Efecto de Carayá (Alouatta caraya) en la dinámica y ... · Tesis presentada para obtener el grado...
111
Dirección: Dirección: Biblioteca Central Dr. Luis F. Leloir, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires. Intendente Güiraldes 2160 - C1428EGA - Tel. (++54 +11) 4789-9293 Contacto: Contacto: [email protected] Tesis de Posgrado Efecto de Carayá (Alouatta caraya) Efecto de Carayá (Alouatta caraya) en la dinámica y regeneración de en la dinámica y regeneración de las selvas de inundación del las selvas de inundación del Paraná medio Paraná medio Bravo, Susana Patricia 2003 Tesis presentada para obtener el grado de Doctor en Ciencias Biológicas de la Universidad de Buenos Aires Este documento forma parte de la colección de tesis doctorales y de maestría de la Biblioteca Central Dr. Luis Federico Leloir, disponible en digital.bl.fcen.uba.ar. Su utilización debe ser acompañada por la cita bibliográfica con reconocimiento de la fuente. This document is part of the doctoral theses collection of the Central Library Dr. Luis Federico Leloir, available in digital.bl.fcen.uba.ar. It should be used accompanied by the corresponding citation acknowledging the source. Cita tipo APA: Bravo, Susana Patricia. (2003). Efecto de Carayá (Alouatta caraya) en la dinámica y regeneración de las selvas de inundación del Paraná medio. Facultad de Ciencias Exactas y Naturales. Universidad de Buenos Aires. http://digital.bl.fcen.uba.ar/Download/Tesis/Tesis_3576_Bravo.pdf Cita tipo Chicago: Bravo, Susana Patricia. "Efecto de Carayá (Alouatta caraya) en la dinámica y regeneración de las selvas de inundación del Paraná medio". Tesis de Doctor. Facultad de Ciencias Exactas y Naturales. Universidad de Buenos Aires. 2003. http://digital.bl.fcen.uba.ar/Download/Tesis/Tesis_3576_Bravo.pdf
Transcript of Efecto de Carayá (Alouatta caraya) en la dinámica y ... · Tesis presentada para obtener el grado...

Di r ecci ó n:Di r ecci ó n: Biblioteca Central Dr. Luis F. Leloir, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires. Intendente Güiraldes 2160 - C1428EGA - Tel. (++54 +11) 4789-9293
Co nta cto :Co nta cto : [email protected]
Tesis de Posgrado
Efecto de Carayá (Alouatta caraya)Efecto de Carayá (Alouatta caraya)en la dinámica y regeneración deen la dinámica y regeneración de
las selvas de inundación dellas selvas de inundación delParaná medioParaná medio
Bravo, Susana Patricia
2003
Tesis presentada para obtener el grado de Doctor en CienciasBiológicas de la Universidad de Buenos Aires
Este documento forma parte de la colección de tesis doctorales y de maestría de la BibliotecaCentral Dr. Luis Federico Leloir, disponible en digital.bl.fcen.uba.ar. Su utilización debe seracompañada por la cita bibliográfica con reconocimiento de la fuente.
This document is part of the doctoral theses collection of the Central Library Dr. Luis FedericoLeloir, available in digital.bl.fcen.uba.ar. It should be used accompanied by the correspondingcitation acknowledging the source.
Cita tipo APA:Bravo, Susana Patricia. (2003). Efecto de Carayá (Alouatta caraya) en la dinámica y regeneraciónde las selvas de inundación del Paraná medio. Facultad de Ciencias Exactas y Naturales.Universidad de Buenos Aires.http://digital.bl.fcen.uba.ar/Download/Tesis/Tesis_3576_Bravo.pdf
Cita tipo Chicago:Bravo, Susana Patricia. "Efecto de Carayá (Alouatta caraya) en la dinámica y regeneración de lasselvas de inundación del Paraná medio". Tesis de Doctor. Facultad de Ciencias Exactas yNaturales. Universidad de Buenos Aires. 2003.http://digital.bl.fcen.uba.ar/Download/Tesis/Tesis_3576_Bravo.pdf

TESIS
CS. BIOLOGICAS
3576 IIIl
)
Bravo ¿Unl
N039 AD DE BUENOS AIRESCIAS EXACTAS Y NATURALES
Susana Patricia(l
f“
03576
—WAYÁ (A’louattacaraya)ENLADINAMICA Y REGENERACION DE'LAS SELVAS DE
INUNDACION DEL PARANA MEDIO
SUSANA PA'I'RK ‘lA BRAVO
DIRECTOR: GABRIEL EDUARDO ZUNINO
LUGAR DE TRABAJO:MUSEO ARGENTINO DE CIENCIAS NATURALES E INSTI'I‘U'I‘ONACIONAL
DE INVESTIGACIONES EN CIENCIAS NATURALES
TESIS PRESENTADA EN CUMPLIMIENTO PARCIAL DE LOS REQUISITOSPARA OPTAR AL TITULO DE DOCTORA EN CIENCIAS BIOLÓGICAS
H;2003 -—

RESUMEN
Entre Julio de l996 y Mayo de 2002 estudié el comportamiento de carayá (Alouatra caraya, Primate,Cebidae) y su papel como dispersor de semillas en la selva de inundación de la Isla Brasilera (Prov. delChaco, Argentina), una de las tantas islas ubicadas a lo largo del Río Paraná. Comencé describiendo los trestipos de bosques que conforrnaban la selva, y estudiando la fenología de las especies arbóreas con fi'utocamoso, potencialmente dispersables por carayá. El bosque que encontré en los albardones más antiguos fueel Bosque de Laureles, tuvo la mayor diversidad y abundancia de especies con fruto camoso y tres estratos,dos arbóreos y uno herbáceo. El Bosque de Sangre de Drago y Timbó crecía en albardones de similar altura alanterior pero de más reciente formación y el Bosque de Palo itá crecía en los albardones más bajos y eraprácticamente monoespecífico. En cuanto a la fenología pude distinguir dos grupos de especies, las queproducían frutos en mayor o menor cantidad todo el año y aquellas que tenían la fructificación temporalmenteacotada. El segundo paso fue estudiar la dieta, el comportamiento y el uso del espacio que realizaba carayá encl Bosque de Laureles. La dieta fue rica en frutos flores y hojas nuevas, además un cuarto de los registros dealimentación fueron sobre estructuras vegetales infestadas por artrópodos. Todo esto hizo que carayá noestuviese limitado energéticamente para desplazarse y pudiera utilizar con alta frecuencia todo su territorio.Luego estudié mediante ensayos de laboratorio la germinación de las semillas dispersadas, y en el campo elpatrón espacial de Ia dispersión. Encontré que el pasaje de las semillas por el tracto digestivo de carayá noafectaba la capacidad de germinación y para algunas especies aumentó la velocidad de germinación. Carayádepositó semillas en las zonas de sus dorrnideros y en los bosteaderos puntuales, que a causa de lasconfrontaciones entre grupos se ubicaban principalmente en los límites de los territorios. Continue estudiandoel impacto de la dispersión en este bosque (i. e. escala local) a Io largo de los años, para ello realicé conteos dejuveniles y renovales arbóreos en los sitios donde carayá deposita las semillas y fuera de ellos. Así encontréque la mayoría de las especies dispersadas se establecían tanto en los dorrnideros como en los bosteaderospuntuales, y no lograban hacerlo fuera de ellos. Con muestreos similares, evalue el impacto de la dispersiónde larga distancia en los tres tipos de bosque de la isla y el crecimiento de los juveniles con medicionessucesivas a lo largo de dos años. Observe que la dispersión de semillas que realiza carayá es fundamental parala colonización de los bosques jóvenes por parte de las especies del Bosque de Laureles, principalmenteOcolea diospyrrfolia, Eugenia punicrfolia y Psycholria carthagenensis. Mientras que estas mismas especiesno pueden establecerse en el Bosque de Palo itá, posiblemente por la mayor frecuencia de inundación quepresenta. Por último, para conocer si el reclutamiento en los bosteaderos se debía a la ausencia de predadoresy patógenos, a la saturación de los mismos debido al gran número de semillas que arribaban, o a una sen'e deinterrelaciones más complejas entre distintos factores. Evalué los daños que sufrían los juveniles, determineque artrópodos visitaban los bosteaderos, la disponibilidad de nitrógeno y fósforo, y la remoción de semillas.Esto me permitió arribar a la conclusión de que sumado al alto número de semillas que arribaban, elmecanismo por el cual se produce el establecimiento de los renovales a lo largo de los años se debería a unabaja tasa de remoción de semillas, una disminución de la herbivoría por la presencia de artrópodos predadoresy un aumento en la velocidad de crecimiento por una mayor disponibilidad de nitrógeno que facilitaría elescape a la mortalidad por ataque de los patógenos.
El efecto de carayá como dispersor de semillas no sólo acelera la regeneración del Bosque deLaureles y la maduración de los bosques jóvenes de la isla, sino que esta aceleración va en aumento debido aque se genera una retroalimentación positiva entre el aumento en la densidad de las especies dispersadas y ladensidad del dispersor. Esta puede ser una explicación para la rápida recuperación de estos bosques que seobserva luego de las inundaciones extraordinarias.
Palabras claves: Aloualta caraya, Río Paraná, dispersión de semillas, regeneración de bosques, selva deinundación, Argentina

ABSTRACT
l studied black howler (Alouaua curayu. Primate, Cebidae) behavior and its role as seed disperscr inthe flooded forest of Brasilera Island (Chaco, Argentina), one of Paraná River‘s islands, between July l996and May 2002. l began by describing of the three kinds of forests which make up the flooded forest and bystudying the phenology of trees with fleshy fruits. ln the oldest and highest areas of Brasilera Island l foundthe “Laurel Forest", which was the most diverse and had the greatest fleshy fruits species abundance. Thisforest had three strata. two of them arboreal and one herbaceous. The “Sangre de Drago and Timbó Forest"grew on areas of similar height which had recently been incorporated to the island. The “Palo ilá Forest" grewon low areas and had the lowest diversity. l could found two phenological sets of species, one set offeredfruits throughout the year, while the other set concentrated its fruit production in only in one or two months.The second step was the study of the black howler's diet, behavior and spatial use, in the Laurel Forest. Fruits,flowers and new leaves represented a high proportion of the diet all year long, and 25 % of feeding recordswere on arthropods infested items. Thus, black howler's travel distances were not energetically constrained,and groups could use their whole territory with high frequency. Then, l studied dispersed seed genuination ¡nthe laboratory and the spatial pattem of seed dispersal in the field. l found that passage through black howler'sdigestivo tract did not affect seed germination, but ingested seeds of some species gcrminated faster than notingested seeds. Black howlers transported seeds from the parent tree to sleeping trees and to latrines. As blackhowler groups confronted and defecated with high frequency within their territory limíts, latrines were moreabundant in these areas. l evaluated the impact of seed dispersal over several years in the Laurel Forest (i. e.,local scale) by sampling tree juveniles and renewals in latrines, and around sleeping trees, and outside theseareas. ln general, trees could grow in the latrines, and around the sleeping trees, but did not grow outsidethem. l evaluated long distance dispersion between the three kinds of forests with similar methods as the onesused in the Laurel Forest, and the growth of juveniles for two years. Seed dispersal by black howler isfundamental for the colonization of new forests by Laurel Forest species, specially Ocolea diospyrifolia,Eugenia punicifolia y Psycholria carlhagenensis. These species could not grow in Palo itá Forest, possinybecause of a higher flood frequency. Finally, l studied whether the recruitment of trees was due to the absenceof predation and pathogens in the latrines, the saturation by the great number of seeds that arrive, or an grthrelated to a higher nitrogen availability in the latrines, are important latrines, nitrogen and phosphorusavailability, and Seed removal in the latrines. l concluded that added to the great number of seeds that arrivedto the latrines, the low rate of seed removal, the attenuated herbivory by the presence of arthropods predators,and the faster for trees recruitment are important as well for trees’ recruitment. Fast growth could helpjuveniles to escape at mortality by pathogens.
Acceleration in Laurel Forest regeneration and in maturation of new forests is an effect of blackhowler seed dispersal, Also, as ¡n Brasilera lsland there are a positive feedback between the increase indispersed species density and disperser density, the acceleration is increasing. lt could be an explication forthe highest restoration of flooded forest after an extraordinary flood.
Key words: Alouaua caraya, Paraná River, seed dispersal, forest regeneration, flooded forest, Argentina

ÍNDICE
Ag! ' ' ' ‘
Introducción General 1
CAPÍTULO llnt. ' 7Materiales y '” " ' Rn IA I l l
|"' ¡7
CAPÍTULO ulntr ' ' ' ,1Materiales y '” " ' 7‘)Il A |l“ ‘m
CAPÍTULO lllInti J . ' 17Materialesy " " ' ¡Rn 'L .IIv 42
CAPÍTULO lVlnh J 40Materiales y ‘” " ' finn lAJn- “
CAPÍTULO vIntl I . ' 6‘Materialesy " " ' 64I) lA _IP. 71
CAPÍTULO v1lnti l ' 'Malerinles y '”“ ' 78I| 'A II" R4
CAPÍTULOvnlntegración y Discusión General (n
BIBLIOGRAFÍA 101
APándíon

AGRADECIMIENTOS
Mi primer agradecimiento debe sernecesariamente a la educación públicaargentina, porque si bien tuvo y tieneimportantes falencias que muchas veces metornaron dificultoso el camino, sin ella nohubiera podido siquiera soñar con alcanzarestudios universitarios. A mis padres querealizaron el esfuerzo durante tantos años, sinesperar la recompensa de una hija“adinerada”, aunque si pueden estar segurosde tener una hija feliz. A mi abuela que meapoyó en cada etapa y me enseñó a formar elcarácter para arremeter contra las dificultadesy perseverar ante quienes nos quierenentorpecer el camino, siento en el alma queno haya podido ver terminado “mi trabajocon los monos”. A mi amor, mi colega, mimarido y “director suplente”, gracias portantas horas compartidas y sufridasdiscutiendo ideas durante todas y cada una delas etapas de esta Tesis, ya fuera en el Museo,la Facultad, el campo, el auto, la playa o laclinica (y seguro que olvido algún lugar). Ami hijo Francisco por robarle cada día tantashoras de atención y cuidados. A mi hermanopor su asistencia técnica en la última etapa ya mi hermana por hacerse cargo de su sobrinodurante los extensos fines de semana que lospadres dedicamos al trabajo. A todos ellos vadedicado este trabajo.
A aquellos machistas, asumidos o no,que creyeron que debían hacerme las cosasdificiles para que comprendiera que haytrabajos que no son para las mujeres. Graciassinceramente, fueron un desafio que me diocada vez más motivos para llevar el trabajoadelante, lo mismo digo a aquellos que meconsideraron demasiado joven o demasiadodesafiante.
Una mención más que especial a AnaSallenave, que comenzó siendo mi mejorayudante en el campo para finalizar siendouna gran amiga y una ayuda fundamental, sinella el cansancio me hubiera vencido en másde una oportunidad. Muchos fueron losestudiantes y amigos a los que deboagradecer por colaborar en el trabajo decampo, compartiendo calor, mosquitos, frío,
mosquitos, “largas calmas chichas”,mosquitos, tormentas furiosas, mosquitos,vacas destructoras de campamentos,mosquitos, falta de agua y ...mosquitos, ¡Yestaban felicesl, Candelaria, Emilio, PabloRodríguez, María Alejandra, María Soledad,Boris, Florencia, Lorena, Lula, PabloSalomón, Silvana, Sebastián, Lola, Verónica,Mariel, Ignacio, Martín, Sonia, Silvina, JuanManuel, Johana, Florence, Laurence, Nancy,Severine, JP, Huges y Frederic (sólo esperono olvidar a ninguno). A Silvana Capria porsu ayuda con las experiencias de germinacióny a Gabriela Pirk por su ayuda para pasar enlimpio datos y sus correcciones del idiomainglés.
Gracias a Alejandra Ribichich por suvaliosa ayuda con la nomenclatura botánica,al Dr. Krapovikas y la Dra. Mirta Arriaga porsus determinaciones, a la Dra. Laura de Caboy la Dra. Silvana por los análisis químicos delos eamalotes, al Dr. Arturo Roig, al Dr.Bachman, al Dr. Alejandro Farji, a la DraCristina Sciocsia y al Dr. Di lorio, por suayuda en la identificación de los artrópodos.
Al Dr. Juan Jose Neiff por suimportante ayuda, sobre todo en las primerasetapas del trabajo, y por ser el responsable demi primer visita al norte y mi enamoramientodesde aquel momento con el Paraná.
‘ Debo agradecer también a todosaquellos que me posibilitaron logísticamentellevar adelante el trabajo, al Museo Argentinode Ciencias Naturales, a la Dra. MarthaPiantanida que tanto me apoyó y toleró, aOlga Vaccaro, Esperanza Varela y JuliánFaivovich. A la comunidad de lsla delCerrito, al Destacamento de la PrefecturaNaval Argentina de Isla del Cerrito, al Centrode Capacitación Comunitaria, a Gustavo, aSencio, al señor Gallo, a Titina, a toda lafamilia Bobis y al Sr. Felipe por susriquísimas hamburguesas y empanadas quenos acercaban a la civilización. Muyespecialmente al Dr. Cao, Liliana y sus hijosque tan amablemente nos recibieron cada vezque fue necesario, haciéndonos sentir parte dela familia.

Susana I’ulriciu Bram
No puedo olvidarme de agradecer aOrejas por aceptamie en el grupo yemocionanne hasta las lágrimas cada vez quedecidía bajar mirándome a los ojos, parasentarse, comer, caminar 0 simplementedescansar a mi lado.
A aquellos que posibilitaroneconómicamente la realización de estetrabajo, Conicet, lFS, A Pas de Loup,
Museo Argentino de Ciencias Naturales"Bernardino Rivadavia”.
A quienes leyeron críticamente losprimeras, segundas, terceras,...versiones dealguno o todos los capítulos del trabajo, Dr.Luis Marone, Dra. Bette Loiselle, Dr. VictorCueto, Dra. Carolin Crockett.

Almmna mraya como dispersor de semillas
INTRODUCCIÓN GENERAL
Las que creen que la creación de lasvegetalesha sida simultánea, y par consecuencia,lada planta viene de semilla o renueva, eslánpersuadidas de que cuando se ve'naeer una plantaen un paraje donde na existia ama se debea lasvientosa las aves. que han llevada la semilla; peroya quisieraquerefluianaran...
Félix de Azara (Viaja par Ia América meridional)
Dispersión es un término que seutiliza para denominar el proceso por el cuallos individuos se alejan del ambienteinmediatamente próximo a sus padres yvecinos, por este proceso la distribución delas especies logra ser menos agrupada(Begon et al., ¡996).
Al estudiar las plantas ya sea desdeuna perspectiva anatómica, ecológica oevolutiva, se ha encontrado una grandiversidad de estructuras y formas quefacilitan la dispersión de las semillas. Entrelos factores ecológicos que las plantaspueden utilizar como dispersores seencuentran tanto factores abióticos, el viento,el agua. e incluso el mismo calor del sol quefacilita la explosión de ciertos frutos, comofactores bióticos, los animales, que puedentransportar semillas ya sea adheridas a sucuerpo o dentro del tracto digestivo alalimentarse de los frutos (Renner, ¡992).
La relación que existe entre losanimales dispersores de semillas y las plantascon frutos puede ser considerada como unmutualismo dinámico, donde los animalesutilizan los frutos como alimento, mientraslas plantas logran movilidad para sus semillas(Howe ¡986). Sin embargo, analizando elcambio de dispersores que muestran lasplantas a lo largo de su distribucióngeográfica o la baja superposición temporalde plantas y dispersores en el registro fósil,se considera que las relaciones entre losmamíferos dispersores y las plantas no debentomarse como relaciones co.evolutivas sinomás bien como relaciones ecológicas que segeneran por la co-ocurrencia espaciotemporalde las especies (Herrera, l985; Howe, 1986;Chapman y Chapman, 2002).
Se han elaborado tres hipótesis paraexplicar cual seria la importancia deldesarrollo de estrategias de dispersión en lasplantas: l) Hipótesis de escape: escape a lamortalidad denso dependiente bajo el árbolparental; 2) Hipótesis de disturbios:ocupación de un espacio abierto; 3) Hipótesisde la dispersión directa: ocupación demicrohábitats especiales necesarios para elestablecimiento de renuevos. Por supuestoestas hipótesis no son mutuamenteexcluyentes (Howe, l986).
Al estudiar la dispersión de semillasno sólo debe considerarse la movilidad comoel único factor a tener en cuenta paradeterminar la eficacia del proceso, también sedebe considerar que no se reduzca lacapacidad para genninar de la semillatransportada y que esta sea depositada en unlugar apropiado para el crecimiento de unnuevo individuo (Herrera y Jordano, 1981;Howe y Smallwood, ¡982; Schupp, 1993).La eficacia del dispersor entonces, incluyedos aspectos a evaluar "cantidad" y "calidad"(Schupp, l993).
Las semillas pueden ser arrojadas sinser ingeridas, regurgitadas, ingeridas ydigeridas o bien ser ingeridas pasar por eltracto digestivo y luego ser defecadas. Esteúltimo mecanismo puede llevar minutos,horas, dias o semanas (Howe l986). Claroestá, que el tratamiento de mejor calidad nolo podemos definir a priori pues dependeráde los requerimientos de la especie botánicaque el animal esté manipulando. Así, untrabajo sobre dispersión de semillas deberesponder tanto preguntas sobre las especiesanimales como sobre las especies vegetales ydel ambiente, ¿Transporta este animal lassemillas de las frutas que consume7, ¿Dónde

son depositadas estas semillas?, ¿Quécaracterísticas tienen los microhábitat donde
son depositadas las semillas?, ¿Que' especiesson transportadas?. ¿Pueden estas semillasgenninar? (Jordano y Godoy, 2002).
Si el trabajo de investigación tienecomo meta, conocer el impacto que ladispersión de semillas producida por unfrugívoro tiene sobre una selva y sudinámica, debe ir más allá de la relaciónmutualista de dos especies, abarcando yrespondiendo preguntas tales como ¿Eshomogénea la selva, en cuanto a sucomposición y estructura?, ya que el impactopodría ser mayor si existen zonas de la selvadonde no se encuentran árboles adultos de lasespecies dispersadas. ¿Cómo es la oferta defrutos comestibles para el frugívoro a lo largodel añ0?, así sabremos si el impactó delfrugívoro es constante en el año o no. ¿Cómoutiliza el espacio el fruglvoro?, puessolamente podría estar influyendo en laregeneración de un sector o tipo de selva.¿Cuál es la densidad del frugívoro?, unfrugivoro con una baja densidad es de esperarque tenga un impacto muy leve.
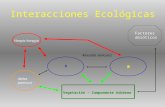
Garber y Lambert ( l998),propusieron un modelo que representa ladispersión de semillas por primates y suimpacto en el ambiente, en el témiino de trescomponentes de interacciones animal-plantaambiente. Estos componentes fueronrepresentados como una fase predispersiva,donde los primates son atraídos hacia la fruta,una fase dispersiva donde los primatesremueven la fruta de la planta y por últimouna fase posdispersiva donde las semillasviables pueden ser predadas, o gerrninar yluego sobrevivir durante los distintos estadioshasta llegar a ser reproductivos. Esta tesispuede enmarcarse dentro de los términos deeste modelo (Fig. l).
El Capítulo l se centra en el estudiode la fase predispersiva, que comprendetodas aquellas características de la vegetaciónque pueden influir en el establecimiento delas plántulas y en la decisión de los monos dealimentarse de tal o cual especie, en estecapítulo intento describir la estructura de lavegetación dentro de la isla centrándome enlos aspectos que más relevancia pueden tener
para carayá (Alouatla caraya), por ejemploidentifico los distintos tipos de bosque queexisten dentro de la isla, la composicióntloristica de los distintos estratos en cada unode ellos, la densidad de las especies arbóreasy la altura del albardón sobre los cualescrecen estos bosques; también describo lascaracteristicas fenológicas de las distintasespecies utilizadas por carayá, en especialaquellas con fruto carnoso potencialmentedispersables por los monos.
Una vez descripto el ambiente, elCapítulo ll se centra en el estudio de la fasedispersiva, es decir el estudio de aquellosaspectos del comportamiento y uso delambiente de carayá que pueden relacionarsecon la remoción de semillas de los árboles.
Estos aspectos van desde el estudio del usodel espacio por parte de los grupos. paradeterminar si existen áreas preferidas para laalimentación, hasta aspectos más puntualescomo el tipo de manipulación que carayáhace de los frutos y la proporción de estos enla dieta. Además en este capitulo pondré aprueba una serie de hipótesis ampliamenteaceptadas en la bibliografia sobre aulladores.
Conociendo ahora el ambiente, cómocarayá lo utiliza y de que especies remuevesemillas, comienzo el estudio de la faseposdispersiva la cual es descripta por losCapítulos Ill, lV, V y Vl. El Capítulo lll lodedico al estudio de los patrones dedispersión que genera carayá y a laevaluación de Ia germinación de las semillasdispersadas mediante pruebas de laboratorio.En el Capítulo IV describo el papel de carayácomo dispersor local, evaluando el éxito deestablecimiento en los distintos sitios cn quecarayá deposita semillas. En el Capítulo Vevalúo a carayá como dispersor de largadistancia, centrándome en el estudio de lallegada y establecimiento de las especies deuno de los bosques de la isla a los otrosbosques. En el Capítqu VI describo algunasde las caracteristicas bióticas y abióticas delos sitios donde carayá deposita las semillas ycomo estas caracteristicas pueden afectar elestablecimiento de los juveniles arbóreos.Finalmente en el Capítulo Vll integro ydiscuto los resultados de los capítulosprecedentes.

FASE PREDISPERSIVA (Capitulo l)
D. l .b ., Posibilidad de¡s l'l ucron . .. E
de los árboles dispersmn por FenologiaA. caraya
Tamaño de la
copaAtributos de
frutos y semillas
FASE DISPERSIVA (Capitulo II)
Uso delespacio y
alimentación
Efectividad de—) A.caraya
como dispersor
\ Manipulacióndefrutos / semillas
Tiempo deretención delas semillas
(Capitulo lll) FASE POSDISPERSIVA (Capítulo IV)
Destino de las ‘. , , . Dispersión local:
Germinacwn semlllas E Credmemo yde sem'uas dispersadas P0r A- establecimient
caraya
/ (CapituloV)Distanciay \patrón de Dispersión dedispersión larga distancia:
Crecimiento y(Capitulo VI) stablecimient
Predadoresy patógenos
Figura l: Estructura de la tesis siguiendo el modelo propuesto por Garber y Lambert (¡998) para ladispersión de semillas por primates.

.-IIOuaIIacarayu como dispersor de semillas
Capítulo l: Área de estudioLas islas del Paraná Medio
INTRODUCCIÓN
Comparado con el Orinoco o elAmazonas el Paraná tiene un régimenhidrológico irregular, pudiendo existir hasta3 inundaciones por año con una duración deentre 2 semanas a 3 meses y una fluctuacióndel nivel del agua entre los 2 y 6 m (Carignany Neiff. ¡992), son frecuentes las crecientesentre Febrero y Marzo y las bajantes entreAgosto y Septiembre (Franceschi y Lewis.l979). Existen también crecientesextraordinarias que pueden durar más de l2meses. las cuales producen un gran impactosobre la vegetación y la fauna, pudiendomorir hasta el 50 % de los árboles (Neiff.¡978. 1999). El último registro de este tipode creciente fue en 1982/83 y las anterioreshabrian ocurrido en ¡905, l9l2, ¡946, |95|,¡966, ¡973 y ¡977 (Neiff, ¡978). Lascrecientes son provocadas fundamentalmentepor las lluvias ocurridas en las nacientes delrío en Brasil (Franceschi y Lewis, ¡979).
En la actualidad el régimenhidrológico estaría siendo alterado por lasdistintas modificaciones que ha sufrido lacuenca, como ser la deforestación masiva yla reestructuración del río (Bucher y Chani,¡999. Neiff, ¡999). Los cambios más fuertesen el pulso anual se atribuyen a los efectos dela represa ltaipú, ubicada en el límite entreBrasil y Paraguay (25° 30' S y 54° 20' O)(Bonetto y Hurtado, 1999).
Las islas del Río Paraná medio seforman por la acumulación de sedimentosque transporta el río, la mayor parte de ellosaportados por su afluente el Bermejo, por lotanto son mayoritariamente de origen andino(Popolozio, 1977). Durante la fase final delas crecientes se depositan los primerossedimentos "depósitos de cauce" que daránorigen a futuras islas, estos son sedimentosde textura gruesa y forman bancos de caucede forma eliptica (Franceschi y Lewis, 1979).Posteriormente en sucesivas inundaciones.debido a la disminución de la velocidad delagua sobre los bancos y luego por la
retención que realizará la vegetación,sedimentan los "depósitos de inundación"que son de textura más fina (Franceschi yLewis, 1979).
Los suelos son semipantanosos, enlas partes más altas son gley húmicos y en losalbardones gley subhúmicos (Franceschi yLewis, ¡979).
Estas islas pueden ser removidas porel mismo rio o bien afirmarse y unirse aotras, asi se conforman las grandes islas quecon el tiempo se adosan a tierra firme eintegran lo que se conoce como valle aluvialdel Río Paraná (Bonetto y Hurtado, 1999).
Dentro de las islas nos encontramoscon bosques que crecen en albardones dedistintas alturas y pastizales con diferentecomposición florística, los albardones estánseparados por lagunas que en algunos casosse conectan entre sí y con el río (obs.personal).
Desde un punto de vistafitogeográftco no hay acuerdo a queprovincia pertenece la vegetación de las isladel Paraná. El primer estudio fitogeográfrcode la Argentina (Cabrera, l976) determinó ¡3provincias y si bien no menciona a las islasdel Paraná específicamente, estas estarianconsideradas como "selva marginal" dentrode la provincia Paranaense, distrito de lasselvas mixtas. Estas selvas producirían unaintrusión de la provincia Paranaense dentrode la provincia Chaqueña aprovechando elmicroclima ribereño de los ríos Paraná yUruguay (Cabrera, ¡976). Sin embargo,Morello y Adámoli (l974) al describir lavegetación del Chaco Argentino, incorporanIa vegetación del valle de inundación delParaná, y definen una subregión ParanáParaguay. Pero admiten que estos bosquesson poco conocidos y contendrían especiesno pertenecientes al Chaco sino ligadas a lasYungas. Martinez-Crovetto (¡963) al realizarel esquema fitogeográfrco de Misionesconsidera la vegetación de las islas del alto

Susana Patricia llruvo
Paraná cómo parte de la "selva higrólilaribereña" incorporando junto a ella tanto a laselva de las zonas más altas de las riberas,como a la selva ribereña que es alcanzadafrecuentemente por las crecidas. El mismocriterio fue seguido por Camevali (1994) alrealizar la fitogeografia de la provincia deCorrientes, quien llamó al conjunto "selvamarginal o ribereña". Sin embargo, ya en elbajo Paraná Burkart (l957) distinguió para elDelta un "bosque fluvial" creciendo en laszonas más bajas e inundables y el “monteblanco" creciendo sobre los albardones másaltos. Franceschi y Lewis (1979) al estudiarlos bosques de las islas de la Provincia deSanta Fe, denominan a estos bosques como"Foreslum nigrum" o "Foresmm insularis"dependiendo de su composición florística,pero no incluyen estas comunidades dentrode una u otra provincia fitogeográficadefinida. Desde una perspectiva ecológica, enel libro "Humedales de la Argentina"(Canevari et aI., l999) se hace referencia a lasislas del Paraná tanto en el capítulocorrespondiente a la región Chaqueña(Bucher y Chani, l999), como en el de laregión de la Cuenca del Plata que incluye alas provincias de Entre Rios, Corrientes yMisiones (Bonetto y Hurtado, l999). SegúnAdámoli (1999) existirían variadas razonespara establecer una unidad funcional entre lasplanicies de inundación de los ríos Paraná yParaguay y con este criterio establece unaregión a la que llama eje Paraguay-ParanáDelta, en la cuál quedarían incluidas las islas.En cuanto al origen de las especies botánicasse encuentra influencia amazónica.paranaense y en algunos casos elementosvinculados con las Yungas (Adámoli, ¡999).
Puesto que las características de laspoblaciones de carayá de uno y otro ambienteresultan diferentes, para los trabajos deprimatología (Rumiz et al., l986; Rumiz,l990; Brown y Zunino, |994; Zunino et a|.,¡996) las islas de los ríos Paraná y Paraguayrepresentaron siempre un ambienteclaramente distinguible de las selvas que seencuentran en tien'a fimte refiriéndose a ellascomo "selva de inundación".
En conclusión existirían criteriosdispares para ubicar las islas del Paranádentro de una región u otra, sin imponar quepauta de clasificación se utilice. Sin
embargo, la mayoría de los autores coincidenen señalar que se trata de humedales muydinámicos, poco estudiados, sin protección yque han estado recibiendo un fuerte impactoambiental en las últimas décadas(Franceschiy y Lewis, |979; Bucher y Chani,l999; Bonetto y Hurtado. 1999; Neiff. l999).
En este capítulo describiré el sitio deestudio prestando especial interés a aquellosaspectos que puedan influir en elcomportamiento del carayá como dispersorde semillas. Primero describiré ladistribución de la vegetación dentro de la IslaBrasilera, luego estudiaré las diferenciasentre los distintos tipos de bosques en la islay finalmente la fenología y las característicasde los frutos que consume carayá.
MATERIALES Y MÉTODOS
El estudio lo llevé a cabo en la lslaBrasilera, Provincia del Chaco, la cual seencuentra en las proximidades de laconfluencia de los ríos Paraná y Paraguay(Fig. 2).
El clima es subtropical templado conuna temperatura media anual de 2|,0 °C yuna amplitud térmica anual de 12,3 °C.Dentro del valle de inundación del Paraná se
registra una menor amplitud térmica diaria yanual que para el resto de la región. Un claroejemplo de esto es la rara ocurrencia deheladas. En la lsla del Cerrito el últimoregistro se produjo en Julio de 2000 (obs.pers.) y de acuerdo con los pobladores elanterior habría ocurrido en la década del 60.
La precipitación media anual de laregión es de l556 mm, las lluviasdisminuyen en invierno sin llegar adeterminarse una verdadera estación seca(Apéndice). En lo que respecta a las islas losposibles déficit hídricos se ven atenuados porlas inundaciones, sin embargo si la sequía seprolonga al terminar la inundación las plantascomienmn a sufrir el déficit rápidamentepues los suelos tienen una baja capacidadpara retener agua útil (Placci, ¡995).
Caracterización de los bosgues de la selvade inundación
Utilicé fotografias aéreas con escalal:3().()()0, para realizar un nmpu de la.

lOOOOOOOOOOOOO0.000000000000000000...0.0.0.0...
Alarm/m camya como dispersor de semillas
27° 18' S
z
59° 40' O
O
¿t ¡stack/ama, . ilN
Bosque de Sangre de Drago y Timbó
D BosquedePaloirá
La a iznlet El 8mm“ s27°2o's59° 37' O
Figura 2: Ubicación de la lsla Brasilera (Provincia del Chaco, Argentina) cn América del Sur. Detalle de la isladonde muestro la distribución de los distintos tipos de bosque.
vegetación de la isla. Digitalicé y analicé lasfotografias utilizando una tabletadigitalizadora, New Sketch 1812 D (Genius)y el programa ldrisi. Así obtuve la superficietotal de la isla y la superficie boscosa. Luegoconfirmé la distribución de los mismos en elcampo.
Las observaciones en el campo merevelaron diferencias en la estructura ycomposición florística de los distintos
parches de bosque. Para caracterizar estosbosques realicé muestreos de vegetación enuna superficie total de l ha en cada uno. A lolargo de los albardones instalé 25 parcelas de20 x 20 m y en ellas registre para todos losárboles con un DAP (diámetro a la altura delpecho) mayor a los lO cm, la especie, sualtura y el valor del DAP. También registré lapresencia de enredaderas, lianas e hierbas.

Susana Patricia Bravo
Para tener una descripción másdetallada de Ia vegetación del bosque donderealicé los estudios que describire’ en elCapitulo ll, incorpore' en los registros de estebosque los árboles con DAP mayor a 5 cm.
Las especies fueron identificadasutilizando la "Flora arbórea del ParqueNacional lguazú" (Dimitri et al., l974),"Flora Ilustrada de Entre Rios" (Burkart,1969), "llustraciones de hojas de especiesarbóreas del Paraguay para la identificaciónde género o especie" (Nakamura, 1983) y"Plantas de la Costa" (Lahitte et al., ¡997).La nomenclatura seguida fue la propuesta porel "Catálogo de Plantas Vasculares de laRepública Argentina“ (Zuloaga y Morrone,l999).
Realice una descripción gráfica delperfil de estos bosques con una transecta delOOmetros de longitud y lO metros de anchoen cada bosque, donde registré para todos losárboles y arbustos, la especie, altura total,altura del inicio de la copa, diámetro de lacopa, forma de la copa
Utilizando las marcas que el aguadejó en los árboles luego de la inundación de200] y con los registros de altura del rio de laciudad de Paso de la Patria (Apéndice),Provincia de Corrientes (obtenidos en eldestacamento de la Prefectura NavalArgentina de isla del Cerrito), calculé laaltura aproximada de los albardones dondecrecía cada tipo de bosque. Utilice dichosregistros pues esta ciudad se encuentra frentea la isla Brasilera sobre la margen derechadel Río Paraná.
Caracterización de los bosques“ variables de importancia
para carayá
Con los datos obtenidos en elrelevamiento de vegetación calcule lassiguientes variables que consideré podríanser importantes en la relación de carayá conla vegetación (Zunino et al., 1996).Variables que se relacionarian con la calidady cantidad de soportes para eldesplazamiento de caraya:Número de individuos (Nl)Altura media (AM)Área basal total (ABT)
Variables que se relacionarian con lacantidad de alimento de alta calidad paracaraya:Número de individuos con fruto camoso(NIFC)Número de especies (NSP)Número de especies con fruto camoso(SPFC)Área basal de los individuos con frutocarnoso (ABlFC)Número de individuos por especie (NI/SP)
Variables que se relacionarian con ladificultad en la búsqueda y obtención delalimento de alta calidad para caraya:Número de individuos con fruto camoso /Número de individuos (NlFC/Nl)Número de especies con fruto camoso /Número de especies (SPFC/NSP)Área basal de individuos con fruto carnoso /Área basal total (ABIFC/ABT)Área basal total / Número de individuos(ABT/Nl)Número de individuos con fruto carnoso /Número de especies (NlFC/NSP)
Con estas variables realicé unanálisis de componentes principales.
Fenología de las especies utilizadas pormuiCabe aclarar que si bien por una
razón de simplicidad hablaré de frutos. encasos como Cecropia pachystachya se tratamás precisamente de infrutescencias. Estecriterio será utilizado en toda la tesis.
Para evaluar la dinámica de la ofertade l'rutos, asi como de las demís estructurasvegetales utilizadas por carayá comoalimento (ver Capitulo ll) realicé registrosfenológicos a lo largo de l año. Lasestructuras vegetales consideradas, ordenadaspor su valor energético de maneradecreciente (Milton, 1980) fueron, Frutas,Flores, Brotes, Hojas nuevas, Hojas maduras.
Cada mes seleccione lO árboles alazar de las especies más importantes en ladieta de car-aya, evalué la abundancia de lasestructuras en cada individuo con un indicesubjetivo que varió entre 0 y 4 (0 = ninguna,4=máxima abundancia), obtuve un valorpromedio cada mes para las distintas

Alaualla caraya como dispersor de semillas ll
estructuras y las distintas especies al cualllamaré "Índice fenológico" (lF). Cuantifiquéla variación mensual de cada estructuraconsiderando todas las especies utilizadascomo:
IFTsp = (ZIF¡)x100/N,lF¡= indice Fenológico de la especie i.N = número de especies consideradas
Descripción de los frutos consumidos mrcarayá
De las principales especies cuyosfrutos son consumidos por car-aya (verCapítulo ll), seleccioné ¡0 individuos al azary colecté entre 10 y lS frutos de cada uno deellos. De cada fruto tomé los siguientesdatos, medida del eje mayor, medida del ejemenor, color, peso fresco, peso seco, tipo defruto y número de semillas. Calculé elporcentaje de agua utilizando la diferenciaentre el peso fresco y el peso seco. El tipo defruto fue establecido consultando las guías deidentificación utilizadas para determinar lasespecies. Para conocer las características delresto de los frutos del bosque no consumidospor carayá colecté bimensualmente a lo largode 2 años todos los frutos que encontraba yrealicé las mismas mediciones.
RESULTADOS
Caracterización de los bosgues de la selvade inundación
La isla Brasilera tiene una superficietotal de 280 ha, de las cuales ¡51,89 ha estáncubiertas por bosques, esto representa másdel S4 % de su superficie. Detecté tres tiposdiferentes de bosque (Fig 2). La superficieque informo para los bosques tiene ciertoerror acan'eado del hecho de que en algunoscasos los albardones se angostan y sobreellos sólo crece una única hilera de árboles,cómo ya lo han descripto (Franceschi yLewis, l979), lo que hace difícil detectarlo através de la fotografia aérea por un lado ydificil de determinar su ancho en el campopor el otro.Bosgue de Laureles (Foto l) (Ocoleadiospyrrfilia): 65.9 ha. Se encuentra en losalbardones más altos y anchos de la isla, conuna altura que varía entre los 5,5 m y los 7 m
sobre el nivel 0 del hidrómetro de Paso de laPatria, estos albardones son los más antiguosy se encuentran mayoritariamente en elcentro de la isla (Fig 2). En la hectárea devegetación que relevé encontré un total de55| árboles con un DAP mayor a los lO cm,donde el ¡6.5| % de los individuospertenecieron a la especie 0. diospyrrfolia.
Al incluir los árboles con un DAPentre 5 y lO cm, se agregaron 452 individuosmás y obtuve la composición florr'stica de losdos estratos arbóreos que caracteriza estebosque (Tabla l y Figura 3 a). El estrato bajoestá compuesto por individuos de entre 2 alO metros de altura, con el 72,28 % de losindividuos entre los 4 y 8 metros. El DAPmedio del estrato es de ll d:8 cm. Está
dominado por Euguem'a punicifolia yPsycholrr‘a carihagenensís, ambas especiesson exclusivas de este estrato y debo destacarque P. carlhagenensr's nunca superó los 6metros, teniendo la mayor parte de losindividuos entre 2 y 3 metros de altura.Otrocomponente importante del estrato, 8,9! %de los individuos relevados, fueron losrenovalcs de O. diospyrrfolia.
El estrato alto está compuesto porindividuos de entre lO y 23 metros de altura,con el 65,00 % de los individuos entre los l Iy I3 metros. El DAP medio del estrato es de30 :t 18 cm. El dosel está dominado por 0.diospyrifolr'a entre los ll y los |8 metros ypor Albizia inundata entre los l5 y 20 metrosde altura.
Otra especie importante por suabundancia es Cecropia pachystachya, perono caracteriza a ninguno de los estratos y sela encuentra ocupando los claros del bosque,ya sean estos de origen natural o antrópico.
El dosel es cerrado y muy poca luzllega al interior puesto que 0. diospyrifoliaE. punicrfolia y P. carrhagenensís sonespecies peremnifolias, con abundante follajey hojas ellpticas u oblongo-lanceoladas.
El estrato herbáceo, fue nulo bajo loslaureles. Sin embargo fue muy abundante enlos claros dominados por C. pachyslachya,donde es común encontrar el suelocompletamente cubierto por Tradescandr'aflumr'nensis (Comelinaceae). Otra especiecomún en los claros es Cleome hassleriana(Caparaceae).

