas (IIAP), ubicado a 5o 56’ Latitud Sur, 76o 5’ Longitud Oeste, a una elevación de 184 m.s.n.m. El clima es húmedo tropical, con una temperatura promedio de 26oC. con precipitación promedio anual de 2200 mm. Sin embargo entre los meses de Junio a Setiembre, la precipitación está por debajo de los 100 mm mensuales.
El experimento se desarrolló sobre un suelo franco arenoso, ácido e infértil, clasificado como Ultisols (Tyler et al., 1978) sobre el cual crecía una plantación de pijuayo de 4 años de edad infestada de gramíneas y leguminosas. Estas fueron identificadas como rabo de zorro (Andropogon bicornis L.), mezcla de braquiarias como: Brachiaria humidicola y Brachiaria decumbens Stapf., Kudzu (Pueraria phaseoloides (Roxb.) Benth), Stylosantes guianensis (Aub.) Sev. y otras especies no determinadas. La densidad de siembra de las plantas de pijuayo fue de 400 por hectárea, es decir a un distanciamiento de 5m x 5m. Establecimiento de los tratamientos Para la aplicación de las dosis crecientes de P y la posterior siembra del centrosoma (C. macrocarpum) tanto las gramíneas como las leguminosas existentes en el campo fueron cortadas y eliminadas en forma manual. Después se realizó el muestreo inicial del suelo a una profundidad de 0 a 15 cm, con un muestreador tipo barreno de 2.5 cm de diámetro interior. Después de ello se adicionó el P como Roca Fosfórica (RF) y las dosis aplicadas fueron 0, 20, 40 y 80 Kg de P/ha. Estas se aplicaron manualmente en forma uniforme al voleo e incorporándose con azadón hasta una profundidad de 5 cm. En la parcela control se removió el suelo de manera similar a los otros tratamientos, pero sin la adición de P. Después se sembró el centrosema en forma tradicional (manual) y con semilla botánica germinada a un distanciamiento de 0.50m x 0.50m utilizándose en total 2 Kg de semilla (Ara & Schaus, 1980). Durante el ensayo se dio una infestación severa de hormigas cortadoras de hojas curuhuinsi (Atta sp.) siendo necesario realizar dos resiembras del centrosema, y para controlar el ataque de estas hormigas se aplicó Fenkil al 3% con un insuflador (aplicador con presión de aire) en forma localizada. El área de cada una de las parcelas fue de 225 m2 con 9 plantas de pijuayo por parcela. Para medir la respuesta a las dosis crecientes de P se utilizó el Diseño de Bloques Completamente Randomizados (BCR), con cuatro tratamientos y tres repeticiones, realizándose la prueba de DLS al 5 % de probabilidades para comparar la diferencia entre los promedios. Evaluaciones a) Suelo. Durante el tiempo de ejecución del ensayo, se efectuaron 3 muestreos de suelos a una sola profundidad de 0-15 cm. El primer muestreo se realizó antes de la aplicación de las dosis de P; el segundo y tercer muestreo se realizaron a los 60 y 150 días después de la siembra del centrosema. Las muestras de suelo fueron secadas al aire y molidos en un molino eléctrico, envasadas, etiquetadas y entregadas al laboratorio. El P disponible fue extraído con la

LUIS A. ARÉVALO, JULIO C. ALEGRE Y RAÚL FASABI
Ecología Aplicada Vol. 2 Nº 1, pp. 93-97 __________________________________________________________________________________________
95
solución de Olsen modificado (Hunter, 1974) y determinado por colorimetría. b) Porcentaje de Cobertura y forraje disponible. El porcentaje de cobertura se midió con un cuadrante de 1 m2 dividido en 100 partes iguales de 10 cm2 cada uno de ellos (Pérez et al., 1993) ). La primera evaluación se realizó 30 días después de la siembra, y las evaluaciones posteriores se realizaron con un intervalo de 15 días hasta cuando se alcanzó el 100% de cobertura. Tanto el muestreo de forraje disponible como el de tejidos del centrosema se realizaron en forma simultánea a 30, 120 y 150 días después de la siembra. Para la evaluación del forraje disponible (materia verde fresca) se utilizó un marco de madera de 0.25 m2; evaluándose en cuatro puntos al azar dentro del área de cada parcela, por cada tratamiento y repetición. El peso fresco de esta biomasa, se determinó en una balanza analítica y luego se colocaron las muestras en una estufa de aire caliente por 78 horas a una temperatura de 70oC, después se determinó el peso seco, las muestras fueron molidas envasadas, etiquetadas y enviadas al laboratorio para su análisis. Las muestras de tejidos fueron analizados por vía húmeda con H2SO4 y H2O2 en una plancha de calentamiento y luego analizados por absorción atómica (Hunter, 1979). Las muestras de hojas de pijuayo fueron colectadas en dos épocas durante el ensayo, la primera de ellas antes de la aplicación del P y la segunda cinco meses después de su aplicación. Para ellos se utilizó la metodología descrita por Grau (1986); es decir se tomo la hoja número ocho (8) del total de hojas de la planta y de esa hoja se colectaron los foliolos del tercio medio a ambos lados del raquis. Para su análisis en el laboratorio se siguió el mismo procedimiento descrito para el caso del análisis de los tejidos del centrosema. Resultados y discusión Los cambios en la concentración del P disponible del suelo por efecto de la aplicación de dosis crecientes de P para el establecimiento de centrosema (C. macrocarpum) dentro de una plantación de pijuayo (B. gasipaes) se presentan en la Tabla 1. Los niveles iniciales de P en el suelo fueron bajos; pero a medida que las dosis adicionadas al suelo fueron mayores, su concentración se incrementó significativamente comparado con el control. El incremento de la concentración de P fue muy alto (hasta 374%) comparado con el control. La alta acidez del suelo (1.21 cmol(+)/L) con 85% de saturación de aluminio ayudó a la rápida solubilidad de la roca fosfórica (Alegre et al., 1994) y por lo tanto aumentó los niveles de P disponible hasta 18.7 mg/Kg después de cinco de su aplicación con la dosis más alta. Los requerimientos de P por la cobertura de centrosema, son más bajos que para los cultivos
anuales; el centrosema responde a bajos niveles en el suelo ya que tiene otros medios de absorción o exploración de un mayor volumen del suelo mediante la asociación con hongos tipo micorrizas (Ruíz, 1994; Ydrogo, 1995). Esta podría ser la causa del incremento de la concentración del P tanto para los tejidos del centrosema como para los tejidos de pijuayo, en el tratamiento control (Tabla 2) comparado con los otros tratamientos. Hubo un mayor incremento en la concentración de P en los tejidos del centrosema para todos los tratamientos que recibieron P.
Tabla 1. Cambios en la concentración del fósforo disponible del suelo, por efecto de la aplicación de dosis crecientes de P. Yurimaguas, Loreto.
Dosis de P Concentración de P en el suelo
AP* 5 MDA** Kg/Ha mg/Kg 0 5.4 5.0 20 5.6 14.0 40 5.3 13.7 80 6.2 18.7 DLS.05 0.71 2.71 * Antes de Aplicar la Roca Fosfórica (RF) ** 5 Meses después de la aplicación de la RF
Tabla 2. Cambios en la concentración de P en los tejidos del centrosema y pijuayo en función al tiempo y dosis de P aplicados al suelo. Yurimaguas, Perú.
Concentración de P Centrosema Pijuayo
Dosis de P
1 MDA* 5 MDA AP** 5 MDAKg/Ha G/Kg 0 0.47 1.02 1.01 1.22 20 0.53 1.72 1.06 2.06 40 0.63 1.96 1.05 1.95 80 0.69 2.16 1.05 2.00 DLS .05 0.52 0.9 0.04 0.1 * Un mes después de la aplicación de roca fosfórica ** Antes de la aplicación de roca fosfórica
La concentración de P en los tejidos del pijuayo siguió la misma tendencia que para el caso del centrosema después de la aplicación de las diferentes dosis crecientes de P al suelo (Tabla 2). Antes de la aplicación de las dosis de P al suelo las concentraciones en los tejidos de pijuayo fue bajo y cuando se hizo las comparaciones iniciales, estas fueron significativamente diferentes que podría deberse a la variabilidad genética innata que presenta esta palmera. Cinco meses después de aplicada la roca

EFECTO DEL FÓSFORO SOBRE Centrosema macrocarpum Benth DENTRO DE UNA PLANTACIÓN DE PIJUAYO (Bactris gasipaes H.B.K.)
Diciembre 2003 __________________________________________________________________________________________
96
fosfórica en sus diferentes niveles hubo un fuerte incremento en función a las dosis crecientes de P, dándose la mayor concentración con la primera dosis (20 kg P/ha) y que también se debería a esta variabilidad y a la mayor eficiencia de absorción de las palmeras evaluadas dentro de esta parcela. b. Porcentaje de cobertura y forraje disponible El porcentaje de cobertura del centrosema fue menor a 20% en los primeros 60 días sin importar la dosis de P adicionada al suelo (Tabla 3). A 90 días hubo un alto incremento en el porcentaje de cobertura alcanzando valores entre 55 a 62% con diferencias estadísticamente significativas entre los tratamientos. Ciento veinte (120) días después de la siembra del centrosema, el tratamiento que recibió 20 Kg de P alcanzó el 100% de cobertura; mientras que los demás tratamientos alcanzaron entre 91 a 98% de cobertura. Tanto el tratamiento control como el tratamiento con mayor dosis de P (80 kg/ha) presentaron los valores más bajos en cobertura. Estas diferencias fueron estadísticamente significativas. Ciento cincuenta días después del establecimiento del centrosema, el porcentaje de cobertura fue del 100% sin importar la dosis de P adicionada al suelo. Tabla 3. Porcentaje de cobertura y producción demateria seca del Centrosema macrocarpum por efecto de la aplicación de dosis crecientes de P.Yurimaguas, Loreto.
Porcentaje de Cobertura (dds*) Materia Seca (dds) Dosis de P 60 90 120 150 120 150
Kg/Ha % T/ha 0 13.9 59.2 91.0 100 1.57 2.90 20 18.3 68.4 100.0 100 2.00 3.95 40 14.8 61.2 98.0 100 1.76 3.57 80 13.0 56.8 93.0 100 1.62 3.90
DLS .05 4.81 11 3.72 1 1.85 2.27
* dds = días después de la siembra Por otro lado, el porcentaje de cobertura del centrosema a través del tiempo parece estar ligado a la fisiología misma de la planta, debido a la tendencia de crecimiento que sigue, similar a los tratamientos que recibieron P. La producción de materia seca fue mayor en todos los tratamientos que recibieron P (Tabla 3) comparado con el testigo, tanto a 120 como a 150 días después de la siembra. Esto sugiere que la cantidad de materia seca (hojas y tallos), estuvo en función a los niveles de P aplicado al suelo; siendo las diferencias más saltantes a los 150 días. Los tratamientos que recibieron 20 y 80 Kg de P/ha tuvieron una producción mayor en 1.05 y 1.0 T/ha que el tratamiento control (Tabla 3). Por otro lado, hubo un incremento en la
producción de materia seca, por efecto fisiológico en el crecimiento del centrosema, entre 120 a 150 días después de sembrado. Este incremento fue de 184, 198, 203 y 241% para las dosis de 0, 20, 40 y 80 kg de P/ha, confirmándose el efecto que tiene el P sobre el crecimiento del centrosema. Conclusiones El presente trabajo de investigación sólo comprendió la fase del establecimiento de la cobertura, la cual nos llevó a las siguientes conclusiones: 1. La adición de P como roca fosfórica, incrementó su concentración tanto en el suelo como en los tejidos del Centrosema macrocarpum, así como también en los tejidos de las plantas de pijuayo. 2. Tanto el porcentaje de cobertura, como la producción de materia seca por el Centrosema macrocarpum se incrementaron por la adición de P, como roca fosfórica. 3. La falta de respuesta entre las dosis de P aplicadas al suelo, sugieren que sólo es necesario la adición de 20 kg de P/ha para el establecimiento y posterior crecimiento del Centrosema macrocarpum Benth; sin embargo al mismo tiempo mejora la concentración de P tanto en el suelo como en los tejidos de pijuayo, sugiriendo una mejor y mayor sostenibilidad del sistema a través del tiempo. Literatura citada Alegre J.C. & Chumbimune R. 1992. Investigaciones
y usos de la roca fosfórica en el Perú. In: Memorias de la II Reunión de la Red Latinoamericana de Roca Fosfórica, San Cristóbal, Edo. de Tachira, Venezuela.
Alegre J.C., Gighuru M. & Sánchez P. 1994. Fertilización Fosfórica en Sistemas con Bajos Insumos en los trópicos húmedos del Perú. Terra. 12 1.
Ara M. & Schaus R. 1980. Establecimiento y producción de gramíneas y leguminosas forrajeras en Yurimaguas. En II Reunión de la Red Internacional de Evaluación de pastos tropicales. Cali, COLOMBIA. : 433-443.
Arévalo L. 1992. Cobertura de leguminosas en plantación de pijuayo para la producción de frutos, Trop Soils Technical Report, 1988 - 1989. North Carolina State University Raleigh, N.C. : 184-196.
Arévalo L.A., Alegre J.C., Bandy D.E. & Szott L.T. 1998. The effect of cattle grazing on soil physical and chemical properties in a silvopastoral system in the Peruvian amazon. Agroforestry System. 40: 109-124.
Ensminger L.E., Pearson R.W. & Arminger W.H. 1967. Effectiveness of Rock phosphate as a Source of phosphorus for plants”. USD Agricultural Research Service, Bulletin.: 41-125.
Fenster W.E. & Leon L.A. 1979. Manejo de la fertilización con fósforo para el establecimiento de

LUIS A. ARÉVALO, JULIO C. ALEGRE Y RAÚL FASABI
Ecología Aplicada Vol. 2 Nº 1, pp. 93-97 __________________________________________________________________________________________
97
pastos mejorados en suelos ácidos e infértiles de América Tropical. In Producción de Pastos en suelos Acidos de los Trópicos. Tergas,L.E.y Sanchez, P. A.(eds) (CIAT, Serie 03SG-5). : 119-133.
Gighuru M.P. & Sánchez P.A. 1988. Phosphate rock fertilization in tilled and no-till low input system in the humid tropics. Agron. J. 80: 943-947.
Hunter A.H. 1974. International soil fertility evaluation and improvement laboratory procedures. Dep. Of Soil Science, North Carolina State Univ., Raleigh.
Hunter A.H. 1979. Suggested soil plant analytical techniques for tropical soils research program laboratories. Agro Services International, Orange City, Fl.
Grau M.G. 1986. Determinación de la hoja más indicativa para el análisis foliar del peach palm (Bactris gasipaes H.B.K.). Tesis Ing. Agrónomo. Universidad Nacional Agraria la Molina, Lima, Perú.
Nair P.K.R. 1993. An Introduction to Agroforestry. Kluwer Academic Publisher.
Perez J.M., Szott L. & Arévalo L.A. 1993. Pijuayo con coberturas de leguminosas. In: Mora Urpi, J., L.T.Szott, Murillo, M. y V.M. Patiño (eds). IV Congreso Internacional sobre Biología, Agronomía e Industrialización del pijuayo. 1991 Iquitos, Perú. Editorial de la Universidad de Costa Rica, San José, Costa Rica. : 209-224.
Ruiz P.O. 1994. Effects of Soil Management on
Vesicular-arbuscular Mycorrhizal Fungi and on Soil Phosphorus Fractions in Ultisols of the Peruvian Amazon. Ph.D. degree dissertation thesis. Soil Science Department. North Carolina State University. Raleigh, N.C., U.S.A.
Sánchez P.A. 1982. Estrategia de producción de pasturas a base de leguminosas en América Tropical. Primer Congreso Nacional de la Ciencia del suelo. Lima - Perú. : 1-37.
Toledo J. & Morales F. 1979. Fertilización de gramíneas y leguminosas forrajeras tropicales con Roca Fosfórica. Producción de Pastos en Suelos Acidos de los Trópicos. CIAT, Cali, Colombia. : 191-210.
Tyler E.J., Buol S.W. & Sánchez P.A. 1978. Genetic association of soil properties encountered in a detailed soil survey in the upper Amazon Basin of Peru. Soil Sci.Soc.Am.J. 42: 771-776.
Vela J. 1991. Establecimiento de tres asociaciones de gramíneas y leguminosas. Investigación sobre pastos tropicales en la Amazonía INIAA - Pucallpa - Perú 1991. Informe Técnico # STO3.
Ydrogo H.F. 1995. Inoculación de lombrices de tierra Pontoscolex corethrurus y presencia de micorrizas vesículo arbusculares en plántulas de arazá (Eugenia stipitata) achiote (Bixa orellana) y pijuayo (Bactris gasipaes) y sus efectos en el crecimiento. Tesis Ing. Agrónomo. Univ. Nacional de San Martín, Tarapoto.

Ecología Aplicada, 2(1), 2003 ISSN 1726-2216 Depósito legal 2002-5474
EFECTOS DEL PARAQUAT SOBRE EL CRECIMIENTO Y LA MORFOLOGÍA DE LA MICROALGA Dunaliella tertiolecta.
Abelardo Calderón Rodríguez1 y Román Felipe Serpa Ibáñez2
Resumen Se estudió el efecto de diferentes dosis del herbicida metilviológeno paraquat, sobre el crecimiento y la morfología de la microalga Dunaliella tertiolecta bajo condiciones no controladas. Se encontró que la mejor forma de evaluar el crecimiento en este caso es la densidad celular. A las dosis menores de 0.008 ppm. se observó una tendencia creciente en la densidad celular, a dosis mayores hasta 0.04 ppm. la tendencia es decreciente. La forma y el movimiento se ven profundamente alterados a concentraciones altas, lo cual los lleva a la muerte aguda (no programada). En concentraciones bajas parece funcionar bien el sistema detoxificante SOD y carotenos, llegándose a determinar una dosis letal media de 0.006 ppm. Palabras Clave: Dunaliella, paraquat, detoxificación.
Abstract
Different doses of the metilviologen herbicide paraquat were applied on the microalgae Dunaliella tertiolecta under non controlled conditions to determine the effects on the growth and the morphology. We found out that, the best way to evaluate the growth in this case is the cellular density. Doses lower than 0.008 ppm had an increasing tendency in cellular density while greater doses up to 0.04 ppm had a declining tendency. The morphology and the movement of the microalgae are profusely disturbed at high concentrations. Finally they die abruptly (non programmed death). At low concentrations, the detoxificant SOD and carotene system seem to function well. A lethal dose of 0.006 ppm was found for this case. Key words: Dunaliella, paraquat, detoxification.
1 Biólogo, M.Sc. Profesor Principal de Fisiología Vegetal. Departamento de Biología. Universidad Nacional Agraria La Molina, Lima – Perú. Apartado Postal 456 Lima 100 Perú. [email protected] 2 Biólogo. Asistente de Investigación del Laboratorio de Fisiología Vegetal. Universidad Nacional Agraria La Molina, Lima – Perú.
Introducción El paraquat (Dicloruro de 1,1-Dimetil-4,4bipiridilo) es un reactivo que promueve la generación de radicales libres (FR) tales como el anión súperoxido O-
2 y el altamente reactivo hidroxilo OH. que reacciona casi con cualquier clase de molécula orgánica. Estos radicales, conjuntamente con el peroxido de hidrógeno (H2O2) son formas incompletamente reducidas del O2 y se conocen como especies de oxígeno reactivo (ROS) y tienen una alta reactividad aun bajo condiciones moderadas que facilitan la vida. Ellos oxidan a una gran variedad de biomoléculas. El ROS inactiva enzimas, oxida a los ácidos grasos insaturados de las membranas, y causa mella en los ácidos nucleicos con el riesgo de provocar mutaciones. A altas concentraciones, el ROS inicia la degradación de las proteínas, altera fuertemente la permeabilidad de las membranas celulares e induce la muerte celular programada o aguda. Estos efectos califican al paraquat como un herbicida de alta toxicidad (Dietz et al.,1998). En las plantas, el paraquat induce alteraciones en la ruta del transporte electrónico a nivel de reductor de la ferredoxina, aumentando el estrés oxidativo (Halliwel et al., 1987; Ascon – Bieto & Talon, 1993).
Se ha reportado que el mecanismo de tolerancia implica elevados niveles de enzimas detoxificantes. Las enzimas antioxidantes mas prominentes son las ROS detoxificantes como la súperoxido dismutasa (SOD), Cu /Zn-SOD, Mn-SOD, Fe-SOD, catalasas (CAT) y un gran numero de peroxidasas reductoras del H2O2. Otros antioxidantes no enzimáticos, como el glutation, el ascorbato, los carotenoídes, los flavonoides, el tocoferol , los ácidos ureíco y lipoíco ,varios aminoácidos ,poliaminas, y una variedad de compuestos fenólicos también cumplen un rol importante en la red de antioxidantes que protegen a las células ( Shannon et al., 1966; Larson, 1988). Existen sin embargo estudios antagónicos (Kao & Hassan, 1985; Bray et al., 1986) en los que haciendo uso de biotipos resistentes al paraquat no se ha encontrado incrementos en los niveles de enzimas detoxificantes. Por el contrario, se ha propuesto que la resistencia en estos biotipos se basa en un mecanismo de captura que evita que el paraquat alcance su sitio de acción en el cloroplasto o que se reduzca la translocación. El mecanismo de captura se da mas bien en organismos multicelulares; la tolerancia a la acción del paraquat a nivel de organismos unicelulares como E. coli y Chlorella sería mas bien atribuible a una disminución en la absorción del producto por

EFECTOS DEL PARAQUAT SOBRE LA MICROALGA Dunaliella tertiolecta
Diciembre 2003 __________________________________________________________________________________________
100
parte de las células antes que a niveles endógenos de enzimas detoxificantes. Esta sería una clase de resistencia al estrés iónico llamada evitación (Levitt, 1980). El resultado del tratamiento con paraquat es un progresivo deterioro de la estructura celular (células hinchadas, citoplasma disgregado, alteración de la permeabilidad, las organelas no se hacen notorias y finalmente la muerte). Estudios ultra estructurales en algas unicelulares tales como la Chlorela y Clamydomonas (Bray et al., 1986) se han llevado a cabo en condiciones de laboratorio, pero no existen trabajos bajo condiciones no controladas, por tal motivo, el objetivo del presente trabajo, ha sido evaluar bajo condiciones no controladas de temperatura e irradiancia, el efecto del paraquat sobre la fisiología (crecimiento y motilidad) y ciertas características morfológicas del alga unicelular Dunaliella tertiolecta. Materiales y Métodos El experimento se realizó en un ambiente externo del Laboratorio de Fisiología Vegetal de la Universidad Nacional Agraria La Molina, durante fines de otoño y principios de primavera. La temperatura diurna varió entre los 20.8 oC y 22.9 oC. El cielo se mostró totalmente cubierto durante los días que duró el experimento. Se inoculó una alícuota de 5 ml. de una cepa de Dunaliella tertiolecta procedente del Instituto del Mar del Perú (IMARPE). a un litro de medio de cultivo agua de mar, envejecida , filtrada y enriquecida con las siguientes sales: NO3 Na (75mg), NaH2PO4 H2O (5mg), Na2EDTA (4.36mg), Cl3 Fe 6H2O (3.15mg), SO4Cu 5H2O (0.01mg), SO4Zn 7H2O (0.022mg), Cl2Co 6H2O (0.01mg), Cl2 Mn 4H2O (0.18 mg), MoO4 Na2 2H2O (0.006mg), Tiamina HCl (0.1mg), Biotina(0.5ug) y Vitamina B12 (0.5ug). El cultivo madre se mantuvo durante nueve horas bajo luz artificial. Prueba de Crecimiento control Se puso 10 ml. del medio de cultivo agua de mar enriquecida en cada uno de 20 viales. Estos medios fueron autoclavados durante 15 minutos a 120 C y 15 libras de presión. Luego de enfriados fueron inoculados con 2 ml. del cultivo madre. La mitad de los viales se mantuvo en la cámara de crecimiento (laboratorio) y la otra mitad se cultivo en un ambiente externo del laboratorio. El crecimiento en cada ambiente se determinó por medio de la densidad celular y por la absorbancia del cultivo a través del tiempo. Para cuantificar la densidad celular en cada vial, se traslado una alícuota de 0.9 ml a otro vial y se le adicionó 0.1 ml. de lugol. De aquí se tomó una pequeña muestra para montarla en la cámara Newbauer y efectuar el conteo. En los casos de mucha densidad celular, la muestra se diluyó
a razón de 1 en 10. Para cuantificar la absorbancia se usó la longitud de onda 640 nm. que corresponde a la clorofila. Tratamiento con Paraquat Se prepararon 24 viales con 9 ml. de medio agua de mar enriquecido y esterilizado al autoclave a 120 C durante 15 minutos. Los viales fueron luego inoculados con 1 ml. del cultivo madre y se incuban en la cámara de crecimiento. En los siguientes días se evalúa el crecimiento por absorbancia de clorofila. Después de seis días de crecimiento, 21 viales son agrupados de tres en tres para someterlos a las concentraciones siguientes del paraquat grado comercial; (0.001, 0.002, 0.004, 0.008, 0.01, 0.02, 0.04 ppm), los 3 viales restantes sirvieron como controles. Todos los cultivos fueron mantenidos en condiciones no controladas (exterior del laboratorio) la irradiancía promedio registrada fue de 1560 umol.m-2 s-1 con cielo nublado. Estos cultivos fueron analizados al día siguiente de haber recibido el tratamiento (séptimo día). Resultados y Discusión En la Figura 1 se observa que el crecimiento de Dunaliella tertiolecta es más pronunciado en el ambiente exterior donde la temperatura diurna varió entre 20.5oC y 23.5 oC mientras que en condiciones del Laboratorio donde temperatura varió entre 18.25oC y 23oC es menor. Estas curvas se hicieron en función de la absorbancia de la clorofila (indicador de crecimiento), sin embargo no parece ser un indicador muy confiable, por cuanto, el cultivo desde la cepa madre al parecer no estuvo axénico. Una mejor forma de evaluación se presenta en la figura 2, en donde las curvas están en función de la densidad celular en el transcurrir de los días. En ella se observa una tendencia similar al encontrado con las absorbancias, a excepción de la disminución en la densidad en los últimos dos días que no se observa en el gráfico de absorbancias vs. días. El análisis cuantitativo del crecimiento establece un intervalo de aproximadamente 10 a 11 días para llegar a un crecimiento máximo, lo cual corrobora lo encontrado por Aguilar (1995). Estas curvas, conjuntamente con otra estándar elaborada al momento del tratamiento con las dosis del paraquat, nos sirvieron para comparar los efectos a partir del sétimo día de crecimiento (primer día de tratamiento) hasta el día onceavo (sexto día de tratamiento) De las Figuras 3.1 y 3.2 densidad celular vs. días (para cada dosis del paraquat) se observa que hay una tendencia al incremento en la densidad celular (crecimiento) en las dosis siguientes 0.001, 0.002, 0.004, 0.008 ppm, mientras que existe una tendencia a la disminución en la densidad celular en las dosis 0.01, 0.02, 0.04 ppm.
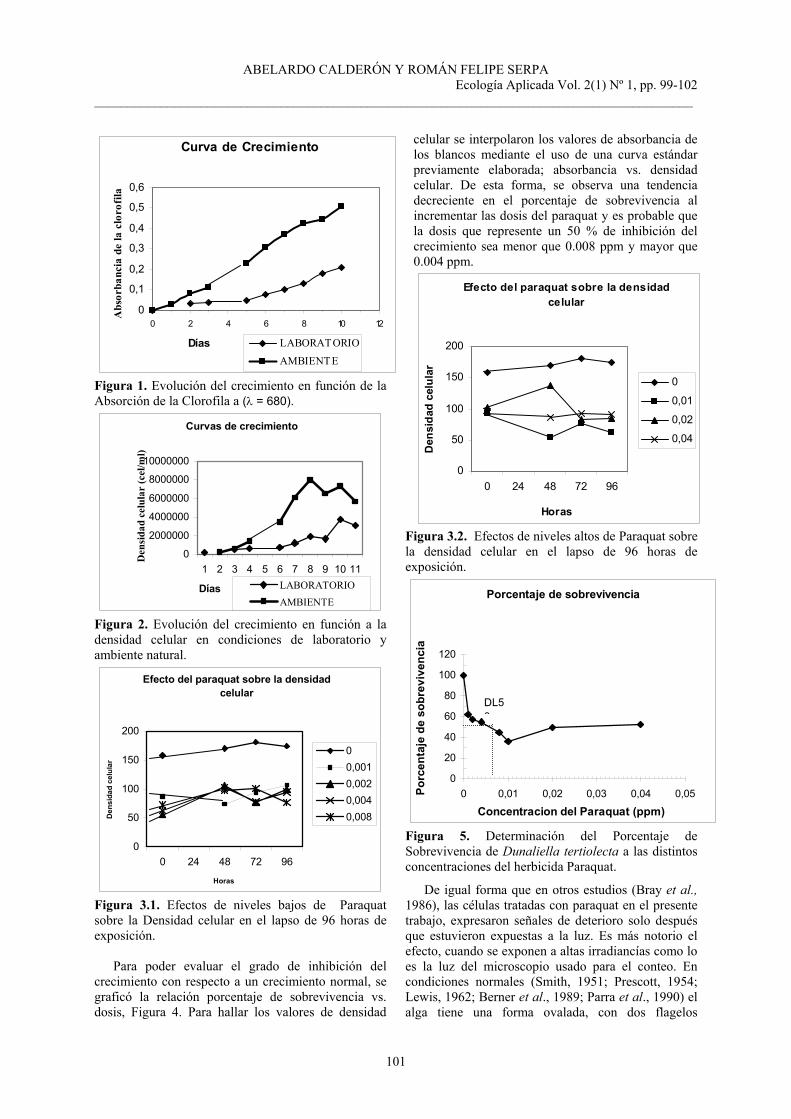
ABELARDO CALDERÓN Y ROMÁN FELIPE SERPA
Ecología Aplicada Vol. 2(1) Nº 1, pp. 99-102 __________________________________________________________________________________________
101
Figura 1. Evolución del crecimiento en función de la Absorción de la Clorofila a (λ = 680).
Curvas de crecimiento
0
2000000
4000000
6000000
8000000
10000000
1 2 3 4 5 6 7 8 9 10 11
Días
Den
sida
d ce
lula
r (c
el/m
l)
LABORATORIOAMBIENTE
Figura 2. Evolución del crecimiento en función a la densidad celular en condiciones de laboratorio y ambiente natural.
Efecto del paraquat sobre la densidad celular
0
50
100
150
200
0 24 48 72 96
Horas
Den
sida
d ce
lula
r
00,0010,0020,0040,008
Figura 3.1. Efectos de niveles bajos de Paraquat sobre la Densidad celular en el lapso de 96 horas de exposición. Para poder evaluar el grado de inhibición del crecimiento con respecto a un crecimiento normal, se graficó la relación porcentaje de sobrevivencia vs. dosis, Figura 4. Para hallar los valores de densidad
celular se interpolaron los valores de absorbancia de los blancos mediante el uso de una curva estándar previamente elaborada; absorbancia vs. densidad celular. De esta forma, se observa una tendencia decreciente en el porcentaje de sobrevivencia al incrementar las dosis del paraquat y es probable que la dosis que represente un 50 % de inhibición del crecimiento sea menor que 0.008 ppm y mayor que 0.004 ppm.
Efecto del paraquat sobre la densidad celular
0
50
100
150
200
0 24 48 72 96
Horas
Den
sida
d ce
lula
r
0
0,01
0,02
0,04
Figura 3.2. Efectos de niveles altos de Paraquat sobre la densidad celular en el lapso de 96 horas de exposición.
Porcentaje de sobrevivencia
0
20
40
60
80
100
120
0 0,01 0,02 0,03 0,04 0,05
Concentracion del Paraquat (ppm)
Porc
enta
je d
e so
brev
iven
cia
DL50
Figura 5. Determinación del Porcentaje de Sobrevivencia de Dunaliella tertiolecta a las distintos concentraciones del herbicida Paraquat.
De igual forma que en otros estudios (Bray et al., 1986), las células tratadas con paraquat en el presente trabajo, expresaron señales de deterioro solo después que estuvieron expuestas a la luz. Es más notorio el efecto, cuando se exponen a altas irradiancías como lo es la luz del microscopio usado para el conteo. En condiciones normales (Smith, 1951; Prescott, 1954; Lewis, 1962; Berner et al., 1989; Parra et al., 1990) el alga tiene una forma ovalada, con dos flagelos
Curva de Crecimiento
0
0,1
0,2
0,3
0,4
0,5
0,6
0 2 4 6 8 10 12
Días
Abs
orba
ncia
de
la c
loro
fila
LABORATORIOAMBIENTE

EFECTOS DEL PARAQUAT SOBRE LA MICROALGA Dunaliella tertiolecta
Diciembre 2003 __________________________________________________________________________________________
102
visibles, cloroplastos en forma de copa con un pirenoide en posición central penetrado por láminas tilakoidales, gránulos de almidón rodeando al pirenoide, un núcleo y tiene una tendencia a movilizarse hacia los bordes del cubreobjetos (fotofobia). Por otro lado, en un primer momento del tratamiento, en los niveles bajos se observa un movimiento normal, es decir rectilíneo, con un giro del alga sobre su eje dorsoventral, asemejando un tirabuzón; después de un cierto tiempo (aproximadamente media hora) se observa una agitación anómala (vibración) en algunas células. Al día siguiente, muchas células tienen un movimiento muy lento pero aun son ovoides, pero otros tantos están inmóviles y son esféricos, tal vez están muertos. Normalmente, los cloroplastos y otros organelos producen niveles tolerables de oxígeno activo a través de la autooxidación del componente transportador de electrones del Fotosistema I. En la presencia de luz, el paraquat acelera la producción de superoxidos según Audus (1981). En niveles bajos del producto, el sistema antioxidante (SOD y otros) reforzado en este caso por la presencia del beta caroteno garantiza la sobrevivencia de los individuos más eficientes en su biosíntesis. Debemos anotar que Dunaliella tertiolecta es un buen productor de beta caroteno cuando se halla bajo estrés salino o de luz, siempre que la célula esté intacta. En niveles superiores al 0.008 ppm los radicales libres (ROS) generan peroxidación de los lípidos de las membranas y por ello es que la permeabilidad se altera y entonces vemos células hinchadas o con incremento del volumen celular, tilakoides disgregados y una serie de daños morfológicos. En las dosis mayores, los procesos descritos son similares pero más violentos y aparecen 15 minutos después de iniciado el tratamiento y a los 30 minutos la mayoría de las células están hinchadas e inmóviles. Las enzimas antioxidantes parecen no funcionar porque a nivel ultraestructural, todas las membranas dejan de ser funcionales, se pierde la compartimentación, se ven corrugadas, disgregadas, similares a los reportados para Chlamydomonas Se sabe que la efectividad del herbicida en Chlorela está relacionada a la presencia en el medio de ciertos iones como el Cu (+2), que inhibe la actividad del paraquat a una concentración de 0.5 ppm. Audus (1981). El Cobre es cofactor de una enzima antioxidante Cu/Zn SOD. Para Dunaliella no se han descrito mecanismos de resistencia, pero al parecer los carotenos juegan un rol importante hasta cierto punto. Literatura citada Aguilar C. 1995. Crecimiento y ciclo de vida de la
microalga Dunaliella salina Teodoresco (Chlorophyta, Volvacales) de las salinas de Los
Chimus (Ancash) y las salinas de Chilca (Lima).Tesis para obtener el titulo de Licenciado en Biología. Universidad Ricardo Palma. Lima-Perú.
Ascon – Bieto J. & Talon M. 1993. Fisiología y Bioquímica Vegetal. Interamericana Mc Graw – Hill –España.
Audus J. 1981. Herbicides. Academic Press, Inc., New York.
Ben-Amots A., Shaish A. & Avron M. 1989. The mode of action of the massively accumulated beta-carotene of Dunaliella bardawil in protecting the alga against the damage by excess irradiation. Plant Physiology. 9: 1040-1043.
Baier M. & Dietz J. 1999. The Costs and Benefits of Oxygen for Photosynthesizing Plant Cells. Progress in Botany Vol 60. Springer – Verlag Berlin Heidelberg.
Berner T., Dubinski Z., Wyman K. & Falkowski P. 1989. Photo adaptation and the package effect in Dunaliella tertiolecta. Journal Phicology. 25: 141-146.
Bray D., Bagu J. & Nakamura K. 1986. Ultra structure of Chlamydomonas reinhardtii following exposure to paraquat. Comparison of wild type and paraquat –resistant mutant. Canadian Journal Bot. 71: 174–182.
Dietz J., Baier M. & Kramer U. 1998 Free Radicals and Reactive Oxygen Species as Mediators of Heavy Metal Toxicity in Plants. University of Bielefeld. D-33615. Germany.
Halliwell E. 1987. Oxidative damage, lipid peroxidation and antioxidant protection in Chloroplasts. Chemistry and Physics of lipids. 44: 327 -340.
Kao S.M. & Hassan H.M. 1985. Biochemical Characterization of a Paraquat – Tolerant Mutant of E coli. J. Biol. Chem. 260: 10478 – 10481.
Larson R.A. 1988. The antioxidants of higher plants. Phytochemistry. 27: 969 – 978.
Levitt J. 1980. Responses of Plants to Environmental Stress. Academic Press, Inc New York Vol 1. Second Edition.
Lewis R.A. 1962 Physiology and Biochemistry of alga. Academic Press. New York – London.
Parra O., Gonzales M. & De La Rosa V. 1990. Caracterización Biológica de una Cepa Chilena de Dunaliella salina Potencialmente Comerciable. Arch. Biol.Med.Exp. 23: 141 -146.
Prescott G.W. 1954. How to Know the Fresh Water Algae. Dubuque, Iowa, W.C Brown.
Shannon L., Kay E. & Lewj Y. 1966. Peroxidase isozymes from horseradish roots. Isolation and Physical Properties. J.Biol Chem. 241: 2166–2172.
Smith G.M. 1951. Manual of Phycology. Waltham, Mass. Chronica Botanical.

