Fósiles -...
24
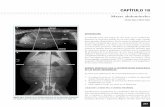
316 Placoparia (Coplacoparia) tournemini (Rouault). Grupo de ejemplares de este trilobites queirúrido, del Ordovícico Medio de Alía (Cáceres). Placoparia (Coplacoparia) tournemini (Rouaul)t. Grupo de ejemplares de este trilobites queirúrido, del Ordovícico Medio de Alía (Cáceres).
Transcript of Fósiles -...

316
Placoparia
(Coplacoparia)
tournemini (Rouault).
Grupo de ejemplares
de este trilobites
queirúrido, del
Ordovícico Medio de
Alía (Cáceres).
Placoparia
(Coplacoparia)
tournemini (Rouaul)t.
Grupo de ejemplares
de este trilobites
queirúrido, del
Ordovícico Medio de
Alía (Cáceres).

317
Fósiles

318
Concentración
de valvas ventrales
(aplanadas, con gran
campo muscular) y
dorsales (abombadas)
del braquiópodo
Eorhipidomella
musculosa (Mélou),
del Ordovícico Medio
de Fuenlabrada de los
Montes (Badajoz).
Moldes internos de
15-16 mm de anchura.
Concentración
de valvas ventrales
(aplanadas, con gran
campo muscular) y
dorsales (abombadas)
del braquiópodo
Eorhipidomella
musculosa (Mélou),
del Ordovícico Medio
de Fuenlabrada de los
Montes (Badajoz).
Moldes internos de
15-16 mm de anchura.

319
xtremadura, espacio abierto a la meseta
castellana por el este y a la infl uencia at-
lántica y portuguesa al oeste, no siempre
presentó la misma arquitectura geográfi -
ca, ni cobijó sobre sus antiguas tierras y
mares los mismos organismos que hoy la pueblan. A
través de los tiempos geológicos su diseño, estructu-
ra y habitantes han sufrido numerosas e importantes
transformaciones que también constituyen parte de su
historia.
Aquellos acontecimientos y seres vivos que no re-
cogen los libros de historia clásicos, están registrados
en forma de fósiles en los diferentes tipos de rocas que
constituyen el sustrato o terruño que hoy denomina-
mos Extremadura. Lugar para el que, en 1931, un exi-
mio geólogo y naturalista extremeño, Eduardo Hernán-
dez Pacheco, establecía sus límites entre “las nevadas
cumbres de Almanzor hasta los fructíferos castañares
y olivares de la alegre y pintoresca Sierra de Aracena,
límite natural y meridional de Extremadura, y desde el
Alentejo portugués a las vertientes de la Sierra Morena,
que dan vista a la llanura Bética”.
El interés e importancia de los fósiles de Extrema-
dura es clave para conocer la historia y la evolución
paleogeográfi ca de la Península Ibérica durante los
tiempos neoproterozoicos y paleozoicos. Los traba-
jos publicados reúnen datos procedentes de unos 400
yacimientos o secciones fosilíferas extremeñas, en los
que se llevan inventariados más de un millar de espe-
cies fósiles de invertebrados marinos, registros micro-
paleontológicos diversos, y señales de actividad en el
sedimento (icnofósiles).
Además de la contribución general de las locali-
dades paleontológicas extremeñas a la Geología de la
Península Ibérica y a la historia de su paleobiodiversi-
dad, algunos yacimientos son de interés especial por no
encontrar parangón en otras regiones ibéricas, o bien
por servir de localidad tipo a varias decenas de fósiles
nuevos para la Ciencia. El descubrimiento y la primera
caracterización de estos últimos en Extremadura moti-
vó que parte de ellos tomasen nombres científi cos rela-
cionados con la región, como por ejemplo ocurre con
E

320
los arqueociatos cámbricos Chouberti-
cyathus extremadurensis, Alconeracya-
thus andalusicus (género defi nido en Al-
conera), Rasetticyathus alconeri (especie
dedicada a Alconera), Erismacoscinus
zafrensis (de Zafra), E. badajocensis (de
la provincia de Badajoz), Antoniocosci-
nus segedanensis (de Segeda, nombre ro-
mano de Zafra). Es también el caso de los
braquiópodos Yorkia zafrensis (del Cám-
brico de Zafra) o Productina pacensis (del
Carbonífero pacense); de los trilobites
Calymenella alcantarae (del Ordovícico
de Valencia de Alcántara), Linguaphillip-
sia sanpedroensis (del Carbonífero de
la Sierra de San Pedro); y de los corales
Disphyllum pedrosensis (Devónico de la
Sierra del Pedroso).
La historia de las investigaciones pa-
leontológicas en Extremadura se inicia a
mediados del siglo XIX de la mano de
Casiano de Prado y progresa, en el úl-
timo cuarto de aquel siglo, merced al
trabajo de otros ilustres miembros de la
Comisión del Mapa Geológico de Espa-
ña, como Lucas Mallada y Justo Egoz-
cue. En la primera mitad del siglo XX, las
principales aportaciones corren a car-
go de geólogos como Ismael Rosso de
Luna, Francisco Hernández Pacheco y
Primitivo Hernández Sampelayo. A par-
tir de 1930 y hasta nuestros días, se mul-
tiplican los hallazgos paleontológicos
por toda Extremadura, primero a cargo
de geólogos y paleontólogos de otros
países europeos (alemanes, holandeses,
franceses), que progresivamente son re-
levados por la generación contemporá-
nea de paleontólogos españoles de las
universidades de Extremadura, Madrid,
Zaragoza, Valencia y Oviedo, junto a
investigadores del Instituto Geológico y
Minero de España y del Consejo Supe-
rior de Investigaciones Científi cas.
Fósiles del Neoproterozoico
Las rocas sedimentarias neoprotero-
zoicas afl oran extensamente en el cen-
tro y oeste de Extremadura, presentando
un contenido fósil de escasa diversidad
y simplicidad orgánica, en la que mu-
chos organismos no presentan partes
duras o susceptibles de conservación.
Por lo general, en las litologías pizarro-
sas se documentan microfósiles de pa-
red orgánica (acritarcos) y vesículas de
cianobacterias coloniales, mientras que
las calizas reúnen fósiles más variados e
importantes.
Los fósiles neoproterozoicos precisan
de técnicas especiales para su estudio,
láminas delgadas en el caso de los restos
de la actividad de las cianobacterias, y
disgregación de las rocas y ataque con
ácidos fuertes para los acritarcos. En al-
gunas circunstancias, por silicifi cación
de las calizas, se ha preservado la es-
tructura de las cianobacterias, las cuales
presentan diseños orgánicos similares a
las actuales, con estructuras segmenta-
das o en forma de fi lamento.
Los acritarcos constituyen la parte
fundamental del plancton de los mares
del Neoproterozoico y del Cámbrico.
Dentro de esta denominación se inclu-
ye una amplia gama de microfósiles de
pared orgánica con formas muy diver-
sas, cuyas vesículas pueden ser globo-
sas o de contorno poligonal, con o sin
procesos espinosos o estructuras de de-
senquistamiento, que se relacionan con
restos de las posibles algas unicelulares
planctónicas que originan estos quistes.
Las cianobacterias bentónicas fl ore-
cieron en los ambientes marinos some-
ros de hace más de 540 millones de años
y, debido a su actividad biológica, con-

