ESTUDIO DE NUEVAS DIANAS TERAPÉUTICAS PARA...
20
1 ESTUDIO DE NUEVAS DIANAS TERAPÉUTICAS PARA EL DESARROLLO DE COMPUESTOS NEUROPROTECTORES Investigadores principales: Dr. Rafael Maldonado López Facultat de Ciències de la Salut i de la Vida UPF Dr. Fernando Rodríguez de Fonseca Hospital Carlos Haya Fundación IMABIS. Málaga Dr. Emilio Fernández Espejo Facultad de Medicina de Sevilla Duración: 3 años
Transcript of ESTUDIO DE NUEVAS DIANAS TERAPÉUTICAS PARA...

1
ESTUDIO DE NUEVAS DIANAS TERAPÉUTICAS PARA EL
DESARROLLO DE COMPUESTOS NEUROPROTECTORES
Investigadores principales:
Dr. Rafael Maldonado López
Facultat de Ciències de la Salut i de la Vida UPF
Dr. Fernando Rodríguez de Fonseca
Hospital Carlos Haya Fundación IMABIS. Málaga
Dr. Emilio Fernández Espejo
Facultad de Medicina de Sevilla
Duración: 3 años

2

3
1. Resumen
Las acil-etanolamidas, anandamida (AEA), oleil-etanolamida (OEA) y
palmitoil-etanolamida (PEA) son endocannabinoides que actúan en los
receptores cannabinoides CB1 (AEA) o del peroxisoma activado por
proliferadores alfa (PPARα) (OEA, PEA). Se han sugerido efectos
neuroprotectores para estos compuestos, dado que se acumulan en el
cerebro después de isquemia o neurotoxicidad, por cuya razón serían
importantes en el control de la supervivencia y plasticidad neuronales. La
neuroprotección inducida por AEA ya ha sido publicada, pero no por OEA o
por PEA. Este proyecto estudiará si estos transmisores modifican la
neurotoxicidad causada por 6-hidroxidopamina en ratas y por MDMA en
ratones normales y genéticamente modificados para anular el gen que
codifica para el receptor PPARα. Específicamente investigaremos si la
administración de OEA/PEA modifica la duración e intensidad de la
neurotoxicidad inducida por 6-hidroxidopamina y MDMA. Se utilizará una
estrategia multidisciplinar para medir diferentes parámetros de toxicidad.
En los estudios comportamentales se evaluarán efectos cognitivos y
motores. Se realizarán correlaciones anatómicas de la neurodegeneración
dopaminérgica para evaluar la expresión de tirosina hidroxilasa y la
inducción de apoptosis. La integridad de la transmisión dopaminérgica se
estudiara midiendo cambios en los transportadores de dopamina y la
inducción de genes pro- o antidegeneración, incluidos el estrés oxidativo y
las enzimas antioxidantes. Los estudios in vitro con cultivos de neuronas
dopaminérgicas se llevarán a cabo para evaluar los efectos protectores de
las acil-etanolamidas. La participación de estos transmisores en los
mecanismos mencionados se verificará mediante métodos farmacológicos.
Así, se desarrollarán nuevos análogos de OEA y PEA, más potentes y
duraderos. El objetivo final de este proyecto es evaluar si estos
intermediarios lipídicos podrían ofrecer nuevas estrategias terapéuticas para
aliviar el daño neuronal.
Los objetivos del proyecto son los siguientes:

4
1. La primera hipótesis es que la oleoil-etanolamida (OEA) y la palmitoil-
etanolamida (PEA) administradas antes de que la neurotoxina 6-
hidroxidopamina (6-OHDA) pueden favorecer la viabilidad o proteger las
neuronas dopaminérgicas de los efectos neurotóxicos en ratas, así como
proteger de la toxicidad cultivos de neuronas mesencefálicas
dopaminérgicas. Los agonistas sintéticos del PPAR-α deberían tener efectos
neuroprotectores similares a los de la OEA y/o la PEA.
2. La segunda hipótesis es que la OEA y la PEA administradas antes que la
neurotoxina 3,4-metilen-dioximetanfetamina (MDMA, éxtasis) pueden
facilitar la viabilidad o proteger las neuronas dopaminérgicas de los efectos
neurotóxicos en ratones. Los agonistas sintéticos del PPAR-α deberían tener
efectos neuroprotectores similares a los de la OEA y/o la PEA. Además, los
animales PPAR-α knock-out deberían presentar una mayor sensibilidad a los
efectos neurotóxicos del MDMA que los ratones de tipo salvaje, mientras
que los efectos neuroprotectores de la OEA y/o la PEA deberían desaparecer
en estos animales modificados genéticamente. Los ratones PPARα knock-out
también serán útiles para identificar el papel de este factor de transcripción
en los efectos neuroprotectores de la AEA, que actúa principalmente en una
diana diferente, el receptor cannabinoide CB1.
3. El perfil farmacológico de la OEA y la PEA se analizará en la expresión
de genes relacionados con la neurodegeneración en el estriado dorsal y el
mesencéfalo ventral de ratas y ratones tratados con OEA/PEA antes de la
administración de una dosis neurotóxica de 6-OHDA o de MDMA. El
estudio se llevará a cabo mediante análisis del genoma completo seguido
por PCR cuantitativa en tiempo real de los genes candidatos. Además, la
expresión de proteínas de los genes candidatos seleccionados será
estudiada en el estriado dorsal y el mesencéfalo ventral de ratas o ratones
tratados con OEA/PEA antes de la administración de una dosis neurotóxica
de 6-OHDA o MDMA. Si fuese necesario, también se realizará un Western
blot, así como immunohistoquímica.
4. Se diseñarán y sintetizarán nuevos análogos de la acil-etanolamida
basados en estructuras de sulfamoil. La capacidad de estos nuevos

