
Productividad Secundaria, Utilización del Hábitat y ... · funciones de alta productividad y...
14
Lara-Domínguez, A. L. y A. Yáñez-Arancibia, 1999. Productividad secundaria, utilización del hábitat y estructura trófica, p. 153-166. In: A. Yáñez-Arancibia y A. L. Lara-Domínguez (eds.). Ecosistemas de Manglar en América Tropical. Instituto de Ecología A.C. México, UICN/ORMA, Costa Rica, NOAA/NMFS Silver Spring MD USA. 380 p. 11 Productividad Secundaria, Utilización del Hábitat y Estructura Trófica Ana Laura Lara-Domínguez 1* , Alejandro Yáñez-Arancibia 2 1 Centro EPOMEX, Universidad Autónoma de Campeche, México 2, Instituto de Ecología, A.C., México * dirección actual del primer autor Resumen Los organismos marinos, en el transcurso de su ciclo de vida, pasan por distintas etapas biológicas, desde huevo hasta adultos reproductores, acoplando cada etapa con características físico-ambientales que son propias para los diferentes ambientes y hábitats. Cada uno de estos momentos son críticos para el óptimo desarrollo de las distintas etapas biológicas. En escalas espaciales y temporales, esto se traduce en una clara separación de ambientes físicos acoplados con cada una de las etapas biológicas de las especies. Dentro de la complejidad de la zona costera, ésta se caracterizada por diferentes subsistemas entre los que se incluyen lagunas costeras y estuarios, bocas de conexión entre aguas protegidas y el mar, plataforma continental adyacente. En latitudes tropicales, estos subsistemas están caracterizados por una gran diversidad de ambientes entre los que se pueden mencionar a los manglares, pastos marinos y zonas palustres entre otros, los cuales son áreas idóneas de reproducción, crianza y alimentación de diferentes organismos. En éste capitulo se describe a los manglares como hábitat critico que contribuye significativamente a la producción secundaria. Se destacan dos importantes funciones ecológicas del ecosistema: 1. Como una área de protección, debido la estructura de sus raíces especializadas y adaptadas a periodos de inundación, lo cual le permite al ecosistema mantener una importante biodiversidad, y sostener poblaciones de importancia ecológica y comercial; 2. Como fuente de carbono (a través del detritus) para muchos de los organismos que residen en los manglares a través de la trama trófica. Esto hace que el ecosistema de manglar funcione como un área de alimentación para juveniles y adultos, crianza para larvas y juveniles de diferentes especies. Abstract The marine organisms, in the course of their life cycle, go through different biological stages from egg adult reproductive, coupling each stage with environmental characteristics for different ambient and habitats. Each one is critical for the optimum development of the different biological stages. In spatial and temporary scales, this is observed as a clear physical environment coupled with each one of the biological stages of the fish species. The coastal zone is a complex environment characterized by different subsystems which are coastal lagoons, estuaries, connection inlets protected waters and the sea, adjacent continental shelf. In tropical latitudes, these subsystems are characterized by a great diversity of environments such mangroves, sea grass, swamp zones. They are suitable reproduction areas, feeding and nursery areas of different organisms. In this chapter we are described the mangroves as critical habitat that contributes significantly to the secondary 153
Transcript of Productividad Secundaria, Utilización del Hábitat y ... · funciones de alta productividad y...

Lara-Domínguez, A. L. y A. Yáñez-Arancibia, 1999. Productividad secundaria, utilización del hábitat y estructura trófica, p. 153-166. In: A. Yáñez-Arancibia y A. L. Lara-Domínguez (eds.). Ecosistemas de Manglar en América Tropical. Instituto de Ecología A.C. México, UICN/ORMA, Costa Rica, NOAA/NMFS Silver Spring MD USA. 380 p. 11
Productividad Secundaria, Utilización del Hábitat
y Estructura Trófica
Ana Laura Lara-Domínguez1*, Alejandro Yáñez-Arancibia2
1 Centro EPOMEX, Universidad Autónoma de Campeche, México 2, Instituto de Ecología, A.C., México * dirección actual del primer autor
Resumen
Los organismos marinos, en el transcurso de su ciclo de vida, pasan por distintas etapas biológicas, desde huevo hasta adultos reproductores, acoplando cada etapa con características físico-ambientales que son propias para los diferentes ambientes y hábitats. Cada uno de estos momentos son críticos para el óptimo desarrollo de las distintas etapas biológicas. En escalas espaciales y temporales, esto se traduce en una clara separación de ambientes físicos acoplados con cada una de las etapas biológicas de las especies. Dentro de la complejidad de la zona costera, ésta se caracterizada por diferentes subsistemas entre los que se incluyen lagunas costeras y estuarios, bocas de conexión entre aguas protegidas y el mar, plataforma continental adyacente. En latitudes tropicales, estos subsistemas están caracterizados por una gran diversidad de ambientes entre los que se pueden mencionar a los manglares, pastos marinos y zonas
palustres entre otros, los cuales son áreas idóneas de reproducción, crianza y alimentación de diferentes organismos. En éste capitulo se describe a los manglares como hábitat critico que contribuye significativamente a la producción secundaria. Se destacan dos importantes funciones ecológicas del ecosistema: 1. Como una área de protección, debido la estructura de sus raíces especializadas y adaptadas a periodos de inundación, lo cual le permite al ecosistema mantener una importante biodiversidad, y sostener poblaciones de importancia ecológica y comercial; 2. Como fuente de carbono (a través del detritus) para muchos de los organismos que residen en los manglares a través de la trama trófica. Esto hace que el ecosistema de manglar funcione como un área de alimentación para juveniles y adultos, crianza para larvas y juveniles de diferentes especies.
Abstract
The marine organisms, in the course of their life cycle, go through different biological stages from egg adult reproductive, coupling each stage with environmental characteristics for different ambient and habitats. Each one is critical for the optimum development of the different biological stages. In spatial and temporary scales, this is observed as a clear physical environment coupled with each one of the biological stages of the fish species. The coastal zone is a complex environment characterized by
different subsystems which are coastal lagoons, estuaries, connection inlets protected waters and the sea, adjacent continental shelf. In tropical latitudes, these subsystems are characterized by a great diversity of environments such mangroves, sea grass, swamp zones. They are suitable reproduction areas, feeding and nursery areas of different organisms. In this chapter we are described the mangroves as critical habitat that contributes significantly to the secondary
153

Ecosistemas de Manglar A. L. Lara-Domínguez & A. Yáñez-Arancibia
154
production. This point of view emphasized In two Important ecological functions of the ecosystem: 1. As a protection area, due to the structure of their specialized and adapted roots to flood periods. This function permits to maintain an important biodiversity and support important commercial and ecological populations; 2. As source of carbon (detritus) for
many animal species that live in the mangroves, through the trophic chain. The above causes make that the mangrove ecosystems operate as a feeding area for juveniles and adults, as a nursery area for larvae and juveniles, protection for eggs, larvae and juveniles of different species.
Introducción
Se ha discutido ampliamente que los ecosistemas de manglar en la zona costera tropical, conforman un hábitat crítico para numerosas especies de Invertebrados (moluscos y crustáceos), y de vertebrados como aves, mamíferos y particularmente peces que utilizan sus recursos en alguna etapa de su ciclo de vida. Considerando a los ecosistemas de manglar como un hábitat particular, Day y Yáñez-Arancibia (1988) establecen, desde el punto de vista teórico, que un hábitat representa una unidad morfofuncional de un todo mayor. Es decir, es el mínimo nivel de información que define un paquete de elementos comunes, normalmente Identificados como una "región" especifica dentro de un ecosistema.
Este componente del sistema costero en la banda subtropical y tropical es de suma importancia por las diferentes funciones que desempeña, que van desde el mantenimiento de las características físicas de la costa, ya que prevén la erosión de la línea de costa por su ubicación y estructura. Así como importantes áreas de alta diversidad biológica por sus funciones de alta productividad y protección de diferentes especies (Szelistowski, 1990; Yáñez-Arancibia et al., 1988; 1991; 1993).
