
EVALUACIÓN DE LA GENOTOXICIDAD DE FITOSTEROLES … · FITOSTEROLES PRESENTES EN EFLUENTES DE LA...
88
UNIVERSIDAD CATÓLICA DE LA SANTÍSIMA CONCEPCIÓN FACULTAD DE CIENCIAS EVALUACIÓN DE LA GENOTOXICIDAD DE FITOSTEROLES PRESENTES EN EFLUENTES DE LA INDUSTRIA DE CELULOSA KRAFT BLANQUEADA ELIZABETH DEL PILAR OÑATE CEA Seminario de título entregado a la Facultad de Ciencias de la Universidad Católica de la Santísima Concepción para optar al grado académico de Licenciado en Química, y al título profesional de Químico Marino Mención manejo medio ambiental Profesor Guía: Dra. María Angélica Mondaca Concepción, 2005
Transcript of EVALUACIÓN DE LA GENOTOXICIDAD DE FITOSTEROLES … · FITOSTEROLES PRESENTES EN EFLUENTES DE LA...

UNIVERSIDAD CATÓLICA DE LA SANTÍSIMA CONCEPCIÓN
FACULTAD DE CIENCIAS
EVALUACIÓN DE LA GENOTOXICIDAD DE
FITOSTEROLES PRESENTES EN EFLUENTES DE LA
INDUSTRIA DE CELULOSA KRAFT BLANQUEADA
ELIZABETH DEL PILAR OÑATE CEA
Seminario de título entregado a la Facultad de Ciencias de la Universidad Católica de la Santísima Concepción para optar al grado académico de
Licenciado en Química, y al título profesional de Químico Marino Mención manejo medio ambiental
Profesor Guía: Dra. María Angélica Mondaca
Concepción, 2005

A mi amada y adorada madre, María del Rosario,
por su inmenso amor, apoyo y paciencia,
entregados incondicionalmente en todas las etapas de mi vida,
especialmente en mi educación superior.
Mamá gracias por todo,
te AMO con todas mis fuerzas y corazón.

AGRADECIMIENTOS
A mis queridos padres, Santiago y Maria, y hermanos por el cariñoso apoyo que me han
brindado incondicionalmente con todo su esfuerzo y dedicación, a familia Rodríguez Cea
por su amor y compañía, así también a mis tíos Joel y Carol, y a mi negro regalón, Matías.
A Sandra, mi gran amiga.
A mi profesora guía Dra. María Angélica Mondaca por haberme acogido amorosamente en
su laboratorio entregándome mucha dedicación y paciencia en la ejecución de la tesis.
Agradezco al proyecto FONDECYT 1040987, “Influencia de la tecnología de tratamiento
en la eliminación biológica de compuestos hormonales activos, contenidos en los efluentes
de la industria de celulosa Kraft", el cual proporcionó los recursos económicos para la
realización de esta tesis, y a la Dra. Gladys Vidal, investigadora titular del proyecto, por su
constante preocupación y apoyo.
No puedo dejar de agradecer al grupo de biotecnología, pues en ellos encontré la amistad y
compañerismo altruistas.
Finalmente y por sobre todo, agradezco a Dios por su infinita fidelidad, pues a ÉL le debo
todo lo que tengo y soy. Gracias Dios mío, por tanto amor entregado aún cuando no lo
merezco.

ÍNDICE
CAPÍTULO I
I.1. Introducción......................................................................................................................2
I.2. Hipótesis...........................................................................................................................4
I.3. Objetivos...........................................................................................................................4
CAPÍTULO II
II.1. Antecedentes bibliográficos............................................................................................6
II.1.1. Los extractivos de la madera........................................................................................6
II.1.2. Proceso productivo de la industria de celulosa Kraft.................................................11
II.1.2.1. Tratamiento de los efluentes y su caracterización fisicoquímica............................16
II.1.3. Principales contaminantes presentes en los efluentes y su composición química......20
II.1.4. Toxicidad de los efluentes y sus efectos sobre los organismos acuáticos..................26
II.1.5. Genotoxicidad de los efluentes de la industria celulósica..........................................30
CAPÍTULO III
III.1. Materiales y métodos...................................................................................................39
III.1.1. Diseño experimental..................................................................................................39
III.1.2. Muestras de efluentes de la industria de celulosa Kraft............................................39
III.1.3. Reactivos...................................................................................................................39
III.1.4. Ensayo “rec” de Bacillus subtilis..............................................................................40
III.1.5. Test de Ames.....................................................................................................…...41
III.1.6. Análisis estadístico...................................................................................................43
i

CAPÍTULO IV
IV.1. Resultados....................................................................................................................45
IV.1.1. Ensayo “rec” de Bacillus subtilis..............................................................................45
IV.1.1.1. β-sitosterol..............................................................................................................45
IV.1.1.2. Estigmasterol..........................................................................................................46
IV.1.1.3. Influentes primarios...............................................................................................48
IV.1.1.4. Efluente secundario anaerobio...............................................................................54
IV.1.1.5. Efluente secundario aerobio...................................................................................54
IV.1.2. Test de Ames.............................................................................................................56
IV.1.2.1. β-sitosterol.............................................................................................................56
IV.1.2.2. Estigmasterol..........................................................................................................57
IV.1.2.3.Estigmasterol con activación metabólica................................................................58
IV.1.2.4. Influentes primarios...............................................................................................59
IV.1.2.5. Efluente secundario anaerobio...............................................................................61
IV.1.2.6. Efluente secundario aerobio...................................................................................62
CAPÍTULO V
V.1.Discusión........................................................................................................................65
CAPÍTULO VI
VI.1. Conclusión...................................................................................................................74
CAPÍTULO VII
VII.1. Bibliografía ................................................................................................................76
ANEXO: Publicaciones y congresos ...................................................................................82
ii

RESUMEN
Los efluentes de la industria de celulosa y papel contienen disruptores endocrinos de
naturaleza química originados naturalmente, tales como los fitosteroles o esteroles
vegetales, los cuales están relacionados con una variedad de anormalidades fisiológicas y
morfológicas de los peces habitantes en aguas receptoras de estos efluentes. Así también,
estos constituyentes de los extractivos de la madera serían responsables de los efectos
genotóxicos en los peces expuestos a residuos de la industria de celulosa Kraft, lo cual
sugiere que tales efectos no tienen relación alguna con las tecnologías de blanqueo TCF
(Total Chlorine Free) o ECF (Elementary Chlorine Free).
El objetivo del presente trabajo fue evaluar la genotoxicidad de los efluentes de la industria
de celulosa Kraft y relacionar el efecto genotóxico de dichos efluentes con la presencia de
fitosteroles en los mismos, utilizando el ensayo “rec” con Bacillus subtilis y el test de Ames
con Salmonella typhimurium.
Los resultados obtenidos indican que el β-sitosterol y estigmasterol presentan efecto
genotóxico demostrado por la diferencia relativa de sobrevivencia entre las cepas B. subtilis
rec(+) y B. subtilis rec(-), siendo estos fitosteroles responsables, en parte, de la
genotoxicidad de los efluentes de la industria de celulosa Kraft blanqueada, la que es
reducida luego del tratamiento biológico tanto aerobio como anaerobio. Los ensayos con el
test de Ames no demostraron mutagenicidad evidente de los esteroles vegetales, así mismo
en los efluentes con y sin tratamiento biológico, lo cual significa que las muestras
ensayadas no son mutagénicas. Estos resultados señalan que otros ensayos de
genotoxicidad/mutagenicidad son necesarios para la confirmación de los mismos.
iii

CAPÍTULO I
INTRODUCCIÓN

2
CAPÍTULO I
I.1. INTRODUCCIÓN
Las nuevas tecnologías de blanqueo, libres de cloro elemental (ECF) y totalmente libres de
cloro (TCF), que han realizado una eliminación parcial y total de cloro como agente
blanqueante, han resultado en la disminución de los compuestos organoclorados emitidos al
ambiente (AOX) (Strömberg et al., 1996), los que han sido considerados, tradicionalmente,
responsables del mayor porcentaje de la toxicidad de los efluentes de la industria de
celulosa Kraft blanqueada. Sin embargo, existe evidencia que la toxicidad de estos
efluentes se debe no solamente a compuestos AOX, sino también, a variados compuestos
no clorados provenientes de la fracción extractiva de la madera, tales como los fitosteroles
y ácidos resínicos, los que durante la etapa de pulpaje de la madera (separación de la
lignina de los otros componentes de la madera) quedan parcialmente absorbidos en la
pulpa, la que posteriormente es lavada. Estas aguas de lavado, llevan consigo, entre otros
compuestos, fitosteroles. Estos son finalmente arrastrados por dichas aguas hacia el
efluente total, contribuyendo, por lo tanto, a la toxicidad crónica de los peces expuestos a
estos efluentes. La toxicidad crónica se manifiesta en efectos biológicos tales como:
alteración en el nivel de esteroides sexuales en el plasma de peces y disminución de su
capacidad reproductiva. Estos disruptores endocrinos, llamados también fitoestrógenos, son
conocidos como “precursores de compuestos con actividad hormonal”. En los efluentes de
celulosa Kraft blanqueada existe una cantidad detectable de fitosteroles, siendo el β-
sitosterol el compuesto predominante del total de esteroles detectados (25-47%)
(Mahmood-Khan & Hall, 2003). Una vía de eliminación de los fitosteroles presentes en los
efluentes es mediante los tratamientos biológicos, sin embargo el porcentaje de remoción
puede ser variable (Kostamo & Kukonen, 2003).
De los contaminantes identificados en los efluentes de la industria de celulosa Kraft
blanqueada, un número significativo son clasificados como clastogénicos, carcinogénicos y
genotóxicos.

3
Las sustancias genotóxicas presentes en los efluentes no tendrían relación alguna con el
cloro o tipos de procesos de blanqueo. Esto sugiere que los derivados provenientes de los
extractivos de la madera serían los responsables de dichos efectos genotóxicos (Ericson &
Larsson, 2000). Los ácidos resínicos son conocidos por su genotoxicidad (Maria et al.,
2004; Pacheco & Santos, 2002), mientras que la escasa información sobre los fitosteroles
señala que estos no muestran actividad mutagénica evidente (Wolfreys & Hepburn, 2002).
No obstante, se ha establecido la actividad genotóxica de metabolitos de estradiol y estrona
(estrógenos).
Los tratamientos secundarios podrían reducir o eliminar la genotoxicidad de los efluentes,
sin embargo el tratamiento no siempre reduce la genotoxicidad, sino que, en algunas
instancias, puede incrementarla. Por lo tanto, los ensayos de genotoxicidad constituyen una
medida efectiva para el monitoreo medio ambiental y para seleccionar procesos de
tratamientos apropiados (Claxton et al., 1998).

4
I.2. HIPÓTESIS
Los fitosteroles, constituyentes de la fracción extractiva de la madera, son responsables de
la genotoxicidad presente en los efluentes de la industria de celulosa Kraft. Dicha
genotoxicidad es reducida por los tratamientos biológicos aerobios y anaerobios.
I.3. OBJETIVOS
I.3.1 Objetivo general
Evaluar la genotoxicidad de los efluentes de la industria de celulosa Kraft y relacionar el
efecto genotóxico de dichos efluentes con la presencia de los fitosteroles en los mismos.
I.3.2. Objetivos específicos
• Evaluar la genotoxicidad de β-sitosterol y estigmasterol.
• Evaluar la genotoxicidad de los efluentes provenientes de la industria de celulosa Kraft
blanqueada sin previo tratamiento biológico.
• Evaluar la genotoxicidad de los efluentes tratados aeróbica y anaeróbicamente.

CAPÍTULO II
ANTECEDENTES BIBLIOGRÁFICOS

6
CAPÍTULO II
II.1. ANTECEDENTES BIBLIOGRÁFICOS
II.1.1. Los extractivos de la madera
Los principales componentes de la madera son: celulosa, hemicelulosa, lignina y
extractivos apolares de la madera.
La celulosa es un polisacárido resultante de la polimerización lineal de grupos anhidro-
glucosa, con enlaces 1-4 β glucosídicos, insoluble en agua y álcalis o en ácidos diluidos a
temperatura ambiente (Vidal, 1994).
La hemicelulosa está formada por polisacáridos no celulósicos, producto de la
polimerización heterogénea de varios azúcares de cinco y seis átomos de carbono, pentosas
(xylosa, arabinosa) y hexosas (manosa, glucosa, galactosa) respectivamente, y de ácidos de
azúcares. La hemicelulosa de la madera dura contiene principalmente xylanos, mientras que
la hemicelulosa de la madera blanda contiene principalmente glucomananos, de aquí su
heterogeneidad química. La hemicelulosa, es el segundo polisacárido más común en la
naturaleza, representa sobre el 20-35% de la biomasa lignocelulósica (Saha, 2003). El
grado de polimerización es bajo por lo que su solubilidad en agua, álcalis y ácidos diluidos
a temperatura ambiente es algo superior (Vidal, 1994).
La lignina es una sustancia mucho más compleja, resultante de la polimerización reticulada
en tres dimensiones de monómeros aromáticos, con un grado de polimerización aún no
definido (Vidal, 1994). Es un polímero formado esencialmente por unidades de
fenilpropano. Los grupos funcionales de la lignina, asociados a estas unidades de
fenilpropano, varían según la especie, los más importantes son: metóxilo, hidróxilo
fenólico, hidróxilo alifático, carbonilo. Las principales unidades estructurales presentes en
la lignina son el alcohol trans-coniferílico, alcohol trans-sinalpílico y el alcohol trans-p-
cumarílico (Guiñez, 2002).

7
Por otra parte, los extractivos son sustancias lipofílicas constituyentes de la madera,
solubles en solventes orgánicos o apolares, comúnmente conocidos como resinas de pino
(pich), constituídos principalmente de: triglicéridos, ácidos grasos y resínicos, ceras,
alcoholes grasos, esteroles y esteril ésteres (Dorado et al., 2001). La mayor parte de la
fracción no polar de la madera, está compuesta por ácidos resínicos y grasos (Ucar &
Balaban 2002), no obstante, esto puede cambiar según sea la especie. En la Tabla II.1 se
muestra la composición de los extractivos en dos maderas distintas, pino y eucaliptos.
Tabla II.1. Principales constituyentes de los extractivos provenientes de diferentes especies
de madera.
Constituyentes de la fracción lipofílica de la madera
Pinus sylvestris (mg/g madera)
Eucaliptus globulus ( mg/g madera)
Ácidos grasos
4,0
0,28
Ácidos resínicos
8,9
N.E.
Esteroles (*)
0,2
0,49
Ceras
1,6
0,06
Ésteres de esteroles
1,2
0,52
Triglicéridos
7,3
0,13
N.E.: no estudiado (*) el valor dado corresponde sólo a la concentración de β-sitosterol.
La variabilidad de las especies de árboles, no sólo está dada por sus diferencias
morfológicas, sino también por su composición química especialmente de los extractivos,
los cuales son responsables del color y olor de la madera y en el mismo alcance, de sus
propiedades físicas.

8
Estos extractivos juegan un rol importante en la protección de la madera hacia ataques
biológicos, tanto del árbol viviente como de los productos de la madera. Sin embargo,
pueden tener efectos adversos en el procesamiento de la madera, causando problemas
técnicos y ambientales (Dorado et al., 2001).
Las composiciones en promedio, sobre la base de peso seco, de la madera son: celulosa
(43%), hemicelulosa (25%) y lignina (28%) y en menor extensión los extractivos de madera
(4%). No obstante, estos valores pueden variar según el tipo de madera, de éste modo, la
madera blanda (MB) como el Pinus pinaster o Pinus radiata y madera dura (MD) como
Eucaliptus globulus o Populus nigra se diferencian fundamentalmente en su composición
química, tal como se muestra en la Figura 2-1.
Figura 2-1. Composición química de la madera.
Hemicelulosa Celulosa
Carbohidratos
Extractivos
MADERA
Lignina
MD: 21% MB: 25%
45%
2-8%
MD: 35% MB: 25%

9
Por otro lado, con relación a los extractivos de la madera, se observa que las maderas
blandas, como el pino, presentan mayor cantidad de extractivos (0,5- 7,0%) que las
maderas duras, como es el caso del eucaliptos (0,2-3,5%) (La Fleur, 1996). Así también, el
contenido de los extractivos fluctúa según el tipo de crecimiento de la madera, ya sea éste
rápido o lento. De este modo, el total de los extractivos es de 4,2 y 5,1% para árboles de
crecimiento rápido y lento, respectivamente (Ucar & Balaban, 2002). Por otra parte, las
maderas blandas son ricas en ácidos resínicos (La Fleur, 1996; Mellanen et al., 1996),
mientras que en las maderas duras abundan los fitosteroles (La Fleur, 1996; Verta et al.,
1996). No obstante, en el Pinus nigra var. pallasiana (MB) se ha encontrado altas
cantidades de esteroles y menores de ácidos resínicos , mientras que en el pino Tracian
black (MD) la concentración de los ácidos resínicos fue mayor que la de esteroles, siendo
esto considerado como un “ecotipo geográfico” (Ucar & Balaban, 2002), atribuyéndose
éste incremento a condiciones ecológicas y climáticas de la región. Cualquiera sea la
especie de madera blandas, duras, de crecimiento rápido o lento, éstas contienen extractivos
en variadas concentraciones.
En la madera de Eucaliptus globulus, los esteroles y ésteres de esteroles encontrados y sus
respectivas concentraciones en (g/g madera) fueron: campesterol (1,15), β-sitosterol (48,1),
estigmastanol (5,8), fucosterol (2,3), cicloartenol (1,7), 24-metilenecicloartenol (0,96),
citrostadienol (1,54), ésteres de β-sitosterol (33,3) y ésteres de estigmastanol (6,7) (del Río
et al., 1999).
Las concentraciones de esteroles encontradas en dos estaciones muestreadas del río
Lanyang en el noreste de Taiwan, y sus respectivas concentraciones en µg/g de peso seco
del material suspendido total, para una estación cercana a una reserva natural, fueron
(µg/g): campesterol (14,8), estigmasterol (45,2) y β-sitosterol (101,0) ( Jeng & Kao, 2002),
mientras que para la estación que está lejos de la reserva natural, las concentraciones fueron
menores: campesterol (4,13), estigmasterol (1,69) y β-sitosterol (6,39) ( Jeng & Kao, 2002),
esto deja en evidencia que las plantas mayores son fuentes predominantes de esteroles, pues
se supone que producto del aumento de las perturbaciones antropogénicas, la erosión de los

