Metabolismo de amino ácidos.
51
1 Metabolismo de amino ácidos • Araiza German Karen • Armenta Medina Priscila • Camacho Badilla Roberto
-
Upload
erik-davila -
Category
Healthcare
-
view
192 -
download
0
Transcript of Metabolismo de amino ácidos.

1
Metabolismo de amino ácidos • Araiza German Karen• Armenta Medina Priscila• Camacho Badilla Roberto

Síntesis de Urea• Elimina el 95% del Nitrógeno sobrante.• La urea se forma apartar de amoniaco, CO2 y aspartato en una
ruta cíclica llamada ciclo de la urea (de Krebs o ciclo de Krebs-Henseleit).
• Tiene lugar en los hepatocitos
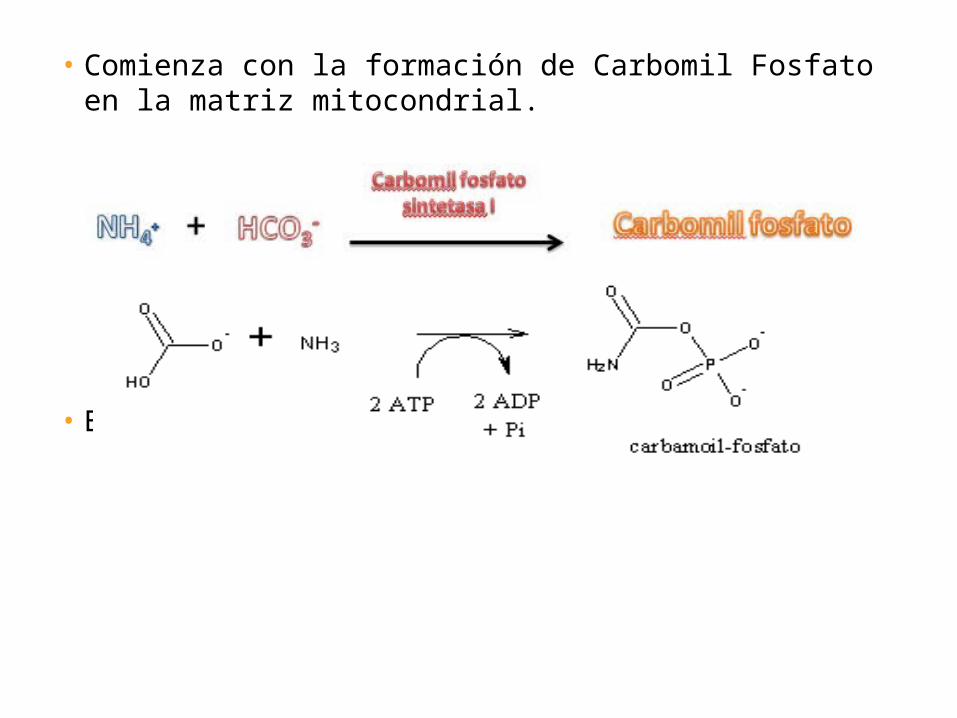
• Comienza con la formación de Carbomil Fosfato en la matriz mitocondrial.
• Es una reacción irreversible.
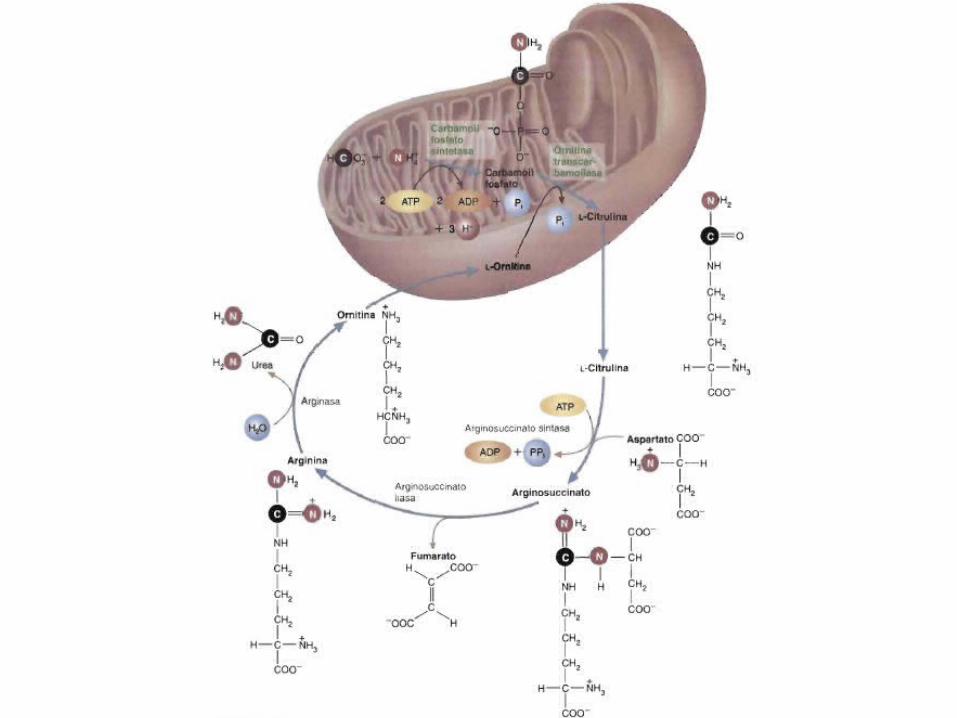

• Una vez formada, la urea difunde fuera de los hepatocitos al torrente sanguíneo.
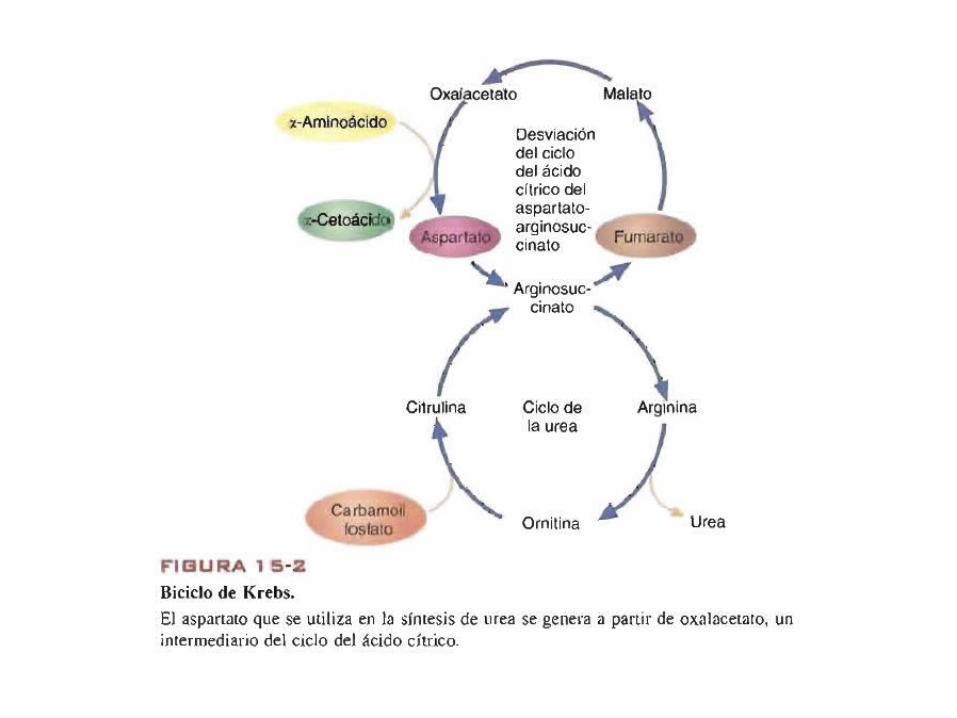
• Finalmente se elimina en la orina por el riñón.• La ornitina vuelve a las mitocondrias para iniciar un nuevo ciclo.• El fumarato se hidrata para formar malato y entrar al ciclo de
acido cítrico.