5
compuestos de regular la expresión génica mediante la activación de los
receptores PPAR-α será validada in vitro y serán ensayados como
neuroprotectores utilizando las técnicas descritas en los objetivos 1 y 2.
2. Resultados
Los principales resultados obtenidos para alcanzar los diferentes objetivos
se describen a continuación.
Objetivo 1. Efectos neuroprotectores de la oleoil-etanolamida y la
palmitoil-etanolamida frente a la 6-OHDA (subproyectos 1 y 3 )
La eficacia neuroprotectora potencial de la administración sistémica de
OEA ha sido ensayada in vivo y en modelos de Parkinson con 6-OHDA
(ratas) y de degeneración de las neuronas dopaminérgicas de la substantia
nigra inducida por MPTP (ratones). Los resultados de los estudios in vivo en
ratas revelaron que la administración periférica de la acil-etanolamida OEA
(0,5, 1 y 5 mg/kg) viene seguida de un rápido paso de la barrera
hematoencefálica y por una rápida eliminación del compuesto. Los estudios
funcionales indicaron que todos los déficits funcionales se redujeron en los
animales tratados con la OEA (5 mg/kg). Además, la reducción de la
densidad inmuno-rreactiva de la tirosina-hidroxilasa del estriado y de la
substantia nigra inducida por 6-OHDA (TH-ir), así como la expresión de
sinaptofisina en el estriado, fueron significativamente antagonizados por la
OEA (5 mg/kg). La respuesta oxidativa a la lesión de la toxina en el
estriado, medida como expresión de la hemooxigenasa-1, también se
redujo tras la administración de la OEA (5 mg/kg). En resumen, la OEA
atraviesa la barrera hematoencefálica tras la administración sistémica y,
dependiendo de la dosis, protege el circuito estriado de la toxicidad inducida
por 6-OHDA. Los ratones C57BL/6 de tipo salvaje y PPAR-α knock-out
convertidos en parkinsonianos mediante una dosis tóxica de MPTP (40
mg/kg) que induce la muerte celular del 70-80% en la substantia nigra,
fueron tratados con OEA sistémica (0, 0,5, 1, y 5 mg/kg) antes y después
de la administración de MPTP. La OEA (5 mg/kg) redujo significativamente
la pérdida de TH-ir en el estriado y la muerte celular nigral. Curiosamente,

6
este efecto neuroprotector de la densidad de TH en estriado, pero no del
número de células dopaminérgicas nigrales, también se encontró en ratones
PPAR-α knock-out, sugiriendo por tanto que los efectos protectores de la
OEA en el estriado dorsal no están mediados por su acción agonista sobre
los receptores PPAR-α. En resumen, la OEA (5 mg/kg) sistémica ejerce
efectos neuroprotectores parciales en los animales parkinsonianos, tanto en
modelos con 6-OHDA como con MPTP, y el efecto en el estriado no parece
estar mediado por los receptores PPAR-α (Galán-Rodríguez et al.,
Neuropharmacology, 2009).
También se llevaron a cabo estudios in vitro de los efectos de la acil-
etanolamida OEA en células cultivadas después de la inducción de estrés
oxidativo mediante 6-OHDA en las neuronas dopaminérgicas primarias y
células cromafinas extraadrenales. Los efectos protectores fueron medidos
por los ensayos de la lactato-deshidrogenasa (LDH) y el bromuro de 3-(4,5-
dimetiltiazol-2-il)-2,5-difeniltetrazólico (MTT). La OEA (0,5 y 1 µM)
administrada antes y después de la toxina ejerce, de forma significativa,
efectos neuroprotectores sobre las neuronas cultivadas. Sin embargo, la
OEA administrada antes de la 6-OHDA fue ineficaz, pero suministrada
después de la 6-OHDA (OEA 1 µM) resultó efectiva. Los efectos de la OEA
siguieron unas curvas de dosis-respuesta en forma de U. Este tipo de
respuesta podría indicar una toxicidad debida a la elevada concentración del
fármaco o que se activan vías intracelulares opuestas con dosis de los
fármacos diferentes. Los efectos protectores inducidos por la OEA fueron
abolidos después de la adición de dosis bajas de capsaicina, agonista de los
receptores TRPV1 (0,1 a 1 µM), o del inhibidor de la adenilato ciclasa MDL-
12,330A (1 y 10 µM), pero no mediante la adición de GW6471, bloqueador
del receptor PPAR-α. Estos resultados sugieren que los efectos protectores
de la OEA sobre las neuronas dopaminérgicas son mayoritariamente
mediados por el bloqueo de los receptores TRPV1 y la activación de la vía
del AMPc. En otra serie experimental, las propiedades de protección in vitro
de la PEA y el WY14643, un agonista selectivo de los receptores PPAR-α, se
evaluó en neuronas dopaminérgicas de la substantia nigra y células
cromafinas extraadrenales noradrenérgicas utilizando un método similar al
descrito anteriormente. La PEA tiene efectos neuroprotectores en las células
de la substantia nigra en el ensayo de la LDH en todas las dosis probadas

