Revista Peruana de Biología v20n1
124
Rev. peru. biol. ISSN 1561-0837 UNIVERSIDAD NACIONAL MAYOR DE SAN MARCOS FACULTAD DE CIENCIAS BIOLÓGICAS V OLUMEN 20 SEPTIEMBRE, 2013 NÚMERO 1 LIMA, PERÚ REVISTA PERUANA DE BIOLOGÍA ECOLOGÍA, PESQUERÍA Y CONSERVACIÓN DEL JUREL (TRACHURUS MURPHYI) EN EL PERÚ NÚMERO ESPECIAL: ECOLOGÍA, PESQUERÍA Y CONSERVACIÓN DEL JUREL (TRACHURUS MURPHYI) EN EL PERÚ EDITORES: JORGE CSIRKE RENATO GUEVARA-CARRASCO MARCO ESPINO
-
Upload
leonardo-romero -
Category
Documents
-
view
396 -
download
0
description
REVISTA PERUANA DE BIOLOGÍA is scientific journal, peer review and published for Facultad de Ciencias Biológicas, Universidad Nacional Mayor de San Marcos, Lima, Peru. It has three numbers, in April, August and December. It publishes articles complete, and original in English or Spanish, on biodiversity, biotechnology, ecology, environmental management and biomedicine themes, according to international standards.
Transcript of Revista Peruana de Biología v20n1

Rev. peru. biol. ISSN 1561-0837
Universidad nacional Mayor de san Marcos
FacUltad de ciencias Biológicas
volUMen 20 septieMBre, 2013 núMero 1
LIMA, PERÚ
revista
perUana de
Biología
ecología, pesqUería y conservación del jUrel (Trachurus murphyi) en
el perú
núMero especial:ecología, pesqUería y
conservación del jUrel (Trachurus murphyi) en
el perúeditores: jorge csirke renato gUevara-carrasco Marco espino

2
RectorDr. Pedro Atilio Cotillo Zegarra Vicerrector de Investigación Dr. Bernardino Ramírez BautistaConsejo Superior de InvestigaciónDr. Manuel Gongora Prado Decana de la Facultad de Ciencias BiológicasMag. Olga Bracamonte GuevaraInstituto de Investigación en Ciencias Biológicas Antonio RaimondiMag. Inés Miriam Gárate Camacho
La Revista Peruana de Biología es una publicación científica arbitrada, producida por el Instituto de Ciencias Biológicas Antonio Raimondi, Facultad de Ciencias Biológicas de la Universidad Nacional Mayor de San Marcos, Lima, Perú, y auspiciada por el Vicerrectorado de Investigación. La Revista es publicada tres veces al año (abril, agosto y diciembre) y esta dedicada a la publicación de artículos científicos originales e inéditos de las áreas de biodiversidad, biotecnología, ecología y biomedicina. La Revista publica los trabajos realizados por académicos e investigadores nacionales y extranjeros, en idioma español o inglés. Los trabajos recepcionados son evaluados por árbitros según criterios internacionales de calidad, creatividad, originalidad y contribución al conocimiento. La Revista es publicada simultáneamente en la página web de la Universidad.