7
• La función terminal de la síntesis de amino ácidos, es la síntesis de proteínas.
• El acervo de aminoácidos es el conjunto de moléculas de aminoácidos de disposición inmediata para su uso en los procesos metabólicos.( alimentos y tejidos)
• Metabolismo de amino ácidos: conjunto complejo de Rx en las que las moléculas de aminoácidos que se requieren para la síntesis de proteínas y otro metabolitos se producen y se degradan de forma continua.
8
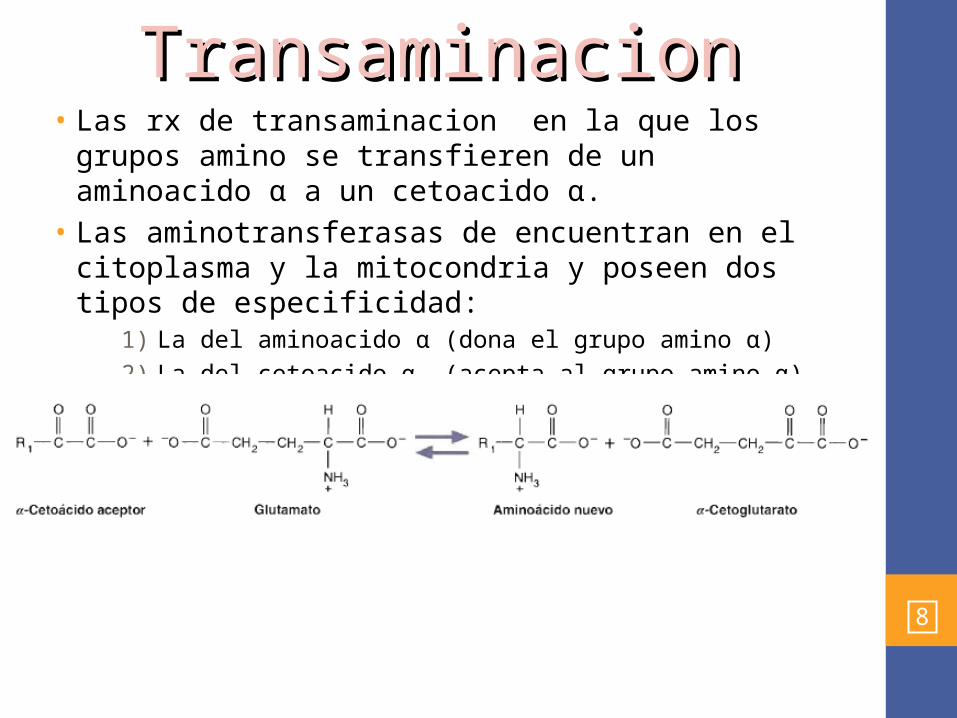
• Las rx de transaminacion en la que los grupos amino se transfieren de un aminoacido α a un cetoacido α.
• Las aminotransferasas de encuentran en el citoplasma y la mitocondria y poseen dos tipos de especificidad:
1) La del aminoacido α (dona el grupo amino α)2) La del cetoacido α (acepta al grupo amino α)
• La mayoria de las veces se utiliza el glutamato como donador del grupo amino:
TransaminacionTransaminacion

9
• Se produce glutamato cuando el α-cetoglutarato acepta un grupo amino, el α-cetoglutarato/glutamato tiene funciones muy importantes en el metabolismo en general.
• El otro par, oxalacetato/aspartato, participa en la eliminacion del nitrogeno en el ciclo de la urea.
• El par, piruvato/alanina participa en el ciclo de la alanina.• Ya que el α-cetoglutarato y el oxalacetato participan en el ciclo
de Krebs, tienen una participacion importante ofrecer energia a la celula.
• Se utiliza la coenzima piridoxal-5´-fosfato (PLP), este se une al sitio activo de la enzima , por interacciones no covalentes y una base de Schiff (R'-CH=N-R, una aldimina) que se forma por la condensación del grupo aldehído del PLP y el gpo 8-amino de un residuo de lisina .
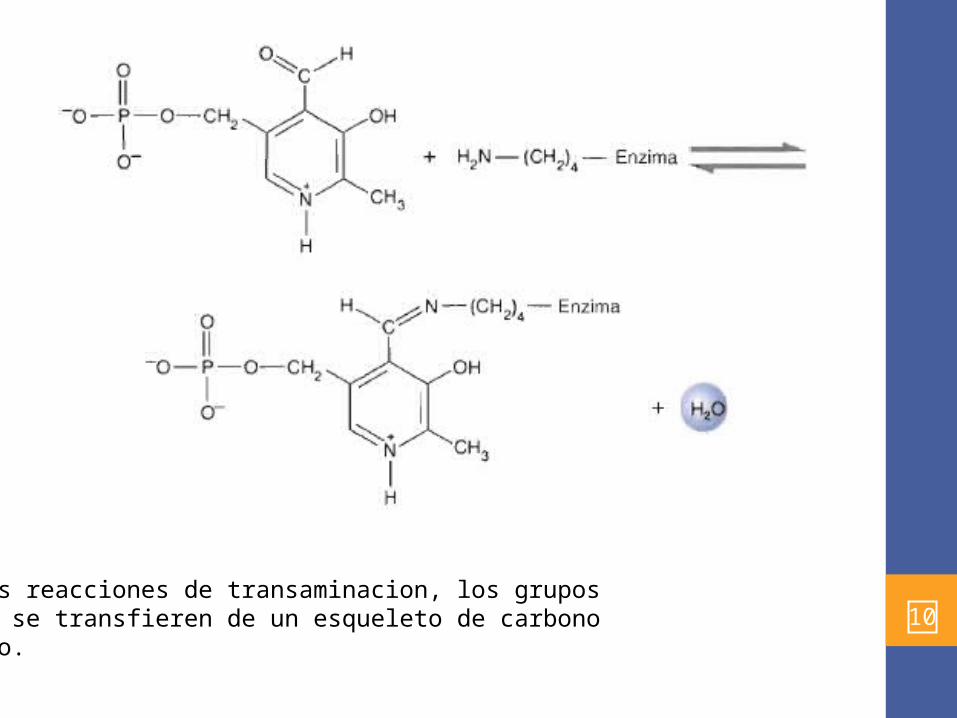
10En las reacciones de transaminacion, los grupos amino se transfieren de un esqueleto de carbonoa otro.