l’) Susana Patricia llmvo
'l‘abla l: Composición florísticas de los bosques de la isla Brasilera, Prov. del Chaco. Los números indican lacantidad de individuos de cada especie que registre dentro de l ha para cada bosque. Todos los individuos tienenun DAP mayor a 5 cm en el bosque de Laureles y mayor a lO cm en los otros dos bosques. El 72,28% de losindividuos del estrato bajo del bosque de Laureles miden entre 4 y 8 m. mientras que el 65 % de los individuosdel estrato alto miden entre l l y l3 m.
. . Laureles Sangre deFam'l'a Es cies —'— Paloitá DraoSubfamflh Pe Estrato bajo Estrato alto g Y
CurumyLauraceae Ocelaa diospyrifolia 68 65 25 14
Nectandra angusnfolia 4 3 7Sp l 4 6 l
Fabaceae Sp4 ISpS l
Mimosoideae Albizía inundan: 8 22 27 lS lInga vera 23 7 18 36
Caesalpinoideae Pellophorum dubium l l
Papilionoideae Geofli‘oea slriala l 3 2
Myrtaceae Eugenia burkarlíana 49 l7Eugenia punicifolía 2 | 7 2Eugenia pyn'fonnis, var. 2pyriformisSp2 8Sp3 3
Euforbíaceae Croton urucurana ¡4 6 18 153Sapium haematosperm um 2 |2 78
Flacourtiaceae Banara argula 8 lO 216 20
Sapotaceae Pauteria salicifolía l
Cecropiaceae Cecropia pachyslachya 270 87 42 l9
Meliaceae Guarea macrophylla 44 lOSubsp. spícaeflora
Tríchília culigua 2
Rubiaceae Psycholría carlhagenensis 30
Salicaceae Salix humboldlíana l l9 12
Asteraceae Tessaria inlegnfolía l l2 l
lndeterminadas
2

Alouaua caraya como dispersor dc semillas 13
En relación con las lianas yenredaderas fueron muy abundantes en elestrato alto del bosque, y en algunos casoscubrían casi por completo la copa de losárboles, incluso de individuos de C.pachyslachya. En los bordes lindantes con laslagunas se formaba una cortina entre lasenredaderas y las ramas de los árboles quecaían hacia el agua. Entre las especies máscomunes de enredaderas están lpomoeindica. Ipomoea cairíca e Ipomoea alba(Convolvulaceae), Cissus palmala(Vitaccae), Paullinia sp (Sapindaceae),Clyroswma callistegioides (Bignoniaceae) yCanavalia bonariensis (Leguminosa).También se pueden encontrar individuos de[sería quuriams (Convolvulaccac) yHoussayanlhus incanus (Sapindaceae).
Bosgue de Sangre de Drago y Timbó (Foto2) (Croton urucurana y AIbízia inundata):24,2 ha. Crece en albardones de la mismaaltura que el bosque anterior pero másangostos, se encuentra mayoritariamente enlos bordes de la isla y existen pequeñosparches en medio de algunas lagunas (Fig. 2).En la hectárea de vegetación que relevéencontré un total de 607 árboles con un DAPmayor a los lO cm. El DAP medio de estebosque fue de 16i 7 cm, y la altura mediafiJe de 12,2 i 3,2 m. Con el 80,88 % de losindividuos entre los 10 y |6 metros.
El 25,20 % de los individuospertenecieron a la especie C. urucurana y el24.88 % a la especie A. inundata. Otraespecie importante fue Tessaria ¡megrrfoliaque se encontraba agrupada en una franjaangosta sobre la barranca del rio, entre losindividuos de esta especie se encontraban losde Salix humboldtíana, la gran densidad deindividuos de esta franja sumada a lapendiente hacía díficultoso moverse en estazona. Los parches pequeños que seencuentran en las lagunas también estáncompuestos por sólo 'estas dos especies(Tabla l Fig 3 b).
Banara argula si bien no fuedominante en el bosque, llamó mi atenciónpor su distribución espacial, todos losindividuos ubicados a lo largo de unaestrecha franja de entre lO y 20 metros deancho donde el terreno tenía menor altura, 5
rn respecto al 0 del hidrómetro de Paso de laPatria.
Debo destacar que el dosel de estebosque es extremadamente abierto, pues lascopas de los árboles en la mayoría de loscasos no llegan a tocarse, su follaje no esabundante, varias especies son caducifolias ysus hojas están compuestas por pequeñosfollolos.
A pesar de no haber relevado losindividuos entre S y lO cm de DAP no existeen este tipo de bosque un estrato inferiordesarrollado (Fig. 3 b). SÍ observé un estratoherbáceo importante. Particularmente la zonamás baja donde se encontraba B. argutaestaba cubierta por Panicum elephamipes(Gmmineae), superando el metro de alturaaunque no crecía debajo de la copa de losárboles.
En cuanto a las enredaderas, si bienalgunas llegaban a la copa de los árboles, novi copas cubiertas por completo como en elbosque de Laureles y la mayoría de losindividuos, por el aspecto de sus tallos,parecian jóvenes.
Bogug de Palo ita (Foto 3) (Banara arguta):6|,7 ha (Fig. 2). Se encuentra en albardonesbajos con una altura de aproximada de entre5 m y 5,5 m respecto al 0 del hidrómetro dePaso de la Patria, analizando los niveles delrío durante el 200! encontré que el suelo deeste bosque estuvo al menos 26 diasinundado, mientras los dos tipos de bosqueanteriores tuvieron agua sobre su suelo comomáximo l5 días (Apéndice). En la hectáreade vegetación que relevé encontré un total de387 árboles con un DAP mayor a los lO cm.El DAP medio de este bosque fue de28 :l: l4 cm, y la altura media fue del2,| :1:4,l m. El 7l,58 % de los individuospresentaron una altura entre los 8 y 17 m.
El 55.8 % de los individuospertenecieron a la especie B. arguta. El restode las especies que completan el bosque seagrupan en zonas en que el suelo está máselevado, y son las mismas especies quedominaban en el estrato alto del bosque deLaureles (Tabla l). El estrato arbustivo esmonoespecíñco con una altura variable entrel y ¡,5 m, formado por una especie espinosaque no he podido identificar hasta elmomento (Fig. 3 c). He visto que en algunos

14 Susana Patricia Bravo
sz» Bosque de Laurelesg H
A 3 ‘ aé, a»
g
< g ..Ñ " 3
g ‘2
.‘I
:3 _o ¡.no _., ;
Altura(m)
101214161820222426283032343638
Altura(m)
101214161820222426283032343638
2468 llll
iililllii'lltl ¿ii‘l101214161820222426283032343638404244464850525456586062
Distancia en la transecta (m)
Bosque de Sangre de Drago y TimbóY b i
__L,l1_.L._l_sl_.l_s.i__.L__L_.L__1__ \ ,.i}, ‘
Illll-l\1‘I-i:’lïi'"""7'nl!‘lllellll2 4 6 81012141618202 2426283032343638 404244464850525456 5860626466687072747678 80828486889092949698100
Distancia en la transecta (m)
V Bosque de Palo itáC
_ r“ » .s »» _ s r I "'"‘*"Ï‘Ï2\\ 3 1 q_ ,, 2 n \ y ll
__ I! fin 2 x 2 ‘ i 2“ ‘ 2 ,Ï ‘ ," l ¡l ’
\ / \ ‘ 7/ ,.4. _€lxj : y 1 r.‘ V
1 11 l , i Uli‘il '15 l l._l._l.'4 ' T i ,; l l 4 1 TT" l I i Á'Ï" mlhi 'i' "'T Ï'Ï l I Ï' v i i2 4 6 81D1214161820222426283032343638404244464850525456586062646668707274767880828466889092949698100
Distancia en la transecta (m)
Figura 3: Perfiles de los distintos tipos de bosque de la isla Brasilera, Provincia del Chaco.l= Ocotea díospyrzfolia, 2 = Banara arguta, 3 = Albizia inundata, 4 = Cecropia pachyslachya,5 = Croton urucurana, 6 = Eugenia punicifolia, 7 = Psychotria carthagenensis, 8 = Arbusto noidentificado, 9 = Sapium haematospermum, lO = Tessaria integrifolia, l l = Inga vera.

Almunia carqr-a como dispersor de semillas l5
casos apoyándose sobre los árboles estearbusto puede superar los 2,50 m.
El dosel es continuo aunque muyabierto, pues si bien B. arguta tiene grandescopas, su follaje es poco abundante ycaducifolio, sus hojas no son grandes ysufren una alta tasa de herbivorla, que enalgunos años afecta a más del 50% de lalámina de sus hojas (obs. personal).
Prácticamente no se encuentranenredaderas y sólo observé P. elephamr‘pesen algunas áreas cercanas al río.
Caracterización de los bosguesconsiderando variables de importanciapara carayá
Al realizar el análisis decomponentes principales obtuve tres factoresque explican el 79,6 % de la varianza entreparcelas. Teniendo en cuenta los dos factoresmás importantes pude ver que el pn'merfactor diferencia al bosque de Sangre deDrago y Timbó de los otros dos por variablesrelacionadas con la cantidad de alimento dealta calidad y la posibilidad de ser encontrado(Fig. 4, Tabla 2), así este bosque es el queposee menor disponibilidad de frutas paracaraya.
Por otra parte el segundo factordiferencia al bosque de Laureles del bosquede Palo itá por la cantidad de especies, siendoinferior el número en el bosque de Palo itá(Fig. 4, Tabla 2).
Por lo tanto al evaluar estos bosquesconsiderando variables de posibleimportancia para caraya., estos bosquestambién son diferenciables, por su oferta dealimento y puede ordenárselos en ordendecreciente de calidad, siendo el bosque deLaureles el de mejor calidad y el bosque deSangre de Drago y Timbó el de peor calidad.
El tercer factor explica sólo un¡2,26 % de Ia varianza entre parcelas y serelacionaría con los soportes para lalocomoción, siendo la variable másimportante el número de individuos(Tabla 2). Sin embargo, no diferencia entrelos tipos de bosque, por lo tanto nopresentarían diferencias para carayá encuanto a las posibilidades para lalocomoción.
Tabla 2: Peso de las distintas variables queconl‘onnan los 3 factores del análisis decomponentes principales que explica el 79,59 %de la varianza de las parcelas de vegetación de laisla Brasilera, Provincia del Chaco.
Factor1 Factor2 FactorsNI ' -0.276 0.299 0.875NIFC 0.808 0.351 0.292NSP -0.032 0.925 0.168SPFC 0.639 0.699 0.006ABlFC 0.925 -0.069 0.050AM -0.002 -0.584 0.072ABT 0.702 -0.026 0.254NI_SP -0.063 -0.665 0.686NIFC_NI 0.948 0.036 -0.068SPFC_NSP 0.886 -0.041 -0.007ABIFC_AB 0.934 0.004 -0.086ABT_NI 0.677 -0.344 -0.304NlFC_NSP 0.738 -0.471 0.250
NI: Número de individuos. NIFC: Número de individuos confruto curiosa. NSP: Número de especies. SPFC: Número deespecies con fruto camoso. ABIFC: Área basal de losindividuos con ll'ulo camoso. AM: Altura media. Área basaltotal. Nl/SP: Número de individuos por especie. NlFC/Nl:Número de individuos con fruto camoso l Número de
individuos totales. SPFC/NSP: Número dc especies con frutocamoso / Número de especies. ABIFC/ABT: Area basal de losindividuos con li'uto eamoso I Área basal total. ABT/NI: Áreabasal total l Número de individuos. NlFC/NSP: Número deindividuos con fruto camoso l Número de especies.
Figura 4: Ubicación de las parcelas de vegetaciónde los distintos bosques de la isla Brasilera en clespacio definido por los factores l y 2 del análisisde componentes principales .0 Representa a las parcelas ubicadas en elBosque de Laureles, 0 las ubicadas en el Bosquede Palo itá y A las ubicadas en el Bosque deSangre de Drago y Timbó. Las elipses ¡destacanel agrupamiento de las distintas parcelas.

Fenología de las especies utilizadas porcarayá
Con los registros fenológicos vi quedurante el invierno (Junio, Julio y Agosto). elnivel de recursos para carayá decrecía(Fig. 5 a). No hubo especies floreciendo eneste período y sólo algunos individuos de ().diospynfolia, B. argma y P. carlhagenensistuvieron frutos (Fig. S b y c). Las hojasnuevas tuvieron un nivel baal producto de larenovación de hojas en las especies perennes,y las hojas maduras mostraron un nivelminimo por la pérdida de hojas que sufrieronlas eSpecies caducifolias.
En la primavera (Septiembre,Octubre y Noviembre) observé el mayornivel de recursos (Fig. S a), es la estación decrecimiento y muestra altos niveles de hojasnuevas, flores y frutos.
Respecto a los frutos, pude detectarentre las especies dos estrategias, unasofrecen fruta en mayor o menor cantidadtodo el año (B. argula, P. carlhagenensis y0. diospyrrfolia), mientras que otrasconcentran la producción en uno o dos meses(C. pachyslachya y E. punicrfolia)(Fig. 5 b y c).
Escrinción de los fïos consumidosp_orcarayá
Los frutos consumidos por carayatienen todos un alto porcentaje de agua, peroson variables en cuanto a tamaño, pesos colory número de semillas (Tabla 3). Al recorrerel bosque y al analizar su composiciónflorística observé que el resto (le los frutoseran cápsulas 0 legumbres secas, por ejemplolos frutos de enredaderas como [pomer albao árboles como Pellophorum dubium, y queen el resto de las variables medidas eran tandiversos como los frutos consumidos. Sólolos frutos de Inga vera tuvieron un ciertocontenido de agua pero inferior al de losfrutos consumidos
lFT
e
100W80
70 +Flous
:3 -I- Hojasnuevas40 + Hojumaduras30 -K-Frulas20100 123460780101112
Meses
b 4 +Eanara algun
-¡.- 0:0!“ dbspyribüa3
3 +Fbycholri'Q 2 carthaqeaensisE
1
0 123456789101112Mesas
c +Cecm a4 pachyslachyn
-)<—Eugenia puntiloh‘a3
lA
.°..
2 2
1."
1
O 123456789101l12Mess
l-‘igura 5: Con el índice l'enológieo total (ll’l‘)represento la abundancia de cada estructuravegetal en la islu Brasilera a lo largo del año (verdetalle cn materiales y métodos). l-ln las liguras by c muestro en detalle la abundancia de losdistintos frutos utiliïados por carayá mediante elíndice fcnológico (ll’).

Alouana caraya como dispersor de semillas 17
Tabla 3: Característica de los frutos consumidos por car-ayacn la ¡sal Brasilera. Luego dc la especie indico eltipo de fruto, A = Aquenios, B = baya, D = drupa. Las referencias de los colores son las siguientes, B/N = blancoy negro, N = negro, 0 = ocre, R ='Rojo y V = verde.
Peso Eje mayor JEje menor Agua No deEspecie (tipo de fruto) Media á: sd Media i sd Media i sd Media :1:sd Color .
o semillas(gr) (mm) (mm) (/0)
Banara argura (B) 0,96 i 0,31 l 1,53 :t 2.6 9,41 :t 1,40 82,36 i 4,59 V > 10
Cecropia pachyslachya (A) 50 "' B/N >100
Císsus palmala (B) ¡,54 :l:0,33 ' ¡{951? “1’15; 88,25 :1:4,03 N l
Eugenia punicifolia (B) 1,47 :t 0,2 9.00 :1:2,1 8,02 i 1.3 86,31 :l:3,02 N 1. . 10,37 :1: 37,75 :t 24,24 :1: 58,56 d:
Cu.ofroea slnalla (D) L9] 3‘67 ¡’47 l ¡‘26 V 1
Ocolea diospyrlfolía (B) 0,94 :1:0,22 ¡Tai 9,03 :t 0,62 55,02 :1:0,91 v 1. . . 18,31 :t 16,46 d: 73,47 :1:
Psuimm guajaba (B) 3,48 :t:2,00 3,48 3’17 17,5] O >10
“Www” Mmmm” 0,30 :t 0.07 7,45 :t 0,74 6,63 i 0,76 84,48:1:3,1l R 2
Urera auramiaca (B) 0,013 """ 2,83 i 0,46 2,82 :l:0.35 R >10' dato obtenido de la bibliografia. “ el peso se obtuvo con los frutos de un único individuo, y dado su tamano debieron ser pesados enconjunto.
DISCUSIÓN escasa luz que llega y también la luzpodría ser la responsable de 1a presencia
La distribución de bosques que muestro de este estrato en el Bosque de Sangre deen el mapa puede perder validez en poco Drago y Timbó. Pero la ausencia detiempo pues la dinámica de la isla es muy herbáceas en el Bosque de Palo ita ya noveloz, y así como una gran superficie puede atribuirsele a la falta de luz sinopuede ser removida por el rlo en pocos que podría relacionarse con la baja alturadias, nuevos bosques pueden crecer en del albardón, que produciría una altaalgunos meses bajo las condiciones frecuenda de inundaciones.adecuadas, por ejemplo durante los años Luego de las tormentas con2000 y 2001, luego de 18 meses con fuertes vientos comunes en la zona,ausencia de crecientes y por lo tanto un observé en el Bosque de Laureles y el debajo nivel de agua en las lagunas observé P810 itá una gran cantidad de claros decomo velozmente un bosque de T_ origen natural provocados por la caida de¡ntegnfolía creció en el medio de una árboles o ramas grandes, supongo quelaguna interna llegando a superar los 2 puede deberse al importante efecto demetros de altura borde que deben tener estos bosques a1
Los distintos bosques tienen encontrarse distribuidos a lo largo de loscienas diferencias y características en albardones. Esto puede explicar en partecomún que vale la pena discutir, por la gran abundancia de C. pachystachyaejemplo la presencia o no de estrato que encontré en ambos bosqueS, pues seherbáceo o la gran cantidad de C. considera a esta especie tlpica de lapachyslachya. cicatrización.
La auscncia de estrato herbáceo Por otra parte si bien encontrédebajo de los laureles puede deberse a la rastros de talado reciente en la isla, se

l8 Susana Patricia llravo
trato mayoritariamente de individuos deE. punicífolia y en algunas ocasionesárboles mayores como O. diospynfolia oInga vera eran tirados para alcanzarpanales. Pero esto no fue lo más común yno creo que la abundancia de C.pachyslachya se deba simplemente aefectos antrópicos. El bosque parece nohaber sido talado masivamente, aexcepción de pequeñas áreas donde seubicaron viviendas con anterioridad a lainundación de los años 82 y 83, lamayoria se encontraban sobre la margendel río frente a la isla del Cerrito,afectando mayoritariamente el Bosque deSangre de Drago y Timbó. Estos lugaresse distinguen claramente por la presenciade árboles de mango.
Tanto el Bosque de Sangre deDrago y Timbó como el de Laureles hansido descriptos anteriormente por otrosautores que le han otorgado distintasnomenclaturas (Franceschi y Lewis, l979;Lewis y Franseschi, |979; Esckuche yFontana, ¡996), sin embargo el Bosque dePalo itá no lo he visto descripto entrabajos anteriores, aunque Esckuche yFontana (1996) citan la especie B. argutaen sus censos dentro de lo que llaman"Selva Riparia del Alto Paraná", estaselva sería el equivalente al bosque deLaureles. Dado que en el Alto Paraná lasislas son originadas por la erosión deantiguas terrazas fluviales y que ladeposición de sedimentos es escasa, no serellenan en su interior grandes áreas comoocurre en el Paraná medio (Eskuche yFontana, l996). Es posible que estas islaspresenten una mayor altura y nocontengan zonas que se inundenfrecuentemente, lo que al parecer sería lacondición apropiada para el desarrollo debosques dominados por esta especie. Eltrabajo de Franceschi y Lewis (1979) hasido realizado varios kilómetros río abajo,en la Provincia de Santa Fe, y esta podríaser la causa de que no hayan encontrado aesta especie en sus censos, aunque, hasido observada en las islas del Paranáubicadas frente a Reconquista (Santa Fe)(G. Zuníno, com. pers.).
Respecto al Bosque de Sangre deDrago y Timbó incluye lo que muchos
autores llaman "Alisal" o "Bosque deAlisos" (Franceschi y Lewis, l979; Lewisy Franseschi, l979; Neiff y Reboratti,l989; Neiff y Poi de Neiff, l990;Esckuche y Fontana, l996;), haciendoreferencia a la abundancia de T.
integnfolia, sin embargo a los fines deeste trabajo no considere correcto utilizaresta nomenclatura pues T. inlegrijblia enel sitio de estudio ocupa sólo una densa yestrecha franja de terreno sobre cl río opequeñas áreas entre bosques, que norepresentan en sí mismos un ambienteutilizable por caraya.
A pesar de que el nivel derecursos disminuye en invierno estosseguirían siendo mayores que en losbosques de tierra firme (Rumiz et al,1986). En tierra firme sólo algunosindividuos de Ficus Iuschnalhitmaofrecen frutos como alimento para carayadurante el invierno (Zuníno, 1989), siendoesta la única especie dispersada porcaraya en esa estación.(Bravo y Zuníno,2000). Los primeros trabajosprimatológicos realizados en el área de lasislas del Paraná al no observar F.Iuschnathiana, consideraron que duranteel invierno caraya se debía alimentar sólode hojas maduras, y que esto era posiblegracias a que estas hojas, por tratarse deespecies de rápido crecimiento, tendrianbajo contenido de compuestossecundarios siendo así más fáciles de
digerir, de esta manera explicaban la grandensidad de carayá que encontraban en lasislas (Rumiz et al., l986; Brown yZuníno, ¡994). Considerando laabundancia de las especies que puedenofrecer frutos todo el año en los distintosbosques, es obvio que el Bosque deLaureles es el que más posibilidadesofrece a carayá de disponer de frutos eninvierno y el Bosque de Sangre de Dragoy Timbó es el más limitante en estesentido.
El patrón de oferta de frutos quedescribe, por el cual existe un grupo derecursos de alta calidad "constante" y otrogrupo disponible sólo estacionalmente essimilar a lo señalado por Estrada (l984)para "Los Tuxtlas", México, limite nortede la distribución del género Aloualla. De

Aloualla caraya corno dispersor de semillas ¡9
todas maneras trabajos realizados en elBajo Paraná indiearian que para algunasespecies la producción de frutas esinfluida más por el régimen hidrológicoque por la estacionalidad climática local(Neiff y Reboratti, 1989; Neiff, 1990).Debido a esta interacción de variablesclimáticas e inundaciones, en Formosa seencontró que podia haber grandesdiferencias entre años en cuanto a laabundancia de las distintas estructurasvegetales, por ejemplo para algunasespecies los pulsos de inundación podíanestimular la expansión foliar y laformación de pimpollos, pero luego eldesarrollo de los frutos podía serinterrumpido si se producía una fuertesequía (Plaeci, l995). Asl en la formacióny maduración de los frutos dc ().diospyrilblía influian las condicionesclimáticas de hasta 8 meses antes (Placei,¡995).
El ordenamiento de los bosquessegún su calidad para carayá llevaimplícito una diferencia forzada en cuantoa cantidad de frutos en la dieta de losgrupos qUe utiliian uno u otro bosque, porende es de esperar que la función decarayá como dispersor de semillas seadiferente en cada uno de ellos. Dada ladiferencia en el número de especies entreel Bosque de Laureles y el de Palo itá,puedo suponer que los grupos de carayaque utilizan este último bosque tienen unadieta menos diversa lo que puedesignificar escasez de alimento en ciertosmeses. Cabe pensar también que en elBosque de Laureles carayá funcione comodispersor para un número mayor deespecies y que su efecto no se limite a unaépoca en particular del año. Para poderconfirmar esto es necesario realizarestudios de dieta y movilidad de losgrupos.

Foto 1: Bosque de Laureles
Foto 2: Bosque de Sangre de Drago y Timbó

Alouana caraya como dispersor de semillas 2!
Capítulo Il: El dispersorUso del espacio, comportamiento de alimentación y patrón de actividad
de los grupos de carayá que habitan en la Isla Brasilera.
“...Pasan de un árbol a otra sin saltar y sinbalancearse slno muy lentamente.parque son pesadas,tristes y serios. ”“...Nadlela dentada, sin duda a cursa desu seriedad.Se aye a más de una nu'llade rflstancla su grito, que esfuerte triste, ronca e insoportable. ’-’
FelixdeAma (Viaja por la Américameridional)
INTRODUCCIÓN
Los monos aulladores (Alouarta,Primate: Cebidae) son unos de los primatesneotropicales de mayor tamaño, tienenhábitos diurnos, son arborlcolas, la dieta esfolívoro-fi'uglvora, y pueden vivir en gruposunimacho o multimachos (Crockett yEisenberg, l987). Son colonimdores deambientes disturbados y muestran una altaplasticidad en su comportamiento, que lespermite utilizar un amplio rango de hábitats(Eisenberg et al, l972). Se caracterizan poremitir aullidos, probablemente como unaforma de marcar el territorio, y por poseeruna cola prensil muy desarrollada que lesfacilita los desplazamientos por el estratoarbóreo del bosque; generalmente se losconsidera estrictamente arbóreos aunque enciertos ambientes o circunstancias puedendesplamrse por el suelo. En los bosques queforman el dosel a gran altura, utilizan casicon exclusividad el estrato medio y alto de lavegetación. Dada su dieta baja en energiamuestran escasa actividad pudiendo dedicaral descanso casi un 80% del tiempo(Croekett y Eisenberg, 1987).
Diariamente los grupos de aulladoresutilizan pequeñas zonas de sus áreas deacción y tardan un tiempo variable enutilizarla por completo. Se ha visto queutilizan diferencialmente la superficie quehabitan, teniendo áreas muy frecuentadas,otras con frecuencia media y áreas con bajafrecuencia de uso. En general la utilizacióndiferencial del espacio se relaciona con la
distribución de los recursos alimenticiosdentro del área (Milton, 1980; Estrada, 1984;Zunino, ¡986; Chapman, 1988; LucenaMendes, 1989; Julliot, 1992; Stoner, 1996).
Las poblaciones de aulladores se veninfluidas por factores tales como la predacióno la mortalidad producida por plagas yenfermedades, pero la disponibilidad dealimento es asumidafi'ecucntemente como elfactor crítico que determina la abundancia deprimates (Milton, 1996; Chapman et aL,1999). En ausencia de cacería es razonableasumir que los recursos alimenticios sean losque limitan la densidad de aulladores(Balcomb et al., 2000).Varios autoresconcluyen que el factor más importante quedetermina la calidad de hábitat para losaulladores es la densidad de las especies máscomunes en su dieta (Gaulin et al., 1980;Stoner, l996).
Estudios comparativos de distintaspoblaciones de aulladores han mostrado quela distribución espacial y temporal de losrecursos tienen influencia sobre el recorridodiario, el área de acción y la intensidad conque los grupos usan el espacio (Milton, l980;Chapman, 1988). La literatura revela una
.serie de hipótesis que inter-relacionan estosfactores. Crockett y Eisenberg (l987)sostienen que el tamaño de las áreas deacción está negativamente conclacionadocon la densidad de la población de aulladores(Hipótesis l). Esta con'elación no seobservada si los aulladores utilizaran el

22 Susana Pauicia Bravo
espacio sólo en función de los recursos puesen el caso de una población con bajadensidad que no ha alcanzado la capacidadde carga del ambiente, deberian encontrarseáreas sin ser utilizadas por ningún grupo quecon el tiempo serían ocupadas por nuevosgrupos; por el contrario y en apoyo de laHipótesis l existen evidencias de que el áreade acción de los aulladores decrece al mismotiempo que la densidad de la población en unsitio aumenta (Chivers, l969), es decir quelos grupos de aulladores tienden a utilizarsuperficies mayores a las que necesitansimplemente porque está disponible. Otrosautores, asocian las pequeñas áreas de acciónen aulladores con las dietas dominadas porhojas (Estrada, 1984; Julliot y Sabatier, l993)(Hipótesis 2), asumiendo que en las selvas lashojas presentan una distribución uniforme enel tiempo y el espacio. Por último, Milton(1980) sugiere que los aulladores minimizanlos gastos energéticos que implican losdesplazamientos dado que sus dietasbásicamente follvoras los limitaenergéticamente (Hipótesis 3). De estahipótesis se deriva que el porcentaje de hojasen la dieta estaría negativamentecorrelacionado con la distancia diariarecorrida por un grupo de aulladores (Milton,1980; Chiarello, ¡993).
La densidad de carayá en la selva deinundación es la mayor reportada para el
Igénero Alouatta (Zunino y Rumiz, 1986;Rumiz,- ¡990; Crockett, 1998). Por ejemplo,en el Bosque de Laureles de la isla Brasilera,Kowalewski (2000) registró una densidad de4,25 ind/ha. De acuerdo con las distintashipótesis mencionadas y considerando la altadensidad de carayá en la Isla Brasilera esperoencontrar que los grupos de carayá tenganpequeñas áreas de acción, posean una dietadominada por hojas maduras y que ladistancia que reconan diariamente estecorrelacionada inversamente con elporcentaje de hojas maduras en la dieta. Siesto se cumple en la lsla Brasilera la calidadde carayá como dispersor de semillas se veríaafectada tanto por la baja cantidad desemillas transportadas como por la cortadistancia a la que serían depositadas.
En este capítulo pondré a pruebaestas hipótesis y trataré de determinar que
factores posibilitan que en la lsla Brasilera lapoblación de carayá alcance una densidad tanelevada. Para ello tendré en cuenta losresultados del capitulo anterior y describirélos patrones básicos de comportamiento, usodel espacio y dieta de carayá en la islaBrasilera, muchos de los cuales se relacionancon la dispersión de semillas.
MATERIALES Y MÉTODOS
Realicé las observaciones sobre dosgrupos de carayá que utilizaban parte delBosque de Laureles en la Isla Brasilera(Fig. 6). Decidí centrar los estudios en estebosque por ser el de mayor oferta de frutospara carayá (ver Capítulo l) y donde ladensidad es 4 veces superior a la mayordensidad reportada para el género (Crockett,¡998).
Utilizando las categorias de edad ysexo definidas .por Rumiz (1990) para estaespecie, que relacionan el tamaño de losindividuos y el color del pelaje con la edad(Foto 4), puedo decir que ambos grupostenían una composición similar. El grupo delos "Gringos" tenía al iniciar el estudio untotal de 16 individuos, 2 machos adultos, lmacho subadulto, 5 hembras adultas, lmacho juvenil, 4 hembrasjuveniles y 3infantes. El grupo de los "Enfennitos" estabacompuesto por 2l individuos, 2 machosadultos, l macho subadulto, 6 hembrasadultas, l hembra subadulta, 3 machosjuveniles, 4 hembras juveniles y 4 infantes.ldentifiqué los individuos por sus marcasnaturales. variaciones en el pelaje y tamañorelativo de sus cuerpos, con estos datosrealice una ficha para cada uno de ellos quefue actualizada a lo largo del estudio.
Seguí los grupos durante períodos de5 a 7 dias consecutivos, bimensualmenteentre mayo de ¡998 y febrero de 2000.Debido a inundaciones prolongadas debíinterrumpir las observaciones durante 2meses (Septiembre y octubre) en ¡998 ydurante 3 meses en 1999 (enero, febrero ymarzo). Cada dla segui los grupos desde lasalida hasta la puesta de sol, es decir desdeque comenzaron hasta que finalizaron superiodo de actividad. Totalicé |680 hs deobservación.

Alouarla earaya como dispersor de semillas 23
Alimentación 1 patrón de actividad
Utilice el método de muestreo porbarn'do (Altmann, 1974; Martin y Bateson,l993) de todos los individuos del grupo queestuvieran visibles. Este es el métodorecomendado para realizar los primerosestudios de patrones de actividad diarios yanuales en un sitio del cual no se poseenconocimientos previos, pues permite tornaruna muestra amplia y representativa delcomportamiento básico de los grupos deprimates (Fragaszi et al., ¡992; Martin yBateson, ¡993). Cada 15 minutos, registre demanera instantánea a que altura se encontrabacada individuo y cual de los siguientescomportamientos estaba realizando,locomoción, movimiento, alimentación,descanso, aullidos e interacciones sociales(juego, acicalado y agresión). La locomociónimplicó cualquier desplazamientosincronizado del grupo de .un sitio a otro ensu área de acción, mientras que elmovimiento era el cambio de posición de losindividuos dentro del grupo. Cuando observéindividuos alimentándose registre queestructura vegetal ingerlan (hoja madura,hoja nueva, pecíolo, flor, fruto verde o frutomaduro) y a que especie pertenecía.Consideró descanso toda aparente inactividadde los animales. Cada vez que fueronobservadas interacciones sociales registre queindividuos se encontraban involucrados.
Registre Ad libr'lum todas lasconfrontaciones (persecuciones, aullidos yagresiones fisicas) entre los grupos focales ycualquier grupo o individuo ajeno a ellos.
Para determinar el tiempo de pasajepor el tracto digestivo de las semillas utilicélos períodos en que al comenzar lasobservaciones no hubo frutos disponibles enel bosque, pero que luego aparecieron yfueron consumidos. De esta forma al nohaber en un principio semillas en las heces,pude tomar registro de cuanto tiempotardaban en aparecer luego de que losanimales comenmr'an a alimentarse de losfrutos.
Algunas de las estructuras vegetalesingeridas por carayá estuvieron infestadas
por artrópodos, para evaluar esta infestaciónrealicé las siguientes mediciones:
- Ocotea diospyrr'foli'a, en julio de1998 colecté 50 frutos de los 8árboles que utilizaron con mayorfrecuencia para alimentarse yotros 50 de 7 árboles con fruta nousados o usados en bajafi'ecuencia.
- Albr'zia inundara, en septiembrede l999 colecté hojas nuevas delos 2 árboles utilizados por losSNPOS'
- Inga vera, en noviembre de 1999colecté 20 flores de 6 árboles, 3dentro del territorio de los gruposobservados y utilizados comofuente de alimento por ellos.
— Banara argura, en mayo de 1999colecté 20 frutos de lO árbolesutilizados por los dos grupos. Enseptiembre de 1999 colecté 20hojas nuevas de l0 árbolesdentro del territorio de los
grupos.- Eugenia pum'cifolr'a, en febrero
de 2000 colecté 20 frutos de lOárboles utilizados por los dosgrupos observados como fuentede alimento.
Registré el porcentaje de unidadesinfestadas en la muestra, el número deinsectos por unidad y la ubicación de losmismos dentro de la estructura vegetal. Sibien las estructuras fueron utilizadas porcar-ayaen más de un periodo de muestreo losporcentajes de infestación los evaluó duranteun único periodo, en los otros periodosconfirmó que estuvieran infestadas (exceptopara Albizr'a inundara).
Colecté frutos infestados con distintogrado de madurez de cada especie y losobservé bajo lupa, realizando cortes paraconocer la ubicación de las larvas dentro delfruto y el daño que iban causando a lassemillas a lo largo de su desarrollo.
Durante un periodo de sequía queduró unos pocos meses los animales sealimentaron de pecíolos de camalotes(Er'chhorm'a crassipes, Pontederiaceae), paratratar de dilucidar que recurso podian estaraportando estas estructuras, colecté

24 Susana Patricia Bmvo
ejemplares y realicé análisis de contenido deagua, fósforo, calcio y magnesio en hoja,pecíolo y raíz. Evalué los resultadosmediante una pmeba de t de Student.
Todas las pruebas estadísticas queutilicé en la Tesis están basadas en Zar(l996). Las pruebas no paramétricas lasutilicé cuado no se cumplían los supuestospara realizar pruebas paramétricas. Losgráficos de cajas, indican la ubicación delvalor promedio con una línea horizontal, losextremos de las cajas indican los valores delpromedio :t el error estándar y los extremosde las barras indican los valores del promedio:1:el desvío estándar.
Uso del espacio
Organicé un sistema de grilla‘concuadrados de 20 m de lado, en toda el áreaque utilizaron los grupos, identificando lasesquinas de cada cuadrado con rótulos dealuminio sobre el árbol más cercano. Losárboles de mayor porte (aprox. DAP > 20cm) fueron identificados hasta especie,numerados y ubicados en un mapa de lagrilla.
El recorrido diario (RD) de losgrupos lo estime dibujando cada dia losdesplazamientos en el. mapa de la grilla ymidiendo luego la longitud del mismo. Pararealizar el dibujo del recon'ido tuve en cuentalos árboles numerados que los animalesutilizaban para desplazarse. Cuando el grupose encontraba avanzando de manera dispersaabarcando grandes áreas registraba en elmapa el punto central del grupo.
Teniendo en cuenta elcomportamiento de los grupos y la forma enque las distintas áreas del bosque fueronusadas por los grupos distinguí doscategorías, territorio y áreas desuperposición.
Considere territorio de un grupo todael área que defendió activamente (Jolly,1985), en estas áreas cada vez que detectabanla presencia de individuos ajenos al grupoiniciaban una persecución corriendo al/losindividuos hasta un cierto punto dondecomenzaban los aullidos. Esa zona fue la queconsideré límite del territorio. El área desuperposición aquellas área que el grupo
usaba fuera de su territorio y de las cuales alser detectado era desplazado. El territoriomás las áreas de superposición de un grupoconstituyen lo que llamaré área de acción.
Dado que el bosque forma unaestrecha franja cuyo ancho varia entre los 20y lOOrn y los grupos lo podían cubrir porcompleto al desplazarse, para determinar lassuperficies de todas las áreas definidas, sólodebí determinar sus extremos en la franja debosque (Fig. 6). Cómo los animales utilimrondiariamente a Io largo del año una mismaárea y las confrontaciones entre gruposfueron frecuentes, no necesite utilizar ningúnmétodo especifico (e.g., poligono convexo),sólo ubicar en el . mapa los sitios queutilizaban, los sitios de donde erandesplazados, los sitios de donde desplazabany los sitio donde aullaban.
Evalué la relación entre el RDpromedio por grupo en cada periodo demuestreo y el porcentaje de hojas maduras enla dieta de ese periodo, y la relación entre elRD promedio y el número de confrontacionesdel grupo, con-una correlación de Spearman.
RESULTADOS
Alimentación
Como ambos grupos se alimentaronde forma similar voy a presentar elporcentaje promedio. A lo largo del año loscarayá consumieron estructuras vegetales de22 especies pertenecientes a IS familias, el63,6% fueron especies arbóreas (Tabla 4).Las especies de árboles de las cualesconsumieron frutos representan el 76,9 % delos individuos del Bosque de Laureles.Caraya' consumió también frutos de dosenredaderas (Tabla 4) y de al menos unaplanta epífita. Los frutos de Psycholrr'acarrhagenensis en general eran comidas sólopor los juveniles. Las semillas de Geofroeastriaua no fueron ingeridas porque al tenerfrutos del tipo drupa, sus semillas seencuentran dentro de un endocarpiolignificado de gran tamaño. entoncesmanipulaban el fruto alimentándose delmesocarpio camoso y arrojaba el resto debajodel árbol parental. El resto de las semillasfueron ingeridas junto con los frutos y

Alouatta cama comodispersorde semillas 25
N27°20's l, \/ >K
59° 37' o x.
27° l8‘S
Arr; “¿k ,.\
—-— Limite del bosque
—- - Limite de los territorios
G o Gringos: Tia-diodo de los Gringos
Enfermitos: Territorio de los Enfermitos
Fig. 6: Ubicación de la grilla en la Isla Brasilera. Territorios de los dos grupos observados.
aparecieron en las heces aproximadamente alas 24 hs de haber sido ingeridas, pero podiancontinuar apareciendo en las heces en menorcantidad hasta 48 hs después de que dejarande alimentarse de fi'utos.
El período de muestreo en el cual sealimentaron de una mayor diversidad defrutos fue mayo de 1999, cuandoconsumieron frutos de Ocotea diospyrifolía,Banara arguta, Cecropia pachystachya yPsychotria carthagenensis.
Los frutos maduros representaron el17.6 i 1.1% del total de los registros
(Fig. 7 a), pero estacionalmente llegaron arepresentar más del 60 % (Fig. 7 b), ei mayorporcentaje de frutos maduros lo ingirieron iosEnfermitos durante julio de 1998 (71 %).
Anualmente las flores representaronel 12,0 :t 3,5 % del total de los registros(Fig. 7 a), sin embargo en algunos mesesllegaron a representar más del 43 %(Fig. 7 b), las flores fueron ingeridas ensimilar proporción a las frutas cuando estasno estuvieron disponibles. Las flores de G.striatta (24,26%; 32,45 %; 29,55 %), Ingavera (42,19 %), y Papilionáceae l (12,66 %)