Revista Peruana de BiologíaHecho el Depósito Legal 98-3017Rev. peru. biol. - ISSN-L 1561-0837Rev. peru. biol. - ISSN 1561-0837Rev. peru. biol. - ISSN 1727-9933 (on line)http://revistasinvestigacion.unmsm.edu.pe/index.php/rpb/http://sisbib.unmsm.edu.pe/BVRevistas/biologia/biologiaNEW.htmhttp://redalyc.uaemex.mx/
©Facultad de Ciencias Biológicas, UNMSM- La Revista Peruana de Biología publica artículos de acceso abierto, distribuido bajo los términos de la Licencia de Atribución Reconocimiento-NoComercial-SinObraDerivada 3.0 de Creative Commons (http://creativecommons.org/licenses/by-nc-nd/3.0/deed.es_ES), que permite el uso no comercial, distribución y reproducción en cualquier medio, siempre que la obra original sea debidamente citadas. Para uso comercial, por favor póngase en contacto con [email protected]@gmail.com
Información adicional a: Revista Peruana de BiologíaFacultad de Ciencias Biológicas UNMSMCiudad Universitaria, Av. Venezuela Cdra. 34 s/n. LimaCasilla Postal: 11-0058 Lima-11, Perú.Teléfono 619-7000-1502 / Telefax 619-7000-1509Editor Jefe, email: [email protected]
revista perUana de BiologíaPublicación científica de la Facultad de Ciencias Biológicas de la
Universidad Nacional Mayor de San Marcos
Resumida/Indizada (Abstracted/Indexed) en:Periódica (Índice de Revistas Latinoamericanas en Ciencias), LIPECS (Literatura Peruana en Ciencias de la Salud), Zoological Record (BIOSIS), Scielo (Scientific Electronic Library Online), Index to American Botanical Literature (The New York Botanical Garden), BIOSIS Previews, Biological Abstracts (BIOSIS), ProQuest (Biological Science Journals), Redalyc, CABI, AGRICOLA.
Foto en carátula: cortesía de Pesquera Diamante.
Comité EditorEditor JefeLeonardo Romero, Universidad Nacional Mayor de San Marcos.Editores asociadosDra. Rina Ramírez, Universidad Nacional Mayor de San Marcos.Dra. Mónica Arakaki-Makishi, Universidad Nacional Mayor de San Marcos.Dra. Diana Silva Dávila, Universidad Nacional Mayor de San Marcos.Dra. Blanca R. León, Profesora Honoraria, Universidad Nacional
Mayor de San Marcos. The University of Texas at Austin, Geography and the Environment, Faculty Member, Estados Unidos.
Dr. Carlos Peña, Laboratory of Genetics, Department of Biology, University of Turku, Finlandia.
Cesar Arana, Universidad Nacional Mayor de San Marcos.José Roque, Universidad Nacional Mayor de San Marcos.
Número especial 20(1): Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el PerúBasado en más de cinco décadas de investigaciones realizadas por el Instituto del Mar del Perú (IMARPE), sobre los aspectos de biología, ecología y pesquería del Jurel Trachurus murphyi en el mar peruano.EditoresJorge Csirke, Comité Consultivo del IMARPE, Perú.Renato Guevara-Carrasco, IMARPE, Perú.Marco Espino, IMARPE, Perú.Editor AsistentePedro Trillo, Instituto de Recursos Acuáticos. Perú.Comité Consultivo del número especialDr. Arnaud Bertrand, Institut de Recherche pour le Développement
(IRD), France. E-mail: [email protected]. François Gerlotto, Director, Instituto de Recursos Acuáticos,
Francia. E-mail: [email protected]. Leo Walter González, Universidad de Oriente, Núcleo de Nueva
Esparta,Venezuela. E-mail: [email protected]. Omar DeFeo, Unidad de Ciencias del Mar, Universidad de la
República, Uruguay. E-mail: [email protected]. Takahisa Mituhasi, Tokyo University of Marine Science and
Technology, Japan. E-mail: [email protected]. Orlando Mora, Consultor independiente, Bogotá, Colombia.