Los estudios sobre los ecosistemas de manglar y su ¡interacción con las comunidades de animales se enfatizan desde que Odum (1970) y Odum y Heald (1972, 1975) establecen la hipótesis de que el carbón de los manglares (hojas, propágulos, y tejidos de la madera) pueden contribuir a la trama trófica basada en el detritus, de las aguas costeras adyacentes. Sin embargo, las diferentes Investigaciones que se han llevado a cabos desde entonces aún no han podido clarificar esta relación y aun permanece mucha Incertidumbre en relación con la Importancia de este material en la producción secundaria de invertebrados, peces, pájaros entre otros.
El presente capitulo, tiene por objetivo sintetizar diferentes investigaciones sobre el papel ecológico de los bosques de manglar en cuanto a su función como hábitat critico que proporciona refugio y protección a las especies animales de importancia ecológica y comercial. Como fuente de alimento a través de la trama trófica y finalmente en la producción secundaria relacionada con las pesquerías comerciales. Este enfoque se enfatiza en los estudios realizados en el sur del Golfo de México.
Utilización del Hábitat Los ecosistemas de manglar en la zona costera
por la forma de las raíces aéreas de sostén y los neumatóforos, se consideran que proporcionan un hábitat critico que puede ser particularmente propicio para ser usado por los juveniles y adultos de especies nectónicas como áreas de protección y alimentación, así como áreas de crianza y desove (Jeyaseelan y Krishnamurthy, 1980; Odum et al., 1982; Bell et al., 1984; Thayer et al., 1987; Robertson y Duke, 1987; Yáñez-Arancibia et al., 1988,1991, 1993; Vega-Cendejas et al., 1994).
En el sur del Golfo de México Yáñez-Arancibia et al. (1988, 1991) evalúan la estacionalidad de la biomasa y diversidad de los peces estuarinos vinculados a la heterogeneidad espacial de la Laguna de Términos. Para lo cual, estudian los patrones de biomasa, densidad, número de especies y talla de la comunidad de peces.
Para establecer el patrón de utilización secuencial de los hábitats de la laguna por las comunidades de peces, se tomaron en cuenta las siguientes consideraciones: 1) el patrón de comportamiento individual de ciertas especies que son definidas como especies dominantes con base a tres parámetros a) por su alta frecuencia de aparición en las capturas y sus valores altos de b) biomasa y c) densidad. 2) Para determinar el uso secuencial de los diferentes hábitats de la laguna se consideró la acoplación de las estrategias biológicas del ciclo de vida de las especies (juvenil, adulto madurando, adulto en reproducción, adulto en descanso) con la estacionalidad ambiental (época de secas, lluvias y nortes), cuantificado por el índice de Abundancia de Rogers y Herke (1985). Con ésta información se infiere el uso programado estacional de las especies.

Ecosistemas de Manglar A. L. Lara-Domínguez & A. Yáñez-Arancibia
155
Figura 1. Localización de los sitios experimentales analizados en la Laguna de Términos. SFL ubicado en el Río Palizada se refiere a los manglares con influencia fluvial; SMS, EP, SM y MP corresponden al área de interacción pastos marinos y manglar; PR, P ubicado en Puerto Real se refiere al hábitat de pastos marinos (modificado de Yáñez-Arancibla et a/., 1993)
Los diferentes ambientes de la Laguna de
Términos que Yáñez-Arancibia et al. (1988, 1991, 1993) analizan, son los hábitats de manglar y de pastos marinos, que generalmente son compo-nentes importantes en los sistemas costeros (Fig. 1). El hábitat de manglar, como área de estudio, se localiza en los sistemas fluvio-Iagunares (SFL) que corresponde a los ríos asociados a la laguna, presentando manglares ribereños; mientras que el hábitat de pastos marinos se ubica tanto en un canal de marea de la Isla del Carmen denominado Estero pargo (SMS. EP, SM. MP) con la presencia de praderas de Thalassia testudinum y asociados a manglares de franja; y en la Boca de Puerto Real, que es un ambiente típicamente marino con la presencia de pastos marinos (PR, P) -[las iniciales dentro de los paréntesis corresponden a la nomenclatura utilizada en las figuras para identificar a cada hábitat]-.
Este último hábitat, también constituye una de las dos bocas que conecta a la Laguna de Términos con el Golfo de México, que comprende la principal ruta de inmigración de peces a la laguna. Dentro de la laguna, los movimientos migratorios de los peces son favorecidos por las corrientes hacia el canal de marea o Estero pargo siguiendo hacia la Boca del Carmen. Esta última boca en un ambiente de agua salobre y turbia que funciona como una ruta de migración de algunas especies de peces que se dirigen a la Sonda de Campeche (Fig. 1). No obstante, esta boca también constituye una ruta de inmigración de los individuos a la laguna por la capa inferior del agua, distribuyéndose principalmente en áreas de baja salinidad y alta turbidez (Yáñez-Arancibia et al., 1991).
Ecosistemas de Manglar A. L. Lara-Domínguez & A. Yáñez-Arancibia
156
Esta inmigración de individuos a la laguna es constante a lo largo de año (Yáñez-Arancibia y Day 1982; Yáñez-Arancibia et al., 1991). Sin embargo, en septiembre y octubre se registra la mayor inmigración de Juveniles relacionada con los cambios climáticos entre las épocas de lluvias y nortes, cuando se manifiesta la mayor descarga de ríos, con valores altos de productividad primaria en las aguas abiertas de la laguna, principalmente cerca de las bocas de los nos. Asimismo, durante este interperiodo, se asocia un incremento de la diversidad de peces y de tallas pequeñas. Similarmente, Febrero constituye el interperiodo entre la época de nortes y la de secas. Durante éste, se registra baja diversidad de especies, individuos de tallas grandes y alta biomasa.
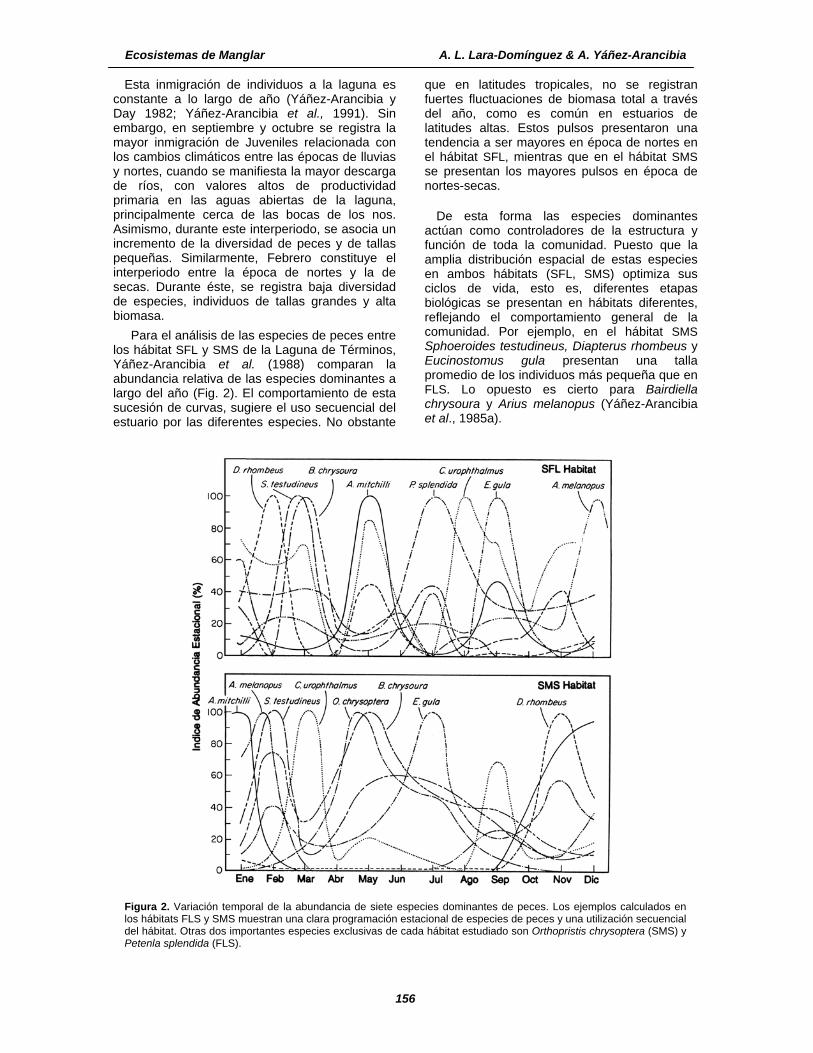
Para el análisis de las especies de peces entre los hábitat SFL y SMS de la Laguna de Términos, Yáñez-Arancibia et al. (1988) comparan la abundancia relativa de las especies dominantes a largo del año (Fig. 2). El comportamiento de esta sucesión de curvas, sugiere el uso secuencial del estuario por las diferentes especies. No obstante
que en latitudes tropicales, no se registran fuertes fluctuaciones de biomasa total a través del año, como es común en estuarios de latitudes altas. Estos pulsos presentaron una tendencia a ser mayores en época de nortes en el hábitat SFL, mientras que en el hábitat SMS se presentan los mayores pulsos en época de nortes-secas.