321
tribuyeron decisivamente a la formación
de rocas carbonatadas con estructura
laminar, bien estromatolítica, bien ma-
siva, debido a su capacidad de depositar
el carbonato cálcico. Las rocas calizas
de este tipo abundan en el entorno del
embalse de Valdecañas y en las trinche-
ras de la antigua carretera nacional V, en
las proximidades del puerto de Miravete
(Cáceres).
En otras áreas de la plataforma mari-
na, además de calizas estromatolíticas,
se desarrolló otro grupo de organismos
tubícolas de pequeño tamaño, próximos
a los cnidarios, cuyos restos fósiles se
conocen con el nombre de Cloudina. Se
trata de los primeros metazoos con es-
queleto calcáreo y también los primeros
bioconstructores, pues normalmente se
agrupan por millones para formar ma-
sas biogénicas de gran porte, parecidas
a arrecifes. Las dimensiones de cada es-
queleto individual varían entre 0,3 a 6,5
mm de diámetro y más de 55 mm de
longitud. Internamente están formados
por una serie de láminas cónicas embu-
tidas unas dentro de otras y curvadas en
su parte distal, presentando una estruc-
tura “cono en cono” multilaminada. La
distribución de los conos embutidos es
siempre asimétrica y cada nueva lámina
se dispone más elevada que la anterior,
estructura que los diferencia de todos
los demás grupos fósiles conocidos. En
Extremadura se encuentran excelentes
ejemplos de bioconstrucciones formadas
por Cloudina en los alrededores de Cas-
tañar de Ibor (Cáceres) y en el embalse
de Cíjara, próximo a Bohonal (Badajoz).
Afl oramiento de calizas
neoproterozoicas
con estructuras
estromatolíticas en
Castañar de Ibor
(Cáceres).
Vendotaénidos,
fi lamentos carbonosos
de apariencia algal.
Pizarras neoproterozoi-
cas de Navalvillar de
Ibor (Cáceres).
La anchura media de
cada cinta es de 1 mm.

322
En los niveles de pizarras que al-
ternan con los carbonatos se registran
restos carbonosos fi lamentosos sin es-
tructura interna, estos reciben el nom-
bre de “vendotaénidos”. Abundan en
las mismas áreas que Cloudina. Las pi-
zarras suelen mostrar, además, diversas
huellas de organismos de cuerpo blando
(icnofósiles), como por ejemplo Nimbia,
Neonereites (sucesión del Estenilla, Ba-
dajoz); Torrowangea (Coria, Cáceres);
Phycodes (Arrocampo, Cáceres), etc.
Fósiles del Cámbrico
La aparición de nuevas formas de
vida a comienzos del Fanerozoico que-
da refl ejada, en Extremadura, por la apa-
rición de multitud de fósiles con partes
duras conservadas, así como de icnofó-
siles que evidencian unas estrategias de
alimentación sedimentívora o marcha
ciertamente depuradas. Entre ellos cabe
citar a Phycodes, Arthrophycus, Ruso-
phycus, etc., en la localidad de Medina
de las Torres (Badajoz).
En las plataformas someras de los
mares del Cámbrico Inferior extreme-
ño, frente a las costas de Gondwana, se
desarrollaron nuevas bioconstrucciones
pero esta vez formadas por arqueociatos
y calcimicrobios, producto de la activi-
dad biológica de cianobacterias que for-
maron importantes depósitos de rocas
calcáreas. Los arqueociatos son los res-
tos fósiles de un grupo de poríferos sin
espículas, con esqueleto calcáreo lami-
Deslizamientos
sinsedimentarios en
las bioconstrucciones
calcáreas del Cámbrico
Inferior. Canteras de
Alconera. (Badajoz)
Montículo carbonático
de calcimicrobios y
arqueociatos.
Detalle de una
superfi cie erosionada
en el Cámbrico Inferior
de Alconera (Badajoz),
mostrando en relieve
numerosos cálices de
arqueociatos. Moneda
de 23 mm de diámetro.

323
nar y perforado, que tuvieron un fl oreci-
miento rápido y se extinguieron al fi nal
del Cámbrico. Las formaciones carbona-
tadas, constituidas en una gran propor-
ción por restos fósiles de cianobacterias
como Epiphyton y Renalcis, junto con
los géneros de arqueociatos ya citados
y Nochoroicyathus, Coscinocyathus, Ra-
setticyathus, Mennericyathus y Protopha-
retra, son abundantes en Extremadura y
afl oran de forma discontinua desde el
límite con la provincia de Sevilla hasta
Mérida, destacando los afl oramientos de
Llerena y Usagre (Badajoz), y con una
exposición espectacular en las canteras
de Alconera (Badajoz), donde puede ob-
servarse el desarrollo superpuesto de nu-
merosas bioconstrucciones. Aunque no
se tiene constancia directa de la explo-
tación de estas canteras por los romanos
podemos deducir que éstos conocían su
existencia, ya que muchos de los silla-
res del teatro romano de Mérida están
tallados en calizas con arqueociatos (el
teatro fue construido en el año 16 a.C.,
como un regalo de Agrippa a la recién
fundada ciudad).
Otros fósiles muy comunes a partir
del Cámbrico Inferior son los trilobites,
un grupo de artrópodos de caparazón
trilobulado y articulado, formado por
un número variable de piezas y segmen-
tos. Sus caparazones y restos de muda
se conservan abundantemente en las
rocas detríticas de grano fi no de los ya-
cimientos pacenses del Cámbrico Infe-
rior próximos a Zafra, Alconera, Llerena,
y también en las pizarras del Cámbrico
Medio de la Albuera del Castellar (Zafra).
De las asociaciones de trilobites destaca
Serrania, un género del Ovetiense, y for-
mas representativas de niveles más ele-
vados del Cámbrico Inferior (Marianien-
se) con Hicksia, Strenuaeva, Strenuella,
Gygantopygus, Protaldonaia, Delga-
della y Serrodiscus; así como trilobites
del Cámbrico Medio (Leoniense-Caesa-
raugustiense) como Paradoxides, Con-
dylopyge, Ctenocephalus, Jincella, Sao,
Holocephalina, etc. En los afl oramientos
antes citados, junto a los trilobites sue-
Alconeracyathus
andalusicus (Perejón).
Sección transversal de
un cáliz de arqueociato
irregular, del Cámbrico
Inferior de Alconera
(Badajoz).