26 Susana Pau-¡ciaBravo
fueron consumidas siempre que estuvierondisponibles. En algunos meses, ingirieronflores de dos especies de las cualescomúnmente consumían frutos (0.díospynfolia representó el l0,l7% de losregistros de alimentación del grupo de losEnfermitos durante noviembre de 1999 y B.argula representó el 10,23 % de los registrosde alimentación del grupo de los Gringosdurante febrero de 2000).
Cuando las estructuras de alto valorenergético (frutas, flores y hojas nuevas) noestuvieron disponibles, cómo ocurrió en Juliode l999, la dieta estuvo dominada por hojasmaduras (Fig. 7 b) y los grupos se dividieronen 2 subgrupos, los cuales se alimentaban enlos extremos opuestos de sus territorios.Durante el dia se reunían sólo para a‘ullarjuntos en las confi'ontaciones. Al atardecer sereunían nuevamente y se alimentaban juntos,
para terminar descansando el grupo completoen el árbol dormidero durante la noche. Lacomposición de estos subgrupos cambió cadadía e incluso durante el dla algún individuopodía cambiar de subgrupo.
Carayá ingirió proteina animalindirectamente con las hojas nuevas, losfrutos y las flores. Cada mes uno de losprincipales componentes de la dieta estuvoinfestado por artrópodos en gran proporción(Tabla 5). El 25,5 % del total de los registrosde alimentación correspondió a la ingestiónde estas estructuras infestadas. El mayorporcentaje, 66,7 %, lo obtuve durante elperiodo de muestreo dejulio de l998 para losEnfermitos.
0. diospyrifolia tenía una altafrecuencia en el ambiente (ver Capítulo l),sin embargo cuando la mayor parte de losárboles de esta especie ofrecieron frutos,
Tabla 4: Especies utilizadas por carayá en la Isla Brasilera como alimento, con la X indico la estructuravegetal de la cual se alimentamn. Fl=Flores, Fr=Frutos, lln=llojas nuevas, llm=l-lojas maduras,Pec = Peclolos.
F0231“ Familia Especie Fl Fr Hn Hm Pec
Árboles Lauráceae Ocolea diospyrifolia X X X XNectandra anguslifolia X X
Mimosácege Albizia inundara X XInga vera X X
Papilionáceas Geofroea strialla X X X XSpl X X X
Mirtáceae Eugenia pum‘cifolia X X X XPsídium guajaba XEugenia pyriformis X
Flgcurtiággae Banara argula X XSapotáceae Pouleria salicrfolia X XMoráceae Cecropia pachyslachya X X XRubiáceae Psycholria carlhagenensis X
Enredaderas Bignoniáceae Clylostoma callistegioides X X XConvolvuláceae Ipomoe alba X X
Ipomoea indica X Xlpomoea cairíca X X
Vitáceae Cissus palmala X X XSapindáceae Paullinia sp. X X
Arbustos Urticáceae Urera aurann'aca XFlia l Sp 2 X X
Plantas Pontederiáceae Eichhornia crassipes Xacuáticas

Alauana caraya como dispersor de semillas 27
car-aya se alimentó sólo de algunos de ellos.Por ejemplo, durante julio de ¡998 losGringos entraban diariamente al territorio delos Enfennitos tratando de alimentarse de losfrutos de ciertos árboles dentro de esteterritorio, generando confrontaciones entreambos grupos. Cuando evaluó la infestaciónde los árboles elegidos, encontré que el 80 %de sus frutos estaba ¡nfestado por larvas deHeilipus spp. (Curculionidae), mientras quelos árboles no utilizados o usados en bajafrecuencia tenían una infestación del 35 %.Encontré al menos 2 especies decurculionidos infestando las semillas de 0.díospyrifolia ambas pertenecientes al géneroHeílipus. Los adultos de Heilipus spp.colocaban un único huevo directamentedentro de la semilla de 0. diospynfolia. Todoel desarrollo de la larva se produjo dentro deesta. la larva se alimentó de los cotiledones ydel embrión vegetal hasta consumirlos porcompleto, luego formó la pupa dentro de lascubiertas de la semilla (Foto S), la cualexternamente conservó intacta su apariencia,sólo era visible el pequeño orificio producidopor la oviposición. Algunas larvas dcHeilipus spp. estaban a su vez infestadas por
una avispa parasitoide de la familiaBrachonidae.
Aproximadamente el 50% de. lassemillas de todas las frutas de B. arguraestaban infestados por avispas. Los huevosfueron colocados dentro de las semillascuando los fi-utos aun estaban inmaduros, lassemillas que se encontraban con larvas tenianforma diferente a las semillas sanas. Cuandolos frutos maduraban el desarrollo de lasavispas ya estaba completo y dentro de estassemillas no quedaban restos del cotiledón nidel embrión vegetal.
En el caso de E. punicifolia los frutoseran infestados en el momento en quemaduraban. los frutos maduros permanecenen la planta sólo 2 o 3 dias y la oferta defrutos maduros en el bosque no dura más deuna semana. Los huevos eran depositados enel pericarpio camoso y luego las larvaspasaban a la semilla, cada fruta tenia enpromedio l,9 d:0,6 larvas. No pude conservarlarvas vivas para observar su desarrollo eidentificar los adultos. Sin embargo observéen el campo que el endospenna de estassemillas podia ser eliminado por completo.
Tabla 5: Estmcturas vegetales infestadas consumidas por caraya en la lsla Brasilera. Los valores deinfestación representan cl porcentaje promedio (zi:desvío estándar) de unidades de la estructura infcstada porárbol para cada especie. La última columna indica Ia frecuencia de consumo de cada estructura durante losperiodos en que la consumieron (valor promedio de ambos grupos :l:desvio estándar).
Especie Estructura
E. punicrfolia Frutos
O. diospyrifolía Frutos
I. vera Flores
A. inundata Hojas nuevas
B. argwa Frutos
B. argura Hojas nuevas
Consumo pormuestreo
20.28 :l:0.0410.68 :l: 14.2929.16 :l:0.] l58.78 :t ll.15
1.63 :l:2.3|0.29 :t 0.40
22.23i 28.l90.61 i 0.86
100 ll.84 :t 2.82¡0.81 1-3.5|15.43 i 7.10
100 ¡1.44 i 2.530.73 i 1.030.26 á: 0.4
100 ¡1.621921ll.4l i 7.98
Infestación
80.42 :t l2.28
82.00 :l: ¡0.30
89.81 :l: ll.36

28 Susana Patricia Bravo
a b
1005“ -5:. ÜPlAz.8059-13.. ‘. un
40%' IFrv
20's.- . Huw una:
Jul. Nw.'Mny. Jul Sep. Nov. Feb.
'98 ‘99 ‘ÜÜ
Figura 7: a —.Porcentajes de registros de alimentación sobre las distintas estructuras vegetalesutilizadas por carayá a lo largo del estudio en la Isla Brasilera. b - Porcentaje de registros dealimentación sobre las distintas estructuras vegetales en cada período de muestreo. Los valores queindico son el promedio de los dos grupos observados. Hm = Hojas maduras, Hn = Hojas nuevas,Frm = Frutos maduros, Frv = Frutos verdes, Fl = Flores, Br = Brotes, Pl. Ac.= Plantas acuáticas.
El consumo de miel, musgo y Tanto las hojas como los pecíolos de estasAliquenes de troncos y ramas lo registreraramente, pero el consumo de agua fuefrecuente. El agua la obtenían de los huecosde los árboles y de las lagunas. Al final delperiodo de sequía ambos grupos seaiimentaron de plantas acuáticas, másexactamente de peciolos de E. crassípes.
plantas tenían 'un alto porcentaje de agua,pero al realizar los análisis demicronutrientes encontré que en los pecíolosse acumulaba calcio en mayor concentraciónque en las hojas (t = 3,48; P = 0,03; gl = 4;Tabla 6).
Tabla 6: Contenido de agua, fósforo, cálcio y magnesio en hojas, pecíolo y raíz de Eichhornia azurea. Losvalores están expresados en porcentaje promedio de peso seco (á:desvio estándar). lnformo un único valor paralas raices pues las muestras que obtuve fueron pequeñas y debieron ser analizadas en conjunto. * P = 0,03según la prueba de t de Student.
E. azurea % de agua Fósforo Cálcio * Magnesio
Hoja 91,59 i 1,5 0,2 l4 al:0,055 0,453 :l:0,453 0,554 :t 0,093Pecíolo 94,44 :l:2,3 0,240 at 0,038 1,199 i 0,305 0,734 i 0, l 47Raíz Sin datos 0,480 0,937 0,803
Patrones de actividad
Al iguul que puro ln uliincntución, clptlil‘ótl dc actividad de ninlios grupos dccnrnyil t'uc similar por lo que nncvnmcntcpresentará los ¡mrccntnjcx promedios dc cndnamixidad.
El descanso representó el 56,5 % deltotal de los registros, algunos meses el 50 %
de los registros mostraron a carayá enactividad (Fig. 8). No encontró ningún patrónen cl ritmo diario de actividad y descanso. Sihicn existió unn disminucii'm dc ln uctividnddumntc ln mcdin nmltnnn y ln incdin tni'dc.ulgunos individuos contintmlmualimentimdose, otros jugaban o se aricalaban.Frecuentemente estos períodos de bajaactividad eran interrumpidos porque los

’/oActividad
Alouarra caraya como dispersor de semillas 29
socialesI aullidos
mow'miento
locomoción
Eldescanso
o W , r ,
es}:‘b‘s‘.\&-fi. +04.___.____.__._.__.___...__.’98 ’99 ‘00
Figura 8: Porcentaje de las distintas actividadesdesarrolladas por carayá durante cada periodo deobservación. El valor que indico es el promediode los dos grupos observados.
grupos detectaban la presencia de otro grupoo individuo dentro de su territorio.
Las interacciones sociales incluyeronprincipalmente el acicalamiento entreindividuos (Foto 6) y el juego. Ambos gruposdesarrollaron largas sesiones deacicalamiento durante los periodos de bajaactividad y luego de las confrontaciones.Usualmente las sesiones consistian en un parde individuos que interactuaban alrededor delS minutos, pero existieron sesiones con unaduración mayor a los 45 minutos. Eljuego loregistre principalmente (más del 70 %), entreinfantes e individuos juveniles o estos conindividuos adultos, ya fueran machos ohembras. Sin embargo, en algunaoportunidad pude ver a los machos adultosjugando entre sí y también al machosubadulto jugar con uno de los machosadultos.
El cuidado aloparental fue común,los infantes podian ser amamantados porhembras que no fueran sus madres e inclusoalgunos juveniles ya completamenteindependientes también podian serlo. En elgrupo de los Enfermitos, una hembra (Nadia)presentó signos de enfermedad que leimpidieron seguir al grupo e incluso moversepor las copa de los árboles y su cría fueadoptada por otra hembra (Monalisa) que
v El interacciones
I alimentación
poseía una cría de similar edad (de 2 o 3meses). Nadia murió pero su cría sobrevivióal menos hasta ser independiente. En cuantoal acarreo de los infantes al comenzar eldestete podían ser transportados porcualquier individuo del grupo, incluidos losmachos y los juveniles de l o 2 años de edad.
Las alturas en las que los grupos sealimentaron, descansaron o se desplazaronfueron similares (Fig. 9). Si bien ambosgrupos tuvieron similar dieta y patrón deactividad, los Enfermitos utilizaron una alturamayor para desarrollar todas las actividades.En general, el territorio de los Enfermitosposeía árboles más altos que el territorio delos Gringos y este fue el motivo por el que laaltura promedio de descanso para losEnfermitos fue mayor. La diferencia en laaltura de alimentación estuvo relacionada conel consumo de enredaderas, mientras losGringos se alimentaban de estas sobre losarbustos, los enfermitos lo hacían sobreárboles altos. La altura promedio delocomoción. de los Gringos fue menor pueseste grupo utjlizaba el suelo cuando 'setrasladaba entre parches de bosque en elextremo del albardón, mientras losEnfermitos raramente utilizaron el suelo ensus traslados.
metros
v a un
zo
r. %
y'o % Ï¡mmm (E) Mmm)
DammLonamccion (E)
(E) Alimentacion (G) mmm)
Figura 9: Alturas a las cuales los dos grupos-decarayá desarrollaron las principales actividades.(E) = Enfermitos, (G) = Gringos.
Uso del espacio
Más del 70 % de los registros losobtuve dentro de los territorios. El territoriodefendido por los Gringos fue de 1,3 ha y eldefendido por los Enfermitos fue de 1,8 ha(Fig. 6), Durante febrero de 2000 losEnfermitos desplazaron a los Gringos y el

30 Susana Patricia Bravo
área defendida en este periodo fue 0,9 hapara los Gringos y 2,8 ha para losEnfermitos.
Las áreas de superposición fueronusadas con baja frecuencia, y los gruposintrusos corrían hasta el limite de su territoriocuando eran detectados y perseguidos porotro grupo; una vez en su ten’itorio los gruposse aullaban a unos pocos metros de distanciauno de otro. Algunas de estasconfrontaciones terminaron con agresionesfisicas, pero lo más común fue que ambosgrupos dejaran de aullar bruscamente,comenzando algunos individuos a comer,mientras los machos adultos en generaldescansaban y algunas hembras losacicalaban y los juveniles de ambos gruposjugaban.- juntos. Cada día los- gruposutilizaban toda la superficie de su territorio,por lo que entraban en contacto visualmentecon los grupos vecinos lo que producíaconfrontaciones entre ellos, en un mismo día(Febrero 2000) el grupo de los Enfennitosllegó a tener hasta 6 confrontaciones congrupos vecinos e individuos dispersantes. Elárea de superposición de los Gringos sobrelos Enfennitos fue de 0,44 ha, los Enfermitosse superpusieron tanto en el territorio de losGringos como en el de un tercer grupo los"Vecinos" sumando en total 0,40 ha.
El área de acción de los Gringos fuede 1,7 ha y el área de acción de losEnfermitos fue de 2,2 ha. El área de acciónde los Enfennitos estuvo incluida en elBosque de Laureles continuo sobre elalbardón, mientras el área de acción de losGringos al ubicarse al final del albardónincluyó este bosque continuo y tresfragmentos pequeños ubicados entre laslagunas (Fig. 6).
El RD promedio fue de 513 :t 253 m.El RD máximo fue de l.515 m (Enfennitos)y el minimo fue de 85 m (Gringos), estos RDlos registre en Febrero de 2000 cuando losGringos fueron desplazados por losEnfermitos. El RD medio de cada período demuestreo estuvo positivamentecorrelacionado con el número deconfrontaciones del grupo en cada periodo-demuestreo (Gringos: r, = 0.77; P < 0.05; N =7; Enfermitos: r, = 0,75; P < 0,05; N = 7;Fig. 10 a), pero no estuvo cor-relacionado con
el porcentaje de hojas maduras ingeridas enese periodo (r = 0.46; P = 0.2|; N = 14; Fig.lO b).
a
RD
ñ
RD
sl8
u so 50 1o 90 no
-/.HojasMain:
Figura lO: Correlación de los recorridos diariospromedio en cada período de observación con elnúmero de confrontaciones del período pam cadagrupos (a) y con el porcentaje de hojas madurasen la dieta de cada período (b).Los círculos blancos identifican los valorescorrespondientes al grupo Gringos. Los circulosnegros identifican los valores correspondientes algrupo Enfennitos.
DISCUSIÓN
El área de acción que detemtine paracarayá en la isla es el area de acción máspequeña informada para el género en unambiente natural (Tabla 7), lo que estaría deacuerdo con la Hipótesis l acerca de larelación inversa entre área de acción ydensidad (Crockett y Eisenberg, l987), yaque la isla posee Ia densidad de aulladoresmás alta que se conoce (Rumiz y Zunino.¡986; Rumiz, 1990; Crockett, 1998; Zuninoet al., 2001). Estudios en el sur de Brasilsobre un grupo de carayá reportaron tambiénun área de acción de 2 ha pero el ambiente noera natural sino que se trataba de una

Alouaua caraya como dispersor de semillas 31
Tabla 7: Áreas de acción expresadas en hectáreasinformada para diferentes especies de aulladores.
Especies Área de Referenciaacción
(El)A. caraya l.7-2.2 a Este esudio
A. caraya 2 Bicca-Marques,1994
A. caraya 5-6 " Zunino, l986
A. seniculus 4-7" Sekulic, 1982
A. palliala 9.9 Glander, ¡980
. seniculus |0-20b Terborgh enCrockett yEisenberg, 1987
\nn
A. seniculus |3-24 a Defler en Crocketty Eisenberg, 1987
A. sem'culus 22 Gaulin y Gaulin,1982
A. paIIiara 3| Milton, 1980
A. pigra 25-50 " Silver et al. ¡998
A. seniculus 45 Julliot, l992
A. Stoner, l996
Estrada, ¡984
Chapman, ¡988Schlichte enCrockett yEisenberg, 1987Palacios yRodri uez, 2001
a Area de acción de dos grupos.rango en base a 4 grupos
pal/¡ata 54¡s . palliala 60
A.paIliala ¡08
A. pigra 125
A. seniculus 182
plantación abandonada de Citrus sinenesísenriquecida con especies nativas (BiccaMarquez, 1994).
Contrariando la Hipótesis 2, a pesarde la extremadamente pequeña área de acciónde caraya en la lsla Brasilera, la dieta noestuvo dominada por las hojas. En tienafirme donde el área de acción promedio es de5,5 ha, las hojas representan el 70 % de ladieta de caraya (Zunino y Rumiz, ¡986),mientras que en la lsla Brasilera dondeencontré un área de acción de alrededor de 2ha, las hojas representaron sólo el Sl % de ladieta. Analizando mensualmente el consumode hojas se observa que en julio de 1999 elporcentaje fue extremadamente alto (Fig.
7 b), ya que durante la semana en queobservé a los grupos no hubo frutos ni floresdisponibles dentro de sus áreas de acción, Sirecalculo el porcentaje anual del consumo dehojas sin tener en cuenta este mes, elporcentaje es menor al 40 %. En “LosTuxtlas” (México) se encontró exactamentelo opuesto para Alouarta palliata, un área deacción inusualmente extensa y una dietadominada por hojas. Estrada (¡984) explicóesto diciendo que las hojas en este sitiotenían la “apariencia” de frutos ya que sudisponibilidad era'impredecible en el tiempoy en el espacio. Del mismo modo podríaexplicar las pequeñas áreas de acción decarayá en la lsla Brasilera diciendo que losfrutos poseen “apariencia” de hojas, ya quelos árboles con fruto camoso representan másdel 70 % de los árboles del bosque y estándisponibles a lo largo de todo el año (verCapítulo l). Por lo tanto podría refonnular laHipótesis 2 como, “los aulladores poseenpequeñas áreas de acción en sitios donde ladieta está dominada por recursos abundantesy de distribución uniforme en el tiempo y elespacio”.
No encontré ninguna correlaciónentre el porcentaje de hojas en la dieta y elrecon'ido diario de los grupos (Hipótesis 3),lo que me permite suponer que en la IslaBrasilera carayá no está restringidoenergéticamente. Por otro lado el recon'idodiario estuvo correlacionado positivamentecon las confrontaciones, este podria ser unefecto indirecto de la alta densidad en la isla,de tal forma que el área de acción no sería elúnico parámetro que se ve afectado por laalta densidad poblacional. Otra evidenciasobre la falta de restricciones energéticas decaraya en la Isla Brasilera serian las horasdedicadas al descanso. Los aulladoresdedican al descanso entre el 63 % y el 80 %del dla (Crockett y Eisenberg, 1987; Estrada,¡984), y en tierra firme carayá descansadurante el 77,4 % del día (Zunino, 1989). Sinembargo, en la lsla Brasilera yo encontré unporcentaje menor, similar al tiempo dedicadoal descanso por Arelesgeofioyi (40 al 54 %),un primate de similar tamaño a carayá perocon una dieta más frugívora (Crockett yEisenberg, ¡987). Para explicar esta falta derestricción energética en la isla yo

32 Susana l’alncia Bravo
considerarla nuevamente la abundancia ydistribución uniforme de los recursos de altacalidad.
A lo largo del año los recursos de altacalidad se vieron suplementadosenergéticamente debido a una alta infestaciónpor insectos, estas estructuras infestadasrepresentaron más del 25 % de la dieta. Otrasespecies de aulladores incorporan artrópodosindirectamente en su dieta, generalmente conlos higos, pero siempre se ha considerado quelo hacen en baja proporción (Crockett yEisenberg,- 1987; Prates et al., 1990). En lashojas nuevas de Albizía inundara encontre’ 2o más áfidos en cada folíolo y toda lasuperficie se encontraba cubierta por lasustancia azucar-ada que los áfrdossegregaban. Esta sustancia consiste en un90-95% de carbohidratos y en alg‘unasocasiones puede contener amino acidos(Holldobler y Wilson, ¡990). Todas lasestructuras infestadas representaron paracarayá una ingestión de azúcares (frutos,néctar o la sustancia segregada por losáfrdos) más proteínas (larvas o insectosadultos). Cuando las proteínas son ingeridasconjuntamente con carbohidratos sonutilizadas para las funciones vitales delorganismo, mientras que si son consumidassolas son metabolizadas para obtener energía.Esta es una de las razones por la que algunasespecies omnívoras prefieren alimentarse defrutos infestados (e.g. Cebus capucinus, Panpaniscus) obteniendo alimentos de altacalidad con mayor facilidad que la queimplica la búsqueda y obtención de insectoslibres (Redford et al.. 1984). Otra posiblerazón por la que carayá puede estaringiriendo estructuras infestadas es que estéutilizando la presencia de los artrópodoscomo un indicio de baja concentración decompuestos secundarios. Sin embargo, segúnestudios realizados en Costa Rica conAlouarra palliara el principal factor paradeterminar la selección de alimento por partede esta especie es el contenido proteico, yluego como factor secundario laconcentración de compuestos secundarios(Bilgener, ¡995). Para discernir cual es elpapel que los insectos puedan tener en ladieta sería necesario determinar cual es laeficiencia digestiva de carayá sobre ellos.
Los árboles de las 9 especies de loscuales se alimentó carayá dentro de sus 2 hade territorio, representaban el 89 % de losárboles registrados en los muestreos devegetación (Ver Capítulo l), esto esta'indicando una distribución espacialmenteuniforme de los recursos para carayá. La bajadiversidad arbórea del Bosque de Laureles dela lsla Brasilera se vio reflejada en la bajadiversidad de la dieta de carayá encomparación con la dieta de otros aulladores(Tabla 8). De acuerdo con algunos autores(Gaulin et al., 1980; Stoner, l996) un factorimportante para determinar la calidad dehábitat de un ambiente para los aulladores esla densidad de las especies más comúnmenteconsumidas, siguiendo esta interpretación laselva de inundación representaría unambiente de excelente calidad para car-ayá yla cual contribuiría a explicar la falta derestricciones energéticas en los gruposestudiados.
’l‘abla 8: Número de especies utilizadas comoalimento por los aulladores de distintas especies.
Número deEspecie . ‘ Referenciaes res usadas
.4.pal/¡ala 96" Glander, l980A.pal/¡ala 73 Milton, 1980A. caraya l8 Zunino, ¡987A.pal/¡ara 22 Chapman, 1988A.fusca 27 Prates el a|.. l990Ajusta 52 Galetti et aL, l994A.'.reniculus l95 Julliot, ¡994A.pal/¡ala 95 - 65 ” Stoner, 1996.4.pigra 74 Silver et aL, l998xl. caraza 22 Este estudio" es sólo el número de especies urbórczis."" los dos grupos estudiados utilimron distintonúmem de especies.
La abundancia y distribuciónuniforme de los recursos de alta calidadtambién sería responsable de la conformaciónde grandes grupos que utilimn pequeñasáreas de acción, puesto que durante los cortosperíodos en que sólo tuvieron hojas madurasdisponibles en sus territorios, los grupos sesubdividieron en 2 subgrupos. Estecomportamiento también fue observado porZunino y Rumiz (1986), durante un breveestudio en la Isla Guascara, próxima a la lslaBrasilera. En tierra firme donde la dieta de

Alouaua caraya como dispersor de semillas 33
carayá está dominada por hojas maduras lasubdivisión de los grupos no fue observada(Zunino, l987), pero en este caso los gruposson de menor tamaño a los encontrados en laisla y en ocasiones aun menores a lossubgrupos que observé en la lsla Brasilera. Sibien los grupos de aulladores han sidocaracterizados generalmente como cohesivosy estables (Chivers, ¡969; Glander, 1980;Gaulin et aL, l980; Mllton, 1980; Gaulin yGaulin, l982; Estrada, 1984;.lulliot, l992), eldividirse en subgrupos no sería exclusivo decarayá, pues fue reportado para Alouatrapal/¡ata cn Costa Rica. donde los subgrupospodían permanecer hasta 4 semanasseparados (Chapman, l988).
En la Fig. l l presento un modelo queresume e integra los resultados discutidoshasta el momento, mostrando que laabundancia temporal y espacial de losrecursos de alta calidad favorecería una altadensidad de la población, mientras queminimizar-ía las restricciones energéticas delos individuos. La alta densidad disminuiríael tamaño de las areas de acción, mientrasque la falta de restricciones energéticas de losindividuos aumentarlan su actividad. Losaltos niveles de actividad conjuntamente conlas pequeñas áreas de acción aumentaría laprobabilidad de encuentros entre grupos, yteniendo en cuenta una buena condiciónnutricional de los individuos estos encuentrosse traduciría en una alta frecuencia deconfrontaciones. Así existiría unaretroalimentación positiva entre el nivel deactividad y la probabilidad de encuentros,porque cuando un individuo de un grupodetecta la presencia de un individuo o grupoextraño, vocaliza y el resto del grupo sedesplaza, por esta razón es que el recon'idodiario correlaciona positivamente con elnúmero de confrontaciones. Por último, laalta densidad y actividad de los grupos haceque no existan áreas de bosque con bajafrecuencia de uso. En los capítulossubsiguientes mostraré como la altafrecuencia de confrontaciones y el uso detoda la superficie del bosque es deimportancia a la hora de evaluar a car-ayacomo dispersor de semillas. Una fuena capazde disminuir considerablemente los recursosalimenticios para carayá dentro de la isla y
por ende afectar toda la cascada de relacionesdescritas serían las inundacionesextraordinarias (e.g. según Neiff (l9_99)
pueden morir hasta el 50 % de los arboles).Así, si se acepta el modelo que propongo,todo el sistema quedaría regulado por estasinundaciones y la selva de inundación seríaentonces un ambiente de calidadtemporalmente variable calidad para carayá.Como sugiere Van Home (1983), no escorrecto utilizar sólo la densidad de unaespecie en un ambiente como indicador de lacalidad de hábitat del mismo.
ac ore u 'nflu e sobr la ls lónde gmlllas
Varios frutos consumidos por carayátuvieron infestadas sus semillas ycontrariamente a lo observado para A.seniculus (Julliot,l996), (por lo menos paraO. diospyrrfalia) earayá estaríaalimentándose de aquellos arboles con mayorinfestación. Conocer si los árboles de unaespecie difieren en la tasa o proporción deremoción de sus semillas y conocer lascausas, es sumamente importante a la hora deevaluar los efectos de la dispersión (Jordanoy Schupp, 2000).Algunos autores sugieren que los mamíferosdispersores pueden alimentarsepreferentemente de frutos infestados(Redford et al., 1984; Valburg, 1992 a,b) y siel insecto se aloja en la semilla, el dispersorpuede actuar de diseminador de este insectotransportándolo en su tracto digestivo(Traveset et al., ¡995). Las semillas delgénero Acacia suelen estar infestadas pordistintas especies de bruchidos que mueren alser ingeridas por ungulados o por el ganado,sin embargo, los estudios sobre lasupervivencia y posterior germinación de lassemillas son contradictorios (Coe y Coe,¡987; Miller y Coe, ¡993; Hauser, ¡994). Porlo tanto será necesario estudiar la respuesta alpasaje por el tracto de cada insecto queinfesta los frutos consumidos por carayá, y lagerminación de las semillas para determinarel efecto indirecto que carayá pudiera estarprovocando en esta interacción. En el caso deO. diospyrrfolia las larvas mueren al ser

34 Susana Patricia Bravo
INUNDACI ONES EXTRAORDINARIAS
de alta calidada
Densidad de
Alouattacama
Áreas de bosque “Probabilidadcon baja de
frecuencia de uso encuentros
Restriec ión
energética
Frecuencia deconfrontaciones
Figura ll: Modelo que integra los resultados de disponibilidad de recursos, comportamiento y uso delespacio de carayá en la lsla Brasilera las flechas indican la dirección de la relación entre variables. El signo +indica una relación positiva entre las variables conectadas por la flecha. El signo - indica una relaciónnegativa entre las variables conectadas por la flecha.
ingeridas por carayá (Bravo y Zunino, 1998)pero aun resta evaluar que sucede con lacapacidad de germinación. En Costa Rica seha observado que Heilipus sp. y al menosotra especie de curculionido son el principalfactor de predación de las semillas de O.endresíana no dispersadas, pero a diferenciade lo que ocurre en la lsla Brasilera lainfestación se daria sólo en el suelo del
bosque y no mientras los frutos están en elárbol (Wenny, 2000).
Es importante desde el punto de vistade la dispersión de semillas el uso diario detodo el territorio y la incorporación de losparches de bosque dentro del área de acciónde los grupos, pues esto implica que lassemillas de todos los árboles del bosquepotencialmente pueden ser trasladadas a

Aloualla caraya como dispersor de semillas 35
cualquier área del albardón o a los parches yquizas también a otros albardones con otrotipo de bosque. Es decir, la movilidad decaraya dentro de la isla no estaría limitadapor la discontinuidad del bosque comopudiera pensarse a priori y diariamente elefecto de carayá como dispersor de semillasestaria actuando sobre toda la superficieboscosa de la isla, lo que aumenta supotencial como dispersor.
Como en la isla Brasilera la distanciarecorrida diariamente por cada grupo noestuvo correlacionada con la calidad de ladieta sino con los encuentros con otro grupoo con individuos solitarios, la alta densidadno estaria afectando sólo el tamaño del áreade acción, sino que también afectaríaindirectamente las distancias recorridasdiariamente por los grupos, y podría estarcondicionando en última instancia ladistancia de dispersión de las semillas.
El consumo de flores por parte decarayá, cuantitativamente más relevante queel consumo de fmtos verdes, puede serconsiderado como predación predispersiva.La predación de flores por primates puede sertan Severa que hasta ha sido hipotetizadacomo la mayor presión de selección sobre lasincronla de la floración para varias especiesde árboles en las selvas tropicales(Struhsaker, ¡978 en Heymann, 1992). Sianalizo en particular la relación de carayácon cada una de las principales especies quesufrieron este tipo de predación, debodiferenciar tres grupos. Uno estaria formadopor aquellas especies de las cuales carayáingirió sus flores cada vez que estasestuvieron disponibles y sólo dejó de hacerlocuando una estructura más ricaenergéticamente (e.g. frutos) estuvodisponible o bien cuando la cantidad deflores en los árboles disminulaconsiderablemente (este sería el caso de Ingavera y la Papilionáceae l), Por otro ladoestaria el grupo compuesto por 0.diospyrifolia y B. argura, cuyas flores fueron
generalmente ignoradas y cuyos frutos sífueron ingeridos siempre que estuvierondisponibles. Para el primer grupo carayárepresentaría un verdadero predador deestructuras reproductivas, mientras que parael segundo grupo de especies seriaprincipalmente dispersor de sus semillas(asumiendo que sus semillas son viablesluego de pasar por el tracto digestivo) y soloactuaria como predador ocasionalmente,cuando no hubiera frutos y fueran los únicosárboles ofreciendo flores. Por último, está elcaso de G. strialla cuyas flores fueron lasmás consumidas, lo cual representaría unaimportante predación, pero no me es posibleincorporar a esta especie en el primer grupo,pues si bien sus semillas no son transportadaspor carayá no he evaluado si la remoción delpericarpio tiene algún efecto favorable parasu germinación y posterior establecimiento.
En resumen al estudiar el uso delespacio y los hábitos alimenticios de carayáen la lsla Brasilera, encontré factores quepueden influir positivamente a la hora deevaluar la calidad de carayá como dispersorde semillas: la visita a los distintos parchesde bosque donde podria depositar semillas, eluso diario de todo el territorio sin que quedenáreas de bosque sin visitar por los grupos, laalta proporción de frutos en su dieta, laingestión de las semillas de todas las especiescon frutos carnosos del bosque, en el casoque su germinación no se vea perjudicada.Sin embargo otros factores pueden influirnegativamente: la alta proporción de floresingeridas, la ingestión y transporte desemillas infestadas y la ingestión de lassemillas de todas las especies con frutocamoso del bosque en el caso de que sugerminación no sea posible luego del pasajepor el tracto digestivo de carayá. Por lo tantoes necesario que estudie ahora donde sondepositadas estas semillas, que ocurre con sugerminación y que ocurre con los insectosque infestan las semillas al pasar por el tractodigestivo.

Foto 4: Diferencias de color en el pelaje
Foto 5: Estadios en el desarrollo de Heilipus sp.
Foto 6: Acicalamiento entre dos hembras de carayá

¡llauaua caraya como dispersor de semillas
Capítulo III: Efectos del dispersorPatrón de dispersión y efectos del pasaje por el tracto digestivo
“mala ave tuvo que haber voladomuchas veces de este moda con suestómagobien llena a lagunas distanm yal conseguir cantones una buena comidade peca, la analogía me hace creer quedevolverlalas semillas en una pelorilla enun atada adecuada para lagerminación. "
Charla Darwin (El origen de las especia)
INTRODUCCIÓN
Teniendo en cuenta la hipótesis dedispersión clásica propuesta por Janzen(¡970). las semillas aumentarlan susprobabilidades de sobrevivir a medida queson alejadas del árbol parental o, más bien,cuando son llevadas a una cierta distancia deeste. Sin embargo, la sombra de semillasgenerada por los animales no suele serhomogénea, es decir no declina gradualmentea medida que nos alejamos del árbol parentalcomo predicen los modelos teón'cos. Laheterogeneidad en la distribución de lassemillas en el suelo es conocida como“limitación en la diseminación” (Jordano yGodoy, 2002; Schupp et al., 2002) y esgenerada por diferentes comportamientos delos animales (e.g. conformación de leks, usode perchas) que determinan distintos patronesde distribución de las semillas (Janzen, 1983;Wenny, 2000).
Los aulladores dispersan las semillasmayormente por endozoocoria, por lo queestas serán depositadas en los sitios dondeellos defequen. Los grupos de aulladorestienen un comportamiento particular, ya quetodos los individuos de un grupo defecansecuencialmente utilizando una misma rama(Milton, ¡980; Crockett y Eisenberg, 1987;Alves y Guix, [992; Julliot, ¡996; Yumoto etal., 1999; Bravo y Zunino, 2000), cuando losgrupos son numerosos suelen utilizar más deuna rama, de modo que entre dos y cincoindividuos defecan secuencialmente en cadauna de ellas (obs. pers.). Dependiendo decaracteristicas ambientales como pueden serla cantidad de lluvia o la estructura de la
vegetación (obs. pers.) las heces, y junto conellas las semillas, pueden o no acumularse engran cantidad debajo de estas ramas(Crockett y Eisenberg, [987; Alves y Guix,1992).
En todos los casos se ha visto que elpatrón de dispersión de semillas que realizanlos aulladores es agrupado (Julliot, 1996;Yumoto et al., 1999; Bravo y Zunino, 2000)a diferencia del patrón azaroso generado porotros primates (e.g. Saguinus mystax y S.fuscicollis, Garber, 1986; Cebus capucinus,Rowell y Mitchell, 1991; C. apeIIa y Atelespaniscus, Zhang y Wang, 1995).
Respecto a la ubicación de los sitiosde defecación en el bosque, la mayoría de lostrabajos coinciden en indicar que seencuentran asociados a los árboles utilizadospor los grupos para pasar la noche("donnideros") (Crockett y Eisenberg, 1987;Julliot, 1996; Bravo y Zunino,‘ 2000).Algunos autores señalan que estos árbolespueden ser los mismos que los grupos deaulladores utilizan para alimentarse de frutos,por lo que en estos casos estaríandepositando semillas debajo de los árbolesparentales y podrían, entonces, no contribuirsignificativamente a su dispersión (Yumotoet al., ¡999). Los aulladores pueden tener unnúmero variable de donnideros, pero notodos son utilizados con igual frecuencia.Julliot (¡996), por ejemplo, encontró a lolargo de dos años de estudio que un grupo deAlouana seniculus tenia 34 donnideros, peroel 35 % de las heces hablan sido depositadas

38 Susana Patricia Bravo
alrededor de los cinco dormideros masusados.
Una vez determinado dónde y cómoson depositadas las semillas, uno de lospuntos que se hace necesario estudiar es enqué condiciones llegan estas, es decir si aúnmantienen su capacidad para germinar y, enese caso, que ocurre con la infestacióndeseripta en el Capítulo ll. Al evaluar lacapacidad de germinación es importantetener en cuenta dos factores: el número deSemillas que germinan y la velocidad a la quelo hacen (Lieberman y Lieberman, l986). Elefecto que tiene el pasaje de las semillas porel tracto digestivo de los aulladores sobre lagerminación es variable. La capacidad degerminación suele no verse afectada, aunquedependiendo de la especie la velocidad sípuede aumentar o disminuir por efecto de losaulladores (Estrada y Comes-Estrada, [984;Julliot, ¡996; Bravo y Zunino, 2000). Porotra parte, el efecto del pasaje por el tractodigestivo de carayá sobre las larvas queinfestan las semillas es fundamental a la horade evaluar en qué condiciones llegan lassemillas al suelo, pues si las larvassobreviven pueden continuar consumiendo elendospenna y el embrión vegetal anulandocualquier posibilidad de supervivencia parala semilla.
La efectividad de un dispersor debeser medida teniendo en cuenta doscomponentes "cantidad" de semillasdispersadas y "calidad" de la dispersión(Schupp, 1993). En el capítulo anterior, aldescribir el consumo de frutos que realizacarayá, la manipulación de los mismos y eluso del espacio por los grupos obtuve unadescripción referida a la cantidad de frutosque pueden ser removidos en el tiempo y elespacio. En este capítulo me referiré adistintos aspectos que hacen a la calidad de ladispersión de semillas realizada por carayá.Explicaré dónde son depositadas y en quécondiciones llegan al suelo. Al igual que loencontrado en otros trabajos (Estrada yComes-Estrada, 1984; Julliot. ¡996; Bravo yZunino, 2000), espero que el pasaje por eltracto digestivo de carayá no perjudique lacapacidad germinativa de las semillas. Dadoque la infestación de 0. diospyrifolía eseliminada al pasar por el tracto digestivo decarayá (Bravo y Zunino, |998). espero que
efectos similares ocunan en el resto de laespecies infestadas. Finalmente. y conrespecto al patrón de dispersión, debería seragrupado de acuerdo a su comportamiento dedefecación y a lo que he visto que ocurría entierra firme (Bravo y Zunino, 2000).
MATERIALES Y MÉTODOS
Patrón de dispersión
En los mismos mapas donde registreel recorrido diario durante los seguimientosde los grupos de carayá, ubique diariamentelos dormideros, los sitios donde defecaban ydonde consumlan frutos. Señalando el lugardonde los frutos de cada especie eraningeridos (l) y el lugar de defecación de lassemillas de esa especie (2), y considerandoque el tiempo de pasaje del alimento por eltracto fue aproximadamente de 24 horas (verCapitulo ll), consideré como distancia dedispersión la distancia entre el registro (2) y elregistro (l) del día anterior.
Como los datos de dispersiónobtenidos para cada período de muestreo noserían independientes, pues las fuentes dealimento y los dormideros usados un díapueden determinar los que serán usados alsiguiente. Asi que sólo describiré el rango enque fluctuaron las distancias y cuál o cuálesfueron las distancias de dispersión másfrecuentes.
Dado que durante estos muestreosobservé que algunos individuos no utilizabanlas misma ramas que el resto para defecar.decidí realizar un segundo muestreo paradeterminar la frecuencia de esecomportamiento y si el no uso de la rama encomún (bosteadero) dependía de la edad ysexo de los individuos o de la actividad queestuviera realizando el grupo. Durante julio de2000 realicé el seguimiento de 3 grupos("Labio Rosa", "Vecinos", "lgnacios"),durante 5 dias cada uno, desde la salida hastala puesta del sol, registrando y numerando enel campo el sitio donde defecaban losindividuos (bosteadero / no bosteadero)teniendo en cuenta si eran hembras, machos ojuveniles y cuál era la actividad del resto delgrupo cuando esto ocurrla (defecaban / nodefecaban). Considere que el grupo defecabaen conjunto cuando en un lapso de l minuto

Alouatla :araya como dispersor de semillas 39
más del 50 % de los individuos del grupo seencontraban defccando. Para cada grupoobservado calcule cuál fue la proporción dedeyecciones fiJera de los bosteaderos y, deestas, cuál era la proporción que se producíacuando el resto del grupo defecaba. Evalue’ laproporción de deyecciones fuera delbostcadero para las distintas categorías deedad y sexo con una prueba de Kruskal Wallis considerando“ a cada grupo como unaréplica
Efectos de la ingestión de semillas sobre lagerminación
Evalué la germinación de las semillasde las especies más frecuentemente ingeridaspor camyá, Banara argula, Cecropiapachyslachya, Cissus palmala, Eugenia¡”miei/olía. Ocorea diospyrifolia (Foto 7) yI’xychorria carlhagenensis (Foto 8).
Para obtener semillas que hubieranpasado por el tracto digestivo de los animales("semillas ingeridas") colecté heces de losindividuos adultos de los dos gruposobservados cada vez que consumieron frutos.En el caso de P. carrhagenensis, colectéheces de juveniles, pues fueron los que laconsumieron mayoritariamente.
Para los controles colecté frutos delos árboles más utilizados por carayá paraalimentarse, teniendorespecial cuidado entomar frutos que estuvieran en igual estadode maduración que los consumidos porcarayá y les extraje sus semillas ("semillascontrol").
Sembré las semillas en cajas de Petrisobre papel de filtro el cuál mantuve húmedocon agua destilada. Las cajas de Petriestuvieron bajo un régimen de 12 horas deluz y l2 horas de oscuridad, y las semillasfueron observadas cada 2 - 3 dias pararegistrar las genninaciones (aparición de laradicula) (Lieberman y Lieberman, ¡936). Elnúmero de cajas de Petri que sembré no fueel mismo para todas las especies y estuvo enrelación con la abundancia de semillas en lasheces (Tabla ll). En cada caja de Petricoloque entre 4 y 6 semillas dependiendo deltamaño dc las mismas. Las especies cuyagerminación ocurre naturalmente enprimavera / verano estuvieron a unatemperatura entre 25 —30 °C, mientras que
aquellas que genninan en otoño / inviemofueron mantenidas a 20 —25 °C. Finalicé lasexperiencias luego de transcurridos 30 díassin registrar nuevas germinaciones.
Consideré cada caja de Petri comouna réplica y evalué la germinación de lasiguiente manera (basada en Lieberman yLieberman, 1986):(l) El número de semillas control germinadas
respecto del número de semillas ingeridasgerminadas al finalizar las experiencias,usando una prueba de U de MannWhitney.
(2) El tiempo que tardó en genninar la mitadde las semillas control respecto del tiempoque tardó en genninar la mitad de lassemillas ingeridas, usando una prueba de tde Student o una prueba de U de MannWhitney.
Efectos de la ingestión sobre los insectosgue infestan las semillas
Para determinar el efecto que elpasaje de las semillas por el tracto digestivode carayá pudiera tener sobre los insectos quelas infestaban, extraje semillas de las hecesde carayá de los dos grupos. Las observé bajolupa, poniendo especial atención paradetectar la condición en que se encontrabanlos insectos (vivos o muertos), si el embriónvegetal habia sido atacado por la infestacióny que porcentaje del endosperrna había sidoconsumido.
RESULTADOS
Patrón de dispersión
La distancia de dispersión de lassemillas fue uniforme a lo largo del añovariando entne 0 y 360 metros. Los 360 m dedispersión los registre para C. pachystachya(Enfennitos, Febrero 2000), la. distancia de0 m (i.e. no dispersión) la encontré tanto paraO. diospyrrfolia (Enfennitos, Julio l998)como para E. punicrfolr'a (Enfennitos,Noviembre ¡998) y representó sólo el l,l %del total de los registros. Las distancias dedispersión más frecuentes estuvieron entre 20y 40 metros (Tabla 9).

