
ESPERMINA SOBRE EL DESARROLLO DE EMBRIONES SOMÁTICOS DE … · 2018-09-26 · Figura 3.2 Perfil...
128
-:. -:s- CICY S OSGRADOEN CIENCIAS ( BIOLÓGICAS Centro de Investigación Científica de Yucatán, A.C. Posgrado en Ciencias Biológicas EFECTO DE ASPARAGINA, GLUTAMINA y ESPERMINA SOBRE EL DESARROLLO DE EMBRIONES SOMÁTICOS DE CHILE HABANERO Capsicum chinense Jacq. Tesis que presenta KARIME ZUBIETA SÁNCHEZ En opción al título de MAESTRO EN CIENCIAS (Ciencias Biológicas: Opción Bioquímica y Biología Molecular) Mérida, Yucatán, México 2014
Transcript of ESPERMINA SOBRE EL DESARROLLO DE EMBRIONES SOMÁTICOS DE … · 2018-09-26 · Figura 3.2 Perfil...

-:. -:s-CICY
SOSGRADOEN
CIENCIAS ( BIOLÓGICAS
Centro de Investigación Científica de Yucatán, A.C.
Posgrado en Ciencias Biológicas
EFECTO DE ASPARAGINA, GLUTAMINA y
ESPERMINA SOBRE EL DESARROLLO DE
EMBRIONES SOMÁTICOS DE CHILE HABANERO
Capsicum chinense Jacq.
Tesis que presenta
KARIME ZUBIETA SÁNCHEZ
En opción al título de
MAESTRO EN CIENCIAS
(Ciencias Biológicas: Opción Bioquímica y Biología Molecular)
Mérida, Yucatán, México
2014


CENTRO DE INVESTIGACIÓN CIENTíFICA DE YUCATÁN, A. C.
POSGRADO EN CIENCIAS BIOLÓGICAS
RECONOCIMIENTO
S· POSGRADO EN
CIENCIAS ( BIOLÓGICAS
Por medio de la presente , hago constar que el trabajo de tesis titulado EFECTO DE
ASPARAGINA, GLUTAMINA y ESPERMINA SOBRE EL DESARROLLO DE
EMBRIONES SOMÁTICOS DE CHILE HABANERO Capsicum chinense Jacq. fue
realizado en el laboratorio BQ09 de la Unidad de Bioquímica y Biología Molecular de
Plantas del Centro de Investigación Científica de Yucatán , A.C. bajo la dirección de la Dra .
Nancy Santana Buzzy, dentro de la opció de Bioquímica y Biología Molecular,
perteneciente al Programa de Posgrado en Cie ias Biológicas de este Centro.
Atentamente,
Director de Docencia
Mérida , Yucatán , México. Octubre de 2014


DECLARACiÓN DE PROPIEDAD
Declaro que la información contenida en la sección de Materiales y Métodos
Experimentales, los Resultados y Discusión de este documento proviene de las
actividades de experimentación realizadas durante el período que se me asignó para
desarrollar mi trabajo de tesis, en las Unidades y Laboratorios del Centro de Investigación
Científica de Yucatán, A.C., y que a razón de lo anterior y en contraprestación de los
servicios educativos o de apoyo que me fueron brindados, dicha información, en términos
de la Ley Federal del Derecho de Autor y la Ley de la Propiedad Industrial, le pertenece
patrimonialmente a dicho Centro de Investigación. Por otra parte, en virtud de lo ya
manifestado, reconozco que de igual manera los productos intelectuales o desarrollos
tecnológicos que deriven o pudieran derivar de lo correspondiente a dicha información, le
pertenecen patrimonialmente al Centro de Investigación Científica de Yucatán, A.C. , y en
el mismo tenor, reconozco que si derivaren de este trabajo productos intelectuales o
desarrollos tecnológicos, en lo especial, estos se regirán en todo caso por lo dispuesto por
la Ley Federal del Derecho de Autor y la Ley de la Propiedad Industrial, en el tenor de lo
expuesto en la presente Declaración .
Firma: __ ~ __ if+-_-___ _
Nombre: IBQ. KARIME ZUBIETA SÁNCHEZ


Este trabajo se llevó a cabo en la Unidad de Bioquímica y Biología Molecular de Plantas
del Centro de Investigación Científica de Yucatán , y forma parte del proyecto titulado
"Recalcitrancia del género Capsicum: estud ios morfológicos, bioquímicos y moleculares
relacionados con los factores que afectan la capacidad de regeneración de plantas de
chile in vitro " (Ciencia básica , CONACYT) y "Obtención de variedades más productivas y
más tolerantes de ch ile habanero a partir de la diversidad genética de la especie en
Yucatán" (FOMIX) bajo la dirección de la Dra. Nancy Santana Buzzy.


AGRADECIMIENTOS
• A Dios, por darme vida e iluminarme cada día
• A mi mamá Leticia del C. Sánchez Rebolledo y a mi papá Víctor J. Zubieta
Escalante por su amor, apoyo e impulso en mi desarrollo personal y profesional
• A mi novio Marco Antonio Ávila Cantarell
• A la Dra. Nancy Santana Buzzy por la dirección de mi proyecto de Maestría, su
asesoría, apoyo y comprensión durante estos dos años de desarrollo profesional
• Al Dr. Ignacio R. Islas Flores por su valiosa y entusiasta asesoría y enseñanza en
la realización de este proyecto
• A los técnicos : M. en C. Adriana Canto Flick y M. en C. Eduardo Balam Uc en el
trabajo técnico
• Al M. en C. Jacobo Pérez Pastrana por su apoyo técnico en el área de histología
• A los miembros de mi comité tutorial : Dra . Sara E. Solís Pereira, Dr. Ignacio R.
Islas Flores por sus sugerencias y críticas para el enriquecimiento de este trabajo
• A los miembros de mi comité evaluador: Dra . Sara E. Solís Pereira, Dr. Ignacio R.
Islas Flores, Dra. Lourdes Iglesias Andreu, Dr. Luis Saénz Carbonell
• A mis amigos de laboratorio: M. en C. Raúl E. Valle Gough, M. en C. Carlos F.
Regla Márquez, IBQ. Mario Puc Chan, M. en C. Jacobo Pérez Pastrana, Dra.
Susana Avilés Viñas, Biol. Mariela Vergara Olivares, Agr. Eduardo Ballina
Rodríguez, IBQ. Pedro Oserio Montalvo y B.E. Maximino Gómez Aguilar por su
amistad, apoyo y consejo . En especial a la Dra . Eunice Gómez Uc por su asesoría
durante el desarrollo de mi proyecto de Tesis
• Al Consejo Nacional de Ciencia y Tecnología (CONACYT) por otorgarme la beca
para realizar mis estudios de maestría (Becario: 280804)


DEDICATORIAS
• A mi mamá, Leticia del C. Sánchez Rebolledo:
A quien debo gran parte de lo que soy ahora. Por ser una madre extraordinaria y por
su inmenso amor
• A mi papá, Víctor J. Zubieta Escalante:
Por su apoyo, cariño y consejo
• A mi novio Marco Antonio Ávila Cantarell:
Mi mejor amigo y compañero. Por su amor y comprensión, por cuidarme, apoyarme y
motivarme cada día
Los amo


íNDICE
INTRODUCCI6N ... .. .......... .. ........ .... .... ........ ...... .. ... .. .... ... ..... ..... ... ... .. ...................... ....... .... . ............... 1
CAPiTULO I .... ..... .. ...................... ... ... ... ....... ... ........ ........ .. ..... .. ............... ... ..... ...... ...... .... .. ... ..... ......... . 3
ANTECEDENTES ... ... .............. ...... ........... .. .. ..... .... ...... .. .... ....... ....... ... .. ............ .. .......... ...... .... ... .. ... 3
1 .1 CULTIVO DE TEJIDOS VEGETALES (CTV) .... .......... .. ... . .. ... ... ........... ........ . ........... .. .. ... .. .... .. .. 3
1.1 .1 EMBRIOGÉNESIS SOMÁTiCA ... ... ....... .... .... ... ............. .. ........ ... ... .. ... .. ......... . ....................... ..... .. . . .4
1.1.1.1 ETAPAS DE LA EMBRIOGÉNESIS SOMÁTICA .. ........................................... .. .. .............. ... ... .... .... . 5
1.1.1.1.1 INDUCCiÓN .. .......... ... ....... ...... ... ..... .... ........ ................ .. .... .. .... ... ... ... ........ .... ......... ... .... .. . .... . 6
1.1.1.1.2 HISTODIFERENCIACIÓN .. . .... ....... .. .. ......... .. ...... ....... ..... .. ......... ... ..... ...... . .... ... ... . .... ..... .. .... . ... 7
1.1 .1.1.3 MADURACiÓN ...... ............. ...... ..... ... ... .... ... ... .. . ........ . .... .... .. .. .......... ... .................. .... .... ... ..... 8
1.1.1.1.4 GERMINACiÓN .... ..... .... ..... ....... .. ... ..... ....... ... ... ....... . ... .. .... ... .. .. ........ ... ........ ........ .. ......... . ..... 9
1.1.2 REGENERACiÓN DE PLANTAS VIA ES ............. .. ........ ... .... .... ........ ... ............ ............... .. .... . ...... . . 10
1.2 GÉNERO CAPS/CUM .... .. ........ ...... .... ....... ...... .......... ....................... ....... ...... ...... ... ... ..... .. ....... 11
1.2.1 CHILE HABANERO (CAPSICUM CHINENSE JAca.) .... .. ...... .. .............. .. ...... .. .......... .. .. .. .. ........ .. .. ... 11
1.2.1 .1 TAXONOMIA .... ...... ........... .... ....... .... ...... .. .. .. .. .. .... .... .... .. ...... .... ................ .. ...................... ... . 12
1.2.1.2 PROBLEMÁTICA DEL CULTIVO .. .... ......................................................................... ... .... .... .. .. . 12
1.3 PROTEINAS EN EMBRIONES CIG6TICOS ...... .... ........ .. ........ .. .. .. .... .. ............................ .... .. 14
1.3.1 PROTEINAS DE RESERVA ................ .. . .. ................................................. .. ........ .. .. ........... .. .... .... 14
1.3.2 PROTEINAS LEA ....... .. ............................ ...... ........ .. ...... .... ...... .. ...... .. ................................ ... .. 15
1.3.3 ACUMULACiÓN DE PROTEINAS EN EMBRIONES SOMÁTICOS ..... ........... .. .. .... .... .. .......................... 16
1.4 POLlAMINAS ...... ... ... ..... ..... .. ............. ............... .. .... .. ............. .. ..... ...... ... .... .. ............. .. ..... .. . .... 18

1.4.1 GENERALIDADES EN PLANTAS ...................................... .... .... .... .. .. .......... ........ .... ................ .... . 18
1.4.2 TIPos DE POLlAMINAS ...... .... ........ ... ................ .... ........ ... .......... ..... ...... .. .......... .. ...... .... ....... .. ... 19
1.4.3 FUNCIONES DE LAS POLlAMINAS ............. .. .. .. .............. .. ............... .......... ..... : .......... .... .............. 20
1.4.4 ANTECEDENTES DE POLlAMINAS EN LA EMBRIOGÉNESIS SOMÁTICA ............. .. ....................... .. .... 22
HiPÓTESiS .... ............ ... .... ... .. ......................... ... ....................... ................... .. .... .. .... ... .................. 25
OBJETIVO GENERAL .. .......... .. ....... .. .... ...................... ............. ......... ...... .... ... .. ....... ..... ..... ........... 25
OBJETIVOS ESPEC[FICOS ................. ............................. ... ........................... ... .. .. .. .... ... ..... ........ 25
JUSTIFiCACiÓN ............. ..... ...... .. ... ........ ....... .... ... .. ..................... ............. ... ........ ...... ...... ...... ..... .. 26
ESTRATEGIA EXPERIMENTAL. ...... ....... .. ......... .... .. ... ..... ... .............. ..... ...................................... 27
BIBLlOGRAF[A .. .... .. ....... ............ .... ............. .... ... .. ... .. ..... ......................... ..... ... ....... ......... ... .......... 28
CAPITULO 11 ......... .. ....... ..... .......... ....... ..... ....... ...... ..... .. ........ .......................................... .................. 39
EFECTO DE ASPARAGINA, GLUTAMINA y ESPERMINA SOBRE LA EFICIENCIA, CALIDAD Y
DESARROLLO DE LOS ESs DE CHILE HABANERO .. .. ...................... .. ...................................... 39
2.1 INTRODUCCiÓN ... ..... ....... ......... ... .... ... ............ ............. ......................... ..... .... ....... ........ ....... . 39
2.2 MATERIALES Y MÉTODOS .. .. ........ ....... ... .... .. ...... ... ... .... .. ... ... ............ .................... ............ .. .42
2.2.1 MATERIAL VEGETAL .................................................................... ................... ........ .......... ... .. .. 42
2.2.2 DESINFECCiÓN y GERMINACiÓN DE SEMILLAS .......................................................................... .43
2.2.3 INDUCCiÓN DE EMBRIOGÉNESIS SOMÁTiCA .............................. ....... ..... ..... ... ........... .... .... ... .... .. .43
2.2.4 ADICiÓN DE ASPARAGINA (ASN), GLUTAMINA (GLN) y ESPERMINA (SPM) EN LA
HISTODIFERENCIACIÓN .. ... ... ... .. .... .. ...... ... .......................... ... ....................... ............... ... .. ...... ........ .. 43
2.2.5 EVALUACiÓN DE LOS TRATAMIENTOS .......... ...... ....................... .. ................ ...... ........ .. ......... .. ... 44
2.2.6 ANÁLISIS HISTOLÓGICO ........................ .... ....................... .... ........ ....... ...... ....................... ........ 44
ii

2.3 RESUL TADOS .................. . ................................. ................ ...... ...... ... ..... ... .. ..... ..... . ... ..... . .... ... 46
2 .3 .1 COMPORTAMIENTO DE LA EFICIENCIA EN LA ES DE CHILE HABANERO SOMETIDA A DIFERENTES
TRATAMIENTOS CON ASN, G LN y SPM ...... ..... ..... ... ... ... ... ... ............. .. . ............. ....... ........... ............... .46
2 .3 .2 EFECTOS DE ASN, GLN y SPM SOBRE LA CALIDAD Y DESARROLLO DE LOS ESs DE CHILE
HABANERO .... ..... ...... .. .... .... ..... .............. ... .. .. .... ..... ........ .. .. ......... ..... ...... ........ ..... ... ........ .. .. . ... ...... .... 48
2.3.3 EFECTOS DE SPM SOBRE LA MORFOLOGIA DE LOS ESs DE CHILE HABANERO ... .. ... .... ...... ........... 52
2.4 DISCUSiÓN ................ ... ......... .... ..... ....... . ......................... ........................ ............... ........ .... ... 54
BIBLlOGRAFIA .... ... . .......................... . ............ ...... ...... .. . ... .... ......... ............... ... .. .. ..... ...... ... ... ..... ... 59
CAPiTULO III .. .... .. ........... .......... ...... ... ..................... ..... ................... .. ... ... ... ..... .. .... ....... ...... .. .... .... . ... 67
EFECTO DE ESPERMINA EN LA ACUMULACiÓN Y COMPOSICiÓN DE PROTEINAS DE LOS
ESs DE C. CHINENSE .. ..... .. ....... ... .. ...... ..... ... ... .. .............. ... ........ .... ........ .. .. .... .. .. ... ..... .... ......... .. .. ... 67
3.1 INTRODUCCiÓN .. .. ........ ..... ..... ... ..... .. ...... ........ .. .. ... .... ... .. ................... .. ....... ..... .. . .... .......... .... 67
3.2 MATERIALES Y MÉTODOS ...... .. ..... .. ... .... ....... ...... ........... .. .................. ... .................. ............ 68
3 .2 .1 EXTRACCiÓN DE PROTEINAS TOTALES .... ........ . ........ .. .... .. .... .. ................... . ................... .. ..... . .. . . 68
3.2.2 DETERMINACiÓN DEL CONTENIDO DE PROTEINAS .... ...... ...... ........ .... ..... ...... ... .................... . , ...... 69
3 .2 .3 DETERMINACiÓN DE LA COMPOSICiÓN DE PROTEINAS POR SDS-PAGE ............ ............... ... ...... . 69
3 .2.4 DETERMINACiÓN DE LA COMPOSICiÓN DE PROTEINAS POR 2D-PAGE ...... ... ........ .. ......... . .... ...... . 70
3 .3 RESULTADOS ...... ..... .......... ... ...... ... . .. . .... ........ .. .. ....... ...... .... ..... ..... .... ..... .. ..... .... ... ... .... ....... .. . 71
3.3.1 EFECTO DE SPM EN EL CONTENIDO PROTEICO DE LOS ESs DE CAPSICUM CHINENSE .................. 71
3.3.2 EFECTO DE SPM EN EL PERFIL PROTEICO DE LOS ESs DE C . CHINENSE. .. ...... ....... .. ....... ... ... .. ... .73
3 .3.3 ANÁLISIS DE LA COMPOSICiÓN DE PROTEINAS POR 2D-PAGE ................................. .. ....... .. ... .... 75
3.4 DISCUSiÓN ..... ... .. .. .... .... . ...... ... ..................... ......... ...................... ..... ...... ............... ... ...... ... .. .. 92
BIBLlOGRAFIA .. ........... ...................... ... . .... .. .... .. .... .... .... .. .......... ....... .. ....... ... ....... ............. ... .. . ..... 97

CAPITULO IV .... ... .. .... ..... .. ........ ......... .. ..... .. ...... ... : ............................. ..... ........................................ 1 01
CONCLUSIONES Y PERSPECTiVAS ............ ....................... ... ..... ... ... ... ............. .... ........ .... ..... .. 101
4.1 CONCLUSIONES ... .. ... .... ........ ..... ................... ........... .................. .................................. ...... 101
4.2 PERSPECTiVAS .............. ............... .... ...... .................... ...................................... .................. 102
iv

íNDICE DE FIGURAS
Figura 1.1 Sistemas de regeneración in vitro . ....... .... ........ .... ............................ .................. 4
Figura 1.2 Proceso de embriogénesis somática indirecta ... ..... ........................... ... ..... ........ 6
Figura 1.3 Principales estados productores de variedades de chile ..................... ....... ..... 11
Figura 1.4 Principales PAs que se encuentran en los seres vivos ... ................. .. .. .... ........ 18
Figura 1.5 Diagrama de flujo de la estrategia experimental general ................................. 27
Figura 2.1 Efecto de Asn , Gln y Spm exógena en el número de embriones somáticos de
C. chinense producidos por explante ... .......................................... ..... .... .. .. .. ..................... 47
Figura 2.2 Respuesta de segmentos de hipocotilos cultivados en medio MS + 4.5 IJM de
2,4-0 suplementado con diferentes concentraciones de Asn, Gln y Spm (30 días de
cultivo) .. ............ ................ ... .. ..... .. .............. ...... ... ..... .... .... ................... ....... .. ....... ... ..... ....... 49
Figura 2.3 Efecto de diferentes concentraciones de Asn , Gln y Spm sobre el desarrollo de
los embriones somáticos de C. Chinense a los 45 días de cultivo .............. .......... .... ... .. .... 50
Figura 2.4 Estadías de desarrollo y morfología de los embriones somáticos de chile
habanero formados en diferentes concentraciones de Asn , Gln y Spm ..... .................... ... 52
Figura 2.5 Análisis histológico de ESs de C. chinense tratados con Spm O, 0.01 , 0.1 Y 1
mM en diferentes estadías de desarrollo . .... .... .... ........................................ ..... .. .. .... ..... .... 53
Figura 3.1 Contenido de proteína total en los ESs de C. chinense tratados con 0.1 mM
Spm ....... ... ... .................. ... ... .. ... .. ... .... .. ... .... .............................. ........... ........................ .. .... . 73
Figura 3.2 Perfil protéico en 12 % SDS-PAGE de embriones somáticos a los 45 dias de
cultivo en medio líquido ............... ......................... ........................................ ...................... 74
Figura 3.4 Isoelectroenfoque y 20 en SDS-PAGE (IEF/SDS-PAGE) de embrión cigótico
maduro de C. chinense .. ........ ... ... .......... ............... ...................................... ....................... 76

Figura 3.5 Isoelectroenfoque y anális is en 20 (SOS-PAGE) de semilla de C. chinense . . 77
Figura 3.6 Isoelectroenfoque y 20 en gel de poliacrilamida con dodecil sulfato de sodio
(IEF/SOS-PAGE) de proteínas de EC maduro de C. chinense .......... .... ............................ 80
Figura 3.7 Comparación por 20 del perfil de proteínas y de su abundancia en la semilla y
el EC de C. chinense ................................................................................... ..... .......... .. ...... 81
Figura 3.8 Isoelectroenfoque y 20 (IEF/SOS-PAGE) de las proteínas de ESs avanzados
de C. chinense tratado con 4.5 IJM de 2,4-0 (TO) ............... .... .. ...... ...... ................ ............. 84
Figura 3.9 Isoelectroenfoque y 20 de las proteínas de ESs avanzados de C. chinense
tratados con 0.1 mM de Spm .. .. .... .. ............................................................ .. ..................... 86
Figura 3.10 Isoelectroenfoque y 20 de las proteínas de A) Ess de C. chinense tratados
con Spm 0.1 mM . B) Ess del tratamiento testigo ..................................................... ...... .... 89
Figura 3.11 Isoelectroenfoque y 20 en gel de poliacrilamida con dodecil sulfato de sodio
(IEF/SOS-PAGE) de ESs avanzado de C. chinense tratado con 0.1 mM de Spm ............ 90
Figura 3.12 Isoelectroenfoque y 20 de proteínas del A) EC de C. chinense. B) ESs de C.
chinense tratado con Spm 0.1 mM y C) ESs de C. chinense del TO .. .. .. ............................ 91
vi

íNDICE DE CUADROS
Cuadro 2.1 Porcentaje de ESs tempranos y avanzados de C. chinense obtenidos con diferentes
concentraciones de Asn , Gln y Spm ............. ........ .. ............................................ .. ...... ... ......... .. ..... .. . 51
Cuadro 3.1 Peso molecular (PM) y punto isoeléctrico (pi) de proteínas específicas del perfil
proteico del EC de C. chinense ...... .. .. .. ................................................... .. .. ... .. .. .................. .. ........... 78
Cuadro 3.2 Peso molecular (PM) y punto isoeléctrico (pi) de proteínas que muestran cambios en
su abundancia en el perfil proteico de la semilla y el EC de C. chinense . ............ .. ............ .. ........... . 82
Cuadro 3.3 Peso molecular (PM) y punto isoeléctrico (pi) de proteínas específicas del perfil
proteico de ESs de C. chinense tratados con Spm 0.1 mM .... .. .............. .. ..... .. .................................. 85
Cuadro 3.4 Peso molecular (PM) y punto isoeléctrico (pi) de proteínas específicas del perfil
proteico de ESs de C. chinense tratados y sin tratar con Spm 0.1mM ........................ .. ........ .. ......... 87


ABREVIATURAS
Asn
ABA
BSA
CTV
EC
ECs
ES
ESO
ESI
ESs
Gln
MS
Spm
2,4-0
Asparag ina
Ácido abscísico
Ácido giberélico
Albúmina de
bovino
Cultivo
vegetales
de
suero
tejidos
Embriogénesis cigótica
Embriones cigóticos
Embriogénesis somática
Embriogénesis somática
directa .
Embriogénesis somática
indirecta.
Embriones somáticos
Glutamina
Murashige y Skoog
Espermina
Ácido 2,4-
Oiclorofenoxiacético


RESUMEN
Tomando en cuenta la incapacidad de los embriones somáticos de chile habanero
(Capsicum chinense Jacq .) para convertirse en planta, en este estudio se realizó la
comparación de diferentes tratamientos con los aminoácidos asparagina (Asn) y
glutamina (Gln) y con la poliamina espermina (Spm) sobre la eficiencia, calidad y
desarrollo de los embriones somáticos . Los resultados mostraron que la espermina fue
favorable en los ESs . El tratamiento de 0.1 mM Spm resultó de mayor interés pues no
disminuyó la eficiencia y elevó notablemente la calidad, mejoró el desarrollo y morfología
de los embriones somáticos. Con este tratamiento se observó la formación de dos
pequeños cotiledones en embriones avanzados, a diferencia de los demás incluyendo al
tratamiento testigo. Para conocer el efecto de esta poliamina sobre el contenido y
composición de proteínas totales de los embriones somáticos, se realizó su cuantificación
y separación (SOS-PAGE y 20-PAGE) en muestras de embriones tratados con 0.1 mM
Spm; en estos análisis se incluyeron como testigos embriones somáticos sin tratar, así
como la semilla y el embrión cigótico maduro de C.chinense. Los análisis proteicos
corroboraron que existe una gran deficiencia en contenido y composición de proteínas en
los embriones somáticos en comparación con los cigóticos de la especie, en
aproximadamente un 90%. Análisis de 20 mostraron que la Spm modificó el contenido y
composición proteica de los embriones propiciando la presencia de 28 puntos de proteína,
que no se encontraron en los embriones sin tratar; de los cuales, cinco se asemejan en
peso molecular y punto isoeléctrico a los reportados como proteínas de reserva para
diferentes especies vegetales . Así mismo, mostró la presencia de 4 puntos de proteína
que también se encontraron en los embriones cigóticos de chile habanero pero no en el
tratamiento testigo. Los resultados permiten inferir que la modificación observada en el
patrón de proteínas de los embriones somáticos con morfología like-normal por la
aplicación de espermina en el medio de cultivo, pudiera estar involucrada en los efectos
favorables sobre el desarrollo y morfología de los embriones. Sin embargo, aún se
requiere otros estudios que permitan solucionar el problema de la conversión de estos
embriones en plantas normales.


ABSTRAeT
Considering the inability of somatic embryos of habanero pepper (Capsicum chinense
Jacq.) to conversion in plant, in this study a comparison with different treatments of amino
acids asparagine (Asn) and glutamine (Gln) and the polyamine spermine (Spm) was
performed on the efficiency, quality and development of somatic embryos. Results showed
that spermine was favorable in the ESs. Spm 0.1 mM treatment was of interest because it
did not decrease the efficiency and significantly increased quality, improved development
and morphology of the embryos. With this treatment the formation of two small cotyledons
in advanced embryos was observed, unlike the others including the control treatment. To
know the effect of this polyamine on the content and composition of total proteins of
somatic embryos, quantification and separation (SDS-PAGE and 2D-PAGE) in samples of
treated embryos with 0.1 mM Spm was performed; in these analyzes were included
untreated somatic embryos, seed and mature zygotic embryo of C.chinense as controls .
Protein analyzes corroborated that there is a high deficiency in protein content and
composition in somatic embryos compared with zygotic species, about 90%. 2D analysis
showed that Spm modified the protein content and composition of embryos favoring the
presence of 28 protein spots which were not found in untreated embryos; five of these
spots are similar in molecular weigth and isoelectric point to the reported as storage
proteins for different plant species. It also showed the presence of 4 protein spots that
were also found in zygotic embryos of habanero pepper but not in the control treatment.
The results allow us to infer that the change observed in the protein pattern of somatic
embryos with like-normal morphology by the application of spermine in the culture
medium, could be involved in the beneficial effects on the development and morphology of
embryos. However, still other studies to solve the problem of the conversion of these
embryos into normal plants are required .


