
Inmunologia-Apunte El Vikingo
57
APUNTE DE INMUNOLOGIA (Actualización 2006) “EL VIKINGO”
-
Upload
apuntesdelcev -
Category
Documents
-
view
217 -
download
0
Transcript of Inmunologia-Apunte El Vikingo
8/7/2019 Inmunologia-Apunte El Vikingo
http://slidepdf.com/reader/full/inmunologia-apunte-el-vikingo 1/53
APUNTE DE
INMUNOLOGIA(Actualización 2006)
“EL VIKINGO”
1

8/7/2019 Inmunologia-Apunte El Vikingo
http://slidepdf.com/reader/full/inmunologia-apunte-el-vikingo 2/53
INDICEINDICEINTRODUCCION………………………………………………………………..…………………………………………………PAG. 3GENERALIDADES……………………………………………………………...........................................................PAG. 3INMUNIDAD INNATA……………………………….................................................................................... PAG. 4
INFLAMACION………………………………………………………………………………………………………PAG. 5o VASODILATACION………………………………………………………………………………………PAG. 6
o ACONTECIMIENTOS CELULARES: TRAFICO LINFOCITARIO………………………………PAG. 6o FACTORES QUE CONDICIONAN EL TRAFICO LINFOCITARIO…………………………….PAG. 6
MOLECULAS DE ADHESION…………………………………………………………..……PAG. 6 QUIMIOTAXIS………………………………………………………………………………….PAG. 7 QUEMOQUINAS………………………………………………………………………………..PAG. 7
o COMPONENTES HUMORALES……………………………………………………………………..…PAG. 8
SISTEMA DE LA COAGULACION…………………………………………………………. PAG. 8 PROTEINAS DE FASE AGUDA………………………………………………………..…… PAG. 8 IFNALFA Y BETA……………………………………………………………………………….PAG. 8 SISTEMA DEL COMPLEMENTO…………………………………………………………….PAG. 8 SISTEMA DE LAS CININAS……………………………………………………………..….PAG. 9
o COMPONENTES CELULARES…………………………………………………………………….……PAG. 10 CELULAS FAGOCITICAS………………………………………………………………….….PAG. 10
• EL NEUTROFILO……………………………………………………………………..PAG. 11• EL MACROFAGO…………………………………………………………………….. PAG. 11• LOS EOSINOFILOS………………………………………………………………….PAG. 12
MASTOCITOS Y BASOFILOS……………………………………………………………….PAG. 12 PLAQUETAS……………………………………………………………………………………..PAG. 13 CELULAS NATURAL KILLER………………………………………………………………..PAG. 13
INMUNIDAD ADAPTATIVA…………………………………………………………………………………………………….PAG. 13 CELULAS DENDRITICAS…………………………………………………………………………………………PAG. 13 ANTIGENOS DE HISTOCOMPATIBILIDAD…………………………………………………………………PAG. 14
PROCESAMIENTO Y PRESENTACION ANTIGENICA…………………………………………………....PAG. 15 LINFOCITOS T Y B…………………………………………………………………………………………………PAG. 16 LINFOCITOS T………………………………………………………………………………………………………PAG. 17
o ONTOGENIA T…………………………………………………………………………………………….PAG. 18
o ACTIVACION T……………………………………………………………………………………………PAG. 19
o LINFOCITOS T CD4+: FENOTIPOS TH1 Y TH2………………………………………………...PAG. 19
o LINFOCITOS T CD8+……………………………………………………………………………………PAG. 20
o LINFOCITOS T REGULATORIOS Y DE MEMORIA....................................................PAG. 21
LINFOCITOS B………………………………………………………………………………………………………PAG. 21o LINFOCITOS B1 Y DE LA ZONA MARGINAL DEL BAZO……………………………………. PAG. 22
o ONTOGENIA B…………………………………………………………………………………………….PAG. 22
o ACTIVACION B……………………………………………………………………………………………PAG. 22 LOS ANTICUERPOS………………………………………………………………………………………………. PAG. 25
o DIVERSIDAD DE INMUNOGLOBULINAS: LOS DIFERENTES ISOTIPOS………………..PAG. 26
o LOS ANTICUERPOS PUEDEN TENER CADENAS LIVIANAS KAPPA O LAMBDAPERO NO AMBOS TIPOS……………………………………………………………………………… PAG. 26
o INMUNOGLOBULINA M………………………………………………………………………………..PAG. 26
o INMUNOGLOBULINA A……………………………………………………………………………..… PAG. 26
o INMUNOGLOBULINA G……………………………………………………………………………….. PAG. 26o INMUNOGLOBULINA D…………………………………………………………………………………PAG. 26o INMUNOGLOBULINA E………………………………………………………………………………… PAG.
27o TEORIA DE LA SELECCIÓN CLONAL……………………………………………………………….PAG. 28
o LA RESPUESTA INMUNE PRIMARIA Y SECUNDARIA EN LA PRODUCCIONDE ANTICUERPOS……………………………………………………………………………………….PAG. 28
ORGANOS LINFATICOS………………………………………………………………………………………….PAG. 29o EL TIMO…………………………………………………………………………………………………….PAG. 29
o LA MEDULA OSEA………………………………………………………………………………………. PAG. 30
o LOS GANGLIOS LINFATICOS………………………………………………………………………..PAG. 30
o EL BAZO…………………………………………………………………………………………………….PAG. 30
INTEGRACION DE LOS PROCESOS: INGRESO DEUN ANTIGENO Y SECUENCIA DE EVENTOS……………………………………………………………………………...PAG. 31MECANISMOS DE LESION INMUNITARIA: REACCIONES DE HIPERSENSIBILIDAD………………………. PAG. 33
HIPERSENSIBILIDAD DE TIPO I……………………………………………………………………………..PAG. 33 HIPERSENSIBILIDAD DE TIPO II…………………………………………………………………………… PAG. 34 HIPERSENSIBILIDAD DE TIPO III……………………………………………………………………….….PAG. 35 HIPERSENSIBILIDAD DE TIPO IV………………………………………………………………………..... PAG. 36
2

8/7/2019 Inmunologia-Apunte El Vikingo
http://slidepdf.com/reader/full/inmunologia-apunte-el-vikingo 3/53
RECHAZO DE TRANSPLANTES………………………………………………………………………………………………..PAG. 37TOLERANCIA INMUNITARIA………………………………………………………………………………………………….PAG. 38
SUPERANTIGENOS……………………………………………………………………………………………………………….PAG. 39AGENTES INFECCIOSOS Y MECANISMOS DE EVASION…………………………………………………………….. PAG. 39INMUNODEFICIENCIAS…………………………………………………………………………………………………………PAG. 40
AGAMMAGLOBULINEMIA DE BRUTON LIGADA AL CROMOSOMA X……………………….…….. PAG. 40 INMUNODEFICIENCIA VARIABLE COMUN……………………………………………………..…………PAG. 40
.
DEFICIT AISLADO DE IgA………………………………………………………………………………………PAG. 40
SINDROME DE HIPER IgM………………………………………………………………………………………PAG. 40 INMUNODEFICIENCIA COMBINADA SEVERA……………………………………………………….……PAG. 41 SINDROME DE WISKOTT-ALDRICH………………………………………………………………………….PAG. 41 DEFICIENCIAS GENETICAS DEL COMPLEMENTO………………………………………………………. PAG. 41 DEFECTOS DE LA FUNCION LEUCOCITARIA…………………………………………………..…………. PAG.
41ASPECTOS GENETICOS EN INMUNOLOGIA………………………………………………………………………………PAG. 43
INTRODUCCIONAntes que nada me gustaría darles la bienvenida a 3er año, lo que no es poco. Muchos de Uds. han sorteadoinfinidad de obstáculos para llegar a esta instancia. Microbiología es una asignatura muy interesante, muylarga y en algunos casos se complica, pero en líneas generales es divertida, entretenida y para nada aburrida.
En esta oportunidad, tienen en sus manos el APUNTE DE INMUNOLOGIA. Existe también el dePARASITOLOGIA MÉDICA y el de VIROLOGIA, conformando en conjunto una suerte de trilogía.¿Por qué redacto este apunte? Básicamente por que me costó entender el libro INMUNOLOGIA DE FAINBOIMa la primera lectura. Esto derivó en histeria y terror: tenía menos de un mes para preparar el parcial, nopodía distinguir que era o no importante, algunas cosas nunca fueron aclaradas y otras simplemente fuerondifíciles de comprender con la lectura sin previa ni posterior explicación. No se asusten…… MICROBIOLOGIAno es una materia tan autodidacta como en los años anteriores, por el contrario los profesores se desvivenpor explicar. En fin, con ayuda de mi novia (estudiante de Ciencias Biológicas) intentamos darles una suertede introducción a la inmunología. El principal objetivo de este texto es que pueda facilitar la comprensión dellibro de FAINBOIM, por lo que recomendamos su lectura luego de inspeccionar estas hojas. Insisto, no dejende leer la bibliografía recomendada!!!! Como siempre esto es solo un consejo y Uds. son los que deciden.Además, les recomiendo que lean el apunte en el orden en que está escrito. Creo que es la única manera depoder comprender todos los contenidos……..en fin……..DISFRUTEN DE LA CURSADA!!!!!
GENERALIDADESBueno.....…para comenzar analizaremos algunos conceptos básicos que nos permitirán entender los procesosque se llevan a cabo durante el curso de una infección. Muchas de los temas que serán analizados acontinuación ya han sido aprendidos tanto en histología como en fisiología, pero como somos humanos ysabemos que nuestra memoria suele jugarnos en contra, haremos una revisión de los mismos. A pesar deque los contenidos son un poco extensos, este capitulo no presentará mayores problemas.
Nuestro sistema inmune nos salva de la muerte segura por una infección constituyendo un proceso defensivocuyo principal objetivo es erradicar al microorganismo que ha invadido al huésped. Las sustancias capaces dedesencadenar una respuesta inmunitaria reciben el nombre de antigenos (antibody generator) oinmunógenos.Si quisiéramos estudiar el orden cronológico en que el ser humano desarrolla una respuesta inmune frente a
un determinado agente (antigeno), deberíamos destacar la: Inmunidad innata o inespecífica que, como su nombre lo indica, constituye una primer barrera de defensa
que actúa de la misma manera independientemente del microorganismo involucrado.
Inmunidad específica o adaptativa, que actúa en una etapa posterior a la inespecífica y está constituidapor células con receptores de alta especificidad (linfocitos T y B) que son capaces de reconocer un ampliorango de microorganismos.
Ya se que el siguiente cuadro parecerá chino básico en un primer momento, pero el mismo destaca en formaconcisa las principales características de ambos tipos de inmunidad. Igualmente cada una de ellas seráanalizada posteriormente en profundidad. Lo importante es que las sepan distinguir y puedan saber cualesson sus componentes………el resto sale por lógica (por supuesto que la lógica viene cuando terminemos deleer tooooooooooddddooooooo el apunte, no? JE-JE…..no desesperen que es fácil!!!!).
FASE I BARRERAS FISICAS Y ANATOMICAS NATURALES
3

8/7/2019 Inmunologia-Apunte El Vikingo
http://slidepdf.com/reader/full/inmunologia-apunte-el-vikingo 4/53
INMUNIDAD INNATA OINMUNIDAD INNATA O INESPECIFICAINESPECIFICA FASE II
(INFLAMACION)
COMPONENTES CELULARES:- NEUTROFILOS- CELULAS NK- MACROFAGOS- MASTOCITOS Y BASOFILOS- EOSINOFILOS- PLAQUETAS- CELULAS ENDOTELIALES- CELULAS DENDRITICAS- MASTOCITOS
COMPONENTES HUMORALES:- COMPLEMENTO- COAGULACION- SISTEMA DE LAS CININAS- PROTEINAS DE FASE AGUDA- IFN ALFA Y BETA
INMUNIDAD ADAPTATIVA OINMUNIDAD ADAPTATIVA O ESPECIFICAESPECIFICA
LINFOCITOS T Y B CON RECEPTORES DE ALTA ESPECIFICIDADRECONOCEN EL ANTIGENO PRESENTADO POR UNA CPA
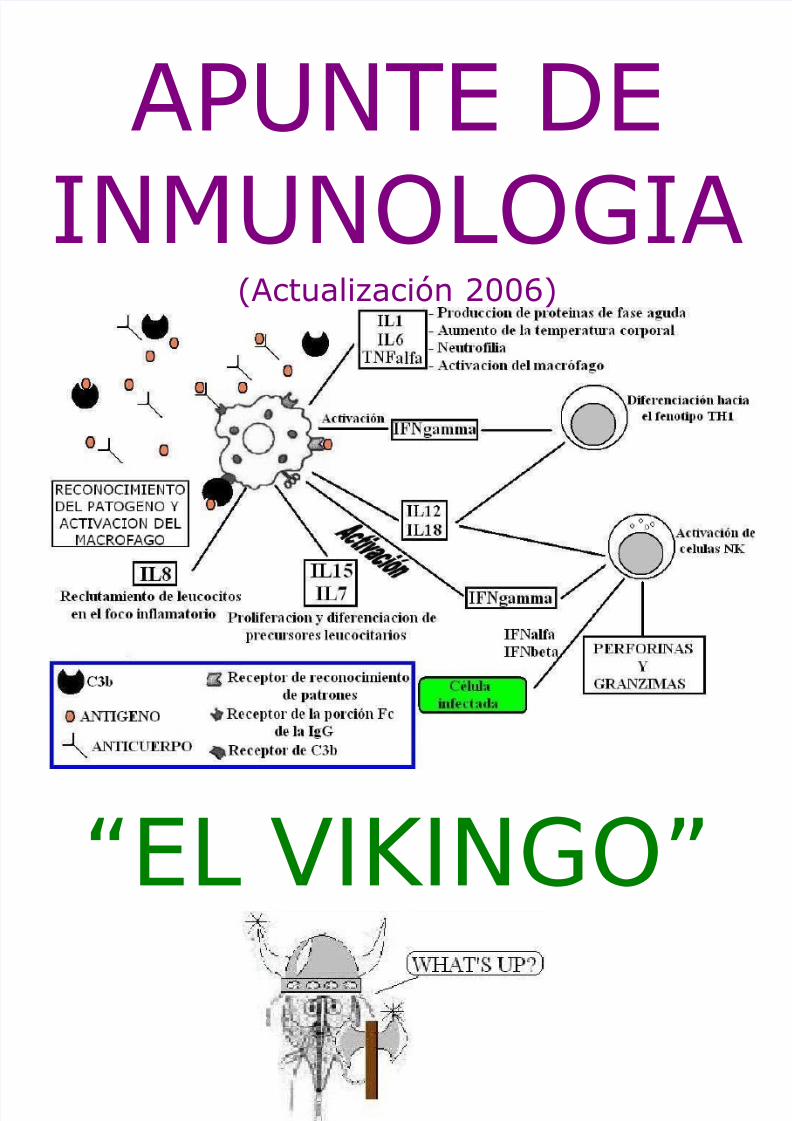
1) INMUNIDAD INNATALas defensas constitutivas y las inducibles son inespecíficas, porque si bien requieren un reconocimientoespecífico entre moléculas y receptores del microorganismo y el huésped, no necesitan un reconocimientoespecifico tipo antigeno-anticuerpo, a excepción de la IgA secretoria. Como se aclaró en el cuadro anterior, lainmunidad innata consta de dos fases:- Fase I: La barreras físicas naturales están representadas por los diferentes epitelios intactos,
siendo la piel la barrera mecánica más efectiva ya que la misma se encuentra en continua descamación, loque contribuye a la eliminación de los microorganismos adheridos a ella. La piel presenta además ciertas
propiedades que inhiben el crecimiento bacteriano, como por ejemplo la baja humedad y Ph, la presenciade sustancias con actividad antibacteriana y antifúngica como la lisozima (que degrada la pared celular dealgunas bacterias), la lactoferrina (que compite con la bacteria por el hierro el cual es esencial para lamultiplicación bacteriana), el ácido láctico secretado por las glándulas sudoríparas y los ácidos grasoslibres liberados por degradación enzimatica de los componentes de las secreciones sebáceas que impiden eldesarrollo de Tinea Capitis (hongo) en el adulto. Cuando el grado de humedad de la piel aumenta, porejemplo a causa del uso de vestimentas muy ajustadas que no permiten la aireación, se presenta un mediopropio para el desarrollo de TRICOFICIAS (hongos) que colonizan diferentes pliegues cutáneos. En el casode quemaduras severas, al no encontrarse la piel intacta, no puede protegerse el tejido celular subcutáneofrente a una eventual infección por Pseudomona aeruginosa (bacteria). Tambien existen aglutininas (queinteractuan con receptores inhibiendo la adherencia de microorganismos al epitelio) y defensinas (quealteran la permeabilidad de las membranas microbianas). La conjuntiva está protegida por el efecto debarrido continuo de las lágrimas y por la presencia de altas concentraciones de lisozima en la secreciónlacrimal. El epitelio de los senos paranasales y del tracto respiratorio, desde la laringe hasta los alvéolos, esuna importante barrera mecánica protegida por una cubierta de mucus, las células ciliadas que barrenel mismo y la IgA secretoria. De esta manera las partículas que toman contacto con el epitelio respiratorioson capturadas por las secreciones y transportadas hacia la laringe, para ser luego deglutidas y destruidaspor los mecanismos efectores del tracto gastrointestinal. Muchas de las bacterias ingeridas se inactivan porla elevada concentración de ácido clorhídrico y por las enzimas gástricas. Otros patógenos como losvirus envueltos son susceptibles a la acción detergente de las sales biliares. Las defensas primarias deltracto urinario están representadas por el mecanismo de arrastre que produce la micción, el Phrelativamente bajo de la orina que inhibe en cierta medida el crecimiento bacteriano y la secreción deproteínas (como la de Tamm y Horsfall) que poseen afinidad por los receptores de E. Coli uropatógenaimpidiendo que los mismos se adhieran al epitelio de las vías urinarias. Un caso muy interesante es el de lavagina, que esta protegida tanto por el bajo Ph de sus fluidos como por el fenómeno de exclusión que
ejerce la flora bacteriana normal compuesta fundamentalmente por Lactobacillus. Esta compite pornutrientes y receptores presentes en el epitelio que permiten la colonizacion de las superficies mucosas yademas producen sustancias con actividad antimicrobiana.- Fase II: En esta etapa, el microorganismo ha logrado evadir las barreras naturales enunciadas
anteriormente y el individuo desarrolla una respuesta inflamatoria al verse afectados los tejidosprofundos. La inflamación se define como una reacción compleja del tejido conjuntivo vascularizado frente alos estímulos nocivos locales. Este proceso se caracteriza por una reacción de los vasos sanguíneos que dalugar a la acumulación de líquido y leucocitos en los tejidos extravasculares. Es útil para destruir, atenuar omantener localizado al agente patógeno y al mismo tiempo iniciar una cadena de acontecimientos que
4

8/7/2019 Inmunologia-Apunte El Vikingo
http://slidepdf.com/reader/full/inmunologia-apunte-el-vikingo 5/53
dentro de lo posible curan y reconstruyen el tejido lesionado. Sino existiera la inflamación, las infecciones sepropagarían de forma incontrolada, las heridas no se curarían nunca y los órganos lesionados presentaríanlesiones supurativas de forma permanente. No obstante los proceso de inflamación y reparación pueden serperjudiciales (ver HIPERSENSIBILIDAD). La inflamación termina cuando se elimina el estimulo nocivo ydesaparecen o quedan inhibidos los mediadores de la misma. La respuesta inflamatoria tiene lugar en eltejido conjuntivo vascularizado, e involucra la participación de elementos tales como el plasma, las célulascirculantes, los vasos sanguíneos y los constituyentes celulares y extracelulares del tejido conjuntivo:
CELULAS CIRCULANTES CELULAS DEL TEJIDO CONJUNTIVO MATRIZ EXTRACELULARNEUTROFILOS MASTOCITOS COLAGENOMONOCITOS FIBROBLASTOS ELASTINA
EOSINOFILOS MACROFAGOS FIBRONECTINALINFOCITOS LINFOCITOS RESIDENTES LAMININABASOFILOS PROTEOGLICANOSPLAQUETAS
La inflamación presenta dos fases bien diferenciadas AGUDA y CRONICA. La inflamación aguda tiene unaevolución relativamente breve, con una duración que oscila entre minutos, horas y pocos días. Suscaracterísticas principales son la exudación de liquido y de proteínas plasmáticas (EDEMA), la modificación enel calibre de los vasos (vasodilatación) que da lugar al aumento del flujo sanguíneo y la migración deleucocitos (predominantemente NEUTROFILOS) desde el punto en el que abandonan la microcirculación hastael foco de la lesión, donde se acumulan. La inflamación crónica tiene una duración mayor y se caracterizahistológicamente por la presencia de linfocitos y macrófagos, la proliferación de vasos sanguíneos, fibrosis(intentos de curación) y necrosis tisular, siendo el macrófago el actor principal de dicho proceso.Los cuatro signos cardinales de la inflamación son: RUBOR (enrojecimiento), CALOR, TUMOR (tumefaccióno induración) y DOLOR. Estos eventos son producidos por el aumento de la permeabilidad vascular y deldiámetro de los vasos sanguíneos (vasodilatación) produciéndose un incremento del flujo sanguíneo local conaumento de la temperatura en la zona de la lesión (lo que explica el calor) que a su vez es responsable delenrojecimiento y disminución de la velocidad de flujo con acumulación local de fluidos (ocasionando eledema). Por otro lado, la liberación de mediadores químicos y la compresión mecánica de los nervios seránresponsables del dolor. Además, el pasaje de fluidos y células producirán la hinchazón.Los eventos producidos durante la inflamación son numerosos. Para realizar una descripción en formaordenada y facilitar su comprensión dichos procesos serán analizados por separado, sin embargo debetenerse en cuenta que la secuencia descripta es arbitraria, ya que todos se inician casi simultáneamente. Lainmunidad innata suele resolver el proceso infeccioso naciente, o al menos controlarlo hasta desarrollar larespuesta inmune adaptativa y ademas puede condicionar el rumbo de esta ultima.
INFLAMACION
5
8/7/2019 Inmunologia-Apunte El Vikingo
http://slidepdf.com/reader/full/inmunologia-apunte-el-vikingo 6/53
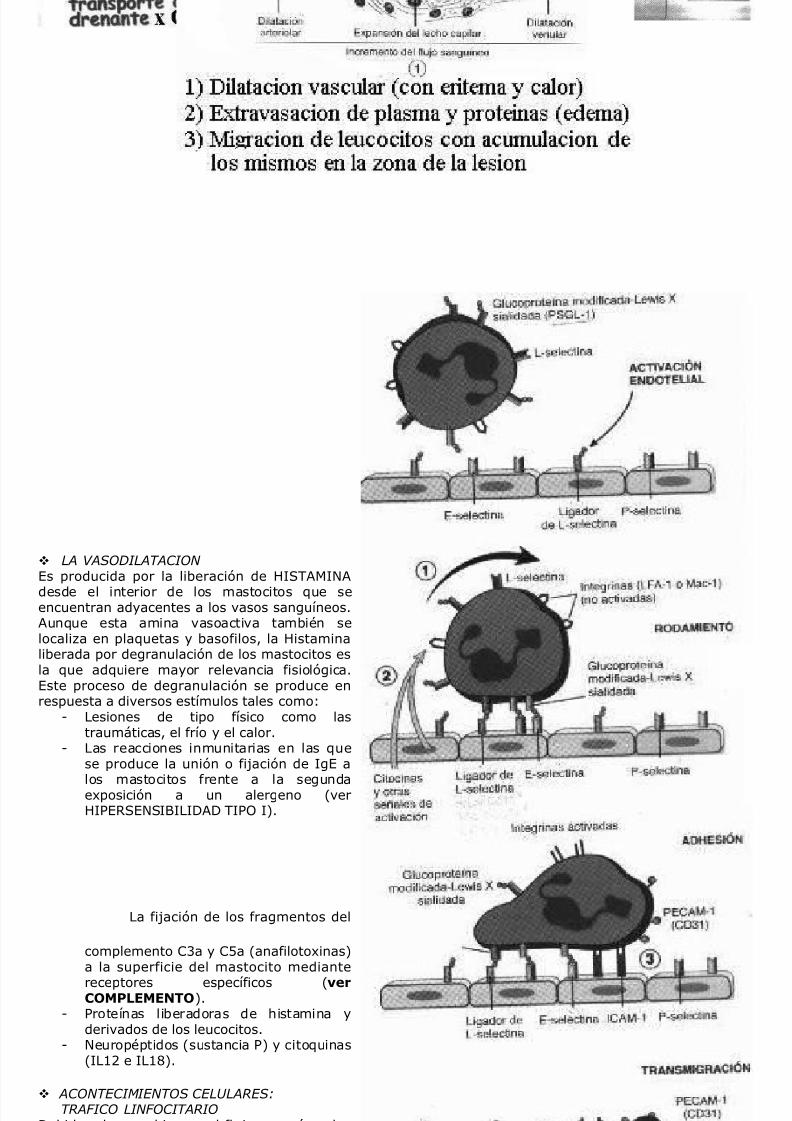
LA VASODILATACION Es producida por la liberación de HISTAMINA
desde el interior de los mastocitos que seencuentran adyacentes a los vasos sanguíneos.Aunque esta amina vasoactiva también selocaliza en plaquetas y basofilos, la Histaminaliberada por degranulación de los mastocitos esla que adquiere mayor relevancia fisiológica.Este proceso de degranulación se produce enrespuesta a diversos estímulos tales como:
- Lesiones de tipo físico como lastraumáticas, el frío y el calor.
- Las reacciones inmunitarias en las quese produce la unión o fijación de IgE a
los mastocitos frente a la segundaexposición a un alergeno (verHIPERSENSIBILIDAD TIPO I).- La fijación de los fragmentos del
complemento C3a y C5a (anafilotoxinas)a la superficie del mastocito mediante
receptores específicos (verCOMPLEMENTO).- Proteínas liberadoras de histamina y
derivados de los leucocitos.- Neuropéptidos (sustancia P) y citoquinas
(IL12 e IL18).
ACONTECIMIENTOS CELULARES:TRAFICO LINFOCITARIO
Debido a los cambios en el flujo sanguíneo losleucocitos se sitúan en la periferia del mismo(marginación), pudiendo contactar directamen-
te con el endotelio activado en forma transi-toria por medio de selectinas, llevando a caboel rodamiento sobre dicha superficie. Mastarde, la acción de diversas quemoquinas pro-
6