324
len encontrarse también restos más raros
de braquiópodos (Yorkia y otros géneros
organofosfáticos) y también equinoder-
mos (eocrinoideos, homalozoos).
Fósiles del Ordovícico
Los fósiles ordovícicos más antiguos
de Extremadura corresponden a las seña-
les dejadas en el sustrato arenoso por un
gran número de invertebrados marinos,
muchos de ellos de cuerpo blando y sin
partes fosilizables. Sus icnofósiles o hue-
llas de actividad son muy frecuentes en
todas las sierras formadas por la “Cuar-
cita Armoricana” del Arenigiense, tanto
en la provincia de Cáceres (Serradilla
y Monfragüe, Las Villuercas, Altamira,
Hospital del Obispo, norte de Ladrillar,
Cranidios de dos
trilobites del Cámbrico
Medio de Zafra
(Badajoz): Conocoryphe
(Parabailiella) sp.
(izquierda, 25 mm de
anchura) y Jincella
sp. (dcha.)

325
sur de Villanueva de la Sierra, Cáceres
ciudad y Puerto Señorina, Alcuéscar,
Sierra de San Pedro al noreste de Valen-
cia de Alcántara o sureste de Herreruela,
Pino de Valencia, etc.), como en la pro-
vincia de Badajoz (norte de San Vicente
de Alcántara y La Codosera, Alburquer-
que, sierras de La Rinconada y El Alji-
be en torno a Helechosa de los Montes,
sierras de Herrera del Duque, Rena,
Siruela, Benquerencia de la Serena-He-
lechal, Cabeza del Buey, Zarza Capilla,
Peñalsordo, Alange, etc.). En general,
las formas dominantes de estas icnoaso-
ciaciones son las pistas de reptación de
artrópodos denominadas Cruziana, co-
rrespondientes al moldeado en arena de
dos surcos paralelos (ahora lóbulos en
relieve), con marcas de estrías oblicuas
dejadas por los apéndices locomotores.
Cruziana se suele atribuir a la actividad
excavadora de ciertos trilobites, respon-
sables también de otras marcas orgáni-
cas en las cuarcitas, como las llamadas
Monomorphichnus (de marcha sobre el
fondo) o Rusophycus (marcas ovaladas
de reposo). Otras huellas frecuentes tam-
bién en el plano de estratifi cación son
Gordia, Palaeophycus, “Rhizomorpha”,
Planolites, Didymaulichnus (pista bilo-
bulada lisa) y Arthrophycus (con sección
cuadrangular y marcas transversas: todas
ellas producidas por organismos desco-
nocidos. Entre los icnofósiles orientados
perpendicularmente a los estratos de
cuarcita, destacan las galerías vertica-
les llamadas Skolithos (tubos simples),
Monocraterion y Diplocraterion (tubo
doble, con entrada y salida), así como
las estructuras complejas tridimensio-
nales llamadas Daedalus (una lámina
en forma cónica). Todas ellas ofrecen
secciones características al atravesar los
planos de estratifi cación, y se atribuyen
a animales de cuerpo blando dotados de
gran movilidad (gusanos marinos).
Los restos fósiles propiamente di-
chos (conchas y caparazones), son poco
frecuentes en los materiales arenoso-
cuarcíticos que componen la “Cuarci-
ta Armoricana”. Sin embargo, hacia la
parte superior de la misma, se cono-
cen algunos niveles de concentración
de grandes lingúlidos (braquiópodos
quitinofosfáticos) y bivalvos, como por
Hicksia hispanica
(Richter y Richter).
Cranidio (región
cefálica) de un trilobites
del Cámbrico Inferior
de Zafra (Badajoz).
Longitud, 25 mm.
Paradoxides
(Eccaparadoxides) sp.
Cranidio (región
cefálica) de un trilobites
del Cámbrico Medio
de Zafra (Badajoz).
Longitud, 25 mm.
Cruziana.
Pista de reptación
atribuida a trilobites,
moldeada en una
cuarcita del Ordovícico
de Las Hurdes.

326
ejemplo ocurre en la vertiente oriental
de la Sierra del Hospital del Obispo y
en la opuesta de la Sierra de Altamira (al
este de los Ibores), en lo que se interpre-
ta como una acumulación catastrófi ca
derivada de tsunamis provocados por
explosiones volcánicas en la plataforma
gondwánica.
Las pizarras que suceden geológi-
camente a estas sierras formadas por la
“Cuarcita Armoricana”, suelen formar
valles deprimidos al pie de las mismas,
donde los yacimientos de fósiles del
Ordovícico Medio son prácticamente
continuos y presentan una diversidad
paleontológica variable, dependiendo
Aspecto del Sinclinal
de Guadarranque al no-
reste de Alía (Cáceres),
mostrando alineaciones
cuarcíticas del Ordoví-
cico Medio y Superior.
En primer término
destaca el “valle de los
trilobites”, con afl o-
ramientos de pizarras
fosilíferas en el fl anco
norte de la estructura.
Derrumbes en el traza-
do del “ferrocarril de
La Jara” entre Puerto
de San Vicente y Alía
(Cáceres). Las rocas son
pizarras fosilíferas del
Ordovícico Medio, en
una de las localidades
paleontológicas más
conocidas de Extrema-
dura.