11
Aminoacidos que forman el cetoglutaratoα
• Arginina• Prolina• Histidina• Glutamato• Glutamina
12
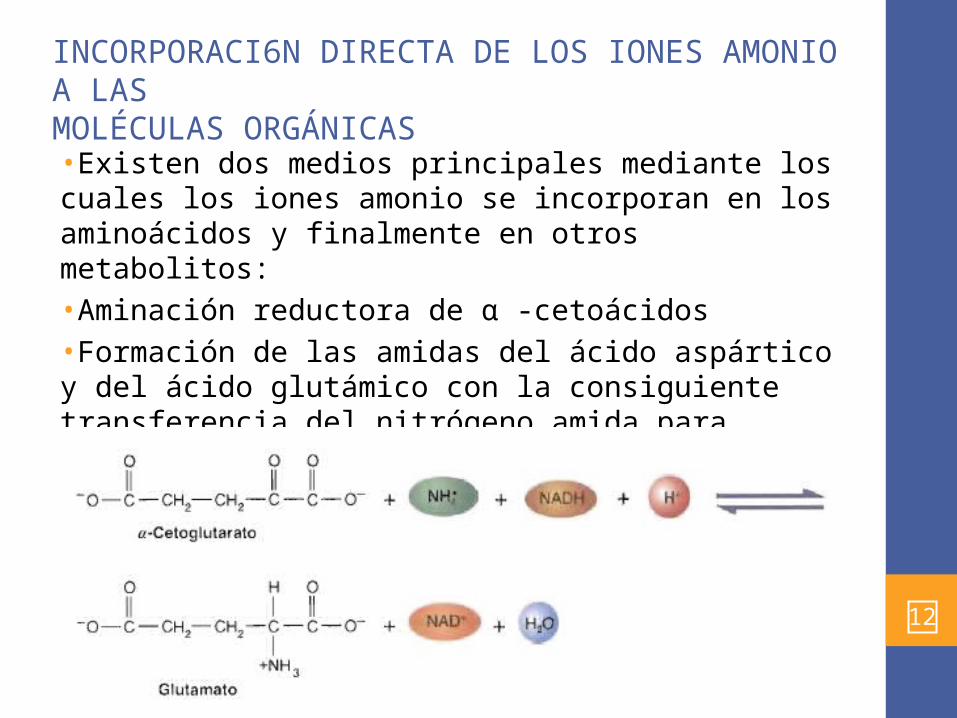
INCORPORACI6N DIRECTA DE LOS IONES AMONIO A LASMOLÉCULAS ORGÁNICAS•Existen dos medios principales mediante los cuales los iones amonio se incorporan en los aminoácidos y finalmente en otros metabolitos: •Aminación reductora de α -cetoácidos •Formación de las amidas del ácido aspártico y del ácido glutámico con la consiguiente transferencia del nitrógeno amida para formar otros aminoácidos.•La deshidrogenasa de glutamato , cataliza la aminación directa del α -cetoglutarato:
13
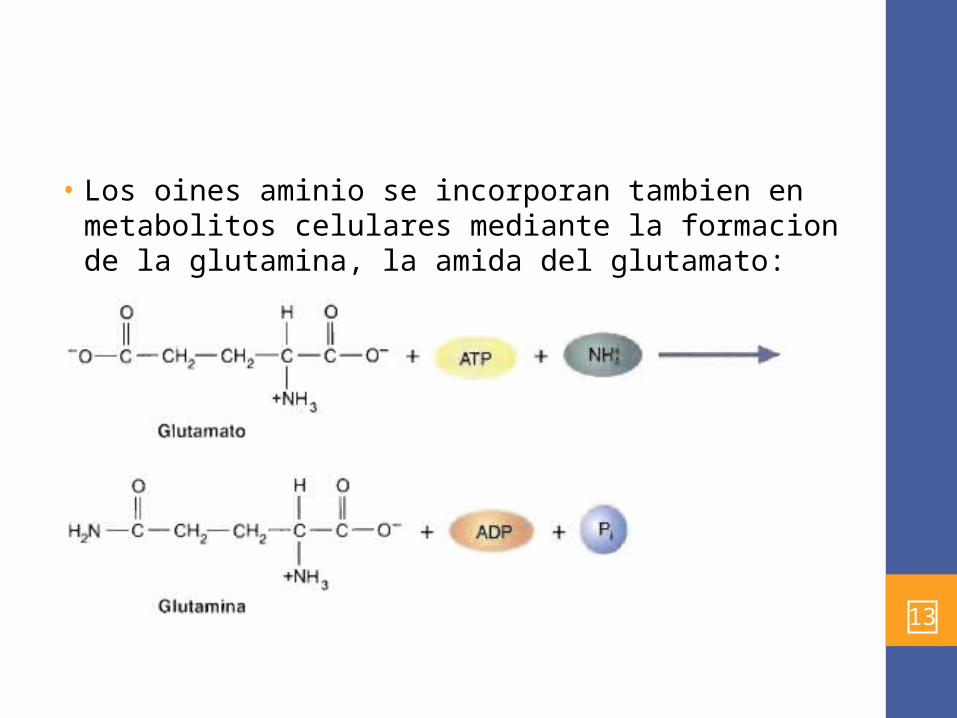
• Los oines aminio se incorporan tambien en metabolitos celulares mediante la formacion de la glutamina, la amida del glutamato:

14
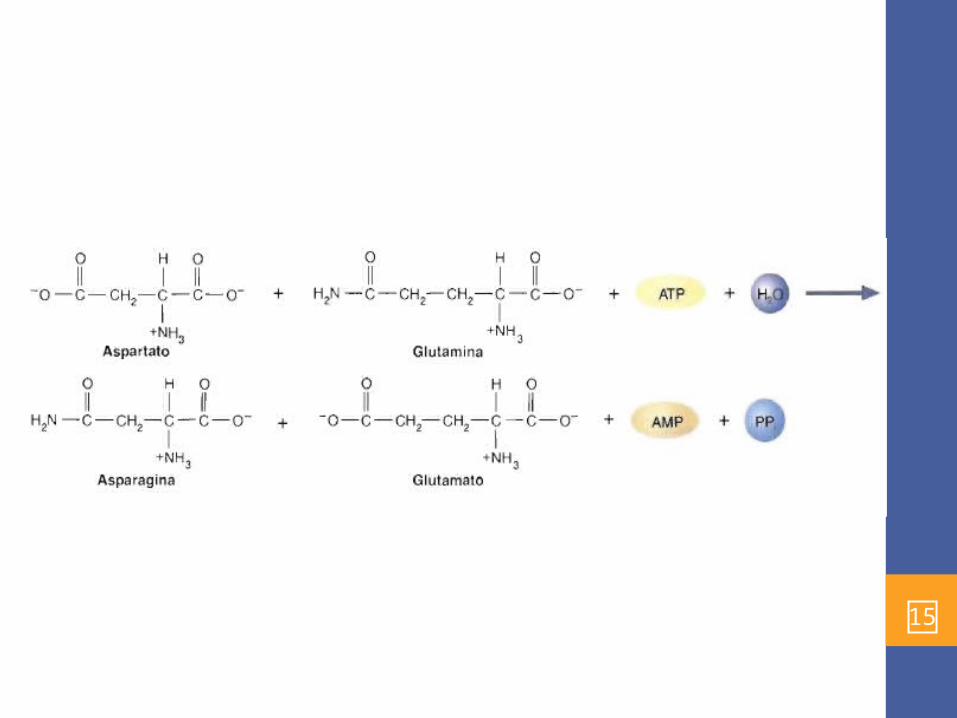
Asparagina• El espartato tambien es un percusor de importante de la
sisntesis de nucleotidos.• La familia del aspartato también contiene asparagina, lisina,
metionina y treonina. Esta última contribuye a la ruta de reacción en la que se sintetiza la isoleucina. La síntesis de isoleucina, que suele considerarse un miembro de la familia del piruvato.
• La asparagina, la amida del aspartato, no se forma directamente a partir del aspartato y el NH, sino que el grupo amida de la glutamina se transfiere mediante transferencia del grupo amida durante una reacción que requiere ATP, catalizada por la asparagina sintetasa:
15
16
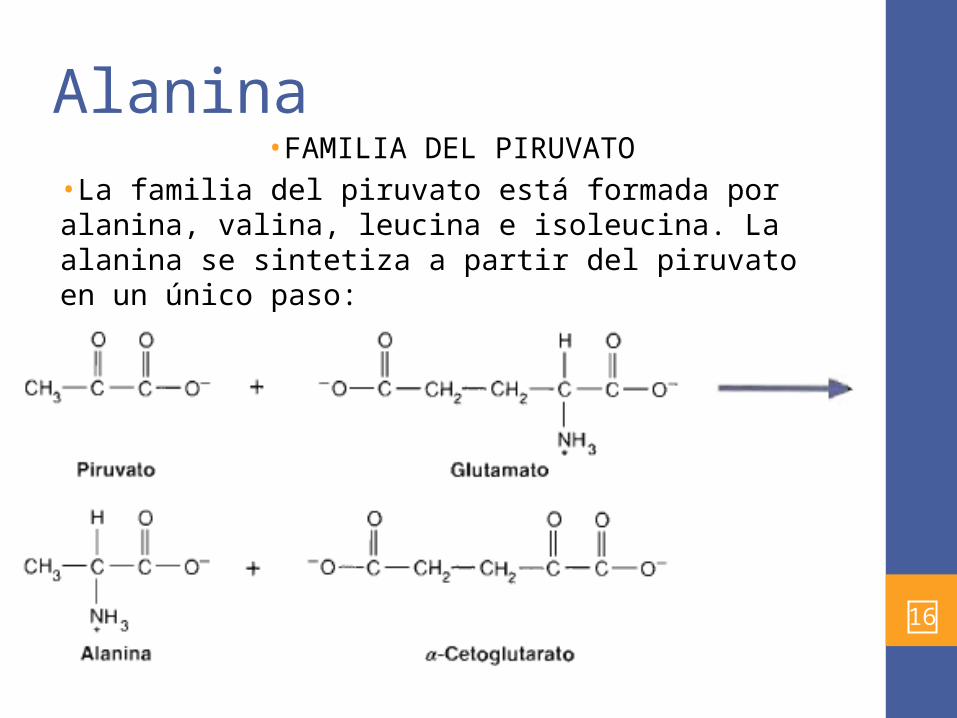
Alanina •FAMILIA DEL PIRUVATO
•La familia del piruvato está formada por alanina, valina, leucina e isoleucina. La alanina se sintetiza a partir del piruvato en un único paso:

17
•El ciclo de la alanina contribuye al mantenimiento de la glucosa sanguínea y la enzima que cataliza esta rx es la alanina aminotransferasa.

18
Acido aspártico• Su biosintesis tiene lugar en la transaminacion del acido
oxalacetico (metabolito intermedario en el ciclo de Krebs)

Sintesis de Serina, Glicina y Cisteina.
•Los miembros de la familia de la serina, que son serina, glicina y cisteína, obtienen sus esqueletos carbonados a partir del intermediario glucolítico glicerato-3-fosfato.
•Los miembros de este grupo desempeñan funciones importantes en numerosas rutas anabólicas.
•La serina es precursora de la etanolamina y la esfingosina.
•La glicina se utiliza en las rutas de síntesis de purinas, porfirinas y glutatión.
Juntas, la serina y la glicina contribuyen a un conjunto de rutas de biosíntesis que se denominan, en conjunto, metabolismo de un carbono. La cistina desempaña un papel significativo en el metabolismo del azufre.