8/7/2019 Inmunologia-Apunte El Vikingo
http://slidepdf.com/reader/full/inmunologia-apunte-el-vikingo 7/53
duce un cambio conformacional de las inte-grinas presentes en los leucocitos, los que aho-ra se adhierenfirmemente al endotelio que-dando éste literalmente revestido (pavimen-tación) por los mismos.Finalmente los leu-cocitos dirigen sus pseudópodos hacia las unio-nes que existen entre las célulasendoteliales, se introducen a través de las mismas y quedan situadas entre las células endoteliales y lamembrana basal. Este evento de diapedesis o trasmigración se produce predominantemente en lasvenulas de endotelio alto (HEV). Poste-riormente atraviesan la membrana basal ingre-sando al espacioextravascular migrando ha-cia un estimulo quimiotáctico.Este mecanismo de salida es utilizado por neutrófilos, monocitos, linfocitos, eosinófilos y macrófagos. Loseventos producidos son bási-camente los mismos en los diversos tipos celulares mencionados, la diferenciaradica en las moléculas de adhesión involucradas. Ade-más, debe quedar bien claro que el cambio en laexpresión de dichas moléculas y en los receptores de quemoquinas determina la amplia gama de destinos alos cuales migrar . Debido a que estos patrones no se modifican en gran medida en los linfocitos, losmismos son capaces de recircular continuamente para aumentar la chance de encontrar suantigeno específico.
No se vuelvan locos tratando de memorizar moléculas específicas, solo sepan describir y enumerar loseventos principales:
EVENTOS QUE OCURRENEN LA LUZ VASCULAR
OTROS EVENTOS
MARGINACION TRANSMIGRACIONO DIAPEDESIS
RODAMIENTOMIGRACION HACIA EL
ESTIMULOQUIMIOTACTICO
ADHERENCIA FIRME
Factores que condicionan el tráfico linfocitarioEl mismo esta determinado por el patrón de expresión de moléculas de adhesión sobre los leucocitos y elendotelio y por las quemoquinas producidas localmente y sus receptores expresados sobre la superficie de laspo-blaciones leucocitarias.- Moléculas de adhesión: están pre-sentes en la superficie de todas las células del organismo e
involucradas en la interacción celular y de las células con moléculas de la matriz extracelular (MEC).Existen 5 familias principales:
Moléculas que establecen interaccionesdébiles (unión transitoria)
Moléculas que permiten sostener laintegridad funcional de los tejidos (uniones
firmes)
Moléculas que establecenuniones relativamente estables
SELECTINAS SIALOMUCINAS CADHERINAS INTEGRINAS SUPERFAMILIA DE IGL SELECTINAE SELECTINAP SELECTINA
CD 34GLYCAM 1MADCAM 1
E CADHERINA CD11/CD 18VLA4
LFA-1 ó MAC-1
ICAM 1, 2 y 3V CAM 1PECAM 1
Las selectinas se expresan en células endoteliales, leucocitos y plaquetas y se adhieren a SIALOMUCINAS.Las sialomucinas se expresan en células endoteliales y leucocitos y se adhieren a L- SELECTINAS.Las cadherinas NUNCA se expresan en leucocitos pero si en las células de Langherans, que al adquirirmovilidad dejan de expresarlas. Las cadherinas se adhieren a la MEC y mantienen la integridad de los tejidos.Las integrinas se expresan en leucocitos y se adhieren tanto a componentes de la MEC como a moléculas dela superfamilia de Ig y complemento.
La superfamilia de Igs se expresan sobre células endoteliales y células presentadoras de antigenos (CPA) y seadhieren a integrinas.
Muchas de estas moléculas se encuentran presentes en forma constitutiva. Sin embargo, al desencadenarsela inflamación se produce una inducción de su síntesis por parte de las células endoteliales, lo que permiteun aumento en la expresión de los mismos, fenómeno conocido con el nombre de ACTIVACION ENDOTELIAL.Este proceso es facilitado por la producción de citoquinas tales como IL6, IL1 y TNFalfa. Estas son proteínasproducidas por muchos tipos celulares entre los cuales se destaca el macrófago activado.Además de esta acción, dichas citoquinas:
- Producen un aumento de la permeabilidad vascular- Desencadenan la respuesta de fase aguda (ver mas adelante)- Facilitan la producción de PGE (prostaglandina E), al inducir la actividad de la ciclooxigenasa a nivel
hipotalámico, con el consecuente aumento de la temperatura corporal (fiebre)- Producen neutrofilia al actuar a nivel de la medula ósea- Activan al macrófago, neutrofilo, reclutan celulas NK, etc.
Existe una gran variedad de citoquinas cuya actividad se destaca por ser pleotrópica (actúa sobre muchostipos celulares) y multifuncional (una determinada citoquina da lugar a acciones reguladoras positivas ynegativas). Sus efectos se producen mediante la fijación a receptores específicos situados en células diana.
Citoquinas queregulan la función
leucocitaria
Citoquinas implicadas en la inmunidadnatural
Citoquinas queactivan célulasinflamatorias
Quemoquinas Citoquinas que estimulan lahematopoyesis
Regulan laactivación,
crecimiento ydiferenciación de
los leucocitos
Producen aumento de la permeabilidadvascular, incremento en la expresión de
moléculas de adhesión, inducen lasíntesis de proteínas de fase aguda,
aumento de la Tº corporal y neutrofilia
Activan almacrófago durante
las respuestasinmunes mediadas
por células
Llevan a caboactividad
quimiotáctica
Actúan como mediadoras delcrecimiento y diferenciación de
leucocitos inmaduros
IL2
IL4IL10TGFbeta
TNFalfa
IL1IL6IFNalfa y beta
IFNgamma
TNFalfaTNFbetaIL5IL10IL18
IL8 IL3
IL7CSF
7

8/7/2019 Inmunologia-Apunte El Vikingo
http://slidepdf.com/reader/full/inmunologia-apunte-el-vikingo 8/53
IL12- Quimiotaxis: es la locomoción orientada según un gradiente químico que permite a los
leucocitos migrar en los tejidos hasta alcanzar la zona de la lesión. Diversas sustancias exógenas yendógenas pueden actuar como factores quimiotácticos.
SUSTANCIAS EXOGENAS SUSTANCIAS ENDOGENASPRODUCTOS BACTERIANOS(PEPTIDOS FORMILADOS)
COMPONENTES DEL SISTEMA DEL COMPLEMENTO(ESPECIALMENTE C5a)
PRODUCTOS DE LA VIA DE LA LIPOOXIGENASA(PRINCIPALMENTE LTB4)
CITOQUINAS(EN PARTICULAR IL8)
La fijación de estas sustancias a sus receptores específicos en el leucocito desencadena un proceso detransducción de señales que culmina con un incremento de las concentraciones de calcio intracelular, lo queposibilita el ensamblaje de sus elementos contráctiles presentes en el citoesqueleto responsables delmovimiento celular (pseudópodos). Además de estimular la locomoción, muchos factores quimiotácticosproducen la activación leucocitaria que es desencadenada mas comúnmente por la fagocitosis y la formación
de complejos antigeno-anticuerpo (ver COMPONENTES CELULARES).- Quemoquinas: son un superfamilia de proteínas cuya acción principal es activar e inducir la
quimiotaxis de tipos específicos de leucocitos, que al poseer receptores para los mismos, migran a lossitios anatómicos donde estas se encuentran y desempeñan sus funciones. Todas las quemoquinasposeen residuos de cisteína conservados formando puentes disulfuro que son esenciales para su
función. La posición relativa de las cisteínas es usada para clasificarlas en cuatro familias: CXC, CC,CX3C y C, donde “C” representa un residuo de cisteína y “X” un aminoácido no conservado.QUEMOQUINAS CXC QUEMOQUINAS CC QUEMOQUINAS C QUEMOQUINAS CX3C
Presenta un residuo deaminoácidos que separa los
dos primeros residuosconservados de cisteína
Tiene adyacentes los dosprimeros residuos
conservados de cisteína
Carecen de dos de loscuatro residuos
conservados de cisteína
Las cisteínas están separadaspor tres aminoácidos no
conservados
IL8 MCP1 (proteínaquimiotáctica de
monocitos) y RANTES
LINFOTAXINA FRACTALTINA
Como ya se habrán dado cuenta, hay algunas cosas que no se sabe bien de donde salen y son un misterio. Labuena noticia es que estas dudas serán aclaradas a continuación cuando desarrollemos los componenteshumorales y celulares de la inflamación.
COMPONENTES HUMORALESA) Sistema de la coagulación
Al formarse FACTOR XIIa, se activa la vía intrínseca de la coagulación con generación de:- TROMBINA (FACTOR IIa): que fragmenta el fibrinógeno soluble circulante para generar un
coagulo insoluble de fibrina. Durante esta conversión, se forman fibrinopéptidos que inducen un
incremento en la permeabilidad vascular y estimulan la actividad quimiotáctica de los leucocitos.También posee propiedades inflamatorias al aumentar la expresión de moléculas de adhesiónleucocitaria en el endotelio vascular.- FACTOR Xa: que actúa como mediador de la inflamación aguda y causa un aumento de la
permeabilidad vascular y de la exudación leucocitaria.
B) Proteínas de fase agudaLas proteínas de fase aguda conforman un conjunto de proteínas, sintetizadas fundamentalmente por elhepatocito, cuya producción y niveles séricos se incrementan dramáticamente en respuesta a la injuriacelular. Esta producción exacerbada responde principalmente a la acción de citoquinas producidas por
8

8/7/2019 Inmunologia-Apunte El Vikingo
http://slidepdf.com/reader/full/inmunologia-apunte-el-vikingo 9/53
monocitos y macrófagos tales como IL1, IL6 y TNFalfa. Constituyen ejemplos de estas sustancias la proteínaC reactiva y la proteína de unión a manosa. Ambas actúan opsonizando (recubriendo) a los microorganismospara que estos sean posteriormente reconocidos por monocitos, macrófagos o neutrófilos.
C) IFNalfa y beta
Estas citoquinas son de relevancia en la actividad antiviral (ver apunte de virología) produciendo la síntesis desustancias que degradan los ácidos nucleicos del virión. Presentan además una notable capacidad depotenciar la actividad citotóxica de las células natural killer (NK) e inducen ciertas actividades proinflamatoriasen el macrófago.
D) Sistema del complemento
El sistema del complemento esta compuesto por 20 proteínas(junto a sus productos de fragmentación), cuya concentraciónmayor se observa en plasma. Este sistema actúa en losprocesos inmunitarios innatos y adaptativos de defensa frente amicroorganismos y su objetivo final es la lisis de los mismos através del denominado complejo lítico de ataque de membrana(CLAM). En el proceso se elaboran diversos componentes delcomplemento que producen aumento de la permeabilidadvascular, quimiotaxis y opsonización. Dichos componentes seencuentran en un principio en forma inactiva en el plasma y senumeran de C1 a C9. El paso mas importante para larealización de las funciones biológicas del complemento es la
activación de su tercer componente o C3. La fragmentación delC3 se puede producir a través de la denominada vía clásica, que se inicia por la fijación del C1 a unanticuerpo (IgM o IgG) unido a un antigeno, es decir, a un complejo inmune, o a través de la vía alternativa,que se puede activar por las superficies de los microorganismos (LPS bacteriano, Igs agregadas yendotoxinas). La vía alternativa implica la participación de un grupo especifico de componentes séricosdenominado sistema de properdina (Properdina P, factores B y D). Además, la vía de las lectinas activa uncomplejo con actividad serinoproteasa que al activarse escinde a los componentes C4 y C2 y da lugar a laformación de C3 convertasa en un proceso que no involucra la participacion del componente C1. En dichasvías, la C3 convertasa divide el C3 en dosfragmentos importantes: el C3a, que es liberado y elC3b. Mas adelante, este ultimo se une a losfragmentos generados previamente para formar C5convertasa que a su vez interactúa con C5 paraliberar C5a e iniciar de este modo la formación deCLAM (C5 a C9). El CLAM produce lisis mediante launión hidrofóbica inicial a la bicapa lipídica de lascélulas diana, formando finalmente canalescilíndricos transmembrana.Las funciones biológicas del sistema delcomplemento se incluyen en dos categorías generales: lisis celular por CLAM y los efectos biológicos de losfragmentos proteolíticos del complemento. Los factoresderivados del complemento afectan a diversos fenómenosde la inflamación aguda:
- Fenómenos vasculares: C3a y C5a
(anafilotoxinas) son los productos de fragmentaciónde los componentes correspondientes delcomplemento. Estos productos incrementan lapermeabilidad vascular y producen vasodilataciónprincipalmente mediante la liberación de histaminadesde los mastocitos. C5a también activa la vía de lalipooxigenasa (metabolismo del ácido araquidónico)en los neutrófilos y monocitos, dando lugar a unincremento en la liberación de mediadoresinflamatorios.- Adhesión, quimiotaxis y activación de los leucocitos: C5a es un potente agente
quimiotáctico para neutrófilos, monocitos, eosinófilos y basofilos. También incrementa la adhesión de
los leucocitos al endotelio mediante la activación de los propios leucocitos y el aumento de laintensidad de unión de las integrinas de superficie a su ligando endotelial.
9

8/7/2019 Inmunologia-Apunte El Vikingo
http://slidepdf.com/reader/full/inmunologia-apunte-el-vikingo 10/53
- Fagocitosis: C3b y C3bi, cuando se fijan a la pared celular bacteriana, actúan como opsoninas
y favorecen la fagocitosis por parte de neutrófilos y macrófagos que presentan receptores para C3b ensu superficie.- Potenciación de la respuesta B: los productos de degradación del C3b (C3bi, C3dg y C3d)
poseen un receptor específico sobre la superficie del LB denominado CR2. El mismo forma parte delcomplejo integrado además por CD19 y CD81. CD19 se encarga de desencadenar un evento detransducción de señales que culmina con la activación del LB, la que es potenciada por losmencionados productos de degradación del complemento mediante la producción de un
entrecruzamiento de las señales, incrementando de esta manera la producción de anticuerposespecíficos.
En síntesis, las funciones del complemento son básicamente:
INFLAMACION OPSONIZACION CITOTOXICIDAD POTENCIACION DE LA RESPUESTA BC3a y C5a C3b C5-C9 C3bi, C3dg y C3d
El mecanismo de ensamblaje del complemento esta fuertemente controlado por proteínas inhibitorias. Lapresencia de estos inhibidores en la membrana celular del huésped es útil para distinguirlo de la mayor parte
de los microorganismos y lo protege frente a una lisis celular inadecuada. Debido a estos mecanismosreguladores, se alcanza un equilibrio que da lugar a la destrucción efectiva de organismos extraños,impidiendo la lesión de las células del huésped. Estos mecanismos reguladores son los siguientes:- Regulación de las convertasa de C3 y C5: el desencadenamiento de la vía alterna en forma
independiente de la clásica presenta un gran dilema, existiendo la posibilidad de que C3b actué sobrela superficie de células extrañas y sobre las propias. Este inconveniente no se presenta con la víaclásica, dado que sus activadores (ANTICUERPOS) no reconocen normalmente células propias. Existeuna gran diversidad de moléculas que inhiben la interacción de C3b con el FACTOR B o incrementan lasusceptibilidad a la degradación del C3b a través de la acción del FACTOR I. El objetivo en ambos
10

8/7/2019 Inmunologia-Apunte El Vikingo
http://slidepdf.com/reader/full/inmunologia-apunte-el-vikingo 11/53
casos es inhibir la formación de C3 convertasa de la vía alterna . Entre dichas moléculas se encuentranCR1, DAF (factor acelerador de decaimiento), MCP (proteína cofactor de membrana) y FACTOR H. Esteultimo es una proteína plasmática que interactúa con la superficie celular (mientras las demás seencuentran sobre la superficie de células propias) uniéndose al ácido siálico. Las células de losmamíferos expresan un alto contenido de este ácido, por lo tanto unen FACTOR H y se ven protegidasde la C3 convertasa de la vía alterna. Los microorganismos patógenos que expresan bajo contenido deácido siálico NO unirán FACTOR H, permitiendo el ensamblado de la C3 convertasa de la vía alternacon la posterior activación del complemento.
- Fijación de los componentes activos del complemento por proteínas plasmáticas
específicas: el primer paso de la vía clásica, en el que C1 se une a un complejo inmune quedabloqueado por un inhibidor específico denominado inhibidor C1. La activación excesiva delcomplemento también se impide por diversas proteínas que actúan sobre la formación de CLAM, comopor ejemplo CD59 (inhibidor de membrana de la lisis reactiva).
E) Sistema de la cininas
El sistema de las cininas genera peptidos vasoactivos a partir de proteínas plasmáticas denominadasquininógenos y mediante proteasas especificas denominadas calicreínas. El sistema de las cininas produce laliberación de bradicinina, un potente agente que incrementa la permeabilidad vascular. La cascada quefinalmente da lugar a cininas se inicia por la activación del FACTOR XII (HAGEMAN) de la vía intrínseca de lacoagulación debido a su contacto con superficies que presentan carga negativa, como el colágeno y lasmembranas basales. Se produce un fragmento del FACTOR XII denominado FACTOR XIIa o activador deprecalicreína, que convierte la precalicreína plasmática en una forma proteolítica activa conocida comocalicreína. Esta última fragmenta una proteína plasmática precursora (cininógeno de alto peso molecular oHMWK) para producir bradicinina.
COMPONENTES CELULARESTodos los elementos celulares que serán analizados a continuación, derivan de un precursor común en medulaósea, el cual posteriormente dará origen a otros precursores de la serie mieloide y eritroide:
11

8/7/2019 Inmunologia-Apunte El Vikingo
http://slidepdf.com/reader/full/inmunologia-apunte-el-vikingo 12/53
A) Células fagocíticas
Las defensas del organismo poseen un número de células diferenciadas para ejecutar funciones específicas.Todas ellas derivan de un precursor común ubicado en la medula ósea denominado célula madre o STEM CELL(recuerdan HISTOLOGIA? Digan si o…….jódanse!!!). Las células que poseen actividad antimicrobiana directason las células fagocíticas, que incluyen a los linfocitos polimorfonuclerares (particularmente neutrófilos), losmonocitos sanguíneos, los histiocitos y las células fagocíticas alojadas en tejidos específicos. A estas debensumarse las células dendríticas, que juegan un papel crucial en la presentación antigénica. Tanto losneutrófilos como los macrófagos deben reconocer y fijar las partículas extrañas para posteriormente
internalizar dichos elementos e inactivarlos mediante diversos mecanismos. Algunos patógenos sonreconocidos en forma directa, ya que conservan sobre su superficie estructuras invariantes compartidas pormuchas clases de microorganismos. Este mecanismo es mediado por una amplia variedad de receptores deRECONOCIMIENTO DE PATRONES (RRP). Los mismos:
- Se encuentran presentes en los microorganismos pero no en sus huespedes- Son esenciales para la supervivencia o patogenicidad de los mismos- Son compartidas por clases enteras de microorganismos
Por lo tanto, aun cuando el sistema inmune innato carece de la especificidad del adaptativo, es capaz dediscriminar lo propio respecto de lo no propio (infeccioso).
RECEPTORES DE RECONOCIMIENTO DE PATRONESRECEPTORES TIPO TOLL RECEPTORES DE LECTINA TIPO C RECEPTOR SCAVENGER
No induce la fagocitosisMedia la internalizacion de microorganismoscon la posterior degradacion, procesamiento
y presentacion de peptidos antigenicos Reconocen lipoproteinasbacterianas, polirribonucleotidos
y ADN bacterianoActiva vias que conducen a la produccion de
mediadores inflamatoriosSu activacion conduce a la secrecion de
numerosas quimocinas y citoquinasReconoce LPS, peptidoglicano y ARN simple y doble
cadenaReconocen residuos de manosa, galactosa y
fucosaRRP SOLUBLES
PROTEINA DE UNION A MANOSA FICOLINA H y L PROTEINA C REACTIVA PROTEINAS SURFACTANTESSon producidas por el higado en etapas tempranas del proceso infeccioso durante la respuesta de
fase aguda Son producidas por celulasalveolares tipo I y secretadassobre el epitelio respiratorioReconocen a sus ligandos y activan el complemento por la via de
las lectinas
Reconoce patogenos y celulaspropias dañadas mediando su
eliminacionTodos estos tipos están presentes en NEUTROFILOS, CELULAS DENDRITICAS y MACROFAGOS, reconociendo los mismos ciertos
patrones moleculares asociados a patógenos (PMAP)
Sin embargo, la mayor parte de los microorganismos no son reconocidos hasta que son recubiertos por
elementos denominados opsoninas, que se unen a receptores específicos situados en los leucocitos y de estamanera facilitan su fagocitosis por los mismosLas opsoninas más importantes son:
C3b y C3biSe generan por la actividad del complemento y son reconocidos porReceptores CCR1, CCR2 y CCR3
ANTICUERPOSSu porción Fc es reconocida por receptores de este fragmentollevando a cabo un proceso denominado citotoxicidad dependiente deanticuerpos (CCDA)
PROTEINAS DE FASE AGUDA Son producidas por el hepatocito en respuesta a la acción de IL1, IL6y TNFalfa
El neutrófiloEs el leucocito polimorfonuclear más abundante. Tiene un vida media corta en la circulación, deaproximadamente 10 horas. Normalmente estas células son marginadas en distintos tejidos, donde completan
su ciclo de vida, de unos pocos días de duración. Pertenecen a la serie medular mieloide y cuando alcanzan sumaduración ya no pueden replicarse. Presentan núcleo multilobulado y numerosos gránulos que contienenenzimas y otras sustancias con acción bactericida. Los neutrófilos son atraídos al sitio de infección porsustancias quimiotácticas como el componente C5a del complemento y otros intermediarios del proceso
12