INTRODUCCiÓN
INTRODUCCiÓN
En los últimos años se ha desarollado un gran interés en establecer los protocolos de
regeneración in vitro, de especies vegetales de importancia agronómica, que permitan su
utilización en los trabajos de mejoramiento y de micropropagación . El chile habanero es
un cultivo reconocido por ser uno de los productos agrícolas de mayor importancia
comercial a nivel internacional y nacional; es altamente demandado por la industria
alimenticia, farmacéutica, cosmética, química y militar debido a sus altos contenidos de
capsaicina (Curry et al., 1999). Además, su explotación constituye una fuente de trabajo
permanente durante el año para los productores que se dedican a su cultivo (Paran et al.,
2007).
Dentro de las vías de regeneración de plantas, la embriogénesis somática (ES) es
considerada el método más eficiente para la propagación masiva de plantas (Merkle y
Dean, 2000). Sin embargo, la mayoría de los reportes (Franck-Duchenne et al., 1998;;
Buyucalaka y Mavituna et al. 1996; Steinz et al., 2003; Ochoa y Ramírez, 2001; Kothari ,
2010) reconocen al género Capsicum como recalcitrante a la morfogénesis in vitro debido
a la baja eficiencia y reproducibilidad de los sistemas de embriogénesis somática
establecidos, así como por el alto indice de deformaciones y en consecuencia, la baja
tasa de germinación de los embriones formados. En el caso de C. chinense, en el año
2013, se reportó un protocolo altamente eficiente y reproducible de ES directa en medio
líquido por Avilés-Viñas, con el cual es posible obtener 1.77 x 104 embriones somáticos
(ESs)/L de medio. No obstante, aún persisten los problemas de un alto índice de
embriones somáticos (ESs) deformados y su incapacidad para germinar y convertirse en
plantas . Es por ello que, actualmente se llevan a cabo diversos estudios histológicos,
bioquímicos y moleculares para entender esta problemática a fin de establecer estrategias
efectivas que contribuyan a su solución . Dentro de estos estudios, se encuentra el
conocer el papel de las poliaminas endógenas y exógenas , así como el de las proteínas
en este proceso.
Lecona et al., 2012 reportaron que el contenido de proteinas totales presentes en los ESs
de C. Chinense, disminuye drásticamente a medida que el embrión somático avanza en
sus estad íos de desarrollo; pasando de contener 2.9 ¡.t.g .mg-1 de proteína en el estadio

INTRODUCCiÓN
globular a 0.4 f!g.mg-1 en el estadía cotiledonar. Los bajos contenidos de proteína en esta
etapa crítica de desarrollo, podría ser una de las posibles causas que afectan el correcto
desarrollo y germinación de los ESs. Por este motivo, en el grupo de trabajo se han
estado implementando estrategias que pudieran favorecer la acumulación y/o
composición de proteínas en los ESs de ch ile habanero.
La adición de compuestos orgánicos nitrogenados, como los aminoácidos y las palia minas
(PAs) al medio de cultivo de especies vegetales, han mostrado que pueden estimular la
ES, así como favorecer el desarrollo, calidad y germinación de los ESs. La aplicación
exógena de aminoácidos tales como L-Prolina (Pro), L-Serina (Ser), L-Glutamina (Gln), L
Asparag ina (Asn); así como de las poliaminas Espermina (Spm), Putrescina (Put) y
Espermidina (Spd) ha permitido aumentar la frecuencia de formación de ESs por explante,
así como el número de ESs germinados y convertidos en plantas (Minocha y Minocha,
1995; Carmona , 1997; Chia and Saunders, 1999; Witjaksono and Litz, 1999; Ronchi et al.,
1984; Claparols et al., 1993). En C.chinense, Lecona (2012), observó una notable mejoría
en los ESs al adicionar L-Asparagina en el medio de cultivo de esta especie. Así también,
se ha indicado que el empleo de poliaminas en cultivos embriogénicos, permite la
regeneración de Capsicum frutescens Mili (Kumar et al. , 2007). López-Erosa (2012),
realizó un estudio de las PAs endógenas y exógenas en los embriones somáticos de chile
habanero, en el cual observó mejoría en su desarrollo y germinación al adicionar
espermina (Spm) al medio de cultivo.
Tanto los aminoácidos como las poliaminas, son utilizados por las células para modular
numerosos procesos celulares; por lo tanto, la adición de alguno de estos compuestos
pudiera resultar en la mejora de la calidad y desarrollo de los ESs de esta especie . Por lo
consiguiente, en el presente trabajo, se llevó a cabo la evaluación de los efectos de los
aminoácidos L-glutamina, L-asparagina y de la poliamina espermina sobre la eficiencia ,
calidad y desarrollo de los ESs de C. chinense. Así como el efecto en la acumulación y
composición de proteínas endógenas de los ESs provenientes del mejor tratamiento
evaluado, esto como un posible indicador del buen desarrollo y calidad de los ESs de esta
especie.
2

CAPíTULO I
ANTECEDENTES
1.1 CULTIVO DE TEJIDOS VEGETALES (CTV)
CAPiTULO I
El cultivo de tej idos vegetales (CTV), es el conjunto de técnicas que permiten bajo
condiciones artificiales , axénicas y controladas, el establecimiento, mantenimiento y
desarrollo de cualquier parte de una planta , desde una célula hasta un organismo
completo (Pérez-Molphe et al. , 1999).
El CTV es una herramienta invaluable para la resolución de problemas básicos y
aplicados en la biotecnología vegetal , ya que por una parte ofrece una serie de sistemas
modelo ideales para la investigación fisiológica, bioquímica , genética y estructural , y por
otro lado tiene una aplicacíón práctica en la clonación , conservación y manipulación in
vitro de cualquier material vegetal (pérez-Molphe et al., 1999).
Pérez-Molphe et al. , 1999, describen tres principios básicos cuya comprensión y
manipulación son determinantes para el éxito o fracaso de cualquier trabajo en el campo
del CTV: la elección del explante, elección del medio y condiciones de cultivo y
condiciones asépticas. La interacción de estos tres principios básicos determinará la
respuesta del tejido cultivado in vitro. Esta respuesta puede presentarse como:
organogénesis (formación de órganos adventicios como raíces y brotes) , ES (formación
de embriones a partir de células somáticas) , formación de tejido calloso o simplemente
desarrollo del tejido .
3
CAPiTULO I
'.
a. el lO
P t
1'91 ag."." ecta
~
,rece / ,,, Pen I
em non., some .cos
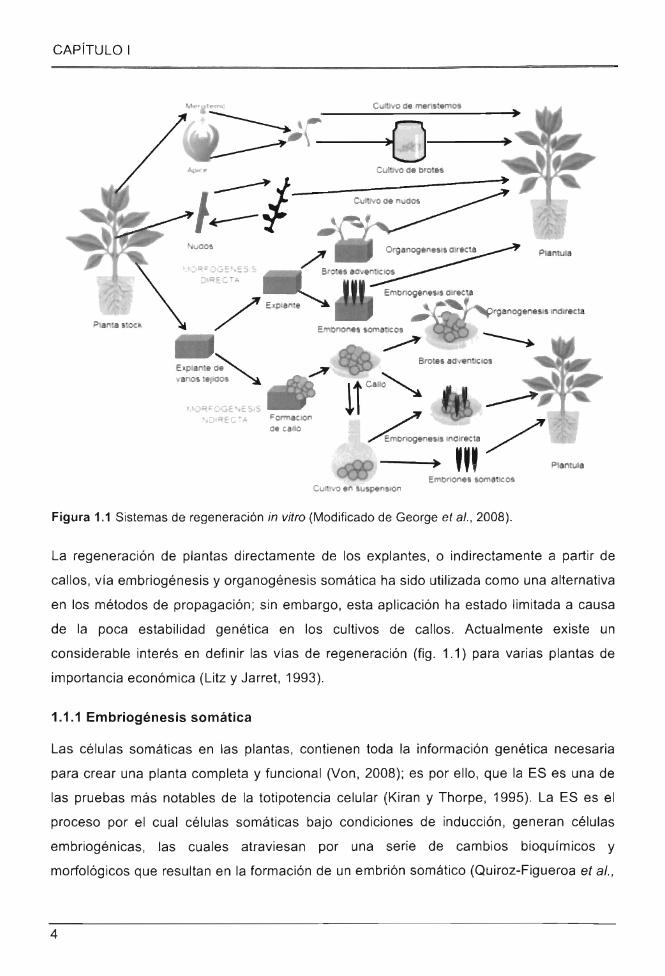
Figura 1.1 Sistemas de regeneración in vitro (Modificado de George et al., 2008).
La regeneración de plantas directamente de los explantes, o indirectamente a partir de
callos , vía embriogénesis y organogénesis somática ha sido utilizada como una alternativa
en los métodos de propagación; sin embargo, esta aplicación ha estado limitada a causa
de la poca estabilidad genética en los cultivos de callos. Actualmente existe un
considerable interés en definir las vías de regeneración (fig. 1.1) para varias plantas de
importancia económica (Litz y Jarret, 1993).
1.1.1 Embriogénesis somática
Las células somáticas en las plantas, contienen toda la información genética necesaria
para crear una planta completa y funcional (Von, 2008); es por ello, que la ES es una de
las pruebas más notables de la totipotencia celular (Kiran y Thorpe, 1995). La ES es el
proceso por el cual células somáticas bajo condiciones de inducción , generan células
embriogénicas, las cuales atraviesan por una serie de cambios bioquímicos y
morfológicos que resultan en la formación de un embrión somático (Quiroz-Figueroa et al. ,
4

CAPíTULO I
2006).
El desarrollo de los embriones somáticos ocurre de manera muy similar al de los
embriones cigóticos (ECs) (Kiran y Thorpe, 1995). Poseen una estructura con dos polos
opuestos, un eje radical-apical , y no poseen conexión vascular con el tejido materno .
Estas estructuras deben ser capaces de crecer y formar plantas normales (Litz y Jarret,
1993). Mediante el proceso de ES, se pueden obtener fácilmente una gran cantidad de
ESs en diferentes y definidas etapas de desarrollo (Van y Clapham, 2007).
Debido a que los ESs son generados a partir de un proceso asexual, la nueva planta será
exactamente igual a la donadora de la célula inicial (Kiran y Thorpe, 1995). Aunque en
ciertos aspectos, los ESs mantienen una similitud con los ECs tanto in vivo como in vitro ,
pueden ocurrir algunas anormalidades en su desarrollo (Litz y Jarret, 1993).
1.1.1.1 Etapas de la embriogénesis somática
Existen dos tipos de ES in vitro, la directa (ESO) y la indirecta (ESI) . En la ESO, los
embriones se producen directamente del explante original ; mientras que en la ESI , es
indispensable obtener un tejido calloso o una suspensión celular embriogénica a partir de
la cual se obtendrá la diferenciación de los ESs en un segundo paso (Pérez-Molphe et al.,
1999).
El proceso de ES consta de diferentes etapas , en cada una de las cuales se requieren
diferentes estímulos y ocurren diferentes fenómenos (Merkele et al. 1995). Estas etapas
son : inducción, histodiferenciación, maduración y germinación (Fig . 1.2; Pérez-Molphe et
al., 1999):
5

CAPITULO I
TEJIDO CALLOSO
I INDUCCiÓN
MASA DE CÉLULAS + AUXINAS PROEMBRIONICAS
EMBRIONES EN ESTADIO GLOBULAR
HISTODIFERENCIACIÓN EMBRIONES EN ESTADIO CORAZÓN -AUXINAS
/1/ EMBRIONES EN ESTADIO TORPEDO
~ 1 MADURACiÓN
EMBRIONES EN ESTADO COTILEDONARIO ABA
DESECACiÓN
~1'!- PLÁNTULAS l GERMINACiÓN
Figura 1.2 Proceso de embriogénesis somática indirecta (Modificado de Pérez-Molphe et al., 1999).
1.1.1.1.1 Inducción
En esta etapa , las células somáticas se convierten en proembriogénicas (capaces de
formar ESs). Para que este proceso suceda , se consideran ciertos factores determinantes
(Pérez-Molphe et al., 1999) :
6
a. Genotipo. Se sabe que diferentes genotipos aún de una misma especie
muestran diferencias notables en la capacidad para iniciar el proceso de
ES. Por lo que ciertos genotipos pueden ser más propensos a ella y en
otros puede ser prácticamente imposible.
b. Grado de diferenciación de las células del explante. Es mucho más
factible obtener ES de tejidos jóvenes y muy poco diferenciados que de
tejidos maduros completamente diferenciados.
C. Auxinas. Entre los efectos que ocasionan este tipo de reguladores del

CAPíTULO I
crecimiento, se encuentra la metilación del ADN que puede favorecer el
cambio del patrón de expresión genética que se requiere para que ocurra
la inducción . Entre las auxinas más efectivas para la inducción de la ES,
se encuentran las sintéticas como los ácidos fenoxiacéticos (ácido 2,4-
diclorofenoxiacético (2,4-0), y ácido 2,4,5- triclorofenoxiacético (2,4,5-T))
de los cuales el que usualmente se emplea es el 2,4-0 . Esto debido a que
las células vegetales son incapaces de degradar o inactivar eficientemente
las auxinas sintéticas como lo hacen con las naturales, por lo que el efecto
de las primeras, son más intensos y duraderos.
d. Aislamiento celular. El aislamiento de una célula o de un grupo de células
respecto al resto del tejido, estimula la conversión de una célula somática
en una proembriogénica . Esto sucede cuando hay necrosis en el tejido
adyacente o cuando se aplican tratamientos físicos para separar las
células.
1.1.1.1.2 Histodiferenciación
Cuando los cultivos son transferidos de un medio de proliferación a un medio de
estimulación del desarrollo embrionario, estos cultivos consisten en una mezcla de células
individuales y agregados celulares (Van, 2008). Durante la histodiferenciación, las masas
de células proembriogénicas se diferencían formando ESs, mediante una división y
diferenciación celular simultáneas. Para que estas células dejen de multiplicarse y pasen
a una fase de diferenciación, se requiere remover las auxinas exógenas o utilizarlas en
concentraciones más bajas (Litz y Jarret, 1993), debido a que resultan ser inhibitorias
para el desarrollo de las masas proembriogénicas en ESs. El grado de diferenciación de
los embriones cultivados en presencia de auxina, varía para diferentes especies (Van,
2008). Uno de los fenómenos iniciales en esta etapa es el establecimiento de la polaridad
en las masas de células proembriogénicas, así como la formación de los tejidos
vasculares ; los cuales se mantendrán durante todo el desarrollo y vida de la planta .
Durante esta etapa, los ESs pasan por una serie de estadías intermedios muy similares a
los que ocurren en la EC. En las dicotiledóneas los estadías son : globular, corazón y
torpedo; y en las monocotiledóneas son : globular, corazón y escutelar (Pérez-Molphe et
al. , 1999).
7

CAPiTULO I
1.1.1.1.3 Maduración
Para que un embrión somático adquiera la capacidad de germinar requiere pasar por una
fase de maduración, durante la cual, básicamente ocurre elongación celular sin división
(Pérez-Molphe et al., 1999). En esta etapa, los ESs sufren cambios bioquímicos y
morfológicos: formación de los órganos de reserva y cotiledones, la expansión coordinada
con la deposición de materiales de reserva , la represión de la germinación y la adquisición
de tolerancia a la desecación (Thomas, 1993). En el caso de las dicotiledóneas, al
embrión somático maduro se le da el nombre de estadía dicotiledonario (Pérez-Molphe et
al. , 1999) .
los ESs acumulan sustancias de reserva con las mismas características que las
presentes en los ECs; las cuales son también dirigidas a los compartimentos subcelulares
correctos (Merkele et al., 1995). Sin embargo, tanto la cantidad como la sincronización de
una sustancia de reserva en particular, puede diferir entre embriones somáticos y
cigóticos (Merkele et al., 1995; Yeung, 1995).
la síntesis y acumulación de las proteínas de reserva y proteínas lEA (late
embryogenesis abundant proteins) durante la embriogénesis cigótica y somática, para la
mayoría de las especies, son reguladas a través de ácido abscícico (ABA) y la desecación
(Dodeman et al., 1997). los cultivos embriogénicos de algunas especies requieren ser
tratados con ABA a concentraciones entre 10 Y 50 ¡,tM, para estimular la maduración . En
otros casos, este regulador de crecimiento es utilizado para reducir el proceso de la
embriogénesis secundaria o para inhibir la germinación precoz. Se han reportado muchos
otros factores que tienen influencia en la maduración de los ESs en diferentes especies,
tales como el etileno, el estrés osmótico, el pH y el fotoperíodo (Van, 2008). Sin embargo,
los embriones no deben ser expuestos a tratamientos de maduración hasta que hayan
alcanzado una apropiada etapa de desarrollo (Zimmerman , 1993).
Generalmente, la maduración de los ECs termina en algún grado de deshidratación,
resultando en una reducción gradual del metabolismo a medida en que el tejido de la
semilla va perdiendo agua y el embrión pasa a un estado metabólicamente inactivo o
quiescente. Este proceso es muy importante, ya que le confiere a la semilla tolerancia a
diferentes tipos de estrés ambientales extremos y le permite conservarse en el tiempo
8

CAPiTULO I
(Kermode, 1990). En embriones somáticos de plantas de semilla ortodoxa es posible
inducir este estado de quiescencia a través de un tratam iento de deshidratación . En
numerosos intentos para mejorar la cal idad de los ESs, se ha observado el papel
estimulante de un bajo potencial osmótico en el medio de maduración en gimnospermas
(Attree y Fowke , 1993), como en angiospermas (McKersie y Brown , 1996). Este efecto
permite imitar el estrés hídrico impuesto naturalmente en sem illas ortodoxas durante
largas etapas de maduración, permitiendo la sobrevivencia de los embriones a la
deshidratación (Von , 2008). En algunas especies, después del tratamiento de
maduración, se induce una desecación parcial la cual aumenta la frecuencia de
germinación de los ESs. Así mismo, se ha sugerido que la desecación disminuye el
contenido endógeno de ABA (Kermode et al., 1989) o cambia la sensibilidad a este
regulador (Finkelstein et al. , 1985) .
Los ESs comparados con los cigóticos de la misma especie, con frecuencia se desarrollan
de manera anormal o son inmaduros . La anomalía más frecuente es la formación doble o
triple del sistema vascular, causada por la pobre polarización del transporte de auxinas .
También se puede encontrar un conten ido excesivamente alto, reducido o ausente de las
reservas de almidón y/o proteicas; que el meristemo apical o rad ical no estén
desarrollados o que la embriogénesis sea repetitiva o secundaria . El hecho de observar
en los ESs de algunas especies, la falta del meristemo apical o de una escasa
acumulación de sustancias lipoproteicas, no debe considerarse como una anormalidad ,
sino que puede ser síntoma de inmadurez debido a la rápida formación de los mismos. En
este caso, una etapa intermedia que permita la maduración de las estructuras formadas
permitirá incrementar el porcentaje de embriones somáticos capaces de germinar
(Radice, 2010).
1.1.1.1.4 Germinación
En esta etapa se lleva a cabo la elongación y reactivación metabólica de un embrión
somático maduro para convertirse en plántula . Para que esto ocurra , se requ ieren
estímulos como la luz, el ácido giberélico o citocininas . Debido a que los ESs carecen de
tej idos de reserva , su germinación sólo ocurre in vitro en donde el medio de cultivo aporta
los nutrientes , o bien , cuando se les proporcionan depósitos artificiales de nutrientes
conocidos como sem illas artificiales (Pérez-molphe et al., 1999).
9

CAPiTULO I
1.1.2 Regeneración de plantas vía ES
La ES es un proceso complejo en el cual la calidad del producto final , es decir, la
sobrevivencia y crecimiento de las plantas regeneradas depende de las condiciones
provistas en las etapas tempranas, cuando los ESs son madurados y germinados.
Solamente aquellos embriones que han acumulado suficiente material de reserva y han
adquirido tolerancia a la desecación al final de la maduración, desarrollan en plantas
normales. Usualmente, los ESs desarrollan en plantas pequeñas comparadas con las
plántulas cultivadas en medio de cultivo libre de reguladores de crecimiento; sin embargo,
existen casos en los que las auxinas y citocininas, estimulan la germinación . Además de
que para algunas especies se requiere la inclusión de componentes extras como la
glutamina y la caseína hidrolizada. Cuando las plantas han alcanzado un tamaño
adecuado, pueden ser transferidas a condiciones ex vitro (Von , 2008).
Las plantas provenientes de ESs crecen de manera similar a las plantas derivadas de
semillas; sin embargo, para muchas especies la variación somaclonal es un problema. En
general , el uso de 2,4-0 y/o una fase prolongada de callo es responsable de inducir
variación genética y epigenética (Von , 2008).
La ES muestra gran cantidad de ventajas comparado con otros sistemas de propagación
in vitro , incluyendo sus altas tasas de multiplicación, caso de crioconservación de callos
embriogénicos, el potencial para su escalamiento en cultivos de suspensiones celulares,
el uso de bioreactores, tecnologías de semillas sintéticas y el hecho de que los cultivos
embriogénicos son blancos factibles para la transferencia genética (Merkle y Oean, 2000).
La regeneración de plantas vía ES, tiene un significado especial debido a que se obtienen
regenerantes genéticamente más estables. Así mismo, posee un considerable potencial
para mejorar el entendimiento de diversos aspectos relacionados al desarrollo
embrionario (Roja et al., 2005). A pesar que la ES se ha obtenido con éxito en menor
número de especies que la organogénesis; cuando ésta es posibe, su productividad en
cuanto al número de plantas que se pueden generar es mayor (Pérez-molphe et al.,
1999). La embriogénesis somática juega un papel importante en la propagación clona!.
Cuando se íntegra con programas de mejoramiento convencional y técnicas de biología
celular, la ES provee una invaluable herramienta para mejorar el ritmo del mejoramiento
genético del cultivo de especies de importancia (Stasolla y Yeung , 2003).
10

CAPITULO I
1.2 GÉNERO CAPSICUM
El género Capsicum pertenece a la familia de las Solanaceaes. Consta de
aproximadamente 20-27 especies, de las cuales cinco son domesticadas (Pickersgill ,
1997): C. annuum, C. baccatum, C. chinense, C. Frutescens y C. pubescens. La
propiedad química que separa al género Capsicum de otros grupos vegetales, es un
alcaloide denominado capsaicina, una sustancia cristalina potente y acre , que no existe
en ningún otro género. La cantidad de capsaicinoides presentes en los chiles varía
considerablemente de especie a especie (Paran et al., 2007)
México es el país con más variedades de chile en el mundo (Inforural, 2012-fuente
Sistema Producto Chile habanero).
Océano Pacifico
Figura 1.3 Principales estados productores de variedades de chile (Inforural, 2012).
1.2.1 Chile habanero (Capsicum chinense Jacq.)
El chile habanero es uno de los productos hortícolas más importantes debido a su
demanda local, nacional e internacional. Yucatán es considerado como centro de
diversidad genética de esta especie por la diversidad de colores, formas, pungencia,
aroma y color de sus chiles (Terán y Rasmussen, 1998); y ocupa el primer lugar en
producción de fruto con 2,574.12 toneladas anuales (SIAP, 2014).
El chile habanero es un producto muy apreciado en el mundo debido que es uno de los
chiles de mayor pungencia o picor (registra de 200,000 a 500,000 unidades Scoville) por
11

CAPiTULO I
su alto contenido de capsaicina (Curry et al., 1999). Además el contenido nutricional del
chile es alto; es fuente de vitaminas, particularmente de la C, y en los tipos pungentes
secos, de vitamina A (Tun Dzul , 2001).
1.2.1.1 Taxonomía
De acuerdo al Código Internacional de Nomenclatura Botánica la taxonomía de chile
habanero se describe como sigue:
1.2.1.2 Problemática del cultivo
Reino: Plantae
Subreino: Embriophyta
Clase: Angiosperma
Subclase: Dicotiledónea
Superorden: Sympetala
Orden: Tubiflorae
Familia: Solanaceae
Género: Capsicum
Especie: C. chinense Jacq.
A pesar de la riqueza genética de chile habanero en la península de Yucatán, este cultivo
ha sufrido una fuerte erosión como consecuencia de la pérdida acelerada de las
variedades criollas ó autóctonas, por la sustitución de éstas por variedades foráneas.
Aunado a esto, la producción de chile habanero se ve afectada por una serie de
situaciones adversas, tales como problemas al establecer cultivos de calidad homogénea,
problemas fitosanitarios , así como polinización cruzada , factores medioambientales
adversos, falta de asesoría técnica , etc. (latounerie et al., 2006).
El mejoramiento y propagación convencional han contribuido significativamente al
mejoramiento genético de plantas; sin embargo, uno de los principales retos que se tienen
es, por un lado, la necesidad de incrementar la productividad y por otro disminuir los
costos de producción (Mondal et al., 2004).
12

CAPíTULO I
Estos problemas podrían ser resueltos por mejoramiento genético a partir de la aplicación
de sistemas de regeneración de plantas; permitiendo la posibilidad de propagar plántulas
en forma masiva a través de la micropropagación . El método considerado como el más
eficiente en regeneración de plantas in vitro, es la embriogénesis somática (ES). Técnica
que como todas las demás en el CTV, requiere sistemas óptimos de regeneración . Sin
embargo, a pesar que otros miembros de la familia de las Solanáceas (papa , tomate,
tabaco y petunia) se utilizan como sistemas modelo de regeneración in vitro; el género
Capsicum es reconocido por su recalcitrancia (incapacidad de las células, tejidos y
órganos vegetales a responder al cultivo in vitro en cualquiera de sus etapas) a la
morfogénesis in vitro; motivo por el cual representa un reto para la aplicación de dichas
técnicas. Esto se refleja por el hecho de que todos los protocolos establecidos para este
género tienen varios problemas en común : baja eficiencia, baja reproducibilidad, baja
capacidad de germinación y alto índice de embriones deformados.
A pesar que la gran mayoría de los protocolos de ES se han establecido para C. annuum,
recientemente se han generado los primeros reportes de embriogénesis somática directa
(López-Puc et al., 2006) e indirecta (Zapata-Castillo et al., 2007) para Capsicum chinense.
Actualmente, se cuenta con un protocolo de ES directa de alta eficiencia en medio líquido
para la regeneración in vitro de chile habanero (Avilés et al. , 2013); protocolo en el cual se
resolvieron los problemas de dependencia del genotipo y del explante. Aún así , este
sistema se ve limitado por la alta frecuencia de embriones con deformaciones en la región
apical, baja capacidad de germinación y por ende la baja conversión de los ESs a plantas
completas. Esto ha motivado diversos estudios para poder entender la(s) causa(s) de la
recalcitrancia del género Capsicum; entre dichos estudios se encuentra la evaluación del
papel de las proteínas en los embriones somáticos y el de las poliaminas (PAs), tanto
endógenas como adicionadas al medio de cultivo.
13

CAPITULO I
1.3 PROTEíNAS EN EMBRIONES CIGÓTICOS
1.3.1 Proteínas de reserva
Las plantas almacenan proteínas en sus células embrionarias y vegetativas para proveer
fuente de carbono, nitrógeno y azufre para su posterior crecimiento y desarrollo. El
almacenamiento y movilización de los aminoácidos que componen a estas proteínas son
aspectos críticos para el ciclo de la vida de las plantas (Herman y Larkins, 1999).
Las semillas contienen muchas proteínas metabólicamente inactivas que funcionan como
reservas ; las cuales varían dependiendo de la especie. De igual manera se presentan
proteínas metabólicamente activas, como las enzimas (Osborne, 1907).
Se han reportado cuatro tipos de proteínas: prolaminas, glutelinas, globulinas y albúminas
predominando las dos primeras (Osborne, 1907). El total de proteínas activas en trigo sólo
alcanza el 15% del total, siendo las principales las albúminas. En la avena (Avena sativa),
el 80% del porcentaje de proteína total corresponde a la globulina, 15% a la prolamina y
5% a la glutelina; en el pepino (Cucurbita pepo) también predomina la globulina . En
cambio en el arroz (Oryza sativa) , 80% corresponde a la glutelina, 10% a la globulina y
5% a la albúmina y a la prolamina . En el maíz (Zea mays) los valores son 31, O, 14 Y 48%
respectiva mente .
Las proteínas de reserva se acumulan principalmente en vacuo las de células
diferenciadas del embrión y del endospermo; y como cuerpos proteicos ensamblados
directamente dentro del retículo endoplásmico rugoso. La síntesis de proteínas de reserva
y la formación de vacuolas especializadas ocurre después de completarse la división
celular, cuando todo el crecimiento adicional ocurre sólo por expansión celular y por
acumulación de sustancias de reserva (Morcillo et al., 2001). Las proteínas de reserva en
semillas que se encuentran en mayor cantidad en las vacuolas son las globulinas 7S y
11 S (Herman y Larkins, 1999). Las proteínas 11 S se encuentran en semillas de coníferas ,
monocotiledóneas y dicotiledóneas (Higuchi y Fukazawa, 1987). Mientras que las
proteínas 7S son predominantes en monocotiledóneas (Morcillo et al., 2001).
14

CAPiTULO I
Otras proteínas como las lectinas, son secuestradas con las proteínas de reseNa, y
posiblemente se encuentren en suficiente concentración para constituir proteínas de
reseNa auxiliares (Herman y Larkins, 1999).
En general, las semillas presentan alto contenido de nitrógeno y de prolina; así como una
baja cantidad de lisina, triptófano y metionina. Contienen algunos minerales cuya
composición es similar a la del resto de la planta; y presentan además, otros compuestos
nitrogenados como aminoácidos (glutamina y asparagina en las nueces), amidas y
alcaloides (Higuchi y Fukazawa, 1987).
1.3.2 Proteínas LEA
Las proteínas LEA (Late embryogenesis abundant proteins), fueron identificadas por
primera vez en semillas de algodón (Gossypium hirsutum), en etapas tardías de la
embriogénesis (Dure et al., 1981). Posteriormente fueron encontradas también en semillas
y órganos vegetativos de diferentes plantas, especialmente bajo condiciones de estrés por
frío, sequedad o alta salinidad , entre otros (Hundertmark y Hincha, 2008). Estas proteínas
se producen abundantemente durante el desarrollo de la semilla, comprendiendo arriba
del 4% de las proteínas celulares (Roberts et al., 1993).
Las proteínas LEA se encuentran conseNadas en una gran variedad de especies (Barceló
et al., 2001; Colorado et al., 1995; Rock y Quatrano, 1995; Delseny et al., 2001). Una
característica común de estas proteínas, es su composición parcial de aminoácidos
cargados y polares; en particular glicina y lisina (Dure et al., 1989), que les confiere su alta
hidrofilicidad (Garay et al., 2000) y estabilidad déficit hídrico (Hundertmark y Hincha,
2008). Por lo regular carecen de algunos aminoácidos como triptófano y muy a menudo
de cisteína. Además, contienen escasos residuos básicos e hidrofóbicos. Estas
características les confieren una función importante: proteger a las células ante los daños
producidos por la brusca deshidratación que sufren las semillas en las últimas fases de
desarrollo (Dure et al., 1989). Estas proteínas son reguladas transcripcionalmente en
respuesta a ABA (Ingram y Bartels, 1996) durante la fase de maduración de la semilla .
Dentro de las proteínas LEA, destacan las dehidrinas (DHN); llamadas también proteínas
RAB (Responsive to ABA) o LEA 11. Estas proteínas se inducen por deshidratación (por
15

CAPiTULO I
calor, sequ ía o frío) acumulándose en grandes cantidades en semillas maduras . Ejemplo
de éstas son las proteínas RAB21 o RAB16A-C de arroz (Mundy and Chua , 1988;
Yamaguch i-Shinozaki et al., 1989) o la proteína RAB18 de Arabidopsis (Goshi et al. ,
1995). En cond iciones normales se ha observado la acumulación dife rencial de las
dehidrinas en diferentes tejidos; sin embargo, en condiciones de estrés o después de la
apl icación de ABA exógeno, se produce un aumento de dichas proteínas en todos los
tejidos, lo que indica su importancia en las respuestas de las plantas a cond iciones
adversas a estrés abiótico (Nylander et al., 2001).
Las proteínas LEA no son específicas de plantas, ya que han sido encontradas en
diferentes organismos como bacterias , cianobacterias, levaduras y nematodos. Se
encuentran asociadas con la tolerancia a la desh idratación ; permitiendo la sobrevivencia
de organismos inclusive en cond iciones de pérd ida completa de agua (Hundertmark y
Hincha, 2008).
1.3.3 Acumulación de proteinas en embriones somáticos
Los ESs expresan a nivel molecular, una serie de proteínas propias del proceso de
embriogénesis cigótica, las cuales nunca aparecen en los brotes adventicios. A estas
proteínas se les considera como marcadores moleculares del proceso de embriogénesis
(Pérez-Molphe et al. , 1999). Sin embargo, frecuentemente se observa , que los ESs no
son capaces de acumular varios tipos de proteínas, asociadas con la madurez de los ECs
en cantidades suficientes (Zimmerman,1993). Un ejemplo de ello son los ESs de alfalfa,
en los cuales se han detectado un 10% de las proteínas que se encuentran almacenadas
en ECs (Stuart et al., 1988).
Se han reportado estudios en los que el contenido de proteínas totales en ESs de
diferentes especies se ve incrementado durante su desarrollo ; tal es el caso en alfalfa (Lai
et al. , 1992), en soya (Stejskal , 1994) Y en Pisum sativum (Griga et al., 2007). Un
comportamiento opuesto se observó en estudios realizados por Lecona et al. (2012) para
Capsicum chinense; en la cual el conten ido de proteínas totales presentes en los ESs no
solamente fue menor a comparación del embrión cigótico, sino que además, su contenido
se vio disminuido a medida que el embrión somático avanzaba en sus estad íos de
desarrollo; pasando de contener 2.5 f-lg.mg-1 de proteínas en el estad ío globular a 0.4
16