40 Susana Putricm Bravo
Tabla 9: Distancias de dispersión para las semillas de especies arbórcas dispersadas por dos grupos de camyáen la lsla Brasilera. Rango = distancia minima - distancia máxima registrada, ‘ informo el valor promedioporque no existió una moda para esta especie en ese pen'odo.
Especie Mes grupo Rango (m) Moda (m)
Ocolea diospyrifolia Julio ¡998 Gringos l00 —160 100Enfennitos 0 —220 180 y 20
Mayo 1999 Gringos l0 —l40 30Enfermitos 20 - 230 80
Cecropia pachysrachya Mayo 1999 Gringos ¡0 —80 42 "
Febrero 2000 Gringos IO —80 lOEnfennitos 20 —360 60
Psychotria carthagenensis Mayo 1999 Gringos 20 —70 30 y 20
Julio l999 Gringos lO —l20 40, Enfennitos lO —80 20
Eugenia punicrfolia Nov. l998 Gringos 20 —80 40Ent‘ennitos 0 - 230 200
Febrero 2000
Enfermitos IO -- 200 20, 40 y l lOBanara argula Mayo l999 Gringos lO- l40 30 y 40
Enfermitos 20 —200 50
Los grupos defecaron básicamente endos situaciones: al finalizar un período dedescanso y antes o después de lasconfrontaciones del grupo. Como resultadode ello, las semillas dispersadas fuerondepositadas alrededor de los donnideros y enlos bordes de los territorios (Fig. l2). LOSgrupos utilizaron los mismos bosteaderos a lolargo de todo el estudio. En las zonas desuperposición o cuando los Enfermitosdesplazaron a los Gringos (ver Capítqu ll)ambos grupos utilimron los mismosbosteaderos.
Los Gringos utilizaron 7 donniderosy los Enfennitos utilizaron 8. Sin embargo,no todos fueron utilizados con igualfrecuencia; ambos grupos tuvieron un"dormidero principal" (Fig. ¡2) dentro de suterritorio donde descansaron el 47,4 % de lasnoches los Gringos y el 52,7 % de las nocheslos Enfennitos (Tabla 10). Observe el mismopatrón en los otros 3 grupos estudiadosdurante el 2000. El grupo de los Gringostambién utilizó con frecuencia (13,2 %) undormidero ubicado en un pequeño fragmentode bosque (Fig. l2). En la lsla Brasilera se
produjo acumulación de las heces en el sueloa lo largo del tiempo, por lo tanto estosdonnideros con alta frecuencia de uso eran elcentro de un área aproximadamente circularde unos 20 metros de diámetro donde el sueloestaba cubierto por una capa de varioscentímetros de heces y semillas, y donde sepodía observar una gran cantidad de árbolesjuveniles (ver Capítulo IV). Por otra parte,pude definir "bosteaderos puntuales" (Foto 9)asociados a las zonas de confrontación (Fig.l2) y a los donnideros utilizados con menorfrecuencia. Estos tienen igualescaracterísticas que el asociado al dormideroprincipal, pero con un diámetro promedio de5 m.
De las 337 deyecciones que registrédurante las observaciones de Julio del 2000el 35 % estuvieron fuera de los bosteaderos("deyecciones independientes") (Fig. 13 a).De estas, el 94 % se produjeron cuando elresto del grupo no estaba defecando. es decirsólo en 7 oportunidades hubo individuos quedefecaron fuera de los bosteaderos al mismotiempo que el resto del grupo lo estabahaciendo en los bosteaderos (Fig. l3b).

lOOOOOOOOOOOOO0.0.0.000COOOOOOOOOOOOOOOOOOOOOO.
Alouarra caraya como dispersor de semillas 41
Respecto a la frecuencia con que losindividuos defecaban fuera de losbosteaderos teniendo en cuenta su edad ysexo no encontré diferencias significativas(H = l,39; P = 0,5; gl = 2; Fig. 13 c).
Tabla lO: Los valores indican el porcentaje denoches que cada grupo de carayá pasó en cadauno de sus árboles dormideros.
Dormidero Gringos Enfermitos
l 47,4% 52,7%
2 18,4% . 14,5%3 13,2% ¡2,7%4 10,5% 7,3%5 5,3% 7,3%6 2,6% 1,8%7 2,6% 1,8%8 1,8%
Efectos de la ingestión de semillas sobre lagerminación
Para la mayoria de las especies elporcentaje de semillas control gerrninadasfue similar al número de semillas ingeridasgerminadas (Tabla ll), exceptuando a B.arguta y Cissus sp. la germinación fue alta(mayor al 50 %) en ambas condiciones.Aunque, las semillas de C. pachystachya enel grupo de los Gringos y E. punicifolíatuvieron un mayor número de semillascontrol germinadas (Tabla ll).
En cuanto a la velocidad degerminación, fue similar para las semillascontrol y las ingeridas en C. pachystachya, C.palmata y P. carthagenensís (Tabla ll). Lassemillas ingeridas germinaron más rápidoque las control en B, arguta y E. punícífolia(Tabla ll). Las semillasde O. diospyrifolr'aingeridas por los Enfermitos germinaron amayor velocidad que sus controles, pero estono sucedió con las semillas ingeridas por losGringos (Tabla 11).ÁAl analizar de maneracruzada las velocidades de germinación deambos grupos, encuentro que las semillasingeridas por los Enfermitos no genninaronmás rápido que las semillas ingeridas por losGringos (t = 0,11; P = 0,92; gl = 79), sinembargo si germinarona mayor velocidad lassemillas control de los Gringos respecto a las
semillas control de los Enferrnitos (t = 2,64;P = 0,02; gl = 79). ‘
Borde del Bosque
Limite de los territorios
D Territoriode los GringosE Territoriodelos Enfemritos
' Sitios de las confrontaciones- Dormiderosprincipales. Donnidero en un fiagmento
de bosque
Figura 12: Ubicación de los dormideros másusados por los dos grupos y sitios donde seprodujeron las confrontaciones entre grupos.
Efectos de la ingestión sobre los insectosgue infestan las semillas
Las observaciones bajo la lupa mepermitieron determinar que el impacto de laingestión de carayá sobre cada una de lasespecies infestadas fue diferente. En 0.diospyrr’folia carayá elimina la mayor partede las larvas de Heílipus sp sin afectar alembrión de la semilla. Sólo un 8 % de laslarvas sobrevivieron al pasaje por el tractodigestivo. El porcentaje de endospermaafectado depende del momento en que esingerida la semilla. En las semillasconsumidas cuando recién maduran sólo hayun huevo y el pequeño canal de laoviposición, a medida que pasa el tiempo eltamaño de la larva es mayor y por endemenor la cantidad de endosperma que queda.
Las avispas que infestan las semillasde B. arguta mueren al pasar por el tractodigestivo de carayá, pero cuando estosingieren los frutos las avispas estándesarrolladas y prontas a nacer. Asi duranteel estadio. de larvas se comen todo elendosperma y embrión de las semillas queinfestan.

42 Susana Pamela Bravo
Tabla 11: Resultados de las pruebas de germinación donde compare el porcentaje de semillas germinadas porcaja de Petri y el tiempo que tardó en gerrninar el 50 % de las semillas (velocidad de germinación) entre lassemillas ingeridas (l) por ambos grupos de carayá y sus respectivos controles (C). Identifique a los gruposcomo G = Gringos y E = Enfennitos. N.= número de cajas de Petri con semillas ingeridas, Nc= número decajas de Petri con semillas control.
. N, Porcentaje de semillas . . .,Especne Grupo NC germinadas Velocrdad de germmacnon
Banara argula G 78 l = 25,83 :1:27,50 l = 22,95 :1:7,1490 C = 26,33 :t 20,83 C = 23,24 :1:7,47
U = 3388,5 P = 0,7 t = 6,60 P < 0.00001
Cecropia pachysrachya G 30 l = 83,83 i 12,66 l = 5,07 i 0,2530 C=93.83:t 10,16 C=5,03 i0,18
U=255P=0,004 U=435P=0,82
E 30 1= 82,16 :t 17,33 1= 5,5 :1:0,9730 C = 78,66 i 20,83 C = 5,27 i 1,31
‘ U=402P=0,61 U=350,5 P=0,2
Cissus sp. E 15 1= 31,00 :t 34,33 l= 53,70 :1:18,0115 C = 33,33 :t 31,00 C = 38,33 i 22,48
U=106P=0,79 U=63P=0,066
Ocotea diospynfolia G 45 l = 55,33 :1:31,66 l = 24,43 :1:14,1445 C = 53,33 :t 30,33 C = 28,41 :t 13,56
U=972,5P=0,77 U=640P=0,2
E 45 I = 64,33 :t 28,00 1 = 24,09 d: 14,9245 C=6l,33:t30,00_ C=35,85:1:13,15
U = 989,5 P = 0,85 U = 441 P = 0,0003
Eugenia punicrfolía G 35 l =77,00 i 22,66 1=27,2| :t 8,8735 C = 98,00 :1:8,00 C = 33,51 i 9,49
U = 417,5 P = 0,022 U = 366,5 P = 0,004
E 35 l =79 i 21,66 1=22,63 :t 6,2535 C = 91,33 i 17,00 C = 28,23 :t:8,24
U=293,5P<0,001 U=208P<0,0001
Psychorria carthagenensis G 17 1=76,50 :t 21,50 l =45,76 i 10,7917 C = 87,50 :t 20,50 C = 48,76 :t 12,80
U =94,5 P= 0,21 U = 135,5 P = 0,76
No encontré larvas en las semillas de E.punicrfoh‘a ingeridas por carayá, sinembargo he observado en el campo que elendosperma de las semillas que seencuentran bajo los árboles parentales y elde las que están en algunas de las hecesdesaparece en poco tiempo quedando sólosus cubiertas.
DISCUSIÓN
Patrón de dismrsión
Habitualmente se considera que elpapel de los dispersores de semillas esalejarlas del árbol parental y permitirles así

Alouaua caraya como dispersor de semillas 43
Deyeccrona('26)
ho
Blue-dem Habana-¡tem
b
'°° cs:A ao:2¡i 60
5 .0
É zo
o ISI
" mui-¡Ad aimAdmddulmddm
Deyucione(Vo)
8
Madina Hembras Juvmilncm kh ¡riñón
Figura 13: a - Porcentaje promedio dedeyecciones fuera y dentro de los bosteaderos. b- Porcentajes de deyecciones fuera de losbosteaderos cuando el resto del grupo defecabao cuando realizaban otra actividad. c Porcentaje de deyecciones fuera de bosteaderoconsiderando edad y sexo de los individuos
escapar de la mortalidad densodependienteque puede existir en sus cercanías("hipótesis de escape"; Janzen, 1970). Sibien en este estudio, una baja proporción desemillas de 0. díospyrr‘folíay E. punicrfoliafueron depositadas debajo del árbolparental, observando las distancias dedispersión (Tabla 9), y considerando lahomogeneidad del bosque (ver Capltulo l)
se puede deducir que, de forma similar a loque ocurre con Ocorea endresíana en CostaRica (Wenny, 2000), las semillas pudieronser depositadas debajo de otros árbolesadultos de la misma especie, por lo tanto enmuchos casos carayá no cumpliría con elrequisito clásico impuesto por Janzen(l970) para definir a un buen dispersor. Sinembargo, es necesario considerar otras dosposibles funciones de la dispersión desemillas. Por un lado, la facilitación delflujo génico dentro de las poblacionesvegetales (Renner, 1992) y, por otro, laposibilidad de llegar a nuevos sitiosfavorables para la germinación ysupervivencia de las plántulas sin unadisminución en la capacidad genninativa(“hipótesis de la dispersión directa") (Howey Smallwood, l982). Considerando elincremento del flujo genico, las semillasdepositadas debajo de otro árbol adulto dela misma especie deberian ser consideradascomo dispersadas. Desde el punto de vistade la hipótesis de la dispersión directa,carayá llevarla semillas aún hasta lospequeños fragmentos de bosque y noperjudicarla la capacidad de germinación delas semillas, incluso la estaría acelerando enalgunos casos. Pero, me resta conocer si sonlos bosteaderos sitios favorables para elestablecimiento y si esto es independientede que se encuentren debajo de un árbolparental. Pero. se han encontrado casos enlos que el alejamiento del árbol pareanparecería no representar una ventaja paralas semillas (Howe y Smallwood, ¡982;Claik y Clark, 1984). Entonces es válidopreguntarse, ¿Es baja la supervivencia desemillas y plántulas debajo del árbolparental para estas especies?.
Carayá utiliza los bosteaderossolamente cuando es todo el grupo quienfinaliza un periodo de descanso y defeca,pero no lo utiliza cuando un únicoindividuo lo hace. Posiblemente el mismocomportamiento ocurra en otras especies de“aulladores, aunque la complejidad y alturade los ambientes tropicales impidanobservarlo con claridad. Puesto que en lalsla Brasilera las heces llegan al sueloprácticamente intactas y alll se acumulan,pude observar cuantificar estecomportamiento. El hecho que el 35 % de

44 Susana Patricia Bravo
las deyecciones se produjeran fuera de losbosteaderos implica que, superpuesto alclásico patrón de dispersión de semillasagrupado producido por los aulladores(Julliot, 1996; Yumoto et al., l999; Bravo yZunino, 2000), carayá estaría produciendoun patrón secundario probablementeazaroso similar al de otros primates.Distintos autores han cuestionado laefectividad de aquellos dispersores quegeneran patrones agrupados donde lassemillas se encuentran en alta densidad.Obviamente, muchos factoresdensodependientes podrian actuar bajoestas condiciones, perjudicando elreclutamiento, al igual que ocurre en lascercanía de los árboles parentales (Howe,1986; Dino y Dominguez, 1986). Por esoconsidero que el patrón secundario azarosode dispersión que carayá produce podría serimportante para la reproducción de algunasespecies vegetales. Sin embargo, algunasobservaciones me llevan a pensar que estepatrón que describo como azaroso no es tal,pues la definición de azar implica que laocurrencia de un hecho en un puntodeterminado no debe afectar la probabilidadde ocurrencia a futuro en este u otro punto.En este sentido, a lo largo de los añosobservé como los sitios donde hablaregistrado la deyección de un animalvolvían a ser usados, es decir que enaquellos sitios donde registre’ la deyecciónde un animal existiría una mayorprobabilidad de que vuelva a ocurrir otroregistro que en un punto cualquiera delbosque. Es probable que gran parte de lospatrones de dispersión azarosa descriptospara distintas especies en realidad no seantales si tenemos estrictamente en cuenta ladefinición de azar. Por ejemplo ¿Podemosconsiderar que un zonal (Turdusrufiventris) o un benteveo (Pitangussulfuratus) que retira frutos de un árbol deP. carthagenensis para luego ingerirlosjunto con la semilla en una percha adeterminada distancia del árbol parentalproduce una dispersión azarosa? Es ciertoque la deyección puede ocurrirazarosamente, pero siempre será sobre laruta que el ave utiliza para ir del árbol confruta a la percha y viceversa. ¿Podemosconsiderar amrosa la dispersión de semillas
producida por animales que se desplazansiguiendo determinadas rutas dentro de susáreas de acción o que usan con distintafrecuencia las distintas zonas? Seguramenteno, ya que las deyecciones se produciráncon mayor probabilidad en las rutas y en laszonas del área de acción más utilizadas(Jordano y Schupp, 2000; Schupp et a|.,2002). De todas formas, para una semillatanto esta situación como las deyeccionesindependientes de carayá son situacionesmuy diferentes a las de un bosteadero, puesni la materia fecal ni las semillas se
acumulan y la cantidad de plántulaspresentes sera' escasa. Sin embargo, desdeel punto de vista del bosque ninguna deestas situaciones representan una dispersiónazarosa de semillas y ambos casos,dispersión en bosteaderos y dispersión endeyecciones independientes, seríanfenómenos que limitarian la diseminaciónpotencial de las semillas.
Efectos de la ingestión de semillas sobrela germinación
Las semillas ingeridas por carayáen general gerrninaron en similarproporción a las semillas control aexcepción de E. punícifolia y C.pachystachya (grupo Gringos) quedisminuyeron el porcentaje de germinación.En cuanto a la velocidad de germinaciónaproximadamente en la mitad de lasexperiencias observé un aumento develocidad de las semillas ingeridas mientrasque la otra mitad no varió la velocidadrespecto a los controles. Si bien losexperimentos de germinación mostraron unmenor porcentaje de semillas germinadasde E. punicrfoli'a cuando provenían defrutos ingeridos, el porcentaje de semillasgerminado fue elevado en los dostratamientos. Además existió un aumentoen la velocidad de germinación cuandopasaban por el tracto digestivo, por lo tantono puedo considerar que estén siendoperjudicadas al ser ingeridas. Las semillasde C. pachystachya ingeridas por el grupoGringos también genninaron menos que suscontroles. Nuevamente el porcentaje desemillas gerrninadas en ambos tratamientosfue elevado. Tampoco puedo concluir un

Aloha/Ia caraya como dispersor de semillas 45
perjuicio de carayá sobre esta especie puesla diferencia entre tratamientos en el otrogrupo no fue significativa y la tendencia fueopuesta, las semillas ingeridas genninaronen mayor proporción que las control.
El hecho de que las semillas controlde 0. diosmrifolia colectadas en losterritorios de ambos grupos tenganvelocidades de germinación diferente indicanuevamente que existen diferenciasíntraespecífrcas entre los árboles de estaespecie (ver Capitulo ll sobre diferencias enla infestación por curculiónidos). Sin.embargo, al comparar la velocidad degerminación de las semillas luego de pasarpor el tracto digestivo de los individuos deuno y otro grupo, encuentro que lasdiferencias desaparecen por lo que el efectodel pasaje por el tracto digestivo estaríainteractuando con estas diferenciasintraespeclcifas de 0. diospyrifolia. Dichode otra manera, dentro de esta especieexistirlan dos tipos de individuos, aquellosque producen semillas con germinación"lenta" y los que producen semillas congerminación "rápida". La escarifrcación quecarayá produce puede acelerar lagerminación de las semillas "lentas" perono influiria sobre aquellas de germinación"rápida". Si integro estos resultados con losdel capítulo anterior encuentro que lassemillas de germinación lenta fueron, a suvez, las más infestadas. ¿Puede ser lagerminación lenta una ventaja para eldesarrollo de los curculiónidos? Tambiénfueron esas semillas las más ingeridas porcar'ayá. Se ha encontrado que lagerminación tardía puede traer desventajaspara algunas especies pues aumentan lasprobabilidades de que se produzcapredación de semillas y patógenos yherblvoros atacar-¡an selectivamente a lasplántulas producto de una germinacióntardía (Seiwa, 1998) ¿El efecto de carayásobre la infestación y la germinacióncontrarresta estas desventajas y permite queestos individuos de O. diospyrr'foliapermanezcan en el sistema?
El aumento en la velocidaddegcrrninación como efecto del pasaje porel tracto digestivo de los dispersores podríaser un efecto bastante
frecuente (Hladik y Hladik, ¡969;Lieberman y Lieberman, 1986;. Nunes,1995; Julliot, 1996; Traveset y Verdú,2002). Sin embargo, como muchos trabajosde dispersión no lo evalúan en susexperiencias de germinación no es posiblesaber que tan generalizado se encuentra(Garber, l986; De Figueiredo, 1993;Ellison et al., 1993; Foord et aL, 1994;Overdorff y Strait, l998). Uno de losfactores citado como regulador de lavelocidad de germinación es lapermeabilidad de la cubierta de la semilla,pues esta actúa sobre la tasa de hidratación(Cronquist, 1984). Es plausible que laescarificación química y mecánica sufridapor las semillas al pasar por el tractodigestivo de carayá aumente lapermeabilidad y. por ende, acelere lagerminación (Traveset y Verdú, 2002). Dela misma forma, la. diferencia entre losindividuos de 0. diospynfolr'a podria estardada por la permeabilidad de las cubiertasde las semillas de cada individuo. Losindividuos de germinación lenta podriantener cubiertas poco penneables queretrasan la hidratación respecto de los degerminación rápida, pero esta diferencia depermeabilidad se verla eliminada al pasarpor el tracto digestivo de carayá. De estemodo, los tiempos de germinación seigualan'an y pasarían a estar regulados porotro factor.
Las semillas de 0. diospyrrfaliadispersadas por carayá pierden la pulpa enmayor o menor medida, trabajos conOcórea endresr'ana han observado que laremoción de la pulpa es fundamental para lagerminación y establecimiento de lasplántulas (Wenny, 2000).
Las experiencias de germinación enel laboratorio me permitieron conocer elefecto del pasaje por el tracto digestivosobre las semillas de las distintas especies ycomprobar que en ningún caso la capacidadde germinación se vio afectada de maneraimportante. Sin embargo esto no significaque en la naturaleza las semillas que llegana los bosteadenos puedan genninar de igualmanera pues muchos otros factores entranen juego (e.g., predación, ataque porpatógenos).

46 Susana Patricia Bravo
Efectos de la ingestión sobre los insectosgue infestan las semillas
Algunos estudios han encontradoque los dispersores evitaban la ingestión defrutos o semillas infestados, como hacen losmurciélagos de Filipinas (Utzurrum yHeideman, |99|). algunas aves de Alaska(Traveset et a|., ¡995) o A. seniculus en laGuyana Francesa (Julliot, ¡996). Sinembargo, otros encuentran que losdispersores pueden ser atraídos por lasfrutas infestadas, como es el cm de losvertebrados que remueven frutas deAcníslus arborescens en Costa Rica(Valburg, l992a), o de varias especies demonos en todo el mundo (Redford et al.,1984). Car-aya parece estar dentro de estesegundo grupo, pues varias de las especiesque consumió tuvieron infestadas sussemillas. Esto puede significar la dispersiónde la infestación conjuntamente con lassemillas como se vio que ocurre con lassemillas de la palmera Syagrusromanzojfiana y de Eugenia sp dispersadaspor tucanes y zorzales en el sudeste deBrasil (Guix y RuiL 1995) y con lassemillas ingeridas por Mimus polyglottosinfestadas con avispas (Nalepa y Piper,1994) o directamente podría implicar Iadestrucción de las semillas infestadas (Coey Coe, ¡987; Hauser, l994). Si bien elimpacto de carayá sobre cada una de lasespecies que consume es diferente, noobservé ni la dispersión de la infestación nila destrucción total de las semillas, sinoefectos indirectos más complejos.
Carayá provocaría un efectopositivo sobre la mayoría de las semillasinfestadas de 0. diospyrifolia. Pues nodispersa al insecto y, más aún, lo eliminamanteniendo la capacidad de germinaciónde las semillas. Ese patrón es similar al quereportan Coe y Coe (1987) para las semillasde acacias ingeridas por ungulados cuandola infestación por bruquidos es reciente. Detodas formas, el beneficio que el pasaje porel tracto de carayá pueda representar paraestas semillas sera inversamenteproporcional al tiempo que carayá tarde eningerir los fi'utos maduros, pues cuanto mástiempo pasen estos en el árbol mayor será eldesarrollo de la larva y, por ende, la
cantidad de endospenna que esta habráconsumido. Por el mismo motivo el aporteenergético que la larva pueda significar enla dieta de carayá será directamenteproporcional al tiempo que carayá tarde eningerir los frutos una vez maduros. Comocarayá consume los frutos desde quemaduran hasta que desaparecen de laplanta, el balance de esta relación particularentre 0. diospyrifolia y carayá comenzaríasiendo positivo para la planta, se volveríapositivo para ambos en algún momento delperíodo de madurez del fruto y pasaria a serpositivo sólo para carayá al estarfinalizando el periodo del fruto en la plantacuando la larva será de gran tamaño perolos restos de los cotiledones seránpequeños. El consumo de frutos en estaúltima etapa incluso podría volversenegativo para la planta pues se estariandispersando semillas con pocasprobabilidades de supervivencia una vezgerrninadas.
Sobre las semillas de B. argulacarayá tendría un efecto neutro. puesto quesi bien las avispas mueren al pasar por eltracto digestivo, cuando carayá ingiere losfrutos las avispas ya se han comido todo elendosperrna y al embrión de las semillasinfestadas. La dispersión de estas semillasno seria una desventaja para la planta puesno reducen la probabilidad de serdispersadas a las‘semillas sanas puesto quedentro de un mismo fruto hay variassemillas algunas infestadas y otras sanas.Más aún, la presencia de semillas infestadaspodría funcionar como un atractivo extrapara que carayá consuma sus frutos. Detodas formas, en esta especie la proporciónde frutos removidos por carayá no fue tanimportante como por ejemplo en 0.diospyrífolia o E. punicifolia, pues cuandoestaba en fruto no era visitada a diario, ni senotaba una merma considerable en laabundancia de frutos de una copa cuandoun grupo de carayá dejaba el árbol luego dealimentarse. Por otra parte fue en la quemas aves observé alimentarse.
De la infestación en E. pum'crfoliapuedo decir que mayormente las larvas soneliminadas luego del pasaje de los frutospor el tracto digestivo de carayá, pues seencuentran en la pulpa (ver Capítulo ll) y

Alouaua caraya como dispersor dc semillas 47
serian digeridas junto con esta. Sinembargo, las observaciones en el campoindican que esto no siempre sería asi,posiblemente en los últimos frutosconsumidos las larvas ya hayan alcanzadola semilla y, a diferencia de lo que ocurreen las otras especies, no serian dañadas porla digestión alli dentro. De ser cierta estahipótesis, al final de la fructificación carayáestaria dispersando ya no las semillas sinoal insecto que las infesta, pero ¿Se debeconsiderar a esto cómo un efecto negativo?Antes de responder debo considerar quecarayá parece estar teniendo ciertainclinación a incorporar a su dietaespecialmente estructuras infestadas (verCapitulo ll). Por otra parte, contrariamentea lo que ocurre con B. argula la mayorparte de los frutos de E. punicífolia sonremovidos por carayá cuando abandonan ungrupo de árboles del cual se alimentaron(obs. pers.). Esto podria implicar un altoimpacto sobre la población de insectos queinfestan esta especie, como se ha vistoocurre con la predación indirecta debrúquidos por parte de vertebradosherbivoros en Costa Rica (Herrera, 1989).Si carayá es un dispersor efectivo de E.punicifolía y la infestación alta de sus frutoses útil para atraerlo, es importante que estosinsectos continúen siendo abundantes
dentro del ecosistema. Por lo tanto, estaproporción de semillas "perdidas" no loseria tanto ya que podria considerarse una"inversión" a futuro que permitiría ladispersión efectiva de semillas en lapróxima fructificación.
En resumen, las semillasdispersadas por carayá en la selva deinundación llegan con capacidad paragerminar al suelo. En algunos casos lohacen con mayor rapidez y en aquellasespecies cuyas semillas estaban infestadas,la infestación es eliminada en gran partedando lugar a una serie de efectosindirectos y relaciones interespeclflcascomplejas. Las semillas son depositadas entres condiciones, heces independientes conun número variable de semillas, dormiderosy bosteaderos donde se acumulan engrandes densidades y donde lascaracteristicas ambientales para una semillaserian muy diferentes a cualquier área delsuelo del bosque. Ahora me resta estudiar silos bosteaderos y donnideros son sitiosfavorables para el establecimiento de estasespecies, qué ocurre debajo de los árbolesparentales de cada una de ellas, si laeliminación de la infestación es efectiva enel campo y se Ve reflejada en laregeneración de las distintas especies. Deesto tratará el siguiente capítulo.

Foto 7: Frutos de Ocotea diospyrifolia
Foto 9: Bosteadero puntual

Alarm/Iacaraya como dispersor de semillas 49
Capítulo IV:Impacto de carayá como dispersor local de semillas en el Bosque de
Laureles
INTRODUCCIÓN
Varios autores han destacado laimportancia de los frugívoros paramantener la heterogeneidad floristica de lasselvas (Lieberman et al., 1979; Harstshom,l980; Howe y Vande Kerckhove. |98|;Bourliere, ¡985; Heymann, l992; Julliot,[997). Lieberman et al. (1979) sugieren queel aumento en la densidad de las principalesespecies vegetales consumidas‘pór Papioanubr's en el área más utilizada por estos, sedebe precisamente a la dispersión desemillas que ellos generan, Milton (¡980)indica que algo similar ocurre con ladistribución agrupada del alimento deAlouarra paIIr'ara en Barro Colorado y a lamisma conclusión llega Julliot (l997)respecto a las especies dispersadas porAlouatla seniculus en la Guayana Francesa.
Sin embargo la mayoria de lostrabajos sobre dispersión de semillas poranimales, sólo describen patrones dedistribución de plántulas producido por eldispersor o los dispersores bajo estudio (e.g., Dinerstein y Wemmer, 1988; Tutin etal., l99|;- Valido y Nogales, 1994;Kobayashi e lzawa, 1997; Kobayashi et al,1999). Pocos trabajos evalúan la posibilidadde que estas plántulas - lleguen a serjuveniles de algunos años de edad, entreellos estarian los de Julliot (¡997) con A.seniculus en la Guayana Francesa, deForget ( ¡997) en el mismo sitio trabajandocon las semillas de Campa procera(Meliaceae) y Vouacapoua americana(Caesalpiniaceae) dispersadas por roedores,Fragoso (l997) estudiando la dispersión deMaximiliana maripo (Palmae) por Tapr'ruslerrestris en Brasil, y Rey y Alcántara(2000) trabajando sobre la dispersión deOlea europea que la aves fruglvorasproducen en España. Pero las característicasbiológicas de las especies dispersadas yproblemas logísticos derivados de lacomplejidad de los distintos ecosistemashacen prácticamente imposible evaluar el
efecto de un frugívoro para el conjunto deespecies que pueda dispersar, es decirevaluar el impacto de un determinadodispersor en la dinámica de un ecosistema.En este sentido el trabajo de Julliot (¡997)es el que más se acerca a ese objetivo,tratando de evaluar el establecimiento de Iamayor cantidad de especies dentro de lasdispersadas por A. seniculus. sin embargodebe limitarse a trabajar con especiesarbóreas, cuyas semillas no presentenmecanismos de dorrnición que le impidangerminar a la sombra, cuyas plántulas yjuveniles puedan '-ser fácilmenteidentificables en el campo y que estuvieranpresentes en al menos el 2% de las heces.De esta manera el número de especies se velimitado a 6 de un total de 39 que seencuentran en las heces de A. sem'culus.
El Bosque de Laureles de las islasdel Río Paraná es un' sistema simple con unbajo número de especies (Capítulo l), por locual pude trabajar con todas las especiesdispersadas por carayá, aunque no pudehacerlo con igual detalle para todas ellas.Esto me permitió realizar una evaluaciónmás realista acerca del pa‘pel que car-ayapue_da tener dentro del bosque comodispersor de semillas. Car-aya dispersa unagran cantidad de semillas con capacidadpara germinar y la mayor parte esdepositada en los bosteaderos, los cualesson utilizados por varios años (Capitulolll). Se ha observado que un patrón dedispersión de este tipo puede estaracumulando semillas principalmente ensitios favorables para la germinación,aumentando en gran medida la tasa dereclutamiento de las especies vegetalesdispersadas. Pero, también podriaobservarse lo contrario, que las semillasfueran acumuladas en áreas con escasasposibilidades de supervivencia (Wenny,2000; Schupp et al., 2002). En losbosteaderos varios factores pueden influir

50 Susana Patricia Bravo
positiva o negativamente en elestablecimiento de los juveniles (CapítuloVl). Por lo tanto, si el balance final de estosfactores favorece el establecimiento.debería encontrar en los bosteaderos grancantidad de juveniles y renovales dedistintas edades.
En el Capítulo lll encontré que enla isla Brasilera las semillas dispersadas porcarayá pueden ser depositadas debajo de losdormideros principales, en bosteaderospuntuales o en deyecciones independientes,y cada uno de estos sitios presentócaracterísticas particulares en cuanto atamaño y abundancia dentro de la isla. Porlo tanto para estudiar el establecimiento delos juveniles en cada uno de ellos deberíautilizar distintos métodos, así que en estecapítulo me centraré en el estudio delestablecimiento de las especies que fueroningeridas por carayá y que se encuentrandebajo de los dormideros principales y enlos bosteaderos puntuales del Bosque deLaureles, aunque también discutiréparcialmente que ocurre con los juvenilesque se desarrollan en las deyeccionesindependientes.
MATERIALES v MÉTODOS
Para determinar categorías deedades establecí los siguientes criterios:consideré árbol o enredadera juvenil a losindividuos menores a 25 cm de altura y sincotiledones. Considere renovales a losárboles mayores de 25 cm de altura noreproductivos, por lo tanto su alturamáxima y DAP serán distinta para cadaespecie (ver más adelante). Consideré comorenoval de enredadera aquellos individuosque teniendo un largo mayor a los 25 cm notenían el tallo lignifrcado.
Dado que el objetivo de la tesis esconocer el impacto que puede tener carayáen la dinámica y regeneración de la selva deinundación y no describir en que etapas desu vida los individuos de cada especietienen las mayores probabilidades demortalidad. El hecho de no trabajar conplántulü, tanto en este capítulo como en elCapítulo V, me facilitó alcanzar el objetivocon un menor esfiJerzo de muestreo sin por
eso afectar la calidad de la informaciónobtenida.
Establecimiento en las áreas dormidero
Coloqué 2 transectas a cada ladodel dormidero principal de cada uno de losgrupos observados durante este estudio y del grupo observada durante el año 2000 (P.Salomón, com. pers.). Las transectas partíandel tronco en direcciones opuestasseleccionadas al azar, tenían 9 m de largo y2 m de ancho (Fig. 14). En ellas conté atodos los individuosjuveniles y determiné aque especie pertenecía cada uno.
Para evaluar el impacto de ladispersión de semillas producida por carayáa lo largo de los años, utilicé transectassimilares pero de 4 m de ancho donde contéy determine específicamente los renovalesconsiderando las siguientes categorías dealtura: entre 0,25 y 0,50 m, entre los 0,5! my 1,00 rn, entre l,Ol y 3,00 m, mayores a3,00 m. Para Psychorria carrhagenensissólo consideré los individuos menores a 0,5m y para Eugenia punicífolia sólo aquellosque tuvieran un DAP menor a 3 cm. Losindividuos que superaban estas dimensionesya producían frutos.
Dado que los dormideroscorrespondían a distintas especies (2 B.argula, 2 Geofi'oea slrialla, l Albiziainundara) realice conteos de juveniles yrenovales con el mismo método debajo deotros 5 árboles de estas mismas especiesque no eran utilizados por los grupos decarayá para pasar la noche ("controles dedormidero“).
Con el mismo método evalué ladensidad de juveniles y renovales debajo de6 árboles adultos de Banara arguta y 6árboles adultos de Ocotea diospyrrfolia("control de parentales"), estas eSpeciesfueron seleccionadas por ser dispersadaspor carayá en gran cantidad (ver Capítulo lly lll) y tener un diámetro de copa similar alde los árboles dormideros. Para evaluar alas otras 2 especies dispersadas en grancantidad por carayá e importantes en elbosque por su abundancia (P.carthagenensis y E. punícifolía) utilicé lastransectas que se hallaban debajo de 0.diospyrífolia como controles de parental,

Alouana caraya como dispersor de semillas
debido a que pertenecen al estrato bajo delBosque de Laureles y son muy abundantesdebajo de las copas de esta especie (verCapitulo l). En este caso ya no se trataba deun único árbol parental del cual lastransectas se alejaban, sino transectascubiertas por la copa de distintosindividuos.
Tanto para juveniles como pararenovales evalué la riquem de cadadormidero y control de dormidero, con esosvalores calcule la riqueza promedio.También estimé la riqueza acumulada comoel número total de especies que encontrédebajo de los dormideros y de los controles,considerando tanto a los juveniles como alos renovales. Esta última es un reflejo delefecto en el Bosque de Laureles de ladispersión de semillas que realiza carayá.mientras que la primera me permitedescribir con más detalle este efecto.
Evalué la densidad de juveniles yrenovales en los dormideros y en loscontroles de dormidero considerando elconjunto de especies encontradas, y alcomparar con los árboles parentales sóloconsidere los de la especie bajo evaluación.En todos los casos promedié las 2 transectasde cada árbol. Evalué las diferencias en ladensidad de juveniles mediante una pruebade t’ de Student. Para evaluar lasdiferencias en la densidad de renovalesentre dormideros y controles considerandolas distintas categorias de altura utilicé unaprueba de Anova de 2 factores,transformando los datos con raiz cuadrada.Los contrastes a posteriori los realicé conpruebas de Tukey. Para evaluarestadísticamente la riqueza promediodebajo de los dormideros y de los controlesde dormidero utilicé una prueba de t’ deStudent para losjuveniles y una prueba detde Student para los renovales. Para P.carthagenensis utilicé una prueba de U deMann-Whitney, ya que sólo consideré unacategoría de altura para esta especie. Nopude comparar estadísticamente lasdensidades de renovales de B. argutadebajo de los dormideros con lasdensidades debajo de los parentales debidoal escaso número de individuos.
Figura 14: Ubicación de las transectas queutilicé para el conteo de los juveniles y losrenovales que creclan debajo de los árbolesdormideros, los árboles controles y los árbolesparentales. El circulo pequeño representa lasección del tronco y el circulo mayor laproyección de la copa.
Para evaluar el impacto de ladispersión en los bosteaderos puntualesconté todos los juveniles presentes en l7.bosteaderos de 3 grupos distintos de carayáy en l7 áreas control. Para evaluar ladensidad de renovales realicé los conteos en¡6 bosteaderos y lO áreas control.
Establ i lento lo steadepuntuales
Determine y marque un centro encada bosteadero (donde se observaba lamayor concentración de heces) y medi ladistancia de cada individuo a este centro.Prómediando las distancias del individuomás distante de cada bosteadero obtuve undiámetro de 5 m, que utilicé paradeterminar el diámetro de las áreas controlubicadas al azar en el bosque, con la únicacondición de no contener bosteaderos.
Para los juveniles y renovales decada especie calcule dos parametros, elporcentaje de bosteaderos o áreas control enque estaba presente (Presencia) y el númeropromedio de individuos en cada una deestas áreas (Abundancia). Al igual que paralos dormideros evaluó la riqueza'de cadabosteadero y área control y calcule lariqueza acumulada y la riquem promedio.
Dado que la luz puede determinarla distribución de los juveniles dentro delbosque (Clark y Clark, 1992; Nicotra et aL,¡999), estime la abertura del dose] en lO

52 Susana Patricia Bravo
bosteaderos puntuales y en 6 controlestomados al azar utilizando el "Indice deiluminación de la copa" (Clark y Clark,l992) y el método del "canopy-scope"("Moosehom" modificado, Brown et al.,2000). También estime’ en cada sitio lacobertura del suelo por parte de lasherbáceas, considerando 5 categorías,100%, 75%, 50%, 25%, 0% decobertura.
Para caracterizar a losjuveniles quecreclan en cada sitio medi la altura, conté elnúmero de hojas y medi el largo de la hojamayor de los juveniles que creclan en 6bosteaderos y 6 áreas control seleccionadosal azar.
Evalué la diferencia del número dejuveniles, renovales y riqueza promedioentre bosteaderos y áreas control conpruebas de U de Mann-Whitney. Realicecorrelaciones de Speannan entre el númerode juveniles y la cobertura de doselestimada por cada método, y entre elnúmero de juveniles y la cobertura deherbáceu. Realicé las mismas correlacionescon la riqueza de cada bosteadero.
Para evaluar la altura de losjuveniles entre sitios (bosteadero y control)y especies (O. diopsyrifalía, E. punicifoliay P. carlhagenensis) utilicé una prueba deAnova de dos factores. Los datos fuerontransformados aplicando logaritmo.
El número de hojas de los juvenilesde cada especie que crecía en losbosteaderos o en las áreas control lo analicepor medio de pruebas de t de Student o deU de Mann-Whitney.
El largo de la hoja mayor de losjuveniles de las distintas especies que seencontraban creciendo en los bosteaderos oen las áreas control lo evalué con unaprueba de Anova de dos factores.
RESULTADOS
Establecimiento en Igs áreas dormidero
La densidad de árboles juvenilesdebajo de los dormideros fue mayorrespecto a la densidad debajo de loscontroles de dormidero (t’ = 2,74; gl = 5,44;
lndividuos’m2
ÉDonnidero Control
n
Figura 15: Densidad promedio de juvenilesdebajo de los árboles donnidero y debajo de losárboles control.
Indivrduos'm:
o
...1151-11!) 1.0! - 3.00 > 3.00
I Allun(m)
Figura l6: Densidad promedio de ren0valespertenecientes a cada categoria de altura debajode los árboles donnidero (gris) y debajo de losárboles control (blando).
o .. ,DJS-0.50
P= 0,04; Fig. ¡5). Al evaluar la densidadde renovales considerando las distintascategorías de altura encuentro diferenciassignificativas tanto entre sitios como entrelas alturas pero no en la interacción (Tabla¡2, Fig. l6). Respecto a la riquezaacumulada encontré un total de ¡2 especiesen los dormideros mientras que 9 especiesestuvieron bajo los árboles control (Tablal3). Pero, como no todas ellas estaban entodos los dormideros, no hubo diferenciassignificativas en cuanto a la riquezapromedio de juveniles (t’ = 0,69; gl = 6, [2;P= 0,5l; Fig. l7). Sin embargo, mientraslos dormideros mantienen su riquezapromedio cuando evalúo los ren0vales, enlos controles de donnidero la riquezapromedio disminuye marcadamente,difiriendo (t = 4,15; gl = lO; P = 0,002; Fig.l7).
Los juveniles de B. arguta tuvierondensidades muy bajas con una gran

