
Redalyc.DINÁMICA DEL CARBONO Y NITRÓGENO DEL SUELO …observó que la mineralización de la...
16
Tropical and Subtropical Agroecosystems E-ISSN: 1870-0462 [email protected] Universidad Autónoma de Yucatán México Suárez M., Gabriela; Campos C., Adolfo; Cruz H., Lourdes DINÁMICA DEL CARBONO Y NITRÓGENO DEL SUELO EN ECOSISTEMAS DE LA COSTA TROPICAL SECA, EN LA MANCHA (CICOLMA), VERACRUZ, MÉXICO Tropical and Subtropical Agroecosystems, vol. 18, núm. 3, 2015, pp. 347-361 Universidad Autónoma de Yucatán Mérida, Yucatán, México Disponible en: http://www.redalyc.org/articulo.oa?id=93944043009 Cómo citar el artículo Número completo Más información del artículo Página de la revista en redalyc.org Sistema de Información Científica Red de Revistas Científicas de América Latina, el Caribe, España y Portugal Proyecto académico sin fines de lucro, desarrollado bajo la iniciativa de acceso abierto
Transcript of Redalyc.DINÁMICA DEL CARBONO Y NITRÓGENO DEL SUELO …observó que la mineralización de la...

Tropical and Subtropical Agroecosystems
E-ISSN: 1870-0462
Universidad Autónoma de Yucatán
México
Suárez M., Gabriela; Campos C., Adolfo; Cruz H., Lourdes
DINÁMICA DEL CARBONO Y NITRÓGENO DEL SUELO EN ECOSISTEMAS DE LA
COSTA TROPICAL SECA, EN LA MANCHA (CICOLMA), VERACRUZ, MÉXICO
Tropical and Subtropical Agroecosystems, vol. 18, núm. 3, 2015, pp. 347-361
Universidad Autónoma de Yucatán
Mérida, Yucatán, México
Disponible en: http://www.redalyc.org/articulo.oa?id=93944043009
Cómo citar el artículo
Número completo
Más información del artículo
Página de la revista en redalyc.org
Sistema de Información Científica
Red de Revistas Científicas de América Latina, el Caribe, España y Portugal
Proyecto académico sin fines de lucro, desarrollado bajo la iniciativa de acceso abierto

Tropical and Subtropical Agroecosystems, 18 (2015): 347 - 361
347
DINÁMICA DEL CARBONO Y NITRÓGENO DEL SUELO EN
ECOSISTEMAS DE LA COSTA TROPICAL SECA, EN LA
MANCHA (CICOLMA), VERACRUZ, MÉXICO
[SOIL CARBON AND NITROGEN DYNAMICS IN TROPICAL
DRY COASTAL ECOSYSTEMS IN LA MANCHA, VERACRUZ,
MEXICO]
Gabriela Suárez M.1 Adolfo Campos C.2*, Lourdes Cruz H.2
1Alumna de Maestría, Universidad Veracruzana, México
e-mail: [email protected] 2Instituto de Ecología, A. C., Carretera antigua a Coatepec, No. 351,
El Haya, Xalapa, Ver., C. P. 91070, México.
e-mail: [email protected], [email protected] *Corresponding author
RESUMEN
Esta investigación se realizó en La Mancha
(CICOLMA), en los sitios de Selva mediana sub-
caducifolia, Acahual y en Matorral sobre dunas. El
objetivo fue estudiar la dinámica del carbono y del
nitrógeno orgánico del suelo para cuantificar la
mineralización de la materia orgánica y el suministro
de nitrógeno. En laboratorio, muestras de suelo se
incubaron a 25 °C por un período de 97 días. En ese
período de incubación se realizaron 10 muestreos para
monitorear la dinámica del carbono (CO2-C) y del
nitrógeno (𝑁𝐻4+, 𝑁𝑂3
−) del suelo. Los resultados
mostraron que, los contenidos de carbono orgánico
acumulativo siguió la siguiente tendencia:
Matorral_Hondonada (2.70 mg g-1) > Acahual (2.0 mg
g-1) > Selva (1.9 mg g-1) > Matorral_Cima (1.2 mg g-
1). La comparación de medias por Tukey (α = 0.05)
reveló diferencias sólo entre Matorral_Hondonada y
Matorral_Cima. Los contenidos de nitrógeno
inorgánico acumulativo presentó el siguiente orden:
Acahual (1284 mg kg-1) > Matorral_Hondonada (1140
mg kg-1) > Selva (1021 mg kg-1) > Matorral_Cima (302
mg kg-1). El análisis estadístico mostró diferencias sólo
entre Matorral_Cima y los demás tratamientos. Se
observó que la mineralización de la materia orgánica y
el suministro de nitrógeno fueron mayores en
Matorral_Hondonada y Acahual, respectivamente. Se
concluye que la mineralización de la materia orgánica
y el suministro de nitrógeno están en función del tipo
de ecosistema y de las propiedades del suelo.
Palabras clave: mineralización de la materia orgánica
del suelo; selva mediana sub-caducifolia; Acahual;
Matorral; duna.
SUMMARY
This study was carried out at the La Mancha Coastal
Research Center (CICOLMA) in tropical semi-
deciduous forest, secondary forest and two locations in
coastal dune scrub (crest and slack). The aim was to
study the carbon and organic nitrogen dynamics of the
soil to quantify organic matter mineralization and
nitrogen supply. In the laboratory, soil samples were
incubated at 25 °C for 97 days and over this time 10
samples were analyzed to monitor the soil’s carbon
(CO2-C) and nitrogen (𝑁𝐻4+, 𝑁𝑂3
−) dynamics. The
results reveal that mean cumulative organic carbon
content was: dune slack (2.70 mg g-1) > secondary
forest (2.0 mg g-1) > semi-deciduous forest (1.9 mg g-
1) > dune crest (1.2 mg g-1). A comparison of the means
using Tukey’s test (α = 0.05) detected differences only
between the dune slack and the crest. Mean cumulative
inorganic nitrogen content was: secondary forest (1284
mg kg-1) > dune slack (1140 mg kg-1) > semi-deciduous
forest (1021 mg kg-1) > dune crest (302 mg kg-1), with
significant differences only between the dune crest and
the other treatments. In general, organic matter
mineralization was higher in the dune slack and
nitrogen supply was higher in the secondary forest. We
conclude that organic matter mineralization and
nitrogen supply are depending on the type of
ecosystem and soil properties.
Key words: soil organic matter mineralization;
tropical semi-deciduous forest; secondary forest;
scrub; dune.

