Gastrulacion y Neurulacion - Embriologia
36
Márquez-Orozco MC Biología del Desarrollo Fascículo 10 1 Biología del Desarrollo _______________________________________________________________________________________________ __________________________________________ Gastrulación Neurulación ___________________________________________________________________ __________________________________________ Dra. María Cristina Márquez Orozco Profesor Titular de Carrera Departamento de Embriología Facultad de Medicina UNAM Fascículo 10 _______________________________________________________
-
Upload
omar-gutierrez -
Category
Documents
-
view
1.773 -
download
11
Transcript of Gastrulacion y Neurulacion - Embriologia

Márquez-Orozco MC
Biología del Desarrollo Fascículo 10
1
Biología del Desarrollo _______________________________________________________________________________________________
__________________________________________
Gastrulación
Neurulación
___________________________________________________________________
__________________________________________
Dra. María Cristina Márquez Orozco
Profesor Titular de Carrera
Departamento de Embriología
Facultad de Medicina
UNAM
Fascículo 10 _______________________________________________________

Márquez-Orozco MC
Biología del Desarrollo Fascículo 10
2
RESPETE EL DERECHO DE AUTOR
APOYE LA CREATIVIDAD Y
LA PRODUCCIÓN DE MATERIAL DIDÁCTICO
Este fascículo o cualquiera de sus partes no deberá reproducirse, ni
archivarse en sistemas recuperables, ni transmitirse en ninguna
forma, ni por ningún medio electrónico, de fotocopiado, grabado o
cualquier otro sin permiso escrito del autor.
BIOLOGÍA DEL DESARROLLO
Tercera edición 2005
4ª reimpresión
Derechos reservados
Edición: Amalia Márquez Orozco
Edición literaria: María Refugio Puente Anguiano
Obra general:ISBN968-7785-00-4
GASTRULACIÓN
NEURULACIÓN:ISBN968-7785-10-1
Impreso en: Gounod 129
México, 06250 D.F.
IMPRESO EN MÉXICO

Márquez-Orozco MC
Biología del Desarrollo Fascículo 10
3
Gastrulación Neurulación
Una de las consecuencias inmediatas de la segmentación es el inicio de la gastrulación que es la formación de las tres hojas blastodérmicas: el ectodermo, el mesodermo y el endodermo.
De nuevo estás ante un proceso biológico que tiene una enorme repercusión clínica, como es el poder identificar los tejidos con un origen común. Este conocimiento ha permitido, en desarrollo de diferentes técnicas, para resolver problemas de salud, por ejemplo por ejemplo, el transplantar células de la médula suprarrenal, derivadas de las crestas neurales, al cuerpo estriado, uno de los núcleos basales del cerebro, que se altera en las personas que padecen; el mal de Parkinson, enfermedad que invalida a los pacientes, debido a que no pueden controlar el temblor de sus músculos voluntarios de sus músculos voluntarios, causado temblor por falta de neurotransmisores, que en este caso son proporcionados por las células de las crestas neurales, que derivan del neuroectodermo nervioso central, del que forma parte el cuerpo estriado.
También esta información ha servido para explicar el origen de algunos síndromes, como la displasia ectodérmica anhidrótica , en la que los genes alterados afectan al ectodermo y sus
derivados: el pelo, los dientes y las glándulas sudoríparas, entre otros.
Te recomiendo que continúes utilizando un marcador para señalar los datos importantes, y con tu material de moldeable, trata de crear modelos tridimensionales. Colorea los esquemas asignando un color a los tejidos del mismo origen y elabora tus propios resúmenes. Es frecuente que el verde se emplee para representar el ectodermo superficial, el azul para el neuroectodermo del sistema nervioso central, el morado para de neuroectodermo de las crestas neurales, el rosa o rojo para el mesodermo y el amarillo para el endodermo. Con esta clave será más fácil que identifique el origen de los tejidos y sus derivados.
Espero que en el primer examen parcial haya obtenido una buena calificación, si no fue así, trata de analizar las causas por las que su rendimiento fue menor al esperado. Procura corregirlas, ya que tú eres el único responsable de tu destino y lo más importante es que te prepares bien para ser un excelente profesional. Sigue adelante con la seguridad de que tu esfuerzo se verá coronado por el éxito, ¡Razona la información! ¡Evita memorizar los datos sin entenderlos! ¡Busca ayuda si la necesitas! ¡Confía en ti mismo Y CONVÉNCETE DE QUE TU
MENTE ES MARAVILLOSA! ¡YO PUEDO! ¡ÉXITO!
Movimientos morfogenéticos
La gastrulación* se inicia cuando las células del macizo celular interno han alcanzado un grado de diferenciación suficiente, para formar las áreas morfogenéticas u órganoformadoras, de las que derivan las hojas blastodérmicas.
*(de gaster, estómago).
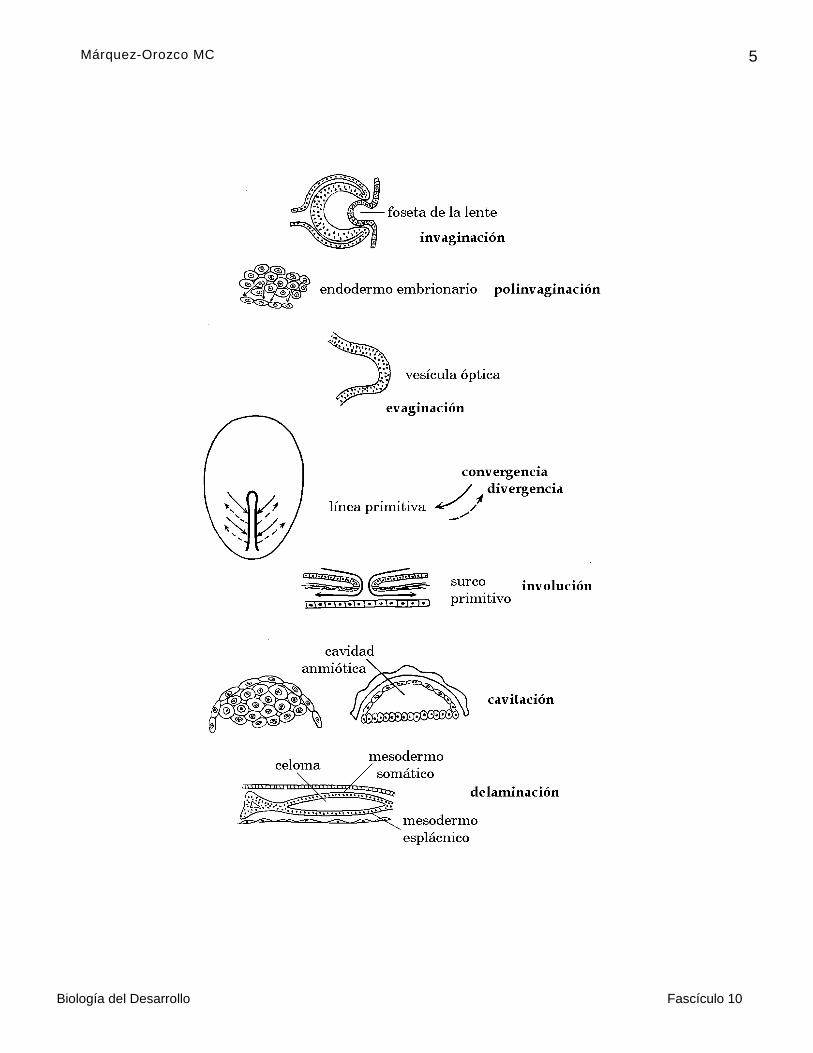
Durante la gastrulación, que empieza en el séptimo día (6.5 días) posterior a la fecundación, las células migran, y lo hacen mediante los llamados movimientos morfogenéticos, que son típicos de grupos específicos de células, en los que funcionan los mismos genes. Los principales movimientos morfogenéticos (figura 1) son:
Invaginación. Hundimiento de un grupo de células que penetran en otro tejido o en la luz de una estructura, para formar un esbozo hueco o uno sólido. Por este se proceso se forma el surco primitivo del embrión , la copa óptica, la foseta de la lente o cristalino, los
folículos pilosos que penetran en la dermis y el surco neural.
Polinvaginación. Es el mismo tipo de movimiento de invaginación, sólo que cada célula se hunde de manera independiente . Lo presentan tanto las células del endodermo, embrionario como las del extraembrionario.
Evaginación. Consiste en la formación de una saliente a partir de un grupo de células, que se proyectan hacia el exterior de una estructura u órgano. Las células evaginadas pueden formar esbozos huecos o sólidos. Por este movimiento se forman la alantoides, el hígado, el páncreas y las vesículas ópticas, éstas le dan al cerebro del embrión aspecto de pez martillo.
Convergencia. Migración de las células para reunirse en un punto determinado , por ejemplo, la línea primitiva de la que se forman el mesodermo y el endodermo embrionarios. Los gonocitos migran desde la pared del saco

Márquez-Orozco MC
Biología del Desarrollo Fascículo 10
4
vitelino hasta las crestas urogenitales, en donde convergen.
Divergencia. Las células se alejan de un punto determinado; y se dispersan o se reúnen en otro; por ejemplo, las del mesodermo, que después de converger en la línea primitiva, penetran por el surco primitivo y se dirigen hacia los bordes del embrión , o las células de las crestas neurales que migran a diferentes partes del cuerpo del embrión.*
Involución. Es equivalente a que las células den vuelta en U para colocarse en un plano distinto al original; se produce por movimientos del mesodermo de la línea primitiva. En un principio, las células convergen en un montículo longitudinal, que es la línea primitiva, después se invaginan para formar en el surco primitivo, y divergen, al penetrar entre el ectodermo y el endodermo embrionarios y sus células diferenciadas, forman el mesodermo que avanzan hasta los bordes del embrión trilaminar.
Cavitación. Consiste en la formación de un espacio entre las células, que en un principio, constituían estructuras sólidas . En algunos casos, además del reacomodo de las células en una pared, como sucede en el desarrollo del blastocele, de la cavidad amniótica o del antro folicular, algunas células degeneran.
Delaminación. Consiste en que un grupo sólido de células se reacomoden para forman hojas o láminas, separadas por un espacio o cavidad. Este movimiento es característico del mesodermo lateral del embrión, que por estas razones también se llama laminar que se divide en una hoja somática que, junto con el ectodermo, constituye la somatopleura, y en una hoja esplácnica que, con el ectodermo, origina la esplacnopleura; entre las dos aparece la cavidad celómica o cavidad general del cuerpo. El mesodermo somático extraembrionario se delamina en una hoja somática y una esplácnica, separadas por el celoma extraembrionario.
Elongación. Aumento de la longitud del eje céfalocaudal de una estructura. Por ejemplo, el alargamiento del embrión, el del sistema nervioso central o el del esófago.
Plegamiento. Consiste en que una o varias hojas de tejido realicen un movimiento envolvente o se flexionen. Entre los tejidos que se pliegan, están los bordes del surco neural durante la formación del tubo neural, y el movimiento que efectúa la pared del cuerpo
y la esplacnopleura para originar un embrión cilíndrico y el tubo digestivo, respectivamente.
Epibolia. Movimiento que consiste en que las células mientras se multiplican, van cubriendo la superficie de una estructura u órgano. Es característico del ectodermo superficial.
Endobolia. Revestimiento de una cavidad . Se debe a que las células, al irse dividiendo, se desplazan en la superficie interna de una estructura hueca, como lo hace el endodermo del saco vitelino primitivo, al tapizar el trofoblasto.
Movimiento amiboide o ameboide. Desplazamiento individual de las células por medio de pseudópodos, como los de las amibas. Se puede observar en las células sexuales primordiales o gonocitos, precursores de las espermatogonias o las ovogonias, que tienen que moverse desde la pared del saco vitelino cercana a la alantoides, al intestino primitivo, y a la raíz del mesenterio dorsal hasta llegar a las crestas urogenitales. Para llevar a cabo este movimiento
Márquez-Orozco MC
Biología del Desarrollo Fascículo 10
5

Márquez-Orozco MC
Biología del Desarrollo Fascículo 10
6
FIGURA 1. MOVIMIENTOS MORFOGENÉTICOS.
Te sugiero que con ayuda de la figura 1 y tu material moldeable las reproduzcas
tridimensionalmente y, además, las colorees para retenerlas con más facilidad.