CAPíTULO I
Ilg.mg-1 en el estadía cotiledonar. Aunado a esto, se encontró la existencia de 9 bandas
(4.0, 5.2, 8.1, 13.7, 20.9, 23.7, 41 , 50 Y 69.3 kOa) en el perfil proteico de los ECs de esta
especie, que no se presentaron en el perfil proteico de los ESs. Cinco de estas bandas
coinciden por su peso molecular con los reportados para proteínas de reserva en otras
especies (Oerbyshire et aL , 1976; Higgins, 1984, Shewry et aL, 1995; Griga et aL, 2007;
Brownfield et aL , 2007; Vladova et aL, 2004). Aunado a ésto, mediante 20 PAGE, este
autor observó en chile habanero marcadas diferencias entre los grupos de proteínas
expresadas en los ECs maduros y las de los ESs en estadía cotiledonar; particularmente,
encontró que un grupo de proteínas que sólo se expresaron en los ECs y no así en los
ESs con un rango de peso molecular de 40-45 KOa y punto isoeléctrico entre 7 y 10, se
encontraba en los ECs y no así en los ESs, las cuales por ambas propiedades pudieran
tratarse de proteínas de reserva (tipo globulinas).
En un estudio realizado en abeto, Flinn et al., (1993) observaron que existía similitud en la
regulación de la expresión de un gen que codifica proteínas de reserva durante los
estadías tempranos e intermedios de la embriogénesis somática y cigótica . Sin embargo,
existieron diferencias en la expresión de genes en los estados posteriores durante la
maduración , lo cual puede ser atribuido a la influencia de la desecación de los embriones
cigóticos, la cual no ocurre en embriones somáticos.
los ESs son menos vigorosos que los cigóticos; por lo que se esperaría que el
incremento en la acumulación de proteínas en los ESs, resultara en un mayor vigor de las
plántulas, así como que el incremento en la disponibilidad de reservas endógenas
permitiera una transición más exitosa a la autotrofia (Morcillo et al., 2001).
No obstante estos estudios , es claro que en el área de las proteínas relacionadas con la
embriogénesis cigótica y somática aún hay un gran desconocimiento acerca de las
proteínas que las regulan . En general , para la mayoría de las especies hortícolas de
importancia comercial como el chile habanero, se han realizado pocos estudios
proteomicos existiendo poca información disponible sobre la secuenciación de estas
proteínas.
17

CAPiTULO I
1.4 POLlAMINAS
1.4.1 Generalidades en plantas
Las poliaminas (PAs) son cationes orgánicos nitrogenados de bajo peso molecular
presentes en todas las células eucarióticas e íntimamente involucrados y requeridos para
distintas funciones biológicas (Tabor y Tabor, 1984; Jian-Ying y Robert, 2006). Su
principal característica es la de poseer a lo largo de su esqueleto carbonado varios grupos
amino (Carmona , 1997). Por su carácter policatiónico pueden unirse y formar complejos
con moléculas polianiónicas, como proteínas , fosfolípidos , pectinas, ADN , ARN, entre
otras (Galston 1983; Galston y Kaur-Shawney, 1987). Debido a estas características, las
PAs afectan la actividad celular, y como consecuencia están involucradas en una amplia
gama de procesos fisiólogicos (Guye et al., 1986; Evans y Malmberg, 1989; Faust y Wang
1982; Bais y Ravishankar, 2002).
Las principales PAs que se encuentran en los seres vivos son la putrescina (Put) (butano-
1, 4 diamina), espermidina (Spd) [N-(3-aminopropil) butano 1-4 diamina] y espermina
(Spm) [NN' -bis- (3-aminopropil) butano 1,4 diamina] (Evans y Malmberg, 1989; Galston
Kaur-Shawney, 1990); figura 1.4.
18
H H 2N~N~N~NH2
H
Espermina
H 2N~NH2 H2N~N~NH2 H
Putrescina Espermidina
Figura 1.4 Principales PAs que se encuentran en los seres vivos

CAPíTULO I
Investigaciones realizadas in vitro e in vivo con Helianthus tuberosus l. han demostrado
que el máximo nivel de síntesis de PAs se encuentra justo antes de la replicación del ADN
y la división celular, por tal motivo, se propone que existe una alta correlación entre los
niveles de poliaminas y ciertos eventos del ciclo celular en plantas (Serafini et al., 1989).
los procesos responsables de regular los niveles de poliaminas libres en la célula son :
nivel de síntesis y degradación de las mismas, su compartamentalización, si se transporta
o no a través de la planta, así como su conjugación con otros compuestos en la célula.
los niveles de PAs libres en la célula a su vez regulan el crecimiento y desarrollo vegetal
(Echevarría, 2003) .
1.4.2 Tipos de poliaminas
las PAs pueden presentarse en forma libre o conjugadas a compuestos fenólicos del tipo
del ácido cinámico y sus derivados (Martin-Tanguy, 1985), a otros compuestos de bajo
peso molecular (péptidos, ácidos grasos) o a macromoléculas tales como proteínas,
ligninas y ácidos nucleicos (Galston y Sawhney, 1990). A continuación se describen
ambos tipos:
• Libres. las principales poliaminas celulares, las PAs alifáticas, adoptan una
estructura lineal pero flexible, lo que es la base para algunas de sus propiedades.
• Conjugadas. En vegetales las formas conjugadas mejor conocidas son las
asociadas a péptidos o a moléculas de bajo peso molecular como ácidos fenólicos,
amidas de ácidos hidroxicinámicos, las cuales son solubles en el citoplasma
(Galston y Kaur-Sawhney, 1990).
En función de su solubilidad se distinguen dos tipos principales de conjugados :
1. Solubles en agua con al menos un grupo amino libre. Se incluyen las PAs alifáticas
(Put, Spd y Spm), las cuales forman complejos con compuestos de bajo peso
molecular y son solubles en el citoplasma de la célula y se encuentran protegidas
contra la degradación durante su transporte a través de la planta (Galston y Kaur
Sawhney, 1990).
2. Insolubles en agua en las cuales todos los grupos amino primarios o secundarios han
sido combinados. Se incluyen aminas alifáticas, aromáticas (tiramina, dopamina,
19

CAPiTULO I
seroton ina, octopamina , triptamina, etc.), y macromoléculas como la celulosa de la
pared celular y sitios anión icos de las membranas. Debido al alto peso molecular de
este complejo , las PAs en este estado no se presentan en el citoplasma celular
(Galston y Kaur-Sawhney, 1990).
Además de los tipos de poliaminas ya mencionados, también existen poliaminas poco
comunes cuya distribución en la naturaleza está más limitada con respecto a las
poliaminas típicas. Estas son : 1,3-diaminopropano, cadaverina (Cad), nor-espermidina (o
termina) , homo-espermidina , termoespermina , nor espermina (o cald ina), homo
espermina , canavalmina, caldopentamina y caldohexamina (Carmona, 1997). La función
de estas pol iaminas es muy poco conocida, si bien , se les han atribuido papeles de
protección específicos, tanto en bacterias como en las plantas adaptadas a ambientes
extremos (Galston y Kaur-Sawhney, 1990).
1.4.3 Funciones de las poliaminas
Las poliaminas juegan diversos roles en el crecimiento y desarrollo vegetal ; son mucho
más abundantes en las plantas que los fitorreguladores como giberelinas y citocininas, y
se requieren en cantidades del orden milimolar para que se induzca una respuesta
biológica (De la Peña, 2003). En las diversas respuestas fisiológicas que inducen, se
incluyen la estimulación y regulación de la división celular, la formación de microtúbulos,
el aumento de longevidad de los óvulos y del polen , la iniciación de la raíz, la in iciación y
desarrollo de flores, la maduración de frutos , la embriogénesis somática y respuestas de
tipo defensivo a estrés biótico y abiótico (Carmona , 1997). Además retardan la
destrucción de clorofilas (Slocum et al., 1984) y previenen la senescencia en hojas
cortadas (Galston y Kaur-Sawhney, 1990); estabilizan las membranas celulares vegetales,
las estructuras de la pared celular y el pH celular (Evans y Malmberg, 1989, Berta et al.,
1997). De igual manera , debido a su carácter de ácido débil , podrían amortiguar el
citoplasma en condiciones de estrés ácido (Rodríguez y Domínguez, 2007).
La aplicación exógena de poliam inas induce la síntesis de ADN . Las PAs aumentan la
estabilidad del complejo de los nucleosomas y participan en la condensación y
remodelación de la estructura de la cromatina y por lo tanto , en los procesos de
replicación y transcripción del ADN. Se sabe que procesos de acetilación que modifican a
20

CAPíTULO I
las histonas pueden acetilar también a las PAs (Carmona, 1997). Estos compuestos
protegen al ADN de la degradación enzimática, degradación por rayos X y gamma (y) , de
las especies reactivas de oxígeno y del calor. En este caso , la capacidad de
compactación del ADN y la capacidad de secuestrar radicales libres, son los atributos
principales de las PAs que brindan estabilidad y protección del ADN (Rodríguez y
Domínguez, 2007).
las poliaminas han mostrado regular también la síntesis de proteínas. la primera
evidencia de que las PAs están involucradas funcionalmente en este proceso biológico,
viene de estudios realizados tanto en procariontes como eucariontes. A partir de estas
investigaciones, se puede concluir que las PAs son capaces de incrementar la tasa de
síntesis de proteínas en concentraciones subóptimas de Mg+2 (Tabor y Tabor, 1984;
Marton y Morris, 1987). Aunado a esto, se ha demostrado una disminución en la tasa de
síntesis de proteínas y en el crecimiento en mutantes de E. coli deficientes de PAs,
cultivadas en ausencia de estos compuestos (Morris y Jorstad, 1973). Se demostró
también que la adición de Put causa un aumento casi inmediato en la síntesis de
proteínas (Young y Srinivasan, 1972). Existen varios sitios específicos en la maquinaria
traduccional donde las poliaminas pudieran actuar. Se sabe que estos compuestos se
unen a los ribosomas facilitando la asociación y estabilidad de las subunidades
ribosomales . Existe también evidencia acerca de la influencia de las PAs en la fidelidad
traduccional (la frecuencia con la que los aminoácidos son incorporados correctamente en
un polipéptido en respuesta a un codón dado) y en la terminación de la cadena
polipeptídica. Estas aminas pueden afectar también ya sea el paso de iniciación o de
enlongación de las cadenas polipeptídicas nacientes (Tabor y Tabor, 1984). De igual
manera , se ha reportado un número de aminoacil ARNt sintetasas que han sido
estimuladas por las PAs; en adición a la influencia en la unión del ARNt a su sintetasa
homóloga, transiciones estructurales inducidas por las poliaminas pueden también
influenciar la interacción codón-anticodón con el ribosoma (Tabor y Tabor, 1984; Marton y
Morris, 1987). En general, se ha encontrado que las PAs estimulan en mayor medida la
síntesis de proteínas de alto peso molecular que aquellas de bajo peso (Tiburcio et al. ,
1993).
De igual manera, se sabe que las PAs están involucradas en las modificaciones
21

CAPITULO I
postraduccionales. Entre los mecanismos más estud iados se encuentra aquel mediante el
cual las PAs pueden unirse a las proteínas por la acción de transglutaminasas (Pegg ,
1986). Otro mecanismo postraduccional en el que actúan las poliaminas, es la
fosforilación de proteínas nucleares, en el cual las PAs tienen un efecto estimulante sobre
proteínas cinasas independientes de AMP cíclico (Tabor y Tabor, 1984).
Cuando se presentan bajas tasas de síntesis de estas moléculas y de división celular, el
conten ido endógeno de poliamínas disminuye (Egea y Mizrah i, 1991); en consecuencia , el
proceso de crecimiento está afectado positivamente por la presencia y actividad de las
PAs (Faust y Wang, 1982), esto evidenciado en un gran número de plantas en las que se
ha relacionado la presencia de estas moléculas con órganos y tejidos en crecimiento
activo. Además de ser esenciales para el crecimiento, bajo condiciones apropiadas,
pueden ejercer funciones específicas de control de diferentes procesos de diferenciación
y morfogénesis; por ejemplo, la formación de nódulos y raíces en Phaseolus aureus Roxb,
la determinación del desarrollo vegetativo o reproductivo en el meristemo del tallo de
Sinapsis alba L. (Havelange et al., 1996), la iniciación y el desarrollo floral , la formación
del polen , el desarrollo del fruto en el chícharo (Pisum sativum L.) (Evans y Malmberg ,
1989; Marquínez et al., 2001 ;), la formación del embrión en zanahoria (D . carota) (Bastola
y Minocha, 1995), la germinación de semillas (Galston y Flores , 1991 ; Bais y Ravishankar,
2002), así como la restauración del potencial morfogenético y el incremento del porcentaje
de respuesta de los explantes en cuanto a formación de brotes en C. frutescens (Kumar et
al., 2007) .
1.4.4 Antecedentes de poliaminas en la embriogénesis somática
Los eventos celulares y morfogenéticos durante la ES están controlados por una serie de
cond iciones de cultivo y efectos genéticos. Montague et al. (1978, 1979) reportaron un
doble incremento en los contenidos de Put en cultivos embriogénicos de zanahoria sobre
los cultivos no embriógenicos; más tarde, Feirer et al. (1984) reportaron en esa misma
especie, que las PAs promovieron el proceso de ES. De igual manera, Fienberg et al.
(1984) observaron un incremento en la producción de Spd y Spm en cultivos
embriogénicos, en comparación con las células no diferenciadas (Kakkar et al., 2000) .
Minocha et al. (1999) realizaron un estudio sobre los niveles de PAs durante el desarrollo
de embriones somáticos y cigóticos en Pinus radiata , en donde altas proporciones de Spd
22

CAPíTULO I
y Spm fueron observados en embriones capaces de germinar y formar plántulas (Steiner
et al., 2007) .
Contrario a lo anterior, Calheiros et al. (1994) demostraron que la aplicación de PAs
exógenas (Put, Spd y Spm) disminuyeron el número de embriones producidos en café.
Sin embargo, se ha demostrado que altos contenidos de PAs en un medio suplementado
con 2,4-0 y BAP es necesario para superar la naturaleza recalcitrante de Hevea
brasiliensis con respecto a la embriogénesis somática (EI-Hadrami y O'Auzac, 1992).
Estos mismos autores, adicionaron PAs exógenas y observaron que hubo una mejora en
la producción de callo embriogénico y un incremento en el número de embriones
somáticos producidos por callo (Kakkar et al" 2000).
En estudios recientes, Steiner et al. , (2007) evaluaron el efecto de las PAs en el
crecimiento, asi como los niveles endógenos en cultivo embriogénico de Araucaria
angustífolia, y encontraron que la adición de PAs al medio de cultivo mejoró el crecimiento
embriogénico, y de igual forma hubo un incremento en los niveles endógenos de otras
hormonas vegetales. Paul et al., (2009) concluyeron que la aplicación de las PAs al medio
embriogénico redujo los contenidos endógenos de las mismas, pero también tuvo un
efecto en el crecimiento de callo embriogénico, en términos de peso fresco y número de
ESs por callo durante estad íos tempranos de la ES en Momordica charantia. Aunado a
esto, Mauri y Manzanera (2011) demostraron que el contenido endógeno de poliaminas
en callo embriogénico, embriones somáticos y cigóticos inmaduros de encino (Quercus
ilex l.), es más abundante que en la fase de maduración y germinación .
En resumen , las poliaminas participan en el proceso embriógenico de muchas especies
vegetales, ya que se ha visto que las estructuras embriogénicas contienen mayor cantidad
de estos compuestos en comparación con las no embriogénicas (Mauri y Manzanera,
2011). Por otro lado, la importancia de las PAs exógenas radica en el hecho de que
mejoran los procesos in vítro , tales como la ES la cual puede ser regulada mediante la
manipulación de los niveles y el metabolismo de las PAs (paul et al., 2009). No obstante,
también se sabe que no todas las especies vegetales responden de igual forma a la ES,
así como al tratamiento in vitro con PAso
Tomando en cuenta estos antecedentes, en este estudio se explorará el efecto de la
23

CAPITULO I
adición exógena de poliaminas y aminoácidos sobre la embriogénesis somática de C.
chinense dado que esta especie hasta ahora ha mostrado una alta recalcitrancia a la
regeneración in vitro a través de tal proceso celular.
24

CAPíTULO I
HIPÓTESIS
Si los aminoácidos y las poliaminas están involucrados en la modulación de diversos
procesos celulares, entonces es posible que la adición de asparagina , glutamina y/o
espermina a cultivos embriogénicos de Capsicum chinense, mejorará la calidad ,
desarrollo y morfología de los embriones somáticos y favorecerá su composición proteica.
OBJETIVO GENERAL
Mejorar la respuesta bioquímica-estructural de los embriones somáticos de chile habanero
adicionando al medio de cultivo los compuestos asparagina, glutamina (aminoácidos) y
espermina (poliamina).
OBJETIVOS ESPECíFICOS
1. Evaluar diferentes concentraciones de asparagina, glutamina y espermina sobre la
eficiencia, calidad y desarrollo de los embriones somáticos de Capsicum chinense.
2. Seleccionar el tratamiento que favorezca el desarrollo de los ESs de chile
habanero para analizar su contenido proteico comparado con el EC de la especie .
3. Analizar el perfil proteico de los ESs y ECs mediante electroforesis SDS-PAGE y
2D-PAGE.
25

CAPíTULO I
JUSTIFICACiÓN
A pesar de que actualmente se cuenta con un protocolo altamente eficiente y reproducible
de embriogénesis somática para la regeneración in vitro de C. chinense, y de la notable
mejora de los embriones somáticos obtenidos, aún persiste su incapacidad para germinar
y convertirse en plantas . Tomando en cuenta la deficiencia en contenido proteico
reportada en los ESs de la especie , y los antecedentes que existen sobre los beneficios
de la adición exógena de aminoácidos y poliaminas durante la ES de diferentes especies
vegetales, en el presente trabajo se evaluó el comportamiento de los ESs de C. chinense
con la aplicación de los aminoácidos asparagina y glutamina y de la poliamina espermina,
así como el efecto del mejor tratamiento evaluado en el contenido y composición proteico
en los ESs de C. chinense.
26
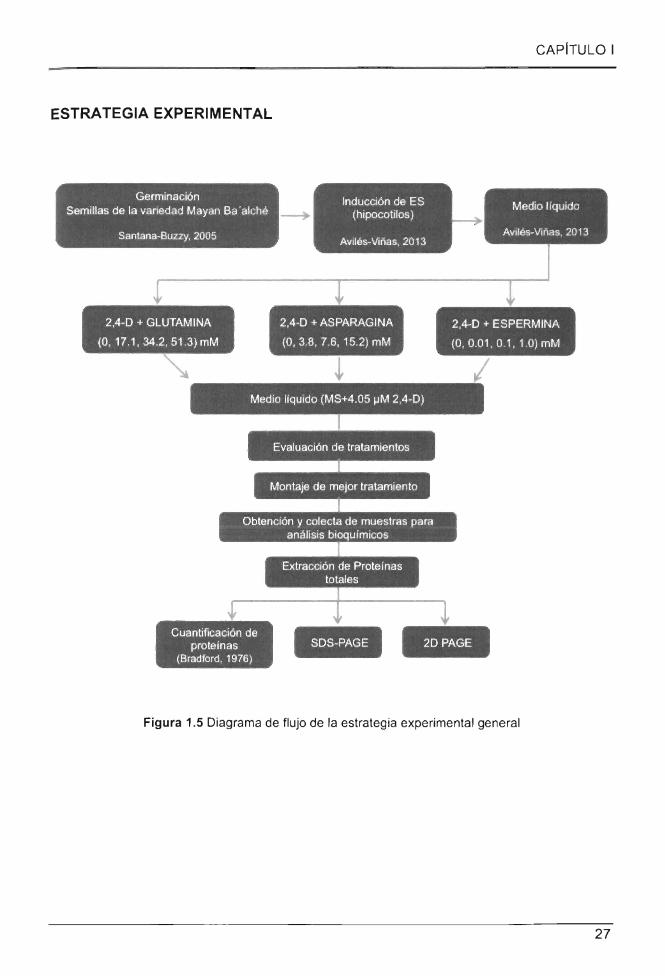
ESTRATEGIA EXPERIMENTAL
Germinación Semillas de la vanedad Mayan Ba 'alché
Santana-Buzzy, 2005
2,4-0 + GLUTAMINA
(O, 17.1, 34.2, 51 .3) mM
] 2,4-0 + ASPARAGINA
(0,3.8,7.6,15.2) mM
Medio liquido (MS+4.05 11M 2,4-0)
Cuantificación de proteínas
(Bradford, 1976) SOS-PAGE
CAPiTULO I
Medio liquido
Avilés-Viñas. 2013
2,4-0 + ESPERMINA
(0,0.01,0.1,1.0) mM
20 PAGE
Figura 1.5 Diagrama de flujo de la estrategia experimental general
27

CAPITULO I
BIBLIOGRAFíA
Attree S.M. Y Fowke L.C. (1993) . Embryogeny of gymnosperms: Advances in synthetic
seed technology of conifers . Plant Cell Tissue Organ Cult. 35, 1-35.
Avilés-Viñas S. , Lecona-Guzmán C., Canto Flick A., López-Erosa S.,Santana-Buzzy N.
(2013) . Morpho-histological and ultrastructural study on direct somatic
embryogenesis of Capsicum chinense Jacq. in liquid medium. Plant Biotechnol
Rep. 7: 277-286.
Bais H.P. Y GA Ravishankar (2002) . Role of polyamines in the ontogeny of plants and
their biotechnological applications. Plant Cell Tiss . Org . Cult., 69, 1-34.
Barceló J., Nicolás G., Sabater B. and Sánchez R. (2001) . Fisiología Vegetal. Ed .
Pirámide, Madrid, España. 568 p.
Berta G, Altamura MM, Fusconi A, Cerruti F, Capitaní F, Bagni N. (1997). The plant cell
wall is altered by inhibition of polyamine biosynthesis . New Phytol 137: 569-577 .
Brownfield DL, Todd, Stone SL, Deyholos MK, Gifford DJ (2007) . Patterns of storage
protein and triacylglycerol accumulation during loblolly pine somatic embryo
maturation . Plant Cell Tissue Organ Cult. 88(2) : 217-223 .
Buyucalaca S. y Mavituna F. (1996). Somatic embryogenesis and plant regeneration of
pepper in liquíd media . Plant Cell Tiss. Org. Cult. 46:227-235.
Calheiros M.B.P., L.G.E. Vieira y S.R.L . Fuentes (1994) . Effects of exogenous polyamines
on direct somatic embryogenesis in coffee. Rev. Bras. Fisiol. Veg ., 6,109-114.
Carmona A.J . (1997) . Contribución al conocimiento del papel de las poliaminas en el
desarrollo del fruto de la platanera . Estudio de las implicaciones fisiológicas de
estas sustancias sobre el crecimiento in vitro de los tejidos del ovario . Tesis de
doctorado. Universidad de la Laguna . 226 p.
Chia JT, Saunders JW. (1999) . Evaluation of sole nitrogen sources for shoot and leaf disc
culture of sugarbeet. Plant Cell Tissue Organ Cult. 59: 47-56 .
28

CAPiTULO I
Claparols 1, Santos MA and Torne ' JM (1993). Influence of some exogenous amino acids
on the production of maize embryogen ic callus and on endogenous amino acid
content. Plant Cell , Tissue and Org. Cult. 34 : 1- 11 .
Colorado P., Nicolás C., Nicolás G. and Rodríguez D. (1995). Convergent effects of stress
and ABA on gene-expression during germination of chick-pea seeds. J. Plant
Physiol. 146: 535-540.
Curry J., M. Alu ru, M. Mendoza, J. Nevarez, M. Melendrez y A. O'Connell (1999).
Transcripsts for possible capsaicinoid biosynthetic genes are diferentially
accumulated in pungent and non-pungent Capsicum spp. Plant Sci., 148, 47-57 .
De la Peña Seaman C. (2003). El efecto de la luz, el pH y la benciladenina en la
biosíntesis de las pol iaminas en suspensiones celulares y su variación durante la
embryogenesis somática de Coffea canephora. Tesis de maestría . Centro de
Investigación Científica de Yucatán . 103 p.
Delseny M. , Bies-Etheve N., Caries C., Hull G., Vicient C., Raynal M., Grellet F. and
Aspart L. (2001) . Late embryogenesis abundant (LEA) protein gene regulation
during Arabidopsis seed maturation . J. Plant Physiol. 158: 419-427.
Derbyshire E., Wright D. and Boulter D. (1976). Legumin and vicilina storage proteins in
legume seeds. Phytochemistry. 15: 3- 24.
Dodeman V.L. , Ducreux G. & Kreis M. (1997). Zygotic embryogenesis versus somatic
embryogenesis . J. Exp. Bot. 48 , 1493-1509.
Dure L 111 , Greenway SC, Galau GA (1981). Developmental biochemistry of cotton seed
embryogenesis and germination : changing messenger ribonucle ic acid
populations as shown by in vitro and in vivo protein synthesis. Biochemistry
20:4162-4168.
Dure L.S., Crouch M., Harada J., Ho T.H.D., Mundy J., Quatrano R., Thomas T. and Sung
Z.R. (1989). Common amino acid sequence domains among the LEA proteins of
higher plants. Plant Mol. Biol. 12: 475-485.
29

CAPíTULO I
Echevarría M. 1. (2003). Función de las poliaminas en el proceso de crecimiento vegetal.
Posibles mecanismos de transducción de señales involucrados. Tesis de
doctorado. Centro de Investigación Científica de Yucatán . 161 p.
Egea-Cortines M. Y Y. Mizrahi (1991). Polyamines in cell división , fruit set and
development, and seed germination in: biochemistry and physiology of
polyamines in plants . Slocum R.o . y H.E. Flores (eds) . CRC Press, Boca Raton ,
pp. 143-158.
EI-Hadrami , l . Y J. D'Auzac (1992). Effects of polyamine biosynthetic inhibitors on somatic
embryogenesis and cellular polyamines in Hevea brasiliensis. J. Plant Physiol.,
140,33-36.
Evans P.T. Y R.l . Malmberg (1989). Do polyamines have roles in plant development?
Plant Physiol. , Annu Rev., Plant Mol. Biol. , 40 , 235-269.
Faust M. Y S.Wang (1982). Polyamines in horticulturally important plants . Horticult. Rev.,
14,333-355.
Feirer RP , Mignon G, Litvay JD (1984). Argin ine decarboxylase and polyam ines required
for embryogenesis in wild carrot. Science 223 :1433-1435.
Fienberg A.A ., J.H. Choi , W.P. lubich y Z.R. Sung (1984). Development regulation of
polyamine metabolism in growth and differentation of carrot cultura . Planta , 162,
532-539 .
Finkelstein R.R., Tenbarge K.M . y Shumway J.E. (1985). Role of ABA in maturation of
rapeseed embryos. Plant Physiol. 78, 630-636 .
Flinn , B.S ., Roberts , D.R., Newton C.H., Cry, D.R. and Taylor, E.P. (1993). Storage protein
gene expression in zygotic and somatic embryos of interior spruce. Physiologia
Plantarum . 89 : 719-730.
Franck-Duchenne , M. , Wang , S. Ben Tahar, y R.N. Beachy. (1998). In vitro stem
elongation of sweet pepper in media containing 2,4-epibrassinolide. Plant Cell.
Tiss. Org . Cult. 53 :79-84 .
30

CAPíTULO I
Galston A.W. (1983). Polyamines as modulators of plant development. Bioscience, 33,
382-388 .
Galston A.W. y H.E. Flores (1991). Polyamines and plant morphogenesis in: biochemistry
and physiology of polyamines in plants . Slocum, R.D. y H.E. Flores (eds) , pp.
175-186.
Galston A.W. y R. Kaur-Sawhney (1987). Polyamines as endogenous growth regulators in:
plant hormone and their role in plant growth and development. Davies , P.J. (ed) .
pp. 280-295.
Galston A.W. y R. Kaur-Sawhney (1990) . Polyamines in plant physiology. Plant Physiol. ,
94,406-410 .
Garay-Arroyo A, Colmenero-Flores JM , Garciarrubio A, Covarrubias (2000). AA: Highly
hydrophilic proteins in prokaryotes and eukaryotes are common during conditions
of water deficit. J Biol Chem, 275:5668-5674.
George E. F., HallGeert Michael, Klerk bGeert-Jan,(2008). Plant Propagation by Tissue
Culture 3rd Edition .Springer. 501 pp.
Goshi F., Bertauche N., Vartanian N. and Giraudat J. (1995). Abcisic acid-dependent and
independet regulation of gene expression by progressive drought in Arabidopsis
thaliana . Mol. Gen. Genet. 246: 10-18.
Griga M, Horacek, J. and Klenoticova H. (2007) . Protein patterns associated with Pisum
sativum somatic embryogenesis. Biologia plantarum 51 (2): 201-211 .
Guye M.G., l. Vigh y M. Wilson (1986) . Polyamine titre in relation to chill-sensitivity in
Phaseolus sp. J. Exp . Bot. 37, 1036-1043.
Havelange A. , P. lejeune, G. Bernier, R. Kaur-Shawney y A.W. Galston (1996) .
Putrescine export from leaves in relation to floral transition in Stropsis alba.
Physiol. Plant. 96 , 59-65.
Herman E. Y larkins B. (1999) . Protein Storage Bodies and Vacuoles . The Plant Cell , Vol.
31

CAPiTULO I
11,601-613.
Higgins T. (1984). Synthesis and regulation of major proteins in seeds. Annu . Rev. Plant
Physiol. , 35 :191-221 . Hvoslef-Eide.
Higuchi, W., y Fukazawa, C. (1987). A rice glutelin and a soybean glycinin have evolved
from a common ancestral gene. Gene 55, 245-253.
Hundertmark M. Y Hincha D. (2008). LEA (Late Embryogenesis Abundant) proteins and
their encoding genes in Arabidopsis thaliana . BMC Genomics. 9:118
doi :10.1186/1471-2164-9-118.
Inforural (2012). Fuente : Sistema producto ch ile habanero-México 2014. Lanzamiento de
la iniciativa "Tsaay Yucatán" 1 a Reun ión : Identificando los desafíos. [Online) .
(Actualizado 2014)
Dispon ible en : htt ://www .tsaaucatan.com
Consulta: 21 de agosto de 2014.
Ingram J. Y Bartels D. (1996). The molecular basis of dehydration tolerance in plants.
Annu . Rev. Plant Physiol. Plant Mol. Biol. 47:377-403 .
Jian-Ying W. y A.C. Robert (2006). Polyamine cell signaling . Human Press. Totowa , New
Jersey. 479 p.
Kakkar R.K., P.K. Nagar, P.S. Ahuja y V.K. Rai (2000). Polyamines and plant
morphogenesis . Biol. Plant., 43 , 1-11 .
Kermode A.R . (1990). Regulatory mechanisms involved in the transition from seed
development to germination . Crit. Rev. Plant Sci. 9, 155-195.
Kermode A.R ., Dumbroff E.B. & Bewley J.D. (1989). The role of maturation drying in the
transition from seed to germination . VII Effects of partial and complete desiccation
on abscisic acid levels and sensistivity in Ricinus communis L. seeds. J. Exp . Bot.
40 , 303-313 .
Kiran K.S y Thorpe, T.A. (1995). Asexual embryogenesis in vascular plants in nature. In
32