Suarez-M., et al 2015
348
INTRODUCCIÓN
Los ecosistemas tropicales se encuentran en constante
transformación y en pérdida de su vegetación primaria.
Esto puede ser por disturbios naturales como los
huracanes o la dinámica de claros, así como por
disturbios de origen antrópico dados por la
proliferación de actividades humanas de alto impacto
e intensidad como la ganadería y la agricultura
(Martínez-Ramos, 1985; Capers et al. 2005; Martínez-
Ramos y García-Orth 2007). Este cambio en la
conformación de la vegetación lleva a continuos
procesos de regeneración forestal. Estos procesos son
los encargados de regresar la composición estructural
y funcional de un sistema a un estado similar al que
tenía antes del disturbio (Martínez-Ramos y García-
Orth 2007). En ecosistemas naturales, sin la
intervención humana, la transformación y movimiento
de materiales dentro del reservorio de materia orgánica
del suelo es un proceso dinámico controlado por el
clima, el tipo de suelo, la vegetación y los organismos
del suelo (Bot y Benítez, 2005). El mantillo que se
acumula en la superficie del suelo y la producción de
biomasa, generalmente conducen a una actividad
biológica alta en el suelo y en la superficie del suelo
(Quideau, 2005). Este equilibrio produce casi un ciclo
cerrado en la transferencia de nutrientes entre el suelo
y la vegetación adaptada a las condiciones del sitio
geográfico, resultando en condiciones casi perfectas
para el crecimiento de las plantas, por ejemplo
condiciones micro climáticas con adecuada humedad
en el suelo (Gregorich and Janzen, 2000; Bot y
Benítez, 2005). Contrario a lo anterior, en sistemas
antrópicos, la actividad biológica del suelo es afectada
por los sistemas de manejo y por los tipos de plantas
que se establecen (Bot y Benítez, 2005). La
simplificación de la vegetación y la pérdida de la capa
de mantillo en los sistemas cultivados, conduce a una
disminución en la cantidad de materia orgánica que
anualmente se incorpora al suelo, y los organismos que
habitan en la superficie del suelo y en la capa de
mantillo también son eliminados (Lal et al. 2000; Bot
y Benítez, 2005). La labranza es una de las prácticas
agrícolas que más reduce el contenido de materia
orgánica del suelo, porque cada vez que se labra es
aireado el suelo. Así que, como la mineralización de la
materia orgánica es un proceso aeróbico, el oxígeno
estimula la actividad de los microorganismos del suelo,
que se alimentan de materia orgánica, acelerando de
esta manera el ciclo del carbono (Magdoff y Weil,
2004; Bot y Benítez, 2005).
La materia orgánica del suelo (MOS) es uno de los
recursos naturales más importantes, y es la base de la
fertilidad del suelo (Sparks, 1999). El término materia
orgánica del suelo se utiliza para describir los
componentes orgánicos y generalmente se refiere al
carbono orgánico total contenido en el suelo (Baldock
y Skjemstad, 2000). La materia orgánica del suelo
puede ser de origen vegetal, animal y microbiano, y
consiste de un continuum de materiales en varios
estados de descomposición por la acción de procesos
bióticos y abióticos (Baldock y Skjemstad, 2000;
Curtin and Wen, 1999). El proceso de mineralización
de la MOS libera carbono a la atmósfera como CO2,
mientras algo de carbono es asimilado en los tejidos
microbianos, por ejemplo en la biomasa microbiana, y
otra parte se convierte en sustancias húmicas a través
del proceso de humificación (Sparks, 1999). La
función más importante de la materia orgánica del
suelo es que proporciona la energía metabólica que
dirige los procesos biológicos del suelo y los efectos
directos e indirectos que esto tiene en otras
propiedades y procesos del suelo (Sparks, 1999; Tan,
2003). El carbono fijado en el proceso de la fotosíntesis
es depositado en el suelo durante el crecimiento de las
plantas, y por este medio se proporciona el sustrato de
carbono a los organismos del suelo que descomponen
física y químicamente la estructura orgánica (Sparks,
1999; Tan, 2003). Los organismos del suelo (biota),
usan los residuos de las plantas y los animales y los
derivados de la materia orgánica como alimentos (Tan,
2003). A medida que descomponen los residuos y la
materia orgánica, los nutrientes son liberados dentro
del suelo en formas que pueden ser absorbidas por las
plantas (Sparks, 1999; Tan, 2003).
La mineralización de la materia orgánica del suelo en
los ecosistemas naturales y perturbados, es una fase de
alta prioridad para el suministro de nitrógeno
inorgánico (Curtin and Campbell, 2008; Jarvis et al.
1996). El amonio (𝑁𝐻4+) es el producto inicial de la
mineralización heterótrofa y después por oxidación
(realizada por microorganismos autótrofos) se
transforman a nitrato (𝑁𝑂3−) a través del proceso
conocido como nitrificación (Jansson and Persson,
1982; Havlin et al. 1999). Las concentraciones de
amonio y de nitratos en el suelo, son de gran
importancia durante el período de crecimiento de las
plantas (Jansson, 1982; Havlin et al. 1999). De los
nutrimentos que las plantas necesitan para su
crecimiento, el nitrógeno es requerido en mayor
cantidad a la que habitualmente se encuentra en forma
disponible en el suelo (Havlin et al. 1999). Las formas
de nitrógeno asimilables por la planta son 𝑁𝐻4+ y 𝑁𝑂3
−
(Havlin et al. 1999). El suministro de nitrógeno al
suelo es sensible a los factores que controlan la
mineralización potencial del nitrógeno (por ejemplo
las propiedades del suelo y las prácticas de manejo) y
a condiciones ambientales (por ejemplo, humedad y
temperatura) que regulan la actividad biótica (Griffin,
2008; Zebarth et al. 2009). En la evaluación del
suministro de nitrógeno por el suelo se utilizan
métodos biológicos que implican la incubación del
suelo en condiciones óptimas para promover la
mineralización de N y medir N inorgánico que se
produce durante un período de tiempo específico

Tropical and Subtropical Agroecosystems, 18 (2015): 347 - 361
349
(Campbell et al. 1998; Parkin et al. 2002). Estos
métodos se basan en el hecho de que la transformación
del N del suelo bajo condiciones de campo involucra
procesos basados en microorganismos (Parkin et al.
2002). Los métodos biológicos, particularmente la
incubación aeróbica, simulan la mineralización de N
en condiciones de campo (Sharifi et al. 2007). Stanford
y Smith (1972) fueron los primeros que propusieron un
método de incubación para evaluar la mineralización
del nitrógeno, incubando muestras de suelo por más de
20 semanas bajo óptimas condiciones de temperatura
y humedad.
En esta investigación se analiza la dinámica del
carbono y nitrógeno orgánico del suelo en parcelas
permanentes de selva mediana sub-caducifolia
conservada, acahual de 17 años (en potrero
abandonado) y en matorral sobre dunas, para generar
conocimiento científico sobre la mineralización de la
materia orgánica del suelo y del suministro de
nitrógeno mineral. Para ello se realizó un experimento
de incubación de muestras de suelo para analizar
específicamente, cómo se comporta la producción de
CO2, y de 𝑁𝐻4+ y 𝑁𝑂3
−, que son las formas principales
de nitrógeno disponible para las plantas, provenientes
de la mineralización de la materia orgánica; y para
identificar si la producción de CO2, 𝑁𝐻4+ y 𝑁𝑂3
− varía
con el tipo de ecosistema bajo estudio. Nuestra
hipótesis fue que el proceso de restauración estimula la
mineralización del carbono y nitrógeno orgánico
debido al efecto regulatorio de la calidad de la materia
orgánica del suelo.
MATERIALES Y MÉTODOS
Área de estudio
El estudio se llevó a cabo en el Centro de
Investigaciones Costeras La Mancha (CICOLMA),
que pertenece al Instituto de Ecología A.C (INECOL).
Está ubicado a 19º 35’ lat. N y 96º 22’ long. W, sobre
la planicie costera del Golfo de México al centro de
Veracruz, 25 km al N de la Ciudad José Cárdel, en el
municipio de Actopan (Figura 1). CICOLMA tiene una
superficie de 83 ha con una gran variedad de
vegetación. Es un mosaico heterogéneo y dinámico,
que cuenta con vegetación en diferentes fases de
sucesión ya sea primaria o secundaria que crece muy
cerca uno de otro y son sitios con condiciones edáficas
muy relacionadas.
El clima de la región se caracteriza por ser estacional,
con una temperatura anual promedio que puede variar
entre los 21.1 ºC en enero y 27.3 ºC en junio. Los
valores máximos y mínimos de temperatura extrema
registrados en la zona son de 40.5 ºC y 6 ºC. El clima
de tipo Aw2 cálido sub-húmedo de acuerdo a la
clasificación de Köeppen (García, 1981). La
precipitación total anual oscila de 899 a 1829 mm, el
mes más lluvioso varia de un año a otro, entre junio y
septiembre. Los registros de precipitación y
temperatura que se tienen para la estación permiten
reconocer claramente dos épocas, la primera
corresponde al periodo de lluvias que se presenta entre
junio y septiembre, donde se tiene una precipitación
total anual del 78%, y la segunda es la época seca que
se registra entre los meses de octubre a mayo
(Travieso-Bello y Campos, 2006). Adicionalmente la
región de La Mancha se encuentra influenciada por
vientos provenientes del noreste y norte, siendo estos
últimos los que permiten diferenciar una tercera época
(“nortes”) con vientos muy fuertes y húmedos que
aportan un porcentaje importante de la precipitación
anual en los meses de noviembre a febrero (Travieso-
Bello y Campos, 2006).
En cuanto a los tipos de suelo que se encuentran en los
terrenos de la CICOLMA, según la clasificación
establecida por FAO/UNESCO, corresponden los
tipos: arenosol cámbico, arenosol lúvico, arenosol
calcárico poco humificado y arenosol calcárico
ligeramente humificado (Travieso-Bello y Campos,
2006). Esto permite reconocer que los suelos de la zona
son principalmente arenosos y pueden variar en su
permeabilidad, fertilidad y concentración de materia
orgánica dependiendo de la topografía de los terrenos
(Travieso-Bello y Campos, 2006).
Este trabajo se centró en el mosaico de vegetación
compuesto por: a) la selva mediana sub-caducifolia, b)
acahual de 17 años (en potrero abandonado) y c)
matorral sobre dunas costera, todos incluidos dentro de
los terrenos pertenecientes a CICOLMA (Figura 2). A
continuación se describen brevemente cada uno de
estos tres tipos de vegetación:
a) La selva mediana sub-caducifolia ubicada en La
Mancha crece sobre dunas costeras, tiene en el área de
conservación su último remanente sobre dunas
(Castillo-Campos, 2006). El paisaje de La Mancha está
muy humanizado, sin embargo, conserva una
importante flora nativa a la que se han incorporado
numerosas plantas introducidas involuntaria o
voluntariamente (Castillo-Campos, 2006). Existe una
gran diversidad de árboles que forman parte de la flora
silvestre de dicha selva (Castillo-Campos, 2006). Es
mediana porque los árboles con los que cuenta no
rebasan los 30 metros de altura, y es subcaducifolia
porque algunos árboles son de hoja caduca y otros de
hoja perenne (Castillo-Campos, 2006). Aquí habita
una cantidad más amplia de animales y vegetales como
mapaches, tlacuaches, armadillos, iguanas, cangrejos,
y dentro de los árboles: nacatles, mulatos, cedros,
caobas, quiebraches, rabia, ficus y ojites, entre otros
(Castillo-Campos, 2006).