Márquez-Orozco MC
Biología del Desarrollo Fascículo 10
7
Matriz extracelular
Para que los movimientos morfogenéticos se lleven a cabo, las células del embrión sintetizan una matriz extracelular que las guía hacia su destino final.
La matriz extracelular se compone de sustancias sintetizadas por las células, que se depositan por fuera de sus membranas celulares en forma de láminas basales, como las de los epitelios, o entre células, como las mesodérmicas, que son estrelladas.
La matriz extracelular es una especie de empaque que puede servir como medio de transmisión mecánica, por ejemplo, en los tendones que soportan fuerzas poderosas; en otros casos, promueven la fijación de sales de calcio, como sucede en el hueso o el depósito de ácido condroitinsulfúrico en el cartílago, que ayudan a que los tejidos soporten grandes pesos y fuerzas.
Entre los componentes de la matriz extracelular que son importantes en la migración de las células están:
El ácido hialurónico, un proteoglicano que forma parte de las láminas basales de las células epiteliales, ávido por el agua. Está compuesto por ácido D-glucurónico y N-acetil-glucosamina que se interpone entre las dos capas de un embrión bilaminar o epiblasto y el hipoblasto, que veremos un poco más adelante y que es fundamental para orientar a las células mesodérmicas derivadas de la línea primitiva.
La fibronectina, una variedad de colágena, es muy importante para dirigir a las células que migran, no sólo orientadas por las láminas basales, sino también por otras células, como lo hacen las del sistema nervioso o los gonocitos. En este caso, tienen sitios de reconocimiento que favorecen la migración celular.
Son muy numerosos los componentes de la matriz extracelular, y su importancia funcional es enorme, tanto para el desarrollo normal del embrión, como para el funcionamiento de los tejidos adultos. Se clasifican en dos grandes grupos, los de tipo colágena y los glucosaminoglicanos (mucopolisacáridos).
Entre los diversos tipos de colágena, que se identifica con números romanos, están:
El tipo I o fibronectina de la piel, del hueso, de los tendones y del tejido conectivo intersticial, que forma el 90% de la colágena del cuerpo.
El tipo II o condronectina, característica de la notocorda, del cartílago, del cuerpo vítreo y de la córnea.
El tipo III, que es otra variante de fibronectina, presente en la piel, en el músculo esquelético y en los vasos sanguíneos.
El tipo IV o laminina, característica de las láminas basales, importante en el crecimiento de las fibras nerviosas y en la cicatrización.
El tipo V o fibronectina, característica de la placenta, del músculo liso y de los vasos sanguíneos.
El tipo X o condronectina, presente en el cartílago hipertrofiado.
Los glucosaminoglucanos en general están mezclados con la colágena, y se distribuyen de manera similar. Los principales son: el ácido hialurónico, el dermatán sulfato o condroitín sulfato B, el condroitín sulfato A y C, el queratán sulfato, el heparán sulfato y el heparín sulfato.
Embrión bilaminar o disco embrionario (segunda semana del desarrollo)
Ahora que sabes qué sustancias guían a las células que deben migrar, te explicaré cómo se realiza la gastrulación.
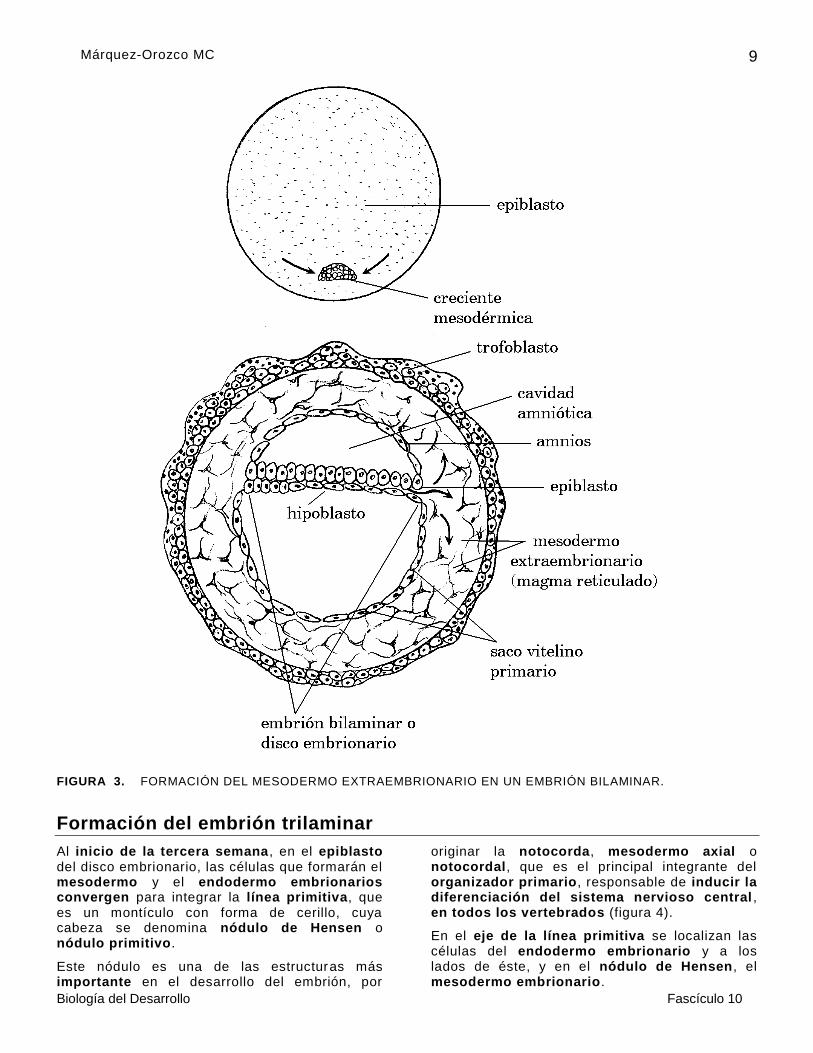
En el séptimo día del desarrollo de un embrión, las células del macizo celular interno , que morfológicamente son iguales, pero químicamente distintas, empiezan a formar una población de células que por polinvaginación se colocan en la parte más profunda del macizo celular interno, y constituyen el hipoblasto o capa interna del embrión, que más adelante se diferenciará en el endodermo extraembrionario . Poco a poco, sus células revisten por endobolia al trofoblasto y forman el saco vitelino primario, que se cierra en la parte ventral.
Algunas células del hipoblasto son un poco más altas y constituyen la placa precordal, que es el primer indicio de la aparición de los ejes de simetría, ya que señalan la posición de la región cefálica. El extremo contrario es la región caudal. En donde se localiza el hipoblasto, es la región ventral, y la dorsal corresponde a las células que están sobre el hipoblasto. Desde luego, también se pueden identificar el lado derecho y el izquierdo, que estarán hacia atrás de la hoja, el derecho, y hacia adelante, el izquierdo (figura 2).
Las demás células del macizo celular interno también se reorganizan y, por cavitación, forman