10
suelos se ve agravada y en consecuencia, estos compuestos presentes en los árboles llegan a
las aguas de dicho río.
Dado que los esteroles, al igual que los ésteres de esteroles, son los constituyentes más
abundantes de la fracción lipofílica de la madera (Tabla II.1), son los principales
responsables de los problemas generados sobre la calidad del producto y operacionales, así
también de los impactos ambientales. De esta forma, los esteroles vegetales o fitosteroles
juegan un papel importante como constituyentes de los extractivos de la madera, pues son
responsables de la toxicidad de los efluentes generados en la industria de celulosa Kraft.
Algunos fitosteroles comúnmente encontrados en los efluentes de la industria de celulosa
Kraft son: β-sitosterol, estigmasterol, campesterol y estigmastanol. La Figura 2-2 muestra
la estructura química de los esteroles anteriormente mencionados.
Figura 2-2: Estructura molecular de esteroles vegetales (fitosteroles).
HO
Sitosterol
HO
Estigmasterol
HO
Campesterol
HO
Estigmastanol

11
II.1.2. Proceso productivo de la industria de celulosa Kraft
La celulosa constituye el principal producto chileno de exportación forestal. El tipo de
celulosa que más se produce en Chile es la blanca de fibra larga BSKP (Bleached Softwood
Kraft Pulp) de Pinus radiata, destinada principalmente a la exportación. También se
produce celulosa de fibra larga no blanqueada UNKP (Unbleached Kraft Pulp) y la celulosa
blanca de fibra corta BEKP (Bleached Eucalyptus Kraft Pulp). Chile produce actualmente,
cerca de 2,8 millones de toneladas de celulosa Kraft, tanto cruda (15%) como blanqueada
(85%). El país exporta el mayor porcentaje de su producción (82%), a países de la
comunidad europea, norteamericana y secundariamente a Asia.
La pulpa es un producto que se obtiene de la madera después de separar las fibras de
celulosa de los otros compuestos orgánicos (hemicelulosa, lignina, fitosteroles, extractos
resinosos, entre otros) presentes en éste recurso.
En general y tal como se ilustra en la Figura 2-3, el proceso de producción de la celulosa se
puede separar en cinco operaciones unitarias claramente definidas: preparación de la
madera, etapa de pulpaje, separación y lavado de la pasta, blanqueo, secado y embalado.
También se identifica a la etapa de pulpaje como una fase crítica, pues compuestos como
los fitosteroles, liberados en esta etapa desde la fracción extractiva de la madera, serían
probablemente responsables de la genotoxicidad del efluente final de la industria de
celulosa Kraft.
A continuación, se hace una descripción sucinta de los principales procesos que intervienen
en la producción de celulosa Kraft y se identifican los principales contaminantes generados
en la etapa de reducción de la madera o etapa de pulpaje.

13
En una primera etapa la madera es descortezada y luego reducida a astillas. La corteza
desprendida es usada como combustible en las calderas de poder para generar vapor y
energía eléctrica, insumos requeridos en distintas fases de la planta. Próximamente, las
astillas ingresan a un digestor continuo dentro del cual son sometidas a cocción junto con el
licor blanco, compuesto por hidróxido de sodio (NaOH) y sulfuro de sodio (Na2S), a
condiciones controladas de temperatura (130-170°C) y presión. El propósito de la etapa de
cocción es liberar las fibras de celulosa de los otros componentes de la madera, mediante la
disolución de la lignina que las mantiene unidas. Continuamente, se agregan y retiran
líquidos de cocción, y en la medida que la mezcla de astillas va descendiendo dentro del
digestor, se transforma lentamente en una pasta compuesta por fibras de celulosa, lignina y
licor de cocción, mientras que una parte importante de la lignina disuelta es retirada en los
líquidos de cocción en forma de licor negro (mezcla de licor blanco y lignina disuelta). En
esta fase se genera una gran cantidad de residuos de fibra de celulosa, los que al ser
depositados en los cuerpos de agua, generan un exceso de materia orgánica. En
consecuencia, el efluente líquido generado está compuesto por sólidos suspendidos (fibras)
y productos de degradación de la lignina. Es de crucial importancia destacar que en la etapa
de pulpaje se liberan también compuestos que forman parte de la fracción extractiva de la
madera tales como fitosteroles (β-sitosterol, estigmasterol, campesterol, entre otros) y
ácidos resínicos ( ácido abiético, ácido dehidroabiético, etc).
La pasta fibrosa proveniente de la etapa de reducción kraft, sigue una etapa de lavado a
altas temperaturas dentro del digestor, en el cual flujos de agua a contracorriente van
eliminando el licor negro de la pasta, el que es posteriormente tratado para regenerar el
licor blanco, a través de un proceso de caustificación. Sin embargo, esto no significa la
ausencia de residuos líquidos con alto contenido de sustancias orgánicas (lignina, ácidos
resínicos, fitosteroles, entre otros), tanto para la fase de pulpaje de la madera como de
separación y lavado.

14
Más tarde, la pasta continúa varias etapas de lavado para eliminar el resto de licor negro,
luego pasa al clasificador de nudos (astillas que no alcanzaron una cocción completa). Estos
son devueltos al digestor continuo y las fibras clasificadas pasan a sucesivas etapas de
separación y lavado, obteniéndose finalmente la celulosa Kraft sin blanquear (pasta café)
que posee todavía altos niveles de lignina.
Los extractivos de la madera tales como los fitosteroles, originados previamente en la etapa
de pulpaje, son liberados en las aguas de lavado de la madera y arrastrados finalmente por
estas aguas hacia el efluente total, así estos compuestos causan serios disturbios fisiológicos
y bioquímicos sobre los peces expuestos a los efluentes de la industria de celulosa Kraft,
pues actúan como disruptores endocrinos en organismos habitantes de aguas receptoras de
estos residuos industriales. Por lo que, los fitosteroles son conocidos como fitoestrógenos o
pueden actuar como precursores de compuestos con actividad hormonal. Es sabido que los
esteroides hormonales poseen propiedades genotóxicas, no obstante, no se conoce
información respecto de la genotoxicidad de los fitosteroles, como constituyentes de estos
efluentes y contribuyentes de la toxicidad de los tales.
La pasta clasificada y lavada, prosigue varias etapas de blanqueo para eliminar el
remanente de lignina contenida en la pasta. Para ello, se utilizan diferentes productos
químicos, tales como, cloro elemental (Cl2), dióxido de cloro (ClO2), oxígeno (O2),
peróxido de hidrógeno (H2O2) y soda cáustica (NaOH). El proceso de blanqueo se lleva a
cabo a través de una serie de etapas, las que constituyen secuencias de blanqueo, usadas
alternativamente. Estas secuencias se clasifican en tres grupos: estándar, se utiliza cloro
elemental; ECF, libre de cloro elemental, se utiliza ClO2; TCF, totalmente libre de cloro, es
decir, no se utiliza ningún agente clorado durante la secuencia.
Las plantas de celulosas más modernas cuentan con un proceso adicional de pre-
deslignificación con oxígeno, reduciendo sustancialmente el consumo de químicos clorados
de blanqueo. Algunas plantas usan ozono (O3) y enzimas como agentes blanqueantes. En la
fase de blanqueo se producen reacciones que solubilizan la lignina residual y se forman
grupos cromóforos, los que dan el color café rojizo a la salida de la etapa de cocción (licor
negro), lográndose una pulpa blanqueada.

15
La pasta procedente de la planta de blanqueo es distribuida uniformemente sobre el
fourdrinier para extraer el exceso de agua produciéndose las primeras uniones entre las
fibras. A continuación, es conducida hacia las prensas de rodillos, donde se le extrae gran
cantidad de agua. La hoja, que a esta altura posee una consistencia de aproximadamente un
46%, entra a los presecadores y luego a los secadores principales. A la salida de esta área,
la hoja posee una consistencia de 87-92% seco.
Finalmente, la hoja pasa por la cortadora dejándola en forma de pliegos, se embalan en
forma de units y se pesan antes de ser guardados en las bodegas. También existe la
posibilidad de enrollar la hoja de celulosa sin usar la cortadora, formándose las bobinas.

16
II.1.2.1. Tratamiento de los efluentes y su caracterización fisicoquímica
Algunas variables relevantes para medir el nivel de toxicidad de las descargas de los
efluentes son: sólidos suspendidos totales (SST), demanda bioquímica de oxígeno,
usualmente medida después de 5 días, por lo que se conoce como DBO5, y los AOX, que
mide la concentración de compuestos organoclorados. Finalmente, el pH, debe aproximarse
a neutro. La DBO5 es originada por la presencia de compuestos fácilmente biodegradables,
tales como los carbohidratos y los ácidos orgánicos. La toxicidad se atribuye a compuestos
extractivos de la madera, tales como terpenos volátiles, ácidos resínicos y fitosteroles, y a
los efluentes de blanqueo, que contienen compuestos fenólicos clorados.
A continuación en la Figura 2-4, se muestra un esquema explicativo de los tratamientos
aplicados a residuos de la industria de celulosa Kraft.
Figura 2-4. Tratamientos aplicados sobre residuos líquidos de la industria de celulosa
Kraft.
Tratamiento terciario
TRATAMIENTOS
Tratamiento primario
Tratamiento secundario
anaerobio aeróbio

17
-Tratamiento primario
Los residuos líquidos procedentes de los diversos procesos de la planta son llevados a la
planta de tratamiento primario, donde existe un clarificador o piscina de decantación, cuya
función es retirar los sólidos suspendidos. Las fibras son llevadas a la superficie del agua
con la ayuda de burbujas de aire inyectadas en el fondo y retiradas por rebalse a través de
los bordes superiores de la piscina. Los sólidos más pesados, una vez decantados, son
retirados desde el fondo por rastrillos rotatorios, los que posteriormente, junto con las
fibras, son prensados para retirarle el agua excedente y depositados en vertederos
especialmente habilitados o para ser quemados en calderas de poder.
El efluente prosigue a una etapa de neutralización, donde se le agregan aditivos químicos
neutralizantes para que los residuos finales no sean ácidos ni alcalinos. Posteriormente, el
efluente está en condiciones para ser devuelto al río. Algunas plantas más modernas, como
las recientemente construidas en Chile, poseen un tratamiento secundario para estos
efluentes, cuyo objetivo principal es reducir la carga orgánica.
-Tratamiento secundario
Tratamiento biológico aerobio: los tratamientos aerobios han sido los primeros
tratamientos biológicos secundarios aplicados, con el objetivo de reducir la carga orgánica
biodegradable (DBO5), siendo habituales tanto en las industrias de pulpa como en las
integradas de pulpa y papel. En los procesos de lodos activos se alcanzan elevados
porcentajes de eliminación de la carga en DBO5, generalmente en el rango 65-99% (Vidal
et al., 1996). La eficacia de eliminación de la DQO varía en función de la relación
DBO/DQO, siendo inferior en el caso de los efluentes procedentes de los procesos de
pulpado químico, en los que dicha relación es baja. La eliminación de color es
prácticamente nula.

18
El tratamiento aerobio reduce fuertemente la toxicidad acuática de los efluentes de
digestión y o cocción de las pulpas mecánicas o químicas, al degradarse aquellas sustancias
como los ácidos grasos, ácidos orgánicos y ciertos extractivos de la madera. Durante el
tratamiento de aguas residuales, las concentraciones de extractivos fueron reducidas sobre
un 97%. En el tratamiento con lodo activado sobre un 94% de los ácidos resínicos y sobre
un 41 % de los esteroles fueron degradados o transformados en otros compuestos (Kostamo
& Kukonen, 2003). No obstante, estos porcentajes de remoción pueden ser variables.
Tratamiento biológico anaerobio: la aplicación de la digestión anaerobia al tratamiento de
los efluentes de la industria de la pulpa y papel se inició a mediados de la década de los
ochenta, se incrementó fuertemente su presencia en los últimos años de la misma. La
digestión anaerobia presenta diferentes ventajas frente al tratamiento aerobio (Tabla II.2),
principalmente cuando los efluentes están fuertemente contaminados por la materia
orgánica.
Tabla II.2. Ventajas de los tratamientos aerobio y anaerobio.
Tratamiento aerobio
Tratamiento anaerobio
Mayor estabilidad del proceso Mayor capacidad de eliminación de la DBO5
Proceso de degradación más rápido Menor tiempo de retención hidráulico Menor volumen de muestra
Menor tasa de producción de fangos Bajo consumo energético Operatividad con mayor DBO del influente Generación de metano como producto secundario utilizable
Por tanto, la aplicación del tratamiento anaerobio es preferentemente indicada en los
efluentes de pulpado mecánico, termomecánico o químico-mecánico y a los condensados
de los evaporadores de los sistemas de recuperación en los procesos Kraft.

19
Otro aspecto a considerar, a favor de la digestión anaerobia de este tipo de efluentes, es el
menor requerimiento de nutrientes que en el caso de los tratamientos aerobios, tanto desde
un punto de vista económico como ambiental. Con aquellos efluentes de mayor
biodegradabilidad y baja toxicidad los porcentajes de depuración varían entre el 60-99%.
Como ya se ha comentado, parte de los efluentes generados por los procesos de pulpa de
celulosa y papel (principalmente aquellos procedentes de los procesos de blanqueo, pulpado
semiquímico y pulpado químico) son de difícil biodegradabilidad y presentan toxicidad
para la biomasa implicada en los sistemas biológicos de tratamientos. Sin embargo, una
dilución apropiada de los efluentes hace posible la puesta en marcha adecuada de los
digestores para que los microorganismos anaerobios tengan una máxima adaptación.
-Tratamiento terciario
Si la aplicación de tratamientos secundarios biológicos no ha sido suficiente, se puede
pensar en aplicar algunos de los pre-tratamientos de detoxificación, tales como adsorción,
precipitación u oxidación de los tóxicos orgánicos. Otros procesos como, la filtración con
arena y osmosis inversa se pueden utilizar para separar cualquier contaminante residual que
no haya sido eliminado durante los procesos de tratamiento biológico previos. No obstante,
la aplicación de un tratamiento anaerobio-aerobio puede evitar en gran medida la toxicidad
y viabilizar la eliminación de la DBO5.

20
II.1.3. Principales contaminantes presentes en los efluentes y su composición química
Los agentes blanqueadores más comúnmente usados en la industria de celulosa y papel son
los químicos clorados. Sin embargo, el uso de estos reactivos resulta en la formación de
compuestos organoclorados prioritariamente polutantes (del Río et al., 2000). El uso de
métodos de blanqueo con oxígeno, ozono o peróxido de hidrógeno (TCF), saludables para
el ambiente en la producción de la pulpa, están reemplazando los químicos clorados de
blanqueo. No obstante, nuevos problemas respecto de la toxicidad de los efluentes tienen
origen con la introducción de los procesos TCF. Algunos de ellos, dicen relación con los
compuestos constituyentes de los extractivos de la madera, los cuales causan problemas
ambientales en la industria de celulosa y papel. Así, durante la etapa de pulpaje, los
extractivos de la madera forman partículas coloidales de resinas. Los fitosteroles, especies
químicas constituyentes de estos coloides, sobreviven a la etapa de pulpaje siendo liberados
en las aguas de lavado durante la etapa de cocción y de blanqueo, pudiendo quedar
almacenados en diferentes partes de la industria o suspendidos en estas aguas hasta
finalmente ser descargados en los efluentes, aún cuando la cantidad de los extractivos de la
madera en las aguas de lavado disminuye dramáticamente a través de los procesos (cerca
del 99% de reducción)(Gutiérrez et al., 2001).
Un estudio realizado por la empresa nacional de celulosa de Pontavedra, España, la cual usa
madera de eucaliptos en un proceso Kraft con blanqueo TCF, caracterizó extractos
coloidales y aguas de procesos, y fueron comparados con la composición lipofílica de la
madera. La Tabla II.3 muestra la composición de la fracción extractiva de la madera, y la
Tabla II.4 los principales esteroles libres y esterificados. Presentes en las muestras antes
mencionadas.
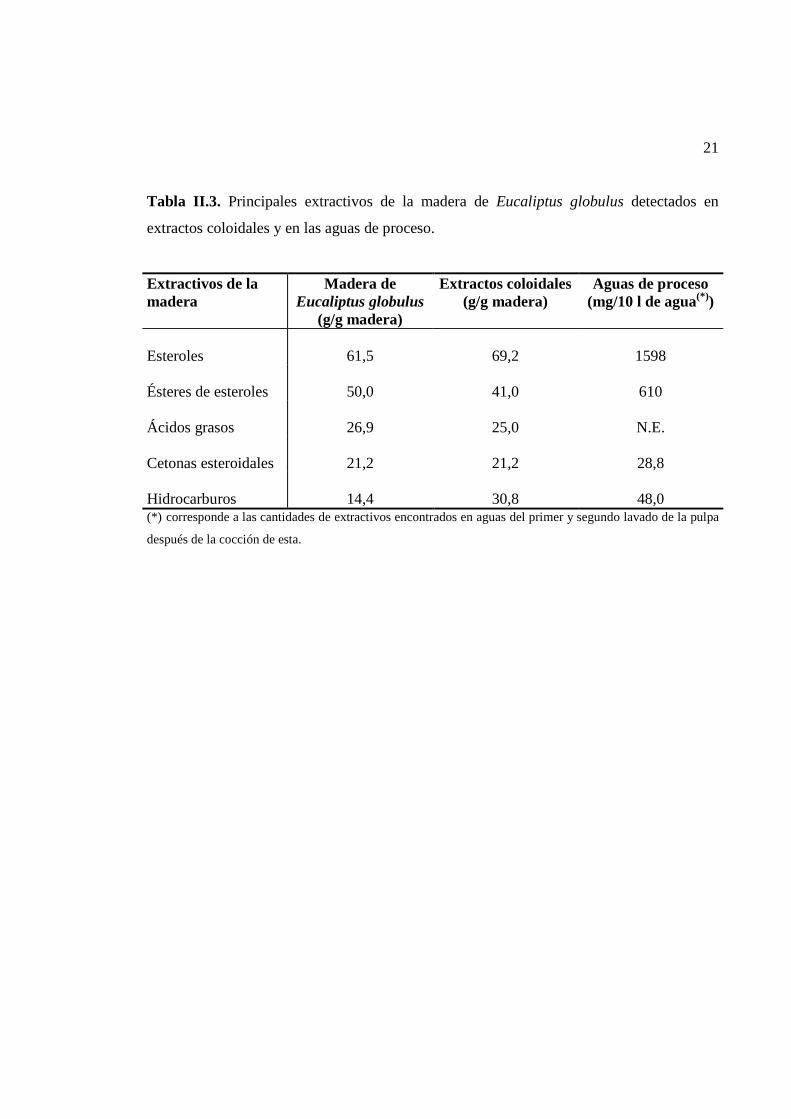
21
Tabla II.3. Principales extractivos de la madera de Eucaliptus globulus detectados en
extractos coloidales y en las aguas de proceso.
Extractivos de la madera
Madera de Eucaliptus globulus
(g/g madera)
Extractos coloidales (g/g madera)
Aguas de proceso (mg/10 l de agua(*))
Esteroles
61,5
69,2
1598
Ésteres de esteroles
50,0
41,0
610
Ácidos grasos
26,9
25,0
N.E.
Cetonas esteroidales
21,2
21,2
28,8
Hidrocarburos
14,4
30,8
48,0
(*) corresponde a las cantidades de extractivos encontrados en aguas del primer y segundo lavado de la pulpa
después de la cocción de esta.