CAPíTULO I
vitro embryogenesis in plants. Kluwer Academic Publishers. Dordrecht. Pp. 17-72.
Kothari S.L., A. Joshi , S. Kachhwaha and N. Ochoa-Alejo (2010). Chilli peppers - A
review on tissue culture and transgenesis Biotechnology Advances, 28(1): 35-48.
Kumar V., A. Sharma, B.C Narasimha Prasad, H.B. Gururaj, P. Giridhar y G. Ravishankar
(2007). Direct shoot bud induction and plant regeneration in Capsicum frufescens
MilI. : influence of polyamines and polarity. Acta Physiol Plant. 29:11-18.
Lai , F.M., Senaratna, T. and McKersie, B.D. (1992). Glutamine exchanges storage protein
synthesis in Medicago sativa L. Somatic embryos. Plant Science. 87:6977.
Latournerie L.M ., M.E. Yupit, S.J. Tun, M.A.M Pech y C.D. Lozada (2006). Fundación
Produce Yucatán, A.C . 1: 15-17.
Lecona-Guzmán C. (2012) . Estudio de las proteínas en la embriogénesis somática de
chile habanero (Capsicum chinense Jacq.). Tesis de Doctorado. 81 p.
Lecona-Guzmán C.A., Solís Marroquín D., Avilés-Viñas S., Santos-Briones C., Santana
Buzzy N. (2012). Changes in the protein profile of habanero pepper (Capsicum
chinense J.) somatic embryos during development. African Journal of
Biotechnology. En edición .
Litz R.E. Y Jarret R.L. (1993). Regeneración de plantas en el cultivo de tejidos:
embriogénesis somática y organogénesis. Cultivo de tejidos en la agricultura
fundamentos y aplicaciones. Centro Internacional de Agricultura Tropical. pp.
143-172.
López-Erosa S. (2012). Efecto de poliaminas exógenas en la embriogénesis somática de
chile habanero (Capsicum chinense Jacq.). Tesis de maestría . Centro de
Investigación Científica de Yucatán . 89 p.
Lopez-Puc G.; Canto-Flick A., Barredo-Pool F., Zapata-Castillo P., Montalvo-Peniche M.,
Barahona-Perez F., Santana-Buzzy N. and Iglesias-Andreu L. (2006). Direct
Somatic Embryogenesis: A Highly Efficient Protocol for In Vitro Regeneration of
Habanero Pepper (Capsicum chinense Jacq.). Hort Science 41 (7) :1645-1650.
33

CAPiTULO I
Marquínez X., J. Norato, L. Reyes, G. García, y A. Caro (2001). Mecanismos fis iológicos
de respuestas a bajas temperaturas y a la aplicación de putrescina en Kikuyo y
Falsa poa . Comafi , 28 , 34-39 .
Martin-Tanguy J. (1985). The occurrence and possible function of hydroxycinnamoyl acid
amides in plant. Plant. Growth Regul. , 3, 381-399 .
Marton L y Morris D (1987). Molecular y cellular functions of the polyam ines . En : PP Mc
Cann, A Pegg y A Sjoerdsma, eds. Inhibition of polyamine Metabolism , pp. 79-
105. San Diego: Academic Press.
Mauri p.v. y J.A. Manzanera (2011). Somatic embryogenesis of holm oak (Quercus ilex
L.): ethylene production and polyamine content. Acta Physiol. Plant. , 33 , 717-723.
McKersie B.D. & Brown D.C.W. (1996) Somatic embryogenesis and artificial seeds in
forage legumes. Seed Sci. Res. 6, 109-126.
Merkele SA, Parrott WA y Flinn B.S. (1995). Morphogenic aspects of somatic
embryogenesis . pp. 155-203 in Thorpe TA (ed .) In Vitro Embryogenesis in
Plants . Kluwer Academic Publishers, Dordrecht, Boston , London .
Merkle SA y Dean J.F.D. (2000) Forest biotechnology. Curro Opin o Biotechnol. 11: 298-
302 .
Minocha S.C. y R. Minocha (1995). Role of polyamines in somatic embryogenesis in :
biotechnology in agriculture and forestry. Somatic embryogenesis and synthetic
seed. Bajaj, Y.P.S . (eds) . Springer - Verlag , Berlin , 30, 53-70 .
Minocha R., D.R. Smith , C. Reeves, K.D. Steele y S.C. Minocha (1999). Polyamine levels
during the development of zygotic and somatic embryos of Pinus radiata. Physiol.
Plant. , 105,155-254.
Mondal T.K. A. Bhattacharya M. Laxmikumaran y P.S. Ahuja (2004). Recent advances of
tea (Camelia sinensis) biotechnology. Plant Cell Tiss . Org . Cult. , 76, 195-254.
Montague M.J ., J.W. Koppenbrink y E.G. Jaworaski (1978). Polyam ine metabolism in
34

CAPiTULO I
embriogenic cells of Daucus carota. 1. Changes in arginine decarboxylase activity.
Plant Physiol., 62,430-433 .
Montague M.J., TA Armstrong y E.G. Jaworski (1979). Polyamines metabolism in
embryogenic cells of Daucus carota . 11. Changes in arginina decarboxylase
activity. Plant Physiol. , 63, 341-345.
Morcillo F., Hartmann C. Duval Y. , y Tregear J. (2001). Regulation of 7S globulin gene
expression in zygotic and somatic embryos of oil palm . Physiologia Plantarum
112: 233-243.
Morris O Y Jorstad C (1973). Growth and macromolecular composition of a mutant of
Escherichia Coli during polyamine limitation. J Bacteriol 113: 271-277
Mundy J. and Chua N.H. (1988). Abscisic acid and water-stress induce the expression of a
novel rice gene. EMBO J. 7: 2279-2286.
Nylander M., Svensson J., Palva E.T. and Welin B.v. (2001). Stress induced accumulation
and tissue- especific localization of dehydrins in Arabidopsis thaliana Plant Mol.
Biol. 45: 263-279 .
Ochoa-Alejo N. and Ramírez-Malagón R. (2001). In vitro chili pepper Biotechnology. In
Vitro Cell. Dev. Biol-Plant. 37(6) : 701-729.
Osborne, T. B. (1907). The proteins of the wheat kernel. Carnegie Institute. Whashington,
whashington D. C., Publ : 84 : 1-119.
Paran l. , A. Ben-Chaim, K. Byoung-Cheorl y M. Jahn (2007). Capsicums. Genome
mapping and molecular breeding in plants. Springer-Verlag Berlin Heidelberg, 5,
209-226 .
Pegg A. (1986). Recent advances in the biochemistry of polyamines in eukaryotes.
Biochem J 234 : 249-262.
Pérez-Molphe, E.M., Ramírez- Malagon R., Núñez-Palenius H.G. y Ochoa-Alejo N. (1999).
Introducción al cultivo de tej idos vegetales . Universidad Autónoma de
35

CAPíTULO I
Aguascalientes , México, pp: 9-11, 16-20, 27-35, 57-60, 81-84 .
Pickergill B. (1997). Genetic resources and breeding of Capsicum spp. Euphytica 96 : 129-
133.
Pickersgill A., K. Mitter y S. Sen-Raychaudhuri (2009). Effect of polyamines on in vitro
somatic embryogenesis in Momordica charantia L. Plant Cell Tiss. Organ. Cult. ,
97, 303-311 .
Quiroz-Figueroa F., Rojas-Herrera R, Galaz-Avalos RM , loyola-Vargas VM. (2006).
Embryo production through somatic embryogenesis can be used tu study cell
differentiation in plants. Review paper. Plant Cell Tiss Organ Cult. 86 : 285-301 .
Radice , S. (2010). Morfogénesis . Biotecnología y Mejoramiento Vegetal 11. Instituto
Nacional de Tecnología Agropecuaria . pp.26-33
Roberts, J. K., Desimone, N. A., Lingle, W. l . and Dure, 111, l . (1993) Cellular
concentrations and uniformity of cell-type accumulation of two lEA proteins in
cotton embryos. Plant Cell 5, 769-780.
Rock C.D . and Quatrano R.S., (1995) . The role of hormones during seed development En :
"Plant Hormones: Physiology, Biochemistry and Molecular Biology" (Davies PJ .
Ed). Kluwer Academic Publishers, Dordrecht pp: 671-697.
Rodríguez M. Y D. Kessler (2007) . Análisis del metabolismo de poliaminas de maíz bajo
condiciones de estrés biótico y abiótico . Tesis de doctorado. Instituto Potosino de
Investigación Científica y Tecnológica , A.C. 208 p.
Roja R., Reddy V.O., Prakash Babu y Padmaja G. (2005). Changes in protein profiles
associated with somatic embryogenesis in peanut. Biologia Plantarum 49 (3) :
347-354 .
Ronchi, V. N., Caligo, M. A., Nozzolini , M. and luccarini , G. (1984) . Stimulation of carrot
somatic embryogenesis by proline and serine. Plant Cell Rep., 1984, 3, 210.
Serafini-Fracassini D., S. Del Duca y P. Torrigiani (1989). Polyamine conjugation during
36

CAPíTULO I
the cell cycie of Helianfhus fuberosus: non-enzymatic and transglutam inase-like
binding activity. Plant Physiol. Siochem ., 27, 659-668 .
Servicio de información agroalimentaria y pesquera , México-2014. Anuario estadístico de
la producción agrícola [Online] .
(Actualizado 2014) .
Disponible en : http://www.siap. ob .mx/cierre-de-Ia- roduccion-a ricola- or-
cu ltivo/
Consulta : 21 de agosto de 2014.
Shewry P., Napier J. and Tatham A. (1995). Seed storage proteins : structure
and biosynthesis. Plant Cell, 7: 945-956
Slocum R.D ., R. Kaur-Shawhne y A.W. Galston (1984). The phyisiology and biochemistry
of polyamines in plants. Arch . Siochem. Siophys ., 235, 283-303.
Stasolla C. y Yeung E.C (2003). Recent advances in conifer somatic embryogenesis:
improving somatic embryo qualityt. Plant Cell , Tissue and Organ Culture 74: 15-
35.
Steiner N., C. Santa-Catarina , V. Silveira , E.I.S . Floh y M.P. Guerra (2007). Polyamine
effect on growth and endogenous hormones levels in Araucaria angustifo/ia
embryogen ic cultures. Plant Cell Tiss. Organ . Cult., 89, 55-62.
Stejskal, J. (1994). Comparative analysis of proteins and isozymes in somatic and zygotic
embryos of soybean (G/ycine max (l.) Merr). Ph D. Thesis , lES Prage .
Stuart, DA, Nelse, J. and Nichol, J.W. (1988). Expression of 7S and 11S alfalfa seed
storage proteins in somatic embryos . J. Plant Physiol. 132: 134-139.
Tabor C.W. y H. Tabor (1984). Polyamines. Ann . Rev. Siochem ., 53, 749-790 .
Terán S. y C.H. Rasmussen (1998). las plantas de la Milpa entre los Mayas. Fundación
Tun Sen Kin. A. C. Yucatán , México . 349 p
Thomas T.l . (1993) Gene express ion during plant embryogenesis and germination : An
37

CAPíTULO I
overview. Plant Cell 5,1401-1410.
Tiburcio A., Campos J. Figueras X. y Besford R. (1993). Recent advances in the
understanding of polyamine functions during plant development. Plant Growth
Regulation 12: 331-340 .
Tun Dzul, J. (2001). Chile habanero, características y tecnología de producción .
SAGARPA, INIFAP. Mocochá, Yucatán, México. 74 p.
Vladova R., Tsanev V. and Petcolicheva K. (2004). Seed storage proteins in Solanaceae
and Cucurbitaceae species. Biologia Plantarum, 48 (4) : 601-603 .
Von Arnold S. y Clapham D. (2007). Spruce Embryogenesis. Plant Embryogenesis. Pp.
31-51
Von Arnold S. (2008). Somatic embryogenesis Plant Propagation by Tissue Culture, vol 1.
Springer. pp . 335.
Witjaksono, Litz RE . (1999). Maturation of avocado somatic embryos and plant recovery.
Plant Cell Tissue Organ Cult. 58 : 141-148.
Yamaguchi-Shinozaki K., Mundy J. and Chua N.H ., (1989). Four tightly linked rab genes
are diferentially expressed in rice. Plant Mol. Biol. 14: 29-39.
Yeung E.C. (1995) Structural and developmental patterns in somatic embryogenesis . pp.
205-247 in Thorpe TA (ed .) In vitro Embryogenesis in Plants . Kluwer Academic
Publishers Dordrecht, Boston , london. 544 pp.
Young D. and Srinivasan P. (1972). Regulation of macromolecular synthesis by putrescine
in a conditional Escherichia coli putrescine auxotroph . J. Bacteriol 112: 30-39 .
Zapata-Castillo P., Canto-Flick A. , lopez-Puc G. , Solís-Ruiz A. Barahona-Pérez F. and
Santana-Buzzy N. (2007). Somatic Embryogenesis in Habanero Pepper
(Capsicum chinense Jacq.) From Cell Suspensions . Hortscience 42(2): 329-333 .
Zimmerman J. l ., (1993) Somatic Embryogenesis: A Model for Early Development in
Higher Plants The Plant Cell, 5, 1411-1423
38

CAPíTULO II
CAPíTULO 11
EFECTO DE ASPARAGINA, GlUTAMINA y ESPERMINA SOBRE lA
EFICIENCIA, CALIDAD Y DESARROllO DE lOS ESs DE CHilE HABANERO
2.1 INTRODUCCiÓN
Uno de los factores de importancia tanto en la inducción como en el desarrollo de la ES,
es la composición de nutrientes del medio de cultivo. La concentración de nitrógeno (N) y
las cantidades relativas de amonio y nitrato pueden ser críticas para el crecimiento y
morfogénesis de las células vegetales (Halperin , 1966; Reinert et al., 1967, Heimer y
Filner 1970; Armstrong y Green 1985; Wilkinson y Thompson , 1987). En especies
vegetales como zanahoria, alfalfa (Lai et al., 1992); arroz (Ozawa et al., 1996), abeto
blanco (Barret et al., 1997) y algodón (González-Benito et al., 1997) se ha promovido la
inducción y el desarrollo de la ES usando diferentes balances y fuentes de N en el medio
de cultivo. Este elemento ha sido considerado como uno de los factores clave para la
maduración de los ESs (Davidonis y Hamilton , 1983; Trolinder y Goodin, 1988b;
Witjaksono y Litz, 1999).
Los compuestos orgánicos nitrogenados tales como los aminoácidos y las poliaminas,
ayudan a las células en su proceso de diferenciación. Los aminoácidos sirven como
fuente primaria de N orgánico para el crecimiento de muchas células eucariotas.
Adicionalmente , éstos promueven la comunicación entre células y tejidos dentro de
organismos pluricelulares (Young et al., 1999). Los aminoácidos como moduladores de
crecimiento y diferenciación celular pueden afectar el metabolismo en general y
consecuentemente la morfogénesis (Basu et al., 1989). Han probado ser efectivos
reguladores morfogenéticos en numerosas especies (Tupy et al., 1983; Ronchi et al.,
1984; Trigiano y Conger, 1987; Basu et al., 1989; John y Guha-Mukherjee, 1997; Nanjo et
al., 1999); y se ha encontrado que el tipo de aminoácido y su concentración desempeñan
papeles importantes en diferentes etapas del proceso de ES (Ronchi et al. , 1984;
Gerdakaneh et al., 2011). Se ha realizado la aplicación exógena de L-Prolina (Pro) , D
Prol ina , L-Serina (Ser) , hidroxiprolina , L-Arginina (Arg) , L-Alanina (Ala), L-Glutamina (Gln) ,
L-Asparagina (Asn); los cuales han demostrado estimular la ES (Ronchi et al., 1984;
39

CAPíTULO 11
Claparols et al., 1993; Buyucalaca y Mavituna , 1996; Guerra et al., 1997 ; Dal Vesco y
Guerra , 2001 ; Sen et al., 2002 ; Gerdakaneh et al., 2011).
La ad ición de aminoácidos como Asn y Gln al medio de cultivo se ha utilizado como una
estrategia para incrementar el contenido de proteínas; y mejorar el desarrollo de los ESs ,
en diferentes especies vegetales (Ch ia and Saunders, 1999; Witjaksono and Litz, 1999),
En el medio de regeneración de alfalfa (M. sativa) , la adición de Gln, alanina (Ala) , Pro y
Arg en concentración de 30 mM incrementa la producción de ESs, su peso seco, la
síntesis de proteínas almacenadas, al igual que el promedio de conversión a planta.
Particularmente la adición de Gln 50 mM produjo un incremento significativo en la síntesis
de proteínas y en el peso seco de los ESs (Lai et al., 1992). La combinación de caseína
hidrolizada y Gln incrementó el número de ESs en Eucalipto (Muralidharan y
Mascarenhas, 1995). En algodón , la adición de 13.7 mM de Gln permitió que los ESs
fueran capaces de convertirse en plántulas (González-Benito et al., 1997). Nuutila et al.,
(2000) describieron que los estad íos embriogénicos tempranos de ESs de cebada ,
requieren la presencia de Gln u otro aminoácido. Morcillo et al. (2001) encontraron que la
adición de Gln y Arg al medio de cultivo de la palma aceitera incrementó la cantidad de
proteinas en los ESs. Schmidt et al., 2005, reportaron que Gln 30 mM promovió el estadío
cotiledonar en ESs de soya e incrementó el contenido de proteínas totales . En Paspalum
scrobiculatum, la adición de Ser, Pro y Gln promovió la maduración de los ESs y su
conversión en plantas (Ceasar e Ignacimuthu, 2010).
Wu et al., 2004, desarrollaron un protocolo para la inducción de la ES y regeneración de
plantas de alta eficiencia en 10 cultivares recalcitrantes de algodón (Gossypum hirsutum
L.) util izando un medio de cultivo con Asn/Gln (7 .6 mM/13.6 mM). En trigo (Triticum
aestivum L.), Sarker et al., 2007, encontraron que la aplicación de Asn 0.75 mM promovió
el proceso de ES así como su regeneración en plantas completas . En C. chinense, la
adición de Asn 3.8, 7.6 Y 15.2 mM favoreció el desarrollo de los ESs e indujo la expresión
de 7 péptidos que en un principio no se encontraban en el perfil proteico de los ESs de
esta especie y que sí están presentes en el perfil proteico de los ECs (Lecona-Guzmán ,
2012).
Las PAs son compuestos orgánicos nitrogenados muy importantes para las plantas , dado
que participan en muchos de sus procesos fisiológicos y de desarrollo (Evans y
40

CAPíTULO 11
Malmberg, 1989; Galston y Kaur-Sawhney, 1990; Carmona, 1997; Berta et al., 1997). Son
consideradas como una clase de reguladores del crecimiento , como segundos
mensajeros hormonales de la prolife ración y división celular en muchos procesos
incluyendo la embriogénesis y como una de las reservas de carbono y nitrógeno por lo
menos en tej idos cultivados (Kakkar et al., 2000 ; Bais y Ravishankar, 2002; Silveira et al.,
2004 a,b; Jimenez, 2005). Existen diversos estud ios que muestran la importancia de las
PAs exógenas para mejorar los procesos in vitro como la ES. Se sabe que las PAs
exógenas pueden inducir la división celular e incrementar la regeneración en cultivos de
células vegetales (Kakkar et al., 2000). Kevers et al., (2000) observaron que ad icionando
PAs al medio de cultivo de Panax ginseng promovieron el proceso de ES; particularmente,
con la ad ición de Spm 0.1 mM lograron incrementar el número de ESs que desarrollaron
tanto el área apical como radical , obten iendo un 81 % de ESs convertidos en plantas.
Steiner et al., (2007) estimularon el crecimiento embriogénico de Araucaria angustifolia
suplementando el medio de cultivo con Put (1 y 0.01 mM) y Spm (0.01 y 0.1 mM). Paul et
al. (2009), concluyeron que la aplicación de PA's al medio embriogénico de Momordica
charantia provocó un incremento en peso fresco y en el número de ESs durante estad íos
tempranos. En el género Capsicum se han realizado estud ios sobre la caracterización de
PAs en la ES (Regla-Márquez, 2011), así como el efecto de la adición de las mismas y de
inhibidores de las enzimas de su biosíntesis en el desarrollo de brotes de C. frutescens M.
(Kumar et.a/. , 2007). En C. chinense , lópez-Erosa (2012) , tras aplicar Put, Spm y Spd en
concentraciones de 0.01 , 0.1 Y 0.1 mM, en el medio de cultivo de los ESs ; obtuvo mejoría
en el desarrollo de los ESs solamente con Spm; detectando en la germinación de los ESs
tratados con Spm 0.1 mM, armonía en la zona apical y radical así como un cambio en su
coloración tornándose verde .
Se sabe que las PAs regulan la síntesis de proteínas (Young y Srinivasan , 1972; Morris y
Jorstad, 1973; Tabor y Tabor, 1984; Marton y Morris, 1987; Evans y Malmberg, 1989) y
están involucradas en las modificaciones postraduccionales (Tabor y Tabor, 1984; Pegg,
1986). En estudios realizados en avena por Tiburcio et al. (1993) , fue possible mantener
alto contenido de proteína en las membranas tilacoides de hojas bajo estrés osmótico
cuando fueron tratadas con DFMA (el cual incrementa los niveles endógenos de Spd y
Spm después de un tratamiento osmótico) . Helleboid et al. (1995) reportaron que la
adición de Put a cultivos embriogénicos de un híbrido de Cichorium intybus l . pretratado
41

CAPíTULO 11
con un inhibidor de PAs, restauró el contenido inicial de Spd , la ES, así como los niveles
de polipéptidos extracelulares relacionados con la ES (SER). Santanen y Simola (1999),
tras estudios realizados en ECs y megametofitos de Picea abies, sugirierion que las PAs
pueden tener un rol en la acumulación de proteínas de reserva , así como en la
maduración de las semillas de dicha especie. Estos autores también sugieren que las
interacciones de las PAs y sus productos con las proteínas, pueden ser críticas para la
división y diferenciación celular.
La regeneración de plantas in vitro de C. chinense vía ES, se ha visto truncada por el alto
índice de ESs deformados, la baja tasa de germinación y por ende de conversión a planta .
La acumulación de proteínas en la ES es de suma importancia para que los embriones
adquieran la capacidad de germinar adecuadamente. En e, chinense, Lecona et al.
(2012) ; reportaron que contrario a lo reportado para ESs de otras especies; el contenido
de proteínas en éstos disminuye con el avance de su desarrollo, llegando a presentar una
cantidad crítica de proteínas en el estadío cotiledonar. Esto permite inferir que dicho
fenómeno pudiera estar relacionado con la baja capacidad de germinación y conversión a
planta de los ESs de esta especie . En atención a estas problemáticas, y con base en los
efectos positivos reportados , de la aplicación de Asn , Gln (aminoácidos) y Spm
(poliamina) sobre la ES de diferentes especies vegetales (particularmente en C. chinense)
se propuso en el presente trabajo evaluar comparativamente el efecto de la aplicación
exógena de Asn, Gln y Spm sobre la eficiencia, calidad y desarrollo de los ESs de chile
habanero con el fin de determinar cual o cuales tratamientos , contribuyen a mejorar
dichas características y analizar el contenido y composición de proteínas totales de los
ESs tratados con el o los mejores tratamientos en relación con el testigo.
2.2 MATERIALES Y MÉTODOS
2.2.1 Material vegetal
Se utilizaron semillas de chile habanero de la variedad Mayan Ba'alché, proveniente del
banco de germoplasma de la Unidad de Bioquímica y Biología Molecular de plantas, del
Centro de Investigación Científica de Yucatán. Esta variedad fue elegida por presentar el
típico fruto naranja , siendo una de las variedades más conocidas y demandadas a nivel
local y nacional. Además de ser una de las variedades con registro de obtentor.
42

CAPíTULO 11
2.2.2 Desinfección y germinación de semillas
la desinfección de las semillas , se realizó de acuerdo con Santana et al. (2005). las
semillas fueron sumergidas en una solución de etanol al 70% por 5 minutos ; se
enjuagaron tres veces con agua destilada estéril y posteriormente se colocaron en una
solución de hipoclorito de sodio comercial (Cloralex: 6% de cloro activo) diluido 3.3 veces
(v/v) = [1.8%] durante 15 minutos. Finalmente las semillas se enjuagaron 3 veces con
agua destilada estéril. Todo el procedimiento se realizó en agitación .
Con el fin de establecer un stock de plantas asépticas para la toma de explantes, se
indujo la germinación de las semillas en condiciones in vitro. Para este propósito, el medio
de cultivo utilizado fue el MS (Murashige y Skoog, 1962), suplementado con 1.15 iJM de
ácido giberélico (AG3), 3% de sacarosa, 0.2% de gelrite, y con pH 5.8. las semillas se
incubaron en oscuridad a una temperatura de 25 ± 2 OC por 10 días. Una vez germinadas,
las plántulas fueron transferidas a un cuarto de fotoperíodo (16 horas luz/8 oscuridad) a
una intensidad lumínica de 40-50 I-lmol.m2.s·\ y a una temperatura de 25 ± 2 oC durante
15 días.
2.2.3 Inducción de embriogénesis somática
Para inducir el proceso de embriogénesis somática, se utilizó el protocolo reportado por
Avilés , et al. (2013) . Se tomaron hipocotilos de plántulas de 30 días en condiciones
asépticas . El explante se colocó en medio MS, suplementado con 9.05 iJM de Acido 2,4-
Oiclorofenoxiacético (2,4-0), 3% de Sacarosa y 0.2% de gelrite. Se colocaron diez
explantes por frasco , con cinco réplicas para cada tratamiento y se mantuvieron a 25 ± 2
oC, en condiciones de fotoperíodo (16 horas luz/8 oscuridad) a una intensidad lumínica de
40-50 I-lmol.m2.s·1 durante 30 días.
2.2.4 Adición de asparagina (asn), glutamina (gln) y espermina (spm) en la
histodiferenciación
Una vez transcurridos los 30 días de inducción, los explantes fueron transferidos a medio
MS liquido como fue reportado por Avilés et al., (2013) ; constituido por 2,4-0 (4 .5 I-lM) Y
sacarosa (30 g.l-1), a un pH de 5.8 por un periodo de 7 días . Posteriormente los
explantes fueron transferidos al mismo medio de cultivo ahora suplementado de manera
43

CAPíTULO 11
individual con diferentes concentraciones de Asn (3.8 mM, 7.6 mM y 15.2 mM), Gln (17.1
mM , 34.2 mM , 51 .3 mM) y Spm (0.01 mM , 0.1 mM y 1 Mm); manteniendo como testigo un
cultivo libre de estos compuestos durante 7 días. Transcurrido este tiempo, los explantes
fueron transferidos a medio MS líquido nuevo libre de aminoácidos y la poliamina y
cultivados durante 14 días. Durante todo este periodo (30 días) los cultivos se
mantuvieron a una temperatura de 25 ± 2°C, con luz continua (24 hrs. luz) a una
intensidad lumínica de 40-50 IJM 'm-2 's-1 en agitación (100 rpm) . Para este procedimiento
se utilizaron matraces Erlenmeyer de 250 mi con 50 mi de medio de cultivo, empleando 5
matraces por cada tratamiento.
2.2.5 Evaluación de los tratamientos
Para evaluar el efecto de la adición de Asn , Gln y Spm sobre el desarrollo de ESs de chile
habanero, se evaluó el número de embriones totales, el porcentaje de embriones
tempranos y tardíos; así como la morfología y calidad de los embriones obtenidos de
dichos tratamientos, comparándolos con los del tratamiento testigo. Esto se realizó
utilizando un estereoscopio marca Leica modelo DFC 320 . Los resultados fueron
analizados empleando análisis de varianza y comparación de medias por la prueba de
Tukey (Ps 0.05). Para ello se utilizó la herramienta informática SPSS Statistics Standard
20 .0.0 de 18M .
2.2.6 Análisis histológico
Con el fin de realizar una mejor evaluación del efecto de los mejores tratamientos sobre la
morfología de los ESs, se recolectaron durante el proceso de ES, muestras de los
embriones en diferentes estad íos de desarrollo para efectuar su análisis histológico.
Las muestras se sumergieron en una solución fijadora (10 mL de formaldehido al 37%, 50
mi de alcohol al 96%, 5 mi de ácido acético al 100%, (FAA) aforada a 100 mi con agua
destilada), posteriormente, se colocaron en un desecador al cual se aplicó vacío por 15
minutos y en donde se mantuvieron con la solución por 48 horas .
Después de fijadas, las muestras fueron deshidratadas mediante la exposición gradual a
etanol (30%, 50%, 70%) durante una hora, (85%, 96%) por 2 horas y (100%) por una
hora. Finalmente las muestras se colocaron en butanol (Sigma) al 100% pasando por dos
44

CAPíTULO 11
cambios cada 24 horas. Durante el segundo cambio se adicionaron hojuelas de parafina
(Paraplast plus, Sigma) y se colocaron en agitación toda la noche.
Para la inclusión, las muestras se colocaron en un horno de circulación forzada de aire
(HCF-71, BG) a 60°C por aproximadamente 2 horas. Una vez derretida la parafina, se
agregaron más hojuelas de ésta cada 2 horas por 24 horas . Posteriormente se realizó un
cambio con parafina líquida, desechando la mitad del volumen y adicionando el mismo
volumen con parafina líquida pura (previamente colocada en el horno). Se realizaron 2
cambios por día durante 3 días.
Una vez realizada la inclusión, las muestras se colocaron en el centro del molde de
aluminio (histo base mold 15 x 15 x 5 mm, Fisher) el cual fue precalentado en la estufa 1
hora antes de utilizarlo. Las muestras se colocaron utilizando pinzas y fueron cubiertas
con parafina líquida pura colocando un cassette (Embedding cassettes M490-2, Simport)
sobre el molde . Posteriormente, los moldes con muestra fueron enfriadas a temperatura
ambiente. Una vez endurecida la parafina, se retiró el molde a fin de realizar cortes de 5-8
IJm en el micrótomo de rotación (Microtom HM 340E, MICROM International GmbH)
utilizando la navajas Leica 818 High Profile (Leica Biosystems). Los cortes se montaron
en portaobjetos 25 x 75 (Corning) y se colocaron a 42°C por 24 horas para su adhesión .
Enseguida , éstos se trataron con butanol al 100% por 10 mino y se desparafinaron con
ultraclear histograde (J .T. Baker) por 15 mino Se continuó con un proceso de hidratación
mediante una exposición gradual de etanol (96%, 50% Y 30% por 2 min) y se hornearon
nuevamente a 3rC por 3 días. Pasado este tiempo, se tiñeron con azul de Toluidina al
0.05%.
Los cortes histológicos se observaron en un estereoscopio marca Leica (MZ FLlII , Leica).
Las fotos fueron tomadas con la cámara del mismo (Leica DFC320 de 3.45 IJm pixel,
Leica Microsystems).
45