Márquez-Orozco MC
Biología del Desarrollo Fascículo 10
8
la cavidad amniótica, el amnios y el epiblasto, que es la capa superficial, que origina:
El epitelio amniótico, que es ectodermo extraembrionario.
Las células del ectodermo embrionario.
La línea primitiva, de la que derivan el mesodermo embrionario, el extraembrionario y el endodermo embrionario.
De esta manera, el embrión es un disco embrionario o un embrión bilaminar, que está bien constituido hacia el día nueve ó 10 del desarrollo, es decir, en la segunda semana, aunque en la actualidad le llaman pre-embrión (figura 2 ).
Antes de que concluya la segunda semana del desarrollo, en la región caudal del embrión aparece una estructura en forma de media luna, que recibe el nombre de “creciente mesodérmica”, y corresponde a la parte caudal de la línea primitiva. De esta estructura migran células que se colocan entre el trofoblasto y el amnios y entre el trofoblasto y el saco vitelino , que son las del mesodermo extraembrionario o magma reticulado, constituido por células estrelladas, que son una especie de empaque. Algunas células de este tejido probablemente derivan de las del saco vitelino primario (figura 3).
FIGURA 2. DESARROLLO DE UN EMBRIÓN BILAMINAR O DISCO EMBRIONARIO.
Márquez-Orozco MC
Biología del Desarrollo Fascículo 10
9
FIGURA 3. FORMACIÓN DEL MESODERMO EXTRAEMBRIONARIO EN UN EMBRIÓN BILAMINAR.
Formación del embrión trilaminar
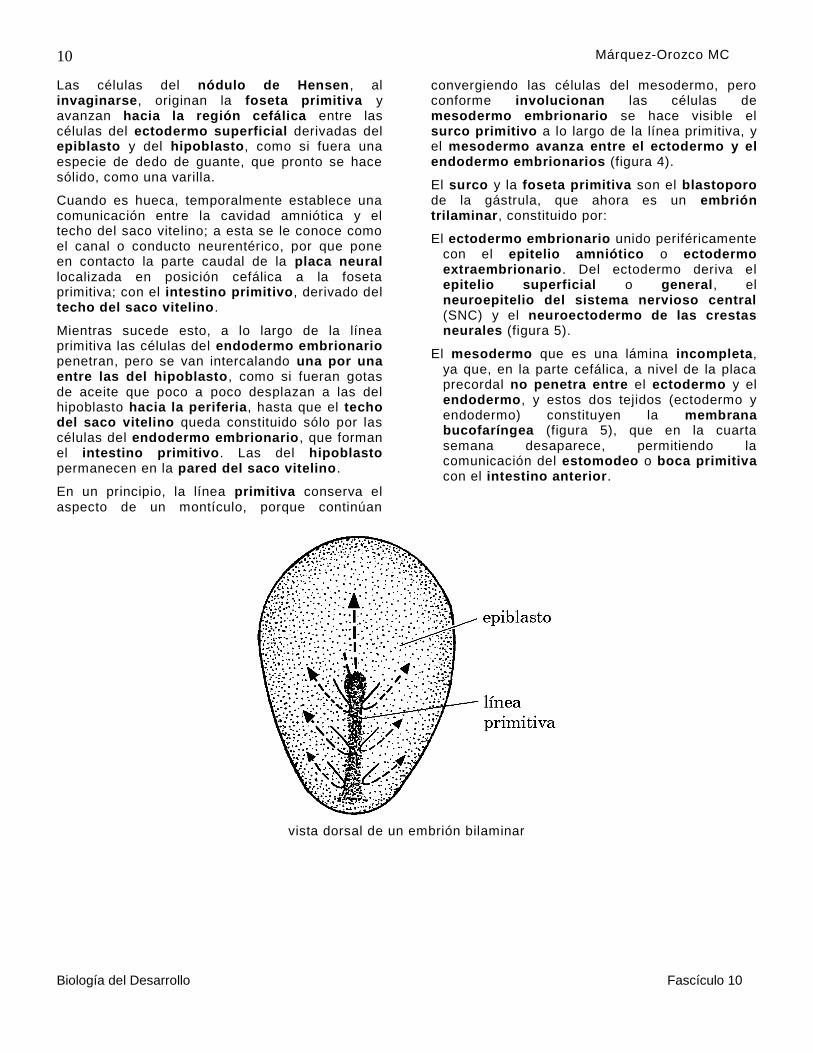
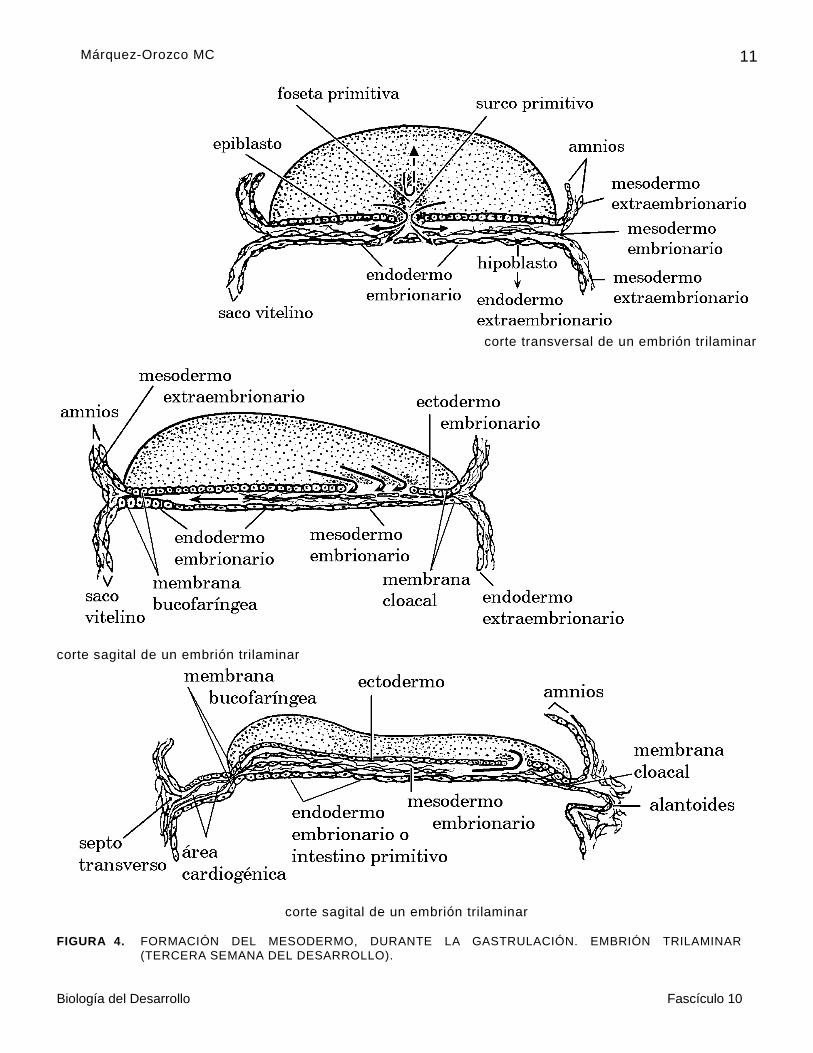
Al inicio de la tercera semana , en el epiblasto del disco embrionario, las células que formarán el mesodermo y el endodermo embrionarios convergen para integrar la línea primitiva, que es un montículo con forma de cerillo, cuya cabeza se denomina nódulo de Hensen o nódulo primitivo.
Este nódulo es una de las estructuras más importante en el desarrollo del embrión, por
originar la notocorda, mesodermo axial o notocordal, que es el principal integrante del organizador primario, responsable de inducir la diferenciación del sistema nervioso central , en todos los vertebrados (figura 4).
En el eje de la línea primitiva se localizan las células del endodermo embrionario y a los lados de éste, y en el nódulo de Hensen, el mesodermo embrionario.
Márquez-Orozco MC
Biología del Desarrollo Fascículo 10
10
Las células del nódulo de Hensen, al invaginarse, originan la foseta primitiva y avanzan hacia la región cefálica entre las células del ectodermo superficial derivadas del epiblasto y del hipoblasto, como si fuera una especie de dedo de guante, que pronto se hace sólido, como una varilla.
Cuando es hueca, temporalmente establece una comunicación entre la cavidad amniótica y el techo del saco vitelino; a esta se le conoce como el canal o conducto neurentérico, por que pone en contacto la parte caudal de la placa neural localizada en posición cefálica a la foseta primitiva; con el intestino primitivo, derivado del techo del saco vitelino.
Mientras sucede esto, a lo largo de la línea primitiva las células del endodermo embrionario penetran, pero se van intercalando una por una entre las del hipoblasto, como si fueran gotas de aceite que poco a poco desplazan a las del hipoblasto hacia la periferia, hasta que el techo del saco vitelino queda constituido sólo por las células del endodermo embrionario , que forman el intestino primitivo. Las del hipoblasto permanecen en la pared del saco vitelino.
En un principio, la línea primitiva conserva el aspecto de un montículo, porque continúan
convergiendo las células del mesodermo, pero conforme involucionan las células de mesodermo embrionario se hace visible el surco primitivo a lo largo de la línea primitiva, y el mesodermo avanza entre el ectodermo y el endodermo embrionarios (figura 4).
El surco y la foseta primitiva son el blastoporo de la gástrula, que ahora es un embrión trilaminar, constituido por:
El ectodermo embrionario unido periféricamente con el epitelio amniótico o ectodermo extraembrionario. Del ectodermo deriva el epitelio superficial o general, el neuroepitelio del sistema nervioso central (SNC) y el neuroectodermo de las crestas neurales (figura 5).
El mesodermo que es una lámina incompleta, ya que, en la parte cefálica, a nivel de la placa precordal no penetra entre el ectodermo y el endodermo, y estos dos tejidos (ectodermo y endodermo) constituyen la membrana bucofaríngea (figura 5), que en la cuarta semana desaparece, permitiendo la comunicación del estomodeo o boca primitiva con el intestino anterior.
vista dorsal de un embrión bilaminar
Márquez-Orozco MC
Biología del Desarrollo Fascículo 10
11
corte transversal de un embrión trilaminar
corte sagital de un embrión trilaminar
corte sagital de un embrión trilaminar FIGURA 4. FORMACIÓN DEL MESODERMO, DURANTE LA GASTRULACIÓN. EMBRIÓN TRILAMINAR
(TERCERA SEMANA DEL DESARROLLO).

Márquez-Orozco MC
Biología del Desarrollo Fascículo 10
12
Tampoco en la región caudal penetra el mesodermo, y el ectodermo junto con el endodermo forman la membrana cloacal (figura 5), que en este caso también degenera y aparecen el orificio anal y el urogenital, si es un embrión masculino, o el anal, el meato urinario y el orificio vaginal, si es femenino.
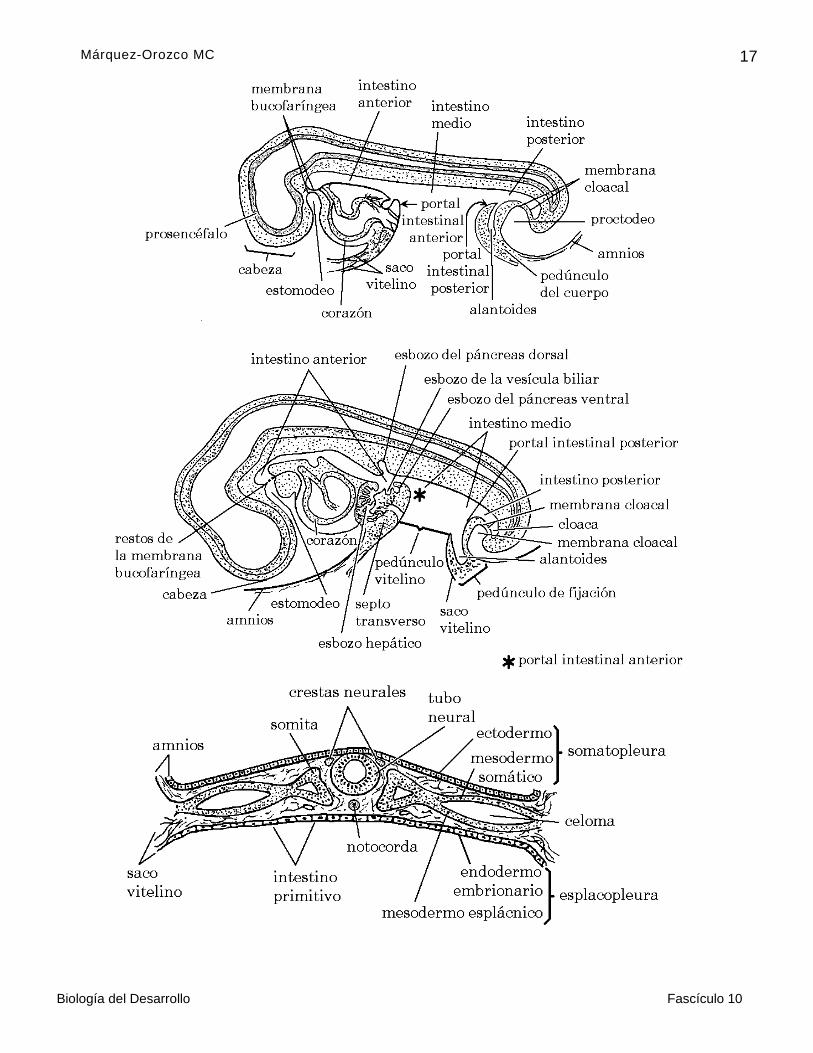
Del mesodermo derivan la notocorda, llamada también mesodermo axial o notocordal, el mesodermo paraxial o somítico, el mesodermo intermedio o nefrotomos y el mesodermo lateral o laminar (figura 5).
El endodermo embrionario, que se continúa con las paredes del saco vitelino y sólo forma endodermo embrionario (figura 5).
Si se superponen los tres discos y después se cortan en sentido transversal y longitudinal, te darás cuenta de la relación que existe entre los tejidos de las tres hojas (figuras 4 y 5), y es posible comprender el porqué pueden interactuar unos con otros.
A modo de catálogo te voy a mencionar las principales estructuras que derivan de cada tejido que forma las hojas blastodérmicas, aunque más adelante, al entrar al estudio de la organogénesis o embriología especial, veremos cómo se diferencia cada una de éstas.
FIGURA 5. TEJIDOS QUE DERIVAN DE LAS HOJAS BLASTODÉRMICAS.