7
(0,5, 1 y 5 µM), aunque sólo la dosis de 5 µM de PEA fue eficaz en el
ensayo del MTT. El WY14643 no induce efectos neuroprotectores en estos
ensayos cuando se administra a dosis de 1, 5 y 10 µM. Estos datos indican
que la PEA induce efectos neuroprotectores sobre las neuronas
dopaminérgicas como lo hace la OEA, aunque el agonista del PPAR-α
WY14643 no tiene estos efectos protectores (Galán-Rodríguez et al.,
Neuropharmacology, 2009).
Además, estábamos interesados en discernir si las neuronas
dopaminérgicas de la substantia nigra pars compacta (SNpc) se ven
afectadas en los ratones PPAR-α knock-out y si los efectos tóxicos inducidos
por MPTP se han modificado en estos ratones mutantes. En primer lugar, los
datos revelaron que los ratones PPAR-α knock-out presentaron un SNpc
reducido en comparación con los ratones de tipo salvaje, tanto en volumen
como en número de neuronas dopaminérgicas. Este resultado sugiere que
los receptores PPAR-α son necesarios para proteger las neuronas
dopaminérgicas nigrales del daño oxidativo normal. La parte lateral de la
SNpc fue la más dañada en los ratones PPAR-α knock-out. A nivel
bioquímico se encontró una disminución significativa de las moléculas
antioxidantes en el SNpc de ratones knock-out, lo que podría explicar la
reducción de la protección contra el daño oxidativo normal. Como era de
esperar, el MPTP causó un rápido inicio de Parkinson en ratones de tipo
salvaje, vinculado a una fuerte reducción en el número de neuronas
dopaminérgicas de la substantia nigra (74%). Sin embargo, los ratones
PPAR-α knock-out eran más resistentes que los ratones de tipo salvaje al
MPTP, ya que este tóxico indujo una reducción menos intensa de las
neuronas dopaminérgicas nigrales (36%). El MPTP indujo un aumento del
óxido nítrico en la substantia nigra pars compacta en ratones de tipo
salvaje, pero la producción de óxido nítrico se redujo en los ratones knock-
out, lo que sugiere que la falta de receptores PPAR-α baja la actividad de la
sintasa del óxido nítrico y la producción de óxido nítrico después del
tratamiento con MPTP (González-Aparicio et al., Neuroscience, 2011).
La participación de los receptores PPAR-α en la sensibilización motora
producida por la morfina y la cocaína también fue investigada en ratones
PPAR-α knock-out y mediante el uso de un agonista selectivo de PPAR-α,

8
WY14643. Los resultados encontrados en ratones indican que el PPAR-α
desempeña un papel inhibidor en la expresión, pero no en la inducción, de
la sensibilización motora a la morfina. Sin embargo, la sensibilización a la
cocaína no se modificó en estos ratones mutantes, lo que sugiere que este
receptor nuclear participa en la activación motora de los opiáceos, pero no
de los psicoestimulantes. Además, la expresión del PPAR-α en cerebro fue
aumentada con la dosis más alta de la administración repetida de morfina,
pero no crónica de la cocaína, lo que sugiere que este receptor podría
desempeñar un papel compensatorio para mantener la homeostasis en la
administración crónica de opioides. Conforme a esto, el WY14643 sistémico
bloqueó la sensibilización a la morfina, confirmando que el PPAR-α
desempeña un papel compensatorio oponiéndose a la sensibilización motora
inducida por la morfina, probablemente a través de una reducción de los
cambios asociados con la inflamación (Fernández-Espejo et al.
Neuroscience, 2009)
También se ha estudiado la distribución anatómica de los receptores
PPRA-α en áreas del cerebro de rata relacionadas con la transmisión
dopaminérgica, así como la expresión de genes específicos relacionados con
la transmisión dopaminérgica y la neuromodulación para acil-etanolamidas
por PCR en tiempo real. El análisis de los efectos de la OEA en el cerebro de
rata se está realizando y comparando con los de las ratas tratadas con 6-
OHDA. Se ha demostrado la permeabilidad de la barrera hematoencefálica
en la PEA y la OEA (Plaza-Zabala et al., Synapse 2010). Finalmente se han
estudiado los posibles cambios inducidos por la administración sistémica de
la OEA en las redes de neuropéptidos diencefálicos (Serrano et al.
Neurofarmacología 2011). Este estudio reveló que la OEA induce cambios
sustanciales en las señales periféricas que se dirigen al hipotálamo, que
parecen desempeñar un papel más importante que el efecto directo sobre
las neuronas peptidérgicas del hipotálamo.
Objetivo 2. Neuroprotección frente a la neurotoxicidad inducida por
3,4-metilen-dioxi-metanfetamina (MDMA) (Subproyecto 1)
La funcionalidad del transportador de dopamina (DAT), se ha evaluado
siguiendo un régimen neurotóxico de MDMA en ratones y se han investigado