Ecología Aplicada, 2(1), 2003 ISSN 1726-2216 Depósito legal 2002-5474
VARIABILIDAD DEL ENSAYO ECOTOXICOLÓGICO CON Chironomus calligraphus Goeldi (DIPTERA: CHIRONOMIDAE) PARA EVALUAR CADMIO, MERCURIO Y PLOMO
José Alberto Iannacone Oliver1, Neil Salazar Capcha1 y Lorena Alvariño Flores1
Resumen
En la evaluación del riesgo ambiental (ERA) de los ambientes acuáticos contaminados por relaves mineros en el Perú se requiere del empleo de organismos biológicos estandarizados como la larva del insecto Chironomus calligraphus Goeldi. Los organismos biológicos para ser usados como herramientas ecotoxicológicas requieren ser sencillos, prácticos, sensibles y repetibles. Por lo que el objetivo de este trabajo fue evaluar el Cd+2, Hg2+ y Pb2+ en forma de cloruro de cadmio (CdCl2), cloruro de mercurio (HgCl2) y nitrato de plomo (Pb3(NO)2) respectivamente, sobre la mortalidad de las larvas de primer estadio del díptero C. calligraphus a 48 h de exposición y así determinar su variabilidad. La especie fue aclimatada y criada en condiciones estandarizadas de laboratorio. Se realizaron seis bioensayos de toxicidad aguda estáticos para cada metal, en cada bioensayo se siguió un diseño experimental en bloques completamente randomizados (DBCR): cuatro repeticiones con seis concentraciones y empleándose 240 larvas por bioensayo. La Concentración Letal media (CL50) en µg del metal L-1 a 48 h de exposición y el coeficiente de variación (CV %) fueron: 132 µg L-1 y 45,45 % para Cd+2; 127 µg L-1 y 22,83 % para Hg2+; y finalmente 18.730 µg L-1 y 33 % para Pb2+. La variabilidad del ensayo con C. calligraphus es comparada con otros organismos biológicos dulceacuícolas al nivel mundial. Palabras clave: Bioensayo, Chironomus, cadmio, ecotoxicología, macroinvertebrado, mercurio, plomo, repetibilidad.
Abstract Environmental Risk Assessment (ERA) of freshwater environment in Peru needs the employment of standardized biological organisms such as the larval midge Chironomus calligraphus Goeldi. The biological organisms to be employed as ecotoxicological tools should be easy, practical, sensitive and reproductible. The aim of this research was to evaluate Cd+2, Hg+2 and Pb+2 as cadmium chlorure (CdCl2), mercury chlorure (HgCl2) and lead nitrate (Pb3(NO)2) forms, on the mortality of first instar larva of diptera C. calligraphus at 48 h exposure and in this way determinate its variability. The species was acclimated and reared under standardized laboratory conditions. Six bioassays of acute static toxicity for each metal were done, in each one a randomized block experimental design (RBED) was used with four repetitions of five concentrations and employing 240 larvae for each bioassay. The median lethal concentration (LC50) in µg metal L-1 at 48 h exposure and the coefficient of variation (CV %) were: 132 µg L-1 and 45,45 % for Cd+2; 127 µg L-1 and 22,83 % for Hg2+; and finally 18.730 µg L-1 and 33 % for Pb2+. The variability of the essay of C. calligraphus is compared with other freshwater biological organisms all over the world. Key words: Bioassay, Chironomus, cadmium, ecotoxicology, lead, macroinvertebrate, mercury, repeatability.
1 Laboratorio de Ecofisiología Animal. Facultad de Ciencias Naturales. Universidad Nacional Federico Villarreal (FCCNM-UNFV). Calle San Marcos 383, Lima 21, Perú. Correo electrónico: [email protected] / [email protected].
Introducción La ecotoxicología es la ciencia que describe y predice el comportamiento de las sustancias en el medio ambiente y las respuestas biológicas del sistema para así, finalmente evaluar el riesgo asociado con estas emisiones (Delvalls & Conradi, 2000). Los bioensayos ecotoxicológicos en el laboratorio con un determinado organismo biológico y el uso de controles permiten predecir el efecto de las sustancias químicas tóxicas (Calow, 1993). Los organismos biológicos para ser usados como herramientas
ecotoxicológicas requieren ser sencillos, prácticos, sensibles y repetibles (Iannacone et al., 1998; Alayo & Iannacone, 2002). La APHA (1989) señala como protocolos estandarizados para la realización de bioensayos de toxicidad aguda y crónica acuática a las larvas de los insectos quironómidos, principalmente Chironomus tentans Fabricius, Chironomus riparius Meigen y Chironomus plumosus (Linnaeus) (Mereggalli et al., 2002; Lahr et al., 2003; Martinez et al., 2003). Estos mosquitos quironómidos son abundantes en los ambientes dulceacuícolas y

ENSAYO ECOTOXICOLÓGICO CON Chironomus calligraphus PARA EVALUAR CADMIO, MERCURIO Y PLOMO
Diciembre 2003 __________________________________________________________________________________________
104
componente importante en las comunidades acuáticas detritívoras (Iannacone et al., 2000; Crane et al., 2002; Sánchez & Tarazona, 2002). Chironomus calligraphus Goeldi, es un insecto Pan-Americano, de distribución predominantemente Neotropical, y que también ocurre en la región Neártica Sur, muy común en los ambientes acuáticos epicontinentales de la ciudad de Lima, Perú (Iannacone et al., 1999, 2000; Spies et al., 2002). C. calligraphus ha demostrado una alta sensibilidad a metales pesados (Iannacone et al., 1998; Iannacone & Dale, 1999; Arrascue et al., 2001) y a plaguicidas (Iannacone & Alvariño, 1998, 2000). Para la evaluación del riesgo ambiental (ERA) de los ambientes acuáticos epicontinentales en el Perú se requiere del empleo de organismos biológicos estandarizados como la larva del insecto C. calligraphus. Los metales pesados, como el cadmio, mercurio y plomo, producen efectos deletéreos en las aguas naturales (Weber, 1996; Iannacone & Alvariño, 1999; Fernández & Beiras, 2001). En Latinoamérica existen algunas experiencias de ejercicios de intra e intercalibración empleando baterías de bioensayos, que incluyen artrópodos, para la evaluación de tóxicos de referencias con el fin de determinar el grado de repetibilidad, precisión o variabilidad de estas pruebas ecotoxicológicas (Forget et al., 2000). Así se han realizado en países como Argentina (Ronco et al., 2000), Chile (Castillo & Schäfer, 2000; Castillo et al., 2000a), Colombia (Díaz-Baez & Perez, 2000), Costa Rica (Castillo et al,. 2000b) y México (Pica et al., 2000). En el Perú, aún no se tienen este tipo de experiencias para los bioensayos ecotoxicológicos. Por lo que, el objetivo de este trabajo fue evaluar el grado de variabilidad o repetibilidad del bioensayo que emplea a C. calligraphus usando al cadmio, mercurio y plomo como tóxicos de referencia. Materiales y métodos Chironomus calligraphus La colección, identificación y cría estandarizada de C. calligraphus, así como el protocolo de bioensayo siguió el procedimiento detallado descrito por Iannacone et al. (2002a). Los ensayos ecotoxicológicos con C. calligraphus se iniciaron con larvas de primer estadio dentro de 24 h de haber eclosionado de las masas de huevos procedente de la Planta de Tratamiento de Aguas residuales domésticas de Villa El Salvador, Lima, Perú durante el 2000 al 2001. Los ejemplares se separaron al azar de los frascos de eclosión y recuperación de las larvas de primer estadio. Diez larvas de primer estadio se distribuyeron al azar en cada concentración de las muestras con metales evaluadas en cada una de las cuatro repeticiones del ensayo experimental. Cada repetición incluyó cinco concentraciones ensayadas y
el control. Las larvas se consideraron muertas si no fueron capaces de moverse coordinada y normalmente cuando fueron pinchadas ligeramente con un alfiler luego de 48 h de exposición. Se emplearon un total de 240 larvas por ensayo. Los bioensayos fueron repetidos seis veces secuencialmente. El pH se midió mediante un potenciómetro. El pH se midió en dos réplicas al inicio del ensayo y a las 48 h de exposición. La temperatura se mantuvo regulada a 20 oC ± 3 oC. Metales y Físico-química del agua Las soluciones madre para los iones tóxicos de cadmio, mercurio y plomo fueron preparadas de la sal metálica a base del cloruro de cadmio (CdCl2), cloruro de mercurio (HgCl2) y nitrato de plomo (Pb3(NO)2) a una concentración de 100 mg L-1. Todas las concentraciones se registraron en concentraciones nominales de ión de metálico. Los matraces aforados previamente fueron lavados con ácido nítrico al 10 % y luego enjuagados en agua destilada y mantenidos a 4 oC (Iannacone et al., 2000). El pH fue ajustado al inicio del ensayo a 7 con una solución NaOH 0,1M o con H2SO4 0,1M. Los datos de oxígeno disuelto se midieron mediante el método de Winkler (APHA, 1989). En el ensayo con Cd2+ se emplearon las siguientes cinco concentraciones 125 µg L-1, 250 µg L-1, 500 µg L-1, 1.000 µg L-1 y 2.000 µg L-1. En el ensayo con Hg2+ se emplearon las siguientes cinco concentraciones 62,5 µg L-1, 125 µg L-1, 250 µg L-1, 500 µg L-1 y 1.000 µg L-1. En el ensayo con Pb2+ se emplearon las siguientes cinco concentraciones 6.250 µg L-1, 12.500 µg L-1, 25.000 µg L-1, 50.000 µg L-1 y 100.000 µg L-1. Para el Cd2+,Hg2+, Pb2+ las cinco concentraciones siguieron un factor de incremento de 2. Las principales características fisicoquímicas del agua destilada son las indicadas por Iannacone et al. (2002b). Diseño Experimental y Análisis de datos Se determinaron la Concentración Letal media (CL50) promedio, su desviación estándar y su Coeficiente de Variación (CV %) para los bioensayos de C. calligraphus expuestos al cadmio, mercurio y plomo utilizando los resultados de las sumas de las cuatro repeticiones de cada concentración. Las diferencias entre las concentraciones se evaluaron a través de un Análisis de Varianza (ANDEVA), previa transformación de los datos a raíz cuadrada del arcoseno, con el fin de ajustar los datos a la distribución normal (Zar, 1996). Se utilizaron los bioensayos como bloques y las variables respuestas provinieron de las sumas de las cuatro repeticiones de cada concentración de cada ensayo. En el caso de existir diferencias significativas entre las concentraciones se realizó una Prueba de Significación DVS (Diferencia Verdaderamente Significativa) de Tukey (Daniel, 1993). Las CL50s se calcularon usando un programa computarizado de la EPA versión 1,5 –1993 (USEPA,

JOSÉ ALBERTO IANNACONE OLIVER, NEIL SALAZAR CAPCHA Y LORENA ALVARIÑO FLORES
Ecología Aplicada Vol. 2 Nº 1, pp. 103-110 __________________________________________________________________________________________
105
1993). Estos valores fueron obtenidos a partir del análisis de regresión lineal entre el logaritmo de la concentración de las muestras con metales y la mortalidad Probit (porcentaje transformado). Estos modelos de regresión lineal fueron verificados usando el estadístico χ2 (Martín & Holdich, 1986). Se calcularon los valores de NOEC (Concentración de efectos no observables) y LOEC (Concentración más baja de efectos observables) para cada metal. Resultados La secuencia en orden de toxicidad de C. calligraphus expuesto a los tres metales fue: Hg2+ > Cd2+ > Pb2+ (Tabla 1). Las Tablas 2, 3 y 4 señalan los porcentajes de mortalidad para cada una de las cinco concentraciones y el control en cada una de los seis bioensayos para el cadmio, mercurio y plomo, respectivamente. Para todos los casos el CV (%) fue menor del 50 %, a excepción del control del ensayo con cadmio que fue del 75,42 %. Sin embargo, en ninguno de los casos la mortalidad en el control sobrepasó el 20 %. La prueba de Tukey mostró que el LOEC para el Cd2+, Hg2+ y Pb2+ fue de 62,5, 125 y 6.250 ug L-1, respectivamente. En cambio, el NOEC para el Cd2+, Hg2+ y Pb2+ fue de < 62,5 < 125 y < 6.250 ug L-1, respectivamente. Para C. calligraphus, el valor promedio de la CL50 al cadmio de 132 µg L-1 ocupa la posición tercera en comparación a otras diez especies del género Chironomus (Tabla 5). Para C. calligraphus, el valor promedio de la CL50 al mercurio de 130 µg L-1 ocupa la posición cuarta en comparación a otras siete especies del género Chironomus (Tabla 6). Para C. calligraphus, el valor promedio de la CL50 al plomo de 18.732 µg L-1 ocupa la posición primera en comparación a otras siete especies del género Chironomus (Tabla 7). La Tabla 8, señala que C. calligraphus ocupa la posición 12va (para el Hg2+), 15va (para el Pb2+) y 18va (para el Cd2+) entre 29 bioensayos ecotoxicológicos empleando diferentes componentes de la biota animal. Discusión Las especies propuestas como herramientas para bioensayos ecotoxicológicos, requieren como requisitos para uso no solo que sean sensibles, sino que los ensayos sean reproducibles con poca variabilidad en su respuesta (Magdaleno et al., 1997; Steevens & Benson, 1998). C. calligraphus es una especie con alta sensibilidad a metales pesados, plaguicidas y a muestras ambientales (Iannacone et al., 2000; Arrascue et al, 2001). Sus valores de CL50 para el cadmio, mercurio y plomo tienen en general el mismo orden de magnitud que otras especies del género (Tablas 5, 6 y 7). En un estudio previo, Iannacone & Dale (1999) encontraron una sensibilidad de C. calligraphus al mercurio (seis veces mayor); al plomo (cinco veces menor) y al mercurio (dos veces
menor que la encontrada en el presente estudio). Sin embargo estos resultados correspondieron para cada metal a un bioensayo sin repeticiones. Entre los numerosos factores bióticos que pudieran influir en la toxicidad del cadmio, mercurio y plomo sobre las especies de la familia Chironomidae, tenemos a la especie, estado de desarrollo y edad. Los valores comparativos de CL50 expuestos al cadmio, mercurio y plomo para las diferentes especies del Chironomus señalados en la Tablas 5, 6 y 7 señalan diferencias por ser especies diferentes, así como estados de desarrollo y edades diferentes por ser utilizados formas larvarias del primer al cuarto estadio. En general se considera que las formas larvarias de estadios tempranos son más sensibles que los estadios superiores (Tablas 5, 6 y 7). Las larvas de primer estadio (plantónicas) son más sensibles que las de segundo estadio (bentónicas) de C. riparius y de las de cuarto estadio (bentónicas) de C. decorus. Además, entre los factores abióticos se señala al tiempo de exposición como un factor de importancia que influiría en la ecotoxicidad de los tres metales pesados (Peck et al., 2002). Los coeficientes de variación (CV %) obtenidos para los 29 ensayos empleando especies biológicas acuáticas pertenecientes a diferentes taxas, variaron entre 2,1 a más de 100 % (Tabla 8). El ensayo con C. calligraphus ocupa la posición 12va, 15va y 18va para Hg2+, Pb2+ y Cd2+, respectivamente, el cual fue aceptable al ser menor al 50 %. Uno de los factores que estaría influyendo en la variabilidad encontrada en el ensayo estático de mortalidad con C. calligraphus expuestos a los tres metales a 48 h de exposición, es la estación del año y la calidad del agua (contenido de oxígeno, pH, sólidos suspendidos, etc.) del lugar colecta de los organismos, ya que los quironómidos en la fase de huevos son obtenidos de cuerpos de agua artificiales en forma periódica a través de las diferentes estaciones del año y luego criados artificial y parcialmente en condiciones estandarizadas de laboratorio (Milani et al., 2003). Las especies criadas artificialmente en forma continua durante todo su ciclo vital tienden a presentar menor variabilidad en su respuesta que las especies criadas parcialmente y obtenidas en forma periódica del campo (APHA, 1989; Calow, 1993). Agradecimientos Al Laboratorio de Ecofisiología Animal de la Universidad Nacional Federico Villarreal, Lima, Perú. Literatura citada Alayo M. & Iannacone J. 2002. Ensayos
ecotoxicológicos con petroleo crudo, Diesel 2 y Diesel 6 con dos subespecies de Brachionus plicatilis Müller 1786 (Rotifera: Monogononta). Gayana. 66: 45-58.

ENSAYO ECOTOXICOLÓGICO CON Chironomus calligraphus PARA EVALUAR CADMIO, MERCURIO Y PLOMO
Diciembre 2003 __________________________________________________________________________________________
106
APHA (AMERICAN PUBLIC HEALTH ASSOCIATION), AWWA (AMERICAN WATER WORKS ASSOCIATION), WPCF (WATER POLLUTION CONTROL FEDERATION). 1989. Standard methods for examination of water and wastewater. 17th. Ed. American Health Association. Washington, D.C.
Arrascue A., Iannacone J., Alvariño L., Basilio S. & Lazcano C. 2001. El insecto Chironomus calligraphus Goeldi y la bacteria Escherichia coli como ensayos ecotoxicológicos para evaluar sedimentos elutriados dulceacuícolas. Rev. per. Ent. 42: 159-173.
Calow P. 1993. Handbook of ecotoxicology. 478 p. Vol. I. Blackwell Science, Sheffield, UK.
Castillo G.C. & Schäfer L. 2000. Evaluation of a bioassay battery for water toxicity testing: A Chilean experience. Environm. Toxicol. 15: 331-337.
Castillo G.C., Vila I.C. & Neild E. 2000a. Ecotoxicity assessment of metals and wastewater using multitrophic assays. Environm. Toxicol. 15: 370-375.
Castillo L.E., Pinnock N. & Martinez E. 2000b. Evaluation of a battery of toxicity tests for use in the assessment of water quality in a Costa Rican laboratory. Environm. Toxicol. 15: 312-321.
Crane M., Sildanchandra W., Kheir R. & Callaghan R. 2002. Relationships between biomarker activity and development endpoints in Chironomus riparius Meigen exposed to an organophosphate insecticide. Ecotoxicol. Environm. Saf. 53: 361-369.
Daniel W.W. 1993. Bioestadística. Tercera Edición. Editorial Limusa (México).
Delvalls T.A. & Conradi M. 2000. Advances in marine ecotoxicology: laboratory tests versus field assessment data on sediment quality studies. Ciencias Marinas. 26: 39-64.
Díaz-Baez M.C. & Perez J.B. 2000. Intralaboratory experience with a battery of bioassays: Colombia experience. Environm. Toxicol.15: 297-303.
Espiritu E.Q., Janssen C.R. & Persoone G. 1995. Cyst-based toxicity tests. VII. Evaluation of the 1-h enzymatic inhibition test (Fluotox) with Artemia nauplii. Environm. Toxicol. Water Qual. 10: 25-34.
Fargasova A. 2001. Winter third- to fourth- instar larvae of Chironomus plumosus as bioassay tools for assessment of acute toxicity of metals and their binary combinations. Ecotoxicol. Environm. Saf. 48: 1-5.
Fernández N. & Beiras N. 2001. Combined toxicity of dissolved mercury with Copper, lead and cadmium on embryogenesis and early larval growth of the Paracentrotus lividus sea-urchin. Ecotoxicol. 10: 263-271.
Forget G., Sánchez A., Arkhipchuk V., Beauregard T., Blaise C., Castillo G., Castillo L.E., Diaz-Baez M.C., Pica Y.G., Ronco A., Srivastava R.C. & Dutka B.J. 2000. Preliminary data of a single-blind, multicountry trial of six bioassays for water toxicity monitoring. Environm. Toxicol. 15: 362-369.
Greene J.C., Miller W.E., De Bacon M.K., Long M. A. & Bartel S. 1985. A comparison of three microbial assay procedures for measuring toxicity of chemical residues. Arch. Environm. Contam. Toxicol. 14: 659-667.
Iannacone J., Dale W. & Alvariño L. 2000. Monitoreo ecotoxicológico del río Rímac (Lima-Perú) empleando a Chironomus calligraphus Goeldi (Diptera: Chironomidae). Rev. Chil. Ent. 27: 25-34.
Iannacone J., Alvariño L. & Gutierrez A. 1999. Cinco ensayos ecotoxicológicos para evaluar metales pesados en el agua dulce. Bol. Soc. Quím. Perú 65: 30-45.
Iannacone J. & Alvariño L. 1998. Ecotoxicidad aguda del insecticida organofosforado temephos sobre Chironomus calligraphus Goeldi (Diptera: chironomidae). Acta Ent. Chilena. 22: 51-53.
Iannacone J. & Alvariño L. 1999. Acute ecotoxicity of heavy metals using juveniles of freshwater snail Physa venustula (Gould, 1847) (Mollusca). Gayana. 63: 101-110.
Iannacone J. & Alvariño L. 2000. Chironomus calligraphus Goeldi y Moina macrocopa (Sars) como herramientas ecotoxicológicas para la evaluación del lindano y clorpirifos. Bol. Soc. Biol. Concepción (Chile). 71: 33-39.
Iannacone J. & Dale W.E. 1999. Protocolo de bioensayo ecotoxicológico para evaluar metales pesados contaminantes de agua dulce con Chironomus calligraphus (Diptera: Chironomidae) y el Crustáceo Moina macrocopa (Crustácea: Cladócera), en el río rímac. Rev. per. Ent. 41: 111-120.
Iannacone J., Alvariño L. & Dale W.E. 1998. Pruebas ecotoxicológicas como una herramienta para la evaluación del impacto ambiental de los ecosistemas acuáticos. Bol. Lima (Perú). 113: 53-68.
Iannacone J., Alvariño L. & Mansilla J. 2002a. Actividad insecticida de cuatro extractos botánicos sobre larvas de los mosquitos Culex quinquefasciatus (Diptera: Culicidae) y Chironomus calligraphus (Diptera: Chironomidae). Wiñay Yachay. 6: 59-74.
Iannacone J., Caballero C. & Alvariño L. 2002b. Employing the freshwater snail Physa venustula Gould as an ecotoxicological tool for environmental risk assessment by pesticides. Agri. Téc. (Chile). 62: 212-225.

JOSÉ ALBERTO IANNACONE OLIVER, NEIL SALAZAR CAPCHA Y LORENA ALVARIÑO FLORES
Ecología Aplicada Vol. 2 Nº 1, pp. 103-110 __________________________________________________________________________________________
107
Janssen C. 1998. Alternative assay for routine toxicity assessments. A review. Chapter 26. pp. 813-839. In: Ecotoxicology. G. Schüürmann, Markert B. Eds.
Khangarot B.S. & Ray P.K. 1989. Sensitivity of midge larvae of Chironomus tentans Fabricius (Diptera Chironomidae) to heavy metals. Bull. Environm. Contam. Toxicol. 42: 325-330.
Lahr J., Maas-Diepeveen J.L., Stuijfzand S.C., Leonards P.E., Druke J.M., Lucker S., Espeldoorn A., Kerkum L.C., van Stee L.L. & Hemdriks A.J. 2003. Response in sediment bioassays used in the Netherlands: can observed toxicity be explained by routinely priority pollutants?. Water Res. 37: 1691-1710.
Magdaleno A., Gomez C.E., Velez C.G. & Accorinti J. 1997. Preliminary toxicity test using the green alga Ankistrodesmus falcatus. Environm. Toxicol. Water Qual. 12: 11-14.
Martin T.R. & Holdich D.M. 1986. The acute lethal toxicity of heavy metals to peracarid crustaceans (with particular reference to freshwater asellids and gammarids). Water Res. 20: 1137-1147.
Martinez E.A., Moore B.C., Schaumloffel J. & Dasgupta N. 2003. Morphological abnormalities in Chironomus tentans exposed to cadmium and copper spiked sediments. Ecotoxicol. Environm. Saf. 55: 204-212.
Mereggalli G., Bettinetti R., Pluymers L., Vermeulen A.C., Rossaro B. & Ollevier F. 2002. Mouthpart deformities and nucleolus activity in field-collected Chironomus riparius larvae. Arch. Environm. Contam. Toxicol. 42: 405-409.
Milani D., Reynoldson T.B., Borgmann U. & Kolasa J. 2003. The relative sensitivity of four benthic invertebrates to metals in spiked-sediment exposures and application to contaminated field sediment. Environ. Toxicol. Chem. 22: 845-854.
Postma J.F., Buckert de Jong M.C., Staats N. & Davids C. 1994. Chronic toxicity of cadmium to Chironomus riparius (Diptera: Chironomidae) at different food levels. Arch. Envrion. Contam. Toxicol. 26: 143-148.
Peck M.R., Klessa D.A. & Baird D.J. 2002. A tropical sediment toxicity test using the dipteran Chironomus crassiforceps to test metal bioavailability with sediment pH change in tropical acid-sulfate sediments. Environm. Toxicol. Chem. 21: 720-728.
Persoone G. 1996. Development and first validation of a “culture free” algal microbiotest: the algaltoxkit. In: Microscale toxicology, advances, techniques and practice, Wells, P.G., Lee K., Blaise, C. (eds.). CRC Publishers.
Pica G.Y., Trujillo G.D. & Hernández H.S. 2000. Bioassay standardization for water quality monitoring in Mexico. Environm. Toxicol. 15: 322-330.
Qureshi S.A., Sakesena A.B. & Singh V.P. 1980. Acute toxicity of four heavy metals to benthic fish food organims for the River Khan Ujjain. Int. J. Environm. Stu. 15 : 59-61.
Rao D.S. & Saxena A.B. 1981. Acute toxicity of mercury, zinc, lead, cadmium and manganese to Chironomus sp. Int. J. Environ. Stud. 16 : 226-226.
Reish D.J. 1993. Effect of metals and organic compounds on survival and bioaccumulation in two species of marine gammaridean amphipod, together with a summary of toxicological research on this group. Jour. Nat. History. 27: 781-794.
Ronco A., Sobrero C., Grassi V., Kaminski L., Massolo L. & Mina L. 2000. WaterTox bioassay intercalibration network: results from Argentina. Environm. Toxicol. 15: 287-296.
Rossino G.B. & Ronco A.E. 1996. Acute toxicity bioassay using Daphnia obtusa as a test organism. Environm. Toxicol.Water Qual. 11: 255-258.
Sánchez P. & Tarazona J.V. 2002. Development of a multispecies system for testing reproductive effects on aquatic invertebrates. Experience with Daphnia magna, Chironomus prasinus and Lymnaea peregra. Aquatic Toxicol. 60: 249-256.
Snell T.W., Moffat B.D., Janssen C. & Persoone G. 1991. Acute toxicity tests using rotifers: III. Effects of temperature, strain and exposure time on the sensitivity of Brachionus plicatilis. Environm. Toxicol. Water Qual. 6: 63-75.
Spies M., Sublette J.E., Sublette M.F., Wülker W.F., Martin J., Hille A., Miller M.A. & Witt K. 2002. Pan-american Chironomus calligraphus Goeldi, 1905 (Diptera: Chironomidae): Species or Complex?. Evidence from external morphology, karyology and DNA sequencing. Aquat. Insects. 24: 91-113.
Steevens J.A. & Benson W.H. 1998. Hyalella azteca 10-day sediment toxicity test: Comparison of growth measurement endpoints. Environm.Toxicol. Water Qual. 13: 243-248.
USEPA. 1993. Probit analysis program. Version 1,5. U.S. Environmental Protection Agency. Washington D.C. U.S.A.
Watts M.M. & Pascoe D. 2000. A comparative study of Chironomus riparius Meigen and Chironomus tentans Fabricius (Diptera: Chironomidae) in aquatic toxicity tests. Arch. Environm. Toxicol. 39: 299-306.
Weber D.N. 1996. Lead-induced metabolic imbalances and feeding alterations in juvenile fathead minnows (Pimephales promelas). Environm. Toxicol. Water Qual. 11: 45-51.
Willemsen A., Vaal M.A. & De Zwart D. 1995. Microbiotests as tools for environmental monitoring. National Institute of Public Health and Environmental Planning (RIVM). The Netherlands, report No 9. 607042005.

ENSAYO ECOTOXICOLÓGICO CON Chironomus calligraphus PARA EVALUAR CADMIO, MERCURIO Y PLOMO
Diciembre 2003 __________________________________________________________________________________________
108
Zar J.H. 1996. Bioestatistical Analysis. 3th Ed. Prentice –Hall. Inc. Upper Saddle River, Jew Jersey.
Tablas citadas en el texto
Tabla 1. Valores de la CL50 y sus límites de confianza para los bioensayos realizados con los tres metales pesados.
Hg2+ Cd2+ Pb2+
Repetición L. Inf. CL50 L. Sup. L. Inf. CL50 L. Sup. L. Inf. CL50 L. Sup.
1 71 113 154 152 210 264 11.490 19.515 27.782
2 45 83 121 138 179 217 * 19.474 *
3 100 150 200 150 193 232 15.460 22.329 29.062
4 102 152 198 37 86 120 19.620 27.593 34.974
5 86 125 162 15 67 104 9.219 12.748 16.257
6 110 154 197 11 59 99 7.485 10.735 13.915
Promedio 86 130 172 84 132 173 12.660 18.732 24.398
D.E 24 28 32 70 69 73 4.906 6.203 8.961
L. inf. = Límite inferior; CL50 = Concentración Letal media; L. sup. = Límite superior; D.E. = Desviación estandar; ( * ) = No determinado.
Tabla 2. Porcentaje de Mortalidad de C. calligraphus expuestas al cadmio a 48 h Repeticiones (% mortalidad)
µg L-1 1 2 3 4 5 6 Promedio ± DESig. CV
control 15 10 10 2,5 2,5 2,5 7,08 ± 5,34 a 75,42 125 32,5 32,5 32,5 67,5 75 77,5 52,92 ± 22,61 b 42,72 250 65 77,5 67,5 92,5 97,5 92,5 82,08 ± 14 c 17,06 500 95 95 97,5 97,5 97,5 97,5 96,67 ± 1,29 d 1,34 1.000 97,5 100 100 100 100 100 99,58 ± 1,02 d 1,02 2.000 100 100 100 100 100 100 100 ± 0 d 0
DE = Desviación Estándar; CV = Coeficiente de Variación; Sig. = Significancia.
Tabla 3. Porcentaje de mortalidad de C. calligraphus expuestas al mercurio a 48h. Repeticiones (% mortalidad)
ug/L 1 2 3 4 5 6 Promedio ± DE Sig. CV
control 15 12,5 15 17,5 15 12,514,58 ± 1,88 a 12,9062,5 35 47,5 32,5 30 35 25 34,17 ± 7,53 b 22,03125 70 67,5 52,5 52,5 50 45 56,25 ± 10,09 c 17,94250 72,5 77,5 67,5 72,5 87,5 75 75,42 ± 6,79 d 9 500 92,5 87,5 95 97,5 95 95 93,75± 3,45 e 3,68 1.000 100 100 100 100 100 100 100 ± 0 f 0 DE = Desviación Estándar; CV = Coeficiente de variación; Sig. = Significancia.
Tabla 4. Porcentaje de mortalidad de C. calligraphus expuestas al plomo a 48 h. Repeticiones (% mortalidad)
ug L-1 1 2 3 4 5 6 Promedio +/- DE Sig. CV
Control 17,5 20 10 12,5 10 10 13,33 ± 4,38 a 32,83 6.250 37,5 42,5 25 20 27,5 35 31,25 ± 8,48 ab 27,13 12.500 47.5 55 35 32,5 57,5 62,5 48,33 ± 12,32 bc 25,48 25.000 65 62,5 57,5 47,5 77,5 82,5 65,42 ± 12,89 c 19,70 50.000 75 67,5 80 77,5 95 95 81,67 ± 11,14 d 13,64 100.000 100 100 100 100 100 100 100 ± 0 e 0
DE = Desviación estándar; CV = Coeficiente de variación; Sig. = Significancia.