Márquez-Orozco MC
Biología del Desarrollo Fascículo 10
13
Ectodermo superficial
Epidermis de la piel. Pelo, pestañas, cejas, uñas.
Olfatorias-epitelio olfatorio. Placodas Ópticas-cristalino o lente. Óticas-laberinto membranoso del oído.
Sudoríparas. Glándulas Mamarias. Sebáceas. Lagrimales.
Epitelio de la cavidad bucal hasta antes de la “V” lingual. Órganos del esmalte y esmalte. Epitelio de la parte final del conducto recto-anal. Bolsa de Rathke-adenohipófisis. Neuroectodermo del sistema nervioso central (cuadro 1).
CUADRO 1
DESARROLLO DEL SISTEMA NERVIOSO CENTRAL
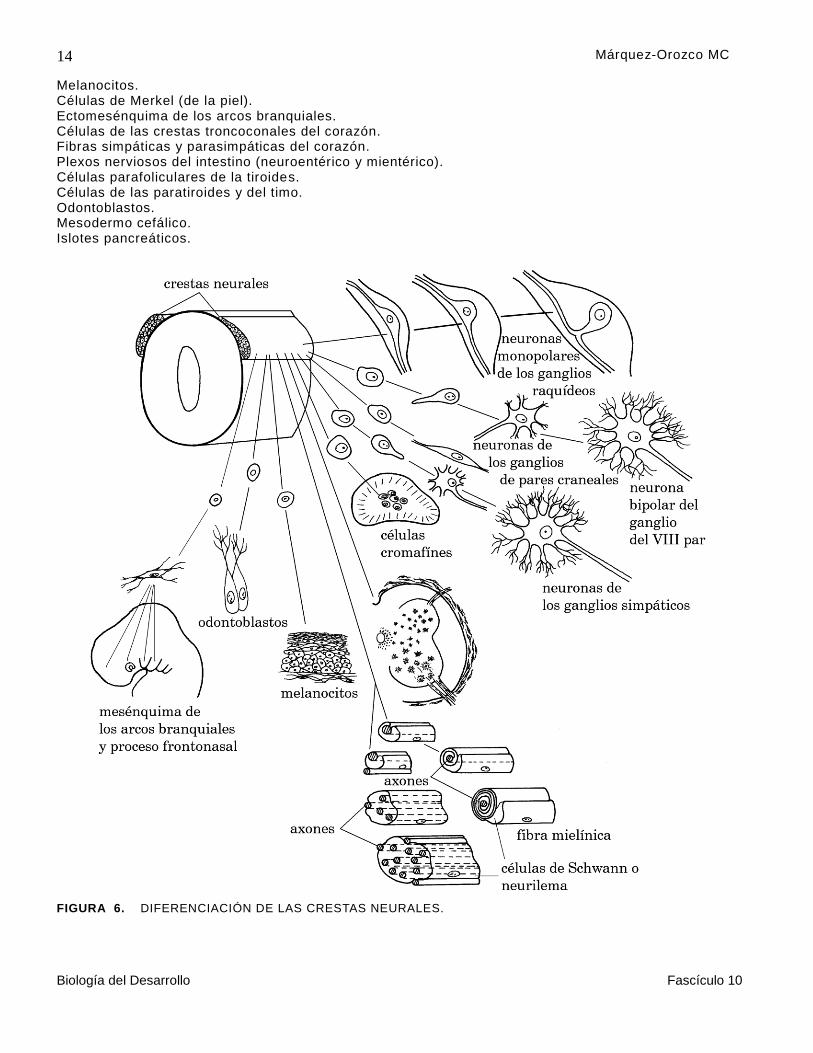
Neuroectodermo de las crestas neurales (figura 6).
Raquídeos. Ganglios Simpáticos. Parasimpáticos de algunos pares craneales.
Las arterias. Células del sistema cromafín de Los cuerpos carotídeos. La médula suprarrenal.
Células de la piamadre y de la aracnoides. Células de Schwann o neurilema que mielinizan el sistema nervioso periférico. Endotelio y estroma de la córnea. Constrictor de la pupila. Coroides. Músculo ciliar.
Márquez-Orozco MC
Biología del Desarrollo Fascículo 10
14
Melanocitos. Células de Merkel (de la piel). Ectomesénquima de los arcos branquiales. Células de las crestas troncoconales del corazón. Fibras simpáticas y parasimpáticas del corazón. Plexos nerviosos del intestino (neuroentérico y mientérico). Células parafoliculares de la tiroides. Células de las paratiroides y del timo. Odontoblastos. Mesodermo cefálico. Islotes pancreáticos.
FIGURA 6. DIFERENCIACIÓN DE LAS CRESTAS NEURALES.

Márquez-Orozco MC
Biología del Desarrollo Fascículo 10
15
FIGURA 7. DESARROLLO DEL MESODERMO PARAXIAL.

Márquez-Orozco MC
Biología del Desarrollo Fascículo 10
16
Mesodermo axial o notocordal (figuras 8 y 9).
Organizador primario induce la diferenciación del sistema nervioso central.
Sólo se conserva como algunas células de los núcleos pulposos de los discos intervertebrales.
Mesodermo paraxial o somítico (figuras 7 y 8).
En la región cefálica este mesodermo se divide en posición caudal a la placa precordal para formar siete pares de esferas sin cavidad, constituidas por células mesenquimáticas, estas estructuras son las somitómeras, de las que se derivan:
Músculos extraoculares.
Músculos masticadores.
Músculos de la expresión facial.
Músculo estilofaríngeo.
Caudal a estas somitómeras esféricas que sólo forman músculos se diferencian 42 a 44 somitómeras ovaladas que se transforman en 42 a 44 pares de somitas típicas, con células epiteliales, que en corte transversal son
triangulares, y desde la región dorsal son cuadrangulares (figuras 7 y 8). Cada somita está constituida por una parte dorsolateral o dermatomo, una ventromedial o esclerotomo y una medial o miotomo.
Dermatomo
Dermis de la piel.
Miotomo
Cuello.
Músculo esquelético de Tronco. Miembros escapulares y pélvicos.
Márquez-Orozco MC
Biología del Desarrollo Fascículo 10
17

Márquez-Orozco MC
Biología del Desarrollo Fascículo 10
18
FIGURA 8. FLEXIÓN Y PLEGAMIENTO DEL EMBRIÓN HUMANO, EN LA TERCERA, CUARTA Y QUINTA
SEMANAS.
Esclerotomo
Vértebras.
Esqueleto axial Costillas. Base del occipital.
Esqueleto de los miembros.
Mesodermo intermedio o nefrotomos (figura 8)
Pronefros riñón transitorio.
Crestas urogenitales(mesonefros, metanefros y gónadas).
Deferentes.
Del epidídimo.
Conductos Conductos Eyaculadores.
mesonéfricos Vesículas seminales.
Mesonefros o de Wolff Conductos de Gartner.
(riñón
transitorio) Conductos Utrículo prostático.
paramesonéfricos Tubas uterinas.
o de Müller. Útero.
Parte superior de la vagina.
MetanefrosRiñón definitivo.

Márquez-Orozco MC
Biología del Desarrollo Fascículo 10
19
Estroma testicular.
Testículos Células de LeydigTestosterona.
Túnica albugínea.
Estroma ovárico.
Ovarios Células esteroidogénicasTestosterona Esteroides
De la teca interna
Túnica albugínea.
Mesodermo lateral o laminar (figura 8).
Por delaminación origina dos hojas, una de mesodermo somático y otra de mesodermo esplácnico. La de mesodermo somático, junto con el ectodermo, forma la somatopleura y la
de mesodermo esplácnico, junto con el endodermo, forma la esplacnopleura; entre las dos hojas aparece el CELOMA o cavidad general del cuerpo, que en esta etapa es doble (figura 8)
Mesodermo somático (figura 8)
Pleura.
Peritoneo. Parietales.
Pericardio.
Esternón.
Parte del blastema de los miembros.
Arteriales.
Vasos sanguíneos Venosos. Linfáticos.
Mesodermo esplácnico (figura 8)
Músculo cardiaco, endocardio.
Arteriales.
Vasos sanguíneos Venosos. Linfáticos.
Pleura.
Peritoneo viscerales
Pericardio.
Mesenterios Dorsal. Ventral.
Submucosa. Tubo digestivo y del aparato respiratorio.
Músculo liso.
Epitelio celómico. Células de Sertoli. Células foliculares.
Endodermo (figura 8)
Mucosa del tubo digestivo y del árbol respiratorio.
Glándulas del tubo digestivo y salivales.

Márquez-Orozco MC
Biología del Desarrollo Fascículo 10
20
Hígado.
Páncreas.
Timo.
Tiroides.
Paratiroides.
De las amígdalas.
Epitelio De la caja timpánica. De la tuba timpánica.
Epitelio de la vejiga urinaria.
Plasma germinal (figura 8)
Ovogonias “óvulos”.
Células sexuales primordiales o gonocitos Espermatogonias.
Espermatozoides.
Moléculas de adhesión celular o CAMs
Como te habrás dado cuenta las células derivadas del cigoto, en unos cuantos días se diferencian, migran y se organizan para formar los tejidos y los órganos primitivos del embrión. Para que se lleven a cabo estos cambios es necesaria la intervención de diferentes tipos de sustancias, entre las que se encuentran las llamadas moléculas de adhesión de las células o CAMs, a las que se les ha dado mucha importancia en los últimos años.
Las propiedades de los CAMs fueron descubiertas por J. Holtfreter, desde el inicio del siglo XX, quién experimentalmente observó que, cuando colocaban en un cultivo de tejidos células de diferente origen como las del ectodermo, las del mesodermo y las del endodermo, mezcladas; se reunían de acuerdo a su origen y tenían un comportamiento similar de acuerdo a éste. Por ejemplo el ectodermo se colocaba en el exterior, el endodermo en el interior y el mesodermo entre los dos.
A pesar de haber descubierto esta propiedad de las células, no lograron identificar las moléculas que unían a las células.
Al final de los 60s y principios de los 70s S. Roth y colaboradores lograron identificar la presencia de diferentes tipos de moléculas de adhesión celular o CAMs en la pared de las células, las cuales eran responsables de ponerlas en contacto y, demostraron que son de dos tipos:
Homofílicas si ponen en contacto células del mismo tipo.
Heterofílicas si unen células de diferente tipo.
En la actualidad, se han aislado numerosos tipos de CAMs, que se han clasificado en dos grandes grupos:
Las cadherinas, que son glucoproteínas dependientes de iones de calcio .
Las inmunoglobulinas de la superfamilia de los CAMs que son semejantes a las moléculas de los anticuerpos y no son dependientes de iones calcio.
Las cadherinas son necesarias para mantener las conexiones intracelulares. Las células con pocas moléculas de cadherinas tienen menor adhesividad que las que tienen muchas de éstas moléculas en sus membranas.
En los embriones de mamíferos se han encontrado tres clases de cadherinas:
La E-cadherina o L-CAM.
La P-cadherina.
La N-cadherina.
La E-cadherina o cadherina epitelial que también se llama L-CAM o uvomorulina es homofílica, se expresa tempranamente en el embrión y después en todas las células de tipo epitelial, tanto embrionarias como adultas, como en las de ectodermo superficial y en sus