•La serina es la fuente principal de glicina.
•Pueden obtenerse cantidades menores de glicina a partir de colina, cuando esta última molécula
se encuentra en exceso.
•La síntesis de glicina a partir de colina consta de dos desbidrogenaciones y un conjunto de
desmetilaciones. La glicina actúa como neurotransmisor inhibidor en el sistema nervioso central.
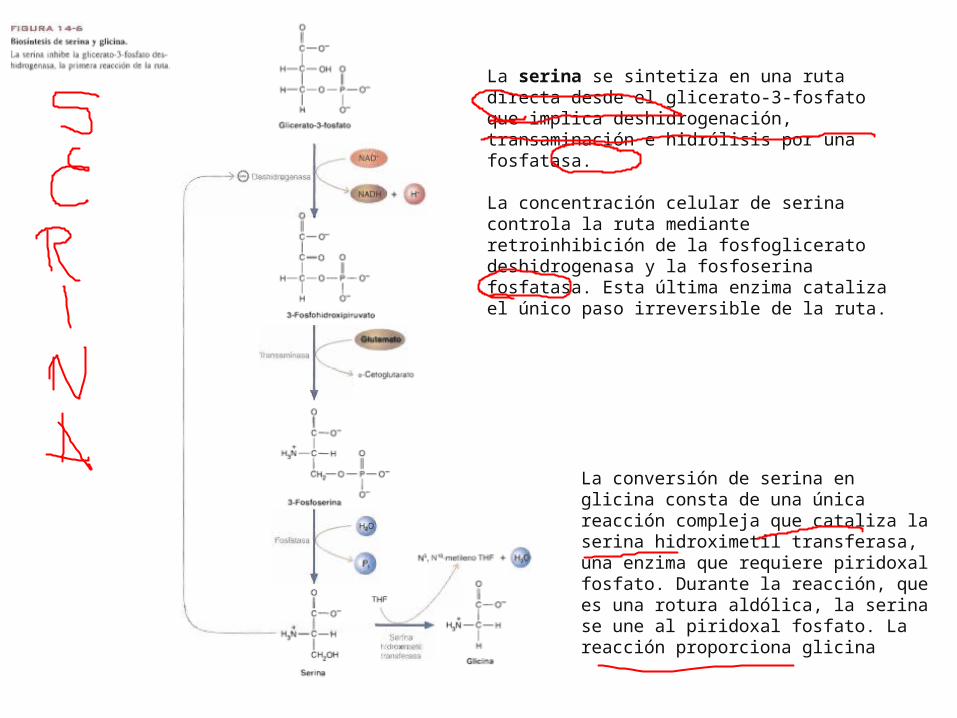
La serina se sintetiza en una ruta directa desde el glicerato-3-fosfato que implica deshidrogenación, transaminación e hidrólisis por una fosfatasa.
La concentración celular de serina controla la ruta mediante retroinhibición de la fosfoglicerato deshidrogenasa y la fosfoserina fosfatasa. Esta última enzima cataliza el único paso irreversible de la ruta.
La conversión de serina en glicina consta de una única reacción compleja que cataliza la serina hidroximetil transferasa, una enzima que requiere piridoxal fosfato. Durante la reacción, que es una rotura aldólica, la serina se une al piridoxal fosfato. La reacción proporciona glicina
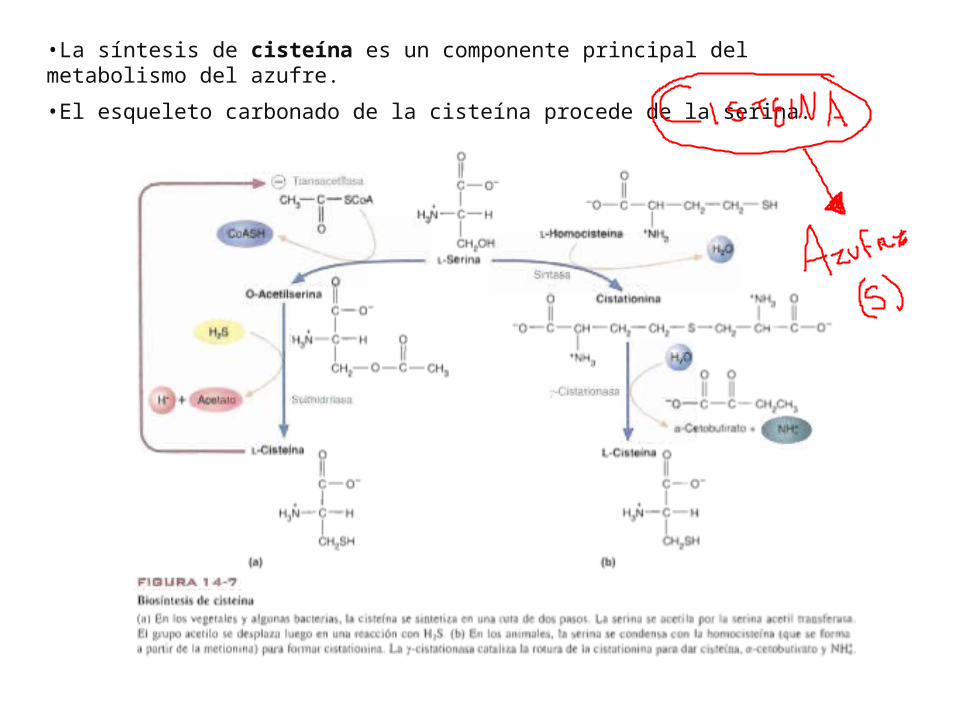
•La síntesis de cisteína es un componente principal del metabolismo del azufre.
•El esqueleto carbonado de la cisteína procede de la serina.
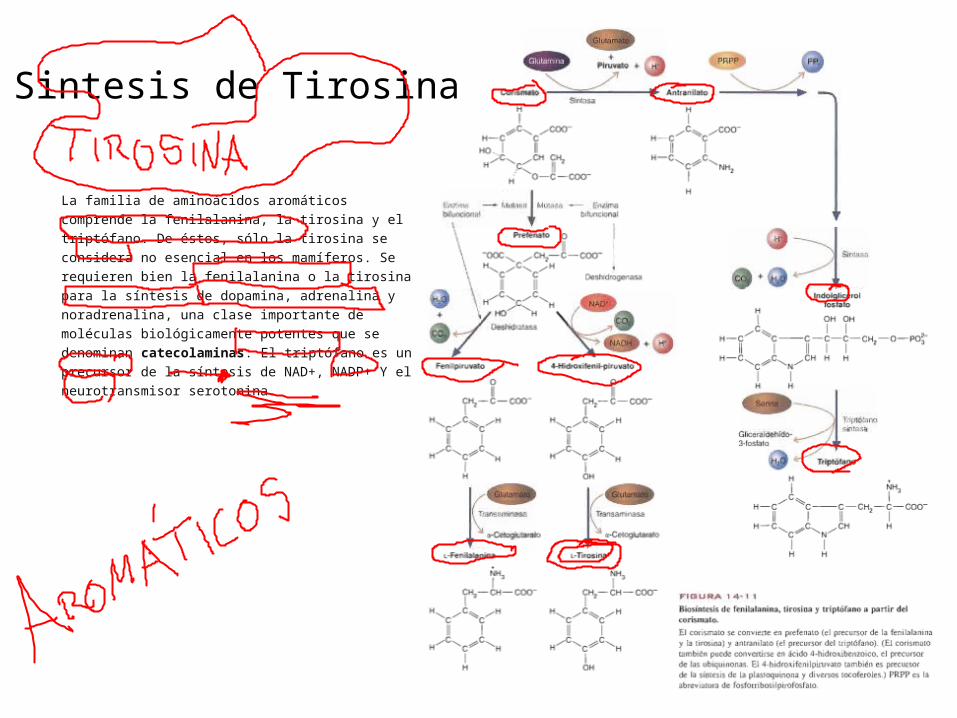
Sintesis de Tirosina
La familia de aminoácidos aromáticos comprende la
fenilalanina, la tirosina y el triptófano. De éstos, sólo la
tirosina se considera no esencial en los mamíferos. Se
requieren bien la fenilalanina o la tirosina para la
síntesis de dopamina, adrenalina y noradrenalina, una
clase importante de moléculas biológicamente potentes
que se denominan catecolaminas. El triptófano es un
precursor de la síntesis de NAD+, NADP+ Y el
neurotransmisor serotonina.

La tirosina no es un aminoácido esencial en los animales debido a que se sintetiza a partir de la fenilalanina en una reacción de hidroxilación. La enzima que participa, la fenilalaina-4-monooxigenasa, requiere la coenzima tetrahidrobiopterina, una molécula semejante al ácido fólico que procede del GTP. Debido a que esta reacción es un primer paso del catabolismo de la fenilalanina.