De esta forma las especies dominantes actúan como controladores de la estructura y función de toda la comunidad. Puesto que la amplia distribución espacial de estas especies en ambos hábitats (SFL, SMS) optimiza sus ciclos de vida, esto es, diferentes etapas biológicas se presentan en hábitats diferentes, reflejando el comportamiento general de la comunidad. Por ejemplo, en el hábitat SMS Sphoeroides testudineus, Diapterus rhombeus y Eucinostomus gula presentan una talla promedio de los individuos más pequeña que en FLS. Lo opuesto es cierto para Bairdiella chrysoura y Arius melanopus (Yáñez-Arancibia et al., 1985a).
Figura 2. Variación temporal de la abundancia de siete especies dominantes de peces. Los ejemplos calculados en los hábitats FLS y SMS muestran una clara programación estacional de especies de peces y una utilización secuencial del hábitat. Otras dos importantes especies exclusivas de cada hábitat estudiado son Orthopristis chrysoptera (SMS) y Petenla splendida (FLS).
Ecosistemas de Manglar A. L. Lara-Domínguez & A. Yáñez-Arancibia
157
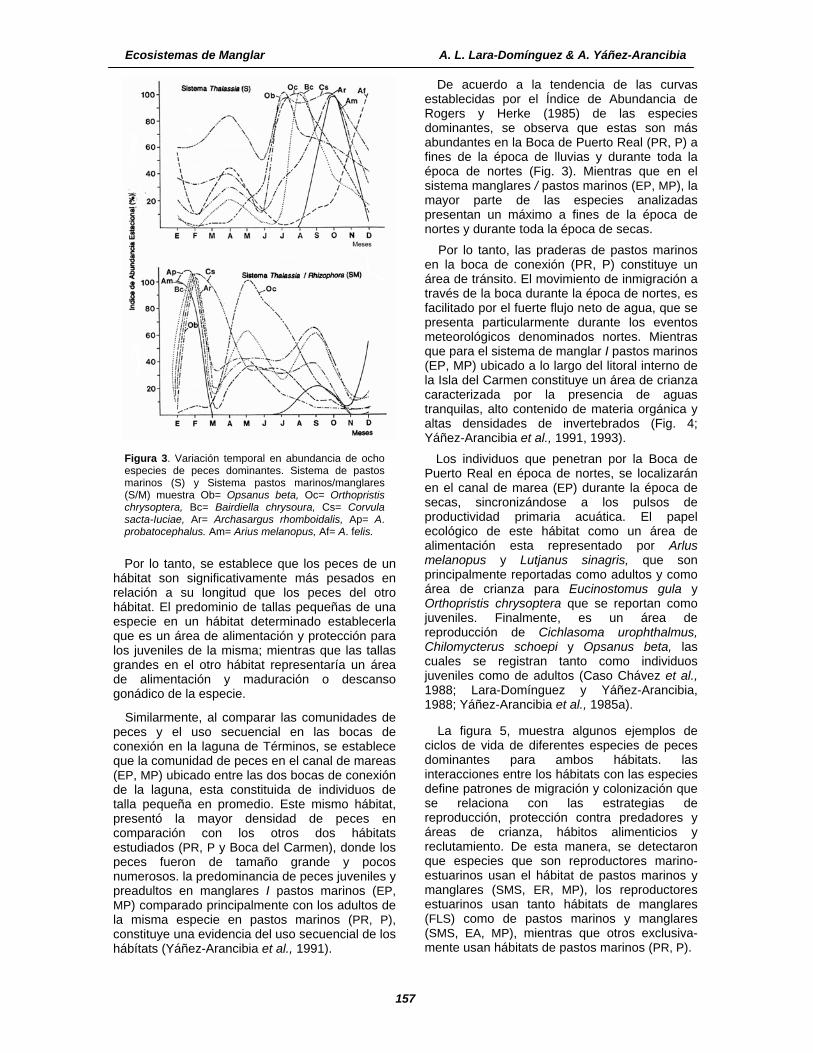
Figura 3. Variación temporal en abundancia de ocho especies de peces dominantes. Sistema de pastos marinos (S) y Sistema pastos marinos/manglares (S/M) muestra Ob= Opsanus beta, Oc= Orthopristis chrysoptera, Bc= Bairdiella chrysoura, Cs= Corvula sacta-Iuciae, Ar= Archasargus rhomboidalis, Ap= A. probatocephalus. Am= Arius melanopus, Af= A. felis.
Por lo tanto, se establece que los peces de un hábitat son significativamente más pesados en relación a su longitud que los peces del otro hábitat. El predominio de tallas pequeñas de una especie en un hábitat determinado establecerla que es un área de alimentación y protección para los juveniles de la misma; mientras que las tallas grandes en el otro hábitat representaría un área de alimentación y maduración o descanso gonádico de la especie.
Similarmente, al comparar las comunidades de peces y el uso secuencial en las bocas de conexión en la laguna de Términos, se establece que la comunidad de peces en el canal de mareas (EP, MP) ubicado entre las dos bocas de conexión de la laguna, esta constituida de individuos de talla pequeña en promedio. Este mismo hábitat, presentó la mayor densidad de peces en comparación con los otros dos hábitats estudiados (PR, P y Boca del Carmen), donde los peces fueron de tamaño grande y pocos numerosos. la predominancia de peces juveniles y preadultos en manglares I pastos marinos (EP, MP) comparado principalmente con los adultos de la misma especie en pastos marinos (PR, P), constituye una evidencia del uso secuencial de los hábítats (Yáñez-Arancibia et al., 1991).
De acuerdo a la tendencia de las curvas establecidas por el Índice de Abundancia de Rogers y Herke (1985) de las especies dominantes, se observa que estas son más abundantes en la Boca de Puerto Real (PR, P) a fines de la época de lluvias y durante toda la época de nortes (Fig. 3). Mientras que en el sistema manglares / pastos marinos (EP, MP), la mayor parte de las especies analizadas presentan un máximo a fines de la época de nortes y durante toda la época de secas.
Por lo tanto, las praderas de pastos marinos en la boca de conexión (PR, P) constituye un área de tránsito. El movimiento de inmigración a través de la boca durante la época de nortes, es facilitado por el fuerte flujo neto de agua, que se presenta particularmente durante los eventos meteorológicos denominados nortes. Mientras que para el sistema de manglar I pastos marinos (EP, MP) ubicado a lo largo del litoral interno de la Isla del Carmen constituye un área de crianza caracterizada por la presencia de aguas tranquilas, alto contenido de materia orgánica y altas densidades de invertebrados (Fig. 4; Yáñez-Arancibia et al., 1991, 1993).