9
los posibles efectos neuroprotectores de la OEA. La administración aguda de
MDMA a la dosis de 10 mg/kg aumentó los niveles extracelulares de
dopamina del estriado con respecto al valor basal en los ratones tratados
con solución salina o con 3 mg/kg de MDMA. Por el contrario, los niveles de
dopamina no aumentaron en los ratones tratados con la dosis de 30 mg/kg
de MDMA. Estos resultados muestran que el tratamiento repetido con MDMA
a la dosis de 30 mg/kg disminuye la actividad funcional del DAT, sugiriendo
posibles efectos neurotóxicos. También se ha estudiado la reacción
apoptótica después de un régimen neurotóxico de MDMA en ratones. La
administración repetida de altas dosis de MDMA (30 mg/kg) en ratones no
indujo la apoptosis en el hipocampo, la corteza frontal y el estriado, medida
24 horas después de la última inyección. De hecho, no hay muerte celular
programada en estas estructuras cerebrales 24 horas después de la última
inyección siguiendo un régimen de altas dosis repetidas de MDMA. Por otra
parte, la administración de MDMA disminuye DAT en el estriado y produce
una disminución en el aprendizaje de la tarea de evitación activa.
Curiosamente, el tratamiento previo con la OEA (5 mg/kg) impide la
reducción de DAT y los retrasos de los déficits cognitivos inducidos por 5
mg/kg de MDMA (Plaza-Zabala et al., Synapse, 2010).
En un segundo experimento, se halló que las diferentes dosis de MDMA
ensayadas (0, 0,03, 0,06, 0,125 y 0,25 mg/kg/infusión) fueron incapaces
de mantener una conducta de autoadministración en ratones deficientes en
el transportador de la serotonina (SERT), mientras que los animales de
tipo salvaje adquirieron este comportamiento. La administración de MDMA
aumentó los niveles extracelulares de dopamina del estriado, tanto en los
animales de tipo salvaje como en los ratones SERT knock-out. Sin embargo,
la concentración extracelular de 5-HT se incrementó sólo en el genotipo de
tipo salvaje, lo que indica una función específica de SERT en las propiedades
reforzantes del MDMA (Trigo et al. Biological Psychiatry, 2007).
Por otra parte, los efectos hipertérmicos del MDMA (1, 5 y 25 mg/kg) se
abolieron en ratones PPAR-α knock-out, lo que indica un papel crucial de
estos receptores en las alteraciones de la temperatura corporal producidas
por MDMA. Por otro lado, hemos obtenido datos que muestran que los
efectos hiperlocomotores del MDMA (20 mg/kg) también disminuyeron en

10
ratones PPAR-α knock-out, sugiriendo una implicación de estos receptores
nucleares en las propiedades psicoestimulantes del MDMA. También hemos
observado que otro cannabinoide, el THC, interactúa con el MDMA en
términos de aprendizaje y procesamiento de la memoria de una forma
dosis-específica. De hecho, el THC (0,3 y 1 mg/kg) puede aumentar los
efectos de bajas dosis de MDMA (3 mg/kg) para producir un deterioro
cognitivo, pero puede tener efectos protectivos sólo a una dosis de THC de
1 mg/kg, frente a las alteraciones inducidas por MDMA a dosis altas (10
mg/kg) (Tschon et al., IVth European Workshop on Cannabinoides Research,
Madrid, 2009).
La descripción anatómica de los receptores PPRA-α en las neuronas
dopaminérgicas del ratón también se ha llevado a cabo y se está evaluando
su papel en la neurogénesis en animales jóvenes y adultos.
Objetivo 3. Estudios moleculares y genéticos (Subproyectos 1 y 2)
Con el uso de un control yoke en el paradigma operante de
autoadministración intravenosa, que es el modelo animal más relevante
para estudiar el potencial adictivo de las drogas de abuso en humanos,
hemos demostrado que la exposición repetida al MDMA induce la expresión
de genes relacionados con las respuestas inflamatorias e inmunológicas en
varias estructuras cerebrales, incluyendo el estriado ventral, la corteza
frontal, el núcleo dorsal del rafe y el hipocampo. Entre ellos, los cuatro
genes para los que la expresión diferencial se validó por qRT-PCR, Camk2a,
Kalrn, DDN y EGR3, han pronosticado sitios de unión para homeobox-1
pancreático duodenal (Pdx1). Además, los cambios de expresión de los
genes identificados en el núcleo dorsal del rafe después de la
autoadministración de MDMA indican que esta región del cerebro puede
estar implicada en el aprendizaje por motivos relacionados con el
comportamiento de búsqueda activa del MDMA (Fernández-Castillo et al.,
Genes Brain and Behavior, 2011).
Por otra parte, se ha estudiado la expresión de genes relacionados con la
neurodegeneración en el estriado dorsal y el mesencéfalo ventral de ratas y
ratones tratados con OEA o PEA antes de una dosis neurotóxica de 6-OHDA

