
Principios de Nueroanatomia Funcional
109
Pontificia Universidad Católica de Valparaíso Principios Neuroanatomía Funcional Cátedra Anatomía Humana II PRINCIPIOS NEUROANATOMÍA FUNCIONAL “Unidad 2: Anatomía Externa e Interna Sistema Nervioso Central” Laboratorio Antropología Física y Anatomía Humana
-
Upload
ysa03048149 -
Category
Documents
-
view
2.952 -
download
3
Transcript of Principios de Nueroanatomia Funcional

Pontificia Universidad Católica de Valparaíso Principios Neuroanatomía Funcional Cátedra Anatomía Humana II
PRINCIPIOS NEUROANATOMÍA FUNCIONAL “Unidad 2: Anatomía Externa e
Interna Sistema Nervioso Central”
Laboratorio Antropología Física y Anatomía Humana

Pontificia Universidad Católica de Valparaíso Principios Neuroanatomía Funcional
EMBRIOLOGÍA DE LAS REGIONES DE SNC
Formación de la Médula Espinal
Las placas basales incluyen los somas de las motoneuronas que posteriormente constituirán las astas anteriores de la médula espinal. Las placas alares corresponden a regiones sensitivas que se diferenciarán en las astas posteriores de la médula espinal. El crecimiento de las placas alares origina el tabique medio posterior. Al sobresalir ventralmente las placas basales se forma el tabique medio anterior, mientras tanto se desarrolla la fisura mediana en la superficie anterior de la médula espinal. El surco limitante delimita ambas placas, y de esta manera también separa las regiones motoras de las sensitivas. Las regiones dorsal (placa del techo) y ventral (placa del piso) en la línea media del tubo neural no poseen neuroblastos y constituyen vías para fibras nerviosas que cruzan la médula espinal de un lado al otro.
Entre las astas ventral y dorsal de los segmentos torácicos hasta el
segundo o tercero lumbar de la médula espinal se acumulan neuronas que formarán el asta lateral o intermedia, que contiene neuronas del sistema nervioso autónomo.
Al plegarse la placa neural aparece un grupo de células a cada lado del surco neural. Estas células tienen un origen ectodérmico y se denominan células de la cresta neural y forman temporalmente una zona intermedia entre el tubo neural y el ectodermo superficial. Esta zona se extenderá desde el prosencéfalo hasta el nivel de los somitas caudales y de cada lado emigra a la cara dorsolateral del tubo neural. Algunas células originan los ganglios de la raíz dorsal de los nervios raquídeos. Las células de la cresta neural se diferenciarán también en neuroblastos simpáticos, células de Schwann, células pigmentarias, odontoblastos, meninges y mesénquima de los arcos faríngeos.
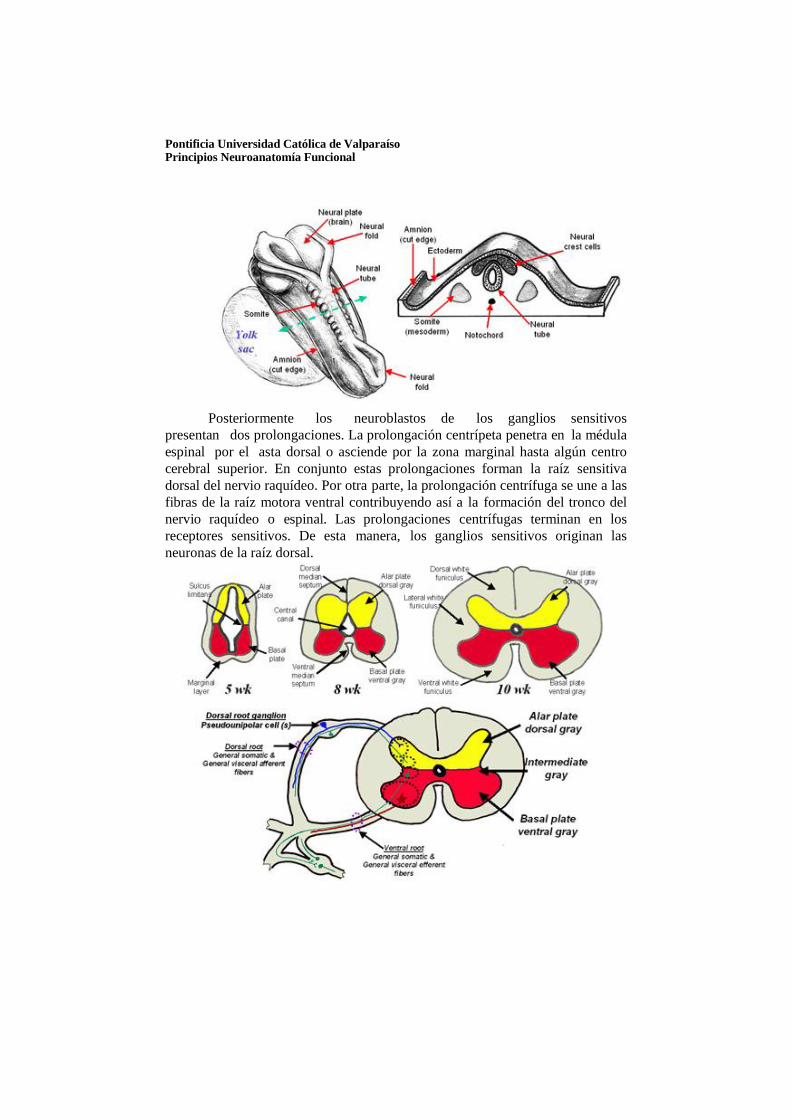
Pontificia Universidad Católica de Valparaíso Principios Neuroanatomía Funcional
Posteriormente los neuroblastos de los ganglios sensitivos presentan dos prolongaciones. La prolongación centrípeta penetra en la médula espinal por el asta dorsal o asciende por la zona marginal hasta algún centro cerebral superior. En conjunto estas prolongaciones forman la raíz sensitiva dorsal del nervio raquídeo. Por otra parte, la prolongación centrífuga se une a las fibras de la raíz motora ventral contribuyendo así a la formación del tronco del nervio raquídeo o espinal. Las prolongaciones centrífugas terminan en los receptores sensitivos. De esta manera, los ganglios sensitivos originan las neuronas de la raíz dorsal.

Pontificia Universidad Católica de Valparaíso Principios Neuroanatomía Funcional
Las fibras nerviosas motoras comienzan a originarse a partir de células situadas en las placas basales de la médula espinal. Se sitúan en haces y forman las raíces nerviosas ventrales. Las raíces nerviosas dorsales se originan en células de los ganglios de la raíz dorsal. Las prolongaciones distales (centrífugas) se unen a la raíz motora y forman el nervio espinal. Casi inmediatamente los nervios se dividen en ramos primarios dorsales y ventrales. Los dorsales inervan la musculatura axial dorsal, las articulaciones intervertebrales y la piel de la espalda. Los ramos ventrales inervan los miembros y la pared anterior del cuerpo, y forman los principales plexos nerviosos.
El proceso es llevado a cabo por las células de Schwann. Estas células se originan en la cresta neural, emigran a la periferia y se disponen alrededor de los axones formando la vaina de Schwann o neurilema. Las fibras en las que se deposita la mielina, adquieren un aspecto blanquecino. La mielina no es sino un enrollamiento de la membrana de la célula de Schwann alrededor del axón.
En el tercer mes de desarrollo, la médula espinal se extiende en toda la
longitud del embrión y los nervios raquídeos atraviesan los forámenes intervertebrales en su nivel de origen. Sin embargo, al aumentar la edad del embrión, el raquis y la duramadre se alargan más rápidamente que el tubo neural y el extremo terminal de la médula se desplaza a niveles cada vez más altos. En el recién nacido el extremo está situado a la altura de L3. Como consecuencia de este crecimiento desproporcionado, los nervios espinales tienen una dirección oblicua desde su segmento de origen en la médula hasta el nivel correspondiente de la columna vertebral. La duramadre permanece unida a la columna vertebral a nivel coccígeo.

En el adulto, la médula espinal termina a la altura de L2-L3. Por debajo
de este sitio, una prolongación filiforme de la piamadre forma el filum terminale y está unido al periostio de la primera vértebra coccígea. Las fibras nerviosas que se encuentran por debajo del extremo terminal de la médula forman la cola de caballo (cauda equina).
Formación del Encéfalo
Mielencéfalo
El mielencéfalo da origen a la médula oblonga (bulbo raquídeo). Difiere de la médula espinal porque sus paredes laterales experimentan un movimiento de eversión, es decir, las placas alares se separan. Se distinguen las placas alares y basales separadas por el surco limitante.
La placa basal, semejante a la de la médula espinal, contiene los
núcleos motores, que se dividen en tres grupos:
Grupo medial o eferente somático: El grupo medial o eferente somático contiene las neuronas motoras que forman la continuación cefálica de las células del asta anterior. Dado que este grupo eferente somático continúa en dirección rostral hacia el mesencéfalo, se denomina columna motora eferente somática. En el mielencéfalo representa las neuronas del nervio hipogloso que se distribuye en los músculos de la lengua. En el metencéfalo y el mesencéfalo la columna está representada por las neuronas de los nervios abducens o motor ocular externo, patético o troclear y oculomotor o motor ocular común, respectivamente. Estos nervios se distribuyen en los músculos del ojo.

Grupo intermedio o eferente visceral especial: El grupo eferente visceral especial se extiende hasta el metencéfalo y forma la columna motora eferente visceral especial. Sus neuronas motoras se distribuyen en los músculos estriados de los arcos faríngeos. En el mielencéfalo la columna está representada por las neuronas de los nervios espinal accesorio, vago y glosofaríngeo.
Grupo lateral o eferente visceral general: El grupo eferente visceral general contiene las neuronas motoras que se distribuyen en los músculos lisos del aparato respiratorio, tracto intestinal y corazón.
La placa alar incluye tres grupos de núcleos sensitivos de relevo.
Grupo lateral o aferente somático: recibe impulsos del oído y la superficie de la cabeza por medio de los nervios vestibulococlear y trigémino.
Grupo intermedio o aferente visceral especial: recibe impulsos de los botones gustativos de la lengua y del paladar, bucofaringe y epiglotis.
Grupo medial, aferente visceral general: recibe información interoceptiva del aparato gastrointestinal y del corazón.
La placa del techo del mielencéfalo consiste en una capa única de células ependimarias cubierta por mesénquima vascularizado, la piamadre. Ambas forman la tela coroidea. Debido a la activa proliferación del mesénquima vascularizado, un cierto número de invaginaciones sacciformes se introducen en la cavidad ventricular subyacente. Estas invaginaciones forman el plexo coroideo, que elabora el líquido cerebro-espinal.

Metencéfalo
El metencéfalo, parecido al mielencéfalo, se caracteriza por sus placas
basales y alares. Sin embargo, se forman dos nuevos componentes: a) el cerebelo y b) la protuberancia (puente).
Las placas basales del metencéfalo contienen tres grupos de neuronas motoras:
El grupo eferente somático medial: que da origen al nervio abducens
El grupo eferente visceral especial: que contiene los núcleos de los nervios trigémino y facial, que inervan los músculos del primero y el segundo arcos faríngeos.
El grupo eferente visceral general: cuyos axones se distribuyen en las glándulas submaxilares y sublinguales.
La capa marginal de las placas basales se expanden considerablemente ya que sirven de puente para las fibras nerviosas que conectan la corteza cerebral y la cerebelosa con la médula espinal. Es por esto que a esta porción del metencéfalo se le denomina puente o protuberancia. Además de fibras nerviosas, la protuberancia contiene los núcleos pontinos o protuberanciales, que se originan en las placas alares del metencéfalo y el mielencéfalo.
Las placas alares de esta vesícula poseen tres grupos de núcleos sensitivos:
Grupo aferente somático (lateral): que contiene neuronas del nervio trigémino y una pequeña porción del complejo vestibulococlear
Grupo aferente visceral especial

Pontificia Universidad Católica de Valparaíso Principios Neuroanatomía Funcional Grupo aferente visceral general.
Cerebelo
Las porciones dorsolaterales de las placas alares se curvan en sentido
medial y forman los labios rómbicos. En la porción caudal del metencéfalo los labios rómbicos están muy separados, pero inmediatamente por debajo del mesencéfalo se acercan en la línea media. Al profundizarse el pliegue protuberancial, los labios rómbicos quedan comprimidos en dirección cefalocaudal y forman la placa cerebelosa. En el embrión de 12 semanas esta placa presenta una pequeña porción en la línea media, el vermis, y dos porciones laterales, los hemisferios. Poco después una cisura transversal separa el nódulo del vermis, y el flóculo lateral de los hemisferios. Desde el punto de vista filogenético este lóbulo floculonodular es la parte más primitiva del cerebelo.


Pontificia Universidad Católica de Valparaíso Principios Neuroanatomía Funcional
En una etapa inicial la placa cerebelosa está formada por las capas neuroepitelial, del manto y marginal. Durante el desarrollo ulterior algunas células formadas por el neuroepitelio emigran hacia la superficie del cerebelo y forman la capa germinal externa. Estas células conservan su capacidad mitótica y proliferan en una zona de la superficie del cerebelo.
Existen dos zonas de histogénesis del cerebelo, una zona ventricular situada por debajo de la placa cerebelosa en desarrollo, la capa marginal interna, y la segunda que termina por quedar en posición superficial a la zona del manto, la capa germinal

Pontificia Universidad Católica de Valparaíso Principios Neuroanatomía Funcional
externa. Esta capa deriva de la células que emigran desde el labio
romboidal. Ambas
capas muestras mitosis proliferativas y producen una progenie de varias capas. Esta fase disminuye al iniciar la fase de neurogénesis y seguida de una gliogénesis. De la placa alar además migran células que forman las futuras neuronas de Purkinje y los núcleos profundos del cerebelo. Las neuronas de Purkinje formadas migran y se ubican bajo la capa germinal externa
En el sexto mes del desarrollo la capa granulosa externa comienza a
producir diversos tipos celulares, los cuales emigran hacia las zonas bajo las células de Purkinje en proceso de diferenciación y dan origen a las células granulosas, células en cesta y células estrelladas. La corteza del cerebelo, que consiste en células de Purkinje, neuronas de Golgi tipo II y neuronas producidas por la capa granulosa externa, alcanza sus dimensiones definitivas después del nacimiento. Gran parte de las células de la capa germinal externa forma neuronas estrelladas, originando la capa molecular. Los núcleos cerebelosos profundos, llegan a su situación definitiva antes del nacimiento. Finalmente la corteza cerebelosa queda formada por tres capas: Molecular, Purkinje y Granulosa. Se establecen contactos sinápticos entre las células de la capas. Axones provenientes de médula, tronco y cerebro ingresan a cerebelo formando los pedúnculos cerebelosos, sinaptando con núcleos profundos del cerebelo y con las células granulosas (fibras musgosas). Axones provenientes de la oliva sinaptan con las células de Purkinje (fibras trepadoras). Los axones de las neuronas de Purkinje son los únicos que salen de corteza para sinaptar con los núcleos profundos. Los axones originados en los núcleos profundos salen por los pedúnculos cerebelosos superiores o inferiores para sinaptar con otros sitios de relevo.

Pontificia Universidad Católica de Valparaíso Principios Neuroanatomía Funcional
Mesencéfalo
Desde el punto de vista morfológico, el mesencéfalo es la más primitiva de las vesículas encefálicas. Cada placa basal posee dos grupos de núcleos motores:
Grupo medial eferente somático: representado por los nervios oculomotor y troclear, que inervan a los músculos del ojo.
Grupo eferente visceral general: pequeño grupo representado por el núcleo de Edinger-Westphal, que inerva el esfínter de la pupila.
La capa marginal de cada placa basal se engrosa y forma el pie
de los pedúnculos cerebrales, estructuras que sirven para el paso de fibras que descienden desde el cerebro hacia los centros inferiores de la médula y de la protuberancia. Las placas alares del mesencéfalo aparecen en un principio en forma de dos elevaciones longitudinales separadas por una depresión poco profunda en la línea media. Al continuar el desarrollo aparece un surco transversal que divide a cada elevación longitudinal en un colículo anterior (superior) (tubérculo cuadrigémino superior) y otro posterior (inferior) (tubérculo cuadrigémino inferior).

Pontificia Universidad Católica de Valparaíso Principios Neuroanatomía Funcional
Los colículos se forman por oleadas de neuroblastos que emigran hacia
la zona marginal suprayacente, donde se disponen en capas estratificadas.
Diencéfalo
Placa del techo y epífisis
El diencéfalo se desarrolla a partir de la porción mediana del prosencéfalo, y se considera que consiste en una placa del techo y dos placas alares, pero que carece de placas del piso y las basales. La placa del techo del diencéfalo está formada por una sola capa de células ependimarias cubiertas por mesénquima vascularizado, las cuales, al combinarse, originan el plexo coroideo del tercer ventrículo. La porción más caudal de la placa del techo se convierte en el cuerpo pineal o epífisis. La primera manifestación de esta glándula es un engrosamiento epitelial de la línea media, pero que hacia la séptima semana comienza a evaginarse. Por último, se convierte en un órgano macizo situado en el techo del mesencéfalo y sirve de intermediario de la acción de la luz y la oscuridad sobre los ritmos endocrinos y del comportamiento.
Placa alar, tálamo e hipotálamo
Las placas basales y alares forman las paredes laterales del diencéfalo. Una hendidura, el surco hipotalámico, divide dichas placas en porciones dorsal y ventral, el tálamo y el hipotálamo, respectivamente.
Como consecuencia de la actividad de proliferación, poco a poco el tálamo sobresale en la luz del diencéfalo. Con frecuencia esta expansión es muy grande, tanto que las regiones talámicas derecha e izquierda se fusionan en la línea media y forman la masa intermedia o comisura gris intertalámica.
El hipotálamo, que deriva de la placa alar, se diferencia en varios grupos de núcleos, que sirven como centros de regulación de funciones viscerales tales como el sueño, la digestión, la temperatura corporal y la conducta emocional. Uno de estos grupos, el tubérculo mamilar, forma una eminencia definida sobre la cara ventral del hipotálamo, a cada lado de la línea media. Algunos autores señalan que hipotálamo y tálamo derivan de la placa alar.

Pontificia Universidad Católica de Valparaíso Principios Neuroanatomía Funcional
Hipófisis o glándula pituitaria
La hipófisis o glándula pituitaria se desarrolla en dos partes
completamente distintas: a) una evaginación ectodérmica llamada bolsa de Rathke y b) una prolongación del diencéfalo hacia abajo, el infundíbulo.
Cuando el embrión tiene tres semanas de edad, aproximadamente, la
bolsa de Rathke se observa como una evaginación de la cavidad bucal, y ulteriormente crece en sentido dorsal hacia el infundíbulo. Hacia el final del segundo mes pierde su conexión con la cavidad bucal y se halla en contacto con el infundíbulo.
En el curso del desarrollo ulterior las células de la pared anterior de la
bolsa de Rathke aumentan rápidamente de número y forman el lóbulo anterior de la hipófisis o adenohipófisis. Una pequeña prolongación de este lóbulo, la pars tuberalis, crece a lo largo del tallo del infundíbulo y por último lo rodea. La pared posterior de la bolsa de Rathke se convierte en la pars intermedia, que en el ser humano parece tener poca importancia.
Pontificia Universidad Católica de Valparaíso Principios Neuroanatomía Funcional
El infundíbulo da origen al tallo y a la pars nervosa o lóbulo posterior
de la hipófisis (neurohipófisis). Está formado por células de neuroglia y posee, además, cierta cantidad de fibras nerviosas provenientes de la región hipotalámica.
Telencéfalo
El telencéfalo, la vesícula encefálica más rostral, consiste en dos evaginaciones laterales, los hemisferios cerebrales, y una porción mediana, la lámina terminal. Las cavidades de los hemisferios, los ventrículos laterales, comunican con la luz del diencéfalo por medio de los agujeros interventriculares (de Monroe).
Hemisferios cerebrales
Los hemisferios cerebrales comienzan a desarrollarse al principio de la quinta semana de vida intrauterina, en forma de evaginaciones bilaterales de la pared lateral del prosencéfalo. Hacia la mitad del segundo mes la porción basal de los hemisferios

Pontificia Universidad Católica de Valparaíso Principios Neuroanatomía Funcional
(porción que inicialmente formó la prolongación del tálamo hacia adelante) comienza a aumentar de tamaño. Como consecuencia de ello, sobresale hacia el interior del ventrículo lateral y también en el piso del agujero interventricular. En cortes transversales esta región de crecimiento rápido tiene aspecto estriado y, por lo mismo, se denomina cuerpo estriado.
En la región donde la pared del hemisferio está unida al techo del diencéfalo no se producen neuroblastos y la zona es muy delgada. En este sitio la pared del hemisferio tiene una sola capa de células ependimarias cubiertas por mesénquima vascularizado que en conjunto forman el plexo coroideo. El plexo coroideo debía formar el techo del hemisferio, pero como consecuencia del crecimiento desproporcionado de varias partes del hemisferio, sobresale en el ventrículo lateral siguiendo una línea llamada fisura coroidea. Inmediatamente por arriba de la fisura coroidea la pared del hemisferio está engrosada, lo cual forma el hipocampo. Esta estructura, que tiene una función relacionada al olfato, sobresale gradualmente en el ventrículo lateral
Al producirse la ulterior expansión de los hemisferios, poco a poco
cubren la cara lateral del diencéfalo, el mesencéfalo y la porción cefálica del metencéfalo, que forma parte de la pared del hemisferio, también se expande hacia atrás y se divide en dos partes: a) una porción dorsomedial, el núcleo caudado, y b) una porción ventrolateral, el núcleo lenticular. Esta división se efectúa por los axones que van hacia la corteza del hemisferio y que provienen de ella y se abren paso por la masa nuclear del cuerpo estriado. El fascículo fibroso formado de esta manera se denomina cápsula interna.