JOSÉ ALBERTO IANNACONE OLIVER, NEIL SALAZAR CAPCHA Y LORENA ALVARIÑO FLORES
Ecología Aplicada Vol. 2 Nº 1, pp. 103-110 __________________________________________________________________________________________
109
Tabla 5. Valores comparativos de CL50 con otras especies de Chironomus expuestas al cadmio.
Especie de Chironomus CL50 Tiempo de EstadioReferencia
µg L-1 Exposición (h)
Chironomus riparius Meigen 13 48 primer Milani et al. (2003) Chironomus riparius Meigen 16,2 72 cuarto Postma et al. (1994) Chironomus calligraphus Goeldi 132 48 primer Original Chironomus calligraphus Goeldi 280 48 primer Iannacone & Dale (1999)Chironomus riparius Meigen 1.200 240 cuarto Watts & Pascoe (2000) Chironomus tentans Fabricius 1.200 240 cuarto Watts & Pascoe (2000) Chironomus calligraphus Goeldi 3.670 24 primer Iannacone & Dale (1999)Chironomus tentans Fabricius 8.050 48 tercero Khangarot & Ray (1989) Chironomus plumosus (Linnaeus) 10.000 96 tercero Fargasova (2001) Chironomus tentans Fabricius 23.250 24 tercero Khangarot & Ray (1989)
Tabla 6. Valores comparativos de CL50 con otras especies de Chironomus expuestas al mercurio.
Especie de Chironomus CL50 Tiempo de Estadio Referencia
µg L-1 Exposición (h)
Chironomus calligraphus Goeldi 20 48 primer Iannacone & Dale (1999) Chironomus tentans Fabricius 29 48 tercero Khangarot & Ray (1989) Chironomus tentans Fabricius 119 24 tercero Khangarot & Ray (1989) Chironomus calligraphus Goeldi 127 48 primer Original
Chironomus calligraphus Goeldi 130 24 primer Iannacone & Dale (1999) Chironomus tendipes 664 48 tercero Rao & Saxena (1981) Chironomus sp. 1.800 40 tercero Qureshi et al. (1980)
Tabla 7. Valores comparativos de CL50 con otras especies de Chironomus expuestas al plomo.
Especie de Chironomus
CL50 µg L-1
Tiempo de Exposición (h)
Estadio
Referencia
Chironomus calligraphus Goeldi 18.730 48 primer Original Chironomus tentans Fabricius 34.670 48 tercero Khangarot & Ray (1989) Chironomus tendipes 50.000 48 tercero Rao & Saxena (1981) Chironomus tentans Fabricius 52.870 24 tercero Khangarot & Ray (1989) Chironomus calligraphus Goeldi 77.180 48 primer Iannacone & Dale (1999)
Chironomus calligraphus Goeldi 13.030 24 primer Iannacone & Dale (1999) Chironomus sp. 200.000 40 tercero Qureshi et al. (1980)

ENSAYO ECOTOXICOLÓGICO CON Chironomus calligraphus PARA EVALUAR CADMIO, MERCURIO Y PLOMO
Diciembre 2003 __________________________________________________________________________________________
110
Tabla 8. Variabilidad o repetibilidad de los bioensayos empleando diferentes especies acuáticas.
Taxa Especie CV % Referencia
Pez Onchorhynchus mykiss 2,1 Castillo et al. (2000)
Microalga Raphidocelis subcapitata 8 Persoone (1996) Echinodermatha Paracentrotus lividus 11 Fernández & Beiras (2001) Bacteria Vibrio fisheri (Microtox) 12 Janssen (1998) Celenterado Hydra attenuata 13 Diaz-Baez & Perez (2000)
Planta Lactuca sativa 14,9 Forget et al. (2000) Rotifera Brachionus plicatilis 15 Snell et al. (1991) Microcrustáceo Artemia franciscana 18 Espiritu et al. (1995) Microalga Raphidocelis subcapitata 18 Janssen (1998)
Planta Allium cepa 20 Forget et al. (2000) Microcrustáceo Daphnia magna 20 Janssen (1998) Insecta Chironomus calligraphus 22,83 Original (Hg2+) Microcrustáceo Daphnia magna 26 Willemsen et al. (1995) Microcrustáceo Daphnia obtusa 30 Rossini & Ronco (1996) Insecta Chironomus calligraphus 33 Original (Pb2+) Microalga Ankistrodesmus falcatus 40 Magdaleno et al. (1997) Microcrustáceo Hyalella azteca 40 Steevens & Benson (1998) Insecta Chironomus calligraphus 45,45 Original (Cd2+)
Planta Lactuca sativa 46 Forget et al. (2000) Microcrustáceo Artemia franciscana 47 Janssen (1998) Microcrustáceo Corophium insidiosum 56 Reish ( 1993) Bacteria Muta-Cromo Placa 60 Pica et al. (2000) Bacteria Vibrio fisheri (Microtox) 65 Greene et al. (1985) Nemátoda Panagrellus redivivus 67 Diaz-Baez & Perez (2000)
Microrganismo Bacillus cereus 70 Forget et al. (2000)
Microcrustáceo Daphnia magna 71 Forget et al. (2000) Microcrustáceo Elasmopus bampo 72 Reish ( 1993) Microcrustáceo Moina macrocopa 77 Iannacone & Dale (1999) Nemátoda Panagrellus redivivus > 100 Pica et al. (2000)
CV % = Coeficiente de Variación

Ecología Aplicada, 2(1), 2003 ISSN 1726-2216 Depósito legal 2002-5474
EFECTO ECOTOXICOLÓGICO AGUDO DEL MERCURIO SOBRE LARVAS DEL “MUY MUY” Emerita analoga (STIMPSON) (DECAPODA: HIPPIDAE) PROCEDENTES DE CUATRO
LOCALIDADES DE LIMA José Alberto Iannacone Oliver1 y Lorena Alvariño Flores1
Resumen
La evaluación de Riesgos Ambientales (ERA) por el mercurio en los ambientes marinos requiere del empleo de especies bioindicadoras. Los organismos biológicos procedentes de localidades con diferentes influencias ambientales presentan respuestas variables en bioensayos ecotoxicológicos. Emerita analoga (Stimpson) es un invertebrado componente del macrozoobentos arenoso marino en la Costa Central del Perú. Se determinó el efecto ecotoxicológico agudo del mercurio (Hg2+) sobre larvas de E. analoga procedentes de cuatro playas de Lima, Perú: Santa María del Mar, Conchán, El Silencio y Puerto Viejo. Las larvas con menos de 24 h fueron obtenidas de hembras ovígeras en octubre y noviembre de 1999. Se alimentaron con la microalga Nannochloris sp. y se evaluaron en agua de mar con luz UV. El Diseño en bloques completamente aleatorizado empleado fue de 6 concentraciones x 4 repeticiones. Los ensayos ecotoxicológicos a 48 h y 96 h de exposición mostraron un valor de Concentración Letal media (CL50) promedio de 14,20 µg L-1 y 7,48 µg L-1, respectivamente. Los parámetros de Concentración de efectos no observables (NOEC) y de Concentración más baja de efectos observables (LOEC) fueron 0,65 µg L-1 y 1,63 µg L-1 a 96 h de exposición, respectivamente. No se encontraron diferencias significativas en los valores de CL50 entre las cuatro localidades evaluadas. El coeficiente de variabilidad (CV) para la CL50 fue aceptable (CV48h = 26,51 % y CV96h = 40,94 %). Los resultados son comparados con otras especies de crustáceos acuáticos al nivel mundial. Se analizan las perspectivas del uso de E. analoga como organismo bioindicador de toxicidad. Palabras claves: Crustacea, Emerita, macrozoobentos, mercurio, toxicidad.
Abstract Environmental Risk Assessment (ERA) for mercury in marine environment needs the use of
bioindicator species. Biological organisms coming from localities with different environmental influences are known to exhibit variable responses in ecotoxicological bioassays. Emerita analoga (Stimpson) is an invertebrate component of marine sand macrozoobenthos in the Peruvian Central coast. The ecotoxicological acute effects of mercury (Hg2+) on larvae of E. analoga from four beaches of Lima, Peru: Santa María del Mar, Conchan, El Silencio and Puerto Viejo were determined. Less than 24 h larvae were obtained from ovigerous females in October and November of 1999. They were fed with the Nannochloris sp. microalga and were evaluated using marine water with UV light. A completely randomized block design was employed with 6 concentrations x 4 repetitions. Ecotoxicological assays at 48 h and 96 h exposure showed a value of average Lethal medium concentration (LC50) of 14,20 µg L-1 and 7,48 µg L-1, respectively. Non Observed Effect Concentration (NOEC) and Lowest Observed Effect Concentration (LOEC) were 0,65 µg L-1 and 1,63 µg L-1 at 96 h exposure, respectively. Non significant differences in values of LC50 among the four localities evaluated were observed. Variability coefficient (VC) of LC50 was acceptable (VC48h = 26.51 % and VC96h = 40,94 %). The results were compared with other species of aquatic crustaceans at the global level. Perspectives of use of E. analoga as a toxicity bioindicador organism were analyzed. Key word: Crustacea, Emerita, macrozoobenthos, mercury, toxicity.
1 Laboratorio de Ecofisiología Animal. Facultad de Ciencias Naturales. Universidad Nacional Federico Villarreal (FCCNM-UNFV). Calle San Marcos 383, Lima 21, Perú. Correo electrónico: [email protected] / [email protected]
Introducción Emerita analoga (Stimpson 1857) “Muy Muy” es un crustáceo macrozoobentónico característico del intermareal e infralitoral de las playas de arena de las Costas del Pacífico Sur de América, a la vez que es un
eslabón importante en las tramas tróficas de este sistema (Oliva et al., 1992; Alvitrés et al., 1999; Contreras et al., 2000). Pese a su gran importancia pocos son los estudios tendientes a conocer aspectos de su ecología (Paul, 1976; Iannacone et al., 1996;

EFECTO ECOTOXICOLÓGICO DEL MERCURIO SOBRE LARVAS DEL Emerita analoga EN LIMA
Diciembre 2003 __________________________________________________________________________________________
112
Homola & Chang, 1997; Faulkes & Paul, 1998; Paul & Bruner, 1999). E. analoga ha sido empleada como especie bioindicadora de importancia ecotoxicológica para el monitoreo de algunos metales pesados (Hernández et al., 2000; Valdovinos & Zúñiga, 2002), de efluentes pesqueros y de ficotoxinas (Bretz et al., 2002; Ferdin et al., 2002; Powell et al., 2002) en el ecosistema marino. Los organismos biológicos procedentes de localidades con diferentes características ambientales podrían presentan diferentes respuestas en bioensayos ecotoxicológicos (Forbes, 1998; Repetto et al., 2000; Alayo & Iannacone, 2002). La contaminación por mercurio produce efectos deletéreos en diferentes componentes de la biota acuática (Pritchard, 1993; Rainbow, 1997; Iannacone & Alvariño, 1999). Se han hecho evaluaciones del efecto del mercurio (Hg2+) y del metilmercurio (MeHg2+) en el ambiente acuático (Baker et al., 1985; Pritchard, 1993; Andres et al., 2002). Según la Ley General de Aguas en el Perú, los límites máximos permisibles para el Hg2+ en aguas de Clase V para el cultivo de moluscos es de 0,1 µg L-1, y según la Agencia de Protección ambiental de los Estados Unidos (USEPA), es de 2 µg L-1 para la protección de los ambientes acuáticos naturales. En el Perú, se han evaluado del ambiente litoral marino a las siguientes tres especies: Tetrapigus niger (Molina) “erizo negro”, Arbacia spatuligera (Valenciennes) “erizo marrón” y Perumytilus purpuratus (Lamarck) (Iannacone et al., 1999; 2001). Muchas formas larvales y postlarvales de crustáceos marinos han sido empleadas para evaluar la ecotoxicidad de metales pesados, por constituir los estadios más sensibles de su ciclo de vida (Marino-Balsa et al., 2000). El objetivo fue determinar el efecto ecotoxicológico agudo del mercurio (Hg2+) sobre zoeas de E. analoga procedentes de cuatro Playas de la localidad de Lima: El silencio, Santa María del Mar, Conchán y Puerto Viejo. Materiales y métodos Emerita analoga Se procedió a la colección, identificación y cría estandarizada de E. analoga. Cincuenta hembras de E. analoga con presencia de masas ovígeras fueron colectadas de cada una de las cuatro playas arenosas: El Silencio, Santa María del Mar, Conchán y Puerto Viejo de Lima, Perú durante octubre y noviembre en 1999. Los ensayos ecotoxicológicos con E. analoga se iniciaron con larvas o “zoeas” de primer estadio dentro de 24 h de haber eclosionado de las masas ovígeras procedentes de cada una de las cuatro playas. Diez larvas de primer estadio se distribuyeron al azar en cada concentración nominal de Hg2+ evaluada en cada una de las cuatro repeticiones del ensayo
experimental. Las larvas se consideraron muertas si no fueron capaces de moverse coordinada y normalmente cuando fueron pinchadas ligeramente con un alfiler luego de 48 h y 96 h de exposición. Se emplearon un total de 240 larvas por ensayo. Los bioensayos semi-estáticos con renovación de la solución a 48 h de exposición fueron repetidos 2 veces en envases plásticos de 0,5 L. La temperatura se mantuvo en una incubadora fluctuante entre 19 oC ± 3 oC. Mercurio y Físico-química del agua La solución madre para el ión tóxico de mercurio fue preparada de la sal metálica a base del cloruro de mercurio (HgCl2) (Iannacone & Alvariño, 1999) a una concentración de 100 mg L-1. Todas las concentraciones se registraron en concentraciones nominales de ión de mercurio. Los matraces aforados previamente fueron lavados con ácido nítrico al 10 % y luego enjuagados en agua destilada y mantenidos a 4 oC. (Iannacone et al., 2000). Los datos de oxígeno disuelto se midieron mediante el método de Winkler (APHA, 1995). En el ensayo con Hg2+ se emplearon las siguientes cinco concentraciones 0,65 µg L-1, 1,63 µg L-1, 4,09 µg L-1, 10,24 µg L-1 y 25,60 µg L-1 (Iannacone & Alvariño, 2002). Para el Hg2+ las cinco concentraciones siguieron un factor de incremento en 2,5. Diseño experimental y Análisis de datos Se determinaron la Concentración Letal media (CL50) promedio, su desviación estándar y su Coeficiente de Variación (CV %) para todos los bioensayos con E. analoga expuestos al mercurio a 48 y 96 h de exposición para las cuatro localidades evaluadas. Las diferencias entre las concentraciones y entre las repeticiones se evaluaron a través de un Análisis de Varianza (ANDEVA) de dos vías, previa transformación de los datos a raíz cuadrada del arcoseno, con el fin de ajustar los datos a la distribución normal (Zar, 1996). En el caso de existir diferencias significativas entre las concentraciones y entre las repeticiones se realizó una Prueba de Significación DVS (Diferencia Verdaderamente Significativa) de Tukey (Daniel, 1993). Las CL50s se calcularon usando un programa computarizado de la EPA versión 1,5 (USEPA, 1993). Estos valores fueron obtenidos a partir del análisis de regresión lineal entre el logaritmo de la concentración de dilución de las muestras de agua (en porcentaje) y la mortalidad Probit (porcentaje transformado). Los porcentajes de mortalidad de las repeticiones para cada concentración fueron sumados. El modelo de regresión lineal fue verificado usando el estadístico χ2 (Martín & Holdich, 1986). Se determinó además el NOEC y el LOEC a 48 h y 96 h de exposición. Resultados y discusión Las variaciones de tres parámetros ecotoxicológicos y el CV respectivo para el crustáceo de E. analoga expuesto al mercurio son señalados en la Tabla 1.

JOSÉ IANNACONE Y LORENA ALVARIÑO
Ecología Aplicada Vol. 2 Nº 1, pp. 111-115 __________________________________________________________________________________________
113
Los porcentajes de mortalidad de las larvas de E. analoga aumentaron con cada una de las concentraciones crecientes de Hg2+ a 48 y 96 h de exposición (Tablas 2 y 3). Tabla 2. Porcentaje de Mortalidad de E. analoga expuestas al mercurio a 48 h. (% mortalidad) Μg L-1 1 2 3 4 Promedio ± DE Sig.
Control 0 a 0 a 0 a 0 a 0 ± 0 a 0,65 0 a 0 a 5 a 5 a 2,5 ± 2,88 a 1,63 1,13 a 0 a 13,33 b 5 a 4,86 ± 6,03 ab 4,09 8,88 b 2,50 a 15 b 10 ab 9,09 ± 5,13 b
10,24 50,60 c 37,50 b 26,19 c 17,50 b 32,94 ± 14,33 c 25,60 87,31 d 100 c 82,50 d 77,50 c 86,82 ± 9,65 d
1 = El Silencio; 2 = Santa María; 3 = Conchán; 4 = Puerto Viejo. DE = Desviación estándar; Sig. = Significancia. Letras iguales en una misma columna indican que los promedios son estadísticamente iguales. La secuencia ecotoxicológica en términos de CL50 de mayor a menor toxicidad fue: Daphnia obtusa > Daphnia pulex > Daphnia cucullata > Daphnia obtusa > Pagurus longicarpus > Daphnia magna > Emerita analoga (96 h) > Daphnia magna > Emerita analoga (48 h) > Corophium insidiosum > Elasmopus bampo > Homarus gammarus > Streptocephalus proboscideus > Thamnocephalus platyurus > Maja squinado > Palaemon serratus (Tabla 4). E. analoga ocupa la posición 7ma entre 16 bioensayos ecotoxicológicos empleando diferentes especies carcinológicas, componentes de la biota animal acuática (Tabla 4). McKinney & Rogers (1992) han señalado al mercurio como uno de los metales de mayor interés por la USEPA (Agencia de Protección Ambiental de los Estados Unidos), por lo que requiere colaboración entre toxicólogos y químicos para coordinar investigaciones en la química de la especiación de metales y en los efectos toxicológicos de la especiación de metales (Wright, 1995). Alayo & Iannacone (2002) presentan una evaluación comparativa global entre E. analoga,
Brachionus plicatilis Müller 1786 (Rotifera) y Artemia salina (L.) (Crustacea) para su eventual empleo en bioensayos ecotoxicológicos. Entre los criterios usados para validar un bioensayo tenemos: “facilidad de uso” que incluye rapidez del bioensayo, simplicidad en la manipulación, disponibilidad del material biológico, poco volumen de muestra y facilidad en la observación y conteo de los organismos afectados (muertos o inmóviles). Los decápodos como E. analoga permiten realizar pruebas de fácil lectura, pero con manipulación simple. La simplicidad de manipulación y lectura dependen esencialmente del tamaño de los organismos;
a mayor tamaño más simple será el método. Por ello, Brachionus al poseer menor tamaño que Emerita, presenta mayor dificultad en su manejo. E. analoga es una especie con alta disponibilidad en el Perú, como material biológico para bioensayos. El criterio de “costo” incluye lo invertido en materiales y los gastos del trabajo, tanto en el cultivo como en la prueba de toxicidad. Los bioensayos con Brachionus son menos costosos que utilizar Emerita o Artemia. La “concordancia ecológica” en el ambiente marino favorece principalmente a E. analoga por su importancia en las redes tróficas. Tabla 3. Porcentaje de Mortalidad de E. analoga expuestas al mercurio a 96 h. (% mortalidad)
µg L-1 1 2 3 4 Promedio ± DE Sig.
control 0 a 0 a 0 a 0 a 0 ± 0 a
0,65 21,22 b 2,29 a 5 a 3,87 a 8,09 ± 8,82 b
1,63 39,40 c 1,30 a 33,33 b 10,73 b 28,69 ± 12,45 c
4,09 33,34 c 11,92 b 25,64 b 45 c 28,97 ± 13,87 c
10,24 84,85 d 57,38 c 52,38 c 72,23 d 66,71 ± 14,74 d
25,60 100 e 100 d 100 d 100 e 100 ± 0 e 1 = El Silencio; 2 = Santa María; 3 = Conchán; 4 = Puerto Viejo. DE = Desviación estándar; Sig. = Significancia. Letras iguales en una misma columna indican que los promedios son estadísticamente iguales. Muchas especies propuestas como herramientas para bioensayos ecotoxicológicos, requieren como requisitos para uso no solo que sean sensibles, sino que sean ensayos reproducibles con poca variabilidad en su respuesta. En el caso de E. analoga al provenir de cuatro localidades diferentes de la costa central de Lima, Perú, no presentó diferencias significativas entre los valores de CL50, NOEC y LOEC para el Hg2+ a 48 y 96 h de exposición (Tabla 1). Finalmente, E. analoga en su fase larvaria en el ciclo de vida de esta especie es apropiada para su empleo en bioensayos ecotoxicológicos para evaluar el metal Hg2+ en el ambiente acuáticos marino.
Tabla 1. Valores de CL50, NOEC y LOEC del mercurio sobre larvas de Emerita analoga procedentes de cuatro playas de Lima, Perú.
Parámetros (ug L-1) El Silencio Santa MaríaConchánPuerto Viejo
Promedio
± DE Coeficiente de variación
CL50 48 h 10,60 10,96 12,92 18,18 13,16 ± 3,49 26,51 CL50 96 h 3,16 8,69 5,40 8,37 6,40 ± 2,62 40,94 NOEC 48 h 1,63 4,09 0,65 4,09 2,61 ± 1,74 66,66 NOEC 96 h < 0,65 1,63 0,65 0,65 0,89 ± 0,49 55,05 LOEC 48 h 4,09 10,24 1,63 10,24 6,55 ± 4,37 66,71 LOEC 96 h 0,65 4,09 1,63 1,63 2,00 ± 1,46 73 NOEC = Concentración de efectos no observables. LOEC = Concentración más baja de efectos observables. DE = Desviación estándar.

EFECTO ECOTOXICOLÓGICO DEL MERCURIO SOBRE LARVAS DEL Emerita analoga EN LIMA
Diciembre 2003 __________________________________________________________________________________________
114
Literatura citada Alayo M. & Iannacone J. 2002. Ensayos
ecotoxicológicos con petróleo crudo, Diesel 2 y Diesel 6 con dos subespecies de Brachionus plicatilis Müller 1786 (Rotifera: Monogononta). Gayana. 66: 45-58.
Alvitres V., Chanamé J., Fupuy J., Chambergo A. & Cortez M. 1999. Cambios en la prevalencia de los helmintos parásitos de Emerita analoga por efecto de “El Niño 1997-97”. Rev. per. Biol. Vol. Extraordinario.: 69-76.
Andres S., Laporte J.M. & Mason R.P. 2002. Mercury accumulation and flux across the gills and the intestine of the blue crab (Callinectes sapidus). Aquat. Toxicol. 56:303-20.
APHA (AMERICAN PUBLIC HEALTH ASSOCIATION), AWWA (AMERICAN WATER WORKS ASSOCIATION), WPCF (WATER POLLUTION CONTROL FEDERATION). 1995. Standard methods for examination of water and wastewater. 19th. Ed. American Health Association. Washington, D.C.
Baker R., Lavie B. & Nevo E. 1985. Natural selection for resistance to mercury pollution. Experientia. 41: 697-699.
Brkovic-Popovic I. 1990. Effect of mercury on the survival of Daphnia magna. Wat. Sci. Tech. 22: 241-246.
Bretz C.K., Manouki T.J. & Kvitek R.G. 2002. Emerita analoga (Stimpson) as an indicator species for paralytic shellfish poisoning toxicity along the California Coast. Toxicon. 40: 1189-1196.
Centeno M.D.F., Perssone G. & Goyvaerts M.P. 1995.
Cyst-based toxicity test. IX. The potential of Thamnocephalus platyurus as test species in comparison with Streptocephalus proboscideus (Crustacea: Brachipoda: Anostraca). Environm. Toxicol. Water Qual. 10: 275-282.
Contreras H., Jaramillo E. & Quijón P. 2000. Natural history of Emerita analoga (Stimpson) (Anomura, Hippidae) in a sandy beach of northern Chile. Rev. chil. Hist. nat. 73: 705-715.
Daniel W.W. 1993. Bioestadística. Tercera Edición. Editorial Limusa (México).
Faulkes Z. & Paul D.H. 1998. Digging in sand crabs: coordination of joints in individuals legs. J. Exp. Biol. 201: 2139-2149.
Ferdin M.E., Kvitek R.G., Bretz C.K., Powell C.L., Doucette G.J., Lefebvre K.A. & Coale S. 2002. Emerita analoga (Stimpson) – possible new indicator species for the phycotoxin domoic acid in California coastal waters. Toxicon. 40: 1259-1265.
Forbes V.E. 1998. Sources and implications of variability in sensitivity to chemicals for ecotoxicological risk assessment. Arch. Toxicol. Suppl. 20: 407-418.
Hernández C., Yánez R. & Rudolph A. 2000. Toxicity response of Emerita analoga (Stimpson, 1857) collected from beaches of South Central Chile. Bull. Environ. Contam. Toxicol. 65: 567-572.
Homola E. & Chang E.S. 1997. Distribution and regulation of esterases that hydrolyze methyl farnesoate in Homarus americanus and other crustaceans. Gen. Comp. Endocrinol. 106: 62-72.
Tabla 4. Valores comparativos de CL50 con especies de crustáceos marinos y dulceacuícolas expuestos al mercurio. Especie de Crustáceos
CL50 µg L-1
Tiempo de
exposición(h) Estadio
Ambiente Referencia
Daphnia obtusa 2 48 Neonato D Rossini & Romo (1996) Daphnia pulex 2,2 48 Neonato D Rossini & Romo (1996) Daphnia cucullata 2,4 48 Neonato D Rossini & Romo (1996) Daphnia obtusa 4 24 Neonato D Rossini & Romo (1996) Pagurus longicarpus 4 168 Adultos M Eisler & Hennekey (1977) Daphnia magna 1,5 – 3,2 48 Neonato D Rossini & Romo (1996) Emerita analoga 6,40 96 Primero M Original Daphnia magna 9,2 48 Neonato D Brkovic-Popovic (1990) Emerita analoga 13,16 48 Primero M Original Corophium insidiosum 20 96 Adultos M Reish (1993) Elasmopus bampo 20 96 Adultos M Reish (1993) Homarus gammarus 48 48 Primero M Marino-Balsa et al. (2000) Streptocephalus proboscideus 60 24 Nauplio D Centeno et al. (1995) Thamnocephalus platyurus 60 24 Nauplio D Centeno et al. (1995) Maja squinado 72 72 Primero M Marino-Balsa et al. (2000) Palaemon serratus 74 72 Primero M Marino-Balsa et al. (2000) M = marino; D = dulceacuícola.

JOSÉ IANNACONE Y LORENA ALVARIÑO
Ecología Aplicada Vol. 2 Nº 1, pp. 111-115 __________________________________________________________________________________________
115
Iannacone J. & Alvariño L. 1999. Acute ecotoxicity of heavy metals using juveniles of freshwater snail Physa venustula (Gould, 1847) (Mollusca). Gayana. 63: 101-110.
Iannacone J. & Alvariño L. 2002. Efecto del detergente doméstico alquil aril sulfonato de sodio lineal (LAS) sobre la mortalidad de tres caracoles dulceacuícolas en el Perú. Ecol. Apl. 1: 81-87.
Iannacone J., Alvariño L., Dionisio M. & Sánchez J. 1999. Efecto del Cu2+ en la embrionación del erizo negro Tetrapigus niger (Molina) y en la fertilización del erizo marrón, Arbacia spatuligera (Valenciennes) (Echinodermatha). Bol. Soc. Quim. Peru. 65: 183-190.
Iannacone J., Dale W. & Alvariño L. 2000. Monitoreo ecotoxicológico del río Rímac (Lima-Perú) empleando a Chironomus calligraphus Goeldi (Diptera: Chironomidae). Rev. Chil. Ent. 27: 25-34.
Iannacone J., García J., Vela H., Ticona C., Torres E., Quinte G. & Vidarte K. 2001. Toxicidad y bioacumulación de plomo en Perumytilus purpuratus (Lamarck, 1819) “chorito” (Bivalvia). Bol. Soc. Quim. Peru. 67: 89-98.
Iannacone J., Villaseca A. & Mari C. 1996. Algunos aspectos ecológicos de la fauna parasitaria (metazoa) de Callianassa garthi “marucha” y Emerita analoga “muy muy” en la costa central del Perú. Congreso Nacional de Ecología. 25-28 de marzo 1996. Lima, Perú.
Lanston W.J. 1990. Toxic effects of metals and the incidence of metal pollution in marine ecosystems. pp. 101- 119. In: Heavy metals in the marine environment (Furness, R.W. & Rainbow, P. S., eds.). CRC Press, Inc. Boca Raton, Florida, USA.
Marino-Balsa J.C., Poza. E., Vazquez. E. & Beiras, R. 2000. Comparative toxicity of dissolved metals to early larval stages of Palaemon serratus, Maja squinado, and Homarus gammarus (Crustacea: Decapoda). Arch. Environ. Contam. Toxicol. 39: 345-351.
Martin T.R. & Holdich D.M. 1986. The acute lethal toxicity of heavy metals to peracarid crustaceans (with particular reference to freshwater asellids and gammarids). Water Res. 20: 1137-1147.
McKinney J. & Rogers R. 1992. Metal bioavailability: EPA workshop identified research needs. Environ. Sci. Technol. 26: 1298-1299.
Oliva M.E., Luque J.L. & Cevallos A. 1992. Parásitos de Emerita analoga (Stimpson) (Crustacea): Implicancias ecológicas. Bol. Lima (Perú). 79: 77-80.
Paul D.H. 1976. Role of proprioceptive feedback from nonspiking mechanosensory cells in the sand crab, Emerita analoga. J. Exp. Biol. 65: 243-258.
Paul D.H. & Bruner J. 1999. Receptor potentials and electrical properties of nonspiking strech-receptive neurons in the sand crab Emerita analoga (Anomura : Hippidae). J. Neurophysiol. 81: 2493-2500.
Powell C.L., Ferdin M.E., Busman M., Kvitek R.G. & Doucette G.J. 2002. Development of a protocol for determination of domoic acid in the sand crab (Emerita analoga): a possible new indicator species. Toxicon 40: 485-492. Pritchard J.B. 1993. Aquatic Toxicology: past,
present, and prospects. Environ. Health Perspectives. 100: 249-257.
Rainbow P.S. 1997. Trace metals accumulation in marine invertebrates: marine biology or marine chemistry. J. mar. biol. Ass. U.K. 77: 195-210.
Reish D.J. 1993. Effect of metals and organic compounds on survival and bioaccumulation in two species of marine gammaridean amphipod, together with a summary of toxicological research on this group. Jour. Nat. History. 27: 781-794.
Repetto G., Peso A. & Repetto M. 2000. Alternative ecotoxicological methods for the evaluation, control and monitoring for environmental pollutants. Ecotoxicol. Environm. Res. 3: 47-51.
Rossino G.B. & Ronco A.E. 1996. Acute toxicity bioassay using Daphnia obtusa as a test organism. Environm. Toxicol. Water Qual. 11: 255-258.
USEPA. 1993. Probit analysis program. Version 1,5. U.S. Environmental Protection Agency. Washington D.C. U.S.A.
Valdovinos C. & Zuñiga M. 2002. Copper acute toxicity tests with the sand crab Emerita analoga (Decapoda: Hippidae): a biomonitor of heavy metal pollution in Chilean coastal seawater?. Bull. Environ. Contam. Toxicol. 69: 393-400.
Wright D.A. 1995. Trace metals and major ion interactions in aquatic animals. Mar. Poll. Bull. 34: 8-17.
Zar J.H. 1996. Bioestatistical Analysis. 3th Ed. Prentice –Hall. Inc. Upper Saddle River, Jew Jersey.