8/7/2019 Inmunologia-Apunte El Vikingo
http://slidepdf.com/reader/full/inmunologia-apunte-el-vikingo 13/53
inflamatorio. Partículas como las bacterias se adhieren a la superficie del neutrófilo y soningeridas en una vacuola llamada fagosoma. Una vez dentro de la célula, la bacteria puedeser inactivada por varios mecanismos. Algunos de ellos pueden ser dependientes del oxigeno,como los que se originan con el estallido respiratorio. Al aumentar la incorporación deoxigeno dentro del neutrófilo, disminuye el Ph dentro del fagosoma y se originan productostóxicos para la bacteria como el anión superóxido, el peroxido de hidrogeno y los radicaleshidroxilo, entre otras moléculas activas.Existe un segundo mecanismo dependiente del oxigeno que requiere además la acción de la
enzima mieloperoxidasa. En presencia de peroxido y de un haluro (cloruro o ioduro) la enzima cataliza lareacción de halogenación del peroxido. Como resultado se forma, por ejemplo en el caso del cloruro, el aniónhipoclorito que es altamente bactericida. La fusión de los gránulos citoplasmáticos con el fagosoma es unfenómeno independiente del oxigeno. Al fusionarse los gránulos liberan las enzimas y otras sustanciasantibacterianas y forman el fagolisosoma y endosoma dentro del cual la bacteria queda sometida a un Ph de3-4 y a la acción de la lisozima, fosfatasas, nucleasas, lactoferrina y proteínas catiónicas antibacterianas.Todo esto hace que el entorno se vuelva altamente hostil para la mayoría de las bacterias. Debe tambiénagregarse, que las enzimas lisosomales como las colagenasas y elastasas son también deletéreas para lostejidos del huésped cuando se liberan y contribuyen a intensificar el proceso inflamatorio y el daño del tejido.
MECANISMOS BACTERICIDAS DEPENDIENTESDEL OXIGENO
MECANISMOS BACTERICIDAS NO DEPENDIENTES DELOXIGENO
ESTALLIDO RESPIRATORIOPRODUCCION DE:
- SUPEROXIDO- OXIGENO SIMPLE
- OTROS RADICALES ACTIVOSSISTMA MIELOPEROXIDASASISTEMA DEL OXIDO NITRICO
LIBERACION EN EL FAGOSOMA DEL CONTENIDOLISOSOMAL:
- PROTEINAS CATIONICAS- HIDROLASAS ACIDAS
- LISOZIMA- LACTOFERRINA- PROTEASAS
El macrófagoEstas células se hallan distribuidas ampliamente en los tejidos, derivan de monocitossanguíneos y su función en la defensa del huésped es más amplia y duradera que la de losneutrófilos. Los macrófagos son también atraídos por C5a y otras sustancias quimiotácticas ytienen la capacidad de fagocitar bacterias y detritus celulares originados durante la infección.A diferencia de los neutrófilos, los macrófagos presentan propiedades particularessumamente relevantes:
- Establecen poblaciones estables en los diversos tejidos, asumiendo fenotipos especializados (células deKupffer en hígado, macrófagos alveolares en pulmón, etc.)
- Actúan como células presentadoras de antigenos (CPA) a células T (los otros dos tipos de CPA son las
células dendríticas y los LB……NO DESESPEREN, LO VEREMOS MAS ADELANTE!!!!).- Presentan una marcada capacidad para producir quemoquinas en respuesta a estímulos de naturaleza
microbiana, citoquinas que inducen una respuesta inflamatoria local y sistémica (IL1, IL6 y TNFalfa),citoquinas que orientan el curso futuro de la respuesta inmune adaptativa (IL12 e IL18), citoquinasque median el reclutamiento de leucocitos en el tejido inflamatorio (IL8), citoquinas que inducen laproliferación y diferenciación de precursores leucocitarios a nivel de la medula ósea (IL7, IL15 y CSF[factores estimulantes de formación de colonias])
- Pueden resintetizar los gránulos lisosomales con su enzimas- Pueden volver a fagocitar- Carecen del sistema mieloperoxidasa y de lactoferrina- Presentan un estallido respiratorio menos espectacular- Pueden ser activados por IFNgamma para aumentar su capacidad bactericida- Producen oxido nítrico, que además puede combinarse con anión superóxido para formar peroxinitrilo,
molécula tóxica para las bacterias y las células humanasEl macrófago suele modificar su patrón de comportamiento al reconocer una gran concentración dequemoquinas, complejos antigeno-anticuerpo y patógenos opsonizados. Sin embargo, el estimulo masimportante lo constituye el reconocimiento del patógeno oposonizado con componentes del complemento oanticuerpos. El cambio producido se denomina ACTIVACION (ver grafico). Al activarse, el macrófago presenta:
- Capacidad incrementada para producir IROs (intermediarios reactivos del oxigeno)- Aumento de la capacidad endocítica- Aumento en la expresión de moléculas HLA de clase I y II- Aumento en la expresión de moléculas coestimulatorias (B7)- Aumento en la expresión de moléculas de adhesión- Aumento de la secreción de citoquinas y quemoquinas proinflamatorias que contribuyen al
reclutamiento y activación de diferentes poblaciones leucocitarias en el sitio de la infección.
13
8/7/2019 Inmunologia-Apunte El Vikingo
http://slidepdf.com/reader/full/inmunologia-apunte-el-vikingo 14/53
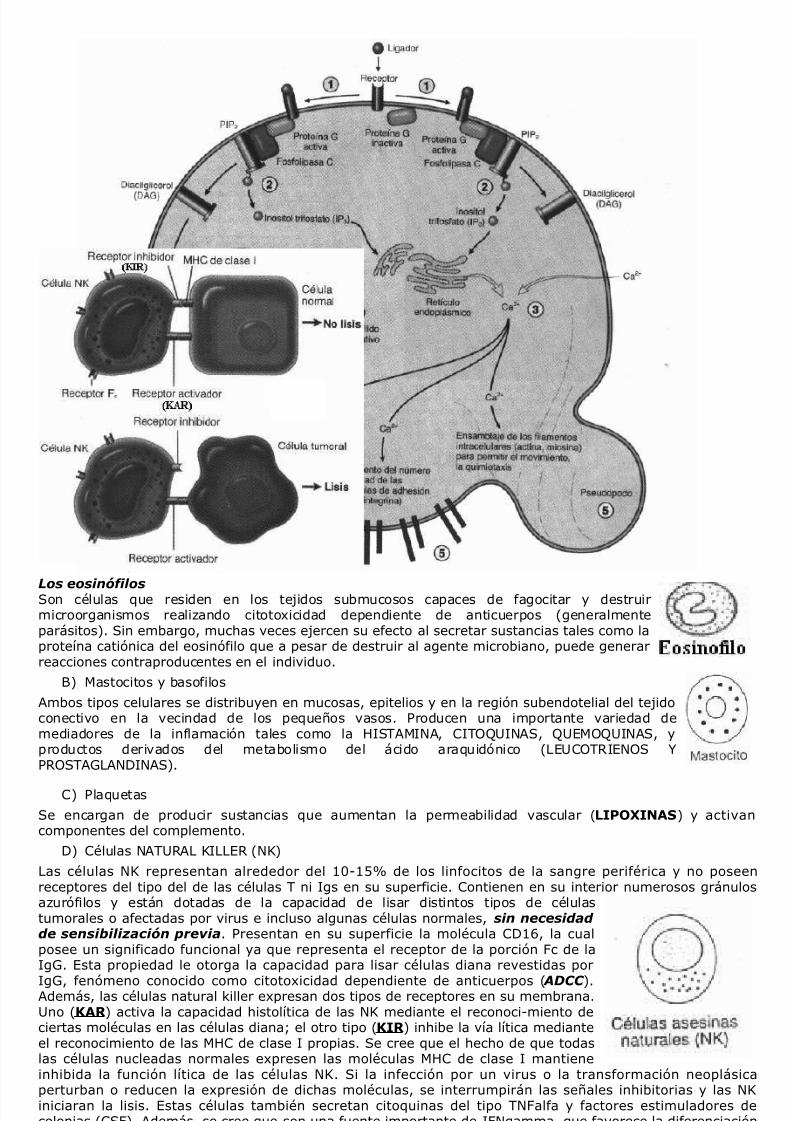
Los eosinófilosSon células que residen en los tejidos submucosos capaces de fagocitar y destruirmicroorganismos realizando citotoxicidad dependiente de anticuerpos (generalmenteparásitos). Sin embargo, muchas veces ejercen su efecto al secretar sustancias tales como laproteína catiónica del eosinófilo que a pesar de destruir al agente microbiano, puede generarreacciones contraproducentes en el individuo.
B) Mastocitos y basofilos
Ambos tipos celulares se distribuyen en mucosas, epitelios y en la región subendotelial del tejidoconectivo en la vecindad de los pequeños vasos. Producen una importante variedad demediadores de la inflamación tales como la HISTAMINA, CITOQUINAS, QUEMOQUINAS, yproductos derivados del metabolismo del ácido araquidónico (LEUCOTRIENOS YPROSTAGLANDINAS).
C) Plaquetas
Se encargan de producir sustancias que aumentan la permeabilidad vascular (LIPOXINAS) y activancomponentes del complemento.
D) Células NATURAL KILLER (NK)
Las células NK representan alrededor del 10-15% de los linfocitos de la sangre periférica y no poseenreceptores del tipo del de las células T ni Igs en su superficie. Contienen en su interior numerosos gránulosazurófilos y están dotadas de la capacidad de lisar distintos tipos de célulastumorales o afectadas por virus e incluso algunas células normales, sin necesidad de sensibilización previa. Presentan en su superficie la molécula CD16, la cualposee un significado funcional ya que representa el receptor de la porción Fc de laIgG. Esta propiedad le otorga la capacidad para lisar células diana revestidas por
IgG, fenómeno conocido como citotoxicidad dependiente de anticuerpos ( ADCC ).Además, las células natural killer expresan dos tipos de receptores en su membrana.Uno (KAR ) activa la capacidad histolítica de las NK mediante el reconoci-miento deciertas moléculas en las células diana; el otro tipo (KIR ) inhibe la vía lítica medianteel reconocimiento de las MHC de clase I propias. Se cree que el hecho de que todaslas células nucleadas normales expresen las moléculas MHC de clase I mantieneinhibida la función lítica de las células NK. Si la infección por un virus o la transformación neoplásicaperturban o reducen la expresión de dichas moléculas, se interrumpirán las señales inhibitorias y las NKiniciaran la lisis. Estas células también secretan citoquinas del tipo TNFalfa y factores estimuladores decolonias (CSF). Además, se cree que son una fuente importante de IFNgamma, que favorece la diferenciaciónde las células TH1 y cumple un papel importante en la respuesta inmune antiviral.El mecanismo citotóxico mediado por las células NK desencadena la apoptosis de la célula blanco a través dela liberación de proteínas contenidas en los gránulos azurófilos especializados presentes en el citoplasma queson liberados con la activación de la célula NK. Los gránulos son lisosomas modificados que contienen dosclases distintas de proteínas denominadas perforinas y granzimas. La perforina es una glicoproteina que alintegrarse a la membrana de la célula blanco forma poros sobre la misma permitiendo el ingreso de lasgranzimas, que a su vez activan una cascada enzimatica que conduce a la fragmentación del ADN celular. La
14
8/7/2019 Inmunologia-Apunte El Vikingo
http://slidepdf.com/reader/full/inmunologia-apunte-el-vikingo 15/53
direccionalidad de la secrecion evita que las mencionadas proteínas destruyan células no infectadas. Ademas,puede mediar la apoptosis de la celula blanco mediante la interaccion FAS- FASL (ver LTCD8+).
2) INMUNIDAD ADAPTATIVALa inmunidad específica está estructurada sobre tres tipos de células: células presentadoras de antigenos(CPAs), Linfocitos T (LT) y Linfocitos B (LB). Los antigenos son capturados por las CPA y procesados para supresentación a los LT recirculantes. Ya en la superficie de la CPA, el antigeno espera el encuentro con un LTportador de un receptor específico del mismo. Al mismo tiempo, los LB reconocen el antigeno libre(conformación nativa sin procesamiento previo) y lo presentan a los LT que ya han reconocido el mismo
antigeno mediante su receptor. Como resultado se genera una cascada de señales que culmina con laproducción de anticuerpos específicos del patógeno y células de memoria. Esta es la forma de presentaciónque tiene lugar ante el primer contacto con un antigeno y constituye la denominada respuesta primaria;mientras que la respuesta secundaria se desencadena ante una segunda exposición al mismo antigeno.Las CPA son un grupo de células especializadas con capacidad para presentar antigenos a los LINFOCITOS T.Existen tres tipos de CPA: MACROFAGOS, LINFOCITOS B y CELULAS DENDRITICAS, de las cuales lasultimas constituyen las CPA mas eficientes en una respuesta primaria al poseer una mayor concentración demoléculas coestimulatorias (B7) sobre su superficie y por lo tanto las unicas capaces de activar LT naive ovirgenes. Sin embargo, las otros dos tipos actúan eficientemente cuando el antigeno es presentado a unlinfocito T de memoria que previamente haya interactuado con el mismo (respuesta secundaria).Que es la presentación de antigenos? ¿Que tipo de interacciones están involucradas? Todo será aclarado a sudebido tiempo, pero para ello debemos primero analizar ciertos conceptos. Las características del macrófagono serán expuestas nuevamente.
Células dendríticasExisten dos tipos de células de morfología dendrítica con funciones muy distintas. Ambosposeen numerosas prolongaciones citoplasmáticas dendríticas finas, a las que deben sunombre. Uno de los tipos son las denominadas células interdigitadas dendríticas (CID) osimplemente células dendríticas. No son células fagocitarias y expresan moléculas MHCclase II en grades cantidades, así como moléculas coestimulatorias B7 (CD80 yCD86). De esta forma están perfectamente preparadas para presentar los antigenos alas células T CD4+. Las CID están ampliamente distribuidas en el organismo. En la pielexisten unas células similares denominadas células de Langerhans. El otro tipo decélulas con morfología dendrítica se encuentra en los centros germinales del bazo y losganglios linfáticos, por lo que se les da el nombre de células foliculares dendríticas(CFD). Poseen receptores para el fragmento Fc de la IgG, por lo que pueden capturar complejos inmunes
(complejos antigeno-anticuerpo).
ANTIGENOS DE HISTOCOMPATIBILIDAD (MHC clase I y II)En los seres humanos, lasmoléculas codificadas porlos genes MHC (complejomayor de histocompatibili-dad se denominan antige-nos HLA (Human-leucocy-te Antigen), debido a quese descubrieron por pri-mera vez en linfocitos. La
función fundamental deestas moléculas que seencuentran en la super-ficie celular, consiste encaptar fragmentos de pép-tidos de proteínas extra-ñas para presentarlos alas células T específicasdel antígeno. Los pro-ductos de los genes MHCse clasifican en tres gru-pos. Los genes de clase I
y II codifican glucoprote-inas de la superficie celu-lar y los genes de clase IIIlo hacen con componentesdel complemento.
Las moléculas MHC se destacan por:
15

8/7/2019 Inmunologia-Apunte El Vikingo
http://slidepdf.com/reader/full/inmunologia-apunte-el-vikingo 16/53
- Su polimorfismo, ya que la secuencia de estos genes (y por ende la secuencia de las proteinascodificadas) difieren entre los individuos de una misma especie.
- Su poligenismo, ya que esta region posee varios genes y moleculas de clase I y II dentro de MHC.
- Su codominancia, ya que al ser individuos diploides tenemos un juego de cromosomas maternos yotros paternos con expresion de ambos para cada uno de los genes del MHC.
La importancia estas tres caracteristicas radica en que disponer de varios genes en cada individuo de lapoblacion que codifican moleculas del MHC con capacidad de unir diferentes peptidos constituye unmecanismo de reaseguro de que al menos algun peptido de cualquier patogeno sera presentadoeficientemente por una molecula de MHC de ese individuo, lo que permitira una excelente activacion de la
respuesta inmune adaptativa y la eliminacion de cualquier agente infeccioso. Este fenomeno se ve potenciadopor el polimorfismo porque aunque existiera un patogeno cuyos peptidos no se unan a las moleculas de claseI o II de un determinado individuo, esta cualidad asegura que en otros individuos eso no ocurrira debido aque el conjunto de moleculas que expresan difiere del conjunto de la persona que es incapaz de presentar lospeptidos del patogeno hipotetico. De esta manera el se asegura la subsistencia de la especie en su luchacontra los distintos agentes microbianos. Por ultimo hay que considerar que las posibilidades de presentardiferentes peptidos derivados de multiples microorganismos aumenta con la codominancia ("dos moleculas –la materna y la paterna – presentan mejor que una”).
- Ser los antigenos diana preferidos en el rechazo de transplantes mediado por células T (ver masadelante).
Por esta razón, y debido a que cada individuo tiene 5 o mas loci codificadores demoléculas MHC, es raro que dos personas posean
conjuntos idénticos de proteínas MHC, lo cual hacemuy difícil el emparejamiento de un donante y unreceptor para el transplante de órganos enhumanos, excepto en el caso de gemelosgenéticamente idénticos.
Los antigenos de clase I se expresan en lasuperficie de todas las células nucleadas y en lasplaquetas. Están codificados por tres lociestrechamente relacionados y designados comoHLA-A, HLA-B y HLA-C. Cada una de estasmoléculas es un heterodímero formado por unacadena polimórfica ALFA, unida mediante un enlace
covalente a un péptido NO polimórfico denominado BETA 2 MICROGLOBULINA,cuya codificación no se hace en el MHC. La región extracelular de la cadenapesada esta dividida en tres dominios: alfa 1, alfa 2 y alfa 3. Los dominios alfa 1 y alfa 2 contienen unahendidura o surco en el que los peptidos se unen a la molécula MHC. Las distintas clases de alelos de clase Ise unen a fragmentos peptídicos diferentes.Los antigenos de clase II están codificados por una región denominada HLA-D, que posee tres subregiones:HLA-DP, HLA-DQ y HLA-DR. Cada molécula de clase II es un heterodímero formado por una cadena alfa yotra beta unidas por enlaces no covalentes. Las dos cadenas son polimórficas y cada una de las tressubregiones HLA-D codifica una o varias cadenas alfa y beta. Las porciones extracelulares de las cadenas alfay beta tienen dos dominios: alfa 1, alfa 2, beta 1 y beta 2. Las moléculas MHC de clase II poseen unahendidura para unirse al antigeno constituida por una interacción de los dominios alfa 1 y beta 1 de las doscadenas y es en esta porción en la que difieren la mayoría de los alelos de clase II. La distribución de lasmoléculas de clase II se limita en gran medida a las CPA.
FUNCIONESMHC clase I MHC Clase
Presentan peptidos de origen endogeno a los LTCD8+Actuan como ligando de receptores de celulas NK
Presentan peptidos de origen exogeno a los LTCD4+
Cada molecula del MHC tiene un solo sitio de union al peptido pero este es degenerado en cuanto a laespecificidad siendo capaz de unir diferentes peptidos siempre y cuando tengan los motivos de unionadecuados, pero solo puede hacerlo de uno a la vez.
PROCESAMIENTO Y PRESENTACION ANTIGENICAYa se ha mencionado que las CPA incluyen a losLB, MACROFAGOS y CELULAS DENDRITICAS.Estos tres tipos celulares poseen receptoresespecíficos que le permiten reconocer antigenosopsonizados, que serán internalizados yprocesados para su posterior presentación alinfocitos T vírgenes. Por otra parte se debe teneren cuenta que ciertos microorganismos como losvirus ingresan a diversas células, que pueden ono ser CPA, mediante la interacción de unreceptor especifico presente en el virión sin lanecesidad de ser opsonizado. El resultado delprocesamiento antigénico es la generación depeptidos que serán presentados por moléculasMHC de clase I o II. Este complejo interaccionarácon el TCR del LT y producirá la primera señal
necesaria para su activación (ver mas adelante).Los peptidos se presentan por moléculas MHC de clase I o II? Para contestar este interrogante debemosanalizar dos vías de procesamiento antigénico denominadas endógena (biosintética) y exógena (endocítica).
16

8/7/2019 Inmunologia-Apunte El Vikingo
http://slidepdf.com/reader/full/inmunologia-apunte-el-vikingo 17/53
- VIA ENDOGENA: mediante esta vía se produce la degradación de proteínas en el citosol celular
mediante el PROTEASOMA (proteasa multicatalítica) que las reconoce al encontrarse estas modificadaspor la unión a UBIQUITINA. Los peptidos así generados deben ser traslocados al RER mediantetransportadores transmembrana dependientes de ATP denominados TAP (transportador asociado a lapresentación antigénica) que forma un heterodímero TAP1/TAP2 presente en la membrana del RER,requiriéndose para el transporte la hidrólisis de ATP. La cadena alfa del MHC I sintetizada se traslada alRER, donde se une temporalmente a la CALNEXINA que se disocia de la misma una vez que la beta 2microglobulina se une a la primera formando el dímero alfa-beta 2 m. Este complejo se asocia a suvez a las proteínas CALRETICULINA Y TAPASINA, que cumplen la función de mantener al heterodímeroparcialmente plegado. Finalmente la TAPASINA del complejo se une a TAP1 y permanece fijada poresta porción al heterodímero TAP1/TAP2 a la espera del ingreso al RER del péptido, que al unirse alMHC de clase I lo estabiliza permitiendo su pasaje a la membrana; siendo este un evento crucial.
Al llegar a la superficie celular, este heterodímero es reconocido por el TCR de los LT CD8+,llevandose a cabo la presentación antigénica.
- VIA EXOGENA: esta vía de procesamiento requiere la endocitosis de antigenos mediada por
receptores presentes en las CPA profesionales (BCR en LB y receptores de manosa, scavenger y defragmentos Fc de Igs en células dendríticas y macrófagos). En lugar de obtenerse a partir de proteínasextrañas sintetizadas en el interior de las células diana, los peptidos presentados a las célulascolaboradoras se obtienen a partir de microorganismos extracelulares o de sus productos, que hansido ingeridos por las CPA y degradados en el entorno acídico de los endosomas. Estos antigenos soninternalizados e ingresan inicialmente a un endosoma que luego deberá fusionarse con los lisosomas
para así degradar las proteínas microbianas y generar los peptidos que posteriormente se unirán aMHC de clase II. Las moléculas MHC de clase II se sintetizan en el citosol y son translocadas al RER.Para evitar que esta molécula se una a un péptido erróneo, la misma se ensambla con la denominadaCADENA INVARIANTE, ocupando esta ultima una parte de la hendidura a la que deben unirse lospeptidos antigénicos. Además cumple la función de dirigirla hasta el endosoma, donde la cadenainvariante es degradada por proteasas, generándose CLIP (péptido invariante asociado a Clase II) quepermanece unido a la molécula MHC de clase II cubriendo la hendidura hasta que otra molécula declase II llamada HLA-DM presente en el endosoma disocia CLIP del MHC de clase II. Mas tarde el péptido se une a esta ultima y el complejo formado es transportado a la superficie celular,donde interactúa con el TCR del LT CD4+.
17

8/7/2019 Inmunologia-Apunte El Vikingo
http://slidepdf.com/reader/full/inmunologia-apunte-el-vikingo 18/53
- VIA DEL CD1: la funcion principal de la molecula CD1 es presentar glucolipidos a Linfocitos T
con TCR alfa/beta, gamma/delta y celulas NK.
Es necesario remarcar que a pesar de que ambas vías se han descrito por separado pareciendo totalmenteindependientes la una de la otra, existe entrecruzamiento entre ellas, por lo que un péptido puede serpresentado tanto por moléculas de clase I como de clase II. Además, muchas de las moléculas MHC de claseI y II de la superficie celular de una célula diana poseen peptidos derivados de las propias proteínas en elsurco de unión (para las moléculas de clase I, fragmentos de proteínas citosólicas y nucleares degradadas; ypara las moléculas de clase II, fragmentos degradados de membrana y proteínas séricas que pasan a travésdel sistema endosoma-lisosoma). Solo una pequeña fracción de las moléculas MHC de clase II de la superficiede una CPA estarán unidas a peptidos. Sin embargo, esto es suficiente como para iniciar una respuestainmunitaria, debido a que solo unos cuantos centenares de dichas moléculas son suficientes para activar unacélula T colaboradora o una T citotóxica.La obligatoriedad de focalizar el reconocimiento T sobre las superficies celulares en cierta forma garantiza laeficacia de su funcion. Seria un desperdicio que un LT helper se active por un antigeno libre, diseminando acualquier parte las citoquinas producidas. Lo mismo se aplica a un T citotoxico. La necesidad de dos clases de
moleculas MHC distintas para una misma funcion (presentar peptidos) pudo haber surgido como unaespecializacion funcional. Las celulas T citotoxicas (CD8) no pueden eliminar y atacar bacterias ni sus toxinas,pero si pueden destruir cualquier celula que lleve epitopes extraños en su superficie presentados pormoleculas MHC clase I. En cambio, el sistema de reconocimiento de peptidos presentados por moleculas declase II que se expresan solo en celulas del sistema inmune, garantiza que los LTCD4+ monten una respuestaactivando macrofagos e induciendo la produccion de anticuerpos por parte de los LB sin afectar otros tejidos.
LINFOCITOS T Y BSon células morfológicamente indistinguibles entre si originadas de un precursor mieloide común en la medulaósea, sin embargo solo los LB maduran en ella. En cambio, los LT salen de la medula ósea como célulasinmaduras y migran al timo, donde completan su maduración. Por ello la medula ósea y el timo se denominanórganos linfáticos primarios, pues de ellos egresan linfocitos con capacidad de reconocer un determinadoantigeno. El resto de los órganos linfáticos de nuestro cuerpo como el bazo, las placas de Peyer en el in-
testino, los ganglios linfáticos, el apéndice, las amígdalas, etc. son llamados órganos linfáticos secundarios yconstituyen los sitios en que se inician las respuestas inmunes adaptativas, dado que es allí donde loslinfocitos son activados por el antigeno. Ambos linfocitos están encargados de llevar a cabo las accionesefectoras de la respuesta inmune adaptativa y son responsables de la presencia de memoria inmunológicafrente a una infección. Por lo tanto son las células encargadas de erradicar un agente que ha podido evadir larespuesta innata y se ha diseminado en el organismo mediante la vía sanguínea; como también de impedir elreingreso o efectuar un clearence mas rápido y efectivo si un agente con el cual ya se ha tenido contactoprevio ingresa al torrente sanguíneo de todas formas.
18

8/7/2019 Inmunologia-Apunte El Vikingo
http://slidepdf.com/reader/full/inmunologia-apunte-el-vikingo 19/53
Los linfocitos son los encargados dereconocer los antigenos circulantes.Los antigenos son sustancias prove-nientes de organismos microbianos,pudiendo pertenecer a la estructurade dicho microorganismo o repre-sentar elementos secretados por losmismos (exotoxinas). Los antigenosson capaces de inducir la activaciónlinfocitaria luego de unirse a losreceptores específicos presentestanto en linfocitos T como en los B.A diferencia de estos, los haptenosNO pueden desencadenar unarespuesta inmunológica por simismos, necesitando estar unidoscovalentemente a una molécula
carrier.Las proteínas constituyen los antigenos de mayor inmunogenicidad, definiéndose esta ultima como unamedida de la capacidad del antigeno para la activación linfocitaria. Sin embargo, los polisacáridos puedentambién actuar como antigenos pero debido a la incapacidad de ser procesados por una CPA, no suelenmediar la activación T (antigenos T independientes). Los linfocitos T necesitan reconocer antigenos
presentados por moléculas de histocompatibilidad (MHC) sobre la superficie de una CPA y al no poderactivarse los LT, no se producirá la colaboración T-B (ver mas adelante).Por el contrario, estas moléculas serán reconocidas por Linfocitos B1 y el resultado será la generación de IgMde baja afinidad y una baja producción de anticuerpos frente a la reinfección (carencia de memoria), eventospuramente dependientes de la activación T.Las zonas de un antigeno que se combinan con la porción Fab de una Ig o con el receptor de los linfocitos(TCR o BCR) reciben el nombre de determinantes antigénicos o epitopos.Se concluye por lo tanto, que el reconocimiento antigénico T es diferente del B. El LB es capaz dereconocer antigenos nativos (presentes en la superficie del microorganismo) y libres (sustancias solublesen plasma como las toxinas bacterianas) sin que medie un procesamiento antigénico previo por lasCPA; mientras que los LT solo pueden reconocer peptidos procesados por estas ultimas.
Linfocitos T Estos linfocitos derivan del TIMO y representan el 60-70% de los linfocitos totales en la sangreperiférica. Se encuentran además en las áreas paracorticales de los ganglios linfáticos y en losmanguitos periarteriolares del bazo. Cada célula T esta programada genéticamente parareconocer a un antigeno especifico unido a la CPA mediante un receptor específico del antigenodenominado TCR. En alrededor del 95% de las células T, el TCR consiste en un heterodímerounido por puentes disulfuro y formado por una cadena polipeptídica alfa y otra beta, cada unade las cuales posee una región variable y otra constante. En una minoría de las células Texistentes en la sangre periférica se encuentra otro tipo de TCR, formado por cadenas polipeptídicas gamma y
delta. Las células con TCR gamma/delta tienden acongregarse en las interfases epiteliales (mucosas). Tantolos TCR alfa/beta como los gamma/delta están unidos aun grupo de cinco cadenas de polipéptidos por un enlacecovalente, formando el llamado complejo molecular CD3.
Las proteínas CD3 no son variables. No se unen alantigeno pero intervienen en la transducción de señaleshacia la célula T, una vez que esta ha reconocido elantigeno. La diversidad de los TCR se genera gracias a losreordenamientos somáticos de los genes que codifican asus cadenas alfa, beta, gamma y delta. Las células Ttambién expresan otras moléculas sobre su superficie. Lasde mayor importancia incluyen CD4, CD8 y CD40 ligando(CD40L). Las dos primeras se expresan en dos subgruposcelulares T excluyentes entre si. Alrededor del 60% de loslinfocitos T expresan CD4 mientras que solo el 30%expresan CD8. Ambas glucoproteinas actúan como
correceptores en la activación de las células T. Durante la presentación antigénica, las moléculas CD4 se unena porciones no polimórficas de las moléculas del complejo de histocompatibilidad de clase II (MHC II)expresadas por las células que presentan el antigeno. Por el contrario, las moléculas CD8 se unen a MHC declase I. Gracias a estas propiedades, las células T colaboradoras (helper) CD4+ solo pueden reconocer a unantigeno en el contexto de los MHC clase II, mientras que las células T citotóxicas CD8+ reconocen a losantigenos unidos a las células solo cuando están asociados a los MHC de clase I. Las células T CD4+ y CD8+llevan a cabo funciones distintas pero algo superpuestas. Puede considerarse que las primeras son unregulador principal ya que a través de la secrecion de ciertas citoquinas las células CD4+ influyen en lafunción de la práctica totalidad de las demás células del sistema inmune, incluidas las otras células T, lascélulas B, los macrófagos y las NK. Existen dos poblaciones funcionalmente distintas de células colaboradorasCD4+. El subgrupo TH1 sintetiza y secreta IL2 e IFN gamma, pero no IL4 ni IL5, mientras que las célulasTH2 producen estos últimos pero no los primeros. Esta distinción es importante porque las citoquinassecretadas por estos dos subgrupos ejercen efectos distintos sobre el resto de las células del sistema inmune.
En general el subgrupo TH1 facilita la hipersensibilidad retardada, la activación de los macrófagos y la síntesisde anticuerpos IgG. Por su parte, el subgrupo TH2 ayuda en la síntesis de otras clases de anticuerpos, entreellos la IgE. Las células T CD8+ pueden secretar citoquinas, sobre todo las del subgrupo TH1, pero ejercensus funciones actuando principalmente como células citotóxicas.
19