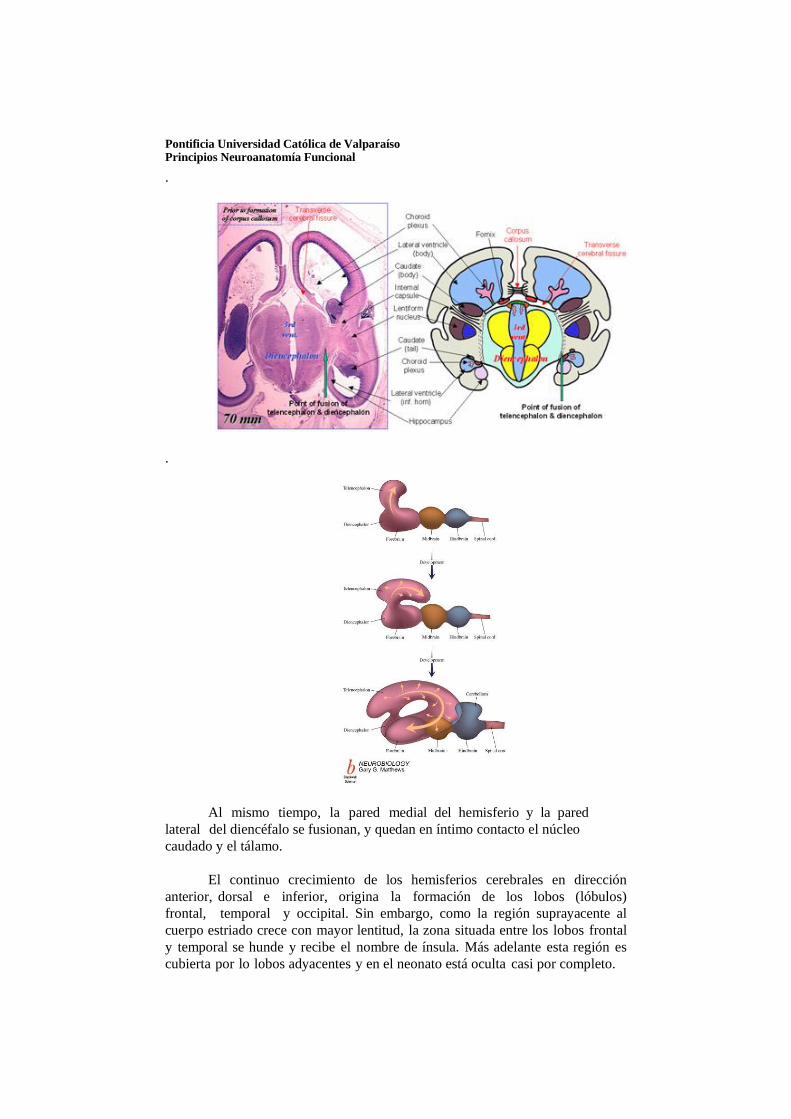
Pontificia Universidad Católica de Valparaíso Principios Neuroanatomía Funcional
.
.
Al mismo tiempo, la pared medial del hemisferio y la pared lateral del diencéfalo se fusionan, y quedan en íntimo contacto el núcleo caudado y el tálamo.
El continuo crecimiento de los hemisferios cerebrales en dirección
anterior, dorsal e inferior, origina la formación de los lobos (lóbulos) frontal, temporal y occipital. Sin embargo, como la región suprayacente al cuerpo estriado crece con mayor lentitud, la zona situada entre los lobos frontal y temporal se hunde y recibe el nombre de ínsula. Más adelante esta región es cubierta por lo lobos adyacentes y en el neonato está oculta casi por completo.

Pontificia Universidad Católica de Valparaíso Principios Neuroanatomía Funcional
En el período final de la vida fetal la superficie de los hemisferios
cerebrales crece con tal rapidez que presenta muchos giros (circunvoluciones) separadas por cisuras y surcos.
Desarrollo de la corteza
La corteza cerebral se desarrolla a partir del palio, que puede dividirse en dos regiones: a) el paleocórtex o arquipalio, zona situada inmediatamente lateral al cuerpo estriado, y b) neocórtex o neopalio, entre el hipocampo y el paleocórtex.
En el neocórtex, grupos de neuroblastos emigran a un sitio por debajo de la piamadre y luego se diferencian en neuronas completamente maduras. Cuando llega la siguiente oleada de neuroblastos, emigran por las capas de células antes formadas hasta llegar a la situación subpial. En consecuencia, los neuroblastos formados en la etapa inicial toman una situación profunda en la corteza, en tanto que los formados más tarde tienen una posición más superficial.
En el recién nacido la corteza tiene aspecto estratificado debido
a la diferenciación de las células en distintas capas. La corteza motora posee abundantes células piramidales, y las áreas sensitivas se caracterizan por células granulosas.

Pontificia Universidad Católica de Valparaíso Principios Neuroanatomía Funcional
Comisuras
En el adulto, los hemisferios derecho e izquierdo están conectados por varios haces de fibras, las comisuras, que atraviesan la línea media. El más importante de estos haces utiliza la lámina terminal. El primero de los fascículos cruzados que aparece es la comisura anterior, y consiste en fibras que conectan el bulbo olfatorio y el área cerebral correspondiente de un hemisferio con los del lado opuesto.
La segunda comisura que aparece es la comisura del hipocampo o del
trígono. Sus fibras nacen en el hipocampo y convergen en la lámina terminal cerca de la placa del techo del diencéfalo. Desde allí continúan y forman un sistema arqueado inmediatamente por fuera de la fisura coroidea, hasta llegar al tubérculo mamilar y al hipotálamo.
La comisura más importante es el cuerpo calloso. Aparece hacia la
décima semana de desarrollo y comunica las áreas no olfatorias de la corteza cerebral derecha e izquierda. En un principio forma un pequeño fascículo en la lámina terminal. Sin embargo, como consecuencia de la expansión continua del neocórtex, rápidamente se extiende primero hacia ventral y después a dorsal, y se dispone de manera de arco sobre el delgado techo del diencéfalo.
Además de las tres comisuras que se desarrollan en la lámina terminal,
aparecen otras tres. Dos de ellas, las comisuras posterior y habenular, se encuentran inmediatamente por delante y por debajo del tallo de la epífisis. La tercera, el quiasma óptico, aparece en la pared rostral del diencéfalo y contiene fibras de las mitades mediales de las retinas.

Pontificia Universidad Católica de Valparaíso Principios Neuroanatomía Funcional
ANATOMÍA EXTERNA E INTERNA DE SNC
I.- Anatomía Externa y Relaciones de Médula Espinal
La médula espinal es una porción alargada del sistema nervioso central que ocupa los dos tercios superiores del canal vertebral. Se extiende desde el borde superior del atlas hasta aproximadamente el disco intervertebral entre la primera y la segunda vértebra lumbar, aunque este límite puede tener variaciones. Se encuentra rodeada por las meninges (duramadre, aracnoides y piamadre). Se continúa cranealmente con la médula oblonga, y caudalmente se estrecha formando el cono medular. El diámetro transversal de la médula no es constante sino que disminuye en dirección cráneo-caudal, presentando dos engrosamientos: cervical y lumbar.
El engrosamiento cervical da origen a grandes nervios espinales para la inervación del miembro superior, y se extiende desde el tercer segmento cervical hasta el segundo segmento dorsal. El engrosamiento lumbar da origen a los nervios espinales que inervan el miembro inferior y se extiende entre el primer segmento sacro, cuyos niveles vertebrales son la IX a XII vértebras torácicas.
A lo largo de la médula pueden observarse los siguientes surcos: un
surco anterior y un surco o tabique posterior dividen a la médula en dos mitades: derecha e izquierda, unidas por una banda comisural de tejido nervioso que contiene el conducto central.
El surco medio anterior o fisura media anterior se ubica a lo largo de toda la superficie ventral de la médula. Su profundidad varía haciéndose más profundo a nivel caudal. Dorsal a él se encuentra la comisura blanca anterior.
El surco medio posterior es menos extenso que el anterior y de él
surge el tabique posterior que penetra en la médula hasta casi llegar al canal central. La longitud del tabique disminuye hacia caudal a medida que el canal se ubica más posterior y la médula se estrecha.
Lateral a ambos lados del surco medio posterior se encuentra el surco
lateral posterior por donde entran las fibras dorsales de los nervios espinales. Entre los dos surcos, el medio posterior y el posterior lateral, se encuentra el fascículo posterior. En las regiones cervical y dorsal superior existe un nuevo surco que divide el fascículo posterior en un fascículo medial (grácil o de Goll) y otro lateral (cuneatus o de Burdach). Este surco recibe el nombre de surco paramedio posterior.
Entre el surco lateral posterior y el surco medio anterior se encuentra el
fascículo anterolateral, dividido por el origen de las raíces espinales ventrales en una porción anterior y otra lateral.

Pontificia Universidad Católica de Valparaíso Principios Neuroanatomía Funcional
El filum terminale es tejido conjuntivo (piamadre) que desciende desde el cono medular. Posee dos partes: el filum terminale interno y el externo. El interno (15cm) está rodeado por duramadre y aracnoides y termina a nivel de S2. El filum terminale externo (5cm) se fusiona con la duramadre y desciende hasta la parte posterior de la primera vértebra coccígea. A ambos lados de la médula espinal se continúan las raíces de los nervios espinales, tanto ventrales como dorsales, que se unen a nivel de los forámenes intervertebrales. Debido a que la longitud del conducto vertebral es mayor que la médula espinal los últimos nervios espinales deben recorrer una distancia apreciable hasta alcanzar sus forámenes de salida formando lo que se conoce como la cola de caballo o cauda equina. La cola de caballo se define como el conjunto de raíces nerviosas que penden del cono medular.
Las raíces espinales ventrales emergen de la médula formando radículas
(filetes) que luego se unen para formar el nervio espinal. Lo mismo sucede con las raíces espinales dorsales, con la diferencia de que antes de dividirse e ingresar a la médula presentan unas dilataciones ovoides llamadas ganglios espinales.

Pontificia Universidad Católica de Valparaíso Principios Neuroanatomía Funcional

Anatomía Interna de la Médula Espinal.
La sustancia gris de la médula espinal tiene una ubicación central y una
forma de columna acanalada. En cortes transversales presenta dos masas simétricas derecha e izquierda conectadas por una comisura gris transversal estrecha. La comisura está atravesada por el canal central. Cada masa lateral presenta una concavidad lateral desde la cual se proyecta una columna lateral en algunos niveles. En cortes transversales las masas laterales parecen tener proyecciones que se denominan astas. Toda esta sustancia gris central se encuentra rodeada por sustancia blanca que mayormente corresponde a fibras longitudinales agrupadas en fascículos o cordones, que en conjunto forman una columna.
La columna gris anterior se proyecta desde la comisura gris en sentido ventrolateral. Es corta y ancha y se encuentra separada de la superficie por el fascículo anterior. Pueden distinguirse en esta asta ventral una cabeza y una base. Entre los segmentos T2 y L1 aparece una columna gris lateral.
La columna gris posterior se proyecta en sentido dorsolateral. Es
estrecha y se aproxima bastante a la superficie, pero se encuentra separada de ella por el fascículo dorsolateral (de Lissauer). Se considera una base, un cuello y un vértice cubierto por una sustancia gelatinosa (de Rolando).

El límite entre la sustancia gris y blanca no siempre es nítido. A nivel
cervical el fascículo lateral se encuentra invadido por bandas de sustancia gris procedente del asta posterior separadas por fibras nerviosas que se entrelazan formando una red, de ahí su nombre de formación reticular. Puesto que pueden encontrarse regiones similares en distintos niveles del sistema nervioso central se ha denominado a estas estructuras en conjunto como sistema reticular.
Existe una relación directa entre la masa de sustancia gris de la médula
espinal y el número de estructuras a las que inerva. Por eso se aprecian los engrosamientos en los niveles cervical y lumbar para la inervación de los músculos del miembro superior e inferior respectivamente.
El conducto central atraviesa toda la médula espinal y la mitad caudal de médula oblonga, abriéndose en el cuarto ventrículo. En el cono medular se expande como un ventrículo terminal fusiforme, transversalmente triangular con base anterior; esta cavidad se oblitera aproximadamente a los 40 años. A nivel cervical y dorsal, el conducto es ligeramente ventral respecto al punto medio de la medula espinal y central en el engrosamiento lumbar. Está ocupado por el líquido cerebro espinal (LCE) y revestido por un epitelio ciliado, el epéndimo. La sustancia gris que rodea al canal central corresponde a la comisura gris, más delgada ventralmente. Anterior a la comisura gris se encuentra la comisura blanca anterior y posterior se encuentra en relación con la comisura blanca posterior. Ésta última está en relación con el tabique medio posterior.

Organización de la Sustancia Gris.
Como se mencionó anteriormente la sustancia gris ocupa en la médula
espinal una ubicación central. De las masas laterales (derecha e izquierda) de sustancia gris se proyectan las astas anteriores y las posteriores. Muchas neuritas cruzan la línea media de las comisuras formando una estructura continua desde el punto de vista funcional. Las neuronas de la sustancia gris son multipolares y presentan características variables como la longitud y el tamaño o la disposición de las dendritas y de los axones. La mayoría son neuronas Golgi I y II. La neuronas con axón largo salen de la sustancia gris y se incorporan a las raíces espinales ventrales o a fascículos ventrales. Los axones cortos de las neuronas Golgi están limitados a la sustancia gris cercana. Algunas neuronas son intrasegmentarias, distribuidas dentro de un solo segmento; otras se extienden a través de varios segmentos, por lo que tienen una distribución intersegmentaria.
En gran parte del sistema nervioso central, los somas neuronales se encuentran agrupados, a menudo en grandes números, lo que indicaría una función común. En ocasiones estos grupos pueden dividirse en grupos más pequeños, lo que implicaría una función particular. En médula espinal existen núcleos anteriores, posteriores y laterales.
Las astas anteriores poseen neuronas de motricidad voluntaria en su
cabeza y de motricidad visceral en la base. En las astas posteriores se encuentra en la cabeza la sensibilidad exteroceptiva, en el cuello la sensibilidad propioceptiva y en la base la sensibilidad visceral.
Brod Rexed reconoció que las neuronas del asta dorsal se encuentran
dispuestas en láminas aplastadas que corren paralelas al eje longitudinal de médula espinal. Cada lámina puede distinguirse de otra por la morfología neuronal y densidad de su unión, como ocurre también en la citoarquitectura del córtex. Cada lámina de Rexed contiene neuronas con distintas conexiones y funciones anatómicas.
En la comisura gris o región intermedia y en el asta posterior las
láminas de Rexed tienen un parecido mas a columnas que a láminas aplastadas. En cualquier segmento espinal existen 10 láminas de Rexed. Algunas láminas se corresponden con los núcleos de la médula espinal.

Pontificia Universidad Católica de Valparaíso Principios Neuroanatomía Funcional Lámina I: Son neuronas que residen en la parte posterior del asta dorsal (lámina marginal o capa de Waldeyer) y se relacionan con el dolor y frío.
Lámina II: Corresponde a la sustancia gelatinosa; son neuronas pequeñas que reciben aferencias en relación al dolor y recepción térmica. Estas neuronas son denominadas interneuronas ya que sus axones no ascienden formando los tractos ascendentes.
Lámina III y IV: Corresponde al núcleo propio, esta región posee muchas fibras mielínicas y de proyección. Una categoría de las neuronas de esta región se relaciona con el tacto.
Lámina V: Capa gruesa que incluye el cuello de la columna dorsal, y puede dividirse en un tercio lateral y dos tercios mediales. Ambas subdivisiones están formadas por una población mixta, pero la primera contiene somas prominentes entremezclados con numerosos haces de fibras. Recibe el nombre de formación reticular y es particularmente prominente a nivel cervical.
Lámina VI: Muy prominente en los engrosamientos cervical y lumbar. Presenta un tercio medial de neuronas d densamente agrupadas y dos tercios laterales con somas de forma triangular de sensibilidad propioceptiva de mayor tamaño y agrupados de forma más laxa. Corresponde a la base del asta dorsal.
Región posteromedial de la lámina VII o Núcleo Dorsal (Clarcke): Este núcleo está presente entre los segmentos C8 y L3, contiene neuronas de segundo orden relacionadas con la propiocepción desde los miembros inferiores. Este núcleo es el lugar de la proyección del fascículo espinocerebeloso dorsal.
Región lateral de la lámina VII o Núcleo Intermediolateral: se localiza en los niveles de T1 a L2, y contiene neuronas preganglionares simpáticas que inervan a los ganglios paravertebrales.
Lámina VIII: Contiene neuronas relacionadas con el control motor, esta se integran con las de la lámina IX. Son las neuronas donde terminan principalmente las vías corticoespinal y extrapiramidales propiamente dichas
Lámina IX: Son neuronas del asta ventral. Se encuentran las neuronas del grupo nuclear lateral de los engrosamientos medulares y el grupo nuclear medial que se extiende a los largo de la médula. Son neuronas largas que forman los plexos nerviosos. Las motoneuronas alfa son la vía final común de Sherrington de la vía piramidal (son los axones que van a formar el componente motor del nervio periférico) y las motoneuronas gamma forman parte de la regulación de la contracción muscular.

Pontificia Universidad Católica de Valparaíso Principios Neuroanatomía Funcional Lámina X: Pequeñas neuronas y fibras amielínicas que rodean al canal central. Estudios fisiológicos indican que se relacionan con temperatura, dolor y sensaciones viscerales.
Pontificia Universidad Católica de Valparaíso Principios Neuroanatomía Funcional Organización de la Sustancia Blanca
La sustancia blanca de la médula contiene fibras nerviosas, neuroglia y
vasos sanguíneos; rodea la sustancia gris y debe su color a la presencia de un gran número de fibras nerviosas mielínicas. Las fibras nerviosas se distribuyen en cinco grupos:
Fibras aferentes de neuronas localizadas en los ganglios espinales y que
penetran por las raíces dorsales y recorren distancias medulares variables Fibras largas ascendentes, derivadas de neuronas espinales que
conducen impulsos a niveles superiores Fibras largas descendentes desde niveles superiores del SNC para
establecer sinapsis con neuronas espinales Fibras que establecen conexiones intrasegmentarias e intersegmentarias. Fibras procedentes de motoneuronas de las columnas grises ventral y
lateral que se incorporan a las raíces ventrales.
Estas fibras forman, en todos los casos excepto el último, haces o fascículos longitudinales. Sin embargo, en mucho niveles puede verse que hay fibras que siguen direcciones oblicuas o incluso horizontales, en particular a través de la línea media de las comisuras gris y blanca, gran parte de las cuales son fibras de decusación. Muchas otras son conexiones comisurales intrasegmentarias que conectan neuronas de las columnas grises con neuronas contralaterales.
Vías ascendentes:
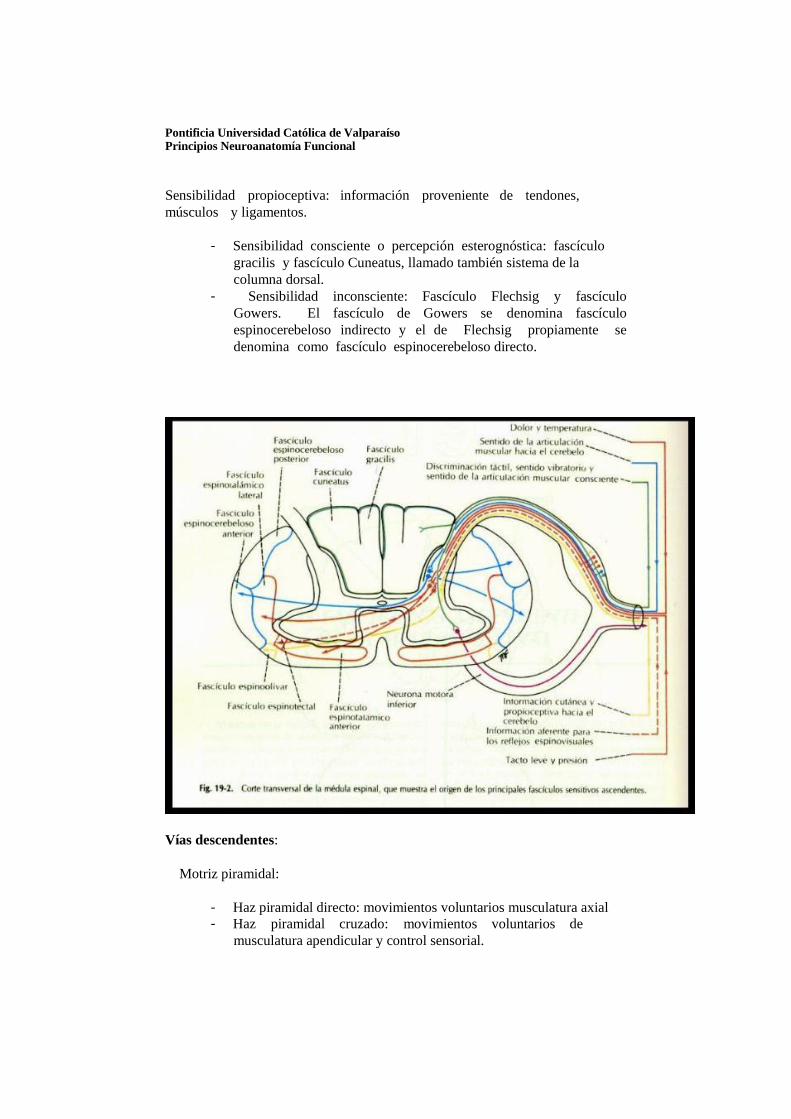
Sensibilidad exteroceptiva: llamado también llamado sistema ventrolateral
- Sensibilidad termoalgésica (dolor y temperatura): fascículo espinotalámico dorsal (cordón lateral).
- Sensibilidad protopática (tacto grosero): fascículo espinotalámico ventral (cordón ventral).
- Sensibilidad epicrítica (tacto fino o presión): región del cordón posterior.
Pontificia Universidad Católica de Valparaíso Principios Neuroanatomía Funcional
Sensibilidad propioceptiva: información proveniente de tendones, músculos y ligamentos.
- Sensibilidad consciente o percepción esterognóstica: fascículo
gracilis y fascículo Cuneatus, llamado también sistema de la columna dorsal.
- Sensibilidad inconsciente: Fascículo Flechsig y fascículo Gowers. El fascículo de Gowers se denomina fascículo espinocerebeloso indirecto y el de Flechsig propiamente se denomina como fascículo espinocerebeloso directo.
Vías descendentes:
Motriz piramidal:
- Haz piramidal directo: movimientos voluntarios musculatura axial - Haz piramidal cruzado: movimientos voluntarios de
musculatura apendicular y control sensorial.

Pontificia Universidad Católica de Valparaíso Principios Neuroanatomía Funcional
Motriz extrapiramidal:
- Fascículo rubroespinal: Movimientos voluntarios de los miembros - Fascículo reticuloespinal: Movimientos automáticos, músculos
axiales y miembros. - Fascículo tectoespinal: Coordina el cuello con los movimientos oculares. - Fascículo vestibuloespinal lateral y vestibuloespinal medial:
equilibrio y posición de la cabeza/músculos del cuello respectivamente.
El tronco del encéfalo está ubicado inferior al cerebro y superior de la médula espinal, relacionándose por dorsal con el cerebelo. Se divide en tres partes, que de arriba hacia abajo son:
- Mesencéfalo - Puente - Médula Oblongada o bulbo raquídeo.

Pontificia Universidad Católica de Valparaíso Principios Neuroanatomía Funcional
II.- Anatomía Externa de Médula Oblongada
La médula oblonga se extiende desde el borde inferior del puente o protuberancia hasta un plano transversal que pasa por encima del primer par de nervios espinales y que corta el borde superior del atlas (C1) dorsalmente y el centro del proceso odontoides ventralmente. En este punto su anatomía interna es semejante a la de la médula espinal pero cambia rostralmente. Se encuentra separada del hueso occipital y del atlas por las meninges y los ligamentos occipitoaxiales. En dirección caudal, la superficie dorsal se encuentra en la hendidura formada en la línea media entre los hemisferios cerebelosos; la porción superior forma el piso del cuarto ventrículo (fosa romboidal).
El canal central medular se prolonga en su mitad inferior, expandiéndose en dirección craneal formando el IV ventrículo; así puede dividirse la médula oblonga en una porción cerrada que contiene el conducto central y una porción abierta que contiene la mitad inferior del cuarto ventrículo.