11
(ratas) o MDMA (ratones) para llevar a cabo el perfil farmacológico de la
OEA y la PEA en términos de expresión génica. Los estudios se han puesto
en marcha y los resultados estarán disponibles en breve. El perfil
farmacológico de la OEA se encuentra en evaluación para analizar la
expresión de genes en el estriado dorsal, mesencéfalo ventral y el
hipotálamo de ratas y ratones. Además, se ha conseguido la expresión de
genes seleccionados en el cerebro de los ratones después del tratamiento
con OEA +/- la neurotoxina MPTP, aunque los resultados finales aún no
están disponibles. Se han realizado y publicado estudios específicos sobre
los efectos de la OEA en péptidos y neurotransmisores del hipotálamo
(Serrano et al., Neuropharmacology, 2011). Un estudio piloto de los efectos
protectores de la OEA contra el estrés oxidativo asociado al daño en el
hígado también se ha completado y publicado (González-Aparicio et al.,
Psychoparmacology, 2011). Además, ha sido desarrollado un método para
evaluar los niveles de la OEA en el tejido cerebral a través de microdiálisis
acoplada a espectrometría de masas en colaboración con el Instituto de
Investigación Scripps (La Jolla, California, EEUU).
Objetivo 4. Síntesis de nuevos análogos de las acil-etanolamidas
(Subproyecto 2)
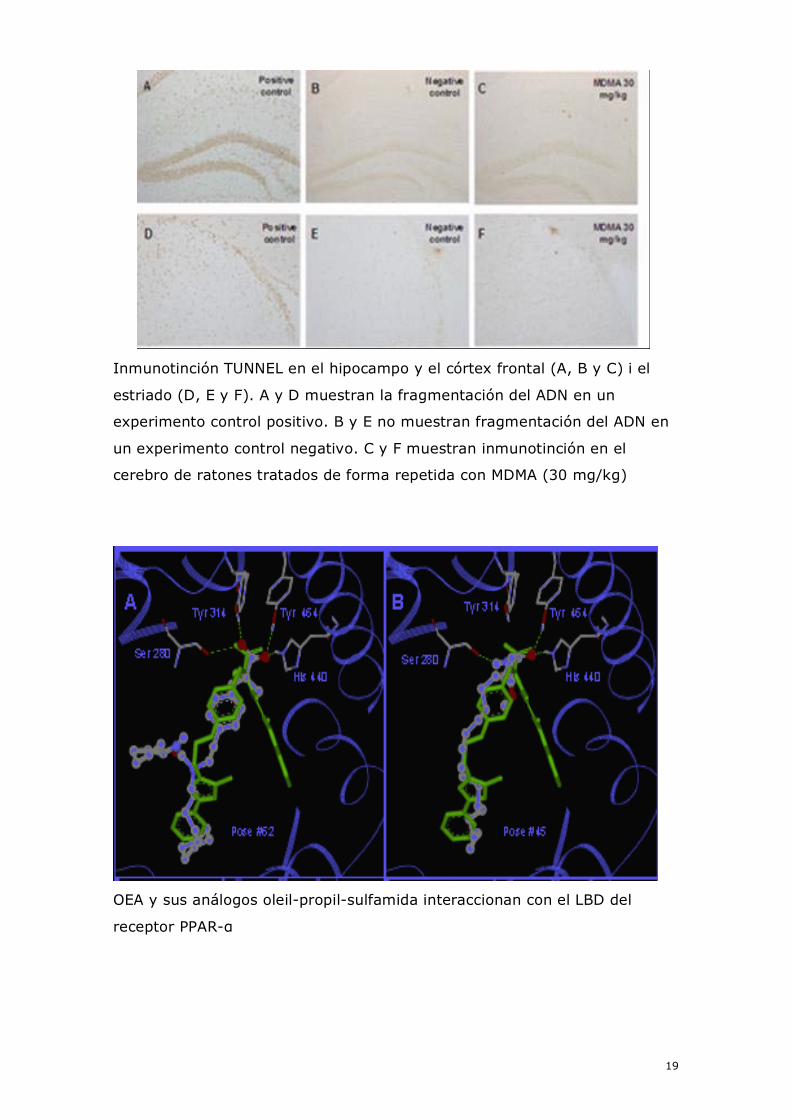
Los requisitos estructurales para la unión de la OEA con el receptor
PPAR-α se han estudiado mediante el análisis de docking o acoplamiento
(Moreno-Santos et al., Journal of Biological Chemistry, presentado). Así, se
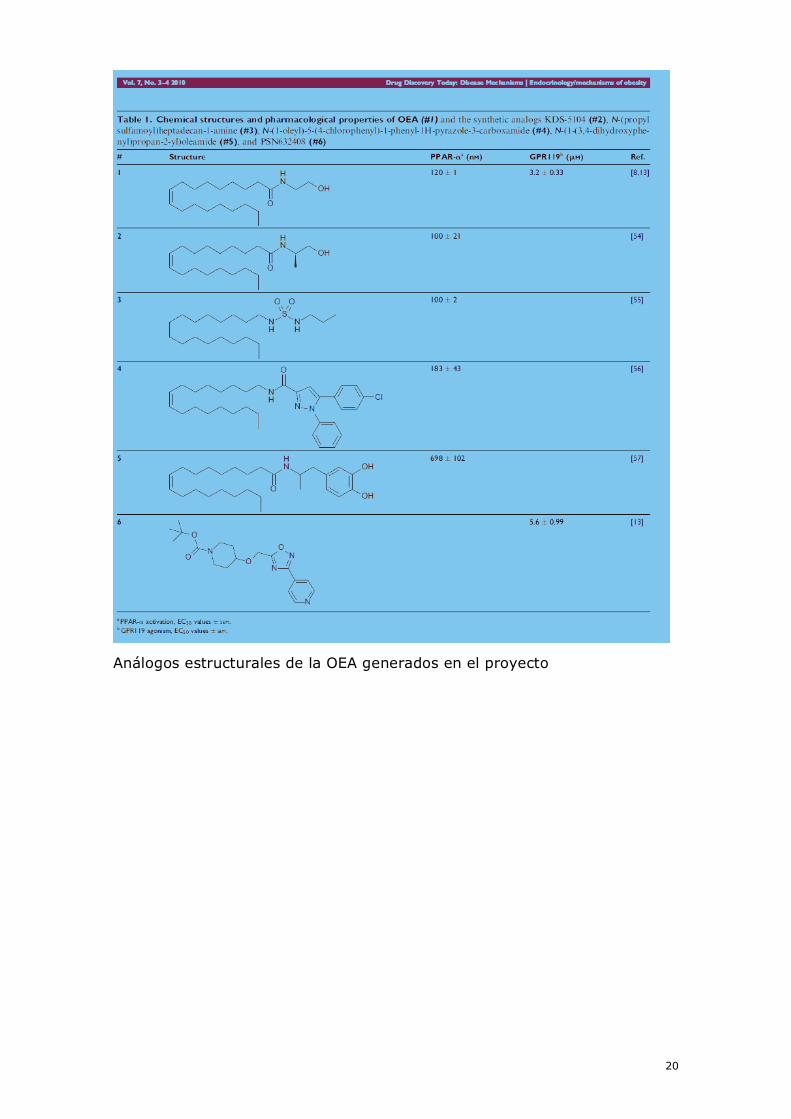
han diseñado nuevos análogos de la acil-etanolamida como
neuroprotectores potenciales. De estos, se ha hallado que los basados en
estructuras de sulfamoil son neuroprotectores in vitro. También se han
sintetizado otros agonistas del PPAR-α procedentes de compuestos
derivados de oleil pirazol y triazoles. La capacidad de estos nuevos
compuestos para la regulación de la expresión génica, a través de la
activación de los receptores PPAR-α, ha sido validada in vitro, también han
sido validados y probados como neuroprotectores utilizando las técnicas
descritas en los objetivos 1 y 2. Se ha generado una patente (Rodríguez de
Fonseca et al., WO2009/050318 A1) y tres publicaciones (Alvarado et al.,
Bioorganic & Medicine Chemistry, 2008; Cano et al., European Journal of
Medicinal Chemistry, 2009; Almeida et al., ChemMedChem., 2010) y están