327
del nivel estratigráfi co. En estas pizarras
se intercalan niveles arenosos y suelen
terminar en una segunda unidad cuar-
cítica importante (la Cuarcita Canteras,
La Cierva o del Torrico), en cuya base
existen también fósiles del Ordovícico
Medio. Las principales áreas donde se
han citado yacimientos de edad Ore-
taniense-Dobrotiviense (antiguo “Llan-
virn-Llandeilo” en su uso ibérico) son,
dentro de la provincia de Cáceres, el sur
de Mirabel, Serradilla, parque de Mon-
fragüe, sureste de Santiago de Alcán-
tara, sur y sureste de Herreruela, este de
Aliseda, este de la ciudad de Cáceres y
suroeste de su término municipal, norte
de Robledollano, norte y este de Caña-
mero, oeste de Solana, norte y este de
Castañar de Ibor, Navatrasierra –entre
Navatrasierra y Navalvillar de Ibor– y
noreste de Alía, entre otras áreas impor-
tantes. En la provincia de Badajoz, los
principales yacimientos paleontológicos
del Ordovícico Medio se sitúan al norte
de San Vicente de Alcántara, Helechosa
de los Montes, noreste y sur de Herrera
del Duque, sur y sureste de Fuenlabra-
da de los Montes, noreste de Casas de
Don Pedro, norte de Talarrubias, sureste
de Navalvillar de Pela, norte y este de
Garlitos, sur de Peñalsordo, sureste de
Capilla y oeste de Cabeza del Buey.
Los fósiles del Ordovícico Medio
extremeño comprenden numerosas
especies de trilobites pertenecientes a
35 géneros, entre los cuales destacan
algunos calymenáceos y homalonóti-
dos como Neseuretus, Colpocoryphe,
Prionocheilus, Eohomalonotus e Ibero-
coryphe; facópidos como Eodalmaniti-
na, Zeliszkella o Crozonaspis; ilénidos
como Ectillaenus; queirúridos como
Placoparia y Eccoptochile; asáfi dos
como Asaphellus, Nobiliasaphus e Isa-
belinia, lichidos (Uralichas) y odonto-
pléuridos (Selenopeltis). Los moluscos
son el siguiente grupo en orden de
abundancia y diversidad, en especial
Excavación temporal
en las pizarras del
Ordovícico Medio de
Fuenlabrada de los
Montes (Badajoz),
emplazada en niveles
particularmente ricos en
trilobites completos.

328
A B
C D
E F

329
los bivalvos como Redonia, Coxicon-
chia, Hemiprionodonta, Cardiolaria,
Praenucula, etc., gasterópodos (Sinui-
tes, Ptychonema), cefalópodos, rostro-
conchas, etc. A continuación se sitúan
los braquiópodos, con una decena
de formas ubicuas entre las que cabe
mencionar los géneros Cacemia, Hete-
rorthina, Eorhipidomella y Aegirome-
na. Los equinodermos más frecuentes
son los cistoideos diplopóridos (Calix,
Oretanocalix, Phlyctocystis) y los res-
tos de crinoideos. Hay también algu-
nos cnidarios (conuláridos) y briozo-
os, así como representantes de grupos
problemáticos, como los macaeridios
y los hiolites. Los ostrácodos (artrópo-
dos diminutos de caparazón bivalvo)
abundan en algunos niveles. Entre las
formas pelágicas, el microplancton de
pared orgánica (acritarcos y quitinozo-
os) suele presentarse mal conservado
por causa de metamorfi smo regional y
de contacto, pero el zooplancton co-
lonial (graptolitos) abunda en la parte
inferior de las sucesiones pizarrosas,
A. Neseuretus tristani (Brongniart in Desmarest). Vista lateral de un capara-zón completo, procedente del Ordovícico Medio de Helechosa de los Montes (Badajoz). Longitud, 43 mm.
B. Eodalmanitina destom-besi (Henry). Agrupación de caparazones juveniles completos (de alrededor de 10 mm de longitud), encontrada en el Ordovícico Medio de Fuenlabrada de los Montes (Badajoz).
C. Zeliszkella torrubiae (Ver-neuil y Barrande). Trilobites facópido del Ordovícico Medio de Fuenlabrada de los Montes (Badajoz). Longitud del ejemplar, 50 mm.
D. Isabelinia glabrata (Salter), un trilobites asáfi do del Ordovícico Medio de Fuenlabrada de los Montes (Badajoz). Longitud del ejemplar, 60 mm.
E. Ectillaenus giganteus (Bur-meister). Trilobites ilénido del Ordovícico Medio de Fuenlabrada de los Montes (Badajoz). Longitud, 80 mm.
F. Eccoptochile almadenen-sis (Romano) (izquierda), un trilobites queirúrido asociado en esta muestra al ilénido Panderia beaumonti (Rouault). Ordovícico Medio de Fuenlabrada de los Mon-tes (Badajoz). Longitud del ejemplar mayor, 50 mm.
G. Agrupación de colonias en forma de diapasón del graptolito Didymograptus (Jenkinsograptus) spinulosus (Perner), procedentes de las pizarras del Ordovícico Medio del Puerto del Clavín (Cáceres). Los ejemplares de mayor tamaño alcanzan 40 mm de longitud.
G

330
A. Uralichas hispanicus (Verneuil y Barrande). Cranidio (parte central cefálica) de un ejemplar en el que se distinguen los surcos y lóbulos, combinados con una ornamentación de grá-nulos, tan característicos de los trilobites Lichida. A la especie ilustrada pertenecen los trilobites más grandes encontrados en Gondwana, con una longitud axial de más de 60 cm. El fragmento de muda fi gurado mide 50 mm de longitud antero-posterior y procede del Ordovícico Medio de Alía (Cáceres).
B. Redonia deshayesi (Rouault). Moldes inter-nos de la valva derecha (arriba) e izquierda (aba-jo), mostrando las ranuras correspondientes al septo anterior. Este molusco bivalvo abunda en los yacimientos del Ordoví-cico Medio extremeño; los ejemplares ilustrados proceden de Herrera del Duque (Badajoz) y miden 20 mm.
C. Cacemia ribeiroi (Shar-pe). Valvas de un órtido (braquiópodo) adaptado a la vida en fondos fango-sos, con la línea cardinal alargada (40 mm) como en los espiriféridos. Ordovícico Medio de Helechosa de los Montes (Badajoz).
D. Selenopeltis gallica (Bruton), un trilobites odontopléurido nectoben-tónico en el que destacan las largas espinas pleura-les. Ejemplar juvenil de 10 mm de longitud. Ordo-vícico Medio de Fuen-labrada de los Montes (Badajoz).
E. Eoglyptograptus sp., un graptolito biseriado del Ordovícico Medio de Alía (Cáceres). Longitud, 32 mm.
donde son frecuentes las colonias en
forma de horquilla del género Didy-
mograptus o las biseriadas de Huste-
dograptus y Eoglyptograptus.
En su conjunto, los fósiles del Ordo-
vícico Medio extremeño reúnen más de
Trocholites fugax (Babin y Gutié-
rrez-Marco), un cefalópodo de con-
cha espiral del Ordovícico Medio de
Garlitos (Badajoz). Diámetro mayor
del ejemplar, 33 mm.
dos centenares de especies, cuya distri-
bución vertical permite establecer divi-
siones y correlaciones muy fi nas para
estos materiales, merced a la veintena
de biozonas defi nidas por la selección
evolutiva y registrática de todos estos in-
vertebrados fósiles.