Pontificia Universidad Católica de Valparaíso Principios Neuroanatomía Funcional
Las arterias vertebrales discurren laterales al bulbo, dando origen a las
pequeñas arterias espinales posteriores. Las superficies anterior y posterior presentan los surcos medio anterior y posterior, respectivamente.
Surco medio anterior. Se extiende por todo la médula oblonga y se continúa caudalmente con el surco medio anterior de la médula espinal. Termina por encima del borde inferior del puente en un foramen ciego triangular. A nivel inferior queda interrumpido por los haces de la decusación piramidal.
Surco medio posterior. Se encuentra solo en la porción cerrada del bulbo. Se continúa caudalmente con el surco medio posterior de la médula espinal. Pierde profundidad hacia craneal y desaparece en el punto donde el canal central se expande para formar el cuarto ventrículo.
Región anterior. Entre el surco medio anterior y el lateral presenta una elevación alargada que corresponde a las pirámides. En su extremo superior, a nivel de la unión con el puente, surge el nervio abducens (VI); en dirección caudal se continúa con el cordón medular anterior. Cada pirámide contiene fibras corticospinales homolaterales, de las que aproximadamente el 70-90% cruzan el surco medio y forman la decusación piramidal. Luego descienden por el cordón lateral formando el fascículo piramidal cruzado. Las fibras que no se decusan continúan por el cordón anterior formando el fascículo piramidal directo. Existe también un grupo de fibras corticospinales que no decusan pero se inclinan posterolateralmente uniéndose a los haces piramidales cruzados constituyendo pequeños componentes no cruzados de éstos.

Los cuatro últimos nervios craneales se originan en médula oblonga. Las radículas del nervio hipogloso se encuentran alineadas con las radículas anteriores de los nervios espinales. Las radículas de los nervios accesorio, vago y glosofaríngeo están alineadas

Pontificia Universidad Católica de Valparaíso Principios Neuroanatomía Funcional
con las radículas dorsales de los nervios espinales y penetran a través de un surco lateral posterior.
Región lateral. Entre el surco anterolateral y el surco posterolateral. Su porción superior presenta un abultamiento que corresponde a la oliva.
Oliva. Elevación oval situada entre los surcos lateral anterior y lateral posterior; está formada por el núcleo olivar inferior. Está localizada lateral a la pirámide, separada de ella por el surco lateral anterior y fibras del nervio hipogloso. Las raíces del nervio facial emergen entre su extremo rostral y el borde inferior de la protuberancia, en el ángulo pontocerebeloso.
Región posterior. Esta región, dorsal a surco lateral posterior, puede dividirse en dos partes, caudal y rostral. La porción caudal constituye una continuación superior de los haces grácil y cuneiforme. El primero linda con el surco medio posterior, y está separado del fascículo cuneiforme por la continuación craneal del surco y el tabique intermedio posterior de la médula espinal. Ambos haces son inicialmente verticales y paralelos, pero a nivel del límite inferior del IV ventrículo divergen, formando ambos una elevación, los tubérculos grácil y cuneiforme formados por los núcleos grácil y cuneiforme, respectivamente.

Pontificia Universidad Católica de Valparaíso Principios Neuroanatomía Funcional
Los pedúnculos cerebelosos inferiores son las elevaciones redondeadas situadas entre el IV ventrículo y las raíces de los nervios glosofaríngeo y vago. Los dos pedúnculos divergen y se inclinan para entrar en los hemisferios cerebelosos; se encuentran cruzados por las estrías medulares que se dirigen al surco medio del IV ventrículo.
Anatomía Interna de Médula Oblongada.
En el límite inferior la estructura interna muestra elementos que son continuación con los presentes en médula espinal.
Decusación piramidal: Las pirámides están formadas por las fibras corticoespinales y corticobulbares. A nivel de la porción inferior de la médula oblonga la mayoría de las fibras corticoespinales pasan al cordón lateral medular del lado opuesto, formando la decusación de las pirámides. Las fibras que no se cruzan descienden por el cordón anterior medular formando el fascículo corticoespinal directo.

Pontificia Universidad Católica de Valparaíso Principios Neuroanatomía Funcional Núcleos delgado y cuneiforme: Forman unos tubérculos en la cara dorsal de la médula oblonga. En ellos hacen sinapsis las neuronas provenientes de los fascículos medulares del mismo nombre que conducen la sensibilidad propioceptiva y táctil. Los axones de están neuronas se arquean (fibras arciformes) hacia el lado opuesto (decusación del lemnisco) detrás a la pirámides y cercanos a la línea media formando el lemnisco medio, que termina en los núcleos ventrolateral y posterior del tálamo.
Núcleo cuneiforme lateral: lateral al núcleo cuneiforme existe un acumulo de neuronas cuyos axones forman las fibras arciformes laterales (fibras arciformes externas) que se dirigen al cerebelo por el pedúnculo cerebelar inferior ipsilateral. Retransmite al cerebelo información sensorial somática desde el tronco superior y el miembro superior, no para la percepción sino para el control de movimientos.
Núcleo olivar inferior y olivares accesorios: El olivar inferior es una gran lámina dentada que protruye formando la oliva. Dorsal y medial a este núcleo se ubican los olivares dorsal y medial. Los axones de estas neuronas se dirigen al cerebelo por el pedúnculo cerebelar inferior heterolateral. El núcleo olivar envía una de las señales excitadoras más fuertes del SNC; es una vía de control sobre la precisión del movimiento.
Formación reticular: Fibras dispersas y células de diverso tamaño que son disgregadas por los entrecruzamientos de fibras. Se ubican en la porción central y lateral de la médula oblonga. El sistema reticular es responsable de los estados de vigilia y sueño. Este sistema participa en fenómenos relacionados con la atención, es decir, cuando se selecciona información, o inversamente, cuando se inhiben señales consideradas irrelevantes.
Núcleo hipogloso: columna larga y delgada que ocupa toda la extensión de la médula oblonga, en la porción inferior se corresponden a las astas dorsales medulares. En la región superior se forma una proyección en el piso del IV ventrículo denominada trígono del hipogloso. Inervación musculatura de la lengua.
Núcleo motor dorsal del vago: lateral al del hipogloso y su posición corresponde en el piso del IV ventrículo al ala gris. Inervación musculatura lisa y glándulas.
Núcleo ambiguo: situado lateral de la sustancia reticular. Se extiende en una longitud similar a la del hipogloso. Sus células dan fibras motoras para el IX, X y XI. Inervación faríngea y laríngea.

Núcleo salivar inferior: Situado medial al fascículo solitario en el límite pontomedular. Motor visceral.
Núcleo del fascículo solitario: Las ramas sensitivas de los nervios VII, IX y X al ingresar en el tallo forman el fascículo descendente, fascículo solitario, situado dorsolateral en la médula oblonga lateral al sulcus limitans. Rodeando al fascículo en su porción inferior están las neuronas del núcleo del fascículo solitario. Inferiormente los fascículos solitarios están cercanos a la línea media formando el núcleo cardiorrespiratorio y rostralmente se relaciona con las vías gustativas por lo que se denomina esta porción como núcleo gustativo.
Raíz descendente del trigémino y núcleo (espinal) de la raíz descendente: homóloga a la sustancia gelatinosa de la médula espinal (Rolando). Ubicados posterolateralmente conducen información sensitiva.
Secciones Transversales de Médula Oblongada

A nivel del foramen magno donde la médula espinal termina y la
médula oblonga comienza, la sustancia gris posee una marcada reorganización. En la decusación piramidal las fibras corticoespinales se interponen entre la sustancia gris ventral y la sustancia gris central. El canal central y la sustancia gris que lo rodea aparecen desplazados por las fibras de la decusación piramidal. La mayoría de las fibras corticoespinales que descienden por la región ventral del tronco cerebral se cruzan originando en médula espinal el fascículo corticoespinal cruzado, mientras que otro grupo de fibras no se cruza formando el fascículo corticoespinal directo.
Adyacente al surco mediano dorsal se encuentra el núcleo grácil que se
rodea del fascículo gracilis el cuál contiene los axones sensoriales primarios que terminan en el núcleo del mismo nombre. Proyectándose posteriormente del asta dorsal se encuentra el núcleo cuneiforme que se hace más prominente en niveles superiores, el fascículo cuneiforme cubre al núcleo y se ubica lateral al fascículo grácil. El núcleo espinal del trigémino se extiende desde la porción superior de médula espinal, pasando a lo largo de médula oblongada y terminando en puente, este núcleo es análogo a la sustancia gelatinosa que se encuentra rodeando el asta posterior de médula espinal. El tracto espinal del nervio trigémino es una banda angosta de fibras nerviosas sensoriales que se interponen entre el núcleo espinal del nervio trigémino y la periferia de la médula oblonga.

Pontificia Universidad Católica de Valparaíso Principios Neuroanatomía Funcional
El núcleo grácil se ubica en el tubérculo grácil, medial al septo mediano dorsal y rodeado por fibras que forman el fascículo grácil, que terminan en el núcleo. El núcleo cuneiforme se encuentra rodeado por las fibras del fascículo cuneiforme.
La decusación sensitiva es la clave de este corte. Numerosas fibras arqueadas
(arciformes) internas se originan de los núcleos grácil y cuneiforme, tomando un curso semicircular que atraviesan la formación reticular y sustancia gris que rodea al conducto central para decusar formando la decusación sensitiva. Las fibras arqueadas internas pasan entre el núcleo espinal del trigémino y la sustancia gris central. Después de decusarse las fibras ascienden como un tracto denominado lemnisco medial, este tracto termina en el tálamo. En la parte ventral de la sustancia gris central se ubica el núcleo hipogloso, que origina fibras para los músculos de la lengua.

Pontificia Universidad Católica de Valparaíso Principios Neuroanatomía Funcional
El núcleo olivar inferior es notorio y los núcleos accesorios olivares medial y posterior también son visibles. El núcleo olivar inferior se asemeja a un engranaje de sustancia gris que origina una gran cantidad de fibras olivocerebelares.
Se reconoce muchos núcleos en el piso del cuarto ventrículo. Más
medial se encuentra el núcleo hipogloso, representando la columna somatomotora de los núcleos motores del tronco. Fibras del hipogloso toman un curso ventrolateral entre la oliva y la pirámide. Mas lateral se encuentra el núcleo motor dorsal del vago (núcleo motor visceral) y medial e inferior el núcleo vestibular (núcleo aferente somático especial). Ventrolateral al núcleo motor dorsal del vago está el núcleo solitario del tracto solitario, en relación con la sensación visceral y el gusto. Algunos axones del núcleo solitario se proyectan al tálamo. El núcleo espinal del trigémino con el tracto ubicado lateral a él se considera como núcleo aferente somático general. El fascículo longitudinal medial y el tracto tectoespinal se ubican cerca del rafe medio y ventromedial al núcleo hipogloso Dorsal a la oliva se encuentra la formación reticular compuesta de neuronas dispersadas por axones mielinizados ascendentes y descendente generando una variedad de vías.

Pontificia Universidad Católica de Valparaíso Principios Neuroanatomía Funcional III.- Anatomía Externa de Puente
El puente se encuentra ventral al cerebelo, inferior al mesencéfalo y
superior a la médula oblonga. La transición está marcada superficialmente por el surco pontomedular, origen aparente de los nervios abductor, facial y auditivo. La superficie ventral es convexa en sentido transversal y excava un surco en la porción petrosa del hueso temporal en sentido lateral hasta el meato acústico interno. Las neuronas de los núcleos pontinos dan origen a haces de fibras transversas que cruzan la línea media y se dirigen contralateralmente al cerebelo, convergiendo a cada lado en el pedúnculo cerebeloso medio. La superficie presenta un surco medio vertical poco profundo, el surco basilar, que suele contener la arteria basilar.
El nervio auditivo está situado anterior a los nervios vestibular y facial al salir del meato auditivo interno. En el ángulo pontocerebeloso, se desplaza en sentido posterior para alinearse con el nervio vestibular; de esta manera, ambos nervios poseen una ubicación dorsal respecto al nervio facial al entrar en el tronco encefálico. Aproximadamente en el nivel mediopontino emerge el nervio trigémino, con una pequeña raíz motora superomedial y una gran raíz sensitiva inferolateral. El nervio motor ocular externo (abducens) emerge en la unión pontobulbar cerca de la línea media y asciende sobre la superficie anterior del puente antes de perforar la duramadre por encima de la parte posterior del cuerpo del esfenoides.
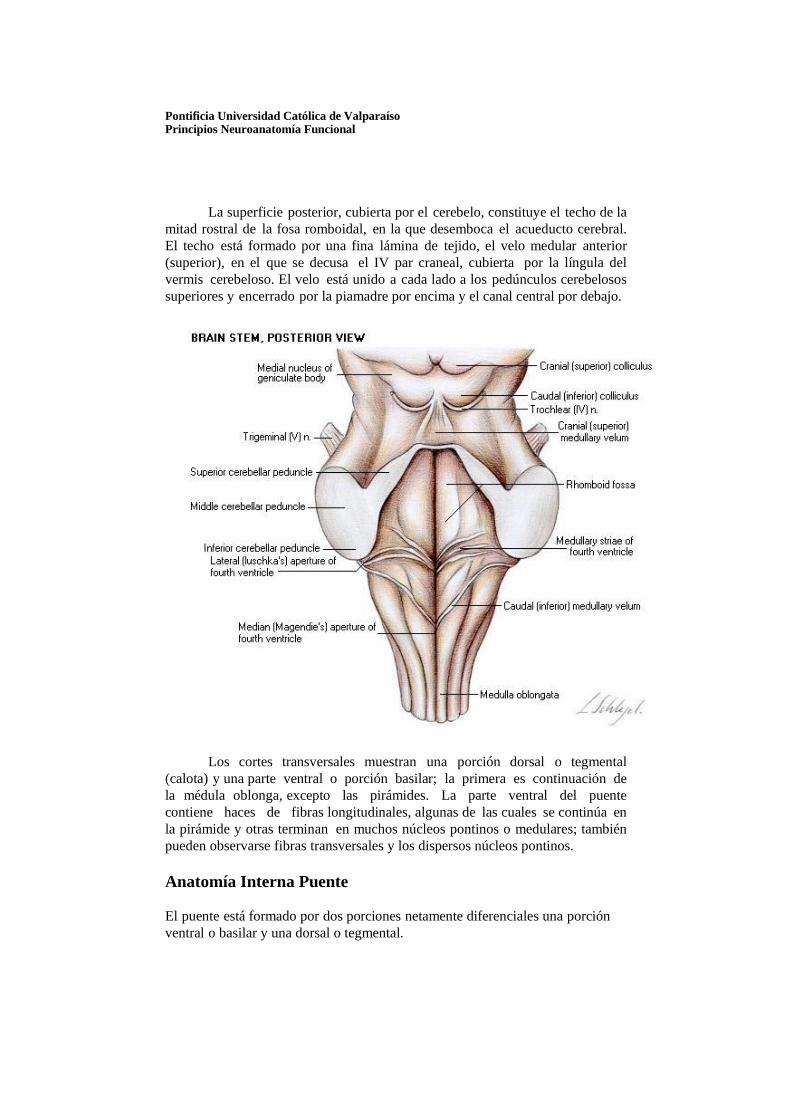
Pontificia Universidad Católica de Valparaíso Principios Neuroanatomía Funcional
La superficie posterior, cubierta por el cerebelo, constituye el techo de la
mitad rostral de la fosa romboidal, en la que desemboca el acueducto cerebral. El techo está formado por una fina lámina de tejido, el velo medular anterior (superior), en el que se decusa el IV par craneal, cubierta por la língula del vermis cerebeloso. El velo está unido a cada lado a los pedúnculos cerebelosos superiores y encerrado por la piamadre por encima y el canal central por debajo.
Los cortes transversales muestran una porción dorsal o tegmental (calota) y una parte ventral o porción basilar; la primera es continuación de la médula oblonga, excepto las pirámides. La parte ventral del puente contiene haces de fibras longitudinales, algunas de las cuales se continúa en la pirámide y otras terminan en muchos núcleos pontinos o medulares; también pueden observarse fibras transversales y los dispersos núcleos pontinos.
Anatomía Interna Puente
El puente está formado por dos porciones netamente diferenciales una porción ventral o basilar y una dorsal o tegmental.

Pontificia Universidad Católica de Valparaíso Principios Neuroanatomía Funcional Porción basilar
Formada por fibras verticales y horizontales entre las cuales se
encuentran los núcleos pontinos. Las fibras verticales corresponden a las corticoespinales; corticoestibares que provienen de la corteza y terminan en núcleos motores del tronco cerebral; y corticopónticas, que provienen de corteza y terminan en los núcleos pónticos. Las fibras horizontales son las que forman los pedúnculos cerebelares medios y nacen, la mayoría, en los núcleos pónticos contralaterales.
Los núcleos pónticos son numerosas neuronas que se ubican entre los
fascículos transversales y verticales de la porción basilar del puente. Fibras temporopontinas,frontopontinas o colaterales corticoespinales hacen sinapsis en los núcleos para conectarse con cerebelo y así regular los mecanismos motores.

Porción tegmental
Se presenta una continuidad de estructuras desde médula oblonga, como formación reticular y lemnisco medial, que en la porción inferior del puente se ubica anteroposterior y medial, apareciendo en la región más rostral del puente como una banda transversal que marca el límite entre la porción tegmental y la porción basilar.
Núcleo del abductor: se encuentra situado a nivel del colículo facial, que es el engrosamiento de la eminencia media del abductor y la rodilla del facial. Ocupa la misma posición que en la médula oblonga. Tiene el núcleo del hipogloso. Inerva el músculo recto lateral del ojo.
Núcleo motor del facial: situado ventrolateral en la mitad inferior de puente. Recibe fibras corticonucleares cruzadas y un pequeño número de fibras homoloaterales, además de unas fibras del haz rubrorreticular homololateral. Los axones de sus neuronas grandes forman el nervio facial, cuyas fibras rodean dorsolateralmente el núcleo del abductor a nivel del piso del IV ventrículo, formando la rodilla del nervio facial. Recibe fibras corticonucleares para el control voluntario y fibras aferentes de su propia raíz sensitiva (a través del núcleo solitario) y el núcleo de la raíz descendente del trigémino. Estas aferencias completan arcos reflejos locales, tal como sucede en los segmentos espinales.
Núcleos y raíces del trigémino: los núcleos motor y principal sensitivo se ubican dorsalmente, y a la altura del tercio superior del puente. El sensitivo queda lateral al motor. Las fibras exteroceptivas se originan en el núcleo del trigémino. La vía termoalgésica de cara desciende para hacer sinapsis en el núcleo espinal (médula oblonga); la vía táctil hace sinapsis en el núcleo principal; mientras que la vía propioceptiva asciende y hace sinapsis en el núcleo mesencefálico. La vía propioceptiva no se origina en el ganglio del trigémino sino que desde el mismo núcleo. Los núcleos sensitivos reciben sensibilidad de cara y cabeza, mientras que el motor da inervación a los músculos masticadores.
Lemnisco trigeminal (trigeminotalámico): se origina en neuronas del núcleo de la raíz descendente contralateral, conduciendo información sobre la sensibilidad dolorosa y térmica en al piel facial y las mucosas de la conjuntiva, la lengua, la boca, la nariz, etc. Los lemniscos juntos forma una banda transversal compuesta, en orden lateral desde la línea media, por los lemniscos medial y trigeminal, el haz espinotalámico lateral y el lemnisco lateral. Luego asciende terminar en el núcleo ventromedial del tálamo.
Núcleos cocleares: El cuerpo restiforme (inicio del pedúnculo cerebelar inferior) está rodeado por una eminencia que corresponde al colículo acústico formado por los núcleos cocleares dorsal y ventral.
Nervios coclear y vestibular: El nervio coclear transmite las impresiones sonoras del órgano sensorial de la audición hasta el núcleo coclear. Luego los axones, homo o contralaterales, forman el lemnisco lateral que asciende lateral al lemnisco medial y termina en el colículo cuadrigémino inferior. Para llegar

al lado opuesto las fibras provenientes de los núcleos cocleares se dirigen hacia la zona límite entre la porción basilar y tegmental en donde forman una banda transversal, denominada cuerpo trapezoide, cuyas fibras se cruzan en ángulo recto con las del lemnisco medial.
Núcleo olivar superior: Corresponde a tres masas nucleadas ubicadas lateral al cuerpo trapezoide. Algunas fibras eferentes terminan en el lemnisco lateral, y otras lo hacen en el núcleo abductor.
Núcleos vestibulares: Las fibras vestibulares en el tallo cerebral hacen sinapsis con algunos de los cuatro núcleos vestibulares: medial o principal, lateral o Deiters, superior o Bechtterew y espinal o descendente. Ocupan el área acústica del piso del IV ventrículo. Otras fibras vestibulares no realizan sinapsis en los núcleos sino que se dirigen al cerebelo junto con las fibras eferentes provenientes de los núcleos vestibulares, formando así el fascículo vestibulocerebeloso. Otras fibras eferentes de los núcleos vestibulares contribuyen a formar el fascículo longitudinal que se extiende dorsalmente a lo largo de todo el tronco cerebral. Este fascículo conecta los núcleos vestibulares (movimientos coordinados oculocefalógiros) a los núcleos motores oculares (III, IV y VI) y a los núcleos encargados de los movimientos de cabeza.
Fascículo espinocerebeloso ventral: en la parte superior del puente, se dirige dorsalmente a través de la raíces del trigémino para alcanzar el pedúnculo cerebelar superior y llegar al vermis del cerebelo.
Secciones Transversales Puente

El colículo facial marca una prominencia en el piso del IV ventrículo. Cada colículo facial se produce porque fibras del núcleo facial rodean (rodilla) al núcleo abductor. El tracto longitudinal medial ocupa una posición paramediana. Las fibras ascendentes del lemnisco medial se intersectan con las fibras transversales del cuerpo trapezoide que se agrega y forma el lemnisco lateral. El lemnisco espinal y trigeminal se localizan en la parte ventral del tegmento póntico.
Lateral al núcleo abductor se ubica el núcleo vestibular que origina el
tracto vestibuloespinal. En la porción ventral del puente grupos celulares forman los núcleos pontinos, junto con fibras transversas las cuales se dirigen al pedúnculo cerebelar medio. Fibras corticoespinales y coricobulbares descienden notoriamente en este nivel destinadas a las pirámides de médula oblonga.
En la parte ventral del puente las fibras motoras corticopontinas, corticobulbares y corticoespinales continúan descendiendo desde la base del pedúnculo. En adición a estas fibras corticofugas el puente contiene transversalmente las fibras pontocerebelares que se originan de los núcleos pónticos para cruzar la línea media e ingresar en el pedúnculo cerebelar medio contralateral. El tegmento varía su estructura interna a diferentes niveles. Este nivel muestra el núcleo motor y sensitivo del trigémino; el núcleo motor es medial al núcleo sensitivo.