22
Tabla II.4. Principales esteroles y esteres de esteroles provenientes de la fracción
extractiva de la madera de Eucaliptus globulus presentes en extractos coloidales y en las
aguas de proceso.
Compuestos Madera de Eucaliptus globulus
(g/g madera)
Extractos coloidales
(g/g madera)
Aguas de proceso (mg/10 l de agua(*))
β-sitosterol
48,1
51,9
1134
Estigmastanol
5,80
13,5
236
Fucosterol
2,30
1,54
145
Campesterol
1,15
0,19
N.E.
Otros esteroles
4,23
2,12
83
Ésteres de β-sitosterol
33,7
27,5
243
Ésteres de stigmastanol
6,70
9,60
85
Otros ésteres de esteroles
10,0
3,80
282
(*) corresponde a las cantidades de extractivos encontrados en aguas del primer y segundo lavado de la pulpa
después de la cocción de esta.
En consecuencia, la composición de los extractivos encontrados en las partículas coloidales
y en las aguas de procesos, fue similar a la composición lipofílica de la madera de
eucaliptos. Del mismo modo, la composición de los esteroles libres y esterificados fue
proporcional a la encontrada en la madera, siendo el β-sitosterol y ésteres de β-sitosterol y
de estigmastanol los más predominantes (del Río et al., 1999).
Aunque los extractivos de la madera corresponden a una pequeña fracción (0.26%) de esta,
muchos de sus constituyentes, como ya se ha dicho, permanecen aún después del pulpaje
contribuyendo a la formación de los extractos resinosos constituidos especialmente de
esteroles y ésteres de esteroles, los que son posteriormente dispersados en la fase acuosa.

23
Esto indica, que bajo condiciones alcalinas en la cocción Kraft, los esteroles y ésteres de
esteroles forman jabones, no ocurriendo mayores cambios estructurales de estos
compuestos, por lo tanto, son los principales constituyentes de los residuos líquidos
generados (del Río et al., 1999).
La Tabla II.5 muestra las concentraciones de los esteroles totales encontrados en los
efluentes tratados y no tratados de la industria de celulosa y papel, las cuales poseen
distintos procesos de reducción de la madera (reducción mecánica o química) y sistemas de
tratamiento de sus efluentes (lagunas aireadas, lodo activado). Mientras que la Tabla II.6
muestra las concentraciones de los fitosteroles particulares presentes en los efluentes
tratados y no tratados.

24
Tabla II.5 . Concentración de esteroles totales presentes en los efluentes tratados y no
tratados.
Madera
Esteroles totales a (µg / l)
Antes Después
Referencia
M b
361-436
353-131
Mahmood-Khan & Hall, 2003
MB
NM
255
Van der Heuvel et al., 2002
MB/MD
1027
100
Mattson et al., 2001
MB/MD
NM c
72-1056
Mellanen et al., 1999
MB
350-1734 d
77-555
Cook et al., 1997
MB
66,67-166,67 e
10-55
Stromberg et al., 1996
MD
37-755,6
3,4-282
Stromberg et al., 1996
MB
20-40
<10-220
Verta et al., 1996
MB/MD
1060-3420
50-210
Verta et al., 1996
MD
610-680
400
Verta et al., 1996
M b
1200 f
280 f
MacLatchy and van der Kraak, 1995
(a) esteroles totales antes y después del tratamiento biológico para distintos procesos y plantas, (b) no está
claro el tipo de madera empleada, (c) no medido, (d) valores considerando remoción media de 78% de la
cantidad mínima emitida y 68% de remoción de la mayor cantidad emitida, (e) menor y mayor valor medidos
por el autor para distintas plantas que procesan el mismo tipo de madera, (f) exclusivamente β-sitosterol.

25
Tabla II.6. Concentración de los fitosteroles presentes en los efluentes tratados y no
tratados.
Madera
Fitosteroles
Concentración de Fitosteroles (µg /l)
Antes Después
Referencia
MB
Campesterol
Estigmasterol
β-sitosterol
Estigmastanol
16,550 – 87,750
23,640 – 105,34
283,77 – 1422,0
26,000 – 05,340
3,64 –28,10
5,20 – 33,71
62,43 – 455,10
5,72 – 33,70
Cook et al., 1997 a
MB/MD
β-sitosterol
-----------
44 – 711
Mellanen et al.,
1999
MB
Campesterol
Estigmasterol
β-sitosterol
β-sitostanol
----------
----------
----------
----------
7,6
21,2
165,4
61,2
Van der Heuvel et al., 2002
M
Campesterol
Estigmasterol
β-sitosterol
β-sitostanol
85 - 67
38 - 30
126 - 240
112 – 98
52 - 36
18 - 20
17 - 37
21 - 38
Mahmood-Khan &
Hall, 2003
(a) estimaciones realizadas por Cook et al., 1997, considerando un caudal de 192.21 m 3/ ton. y 17.84 m 3/ ton.
de pulpa producida para el mínimo y máximo valor dado, respectivamente.

26
II.1.4. Toxicidad de los efluentes y sus efectos sobre los organismos acuáticos
A partir de los estudios realizados sobre Perca fluvialitis (1984/1985), se ha demostrado
que los efluentes provenientes de las industrias de celulosa Kraft con blanqueo
convencional causan serias alteraciones sobre las funciones bioquímicas y fisiológicas
vitales, las cuales pueden afectar la sobrevivencia de los peces. Los resultados revelan que
los peces son fuertemente afectados por estos efluentes. Algunos síntomas son: reducción
del crecimiento gonadal, dilatación del hígado, fuerte inducción de la actividad de
ethoxyresorufin-O-deethylase (EROD) en el hígado, alteración en el metabolismo de
carbohidratos, disturbios en el balance iónico del plasma, estimulación en la producción de
células rojas y una alterada capa de células blancas indicando un debilitado sistema inmune.
La disminución en la talla de las gónadas indica inhibición del crecimiento gonadal y un
subsiguiente daño o retardo en el desarrollo de las células germinales. Esto va acompañado
por reducción de los niveles esteroidales, lo cual fortalece las sospechas de que la
exposición de los peces a los efluentes de la industria de celulosa Kraft blanqueada produce
serios disturbios en su reproductividad fisiológica. En consecuencia, los parámetros
bioquímicos y fisiológicos (ej. Actividad de EROD, tamaño de gónadas, cantidad de
linfocitos, metabolitos de carbohidratos y composición ionico del plasma), pueden ser
usados como indicadores sensitivos y confiables de los efectos tóxicos sobre las
poblaciones naturales de peces expuestos a estos efluentes (Förlin et al., 1995). Aunque la
toxicidad de los efluentes en general ha disminuido durante las últimas décadas,
alteraciones reproductivas son comunes en los peces receptores y se han asociado con un
estado hormonal alterado.
La disrupción endocrina observada en los peces expuestos a los efluentes de las industrias
de celulosa Kraft, dice relación con los compuestos organoclorados como las dioxinas y
furanos, que se generan durante el proceso de blanqueo con cloro elemental o estándar. Sin
embargo, los efectos sobre la reproducción de los organismos acuáticos, en análisis de
terreno y laboratorio, para los efluentes provenientes de los procesos de blanqueo ECF y
TCF, se siguen observando.

27
Esto sugiere que los compuestos derivados de la madera, como los fitosteroles, presentes en
los efluentes de la industria de celulosa Kraft, pueden causar cambios hormonales en peces,
estos compuestos hormonalmente activos llamados fitoestrógenos, han sido identificados
en numerosas plantas comestibles y asociados con efectos adversos sobre la reproducción
del ganado y de la vida silvestre. La madera contiene fitoestrógenos tales como el β-
sitosterol. Estos estrógenos derivados de la madera incluyendo el β-sitosterol pueden
suprimir la producción de esteroides testiculares y consecuentemente impedir la capacidad
de reproducción en el pez macho (Mellanen et al., 1999). Además, se ha encontrado que el
poliqueto Mustela putorius expuesto a dosis de fitosteroles, además de sufrir disrupción
endocrina, cambios en los niveles de testosterona y estradiol, su metabolismo intermediario
también se ve afectado con un incremento en el contenido de glicógeno en el hígado, así
como una aumentada actividad de glucosa 6-fosfatasa del riñón y disminución en los
niveles de colesterol “malo” o LDL (Lipoprotein Density Light) (Nieminen et al., 2002).
Por tanto, se ha demostrado que los efluentes de la industria de celulosa Kraft pueden ser
una fuente de fitosteroles (fitoestrógenos) para el medio acuático. Así, el β-sitosterol reduce
la capacidad reproductora en el pez Carassius auratus por disminución de niveles de
testosterona y 11-Ketotestosterona en los machos, y niveles de testosterona y 17 β-estradiol
en las hembras. Además, el β-sitosterol induce la expresión del gen vitelogenino en trucha
Oncorhynchus mykiss (Nieminen et al., 2002).
El hígado de pez macho o juvenil puede ser inducido por estrógenos para sintetizar y
secretar grandes cantidades de ésta proteína. Esta inducción conduce a la disminución del
tamaño de huevo y gónada, decrecimiento de las características sexuales secundarias y
reducción de los niveles de esteroides sexuales del plasma luego de la exposición a β-
sitosterol, pudiendo ser, tales síntomas, independientes de los efectos en la gónada
pituitaria. De esta manera, el nivel de vitelogenina en pez macho, puede ser usado como un
biomarcador ecotoxicológico específico y confiable de contaminación estrogénica de los
efluentes (Mellanen et al., 1999).

28
Algunos resultados indican que el efecto estrogénico está asociado con la exposición hacia
los efluentes de la industria de celulosa Kraft, pues el incremento de la expresión del gen
vitelogenino en peces machos, visto en la cercanía de la industria en estudio, es inducido
por contaminantes estrogénicos derivados de la madera o sus semejantes presentes en los
efluentes. La industria de celulosa Kraft blanqueada es responsable de las grandes
descargas de compuestos derivados de la madera tales como, esteroles y ácidos resínicos,
en fracciones solubles y particuladas, dentro del lago. Compuestos estrogénicos
descargados en el licor negro y en los efluentes pueden causar continuos problemas de
reproducción en los peces, ya sea en ausencia de blanqueo con cloro o en presencia de
tratamiento secundario.
Por otra parte, un fuerte efecto de algunos efluentes de la industria de celulosa y papel es el
desarrollo de características sexuales secundarias masculinas (disrupción endocrina
androgénica) en pez hembra Poecilia reticulata (Larsson et al., 2002). Los fitosteroles
presentes en el licor negro no pueden ser eliminados como una fuente de masculinización
porque bajos niveles de éste licor están presentes en los efluentes. La escasez de hembras
altamente masculinizadas permite predecir que una reducción total en el grado de
masculinización está asociado con la recuperación del tall-oil (licor negro). Las industrias
ahora recuperan aproximadamente el 97% del tall-oil y terpenos (productos
complementarios a los procesos de deslignificación de la pulpa de madera) (Cody, 1997).
Otros estudios indican que los fitosteroles, tales como el β-sitosterol, campesterol y
estigmasterol contienen anillos esteroidales y podrían ser potencialmente convertidos por
mycobacterium sp. en esteroides, incluyendo androstadienediona y androstenediona, los
cuales pueden actuar como andrógenos (Larsson et al., 2002; Jenkins et al., 2001). La
modificación bacteriana no es la única fuente posible de compuestos androgénicos
presentes en los efluentes de la industria de celulosa y papel, sino que también pueden ser
suplidos directamente, al menos en pequeñas cantidades, por los productos vegetales. Por
ejemplo, la androstenediona se ha identificado en el polen del pino scotch (Pinus
sylvestris).

29
Los esteroides presentes en estos efluentes pueden tener efectos masculinizantes como
andrógenos o como precursores de andrógenos o por la interferencia con esteroides
biosintéticos o por los procesos de biodegradación en los ovarios, tejidos renales o hígado
(Jenkins et al., 2001). El mecanismo de acción de los disruptores endocrinos, provenientes
de los efluentes de la industria de celulosa, aún no está dilucidado (Larsson et al., 2002).
Por otro lado, la inducción de la actividad de glutation reductasa (GR) por los efluentes de
la industria de celulosa en experimentos, “in vitro” y “in vivo”, es una respuesta al estrés
oxidativo que puede indicar la presencia de compuestos en los efluentes que causan
incremento en la demanda de glutation reducido (GSH). Muchos compuestos o sus
metabolitos pueden sufrir ciclos redox y generar, por lo tanto, oxiradicales causando daño
oxidativo.
Aunque haya una gran cantidad de fitosteroles detectables en los extractos coloidales (del
Río et al., 1998), en las aguas de proceso (Gutíerrez et al., 2001) y consecuentemente en los
efluentes de la industria de celulosa Kraft (Cook et al, 1997), el β-sitosterol es el
compuesto dominante en la fracción de esteroles detectados. Debido a los efectos adversos
de estos compuestos, se hace necesario reducir la emisión de ellos hacia el medio acuático,
empleando tecnologías de tratamientos de efluentes como lagunas de estabilización aireada
y lodos activados. Estudios ya realizados con 22 plantas de pulpaje en EE UU muestran
eficiencia de remoción de los fitosteroles en el rango de 64-79% (Cook et al., 1997). En el
mismo contexto, en la industria de celulosa Kraft con blanqueo ECF localizada al este de
Finlandia, durante el tratamiento de sus aguas residuales, las concentraciones de los
extractivos de la madera fueron reducidas sobre un 97%. En el tratamiento de lodo
activado, sobre un 94% de los ácidos resínicos y sobre un 41% de los esteroles fueron
degradados o transformados en otros compuestos (Kostamo & Kukkonen, 2003). Esteroles
totales medidos en los efluentes de la industria de celulosa fueron removidos, mediante el
sistema de tratamiento de lodo activado, con un promedio de eficiencia de remoción de un
72 y 66% para las dos industrias inspeccionadas ( Mahmood-Khan & Hall, 2003).

30
II.1.5. Genotoxicidad de los efluentes de la industria celulósica
La genotoxicidad es el efecto que se produce sobre el ADN y otros blancos celulares que
controlan la integridad del material genético. Esto incluye inducción de aductos de ADN,
ruptura de la doble hélice, mutaciones puntuales y alteraciones cromosomales (estructurales
y numéricos) (Gollapudi & Krishna, 2000). En otras palabras, la genotoxicidad es el daño a
nivel del ADN producto de las interacciones entre las moléculas de ADN y compuestos
químicos tóxicos.
La modificación química de ADN es generalmente aceptada como una etapa de iniciación
de la carcinogénesis, por lo tanto, futuras generaciones podrían ser afectadas por una
reducida viabilidad embriónica y desordenes genéticos, siendo de crucial importancia la
evaluación de la genotoxicidad en los estudios ambientales (Ericson & Larsson, 2000).
Se han identificado varias industrias, tales como manufactureras de tinturas, refinerías de
petróleo, de metales e industrias de celulosa y papel, como emisoras de efluentes
genotóxicos y mutagénicos (Claxton et al., 1998).
Aunque los análisis químicos son los métodos principales para evaluar el potencial de
toxicidad de los efluentes, está claro que tal aproximación contiene limitaciones inherentes.
En contraste a los análisis químicos, los bioensayos proveen un medio de evaluación de
toxicidad de mezclas complejas sin un previo conocimiento sobre la composición química
de la mezcla. Así, la genotoxicidad de los efluentes puede ser efectivamente evaluada, a
través de bioensayos sin requerir información previa sobre su composición química. La
evaluación biológica, toxicidad aguda y test de genotoxicidad, para las descargas de los
efluentes es obligatoria en algunos campos, mientras en otros los efluentes son evaluados
sólo por sus propiedades químicas y físicas.
En los efluentes de la industria de celulosa Kraft blanqueada, que son descargados a partir
de las eficientes plantas de tratamiento secundario, se ha detectado baja toxicidad aguda.
Sin embargo, tales efluentes pueden incluir compuestos que causan efectos a largo plazo,
siendo, por ejemplo, genotóxicos, hormonalmente activos o bioacumulativos.