E-mail: [email protected]. Jorge Oliva Nuñez, Universidad Nacional Pedro Ruiz Gallo –
Lambayeque, Perú. E-mail: [email protected]

1
Número especial: Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú
Special issue: Ecology, fisheries and conservation of Jack mackerel (Trachurus murphyi) in Peru
CONTENIDO
Presentación3 El IMARPE y el número especial Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú
The IMARPE and the special issue Ecology, fisheries and conservation of jack mackerel (Trachurus murphyi) in Peru
Germán Vásquez Solís Talavera
4 Agradecimientos
introducción5 El Jurel Trachurus murphy en el Perú The Jack mackerel Trachurus murphy in Peru Jorge Csirke
el medioambiente del jurel (Trachurus murphyi) en el Perú 9 El jurel Trachurus murphyi y las variables ambientales de macroescala The Jack mackerel Trachurus murphyi and the environmental macro-scale variables Marco Espino
21 Patrones de variabilidad ambiental en el mar peruano Environmental variability patterns in the Peruvian waters Roberto Flores, Marco Espino, Gertrudes Luque y Jorge Quispe
biología del jurel (Trachurus murphyi) en el Perú29 Aspectos reproductivos del jurel Trachurus murphyi Reproductive aspects of Jack mackerel Trachurus murphy Ángel Perea, Julio Mori, Betsy Buitrón y Javier Sánchez
35 Escala de madurez gonadal del jurel Trachurus murphyi Nichols 1920 Scale of gonad maturity stages of Jack mackerel Trachurus murphyi Nichols 1920 Javier Sánchez, Ángel Perea, Betsy Buitrón y Liz Romero
45 Edad y crecimiento del jurel Trachurus murphyi en el Perú Age and growth of Jack mackerel Trachurus murphyi in Peru Teobaldo Dioses
53 Validación del ritmo de formación de los anillos de crecimiento en otolitos del jurel Trachurus murphyi Nichols 1920 Validation of the formation rate of growth rings in otoliths of Jack mackerel Trachurus murphyi Nichols 1920 Carlos Goicochea, Jorge Mostacero, Patricia Moquillaza, Teobaldo Dioses, Yeny Topiño y Renato Guevara-Carrasco
61 Estimación del crecimiento del jurel Trachurus murphyi capturado en el Perú, a partir del análisis de las frecuencias por tallas Estimation of growth parameters of Jack mackerel Trachurus murphyi caught in Peru, from length frequency analysis
Erich Díaz
ecología del jurel (Trachurus murphyi) en el Perú67 Patrones de distribución y abundancia del jurel Trachurus murphyi en el Perú Abundance and distribution patterns of Jack mackerel Trachurus murphyi in Peru Teobaldo Dioses
Revista PeRuana de Biología
Volumen 20 Septiembre, 2013 Número 1Rev. peru. biol. ISSN-L 1561-0837

2
75 Ecología trófica del jurel Trachurus murphyi en el Perú entre 1977 – 2011 Trophic ecology of Jack mackerel Trachurus murphyi in Peru, 1977 – 2011 Ana Alegre, Pepe Espinoza y Marco Espino
83 Variabilidad espacial y temporal de larvas de jurel Trachurus murphyi en el Perú entre 1966 – 2010 Spatial and temporal variability of Jack mackerel Trachurus murphyi larvae in Peru between 1966 – 2010 Patricia Ayón y Jonathan Correa
87 Biomasa acústica y distribución del jurel Trachurus murphyi en el Perú Acoustic biomass and distribution of Jack mackerel Trachurus murphyi in Peru Marceliano Segura y Aníbal Aliaga
Pesquería, manejo y conservación del jurel (Trachurus murphyi) en el Perú97 Análisis de la pesquería del jurel Trachurus murphyi en el Perú Analysis of the Jack mackerel Trachurus murphyi fishery in Peru Miguel Ñiquen, Marilú Bouchon, Danny Ulloa y Ana Medina
107 Medidas de conservación y ordenación pesquera del jurel Trachurus murphyi en el Perú Conservation and fishery management regulations of Jack mackerel Trachurus murphyi in Peru Jorge Zuzunaga

3
El IMARPE y el número especial Ecología, pesquería y conservación del jurel
Rev. peru. biol. special issue 20(1): 003 - 004 (September 2013)Ecology, fisheries and conservation of Jack mackerel (Trachurus murphyi) in Peru. Editors: Csirke J., R. Guevara-Carrasco & M. Espino
Rev. peru. biol. número especial 20(1): 003 - 004 (Septiembre 2013)Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú. Editores: Csirke J., R. Guevara-Carrasco & M. Espino
Facultad de ciencias Biológicas unMsMISSN-L 1561-0837
El IMARPE y el número especial Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú
Germán Vásquez Solís Talavera
The IMARPE and the special issue Ecology, fisheries and conservation of Jack mackerel (Trachurus murphyi) in Peru
Instituto del Mar del Perú, esquina Gamarra y General Valle S/N Chucuito, Callao, Perú.