8/7/2019 Inmunologia-Apunte El Vikingo
http://slidepdf.com/reader/full/inmunologia-apunte-el-vikingo 20/53
ONTOGENIA T Los precursores de LT provienen de la medula ósea. Con la llegada al timo sufren un proceso de maduraciónque finaliza con la exportación de LT funcionalmente maduros a la sangre periférica y a los órganos linfáticossecundarios. Esta madurez se alcanza con la expresión en la membrana del receptor de células T (TCR) quereconoce antigenos expresados sobre moléculas MHC. Las células T que ingresan al timo (timocitos)provenientes de medula ósea no expresan TCR, ni tampoco las moléculas correceptoras CD4 o CD8 (timocitosdoble negativos). El proceso de maduracion centra primeramente su atencion en la generacion de un receptorfuncional (TCR). El rearreglo de la cadena beta se produce en dos etapas: 1) se reordenan los fragmentos D-Jen ambos cromosomas y 2) se asocia un fragmento V al DJ ya arreglado. Se intenta en un cromosoma y encaso de no lograrse el reordenamiento exitoso se sigue en el segundo (exclusion alelica). El reordenamientoexitoso se visualiza con la expresion en la membrana de la cadena beta, junto a una alfa sustituta y elcomplejo CD3 (PRE-TCR). La expresion en la membrana de este ultimo induce la expresion de las moleculascorreceptoras CD4 y CD8, transformandose en timocitos DOBLE POSITIVOS. Estos expresan el TCR funcionalluego de efectuarse un rearreglo productivo en la cadena alfa, que no sufre exclusion alelica. Las células Treconocen el antigeno en asociación con las propias moléculas MHC pero no asociados con moléculas MHCextrañas, es decir, presenta un restricción MHC , lo que refleja un proceso de selección positiva durante eldesarrollo en el timo, por lo cual las células inmaduras que sean capaces de reconocer peptidos extrañospresentados por moléculas MHC propias son seleccionados para sobrevivir, mientras que los restantes que nopodrían ser utilizados por el organismo, mueren.En la corteza tímica, más del 95% de los timocitos muere, al no ser rescatados de la apoptosis por señalestraducidas a través de su receptor T. Estas señales son generadas como producto del reconocimiento de MHCpropias expresadas en las células epiteliales de la corteza. Por lo tanto, serán seleccionados positivamente
aquellos clones T cuyos receptores puedan reconocer MHC propias. Luego, principalmente a nivel del áreacorticomedular, transcurrirá el proceso de selección negativa. A través del mismo, aquellos clones T capacesde reconocer con alta afinidad oavidez MHC propias presentes en lasuperficie de células dendríticas ymacrófagos, mueren por apoptosis.De este modo se tiende a eliminarclones autorreactivos, garantizandola tolerancia a los tejidos propios.Los clones T así seleccionados ex-presando el TCR funcional y un únicocorreceptor CD8 o CD4, son expor-tados a la circulación. Desde allí po-
drán acceder a los órganos linfáticossecundarios, lugar donde transcurreel proceso de presentación antigé-nica. La selección de un fenotipoCD4 o CD8 es puramente azarosa,ya que si los linfocitos T reconocencon su TCR un péptido presentadopor moléculas MHC de clase II seorienta a la clase CD4, mientras quesi interacciona con un péptido aso-ciado a una molécula MHC de clase Ise diferencia al fenotipo CD8.
Activación T
20
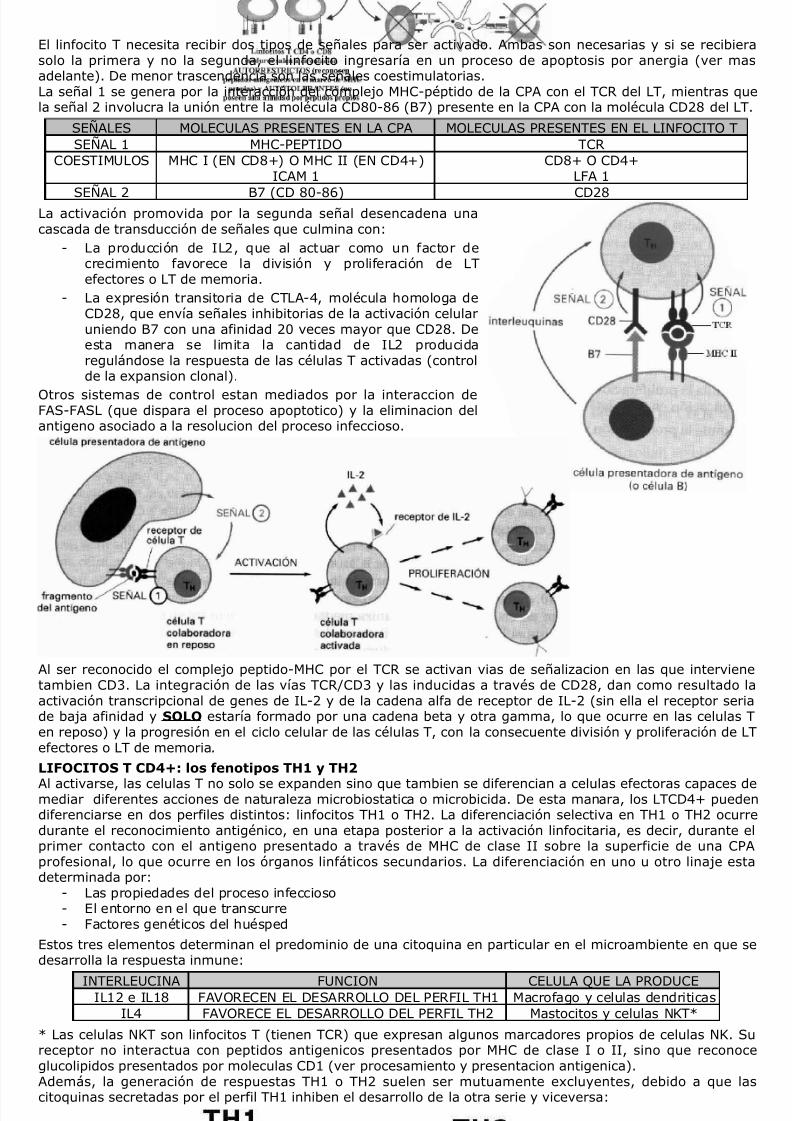
8/7/2019 Inmunologia-Apunte El Vikingo
http://slidepdf.com/reader/full/inmunologia-apunte-el-vikingo 21/53
El linfocito T necesita recibir dos tipos de señales para ser activado. Ambas son necesarias y si se recibierasolo la primera y no la segunda, el linfocito ingresaría en un proceso de apoptosis por anergia (ver masadelante). De menor trascendencia son las señales coestimulatorias.La señal 1 se genera por la interacción del complejo MHC-péptido de la CPA con el TCR del LT, mientras quela señal 2 involucra la unión entre la molécula CD80-86 (B7) presente en la CPA con la molécula CD28 del LT.
SEÑALES MOLECULAS PRESENTES EN LA CPA MOLECULAS PRESENTES EN EL LINFOCITO TSEÑAL 1 MHC-PEPTIDO TCR
COESTIMULOS MHC I (EN CD8+) O MHC II (EN CD4+)ICAM 1
CD8+ O CD4+LFA 1
SEÑAL 2 B7 (CD 80-86) CD28La activación promovida por la segunda señal desencadena unacascada de transducción de señales que culmina con:
- La producción de IL2, que al actuar como un factor decrecimiento favorece la división y proliferación de LTefectores o LT de memoria.
- La expresión transitoria de CTLA-4, molécula homologa deCD28, que envía señales inhibitorias de la activación celularuniendo B7 con una afinidad 20 veces mayor que CD28. Deesta manera se limita la cantidad de IL2 producidaregulándose la respuesta de las células T activadas (controlde la expansion clonal).
Otros sistemas de control estan mediados por la interaccion deFAS-FASL (que dispara el proceso apoptotico) y la eliminacion delantigeno asociado a la resolucion del proceso infeccioso.
Al ser reconocido el complejo peptido-MHC por el TCR se activan vias de señalizacion en las que intervienetambien CD3. La integración de las vías TCR/CD3 y las inducidas a través de CD28, dan como resultado laactivación transcripcional de genes de IL-2 y de la cadena alfa de receptor de IL-2 (sin ella el receptor seriade baja afinidad y SOLO estaría formado por una cadena beta y otra gamma, lo que ocurre en las celulas Ten reposo) y la progresión en el ciclo celular de las células T, con la consecuente división y proliferación de LTefectores o LT de memoria.
LIFOCITOS T CD4+: los fenotipos TH1 y TH2
Al activarse, las celulas T no solo se expanden sino que tambien se diferencian a celulas efectoras capaces demediar diferentes acciones de naturaleza microbiostatica o microbicida. De esta manara, los LTCD4+ puedendiferenciarse en dos perfiles distintos: linfocitos TH1 o TH2. La diferenciación selectiva en TH1 o TH2 ocurredurante el reconocimiento antigénico, en una etapa posterior a la activación linfocitaria, es decir, durante elprimer contacto con el antigeno presentado a través de MHC de clase II sobre la superficie de una CPAprofesional, lo que ocurre en los órganos linfáticos secundarios. La diferenciación en uno u otro linaje estadeterminada por:
- Las propiedades del proceso infeccioso- El entorno en el que transcurre- Factores genéticos del huésped
Estos tres elementos determinan el predominio de una citoquina en particular en el microambiente en que sedesarrolla la respuesta inmune:
INTERLEUCINA FUNCION CELULA QUE LA PRODUCEIL12 e IL18 FAVORECEN EL DESARROLLO DEL PERFIL TH1 Macrofago y celulas dendriticasIL4 FAVORECE EL DESARROLLO DEL PERFIL TH2 Mastocitos y celulas NKT*
* Las celulas NKT son linfocitos T (tienen TCR) que expresan algunos marcadores propios de celulas NK. Sureceptor no interactua con peptidos antigenicos presentados por MHC de clase I o II, sino que reconoceglucolipidos presentados por moleculas CD1 (ver procesamiento y presentacion antigenica).Además, la generación de respuestas TH1 o TH2 suelen ser mutuamente excluyentes, debido a que lascitoquinas secretadas por el perfil TH1 inhiben el desarrollo de la otra serie y viceversa:
21

8/7/2019 Inmunologia-Apunte El Vikingo
http://slidepdf.com/reader/full/inmunologia-apunte-el-vikingo 22/53
Tanto las celulas TH1 como TH2 no requieren la existencia de señales coestimulatorias para activarse en lostejidos perifericos. Solo precisan reconocer el mismo complejo peptido-MHC que reconocieron originalmenteen el organo linfatico secundario.
Debe recalcarse que el patrón de citoquinas de cada perfil es sumamente variado y por ende sus funcionestambién lo son:
De lo anterior puede deducirse que mientras el fenotipo TH1 tiende a originar acciones contra organismos
intracelulares, el TH2 hace lo propio frente a los extracelulares. Ademas, la funcion central de este ultimo escolaborar con los LB en los organos linfaticos secundarios a fin de permitirles montar una produccion efectivade anticuerpos, como se vera al analizar la activacion del LB.
LINFOCITOS T CD8+La activación de los LT CD8+ vírgenes requiere mayor actividad coestimulatoria que los LT CD4+, por lo quese requiere la presencia de estos últimos, los que deben reconocer previamente el antigeno sobre la mismaCPA. La interacción CD40-CD40L induce en la CPA una mayor expresión de moléculascoestimulatorias B7, alcanzándose de esta manera los niveles requeridos para activar a los precursores citotóxicos.
SEÑALES MOLECULAS PRESENTES EN LA CPA MOLECULAS PRESENTES EN EL LT CD8+SEÑAL 1 MHC I-PEPTIDO TCRSEÑAL 2 B7 CD28
COESTIMULATORIAS MHC I CD8No es necesario el encuentro simultáneo de los LT CD4+ y LT CD8+ sobre las CPA. Luego de este proceso, elLT CD8+ abandona el órgano linfático secundario en busca de las células infectadas por el mismo patógenoque dio lugar a su activación, contactando con ella mediante interacciones inespecíficas transitorias queestabilizan la unión, adquiriendo mayor afinidad si el péptido antigénico presentado por MHC I sobre lamembrana de la célula es reconocido por el LT CD8+.
La citotoxicidad se lleva a cabo por un proceso similar al realizado por las células NK (secrecion de perforinas
y granzimas) y además induce la apoptosis celular mediante la interacción FAS-FASL al producirse latrimerizacion del FAS sobre la membrana de la celula blanco y reclutarse proteinas adaptadoras al dominio demuerte de FAS, con la posterior activacion de la CASPASA 8 y 3.Una vez que el LT citotoxico descarga el contenido de sus granulos sobre la membrana de la celula diana, sesepara de esta y puede volver a reconocer y matar otras celulas infectadas. De hecho, el reconocimiento delantigeno a traves del TCR no solo induce la descarga de los granulos, sino que ademas estimula la sintesis deperforina y granzima para la formacion de nuevos granulos. En consecuencia, la citotoxicidad T es muyeficiente ya una sola celula efectora puede inducir la apoptosis de muchas celulas infectadas.Los LTCD8+ efectores no requieren señal 2 para activarse, lo que les permite destruir celulasparenquimatosas infectadas de diferenes tejidos que no suelen presentar moleculas coestimulatorias.
22

8/7/2019 Inmunologia-Apunte El Vikingo
http://slidepdf.com/reader/full/inmunologia-apunte-el-vikingo 23/53
PRODUCCION DE CITOQUINAS POR LTCD8+
LINFOCITOS T REGULADORESLa actividad de los LTH1, LTH2 y LTCD8+ citotoxicos esta regulada por celulas T reguladoras con capacidadde inhibir la expansion clonal T y la produccion de citoquinas por parte de los 3 tipos de celulas efectorasmencionadas mediante la elaboracion de citoquinas inhibitorias (IL10 y TGFbeta) y/o el establecimiento deinteracciones directas (celula-celula) con las mismas.
LINFOCITOS T DE MEMORIASe mantienen en funcion de una proliferacion homeostatica que tiende a reemplazar a las celulas quemueren. Esta renovacion no requeriria que sean nuevamente estimuladas por el antigeno. Existen 2subpoblaciones: 1) LT de memoria efectores (ingresan a los tejidos perifericos) y 2) LT de memoria centrales(ingresan al area T de los organos linfaticos secundarios.
Linfocitos B Los linfocitos B constituyen el 10-20% de la población de linfocitoscirculantes en la sangre peri-férica. Se encuentran también enla medula ósea, en los tejidoslinfoides periféricos como losganglios linfáticos, el bazo o lasamígdalas y en órganos extra-linfáticos como el tracto gastro-
intestinal. En los ganglios linfáticos, ocupan lacorteza superficial, mientras que en el bazo seencuentran en la pulpa blanca. En ambas
localizaciones se congregan formando folículoslinfoides que al activarse desarrollan centrosgerminales.Al recibir una estimulación antigénica, las célulasB se transforman en células plasmáticassecretoras de inmunoglobulinas, que son lassustancias mediadoras de la inmunidad humoral.Las células B reconocen a los antigenos a travésdel complejo receptor del antigeno. Esta es unaIgM (BCR) presente en la superficie de todas ellasque posee una única especificidad antigénica. Además de la IgM de membrana, el complejo posee unheterodímero de proteínas no polimórficas transmembrana: Igalfa e Igbeta. Estas no se unen al antigeno pero
son imprescindibles para la transducción de señales a través del receptor. Los LB también expresan otrasmoléculas esenciales para su función, como receptores para el complemento, para fragmentos Fc de Ig yCD40. Debemos además recordar que a esta Ig se asocian tanto el complejo CD19-CD81 como CR2 (verPOTENCIACION DE LA RESPUESTA B).
23

8/7/2019 Inmunologia-Apunte El Vikingo
http://slidepdf.com/reader/full/inmunologia-apunte-el-vikingo 24/53
Linfocitos B1 y de la zona marginal del bazoSon linfocitos ubicados en la zona marginal del bazo (por lo queresponden en forma eficiente contra patogenos que se hallan ensangre) y cavidades internas (peritoneal y pleural). Reconocenpolisacáridos bacterianos con mayor frecuencia que los linfocitos B2,por lo que se consideran una primer línea de defensa contra lasbacterias. Son capaces de responder rápidamente diferenciándose enplasmocitos aun sin colaboración T. Como el reconocimiento de lospatógenos se realiza por un mecanismo T-independiente, losanticuerpos generados (denominados anticuerpos naturales) poseenbaja afinidad (no desarrollan hipermutación somática) y no presentancambio de isotipo (solo se generará IgM). Además no se producirámemoria inmunológica. Los antigenos T independientes son confrecuencia grandes polímeros (LPS de bacterias capsuladas) condeterminantes antigénicos idénticos repetidos, que se unen a múltiplespuntos de las moléculas de anticuerpos unidos a la membrana de lacélula B actuando como receptores para los antigenos y pudiendogenerar una señal suficientemente intensa como para activardirectamente a las células B.
Linfocitos B2Constituyen mas del 90% de los LB presentes en la sangre y tejidos linfaticos secundarios. Su funcionprincipal es reconocer antigenos proteicos y diferenciarse en celulas productoras de anticuerpos especificos
gracias a la colaboracion que reciben de los LTH2.ONTOGENIA BA semejanza de los linfocitos T, los LB alcanzan la madurez funcional con la expresión en la membrana de losreceptores de reconocimiento antigénico (BCR). En los LB, estos receptores son las inmunoglobulinas desuperficie. Los reordenamientos de las cadenas pesadas y livianas identifican a los diferentes estadios demaduración de los LB. Uno de los primeros eventos lo constituye el reordenamiento de la cadena pesada. Elreordenamiento de un gen D próximo a uno de los genes J (D- J) identifica al estadio temprano pro-Btemprano. Este reordenamiento se produce en uno solo de los cromosomas. Si este no es productivo, seintenta el reordenamiento en el otro cromosoma. Cuando se alcanza un reordenamiento productivo seexpresa una cadena MU en el citoplasma que identifica el estadio PRE-B. Estas células comienzan elreordenamiento de las cadenas livianas. Un cromosoma intenta ordenar la cadena KAPPA, acercando un gen V junto a otro J, realizando varios intentos en ese cromosoma. Si fracasa en generar una cadena liviana
productiva, intenta con el loci KAPPA del otro cromosoma. Si este también falla, recién ahora se intentareordenar los genes LAMBDA (un cromosoma por intento). En otras palabras, un clon que expresa una cadenaLAMBDA, posee ambos loci KAPPA abortivamente reordenados. Estos fenómenos explican la exclusión alélica,por el cual una célula B solo expresa uno de los alelos posibles de las cadenas liviana y pesada (el materno oel paterno) y solo una de las cadenas livianas posibles. Para comprender este tema se recomienda leerprimero la sección GENETICA EN INMUNOLOGIA.Una vez que la cadena liviana es ensamblada con la cadena pesada y expresada en la membrana se haalcanzado el estadio linfocitario B inmaduro. Este es el estadio seleccionado para generar la tolerancia conautoantígenos.
TOLERANCIA CENTRAL:Los LB inmaduros pueden percibir a traves de su BCR
SEÑAL INTENSA SEÑAL LEVE AUSENCIA DE SEÑALConduce a la APOPTOSIS o EDICION (reemplazo
del BCR por uno no reactivo)
Conduce a la ANERGIA
(ausencia de respuesta) Conduce a la MADURACIONLos linfocitos B con IgM en su superficie que sobrevivan a la induccion de tolerancia central, abandonan lamedula ósea y completan su maduración en el bazo, expresando IgM e IgD. En este organo, sufren unproceso de selección negativa si reciben señales provenientes de moleculas propias (induccion de toleranciaperiferica). La población mayoritaria de LB en sangre y tejidos linfáticos secundarios son los denominadosLB2, que abandonan el bazo y comienzan a circular en busca del antigeno hacia el cual expresan un receptorespecífico.
Activación BLa producción de anticuerpos frente a antigenos proteicos requiere que el LB reciba dos tipos de señales. Laprimera a través de la interacción entre BCR y el antigeno; mientras que la segunda es provista por lainteracción con el linfocito T CD4+ helper 2. Esta segunda señal es generada por linfocitos T previamenteactivados por epitopes antigenicos en el mismo antigeno (RECONOCIMIENTO LIGADO). A esta interacción se
la conoce como cooperación T-B. Como los antigenos proteicos requieren de una segunda señal, se losdenomina antigenos T-dependientes. Existe sin embargo (como se mencionó anteriormente), un numero demoléculas capaces de activar LB en ausencia de la colaboración T. A estos se los denomina antigenos T-independientes y son sustancias (LPS bacteriano) que poseen numerosos determinantes antigénicosrepetidos, lo que provoca la activación B por producir un entrelazamiento de las Igs de superficie (CROSS-LINKING).
SEÑALES LINFOCITO B ANTIGENO LINFOCITO TH2SEÑAL 1 BCR EPITOPE ANTIGENICO -SEÑAL 2 CD40 - CD40L
El reconocimiento antigénico a través del BCR media dos procesos diferentes:- Transducción de señales de activación hacia el interior de la célula B- Endocitosis del antigeno con posterior procesamiento y presentación de los peptidos resultantes
asociados a MHC de clase II sobre sus superficie (no se olviden que es una CPA), los que seranreconocidos por LTH2 especificos. El reconocimiento por parte del TCR del peptido antigenicopresentado por la molecula de clase II induce en el LTH2 la expresion de la molecula CD40L quereconoce en la membrana del LB a la proteina CD40.
24

8/7/2019 Inmunologia-Apunte El Vikingo
http://slidepdf.com/reader/full/inmunologia-apunte-el-vikingo 25/53
La activación del LBinduce un incremento delas moléculas coestimu-latorias B7, adquiriéndo-se de esta forma la se-gunda señal de la activa-ción T (la primera se ha-bía originado en el reco-nocimiento por parte delTCR del complejo MHC-péptido ubicado sobre laCPA). La activación Bpermite la proliferación ydiferenciación en célulasplasmáticas productorasde anticuerpos o LB dememoria. Los plasmoci-tos (células productorasde anticuerpos) comien-
zan a proliferar constituyendo el centro germinal, que se origina a partir deunas pocas células B fundadoras específicas. A las 72 horas comienza a
observarse la proliferación, en el centro del folículo, de linfoblastos que reciben el nombre de centroblastos.Estos últimos son células grandes que presentan un citoplasma muy basófilo, carecen de Igs de superficie yse ubican en los espacios que dejan las prolongaciones de las células foliculares dendríticas en el gangliolinfático (zona oscura), donde también seobservan linfocitos T. Una vez diferenciadas acentrocitos, reexpresan las Igs y se localizan enla zona clara basal del centro germinal, donde in-teraccionan con las células foliculares dendríticasy algunos linfocitos T que corresponden al mismoclon que se activó originalmente en la zonaparacortical y continúan aquí su expansión clonalpara colaborar con los LB.Cada clon de LB produce un solo tipo de
anticuerpos y por lo tanto una sola especificidad.El rol del antigeno consiste en seleccionaraquellas células productoras de anticuerposreactivos frente al mismo. Aquellos clones queposeen alta especificidad para el antigeno en suBCR proliferarán, originándose de esta maneraplasmocitos productores de anticuerpos queposeen la misma afinidad que el BCR original.
25

8/7/2019 Inmunologia-Apunte El Vikingo
http://slidepdf.com/reader/full/inmunologia-apunte-el-vikingo 26/53
Los centrocitos están programados para morir en un determinado periodo, a menos que contacten con elantigeno. Se produce entonces una competencia de tipo darwiniana en la cual aquellos centrocitos queexpresen BCR de mayor afinidad podrán contactar con al antigeno y salvarse de morir. La selección de loscentrocitos involucra dos etapas:
- Contacto y endocitosis del antigenos: los centrocitos pueden contactar con el antigeno
soluble o presentado sobre la membrana de las células foliculares dendríticas. En este ultimo caso losanticuerpos generados pueden unirse al antigeno remanente formando complejos inmunes que puedenser captados por las células foliculares dendríticas a través de sus receptores para Fc de IgG y paraC3b y sus productos de degradación. Como la disponibilidad del antigeno en esta etapa esgeneralmente muy baja, serán seleccionados aquellos clones cuyas inmunoglobulinas de superficietengan la mayor afinidad por el antigeno. Este proceso ocurre a los pocos días de la estimulaciónantigénica.
- Migración hacia la parte más externa de la zona clara: donde se concentran las TH2
específicas (que migraron con los linfocitos B activados y sufrieron también una serie detransformaciones celulares). Allí los centrocitos vuelven a interactuar con las células TH2 de maneraparecida al primer contacto. Esta participación de las células T para que continué el proceso deselección sirve para impedir que los centrocitos que mutaron su BCR y puedan reconocerautoantígenos sigan diferenciándose, ya que no van a encontrarse LTH2 específicos para peptidosderivados de autoantígenos.
En el centro germinal se produce también el cambio de isotipo de cadena pesada, que depende de laproducción de citoquinas y de señales desencadenadas por la interacción de las moléculas CD40 y CD40 L.Así, la célula deja de producir IgM y comienza a producir IgG, IgA o IgE. Al dejar el centro germinal, lamayoría de los linfocitos B de memoria han cambiado el isotipo que expresan.
26