Márquez-Orozco MC
Biología del Desarrollo Fascículo 10
21
derivados; en el epitelio urogenital, en el parénquima hepático y pancreático; en el epitelio del árbol respiratorio, en el del timo, en la tiroides y en el de las paratiroides.
La P-cadherina o cadherina placentaria, es homofílica se expresa por primera vez en las células del trofoblasto que se ponen en contacto con el epitelio uterino . Probablemente facilita el contacto de estos dos tejidos. También se ha detectado en el ectodermo extraembrionario, en el endodermo, en la notocorda, en el mesodermo lateral y en la placenta. En el adulto se expresa en la epidermis y en el epitelio pigmentario de la retina.
La N-cadherina o cadherina neural, llamada también C-Cal-CAM o A-CAM, es homofílica. En un principio se expresa en las células mesodérmicas y notocordales, de la gástrula; cuando dejan de expresarse la E-cadherina. Se encuentra en las células de las crestas neurales antes de que inicien su migración , los pierden durante la migración y se vuelven a expresar cuando se agrupan para formar los ganglios espinales, simpáticos y parasimpáticos. También se encuentran en las células del sistema nervioso, en la lente, el músculo estriado, en los gonocitos o células sexuales primordiales y en el esbozo renal.
Las inmunoglobulinas de la superfamilia de CAMs son de dos clases:
La N-CAM.
La Ng-CAM.
La N-CAM o molécula de adhesión de las células neurales, es homofílica, no dependiente de iones de calcio, es una cadena simple de polipéptidos anclada a la membrana celular por una secuencia de aminoácidos transmembranales unidos a las proteínas o a los lípidos. El sitio de reconocimiento está en la cadena externa. La molécula de N-CAM es rica en ácido siálico, que es un sacárido de 10 carbonos, derivado de la unión de D-manosa y ácido pirúvico, tiene una carga altamente negativa.
La N-CAM de las células embrionarias tienen 30% de ácido siálico y la adulta 10%. La adhesión de las células es mayor cuando hay menos ácido siálico y no se pueden adherir si existen altas concentraciones. Esto podría explicar la probabilidad de que se altere el sistema nervioso, si la concentración de ácido siálico es anormal y que su desarrollo sea normal si se produce en cantidad adecuada.
A medida que avanza el desarrollo disminuye la cantidad de ácido siálico y aumenta la adhesión celular.
La N-CAM interviene en el reconocimiento de los axones con las células de los órganos blanco, en la formación de las placas neuromusculares y en la diferenciación de los nervios, llamada fasciculación que se debe a la unión de las fibras nerviosas en haces.
La N-CAM y la N-cadherina se encuentran en las mismas células del sistema nervioso pero en diferentes sitios. En el tubo neural la N-CAM se concentra en la parte externa y la N-cadherina cerca de la luz del tubo . En etapa embrionaria, la N-CAM se expresa en el epiblasto, en la placa y en el tubo neural, en las placodas y en el mesodermo y en el adulto en el sistema nervioso, el músculo esquelético, en las placas neuromusculares, en el músculo cardiaco y el liso, en la corteza suprarrenal, en los ovarios y en los testículos, en el epitelio del riñón y en el del intestino, en los mesenterios y en el bazo.
La Ng-CAM o molécula de adhesión de las células gliales, es heterofílica, y no dependiente de iones calcio . Al igual que la N-CAM tiene una cadena polipeptídica con un sitio de reconocimiento externo y está anclada a la membrana celular por aminoácidos transmembranales. Se expresa en etapas tardías en las neuronas y en las células gliales.
Tanto la N-CAM como la L-CAM o E-cadherina se expresan en el epiblasto y en el hipoblasto, dejan de expresarse durante la migración de las células del mesodermo de la línea primitiva y se re-expresan en los derivados mesodérmicos que ya conoces.
Antes de la inducción primaria se expresan en el epiblasto N-CAM y L-CAM, en cuanto inicia la diferenciación del sistema nervioso central, en la placa neural se sigue expresando N-CAM y desaparece L-CAM, en cambio en las células del ectodermo superficial se sigue expresando L-CAM y deja de hacerlo N-CAM.
Además de las moléculas de adhesión celular descritas se han descubierto otras que intervienen en el reconocimiento, migración e integración de los tejidos de un organismo de manera armónica, por lo tanto, son muy importantes en el desarrollo normal y podrían explicar distintos tipos de patología como los debidos a falta de migración o la migración anormal de algunas células , por ejemplo las neuronas o los gonocitos.

Márquez-Orozco MC
Biología del Desarrollo Fascículo 10
22
Plegamiento del embrión
En la figura 8 observa cómo el embrión trilaminar, que en la tercera semana era plano, al inicio de la cuarta semana se ha flexionado, ya que el sistema nervioso central por crecimiento diferencial también lo ha hecho, y va tomando la forma de “C”.
Mientras esto sucede, la pared del embrión, o somatopleura, y la esplacnopleura se pliegan, se cierran ventralmente, y el embrión se vuelve cilíndrico. En el cuerpo queda una cicatriz en la región ventral, la línea alba (figura 8), que en las
mujeres embarazadas se pigmenta por acción de las hormonas y es la línea morena.
También te puedes dar cuenta que el aspecto del embrión va cambiando de la tercera a la quinta semana de desarrollo, época en que ya se han diferenciado el corazón, el hígado, el pulmón, el intestino, el páncreas, las crestas urogenitales y el sistema nervioso ha modificado su aspecto. Recuerda que desde el final de la tercera semana o principio de la cuarta, el corazón empieza a latir, aunque todavía no tiene la forma que conoces ya sólo es un tubo flexionado.
Neurulación
El esbozo del sistema nervioso central (SNC) humano aparece en el embrión presomítico, durante la tercera semana del desarrollo, debido a que el complejo cordomesodérmico u organizador primario, constituido por tres categorías de mesodermo, el axial o notocordal, el cefálico y el somítico o paraxial, liberan evocadores.
Los evocadores son sustancias químicas que actúan como hormonas morfogenéticas, y son las responsables de inducir al neuroectodermo, situado en el centro del ectodermo superficial, sobre el complejo cordomesodérmico , a diferenciarse en la placa neural.
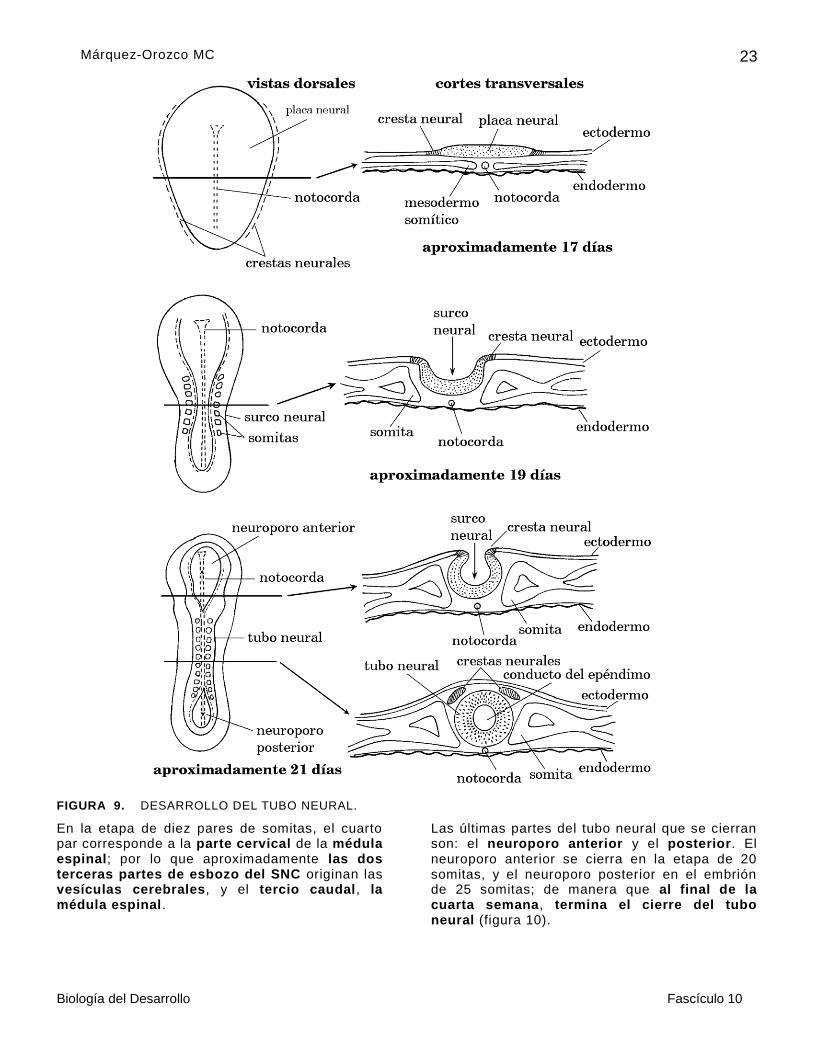
La placa neural tiene contorno piriforme, debido a que la región cefálica es más amplia que la caudal. A los lados de la placa neural hay dos bandas de neuroectodermo, que darán origen a las crestas neurales, que hacia el lado externo están en contacto con el ectodermo superficial (figura 9).
La placa neural inducida por el complejo cordomesodérmico u organizador primario es
un epitelio engrosado, constituido por células columnares, que tienen su núcleo colocado a diferentes niveles y sus membranas son muy delgadas. Por esta razón, durante muchos años los investigadores pensaron que se trataba de un epitelio multinucleado, es decir, un sincicio. Con el perfeccionamiento de las técnicas histológicas y el uso del microscopio electrónico, se precisó su estructura real.
Cuando la placa neural, engrosada, se invagina en sentido longitudinal, desde la región dorsal del embrión y toma el aspecto de una zapatilla, ya que el surco neural se profundiza, sobre todo en la parte media del embrión, y sus bordes se acercan. Al principio de la cuarta semana, en el embrión con siete pares de somitas, sus bordes se unen y empiezan a formar el tubo neural a la altura del cuarto al sexto par de somitas.
El tubo neural continúa su cierre, al mismo tiempo, hacia la región cefálica y a la caudal, como si dos cierres de cremallera avanzaran en sentido contrario (figura 9).
Márquez-Orozco MC
Biología del Desarrollo Fascículo 10
23
FIGURA 9. DESARROLLO DEL TUBO NEURAL.
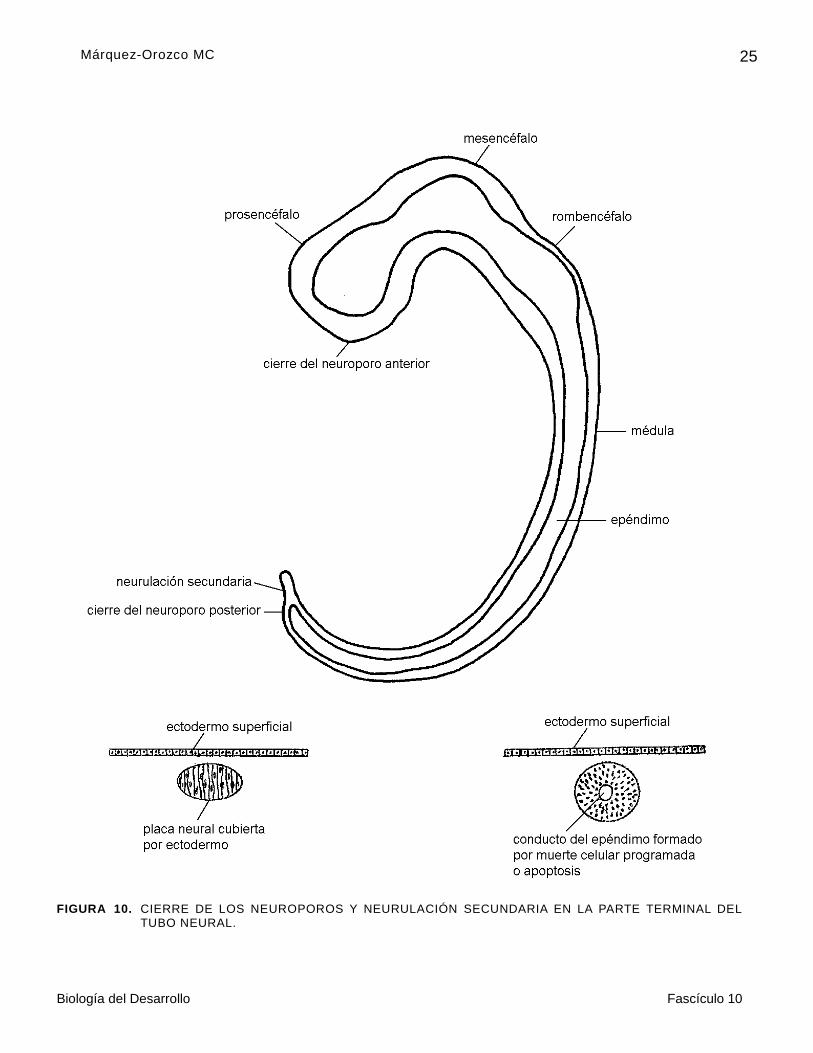
En la etapa de diez pares de somitas, el cuarto par corresponde a la parte cervical de la médula espinal; por lo que aproximadamente las dos terceras partes de esbozo del SNC originan las vesículas cerebrales, y el tercio caudal, la médula espinal.
Las últimas partes del tubo neural que se cierran son: el neuroporo anterior y el posterior. El neuroporo anterior se cierra en la etapa de 20 somitas, y el neuroporo posterior en el embrión de 25 somitas; de manera que al final de la cuarta semana, termina el cierre del tubo neural (figura 10).
Márquez-Orozco MC
Biología del Desarrollo Fascículo 10
24
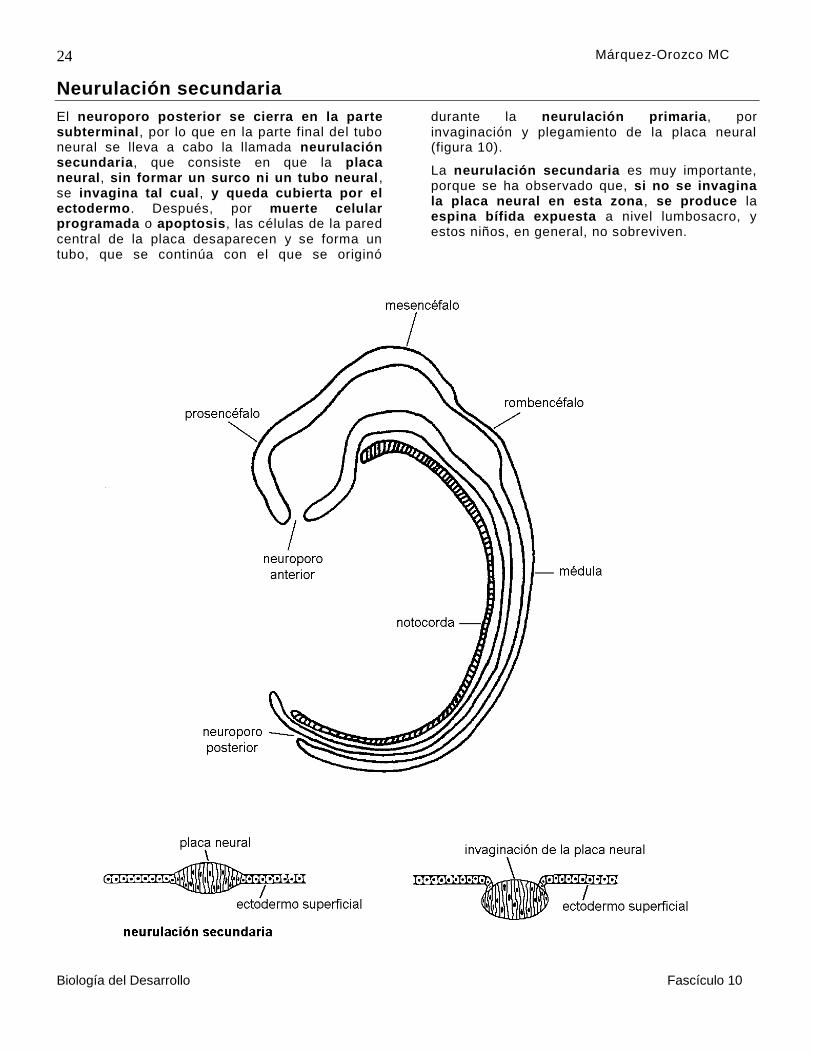
Neurulación secundaria
El neuroporo posterior se cierra en la parte subterminal, por lo que en la parte final del tubo neural se lleva a cabo la llamada neurulación secundaria, que consiste en que la placa neural, sin formar un surco ni un tubo neural , se invagina tal cual, y queda cubierta por el ectodermo. Después, por muerte celular programada o apoptosis, las células de la pared central de la placa desaparecen y se forma un tubo, que se continúa con el que se originó
durante la neurulación primaria, por invaginación y plegamiento de la placa neural (figura 10).
La neurulación secundaria es muy importante, porque se ha observado que, si no se invagina la placa neural en esta zona , se produce la espina bífida expuesta a nivel lumbosacro, y estos niños, en general, no sobreviven.
Márquez-Orozco MC
Biología del Desarrollo Fascículo 10
25
FIGURA 10. CIERRE DE LOS NEUROPOROS Y NEURULACIÓN SECUNDARIA EN LA PARTE TERMINAL DEL
TUBO NEURAL.