Pontificia Universidad Católica de Valparaíso Principios Neuroanatomía Funcional
El IV ventrículo es menor en esta sección, dado que cranealmente se
continúa como acueducto cerebral del mesencéfalo. El velo medular superior constituye el techo del IV ventrículo el cual se ubica entre los dos pedúnculos cerebelares superiores. En la parte lateral del tegmento del puente están los 4 lemniscos: el lemnisco espinal se ubica lateral al lemnisco medial y el lemnisco trigeminal dorsolateral a éste. El lemnisco lateral se orienta verticalmente ascendiendo cerca del margen lateral del tegmento. Ventralmente se encuentran los núcleo pontinos y las fibras pontocerebelares que se intersectan con las fibras corticoespinales, corticoponticas y corticobulbares. La fibras pontocerebelares conectan los núcleos pónticos con el corte cerebelar contralateral vía pedúnculos cerebelares.
IV.- Anatomía Externa Mesencéfalo
El mesencéfalo atraviesa el hiato de la tienda del cerebelo y conecta el puente y el cerebelo con el encéfalo anterior. Es el segmento tronco encefálico más corto, con no más de 2cm de longitud, y en su mayor parte está situado dentro de la fosa craneal posterior. Lateral a él se encuentran los giros del hipocampo, que ocultan sus lados cuando se examina la superficie inferior del cerebro. Con fines descriptivos, se divide en tres regiones, pasando de dorsal a ventral: el techo, el tegmento y la base del pedúnculo. El tegmento y la base del pedúnculo conforman el pedúnculo cerebral. El techo y el tegmento se encuentran separados por el acueducto cerebral que conecta el tercer y cuarto ventrículos. El acueducto se encuentra rodeado por la sustancia gris periacueductal que contiene neuronas que forman parte del circuito para supresión del dolor endógeno. En el techo se encuentran los tubérculos cuadrigéminos, cuatro elevaciones redondeadas, un par superior y otro inferior.
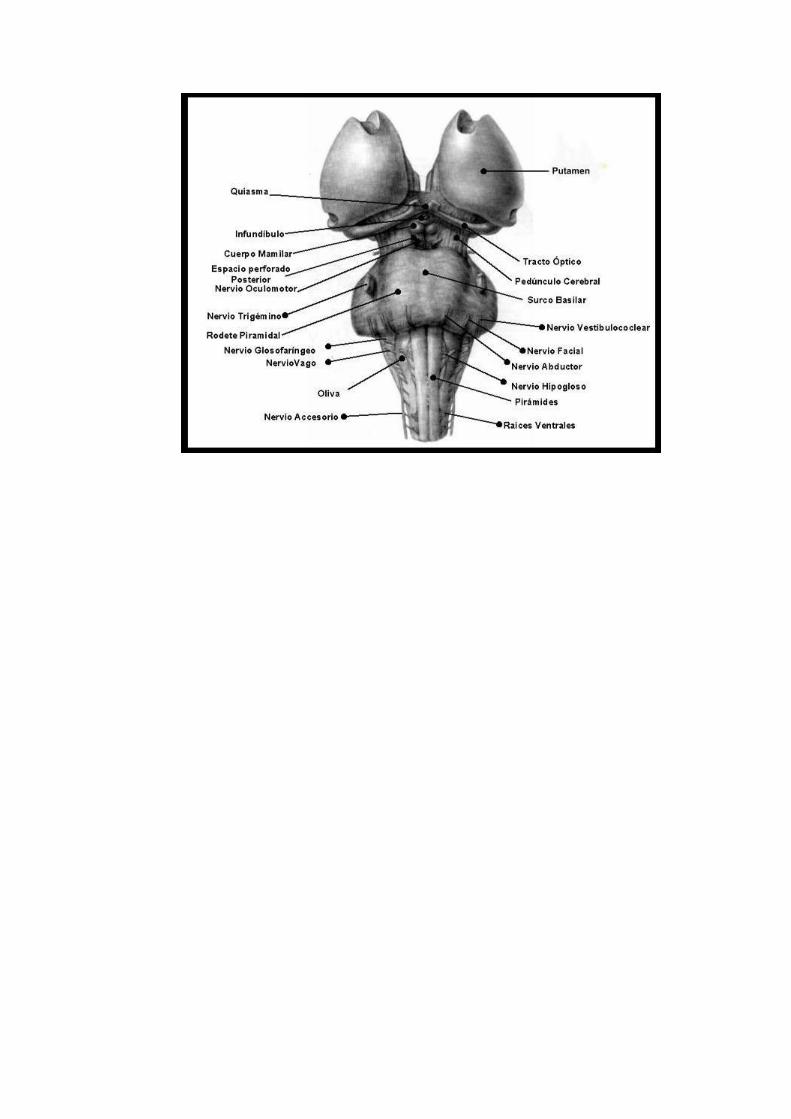
Pontificia Universidad Católica de Valparaíso Principios Neuroanatomía Funcional
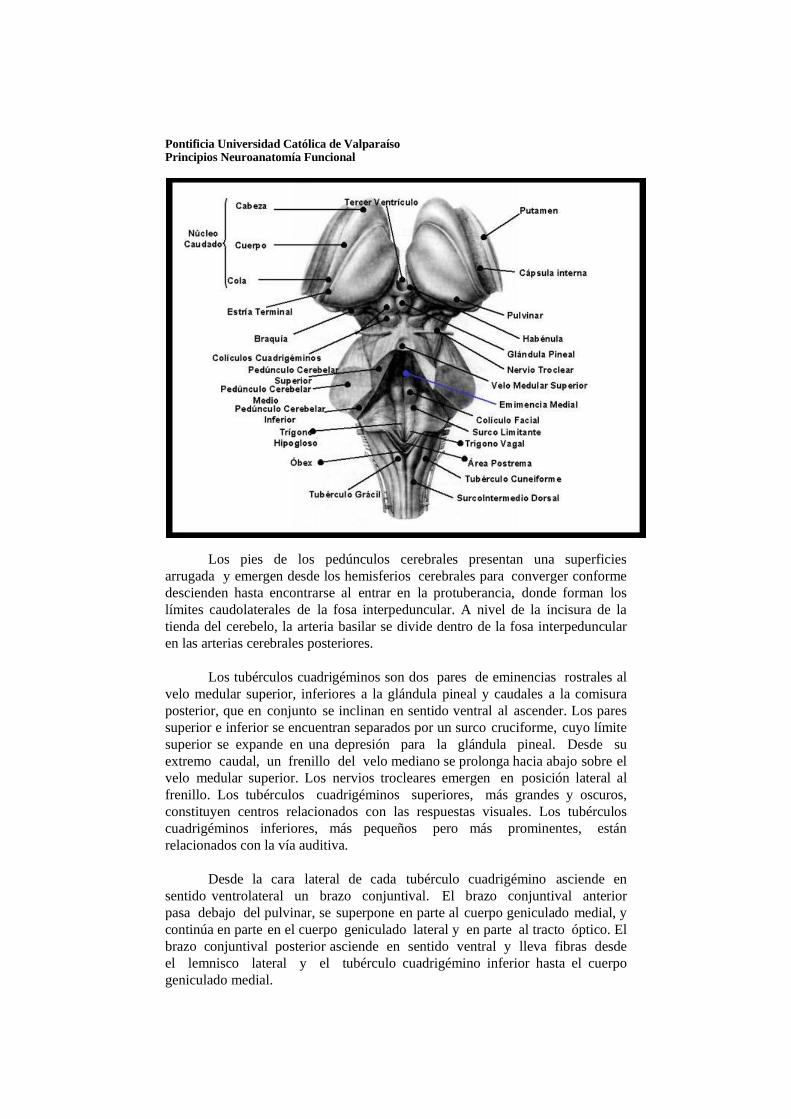
Los pies de los pedúnculos cerebrales presentan una superficies arrugada y emergen desde los hemisferios cerebrales para converger conforme descienden hasta encontrarse al entrar en la protuberancia, donde forman los límites caudolaterales de la fosa interpeduncular. A nivel de la incisura de la tienda del cerebelo, la arteria basilar se divide dentro de la fosa interpeduncular en las arterias cerebrales posteriores.
Los tubérculos cuadrigéminos son dos pares de eminencias rostrales al
velo medular superior, inferiores a la glándula pineal y caudales a la comisura posterior, que en conjunto se inclinan en sentido ventral al ascender. Los pares superior e inferior se encuentran separados por un surco cruciforme, cuyo límite superior se expande en una depresión para la glándula pineal. Desde su extremo caudal, un frenillo del velo mediano se prolonga hacia abajo sobre el velo medular superior. Los nervios trocleares emergen en posición lateral al frenillo. Los tubérculos cuadrigéminos superiores, más grandes y oscuros, constituyen centros relacionados con las respuestas visuales. Los tubérculos cuadrigéminos inferiores, más pequeños pero más prominentes, están relacionados con la vía auditiva.
Desde la cara lateral de cada tubérculo cuadrigémino asciende en
sentido ventrolateral un brazo conjuntival. El brazo conjuntival anterior pasa debajo del pulvinar, se superpone en parte al cuerpo geniculado medial, y continúa en parte en el cuerpo geniculado lateral y en parte al tracto óptico. El brazo conjuntival posterior asciende en sentido ventral y lleva fibras desde el lemnisco lateral y el tubérculo cuadrigémino inferior hasta el cuerpo geniculado medial.

Pontificia Universidad Católica de Valparaíso Principios Neuroanatomía Funcional
Anatomía Interna Mesencéfalo
En el se consideran dos porciones: una basal pedúnculo cerebral y una dorsal el tectum o lámina cuadrigémina.
Acueducto cerebral (Silvio): es un conducto ubicado ventral a la lámina cuadrigémina que comunica los ventrículos III y IV. Las sustancia gris periacueductal (regulación dolor) rodea a este conducto.
Pedúnculo cerebral: posee dos porciones: la base o pie del pedúnculo con una espacio medio que corresponde a la fosa interpeduncular (emergencia del III par craneal) y el tegmento separado de la base por la sustancia nigra. La base está formada por fibras de origen descendentes corticales (temporopóntico, frontopóntico, corticobulbar lateral, corticobulbar medio y corticoespinal).
La sustancia nigra: es una lámina que se extiende hasta el diencéfalo y está formada por neuronas con melanina. Las fibras aferentes de la sustancia nigra son corticonigrales, estrionigrales y talamonigrales. Las fibras eferentes son nigroreticulares y nigrorúbricas. La sustancia nigra forma parte de los ganglios basales.

Pontificia Universidad Católica de Valparaíso Principios Neuroanatomía Funcional Núcleo ruber o rojo: dorsal y medial a la sustancia nigra y se extiende desde el borde inferior del colículo inferior hasta el subtálamo. Está formado por células pequeñas y grandes. Las fibras eferentes son el fascículo rubroespinal al lado opuesto y rubrotalámico que conecta con el núcleo ventrolateral. El núcleo rojo se conecta con sustancia reticular, oliva bulbar y núcleos motores del tronco cerebral. Las fibras aferentes son las provenientes de los núcleos cerebelares heterolaterales, las frontorubricas, las provenientes del subtálamo por medio del tracto prerrubral y por último las provenientes del globo pallidus.
Decusación de los pedúnculos cerebelares superiores (braquia conjuntiva): en la zona de unión del puente con el mesencéfalo las fibras provenientes del cerebelo se cruzan (decusación) para terminar principalmente en el núcleo rojo. Otras fibras sinaptan en núcleos ventrolateral y centro mediano del tálamo y núcleos del tronco cerebral.
Núcleo interpeduncular: se ubica en el piso de la fosa interpeduncular. Recibe el fascículo habenulopeduncular.
Núcleo troclear: se ubica a la altura del tubérculo cuadrigémino inferior y da inervación al músculo oblicuo superior del ojo.
Núcleo oculomotor: se ubica rostral al núcleo troclear. Inervación de los músculos del ojo, excepto recto lateral y oblicuo superior.
Núcleo visceral del oculomotor (Edinger-Westphal): rostral al anterior y es el origen de las fibras preganglionares del ganglio ciliar que inerva al músculo ciliar y constrictor de la pupila.
Núcleo mesencefálico del trigémino: Rodea la sustancia periacueductal y forma las fibras de la porción mesencefálica del trigémino que desciende a la parte caudal para reunirse con los otros componentes y formar el lemnisco trigeminal. Da inervación propioceptiva de los músculos masticadores y articulación temporomandibular.
Lemnisco medial: Acompañado de los fascículos espinotalámicos.
Fascículo longitudinal dorsal: si sitúa la lateral en la sustancia gris periacuductal que es una importante vía eferente hipotalámica por la cual descienden impulsos viscerales motores hasta los centros vegetativos y sustancia reticular.

Pontificia Universidad Católica de Valparaíso Principios Neuroanatomía Funcional Fascículo longitudinal medio: se ubica ventral adyacente a los núcleos del oculomotor y troclear con lo cuales establece conexiones.
Núcleo intersticial (Cajal): Se sitúa lateral y adyacente al fascículo longitudinal medio al nivel de la extremidad superior del núcleo del tercer par. Posee fibras aferentes vestibulares a través del fascículo longitudinal medio, colículo cuadrigémino superior y globus pallidus. Las vías eferentes contribuyen a formar el fascículo longitudinal medio.
El tectum y tegmento se continúan rostralmente con el diencéfalo, entre
ellos se ubica una zona difusa que es la región pretectal que contiene al núcleo pretectal, que recibe fibras del tracto óptico provenientes de la retina ipsilateral que a su vez hacen sinapsis con el núcleo visceromotor del III par. La región pretectal además es atravesada por fascículos aferentes somáticos (lemniscos y espinotalámicos) y aferentes viscerales (pedúnculo mamilar).
Colículo cuadrigémino inferior: contiene en su interior el núcleo cuadrigemino inferior. La mayoría de las fibras del lemnisco lateral terminan en este núcleo (ipsilateral y heterolateral). El núcleo sinapsa con el cuerpo geniculado medial heterolateral.
Colículo cuadrigémino superior: Son centros de coordinación refleja en relación con las vías ópticas. Por su braquia llegan fibras desde cuerpo geniculado lateral, corteza visual y tractos ópticos. Posee un estrato zonal superficial que recibe fibras corticales, un estrato gris que recibe fibras del tracto y corticales, un estrato óptico que recibe fibras de retina y cuerpo geniculado lateral y estrato lemniscal que recibe fibras aferentes somáticas.

Secciones Transversales Mesencéfalo
El colículo inferior constituye una masa homogénea ovoide de sustancia gris que protruye en la superficie posterior del mesencéfalo. Es un importante sitio de procesamiento de las vías auditivas: sus aferencias provienen del lemnisco lateral y de fibras del cuerpo geniculado medial; sus eferencias se dirigen al cuerpo geniculado medial a través del brazo conjuntival inferior. La comisura del colículo inferior permite la comunicación entre ambos colículos. El desplazamiento posterolateral del lemnisco lateral, produce un acercamiento de éste al colículo inferior.
El núcleo troclear se ubica en la sustancia gris periacueductal cercano a
la línea media, posterior al fascículo longitudinal medial (FLM). Sus fibras se dirigen posterolateralmente para luego rodear la sustancia gris periacueductal, se decusan

Pontificia Universidad Católica de Valparaíso Principios Neuroanatomía Funcional totalmente a nivel del velo medular superior y emergen inferiormente al colículo inferior en la superficie posterior del mesencéfalo. El nervio troclear inerva solamente al músculo oblicuo mayor o superior del ojo.
El núcleo mesencefálico del trigémino está lateral al acueducto
cerebral. El tracto trigeminotalámico anterior se origina contralateralmente de los núcleos espinal y principal del V par y se ubica posterior al tracto tegmental central, mientras que el tracto trigéminotalámico posterior se origina del núcleo principal del V par ipsilateral y se ubica entre el lemnisco medio y el tracto espinotalámico. La decusación de los pedúnculos cerebelosos superiores se ubican en la porción central del tegmento, anterior al acueducto cerebral. El lemnisco medial asciende posterior a la sustancia negra; los lemniscos trigeminal y espinal se sitúan lateralmente al lemnisco medial. El lemnisco lateral está posterior al lemnisco trigeminal.
La sustancia negra es un núcleo motor situado a lo largo del mesencéfalo
entre el pie peduncular y el tegmento. Está compuesto por una pars compacta y una pars reticularis. Las neuronas multipolares de la pars compacta contienen en su citoplasma gran cantidad de gránulos de melanina y las principales catecolaminas, mientras que la pars reticularis no tiene melanina y presenta un color café rojizo debido a la presencia de un pigmento que contiene hierro. Este núcleo establece conexiones dopaminérgicas con la corteza cerebral, hipotálamo, núcleos basales (son la fuente dopaminérgica más importante del neostriatum, o sea, del núcleo caudado y putamen) y médula espinal. Se relaciona funcionalmente con el tono muscular. La degeneración de gran parte de este núcleo y, por consiguiente, la deprivación dopaminérgica del núcleo caudado a través del tracto nigroestriatal explican la enfermedad de Parkinson.
El pie peduncular constituye la porción basilar del mesencéfalo. Es una
columna de sustancia blanca ubicada anteriormente a la substancia nigra, cuyas fibras corticofugales comunican la corteza cerebral con las astas anteriores de la médula espinal, núcleos de pares craneales y núcleos pontinos. En su quinto medial se ubican las fibras frontopontinas. El quinto lateral está ocupada por fibras parieto, occipito y temporopontinas. La porción media está constituida por fibras corticoconucleares y corticoespinales organizadas somatotópicamente para la musculatura de la cabeza y extremidades (ordenadas de medial a lateral).
La formación reticular a este nivel está constituida por: (1) el núcleo
tegmental dorsal de la sustancia gris periacueductal, recibe aferencias del cuerpo mamilar. (2) el núcleo tegmental ventral se ubica anterior al FLM y parece ser una continuación del núcleo central superior. (3) parte del núcleo reticular pontino oral. (4) el locus ceruleus del grupo reticular central, en la profundidad del colículo inferior. (5) el núcleo pedunculopontino del grupo reticular lateral es el único que recibe aferencias directamente del globo pálido. (6) núcleo cuneiforme .

Pontificia Universidad Católica de Valparaíso Principios Neuroanatomía Funcional
El colículo superior conforma un elaborado centro reflejo de estructura laminar con una organización retinotópica que se corresponde con la corteza visual. Las capas superficiales reciben aferencias directas de la retina, corteza visual y tracto espino tectal, mientras las más profundas reciben aferencias de los sistemas somatosensorial y auditivos. Las eferencias conforman los tractos tectoespinal y tectobulbar, los cuales se relacionan con respuestas reflejas de movimientos oculares, cabeza y cuello ante estímulos visuales. Cercano a la porción anterior de los colículos superiores existe una pequeña agrupación neuronal denominada núcleo pretectal, cuyas aferencias se relacionan con el reflejo pupilar (fotomotor). La luz sobre la retina excita a fibras que viajan por los nervios y tracto ópticos hasta sinaptar con neuronas del núcleo pretectal que proyectan sus fibras al núcleo Edinger-Westphal del III par (parasimpático). De aquí emergen fibras que viajan por el III par hasta el ganglio ciliar donde sinaptan con neuronas postganglionares que envían sus axones al esfínter pupilar para producir miosis (disminución del diámetro pupilar). El núcleo pretectal también tiene que ver con el reflejo de acomodación del cristalino (aumento de la curvatura del cristalino).
Las fibras somatomotoras y visceromotoras del nervio oculomotor se
originan en neuronas que conforman una serie de agrupaciones celulares denominadas complejo nuclear del oculomotor, el cual se ubica anterolateralmente al extremo superior del acueducto cerebral en la sustancia gris periacueductal posterior al FLM. El III par tiene 2 núcleos motores:

El núcleo motor principal, cuyas fibras se dirigen anteriormente a
través del tegmento medial atravesando el núcleo rojo para luego emerger en la fosa interpeduncular. Recibe aferencias de ambos hemisferios cerebrales mediante fibras corticonucleares, de la corteza cerebral . Además contribuye con el FLM que permite la conexión de este núcleo con los núcleos del IV, VI y vestibulares del VIII par craneal.
Las fibras somatomotoras del III par inervan a todos los músculos extrínsecos del ojo excepto el oblicuo superior y el recto lateral. El núcleo accesorio parasimpático (núcleo Edinger-Westphal) se ubica posterior al núcleo motor principal. Estas neuronas preganglionares emiten axones que viajan junto a las otras fibras del III par hasta sinaptar en el ganglio ciliar, del cual surgen los nervios ciliares que inervan las fibras circulares del iris y músculos ciliares. A este núcleo parasimpático llegan fibras corticonucleares involucradas en el reflejo de acomodación y fibras desde los núcleos pretectales relacionadas con reflejos visuales.
A cada lado de la línea media, en el centro del tegmento, se
observa una agrupación neuronal bien delimitada de forma oval: el núcleo rojo. En preparaciones frescas este núcleo presenta una coloración rojiza debido a su gran vascularización y a la presencia de un pigmento citoplasmático que contiene hierro. El núcleo rojo está compuesto por una parte caudal magnocelular y una rostral parvicelular. Recibe aferencias desde: (1) corteza cerebral mediante fibras cortico-rubrales (2) de cerebelo mediante el pedúnculo cerebeloso superior. Algunas fibras de este pedúnculo pasan sin sinaptar formando una cápsula alrededor del núcleo rojo o lo atraviesan en dirección hacia los núcleos talámicos ventrales lateral, y ventral anterior, (3) de núcleos hipotalámicos, subtalámicos, lenticular, substantia nigra y médula espinal. Las eferencias del núcleo rojo son (1) el tracto rubroespinal que se origina en las células de la parte caudal de este núcleo y cuyas fibras se decusan antes de descender por la médula espinal (2) el tracto rubrorreticular que conecta el núcleo rojo con la formación reticular (3) conecciones con substantia nigra.
Entre el pie peduncular y el tegmento se observa la substantia
nigra. Los lemniscos medial, espinal y trigeminal forman una banda curva en la región posterior del tegmento. El brazo conjuntival inferior va en dirección del núcleo geniculado medial y se ubica posterolateralmente al tracto espinotalámico en la región tegmental dorsal. Inmediatamente posterior a la fosa interpeduncular y en la línea media se encuentra el núcleo interpeduncular. Posterior a este núcleo está el área tegmental ventral (ATV) cuyos axones se proyectan por el sistema dopaminérgico mesolímbico al cuerpo estriado (núcleo acumbens), amígdala, área septal y lóbulo frontal. El AVP se relaciona con la recompensa en casos de adicción a cocaína y sus disfunciones se relacionan con la esquizofrenia.