Email Germán Vásquez Solís Talavera: [email protected]
Publicado online: 11/10/2013Publicado impreso: 15/10/2013
Presentación
© Los autores. Publicado por la Revista Peruana de Biología de la Facultad de Ciencias Biológicas, Universidad Nacional Mayor de San Marcos. Este es un artículo de acceso abierto, distribuido bajo los términos de la Licencia de Atribución Reconocimiento-NoComercial-SinObraDerivada 3.0 de Creative Commons (http://creativecommons.org/licenses/by-nc-nd/3.0/deed.es_ES), que permite el uso no comercial, distribución y reproducción en cualquier medio, siempre que la obra original sea debidamente citadas. Para uso comercial, por favor póngase en contacto con [email protected].
Citación:Vásquez Solís Talavera G. 2013. El IMARPE y el número especial Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú. En: Csirke J., R. Guevara-Carrasco & M. Espino (Eds.). Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú. Rev. peru. biol. número especial 20(1): 003- 004 (Septiembre 2013)
El mar peruano es la cuarta región natural del Perú y es considerado como uno de los sistemas más productivos del planeta, si no el más. Su particular situación geográfica le permite contar con características muy propias que lo convierten en un inmenso sistema de producción de biomasa, que sustenta una compleja malla trófica de comunidades biológicas que se desarrollan en condiciones de variabilidad extrema.
Dentro de este escenario de alta complejidad, el mar peruano ha sido una fuente de alimento para el poblador peruano, desde tiempos inmemoriales. Así lo indican múltiples testimonios que se encuentran en restos arqueológicos de nuestras ancestrales culturas afincadas en la costa peruana desde hace más de 6000 años, tales como Caral, Bandurria y Áspero, donde se reportan diferentes especies de peces y moluscos, entre las cuales destaca el jurel como parte de la dieta de nuestros ancestros.
El Estado peruano inicia los estudios formales de los recursos del mar peruano desde fines del siglo XIX e inicios del siglo XX, en la época de la explotación del guano de isla, producto natural que marcó una importante etapa en la economía y la historia nacional. Luego, las investigaciones fueron continuadas por el Instituto del Mar del Perú (IMARPE) creado en 1964; una década en la cual el Perú comenzó a convertirse en uno de los países más importantes del mundo en materia de producción pesquera. Gracias a la actitud visionaria de las autoridades y actores principales de la pesca nacio-nal de aquella época y al efectivo aporte de la cooperación internacional, el IMARPE pudo amalgamar un notable grupo de trabajo conformado por profesionales, técnicos y administrativos que han logrado constituir y mantener un importante sistema de investigación del mar peruano y sus pesquerías. Esto ha permitido que hoy en día el Perú cuente con un buen y creciente conocimiento sobre la compleja dinámica de su mar y sus recursos vivos, así como con una extraordinaria base de datos científicos, considerada por muchos expertos como una de las más extensas y completas del mun-do. Esto se manifiesta en el creciente interés de entidades nacionales e internacionales por cooperar con el IMARPE en la investigación de un mar de características únicas.
Desde que asumí la Presidencia del Consejo Directivo del IMARPE en septiembre de 2011, pude constatar la complejidad de las labores que realizan sus científicos y de las dificultades que enfrentan con mucho profesionalismo para cumplir con su misión, muchas veces en condiciones muy difíciles generadas por las carencias propias de un país en desarrollo como el nuestro.
Gracias al denodado esfuerzo de quienes laboran y han laborado en el IMARPE desde su creación, es que hoy podemos contar con un acervo de conocimiento cien-tífico que respalda la toma de decisiones para la gestión del uso de los recursos vivos del mar. Dicho conocimiento científico se manifiesta en las publicaciones científicas periódicas que el IMARPE ha venido produciendo a lo largo de su historia en sus labores de evaluación y conocimiento de nuestros recursos marinos y que, desde este año 2013 tienen visibilidad y un uso abierto mayor a través del Repositorio Digital inserto en la página web institucional.

4
Vásquez Solís Talavera
Rev. peru. biol. número especial 20(1): 003 - 004 (Septiembre 2013)Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú.