Márquez-Orozco MC
Biología del Desarrollo Fascículo 10
26
Mecanismos que intervienen en el cierre del tubo neural
En la actualidad, se sabe que las células neuroepiteliales, que al principio del desarrollo eran cuboides, tienen en su citoplasma microtúbulos y microfilamentos dispuestos al azar. Cuando son inducidos por el organizador primario para formar la placa neural, las células se vuelven columnares, porque los microtúbulos
se vuelven paralelos al eje mayor de las células neuroepiteliales (figura 11).
Los microfilamentos de actina se colocan en el ápice de las células, y se unen a desmosomas por medio de una proteína llamada espectrina, que es característica de su membrana plasmática (figura 11).
FIGURA 11. DESARROLLO DEL NEUROEPITELIO.
Por contracción de los microfilamentos , se reduce poco a poco el ápice de las células, como si se tratara de la jareta de una bolsa de tabaco, y se forma el surco neural y del tubo neural (figura 11).
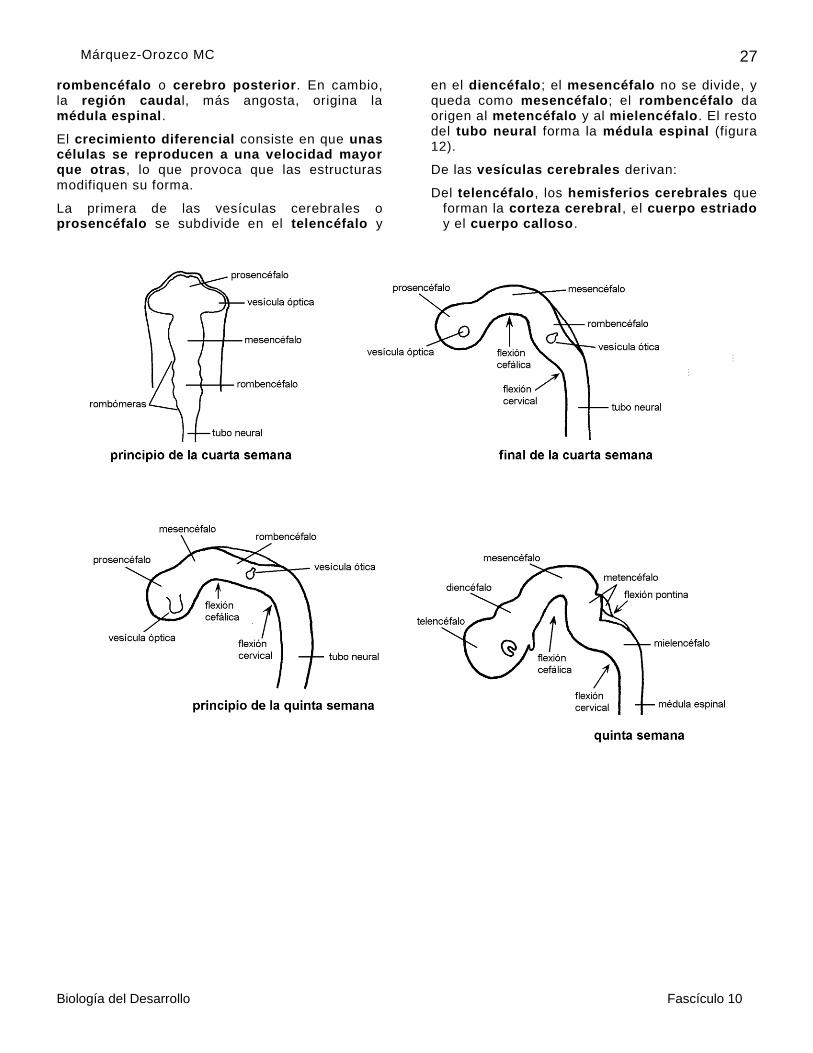
La región cefálica de la placa neural , más amplia desde el inicio del desarrollo, por crecimiento diferencial de sus paredes, forma el esbozo de las futuras vesículas cerebrales primitivas, el prosencéfalo o cerebro anterior, el mesencéfalo o cerebro medio y el
Márquez-Orozco MC
Biología del Desarrollo Fascículo 10
27
rombencéfalo o cerebro posterior. En cambio, la región caudal, más angosta, origina la médula espinal.
El crecimiento diferencial consiste en que unas células se reproducen a una velocidad mayor que otras, lo que provoca que las estructuras modifiquen su forma.
La primera de las vesículas cerebrales o prosencéfalo se subdivide en el telencéfalo y
en el diencéfalo; el mesencéfalo no se divide, y queda como mesencéfalo; el rombencéfalo da origen al metencéfalo y al mielencéfalo. El resto del tubo neural forma la médula espinal (figura 12).
De las vesículas cerebrales derivan:
Del telencéfalo, los hemisferios cerebrales que forman la corteza cerebral, el cuerpo estriado y el cuerpo calloso.

Márquez-Orozco MC
Biología del Desarrollo Fascículo 10
28
FIGURA 12. DESARROLLO DE LAS FLEXIONES Y VESÍCULAS CEREBRALES.