Pontificia Universidad Católica de Valparaíso Principios Neuroanatomía Funcional Lateral y posteriormente al núcleo rojo se ubica la formación reticular del tegmento mesencefálico.
V.- Columna de Núcleos del Tronco Cerebral
En el desarrollo embrionario el tubo neural se compone de dos paredes laterales que están divididas por el surco limitante en dos porciones: las láminas basal y alar. De éstas últimas se originan las estructuras sensitivas, mientras que de las primeras se originan las motoras. A su vez el sistema nervioso se divide en componentes: uno somático por lo cual reaccionamos frentes al medio recibiendo estímulos (somático aferente) y respondiendo a ellos (somático eferente) y el que se relaciona con el funcionamiento vísceras (aferente y eferente visceral). Existe además un grupo de estructuras que derivan de los arcos branquiales que poseen características anatómicas especiales (músculo estríado y control voluntario) y desempeñan funciones vegetativas (masticación y deglución). Estos componentes funcionales están ordenados en columnas de núcleos que se extienden de la médula al mesencéfalo. En el tronco encefálico las astas medulares son sustituidas por los nervios craneales. Los núcleos sensoriales de estos nervios contienen neuronas que reciben información aferente de las estructuras craneales, mientras que los núcleos motores contienen las neuronas motoras que inervarán diversas estructuras.
Existe una diferencia en cuanto al orden adoptado por las columnas en el tronco encefálico: en este segmento las columnas adoptan un orden latero-medial, a diferencia de médula espinal donde el orden era dorso-ventral. Otra diferencia es la diferenciación entre las columnas de células viscerales y somáticas. Además, como se menciono anteriormente, al existir estructuras especializadas en la cabeza tanto sensoriales como motoras, existe una diferenciación adicional de los núcleos de los nervios craneales: las estructuras motoras especiales son los músculos de origen branquiomérico, mientras que las sensoriales incluyen el epitelio olfatorio, la retina, la cóclea, el laberinto y los botones del gusto. Estas estructuras están inervadas por axones aferentes y motores especiales.

Pontificia Universidad Católica de Valparaíso Principios Neuroanatomía Funcional
La clasificación de las columnas sería:
Columna Somática Aferente Especial: audición y equilibrio. Columna Somática Aferente General: sensibilidad exteroceptiva
(dolor, calor, frío, tacto) y sensibilidad propioceptiva (músculos y articulaciones).
Columna Visceral Aferente General y Especial: Gusto y olfato. Sensación de órganos viscerales y procesos de estímulos químicos.
Columna Visceral Eferente General: inervación motora
Columna Somática Eferente Especial: Son viscerales porque desempeñan función autónoma tales como deglución, masticación y especiales porque inervan músculos de los arcos branquiales.
Columna Somática Eferente General: Inervación musculatura esquelética.
Núcleos nervios craneales
Existe una relación funcional en cuanto a la ubicación de las columnas con respecto a la línea media. Las columnas motoras están situadas junto a la línea media, es decir, mediales a las columnas sensitivas. Estos dos tipos de columnas se encuentran separadas por el sulcus limitans.

Pontificia Universidad Católica de Valparaíso Principios Neuroanatomía Funcional
Columna Somática Eferente General: situada cerca de la línea media de médula oblonga, puente y mesencéfalo e inervan a las estructuras derivadas de los somites (músculos motores de ojo y lengua). Al nivel de mesencéfalo se encuentran los núcleos oculomotor (III) y troclear (IV), a nivel del piso de IV ventrículo los núcleos del abductor (VI) e hipogloso (XII).
Columna Somática Eferente Especial: situada en médula oblonga y protuberancia, se encuentra desplazada del IV ventrículo. Inerva musculatura derivada de arcos branquiales. El ambiguo da inervación a los músculos faríngeos y laríngeos (tercer y cuarto arco). El motor del facial inerva los músculos faciales (segundo arco) y el núcleo motor del trigémino que inerva a los músculos masticadores (primer arco). A nivel de la protuberancia se encuentran los núcleos motor del trigémino (V) que inerva a los músculos masticadores y el núcleo del facial (VII) para los músculos de la mímica. A nivel de médula oblonga se encuentra el núcleo ambiguo que envía fibras al glosofaringeo (IX), vago (X) y accesorio (XI).
Columna Visceral Eferentes General: ubicada en el piso del IV ventrículo de médula oblonga y puente. Las neuronas de estos núcleos forman las fibras preganglionares que dan inervación a músculo cardiaco, músculo liso y glándulas. El vago se distribuye en vísceras torácicas y abdominales. Los núcleos mucolacrimonasal y salivales terminan en los núcleos submaxilar otico y esfenopalatino. El núcleo visceral oculomotor en el ganglio ciliar. A nivel del mesencéfalo se encuentra el núcleo oculomotor (III). En el puente encontramos el núcleo salival superior (VII). En médula oblonga, el salival inferior (IX) y el núcleo motor dorsal del vago o cardioneumoentérico (IX).
Columna Aferente Visceral General y Especial: Posee solo un núcleo, el núcleo solitario (VII), que posee una porción rostral que recibe información gustativa y una caudal de sensibilidad visceral y procesos de estímulos químicos. Se encuentra ubicado prácticamente en médula oblonga.
Columna Somática Aferente General: se extiende desde médula espinal rostral al mesencéfalo; posee los tres núcleos trigéminos. En mesencéfalo y la porción rostral del puente se encuentra el núcleo trigémino mesencefálico que media la sensibilidad propioceptiva. En la porción media del puente se ubica el núcleo trigémino principal que media la sensibilidad exteroceptiva junto con el núcleo trigémino espinal, ubicado este último en la porción caudal del puente y a lo largo de médula oblonga y la porción rostral de médula espinal.
Columna Somática Especial: ubicada en la protuberancia, contiene los núcleos vestibular y coclear (VIII)
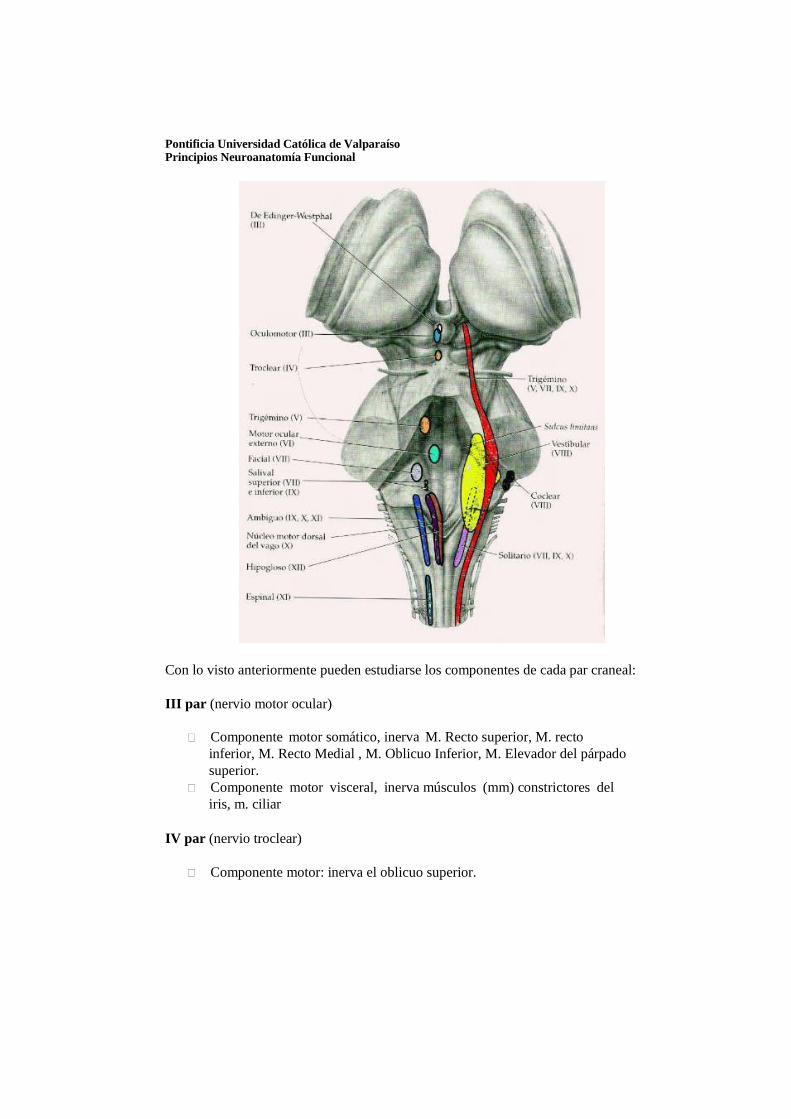
Pontificia Universidad Católica de Valparaíso Principios Neuroanatomía Funcional
Con lo visto anteriormente pueden estudiarse los componentes de cada par craneal:
III par (nervio motor ocular)
Componente motor somático, inerva M. Recto superior, M. recto inferior, M. Recto Medial , M. Oblicuo Inferior, M. Elevador del párpado superior.
Componente motor visceral, inerva músculos (mm) constrictores del iris, m. ciliar
IV par (nervio troclear)
Componente motor: inerva el oblicuo superior.

V par (nervio trigémino)
Componente sensorial o mesencefálico: propiocepción o principal: tacto o espinal: presión y termoalgesia
Componente motor: inerva a los mm. masticadores
VI par (motor ocular externo)
Componente motor: inerva el recto lateral
VII par (nervio facial)
Componente motor: o Motor del facial. inerva los músculos de la mímica. o Núcleo salival superior: glándulas lacrimales, nasales y salivales.
Componente sensitivo: o Núcleo solitario: gusto dos tercios anteriores de la lengua.
VIII par (vestíbulococlear)
Cocleares: audición Vestibulares: equilibrio
IX par (nervio glosofaríngeo)
Componente motor: o Núcleo ambiguo: inerva el estilofaríngeo o Núcleo salivar inferior: glándulas salivales
Componente sensitivo: o Núcleo solitario: gusto del tercio posterior de la lengua
X par (nervio vago)
Componente motor: o Núcleo ambiguo: mm. laríngeos y faríngeos o Núcleo motor dorsal: actividad cardiaca, pulmonar y entérica
Componente sensitivo: o núcleo solitario: sensibilidad visceral, cuerpos carotídeos y el
gusto del paladar

Pontificia Universidad Católica de Valparaíso Principios Neuroanatomía Funcional XI par (nervio accesorio espinal)
Componente motor: inerva el m. esternocleidomastoideo; sus fibras
superiores ayudan a la deglución.
XII par (nervio hipogloso)
Componente motor: inervación de los músculos de la lengua.
VI.- Anatomía Externa Cerebelo
El cerebelo, la parte de mayor tamaño del rombencéfalo, se encuentra dorsal al puente y a la médula oblonga, y su región media está separada de estas regiones por el IV ventrículo. Se encuentra ubicado en la fosa craneal posterior, está cubierto por la tienda del cerebelo (pliegue de la duramadre) y es prácticamente esférico, pero está ligeramente comprimido en su región media y aplanado, siendo su diámetro mayor el transversal. Su peso promedio en varones es de uno 150g. En los adultos, la relación del cerebelo con el cerebro es de aproximadamente 1:10, y en niños 1:20.
Relaciones Anatómicas
Cara superior: plana y cubierta por la tienda del cerebelo. Cara inferior: convexa y se aloja en la fosa cerebelosa (hueso occipital). Cara anterior: se relaciona con el techo del IV ventrículo.
El cerebelo está cubierto por una corteza laminada de neuronas y células
gliales. Internamente el cerebelo está formado por sustancia blanca, que contiene los núcleos cerebelosos grises. El cerebelo puede subdividirse en divisiones medial-lateral o anterior-posterior. El cerebelo en la línea media recibe el nombre de vermis. El resto de la corteza forma los hemisferios cerebelosos.

La superficie cerebelosa está dividida por numerosos surcos transversos curvos, que le proporcionana un aspecto laminado y la separan en giros. Los surcos más profundos la dividen en lóbulos o lobos. El cerebelo se encuentra dividido en tres lobos: anterior, medio y floculonodular. El lobo anterior está separado del lobo medio por la surco primario o primo. El lobo medio (posterior) es el mayor y se ubica entre los surcos prima y uvulonodular. El lobo floculonodular se ubica dorsal al surco uvulonodular. Un surco cincunferencial se encuentra a lo largo del borde del cerebelo y lo separa anatómicamente en superior e inferior.
Funcionalmente el cerebelo se divide en regiones las cuales difieren
entre los anatomistas. El lobo anterior, úvula y pirámide constituyen el paleocerebelo cuya función es la coordinación muscular. El lobo medio a excepción de úvula y pirámide constituye el neocerebelo y está situado con los movimientos finos. El lobo floculonodular constituye el arquicerebelo relacionado con el sistema vestibular. Actualmente aplicamos otro tipo de clasificación que divide funcionalmente al cerebelo en tres regiones: espinocerebelo, cerebrocerebelo y vestibulocerebelo. Esta clasificación permite una mayor correlación anatómica funcional del cerebelo.


Pontificia Universidad Católica de Valparaíso Principios Neuroanatomía Funcional

Pontificia Universidad Católica de Valparaíso Principios Neuroanatomía Funcional
Anatomía Interna Cerebelo
La sustancia gris se distribuye externamente en el cerebelo formando la corteza cerebelar, mientras que la sustancia blanca se distribuye internamente formando el árbol de la vida. Existen además en la sustancia blanca 4 pares de núcleos grises.
Corteza Cerebelar
Está plegada por muchos surcos transversales paralelos en folias cerebelosas. Las capas de la corteza cerebelar son:
Capa molecular: Está formada por las fibras paralelas, las dendritas
de las células de Purkinje, las fibras trepadoras que con ellas hacen sinapsis las células en cesto y células estrelladas. Las células en cesto envía su axón transversalmente al eje de la laminilla; de él parten ramificaciones que sinaptan con las células de Purkinje en forma de cesto.

Pontificia Universidad Católica de Valparaíso Principios Neuroanatomía Funcional
Capa células de Purkinje: Son grandes células. De lo alto de su cuerpo parten uno o varios tallos que penetran la zona molecular arborizándose y prolongándose hasta la misma superficie del cerebelo. Del otro lado de la célula emerge un axón que ingresa a la sustancia blanca para sinaptar en los núcleos
grises que se ubican en la sustancia blanca del cerebelo. El axón emite prolongaciones que sinaptan con otras células de Purkinje y elementos de la capa molecular. En esta capa existen una neuroglia (Epitelial de Golgi) y las células de Bergman.
Capa granular: Presenta células con aspecto granular, su axón se dirige a la capa molecular. Las dendritas varían de 3 a 5 y permiten formar los glomérulos donde sinaptan las fibras musgosas. Existen también células tipo Golgi II, cuyas dendritas ascienden a la capa molecular y su axón sinapta en el glomérulo.
Los distintos tractos llegan al cerebelo a través de las fibras musgosas y trepadoras. Las fibras trepadoras provienen de los núcleos olivares inferiores y según se ha descrito cada una de ellas sinapta con una sola célula de purkinje. Las fibras musgosas provienen de los restantes tractos aferentes y sinaptan en los glomérulos.

Pontificia Universidad Católica de Valparaíso Principios Neuroanatomía Funcional Núcleos Grises
Núcleos grises, desde la parte lateral a la línea media se encuentran: el núcleo dentado (oliva cerebelosa), núcleo emboliforme, núcleo globoso y núcleo del techo (fastigial). El núcleo más grande es el dentado. Cada núcleo está compuesto de neuronas grandes y multipolares, con dendritas de ramificación simple. Los axones forman la eferencia cerebelosa por los pedúnculos cerebelares.
Núcleo fastigial: ubicado en el vermis, dorsal y superior del techo del
cuarto ventrículo. Núcleo globoso: se ubica lateral al fastigial y de menor tamaño.
Núcleo dentado: se ubica en la parte interna de la sustancia blanca hemisférica. Es una lámina de sustancia gris, plegada e irregular.
Emboliforme: se ubica medial al dentado, dorsal y lateral del núcleo fastigial

Pontificia Universidad Católica de Valparaíso Principios Neuroanatomía Funcional Pedúnculos cerebelares
Son tres estructuras de sustancia blanca que conectan el cerebelo con
otros centros nerviosos. Son tres pares de pedúnculos: superior, medio e inferior.
Inferior: Se forma en la superficie posterolateral de la mitad superior de la médula oblonga. Consta de un haz externo de fibras compactas localizado en la cara dorsolateral de la porción superior de médula oblonga (cuerpo restiforme) y una porción medial formada por haces dispersos de fibras que entran en el cerebelo pasando a través de los núcleos vestibulares (cuerpo yuxtarestiforme).

Pontificia Universidad Católica de Valparaíso Principios Neuroanatomía Funcional
Aferentes (cuerpo restiforme)
o Fascículo espinocerebloso posterior (directo), viene de médula espinal
(núcleo de Clarck) y se distribuye en el paleocerebelo: lóbulo anterior, úvula y pirámide.
o Fascículo cuneocerebeloso, desde el núcleo cuneiforme lateral (estímulos propioceptivos y tactiles del cuello y miembro superior) al vermis.
o Fascículo olivocerebeloso, desde los núcleos olivares inferiores hasta la corteza del neocerebelo
o Fascículo reticulocerebeloso, desde la formación reticular de la médula oblonga hasta el vermis.
o Fascículo vestibulocerebeloso, desde núcleos vestibulares y el nervio vestibulococlear hasta al arquicerebelo.
Eferentes (cuerpo yuxtarestiforme)
o Fibras cerebelovestibulares, que van a los núcleos vestibulares. o Fibras cerebeloreticulares, que terminan en la sustancia
reticular de puente y bulbo.
Medio: es el más grande de los tres pedúnculos. Nace en la región posterolateral del puente y se continúa con la sustancia blanca del hemisferio cerebelar. Está formado por la vía corticopontocerebelosa que son fibras transversales que van de un hemisferio cerebelar a otro atravesando al puente. Sus fibras se originan en los núcleos pónticos, que reciben sinapsis desde corteza frontal, parietal y temporal. Las fibras ponticocerebelosas terminan en cerebelo.
Superior: sale de la porción superior de la valécula (escotadura anterior
y media del cerebelo), para llegar al sector inferior del mesencefalo. La mayoría de las fibras son eferentes y provienen de los núcleos, penetran en el tegmento mesencefálico para cruzar y terminar en el núcleo rojo y núcleo ventrolateral talámico. De esta manera queda configurada la vía cerebelo-rubro-talámica que completada con las conexiones del tálamo y el área 4 de corteza constituye el modo en que el cerebelo modifica la actividad motora voluntaria. Existen fibras eferentes descendentes a los núcleos olivares y reticulares. Hay fibras aferentes como: fascículo espinocerebeloso anterior cruzado y fibras rubrocerebelosas

Pontificia Universidad Católica de Valparaíso Principios Neuroanatomía Funcional
Las fibras eferentes del cerebelo llevan impulsos excitadores originados en los núcleos grises hasta tálamo, núcleo rojo, formación reticular y núcleos vestibulares. La actividad de los núcleos está modulada por las células de Purkinje. La acción de células de Purkinje en los núcleos cerebelares y vestibulares es inhibidora. La actividad de las células de Pukinje a su vez esta condicionada por las fibras trepadoras (excitadora), fibras paralelas y células en cesto. Las células en grano son excitadoras produciendo depolarización de las células de Purkinje. Las células en cesto son inhibidoras, un estimulo de una fibras musgosa se traduce en la excitación de un grupo de células de Purkinje y en la inhibición de las células contiguas.
VII.- Anatomía del Diencéfalo
El diencéfalo forma parte del prosencéfalo, estructura que se desarrolla a partir de la vesícula cerebral primaria anterior. Esta vesícula prosencefálica y su cavidad, el futuro III ventrículo, se diferencian pronto en un diencéfalo caudal y un telencéfalo rostral. A partir de los lados del telencéfalo se desarrollan dos divertículos, derecho e izquierdo, que formarán los hemisferios cerebrales, cada uno con un ventrículo interno. Los puntos de evaginación se convierten en los forámenes interventriculares, a través de los cuales se comunican los tres ventrículos. El diencéfalo corresponde en gran parte al III ventrículo y las estructuras adyacentes; el telencéfalo comprende un telencéfalo medio, que contiene una limitada extensión hacia adelante del III ventrículo, y los grandes hemisferios telencefálicos lateras cada uno con un ventrículo lateral.
El prosencéfalo en desarrollo da lugar al telencéfalo por ventral y al
diencéfalo por dorsal. Las paredes laterales del diencéfalo en desarrollo muestran cuatro regiones de crecimiento algo diferenciadas, que forman por separado el epitálamo en la parte más superior, los tálamos dorsal y ventral en el centro y el hipotálamo en la porción inferior. El epitálamo comprende en el cerebro maduro los núcleos paraventriculares anterior y posterior, los núcleos habenulares medial y lateral, la estría medular del tálamo y el cuerpo pineal. El tálamo dorsal se subdivide en masas pronucleares, que se transformarán en los grandes núcleos principales del tálamo. El esbozo talámico ventral da lugar a los núcleos geniculados reticular y ventral del tálamo, entre otros. El término

Pontificia Universidad Católica de Valparaíso Principios Neuroanatomía Funcional
subtalámico es usado a veces como sinónimo del tálamo ventral e incluye el núcleo subtalámico, el cual se encuentra íntimamente relacionado con los núcleos basales y la sustancia nigra.
Superficie inferior. Está formada por estructuras hipotalámicas, de adelante hacia atrás: el quiasma óptico, con la cintilla óptica a cada lado, el infundíbulo con el túber cinereum, y los cuerpos mamilares.
Superficie superior. Se halla debajo del trígono cerebral, que es un grueso haz de fibras que se origina en el hipocampo del lóbulo temporal y se arquea posteriormente sobre el tálamo para unirse al cuerpo mamilar. La verdadera pared superior del diencéfalo es el techo del tercer ventrículo, que posee el epitelio ependimario y tela coroidea del tercer ventrículo con pliegues vasculares denominadas plexos coroides.
Superficie externa. Está limitada por la cápsula interna de sustancia blanca, que consta de fibras nerviosas que unen la corteza cerebral con partes del tronco cerebral y médula espinal. Por la cápsula interna descienden fibras piramidales y corticoponticas.
Superficie interna, o pared lateral del tercer ventrículo está formada en la parte superior, por la superficie interna del tálamo y, en la parte inferior, por el hipotálamo. Estas dos áreas están separadas por el surco hipotalámico.
El diencéfalo se compone de cuatro regiones: tálamo, hipotálamo,
epitálamo y subtálamo.
Tálamo: Los tálamos están separados por el III ventrículo y en algunos casos comunican entre si por un comisura gris intertalámica. Lateral al tálamo se ubica la cápsula interna que lo separa del núcleo lenticular. Dorsal se relaciona con parte del sistema ventricular y ventral con el trígono y cabeza del núcleo caudado. Inferiormente el tálamo se relaciona con el hipotálamo, región subtalámica y mesencéfalo.
Hipotálamo: Se exterioriza en la cara inferior del cerebero presentando una cara anterior constituida por la lámina supraóptica y una cara inferior constituida por el túber cinereum y los tubérculos mamilares. La cara interna o profunda del hipotálamo forma el vértice del III ventrículo.