Aloualla caraya como dispersor de semillas 53
Tabla ¡2: Resultados del Anova de dos factorescomparando la densidad de renovales debajo delos dormideros con la densidad de renovalesdebajo de los controles para las distintascategorías de altura (0,25 - 0,50 m, 0.5| 1,00 m, |,0l —3,00 y > 3 m). CM = Cuadrados
medios.
Factores gl CM F
Sitio (S) l 7,56 56,62 <0,0000|Altura (A) 3 0,96 7,20 0,0007
S x A 3 0,l4 ¡,03 0,37Error 36 ¡3,35
variabilidad, tanto debajo de los dormideroscomo debajo de los árboles parentales, perola densidad fue mayor debajo de los árbolesparentales (Fig. ¡8 a). Al evaluar ladensidad de renovales observé queraramente se los encontraba,indistintamente que muestreara debajo delos árboles parentales o debajo de losdormideros (Fig. ¡8 a).
Encontré juveniles de 0.díospyrrfolia (Foto lO) en mayor densidaddebajo de los dormideros que de losparentales (t’ = 3,09; gl = 18,60; P = 0,006;Fig. l8 b). Respecto a los renovales de estaespecie fue significativa la interacción entrelas densidades de renovales de distintasalturas y el lugar donde se encontrabancreciendo (dorrnidero o árbol parental),(F = 3,|4; gl = 3; P = 0,037), los renovalesentre l - 3 m y los mayores de 3 rn fueronsignificativamente más abundantes debajode los dormideros (Prueba de Tukey,P < 0,05; Fig. 18 b).
Los juveniles de E. punicrfolia(Foto ll) tuvieron una densidad promediomayor debajo de los árboles donnidero,pero las diferencias fueron no significativas(t' = 1,72; gl = 5,31; P =0,l4; Fig. 19 a).En cuanto a los renovales, hubo diferenciassignificativas entre las distintas categoriasde altura (F = 5,19; gl = 3; P = 0,004), losrenovales mayores de 3 metros fueronmenos abundantes respecto al resto de lascategorías (Prueba de Tukey, P < 0,05; Fig.l8 a). Si bien la densidad de renovales fuemayor en los dormideros no detecte
lndellDi/I'llUO oo
Figura l7: Riqueza promedio de juveniles yrenovales en los dormideros (gris) y loscontroles de donnidero (blanco)." P = 0,002 (Prueba de t de Student).
diferencias significativas (F = 1,26; gl = l;P = 0,27). La interacción entre la altura y elsitio de crecimiento fue no significativa(F = 0,78; gl = 3; P = 0,51).
Las densidades de juveniles de P.carlhagenensis (Foto l2) debajo de losdormideros fueron mayores a las queencontré debajo de los parentales pero nohubo diferencia estadísticamentesignificativas (U = 5,00; N = S; N = 5;P = 0,12; Fig. l9 b), en cuanto a losrenovales tuvieron densidades similaresdebajo de los dormideros y debajo de loscontroles (U = 3,00; N = 4; N = 5; P = 0,08;Fig. l9 b).
Establecimiento en los bosteaderospuntuales
El -número de individuos porbosteaderos fue mayor al número deindividuos por áreas control tanto para losjuveniles (U = 23,5; N = l7; N = 17;P = 0,00003; Fig. 20 a) como para losrenovales (U = 5,5; N = lO; N = 17;P = 0,00008; Fig. 20 a). En los bosteaderosencontré una riqueza acumulada de l4especies, mientras que para las areas controlfue de l0 especies (Tabla l4). La riquezapromedio fue significativamente mayor enlos bosteaderos que en las área control paralos juveniles (U = 52,5; N = l7; N = l7;P= 0,001; Fig. 20 b) y para los renovales(U = l4,00; N = IO; N = l7; P= 0,0005;Fig. 20 b). Considerando juveniles yrenovales en conjunto, en los bosteaderos

54 Susana Patricia Bravo
Tabla 13: Números dejuvenilcs (J) y renovales (R) de cada especie conmbilizndos bajo los árboles dormideroy bajo los árboles control de donnidero. La última fila conesponde al número total de especies en cada sitio(Riqueza acumulada).
Dormideros ControlFami Especies J R J R
Lauraceae Ocotea diospyrifolia 238 107 77 ¡2Nectandra angusnfolia 2 3 _ _
Fabaceae Inga vera 3 13 _ 2Myrtaeeae Eugenia punicifolia 317 l90 9] IS
Eugenia pyriformis Psidium 9 _ 4 lguajaba 3 _ _ _
Flacourtiaceae Banara argula 2 5 83 4Sapotaceae Pouteria salicifolia 2 2 _ _CecrOpiaceae Cecropia pachysrachya 23 ¡5 lO 4Rubiaceae Psycholria carthagenensis ¡065 87 66 4Vitáceae Cissus palmata 6 4 6 2
Sapindáceae Paullím'a spp'. l l 7 3| lRigueza
l2 9acumulada
Tabla 14: Números de juveniles (J) y renovales (R) de cada especie contabilizados en los bosteaderospuntuales y en las áreas control. La última fila corresponde al número total de especies en cada sitio (Riquezaacumulada).
Dormideros ControlFamilia EW.“ J R J R
Lauraceae Ocotea diospyrifolia 943 43 24 8Nectandra angustifolïa 2 3 0 l
Fabaceae Inga vera 12 5 6 2Myrtaceae Eugenia pum'cifolía 53l 67 26| 40
Eugenia pyriformis 2 I 4 0Eugenia burkarlíana 5 24 2 0Psídium guajaba 2 l l lSpl l 0 0 0
Flacouniaceae Banara arguta 0 I l OSapotaceae Pouleria salicifolía 7 4 l 0Cecropiaceae Cecropia pachyslachya 0 IO 0 0Rubiaceae Psychorria carlhagenensis 629 IOO 319 40
Vitáceae Cissus palmala 4 0 0 0
Sapindáceae Paullínia spp. 60 0 0 0nguezaacumulada

Aloualla caraya como dispersor de semillas 55
RENOVALES
a Banara argula
MMM .0.8B m
0.40 %0 oo Demideml’mmd
JUVENILES
0,1540” 0.5l-l .00 Mil-LN >3Alan (In)
MM"?
OO R8
0.30.50 0.5l- I.W LoI4.00 >3
Ahull (In)
Figura ¡8:Densidad promedio de juveniles y renovales de las distintas alturas, debajo de los árbolesdonnidero (gris) y de los árboles parentales (blanco), para Banara arguta (a) y Ocotea diospyrifolia
de las l4 especies 8 tuvieron una presenciamayor al 20 % (Fig. 21 a), mientras que enlas áreas control de las lO especies sólo 3superaron el 20 % de presencia y suabundancia fue notoriamente inferior a la quetuvieron en los bosteaderos (Fig. 2] b). Losjuveniles de E. puniclfolia y P.carrhagenensís estuvieron presentes en el100 % de las áreas bosteadero y control. Losjuveniles de 0. diospynfolia estuvieronpresentes en el l00 % de las áreas bosteadero(Fig. 2| a) y en el 56,7 % de las áreas control(Fig. 2| b).
La altura de los juveniles de E.punícifalia, 0. diospyrifolía y P.carrhagenesís fue mayor en los bosteaderospuntuales (F = 12,12; gl = l; P = 0,002; Fig.22 a), no hubo diferencias significativas entrelas especies (F= 1,87; gl = 2; P=0,l7), ytampoco en la interacción (F.= 0,36; gl =2;
P= 0,70). La hoja mayor fue más larga enaquellos individuos que crecían en losbosteaderos (F = 23,77; gl = l; P < 0,00001;Fíg. 22 b), También hubo diferencias en ellargo de la hoja mayor entre las distintasespecies (F = 12,22; gl = 2; P = 0,0002). Sinembargo. la interacción no fue significativa(F = 0,46; gl = 2; P = 0,6). Los juveniles delas tres especies que crecian en ambos sitiosno difirieron significativamente en el númerode hojas_(Fig. 22 c). Además, debo destacarque tanto la altura como el largo de la hojamayor fueron menos variables para losjuveniles que crecian en los bosteaderos (Fig.22 a y b). '
La densidad de juveniles no estuvocorrelacionada con la abertura del doselestimada por los dos métodos (Índice deiluminación R, = 0,049; P = 0,85; Canopy

Susana I’alncna Bravo
JUVENILES
a Eugenia punicifolia
Individuos’m:
b Psychotria carthagenensis
¡000
lI'IdIVIduosliInz
É .1¡manila ¡anda
scope R, = 0,30; p = 0,24; Fig. 23 a) ni con lacobertura del sotobosque mi = 0,36;P = 0,20; Fig. 23 a). La riqueza sl estuvocorrelacionadas con la abertura del dose](Índice de iluminación R. = 0,45; p = 0,07;Canopy-scope Rs= 0,58; P = 0,02; Fig.23 b), aunque no con la cobertura delsotobosque (R. = 0.42; P = 0.13; Fig. 23 b).
DISCUSIÓN
Dentro del Bosque de Laureles tantolos donnideros como los bosteaderospuntuales representaron áreas con un altonúmero de juveniles y renovales, y una altariqueza. Es decir que carayá estaria ayudandoa mantener la diversidad del bosqueposibilitando el establecimiento de gran partede las especies que lo componen. Losbosteaderos de carayá parecen ser sitiosfavorables para el crecimiento de losjuveniles de al menos las tres especies másabundantes, 0. diospyrifolia, E. punicrfolia yP. carrhagenensis, pues los individuos quealll crecen tienen mayor altura y hojas demayor tamaño.
RENOVALES
Individuos/mi
Alu. (m)
Figura l9: Densidad promedio de juveniles yrenovales de distintas alturas debajo de losárboles donnidero (gris) y debajo de los árbolesparentalcs (blanco) para Eugenia punic'ifolia (a) yPsycholria carlhagenensis (b). Para esta últimaconsideré únicamente la primer categoría deren0vales pues los individuos más altos yaproducían frutos.
Si bien los juveniles ya no pueden serconsiderados plántulas, de todas formasrepresentan las primeras etapas delcrecimiento y a eso se debe tanto su altadensidad promedio como la granvariabilidad, pues son más sensibles a laherbivoría y están más ligados que losrenovales al número de semillas que llegarona cada sitio durante las últimasfructificaciones. Los renovales representan alos sobrevivientes de varios años, por eso sumenor variabilidad ya que no existe unarelación tan directa con la cantidad desemillas que llegaron a cada sitio, ni sondemasiado sensibles a la herbivoría. Esto eslo que se conoce como desacople de losestadios de la dispersión de semillasproducida por los vertebrados, es decir dadoque hay distintos procesos operando en cadaestadio, a medida que se alejan en el tiempoaumenta la independencia entre ellos,pudiendo volverse completamenteindependiente uno de otro (Jordano yHerrera, 1995).
La alta varianza de la densidad dejuveniles en los donnideros podría estar dadaen parte por una distribución contagiosa

Alouaua caraya como dispersor de semillas S7
dentro del agrupamiento que ellosrepresentan dentro del bosque. Comodescribe Julliot (1997), la dispersión de losaulladores se caracteriza por un patrónagrupado a distintas escalas, en el bosquezonas de alta concentración de
Jumla ¡ande!
Instala Ronda
Figura 20: Número promedio de individuos (a) yriqueza promedio (b) que registró en los conteosdc juveniles y renovales en los bosteaderospuntuales (gris) y en las áreas control (blanco).
semillas alrededor de los donnideros, en eldonnidero agrupaciones menores formadasdebajo de las ramas desde las que los monosdefecan (bosteaderos) y dentro de estos lasagrupaciones más pequeñas que representanlas heces. Asi alrededor de los donniderosexisten zonas con densidades muy altas yotras con densidades más bajas, por lo tantoal colocar las transectas al azar estas podíanincorporar zonas con distinto número dejuveniles. Sin embargo, en los bosteaderospuntuales la alta varianza que registró nopuede deberse a un efecto del muestreo, puesen ellos conté todos los juveniles'presentes.En este caso la variabilidad se deberiaexclusivamente a factores tales como unallegada diferente de semillas a cadabosteadero dada por su frecuencia de uso,una diferencia en la tasa de remoción desemillas o en la tasa de mortalidad de las
plántulas. En el caso de los bosteaderospuntuales la llegada de luz y la posiblecompetencia con las herbáceas, no han sidofactores importantes en la determinación delnúmero de juveniles, a diferencia de loencontrado en Gabón, donde la cobertura dela vegetación y la desecación pueden serfundamentales para el éxito de lagerminación de las semillas dispersadas porlos gorilas (Rogers et al., 1998). Sinembargo, el número de juveniles puede serafectado por otros factores bióticos yabióticos como la herbivorla (Schupp, 1988),los patógenos (Augspurger, l984) o ladisponibilidad de nutrientes (Burslem et al.,1996) que sl podrían en este caso estarinfluyendo sobre el número de juvenilespresente en los bosteaderos puntuales decarayá (ver Capítulo Vl). La abertura deldosel fue importante en relación con lariqueza que podia encontrar tanto en losbosteaderos como en las áreas control,estando positivamente correlacionadas.integrando los resultados del número deindividuos y de la riqueza respecto a lallegada de luz puedo suponer que bosteaderosque difieran sólo por la llegada de luz tendránun número similar de individuos pero lariqueza será menor en aquellos con menorluminosidad, esto no es necesariamente asien otros sistemas donde la composiciónespecífica de juveniles en un área dada puedevariar con la luz pero esto no implica que lohaga su riqueza (Dalling et al., 1998; Nicotraet al., ¡999).
La baja varianza en el númeropr'omedio de los renovales indica que entodos los bosteaderos puntuales y donniderosel número de individuos que logranestablecerse es similar, y lo más importantees que siempre este número es mayor al queencuentro en los controles. Al analizar ladensidad de los renovales de los donniderosteniendo en cuenta su altura, para todas lascategorías las densidades son mayores que enlos controles, lo que implica que elestablecimiento se ha'ido produciendo a lolargo de los años y no que se ha dado poralgún evento particular. Es.

C-awhwww
58 Susana Patricia Bravo
¡amm
PIM l”
NW ¡mmMmm 1
¡nos veuParfumpam
CluulpumEugeniahumana
Euoulamm.
N ¡a
Presencia (‘36)
WI ¡"Bum
20 Ao no oo 130 ¿o no
Abundana‘a (indlbosteadero)
s {o ¡o
Abundancia (Ind/área)
Figura 2|: Presencia y abundancia de juveniles (lisos) y renovales (rayas) en los bosteaderos puntuales delBosque de Laureles (a) y en las áreas control (b).Presencia = porcentaje debosteaderos en donde encontré individuos de una especies.Abundancia = Número promedio de individuos por bosteadero.Observar el mayor número de especies, con mayor presencia cada una de ellas y mayor abundancia cn losbostmderos puntuales.Nota: tener en cuenta el cambio de escala cn cada grafico.
decir, la dispersión de semillas que camyá harealizado en los últimos años ha sidoefectiva, permitiendo el establecimiento deespecies que no logran establecerse en otrmzonas del bosque. Sin embargo, no se debeconfundir este patrón con una mayorprobabilidad de éxito para las semillas quearriban o con una menor mortalidad, puespuede deberse simplemente al mayor númerode semillas que llegan
La abundancia de juveniles yrenovales de cada especie que creclan comoefecto de la dispersión de semillas producidapor carayá, fue similar a la que mantenían losadultos de esas especies en el Bosque deLaureles (ver Capítulo l). Al encontrarpatrones similares algunos autores los
consideraron como una prueba del efecto dcldispersor sobre la estructura de la vegetación(Liebennan et al., l979). Sin embargo.considero que esta asociación no puededeterminar una relación de causa efecto puessimplemente puede representar que carayáestá alimentandose de los fi'utos en igualproporción a la que estos son ofrecidos por elambiente y de esta manera su efecto comodispersor es simplemente mantener laestructura y composición florística delbosque y no generarla. Para con0cer como segenera esta estructura debo realizar estudiosdurante la formación del bosque por ejemploen la isla sobre los albardondes más nuevos oen islas de reciente formación, y determinarallí el impacto de carayá (ver Capítulo V).

Alaualla caraya como dispersor de semillas 59
El impacto de carayá en laregeneración de la selva de inundación puedeser aun mayor al que observé, pues por unlado debe sumarse el efecto de lasdeyecciones independientes que no evalué(ver Capitulo lll), y por otro los juveniles yrenovales que pudieran originarse a partir deestas deyecciones podrian estar incorporadosa los distintos controles de
a
_z; l i i5.. ag .
eL
2
l l; ¡mnlrlfinlln I'.eminem”! 0. ¡[hannfilra
bll
A ro
É l
Éó2 III
31 4 II' e
É z
L'punklfinllal f. (WW! ll hpplfidlo
O
unaun
e É É
ti i
P."Wikun I'.WII!!! 0.dannfipha
Nin-edeth
a
Nu
nv
Figura 22: Altura, número de hojas y largo de la hojamayor de los juveniles de E. punicifolia, P.carthagenensis y O. diospyrifolia que creclan en losbosteaderos puntuales (gris) yen las áreas control(blanco).
mis mediciones. Al Seleccionar las áreascontrol pude observar que juveniles aisladosde dos de las especies presentes en los
controles (0. diospyrifolr‘a y E. punicr'folia)muchas veces provenían clararhente dedeyecciones independientes, pues seencontraban creciendo entre restos de materiafecal, por supuesto en ese caso los controleseran desechados. De la observación yseguimiento de 3 deyecciones independientesencontré que mientras la materia fecaldesaparecla por completo en pocos meses, lassemillas germinaban, y las plántulassobrevivían transformándose en juveniles queal menos alcanzaron los 3 años de edad. Lascaracteristicas de estos juveniles "fueronsimilares a las de los juveniles de las áreascontrol (P. carthagenensís (l individuo):altura 7 cm, número de hojas 3 y largo de lahoja mayor 4 cm; E. punicr'folr'a: altura6,4 cm, número de hojas promedio 3,4 ylargo de la hoja mayor 2,2 cm). Estasobservaciones concuerdan con la idea de queparte de los juveniles contabilizados en lasáreas control se encuentren alli también porefecto de la dispersión de semillas producidapor carayá, pues al no existir restos demateria fecal las áreas no serian rechazadascomo controles. Para determinar con mayorexactitud el impacto de carayá en laregeneración del la selva de inundacióndebería realizar un seguimiento en el tiempode las semillas que llegan al suelo en lasdeyecciones independientes.
Si bien la riqueza acumulada en losdonnideros y en los bosteaderos puntualesfue levemente mayor al número total deespecies presente en los controles, existengrandes diferencias entre estos sitios. Pues, elnúmero de especies que puede encontrarse enun único bosteadero o dormidero sl essignificativamente superior al que puedeencontrarse en una única área control, peromás notorio aún es que, como indican losvalores de presencia, cada especie .esencontrada con mayor frecuencia en losbosteaderos que en las áreas control y, tantoen los bosteaderos puntuales como en losdonnideros, están representadas por _unmayor número de individuos. Todo esto hacede los bosteaderos puntuales y donniderosáreas de alta diversidad dentro del Bosque deLaureles.
Hubo especies para las cuales en losbosteaderos sólo registre la presencia derenovales (e. g. C. pachystachya y.B. argura,

60 Susana Patn’ciu Bravo
Fig. 21 b). Estos estuvieron en bajo número ytodos tenían aproximadamente la mismaaltura, lo que hace suponer que para estasespecies la dispersión y/o el establecimiento
a
omo
son
540072:00
amoIz o 0
loa o o 9 °n ' 9 ° ’ °o) H 25 U 43 3\
Madedunm
wo 4o
sm
¿400l 1
im" 'i¿200 ° o
¡oo .0 o o ° 0 o 4' oo. on .o n n ¡2 u. zu(mw
oooa
sao
34002300azen)Iz o o
tuo o| . an
0 1|.) JU 60 IO lo“ l0
Columndemas: (’/.)
están asociados a eventos particulares, ya seapor factores abioticos como podrian ser añosespecialmente
1.1
O65
055
0 O549E 0 O 0
\5O O 0 0
25O Oli
U.‘ l‘ 23 .‘3 43 ‘1¡Mandequ
150
65O
.‘5
. o ognx o oo oH
¡o o o o15
i oIS
u 4 l l! lo 10
(Iron-mv:15
O65
0.13
.. o15
3 ..¡5
01.1
lS Lu zu ¡u oo no Iuo lu
Cuban-I de¡[aunque 19-)
Figura 23: Correlación entre el número de individuos juveniles (a) y la riquen (b) de los bosteaderos(círculos blancos) y de las áreas control (círculos negros) con la cobertura de dosel evaluada con el Indice deiluminación. el método Canopy-scope y con la cobertura del sotobosque.
lluviosos o secos. o por factores bioticos.como una fi'uctificación extraordinaria de laespecie o una baja oferta de frutos del restode las especies del bosque. Un caso similar aestos es el de Tetragaslris panamensis en lalsla de Barro Colorado, ya que Alouatlapalliata fue el visitante más frecuente de laespecie y quien más semillas trasladó,durante un estudio de dispersión de semillasllevado a cabo durante 1978 (Howe, ¡980).Sin embargo según los estudios de dietallevados a cabo por Milton durante variosaños en ese sitio (Milton, l980) los frutos deT. panamensis no estaban incluidos en la
dieta de A. palh'ata, al parecer esta especieera consumida sólo en aquellos años en losque producla gran cantidad de frutos.
Cecropía pachystachya como todaslas especies de su género se encuentrageneralmente asociada a los claros pues sussemillas presentan mecanismos de dormiciónque son desactivados por la luz directa(Vazquez-Yanes y Smith, l982). Esta puedeser la razón por la cual no encuentrojuveniles y sus renovales los observé en unbajo porcentaje de bosteaderos a pesar de quesus semillas llegan a todos ellos manteniendo

Alouaua caraya como dispersor de semillas 6|
su capacidad de germinación (ver Capítulolll).
Para 0. diospyrifolia la dispersiónproducida por carayá tendría un efectosumamente importante. De los 1503juvenilescontabilizados el 87,3% crecían en losbosteaderos y dormideros de carayá, y de los188 renovales el 80,8%. Los juveniles yrenovales contabilizados debajo de losárboles parentales representaron apenas el8% y el 3 % respectivamente. El resto,aproximadamente un l0 % de losjuveniles yl7 % de los rcnovales, fue contabilizado enlos distintos controles, los cuales puedenestar incluyendo tanto individuos creciendoen deyecciones independientes de carayá,individuos creciendo a partir de semillasdispersadas por otros medios o bien desemillas caidas de algún árbol parental. Porlo tanto es claro que cualquier modificaciónen la abundancia de carayá dentro del Bosquede Laureles implicaría para 0. diospynfoliauna importante reducción en el reclutamientode individuos pudiendo llegar a perder ladominancia dentro del bosque. Posiblementeparte de este aumento en el reclutamientocomo efecto de la dispersión de semillas querealiza carayá se deba a los efectos deeliminación de larvas descriptos en elCapítulo lll. En Costa Rica para Ocoteaendresiana, observaron que una proporciónimportante de las semillas eran predadas bajolos árboles parentales por Heilipus sp. y otraespecie de curculiónido, y para elreclutamiento de nuevos individuos seríafundamental la dispersión de semillas
realizada por los pájaros campana (Prom'asrricarunculara) que les permite escapar deesta predación (Wenny, 2000).
Al analizar las caracteristicas flsicasde los juveniles de 0. diospyrifolia, E.punicifolla y P. carthagenensis que crecíanen los bosteaderos puntuales con aquellosque crecían en las áreas control, encontré quelos juveniles de los controles eran más bajosy tenian hojas más pequeñas. Esto puederelacionarse con un mayor vigor de losindividuos que crecen a partir de las semillasdispersadas por carayá, como ha sidoreportado para las semillas dispersadas porlos rinocerontes en Nepal (Dinerstein yWemmer, ¡988) o para las semillas deMyrtus commum'sdispersadas por mamíferos(Traveset et aL, 200l, en Traveset y Verdú,2002). Esto se deberia a que crecen con unadisponibilidad mayor de nutrientes aportadospor las heces (Traveset y Verdú, 2002).
Los resultados indican un incrementode la diversidad por un aumento del númerode especies de juveniles que pueden estarcreciendo en el suelo de un bosque. Enconclusión, carayá es un dispersor efectivoen lo que hace a la dispersión local de lamayoria de las especies que componen elBosque de Laureles. Esto no significa quepueda tener un papel importante dentro de laisla como dispersor de larga distancia,facilitando la llegada de especies del Bosquede Laureles a los otros tipos de bosque menosdiversos. Esto es lo que evaluará en elsiguiente capitulo.
Yoya sabía aa que nos contó,que las laureles crecen dela bostade los monos
Rali(7 años, l. del Canita)

Foto 10:Juveniles de Ocotea diospyrifolia
Foto ll :Juvenile de Eugeniapunicifolia
Foto 12:Juvenile de Psychotria carthagenensis

AIauaIIa car-aya como dispersor de semillas 63
Capítulo V:Impacto de carayá como dispersor de semillas de larga distancia
"...reflexianen, ningún ave come las semillasdemasiado pequeñas. que aunque las comierano las transportaría a distancias muylejanas... "
Felix de Azara (Viajespor la América meridional)
INTRODUCIÓN
La dispersión de larga distancia esdefinida como aquella que permite lacolonización de nuevos hábitats y Iaexpansión en la distribución geográfica deuna especie (Augspurger y Kitajima, 1992;Clark et a|., l999). Si bien se considera quelos eventos de dispersión de larga distanciason raros y están asociados a bajasdensidades de semillas, la dispersión desemillas de larga distancia sería un factordominante en el control de la dinámicaespacial de las poblaciones vegetales(Bullock et a|., 2002). Este tipo de dispersióndeterminaría la velocidad y el patrón deexpansión en la distribución de una especie(Shigesada y Kawasaki, 2002) ya sea a escalalocal, de paisaje o geográfica (Bullock et al.,2002). Por ejemplo, uno de los pocos trabajosque registró y cuantificó la dispersión delarga distancia (Fragoso, 1997), encontró quelos renovales de entre l y 5 años de lapalmera Maximiliana maripa se agrupabanprincipalmente en bosteaderos de tapires quese encontraban aproximadamente a 2 Km dedistancia de los árboles adultos de la especie,a pesar de que las semillas eran dispersadasen igual o mayor proporción por otrosdispersores a sitios próximos de los árbolesparentales.
La expansión de la distribución, acualquier escala, de una especie mayormentees estimada mediante simulaciones queconjugan modelos de dinámica poblacionalcon modelos de dispersión (Shigesada yKawasaki, 2002). Por ejemplo, Parker (2000)genera un modelo para simular la invasión
del arbusto Cystisus scoparius en pastizalesde Festuca spp., resultando que de todos losparámetros demográficos que incluye, el queejerce mayor influencia sobre la tasa deexpansión de la población es la distanciamáxima de dispersión. Lo mismo encuentranNeubert y Caswell (2000) con modelosdesarrollados para dos herbáceas (Calatheaovandensis y Dipsacus sylveslrís), y similaresson los resultados de Woolcock y Cousens(2000, en Bullock et al., 2002).
Existen diversas técnicas para elestudio de la dispersión en el campo, perotodas ellas dificilmente se pueden aplicar a ladispersión de larga distancia y cuando se hacesuelen tener grandes errores (Shigesada yKawasaki, 2002). Los métodos más comunespara construir las curvas de dispersión son lastrampas de semillas y el conteo de plántulasen el campo, a distintas distancias de lafuente de semillas. La distancia máxima dedispersión en general es subestimada, pueslos muestreos se realizan desde la fuente desemillas, pero ¿Hasta que distancia se debencolocar trampas? ¿Cuál es el origen de lassemillas o plántulas que se encuentran a lasdistancias más largas? ¿Cuál fue el dispersorque las depositó allí? (Greene yCalegoropoulos, 2002). Probablemente, ladispersión de larga distancia no sea un eventotan extraño como se cree, sino que está pocoestudiado y en muchos casos con métodosincorrectos, pues las trampas de semillas nologran capturar la mayor parte de la.dispersión provocada por vertebrados, porquees discontinua en tiempo y espacio, aunque.

64 Susana Patricia Bravo
este grupo suele ser el responsable de ladispersión a mayores distancias (Clark et al.,l999). Por lo tanto, las estimaciones sobre lacolonización de nuevos hábitats por parte deuna especie estarán influenciadasmarcadamente por el diseño de muestreoutilizado para estimar la maxima distancia dedispersión (Bullok et al., 2002). Paradisminuir los errores se recomienda estudiar
este tipo de dispersión mediante laobservación directa y seguimiento de losdispersores, habituando a estos a la presenciadel observador, conociendo susdesplazamientos, el uso del espacio, suspatrones de alimentación y defecación(Greene y Calegoropoulus, 2002; Schupp eta|., 2002; Williamson, 2002).
Dados los problemas de medición‘ yel hecho de que las distancias de dispersiónvarían ampliamente dependiendo de cadasistema planta-dispersor-ambiente, no se hafijado una distancia crítica mínima paraconsiderar que se está ante un evento dedispersión de corta o larga distancia (Greeney Calegoropoulos, 2002). Por ejemplo,algunos autores consideran distancias largasde dispersión a partir de los 100 m(Augspurger y Kitajima, 1992), mientrasotros ya lo hacen a partir de los 30 m (Clarket aL, 1999). Dada la particular estructurafisonómica de la Islas Brasilera (Capitulo l),consideraré dÍSpersión de corta distancia a laproducida dentro de un mismo bosque(Capitulo lV) y dispersión dc larga distanciaaquella que penniten la llegada de especiesde un tipo de bosque a otro, sin tener encuanta la distancia real en metros, sino lasconsecuencias ecológicas que implica. Dadoque observé que carayá puede desplazarseentre fragmentos de bosque y que es undispersor efectivo a escala local, evaluaré eneste capítulo el papel de carayá comodispersor entre los distintos tipos de bosquepresentes en la Isla Brasilera y establecere’ sirealiza una dispersión efectiva de largadistancia. Consideraré dispersor de largadistancia a carayá si encuentro en losbosteaderos de un tipo de bosque unacomposición específica de juveniles yrenovales similar a la de árboles adultos deotro de los tipos de bosques presentes en la
isla. Espero encontrar una composición dejuveniles similar a la de los bosteaderos delBosque de Laureles, tanto en los del Bosquede Sangre de Drago y Timbó como en los delBosque de Palo itá. En aquellos bosquesdonde la composición especifica de losrenovales en los bosteaderos difiera de la
composición especifica de juveniles, esperoencontrar que la mortalidad de juveniles seamayor a la del Bosque de Laureles y elcrecimiento de losjuveniles sea menor.
MATERIALES Y MÉTODOS
Al igual que en el capítulo anterior notrabajé con el estadío de plántulas pues ladensidad en los bosteaderos era tan elevadaque la identificación de los individuos en eltiempo era prácticamente imposible. Por otraparte, sea cual fuere el factor que produzca lamortalidad en esa etapa, siempre es muyelevada (Augspurger, 1984; De Steven,l99l). Se ha llegado a observar hasta un95 % de mortalidad entre las plántulas (Rey yAlcántara, 2000). Por lo tanto para realizarseguimientos en el tiempo es necesariotrabajar con un número de individuos variasveces mayor al que utilicé en este estudio.
Caravá como dispersor de semillas en losdistintos tipos de bosgue
Siguiendo el mismo procedimientodescrito en el Capítulo lV para losbosteaderos puntuales del Bosque deLaureles, realicé el conteo de juveniles yrenovales en bosteaderos puntuales de los dostipos de bosque restantes determinados en elCapítulo l, Bosque de Palo itá y Bosque deSangre de Drago y Timbó.
Dado que la composición de especiesde juveniles y renovales de los bosteaderosdel Bosque de Laureles resultó similar a la delos bosteaderos del Bosque de Sangre deDrago y Timbó, pero este último tiene unacomposición de individuos adultos muydiferente (ver Capítulo l), fonnulé lasiguiente hipótesis a posteriori, “como efectode la dispersión de semillas producida porcarayá el Bosque de Sangre de Drago yTimbó en algunos años tendrá una

Aloualla caraya como dispersor de semillas 65
composición florística similar al Bosque deLaureles”, es decir dominarán en el bosque0. diospyrifolia y E. punicrfolia. De estahipótesis se desprenden las siguientespredicciones, la comunidad de renovales delbosque deberá estar dominada por 0.diospyrifolia y E. punicifolia y los individuosdeben estar creciendo desde los bosteaderosde carayá. Para poner a prueba estaspredicciones realicé un muestreo de renovalesmayores al metro de altura en una hectáreagrillada del bosque, y registre para cadaindividuo su ubicación espacial según lagrilla, si se encontraba creciendo dentro ofuera de un bosteadero, su altura y DAP, ladistancia al individuo adulto más cercano,especie y DAP de este individuo. Con estosdatos obtuve la pr0porción de individuos ‘decada especie y su distribución espacial paraobservar Si seguía el mismo patrón descritopara la dispersión generada por carayá en elBosque. de Laureles (ver Capítulo lll). Notuve en cuenta para este estudio a .P.carthagenensis pues los individuos de estaespecie llegan muy rápido a la madurez,produciendo frutos, por lo qúe nopresentarian patrones de distribución tanclaros. j
Para obtener una evaluación del éxitoen el establecimiento dentro de cada bosqueevalué la diferencia entre el número dejuveniles y el número de renovales porbosteaderos. con pruebas de U de MannWhitney. Para evaluar posibles diferencias enel establecimiento en los distintos bosquescompare el número de I renovales porbosteadero en cada bosque con una prueba deAnova de un factor. Para analizar la riquezapromedio en los bosteaderos puntualescomparé el _ número de especies porbosteadero parajuveniles y renovales en cadabosque con pruebas de.U de Mann-Whitney yla riqueza promedio de los renovales entrebosques con una prueba de Anova de unfactor.
Mortalidad y crecimiento de los árbolesjuveniles .
“Dado que las mayores diferencias encuanto al establecimiento de las especies
dispersadas lo obtuve entre los Bosques deLaureles y Palo ita (resultados de la secciónanterior), decidi comparar el crecimiento y lasupervivencia de los juveniles en ambosbosques. Enjulio de 2000 realicé un muestreoen 6 bosteaderos puntuales de cada bosquecontando e identificando todos los juveniles,midiendo su altura, contando el número dehojas y midiendo el largo de la hoja mayor.Estas mediciones las repetí a los 2, IO, 18 y23 meses. Debí suspender los conteos en unode los bosteaderos del Bosque de Laurelesluego de la segunda medición porque lasmarcas se perdían entre mediciones.
Dado que durante marzo de 2001hubo una inundación de aproximadamente 15días (Apéndice), pude describir los efectos debreves periodos de inundación sobre losjuveniles de 0. diáspjn'ifolia, E. punicifolia yP.' carthagenensis. Considere crecimientopreinundación aquellas áreas de los juvenilescubiertos por arcilla y crecimientopostinundación aquellas áreas de losjuvenilesque estando por debajo de los niveles de lainundación no estaban cubiertas por arcilla.
Calcule' la proporción de individuosdesaparecidos entre mediciones, sindistinción por especies, y la dividí por elnúmero de meses transcurridos, de estamanera obtuve" una tasa de mortalidadmensual para los distintos períodos. Comparéla mortalidad entre periodos y entre bosquescon un Anova de dos factores de medidasrepetidas, donde cada bosteadero representóun bloque. Para realizar las comparaciones aposteriori utilicé la pruebade Tukey.
Comparé gráficamente el crecimientoen altura, número de hojas y largo de la hojamayor de las especies más abundantes encada bosque, 0. diospynjfolia, E. punicifolía yP. carlhagenensis en el Bosque de Laureles y0. dioSpyrifolia y E. punicifolia en el Bosquede Palo itá.
RESULTADOS
Carayá como dispersor de semillas en losdistintos tipos de bosgue
Tanto en el Boscjuede Laureles comoen el Bosque de Sangre de Drago y Timbó el

66 Susana Patricia Bravo
Tabla 15: Número promedio de juveniles y renovales i desvío estandar en los bosteaderos de cada bosque,entre paréntesis indico el número de bosteaderos muestreado. En 1a última columna indico el valor de laprueba U de Mann-Whitney y la probabilidad asociada.
Bosque Juveniles Renovales
Laureles 141,8 d: 136,0 25,1 i 11,9 U = 10,0(17) (10) P = 0,0002
Palo ¡tá 18,2 d: 9,2 13,1 i 12,3 U = 24,5(10) (7) P = 0,3
SangredeDragoy Timbó 37’?¡inzó‘s “¿3389 g;
número promedio de renovales por 10bosteadero fue significativamente menor quecl número dejuveniles, pero por otro lado fue amenos vnriable (Tabla 15). Por el contrario, fi 6cn el Bosque de Palo itá el número promedio S
de juveniles por bosteadero fue similar a1 de g 4renovales, y este último fue más variable. No Eobservé diferencias estadísticamente 2
significativas entre el número promedio de orenovales entre bosques (F = 2,96; P = 0,07; Imih mtbgl =24). Pero dado la significancia fuemarginalmente no significativa no puedotenerla en cuenta para arribar a unaconclusión ya que la prueba pudo no sersuficientemente potente. En los bosteaderosdel Bosque de Laureles el número promediode renovales fue el mayor, mientras en elBosque de Palo itá el número promedio fue elmenor y similar al promedio del Bosque deSangre de Drago y Timbó. Sin embargo,mientras en los bosques de Laureles y Sangrede Drago y Timbó la varianza fue deaproximadamente el 50 % del promedio, en elBosque de Palo itá varianza y promediofueron similares (Tabla 15).
Mientras la riqueza promedio fuesimilar para juveniles y renovales en elBosque de Laureles y en el de Sangre deDrago y Timbó, en el Bosque de Palo itá lariqueza promedio de los renovales fuesignificativamente menor que Ia de juveniles(U = 8; N = 10; N = 7; P=0,004; Fig. 23).Comparando la riqueza promedio de losrenovales entre bosques encontré que existíandiferencias significativas (F=5,04; gl=24;P = 0,01) siendo menor en el bosque de Paloitá con respecto a los otros dos tipos de
Figura 24:Riqueza promedio por bosteadero parajuveniles y renovales en el Bosque de Laureles(gris), en el Bosque de Sangre de Drago y Timbó(negro) y en el Bosque de Palo itá (blanco).
bosques (Prueba de Scheffé, P=0,03; Fig.24).
El número total de especies presentesen los bosteaderos fue de 12 para losjuveniles y l l para los renovales en elBosque de Laureles (Fig 25 a). En el Bosquede Sangre de Drago y Timbó encontré lasmismas 10 especies tanto entre los juvenilescomo entre los renovales (Fig 25 b), mientrasque en el Bosque de Palo itá encontré 10especies en los conteos de juveniles y 5 en losde renovales (Fig 25 c)
Como señalé en el Capítulo 1V losjuveniles de E. punicg'folia,P. carthagenensisy 0. diospyrifolia estuvieron presentes entodos los bosteaderos del Bosque de Laureles.Algo similar ocurrió en el Bosque de Sangrede Drago y Timbó, donde estuvieronpresentes en el 91 % de los bosteaderos. Losrenovales de estas especies también tuvieronlos mayores porcentajes de presencia, 90 %para O. diospyrifolia, 80 % E. punicrfolia y

lOOOOOOOOOOOOOOOOOOOOOOOOOOOOOOOOOOOOO000......
Aloualta caraya como dispersor de semillas 67
a
amor: avvuia
Comp" pachyshchyn
Pouterln solicito":
Pnuflinia app.
n .r .2 ¡ra
Noaandra IngustllollnMimosa 1
¡ng! ver:Psldlum quejaba
Eugeniubunurtnne
Eugenia pymormis
Psydmlrínanhogomnsis
120 140 100
Paulllnla spp.
Quotes diowyn'olh
Nodnndm nngusflíolla
lngl ver.
Psidlumqual-ba
Eugenia punidfolia
Oecropiapadiyfladvyl
Banlra ¡Igual
eo lo zo
Presencia (°/o) Abundancia (ind./bosteadem)
C
Desconocid- 2
Cecropla paehysiaehye
Paullinia spp.
Ooolen diospynfolia
Nedandra angusfiloliaMlnnoon 1
Ingaver.Eugenia punícifolín
Citan palmasEugenia pyri'ormis
Psychotría cannauenensis
y 5 10 . a5
Presencia (%) Abundancia (ind/bosteadero)
Figura 25: Presencia y abundancia de juveniles (lisos) y renovales (rayas) en los bosteaderos del Bosque deLaureles (a), el Bosque de Sangre de Drago y Timbó (b) y el Bosque de Palo itá (c). Presencia = porcentaje debosteaderos en donde encontré individuos de una especies. Abundancia = Número promedio de individuospor bosteadero.Notar las riquezas similares en el Bosque de Laureles y el Bosque de Sangre de Drago y Timbó, pero conmenores abundancias en este último. Notar también la pérdida de riqueza en la composición de renovales delBosque de Palo itá. ' ' ‘- yNota: observar los cambios de escala entre gráficos.

