
INSTITUTO POLITÉCNICO NACIONAL CENTRO …cinética geostrófica (J/m3) y (f) anomalía de la...
83
INSTITUTO POLITÉCNICO NACIONAL CENTRO INTERDISCIPLINARIO DE CIENCIAS MARINAS EFECTOS DE LA ENERGÍA CINÉTICA Y POTENCIAL SOBRE LA DISTRIBUCIÓN DE LARVAS DE PECES MESOPELÁGICAS EN EL SUR DEL GOLFO DE CALIFORNIA. TESIS QUE PARA OBTENER EL GRADO DE DOCTORADO EN CIENCIAS MARINAS PRESENTA FERNANDO CONTRERAS CATALA LA PAZ, B.C.S., JUNIO DE 2016.
Transcript of INSTITUTO POLITÉCNICO NACIONAL CENTRO …cinética geostrófica (J/m3) y (f) anomalía de la...

INSTITUTO POLITÉCNICO NACIONAL
CENTRO INTERDISCIPLINARIO DE CIENCIAS MARINAS
EFECTOS DE LA ENERGÍA CINÉTICA Y POTENCIAL SOBRE LA
DISTRIBUCIÓN DE LARVAS DE PECES MESOPELÁGICAS EN EL SUR DEL
GOLFO DE CALIFORNIA.
TESIS
QUE PARA OBTENER EL GRADO DE
DOCTORADO EN CIENCIAS MARINAS
PRESENTA
FERNANDO CONTRERAS CATALA
LA PAZ, B.C.S., JUNIO DE 2016.



DEDICATORIA
A mi queridísima madre por todo.
A mi hermano Jaime Contreras
A la pequeñita que me hace llorar, sonreír
y sobre todo que ha sabido aguantar mis
ausencias (Sofía A. Contreras A.). ¡Te
adoro hija!
En especial al Dr. Miguel F. Lavín y al M.
en C. Alejandro Zarate quienes se nos
adelantaron en este ciclo natural por el
cual cada ser vivo tiene que pasar. ¡Se les
extraña!
A todos aquellos que aprecien el trabajo en
equipo y muestren interés por el presente
trabajo.

AGRADECIMIENTOS
Al Centro Interdisciplinario de Ciencias Marinas del Instituto Politécnico Nacional.
Al Consejo Nacional de Ciencia y Tecnología (CONACyT) por la beca otorgada
durante mis estudios de doctorado.
Al comité tutorial (Dra. Laura Sánchez Velasco, Dr. Emilio Beier, Dr. Jaime Gómez
Gutiérrez, Dr. David Aurioles Gamboa, Dr. Eduardo Santamaría del Ángel y S. A.
Patricia Jiménez Rosenberg) por los comentarios y sugerencias realizadas al
presente estudio.
A los proyectos: SEP-CONACyT (contrato 2014- 236864) por el Instituto Politécnico
Nacional (proyecto multidisciplinario 2015-0176) y CONACyT (SEP-2011-168034-
T).
Al Centro de Investigación Científica y de Estudios Superiores de Ensenada
(CICESE) en especial al laboratorio de Circulación y dispersión costera (CIRCOS).
Al Centro de Investigación Científica y de Estudios Superiores de Ensenada Unidad
La Paz (CICESE-ULP) por lo apoyos otorgados durante mis estancias y congresos.
Al Dr. Víctor M. Godínez Sandoval (CICESE) por los comentarios vertidos al
presente manuscrito
Al laboratorio del cuerpo académico Ecología de fitoplancton (CA-UABC-158)
dirigido por el Dr. Eduardo Santamaría del Ángel por permitirme realizar una
estancia de investigación.
Al laboratorio de Ecología de Pinnípedos dirigido por el Dr. David Aurioles.
Al Consejo Sudcaliforniano de Ciencia y Tecnología (COSCYT) por apoyarme
económicamente para asistir al Larval Fish Conference Quebec, Canadá.
Al personal técnico y científico que participó en los cruceros oceanográficos a bordo
de los buques oceanográficos Francisco de Ulloa (CICESE) y El Puma (UNAM).
A tod@s mis compañer@s del grupo LEGOZ del departamento de Plancton y
Ecología Marina del CICIMAR.

AGRADECIMIENTOS ESPECIALES
Debo dar mil gracias a la Dra. Laura Sánchez por confiar en mí desde el inicio de
esta aventura que me acercó al conocimiento de las larvas de peces y las
interacciones físico-biológicas, así mismo gracias por el apoyo brindado y sobre
todo por la amistad mostrada hacia mi persona y familia.
Al Dr. Emilio Beier por sus valiosos consejos y la amistad prestada a mi familia.
Al Dr. Víctor Godínez por todos esos aprendizajes durante este episodio y sobre
todo por su amistad.
Al Dr. Miguel F. Lavín † (1951-2014) por haberme dejado enseñanzas para mi vida
personal y como profesional.
Al Dr. Eduardo Santamaría por sus consejos siempre atinados y sobre todo por su
amistad y confianza.
Al Sr. Francisco Becerra por su apoyo
A toda mi familia en Colima y en la Ciudad de México por su apoyo.
A tod@s mis amig@s en La Paz y en la Ciudad de México por sus porras durante
este proceso difícil pero a la vez emotivo.

ÍNDICE
Relación de figuras------------------------------------------------------------------------------------I
Relación de tablas-----------------------------------------------------------------------------------V
Glosario------------------------------------------------------------------------------------------------VI
Resumen----------------------------------------------------------------------------------------------XI
Abstract-----------------------------------------------------------------------------------------------XII
1. Introducción-----------------------------------------------------------------------------------1
2. Antecedentes---------------------------------------------------------------------------------3
2.1. Área de estudio------------------------------------------------------------------------------4
3. Hipótesis----------------------------------------------------------------------------------------9
4. Objetivo general------------------------------------------------------------------------------10
4.1. Objetivos particulares---------------------------------------------------------------------10
5. Métodos--------------------------------------------------------------------------------------11
5.1 Análisis de datos----------------------------------------------------------------------------13
6. Resultados-----------------------------------------------------------------------------------20
6.1 Estructura hidrográfica y circulación---------------------------------------------------20
Transectos con poca evidencia de remolinos de mesoescala ------------------20
Transectos con evidencia de remolinos de mesoescala -------------------------28

6.2 Distribución tridimensional de larvas de peces mesopelágicas-----------------34
Transectos con poca evidencia de remolinos de mesoescala ------------------34
Transectos con evidencia de remolinos de mesoescala -------------------------41
7. Discusión ------------------------------------------------------------------------------------45
8. Conclusiones---------------------------------------------------------------------------------50
9. Consideraciones-----------------------------------------------------------------------------45
Literatura citada-----------------------------------------------------------------------------52
Apéndice I ------------------------------------------------------------------------------------63

I
RELACIÓN DE FIGURAS
Pág.
Figura 1. Localización del área de estudio (batimetría del Golfo de California y
Pacífico adyacente
5
Figura 2. Representación de las principales corrientes en la región del Pacífico
Central Mexicano (PCM): CC, Corriente de California; CGC, Corriente del Golfo
de California; y CCM, Corriente Costera Mexicana (Figura tomada de Pantoja et
al., 2012).
8
Figura 3. Localización del área de estudio y de las estaciones de muestreo
durante los cinco cruceros. (a) Estaciones con baja energía cinética geostrófica y
(b) estaciones con alta energía cinética geostrófica en el sur del Golfo de
California. Estrellas negras, solo lances de CTD. Círculos rojos lances de CTD y
muestreos de zooplancton
17
Figura 4. Distribución tridimensional de larvas de peces sobre la línea con baja
energía (L1). (a) Imagen de la concentración de clorofila a superficial (MODIS-
AQUA LAC) durante el 17 de Agosto de 2005. Los puntos representan las
estaciones de muestreo. (b) Sección vertical de las velocidades geostróficas (m/s)
en L1. (c) Distribución vertical de larvas de Vinciguerria lucetia (larvas/10m2) sobre
la anomalía de la densidad (kg/m3). (d) larvas de Diogenichthys laternatus sobre
la estructura térmica (Θ oC). (e) Gráficos de la distribución del flujo de energía
cinética geostrófica (J/m3) y (f) anomalía de la energía potencial (J/m3). La línea
roja en las Figuras 4c y d representa la profundidad de la capa de mezcla
superficial.
21
Figura 5. Distribución tridimensional de larvas de peces sobre la línea con baja
energía (L2). (a) Imagen de la concentración de clorofila a superficial (MODIS-
AQUA LAC) durante el 11 de Julio de 2011. Los puntos representan las estaciones
de muestreo. (b) Sección vertical de las velocidades geostróficas (m/s) en L2. (c)
Distribución vertical de larvas de Vinciguerria lucetia (larvas/10m2) sobre la
anomalía de la densidad (kg/m3). (d) larvas de Diogenichthys laternatus sobre la
estructura térmica (Θ oC). (e) Gráficos de la distribución del flujo de energía
cinética geostrófica (J/m3) y (f) anomalía de la energía potencial (J/m3). La línea
roja en las Figuras 5c y d representa la profundidad de la capa de mezcla
superficial.
24

II
Figura 6. Distribución tridimensional de larvas de peces sobre la línea con baja
energía (L3). (a) Imagen de la concentración de clorofila a superficial (MODIS-
AQUA LAC) durante el 11 de Julio de 2011. Los puntos representan las estaciones
de muestreo. (b) Sección vertical de las velocidades geostróficas (m/s) en L3. (c)
Distribución vertical de larvas de Vinciguerria lucetia (larvas/10m2) sobre la
anomalía de la densidad (kg/m3). (d) larvas de Diogenichthys laternatus sobre la
estructura térmica (Θ oC). (e) Gráficos de la distribución del flujo de energía
cinética geostrófica (J/m3) y (f) anomalía de la energía potencial (J/m3). La línea
roja en las Figuras 6c y d representa la profundidad de la capa de mezcla
superficial.
25
Figura 7. Distribución tridimensional de larvas de peces sobre la línea L4 (con baja
energía). (a) Imagen de la concentración de clorofila a superficial (MODIS-AQUA
LAC) durante el 3 de Mayo de 2012. Los puntos representan las estaciones de
muestreo. (b) Sección vertical de las velocidades geostróficas (m/s) en L4. (c)
Distribución vertical de larvas de Vinciguerria lucetia (larvas/10m2) sobre la
anomalía de la densidad (kg/m3). (d) larvas de Diogenichthys laternatus sobre la
estructura térmica (Θ oC). (e) Gráficos de la distribución del flujo de energía
cinética geostrófica (J/m3) y (f) anomalía de la energía potencial (J/m3). La línea
roja en las Figuras 7c y d representa la profundidad de la capa de mezcla
superficial.
27
Figura 8. Distribución tridimensional de larvas de peces sobre la línea con alta
energía (H1). (a) Imagen de la concentración de clorofila a superficial (MODIS-
AQUA LAC) durante el 29 de Julio de 2011. Los puntos representan las estaciones
de muestreo. (b) Sección vertical de las velocidades geostróficas (m/s) en H1. (c)
Distribución vertical de larvas de Vinciguerria lucetia (larvas/10m2) sobre la
anomalía de la densidad (kg/m3). (d) larvas de Triphoturus mexicanus sobre la
estructura térmica (Θ oC). (e) Gráficos de la distribución del flujo de energía
cinética geostrófica (J/m3) y (f) anomalía de la energía potencial (J/m3). La línea
roja en las Figuras 8c y d representa la profundidad de la capa de mezcla
superficial.
29
Figura 9. Distribución tridimensional de larvas de peces sobre la línea con baja
energía (H2). (a) Imagen de la concentración de clorofila a superficial (MODIS-
AQUA LAC) durante el 11 de Julio de 2010. Los puntos representan las estaciones
31

III
de muestreo. (b) Sección vertical de las velocidades geostróficas (m/s) en H2. (c)
Distribución vertical de larvas de Vinciguerria lucetia (larvas/10m2) sobre la
anomalía de la densidad (kg/m3) y (d) larvas de Diogenichthys laternatus sobre la
estructura térmica (Θ oC). (e) Gráficos de la distribución del flujo de energía
cinética geostrófica (J/m3) y (f) anomalía de la energía potencial (J/m3). La línea
roja en las Figuras 9c y d representa la profundidad de la capa de mezcla
superficial.
Figura 10. Distribución tridimensional de larvas de peces sobre la línea con baja
energía (H3). (a) Imagen de la concentración de clorofila a superficial (MODIS-
AQUA LAC) durante el 15 de Octubre de 2007. Los puntos representan las
estaciones de muestreo. (b) Sección vertical de las velocidades geostróficas (m/s)
en H3. (c) Distribución vertical de larvas de Vinciguerria lucetia (larvas/10m2) sobre
la anomalía de la densidad (kg/m3) y (d) larvas de Triphoturus mexicanus sobre la
estructura térmica (Θ oC). (e) Gráficos de la distribución del flujo de energía
cinética geostrófica (J/m3) y (f) anomalía de la energía potencial (J/m3). La línea
roja en las Figuras 10c y d representa la profundidad de la capa de mezcla
superficial.
33
Figura 11. Dendrograma de los hábitats de larvas de peces definidos por la
medida de disimilitud de Bray-Curtis de las líneas con baja energía cinética
geostrófica. Larvas de peces recolectadas en el sur del Golfo de California.
35
Figure 12. Triplot basado en el diagrama de ordenación del análisis canónico de
correspondencias (ACC) sobre líneas con baja energía cinética geostrófica. Las
muestra biológicas (cuadros y círculos), variables ambientales (flechas),
centroides de los hábitats (símbolos rojos) y de las especies (estrellas); primer eje
horizontal y segundo eje vertical. Datos recolectados en 5 cruceros en el sur del
Golfo de California y Pacífico adyacente. Sal, Salinidad absoluta; BZ, Volumen
desplazado del zooplancton; Fluo, fluorescencia; Temp, Temperatura
conservativa; OD, Oxígeno disuelto; HLP, Hábitat de larvas de peces.
39
Figura 13. Dendrograma de los hábitats de larvas de peces definidos por la
medida de disimilitud de Bray-Curtis en transectos con alta energía cinética
geostrófica. Muestras de larvas de peces recolectadas en zonas con alta energía
cinética geostrófica en el sur del Golfo de California.
43
Figura 14. Triplot basado en el diagrama de ordenación del análisis canónico de
correspondencias (ACC) sobre líneas con alta energía. Las muestras biológicas
44

IV
(cuadros y círculos), variables ambientales (flechas), centroides de los hábitats
(símbolos rojos) y de las especies (estrellas); primer eje horizontal y segundo eje
vertical. Datos recolectados en transectos con alta energía en el sur del Golfo de
California. Sal, Salinidad absoluta; BZ, Volumen desplazado del zooplancton;
Fluo, fluorescencia; Temp, Temperatura conservativa; OD, Oxígeno disuelto; HLP,
Hábitat de larvas de peces.