Pontificia Universidad Católica de Valparaíso Principios Neuroanatomía Funcional
Epitálamo: Corresponde al conjunto formado por los trígonos de la habénula, comisura interhabenular, comisura blanca posterior y el cuerpo pineal.
Subtálamo: Corresponde a la unión mesodiencéfalica
Anatomía del Tálamo
Masa grande y ovalada de sustancia gris que forma la mayor parte del diencéfalo. Está ubicado a cada lado del tercer ventrículo. Está formado por un grupo de núcleos interpuestos en la vía sensitiva. En él se integran los distintos tipos de sensibilidad para ser proyectados luego a corteza cerebral.
La sustancia gris contiene también sustancia reticulada y sustancia
blanca. Está formada por láminas medulares, una lateral y otra medial, bifurcada hacia craneal. Anatómicamente existen seis grupos nucleares de acuerdo a su posición en relación a la lámina medular medial.

Los núcleos tálamicos se dividen funcionalmente en núcleos de relevo y
núcleos de proyección difusa. Los anteriores, mediales y laterales son de relevo y los restante de proyección difusa. Los núcleos de relevo son esenciales para las funciones cerebrales y cada uno desempeña un papel en la percepción, volición y cognición. Para cada modalidad sensorial existe un núcleo de relevo (excepto el olfato). Los núcleos de relevo se proyectan a la corteza asociativa: asociativa parietotemporooccipital, prefrontal y asociación límbica, para la percepción, funciones cognitivas y aprendizaje/memoria respectivamente. Los núcleos de proyección difusa actúan en el despertar y en la regulación de la excitabilidad de las regiones más amplias del córtex. Los núcleos de proyección difusa conectan varias áreas corticales, a diferencia de los de relevo que lo hacen a una sola área.
Núcleos de Relevo
1.- Grupo Anterior: Formado por los núcleos talámicos anterior y dorsolateral que reciben conexiones de los cuerpos mamilares y formación hipocampal y se conecta con el giro cingulado. Está encargado del tono emocional, mecanismo de memoria reciente y aprendizaje.
2.- Grupo Medial: Contiene al núcleo dorsomedial. El tálamo medial se encarga de integrar una variedad de información sensitiva, que incluye información somática, visceral, olfatoria y de relacionarla con los sentimientos emocionales y los estados subjetivos.
3.- Grupo Lateral: Se compone de los siguientes núcleos: anteriorventral, lateralventral, posterioventral, geniculado lateral, geniculado medial, pulvinar y posterior lateral.
El anterior ventral recibe aferencia de los ganglios basales y da
eferencia a la corteza suplementaria, planifica el movimiento. El lateral ventral recibe aferencia del cerebelo y da eferencia a corteza
premotora y motora primaria, planifica y controla movimientos. El posterior ventral recibe aferencia de médula espinal, tronco cerebral,
lemnisco medial y trigeminal y da eferencia a la corteza sensorial somática primaria, tacto, propiocepción, dolor y temperatura.
Geniculado lateral recibe aferencia de retina y da eferencia a corteza visual primaria, visión.
Geniculado medial recibe aferencia de colículo inferior y da eferencia a corteza auditiva primaria, audición.
Pulvinar, recibe aferencia de colículo superior, lobo parietal, temporal y occipital y da eferencia a la corteza asociativa parietal, temporal y occipital, integración sensorial, percepción y lenguaje.
Posteriorlateral, recibe aferencia de colículo superior, lobo occipital y da eferencia a la corteza asociativa parietal posterior, integración sensorial.

Núcleos de Proyección Difusa
4.- Intralaminares: reciben aferencia de tronco, y médula espinal y dan eferencia a cortex y ganglios basales, para la regulación de la actividad cortical.
5.- Núcleos de la línea media, recibe aferencia de la formación reticular e hipotálamo y da eferencia a cortex y alocorteza, para la regulación de la excitabilidad neuronal.
6.- Núcleo reticular, recibe aferencias de tálamo y cortex y da eferencia a tálamo, para la regulación de la actividad neuronal talámica.
Núcleo Ventral Anterior: Se comunica con la formación reticular,
cuerpo estríado y corteza premotora, sustancia nigra y núcleos talámicos. Parece influir en las actividades de corteza motora.
Núcleo Ventral Intermedio: Comunicaciones similares que el anterior y además con el núcleo rojo y cerebelo.
Núcleo Ventral Lateral: Se divide en posteromedial y posterolateral. El núcleo posteromedial recibe las vías gustatoria y trigeminal ascendente, mientras que el núcleo ventral posterolateral recibe los lemniscos medial y espinal ascendentes.
A considerar.
1.- El tálamo está formado por un intrincado grupo de neuronas que se comunican entre si.
2.- Una gran cantidad de información sensitiva excepto la olfatoria converge en el tálamo para su integración.
3.- La via olfatoria se conecta con el sistema límbico y ahí podría relacionarse con el tálamo a través del fascículo mamilotalámico.
4.- Existe estrecha relación entre tálamo y corteza cerebral. Se sabe que al eliminar corteza es posible apreciar sensasiones imperfectamente.

Pontificia Universidad Católica de Valparaíso Principios Neuroanatomía Funcional

Anatomía del Epitálamo
El epitálamo comprende los núcleos paraventriculares anterior y
posterior, los núcleos habenulares medial y lateral, la estría medular del tálamo y la epífisis o glándula pineal.
Núcleos paraventriculares. Se encuentran inmediatamente por debajo del epéndimo del III ventrículo, en posición dorsal. Reciben aferencias del hipotálamo, en particular de las áreas hipotalámicas anterior y lateral, los núcleos septales, el núcleo del lecho de la estría terminal y la formación del hipocampo. Presenta aferencias adicionales del tronco encefálico y pueden incluir fibras noradrenérgicas desde el locus coeruleus (puente). Estos núcleos se proyectan sobre el núcleo auditivo, la amígdala y la formación del hipocampo.
Núcleos habenulares. Los núcleos habenulares ocupan una posición posterior en el ángulo dorsomedial del tálamo, inmediatamente por debajo del epéndimo del tercer ventrículo, situándose la estría del tálamo superior y lateralmente. Desde el margen ventral de los núcleos emerge un fascículo de sustancia blanca, el haz habenulopeduncular, que progresa en dirección caudal e inferior hacia la región interpeduncular del mesencéfalo. El núcleo habenular medial es colinérgico y envía eferencias hasta el núcleo interpeduncular del mesencéfalo. Desde el núcleo habenular lateral, pasan fibras a los núcleos del rafe y la formación reticular mesencefálica adyacente, a la parte de la sustancia negra y el área tegmetaria ventral, y al hipotálamo y el prosencéfalo basal.
Estría medular. La estría medular cruza la cara superomedial del tálamo, corre medial al triángulo de la habénula y envía abundantes fibras al núcleo habenular ipsilateral. Otras fibras de la estría cruzan en la lámina pineal anterior y se decusan formando la comisura habenular para llegar al núcleo habenular contralateral.

Anatomía del Hipotálamo
Pese a ser una estructura muy pequeña en el sistema nervioso,
contiene el sistema integrador que controla el equilibrio de líquidos y electrolitos, la ingestión de alimentos y el equilibrio de energía, la reproducción, la termorregulación, las respuestas inmunes y muchas respuestas emocionales, a través de sistemas efectores autónomos y endocrinos.
Se extiende en sentido posterior desde la lámina terminal hasta un plano
vertical que pasa por detrás de los cuerpos mamilares, y desde el surco hipotalámico hasta la base del cerebro, debajo del III ventrículo. Esta parte anterior de la región, el área preóptica, pertenece en sentido estricto al telencéfalo impar, pero se incluye aquí por razones funcionales.
El hipotálamo se encuentra bordeado a los lados por la parte
anterior del subtálamo, la cápsula interna y las cintillas ópticas; por detrás se sitúa la porción tegmentaria del subtálamo y el tegmento mesencefálico; dorsalmente se relaciona con los núcleos talámicos dorsales. En la parte anterior, el quiasma óptico, la lámina terminal y la comisura anterior separan el área preóptica del tabique precomisural.
Las estructuras presentes en el suelo del III ventrículo alcanzan la
superficie pial en la fosa interpeduncular. Desde delante hacia atrás se encuentran: quiasma óptica; el tuber cinereum, las eminencias tuberales y el tallo infundibular; los cuerpos mamilares; y la sustancia perforada posterior (espacio perforado posterior), localizada en la divergencia de los pedúnculos cerebrales y está atravesada por ramas centrales pequeñas de las arterias cerebrales posteriores.
Los cuerpos mamilares son eminencias lisas, hemisféricas, que contactan
entre sí y se sitúan por delante de la sustancia perforada posterior. Cada uno de los tubérculos mamilares contiene núcleos encerrados por fascículos blancos procedentes en gran parte del fórnix.
El tuber cinereum, situado entre los cuerpos mamilares y el quiasma
óptico, es una masa convexa de sustancia gris; su cara ventral presenta una serie de eminencias con surcos intermedios de profundidad variable. A partir de él, el infundíbulo hueco y cónico se continúa en sentido ventral con el lóbulo posterior de la hipófisis. Alrededor de la base del infundíbulo se encuentra la eminencia media, delimitada por un surco tuberoinfundibular superficial. El tuber cinereum presenta también un par de eminencias laterales que marcan los núcleos tuberales laterales, y una pequeña eminencia pos- infundibular media.

Pontificia Universidad Católica de Valparaíso Principios Neuroanatomía Funcional
Divisiones y núcleos del hipotálamo
El hipotálamo se puede dividir en sentido anteroposterior en las regiones quiasmáticas (supraóptica), tuberal (infundibulotuberal) y posterior (mamilar), y en sentido mediolateral en zonas periventricular, intermedia (medial) y lateral.

Pontificia Universidad Católica de Valparaíso Principios Neuroanatomía Funcional Zona periventricular. Bordea el III ventrículo. En la pared anterior del ventrículo se encuentra el órgano vascular de la lámina terminal, que se continúan en dirección dorsal con el núcleo preóptico medio y el órgano subfornical. A cada lado de la región quiasmático se encuentran una parte del núcleo preóptico, el pequeño núcleo supraquiasmático y neuronas periventriculares mediales al núcleo periventricular que se funden con él. En la región tuberal, el grupo de células periventriculares se extiende alrededor de la base del III ventrículo para formar el núcleo arcuato sobre la eminencia media. En la región posterior, la estrecha zona periventricular se continúa hacia los lados con área hipotalámica posterior, y por detrás de la con la sustancia gris periacueductal mesencefálica.
Zona intermedia. Esta zona contiene los núcleos paraventricular y supraóptico, los grupos nucleares intermedios, los núcleos ventromedial y dorsomedial, y el tubérculo mamilar y los núcleos tuberomamilares.
Zona lateral. En la región tuberal, los núcleos laterales del tuber están bien definidos en el hombre. Los núcleos preóptico, anterior, dorsomedial y ventromedial comprenden todos ellos neuronas de tamaño pequeño o medio, mientras que el hipotálamo posterior se componen de escasas neuronas grandes diseminadas entre otras más pequeñas.
VIII.- Anatomía Externa de los Hemisferios
En los vertebrados más primitivos ambos hemisferios participan predominantemente en el procesamiento de señales olfatorias, que llegan en sentido rostral al lóbulo olfatorio. Dicho lóbulo se alarga formando un bulbo olfatorio, conectado a los hemisferios a través del tracto olfatorio. En la base de cada hemisferio, unas masas de sustancia gris, los núcleos basales, forman centros motores precoces. La pared del hemisferio es el palio, donde presumiblemente se integran la información olfatoria y otros tipos de información. Durante la evolución visual, las vías auditivas, entre otras, se han extendido a través del tálamo hasta el palio cerebral, lo que constituye un ejemplo de encefalización. Así pues, cada hemisferio se ve agrandado por un neopalio adicional. La pared medial del hemisferio se especializa convirtiéndose en la formación hipocámpica, considerada durante mucho tiempo como primariamente olfatoria, concepto insostenible en la actualidad.
En los mamíferos superiores el neopalio está muy agrandado y el
lóbulo piriforme aparece relativamente reducido; las vías motoras neopaliales se desarrollan desde él, pero los ganglios basales siguen siendo partes esenciales del control motor. La expansión del neopalio de los mamíferos se debe en gran parte al crecimiento de áreas de asociación relacionadas con la interacción entre conexiones aferentes y eferentes. La formación hipocámpica se denomina con frecuencia arquepalio o corteza primaria, y el lóbulo piriforme se conoce como paleopalio o corteza antigua; sin embargo, algunos autores agrupan ambas estructuras bajo el nombre de arqueopalio.

Pontificia Universidad Católica de Valparaíso Principios Neuroanatomía Funcional
El telencéfalo comprende:
Los hemisferios cerebrales, sus comisuras y cavidades. La parte anterior del III ventrículo
Cada hemisferio cerebral tiene un estrato externo de neuronas, la corteza o palio, una masa interna de prolongaciones neurales, los núcleos basales de situación profunda y un ventrículo lateral. Los hemisferios constituyen la parte de mayor tamaño del cerebro y, vistos desde arriba, tienen forma oval, más anchos por detrás que por delante, su diámetro mayor transverso se sitúa en la línea media que une las dos eminencias parietales. Los hemisferios están separados incompletamente por una profunda hendidura media, el surco interhemisférico, y cada hemisferio contiene un ventrículo lateral.
La anatomía externa de los hemisferios cerebrales se describe en la
guía 1 de laboratorio.

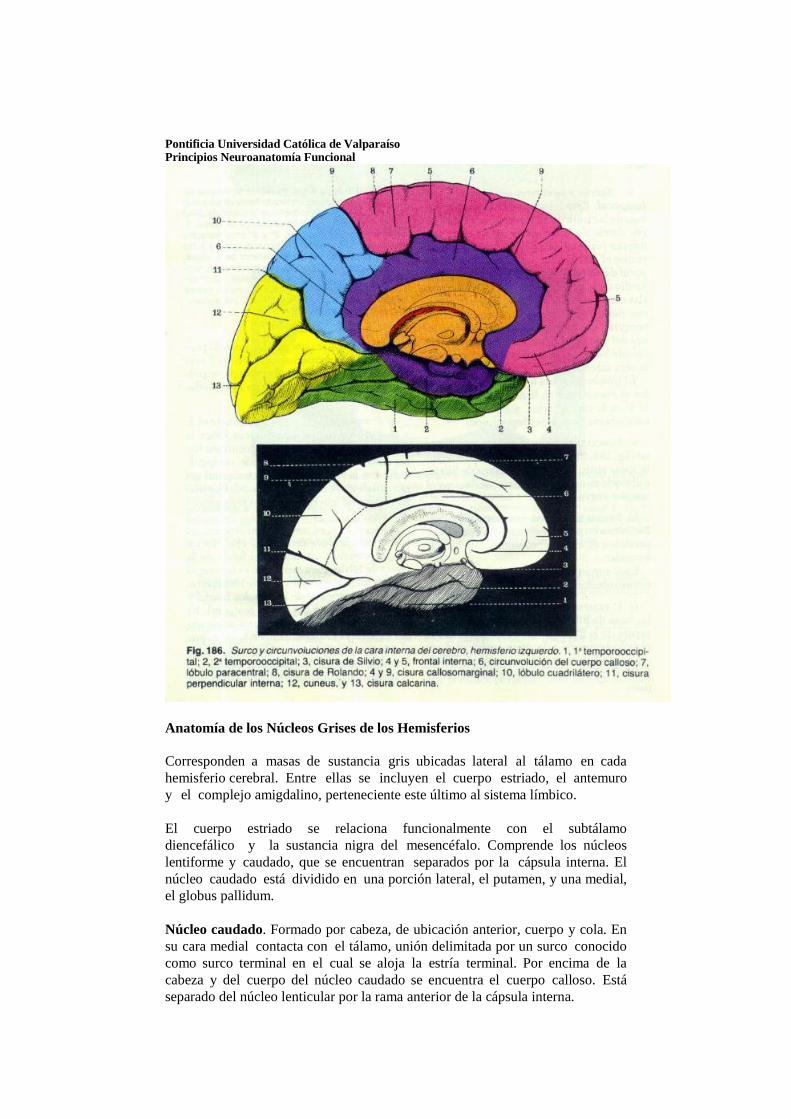
Pontificia Universidad Católica de Valparaíso Principios Neuroanatomía Funcional
Anatomía de los Núcleos Grises de los Hemisferios
Corresponden a masas de sustancia gris ubicadas lateral al tálamo en cada hemisferio cerebral. Entre ellas se incluyen el cuerpo estriado, el antemuro y el complejo amigdalino, perteneciente este último al sistema límbico.
El cuerpo estriado se relaciona funcionalmente con el subtálamo diencefálico y la sustancia nigra del mesencéfalo. Comprende los núcleos lentiforme y caudado, que se encuentran separados por la cápsula interna. El núcleo caudado está dividido en una porción lateral, el putamen, y una medial, el globus pallidum.
Núcleo caudado. Formado por cabeza, de ubicación anterior, cuerpo y cola. En su cara medial contacta con el tálamo, unión delimitada por un surco conocido como surco terminal en el cual se aloja la estría terminal. Por encima de la cabeza y del cuerpo del núcleo caudado se encuentra el cuerpo calloso. Está separado del núcleo lenticular por la rama anterior de la cápsula interna.

Pontificia Universidad Católica de Valparaíso Principios Neuroanatomía Funcional
Núcleo lenticular. Situado bajo la corteza de la ínsula, de la que se encuentra separado por la sustancia blanca y por la sustancia gris del antemuro. El antemuro o claustro divide la sustancia blanca subcortical insular para crear las cápsulas extrema y externa del núcleo lenticular. El putamen, en posición lateral, está separado del globus pallidus por una lámina de axones mielínicos, conocida como lámina medular externa.

Pontificia Universidad Católica de Valparaíso Principios Neuroanatomía Funcional
IX.- Anatomía de la Corteza Cerebral
Desde la etapa prenatal y a lo largo de la vida existen fenómenos persistentes de remodelación neuronal. Durante los primeros meses de gestación se forman las seis capas fundamentales de la corteza cerebral, proceso que depende en gran medida de la migración de las neuronas a lo largo de prolongaciones citoplasmáticas provenientes de las células de Cajal-Retzius, glía radial y migración tangencial por substrato.

Pontificia Universidad Católica de Valparaíso Principios Neuroanatomía Funcional
En etapas posteriores los axones son guiados hacia sus blancos
siguiendo los gradientes de concentración de diversas sustancias químicas presentes en el microambiente tisular desarrollado por la neuroglía.
Después del nacimiento se forman circuitos sinápticos maduros a través de mecanismos complejos que involucran la potenciación a largo plazo. Tales fenómenos permiten la adquisición de destrezas y el almacenamiento de memoria, y constituyen las bases moleculares del aprendizaje.
Organización Laminar de la Corteza Cerebral
La corteza cerebral tiene una organización de 6 capas celulares características en sus linderos neopaleales y se conocen como neocorteza. La alocorteza que tiene un menor número de capas celulares esta constituida por el paleocortex (corteza olfatoria) y el arquicortex (hipocampo y giro dentado del hipocampo).
La corteza cerebral es la capa más externa de la substancia gris que
cubre a los giros y a los surcos de los hemisferios cerebrales. Su superficie es de 2,200 centímetros cuadrados, con un espesor promedio de 2.5 mm. que va desde los 4.5 mm en la circunvolución pre-central (área premotora), a un espesor de 1.5 mm en el piso de la cisura calcarina. El grosor de la corteza es mayor siempre en la cima del giro y menor en el piso del surco.
Existen entre 14 y 20 billones de neuronas entre las que se encuentran
las piramidales, las estrelladas, las fusiformes, y se reparten como neuronas aferentes, eferentes y de asociación. La mayoría de las células corticales tienen función asociativa.
Células de la Corteza Cerebral
Células piramidales. Llevan ese nombre por su forma. La mayoría tienen un diámetro de 10 a 50 mm pero también hay células piramidales gigantes conocidas como células de Betz cuyo diámetro puede ser hasta de 120 mm. Se encuentran en la giro precentral motor. Los vértices están orientados hacia la superficie pial de la corteza. Una gruesa dendrita va hasta la piamadre y emite ramas colaterales. Las neuritas poseen espinas dendríticas para las sinapsis con otras neuronas. El axón nace de la base del cuerpo celular y termina en las capas más profundas o entra en la sustancia blanca como fibra de proyección, asociación o comisural.

Pontificia Universidad Católica de Valparaíso Principios Neuroanatomía Funcional Células estrelladas o granulosas. Son pequeñas, 8 mm y tienen forma poligonal. Poseen múltiples dendritas y un axón relativamente corto que termina en una neurona cercana.
Células fusiformes. Tienen su eje longitudinal vertical a la superficie y están concentrados principalmente en las capas corticales más profundas. Las dendritas se originan en cada polo del cuerpo celular, mientras que la dendrita superior asciende hacia la superficie de la corteza y se ramifica en las capas superficiales. El axón se origina en la parte inferior del cuerpo celular y entra en la sustancia blanca como fibra de proyección, asociación o comisural.
Células horizontales de Cajal. Son pequeñas células fusiformes orientadas horizontalmente que se hallan en las capas más superficiales de la corteza. Se origina una dendrita a cada lado del axón que corre paralelamente a la superficie de la corteza haciendo contacto con las dendritas de las células piramidales.
Células de Martinotti. Son pequeñas células multiformes presentes en todos los niveles de la corteza. La célula tiene dendritas cortas pero el axón se dirige hacia la piamadre de la corteza, donde termina en una capa más superficial, en general, en la más superficial.
Las capas o láminas de la corteza se dividen por densidad y disposición de las células en:
Capa molecular (capa plexiforme). Es la más superficial. Consiste en una red densa de fibras nerviosas orientadas tangencialmente. Estas derivan de dendritas de células piramidales y fusiformes, los axones de células estrelladas y de Martinotti. También hay fibras aferentes que se originan en el tálamo, de asociación y comisurales. Entre las fibras nerviosas hay algunas células de Cajal. Por ser la capa más superficial se establecen gran cantidad de sinapsis entre diferentes neuronas.
Capa granular externa. Contiene un gran número de pequeñas células piramidales y estrelladas. Las dendritas de estas células terminan en la capa molecular y los axones entran en las capas más profundas.
Capa piramidal externa. Esta capa está compuesta por células piramidales. Su tamaño aumenta desde el límite superficial hasta el límite más profundo. Las dendritas pasan hasta la capa molecular y los axones hasta la sustancia blanca como fibras de proyección, asociación o comisurales.
Capa granular interna. Esta capa está compuesta por células estrelladas dispuestas en forma muy compacta. Hay una gran concentración de fibras dispuestas horizontalmente conocidas en conjunto como la banda externa de Baillarger.