CAPíTULO II
2.3 RESULTADOS
2.3.1 Comportamiento de la eficiencia en la ES de chile habanero sometida a
diferentes tratamientos con Asn, Gln y Spm
Al analizar el efecto de Asn , Gln y Spm sobre la eficiencia de la ES de C.chínense
(número de ESs totales obtenidos por explante), se observó que en todos los tratamientos
se formaron entre 33 y 64 ESs/explante a los 30 días de cultivo en medio líquido. El
tratamiento que mostró el mayor número de ESs, fue el tratamiento testigo (MS + 2,4-0
4.5 ¡JM) , con 64±4.2 ESs/Exp, respuesta que fue significativamente diferente sólo en los
tratamientos con Asn 7.6 mM, Gln 34.2 mM, Gln 51 .3 mM y Spm 1 mM , afectando
negativamente la eficiencia del sistema al presentar los menores valores de embriones
formados (43±4.8, 39±4, 33±1 y 38±6 ESs/Exp respectivamente) . Los tratamientos
restantes aunque mostraron menor número de ESs con respecto al tratamiento testigo, no
variaron significativamente de éste (Fig . 2.1 Y 2.2). Particularmente, se observó que a
medida que se aumentó la concentración de Gln, la respuesta de formación de ESs fue
disminuyendo (Fig . 2.1).
46

80
~ 70 = j 60 1 .. 50
~ 40 J
~ 30 j ~ 20
~ 10
O
~ ~~ 1 -[ 60 1 II so ~ 40 j
il 30 j ~ 20 j ~ 10
a 1
TO
1
0 +----
L r
80
! 70 = % 60
II SO
l40 ~ 30
~ 20
~ 10 I
I L_
TO
1
TO
CAPiTULO 11
Asparagina
.b
ab b
ASN 3.8 mM ASN 7.6mM ASN 15.2 mM
Tratamientos
Glutamina
b b
GLN 17.1 mM GLN 34.2 mM GLN 51.3 mM
Tratamientos
Espermina
ab ab
b
SPM 0.01 mM SPM 0.1 mM SPM 1 mM
Tratamientos
Figura 2.1 Efecto de Asn , Gln y Spm exógena en el número de embriones somáticos de C.
chinense producidos por explante. TO: tratamiento testigo. Cada valor es el promedio de cinco
réplicas independientes con el error estándar. Letras diferentes indican diferencia significativa por la
prueba de Tukey (p<0.05).
47

CAPíTULO 11
2.3.2 Efectos de Asn, Gln y Spm sobre la calidad y desarrollo de los ESs de chile
habanero
En la figura 2.2 se observa la respuesta de los explantes expuestos a los diferentes
tratamientos (30 días de cultivo en medio liquido). Todos respondieron con la formación
de ESs. Los embriones obtenidos bajo tratamiento con Spm en las concentraciones de
0.01 y 0.1 mM fueron los que mostraron mejoría en cuanto a calidad de tejido (mejor
diferenciaciado y con mejor consistencia) comparado con el tratamiento testigo. En
particular, los ESs tratados con Spm 0.1 mM mostraron morfologías mejor delimitadas,
con mayor calidad, avanzando en su desarrollo con mayor rapidez y sincronía en
comparación con los demás tratamientos . Aquellos que fueron expuestos a Spm 1 mM
mostraron tejidos fenolizados . Por otro lado, los ESs tratados con glutamina, fueron los
que presentaron la más baja calidad; al aplicar Gln 17.1 mM, se inhibió el desarrollo de los
embriones y aunque con las concentraciones más altas de este aminoácido, se
apreciaron ESs de mayor tamaño, éstos se observaron con cierta desdiferenciación de
tejido. En cuanto a los efectos producidos por Asn, a la concentración de 3.8 mM, hubo un
atraso en el desarrollo de los ESs, mientras que con las concentraciones de 7.6 y 15.2
mM , el proceso de ES mostró la mayor asincronía entre tratamientos.
48

CAPíTULO 11
Figura 2.2 Respuesta de segmentos de hipocotilos cultivados en medio MS + 4.5 IJM de 2,4-D
suplementado con diferentes concentraciones de Asn , Gln y Spm (30 días de cultivo).
Transcurridos 45 días de cultivo, se observó en todos los tratamientos, la disgregación de
los ESs del tej ido que les dio origen . Se obtuvo mayor cantidad de embriones disgregados
con los tratamientos de Asn 3.8 mM , Spm 0.01 mM y Spm 0.1 mM. Estos dos últimos
tratamientos mostraron ESs mejor desarrollados y con menos deformaciones en
comparación con los demás tratamientos. Por el contrario, se observó que la menor
cantidad de embriones disgregados se obtuvo con los tratamientos Spm 1 mM, y todos los
tratamientos con glutamina. El empleo de Asn no favoreció la morfología de los
embriones, observando embriones poco definidos. los ESs tratados con Gln se
observaron con la más baja calidad y desarrollo de todas las condiciones evaluadas (Fig .
2.3).
49

CAPíTULO 11
Figura 2.3 Efecto de diferentes concentraciones de Asn, Gln y Spm sobre el desarrollo de los
embriones somáticos de C. Chinense a los 45 días de cultivo.
En la cuadro 2 .1 se muestra que, a los 45 días de cultivo , el tratamiento con 0.1 mM de
Spm alcanzó el mayor porcentaje (71 .9 ± 1.4%) de ESs en estadíos avanzados de
desarrollo (torpedo y cotiledonar) , difiriendo significativamente del tratamiento testigo
(54.7 ± 1.5%), y de los tres tratamientos con Gln . Estos últimos propiciaron los menores
porcentajes de ESs en estadíos avanzados (32 .8 ± 4.2, 33 .2 ± 3.5 y 27.3 ± 4 .6 %); siendo
significativamente diferentes de los demás tratamientos. En lo que concierne a los ESs
tratados con Asn , arriba del 60% de los ESs formados alcanzaron estad íos avanzados de
desarrollo.
50
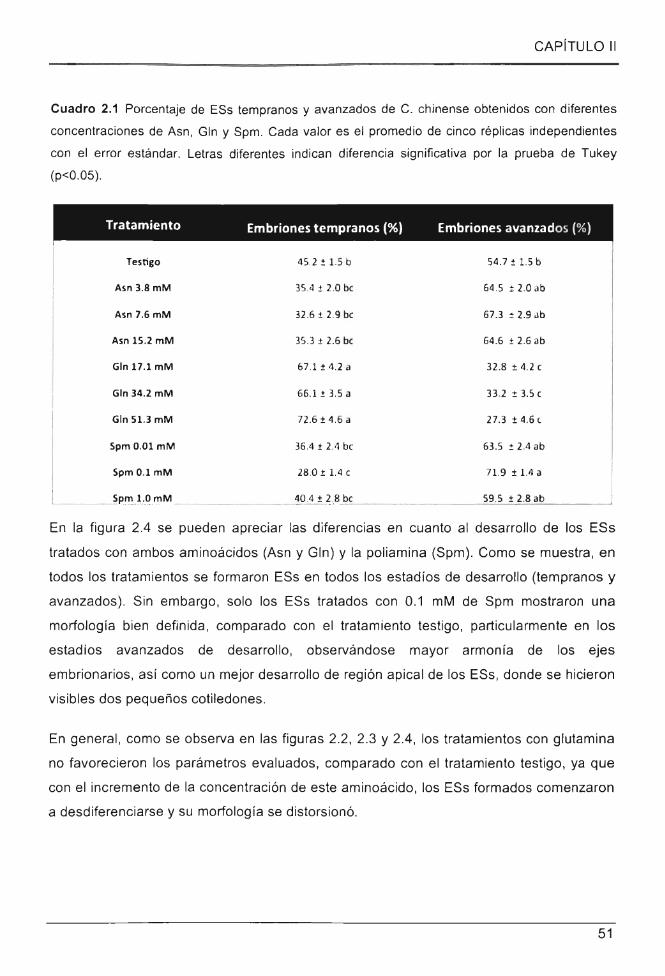
CAPiTULO II
Cuadro 2.1 Porcentaje de ESs tempranos y avanzados de C. chinense obtenidos con diferentes
concentraciones de Asn , Gln y Spm. Cada valor es el promedio de cinco réplicas independientes
con el error estándar. Letras diferentes indican diferencia significativa por la prueba de Tukey
(p<O.05).
Tratamiento Embriones tempranos (%) Embriones avanzados (%)
Testigo 45.2 ± 1.5 b 54 .7 ± 1.5 b
Asn 3.8 mM 35.4 ± 2.0 be 64 .5 :!: 2.0 ab
Asn 7.6 mM 32.6 ± 2.9 be 67.3 :!: 2.9 ab
Asn 15.2 mM 35.3 ± 2.6 be 64 .6 ± 2.6 ab
Gln 17.1 mM 67 .1 ± 4.2 a 32.8 ± 4.2 e
Gln 34.2 mM 66.1 ± 3.5 a 33.2 ± 3.5 e
Gln 51.3 mM 72 .6 ± 4.6 a 27.3 ± 4.6 e
5pmO.01 mM 36.4 ± 2.4 be 63 .5 ! 2.4 ab
Spm 0.1 mM 28.0 ± 1.4 e 71.9 ± 1.4 a
S m 1.0 mM 40.4 ± 2.8 be 59.5 ± 2.8 ab
En la figura 2.4 se pueden apreciar las diferencias en cuanto al desarrollo de los ESs
tratados con ambos aminoácidos (Asn y Gln) y la poliamina (Spm). Como se muestra, en
todos los tratamientos se formaron ESs en todos los estad íos de desarrollo (tempranos y
avanzados) . Sin embargo , solo los ESs tratados con 0.1 mM de Spm mostraron una
morfología bien definida , comparado con el tratamiento testigo, particularmente en los
estadíos avanzados de desarrollo, observándose mayor armonía de los ejes
embrionarios, así como un mejor desarrollo de región apical de los ESs, donde se hicieron
visibles dos pequeños cotiledones.
En general, como se observa en las figuras 2.2, 2.3 Y 2.4, los tratamientos con glutamina
no favorecieron los parámetros evaluados, comparado con el tratamiento testigo, ya que
con el incremento de la concentración de este aminoácido, los ESs formados comenzaron
a desdiferenciarse y su morfología se distorsionó.
51
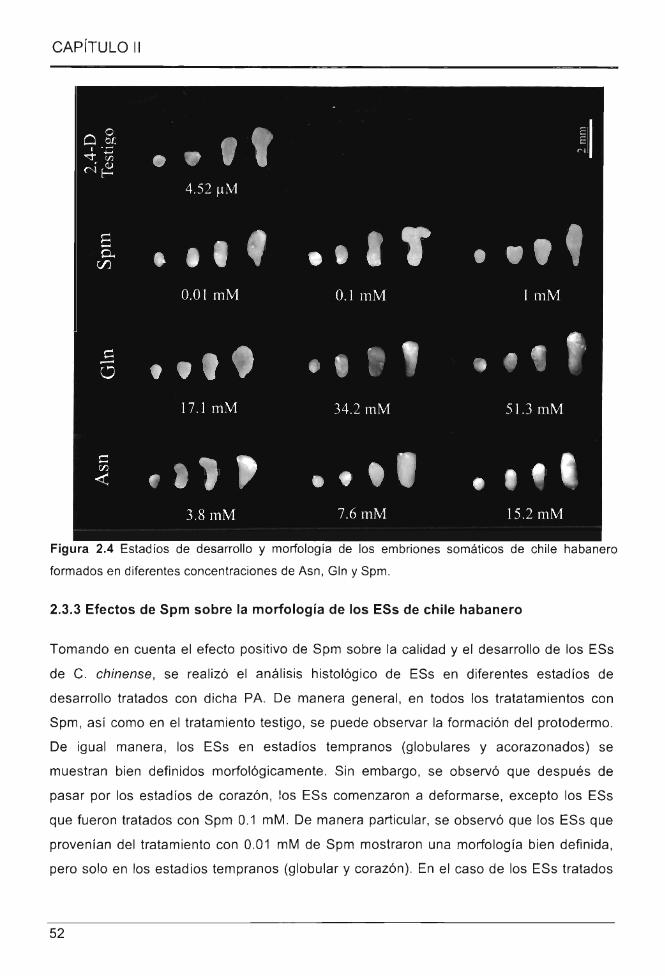
CAPíTULO 11
~ t ~ I O ~. , 1'--
-r7 • .. r I
• 'J ('1 f-
-lS2 p\l
,.... , I , , = I , c... • • • • • • C/)
0.01 m\l () . 1 III \1 ImM
e , t ~ ... ~ " • • f~' - • " , ~ lJ •
17.1 m\l 1-l .~ m!\l 51.l filM
:::
1 , , I 'f) I t • « t • • • • 3.X mM 7.() mM 15.2 111\1
Figura 2.4 Estadíos de desarrollo y morfología de los embriones somáticos de chile habanero
formados en diferentes concentraciones de Asn, Gln y Spm.
2.3.3 Efectos de Spm sobre la morfología de los ESs de chile habanero
Tomando en cuenta el efecto positivo de Spm sobre la calidad y el desarrollo de los ESs
de C. chinense, se realizó el análisis histológico de ESs en diferentes estadíos de
desarrollo tratados con dicha PA. De manera general , en todos los tratatamientos con
Spm , así como en el tratamiento testigo, se puede observar la formación del protodermo.
De igual manera , los ESs en estadíos tempranos (globulares y acorazonados) se
muestran bien definidos morfológicamente. Sin embargo, se observó que después de
pasar por los estad íos de corazón, los ESs comenzaron a deformarse, excepto los ESs
que fueron tratados con Spm 0.1 mM. De manera particular, se observó que los ESs que
provenían del tratamiento con 0.01 mM de Spm mostraron una morfología bien definida ,
pero solo en los estad íos tempranos (globular y corazón) . En el caso de los ESs tratados
52
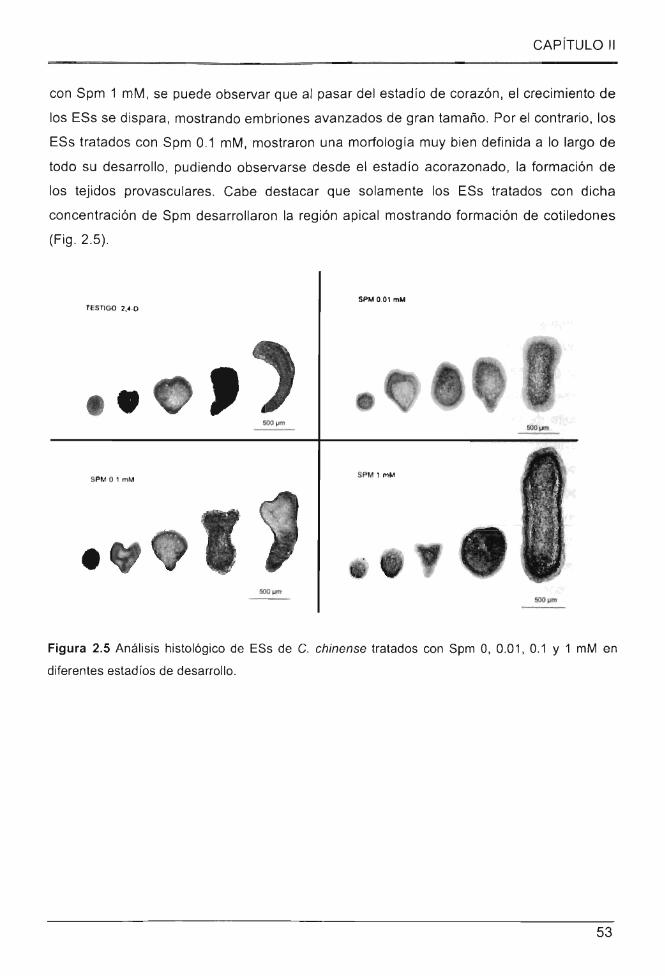
CAPITULO II
con Spm 1 mM , se puede observar que al pasar del estadía de corazón, el crecimiento de
los ESs se dispara, mostrando embriones avanzados de gran tamaño. Por el contrario, los
ESs tratados con Spm 0.1 mM , mostraron una morfología muy bien definida a lo largo de
todo su desarrollo, pudiendo observarse desde el estadía acorazonado, la formación de
los tejidos provasculares. Cabe destacar que solamente los ESs tratados con dicha
concentración de Spm desarrollaron la reg ión apical mostrando formación de cotiledones
(Fig . 2.5) .
SPMOOl mM TESTIGO 2.' o
SPMO 1 mM SPM 1 mM
Figura 2.5 Análisis histológico de ESs de C. chinense tratados con Spm 0, 0.01 , 0.1 Y 1 mM en
diferentes estad íos de desarrol lo.
53

CAPíTULO 11
2.4 DISCUSiÓN
Al evaluar la eficiencia de la ES de C. chinense, bajo diferentes concentraciones de los
aminoácidos Asn y Gln; y de la poliamina Spm, se corroboró el alto potencial
embriogénico de la especie previamente reportado por Avilés-Viñas en 2013. Los
resultados mostraron que el tratamiento testigo no fue superado en eficiencia por ninguno
de los tratamientos evaluados. Bajo la mayoría de los tratamientos, la eficiencia no varió
significativamente respecto al TO; excepto por Asn 7.6 mM, Gln 34.2, Gln 51.3 mM y Spm
1 mM , con los cuales fue afectada negativamente.
Los efectos positivos de aminoácidos sobre los embriones somáticos, particularmente al
aumentar el número de ESs/explante, acelerando el desarrollo, incrementando el número
de ESs germinados, incluso la conversión en plantas, han sido reportados en diferentes
especies vegetales. En este estudio, la adición de Asn 7.6 y 15.2 mM favoreció el
desarrollo de los ESs, alcanzando los estad íos más avanzados en menor tiempo. Esto
evidenciado por la presencia de ESs en estadíos avanzados a los 30 días de cultivo
(medio líquido), a diferencia de los demás tratamientos. Esto coincide con estudios
realizados por Lecona-Guzmán (2012), en los que observó que en general, la aplicación
de Asn en la ES de C. chinense favorece la velocidad del desarrollo de los ESs,
observándose embriones avanzados en su desarrollo a los 65 días de cultivo . En este
estudio, la adición de este aminoácido al medio de cultivo propició la obtención de un
mayor porcentaje de ESs avanzados, de manera similar al tratamiento con Spm. La
adición de Asn ha promovido la ES en otras especies como trigo (Sarker et al., 2007) y
algodón (Wu et al. , 2004) llegando inclusive a lograr su conversión en planta. La
presencia de aminoácidos en el medio de cultivo resulta favorable debido a que estos
sustituyen parcialmente el amonio (NH 4 +) e incrementan los niveles de N reducido , el cual
es metabolizado rápidamente por las células vegetales, estimulando el rápido crecimiento
y el desarrollo celular (Gamborg , 1970; Grimes y Hodges, 1990). Este efecto estimula el
desarrollo de los ESs en diversas especies vegetales y permiten la adaptación a cambios
ambientales, especialmente aquellos que ocasionan estrés en el organismo (Young et al. ,
1999), como es el caso de la morfogénesis in vitro .
A pesar de los efectos positivos provocados por Asn , este aminoácido no favoreció la
calidad ni la morfología de los ESs respecto al tratamíento testigo (TO). Cabe mencionar
54

CAPíTULO II
que bajo la concentración de 3.8 mM Asn, los ESs mostraron retraso en su desarrollo; sin
embargo, a los 45 dias de cultivo, estos embriones mostraron un mejor desarrollo en
comparación con las concentraciones restantes de Asn .
En lo que respecta a Gln, todas las concentraciones empleadas tuvieron efectos
negativos sobre el proceso embriogénico. En cuanto a eficiencia, a medida que se
incrementó la concentración de este aminoácido, disminuyó el número de ESs formados
por explante e inhibió su desarrollo al mostrar el menor porcentaje de éstos en estadios
avanzados a los 45 dias de cultivo. Aunado a esto, desde las primeras etapas de
desarrollo, los embriones formados bajo Gln, fueron los más deficientes en calidad,
desarrollo y morfologia; además de mostrar menor capacidad para disgregarse del tejido
de origen. Diferentes estudios han mostrado los efectos positivos de Gln en la ES de
numerosas especies vegetales; sin embargo los aminoácidos también pueden inhibir el
crecimiento en cultivos vegetales (Gamborg, 1970; Heimer y Filnes, 1970; Fukunaga y
King, 1982; Tupy et al., 1983; Ronchi et al., 1984; John and Mukherjee, 1997). la falta de
crecimiento en presencia de aminoácidos individuales puede deberse a la incapacidad de
los compuestos para sustituir al amonio ó por la inhibición de reacciones metabólicas en
las células (Gamborg , 1970). los efectos negativos de Gln sobre el desarrollo de la ES
puede deberse a la inhibición de auxinas (Jacques et al., 1995). la necesidad de
aminoácidos en el medio de cultivo depende de la especie, explante o de la respuesta
morfogenética deseada. Merkle et al., (1995) reportó que la adición de aminoácidos al
medio de cultivo puede inhibir ó promover el desarrollo y conversión de los ESs.
El uso de Spm (poliamina), en concentraciones de 0.01 y 0.1 mM, no afectó la eficiencia
de la ES al no mostrar diferencia significativa en el número de ESs/explante respecto al
TO. Esto coincide con lo reportado en 2012 por lópez-Erosa, quien tampoco encontró
diferencia significativa en la eficiencia al utilizar las mismas concentraciones de Spm
durante los primeros 15 dias de histodiferenciación de los ESs de C. chinense. A pesar de
la disminución significativa del número de ESs obtenido en presencia de 1 mM Spm, en
este estudio, contrario a lo reportado por lópez-Erosa (2012), en ambos estudios se
observó gran coincidencia en el patrón de comportamiento de la eficiencia tras el efecto
de las tres concentraciones evaluadas (0 .01 , 0.1 Y 1 mM), en los cuales el número de ESs
formados resultó mayor bajo la concentración de 0.1 mM . En otras especies vegetales;
55

CAPíTULO 11
bajo tratamiento con PAs , se ha observado el incremento del número de ESs formados;
tal es el caso de Hevea brasiliensis (EI-Hadrami et al. 1989a, 1989b), Panax ginseng
(Kevers et al. , 2000) quienes lograron incrementar cuatro veces el No. de ESs producidos
con la ad ición de PAs (0 .01, 0.1 Y 1.0 mM); yen Momordica charantia (Paul et al. , 2009),
al adicionar Put 0.1, 0.5 Y 1.0 mM ; Spd y Spm 0.1, 0.5 Y 1.0 flM. En este estudio, además
de mantener la eficiencia de la ES, Spm 0.01 y 0.1 mM favorecieron la calidad de los ESs
de C.chinense a diferencia de los demás tratamientos a los 30 días de cultivo (medio
líquido). Así mismo, bajo estas dos condiciones (como con Asn 3.8mM), hubo mayor
cantidad de ESs que fueron capaces de separarse del explante a los 45 días de cultivo.
En general, el uso de Spm (0 .01, 0.1 Y 1 mM) propició la disminución de deformaciones
en los ESs, observando a los 45 días de cultivo, estructuras mejor definidas en
comparación con los demás tratamientos, incluido el TO. Estos resultados coinciden con
López-Erosa (2012), quien evaluó el efecto de Put, Spd y Spm sobre la ES de esta
especie y encontró que sólo con la aplicación de Spm se logró mejorar el desarrollo y
disminuir la frecuencia de anormalidades de los ESs de chile habanero.
Contrario a lo anterior, con Spm 1 mM, además de haber disminuido la producción ESs,
éstos mostraron fenolización en su tejido transcurridos 30 días de cultivo. Y se encontró
menor cantidad de ESs disgregados en el medio de cultivo a las 45 días. Respecto a esto,
se han reportado efectos negativos de las PAs sobre cultivos embriogénicos; tal es el
caso de Calheiros et al. (1994), quienes al adicionar PAs en el medio de cultivo para ES
directa de Coffea canephora y arabica, observaron que tratamientos con Spd (50 y 100
flM) redujeron el No. de ESs en dos genotipos de Coffea canephora (N90-6 y N84-3). Así
mismo, Cuando Spd y Spm (20 flM) fueron aplicadas en combinación , el efecto negativo
de la Spd fue significativo en ambas especies; Spm inhibió el proceso de ES en las dos
especies, pero con diferencias estadísticas no significativas. Patil et al., (1998) observaron
una reducción en la inducción de ESs por Put (0.1 , 0.2, 0.5 mM), Spd (0.1 y 0.2 mM) en
garbanzo. Rastogi y Davies (2000) y Silveira et al. (2004) reportaron que en cultivos en
suspensión de Pinus taeda y callos de Nicotiana tabacum, altos niveles de Put estaban
asociados con la reducción del crecimiento celular. Asimismo, Silveira et al. (2006)
observaron una reducción en el crecimiento de cultivos en suspensión de Araucaria
angustifolia aplicando Spd y Spm (1 .0 mM) al medio de cultivo . El efecto negativo de Spm
1 mM en C. chinense puede deberse a que resulta ser una concentración muy alta que
56

CAPíTULO 11
pudiera desbalancear los niveles endógenos de PAs en los ESs. Oe igual manera, pudiera
deberse en parte a un mecanismo de inhibición por retroalimentación aumentando la
síntesis de etileno. Existe una fuerte evidencia de la competencia entre las vías
biosintéticas de las PAs y el etileno (Minocha et al., 1990).
En este estudio, Spm 0.1 mM, benefició a los ESs de C. chinense en diferentes aspectos .
Por una parte, no afectó la eficiencia de la ES; mejoró la calidad de los ESs al mostrarse
mejor definidos y delimitados en cuanto a estructura. Así mismo, el crecimiento y
desarrollo de éstos, presentó mayor sincronía entre los diferentes tratamientos a los 30
días de cultivo . Transcurridos 45 días, permitió obtener el mayor porcentaje de ESs en
estadías avanzados de desarrollo, difiriendo significativamente del TO. En cuanto a
morfología, favoreció notablemente a los ESs al mostrar estadías mejor definidos;
particularmente en los estadías avanzados, se observó la mejoría de la región apical de
los embriones mostrando la formación de cotiledones, no así en los demás tratamientos.
Esto coincide con lo reportado por Kevers et al., (2000) y López-Erosa (2012) quienes al
adicionar 0.1 mM Spm al medio de cultivo de Panax ginseng y C. chinense
respectivamente, lograron incrementar el número de ESs que desarrollaron de manera
armoniosa el área apical como radical; así como la emisión de hojas cotiledonares en la
etapa de germinación en el caso de C.chinense . Las PAs mejoran el proceso
embriogénico de muchas especies vegetales (Feirer et al., 1984; Steiner et al., 2007; Paul
et al., 2009). En Hevea brasiliensis, ha sido necesario un alto contenido de PAs en un
medio suplementado con 2,4-0 y BAP para superar su naturaleza recalcitrante a la ES
(EI-Hadrami y O'Auzac, 1992). Las PAs desempeñan diversos papeles en muchos
procesos fisiológicos, de crecimiento y de desarrollo vegetal (Kakkar et al., 2000; Basis y
Ravishankar, 2002; Silveira et al. , 2004 a,b; Jimenez, 2005). Existe una alta correlación
entre los niveles de PAs y ciertos eventos de la división celular en plantas (Serafini et al.,
1989). la ES puede ser regulada mediante la manipulación de los niveles y el
metabolismo de las PAso Las PAs exógenas pueden inducir la división celular e
incrementar la regeneración en cultivos de células vegetales (Kakkar et al., 2000). Steiner
et al., 2007 sugieren que los resultados positivos observados en el crecimiento de cultivos
embriogénicos de araucaria angustifolia por la adición de PAs puede ser el resultado de
su interacción con otras hormonas.
57

CAPíTULO 11
Respuestas diferentes de las diferentes fuentes de N orgánico, indican el requerimento de
aminoácidos y PAs específicas para eventos específicos durante la morfogénesis (Sarker
et al. , 2007). El mejor tratamiento aplicado en este estud io resultó ser el de Spm 0.1 mM ,
el cual repercutió favorablemente en el desarrollo , morfología y la calidad de ESs ,
superando al tratamiento testigo (TO).
58

CAPíTULO II
BIBLIOGRAFíA
Armstrong C.L. y Green C.E. (1985). Establishment and maintenance of friable ,
embryogenic maize callus and the involvement of l-proline . Planta 164: 207-214.
Avilés-Viñas S., lecona-Guzmán C. , Canto Flick A. , lópez-Erosa S.,Santana-Buzzy N.
(2013). Morpho-histological and ultrastructural study on direct somatic
embryogenesis of Capsicum chinense Jacq. in liquid medium. Plant Biotechnol
Rep. 7: 277-286.
Bais H.P. Y Ravishankar GA (2002). Role of polyamines in the ontogeny of plants and
their biotechnological applications. Plant Tiss. Org . Cult. 69: 1-34.
Barrett JD, Park YS y Bonga JM . (1997). The effectiveness of various nitrogen sources
white spruce (Picea glauca (Moench) Voss) somatic embryogenesis . Plant Cell
Rep. 16:411-415.
Basu A.; Sethi U.; Guha-Mukherjee S. (1989) Regulation of cell proliferation and
morphogenesis by aminoacids in Brassica tissue cultures and its correlation with
threonine deaminase. Plant Cell Rep . 8: 333- 335; 1989.
Berta G, Altamura MM, Fusconi A, Cerruti F, Capitani F, Bagni N. (1997). The plant cell
wall is altered by inhibition of polyamine biosynthesis. New Phytol137 : 569-577 .
Buyucalaca S. y Mavituna F. (1996). Somatic embryogenesis and plant regeneration of
pepper in liquid media. Plant Cell Tiss. Org . Cult. 46 :227-235.
Calheiros M.B.P., l .G.E. Vieira y S.R.L. Fuentes (1994). Effects of exogenous polyamines
on direct somatic embryogenesis in coffee . Rev. Bras. Fisiol. Veg ., 6,109-114.
Carmona A.J . (1997). Contribución al conocimiento del papel de las poliaminas en el
desarrollo del fruto de la platanera. Estudio de las implicaciones fisiológicas de
estas sustancias sobre el crecimiento in vitro de los tejidos del ovario. Tesis de
doctorado. Universidad de la laguna. 226 p
Ceasar S.A., Ignacimuthu S. (2010). Effects of cytokinins , carbohydrates and amino acids
59