31
Muchos de estos compuestos aparecen a bajas concentraciones o son de características
químicas desconocidas y esto no sólo hace que los análisis químicos requieran tiempo, sino
también métodos de alta tecnología. Por ésta razón, el uso de ensayos biológicos en el
monitoreo de los efluentes puede ser útil. Resultados muestran que los efluentes o aguas
residuales pueden tener diversas propiedades y debido a esto su evaluación podría no ser
limitada sólo análisis químico o a uno o dos ensayos. Por tanto, una rápida batería de
bioensayos para la evaluación de la toxicidad aguda y crónica, la genotoxicidad y los
efectos hormonales podría ser conveniente para el monitoreo biológico del efluente total
(Piia Pessala et al. , 2004).
De los procesos de pulpaje resulta una compleja descarga conteniendo lignina disuelta,
productos de degradación de la celulosa y extractivos de la madera tales como terpenoides,
ácidos resínicos, fitosteroles y constituyentes fenólicos. Además, compuestos derivados del
proceso de blanqueo. De los contaminantes identificados en los efluentes de la industria de
celulosa Kraft un número significativo de químicos (sobre 300) son clasificados como
clastogénicos, mutagénicos y carcinogénicos. Houk (1992) enumeró 31 compuestos
genotóxicos, originados naturalmente de la pulpa o formados en la etapa de cloración. Los
efluentes totales provenientes de la industria procesadora de la madera son una de las
mayores fuentes de contaminación acuática, pues inducen activación de biotransformación
y genotoxicidad a diferentes niveles biológicos. En estudios de genotoxicidad
(mutagenenicidad) con bacterias se ha demostrado que los efluentes de la industria
celulósica poseen un potencial daño sobre el ADN. Utilizando el test de Ames, se ha
descrito la inducción de mutación, principalmente, por sustitución de bases. Estudios
recientes han demostrado que los efluentes de la industria de celulosa Kraft blanqueada
podrían ser una fuente de compuestos genotóxicos, pues se ha observado un aumento en la
concentración de aductos de ADN dosis-dependiente en Onchorhynchus tshawytscha y
Perca fluvialitis, indicando que los peces fueron expuestos a contaminantes potencialmente
genotóxicos.

32
Por otra parte, se demostró elevados niveles de micronúcleos hepáticos en trucha rainbow
(Onchorhynchus mykiss) después de la inyección de una fracción mutagénica proveniente
de extractos de efluente de industria de pulpa, a pesar de la ausencia de respuesta
mutagénica usando la cepa S. typhimurium TA-100 sin activación microsomal. En otros
trabajos, mediante el test de Ames, se ha evidenciado genotóxicidad en trucha, debido al
contenido de mutágenos en dichos efluentes. (Maria et al., 2003; Pirjo Sipi et al., 1997;
Ericson & Larsson, 2000).
Los estudios de genotoxicidad, utilizando el test de Ames, han mostrado que licores
provenientes de la etapa de cloración son significativamente más genotóxicos que otros
procesos de extracción de lignina (extracción alcalina, etapas de dióxido de cloro o
hipoclorito), y que los efluentes liofilizados son genotóxicos en cultivos de células de
mamíferos. Ensayos “in vivo” e “in vitro” han demostrado que químicos principalmente
clorados de bajo peso molecular son responsables de la actividad genotóxica (Maria et al.,
2004). No obstante, se ha demostrado el potencial genotóxico de ácido dehidroabiético
(DDHA) y ácido abiético (AA) (Pacheco & Santos, 2002; Maria et al., 2003). Otros
estudios, indican que los efluentes bioblanqueados (hemicelulasa) poseen leve efecto
genotóxico, señalando que compuestos clorofenólicos presentes, en el efluente derivado del
proceso de blanqueo convencional Kraft, están también presentes en el efluente del proceso
de blanqueo enzimático (Pérez-Alzola & Santos, 1997). No así, en un experimento
recientemente realizado en el laboratorio, trucha rainbow (Oncorhynchus mykiss) expuestas
a un efluente de industria de celulosa kraft proveniente de un proceso totalmente libre de
cloro, exhibieron formación de aductos de ADN de hígado, sugiriendo que, al menos parte
de sustancias genotóxicas presentes en los efluentes de la industria no tienen relación
alguna con el cloro o tipos de procesos de blanqueo, debido que, al parecer derivados
provenientes de los extractivos de la madera serían los responsables de dichos efectos
genotóxicos (Ericson & Larsson, 2000).

33
Los ácidos resínicos son conocidos por su mutagenicidad (genotoxicidad), mientras que la
escasa información sobre los fitosteroles (fitoestrógenos) señalan que estos y ésteres de
fitosteroles no muestran actividad mutagénica evidente en ninguno de los ensayos
utilizados (ensayo de mutación bacteriana y ensayo de aberración cromosomal “in vitro”)
cuando es evaluado de acuerdo a las pautas OECD (Organisation for Economic Co-
operation and Development) para la evaluación de la genotoxicidad de químicos (Wolfreys
& Hepburn, 2002). Otros resultados muestran que un concentrado de oxido de fitosteroles
conteniendo aproximadamente un 30% de oxido de fitosteroles no posee potencial
genotóxico ni evidencia de toxicidad cuando es administrado en la dieta de rata por 90 días
consecutivos (Lea et al., 2004). En contraste, se ha establecido la actividad genotóxica de
los metabolitos de estradiol y estrona. El potencial genotóxico de estradiol, estrona y
acetato de ciproterona con sus análogos puede jugar un rol anormal bajo condiciones
fisiológicas y terapéuticas (Joosten et al. , 2004).
La activación de ciertos xenobióticos resulta en la producción de oxiradicales, los cuales
reaccionan con muchas moléculas biológicas, incluyendo el ADN cuya integridad puede ser
afectada. Considerando los riesgos asociados con el ADN, la implementación de
biomarcadores de genotoxicidad tales como aducto de ADN, ruptura de la doble hélice de
ADN, micronúcleo y anormalidades nucleares eritrocíticas (ENAs; Erythocytic Nuclear
Abnormalities) es de particular importancia.
La predominancia de los metabolitos epóxidos, junto con los metabolitos de saturación y/o
conjugación de la fase II, contribuyen a la formación de enlaces covalentes con el ADN.
Así, efectos genotóxicos pueden ser esperados en los peces expuestos hacia xenobióticos, si
la fase II (conjugación) es estresada y el mecanismo de reparación de ADN es inhibido. No
obstante, no ha sido posible establecer una relación entre actividad de biotransformación y
genotoxicidad, puesto que los resultados obtenidos son variables, así por ejemplo, en A.
anguilla expuesta a ácido abiético, una baja actividad de EROD hepático parece ser capaz
de inducir un incremento en la ruptura de las hebras de ADN, mientras que anormalidades
nucleares eritrocíticas (ENAs) fueron observadas sólo después de una larga exposición
hacia mayores concentraciones de ácido Abiético (Maria et al., 2004).

34
En otros estudios, la inducción de ENA en A. anguilla expuesta a los efluentes de la
industria celulósica no fue asociada con una inducción precedente o simultanea de EROD,
señalando que ésta respuesta genotóxica no depende de una alta variedad de actividad de
biotransformación. Sin embargo, los mayores niveles de ENA fueron observados cuando
los peces exhiben los mayores niveles de actividad de EROD, sugiriendo que,
particularmente para ácido dehidroabiético (DHAA), una fuerte actividad de
biotransformación podría potenciar la expresión de la genotoxicidad. Por lo tanto, la
eventual interdependencia entre las respuestas previas puede variar con el contaminante.
Algunos autores sugieren que la actividad enzimática aumentada de la fase I, tal como la
actividad de EROD, es una importante alteración fisiológica que puede promover un efecto
genotóxico adverso, causado por la biotransformación de algunos compuestos de los
efluentes, tales como DHAA, AA, fitosteroles y reteno en metabolitos activos. Sin
embargo, la relación entre la inducción de las oxigenasas de función mixta (MFO; Mixed
Function Oxigenase) y la genotoxicidad en peces no está completamente clarificada (Maria
et al., 2003).
Los tests de genotoxicidad de complejos efluentes industriales y sitios medio ambientales
contaminados demostraron que éstas mezclas ambientales contienen tóxicos potencialmente
cancerígenos no identificados, y por lo tanto, no regulados. Se ha demostrado que extractos
de los efluentes de la industria celulósica contienen actividad mutagénica e inducen
respuestas bioquímicas en los peces, tal como una actividad aumentada del sistema
enzimático MFO.
Los tratamientos de los efluentes de la industria de celulosa Kraft han mostrado reducción o
eliminación de la genotoxicidad. Sin embargo, resultados demuestran que los compuestos
genotóxicos están aún presentes luego de ser tratados con lodo activado, pues las rupturas
de las hebras de ADN, en muestras de sangre y homogeneizado de hígado de peces, ocurrió
a las 72 horas de exposición. En otros casos, tratamiento y remediación, han mostrado
aumento de la genotoxicidad, tal como el tratamiento de aceite crudo mediante degradación
por hongos (Claxton et al., 1998).

35
Debido a que, muchas veces la genotoxicidad de las aguas residuales industriales o
efluentes permanece aún después de ser tratadas, surge la siguiente interrogante ¿son las
sustancias genotóxicas no biodegradables? la respuesta es probablemente no. Los
compuestos genotóxicos son, en general hidrofóbicos, fácilmente absorbidos dentro del
lodo. Por lo tanto, una vez que la interacción xenobiótico-ADN toma lugar, estos tóxicos
podrían ser modificados biológicamente tornándose en metabolitos aún más tóxicos. Esto
debe ser otro factor desconocido involucrado en los bajos rangos de remoción de la
genotoxicidad en los procesos de lodo activado (Takigami et al., 2002). La genotoxicidad
remanente en los efluentes tratados sugiere que muchos micropolutantes genotóxicos
desconocidos pueden pasar a través de los procesos de tratamiento de aguas residuales, tal
como sucede en el tratamiento de lodo activado. Resultados del ensayo “rec” de B. subtillis
muestran que la contaminación genotóxica es ampliamente extendida en las aguas
ambientales y éste tipo de contaminación no puede ser efectivamente removida por los
procesos de tratamientos convencionales. En consecuencia, para proteger los ecosistemas,
se hace necesario desarrollar nuevas estructuras de tecnología ambiental (Takigami et al.,
2002). Estrategias de tratamiento han fluctuado desde simples hasta sofisticados esfuerzos
de ingeniería que incluyen el aumento del pH del efluente, substitución de cloro por
dióxido de cloro, tratamiento de los efluentes con dióxido de sulfuro, uso de lagunas
aireadas, y el uso de métodos de intercambio iónico.
Los procesos de tratamiento podrían reducir o eliminar la genotoxicidad de un efluente. Un
ejemplo donde el tratamiento puede reducir efectivamente la genotoxicidad del efluente
industrial en varios órdenes de magnitud es ilustrado por el uso de tratamiento secundario
de aguas biotratadas (lagunas de aireación) de la industria de celulosa. Sin embargo, el
tratamiento de aguas no siempre reduce la genotoxicidad de los efluentes y puede en
algunas instancias incrementar la genotoxicidad. Por lo tanto, los ensayos de genotoxicidad
proveen un medio efectivo para el monitoreo medio ambiental y para seleccionar procesos
de tratamientos apropiados (Claxton et al., 1998).

36
Los estudios sobre la genotoxicidad de los efluentes industriales, evaluados principalmente
a través del bioensayo de mutagenicidad en S. typhimurium, indican que estos presentan un
relativo potencial mutagénico.
Cerca del 60% de los estudios han usado el ensayo con S. typhimurium o test de Ames, el
22% han utilizado otros ensayos de mutación, 10% usó ensayos cromosomales, y el 7%, el
ensayo “rec” de B. subtillis, para determinar daño al ADN; Aproximadamente el 2,5%
utilizaron ensayos con animales vivos (Claxton et al., 1998). La razón por la que se ha
usado mayormente el ensayo con S. typhimurium y el uso limitado de otros ensayos es que
este (Ames) representa mejor lo que sucedería en las células eucarióticas (organismos
superiores).
Aunque los bioensayos comúnmente disponibles no pueden proveer información precisa y
detallada sobre la naturaleza química de los mutágenos detectados, tales ensayos proveen
una posible respuesta a la interrogante si las muestras examinadas contienen mutágenos a
niveles potencialmente dañinos para los organismos. Por lo tanto, se ha visto que la
estrategia más razonable para la evaluación de las muestras ambientales es el uso de
ensayos biológicos como un test preliminar para detectar la presencia de compuestos
mutagénicos y genotóxicos.
El ensayo “rec” de B. subtilis fue especialmente diseñado para evaluar la genotoxicidad de
varios tipos de aguas. Este ensayo puede ser una herramienta muy poderosa, gracias a su
mayor sensibilidad para la detección de mutágenos en las aguas altamente poluídas,
comparado con otros ensayos de mutagenicidad (Takigami et al., 2002).
Aunque el ensayo “rec” de B. subtilis, ensayo de daño a nivel de ADN, no es un ensayo de
mutagenicidad, este es muy útil, sumado a los ensayos mutagénicos para programas de
screening preliminares (Mazza, 1982). El acoplamiento de ensayos utilizando bacterias (test
de Ames) con ensayos químicos amplía la utilidad de estos bioensayos y permite el
aislamiento e identificación de fracciones químicas definidas que contienen actividad
genotóxica (Claxton et al., 1998).

37
Estos bioensayos son complementarios cuando son aplicados a un screening de los
efluentes de la industria de celulosa Kraft blanqueada por su potencial genotóxico (Rao et
al., 1995).
Los bioensayos de genotoxicidad constituyen una herramienta integral en la evaluación de
compuestos genotóxicos presentes en el ambiente suministrando información útil sobre los
posibles riesgos hacia el medio ambiente y su ecosistema.

CAPÍTULO III
MATERIALES Y MÉTODOS

39
CAPÍTULO III
III.1. MATERIALES Y MÉTODOS
III.1.1. Diseño Experimental
Se evalúo la genotoxicidad mediante el ensayo “rec” de Bacillus subtilis y la mutagenicidad
con Salmonella typhimurium o test de Ames sin activación metabólica, sobre muestras de
fitosteroles que correspondieron a soluciones estándares de β-sitosterol y estigmasterol
cuyas concentraciones fluctuaron entre 1 y 32 mgL-1. Así también, muestras de efluentes de
la industria de celulosa Kraft con y sin tratamiento biológico, en dosis de 0,1 a 0,5 mL. Para
estigmasterol se realizó el ensayo de mutagenicidad en presencia de activación metabólica
(mezcla S9).
III.1.2. Muestras de efluentes de la industria de celulosa Kraft blanqueada
Las muestras fueron obtenidas de industrias de celulosa Kraft blanqueada que utilizan como
materia prima el Pinus radiata. Las etapas de blanqueo de éstas industrias están sujetas a
tecnologías libre de cloro elemental (ECF), con secuencias de Blanqueo D-Eop-D-D,
previo a una etapa de deslignificación con oxígeno.
Las muestras de efluente de la industria celulósica fueron previamente sometidas a
tratamiento biológico aerobio y anaerobio en el laboratorio de biotecnología ambiental del
Centro de Ciencias Ambientales EULA, Chile. Dichos tratamientos corresponden a laguna
aireada y a un biorreactor anaerobio con biomasa inmovilizada.
III.1.3. Reactivos
Los fitosteroles, β-sitosterol (22,23-Dihydrofucosterol) cuyo peso molecular es 414,7
g/gmol con un 95% de pureza, constituido por un 75,5% de β-sitosterol y un 13,0% de β-
sitostanol, y el estigmasterol, 3β-Hydroxy-24-ethyl-5,22-Cholestadiene, con peso molecular

40
412,7 g/gmol y un 95% de pureza fueron proveídos por Calbiochem y Sigma,
respectivamente.
Araclor 1254 inducido en KCl, preparaciones de homogeneizado de hígado de rata
(fracción microsomal S9) y nicotinamín adenín dinucleótido fosfato reducido, NADPH
Regensys A, fueron obtenidos de Gene X Press Life Science Business.
III.1.4. Ensayo “rec” de Bacillus subtilis
El ensayo “rec” de B. subtilis se llevó a cabo de acuerdo a lo descrito por Mazza (1982),
utilizándose las cepas B. subtilis 1652 rec(+) y B. subtilis 1791 rec(-).
Se preparó una solución estándar de 100 mgL-1 de β-sitosterol y estigmasterol. A partir de
ellas se realizaron diluciones crecientes de 1, 2, 4, 8, 16 y 32 mgL-1. Luego desde un cultivo
de B. subtilis de toda la noche se efectúo una dilución seriada de la cepa rec(+) y rec(-)
hasta 10-4 y ésta ultima diluida a la mitad. A continuación se adicionó 0,1 mL de cada una
de las concentraciones, anteriormente mencionadas, del fitosterol correspondiente y 0,1 mL
de cada cepa previamente diluida, en tubos que contenían agar soft mantenidos en baño
maría en seco a una temperatura de 45°C. Finalmente, el contenido de estos tubos fueron
vertidos sobre placas petri rotuladas y preparadas con agar nutritivo, agitando cada vez los
tubos antes de ser vertidos en un vortex modelo M-16710-12-26 y distribuyendo la mezcla
homogéneamente sobre la superficie de las placas. Una vez solidificado el agar, las placas
se incubaron a 37°C durante 19 a 24 horas. El control se realizó del mismo modo pero en
ausencia de las soluciones de β-sitosterol y estigmasterol. Después de incubar se realizó el
recuento de las colonias desarrolladas en las placas y los resultados fueron expresados
como unidades formadoras de colonias (UFC).
Las muestras de efluentes ajustadas a pH 7,0 fueron previamente filtradas a través de filtros
millipore de 0,25 µm y se realizó, posteriormente, el ensayo “rec” de B. subtilis del mismo
modo descrito para los fitosteroles. En dosis de 0,1, 0,3 y 0,5 mL de efluente. Todos los
ensayos se realizaron en duplicado.