Pontificia Universidad Católica de Valparaíso Principios Neuroanatomía Funcional
Capa ganglionar (capa piramidal interna). Esta capa contiene células piramidales muy grandes y de tamaño mediano. Entre las células piramidales hay células estrelladas y de Martinotti. Además hay un gran número de fibras dispuestas horizontalmente que forman la banda interna de Baillarger. En las zonas motoras del giro precentral, las células de proyección de Betz dan origen aproximadamente al 3% de las fibras de proyección del haz corticoespinal.
Capa multiforme (capa de células polimórficas). Aunque la mayoría de las células son fusiformes, muchas son células piramidales modificadas cuyo cuerpo celular es triangular u ovoide. Las células de Martinotti también son conspicuas en esta capa. Hay muchas fibras nerviosas que entran en la sustancia blanca subyacente.
Organización Columnar de la Corteza Cerebral
Numerosos estudios han demostrado que la corteza funciona en verdaderos módulos de columnas verticales que analizan aspectos diversos de la información que llega a la corteza. Esta organización columnar vertical es difícil de visualizar morfológicamente.
Un estudio que combina los registros neurofisiológicos (microelectrodos) con la histología de la corteza cerebral, sugiere que la corteza esta organizada en unidades verticales de actividad funcional. A continuación se adjunta un artículo de lectura obligatoria sobre la morfofisiología de las columnas corticales.

Pontificia Universidad Católica de Valparaíso Principios Neuroanatomía Funcional Hubel y Wiesel estudiando la vía visual al igual que Mountcastle en las regiones parietales comprobaron que la corteza cerebral está organizada de una forma curiosa. Existen columnas de células que tienen todas ellas la misma función. Así existen columnas que solo responden a cambios de orientación del estímulo visual. Otras columnas son de binocularidad. Otras son de visión de colores o de identificación del objeto visual. Estas columnas están conectadas entre si y funcionan como microunidades. Cuando se integran el conjunto de información de las diferentes columnas uno extrae la información global de lo que está viendo. Parece que toda la corteza funciona de esta forma, y los impulsos sensitivos o auditivos son integrados de la misma forma.
Fibras de los Hemisferios Cerebrales
Fibras de asociación
Las fibras de asociación corresponden a axones de neuronas que proyectan a otras áreas corticales tanto ipsi como contralateralmente. Algunas de estas fibras conectan circunvoluciones vecinas, o áreas adyacentes de la misma circunvolución. Estas fibras cortas de asociación pueden proyectarse sin salir de la corteza (son intracorticales), otras de hecho salen a la sustancia blanca (sub corticales) para luego llegar al sitio de destino. Existen también largos fascículos de asociación para conectar áreas mas lejanas. Entre éstos estan los conocidos fascículos : longitudinal superior , arqueado, occipitofrontal inferior, longitudinal inferior , uncinado, occipital verticales. Existe otro grupo de fibras de asociación que conectan ambos hemisferios recíprocamente. Estas son las fibras comisurales, entre ellas tenemos aquellas que forman el cuerpo calloso, el fórnix, y la comisura blanca anterior.

Pontificia Universidad Católica de Valparaíso Principios Neuroanatomía Funcional
Fibras de proyección
Estas fibras incluyen tanto fibras que se originan en la corteza cerebral y que se dirigen a estructuras subcorticales, como aquellas que se originan en estructuras sub corticales y terminan en la corteza cerebral.
Las fibras de proyección corticofugas se dirigen al tálamo, cuerpo estriado, hipotálamo, sustancia negra, núcleo rojo, formación reticular, colículos, núcleos motores del tronco encefálico y médula espinal. Estas fibras de proyección transcurren por la cápsula interna, la cápsula externa y los pedúnculos cerebrales.
Cápsula interna
Es la principal estructura de sustancia blanca del cerebro, que contiene las fibras de proyección. Tiene la forma de un abanico el que en un corte horizontal muestra claramente las regiones que se distinguen en ella, estas son la extremidad anterior, la rodilla y la extremidad posterior.
La extremidad anterior contiene las fibras del lóbulo frontal hacia: a) tálamo (núcleo dorso-mediano) y viceversa, b) desde corteza cingulada a núcleo anterior del tálamo y viceversa, c) núcleos pontinos, d) hipotálamo, e) cuerpo estriado (cabeza n. caudado).
La rodilla de la cápsula interna contiene las fibras: a) córtico-nucleares, b ) algunas fibras córtico-reticulares y c) fibras tálamo-corticales hacia corteza motora y premotora.
La extremidad posterior de la cápsula interna contiene: a) via córtico espinal, ordenada somatotópicamente (miembro superior representado en la zona anterior, tronco en la zona media y miembro inferior representado en la zona posterior), b) vía córtico rubral, c) la radiación talámica superior (hacia la corteza parietal), d) radiación talámica posterior que compromete al núcleo pulvinar. Por último las porciones e) retrolenticular y f) sublenticular de la cápsula interna proyectan hacia las cortezas visual primaria y auditiva primaria respectivamente.
Estudio Citoarquitectónico de la Corteza
Aunque la corteza cerebral no tiene una estructura uniforme, ha sido factible dividirla en diferentes áreas por su espesor y características celulares. Campbell en 1905 describió 22 áreas corticales. Brodman en 1909 las aumentó a 47 y que por su configuración siguen vigentes sobre todo para la relación entre estructura y función. Von Economo en 1929 delimita 109 áreas. Vogt en 1919 parceló el cerebro en 200 áreas.
Economo y Koskinas(1925) describen cinco tipos distintos de corteza arquitectural cerebral en orden a su constitución, a saber:

El piramidal agranular (1) la corteza es gruesa con seis capas bien
diferenciadas, células granulares en las capas II y IV y células piramidales en las capas III y V.
El polar parietal (3) que tiene más células granulares y células piramidales más pequeñas.
El polar frontal (2) y granular (4) de corteza delgada muy bien estratificada, con mayor número de células piramidales en el polar frontal que en el granular.
El coniocortex o polar (5) en el que las células granulosas se desarrollan hasta en las capas de células piramidales.
Brodmann (1868-1918) descubrió que la corteza cerebral humana esta estructurada siguiendo los mismos principios generales que en los demás mamíferos y en 1909 publicó su celebre obra “Vergleichende Lokalisationslehre der Grosshirnrinde in ihren Prinzipien dargestellt auf Grund des Zellenbaues” que trata sobre la localización de las distintas áreas cerebrales, de acuerdo a sus estructuras citoarquitectónicas. Esta obra establece la división de la corteza en 51 áreas y estudia su localización comparativa en diferentes mamíferos. De plena vigencia, constituye la localización topográfica por excelencia cuando nos referimos a un área cortical en cualquier especie animal.
La Corteza Cerebral está colonizada por multitud de poblaciones neuronales distintas; sin embargo, estas poblaciones se pueden reunir en dos grandes grupos, interneuronas o células de proyección intracortical y células piramidales (dada la morfología de su cuerpo celular) de proyección a larga distancia. Esta diversidad celular sirvió a Brodmann de base para subdividir esta estructura en seis capas horizontales contadas desde la superficie pial (en íntimo contacto con la piamadre) a la sustancia blanca (porción interna de la Corteza Cerebral por donde transcurren los axones que entran y salen de esta estructura).
Cada una de estas capas posee una especialización, de tal modo que podemos clasificarlas utilizando diversos parámetros. Por ejemplo, morfología neuronal predominante, estructuras sobre las que proyectan o bien proyecciones subcorticales que reciben. De este modo, la capa cuatro es importante por recibir información proveniente del tálamo y estar colonizada por neuronas de pequeño tamaño (capa granular). Las capas superiores (capas supragranulares) poseen células de pequeño y mediano tamaño cuya principal función es conectar distintas áreas corticales homo y contralaterales. Las capas inferiores o infragranulares poseen neuronas de tamaño mediano y grande que proyectan principalmente a estructuras subcorticales. Así, mientras la capa seis va a proyectar básicamente al tálamo, la capa cinco va a dar lugar a proyecciones a estructuras colocadas en el cerebro medio, posterior y médula espinal.


Pontificia Universidad Católica de Valparaíso Principios Neuroanatomía Funcional

Pontificia Universidad Católica de Valparaíso Principios Neuroanatomía Funcional
X.- Localizaciones Funcionales en Corteza
La neurociencia intenta establecer una correlación entre función y estructura a distintos niveles. En este capítulo nos interesaremos por la confección de una cartografía a diferentes niveles, incluyendo sistemas como las redes corticales y sus conexiones. Es difícil y arbitrario, en el mejor de los casos, dar la definición de qué es una función. Los clínicos, los fisiólogos y los psicólogos tienen diferentes conceptos sobre el mismo comportamiento
Ciertas formulaciones psicológicas pueden no ser apropiadas para
describir la función cerebral o las conexiones reales. Por otra parte, la anatomía y la fisiología por sí solas puede que no proporcionen incluso las preguntas, y mucho menos las respuestas, sobre el comportamiento. Los avances recientes en la confección de modelos cognitivos representan un paso importante en la interpretación de los déficits funcionales. Estos modelos, a menudo representados mediante diagramas de cajas y flechas, se basan en los principios del procesamiento de la información y en la metáfora del ordenador. Sin embargo, el fraccionamiento o las distintas fases de la función presentan muchos problemas metodológicos y conceptuales. Descomponer un comportamiento complejo en sus componentes conlleva el riesgo de perder el significado y la significación biológica del comportamiento para el organismo. Desde el punto de vista de localización, es probable que los comportamientos más complejos, tales como la comprensión, tengan un input amplio y se vean afectados por lesiones en muchas áreas. Las funciones fraccionadas minuciosamente, por ejemplo "buffer de memoria a corto plazo", son, con frecuencia, el componente de muchos comportamientos complejos e, incluso, es menos probable que puedan ser localizados, si es que existe dicha posibilidad.
El fraccionamiento de la función implica, a menudo, que tenga
lugar una activación en serie de los procesos. Sin embargo, estudios fisiológicos y la confección de modelos informáticos recientes han dado énfasis al hecho que los procesos mentales operen tanto en paralelo como en serie. (Hinton, Anderson, 1981; McClelland, Rumelhart, 1986).
Existen unidades discretas de cognición que tienen,
presumiblemente, equivalentes similares en una organización neural, pero hasta el momento hay pocas pruebas de que la mayor parte del fraccionamiento de las funciones mediante modelos cognitivos tenga su correspondiente organización fisiológica. Se ha expresado la duda referente a si el mundo humano de la percepción y del procesamiento del input puede ser analizado en elementos independientes. A menudo se ha probado el fracaso de los intentos de segregar la percepción del procesamiento central. La expectativa y el conocimiento previo, en otras palabras el precepto, influyen mucho en cómo se perciben los estímulos. El procesamiento "de arriba hacia abajo" o la influencia de la memoria y las asociaciones en el momento de la percepción continúan siendo más válidos que lo que proporcionan muchos modelos funcionales modulares.

Algunas funciones complejas comunes, como la evocación de palabras,
pueden estar distribuidas de forma difusa por el área del lenguaje, ya que parecen verse afectadas por lesiones localizadas en distintas regiones. Ello puede atribuirse, probablemente, a las múltiples maneras en las que las palabras están relacionadas con los objetos que representan. Por ejemplo, la palabra "manzana" evoca color, forma, sabor, textura y el concepto de Adán y Eva comiéndola en el pecado original. La amplia distribución de ciertas funciones cognitivas, como la atención dirigida y la memoria, es biológicamente predecible, si consideramos sus conexiones con muchos otros procesos. Abastecen a otros procesos cognitivos y forman el núcleo de la cognición.
Otras funciones integradoras, como la resolución de problemas, la
inteligencia o el juicio, desafían los esfuerzos destinados a localizarlas. Algunas de estas funciones cognitivas aún continúan esperando definiciones ampliamente consensuadas. Las grandes redes neuronales son necesarias para que puedan llevarse a cabo las tareas cognitivas complejas, las cuales no es probable que estén restringidas a las circunvoluciones o lóbulos cerebrales.
En el pasado, los mapas corticales eran anatómicos, basados
en la citoarquitectura, como el conocido mapa de Broadmann de 1909. Éste indicó las fronteras de la citoarquitectura usando distintos gráficos, pero en realidad hay relativamente poco consenso entre los investigadores acerca del número de áreas que pueden distinguirse con seguridad. La confección del mapa arquitectónico completo de un cerebro es un proceso muy difícil y que requiere mucho tiempo y puede que existan más variaciones individuales de las que se aprecian en las pocas muestras disponibles en cada estudio.
Además, existen varias tinciones y técnicas que dan como resultado
mapas diferentes. Un ejemplo de ello es la arquitectura de la mielina de Flechsig. La técnica reciente más usada son los mapas arquitectónicos pigmentados de Braak. Se considera que son útiles para distinguir las áreas de importancia funcional en la corteza temporoparietal. El futuro augura nuevos mapas con tinciones funcionales, como la citocromo oxidasa y los marcadores de neurotransmisores. Sin embargo, actualmente, la relación entre los mapas citoarquitectónicos y la función sólo es provisional, con la excepción de las áreas corticales primarias, como las cortezas estriadas y piramidales. Los mapas funcionales han evolucionado a través de varias técnicas fisiológicas, desde la ablación cortical, pasando por la degeneración retrógrada, la estimulación cortical hasta la activación funcional. Uno de los mejores ejemplos de un mapa funcional es el de la corteza del lenguaje de Penfield que procede de la estimulación cortical. Algunas de las pruebas recientes procedentes de experimentos fisiológicos indican una gran cantidad de plasticidad de los mapas corticales, lo cual modifica el concepto de correspondencia exacta de la función y las áreas corticales.

La corteza del lenguaje es anatómicamente contigua desde el área de
Wernicke hasta el área de Broca, pasando por el opérculo temporal, la ínsula y el opérculo frontal. Los anatomistas también han demostrado la contigüidad de la corteza del lenguaje, la cual se hace evidente cuando se despliega la cisura de Silvio (Krieg, 1963).
La organización de la corteza del lenguaje se conceptualiza más como
una red dinámica que como una conexión de centros fijos. Los modelos de inteligencia computacional y artificial modernos proporcionaron términos como proyecciones "feed-forward" y "feed-back" y la operación de redes en paralelo, en serie y de manera recurrente. Algunos de los estudios de la estimulación de Ojemann sugieren más una organización más concéntrica que el modelo clásico conexionista posterior anterior. La percepción y el ensamblaje fonémicos ocupan las regiones perisilvianas centrales, mientras que la memoria y la selección léxica tienen lugar de una forma más periférica en las cortezas
de asociación. De una manera general, este modelo también es compatible con los estudios de localización de una lesión.
La localización de una lesión ha sido una de las técnicas más
importantes de correlación entre la estructura y la función. La observación de que las lesiones ubicadas en el mismo lugar se asociaban a déficit similares o a un mismo comportamiento alentó a los clínicos y a los anatomistas a extraer conclusiones de las lesiones acerca de la manera cómo funciona el cerebro.
Los principios generales desarrollados por muchos investigadores
en las correlaciones anatómicas de las funciones cognitivas complejas pueden resumirse de la siguiente manera:
1.- Los componentes de una única función simple están representados dentro de múltiples zonas interconectadas, que colectivamente constituyen una red integrada para esa función.
2.- Ciertas áreas corticales contienen el substrato neural para los componentes de varias funciones complejas y pueden, por lo tanto, pertenecer a varias redes que se solapan parcialmente.
3.- Es probable que las lesiones que afectan a cualquier región cortical causen múltiples déficit.
4.- La misma función puede verse dañada como consecuencia de una lesión en una de las varias áreas corticales, cada una de las cuales es un componente de una red integrada para esa función.
5.- Por regla general, tiene lugar una compensación (restablecimiento del déficit) sólo si en la lesión están involucrados uno o pocos componentes corticales.

6.- Los daños graves y permanentes que afectan sólo a una función compleja requieren, normalmente, que varios o todos los componentes de la red relevante se involucren de forma simultánea. Estos principios son lo suficientemente generales para explicar muchos de los hallazgos en los estudios sobre la localización de las lesiones y también pueden aplicarse a los estudios sobre estimulación.
Durante un siglo, la correlación clinicoanatómica ha sido el patrón oro
de la localización de las lesiones cerebrales. Sin embargo, sus puntos débiles eran la variabilidad de las descripciones del déficit y la diferencia de tiempo entre los datos clínicos y los de la autopsia. Antes, los pacientes a los que se les practicaba una autopsia tenían, con frecuencia, déficit cognitivos poco documentados y un gran intervalo entre los resultados anatómicos y clínicos.
Los métodos modernos de localización de las lesiones in vivo, en
especial la visualización por medio de la TAC y la RM, permitieron aplicar pruebas estandarizadas a un grupo de pacientes similares en un estadio bien definido de un accidente vascular, con una localización de la lesión in vivo simultánea. La descripción del déficit, influenciada por la neuropsicología cognitiva, ha evolucionado notablemente.
Cada caso requiere una gran cantidad de pruebas cognitivas complejas, lo cual dificulta la estandarización, sumando un gran número de casos clínicos que son difíciles de integrar. Las definiciones y las descripciones de función se continúan modificando. El análisis de lesiones realizado in vivo también se ha convertido en complejo con la evolución de nueva tecnología que ofrece cada vez más resolución anatómica y que es sensible a las alteraciones in vivo de la estructura y la química. Actualmente, se pueden medir las alteraciones en el flujo sanguíneo cerebral mediante varias técnicas tales como el xenón, el SPECT, el PET y la RMf. Estos cambios representan, en realidad, modificaciones fisiológicas relacionadas con la función y ello da lugar a la imagen funcional. Desde que la imagen funcional ha adquirido importancia, el método lesional se ha dejado en un segundo término hasta cierto punto, incluso algunos lo consideran obsoleto. Sin embargo, se ha avanzado sustancialmente tanto en la metodología como en el conocimiento e intentaré resumir estos avances, especialmente en el ámbito del lenguaje del procesamiento y visuoespacial y discutir sobre los estudios de RM que ofrecen una alta resolución anatómica.

Visión general de la función de cada lobo
El lobo frontal está situado en la porción anterior del cerebro, separado
por el surco central del lobo parietal, y por el surco lateral del lobo temporal. La porción postcentral es la que lleva en sí el homúnculo de Penfield y
el córtex motor. La porción anterior que se denomina región prefrontal tiene una gran importancia en el hombre y constituye una de las principales áreas asociativas. Durante años se conoció como el cerebro mudo pero más adelante se verá su enorme importancia.
El lobo parietal, es el lobo peor demarcado en comparación al resto.
Detrás del surco central y superior al surco central no tiene límites con lobos temporal y occipital. Se consideró la corteza parietal como un área silente, pero ahora no hay duda para concluir que la corteza parietal anterior contiene el mecanismo para la percepción táctil. Pero la parte más importante de las funciones del lobo parietal componen un centro para la integración de la información somatosensorial, con la auditiva y la visual, con el fin de construir lo que se denomina el esquema corporal y su relación con el espacio exterior. Las actividades construccionales implican fundamentalmente el lobo parietal izquierdo. La comprensión de los aspectos gramaticales y sintácticos del lenguaje es una función del lobo parietal dominante. En cambio el reconocimiento y utilización de números, principios aritméticos y cálculo se sitúa en el lobo parietal no dominante. Las alteraciones más curiosas del lobo parietal son las agnosias, que constituyen la incapacidad para reconocer determinados aspectos de la visión, la audición o la percepción corporal.
El lobo temporal se separa del frontal por el gran surco lateral. En él se
sitúa el lenguaje auditivo, la memoria, y por sus grandes conexiones con el sistema límbico las funciones emocionales.
Los lobos occipitales son el término de las vías geniculocalcarinas (del
cuerpo geniculado a la corteza visual) y son esenciales para la percepción y reconocimiento visual.
Lobo de la ínsula, situado en la profundidad del surco lateral, posee una función vegetativa, controla funciones autonómicas como la temperatura, la tensión arterial, la frecuencia cardiaca, el grado de dilatación de los vasos sanguíneos que va a guardar relación con la expresión de las emociones en la cara.
Sistema límbico es el gran cerebro asociativo que controla la memoria
y las emociones, es un haz oval que conecta entre sí las distintos giros y lobos.

Áreas Funcionales Corticales
La organización de las conexiones del encéfalo permite que múltiples
impulsos excitatorios e inhibitorios sean integrados en una única experiencia mental. Cuando uno experimenta un objeto del entorno, por ejemplo, la imagen de un café de noche. Existe una imagen con visual con el color, forma de las mesas y yuxtaposición de las figuras, se capta el movimiento de las personas y el ruido de los coches.
El complejo de olores de las cervezas en las mesas, y la sensación de frío
o calor ambiental. No sólo hay estas percepciones inmediatas sin un surtido de asociaciones de recuerdos que esta imagen suscita.
De alguna forma, nosotros integramos todas estas múltiples aferencias
en una experiencia con sentido y nos movemos hacia la mesa para saludar a la persona sentada o para pedir algo de comer porque estamos con apetito.
Cada aspecto del entorno excita distintas poblaciones de neuronas en
diferentes áreas corticales del encéfalo, de este modo el cerebro organiza en una experiencia mental única y controla la ejecución de la acción apropiada. Para comprender cómo tiene lugar la integración se debe tener una apreciación de las vías neuroanatómicas a través de las cuáles se procesa la información.
En las cortezas de asociación visuales más primarias (área 18 y 19) se
aprecian el color básico, la forma, la profundidad y la textura. A medida que los impulsos excitan otras áreas de asociación de orden superior más anteriores, en las vías de conexión hacia el lóbulo temporal y la región hipocampal (áreas 37, 20 y 21), se empieza a atribuir significado a la percepción visual.
Las lesiones en la región occipitotemporal pueden causar una agnosia
visual, en la que el individuo puede ser capaz de ver un objeto pero no reconocerlo. Por ejemplo, uno puede no ser capaz de distinguir visualmente un cerdo de un perro, o a menudo ser incapaz de reconocer visualmente la identidad específica de su propia cara en el espejo.
Cada modalidad sensitiva tiene un área cortical primaria y un
sistema de conexiones para el procesamiento sensorial. Una característica importante de las cortezas primarias visual, auditiva y somatosensorial es su organización topográfica. En el sistema somatosensorial, el córtex primario está somatotópicamente organizado, de tal forma que partes específicas de la corteza cerebral reciben las aferencias de partes concretas del organismo, mientras que el córtex calcarino está retinotópicamente organizado, y el córtex auditivo está tonotópicamente organizado.