8/7/2019 Inmunologia-Apunte El Vikingo
http://slidepdf.com/reader/full/inmunologia-apunte-el-vikingo 27/53
Como ya dijimos,el BCR posee una estructura similar a la de los anticuerpos. El mismo esta conformado por una cadenapesada y otra liviana, las que presentan dominios constantes y variables. Estos últimos proveen a losanticuerpos de la diversidad necesaria para reconocer los diferentes agentes microbianos por medio de dosmecanismos:
- Recombinación génica: es un evento que ocurre previo al contacto anti-génico y se
caracteriza por la asociación de segmentos génicos que codifican para las regiones variables de lascadenas pesadas (V, D y J) y cadenas livianas (V y J) que se rearreglan y ensamblan dando lugar a unaltísimo numero de anticuerpos diferentes; mientras que los rearreglos de las cadenas constantesdeterminan el isotipo particular del anticuerpo (ver más adelante).
- Hipermutación somática: es un evento que opera luego de producido el ensamble de los
genes funcionales para Ig (recombinación génica) y en forma posterior al contacto antigénico. Es unproceso que ocurre en el centro germinal y consiste en introducir mutaciones puntuales en losdominios variables de las cadenas pesada y liviana contribuyendo notablemente al incremento de laafinidad de los anticuerpos producidos a medida que progresa la respuesta inmune humoral. Algunasmutaciones disminuyen la capacidad de reconocimiento y provocan la apoptosis pero otras laincrementan y seran los LB que han sufrido estas mutaciones los que seran seleccionados en el centrogerminal ya que tendran mayor chance de competir con éxito en la captura del antigeno que loscentrocitos que poseen Ig de baja afinidad. La selección esta dada por la señales de supervivenciaimpartidas por LTCD4+.
Los centrocitos que han sobrevivido al proceso de selección dan lugar a dos tipos celulares: 1)
plasmoblastos, que abandonan el centro germinal para completar su diferenciacion a celulas plasmaticasproductoas de inmunoglobulinas de alta afinidad y 2) LB de memoria. Esto depende de varios factores:
DIFERENCIACION A PLASMOBLASTOS DIFERENCIACION A LB MEMORIACentrocitos con receptores de alta afinidad
IL10 producida por LTInteraccion CD40-CD40L
IL4 producida por LT
Una vez que abandonan el centro germinal los LB de memoria recirculan por los tejidos linfaticos secundarioso hacen homing (migracion sitio-especifico) y quedan retenidos transitoriamente. La reexposicion al antigenoinduce una rapida y masiva expansion clonal de los LB de memoria, generandose entre 8 y 10 veces masplasmocitos que en la respuesta primaria. Los LB de memoria expresan BCR de alta afinidad por el antigenocomo consecuencia de su paso por el centro germinal y niveles elevados de moleculas de clase II. Ambascaracteristicas contribuyen a que estos contacten mas rapidamente con LTH2 activados que los LB virgenesde la respuesta primaria y la produccion de anticuerpos sea mas rapida en la respuesta secundaria. Si los
niveles de anticuerpos que persisten en la circulacion son suficientemente altos, el reingreso del antigeno enel organismo no genera una respuesta secundaria ya que los mismos son capaces de neutralizar el antigenofavoreciendo su inmediata depuracion por el sistema fagocitico mononuclear. En cambio, si los niveles deanticuerpos no son suficientes para neutralizar por completo el antigeno, los LB de memoria que expresen
27

8/7/2019 Inmunologia-Apunte El Vikingo
http://slidepdf.com/reader/full/inmunologia-apunte-el-vikingo 28/53
BCR de mayor afinidad son los primeros en ser activados por el antigeno libre nuevamente. Esta activacionpuede dar lugar a una nueva respuesta del centro germinal.
LOS ANTICUERPOS Las moléculas mas sencillas de anticuerpo tienen forma de Y con dos lugares idénticos de unión al antigeno,uno en cada punta de los dos brazos de la Y. Como tienen dos lugares de unión al antigeno, se dice que son
bivalentes. Cuando las moléculas de antigeno tienen treso más determinantes, las moléculas de anticuerposbivalentes pueden establecer enlaces cruzados entremoléculas de antigeno, formando amplias redes que
pueden ser fagocitadas y degradadas rápidamente por losmacrófagos. La eficiencia de la unión del antigeno y laformación de enlaces cruzados se ve altamenteincrementada por la presencia en los anticuerpos de unaregión bisagra flexible, que permite variar la distancia
entre los dos lugares de unión al antigeno. El efecto protector de los anticuerpos nose debe solamente a su capacidad de unión al antigeno, sino que tambiénparticipan en diversas actividades mediadas por su porción Fc de la molécula enforma de Y. Es esta parte la que determina lo que le sucederá al antigeno despuésde unirse al anticuerpo (ver mas adelante).Existe una gran diversidad de inmunoglobulinas altamente específicas frente a un
determinado antigeno. Losanticuerpos sonproteínas y estasúltimas son codificadas porgenes. Por lo tanto, ladiversidad de
anticuerpos plantea unproblema genéticoparticular: ¿Como puede unanimal producir másanti-cuerpos que losgenes que existen en su genoma? El sistema inmunitario de los mamíferos ha desarrollado mecanismosgenéticos únicos que lo capacitan para generar un número casi ilimitado de cadenas livianas y pesadasdiferentes de una forma altamente económica: fusionando segmentos genéticos separados antes deque sean transcritos (ver aspectos genéticos en inmu-nologia). Los extremos ami-noterminales de lascadenas livianas y pesadas son los que se unen entre si formando el lugar de unión del antigeno, y lavariabilidad de sus sec-uencias de aminoácidos cons-tituye la base estructural de la diversidad de estoslugares de unión. La variabilidad de las regiones variables esta restringida en su mayor parte a tres pequeñas
regiones hi-pervariables de cada cadena. Los anticuerpos o Igs, son secretado por las células plas-máticasuna vez que su precursor, el LB, es activado por una antigeno especifico. Están conformados por cuatrocadenas peptídicas: dos pesadas (H) y dos livianas (L). Ambas cadenas pesadas se encuentran unidas entre simediante puentes disulfuro. Además, las cadenas livianas se asocian a las pesadas a través de dichospuentes. Tanto la cadena pesada como la liviana poseen una porción variable (de unión al antigeno) y unaporción constante (Fc).
Diversidad de inmunoglobulinas (Igs): los diferentes isotiposEl analisis de las secuencias de aminoacidos de los dominios constantes de las cadenas pesadas permitiodistinguir en el hombre 5 clases de anticuerpos: IgM, IgG, IgA, IgE e IgD con cadena H mu, gamma, alfa,epsilon y delta respectivamente. Como se mencionará mas adelante, el primer isotipo en aparecer es la IgM.Posteriormente se producirá el cambio de isotipo. El objetivo es presentar una variación en la porción Fc(cadena pesada) de los anticuerpos para que estos sean reconocidos por diferentes tipos celulares encargadosde una respuesta efectiva específica contra un determinado antigeno.Es por esta razón que frente a diversos alergenos se genera el isotipo IgE produciendo eventos quedeterminan la rápida desaparición del mismo (ver HIPERSENSIBILIDAD TIPO I), que en respuesta a unantigeno presente en la circulación sanguínea se elabore IgG activadora del sistema del complemento y de
28

8/7/2019 Inmunologia-Apunte El Vikingo
http://slidepdf.com/reader/full/inmunologia-apunte-el-vikingo 29/53
citotoxicidad dependiente de anticuerpos o que ante el contacto de un organismo con las mucosas se genereIgA que impedirá su fijación a los epitelios. ¿Como distingue el organismo cual es la mejor opción frente a undeterminado agente? Quien sabe…..la inteligencia de este minucioso proceso por momentos desborda nuestracapacidad de comprensión.A continuación se analizarán las principales características de los diferentes isotipos de INMUNOGLOBULINAS.
Los anticuerpos pueden tener cadenas livianas KAPPA o LAMBDA, pero no ambos tiposAdemás de las cinco cadenas pesadas (alfa, gamma, epsilon, delta y mu), los vertebrados superiores tienendos tipos de cadenas ligeras: KAPPA y LAMBDA, que pueden estar asociadas con cualquiera de las cadenaspesadas. Una determinada molécula de anticuerpo contiene siempre cadenas livianas idénticas y cadenas
pesadas idénticas; por consiguiente, sus dos lugares de unión al antigeno siempre son idénticos. Esta simetríaes crucial para la función de formación de enlaces cruzados que tienen los anticuerpos secretados. Porconsiguiente, una molécula de IgG puede tener cadenas ligeras KAPPA o LAMBDA pero no ambas a la vez.
INMUNOGLOBULINA M La IgM, cuya cadena pesada es siempre MU, es la primera clase de anticuerpo que producen las células B endesarrollo, aunque posteriormente muchas células B pasan a producir otras clases de anticuerpo.Inicialmente, el precursor inmediato de una célula B (célula PRE B), solo fabrica cadenas MU, las cuales seasocian con cadenas polipeptídicas no livianas (cadenas sustitutas) y situadas en la membrana plasmática. Encuanto incrementa la síntesis de las cadenas livianas, estas se combinan con las MU, desplazando las cadenassustitutas y formando una molécula de IgM de cuatro cadenas que se inserta en la membrana plasmática.Ahora la célula tiene receptores a los que puede unirse el antigeno en su superficie; en este momento la
célula recibe el nombre de célula B virgen.Muchas células B vírgenes empiezan también aproducir moléculas IgD de superficie, con elmismo lugar de unión al antigeno que lasmoléculas IgM.La IgM no es solo la primera clase de anticuerpoen aparecer en la superficie celular de las célulasB en desarrollo, sino que también es la claseprincipal de Ig secretada a la sangre en lasprimeras fases de una respuesta primaria deanticuerpos. La forma de IgM de secrecion secompone de cinco unidades de 4 cadenas,teniendo así un total de 10 lugares de unión parael antigeno y presentando de esta manera diez
cadenas pesadas MU y diez livianas. Cadapentámero contiene una copia de otra cadenapolipeptídica denominada cadena J. Las célulassecretoras de IgM producen la cadena J y lainsertan covalentemente entre dos regionesadyacente de la porción Fc. Los anticuerpos IgMsuelen presentar baja afinidad por el antigeno, yaque son producidos en forma previa al desarrollode hipermutación somática en las células B. LaIgM se expresa de forma monomérica sobre la
superficie de los LB, actuando como receptor antigénico (BCR).Pese a que existen receptores de la porción Fc de esta Ig sobre la superficie de ciertos leucocitos, los mismosno parecen mediar fenómenos de activación celular. Por lo tanto NO PRODUCEN EL RECLUTAMIENTO DE
ORGANISMOS EFECTORES COMO LA IgG . Por el contrario, son eficientes a la hora de cumplir una funciónneutralizante y posee una notable capacidad para activar la via clasica del complemento.
INMUNOGLOBULINA AEste isotipo se encuentra presente a nivel circulante (forma monomérica) y en secreciones (saliva, lagrimas,secreciones del tracto respiratorio, genitourinario e intestinal) seencuentra en forma dimérica, cumpliendo un papel critico en losmecanismos de defensa de mucosas. Es producida a nivel local porplasmocitos presentes en la lámina propia, debiendo atravesar el epiteliopara ejercer su acción. La IgA no activa la vía clásica del complemento
pero es capaz de formar com-
29

8/7/2019 Inmunologia-Apunte El Vikingo
http://slidepdf.com/reader/full/inmunologia-apunte-el-vikingo 30/53
plejos inmunes y activar la vía alterna. Su función principal reside en su capacidad de prevenir la adhesiónde los microorganismos a las superficies epiteliales que recubren los tractos respiratorio, digestivo ygenitourinario. Por lo tanto actúan sobre superficies comunicadas con el exterior como anticuerponeutralizante.
INMUNOGLOBULINA G Es la Ig presente en mayores concentraciones plasmáticas, encontrándose siempre en forma monomérica.Expresa cuatro diferentes subclases: IgG1, IgG2, IgG3 e IgG4. Todas presentan una cadena pesada GAMMA.Esta inmunoglobulina representa el único isotipo capaz de atravesar la placenta. Presenta ciertas propiedadesque son compartidas con las IgM.
Estas Igs son las encargadas de mediar el reconocimiento antigénico y reclutar mecanismos efectores dediferente naturaleza tendientes a eliminar el agente microbiano y/o sus productos tóxicos.- Función neutralizante: los microorganismo poseen sobre su superficie contrarreceptores de
moléculas presentes en una determinada célula blanco del huésped. Dicha interacción permitiría elingreso del agente a la célula. La función neutralizante de los anticuerpos impediría este proceso alunirse a los contrarreceptores presentes en el patógeno. Un mecanismo similar podría operar en lainmunidad hacia las toxinas producidas por los microorganismos, neutralizando su actividad toxica.- Reclutamiento de mecanismos efectores: al unirse al agente, los anticuerpos actúan como
opsoninas produciendo dos eventos de gran importancia. Por un lado el fragmento Fc activa el sistemadel complemento. Por otro, los receptores leucocitarios reconocen la porción Fc de la Ig permitiendo lainternalización de los microorganismos.
Debe recalcarse que son los anticuerpos integrados en complejos inmunes y no los anticuerpos libres los quedesencadenan los procesos enunciados anteriormente.
INMUNOGLOBULINA DSe presenta en forma monomérica y constituye menos del 1% del total de Igs circulantes. Se expresa juntocon la IgM sobre la superficie de los linfocitos B maduros. A nivel circulante se desconoce su significadofuncional.
INMUNOGLOBULINA E Presenta una estructura monomérica y constituye el isotipo de Ig presente en menor concentraciónplasmática. No muestra capacidad para activar el sistema del complemento. Su función consiste en inducir ladegranulación de los mastocitos, con liberación de sustancias mediadoras de diferentes procesos.La IgE se produce como resultado de un primer contacto con el antigeno. Su gran afinidad por los receptoresFc de esta Ig en los mastocitos, provoca que los mismos se adhieran a estos últimos. Esta unión no producela activación celular, que si se efectúa ante una reexposición al mismo antigeno. En esta ocasión, se produceun entrecruzamiento de la IgE unida a la superficie del mastocito, con la posterior liberación de HISTAMINA yotras sustancias que ya han sido analizadas.Los anticuerpos IgE inducen además mecanismos citotóxicos contra ciertos helmintos (parásitos) mediados através de la interacción de los anticuerpos IgE que recubren el parásito, con los receptores de la porción Fcexpresados predominantemente en la superficie de granulocitos eosinófilos, que al activarse liberan potentesagentes citotóxicos capaces de mediar la destrucción del parásito.
30

8/7/2019 Inmunologia-Apunte El Vikingo
http://slidepdf.com/reader/full/inmunologia-apunte-el-vikingo 31/53
Teoría de la selección clonal El sistema inmune puede responder a millones de antigenos diferentes deuna manera altamente específica debido que durante el desarrollo, cadalinfocito queda destinado a reaccionar con un antigeno concreto antes dehaber sido expuesto a este antigeno. Por lo tanto, un antigeno extrañoestimula selectivamente aquellas células que expresen receptorescomplementarios y específicos del antigeno y que por ende ya estándestinados a responder ante este. La teoría de la selección clonalproporciona una trama conceptual útil para comprender la base celular de la
memoria inmunológica. En un animal adulto, las células T y B de los órganoslinfoides secundarios son una mezcla de células que se hayan por lo menosen tres fases distintas de maduración: células vírgenes o naive, células dememoria y células activadas. Cuando las células vírgenes se encuentran conun antigeno por primera vez, algunas de ellas se estimulan paramultiplicarse y se transforman en células activadas, es decir, en célulasdedicadas activamente a producir una respuesta. Otras células vírgenes seestimulan para multiplicarse y diferenciarse en células de memoria, es decir,en células que no producen respuesta pero que fácilmente se inducen atransformarse en células activadas ante un encuentro posterior con elmismo antigeno. Mientras que las células vírgenes y las células dememoria pueden vivir durante meses o incluso años, las célulasefectoras mueren por apoptosis en unos cuantos días. Según esteesquema, la memoria inmunológica se genera durante la respuestaprimaria, en parte porque la proliferación de la célula virgen desencadenadapor el antigeno genera muchas células de memoria (expansión clonal) y enparte por que las células vírgenes se diferencian en células de memoria queson capaces de responder mas fácilmente al antigeno que una célula virgen.Los antigenos pueden persistir en los órganos linfoides durante largo tiempoconstituyendo una respuesta primaria y se cree que una estimulacióncontinua mediante un antigeno contribuye al mantenimiento de dichamemoria a largo plazo. Tambien se postula que puede mantenerse la produccion de anticuerpos sin mediarreexposicion al antigeno por dos mecanismos no mutuamente excluyentes: 1) que las celulas plasmaticashumanas presenten una vida media extremadamente larga; 2) que se produzca la diferenciacion continua decelulas B de memoria a celulas plasmaticas de vida media larga.
La respuesta inmune primaria y secundaria en la producción de anticuerposLa respuesta inmune primaria se genera ante un primer contacto con el antigeno. Se caracteriza por un lentoincremento de las concentraciones plasmáticas de Igs correspondientes al isotipo IgM en una etapa temprana(4-5 días). Los anticuerpos IgG son detectados más tardíamente.La reexposición de un individuo al mismo antigeno, genera la denominada respuesta inmune secundaria en laque la producción de Igs es más rápida, el isotipo predominante es IgG y existe un incremento en la afinidadhacia el antigeno inductor. Por lo tanto, permite montar una respuesta inmune más veloz y efectiva hacia lospatógenos con los que se ha tenido contacto previo.La respuesta secundaria requiere la presencia de memoria inmunológica establecida durante el primercontacto con el antigeno y refleja la existencia de clones linfocitarios antigeno-específicos previamenteexpandidos. Esta memoria es facilitada por el depósito de antigenos en las CFD, que podrían así desencadenar estímulos permanentes, permitiendo que el reconocimiento T perdure en el tiempo. Nosreferimos a la memoria T debido a que el desarrollo de memoria en las células B es un proceso que requierela colaboración de linfocitos T.
La hipótesis aceptada, es que las células de memoriase originan a partir de células efectorassobrevivientes que después de un encuentro con elantigeno han retornado al estado de reposo,circulando por el organismo listas para responderante una nueva exposición al mismo antigeno que lageneró. Este trafico, que de alguna u otra forma esde características muy permisivas, se realiza graciasa la perdida del patrón de expresión de selectinaspropias de Linfocitos vírgenes. Se debe tener encuenta que al diferenciarse, cada tipo celular
adquiere un nuevo patrón de expresión que lepermite adquirir nuevas propiedades y la capacidad de migrar hacia diferentes destinos (ver tráficolinfocitario).
ORGANOS LINFATICOS El timo y la medula ósea constituyen órganos linfáticoscentrales donde se originan y maduran los linfocitos T(por ser derivados del timo) y B (derivados de lamedula ósea o bone marrow) respectivamente. Unavez que los mismos maduran tanto en la medula óseacomo en el timo, pasan a la sangre donde recirculanpasando a través de los órganos linfáticos periféricos.Ellos constituyen el sitio donde convergen las CPA y los
linfocitos y estos últimos se activan para convertirseen células efectoras o células de memoria. Las CPAtransportan hasta estos órganos la carga de antigenoscapturada en la periferia. Allí residen además, células
31
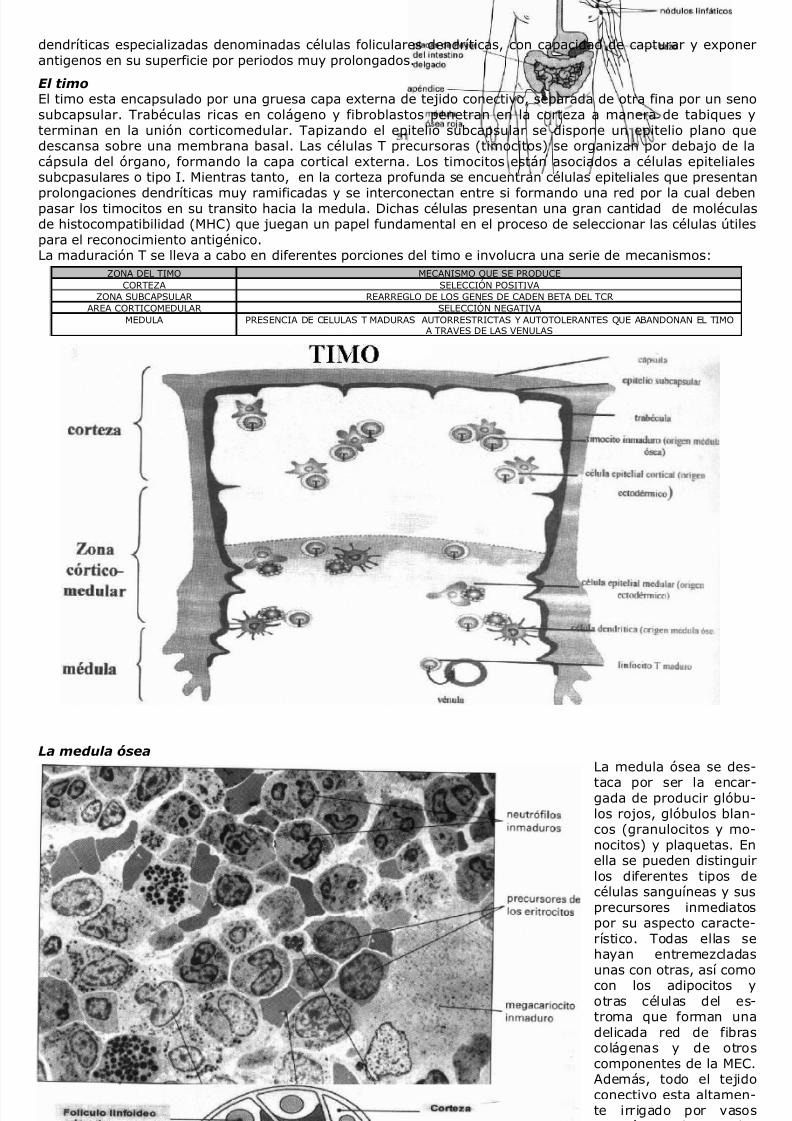
8/7/2019 Inmunologia-Apunte El Vikingo
http://slidepdf.com/reader/full/inmunologia-apunte-el-vikingo 32/53
dendríticas especializadas denominadas células foliculares dendríticas, con capacidad de capturar y exponerantigenos en su superficie por periodos muy prolongados.
El timoEl timo esta encapsulado por una gruesa capa externa de tejido conectivo, separada de otra fina por un senosubcapsular. Trabéculas ricas en colágeno y fibroblastos penetran en la corteza a manera de tabiques yterminan en la unión corticomedular. Tapizando el epitelio subcapsular se dispone un epitelio plano quedescansa sobre una membrana basal. Las células T precursoras (timocitos) se organizan por debajo de lacápsula del órgano, formando la capa cortical externa. Los timocitos están asociados a células epitelialessubcpasulares o tipo I. Mientras tanto, en la corteza profunda se encuentran células epiteliales que presentan
prolongaciones dendríticas muy ramificadas y se interconectan entre si formando una red por la cual debenpasar los timocitos en su transito hacia la medula. Dichas células presentan una gran cantidad de moléculasde histocompatibilidad (MHC) que juegan un papel fundamental en el proceso de seleccionar las células útilespara el reconocimiento antigénico.La maduración T se lleva a cabo en diferentes porciones del timo e involucra una serie de mecanismos:
ZONA DEL TIMO MECANISMO QUE SE PRODUCECORTEZA SELECCIÓN POSITIVA
ZONA SUBCAPSULAR REARREGLO DE LOS GENES DE CADEN BETA DEL TCRAREA CORTICOMEDULAR SELECCIÓN NEGATIVA
MEDULA PRESENCIA DE CELULAS T MADURAS AUTORRESTRICTAS Y AUTOTOLERANTES QUE ABANDONAN EL TIMOA TRAVES DE LAS VENULAS
La medula óseaLa medula ósea se des-
taca por ser la encar-gada de producir glóbu-los rojos, glóbulos blan-cos (granulocitos y mo-nocitos) y plaquetas. Enella se pueden distinguirlos diferentes tipos decélulas sanguíneas y susprecursores inmediatospor su aspecto caracte-rístico. Todas ellas sehayan entremezcladasunas con otras, así como
con los adipocitos yotras células del es-troma que forman unadelicada red de fibrascolágenas y de otroscomponentes de la MEC.Además, todo el tejidoconectivo esta altamen-te irrigado por vasossanguíneos de paredesmuy finas, denominados
sinusoides, en cuyo interior van a
parar las nuevas células.Los ganglios linfáticosLa estructura de un ganglio linfáticoesta constituida por una capa
32