Suarez-M., et al 2015
350
Figura 1. Localización de los sitios de estudio.
Figura 2. Parcelas del estudio a largo plazo de la vegetación y dinámica de regeneración del paisaje, dentro del
proyecto: “Estructura de la vegetación y dinámica de la regeneración forestal en un mosaico heterogéneo en el trópico
seco del centro de Veracruz” (Laborde, CB2010/152457 CONACYT).
El símbolo de esta selva mediana subcaducifolia, es el
Ficus secapalos o matapalos. Aquí habita un cangrejo
rojo, el cual influye grandemente en la conservación de
esta selva, porque con sus madrigueras además de que
oxigenan la tierra, la proveen de una gran cantidad de
materia orgánica que sirve como fertilizante a los
árboles de la zona (Castillo-Campos, 2006).
Adicionalmente se pueden identificar otro tipo de
dinámicas en esta selva como lo es la dinámica de
claros, pues debido a la gran cantidad de elementos

Tropical and Subtropical Agroecosystems, 18 (2015): 347 - 361
351
involucrados (variables ambientales, variables
estructurales) la selva pueda presentar procesos de
“mini sucesiones” en respuesta a este tipo de disturbio
cuya intensidad y duración son variables para cada
caso (Begon et al. 2006).
b) Acahual. En La Mancha se tienen diferentes parches
de vegetación secundaria que se han clasificado en tres
categorías: pastizal, matorral y bosque secundario
(Travieso-Bello et al. 2006). Este sitio ha pasado por
transformaciones derivadas de la perturbación de la
selva baja caducifolia y la introducción de pastizales
cultivados con fines ganaderos y que presentan
diversos grados de perturbación es por eso que las
comunidades de plantas que se han establecido ahora
pasan por procesos para poder establecerse en el lugar
(Travieso-Bello et al. 2006). La vegetación en este
sitio no suele conservarse como tal, sino que al
detenerse el disturbio agropecuario que la generó se
inician procesos sucesionales que dan lugar a
comunidades con distinta estructura y composición
florística (Travieso-Bello et al. 2006). Cabe mencionar
que en 1995 durante 20 años esta tierra fue usada para
la ganadería, usando el suelo para cultivar pastos que
hasta estas fechas siguen tomando parte del terreno
(Travieso-Bello et al. 2006). Actualmente, a 17 años
de haber detenido el disturbio antrópico, se tiene un
acahual con dosel arbóreo de entre 4 y 10 m de altura,
donde la riqueza de herbáceas frecuentemente supera
el 50% del total de especies registradas para la región
(Mesa, 2013).
c) Matorral. Las dunas costeras constituyen un tipo de
ecosistema formado a partir de la acumulación de
arena por el viento. Estas son modeladas por procesos
geomórficos como la acción del agua, del viento y
procesos biológicos (Moreno-Casasola y Vázquez,
2006; Moreno-Casasola y Travieso-Bello, 2006;
López-Ramírez, 2007). Este es un ecosistema que
presta numerosos servicios ambientales tales como:
atrapar y almacenar arena (zona de amortiguamiento
entre el mar y la tierra), proteger otros ecosistemas de
las fuerzas de la naturaleza (vientos huracanados),
entre otros (Moreno-Casasola y Travieso-Bello, 2006).
Por sus características propias las dunas exhiben una
serie de gradientes relacionados con los factores
ambientales que se presentan en cada una de sus zonas
(Moreno-Casasola y Travieso-Bello, 2006). En
algunas zonas se pueden observar las coberturas
vegetales abiertas compuestas de herbáceas y arbustos
bajos, que con el tiempo se van transformando en
pastizales con algunos arbustos (Moreno-Casasola y
Vázquez, 2006; Moreno-Casasola y Travieso-Bello,
2006). Se considera que estos matorrales pueden crecer
en tal medida que se encaminen a convertirse en una
selva baja (Moreno-Casasola y Travieso-Bello, 2006),
así como el establecimiento de manchones de
vegetación, hacen menos posible la retención de agua
de lluvia necesaria para el crecimiento de otro tipo de
especies. En este sitio los nutrimentos suelen estar
menos disponibles por la densidad menor de
microorganismos que descompongan la materia
orgánica del suelo (Mesa, 2013).
Muestreo de suelos en las áreas de estudio
Fase de campo
Para el muestreo de suelos, cada parcela experimental
se dividió en tres secciones. En cada sección se aplicó
un muestreo sistemático que consistió en distribuir
cinco puntos espaciados cada cinco metros,
correspondiendo a cinco sub-muestras de suelo. Las
cinco sub-muestras de cada sección se mezclaron para
formar una muestra compuesta. Por las características
topográficas de la parcela de Matorral sobre duna, el
esquema de muestreo de suelos se aplicó en la Cima y
en la Hondonada de la duna. En todos los casos, las
muestras de suelo se obtuvieron de 0 - 10 cm de
profundidad del suelo. Las muestras de suelo se
colocaron en bolsas de plástico y se trasladaron al
laboratorio del INECOL, en Xalapa. En el laboratorio,
las muestras de suelo se colocaron en el área de secado.
Las muestras de suelo con la estructura de campo se
secaron al aire en charolas de 20 x 20 cm.
Experimento de incubación
Las muestras secas y sin tamizar se llevaron al
laboratorio para instalar el experimento de incubación.
Otra porción de muestra se tamizó para la realización
del análisis físico y químico de suelos.
Previamente a la incubación se pesaron 20 g de suelo
seco al aire (sin tamizar) y se colocaron en frascos de
polipropileno de 100 mL. Posteriormente, a las
muestras de suelo se les agregó agua destilada hasta
55% de su capacidad de almacenamiento de agua,
enseguida se mezclaron cuidadosamente el suelo con
el agua con una espátula para homogeneizar la
humedad en toda la muestra de suelo. El frasco que
contenía el suelo, se colocó dentro de un frasco de
polipropileno de 1.2 L (de boca ancha), se aplicó de 1
a 2 mL de agua destilada en el fondo de dicho frasco
(1.2 L) para mantener una atmósfera saturada de
humedad, además se colocó dentro otro frasco de
polipropileno de 100 mL, que contenía 20 mL de una
solución de NaOH 0.25 M, para capturar el CO2
respirado durante el período de incubación, finalmente
se selló el frasco de 1.2 L con masking tape para evitar
fugas de CO2. El experimento de incubación
contempló tres réplicas de cada muestra y dos réplicas
de blancos (frascos que no tuvieron suelo, sólo el agua
y el vial con el NaOH) para cuantificar la
carbonatación ambiental del NaOH, que se podría
producir al abrir la incubadora y los frascos. Para
mantener la humedad del suelo constante (55% de
espacio poroso ocupado por agua), durante el período