V
RELACIÓN DE TABLAS
Pág.
Tabla 1. Masas de agua en el Golfo de California y Pacífico Transicional Tropical-
Subtropical de acuerdo a Durazo & Baumgartner (2002), Castro et al. (2006),
Lavín et al. (2009).
6
Tabla 2. Información general de los cruceros oceanográficos realizados en el sur
del Golfo de California.
18
Tabla 3. Valores promedio de la energía cinética geostrófica y anomalía de la
energía potencial (J/m3) referidos a 300 m de profundidad (ver la posición de los
transectos en Figura 3).
22
Tabla 4. Prueba de Olmstead–Tukey, ANOSIM (de una vía) y el análisis de
SIMPER entre la abundancia larval de los hábitats (HLP), clasificada de acuerdo
a la medida de disimilitud de Bray-Curtis. J: jerarquía, X: promedio de la
abundancia, %F: frecuencia de ocurrencia, %S: Porcentaje de similitud (%
Contribución). D: dominante, C: constante, F: frecuente y R: rara.
36
Tabla 5. Porcentaje de contribución de las variables exploratorias por el análisis
canónico de correspondencia
40

VI
GLOSARIO
Anticiclónico. Se usa para definir el sentido de rotación de fenómenos oceánicos
y atmosféricos contrario a la dirección de giro de los ciclones y al de la Tierra, en el
mismo sentido de las manecillas del reloj en el hemisferio norte y en sentido
contrario en el hemisferio sur.
Batimetría. Proviene del griego βαθυς, profundo, y μετρον, medida. Se refiere a la
configuración del suelo marino o a un mapa donde este delineado el perfil del fondo
de un cuerpo de agua por medio de contornos de igual profundidad (isóbatas).
Biomasa. Es la cantidad de materia viva producida en un área determinada de la
superficie terrestre, o por organismos de un tipo específico, expresada en peso por
unidad de área o volumen.
Capa de mezcla. Estrato donde existe activa turbulencia homogeneizando algunos
rangos de profundidades.
Ciclónico. Es el sentido de giro de un fluido igual al de los ciclones y de la Tierra;
en sentido contrario al de las manecillas del reloj en el hemisferio norte y en el mismo
sentido en el hemisferio sur.
Clorofila. Molécula con un anillo de porfirina y un núcleo formado por un átomo de
magnesio. El anillo de porfirina es un tetrapirrol con cuatro anillos pentagonales de
pirrol enlazados para formar un anillo mayor que es la porfirina y una cadena larga
llamada fitol. Su función en el proceso fotosintético es la absorción de energía
luminosa en la banda de los azules.

VII
Corriente geostrófica. Es la corriente definida al resolver las ecuaciones
hidrodinámicas asumiendo que la componente horizontal de la fuerza de Coriolis
está en balance con la componente del gradiente de presión:
CTD. Instrumento oceanográfico para obtener un perfil de temperatura y la
conductividad en función de la profundidad. Con estos datos se puede generar las
características T-S (Temperatura-salinidad) de la columna de agua. Ese
instrumento se puede bajar al agua desde un barco con cable cuyo núcleo contenga
conectores eléctricos por lo tanto recibir, observar y almacenar los datos en tiempo
real. Las siglas CTD provienen del inglés Conductivity, Temperature, Depth.
Decibar. Es la unidad práctica de presión para trabajos oceanográficos, es la
décima parte de un bar. Para fines prácticos equivale a un metro de profundidad.
Densidad. Es una medida de la masa de una sustancia por unidad de volumen ρ
= m/V. El recíproco del volumen específico. En Oceanografía, la densidad del agua
de mar es numéricamente equivalente a la gravedad específica y es una función de
la salinidad, temperatura y la presión. Densidad ρ = (ρs,t,p)
Demersal. El término se aplica para los organismos que vive muy próximo al fondo
de la masa de agua que es su hábitat.
Energía cinética. La energía cinética es la energía del movimiento. Cuando el agua
marina tiene un movimiento vertical u horizontal - tiene una energía cinética. Existen
al menos tres formas de energía cinética - vibratorio (la energía debido a movimiento
de vibración), de rotación (la energía debido a movimiento de rotación), y de
traslación (la energía debido al movimiento de un lugar a otro). La energía cinética
depende de dos variables: la masa (m) del objeto y la velocidad (v) del objeto. La
siguiente ecuación se utiliza para representar la energía cinética (EC) de un objeto.

VIII
0.5 2EC m v
donde m es la masa del objeto y v es la velocidad del objeto.
Energía potencial. Es la capacidad que tiene un cuerpo para realizar un trabajo de
acuerdo a la configuración que ostente en el sistema de cuerpos que ejercen fuerzas
entre sí, es decir, la energía potencial es la energía que es capaz de generar un
trabajo como consecuencia de la posición de un cuerpo. A la misma puede
considerársela como la energía almacenada en el sistema o la medida de un trabajo
que el sistema puede ofrecer.
Estratificación. Condición del fluido que implica la existencia de dos o más capas
horizontales arregladas según su densidad, de tal manera que las capas menos
densas están sobre las más densas
Fisiografía. Está definida como la descripción de la naturaleza a partir del estudio
del relieve y la litosfera, en conjunto con el estudio de la hidrosfera, la atmósfera y
la biosfera.
Flexión. Inicia con la flexión del notocordio y termina con la formación de la placa
hipúrica de una larva de pez.
Geostrofía. Método para analizar las corrientes en una zona, que consiste en
asumir que existe un balance entre la fuerza de Coriolis y el gradiente horizontal de
presión.
Isotaca. La línea o superficie que une todos los puntos de magnitud de velocidad
sobre una gráfica en el espacio o en el tiempo; por ejemplo la magnitud del flujo de
una corriente.

IX
Isopicna. Línea o superficie que une todos los puntos de densidad constante sobre
una gráfica en el espacio o en el tiempo.
Isoterma. Una línea o superficie que conecta en una gráfica todos los puntos de
igual valor de la temperatura, ya sea en el tiempo o en el espacio.
Larva. (Del latín Larva, fantasma). En los animales con desarrollo indirecto (con
metamorfosis) y se define como el estadio que precede a la eclosión del huevo; es
diferente en forma y pigmentación al juvenil y al adulto, y debe de sufrir una etapa
de transformación antes de asumir las características del adulto.
Masas de Agua. Volumen de agua usualmente identificado por rangos de
temperatura y salinidad que permiten distinguirlo de las aguas circundantes. Su
formación ocurre en contacto con la atmósfera y por la mezcla de dos o más tipos
de agua.
Meandro. Son las curvaturas muy pronunciadas (algunas en forma de “u”) en la
trayectoria de un río o de una corriente marina.
Meroplancton. Grupo representado por organismos que forman parte del plancton,
solamente durante una parte de su ciclo de vida.
Mesoescala. Se refiere a escala en espacio y tiempo de fenómenos
oceanográficos, la cual va de decenas hasta cientos de kilómetros y de semanas a
meses.
Mesopelágico. Zona en que uno de los niveles en los que está dividido el océano
según su profundidad se sitúa entre 200 y 1000 metros.
Oxígeno disuelto. Concentración de oxígeno presente en el agua.

X
Pelágico. (Del griego (pélagos), "mar abierto") es la parte de la columna de agua
del océano que no está sobre la plataforma continental. Los organismos que habitan
esta área se denominan pelágicos.
Picnoclina. Son capas de agua en donde la densidad del agua cambia rápidamente
con la profundidad. La mayoría de las veces coincide con la termoclina y como esta
tiene gran variación estacional en latitudes medias y casi nulas en latitudes altas.
Plancton. Organismos que no pueden desplazarse de manera autónoma en el agua
y que son arrastrados por las corrientes “viven a la deriva”.
Postflexión. Inicia con la osificación de la placa hipúrica y termina antes de que se
complete el número de elementos de aletas pares e impares.
Preflexión. Fase en la que se da la absorción del saco vitelino, la apertura de la
boca y el ano, la pigmentación de los ojos y el notocordio está recta.
Taxón. Cualquier unidad taxonómica definible, por ejemplo la subespecie, la
especie, el género, la familia.
Termoclina. Capa que presenta un cambio abrupto de temperatura con la
profundidad. Se clasifican en termoclina estacional y termoclina permanente.
(Véase estratificación).
Transecto. Es la línea que resulta de unir a varios sitios de muestreo a lo largo de
la cual se quieren graficar y analizar los datos en forma independiente.
Tridimensional. Se refiere a la distribución vertical de las especies

XI
RESUMEN
Se analizaron los efectos de la energía cinética geostrófica y anomalía de la energía
potencial en remolinos sobre la distribución tridimensional de las larvas de peces de
especies mesopelágicas (Vinciguerria lucetia, Diogenichthys laternatus,
Benthosema panamense y Triphoturus mexicanus) en el sur del Golfo de California
durante estaciones con fuerte estratificación (verano y otoño). La mayor abundancia
de larvas fue encontrada en las estaciones de muestreo en las zonas con baja
energía cinética geostrófica (< 7.5 J/m3) donde la distribución de las especies
dominantes tendía a ser estratificada. Las larvas de V. lucetia (con abundancia
media de 318 larvas/10m2) y B. panamense (174 larvas/10m2) se encuentra
principalmente en y por encima de la picnoclina (típicamente ~ 40 m de
profundidad). Por el contrario, las larvas de D. laternatus (60 larvas /10m2) se
localizaron principalmente en y por debajo del picnoclina. Por otro lado, en las
estaciones de muestreo de las áreas con alta energía cinética geostrófica (> 21
J/m3), donde se detectaron anteriormente remolinos de mesoescala, las larvas de
la especie dominante tenían baja abundancia y se distribuyeron de manera más
uniforme a través de la columna de agua. Por ejemplo, en un remolino ciclónico, las
larvas de V. lucetia (34 larvas/10m2) extendieron su distribución hasta límite de
muestreo 200 m de profundidad por debajo de la picnoclina, mientras que larvas de
D. laternatus (29 larvas/10m2) se encontraron hasta la superficie, esto
probablemente como consecuencia de los efectos de mezcla del remolino. Los
resultados mostraron que el nivel del flujo de energía cinética geostrófica afecta a
la distribución tridimensional y la abundancia de las larvas de peces mesopelágicas
durante los periodos de estratificación fuerte, lo que indica que las áreas con menor
energía cinética geostrófica pueden ser ventajosas a la estabilidad de la columna
de agua, para la alimentación y el desarrollo de las larvas de peces mesopelágicas.
Palabras clave: Flujo de energía cinética geostrófica, estratificación, remolinos de
mesoescala, larvas de peces mesopelágicas, distribución vertical, Golfo de
California, Pacífico mexicano.

XII
ABSTRACT Effects of geostrophic kinetic energy flux on the three-dimensional distribution of fish
larvae of mesopelagic species (Vinciguerria lucetia, Diogenichthys laternatus,
Benthosema panamense and Triphoturus mexicanus) in the southern Gulf of
California during summer and fall seasons of stronger stratification were analyzed.
The greatest larval abundance was found at sampling stations in geostrophic kinetic
energy-poor areas (<7.5 J/m3), where the distribution of the dominant species tended
to be stratified. Larvae of V. lucetia (average abundance of 318 larvae/10m2) and B.
panamense (174 larvae/10m2) were mostly located in and above the pycnocline
(typically ~ 40 m depth). In contrast, larvae of D. laternatus (60 larvae/10m2) were
mainly located in and below the pycnocline. On the other hand, in sampling stations
from geostrophic kinetic energy-rich areas (> 21 J/m3), where mesoscale eddies
were present, the larvae of the dominant species had low abundance and were
spread more evenly through the water column, in spite of the water column
stratification. For example, in a cyclonic eddy, V. lucetia larvae (34 larvae/10m2)
extended their distribution to, at least, the limit of sampling 200 m depth below the
pycnocline, while D. laternatus larvae (29 larvae/10m2) were found right up to the
surface, both probably as a consequence mixing and secondary circulation in the
eddy. Results showed that the level of the geostrophic kinetic energy flux affects the
abundance and the three-dimensional distribution of mesopelagic fish larvae during
the seasons of stronger stratification, indicating that the areas with low geostrophic
kinetic energy may be advantageous for the feeding and development of
mesopelagic fish larvae for the water column stability.
Keywords: Geostrophic kinetic energy flux, stratification, mesoscale eddies,
mesopelagic fish larvae, vertical distribution, Gulf of California, Pacific Mexican.