68 Susana Patricia Bravo
o Árbol adulto de Sangre de Drago o Timbó
- Renovales de Ocolea diospyrifolia yEugenia punicl'folia
° Árbol adulto de Banara argula
A Alisos
Figura 26: Distribución de los renovales de Ocolea dlospyrififlia y Eugenia punicifolia en l ha del Bosque deSangre de Drago y Timbó donde no haba adultos de estas especies. Un único individuo crecía fuera de losbosteaderos de carayá.
algo menor, 60 %, para P. carlhagenensis. Encuanto a la abundancia promedio porbosteadero, nuevamente fueron estas tresespecies las más abundantes en ambosbosques, tanto en los conteos de juvenilescomo de renovales (Fig. 25ayb). Losbosteaderos del Bosque de Sangre de Drago yTimbó tuvieron una mayor presencia deCecropia pachyslachya y de Inga vera quelos bosteaderos del Bosque de Laureles,mientras que en estos hubo una mayorpresencia de Eugenia burkartiana (Fig. 25
a y b). En el Bosque de Sangre de Drago yTimbó fue en el único donde encontrérenovales de enredaderas (Pau/¡nio spp. yCissus palmala). En los bosteaderos delBosque de Palo itá los juveniles de O.diospyrifolia estuvieron presentes en el 80 %de ellos (Fíg. 25 c). Igual porcentaje registrepara los juveniles de Nectandra anguslifoliapero no encontré renovales, en general es loque ocurre con la mayoría de las especies eneste bosque. E. punicifolia tuvo una presenciadel 60 % para los juveniles y P.

Áloualla caraya como dispersor de semillas 69
carlhagenensis del 40 %, pero ambastuvieron una abundancia muy baja con menosde 5 individuos por bosteadero, y no encontrérenovales de ninguna de las dos especies (Fig.25).
En el Bosque de Sangre de Drago yTimbó no encontré renovales de las especiesdominantes (ver Capítulo l), de los 78renovales contados el 81 % pertenecieron aO. diospyrifolia, el 10 % a Inga vera y el 9 %a E. puniczfolía. Sólo un individuo de 0.diospyrifolia no estaba creciendo en unbosteadero. el resto de los renovales creciandesde los bosteaderos y al ubicarlos en elmapa del bosque observé un patrón depequeños parches cercanos a arboles adultosde Crolon urucurana y AIbízia inundata, y endos casos estas agrupaciones de renovalesrodeaban por completo individuos de B.argula de gran porte (Fig 26); Ocho de lasparcelas se encontraban dentro de la franja deTessaria integnfolia que bordea el rio y allino encontré bosteaderos (Fig. 26). Laestructura de edades de los renovales de O.diospynfolia se puede ver reflejada tanto enla altura como en el DAP (Fig. 27 a y b). Losrenovales más altos alcanzaron los lS m,llegando la copa al dosel del bosque (verCapítulo l) pero con un DAP de |7 cm lo queindica que se trata de individuos de pocosaños de edad. Todas las categorías tuvieronun porcentaje importante de individuos. Paralas otras dos especies, también encontréindividuos de distintas alturas y DAP pero nome fue posible armar una estructura deedades completa por el menor número derenovales.
Mortalidad 1 crecimiento de los árbolesjuveniles
La proporción de mortalidad mensualdisminuye en el tiempo (Tabla 16), siendomuy alta y significativamente distinta al restode los periodos durante los 2 primeros mesespara ambos bosques (Fig. 28). El'número deindividuos por bosteadero que 'sobrevivenluego de los 23 meses de seguimientos seestabiliza en el tiempo alrededor de los 40individuos en el Bosque de'Laureles (Fig.
29 a) y alrededor de S en el Bosque de Paloitá (Fig. 29 b).
Al analizar la 5¿mortalidad de losjuveniles de las 3 especies más abundantes ena
lndivvdinsiv.)
a 'H ' lluml2345618910lll2lll4l5Alla-(m)
lnfividuns(56)
8
3.ÜÜÜ,6,.ÜWÜÚÜÜÚQÜDAP(crn)
Figura 27: Estructura de edades de los renovalesque crecen en los bosteaderos del Bosque deSangre de Drago y Timbó, al estimar la edadutilizando la altura (a) o el DAP (b).
los . bosteaderos, 0. diaspyrifolia, E."punícifolia y P. carthagenensis, encontrédistintos patrones. El número de juveniles de0. diospynfolia en el Bosque de Laureles seestabilizó alrededor de 10 individuos porbosteadero, mientras que en el Bosque dePalo itá se estabilizó alrededor de 2individuos por bosteadero. E. punicifolia enel Bosque de Laureles no llegó a estabilizar elnúmero de juveniles por bosteadero a lo largo'de los 23 meses de Seguimiento, mientras enel Bosque de Palo itá desaparecieronprácticamente todos los individuos. De los 6individuos que creclan en 3 bosteaderos altiempo 0,’ sólo 2 sobrevivieron en uno deellos. Los juveniles de P. carthagenensis seencuentran en el orden de los 16 individuospor bosteadero en los últimos 2 períodos de

70 Susana l’anicia Bravo
Factores gl CM F P
Entre Bosques
Bosque l 0,0004 0,12 0,74Bosteadero 9 090038
Dentro del Bosque
T' 3 0,0595 14,96 < 0,00001lempo 0’63
' , 3 0,0020 0,59Tiempo x Bosque 0,0040
Tiempo x bosteadero 27
Tabla l6: Resultados del Anova de dos factores de medidas repetidas comparando la proporción demortalidad mensual entre período de observación, y entre los bosques de Palo ita y de Laureles.
muestreo en el Bosque de Laureles, mientrasque prácticamente desaparecen de losbosteaderos del Bosque de Palo itá dondecomenzaron siendo 6 individuos localizadosen 3 bosteaderos y al finalizar el seguimientosólo sobrevivía l de ellos.
En cuanto al crecimiento en elBosque de Laureles, los juveniles de 0.diospyrrfolía al iniciar el estudio tuvieron unaaltura promedio de ll cm llegando a unaaltura promedio de l6 cm al cabo de los dosaños de mediciones. Crecieron con mayoraceleración en los períodos que incluyeron alverano (Fig. 30 a). El número de hojas fuemuy variable, disminuyendo durante losperíodos en que los juveniles no crecían enaltura y aumentando cuando se aceleraba elcrecimiento, el largo de la hoja mayor fuecreciendo a lo largo tiempo (Fig. 30 a). En elBosque de Palo itá los juveniles de estaespecie tuvieron una altura promedio inicialsimilar a la del Bosque de Laureles, ll,9 cmpero crecieron a mayor velocidad y en todoslos períodos menos durante el primerinvierno, alcanzando una altura promedio de24 cm (Fig. 30 a), sin perder hojas y con unarelación aproximada de l hoja nueva por cadanuevo centímetro ganado en altura. Las hojasfueron de mayor tamaño que en el Bosque deLaureles y crecieron de manera constantedurante el primer verano y el inviernosubsiguiente, la velocidad de crecimientodecayó hacia el segundo verano pero hay quetener en cuenta que las hojas ya tenían untamaño promedio de 9,5 cm lo que las acerca
en tamaño a las hojas de los individuosadultos (Fig. 30 a).
Los juveniles de E. punicifoliacrecieron de manera continua en ambos
9 IJ <>
É
'g'. o lS
É a
g oro !á .E3, 0.058' II l
nm ' i"'W 2. A lOb ¡ab nt,
HH l Mus
Figura 28:Proporción de individuos muertos pormes en los bosteaderos del Bosque de Laureles(linea continua) y del Bosque de Palo ita (lineadiscontinua) en cada conteo. Considerando todaslas especies en conjunto. La flecha indica elmomento de la inundación que afectó ambosbosques y las lineas horizontales indican losmeses de invierno.
Los meses que no companen la misma letradifieren significativamente (Prueba de Tukey P <0,001).
bosques, a excepción del primer invierno enel Bosque de Palo ita. A pesar de esto, alinicio de las mediciones la altura promedio delos individuos fue similar en ambos bosques,8,4 cm en el Bosque de Laureles y 8,7 cm enel Bosque de Palo itá. Pero en la últimamedición los individuos del Bosque deLaureles tuvieron una altura promedio de

¡Iloualla caraya como dispersor dc semillas 7|
¡9,2 cm y los del Bosque de Palo ita 3 l ,0 cm.Las hojas aumentaron en número y tamaño demanera similar en ambos bosques (Fig. 30 b).a
N”IndIBosleadcro
3 I
o + 4.a no 4 a ¡2 l6 20 24
T Meu-s
b20
g IS l
3 lO . .3a 5 - .
0 AL +o 4 a ¡2 l6 zo 24
f “mi
Figura 29: Número promedio de individuos en losbosteaderos del Bosque de Laureles (a) y delBosque de Palo ita (b) en cada conteo. La flechaindica el momento de la inundación que afectóambos bosques (marzo 200]) y las lineashorizontales indican los meses de invierno.Observar la diferencia de escala entre ambosgráficos.
En cuanto al crecimiento de losjuveniles de P. carthagenensis (considerandosólo el Bosque de Laureles), los individuoscrecieron a lo largo de todo el año con unanotoria pérdida de altura durante el últimoperíodo, de 24,30 cm a ¡8,74 cm ( Fig. 30 c).Este cambio se debió a la rotura de muchosindividuos afectados por una importantecaída de árboles. El número de hojas si bienvarió se mantuvo siempre alrededor de las 6hojas promedio por individuo, observé quedistintos factores generaban mortalidad de lashojas (helada, inundación, sequía) pero estasbrotaban y creclan rápidamente, reponiendoel número una vez pasado el disturbio,aunque tardando algunos meses en alcanzar el
tamaño definitivo el cual solía ser mayor altamaño de la hoja perdida (e.g., tercer períodode muestreo) de ahí que el tamaño de lashojas de esta especie tenga crecimientonegativo en la mayor parte de los muestreos(Fig. 30 c).
Los juveniles de las tres especies másabundantes tuvieron diferentes estrategiaspara superar la inundación de l5 días a la quese vieron sometidos durante marzo de 200].Los juveniles de P. carlhagenensis (Foto l3)perdieron casi por completo las hojas (Fig.30 c) y la parte distal del vástago, tomando elaspecto de estar completamente muertoshasta que comenzaron a rebrotar, los de O.diospyrífolia (Foto 14), si bien no perdieronlas hojas durante la inundación, estas fueronreemplazadas durante el períodopostinundación (Fig. 30 a), en algunosindividuos cayeron todas las hojas ._y lasnuevas crecieron todas a partir del nuevosegmento de vástago desarrollado luego de lainundación; los de E. punicífolja (Foto‘lS) noperdieron hojas, sacando hojas nuevas y demayor tamaño en las áreas del vástago decrecimiento postinundación. Este aumento dealtura postinundación, fácilmente medible enE. Punicg'foh‘a,fue de aproximadamente lcmpor mes, indicando que la velocidad decrecimiento no fue constante durante los 8meses del segundo perlodo de muestreo, sinoque se aceleró notoriamente durante’ losúltimos 2 meses. Por otra parte, analizandoport separado el crecimiento promedio detodos los individuos en cada bosteadero delBosque de Laureles, el único bosteadero enque el crecimiento de los juveniles no siguióla tendencia promedio fue aquel que por estara mayor altura en el albardón no fuealcanzado por la inundación (Fig. 3]). Estemismo bosteadero si mostró crecimientocuando el agua provino de las lluvias comoen el último período de muestreo.
DISCUSIÓN
El número de renovales fue menor alde juveniles en los tres bosques, esto eraesperable dada la alta mortalidad de losárboles en sus primeros meses de vida(Augspurger, l984). La gran variabilidad del

72 Susana Patricia Bravo
BOSQUE DE LAURELESa
IB 5,7'Í' Altura
'6 -o- Luaodonola me: 3.5¡4 -a- Num.de han:
¡2 .13 3‘E 2U
'° .31 fa - z
4.96
4 num-uneuau 470 1 ' * I0 ll 13l-l l—-1
Mens
b
24 . 5°22 b -.- Amara20 - + Largodo hojama) "0
1:)- Num. de hojas ‘30 a
E fi20 ¡vz|0
0
C
32 7.2All
29 +- "m . <6.826 -O- Largon hoyama)
—o- Nunmmj'a- « 6.4
4 6.0 1E - 5-6 É
. 5.2 2°z< 4.3
4.4
4.0
Bosque de Palo itá respecto al número derenovales podría estar dada por pequeñasdiferencias topográficas del suelo queproducirían diferencias en el número de diasde anegamiento, sobre todo considerandoque, con diferencias que van de los 50 cm y2 rn en la altura de los albardones, se llegarona formar bosques tan disímiles como elBosque de Palo itá’y el Bosque de Laureles(ver Capítulo l).
La composición especifica de loshosteaderos de los tres bosques es similnrcuando analizo a los juveniles. pero esto yuno es así cuando nnalizo la composiciónespecífica de los renovales. En el Bosque de
BOSQUE DE PALO ITA
2024 .
22 L —¡— Altura IB
zo + Lugodohojama) 4 ¡6
¡3 -o- Num.de hoja! J Nlbe ¡4. t 12 .2° á
l2 |0 E7IO - 1 s _s .ú . o
4 A - 4
o 2 + ¡o u nD-l l-IMacs
50
40
30 E2
20 É‘z.
i .0
o
Figura 30: Altura, largo de la hoja mayor ynúmero de hojas promedios de los juveniles de O.diospyrífolia (a), E. punicifolia (b) y P.carthagenensis (c) que crecen en los bosteaderosde cada bosque. Para la última especie muestroúnicamente el gráfico del Bosque de laureles dadala casi completa desaparición de los individuos enel Bosque de Palo itá.La flecha-indica el momento de la inundación queafectó ambos bosques (marzo 200i) y las lineashorizontales señalan los meses de invierno.
Palo ita' el número de especies disminuyenotoriamente en el paso de juveniles arenovales. Por lo tanto carayá seria undispersor de larga distancia efectivo para lasespecies que componen el Bosque deLaureles sólo Cuando son llevadas al Bosquede Sangre de Drago y Timbó. En contra de mihipótesis la mortalidad de juveniles en losbosteaderos del Bosque de Laureles y los delBosque de Palo itá fue similar y elcrecimiento (le'lus principales especies fuemayor en el último bosque.
De los conteos de juveniles se puedeinferir que un mayor número de semillas es

Alouarla canoa como dispersor de semillas 73
dispersado por carayá en el Bosque deLaureles, lo cual es lógico pues se trata de unproceso de dispersión local, cercano a lasfuentes de semillas. Semillas de las mismasespecies son transportadas a los Bosques deSangre de Drago y Timbó y de Palo ita, peroen un número menor, lo cual también eslógico pues se trata de eventos de dispersiónde larga distancia. Considerando que elBosque de Palo itá difiere de los otros dos porsn bajo nivel de suelo (ver Capitulo l), ycomo carayá hace llegar semillas de lasmismas especies a los tres bosques. lafrecuencia de inundación en el Bosque dePalo itá sería lo que impide que varias deestas especies se establezcan. Así, ladinámica del rÍo sería responsable demantener la diversidad de ambientes dentrode la isla. De forma similar se comportan lospastizales inundables del río NieuweMerwede en los Países bajos, donde lassemillas de las especies de las zonas más altasllegan y genninan en los pastizales máscercanos al río pero no logran sobrevivir másde un año bajo el régimen de alta frecuenciade inundaciones (Lenssen et al., 1999). Estafalta de correspondencia entre la composiciónespecífica de las semillas que arriban y lacomposición específica de la vegetación quelogra establecerse es muy común en loshumedales (Keddy y Reznicek, l982;Schneider y Sharitz, 1988; Welling et al.,l988). También estudios experimentales hanencontrado que la hidrología es el principalfactor para determinar la composición de lascomunidades vegetales de los humedales(Weither y Keddy, 1995).
El caso en el Bosque de Sangre deDrago y Timbó es diferente, este bosquecrece sobre un albardón de igual altura alBosque de Laureles, pero es de formaciónmás reciente (ver Capítulo l). Como reflejanlos conteos de juveniles y renovales, ladiversidad específica se mantiene en ellos,pero a diferencia de lo que ocurre en elBosque de Laureles esta composición no secorrelaciona con la composición del bosqueadulto de Sangre de Drago y Timbó, sino querefleja la composición del Bosque deLaureles. Asi el efecto de carayá comodispersor de larga distancia permitiría el
aumento de la riqueza y de mantenerse lascondiciones podria llevar al Bosque deSangre de Drago y Timbó a una fisonomía
Figura 3l: Altura promedio general en cadaconteo de los juveniles de O. piospyrifolia, E.punicifolia y P. carthagenensis que crecen en losbosteaderos del Bosque de Laureles. La lineacontinua indica al bosteadero que no fuealcanzado por la inundación. La flecha indica elmomento de la inundación que afectó ambosbosques (marzo 200l) y las lineas horizontales losmeses de invierno.
similar a la del Bosque de Laureles. Algosimilar se ha observado en el ParqueNacional Manu (Perú) donde las especies delbosque maduro dispersadas por vertebrados,estarían lentamente arribando yestableciéndose en el bosque más jovencercano al río y dominado por especiesanemófilas (Foster et al., 1986). Es dedestacar que en el Bosque de Sangre deDrago y Timbó fue frecuente elestablecimiento de C. pachystachya, cuyosrenovales raramente los encontre en elBosque de Laureles, pero cuyos individuosadultos fueron una de las especiesdominantes. También Cecropr'a sp es una delas primeras especies en llegar a los bosquesjóvenes en el Parque Naciónal Manu, ytambién en este caso domina en bosques unpoco más antiguos (Foster et al., 1986).
Por otra parte al muestrear losrenovales en el Bosque de Sangre de Drago yTimbó no encontré renovales de las especiesdominantes, sólo de Tessarr'a integ'rr'foliaperocomo en todos los casos estos estaban dentro .de la estrecha franja cercana al margen del rloocupada por los adultos de esta especie (verCapítulo l) decidí no considerarlos al analizarlos datos. El resto de los renovales crecían en

74 Susana Palricia Bravo
los bosteaderos y aproximadamente el 8l %perteneció a 0. diospyrifolia. El patrón dedispersión que generó carayá en este bosquees similar al que describí para el Bosque deLaureles (ver Capítulo lll). Los bosteaderospuntuales y los grandes bosteaderos queprodujeron la acumulación de renovalesalrededor de las 2 B. arguta en este bosque(Fig. 26), corresponderían a los donniderosprincipales de los 2 grupos de carayá queutiliun la zona de bosque relevada (para unode los casos esto fue confirmado). Así lahipótesis acerca de que el efecto de carayácomo dispersor de semillas llevaría con eltiempo al Bosque de Sangre de Drago yTimbó a un bosque de similar composición alBosque de Laureles fue avalada por ladistribución espacial de los renovales. '
El caso de Inga vera es particularpues si bien en el Capitulo ll la considerécomo una especie sobre la cual carayá teníaun efecto predador por alimentarse de susflores y no dispersar sus semillas, las semillasde esta especie aparecieron en_-las heces delos carayá que habitaban el bosque de Palo itáy el Bosque de Sangre de Drago y Timbó.Estas semillas germinaban originandojuveniles y renovales ante lo cual deboconcluir que la relación de carayá con estaespecie depende de la composición florísticadel bosque y de la oferta de alimento que estépresente, pudiendo pasar de ser un claropredador a un dispersor efectivo.
La marcada mortalidad -de juvenilesdel primer periodo se debería a la altamortalidad común entre los más pequeños. Lamayor proporción de árboles jóvenes mueredurante los 2 primeros meses, en parte debidoa factores densodependientes, pero paraalgunas especies las causas de mortalidad nodependen de la densidad, y probablementevaríen no solo interespecíflcamente sinotambién entre años (Augspurger,- 1984;Howe, ¡993). En este caso, ,la densidadpodría ser un factor importante de lamortalidad para los individuos más jóvenes,pero no para el resto de los juveniles, dadoque si bien el número de individuosque yomido va disminuyendo en el tiempo, esto nosignifica que el número total de individuos enel bosteadero lo haga, por el contrario en un
bosteadero que está siendo utilimdo confrecuencia el número tiende a mantenerseconstante pues nuevos individuos nacen todoel tiempo.
Se observa claramente que los mesesdel primer invierno fueron un período dificiltanto para los individuos de O. diopyrifolia.que crecieron de manera más lenta tambiénen el segundo invierno (tercer período demuestreo), como para aquellas especies quecrecieron prácticamente de manera continuaen el resto de los períodos. Existen 2 factoresque podrían explicar esto, la crudeza de esteprimer invierno, atipico en las islas y elaumento en la disponibilidad de agua a partirdel segundo período (Apéndice). El primerinvierno estuvo caracterizado por unaimportante sequía, agudizada por el bajonivel del río durante casi todo el año previo,lo que llevó a una desaparición completa delagua de las lagunas internas, bajando elefecto atemperante del Río Paraná ypermitiendo el registro de fuertes heladas.Estas heladas fueron responsables directas dela pérdida de hojas de los juveniles de P.carthagenensis, mientras la falta de aguapareció ser lo que más afectó la pérdida dealtura y hojas de los juveniles de 0.diospynfolia a los cuales se les secaba laporción apical progresivamente y perdíanhojas de distintos sectores. Si bien en elBosque de Laureles, durante el segundoinvierno (tercer_ período de muestreo),observé un aumento de la mortalidad generalrespecto al segundo período (primavera verano) (Fig. 28 y 29 a), hubo un efectomenos marcado que en el primer invierno.sobre el crecimiento de los juveniles de 0.diospyrifolia (Fig. 30 a). Esto podria deberSea diversas causas, que este periodo incluyetambién los primeros meses de la primavera,que no hubo una marcada sequía durante esteinvierno, ni se produjeron heladas(Apéndice). En el Bosque de Palo itá losjuveniles continuaron creciendo a igual ritmoque en el período anterior (Fig. 30 a) y noobservé un aumento de la mortalidad general(Fig. 28 y 29 b). Aquí podría influir que losefectos de la inundación sean más duraderosen este bosque ya que la misma dura más yalcanza mayor altura, lo que permite el aporte

Alouaua caraya como dispersor de semillas 75
de mayor cantidad de nutrientes. El leveaumento en la mortalidad que se observa enel último período para ambos bosques (Fig.28 y 27 a y b) se relacionó con una fuertetormenta que provocó la caída de un grannúmero de árboles y ramas sobre losbosteaderos. Como Weiher y Keddy (1995)han señalado los factores estocásticostambién tiene un papel importante sobre elreclutamiento de individuos en losecosistemas.
Básicamente tres observaciones mellevaron a considerar importante el efecto dela inundación en el crecimiento de losjuveniles, el notorio crecimientopostinundación, que este fuera mayor en elbosque con más días de anegamiento (Bosquede Palo ita) y la ausencia de crecimiento en elbosteadero que no fue alcanzado por lainundación. En otros ecosistemas tropicalesse ha observado que la cantidad de agua y eltiempo que esta permanece en el suelo esimportante para determinar la velocidad decrecimiento de los juveniles de Vouocapouaamericana (Caesalpiniaceae) (Forget, 1994).
Un aumento en la velocidad decrecimiento hace decrecer la tasa demortalidad entre los juveniles, aunque no seconoce cuál es el mecanismo, podría facilitarel escape de los juveniles a los factores quegeneran mortalidad entre los más pequeñossimplemente por su tamaño (e.g. herbivoria),o bien deberse al aumento en el vigor de losjuveniles a medida que crecen por unaumento de sus protecciones químicas yestructurales (Walters y Reich, 2000). Porejemplo, la mortalidad debida al ataque de loshongos disminuye cuando comienza lalignificación (Augspurger, ¡984). En la islaBrasilera los juveniles que crecían en losbosteaderos del Bosque de Palo itá crecieroncon mayor velocidad, esto podria ayudarlos aaumentar las posibilidades de supervivencia
durante las inundaciones. Al igual que otrosinvestigadores (Placcí, 1995), he observadoque los juveniles arbóreos (0. diospyrifqlia yE. punicifolia) de mayor edad sobreviven alas inundaciones sin mostrar daños aparentes,aun cuando hayan sido cubiertos en sutotalidad por el agua.
En conclusión, la dispersión desemillas producida por carayá tiene un efectorelevante tanto en la regeneración de la selvade inundación como en su dinámica,facilitando por un lado el reemplazo deejemplares para la mayoría de las especiesque conforman el Bosque de Laureles de losalbardones altos de mayor edad ymanteniendo la proporción entre las distintasespecies (Capitulo IV). Mientras que por otrolado, posibilita la colonización por parte deespecies de este bosque de los albardonesmás nuevos, ocupados por Bosques de Sangrede Drago y Timbó. Carayá también hacellegar semillas de las especies del Bosque deLaureles a los albardones más bajos, donde seencuentra el Bosque de Palo ita, pero laelevada frecuencia de inundación nopermitiría la colonización. Por lo tanto lainteracción de la dispersión de semillasproducida por carayá con las inundaciones esfundamental para facilitar el crecimiento delos juveniles en los bosteaderos y paramantener la diversidad de bosques dentro dela isla. Mis resultados mostraron que elestablecimiento en los bosteaderos no seproduce porque sean “sitios seguros” dondeexiste una baja tasa de mortalidad, sino quese deberia en parte a una continua yabundante llegada de semillas, a un rápidocrecimiento y a un aumento en el vigor de losjuveniles que crecen en ellos (Capitulo IV).Pero no sé aun que factores estaríanfavoreciendo o afectando el crecimiento delos juveniles, ni cuales provocan sumortalidad, eso es lo que estudiaré en elpróximo capítulo.

Foto 13:Juvenil de P. carthagenensis luego de la inundación
Foto 14: Juveniles de 0. diospyrg'folialuego de la inundación

Alouatta carayn como dispersor de semillas 77
Capítulo VI:
¿Hay factores que facilitan o perjudican el establecimiento de los
juveniles en los bosteaderos?
"El olor basta para hacerles encontrar excrementosy cadáveres. bajo los cuales practican agujerosdonde sus pequeñuelos encuentran el alimento a sualcance. Parece en consecuencia, que estos insectosno educan a sus hijos y no les dan ningunainstrucción. y también que la hembra sola trabajapara asegurar a su progenie una habitación yalimentos"
"Su olfato es tan fino. que antes de que una personahaya terminado de hacer sus necesidades en plenocampo. varios de estos insectos han acudido ya allugar"
Felix de Azara(Viajespor la América meridional)
INTRODUCCIÓN
Las hipótesis propuestas para explicar eldesarrollo de las estrategias dispersivas enlos vegetales llevan implícito que ladispersión significa un aumento en lasprobabilidades de supervivencia de ladescendencia (Howe y Smallwood, ¡982).Por lo tanto será crucial que ocurre con lasemilla una vez que el dispersor primario,en este caso carayá, la deposita en el suelo(Herrera et aL, 1994; Jordano y Herrera,l995; Schupp y Fuentes, 1995). Tantofactores bióticos como abióticos entrarán enel juego durante esta etapa posdispersiva ytendrán gran influencia sobre la estructurade la comunidad (Chambers y MacMahon,¡994)
El comportamiento de losfrugívoros influye directamente en la etapade remoción de semillas de los árboles y suposterior deposición en el suelo, pero seconsidera que los frugívoros pueden afectarsólo indirectamente sobre la fase dereclutamiento, por ejemplo dejando lassemillas en zonas de alta o baja predación oen zonas con mayor o menor disponibilidadde nutrientes, pero no pueden afectardirectamente la predación o disponibilidadde nutrientes del suelo (Augspurger, ¡984;Jordano y Herrera, 1995). Por lo mismoJordano y Godoy (2002) encontraron quelos frugivoros participaban activamente enla determinación de la lluvia de semillas,pero luego otros factores afectaron lacomposición específica del banco desemillas del suelo en el SE de España. Sin
embargo, en el caso de mamíferos de grantamaño y especialmente cuado formanbosteaderos, la materia fecal q'ue acompañaa la lluvia de semillas puede influir demanera importante en la suerte de la semillay de las plántulas (Andresen, 200]). Estetipo de dispersor no sólo estaria dejando lassemillas en un sitio con determinadaspropiedades, sino que también seriaresponsable de generar y mantenercaracterísticas que facilitarán o perjudicaránel establecimiento y crecimiento.
En relación a la materia fecal deotras especies de monos aulladores se hatrabajado principalmente con el conjunto deescarabajos estiercoleros (Estrada et al.,1993; Feer, 1999; Andresen, 200]), que enparte disgrcgan las heces y en parte lasentierran, teniendo por Io tanto distintoefecto sobre las semillas y su germinación(Andresen, 200]). En ambientes tropicalesno llegan a formarse bosteaderos pues lamateria fecal es removida en pocas horaspor escarabajos estichólerós, hormigas yroedores (Estrada y Coates-Estrada, 1991;Levey y Byrne, ¡993; Feer, 1999;Andresen, 2001). Gran parte de estosanimales son atraídos por la propia materiafecal y otros por la abundancia de semillasque pueda haber junto con la materia fecal(Andresen, ¡999), pero en ninguno de losestudios se describe el ensamble completode artrópodos asociado, ni los daños que losherblvoros puedan estar causando dentro delos bosteaderos.

Susana l’alricia Bravo
Se ha descripto en el caso de losgrandes mamíferos que las heces al tardarun tiempo prolongado en descomponersesirven de fertilizante a las plántulas y lashacen crecer más vigorosas (DinersteinWemmer, l988; Traveset et al., 200i). Enel caso de carayá en la selva de inundación,la continua caída de heces y orina podríaademás de fertilizar, tener efectos negativosen las plántulas comó daños fisicos oquemaduras. En la isla Brasilera por lotanto, la supervivencia de los juveniles enlos bosteaderos podría estar afectada porfactores positivos y negativos, como unamayor concentración de nitrógeno yfósforo, o mayor herbivoría por estarcreciendo en altas densidades.
Teniendo en cuenta los resultadosdel Capítulo V donde una importanteproporción de juveniles murió durante losdos años de observaciones, no es posiblepensar en los bosteaderos como sitiosseguros donde las semillas que arribanlogran escapar de la predación y donde lamortalidad de los juveniles es baja porcrecer alejados de los árboles parentales.Pero aún no he descripto cuales son estosfactores que pueden estar actuando y en quegrado lo haría cada uno de ellos. Losresultados del alto establecimientoobservado (Capituo lV y V) podrían sersimplemente el resultado de la saturaciónde depredadores y patógenos por el altonúmero de semillas que arriban, 0 bientratarse de una serie de interrelaciones máscomplejas donde se suman factorespositivos que facilitan el crecimiento ypermiten a los juveniles escapar a lamortalidad. Por ello el objetivo de estecapítulo será obtener una descripción de lascondiciones en las que se encuentran losjuveniles dentro de los bosteaderos ydeterminar si los predadores o posiblesdispersores secundarios reconocen losbosteaderos como parches de oferta desemillas. Para ello realicé unacuantificación de todos los daños quemostraron los juveniles en los bosteaderos,y de la concentración de nitrógeno y fósforode los juveniles que crecen en losbosteaderos respecto de los que crecenfuera. Describí la comunidad de artrópodosque pueda estar visitando los bosteaderos.
Y finalmente, evalué experimentalmente laremoción de semillas de los bosteaderos ydebajo de los árboles parentales pordistintas taxas.
MATERIALES Y MÉTODOS
Tipos de daño gue afectan el crecimiento
Para obtener una descripcióndetallada de los daños que pudieran estarperjudicando el desarrollo de los juvenilesque crecen en los bosteaderos, en Julio de2000 seleccioné al azar 5 bosteaderos delBosque de Laureles y 6 bosteaderos delBosque de Palo itá. La distancia mínimaentre bosteaderos fue de aproximadamente40 metros. En cada uno registré el númerototal de juveniles, el número de hojas decada individuos, el número de individuosdañados, el número de hojas dañadas decada juvenil y en que proporciónaproximada estaba afectada cada una deellas. Las proporciones fueron determinadaspor dos observadores independientes y encaso de divergencias registre el promedio.Con estos datos calcule la proporción deindividuos afectados por cada tipo de daño(ver más abajo) en el bosteadero, laproporción de hojas afectadas por cada tipode daño por individuo y un índice deintensidad de ataque (llA) calculado paracada individuo y luego obtuve un promediopara el bosteadero,
Nllt
llA = (Z (ppaHi)/NHt)xlOOI- l
ppaHi = proporción de la hoja i afectada.NHt = Número total de hojas del individuo
Los tipos de daño que consideré fueron lossiguientes:
Cortes: pérdida de área foliar.Daño por herbivoría: cuando un artrópodoera observado comiendo las hojas o bien sila forma de los cortes permitía inferir quehabia sido producido por un insectoalimentándose.Quemadura por la orina: La hoja seencontraba húmeda, amorfa y con unatonalidad notoriamente más oscura. Engeneral el daño comprometía la zona apical

Aloualla caraya como dispersor de semillas 79
y primeras hojas o bien un lateral de todo elindividuo.
Hojas secas: cuando parte o la totalidad dela hoja moría pero aún permanecíadesplegada.Patógenos: determine su presencia porcambios en la coloración, que implicabanpérdida de intensidad del verde virando alamarillo de manera no uniforme, en generalcon apariencia punteada. Cambios detextura que implicaban un aumento en larugosidad. Y cambios en la forma de lashojas que solían implicar la contracción dealgún sector. Dado que si bien en lamayoría de los casos los patógenos parecianestar atacando el ¡00 % de la hoja, no mefue posible estimar para ellos el IlA. Por lotanto no consideré a los patógenos cuandoevalué la intensidad del ataque utilizandoeste índice.
Compará el número de juvenilesafectado por cada uno de los distintos dañosen cada bosque con una prueba de Anovade dos factores y la prueba paracomparaciones a posteriori de Tukey.Realice una prueba de Anova de un factorpara cada bosque utilizando la proporciónde hojas afectadas como una primeradescripción de la intensidad de ataque queincluyera a todos los daños y luego unaprueba de Anova de un factor para cadabosque utilizando el llA. Para cada especiedetermine diferencias en la proporción dehojas afectadas por medio de una prueba deAnova de un factor. Todos los datos fuerontransformados con raíz cuadrada. Paradeterminar que tipo de daño atacaba conmayor intensidad a las dos especies másafectadas según el análisis anterior, realicéuna prueba de Anova de un factor para O.diospyrifolia realizando comparaciones aposteriori con la prueba de Tukey, estosdatos fueron transformados con raizcuadrada. Para P. carlhagenensis realicéuna prueba de Kruskal-Wallis y pruebas decomparaciones múltiples a posteriori paraanálisis no paramétricos, transforme losdatos sumándoles 0,0001 y aplicandologaritmo en base lO.
Qisponibllldad de nutrientes
Evalué el contenido de nitrógeno yfósforo directamente en los juvenilessiguiendo las recomendaciones de Sollins(1998), dado que en algunos bosques elcontenido de nutrientes del suelo puede sermuy variable temporal y espacialmente, ypor otra parte la simple presencia denitrógeno y fósforo en el suelo no significaque esté disponible para la vegetación. Enagosto de 2000, colecté en el Bosque deLaureles, juveniles de Ocotea diospynfolia,Psychorria carthagenensis y Eugeniapunícifolia que creclan en 10 bosteaderosseleccionados al azar y debajo de l0 árbolesparentales también seleccionados al azar.
El método utilizado para evaluar lasconcentraciones de nitrógeno y fósforo fueel de micro-Kjedahl. Los análisis fueronrealizados en la Cátedra de Ecología de laFacultad de Ciencias Agrarias de laUniversidad de Buenos Aires.
Para cada especie compare lasconcentraciones encontradas en los.individuos que provenían de losbosteaderos y los que crecian debajo de losárboles parentales con pruebas de t deStudent.
Artrómdos en los bosteaderos
En el Bosque de Laurelesseleccionó al azar 20 bosteaderos, con unadistancia mínima entre ellos de 40 m.Seleccione un sitio control dentro de unradio de 20 m de cada bosteadero, con igualcobertura vegetal y altura del suelo. Encada sitio coloqué una trampa de caida paraartrópodos. Las trampas permanecieronabiertas durante 7 dias. Este muestreo loreiteró en cada estación del aflo (febrero,mayo, julio y noviembre).
Las trampas en las que cayó unagran cantidad de hojarasca, materia fecal delos carayá, perdieron el agua o fueronremovidas por algún animal, fueronanuladas junto con su contraparte contmI obosteadero, de manera de mantener undiseño balanceado.

Por observación bajo lupadeterminó morfoespecies, que luego fueroncon'oboradas e identificadas cuando fueposible consultando con distintosespecialistas. Los hábitos alimenticios delas diferentes especies los obtuve de labibliografia y de las consultas conespecialistas.
Para analizar las diferencias enriqueza y número de individuos por trampaentre sitios utilicé pruebas de t de Student opruebas de U de Mann —Whitney para losdatos de cada estación.
Remoción de semillas
En mayo de 2001 realicé en elBosque de Laureles experiencias deremoción de Semillas dentro y fuera de llosbosteaderos, con las semillas de las especiesmás abundantes, Ocotea diospynfolia yPsychotria carthagenensix.
Seleccione al azar lO bosteaderos,con una distancia minima entre ellos de 40m, y lO sitios control ubicados debajo de lacopa de árboles parentales de ambasespecies. En cada uno de estos sitios ofrecísemillas en 3 cajas de Petri. Una de ellascolocada sobre un cilindro plástico a unaaltura de entre 2 y 3 cm, abierta sólo de díaque permitía el acceso a los vertebradosdiurnos, otra en igual condición permanecíaabierta de noche para permitir el acceso alos vertebrados nocturnos, y la terceraubicada en el fondo de un hueco y cubiertacon una rejilla metálica permitía el accesosólo a los invertebrados. Dentro de cadacaja coloque lO semillas de O. diospyrifoliay l0 de P. carlhagenensis. Las semillasfueron ofrecidas durante 4 dlasconsecutivos. Al atardecer y al amanecerregistre el número de semillas removidas yrepuse las faltantes. No pude realizar laexperiencia con las semillas de Eugeniapunicifolia dado que los fi'utos de estaespecie duran en los árboles alrededor de lsemana, y la fructificación puede ocurrir encualquier momento entre noviembre yfebrero (ver Capítulo l).
El número de semillas removido
por los invertebrados lo dividi por 2 paralos análisis, pues las semillas estabandisponibles el doble de tiempo para este
Susam‘ l'atncia Bravo
taxa. Para cada especie de semillas evaluélas diferencias de remoción por cada taxaen cada sitio om un Anova de 2 factores.
RESULTADOS
Tims de daño gue afectan el crecimiento
Al analizar los daños que afectan elcrecimiento de los juveniles en cada bosqueencontré que existían diferenciassignificativas tanto entre bosques (F = 4,45;gl = l; P = 0,04; Fig 32) como entre tiposde daño (F = l0,32; gl = 4; P < 0,0000l)pero no en la interacción (F = 1,65; gl = 4;P = 0,l8). Existe una proporción mayor deindividuos afectados en los bosteaderos delBosque de Laureles y el ataque porpatógenos es el tipo de daño q ie más afecta.a losjuveniles en ambos bosqt es (Fig. 32).
Figura 32:Proporción de individuos afectadospor distintos factores en el Bosque de Laureles(gris) y en el Bosque de Palo ita (blanco). Losfactores que no comparten la misma letradifieren significativamente (Prueba de Tukeyl’ < 0,0005).
Respecto a Ia proporción de hojasafectadas encuentro nuevamente 'que lospatógenos no sólo atacan más individuossino que atacan una mayor proporción dehojas de los individuos tanto en el Bosquede Laureles (F = 7,93; gl = 4; P = 0,0005;Fig.33 a), como en el Bosque de Palo itá(F = 8,08; gl = 4; P = 0,0005; Fig.33 b).Dejando de lado el ataque de los patógenosy evaluando la intensidad del ataque con elindice llA no encontré diferenciassignificativas entre distintos tipo de daño enninguno de los dos bosques (Bosque deLaureles, F = 0,04; gl = 3, P = 0.99; Bosque

Aloualla caraya como dispersor de semillas 3 l
Tabla ¡8: Lista de artrópodos capturados en los bosteadcros (B) y en los sit-ioscontrol (C) durante cadaestación del año. Para cada especie indico el número total de individuos capturados y hábitosalimenticios, estos últimos obtenidos de bibliografia y consultas a especialistas. (E) Estiercoles
WN lnviemo Primavera Verano Otoño _FAMlLlA llábilo
EsEcie B C B C B C B (.‘OPILIONIDA
GONYLEP'I'IDAE
Sp l I l 0 0 0 0 0 0
Sp 2 0 0 0 l 0 0 0 0
ARANEAF.
SALTlCllME
Phiale raburijbliala 0 0 l 0 0 0 0 O Predador
Phiale sp I 0 Ü 0 0 0 l 0 0 Predador
Phiale sp 2 0 0 | 0 0 0 0 0 Predador
Phiale sp 3 0 0 0 0 l 0 0 0 Predador
Lycosidne l 0 0 0 O 0 0 0 Predador
Schizacosa maliliom l 0 0 0 0 0 0 0 l’redndor
Lycosa cfi'. ardalína l 0 0 0 l 0 0 0 Predador
Lycosa cji'. malilíosa 0 0 l 0 0 0 0 0 Predudor
Lycosa sp. 2 0 4 2 O 0 0 0 l’rcdador
ARANEIDAE
Alpaida sp. l O 0 0 0 0 0 i’redador
lSOPODA
Sp l l | 0 0 0 0 0 0
ORTOPTERA
Sp l 0 0 l 0 0 0 0 0 Fitófngo
Sp 2 l 0 0 0 0 0 0 0 Filófago
Sp 3 l 0 O 0 0 0 0 0 Filófago
DERMAPTERA
Sp | 0 0 0 0 2 0 0 0 Predador
COLEOPTERA
Sp l 0 0 0 0 0 l 0 0 Predador
Sp 2 0 0 0 0 0 l 0 0 l’redudor
Sp 3 0 0 l 0 0 0 0 0
Sp 4 0 0 l 0 0 0 0 0
CICINDELIDAE
Sp I 0 0 0 4 0 0 0 l’redmlor
Sp 2 0 0 0 0 3 l 0 0 Predador
Sp 3 0 0 0 0 l 0 0 Predador
SCARABIDAE
Sp l 0 0 l 0 0 0 0 0 Predador
Sp 2 0 0 l 0 0 0 0 0 Predador
Sp 3 0 0 0 0 7 0 l 0 (E)
Odonthophagus hirculus l 0 0 0 3 0 0 0 (E)
Canlhon sp l I 0 0 0 ¡6 0 0 0 (E)
Camhon sp 2 0 0 l 0 0 0 0 0 (E)

82 Susana Patricia Ilmvo
ORDEN Invierno Primavera Verano OtoñoFAMILIA llábilo
M B C B C B C B COmherus sulcalor 0 0 0 0 2 0 0 0
S'l'APHlLlNlDAE 0 0 0 Prcdador
Sp l S 0 0 0 0 0 2 0
CASS! DA F,
Sp | 0 0 0 S l 3 0 0 Filófago
CURCULlONlDAE
Sp l 0 0 0 0 2 0 0 0 Filófago
Sp 2 0 0 0 0 I 0 0 0 Filó [ago
HYMI‘ZNOPTERA
Sp | (avispn) I 0 I 0 0 0 0 Prcdndor
Sp 2 (avispa) 0 | 0 0 0 0 0 Predador
I’ORMICIDAE
Odontomachux haemalodex 2 0 l l 9 l | 5 | 0 Predador
Phcida/c xp. I6 2 2 l I 0 3 l l’rcdndor
Camponolus sericeivemris 0 0 0 0 l 0 0 0 Filófago
Camponolus sp I 2 0 l l l 0 0 0 ?
(."amponomx sp 2 0 l 5 5 2 0 0 0
Campanalus sp 3 0 l 0 0 0 0 0 0
Procn'plocerus sp. l 0 0 0 0 0 0 0
Zacryplocerus clypealus 0 l 0 0 l 0 0 0
Paractyplocenls sp. l 0 l 0 l 0 0 0
Acromirmex cfi‘. hixpidu: 0 0 2 0 2 l 0 0 Cortadora
I’aralrechina xp. 0 2 0 4 0 0 0 0 ?
Sp l 0 0 0 l 0 0 0 0
Sp 2 0 0 0 l 0 0 0 0
Sp 3 0 0 0 0 l 0 0 0
CHALCIDIDAE
Sp I 0 0 2 0 0 0 0 0
Sp 2 0 0 2 0 0 0 0 0
Sp 3 0 0 0 l 0 2 0 0
MUTILLIDAE
Sp l 0 0 0 2 0 0 0 0 Prcdador
HEMlPTEROIDE
Sp | 0 l 0 0 l 0 0 0
HOMOP'I‘ERA
CICADEUDAI'I
Sp | 0 0 0 0 2 0 0 0 Filófago
DERMAPÏEROS
Sp l 0 0 0 0 l l 0 0 Predador
OR'I‘OI’TI-ZRA
5p l 0 o ¡ o o 0 0 0 Filófago
Sp2 l 0 0 0 0 0 0 0 Fitófago
Sp 3 l 0 0 0 0 0 0 0 Filófago
llli'l'líROl’TlïRA

Alouarla caraya como dispersor de semillas 83
Sp i 0 l 0 3 0 0 0 9
Sp 2 0 0 0 2 0 0 0 ?
TOTAL 42 42 34 73 i7 7 l
Dipteros de al menos 2 especies en los bosteaderos con material fecal fresca y al menos 2 especies deácaros en todos los bosteaderos
937m __. I l [i l |Secas Cotton PetogenoebQuem-din" HW.
°' F]M, I:L ..._.Seu-se Conesa Femme-b
Habia varian
Figura 33: Proporción de hojas afectadas por losdistintos factores en los juveniles de losbosteaderos del Bosque de Laureles (a)y en ios'del Bosque de Palo ita (b). Los factores que nocomparten la misma letra difierensignificativamente (Prueba de Tukey P < 0,005).
de Palo itá , F=0,96; gl=3; P=0,43;Fig. 34).
Las tres especies más abundantesfueron afectadas en distinto grado por cadafactor (F = 9,6]; gl = 2; P = 0,003), iosindividuos de E. punicifolia tuvieron unaproporción menor de hojas afectadas quelos de O. diospyrifolia y P. carraghenensis(Prueba de Tukey P < 0,05; Tabla ¡7). Losindividuos de 0. diospyrr'folia fueronafectados más por los patógenos que por elresto de los factores (F = 7,33; gi =4;P = 0,0008; Tabla i7) y los de P.carrhagenensis presentaron una mayorproporción de hojas afectadas por cortes,herbivoria y patógenos que hojas secas o
quemadas (H = ¡0,13; gi = 4; P = 0,04;Tabla 17).
Disponibilidad de nutrientes
La concentración de fósforo nomostró diferencia significativa en las tresespecies estudiadas entre los individuos quecrecian en los bosteaderos y los que creciandebajo de los árboles parentales. Sinembargo la tendencia en ias tres especiesfue ia de una concentración levementemayor en ios controles, entre un 0,02 % yun 0,03 % (Fig. 35 a). Sólo para Psychom'acarrhagenensis encontré diferenciassignificativas en la concentración denitrógeno, siendo un 0,5 % mayor enaquellos individuos que creclan en iosbosteaderos. Para ias otras dos especies lasdiferencias fueron no significativas, pero laconcentración fue levemente mayor en losbosteaderos, 0,i % para 0. diospynfolia y0,02 % para E. pun'r'cífolía (Fig. 35 b).
AgÉQogog en los bostegdeggg
En todas las estaciones la riqueza yel número total de individuos capturadosfue‘mayor en los bosteaderos (Tabla 18).Tanto en el analisis de ia riqueza como enel análisis del número de individuos obtuvediferencias significativas en el verano,siendo mayor en ambos casos para iosbosteaderos. En ei resto de las estacionesesta tendencia de mayor abundancia yriqueza de artrópodos en los bosteaderos semantuvo pero las diferencias fueron‘ nosignificativas (Fig. 36).
Los artrópodos más abundantes enlos bosteaderos fiJeron las hormigas, iosescarabajos estiercoleros y las arañas,mientras que en los controles los másabundantes fueron las hormigas (Tabla 18).Tanto en los controles como en iosbosteaderos en invierno y otoño ia hormiga.