Suarez-M., et al 2015
352
de incubación se pesaron los frascos con el suelo cada
semana y cuando fue el caso se hizo el ajuste de
humedad. Las muestras de suelo se incubaron por
triplicado a 25 oC (+/- 1 oC) en un arreglo aleatorio en
una incubadora (Lab– Line, Imperial III Incubator) por
un período de 97 días. En ese período de incubación se
hicieron 10 muestreos (1, 2, 3, 7, 10, 27, 35, 45, 66 y
97 día) para monitorear la dinámica del carbono y del
nitrógeno del suelo.
Las muestras se mantuvieron destapadas en charolas
de almacenamiento de plástico (60 x 30 x 10 cm), en
cuya base se colocó una película de agua destilada,
finalmente se taparon con un film plástico perforado
con una aguja para tener condiciones aeróbicas. Las
muestras de la charola de almacenamiento fueron
mantenidas en la misma incubadora, a la misma
temperatura y humedad del aire que los frascos de 1.2
L para medición; hasta que fueron transferidas y
mantenidas en los frascos de medición, durante el
periodo de cuantificación. Por ejemplo, para la
medición del día 1, en el momento de instalar el
experimento, se debieron colocar los triplicados de las
muestras y los blancos en sus respectivos frascos de
medición, con sus respectivos viales con NaOH,
sellados al aire. Cumplido este plazo (por ejemplo 1
día) se retiraron de la incubadora, se taparon los viales
con NaOH, y se hizo la determinación de CO2-C por
acidimetría con el método de Anderson (1982). Así
que, el CO2 respirado fue atrapado en solución alcalina
de NaOH 0.25 M (agregando un exceso de BaCl2 1M,
para precipitar el ion carbonato) y se midió en los días
1, 2, 3, 7, 10, 27, 35, 45, 66 y 97 (10 muestreos),
después de haber instalado el experimento. El CO2-C
se calculó de la siguiente manera:
CO2-C (mg) = (mL HCl b – mL HCl m) * M * E
Donde:
HCl b= Volumen de ácido clorhídrico usado para
titular el blanco.
HCl m = Volumen de ácido clorhídrico usado para
titular la muestra.
M = Molaridad del ácido
E = peso equivalente (6 para C).
El N inorgánico se determinó por extracción del suelo
en cada muestra en 11 fechas de muestreo incluyendo
el T0 (tiempo cero, muestra no incubada), 1, 2, 3, 7, 10,
27, 35, 45, 66 y 97. La determinación de nitrógeno
inorgánico (Mulvaney, 1996) se basa en la extracción
del amonio intercambiable por equilibrio de la muestra
de suelo con KCL 2 N y su determinación por
destilación mediante arrastre de vapor en presencia de
MgO. La adición de la aleación de Devarda permite
incluir la determinación de nitratos.
Análisis estadístico
El modelo lineal general (GLM) fue usado para evaluar
las diferencias en los contenidos de carbono orgánico
total acumulado y para el nitrógeno inorgánico
acumulado durante el periodo de la incubación y los
sitios experimentales. Las diferencias significativas
para las medias fueron realizadas por el procedimiento
de Tukey (p ≤ 0.05). Las pruebas estadísticas y los
gráficos fueron realizados usando SAS (2000) y
SigmaPlot 13.0, respectivamente.
RESULTADOS Y DISCUSIÓN
Propiedades del suelo
En la tabla 1 se presentan los datos de las propiedades
químicas y físicas de los suelos en los sitios estudios.
En general, todos los sitios presentaron texturas
gruesas, con valores de arena mayores que 80%. Los
suelos de los sitios de Acahual y Selva presentaron una
textura de tipo arenoso franco, con valores de limo de
10.8 % y 9.5 %, respectivamente; mientras que los
sitios de Matorral presentaron textura de arena (>
93%), con menos de 2.0 % de arcilla. Los suelos de
Acahual y Selva presentaron un pH neutro (7) y
ligeramente alcalino (7.4) respectivamente; mientras
que los sitios del Matorral presentaron un pH
moderadamente alcalino (7.8), siendo el
Matorral_Cima el que presentó el valor más alto (8).
Los valores de pH altos (Acahual y Selva) y muy altos
(Matorral_Cima y Hondonada) podrían ser atribuidos
a la presencia de carbonatos en el suelo. En el campo,
estos suelos mostraron fuerte efervescencia a la
aplicación de HCl (10%). En todos los sitios, los
niveles de carbono orgánico del suelo fueron bajos,
destacándose el contenido más bajo en el sitio de
Matorral_Cima y el valor más alto en el sitio de
Matorral_Hondonada. Los niveles de nitrógeno total
siguieron un patrón similar a los del carbono orgánico.
En los sitios de Acahual y Selva, la CIC se ubica en un
rango medio (12.11 cmol kg-1 y 15.10 cmol kg-1
respectivamente), mientras que en
Matorral_Hondonada está en un nivel bajo (8.05 cmol
kg-1) y en Matorral_Cima la CIC está en un nivel muy
bajo (2.50 cmol kg-1). Las concentraciones de calcio
intercambiable están en un nivel alto en los sitios de
Acahual y Selva (10.54 cmol kg-1 y 12.76 cmol kg-1
respectivamente), mientras que en los sitios de
Matorral_Cima y Matorral_ Hondonada están en un
nivel medio (5.45 cmol kg-1 y 8 cmol kg-1). Las
concentraciones de magnesio se ubican en un nivel
medio en el sitio de Selva (2.63 cmol kg-1), bajo en los
sitios de Acahual (0.99 cmol kg-1) y
Matorral_Hondonada (0.68 cmol kg-1), y muy bajo en
Matorral_Cima (0.21 cmol kg-1). Los valores
reportados de potasio indican concentraciones bajas en
los sitios de Acahual (0.42 cmol kg-1) y Selva (0.32
cmol kg-1), y concentraciones muy bajas en los sitios

Tropical and Subtropical Agroecosystems, 18 (2015): 347 - 361
353
de Matorral_Cima (0.03 cmol kg-1) y Matorral_
Hondonada (0.24 cmol kg-1). Finalmente, las
concentraciones de sodio son altas en el sitio de
Acahual (0.89 cmol kg-1), medias en los sitios de
Matorral_Hondonada y Selva (0.60 cmol kg-1 y 0.65
cmol kg-1 respectivamente), y bajas en Matorral_Cima
(0.23 cmol kg-1).
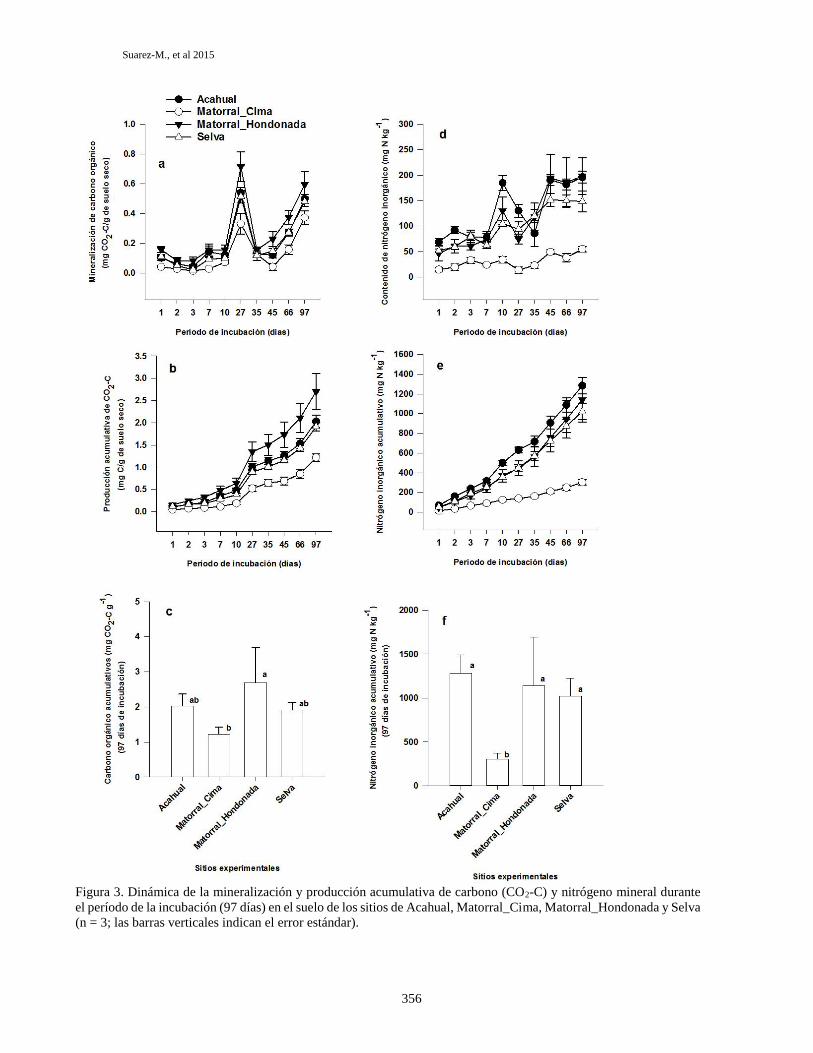
Dinámica de la mineralización del carbono
orgánico de suelo
En la Figura 3a se presenta la dinámica de la
mineralización de carbono orgánico durante el período
de la incubación para los sitios estudiados. Como
puede verse en la Figura 3a, durante el período de la
incubación, el Matorral_Hondonada presentó los
valores promedio más altos y el Matorral_Cima los
valores promedio más bajos, mientras que los sitios de
Acahual y Selva se situaron en un nivel intermedio en
cuanto a la dinámica del carbono orgánico. En
particular, se observa para todos los sitios un pico en el
día 27 de la incubación, y a partir del día 45 comienza
un incremento gradual en la mineralización del
carbono orgánico. Las variaciones observadas en la
mineralización del carbono orgánico durante el
período de la incubación podrían estar relacionadas
con distintos reservorios de carbono presentes en el
suelo, que difieren en su biodisponibilidad (Whalen y
Sampedro, 2010). El tipo de vegetación y las especies
de árboles tienen un efecto importante en la dinámica
del carbono orgánico de la capa superficial del suelo
(Berg and Laskowski, 2005).
La Figura 3b presenta las curvas de la mineralización
acumulativa de CO2-C, como una función del tiempo
de incubación. En el sitio de Acahual, el valor
promedio de CO2-C total acumulado al final del
período de la incubación fue de 2.0 mg g-1 de suelo
seco, con un valor mínimo 1.82 y un valor máximo de
4.0 mg CO2-C g-1 de suelo seco; en el sitio de
Matorral_Cima, el valor promedio de CO2-C total
acumulado fue de 1.21mg g-1, con un valor mínimo de
1.0 y un valor máximo de 1.5 mg CO2-C g-1;en el sitio
de Matorral_Hondonada el valor promedio fue de 2.7
mg CO2-C g-1, con un valor mínimo de 1.8 y un valor
máximo de 4.0 mg g-1; el sitio de Selva presentó un
valor promedio de 1.9 mg CO2-C g-1, con un valor
mínimo de 1.64 y un valor máximo de 2.29 mg CO2-C
g-1.El patrón de evolución de CO2-C acumulativo
(Figura 3b) revela que el sitio Matorral_Hondonada
presenta el valor más alto y el sitio Matorral_Cima el
valor más bajo. La cantidad de carbono orgánico del
suelo mineralizado es una señal del estado funcional de
un ecosistema (Berg y Laskowski, 2005).
Las diferencias observadas en el flujo de carbono
acumulado en el día 27, podrían estar relacionadas con
la fracción orgánica lábil, que es la más disponible para
los microorganismos que participan en la
mineralización de la materia orgánica del suelo (Curtin
et al. 1998; Franzluebbers et al. 2000; Huang et al.
2005).
El análisis de varianza (ANOVA) mostró que el
contenido de carbono orgánico acumulado en los 97
días de incubación fue estadísticamente diferente en
alguno de los tratamientos bajo estudio (Tabla 2). El
promedio de los contenidos de carbono orgánico
acumulativo siguió la siguiente tendencia:
Matorral_Hondonada > Acahual > Selva >
Matorral_Cima. Sin embargo, la comparación de
medias por el procedimiento de Tukey (α = 0.05)
reveló diferencias significativas sólo entre
Matorral_Hondonada y Matorral_Cima (Figura 2c).
Aunque la prueba estadística haya resultado no
significativa para la mayoría de los casos, es de resaltar
los valores promedio de carbono orgánico en los sitios
de Acahual (2.03 mg CO2-C g-1) y de Selva (1.90 mg
CO2-C g-1) con respecto al Matorral_Cima (1.21 mg
CO2-C g-1) y al Matorral_Hondonada (2.70 mg CO2-C
g-1). Los valores altos de carbono orgánico registrados
en Matorral_Hondonada podrían estar relacionados a
la forma topográfica del sitio (Hondonada) y a la
presencia de árboles y arbustos, que favorecen la
acumulación de materia orgánica (por ejemplo
cantidad y calidad del matillo) en el suelo, así como
por la humedad que podría acumularse en el suelo,
debido al efecto del micro-relieve que recibe
escorrentía de la parte superior del terreno
(Schoeneberger y Wysocki, 2005).
La mineralización de la materia orgánica con
frecuencia está relacionada con el tipo de vegetación,
las propiedades del suelo y la posición topográfica
(Berg y Laskowski, 2005). Las diferencias en la
calidad de la materia orgánica del suelo entre los sitios
de estudio podría también haber influenciado el
proceso de mineralización. Al respecto se señala que la
fracción lábil de la materia orgánica tiene
concentraciones más altas de carbono y de nitrógeno
que las fracciones estabilizadas (Gregorich et al.
1996). Estos resultados son, por lo tanto un reflejo de
la actividad biológica y de las propiedades del suelo de
los ecosistemas estudiados.