Ecología Aplicada, 2(1), 2003 ISSN 1726-2216 Depósito legal 2002-5474
MACROINVERTEBRADOS EN LAS LAGUNAS DE PUERTO VIEJO, LIMA – PERÚ José Iannacone1, Jaime Mansilla2 y Karen Ventura3
Resumen
Se realizó un estudio para evaluar la biodiversidad y similaridad de los macroinvertebrados del bentos de las lagunas de Puerto Viejo del 15 de Julio al 15 de Octubre del 2001. Se realizaron siete muestreos (M) temporales en ocho estaciones (E) espaciales en sus lagunas naturales y artificiales. Se calculó la diversidad alfa usando el Indice de Diversidad de Shannon-Wienner (H´), el de Equidad de Pielou (J´) y el Indice de Dominancia de Simpson (C). Para la determinación de la diversidad beta se empleó los Indices cuantitativos de Sörensen y Morisita-Horn. Un total de 5519 especímenes, tres filas, seis órdenes, nueve familias y 10 especies fueron colectadas en el macrozoobentos durante todo este estudio. Las taxas más dominantes durante este estudio fueron: Melanoides tuberculata (Müller) (Mesogastropoda: Thiaridae) con 3276 individuos (59,36 %), Chironomus sp. (Diptera: Chironomidae) con 1281 individuos (23,21 %) y Heleobia cumingi (Orbigny) (Mesogastropoda: Hydrobiidae) con 891 individuos (16,14 %). Estas tres especies presentaron una distribución espacial amontonada a lo largo de todos los muestreos realizados. Los valores de diversidad alfa mediante los Indices de diversidad de Shannon-Wiener, de Equidad de Pielou y de Simpson no variaron significativamente entre los siete muestreos realizados y entre las ocho estaciones censadas. Palabras claves: Chironomus, Heleobia, humedales, macrozoobentos, Melanoides, Perú
Abstract A study was performed to evaluate the biodiversity and similarity of benthic macroinvertebrates of the Puerto Viejo Lagoons from15 july to 15 october, 2001. Seven samplings (Sa) were conducted in eight spatial stations (St) in their natural and artificial lagoons. The alpha diversity was calculated using the Shannon-Wiener (H´), evenness of Pielou and dominance of Simpson (C) indexes. Beta diversity was determined employing quantitative Sörensen and Morisita-Horn indexes. A total of 5519 specimens belonging to three phyla, six orders, nine families and ten species were collected from macrozoobenthos during all the study. The most abundant taxa were Melanoides tuberculata (Müller) (Mesogastropoda: Thiaridae) with 3276 individuals (59,36 %), Chironomus sp. (Diptera: Chironomidae) with 1281 individuals (23,21 %) and Heleobia cumingii (Orbigny) (Mesogastropoda: Hydrobiidae) with 891 specimens (16,14 %). These three species had a crowded spatial distribution in all the samplings. The values of alpha diversity: the Shannon-Wiener, equableness of Pielou and Simpson indexes did not change significantly among the seven samplings and in the eight surveyed stations. The values of beta diversity (quantitative indexes of Sörensen and Morisita- Horn) were higher between Sa3 and SA4; and also in Sa5, Sa6 and Sa7 between each other. For the eight spatial stations Sörensen and Morisita- Horn cuantitative indexes showed a similarity over 70% between St3- St4; St4- St6 and St5- St7. St2- St6 showed the highest disssimilarity employing both cuantitative indexes. Key words: Chironomus, Heleobia, macrozoobentos, Melanoides, Peru, wetlands.
11 Laboratorio de Ecofisiología. Facultad de Ciencias Naturales y Matemáticas. Universidad Nacional Federico Villarreal. Calle San Marcos 383, Pueblo Libre, Lima 21, Perú. Correo electrónico: [email protected] 2 Jaime Mansilla Correo electrónico: [email protected] 3 Karen Ventura Correo electrónico: [email protected]
Introducción Los humedales comprenden diversos cuerpos de agua, temporales o permanentes, ámbitos que son hábitats para microorganismos, flora y fauna características (Brown et al., 1997; Ugarte, 1998; Franco et al., 2000; Pulido 2000). Sin embargo, a pesar que se ha logrado algunos avances encaminados a la conservación de estos ecosistemas sobre todo en la costa, son los primeros que sufren presión antropogénica (Obando et al., 1998), por lo que aún queda mucho por hacer en el campo legal (CONAM,
1999), político y económico (Reaño & Guardia, 2001). Las comunidades bentónicas sean marinas o epicontinentales son todas las especies que viven en relación íntima con el fondo, ya sea para fijarse en él, para excavarlo, o para movilizarse sobre su superficie, siendo el factor básico para su establecimiento, “su relación íntima con el fondo” (Salazar et al., 2001). Las especies bentónicas mayores de 1 mm de longitud, se consideran macrozoobentos (Jaramillo et al., 1998).

JOSÉ IANNACONE, JAIME MANSILLA Y KAREN VENTURA
Ecología Aplicada Vol. 2 Nº 1, pp. 116-124 __________________________________________________________________________________________
117
La determinación cualitativa y cuantitativa de la comunidad de invertebrados nos sirven como indicadores biológicos de la calidad del agua (Prat et al., 1999; Cauchie et al., 2000; Gaete et al., 2000); así como su rol como transportadores mecánicos de bacterias u otros patógenos por vía externa e interna (Angeler et al., 2000; Kim et al., 2000). En el departamento de Lima, Perú, se encuentran los humedales de las lagunas de Puerto Viejo, un conjunto de cuerpos de agua con altas salinidades de diferentes profundidades en medio de un extenso gramadal y juncal, que recoge las afloraciones hídricas del acuífero del Río Mala, con un área de doscientas has. Estas lagunas, conforman junto a otros humedales de la Costa, una red de afloramientos casi continuos que son una fuente de vida para muchísimas especies (Reaño & Guardia, 2001; Pulido 2000). A la fecha existen muy escasos trabajos publicados sobre este ecosistema natural con relación a inventarios y censos de su biota (Ciriaco et al., 1997; León, 1997). Los estudios sobre lagunas costeras naturales son de urgente necesidad (Arias & Sierra, 2001), porque se sabe que la destrucción de los hábitats y la extinción de la vida silvestre ocasiona la ruptura de una cadena ecológica (Obando et al., 1998), es por ello el interés de realizar el presente trabajo durante el 2001. La importancia del estudio de este humedal de Puerto Viejo radica en que sirve de refugio, nidificación y alimentación de aves continentales y locales, provenientes del Hemisferio Norte y de zonas altoandinas respectivamente. Muchas de estas aves residentes o migratorias se alimentan del macrozoobentos de estos humedales. El objetivo de presente trabajo fue evaluar la biodiversidad alfa y beta del macrozoobentos de las Lagunas de Puerto Viejo durante el 15 de Julio al 15 de Octubre del 2001. Materiales y métodos Los pantanos de Puerto Viejo, se localizan en la provincia de Cañete en el distrito de San Antonio de Mala (Km. 71 de la panamericana Sur) (12° 33-34´ Lat. Sur 76° 42-43´ Long. Oeste). En estas lagunas recorren las afloraciones hídricas del acuífero de Mala que están ubicadas paralelamente al océano, sus profundidades oscilan entre 0,3 m a 2,4 m y se encuentran a 20 msnm, la temperatura fluctúa de 17 y 22 °C. Las plantas vasculares dominantes de éste ecosistema son Distichlis spicata (L.) Green (Poaceae), Salicornia fruticosa, Cyperus laevigatus L. (Cyperaceae), el junquillo Scirpus americanus (Cyperaceae), Potamogeton striatus R & P. (Potamogetonaceae) y Bacopa monnieri (L.) Pennell (Scrophulariaceae) (León et al., 1995; Reaño & Guardia, 2001). Los siete muestreos se realizaron quincenalmente desde el 15 de julio al 15 de octubre de 2001. Se establecieron 8 estaciones de muestreo en las
diferentes lagunas (Figura 1). Las características de las ocho estaciones se presentan en Tabla 1.
Figura 1. Puntos de muestreo de macrozoobentos en las Lagunas de Puerto Viejo. Se colectó 1 L de sustrato bentónico de cada estación a 1m de la orilla empleando un envase de plástico de 2 L de capacidad, el cual fue introducido manualmente al sustrato y posteriormente fueron tamizadas con un colador de plástico de 1000 µ, para separar el macrozoobentos, siguiendo las recomendaciones de Wetzel & Likens (2000). Se determinó la temperatura (T°), pH y Oxígeno disuelto (estos tres parámetros fueron tomados en todas las estaciones solo en los dos últimos muestreos), presencia de vegetación vascular acuática, tipo de gránulo del sustrato predominante y rango de profundidad de cada estación (Tabla 1). La conductividad eléctrica para todas las estaciones de muestreo fue de 1mS. Además mediante la transparencia del agua se calificó a cada estación en dos categorías: clara o turbia, si es que se observaba o no el disco de Secchi a 10 cm de profundidad. En el laboratorio, cada muestra fue separada en morfoespecies y se fijaron en viales de vidrio de 5mL, con alcohol etílico al 70%, debidamente rotulados para su posterior identificación y clasificación. Para los moluscos se siguió el procedimiento descrito por Thiengo et al. (2001) y para la identificación de los invertebrados se usó claves taxonómicas especializadas (Merritt & Cummins, 1984; Borror et al., 1995; Vivar et al., 1998; Mazzucconi, 2000). Muestras de importancia taxonómica fueron depositadas en la colección malacológica del Museo de Historia Natural- Universidad Nacional Mayor de San Marcos (MHN, UNMSM) y en la colección entomológica del Programa Nacional de Control Biológico, Servicio Nacional de Sanidad Agraria (PNCB, SENASA), Lima, Perú. Se catalogaron las morfoespecies en tres gremios alimentarios: depredadores, fitófagos y detritívoros empleando los
GGaarriittaa ddee CCoonnttrrooll
CCaasseettaa
MMiirraaddoorr
CCaarrrreetteerraa PPaannaammeerriiccaannaa SSuurr
== EEssttaacciioonneess ddee mmuueessttrreeoo
ZZoonnaa NNoorrttee
ZZoonnaa SSuurr 11
22
== CCuueerrppoo ddee aagguuaa
33
44 88
77
55
66
ZZoonnaa CCeennttrraall
NN

MACROINVERTEBRADOS EN LAS LAGUNAS DE PUERTO VIEJO, LIMA – PERÚ
Diciembre 2003 __________________________________________________________________________________________
118
criterios propuestos por Iannacone et al. (2000a) y Salazar & Iannacone (2001). Con la finalidad de evaluar la diversidad alfa en las comunidades macrozoobentónicas de cada estación y muestreo, fueron utilizados los siguientes índices: Shannon- Wiener, que varía de 0 a log base 2 del número de morfoespecies censadas, determinadas por el número de morfoespecies presentes en la comunidad y basándose en la escala logarítmica escogida H’ = -Σpilogpi,donde , H’ = índice de diversidad de Shannon-Wiener, (log) = logaritmo base 2; pi = ni/N; donde ni = al número de individuos de cada morfoespecie, N = número total de individuos (Iannacone et al., 2000a). El otro índice utilizado fue el de Simpson (C) que varía entre 0 y 1, cuya fórmula es: C =1 -Σ (ni/N)2, donde ni = al número de individuos en la muestra o estación, siendo el valor equivalente a 1 como el de máxima diversidad. Se calculó el índice de equidad de Pielou, empleando J´= H’/log2S, donde S = es el número máximo de especies en la muestra. Además, se utilizaron dos procedimientos no paramétricos para estimar la riqueza de especies de este ecosistema: Chao 2 y Jacknife de primer orden (Jack 1) (Moreno, 2001).
Para la diversidad beta de similaridad, se uso el índice de Sörensen cuantitativo (IScuant) = 2pn/ an+bn, donde, an = número total de individuos en el sitio a (muestreo o estación), bn = número total de individuos en el sitio b (muestreo o estación), pn = sumatoria de la abundancia mas baja de cada una de las morfoespecies compartidas entre ambos sitios; y el índice de Morisita –Horn, que varía de 0 a un valor máximo próximo a uno. IM-H =2Σ(anixbnj)/ (da+db)aNxbN, donde: ani = número de individuos de la i-ésima morfoespecie en el sitio A (muestreo o estación), bnj =número de individuos de la - j ésima
morfoespecie en el sitio B (muestreo o estación), da = Σ(ani)2/ aN2 y db = Σ(bnj)2/bN2 (Moreno, 2001). Se empleó el coeficiente de correlación de Pearson para determinar si existía algún tipo de correlación lineal entre las poblaciones de los tres macroinvertebrados más abundantes tanto por muestreo como por estaciones. El ANDEVA se utilizó para determinar si el número de individuos promedio entre los macroinvertebrados más abundantes, H´, J y C por muestreo y estación por separado, fueron estadísticamente iguales. Para las especies más abundantes se determinó la distribución espacial (al azar, amontonada o uniforme), utilizando el criterio de S2/X (Varianza/ Promedio de individuos por estación), realizándose la determinación en cada uno de los siete muestreos. Solo se incluyeron en el análisis las estaciones que presentaron a la especie evaluada. Para todos los casos se empleó un nivel de significancia de alfa = 0,05. Resultados Se colectaron un total de 5519 ejemplares distribuidos en 10 especies, 9 familias en 6 órdenes. El número total de familias y el número de individuos
colectados en cada uno de los 7 muestreos y 8 estaciones realizados durante los tres meses de monitoreo, se observa en la Tabla 2. El orden Mesogastropoda con dos especies representó el 75,77 % de toda la muestra censada. La familia con mayor número de especímenes fue Thiaridae, con la especie Melanoides tuberculata (Muller). Las larvas de la familia Chironomidae con Chironomus sp. fue entre los insectos la más predominante (Tabla 2). La familia Hydrobiidae con Heleobia cumingii (Orbigny) con 891 individuos fue el tercer invertebrado en orden de importancia. Con relación a la posición trófica, el gremio alimentario cualitativamente dominante fueron depredadores (60 %), luego los detritívoros con 20 %
Tabla 1. Principales características de las ocho estaciones de los humedales de Puerto Viejo, Lima, Perú. Estación (E)
Tipo de laguna
Tipo de agua
Profundidad a 1 m de la orilla (Rango en cm)
Sustrato Predominante del fondo
Vegetación
Temperatura T°C
PH
O2 disuelto
E1 A Clara 52 – 78 Arenoso Sí 23 8 10 E2 N Clara 37 – 54 Arcilloso Sí 23 8 8 E3 N Turbia 15- 28 Arenoso Sí 26 8 3 E4 N Clara 40 – 57 Arenoso No 25 9 2 E5 N Clara 20 – 45 Arenoso No 24 9 3 E6 A Turbia 19 – 43 Arenoso No 24 9 3 E7 N Clara 2 – 30 Arenoso Sí 24 9 3 E8 A Clara 30 – 45 Arenoso No 23 8 1 A = Artificial. N = Natural. V = Vegetación = Sí (incluye Cyperus laevigatus, Scirpus americanus, Potamogeton striatus, Distichlis spicata, Bacopa monnieri).

JOSÉ IANNACONE, JAIME MANSILLA Y KAREN VENTURA
Ecología Aplicada Vol. 2 Nº 1, pp. 116-124 __________________________________________________________________________________________
119
y los fitófagos con 20 %; en cambio cuantitativamente, los fitófagos dominaron con 75,5 %, luego los detritívoros con 23,34 % y finalmente con un menor porcentaje estuvieron los depredadores (1,12 %) (Tabla 2).
Figura 2. Variaciones poblacionales de tres macroinvertebrados en las Lagunas de Puerto Viejo, Lima, Perú. La Figura 2 nos indica la variación del número de individuos totales de M. tuberculata, Chironomus sp. y H. cumingii con relación a los siete muestreos realizados en los humedales de Puerto Viejo. Se puede observar una tendencia a disminución del número total de larvas de III y IV estadio de Chironomus a través de los muestreos realizados; en cambio Heleobia, a excepción del primer muestreo presenta una tendencia a incrementar sus poblaciones. Sin embargo, el análisis de correlación lineal mostró que no existe ningún tipo de relación significativa entre las poblaciones de estos tres invertebrados por muestreo (Tabla 3). Además el ANDEVA mostró que el número promedio de individuos por muestreo para estas tres especies varió significativamente entre sí (M. tuberculata = 468 ± 413,73 (127 – 1311); Chironomus sp. = 183 ± 130,64 (52 – 409) y H.
cumingii = 127,28 ± 122,06 ( 0 – 257) (F = 3,45; g. l. = 2 y 18; P = 0,05). La Figura 3 nos indica la variación del número de individuos totales de M. tuberculata, Chironomus sp. y H. cumingii con relación a las ocho estaciones realizadas en los humedales de Puerto Viejo. Sin
embargo, el análisis de correlación lineal mostró que solo existe relación negativa y significativa entre las poblaciones de Chironomus y Heleobia (Tabla 3). Además el ANDEVA mostró que el número promedio de individuos por estación para estas tres especies no varió significativamente entre sí (M. tuberculata = 409,50 ± 548,05 (0 – 1421); Chironomus sp. = 160,12 ± 35,87 (95 – 202) y H. cumingii = 111,37 ± 95,89 (5 – 294); F = 1,97; g. l. = 2 y 21; P = 0,16).
Figura 3. Variaciones poblacionales espaciales de tres macroinvertebrados en las Lagunas de Puerto Viejo, Lima, Perú. Para casi todos los casos la distribución espacial de M. tuberculata, Chironomus sp. y H. cumingii fue amontonada, debido a que presentaron valores mayores a 1 a la relación Varianza/ Promedio del número de individuos (Tabla 4). Solo en el segundo
Tabla 2. Macrozoobentos de la Laguna de Puerto Viejo, Lima, Perú durante el 15 de julio Al 15 de octubre del 2001.
Orden Familia Especie Posición Trófica Número (Abundancia porcentual)
Mesogastropoda Thiaridae Melanoides tuberculata F 3276 (59,36) Diptera Chironomidae Chironomus sp. (l) D 1281 (23,21) Mesogastropoda Hydrobiidae Heleobia cumingii F 891 (16,14) Hemiptera Notonectidae* Notonecta peruviana (a, ni) P 30 (0,54) Odonata Coenagrionidae sp. no identificada (na) P 24 (0,43) Diptera Culicidae* Aedes sp. (l, p) D 10 (0,18) Coleoptera Hydrophilidae Berosus sp. (l) P 3 (0,06) Tricladida Planariidae Girardia festai P 2 (0,04) Odonata Libellulidae sp. no identificada (na) P 1 (0,02) Coleoptera Hydrophilidae Tropisternus sp. (l) P 1 (0,02) P= Depredador D = Detritívoro F = Fitófago a = adulto ni = ninfa l = larva na = nayade p = pupa * Organismos no típicamente considerados del macrozoobentos.
0
500
1000
1500
0 1 2 3 4 5 6 7Muestreos
No
de in
divi
duos
to
tale
s
MelanoidestuberculataChironomus sp.
0
200
400
600
800
1000
1200
1400
1600
0 1 2 3 4 5 6 7 8Es t ac io ne s
M elano idestuberculataChirono mus sp .
Heleob ia cuming ii

MACROINVERTEBRADOS EN LAS LAGUNAS DE PUERTO VIEJO, LIMA – PERÚ
Diciembre 2003 __________________________________________________________________________________________
120
muestreo para M. tuberculata se observó una distribución al azar, quizás influenciado por ser solo dos estaciones las que presentaron este caracol.
Tabla 4. Valores de distribución espacial (Varianza / Promedio de individuos) de tres macroinvertebrados de las lagunas de Puerto Viejo, Lima, Perú.
Mue
stre
os
Mel
anoi
des
tube
rcul
ata
Tipo
de
dist
ribuc
ión
Chi
rono
mus
sp
.
Tipo
de
dist
ribuc
ión
Hel
eobi
a cu
min
gii
Tipo
de
dist
ribuc
ión
M1 436,57 A 27,14 A 22,77 A M2 0,18 Az 11,80 A -* - M3 4,16 A 11,20 A - - M4 18,36 A 14,53 A - - M5 36,20 A 3,56 A 13,09 A M6 23,81 A 6,46 A 42,61 A M7 52,84 A 6,54 A 37,04 A * = se presentó esta especie en ≤ a una estación. A = distribución amontonada. Az = distribución al azar.
Los resultados de los Indices de diversidad de Shannon-Wiener (H´) y de equidad (J´), así como el de Simpson (C), calculados para los siete muestreos temporales y para las ocho estaciones espaciales, muestran que el H’ con mayor valor promedio fue en el segundo muestreo (0,99) y con el menor valor el cuarto muestreo (0,69). C muestra valores más bajos, siendo el mayor valor el segundo y tercer muestreo (0,43) y con el menor valor en el primer muestreo (0,35). J en el primer muestreo alcanzó el mayor valor
(0,89) y el menor valor lo presentó en el quinto muestreo temporal (0,49). Con relación a las estaciones de muestreo (E), H’ presentó el mayor valor promedio en la segunda estación (0,99) y con el menor valor la sexta estación (0,55). C muestra valores más bajos, siendo el mayor valor la tercera estación (0,49) y con el menor valor en la primera estación (0,26). J en la cuarta estación alcanzó el mayor valor (0,87) y el menor valor lo presentó en la sexta estación espacial (0,49). Sin embargo, no se encontraron diferencias significativas en los valores de H´, J´ y C, ni por muestreo, ni tampoco por estaciones para los macroinvertebrados de los humedales de Puerto Viejo (Tabla 5).
Tabla 5. Valores comparativos del estadístico F y sus respectivos valores de probabilidad entre H´, J´ y C para los muestreos temporales y estaciones espaciales del macrozoobentos de las Lagunas de Puerto Viejo, Lima, Perú (F = Prueba de Fisher del ANDEVA; P = Probabilidad; g.l.= grados de libertad).
Indi
ces
F m
uest
reos
P m
uest
reos
g.l.
F Es
taci
ones
P est
acio
nes
g.l.
H´ 0,86 0,52 5 y 33 1,43 0,23 7 y 31J´ 0,65 0,66 5 y 33 1,91 0,10 7 y 31 C 0,16 0,97 5 y 33 1,21 0,33 7 y 31
Los dos procedimientos no paramétricos para estimar la riqueza específica de este humedal: Chao 2 y Jacknife de primer orden (Jack 1), para la comunidad de macroinvertebrados de Puerto Viejo, obtuvieron valores de 18 y 13,5 respectivamente. Lo cual indica en comparación a la riqueza de especies de 10, obtenida a lo largo de todo el muestreo, que aún se requiere un mayor monitoreo de los macroinvertebrados bentónicos durante otras épocas del año. Para evaluar la similaridad por muestreos temporales los índices cuantitativos de Sörensen y Morisita-Horn mostraron coincidencia entre los M3 y M4; así como entre M5, M6, M7 entre sí. Sin embargo, solo Sörensen indicó una alta similaridad entre M2 y M4 (Tabla 6). En contraste, solo Morisita indicó una similaridad entre M1 con M5, M6 y M7. Por ambos índices en los muestreos M1 y M3; así como M1 y M4 presentaron la más alta disimilaridad en todo el monitoreo. Para las ocho estaciones espaciales, el índice de Sörensen cuantitativo mostró similaridades sobre el 70% entre E3 y E4; E4 y E6; E5 y E7, y E2 y E8. En cambio para Morisita- Horn E3 y E4; E3 y E6; E4 y E6; E5 y E7; E5 y E8, y entre E7 y E8 presentaron las mayores similaridades. Las estaciones E2 y E6 presentaron la mayor disimilaridad empleando ambos índices cuantitativos (Tabla 7).
Tabla 3. Matriz de correlación entre el número de individuos totales, según fecha de muestreo y según estación de muestreo, para tres especies de macroinvertebrados de las Lagunas de Puerto Viejo, Lima, Perú.
Por Muestreo Melanoides
tuberculata Chironomus
sp. Heleobia cumingii
Melanoides tuberculata
- 0,46* (0,29)**
0,67 (0,09)
Chironomus sp.
0,15 (0,71)
- -0,20 (0,66)
Por E
stac
ión
Heleobia cumingii
0,17 (0,67)
-0,71 (0,048)
-
* = Coeficiente de correlación de Pearson ** = Significancia o significación observada (valor P) Valores sobre la diagonal corresponden a correlaciones entre especies según fechas de muestreo (n = 7). Valores debajo de la diagonal corresponden a correlaciones entre especies según estaciones de muestreo (n = 8).
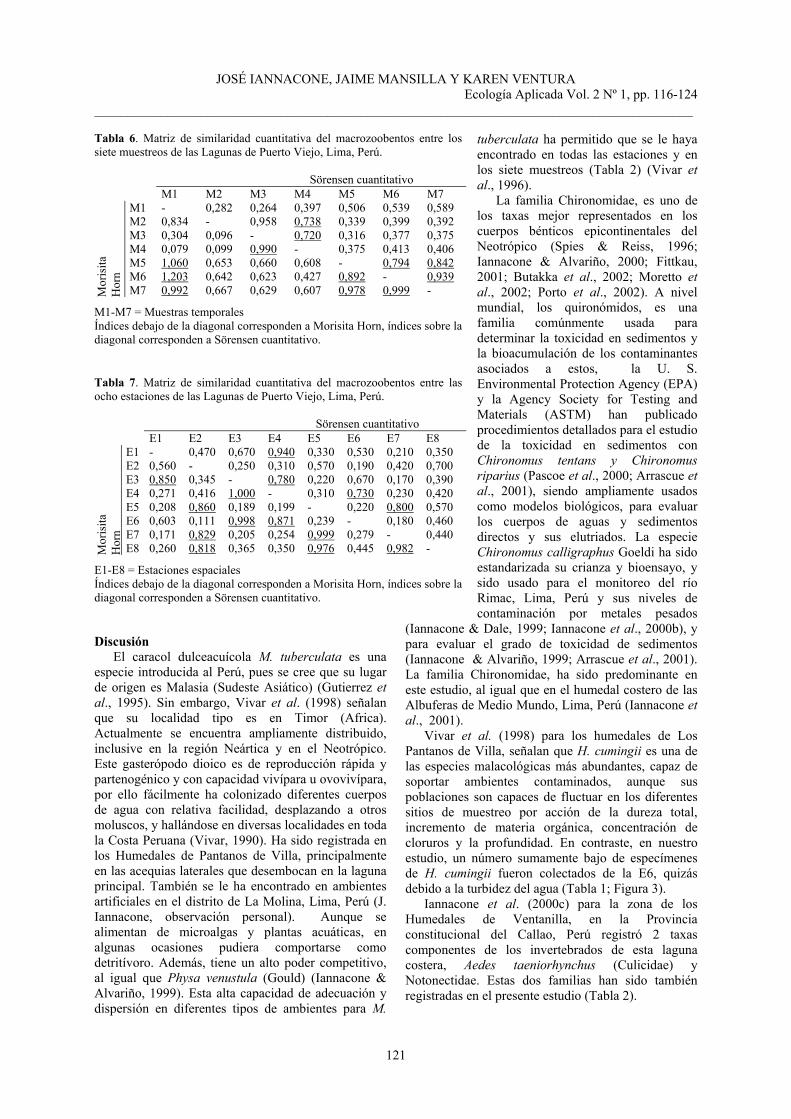
JOSÉ IANNACONE, JAIME MANSILLA Y KAREN VENTURA
Ecología Aplicada Vol. 2 Nº 1, pp. 116-124 __________________________________________________________________________________________
121
Discusión El caracol dulceacuícola M. tuberculata es una especie introducida al Perú, pues se cree que su lugar de origen es Malasia (Sudeste Asiático) (Gutierrez et al., 1995). Sin embargo, Vivar et al. (1998) señalan que su localidad tipo es en Timor (Africa). Actualmente se encuentra ampliamente distribuido, inclusive en la región Neártica y en el Neotrópico. Este gasterópodo dioico es de reproducción rápida y partenogénico y con capacidad vivípara u ovovivípara, por ello fácilmente ha colonizado diferentes cuerpos de agua con relativa facilidad, desplazando a otros moluscos, y hallándose en diversas localidades en toda la Costa Peruana (Vivar, 1990). Ha sido registrada en los Humedales de Pantanos de Villa, principalmente en las acequias laterales que desembocan en la laguna principal. También se le ha encontrado en ambientes artificiales en el distrito de La Molina, Lima, Perú (J. Iannacone, observación personal). Aunque se alimentan de microalgas y plantas acuáticas, en algunas ocasiones pudiera comportarse como detritívoro. Además, tiene un alto poder competitivo, al igual que Physa venustula (Gould) (Iannacone & Alvariño, 1999). Esta alta capacidad de adecuación y dispersión en diferentes tipos de ambientes para M.
tuberculata ha permitido que se le haya encontrado en todas las estaciones y en los siete muestreos (Tabla 2) (Vivar et al., 1996). La familia Chironomidae, es uno de los taxas mejor representados en los cuerpos bénticos epicontinentales del Neotrópico (Spies & Reiss, 1996; Iannacone & Alvariño, 2000; Fittkau, 2001; Butakka et al., 2002; Moretto et al., 2002; Porto et al., 2002). A nivel mundial, los quironómidos, es una familia comúnmente usada para determinar la toxicidad en sedimentos y la bioacumulación de los contaminantes asociados a estos, la U. S. Environmental Protection Agency (EPA) y la Agency Society for Testing and Materials (ASTM) han publicado procedimientos detallados para el estudio de la toxicidad en sedimentos con Chironomus tentans y Chironomus riparius (Pascoe et al., 2000; Arrascue et al., 2001), siendo ampliamente usados como modelos biológicos, para evaluar los cuerpos de aguas y sedimentos directos y sus elutriados. La especie Chironomus calligraphus Goeldi ha sido estandarizada su crianza y bioensayo, y sido usado para el monitoreo del río Rimac, Lima, Perú y sus niveles de contaminación por metales pesados
(Iannacone & Dale, 1999; Iannacone et al., 2000b), y para evaluar el grado de toxicidad de sedimentos (Iannacone & Alvariño, 1999; Arrascue et al., 2001). La familia Chironomidae, ha sido predominante en este estudio, al igual que en el humedal costero de las Albuferas de Medio Mundo, Lima, Perú (Iannacone et al., 2001). Vivar et al. (1998) para los humedales de Los Pantanos de Villa, señalan que H. cumingii es una de las especies malacológicas más abundantes, capaz de soportar ambientes contaminados, aunque sus poblaciones son capaces de fluctuar en los diferentes sitios de muestreo por acción de la dureza total, incremento de materia orgánica, concentración de cloruros y la profundidad. En contraste, en nuestro estudio, un número sumamente bajo de especímenes de H. cumingii fueron colectados de la E6, quizás debido a la turbidez del agua (Tabla 1; Figura 3). Iannacone et al. (2000c) para la zona de los Humedales de Ventanilla, en la Provincia constitucional del Callao, Perú registró 2 taxas componentes de los invertebrados de esta laguna costera, Aedes taeniorhynchus (Culicidae) y Notonectidae. Estas dos familias han sido también registradas en el presente estudio (Tabla 2).
Tabla 6. Matriz de similaridad cuantitativa del macrozoobentos entre los siete muestreos de las Lagunas de Puerto Viejo, Lima, Perú. Sörensen cuantitativo M1 M2 M3 M4 M5 M6 M7
M1 - 0,282 0,264 0,397 0,506 0,539 0,589 M2 0,834 - 0,958 0,738 0,339 0,399 0,392 M3 0,304 0,096 - 0,720 0,316 0,377 0,375 M4 0,079 0,099 0,990 - 0,375 0,413 0,406 M5 1,060 0,653 0,660 0,608 - 0,794 0,842 M6 1,203 0,642 0,623 0,427 0,892 - 0,939
Mor
isita
H
orn
M7 0,992 0,667 0,629 0,607 0,978 0,999 -
M1-M7 = Muestras temporales Índices debajo de la diagonal corresponden a Morisita Horn, índices sobre la diagonal corresponden a Sörensen cuantitativo.
Tabla 7. Matriz de similaridad cuantitativa del macrozoobentos entre las ocho estaciones de las Lagunas de Puerto Viejo, Lima, Perú. Sörensen cuantitativo E1 E2 E3 E4 E5 E6 E7 E8
E1 - 0,470 0,670 0,940 0,330 0,530 0,210 0,350 E2 0,560 - 0,250 0,310 0,570 0,190 0,420 0,700 E3 0,850 0,345 - 0,780 0,220 0,670 0,170 0,390 E4 0,271 0,416 1,000 - 0,310 0,730 0,230 0,420 E5 0,208 0,860 0,189 0,199 - 0,220 0,800 0,570 E6 0,603 0,111 0,998 0,871 0,239 - 0,180 0,460 E7 0,171 0,829 0,205 0,254 0,999 0,279 - 0,440
Mor
isita
H
orn
E8 0,260 0,818 0,365 0,350 0,976 0,445 0,982 -
E1-E8 = Estaciones espaciales Índices debajo de la diagonal corresponden a Morisita Horn, índices sobre la diagonal corresponden a Sörensen cuantitativo.