331
A B
C
D E

332
Los materiales del Ordovícico Su-
perior extremeño no son tan fosilíferos
como los precedentes, restringiéndose
a hallazgos bastante puntuales en la
provincia de Cáceres (noreste de Va-
lencia de Alcántara, sur de Herreruela,
noroeste de Cañamero, oeste de Na-
vatrasierra, noreste de Alía) y en la de
Badajoz (norte de Talarrubias, Garlitos,
sur y sureste de Capilla). Los fósiles se
presentan tanto en pizarras como en
cuarcitas, y son principalmente braquió-
podos (Gelidorthis, Svobodaina, Dra-
bovia, Reuschella, Aegiromena, etc.) y
trilobites (Calymenella, Crozonaspis,
Dalmanitina, Scotiella, Deanaspis, Zeti-
llaenus, Actinopeltis, etc.). Se conocen
también algunos equinodermos (cistoi-
deos, crinoideos) y briozoos, que están
en estudio. Casi todas las asociaciones
mencionadas son de edad Berounien-
se (Caradociense a Ashgilliense basal
como máximo). Las litofacies calcáreas
(Caliza Urbana), de edad Kralodvorien-
se (Ashgilliense “medio”), se presentan
únicamente al sureste de Capilla y en
Garlitos (Badajoz), donde brindaron
asociaciones de conodontos y raros
graptolitos. Los materiales del Ordoví-
cico terminal (Kosoviense o Ashgillien-
se/Hirnantiense), coetáneos con la gla-
ciación con la que culmina el periodo
Ordovícico, libraron un único fósil (un
equinodermo homalozoo) al oeste de
Navatrasierra (Cáceres), que no obstan-
te representa el quinto yacimiento de
esta edad conocido en todo el suroeste
de Europa.
A pesar de la aparente abundancia
de localidades paleontológicas extre-
meñas del periodo considerado, restan
por investigar aún extensos afl oramien-
tos pacenses de pizarras ordovícicas. De
ellos destacamos los ubicados entre Al-
burquerque y Puebla de Obando, entre
Cheles y Valencia de Mombuey y, sobre
todo, en La Serena, Sierra de Hornachos
y Sierra del Pedroso donde, por el mo-
mento, no se conoce ningún fósil ordo-
vícico de grupos clave como trilobites,
braquiópodos o graptolitos.
Svobodaina havliceki
Villas. Braquiópodo del
Ordovícico Superior
conservado en arenisca.
Moldes internos de la
valva ventral (izquier-
da) y dorsal (derecha),
procedentes de Garlitos
(Badajoz). La longitud
mayor es de 20 mm.

333
Fósiles del Silúrico
El Silúrico de Extremadura compren-
de afl oramientos poco extensos, restrin-
gidos al núcleo de los sinclinales hercíni-
cos de mayor continuidad lateral donde
la sucesión de este periodo se inicia con
una formación extraordinariamente fosi-
lífera. Se trata de pizarras negras grap-
tolíticas, también llamadas “ampelitas”
por su utilización antigua para el abo-
nado de viñedos, donde se acumulan
por millones los fósiles de las colonias
planctónicas macroscópicas de los ya
citados graptolitos. Cada una de ellas
suele estar compuesta por una sola rama
o estipe (colonias uniseriales), pero hay
también colonias con dos estipes unidas
dorsalmente (formas biseriales), e inclu-
so colonias ramifi cadas radialmente. Los
graptolitos son excepcionalmente preci-
sos desde el punto de vista estratigráfi co,
de modo que a partir de los yacimientos
extremeños han podido caracterizarse
numerosas biozonas de las épocas y
edades Llandovery (Telychiense), Wen-
lock (Sheinwoodiense, Homeriense) y
Ludlow (Ludfordiense). Las rocas del
Pridoli no contienen fósiles en ninguna
localidad, pero deben de estar repre-
sentadas en las potentes alternancias de
areniscas y pizarras en las que se registra
el paso al Devónico, materializadas por
los niveles previos a los que libraron los
fósiles devónicos más antiguos.
Los principales afl oramientos de piza-
rras negras graptolíticas en Extremadura,
donde se llevan localizados numerosos
yacimientos con graptolitos silúricos, se
sitúan, dentro de la provincia de Cáce-
res, al norte de Serradilla y parque de
Monfragüe, al norte de Castañar de Ibor,
oeste de Navatrasierra, noreste de Alía,
embalse de Cijara, Cáceres ciudad y su-
roeste de su término municipal, y sureste
Monograptus fl emingii
(Salter). Colonia con-
servada en tres dimen-
siones en el interior de
un nódulo, y vaciado
en látex (centro) de una
de sus mitades. En este
último se distinguen con
claridad las celdillas
ganchudas donde vivie-
ron los zooides. Silúrico
Superior de Herrera del
Duque (Badajoz). Di-
mensiones del fragmen-
to colonial: 30 x 3 mm.
Spirograptus turricu-
latus (Barrande), un
graptolito de esqueleto
enrollado en espiral
turriculada, del Silúrico
Inferior de Alía (Cáce-
res). El diferente aspecto
de las dos colonias de
la fotografía obedece a
que fueron aplastadas
con dos orientaciones
distintas. La colonia de
la derecha mide 17 mm.
Opérculo bilobulado
(“peltocárido”) de un
cefalópodo nautiloideo,
conservado junto a
restos de braquiópodos
epiplanctónicos en
pizarras negras grapto-
líticas. Silúrico Superior
de Herrera del Duque
(Badajoz).
Anchura, 24 mm.
Colonias uniseriadas
de Monograptus sp., el
género más representa-
tivo de entre todos los
graptolitos silúricos. Los
ejemplares proceden
de Pino de Valencia
(Badajoz), y su anchura
máxima varía entre 3 y
3,5 mm.