CAPiTULO 11
on induction and maturation of somatic embryos in kodo millet (Paspalum
scorbiculatum Linn) . Plant Cell Tiss Organ Cult. 102: 153-162.
Chia JT, Saunders JW. (1999) . Evaluation of sole nitrogen sources for shoot and leaf disc
culture of sugarbeet. Plant Cell Tissue Organ Cult. 59 : 47-56 .
Claparols 1, Santos MA and Torne ' JM (1993) . Influence of some exogenous amino acids
on the production of maize embryogenic callus and on endogenous amino acid
content. Plant Cell, Tissue and Org . Cult. 34: 1-11 .
Dal Vesco L.L. y Guerra M. P. (2001). The effectiveness of nitrogen sources in Feijoa
somatic embryogenesis . Plant Cell , Tissue and Organ Culture 64: 19-25.
Davidonis , G.H. Y Hamilton, R.H. (1983). Plant regeneration from callus tissues of
Gossypium hirsutum L. Plant Sci. Lett. 32:89-93.
EI-Hadrami l., M.P. Y J . D'Azauc (1989a) . Clonal variability of embryogenic potential in
Hevea brasiliensis: relations with polyamines and peroxidases in calli. Compt.
Ren . Acad. Sci ., 308, 299-305.
EI-Hadrami l. , N. Michaux-Ferriere, M.P. Carron , y J. D'Azauc (1989b) . Polyamines, a
posible limiting factor in somatic embryogenesis of Hevea brasiliensis. Compt.
Ren . Acad . Sci., 308, 205-211 .
EI-Hadrami, L. Y J. D'Auzac (1992). Effects of polyamine biosynthetic inhibitors on somatic ,
embryogenesis and cellular polyamines in Hevea brasiliensis. J. Plant Physiol.,
140, 33-36.
Evans P.T. Y R.L. Malmberg (1989) . Do polyamines have roles in plant development?
Plant Physiol., Annu Rev., Plant Mol. Biol., 40 , 235-269.
Feirer RP, Mignon G, Litvay JO (1984) . Arginine decarboxylase and polyamines required
for embryogenesis in wild carrot. Science 223:1433-1435
Fukunaga Y. y King J. (1982) . The effect of L-amino acids on the growth of and nitrate
reductase activity in cultured cells of Datura innoxia . Plant Science Letters . 24 :45-
60

CAPíTULO II
54 .
Galston A.W. y R. Kaur-Sawhney (1990). Polyamines in plant physiology. Plant Physiol. ,
94,406-410.
Gamborg , O. l ., Plant Physiol. , (1970) . The Effects of Aminoacids and Ammoniumon the
Growthof Plant Cells in Suspension Culture 1970, 45, 372-375 .
Gerdakaneh M., Mozafari A. y Sarabi B. (2011). Effects of different amino acids on
somatic embryogenesis of strawberry (Fragaria x ananassa Duch). Acta Phyiol
Plant . 33: 1847-1852.
González-Benito ME, Carvalho JMF-C y Pérez C. (1997) . Somatic embryogenesis of an
early cotton cultivar. Pesq. Agrop. Bras. 32(5) : 485-488.
Grimes H.D. Y Hodges, T.K. (1990). The inorganic N03': NH/ ratio influences plant
regeneration and auxin sensitivity in primary callus derived from immature
embryos of indica rice (Oryza sativa l.) . J . Plant Physiol.136: 362-367.
Guerra MP, Pescador R, Dal Vesco ll, Nodari RO y Ducroquet JPHJ. (1997). In vitro
morphogenesis in Feijoa sellowiana: Somatic embryogenesis and plant
regeneration . Acta Hort. 452: 27-35.
Halperin W. (1966). Alternative morphogenetic events in cell suspensions. Amer. J. Bot.
53(5) : 443-453.
Heimer Y.M. Y Filner P. (1970) . Regulation of the nitrate assimilation pathway of cultures
tobacco cells 11. Properties of a variant cell line. Bioch . et Biophysica Act. (BBA)
215 : 152-165.
Helleboid S., Couille J., Hilbert, J. y Vasseur J. (1995). Inhibition of direct somatic
embryogenesis by a-difluoromethylarginina in a Cichorium hybrid: effects on
polyamine content and protein patterns . Planta 196: 571-576 .
Jiménez VM . (2005) . Involvement of plant hormones and plant growth regulators on in
vitro somatic embryogenesis. Plant Growth Regul. 47 : 91-110.
61

CAPiTULO 11
John, S. J. y Guha-Mukherjee, S. (1997) en Plant Molecular Biology and Biotechnology
(eds Tewary, K. K. and Singhal , G. S.), Narosa Publishing House, New Delh i,
1997, pp. 17- 28 .
Kakkar R.K., Nagar P.K., Ahuja P.S . y Rai V.K. (2000). Polyamines and plant
morphogenesis. Biologia plantarum 43(1): 1-11 .
Kevers C., N. Le Gal, M. Monteiro, J. Dommes y T. Gaspar (2000) Somatic
embryogenesis on Panax ginseng in liquid cultures : a role for polyamine and their
metabolic pathways. Plant Growth Regul. , 31 , 209-214 .
Kumar V., A. Sharma, B.C Narasimha Prasad, H.B. Gururaj, P. Giridhar y G. Ravishankar
(2007). Direct shoot bud induction and plant regeneration in eapsicum frutescens
MilI. : influence of polyamines and polarity. Acta
Kumar V., P. Giridhar, A. Chandrashekar y G.A. Ravishankar (2008). Polyamines
influence morphogenesis and caffeine biosynthesis in in vitro cultures of eoffea
canephora P. es Fr. Acta Physiol . Plant., 30,217-223 .
Lai , F.M., Senaratna, T. and McKersie, B.O. (1992). Glutamine enchanges storage protein
synthesis in Medicago sativa L. Somatic embryos. Plant Science. 87 :6977.
Lecona-Guzmán C. (2012). Estudio de las proteínas en la embriogénesis somática de
chile habanero (eapsicum chinense Jacq.). Tesis de Doctorado. 81 p.
Lecona-Guzmán C.A., Solís Marroquín D., Avilés-Viñas S., Santos-Briones e. , Santana
Buzzy N. (2012). Changes in the protein profile of habanero pepper (eapsicum
chinense J.) somatic embryos during development. AJB . 11 (47); 10761-10768.
López-Erosa S. (2012). Efecto de poliaminas exógenas en la embriogénesis somática de
chile habanero (eapsicum chinense Jacq.). Tesis de maestría . Centro de
Investigación Científica de Yucatán . 89 p.
Marton L Y Morris O (1987). Molecular y cellular functions of the polyamines. En : PP Mc
Cann, A Pegg y A Sjoerdsma, eds . Inhibition of polyamine Metabolism , pp. 79-
105. San Diego: Academic Press.
62

CAPíTULO II
Merkle SA, Parrot WA y Flinn BS (1995). Morphogenic aspects of somatic embryogenesis.
En : Thorpe TA (ed). In vitro Embryogenesis in Plants (pp. 155-203). Kluwer
Academic Publishers, Dordrecht.
Minocha S.C., Minocha R. y Robie C.A. (1990). High-performance liquid chromatograph ic
method for the determination of dansyl-polyamines. J. Chromatogr 511 : 177 -183 .
Morcillo F., Hartmann C. Duval Y., y Tregear J. (2001). Regulation of 7S globulin gene
expression in zygotic and somatic embryos of oil palm. Physiologia Plantarum
112: 233-243.
Morris D.R. Y Jorstad C.M. (1973). Growth and macromolecular composition of a mutant of
Eseherieia eoliduring polyamine limitation. JB. 113: 271 -277 .
Muralidharan EM. Y Mascarenhas AF. (1995). Somatic embryogenesis in Eucalyptus. In :
Jain SM, Gupta PK. Y Newton RJ . (eds) . Somatic embryogenesis in woody
plants, Vol. 2, Angiosperms. Kluwer Academic Publishers, Dordrecht. 23-40 pp .
Murashige, T. and Skoog, F. (1962). A revised medium for rapid growth and bioassays
with tobacco cultures. Physiol. Plant. 15:473-479.
Nanjo T., Kobayashi M., Yoshiba Y., Sanada Y., Wada K., Tsukaya H., Kakubari Y.,
Yamaguchi-Shinozki K. y Shinozaki K., (1999) Biological functions of proline in
morphogenesis and osmotolerance revealed in antisense transgenic Arabidopsis
tha/iana. Plant J. Cell Mol. Biol. , 18, 185- 193.
Nuutila, A.M ., Hamalainen , J. and Mannonen, l . (2000). Optimization of media nitrogen
and copper concentrations for regeneration of green plants from polyembryogenic
cultures of barley (Hordeum vulgare l.) . Plant Science. 151 : 85-92 .
Ozawa K, Ling D-H y Komamine A. (1996). High-frecuency somatic embryogenesis from
suspension- cultu red clusters of cells of an interspecific hybrid of Oryza. Plant Cell
Tiss. Org. Cult. 46: 157-159.
Patil P. , R. Chandra , S. Khetarpal , y R. Plisetty (1998). Influence of polyamine and
ethylene inhibitors on somatic embryo induction in ch ick pea (Cieer arietinum L.)
63

CAPiTULO "
Plant Physiol. , 101 , 213-219.
Paul A., K. Mitter y S. Sen-Raychaudhuri (2009) . Effect of polyamines on in vitre somatic
embryogenesis in Momordica charantia L. Plant Cell Tiss . Organ . Cult. , 97 , 303-
311 .
Pegg A (1986). Recent advances in the biochemistry of polyamines in eukaryotes .
Biochem J 234: 249-262.
Rastogi R. Y Davies P.J. (2000) . Effects of light and plant growth regulators on polyamine
metabolism in higher plants. In : Slocum R.D . y Flores H.E. (eds) . Biochemistry
and physiology of polyamines in plants (pp. 187-199). CRC Press, Boca Raton .
Regla-Márquez C.F (2011) . Estudio de las poliaminas durante el proceso de
embriogenesis somática de ch ile habanero (Capsicum chinense Jacq). Tesis de
maestria . Centro de Investigación Científica de Yucatán . México. 59 p.
Reinert J. , Tozawa M. y Semeroff S. (1967) . Nitrogen compounds as factors of
embryogenesis in vitro. Nature 216 : 1215-1216.
Ronch i, V. N., Caligo, M. A , Nozzolin i, M. and Luccarini, G. (1984). Stimulation of carrot
somatic embryogenesis by proline and serine. Plant Cell Rep. , 1984, 3, 210.
Santana-Buzzy N.,Canto-Flick A., Barahona-Pérez F., Montalvo-Peniche M., la pata
Castillo P., Solís-Ruiz A., laldívar-ColIí A , Gutiérrez-Alonso O., and Miranda
Ham M. 2005 . Regeneration of Habanero Pepper (Capsicum chinense Jacq .) Via
Organogenesis Hortscience 40(6) : 829-1831 .
Santanen A Y Simola L. (1999). Metabolism of L(U-14C)-arg inine and L(U_14C)-orn ith ine in
maturing and vernalized embryos and megagametophytes of Picea abies.
Physiolog ia Plantarum 107: 433-440.
Sarker, K. K., Kabir, A H., Sharm in, S. A , Nasir, l ., & Alam, M. F. (2007) . Improved
somatic embryogenesis using L-asparagine in wheat (Triticum aestivum L.).
Sjemenarstvo, 24(3-4) , 187-196.
64

CAPiTULO 11
Schmidt MA, Tucker D.M., Cahoon E.B. y Parrot W.A. (2005) . Towards normalization of
soybean somatic embryo maturation . Plant Cell Rep. 24: 383-391.
Sen J., Kalia S. y Guha-Mukherjee S. (2002). Level of endogenous free amino acids
during various stages of culture of Vigna mungo (L.) Hepper-somatic
embryogenesis , organogenesis and plant regeneration . Curro Sci. 82: 429-433 .
Serafini-Fracassini D., S. Del Duca y P. Torrigiani (1989) . Polyamine conjugation during
the cell cycle of He/ianthus tuberosus: non-enzymatic and transglutaminase-like
binding activity . Plant Physiol. Biochem ., 27 , 659-668.
Silveira V., Floh E. 1. S., Handro W. y Guerra M.P. (2004). Effect of plant growth regulators
on the cellular growth and levels of intracellular protein, starch and polyamines in
embryogenic suspension cultures of Pinus taeda. Plant Cell, Tissue and Organ
Culture 76: 53-60 .
Silveira V., Santa-Catarina C., Tun N.N., Scherer G.F .E., Handro W. y Guerra M.P. et.
(2006). Polyamine effects on the endogenous polyamine contents, nitric oxide
release, growth and differentiation of embryogenic suspension cultures of
Araucaria angustifo/ia (Bert .) O. Ktze. Plant Sci. 171 : 91-98.
Steiner N., C. Santa-Catarina, V. Silveira, E.I.S . Floh y M.P. Guerra (2007). Polyamine
effects on growth and endogenous hormones levels in Araucaria angustifolia
embryogenic cultures . Plant Cell Tiss. Organ . Cult., 89, 55-62.
Tabor C.W. y H. Tabor (1984). Polyamines. Ann. Rev. Biochem ., 53, 749-790 .
Tiburcio A., Campos J. Figueras X. y Besford R. (1993) . Recent advances in the
understanding of polyamine functions during plant development. Plant Growth
Regulation 12: 331-340.
Trigiano R.N. Y Conger B.v. (1987). Regulation of Growth and somatic embryogenesis by
proline and serine in suspension cultures of Dacty/is g/omerata. Journal of Plant
Physiology. 130: 49-55.
Trolinder N.L., Goodin J. R. (1988b). Somatic embryogenesis in cotton . 11. Requirements
65

CAPíTULO 11
for embryo development and plant regeneration. Plant Cell Tiss . Organ Cult.
12:43-53.
Tupy, J., Hrabetova, E. and Capkova , V. (1983). Amino acids and bivalent cations in the
growth of tobacco pollen in mass culture . , Plant Sci . Lett., ,30,91-98.
Wilkinson T.C. y Thomson S.A. (1987). Genotype, medium and genotype X medium
effects on the establishment of regenerable maize callus. Madyca 32: 89-105.
Witjaksono y Litz RE . (1999) . Maturation of avocado somatic embryos and plant recovery .
Plant Cell Tissue Organ Cult. 58: 141-148.
Wu J., Zhang X., Nie Y., Jin S. y Liang S. (2004). Factors affecting somatic embryogenesis
and plant regeneration from a range of recalcitrant genotypes of chinese cottons
(Gossypium hirsutum L.). In vitro Cell. Dev. Biol.-Plant 40: 371-375.
Young D. Y Srinivasan P. (1972) . Regulation of macromolecular synthesis by putrescine in
a conditional Escherichia coli putrescine auxotroph . J. Bacteriol 112: 30-39.
Young BG, Jack DL, Smith DW y Saier Jr. MH . (1999) . The amino acid/auxin : proton
symport permease family. Biochim. Biophys . Acta 1415: 306-322.
66

CAPíTULO 111
CAPíTULO 111
EFECTO DE ESPERMINA EN lA ACUMULACiÓN Y COMPOSICiÓN DE
PROTEíNAS DE lOS ESs DE C. chinense
3.1 INTRODUCCiÓN
El crecimiento de los cultivos embriogénicos está asociado generalmente con cambios en
la síntesis y movilización de proteínas, carbohidratos y lípidos . los niveles de estas
moléculas depende de las diferentes fases de crecimiento (lulsdorf et al., 1992), y
funcionan como moléculas regulatorias o como suplemento de energía para el crecimiento
celular (Nomura y Komamine, 1995).
Durante las etapas de histodiferenciación-maduración, los ESs acumulan moléculas de
reserva (Von et al. , 2002) similares a las de los ECs. Entre éstas, las proteínas son unas
de las más importantes y por ello , unas de las más estudiadas en términos de niveles y
patrones de expresión (Roja et al., 2005; Brownfield et al., 2006) .
las proteínas influencian directamente la bioquímica celular y proveen un análisis más
preciso de los cambios ocurridos durante el crecimiento y desarrollo celular (Chen y
Harmon, 2006). los primeros estudios sobre la acumulación y composición de proteínas
endógenas de ESs de chile habanero realizados por Lecona et al., 2012 , revelaron que
hay una disminución crítica en los contenidos de proteínas totales a medida que los
embriones avanzan en su desarrollo, pasando de contener 2.9 f..lg .mg-1 (PF) en el estadío
globular a 0.4 g.mg-1 (PF) de proteínas en el estadío cotiledonar. Este comportamiento es
contrario a lo reportado para diferentes especies vegetales en los que el contenido de
proteínas se incrementa durante su desarrollo (Lai et al., 1992; Stejskal, 1994; y Griga et
al., 2007). De igual manera, Lecona et al. (2012) encontraron 9 bandas en el perfil
protéico de los ECs que no se observaron en el perfil de los ESs. Cinco de estas bandas,
tuvieron idéntico peso molecular (PM) al de proteínas de reserva reportadas en otras
especies vegetales. Así mismo, el análisis proteico en 2D mostró marcadas diferencias
entre los grupos de proteínas expresadas en los ECs maduros y ESs cotiledonares de C.
chinense. La deficiencia en el conten ido y composición de proteínas en la etapa
67

CAPíTULO 111
cotiledonar de los ESs, sug iere que este factor es una de las posibles causas que impiden
el correcto desarrollo y germinación de los embriones somáticos de ch ile habanero.
las PAs regulan la síntesis de proteínas (Young y Srinivasan , 1972; Morris y Jorstad ,
1973; Tabor y Tabor, 1984; Marton y Morris, 1987; Evans y Malmberg , 1989) y están
involucradas en las mod ificaciones postraduccionales (Tabor y Tabor, 1984; Pegg , 1986).
Así mismo, las interacciones de las PAs y sus derivados con las proteínas, pueden ser
críticas para la división y diferenciación celular (Santanen y Simola , 1999). Esto permite
suponer que la adición de Spm 0.1 mM (tratamiento utilizado en este estudio) pudiera
estar provocando cambios en el conten ido y composición de proteínas totales en los ESs
de ch ile habanero. Por consiguiente, en este estudio se realizó el anális is del conten ido y
composición de proteínas totales de los ESs de esta especie en estad íos avanzados de
desarrollo adicionados con Spm 0.1 mM y se comparó con los ESs avanzados
provenientes del tratam iento testigo (TO), EC maduro y la semilla de chile habanero.
3.2 MATERIALES Y MÉTODOS
la técnica de isoelectroenfoque (pi) para los análisis proteicos en pl-SDS-PAGE se
realizó siguiendo una modificación del protocolo descrito en el manual ZOOM IPGRunner
System for isoelectric focusing of ZOOM Strips, catálogos ZM0001 , ZM0002, ZM0004 . la
modificación ha sido descrita por Islas-Flores y Brito-Argáez (2014).
3.2.1 Extracción de proteínas totales
las muestras de sem illa, embrión cigótico maduro, ESs avanzados tratados con 0.1 mM
Spm y ESs avanzados del tratamiento testigo de chile habanero, se liofilizaron y
posteriormente se maceraron en presencia de nitrógeno líquido hasta obtener un polvo
fino. Inmediatamente se añadió amortiguador de extracción Tris 50 mM pH 7.4 adicionado
con 10 mM de ácido ascórbico, 50 mM de NaCI, 10% de Glicerol, 1 mM de EDTA, 1 mM
de EGTA e inhibidores de proteasas (Cocktail Complete TM-Roche) hasta formar una
pasta. l a relación de extracción fue 1: 1 (1 g de tej ido por mi de amortiguador de
extracción); los extractos se centrifugaron a 14,000 x g durante 15 minutos a 4 oC. El
sobrenadante se transifirió a un nuevo tubo Eppendorf y la pastilla se resuspend ió con
amortiguador de extracción ; posteriormente se centrifugó a 14,000 x g durante 15 mino a 4
68

CAPíTULO 111
OC . El sobrenadante se recuperó y se mezcló con el recuperado en la primera
centrifugación. Se tomaron 40 11-1 del sobrenadante para cuantificar la concentración de
proteína en cada una de las cuatro muestras por triplicado. El extracto restante se
conservó a - 20 oC hasta su uso.
3.2.2 Determinación del contenido de proteínas
la proteína soluble presente en las diferentes muestras se determinó siguiendo el método
de Bradford (1976). Para realizar la curva patrón de proteínas se utilizó como estándar
albúmina de suero bovino (BSA) en concentraciones de: O, 1, 2.5, 5.0, 7.5, 10, 15 Y 20 f.l.g
Y para preparar las muestras, se utilizó 5 11-1 de cada extracto protéico, el volumen se
ajustó con agua destilada a 800 11-1 Y se adicionó 200 11-1 del reactivo de Bradford comercial
marca Bio-Rad . Cada muestra se analizó por triplicado. la absorbancia de la curva patrón
y de las muestras se determinó a 595 nm; utilizando un espectrofotómetro Genesys 5 UV
VIS.
3.2.3 Determinación de la composición de proteínas por SDS-PAGE
Para analizar el perfil de proteína total presente en los ESs de chile habanero tratados con
y sin Spm 0.1 mM; en la semilla y el EC maduro de esta especie ; las proteínas se
separaron mediante SDS-PAGE (electroforesis en geles de poliacrilamida con dodecil
sulfato de sodio). Brevemente, se prepararon geles de poliacrilamida al 12% (separación)
y 4% (concentrador) . Los geles se colocaron en un equipo de electroforesis Bio-Rad, el
voltaje se proporcionó mediante una fuente Power Pac 3000. las muestras de proteína
(30 I1-g) se adicionaron con 8 11-1 de amortiguador de laemmli 5x, el volumen se completó
hasta 40 ~d con agua destilada. Se mezcló generosamente y enseguida, a la mezcla se le
dió un pulso de centrífuga a 14,000 x g, se calentó a 95 oC por 5 min o Inmediatamente las
muestras se colocaron en hielo por 3 min y se les dió un pulso de centrifuga a 14,000 x g.
El marcador de peso molecular fue del kit Mark 12™ (Invitrogen Cat. lC5677), se
aplicaron 6 I1-I/gel.
El marcador y las muestras se aplicaron por separado en los pozos del gel de apilamiento.
Se utilizó como amortiguador de corrida 1x Trizma base 0.025 M, Glicina 0.19 M, SDS
0.05% . La electrofores is se realizó a 100 volts por 1:45 h. a temperatura ambiente.
69

CAPíTULO 111
Finalizada la electroforesis, el gel se tiñó con una solución 0.25% de azul brillante de
Coomasie en 20% metanol y 10% Ácido acético, en agitación toda la noche. Para desteñir
el gel , éste se adicionó con una solución compuesta de 25% de metanol y 10% de ácido
acético . El perfil electroforético se fotodocumentó con una cámara Nikon COOLPIX S3000
para su posterior análisis.
3.2.4 Determinación de la composición de proteínas por 2D-PAGE
La variación en la composición de proteínas también fue anal izada mediante la técnica de
Isoelectroenfoque (pi) y electroforesis en gel de poliacrilamida con dodecil sulfato de sodio
(IEF/SDS-PAGE), utilizando tiras de isoelectroenfoque y la cámara de electroforesis
ZOOM® IGPRUNNER n, System .
La muestra restante del proceso de extracción (1 mi) almacenada a -80 oC , se descongeló
y se precipitó adicionándole 500 ~d de ácido tricloroacético (10%) y 3 ¡.,t.1 de ~
mercaptoetanol (0.07%), enseguida la mezcla se aforó a 5 mi con acetona al 100%. La
muestra se incubó durante 2 h. a - 20 oC. La mezcla se centrifugó a 16,000 x g durante 20
min a 4 oC. Se descartó el sobrenadante, la pastilla se lavó tres veces con 1 mi de
solución de lavado (acetona al 100%, adicionada con 0.07% de ~-mercaptoetanol)
centrifugando a 16,000 x g durante 20 min a 4 oC cada vez. El sobrenadante se desechó.
La pastilla se dejó secar por 2 min a 25 oC y se resuspendió con agua ultrapura; de tal
manera que al cargar 10 ¡.,t.1 de muestra en los geles este volumen debía contener 15 ¡.,t.g ó
20 ¡.,t.g de proteína total según el caso. Las muestras se sonicaron por 5 min a temperatura
ambiente.
Las tiras de isoelectroenfoque se colocaron en el cassette (ZOOM- IPGRUNNER) y se
hidrataron en una mezcla de proteína (1 O ~d) adicionada con 140 ¡.,t.1 de amortiguador de
lisis (8M Urea, 2% de IGPAL CA-640, 2% anfolitos (1 .6% del rango 3-10) más (0.4% del
rango 5-7) , 4 ¡.,t.1 de 10 mM de ditiotreitol y 1.5 mg de azul de bromofenol). Las tiras se
dejaron hidratar toda la noche.
Los cassettes IGPRUNNER conteniendo las tiras hidratadas, se colocaron en la cámara
de isoelectroenfoque sobre la base de corrida (ZOOM® IGPRUNNER Core) y se
introdujeron en la cámara Mini-Cell Chamber. Armada la cámara, en la misma se vertieron
70

CAPíTULO III
600 mi de agua desionizada. Posteriormente se realizó el isoelectroenfoque
incrementando el voltaje de la siguiente manera : 200 volts por 20 min, 450 volts por 15
min, 750 volts por 20 min y 2000 volts por 2 h. Concluido el isoelectroenfoque, las tiras se
introdujeron en tubos de ensaye con 1 mi de amortiguador de laemmli 1X y se
equilibraron en agitación suave durante 15 min a 25°C.
la segunda dimensión se realizó en geles de poliacrilamida al 12%. Brevemente, las tiras
conteniendo a las proteínas isoelectroenfocadas, se colocaron sobre el gel de
apilamiento, orientando el lado ácido (+) de la tira hacia la posición del carril donde fue
aplicado el marcador de peso molecular de proteínas (200,000-21,500 da). Se vertió
agarosa líquida al 0.5 % con amortiguador de corrida 1 X (Trizma base 0.025 M, Glicina
0.19 M, SOS 0.05 %) sobre la tira y se dejó solidificar. Una vez que la tira quedó
inmovilizada, se agregó amortiguador de laemmli 1X (Tris-HCI 60 mM pH 6.8, 2% SOS,
10% glicerol , 5% p-mercaptoetanol , 0.01 % Azul de bromofenol), sobre la agarosa. En una
poceta separada del resto del gel , se adicionó el marcador de peso molecular y se
cubrieron los carriles con amortiguador de corrida, la electroforesis se desarrolló a 100
volts durante 2 h. a temperatura ambiente. Concluida la electroforesis, los geles fueron
teñidos con nitrato de plata según el procedimiento reportado en Islas-Flores y Brito
Argáez (2014).
los patrones de proteína en los geles fueron registrados fotográficamente con una
cámara Nikon COOlPIX S3000 y evaluados comparativamente entre sí.
3.3 RESULTADOS
3.3.1 Efecto de Spm en el contenido proteico de los ESs de Capsicum chinense
En la figura 3.1, se muestra la cuantificación de proteínas extraídas de los ESs tratados
con y sin 0.1 mM Spm, y del embrión cigótico maduro de la misma especie. El EC
presentó el mayor contenido de proteínas con 57.71 ± 4.7 f.lg/mg peso seco (PS),
superando significativamente a los ESs tratados con y sin Spm en un 90.6% y 86 .3%,
respectivamente. No se encontró diferencia significativa en la acumulación de proteínas
entre los ESs del TO y los ESs tratados con Spm . Debido a la diferencia en el contenido
proteico entre el EC y los ESs de chile habanero , se decidió comparar la cantidad de
71

CAPíTULO 111
proteína de los ESs respecto a la del hipocotilo en inducción. El resultado mostró que los
ESs tratados con Spm tuvieron un decremento en contenido de proteína (5.4 ± 0.4 !!g/mg
PS) respecto a los ESs sin tratar (ESs TO), de los cuales se obtuvo la cantidad de 7.92 ±
0.2 !!g/mg PS, difiriendo significativamente entre sí.
72
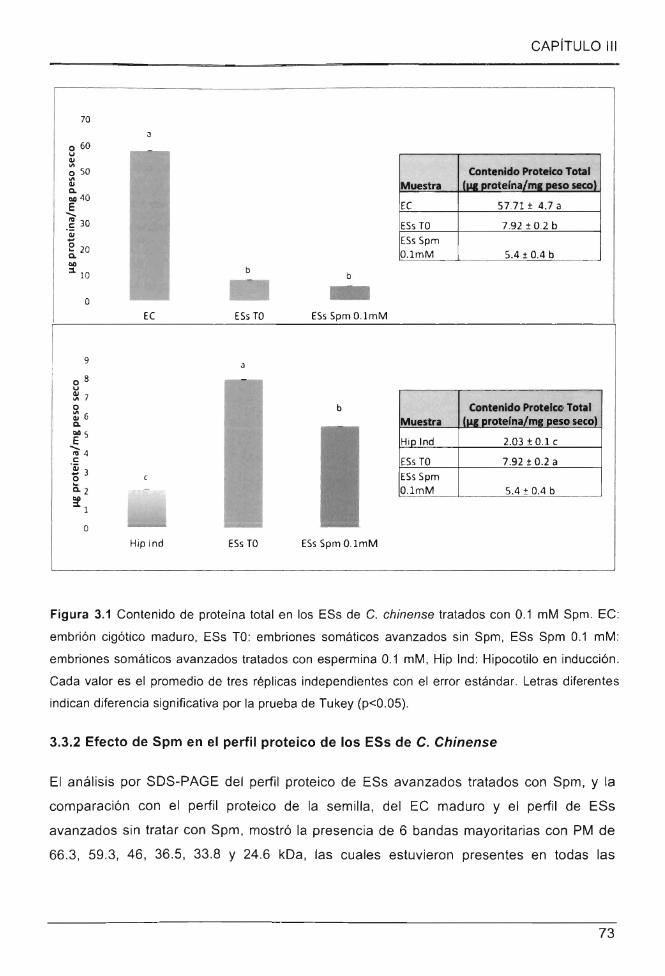
CAPiTULO 111
70
O 60 ... 111 VI
O 50 Contenido Proteico Total VI 111 o. Muestra (IJ,I protelna/ml peso seco) 1>0 40 E EC S7.71 ± 4 .7 a ..... ~ 30 .~ ... ~ 20 o.
ESs ro 7.92 ± 0.2 b ESs Spm O.lmM 5.4 ± 0.4 b
1>0 b :1 10 b
O
EC ESs ro ESs Spm O.lmM
9
0 8 ... ~ 7 O b ~ 6 o.
Contenido Proteico Total Muestra I (IJ,I protelna/ml peso seco)
e5 ..... Hip Ind 2.03 ± 0.1 e
1\1 4 e
] 3 O ...
ESs ro 7 .92 ± 0.2 a
ESs Spm 0. 2 O.lm M 5.4 ± 0.4 b
~ l
O
Hip Ind ESs ro ESs Spm O.lm M
Figura 3.1 Contenido de proteína total en los ESs de C. chinense tratados con 0.1 mM Spm. EC:
embrión cigótico maduro, ESs TO: embriones somáticos avanzados sin Spm, ESs Spm 0.1 mM:
embriones somáticos avanzados tratados con espermina 0.1 mM, Hip Ind : Hipocotilo en inducción.
Cada valor es el promedio de tres réplicas independientes con el error estándar. Letras diferentes
indican diferencia significativa por la prueba de Tukey (p<0.05).
3.3.2 Efecto de Spm en el perfil proteico de los ESs de C. Chinense
El análisis por SDS-PAGE del perfil proteico de ESs avanzados tratados con Spm, y la
comparación con el perfil proteico de la semilla , del EC maduro y el perfil de ESs
avanzados sin tratar con Spm, mostró la presencia de 6 bandas mayoritarias con PM de
66.3, 59.3, 46 , 36 .5, 33.8 Y 24.6 kDa, las cuales estuvieron presentes en todas las
73