MACROINVERTEBRADOS EN LAS LAGUNAS DE PUERTO VIEJO, LIMA – PERÚ
Diciembre 2003 __________________________________________________________________________________________
122
Melo et al. (2002) y Santos et al. (2002) han evaluado la fauna de coleópteros acuáticos en el neotrópico de Brazil, registrando a las larvas de la familia Hydrophilidae, con Tropisternus y Berosus como los géneros más abundantes, dentro de los coleópteros. En el presente estudio también se registró a ambos géneros en las Lagunas costeras de Puerto Viejo, Lima, Perú. El turbelario Girardia festai (= Dugesia festai) registrado para el humedal de Pantanos de Villa, se encuentran presente solo en la séptima estación de muestreo (Sarmiento & Morales, 1998). No han sido registrados en Puerto Viejo, moscas dipteras ciclorrafas muy comunes en humedales dulceacuícolas (Keiper et al., 2000). Como señaló Vivar et al. (1998) el estudio de los macroinvertebrados de Los Pantanos de Villa, Lima, Perú constituyen un grupo de especies claves para el monitoreo y conservación de ese ecosistema particular, pues esta biota acuática, puede actuar como hospederos intermediarios, controladores biológicos, indicadores de la calidad del agua y parte del flujo energético para algunos vertebrados. Algunos muestreos temporales son más similares entre sí, como es el caso de M5, M6 y M7; así como entre M3 y M4 (Tabla 6). Los parámetros ecológicos evaluados en la Tabla 1, durante el periodo de estudio de 90 días, no fueron muy fluctuantes. Por lo que se requeriría mayores estudios para determinar si la insolación, salinidad u otros factores fisicoquímicos serían los responsables de explicar las altas similaridades encontradas en dichos muestreos. Las estaciones de muestreo que mostraron las mayores consistencias por ambos índices de similaridad fueron E3 y E4; E4 y E6, y finalmente E5 y E7 (Tabla 7). E3 y E4 fueron bastante similares debido a que fueron lagunas naturales, presentaron sustrato arenoso, temperaturas semejantes, pero ligeramente más altas que el resto de las estaciones y oxígeno disuelto similares (Tabla 1). Así como la cercanía espacial de ambos puntos de muestreo (Figura 1). E4 y E6 son coincidentes en la ausencia de vegetación, fondo arenoso, temperatura, pH y oxígeno disuelto (Tabla 1). Finalmente para E5 y E7 su similaridad se relaciona a que ambas son lagunas naturales, presentan aguas claras, sustrato arenoso, temperatura, pH y oxígeno disuelto (Tabla 1). Así como la cercanía espacial de ambos puntos de muestreo (Figura 1). Los gremios de fitófagos y detritívoros incluyeron pocas especies con un número altos de especímenes con amplia distribución espacial y temporal. En contraste los depredadores fueron numéricamente en bajo número, pero con mayor riqueza de taxas (Tabla 2). Este mismo patrón fue encontrado en la fauna de invertebrados en los humedales de Ventanilla, Callao, Perú (Iannacone et al., 2000b).
El censo de la fauna de invertebrados en el presente estudio para la Laguna de Puerto Viejo, humedal como Los Pantanos de Villa, permite también justificar el establecimiento de estrategias para la conservación de este ecosistema. Observaciones adicionales deberían ser hechas en estos cuerpos de agua para determinar el cambio espacio –temporal de los componentes de macroinvertebrados bentónicos. Entre las principales conclusiones de este trabajo tenemos: (1) un total de 5519 especímenes, tres filas, seis órdenes, nueve familias y diez especies fueron colectados en el macrozoobentos durante todo este estudio; (2) las tres especies más dominantes durante este estudio fueron: Melanoides tuberculata (Müller) (Mesogastropoda: Thiaridae) con 3276 individuos (59,36 %), Chironomus sp. (Diptera: Chironomidae) con 1281 individuos (23,21 %) y Heleobia cumingii (Mesogastropoda: Hydrobiidae) con 891 individuos (16,14 %). Estas tres especies presentaron una distribución espacial amontonada o contagiosa; (3) los valores de diversidad alfa: los Indices de diversidad de Shannon-Wiener, de Equidad de Pielou y de Simpson no variaron significativamente entre los siete muestreos realizados y entre las ocho estaciones censadas; (4) los valores de diversidad beta (los índices cuantitativos de Sörensen y Morisita-Horn) por muestreo fueron más altos entre los M3 y M4; así como entre M5, M6, M7 entre sí. Por ambos índices en los muestreos M1 y M3; así como M1 y M4 presentaron la más alta disimilaridad en todo el monitoreo y (5) para las ocho estaciones espaciales, los índices de Sörensen y Morisita- Horn cuantitativo mostraron similaridades sobre el 70% entre E3 y E4; E4 y E6; E5 y E7. Las estaciones E2 y E6 presentaron la mayor disimilaridad empleando ambos índices cuantitativos. Agradecimientos A María Isabel La Torre Acuy por la identificación taxonómica de la flora vascular de los humedales de Puerto Viejo. Literatura citada Angeler D.G., Alvarez-Cobelas M., Rojo C. &
Sánchez-Carrillo S. 2000. The significance of water inputs to plankton biomass and trophic relationships in a semi-arid freshwater wetland (central Spain). J. Plankton Res. 22: 2075- 2093.
Arias C.N.F. & Sierra P.C. 2001. La Gestión ambiental marina y costera en Colombia: un reto institucional. Libro de Resúmenes del IX Congreso Latinoamericano sobre Ciencias del Mar. San Andrés Isla, Colombia. Sept. 16 – 20, 2001. Universidad Nacional de Colombia. : 501-504.
Arrascue A., Iannacone J., Alvariño L., Basilio S. & Lazcano C. 2001. El insecto Chironomus calligraphus Goeldi y la bacteria Escherichia coli

JOSÉ IANNACONE, JAIME MANSILLA Y KAREN VENTURA
Ecología Aplicada Vol. 2 Nº 1, pp. 116-124 __________________________________________________________________________________________
123
como ensayos ecotoxicológicos para evaluar sedimentos elutriados dulceacuícolas. Rev. per. Ent. 42: 159-173.
Borror D., De Long D. & Thriplehorn C. 1995. An introduction to the study of insects. Saunders College Publishing. 6th Edition. USA.
Brown S.C., Smith K. & Batzer D. 1997. Macroinvertebrate responses to wetland restoration in Northern New York. Environ. Entomol. 26: 1016-1024.
Butakka C.M.M., Komatsu E.H. & Takeda A.M. 2002. Composição da fauna bentônica da lagoa Ventura, sistema rio Ivinheima, bacia do Alto Rio Parana (MS, PR). XXIV Congresso Brasileiro de Zoologia: A Zoologia e os Ecossistemas Costeiros. 17 a 22 de fevereiro de 2002, Itajaí – Santa Catarina, Brazil. : 256.
Cauchie H.M., Hoffmann L. & Thomé J.P. 2000. Metazooplankton dynamics and secondary production of Daphnia magna (Crustacea) in an aerated waste stabilization pond. J. Plankton Res. 22: 2263 - 2287.
CONAM (Consejo Nacional del Ambiente). 1999. Perú Megadiverso. Prioridades en uso y conservación de la biodiversidad para el desarrollo sostenible. Del Río, M. L. (Ed.). Lima, Perú.
Ciriaco F.J., Campos G., Rodríguez C. & Aguilar C. 1997. Caracterización del ecosistema lagunas de Puerto Viejo. Libro de Resúmenes del Congreso Peruano de Ecología, 25 al 28 marzo de 1997, Lima-Perú. Universidad Nacional Agraria La Molina. : 53.
Fittkau E.J. 2001. Amazonian Chironomidae (Diptera: Chironomidae): A contribution to chironomid research in the neotropics. Amazoniana. 16: 313-324.
Franco P.J., Sulca Q.L. & Cáceres M.C. 2000. Cyanophytas de los Humedales del Valle de Ite Tacna. Libro de Resúmenes del VIII Congreso Nacional de Botánica, Abril 24- 28, 2000. Arequipa. : 51.
Gaete H., Bay-Schmith E., Baeza J. & Rodríguez J. 2000. Ecotoxicological assessment of two pulps mill effluent, Biobio River Basin, Chile. Bull. Environ. Contam. Toxicol. 65: 183-189.
Gutierrez A.A., Perera G.P., Yong M.C., Ferrer J.L. & Sánchez J.N. 1995. Distribución y posible competencia entre Melanoides tuberculata y Tarebia granifera (Prosobranchia: Thiaridae) en el lago Hanabanilla, Cuba. Rev. Cubana Med. Trop. 15: 1-7.
Iannacone J., Alayo M., Arrascue A., Sánchez J. & Abanto M. 2001. Las trampas de luz para evaluaciones rápidas de la biodiversidad de la artropofauna: análisis de tres casos. Wiñay Yachay. 5: 7-20.
Iannacone J., Alayo M. & Sánchez J. 2000a. Biodiversidad de la artropofauna del Bosque
Zárate, Lima, Perú, empleando tres técnicas de censo. Wiñay Yachay. 4(1): 27-46.
Iannacone J. & Alvariño L. 1999. Ecotoxicidad aguda de metales pesados empleando juveniles del caracol de agua dulce Physa venustula (Gould, 1847) (Mollusca). Gayana. 63: 101-110.
Iannacone J. & Alvariño L. 2000. Chironomus calligraphus Goeldi y Moina macrocopa (Sars) como herramientas ecotoxicológicas para la evaluación del lindano y clorpirifos. Bol. Soc. Biol. Concepción. 71: 33-39.
Iannacone J., Alvariño L., Moreno R., Reyes M. & Chauca J. 2000b. Culícidos (Diptera) del río Chillón y sectores adyacentes de la provincia constitucional del Callao, Perú, durante el Niño 1997-1998. Acta Ent. Chilena. 24: 51-60.
Iannacone J. & Dale W. 1999. Protocolo de bioensayos ecotoxicológicos para evaluar metales pesados en el agua dulce con Chironomus calligraphus (Diptera: Chironomidae) y Moina macrocopa (Crustácea: Cladocera), en el río Rímac, Lima, Perú. Rev. per. Ent. 41: 111-120.
Iannacone J., Dale W. & Alvariño L. 2000c. Monitoreo ecotoxicológico del río Rímac (Lima- Perú) empleando a Chironomus calligraphus Goeldi (Diptera: Chironomidae). Rev. Chilena Ent. 27: 23-34.
Jaramillo E., Carrasco F., Quijon P., Pino M. & Contreras H. 1998. Distribución y estructura comunitaria de la macrofauna bentónica en la costa del norte de Chile. Rev. Chilena Hist. Nat. 71: 459-478.
Keiper J.B., Walton W.E. & Foote B.A. 2002. Biology and ecology of the higher diptera from freshwater wetlands. Annu. Rev. Entomol. 47: 207-232.
Kim H.W., Hwang S.J. & Joo G.J. 2000. Zooplankton grazing on bacteria and phytoplankton in a regulated large river (Nakdong River, Korea). J. Plankton Res. 22: 1559 – 1577.
León B. 1997. Uso acertado de los humedales de la Costa Central: Retos y posibilidades. Resúmenes del Congreso Peruano de Ecología, 25 al 28 marzo de 1997, Lima-Perú. Universidad Nacional Agraria La Molina. : 6.
León B., Cano A. & Young K.R. 1995. La flora vascular de los Pantanos de Villa, Lima, Perú: Adiciones y guía para las especies comunes. Pub. Mus. Hist. Nat., UNMSM (B). 38: 1-39.
Mazzucconi S.A. 2000. A review of the South american Notonecta (Paranecta) including N. inca new species from Peru (Heteroptera: Notonectidae). First Abstract Book I. XXI International Congress of Entomology, Brazil.: 267.
Melo M., Mattos M.L.T., Martins J.F.S., Dias R.A., Rodenas P.A., Benetti C.J. & Florentín G.L. 2002. Registro de coleoptera em ecossistema de arroz irrigado tratado com inseticida. XXIV Congresso

MACROINVERTEBRADOS EN LAS LAGUNAS DE PUERTO VIEJO, LIMA – PERÚ
Diciembre 2003 __________________________________________________________________________________________
124
Brasileiro de Zoologia: A Zoologia e os Ecossistemas Costeiros. 17 a 22 de fevereiro de 2002, Itajaí – Santa Catarina, Brazil. : 256.
Merrit R.W. & Cummins K.M. 1984. An introduction to the aquatic insects of North America. Kendall. Hunt Publishing Co. Dubuque, IA. U.S.A.
Moreno C.E. 2001. Métodos para medir la biodiversidad. Manuales y Tesis SEA. Sociedad Entomológica Aragonesa Ed. Madrid, España.
Moretto Y., Takeda A.M. & Butaca C.M.M. 2002. Distribução espacial de larvas de Chironomidae (Diptera) em seis diferentes ambientes da planície aluvial do Rio Paraná. XXIV Congresso Brasileiro de Zoologia: A Zoologia e os Ecossistemas Costeiros. 17 a 22 de fevereiro de 2002, Itajaí – Santa Catarina, Brazil. : 587.
Obando C.M., Campos C.M., García Z.P. & Romero N.M. 1998. Inventario de la diversidad ornitológica del humedal del caucato Pisco durante 1997. Ecología (Perú). 1: 72-75.
Pascoe D., Wenzel A., Janssen C., Girling A.E., Jüttner I., Fliedner A., Blockwell S.J., Maund S.J., Taylor E.J., Diedrich M., Persoone G., Verhelst P., Stephenson R.R., Crossland N.O., Mitchell G.C., Pearson N., Tatterfield L., Lay J.P., Peither A., Neumeier B. & Velletti A.R. 2000. The development of toxicity tests for freshwater pollutants and their validation in stream and pond mesocosms. Wat. Res. 34: 2323-2329.
Porto L.A.C., Ribeiro K.D. & Silva R.C. 2002. Invertebrados bentônicos do alto rio Tocantins- Goiás. XXIV Congresso Brasileiro de Zoologia: A Zoologia e os Ecossistemas Costeiros. 17 a 22 de fevereiro de 2002, Itajaí – Santa Catarina, Brazil. : 584.
Prat N., Toja J., Sola C., Burgos M.D., Plans M. & Rieradevall M. 1999. Effect of dumping and cleaning activities on the aquatic ecosystems of the Guadiamar River following a toxic flood. Sci. Total Environ. 242: 231 - 248.
Pulido V. 2000. Memorias sobre el Taller de Conservación de Humedales de Sitios Ramsar en el Pacífico Sur. 20 –21 de octubre de 1999, Trujillo, Perú.
Reaño G. & Guardia M. 2001. Salvemos Puerto Viejo. Recurso ecoturístico de Lima. Disponible en: http//:www.conam.gob.pe/chm/docs/otros/eco/otros/ecoptoviejo leído el 20 de noviembre del 2001.
Salazar N.C. & Iannacone J. 2001. Censos rápidos empleando la técnica de barber para evaluar la artropofauna del Parque Nacional Yanachaga-Chemillén, Sector Rio Pescado Oxapampa- Pasco. Bol. Lima (Perú). 125: 126-130.
Salazar N.C., Iannacone J., Alvariño L., Orozco R. & Miranda R. 2001 Estructura macrozoobentónica de fondo blando en la bahía de Ilo, Moquegua- Perú. Wiñay Yachay. 5: 179-192.
Santos A.D.D., Ferreira Jr.N. & Nessimian J.L. 2002. Caracterização das formas adultas e larvais de Hydrophiloidea (Insecta, Coleoptera) sensu Hansen 1991, para o estado do Rio de Janeiro e localidades vizinhas. XXIV Congresso Brasileiro de Zoologia: A Zoologia e os Ecossistemas Costeiros. 17 a 22 de fevereiro de 2002, Itajaí – Santa Catarina, Brazil. : 256.
Sarmiento L. & Morales M.E. 1998. Protozoarios, turbelarios y nemátodos de los Pantanos de Villa, Lima, Perú: su importancia en el ecosistema. pp. 41-54. Boletín del Museo de Historia Natural, Universidad Nacional Mayor de San Marcos. Serie de divulgación No 11. Cano, A.; Young, K. R. (Eds.). Lima, Perú.
Spies M. & Reiss F. 1996. Catalog and bibliography of Neotropical and Mexican Chironomidae. Spixiana. 22: 61-119.
Thiengo S.C., Fernández M.A., Boaventura M.F., Grault C.E., Silva H.F.R., Mattos A.C. & Santos S.B. 2001. Freshwater snails and schistosomiasis mansoni in the state of Rio de Janeiro, Brazil: I- Metropolitan Mesoregion. Mem. Inst. Oswaldo Cruz. 96: 177-184.
Ugarte J.A.N. 1998. Los humedales de la Reserva Nacional Salinas y Aguada Blanca: 21 000 Flamencos que proteger. Libro de Resúmenes del IV Congreso Latinoamericano de Ecología. II Congreso Peruano de Ecología. 20 al 25 de Octubre 1998. Arequipa- Perú. Instituto Regional de Ciencias Ambientales. Facultad de Ciencias Biológicas y Agropecuarias. Departamento Académico de Biología. Universidad San Agustín. Arequipa - Perú.
Vivar R. 1990. Un gasterópodo de la familia Thiaridae en el Perú: Melanoides tuberculata (Müller, 1774). Bol. Lima (Perú). 69: 33-34.
Vivar R., Larrea H., Huaman P., Yong M. & Perera G. 1996. Some ecological aspects of the freshwater molluscan fauna of Pantanos de Villa, Lima, Perú. Malacol. Rev. 29: 65- 68.
Vivar G.R., Ramírez M.R. & Huamán P. 1998. Moluscos de los Pantanos de Villa y su aporte a la Conservación. pp. 55-73. Boletín del Museo de Historia Natural, Universidad Nacional Mayor de San Marcos. Serie de divulgación No 11. Cano, A.; Young, K. R. (Eds.). Lima, Perú.
Wetzel R.G. & Likens G.E. 2000. Limnological analyses. 3th Ed. Springer- Verlag, New York.

Ecología Aplicada, 2(1), 2003 ISSN 1726-2216 Depósito legal 2002-5474
REPRODUCCIÓN DE LA TARICAYA Podocnemis unifilis TROSCHEL 1848 (TESTUDINES: PODOCNEMIDIDAE) EN LA VÁRZEA DEL MEDIO SOLIMÕES, AMAZONAS, BRASIL.
Augusto Fachín Terán1 y Eduardo Matheus von Mülhen2
Resumen
Este trabajo fue realizado de julio a noviembre de 1998, en la Reserva de Desarrollo Sustentable Mamirauá. La reserva está localizada en la parte media del río Solimões, cerca de la ciudad de Tefé, Amazonas, Brasil. El período de desove de Podocnemis unifilis ocurrió en los meses de agosto y septiembre, y la eclosión en octubre y noviembre. Los nidos fueron construidos en sustratos de greda (67.5%), arena (25%), y hojarasca (7.5%). La obtención de crías fue mayor en la playa de arena y menor en los barrancos de tierra gredosa. El hombre y la iguana Tupinambis, son los mayores depredadores de huevos. La protección y manejo de los locales de desove, la educación ambiental, la participación comunitaria y la fiscalización permanente del Instituto Brasilero de los Recursos Naturales y del Medio Ambiente – IBAMA son entre las actividades más importantes para viabilizar la recuperación de la población de P. unifilis en esta parte de la Amazonía. Palabras clave: Amazonía, depredación, Podocnemis unifilis, quelonio, reproducción.
Resumo Este trabalho foi realizado de julho a novembro de 1998, na Reserva de Desenvolvimento Sustentável Mamirauá. A reserva está situada no médio Solimões, perto da cidade de Tefé, Amazonas, Brasil. O período de desova de Podocnemis unifilis ocorreu nos meses de agosto e setembro, e a eclosão em outubro e novembro. Os ninhos foram construídos em substratos de argila (67,5%), areia (25%), e folhiço (7,5%). A obtenção de filhotes foi maior na praia de areia e menor nos barrancos de terra argilosa. O homem e o lagarto Tupinambis, foram os maiores predadores de ovos. A proteção e o manejo dos locais de desova, a educação ambiental, a participação comunitária e a fiscalização permanente do Instituto Brasileiro dos Recursos Naturais e do Meio Ambiente-IBAMA estão entre as atividades mais importantes para viabilizar a recuperação da população de P. unifilis nesta parte da Amazônia. Palavras chave: Amazônia, Podocnemis unifilis, predação, quelônio, reprodução.
Abstract I studied the nesting biology of Podocnemis unifilis from July to November 1998 in the Mamirauá Sustainable Development Reserve located in the Solimões River near Tefé, mazonas, Brazil. Podocnemis unifilis nested in August and September, with the hatching event occurring in October and November. Nests were excavated in clay soils (67.5%), sand (25%), and leaf litter (7.5%). Hatching success was highest in the sand beach nests and lowest in the clay banks nests. Humans and the tegu lizard (Tupinambis) were the main egg predators. This turtle population can be recuperated only by nesting beaches protection, educational programs for the inhabitants of the Reserve, participation of the community in the conservation and management program, and permanent guarding of the nesting beaches by the Instituto Brasilero de los Recursos Naturales y del Medio Ambiente personnel. Kew words: Amazonía, predation, Podocnemis unifilis, turtle, reproduction
Introducción La taricaya, Podocnemis unifilis, es un quelonio de tamaño medio dentro de la familia Pelomedusidae. En función de la continua presión de caza a que está siendo sometida, sus poblaciones han disminuido en
las áreas donde está distribuida, y actualmente la especie esta en la lista 1, como especie amenazada (UICN, 1996). En la Reserva de Desarrollo Sustentable Mamirauá –RDSM, esta especie viene siendo permanentemente
____________________________ 1 Universidade do Estado do Amazonas. Escola Normal Superior. Avenida Darcy Vargas 2490 - Bairro Chapada. CEP: 69050-020, Manaus, Amazonas, Brasil. Correo electrónico: [email protected] Universidad Nacional de la Amazonía Peruana. Departamento Académico de Ecología y Manejo. Facultad de Ciencias Biológicas. Casilla Postal 496. Iquitos, Loreto-Perú. 2 Museu Paraense Emílio Goeldi -Campus de Pesquisa - Centro de Zoologia - Laboratório de Herpetologia-Av. Perimetral, 1901/1907 - Terra Firme - Cx. Postal 399- CEP 66017-970 - Belém, Pará, Brasil. Correo elctrónico: [email protected]

REPRODUCCIÓN DE LA TARICAYA Podocnemis unifilis, AMAZONAS, BRASIL
Diciembre 2003 _______________________________________________________________________________________
126
explorada para uso en la alimentación de los comunitarios que viven en la reserva; así como comercializada ilegalmente en las ciudades de Tefé y Alvarães (Fachín-Terán, 1999). En esta reserva, la población de P. unifilis está seriamente amenazada por el hombre que usa este recurso de manera no sustentable. Según Vogt et al. (1994), uno de los primeros pasos para el manejo y la conservación de cualquier especie es el entendimiento de su ecología reproductiva. Podocnemis unifilis tal vez sea la especie más investigada dentro de los Podocnemis en lo que se refiere a su ecología reproductiva. Una serie de trabajos fueron realizados en la Amazonía de Brasil (Vanzolini, 1977; Souza & Vogt, 1994), de Colombia (Medem, 1964; Foote, 1978; Paéz, 1995; Paéz & Bock, 1997, 1998), de Venezuela (Thorbjarnarson et al., 1993), de Bolivia (Caballero, 1996) y del Perú (Ponce, 1979; Fachín-Terán, 1992, 1993; Fachín-Terán et al., 1997; Soini, 1995a, 1995b, 1995c; Soini & Coppula, 1995; Soini & Soini, 1995a, 1995b, 1995c; Soini, 1996, 1997). A pesar de todo este esfuerzo, el único registro sobre reproducción de esta especie en la várzea del medio Solimões es una nota científica sobre nidificación (Thorbjarnarson & Silveira, 1996). Casi todos los aspectos sobre la ecología de esta especie son desconocidos en esta parte de la región amazónica. El propósito de este estudio fue colectar información básica sobre el periodo de nidificación y eclosión, y cuantificar la pérdida de nidos y la sobrevivencia de embriones de P. unifilis en la RDSM. Esto ayudará en el futuro a planificar estudios más detallados sobre la ecología de esta especie. Materiales y métodos Nuestro estudio fue realizado en el Sector Jarauá de la Reserva de Desarrollo Sustentable Mamirauá - RDSM, que está localizada en la Amazonía Occidental Brasilera, entre los ríos Japurá, Solimões y Auti-Paraná, cerca de la ciudad de Tefé, en el Estado de Amazonas (03º08'S, 64º45'W y 2º36'S, 67º13'W). Esta reserva tiene una extensión de 1´124,000 ha y está dividida en dos partes: un Área Focal, con cerca de un sexto del área total (260,000 ha), y un Área Subsidiaria complementaria. El Área Focal esta dividida políticamente en nueve sectores. El sector Jarauá esta situado en el río Japurá, y es el mayor de los sectores, con área de 563 km2, y población estimada en 338 habitantes. La RDSM alberga individuos dispersos de Podocnemis expansa, una pequeña población de P. unifilis y una población relativamente grande de P. sextuberculata. Existen cuatro especies de quelonios menos abundantes: Phrynops raniceps, Platemys platycephala, Chelus fimbriatus y Kinosternon scorpioides. También ocurre la especie terrestre
Geochelone denticulata, adaptada a las condiciones de la várzea (Fachín-Terán, 1999). Los nidos de taricaya fueron estudiados en la playa de Pirapucu (2º53'14”S y 64º52'8”O) y en el Paraná de Jarauá (2º50'52”S y 64º58'29”O). Los locales de reproducción fueron visitados periódicamente de julio a noviembre de 1998. Estas áreas fueron identificadas con la ayuda de dos asistentes de campo de la comunidad de San Raimundo de Jarauá. Durante la temporada de desove de 1998, la Playa de Pirapucu fue protegida en común acuerdo entre los comunitarios y el Proyecto Mamirauá, para realizar estudios sobre ecología reproductiva de los quelonios. Algunos nidos encontrados en el Paraná de Jarauá fueron dejados intactos con la finalidad de obtener información in situ sobre el periodo de incubación y cambios en el hábitat en función de la época de vaciante y creciente. Otros fueron transplantados a la Playa de Pirapucu para evitar la depredación por parte del hombre. Una vez localizado el nido, fueron registradas las siguientes informaciones: fecha de desove, local, distancia del nido a la orilla, tipo de sustrato, diámetro externo del nido, diámetro de la entrada, profundidad y diámetro mayor del nido, número y dimensiones de los huevos. Para determinar el tamaño de los huevos, en nueve nidos se tomó una muestra de 10 unidades, y en dos nidos se tomó la nidada completa. El largo y el ancho de los huevos fue medido con un calibrador vernier cuya precisión era de 1 mm. Dos nidos depredados parcialmente fueron transplantados y usados en las análisis. En 13 nidos eclosionados, se tomó una muestra de 10 crías, y en dos nidos todas las crías fueron medidas. La biometría de los individuos fue realizada tomándose en cuenta las medidas padrón de largo y ancho del caparazón, y largo del plastrón (Medem, 1976). Todas estas medidas fueron realizadas en línea recta en el punto de mayor amplitud. Los huevos y las crías fueron pesados en balanza Pesola con precisión de 1 g. Después de las mediciones, las crías fueron liberadas próximas de la ribera, en el local donde fue realizado el desove. Datos del nivel del agua fueron colectados diariamente a las 6:00 am, mediante el uso de escalas móviles, en una base fija situada en la comunidad de San Raimundo de Jarauá. El ecosistema de várzea alberga una diversidad de tipos de hábitat de los cuales algunos aparecen solo estacionalmente. Algunas denominaciones de estos hábitats solo son usadas localmente y no conseguimos una traducción del portugués al idioma español. A continuación definimos cada uno de estos términos: Creciente: fase de aumento del nivel de agua en el ciclo hidrológico anual. Seca: fase mas baja del nivel de agua en el ciclo hidrológico anual.

AUGUSTO FACHÍN TERÁN Y EDUARDO MATHEUS VON MÜLHEN
Ecología Aplicada Vol. 2 Nº 1, pp. 125-132 _______________________________________________________________________________________
127
Repiquete: aumento temporal del nivel de agua que precede a la creciente principal (oscilaciones del nivel de agua). Poza: cuerpo de agua formado cuando el nivel del agua disminuye. Caños: canales que drenan los lagos en la várzea. Lagos: cochas de la planicie inundada. Resaca: brazo de cualquier cuerpo de agua (usualmente lago o canal) que termina en la floresta, y que seca casi por completo durante la época de vaciante. Paraná: ramal del canal principal de un río que forma un brazo que atraviesa la floresta inundada de la várzea, conectándose aguas abajo al mismo cuerpo de agua. Resultados La nidificación de P. unifilis en la RDSM, ocurre con la disminución del nivel de agua que expone las playas de arena y barrancos de tierra (Figura 2). Los desoves ocurrieron entre el 21 de agosto y 21 de septiembre. El 72,5% de posturas ocurrió entre el 21 y 30 de agosto (Figura 1). En ese periodo también se registraron cinco nidos fuera del área de estudio, en la Playa de Santa Luzia de Horizonte en el río Solimões (Fachín-Terán, obs. pers.).
Figura 1. Período de desove (N=40) y eclosión (N=21) en función del nivel del agua, registrados en Podocnemis unifilis, en el sector Jarauá de la RDSM. Fueron registrados 40 nidos de P. unifilis en diferentes locales del sector Jarauá: Resaca de Tucuxi (N=11), Playa de Pirapucu (N=8), entrada del Paraná de Jarauá (N=8), Resaca de Arthur (N=5), entrada del caño del Lago Baixo (N=4), Resaca de Panema (N=2), Poza de Jaraqui (N=1), y Playa de Putiri (N=1). De los 40 nidos, 27 (67.5%) fueron depositados en sustrato arcilloso, 10 (25%) en arena, y tres (7.5%) en hojarasca. Siete nidos que se encontraban en sustrato arcilloso y que corrían peligro de depredación por parte del hombre fueron transplantados a la playa de Pirapucu en el rió Japura y colocados en sustrato arenoso.
En 23 nidos donde fue registrado el relieve, 16 (69.6%) ocurrieron en la parte alta o “meseta” y siete (30.4%) en terreno inclinado. La localización de los nidos en sustrato arenoso solo fue posible con el suelo recientemente removido donde el sustrato es más oscuro, cubierto por arena húmeda removida por las hembras. Nidos construidos en sustrato arcillo, externamente tienen diferencia en la coloración con relación al sustrato del alrededor y también presentan rastros de las uñas sobre la cobertura externa del nido. Los nidos de taricaya son excavaciones con una entrada más o menos circular y una galería que se extiende hasta una cámara mayor, donde son depositados los huevos. Las dimensiones de los nidos son presentadas en la tabla 1. Fueron medidos 207 huevos, siendo el largo medio de 42.4+0.2 mm, con un rango de 36.2 a 47.8 mm; el ancho medio de 30.7+0.2 mm, con un rango de 26 a 34 mm; el peso medio de 23.2+3.5 g, con un rango de 14 a 28 g. Encontramos dos huevos pequeños que midieron 36 x 23.8 mm y 29.7 x 24.3 mm, y pesaron 10 y 11 gramos respectivamente. Huevos recientes presentaron color rosáceo y forma elíptica, con una secreción pegajosa y transparente. En 16 nidos incubados en sustrato arenoso, el tiempo medio de incubación de los huevos fue mayor
(61.7+3 días), con respecto a cinco nidos incubados en sustrato arcilloso (72.8+0.44 días). Fueron medidas 223 crías, siendo el largo medio del caparazón de 43.1+0.2 mm, con un rango de 37.6 a 48.5 mm; el ancho medio del caparazón fue 38.7+0.2 mm, con un rango de 33.7 a 43.4; el largo medio del plastrón fue 40.2+0.2 mm, cun un rango de 32.8 a 44.6 mm; el peso medio fue 16+2.3 g, rango de 11 a 21 g. Encontramos tres crías pequeñas que midieron entre 32.5 a 33.5 mm, y pesaron de 7 a 8 gramos. En una nidada de 35 huevos que se desarrolló normalmente, todas las crías eran pequeñas.
En la muestra de 10 ejemplares medidos, el tamaño medio del largo del caparazón fue de 37.4+0.08 mm, rango de 35.2 a 38.3 mm; con un peso medio de 7.4+0.52 g, y rango de 7 a 8 g. Del total de huevos incubados, 66.8% eclosionaron. El porcentaje de crías fue mayor en nidos dejados intactos en sustrato arenoso (92.2%), que en nidos transplantados (58.3%) y nidos dejados intactos en sustrato arcilloso (44.6%) (Tabla 2). La prueba de Tukey evidencia diferencia estadísticamente significativa con relación a la cantidad de crías obtenidas en función del tipo de sustrato (p<0.05). De los 40 nidos, 18 (45%) permanecieron intactos, 15 (37.5%) fueron depredados por el hombre, cuatro (10%) por la iguana Tupinambis y tres (7.5%) inundados por un aumento temporal del nivel de las aguas (repiquete).
012345678910
11 15 19 23 27 31 4 8 12 16 20 24 28 2 6 10 14 18 22 26 30 3 7 11 15 19 23Agosto Septiembre Octubre Noviembre
Núm
ero
-40-30-20-10010203040506070
Nív
el d
el a
gua
(cm
)
Desove
Eclosión
Nivel del agua

REPRODUCCIÓN DE LA TARICAYA Podocnemis unifilis, AMAZONAS, BRASIL
Diciembre 2003 _______________________________________________________________________________________
128
Discusión Diversos autores observaron que la reproducción de los Pelomedusidae está relacionada al ciclo anual de vaciante y creciente, siendo que el desove y la incubación es realizada en la época de la seca, y el nacimiento de las crías coincide con el inicio de la creciente (Alho & Pádua, 1982; Fachín-Terán, 1992; Thorbjarnarson et al., 1993; Soini, 1996, 1997). Este mismo patrón fue observado en la RDSM, durante nuestro estudio. El periodo de desove de P. unifilis varía ampliamente a través de la distribución de la especie (Vogt & Soini, in press). Se puede esperar la existencia de variaciones en los periodos de nidificación en función de las diferencias existentes en las épocas de vaciante y creciente a lo largo de la cuenca Amazónica (Pezzuti, 1998), y la cuenca del Orinoco. Podocnemis unifilis desova en enero y febrero en el rió Capanaparo, Venezuela (Thorbjarnarson et al., 1993); octubre a marzo en el río Caquetá, Colombia (Medem, 1964); julio y agosto en la Reserva Nacional Pacaya-Samiria y en la Reserva de Biosfera del Manú, Perú (Fachín-Terán, 1992; Soini & Copula, 1995; Mitchell & Quiñones, 1994); agosto y septiembre en el rio Iténez, Bolivia (Caballero, 1996). En el Brasil el periodo de desove es entre junio y julio en el río Purus, agosto y septiembre en el río negro, río Jaú, río Trombetas, río Tapajós y río de las Muertes (Vogt & Soini, in press). Las actividades de nidificación de P. unifilis en la RDSM, se concentran durante la última semana del
mes de agosto. Esto ocurre luego que surgen las partes más altas de las playas y de los barrancos en las márgenes de los cuerpos de agua. En esta reserva P. sextuberculata concentra sus actividades de nidificación en el inicio del mes de septiembre (Pezzuti, 1998). La especie de mayor tamaño, P. expansa desova solamente después que el agua alcanza su nivel más bajo en los meses de septiembre y octubre. Un patrón similar fue observado
con P. expansa en el río Trombetas, Brasil (Alho, 1982) y en el río Pacaya, Perú (Soini, 1997). Entre los quelonios P. unifilis parece ser la especie en que los procesos reproductivos están más adaptados a la diversidad de habitats de la várzea. Esta especie puede desovar en playas de arena, en barro semi-seco y barrancos a las márgenes de los ríos, lagos, paranas y caños (Fachín-Terán, 1992; Souza & Vogt, 1994; Soini & Soini, 1995; Caballero, 1996). Los suelos en estés ambientes pueden variar desde arenoso puro, areno-arcilloso, arcilloso amarillo a ceniza, y hojarasca (Fachín-Terán, 1992; Soini & Soini 1995a). En la RDSM, Thorbjarnarson e Silveira (1996), encontraron en el lago Sanguessuga (= Lago callucallu), un nido construido sobre un colchón de vegetación fluctuante y materia orgánica. El nido fue construido en sustrato compuesto principalmente por hojas en descomposición. Según Vogt & Flores-Villela (1986), especies con amplia distribución geográfica, tienen la plasticidad de comportamiento materno para compensar diferencias climáticas, escogiendo sitios con características distintas o por la desova en época mas caliente o mas fría. El tamaño medio de la nidada de P. unifilis mencionado por diferentes autores varia con la distribución geográfica de la especie. Son 23.3 huevos en Venezuela (Thorbjarnarson et al., 1993); 22.4 (Medem, 1964), 27.3 (Foote, 1978), y 26.8 en Colombia (Páez, 1995); 21 (Ponce, 1979), 23.6 (Landeo, 1997b), 31.3 (Fachín-Terán, 1993) y 34.5 en el Perú (Soini, 1996); 27 en Bolivia (Caballero, 1996);
Tabla 1. Dimensiones de los nidos, número y tiempo de eclosión de P. Unifilis en la RDSM. X D.E. RANGO N
Diámetro externo del nido (cm) 23,4 9,6 13,5-37,0 10 Diámetro de la entrada del nido (cm) 11,9 2,1 9,5-16,0 8 Profundidad hasta el primer huevo (cm) 8,8 1,4 6,5-11,0 10 Diámetro mayor de la cámara de huevos (cm) 16,3 2,2 13,5-19,0 10 Profundidad total del nido (cm) 18,2 3,1 13,5-23,0 10 Distancia del nido a la orilla (m) 15,9 11,5 3,0-33,0 22 Número de huevos / nido 35,1 5,9 26-49 28 Tiempo de eclosión de los huevos (días) 64,4 5,5 56-73 21
X = Promedio, D.E. = Desviación estándar, N =Tamaño de la muestra
Tabla 2. Sobrevivencia y causas de perdida en nidos de P. unifilis en la RDSM. TIPOS DE SUSTRATO ARCILLA ARENA ARCILLA►ARENA TOTAL GENERAL N=8 % N=8 % N=7 % N=23 % Crías vivas 117 44,6 284 92,2 140 58,3 541 66,8 Huevos sin desarrollo 32 12,2 5 1,6 52 21,7 89 11,0 Huevos comidos por Tupinambis 10 3,8 0 0,0 20 8,3 30 3,7 Embriones atacados por larvas de mosca 1 0,4 0 0,0 19 7,9 20 2,5 Embriones desarrollados – muertos 8 3,1 19 6,2 9 3,8 36 4,4 Embriones muertos por la inundación 94 35,9 0 0,0 0 0,0 94 11,6 TOTAL DE HUEVOS 262 100,0 308 100,0 240 100,0 810 100,0 N= Número de nidos. Arcilla ►Arena = Nidos transplantados de sustrato arcilloso a sustrato arenoso.