1
1. INTRODUCCIÓN
El estudio de los remolinos de mesoescala y su impacto en el ecosistema
biológico es complejo; esto debido a la variabilidad en su origen, dinámica física y
constante evolución, esto se ve reflejado en la variabilidad de la respuesta biológica
ante los cambios físicos de los remolinos (Eden et al. 2009). De manera general, se
ha documentado que los remolinos de mesoescala estimulan la productividad
biológica en los océanos, debido a que advectan, mezclan y redistribuyen las masas
de agua. Estos remolinos generan límites hidrográficos y dinámicos que pueden
definir hábitats plantónicos contiguos que influyen en la distribución y sobrevivencia
de los organismos del zooplancton (Sánchez-Velasco et al. 2013, Contreras-Catala
et al. 2015). Dichos hábitats planctónicos se caracterizan por presentar propiedades
físicas y químicas diferentes, asociadas al origen y evolución de los remolinos, y a
las propiedades de las masas de agua de la región.
La formación de los remolinos de mesoescala puede ser debido por diversas
causas, de los cuales se han detectado al menos estos mecanismos de formación:
i) inestabilidades de corrientes costeras (Zamudio et al. 2008, Pantoja et al. 2012),
donde por interacción de la corriente y la plataforma continental (efectos de punta)
se originan remolinos que atrapan aguas costeras y/u oceánicas; ii) por la
interacción de la circulación de gran escala con la topografía del fondo (Kurian et al.
2011); iii) por el forzamiento del viento local (Pares-Sierra et al. 1993) y iv) por
ondas atrapadas en la costa de origen ecuatorial .
En este contexto las estructuras de mesoescala como los remolinos y las
surgencias, generalmente asociadas a procesos de mezcla, convergencia y

2
divergencia, son áreas donde la energía cinética típicamente es alta (Myers &
Drinkwater 1989, Piontkovski et al. 1995, Omand et al. 2015).
Numerosos estudios han descrito las relaciones cualitativas entre remolinos
de mesoescala y el fitoplancton (Crawford et al. 2007, Moore et al. 2007, Brown et
al. 2008), y los remolinos con el zooplancton (Fernández et al. 2004, Sánchez-
Velasco et al. 2006, Eden et al. 2009).
En el Golfo de California (GC) y el Pacifico adyacente (PA) se ha
documentado la presencia recurrente de remolinos de mesoescala durante todo el
año (Álvarez-Borrego 2012, Lavín et al. 2014). Estos han sido reportados mediante
el uso de imágenes de satélite (Pegau et al. 2002, López-Calderon et al. 2008,
Kurczyn et al. 2012), modelos numéricos inferidos por datos satelitales (Beier 1997,
Martínez & Allen 2004, Zamudio et al. 2007, 2008) y con datos hidrográficos in
situ (Badán-Dangon et al. 1985, Emilsson & Alatorre 1997, Lavín et al. 1997, 2013,
Navarro-Olache et al. 2004, Kurczyn et al. 2013). Sin embargo, pocos estudios han
cuantificado y relacionado la energía cinética y/o potencial con la distribución de los
organismos planctónicos.
2. ANTECEDENTES
Los estudios dirigidos a conocer los efectos de los remolinos de mesoescala
y su impacto en la distribución de las larvas de peces a nivel mundial son aún
insuficientes comprados con la frecuencia e importancia de estos en la productividad
del océano. Los primeros trabajos no se enfocaron al muestreo de un remolino en
particular, algunos de ellos fueron eventos casuales. Podemos ejemplificar unos de

3
los más importantes ya dirigidos con muestreos intensivos y haciendo énfasis en su
origen y evolución.
En el Mar de Arabia y Mar Negro Piontkovski et al. (1995) reportaron que la
heterogeneidad espacial más alta de la biomasa del zooplancton se encontró en
regiones donde la energía potencial disponible era más alta, lo cual se asoció a
campos de remolinos de mesoescala. Ladd et al. (2009) en el Golfo de Alaska
observaron que las mayores concentraciones de clorofila se presentaron en áreas
con alta energía cinética turbulenta, justo donde hay una formación continua de
remolinos, sugiriendo que la energía cinética turbulenta puede ser una variable
predictiva de los florecimientos de fitoplancton en esta región. Nieto et al. (2013)
registraron relaciones entre remolinos, filamentos y la distribución espacial del
hábitat de desove de la sardina del Pacífico, observando que el área con mayor
probabilidad de encontrar huevos de sardina fuera de la costa coincide con altos
valores de la energía cinética turbulenta de los remolinos.
En este contexto diferentes autores (Zamudio et al. 2008, Lavín et al. 2013,
2014) han descrito el mecanismo de formación de los remolinos en el sur del Golfo
de California. Zamudio et al. (2008), Lavín et al. (2013, 2014) reportaron que la
interacción de la Corriente Costera Mexicana hacía el polo con las irregularidades
en la topografía, generan frecuentemente remolinos por inducción de
inestabilidades baroclínicas y que este mecanismo se puede ver reforzado por la
llegada de ondas atrapadas en la costa de origen ecuatorial. Esto sugiere que el sur
del Golfo de California y el Pacífico adyacente son áreas energéticas con elevada
actividad de mesoescala. Sin embargo hasta el momento, no existen estudios

4
multidisciplinarios donde se haya examinado el impacto de la energía cinética
geostrófica de los remolinos sobre los organismos planctónicos.
Un buen indicador biológico para la observación del efecto del flujo de la
energía cinética geostrófica y potencial de los remolinos sobre los organismos del
zooplancton pueden ser las larvas de peces de las especies mesopelágicas más
comunes (Vinciguerria lucetia, Diogenichthys laternatus, Benthosema panamense y
Triphoturus mexicanus), debido a sus altas abundancias y amplias distribuciones,
que son componentes importantes del ecosistema pelágico en el Pacífico
nororiental, incluyendo el Golfo de California (Ahlstrom 1959, Sassa et al. 2007,
Landry et al. 2012). En el GC, estas larvas tienen una distribución heterogénea
desde la región del Archipiélago medio hasta la boca en el sur del golfo (ej. Danell-
Jiménez et al. (2009), Contreras-Catala et al. (2012), Apango-Figueroa et al.
(2015)).
Aunque existen estudios de la abundancia en función de la profundidad, se
ha visto que estas especies tienden a exhibir gradientes verticales en abundancia.
Larvas de V. lucetia y B. panamense tienen mayores abundancias en la capa
superficial, disminuyendo con la profundidad y larvas de D. laternatus y T.
mexicanus tienen una mayor abundancia en la capa de ~ 200 a 100 m de
profundidad, disminuyendo hacia la superficie (Sánchez-Velasco et al. 2007, 2013,
Contreras-Catala, Sanchez-Velasco, et al. 2012, Contreras-Catala et al. 2015).
2.1 ÁREA DE ESTUDIO
La región sur GC y el Pacífico adyacente (Fig. 1); se localizan en una zona
donde convergen diferentes masas de agua (ver Tabla 1), aguas de origen subártico

5
acarreadas por ramificaciones de la Corriente de California, agua superficial tropical
traída por la Corriente Costera Mexicana (CCM) y aguas con alta salinidad
generadas en el Golfo de California.
Figura 1. Localización del área de estudio (batimetría del Golfo de California y Pacífico adyacente), ver estaciones de muestreo en las Figs. 3a y b.

6
Tabla 1. Masas de agua en el Golfo de California y Pacífico Transicional Tropical-Subtropical de acuerdo a Durazo & Baumgartner (2002), Castro et al. (2006), Lavín et al. (2009).
Masas de agua Abreviación Salinidad Temperatura (oC)
Agua del Golfo de
California AGC >34.9 ≥12
Agua Superficial
Tropical AST <34.9 ≥18
Agua de la Corriente
de California ACC 33.8 ≤ S ≤34.5 12 ≤ T ≤ 18
Agua Subsuperficial
Subtropical ASsSt 34.5 ≤ S < 34.9 9 ≤ T ≤ 18
Agua Intermedia del
Pacífico AIP 34.5 ≤ S ≤ 34.8 4 ≤ T < 9
Agua Profunda del
Pacífico APP > 34.5 < 4
Agua subÁrtica ASA ≤ 34.9 8 < T < 21
Por un lado el GC se caracteriza por ser un mar semicerrado y por ser una
cuenca de evaporación (Beier 1997, Lavín et al. 2009), donde se presentan varios
procesos hidrodinámicos que pueden promover el enriquecimiento de las capas
superiores del océano y el transporte (o retención) de huevos y larvas de peces
(Peguero-Icaza et al. 2008). Se caracteriza por presentar una circulación superficial
estacional, ciclónica en verano y anticiclónica en invierno) (Beier 1997), surgencias
inducidas por el viento en el sur del golfo (que se producen en el lado continental

7
durante el invierno y el lado peninsular durante verano) como respuesta al régimen
de vientos monzónico (Badan-Dangon et al. 1991, Lavín et al. 2009).
Castro et al. (2006) y Lavín et al. (2009), mencionan que la estructura
termohalina de las capas superiores del área donde se da la comunicación entre el
GC y el Pacífico (“la boca”) es compleja debido a la confluencia de las aguas
superficiales con características distintas, como es el Agua Superficial tropical
(AST), el Agua de la Corriente de California (ACC) de origen subártico y el Agua del
Golfo de California (AGC). Las diferencias de temperatura y densidad entre estas
masas de agua superficial causan frentes, que tienden a desarrollar estructuras de
mesoescala como remolinos, chorros y meandros que son frecuentemente
formados en la región.
En la región del Pacífico se presenta un cambio en cuanto a la proporción
de las masas debido a la convergencia de diferentes corrientes oceánicas: la
Corriente de California con aguas de origen subártico con flujo hacia el ecuador, la
cual lleva agua relativamente fría y densa, la corriente Costera Mexicana con flujo
hacia el polo (Figura 2), la cual trae consigo agua relativamente cálida y de menor
densidad de origen tropical y en menor proporción aguas con alta salinidad
generadas en el GC (Lavín et al. 2006, Pantoja et al. 2012). Aproximadamente frente
a la punta de la Península de Baja California la Corriente de California se separa
en dos corrientes (dependiendo de la época del año), una alimentando a la corriente
Nor-Ecuatorial con flujo hacia el oeste, mientras que la otra corriente continúa su
flujo hacia el ecuador a lo largo de la costa de México. (Kessler 2006, Godínez et
al. 2010). En contraste, el flujo polar de la corriente Costera Mexicana es

8
particularmente evidente en verano (Beier et al. 2003, Kessler 2006, Lavín et al.
2006).
Figura 2. Representación de las principales corrientes en la región del Pacífico Central Mexicano (PCM): CC, Corriente de California; CGC, Corriente del Golfo de California; y CCM, Corriente Costera Mexicana (Figura tomada de Pantoja et al., 2012).
Adicionalmente a este sistema de Corrientes, la región de estudio se
caracteriza por presentar surgencias costeras intensas, forzadas por la variación
estacional de los vientos provenientes del noroeste. Strub et al. (1987)
caracterizaron este patrón de vientos en dos regímenes estacionales: uno otoño-
invierno, donde los vientos promedio son casi cero, o débilmente hacia el sur, y otro
primavera-verano donde los vientos fluyen hacia el sur y son en gran medida

9
responsables de que en esta región ocurran las surgencias costeras (Kurczyn et al.
2012).
La zona de confluencia entre diferentes masas de agua (Corriente Costera
Mexicana y Corrientes del Golfo) genera remolinos en la entrada del golfo (Kurczyn
et al. 2012, Pantoja et al. 2012). Donde la presencia constante de remolinos
anticiclónicos en la zona está relacionada tanto con la intensificación de la Corriente
Costera Mexicana como con la fuerte circulación que ocurre en la entrada del Golfo
de California (Kurczyn et al., 2012) y la máxima de generación de remolinos
ciclónicos está relacionado con el máximo de surgencias presentes en la zona. El
mayor número de generación de remolinos ocurre donde existe un cambio abrupto
en la línea de costa.
En este contexto, es probable que los remolinos de mesoescala tengan un
papel importante en el transporte y/o retención de las propiedades químicas y de
los organismos planctónicos, así como la redistribución y mezcla de propiedades
físicas en los diferentes niveles de la columna de agua.
3. HIPÓTESIS
Sobre la base de estos estudios de la distribución vertical de larvas de peces
(Sánchez-Velasco et al. 2007, Danell-Jiménez et al. 2009, Contreras-Catala et al.
2012, 2015) y de los procesos físicos como remolinos de mesoescala (Zamudio et
al. 2008, Lavín et al. 2013, 2014), indican que durante los periodos de estratificación
fuerte (con alta anomalía de energía potencial) donde el flujo de la energía cinética
geostrófica es baja (con ausencia de remolinos de mesoescala), la distribución

10
vertical de las larvas de peces serán estratificadas. Por el contrario, en las zonas
donde la energía cinética geostrófica es alta (zonas con influencia de remolinos de
mesoescala), las larvas se extenderá en la columna de agua por procesos de
mezcla y la advección vertical asociados con la circulación secundaria dentro de los
remolinos.
4. OBJETIVO GENERAL
Describir los efectos de las zona más energéticas (mayor energía cinética
geostrófica) distribución tridimensional de las larvas de peces de cuatro
especies mesopelágicas más abundantes (V. lucetia, D. laternatus, B.
panamense y T. mexicanus) en el sur del Golfo de California durante la
temporada de estratificación fuerte de verano/otoño.
4.1 OBJETIVOS PARTICULARES
Monitorear los remolinos de mesoescala mediante el uso de imágenes de
satélite NOAA/AVHRR, GOES SST y Aqua/MODIS LAC
Describir los gradientes ambientales de temperatura, densidad y velocidades
geostróficas en el área de estudio.
Identificar las zonas con baja actividad de mesoescala, es decir, las zonas
con baja energía cinética geostrófica, y sobre una actividad de mesoescala
fuerte, es decir, las zonas con alta energía cinética geostrófica.
Analizar y comparar estadísticamente la distribución tridimensional de las
larvas de peces más abundantes en zonas de baja actividad de mesoescala
(zonas con baja energía cinética geostrófica), y sobre una actividad de

11
mesoescala fuerte (zonas con alta energía cinética geostrófica) durante
periodos de estratificación (con altas anomalías de energía potencial).
5. MÉTODOS
En el presente estudio se analizaron y compararon datos provenientes de 5
cruceros oceanográficos, (Agosto 2005, Octubre 2007, Julio 2010, Julio 2011 y
Abril-Mayo 2012), en el sur del Golfo de California durante periodos cálidos cuando
la estratificación fue alta, excepto en abril-mayo cuando la capa de mezcla
superficial estaba disminuyendo (periodo frío). En 3 de ellos se observaron
remolinos de diversos orígenes y evolución (ver Tabla 2). Los remolinos fueron
monitoreados mediante imágenes de satélite (NOAA/AVHRR, GOES SST y
Aqua/MODIS LAC) meses antes, durante y después de cada crucero. El monitoreo
satelital estuvo a cargo del personal del laboratorio de Circulación y Dispersión
Costera (CIRCOS) del Departamento de Oceanografía Física del CICESE y del
laboratorio de Ecología del fitoplancton de la Facultad de Ciencias Marinas, UABC.
En cada crucero se diseñó una red de estaciones oceanográficas que cubrió
la estructura de la columna de agua, de acuerdo a la disponibilidad e infraestructura
durante el muestreo; los cruceros se realizaron a bordo del B/O Francisco de Ulloa
del CICESE y el B/O El Puma de la UNAM.
Se realizaron arrastres estratificados de zooplancton durante el día y la noche
a diferentes niveles de profundidad los cuales fueron seleccionados de acuerdo a la
estructura hidrográfica y logística disponible en cada crucero (ver Tabla 2). Durante