CAPITULO 111
muestras analizadas (fig . 3.2, marcas rojas). El perfil proteico de la semilla contuvo 17
bandas que corresponden a péptidos con PM ubicados entre 14.4 y 91 .2 kDa. Cinco de
estos péptidos (87, 41.5, 32 .6, 18 Y 14.4 kDa) fueron exclusivos de semilla (fig . 3.2,
flechas naranjas).
El perfil del EC reveló 16 bandas con PM entre 21 .9 y 200 kDa. Tres de estos péptidos
con PM de 80.8, 62.4 Y 29 kDa , fueron específicos del EC (fig. 3.2, flechas verdes) . El
perfil proteico de la semilla y del EC mostraron 6 bandas comunes cuyos PM fueron de
91.2 , 72.5, 64.7,52.9,27.5 Y 21 .9 kDa (fig. 3.2, flechas negras).
KDa M
200
116.3 97.4
66.3
55.4
36.5
31
21.5
14.4
5 EC ro 5pm
100
JI .l ... ,. , 11.5 .. , 64' n. S'l 52,'
.. O
•• '1.1 , .. ns
)]1
JI .4
" , .. n .s
lO.
21..9 lLS
, ..
Figura 3.2 Perfil protéico en 12 % SDS-PAGE de embriones somáticos a los 45 dias de cultivo en
medio líquido. S: Semilla de C. chinense; EC: Embrión cigótico maduro; TO: ESs avanzados del
tratamiento testigo; Spm: ESs avanzados tratados con Spm 0.1 mM; M: Marcador de peso
molecular.
74

CAPíTULO 111
En lo que respecta a los ESs , se encontró el mismo número de bandas (14) tanto en los
ESs tratados con Spm como en ESs sin tratar cuyo rango fue de 21 .5 a 200 kDa. Dos de
esas bandas con PM de 21 .5 y 24.6 kDa fueron más abundantes en los ESs sin tratar (TO)
que en los ESs con Spm. los ESs, tanto tratados con Spm como sin tratar, mostraron 7
péptidos específicos con pesos moleculares de 78 .7, 51 .6, 42.8, 39, 31.4, 28 .6 Y 21.5 kDa
(fig. 3.2, flechas azules) que no estuvieron presentes en EC ni en semilla de C. chinense.
Una banda con PM de 200 kDa, se observó en el EC así como en los ESs con y sin Spm,
pero no en el perfil de la semilla .
3.3.3 Análisis de la composición de proteínas por 2D-PAGE
La composición de proteinas (15 iJg) de las muestras de la sem illa y el EC de chile
med iante Isoelectroenfoque-SDS-PAGE, inicialmente se analizaron en un gradiente lineal
de pH 3-10 ; Y la segunda dimensión en presencia de SOS empleándose para ello un gel
de poliacrilamida al 12% (F ig. 3.3 Y 3.4) . Se observó que las manchas de proteína
resueltas se concentraron en un rango de pH de 5 a 7, razón por la cual se decidió que
para los sigu ientes ensayos se utilizarían tiras de isoelectroenfoque con un rango de pH
4-7, esperando obtener con ello , una mejor separación de las proteínas observadas.
75
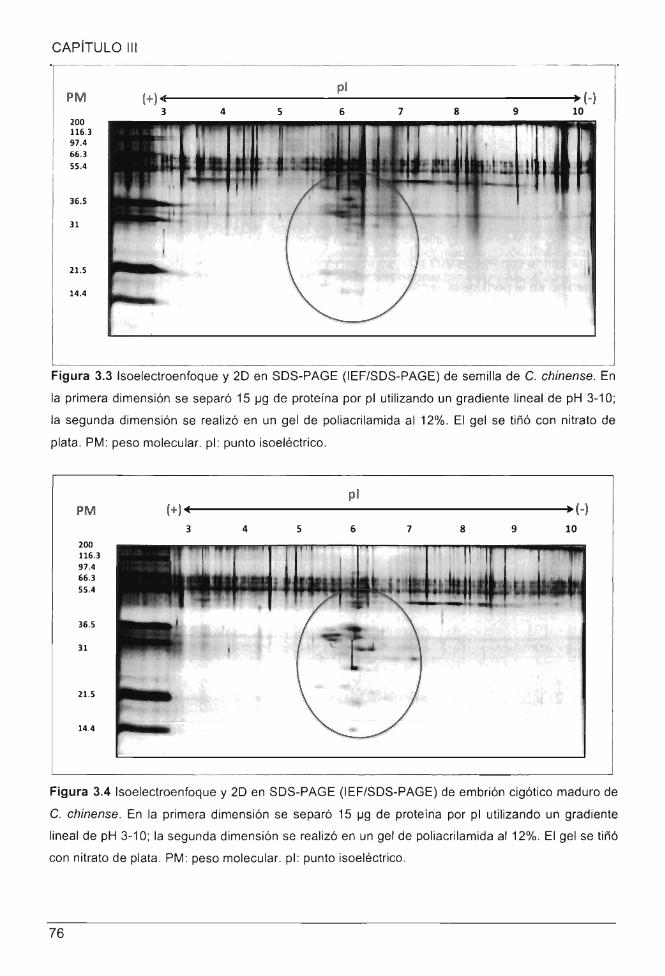
CAPiTULO 111
PM
200 116.3 97.4 66.3 55.4
36.5
31
21 .5
14.4
pi (+)+I(---------------------+~ (-)
3 4 S 6 7 8 9 10
Figura 3.3 Isoelectroenfoque y 2D en SDS-PAGE (IEF/SDS-PAGE) de semilla de C. chinense. En
la primera dimensión se separó 15 IJg de proteína por pi utilizando un gradiente lineal de pH 3-10;
la segunda dimensión se realizó en un gel de poliacrilamida al 12%. El gel se tiñó con nitrato de
plata . PM: peso molecular. pi : punto isoeléctrico.
pi PM (+) 1( ~ (-)
3 4 S 6 7 8 9 10
200 116.3 97.4 66.3 55.4
36.5
31
21.5
14.4
Figura 3.4 Isoelectroenfoque y 2D en SDS-PAGE (IEF/SDS-PAGE) de embrión cigótico maduro de
C. chinense. En la primera dimensión se separó 15 IJg de proteína por pi utilizando un gradiente
lineal de pH 3-10; la segunda dimensión se realizó en un gel de poliacrilamida al 12%. El gel se tiñó
con nitrato de plata . PM : peso molecular. pi : punto isoeléctrico.
76
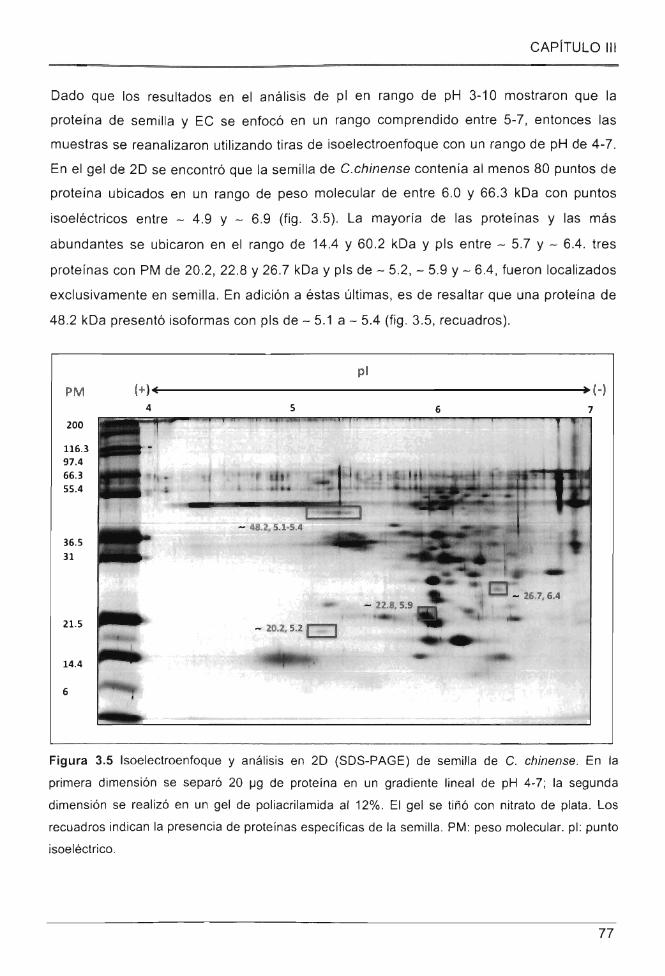
CAPiTULO III
Dado que los resultados en el análisis de pi en rango de pH 3-10 mostraron que la
proteína de semilla y EC se enfocó en un rango comprendido entre 5-7, entonces las
muestras se reanalizaron utilizando tiras de isoelectroenfoque con un rango de pH de 4-7.
En el gel de 2D se encontró que la semilla de C.chinense contenía al menos 80 puntos de
proteína ubicados en un rango de peso molecular de entre 6.0 y 66 .3 kDa con puntos
isoeléctricos entre - 4.9 Y - 6.9 (fig. 3.5) . la mayoría de las proteínas y las más
abundantes se ubicaron en el rango de 14.4 y 60.2 kDa y pis entre - 5.7 Y - 6.4. tres
proteínas con PM de 20.2 , 22 .8 Y 26.7 kDa y pis de - 5.2, - 5.9 Y - 6.4, fueron localizados
exclusivamente en semilla. En adición a éstas últimas, es de resaltar que una proteína de
48.2 kDa presentó isoformas con pis de - 5.1 a - 5.4 (fig . 3.5, recuadros).
PM
200
116.3 97.4 66.3 55.4
36.5 31
21.5
14.4
6
pi
(+)+4----------------------... (-) 4 S 6 7
- 20.2, 5.2 G:J
Figura 3.5 Isoelectroenfoque y análisis en 20 (SOS-PAGE) de semilla de C. chinense. En la
primera dimensión se separó 20 I-Ig de proteína en un gradiente lineal de pH 4-7; la segunda
dimensión se realizó en un gel de poliacrilamida al 12%. El gel se tiñó con nitrato de plata . Los
recuadros indican la presencia de proteínas específicas de la semilla . PM : peso molecular. pi : punto
isoeléctrico.
77

CAPíTULO III
El perfil proteico del EC (fig . 3.6) , mostró un total de 162 puntos de proteína, ubicadas en
un rango de 5.8 y 116.3 kDa y pi entre - 4.1 Y - 7. la mayoría de las proteínas y con
mayor abundancia se localizaron en un rango entre 13.3 y 58 .7 kDa y pi entre - 5.3 Y -
6.7. Así mismo, se observaron 30 puntos de proteína que se presentaron solamente en el
perfil proteico del EC (cuadro 3.1 y fig . 3.6, recuadros) .
78
Cuadro 3.1 Peso molecular (PM) y punto isoeléctrico (p i) de proteínas específicas
del perfil proteico del EC de C. chinense.
Embrión cigótico
PM (kDa) pi
116.3 - 5.58
69.1 - 5.30
66.3 - 5.27
64.1 - 5.27
59.8 - 6.88
69 .1 - 5.30
66 .3 - 5.27
64.1 - 5.27
59.8 - 6.88
57.6 - 6.87
48 .8 - 5.54
46 - 5.67
44 .1 - 5.67
41 .2 - 5.91
39.3 - 4.76
39.3 - 5.60
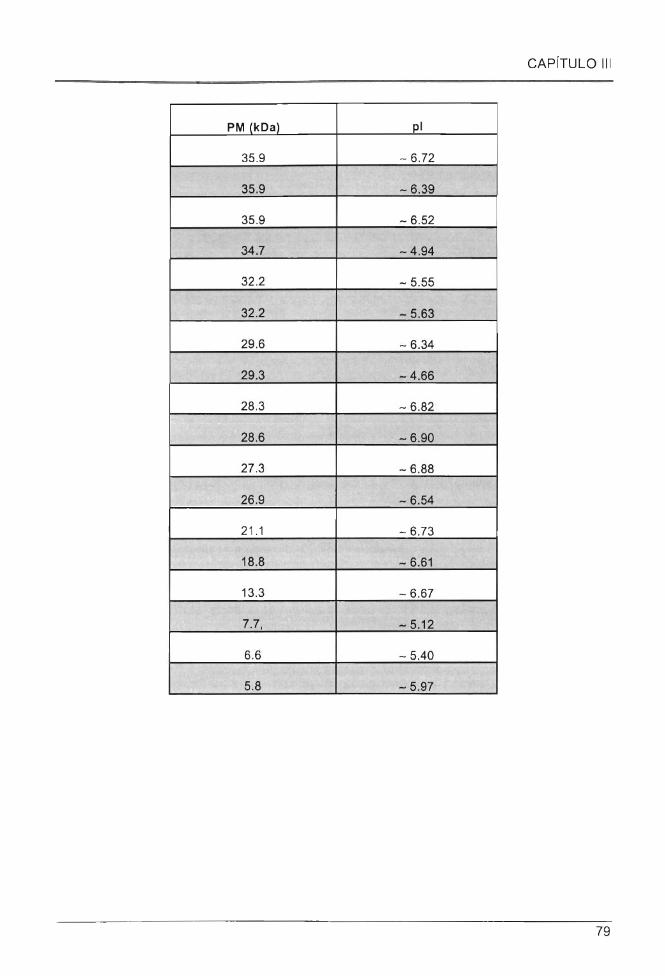
CAPíTULO 111
PM (kDa) ~I
35.9 - 6.72
35.9 - 6.39
35.9 - 6.52
34.7 - 4.94
32.2 - 5.55
32.2 - 5.63
29 .6 - 6.34
29.3 -4.66
28 .3 - 6.82
28.6 - 6.90
27 .3 - 6.88
26.9 - 6.54
21.1 - 6.73
18.8 - 6.61
13.3 - 6.67
7.7, - 5.12
6.6 - 5.40
5.8 - 5.97
79
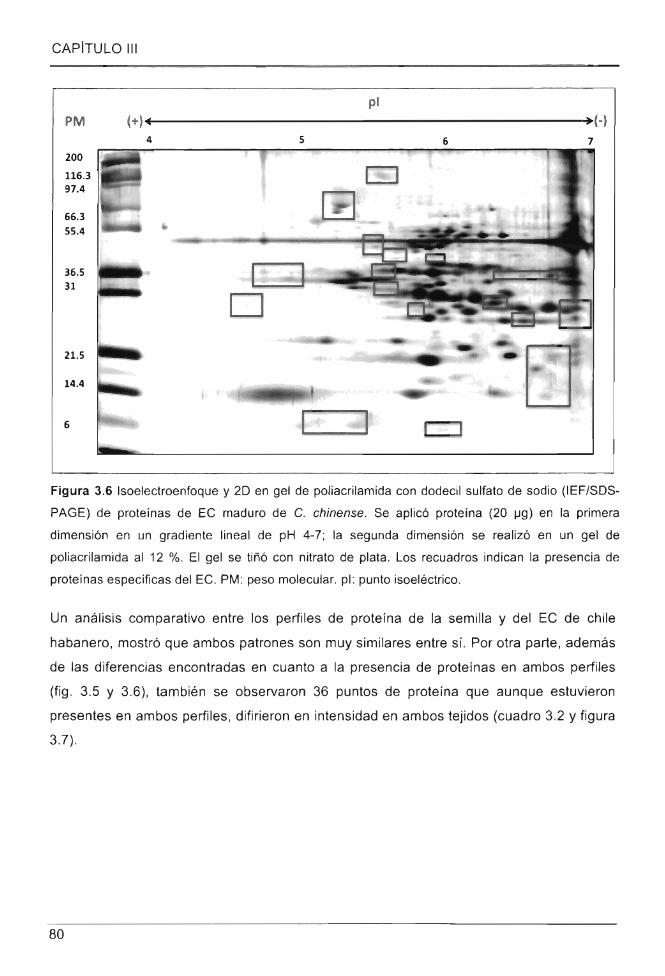
CAPiTULO 111
pi
PM +----------------------+-(-) (+) e 4 5
200
116.3 97.4
66.3 55.4
36.5 I 31
D 21.5
14.4
6
Figura 3.6 Isoelectroenfoque y 20 en gel de poliacrilamida con dodecil sulfato de sodio (IEF/SOS
PAGE) de proteínas de EC maduro de C. chinense. Se aplicó proteína (20 IJg) en la primera
dimensión en un gradiente lineal de pH 4-7; la segunda dimensión se realizó en un gel de
poliacrilamida al 12 %. El gel se tiñó con nitrato de plata . Los recuadros indican la presencia de
proteínas específicas del EC. PM: peso molecular. pi : punto isoeléctrico.
Un análisis comparativo entre los perfiles de proteína de la semilla y del EC de chile
habanero, mostró que ambos patrones son muy similares entre sí. Por otra parte, además
de las diferencias encontradas en cuanto a la presencia de proteínas en ambos perfiles
(fig . 3.5 Y 3.6), también se observaron 36 puntos de proteína que aunque estuvieron
presentes en ambos perfiles, difirieron en intensidad en ambos tejidos (cuadro 3.2 y figura
3.7).
80
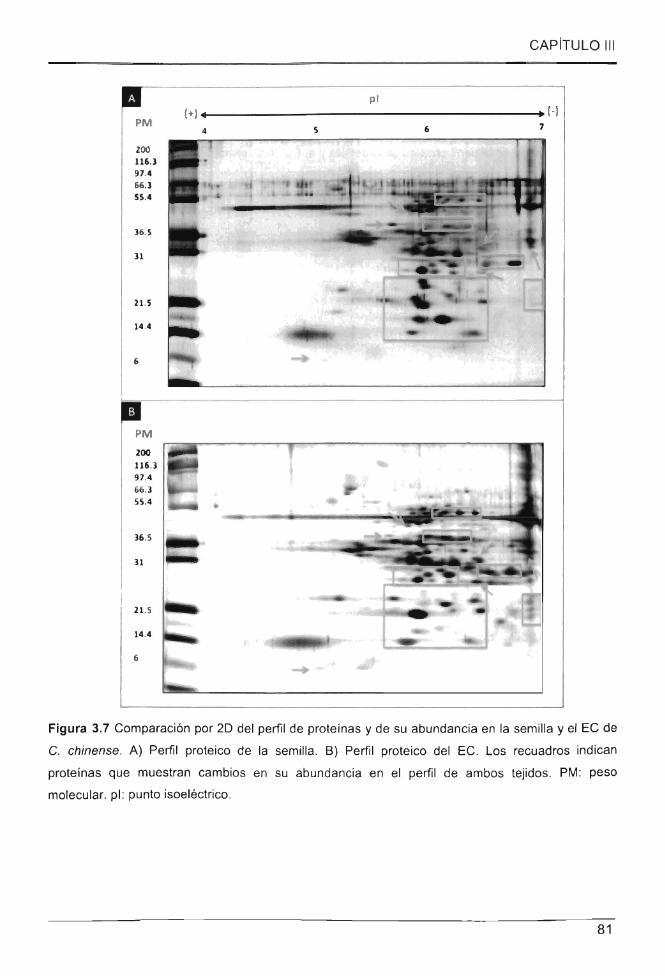
CAPiTULO 111
pi
(+1 4 ~ H PM
6 7 41 5
200 116.3 97.4 66.3 55.4
36.5
31
21 5
144
Figura 3.7 Comparación por 20 del perfil de proteínas y de su abundancia en la semilla y el EC de
C. chinense. A) Perfil proteico de la semilla . B) Perfil proteico del EC. Los recuadros indican
proteínas que muestran cambios en su abundancia en el perfil de ambos tejidos. PM: peso
molecular. pi : punto isoeléctrico.
81
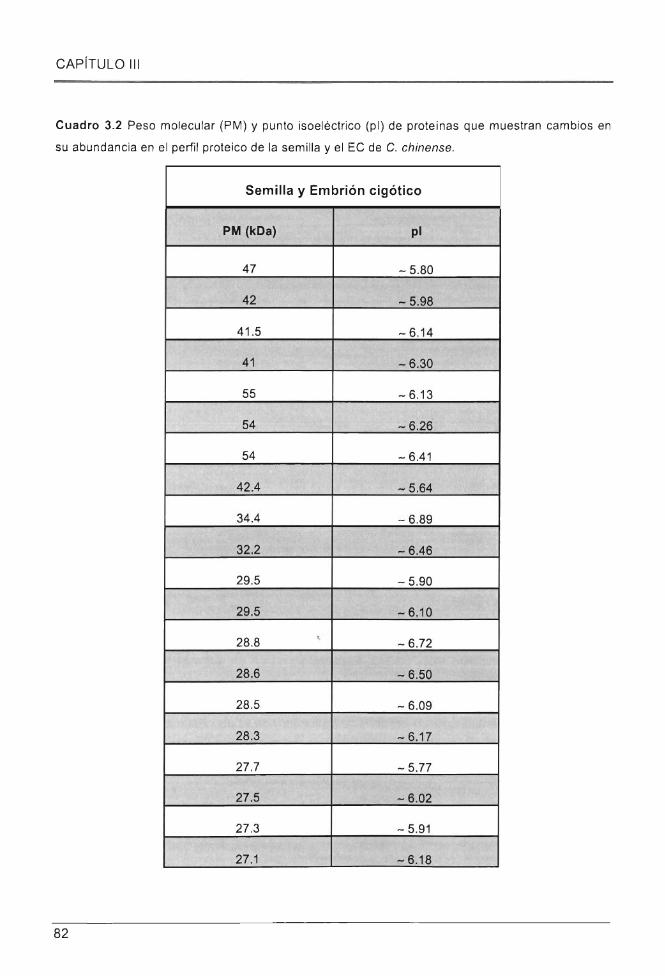
CAPíTULO 111
Cuadro 3.2 Peso molecular (PM) y punto isoeléctrico (pi) de proteínas que muestran cambios en
su abundancia en el perfil proteico de la semilla y el EC de C. chinense.
Semilla y Embrión cigótico
PM (kDa) pi
47 - 5.80
42 - 5.98
41 .5 - 6.14
41 - 6.30
55 - 6.13
54 - 6.26
54 - 6.41
42.4 - 5.64
34.4 - 6.89
32.2 - 6.46
29.5 - 5.90
29.5 - 6.10
28 .8 - 6.72
28.6 - 6.50
28 .5 - 6.09
28 .3 -6.17
27 .7 - 5.77
27 .5 - 6.02
27 .3 - 5.91
27.1 - 6.18
82

CAPiTULO 11 1
PM (kDa) pi
27 .1 - 6.40
25.5 - 5.94
24.7 - 5.95
23.8 - 6.42
23 .2 - 6.88
23 .2 - 6.14
21 .8 - 6.88
21 .8 -6.46
21 .5 - 6.19
20.8 - 6.90
20 .6 - 5.92
17.6 - 6.08
17.1 - 5.89
14.6 - 6.36
14.2 - 5.81
6.3 - 5.05
El perfil proteico de los ESs sin tratar con Spm (TO) (fig . 3.8) , permitió ubicar un total de 33
puntos de proteína en un rango de 13.7 y 87 kDa ; y pi entre - 4.8 Y - 6.3. El mayor
número de proteínas , así como las más abundantes fueron ubicadas en un rango de 21 .5
y 52.1 kDa y rangos de pi de - 5.2 Y - 6.1. Bajo las cond iciones empleadas en este
estudio, no se encontraron proteínas específicas de esta muestra .
83
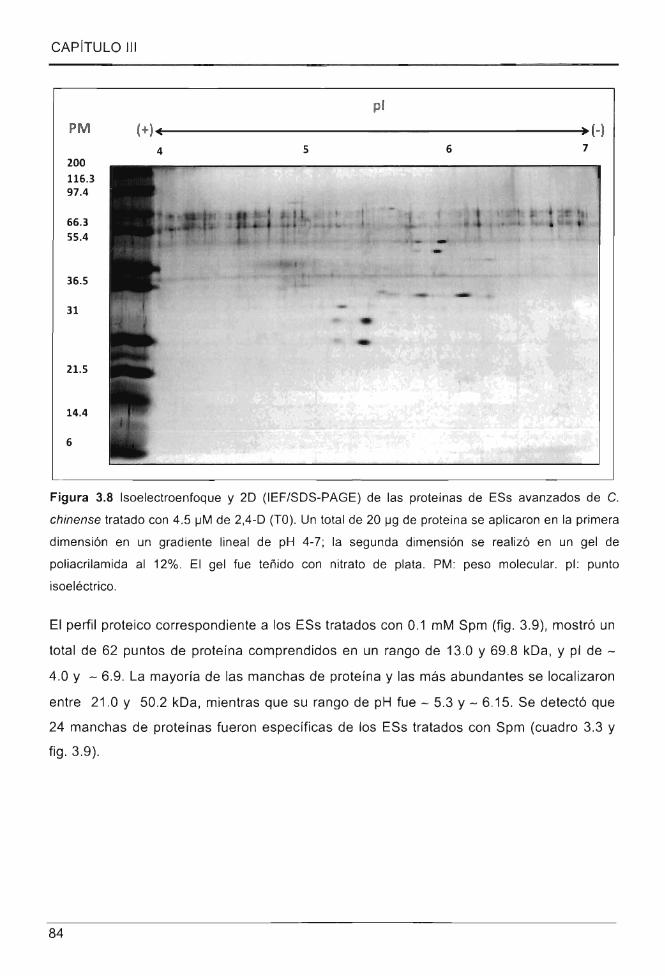
CAPíTULO 111
pi
PM (+) e . (-) 4 5 6 7
200 116.3 97.4
66.3 55.4
36.5
3l
21.5
14.4
6
Figura 3.8 Isoelectroenfoque y 20 (IEF/SOS-PAGE) de las proteínas de ESs avanzados de C.
chinense tratado con 4.5 I-lM de 2,4-0 (TO). Un total de 20 I-lg de proteína se aplicaron en la primera
dimensión en un gradiente lineal de pH 4-7; la segunda dimensión se realizó en un gel de
poliacrilamida al 12%. El gel fue teñido con nitrato de plata . PM : peso molecular. pi: punto
isoeléctrico.
El perfil proteico correspondiente a los ESs tratados con 0.1 mM Spm (fig . 3.9), mostró un
total de 62 puntos de proteína comprendidos' en un rango de 13.0 y 69.8 kDa, y pi de -
4.0 Y - 6.9. la mayoría de las manchas de proteína y las más abundantes se localizaron
entre 21 .0 y 50.2 kDa , mientras que su rango de pH fue - 5.3 Y - 6.15. Se detectó que
24 manchas de proteínas fueron específicas de los ESs tratados con Spm (cuadro 3.3 y
fig . 3.9).
84

CAPíTULO 111
Cuadro 3.3 Peso molecular (PM) y punto isoeléctrico (pi) de prote ínas específicas del perfil
proteico de ESs de C. chinense tratados con Spm 0.1 mM.
Embrión somático tratado con 0.1 mM Spm
PM (kDa) pi
66.3 - 4 .02
66.3 - 5.59
64.9 ñA!'i
59.5 - 5.64
59.5 - 5.69
59.5 - 5.93
59.5 - 588
59.5 - 5.83
58 .1 - 6.73
58.1 - 5.78
58.1 - 6.45
56.8 - 5 .55
56.8 - ñ . R~
55.4 - 6.04
50 .2 - 5.69
50.2 - ñ.1ñ
49.1 - 4.03
46.0 - 5.48
40.7 - 6.86
40.7 - 6 .92
85
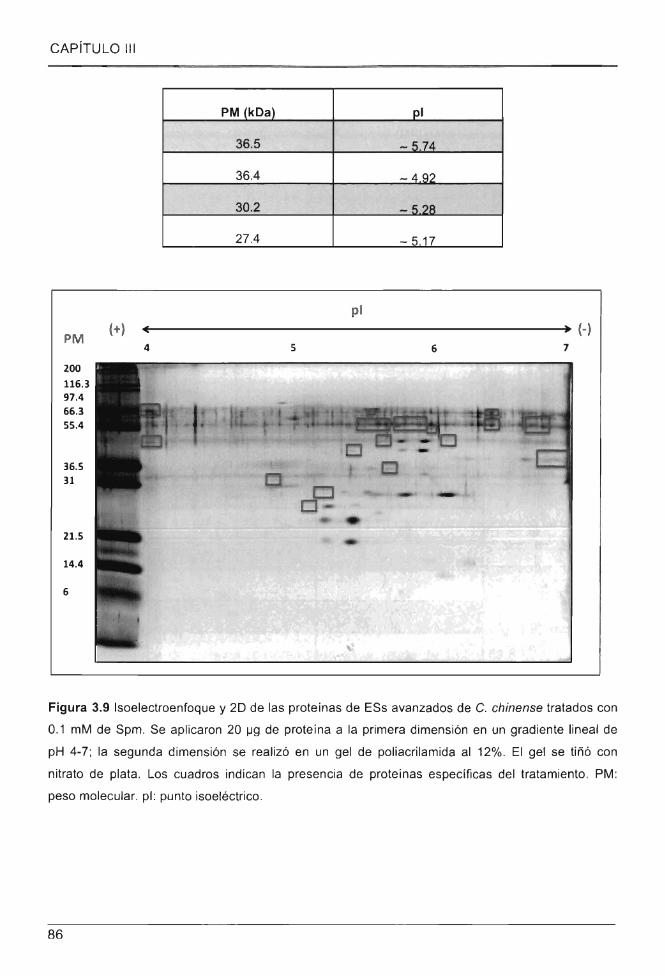
CAPíTULO 111
PM (kDa) pi
36.5 - 5.74
36.4 -4.92
30.2 - 5.28
27.4 - 5.17
pi (+) 4 . (-)
PM 4 5 6 7
200 116.3 97.4 66.3 55.4
36.5 31
21.5
14.4
6
Figura 3.9 Isoelectroenfoque y 20 de las proteínas de ESs avanzados de C. chinense tratados con
0.1 mM de Spm. Se aplicaron 20 I-Ig de proteína a la primera dimensión en un gradiente lineal de
pH 4-7 ; la segunda dimensión se realizó en un gel de poliacrilamida al 12%. El gel se tiñó con
nitrato de plata. Los cuadros indican la presencia de proteínas específicas del tratamiento. PM:
peso molecular. pi : punto isoeléctrico.
86