8/7/2019 Inmunologia-Apunte El Vikingo
http://slidepdf.com/reader/full/inmunologia-apunte-el-vikingo 33/53
externa (corteza) y una interna (mé-dula). Los vasos linfáticos son los en-cargados de recoger el fluido extra-celular que llega desde los tejidos al producirse el edema característico de un proceso inflamatorio. Este fluidotransporta elementos celulares desde la periferia tales como CPAs carga-das de antigenos, hasta el sitio deencuentro con el linfocito T en la zona paracortical. La linfa atraviesa el órgano y en su recorrido colecta a loslinfocitos T y B que llegaron por vía sanguínea. Sale por los vasos linfáticos eferentes y a través del con-ductotoráxico vuelve a la sangre.En la corteza se observan estructuras redondeadas denominadas folículos primarios, que son colonizadospor LB vírgenes procedentes de la medula ósea. Los mismos se encuentran rodeados de LT que se disponenen forma difusa, en lo que constituye la zona paracortical o timo-dependiente.Para ingresar al ganglio linfático, los linfocitos que viajan en el torrente circulatorio deben atravesar venulaspostcapilares (HEV) que se encuentran en la corteza difusa, ya que los linfocitos tienen la capacidad específicade adherirse a este endotelio especializado.
El bazoEste órgano posee dos zonas bien delimitadas,denominadas pulpa roja y pulpa blanca. Lasangre, transportando linfocitos y antigenos,llega por la arteria trabecular y la arteriolacentral. Estas vuelcan su contenido en el senomarginal (vénula especializada) que a su vezdrena en la vena trabecular. La pulpa blancaconsiste en un manguito de linfocitos que rodealas pequeñas arteriolas esplénicas, desde su
origen como ramas arteriales hasta su termi-nación. En el corte longitudinal de una arteriolase observan los linfocitos T rodeando a la arte-riola central, que a su vez presentan una coronade linfocitos B en su periferia. Cuando se activanestos últimos, forman los centros germinales queconstituyen la zona del manto.El bazo se diferencia de los otros órganos lin-fáticos secundarios en que tanto la entrada comola salida de los linfocitos se realiza a través de lacirculación sanguínea. La pulpa roja es el sitiodonde se colectan y eliminan de la circulación los
glóbulos rojos en estado senescente. En la pulpablanca se colectan los antigenos provenientes dela sangre. Ambas zonas están separadas por lazona marginal.
INTEGRACION DE LOS PROCESOS: INGRESO DE UN ANTIGENO Y SECUENCIA DE EVENTOS
Para poder entender todos los acontecimientos hasta aquí analizados, es necesario describir un supuesto casoen que un organismo invade al huésped humano. Solo así se aclararán todos los interrogantes que supongodeberán tener al respecto. Pero no desesperen, una vez finalizada la lectura de esta sección les aseguro quecomprenderán la totalidad de los conceptos desarrollados.Pedrito concurre al colegio como todos los días y al llegar al aula advierte que Lucas (su compañerito debanco) esta desganado, fatigado y se siente mal. No quiere jugar y para colmo tiene unas manchas raras enla cara y en los brazos. Pedrito no le da mucha importancia y trata de resolver un problema de matemáticaque le dictó la profesora, cuando de repente Lucas le estornuda en la cara (QUE ASCO!!!).Pobre Pedrito, unos días después el medico le informa a su mama que no puede ir al colegio por padecerSARAMPION . Además en el examen de rutina le descubren Toxocarosis provocada por un parásitoconocido como larva migrante visceral. QUE MALA LECHE!!!! Pero la desventura de este chico nos ayudaráa describir los eventos que se suceden a continuación.El virus del SARAMPION ingresa por vía aérea y necesita adherirse a las células del epitelio respiratorio. Elmucus propio de dicha vía intenta impedir en un primer momento la interacción virus-célula. Las partículasvirales que toman contacto con el epitelio respiratorio son capturadas por las secreciones y transportadashacia la laringe, para luego ser deglutidas y destruidas por los mecanismos efectores del tractogastrointestinal. Si estos mecanismos no evitan la colonización del tracto respiratorio por el virus, esteingresa en las células para posteriormente replicar y diseminarse por vía hematógena.
33

8/7/2019 Inmunologia-Apunte El Vikingo
http://slidepdf.com/reader/full/inmunologia-apunte-el-vikingo 34/53
Dependiendo de cual fue la vía deentrada al organismo, los antigenos sonatrapados por diferentes órganoslinfáticos secundarios. Antigenos queingresan al espacio intercelular (piel) decualquier tejido, son barridos por losvasos linfáticos hacia el ganglio regional,mientras que los que ingresan por víarespiratoria o digestiva son atrapadospor los ganglios regionales, amígdalas,adenoides, placas de Peyer y apéndice.Los que hacen su ingreso o se diseminanpor vía sanguínea, como el caso delSARAMPION, son recogidos pormacrófagos del bazo, hígado o pulmón.Sin embargo, solo el bazo, con unaarquitectura que permite un íntimocontacto de las CPA con las regiones T yB, es capaz de generar una respuestainmune contra antigenos que seencuentran en la circulación. Así es comolos antigenos son secuestrados por
diferentes CPA que los internalizan,procesan y posteriormente presentansobre su superficie diversos peptidospudiendo además activarse y producirmediadores de la inflamación. Esteproceso facilitará la extravasación deleucocitos hacia la zona de ingreso delpatógeno por el efecto vasodilatador y elaumento de la permeabilidad vascular,contribuyendo al encuentro con elmismo.
Los linfocitos T y B vírgenes (naive) recirculan continuamente entre la sangre los órganos linfáticossecundarios y la linfa, para retornar a la circulación sanguínea a través del conducto toráxico. Este circuitocontinúa en la medida en que el linfocito no se encuentra con su antigeno y eventualmente muere por
apoptosis. En cambio, si los linfocitos reconocen a su antigeno, son secuestrados en los órganos linfáticossecundarios donde se diferencian a efectores capaces de combatir la infección.El TCR solo puede reconocer epitopes de macromoléculas que fueron procesadas y luego presentadas al TCRen la superficie de CPAs. En cambio, el BCR reconoce el epitope en su conformación nativa (no requiereprocesamiento). Si bien existen ciertos antigenos que pueden desencadenar una respuesta humoral sincolaboración de las células T (antigenos T-independientes), la producción de anticuerpos frente a la mayoríade los antigenos microbianos (proteínas), requiere de la colaboración T.
34

8/7/2019 Inmunologia-Apunte El Vikingo
http://slidepdf.com/reader/full/inmunologia-apunte-el-vikingo 35/53
Para que el encuentro entre los linfocitos y su antigeno sea exitoso, debe estar garantizada la llegada de losantigenos a los órganos linfáticos secundarios. El mecanismo mas eficiente e importante para desencadenar
una respuesta primaria esque el antigeno sea trans-formado por CPAs profe-sionales que se ubican enlas áreas paracorticales T,donde presentan péptidosantigénicos asociados amoléculas MHC a los linfo-citos T que se extravasanen esos órganos a travésde HEV. A su vez, en loslinfáticos se expresan cier-tas moléculas de adhesióny se producen quemoqui-nas que son reconocidaspor moléculas presentesen el linfocito vírgen.De esta manera, ciertasquemoquinas producidaspor el ganglio linfático fa-
cilitan el encuentro transi-torio entre el linfocito T y la CPA (monitoreo) permitiendo detectar la presencia de su antigeno especifico.Si el LT no encuentra su antigeno especifico, se separa de la CPA y abandona el ganglio por el linfáticoeferente y luego a través del conducto toráxico pasa a la circulación sanguínea, para continuarextravasándose en otros órganos linfáticos secundarios y eventualmente si nunca los encuentra, morir porapoptosis. En las pocas ocasiones en que el linfocito encuentra su antigeno sobre la superficie de la CPA, elreconocimiento antigénico induce un cambio conformacional en la molécula LFA1, que aumentamarcadamente la afinidad por sus ligandos ICAM, estabilizándose la unión entre ambas células. Estaasociación puede persistir por muchos días mientras la célula T se activa, prolifera y su progenie se diferenciaa células efectoras. Las mismas luego se despegan y excepto aquellas células TH2 que estarán disponiblespara proveer a los linfocitos B la señal coestimulatoria necesaria para su activación (cooperación T-B), el restode los linfocitos T efectores abandona el ganglio por el linfático eferente y luego a través del conductotoráxico, pasando a la circulación sanguínea para ser distribuidas al tejido donde se ha instalado la infección.
Mas tarde, los linfocitos B vírgenes son atraídos hacia los folículos linfoides por diversas quemoquinasproducidas en esta localización. Si en su trayecto hacia los mismos, no contactan con su antigeno especifico y
no encuentran a linfocitos TH2que hayan sido activados porpeptidos derivados del antigenopara el cual son específicas,abandonan el ganglio a través dellinfático eferente para retornar ala sangre a través del conductotoráxico. Si en cambio contactancon su antigeno pero no recibenla señal coestimulatoria aportada
por la célula TH2 especifica acti-vada por el mismo antigeno,morirán dentro de las siguientes24 horas. Si en su trayecto hacialos folículos linfoides, contactancon su antigeno específico y ade-más reciben la señal coestimu-latoria aportada por el LTH2 acti-vado por peptidos derivados delantigeno para el cual son especi-ficas, se activan y proliferan dife-renciándose hacia células produc-toras de anticuerpos (plasmoci-
tos) y células B de memoria. Es-tos acontecimientos tienen lugaren la interfase entre los folículosy el área T (borde del folículo primario). Allí las células B co-mienzan a proliferar generandoun foco primario (expansiónclonal). Luego, algunas de estas
células B migran a los cordones medulares y continúan dividiéndose por 2 o 3 días hasta que se diferencianen células plasmáticas productoras de los anticuerpos mas tempranos en respuesta al antigeno, hasta quemueren por apoptosis. Otras células B del foco migran al centro del folículo para establecer un centrogerminal. Los centros germinales son sitios de intensa proliferación, generándose centroblastos, sobre los queopera la hipermutación somática. Estos posteriormente se diferenciarán a centrocitos que sufrirán switch deisotipo y eventualmente se diferenciarán a plasmoblastos o a células B de memoria. Los plasmoblastosabandonan el centro germinal y vía linfático eferente y conducto toráxico pasan a la circulación que los
35

8/7/2019 Inmunologia-Apunte El Vikingo
http://slidepdf.com/reader/full/inmunologia-apunte-el-vikingo 36/53
transporta a la medula ósea. Allí se diferencian a plasmocitos que persisten secretando anticuerpos poraproximadamente 4 semanas.Por otra parte, Toxocara canis (larva migrante visceral) es un parásito (gusano) que actúa como unmicroorganismo extracelular, por lo tanto se generará frente al mismo una respuesta tendiente a laproducción de anticuerpos (preferentemente IgE). Estos últimos podrán eliminar al patógeno por alguno delos siguientes mecanismos:
- Opsonización: mediante la cual el anticuerpo recubre el patógeno y facilita que este sea
ingerido y destruido por células fagocíticas, ya que las mismas cuentan con receptores que unen elfragmento Fc de las inmunoglobulinas.
- Neutralización: el anticuerpo se une al patógeno, bloqueando sus adhesinas o las toxinas
producidas por el mismo, neutralizando su sitio activo.- Activación del sistema del complemento: a través del fragmento Fc de los anticuerpos IgG
o IgM.
Los complejos inmunes contribuyen a la expansión clonal de los LB por períodos prolongados de tiempo,manteniéndose los LB de memoria. Este clon de memoria expandido responderá ante un segundo contacto(reexposición) con el mismo antígeno, en lo que constituye la respuesta secundaria.La inmunidad mediada por células genera tres tipos diferentes de células T efectoras:- Células CD4+ TH1, que presentan capacidad para activar a los macrófagos y por lo tanto
juegan un papel fundamental en la destrucción de patógenos intracelulares.- Células CD4+ TH2, que colaboran con los LB a fin de producir una adecuada respuesta
humoral .
- Células TCD8+ citotóxicas las cuales median la destrucción de células infectadas por virus.
En el caso del sarampión, se genera inmunidad de por vida, por lo que NO existe reinfección y los anticuerposproducidos actúan por un mecanismo de neutralización. Además, en un principio la respuesta innata esmediada por células NK que lisan las células infectadas prolongándose esta situación durante el desarrollo dela respuesta adaptativa, mediada esta última por linfocitos TCD8+ citotóxicos.
MECANISMOS DE LESION INMUNITARIA: REACCIONES DE HIPERSENSIBILIDAD
36

8/7/2019 Inmunologia-Apunte El Vikingo
http://slidepdf.com/reader/full/inmunologia-apunte-el-vikingo 37/53
El ser humano habita un entorno cargado de sustancias capaces de provocar respuestas inmunitarias. Elcontacto con estos agentes no solo induce una respuesta inmunitaria protectora, sino también reacciones que
pueden ser nocivas para los tejidos. Los antigenos exógenos se encuentran en el polvo, los pólenes, losalimentos, los fármacos, los agentes microbianos, los productos químicos y muchos derivados sanguíneosutilizados en la práctica clínica. Las respuestas inmunitarias provocadas por estos antigenos exógenosadoptan diversas formas, que van desde las molestias triviales, como el prurito cutáneo, a enfermedadespotencialmente mortales como el asma bronquial. Estas respuestas reciben en conjunto el nombre dereacciones de hipersensibilidad y pueden ponerse en marcha por la interacción del antigeno con losanticuerpos humorales, o bien por mecanismos inmunitarios de tipo celular. No solo los antigenos exógenospueden desencadenar reacciones inmunitarias nocivas para los tejidos, sino que también pueden hacerlo losantigenos intrínsecos del organismo. Algunas reacciones inmunitarias contra el propio organismo sondesencadenadas por antigenos homólogos distintos en las personas con carga genética diferente. Lasreacciones transfusionales y el rechazo de transplantes son ejemplos de trastornos causados por antigenoshomólogos. Otro tipo lo constituyen las enfermedades autoinmunes.
HIPERSENSIBILIDAD TIPO I Es una reacción inmunológica de desarrollo rápido, que ocurre poco minutos después de la combinación deuna antigeno con un anticuerpo unido a mastocitos o a basofilos en personas previamente sensibilizadas alantigeno en cuestión. Puede manifestarse como una respuesta generalizada o como una reacción local. Laenfermedad generalizada suele producirse por la inyección intravenosa de un antigeno y provoca prurito,contracción de los bronquios, edema laringeo con obstrucción respiratoria, vómitos, diarrea y shock quepuede llevar a la muerte. Las reacciones locales dependen de la puerta de entrada del alergeno (sustanciasgeneralmente inocuas que son capaces de inducir la producción de cantidades importantes de IgE en ciertosindividuos) y adoptan la forma de tumefacciones cutáneas localizadas (ronchas), exudado nasal y conjuntival(rinitis y conjuntivitis alérgica), asma bronquial o gastroenteritis alérgica. Muchas reacciones locales dehipersensibilidad tipo I tienen dos fases bien definidas: la respuesta inicial, caracterizada por vasodilatación,salida de liquido de los vasos y, dependiendo de la localización, espasmo del músculo liso bronquial osecreciones glandulares. En muchos casos tiene lugar una segunda fase tardía, entre 2 y 8 horas después,sin nueva exposición al antigeno. Esta reacción persiste por varios días y se caracteriza por una infiltraciónmas intensa del tejido por eosinófilos, neutrófilos, basofilos, monocitos y células T CD4+, así como pordestrucción hística de las células epiteliales de las mucosas.
37
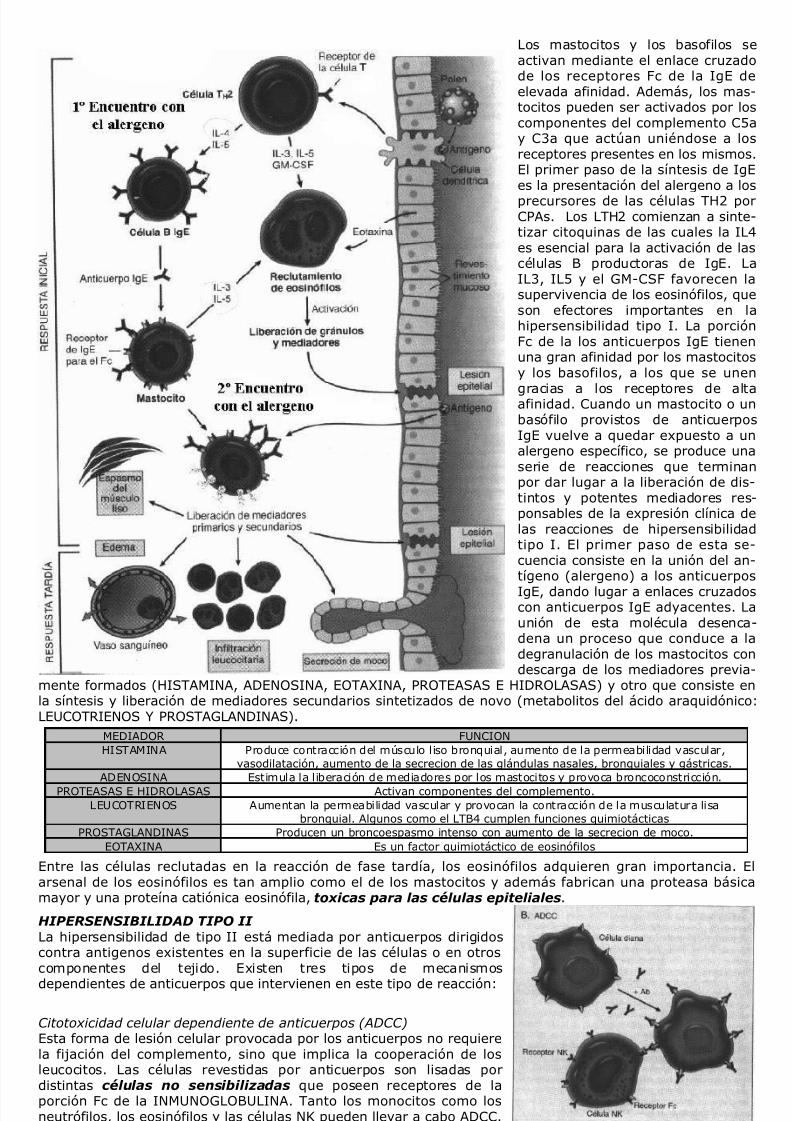
8/7/2019 Inmunologia-Apunte El Vikingo
http://slidepdf.com/reader/full/inmunologia-apunte-el-vikingo 38/53
Los mastocitos y los basofilos seactivan mediante el enlace cruzadode los receptores Fc de la IgE deelevada afinidad. Además, los mas-tocitos pueden ser activados por loscomponentes del complemento C5ay C3a que actúan uniéndose a losreceptores presentes en los mismos.El primer paso de la síntesis de IgEes la presentación del alergeno a losprecursores de las células TH2 porCPAs. Los LTH2 comienzan a sinte-tizar citoquinas de las cuales la IL4es esencial para la activación de lascélulas B productoras de IgE. LaIL3, IL5 y el GM-CSF favorecen lasupervivencia de los eosinófilos, queson efectores importantes en lahipersensibilidad tipo I. La porciónFc de la los anticuerpos IgE tienenuna gran afinidad por los mastocitosy los basofilos, a los que se unen
gracias a los receptores de altaafinidad. Cuando un mastocito o unbasófilo provistos de anticuerposIgE vuelve a quedar expuesto a unalergeno específico, se produce unaserie de reacciones que terminanpor dar lugar a la liberación de dis-tintos y potentes mediadores res-ponsables de la expresión clínica delas reacciones de hipersensibilidadtipo I. El primer paso de esta se-cuencia consiste en la unión del an-tígeno (alergeno) a los anticuerpos
IgE, dando lugar a enlaces cruzadoscon anticuerpos IgE adyacentes. Launión de esta molécula desenca-dena un proceso que conduce a ladegranulación de los mastocitos condescarga de los mediadores previa-
mente formados (HISTAMINA, ADENOSINA, EOTAXINA, PROTEASAS E HIDROLASAS) y otro que consiste enla síntesis y liberación de mediadores secundarios sintetizados de novo (metabolitos del ácido araquidónico:LEUCOTRIENOS Y PROSTAGLANDINAS).
MEDIADOR FUNCIONHISTAMINA Produce contracción del músculo liso bronquial, aumento de la permeabilidad vascular,
vasodilatación, aumento de la secrecion de las glándulas nasales, bronquiales y gástricas.ADENOSINA Estimula la liberación de mediadores por los mastocitos y provoca broncoconstricción.
PROTEASAS E HIDROLASAS Activan componentes del complemento.LEUCOTRIENOS Aumentan la permeabilidad vascular y provocan la contracción de la musculatura lisa
bronquial. Algunos como el LTB4 cumplen funciones quimiotácticasPROSTAGLANDINAS Producen un broncoespasmo intenso con aumento de la secrecion de moco.
EOTAXINA Es un factor quimiotáctico de eosinófilos
Entre las células reclutadas en la reacción de fase tardía, los eosinófilos adquieren gran importancia. Elarsenal de los eosinófilos es tan amplio como el de los mastocitos y además fabrican una proteasa básicamayor y una proteína catiónica eosinófila, toxicas para las células epiteliales.
HIPERSENSIBILIDAD TIPO II La hipersensibilidad de tipo II está mediada por anticuerpos dirigidoscontra antigenos existentes en la superficie de las células o en otroscomponentes del tejido. Existen tres tipos de mecanismos
dependientes de anticuerpos que intervienen en este tipo de reacción:
Citotoxicidad celular dependiente de anticuerpos (ADCC)Esta forma de lesión celular provocada por los anticuerpos no requierela fijación del complemento, sino que implica la cooperación de losleucocitos. Las células revestidas por anticuerpos son lisadas pordistintas células no sensibilizadas que poseen receptores de laporción Fc de la INMUNOGLOBULINA. Tanto los monocitos como losneutrófilos, los eosinófilos y las células NK pueden llevar a cabo ADCC.Intervienen en este proceso tanto la IgG como la IgE.
Reacciones dependientes del complemento
38

8/7/2019 Inmunologia-Apunte El Vikingo
http://slidepdf.com/reader/full/inmunologia-apunte-el-vikingo 39/53
Existen dos mecanismos por los que el anticuerpo y el complemento pueden dar lugar a una hipersensibilidadde tipo II: la lisis directa y la opsonización. En el primer caso, el anticuerpo IgG o IgM reacciona con unantigeno de la superficie celular causando la activación del complemento y provocando la formación del CLAM.
En el segundo, lafijación del anti-cuerpo o del frag-mento C3b sobre lasuperficie de lascélulas (opsoniza-ción) las convierteen susceptibles a lafagocitosis. Clínica-mente, estas situa-ciones se producenen las reaccionestransfusionales, enlas que las célulasde un donante in-compatible reaccio-nan con los anti-cuerpos autóctonos
del receptor; y en la eritroblastocis fetal, en la que existe una incompatibilidad Rh entre la madre y el feto y
un pasaje de IgG anti-eritrocitos fetales por la placenta, causando la destrucción de los mismos.
Disfunción celular mediada por anticuerposEn algunos casos, los anticuerpos dirigidos contra losreceptores de la superficie celular dificultan o alteran laregulación funcional sin provocar lesión celular niinflamación. Por ejemplo, en la MIASTENIA GRAVIS, losanticuerpos reaccionan con los receptores de acetilcolina delas placas motoras terminales de los músculos esqueléticos,alterando la transmisión neuromuscular con la consiguientedebilidad de los músculos. En la enfermedad de GRAVES lasituación es inversa, existiendo una estimulación de la
función celular, ya que los anticuerpos dirigidos contra elreceptor de TSH presentes en las células tiroideas provocanel desarrollo de hipertiroidismo.
HIPERSENSIBILIDAD DE TIPO III (MEDIADA POR INMUNOCOMPLEJOS)
Lasreacciones de hipersensibilidad de tipo III se deben a complejos antigeno-anticuerpo que lesionan los tejidosa causa de su capacidad para activar el sistema del complemento. La reacción toxica se inicia cuando el
39
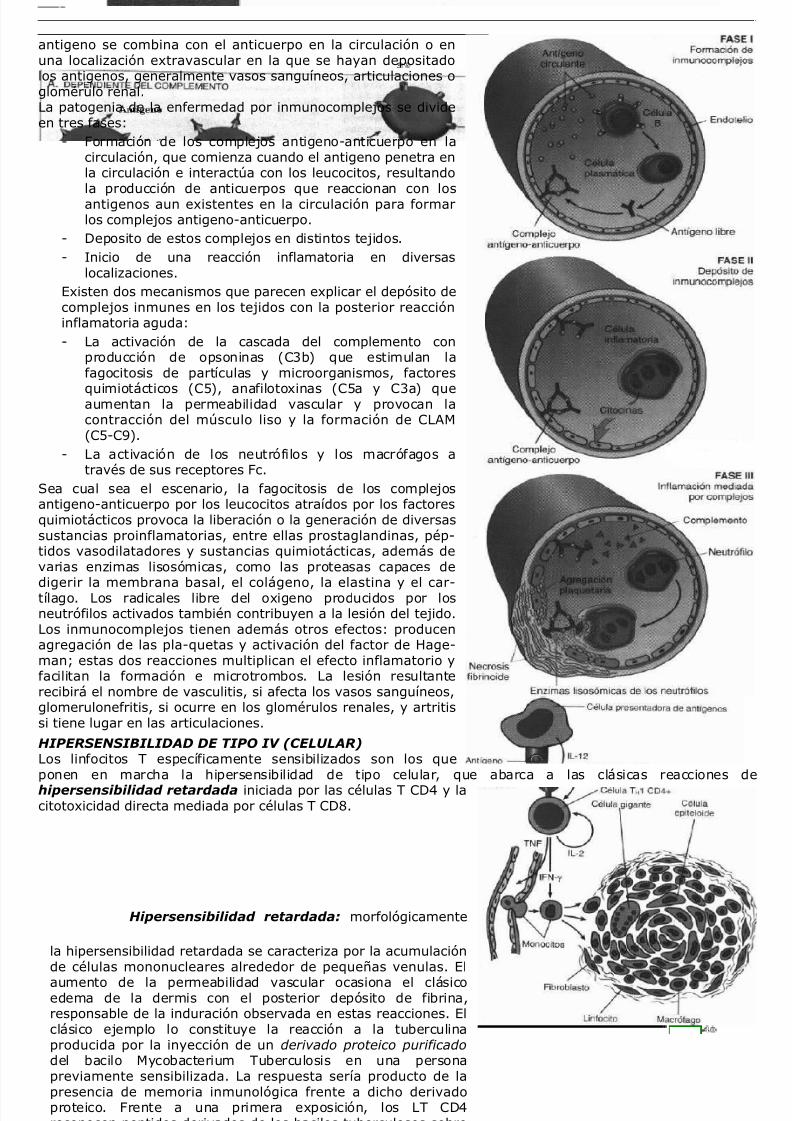
8/7/2019 Inmunologia-Apunte El Vikingo
http://slidepdf.com/reader/full/inmunologia-apunte-el-vikingo 40/53
antigeno se combina con el anticuerpo en la circulación o enuna localización extravascular en la que se hayan depositadolos antigenos, generalmente vasos sanguíneos, articulaciones oglomérulo renal.La patogenia de la enfermedad por inmunocomplejos se divideen tres fases:
- Formación de los complejos antigeno-anticuerpo en lacirculación, que comienza cuando el antigeno penetra enla circulación e interactúa con los leucocitos, resultandola producción de anticuerpos que reaccionan con losantigenos aun existentes en la circulación para formarlos complejos antigeno-anticuerpo.
- Deposito de estos complejos en distintos tejidos.- Inicio de una reacción inflamatoria en diversas
localizaciones.Existen dos mecanismos que parecen explicar el depósito decomplejos inmunes en los tejidos con la posterior reaccióninflamatoria aguda:- La activación de la cascada del complemento con
producción de opsoninas (C3b) que estimulan lafagocitosis de partículas y microorganismos, factores
quimiotácticos (C5), anafilotoxinas (C5a y C3a) queaumentan la permeabilidad vascular y provocan lacontracción del músculo liso y la formación de CLAM(C5-C9).
- La activación de los neutrófilos y los macrófagos através de sus receptores Fc.
Sea cual sea el escenario, la fagocitosis de los complejosantigeno-anticuerpo por los leucocitos atraídos por los factoresquimiotácticos provoca la liberación o la generación de diversassustancias proinflamatorias, entre ellas prostaglandinas, pép-tidos vasodilatadores y sustancias quimiotácticas, además devarias enzimas lisosómicas, como las proteasas capaces de
digerir la membrana basal, el colágeno, la elastina y el car-tílago. Los radicales libre del oxigeno producidos por losneutrófilos activados también contribuyen a la lesión del tejido.Los inmunocomplejos tienen además otros efectos: producenagregación de las pla-quetas y activación del factor de Hage-man; estas dos reacciones multiplican el efecto inflamatorio yfacilitan la formación e microtrombos. La lesión resultanterecibirá el nombre de vasculitis, si afecta los vasos sanguíneos,glomerulonefritis, si ocurre en los glomérulos renales, y artritissi tiene lugar en las articulaciones.
HIPERSENSIBILIDAD DE TIPO IV (CELULAR)Los linfocitos T específicamente sensibilizados son los queponen en marcha la hipersensibilidad de tipo celular, que abarca a las clásicas reacciones dehipersensibilidad retardada iniciada por las células T CD4 y lacitotoxicidad directa mediada por células T CD8.- Hipersensibilidad retardada: morfológicamente
la hipersensibilidad retardada se caracteriza por la acumulaciónde células mononucleares alrededor de pequeñas venulas. El
aumento de la permeabilidad vascular ocasiona el clásicoedema de la dermis con el posterior depósito de fibrina,responsable de la induración observada en estas reacciones. Elclásico ejemplo lo constituye la reacción a la tuberculinaproducida por la inyección de un derivado proteico purificadodel bacilo Mycobacterium Tuberculosis en una personapreviamente sensibilizada. La respuesta sería producto de lapresencia de memoria inmunológica frente a dicho derivadoproteico. Frente a una primera exposición, los LT CD4reconocen peptidos derivados de los bacilos tuberculosos sobrela superficie de CPAs asociados a moléculas MHC de clase II. Este primer encuentro estimula ladiferenciación de las células T CD4 a un perfil TH1. Cuando se administra la inyección intracutánea detuberculina a una persona que estuvo antes expuesta al bacilo tuberculoso, las células TH1 de memoriaentran en contacto con el antigeno existente sobre la superficie de CPAs y se activan. Estos cambios vanacompañados por la secrecion de citoquinas del tipo TH1, responsables de la expresión de lahipersensibilidad retardada. Con determinados antigenos persistentes o no degradables, como es el caso del
40