Los individuos que penetran por la Boca de Puerto Real en época de nortes, se localizarán en el canal de marea (EP) durante la época de secas, sincronizándose a los pulsos de productividad primaria acuática. El papel ecológico de este hábitat como un área de alimentación esta representado por Arlus melanopus y Lutjanus sinagris, que son principalmente reportadas como adultos y como área de crianza para Eucinostomus gula y Orthopristis chrysoptera que se reportan como juveniles. Finalmente, es un área de reproducción de Cichlasoma urophthalmus, Chilomycterus schoepi y Opsanus beta, las cuales se registran tanto como individuos juveniles como de adultos (Caso Chávez et al., 1988; Lara-Domínguez y Yáñez-Arancibia, 1988; Yáñez-Arancibia et al., 1985a).
La figura 5, muestra algunos ejemplos de ciclos de vida de diferentes especies de peces dominantes para ambos hábitats. las interacciones entre los hábitats con las especies define patrones de migración y colonización que se relaciona con las estrategias de reproducción, protección contra predadores y áreas de crianza, hábitos alimenticios y reclutamiento. De esta manera, se detectaron que especies que son reproductores marino-estuarinos usan el hábitat de pastos marinos y manglares (SMS, ER, MP), los reproductores estuarinos usan tanto hábitats de manglares (FLS) como de pastos marinos y manglares (SMS, EA, MP), mientras que otros exclusiva-mente usan hábitats de pastos marinos (PR, P).

Ecosistemas de Manglar A. L. Lara-Domínguez & A. Yáñez-Arancibia
158
Figura 4. Variación de diferentes parámetros ambientales en función de las bocas de conexión (BP= Puerto Real; BC= Boca del Carmen) y del canal de marea ubicado en la Isla del Carmen (EP= Estero Pargo)
El uso de diferentes hábitats por las diferentes estrategias de vida, optimiza las tasas de reclutamiento, por ejemplo, los Juveniles de los reproductores marinos e.g., Archosargus rhomboidalis y Haemulon plumieri migran del hábitat de pastos marinos y manglar (SMS EP. MP) en la época de secas cuando la productividad es alta en este hábitat. Mientras que los reproductores dulceacuicolas-estuarinos como Arius melanopus y Bairdiella chrysoura se alimentan como subadultos en el hábitat de pastos marinos y manglar (SMS EP. MP) durante la época de secas y regresan a FLS para reproducirse durante la época de lluvias, usando ambos hábitats durante los periodos de mayor productividad. Finalmente, Cichlasoma urophthalmus y Urolophus jamaicensis usan exclusivamente SMS durante diferentes etapas de su ciclo de vida, principalmente cuando se registra la productividad más alta en el área (Yáñez-Arancibia et al., 1985a).
Por otro lado, muchas de las especies analizadas son de importancia comercial y el separar sus ambientes de alimentación, reproducción y crianza favorece su permanencia en el ecosistema. De manera que los diferentes mecanismos de reclutamiento de los peces costeros dependientes estuarinos o relacionados a estuarios pueden estar controlados por dos principales tipos de procesos: 1) los procesos
físicos que tiene su principal influencia durante las etapas de huevos y larvas sobre la plataforma continental, las bocas estuarinas y parte del estuario; y 2) procesos biológicos que tiene su principal influencia durante la distribución y abundancia de juveniles dentro del estuario. En el sur del Golfo de México, las especies de peces analizadas ya han sido transportadas, por procesos físicos, desde los ambientes marinos a las aguas costeras de diferentes ambientes como manglares, pastos marinos, lagunas costeras entre otros. Mientras que dentro de las áreas costeras predominan los procesos biológicos determinando el uso secuencial del hábitat acoplado a las estrategias biológicas.
Comparativamente en la costa del Pacifico Tropical Este, Szelistowsky (1990) establece que existen tres variables ecológicas que probablemente son importantes en la constitución de la estructura de las comunidades de peces en los manglares. Estas incluyen la proximidad de los manglares con respecto a otros hábitats, la mejor ruta del flujo de energía y las limitaciones físicas impuestas por el mismo sistema. Para este autor los hábitats de arrecifes de coral y pastos marinos en Punta Morales, Costa Rica, se encuentran pobremente desarrollados y es aquí donde los manglares

Ecosistemas de Manglar A. L. Lara-Domínguez & A. Yáñez-Arancibia
159
Figure 5. Ciclo de vida de seis especies de peces selectas: desovadores marino - estuarinos A. rhomboidalis y H. plumieri; dosovadores estuarinos C. urophthalmus y U. jamaicencis; desovadores agua dulce - estuarlnos A. melanopus y B. chrysoura. Los peces migratorlos usan los hábitats SMS y FLS en los periodos de mayor productividad para alimentación, desove o crianza.
tienen un papel ecológico fuertemente vinculado a los estuarios, bahías y/o grandes sistemas deltáicos (e.g., Golfo de Nicoya, Golfo Dulce). Donde las comunidades de peces están fuertemente dominadas por fauna estuarina típica, siendo abundantes el grupo de los bagres (Ariidae), de las corvinas (Sciaenidae), mojarras (Gerreidae) y lisas (Mugilidae).
Otro aspecto de los ecosistemas de manglar que aún no se discute a fondo, es que sus parámetros físicos son altamente variables tanto dentro como entre el sistema de raíces, y los
estudios ecológicos se han limitado a observa-ciones de peces pasajeros más que dentro de las raíces (Thayer et al., 1987). En general entre las raíces, se pueden registrar amplios .rangos de salinidad, temperatura, fuerza de la marea y turbidez, que pueden limitar la distribución de los peces. Únicamente Thayer et al. (1987) ,en el sur de Florida, Zárate Lomeli (1996), y Yáñez-Arancibia et al., (1994) en Laguna de Términos y Vega Cendejas (1998) en Ría Celestún, México, han caracterizado y cuantificado las comunidad peces en el sistema de raíces adyacente a los hábitats de manglar.

Ecosistemas de Manglar A. L. Lara-Domínguez & A. Yáñez-Arancibia
160
Este último autor encuentra que, los factores más importantes de producción natural que proporcionan las raíces de manglar para las poblaciones de peces fueron la turbidez que les ofrece protección, la materia orgánica como resultado de la dinámica del detritus que asegura la disponibilidad de alimento, y las relaciones biogeogréficas de las especies.
Finalmente, son pocos los estudios sobre los primeros estados de vida de los peces (huevos y larvas) en los manglares, y la supuesta importancia de los manglares como sitios de desove y áreas de crianza de larvas esta basada más sobre especulación que sobre una fuerte base de datos. Ramírez et al. (1989) examinaron localidades de desove y distribución de larvas de anchoas en el sistema de manglar así como
adyacente al mismo en Punta Morales (Golfo de Nicoya). Los resultados obtenidos muestran que, las anchoas parecen desovar próximas pero no dentro de los manglares reclutándose dentro del sistema a una longitud de 12 mm.
Ramírez et al. (1990) conducen una evaluación de ictioplancton en la misma área encontrando que las densidades de huevos planctónicos fueron menores dentro de los manglares que en las localidades adyacentes, pero que la densidad y diversidad de larvas no difiere significativamente entre las estaciones de muestreo. Sin embargo, son necesarias más Investigaciones antes de formular generalizaciones acerca del uso de los sistemas de manglar por los primeros estados de vida de los peces.
Estructura Trófica
En relación de como las principales rutas del flujo de energía pueden influir en la estructura de las comunidades de peces, Odum y Heald (1972, 1975) concluyen que la hojarasca del manglar, vía ruta del detritus, es la principal fuente de energía para los consumidores de la trama trófica, siempre que la biomasa de las algas y del plancton fueran bajas. Consecuentemente, dominarían la comunidad los peces capaces de utilizar directamente el detritus del manglar y los otros que predan sobre los detritívoros y hacen notar que algunas zonas costeras de importancia comercial se localizan próximas a los bosques de manglar y pastos marinos. (Heald y Odum, 1970; Odum, 1971; Heald, 1971; Odum y Heald, 1972, 1975).