41
El ensayo “rec” de B. subtilis se basa en la diferencia relativa de sobrevivencia (N/No) de la
cepa rec(+) capaz de reparar el daño causado sobre el ADN, y su cepa deficiente rec(-). La
diferencia entre sus sobrevivencias es, por lo tanto, interpretado simplemente como
genotoxicidad. Usando este criterio los resultados son expresados en una curva dosis-
respuesta, dónde un claro efecto se manifiesta cuando la sobrevivencia de la cepa rec(-) es
significativamente menor respecto de la sobrevivencia de la cepa rec(+), a medida que
aumenta la dosis o concentración de la muestra a evaluar. Por lo tanto, el valor de eficiencia
de placa (A) menor que 1 indica presencia de efecto genotóxico, Así:
Dónde,
-N/No rec(-) = unidades formadoras de colonias (UFC) de la cepa rec(-) de placa ensayo en
presencia del contaminante o muestra / UFC de la cepa rec(-) en ausencia del contaminante
(control).
-N/No rec(+) = unidades formadoras de colonias (UFC) de la cepa rec(+) de placa ensayo
en presencia del contaminante o muestra / UFC de la cepa rec(+) en ausencia del
contaminante (control).
III.1.5. Test de Ames
Los análisis de mutagenicidad se realizaron utilizando el test de Ames descrito por Ames et
al. (1975) y modificado por Maron & Ames (1983).
A tubos con top agar, suplementado con 0,05 mM de histidina y biotina, y mantenidos en
baño seco a 45°C, se adicionó en duplicado 0,1mL de cada una de las diluciones de las
soluciones estándares, anteriormente preparadas, del fitosterol correspondiente, luego se
agregó por separado 0,1mL de un cultivo de 18-24 h de S. typhimurium, cepas TA-100 y T-
A98. A continuación, se agitó el contenido de cada tubo en un vortex modelo M-16710-12-
26 y se vertió sobre placas petri con agar mínimo glucosa adecuadamente rotuladas y se
A = N/No rec(-) / N/No rec(+) (1)

42
homogenizaron manualmente. Una vez solidificado el agar de superficie (top agar), las
placas se incubaron a 37°C durante 72 h. Se efectuó el recuento y los resultados fueron
expresados como número de revertantes promedio por placas ± desviación estándar (DE)
para tratamientos y controles.
Las muestras de los efluentes ajustadas a pH 7,0 se filtraron a través de filtros millipore de
0,25 µm y se realizó el test de Ames del mismo modo descrito para los fitosteroles. En
dosis de 0,1, 0,3 y 0,5 mL de efluente. Todos los ensayos se realizaron en duplicado.
El test de Ames con activación metabólica, en presencia de fracción S9, fue realizado sólo
para el estigmasterol, agregando 0,5 mL de ésta fracción en los tubos con top agar, antes de
adicionar las diluciones de la solución estándar de estigmasterol.
Para cada ensayo se realizó un control positivo con 0,1mL de 2-Antramina y un control
negativo con 0,1 mL de agua estéril, cada uno de ellos en duplicados.
Previo a los ensayos se confirmaron las características fenotípicas de las cepas utilizadas.
La determinación de la tasa de reversión espontánea, la tasa de reversión por acción de un
agente mutágeno conocido (control positivo) y la viabilidad de la cepa se realizaron en cada
experimento.
Usando éste procedimiento, la mutagenicidad se expresó en términos de la razón de
mutagenicidad (RM), expresada como la relación entre el número de revertantes obtenidas
con la muestra ensayada y el número de revertantes naturales (reversión espontánea).
Se considera una respuesta positiva cuando la RM es mayor o igual a 2 y existe un claro
efecto dosis-respuesta. Así,
Donde,
-E = número de colonias revertantes placa ensayo (revertantes inducidas).
-C = número de colonias revertantes en la placa control negativo ( revertantes espontáneas).
RM = E / C (2)

43
III.1.6. Análisis estadístico
Los resultados del ensayo “rec” de B. subtilis así como también, del test de Ames fueron
tratados con el programa Statistix for Windows 2.2.
Con el propósito de encontrar diferencias significativas (p<0,05) los resultados obtenidos
de eficiencia de placa y razón de mutagenicidad (variables dependientes), fueron sometidos
a análisis de varianza de una y dos vías (ANOVA). Luego de encontrar diferencias se
aplicó la prueba paramétrica Bonferroni para comparación de promedios, ya sea de
eficiencia de placa y/o razón de mutagenicidad, según corresponda. Las variables
independientes utilizadas fueron dosis (o concentración) y muestra, la cual en algunas
instancias posee réplicas, es decir el ensayo se realizó más de una vez con la misma
muestra. En el caso del test de Ames se incluyó además la variable independiente cepa con
dos niveles, TA-100 y TA-98.

CAPÍTULO IV
RESULTADOS

45
CAPÍTULO IV
IV.1. RESULTADOS
IV.1.1. Ensayo “rec” de Bacillus subtilis
En la Figura 4-1 y 4-2 se muestran los resultados de genotoxicidad al ensayar los
fitosteroles, β-sitosterol y estigmasterol, respectivamente con B. subtilis.
IV.1.1.2. β-sitosterol
En la Figura 4-1 se observan los resultados obtenidos al ensayar el β-sitosterol. La
sobrevivencia de ambas cepas se mantiene constante hasta los 16 mgL-1. A partir de esta
concentración la cepa rec(+) tiende a incrementar su sobrevivencia a medida que aumenta
la concentración de β-sitosterol, lo que podría deberse a que dicho compuesto estaría siendo
metabolizado por esta cepa. Mientras que la sobrevivencia de la cepa rec(-) se ve levemente
afectada en presencia de éste compuesto puesto que, dicha cepa se caracteriza por ser
incapaz de reparar el daño causado a nivel del ADN. Por lo tanto, el β-sitosterol muestra un
efecto genotóxico a concentraciones superiores a 16 mgL-1, debido a la diferencia relativa
de sobrevivencia existente entre las cepas.
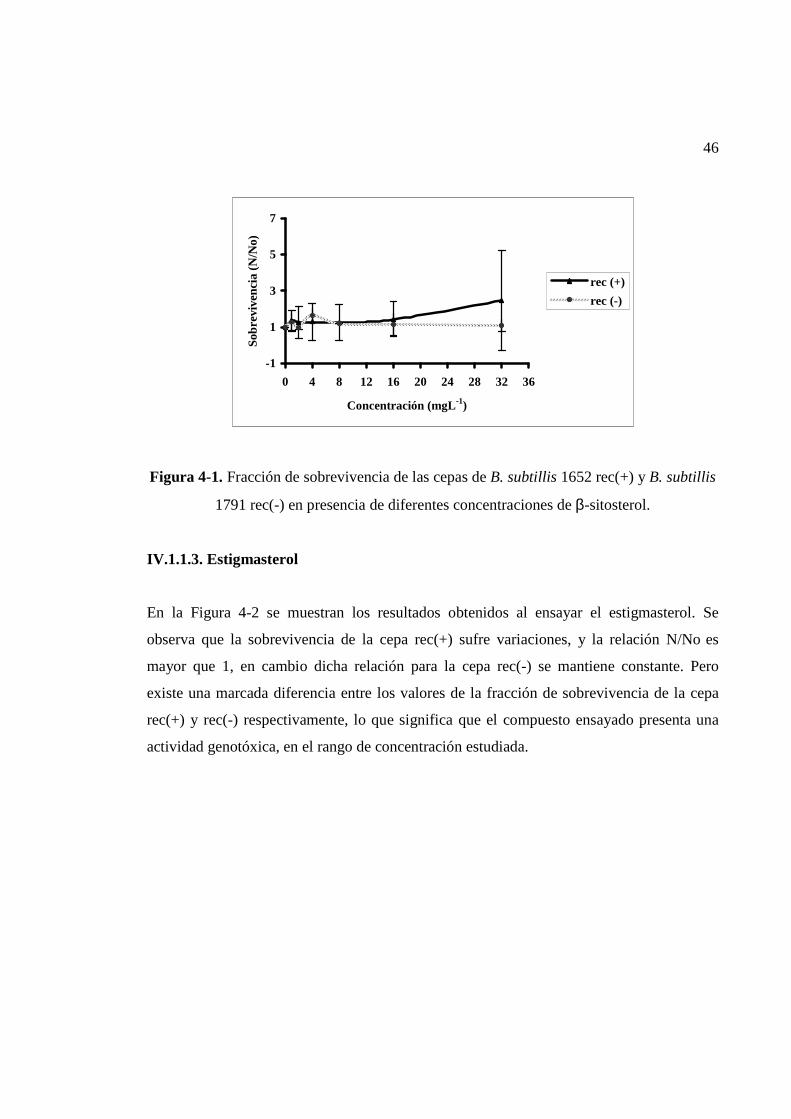
46
-1
1
3
5
7
0 4 8 12 16 20 24 28 32 36
Concentración (mgL-1)
Sob
revi
venc
ia (
N/N
o)
rec (+)
rec (-)
Figura 4-1. Fracción de sobrevivencia de las cepas de B. subtillis 1652 rec(+) y B. subtillis
1791 rec(-) en presencia de diferentes concentraciones de β-sitosterol.
IV.1.1.3. Estigmasterol
En la Figura 4-2 se muestran los resultados obtenidos al ensayar el estigmasterol. Se
observa que la sobrevivencia de la cepa rec(+) sufre variaciones, y la relación N/No es
mayor que 1, en cambio dicha relación para la cepa rec(-) se mantiene constante. Pero
existe una marcada diferencia entre los valores de la fracción de sobrevivencia de la cepa
rec(+) y rec(-) respectivamente, lo que significa que el compuesto ensayado presenta una
actividad genotóxica, en el rango de concentración estudiada.
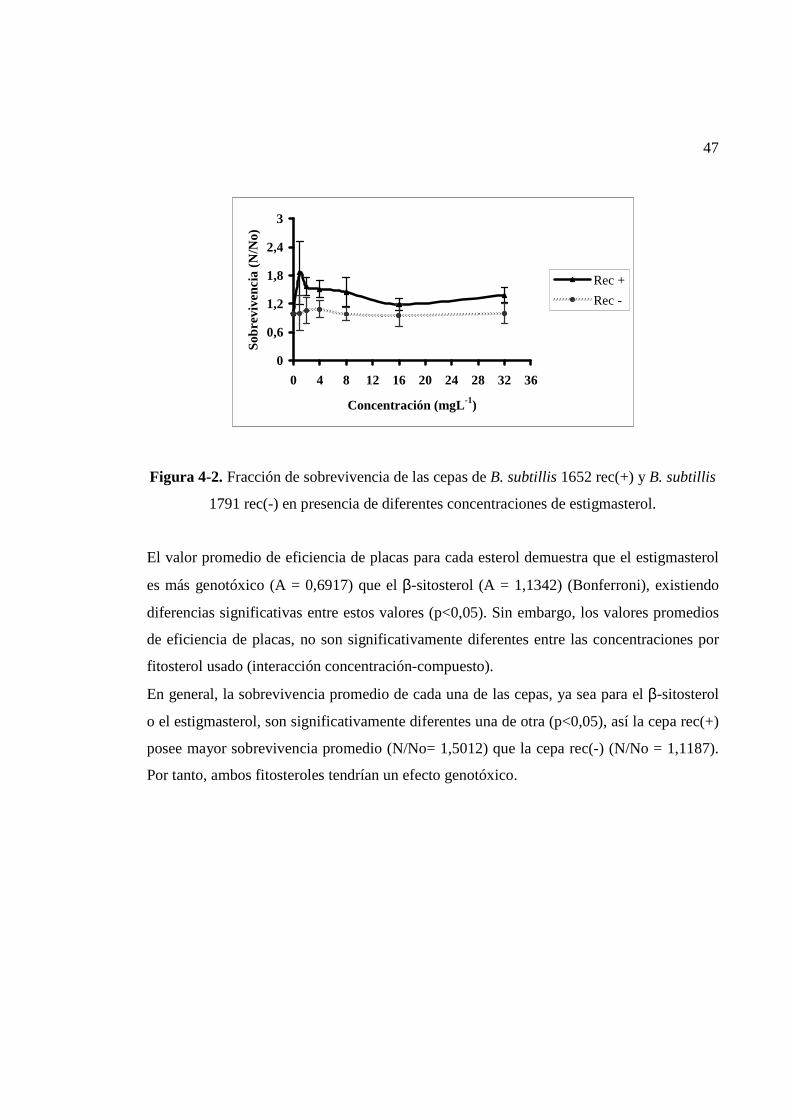
47
0
0,6
1,2
1,8
2,4
3
0 4 8 12 16 20 24 28 32 36
Concentración (mgL-1)
Sob
revi
venc
ia (
N/N
o)
Rec +
Rec -
Figura 4-2. Fracción de sobrevivencia de las cepas de B. subtillis 1652 rec(+) y B. subtillis
1791 rec(-) en presencia de diferentes concentraciones de estigmasterol.
El valor promedio de eficiencia de placas para cada esterol demuestra que el estigmasterol
es más genotóxico (A = 0,6917) que el β-sitosterol (A = 1,1342) (Bonferroni), existiendo
diferencias significativas entre estos valores (p<0,05). Sin embargo, los valores promedios
de eficiencia de placas, no son significativamente diferentes entre las concentraciones por
fitosterol usado (interacción concentración-compuesto).
En general, la sobrevivencia promedio de cada una de las cepas, ya sea para el β-sitosterol
o el estigmasterol, son significativamente diferentes una de otra (p<0,05), así la cepa rec(+)
posee mayor sobrevivencia promedio (N/No= 1,5012) que la cepa rec(-) (N/No = 1,1187).
Por tanto, ambos fitosteroles tendrían un efecto genotóxico.

48
IV.1.1.3. Influentes primario
En la Figura 4-3 se muestra una fotografía del ensayo “rec” donde es posible apreciar la
diferencia relativa de sobrevivencia de las cepas de B. subtilis 1652 rec(+) y B. subtilis
1791 rec(-) en función de la dosis creciente de Influente 1, lo cual representa un efecto
genotóxico, ya que a medida que aumenta la dosis del Influente (de izquierda a derecha) las
unidades formadoras de colonias desarrolladas por la cepa rec(+) son mayores en
comparación con las UFC de la cepa rec(-), y estas a su vez, son menores que las placas del
control. En la figura 4-4, se presentan en la fila superior el crecimiento de colonias de B.
subtilis 1652 rec(+) y en la fila inferior las UFC de la cepa de B. subtilis 1791 rec(-),
desarrolladas en ausencia del contaminante (Influente 1).

49
Figura 4-3. Diferencia relativa de sobrevivencia de las cepas de B. subtilis en función del
aumento de la dosis de influente 1. Donde la fila superior corresponde a UFC de la cepa de
B. subtilis 1652 rec(+) y la fila inferior a UFC de B. subtilis 1791 rec(-).
Figura 4-4. UFC de la cepa de B. subtilis 1652 rec(+) (fila superior) y UFC de B. subtilis
1791 rec(-) (fila inferior) desarrolladas en ausencia del contaminante.

50
En la Figura 4-5 se muestran los resultados obtenidos al ensayar una muestra de efluente de
la industria de celulosa Kraft sin previo tratamiento biológico (Influente 1).
A)
0
0,4
0,8
1,2
1,6
2
0 0,1 0,2 0,3 0,4 0,5 0,6
Dosis (mL)
Sob
revi
venc
ia (
N/N
o)
rec (+)
rec (-)
B)
0
0,4
0,8
1,2
1,6
2
0 0,1 0,2 0,3 0,4 0,5 0,6
Dosis (mL)
Sob
revi
venc
ia (
N/N
o)
rec (+)
rec (-)
C)
0
0,4
0,8
1,2
1,6
2
0 0,1 0,2 0,3 0,4 0,5 0,6
Dosis (mL)
Sob
revi
venc
ia (
N/N
o)
rec (+)
rec (-)
Figura 4-5. Fracción de sobrevivencia de las cepas de B. subtillis 1652 rec(+) y B. subtillis
1791 rec(-) en presencia de diferentes dosis de Influente 1 con tres réplicas (A, B y C).

51
El Influente 1 (Figura 4-5) presenta efecto genotóxico en el rango de dosis usada, ya que
existe diferencia entre las sobrevivencias de ambas cepas, siendo la sobrevivencia de la
cepa rec(-) menor (N/No = 0,9444) respecto de la cepa rec(+)(N/No = 1,0026), exepto en la
réplica A, dónde el valor de eficiencia de placa promedio en la dosis de 0,1 mL es distinto
estadísticamente de la réplica B y C en esta misma dosis. Así también dentro de la misma
réplica (A) el valor de eficiencia de placa promedio a la dosis de 0,1 mL es diferente de las
dosis de 0,3 y 0,5 mL (Bonferroni). Esto último, puede ser atribuido a la elevada desviación
estándar en esta primera dosis respecto de las otras, debido a un error experimental al
ensayar dicha dosis (Figura 4-5-A). De hecho, las DE sufren cambios entre las réplicas,
aunque no significativos estadísticamente, siendo los valores de las desviaciones estándares
de las Figuras 4-5-B y C mayores en comparación con los valores de las DE de la Figura 4-
5-A. De lo contrario, podríamos inferir que las variaciones en la sobrevivencia de la cepa
rec(+) de la réplica A se debe a que las bacterias utilizarían los compuestos presentes en el
influente como sustrato para sus procesos metabólicos, y no a las variaciones de las
desviaciones estándares de la sobrevivencia de la cepa rec(+) entre las dosis ensayadas
(Figura 4-5-A). La sobrevivencia de la cepa rec(-) disminuye a medida que aumenta la
cantidad de influente ensayado (Figura 4-5-A), lo que significa un efecto genotóxico en las
dosis ensayadas. En resumen, el Influente 1 (Figura 4-5) muestra efecto genotóxico en las
dosis ensayadas, no existiendo diferencias significativas entre los valores promedios de
eficiencia de placas de cada una de las réplicas (A, B y C), a pesar de que las Figuras 4-5-B
y C presentan mayores desviaciones estándares en comparación con la Figura 4-5-A. No
obstante, la eficiencia de placa promedio varía significativamente con la dosis ensayada por
réplica realizada (interacción réplica-dosis; p<0,05).
En la Figura 4-6 se presentan los resultados obtenidos al ensayar un efluente sin previo
tratamiento biológico con dos réplicas D y E (Influente 2).

52
D)
0
0,4
0,8
1,2
1,6
2
0 0,1 0,2 0,3 0,4 0,5 0,6
Dosis (mL)
Sob
revi
venc
ia (
N/N
o)
rec (+)
rec (-)
E)
0
0,4
0,8
1,2
1,6
2
0 0,1 0,2 0,3 0,4 0,5 0,6
Dosis (mL)
Sob
revi
venc
ia (
N/N
o)
rec (+)
rec (-)
Figura 4-6. Fracción de sobrevivencia de las cepas de B. subtillis 1652 rec(+) y B. subtillis
1791 rec(-) en presencia de diferentes dosis de Influente 2 con dos réplicas (D y E).
Podemos observar que hay variaciones en las desviaciones estándares de las sobrevivencias
de las cepas en cada una de las réplicas realizadas por dosis evaluada, así por ejemplo, la
desviación estándar de la sobrevivencia de la cepa rec(-) a los 0,3 mL es mayor en la réplica
D, mientras que a los 0,5 mL la desviación estándar de la sobrevivencia de ésta misma cepa
decrece en la réplica E, no así la desviación estándar de la sobrevivencia de la cepa rec(+),
pues se ve aumentada en ésta dosis con respecto a la Figura 4-6-D.