Suarez-M., et al 2015
354
Tabla 1. Propiedades físicas y químicas del suelo en los sitios experimentales.
Sitios Acahual Matorral_Cima Matorral_Hondonada Selva
pH (H2O) 7.0 8.0 7.8 7.4
K (cmol kg-1) 0.42 0.03 0.24 0.32
Na (cmol kg-1) 0.89 0.23 0.60 0.65
Ca (cmol kg-1) 10.54 5.45 8.0 12.76
Mg (cmol kg-1) 0.99 0.21 0.68 2.63
CIC (cmol kg-1) 12.11 2.50 8.05 15.10
P (mg kg-1) 2.2 1.8 1.4 6.9
C. orgánico, % 3.1 1.8 3.8 2.4
N. total, % 0.31 0.17 0.39 0.27
C/N 10.0 10.5 9.7 8.8
Limo, % 10.88 3.53 4.94 9.54
Arcilla, % 4.03 1.59 1.24 5.70
Arena, % 85.09 94.88 93.82 84.76
Dinámica de la mineralización del nitrógeno
orgánico del suelo
En la Figura 3d se presenta los datos de la
mineralización del N del suelo durante el período de la
incubación para los sitios estudiados. El sitio Acahual
presentó valores promedio de nitrógeno inorgánico que
variaron de 68.5 a 195.8 mg N kg-1, con un valor
mínimo de 19.1 y un valor máximo de 231.6 mg N kg-
1; en el sitio Matorral_Cima los valores promedio
variaron de 13.7 a 54.7 mg N kg-1, con un valor mínimo
de -6.8 y un valor máximo de 77.7 mg N kg-1; el
Matorral_Hondonada presentó valores promedio que
variaron de 46.2 a 197.9 mg N kg-1, con un valor
mínimo de 15.0 y un valor máximo de 341.8 mg N kg-
1; en el sitio de Selva los valores promedio variaron de
51.5 a 151.7 mg N kg-1, con un valor mínimo de 42.7
y un valor máximo de 213.8 mg N kg-1. En la mayoría
de los sitios (Figura 3d), el contenido de nitrógeno
inorgánico comenzó a incrementarse a partir del día 7
y también se destaca que después del día 45 de la fecha
de incubación, se presentaron pocos cambios en los
valores del nitrógeno inorgánico.
En general, se observa que la mineralización del
nitrógeno en el Matorral_Cima tuvo pocos cambios
durante el período de la incubación y también fue el
sitio que presentó los valores más bajos de nitrógeno
inorgánico. En la Figura 3d se aprecia que al final del
período de la incubación, los sitos
Matorral_Hondonada y Acahual presentaron los
valores más altos de nitrógeno inorgánico. En
particular, el Acahual es el sitio que presenta las
mayores variaciones en la dinámica del nitrógeno
durante el período de incubación. La mineralización
del nitrógeno orgánico es un componente importante
del suministro de nitrógeno inorgánico para las plantas
y los microorganismos del suelo (Havlin et al. 1999).
El nitrógeno inorgánico usualmente constituye menos
del 5 % del nitrógeno total en los suelos con niveles de
materia orgánica típicos (Magdoff et al. 1997). La
proporción mayor del nitrógeno total del suelo está
físicamente y químicamente protegida de la actividad
microbiana en el reservorio de materia orgánica
estable, mientras que las fracciones lábiles de la
materia orgánica son una fuente importante de
nitrógeno para las plantas y los microorganismos
(Magdoff et al. 1997). La variabilidad observada en la
mineralización del nitrógeno orgánico durante el
período de la incubación podría deberse a diferentes
patrones en la actividad de los microorganismos del
suelo, que controlan la transformación de nitrógeno
orgánico a nitrógeno mineral (Whalen y Sampedro,
2010). Estos resultados demuestran que los residuos
orgánicos aportados por las parcelas de Acahual y
Matorral_Hondonada son más lábiles que los
aportados por la Selva. La calidad de los residuos
orgánicos influye en la mineralización de la materia
orgánica del suelo (Magdoff et al. 1997). Esto sugiere
que la calidad de la materia orgánica más que la
cantidad, controla el proceso de mineralización.
La Figura 3e presenta las curvas de la mineralización
acumulativa del nitrógeno inorgánico, como una
función del tiempo de incubación. Los sitios Acahual
y Matorral_Cima presentaron los valores más altos y
más bajos respectivamente, de nitrógeno inorgánico
acumulativo durante el período de la incubación. En el
Acahual, el valor promedio de nitrógeno inorgánico
acumulado al final del período de la incubación fue de
1284 mg N kg-1, con un valor mínimo de 1026 y un
valor máximo de 1620 mg N kg-1; en el sitio de
Matorral_Cima, el valor promedio de nitrógeno
inorgánico total fue de 302 mg N kg-1, con un valor