Esto ha conducido a generalizaciones acerca de la estructura de la trama trófica en los estuarios con manglares. Por ejemplo, se estableció que en la mayoría de los estuarios la fuente primaria de alimento proviene del detritus del manglar. Esto condujo a estudiar cuantitativamente la hojarasca, así como a estudiar experimentalmente la dinámica de la descomposición de las hoja (Pool et al., 1975; Fell et al., 1975; Malley, 1978; Fell y Master, 1980; Bunt, 1982; Twilley et al., 1986; Flores-Verdugo et al., 1987; González-Farías y Mee, 1988; Woodroffe et al., 1988). No obstante las múltiples investigaciones sobre el tópico, esta afirmación continúa relativamente sin respuesta.
Una forma de establecer el vínculo del detritus en la trama trófica es a través del estudio del contenido estomacal de las especies que conforman la comunidad estuarina asociada a ecosistemas de manglar. Sobre este tópico, se lIevó a cabo un estudio sobre dinámica alimenticia del contenido estomacal de los peces en la Ría Celestún, al norte de la Península de Yucatán. Los resultados obtenidos, muestran que los
micro-crustéceos son la presa más Importante al constituir el 51% de la biomasa total (Vega Cendejas, 1998). Estos autores concluyen que, el consumo de este grupo es característico en la estructura trófica de los peces asociados a las raíces de manglar y representa, través de la dieta del cangrejo, la incorporación indirecta de detritus a su alimentación así como de otros tipos de alimento, tales como el fitoplancton. De tal forma que, la fauna que mantiene a las poblaciones de peces en este ecosistema costero, dependen directa e indirectamente del detritus proveniente del manglar.
Estos estudios han conducido a realizar análisis más complejos como son las caracterizaciones isotópicas de los organismos en pantanos salobres y de pastos marinos de latitudes templadas Esta técnica consiste en identificar la relación de isótopos estables (δ 13C/12C, δ15N/14N) al marcar al detritus en la trama trófica. Diferentes autores concluyen que detritus vascular en los pantanos salobres y de pastos marinos en estas latitudes, puede no ser tan importante como se pensaba previamente (Thayer et al., 1978; Haines y Montague, 1979; Mc Connaughey y McRoy, 1979; Huges y Sherr, 1983).
Rodelli et al. (1984) implementan esta metodología para determinar la proporción de isótopos estables en la biota de los pantanos de manglar, bocas costeras yaguas costeras en Malasia y concluye que los manglares constituyen una significativa fuente de carbón para muchos animales que residen en los manglares y en las bocas, pero no en aguas costeras. Por otra parte, Stoner y Zimmerman (1988) encontraron en una laguna rodeada de manglar en Puerto Rico que; los camarones se alimenten principalmente de organismos detritívoros, la proporción de isótopos de carbón

Ecosistemas de Manglar A. L. Lara-Domínguez & A. Yáñez-Arancibia
161
tanto del camarón como en sus presas sugiere que la mayoría del origen de su carbón proviene de algas bénticas más que de los manglares.
Para la Laguna de Términos, en Campeche Raz Guzmán y de la Lanza (1993) establecen la proporción isotópica del carbono orgánico en camarones, sedimento y vegetación. Determinan que el δ13C de Rhizophora mangle es de -25.6 a -30.2 ‰ registrándose entre los valores más ligeros de la vegetación del sur y centro de la laguna Dicho carbono es posible que se incorpore a las redes tróficas locales e influya en la proporción isotópica de los consumidores. Puesto que su estudio se enfoca principalmente a las praderas de pastos marinos (Thalassia testudinum y Dictyota sp) como fuente de carbono para los macrocrustáceos, principalmente camarones. Los resultados del análisis de δ13C coincide con el patrón de migración de Peneus setiferus dentro de la Laguna de Términos. Existiendo una relación aparente entre el δ13C, la talla de los Juveniles y la localidad de colecta.
Otra forma de determinar el papel ecológico del detritus proveniente de los manglares en la zona costera es a través del uso de modelos ecológicos que constituyen una herramienta útil para el análisis, la integración, la síntesis y consecuentemente la predicción del sistema y sus recursos bióticos. En esta línea. Soberón-Chávez et al. (1988) desarrollaron un modelo ecológico para explicar los cambios en dirección e intensidad de las interacciones entre las aguas protegidas de la Laguna de Términos y la plataforma continental de la Sonda de Campeche y cuales son los factores que producen alteraciones en la diversidad, distribución, abundancia y persistencia de los recursos. En este modelo un compartimiento lo constituye la biomasa de los desechos foliares de los manglares, que dentro de la laguna se clasificó en dos tipos diferentes, que a continuación se describen.
Manglar de Influencia marina, se refiere a los desechos foliares que se localizan en la Isla del Carmen. La principal característica de este manglar es que no presenta influencia fluvial. Por lo cual, se considera que la producción de los desechos foliares en esta área, están relacionados con la precipitación y la iluminación. Manglar de influencia fluvial, que son los desechos foliares de los manglares que se localizan en la desembocadura del Río Palizada. Los procesos que controlan la producción en esta zona son únicamente con la iluminación.
Adicionalmente, en el detritus convergen los flujos de material proveniente de todos los componentes del sistema en estudio. En promedio, el 5.3% del detritus total en la laguna
proviene de los manglares, teniendo su máximo impacto en el sistema fluvio lagunar este (Subsitema III) donde equivale al 14%. Asimismo, se determina el flujo del detritus a través de los diferentes hábitats de la Laguna de Términos. Esto es, la cantidad de detritus proveniente de los manglares de influencia marina presenta una tasa de exportación de 0.083 g/mes (g de desecho). En cuanto a la cantidad de detritus proveniente de los manglares de influencia fluvial, representa una tasa de exportación de 0.04 g/mes (g de desecho) (Soberón-Chávez et al., 1988).
El modelo establece que la exportación del detritus a otros ecosistemas esta en función de los vientos, descarga fluvial y una constante. De tal manera que, el flujo de exportación del detri-tus de la plataforma carbonatada de la Sonda de Campeche (Subsistema B) se divide en dos partes: alrededor del 90% se dirige a la plataforma terrlgena (subsistema A), y el resto entra a la laguna acarreado principalmente por los vientos del norte a través de la Boca de Puerto Real (Subsistema 1). Ya dentro de la laguna, el detritus que se exporta de la Cuenca Central (Subsistema 11) se divide en dos partes, la primera es acarreada por los vientos del norte al sistema fluvio lagunar del Este (Subsistema III), y el resto se dirige al sistema fluvio-Iagunar Oeste y Boca del Carmen (Subsistema IV). Finalmente, la laguna de Términos exporta detritus a través de la Boca del Carmen hacia la plataforma terrígena impulsado por la circulación de la laguna y por la descarga fluvial del Río Palizada. Este flujo es 6.594 veces mayor que el que entra a la laguna por la Boca de Puerto Real (Soberón-Chávez et al., 1988).
Este intercambio de detritus entre los diferentes hábitats, esta vinculado al patrón de migración de los organismos nectónicos y el modelo define 3 pulsos migratorios:
1. Peces básicamente subadultos y adultos, provenientes del Subsistema B que entran a la laguna hacia las praderas de pastos marinos (subsistema 1) en época de secas con fines de alimentación. Regresan al Subsistema B en época de lluvias, moviendo en esta forma entre 60 y 85 ton/año de materia entre los diferentes ambientes.
2. Organismos que entran en lluvias a la laguna como larvas y juveniles a través del Subsistema 1. Se dirigen al Subsistema III a través del Subsistema ll y salen de la Laguna en secas pasando por el Subsistema IV. Estos organismos regresan al Subsistema B a través del Subsistema A. Causando un movimiento de aproximadamente 200 ton/año.