334
de Aliseda. En Badajoz, el Silúrico fosi-
lífero se conoce al noroeste de Villar del
Rey, sur de Fuenlabrada de los Montes
y Herrera del Duque, oeste de Peloche,
Garlitos, oeste de Helechal, sur y sures-
te de Capilla, Zarza de Alange, Alange y
Villanueva del Fresno.
Las numerosas especies de graptoli-
tos contabilizadas hasta ahora, para el
conjunto de yacimientos extremeños,
pertenecen sobre todo a géneros unise-
riales como Monograptus, Stimulograp-
tus, Pristiograptus, Streptograptus, Mo-
noclimacis, Torquigraptus, Spirograptus,
Oktavites y Saetograptus; a formas bise-
riales como Parapetalolithus, Metaclima-
cograptus, Retiolites o Stomatograptus; y
a formas uniseriales ramifi cadas (Cyrto-
graptus). Además de graptolitos, muchos
horizontes de pizarras negras silúricas
contienen también fósiles de otros or-
ganismos, generalmente en forma de
restos desarticulados o transportados,
entre ellos cefalópodos nautiloideos de
concha recta con opérculo, rhyncho-
nelláceos (braquiópodos) epiplanctóni-
cos, raros bivalvos, partes de artrópodos
(mandíbulas de fi locáridos, patas o cutí-
culas de euriptéridos), conodontos (con-
servados delicadamente en las pizarras
negras) y algunos cornulítidos (conchas
tubulares de adscripción problemática).
Fósiles del Devónico
El Devónico extremeño suele ser bas-
tante fosilífero, especialmente aquellas
unidades o afl oramientos con pizarras
arcillosas, areniscas con hierro y lente-
jones calcáreos. Los braquiópodos son
los fósiles más característicos de este
periodo y los que más abundan en toda
la región, seguidos a gran distancia por
los restos de trilobites, moluscos (bival-
vos, cefalópodos, gasterópodos), corales
rugosos solitarios, tabulados, equinoder-
mos (crinoideos), tentaculitoideos, brio-
zoos, conodontos y ostrácodos. También
se conocen algunos hallazgos puntuales
de restos de peces, poríferos, estroma-
tolitos y crustáceos fi locáridos. Pero, a
excepción de los braquiópodos y de al-
gunos conodontos, ostrácodos y corales,
el resto de los grupos mencionados aún
no ha sido objeto de estudio paleontoló-
gico detallado.
Los yacimientos del Devónico In-
ferior son bastante escasos al norte de
la alineación magmática de Los Pedro-
ches, conociéndose algunas secciones
puntuales en la Sierra de San Pedro (al
suroeste del término municipal de Cá-
ceres), sur de Herrera del Duque, nor-
te del término de Talarrubias y este de
Garlitos, las tres últimas de ellas encla-
vadas en la provincia de Badajoz. Los
niveles del Lochkoviense y Praguiense
son poco fosilíferos, destacando algu-
nos braquiópodos rhynchonelláceos y
trilobites homalonótidos pero, en cam-
bio, los materiales del Emsiense reúnen
una gran diversidad de braquiópodos
(una veintena de géneros), e interesan-
tes asociaciones de conodontos, en este
caso restringidas a las intercalaciones
carbonatadas.
Los materiales del Devónico Infe-
rior resultan mucho más fosilíferos al
sur del eje de Los Pedroches, donde se
continúan con horizontes fosilíferos del
Devónico Medio, prácticamente desco-
nocidos en la Extremadura septentrional
(salvo algunos bivalvos del Givetiense
terminal). Sus principales yacimientos
paleontológicos, todos en la provincia
de Badajoz, se ubican en los alrededores
de Alange, sur y sureste de Oliva de Mé-
rida, Campillo de Llerena, suroeste de
Asociación de braquió-
podos estrofoménidos
(Boucotstrophia) y
espiriféridos (Brachys-
pirifer, a la derecha) en
areniscas del Devóni-
co Inferior de Alange
(Badajoz). La valva de
mayor tamaño mide 35
mm de anchura.
Apousiella, un braquió-
podo espiriférido abun-
dante en las areniscas
del Devónico Superior
de Herrera del Duque
(Badajoz). El ejemplar
más grande mide 25 mm
de anchura.
Pizarras del Devónico
Superior de Herrera
del Duque (Badajoz),
con concentración de
valvas de braquiópodos
espiriféridos (Cyrtospi-
rifer) y rinconélidos. El
ejemplar superior mide
24 mm de anchura.
Asociación del braquió-
podo estrofoménido
Douvillina (izquier-
da, longitud: 20 mm)
con un pigidio de un
trilobites asteropigínido.
Areniscas del Devónico
Superior de Herrera del
Duque (Badajoz).

335
Higuera de la Serena y norte de Peraleda
de Zaucejo. En ellos destacan asociacio-
nes del Lochkoviense, Praguiense, Em-
siense Superior y Givetiense, dominadas
en el primer caso por trilobites, y en el
resto por braquiópodos (de una decena
de géneros) y algunos corales. Otra zona
apenas investigada para las raras asocia-
ciones del Devónico Inferior y Medio es
la prolongación en Extremadura del área
portuguesa de Portalegre, donde prolife-
ran estos fósiles, que apenas han sido
entrevistas al oeste de La Codosera y al
norte de Villar del Rey (Badajoz).
Las sucesiones devónicas más fosi-
líferas de Extremadura resultan ser, no
obstante, las unidades detríticas del De-
vónico Superior, que afl oran en la Sierra
de San Pedro (sur de Aliseda y suroeste
del término municipal de Cáceres), la to-
talidad del sinclinal de Herrera del Du-
que, sureste de Capilla y de Cabeza del
Buey, y sureste de Garlitos (Badajoz). La
abundancia y diversidad de las especies
de braquiópodos en estas localidades,
ha permitido diferenciar cuatro biozo-
nas sucesivas dentro del Frasniense, en
las que predominan formas de espiriféri-
dos como Apousiella, Cyrtospirifer y Ri-
gauxia; estrofoménidos como Douvillina
y Pradochonetes; rinconélidos (Cypho-
terorhynchus, Paropamisorhynchus, Por-
thmorhynchus) y prodúctidos (Producte-
lla), muy frecuentemente concentrados
en horizontes lumaquélicos dentro de
las areniscas, o bien cubriendo extensas
superfi cies en las pizarras.
Los fósiles del Frasniense terminal y
del Fameniense están representados en
contadas localidades, entre ellas las ubi-
cadas en un sector de la Sierra de San
Pedro, al suroeste del término municipal
de Cáceres, sureste de Cabeza del Buey