Márquez-Orozco MC
Biología del Desarrollo Fascículo 10
29
Del diencéfalo, el epitálamo, el tálamo, el hipotálamo, el quiasma óptico, la neurohipófisis, el tallo hipofisiario y la glándula pineal.
Del mesencéfalo, que se conserva como mesencéfalo, se diferencian los colículos superiores, visuales, y los inferiores, acústicos; los núcleos del III, IV y V pares craneales, los núcleos rojos, la sustancia nigra y los pedúnculos cerebrales.
Del metencéfalo, el cerebelo y el puente, con los núcleos del V, VI, VII y VIII pares craneales.
Del mielencéfalo, la médula oblongada, con núcleos del V, VIII, IX, X, XI y XII pares craneales.
Cada una de las tres primeras vesículas cerebrales y la médula tiene cavidades primitivas que son: la del prosencéfalo, la del mesencéfalo, la del rombencéfalo y el conducto medular del tubo neural o epéndimo.
La cavidad del prosencéfalo se subdivide en: la del telencéfalo y la del diencéfalo. La del mesencéfalo es única. La del rombencéfalo forma la del metencéfalo y la del mielencéfalo. La de la región medular del tubo neural también es única (figura 13). Más adelante:
La cavidad del telencéfalo origina los ventrículos laterales y la parte rostral del III ventrículo.
La del diencéfalo forma la mayor parte del III ventrículo.
La cavidad mesencefálica constituye el acueducto cerebral.
Las cavidades del metencéfalo y la del mielencéfalo, constituyen el IV ventrículo.
La de la parte medular del tubo neural origina el conducto medular o del epéndimo (figura 13).
FIGURA 13. CAVIDADES DEL TUBO NEURAL.
Flexiones del tubo neural
Antes del cierre de los neuroporos (tercera semana), el esbozo de las tres vesículas cerebrales es recto, igual que la médula
espinal. Entre las vesículas hay constricciones que las delimitan. En el prosencéfalo se desarrollan las vesículas ópticas, que son un

Márquez-Orozco MC
Biología del Desarrollo Fascículo 10
30
par de evaginaciones de sus paredes. El mesencéfalo es una vesícula dilatada, y en el rombencéfalo se adelgaza el techo (figura 12).
Los mensajes químicos, que pasan del organizador primario el neuroectodermo, son particulares para cada parte de éste, y le programan un ritmo de multiplicación celular característico. Por esta razón, durante la cuarta semana, se inicia la diferenciación de las flexiones cerebrales.
Las dos primeras, de concavidad ventral, aparecen primero la cefálica y después la cervical. La primera y más profunda se
diferencia a nivel del mesencéfalo , que por esto se llama mesencefálica o cefálica, y la segunda o cervical, se forma entre el rombencéfalo y la región cervical de la médula espinal (figura 12).
La tercera flexión, es la póntica o pontina, de concavidad dorsal, se diferencia en la quinta semana del desarrollo, entre el metencéfalo y el mielencéfalo. Al acentuarse estas flexiones, la extensión del encéfalo se reduce, y su forma se modifica hasta que se parece a la del adulto en el cuarto mes fetal, cuando las vesículas telencefálicas se colocan en su posición definitiva (figura 12).
Crecimiento
Durante el desarrollo de un nuevo organismo intervienen una gran cantidad de procesos, muy complejos que siguen una secuencia precisa para que el individuo tenga estructuras armónicamente constituidas.
El crecimiento es uno de estos procesos, que; cuando se trata de una célula, se puede definir como el aumento de la cantidad de citoplasma o como un aumento de masa, lo que significa que predomina el anabolismo* sobre el catabolismo**.
*(de anabolé, progresión o síntesis de sustancias). **(de kataballein, echar abajo o degradación de sustancias).
Cuando se trata de un organismo o de un tejido también crece por multiplicación celular, aunque no aumente el tamaño de las células, lo que pasa, durante la segmentación. Otra forma de crecer es por el depósito de matriz extracelular, lo que sucede en el tejido conectivo, el óseo o el cartilaginoso.
Durante el desarrollo, el crecimiento es enorme, si tomamos en cuenta que un ovocito que mide
de 80 a 120 m al ser fecundado, inicia el desarrollo de un ser humano, que al nacimiento pesa de 2,501 a 3,500 g y mide, de pie, aproximadamente 50 cm, y cuando alcanzan la edad adulta una persona de estatura regular
mide 1.60 m y pesa 60 kg.
Existen diferentes patrones de crecimiento , uno de éstos es el crecimiento diferencial , por el que los órganos adquieren su forma. Esto significa que, mientras unas células crecen rápido, otras lo hacen a velocidad media, y
otras, son lentas. Por ejemplo, al ver el desarrollo del tubo neural, originalmente recto, te habrás dado cuenta de que, la pared dorsal y la ventral del tubo son de diferentes longitud, lo que se debe a que sus células se multiplicaron a distintos ritmos, e indujeron la formación de la flexión mesencefálica, de la cervical y de la pontina, que cambian la morfología del tubo neural.
El embrión crece de forma desproporcionada, pues lo hace más rápido a nivel de la cabeza que al nivel del tronco, o los brazos que las piernas, aunque después se compensa; a esto se le llama crecimiento alométrico, y tiene un límite. Es un crecimiento determinado , esto es con límites fijos, dado por la interacción genético-ambiental.
Entre los factores que intervienen para inducir el crecimiento está la hormona de crecimiento o somatotropina, producida por la adenohipófisis. También los tejidos producen factores de crecimiento, como el neural, el epidérmico, el fibroblástico o la eritropoyetina, que actúan sobre tejidos específicos o, por el contrario, producen sustancias que lo inhiben. A pesar de que se conocen numerosas sustancias que regulan el crecimiento, aún falta mucho por investigar al respecto.
Diferenciación
Otro de los procesos importantes en el desarrollo es la diferenciación, que se puede definir como
la aparición de nuevos componentes celulares. Por ejemplo, en una célula pueden

Márquez-Orozco MC
Biología del Desarrollo Fascículo 10
31
sintetizarse sustancias específicas como pigmento, microfilamento, o se pueden modificar algunos organelos citoplásmicos , como pasa en el espermatozoide.
La diferenciación se debe a la interacción nucleocitoplásmica, lo que significa que los genes de una célula se activen o se prendan para hacerlos funcionales, de acuerdo al citoplasma en el que se encuentran. Como recordarás, los cambios en la composición del citoplasma se observan desde que el huevo se divide en células más pequeñas, que tienen diferentes componentes citoplásmicos . Durante la segmentación, las células se diferencian, al principio químicamente, después morfológicamente, y al final lo hacen funcionalmente; por esta razón se considera que hay tres niveles de diferenciación:
La quimiodiferenciación, es la diferenciación química o invisible, que se manifiesta desde la segmentación. En el embrión de ocho blastómeros, las células del casquete animal se tiñen de manera distinta que las del casquete vegetativo, por tener diferente proporción de ribonucleoproteínas y de vitelo, aunque su forma es similar.
Puede detectarse por medio de técnicas bioquímicas, de tinción, inmunocitoquímicas, de fluorescencia, histoquímicas, de radioinmunoanálisis y de cultivo de tejidos, que son en la actualidad herramientas poderosas de trabajo.
La histodiferenciación, es la diferenciación histológica o morfológica, que se observa desde que, en la mórula, las células internas son más voluminosas y forman uniones en hendidura y las externas, menos voluminosas, originan uniones estrechas. Como recordarás, de las células internas derivan el macizo celular interno, y de las externas, el trofoblasto. En embriones más desarrollados se forman tejidos con células de diferente forma. Este grado de diferenciación se puede identificar mediante técnicas histológicas y de microscopia electrónica.
La auxanodiferenciación, es la diferenciación funcional, pero implica el crecimiento diferencial, por el cual los órganos adquieren su morfología, en que los órganos se coloquen en su posición definitiva y que sean funcionales. Es el nivel de mayor complejidad y se puede comprobar mediante técnicas bioquímicas, fisiológicas y por las utilizadas en morfología, como la microdisección o la colocación de marcas con colorantes vitales, tinta china o metales.
El término proviene de auxé, crecimiento. Por ejemplo, en el corazón, durante la auxanodiferenciación, adquiere la forma definitiva, funciona y desciende de la zona del futuro cuello, al tórax. En este proceso también hay células que están programadas para morir, proceso que se llama apoptosis o muerte celular programada. Esto, por ejemplo, se observa en las membranas interdigitales de las manos y de los pies, que normalmente deben degenerar, pero si no lo hacen pueden inducir una sindactilia, que se produce por la falta de separación de los dedos, en lugar de una fusión de los mismos, por falla en la apoptosis.
Márquez-Orozco MC
Biología del Desarrollo Fascículo 10
32
Organización del desarrollo
Otro de los procesos importantes que intervienen en el desarrollo es la organización.

Márquez-Orozco MC
Biología del Desarrollo Fascículo 10
33
FIGURA 14. CADENA, CASCADA O SECUENCIA DE ORGANIZACIÓN.
Los organizadores son tejidos que inducen a otros tejidos a diferenciarse; actúan por medio de evocadores u hormonas morfogenéticas, que son sustancias químicas del tipo de las ribonucleoproteínas, las cuales pasan del organizador al tejido al que inducen a diferenciarse.
Los evocadores son señales químicas para que los genes de las células inducidas a diferenciarse se activen o se prendan y dirijan la síntesis de proteínas específicas .
Los evocadores pueden pasar al tejido inducido de tres maneras:
Por difusión simple.
A través de una matriz extracelular .
Por medio de uniones celulares específicas.
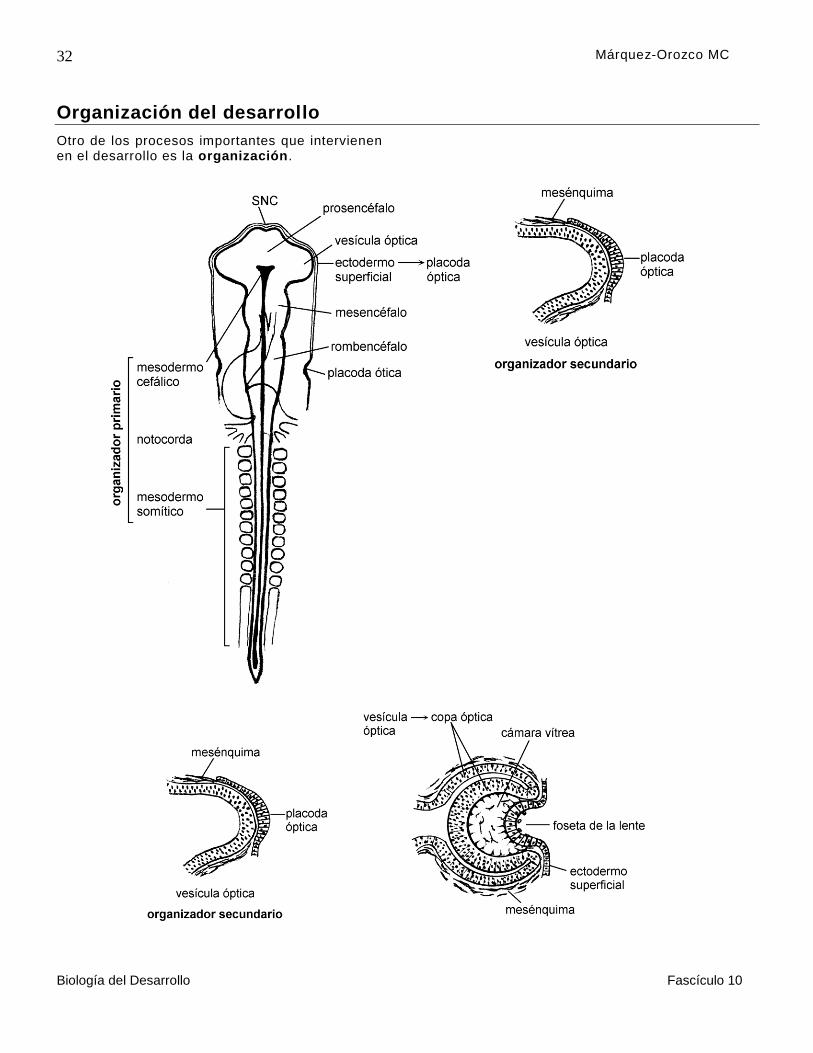
El primero de los organizadores que actúa es el organizador primario o complejo cordomesodérmico, que induce la diferenciación del sistema nervioso central . El complejo cordomesodérmico está constituido por
la notocorda, el mesodermo somítico y el mesodermo cefálico . A este proceso se le conoce como inducción primaria.
Al complejo cordomesodérmico se llama organizador primario porque es el primero que actúa sobre otro tejido, aunque en la actualidad, se sabe que sobre este organizador influyen las células del hipoblasto , pero de todas maneras su función es tan importante; que un investigador, Hans Spemann, en 1935, obtuvo el premio Nobel por descubrir su acción.
Spemann describió que cuando retiraba el tejido del labio dorsal del blastoporo, de la gástrula de anfibios, que equivale a la línea primitiva, se detenía el desarrollo; en cambio, cuando lo transplantaba a otra gástrula, inducía la diferenciación de un segundo tubo neural; lo llamó organizador primario.
El organizador primario tiene tres características fundamentales:
Universalidad. Porque es el mismo en todos los vertebrados.