12
julio de 2010 (L2, H1 y H2) y julio de 2011 (H1), los arrastres se realizaron cada 17
m hasta la termoclina, y en cada capa de la termoclina 50 m hasta 200 m de
profundidad. En agosto de 2005 (L1), octubre de 2007 (H3) y abril-mayo de 2012
(L4), los arrastres fueron cada 50 m desde la superficie hasta 200 m de profundidad.
Se utilizaron redes de zooplancton cónicas de apertura-cierre, con un tamaño
de 50 cm de diámetro boca, con una longitud 250 cm y malla de 505μm
(http://www.generaloceanics.com). Esta red se ubica cerrada en la parte inferior del
estrato donde se tomen las muestras, a continuación, esta se abrió manualmente
con un mensajero de bronce, y se inició la operación de arrastre. Cuando se llegó a
la parte superior del estrato a muestrear, la red se cerró con otro mensajero y el
lance es terminado. Este sistema evita la captura de muestra de otros organismos
de otras capas. Para garantizar un muestreo preciso de cada estrato de
profundidad, la profundidad de la red se calculó por el método del coseno del ángulo
del cable, siguiendo las especificaciones estándar de Smith y Richardson (Smith &
Richardson 1979). Esta técnica de muestreo estratificado ha sido utilizado con éxito
en estudios previos (Sánchez-Velasco et al. 2007, Danell-Jiménez et al. 2009,
Contreras-Catala et al. 2012, 2015).
El volumen de agua filtrada se calculó utilizando flujómetros calibrados
colocados en la boca de cada red. Las muestras se fijaron con formaldehido al 5%
neutralizado con borato de sodio. La biomasa del zooplancton, se estimó mediante
el método del volumen desplazado (Kramer et al. 1972) y se estandarizaron a
ml/1000 m3. Las larvas de peces se separaron de todas las muestras y V. lucetia,
D. laternatus, B. panamense y T. mexicanus se identificaron utilizando el atlas

13
CALCOFI (Moser 1996). La etapa de desarrollo se determinó en relación con la
flexión de la notocorda siguiendo los criterios de Kendall et al. (Kendall et al. 1981)
y sólo se seleccionaron las larvas de peces en pre-flexión. La abundancia de larvas
de peces se estandarizó a número de larvas por cada 10 m2 (Smith & Richardson
1979).
5.1 Análisis de datos
Con los datos de CTD se calcularon las velocidades geostróficas a la
profundidad mínima común entre pares de estaciones (usualmente 1000 m) a partir
de las distribuciones de la Θ (°C) y SA utilizando el software de TEOS-10. Se empleó
una interpolación de mapeo objetivo estándar utilizando la correlación Gaussiana
clásica con errores relativos de 0,1 a 70 km en la escala horizontal de longitud y de
30 m en la escala vertical (Chelton et al. 1998). Adicionalmente se calculó la
profundidad de la capa de mezcla superficial de acuerdo a la metodología de Kara
et al. (2000). Esta emplea un criterio basado en la densidad que tiene una diferencia
de temperatura fija de 0.8 °C.
Con el objetivo de identificar y diferenciar regiones con baja y alta actividad
de mesoescala. Se calculó la energía cinética geostrófica (J/m3)
01 z
rg hz dz
hEC

14
donde g es la velocidad geostrófica normal en cada sección, h es la
profundidad como se definió anteriormente, ρ es la densidad del agua y dz
representan el área diferencial de cada celda. ECg se puede entender como la
energía cinética geostrófica media a lo largo de cada transecto en referencia al nivel
de 300 m de profundidad. El flujo de energía cinética geostrófica incluye la
contribución de los remolinos, filamentos y otros procesos de mesoescala, pero se
consideró que cuando un remolino está presente, la mayor parte del flujo de energía
cinética geostrófica resulta de la dinámica del remolino.
Así mimo se calculó la anomalía de la energía potencial (φ) la cual es definida
como la cantidad de trabajo por unidad de volumen requerida para distribuir la
columna de agua completamente a una profundidad específica (Simpson et al.
1981) la cual se expresa en J/m3.
donde z es la coordenada vertical (positivos hacia arriba), ρ (z) es el perfil de
densidad en una columna de agua de la profundidad h. Esta anomalía de energía
potencial es cero para una columna de agua completamente mezclada. Todas las
situaciones estratificadas con diferencias en ρ dan un cambio en φ, ponderada con
la profundidad del agua. La energía potencial tiene anomalía positiva para una
columna de agua estratificada estable y se vuelve negativa para una columna de
agua estratificada de forma inestable. Físicamente, φ estima la cantidad de energía
por unidad de volumen que es necesaria para mezclar toda la columna de agua,
0 01 1( ) ; ;
h hgz dz dz
h h

15
dando lugar a la mezcla vertical completa (Fiedler et al. 2013). La profundidad
especificada h=300 m fue seleccionada para cubrir la picnoclina en todos los lugares
y épocas del año dentro de la zona de estudio.
La anomalía de la energía potencial de Simpson et al. (Simpson et al. 1981)
se puede entender como una medida de la estratificación de columna de agua.
Los valores en los transectos promediados para la energía cinética
geostrófica varían 5-30 J/m3 y de 400-1000 J/m3 para la anomalía de la energía
potencial (Tabla 3). Por lo tanto "bajo" y "alto", son muy diferentes en ambos casos,
y no son directamente comparables.
Para cada conjunto de datos de los transectos se construyeron dos matrices
biológicas, (Fig. 3; Tabla 2). Para evaluar la significancia estadística y observar si
existen diferencias significativas de la abundancia de larvas total entre el día y la
noche, se utilizó una prueba no paramétrica de Mann-Whitney (Sokal & Rohlf 1985,
Siegel & Castellón 1988). Posteriormente se aplicó un análisis de agrupamiento
basado en una matriz de abundancia de especies vs muestras (Clarke & Ainsworth
1993) para determinar grupos de muestras de larvas de peces o con semejanza en
la abundancia y distribución de las especies colectadas en estas. Con una matriz
de datos triangular se construyó un dendrograma con la medida de similitud de Bray-
Curtis; (Sokal & Rohlf 1985) utilizando el ligamiento beta flexible (β = -0.25) sobre la
base de datos transformada a raíz cuarta con el objetivo de reducir al mínimo el
efecto de valores atípicos (Field et al. 1982). El escalamiento del dendrograma se
realizó en base a la función de Wishart´s (Wishart 1969). Esta función calcula la
pérdida de información en cada paso del análisis de agrupamiento jerárquico. A

16
medida que se aglomeran los grupos, la cantidad de información disminuye hasta
que todos los grupos se aglomeran y no se mantiene ninguna información. Esta
función objetiva se re-escala desde 0% a 100% de información (McCune et al.
2002).
Para detectar si existen diferencias significativas entre los hábitats de larvas
de peces definidos a priori, se aplicó un análisis de similitud unidireccional
(ANOSIM). Este procedimiento utiliza la diferencia entre los rangos de los valores
promedio de la disimilitud de Bray-Curtis en las abundancias y tipos de organismos,
entre replicas entre las muestras (rb) y dentro de las muestras (rw) para dar un
estadístico de prueba,
1
14
b wr rR
n n
Donde n es el número total de réplicas sumadas de dos muestras. R es
escalada entre -1y 1 y el valor de cero representa la hipótesis nula de que no existen
diferencias significativas entre el conjunto de muestras o asociaciones (Clarke 1993,
Clarke & Ainsworth 1993).

17
Figura 3. Localización del área de estudio y de las estaciones de muestreo durante los cinco cruceros. (a) Estaciones con baja energía cinética geostrófica y (b) estaciones con alta energía cinética geostrófica en el sur del Golfo de California. Estrellas negras, solo lances de CTD. Círculos rojos lances de CTD y muestreos de zooplancton.

18
Tabla 2. Información general de los cruceros oceanográficos realizados en el sur del Golfo de California.
Crucero
Agosto
de
2005
Julio
de
2010
Julio
de
2010
Abril
de
2012
Julio de
2011
Julio de
2010
Octubre de
2007
Línea L1 L2 L3 L4 H1 H2 H3
Días de
muestreo 2 2 3 3 4 2 4
Estratos de
muestreo de
zooplancton
(m)
0-50;
50-100;
100-
150
y 150-
200
0-15;
15-30;
30-45;
50-
100;
100-
150
y 150-
200
0-50;
50-
100;
100-
150
y 150-
200
0-15
15-50
50-200
0-17;
17-34;
34-51;
50-100;
100-150
y 150-
200
0-15;
15-30;
30-45;
50-100;
100-150
y 150-
200
0-50;
50-100;
100-150
y 150-200
Estaciones
de muestreo
biológicas
7 9 10 16 16 8 13
Estructura
de
mesoescala
Remolino
ciclónico
Remolino
ciclónico
Remolino
ciclónico
Para determinar la jerarquía de las especies en cada hábitat como especies
dominantes, frecuentes, constantes y poco comunes, se aplicó la prueba de
Olmstead-Tukey. Esta prueba considera la abundancia relativa media contra el
porcentaje de la frecuencia de aparición de cada especie (Sokal & Rohlf 1985). Así
mismo se determinó la contribución de las especies en cada hábitat , utilizando el
porcentaje de similitud (SIMPER) utilizando el programa computacional PRIMER v7;
(Clarke & Warwick 2001). Este análisis descompone la contribución de cada especie
(u otra variable) a la similitud observada entre las muestras. Permite la identificación
de las especies que son más importantes en el patrón observado de similitud. El

19
método utiliza la medida de similitud de Bray-Curtis, comparando a su vez, cada
muestra en el Grupo i con cada muestra en el Grupo ii. El método Bray-Curtis opera
a nivel de especie, y por lo tanto la similitud media entre los grupos i y ii se puede
obtener para cada especie (Clarke & Warwick 2001).
Para definir la relación entre las variables ambientales y la distribución
tridimensional de larvas de peces, se aplicó un análisis canónico de
correspondencia (Ter Braak 1986), Esto se obtiene mediante el uso de la matriz
estandarizada de abundancia de larvas de peces y con las variables ambientales
tomando los promedios por estrato en la columna de agua de: volumen desplazado
del zooplancton (ml/1000 m3), temperatura conservativa (Θ °C), salinidad absoluta
(SA, g/kg), concentración de clorofila a (mg/m3) y oxígeno disuelto (mL/L).

20
6. RESULTADOS
6.1 Estructura hidrográfica y circulación
6.1.1 Transectos con poca evidencia de remolinos de mesoescala.
Las secciones de todas las propiedades hidrográficas se representan a la
profundidad de 220 m de profundidad, justo por debajo del máximo de muestreo de
larvas de peces.
En Agosto de 2005 en la sección L1 (Fig. 4a), las velocidades geostróficas
fueron < 0.1 m/s excepto cerca de la península de Baja California donde
aparentemente fluyó una corriente costera (Fig. 4b). La picnoclina se definió como
la capa con un fuerte gradiente de densidad, entre las isopicnas de 25 y 23 kg/m3 a
~ 50 m de profundidad debajo de la capa de mezcla superficial que fluctuó entre
~ 10 y 25 m de profundidad (Fig. 4c). La estructura térmica (Fig. 4d) mostró una
termoclina somera entre las isotermas de 25 y 20 oC, coincidiendo con la picnoclina.
El flujo de energía cinética geostrófica tuvo un valor promedio de 4.8 J/m3 (Tabla
3), mostrando bajos valores a lo largo del transecto (Fig. 4e), excepto cerca de la
costa peninsular (~ 10 J/m3). El promedio de la anomalía de la energía potencial fue
de 1070 J/m3 (Tabla 3) siendo alta a lo largo del transecto (Fig. 4f).

21
Figura 4. Distribución tridimensional de larvas de peces sobre la línea con baja energía (L1). (a) Imagen de la concentración de clorofila a superficial (MODIS-AQUA LAC) durante el 17 de Agosto de 2005. Los puntos representan las estaciones de muestreo. (b) Sección vertical de las velocidades geostróficas (m/s) en L1. (c) Distribución vertical de larvas de Vinciguerria lucetia (larvas/10m2) sobre la anomalía de la densidad (kg/m3). (d) larvas de Diogenichthys laternatus sobre la estructura térmica (Θ oC). (e) Gráficos de la distribución del flujo de energía cinética geostrófica (J/m3) y (f) anomalía de la energía potencial (J/m3). La línea roja en las Figuras 4c y d representa la profundidad de la capa de mezcla superficial.

22
Tabla 3. Valores promedio del flujo de energía cinética geostrófica y anomalía de la energía potencial (J/m3) referidos a 300 m de profundidad (ver la posición de los transectos en Fig. 3).
Crucero Línea
Estructura
de
mesoescala
Flujo de
energía
cinética
geostrófica
(J/m3)
Anomalía de
la energía
potencial
(J/m3)
Agosto de 2005 L1 4.8 1074.0
Julio de 2010 L2 5.2 775.8
Julio de 2010 L3 7.2 799.4
Abril-Mayo de 2012 L4 6.9 422.64
Julio de 2011 H1 Remolino ciclónico
33.2 928.03
Julio de 2010 H2 Remolino ciclónico
21.3 744.19
Octubre de 2007 H3 Remolino anticiclónico
13.6 1097.68
Durante Julio de 2010 en ambos transectos, L2 y L3 la picnoclina se observó
a ~ 40 m de profundidad entre las isopicnas de 25 y 23 kg/m3, la capa de mezcla
superficial se encontró entre ~ 15 y 20 m de profundidad (Figs. 5c y 6c). Sin embargo
en L3 la picnoclina y la capa de mezcla superficial se profundizó a ~ 50 m cerca de
la costa continental. La termoclina se ubicó en las isotermas de 26 y 18 oC
coincidiendo con la picnoclina (Figs. 5d y 6d). Las velocidades geostróficas fueron
≤ 0.1 m/s, excepto en la línea L3 cerca de la costa continental, donde las velocidades
llegan hasta 0.2 m/s, posiblemente asociadas a un chorro con surgencias cerca de
las estaciones costeras (Fig. 6b). Los valores de la energía cinética geostrófica
fueron bajos a lo largo de las dos líneas, con un promedio de 5,2 y 7,2 J/m3

23
respectivamente (Tabla 3), con un ligero aumento en las velocidades geostróficas
que fueron ≤ 0,1 m/s (Figs. 5e y 6e). Por el contrario, los valores de anomalías de
energía potencial fueron altos a lo largo de dos transectos, en su mayoría > 700
J/m3 (Figs. 5f y 6f).