53
Probablemente la variación de las desviaciones estándares de las sobrevivencias de las
cepas entre las dosis por réplicas efectuadas, sea la razón por la cual no es posible encontrar
un efecto genotóxico constante en cada réplica en el rango de dosis evaluado. Sin embargo,
es posible apreciar la presencia de genotoxicidad en el Influente 2 (Figura 4-6),
simplemente por la existencia de diferencia entre las sobrevivencias de la cepa rec(+) en
relación a la cepa rec(-) no encontrándose diferencias significativas de los valores
promedios de eficiencia de placas entre las réplicas ni en su interacción con las dosis
ensayadas. Por otra parte es posible señalar la existencia de un efecto citotóxico,
demostrado por la disminución de la sobrevivencia de la cepa rec(+)(Figura 4-6-E).
Ambos efluentes de celulosa Kraft sin previo tratamiento biológico, Influentes 1 y 2,
muestran efecto genotóxico (Figura 4-5 y Figura 4-6), encontrándose diferencias
significativas en sus promedios de eficiencias de placas entre las réplicas de ambos
Influentes por dosis ensayada (p<0,05). Por tanto, la genotoxicidad se hace evidente en
cada una de las réplicas de los influentes, por la notoria diferencia existente entre la
sobrevivencia de la cepa rec(+) respecto de la cepa rec(-) en el rango de dosis empleada,
siendo la fracción de sobrevivencia de la cepa rec(+) mayor respecto de la cepa rec(-) en
cada una de las réplicas de los Influentes ensayados ( 1y 2).
Las Figuras 4-7 y 4-8 muestran los resultados de Influente 2 sometido a tratamiento
anaeróbico y aeróbico, respectivamente (efluentes secundarios).

54
IV.1.1.4. Efluente secundario anaerobio
0
0,4
0,8
1,2
1,6
2
0 0,1 0,2 0,3 0,4 0,5 0,6
Dosis (mL)
Sob
revi
venc
ia (
N/N
o)
rec (+)
rec (-)
Figura 4-7. Fracción de sobrevivencia de las cepas de B. subtillis 1652 rec(+) y B. subtillis
1791 rec(-) en presencia de diferentes dosis de efluente tratado anaeróbicamente.
IV.1.1.5. Efluente secundario aerobio
0
0,4
0,8
1,2
1,6
2
0 0,1 0,2 0,3 0,4 0,5 0,6
Dosis (mL)
Sob
revi
venc
ia (
N/N
o)
rec (+)
rec (-)
Figura 4-8. Fracción de sobrevivencia de las cepas de B. subtillis 1652 rec(+) y B. subtillis
1791 rec(-) en presencia de diferentes dosis de efluente tratado aeróbicamente.

55
La muestra tratada anaeróbicamente presenta un valor promedio de eficiencia de placa (A =
0,9844) mayor que la obtenida del tratamiento aeróbico (A = 0,9256), indicando que éste
último tratamiento presentaría mayor efecto genotóxico, en tanto que para el tratamiento
anaeróbico casi no hay efecto. No obstante, estos valores promedios de A entre ambos
tratamientos, no son estadísticamente diferentes.
Los tratamientos secundarios, anaerobio y aeróbico, reducirían la genotoxicidad de los
efluentes sin previo tratamiento biológico, pues los A promedio de las réplicas A, E (Figura
4-5 y Figura 4-6) y los tratamientos biológicos (Figura 4-7 y Figura 4-8) varían
significativamente en las dosis por muestra analizada (interacción dosis-muestra; p<0,05).
Tal diferencia no se pudo apreciar con los Figuras 4-5-B, 4-5-C, 4-6-D y los tratamientos
(Figura 4-7 y Figura 4-8), debido probablemente a la mayor desviación estándar que poseen
las réplicas de la Figura 4-5-B y 4-5-C, con relación, a la Figura 4-5-A, y a la variabilidad
de las desviaciones estándares en la Figura 4-6-D, respecto de la Figura 4-6-E, entre las
cepas.
Por lo tanto, la genotoxicidad de los efluentes con y sin tratamiento es significativamente
diferente (p<0,05). Los tratamientos biológicos (aeróbico y anaeróbico), reducen los efectos
genotóxicos de los efluentes sin mostrar diferencias estadísticas entre uno y otro
tratamiento.
Debido a la poca diferencia existente entre los A promedio de los fitosteroles versus los
efluentes e influentes es posible inferir que la presencia de estos compuestos en los
efluentes de la industria celulósica potenciaría el efecto genotóxico de dichos efluentes, el
cual es reducido por los tratamientos biológicos.

56
IV.1.2. Test de Ames
En la Tabla IV.1 y Tabla IV.2 se muestran los resultados de mutagenicidad al ensayar los
fitosteroles, β-sitosterol y estigmasterol, respectivamente con S. typhimurium / Test de
Ames.
IV.1.2.1. β-sitosterol
Tabla IV.1. Detección del efecto mutagénico de β-sitosterol utilizando las cepas de S.
typhimurim TA-100 y TA-98 en ausencia de la fracción S9.
Concentración (mgL-1)
Revertantes por placas ± DE RM
TA-100 TA-98 TA-100 TA-98 1 2 4 8 16 32 0,1 mL de agua destilada estéril 0,1 mL de 2-Antramina
15,5 ± 4,05 29,5 ± 2,12 0,97 1,05 10,5 ± 2,12 31,0 ± 0 0,66 1,11 8,5 ± 2,12 18,5 ± 2,12 0,53 0,66 11,0 ± 2,83 16,0 ± 5,66 0,69 0,57 13,0 ± 5,66 27,5 ± 0,71 0,81 0,98 12,5 ± 2,12 28,0 ± 2,83 0,78 1,0 16,0 ± 5,66 28,0 ± 2,83 7,5 ± 2,12 26,0 ± 2,83

57
IV.1.2.2. Estigmasterol
Tabla IV.2. Detección del efecto mutagénico de estigmasterol utilizando las cepas de S.
typhimurim TA-100 y TA-98 en ausencia de la mezcla S9.
El β-sitosterol y estigmasterol no presentan efecto mutagénico evidente, no encontrándose
diferencias significativas entre ellos. Es decir, la razón de mutagenicidad (RM) promedio
de las cepas no difiere significativamente entre los fitosteroles analizados. Sin embargo, el
estigmasterol presenta una razón de mutagenicidad promedio mayor que el β-sitosterol.
Concentración (mgL-1)
Revertantes por placas ± DE RM TA-100 TA-98 TA-100 TA-98
1 2 4 8 16 32 0,1 mL de agua destilada estéril 0,1 mL de 2-Antramina
07,5 ± 2,12 20,0 ± 4,24 0,47 0,71 12,5 ± 4,95 26,5 ± 13,4 0,78 0,95 10,5 ± 2,12 22,0 ± 11,3 0,66 0,79 13,0 ± 1,41 18,5 ± 2,12 0,81 0,66 18,5 ± 7,78 29,0 ± 8,49 1,16 1,04 10,5 ± 3,54 21,0 ± 11,3 0,66 0,75 16,0 ± 5,66 28,0 ± 2,83 7,5 ± 2,12 26,0 ± 2,83

58
IV.1.2.3. Estigmasterol con activación metabólica
Tabla IV.3. Detección del efecto mutagénico de estigmasterol utilizando las cepas de S.
typhimurim TA-100 y TA-98 en presencia de la fracción S9.
Concentración (mgL-1)
Revertantes por placas ± DE RM
TA-100 TA-98 TA-100 TA-98
1 2 4 8 16 32 0,1 mL de agua destilada estéril 0,1 mL de 2-Antramina
321 ± 30,4 16,5 ± 4,95 1,44 0,63
288 ± 14,8 21,5 ± 14,8 1,29 0,83
295 ± 20,5 15,5 ± 4,95 1,32 0,60
306 ± 39,6 23,5 ± 6,36 1,37 0,90
295 ± 137 27 ± 5,66 1,32 1,04
274 ± 120 21,5 ± 2,12 1,23 0,83
373 ± 84,1 54 ± 14,8 223 ± 14,1 26 ± 12,7
La RM varía con la cepa por estigmasterol con y sin S9 (p<0,05). Es decir, para ambos,
estigmasterol con y sin S9, la razón de mutagenicidad de la cepa TA-100 es mayor respecto
de la RM de la cepa TA-98. En otras palabras, al ensayar el estigmasterol en presencia de
S9 se tiene que la razón de mutagenicidad de la cepa TA-100 incrementa significativamente
(p<0,05) con relación a la RM de esta misma cepa para el estigmasterol ensayado en
ausencia de activación metabólica. Esto sugiere la presencia de un efecto, lo que
significaría que el estigmasterol por acción de las enzimas hepáticas se activaría, por tanto,
actuaría como un promutágeno, siendo necesario realizar el ensayo a mayores
concentraciones para confirmar su comportamiento promutagénico.
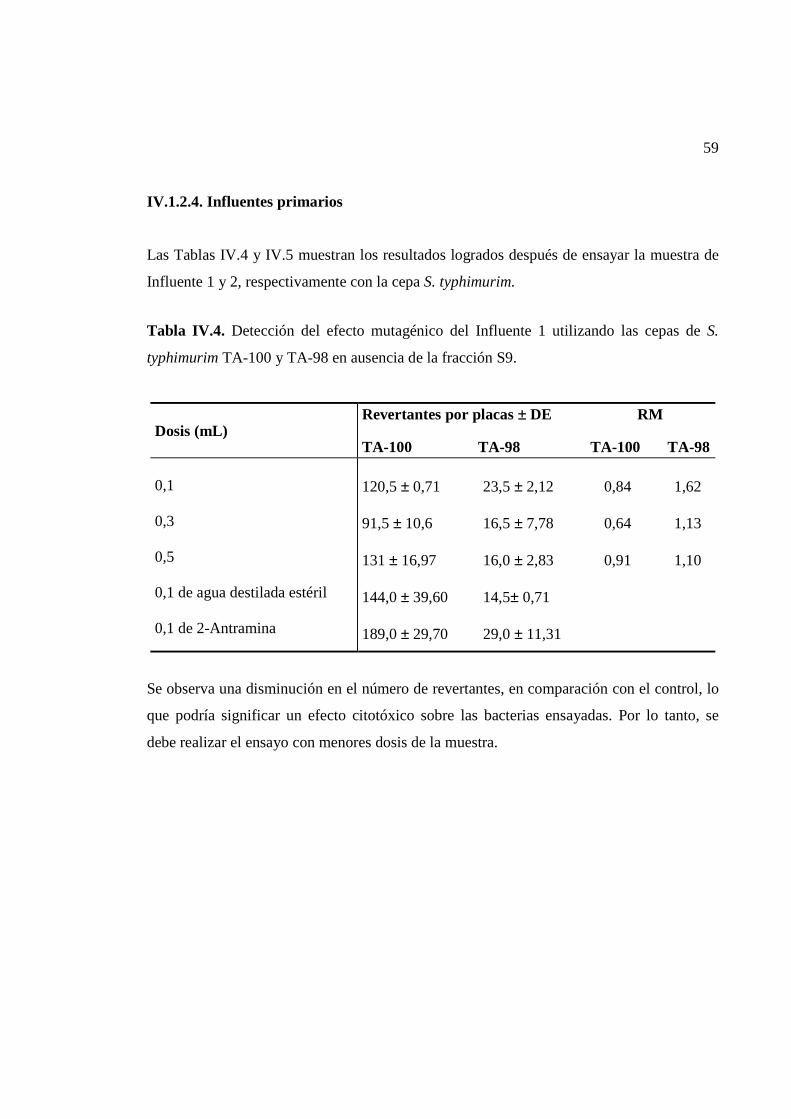
59
IV.1.2.4. Influentes primarios
Las Tablas IV.4 y IV.5 muestran los resultados logrados después de ensayar la muestra de
Influente 1 y 2, respectivamente con la cepa S. typhimurim.
Tabla IV.4. Detección del efecto mutagénico del Influente 1 utilizando las cepas de S.
typhimurim TA-100 y TA-98 en ausencia de la fracción S9.
Dosis (mL)
Revertantes por placas ± DE RM
TA-100 TA-98 TA-100 TA-98 0,1 0,3 0,5 0,1 de agua destilada estéril 0,1 de 2-Antramina
120,5 ± 0,71 23,5 ± 2,12 0,84 1,62
91,5 ± 10,6 16,5 ± 7,78 0,64 1,13
131 ± 16,97 16,0 ± 2,83 0,91 1,10
144,0 ± 39,60 14,5± 0,71
189,0 ± 29,70 29,0 ± 11,31
Se observa una disminución en el número de revertantes, en comparación con el control, lo
que podría significar un efecto citotóxico sobre las bacterias ensayadas. Por lo tanto, se
debe realizar el ensayo con menores dosis de la muestra.

60
Tabla IV.5. Detección del efecto mutagénico del Influente 2 utilizando las cepas de S.
typhimurim TA-100 y TA-98 en ausencia de la fracción S9.
Dosis (mL)
Revertantes por placas ± DE RM
TA-100 TA-98 TA-100 TA-98
0,1 0,3 0,5 0,1 de agua destilada estéril 0,1 de 2-Antramina
129,5 ± 9,19 20,5 ± 2,12 0,90 1,41
142,0 ± 4,24 19,5 ± 7,78 0,99 1,34
143,0 ± 1,41 20,0 ± 2,83 0,99 1,38
144,0 ± 39,60 14,5± 0,71
189,0 ± 29,70 29,0 ± 11,31
Los Influentes 1 y 2 poseen razón de mutagenicidad significativamente diferentes entre las
cepas TA-100 y TA-98 (p<0,05). Siendo la razón de mutagenicidad de la cepa TA-98
mayor que la razón de mutagenicidad de la cepa TA-100 (Bonferroni). Por tanto, se sugiere
la presencia de compuestos que alterarían el marco de lectura en estos influentes. Para
confirmar la presencia de estos compuestos es necesario realizar estos ensayos aumentando
la cantidad o dosis de las muestras.
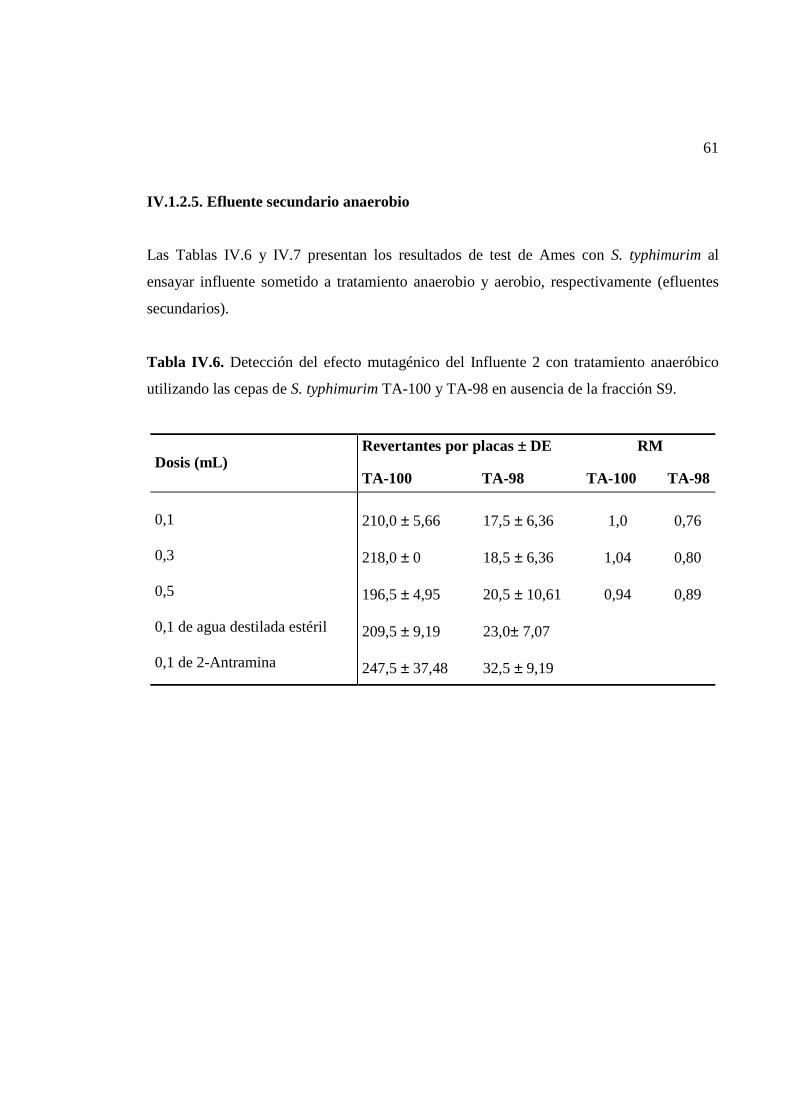
61
IV.1.2.5. Efluente secundario anaerobio
Las Tablas IV.6 y IV.7 presentan los resultados de test de Ames con S. typhimurim al
ensayar influente sometido a tratamiento anaerobio y aerobio, respectivamente (efluentes
secundarios).
Tabla IV.6. Detección del efecto mutagénico del Influente 2 con tratamiento anaeróbico
utilizando las cepas de S. typhimurim TA-100 y TA-98 en ausencia de la fracción S9.
Dosis (mL)
Revertantes por placas ± DE RM TA-100 TA-98 TA-100 TA-98
0,1 0,3 0,5 0,1 de agua destilada estéril 0,1 de 2-Antramina
210,0 ± 5,66 17,5 ± 6,36 1,0 0,76
218,0 ± 0 18,5 ± 6,36 1,04 0,80
196,5 ± 4,95 20,5 ± 10,61 0,94 0,89
209,5 ± 9,19 23,0± 7,07
247,5 ± 37,48 32,5 ± 9,19

62
IV.1.2.6. Efluente secundario aerobio
Tabla IV.7. Detección del efecto mutagénico del Influente 2 con tratamiento aeróbico
utilizando las cepas de S. typhimurim TA-100 y TA-98 en ausencia de la fracción S9.
Dosis (mL)
Revertantes por placas ± DE RM
TA-100 TA-98 TA-100 TA-98 0,1 0,3 0,5 0,1 de agua destilada estéril 0,1 de 2-Antramina
92,0 ± 15,56 18,0 ± 1,41 0,64 1,24
107,0 ± 18,38 16,0 ± 4,24 0,74 1,10
139,0 ± 14,85 17,5 ± 1,21 0,97 1,21
144,0 ± 39,60 14,5± 0,71
189,0 ± 29,70 29,0 ± 11,31
La razón de mutagenicidad varía significativamente con la cepa y los efluentes usados
(p<0,05). Así, La razón de mutagenicidad promedio para el efluente tratado
anaerobicamente es menor que la razón de mutagenicidad del efluente tratado vía aeróbica,
siendo, en consecuencia, más efectivo el tratamiento anaeróbico en la remoción o
eliminación de compuestos genotóxicos de las muestras de influentes. Así también, la razón
de mutagenicidad de la cepa TA-98 del efluente tratado aeróbicamente es mayor que para el
efluente obtenido del tratamiento anaeróbico. En tanto que, la razón de mutagenicidad de la
cepa TA-100 del efluente anaerobio es mayor que del efluente aeróbico.
Para las muestras de los efluentes con y sin tratamiento biológico, la razón de
mutagenicidad de la cepa TA-98 es significativamente mayor que la razón de
mutagenicidad de la cepa TA-100 (p<0,05; Bonferroni).
La razón de mutagenicidad de las cepas es diferente significativamente entre las muestras
de efluentes con y sin tratamiento (p<0,05). Siendo la razón de mutagenicidad promedio de
los influentes mayor que para los efluentes (Bonferroni).