12
bajo evaluación una segunda patente (Oficina Española de Patentes,
201130486) y un manuscrito (Moreno-Santos et al., Journal of Biological
Chemistry, presentado).
3. Relevancia y posibles implicaciones clínicas de los resultados
finales obtenidos
Los enfoques multidisciplinares que se han utilizado en este proyecto de
investigación han permitido la consecución de los diferentes objetivos
incluidos en el mismo, y han proporcionado información de interés para el
desarrollo de nuevas estrategias terapéuticas para la neuroprotección. Los
principales resultados del proyecto que podrían tener aplicaciones clínicas
potenciales pueden resumirse de la siguiente manera:
1. La identificación de los efectos neuroprotectores de dos nuevos
compuestos, oleoil-etanolamida y palmitoil-etanolamida en diferentes
modelos animales. Estos efectos neuroprotectores se han investigado en
dos modelos complementarios de neurotoxicidad: los efectos neurotóxicos
producidos por 6-OHDA en ratas, que promueve una degeneración de las
fibras dopaminérgicas, y la neurotoxicidad promovida por la administración
repetida de MDMA en ratones, que también produce neurotoxicidad
dopaminérgica. Los efectos neuroprotectores en estos modelos animales
para las neuronas dopaminérgicas sugieren un potencial interés terapéutico
de estos nuevos compuestos para la enfermedad de Parkinson y/o para
prevenir las consecuencias perjudiciales del consumo de drogas de abuso
sintéticas. Deberían llevarse a cabo ensayos clínicos con estos compuestos
para confirmar el interés de estos resultados.
2. La identificación de los mecanismos específicos involucrados en
los efectos neuroprotectores de la oleoil-etanolamida y la palmitoil-
etanolamida. La participación selectiva de los receptores PPAR-α en
algunos de los efectos neuroprotectores de estos compuestos identifica una
nueva diana para el desarrollo de nuevos agentes neuroprotectores. La OEA
y la PEA han sido las herramientas farmacológicas utilizadas en este
proyecto para la identificación de este nuevo mecanismo subyacente a los