Cada región sensitiva primaria tiene conexiones con áreas de asociación
de modalidad específica, donde tiene lugar la convergencia e integración de diferentes atributos de la experiencia sensorial. En puntos sucesivos, los axones de diferentes áreas sensitivas de asociación de modalidad específica, empiezan a converger en lo que se denominan áreas de asociación multimodal. En estas áreas se ha demostrado que existen neuronas, que por ejemplo, se activan en respuesta al estímulo visual, encontrándose entremezcladas con neuronas que responden a estímulos auditivos, y con neuronas que responden a estímulos sensitivos múltiples.
Las áreas de asociación multimodal son tres:
El sistema límbico. El área temporoparietal que incluye el lobo temporoparietal inferior y
las zonas laterales del surco temporal superior. El área prefrontal que incluye vastas expansiones del córtex por delante
de las cortezas motoras del lobo frontal.
Área Frontal
Area Motora Primaria: se extiende sobre le limite superior del lobulillo paracentral. Si se estimula produce movimientos aislados en el lado opuesto del cuerpo y contracción de grupos musculares relacionados con la ejecución de un movimiento específico. Las áreas del cuerpo están representadas en forma invertida en el giro precentral (homúnculo). Comenzando desde abajo hacia arriba: deglución, lengua, maxilares, labios, laringe, párpado y cejas, dedos, manos, muñeca, codo, hombro y tronco etc. La función del área motora primaria consiste en llevar a cabo los movimientos individuales de diferentes partes del cuerpo. Como ayuda para esta función recibe numerosas fibras aferentes desde el área premotora, la corteza sensitiva, el tálamo, el cerebelo y los ganglios basales. La corteza motora primaria no es responsable del diseño del patrón de movimiento sino la estación final para la conversión del diseño en la ejecución del movimiento.

Pontificia Universidad Católica de Valparaíso Principios Neuroanatomía Funcional
Area Premotora: no posee células gigantes de Betz. La estimulación eléctrica de esta zona produce movimientos similares a los del área motora primaria pero se necesita estimulación más intensa para producir el mismo grado de movimiento. Recibe numerosas aferencias de la corteza sensitiva, tálamo y ganglios basales. La función de ésta área es almacenar programas de actividad motora reunidos como resultado de la experiencia pasada; es decir programa la actividad motora primaria.
Area Motora Suplementaria: se ubica en el giro frontal medial y por delante del lobulillo paracentral. La estimulación de esta área da como resultado movimientos de las extremidades contralaterales pero es necesario un estímulo más fuerte que el necesario en la zona primaria. La eliminación de ésta área no produce una pérdida permanente de movimiento.
Campo Ocular Frontal: se extiende hacia delante desde el área facial del giro precentral hasta la el giro frontal medial. La estimulación de esta área produce movimientos conjuntos de los ojos en especial en el lado opuesto. Controla los movimientos de seguimiento voluntario de los ojos y es independiente de los estímulos visuales. El seguimiento involuntario ocular de los objetos en movimiento comprende el área visual en la corteza occipital que está conectada al campo visual en la corteza occipital que está conectada al campo ocular frontal por fibras de asociación.
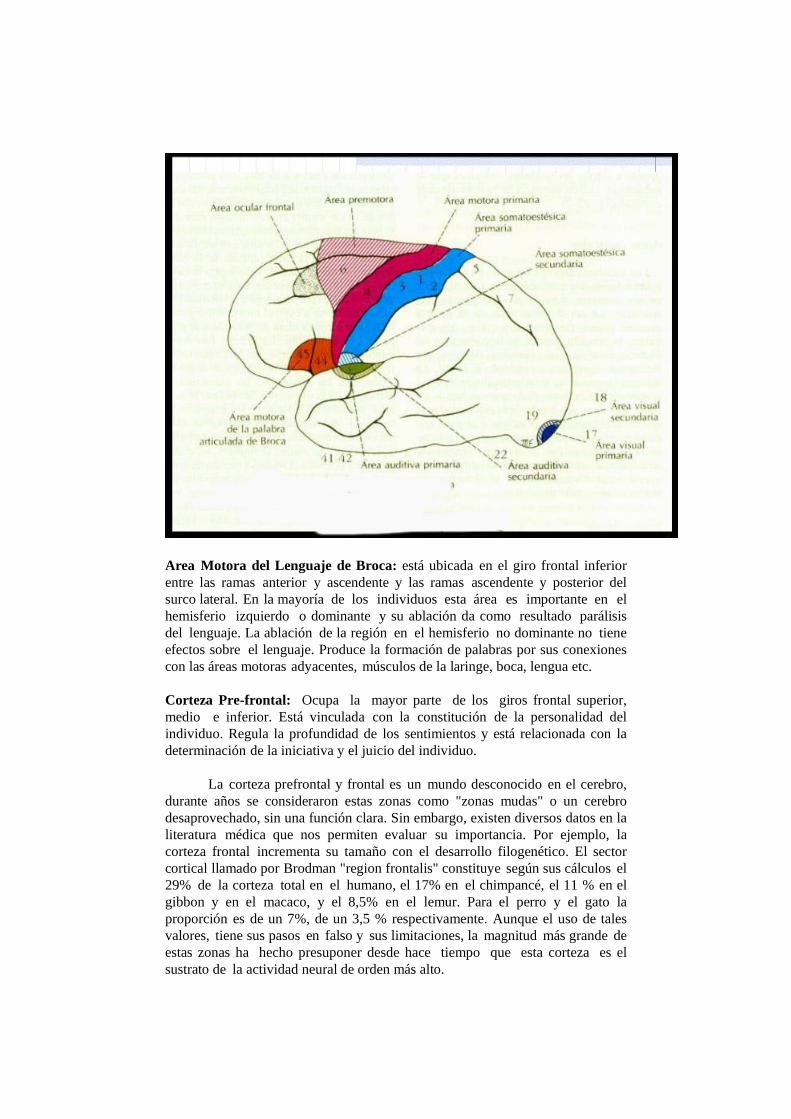
Area Motora del Lenguaje de Broca: está ubicada en el giro frontal inferior entre las ramas anterior y ascendente y las ramas ascendente y posterior del surco lateral. En la mayoría de los individuos esta área es importante en el hemisferio izquierdo o dominante y su ablación da como resultado parálisis del lenguaje. La ablación de la región en el hemisferio no dominante no tiene efectos sobre el lenguaje. Produce la formación de palabras por sus conexiones con las áreas motoras adyacentes, músculos de la laringe, boca, lengua etc.
Corteza Pre-frontal: Ocupa la mayor parte de los giros frontal superior, medio e inferior. Está vinculada con la constitución de la personalidad del individuo. Regula la profundidad de los sentimientos y está relacionada con la determinación de la iniciativa y el juicio del individuo.
La corteza prefrontal y frontal es un mundo desconocido en el cerebro,
durante años se consideraron estas zonas como "zonas mudas" o un cerebro desaprovechado, sin una función clara. Sin embargo, existen diversos datos en la literatura médica que nos permiten evaluar su importancia. Por ejemplo, la corteza frontal incrementa su tamaño con el desarrollo filogenético. El sector cortical llamado por Brodman "region frontalis" constituye según sus cálculos el 29% de la corteza total en el humano, el 17% en el chimpancé, el 11 % en el gibbon y en el macaco, y el 8,5% en el lemur. Para el perro y el gato la proporción es de un 7%, de un 3,5 % respectivamente. Aunque el uso de tales valores, tiene sus pasos en falso y sus limitaciones, la magnitud más grande de estas zonas ha hecho presuponer desde hace tiempo que esta corteza es el sustrato de la actividad neural de orden más alto.

Pontificia Universidad Católica de Valparaíso Principios Neuroanatomía Funcional
Esta noción, que puede ser esencialmente correcta, no ha sido probada.
La historia del estudio clínico del lobo frontal es abundante. Un caso citado frecuentemente en la literatura médica es el de Phineas Gage, un obrero de la construcción de ferrocarriles, quien a mediados del siglo XIX fue herido accidentalmente por una barra de hierro puntiaguda que penetró en su frente de manera oblicua y desde abajo, atravesando la base del cráneo, e infligiéndole un daño masivo en el lobo frontal izquierdo (Harlow 1848). Aparte del aspecto curioso de la herida y de la increíble supervivencia de la víctima, lo interesante del caso es la notable descripción clínica que Harlow realizó describiendo los cambios de personalidad que el paciente presentaba a consecuencia de la herida. Estos cambios eran tan importantes que, tras el accidente y a los ojos de sus amigos, Gage "ya no era Gage". De modo diferente a su conducta previa, Gage mostraba una conducta caprichosa, con pobre capacidad de planear acciones y una impulsividad incontrolada. Veinte años después, en 1868, Harlow escribió: "Un niño en su capacidad intelectual y manifestaciones, pero tiene las pasiones animales de un hombre fuerte".
La corteza frontal es una región cardinal en la convergencia de las tres
vías transcorticales que proceden de las vías visuales, auditivas y somatoestésicas primarias. En resumen, las conexiones del lobo frontal están al frente todas las conexiones sensoriales, y del sistema némonico-emocional límbico.
La función clave que representa el lobo frontal es la unificación
temporal del comportamiento con un único propósito o meta, es decir la estructuración de aquel comportamiento que implica una novedad, que es creativo, la base del comportamiento inteligente. El comportamiento innato e instintivo, que es por definición invariable y común a todas las especies, se excluye de las categorías del comportamiento estructurado hacia un fin, y por tanto, no está regulado por la región prefrontal.
Tres funciones son específicas para la región frontal, las funciones son:
memoria a corto plazo, que permite una función retrospectiva, un set o conjunto preparatorio que permite una función prospectiva o anticipatoria y una función protectora o de control de interferencias, es decir influencias internas y externas, entre las que se incluyen recuerdos y datos de memoria que interfieran con la formación de estructuras de comportamiento. Las dos primeras funciones se localizan ampliamente en la región dorsolateral, mientras que la tercera se localiza en la región orbitaria interna de la corteza frontal. Por eso, las lesiones del lóbulo frontal en humanos se caracterizan por distinta semiología clínica. Los trastornos de atención son una característica prominente de los síndromes clinicos prefrontales. Tales desórdenes son responsables parcialmente de déficits cognitivos, entre los que se incluye distorsión de percepciones. Las lesiones de la corteza prefrontal dorsolatral pueden resultar en apatía y falta de movimiento espontáneo. Las lesiones orbitarias pueden conducir a hipercinesia, euforia y desinhibición.

El resultado más común y característico del daño frontal en el humano es el fallo para organizar nuevos y deliberados esquemas de comportamiento secuencial. Este fallo se acompaña de un concretamente temporal, una cualidad de inmediatez y falta de perspectiva en tiempo. Prospectiva a la vez que retrospectiva que invade en todas las formas de comportamiento y especialmente en el lado izquierdo el lenguaje hablado. El lobo frontal, al guardar una relación tan directa con la capacidad de creación y el manejo de las novedades de conducta puede decirse que está relacionado de una manera indudable con la inteligencia, considerada como capacidad de creación. Esto no quiere decir que la inteligencia sea una función del lóbulo frontal, quiere decir únicamente que para poder realizar una actividad creativa, que en definitiva, es una actividad propiamente humana se necesita de alguna manera el funcionamiento adecuado de las estructuras frontales: la capacidad de orientar la actuación hacia una meta, y de evitar la distorsión por otras actividades que descentran la atención y la capacidad de generar un comportamiento estable.
Área Parietal
Area Somatoestésica Primaria: ocupa el giro postcentral sobre la superficie lateral del hemisferio y la parte posterior del lobulillo paracentral sobre la superficie medial. Histológicamente es un área de tipo granuloso con capa externa de Ballinger muy ancha y obvia. La mitad opuesta del cuerpo está representada de forma invertida: faringe, lengua, cara,..., dedos, mano, brazo, tronco, muslo,.., pierna, pie. La porción de una parte del cuerpo en particular se relaciona con su importancia funcional y no con su tamaño. Por ejemplo superficies grandes ocupan la mano, la cara, labios y el pulgar. Aunque la mayoría de las sensaciones llegan a la corteza desde el lado contralateral del cuerpo, algunas provenientes de la región oral van en el mismo sentido.
Area Parietal de Asociación: Ocupa el lobo parietal superior que se extiende hacia la superficie medial del hemisferio (precuña) y lobo parietal inferior. Tiene muchas conexiones con otras áreas sensitivas de la corteza. Se cree que su principal función consiste en recibir e integrar diferentes modalidades sensitivas. Por ejemplo reconocer objetos colocados en las manos sin ayuda de la vista, es decir maneja información de forma y tamaño relacionándola con experiencias pasadas.
Área Occipital
Area Visual Primaria: ubicada en las paredes de la parte posterior del surco calcarino ocasionalmente alrededor del polo occipital. Histológicamente es un área de corteza delgada, del tipo granuloso con sólo algunas células piramidales. Recibe fibras que vienen de la retina. La mácula lútea, área central de la retina (área de la visión más perfecta) está representada en la corteza en la parte posterior. Las partes periféricas de la retina están representadas por el área anterior.

Area Visual Secundaria: rodea el área visual primaria. Recibe fibras aferentes del área visual primaria y otras áreas corticales y el tálamo. La función consiste en relacionar la información visual recibida por el área visual primaria con experiencias visuales pasadas, lo que permite reconocer y apreciar lo que se está viendo. Se cree que existe un campo ocular occipital en el área visual secundaria cuya estimulación produce la desviación conjugada de los ojos cuando está siguiendo a un objeto, movimientos involuntarios que dependen de los estímulos visuales.
Área Temporal
Area Auditiva Primaria: está ubicada en la pared inferior del surco lateral. Histológicamente de tipo granuloso, es un área de asociación auditiva. La parte anterior del área auditiva primaria está vinculada con la recepción de sonidos de baja frecuencia mientras que la parte posterior con los de alta frecuencia. Una lesión unilateral produce sordera parcial en ambos oídos con mayor pérdida del lado contralateral.
Area Auditiva Secundaria: ubicada detrás del área auditiva primaria. Se cree que esta área es necesaria para la interpretación de los sonidos.
Area Sensitiva del Lenguaje de Wernicke: está ubicada en el hemisferio dominante izquierdo, principalmente, principalmente en la circunvolución temporal superior. Está conectado con el área de Broca por el haz de fibras llamado fascículo arcuato. Recibe fibras de la corteza visual (occipital) y de la corteza auditiva (temporal superior). Permite la compresión del lenguaje hablado y de la escritura, es decir que uno pueda leer una frase, comprenderla y leerla en voz alta.

Otra gran área de asociación desarrollada de modo mucho más importante en humanos que en los animales es el lóbulo temporal. Junto con el área prefrontal, ya descrita, contiene muchas funciones específicamente humanas; en él se sitúa una amplia representación de contenidos de memoria, de lenguaje y, en él se produce una relación directísima con el sistema emocional. Así, los estudios de las conexiones del neocórtex del lobo temporal, realizados en animales, nos han mostrado que esta zona del cerebro está ampliamente conectada con el resto de estructuras cerebrales, pero fundamentalmente con el lóbulo temporal más profundo y el sistema límbico, o cerebro emocional.
En el hombre, conocemos las funciones del lobo temporal humano, gracias al estudio de los pacientes epilépticos. Así, dos tercios de las epilepsias focales - es decir, con un inicio en un punto del cerebro -, tienen su origen en el lobo temporal. Se ha comprobado que las crisis epilépticas del lobo temporal se relacionan con la personal identidad. Tras estimulación eléctrica del lobo temporal se induce la aparición de experiencias previas que, en realidad, forman parte del pasado del paciente.
Entre los fenómenos afectivos que se producen en las crisis del lóbulo
temporal, la sensación de miedo es la más frecuente. Suele asociarse a una sensación visceral, generalmente a nivel del abdomen, que es muy típica. Otros fenómenos afectivos, que pueden aparecer al estimular el lóbulo temporal son: sensación de tristeza, soledad, vergüenza, culpa, enfado o ira, excitación anticipatoria, placer, excitación sexual y sensación de contento o felicidad.

En los fenómenos perceptivos a nivel temporal (generalmente visuales o auditivos) no se produce con un tiempo real, excepto la música: las escenas son estáticas. Los detalles perceptivos pueden ser fragmentarios y les falta la sensación de viveza. El paciente permanece pasivo y no siente que participa en la escena.
La formación de una experiencia depende de una red de neuronas
excitadas o inhibidas en grandes áreas de la corteza temporal y conectada con el sistema límbico, el hipocampo y la amígdala. Lo que lleva una información específica no es la actividad de una única célula, sino un patrón específico de conexiones que enlazan neuronas y crean un patrón específico de excitación e inhibición. Estos circuitos componen la memoria. Las interconecitvidad de esas redes puede ser modificada por las experiencias repetidas, que inducen cambios plásticos sinápticos como el fenómeno de la plasticidad neural, y es la base neurológica de la memoria y el aprendizaje. Todo el sistema temporo-límbico es un sistema tremendamente moldeable a los cambios plásticos sinápticos.
Otras áreas
Área del gusto: está ubicada en el extremo inferior del giro postcentral de la pared superior del surco lateral en el área adyacente de la ínsula.
Área vestibular: está situada cerca de la parte del giro postcentral vinculada con las sensaciones de la cara. Su localización opuesta al área auditiva.
Insula: está enterrada dentro del surco lateral y forma su piso. Histológicamente es granulosa.
Corteza Límbica
Las sensaciones de tipo afectivo son comunes a hombre y animales, y en ambos están situadas en la parte más interna del cerebro, a nivel de la amígdala, el hipocampo y los circuitos límbicos centroencefálicos, es el "paleocórtex" o cerebro de mamífero primitivo, que se denomina también el sistema límbico. En la amígdala se vivencian los afectos, el sistema del hipocampo, inicia los procesos de consolidación de la memoria, que como hemos ya visto guardan una relación muy importante con el neocórtex temporal. El sistema límbico es así una zona profunda situada entre ambos hemisferios cerebrales, rodeando al cuerpo calloso que es un haz de conexión entre ambos hemisferios. Es el área donde se sitúan sentimientos y emociones: el placer, el temor, la tristeza, la alegría, el enfado, la ira, el deseo sexual, etc. Todas estas sensaciones actúan sobre el neocórtex modulando su actividad.
El cerebro es una estructura formada células vivas, las neuronas, que
tienen capacidad de adaptación y cambio. Las neuronas se "comunican" entre sí por medio de conexiones que se denominan sinapsis. Una neurona típica con frecuencia recibe decenas de miles de contactos sinápticos.

Las conexiones entre neuronas dan lugar a circuitos neuronales, son
esos circuitos neuronales, los que dan lugar a la actuación del ser humano. Cambios en el número, tipo y función de las conexiones entre las neuronas son los que dan lugar a procesos tan dispares como la memoria, el aprendizaje y la reparación de funciones tras una lesión. Estos cambios son lo que se llama plasticidad neural. Pongamos algunos ejemplos de lo que es la plasticidad neural. Todos conocemos la gran capacidad que tienen los ciegos de desarrollar otros sentidos con los que valerse para realizar su vida habitual, se ha comprobado como en ellos las áreas cerebrales que corresponden a la visión, están disminuidas; mientras que las que corresponden al tacto o al oído mucho más desarrolladas. Por diversos estudios neurológicos, se conoce que en el cerebro humano está representada el área que controla el movimiento de la mano. Se ha constatado que los violinistas, virtuosos del movimiento manual, tienen mucho más desarrollada el área de la mano (hasta 5 o 10 veces más grande) que sujetos que no se dedican a la interpretación musical. Mediante técnicas de neuroimagen (fundamentalmente, la Tomografía por Emisión de Positrones, PET) se ha comprobado en sujetos obsesivos una mayor actividad en lóbulo frontal. Tras la realización de psicoterapia, sin ningún tratamiento farmacológico, y habiendo mejorado su situación mental, se pudo comprobar que estas áreas disminuyen en su actividad.
Bibliografía
CARPENTER, M. B. Fundamentos de Neuroanatomía. 4a ed. Médica Panamericana, 1994. FRIEDMANN, G.; BÜCHELER, E.; THURN, P. Tomografía Computadorizada del Cuerpo Humano. Salvat, 1986. GERHARDT, P.; FROMMHOLD, W. Atlas de Correlaciones Anatómicas en TC y RMN. Salvat, 1989. LATARJET, M.; RUIZ LIARD, A. Anatomía Humana. 2a ed. Médica Panamericana, 1990. LIPPERT, H. Anatomía. Texto y Atlas. 4ª ed. Marban, 1999. MACKINNON, P. C. B.; MORRIS, J. F. Oxford Anatomía Funcional. Médica Panamericana, 1993-94. MARTIN, J. H. Neuroanatomía. Prentice Hall. Tercera Edición. Texto y Atlas. McMINNM, R. M. H.; HUTCHINGS, R. T.; ABRAHAMS, P. H. Atlas de Anatomía Humana. 2a ed. Océano, 1991. MÖLLER, T. B.; REIF, E.; STARK, P. Atlas de Anatomía Radiológica. Marban, 1994. MOORE, K. L. Anatomía con Orientación Clínica. 3a ed. Médica Panamericana, 1993. MOORE, K. L. Embriología Clínica. 4a ed. Interamericana; McGraw-Hill, 1989. NOBACK, C.R., STROMINGER, N.L.; DEMAREST. R.J. El Sistema Nervioso. Introducción y repaso. 4ª ed. McGrawHill-Interamericana, 1993. NOLTE, J. El Cerebro Humano. Introducción a la Anatomía Funcional. 3ª ed. Mosby/Doyma, 1994- ORTS LLORCA, F. Anatomía Humana. 5a ed. Científico-Médica, 1982-83. PANSKY, B. Anatomía Humana, 6ª ed. McGrawHill-Interamericana, 1998. PUTZ, R.; PABST, R. Sobotta Atlas de Anatomía Humana. 20a ed. Médica Panamericana, 1994.

ROHEN, J. W.; YOKOCHI, C. Atlas Fotográfico de Anatomía Humana. 3a ed. Mosby; Doyma Libros, 1994. ROUVIERE, H.; DELMÁS, A. Anatomía Humana: descriptiva, topogràfica y funcional. 9a ed. Masson, 1988. SADLER, T. W. Langman Embriología Médica. 6a ed. Médica Panamericana, 1993. SNELL, R. S. Neuroanatomía Clínica. 2a ed. Médica Panamericana, 1990. TUCHMANN-DUPLESSIS, H. Embriología. Cuadernos Prácticos. 2a ed. Masson, 1984. WILLIAMS, P. L.; WARWICK, R. Anatomía de Gray. Salvat, 1992. WILSON-PAUWELS, L.; AKESSON, E. J.; STEWART, P. A. Nervios craneanos. Anatomía i Clínica. Panamericana, 1991. YOUNG, P. A. y YOUNG P. H. Neuroanatomía Clínica Funcional. Editorial: Masson, Barcelona. 2001.

















![3 ACOTACIÓN-09.ppt [Modo de compatibilidad] · ACOTACIÓN FUNCIONAL Principios generales UNE 1-039-94; (ISO 129:1973) Definiciones COTA FUNCIONAL (F)COTA FUNCIONAL (F): Ct il l f](https://static.fdocuments.es/doc/165x107/5f828e52ea91ff26d951d845/3-acotacin-09ppt-modo-de-compatibilidad-acotacin-funcional-principios-generales.jpg)