24
Figura 5. Distribución tridimensional de larvas de peces sobre la línea con baja energía (L2). (a) Imagen de la concentración de clorofila a superficial (MODIS-AQUA LAC) durante el 11 de Julio de 2011. Los puntos representan las estaciones de muestreo. (b) Sección vertical de las velocidades geostróficas (m/s) en L2. (c) Distribución vertical de larvas de Vinciguerria lucetia (larvas/10m2) sobre la anomalía de la densidad (kg/m3). (d) larvas de Diogenichthys laternatus sobre la estructura térmica (Θ oC). (e) Gráficos de la distribución del flujo de energía cinética geostrófica (J/m3) y (f) anomalía de la energía potencial (J/m3). La línea roja en las Figuras 5c y d representa la profundidad de la capa de mezcla superficial.

25
Figura 6. Distribución tridimensional de larvas de peces sobre la línea con baja energía (L3). (a) Imagen de la concentración de clorofila a superficial (MODIS-AQUA LAC) durante el 11 de Julio de 2011. Los puntos representan las estaciones de muestreo. (b) Sección vertical de las velocidades geostróficas (m/s) en L3. (c) Distribución vertical de larvas de Vinciguerria lucetia (larvas/10m2) sobre la anomalía de la densidad (kg/m3). (d) larvas de Diogenichthys laternatus sobre la estructura térmica (Θ oC). (e) Gráficos de la distribución del flujo de energía cinética geostrófica (J/m3) y (f) anomalía de la energía potencial (J/m3). La línea roja en las Figuras 6c y d representa la profundidad de la capa de mezcla superficial.

26
En el transecto muestreado durante Abril y Mayo de 2012 L4 (Fig. 7a) las
velocidades geostróficas fueron ≤ 0.1 m/s (Fig. 7b), la picnoclina se ubicó a ~ 40 m
de profundidad (25.5 and 25 kg/m3) y la capa de mezcla superficial se encontró a ~
10 m de profundidad (Fig. 7c). La termoclina se observó entre las isotermas de 22
y 18 oC, similar a la profundidad de la picnoclina (Fig. 7d). El flujo de energía cinética
geostrófica tuvo un promedio de 6.9 J/m3 (Tabla 3), mostrando valores bajos a lo
largo del transecto, excepto en zonas donde la velocidad geostrófica era fuerte (Fig.
7e.). Mientras que la anomalía de la energía potencial fue relativamente baja y poco
variable a lo largo del transecto con valores de ~ 450 J/m3 (Fig. 7f).

27
Figura 7. Distribución tridimensional de larvas de peces sobre la línea L4 (con baja energía). (a) Imagen de la concentración de clorofila a superficial (MODIS-AQUA LAC) durante el 3 de Mayo de 2012. Los puntos representan las estaciones de muestreo. (b) Sección vertical de las velocidades geostróficas (m/s) en L4. (c) Distribución vertical de larvas de Vinciguerria lucetia (larvas/10m2) sobre la anomalía de la densidad (kg/m3). (d) larvas de Diogenichthys laternatus sobre la estructura térmica (Θ oC). (e) Gráficos de la distribución del flujo de energía cinética geostrófica (J/m3) y (f) anomalía de la energía potencial (J/m3). La línea roja en las Figuras 7c y d representa la profundidad de la capa de mezcla superficial.

28
6.1.2 Transectos con evidencia de remolinos de mesoescala
Las secciones de las velocidades geostróficas y estructura hidrográfica que
cruzan los remolinos observados en el Golfo se muestran en las Figs. 8-10.
Las imágenes de satélite definieron la presencia de un remolino durante Julio
de 2011 en la sección H1 (Fig. 8a). Las velocidades geostróficas confirmaron la
presencia de un remolino ciclónico con un diámetro de ~ 150 km extendiéndose a
más de 300 m de profundidad, las velocidades azimutales fueron > 0.35 m/s (Fig.
8b). La profundidad de la picnoclina se observó a ~ 50 m, entre las isopicnas de
~25.5 y 22 kg/m3 comprimiéndose entre las estaciones A08 y A14, donde la capa
de mezcla superficial estuvo a ~ 20 de profundidad (Fig. 8c). Las isopicnas por
debajo de la picnoclina se observaron en forma de domo en la parte central del
transecto (estaciones A08 y A14), en todas las profundidades hasta 300 m. La
termoclina se encontró entre las isotermas de 16 y 18 oC mostrando una distribución
similar a la picnoclina. La descripción detallada de este remolino se presentan en
Sánchez-Velasco et al. (2013). El promedio del flujo de energía cinética geostrófica
fue la más alta (33,2 J/m3) (Tabla 3), justo donde se observaron los mayores valores
de las velocidades geostróficas > 0,3 m/s (Fig. 8e). Si bien la anomalía de la energía
potencial tuvo un promedio de 928 J/m3, esta fue alta a lo largo del transecto (Fig.
8f).

29
Figura 8. Distribución tridimensional de larvas de peces sobre la línea con alta energía (H1). (a) Imagen de la concentración de clorofila a superficial (MODIS-AQUA LAC) durante el 29 de Julio de 2011. Los puntos representan las estaciones de muestreo. (b) Sección vertical de las velocidades geostróficas (m/s) en H1. (c) Distribución vertical de larvas de Vinciguerria lucetia (larvas/10m2) sobre la anomalía de la densidad (kg/m3). (d) larvas de Triphoturus mexicanus sobre la estructura térmica (Θ oC). (e) Gráficos de la distribución del flujo de energía cinética geostrófica (J/m3) y (f) anomalía de la energía potencial (J/m3). La línea roja en las Figuras 8c y d representa la profundidad de la capa de mezcla superficial.

30
En la línea H2 durante Julio de 2010, se muestreo parte de un remolino
ciclónico el cual es evidente en la Fig. 9a. Las velocidades geostróficas (Fig. 9b)
demuestran esta rotación con velocidades azimutales > 0.1 m/s indicando la
presencia de un remolino ciclónico débil con una tendencia superficial anticiclónica.
El diámetro del remolino fue de ~ 60 km (entre C04 y C07) con una profundidad >
300 m. La picnoclina se observó hundida entre las isopicnas de 25 y 23 kg/m3, la
capa de mezcla superficial se engrosó de ~ 25 m (C01 y C02) a ~ 50 m de
profundidad (C06 y C08), lo que indica una tendencia a una circulación anticiclónica
superficial. Sin embargo las isopicnas por debajo de la picnoclina formaron un domo
entre las estaciones C05 y C07, mostrando una larga intrusión de las isopicnas
(26.5) desde los 290 m en C01 hasta 190 m en C06 (Fig. 9c). La estructura térmica
en la Fig. 9d fue muy similar a la distribución de las isopicnas, la isoterma subió
desde los 260 m hasta los 180 m. La descripción detallada de este remolino se
puede consultar en Contreras-Catala et al. (2015). El flujo de energía cinética
geostrófica presenta un valor promedio alto (21,3 J/m3) (Tabla 3), con valores
máximos donde las velocidades geostróficas eran altas (Fig. 9e). La anomalía de la
energía potencial fue alta, presentando un promedio de 744,19 J/m3 (Tabla 3),
mostrando un ligero aumento en la costa continental (Fig. 9f).

31
Figura 9. Distribución tridimensional de larvas de peces sobre la línea con baja energía (H2). (a) Imagen de la concentración de clorofila a superficial (MODIS-AQUA LAC) durante el 11 de Julio de 2010. Los puntos representan las estaciones de muestreo. (b) Sección vertical de las velocidades geostróficas (m/s) en H2. (c) Distribución vertical de larvas de Vinciguerria lucetia (larvas/10m2) sobre la anomalía de la densidad (kg/m3) y (d) larvas de Diogenichthys laternatus sobre la estructura térmica (Θ oC). (e) Gráficos de la distribución del flujo de energía cinética geostrófica (J/m3) y (f) anomalía de la energía potencial (J/m3). La línea roja en las Figuras 9c y d representa la profundidad de la capa de mezcla superficial.

32
Un remolino más pequeño fue detectado con las imágenes de satélite (Fig.
10a) durante Octubre de 2007 en la sección H3. Las velocidades geostróficas (Fig.
10b) mostraron una rotación anticiclónica con grandes velocidades azimutales >
0.25 m/s con un diámetro de ~ 90 km y una profundidad de 70 m en su centro. La
picnoclina se extendió desde las isopicnas de 25 a 23 kg/m3 (Fig. 10c), la termoclina
se observó entre las isotermas de 26 y 18 oC (Fig. 10d). Tanto la picnoclina como la
termoclina se deprimen en la parte central (entre A02 y A11) justo donde la capa de
mezcla superficial se incrementa desde 20 m en su margen hasta ~ 70 m de
profundidad en el centro. La descripción de este remolino se puede consultar en
Contreras-Catala et al. (2012). El flujo de energía cinética geostrófica tuvo un valor
promedio de 13.6 J/m3 (Tabla 3), los valores más bajos se presentaron en el centro
del remolino (Fig. 10e). Por el contrario, la anomalía de la energía potencial mostró
los valores más altos en el centro del remolino ~ 1,100 J/m3, disminuyendo hacía
los márgenes, principalmente del lado de la costa continental (Fig. 10f).

33
Figura 10. Distribución tridimensional de larvas de peces sobre la línea con baja energía (H3). (a) Imagen de la concentración de clorofila a superficial (MODIS-AQUA LAC) durante el 15 de Octubre de 2007. Los puntos representan las estaciones de muestreo. (b) Sección vertical de las velocidades geostróficas (m/s) en H3. (c) Distribución vertical de larvas de Vinciguerria lucetia (larvas/10m2) sobre la anomalía de la densidad (kg/m3) y (d) larvas de Triphoturus mexicanus sobre la estructura térmica (Θ oC). (e) Gráficos de la distribución del flujo de energía cinética geostrófica (J/m3) y (f) anomalía de la energía potencial (J/m3). La línea roja en las Figuras 10c y d representa la profundidad de la capa de mezcla superficial.

34
6.2 Distribución tridimensional de larvas de peces mesopelágicas
6.2.1 Transectos sin influencia de remolinos de mesoescala.
En la presente sección se presentan los resultados estadísticos que se
aplicaron con la matriz de la abundancia de larvas de peces en los transectos con
poca evidencia de remolinos de mesoescala.
La prueba de Mann-Whitney mostró que no existen diferencias significativas
entre los muestreos realizados durante el día y la noche (con nivel de confianza del
95%). Con un nivel de corte del 13% de la información retenida la medida de
disimilitud de Bray-Curtis definió dos grupos de muestras a los que se les denominó
hábitats de larvas de peces (Fig. 11) los cuales fueron significativamente diferentes
(ANOSIM: R=0.4, con un nivel de confianza del 95%) y se nombraron de acuerdo a
su ubicación en la columna de agua como: “hábitat superficial”, localizado de la
picnoclina hacia la superficie principalmente (área sombreada en Fig. 11) y “hábitat
subsuperficial”, ubicado principalmente por debajo de la picnoclina (área blanca en
Fig. 11).

35
Figura 11. Dendrograma de los hábitats de larvas de peces definidos por la medida de disimilitud de Bray-Curtis de las líneas con baja energía cinética geostrófica. Larvas de peces recolectadas en el sur del Golfo de California.
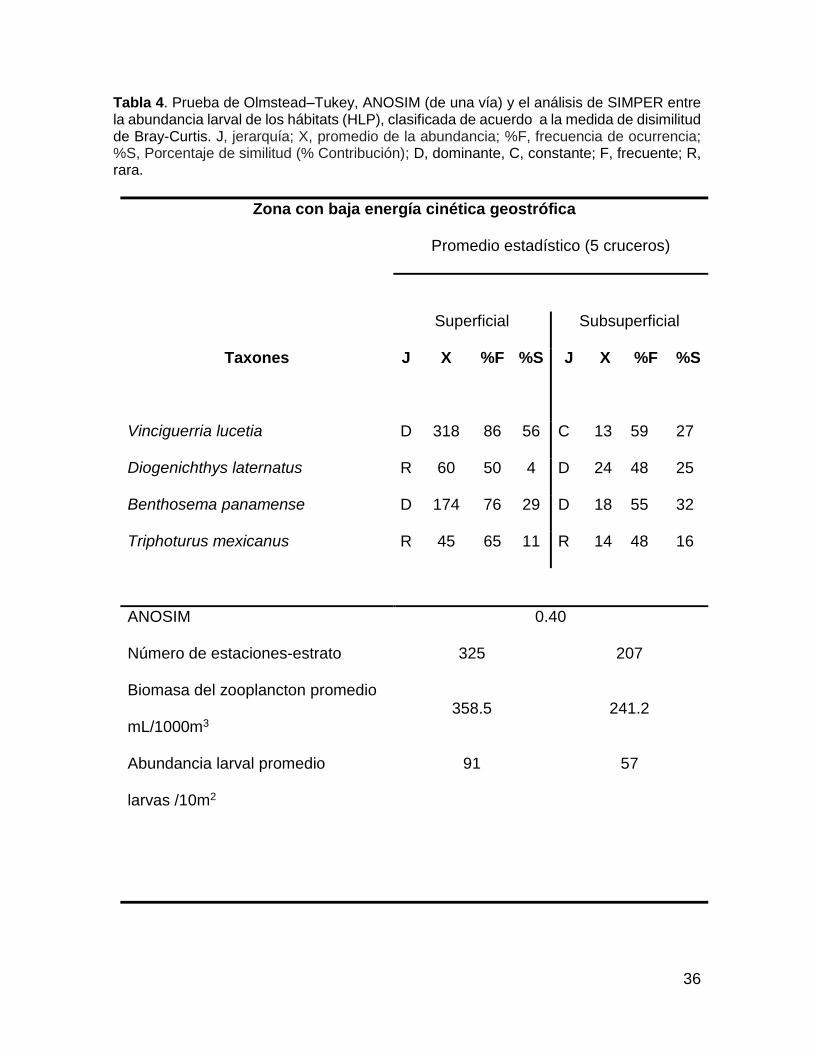
El “hábitat superficial” se localizó en y sobre la termoclina, se conformó por
325 muestras y presentó altas abundancias larvales promedio (91 larvas/10 m2).
Las especies dominantes fueron V. lucetia y B. panamense, especies con afinidad
a la capa superficial. La contribución de estas dos especies para la conformación
de este hábitat fue de 56 y 29% respectivamente (Tabla 4). La distribución de V.
lucetia se observa en las Figs. 4c, 5c, 6c y 7c. Sus mayores abundancias se
observaron desde la termoclina hasta la superficie, asociándose sobre todo con las
estaciones de baja energía cinética geostrófica, excepto durante Agosto de 2005
(Fig. 3c), donde su distribución es homogénea en la columna de agua. Las larvas
de B. panamense mostraron una distribución similar a las larvas de V. lucetia, para
Agosto de 2005 presentaron altas abundancias en toda la columna de agua.
36
Tabla 4. Prueba de Olmstead–Tukey, ANOSIM (de una vía) y el análisis de SIMPER entre la abundancia larval de los hábitats (HLP), clasificada de acuerdo a la medida de disimilitud de Bray-Curtis. J, jerarquía; X, promedio de la abundancia; %F, frecuencia de ocurrencia; %S, Porcentaje de similitud (% Contribución); D, dominante, C, constante; F, frecuente; R, rara.
Zona con baja energía cinética geostrófica
Promedio estadístico (5 cruceros)
Superficial Subsuperficial
Taxones J X %F %S J X %F %S
Vinciguerria lucetia D 318 86 56 C 13 59 27
Diogenichthys laternatus R 60 50 4 D 24 48 25
Benthosema panamense D 174 76 29 D 18 55 32
Triphoturus mexicanus R 45 65 11 R 14 48 16
ANOSIM 0.40
Número de estaciones-estrato 325 207
Biomasa del zooplancton promedio
mL/1000m3 358.5 241.2
Abundancia larval promedio
larvas /10m2
91 57