Tropical and Subtropical Agroecosystems, 18 (2015): 347 - 361
355
mínimo de 237 y un valor máximo de 390 mg N kg-1;
el sitio Matorral_Hondonada presentó un valor
promedio de 1140 mg N kg-1 al final del período de la
incubación, con un valor mínimo de 543 y un valor
máximo de 1861 mg N kg-1; en el sitio de Selva, el
valor promedio de nitrógeno inorgánico total fue de
1021 mg N kg-1, con un valor mínimo de 833 y un valor
máximo de 1337 mg N kg-1.
El comportamiento del N en los suelos a lo largo de la
incubación, sobre todo en el caso del Acahual y la
Selva confirmaron la importancia de la textura y tipo
de suelo en el proceso de la mineralización, lo que
concuerda con otros autores (por ejemplo, Seneviratne
et al. 1999), que observaron que los parámetros que
determinan la liberación de nitrógeno son muy
dependientes del tipo de suelo y de la composición de
su materia orgánica. La estructura y las especies de la
vegetación afectan los procesos del ecosistema como
la producción de mantillo (Berg y Laskowski, 2005).
La calidad del mantillo que entra al suelo se ha
propuesto como un importante controlador de la
dinámica de mineralización y liberación de nutrientes
(Berg y Laskowski, 2005). La composición del
mantillo varía mucho entre las especies de vegetación,
incluso cuando se comparan especies estrechamente
relacionadas, lo que da lugar a variaciones en las
características químicas de la materia orgánica del
suelo y a la colonización por los microorganismos
(Stevenson y Cole, 1999).
El análisis de varianza (ANOVA) (Tabla 2) mostró que
el contenido de nitrógeno inorgánico acumulado en los
97 días de incubación fue estadísticamente diferente en
alguno de los tratamientos bajo estudio. El promedio
de los contenidos de nitrógeno inorgánico acumulativo
presentó el siguiente orden (Figura 3f): Acahual >
Matorral_Hondonada > Selva >Matorral_Cima. Sin
embargo, la comparación de medias por el
procedimiento de Tukey (α = 0.05) mostró diferencias
significativas sólo entre Matorral_Cima y los demás
tratamientos (Figura 3f). Es de resaltar el valor
promedio de nitrógeno inorgánico en el sitio de
Acahual (1284.66 mg kg-1), que podría estar
relacionado a la presencia de leguminosas en la
composición florística de este sitio (comunicación
personal, Javier Laborde, CONACYT
CB2010/152457). También, el valor promedio de
nitrógeno inorgánico (1140.79 mg kg-1) registrado en
el sitio de Matorral_Hondonada es de tomarse en
cuenta, y podría estar igualmente relacionado con la
acumulación de materia orgánica que se favorece por
la forma topográfica del sito (hondonada) y por la
presencia de árboles y arbustos que aportan residuos
orgánicos a la superficie del suelo. Por lo que, este
estudio sugiere que la intensidad de la mineralización
del nitrógeno orgánico del suelo podría estar muy
ligada a la calidad del mantillo de los diferentes
ecosistemas estudiados y al efecto de la posición del
suelo en el micro-relieve (Schoeneberger y Wysocki,
2005).
En la Figura 4a se presentan los resultados de la
dinámica de la concentración del amonio (𝑁𝐻4+) del
suelo durante el período de la incubación para los sitios
estudiados. En el sitio de Acahual, los valores
promedio de amonio variaron de 5.0 a 45 mg 𝑁𝐻4+ kg-
1, con un valor mínimo de -7.2 y un valor máximo de
80.8 mg 𝑁𝐻4+ kg-1; en el sitio de Matorral_Cima los
valores promedio variaron de 5.0 a 16 mg 𝑁𝐻4+ kg-1,
registrándose un valor mínimo de -3.5 y un valor
máximo de 35 mg 𝑁𝐻4+ kg-1; en el sitio de
Matorral_Hondonada los valores promedio variaron de
21 a 57 mg 𝑁𝐻4+ kg-1, presentándose un valor mínimo
de 6.1 y un valor máximo de 113 mg 𝑁𝐻4+ kg-1; en el
sitio de Selva los valores promedio variaron de 13 a 53
mg 𝑁𝐻4+ kg-1, con un valor mínimo de 3.8 y un valor
máximo de 70 mg 𝑁𝐻4+ kg-1.
El patrón general de la dinámica del amonio muestra
que a partir del día 7 y para la mayoría de los sitios, la
variabilidad en la concentración de amonio fue alta. Se
puede ver que en la mayoría de los sitios existen picos
en la producción de amonio. La cantidad y la calidad
de la materia orgánica del suelo determinan la relación
de liberación de nitrógeno por el proceso de
amonificación (Rice, 2005). La riqueza de especies de
plantas puede influir en el proceso de amonificación
por alterar la calidad de la materia orgánica del suelo
(Rice, 2005).
La Figura 4b presenta la dinámica total de la
concentración de amonio (𝑁𝐻4+) durante el período de
la incubación. En el sitio de Acahual, el valor promedio
de 𝑁𝐻4+ total acumulado al final del experimento fue
de 200 mg 𝑁𝐻4+ kg-1, con un valor mínimo de 148 y un
valor máximo de 294 mg 𝑁𝐻4+ kg-1; en el sitio de
Matorral_Cima, el valor promedio de 𝑁𝐻4+ total
acumulado fue de 110 mg 𝑁𝐻4+ kg-1, con un valor
mínimo de 54 y un valor máximo de 170 mg 𝑁𝐻4+ kg-
1; en el sitio de Matorral_Hondonada, el valor
promedio de 𝑁𝐻4+ total acumulado fue de 325 mg 𝑁𝐻4
+
kg-1, con un valor mínimo de 179 y un valor máximo
de 477 mg 𝑁𝐻4+ kg-1; el sitio de Selva presentó un valor
promedio de 𝑁𝐻4+ total acumulado de 261 mg 𝑁𝐻4
+ kg-
1, con un valor mínimo de 195 y un valor máximo de
307 mg 𝑁𝐻4+ kg-1.
Suarez-M., et al 2015
356
Figura 3. Dinámica de la mineralización y producción acumulativa de carbono (CO2-C) y nitrógeno mineral durante
el período de la incubación (97 días) en el suelo de los sitios de Acahual, Matorral_Cima, Matorral_Hondonada y Selva
(n = 3; las barras verticales indican el error estándar).

Tropical and Subtropical Agroecosystems, 18 (2015): 347 - 361
357
El patrón de la concentración acumulativa del amonio
muestra que el Matorral_Hondonada produce las
mayores cantidades de amonio, mientras que el sitio
Matorral_Cima produce las menores cantidades de
amonio.
El análisis de varianza (ANOVA) (Tabla 2) mostró que
el contenido de amonio acumulado en los 97 días de
incubación fue estadísticamente diferente en algunos
de los tratamientos bajo estudio. El promedio de los
contenidos de amonio acumulativo reveló que el
proceso de amonificación decreció en el siguiente
orden: Matorral_Hondonada> Selva >
Acahual>Matorral_Cima. La comparación de medias
por el procedimiento de Tukey (α = 0.05) mostró
diferencias significativas entre Matorral_Hondonada y
Acahual y Matorral_Cima pero no con Selva (Figura
4c). El valor promedio de amonio en el sitio de Selva
fue estadísticamente diferente del registrado en el sitio
de Matorral_Cima pero no del valor promedio
registrado en el sitio de Acahual. En general se observa
que la producción de amonio fue importante en los
sitios de Matorral_Hondonada y de Selva. Rosenkranz
et al. (2012) mencionan que la presencia de
leguminosas reduce la velocidad de amonificación, lo
que significa que una mayor proporción del reservorio
de nitrógeno total estuvo involucrado en el proceso de
amonificación.
En la Figura 4d se presentan los datos de la evolución
de los nitratos (𝑁𝑂3−) del suelo durante el período de la
incubación para los sitios estudiados. En el sitio de
Acahual, los valores promedio de nitratos variaron de
61 a 182 mg 𝑁𝑂3− kg-1, con un valor mínimo de 37 y
un valor máximo de 207 mg 𝑁𝑂3− kg-1; en el sitio de
Matorral_Cima los valores promedio variaron de 7.4 a
42 mg 𝑁𝑂3− kg-1, registrándose un valor mínimo de -
3.2 y un valor máximo de 61 mg 𝑁𝑂3− kg-1; en el sitio
de Matorral_Hondonada los valores promedio variaron
de 23 a 175 mg 𝑁𝑂3− kg-1, presentándose un valor
mínimo de 5.4 y un valor máximo de 290 mg 𝑁𝑂3− kg-
1; en el sitio de Selva los valores promedio variaron de
38 a 125 mg 𝑁𝑂3− kg-1, con un valor mínimo de 15.8 y
un valor máximo de 178 mg 𝑁𝑂3− kg-1.En general se
puede observar que la concentración de nitratos en la
mayoría de los sitios mostró pocos cambios durante los
primeros 27 días, y a partir de esta fecha se observó
una tendencia de incremento en la concentración de
nitratos. En la Figura 4d se destaca que el sitio Acahual
presentó las concentraciones de nitratos más altas,
incluso con la presencia de un pico en el día 10,
mientras que el sitio Matorral_Cima presentó las
concentraciones más bajas y con ligeros cambios en
todo el período del experimento. Estos resultados
podrían estar relacionados a la disponibilidad de
amonio en los sitios estudiados. La nitrificación es un
proceso importante en el ciclo del nitrógeno, y consiste
en un proceso de dos etapas: la oxidación de amoniaco
(NH3) a nitrito (NO2) y posteriormente, nitrito a nitrato
(Jarvis et al. 1996). Este proceso influye en la
producción primaria y controla la retención de
nitrógeno en el ecosistema (Jarvis et al. 1996).
La Figura 4e presenta la evolución total de la
concentración de nitratos (𝑁𝑂3−) como una función del
tiempo de la incubación. En el sitio de Acahual, el
valor promedio de 𝑁𝑂3− total acumulado al final del
experimento fue de 1083 mg 𝑁𝑂3− kg-1, con un valor
mínimo de 841 y un valor máximo de 1325 mg 𝑁𝑂3−
kg-1; en el sitio de Matorral_Cima, el valor promedio
de NO3- total acumulado fue de 192 mg NO3
- kg-1, con
un valor mínimo de 149 y un valor máximo de 230 mg
𝑁𝑂3− kg-1; en el sitio de Matorral_Hondonada, el valor
promedio de 𝑁𝑂3− total acumulado fue de 815 mg 𝑁𝑂3
−
kg-1, con un valor mínimo de 364 y un valor máximo
de 1384 mg 𝑁𝑂3− kg-1; el sitio de Selva presentó un
valor promedio de 𝑁𝑂3− total acumulado de 759 mg
𝑁𝑂3− kg-1, con un valor mínimo de 576 y un valor
máximo de 1029 mg 𝑁𝑂3− kg-1. El patrón de evolución
de la concentración de los nitratos muestra que el
Acahual origina las mayores cantidades de nitratos,
mientras que el sitio Matorral_Cima produce las
menores cantidades de nitratos. Otro estudio (Hamer y
Makeschin, 2009) ha reportado también relaciones
altas de nitrificación en sitios con leguminosas.
El análisis de varianza (ANOVA) (Tabla 2) mostró que
el contenido de nitratos acumulado en los 97 días de
incubación fue estadísticamente diferente en alguno de
los tratamientos bajo estudio. El promedio de los
contenidos de nitratos acumulativos mostró que el
proceso de nitrificación presentó el siguiente orden
(Figura 4f): Acahual > Matorral_Hondonada > Selva >
Matorral_Cima. La comparación de medias por el
procedimiento de Tukey (α = 0.05) mostró diferencias
significativas sólo entre Matorral_Cima y los demás
tratamientos (Figura 4f). Sin embargo es de resaltar la
importancia del proceso de nitrificación en la
producción de nitratos principalmente en el sitio de
Acahual (1083 mg kg-1), pero también en los sitios de
Matorral_Hondonada (815 mg kg-1) y Selva (759 mg
kg-1). En ese sentido, Malchair et al. (2010)
encontraron que la presencia de leguminosas
incrementó la nitrificación, estimulada por la presencia
de bacterias oxidantes del amonio. Es decir, el aumento
de la disponibilidad de amonio puede influir en el
potencial de nitrificación. La disponibilidad de amonio
es generalmente reconocida como un factor limitante
para la nitrificación (Havlin et al. 1999). Este estudio
revela la contribución clave de la parcela de Acahual
de 17 años (en potrero abandonado) al proceso de
nitrificación.