Ecosistemas de Manglar A. L. Lara-Domínguez & A. Yáñez-Arancibia
162
3. Organismos provenientes del Subsistema A que entran en lluvias a la laguna por el Subsistema IV y salen por ésta misma ruta Asimismo existen organismos que salen por el Subsistema IV y que no forman exclusivamente parte del Patrón de Migración 2 y que constituyen parte de este tercer patrón. Este pulso migratorio mueve casi 160 ton/año.
Con este mismo enfoque se llevó a cabo un análisis sobre la ecología trofodinámica de dos hábitat críticos (pastos marinos y manglares) en la Laguna de Términos. Se empleo el Programa ECOPATH que está diseñado para la construcción de modelos del ecosistema a través de las interacciones tróficas y flujos de nutrientes (Christensen y Pauly, 1996). El modelo tiene una entrada total de 9.474 g peso seco/m2/año, con 18.7% de respiración. El detritus proveniente del manglar, es un Importante aporte "extra" al detritus de la laguna, cuyo suministro es de 5797.68 g peso seco/m2/año. En este modelo el detritus constituye el 82% del flujo total, tal como un sistema maduro trabaja. Asimismo, el porcentaje de eficiencia de transferencia de los productores fluctuó de 6.4 a 0.7%.
De acuerdo a los resultados obtenidos se determinó que existen cuatro niveles tróficos para estas comunidades, esto significa casi el 90% de los consumidores de estos hábitat corresponden al segundo y tercer nivel trófico con un amplio espectro y dependientes del aporte de detritus. Subrayando que muchas de las especies de importancia comercial, por ejemplo los camarones, presentan un uso muy Intenso de estas áreas durante alguna etapa de su ciclo de vida (Rivera et al., 1997).
Similarmente, Vega Cendejas (1994, 1998) determina que del total de los flujos del sistema de manglar analizado, el 22.7% se utiliza para los procesos respiratorios, 0.01 % para exportación y 35% se canaliza a detritus. Los niveles tróficos de los consumidores son relativamente bajos, lo que refleja por una parte, que la mayoría de los grupos de peces que lo utilizan son juveniles y por otra, la dependencia directa e indirecta para su producción de los productores primarios como las macrofitas, fitoplancton y el detritus.
Productividad Pesquera
Numerosas investigaciones se han realizado en torno a las variables que controlan la diversidad y abundancia de los recursos pesqueros. Destacan por su Importancia las áreas vegetadas costeras puesto que funcionan como zonas de protección de juveniles de especies de importancia comercial. Así como por la gran disponibilidad de alimento que representan a través del detritus, o bien como fertilizadores de las áreas costeras adyacentes que estimulan la producción primaria en aguas costeras.
No obstante, éstas funciones están controlados por procesos físico-ambientales de la zona costera como son: 1) las condiciones físico-químicas del agua (transparencia, nutrientes, salinidad, temperatura), 2) Latitud geográfica, 3) batimetría y tipos de sedimentos, 4) meteorología y clima, 5) descarga de los ríos, 6) rangos de marea y variación del nivel del mar, 7) lagunas costeras y estuarios adyacentes, 8) dinámica de interacción entre los estuarios y el mar. Que en su conjunto se han denominado mecanismos de producción natural (Soberón Chávez y Yáñez-Arancibia, 1985).
La relevancia pesquera del Estado de Campeche en el sur del Golfo de México, depende de la interacción entre laguna de Términos y la Sonda de Campeche. Por lo que se ha sugerido que ambos sistemas ecológicos tienen interdependencia reciproca. Por lo tanto en los últimos 20 años se ha manejado la hipótesis de que la productividad pesquera de la plataforma
marina de Campeche depende, en parte, de diversos efectos ecológicos de la Laguna de Términos hacia el mar (Sánchez et al., 1981).
Esto hace a la región de la Laguna de Términos particularmente crítica para la Sonda de Campeche por las extensas áreas costeras vegetadas que presenta, con diferentes asociaciones como son manglares, pastos marinos, pantanos fluvio-deltáicos. También, por sus sistemas lagunares-estuarinos y los aportes de agua dulce, sedimentos, nutrientes y materia orgánica que descargan en la Sonda de Campeche. Asimismo, existe un gran intercam-bio de fauna marina y estuarina entre ambas áreas y los peces consumidores secundarios pueden ser importantes en este flujo de energía y nutrientes (Robertson y Duke, 1990; Twilley et al., 1996).
Como resultado el vinculo de la Laguna de Términos junto con la Sonda de Campeche ha sido importantes por las capturas de ostión, peces, pero principalmente de camarones peneidos. Las pesquerías de peneidos en México -por volumen de captura y por divisas- en la Sonda de Campeche (más del 50 % de la captura de camarón del Golfo de México), comprenden para cada una de las tres especies: Peneus duorarum (camaron rosado) contribuye con el volumen de 90.0%, Peneus setiferus (camarón blanco) con 5.2% y Peneus aztecus (camarón café) con 3.9% (Schultz y Chávez, 1976).

Ecosistemas de Manglar A. L. Lara-Domínguez & A. Yáñez-Arancibia
163
De igual forma, la Sonda de Campeche aporta más del 40% de la captura demersal del Golfo de México. Esta es una pesquería multiespecífica estrechamente vinculada a la pesquería del camarón. Su estabilidad, aumento o disminución es el reflejo de la variabilidad natural de los procesos físico-ambientales y biológicos como consecuencia de interacciones ecológicas en la zona costera.
Las características ambientales de mayor implicación ecológica para los peces demersales de la plataforma continental adyacente a la Laguna de Términos dependen fuertemente, del flujo de los ríos Grijalva y San Pedro en el extremo occidental, del río Champotón en el oriente y de la descarga de la Laguna de Términos, a través de su boca occidental (Boca del Carmen), siendo la influencia de dicha laguna determinante en la dinámica ictiológica de la plataforma adyacente (Sánchez et al., 1981 ; Yáñez-Arancibia et al., 1985b A. Yáñez-Arancibia y Sánchez, 1986).
Otro factor determinante en la composición y abundancia de la fauna de la plataforma continental es la influencia de la vegetación costera. El Estado de Campeche presenta la cobertura de manglar más importante en relación con los otros estados del Golfo de México (aproximadamente 40 %), de aquí su relevancia en la producción pesquera en la Sonda de
Campeche. En éste sentido, Turner (1977) ha señalado una correlación positiva entre el cociente de la captura de camarones peneidos entre las áreas de vegetación intermareal, con la latitud para las costas de Texas y Florida. Deegan et al. (1986) concluyen que las capturas pesqueras del Golfo de México se correlacionan con áreas estuarinas y descargas de ríos, fisiografía y vegetación litoral.
La relación de los recursos pesqueros con los sistemas estuarinos, es bien conocida puesto que éstos son fundamentales en el sostenimiento de alguna etapa del ciclo de vida de más del 90% de las especies marinas que tienen valor comercial. En el sur del Golfo de México, más del 75% de las poblaciones de peces dominantes en la plataforma continental son dependientes estuarinas (Yáñez-Arancibia y Sánchez-Gil, 1988). De acuerdo a la producción pesquera de la región y la infraestructura disponible, una extrapolación del valor de los recursos pesqueros que dependen de la Laguna de Términos tiene un "valor intrínseco” de alrededor de 60 x 106 de dólares anuales (en 1988). Esto puede relacionarse con las áreas de manglar y pastos marinos, lo cual plantea el valor económico que representan los hábitats lagunares-estuarinos como consecuencia de su valor ecológico (Yáñez-Arancibia y Aguirre León, 1988).
Literatura Citada
Bell, J. D., D. A. Pollard, J. J. Burchmore, B. C. Pease y M. J. Middleton, 1984. Structure of fish community in temperate tidal mangrove creek in Botany Bay, New South Wales . Aust. J. Mar. Freshwat. Res., 35: 33-48.