37
Continuación Tabla 3
Zona con alta energía cinética geostrófica
HLP I HLP II
Taxones J X %F %S J X %F %S
Vinciguerria lucetia D 34 88 69 D 47 72 26
Diogenichthys laternatus D 29 63 25 R 24 49 10
Benthosema panamense R 18 21 D 29 80 36
Triphoturus mexicanus R 7 44 D 30 75 27
ANOSIM -0.23
Número de estaciones-estrato 50 77
Biomasa del zooplancton promedio
mL/1000m3 467 206
Abundancia larval promedio
larvas /10m2 25 33
El hábitat subsuperficial” se localizó principalmente sobre y por debajo de la
termoclina. Este hábitat se definió por 207 muestras, con una abundancia larval
promedio de 57 larvas/10m2. D. laternatus y B panamense fueron las especies más
dominantes presentando una contribución > 25 % para la conformación de este

38
hábitat (Tabla 3). Las larvas de D. laternatus se localizaron sobre toda la columna
de agua, pero con altas abundancias por debajo de la termoclina, en particular con
el flujo de energía cinética geostrófica baja. Sin embargo durante Agosto de 2015
estas fueron completamente ausentes (Figs. 3d, 4d, 5d y 6d).
La delimitación entre estos dos hábitats se evidenció con el análisis canónico
de correspondencias (Fig. 12) con una correlación de Pearson de 0.72 (Tabla 4). El
“hábitat superficial” presentó una correspondencia con altos valores de oxígeno
disuelto, temperatura, salinidad y volumen desplazado del zooplancton,
asociándose a altas abundancias de V. lucetia y B. panamense estas dos especies
correspondieron especialmente a altos valores de temperatura. Donde el “hábitat
subsuperficial” mostró una correspondencia inversa de los parámetros ambientales
donde: oxígeno disuelto, temperatura, salinidad y volumen desplazado del
zooplancton se asociaron a altas abundancias de D. laternatus.

39
Figure 12. Triplot basado en el diagrama de ordenación del análisis canónico de correspondencias (ACC) sobre líneas con baja energía cinética geostrófica. Las muestras biológicas (cuadros y círculos), variables ambientales (flechas) y centroides de los hábitats (símbolos rojos) y las especies (estrellas); primer eje horizontal y segundo eje vertical. Datos recolectados en 5 cruceros en el sur del Golfo de California. Sal, Salinidad absoluta; BZ, Volumen desplazado del zooplancton; Fluo, fluorescencia; Temp, Temperatura conservativa; OD, Oxígeno disuelto; HLP, Hábitat de larvas de peces.

40
Tabla 5. Porcentaje de contribución de las variables exploratorias por el análisis canónico de correspondencia.
Zona con baja energía cinética geostrófica
Variable % Variable
explicada
Contribución por variable
(%correlación de la variable
por hábitat)
Temperatura conservativa 14.6 39.7
Oxígeno disuelto 17.7 48.3
Salinidad absoluta 2.8 7.5
Volumen desplazado del
zooplancton 0.8 2.3
Fluorescencia 0.8 2.2
Zona con alta energía cinética geostrófica
Temperatura conservativa 19 51.7
Oxígeno disuelto 16.9 46
Salinidad absoluta 10.3 28
Fluorescencia 2.8 7.7
Volumen desplazado del
zooplancton 2.2 6.1

41
Transectos con presencia de remolinos de mesoescala.
En las muestras con alta energía cinética geostrófica, afectados por
remolinos de mesoescala, la abundancia total de larvas demostró una vez más no
existen diferencias estadísticamente significativas entre el día y la noche (con un
nivel de confianza del 95%). En el dendrograma (disimilitud de Bray-Curtis) el corte
se realizó a un nivel en el que se retiene el 13% de la información aproximadamente
con la diferenciación de dos hábitats de larvas de peces (Fig. 13), estos hábitats
fueron significativamente diferentes (ANOSIM: R = -0.23, con un nivel de confianza
del 95%). En contraste con el conjunto de datos de baja energía, el valor negativo
en ANOSIM mostró una gran variabilidad dentro de los hábitats. El primer hábitat se
caracterizó porque la mayoría de las muestras fueron del remolino ciclónico de Julio
de 2010 (sombreado gris en la Fig. 13); y el segundo hábitat, se conformó
principalmente por muestras agrupadas del remolino anticiclónico de Octubre de
2007 y el remolino ciclónico de Junio de 2011 (área en blanco en la Fig. 13).
El “primer hábitat” (remolino ciclónico de 2010) línea H2, presentó la
abundancia promedio de larvas más baja del estudio (25 larvas/10 m2) se conformó
por 50 muestras (Tabla 3). Las especies dominantes fueron V. lucetia (de afinidad
superficial) y D. laternatus (de afinidad profunda), las cuales contribuyeron con
~69% y 25%, respectivamente. Ambas especies fueron relativamente abundantes
en toda la columna de agua y tienden a evitar las estaciones con alta energía
cinética geostrófica (Figs. 9c y d).

42
El “Segundo hábitat” (remolino ciclónico de 2011 y remolino anticiclónico de
2007); Líneas H1 y H3, respectivamente, la abundancia promedio de larvas fue
ligeramente superior a 33 larvas/10 m2 en 77 muestras. A pesar de que las larvas
de V. lucetia fueron dominantes como en el primer hábitat, las larvas de B.
panamense y T. mexicanus también fueron especies características de este hábitat,
marcando la diferencia con el primer hábitat. La contribución de cada una de las tres
especies fue > 26% (Tabla 4). V. lucetia (Figs.8c y 9c) se distribuyó a lo largo de la
columna de agua en todas las muestras, como en el primer hábitat. Del mismo modo
larvas de T. mexicanus (Figs. 8d y 9d) y larvas de B. panamense se observaron en
todas las profundidades muestreadas, sin embargo T. mexicanus fue la especie más
abundante en los primeros 100 m de profundidad. En las Figs. 8c y d, 10c y d, se
observa una tendencia a evitar las estaciones con la energía cinética geostrófica
más alta.
Las definiciones de estos dos hábitats de larvas de peces también fueron
corroborados por el ACC (Fig. 14), con una correlación de Pearson de 0.67 (Tabla
5). El primer hábitat (remolino ciclónico de 2010) tuvo una correspondencia negativa
con la temperatura, oxígeno disuelto y salinidad intermedia, asociándose
principalmente con altas abundancias de larvas de D. laternatus y V. lucetia. Por
otro lado, el segundo hábitat de larvas de peces (remolino ciclónico de 2011 y
remolino anticiclónico de 2007) presentó una correspondencia positiva con la
temperatura, fluorescencia y oxígeno disuelto y se asoció con alta abundancia de
larvas de T. mexicanus y B. panamense.
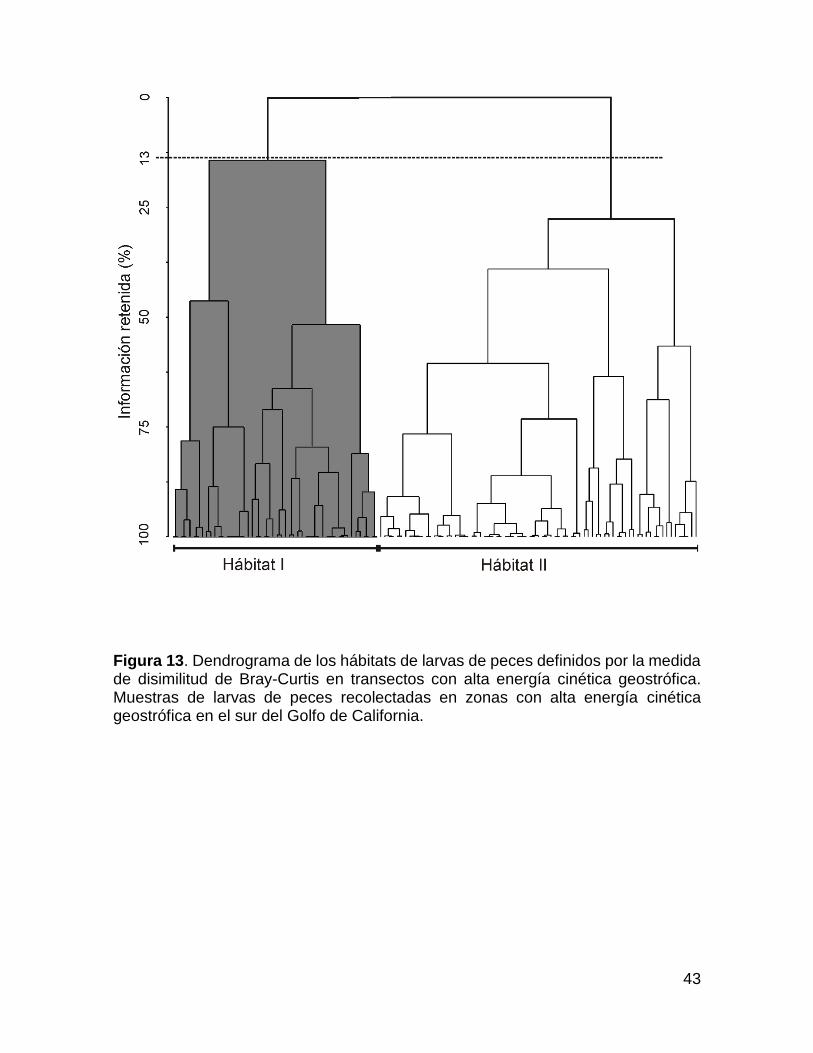
43
Figura 13. Dendrograma de los hábitats de larvas de peces definidos por la medida de disimilitud de Bray-Curtis en transectos con alta energía cinética geostrófica. Muestras de larvas de peces recolectadas en zonas con alta energía cinética geostrófica en el sur del Golfo de California.

44
Figura 14. Triplot basado en el diagrama de ordenación del análisis canónico de correspondencias (ACC) sobre líneas con alta energía cinética geostrófica. Las muestras biológicas (cuadros y círculos), variables ambientales (flechas), centroides de los hábitats (símbolos rojos) y de especies (estrellas); primer eje horizontal y segundo eje vertical. Datos recolectados en transectos con alta energía en el sur del Golfo de California. Sal, Salinidad absoluta; BZ, Volumen desplazado del zooplancton; Fluo, fluorescencia; Temp, Temperatura conservativa; OD, Oxígeno disuelto; HLP, Hábitat de larvas de peces.