Suarez-M., et al 2015
358
Figura 4. Dinámica de la mineralización y producción acumulativa de amonio (𝑁𝐻4
+) y nitratos (𝑁𝑂3−) durante el
período de la incubación (97 días) en el suelo de los sitios de Acahual, Matorral_Cima, Matorral_Hondonada y Selva
(n = 3; las barras verticales indican el error estándar).

Tropical and Subtropical Agroecosystems, 18 (2015): 347 - 361
359
Tabla 2. Análisis de varianza (ANOVA) para los contenidos acumulativos de carbono orgánico, nitrógeno
inorgánico, amonio y nitratos en los sitios experimentales.
Fuente de
variación DF
Suma de
cuadrados
Cuadrados
medios F-Valor Pr >F
Carbono orgánico acumulativo
Modelo 3 6.648975 2.216325 7.48 0.0015
Error 20 5.929516 0.296475
Total 23 12.578492
Nitrógeno inorgánico acumulativo
Modelo 3 3428871.74 1142957.25 11.54 0.0001
Error 20 1981718 99085.9
Total 23 5410589.74
Amonio acumulativo
Modelo 3 150734.78 50244.92 10.17 0.0003
Error 20 98777.65 4938.88
Total 23 249512.44
Nitratos acumulativo
Modelo 3 2526851.87 842283.87 13.12 <0.0001
Error 20 1284356.41 64217.82
Total 23 3811208.04
CONCLUSIONES
En función de la hipótesis y el objetivo planteado, se
pudo llegar a las siguientes conclusiones: El estudio de
incubación permitió cuantificar las formas lábiles del
reservorio de materia orgánica del suelo que
suministran el nitrógeno disponible para las plantas, y
que representan uno de los principales indicadores de
la fertilidad del suelo. La mineralización del carbono y
del nitrógeno orgánico estuvo en función del tipo de
ecosistema y de las propiedades del suelo. Aunque se
presentaron pocas diferencias de textura entre los
suelos de los sitios estudiados, los efectos de esas
diferencias texturales en el proceso de mineralización
podrían ser ecológicamente importantes por el
suministro de nutrientes. La parcela de Acahual, por su
historia de manejo (en potrero abandonado), podría ser
el sitio actualmente con la producción de residuos
orgánicos más lábiles, comparado con los otros sitios.
Los resultados de este estudio sugieren que las
Hondonadas de la parcela de Matorral también son
muy activas en los aportes de materia orgánica lábil,
pero este proceso podría estar acotado por la dinámica
natural del sistema de duna activa que forma este
paisaje. Los resultados de la parcela de Selva muestran
que es un sistema más estable en relación al proceso de
mineralización de la materia orgánica del suelo. En
esta investigación, el estudio de la mineralización de la
materia orgánica se realizó en muestras de suelo de la
capa superficial, así que más investigaciones son
necesarias para evaluar el potencial de mineralización
de la materia orgánica en los diferentes horizontes del
suelo.
Agradecimientos
Los autores agradecen el financiamiento otorgado por
el Consejo Nacional de Ciencia y Tecnología
(Proyecto CONACYT CB2010/152457). Gabriela
Suárez Mandujano contó con el apoyo de una beca
(becaria #19781) que contribuyó a la tesis que para
obtener el grado de Ingeniero Agrónomo presentó en
la Universidad Veracruzana. Agradecemos a Rosario
Landgrave su apoyo para la elaboración del mapa de
ubicación del área de estudio.
REFERENCIAS
Anderson, J.P.E. 1982. Soil respiration. In: Page, A. L.,
Miller, R. H., Keeney, D. R. (eds.). Methods
of Soil Analysis, Part 2, Chemical and
Microbiological Properties. 2nd edition.
Agronomy Monograph, No. 9. ASA.
Madison. WI. pp. 831-871.
Baldock, J.A. and J.O. Skjemstad. 2000. Soil organic
carbon/soil organic matter. In: Peverill, K. I.,
Sparrow, L. A., and Reuter, D. J. (eds.). Soil
Analysis: An Interpretation Manual. CSIRO
Publishing, Collingwood, Australia. pp. 159-
170.
Begon, M. Townsend. C.R. and Harper, J.L. 2006.
Ecology: From individuals to ecosystems.
Blackwell Publishing. Malden. MA. USA. p
469.