•A pesar de la complejidad de las rutas degradativas de los aminoácidos, pueden realizarse las siguientes afirmaciones generales.
•El catabolismo de los aminoácidos normalmente comienza con la eliminación del grupo amino. Los grupos amino pueden luego eliminarse en la síntesis de la urea.
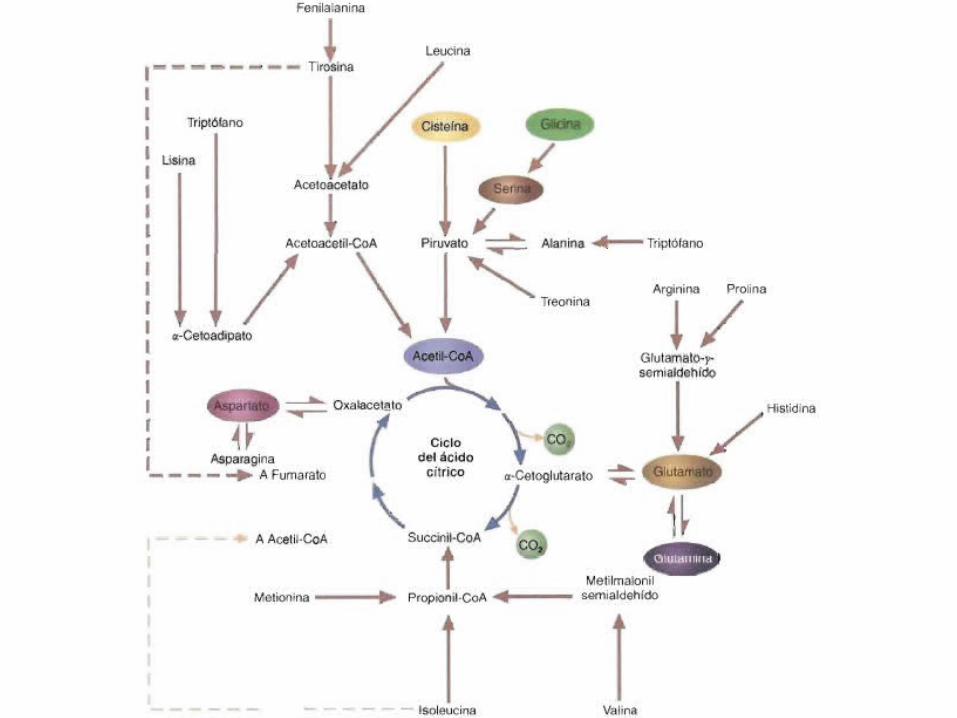
•Los esqueletos carbonados que se producen a partir de los aminoácidos se degradan posteriormente para formar siete productos metabólicos: acetil-CoA, aetoacetil-CoA, piruvato, ct-cetoglutarato, succi- nil-CoA, fumarato y oxalacetato.
Dependiendo de los requerimientos del animal, estas moléculas se utilizan para sintetizar ácidos grasos o glucosa o para generar energía. Los aminoácidos que se degradan para formar acetil-CoA o acetoacetiJ-CA se denominan cetogénicos, debido a que pueden convertirse en ácidos grasos o cuerpos cetónicos. Los esqueletos carbonados de los aminoácidos glucogénicos, que se degradan a piruvato o a un intermediario del ciclo del ácido cítrico, pueden utilizarse a continuación en la gluconeogénesis. Tras considerar las rutas de desaminación y la síntesis de urea, se describen las rutas que degradan los esqueletos carbonados.

La eliminación del grupo a-amino de los aminoácidos incluye dos tipos de reacciones químicas: transaminación y desamioación oxidativa. Ya se han descrito ambas reacciones. (Recuerde que las reacciones de transaminación ocupan posiciones importantes en la síntesis de los aminoácidos no esenciales.) Debido a que estas reacciones son reversibles, los grupos ami no se desvían fácilmente de los aminoácidos abundantes y se utilizan para sintetizar los que son escasos. Los grupos amino quedan disponibles para la síntesis de urea cuando los aminoácidos se en- cuentran en exceso. La urea se sintetiza en cantidades especialmente elevadas cuan- do la alimentación tiene abundantes proteínas o cuando hay una degradación masiva de proteínas, por ejemplo, durante la inanición.

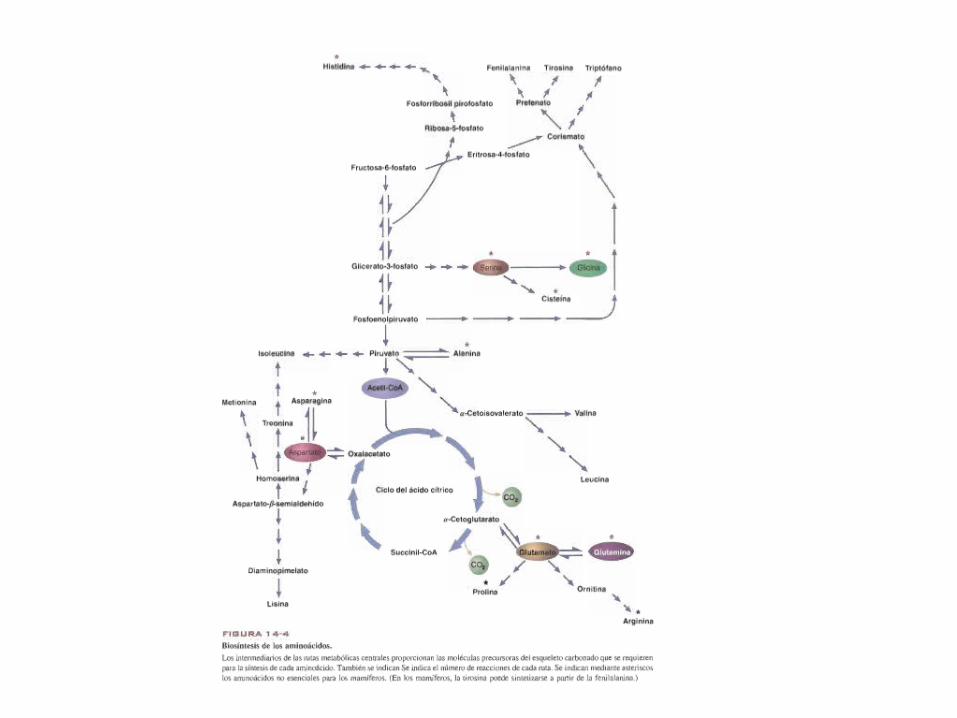

Catabolismo de los aminoácidos• Comienza con la eliminación del grupo amino.• Estos pueden eliminarse en la síntesis de urea.• Los esqueletos de carbono se degradan para formar diferentes
productos metabólicos.• Acetil-CoA, aetoacetil-CoA, piruvato, a-cetoglutarato, succinil-CoA,
fumarato y oxalacetato.
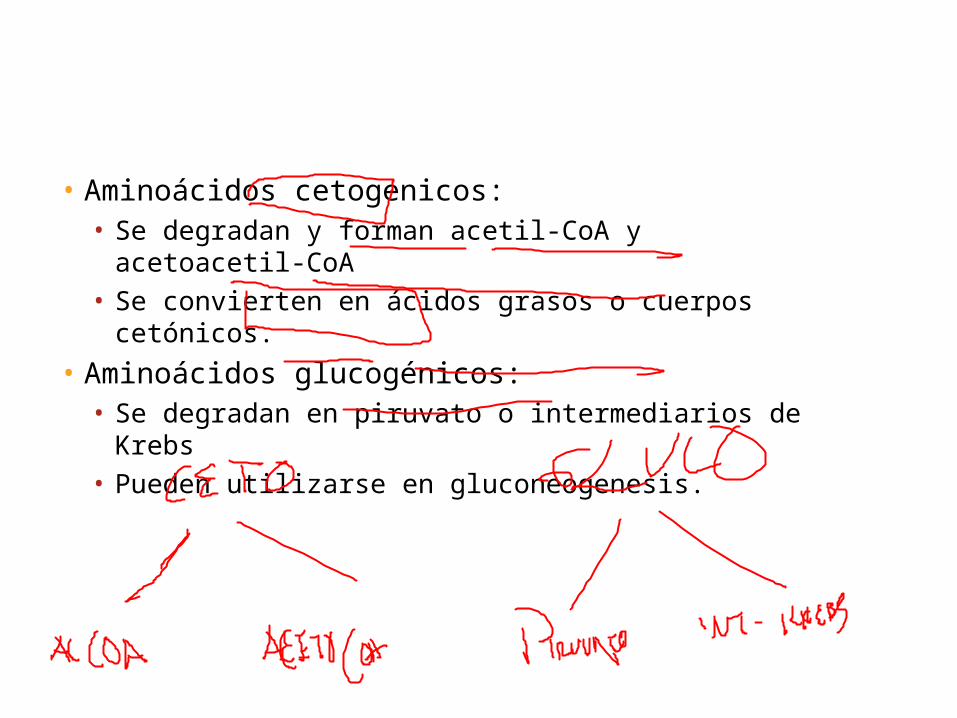
• Aminoácidos cetogenicos:• Se degradan y forman acetil-CoA y acetoacetil-CoA• Se convierten en ácidos grasos o cuerpos cetónicos.
• Aminoácidos glucogénicos:• Se degradan en piruvato o intermediarios de Krebs• Pueden utilizarse en gluconeogenesis.