AUGUSTO FACHÍN TERÁN Y EDUARDO MATHEUS VON MÜLHEN
Ecología Aplicada Vol. 2 Nº 1, pp. 125-132 _______________________________________________________________________________________
129
y en Brasil de 20 (Cantarelli & Herde, 1989), 23 (Valle et al., 1973), 23.7 (Souza & Vogt, 1994), y 24.4 (Vanzolini, 1977). El tamaño medio de 35.1 huevos encontrado en la RDSM, es el mayor registro para esta especie en el Brasil y solo es semejante al encontrado por Soini (1996) en el río Pacaya, Perú. Los dos locales donde el tamaño medio de la nidada es mayor, son áreas inundables de várzea que renuevan anualmente sus nutrientes a través de los ríos que traen sedimentos de origen andina, siendo muy productivas en términos de nutrientes disponibles.
Figura 2. Huevos de Taricaya Podocnemis unifilis en la Várzea del Medio Solimões, Amazonas, Brasil
La revisión de trabajos que reportan periodo de incubación para P. unifilis indican un rango de 55 días (Soini, 1997) a 97 días (Soini & Coppula, 1995). Generalmente los tiempos más cortos de incubación son obtenidos en nidos construidos en playas de arena con mucha exposición al sol (Thorbjarnarson et al., 1993; Fachín-Terán, 1993; Soini, 1997), y los de mayor periodo de incubación son aquellos construidos en barrancos a las orillas de los ríos, lagos y paranas, donde la vegetación rápidamente cubre el nido y como consecuencia la temperatura de incubación es menor (Fachín-Terán, 1993; Soini & Coppula, 1995). En el río Caquetá, Colombia, lluvias frecuentes contribuyeron para bajas temperaturas y periodo de incubación mas prolongado en P. unifilis (Paéz & Bock, 1998). Según Schwarzkopf & Brooks (1987), y Vogt & Bull (1982, 1984), las temperaturas de incubación son resultado del clima en determinado año y de las características físicas del nido y del sustrato. En condiciones de cautiverio, playas de arena construidas sobre sustrato húmedo y vegetación arbórea cercana, disminuyen el número de horas de sol en la playa; aumentando el periodo de incubación de los huevos, siendo registrado en P. unifilis un tiempo medio de incubación de 112 días con rango de 101 a 136 días (Fachín-Terán et al., 1997). Altas temperaturas de incubación aceleran el desarrollo embrionario (Bull et al., 1982; Schwarzkopf & Brooks, 1987; Georges, 1989), por tanto disminuyen el periodo de incubación de los embriones y aumentan la posibilidad de completar su desarrollo (Schwarzkopf & Brooks, 1987). En este estudio, nidos con menor tiempo de incubación fueron aquellos construidos en sustrato arenoso; en estas condiciones solamente un nido fue cubierto por la vegetación herbácea, lo que aumentó el tiempo de
incubación a 69 días. Según Vogt & Bull (1984) la vegetación puede ser el factor más importante en la determinación de la temperatura del sustrato. Los nidos construidos en sustrato arcilloso tuvieron periodo de incubación mayor, y estaban situados cerca de la vegetación arbórea, siendo cubiertos rápidamente por la vegetación herbácea, lo que influenció en el tiempo de exposición del nido a los rayos solares. Sumado a esto, los lugares de desove en el sustrato arcilloso son más húmedos. Esto puede producir temperaturas más bajas y un mayor tiempo de incubación de las crías, con peligro de que el nido sea alcanzado por la creciente, matando los embriones. Crecientes repentinas pueden causar grandes pérdidas en los nidos de los quelonios. Soini (1997) cuantifico una pérdida anual de nidos de P. unifilis de 10 a 50 % por inundación. La pérdida de nidos de esta especie por inundación en el río Caquetá, Colombia, podría haber sido elevada (64%), si los nidos no hubiesen sido transplantados (Paéz & Bock, 1998). Michell & Quiñones (1994) señalan que la mayor tasa de perdida natural de nidos de P. unifilis, exceptuándose la colecta de los huevos por el hombre, se debe a la inundación adelantada de los sitios de nidificación. Por otro lado, la perdida de nidos puede variar entre años y en función del clima. Landeo (1997b) reportó para el río Manú, la pérdida por inundación de 28.68% de nidos de P. unifilis en 1993, y de 0.69% en 1994. Pezzuti (1998) en la RDSM, encontró perdida de 58% de nidos de P. sextuberculata, por el fuerte repiquete durante la seca de 1996. En este estudio, solo tres (7.5%) nidos fueron inundados por el repiquete. P. unifilis desova cuando aparecen las partes superiores de las playas y de los barrancos, lo que puede contribuir para que sus nidos tengan menos posibilidades de ser inundados por la creciente del río. Soini (1995, 1997) reporta que el predador más importante de huevos de P. unifilis es el reptil Tupinambis, que posee extrema facilidad para detectar nidos, aunque hayan sido depositados varios días atrás. Fachín-Terán (1994) cita como principal depredador a las hormigas, y Landeo (1997b) cita una especie de grillo. En la RDSM, Tupinambis fue el mayor predador no humano de nidos de P. unifilis situados en los barrancos próximos a la floresta (Pezzuti, 1996). Esta observación fue confirmada en este estudio, a través de hallazgos de nidos abiertos y de cáscaras de huevos esparcidos cerca del nido y de la vegetación arbustiva. En las playas de arena no encontramos señales que indicasen la presencia de este animal, ni vestigios de nidos depredados. Esto puede ser debido a que la floresta queda un poco lejos de la playa y a la existencia de una barrera natural constituida por una poza de agua que separa estos hábitats. La presencia constante del personal del proyecto, también puede haber contribuido para que este predador no frecuente la playa.

REPRODUCCIÓN DE LA TARICAYA Podocnemis unifilis, AMAZONAS, BRASIL
Diciembre 2003 _______________________________________________________________________________________
130
En nidos transplantados, 7.9% de los embriones fueron atacados por larvas de mosca. Vogt et al. (1994) registraron una baja sobrevivencia en crías de Peltocephalus dumerilianus (39.5%) en función del ataque de larvas de moscas, a las crías que estaban dentro del nido completando su desarrollo. Según Pezzuti (1998) existe la posibilidad de que la manipulación de los huevos pueda facilitar la actividad predatoria de estés dípteros sobre los embriones de quelonios, sugiriendo que seria importante investigar cuando y como la mosca introduce sus huevos en el nido y como ocurre el desarrollo de sus larvas. La colecta de huevos por las poblaciones indígenas y ribereñas de la Amazonía es una actividad tradicional periódica importante y está relacionada al ciclo reproductivo de los quelonios (Bates, 1863; Soini, 1997; Landeo, 1997a, 1997b). Cuando las playas no son preservadas y existen poblaciones humanas próximas, generalmente la depredación es de 100% de los nidos de P. unifilis por el hombre, como ocurre en el bajo río Jau (Rebêlo et al., 1996). Donde existen programas de preservación, la depredación por el hombre disminuye, y puede variar de local para local y de año para año. Así, en la Reserva Nacional Pacaya-Samiria, Perú, la depredación fue 30.7% en el río Samiria (Fachín-Terán, 1994) y 37% en el río Pacaya (Soini & Coppula, 1995). En el río Manú, Perú, en años diferentes fue registrada una depredación de nidos por el hombre de 39.4 y 52.1% (Quiñones, 1991; Landeo, 1997a). La captura de P. unifilis y la colecta de sus huevos para consumo por los comunitarios son una amenaza permanente para esta especie en la RDSM, ya que existe un gran número de pescadores utilizando el Paraná de Jarauá. Esta situación se tiende a agravar en función a la implementación desde 1998, del Proyecto de comercialización de Pescado–PCP, que explora comercialmente el recurso pesquero en esta parte de la reserva. Como consecuencia, aumenta la posibilidad de que los pescadores capturen los quelonios en sus redes malladeras. En esta misma reserva, los resultados encontrados para P. sextuberculata, sugieren que la pesca con redes malladeras para fines comerciales y la captura de hembras adultas en las playas pueden llevar a una drástica reducción de la población de hembras reproductivas (Fachín-Terán et al., 2003). La transferencia de nidos de P. unifilis a locales protegidos en la RDSM podría ser una opción para producir más crías. Entre tanto en especies donde la temperatura de incubación determina el sexo (e.g. P. expansa, Alho et al., 1984; P. unifilis, Souza & Vogt, 1994; Paéz, 1995; P. sextuberculata, Pezzuti, 1998) esta manipulación debe ser realizada con mucha cautela. Experimentos de manipulación de nidos de tortugas marinas mostraron el peligro de masculinizar la población (Mrosovsky, 1982). Paéz & Bock (1998)
recomendaron que en proyectos de manejo de P. unifilis, transferencias de nidos solo deben ser realizados después de pasado el periodo critico de determinación del sexo o transplantados a locales donde la temperatura sea semejante al local inicial. Esto con la finalidad de evitar desvíos para un sexo. La eclosión de P. unifilis en nidos transplantados puede ser alta, pudiendo variar entre 70 a 80% (Soini, 1995). En este estudio a pesar del cuidado en el traslado de los huevos, el porcentaje de eclosión fue menor (58.3%). El relativamente alto número de huevos sin desarrollo (21.7%) puede ser debido a daños ocurridos en el embrión producidos por la vibración durante el transporte en deslizador. Entre tanto no descartamos la posibilidad de elevada ocurrencia de huevos infértiles. A pesar del peligro de desvío para un determinado sexo, la transferencia de nidos para locales seguros, parece ser por el momento la única manera de incrementar la población de esta especie; ya que la nidificación ocurre en áreas de exposición permanente para la depredación por el hombre. Esta estrategia es necesaria a pesar de la posibilidad de modificar la proporción de sexos; siendo la única alternativa hasta la realización de acciones de preservación y de estudios mas completos sobre determinación del sexo y dinámica poblacional de esta especie en la RDSM. La protección y manejo de los locales de desove, la participación comunitaria en la preservación de playas y lagos, la educación ambiental continuada en todos los niveles, la liberación de los ejemplares capturados por los pescadores, y la fiscalización permanente por parte del Instituto Brasilero del Medio Ambiente y de los Recursos Naturales Renovables-IBAMA, en la reserva y en los mercados de las ciudades de Tefé y Alvarães, son acciones importantes para la recuperación de la población de P. unifilis en la Reserva Mamirauá. Agradecimientos A la Sociedad Civil Mamirauá-SCM, por el apoyo logístico y facilidades concedidas para la realización de este estudio. Al Dr. José Márcio Ayres, defensor incansable de la várzea, fallecido en marzo de 2003, por darnos la oportunidad de trabajar en la Reserva de Desarrollo Sustentable Mamirauá. A los Drs. John Thorbjarnarson y Richard C. Vogt por la eficiente gestión y soporte en el campo. Al Dr. Renato Cintra, Maria de Fátima Gomes y Souza Soares y Miriam Elenit Lima de Fachín por sus valiosas sugestiones. A Masidonio Pinho de Carvalho, Mariceudo Pinho de Carvalho, Paulo Roberto Barbosa Pereira y Antônio Martins (Presidente) por su valiosa ayuda durante la colecta de datos. Augusto Fachín Terán, recibió una bolsa de doctorado del CAPES, durante su permanencia en el Brasil en el Departamento de Ecología del INPA. Este estudio fue financiado por el

AUGUSTO FACHÍN TERÁN Y EDUARDO MATHEUS VON MÜLHEN
Ecología Aplicada Vol. 2 Nº 1, pp. 125-132 _______________________________________________________________________________________
131
CNPq/MTC y DFID. Dos revisores dieron valiosas contribuciones al texto final. Literatura citada Alho C.J.R. & Pádua L.F.M. 1982. Sincronia entre
regime de vazante do rio e o comportamento de nidificação da tartaruga da Amazônia Podocnemis expansa (Testudinata, Pelomedusidae). Acta Amazônica. 12: 323-326.
Alho C.J.R., Danni T.M.S. & Padua L.F.M. 1984. Influência da temperatura de incubação na determinação do sexo da tartaruga da Amazônia Podocnemis expansa (Testudinata, Pelomedusidae). Revista Brasileira de Biologia. 44: 305-311.
Bates H.W. 1863. The naturalist on the river Amazon. John Murray, London.
Bull J.J., Vogt R.C. & McCoy J. 1982. Sex determining temperatures in turtles: a geographic comparasion. Evolution. 36(2): 326-332.
Caballero J. 1996. Comparación de cuatro tratamientos de manejo para la protección de la peta de agua (Podocnemis unifilis), Troschel 1840, y estimación de la supervivencia de las crías en las playas del río Itenez. Tesis de Licenciado en Ciencias Biológicas. Universidad Autónoma "Gabriel Rene Moreno", Facultad de Ciencias Agrícolas. Santa Cruz de la Sierra-Bolivia.
Cantarelli V.H. & Herde L.C. 1989. Projeto quelônios da Amazônia 10 anos. Cantarelli, V. H. e L. C. Herde (eds.).Instituto Brasileiro do Meio Ambiente e dos Recursos Naturais Renováveis-IBAMA. Ministério do Interior, Brasília.
Fachín-Terán A. 1992. Desove y uso de playas para nidificación de taricaya (Podocnemis unifilis) en el río Samiria, Loreto-Perú. Boletín de Lima. 79:65-75.
Fachín-Terán A. 1993. Características de Podocnemis unifilis (Reptilia, Testudines) en el río Samiria, Loreto. Boletín de Lima. 87: 69-74.
Fachín-Terán A. 1994. Depredación de la taricaya Podocnemis unifilis en la Reserva Nacional Pacaya-Samiria, Loreto. Boletín de Lima. 16(91-96): 417-423.
Fachín-Terán A., Acosta A., Vilchez I. & Taleixo G. 1997. Reproducción de la taricaya Podocnemis unifilis (Reptilia: Testudines) en cautiverio, Iquitos, Perú. En: Tula G. Fang, Richard E. Bodmer, Rolando Aquino y Michael H. Valqui (eds.). Manejo de Fauna Silvestre en la Amazonía, pp.185-189. La Paz, Bolivia.
Fachín-Terán A. 1999. Ecologia de Podocnemis sextuberculata (Testudines, Pelomedusidae), na Reserva de Desenvolvimento Sustentável Mamirauá, Amazonas, Brasil. Tese de Doutorado. Manaus: INPA/UFAM.
Fachín-Terán A., Vogt R.C. & Thorbjarnarson J.B. 2003. Estrutura populacional, razão sexual e
abundância de Podocnemis sextuberculata (Testudines, Podocnemididae) na Reserva de Desenvolvimento Sustentável Mamirauá, Amazonas, Brasil. Phyllomedusa. 2(1): 43-63.
Foote R.W. 1978. Nesting of Podocnemis unifilis (Testudines: Pelomedusidae) in the Colombia Amazon. Herpetologica. 34: 333-339.
Georges A. 1989. Female turtles from hot nets: is the duration of the development of proportion of development that matters?. Oecologia. 81: 323-328.
Landeo C. 1997a. Usuarios del recurso taricaya (Podocnemis unifilis) en el río Manú. En: Tula G. Fang, Richard E. Bodmer, Rolando Aquino y Michael H. Valqui (eds.). Manejo de Fauna Silvestre en la Amazonía, pp.181-183. La Paz, Bolivia.
Landeo C. 1997b. Factores limitantes de la población pre-eclosional de la taricaya Podocnemis unifilis en el río Manú. En: Tula G. Fang, Richard E. Bodmer, Rolando Aquino y Michael H. Valqui (eds.). Manejo de Fauna Silvestre en la Amazonía, pp.185-189. La Paz, Bolivia.
Medem F. 1964. Morphologie, Oekologie und Verbreitung der Schildkröte, Podocnemis unifilis in Kolumbiem. Senckenb. Biol. 45: 353-368.
Medem F. 1976. Recomendaciones respecto a contar el escamado y tomar las dimensiones de nidos, huevos y ejemplares de los Crocodylia y Testudines. Lozania. 20: 1-17.
Mitchell C.L. & Quiñones L. 1994. Manejo y conservación de la taricaya (Podocnemis unifilis) en la Reserva de Biosfera del Manú, Madre de Dios. Boletín de Lima. 16 (91-96): 425-436.
Mrosovsky N. 1982. Sex ratio bias in hatchling sea turtles from artificially incubated eggs. Biol. Conserv. 23: 309-314.
Páez V.P. 1995. The conservation and nesting ecology of the endangered yellow-spotted Amazonian river turtle, Podocnemis unifilis. Ph. D. Thesis, Ohio University, Athens.
Páez V.P. & Bock B.C. 1997. Nesting ecology of the yellow-spotted river turtle in the Colombian Amazon. In: Van Abemma, J. (Ed.). Proceedings: Conservation, Restoration, and Management of Tortoises and Turtles - An International Conference. New York: New York Turtle and Tortoise Society.: 219-224.
Páez V.P. & Bock B.C. 1998. Temperature effect on incubation period in the Yellow-Spotted River Turtle Podocnemis unifilis, in the Colombian Amazon. Chelonian Conservation and Biology. 3(1): 31-36
Pezzuti Juarez C.B. 1998. Ecologia reprodutiva da iaçá, Podocnemis sextuberculata (Testudines, Pelomedusidae) na RDSM, Amazonas, Brasil. Dissertação de Mestrado. Manaus, INPA/UFAM.

REPRODUCCIÓN DE LA TARICAYA Podocnemis unifilis, AMAZONAS, BRASIL
Diciembre 2003 _______________________________________________________________________________________
132
Ponce M. 1979. Podocnemis unifilis Troschel 1848 “taricaya” (Chelonia, Pleurodira, Pelomedusidae) en el Bosque Nacional “Alexander von Humboldt”, Loreto-Perú. Tesis de Biólogo. Universidad Nacional Agraria. La Molina, Lima.
Rebêlo G.H., Moreira G., Lugli L., Marajó L., Raposo J.C., Queiroz A.L., Cruz R.F. & Reimann C. 1996. Os quelônios do Parque Nacional do Jaú (AM). Reporte técnico para a Fundação Vitória Amazônica. (Não publicado).
Schwarzkopf L. & Brooks J.R. 1987. Nest-site selection and offspring sex ratio in painted turtles, Chrysemys picta. Copeia. 7(1): 55-61.
Soini P. 1995a. Estudio e incubación de los huevos de quelonios acuáticos, 1986. Informe Nº 22. En: Soini, P., A. Tovar y U. Valdez (ed.), Reporte Pacaya-Samiria. Investigaciones en Cahuana: 1980-1994, pp. 247-250. CDC-UNALM/FPCN/TCN. Lima, Perú.
Soini P. 1995b. Estudio y manejo de quelonios acuáticos, 1987. Informe Nº 26. En: Soini, P., A. Tovar y U. Valdez (ed.), Reporte Pacaya-Samiria. Investigaciones en Cahuana: 1980-1994, pp. 279-287. CDC-UNALM/FPCN/TCN. Lima, Perú.
Soini P. 1995c. Manejo de quelonios acuáticos, 1992. Informe Nº 38. In: Soini, P., A. Tovar y U. Valdez (ed.), Reporte Pacaya-Samiria. Investigaciones en Cahuana: 1980-1994, pp. 395-399. CDC-UNALM/FPCN/TCN. Lima, Perú.
Soini P. 1996. Reproducción, abundancia y situación de quelonios acuáticos en la Reserva Nacional Pacaya-Samiria, Perú. Folia Amazonica. 8(1): 147-164.
Soini P. 1997. Ecología y manejo de quelonios acuáticos en la amazonía peruana. En: Tula G. Fang, Richard E. Bodmer, Rolando Aquino y Michael H. Valqui (eds.). Manejo de Fauna Silvestre en la Amazonía, pp.167-173. La Paz, Bolivia.
Soini P. & Coppula M. 1995. Estudio, reproducción y manejo de los quelonios del género Podocnemis (charapa, cupiso y taricaya) en la cuenca del Pacaya, río Pacaya, Loreto-Perú. Informe Nº 2. En: Soini, P., A. Tovar y U. Valdez (ed.), Reporte Pacaya-Samiria. Investigaciones en Cahuana: 1980-1994, pp. 3-30. CDC-UNALM/FPCN/TCN. Lima, Perú.
Soini P. & Soini M. 1995a. Ecología reproductiva de la taricaya (Podocnemis unifilis) y sus implicaciones en el manejo da la especie. Informe Nº 9. En: Soini, P., A. Tovar y U. Valdez (ed.), Reporte Pacaya-Samiria. Investigaciones en Cahuana: 1980-1994, pp. 99-128. CDC-UNALM/FPCN/TCN. Lima, Perú.
Soini P. & Soini M. 1995b. Ensayos de incubación de huevos de los quelonios del género Podocnemis
(charapa, taricaya y cupiso). Informe Nº 12. En: Soini, P., A. Tovar y U. Valdez (ed.), Reporte Pacaya-Samiria. Investigaciones en Cahuana: 1980-1994, pp. 169-176. CDC-UNALM/FPCN/TCN. Lima, Perú.
Soini P. & Soini M. 1995c. Un resumen comparativo de la ecología reproductiva de los quelonios acuáticos. Informe Nº 19. En: Soini, P., A. Tovar y U. Valdez (ed.), Reporte Pacaya-Samiria. Investigaciones en Cahuana: 1980-1994, pp. 215-226. CDC-UNALM/FPCN/TCN. Lima, Perú.
Souza R.R. & Vogt R.C. 1994. Incubation temperature influences sex and hatchling size in the neotropical turtle Podocnemis unifilis. Journal of Herpetology. 28(4): 453-464.
Thorbjarnarson J.B., Perez N. & Escalona T. 1993. Nesting of Podocnemis unifilis. Journal of Herpetology. 27 (3): 344-347.
Thorbjarnarson J.B. & da Silveira R. 1996. Podocnemis unifilis (Yellow-headed Sideneck). Nesting. Herpetological Review. 27(2): 77-78.
IUCN. 1996. Red List of Threatened Animals. Compiled and Edited by Jonathan Baillie and Brian Groombridge. IUCN, Gland, Switzerland.
Valle R.C., Alfinito J. & da Silva M.M.F. 1973. Contribuição ao estudo da tartaruga Amazônica. Em: Preservação da tartaruga Amazônica. Simpósio Internacional sobre manejo de Fauna Silvestre e Pesca Fluvial e Lacustre Amazónica. IBDF/SUDEPE/IICA, Manaus.
Vanzolini P.E. 1977. A brief biometrical note on the reproductive biology of some South American Podocnemis (Testudines, Pelomedusidae). Papeis Avulsos de Zoologia. 31: 79-102.
Vogt R.C. & Bull J. 1982. Temperature controlled sex determination in turtles: ecological and behavioral aspects. Herpetologica. 38(1): 156-164.
Vogt R.C. & Bull J. 1984. Ecology of hatchling sex ratio in map turtles. Ecology. 65(2): 65-74.
Vogt R.C. & Flores-Villela O. 1986. Determinación del sexo en tortugas por la temperatura de incubación de los huevos. Ciencia. 37: 21-32.
Vogt R.C. 1994. Temperature controlled sex determination as a tool for turtle conservation. Chelonian Conservation and Biology. 1(2): 159-162.
Vogt R.C., Cantarelli V.H. & de Carvalho A.G. 1994. Reproduction of the Cabeçudo, Peltocephalus dumerilianus, in the Biological Reserve of rio Trombetas, Pará, Brazil. Chelonian Conservation and Biology. 1(2): 145-148.
Vogt R.C. & Soini P. In Press. Podocnemis unifilis Troschel, 1848. Tracajá, Terecay, Yellow-Spotted Amazon Turtle. IUCN/SSC. Conservation Biology of Freshwater Turtles. Vol. II. Eds. A. Rhodin and P. Pritchard.

Ecología Aplicada, 2(1), 2003 ISSN 1726-2216 Depósito legal 2002-5474
EL “COCODRILO DE TUMBES” (Crocodylus acutus Cuvier 1807): ESTUDIO PRELIMINAR DE SU ESTADO ACTUAL EN EL NORTE DE PERÚ.
Armando H. Escobedo Galván1 y Fernando Mejía Vargas2
Resumen:
De Junio del 2002 a Febrero del 2003, se han realizado recorridos en la zona norte de Perú, específicamente en la región de Tumbes, para conocer el estado actual de Crocodylus acutus, mediante recorridos diurnos y nocturnos, estimando una densidad de 0.18 ind/Km siendo una de las densidades más bajas de C. acutus en su ámbito de distribución. La estructura poblacional es de 82.61% juveniles, 4.35% de sub-adultos y 13.04% de adultos, siendo similar a lo reportado en otras poblaciones. Palabras claves: Crocodylus acutus, Cocodrilo de Tumbes, estructura poblacional, Río Tumbes.
Abstract In order to assess the actual status of the Crocodylus acutus populations from northern Perú, Tumbes Region, from June 2002 to February 2003, diurnal and nocturnal surveys were conducted. A density of 0.19 indviduals / km was estimated. This is the lowest density for C. acutus worldwide. The following population structure was found: 82.61% juveniles, 4.37% subadults and 13.09% adults. This is similar to population structures reported elsewhere. Key words Crocodylus acutus, Crocodile of Tumbes, it structures populational, Tumbes River.
Introducción
Crocodylus acutus, conocido como “Lagarto”, “Cocodrilo Americano” o “Cocodrilo de Tumbes”, está presente en ambas costas de la región neotropical del continente americano, desde el extremo sur de Florida, hasta los llanos del Orinoco en el noreste de Venezuela, en el Atlántico, mientras que en el Pacífico va desde Sinaloa en México, hasta el norte de Perú (Medem, 1983; Thorbjarnarson, 1989).
A pesar de su amplia distribución, sus poblaciones han sufrido una disminución debido principalmente a la pérdida de sus hábitats y la cacería furtiva. Esto ha hecho, que se encuentre en peligro de extinción (Thorbjarnarson, 1988; Ross, 1998).
Por ello, el Cocodrilo de Tumbes, ha sido incluido dentro del Apéndice I del Convenio Internacional para el Tráfico de Especies Amenazadas de Flora y Fauna (CITES, 1995), como consecuencia dentro de su ámbito de distribución se le considera una especie en peligro de extinción que debe ser protegida, razón por la cual no se permite la comercialización de productos y subproductos de esta especie provenientes de su hábitat silvestre. Además, en Perú, desde 1950, bajo la Resolución Suprema Nº 345, se prohíbe la caza de Crocodylus acutus en su hábitat natural.
A pesar de las leyes vigentes tanto internacionales como nacionales para la conservación del “Cocodrilo de Tumbes”, su situación no se ha revertido como debería de esperarse.
En Perú, esta especie tiene una distribución histórica que abarcaba desde el río Zarumilla por el norte hasta en río Chira por el sur (Medem, 1983), sin embargo las últimas evaluaciones han reportado una aparente desaparición de esta especie en las cuencas del Río Chira. Antecedentes
El primer censo de C. acutus, para conocer su situación en la región, se llevó acabo en 1970, registrando un total de 40 individuos para toda la cuenca del Río Tumbes, posteriormente en 1980, solo se registró 5 individuos en un sector del Río Tumbes, para 1985, se registraron cuatro ejemplares en 12 Km. del estero corrales en los manglares de Tumbes (Vásquez & Pickens, 1995) y recientemente, en 1997, miembros del Grupo de Especialistas de Cocodrilos determinaron la presencia de no más de 6 parejas reproductivas en la zona.
El objetivo del presente trabajo fue determinar el estado actual de la población de Crocodylus acutus en la zona de Tumbes.
________________________ 1 Escuela de Ciencias Ambientales, Universidad Latina de Costa Rica y Asociación Costarricense de Investigadores en Crocodilidos. Apartado Postal: 27-2100 Guadalupe, San José, Costa Rica. Correo electrónico: [email protected] 2 Universidad San Luis Gonzaga de Ica. Av. San Martín 465 la Tinguiña Ica-Perú. Correo electrónico: [email protected]

EL “COCODRILO DE TUMBES” (CROCODYLUS ACUTUS CUVIER ) EN EL NORTE DE PERÚ
Diciembre 2003 _____________________________________________________________________________________
134
Material y Métodos: El estudio se realizó desde Junio del 2002 a Febrero del 2003. La evaluación se llevó acabo en el Río Tumbes (límites del Parque Nacional Cerros de Amotape y la Zona Reservada de Tumbes) desde el sector denominado Cóndor Flores hasta el sector Bocana Carrillo incluyendo los sectores de Bocana Murciélago, Cabo Inga y el Guanábano. También se evaluaron los esteros conocidos como La Chepa, Barranco Blanco y La Canela. Se utilizó la metodología descrita por Chabreck (1966), que consiste en el conteo visual nocturno, ya que los ojos de los cocodrilos poseen un tapetum lucidum, refractor de luz, el cual presenta
un color rojizo que permite detectarlos hasta una distancia de 200 metros cuando se les alumbra en la noche (Salas, 1985). Para evaluar la población de esta especie se realizó primero un reconocimiento del área de estudio en el día y luego observaciones directas diurnas y nocturnas en las zonas antes determinadas, en el sector del Río Tumbes, específicamente en el sector denominados El Guanabano, Bocana Carrillo y Quebrada Honda. Los especímenes se estudiaron desde la orilla del río, en el sector de los esteros de manglares y también en forma directa en bote o canoas. Se calculó el tamaño de los individuos observados durante los recorridos, mediante la estimación de la longitud cefálica y distancia interocular. Las estimaciones fueron agrupadas en tres categorías, basadas en la clasificación de Thorbjarnarson (1989): a) juveniles < 90 cm, b) Sub-adultos de 90 a 180 cm, y c) Adultos > de 180 cm. Aquellos individuos que sólo mostraron el
brillo de los ojos fueron designados como “ojos” (King et al., 1990). Resultados y Discusión: Se realizaron 15 recorridos (2 recorridos por sector, excepto el de el estero la Canela), para un total de 121.5 Km, observándose 22 individuos. Se estimó una densidad de 0.18 ind/Km, siendo una de las densidades más bajas para la especie en todo su rango de distribución, este valor puede verse afectado por factores climáticos (principalmente el fenómeno de El Niño y La Niña), la estación, el efecto de la marea y la hora en que se realizó el estudio (Cerrato, 1991), además de ser la zona de
Tumbes el límite sur de distribución, no es precisamente una especie abundante ya que enfrenta la problemática de ser una población limítrofe, al no estar en contacto con otras poblaciones (se desconoce el estado de C. acutus en Ecuador). Ello conduce a que no se vea beneficiada por la recolonización conforme se pierden individuos locales, además de reducir depresiones poblacionales debido a la consanguinidad. La expansión de las diferentes actividades antrópicas en las márgenes del Río Tumbes y los esteros como, la actividad ganadera, expansión de tierras agrícolas y actividades ilegales como la extracción de la madera, la caza furtiva de la fauna silvestre y la extracción de oro en forma artesanal traen como consecuencia la presencia de personas en forma permanente en zonas que antes fueron el hábitat de esta especie quedando sin zonas de reproducción y como consecuencia, se refugian en zonas cada vez más inaccesibles para el hombre.
0102030405060708090
Juveniles Sub-adultos Adultos
Categorias
Porc
enta
je (%
)
Este estudioTumbes (Perú)
Sasa y Chaves1992 Río Tárcoles(C.R.)Chaves 1993 RíoTulín (C.R.)
Gaby et al. 1985Point Turk (Florida)
Thorbjarnarson1989 EtangSaumatre (Haití)
Figura 1. Estructura y tendencia de la población de C. acutus estimada para la zona de Tumbes, en comparación con otras poblaciones estimadas de la especie.