8/7/2019 Inmunologia-Apunte El Vikingo
http://slidepdf.com/reader/full/inmunologia-apunte-el-vikingo 41/53
bacilo tuberculoso, el infiltrado linfocitario perivascular inicial esta constituido por macrófagos, los quesuelen sufrir una transformación morfológica hacia células de aspecto epitelial denominadas célulasepitelioides. El granuloma (ver figura) es una estructura conformada por un agregado de célulasepitelioides, generalmente rodeadas de una corona de células leucocitarias (inflamación granulomatosa).
- Citotoxicidad mediada por células T: en esta variante de hipersensibilidad de tipo IV, las
células T CD8 sensibilizadas destruyen las células diana productoras de antigeno. A estas células efectorasse las denomina linfocitos T citotóxicos (LTC). Los LTC dirigidos contra los antigenos de histocompatibilidadde la superficie celular, desempeñan un papel importante en el rechazo de transplantes. Además intervienenen la resistencia frente a infecciones virales. En las células infectadas por virus, los peptidos virales seasocian a las MHC de clase I y el complejo así formado es transportado a la superficie para ser reconocidopor el TCR de los linfocitos T CD8+. La lisis de las células infectadas antes de que se complete la replicaciónviral, permite eliminar la infección.
RECHAZO DE TRANSPLANTES El rechazo de un transplante depende del reconocimiento que el huésped hace con respecto al tejido extrañotransplantado. En el hombre, los antigenos responsables del rechazo son los del HLA (MHC).En el rechazo participan tanto la activación de los LT CD8+ como las reacciones de hipersensibilidad retardadadesencadenadas por las células T helper CD4+. Se han propuesto dos vías distintas por las cuales se generaesta reacción de rechazo:- Vía directa: las células T del receptor del transplante reconocen las moléculas MHC sobre la
superficie de una CPA del donante. Se cree que los imunógenos más importantes son las célulasdendríticas del donante, ya que no solo expresan grandes cantidades de moléculas MHC de clase I y II,sino que también están dotadas de moléculas coestimulatorias como B7. Una vez activados, los LTCmaduros lisan al tejido injertado.
41
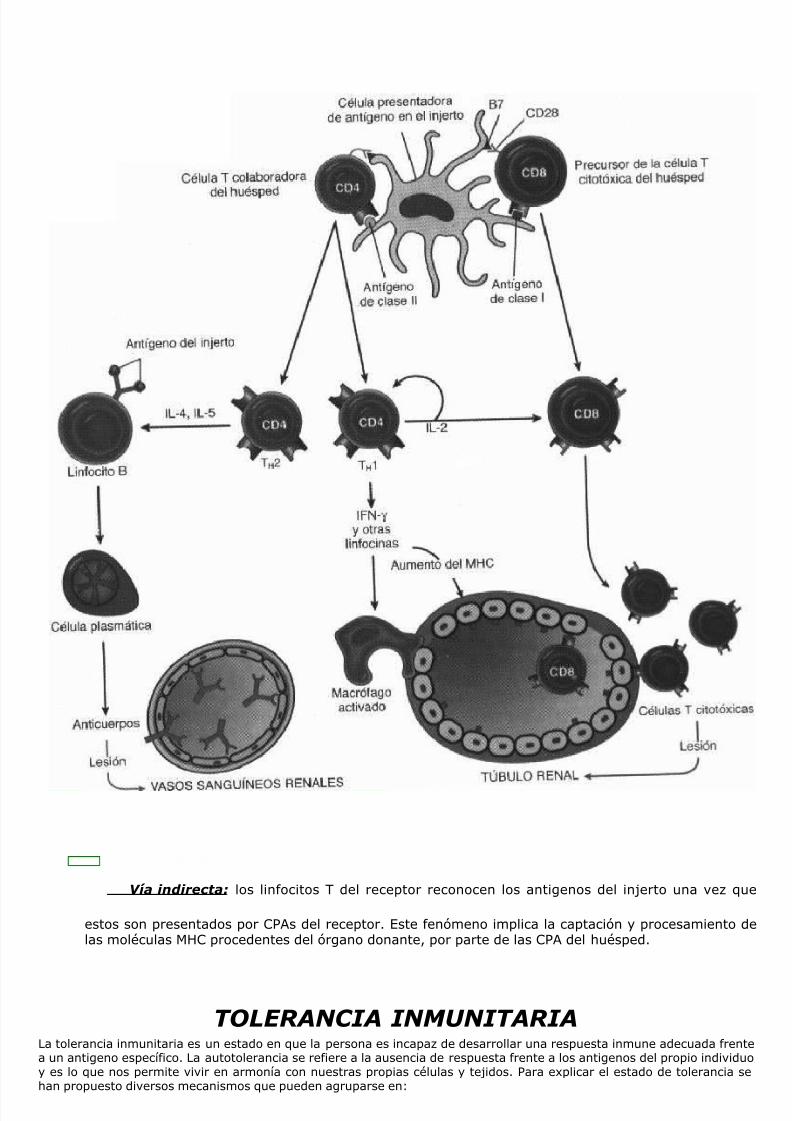
8/7/2019 Inmunologia-Apunte El Vikingo
http://slidepdf.com/reader/full/inmunologia-apunte-el-vikingo 42/53
- Vía indirecta: los linfocitos T del receptor reconocen los antigenos del injerto una vez que
estos son presentados por CPAs del receptor. Este fenómeno implica la captación y procesamiento delas moléculas MHC procedentes del órgano donante, por parte de las CPA del huésped.
TOLERANCIA INMUNITARIALa tolerancia inmunitaria es un estado en que la persona es incapaz de desarrollar una respuesta inmune adecuada frentea un antigeno específico. La autotolerancia se refiere a la ausencia de respuesta frente a los antigenos del propio individuoy es lo que nos permite vivir en armonía con nuestras propias células y tejidos. Para explicar el estado de tolerancia sehan propuesto diversos mecanismos que pueden agruparse en:
- Mecanismos de tolerancia CENTRAL: consiste en la deleción clonal de los linfocitos T y B autorreactivos
durante su maduración en los órganos linfáticos primarios (TIMO Y MEDULA OSEA). Se cree que los timocitos
42

8/7/2019 Inmunologia-Apunte El Vikingo
http://slidepdf.com/reader/full/inmunologia-apunte-el-vikingo 43/53
poseen la capacidad de presentar antigenos autólogos en asociación con MHC propias. Las células T en desarrolloque expresan receptores de alta afinidad para estos autoantígenos sufren una selección negativa y acabansufriendo apoptosis, por lo que en el conjunto de células T periféricas no existen células autorreactivas o si existenson escasas. Cuando un linfocito B en desarrollo encuentra un antigeno unido a la membrana en el interior de lamedula ósea, esa célula sufre apoptosis. No obstante, existen muchos autoantígenos que no son presentados enel timo, por lo que las células T poseedoras de receptores para los mismos escapan del timo y pasan a la periferia.Lo mismo sucede en el sistema de células B, de forma que en la sangre periférica de personas sanas seencuentran células B que poseen receptores para diversos autoantígenos.
- Mecanismos de tolerancia PERIFERICA: las células T autorreactivas que escapan a la selección
negativa intratímica podrían provocar lesiones hísticas, salvo que fueran eliminadas o neutralizadas en los tejidosperiféricos. Existen varios mecanismos que operan a este nivel:
Deleción clonal por muerte celular inducida por la activación: consiste en la
muerte por apoptosis de las células T activadas, a través del sistema FAS-FASL.
Anergia clonal: se trata de una inactivacion funcional prolongada e irreversible de los
linfocitos, que se produce cuando se encuentran con antigenos en determinadas condiciones. Si la célulaque presenta el antigeno no posee CD28 (que interacciona con el B7 de la CPA), la señal liberada seránegativa y la célula se volverá anérgica (inactiva), no pudiendo ser activada ni siquiera ante lapresentación del antigeno por una CPA competente capaz de emitir coestimulación. Como los tejidosnormales no expresan estas moléculas, o lo hacen solo muy débilmente, el encuentro entre las células Thelper específicas y sus correspondientes autoantígenos específicos causará la anergia clonal. Estemecanismo se aplica también a los linfocitos B de los tejidos.
Celulas T reguladoras: la activacion de los clones T autorreactivos esta bajo su control.
La deplecion de estas se asocia con la induccion de autoinmunidad ya que esto elimina el principalmecanismo de control en la periferia de los clones T autorreactivos.
La alteración de los mecanismos de tolerancia periférica contribuye a la patogenia de las enfermedades autoinmunitarias.
43
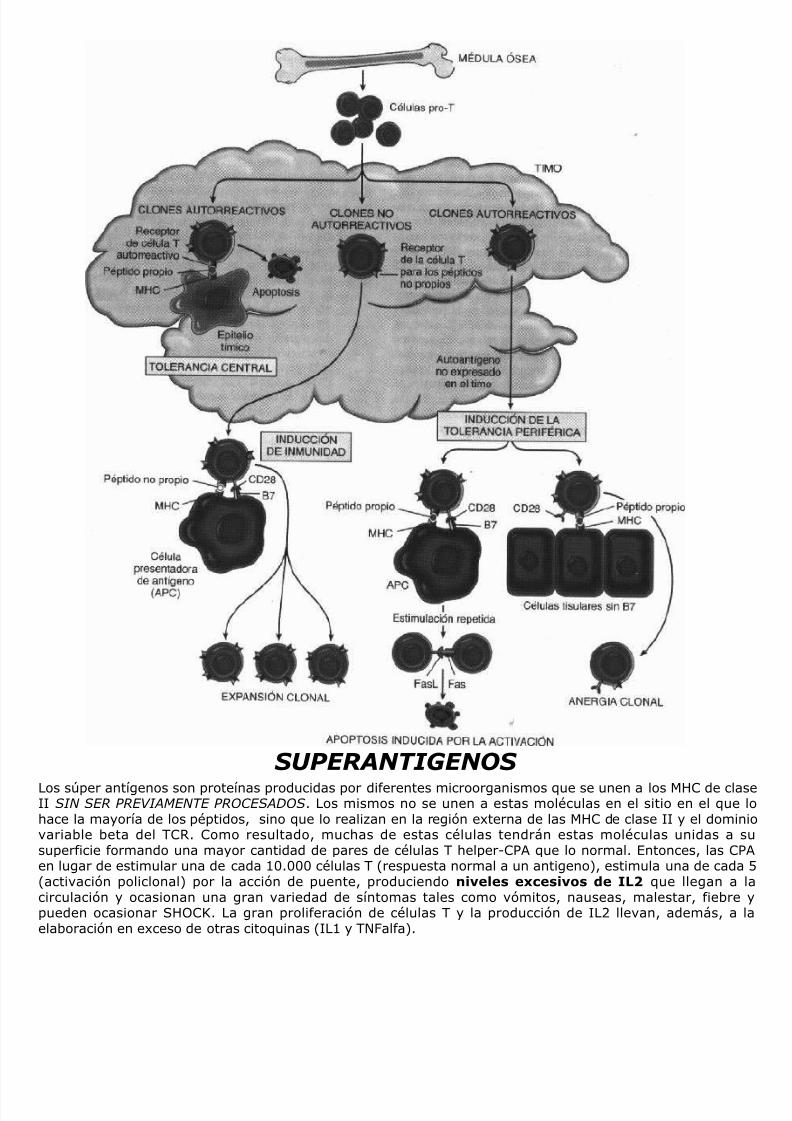
8/7/2019 Inmunologia-Apunte El Vikingo
http://slidepdf.com/reader/full/inmunologia-apunte-el-vikingo 44/53
SUPERANTIGENOS Los súper antígenos son proteínas producidas por diferentes microorganismos que se unen a los MHC de claseII SIN SER PREVIAMENTE PROCESADOS. Los mismos no se unen a estas moléculas en el sitio en el que lohace la mayoría de los péptidos, sino que lo realizan en la región externa de las MHC de clase II y el dominiovariable beta del TCR. Como resultado, muchas de estas células tendrán estas moléculas unidas a susuperficie formando una mayor cantidad de pares de células T helper-CPA que lo normal. Entonces, las CPAen lugar de estimular una de cada 10.000 células T (respuesta normal a un antigeno), estimula una de cada 5(activación policlonal) por la acción de puente, produciendo niveles excesivos de IL2 que llegan a lacirculación y ocasionan una gran variedad de síntomas tales como vómitos, nauseas, malestar, fiebre ypueden ocasionar SHOCK. La gran proliferación de células T y la producción de IL2 llevan, además, a laelaboración en exceso de otras citoquinas (IL1 y TNFalfa).
44
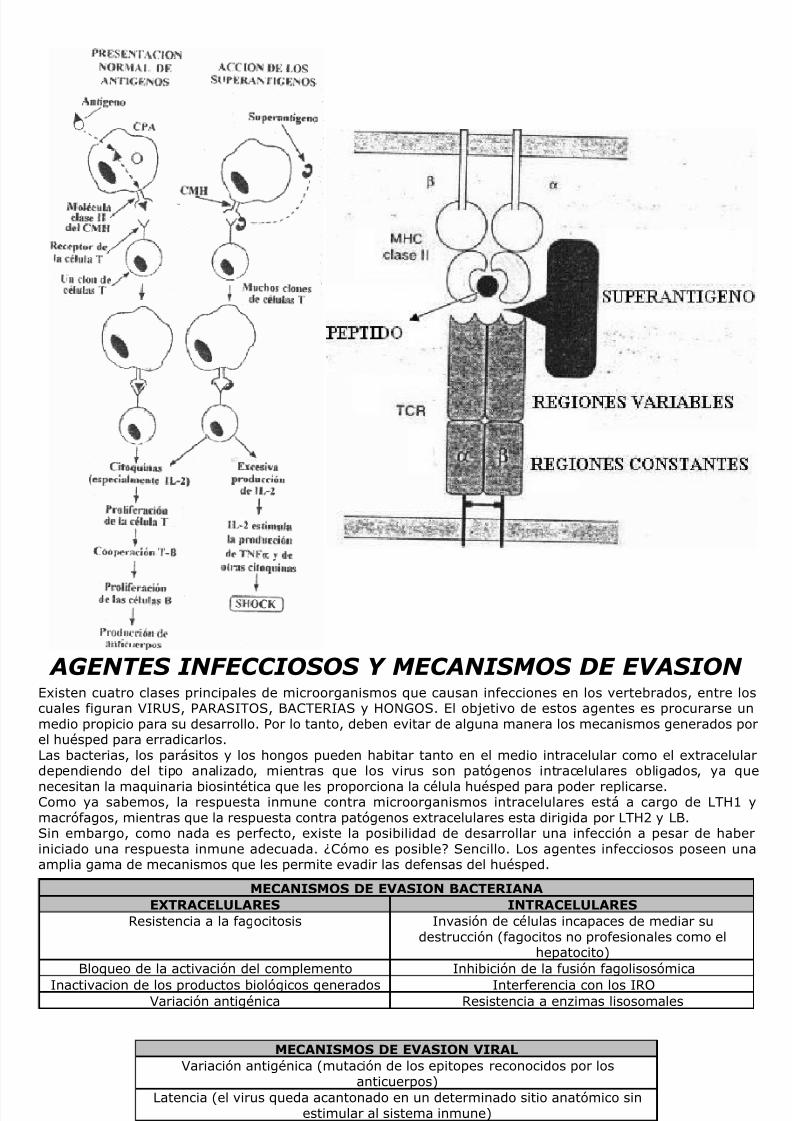
8/7/2019 Inmunologia-Apunte El Vikingo
http://slidepdf.com/reader/full/inmunologia-apunte-el-vikingo 45/53
AGENTES INFECCIOSOS Y MECANISMOS DE EVASION Existen cuatro clases principales de microorganismos que causan infecciones en los vertebrados, entre loscuales figuran VIRUS, PARASITOS, BACTERIAS y HONGOS. El objetivo de estos agentes es procurarse unmedio propicio para su desarrollo. Por lo tanto, deben evitar de alguna manera los mecanismos generados porel huésped para erradicarlos.Las bacterias, los parásitos y los hongos pueden habitar tanto en el medio intracelular como el extracelulardependiendo del tipo analizado, mientras que los virus son patógenos intracelulares obligados, ya que
necesitan la maquinaria biosintética que les proporciona la célula huésped para poder replicarse.Como ya sabemos, la respuesta inmune contra microorganismos intracelulares está a cargo de LTH1 ymacrófagos, mientras que la respuesta contra patógenos extracelulares esta dirigida por LTH2 y LB.Sin embargo, como nada es perfecto, existe la posibilidad de desarrollar una infección a pesar de haberiniciado una respuesta inmune adecuada. ¿Cómo es posible? Sencillo. Los agentes infecciosos poseen unaamplia gama de mecanismos que les permite evadir las defensas del huésped.
MECANISMOS DE EVASION BACTERIANAEXTRACELULARES INTRACELULARES
Resistencia a la fagocitosis Invasión de células incapaces de mediar sudestrucción (fagocitos no profesionales como el
hepatocito)Bloqueo de la activación del complemento Inhibición de la fusión fagolisosómica
Inactivacion de los productos biológicos generados Interferencia con los IROVariación antigénica Resistencia a enzimas lisosomales
MECANISMOS DE EVASION VIRALVariación antigénica (mutación de los epitopes reconocidos por los
anticuerpos)Latencia (el virus queda acantonado en un determinado sitio anatómico sin
estimular al sistema inmune)Alteraciones de la respuesta inmune del huésped
Inhibición de la inmunidad humoral (producción de receptores deanticuerpos y de complemento)
Interferencia a nivel de la presentación antigénica (disminución enla expresión de MHC clase I) Interferencia en la regulación de citoquinas (producción de
homólogos de IL-10 que inhiben la respuesta TH1)
45

8/7/2019 Inmunologia-Apunte El Vikingo
http://slidepdf.com/reader/full/inmunologia-apunte-el-vikingo 46/53
MECANISMOS DE EVASION PARASITARIAVariación antigénica
Liberación al medio extracelular de los anticuerpos unidos a lasuperficie del parásito
Supresión y/o bloqueo de la respuesta inmune del huésped(producción de proteasas que clivan los anticuerpos
INMUNODEFICIENCIAS Las deficiencias inmunitarias primarias casi siempre son de tipo genético y afectan un aspecto especifico de lainmunidad humoral o celular, mecanismos de defensa inespecíficos mediados por proteínas del complementoo por células como los fagotitos o las células NK.Los defectos de células T se asocian casi siempre a defectos en la síntesis de los anticuerpos, por lo que losdéficit aislados de células T suelen ser indistinguibles de los déficit combinados de células T y B.Casi todas las inmunodeficiencias primarias se manifiestan a edades muy tempranas, entre los 6 meses y losdos años, descubriéndose a causa de la propensión de estos lactantes a sufrir infecciones a repetición. Lasinmunodeficiencias secundarias son producidas por complicaciones de una infección (HIV), malnutrición,envejecimiento o efectos secundarios de la inmunosupresión, la radiación o la quimioterapia antineoplásica,quemaduras o por otras enfermedades autoinmunitarias.De más esta decir que ambos tipos de inmunodeficiencias son importantes por igual, pero en esta ocasión noslimitaremos al estudio de las inmunodeficiencias primarias.
AGAMMAGLOBULINEMIA DE BRUTON LIGADA AL CROMOSOMA XLa enfermedad de Bruton es una de las formas mas frecuentes de inmunodeficiencia primaria. Se caracterizapor la ausencia de precursores de las células B (células PRO-B y PRE-B) que puedan diferenciarse haciacélulas B. Durante la diferenciación normal de las células B en la medula ósea, se produce primero unreordenamiento de los genes de cadena pesada de las inmunoglobulinas, que va seguido del reordenamientode los genes de las cadenas livianas. En la agammaglobulinemia de Bruton, la maduración de las células B sedetiene después del reordenamiento de los genes de cadena pesada. Como no se producen cadenas livianas,resulta imposible formar y transportar hacia la membrana celular la molécula completa de inmunoglobulina,que debe contener la cadena liviana y la pesada. En el citoplasma de las células se encuentran cadenaspesada libres. Este bloqueo de la diferenciación se debe a mutaciones en una tirosina kinasa citoplasmáticadenominada tirosina kinasa de Bruton (BTK). Se sabe que las proteínas tirosina kinasas intervienen en latransducción de la señal y se cree que la BTK es esencial para la transducción de señales que dirige las célulasPRO-B hacia las PRE-B y estas a células B maduras. El gen BTK se encuentra en el cromosoma X y es por esta
razón que afecta casi exclusivamente a varones.INMUNODEFICIENCIA VARIABLE COMUN (IDVC)Esta alteración, relativamente frecuente pero mal definida, abarca un heterogéneo grupo de trastornos. Lacaracterística común a todos estos pacientes es la hipogammaglobulinemia que en general abarca a todas lasclases de anticuerpos, aunque a veces solo afecta a la IgG. El diagnostico de inmunodeficiencia variablecomún se hace tras haber excluido las demás causas bien definidas de disminución de la síntesis deanticuerpos. No se conoce bien la base molecular de la diferenciación anormal de las células B, pero segúnalgunos autores lo que sucedería es que las células T serian defectuosas, lo que les impediría enviar señalesadecuadas para la maduración y activación de las células B.
DEFICIT AISLADO DE IgAComo la IgA es la principal inmunoglobulina existente en secreciones externas, las defensas de las mucosasestan debilitadas, lo que facilita la infección de los aparatos respiratorio, digestivo y urogenital. Los pacientessintomáticos suelen presentar infecciones pulmonares a repetición y diarrea. El defecto básico afecta ladiferenciación de los linfocitos B productores de IgA, aunque su base molecular sigue siendo desconocida.
SINDROME DE HIPER IgMOriginalmente se pensaba que el síndrome de hiper IgM se debía a un trastorno de las células B, ya que lospacientes afectados fabrican anticuerpos IgM, pero son incapaces de producir IgG, IgA e IgE. En la actualidadse sabe que se trata de un trastorno de las células T, en el que la alteración de estas les impide activar a lascélulas B con la posterior elaboración de isotipos diferentes a IgM. La capacidad de las células B productorasde IgM para poner en marcha la trascripción de genes que codifican otros isotipos de Igs depende de señalesmediadas por citoquinas y por el contacto de las células T CD4+. En este proceso interviene la interacciónfísica entre las moléculas CD40 de las células B y CD40L expresados por las células T activadas. Por lo tanto,las mutaciones de CD40 o CD40L impiden la interacción necesaria entre las células T y B (cooperación T-B)para que se produzca el cambio de isotipo y el paciente desarrolla esta inmunodeficiencia. En el 70% de los
casos, la mutación afecta al gen CD40L situado en el cromosoma X. Por lo tanto, estos pacientes tienen laforma de la enfermedad ligada al cromosoma X. En los restantes, no se sabe con exactitud cuales son lasmutaciones precisas, aunque se cree que afectan a CD40 o a otras moléculas de las células B en la vía deseñalización CD40.
SINDROME DE DIGEORGE (HIPOPLASIA TIMICA)El síndrome de DIGEORGE es un ejemplo de déficit de células T secundario a la falta de desarrollo de losarcos branquiales tercero y cuarto. Dichos arcos son el origen del timo, las paratiroides, algunas células de latiroides y el cuerpo ultimo branquial. Por lo tanto, estos pacientes sufren una disminución variable de lainmunidad celular (debido a la hipoplasia o ausencia de timo), tetania (a causa de la ausencia de laparatiroides, que regula el metabolismo del calcio) y malformaciones congénitas del corazón y los grandesvasos. Además, pueden presentar anomalías de la boca, los oídos y la cara.
INMUNODEFICIENCIA COMBINADA SEVERA
Las inmunodeficiencias combinadas severas (IDCS) constituyen una constelación de síndromes genéticamentedistintos que tienen en común defectos variables de las respuestas inmunitarias tanto humoral como celular.La llamada forma clásica, inicialmente descrita en lactantes suizos, parece consecuencia de un defecto en lacélula precursora linfoide común y es muy rara. Lo más frecuente es que el defecto se encuentre en el
46