84 Susana Pameia Bravo
Tabla l7: Proporción de hojas afectadas en los individuos de tres especies por los distintos tipos de daño.Las letras diferentes indican diferencias significativas con una probabilidad menor al 0,05 de lu prueba deTukey para 0. diospyrifolia y de comparaciones múltiples entre tratamientos para P. carrhagenensis.
E. pum'crfolr'a 0. diospyrrfolia P. carthagenensr's
Secas 0,05 0,09 a 0,0] c
Quemaduras 0,00 0,0l a 0,02 cCortes 0,02 0,08 a 0,2l d
Herbivoría 0,04 0,05 a 0,l5 d
patógenos 0,l3 0,50 b 0,28 dTotal ' -0,24 0,67 0,66
Tabla l9: Resultados del Anova comparando la remoción de semillas que realizó cada taxa (vertebradosdiurnos, nocturnos e invertebrados) en los bosteaderos y en los controles (Sitio). CM = Cuadradosmedios.
0. diospyrifoliaQ
P. carlhagenensr's
Factores gl CM F P Factores gl CM F P
Taxa (T) 2 0,25 l,63 0,2] Taxa (T) 2 0,55 0,42 0,65Sitio (S) l 0,37 0,24 0,62 Sitio (S) l 0,20 0,16 0,69
T x A 2 0,37 0,24 0,79Error 54 0,]6
más abundante fue Pher'dole sp. y en veranoy primavera Odonromachus haemarodes.pero estas especies siempre las capture enmayor número en los bosteaderos. En loscontroles encontré sólo 3 individuos de 2especies diferentes de arañas mientras queen los bosteaderos fueron 15 individuos de8 especies (Tabla 18). Siempre que se tratóde arácnidos adultos encontré uno portrampa, pero en el caso de individuosjuveniles fue común encontrarlos de a pares
En cuanto al grupo de loscoleopteros, los individuos de las cincoespecies de escarabajos estiercoleros losencontré prácticamente sólo en losbosteaderos y principalmente en verano(Tabla 18). Fue notorio como algunasespecies mostraron cierta especificidad desitio, por ejemplo mientras los Staphilinidosque encontré en Otoño-lnviemo estuvieronpresentes sólo en los bosteaderos, loscoleopteros pertenecientes a la FamiliaCassidae que encontré en PrimaveraVerano estuvieron mayoritariamente en lasáreas control (Tabla 18). Algo similarocun-iria con las avispas parásitas (Flia.
T x A 2 0,70 0,54 0,54Error 54 |,3l
Chalcididae), que si bien estuvieronpresentes en bajo número, 2 especies lascapturé sólo en los bosteaderos y la tercerasólo en las áreas control (Tabla 18).
Remoción de semillas
La remoción de semillas de ambasespecies en los bosteaderos y debajo de losárboles parentales fue muy baja. Tanto para0. diospyrrfolía como para P.carrhagenensis encontré que losvertebrados diurnos y nocturnosremovieron mía semillas de losbosteaderos, mientras que los invertebradosremovieron más semillas de los controles(Fig. 37 a y b), sin embargo las diferenciasno fueron estadísticamente significativas(Tabla ¡9).
DISCUSIÓN
Tipos de daño gue afectan el crecimiento
Losjuveniles de los bosteaderos delBosque de Laureles sufrieron más daños

Alouana caraju como dispersor de semillas 85
por herbivorla y cortes probablementecomo efecto de la alta densidad en que seencuentran. Los patógenos fueron losmayores generadores de daño entre losjuveniles de los bosteaderos de ambosbosques, la mayor parte era claramenteatribuibles a hongos (obs. pers.). Loshongos tienen distintas formas detransmisión, la primaria que se producedesde el suelo cuando este tiene un altonivel de infestación, la secundaria que es laque se produce desde un individuo cercanoenfermo y que se da sólo cuando losindividuos crecen en altas densidades, o lamediada por los insectos que puedentransportar esporas entre plantas (Jenningsy Lysek. 1999). Una alta proporción delataque de los hongos en los bosteaderos esespcrable ya que dada las característicasque estos presentan, permanencia en eltiempo por años, alta densidad de juvenilesy gran cantidad de insectos asociados, lainfestación por los hongos seria factible enlas tres formas conocidas. Según estudiosrealizados en Barro Colorado, Panamá, loshongos fueron el principal factor demortalidad para las plántulas de 9 especiesarbóreas (Augspurger, l984), aunqueexistieron diferencias interespeclficas, losescapes más efectivos estuvieron dados porla dispersión a áreas de baja cobertura o ladispersión en bajas densidades. Una vezinfestadas la muerte se producía en apenasuna semana, pero las plántulas de todas lasespecies lograban escapar a la mortalidad sise encontraban lignificadas (Augspurger,l984). Asi el aumento en'la velocidad degerminación (Capítulo lll) y el rápidocrecimiento de los juveniles en losbosteaderos (Capitulo V) pueden ayudar aque el ataque tan importante de los hongosen los bosteaderos no tenga efectos letalestan generalizados en la población dejuveniles ya que en corto tiempo ellos seencontrarían lignificados.
Si bien los hongos fueron el factormás relevante en ambos bosques, atacandomás individuos y de forma másgeneralizada (mayor número de hojas delindividuo atacado), al diferenciar entreespecies encuentro que existen diferenciasinterespecíficas en cuanto a los tipos dedaño. Estas diferencias han sido descripta
por otros autores tanto para el ataque ajuveniles como a semillas (Chapman, 1989;Dalling et al., 1998; Jordano y Godoy,2002; Lambert, 2002) y no deberiansorprender ya que cada especie poseeriaestrategias propias en cuanto a crecimiento(ver Capitulo lV) y defensa, a la vez quecada herbívoro también puede mostrarpreferencias en su dieta y además lospatógenos podrian tener mayor o menorespecificidad.
Es importante considerar que laforma de detección de los daños no esequivalente para todos los tipos. Porejemplo, las zonas quemadas sólo esposible detectarlas al poco tiempo que seproduce la quemadura, luego pasan a estarsecas o bien desaparecen. En algunos casosllevan a la muerte completa de losindividuos. Este tipo de daño podría ser unimportante factor de mortalidad para losindividuos ubicados en el centro delbosteadero.
alo
9
|1
6
< .1
l: 4
J
1 I—Il
o San! Own-tn Cm Habita-u
b tlo
á
—»uhuowuo DDCauHaüm-ia
Figura 34: Intensidad del ataque por losdistintos factores en evaluada con el lndice llA(ver materiales y métodos) en los bosteaderosdel Bosque de Laureles (a) del Bosque de Palo¡m (b).

86 Susana Pam'cia Bravo
Por lo tanto, este tipo de daño podría estarsiendo subestimado por el muestreo querealicé. Además podría estarsobreestimando los cortes, pues puedenincluir tanto efectos de herbivoría, comozonas quemadas o secas de una hoja que sehayan perdido. A pesar de esto, consideroque el muestreo que realicé es una buenaaproximación para obtener una descripcióngeneral del ambiente en el que seencuentran los juveniles al crecer en unbosteadero.
Los resultados de este capítuloconcuerdan con lo expuesto en el capituloanterior. En los bosteaderos de ambosbosques los juveniles sufren daños queestarían perjudicando su crecimiento, por lotanto no es posible considerarlos comositios seguros libres de herbivoría, ataquepor patógenos u otro tipo de perjuicio, peroa pesar de esto en los bosteaderos seproduce el mayor reclutamiento deindividuos para las principales especies quecomponen el Bosque de Laureles. Existencasos en los cuales los predadores ypatógenos se vuelven irrelevantes al evaluarel reclutamiento, pues no lograrlan dejar alas plantas por debajo de la densidadnecesaria para evitar la competenciaentreellas o para que otros sucesos limiten alas poblaciones en cuanto a número ydistribución (Diizo y Dominguez, 1986).Los bosteaderos de carayá en la selva deinundación serian un ejemplo de ello.
Disponibilidad de nutrientes
La diferencia en el tamaño de losindividuos que crecen dentro y fuera de losbosteaderos (ver Capítulo lV) no podria seratribuido a una diferencia en ladisponibilidad de nitrógeno, pero otrosfactores podrían estar actuandopositivamente en los bosteaderos, como serel contenido de materia orgánica del suelo,la menor compactación y la mayorhumedad del suelo en esos sitios.
Las muestras para los análisis denutrientes las tomé durante el invierno delaño 2000, este fue particularmente duropara el crecimiento de los juveniles de 0.dlospynfolía y E. pum‘cifolia (ver CapituloV), con una baja disponibilidad de agua
tanto de lluvia como del río (Apéndice). Porende es probable que estos juveniles noestuvieran captando nutrientes del suelo engran cantidad y de ahi que las diferenciasde concentración respecto a los juvenilesque crecían fuera de los bosteaderos nofueran elevadas. Por el contrario los juve
ac a
0 4
0 JÍ?C.
°' 0.2
0 |
A c 7P samsunrmi: 0. dmnflfilha E punu'lfillm
5
4 . l
J
J
Ez 2
l
o . 'ÏP ¡Wikun ().Milla!!! E"makina
Figura 351Porcentaje de fósforo (a) y nitrógeno(b) en los juveniles de las tres principalesespecies que crecen en los bosteaderos delBosque de Laureles (gris) y debajo de losparentales (blanco). ' Prueba de t de Studentp<0,05.
Niles de P. carrhagenensis, en los que siobservé diferencias significativas, crecieronde manera continua (ver Capítulo V). Paraconfirmar esta hipótesis sería necesariorealizar un seguimiento en el tiempo de laconcentraciónÉde nutrientes en los juvenileso por lo menos realizar un nuevo muestreoen una época donde se observe uncrecimiento importante de losjuveniles.
Artrópodos en los L ‘ ’ w
En general la materia fecal de losanimales en la naturaleza es consideradacomo un recurso efimero y de aparición

Alornma caraya como dispersor de semillas 87
azarosa en el tiempo y el espacio, lo queimpide la formación de comunidades deartrópodos estructuradas a partir de un usodiferencial del recurso, sino más bien elrecurso es utilizado por quien llega primero(Hanski, l99l). Sin embargo, en la materiafecal del ganado y de otros grandesvertebrados que puede durar varios días sepueden encontrar múltiple especies que sesuceden temporalmente, las más tardías seaprovechan de los efectos de la actividad delas especies más tempranas, por ejemplo lostúneles que producen los escarabajosestiercoleros facilitan la llegada de oxigenoa las larvas coprófagas de algunas moscas,pero también facilitan el acceso a suspredadores (Hanski, 199]). El caso de losbosteaderos de carayá serían un casoextremo en donde es posible encontrar
Propde¡anillasmmm/idas
Figura 37: Proporción de semillas de Ocoleadiospyrifolia (a) y Prychalria carrhagenensis(b) removidas de los bosteaderos (gris) y dc loscontroles (blanco) por cada taxa durante losexperimentos de remoción.
materia fecal de distinta antigüedad y sibien no siempre habrá materia fecal fresca,esto ocurre con bastante frecuencia durantelos años en que los bosteaderos son
utilizados, de ahi que sea posible encontrartodas estas especies a un mismo tiempo.
No solo los hábitos alimenticios delos artrópodos scn importantes para evaluarposibles interacciones entre ellos y elestablecimiento de las semillas dispersadas.Por ejemplo, en el caso de los escarabajosestiercoleros es importante elcomportamiento de oviposición. puesindirectamente pueden acarrear semillas ytransformarse en dispersores secundarios.El acarreo y enterramiento de semillas porparte de los escarabajos estiercoleros esmayor cuanto mayor es la cantidad demateria fecal que rodea las semillas(Andresen, 2002). Este acarreo dependetanto del tamaño de la semilla como delescarabajo, por ejemplo escarabajosmenores a l cm sólo son importantes en laremoción de semillas menores a 0,5 cm(Feer, ¡999). Considerando los resultadosde Feer (¡999), y dado que el tamaño de 4de las 5 especies que capturé fue menor a lcm sólo podrían enterrar semillas pequeñascomo las de Banara arguta y Cecropiapachysrachya, semillas como las de E.punicifolr'a y P. carlhagenensís sólopodrian ser enten'adas en la isla por unaespecie de estiercolero que mide 1,5 cm yque además seria poco abundante. Semillasgrandes como las de 0. diospyrü'olílas nopodrían ser removidas ni enterradas porninguna de las especies que encontré.
Si bien el enterramiento facilita elescape a la remoción por roedores, para lasespecies de los bosques tropicales laeficiencia de la germinación decrece(Vander Wall, 1993; Andresen, 1999), porejemplo las semillas de Micropholísguyanensis (Sapotaceae) de tamañoequivalente a las semillas de 0.diospyrrfolia, si son enterradas a 5 cm deprofundidad sólo la mitad de las plántulaslogran alcanzar la superficie (Andresen,200l), algo similar ocurre con otrasespecies de semillas pequeñas que muestranuna disminución en el establecimientocuando aumenta la profundidad a la que sonenterradas (Dalling et aL, 1994).
Respecto a la hormigas, si bienfueron uno de los grupos más abundantes,ninguna de las especies más abundantes enlos bosteaderos es considerada granlvora o

88 Susana Pau-¡ciaBravo
invierno
g U = 32 g 6 U = 81.5E N = N = IS E N = N = ¡5E = 0.2| (¿l p = 02\ : 4un U)
.9 g3 .12
a I .2 2u‘3 2i - I
0Bosteadero Control Bosteadero Control
Primavera
tu a 6g- U = 122,5 E U = l ¡9m N = N = ¡9 g N = N = 153.-. p - 0‘09 : 4 p = 0,07\ u)a g3 .12 i
.9 I % 2 I0‘ I s
0Bosteadero Control Bosteadero Control
Verano
6g tuE 8g ¡g 3’“ g 4 U= l07\ El=40 : N= N = 2|
g í p = 0.0008 8 p = 0. 43.cÏr g 2m .
I g Ï0
Bosteadero Control Bosteadero Conirol
Otoño
6g tuE Q.m E= e\ : 4fi ena, o3 3.9' 29 2m ..>.'o
E¡ _ o ¡ _
Bosteadero Control Bosteadero Control
l-‘iguru 362Riqucm promedio ( L desvio cslúndur) dc urlrópmius por lrumpu y número prmncdiu du:individuos ( 1 desvio cslúndur) caidos por lnuupu cn cudu cslnción.

AIouaIIa caraya como dispersor de semillas B9
cortadora, son más bien cazadoras,carroñeras u omnivoras, por lo tantorepresentarían una protección para lasplántulas y las semillas, de igual formapodrían estar actuando las arañas. Ambosgrupos podrían estar reduciendo el impactode los herblvoros, de manera similar a loque se observa en las especies vegetales queatraen artrópodos cazadores por medio denectarios extraflorales (Holldobler yWilson, 1990).
Remoción de semillas
El nivel de remoción de semillas enla isla Brasilera fue muy bajo, en ambientestropicales las semiler suelen ser removidasen proporciones muy elevadas, por ejemploel 90 % de las semillas presentes en lasheces de Alouarra palliata en los Tuxtlas,México (Estrada y Coates-Estrada, ¡991),el 68,8 % para Necrandra sp., una Lauraceacon semillas de similar tamaño a las de 0.diospyrrfolia, también en los Tuxtlas(Sánchez-Cordero y Martinez-Gallardo,¡998) y entre el 90 % y 93 % de lassemillas que están en las heces de Alouarraseniculus en la Guayana Francesa (Feer,¡999)
El olor de la materia fecal no sóloatraeria a los escarabajos estiercoleros sinoque puede funcionar como atractor para losroedores (Andresen, 1999; Andresen,2002), este puede ser uno de los motivospor lo que los bosteaderos hayan tenido unamayor remoción de semillas por parte delos vertebrados n0cturnos que las zonascontrol. La baja tasa de remoción podríadeberse a la ausencia de grandes roedoresen el área, aunque en experiencias similaresllevadas a cabo en Costa Rica con Ocoreaendresiana la mayor parte de ladepredación estuvo dada por roedores depequeño tamaño (Wenny, 2000).
La tasa de remoción de semillas delos vertebrados diurnos fue aun menor a latasa de remoción de los vertebradosnocturnos, probablemente esto se deba aque si bien durante el invierno de 2000observamos actividad diurna de losroedores esto coincidió con un considerablecrecimiento poblacional que para el 200l,luego de la inundación. habría decrecido
considerablemente (obs. pers.). Por otrolado, una gran parte de las aves del lugar sealimentan de frutos, pero no serian atraídassimplemente por las semillas limpias comocolocamos en los experimentos y como seencuentran en los bosteaderos. La tendenciacontrapuesta de los vertebrados respecto alos artrópodos acerca del sitio para obtenersemillas resulta llamativa, seria convenienterealizar este mismo experimento con lassemillas más pequeñas de Banara argura yCecropia pachysrachya para conocer si estatendencia se mantiene. En la selva lluviosade Costa Rica se ha encontrado que lashormigas son los más importantesremovedores de semillas de este tipo y quesuelen obtenerlas de las heces de losvertebrados incluso de Alouaua palliala(Levey y Byrne, l993; obs. pers.).
Es de esperar que las hormigas seanlos principales responsables de la remociónpor los invertebrados en los experimentos,entonces una razón la baja remoción desemillas que se encuentran en losbosteaderos podria ser la importantepresencia de hongos que existiría en losmismos, ya que las hormigas evitan condistintas estrategias su contaminación y lade las cámaras de sus honnigueros(Hblldobler y Wilson, ¡990).
En otros ambientes no tropicalestambién se ha encontrado por experienciasde adición de semillas que la aparición deplántqu sólo aumenta cuando se limita lapredación postdispersiva (Jordano y Godoy,2002), sin embargo en la isla Brasilerapareciera que es más importante ladepredación predispersiva. Durante unperiodo de alta densidad de roedores en elinvierno del 2000 realicé un monitoreo deeste tipo de depredación y resultó que 92de 100 árboles de P. carrhagenensisseleccionados a lo largo de una transecta de600 metros mostró signos de predaciónpredispersiva por roedores (restos de frutasy de cubiertas de semillas roldas, ramilletesde frutos cortados acompañados demonticulos con los restos de las cubiertasde las Semillas roldas y heces de roedoressobre y debajo de los árboles). También 0.diospyrifolr'a, Banara arguta y E.puniclfolia sufren predación predispersiva

90 Susana Patricia llmvo
9 7
Avispas parásitas
Hormigas
predadoras Arañas g Moscaspredadoras//Escarabaj os
predadores
\ Moscascon
dieta mixta
ti ‘
estiercoleros
Hormigas cortadoras '
Colémbolos y granívoras Escarabajos estiercoleras\ - ' l
<—-—
I i
' ll l
l l
l l
I l
t I
I ImÑ ' Materia fecal '
Semillas + | de lI - carayá 'l [I’ ’ _ i _ - l dI /z /
zz' /Roedores ' / z ' l/
/ / Ü’
m Plantasjuveniles
Figura 38: Red alimentaria postulada a partir de los resultados del Capítulo V para los bosteaderos decarayá y basada en la red alimentaria que se encuentra en la materia fecal del ganado en Europa (l lanski.|99|). Los recuadros gruesos indican los componentes particulares que fonnnrían parte de la cadenaalimentaria dentro de los bosteaderos. Los flechas con linea puntuada indican los factores con menorimportancia y el signo de pregunta indica que la relación se supone a partir de pocas observaciones.

Aloualla caraya como dispersor de semillas 9|
pero por parte de artrópodos (ver Capítuloll).
Considero que varios son losmotivos por los cuales el impacto de losartrópodos y vertebrados que remuevensemillas es menor al encontrado enambientes tropicales, la menor densidad deartrópodos sumada a su variación en el año,la presencia de gran cantidad de artrópodoscazadores y finalmente la gran cantidad demateria fecal que llega al suelo. esto haceque el reclutamiento de individuos de lasdistintas especies de árboles sea tanefectivo y se produzca de manera tanconcentrada en los bosteaderos. Porejemplo mientras en Los Tuxtlas, México,la producción de materia fecal de losaulladores que llega al suelo fue estimadaen ¡1,2 gr. ha" dia" (Estrada et aL, 1993),en Barro Colorado, Panamá Sl gr. ha'l dla'l(Gill, 1991), en la Guyana Francesa55 gr. ha"dla" y en Manaus, Brasil losmontículos de materia fecal de Alouattaseniculus que llegan al suelo varian entrelos 5 y 30 gr., en la selva de inundación hayuna llegada diaria al suelo de entre 5 ylO Kg por ha de materia fecal, lo queproduce los bosteaderos ya descritos deimportante diámetro y con varioscentímetros de profundidad (ver Capituloslll y lV). Esto se debe a que a diferencia delos otros sitios donde se ha estudiado a losaulladores, en esta selva la densidad esextremadamente alta, y las heccs de cadaindividuo llegan casi completas al suelo sinser interceptadas por la vegetación.
Integración
Considerando los resultados delpresente capítulo y la red alimentariadescripta para la materia fecal del ganadoen Europa, la Figura 38 describeesquemáticamente la red alimentaria que es
posible postular para un bosteadero decar-ayaen la isla Brasilera.
Al modelo original propuesto parala materia fecal del ganado en Europa en laselva de inundación de Argentina se sumanlas arañas y las hormigas carnívoras entreotros. Según algunas observaciones quepude realizar, ciertas aves insectivoras sealimentaron en los bosteaderos (e.g.,Xiphocolaptes major). También los Coatiespodrían estar alimentándose de artrópodosen los bosteaderos, pero dada la fuertepresión de caza que esta sufriendo estaespecie en el lugar no me fue posiblecomprobarlo fehacientemente.
La herbivoría y_granivorla existentanto en los bosteaderos como fuera de ellosy es necesario que los incorpore en elesquema, aunque no serían muyimportantes según la descripción de losfactores adversos para el crecimiento, losexperimentos de remoción y el bajo númerode especies de artrópodos herbivorosencontrados en los bosteaderos. El factornegativo más importante para elcrecimiento de las plántulas es laimportante presencia que tienen lospatógenos, en particular los hongos, y elfactor positivo más importante sería laabundante y continua llegada de semillas.
Los resultados sugieren que lasimple hipótesis de saturación de hcblvorosy patógenos no seria la explicación másapropiada para postular como mecanismode reclutamiento en los bosteaderos. Másbien se trataría de una serie de factoresintel-relacionados, en parte saturación de lospredadores de semillas y patógenos, enparte disminución de la herbivoría por lapresencia de predadores y en parte unamayor disponibilidad de nitrógeno queaceleraría la velocidad de crecimiento yaumentaría el vigor de los juveniles.

¡llouafla caraya como dispersor de semillas 93
Capítulo VII:El papel de Alouatta caraya en
la dinámica y regeneración de la selva de inundación,
“Desde el Río de la Plata hasta las Misiones no seencuentran bosques más que a orillas de los ríos yarroyos. y los ríos, pero estos bosques se destruyen amedida que el país se puebla. "
"Hay un laurel (muydiferente del de España) que seemplea sobre todo para las ensambladuras de losnavíos "
"Se corra también un poco de leña a orillas de losarroyos de la costa septentrional y en las islas delParaná y del Uruguay. Se encuentra también unpocode madera propia para la construcción de carreras,casas y barcos más o menos grandes.... "
Felix de Azara(Viajespor la América meridional)
Integración y discusión general
En la isla Brasilera pude distinguirtres tipos de bosques el de Laureles, el deSangre de Drago y Timbó y el de Palo itá, entodos ellos habitan grupos de caraya aunquede distinto tamaño. En el Bosque de Laureles,carayá remueve las semillas de la mayor partede las especies que componen el bosque.Estas semillas son depositadas en losbosteaderos puntuales, en los donnideros o endeyecciones independientes. El 75 % de lasdeyecoiones se producen en bosteaderos, yasean puntuales o los que se encuentranrodeando los árboles dormidero. Losbosteaderos se encuentran a distancias muyvariables respecto de los árboles parentales eincluso en algunos casos debajo de ellos,provocando que la dispersión de semillasmediada por caraya sea independiente de ladistancia.
El pasaje de las semillas por el tractodigestivo de caraya tiene distintos efectosdependiendo de la especies. Puede ser neutro,puede aumentar la velocidad de germinacióno puede eliminar larvas que se encuentrendentro de la semilla. En O. diospyrifoliaincluso, existen diferencias intraespecificasen cuanto a la velocidad de germinación queson eliminadas por ese mecanismo. Una vezen el suelo, tanto en los bosteaderos comofuera de ellos, las semillas no tuvierongrandes riesgos de ser predadas o removidas,al menos las de 0. diospyrifolia y las de P.
carthagenensis. Sin embargo, no evalué esteriesgo en semillas de tamañopequeño, ni evalue’el ataque de los patógenossobre las semillas. _
Carayá actúa como dispersor local enel Bosque de Laureles, y su efecto se puedeapieciar en los bosteaderos, donde encontréun tercio más de especies y 6 veces másjuveniles que en las zonas control tomadas alazar dentro del bosque. Al evaluar laabundancia de renovales en los bosteaderosencontré que hay en promedio casi S vecesmás renovales que en las áreas control. En losbosteaderos y alrededor de los árbolesdonnideros encontré individuos de distintasedades reclutados a lo largo de varios años,mientras en sus respectivas áreas control paraalgunas especies no había renovales, sólohabia renovales pequeños para otras. En elúnico caso que encontré renovales debajo delos árboles parentales fue para P.carthagenensis y E. punicifolía. Por lo tanto,los bosteaderos de carayá representan zonasde reclutamiento dentro del Bosque deLaureles y cuentan con una mayor diversidadde juveniles y renovales respecto al resto delbosque. En cuanto al crecimiento, losjuveniles de 0. diospyrifolia, E. punicifolia yP. carlhagenensis que crecían en losbosteaderos tenian mayor altura y hojas demayor tamaño que los que crecían en lasáreas control, así los bosteaderos serian nosólo sitios favorables para el establecimiento,

94 Susana Patricia Bravo
sino también para el crecimiento. En losbosteaderos habría una disponibilidad mayorde nitrógeno para el desarrollo de losjuveniles. Por otra parte los bosteaderostendrian asociada toda una red tróñca deartrópodos que interactuaría de distintasformas con las semillas yjuveniles arbóreos.
Como representa la Figura 39, losgrupos de carayá que habitan en el Bosquesde Palo itá y en el de Sangre de Drago yTimbó, también tendrían acceso a zonas delBosque de Laureles. Al llevar las semillas delas especies que componen este bosque a losbosteaderos que se ubican en los otros dos,carayá actúa como dispersor de largadistancia. Las semillas de estas especiesgerminaron en los tres boSques', ya queprácticamente encontré individuos juvenilesde todas ellas. El número de juveniles en losbosteaderos del Bosque de Laureles fueaproximadamente 4 veces superior al de losbosteaderos del Bosque de Sangre de Drago yTimbó, y 6 veces superior al de los delBosque de Palo itá. La diferencia entre elBosque de Laureles y los otros dos se deberíaa que un número menor de semillas estásiendo depositado en ellos pues además detratarse de una dispersión de larga distanciadonde las fuentes de semillas son lejanas, losgrupos de carayá del Bosque de Laureles sonmucho más numerosos. La diferencia entre elBosque de Sangre de Drago y Timbó y el dePalo itá se debería a diferencias en lamortalidad de plántulas y juveniles, odiferencias en la tasa de pnedación yremoción de semillas.
La composición específica de losrenovales en los bosteaderos del Bosque deSangre de Drago y Timbó es similar a la delos juveniles y a la del Bosque de Laureles.Los conteos de renovales muestran que a lolargo de los años un considerable número deindividuos de las especies que caracteriïan alBosque de Laureles han logrado establecerseen los bosteaderos del Bosque de Sangre deDrago y Timbó, por lo que es de esperar quepor efecto de la dispersión de semillasrealizada por carayá la fisonomía de estebosque con los años se asemeje ala del
Bosque de Laureles (Fig. 39).En los conteosde renovales del Bosque de Palo itá tanto elnúmero de especies como la cantidad deindividuos promedio por bosteadero sereduce a la mitad respecto a los otrosbosques. Es decir, que la mayoría de lasespecies no logran establecerse en estebosque, por lo que es de esperar que sufisonomía no cambie por efecto de ladispersión de semillas por parte de carayá(Fig. 39). Las inundaciones serían un factorde mortalidad entre los juveniles máspequeños y dado que este bosque tiene unnivel de suelo algunos metros más bajo quelos otros dos, las inundaciones son másfrecuentes y prolongadas, por lo tanto podríanser las' responsables de la falta dereclutamiento en el Bosque de Palo itá (Fig.39). Si bien el estudio de crecimiento de losjuveniles indicó que la tasa de mortalidad eneste bosque fue similar a la del Bosque deLaureles y que la desaparición de especies delos bosteaderos del Bosque de Palo ita, sedebió al bajo número de juveniles que teníanal principio de las observaciones, hay quetener en cuenta que durante el primer año deobservación no hubo inundaciones, y en elsegundo hubo una inundación de duraciónprolongada (lS días), que afectó a todos lostipos de bosque, y dos inundaciones duranteel 2002 que sólo afectaron parte del Bosquede Palo itá, pero duraron menos de unasemana y apenas alcanzaron algunoscentímetros de altura (Apéndice). Por lotanto, el periodo de muestreo no incluyó elefecto de la frecuencia diferencial deinundaciones prolongadas entre estosbosques. Este efecto estaria reflejado en elmuestreo de renovales, pues los individuostenían más de dos años de edad, ya que susalturas son superiores a la mayor alturaalcanzada por los juveniles que observécrecer durante ese período. Como destaqué enel Capítulo V el número de renovales es muyvariable en el Bosque de Palo itá,probablemente debido a diferenciastopográficas que pueden facilitar lasupervivencia de los juveniles en losbosteaderos ubicados en la zona más alta

Dispersiónlocal
95
'..<Cae
.2"
(Cap.I)
WII
(Clpvun
Alouatla caraya como dispersor de semillas
BosquedeLaurelesJuveniles
(Cap.IV)
\/
O
Renovales
(CapiIV)
(Cap.V)
<':=
dioqueseencuentranenunbosteadero,aexcepción
eroderenovalesenelBosquedePaloitaesmuyvariable.
sugrosorrelativoserelacionaconelnúmerodeindividuosinvolucradosasícomoel
Dispersióndelargadistancia
BosquedePaloitá
,(Cap.l)
Juveniles(Cap.V)Renovales
. (Cap.V)\T E(CapiV),3:m
A“ Inundacióndada
BosquedeSangredeDragoyTimbóprerdadores
(Cap.l)
a
OJuveniles
(Cap.V)
(Cap.V)
Figura39:Integracióndelosresultadosdescribiendoelpapeldecarayácomodispersorlocalydelargadistanciaparalasespeciesquecomponen
elBosquedeLaurelesdelaIslaBrasilera.
Lasflechasgrisesrepresentaneltrasladodesemillasysugrosorserelacionaconlacantidad(sinunaescalareal).
Loscírculosrepresentanelconjuntodeindividuosprome
relativoentreelrestomantienenunaescalareal.Tenerencuentaqueelnúm
Lasflechasqueconectanloscírculosindicanlospasajesdeunoaotroestadio,
largoindicaunmayoromenortiempoinvolucradoenelproceso.
LasflechasquesedirigendesdelosCírculosalosfactoresambientalesindicanlasalidadeindividuoscomoefectodeundeterminadofactoryaquellasconsentido deloscirculos“semillas”eltamaño
opuestoindicanunefectopositivodeundeterminadofactorambientalparalasupervivenciadeindividuososobreelprocesodecreciento.

96 Susana Patricia Bravo
dentro de dicho bosque.Si bien las inundaciones producirían
mortalidad entre losjuveniles, la velocidad decrecimiento en los bosteaderos aumentaría
una vez que estas terminan y los juvenilesque sobrevivir recuperan con creces labiomasa perdida. La influencia de lasinundaciones en el crecimiento fue másnotoria en los escasos individuos quesobreviven en los bosteaderos del Bosque dePalo ita. La diferencia en el crecimientoposinundación entre bosques podría deberseal mayor tiempo y nivel de inundaciónalcanzado en el Bosque de Palo itá que podríarepresentar un mayor aporte de nutrientes, y ala mayor luminosidad de este bosque.
Que tanto los bosteaderos como losdorrnideros de carayá mostraran ser sitiosfavorables para el crecimiento yestablecimiento de los juveniles arbóreos, nosignifica que una semilla que llegue a unbosteadero tenga asegurada su germinación yposterior crecimiento, por el contrarioalgunas semillas serán removidas, otrasgerminarán y sufrirán una gran mortalidad aconsecuencia de diversos factores (sequías,inundaciones y patógenos entre otros), y sóloalgunos pocos juveniles lograránestablecerse. Por lo tanto la probabilidad deque una semilla dispersada por carayá lleguea dejar un individuo adulto es baja, perocuando una semillas no es dispersada porcarayá esta probabilidad es casi nula para lamayoría de las especies. La reproducciónefectiva de los individuos de algunas especiescuando son dispersadas por carayá, selograría debido a la llegada continua de unagran cantidad de semillas que puede saturarpredadores y patógenos, la presencia deartrópodos predadores que pueden limitar alos artrópodos fitófagos y a 'una mayordisponibilidad de nitrógeno que aceleraría elcrecimiento, aumentando las posibilidades deescapar de la mortalidad producida porpredadores y patógenos sobre los individuosmás pequeños. Para responder la preguntaoriginal, ¿Cuál es el papel de carayá en ladinámica y regeneración de la selva deinundación”), no interesa la suerte particular
de una semilla dispersada, sino laprobabilidad de dejar descendientes para elconjunto de árboles que forman esta selva.Por lo tanto, el papel de carayá por medio dela formación de bosteaderos, sería de sumaimportancia en la dinámica de la selva deinundación, pues logra que en ellos seestablezcan individuos de las principalesespecies que la componen. La dispersión desemillas que realiza carayá es además elprincipal factor que permite la llegada de lasespecies de fruto camoso a los bosquesnuevos dominados por especies dispersadaspor factores abióticos (Bosque de Sangre deDrago y Timbó), cambiando de este modo sufisonomía y composición. La dispersión desemillas por carayá no parece ser un factorimportante en la dinámica del Bosque de Paloítá, ni para la reproducción de los individuosde la especie dominante en este bosque(Banara arguta).
Claramente el efecto de carayámantiene o tiende a incrementar la diversidadalfa dentro de los bosques, pero el factor quemantendría la riqueza de ambientes en la isla(i.e., diversidad beta) sería el régimen deinundaciones del río. Las inundaciones seríanlas responsables de generar nuevas áreas en laisla, ofreciendo una superficie apta para elestablecimiento de Bosques de Sangre deDrago y Timbó en las más elevadas, o deBosques de Palo itá en las de menor altura.Carayá transformará mediante la dispersiónde semillas los Bosques de Sangre de Drago yTimbó en nuevos Bosques de Laureles. Miestudio no puede responder ¿Cuál es el origendel primer Bosque de Laureles en una islanueva? ¿Llegan las semillas en el tractodigestivo de carayá cuando este arriba? o¿Carayá arriba a la isla luego que las semillashan llegado por otros medios y ya existenlaureles? Para responder estas preguntas sedebería tener acceso a una isla de recienteformación y monitorear cuando se produce lallegada de carayá y cuando la aparición de losárboles de frutos camosos.
Dentro del proceso de expansión dela distribución de una especie se puedendelimitar tres fases, la de establecimiento, la

Alouarra caraya como dispersor de semillas 97
de expansión y finalmente la fase desaturación (Shigesada y Kawasaki, 2002). Alrealizar simulaciones sobre la expansión deuna especie, basadas en los modelos queconsideran los efectos de la dispersión localmás los efectos de la dispersión de largadistancia, se observa que los límites de ladistribución no son continuos, sino que seproducen poblaciones satélites por fuera de lapoblación original que se van expandiendo,colapsando entre sí y con la poblaciónoriginal, estas poblaciones explicarlan laaceleración en la fase de expansión dealgunas especies (Clark, ¡998; Clark et al.,¡998; Shigesada y Kawasaki, 2002). A unmodelo de este tipo se asemejarlan los efectosde carayá como dispersor. Mientras que elBosque de Laureles se encontraría en la fasede saturación, en el Bosque de Sangre deDrago y Timbó, los bosteaderos y donniderosrepresentarian las colonias satélites, quecuando comiencen a producir frutos seexpandirían, a la vez que generarían nuevascolonias como efecto de la dispersión localque produzca carayá.
En el caso particular de la selva deinundación la aceleración podría ser aunmayor a lo esperado por el simple mecanismode generación y expansión de colonias. Pues,cuando los renovales lleguen a la madurez ycomiencen a producir frutos, representarán unaumento de recursos de alta calidad paracarayá, lo que llevaría a un aumento de lasdensidades y tamaño de los grupos en elBosque de Sangre de Drago y Timbó deacuerdo con el modelo propuesto en elCapítulo ll. Un aumento de densidad en losgrupos de carayá implicaría, mayor densidadde bosteaderos, ya que los territorios serianmás pequeños, un aumento en el número dedonnideros y un aumento en el número debosteaderos puntuales relacionado con elincremento en las confrontaciones entregrupos. Un aumento en el tamaño de losgrupos implicará, bosteaderos de mayortamaño y un mayor número de semillasremovidas por árbol. Se generaría de estaforma una retroalimentación positiva entre ladensidad del dispersor y la de los dispersados,
por lo tanto no sólo la expansión seríaacelerada sino que a su vez esta aceleracióniría en aumento (Fig. 40). Esto podríaexplicar la rápida recuperación de la selva deinundación de las islas del Paraná y de suspoblaciones de carayá luego de lasinundaciones extraordinarias, sólo seríanecesario que sobrevivieran unos pocosárboles de fmto camoso en cada isla.
La fenología de la mayoría de estasespecies es altamente variable, no siguepatrones temporales demasiado fijos y seencuentra asociada a la disponibilidad deagua, ya sea que provenga de los ciclos deinundación o de las lluvias (Placci, 1995). Engeneral, las inundaciones funcionan comodisparadores de la floración, pero el'procesode formación de frutos puede abortarse si seproduce posteriormente una sequía (Placci,1995). Por lo tanto, cualquier modificación dela dinámica del río que suspenda lasinundaciones tendría un efecto sobre lafenologla, disminuyendo la disponibilidad defrutos y por ende produciendo un efectocontrario al descrito en el parrafo anterior,pues disminuirla la disponibilidad de recursosde alta calidad (flores y frutos). Si estasuspensión se vuelve permanente llevaría auna disminución notable en la densidad decarayá acarreando una degradación generalde los bosques, al obstaculizar laregeneración en el Bosque de Laureles y lasmodificaciones en el Bosque de Sangre deDragoyTimbó.
Mi trabajo integra todas las etapas dela dispersión de semillas para evaluar cual esel papel de carayá en la dinámica yregeneración de la selva de inundación,encontrando que la población de esta especieen la lsla Brasilera junto con el régimenpulsátil del Río Paraná son factoresfundamentales para mantener la dinámica yfisonomía actual de estos bosques. Sinembargo, dado el mal estado de conservaciónde la isla, sería importante discutir la validezde estos resultados. El principal impacto quepresenta actualmente la lsla Brasilera es lacontinua y alta presión de caza, que sumadoal pequeño tamaño de la isla a producido la

98 Susana Patricia Bravo
INUNDACIONES EXTRAORDINARIAS
Densidad deAlouarta caraya
+
Abundancia espacial Áreas de bosque Probabilidady temporal de con baja delos recursos frecuencia de uso encuentros
de alta calidada
+
Restricción . _ +energética Actrvrdad v
Frecuencia deconfrontaciones
Figura 40: Modelo que integra los resultados de disponibilidad de recursos, comportamiento y uso del espaciode caraya en la lsla Brasilera con su impacto como dispersor de semillas. Las flechas indican la dirección dela relación entre variables. El signo + indica una relación positiva entre las variables conectadas por la flecha.El signo - indica una relación negativa entre las variables conectadas por la flecha. La linea punteada indica laretroalimentación positiva propuesta.