ARMANDO H. ESCOBEDO Y FERNANDO MEJÍA
Ecología Aplicada Vol. 2 Nº 1, pp. 133-135 _____________________________________________________________________________________
135
En el caso de los esteros la expansión agrícola y la actividad langostinera ha crecido de tal manera que en los sectores del estero la Chepa y Barranco Blanco estas expansiones llegan hasta la orilla de los esteros, y debido a esta expansión se ha talado casi la totalidad del manglar. En algunos casos, los cocodrilos son cazados o capturados muchas veces para ser comercializados vivos en la zona de Puerto Pizarro. La estructura poblacional en la zona estudiada, presenta un gran cantidad de juveniles, los cuales representan un 82.61%, mientras que los sub-adultos y adultos representan un 4.35% y 13.04% respectivamente. Este tipo de estructura poblacional a sido observada en otros estudios (Thorbjarnarson, 1989; Sasa & Chaves, 1992; Piedra, 2000), la cual puede estar asociada al tipo de estrategia poblacional de la especie para su sobre vivencia (Figura 1). El bajo porcentaje de sub-adultos puede deberse a dos causas; a que esta clase es muy difícil de observar, lo cual genera un sesgo en los muestreos y que la etapa de sub-adulto es relativamente corta, llegando a la madurez sexual en poco tiempo (Sasa & Chaves, 1992). Sin embargo, la evaluación de C. acutus, presentada en este trabajo no constituye un censo poblacional completo, sino una indicación del número mínimo de cocodrilos que se encuentran en la zona. Agradecimientos: Agradecemos a Luis Escobedo Cantú, Socorro Galván Carreón, por su apoyo para la realización de este estudio, a Carlos Arias por su hospitalidad y aporte logístico, al personal del Instituto Nacional de Recursos Naturales (INRENA) en Tumbes especialmente a Fernando Cuadros por su ayuda en la realización del estudio, a Karina Ramírez, Pavel Cartagena y a la Lic. Rosario Acero directora del INRENA por su apoyo en cuanto a los permisos, y por último un agradecimiento especial a la Dra. Miryam Venegas de Anaya por el aporte con su valioso conocimiento. Literatura citada: Cerrato C. 1991. Composición y Tamaño de
Poblaciones Silvestres de Caimanes (Caiman crocodilus chiapasus) y Cocodrilos (Crocodylus acutus) de la Costa Caribe de Honduras, Centro América. Tesis de Maestría para optar al grado de Magister Sceientiae en Manejo y Conservación de Vida Silvestre. Programa Regional en Manejo de Vida Silvestre para Mesoamérica y el Caribe. Universidad Nacional. Heredia, Costa Rica.
Chabreck R.H. 1966. Methods of determining the size and composition of Alligátor population in Luisiana. Proc. 20th Ann. Conf. S.E. Assoc. Game Fish Comm. 20: 105-112.
Chaves G. 1993. Situación poblacional del cocodrilo amarillo (Crocodylus acutus Reptilia, Crocodilidae) en el Pacifico Central de Costa Rica. Ecología. Reporte científico Nº 2. Universidad Estatal a Distancia.
Convención Internacional para el Tráfico de Especies Amenazadas de Flora y Fauna Silvestre (CITES). 1995. Convención para el Comercio de Especies Amenazadas. U.S. Departament of Interior. Fish and Wildlife Service. Washington.
King F.W., Espinal M. & Cerrato C. 1990. Distribution and status of the crocodilians of Honduras. Results of a survey conducted for the convention on International Trade in Endangered Species of Wild Fauna and Flora and the Honduras Secretaría de Recursos Naturales.
Medem F. 1983. Los Crocodylia de Colombia. Vol. II. Editorial Carrera. Bogotá, Colombia. Piedra L. 2000. Estado de las poblaciones de
cocodrilos (Crocodylus acutus) (Reptilia: Crocodylidae) en tres ríos del Pacífico Central de Costa Rica. Tesis de Licenciatura. Universidad Nacional de Costa Rica.
Ross J.P. 1998. Crocodiles. Status survery and conservation action plan. 2nd Edition. UICN/SSC. Crocodile Specialist Group. Gland, Switzerland and Cambridge, UK. Viii.
Salas C.E. 1985. Contribución al conocimiento sobre el manejo del Crocodylus acutus Cuvier (Crocodylia, Crocodylidae) en el Refugio Nacional de Fauna Silvestre Dr. Rafael Lucas Rodríguez Caballero. Universidad de Costa Rica, San José, Costa Rica.
Sasa M. & Chaves G. 1992. Tamaño, estructura y distribución de una población de Crocodylus acutus (Crocodylia: Crocodylidae) en Costa Rica. Rev. Biol. Trop. (40): 131-134.
Thorbjarnarson J.B. 1988. Status and ecology of American crocodile in Haiti. Bull. Fla. State Museum. 33(1): 1-86.
Thorbjarnarson J.B. 1989. Ecology of the American Crocodyle, Crocodylus acutus, in Crocodiles international Union for the conservation of the Nature, Gland, Switzerland. : 228-257.
Vásquez P.G. & Pickens C. 1995. Estado de Conservación de los Crocodylia en el Perú. pp. 135-157. En: Larriera A. & Verdade L. (Eds.) La Conservación y el Manejo de caimanes y cocodrilos de América Latina, Vol. 1. Fundación Banco Bica. Santo Tomé, Santa Fe, Argentina. ISBN-950-9632-21-X.

Ecología Aplicada, 2(1), 2003 ISSN 1726-2216 Depósito legal 2002-5474
PERÍODOS DE ACTIVIDAD Y EFECTO DE LAS VARIABLES AMBIENTALES EN COCODRILOS (Crocodylus acutus Cuvier 1807): EVALUANDO LOS MÉTODOS DE DETERMINACIÓN DE LA
FRACCIÓN VISIBLE. Armando H. Escobedo Galván1
Resumen:
Se realizaron un total de 8 muestreos durante los meses de Octubre y Noviembre de 2000, en La Estación Experimental La Rambla ubicada en Río Frío de Sarapiquí en la provincia de Heredia, Costa Rica. Se determinó que los cocodrilos presentan un pico de actividad durante la noche, y que el número de cocodrilos observados se ve afectado por condiciones climáticas previas y durante los muestreos. Se comparó los diferentes métodos estadísticos para calcular la fracción visible, determinando que no existen diferencias entre los métodos y la fracción visible calculada en base al tamaño real de la población cautiva. Palabras claves: Cocodrilo, Crocodylus acutus, fracción visible, Estación Experimental La Rambla.
Abstract During the months of October and November 2000, eight samples were made, at the Station Experimental La Rambla, which is locatated in Rio Frío de Sarapiquí, in Heredia. It was determinate that crocodiles are much more active during night time, and that the number of crocodiles that were seen has been affected with previous climate conditions. Different statistic methods were compared, to calculate the visible fraction, and conclude that there are no difference between the methods, and the visible fraction calculated based on the real captive population. Key words: Crocodile, Crocodylus acutus, Experimental Station La Rambla, visible fraction.
1 Escuela de Ciencias Ambientales, Universidad Latina de Costa Rica y Asociación Costarricense de Investigadores en Crocodilidos. Apartado Postal: 27-2100 Guadalupe, San José, Costa Rica. Correo electrónico: [email protected]
Introducción De las 23 especies de crocodílidos presentes en el planeta, en Costa Rica se encuentra el cocodrilo americano o cocodrilo amarillo (Crocodylus acutus), el cual pertenece a la familia Crocodylidae y se distribuye a lo largo de la costa Pacífica y Caribeña. En el Pacífico Seco se han reportado en el Río Cañas, el Río Tempisque, el Río Bebedero (King et al., 1990; Sánchez et al., 1996; Bolaños et al., 1997) y en playa Nancite (Plotkin & Zanella, 1994). En el Pacífico Central esta presente en los Ríos Grande de Tárcoles, Jesús María y Tusubres (Sassa & Chaves, 1992; Motte, 1994; Piedra, 2000). Bolaños et al. (1997) reportan la presencia de C. acutus en la región Pacífico Sur en los ríos Grande de Térraba, Sierpe y Coto, mientras en la costa Caribeña se encuentra en el Refugio de Vida Silvestre Caño Negro, en la región de Sarapiquí, río San Carlos y en las llanuras de Tortugero; siendo la topografía, la altura (Bolaños et al., 1997) y la temperatura (Kushlan & Mazzotti, 1989) factores que afectan su distribución. Los cocodrilos tienen preferencias por hábitats que presenten una comunicación entre aguas de clasificación bajas y aguas profundas con acceso a zonas secas para asolearse y un lugar para la anidación (Thorbjarnarson, 1989). El apareamiento ocurre entre los meses de diciembre y enero, mientras
que la anidación se da entre enero y febrero (Medem, 1981). C. acutus deposita sus huevos en huecos de unos 40 cm de profundidad por 70 cm de diámetro, esto lo realizan en el suelo en las cercanías de un riachuelo o en la orilla del río. En cuanto a su dieta es exclusivamente carnívoro y en algunas ocasiones se les ha observado consumiendo carroña. Su habilidad para destrozar es poca ya que sus dientes están diseñados para atrapar y retener (Muñoz, 1986). Los juveniles se alimentan principalmente de artrópodos y a medida que se desarrollan se empiezan a alimentar de presas más grandes. A nivel mundial los cocodrilos han sido afectados por la destrucción de su hábitat y la caza comercial. Estos reptiles son cazados por diferentes motivos dentro de las cuales destaca el valor de su piel, uso como mascota, el temor que infunde en las comunidades humanas; por lo que sus poblaciones sufren una gran presión comercial y en general de cacería furtiva (Eltringham, 1984). Sin embargo se ha demostrado que si se dejan de explotar a las especies de cocodrilos por varios años ellas pueden recuperar su nivel de población naturalmente (Chabreck, 1967).

ARMANDO H. ESCOBEDO GALVÁN
Ecología Aplicada Vol. 2 Nº 1, pp. __________________________________________________________________________________________
137
Revisión de Literatura La necesidad de conservar y evaluar las poblaciones de cocodrilos ha sido sólidamente justificada y se fundamenta en: 1) el papel y las funciones ecológicas que desempeñan en los ambientes acuáticos donde habitan; ya que con sus constantes movimientos pueden evitar la proliferación de plantas acuáticas (Kushlan, 1974), 2) los usos medicinales tradicionales de las especies por comunidades autóctonas, 3) en épocas secas favorecen la formación de pozas para el abrevadero de algunas especies silvestres y domésticas (Kushlan, 1974), 4) su importancia económica en términos de productos y subproductos, y 5) su valor como atractivo turístico. La necesidad de evaluar las poblaciones silvestres de cocodrilos se ha sentido en el país desde hace varios años, ya que constituyen un recurso faunístico muy valioso. A diferencia de otras especies de fauna silvestre, los cocodrilos tienen la habilidad de soportar programas de manejo basados en un aprovechamiento económico. De tal manera que constituyen un potencial para integrar la investigación y el manejo de sus poblaciones silvestres con la conservación de su hábitat y el desarrollo rural en humedales tropicales. Por lo cual es necesario determinar el tamaño de población para poder establecer un programa de manejo apropiado para la especie. Para estimar el tamaño de las poblaciones de cocodrilos en el campo se han desarrollado varias metodologías pero la más usada por su facilidad, es el conteo visual nocturno descrita por Chabreck (1966), ya que los ojos de los cocodrilos poseen un tapetum lucidum, refractor de luz, el cual presenta un color rojizo y esto permite detectarlos hasta una distancia de 200 metros cuando se les alumbra en la noche (Salas, 1985). El estudio del tamaño de las poblaciones se analiza empleando fracciones visibles de la población que se han estimado por métodos estadísticos, sin conocer la porción real de la población y para estimar su tamaño se han propuesto modelos relacionados con un tipo de distribución, como el que sugiere King et al. (1990), el cual esta basado en la distribución normal. Otro modelo se basa en la distribución binomial sugerido por Messel et al. (1987), lo cual trae otro problema ya que el tipo de distribución de la fracción visible no ha sido estudiado. Debido a que estas estimaciones se realizan descartando datos, también se puede estimar por medio del número máximo de individuos observados propuesta por King & Messel (en Cerrato, 1991). En estudios anteriores como el de Campos et al. (1994), demostró que el clima y el periodo de luna son factores que influye en el número de individuos observados. El presente estudio pretende, determinar el porcentaje real visible de la población de cocodrilos
cautivos en relación al patrón de actividad y compararlos con los diferentes métodos, para determinar cual metodología es el que se aproxima más a la fracción real. Otro de los objetivos fue el de evaluar si el período de luna tiene algún efecto en cuanto a la actividad de los cocodrilos y determinar el patrón de actividad nocturno de C. acutus. Material y Métodos: Área de estudio: El lugar se encuentra situado en La Rambla, Río Frío de Sarapiquí, Heredia, 10° 22´ 00” latitud N y 83° 55´ 00” longitud O. Este estudio se realizó durante los meses de Octubre y Noviembre del 2000, en la estación experimental de A.C.I.C. (Asociación Costarricense de Investigación de Crocodílidos). El lugar cuenta con una extensión de 2.5 hectáreas, la zona presenta Bosque Tropical Siempre Verde, con una precipitación promedio anual de 3000-4500 mm, con una temperatura entre 27-30°C y con una humedad relativa del 86%. La parte donde se encuentran los cocodrilos es en una pileta redonda que tiene 120m de circunferencia, de una profundidad es de 1m a 1.30m. El lugar presenta playones y algunos zacates y arbustos, por sus características es sencillo determinar el total de individuos presentes en el área. Metodología: Este estudio se realizó basado en la metodología descrita por Chabreck (1966), la cual consiste en la observación nocturna. El conteo se realizó cada 30 minutos en un período de seis de la tarde a cinco de la mañana contando el número de individuos activos usando una linterna para ver su reflejo. Se tomó en cuenta las condiciones climáticas y el período de luna cada vez que se realizó el muestreo. En el área de estudio se determinó que el tamaño de la población es de 78 individuos, mediante la captura de los mismos. Después se determinó la fracción visible como el porcentaje de individuos observados con respecto al total de individuos en la pileta, el cual se comparó con los diferentes métodos ya existentes para determinar la fracción visible. Análisis estadístico: Se analizó el efecto de muestreos y horas sobre el número de cocodrilos observados. También se determino por medio de un X² si los diferentes períodos de luna afectan la actividad de los cocodrilos y el número de individuos observados. También se analizó si los individuos presentan horas de mayor actividad. El porcentaje de la fracción visible se determinó usando el promedio de todos los datos obtenidos durante los ocho muestreos realizados, y se comparó con métodos estimadores para determinar la fracción
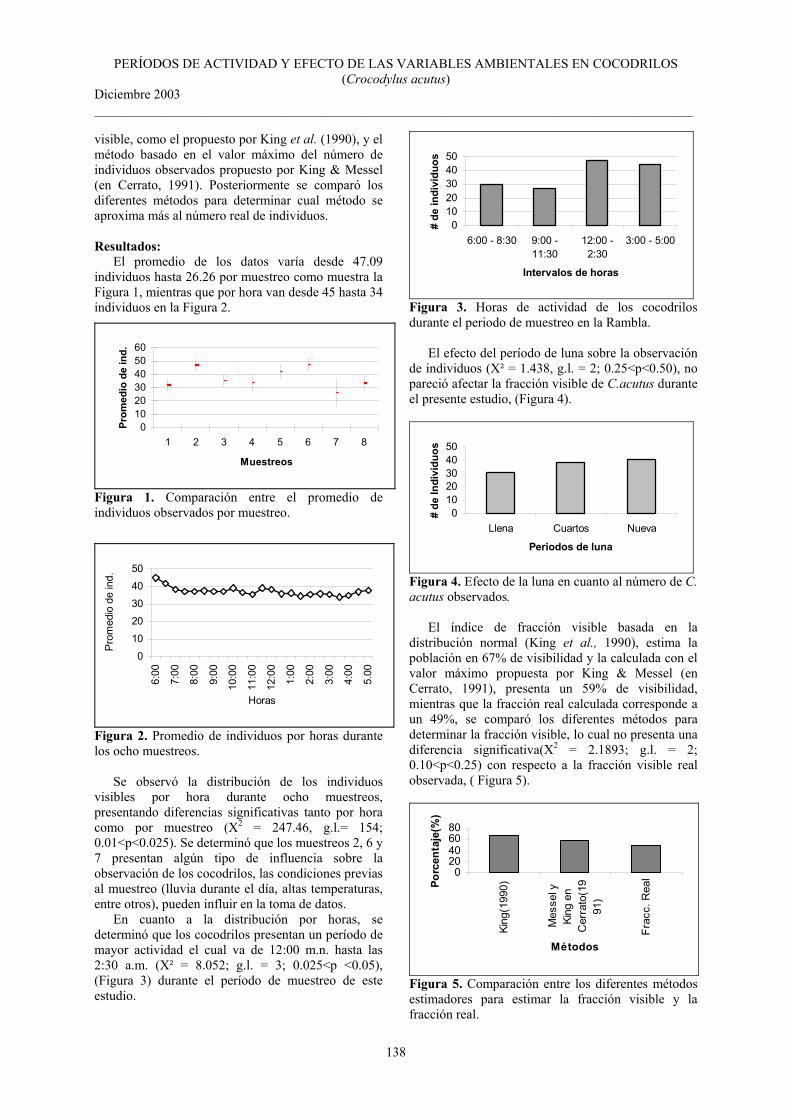
PERÍODOS DE ACTIVIDAD Y EFECTO DE LAS VARIABLES AMBIENTALES EN COCODRILOS (Crocodylus acutus)
Diciembre 2003 __________________________________________________________________________________________
138
visible, como el propuesto por King et al. (1990), y el método basado en el valor máximo del número de individuos observados propuesto por King & Messel (en Cerrato, 1991). Posteriormente se comparó los diferentes métodos para determinar cual método se aproxima más al número real de individuos. Resultados: El promedio de los datos varía desde 47.09 individuos hasta 26.26 por muestreo como muestra la Figura 1, mientras que por hora van desde 45 hasta 34 individuos en la Figura 2.
0102030405060
1 2 3 4 5 6 7 8
Muestreos
Prom
edio
de
ind.
Figura 1. Comparación entre el promedio de individuos observados por muestreo.
0
10
2030
4050
6:00
7:00
8:00
9:00
10:0
0
11:0
0
12:0
0
1:00
2:00
3:00
4:00
5.00
Horas
Pro
med
io d
e in
d.
Figura 2. Promedio de individuos por horas durante los ocho muestreos. Se observó la distribución de los individuos visibles por hora durante ocho muestreos, presentando diferencias significativas tanto por hora como por muestreo (X2 = 247.46, g.l.= 154; 0.01<p<0.025). Se determinó que los muestreos 2, 6 y 7 presentan algún tipo de influencia sobre la observación de los cocodrilos, las condiciones previas al muestreo (lluvia durante el día, altas temperaturas, entre otros), pueden influir en la toma de datos. En cuanto a la distribución por horas, se determinó que los cocodrilos presentan un período de mayor actividad el cual va de 12:00 m.n. hasta las 2:30 a.m. (X² = 8.052; g.l. = 3; 0.025<p <0.05), (Figura 3) durante el período de muestreo de este estudio.
01020304050
6:00 - 8:30 9:00 -11:30
12:00 -2:30
3:00 - 5:00
Intervalos de horas
# de
indi
vidu
os
Figura 3. Horas de actividad de los cocodrilos durante el periodo de muestreo en la Rambla. El efecto del período de luna sobre la observación de individuos (X² = 1.438, g.l. = 2; 0.25<p<0.50), no pareció afectar la fracción visible de C.acutus durante el presente estudio, (Figura 4).
01020304050
Llena Cuartos Nueva
Periodos de luna
# de
Indi
vidu
os
Figura 4. Efecto de la luna en cuanto al número de C. acutus observados. El índice de fracción visible basada en la distribución normal (King et al., 1990), estima la población en 67% de visibilidad y la calculada con el valor máximo propuesta por King & Messel (en Cerrato, 1991), presenta un 59% de visibilidad, mientras que la fracción real calculada corresponde a un 49%, se comparó los diferentes métodos para determinar la fracción visible, lo cual no presenta una diferencia significativa(X2 = 2.1893; g.l. = 2; 0.10<p<0.25) con respecto a la fracción visible real observada, ( Figura 5).
020406080
King
(199
0)
Mes
sel y
King
en
Cer
rato
(19
91)
Frac
c. R
eal
Métodos
Porc
enta
je(%
)
Figura 5. Comparación entre los diferentes métodos estimadores para estimar la fracción visible y la fracción real.

ARMANDO H. ESCOBEDO GALVÁN
Ecología Aplicada Vol. 2 Nº 1, pp. __________________________________________________________________________________________
139
Discusión: Cerrato (1991), determinó que los aspectos ambientales que podrían influir en los conteos están la velocidad del viento, lluvia, el período de luna y neblina. También Motte (1994), determinó que la temperatura del aire y del agua influye en el número de cocodrilos observados. En los resultados se nota que el promedio en los muestreos 2 y 6 es más alto que en los demás (Figura 1). Esto puede estar afectado por la presencia de lluvia o llovizna durante el día. Este autor (Cerrato, 1991), encontró que con la presencia de lluvia hay un aumento en el número de individuos observados, mientras que el promedio del muestreo 7 es el más bajo, él cual puede estar distorsionado ya que ese día se les alimentó. En cuanto al promedio por hora, se observó que las condiciones para cada una fueron homogéneas ya que el estado del tiempo en esa zona presenta gran variación la cual se podía dar en el momento del conteo. Se estableció un intervalo de mayor actividad durante el período de estudio, que va desde las 12:00m.n hasta las 2:30a.m.; en especies como, Caiman crocodilus, se ha determinado que tiene durante la noche algunas horas de actividad y otras de reposo, y que es más activo durante la lluvia y menos en noches de luna (Rivero, 1998), sin embargo en este estudio no se encontró ningún efecto del período de luna sobre la actividad de los individuos, lo cual puede estar justificado por la presencia de neblina en la mayoría de las horas en que se realizaron los muestreos. De acuerdo con los datos obtenidos en los resultados la fracción visible (King et al., 1990) difiere en un 18% con respecto al valor real calculado (49%), lo cual significa que presenta un error de 18% respecto al índice de visibilidad de este estudio en la determinación del tamaño de población. También se determinó la fracción visible basada en el número máximo de cocodrilos observados durante los muestreos (60%), lo cual tiene un 11% de diferencia con respecto al valor calculado, pero en comparación con King et al. (1990) su diferencia es menor. Este error puede estar relacionado con diferentes aspectos tales como el espacio en que se encontraban, ya que el área de acción de esta especie puede ser de 3 a 4 kilómetros, el cual aumenta o disminuye de acuerdo a la dominancia del macho (Acosta, 1998). Pero Sasa & Chaves (1992), no encontraron relación entre la longitud de los playones y el número de individuos en ellos, lo cual indica que su distribución es desigual. Otra de las razones puede ser la disponibilidad de recursos ya que los cocodrilos por naturaleza tienen diferentes tipos de dietas conforme avanzan de edad y tamaño. Los neonatos se alimentan básicamente de insectos, peces pequeños y otras especies de menor tamaño, los juveniles de peces y vertebrados pequeños. Cuando su desarrollo corporal los califica
como adulto se alimentan de peces, vertebrados grandes y carroña (Montero, 1997). Esto puede traer consecuencias en poblaciones silvestres ya que para el diseño de una estrategia de manejo que implica tanto conservación como aprovechamiento sostenible se han calculado el tamaño de la población con la fracción visible, (Cerrato, 1991) lo cual sobre estima el tamaño real de población. Esto representaría un efecto a la hora de cosechar poblaciones de adultos y huevos para el manejo, también en cuento al estatus y situación de las poblaciones de cocodrilos. Sin embargo no se encontraron diferencias entre los dos métodos y la fracción visible real, por lo cual se considera que es mejor emplear el método que más se aproxime al valor real. En este caso se recomendaría el método propuesto por King & Messel en Cerrato (1991), ya que es más simple y no esta relacionado con el tipo de distribución de la fracción visible, la cual no ha sido estudiada. Agradecimientos: En primer lugar agradecerle a la Lic. Liliana Piedra por su tiempo, su cooperación y apoyo en la realización de este estudio, también al Lic. Santos Muñoz por su colaboración, a Carlos Chaves Cavallini, Gabriela Coto y Kessery Kamara por su ayuda en el conteo de los cocodrilos, a Esteban Jiménez Campos por su ayuda en la captura de los individuos en estudio, a Erick Fuchs por su ayuda en cuanto al análisis de los datos, a Marcela Sequeira por su apoyo logístico y por ultimo a la Asociación Costarricense de Investigación de Crocodilos (A.C.I.C) por su colaboración y cooperación para realizar este estudio en La Estación Experimental La Rambla. Literatura citada: Acosta M. & Brenes M. 1998. Cocodrilo Americano
(Crocodylus acutus). III Día de Campo. M.F.V.S. Escuela Centroamericana de Ganadería.
Bolaños J., Sánchez J. & Piedra L. 1997. Inventario y estructura poblacional de crocodílidos en tres zonas de Costa Rica. Rev. Biol. Trop. 44(3)/45(1): 283-287.
Campos Z., Coutinho M. & Abercrombie C. 1994. Night-light, size structures, and sex ratios in wild populations of yacare (Caiman crocodilus yacare) in the Brazilian Pantanal. Vida Silvestre Neotropical. 4(1): 46-50.
Cerrato C. 1991. Composición y Tamaño de Poblaciones Silvestres de Caimanes (Caiman crocodilus chiapasius) y Cocodrilos (Crocodylus acutus) de la Costa Caribe de Honduras, Centro América. Tesis de Maestría para optar al grado de Magister Sceientiae en Manejo y Conservación de Vida Silvestre. Programa Regional en Manejo de

PERÍODOS DE ACTIVIDAD Y EFECTO DE LAS VARIABLES AMBIENTALES EN COCODRILOS (Crocodylus acutus)
Diciembre 2003 __________________________________________________________________________________________
140
Vida Silvestre para Mesoamérica y el Caribe. Universidad Nacional. Heredia, Costa Rica.
Chabreck R.H. 1966. Methods of determining the size and composition of Alligator population in Luisiana. Proc. 20th Ann. Conf. S.E. Assoc. Game Fish Comm. 20: 105-112.
Chabreck R.H. 1967. The American Alligator – Past, present and future. Pro. 21th Ann. Conf. S.E. Assoc. Game Fish Comm. 21: 554-558.
Eltringham S.K. 1984. Wildlife resoures and economic development. Johh Wiley & Sons.
King F.W., Espinal M. & Cerrato C. 1990. Distribution and status of the crocodilians of Honduras. Results of a survey conducted for the convention on International Trade in Endangered Species of Wild Fauna and Flora and the Honduras Secretaría de Recursos Naturales. Mercanogr.
Kushlan J. & Mazzotti F. 1989. Historic and present distribution of the American Crocodile in Florida. Journal of Herpetology. 23(1): 1-7.
Kushlan J.A. 1974. Observations on the role of the American Alligator (Alligator mississsippiensis) in the southern Florida wetlands. 4: 993-996.
Medem F. 1981. Los Crocodylia de Sur de América. Los Crocodylia de Colombia. Vol. I. Editorial Carrera. Bogotá, Colombia.
Messel H., Vorlicek G., Wells A. & Green W. 1981. The Blyth-Cadell River System study and the status of Crocodylus porosus in tidal waterways of Northern Austrilia (Surveys of tidal river system in the Northern Territori and their crocodile populations: Monograph 1). Pergamon press. Brisbane, Australia.
Montero A. 1997. Cocodrilo Americano (Crocodylus acutus). II Versión Día de Campo. Escuela Centroamericana de Ganadería.
Motte M. 1994. Abundancia, distribución e impacto de predación del cocodrilo (Crocodylus acutus Cuvier 1807) sobre el ganado vacuno en las fincas
aledañas al Río Grande de Tárcoles, Costa Rica. Tesis de Maestría para optar al grado de Magister Sceientiae en Manejo y Conservación de Vida Silvestre. Programa Regional en Manejo de Vida Silvestre para Mesoamérica y el Caribe. Universidad Nacional. Heredia, Costa Rica.
Muñoz G. 1986. El caimán de la costa (Crocodylus acutus). Editorial Primicia. Barquisiento, Venezuela.
Piedra L. 2000. Estado de las poblaciones de cocodrilos (Crocodylus acutus) (Reptilia: Crocodylidae) en tres ríos del Pacífico Central de Costa Rica. Tesis de Licenciatura. Universidad Nacional, Costa Rica.
Plotkin P. & Zanella G. 1994. Crocodiles predate on sea turtles. Crocodiles Specialist Group Newsletter. 13(1): 11-12.
Rivero J. 1998. Los Anfibios y Reptiles de Puerto Rico. 2da Edición revisada. Editorial de la universidad de Puerto Rico, San Juan, Puerto Rico.
Salas C.E. 1985. Contribución al conocimiento sobre el manejo del Crocodylus acutus Cuvier ( Crocodylia, Crocodylidae) en el Refugio Nacional de Fauna Silvestre Dr. Rafael Lucas Rodríguez Caballero. Universidad de Costa Rica, San José, Costa Rica.
Sanchez J., Bolaños J. & Piedra L. 1996. Población de Crocodylus acutus (Crocodylia: Crocodylidae) en dos ríos de Costa Rica. Rev. Biol. Trop. 44(2): 835-840.
Sasa M. & Chaves G. 1992. Tamaño, estructura y distribución de una población de Crocodylus acutus (Crocodylia: Crocodylidae) en Costa Rica. Rev. Biol. Trop. (40): 131-134.
Thorbjarnarson J.B. 1989. Ecology of the American Crocodyle, Crocodylus acutus, in Crocodiles nternational Union for the conservation of the Nature, Gland, Switzerland. : 228-257.
.