CAPíTULO III
la comparación de los perfiles protéicos por pi y 2D de los ESs tratados y sin tratar,
permitió observar el efecto de 0.1 mM Spm sobre los ESs de chile habanero. Los dos
perfiles fueron similares dado que ambas muestras, contuvieron 30 manchas de proteína
exclusivas y que no se encontraron en la semilla ni en el EC de la especie (cuadro 3.4).
De igual forma, 7 puntos de proteína de 41 .9 kDa, -6.51; 38.5 kDa, -5.10; 37.6 kDa ,
-5.40; 37.6 kDa , - 5.58 ; 16.6 kDa, -5.99; 16.6 kDa, -6.26 y 13.3 kDa, -6.10 (PM , pi)
variaron en intensidad entre ambos ESs (fig . 3.10). Aunado a ésto, se detectaron 28
puntos de proteína inducidos por el uso de Spm, y que no se encontraron en los ESs sin
tratar (fig . 3.11); dichos puntos de proteína corresponden a los 24 específicos para Spm
(cuadro 3.3), además de 4 proteínas de 66 .3 kDa, -6.39; 57 .8 kDa, -5.42; 42 kDa , -6.28
y 29.3 kDa, -5.62 (PM, pi). Cabe señalar, que no se encontraron proteínas en el perfil
protéico de los ESs del TO que no estuvieran en el perfil de los ESs tratados con 0.1 mM
Spm.
Cuadro 3.4 Peso molecular (PM) y punto isoeléctrico (pi) de proteínas específicas del perfil
proteico de ESs de C. chinense tratados y sin tratar con Spm 0.1 mM .
Embriones somáticos tratados y sin tratar con Spm 0.1 mM
PM (kDa) pi
78.4 - 4.94
73.2 - 5.53
73.2 - 5.61
65.6 - 5.01
59 .5 - 5.72
51.1 - 5.80
46.8 - 5.79
46.8 - 5.94
41 .9 - 6.51
38.5 - 5.10
87
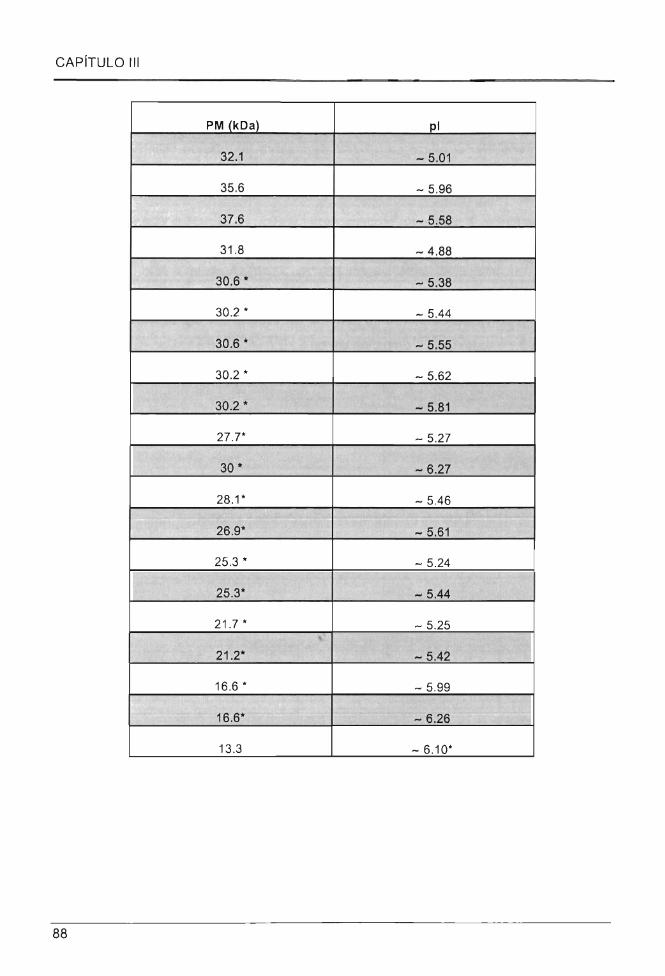
CAPíTULO 111
PMlkD'!l pi
32.1 - 5.01
35 .6 - 5.96
37.6 - 5.58
31.8 - 4 .88
30.6 * - 5.38
30.2 " - 5.44
30.6 * - 5.55
30.2 " - 5.62
30.2 • - 5.81
27.7* - 5.27
30 * - 6.27
28.1" - 5.46
26.9* - 5.61
25.3 " - 5.24
25.3* - 5.44
21 .7 * - 5.25
21 .2* - 5.42
16.6 * - 5.99
16.6* - 6.26
13.3 - 6.10*
88
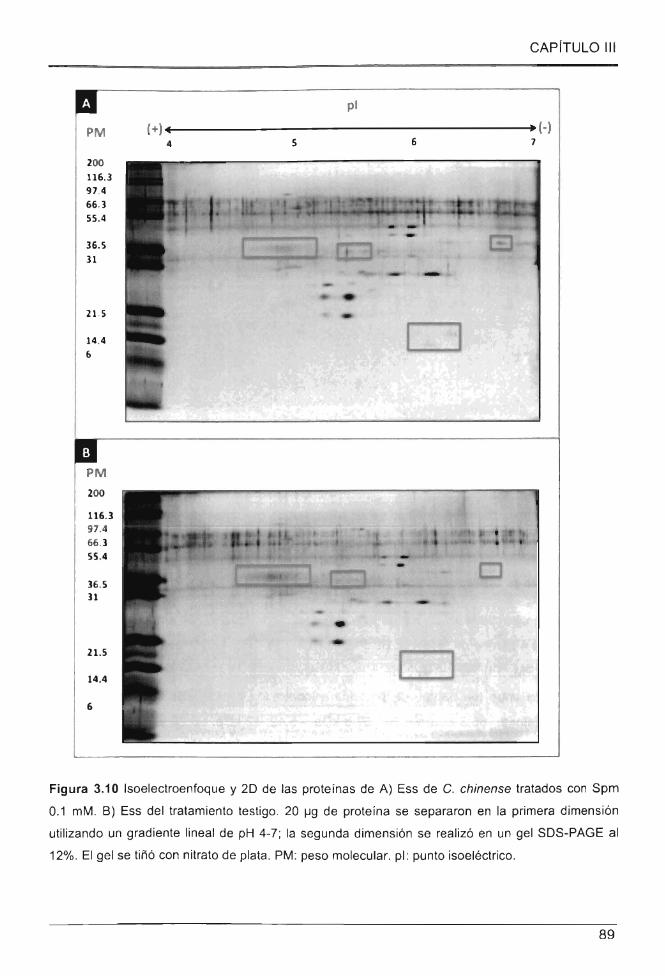
PM
200 116.3 97.4 66.1 55.4
36.5
31
21 .5
14.4 6
PM 200
116.3 97.4 66.3 55.4
36.5 31
21 .5
14.4
6
CAPITULO 111
pi
+------------------+·H (+) • 4 5 6 7
Figura 3.10 Isoelectroenfoque y 20 de las proteínas de A) Ess de C. chinense tratados con Spm
0.1 mM. B) Ess del tratamiento testigo. 20 j.Jg de proteína se separaron en la primera dimensión
utilizando un gradiente lineal de pH 4-7; la segunda dimensión se realizó en un gel SOS-PAGE al
12%. El gel se tiñó con nitrato de plata . PM : peso molecular. pi : punto isoeléctrico.
89

CAPiTULO 111
pi PM +----------------------+. (-) (+) 11
4 s 6 7 200
116.3 97.4 66.3 55.4
36.5 31
21.5
14.4
6
Figura 3.11 Isoelectroenfoque y 20 en gel de poliacrilamida con dodecil sulfato de sodio (IEF/SOS
PAGE) de ESs avanzado de C. chinense tratado con 0.1 mM de Spm. Los recuadros indican 28
proteínas presentes en los ESs tratados con Spm que no se encontraron en los ESs del TO . PM :
peso molecular. pi : punto isoeléctrico.
El perfil de polipéptidos en 2D de los ESs con y sin Spm , mostró que ambos comparten la
expresión de 4 puntos de proteína : 66.3 kDa , ,...5.7; 50.4 kDa , -6.0; 36.2 kDa, -5.4 y 30.2
kDa , -6.1 (PM , PI) con el EC de chile habanero (Fig . 3.12, flechas blancas). Cabe
destacar que entre los puntos de proteína encontrados en los ESs tratados con Spm
había 4 proteínas: 66.3 kDa , - 6.39 ; 57 .8 kDa , -5.42 ; 42 kDa , -6 .28 y 29 .3 kDa, -5.62
(PM , pi) que también estaban presentes en los ECs de la misma especie (fig . 3.12,
flechas amarillas); no obstante , esas proteínas estuvieron ausentes en los ESs del
tratamiento testigo.
90

PM
200 116.3 97.4 66.3
55.4
36.5 31
21.5
14.4
PM
200
116.3 97.4 66.3 55.4
36.5 31
21.5
14.4 6
CAPiTULO 111
pi +-_______________ t (-)
(+) • 4
Figura 3.12 Isoelectroenfoque y 20 de proteínas del A) EC de C. chinense. B) ESs de C. chinense
tratado con Spm 0.1 mM y C) ESs de C. chinense del TO. Un total de 20 I-'g de proteína se
separaron en la primera dimensión en un gradiente lineal de pH 4-7; la segunda dimensión se
realizó en un gel de poliacrilamida al 12%. El gel se tiñó con nitrato de plata . Flechas blancas:
Proteínas presentes en el EC , ESs tratados con Spm 0.1 mM y ESs sin tratar. Flechas amarillas:
proteínas presentes en el EC y en los ESs tratados con Spm. PM: peso molecular. pi: punto
isoeléctrico.
91

CAPíTULO 111
En general se observó que las proteínas que fueron comunes entre la sem illa y el EC, así
como entre los ESs con y sin Spm, son las más abundantes. Mientras que aquellas
proteínas que difieren entre perfiles son de mediana y baja abundancia .
3.4 DISCUSiÓN
los resultados de la cuantificación de proteína mostraron una acumulación
significativamente superior de proteínas totales (90% más) en el EC que en ESs de C.
ehinense, con respecto a las cantidades acumuladas en ESs avanzados de esta especie,
tanto tratados como sin tratar con 0.1 mM Spm . Existen numerosos reportes en los que
trabajando con diferentes especies, se ha descrito que los ESs no son capaces de
acumular varios tipos de proteínas asociadas con la madurez de los ECs en cantidades
suficientes, debido a que a diferencia de los ESs, los embriones cigóticos pasan por
etapas de dormancia y desecación , eventos de tiempo que les permiten acumular
proteínas y otras sustancias de reserva (Zimmerman , 1993). En los ESs de alfalfa, se ha
detectado sólo un 10% del total de las proteínas almacenadas en los ECs (Stuart et a/.,
1988). De igual manera, Winkelmann et al. (2006), obtuvieron bajos rendimientos de
proteína a partir de ESs de Cye/amen persieum MilI. la respuesta obtenida en este
estudio coincide con lo reportado por lecona et al. (2012) , quienes detectaron contenidos
críticos de proteínas en ESs cotiledonares de chile habanero (0.4 ¡,tg/mg PF), comparado
con el contenido registrado en el EC de esta misma especie (14 ¡,tg/mg PF); así mismo en
este estudio se dectaron (57.71 g/mg PF) en el EC. la comparación estadística del
contenido de proteínas del EC (testigo) con los ESs de C.chinense, no mostró diferencia
significativa entre los ESs con y sin Spm. Sin embargo, cuando se compararon
estadísticamente utilizando el Hipocotilo en inducción como testigo, se pudo observar una
disminución significativa en el contenido proteico (aprox. 2.5 ¡,tg/mg PF) de los ESs
tratados con la PA. Cabe señalar que en este análisis estadístico se tomó el hipocotilo en
inducción como testigo debido a que el contenido de proteína presente en esta muestra
(2.03 ± 0.1 ¡,tg/mg PS) y por ser el punto de partida del proceso, permitió realizar una
mejor comparación entre los ESs. El papel de las PAs en la regulación de la síntes is de
proteínas ha sido ampliamente documentado (Young y Srinivasan , 1972; Morris y Jorstad ,
1973; Tabor y Tabor, 1984; Marton y Morris, 1987; Evans y Malmberg , 1989).
92

CAPíTULO 111
El análisis de los perfiles proteicos de la semilla, EC, ESs del TO y ESs tratados con Spm
por SDS-PAGE, mostró 6 bandas con PM de 66.3, 59.3, 46, 36 .5, 33 .8 Y 24.6 kDa , las
cuales estuvieron presentes en todas las muestras analizadas. Dos de estos péptidos
(36 .5 y 46 kDa) presentaron pesos moleculares similares a los reportados para algunas
proteínas de reserva, reportadas en semilla y ESs de P. Taeda (Brownfield et al., 2007) .
lecona et al. (2012), encontraron 7 bandas que estuvieron presentes en todos los
estadías de los ESs y en el EC de chile habanero, una de las bandas reportadas en este
trabajo (46 kDa) , obtuvo un PM similar a una de las reportadas (45.5 kDa) en todos los
estad íos de los ESs así como en el EC de chile habanero por dichos autores. Así mismo,
en este estudio, se encontraron 3 proteínas con PM y pi de 50.4 kDa, -5.97 ; 36.2 kDa ,
-5.43 y 30.2 kDa, -6 .09 respectivamente, que se encontraron tanto en embriones
somáticos como en cigóticos (EC). A nivel molecular, en los ESs se expresan una serie de
proteínas propias del proceso de embriogénesis cigótica, las cuales podrían ser
consideradas como marcadores del proceso de embriogénesis (Pérez-Molphe et al.,
1999). En el perfil proteico de la semilla, se observaron 17 bandas que corresponden a
péptidos con PM ubicado entre 14.4 y 91 .2 kDa. Estos péptidos se encuentran
comprendidos en el mismo rango en el que se han obtenido péptidos en semillas de
diferentes variedades de C. annuum y C. frutescens por Oideigah et al. (1999), quienes
encontraron 12 bandas en un rango de 22-98 kDa. De los cinco péptidos que se
encontraron exclusivamente en el perfil proteico de la semilla de chile habanero (87, 41 .5,
32.6, 18 Y 14.4 kDa), el último péptido (14.4 kDa) pudiera estar asociado con la etapa de
desecación de la semilla, tomando en cuenta lo reportado por Brownfield et al. (2007),
quienes detectaron dos proteínas de 14 y 14.5 kDa en semilla de P. taeda, las cuales
fueron relacionadas con dicho proceso. Dos de las bandas específicas de la semilla que
fueron detectadas en este estudio (14.4 y 41 .5 kDa), poseen similar PM (13.7 Y 41 kDa) a
dos de las bandas reportadas por Lecona et al. (2012), las cuales resultan distintivas del
EC de chile habanero.
En este trabajo se encontraron tres bandas con PM de 80 .8, 62.4 Y 29 kDa, las cuales
fueron específicas del EC. También se observaron otras 6 bandas cuyos PM fueron de
91 .2, 72.5, 64.7, 52 .9, 27 .5 Y 21 .9 kDa, las cuales estuvieron presentes tanto en la
semilla como en el EC. Entre estos péptidos, el que corresponde a un PM de 21.9 kDa
posee un PM similar a una proteína (20 .9 kDa) reportada como específica para EC, por
93

CAPiTULO 111
Lecona et al. (2012). Estos péptidos distintivos de la semilla y el EC, podrían considerarse
como marcadores de embriogénesis cigótica.
En este estudio se reportaron 7 bandas específicas de los ESs con y sin Spm . Lecona et
al. (2012) reportaron 2 bandas específicas para los ESs cotiledonares de C. chinense
(20 .5 y 51 .8 kDa) . Estas últimas tienen un PM sim ilar a dos de los 7 péptidos encontrados
en este estudio cuyos pesos moleculares corresponden a 21 .5 y 51 .6 kDa .
La combinación del isoelectroenfoque y el 2D, permitió una separación más fina de los
extractos proteicos de las diferentes muestras. Obteniendo además de su peso molecular
también su punto isoeléctrico.
Al analizar los perfiles proteicos de la semilla y el EC, se observó que ambos muestran
una composición abundante en proteínas, y una gran similitud entre ellos. La mayor
abundancia de proteínas en ambos perfiles fue en el rango de: 14.4 y 60.2 kDa y pi entre
-5.7 y -6.4 en la semilla , y 13.3 Y 58 .7 kDa y pi entre -5.3 y -6 .7 en el EC. Se conoce
que existe comunicación entre los tejidos propios de la semilla y el EC, en dónde el
embrión puede tomar las proteínas almacenadas en el endospermo; para acumularlas en
los cotiledones (Bewley et al., 2013). Aquellas proteínas que se encuentran en el perfil del
EC y no en la semilla (Cuadro 3.1) , pudieran ser producto de un proceso de estrés al ser
separado de los tejidos propios de la semilla.
El perfil proteico de la semilla , mostró además 3 puntos de proteína con PM y pi de 20 .2
kDa , -5.2; 22 .8 kDa, -5.9 y 26.7 kDa , -6.4; así como un grupo de puntos de proteína , ubicadas con un PM de 48 .2 kDa y un rango de pi de - 5.1 a 5.4, las cuales sólo se
observaron en el perfil de la semilla y no en el de las demás muestras evaluadas. Estas
proteínas pueden considerarse como propias de la testa .
Lecona et al. (2012), reportaron que la mayoría de las proteínas encontradas tanto en
embriones cigóticos como somáticos de C. chinense se encontraron en un rango de PM
de 20 - 130 kDa y pi de - 4 - 7. Puigderrajols et al. (2002) reportaron la acumulación de
proteínas pequeñas (15 a 30 kDa) relacionadas con el choque térmico durante la
maduración de ESs de Quercus suber L. en respuesta a diferentes tratamientos de estrés .
En este estudio, 16 de los 30 puntos de proteína específicos de los embriones tratados y
94

CAPiTULO 111
sin tratar con Spm; mostraron pesos moleculares comprendidos entre 15 y 30 kDa
(cuadro 3.4 , asteriscos). Muchas de las proteínas de choque térmico son chaperonas
moleculares que se forman en respuesta a estrés y también son reguladas durante el
desarrollo. Se ha reportado que la acumulación de este tipo de proteínas está involucrada
en el disparo de la embriogénesis somática (Winkelmann et a/., 2006). En este trabajo, 5
puntos de proteína específicos de los embriones somáticos con pesos moleculares de
46.8,46.8, 51 .1, 73.2 Y 73.2 kDa, y con puntos isoeléctricos de: - 5.94, 5.79, 5.80, 5.61 Y
5.53 respectivamente, presentaron un PM comprendido entre los rangos de 47-50 kDa y
70-75 kDa reportados para proteínas de reserva del tipo globulinas por Griga et a/., 2007.
Es importante señalar que la notable diferencia entre los perfiles detectados antes del
isoelectroenfoque de los ESs tratados y sin tratar con Spm (fig. 3.2) Y los perfiles
obtenidos después de realizar dicha técnica (fig. 3.10) no se debe a una extracción de
proteínas deficiente o a una falta en contenido de proteínas como se observa en la fig .
3.2, si no a que muy probablemente, debido a las condiciones utilizadas en el
isoelectroenfoque, no fue posible enfocar proteínas con pH muy básico (7-10) las cuales
pudieron dirigirse al extremo del electrodo y no entrar al gel.
El análisis por SDS-PAGE no mostró diferencias en cuanto a presencia y ausencia de
bandas de los ESs tratados o no con Spm; sin embargo, el análisis por 2D-PAGE permitió
encontrar diferencias entre los perfiles de ambos embriones . Los ESs tratados con Spm
mostraron 28 puntos de proteína que no se encontraron en los ESs sin tratar; dos de
estos polipéptidos (27.4 kDa, -5 .17 y 5.69; 59.5 kDa, -5.93) poseen un PM y pi similar a
las reportadas por Winkelmann et al. (2006) como proteínas de reserva , tipo globulinas
11 S en embriones somáticos y en el endospermo de Cye/amen persieum MilI. Estos
autores encontraron un grupo de proteínas de aproximadamente 27 kDa y pi entre 5 y 6.
Así como una con PM y pi de 59.12 y 6.46 respectivamente. Además, otros 3 polipéptidos
específicas del tratamiento con Spm en este estudio: 50.2 kDa, -5.69; 50 .2 kDa, -6 .16 y
49.1 kDa, 4.03 (PM,pl) presentaron un PM en el rango reportado por Griga et al. (2007) y
que corresponde a proteínas del tipo globulinas (47-50 kDa). En estudios realizados con
dos variedades de Capsieum annuum, Vladova et al. (2004), encontraron dos
subunidades de globulinas 11 S. Se ha sugerido que las PAs pueden tener un rol en la
acumulación de proteínas de reserva (Santanen y Simola, 1999).
95

CAPíTULO 111
En este estudio la espermina tuvo un efecto en la composición proteica de los embriones
somáticos pues propició la expresión de 28 puntos de proteína que no estaban presentes
en los ESs sin tratar; cuatro de estos polipéptidos estuvieron presentes en el perfil
proteico del EC. Aunado a esto, la Spm causó un ligero aumento en la intensidad de 7
puntos de proteína . Este efecto puede atribuirse a que las poliaminas han demostrado
estar involucradas funcionalmente en la síntesis de proteínas (Young y Srinivasan, 1972;
Morris y Jorstad, 1973; Tabor y Tabor, 1984; Marton y Morris, 1987; Evans y Malmberg,
1989). Las poliaminas pueden estar actuando directa o indirectamente en diferentes
niveles metabólicos, así como en sitios específicos de la maquinaria traduccional ,
regulando la síntesis de proteínas que a su vez, podrían estar involucradas en la
regulación de diferentes eventos celulares como la división y diferenciación celular.
Santanen y Simola (1999) reportaron que las interacciones de las PAs y sus derivados
con las proteínas, pueden ser críticos para ambos procesos celulares. Lo anterior permite
inferir que en este estudio, los cambios observados en el patrón de proteínas de los ESs
por la aplicación de Spm, están involucrados en las mejoras observadas sobre el
desarrollo, calidad y morfología de los embriones somáticos de chile habanero.
96

CAPíTULO III
BIBLIOGRAFíA
Bewley J. D., Bradford K. J., Hilhorst H. W.M . y Nonogaki H. (2013). Seeds. Physiology of
development, germination and dormancy. Springer. 381 p.
Bradford , M.M. (1976). A rapid and sensitive method for the quantification of microgram
quantities of protein utilizing the principie of prote in-dye binding. Anal. Biochem.,
72: 248-254.
Brownfield Dl, Todd, Stone Sl, Deyholos MK, Gifford DJ (2007). Patterns of storage
protein and triacylglycerol accumulation during loblolly pine somatic embryo
maturation . Plant Cell Tissue Organ Cult. 88(2): 217-223.
Chen SX y Harmon AC (2006). Advances in plant proteomics. Proteomics, 6: 5504-5516.
Evans P.T. Y R.l . Malmberg (1989). Do polyamines have roles in plant development?
Plant Physiol., Annu Rev. , Plant Mol. Biol. , 40, 235-269.
Griga M, Horacek, J. and Klenoticova H. (2007). Protein patterns associated with Pisum
sativum somatic embryogenesis. Biologia plantarum 51 (2) : 201-211.
Islas-Flores y Brito-Argáez (2014). Manual de prácticas para electroforésis de proteínas .
México. 77pp .
lai , F.M., Senaratna, T. and McKersie, B.D. (1992) . Glutamine exchanges storage protein
synthesis in Medicago sativa l. Somatic embryos. Plant Science. 87 :6977.
lecona-Guzmán C.A. , Sol ís Marroquín D., Avilés-Viñas S., Santos-Briones C., Santana
Buzzy N. (2012). Changes in the protein profile of habanero pepper (Capsicum
chinense J.) somatic embryos during development. AJB . 11(47); 10761-10768.
lulsdorf MM ., Tautorus TE ., Kikcio SI. y Dunstan DI. (1992). Growth parameters of
embryogenic suspension culture of interior spruce (Picea glauca-engelmannii
complex) and black spruce (Picea mariana Mili.) . Plant Sci. 82 : 227-234.
Marton l Y Morris D (1987). Molecular y cellular functions of the polyamines. En : PP Mc
97

CAPíTULO 111
Cann, A Pegg y A Sjoerdsma, eds. Inhibition of polyamine Metabolism, pp. 79-
105. San Diego: Academic Press.
Morris D.R. Y Jorstad C.M. (1973) . Growth and macromolecular composition of a mutant of
Eschericia coli during polyamine limitation . JB. 113: 271-277.
Nomura K. Y Komamine A. (1995) . Physiological and biochemical aspects of somatic
embryogenesis . In: Thorpe TA (ed) In vitro Embryogenesis in Plants (pp. 249-
266). Klumer Academic Publishers, Dordrecht.
Odeigah P.G.C., Oboh B. y Aghalokpe 1.0. (1999). The characterization of Nigerian
varieties of pepper, Capsicum annuum and Capsicum frutescens by SDS
polyacrylamide gel electrophoresis of seed proteins. Genetic Resources and Crop
Evolution . 46 : 127-131 .
Pegg A. (1986). Recent advances in the biochemistry of polyamines in eukaryotes.
Biochem J 234: 249-262.
Pérez-Molphe E.M., Ramírez-Malagón R. Núñez-Palenius H.G. y Ochoa-Alejo N. (1999) .
Introducción al cultivo de tejidos vegetales . Universidad Autónoma de
Aguascalientes, México, pp: 9-11,16-20,27-35,57-60, 81-84.
Puigderrajols P., Jofré A. , Mir G., Pla M., Verdaguer D., Huguet G. y Molinas M. (2002) .
Developmentally and stress-induced small heat shock proteins in cork oak
somatic embryos. J. Exp . Bot. 53 : 1445-1452.
Roja R. A, Reddy VD, Prakash Babu P, Padmaja G (2005) . Changes in protein profiles
associated with somatic embryogenesis in peanut. Biol Plant 49(3) : 347-354 .
Santanen A. Y Si mola L. (1999). Metabolism of l(U-14C)-arginine and l(U_14C)-ornithine in
maturing and vernalized embryos and megagametophytes of Picea abies.
Physiologia Plantarum 107: 433-440.
Stejskal, J. (1994) . Comparative analysis of proteins and isozymes in somatic and zygotic
embryos of soybean (Glycine max (l .) Merr) . Tesis de doctorado, IEB Prage.
98

CAPíTULO 111
Stuart, DA, Nelse, J. and Nichol, J.W. (1988). Expression of 7S and 11 S alfalfa seed
storage proteins in somatic embryos. J. Plant Physiol. 132: 134-139.
Tabor C.W. y H. Tabor (1984) . Polyamines. Ann. Rev. Biochem ., 53 , 749-790 .
Vladova R., Tsanev V. y Petcolicheva K. (2004). Seed storage proteins in So/anaceae and
Cueurbitaeeae species. Biologia Plantarum 48 (4): 601-603 .
Von Arnold S, Sabala 1, Bozhkovl P, Dyachokl J, Filonoval L. (2002) . Developmental
pathways of somatic embryogenesis . Plant Cell Tissue Organ Cult. 69:233-249.
Winkelmann T., Heintz D., Dorsselaer A. V., Serek M. y Braun H-P. (2006) . Proteomic
analyses of somatic and zygotic embryos of eye/amen persieum MilI. Reveal new
insights into seed and germination physiology. Planta. 224: 508-519.
Young D. Y Srinivasan P. (1972) . Regulation of macromolecular synthesis by putrescine in
a conditional Eseheriehia coli putrescine auxotroph . J. Bacteriol 112: 30-39 .
Zimmerman J.l. (1993) . Somatic embryogenesis: a model for early development in higher
plants. The PlantCel1. 5: 1411 -1423.
99


CAPíTULO IV
CAPíTULO IV
CONCLUSIONES Y PERSPECTIVAS
4.1 CONCLUSIONES
• Asparagina favoreció la velocidad de desarrollo de los ESs, mientras que
Glutamina afectó negativamente la embriogénesis somática en todas las
concentraciones evaluadas.
• Espermina a una concentración de 0.1 mM, resultó el mejor tratamiento evaluado
al mejorar de manera integral el desarrollo y la calidad de los ESs, mostrando un
meristemo caulinar de apariencia normal (formación de cotiledones) .
• Los ESs de C. chinense mostraron un contenido y composición de proteínas
deficiente en comparación con los ECs de la especie.
• Spm 0.1 mM modificó el contenido y la composición de proteínas de los ESs al
permitir la expresión de 28 polipéptidos que se encontraron ausentes en los ESs
sin tratar, así como el cambio en abundancia de 7 polipéptidos.
101

CAPiTULO IV
4.2 PERSPECTIVAS
102
~ Espermina 0.1 mM demostró tener mayores efectos positivos sobre el desarrollo
de los ESs de chile habanero superando al tratamiento testigo; sin embargo, aún
es necesario ajustar las condiciones de cultivo de los ESs. Al observar que Spm
0.01 mM permite que los ESs tempranos tengan un desarrollo más armonioso y,
Spm 0.1 mM favorece particularmente el estadío cotiledonar, una estrategia
pudiera contemplar el uso de dichos tratamientos durante la histodiferenciación de
los ESs (la primera semana con 0.01 mM y la segunda con 0.1 mM de Spm).
~ Aportaría información de mucho interés probablemente, si se realizara un análisis
bidimensional de proteínas totales de los ESs en estad íos tempranos como
avanzados, utilizando la técnica NON-EQUILlBRIUM GEL ELECTROPHORESIS
para enfocar proteínas con puntos isoeléctricos muy básicos con rango de pH de 7
a 10, a fin de observar el comportamiento y los efectos de este tipo de proteínas .
~ Es posible seguir ajustando el método de preparación de las muestras proteicas
para realizar el 2D-PAGE. Una alternativa es cambiar el agua como medio de
resuspensión de las proteínas precipitadas , por el amortiguador de lisis utilizado; el
cual probablemente permita una mejor disolución de las proteínas extraídas. Otra
alternativa es ajustar el porcentaje de anfolitos utilizados en dicho amortiguador
para proteger a las proteínas que no lograron enfocarse en este estudio.
~ Sería idóneo secuenciar algunos de los polipéptidos separados por 20 con el
propósito de identificar a las proteínas que sufren modificaciones en abundancia o
en punto isoeléctrico. Dichas proteínas probablemente desempeñan funciones
regulatorias en la embriogénesis somática .


