45
7. DISCUSIÓN
En el presente estudio se mostraron los efectos de energía cinética
geostrófica sobre la distribución de las larvas de peces mesopelágicas (V. lucetia,
D. laternatus, B. panamense y T. mexicanus) durante años con periodos de
estratificación fuerte, comparando las áreas con alta energía cinética geostrófica
asociados con remolinos de mesoescala con zonas con baja energía donde la
estructura de la columna de agua es más estable. Se elaboró una relación
estadística entre la abundancia larvaria, su distribución tridimensional, y la
estructura de la columna de agua. La mayoría de los estudios anteriores (por
ejemplo, (Contreras-Catala, Sanchez-Velasco, et al. 2012, Sánchez-Velasco et al.
2013, Contreras-Catala et al. 2015)) sugirieron relaciones cualitativas, pero no
realizó una comparación estadística de las zonas con alta y baja energía cinética
geostrófica.
Debido a que la mayor parte de las muestras se encontraban en el verano y
principios de otoño, la columna de agua tenía valores relativamente altos de la
anomalía de energía potencial en todos los transectos analizados, como se
esperaba de acuerdo con los registros anteriores en el golfo durante el verano
(Peguero-Icaza et al. 2008). El presente trabajo se considera una clasificación
previa de los transectos analizados: i) los transectos que no mostraron evidencia de
actividad de mesoescala, donde la energía cinética geostrófica es baja (<10 J/m3),
con isotermas e isopicnas generalmente horizontales así como velocidades
geostróficas < 0,1 m/s; y ii) transectos con evidente actividad de mesoescala con

46
valores de energía cinética geostrófica > 12 J/m3, y la presencia de remolinos
ciclónicos y anticiclónicos, donde las velocidades geostróficas fueron > 0.1 m/s. En
el primer caso (zonas bajas en energía), dos hábitats de larvas de peces se
definieron estadísticamente (superficial y subsuperficial) (Fig. 11). Uno de ellos fue
dominado por larvas de V. lucetia y B. panamense encontrándose principalmente
en y por encima de la picnoclina (~ 40 m de profundidad); mientras que el segundo
hábitat se caracteriza por larvas de D. laternatus ubicadas principalmente en y por
debajo del picnoclina (Figs. 3-6). Este resultado coincide con observaciones
cualitativas anteriores de su distribución vertical (por ejemplo, Danell-Jiménez et al.
2009, Sánchez-Velasco et al. 2013). La corroboración estadística proporcionada por
el presente estudio sugiere que larvas de V. lucetia y B. panamense tienden a
congregarse en la capa superficial, mientras que larvas de D. laternatus se
concentran en la capa subsuperficial. Las distribuciones de profundidad opuestas
de las larvas de estas especies mesopelágicas pueden implicar diferentes
adaptaciones fisiológicas para el ambiente, tales como larvas de V. lucetia y B.
panamense prefieren distribuirse en agua relativamente cálida y “productiva” (> 20
°C y > 1.5 mg/m3), mientras que D. laternatus tolera condiciones hipóxicas (< 1/L
mL), como se ha sugerido en estudios anteriores (Apango-Figueroa et al. 2015,
Davies et al. 2015).
A pesar de esta separación de los hábitats de larvas de peces, en todas las
estaciones de muestreo las mayores densidades de las larvas mesopelágicas
coincidieron en el picnoclina, donde se han registrado los máximos de clorofila en
períodos de estratificación en diferentes océanos (Lasker et al.1975)

47
Lasker et al. (1975) relaciona la capa del máximo clorofila con el éxito de la
primera alimentación de las larvas de especies epipelágicas (larvas de anchoa). Es
posible que la picnoclina y la capa del máximo de clorofila también juegan un papel
importante en la alimentación de las larvas de las especies mesopelágicas, esto se
indica en el presente estudio por su alta abundancia y frecuencia en dicha capa;
larvas asociadas al "hábitat superficial de larvas de peces" migran principalmente
en la capa de mezcla, entre el máximo de clorofila y la superficie, por el contrario, la
especie asociada al " hábitat subsuperficial de larvas de peces", la migración es
principalmente entre la capa profunda y el máximo de clorofila.
Las excepciones a estos patrones de distribución de las larvas se registraron
en agosto de 2005, cuando los niveles de energía cinética geostrófica fueron bajos,
las larvas de B. panamense y V. lucetia se distribuyeron en de toda la columna de
agua muestreada, y no se encontraron larvas de D. laternatus (Fig. 3d). Esta
circunstancia se asoció con anomalías positivas de temperatura que aparentemente
aumentaron el desove de B. panamense y redujeron la de D. laternatus. Aunque no
existe una literatura específica para este problema, ya se ha mencionado que estos
cambios en la abundancia de larvas de peces durante agosto de 2005 se han
observado incidentalmente (Peguero-Icaza et al. 2011).
Dos hábitats de larvas de peces también fueron estadísticamente definidos
(con un nivel de confianza del 95%) en las estaciones de muestreo de áreas con
alta energía cinética geostrófica, donde se detectaron remolinos de mesoescala
(Fig. 13). En ambos hábitats, la abundancia de larvas de la especie dominante fue
menor que en las zonas con baja energía, y las larvas se expandieron en la columna

48
de agua. Uno de los hábitats fue formado por muestras de un remolino ciclónico
(julio de 2010), donde la energía cinética geostrófica era la relatividad alta, las larvas
de V. lucetia y D. laternatus tuvieron la abundancia de larvas más bajo del estudio
(Fig. 7). El segundo hábitat agrupando principalmente muestras de transectos
influenciados por el otro remolino ciclónico (julio de 2011) y el remolino anticiclónico
(octubre de 2007), donde se registró la energía cinética geostrófica más alta del
estudio (> 12 J/m3). En este último grupo, aunque V. lucetia era la especie
dominante, B. panamense y T. mexicanus fueron también importantes, en contraste
con el remolino ciclónico (julio de 2010). Por lo tanto la separación de los dos grupos
de zonas con alta energía cinética geostrófica correspondía más a cambios en la
abundancia de larvas de peces, producto de la intensidad de desove de cada
especie, que a posibles efectos de la rotación de los remolinos sobre la distribución
de las larvas.
La rotación del remolino, independientemente del su sentido de giro, podría
generar mezcla por procesos de convergencia y divergencia, lo que resulta en
condiciones menos estables y favorables tanto para el desove y la alimentación de
las larvas, debido a la dispersión vertical disminuye la tasa de encuentro presa-
depredador.
El efecto de los remolinos en los organismos del zooplancton es compleja
debido a los remolinos evolucionan con el tiempo como resultado de procesos tales
como la difusión y la interacción con el viento. Además, los procesos
submesoescala como circulación secundaria geostrófica y mezcla pueden modular
la distribución/estructura de la comunidad del plancton a través de flujos verticales

49
localizadas en la periferia de los remolinos, por lo tanto se requerirán observaciones
más detalladas y modelos para entender completamente la interacción de los
organismos del zooplancton con la dinámica de los remolinos.
Los fenómenos físicos que afectan zooplancton pueden ocurrir en diferentes
escalas simultáneamente y pueden tener efectos sinérgicos que son difíciles de
medir (Haury et al. 1978, Chelton et al. 2011, León-Chávez et al. 2015). Se
considera que los procesos de mezcla y la convergencia/divergencia que se
producen en los remolinos influyen en la fertilización de las aguas superficiales del
océano, y por lo tanto el posterior enriquecimiento de las redes tróficas (Sangrà et
al. 2005). Sin embargo, los resultados obtenidos en este trabajo, la toma de
muestras obtenidas de estratos de baja resolución, pueden indicar que para los
consumidores primarios de la cadena trófica, por ejemplo, las larvas de peces
mesopelágicas, los remolinos pueden no proporcionar las condiciones óptimas para
el desarrollo de las larvas. Por ejemplo; Holliday et al. (2011), Nieto et al. (2013) y
Song et al. (2012), con diferentes argumentos, han sugerido la posibilidad de que
los remolinos no siempre son favorables para el desarrollo de los huevos y larvas
de peces y estos argumentos deberán aplicarse en estudios multidisciplinarios
futuros. Sin embargo, puede ser que la estratificación de la columna de agua sea
uno de los factores más importantes en la formación y mantenimiento de los hábitats
de algunos organismos zooplanctónicos (Norrbin et al. 1996), incluyendo las larvas
de peces mesopelágicas.
Los resultados de este trabajo en el sur del Golfo de California y Pacífico
adyacente, basados en transectos influenciados por los remolinos de mesoescala

50
(áreas con alta en energía cinética geostrófica) y en transectos con la actividad de
mesoescala débil (zonas bajas en energía), ambas durante periodos de
estratificación fuerte, sugirieron que el nivel del flujo de energía cinética geostrófica
afecta la abundancia y distribución tridimensional de las larvas de peces
mesopelágicas.
A medida que la mayor abundancia de larvas se correlacionó con baja
energía, y la asociación de las larvas fue mayor en las muestras provenientes de
zonas de baja energía, parece que las áreas con alta energía cinética geostrófica,
no proporcionan las mejores condiciones para el desarrollo de las larvas
mesopelágicas.
8. CONCLUSIONES
Los resultados obtenidos en el presente estudio mostraron que la distribución
tridimensional de las larvas de peces mesopelágicas (Vinciguerria lucetia,
Diogenichthys laternatus, Benthosema panamense y Triphoturus mexicanus) en el
sur del Golfo y Pacífico adyacente, depende en gran medida de la energía cinética
geostrófica durante los periodos de estratificación.
Por un lado en regiones donde hubo poca evidencia de procesos de
mesoescala y con baja energía cinética geostrófica < 10 J/m3 la distribución de las
especies dominantes tendieron a ser estratificadas y dependientes de la picnoclina.
Las larvas de V. lucetia (con abundancia media de 318 larvas/10m2) y B. panamense
(174 larvas/10m2) se encuentra principalmente en y por encima de la picnoclina

51
(típicamente ~ 40 m de profundidad). Por el contrario, las larvas de D. laternatus (60
larvas / 10m2) se localizaron principalmente en y por debajo del picnoclina.
Por otro lado, en las estaciones de muestreo donde la energía cinética
geostrófica fue alta y hubo evidencia de remolinos de mesoescala, las larvas de la
especie dominante tenían baja abundancia y se distribuye de manera más uniforme
a través de la columna de agua. Por ejemplo, en un remolino ciclónico, V. lucetia
larvas (34 larvas/10m2) extendieron su distribución a, al menos, el límite de
muestreo 200 m de profundidad por debajo de la picnoclina, mientras que larvas de
D. laternatus (29 larvas/10m2) se encontraron hasta la superficie.
Estos resultados mostraron que el nivel de la energía cinética geostrófica
durante periodos de fuerte estratificación influencia la abundancia y distribución
tridimensional de las larvas de peces mesopelágicas, lo que indica que las áreas
menos energéticas pueden ser ventajosas para el desarrollo de las larvas de peces
mesopelágicas.
9. CONSIDERACIONES
Aunque hay relativamente pocas observaciones con las que se puede
generalizar, se sugiere y son convenientes más muestreos in situ con análisis
estadísticos más amplios y utilizar otras herramientas como el contenido de ácidos
grasos e isótopos estables lo cual nos permitirá complementar un ambiente como
favorable o desfavorable, por lo tanto es tentador especular que este resultado
también podría aplicarse a otras regiones.

52
LITERATURA CITADA
Ahlstrom, E.H. 1959. Vertical distribution of pelagic fish eggs and larvae off
California. Fish. Bull. 60: 107–146.
Álvarez-Borrego, S. 2012. Phytoplankton biomass and production in the Gulf of
California: a review. Bot. Mar. 55: 119–128.
Apango-Figueroa, E., L. Sánchez-Velasco, M.F. Lavín, V.M. Godínez & E.D.
Barton. 2015. Larval fish habitats in a mesoscale dipole eddy in the gulf of
California. Deep Sea Res. Part I Oceanogr. Res. Pap. 103: 1–12.
Badan-Dangon, A., C.E. Dorman, M.A. Merrifield & C.D. Winant. 1991. The Lower
Atmosphere Over the Gulf of California. J. Geophys. Res. 96: 16877–16896.
Badán-Dangon, A., C.J. Koblinsky & T. Baumgartner. 1985. Spring and summer in
the Gulf of California: observations of surface thermal patterns. Ocean. Acta 8:
13–22.
Beier, E. 1997. A Numerical Investigation of the Annual Variability in the Gulf of
California. J. Phys. Oceanogr. 27: 615–632.
Beier, E., M.F. Lavín, J. Gómez, V.M. Godínez & J. García. 2003. La corriente
costera Mexicana. GEOS 23: 152.
Ter Braak, C.J.F. 1986. Canonical correspondence analysis: a new eigenvector
technique for multivariate direct gradient analysis. Ecology 67: 1167–1179.
Brown, S.L., M.R. Landry, K.E. Selph, E. Jin Yang, Y.M. Rii & R.R. Bidigare. 2008.
Diatoms in the desert: Plankton community response to a mesoscale eddy in

53
the subtropical North Pacific. Deep. Res. Part II Top. Stud. Oceanogr. 55:
1321–1333.
Castro, R., V.M. Godínez, M.F. Lavín, E. Beier & C.E. Cabrera-Ramos. 2006.
Hydrography of the southern Gulf of California during the North American
Monsoon Experiment: Campaign NAME-02, August 7-20, 2004.
Chelton, D.B., R.A. deSzoeke, M.G. Schlax, K. El Naggar & N. Siwertz. 1998.
Geographical Variability of the First Baroclinic Rossby Radius of Deformation.
J. Phys. Oceanogr. 28: 433–460.
Chelton, D.B., P. Gaube, M.G. Schlax, J.J. Early & R.M. Samelson. 2011. The
Influence of Nonlinear Mesoscale Eddies on Near-Surface Oceanic
Chlorophyll. Science (80-. ). 334: 328–332.
Clarke, K.R. 1993. Non‐parametric multivariate analyses of changes in community
structure. Aust. J. Ecol. 18: 117–143.
Clarke, K.R. & M. Ainsworth. 1993. A method of linking multivariate community
structure to environmental variables. Mar. Ecol. Ser. 92: 205.
Clarke, K.R. & R.M. Warwick. 2001. Change in marine communities: an approach
to statistical analysis and interpretation. 2nd edn PRIMER-E Plymouth.
Contreras-Catala, F., L. Sánchez-Velasco, E. Beier & V.M. Godínez. 2015. Efectos
de un remolino de mesoescala sobre la distribución de larvas de peces
mesopelágicas en el Golfo de California. Rev. Biol. Mar. Oceanogr. 50: 575–
582.

54
Contreras-Catala, F., L. Sanchez-Velasco, M.F. Lavin & V.M. Godinez. 2012.
Three-dimensional distribution of larval fish assemblages in an anticyclonic
eddy in a semi-enclosed sea (Gulf of California). J. Plankton Res. 34: 548–
562.
Contreras-Catala, F., L. Sánchez-Velasco, M.F. Lavín & V.M. Godínez. 2012.
Three-dimensional distribution of larval fish assemblages in an anticyclonic
eddy in a semi-enclosed sea (Gulf of California). J. Plankton Res. 34: 548–
562.
Crawford, W.R., P.J. Brickley & A.C. Thomas. 2007. Mesoscale eddies dominate
surface phytoplankton in northern Gulf of Alaska. Prog. Oceanogr. 75: 287–
303.
Danell-Jiménez, a., L. Sánchez-Velasco, M.F. Lavín & S.G. Marinone. 2009.
Three-dimensional distribution of larval fish assemblages across a surface
thermal/chlorophyll front in a semienclosed sea. Estuar. Coast. Shelf Sci. 85:
487–496.
Davies, S.M., L. Sánchez-Velasco, E. Beier, V.M. Godínez, E.D. Barton & A.
Tamayo. 2015. Three-dimensional distribution of larval fish habitats in the
shallow oxygen minimum zone in the eastern tropical Pacific Ocean off
Mexico. Deep Sea Res. Part I Oceanogr. Res. Pap. 101: 118–129.
Durazo, R. & T.R. Baumgartner. 2002. Evolution of oceanographic conditions off
Baja California: 1997-1999. Prog. Oceanogr. 54: 7–31.
Eden, B.R., D.K. Steinberg, S. a. Goldthwait & D.J. McGillicuddy. 2009.