336
(Badajoz) y región de Alange (Badajoz).
En sus litologías areniscosas dominan los
braquiópodos (doce géneros en la Sierra
de San Pedro), pero cuando existen li-
tofacies distintas (pizarras con nódulos),
suelen abundar los moluscos (bivalvos
como Buchiola; varios géneros de ce-
falópodos ammonoideos), ostrácodos y
homocténidos.
Fósiles del Carbonífero
El Carbonífero marino de la parte
centroibérica extremeña se encuen-
tra restringido al sinclinal de la ciudad
de Cáceres y al sur y sureste de Aliseda
(Sierra de San Pedro, Cáceres). Los fósi-
les más abundantes (la mayoría del Tour-
naisiense Superior, algunos del Viseense
Medio-Superior), son los braquiópodos
(dieciséis formas distintas de estrofomé-
nidos, espiriféridos, conétidos, prodúc-
tidos y otros), trilobites (ocho proétidos
diferentes), corales rugosos solitarios (al
menos siete formas distintas), briozoos,
bivalvos, cefalópodos (nautiloideos y
ammonoideos), ostrácodos, conodontos,
foraminíferos, crinoideos, dientes de pe-
ces y algas calcáreas. Casi todos ellos,
excepto los trilobites, permanecen aún
sin describir.
El retazo mejor estudiado del Carbo-
nífero marino extremeño es, sin duda, la
cuenca de Los Santos de Maimona, ubi-
cada en la Zona de Ossa Morena y que
se extiende a lo largo de 11 km entre
esa localidad, Fuente del Maestre y Feria
(Badajoz). Allí se conserva una sucesión
del Viseense Superior, discordante sobre
un basamento formado por rocas cám-
bricas que comprende ocho unidades
litoestratigráfi cas. Estas representan des-
de medios litorales hasta ambientes de
aguas profundas en un espacio bastante
reducido. Los fósiles son especialmente
frecuentes en las rocas carbonatadas, de
las que tomaremos dos ejemplos, de-
nominados informalmente “calizas de
Siphonodendron” y “calizas masivas”,
con los que ilustrar las condiciones ex-
cepcionales de preservación y su eleva-
do interés paleontológico.
La “caliza de Siphonodendron” está
constituida por margas y calizas con
abundantes colonias de corales rugosos
del género Siphonodendron, a lo que
alude su nombre. La unidad, de 8 a 35
m de espesor total, representa una lla-
nura arrecifal con varias fases de colo-
nización y desarrollo, en la cual el mar
Schizophoria sp.
Molde interno de la
valva ventral de
un braquiópodo
estrofoménido,
conservado en
materiales
vulcanoclásticos del
Carbonífero Inferior
de Aliseda (Cáceres).
45 mm de anchura.

337
dos y las algas proporcionaban un sus-
trato rígido al que se fi jaban, y sobre el
que se desarrollaron en condiciones óp-
timas, los Siphonodendron, un género
de corales coloniales ramosos, especial-
mente frecuente en el período Carboní-
fero. Otras colonias ramosas de briozo-
os, corales tabulados y algas rodofi tas
solenoporáceas también crecían junto
a los Siphonodendron pero en número
bastante más reducido. Las ramas corali-
nas también proporcionaron nichos am-
bientales locales favorables para el de-
sarrollo de esponjas, corales solitarios,
gasterópodos, trilobites, ostrácodos y un
sinfín de organismos que, tanto sésiles
como vágiles, vivían al amparo de los
corales coloniales.
Los corales del género Siphonoden-
dron crecían hacia arriba con vigor
Cummingella arbizui
(Hahn, Hahn y Rábano).
Trilobites proétido del
Carbonífero Inferior
de Aliseda (Cáceres).
Longitud, 25 mm.
abierto se situaría hacia el Sur y el conti-
nente al Norte (en términos geográfi cos
actuales). En la bioconstrucción de estos
niveles participan corales ramosos, gran-
des braquiópodos gigantoprodúctidos,
algas, tabulados y briozoos. También
son frecuentes los fósiles de corales soli-
tarios, braquiópodos diversos, moluscos
(especialmente gasterópodos), ostráco-
dos y foraminíferos.
Los afl oramientos de la “caliza de Si-
phonodendron” se extienden desde unos
cerros cercanos a Los Santos de Maimo-
na hasta las proximidades de Feria, a lo
largo de las laderas de Sierra Cabrera y
el cerro Armeña, en el término munici-
pal de Fuente del Maestre. La excelente
preservación y la posición horizontal de
la unidad en algunos lugares próximos a
Sierra Cabrera permite observar, en deta-
lle, un ecosistema arrecifal de hace 330
millones de años.
La llanura arrecifal que representa
esta unidad carbonatada se desarrolló
sobre materiales terrígenos cuando la
profundidad del mar era pequeña y los
aportes detríticos fueran lo bastante es-
casos como para no impedir el desarro-
llo de los organismos constructores de
arrecifes. La secuencia de la biocons-
trucción fue, en esencia, la siguiente:
a) en un primer momento, el sustrato
se vio colonizado por crinoideos, cora-
les solitarios, pequeños braquiópodos y
briozoos, cuyos caparazones calcáreos
se acumulaban sobre el fondo al morir;
b) dichos restos permitieron la fi jación
de larvas de otros organismos de ma-
yor tamaño, especialmente algas rojas
y gigantoprodúctidos; estos grandes
braquiópodos (5-15 cm de diámetro) se
situaban sobre las superfi cies tapizadas
por las conchas; c) los gigantoprodúcti-