13
efectos neuroprotectores. Sin embargo, las propiedades farmacocinéticas y
farmacodinámicas de estos lípidos se pueden mejorar con el fin de obtener
compuestos más apropiados para dirigirse in vivo a los receptores PPAR-α y
para obtener nuevos fármacos neuroprotectores. Por otra parte, la
neuroinflamación parece participar en la sensibilización de los efectos de las
drogas de abuso, y el receptor nuclear PPAR-α desempeña un papel
destacado en estas respuestas inflamatorias. Este receptor nuclear participa
en la activación motora provocada por opiáceos y podría ser útil para el
diseño de nuevos fármacos destinados a tratar el abuso de estas drogas.
3. La caracterización de los cambios en la expresión génica en
diferentes estructuras cerebrales durante estos procesos
neurodegenerativos. Hemos identificado los cambios específicos en la
expresión de genes que podrían tener un papel importante en estos efectos
neuroprotectores utilizando técnicas de análisis del genoma completo
seguido por PCR cuantitativa en tiempo real de los genes candidatos. Por lo
tanto, estos genes son de interés para entender mejor los mecanismos
implicados en estos efectos neuroprotectores y para promover futuras
actividades de investigación para identificar nuevos potenciales agentes
neuroprotectores.
4. La generación de nuevos análogos de acil-etanolamida que
podrían ser de interés como nuevos agentes neuroprotectores. Estos
análogos de la acil-etanolamida pueden mejorar las propiedades
farmacocinéticas y farmacodinámicas de la OEA y la PEA y podrían, por
tanto, ser más adecuados que los lípidos originales para posibles
aplicaciones como agentes neuroprotectores, ya que muestran propiedades
neuroprotectoras similares, que son patentables, y dependen de los
receptores PPAR-α.
En resumen, este proyecto ha identificado los efectos neuroprotectores
de la oleoil-etanolamida (OEA) y la palmitoil-etanolamida (PEA), así
como la diana neurobiológica para estas respuestas farmacológicas. Los
nuevos compuestos análogos que se han generado pueden ser de
interés potencial como nuevos agentes neuroprotectores. Además, los

14
genes candidatos identificados pueden ser de interés potencial para el
desarrollo de nuevas estrategias de investigación.
4. Publicaciones
Trigo JM, Renoir T, Lanfumey L, Hamon M, Lesch KP, Robledo P, Maldonado
R.
3,4-methylenedioxymethamphetamine self-administration is abolished in
serotonin transporter knockout mice.
Biological Psychiatry, 2007 Sep 15; 62(6):669-79. Epub 2007 Feb 16.
PMID: 17306775 (IF: 8.926).
Alvarado M, Goya P, Macías-González M, Pavón FJ, Serrano A, Jagerovic N,
Elguero J, Gutiérrez-Rodríguez A, García-Granda S, Suardíaz M, Rodríguez
de Fonseca F.
Antiobesity designed multiple ligands: Synthesis of pyrazole fatty acid
amides and evaluation as hypophagic agents.
Bioorganic & Medicine Chemistry, 2008 Dec 1;16(23):10098-105. Epub
2008 Oct 14. PMID: 18952442 (IF: 2.822).
Galan-Rodriguez B, Suarez J, Gonzalez-Aparicio R, Bermudez-Silva FJ,
Maldonado R, Robledo P, Rodriguez de Fonseca F, Fernandez-Espejo E.
Oleoylethanolamide exerts partial and dose-dependent neuroprotection of
substantia nigra dopamine neurons.
Neuropharmacology, 2009 Mar; 56(3):653-64. Epub 2008 Dec 7. PubMed
PMID: 19070629 (IF: 3.909).
Fernandez-Espejo E, Ramiro-Fuentes S, Rodriguez de Fonseca F.
The absence of a functional peroxisome proliferator-activated receptor-
alpha gene in mice enhances motor sensitizing effects of morphine, but not
cocaine.
Neuroscience, 2009 Dec 1; 164(2):667-75. Epub 2009 Aug 18. PMID:
19698765 (IF: 3.292).