Bunt, J.S., 1982. Studies of litter fall in tropical Australia, p: 223-237. In: Clough, B.F. (Eds.) Mangrove Ecosystems in Australia. Structure, function and management. Australia Institute of Marine Science, Canberra.
Caso-Chávez, M., A. Yáñez-Aranclbla, A. L. Lara-Domlnguez, 1986. Biologla, ecologla y dinámica de poblaciones de Cichlasoma urophthalmus (Günther) en Hábitat de Thalassia testudinum y Rhizophora mangle Laguna de Términos, Campeche (Pisces: Cichlidae). Biotica, 11 (2): 79-111.
Chrlstensen V. y D. Pauly, 1996. ECOPATH for Windows. A users Guide. ICLARM Software. Internatlonal Center Living Aquatic Resources Manag. Makati City, Philippines. 71 p.
Day, J. W., Jr. y A. Yáñez-Arancibia, 1988. Consideraciones ambientales y fundamentos ecológicos para el manejo de la región de Laguna de Términos, sus hábitats y recursos pesqueros, Cap. 23: 453-482. In: A. Yáñez-Arancibia y J.W. Day, Jr. (Eds.) Ecología de los Ecosistemas Costeros en el Sur del Golfo de México: La Reglón de la Laguna de Términos. Inst. Cienc. del Mar y Limnol. Univ. NaI. Autón. México, 518 p.
Deegan, L. A., J. W. Day, Jr., J. G. Gosselink, A. Yáñez-Arancibia, G. Soberón Chávez y P. Sánchez, 1986. Relationships among physical characteristics, vegetation, distribution and fisheries yields in Gulf of Mexico estuaries, p: 83-100. In: A D. Wolfe (Ed.) Estuarine Variability. 509 p.
Fell, J. W., R. C. Cefalu, l. M. Master y A. S. Tallman, 1975. Mlcroblal activitles In the mangrove (Rhizophora mangle) leaf detrital system, p. 681-679. In: G. Walsh., S. Snedaker and H. Teas (Eds.) Proc.lnt. Symp. Biol. Mange. Mangr. Univ. of Florida, Gainesville, Fla.

Ecosistemas de Manglar A. L. Lara-Domínguez & A. Yáñez-Arancibia
164
Flores-Verdugo, F., J. W. Day, Jr. y R. Briceño Dueñas,1987. Structure litterfall, decomposition and detritus dynamics of mangrove in a Mexican coastal lagoon with an ephemeral inlet. Mar. Ecol. Prog. Ser., 35: 83-90.
González-Farías, F. y L.D. Mee, 1988. Effect of mangrove humic-like substances on biodegradation rates of detritus. J. Exp. Mar. Biol. Ecol., 119: 1-13.
Halnes, E.B. y C.L. Montague, 1979. Food sources of estuarine invertebrates analyzed using 13C¡12C ratios. Ecology, 60: 48-56.
Heald, E.J., 1971. The Production of Organic Detritus in a South Florida Estuary. University of Mlami Sea Grant Technical Bulletin, 6. Coral Gables, Fla., 110.
Heald, E.J. y W. E. Odum, 1970. The contribution of mangrove swamps to Florida fisherles. Proc. Gulf Caribb. Fish. Inst., 22: 130-135.
Huges, E. H. Y E. B. Sherr, 1983. Subtidal food webs in a Georgia estuary: 13C¡12C analysls. J. Exp. Mar. Biol. Ecol., 67: 227-242
Jeyaseelan, M.J.P. y K. Krishnamurthy, 1980. Role of mangrove forests of Pichavaram as fish nurseries. Proc.lndian. Nat. Sci Acad. B., 46: 48-53.
Malley, D. F., 1978. Degradation of mangrove leaf litter by the tropical sesarmid crab Chiromanthes onychophorum. Mar. Biol., 49: 377-386.
McConnaughey, T. y C. P. McRoy, 1979. 13C label identifies eelgrass (Zostera marina) carbon in an Alaskan estuarine food web. Mar. Biol., 53: 263-269.
Odum, W. E., 1970. Utilization of the direct grazing and plant detritus food chains by the striped mullet Mugil cephalus, p: 222-240. In: J.H. Steele (Ed.) Marine Food Chains. Univ. Callf. Press, Berkely.
Odum, H. T., 1971. An energy circuit language for ecological and social systems, its physical basis, p: 139-211. In: B. Patten (Ed.) System Analysis and Simulation in Ecology. Vol. 2. New York. Academic Press.
Odum, W. E. y E. J. Heald, 1972. Trophic analysis of an estuarine mangrove community. Bull. Mar. Sci., 22: 671-738.
Odum, W. E. y E. J. Heald, 1975. The detritus-based fod web of an estuarlne mangrove community, p: 265-286. In: L Cronin (Ed.) Estuarine Research, Academlc Press Inc., New York.
Odum, W. E., C.C. Mclvory T.J. Smlth III, 1982. The ecology of the mangrove of South Florida: a Community Profile. U.S. Fish and Wildllfe Services, Technical Report FWS/OBS 81-24, Washington, D.C. 154 p.
Pool, D.J., S. Snedaker y A.E. Lugo, 1975. Litter production in mangrove forests of south Florida and Puerto Rico, p: 213-237. In: G. Walsh, S. Snedaker y H. Teas (Eds.) Proc. Int. Symp. Biol. Mange. Mangr. University of Florida, Gainesville, Florida.
Ramirez, A.R., W.A. Szelistowski y M.I. López, 1989. Spawning pattern and larval recruitment in Gulf of Nicoya anchovies (Pisces: Engraulidae). Rev. Biol. Trop., 37: 55-62.
Ramirez, A.R., M.I. López y W.A. Szelistowski, 1990. Composition and abundance of ichthyoplankton in a Costa Rica mangrove estuary. Rev. Biol. Trop.
Raz Guzmán, A., y G. de la Lanza,1993. δ13C of zooplankton, decapods crustaceans and amphipods from Laguna de Terminos, Campeche (Mexico) with reference to food sources and trophic position. Ciencias Marinas, 19(2): 245-264.
Rivera, E., A. L. Lara-Dominguez, G. J. Vlllalobos Zapata, A. Yáñez-Arancibia, 1997. Trophodynamic ecology of two critical habltats (seagrasses and mangroves) in the Southern Gulf of Mexico, Termlnos Lagoon, Campeche. ICLARM.
Robertson, A. I., y N. C. Duke, 1987. Mangroves as a nursery sites: comparisons of the abundance and species composition of fish and crustaceans in mangroves and other nearshore habitats in tropical Australia. Marine Biology, 96: 193-205.
Robertson, A.I., y N.C. Duke, 1990. Mangroves fish-community in tropical Queensland, Australia: Spatial and temporal patterns in densities, biomass and community structure. Marine Biology, 104: 369-379.
Rodelll, M.R., J.N. Gearing, P .J. Gearing, N. Marshall y A. Sasekumar, 1984. Stable isotope ration as a tracer of mangrove carbon in Malaysian ecosystems. Oecologia, 61: 326-333.
Roger B. D. Y W. H. Herke, 1985. Estuarine-dependent fish and crustacean movements and weir management, p: 201-219. In: C.F. Beyan, P.J. Zurank and R.H. Chabreck (Eds.) Fourth Coastal Marsh and Estuary Management Symposium. Baton Rouge, Louisiana State University Press.

Ecosistemas de Manglar A. L. Lara-Domínguez & A. Yáñez-Arancibia
165
Sánchez, P., A. Yáñez-Aranclbla y F. Amezcua Linares, 1981. Diversidad, distribución y abundancia de las especies y poblaciones de peces demersales de la Sonda de Campeche (Verano 1978). An. Inst. Cienc. del Mar y Limnol. Univ. Nal. Autón. México, 8(1): 209-240.
Schultz, L. y E. Chávez, 1976. Contribución al conocimiento de la biología pesquera del camarón blanco Penaeus setiferus (L.) en el Golfo de México. Mem. Simp. Biología y Dinámica Poblacional de Camarones. Guaymas, Son. 8-13 agosto, 1976.