Márquez-Orozco MC
Biología del Desarrollo Fascículo 10
34
Regionalidad. Porque induce de manera distinta la diferenciación de la región cefálica que de la caudal, ya que el sistema nervioso tiene gradientes de diferenciación céfalocaudales.
Polaridad. Es la capacidad de reconocer el eje axial o céfalocaudal.
Al estudiar el desarrollo del ojo, observó que existen cascadas o secuencia de organizadores, ya que el sistema nervioso, una vez inducido a diferenciarse, se transforma en un organizador secundario. A este proceso se le conoce como inducción secundaria.
Como ejemplo, se puede mencionar a las vesículas ópticas, que son evaginaciones del prosencéfalo, las cuales actúan sobre el ectodermo superficial para que éste forme las placodas ópticas, de las que se origina, a cada lado, una lente o cristalino. Durante la evolución
de la lente, primero se engrosa el ectodermo y forma una placoda óptica, después se invagina en el interior de la copa óptica, y forma la foseta de la lente, que se separa del ectodermo superficial, como una vesícula, que se hace maciza por el crecimiento de las células de su pared posterior y, una vez diferenciada, la lente actúa como un organizador terciario , sobre el ectodermo superficial y sobre el mesodermo que están enfrente de ésta, y los induce a diferenciarse en la córnea transparente (figura 14).
Las cascadas, secuencias o cadenas de organización, son como una carrera de relevos, en la que a un corredor le toca correr un tramo y le pasa la estafeta al siguiente corredor, para que avance otro tramo, y así sucesivamente, hasta que el último llega a la meta.
Organizador primario = complejo cordomesodérmicoinduce a la diferenciación del SNC.
Organizador secundario = vesícula ópticainduce la diferenciación de la lente.
Organizador terciario = lenteinduce la diferenciación de la córnea transparente.
Otros de sus experimentos permitieron observar que aunque un organizador actúe sobre el tejido al que va a inducir, éste puede responder o no a la acción del organizador, lo que depende de la etapa del desarrollo en que se lleva a cabo la inducción. La competencia es la capacidad que tiene un tejido de responder a la acción de un organizador.
Cada tejido tiene un periodo crítico para responder a la acción de los organizadores y, a medida que avanza el desarrollo, disminuye la competencia.
La determinación es la fijación de las nuevas características que se adquieren durante la diferenciación, y, como ya lo expliqué, en los huevos “en mosaico”, se establece desde que se forma el vitelo y las ribonucleoproteínas, en el ovocito de los anfibios; en cambio, en los huevos “de regulación” aparece después de la segmentación, en la cual los componentes celulares como el vitelo y las ribonucleoproteínas, se distribuyen en distintas células y sólo algunos genes actúan en cada tipo de blastómero.
La restricción consiste en que las células pierden la capacidad de activar cualquier gene y, por lo tanto, dejan de ser totipotenciales . Esto no es irreversible pues, como recordarás, al transplantar un núcleo somático al citoplasma del ovocito, aunque ésta es una maniobra
experimental, se reactivan los genes. También se puede producir de manera anormal , cuando el medio en el que se encuentran las células se modifica, por ejemplo, al estar expuestas a fármacos, tóxicos, radiaciones u otros agentes que las alteran.
Los tejidos pueden interactuar de diferente manera y se han descubierto al menos tres tipos de interacciones, que son:
Instructiva. Cuando la respuesta del tejido depende del tipo de tejido que actúa como organizador. Por ejemplo, si la yema pulmonar recibe evocadores del mesénquima del pulmón forma tejido pulmonar, pero si el mesénquima es gástrico, se diferencia en glándulas gástricas; si es hepático, forma trabéculas hepáticas; y si es intestinal, origina vellosidades intestinales.
Permisiva. Cuando la respuesta del tejido inducido es la misma bajo la acción de diferentes organizadores. Por ejemplo, los esbozos pancreáticos forman páncreas, aunque sean inducidos por mesénquima pulmonar, gástrico, hepático o intestinal.
Recíproca. Cuando un tejido que induce a otro a diferenciarse después actúa sobre su organizador, para inducirlo a que realice alguna actividad específica. Por ejemplo, durante el desarrollo de los miembros, el blastema, que es un mesodermo denso, induce al ectodermo a formar un

Márquez-Orozco MC
Biología del Desarrollo Fascículo 10
35
engrosamiento, que es la cresta ectodérmica apical, y, una vez que se diferenció, induce al blastema a formar el factor de mantenimiento de la cresta ectodérmica apical .
El comprender cómo se desarrolla el embrión ha favorecido que se entienda el origen de las estructuras y la manera en que se producen las malformaciones; ha permitido identificar sus causas y, sobre todo, en algunos casos se han podido prevenir.
Sobre este tema se puede decir mucho más, pero he reservado una parte que explicaré a medida que abordemos los temas de la organogénesis.
Procura revisar las referencias bibliográficas incluidas y otras de la biblioteca, para aumentar tus conocimientos. ¡AHORA RECOGE LOS FRUTOS
DE TU DEDICACIÓN Y ESFUERZO! ¡VAS AL TRIUNFO! ¡ADELANTE!
Bibliografía
Carlson BM. Gametogénesis y fecundación En
Embriología básica de Patten. 5 ed. México: Interamericana, McGraw-Hill, 1990:193-258.
Carlson BM. Transport of gametes and fertilization.
En Human embryology and developmental biology. St. Louis, Missouri: Mosby-Year book, 1994:51-64, 65-86.
Diccionario Mosby de medicina y ciencias de la salud. 1 ed Colombia: Mosby-Doyma Libros,
1995:1538 pp.
Diccionario terminológico de ciencias médicas. 10
ed. Salvat Mallorca: Salvat, 1968:1188 pp.
Gilbert SF. The cellular basis of morphogenesis. En
Developmental biology 4th ed. Suderland: Sinauer, 1994:77-117, 234-237, 244-294.
Ham RG, Veomett MJ. Embryology and
morphogenesis. Cell differentiation in the embryo-mosaic and regulative patterns of development. Embryonic induction. Morphogenesis En Mechanisms of development 1th ed. St. Louis, Missouri: Mosby, 1980:35-53, 406-427, 471-564.
Hamilton WJ Mossman HW. Formación de las capas
germinales. Determinación, diferenciación, los mecanismos organizadores, desarrollo anormal y gemelación. En Embriología humana 4 ed. Buenos Aires: Editorial Intermédica, 1973:66-85,195-228.
Larsen WJ. The second week. The third week. The
fourth week. En Human embryology 1th ed. Singapore: Churchill Livingstone, 1993:33-92.
Lodish H. Baltimore D. Multicellularity: cell-cell and
cell-matrix interactions. En Molecular cell biology. 3th ed. New York: Scientific American books, 1995:1123-1169.
Márquez-Orozco MC. Bases morfológicas y
fisiológicas del desarrollo del sistema nervioso En Salas M Ontogenia neural 1 ed. México: Coordinación de la Investigación Científica, Universidad Nacional Autónoma de México, Sociedad Mexicana de Ciencias Fisiológicas, 1991:73-128.
Moore KL Persaud TVN. Inicio del desarrollo humano.
En Embriología clínica. 5 ed. México: Nueva Editorial Interamericana McGraw-Hill de México, 1996:43-82
Pansky B. Segunda semana del desarrollo: embrión
de disco germinativo bilaminar. Mecanismos generales del desarrollo normal. En Embriología Médica. 1 ed. Argentina Editorial Médica Panamericana, 1985:53-67, 76-83.
Patten BM. Formación de las hojas embrionarias.
Diferenciación inicial del cuerpo y constitución de los sistemas orgánicos. En Embriología Médica 5 de. Buenos Aires: El Ateneo, 1969:52-62, 63-73, 74-95.
Sadler TW. De la ovulación a la implantación. En
Langman Embriología Médica. 7 ed. México: Editorial Médica Panamericana, 1996:49-83.
Ejercicios de autoevaluación
Resuelve las siguientes preguntas
¿Qué es la gastrulación? ¿En qué día de la gestación se inicia?
¿Qué son los movimientos morfogenéticos?
Define los siguientes términos y da ejemplos de las estructuras en las que se presentan.
Invaginación.
Polinvaginación.
Evaginación.
Convergencia y divergencia.
Involución.
Cavitación.
Delaminación.
Elongación.
Plegamiento.
Epibolia y endobolia.
Movimiento amiboide.

Márquez-Orozco MC
Biología del Desarrollo Fascículo 10
36
¿Qué es la matriz extracelular? ¿Cuál es su importancia funcional durante el desarrollo embrionario?
¿Qué función tiene el ácido hialurónico en el desarrollo embrionario?
Explica la importancia de la fibronectina en el desarrollo embrionario
¿Cuáles son los dos grandes grupos de matriz extracelular?
Explica el proceso de diferenciación de las estructuras del disco embrionario.
¿Qué es la creciente mesodérmica? ¿Qué deriva de ésta?
Explica la formación de la línea primitiva, los tejidos que la integran y su importancia en el desarrollo
¿Qué es el nódulo de Hensen? ¿Cuál es su importancia en el desarrollo?
Explica la formación del mesodermo embrionario.
¿Qué es el embrión trilaminar?
¿Qué tipos de tejidos derivan del ectodermo superficial?
¿Qué deriva del neuroectodermo que forman la placa neural?
Menciona los derivados de las crestas neurales.
¿Cuál es la función de la notocorda? ¿Qué deriva de la notocorda en el adulto?
¿Qué es una somitómera? ¿Qué deriva de los siete primeros pares?
¿Cuáles son los tres componentes de una somita típica? ¿A qué da origen cada una de sus partes?
Menciona los derivados del mesodermo intermedio.
¿Qué tejidos forman la somatopleura y la esplacnopleura?
¿Qué es el celoma? ¿Cómo se forma?
Menciona los derivados del mesodermo somático.
Menciona los derivados del mesodermo esplácnico.
Enumera los derivados del endodermo.
¿Que deriva de los gonocitos?
¿Qué son los CAMs? ¿Cuál es su función?
¿Qué características tienen los CAMs de acuerdo con su afinidad?
¿Qué son las cadherinas? ¿Qué propiedades y funciones tienen?
¿Qué es la N-CAM y la Ng-CAM? ¿En dónde se expresan? ¿Qué propiedades y funciones tienen?
¿Qué CAMs se expresan en el epiblasto y en el hipoblasto? ¿Qué cambios experimentan durante la migración y la inducción del sistema nervioso la L-CAM y la N-CAM?
¿Qué es la neurulación primaria y la secundaria? ¿Qué flexiones y vesículas cerebrales se diferencian? ¿Qué deriva de cada una?
¿Qué es el organizador primario? ¿Qué propiedades tiene?
¿Qué es un evocador? ¿De qué manera pueden pasar?
¿Qué es el crecimiento? ¿Qué tipos de crecimiento existen?
¿A qué se debe la diferenciación? ¿Cuáles son sus niveles?
¿Qué es la competencia? ¿Cómo cambia durante el desarrollo?
¿Qué es la determinación y la restricción?
De un ejemplo de una cascada de organizadores.
Explique los tres tipos de interacción que existen entre los organizadores y los tejidos inducidos. Da ejemplos.