Proteínas
18
C U R S O: BIOLOGÍA MENCIÓN MÓDULO 3 Unidad I: Organización, Estructura y Actividad Celular Proteínas y Metabolismo.
-
Upload
victor-lillo -
Category
Documents
-
view
213 -
download
1
description
Características químicas, clasificación y función de las proteínas . Mecanismos de acción, propiedades y variables que afectan la actividad enzimática
Transcript of Proteínas

C U R S O: BIOLOGÍA MENCIÓN
MÓDULO 3
Unidad I: Organización, Estructura y Actividad Celular
Proteínas y Metabolismo.
Modelos estructurales de proteínas transportadoras de oxígeno (hemoglobinas) basados en análisis de cristalografía de rayos X.

I) Proteínas.
Las proteínas son las principales macromoléculas de los seres vivos, constituyen más de la mitad del peso seco de una célula y son responsables de una gran cantidad de funciones. Cada uno de nosotros tiene más de diez mil tipos diferentes de proteínas, cada una de ellas con una estructura tridimensional única que le confiere una función específica.
Todas las proteínas están formadas por la misma estructura básica, son polímeros lineales formados por la unión de monómeros llamados aminoácidos. Los aminoácidos son moléculas constituidas por C, H, O, N y en algunos casos poseen también átomos de azufre (S).
1. A minoácidos (TABLA 1)
Son moléculas de bajo peso molecular que se caracterizan por poseer un grupo amino (-NH2), que tiene características básicas, y un grupo carboxilo (-COOH), con propiedades ácidas, ambos grupos unidos al mismo carbono central.
La fórmula general de un aminoácido es:
Átomo de carbono α (quiral)
H
H2N C COOH
Grupo amino R Grupo carboxilo (Básico) (Radical) (Ácido)
R es el radical, cadena lateral que diferencia a los aminoácidos en cuanto a tamaño, forma, carga eléctrica, etc., dando origen a alrededor de 20 aminoácidos diferentes que se encuentran formando parte estructural de las proteínas. Al pH del plasma sanguíneo (7,2) o del líquido intracelular (7,1) el grupo amino y el grupo carboxilo están ionizados.
Figura 1. Los aminoácidos son anfóteros, es decir pueden actuar como ácidos (dadores de protones) o como bases (aceptores de protones) dependiendo del medio.
Tabla 1. Los veinte aminoácidos proteicos y sus características estructurales.
2
C
H
R
COOH+H3N C
H
R
COO-H2NC
H
R
COO-+H3N
pH ácido pH neutro pH básico

C
H
COO-
NH3+
CH2CH2C
-O
O
Grupo Rno polar
Grupo RPolar sin carga
Grupo R polarcon carga negativa o positiva
1.Alanina (Ala)
9.Glicina (Gly)
16.Ácido aspártico (Asp)
2. Valina (Val)
10.Serina (Ser) 17. Ácido glutámico (Glu)
3.Leucina (Leu)
11.Treonina (Thr)
18. Lisina (Lis)
4.Isoleucina (Ile)
12. Cisteína (Cys)
19. Arginina (Arg)
5.Prolina (Pro)
13.Tirosina (Tyr)
20.Histidina (His)
6.Fenilalanina (Phe)
14.Asparagina (Asn)
7.Triptófano (Trp) 15.Glutamina (Gln)
3
Tabla 2. Necesidades diarias de aminoácidos esenciales (g)
Arginina 0(1)Histidina ?(2)Isoleucina 1,30Leucina 2,02Lisina 1,50Metionina 2,02Fenilalanina 2,02Treonina 0,91Triptófano 0,46Valina 1,50
(1) Necesario para bebes y niños en crecimiento
(2) Esencial pero en cantidades inferiores a las expresadas
C
H
COO-CH3
NH3+
C
H
COO-H
NH3+
C
H
COO-CH
NH3+
CH3
CH3
C
H
COO-CH2
NH3+
OH
C
H
COO-CH2
NH3+
CH
CH3
CH3
C
H
COO-CH
NH3+OH
CH3 C
H
COO-
NH3+
CH2CH2CH2CH2+H3N
C
H
COO-
NH3+
CHCH2
CH3
CH3C
H
COO-
NH3+
CH2HS C
H
COO-
NH3+
CH2CH2CH2NHCH2N
NH2+
C
CN
C
C
H
COO-
H2
+
H2
H2
H2
C
H
COO-
NH3+
CH2 C
H
COO-
NH3+
CH2
NH
CHC
HNCH
+
C
H
COO-
NH3+
CH2 C
H
COO-
NH3+
CH2
C
H
COO-CH2
NH3+
C
NH2
O
C
H
COO-
NH3+
CH2
C
C
NH
H
C
H
COO-
NH3+
CH2CH2C
NH2
O

8.Metionina (Met)
Concepto de aminoácido esencial.La mitad de los aminoácidos necesarios
para armar proteínas deben ingerirse en la dieta debido a que los animales no los podemos sintetizar, recibiendo el nombre de aminoácidos esenciales (Tabla 2).
2. Polipéptidos.
Los aminoácidos se unen entre sí mediante enlaces peptídicos para construir los polipéptidos. Estas uniones son enlaces covalentes tipo amida, formados entre el grupo carboxilo de un aminoácido y el grupo amino de otro, liberándose una molécula de agua (síntesis por deshidratación). Los enlaces peptídicos son idénticos a lo largo de las cadenas peptídicas. Al formarse estos enlaces desaparece el carácter anfótero de los aminoácidos.
La unión de dos aminoácidos origina un dipéptido (Figura 2); de tres, un tripéptido y desde 4 o más aminoácidos unidos se les denomina oligopéptidos (Figura 3). A mayores oligomerizaciones se les llama polipéptidos (200 a 300 residuos aminoacídicos). Las proteínas son consideradas cadenas polipeptídicas, que pueden estar conformadas por una o varias cadenas dispuestas en una configuración espacial definida.
La cadena polipeptídica puede mostrar propiedades ácidas o básicas por los grupos presentes en las cadenas laterales de sus aminoácidos. Existe un pH particular para cada proteína en el que su carga neta es cero. Figura 2. Formación de un dipéptido.
4
C
H
COO-
NH3+
CH2CH2SCH3
H2N C C
R
H
O
OH + H N
H
C
R
H
C
O
OH
N
H
C
R
H
C
O
OHH2N C C
R
H
O
C
O
N
HEnlace peptídico
+ H2O

Figura 3. Estructura lineal de un oligopéptido. Por ambos extremos de la molécula se pueden agregar más aminoácidos.
3. P roteínas .
a) Estructura proteica.
Las proteínas son largas cadenas de aminoácidos. Cada una de ellas podría adoptar, en principio, infinidad de formas, pero in vivo sólo presenta una, la más estable y la única que permite el desarrollo de su función específica. Existen cuatro tipos de organización proteica, (Figura 4).
Figura 4. Distintos niveles de organización proteica.
Las proteínas no son simplemente polímeros al azar de longitud variable. Cada tipo de molécula proteica posee una composición química específica (composición cualitativa en aminoácidos), una secuencia ordenada y única de esos aminoácidos, y un determinado peso molecular.
La forma específica que determina la función de una proteína está dada por cuatro niveles sucesivos de estructura, en la cual cada nivel prepara al siguiente.
Estructura primaria, es la secuencia de aminoácidos unidos por enlaces peptídicos. El orden de colocación de estos aminoácidos viene determinado genéticamente, es decir, está “escrito” en el material hereditario.
La estructura primaria es importante porque determina la conformación tridimensional específica de la proteína, necesaria para su función. Además, como es la traducción lineal de la secuencia de nucleótidos de los ácidos nucleicos, proporciona información sobre la contribución genética para la síntesis de proteínas.
Estructura secundaria, es la forma en que la cadena lineal de aminoácidos puede plegarse. Está condicionada por las interacciones entre las cadenas laterales de
5

aminoácidos y por las posibilidades de rotación alrededor de los enlaces. Resulta de la formación de enlaces puentes de hidrógeno de manera regular entre las cadenas de aminoácidos. Existen dos conformaciones más comunes alfa-hélice o beta-plegada.
Estructura terciaria, resulta de las interacciones entre los grupos R de la molécula de proteína y entre los grupos R y el medio circundante. Estas interacciones pueden ser puentes disulfuro, puentes de hidrógeno, e interacciones hidrofílicas o hidrofóbicas.
Se han identificado estructuras terciarias formadas exclusivamente por alfa-hélice, por lámina beta y combinaciones entre ambas. En los dos primeros tipos se forman proteínas fibrosas (muy resistentes e insolubles en agua), como la miosina del músculo, el colágeno y la elastina del tejido conectivo, las que realizan funciones estructurales. La combinación de hélices alfa y láminas beta origina proteínas globulares (solubles en agua).
Estructura cuaternaria, para ser funcionales algunas proteínas requieren de la asociación de varias cadenas polipeptídicas o subunidades; se dice que presentan estructura cuaternaria, refiriéndose a la disposición espacial de los polipéptidos para formar una proteína de mayor complejidad. Se suelen mantener unidos por enlaces químicos débiles (interacciones hidrofóbicas o fuerzas de van der Waals).
El número de polipéptidos que pueden formar parte de una proteína varía en cada caso. Un ejemplo clásico es la hemoglobina, que es un tetrámero formado por cuatro cadenas polipeptídicas relacionadas entre sí, ligadas a un átomo central de hierro.
b) Propiedades de las proteínas.
Cada proteína presenta propiedades físico - químicas derivadas de su composición aminoacídica y su conformación espacial. Son propiedades propias de todas las proteínas:
La especificidad y la desnaturalización.
El hecho que la función de una proteína dependa de su forma específica se hace evidente cuando se altera. En un proceso llamado desnaturalización (Figura 5), las cadenas de los polipéptidos se desenroscan, por lo que pierden su forma específica y, como resultado de ello su función. Por ejemplo, imaginemos lo que ocurre cuando freímos un huevo. El calor desnaturaliza a las proteínas de la clara (Ovoalbúmina), volviéndola de apariencia sólida, blanca y opaca. Los cambios en el pH celular también pueden desnaturalizar a muchas proteínas.
Figura 5. Desnaturalización de una proteína
C) Clasificación de las proteínas
Las proteínas se pueden clasificar según diversos criterios, por ejemplo:
6

c.1. Según su configuración espacial pueden ser:
Fibrosas: Son insolubles en agua; presentan gran resistencia física, por lo cual están generalmente vinculadas con acciones mecánicas o de protección, por ejemplo, colágeno (en tejidos conjuntivos, cartilaginosos, parte orgánica de los huesos), elastina (en tendones y vasos sanguíneos), queratina (piel, pelos, uñas, plumas, pezuñas, etc.).
Globulares: Formadas por cadenas plegadas que pueden adquirir formas esféricas o globulares, generalmente son solubles en agua. Las proteínas globulares desempeñan papeles muy dinámicos en el organismo; pertenecen a esta categoría todas las enzimas, anticuerpos, algunas hormonas (insulina, somatotrofina (GH), prolactina, etc.).
c.2. Según su composición química se pueden clasificar en:
Proteínas simples (holoproteínas): cuando su hidrólisis produce solamente aminoácidos. Por ejemplo; insulina, colágeno, albúminas, histonas, etc.
Proteínas conjugadas (heteroproteínas), presentan un grupo proteico simple y otro
componente, llamado grupo prostético, que pueden ser aniones, cationes, carbohidratos, lípidos, etc.
Ejemplos de heteroproteínas: Glucoproteínas: poseen hidratos de carbono como grupo prostético. Ej.
mucoproteína con función protectora; anticuerpos con función defensiva. Cromoproteínas: poseen un pigmento como grupo prostético. Ej. hemoglobina,
hemocianina y mioglobina, que transportan oxígeno; los citocromos que transportan electrones.
Fosfoproteínas: posee fósforo como grupo prostético. Ej. caseína. Lipoproteínas: tienen un lípido como grupo prostético. Ej. HDL (“colesterol
bueno”), LDL (“colesterol malo”) que transportan lípidos en la sangre. Nucleoproteínas: son proteínas asociadas a ácidos nucleicos. Ej. nucleosomas de
la cromatina; los ribosomas.
d) Funciones de las proteínasLas proteínas dirigen prácticamente la totalidad de los procesos vitales, incluso aquellos
destinados a la producción de ellas mismas. Determinan la forma y la estructura de las células. Sus funciones se relacionan con sus múltiples propiedades, que son el resultado de la composición de aminoácidos, de la secuencia y del modo en que la cadena se pliega en el espacio (Tabla 3).
Tabla 3. Funciones proteicas.FUNCION EJEMPLOS
Estructural
Ciertas Glucoproteínas forman parte de las membranas y participan como receptores o facilitan el transporte de sustancias.
Las proteínas del citoesqueleto, de las fibras del huso, de los cilios, flagelos y de los ribosomas.
Proteínas que confieren resistencia y elasticidad a los tejidos: El colágeno del tejido conjuntivo fibroso. La elastina el tejido conjuntivo elástico. La queratina de la epidermis.
Hormonal La insulina y el Glucagón (que regula la glicemia), la hormona del crecimiento, la Calcitonina (que regula la calcemia)
Homeostática La albúmina (Mantención del equilibrio osmótico).
Defensiva Inmunoglobulinas actúan como anticuerpos. La trombina y el fibrinógeno participan en la formación de coágulos, y
evitan las hemorragias.
7

Transporte
Transportan O2, la hemoglobina, en vertebrados y la mioglobina, en el interior de la célula muscular.
Lipoproteínas transportan lípidos en la sangre. Proteínas transportadoras de la membrana plasmática que regulan
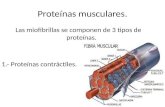
el paso de solutos.Contráctil Actina y miosina son parte de las miofibrillas, responsables de la
contracción muscular.
Reserva La Ovoalbúmina del huevo. la gliadina del grano de trigo, entre otras, son la reserva de aminoácidos
para el desarrollo del embrión.Enzimática Catalizan las reacciones químicas dentro de la célula. La más importante
es la Ribulosa bifosfato carboxilasa.
II. Enzimas y metabolismo.
Las enzimas son proteínas que actúan como catalizadores biológicos que aceleran las reacciones químicas dentro de la célula sin transformarse ellas mismas en una molécula diferente. Las células transforman la energía que toman del entorno en energía química la que les resulta útil para realizar trabajos químicos, mecánicos, etc.
El metabolismo celular, comprende a todas las reacciones químicas que suceden en la célula, permitiendo que ésta pueda:
Obtener energía química del entorno, como por ejemplo: de la luz solar, de sustancias orgánicas, de reacciones exotérmicas, etc.
Convertir los nutrientes ingeridos en precursores de macromoléculas orgánicas. Sintetizar y degradar biomoléculas propias de la fisiología celular.
Mayoritariamente las reacciones químicas que determinan el metabolismo están ligadas a secuencias llamadas rutas metabólicas, así el producto de una es el sustrato de otra; éstas ocurren en un determinado orden, controlado por:
La regulación de cada reacción por proteínas catalizadoras (enzimas). El acoplamiento de las reacciones, donde la energía requerida para las reacciones
endergónicas se desprende de las reacciones exergónicas. La síntesis de transportadores energéticos que capturan la energía de las reacciones
exergónicas y la conducen a las endergónicas.
Existen dos grupos de rutas metabólicas: las anabólicas y las catabólicas.
El catabolismo es la degradación enzimática de moléculas complejas a moléculas sencillas. Principalmente ocurren por reacciones de oxidación donde se libera energía libre de las grandes moléculas orgánicas y parte de esa energía se almacena en los enlaces fosfato del ATP.
El anabolismo es la construcción enzimática de moléculas orgánicas complejas a partir de moléculas precursoras sencillas. Para esto se requiere un aporte de energía aportado por el ATP.Ambos procesos, catabolismo y anabolismo suceden simultáneamente y son interdependientes, pero las etapas de ambos procesos son distintas.
1. Regulación del metabolismo.
La velocidad del catabolismo y del anabolismo está regulada por las necesidades energéticas celulares en cada instante y está controlada principalmente por las enzimas y las hormonas.
Las reacciones químicas para su inicio requieren siempre un aporte de energía llamada Energía de activación. Las enzimas son un tipo especial de proteínas que aceleran las reacciones químicas tanto en el medio intra como en el extra celular. Esta aceleración se debe a la capacidad que tienen las enzimas en disminuir la Energía de activación (Ea) de la reacción
8

química, es decir, permiten que una reacción ocurra en un breve lapso de tiempo. La energía de activación (Ea) representa la energía mínima necesaria que deben alcanzar los reactantes (sustratos) para pasar a productos (Figura 6).
Figura 6: Las enzimas aceleran las reacciones químicas disminuyendo la energía de activación.
2. Propiedades de las enzimas:
Son de naturaleza proteica. Tienen una acción específica (actúan sobre un determinado sustrato).
9
La Ribozima es la única enzima no proteica

Actúan en pequeñísimas cantidades. Permanecen inalteradas al final de la reacción, por lo que son reutilizables. Aceleran las reacciones químicas. Sólo aceleran reacciones favorables, o sea, que pueden ocurrir espontáneamente. No modifican el equilibrio de la reacción. Son sintetizadas en el retículo endoplasmático rugoso.
3. Estructura de las enzimas.
La molécula sobre la cual actúa una enzima recibe el nombre de sustrato, que puede ser uno o más, dependiendo de la enzima y la reacción catalizada. El lugar enzimático al que se une el o los sustratos se denomina sitio o centro activo. El sitio activo no sólo reconoce al sustrato y lo acopla, sino que también lo orienta en una determinada dirección. Las moléculas de sustrato se unen al centro activo de la enzima formando un complejo activado enzima-sustrato. Este sitio contiene los grupos funcionales que se unen al sustrato y efectúan la acción catalítica. Su geometría y su carga eléctrica están relacionadas con la conformación del sustrato y con el tipo de reacción, siendo responsables de la especificidad de la enzima.
Producida la reacción química, la enzima queda libre para efectuar una nueva reacción, así pocas moléculas de enzima procesan muchas de sustrato.
Enzima + Sustrato Complejo Enzima-Sustrato Productos + Enzima
El primer modelo sugerido para explicar la interacción enzima-sustrato fue propuesto por el químico Emil Fisher, denominado modelo llave-cerradura, que supone que la estructura del sustrato y la del sitio activo son exactamente complementarias, de la misma forma que una llave encaja en una cerradura ( Figura 7a).
Estudios posteriores sugirieron que el sitio activo es mucho más flexible que una cerradura. La interacción física entre las moléculas de enzima y sustrato produce un cambio en la geometría del centro activo, mediante la distorsión de las superficies moleculares. Este modelo llamado encaje inducido impondría cierta tensión a las moléculas reaccionantes, facilitando aún más la reacción (Figura 7 b).
10
a)
b)

Figura 7. (a) En el modelo llave-cerradura los sustratos interactúan en forma precisa con el sitio activo. (b) En el encaje inducido, la forma del sitio activo es complementaria del sustrato sólo después que éste se une a la enzima.
4. Clasificación enzimática:a) enzimas simples, están compuestas solamente por proteínas;
b) enzimas conjugadas, tienen una parte proteica llamada Apoenzima, que es inactiva, y una estructura o factor adicional, denominado cofactor enzimático. La proteína unida al cofactor se llama Holoenzima, que es la enzima activa (Figura 8). El cofactor puede ser un ión metálico, como por ejemplo: Mg2+, Mn2+, Cu2+, Zn2+, Cl-, Na+, K+, y otros; o una molécula orgánica (coenzima).La mayor parte de las coenzimas están hechas a partir de vitaminas, o bien son vitaminas en sí.
De la riboflavina (Vit. B2) derivan las coenzimas FMN (flavina mononucleótido) y FAD (Flavina adenina dinucleótido).
Del ácido nicotínico o niacina (Vit. B3) derivan las coenzimas NAD (nicotinamida adenina dinucleótido) y NADP (nicotinamida adenin dinucleótido fosfato).
Del ácido pantoténico (Vit. B5) deriva la coenzima A (CoA).
5. Factores que afectan la actividad enzimática.
11
Figura 8.Componentes y mecanismo de reacción de una enzima. Apoenzima es la enzima inactiva que requiere de un cofactor para activarse a Holoenzima.

La velocidad de la reacción enzimática depende de la concentración de enzimas y sustrato, de la concentración de las coenzimas, del pH, de la temperatura y de la presencia o ausencia de inhibidores.
a) Efecto del pH, las enzimas actúan dentro de límites estrechos de pH (pH óptimo de la reacción). Por ejemplo, la pepsina tiene un pH óptimo de 1,5 a 2.Cuando se grafica la actividad enzimática para valores crecientes de pH, comenzando desde la zona ácida, se obtiene una curva en forma de campana. El peak de la curva corresponde al pH óptimo en el cual la enzima tiene su máxima actividad. En medios muy ácidos o muy alcalinos, la proteína se desnaturaliza y se inactiva la enzima. Algunas enzimas tienen actividad óptima a un pH ácido y otras en un pH alcalino (Figura 9a).
b) Temperatura, la mayoría de las veces la velocidad de las reacciones enzimáticas aumenta con la temperatura, dentro del intervalo en que la enzima es estable y activa. La velocidad por lo general se duplica por cada 10°C de aumento térmico. La actividad enzimática máxima se alcanza a una temperatura óptima, luego la actividad decrece y finalmente cesa por completo; la actividad enzimática disminuye a causa de la desnaturalización progresiva de la enzima por acción de la temperatura. A bajas temperaturas, las reacciones disminuyen mucho o se detienen, pero la acción catalítica reaparece cuando la temperatura se eleva a valores normales para la enzima. (Figura 9b)
Actividad de la enzima ß–Glucosidasa de Penicillium purpurogenum (un hongo filamentoso).
Figura9a: Efecto del pH. Figura 9 b: Efecto de la temperatura.
c) Concentración del sustrato: principalmente la velocidad de la reacción o catálisis varía de acuerdo a la concentración del sustrato.
A baja concentración, la velocidad de la reacción es directamente proporcional a la concentración del sustrato. A concentraciones intermedias, la variación de la concentración del sustrato modifica la velocidad de reacción en una forma que no es proporcional. A altas concentraciones del sustrato, la velocidad de la reacción no depende de la concentración de éste.
12

Cuando las concentraciones del sustrato son bajas, la velocidad aumenta rápidamente. A medida que el sustrato aumenta, la curva se satura y alcanza un punto de equilibrio en el cual la velocidad no depende del sustrato. En este caso, el exceso de sustrato no es atacado por la enzima, ya que toda la enzima se encuentra como complejo enzima-sustrato y se alcanza el máximo de velocidad. (Figura 10)
Figura10: Efecto de la concentración del sustrato sobre la velocidad de una reacción enzimática.
d) Inhibición de la actividad enzimática.
Existen algunos compuestos químicos capaces de bloquear o disminuir la actividad enzimática. Esto ha permitido explicar algunos mecanismos de acción de medicamentos antimicrobianos, así como también dilucidar mecanismos de acción enzimática. Es una forma de control de las reacciones químicas. Puede ser reversible e irreversible.
d.1. Reversible. Las regiones funcionales de la enzima no cambian y sus efectos se pueden eliminar; puede ser a su vez:
Competitiva, el inhibidor (I) y el sustrato (S) se parecen y compiten por unirse al centro activo de la enzima. Se forma un complejo Enzima-inhibidor reversible. Su efecto se puede invertir aumentando la concentración del sustrato.
No competitiva, el inhibidor (I) se une a la enzima o al complejo enzima-sustrato de modo reversible, en un sitio distinto del centro activo, pero esencial para la molécula. Su efecto no puede invertirse por incremento de la concentración del sustrato.
d.2. Irreversible (Acompetitiva). Las regiones funcionales de la enzima sufren cambios permanentes. El inhibidor se une tan estrechamente a ella que se disocian con mucha lentitud y la actividad enzimática disminuye o, incluso, se pierde.
6. Regulación de la actividad enzimática.
Las enzimas suelen formar parte de sistemas multienzimáticos secuenciales, que son la base del control de las vías metabólicas; estas enzimas deben actuar coordinadamente en
13

tiempos adecuados para que las distintas rutas metabólicas cumplan con su función y no alteren los procesos vitales.
Una forma de regulación enzimática está relacionada con la disponibilidad de la enzima para que ocurra una reacción química. Existen tres formas de regular la concentración de las enzimas activas:
Regulación de la expresión génica, que controla la cantidad y velocidad de síntesis enzimática.
Proteolisis enzimática, que modifica la cantidad de enzima activa disponible. Modificación covalente de formas activas e inactivas interconvertibles por acción de
otras enzimas.
7. Control genético.
Las enzimas sintetizadas por una célula están determinadas por la información genética que se expresa en esta célula. Si las enzimas están siempre presentes en cantidades casi constantes, se denominan constitutivas. Aquellas sintetizadas sólo en respuesta a la presencia de ciertos sustratos se llaman inducidas o adaptativas. Los genes que codifican las enzimas inducidas suelen mantenerse reprimidos y se transcriben en respuesta al agente inductor.
El control metabólico en organismos superiores puede también estar regulado por el sistema endocrino. Las hormonas estimulan o inhiben rutas metabólicas de los tejidos blancos.
8. ¿Qué es el aspartamo? El aspartamo es un potente edulcorante. Es cerca de 200 veces más dulce que el azúcar (sacarosa), por lo que sólo se necesitan unas bajas concentraciones para endulzar alimentos y bebidas. Las cantidades utilizadas son tan pequeñas que el aspartamo casi no aporta calorías. El excelente perfil de su gusto lo ha convertido en uno de los principales edulcorantes bajos en calorías de todo el mundo.
El aspartamo es una molécula simple, compuesta por la combinación de dos aminoácidos (ácido aspártico y fenilalanina) y una pequeña cantidad de metanol. ¿Qué sucede cuando se toma aspartamo? Al igual que otros alimentos, el aspartamo se digiere en los intestinos descomponiéndose en sus distintos componentes. Los tres pasan a la corriente sanguínea y se metabolizan de la misma manera que si estos componentes formasen parte de cualquier otro alimento. El aspartamo en sí no puede atravesar intacto la pared intestinal y nunca penetra así en la corriente sanguínea.
La IDA (La Ingesta Diaria Admisible) es una valoración de la cantidad de un aditivo alimentario que se puede ingerir diariamente durante toda una vida sin sufrir un riesgo apreciable para la salud. La IDA de 0-40 mg/kg de peso corporal para el aspartamo significa que se pueden consumir hasta 40 mg de aspartamo por cada kilo de peso corporal de una persona, todos los días de su vida, sin un riesgo apreciable para la salud. Esto significa hasta un total de 2,8 gramos de aspartamo diarios para una persona normal de 70 kg. En términos prácticos, 2,8 gramos de aspartamo equivalen al dulzor de 560 g de azúcar, o la cantidad de edulcorante contenido en catorce latas de 330 ml de una bebida carbonatada endulzada con aspartamo. También es comparable al aspartamo contenido en 125 pastillas edulcorantes normales.
Pero, ¿qué sucede con los componentes del aspartamo? ¿Por qué las etiquetas de productos que contienen aspartamo avisan de que "contiene una fuente de fenilalanina"? La fenilalanina es un aminoácido esencial, o sea, que los seres humanos no pueden producir bastante para mantenerse sanos y deben obtenerla en su dieta. En el cuerpo la fenilalanina no es sólo un componente de las proteínas, sino también el precursor de los neurotransmisores, como la dopamina, la noradrenalina y la adrenalina. Algunas personas sufren un extraño mal heredado, la Fenilcetonuria (PKU), que implica que su capacidad para metabolizar la fenilalanina se ve seriamente perjudicada. Estas personas pueden tener concentraciones muy altas de fenilalanina en el plasma, lo que puede producirles daños neuronales y afectar a su desarrollo cerebral. Por eso en muchos países es un requisito legal hacer un test de fenilcetonuria en los
14

niños poco después de su nacimiento. El tratamiento de esta condición incluye un cuidadoso control de todas las fuentes de fenilalanina en la dieta. La fenilalanina es uno de los componentes del aspartamo y se libera cuando éste es digerido en el intestino. Esta es la razón de que los productos que contienen aspartamo avisen de que contienen una fuente de fenilalanina, información útil para personas que padecen fenilcetonuria.
15