Desaminación• Incluye dos tipos de reacciones: transaminación y desaminación
oxidativa• Reacciones reversibles.• Los grupos amino se desvían de los aa’s abundantes para
formar aa’s escasos.• Pasan al ciclo de la urea cuando los aa’s están en exceso.

Catabolismo de los esqueletos carbonados de los aminoácidos• Aminoácidos que forman Acetil-CoA:• Alanina, Serina, Glicina, Cisteína, Treonina, Lisina, Triptófano, Tirosina,
Fenilalanina y Leucina.• Aminoácidos que forman a-cetoglutarato:• Glutamato y Glutamina, Arginina, Prolina e Histidina.
• Aminoácidos que forman Succinil-CoA:• Metionina, Isoleucina y Valina.
• Aminoácidos que forman Oxalacetato:• Aspartato y Asparagina

Degradación de los Nucleótidos.• En seres vivos, los nucleótidos de purina y pirimidina se están
degradando constantemente.• En animales se debe al recambio normal de los ácidos nucleicos
y de los nucleótidos, y la digestión de los ácidos nucleicos de los alimentos.

• Durante la digestión , los ácidos nucleicos se hidrolizan a oligonucleotidos por las enzimas denominadas nucleasas.
*oligonucleotidos: segmentos cortos de ácidos nucleicos que contienen menos de 50 nucleótidos.
• Las enzimas especificas de la rotura de enlaces internucleotidos en el DNA de denominan Dnasas.
• Las que degradan el RNA se denominan RNasas

• Los oligonucleotidos se siguen degradando por varias fosfodiesterasas, un proceso que da lugar a una mezcla de mononucleótidos.
• Nucleotidasas eliminan los grupos fosfato de los nucleótidos dando nucleosidos.
• Estas ultimas moléculas de hidrolizan por las nucleosidasas a las bases libres y ribosa o desoxirribosa, que posteriormente se absorben.
• Los nucleosidos pueden absorberse por los enterocitos intestinales.

• En contraste con los procesos catabólicos de otras clases principales de biomolecular, el catabolismo de la purinas y las pirimidinas no produce síntesis de ATP.

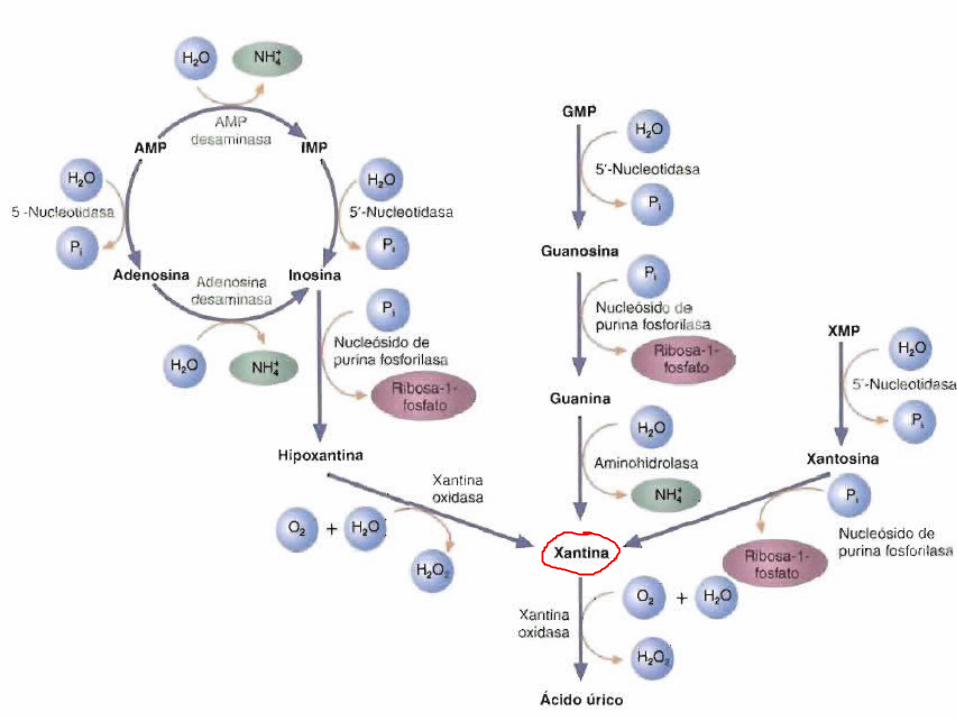
Catabolismo de purinas• Existen variaciones de las rutas especificas que utilizan los
diferentes organismos para degradar el AMP. • En el musculo, el AMP inicialmente se convierte en IMP por la
AMP deasminasa (que también se denomina adenilato aminohidrolasa).
• El IMP se hidroliza a inosina por la 5’-nucleotidasa.

• En la mayoría de los tejidos, el AMP se hidroliza por la 5’- nucleotidasa para formar adenosina.
• Luego la adenosina se desamina por la adenosina desaminasa (adenosina aminohidrolasa) para formar inosina.
• La purina nucleosido fosfohidrolasa convierte la inosina, guanosina y xantocina en hipoxantina, guanina y xantina respectivamente.

• La hipoxantina se oxida a xantina por la xantina oxidasa.• Las reacciones que cataliza la xantina oxidasa producen O2 además
de formar H2O2.• La guanina se desamina a xantina por la guanina desaminasa
(guanina aminohidrolasa).• Las moléculas de xantina se oxidan posteriormente a ácido urico por
la xantina oxidasa.

Enfermedades por defectos de catabolismo de purinas• La gota: se caracteriza por concentraciones sanguíneas
elevadas de acido úrico y ataques recurrentes de artritis. Esta producida por varias anomalías metabólicas.
• Se conoces 2 enfermedades con inmunodeficiencia consecuencias del mal metabolismo de purinas• Deficiencia de adenosina desaminasa• Deficiencia de nucleosido de purina fosforilasa.