8/7/2019 Inmunologia-Apunte El Vikingo
http://slidepdf.com/reader/full/inmunologia-apunte-el-vikingo 47/53
compartimiento de las células T, con alteraciones secundarias de la inmunidad humoral. Los defectos de lascélulas T pueden producirse en cualquiera de las fases de su vía de diferenciación y activación. La forma másfrecuente, que representa el 50-60% de los casos, esta ligada al cromosoma X, lo que hace que la IDCS seamas frecuente en los niños que en las niñas. El defecto genético de la forma ligada al cromosoma X es unamutación en la cadena GAMMA de un receptor común a varias citoquinas. Esta proteína transmembrana formaparte de los componentes de la transducción de señales de los receptores para la IL-2, IL-4, IL-7, IL-9, IL-11e IL-15. Como consecuencia de esta mutación, las células progenitoras linfoides de los pacientes con IDCSligada al sexo no son activadas por estas citoquinas, lo que se traduce en un déficit funcional de todos losfactores de crecimiento. Como muchas de estas citoquinas son necesarias para el desarrollo del sistemalinfoide, la IDCS ligada al cromosoma X se asocia a profundos defectos de las primeras fases del desarrollo delas células T y de las últimas fases del desarrollo de las células B.Los restantes casos de IDCS se heredan en forma autonómica recesiva y la causa mas frecuente es un déficitde la enzima adenosina desaminasa (ADA). Aunque los mecanismos por los que el déficit de ADA produceIDCS son algo oscuros, se ha propuesto que el mismo causa la acumulación de desoxiadenosina y de susderivados (desoxi-ATP), que son especialmente tóxicos para los linfocitos inmaduros.Se han descrito algunas otras causas menos frecuentes de IDCS autosómica recesiva, a saber:
- Las mutaciones de los genes activadores de la recombinasa (RAG1 y RAG2) impiden el reordenamientode genes somáticos esenciales para la organización de losreceptores de la célula T y de los genes de las Igs. Ellobloquea el desarrollo de las células T y B.
- Una kinasa intracelular denomina JAK3 es esencial para latransducción de señales a través de la cadena GAMMA del
receptor de citoquinas. Por lo tanto, las mutaciones deJAK3 tienen los mismos efectos que las de la cadenaGAMMA. La diferencia entre ambas IDCS radica en suspatrones de transmisión hereditaria.
- Las mutaciones que alteran la expresión de las moléculasMHC de clase II impiden el desarrollo de las células TCD4+. Las moléculas MHC de clase II presentan antigenosa los linfocitos T CD4+ y durante el desarrollo de lascélulas T en el timo, la diferenciación de las células CD4+depende de la interacción con las moléculas MHC de claseII expresadas por el epitelio tímico. Las células T CD4+colaboran con el resto de las células T y con las B, por lo que la deficiencia de MHC de clase II se
asocia a una inmunodeficiencia combinada. En estos casos, el defecto genético no radica en los genesMHC, sino en los factores de trascripción que regulan su expresión.
SINDROME DE WISKOTT-ALDRICHSe trata de una enfermedad recesiva ligada al cromosoma X que se caracteriza por trombocitopenia, eccemay una importante tendencia al desarrollo de infecciones a repetición, que provocan la muerte precoz delpaciente. El defecto genético de este síndrome se localiza en el cromosoma X, lugar donde se encuentra elgen de la enfermedad, llamado proteína del síndrome de Wiskott-Aldrich (WASP). Esta proteína multifuncionalinterviene en el mantenimiento de la integridad del citoesqueleto y en la transducción de señales.
DEFICIENCIAS GENETICAS DEL SISTEMA DEL COMPLEMENTOSe han descrito deficiencias hereditarias de la práctica totalidad de los componentes del complemento y dedos de los inhibidores, siendo la que afecta a C2 la más frecuente de ellas.
DEFECTOS EN LA FUNCION LINFOCITARIA
Se han observado casos de alteraciones en cada una de las fases de la función leucocitaria, desde laadherencia al endotelio vascular hasta la actividad microbicida y se ha descrito la existencia de déficitgenéticos clínicos en cada uno de estos pasos críticos del proceso. Son los siguientes:- Defectos en la adhesión leucocitaria: el déficit de adhesión leucocitaria tipo I (LAD1)
consiste en una alteración de la biosíntesis de la cadena BETA 2 (CD18) compartida por las integrinasLFA-1 y MAC-1, produciendo alteraciones en la adhesión, diseminación, fagocitosis y generación delestallido respiratorio de los neutrófilos. Por otro lado, LAD2 es un defecto generalizado delmetabolismo de la fucosa (mutaciones en la fucosil transferasa) provocando la ausencia del ligando dela E selectina (presente en el endotelio activado): sialil Lewis X (presente en la superficie delneutrófilo).- Defectos en la fagocitosis: uno de estos trastornos es el SINDROME DE CHEDIAK-HIGASHI,
una afección autosómica recesiva caracterizada por neutropenia, degranulación alterada y retraso enla destrucción de los microorganismos. Los neutrófilos y otros leucocitos tienen gránulos gigantes, quese pueden observar fácilmente en el frotis de sangre periférica y que se considera el resultado de unafusión aberrante de organelas. En este síndrome, se produce una disminución de la transferencia de
47

8/7/2019 Inmunologia-Apunte El Vikingo
http://slidepdf.com/reader/full/inmunologia-apunte-el-vikingo 48/53
enzimas lisosomales a las vacuolas fagocíticas (con susceptibilidad frente a las infecciones), a losmelanocitos (con albinismo), a las células del sistema nervioso central (con trastornos nerviosos) y alas plaquetas (con cuadros de tipo hemorrágico). La identificación de un gen candidato sugiere que eldefecto básico de este trastorno se encuentra en una proteína asociada a la membrana que, a su vez,esta implicada en el acoplamiento y fusión de las membranas de las organelas. Este mecanismo podríaexplicar la incapacidad para secretar componentes lisosomales hacia los fagosomas o hacia el exteriorde la célula. También se afecta la secrecion de gránulos líticos por las células T citotóxicas, lo queexplica la inmunodeficiencia grave que se observa en este trastorno.
- Defectos de la actividad microbicida: la importancia de los mecanismos bacterianos
dependientes del oxigeno queda demostrada por la existencia de un grupo de trastornos congénitos dela capacidad de destrucción bacteriana, que se denomina enfermedad granulomatosa crónica (EGC) yque hace que los pacientes sean susceptibles a padecer infecciones bacterianas recurrentes. Laenfermedad granulomatosa crónica se debe a defectos hereditarios en los genes que codifican diversoscomponentes de la NADPH oxidasa, que genera el superóxido. Las variantes mas frecuentes son undefecto ligado al cromosoma X en uno de los componentes unidos a la membrana plasmática y los
defectos autonómicos recesivos en los genes que codifican dos de los componentes citoplasmáticos.
MICROORGANISMO PREVALENTE DEFICIENCIA BACTERIA VIRUS HONGOS PARASITOS
LB Si Si No NoLT Si Si Si Si
FAGOCITOS Si Si Si NoCOMPLEMENTO
(C5-C9)Si No No No
48

8/7/2019 Inmunologia-Apunte El Vikingo
http://slidepdf.com/reader/full/inmunologia-apunte-el-vikingo 49/53
ASPECTOS GENETICOS EN INMUNOLOGIALos CONCEPTOS desarrollados a continuación son analizados en un apartado por presentar ciertas dificultadesen su comprensión. Para llegar a entenderlos se necesitan conceptos básicos de genética, biología celular ypor supuesto, de alguien que sepa comunicar los contenidos de tal manera que puedan ser asimilados. Apartir de este momento se realizará una revisión de los aspectos genéticos en inmunología y ya que esta noes un área en la que acostumbre a desenvolverme con soltura, los dejaré en compañía de alguien que si lohace. Espero que el desarrollo de estos temas no les presente mayores inconvenientes.
GENÉTICA DE LA DIVERSIDAD DE ANTICUERPOS
Los anticuerpos tienen una estructura muy particular: cadenas pesadas y livianas, con regiones constantes yvariables (fig. 1). Pero lo más interesante es que éstas se encuentran codificadas en genes diferentes, sesintetizan las cadenas polipeptídicas por separado, y luego se ensamblan para dar lugar a una proteína muycompleja y de gran utilidad, tanto para la defensa del cuerpo humano como para la investigación y diversastécnicas de laboratorio.
Fig. 1. Estructura de un anticuerpo.
ESTRUCTURA DE LOS GENES.
Tomemos primero a la cadena liviana (L). Las regiones constantes de dichas cadenas pueden estarcodificadas por los genes κ (KAPPA) o λ (LAMBDA). Las regiones variables se encuentran upstream o ríoarriba (hacia el extremo 5´) de los genes κ y λ .
Los genes κ . Son 150 Vκ , es decir 150 posibles regiones variables κ para la Cκ (región constante κ ).Más adelante explicaremos cómo las regiones Vκ sufren un proceso conocido como recombinación somática,en el cual se corta y empalma el DNA, quedando elegidas sólo una región Vκ con una única Cκ disponible.
Los genes λ . También presentan amplia gama de Vλ (regiones variables λ ), y sólo una posible Cλ .Además, éstas sufren el proceso de recombinación somática.
Pasemos ahora a las cadenas pesadas (también las llamaremos H, del inglés: Heavy): Tenemos, dentro delpool de genes disponibles para el armado, varias posibles regiones variables, pero también hay variasposibilidades para las constantes. Se presentan en tándem, en el siguiente orden: Cµ , Cδ , Cε, Cy Cα . ELORDEN EN QUE SE PRESENTAN EN EL DNA ES SUMAMENTE IMPORTANTE YA QUE, COMO SE EXPLICARÁLUEGO, EN EL MOMENTO DEL SWITCH DE CLASE O CAMBIO DE ISOTIPO DEL ANTICUERPO, ÉSTE VAOCURRIENDO DE ACUERDO AL ORDEN GENÉTICO DE LAS REGIONES CONSTANTES DE LAS CADENASPESADAS. Es decir: primero se secreta IgM (de la cadena µ ), luego IgD (de la δ ), y posteriormente IgG, IgEe IgA, siguiendo exactamente la misma lógica. Pero no nos compliquemos con esto ahora, porque lo van aentender mejor después... Sólo pretendo que estén alertas cuando aparezca de nuevo el tema.
Un comentario que vale la pena realizar, es que el llamado proceso de recombinación somática es un corte yempalme a nivel de DNA, antes de la transcripción y la posterior traducción. Es uno de los procesos másfascinantes de la biología molecular, sobre todo por el control que se ejerce, dado que un error en éste podríaser muy perjudicial.
Entonces quedamos en que se elige la región constante y esta luego se corta y empalma con la regiónvariable elegida. Así llegamos al gen maduro, el cual se transcribe y traduce, dando lugar a la cadena pesadadel anticuerpo. Si recordamos la primera parte, ésta se une con la cadena liviana que ya sintetizamos alprincipio de la explicación, y voila!!! Tenemos el anticuerpo.
49

8/7/2019 Inmunologia-Apunte El Vikingo
http://slidepdf.com/reader/full/inmunologia-apunte-el-vikingo 50/53
EXCLUSIÓN ALÉLICA
El proceso de exclusión alélica (fig. 2) ocurre con lamaduración del linfocito B (¡¡¡lean con detalle este tema enel apunte, y relaciónenlo con lo que explico acá!!!). Cuando ellinfocito B está maduro, ya para la exhibición en membrana delBCR, en primer lugar sucede el rearreglo de la cadena pesada. ¿Cómo ocurre? Bueno, se selecciona al azar uno delos dos alelos (materno o paterno, recuerden que tenemos losgenes al doble con la misma información ya que somosdiploides) y comienza el “rearreglo”, esto es todo el proceso derecombinación somática, corte y empalme de las regionesconstantes y variables elegidas. Mientras se rearregla uno solode los dos alelos, el otro espera... si el rearreglo es exitoso,
queda ese como gen maduro para ser transcripto, inhibiéndosepara siempre el otro alelo. Pero si no es bueno, se deja esealelo y se pasa al otro, ocurriendo nuevamente un rearreglo detodos los posibles. Si es viable, ya queda con ese gen maduropara la cadena pesada del anticuerpo, sino la célula B entra aapoptosis ya que no quedan más alelos para probar rearreglos.Ahora que ya tenemos la cadena pasada con su región variabley constante seleccionadas , debemos rearreglar las cadenaslivianas. Se empieza con la cadena κ . Si sale bien, ya quedaese rearreglo, de lo contrario, pasa al otro alelo κ . Si nopuede llegar a una cadena liviana normal a través de los genesκ , pasa a los λ . La variabilidad de los genes λ es mayor, ya
que hay 6 genes posibles para la cadena liviana, y recuerdenque se duplica porque hay dos alelos de cada uno. Entonceshay 12 posibilidades para rearreglar exitosamente la cadenaliviana a través de genes λ . Al igual que antes, va probandocon cada uno de las posibles λ , hasta que tiene un genmaduro viable, sino va pasando por los otros alelos.En conclusión, sumando, hay 14 posibilidades para llegar a unrearreglo exitoso de las cadenas livianas.Algo súper importante: las opciones que van probando son alazar y cuando encuentra un rearreglo bueno ¡¡¡no siguenprobando otros!!! Otra cosa: habrán notado que hice hincapiéen que se comienza rearreglando las cadenas pesadas, y ensegundo lugar ocurren los rearreglos de las livianas. Eso es
porque siempre ocurre en ese orden.
Fig. 2. Genética de la maduración del linfocito B. Proceso de exclusión alélica.
PROCESO DE RECOMBINACIÓN SOMÁTICA
¿Cómo ocurre el corte y empalme de genes? ¡¡Qué tema fascinante!! Paso a describir el proceso, sin ahondardemasiado en nombres de enzimas, etc. para no entrar en un grado de detalle tal que se pierdan losconceptos básicos, ya que al fin y al cabo lo que importa es que quede fija la idea. De todos modosrecomiendo que lean el tema de alguno de los libros citados en la bibliografía.
... EN LAS CADENAS LIVIANAS
Resulta que los genes de la cadena V (variable) se encuentran río arriba de los del respectivo gen C(constante) (fig.3). Ahora bien, entre ellos, hay otro grupo de genes: los J (del inglés “joining”). Este grupode genes lo que permite es la unión entre los V y los C seleccionados para realizar el rearreglo. El corte yempalme de DNA es llevado a cabo por un grupo de enzimas (RAG 1 y RAG 2) las cuales en una serie depasos dan como resultado un segmento de DNA con un solo V, un J y un C. Notar que lo que se está diciendocuando se habla de recombinación es que se pierde cierta información de genes C, V y J que no se utilicen.Ya armado el segmento VJC correspondiente, éste se transcribe y traduce generándose de esta manera lacadena liviana.Como se explicó anteriormente, el rearreglo elegido puede no ser viable, y esto puede deberse a variosfactores. Citemos un ejemplo: las enzimas no cortan ni pegan estrictamente, lo cual proporciona ciertavariabilidad a la generación de cadenas livianas; pero al mismo tiempo, esto es poco eficiente ya que loscortes y empalmes no estrictos pueden alterar el marco de lectura en el momento de la traducción.
50

8/7/2019 Inmunologia-Apunte El Vikingo
http://slidepdf.com/reader/full/inmunologia-apunte-el-vikingo 51/53
Fig. 3. Genética de la síntesis dela cadena liviana del anticuerpo.
... EN LAS CADENAS PESADAS:SWITCH DE CLASE O CAMBIO DE ISOTIPO.Como ya lo había dicho, las regiones constantes de la cadena pesada se encuentran en tándem, con cierto
orden: Cµ , Cδ , Cε , C , Cα . (fig.4) Cada una de estas regiones tiene su secuencia promotora yterminadora de la transcripción. Cuando se transcriben cadenas constantes, en el transcripto primarioaparecen las cinco posibles C. Dicho transcripto posee secuencias de splicing alternativo para unir diferentes Ca la V ya seleccionada.
Fig. 4. Orden de las regiones del DNA que codifican para regiones variables, de diversidad, de unión yconstantes de los anticuerpos.
En el caso de las cadenas pesadas, vemos que no haysolamente un segmento J intermediario, sino que aparecentambién los llamados segmentos D (del inglés “diversity”). Así se seleccionan un segmento V de la región variable, uno D yotro J. Ocurre el corte y empalme de los segmento VDJseleccionados, a nivel de DNA. Luego se transcriben los quefueron elegidos (¡¡los otros ya se perdieron!!) junto contodos los C (Cµ , Cδ , Cε , C , Cα ). Posteriormente, porsplicing alternativo del transcripto primario, se selecciona la Ccorrespondiente (fig. 5).Espero que haya quedado claro ya que el resto de laexplicación se basa en esto.
Ahora volvamos sobre el tema de maduración de los linfocitosB.Mientras el linfocito está en estadío de Pre-B ocurren losrearreglos en las cadenas livianas, y expresión de la cadenapesada Cµ . Ojo!! Esta cadena es citoplasmática, no seencuentra en membrana todavía. Ya en el estadío de célula B inmadura, el linfocito expresa enmembrana IgM, con la región variable ya rearreglada y lacadena Cµ como constante.Por la acción de ciertos factores de dife-renciación, la célula Binmadura pasa a ser madura. Ahora no sólo expresa IgM sinoque también tiene en su membrana IgD (¡¡¡recordemos quela cadena C δ se encuentra a continuación de la C µ !!!).
Una vez que encuentra el antígeno, la célula B se activa:prolifera entonces generando plasmocitos, secretores deanticuerpos (luego veremos cómo es que los anticuerpos de
membrana Fig. 5. Switch de clase
51

8/7/2019 Inmunologia-Apunte El Vikingo
http://slidepdf.com/reader/full/inmunologia-apunte-el-vikingo 52/53
pasan a ser secretados), y células B de memoria.Bien, ¿estamos hasta acá? Ok, continuamos la explicación. Ahora veremos qué es lo que pasa genéticamentepara que haya diferentes tipos de inmunoglobulinas.
EL MECANISMO DEL SWITCH DE CLASE O CAMBIO DE ISOTIPO.En primer lugar aclaro que este es un rearreglo a nivel del DNA una vez que la célula B ve al antígeno y seactiva para proliferar.El Switch de clase consiste en que las enzimas corten el DNA, delecionando las zonas que codifican pararegiones constantes de cadenas pesadas. Entonces se van perdiendo C. La segunda observación que debo hacer es que LAS ZONAS VARIABLES YA FUERON SELECCIONADAS
ANTERIORMENTE, NO HAY NUEVA SELECCIÓN DE V, D Y J COMO EXPLICAMOS MÁS ARRIBA. Los V, D, J quese eligieron por exclusión alélica desde el comienzo quedan para siempre porque la otra porción de DNA ya seperdió. Es decir que las regiones variables de unión al antígeno no cambian, aunque cambie el isotipo de la inmunoglobulina que secrete el linfocito B.
Los Plasmocitos: estas células B activas pueden no cortar el DNA, pasando solo a ser secretoras de IgM eIgD. Otras sí cortan la zona de DNA que codifica para Cµ y Cδ , dando como resultado la secreción de IgG. Sicortan más allá de Cε entonces se expresa C y se secreta IgE. Por último, si se corta ésta última se da lugara la transcripción y traducción de la cadena Cα , secretándose entonces IgA.
Fig.6. Corte y empalme de segmentos VDJ.
¡¡El plasmocito no lleva anticuerpos en su membrana!! Secreta las Igs y luego deja de ser funcional para
morir. Para expresar anticuerpos de membrana a solubles, lo único que hacen es delecionar el dominiotransmembrana de la Ig. Pero eso lo veremos después con más detalle.
Las Células B de memoria: TODAS las células B de memoria cortan las cadenas Cµ y Cδ , por lo tanto estetipo de células sólo puede expresar IgG, IgE o IgA. OJO!!! Las células de memoria no secretan anticuerpos,sino que los mantiene en la membrana, es decir que a diferencia de los plasmocitos, no delecionan el dominiotransmembrana, como lo explicaremos en un ratito.
¿Por qué solamente cortando Cµ y Cδ , por ejemplo, se puede expresar la siguiente cadena, Cε, sin expresarC ni Cα ? Esto es porque, como ya se había comentado, cada C tiene su propio iniciador de la transcripcióny su terminador.
La utilidad de las células de memoria es entonces la rapidez con la que van a responder al antígeno si vuelvea aparecer en nuestro organismo. Dado que la selección de V D J ya está realizada, y el anticuerpo solo tiene
que ser secretado, los cambios en el DNA son mucha más rápidos, y en tiempos mucho menores puedenestar secretando las inmunoglobulinas.Es decir que como todos los rearreglos a nivel del DNA ya están hechos, la reacción de estas células dememoria es mucho más rápida. OJO!! Las células de memoria que se convierten en plasmocitos en larespuesta secundaria, nunca secretan IgM o IgD, porque recordemos que las células de memoria siempredelecionan Cµ y Cδ !!!!.
¿ANTICUERPOS DE MEMBRANA O SOLUBLES?Hay una secuencia de aminoácidos, por supuesto codificada en el DNA, que es hidrofóbica y por lo tantointeractúa con la membrana de forma tal que se queda anclada allí . Esa secuencia es la que hace que el anticuerpo sea de membrana y no secretado.Este cambio no es a nivel del DNA sino del transcripto primario, que al sufrir splicing alternativo (existensecuencias dadoras y aceptoras de splicing en sus intrones) va a quitar o dejar el RNA que codifica la cadenita
de aminoácidos hidrofóbicos que se pegan a la membrana.Por otro lado, el transcripto primario de la Ig se poliadenila en forma diferencial (recordemos que lapoliadenilación era una de las tres modificaciones del transcripto primario que luego será mRNA maduro listopara ser captado por los ribosomas y ser traducido a proteínas. Las otras dos modificaciones que sufre eltranscripto primario son splicing y capping, revisen los apuntes de biología celular !!!).Bueno, volviendo a lo que estábamos diciendo, los transcriptos de las Ig se poliadenilan en forma diferencial,es decir que va a haber algunos mRNAs que lleven la cola poli A más adelante, y otros más atrás. Por lotanto, los mRNAs tendrán diferente tamaño: van a haber transcriptos más cortos, y transcriptos largos.Antes debo dejar en claro que para que ocurra el splicing, deben existir en el transcripto primario dossecuencias que permiten el corte: una dadora y una aceptora.
Los transcriptos largos, tienen las secuencias dadoras y aceptoras de splicing, entonces SÍ SUFREN SPLICING y remueven con él el primer codón stop. Luego, la traducción se realiza hasta un segundo codón
stop, incluyendo así a la secuencia de aminoácidos hidrofóbica que ancla el anticuerpo a la membrana. Estasinmunoglobulinas son de membrana.
52

8/7/2019 Inmunologia-Apunte El Vikingo
http://slidepdf.com/reader/full/inmunologia-apunte-el-vikingo 53/53
Los transcriptos cortos, tienen secuencias dadoras de splicing, pero no las aceptoras. NO SUFREN SPLICING, y por lo tanto se traducen hasta el primer codón stop, sin incluir la secuencia hidrofóbica. Como notiene la secuencia hodrofóbica, entonces estos anticuerpos son secretados.
RECEPTORES T.Los receptores de linfocitos T tienen una estructura similar a los anticuerpos, con cadenas livianas y pesadas,con regiones constantes y variables.Los mecanismos de rearreglos que ocurren para la síntesis de receptores T son similares a los explicados paralas inmunoglobulinas de los linfocitos B. La diferencia principal entre los receptores B y los T es que elreconocimiento de los epitopes de antígenos es mucho más “fino” en los anticuerpos que en los receptores T.
# Como resumen final haremos un recorrido por todos los procesos explicados para ver dónde esque se genera diversidad de anticuerpos.
Para comenzar, la variabilidad genética ofrecida se debe a la selección de ciertas regiones de DNA y a ladeleción de otras. Esto es la recombinación somática. La selección de diferentes combinaciones de segmentosVDJ para las cadenas pesadas, y VJ para las livianas, más la flexibilidad de unión en el momento del empalmede los segmentos de DNA cortados por las enzimas. De la mano de este proceso está también la generaciónde regiones hipervariables del anticuerpo (en la zona de reconocimiento del epitope!!!).Otra manera de explicar la diversidad de anticuerpos es por las diferentes combinaciones de cadenas livianascon pesadas.Y a no olvidarnos de que siempre las mutaciones somáticas juegan un papel importante en la generación dediversidad.
Es momento ahora de hablar de la exclusión alélica. El hecho de seleccionar un alelo u otro también es fuentede diversidad. Debe destacarse que una vez que uno de los alelos fue seleccionado y funciona bien, el otroalelo se inactiva para siempre, y no es posible cambiar de alelo!!
Posteriormente, los rearreglos para el cambio de isotipo de inmunoglobulina que ocurren a nivel deltranscripto primario, mediante splicing alternativo, cuando hablamos de un linfocito B maduro con anticuerposIgM e IgD en su membrana. También se efectúan rearreglos a nivel del DNA, con deleción de regiones C, paradar lugar a la secreción de anticuerpos IgG, IgE e IgA.
Para terminar nombraremos otro cambio que explicamos con anterioridad: el paso de inmunoglobulinas demembrana a solubles. Estos cambios se producen por poliadenilación diferencial , derivando en un splicingalternativo que conserva o desecha el dominio transmembrana del anticuerpo.
BIBLIOGRAFIA
BIOLOGIA MOLECULAR DE LA CELULA. ALBERTS. 3ERA EDICION. 2000 INTRODUCCION A LA INMUNOLOGIA HUMANA. FAINBOIM Y GEFFNER. 5TA EDICION. 2005 PATOLOGIA GENERAL Y ESTRUCTURAL. ROBBINS. 6TA EDICION. 2000


