Editores: Csirke J., R. Guevara-Carrasco & M. Espino
Además de las publicaciones periódicas, el IMARPE ha producido notables e importantes publicaciones científicas entre las que podríamos citar las siguientes: Atlas del Instituto del Mar del Perú (1965), Oceanografía de las aguas costeras del Perú (1970), Tempe-ratura promedio de la superficie del mar frente a la costa peruana, periodo 1928-1969 (1972), Clave para identificar los peces marinos del Perú (1974, 1998), La investigación cooperativa de la Anchoveta y su Ecosistema (ICANE) entre Perú y Canadá (1981), El Niño y su impacto en la Fauna Marina (1985), Lista sistemática de moluscos marinos del Perú (1987), The Peruvian Anchoveta and its Upwelling Ecosystem: Three decades of Change (1987), Recursos y dinámica del ecosistema de afloramiento peruano (1988), The Peruvian Upwe-lling Ecosystem: Dynamics and Interactions (1989), Catálogo comentado de los peces marinos del Perú (2001), The Northern Humboldt Current System: Ocean Dynamics, Ecosystem Processes, and Fisheries (2008) y el Libro de peces de aguas profundas del Perú (2009).
Aprovechando la oportunidad que nos brinda la Universidad Nacional Mayor de San Marcos a través de la Revista Peruana de Biología, me corresponde la tarea de presentar un esfuerzo de síntesis sobre una de las especies emblemáticas del mar peruano: el jurel. Ésta es y ha sido una de las especies más importantes del consumo humano directo del poblador nacional desde épocas remotas, por lo cual desde el año 2002 sólo se le puede pescar para ese fin. Su situación ha sido objeto de múltiples controversias en los últimos años, en gran medida por el desconocimiento de su dinámica poblacional en relación a la naturaleza del mar peruano. Por ello, en un esfuerzo estrictamente institucional, 27 investigadores del IMARPE asumieron la tarea de elaborar una síntesis del conocimiento de esta especie, la misma que se presenta ahora como un avance en trece capítulos o artículos de este primer volumen del IMARPE y de la Revista Peruana de Biología dedicado enteramente al jurel.
Con esta obra titulada Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú, se pone a disposición del público nacional e internacional, más de cinco décadas de información sobre sus características biológicas y ecológicas, sobre su pesquería y la complejidad de su manejo, constituyéndose en un referente obligado para aquellos que quieran investigar y conocer sobre este importante recurso.
Antes de finalizar, debo destacar que coincidentemente la publicación de este trabajo científico se produce en el año del jubileo por la conmemoración del quincuagésimo aniversario de creación del IMARPE. Entendemos que esa es una buena señal, pues además hay otros esfuerzos internos en camino, que esperamos vean la luz a los 50 años de creación de una entidad de la cual los peruanos debemos sentirnos orgullosos. Al difundir el conocimiento científico sobre el mar peruano, también estamos cumpliendo con la misión de incorporar a más peruanos al torrente de ciudadanos informados que puedan tener una visión más completa y objetiva sobre la naturaleza de los recursos vivos marinos de esta nuestra todavía poco conocida cuarta región geográfica.
Finalmente, expreso mi agradecimiento personal y calurosa felicitación a todos los actores de dentro y fuera del IMARPE sin cuyo esfuerzo no habría sido posible esta magnífica obra.
Callao, setiembre de 2013.
Calm. (r) Germán Vásquez Solís Talavera
Presidente del Consejo Directivo del IMARPE
Agradecimientos
Jorge Csirke, Renato Guevara-Carrasco y Marco Espino, editores de este número especial de la Revista Peruana de Biología deseamos agradecer a los colaboradores y expertos nacionales y extranjeros, que han contribuido con sus valiosos comentarios, críticas y sugerencias a la realización de esta obra; en especial a: Arnaud Bertrand (Sète Cedex, Francia), Omar DeFeo (Montevideo, Uruguay), Francois Gerlotto (Castelnau-le-Lez, Francia), Jordi Lleonart (Barcelona, España), Takahisa Mituhasi (Tokyo, Japón), