Proteínas: Plegamiento y conformación Proteínas: Plegamiento y ...
Upload
nicky-xikitho-kevinnCategory
view
14download
0
1.1. PROTEÍNAS.Los compuestos nitrogenados de la carne están constituidos por proteínas, también encontramos aminas, compuestos guanidínicos, compuestos de amonio cuaternario, aminoácidos libres y péptidos. Las proteínas representan el componente más abundante de la materia seca del músculo y desempeñan un papel fundamental en las funciones fisiológicas “in vivo”, en los cambios que se originan después de la muerte del animal y en las propiedades de la carne para su consumo en fresco y la industrialización.
EL contenido de proteínas de la carne de vacuno es aproximadamente del 20%, muy parecido también al contenido de proteínas en pollo y cerdo. Las proteínas musculares se pueden clasificar atendiendo a su solubilidad en tres grandes grupos (Primo et al., 1997; Ordoñez et al., 1998): proteínas sarcoplásmicas, proteínas contráctiles o miofibrilares, y proteínas del estroma.
- Proteínas del aparato contráctil (proteínas miofibrilares), extraíbles en su mayor
parte con disoluciones salinas concentradas (miosina, actina, tropomiosina, troponina, etc).
- Proteínas solubles (proteínas sarcoplasmáticas), extraíbles con agua o disoluciones salinas diluidas (mioglobina, enzimas).
- Proteínas insolubles (proteínas del tejido conjuntivo y proteínas de los orgánulos).
Proteínas del aparato contráctil o miofibrilares.Se conocen unas 20 proteínas miofibrilares diferentes. Las más abundantes son miosina y actina, que suponen el 65-70% del total. El resto son las proteínas tropomiosina y troponina, importantes para la contracción, y distintas proteínas del citoesqueleto. Son las responsables de la estructura muscular y de la transformación de la energía química en energía mecánica, durante los fenómenos de contracción y relajación muscular. Tras el sacrificio producen el rigor mortis y la exudación.
La actina es el principal componente de los filamentos delgados. Cuando se encuentra en forma monomérica se denomina actina G que al polimerizarse forma filamentos de actina F. La forma filamentosa de la actina forma el esqueleto del filamento delgado y aloja a la tropomiosina y al complejo troponina. Se combina con la miosina para formar el complejo actomiosina que interviene en el mecanismo de contracción-relajación muscular.
La miosina es una proteína con una parte filamentosa, la cola y el cuello, y una parte globular, las dos cabezas de miosina que tienen capacidad para unirse a la actina y al ATP. Estas cabezas son responsables de la actividad enzimática (ATPasa) y de su capacidad para interaccionar con la actina de los filamentos delgados. En el filamento delgado también se encuentran la tropomiosina y el complejo troponina, ambos regulan la contracción muscular y cada una representa el 5% de las proteínas miofibrilares. Entre las proteínas miofibrilares hay una serie de ellas del citoesqueleto, que contribuyen a mantener el armazón estructural en el que funcionan las proteínas contráctiles de la fibra muscular. Entre estas destacan la conectina o titina (10% de las proteínas miofibrilares), la nebulina (4%) que constituyen la denominada línea N2, la α-actinina o proteína mayoritaria de los discos Z, y las proteínas de los.
Proteínas solubles o sarcoplásmicas.

Las proteínas sarcoplásmicas constituyen un 20-30% del total y en ellas se incluyen enzimas, pigmentos (mioglobina) y albúminas, que se extraen con agua o disoluciones salinas diluidas (0.5%). La mioglobina, que se localiza principalmente en el músculo cardíaco y estriado, es una proteína conjugada hemoglobular monomérica de una masa molecular de 16 kDa. La cantidad de mioglobina muscular se ve afectada por factores genéticos, por la edad, la dieta del animal, el tipo de fibra muscular, la especie y el ejercicio. En cerdos se observa un descenso en el contenido de mioglobina cuando los animales tienen una deficiencia en hierro, también se ha descrito un aumento cuando los animales poseen una deficiencia en vitamina E y por el ejercicio (Belitz et al., 1997).
La mayor parte de las proteínas solubles son enzimas, principalmente enzimas necesarias para la glicolisis y el ciclo de las pentosas-fosfato. También existe una serie de enzimas relacionadas con el metabolismo del ATP, como la creatin-kinasa y la ADP-desaminasa. Además, existen enzimas a las que se les atribuye el envejecimiento de la carne en los fenómenos postmortem (Parreño et al., 1994), como son las proteinasas musculares, especialmente calpaínas y proteinasas lisosomales (catepsinas B, H, L, etc.).
Proteínas insolubles o del estroma.Las proteínas del estroma representan la fracción insoluble proteica de las proteínas musculares. Están constituidas principalmente por proteínas del tejido conjuntivo y se distribuyen por todo el organismo animal, formando parte del esqueleto y de la estructura de órganos, tendones, nervios, resto de membranas y la porción insoluble del aparato contráctil.
Sus principales componentes son el colágeno, la elastina y la reticulina. Son proteínas de valor nutritivo, capacidad de retención de agua y poder emulsionante menores que las anteriormente descritas (Ordoñez et al., 1998).
Proteínas sarcoplásmicas Las proteínas sarcoplásmicas constituyen alrededor del 29 % del total de las proteínas del músculo esquelético (Kauffman, 2001) y corresponden a la fracción soluble a baja fuerza iónica (0,1 o inferior) a pH neutro (Pearson y Young, 1989). Dependiendo de las condiciones usadas para su extracción, esta fracción puede contener entre 100 y 200 proteínas diferentes. Su composición variará según la velocidad y el grado de homogeneización del tejido antes de la extracción, el pH, la naturaleza del solvente utilizado para la extracción y la fuerza centrífuga seleccionada para separar las proteínas sarcoplásmicas (solubles) de la fracción insoluble. La fracción de proteínas sarcoplásmicas contiene las mismas proteínas que se extraen de otras células, es decir, enzimas asociados con la glucólisis y síntesis de carbohidratos y proteínas, junto con otras proteínas exclusivas de la célula muscular, tales como precursores para la síntesis de la miosina y la mioglobina (Whitaker, 1977).
La mioglobina es la proteína más abundante de esta fracción y está formada por una porción proteica, la globina, y una porfirina o grupo hemo que es responsable de su color. Dentro del anillo de porfirina, y en posición central, existe un átomo de hierro que mantiene unido el complejo formado por la globina y el grupo hemo.
El color de la carne dependerá del estado de oxidación de la mioglobina, el tipo de ligando unido al grupo hemo y el estado de la globina. El hierro del anillo de porfirina

puede existir en dos formas: como hierro ferroso reducido (+2) o férrico oxidado (+3). Cuando el hierro del grupo hemo está en estado +2 (ferroso) se denomina mioglobina, de color púrpura y es la forma que se encuentra mayoritariamente en el músculo. Cuando se produce la oxidación de la mioglobina, el átomo de hierro ferroso se convierte en férrico, formándose metamioglobina que es de color marrón. La unión al oxígeno por oxigenación produce oximioglobina dando un color rojo brillante al músculo. La interconversión de las tres formas ocurre fácilmente, dependiendo de las condiciones del medio como la presión parcial de oxígeno, temperatura y pH, produciendo los cambios de color de la carne (Young y West, 2001).
Proteínas miofibrilares.Las proteínas miofibrilares constituyen alrededor del 60 % del total del músculo esquelético (Kauffman, 2001). Son las proteínas estructurales que forman las miofibrillas y poseen una solubilidad intermedia entre las proteínas sarcoplásmicas y las del estroma (Pearson y Young, 1989). Las proteínas miofibrilares, cuya unidad estructural se denomina sarcómero son las responsables de la contracción muscular aunque no todas las proteínas de esta fracción se encuentran directamente relacionadas con la contracción. Por tanto, según su función las podemos dividir en 3 grupos (Pearson y Young, 1989):
- Proteínas contráctiles mayoritarias, incluyen actina y miosina.- Proteínas reguladoras, juegan un importante papel en la iniciación y control de la
contracción y relajación muscular.- Proteínas del citoesqueleto o estroma, aportan soporte estructural y mantienen las
miofibrillas alineadas.
En el Cuadro 17 se muestran las diversas proteínas miofibrilares según los 3 grupos indicados anteriormente, las proporciones relativas de cada proteína en la miofibrilla, su localización en el sarcómero y su función principal. La miosina y la actina combinadas constituyen el 70 % del total de las proteínas miofibrilares siendo la miosina un 50 % y la actina un 20 % del total.
Las proteínas reguladoras, aunque no participan directamente en el proceso de la contracción muscular, juegan un papel importante en la modulación de la contracción e inician el movimiento en el músculo esquelético. En este grupo se incluyen la tropomiosina, las troponinas y las actininas. La tropomiosina supone un 3 % del total de proteínas miofibrilares. En el músculo, la tropomiosina se encuentra siempre asociada con la actina y con el complejo de las troponinas.
Por otra parte, las troponinas constituyen el 4,5 % del total de las proteínas miofibrilares. La troponina consiste en 3 subunidades llamadas troponina C, troponina I y troponina T (1,15 %, 1,35 % y 1,95 % respectivamente, del total de las proteínas miofibrilares). La troponina C se llama así porque tiene alta afinidad por el Ca2+ libre. La troponina I debe su nombre al hecho de inhibir la interacción de la miosina y la actina en la relajación muscular junto con la tropomiosina, la troponina C y la troponina T. La troponina T recibe este nombre porque se une a la tropomiosina y conecta el complejo troponina C-troponina I a la actina-F. Las subunidades de la troponina juegan un papel importante en el ciclo contracción- relajación.
Las actininas actúan en la regulación de la contracción, aunque menos directamente que la tropomiosina y el complejo de troponinas. La α-actinina es probablemente la más importante del grupo de actininas y comprende el 1 % del total de proteínas

miofibrilares. Es el componente mayoritario de la línea-Z y ancla los filamentos delgados de los sarcómeros adyacentes. Las otras actininas presentes en el músculo incluyen las β-actinina, γ-actinina y la Eu-actinina que comprenden menos del 0,01 %, 0,01 % y 0,3 % de las proteínas miofibrilares, respectivamente. La β-actinina se encuentra al final de la banda A de los filamentos delgados y regula su longitud, la γ−actinina se localiza en los filamentos delgados e inhibe la polimerización de la G- actina. La Eu-actinina se encuentra en el disco Z y probablemente no tiene función en la regulación de la contracción pero contribuye a la densidad del disco Z por interacción con la actina y la α-actinina.
Las proteínas del citoesqueleto, el tercer grupo de proteínas miofibrilares aislado e identificado en el músculo, son proteínas estructurales y sirven de soporte a las proteínas contráctiles mayoritarias y a las reguladoras. Este grupo incluye la conectina, proteína-C, miomesina, nebulina, desmina, filamina, vimentina, sinemina, proteína-X, proteína-H, proteína-I, proteína-F y creatinquinasa. La conectina, es un componente de alto peso molecular que constituye el 8 % del total de las proteínas de la miofibrilla. Se encuentra localizada entre los sarcómeros y parece mantener los filamentos gruesos en posición lateral en medio de la banda-A. La proteína-C constituye el 1,5 % de las proteínas miofibrilares y une las moléculas de miosina en los filamentos gruesos, aunque está situada en los filamentos delgados. El resto de proteínas constituyen menos del 1 % de las proteínas miofibrilares. Sus funciones y localización se recogen en la Tabla 1.1.
Cuadro 17.- Proteínas miofibrilares de la carne.

La miosina es el principal constituyente de los filamentos gruesos y está constituida por una cabeza globular con un peso molecular de alrededor de 480 kDa y una longitud de unos 1600 Å. Está formada por dos cadenas idénticas de polipéptidos de unos 225 kDa que pueden separarse tratando la miosina con disoluciones concentradas de urea o guanidina (Pearson y Young, 1989). Cada una de las cadenas posee una estructura de α-hélice y, a su vez, se enrollan formando una doble hélice. Al final de la molécula de miosina, ambas cadenas se pliegan en una estructura globular formada por dichas cadenas más dos cadenas de polipéptidos más pequeñas (Pearson y Young, 1989).
A partir de los estudios de fragmentación enzimática se ha obtenido información muy importante sobre la estructura de la miosina y su actividad ATPasa. La exposición de la miosina a tripsina o quimotripsina da lugar a dos fragmentos llamados meromiosina pesada y meromiosina ligera (HMM y LMM). La meromiosina ligera no tiene actividad ATPasa ni capacidad de unión a la actina. La meromiosina pesada tiene actividad ATPasa y capacidad de unión a la actina pero no forma filamentos. El tratamiento con tripsina de la meromiosina pesada da lugar a dos subfragmentos llamados HMM-S1 y HMM-S2. El subfragmento 1 tiene actividad ATPasa y capacidad de unión a la actina mientras que el fragmento 2 no tiene estas características, lo cual indica que dicha actividad reside enteramente en su cabeza y que hay dos sitios catalíticos por molécula de miosina (Whitaker,1977).
La actina es el mayor constituyente de los filamentos delgados y supone el 20 % de las proteínas miofibrilares (Asghar, 1985). La actina puede encontrarse en dos formas, actina-G (actina globular) y actina-F (actina filamentosa) que se obtiene por la polimerización de la actina-G. La forma monomérica (actina-G) es estable en agua destilada, pero bajo ciertas condiciones polimeriza formando actina-F (Whitaker 1977).

Durante la contracción muscular se produce la unión de los filamentos de actina y de miosina formando el complejo actomiosina. También, en procesos in vitro en los que se ponen juntos miosina y actina se forma dicho complejo. Su formación va acompañada de un aumento de la viscosidad de la solución (Morrissey, 1987). La composición y el peso molecular de la actomiosina dependen mucho de las condiciones experimentales, tales como el pH y las concentraciones de KCl, MgCl2 y proteína (Morrissey, 1987). Los complejos de actomiosina pueden ser extraídos del músculo por exposición prolongada a 0,6 M de KCl. Una propiedad de los complejos de actomiosina es que se disocian en presencia de ATP y Mg2+ y, cuando esto ocurre, la disociación va acompañada por una disminución rápida y elevada de la viscosidad de la solución y por la hidrólisis del ATP. Cuando la hidrólisis se completa, la actina y la miosina se reagregan por la interacción de los grupos sulfhidrilos que parecen ser los responsables tanto de la actividad ATPasa de la miosina como de su capacidad de unión a la actina (Lehninger, 1970).
Proteínas del estroma.Las proteínas del estroma suponen el 10-15 % del total de proteínas en el músculo esquelético (Morissey et al., 1987) y constituyen la fracción insoluble (en solventes acuosos neutros y disoluciones diluidas de sal) de las proteínas de la célula muscular. La mayoría de las proteínas del tejido conectivo se clasifican como fibrosas y consisten en cadenas de polipéptidos dispuestos paralelamente unas a otras formando láminas o fibras. Son las encargadas de aportar dureza, dar forma y proteger el músculo esquelético (Pearson y Young, 1989).
Las principales proteínas del tejido conectivo son el colágeno, la elastina y la queratina. Junto con estas proteínas existen otras, que se encuentran unidas covalentemente a los carbohidratos y se denominan glicoproteínas. Aunque el porcentaje de la composición puede variar ampliamente dependiendo de la fuente del músculo, el colágeno frecuentemente supone el 95 % del total de las proteínas del estroma y la elastina supone el 5 % de la proteína total del estroma (Kauffman, 2001).
Las proteínas son macromoléculas compuestas por carbono, hidrógeno, oxígeno y nitrógeno. La mayoría también contienen azufre y fósforo. Las mismas están formadas por la unión de varios aminoácidos, unidos mediante enlaces peptídicos. El orden y disposición de los aminoácidos en una proteína depende del código genético, ADN, de la persona.
1.1.1. Las proteínas son clasificables según su estructura química en:
Proteínas simples: Producen solo aminoácidos al ser hidrolizados.Albúminas y globulinas: Son solubles en agua y soluciones salinas diluidas (ej.: lactoalbumina de la leche).Glutelinas y prolaninas: Son solubles en ácidos y álcalis, se encuentran en cereales fundamentalmente el trigo. El gluten se forma a partir de una mezcla de gluteninas y gliadinas con agua.Albuminoides: Son insolubles en agua, son fibrosas, incluyen la queratina del cabello, el colágeno del tejido conectivo y la fibrina del coagulo sanguíneo.Proteínas conjugadas: Son las que contienen partes no proteicas. Ej.:

nucleoproteínas.Proteínas derivadas: Son producto de la hidrólisis.
En el metabolismo, el principal producto final de las proteínas es el amoníaco (NH3) que luego se convierte en urea (NH2)2CO2 en el hígado y se excreta a través de la orina.
1.1.2. Las diferencias entre los tipos de proteína:¿de origen animal o vegetal?Las proteínas son macromoléculas formadas por la unión de miles o cientos de aminoácidos. Los aminoácidos se dividen en aminoácidos esenciales y no esenciales. Los esenciales son aquellos que no son elaborados por nuestro organismo y deben incorporarse a través de la dieta. Los no esenciales son sintetizados por nuestro metabolismo.Los aminoácidos son fundamentales para el buen funcionamiento del organismo. Para una persona adulta son ocho los aminoácidos esenciales, mientras que durante el crecimiento se precisan dos más. Aminoácidos esenciales: fenilalanina, leucina, isoleucina, lisina, metionina, treonina,
triptofano y valina. Durante la infancia y adolescencia: arginina e histidina. Aminoácidos no esenciales: alanina, cisteina, cistina, glicina, hidroxiprolina, prolina,
serina, tirosina, ácido aspártico, y glutámico.
La calidad de una proteína depende de su contenido en aminoácidos esenciales. Esa calidad está medida por un índice llamado valor biológico.Por lo tanto, una proteína es de alta calidad o tiene un alto valor biológico cuando es rica en aminoácidos esenciales.
Las proteínas con un valor biológico alto son además de las proteínas de la leche materna, la de los huevos. Le siguen las proteínas de la carne y el pescado y luego los lácteos. Se considera que las proteínas de origen animal son más nutritivas y completas que las de origen vegetal, que son incompletas y de un menor valor biológico. Para que las proteínas vegetales sean completas deben mezclarse entre sí. Por ejemplo: una legumbre + un cereal o un fruto seco + arroz. En un desayuno, al mezclar la leche con los cereales, la proteína del cereal se completa con las de la leche.
Cuadro 18.- Calidad de las proteínas.
Alimento valor biológico Alimento valor biológicoLeche materna 100 Soja 70Huevo 100 Arroz 60Carne 75 Trigo 50Pescado 75 Legumbres 40Leche de vaca 75 Maíz 40
Los alimentos que nos aportan proteínas completas o de alto valor biológico son todos los de origen animal: Todas las carnes, los huevos y el pescado Todos los quesos La leche y todos sus derivados (yogur) Crustáceos y mariscos.
Los alimentos que nos aportan proteínas incompletas, son todos de origen vegetal:

la soja las legumbres (lentejas , garbanzos) los frutos secos los cereales y sus derivados (harinas, arroz. Pan ) hortalizas y frutas
1.1.3. AMINOÁCIDOS Y ENLACE PEPTÍDICO.
1.1.3.1. Estructura de los aminoácidos.Los aminoácidos proteicos tienen una estructura formada por un grupo amino y un grupo carboxilo unidos al mismo carbono, el carbono α, que está unido a su vez a un hidrógeno y a otro grupo característico de cada aminoácido (en la glicina, es otro hidrógeno). Considerando como ejemplo la alanina, se pueden destacar los siguientes aspectos comunes de la estructura de los aminoácidos:
Como se ha indicado, estos aspectos estructurales son comunes a todos los aminoácidos, con la excepción de la glicina, que tiene un hidrógeno unido al carbono α, y de la prolina, que es propiamente un iminoácido, no un aminoácido.
Todos los aminoácidos, con la excepción de la glicina tienen un carbono α asimétrico. Consecuentemente, pueden existir en principio en dos formas, la L y la D.Todos los aminoácidos de las proteínas pertenecen a la serie L.
1.1.3.2. Aminoácidos proteicos.Las proteínas de todos los seres vivos están constituidas por una veintena de aminoácidos distintos, además de algún otro formado por modificaciones posteriores a la síntesis de la cadena polipeptídica.
Desde el punto de vista nutricional, es muy importante tener en cuenta que, aunque la mayoría de los aminoácidos pueden sintetizarse en el organismo siempre que se disponga de suficiente nitrógeno orgánico, esto no sucede con todos, Algunos son "esenciales", es decir, no pueden sintetizarse y tienen que obtenerse ya como tales de las proteínas de la dieta.

1.1.3.3. Aminoácidos alifáticos.Los aminoácidos alifáticos tienen carácter hidrófóbico, tanto más marcado cuanto mayor es la longitud de la cadena. La glicina tiene un tamaño muy pequeño, y permite con su presencia la formación de estructuras particulares, como la triple hélice del colágeno. Todos ellos son muy estables desde el punto de vista químico, y no se ven afectados prácticamente por ningún proceso de los que se llevan a cabo en la industria alimentaria. La valina, leucina e isoleucina son aminoácidos esenciales, pero su abundancia en casi todas las proteínas hace que nunca sean los limitantes de su valor nutritivo.
Glicina Alanina Valina
Leucina Isoleucina
Aminoácidos alifáticos
1.1.3.4. Prolina.La prolina es un iminoácido, es decir, su grupo amino no es un grupo amino primario, como los de los demás aminoácidos, sino secundario. La presencia del anillo impide el giro sobre ese enlace, y consecuentemente la organización de la estructura secundaria de la proteína. Aquellas proteínas con abundante prolina o bien tienen muy poca estructura secundaria (caseínas) o bien tienen una estructura peculiar (colágeno).
1.1.3.5. Aminoácidos aromáticos.Los tres aminoácidos aromáticos son esenciales, fenilalanina y triptófano estrictamente, es decir, en ningún caso se pueden sintetizar, mientras que la tirosina se puede obtener de la dieta o sintetizarla a partir de la fenilalanina. Además de formar parte de las proteínas, son precursores de otros compuestos biológicos. En las proteínas, son responsables de su absorción en el UV próximo. El triptófano es relativamente inestable, mientras que fenilalanina y tirosina son estables.
Fenilalanina Tirosina Triptófano
Aminoácidos aromáticos
1.1.3.6. Aminoácidos con azufre.Los dos aminoácidos azufrados son esenciales, la metionina estrictamente, mientras que la cisteina puede formarse a partir de la metioniona (no al revés). Las dietas basadas en leguminosas pueden ser deficientes en estos aminoácidos. Además, ambos son bastante inestables frente a condiciones de oxidación. La cisteina es muy imporatnte en el mantenimiento de la estructura terciaria y cuaternaria de la mayoría de las proteínas mediante la formación de puentes disulfuro, en dímeros de la cisteina formados por oxidación.
Metionina Cisteina
Aminoácidos con azufre
1.1.3.7. Aminoácidos alifáticos con hidroxilo.

Estos aminoácidos tienen carácter hidrofílico. La treonina es esencial, pero no la serina. Ambos aminoácidos, especialmente la serina, pueden estar modificados por fosforilación, o por glicosilación en el caso de las glicoproteínas. Son relativamente inestables en medio alcalino.
Serina Treonina
Aminoácidos alifáticos con hidroxilo
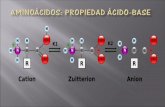
1.1.3.8. Aminoácidos ácidos.El ácido aspártico y el ácido glutámico tienen un grupo carboxilo en la cadena lateral, además del que forma el enlace peptídico. Este grupo carboxilo puede estar o no ionizado en función del pH del medio. Son aminoácidos hidrófilos, y los responsables de las cargas - de la proteína.
Acido aspártico Acido glutámico
Aminoácidos ácidos
1.1.3.9. Aminoácidos con grupos amida.Estos aminoácidos, de carácter hidrófilo, son las amidas del amonio del aspártico y glutámico. Pierden el amonio con relativa facilidad a temperatura elevada, pero esta pérdida es irrelevante desde el punto de vista nutricional.
Asparagina Glutamina
Aminoácidos con grupos amida
1.1.3.10. Aminoácidos básicos.Los aminoácidos básicos son la lisina, arginina e histidina. La lisina es esencial, y además el aminoácido limitante en las dietas basadas en cereales, muy extendidas entre la población mundial. La arginina y la histidina son esenciales para los niños. Estos aminoácidos son hidrofílicos, teniendo o no carga + en función del pH del medio. Son relativamente inestables, especialmente la lisina, pudiendo reaccionar con los carbohidratos a temperaturas elevadas.
Lisina Arginina Histidina
Aminoácidos básicos
1.1.3.11. Selenocisteína.La selenocisteína es el equivalente a la cisteína, pero con átomo de selenio en lugar del átomo de azufre. Se ha encontrado en unas pocas proteínas, la glutatión peroxidasa, la tetraikiodotironina 5' deiodinasa y la formato deshidrogenasa. Precisamente forman parte de este aminoácido es el único papel fisiológico

conocido del selenio. Este aminoácido se incorpora a las proteínas durante su síntesis, mediante una codificación particular.
1.1.3.12. Aminoácidos modificados por hidroxilación.Algunas proteínas contienen aminoácidos hidroxilados. Los más abundantes son la hidroxiprolina y la hidroxilisina, que se producen, después de la síntesis de las cadenas polipeptídicas, a expensas de la prolina y la lisina. Es decir, estos aminoácidos no son insertados como tales en la cadena polipeptídica, y no tienen un código genético propio. Estos aminoácidos son particularmente abundantes en el colágeno.
Hidroxiprolina Hidroxilisina
Aminoácidos hidroxilados
1.1.3.13. Otros aminoácidos.Otros aminoácidos producidos por modificación tras la síntesis de las proteínas son la n-metil-lisina, que se encuentra en la miosina, la metil-arginina y el α -carboxiglutamato, presente en algunas enzimas.
N-metil-lisina α -carboxiglutámico
Otros aminoácidos modificados
1.1.4. Enlace peptídico.Se llama enlace peptídico al enlace amina formado entre el grupo carboxilo de un aminoácido y el grupo amino del siguiente. Nominalmente es un enlace sencillo, pero en realidad tiene un porcentaje significativo de doble enlace, por resonancia con el grupo carbonilo del ácido. La consecuencia es que el enlace da lugar a una unidad rígida planar, con el N del grupo NH en posición trans con respecto al grupo carbonilo.
El enlace peptídico puede romperse con relativa facilidad por hidrólisis, catalizada por ácidos, álcalis y sobre todo por muchas enzimas, las llamadas proteasas o proteinasas.

1.1.5. Puentes disulfuro.Los puentes disulfuro se forman entre los azufres de la cadena lateral de dos restos de cisteina, por una reacción de oxidación. Son esenciales en el mantenimiento de la estructura terciaria y cuaternaria de la mayoría de las proteínas. Pueden formarse y romperse con relativa facilidad en el procesado de los alimentos, dependiendo de las condiciones.
1.1.6. Estructura de las proteínas:

Estructura de las proteínas. Proyecto del Genoma Humano.
1.1.6.1. Estructura primaria.Una cadena polipeptídica consiste en una cadena lineal de aminoácidos unidos por enlaces peptídicos. El primer puesto de la cadena corresponde al grupo amino terminal, y la estructura primaria es la secuencia en la que están situados todos los constituyentes hasta llegar al carboxilo terminal. Esta secuencia está codificada genéticamente.
Existen cadenas polipeptídicas de cualquier número de aminoácidos, sin que exista una solución de continuidad entre péptidos y proteínas. Por convención, se suele considerar proteína a quellos polipéptidos con un peso molecular del orden de 10.000 o más.
1.1.6.2. Estructura secundaria.La estructura secundaria es la forma en la que la cadena polipeptidica se pliega en el espacio. En una proteína, cada tramo de cadena polipeptídica tiene distinta estructura secundaria. Existen varias formas definidas de estructura secundaria, las más importantes de las cuales son las llamadas hélice y hoja plegada.

Las estructuras secundarias definidas están mantenidas por puentes de hidrógeno formados exclusivamente entre los grupos amino y carboxilo que constituyen el esqueleto de la cadena polipeptídica. Consecuentemente, los parámetros estructurales (distancias, ángulos) serán iguales, independientemente de la proteína y de los aminoácidos que formen la estructura.
1.1.6.3. Estructura terciaria.La estructura terciaria de la proteína es la forma en la que se organizan en el espacio los diferentes tramos de la cadena polipeptídica, que pueden tener una estructura secundaria definida, como las hélices u hojas o no tenerla. La estructura terciaria está mantenida por enlaces iónicos y de puentes de hidrógeno entre las cadenas laterales de los aminoáciodos, enlaces hidrofóbicos y eventualmente puentes disulfuro.
1.1.6.4. Estructura cuaternaria.La estructura cuaternaria de una proteína es la forma en la que se asocian las distintas subunidades constituyentes, si es que existen. Es decir, para poder hablar de estructura cuaternaria es necesario que la proteína esté formada por varias subunidades. Como ejemplos de proteínas con estructura cuaternaria se puede considerar la hemoglobina, las inmunoglobulinas o la miosina.
1.2. ENZIMAS.Estructura de la triosafosfato isomerasa. Conformación en forma de diagrama de
cintas rodeado por el modelo de relleno de espacio de la proteína. Esta proteína es una eficiente enzima involucrada en el proceso de transformación de azúcares en energía en las células.
Las enzimas son moléculas de naturaleza proteica que catalizan reacciones químicas, siempre que sean termodinámicamente posibles: Una enzima hace que una reacción química que es energéticamente posible pero que transcurre a una velocidad muy baja, sea cinéticamente favorable, es decir, transcurra a mayor velocidad que sin la presencia de la enzima. En estas reacciones, las enzimas actúan sobre unas moléculas denominadas sustratos, las cuales se convierten en moléculas diferentes denominadas productos. Casi todos los procesos en las células necesitan enzimas para que ocurran a unas tasas significativas. A las reacciones mediadas por enzimas se las denomina reacciones enzimáticas.
Debido a que las enzimas son extremadamente selectivas con sus sustratos y su velocidad crece sólo con algunas reacciones, el conjunto (set) de enzimas sintetizadas en una célula determina el tipo de

metabolismo que tendrá cada célula. A su vez, esta síntesis depende de la regulación de la expresión génica.
1.2.1. Estructuras y mecanismos.Diagrama de cintas que representa la estructura de una anhidrasa carbónica de tipo II. La esfera gris representa al cofactor zinc situado en el centro activo.
Las enzimas son generalmente proteínas globulares que pueden presentar tamaños muy variables, desde 62 aminoácidos como en el caso del monómero de la 4-oxalocrotonato tautomerasa, hasta los 2.500 presentes en la sintasa de ácidos grasos.
Las actividades de las enzimas vienen determinadas por su estructura tridimensional, la cual viene a su vez determinada por la secuencia de aminoácidos. Sin embargo, aunque la estructura determina la función, predecir una nueva actividad enzimática basándose únicamente en la estructura de una proteína es muy difícil, y un problema aún no resuelto.
Casi todas las enzimas son mucho más grandes que los sustratos sobre los que actúan, y solo una pequeña parte de la enzima (alrededor de 3 a 4 aminoácidos) está directamente involucrada en la catálisis. La región que contiene estos residuos encargados de catalizar la reacción es denominada centro activo. Las enzimas también pueden contener sitios con la capacidad de unir cofactores, necesarios a veces en el proceso de catálisis, o de unir pequeñas moléculas, como los sustratos o productos (directos o indirectos) de la reacción catalizada. Estas uniones de la enzima con sus propios sustratos o productos pueden incrementar o disminuir la actividad enzimática, dando lugar así a una regulación por retroalimentación positiva o negativa, según el caso.
1.2.2. Cinética enzimática.Mecanismo para una reacción catalizada por una enzima con un único sustrato. La enzima (E) une un sustrato (S) y genera un producto (P).
La cinética enzimática es el estudio de cómo las enzimas se unen a sus sustratos y los transforman en productos. Los datos de equilibrios utilizados en los estudios cinéticos son obtenidos mediante ensayos enzimáticos.
1.2.3. Clasificación y nomenclatura de enzimas.El nombre de una enzima suele derivarse del sustrato o de la reacción química que cataliza, con la palabra terminada en -asa. Por ejemplo, lactasa proviene de su sustrato lactosa; alcohol deshidrogenasa proviene de la reacción que cataliza que consiste en "deshidrogenar" el alcohol; ADN polimerasa proviene también de la reacción que cataliza que consiste en polimerizar el ADN.
La Unión Internacional de Bioquímica y Biología Molecular ha desarrollado una nomenclatura para identificar a las enzimas basadas en los denominados Números EC. De este modo, cada enzima queda registrada por una secuencia de cuatro números precedidos por las letras "EC". El primer número clasifica a la enzima según su mecanismo de acción. A continuación se indican las seis grandes clases de enzimas existentes en la actualidad:

EC1 Oxidorreductasas: catalizan reacciones de oxidorreducción o redox. Precisan la colaboración de las coenzimas de oxidorreducción (NAD+, NADP+, FAD) que aceptan o ceden los electrones correspondientes. Tras la acción catalítica, estas coenzimas quedan modificadas en su grado de oxidación, por lo que deben ser recicladas antes de volver a efectuar una nueva reacción catalítica. Ejemplos: deshidrogenasas, peroxidasas.
EC2 Transferasas: transfieren grupos activos (obtenidos de la ruptura de ciertas moléculas) a otras sustancias receptoras. Suelen actuar en procesos de interconversión de monosacáridos, aminoácidos, etc. Ejemplos: transaminasas, quinasas.
EC3 Hidrolasas: catalizan reacciones de hidrólisis con la consiguiente obtención de monómeros a partir de polímeros. Actúan en la digestión de los alimentos, previamente a otras fases de su degradación. La palabra hidrólisis se deriva de hidro → 'agua' y lisis → 'disolución'. Ejemplos: glucosidasas, lipasas, esterasas.
EC4 Liasas: catalizan reacciones en las que se eliminan grupos H2O, CO2 y NH3
para formar un doble enlace o añadirse a un doble enlace. Ejemplos: descarboxilasas, liasas.
EC5 Isomerasas: actúan sobre determinadas moléculas obteniendo de ellas sus isómeros funcionales o de posición, es decir, catalizan la racemización y cambios de posición de un grupo en determinada molécula obteniendo formas isoméricas. Suelen actuar en procesos de interconversión. Ejemplo: epimerasas (mutasa).
EC6 Ligasas: catalizan la degradación o síntesis de los enlaces denominados "fuertes" mediante el acoplamiento a moléculas de alto valor energético como el ATP. Ejemplos: sintetasas, carboxilasas.
1.2.4. Aplicaciones industriales.Las enzimas son utilizadas en la industria química, y en otros tipos de industria, en donde se requiere el uso de catalizadores muy especializados. Sin embargo, las enzimas están limitadas tanto por el número de reacciones que pueden llevar a cabo como por su ausencia de

estabilidad en solventes orgánicos y altas temperaturas. Por ello, la ingeniería de proteínas se ha convertido en un área de investigación muy activa donde se intentan crear enzimas con propiedades nuevas, bien mediante diseño racional, bien mediante evolución in vitro. Estos esfuerzos han comenzado a tener algunos éxitos, obteniéndose algunas enzimas que catalizan reacciones no existentes en la naturaleza.
Cuadro 19.- A continuación se muestra una tabla con diversas aplicaciones industriales de las enzimas:
AplicaciónEnzimas utilizadas
Usos
Procesado de alimentos
La amilasa cataliza la degradación del almidón en azúcares sencillos.
Amilasas de hongos y plantas.
Producción de azúcares desde el almidón, como por ejemplo en la producción de jarabe de maíz. En la cocción al horno, cataliza la rotura del almidón de la harina en azúcar. La fermentación del azúcar llevado a cabo por levaduras produce el dióxido de carbono que hace "subir" la masa.
ProteasasLos fabricantes de galletas las utilizan para reducir la cantidad de proteínas en la harina.
Alimentos para bebés TripsinaPara pre-digerir el alimento dirigido a bebés.
Elaboración de cerveza
Cebada germinada utilizada para la elaboración de malta.
Las enzimas de la cebada son liberadas durante la fase de molido en la elaboración de la cerveza.
Las enzimas liberadas degradan el almidón y las proteínas para generar azúcares sencillos, aminoácidos y péptidos que son usados por las levaduras en el proceso de fermentación.
Enzimas de cebada producidas a nivel industrial
Ampliamente usadas en la elaboración de cerveza para sustituir las enzimas naturales de la cebada.
Amilasa, glucanasa y proteasas
Digieren polisacáridos y proteínas en la malta.
Betaglucanasas y arabinoxilanasas
Mejoran la filtración del mosto y la cerveza.
Amiloglucosidasas Producción de cerveza baja en

y pululanasascalorías y ajuste de la capacidad de fermentación.
ProteasasEliminan la turbidez producida durante el almacenamiento de la cerveza.
Acetolactatodecarboxilasa (ALDC)
Incrementa la eficiencia de la fermentación mediante la reducción de la formación de diacetilo.
Zumos de frutasCelulasas, pectinasas
Aclarado de zumos de frutos.
Industria láctea
Queso de Roquefort.
Renina, derivado del estómago de animales rumiantes jóvenes (como terneros y ovejas).
Producción de queso, usada para hidrolizar proteínas.
Enzimas producidas por bacterias
Actualmente, cada vez más usadas en la industria láctea.
LipasasSe introduce durante el proceso de producción del queso Roquefort para favorecer la maduración.
LactasasRotura de la lactosa en glucosa y galactosa.
Digestión de carne PapaínaAblandamiento de la carne utilizada para cocinar.
Industria del almidón
Glucosa.
Fructosa.
Amilasas, amiloglucosidasas y glucoamilasas
Conversión del almidón en glucosa y diversos azúcares invertidos.
Glucosa isomerasa
Conversión de glucosa en fructosa durante la producción de jarabe de maíz partiendo de sustancias ricas en almidón. Estos jarabes potencian las propiedades edulcorantes y reducen las calorías mejor que la sacarosa y manteniendo el mismo nivel de dulzor.
Industria del papel Amilasas, xilanasas, celulasas
Degradación del almidón para reducir su viscosidad, añadiendo

Una fábrica de papel en Carolina del Sur.
y ligninasas
apresto. Las xilanasas reducen el blanqueador necesario para la decoloración; las celulasas alisan las fibras, favorecen el drenaje de agua y promueven la eliminación de tintas; las lipasas reducen la oscuridad y las ligninasas eliminan la lignina para ablandar el papel.
Industria del biofuel
Celulosa en 3D.
CelulasasUtilizadas para degradar la celulosa en azúcares que puedan ser fermentados.
LigninasasUtilizada para eliminar residuos de lignina.
Detergentes biológicos
Principalmente proteasas, producidas de forma extracelular por bacterias.
Utilizadas para ayudar en la eliminación de tintes proteicos de la ropa en las condiciones de prelavado y en las aplicaciones directas de detergente líquido.
AmilasasDetergentes de lavadoras para eliminar residuos resistentes de almidón.
LipasasUtilizadas para facilitar la eliminación de tintes grasos y oleosos.
CelulasasUtilizadas en suavizantes biológicos.
Limpiadores de lentes de contacto
ProteasasPara eliminar restos proteicos de las lentes de contacto y así prevenir infecciones.
Industria del hule CatalasaPara generar oxígeno desde el peróxido, y así convertir el látex en hule espumoso.
Industria fotográfica Proteasa (ficina)
Disolver la gelatina de las películas fotográficas usadas, permitiendo así la recuperación de su contenido en plata.
Biología molecular Enzimas de restricción, ADN ligasa y
Utilizadas para manipular el ADN mediante ingeniería genética. De gran importancia en farmacología,

ADN de doble hélice.
polimerasas
agricultura, medicina y criminalística. Esenciales para digestión de restricción y para la reacción en cadena de la polimerasa.