63
La razón de mutagenicidad varía significativamente con las cepas, TA-100 y TA-98, por
muestras analizadas (p<0,05). No encontrándose diferencias significativas en la razón de
mutagenicidad promedio de la cepa TA-100 entre las muestras de efluente con y sin
tratamiento biológico. En tanto que, la razón de mutagenicidad promedio de la cepa TA-98
de las muestras sin y con tratamiento poseen diferencias significativas. Siendo, la muestra
de efluente sin tratamiento biológico (influente) la que posee una razón de mutagenicidad
promedio, de la cepa TA-98, mayor que las muestras de los efluentes ya tratados
(Bonferroni).
La razón de mutagenicidad varía entre las muestras de influentes, efluentes y fitosteroles
(p<0,05). Ambos fitosteroles estudiados poseen razón de mutagenicidad menor que las
muestras de los efluentes con y sin tratamiento, siguiendo la razón de mutagenicidad de
influentes, efluentes y fitosteroles un orden decreciente, respectivamente. Entre los
fitosteroles, β-sitosterol y estigmasterol no hubo diferencias. Por otra parte se tiene que la
RM varía entre las cepas por muestras (p<0,05), así la razón de mutagenicidad de la cepa
TA-98 es mayor para influentes que para los efluentes, mientras que para la cepa TA-100 la
RM es mayor para los efluentes que influentes.
Aún cuando se han encontrado diferencias significativas en la razón de mutagenicidad entre
las muestras de influentes y efluentes; influentes, efluentes y fitosteroles, no es posible
afirmar que existe mutagenicidad, ya sea de sustitución de pares de base o desplazamiento
del marco de lectura, puesto que no hay ningún valor de razón de mutagenicidad superior o
igual a dos. Pero los resultados sugieren actividad promutagénica para el estigmasterol y la
posible presencia, en los efluentes provenientes de la industria de celulosa Kraft, de
compuestos que causarían mutación por alteración del cuadro de lectura. En consecuencia,
a mayores concentraciones o dosis de las muestras se podría manifestar la mutagenicidad.

CAPÍTULO V
DISCUSIÓN

65
CAPÍTULO V
V.1. DISCUSIÓN
El β-sitosterol presenta una leve disminución de la sobrevivencia de la cepa rec(-) lo que
confirma su incapacidad de reparación genética. Mientras que, la cepa rec(+) tiende a
incrementar su sobrevivencia en presencia de este compuesto, lo cual demuestra que este
esterol vegetal podría estar siendo metabilizado por dicha cepa. Por su parte, el
estigmasterol muestra genotoxicidad en el rango de concentración estudiada, debido a la
relativa diferencia de sobrevivencia entre una cepa y otra, no obstante los valores de
eficiencia de placa varían dentro del rango de concentración, debido a que la fracción de
sobrevivencia de la cepa rec(+) sufre variaciones en tanto que la cepa rec(-) se mantiene
prácticamente constante, resultando esto en un progresivo aumento de los valores de
eficiencia de placa hasta los 16 mgL-1 y un descenso de esta luego de dicha concentración.
Por tanto, el efecto genotóxico mostrado se ve condicionado por las variaciones que
presenta la sobrevivencia de la cepa rec(+) y no al decrecimiento paulatino de la
sobrevivencia de la cepa rec(-) ante un agente dañino, esto indica entonces que
probablemente este fitosterol cause daño sobre el ADN celular, pero ambas cepas serían
capaces de reparar dicho daño, sólo que la cepa rec(+) es más eficiente en reparar en
comparación a la cepa rec(-) cuyo mecanismo de reparación se encuentra alterado.
Aún cuando los valores de eficiencia de placa entre los concentraciones usadas por
fitosterol analizado no son estadísticamente distintos, el valor promedio de eficiencia de
placa para el β-sitosterol es significativamente mayor que para el estigmasterol, indicando
que este último tendría un efecto genotóxico mayor, sin embargo ya hemos señalado que la
disminución de la eficiencia de placa del estigmasterol no se debe principalmente al
decaimiento de la sobrevivencia de la cepa rec(-), sino a las variaciones de la razón N/No
de la cepa rec(+). No obstante, la diferencia entre las sobrevivencias de ambas cepas indica
la presencia de efecto genotóxico.

66
Se sabe que las maderas blandas son ricas en ácidos resínicos en tanto que en las maderas
duras abundan los fitosteroles, pero cuando se utilizan ambos tipos de madera, la
concentración de los fitosteroles descargados en los efluentes se acentúa aún más (Tabla
II.5), lo que no implica la ausencia de estos compuestos en los efluentes provenientes de
procesos en los que se utiliza sólo madera blanda como es el Pinus radiata. Estas
concentraciones pueden ser variables y no estar sujetas únicamente al tipo de madera sino
también a las condiciones geográficas que la afectan (ecotipo geográfico). Además, la
madera blanda de Pinus radiata es de crecimiento rápido y en consecuencia posee mayor
cantidad de extractivos que las maderas de crecimiento lento, por lo tanto el procesamiento
de la madera eliminaría cantidades detectables de esteroles provenientes de la fracción
extractiva de la madera. De hecho, en las dos primeras etapas de lavado de la pulpa se
liberan significativas cantidades de fitosteroles, los que son reducidos a través del proceso
productivo. Sin embargo, cuando la remoción de esteroles totales es considerable, estos
están aún presentes en el efluente final (3,4 mgL-1) (Tabla II.5), por lo tanto, la
genotoxicidad de estos efluentes podría ser atribuida, en cierta medida, a los fitosteroles
como el β-sitosterol y estigmasterol, puesto que ambos presentan efecto genotóxico en el
rango de 1-32 mgL-1, según el ensayo “rec” de B. subtilis realizado. Siendo el β-sitosterol,
el esterol predominante encontrado en los extractivos de la madera, en las aguas de lavado
y consecuentemente en los efluentes, no obstante, este esterol no muestra un claro afecto
genotóxico en las primeras concentraciones ensayadas (1-16 mgL-1), por lo que este podría
causar efectos dañinos sobre el ADN, cuando los organismos son expuestos a los efluentes
en un tiempo prolongado permitiendo así su bioacumulación.
Si bien es cierto, las cantidades de β-sitosterol y estigmasterol encontradas en los efluentes
de la industria de celulosa Kraft son pequeñas (Tabla II.6), cabe señalar que estos pueden
tener efecto genotóxico adverso sobre los organismos acuáticos, puesto que tales
compuestos, como ya se mencionó, podrían bioacumularse y/o biomagnificarse, siendo
posible la presencia de ellos a mayores concentraciones en dichos organismos, causando tal
efecto.

67
En este mismo contexto, se sabe que los fitosteroles son liposolubles, por lo que se
solubilizan fácilmente en las membranas biológicas permitiendo el ingreso de estos a los
organismos por vía dérmica pudiendo ser de este modo, bioacumulados y/o
biomagnificados.
En las tres réplicas de Influente 1 existe presencia de genotoxicidad debido a la diferencia
manifestada de las sobrevivencias entre las cepas, no encontrándose diferencias
significativas entre los promedios de eficiencia placa por réplicas, sin embargo las
diferencias fueron dadas por las dosis ya que para la primera réplica a la dosis de 0,1 mL la
eficiencia de placa promedio es significativamente mayor en comparación a las demás
réplicas a esta misma dosis y dentro de la misma réplica. Esta diferencia se atribuiría a un
error experimental ocurrido, de ahí su mayor desviación estándar, y por tal razón se repite
el ensayo resultando entonces la réplica B, donde se ve claramente una disminución de los
valores de eficiencia de placa a medida que aumenta la dosis de la muestra ensayada,
sugiriendo que la fracción de sobrevivencia de ambas cepas decrecen, siendo la cepa rec(+)
más eficiente en reparar el daño causado sobre su ADN puesto que su mecanismo de
reparación no se encuentra mutado sino en condiciones normales, no así la cepa rec(-).
Debido a las mayores desviaciones estándares encontrada en esta réplica se realizó una
tercera réplica, la cual confirma la presencia de efecto genotóxico para este influente
(Influente 1) y que la variabilidad en las desviaciones estándares se puede deber no tan sólo
a errores experimentales sino también a errores aleatorios.
El Influente 2 muestra genotoxicidad en ambas réplicas no existiendo ningún tipo de
diferencia estadística entre ellas. Sin embargo, por el aumento de la sobrevivencia de la
cepa rec(-) en la dosis de 0,3 mL se efectuó una segunda réplica mejorándose tal
irregularidad, lo cual queda demostrado por la menor desviación estándar lograda, de la
cepa mutada, en esta última réplica. No obstante, es posible apreciar también un efecto
citotóxico, pues la sobrevivencia de la cepa madre tiende a disminuir progresivamente, en
tanto que, la cepa rec(-) se mantiene relativamente constante, esto probablemente ocurra
debido a un daño sobre la membrana de la célula madre causando consecuentemente la
muerte de ella y la declinación en su sobrevivencia respecto de su cepa mutada.

68
Los tratamientos biológicos reducen la genotoxicidad presente en los efluentes de la
industria de celulosa Kraft, puesto que la diferencia entre las sobrevivencias de ambas
cepas de los efluentes biológicamente tratados es menor comparada con los influentes. Esto
indica que la genotoxicidad de los efluentes luego de ser tratados es reducida o
prácticamente eliminada, pues en el tratamiento anaeróbico casi no hay efecto debido a la
muy poca diferencia existente entre las sobrevivencias de las cepas, mientras que en el
efluente tratado aeróbicamente la diferencia entre sus sobrevivencias es un poco mayor,
pero menor respecto de los influentes. No obstante, valores de eficiencia de placa promedio
son menores para los efluentes, aunque no significativamente, en comparación con los
influentes lo que puede ser atribuido a las irregularidades que presenta la fracción de
sobrevivencia de la cepa rec(+) en tanto que la cepa rec(-) se mantiene prácticamente
constante en todas las muestras de efluentes e influentes. Sólo para el caso del tratamiento
anaeróbico el promedio de A es mayor que el influente (Figura 4-4-E), aunque no
significativamente, señalando con esto que el tratamiento anaerobio sería más eficiente en
reducir la genotoxicidad de los efluentes de la industria de celulosa Kraft que el tratamiento
aeróbico ya que el efluente obtenido de este tratamiento posee un A promedio menor que
dicho influente, y que el efluente proveniente del tratamiento anaeróbico. Es conocido que
el tratamiento biológico aerobio presenta ventajas sobre el tratamiento anaerobio tales
como metabolismo celular rápido por tanto una mayor reducción de la materia orgánica,
mayor estabilidad del proceso y facilidad de su control. En contraste, el metabolismo
anaerobio es más lento por lo que requiere mayor volumen y las sustancias tóxicas deben
permanecer más tiempo en el reactor (mayor tiempo de residencia), además implica mayor
dificultad de operación dado que una falla en el proceso significa consumo de tiempo para
que los microorganismos colonicen el reactor y ponerlo nuevamente en funcionamiento.
Sin embargo, el tratamiento anaerobio genera una baja producción de fangos, produce
metano como subproducto utilizable y no necesita energía para su funcionamiento. La
mayoría de las plantas de tratamiento biológicos a nivel industrial son de tipo aerobias,
pues el principal objetivo es lograr una mayor reducción de la DBO5 con facilidad de
operación.

69
En este contexto, según los resultados obtenidos, estos no son suficientes para afirmar la
eficiencia de uno u otro tratamiento, pues cabe la posibilidad que el efluente aeróbico posea
mayor genotoxicidad debido a la transformación de la materia orgánica en otros
compuestos que causen daño sobre el ADN celular (Takigami et al., 2002), que contenga
una mayor DBO5 debido a una mayor cantidad de materia celular, y por otra parte la
posible eliminación de la genotoxicidad por parte del tratamiento anaeróbico se deba a que
la tasa de destrucción de los orgánicos peligrosos se ha maximizado mediante la
inmovilización de la masa sobre un soporte. Además, no se encontraron diferencias
significativas entre estos tratamientos de modo que se pueda asegurar la eficiencia de
eliminación o aumento de la genotoxicidad por parte del tratamiento anaerobio y aerobio
respectivamente (Claxton et al., 1998). Pero sí se observa eliminación de la genotoxicidad
de los efluentes tratados biológicamente, lo que estaría de acuerdo con la reducción de los
fitosteroles contenidos en ellos luego de ser tratados.
De los fitosteroles, analizados con el test de Ames, los valores de razón de mutagenicidad
sugieren que el estigmasterol con y sin S9 induciría mutación por substitución de base CG,
mientras que el β-sitosterol causaría mutación por desplazamiento en el marco de lectura.
Además la marcada diferencia en la RM de la cepa TA-100 para el estigmasterol ensayado
con S9 respecto de la RM del estigmasterol sin S9 indicaría que el sistema de activación
metabólico estaría biotransformando los compuestos en metabolitos más reactivos hacia el
ADN, los que serían responsables de la mutación por desplazamiento del marco de lectura.
Sin embargo, no existen diferencias significativas entre los valores promedio de la razón de
mutagenicidad de cada una de las cepas por compuesto estudiado, ni existe, al menos, un
valor superior a dos, de modo que no se puede afirmar que estos fitosteroles posean una
respuesta positiva, es decir, sean mutagénicos en presencia o ausencia de fracción S9, según
corresponda. No obstante, el estigmasterol muestra actividad promutagénica, puesto que
ensayado con fracción metabólica se activaría. Por lo que se hace necesario profundizar
estos estudios variando las concentraciones, utilizando fracción S9 y otros ensayos, y
realizar bioensayos cuyas respuestas sean dadas en función del tiempo.

70
Las muestras de influentes provenientes de la industria de celulosa Kraft nos indican que
luego de ser tratadas biológicamente estas disminuyen su razón de mutagenicidad, sin
embargo, ambos, influentes y efluentes, inducirían mutaciones por desplazamiento del
cuadro de lectura, esto se explica porque los efluentes son mezclas complejas donde uno de
los contaminates son los fitosteroles, entre otros de los miles de compuestos gentóxicos
existentes en estos efluentes, tales como ácidos resínicos, compuestos clorados, etc.(Houk,
1992; Maria et al., 2004; Pacheco & Santos, 2002; Maria et al, 2003; Pérez-Alzola &
Santos, 1997). Pero esta inducción podría ser significativamente menor luego de que el
Influente (2) es sometido a tratamiento secundario. En el efluente obtenido luego de ser
sometido a tratamiento aeróbico se sugiere la presencia de compuestos que causarían
mutación por desplazamiento del marco de lectura, tales compuestos no serían eliminados
eficientemente por este tratamiento desde la muestra de Influente (2), mientras que en el
efluente resultante del tratamiento anaeróbico estarían presentes compuestos que causarían
mutación por sustitución de bases (GC), esto indica que luego del tratamiento anaeróbico
compuestos orgánicos biodegradables presentes en el Influente (2) serían biotransformados
en compuestos capaces de causar mutación por sustitución de bases. Debido a que la razón
de mutagenicidad promedio de la muestra sometida a tratamiento anaeróbico es menor, se
cree que este tratamiento biológico es más efectivo que el tratamiento aeróbico en la
eliminación de la genotoxicidad.
Los resultados obtenidos con el test de Ames no muestran, al menos un valor superior a 2, y
no existe un claro efecto dosis-respuesta, por tanto, no es posible afirmar con certeza de que
los fitosteroles, influentes y efluentes sean mutagénicos, pero sí muestran o sugieren cierto
efecto mutagénico, haciéndose necesario usar una batería de bioensayos para determinar la
presencia o ausencia efectiva de la mutagenicidad (genotoxicidad), ya que debido a las
diversas propiedades y respuestas bioquímicas de los efluentes de la industria de celulosa
Kraft, la evaluación de estos efluentes podría no solamente estar limitada a uno o dos
bioensayos o sólo a análisis químico, sino a una batería de métodos, puesto que la ausencia
de un efecto específico no siempre refleja la ausencia de contaminación con efecto
genotóxico/mutagénico.