55
Zooplankton community structure in a cyclonic and mode-water eddy in the
Sargasso Sea. Deep. Res. Part I Oceanogr. Res. Pap. 56: 1757–1776.
Emilsson, I. & M.A. Alatorre. 1997. Evidencias de un giro ciclónico de mesoescala
en la parte sur del Golfo de California. In: Contribuciones de la Oceanografía
Física en México. Monogr. No. 3, Unión Geofísica Mex. 173–182.
Fernández, E., F. Álvarez, R. Anadón, S. Barquero, A. Bode, A. García, C. Garca-
Soto, J. Gil, N. González, A. Iriarte, B. Mouriño, F. Rodríguez, R. Sánchez, E.
Teira, S. Torres, L. Valdés, M. Varela, R. Varela & M. Zapata. 2004. The
spatial distribution of plankton communities in a Slope Water anticyclonic
Oceanic eDDY (SWODDY) in the southern Bay of Biscay. J. Mar. Biol. Assoc.
UK 84: 501–517.
Fiedler, P.C., R. Mendelssohn, D.M. Palacios & S.J. Bograd. 2013. Pycnocline
Variations in the Eastern Tropical and North Pacific, 1958–2008. J. Clim. 26:
583–599.
Field, J.G., K.R. Clarke & R.M. Warwick. 1982. A practical strategy for analyzing
multispecies distribution patterns. Mar. Ecol. Prog. Ser 8: 37–52.
Godínez, V.M., E. Beier, M.F. Lavín & J.A. Kurczyn. 2010. Circulation at the
entrance of the Gulf of California from satellite altimeter and hydrographic
observations. J. Geophys. Res. 115: C04007.
Haury, L.R., J.A. McGowan & P.H. Wiebe. 1978. Spatial Pattern in Plankton
Communities, p. 277–327. In J.H. Steele (ed.). . Springer US, Boston, MA.
Holliday, D., L.E. Beckley & M.P. Olivar. 2011. Incorporation of larval fishes into a

56
developing anti-cyclonic eddy of the Leeuwin Current off south-western
Australia. J. Plankton Res. 33: 1696–1708.
Kara, A.B., P.A. Rochford & H.E. Hurlburt. 2000. An optimal definition for ocean
mixed layer depth. J. Geophys. Res. 105: 16,803–816,821.
Kendall, a. W., E.H. Ahlstrom & H.G. Moser. 1981. Early Life Hisfory Stages of
Fishes and Their Characters. Ontog. Syst. Fishes 1–12.
Kessler, W.S. 2006. The circulation of the Eastern Tropical Pacific: A review. Prog.
Oceanogr. 69: 181–217.
Kramer, D., M.J. Kalin, E.G. Stevens, J.R. Thrailkill & J.R. Zwiefel. 1972. Collecting
and processing data on fish eggs and larvae in the California Current region.
NOAA Tech. Rep. NMFS Circ-370 38.
Kurczyn, J.A., E. Beier, M.F. Lavín & A. Chaigneau. 2012. Mesoscale eddies in the
northeastern Pacific tropical-subtropical transition zone: Statistical
characterization from satellite altimetry. J. Geophys. Res. Ocean. 117:
C10021.
Kurczyn, J.A., E. Beier, M.F. Lavín, A. Chaigneau & V.M. Godínez. 2013. Anatomy
and evolution of a cyclonic mesoscale eddy observed in the northeastern
Pacific tropical-subtropical transition zone. J. Geophys. Res. Ocean.
Kurian, J., F. Colas, X. Capet, J.C. McWilliams & D.B. Chelton. 2011. Eddy
properties in the California Current System. J. Geophys. Res. 116: C08027.
Ladd, C., W.R. Crawford, C.E. Harpold, W.K. Johnson, N.B. Kachel, P.J. Stabeno

57
& F. Whitney. 2009. A synoptic survey of young mesoscale eddies in the
Eastern Gulf of Alaska. Deep. Res. Part II Top. Stud. Oceanogr. 56: 2460–
2473.
Landry, M.R., M.D. Ohman, R. Goericke, M.R. Stukel, K.A. Barbeau, R. Bundy &
M. Kahru. 2012. Pelagic community responses to a deep-water front in the
California Current Ecosystem: overview of the A-Front Study. J. Plankton Res.
34: 739–748.
Lavín, M.F., E. Beier, J. Gómez-Valdés, V.M. Godínez & J. García. 2006. On the
summer poleward coastal current off SW México. Geophys. Res. Lett. 33.
Lavín, M.F., R. Castro, E. Beier, C. Cabrera, V.M. Godínez & A. Amador‐
Buenrostro. 2014. Surface circulation in the Gulf of California in summer from
surface drifters and satellite images (2004–2006). J. Geophys. Res. Ocean.
Lavín, M.F., R. Castro, E. Beier & V.M. Godínez. 2013. Mesoscale eddies in the
southern Gulf of California during summer: Characteristics and interaction with
the wind stress. J. Geophys. Res. Ocean. 118: 1–15.
Lavín, M.F., R. Castro, E. Beier, V.M. Godínez, A. Amador & P. Guest. 2009. SST,
thermohaline structure, and circulation in the southern Gulf of California in
June 2004 during the North American Monsoon Experiment. J. Geophys. Res.
114: C02025.
Lavín, M.F., R. Durazo, E. Palacios, M.L. Argote & L. Carrillo. 1997. Lagrangian
Observations of the Circulation in the Northern Gulf of California. J. Phys.
Oceanogr. 27: 2298–2305.

58
León-Chávez, C.A., E. Beier, L. Sánchez-Velasco, E.D. Barton & V.M. Godínez.
2015. Role of circulation scales and water mass distributions on larval fish
habitats in the Eastern Tropical Pacific off Mexico. J. Geophys. Res. Ocean.
n/a–n/a.
Lopez-Calderon, J., A. Martinez, A. Gonzalez-Silvera, E. Santamaria-del-Angel &
R. Millan-Nu??ez. 2008. Mesoscale eddies and wind variability in the northern
Gulf of California. J. Geophys. Res. C Ocean. 113: C10001.
MartÍnez, J. a. & J.S. Allen. 2004. A Modeling Study of Coastal-Trapped Wave
Propagation in the Gulf of California. Part II: Response to Idealized Forcing. J.
Phys. Oceanogr. 34: 1332–1349.
McCune, B., J.B. Grace & D.L. Urban. 2002. Analysis of Ecological Communities.
MjM Software Design.
Moore, T.S., R.J. Matear, J. Marra & L. Clementson. 2007. Phytoplankton
variability off the Western Australian Coast: Mesoscale eddies and their role in
cross-shelf exchange. Deep. Res. Part II Top. Stud. Oceanogr. 54: 943–960.
Moser, H. 1996. The early stages of fishes in the California current region.
California cooperative oceanic fisheries investigations, Atlas No. 33, …
Cooperative Oceanic Fisheries Investigations.
Myers, R.A. & K. Drinkwater. 1989. The influence of Gulf Stream warm core rings
on recruitment of fish in the northwest Atlantic. J. Mar. Res. 47: 635–656.
Navarro-Olache, L.F., M.F. Lavín, L.G. Álvarez-Sánchez & A. Zirino. 2004. Internal
structure of SST features in the central Gulf of California. Deep Sea Res. Part

59
II Top. Stud. Oceanogr. 51: 673–687.
Nieto, K., S. Mcclatchie, E.D. Weber & C.E. Lennert-cody. 2013. Effect of
mesoscale eddies and streamers on sardine spawning habitat and recruitment
success off southern and central California 2–14.
Norrbin, M.F., C.S.D.A. Vis & S.M. Gallager. 1996. Differences in fine-scale
structure and composition of zooplankton between mixed and stratified regions
of Georges Bank. Deep. Res. Part II Top. Stud. Oceanogr. 43: 1905–1924.
Omand, M.M., E.A. D’Asaro, C.M. Lee, M.J. Perry, N. Briggs, I. Cetinić & A.
Mahadevan. 2015. Eddy-driven subduction exports particulate organic carbon
from the spring bloom. Science (80-. ). 348: 222–225.
Pantoja, D.A., S.G. Marinone, A. Parés-Sierra & F. Gómez-Valdivia. 2012.
Numerical modeling of seasonal and mesoscale hydrography and circulation
in the Mexican Central Pacific. Ciencias Mar. 38.
Pares-Sierra, A., W.B. White & C.-K. Tai. 1993. Wind-driven coastal generation of
annual mesoscale eddy activity in the California Current.
Pegau, W.S., E. Boss & A. Martinez. 2002. Ocean color observations of eddies
during the summer in the Gulf of California. Geophys. Res. Lett. 29: 1–3.
Peguero-Icaza, M., L. Sánchez-Velasco, M.F. Lavín & S.G. Marinone. 2008. Larval
fish assemblages, environment and circulation in a semienclosed sea (Gulf of
California, Mexico). Estuar. Coast. Shelf Sci. 79: 277–288.
Peguero-Icaza, M., L. Sánchez-Velasco, M.F. Lavín, S.G. Marinone & E. Beier.

60
2011. Seasonal changes in connectivity routes among larval fish assemblages
in a semi-enclosed sea (Gulf of California). J. Plankton Res. 33: 517–533.
Piontkovski, S., R. Williams, W. Peterson & K. V K. 1995. Relationship between
oceanic mesozooplankton and energy of eddy fields. Mar. Ecol. Prog. Ser.
128: 35–41.
Sánchez-Velasco, L., E. Beier, C. Avalos-García & M.F. Lavín. 2006. Larval fish
assemblages and geostrophic circulation in Bahía de La Paz and the
surrounding southwestern region of the Gulf of California. J. Plankton Res. 28:
1081–1098.
Sánchez-Velasco, L., S.P. a. Jiménez-Rosenberg & M.F. Lavín. 2007. Vertical
Distribution of Fish Larvae and Its Relation to Water Column Structure in the
Southwestern Gulf of California 1. Pacific Sci. 61: 533–548.
Sánchez-Velasco, L., M.F. Lavín, S.P.A. Jiménez-Rosenberg, V.M. Godínez, E.
Santamaría-del-Angel & D.U. Hernández-Becerril. 2013. Three-dimensional
distribution of fish larvae in a cyclonic eddy in the Gulf of California during the
summer. Deep Sea Res. Part I Oceanogr. Res. Pap. 75: 39–51.
Sangrà, P., J.L. Pelegrí, a. Hernández-Guerra, I. Arregui, J.M. Martín, a. Marrero-
Díaz, a. Martínez, A.W. Ratsimandresy & a. Rodríguez-Santana. 2005. Life
history of an anticyclonic eddy. J. Geophys. Res. C Ocean. 110: 1–19.
Sassa, C., K. Kawaguchi, Y. Hirota & M. Ishida. 2007. Distribution depth of the
transforming stage larvae of myctophid fishes in the subtropical-tropical waters
of the western North Pacific. Deep. Res. Part I Oceanogr. Res. Pap. 54: 2181–

61
2193.
Siegel, S. & N.J. Castellón. 1988. Non-parametric Statistics for the Behavioral
Sciences. McGraw-Hill International Editions, Statistics Series.
Simpson, J.H., D.J. Crisp & C. Hearn. 1981. The Shelf-Sea Fronts: Implications of
their Existence and Behaviour [and Discussion]. Philos. Trans. R. Soc. A
Math. Phys. Eng. Sci. 302: 531–546.
Smith, P.E. & S.L. Richardson. 1979. Tecnicas modelo para prospecciones de
huevos y larvas de peces pelágicos. FAO Doc. Tec. Pesca 175.
Sokal, R.R. & F.J. Rohlf. 1985. Biometry. Blume, Barcelona, Spain. 587 pp.
Song, H., A.J. Miller, S. McClatchie, E.D. Weber, K.M. Nieto & D.M. Checkley.
2012. Application of a data-assimilation model to variability of Pacific sardine
spawning and survivor habitats with ENSO in the California Current System. J.
Geophys. Res. Ocean. 117: 1–15.
Strub, P.T., J.S. Allen, A. Huyer, R.L. Smith & R.C. Beardsley. 1987. Seasonal
cycles of currents, temperatures, winds, and sea level over the northeast
Pacific continental shelf: 35°N to 48°N. J. Geophys. Res. Ocean. 92: 1507–
1526.
Wishart, D. 1969. 256. Note: An Algorithm for Hierarchical Classifications.
Biometrics 25: 165–170.
Zamudio, L., P. Hogan & E.J. Metzger. 2008. Summer generation of the Southern
Gulf of Colifornia eddy train. J. Geophys. Res. Ocean. 113: 1–21.

62
ceanic eddies at Cabo Corrientes and the Maria Islands, Mexico. J. Geophys. Res.
112.
Zamudio, L., H.E. Hurlburt, E.J. Metzger & C.E. Tilburg. 2007. Tropical wave-
induced oceanic eddies at Cabo Corrientes and the María Islands, Mexico. J.
Geophys. Res. Ocean. 112: 1–17.

63
Apéndice I
Morfología de los remolinos muestreados en el sur del Golfo de California.
Remolino Anticiclónico
(Octubre de 2007) Ciclónico (Julio de
2010) Ciclónico (Julio de
2011)
Posible origen
Inestabilidades de aguas costeras poco profundas
Inestabilidades de la CCM hacia el polo
Inestabilidades de la CCM hacia el polo.
Fase evolutiva
Inicio de la disipación
Inicio de disipación Maduro
Dimensiones Diámetro / profundidad
~90 km ~70 m ~60 km ~300 m ~147 km ~470 m
Duración ~ 6 semanas ~ 3 semanas ~ 12 semanas
Periodo de rotación
~ 3.5 días ~ 8.4 días ~ 12 días
CCM, Corriente Costera Mexicana


