Soberón-Chávez, G. y Yáñez-Arancibia, A. 1985. Control ecológico de los peces demersales: Variabilidad ambiental de la zona costera y su influencia en la producción natural de los recursos pesqueros, p: 399-486. In: A. Yáñez-Arancibia (Ed.) Recursos Pesqueros Potenciales de México: La Pesca Acompañante del Camarón. Programa Universitario de Alimentos, Instituto de Ciencias del Mar y Limnología, Instituto Nacional de Pesca, UNAM. México D F.
Soberón Chávez, G. A. Yáñez-Arancibia y J.W. Day, Jr., 1988. Fundamentos para un modelo ecológico preliminar de la Laguna de Términos, Cap. 20: 381-414: In: A. Yáñez-Arancíbia.y J.W. Day, Jr. (Eds.) Ecología de los Ecosistemas Costeros en el Sur del Golfo de México: La Reglón de la Laguna de Términos. Inst. Cienc. del Mar y Llmnol. Univ. NaI. Autón. México, Editorial Universitaria, México, 518 p.
Stoner, A. W. and R. J. Zimmerman, 1988. Food pathways associated with penaeid shrimp in a mangrove-fringed estuary. Fishery Bulletin, 86(3), 543-551.
Szelistowski, W.A., 1990. lmportance of mangrove plant litter in fish food webs as temporary, floating habitat in the Gulf of Nicoya, Costa Rica.PhD Dissertation, University of South Carolina, 217 p.
Thayer, G.W.,P.L. Parker, M.L. LaCroix y B. Fry, 1978. The stable carbon isotope ration of some components of an eelgrass (Zostera marina) bed, Oecologia, 35: 1-12.
Thayer, G. W., R. D. Colby y F.W. Hetter, Jr., 1987. Utilization of the red mangrove prop root habitat by fishes in south Florida. Mar. Ecol. Prog. Ser., 35: 25-38.
Turner, E. R., 1977. lntertidal vegetation and commercial yields of penaeid shrimp. Trans. Am. Fish. Soc., 106(5): 411-416.
Twllley, R. R., A. E. Lugo y C. Patterson-Zucca, 1986. Litter production and turnover in basin mangrove forest in southwest Florida. Ecology, 67: 670-683
Twllley, R. R., S. C. Snedaker, A. Yáñez-Arancibia y E. Medina, 1996. Biodiversity and ecosystem processes in tropical estuaries: Perspectives of mangrove ecosystems, p: 327-370. In: H. Mooney, H.J. Cushman, E. Medina, E.O. Salas and E.D. Schulze (Eds.) Biodiversity and Ecosystems Functions: A Global Perspective. John Wiley and Sons, N.Y.
Vega Cendejas, M. E., 1994. Trophic Interrelations in a beach seine fishery from the northwestern coast o, the Yucatan peninsula Mexico. Journal of Fish Biology, 44: 647-659.
Vega Cendejas, M. E., 1998. Trama trófica de la comunidad nectónica asociada al ecosistema de manglar en el liltoral norte de Yucatán. Tesis Doctor en Ciencias (Biología). Univ. NaI. Autón. México, Fac. Ciencias. 170 p.
Vega Cendejas, M. E., U. Ordoñez y M. Hernández, 1994. Day-night variation of fish population in mangroves of Celestun Lagoon Mexico. International Journal Ecology and Environmental Sciences, 20: 99-108.
Woodroffe, C. D., K. N. Bardsley, P. J. Ward y J. R. Hanley, 1988. Production of mangrove litterfall in a macrotidal embayment, Darwin Harbour, N.T., Australia. Est. Coast. Shelf Sci. 26: 581-598.
Yáñez-Aranclbla, A. y A. Aguirre León, 1988. Pesquerías en la región de la Laguna de Términos. Cap. 22: 431-452. In: A. Yáñez-Arancibia y J. W. Day. Jr. (Eds.), Ecología de los Ecosistemas Costeros en el Sur del Golfo de México: La Región de la Laguna de Términos. Inst. Cienc. del Mar y Limnol. UNAM, Coast. Ecol. Inst. LSU. Editorial Universitaria, México DF.
Yáñez-Arancibia, A. y J. W. Day, Jr., 1982. Ecological characterization of Terminos Lagoon, a tropical lagoon-estuarine system in the Southern Gulf of Mexico. ln: P. Lasserre y H. Postma (Eds.) Coastal Lagoons. Oceanologica Acta, Vol. Spec., 5(4): 431-440.
Yáñez-Arancibia, A., A. L. Lara-Domínguez, 1988. Ecology o, three sea catfishes (Arildae) In a tropical coastal ecosystem-southern Gulf o, Mexico. Marine Ecology Progr. Ser., 49: 215-230.
Yáñez-Aranclbla, A. y P. Sánchez-GII, 1988. Ecología de los Recursos Demersales Marinos. Fundamentos en Costas Tropicales. AGT Editor, S.A., 230 p.
Yáñez-Arancibia, A., G. Soberón Chávez y P. Sánchez, 1985a. Ecology of control mechanisms of natural fish production in the coastal zone. Chap. 27: 571-594. In: A. Yáñez-Arancibia (Ed.). Fish Community

Ecosistemas de Manglar A. L. Lara-Domínguez & A. Yáñez-Arancibia
166
Ecology in Estuaries and Coastal Lagoons: Towards an Ecosystem Integration. 654 p.
Yáñez-Arancibia, A., A.L. Lara-Domínguez, A. Aguirre León, S. Díaz-Ruiz, F. Amezcua Linares, P. Chavance y D. Flores, 1985b. Ecología de las poblaciones de peces dominantes en estuarios tropicales. Factores ambientales regulando las estrategias biológicas y poblaciones, Chap. 15: 311-366. In: Yáñez-Arancibia, A. (Ed.) Fish Community Ecology in Estuarles and Coastal Lagoons: Towards an Ecosystem Integratlon. Editorial Universitaria, UNAM-PUAL-ICML. Mexlco, D.F., 654 p.
Yáñez-Aranclbla, A., A. L., Lara-Dominguez, J. L. Rojas Galavlz, P. Sánchez Gil, J.W. Day y C. Madden, 1988. Seasonal biomass and diversity of estuarine fishes coupled with tropical habitat heterogeneity (southern Gulf of Mexico). J. Fish. Biol. 33 (Supplement A): 191-200.
Yáñez-Arancibia, A. P. Sánchez y A. L. Lara-Domínguez, 1991. Interacciones ecológicas estuario-mar: estructura funcional de bocas estuarinas y su efecto en la productividad del ecosistema. In: II Simposio sobre Ecosisstemas da Coasta Sul Brasilerira: Estrutura, Funcao e Manejo. Publ. ACIESP, 71(4): 49-83.
Yáñez-Arancibia, A., A. L. Lara-Domínguez y J. W. Day, Jr., 1993. Interaction between mangrove and seagrass habitat mediated by estuarine nekton assemblages: coupling of primary and secondary production. Hydrobiologia, 264: 1-12
Yáñez-Arancibia, A., A.L. Lara-Domínguez, M. E. Vega Cendejas, G.J. Vlllalobos Zapata, E. Rivera, S. M. Hernández, U. Ordóñez, E. Pérez, H. Álvarez Guillén y F. Vera Herrera.1994. Funcionalidad Ecológica de los Sistemas de Manglar en la Península de Yucatán: Estudio Comparativo de la Laguna de Términos, Campeche y Laguna de Celestún, Yucatán. Informe Técnico Final. Convenio UAC-Programa EPOMEX-CONACyT. Clave F467-19109
Zarate Lomen, D., 1996. El manglar como hábitat critico para las comunidades nectónicas en Estero Pargo, Laguna de Términos, México. Tesis Maestría en Ciencias del Mar. Oceanografía Biológica y pesquera, UACPyP - CCH. Univ. Nal. Autón. México. 107 p.


