Alouana caraya como dispersor de semillas 99
desaparición de los grandes carnívoros (pumay yaguareté) y la casi ausencia de herbivorosde tamaño medio y grande, (comocarpinchos, guasuncho o ciervos de lospantanos). Si, es frecuente encontrarpequeños gatos, rapaces, zorros, víboras ymicroroedores. Se ha hipotetizado que lasinteracciones de arriba hacia abajo (“top down”) podrían estar estructurando ladinámica de la vegetación en los bosquesneotropicales, los grandes predadoresregularían las poblaciones de los herbivorosde tamaño medio, quienes a su vezregularían el reclutamiento de las plántulas(Terborgh, 1992). Los efectos de ladefaunación no son simples de predecir puesdependen del ecosistema en si. Por ejemplo,la depredación de semillas y plántulasaumentó extraordinariamente cuando seeliminaron los mamíferos de más de l Kg depeso en las islas del Lago Gatun en Panamá(Asquith et aL, 1997). Pero en los Tuxtlas,México, con una defaunación similar, sin
z/
mamíferos predadores ni herbivoros mayoresa l Kg de peso, se observó el efecto opuesto,al parecer esto se explicaría por un aumentodesmedido en la densidad de la víboraBothrops spp., que habría ocasionado unaimportante reducción en las poblaciones depequeños roedores del área (Dirzo y Miranda,l99l en Asquith et al, 1997). De todasformas, los resultados de esta tesis deben serinterpretados en el contexto de unadescripción de las relaciones ecológicas quese producen en el marco de la dispersión desemillas que realiza actualmente carayá en laSelva de inundación de las islas del Río
Paraná. Teniendo en cuenta que la granmayoría de estas islas sufren cierta presión decaza, los resultados serán similares en lamayoría de ellas. Es de esperar que en islascon su fauna completa estas relaciones sevuelvan aun más complejas haciéndosequizás menos notorio el impacto de carayá ydisminuyendo la intensidad y velocidad delos cambios en los bosques de las islas.

Alaualla caraya como dispersor de semillas l0l
BIBLIOGRAFÍA
Adámoli, .l. ¡999. Los humedales del Chaco y dell’antanal. En: Tópicos sobre humedalessubtropicales y templados de Sudamérica.Malvárez, Al (Ed). Unesco, Montevideo.Uruguay. Pp. 85 - 93.
Altmann, J. I974. Observational study of behavior:Sampling methods. Behaviour 49: 227 - 267.
Alves, lMSC y Guix, JC. ¡992. Feeding habits ofAlouatra caraya in a semi-natural area (SEBrazil). Mammalia 56: 469 - 472.
Andresen, E. l999. Seed dispersa] by monkeys andthe fate of dispersed seeds in Peruvianruinforcst. Biotropiea 3|: 145 - ¡58.
Andresen. [-2.200l. Effects of dung presence, dungamount and secondary dispersal by dung beetleson the l'ate of Micrapholis guyanensis(Sapotaccae) seeds in Central Amazonia.Joumal of'l‘ropical Ecology l7: 6| - 78.
Andresen, E. 2002. Primary seed dispersal by redhowler monkeys and the effect of defecationpatterns on the fate of dispersed seeds.Biotropica 34: 26] —272.
Asquith, NM; Wright, SJ y Clauss, J. ¡997. Doesmammal community composition controlrecruitment in neotropical forests? Evidencefrom Panama. Eoology 78: 94| - 946.
Augspurger, CK. ¡984. Seedling survival oftropical tree species: Interaction of dispersaldistance, light-gap, and pathogens. Ecology65: [705 - l7l2.
Augspurger, CK y Kitajima, K. ¡992. Experimentalstudies of secdling recruitment from contrastingseed distributions. Eeology 73: l270 - ¡284.
Balcomb, SR; Chapman, CA y Wrangham, RW.2000. Relationship between chimpanzee (Pantrogloafivles) density and large fleshy-li'uit treedensity: Conservation implications. AmericanJournal of Primatology 5]: l97 - 203.
Begon, M; Harper, JL y Townsend, CR. ¡996.Ecology: lndividuals, populations andcommunities. Tercera edición. BlackwellScience, Reino Unido. 1068 p.
Bicca-Marquez, JC. 1994. Padrao de utilizaeao deurna ilha de mata por 'Alouana caraya(Primates: Cebidae). Revista Brasilera deBiologia54: l6| - l7l.
Bilgener, M. l995. Chemical factors influencingfood choice of howler monkey (Alouallapal/iaIa). Tropical Journal of Zoology l9: 291 303
Bonetto, AA y Hurtado, S. 1999. Cuenca del Plata.En: Los humedales de la Argentina:Clasificación, situación actual, conseravción ylegislación. Canevarí, P; Blanco DE; Bucher,
EH; Castro, G y Davidson, l (Eds). Segundaedición. Wetland International. Pp. 31 - 72.
Bourliere, F. ¡985. Primate communities: theirstructure and role in tropical eoosystems.International Journal of Primatology 6: l - 26.
Bravo, SP y Zunino, GE. 1998. Effects of blackhowler monkey (Alouana caraya) seedingestion on ínseet larvae. American Journal ofPrimatology45: 4ll - 415.
Bravo, SP y Zunino, GE. 2000. Gennination ofthree arborcal seeds dispersed by black howlermonkeys (Alouatta caraya). Folia Primatologica7|: 342 - 345.
Brown, A y Zunino, GE. 1994. Habitat,distribución y problemas de conseravción de losprimates de la Argentina. Vida SilvestreNeotropical 3: 30 - 40.
Brown, N; Jennings, S; Wheeler, P y Nabe-Nlelsen,.l. 2000. An improved methods for rapidassessment of forest understorey lightenvironments. Journal of Applied Eoology37: ¡044 - ¡053.
Bucher, EH y Chani, JM. ¡999. Chaco. En: Loshumedales de la Argentina: Clasificación,situación actual, conservación y legislación.Canevarí, P; Blanco DE; Bucher, EH; Castro, Gy Davidson, l (Eds). Segunda edición. Wetlandlntemmional. Buenos Aires. Argentina. Pp. 73 96.
Bullock, JM; Moy, lL; waell, RF; Coulson, SJ;Nolan, AM y Caswell, H. 2002. Plant dispersa!and eolonization processes at local andIandscape scales. En: Díspersal ecology.Bullock, JM; Kenward, RE y Hails, RS (Eds).British Ecological Society, Blackwell ScienceLtd. Reino Unido. Pp. 279 - 302.
Burkart. A. 1957. Ojeada sinóptica sobre lavegetación del delta del Río Paraná. Darwinianal l: 457 - 461.
Burkart, A. 1969. Flora ilustrada de Entre Rlos(Argentina). Colección Científica INTA 6: l 415.
Burslem, DFRPP; Grubb. PJ y Turner, lM. 1996.Responses to simulated drought and elevatednutrients supply among shade-tolerant treeseedlings of lowland tropical forest inSingapore. Biotropica 28: 639 - 648.
Cabrera, AL. 1976. Regiones fitogeográficas deArgentina. En: Enciclopedia Aregntina deAgricultura y Jardinería. Tomo ll, fasclculo l.Segunda edición. ACME. Argentina. 85 p.
Canevarí, P; Blanco DE; Bucher, E; Castro, G yDavidson, l (Eds). 1999. Los humedales de laArgentina, clasificación, situación actual,

102 Susana Patricia Bravo
conservación y legislación. Segunda edición.Wetland International. Argentina. 208 p.
Carignan, RC y Neil‘f, JJ. [992. Nutrient dynamicsin the floodplain ponds of the Paraná River(Argentina) dominated by the water hyaeinthEichornia crassipes. Biogeochemistry l7: 85 12].
Carnevali, R. l994. Fitogeograña de la Provinciade Corrientes. lN'l‘A. Corrientes. Argentina.324 p.
Chambers, .lC y MueMahon, JA. ¡994. A day in thelife of a seed: Movements and fates of seeds andtheir implications for natural and managedsystems. Annual Review of lícology andSystematic 25: 263 - 292.
Chapman, CA. ¡988. Pattern foraging and rangeuse .by three species ol‘ neotropieal primates.Primates 29: l77 - 194.
Chapman, CA. l989. Primate seed dispersa]: thefate of seed dispersed seeds. Biotropiea 2l:l48 - 154. .
Chapman, CA; y Chapman, LJ. 2002. Plant-animalcoevolution: is it thwarted by spatial andtemporal variation in animal foraging? En: Seeddispersal and frugivory: Ecology, Evolution andConservation. Levey, DJ; Silva, WR y Galetti,M (Eds). CABI Publishing. Reino Unido. Pp.275 - 290.
Chapman, CA; (iautiervllion, A; Oates, JF yOnderdonk, DA. 1999. African primatescommunities: Determinants of structure andtretas to survival. En: Primates Communities.Fleagle, JG; Janson, Cl-l y Reed, KE (Eds.).Cambridge University Press. USA. Pp: l - 37.
Chiarello, AG. l993. Activity pattem of the brownhowlcr monkey Aloualla fusca. Geoliroy ¡812,in a forest frugment of southeastcm Brazil.Primates 34: 289 - 293.
Chivers, DJ. ¡969. On the daily behavior andspaeing ol' howling monkey groups. FoliaPrimatologiea ¡0:48 - l02.
Clark, JM. 1998. Why trees migarte so fast:confronting theory with dispersal biology andpaleorecord. The American Naturalist¡52: 204 —224.
Clark, DA y Clark, DB. 1984. Spacing dynamics oftropical rain l'orest tree: Evaluation ol' Janzen Connell Model. American Natumlist 124: 769 788.
Clark, DA y Clark, DB. l992. Life history ol'canopy and emergent trees in a neotropical rainforest. Ecological Monographs 62: 3l5 - 344.
Clark, JS; Fastie, C; llum, G; Jackson, ST;Johnson, C; King, GA; Lewis, M; Lynch, J;Pacala, S; Prentice, C; Schupp, EW; Webb lll,T y Wyckoff, P. ¡998. Reid’s paradox of rapidplant migration: Dispersal theory andinterpretation of paleoecological records.Biosciencc 48: l3- 24.
Clark, JS; Silman, S; Kern, R; Macklin, F. yLambers, JllR. l999. Seed dispersa] near andfar: Pattems across temperate and tropicalforest. Ecology 80: ¡475 - ¡494.
Coe, M y Coe.C. ¡987. Large herbivores. aeaeiatrees and bruchid beetles. South Africa Journalof Science 83: 624 - 635.
Crockett. CM. |998. Conservation biology ol'genusA/ouana. Intemational Journal of Primatology,l9: 549 - S76.
Crockett, CM y JF lïisenbcrg. ¡987. Ilowlers:Variation in group size and demography. En:Primates Societies. Srn'uts. BB; Cheney, DL;Seyfarth, RM; Wrangham. RW y Slruhsaker,'IT (Eds). University ol‘ Chicago Press, USA.Pp: 54 - 68.
Cronquist, A. 1984. Introducción a la botánica.Séptima edición. Harper and Row Publishers.lnc.USA. 848 p.
Dalling, JW; l-lubbell, SP y Silver-a, K. ¡998. Seeddispersal, seedling establishment and gappartitioning among tropical pioneer trees.Joumal of Ecology 86: 674 - 689.
Dalling, JW; Swaine, MD y Ganvood, NC. I994.Ell'ect of soil depth on seedling emergenee intropical soil seed-bank investigations.Functional Ecology 9: ll‘) - l2].
De Figueredo, RA. ¡993. lngestion of Ficusenormis seeds by howler monkeys (Alouatmfusca) in Brazil: ell'ects on seed genuination.Journal of Tropical lïcology 9: 54] - S43.
Dc Stevens, D. |99|. Experiments on mechanismsof tree establishment in old-field suecession:seedling survival and growth. l-Zcology,72: 1076- |088.
Dimitri, MJ; Volkart, C; Ambrosius, C y TiburcioF. ¡974. La flora urborea del Parque NacionalIguazú. Anales de Parques Nacionales l2: l 180.
Dinerstein, E y Wemmer, CM. l988. Frutirhinoeeros eat: dispersa] ol' Trewia nudiflora(Euphorbiaceae) in lowland Nepal. licology69: 1768 - |774.
Dirzo, R y Dominguez, CA. l986. Seed Shadows,seed predation and the advantages ol' dispersa].En: l-‘rugivores and seed dispersal. l-Zstrada,A yFleming, Tl-l. (Eds). Junk Publishers. PaisesBajos. Pp: 237 - 249.
Eisenberg, JF; Muckenhirn, NA y Rudran, R. ¡972.'l'he relation between ecology and socialstructure in primates. Science I76: 863 - 824.
Ellison, AM: Denslow, JS; Loiselle, B y BrenesMD. ¡993. Seed and seedling ecology ofncotropicul melastomutaceae. Ecology74: ¡733 - 1749.
Eskuche, U y Fonatan, JL. ¡996. La vegetación delas islas del alto Paraná. l. Las comunidades deBosque. Folia Botánica et GeobotanieaCorrentesiana ll: l - l3.

Alouuua caraya como dispersor de semillas 103
Estrada, A. 1984. Resource use by howler monkeys(A/oualla paIIiara) in the rain forest of Los'l'uxtlas, Veracruz, Mexico. International JournalofPr-¡matology5: |05 - l3l.
Estrada, A y Coatcs-Estrada, R. ¡984. Fruit eatingand seed dispersal by howling monkeys(Alaualta paIliata) in the tropical rain forest ofLos Tuxtlas, México. International Journal ofPrimatology 6: 77 - 9l.
Estrada. A y Costes-Estrada, R. 199]. Howlermonkeys (Alouarla pal/ima), dung beetles(Scarabaeidae) and seed dispersal: Eoologicalinteractions in the tropical rain forest of Los'l‘uxtlas, Mexico. Journal of Tropical Ecology7: 459 - 474.
Estrada, A: Halffler, G; Costes-Estrada, R y MerittJr, DA. ¡993. Dung betls attracth tomammalian herbivore (Alouatla palliala) andomnivore (Nasua narica) dung in the tropicalrain forest of Los Tuxtlas, Mexico. Journal ofTropical Eoology 9: 45 - 54.
Feer, F. ¡999. Effects of dung beetles (Scarabidae)on seeds dispersed by howler monkeys(Aloualla seniculus) in the French Guianan rainforest. Journal of Tropical Ecology 15: |29l42.
Foord, SH; RJ van Arde y Ferreira, SM. ¡994. Seeddíspcrsal by vervet monkeys in rehabilitatingcoastal dune forest at Richard Bay. South AfricaJoumal of Wildlife Reserve 24: 56 - 59.
Forget. P-M. ¡994. Recruitment pattem ofI'ouacapoua americana (Caesalpiniaceae), arodent-dispersed tree species in French Guiana.Biotropica 26: 408 - 4l9.
Forget, P-M. ¡997. Effect of microhabitat on seedfate and seedling perfonnance in two rodentdispersed tree species ¡n rain forest in FrenchGuiana. Journal of Ecology 85: 693 - 703.
Foster, RB; Arce B., .l y Wachter, TS. l986.Dispersal and sequential plant communities inAmazonian Peru floodplain. En: Frugivores andseed dispersal. Estrada, A y Fleming, TH. (Eds).Junk Publishers. Paises Bajos. Pp: 357 - 384.
Fragaszi, DM; Boinski, S y Whipple, J. l992.Behavioral sampling in the field: comparison ofindividual and group sampling methods.American Journal of Primatology 26: 259 - 275.
Fragoso, JM. 1997. Tapir-generated seed Shadows:Scale-dependent patchiness in-the Amazon rainforest. Joumal of Eeology 85: 5l9 - 529.
Franceschi, EA y Lewis, JP. 1979. Notas sobre lavegetación del valle santafesino del Río Paraná(República Argentina). Eoosur 6: 55 - 82.
Garber, l’A. l986. The eeology of seed dispersal intwo species of callitrichid primates (Saguinusmyslax and Sagulnus flucicollis). AmericanJournal of Primatology IO: ¡55 - l70.
Galetti, M; Pedroni, F y Morellato, LPC. ¡994. Dietof the brown hower monkey Aloualla finca in
forest fragment in southeastem Brazil.Mammalia 58: lll —l13.
Garber, PA y Lambert, JE. 1998. Primates as seeddispersers: Ecological processes and directionsfor future research. American Journal ofPrimatology 45: 3 - 8.
Gaulín, SJC y CK Gaulín. ¡982. Behavioralecology of Alouatra seniculus ¡n Andean cloudforest. International Journal of Primatology3: l - 32.
Gaulin, SJ; Knight, Dl-l y Gaulin, CK. ¡980. Localvariance in Alouarta group size and foodavailability on Barro Colorado Island.Biotropica l2: l37 - 143.
Gill, B. l99l. Dung beetles in tropical Americanforest. En: Dung beetle eeology. Hanski, l yCamberfor't, Y (Eds). Princeton UniversityPress. USA. Pp. 21 l - 229.
Glander, KE. l980. Feeding patterns in mantledhowling monkeys. En: Foraging behavior:Ecological, ethological, and psychologicalapproaches. Kamil, AC y Sargent TD (Eds).Garland Press. USA. Pp: 231 - 257.
Greene, DF y Calegoropoulos, C. 2002. Measuring.and modelling seed dispersalof terrestrial plants.En: Dispersal eeology. Bullock, JM; Kenward,RE y Hails, RS (Eds). British EcologicalSociety, Blackwell Science Ltd.Reino Unido.Pp. 3 - 23.
Guix, JC y.RuiL X. 1995. Toucans and thrushes aspotential dispersers of seed-predatory weevillarvae in southeastem Brazil. Canadian Journalof Zoology 73: 745 - 748.
Hanski, l. ¡991. The dung inseet community. En:Dung beetle ecology. Hanski, l-y Camberfort, Y(Eds), Princeton university Press. USA. Pp. 5 21
llartshom, GS. l980. Neotropical forest dynamics.Biotropica |2: 23 - 30.
Hauser. TP. 1994. Germination, predation anddispersal of Acacia albida seeds. Oikos7|:421-426.
Herrera, CM. 1985. Deterrninants of plant-animalcoevolution: the case of mutualistic dispersal ofseeds by vertebrates. Oikos 44: 132 - l4l.
Herrera, CM. l989. Vertebrate fi'ugívoresand theirintcraction with invertebrate fnrit predators:Supporting evidence from a Costa Rica dryforest. Oikos 54: 185 - 188.
Hen'era, CM y Jordano, P. |93|. Prunus mahaleband birds: The high-efficiency seed dispersalsystem of a temperate fruiting tree. EoologicalMonographs5|: 203 - 22l.
Hen'era, CM; Jordano, P; López Soria, L y Amat.JA. ¡994. Recruitment of a meat-fruiting, birddispersed tree: Bridging frugivore activity andseedling establishment. Ecological Monographs64: 3 |5 - 344.

¡04 Susana Patricia Bravo
Heymann, EW. 1992. The role of primates intropical ecosystems. En: Animal-Plantinteractions in tropical environments.Barthlott,W; Naumann, CM; Schmidt-Loske, Ky Schuchmann, KL (Eds). Alemania. Pp. 103 108.
Hladik, A y Hladik, CM. ¡969. Rappons trophiquesentre vegetation et primates dans la foret deBam) Colorado (Panama). Revista d’Ecologie.(Terre vie) l: 25 - Il7.
Hólldobler, B y Wilson, EO. 1990. The Ants.Harvard University Press. Massachusetts. USA.732 p.
llowe, lli’. ¡980. Monkey dispersal and waste of aneotropical fruit. Ecology 6|: 944 - 959.
Howe, HF. 1986. Seed dispersal by fruit-eatingbirds and mammals. En: Seed dispersa]. DRMurray (Ed). Academic Press. USA. Pp.l23 l90.
Howe, l-lF. l993. Aspects of variation in aneotropical seed dispersal system. VegetatiolO7/l08: ¡49 - ¡62.
Howe, HF y Smallwood, J. 1982. Ecology of seeddispersa]. Annual Review of Ewlogy andSystematics l3: 20] - 223.
llowe, llF y Vande Kerckhove, GA. 198].Removal of wild nutmeg (Virola surinamensis)crops by birds. Ecology 62: |093 - l l06.
Janzen, H. l970. Herbivores and the number oftreespecies in tropical forest. American Naturalist104: 50] - 528.
Janzen, H. l983. Dispersal of seeds by vcrtcbratcguts. En: Coevolution. Futuyma, DJ y Slatkin,M (Eds). Sinaver Associates inc. Sunderland.USA. Pp. 232 - 262.
Jennings, DH y Lysek, G. ¡999. Fungal Biology:Understanding the fungal lifestyle. Segundaedición. Bios Scientific Publishers. ReinoUnido. l65 p.
Jolly, A. 1985. The evolution of primates behavior.Macmillan Publishing Company. USA. 526 p.
Jordano, P y Godoy, JA. 2002. Frugivore-generatedseed Shadows: a landscape view of demographicand genetic effects. En: Seed dispersal andfrugivory: Ecology, Evolution andConservation. Levey, DJ; Silva, WR y (ialetti,M (Eds.). CABI Publishing. Reino Unido. Pp.305 - 321.
Jordano, P y Herrera, M. l995. Shufflíng theOffspring: Uncoupling and spatial discordanceof multiple stage in vertebrate seed dispersal.Eeoscience 2: 230 - 237.
Jordano, P y Schupp, E. 2000. Seed dispersereffectiveness: quantity component and patternsof seed rain for Prunus mahaleb. EcologicalMonographs 70: 591 —615.
Julliot, C.I992. Utilisation des ressourcesalimentaires par le singe hurler roux, Alouanasem'cu/us (Atclidae, Primate), cn Guyanc:
impact de la disseminatíon des gmines sur laregeneration lbrestiere. These de Doctoral,Universite de Tours. Francia.
Julliot, C. l994. Frugivory and seed dispersal byred howler monkeys: evolutionary aspcct.Reviste d’lïcologie (Terre Vie) 49: 33l - 336.
Julliot, C. l996. Seed dispersal by red howlingmonkeys (¡l/omnia seniculus) in the tropicalmin forest of French Guiana. internationalJournal ol'Primatology 17: 239 - 258.
Julliot, C. l997. Impact of seed dispersal by redhowler monkeys Alouana seniculus on theseedling population in the understorey oftropical rainforest. Journal of lícology 85: 43| 440.
Julliot, C. _vSabatier, D.l993. Diet of red howlermonkeys (Alouaua seniculus) in FrenchGuyana. International Journal of Primatologyl4: 527 - 550.
Keddy, PA y Reznicek, AA. |982. 'l‘hc role of seedbanks in the persistence of Ontario’s coastalplain flora. American Journal of Botany 69: l3 22.
Kobayashi, M e lzawa, K.l997. Distributionpatterns of seedlings of Pharus virescens Doell(Poaceae: Bambusoideae) in the Neotropicalrain forest of a Macarena and Tinigua NationalParks. Colombia. Field Studies of Fauna andl-‘lora,La Macarena, Colombia l l: 47 - 56.
Kobayashi. M; Shimooka, Y e l'zawa, K. ¡999.Frugivore-dispersed seedlings of milpeso palm()enocarpus balaua Mart. ln the ncotropicalrain forest of La Macarena, Colombia. FieldStudies of Fauna and Flora, La Macarena,Colombia l3: 4l - 46.
Kowalewski, MM. 2000. Birth seasonality in blackhowler monkeys (Alouaua caraya) in an islandsystem in northern Argentina. Master's thesis,State University of New York at Stony Brook.USA
Lahitte, llB; llurrell, JA; Mehltreter, K; Belgrano,M; Jankowski, LS; Haloua, MP y Canda, (i.¡997. Plantas de la costa. LOLA. Argentina.200 p.
Lambert. JE. 2002. Exploring the link betweenanimal frugivory and plant strategies: The caseof primate fruti processing and post-dispersa]seed fate. En: Seed dispersa] and lrugivory:Eoology, Evolution and Conservation. Levey.DJ; Silva, WR y Galetti, M (Eds.). CAlilPublishing. Reino Unido. Pp. 365 - 379.
Lenssen. J.; Menting, F; van der Putten, W y Blom.K. 1999. Control of plant species richness andzonation of functional groups along a freshwaterflooding gradient. Oikos 86: 523 - 534.
Levey, DJ y Byrne, MM. ¡993. Complex ant-plantinteractions: min forest ants as secondarydispersers and post-dispersal seed predators.Eeology 74: l802 —l812.

Alouana caraya como dispersor de semillas ¡05
Lewis, JP y Fransceschi, EA. 1979. Notas sobre ladinámica de la vegetación del valle del RíoParaná. Ecosur 6: ¡45 - 163.
Liebennan, DJ; may, B; Swaine, MD y Liebennan,M. 1979. Seed dispersa] by baboons in the ShaiHills, Ghana. Eeology 60: 65 - 75.
Lieberman, M y Lieberman, DJ. |986. Anexperimental study of seed ingestión andgermination in plant-animal assemblage inGhana. Journal of Tropical Ecology 2: ll3¡26.
Lucena Mendes, S. ¡989. Estudo ecológico deAllouata fusca.(Primate: Cebidae) na estacáoBiológica de Caratinga, MG. RevistaNordestina Biológica6: 7l-l04.
Martin. P y Bateson, P. [993. Mesuring behaviour.Segunda edición. Cambridge University Press.Reino Unido. 222 p.
Martinez-Crovetto, R. I963.fitogeogrático de MisionesArgentina). Bonplandia l: l7l - 223.
Milton, K. [980. The foraging strategy of howlermonkeys, a study in primate economics.Columbia University Press. USA. 163 p.
Milton, K. |996. Effects of bot fly (Alouallamylabueria) parasitism on a free-ranging howlermonkey (Aloualla palliala) population inPanama. Journal onoology 239: 39 - 63.
Miller, MF y Coe, M. ¡993. ls it advantageous forAcacia seeds to be eaten by ungulates? Oikos66: 364 - 368.
Morello, J y Adámoli, J. ¡974. Las grandesunidades de vegetación y ambiente del ChacoArgentino, segunda parte: Vegetación yambiente de la Provincia del Chaco. SerieFitogeográfiea N° l3. INTA. Buenos Aires.Argentina.l30 p.
Nakamura, G. 1983. ilustraciones de hoju deespecies arbóreas del Paraguay paraidentificación de género o especie. Nota deTecnologia de la Madera 6: l-7l.
Nalepa, CA y Piper, WH. ¡994. Bird dispersal ofthe larval stage of a seed predator. Oecologial00: 200 - 202.
Neiff, JJ. 1978. Fluctuaciones de la vegetaciónacuática en ambientes del valle de inundacióndel Paraná medio. Physis 38: 4| -53.
NeilT, JJ. ¡990. Ideas para la interpretaciónecológica del Paraná. lnterciencia 15: 424 - 441.
NeilT, JJ. 1999. El régimen de pulsos en rios ygrandes humedales de Sudamérica. En: Tópicossobre humedales subtropieales y templados deSudamérica. Malvarez, Al (Ed). Unesco.Uruguay. Pp. 97 - 146. '
NeilT, JJ y Poi de NeilT, A. 1990. Litterl'all, leafdecomposition and litter colonization ofTessaria imegrífolia in the Paraná Riverfloodplain. Hydrobiologia 203 : 45 - 52.
Esquema(República
Neifl', JJ y Reboratti, ¡989. Estructura y dinámicade bosques de Tessaria inlegrifolia. ll: análisisdel crecimiento y la productividad. Boletín de laSociedad Argentina de Botánica 26: 39 -43. _
Ncubert, MG y Caswell, H. 2000. Demography anddispersaI:-calculation and sensitivity analysis ofinvasion speed for stmctured populations.Ecology8|: ¡613 - l628.
Nicotra, AB; Chazdon, RL e Iriarte, SVB. l999.Spatial heterogeneity of light and woodyseedling regeneration in tropical wet forests.Ecology 80: l908 - l926.
Nunes, A. ¡995. Um teste de germinacito emsementcs dispersas por maeacos-Aranha emMaracá, Roraima; Brasil. Studies onNeotropical Fauna and Environment 30: 31 - 36.
OverdortT, DJ y Strait, SG. ¡998. Seed handling bytree prosimian primates in southeastemMadagascar: lmplications for seed dispersal.American Journal of Primatology 45: 69 - 82.
Palacios, E y Rodríguez, A. 2001. Ranging pattemand use of space in a group of red howlersmonkeys (Alauaua seniculus) in theSoutheastern Colombian rainforest. AmericanJournal of Primatology 55: 233 - 251.
Parker, IM. 2000. Invasion dynamics of Cytlsusscoparius: a matrix model approach. EcologiealApplications ¡0: 726 - 743.
Placci, LG. 1995. Estructura y comportamientol‘enológicoen relación a un gradiente hídrico enbosques del este de Formosa. Tesis Doctoral.Universidad de La Plata. Argentina.
Popolizio, E. 1977... Contribución a lageomorfología de la Provincia de Con'ientes.VII Geociencias 7: l -45.
Prates, JC; Pires Gayer, SM; Kunz Jr, LP y Buss,G. 1990. Feeding habits of the brown howlermonkey Alouatra fusea clamitans (Cabrera,l940) (Cebidae, Alouattinae) in the ltapua StatePark: a preliminary report. Acta BiologicaLeopoldensiana ¡2: ¡75 - ¡88.
Redford, KH; Bouchardet da Fonseca, GA yLacher, TE. 1984. The relationship betweenl'rugivory and insectivory in primates. Primates25: 433 - 440.
Renner, SS. |992. Seed dispersal. Progress inBotany 53: 435-453.
Rey, PJ y Alcántara, JM. 2000. Reeruitmentdynamics of fleshy-fruited plant (0Iea_europea): eonnecting patterns of seed dispersalto seedling establishment. Journal of .Ewlogy88: 622 - 633.
Rogers, ME; Voysey, BC; McDonald, KE; Pamell,RJ y Tutin, CEG. ¡998. Lowland gorillas andseed dispersa]: The importance of nest sites.American Journal of Primatology 45: 45 - 68.
Rowell, TE y Mitchell, BJ. |99|. Comparison ofseed dispersa] by guenons in Kenya and

l06 Susana Patricia Bravo
capuchins in Panamá. Journal of TropicalEcology 7: 269 - 274.
Rumiz, Dl. l990. Aloualra caraya: Populationdensity and demography in nonhem Argentina.American Journal of Primatology 21:279-294.
Rumi; DI; Zunino, GE; Obregozo, ML y RuiL JC.¡986. Alouana caraya: Habitat and resourceutilization in northern Argentina. En: Currentperspectives in primates social dynamics. 'l‘aub,DM y King, FA (Eds). Van Nostrand Reinhold.USA. Pp. ¡75 - ¡93.
Sánchez-Cordero, V y Martínez-Gallardo, R. 1998.Postdispersal fruit and seed removal by forestdwelling rodents in lowland rainforest inMéxico. Journal of'l‘ropical Ecology l4: 139 —lSl.
Schneider, Rl, y Seharitz, RR. I988. Hydrochoryand regenemtion in a bald cypress-water tupeloswamp forest. Ecology 69: l055 —1063.
Schupp, EW. 1988. Seed and early seedlingpredation in the forest understory and in treefallgaps. Oikos 5|: 7! - 78.
Schupp, EW. l993. Quantitative, quality andeffectiveness of seed dispersal by animals.Vegetatio l07/108:15-29.
Schupp, EW y Fuentes, M. |995. Spatial patterns ofseed dispersal and the unification of plantpopulation ecology. Ecoscience 2:267 —275.
Schupp, EW; Milleron, T y Russo, SE. 2002.Dissemination limitation and the origin andmaintenance of species-rich tropical forest. En:Seed dispersa] and frugivory: Ecology,Evolution and Conservation. Levey, DJ; Silva,WR y Galetti, M (Eds.). CABl Publishing.Wallingford. Reino Unido. Pp. 305 - 32l.
Seiwa, K. [998. Advantages of early germinationfor growth and survival of seedlings of Hacermono under different overstorey phenologies indeciduous broad-leaved forests. Journal ofEcology 86: 2l9 - 228.
Sekulic, R. ¡982. Daily and seasonal pattem ofroaring and spacing in four red howler Aloualraseniculus troops. Folia Primatologica 39: 22 —4
Shigesada, N y Kawasaki, K. 2002. invasión andthe range expansion of especies: Effects oflong-distance dispersa]. En: Dispersal eoology.Bullock, JM; Kenward, RE y Hails, RS (Eds).British Ecological Society, Blackwell ScienceLtd. Reino Unido. Pp. 350 - 373.
Silver, SC; Ostro, LET; Yeagcr, CP y llorwich, R.¡998. Feoding eoology of black howler monkey(Aloualla pigra) in northern Belize. AmericanJournal of Primatology 45: 263 - 279.
Sollins, P. l998. Factors influencing speciescomposition in tropical lowland rain forest:Does soil matter? Eoology 79: 23 - 30.
Stoner, KE. ¡996. Habitat selection and seasonalpatterns of activity and foraging of mantled
howling monkeys (Aloualla palliata) innortheastern Costa Rica. International Journal ofPrimatology |7: l - 30.
Terborgh. J. |992. Maintenance of diversity intropical forest. Biotropica 24: 283 - 292.
Traveset. A y Verdú, M. 2002. A meta-analysis ofthe effect of gut treatment on seed germination.E : Seed dispersal and frugivory: líeology,Evolution and Conservation. chey, DJ; Silva.WR y Galetti, M (Eds). CAlll Publishing.Reino Unido. Pp. 339 - 350.
Traveset, A; Willson, MF y Gaiter, JC. ¡995.Avoidance by birds of insect-infested fruits ofVaccnium ovalifolium. Oikos 73: 381 - 386.
Tutin, ClíG; Williamson, EA; Rogers, ME yl-‘emández, M. |99|. A case study of plantanimal relationship: Cola Ii:ae and Iowlandgorillas in the Lope Reserve, Gabon. Journal of'I‘ropical Ecology 69: 751-763.
Utzurrum, RCB y Heideman, Pl). |99|.Differential ingestion of viable vs nonviableFicus seeds by fruits bats. Biotropica 23: 3|] 3l2.
Valhurg, LK. ¡992 a. Eating infested fruits:lnteractions in a plant-disperser-pest triad.Oikos 65: 25 - 28.
Valburg, LK. ¡992 b. Feeding preferences ofcommon bush-tangers for insect-infested fruits:Avoidance or attraction? Oikos 65: 29 - 33.
Valido. A y Nogales, M. l-‘rugivory and seeddispersa] by the li'znrd Gal/olía gallori(Lacertidae) in xeric habitat of Canary lsland.Oikos 70: 403 - 4| l.
Vander Wall, SB. 1997. Dispersal of singleleafpiñon pine (Pinus monophylla) by seed-cachingrodents. Journal of Mammalogy 78: l8l —l9l.
Van I-lorne, B. l983. Density as a misleadingindicator of habitat quality. Journal of Wildlifemanagement 47: 893 - 90l.
Vázquez-Yanes, C y Smith, H. ¡982. l’hytochromecontrol of seed germination in the tropical rainforest pioneer trees Cecropia obtusifolia andPiper aurilum and its ecological signiticance.New l’hytology 92: 477 - 485.
Walters. MB y Reich, PB. 2000. Seed size, nitrogensupply. and growth rate affect trcc seedlingsurvival in deep Shade. Ecology 8|: |887|90|.
Weiher, F. y Keddy, PA. |995. The assembly ofexperimental wetland plant communities. Oikos73: 323 - 335.
Welling, Cll; Pederson, Rl. y Van der Valk, AG.l988. Rccruitment from the seed bank and theenvelopment of Innation of cmergent vegetationduring a drawdown in a praire wetland. Journalof Ecology 76: 483 - 496.
Wenny, DG. 2000. Seed dispersal, seed predation,and seedling recruitment of a neotropical

Aloualla caraya como dispersor de semillas 107
montane tree. Ecological Monographs 70: 331 35].
Williamson, M. 2002 Overwiew and synthesis: thetale of the tail. En En: Dispersal ecology.Bullock, JM; Kcnward, RE y Hails, RS (Eds).British Ecological Society, Blackwell ScienceLtd. Reino Unido. Pp. 431 - 443.
Yumoto, T; Kimura, K y Nishimura, A. ¡999.Estimation of the retention times and distancesol' seed dispersed by two monkey species.Alouaua seniculus y Lagolhrix Iagolricha, in aColombian forest. Eoological Researchl4: ¡79 - l9l.
Zar. Jll. l996. Bioestatistical analysis. Terceraedición. Prentice l-lall. USA. 662 p.
Zhang, S-Y y Wang, L-X. 1995. Fruit Consumptionand seed dispersal ol" leiphus cinnamomum(Rhamnaceae) by two sympatric primates(Cebus apella and Ale/es paniscus) in FrenchGuiana. Biotropica 27: 397: 40l.
Zuloaga, F0 y Monone, O. (Eds) 1999. Catálogode las plantas vasculares de la RepúblicaArgentina ll. Tomos l y ll, Vol 74. MissouriBotanical Garden Press. USA.
Zunino, GE. ¡986. Algunos aspectos de la..
ecología y etologla del mono aullador negro(Alouatla caraya) en parches de bosque. TesisDoctoral, Universidad de Buenos Aires.
Zunino, GE. 1987. Nutrición en primates follvoros:la dicta de' Alaualta caraya en vida silvestre.Boletin Latinoamericano de Primatologla5: 78 —87.
Zunino, GE. ¡989. Habitat, dieta y actividad delmono aullador negro (Alcualla caraya) en elnoreste de la Argentina. Boeltln PrimatológicoLatinoamericano l: 74 - 97.
Zunino, GE; Bravo, SP; Reisenman, C y MuradFerreira, F. 1996. Characteristics of two types ofhabitat and the status of the howler monkey(Alouaua caraya). Neotropical Primates 4:4850
Zunino, GE; Gonmlcz, V; Kowalewski, MM yBravo, SP. (2001). Alouatla caraya relationsamong habitat, density and social organization.Primate Report. 61:37 - 46.
Zunino, GE y Rumiz, Dl. 1986. Observaciones sobreel comportamiento ten'itorial del mono aulladornegro (Aloualla etzraya). Boletin PrimatológicoArgentino 4: 36 - 52.

Alouaua caraya como dispersor de semillas 109
Apéndice :Clima e hidrología
Temperaturas 1 precipitaciones promedio históricas (Resistencia, l98l —1990)
23 320
za.1260
24
22L .m
20.
¡e “°
Nh-ID
|4
‘2 ‘ l I ¡o
Datos obtenidos del Servicio Meteorológico Nacional
Desvío res ecto al romedio de la tem eratura las reci itaciones durante el estudio decrecimiento de los juveniles
120 5
80
40
'40 E:
120 5—PredpitadonJJASONDEFMAMJJASONDEFMAM ---'-TempemtureF .L : 12000 2001 2002
Datos regionales, obtenidos del International Research Institute for Clima Prediction, ColumbiaUniversity.

1 lo Susana Patricia Bravo
Períodos de máxima altura del río durante el estudio de crecimiento de los 'uveniles
“A
11IIIIII¡LlullllllllllllIIIIIILILIIIIIIIIAIIIIII
Ul
AlturadelRioParana(rn)
Utu
.- N N°. ‘-? q"P “t‘ °°N to - ó
El Bosque de Palo ¡tá se encuentra sobre albardones entre S y 5,5 m, mientras que elBosque de Laureles se encuentra sobre albardones cuya altura varía entre 5,5 rn y 7 m. La altura delrío corresponde al hidrómetro de Paso de la Patria y los datos fueron facilitados por la PrefecturaNaval Argentina.


