338
pero, como la profundidad era escasa
(normalmente entre 0,5 y 1 m), llega-
ba un momento en que las colonias de
corales alcanzaban el nivel del agua
en bajamar. Entonces el crecimiento
vertical era sustituido por un desarro-
llo preferente en la horizontal, con la
consiguiente extensión lateral de la
llanura arrecifal. Por ello, en la caliza
de Siphonodendron las colonias de co-
rales alcanzan muy raramente medio
metro de altura, aún cuando pueden
llegar a ocupar gran extensión lateral.
Cuando todos los espacios estaban cu-
biertos por las colonias de corales, el
crecimiento se detenía y las colonias
morían. Sin embargo, esto no siempre
llegaba a suceder, debido a frecuentes
movimientos sísmicos que tendían a
producir un hundimiento o subsidencia
de la plataforma en la que se encon-
traban los corales. Cuando se registraba
uno de esos pequeños hundimientos, se
renovaba el crecimiento de los corales.
Si la subsidencia se producía cuando la
llanura arrecifal estaba totalmente col-
matada y los corales habían muerto ya,
podía repetirse de nuevo todo el pro-
ceso.
Con cierta frecuencia, durante cual-
quiera de las fases anteriormente des-
critas, tormentas de excepcional vio-
lencia azotaban la llanura arrecifal. Si
estas tormentas se producían antes de
que los corales estuviesen fi rmemente
fi jados al sustrato proporcionado por
los gigantoprodúctidos, todas las con-
chas de braquiópodos y las colonias de
corales eran removilizadas, quedando
fragmentadas o volcadas. El resultado
es que en esta llanura arrecifal pode-
mos encontrar niveles en los que los
organismos bioconstructores (corales
y gigantoprodúctidos), se conservan
completos y en posición de vida, y ca-
pas en las que sólo se puede observar
una masa de restos fragmentados y re-
sedimentados.
El segundo ejemplo seleccionado de
la cuenca carbonífera es la “caliza ma-
siva”, que alcanza un máximo de 40 m
de espesor y está formada por calizas
masivas con intercalaciones de calizas
tableadas y margas. Vista al microsco-
pio, la caliza se compone sobre todo de
caparazones de organismos unicelulares
denominados foraminíferos que, en al-
gunos niveles, llegan a constituir más del
30% de la roca. Además se encuentran
corales coloniales masivos del género Li-
thostrotion, corales ramosos de los géne-
ros Siphonodendron y Diphyphyllum, y
otros grupos fósiles como algas, briozoos
o microbraquiópodos (adultos de 3 ó 4
mm). Los corales coloniales se conser-
van muchas veces en la posición origi-
nal en la que vivían y son los típicos de
aguas someras y agitadas que, normal-
mente, aparecen a techo de los estratos
compuestos por foraminíferos. Las capas
de caliza masiva se formaban por la acu-
mulación constante de los caparazones
de estos pequeños organismos. Cuando
cesaba el aporte de sus restos, las aguas
en constante movimiento provocaban
Colonia de
Siphonodendron de
gran tamaño, cuyo
crecimiento en abanico
quedó limitado por la
escasa profundidad
del agua, para pasar
a desarrollarse
lateralmente.
Finalmente, al
producirse una
subsidencia pudo
renovar el crecimiento
hacia arriba.

339
la cementación del sustrato calcáreo,
creando la base idónea para la fi jación
de las colonias de corales. En este caso,
éstas no estaban sustentadas por las con-
chas de gigantoprodúctidos, como ocu-
rría en la caliza de Siphonodendron, sino
por un suelo endurecido y por ello aquí
pueden desarrollarse colonias de cora-
les masivos. Como la limitación de la
profundidad era menor, aquí también se
desarrollan colonias de corales ramosos
que alcanzan hasta 2 metros de altura y
más de 3 metros de diámetro, observa-
bles en las localidades de la Peraleda y
el cerro Armeña.
Otras capas de la cuenca de Los
Santos de Maimona contienen también
fósiles de interés, muchas de ellas son
especialmente ricas en microfósiles. Sin
embargo, los dos ejemplos presentados
ilustran acerca de su importancia geoló-
gica y paleontológica general, que atrae
cada año las visitas de numerosos geólo-
gos españoles y extranjeros.
Por último, las series del Carbonífero
continental (o de infl uencia continental),
se reconocen por contener vegetales fó-
siles, como helechos o licofi tas arbores-
centes, los cuales se acumulan muchas
veces como restos previamente “fl ota-
dos” en la cuenca de sedimentación. En-
tre otros afl oramientos mencionaremos
los de Hinojosa, Maguilla, La Lapa, Ca-
sas de Reina y Berlanga (Badajoz), que
contienen formas clásicas como Lepido-
dendron, Archaeocalamites, Asterocala-
mites, Stigmaria, Knorria, Archaeopteris,
Anisopteris, etc., todas ellas del Carboní-
fero Inferior de Ossa Morena. Los afl ora-
mientos del Carbonífero Superior empla-
zado en la Zona Centroibérica brindan
muchos menos restos paleobotánicos,
como una fl ora con Alethopteris del
Westfaliense, localizada en la Sierra de
San Pedro (Cáceres), y diversos palino-
morfos del Estefaniense, encontrados al
sur de la Sierra del Saltillo (Cáceres).
Fósiles post-paleozoicos
Hasta la fecha no se conocen en Ex-
tremadura depósitos fosilíferos del Meso-
zoico (Era Secundaria) o del Paleógeno,
cuyas posibles cuencas continentales
fueron erosionadas completamente y de
las que no queda vestigio alguno.
Los hallazgos paleontológicos en
las cuencas del Neógeno se limitan a
los restos de un rinoceronte del Mioce-
no Medio, Hispanotherium matritensis,
aparecidos cerca de Plasencia (Cáceres).
Los niveles lacustres de la Tierra de Ba-
rros libraron algunos restos micropaleon-
tológicos (escamas de peces, oogonios
de caráceas), y escasos gasterópodos del
Mioceno Superior, que por el momento
permanecen sin estudiar.
Los materiales del Pleistoceno Infe-
rior y Medio han brindado restos poco
importantes de grandes mamíferos en
contados yacimientos por toda la región,
la mayor parte de ellos ubicados en con-
textos arqueológicos tales como cuevas
o terrazas fl uviales.
Superfi cie erosionada
en la “caliza de
Siphonodendron” del
Carbonífero de Los
Santos de Maimona
(Badajoz). Se observan
dos ciclos de
crecimiento de
corales ramosos sobre
los braquiópodos
gigantoprodúctidos.
En la parte superior se
observa una capa
constituida por
colonias fragmentadas
y removilizadas por
una tormenta.