71
Siendo necesario medir varias respuestas biológicas para evaluar los efectos antropogénicos
sobre los ecosistemas acuáticos y particularmente de los efluentes, logrando así una mayor
comprensión de los efectos biológicos de los efluentes.
Por otra parte, los resultados obtenidos en el test de Ames y ensayo “rec” de B. subtilis,
sugieren que una respuesta tiempo dependiente es necesaria y útil de ser evaluada,
considerando que los compuestos contaminantes como los fitosteroles pueden ser
bioacumulados y biomagnificados en el tiempo.
Por otro lado, compuestos con actividad hormonal (estrógenos), derivados de los
fitosteroles, muestran respuestas genotóxicas, siendo entonces los fitosteroles precursores
de compuestos con actividad genotóxica. (Joosten et al. , 2004; Larsson et al., 2002;
Jenkins et al., 2001)
En el test de Ames para un screening general se sugiere realizar una secuencia
metodológica (tier aproach) usando las cepas TA-98 y TA-100 con y sin activación
metabólica. En este caso luego de obtenerse una respuesta negativa con las cepas ya
mencionadas lo recomendable es usar otras cepas tales como TA1535 y TA97 con y sin
activación metabólica, confirmando el experimento sólo si se obtiene una respuesta positiva
usando las condiciones y cepas en la cual se observe la respuesta mutagénica. Si con la
cepa TA97 se obtiene respuestas leve o errada, la cepa TA1537 puede ser usada en una
tentativa por clarificar los resultados (Mortelmans & Zeiger, 2000).
Los resultados obtenidos con el test de Ames estarían en acuerdo con Wolfreys et al. (2002)
que encontró que los fitosteroles y ésteres de fitosteroles no muestran ninguna evidencia de
actividad mutagénica en ninguno de los ensayos, ensayo de mutación bacteriana y ensayo
de aberración cromosomal “in vitro”. Por otro lado, el ensayo “rec” de B. subtilis señala
que los fitosteroles presentan actividad genotóxica, junto con los influentes de la industria
de celulosa Kraft, la cual disminuye luego de tratar los influentes biológicamente. Estando
de acuerdo esto con informes que dicen que los fitosteroles o sus derivados como hormonas
esteroidales presentan genotoxicidad, así también los efluentes de la industria de celulosa
Kraft (Kinae, et al. 1981; Kotelevtsev, et al., 2000; Rao, et al., 1995), los cuales pueden

72
incluir sustancias peligrosas que causan efectos a largo plazo, siendo por ejemplo,
genotóxicas, hormonalmente activos o bioacumulativas, dichas sustancias aparecen a bajas
concentraciones o son de carácter químico desconocido, lo cual hace que su análisis
químico requiera tiempo y métodos de alta tecnología. Por lo tanto, el uso de bioensayos en
el monitoreo de los efluentes tiene una gran útilidad (Piia Pessala et al. , 2004).
En resumen, se puede decir que ambos ensayos son útiles para determinar efecto a nivel del
ADN, sin embargo, dado las diversas propiedades de los efluentes de la industria de
celulosa Kraft, los resultados son variables, siendo conveniente usar una rápida batería de
bioensayos a pequeña escala para la evaluación de la genotoxicidad y los efectos
hormonales, complementado con bioensayos de toxicidad aguda y crónica, con el propósito
de establecer una relación entre toxicidad y genotoxicidad, y esclarecer aún más los
resultados (Rao et al., 1995; Maria et al., 2004; Maria et al., 2003). Además la respuesta
negativa en el test de Ames no necesariamente indica ausencia de genotoxicidad pues sólo
se evalúa un cambio genético en el operón de histidina, mientras que el ensayo “rec” de B.
subtilis, evalúa el daño en el ADN total, por lo tanto el daño encontrado en este ensayo
podría deberse a otro mecanismo o propiedad del ADN y no sólo a un mecanismo de
mutación expresado por la reversión de la mutación en el operón de histidina. Un estudio de
genotoxicidad y/o mutagenicidad no puede evaluarse con un par de ensayos, se sugiere una
batería de ensayos donde se incluyan otros organismos, además de bacterias, tales como
hongos, células animales, entre otros (Gollapudi & Krishna, 2000).

CAPÍTULO VI
CONCLUSIÓN

74
CAPÍTULO VI
VI.1. CONCLUSIÓNES
1. Los resultados obtenidos, de acuerdo al ensayo “rec” de Bacillus subtilis, indican
que los fitosteroles, estigmasterol y β-sitosterol, muestran efecto genotóxico. Estos
no son mutagénicos según el test de Ames.
2. El efecto genotóxico de los efluentes ensayados estaría dado, en parte, por la
presencia de los fitosteroles contenidos en ellos.
3. Los tratamientos biológicos, aerobio y anaerobio disminuyen el efecto genotóxico
de los efluentes provenientes de la industria de celulosa Kraft.
4. Considerando que los resultados presentados son preliminares, se sugiere realizar
otros ensayos para la confirmación de los mismos.
5. Es importante destacar que no existe información en la literatura acerca de las
propiedades genotóxicas de los fitosteroles como constituyentes de los efluentes de
la industria de celulosa Kraft, por lo tanto los resultados obtenidos serían un aporte
en este contexto.

CAPÍTULO VII
BIBLIOGRAFÍA

76
CAPÍTULO VII
VII.1. BIBLIOGRAFÍA
• Ames B., J. McCann, E. Yamasaki (1975) Methods for detecting carcinogens and
mutagens with the Salmonella/mammalian-microsome mutagenicity test. Mutation
Research, 31: 347-364.
• Claxton L., V. Houk, T. Hughes (1998) Genotoxicity of industrial wastes and effluents.
Mutation research, 410: 237-243.
• Cody R., S. Bortone (1997) Masculinization of mosquitofish as an indicator of
exposure to Kraft mill effluent. Bulletin Environmental Contamination & Toxicology,
58: 429-436.
• Cook D., L. La Fleur, A. Parrish, J. Jones, D. Hoy (1997) Characterization of plant
sterols from 22 US pulp and paper mills. Water Science Technology, 35: 297-303.
• del Río J., J. Romero, A. Gutiérrez (2000) Analysis of deposits produced in Kraft pulp
mills using a totally chlorine free bleaching sequence. Journal of Chromatography A,
874: 235-245.
• del Río J., A. Gutierrez, F. Gonzales-Vila (1999) Analisys of impurities occurring in a
totally chlorine free-bleanched Kraft pulp. Journal of Chromatography A, 830: 227-232.
• del Rio J., A. Gutierrez, F. Gonzalez-Vila, F. Martin, J. Romero (1998)
Characterization of organic deposits produced in the Kraft pulping of Eucaliptus
Globulus wood. Journal of Chromatography A, 82: 457-465.
• Dorado J., T. van Beek, F. Claassen, R. Sierra-Alvarez (2001) Degradation of
lipophilic wood extractive constituents in Pinus sylvestris by the white-rot fungi
Bjerkandera sp. And Trametes versicolor. Wood Science and Technology, 35: 117-125.
• Ericson G. & A. Larsson (2000) DNA Adducts in Perch ( Perca Fluvialitis) Living in
coastal water polluted with bleached pulp mill effluents. Ecotoxicology and
Environmental Safety, 46: 167-173.

77
• Espinosa C. (2000) Evaluación de los impactos de la producción de celulosa.
Publicaciones Terram, 4: 1-11.
• Field J., R. Sierra-Alvarez, G. Lettinga, J. Rintala (1992). Environmental
biotechnology for the treatment of forest industry pollutants profiles on biotechnology,
IV Congreso Nacional y I Hispano-Luso de Biotecnología (BIOTEC): 357-395.
• Förlin L., T. Andersson, L. Balk, A. Larsson (1995) Biochemical and physiological
effects in fish exposed to bleached Kraft mill effluents. Ecotoxicology and
Environmental Safety, 30: 164-170.
• Gollapudi B., G. Krishna (2000) Practical aspects of mutagenicity testing strategy: an
industrial perspective. Mutation Research, 455: 21-28.
• Gutiérrez A., J. Romero, J. del Rio (2001) Lipophilic extractives in process waters
during manufacturing of totally chlorine free Kraft from eucalypt wood. Chemosphere,
44: 1237-1242.
• Houk V. (1992) The genotoxicity of industrial wastes and effluents. A review,
Mutation Research, 227: 91-138.
• Jansson M., P. Wormald, O. Dahlman (1995) Reactions of wood extractives during
ECF and TCF bleaching of Kraft pulp Pulp and Paper Canadian, 96: 42-45.
• Jenkins R., R. Angus, H. McNatt, W. Howell, J. Kemppainen, M. Kirk, E. Wilson
(2001) Identification of androstenediona in a river containing paper mill effluent.
Environmental Toxicology and Chemistry, 20: 1325-1331.
• Jeng W.-L. & S.-J. Kao (2002) Lipids in suspended matter from the human-disturbed
Lanyang river, northeastern Taiwan. Environmental Geology, 43: 138-144.
• Joosten H., F. van Acker, D. van den Dobbelsteen, G. Horbach, E. Krajnc (2004)
Genotoxicity of hormonal steroids. Toxicology Letters, 151: 113-134.
• Kinae N., T. Hashizume, T. Makita, I. Tomita, I. Kimura, H. Kanamori (1981) Studies
on the toxicity of pulp and paper mill effluents-II. Mutagenicity of the extracts of the
liver from spotted sea trout (Nibea mitsukurii). Water Research, 15: 25-30.
• Kostamo A. & J. Kukkonen (2003) Removal of resin acids and sterols from pulp mill
effluent by activated sludge treatment. Water Research, 37: 2813-2820.

78
• Kotelevtsev S., O. Hänninen, P. Lindström-Seppä, S. Huuskonen, L. Stepanova, V.
Glaser, A. Beim (2000) Mutagenicity of bleached and unbleached effluents from
Baikalsk pulp and paper mill at lake Baikal, Russia. Aquatic Ecosystem Health and
Management, 3: 95-104.
• Larsson D., K. Kinnberg, J. Sturve, E. Stephensen, M. Skon, L. Forlin (2002) Studies
of masculinization, detoxification, and oxidative stress responses in guppies (Poecilia
reticulata) Exposed to effluent from a pulp mill. Ecotoxicology and Environmental
Safety, 52: 13-20.
• LaFleur L. (1996) Sources of pulping and bleaching derived chemical in effluents.
Environmental fate and effects of pulp and paper mill effluents, M. Servos, K.
Munkittrick, J. Carey, G. van der Kraak, (Eds), St Lucie Press, Delray Press, FL. : 21-
31.
• Lea L., P. Hepburn, A. Wolfreys, P. Baldrick (2004) Safety evaluation of phytosterol
esters. Part 8. Lack of genotoxicity and subchronic toxicity with phytosterol oxides.
Food and Chemical Toxicology, 42: 771-783.
• MacLatchy D. & G. Van der Kraak (1995) The phytoestrogen β-sitosterol alters the
reproductive endocrine status of goldfish. Toxicology and Applied Pharmacology, 134:
305-312.
• Mahmood-khan Z. & E. Hall (2003) Occurrence and removal of plant sterols in pulp an
paper mill effluents. Journal Environmental Engineering Science, 2: 17-26.
• Maria V., A. Correia, M. Santos (2004) Anguilla anguilla L. genotoxic and liver
biotransformation responses to abietic acid exposure. Ecotoxicology and Environmental
Safety, 58: 202-210.
• Maria V., A. Correia, M. Santos (2003) Genotoxic and hepatic biotransformation
responses induced by the overflow of pulp mill and secondary-treated effluents on
Anguilla anguilla L. Ecotoxicology and Environmental Safety, 55: 126-137.
• Maron D., & B. Ames (1983) Revised methods for the Salmonella mutagenicity test.
Mutation Research/Environmental Mutagenesis and Related Subjects, 113: 173-215.

79
• Mattsson K., K-J. Lehtinen, J. Tana, J. Härdig, J. Kukkonen, T. Nakari, C. Engström
(2001) Effects of pulp mill effluents and restricted diet on growth and physiology of
rainbow trout (Oncorhynchus mykiss). Ecotoxicology and Evironmental Safety, 49:
144-154.
• Mazza G. (1982) Bacillus subtilis “rec assay” Test with isogenic strains. Applied and
Environmental Microbiology, 43: 177-184.
• Mellanen P., T. Petänen, J. Lehtimäki, S. Mäkelä , G. Bylund, B. Holmbom, E.
Mannila, A. Oikari, R. Santti (1996) Wood-derived estrogens: studies in vitro with
breast cancer cell lines and in vivo in trout. Toxicology and Applied Pharmacology,
136: 381–388.
• Mellanen P., M. Soimasuo, B. Holmbom, A. Oikari, R. Santti (1999) Expression of the
vitellogenin gene in the liver of juvenile whitefish (Coregonus lavaretus L. s.l.) exposed
to effluents from pulp and paper mills. Ecotoxicology and Environmental Safety, 43:
133-137.
• Guiñez D. (2002) Efecto del porcentaje de sustitución de cloro por dioxido de cloro
sobre las características de los efluentes de blanqueo de pulpa Kraft de Pinus radiata.
Informe de memoria de título para optar al título de Ingeniero Civil Químico. Facultad
de ingeniería, departamento de ingeniería química. Universidad de Concepción.
• Mortelmans K. & E. Zeiger (2000) Test Ames Salmonella / microsome mutagenicity
assay. Mutation Research, 455: 29-60.
• Nieminen P., A. Mustonen, P. Lindström-Seppä, J. Asikainen, H. Mussalo-Rauhamaa,
J. Kukkonen (2002) Phytosterols act as endocrine and metabolic disruptors in the
european polecat (Mustela putorius). Toxicology and Applied Pharmacology, 178: 22–
28.
• Pacheco M. & M. Santos (2002) Biotransformation, genotoxic, and histopathological
effects of environmental contaminants in european eel (Anguilla anguilla L.)
Ecotoxicology and Environmental Safety, 53: 331-347.

80
• Pérez-Alzola L. & M. Santos (1997) In vitro genotoxic evaluation of conventional
bleached and biobleached softwood pulp mill effluents. Mutation Research, 395: 107-
112.
• Pessala P., E. Schultz, T. Nakari, A. Joutti, S. Herve (2004) Evaluation of wastewater
effluents by small-scale biotest and fractionation procedure. Ecotoxicology and
Environmental Safety, 59:263-272.
• Rao S., B. Quinn, B. Burnison, M. Hayes, C. Metcalfe (1995) Assesment of the
genotoxic potential of pulp mill effluent using bacterial, fish and mammalian assays.
Chemosphere, 31: 3553-3566.
• Saha B. (2003) Hemicellulose bioconversion. J Ind Microbiol Biotechnol, 30: 279-291.
• Sipi P., C. Rosenberg, Z. Rudek, P. Jäppinen, H. Vainio, H. Norppa (1997)
Cytogenetic effects of softwood Kraft pulp bleaching effluents and methanesulfonyl
choride in chinese hamster ovary cells. Mutation Research, 390: 105-112.
• Strömberg L., R. Morck, P. Sousa, O. Dahlman (1996) Effects of internal process
changes and external treatment on effluent chemistry. Environmental fate and effects of
pulp and paper mill effluents, M. Servos, K. Munkittrick, J. Carey, G. van der Kraak,
(Eds), St Lucie Press, Delray Press, FL., USA.: 3-19.
• Takigami H., S. Matsui, T. Matsuda, Y. Shimizu (2002) The Bacillus subtilis “rec
assay”: a powerful tool for the detection of genotoxic substances in the water
environment. Prospect for assessing potential impact of pollutants from stabilized
wastes. Waste Management, 22: 209-213.
• Ucar G. & M. Balaban (2002) Ciclohexane extracts of black pine wood naturally
grown in eastern Thrace. Holz als Roh- und Werkstoff, 60: 34-40.
• Vidal G. (1994) Biodegradación de compuestos tóxicos en efluentes de industrias de
madera. Memoria de grado para optar al grado de doctor por la Universidad de Santiago
de Compostela. Departamento de ingeniería química, Universidad de Santiago de
Compostela.
• Vidal G., R. Méndez y J. Lema (1996) La industria celulósica de pasta y papel.
Departamento de ingeniería química, Universidad de Santiago de Compostela, 139-145.

81
• Verta M., J. Ahtiainen, T. Nakari, A. Langi, E. Talka (1996) The effect of waste
constituents on the toxicity of TCF and ECF pulp bleaching effluents. Environmental
fate and effects of pulp and paper mill effluents, M. Servos, K. Munkittrick, J. Carey,
G. van der Kraak, (Eds), St Lucie Press, Delray Press, FL.: 41-51.
• Van den Heuvel M., R. Ellis, L. Tremblay, T. Stuthridge (2002) Exposure of
reproductively maturing rainbow trout to a New Zealand pulp and paper mill effluent.
Ecotoxicology and Environmental Safety, 51: 65-75.
• Wolfreys A. & P. Hepburn (2002) Safety evaluation of phytosterol esters. Part 7.
Assesment of mutagenic activity of phytosterols, phytosterol esters and cholesterol
derivative, 4-cholesten-3-one. Food and Chemical Toxicology, 40: 461-470.
• Información obtenida de las siguientes paginas: www.cmpc.cl, www.papelnet.cl,
www.arauco.cl.

82
ANEXO
Publicaciones
- Xavier, C. R., Oñate, E., Mondaca, M. A. and Vidal, G. 2005. Genotoxicity assess on
kraft pulp mill effluent treated by activated sludge and aerated lagoon. Ecotoxicology and
Environmental Safety (submitted).
Congresos
1. Oñate, E., Decap, J., Chamorro, S., Vidal, G. y Mondaca, M. A. 2004. Evaluación de la
genotoxicidad de fitosteroles presentes en efluentes de fábrica de celulosa Kraft
mediante el ensayo “Rec” de Bacillus subtilis. XXVI Congreso Chileno de
microbiología. Valparaíso, 1 al 3 de Diciembre.
2. Oñate, E., Xavier, C., Belmonte, M., Vidal, G., Mondaca, M. A. 2005. Ensayo “rec”
con Bacillus subtilis usado para evaluar la remoción de la genotoxicidad por el
tratamiento de efluentes de la industria de celulosa Kraft. XXVII Congreso Chileno de
microbiología. Pucón, 13 al 16 de Octubre (Enviado).
3. Oñate, E., Xavier, C., Mondaca, M. A., Belmonte, M., Vidal, G. 2005. La
genotoxicidad como indicador de eficiencia en sistemas de tratamientos aeróbicos. XVI
Congreso de Ingeniería Sanitaria y Ambiental AIDIS-Chile. Viña del Mar, 23 al 26 de
Octubre (Aceptado).
4. Xavier, C. R., Oñate, E., Mondaca, M. A., and Vidal, G. 2006. Kraft pulp mill effluent
treated by activated sludge and aerated lagoon genotoxicity assess. 6° Fate and effects
of pulp and paper mill effluents. Vitoria-Espírito Santo (Brasil), 9 al 12 de Abril
(Enviado).

12
Figura 2-3. Proceso productivo y generación de residuos líquidos en la etapa de pulpaje de la industria de celulosa Kraft.
Madera Etapa de preparación de la madera
Etapa de pulpaje de la madera
Etapa de separación y
lavado
Etapa de secado y embalado
Etapa de blanqueo
RIL ES Fitosteroles
Emisiones
¿Genotoxicidad?
Calderas de
recuperación
Tratamiento de
efluentes
Licor negro


