15
Cano C, Páez JA, Goya P, Serrano A, Pavón J, Rodríguez de Fonseca F,
Suardíaz M, Martín MI.
Synthesis and pharmacological evaluation of sulfamide-based analogues of
anandamide.
European Journal of Medicinal Chemistry, 2009 Dec; 44(12):4889-95. Epub
2009 Aug 12. PMID: 19762126 (IF: 3.269).
Plaza-Zabala A, Berrendero F, Suarez J, Bermudez-Silva FJ, Fernandez-
Espejo E, Serrano A, Pavon FJ, Parsons LH, De Fonseca FR, Maldonado R,
Robledo P.
Effects of the endogenous PPAR-alpha agonist, oleoylethanolamide on
MDMA-induced cognitive deficits in mice.
Synapse, 2010 May; 64(5):379-89. PMID: 20029832 (IF: 2.557).
Orejarena MJ, Lanfumey L, Maldonado R, Robledo P.
Involvement of 5-HT2A receptors in MDMA reinforcement and cue-induced
reinstatement of MDMA-seeking behaviour.
International Journal of Neuropsychopharmacology, 2010 Oct; 14:1-14.
[Epub ahead of print]. PMID:20942998 (IF: 4.874).
Almeida B, Joglar J, Luque Rojas MJ, Decara JM, Bermúdez-Silva FJ, Macias-
González M, Fitó M, Romero-Cuevas M, Farré M, Covas MI, Rodríguez de
Fonseca F, de la Torre R.
Synthesis of fatty acid amides of catechol metabolites that exhibit
antiobesity properties.
ChemMedChem, 2010 Oct 4; 5(10):1781-7. PMID: 20718074 (IF: 3.232).
Serrano A, Pavón FJ, Tovar S, Casanueva F, Señarís R, Diéguez C, de
Fonseca FR.
Oleoylethanolamide: effects on hypothalamic transmitters and gut peptides
regulating food intake.
Neuropharmacology, 2011 Mar; 60(4):593-601. Epub 2010 Dec 21. PMID:
21172362 (IF: 3.909).

16
Gonzalez-Aparicio R, Flores JA, Tasset I, Tunez I, Fernandez-Espejo E.
Mice lacking the PPAR-alpha gene present reduced number of dopamine
neurons in the substantia nigra without altering motor behavior or
dopamine neuron decline over life.
Neuroscience. 2011 [Epub ahead of print] PubMed PMID: 21463665 (IF:
3.292).
Fernández-Castillo N, Orejarena MJ, Ribasés M, Casas M, Robledo P,
Maldonado R, Cormand B.
Active and passive MDMA (‘ecstasy’) intake induces differential
transcriptional changes in the mouse brain.
Genes Brain and Behavior. Under 2nd review (IF: 3.795).
González-Aparicio R, Galán-Rodríguez B, Serrano A, Pavón FJ, Parsons LH,
Maldonado R, Robledo P, Rodríguez de Fonseca F, Fernández-Espejo E.
Systemic oleoylethanolamide exerts dose-dependent neuroprotection of the
nigrostriatal circuit in experimental Parkinsonism.
Psychopharmacology. Submitted (IF: 4.103).
Moreno-Santos I, Romero-Cuevas M, Suardiaz M, Pavón J, Serrano A, Cano
C, Goya P, Rodríguez de Fonseca F, Macías-Gonzalez M.
Sulfamide analogs of oleoylethanolamide as PPARalpha agonist by biological
and computational analyses.
Journal of Biological Chemistry. Submitted (IF: 5.328).
Patentes procedentes de este proyecto
Rodriguez de Fonseca F, Macías-González M, Serrano A, Goya P, Alvarado
M, Helguero J, Jagerovic N.
Pyrazole derivatives of fatty acid amides as PPAR-alpha specific receptor
activators, preparation methods and thereof use of them.
WO2009/050318 A1 (Patente). Fecha de prioridad: 15 Octubre 2007.
Rodríguez de Fonseca F, Suárez Pérez J, Romero-Cuevas M, Fernández-
Espejo E, Goya P, Páez Prósper JA.

17
Uso de derivados de sulfamidas como neuroprotectores. P201130486
(Patente). Fecha de prioridad: 30 Marzo 2011.

18
ANEXO. FIGURAS
Imágenes de inmunofluorescencia de la expresión de OX6 en tejido del
estriado 96 horas después de la infusión bilateral de 6-OHDA y del
pretratamiento con OEA. La OEA se infundió en el estriado derecho a una
dosis de 5µM. Estas imágenes muestran que la reacción de la microglia
estaba presente en ambos estriados dañados, pero el número de células
microgliales reactivas (células ramificadas) era menor en el estriado tratado
con OEA (zona amplificada que muestra microglia OX6+ dispersada con
morfología ramificada). Abreviaturas: str, striatum; cc, corpus callosum
19
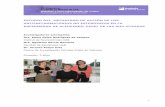
Inmunotinción TUNNEL en el hipocampo y el córtex frontal (A, B y C) i el
estriado (D, E y F). A y D muestran la fragmentación del ADN en un
experimento control positivo. B y E no muestran fragmentación del ADN en
un experimento control negativo. C y F muestran inmunotinción en el
cerebro de ratones tratados de forma repetida con MDMA (30 mg/kg)
OEA y sus análogos oleil-propil-sulfamida interaccionan con el LBD del
receptor PPAR-α
20
Análogos estructurales de la OEA generados en el proyecto