Ecología Aplicada, 2(1), 2003 ISSN 1726-2216 Nota científica Depósito legal 2002-5474
NEW EVIDENCE ON THE LOGGERHEAD SEA TURTLE Caretta caretta (Linnaeus 1758) IN PERU Shaleyla Kelez1, Ximena Vélez-Zuazo1 y Camelia Manrique1
Resumen
La tortuga marina cabezona, Caretta caretta (Linnaeus 1758), tiene amplia distribución en el ámbito mundial. Sin embargo, en el Perú su presencia no estaba confirmada ni documentada adecuadamente. En esta nota, se presentan datos de restos de individuos de esta especie encontrados en caletas y puertos pesqueros, así como de individuos capturados incidentalmente durante faenas pesqueras con espinel. De esta manera, la presencia de esta especie en el mar peruano quedaría confirmada. Los individuos encontrados son juveniles y pertenecerían a poblaciones reproductivas cuyas playas de anidamiento se encuentran en el Pacífico Oeste. Palabras clave: Tortuga marina cabezona, Caretta caretta, Perú,.distribución.
Abstract The loggerhead sea turtle, Caretta caretta (Linnaeus 1758), is widely distributed in the world-wide scope. Nevertheless, in Peru its presence was not confirmed nor documented suitably. In these note, data of rests from this species’ individuals found on inlets and ports as well as of individuals incidentally captured during fishing activities with longline is presented. In this way, the presence of this species in the Peruvian Sea would be confirmed. The individuals found are juveniles and could belong to reproductive populations belonging to West Pacific rookeries. Key words: Loggerhead sea turtle, Caretta caretta, Peru, distribution.
1 Grupo de Tortugas Marinas-Perú – gtm-perú y Asociación Peruana para la Conservación de la Naturaleza - APECO. parque josé de acosta 187, Lima 17, Perú (Correo electrónico: [email protected])
Introduction The loggerhead sea turtle, Caretta caretta (Linnaeus 1758), is widely distributed in tropical and subtropical waters in the Atlantic, Pacific and Indian Oceans, and Mediterranean Sea. In the Pacific, this species has been reported as far north as Alaska and as far south as Chile (Márquez, 1990). In Peru, however, C. caretta has not been officially documented, although there have been anecdotal reports since the 1950’s [Carr, 1952; Koepcke, 1961; Moller-Hergt, 1976; Grimwood & Whitmore, 1978 (in Frazier, 1979 p. 24); Aranda, 1989; Aranda & Chandler, 1989]. Frazier in 1979 mentioned that if the loggerhead turtle is common in northern Chile it could be possible that this species could be present in southern Peru. Later publications neither confirm nor mention that the loggerhead turtle occurs in Peru (Hays-Brown & Brown, 1982; Márquez, 1990; Carrillo & Icochea, 1995; Morales & Vargas, 1996). Despite this lack of documentation, the National Institute of Natural Resources (INRENA) in 1999 classified this species as vulnerable. Materials and methods During January and February 2002, a field survey to determine the current status of sea turtles along the southern coast of Peru was conducted. A total of 22 ports, inlets and beaches were surveyed. In each of these places, fishermen, authorities and members of the community were interviewed. All evidence of sea turtles in the area was recorded. The carapaces found were measured. The Curve Carapace Length notch to
tip (CCLn-t) was taken in all the cases but the Straight Carapace Length notch to tip (SCLn-t) was taken when it was possible. All the carapaces were photographed. From January to May 2003, on board observations during longline fishing activities, targeting “dolphinfish” Coryphaena hippurus and several species of sharks, were conducted. All the sea turtles incidentally captured were identified, measured, weighted, tagged, photographed and returned to the sea. The date, time and geographic position during the incidental capture was recorded. Results and discussion The results presented here are focused in the loggerhead sea turtle. Results on other sea turtles are excluded. In the 2002 survey, 39 individuals were recorded. Later analyses of these carapaces confirmed that three of them were C. caretta. The first carapace was found hanging as an ornament in a restaurant in Boca del Rio, Tacna. The turtle was reported to have been originally landed in the Vila Vila inlet (Figure 1). This carapace measured 57.8 cm SCLn-t and had only four lateral scutes, not common in this species but previously observed (Wyneken, 2001). The second carapace was found in Vila Vila inlet, Tacna, and measured 61.3 cm SCLn-t. According to the fisherman Luis Arce, he accidentally captured it 30-nautic miles west offshore during his longline fishing activities targeting shortfin mako (Isurus oxyrinchus). The third carapace was found in the Coastal Laboratory of the Peruvian Sea Institute (IMARPE) in

NEW EVIDENCE ON THE LOGGERHEAD SEA TURTLE Caretta caretta IN PERU
Diciembre 2003 __________________________________________________________________________________________
142
Ilo, Moquegua, and measured 63.9 cm CCLn-t. This carapace was purchased between 1996-97 in Atico, an inlet located to the north of Arequipa (M. Quiroz per. comm.).
Figure 1. All the turtles (Caretta caretta) were found in central an southern Peruvian Sea. During the longline fishing activities on-board observations, seven Caretta caretta were incidentally captured. All the turtles were found in central an southern Peruvian Sea (Figure 1). The average size of these individuals was 57.0 cm CCLn-t with a range of 48.5 – 62.5, the average weight was 23.1 Kg. with a range of 15 – 28. Five out of seven individuals were found with the hook in their mouths or throats. All the individuals mentioned in these note are juveniles because they are under 70 cm minimum size for adults (Márquez, 1990). These findings confirm the previous anecdotal observations and other recent report (J. Alfaro pers. comm.) that document this species’ presence in the southern coast of Peru. Since there are no known nesting beaches for C. caretta in the Eastern Pacific Ocean, these turtles likely originated from rookeries in the Western Pacific (Resendiz et al., 1998; Nichols et al., 2000). Conclusions On the basis of these findings, the presence of the loggerhead sea turtle in the Peruvian Sea in now confirmed and out of question. According to the size data obtained, the loggerhead turtle individuals are juveniles and would be migrating from rookeries in the Western Pacific, where they were born and will go back as adults to reproduce theirselves. We recommend that further research be conducted to determine the specific rookeries of origin of these individuals, the population status, and the ecological role of the Peruvian marine environment in their life cycle. Acknowledgments: To Dr. Jack Frazier for his asserted observations and to Dr. Pritchard for one individual species
identification. The authors wish to acknowledge use of the Maptool program for analysis and graphics in this paper. Maptool is a product of SEATURTLE.ORG. (Information is available at www.seaturtle.org). Funding came from AWARE Project, Idea Wild and the U.S. National Marine Fisheries Service. References Aranda C. 1989. Marine turtles in Peru. Marine Turtle
Newsletter. 45: 8-9. Aranda C. & Chandler M. 1989. Las Tortugas Marinas
del Perú y su situación actual. Boletín de Lima. 62: 77-86.
Carr A.F. 1952. Handbook of turtles: The turtles of the United States, Canada and Baja California. Cornell University Press, Ithaca.
Carrillo N. & Icochea J. 1995. Lista taxonómica preliminar de los reptiles vivientes del Perú. Publicaciones del Museo de Historia Natural UNMSM (A). 49: 1-27.
Frazier J. 1979. Marine Turtles in Peru and the East Pacific. Office of Zoological Research, National Zoological Park, Smithsonian Institution. Unpublished manuscript.
Hays-Brown C. & Brown W. 1982. Status of sea turtles in the Southeastern Pacific: Emphasis on Peru. In: K.A. Bjorndal (Ed.). Biology and Conservation of Sea Turtles. Smithsonian Institution Press. Washington, D.C. : 235-240.
Koepcke H. 1961. Synökologischen Studien an der Westseite der Peruanischen Anden, Bonner geographische Abhandlungen, Bonn.
Márquez M.R. 1990. FAO Species Catalogue. Vol. 11: Sea turtles of the world. An annotated and illustrated catalogue of sea turtles species known to date. FAO Fisheries Sinopsis Nº 125, Vol. 11. Rome, FAO.
Morales V.R. & Vargas P. 1996. Legislation protecting marine turtles in Peru. Marine Turtle Newsletter. 75: 22-23.
Moller-Hergt G.S. 1976. Análisis de la Situación Actual de los Mamíferos, Aves y Reptiles Silvestres del Perú. Biology Thesis, Universidad Nacional Agraria La Molina (UNALM), Lima, Perú.
Nichols W.J., Resendiz A., Seminoff J.A. & Resendiz B. 2000. Transpacific loggerhead turtle migration monitored with satellite telemetry. Bulletin of Marine Science. 67: 937-947.
Resendiz A., Resendiz B., Nichols W.J., Seminoff J.A. & Kamezaki N. 1998. First confirmation of a trans-Pacific migration of a tagged loggerhead sea turtle (Caretta caretta), released in Baja California. Pacific Science. 52: 151-153.
Wyneken J. 2001. The Anatomy of Sea Turtles. U.S. Department of Commerce NOAA Technical MemorandumNMFS-SEFSC-47.

Ecología Aplicada, 2(1), 2003 ISSN 1726-2216 Nota científica Depósito legal 2002-5474
OBSERVACIONES PRELIMINARES SOBRE COMPORTAMIENTO PRENATAL Y POSTNATAL DE Rattus rattus var. norvegicus (Linneo 1758) EN EL LABORATORIO
Martha Williams de Castro1, Alexandra Castillo2 y Constanza Rosas3
Resumen Con el objeto de lograr entrenamiento en la observación de roedores silvestres traídos al laboratorio, se llevó a cabo un estudio del comportamiento durante la preñez y lactancia de Rattus rattus var. norvegicus (Linneo 1758). Se observó que la agresividad de la madre aumentó conforme iba acercándose el parto, el cual se produjo al décimo séptimo día de gestación. Desde que las crías nacieron, la madre se mostró muy celosa y protectora; y nunca rechazó a sus crías a pesar de que estas eran manipuladas diariamente, para evaluar su peso y sus medidas de longitud cabeza - cuerpo. No se presentó canibalismo. El éxito reproductivo fue del 100% pues todas las crías llegaron a la adultez. Para el peso se encontró que las diferencias sexuales se hacen significativas a partir del día 29 (p = 0.006) mientras que para la longitud cabeza- cuerpo ocurre a partir del día 32 (p = 0.012). El destete se produjo el día 30. Palabras clave: Comportamiento maternal, gestación, parto, lactancia, destete, éxito reproductivo, bioterio.
Abstract The present research work was conducted with the objective of acquiring training in the observation of wild rats brought to laboratory conditions. It consisted in observing pre and post natal behavior in Rattus rattus var. norvegicus (Linneo 1758). The fact that the mother's aggressiveness increased with the arrival of parturition, event that occurred on the 17th day of gestation was observed. Once the pups were born, the mother showed herself protective and careful. She never rejected her pups even when they were manipulated daily to assess their weight and head - body size development. Cannibalism was not observed. The reproductive success was a 100% considering the fact that all pups reached adulthood. In relation to weight and head -body size, it was found that sexual differences became significant around day 29th (p = 0.006) and day 32nd (p = 0.012), respectively. Weaning occurred on day 30th. Key words: Maternal behavior, gestation, parturition, lactation, weaning, reproductive success, rearing center.
1 Laboratorio de Fisiología Animal y Biorremediación. Departamento de Biología, Universidad Nacional Agraria La Molina. Apartado 456. Lima 100. Correo electrónico: [email protected] 2 Departamento de Biología, Universidad Nacional Agraria La Molina. Apartado 456. Lima 100. Correo electrónico: [email protected] 3 Departamento de Biología, Universidad Nacional Agraria La Molina. Apartado 456. Lima 100. Correo electrónico: [email protected]
Introducción La rata blanca de laboratorio Rattus rattus var. norvegicus, goza de general aceptación para el estudio del comportamiento. Entre las ventajas de trabajar con esta especie destacan: su tamaño pequeño, su limpieza, facilidad de manejo y de alojamiento, y el bajo costo de su mantenimiento (Munn, 1950). Por su ciclo de vida corto, es ideal para realizar estudios de comportamiento materno. Esta especie alcanza la madurez sexual entre los 60 y 90 días, y posee un periodo de gestación que dura de 22 a 24 días, al final del cual nacen de 8 a 12 individuos. Puede tener de 8 a 12 nidadas por año y tiene una vida media de dos años (Vega, 2000). En el presente trabajo, realizado en el Laboratorio de Fisiología Animal de la UNALM, se hacen observaciones del comportamiento de una rata madre durante los periodos de gestación, lactancia y destete;
y del comportamiento de sus once crías. Además, se comparan los datos de la evolución del peso y el crecimiento cabeza - cuerpo de las mismas. Con esta investigación se busca establecer patrones de conducta tanto de la madre como de las crías y establecer cómo se va desarrollando el grado de diferencia en peso y tamaño entre hembras y machos. El entrenamiento con Rattus rattus var. norvegicus es de gran importancia, ya que brinda la experiencia para posteriormente realizar estudios con roedores silvestres de los que su biología se sabe muy poco. Para citar unos ejemplos, el entrenamiento posibilita realizar estudios como el de Künkele (2000) que muestra los efectos del tamaño de camada en la energética reproductiva de Cavia porcellus, o el estudio de Veloso & Bozinovic (2000) que examina los efectos de la calidad de alimento en la energética reproductiva de Octodon degus; o finalmente el

COMPORTAMIENTO PRENATAL Y POSTNATAL DE Rattus rattus var. norvegicus
Diciembre 2003 __________________________________________________________________________________________
144
estudio de Sumbera et al. (2003) en el que por primera vez se reporta aspectos de la biología de Heliophobius argenteocinereus. Revisión de Literatura A diferencia de otras especies que requieren experiencias previas para mostrar un cuidado materno adecuado, las ratas hembras de Rattus rattus var. norvegicus bajo condiciones de laboratorio, y sin ninguna experiencia previa con crías, usualmente cuidan de forma adecuada a su primera camada. La madre inexperta construye un nido, alumbra, reúne, limpia, y atiende a sus crías bajo la influencia de los niveles cambiantes de las hormonas (Fleming et al., 1999). Las hormonas juegan un rol muy importante en el desencadenamiento del comportamiento materno. En los últimos días de gestación, los niveles de oxitocina cambian dramáticamente en áreas del cerebro relacionadas con la conducta materna y retornan a la normalidad días después del parto. El contacto físico entre madre y crías, particularmente el que se da en la lactancia, es el que sigue desencadenando el comportamiento materno (Pedersen, 1979, 1992, citado por Zubritsky, 1997). Otras hormonas relacionadas con el comportamiento materno son: la progesterona que disminuye durante los últimos días de gestación, y el estrógeno y la prolactina que incrementan justo antes del parto. Estas hormonas pueden preparar lugares que regulen el aprendizaje y la memoria, y estimular el área préoptica media para responder con cuidado maternal al nacimiento de sus crías (Loy & Gerlach, 1988; Numan, 1991,citados por Kinsley et al., 1999). Además, las hormonas prolactina y la oxitocina son necesarias para la producción de leche. La influencia hormonal y la exposición a las crías, hacen que la madre involucre respuestas motoras ante ellas, como el levantarlas, movilizarlas hacia sitios seguros y colocarse sobre ellas (Fleming et al., 1999); además de evitar el hacerles daño (Peters et al., 1991). La sensibilidad hacia las crías aún se mantiene alta, semanas después del destete. El comportamiento materno también incluye la desaparición de conductas infanticidas que han sido observadas en ratas nulíparas y otras que ya han tenido crías pero que no se encuentran gestando. La desaparición del infanticidio se da tiempo antes del parto, y se presume que también está influenciado por factores fisiológicos (Peters & Cristal, 1983; citados por Peters et al., 1991). Después de un período promedio de gestación de 21 días, poco antes del parto, la madre comienza la construcción del nido. Utiliza de entre todos los materiales que se encuentran a su disposición, los que han estado relacionados con ella en alguna época de su vida (Riess, citado por Barnett, 1963). Barnett (1963) afirma que la construcción del nido es una actividad
homeostática ya que contribuye a mantener constante la temperatura corporal de las crías. Antes del parto, el dolor abdominal en la madre producido por las contracciones uterinas intensas la obliga a ponerse en posición de cúbito abdominal y a estirar las patas del tren posterior (Vega, 2000). El número de crías nacidas por parto fluctúa entre 8 y 12 crías (Shively, 1993; Vega, 2000). La madre ayuda a las crías a salir jalándolas con los dientes. Luego ingiere la placenta, el cordón umbilical y se deshace de las membranas fetales de las crías, con la finalidad de estimularlas y eliminar el contenido líquido de las vías respiratorias de los recién nacidos. Después de nacer, las crías desarrollan una atracción química hacia el vientre materno y se adhieren a los pezones como consecuencia de la experiencia intrauterina con el líquido amniótico. La atracción a menudo es seguida por el desarrollo de preferencias en los pezones. El reconocimiento de la madre se da por el olor que ella emana (Pederson & Blass, 1981; citados por Fleming et al., 1999). La estimulación causada por la lactancia provoca que la leche sea expulsada directamente dentro de la boca de las crías . Las crías recién nacidas tienen un peso de 5-6 gramos, nacen sin pelo, ciegas y sordas (Vega, 2000). Emiten dos tipos de ultrasonidos. El primero en respuesta al frío, desencadena la actitud de búsqueda por parte de la madre. El segundo en respuesta a un trato brusco de la madre, causa que ésta deje de provocar el estímulo que molesta a la cría. Alrededor del día 7 después del nacimiento el pelo comienza a hacerse más visible. Entre los días 14 y 17, los ojos y oídos comienzan a abrirse (Shively, 1993) y las crías se hacen más móviles. La hiperactividad se debe a que las áreas excitatorias del cerebro asociadas con la actividad ya han madurado pero las áreas inhibitorias de la actividad no lo han hecho aún. La maduración recién se da a inicios de la tercera semana (día 21). El destete también se produce alrededor del día 21 ya que la madre experimenta dolor en los pezones y por la presencia de pelo en las crías. Materiales y métodos En el mes de mayo del 2001 se inició el experimento con una rata madre preñada (segunda camada), criada en un bioterio. Antes del parto la madre fue puesta en una jaula (29 x 29 x 29 cm) y todos los días se le administraba agua ad libitum además de 40 g de alimento balanceado Tipo I (Tabla 1) antes del nacimiento de las crías y luego del Tipo II (Tabla 1) después del nacimiento. La madre fue pesada tres veces antes del parto para su control. El noveno día después del parto se trasladaron madre y crías a otro ambiente (54x 29.5x 29 cm) para su seguridad y comodidad, ya que se buscaba proveer a los animales de un ambiente que los enriqueciera (Chmiel &

MARTHA WILLIAMS, ALEXANDRA CASTILLO Y CONSTANZA ROSAS
Ecología Aplicada Vol. 2 Nº 1, pp. 143-148 __________________________________________________________________________________________
145
Noonan, 1996). Por último, en la tercera semana después del nacimiento, se colocaron en el ambiente una serie de trampolines y juegos para la estimulación mental y el desarrollo de conductas específicas de la especie. Tabla 1. Composición de las dietas utilizadas en la experimentación (Tipo I y II) y de dietas que muestran las composiciones nutricionales recomendadas.
Componentes Dieta Tipo I a
Dieta Tipo II b
Composiciones nutricionales recomendadas
Dieta Básica Materia seca (%) 88.433 96.900c Proteína (%) 21.000 17.216 14.700 c Fibra (%) 5.000 3.849 7.400 c Grasa (%) 4.000 5.000 c Ceniza (%) 8.000 3.700 c Extracto etéreo (%) 4.363 6.800 c EM. Aves Mcal/kg 2.880 Aminoácidos Lis (%) 0.920 0.920 d Met (%) 0.622 Met –Cis (%) 0.980 0.980 d Arg (%) 1.075 0.430 d Treo (%) 0.620 0.620 d Trip (%) 0.227 0.200 d Gli- Ser (%) 1.681 His (%) 0.457 0.280 d Ile (%) 0.742 0.620 d Leu (%) 1.411 1.070 d Fen (%) 0.822 Fen-Tir (%) 1.390 1.020 d Val(%) 0.852 0.740 d Minerales Fósforo (%) 0.370 0.300 d Ca(%) 0.630 0.500 d Na(%) 0.050 0.500 d (a) Composición del alimento balanceado Iniciarina Reforzada suministrado a la madre durante el periodo de gestación. (b) Composición del alimento para ratas suministrado a la madre durante el periodo de lactancia. (c) Valores hallados en una ración que cubría los requerimientos nutricionales de las ratas. Fuente: Gutiérrez, 2001. (d) Requerimientos nutricionales estimados para el crecimiento de las ratas. Fuente: National Research Council, 1995. Un día después del nacimiento de las crías (5 hembras y 6 machos) se inició la evaluación diaria del peso y la evaluación interdiaria de la longitud cabeza - cuerpo. Para la identificación de las crías se ideó un sistema de marcación en el cual las uñas de las hembras fueron pintadas con rojo y la de los machos con azul. Se anotaron todos los patrones de comportamiento de la madre y de las crías, así como la evolución de las características físicas, hasta el día en que madre e hijos fueron completamente aislados. Para efectos del análisis estadístico de los datos obtenidos de las crías,
se usó la prueba T tanto para evaluar peso como longitud cabeza-cuerpo. Resultados Comportamiento de madre y crías Desde un comienzo, la rata madre se mostró agresiva y fue difícil su manipulación. Se esperaba que las crías nacieran al día 21 de gestación mas el nacimiento se produjo al día 17. Horas antes del inicio del parto, la madre empezó con la construcción del nido. Para la construcción de éste, la madre mostró preferencia por el papel periódico a pesar de haber tenido al alcance otros materiales, como pedazos de tela y lana. Cuando se trató de incorporar éstos últimos al nido, la madre reaccionaba rápidamente, y los alejaba. Usó todo el papel periódico de la jaula formando una especie de túnel al que le clausuró todas las entradas. La madre parió 11 crías (5 hembras y 6 machos), a las que cuidó y protegió celosamente sin hacer distinción por alguna en particular. La madre trataba de mantener a las crías en el nido todo el tiempo. Cada vez que una cría intentaba acercarse a las experimentadoras la madre la tomaba cuidadosamente con sus dientes y la colocaba dentro del nido rápidamente. Este comportamiento se observó aún después de que las crías se mostraran independientes; es decir cuando los ojos y oídos ya estaban abiertos y podían desplazarse con confianza. En la exploración del ambiente, las hembras se mostraron más curiosas que los machos y en el inicio de los primeros signos de juego, las hembras se mostraron más activas que los machos. En cuanto al alimento y agua consumidos por la madre durante la gestación y lactancia, se observó un mayor consumo en este último periodo. Debido a ello se le administró el alimento Tipo II (Tabla 1), dieta alta en proteínas formulada especialmente para ratas. Desarrollo de las características físicas de las crías En la Tabla 1 se compara la concentración de componentes de las dietas Tipo I y II con las concentraciones de componentes de otras dietas que se sabe satisfacen los requerimientos nutricionales de las ratas (National Research Council, 1995; Gutiérrez, 2001). Se puede observar que la concentración de los componentes de las dietas utilizadas en este experimento, excede a la concentración de las dietas recomendadas, porque en la elaboración de las dietas se busca un margen de seguridad. En la Tabla 2 se muestra un resumen de la aparición de ciertas características físicas y de comportamiento de las crías. Las crías nacen sin pelo, dientes, con los ojos y oídos cerrados, dependiendo completamente del cuidado de la madre. La aparición del pelo en promedio comienza a partir del séptimo día, presentándose como una especie de pelusa la cual

COMPORTAMIENTO PRENATAL Y POSTNATAL DE Rattus rattus var. norvegicus
Diciembre 2003 __________________________________________________________________________________________
146
alrededor del día 11 ya es un pelo grueso y blanco brilloso. Para esta última fecha los dientes ya están formados. Los ojos y oídos se abren alrededor del día 15, siendo éste el comienzo de la exploración de su ambiente, el cual ya reconocen muy bien por el olfato. Empiezan a aproximarse a la comida y al agua como parte de un juego y a partir del día 17 después del nacimiento ya se alimentan solas. Las hembras se muestran más curiosas y avezadas que los machos para explorar lo que se encuentra fuera de su nido y especialmente los juegos y trampolines dispuestos para propiciar la estimulación mental Tabla 2. Características físicas y comportamiento de las crías de Rattus rattus var norvegicus, observaciones efectuadas en el Laboratorio del Fisiología Animal UNALM; del 23 de mayo (día 7) al 12 de junio del 2001 (día 30).
Apa
rició
n de
pe
lo
Apa
rició
n de
di
ente
s
Ape
rtura
de
ojos
y
oíd
os
Inic
io d
e
alim
enta
ción
por
sí
mis
mos
Expl
orac
ión
de
ambi
ente
y
prim
eros
sign
os
de ju
ego
Des
tete
par
cial
Des
tete
to
tal
Día 7 X
Día 11 X
Día 15 X
Día 17 X X
Día 22 X
Día 30 X
El día 22 después del nacimiento se separó la madre de 9 de sus crías. Con este destete parcial se evita problemas de mastitis con la madre. El día 30 se produjo el destete total y hasta esta fecha se observaron cuidados maternales. Como producto de la separación la madre se mostró inapetente y redujo su actividad. A pesar de que el canibalismo es un fenómeno que ocurre muy frecuentemente en roedores criados en bioterios, en este caso no se presentó. Peso: Desde el nacimiento se observó que los machos tienden a tener mayor peso que las hembras. La prueba T utilizada muestra que aproximadamente a fines de la cuarta semana (día 29) se comienzan a expresar significativamente la diferencia de peso entre hembras y machos (p = 0.006). Para el análisis de datos se trabajó con los datos obtenidos a partir del día 5 después del nacimiento pues fue para entonces cuando se pudo realizar el sexado correspondiente. El crecimiento tanto de las hembras como de los machos puede modelarse con una función sigmoidea.
Esto concuerda con lo observado por Zullinger et al. (1984): "...el aumento de peso en los mamíferos no es lineal ... las curvas de crecimiento de muchas de las especies son de forma sigmoidal". Los resultados de peso, ajustados a la curva sigmoidal de Gompertz arrojaron los siguientes valores: y = 449.292*e^(-e^(-0.037(x-45.65))) con un R2 de 0.996279 para los machos y la siguiente ecuación y = 269.292*e^(-e^(-0.045(x-36))) con un R2 de 0.9941 para las hembras, en las cuales e es la base del logaritmo neperiano, la variable dependiente es el peso en gramos y la independiente es el tiempo en días. La Figura 1 muestra las líneas de tendencia, observándose que las diferencias entre los sexos van acentuando conforme va aumentando el peso.
Figura 1.- Línea de tendencia en el aumento de peso dpeso en hembras y machos
y = 4.3945x - 44.791R 2 = 0.9462
y = 3.575x - 32.207R 2 = 0.96138
48
88
128
168
208
248
5 15 25 35 45 55 65 Días después del nacimiento
Peso
pro
med
io (g
)
Hembras
Machos Machos
Hembras
Longitud cabeza - cuerpo: Las dimensiones de longitud cabeza-cuerpo también se ajustan a la ecuación de Gompertz arrojando los siguientes valores: y = 42.12*e^(-e^(-0.0219(x-37.37))) con un R2 de 0.99123 para los machos y la siguiente ecuación y = 30.337*e^(-e^(-0.027(x-25.38))) con un R2 de 0.986398 para las hembras, en las cuales e es la base del logaritmo neperiano, la variable dependiente es la longitud en centímetros y la independiente es el tiempo en días. Al nacer, las crías no muestran diferencias en el tamaño, pero a partir de la segunda semana, los machos muestran un mayor tamaño que las hembras (Figura 2). La línea de tendencia muestra con un R2 de 0.9855 para los machos y 0.9826 para las hembras, que las diferencias en tamaño se van acentuando conforme van adquiriendo mayor tamaño. La prueba T realizada mostró diferencias significativas en las diferencias de longitudes entre hembras y machos alrededor del día 32 (p = 0.012). Esto concuerda con lo ocurrido con el peso, que también se hace significativo alrededor de esta fecha. Discusión Las ratas adquiridas en centros de crianza o bioterios muestran un comportamiento agresivo, ya que se desarrollan en espacios muy pequeños y el

MARTHA WILLIAMS, ALEXANDRA CASTILLO Y CONSTANZA ROSAS
Ecología Aplicada Vol. 2 Nº 1, pp. 143-148 __________________________________________________________________________________________
147
contacto con los humanos es mínimo. Esta agresividad es aún mayor si la rata se encuentra gestando o lactando. Debido a todo ello es comprensible que la rata del experimento se mostrara en extremo agresiva y de difícil manipulación.
La preferencia de la madre por papel periódico en la construcción del nido podría deberse a que éste es uno de los elementos con los que más se ha relacionado durante toda su vida ya que nació, creció y tuvo su primera camada en un ambiente compuesto únicamente por papel periódico. Riess (citado por Barnett, 1963) sugirió que la construcción del nido depende de un tipo de experiencia específico en un estadío de vida temprano. El mencionado autor crió ratas en un ambiente carente de elementos. Cuando estas ratas tuvieron que construir un nido, no lo hicieron a pesar de tener material disponible. En este estudio, la preferencia por el papel periódico puede interpretarse como la preferencia por un elemento con el que la madre se siente relacionada y por tanto le da suficiente seguridad para depositar a sus crías pues ella ya conoce las potencialidades del elemento (imprinting). Según Shively (1993) el parto de las ratas se produce entre el día 21 y día 23 de gestación pero en este caso se produjo en el día 17. Este adelanto pudo deberse a dos factores: (i) mayor actividad física producto de un área más amplia de desenvolvimiento, y (ii) estrés producido por el cambio de ambiente y el traslado continuo hacia diferentes áreas de trabajo. El comportamiento dedicado de la madre hasta días después del destete es coincidente con lo encontrado en la literatura (Steuer et al., 1987; Peters et al., 1991; Fleming et al., 1999), en la que se explica que este comportamiento está regulado por los cambios de niveles hormonales, principalmente por los cambios de cuatro hormonas: la oxitocina, el estrógeno, la progesterona y la prolactina. Muchos estudios de la energética reproductiva en mamíferos han confirmado que la reproducción es el periodo de vida que demanda mayor cantidad de energía (Bronson, 1989; citado por Veloso & Bozinovic, 2000). Especialmente durante la lactancia
el gasto de energía es muy alto (Millar, 1979; Oftedal, 1984; Thompson & Nicoll, 1986; Kenagy, 1987; 1989: McClure, 1987; Hayes et al., 1992; Thompson, 1992; citados por Veloso & Bozinovic, 2000). Una malnutrición durante la gestación y lactancia propicia que las crías nazcan con bajo peso, que estas sean más
propensas a las enfermedades y a la muerte y que las madres produzcan menor cantidad y calidad de leche (Stylianopoulos, 1999). Es por ello que se buscó alimentos enriquecidos para alimentar a la madre y a sus crías. Antes del parto se alimentó a la madre con un alimento formulado para pollos (Tipo I) y en una etapa postnatal se alimento a todas las ratas con el alimento Tipo II. El cambio de dieta se debió a que el segundo era formulado específicamente para ratas. Ambos tipos de alimento tenían un alto contenido proteico (Tabla 1) y cubrían satisfactoriamente los requerimientos nutricionales para el crecimiento de las crías. La calidad de la dieta en la etapa prenatal y
postnatal fue factor muy importante para la obtención de 92% de preñez (si se considera que 12 es la mayor cantidad de crías obtenidas por camada; Shively, 1993; Vega, 2000) y de 100% de éxito reproductivo, ya que la totalidad de crías llegaron a la adultez. La proteína brindada en la dieta sería una de las explicaciones a la ausencia de canibalismo. La madre obtenía suficiente proteína y no necesitaba una cantidad adicional proveniente de sus crías. Otra de las explicaciones sería la reducción en el estrés producido por el ambiente, ya que el área del nuevo ambiente proporcionado triplicaba las dimensiones del área donde la rata madre había vivido. La dieta también habría influido enormemente en el desarrollo del fenotipo, particularmente en la expresión del pelo. Estudios realizados en ratas con tipos diferentes de dietas: pobres y enriquecidas en nutrientes (Stylianopoulos, 1999), mostraron que el pelo de las ratas expuestas al primer tratamiento era áspero y opaco. Mas aún, en algunos casos llegaban a perder el pelo y desarrollaban dermatitis. En cambio las ratas alimentadas con la dieta enriquecida con formulaciones muy parecidas a los alimentos tipo I y II; mostraban el pelo liso y brillante, lo cual concuerda con nuestro alimento Tipo II, ya que las ratas mostraron un pelo muy brilloso. Conclusiones Los elementos con los que la madre estuvo relacionada en algún momento de su vida fueron los que escogió para la construcción de su nido. El alimento usado cubrió los requerimientos alimenticios de la madre durante la preñez y lactancia, así como también los requerimientos nutricionales de las crías durante su desarrollo. Esto se comprueba con el 100 % de supervivencia de las crías. Como en la mayoría de mamíferos, en las ratas el dimorfismo sexual está dirigido a los machos. Estos
Figura 2.- Línea de tendencia en el aumento de la dimensión cabeza- cuerpo en hembras y machos
y = 0.3162x + 3.629R 2 = 0.9855
y = 0.2901x + 3.7944R 2 = 0.98266
10
14
18
22
5 15 25 35 45 Días después del nacimiento
Logi
tud
cabe
za-c
uerp
o
prom
edio
(cm
)
HembrasMachosMachosHembras

COMPORTAMIENTO PRENATAL Y POSTNATAL DE Rattus rattus var. norvegicus
Diciembre 2003 __________________________________________________________________________________________
148
últimos presentan mayor peso y tamaño que las hembras. Las diferencias de dichas magnitudes entre los sexos son muy pequeñas al nacer y se hacen significativas al primer mes de vida. Durante todo el experimento se observaron comportamientos normales tanto de la madre como de la crías, observándose que para el juego, las hembras se mostraban más curiosas y activas que los machos. Mediante este experimento se ha logrado obtener el entrenamiento buscado en la observación del comportamiento prenatal y postnatal de roedores de laboratorio que servirá de base para la observación del comportamiento de roedores silvestres. Agradecimientos A Erika Paliza y a Mirtha Yrivarren por su apoyo y ayuda incondicional. Bibliografía Barnett S.A. 1963. The Rat a study in Behaviour.
Aldine Publishing Company. Chicago - USA. Chmiel D.J. & Noonan M. 1996. Preference of
Laboratory Rats for Potentially Enriching Stimulus Objects. Laboratory Animals. 30: 97-101.
Fleming O.S., O'Day D.H. & Kraemer G.W. 1999. Neurobiology of mother infant interactions: experience and central nervous system plasticity across development and generations. Neuroscience and Biobehavioral Reviews. 23: 673- 685.
Gutierrez C. 2001. Comparación de Tres Métodos en la Determinación de la Digestibilidad de una Dieta Estándar en Rattus norvergicus. Journal Boliviano de Ciencias 3 (6):9-9
Kinsley C.H., Madonia L., Gifford G.W., Tureski K., Griffin G.R., Lowry C., Williams J., Collins J., MClearie H. & Lambert K.G.1999. Motherhood improves learning and memory. Nature. 402 (11): 137 - 138.
Künkele J. 2000. Effects of litter size on the energetics of reproduction in a highly precocial rodent, the Guinea Pig. Journal of Mammalogy. 81 (3): 691-700.
Munn N.L. 1950. Handbook of Psychological Research on the Rat. The Riverside Press. Cambridge Massachusetts. USA.
NATIONAL RESEARCH COUNCIL. 1995. Nutrient Requirements of Laboratory Animals. Fourth
Revised Edition. National Academy Press. Washington DC. http//books.nap.edu/books/0309051266/html/index.html
Peters L.C. , Sist T.C. & Kristal M.B. 1991. Maintenance and Decline of the Suppression of Infanticide in Mother Rats. Physiol & Behav. 50 (2): 451-456
Shively M.J. 1993. Anatomía Veterinaria Básica, Comparativa y Clínica. Editorial El Manual Moderno S.A, México D.F. : 341- 350.
Steuer M.A., Thompson A.C., Doerr J.C., Youakim M. & Kristal M.B. 1987. Induction of Maternal Behavior in Rats: Effects of Pseudopregnancy Termination and Placenta-Smeared Pups. Behavioral Neuroscience. 101 (2): 219-227
Stylianopoulos C. 1999. Efectos de la fortificación y enriquecimiento de tortillas de maíz en el crecimiento y desarrollo cerebral de ratas durante dos generaciones. Tesis para optar el titulo de Master Scientiae. Instituto Tecnológico de Monterrey. Unidad de Postgrado. Maestría en Biotecnología. Monterrey - México. TRANSFERENCIA 48. Año 12. Número 48
http://www.mty.itesm.mx/die/ddre/transferencia/Transferencia48/eep-06.htm
Sumbera R., Burda H. & Chitaukali W. 2003. Reproductive biology of a solitary subterranean Bathyergid Rodent, the Silvery Mole - Rat (Heliophobius argenteocinereus). Journal of Mammalogy. 84 (1): 278 - 287.
Vega M. 2000 . Género Rattus. Portal de Veterinaria. (ISSN 1667-507X)
http://www.portalveterinaria.com/sections.php?op=viewarticle&artid=7
Veloso C. & Bozinovic F. 2000. Effect of food quality on the energetics of reproduction in a precocial rodent, Octodon degus. Journal of Mammalogy. 81 (4): 971-978.
Zubritsky E. 1997. The Limits of Love. What is the biology of motherly love? And why does it sometimes break down? Endeavors Magazine.13 (2).
Zullinger E.M, Ricklefs R., Redford K.H. & Mage G. M. 1984. Fitting sigmoidal equations to mammalian growth curves. Journal of Mammalogy. 65 (4): 607-636.


