Suarez-M., et al 2015
360
Berg, B. y Laskowski. R. 2005. Litter decomposition:
A guide to carbon and nutrient turnover.
Academic Press. NY, USA. p. 448.
Bot, A. and Benites, J. 2005. The importance of soil
organic matter: Key to drought-resistance soil
and sustained food production. FAO. Soil
Bulletin 80. Rome.
Campbell, C.A., Jame, Y. W. and de Jong, R. 1998.
Predicting net nitrogen mineralization over a
growing season: Model verification.
Canadian Journal of Soil Science. 68: 537–
552.
Capers, R.S., Chazdon, R. L., Redondo-Brenes, A. and
Vílchez-Alvarado, B. 2005. Successional
dynamics of woody seedling communities in
wet tropical secondary forests. Journal of
Ecology. 93:1071-1084.
Castillo-Campos, G. 2006. Las Selvas. En: Moreno-
Casasola, P. (ed.). Entornos veracruzanos: la
costa de La Mancha. Instituto de Ecología
A.C. Xalapa, México. pp. 221-230.
Curtin, D. and Wen, G. 1999. Organic matter fractions
contributing to soil nitrogen mineralization
potential. Soil Science Society of America
Journal. 63:410-415
Curtin, D. and Campbell, C. A. 2008. Mineralizable
nitrogen. In: Carter, M. R. and Gregorich, E.
G. (eds.). Soil Sampling and Methods of
Analysis. CRC. Boca Raton, FL. USA. pp.
599-606.
Curtin, D., Campbell, C. A., Jalil, A. 1998. Effects of
acidity on mineralization: pH dependence of
organic matter mineralization in weakly
acidic soils. Soil Biology and Biochemestry.
30: 57–64.
Franzluebbers, A.J., Haney, R. L., Honeycutt, C. W.,
Schonberg, H. H., Hons, F. M. 2000. Flush of
carbon dioxide following rewetting of dried
soil relates to active organic pools. Soil
Science Society of America Journal. 64:613–
623.
García, E. 1981. Modificaciones al Sistema de
Clasificación Climática de Kӧeppen para
Adaptarlo a las Condiciones de la República
Mexicana. Offset Larios. México. p. 246.
Gregorich, E.G. and Janzen, H. H. 2000.
Decomposition. In: Sumner, M. E. (ed.).
Handbook of Soil Science. CRC Press, Boca
Raton, Florida, USA. pp. C-107-C-128.
Gregorich, E.G., Ellert, B. H., Drury, C. F. and Liang,
B. C. 1996. Fertilization effects on soil
organic matter turnover and corn residue C
storage. Soil Science Society of America
Journal. 60:472-476.
Griffin, T.S. 2008. Nitrogen availability. In: Schepers,
J. C. and Ruan, W. R. (eds.). Nitrogen
availability in agricultural systems. Agron.
Monogr. 49. ASA, CSSA, and SSSA,
Madison, WI. pp. 613–646.
Hamer, U., Makeschin, F. 2009. Rhizosphere soil
microbial community structure and microbial
activity in set aside and intensively managed
arable land. Plan Soil. 316: 57-69.
Havlin, J. L., Beaton, J. D., Tisdale, S. L. and Nelson,
W. L. 1999. Soil Fertility and Fertilizers: An
Introduction to Nutrient Management. Sixth
edition. Prentice-Hall, Inc. Upper Saddle
River. New Jersey, USA.
Huang, P. M., Wang, M. K. and Chiu, C. 2005. Soil
mineral-organic matter-microbe interactions:
Impacts on biogeochemical processes and
biodiversity in soils. Pedobiología. 49:609–
635.
Jansson, B. H. 1982. Nitrogen mineralization in
relation of C:N ratio and decomposability of
organic materials. Plant and Soil. 181:39–45.
Jansson, S. L. and Persson, J. 1982. Mineralization and
immobilization of soil nitrogen. In:
Stevenson, F. J. (ed.). Nitrogen in agricultural
soils. Monography Nº 22. American Society
of Agronomy. Madison, Wisconsin, USA. pp.
229-252.
Jarvis, S. C., Stockdale, E. A., Shepherd, M. A. and
Powlson, D. S. 1996. Nitrogen
mineralizations in temperate agricultural
soils: processes and measurement. Advances
in Agronomy 57:187-235.
Lal, R. Kimble. J.M. Stewart. B.A. (eds.). 2000. Global
climate change and tropical ecosystems. CRC
Press, Boca Raton, FL.
López-Ramírez, M. C. 2007. Germinación,
establecimiento y crecimiento de seis
especies en matorrales de dunas costeras.
Tesis de Maestría (Ecología y Ciencias
Ambientales). Universidad Nacional
Autónoma de México, México DF, México.
p.169.
Magdoff, F., Lanyon, L. and Liebhardt, B. 1997.
Nutrient cycling, transformations and flows:
Implications for a more sustainable
agriculture. Advances in Agronomy 60: 1-73.
Magdoff, F. and Weil, R. R. (eds.). 2004. Soil organic
matter in sustainable agriculture. CRC Press,
Boca Raton, FL.
Malchair, S. D., Boeck, H. J., Lemmens, C., Merckx,
R., Nijs, L., Ceulemans, R., Carnol, M. 2010.
Do climate warming and plant species
richness affect potential nitrification, basal
respiration and ammonia-oxidizing bacteria
in experimental grasslands. Soil Biology and
Biochemistry. 42: 1944-1951.
Martínez-Ramos, M. 1985. Claros, ciclos vitales de los
árboles tropicales y regeneración natural de
las selvas altas perennifolias. En: Gómez-
Pompa, A. y Del Amo, S. (eds.).
Investigaciones sobre la Regeneración de
Selvas Altas en Veracruz, México. Instituto
Nacional de Investigaciones sobre Recursos

Tropical and Subtropical Agroecosystems, 18 (2015): 347 - 361
361
Bióticos, Xalapa y Editorial Alhambra
Mexicana, S.A. de C.V. pp. 191-239.
Martínez-Ramos, M. y García-Orth, X. 2007. Sucesión
ecológica y restauración de las selvas
húmedas. Boletín de la Sociedad Botánica de
México. 80: 69-84.
Mesa S. N. 2013. "Estructura de la comunidad de
regeneración de avanzada asociada a un
mosaico de vegetación en el trópico seco de
Veracruz. Tesis de Maestría. Instituto de
Ecología, A.C., Xalapa, Ver. México. p. 105.
Moreno-Casasola, P. y Travieso-Bello, A. C. 2006.
Las Playas y las Dunas. En: Moreno-
Casasola, P. (ed.). Entornos veracruzanos: la
costa de La Mancha. Instituto de Ecología,
A.C., Xalapa, México. pp. 205-220.
Moreno-Casasola, P. y Vásquez, G. 2006. Las
Comunidades de las Dunas. En: Moreno-
Casasola, P. (ed.). Entornos veracruzanos: la
costa de La Mancha. Instituto de Ecología,
A.C., Xalapa, México. pp. 285-310
Mulvaney, R. L. 1996. Nitrogen – Inorganic forms. In:
Sparks, D. L. (ed.) Methods of Soil Analysis,
Part 3, Chemical Methods. Soil Science
Society of America. Madison, WI. pp. 1123-
1184.
Parkin, T. B., Kaspar, T. C. and Cambardella, C. 2002.
Oat plant effects on net nitrogen
mineralization. Plant and Soil 243:187–195.
Quideau, S. A. 2005. Organic matter accumulation. In:
Lal, R. (ed.). Encyclopedia of soil science.
CRC Press. pp. 891-894.
Rice, C. W. 2005. Organic matter and nutrient
dynamics. In: Lal, R. (ed.). Encyclopedia of
soil science. CRC Press. pp. 925-928.
Rosenkranz, S., Wilcke, W., Eisenhauer, N., and
Oelmann, Y. 2012. Net ammonification as
influenced by plant diversity in experimental
grasslands. Soil Biology and Biochemistry
48: 78-87.
SAS Institute Inc. 2000. SAS User´s Guide: SAS
Institute, Inc. Cary. NC. USA.
Schoeneberger, P. J., and Wysocki, D. A. 2005.
Hydrology of soil and deep regolith: A nexus
between soil geography, ecosystems and land
management. Geoderma 126: 117-128.
Sharifi, M. B., Zebarth, D., Burton, C., Grant, G.,
Porter, J., Cooper, Y., Leclerc, G., Moreau,
and Arsenault, W. J. 2007. Evaluation of
laboratory-based measures of soil mineral
nitrogen and potentially mineralizable
nitrogen as predictors of field-based indices
of soil nitrogen supply in potato production.
Plant and Soil 301: 203–214.
SigmaPlot (version 13). 2013.Systat Software Inc.
Chicago IL, USA
Seneviratne, G., van Holm, L. H. J., Balachandra, L.
H. J. and Kulasooriya, S. A. 1999.
Differential effects of soil properties on leaf
nitrogen release. Biology Fertilizant Soils. 28:
238-243.
Sparks, D. L. (Ed.). 1999. Soil Physical Chemistry.
CRC Press, Boca Raton, FL.
Stanford, G. and Smith, S. J. 1972. Nitrogen
mineralization potentials of soils. Soil
Science Society of America Journal. 36: 465-
472.
Stevenson, F. J. and Cole, M. A. 1999. Cycles of soil.
2nd (ed).John Wiley y Sons, New York, USA.
p. 427.
Tan, K. M. 2003. Humic Matter in Soil and the
Environment. Marcel Dekker, Inc, New York.
Travieso-Bello, A. C. y Campos A. 2006. Los
Componentes del Paisaje. En: Moreno-
Casasola, P. (ed.). Entornos veracruzanos: la
costa de La Mancha. Instituto de Ecología,
A.C., Xalapa, México. pp. 139-150
Travieso-Bello, A. C., Gómez R. y Moreno-Casasola,
P. 2006. Los cultivos, los pastizales y los
acahuales. En: Moreno-Casasola, P. (ed.).
Entornos veracruzanos: la costa de La
Mancha. Instituto de Ecología, A.C., Xalapa,
México. pp. 261-272
Whalen, J. K. and Sampedro, L. 2010. Soil Ecology
and Management. CAB International, UK
Zebarth, B. J., Dury, C. F., Tremlay, N. and
Cambouris, A. N. 2009. Opportunities for
improved fertilizer nitrogen management in
production of arable crops in eastern Canada:
A review. Soil Science Society of America
Journal. 89: 113-132.
Submitted September 22, 2015 - Accepted December 07, 2015


