Deficiencia de adenosina desaminasa• Las concentraciones elevadas de dATP inhiben la ribonucleotido
reductasa. Como consecuencia, la sintesis de DNA se deprime.• Ésta distorsión metabólica se observa principalmente en los
linfocitos T y B.• Los niños con deficiencia con este padecimiento normalmente
fallesen antes de los dos años debido a infecciones masivas.

Deficiencia de nucleosido de purina fosforilasa.• Las concentraciones de nucleótidos de purina son elevadas y
disminuyen la síntesis de ácido úrico. • Las concentraciones elevadas de dGTP aparentemente son las
responsables del deterioro de las células T que es característico de esta enfermedad.

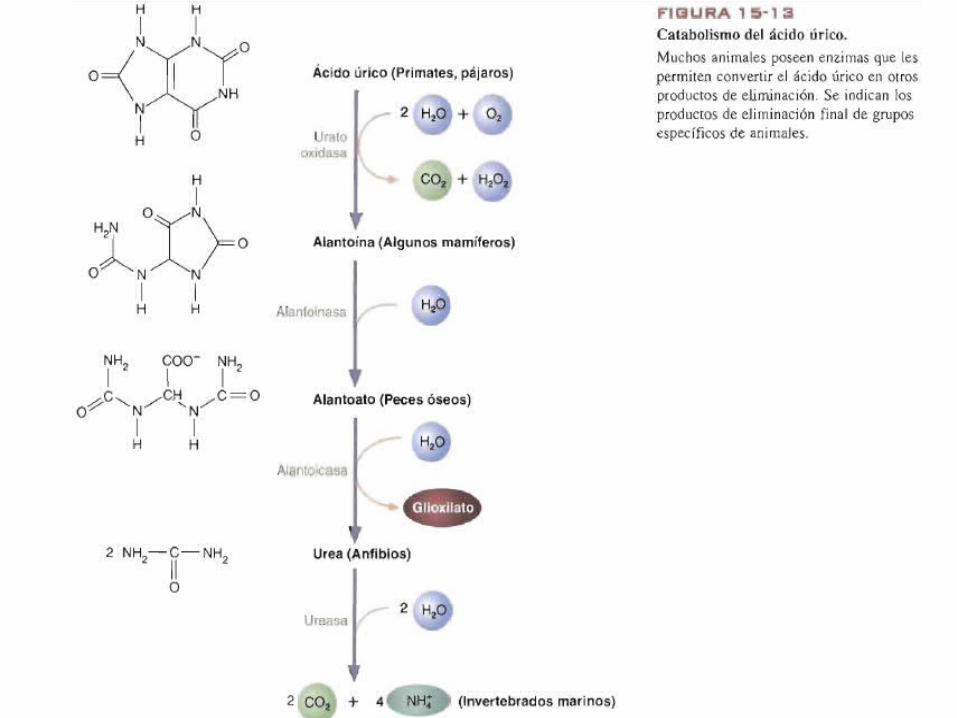
Catabolismo de acido úrico• Muchos animales degradan mas el acido úrico.• La uratooxidasa convierte en acido úrico en alantoina (producto
de eliminación en muchos mamíferos). La alantoinasa cataliza la hidratacion de la alantoina para formar alantoato que eliminan los peces óseos.

• Otros peces así como los anfibios producen alantoicasa, que fracciona el acido alantoico en glioxilato y urea.
• Finalmente los invertebrados marinos degradan la urea a NH4+ y CO2 en una reacción catalizada por la ureasa.

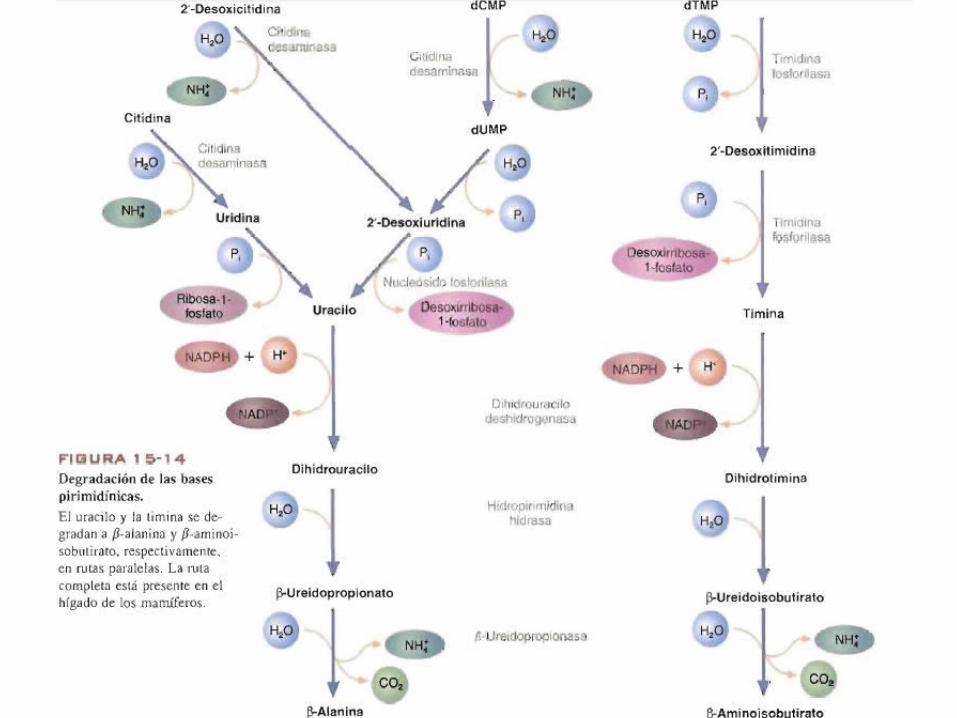
Catabolismo de pirimidina• En los humanos, a diferencia de la purina, la pirimidina si puede
degradarse.• Antes de que pueda degradarse, la citidina y la desoxicitidina se
convierten en uridina y desoxiuridina respectivamente, por reacciones de desaminacion que cataliza la citidina desaminasa.
• El desoxicitidilato( dCMP) se desamina para formar desoxiuridilato (dUMP).

• La ultima molecula se convierte posteriormente en desoxiuridina por la 5’-nucleotidasa.
• La uridina y la desoxiuridina se degradan aun mas por la nucleosido fosforilasa para formar uracilo.
• la timina se forma apartir del timidilato (dTMP) por las acciones secuenciales dela 5’- nucleotidasa y la nucleosido fosforilasa.
• el uracilo y la timina se convierten en sus productos finales, l3-alanina y l3- aminoisobutirato respectivamente en rutas paralelas.

• Se reducen por la dihidrouracilo deshdrogenasa a sus correspondientes derivados dihidro. Al hidrolizarse estas ultimas moleculas se abren los anillos dando l3-ureidopropionato y l3- ureidoisobutirato, respectivamente.
• Finalmente, la l3-alanina y el l3-aminoisobutirato se producen en reacciones de desaminacion catalizadas por la l3-ureidopropionasa.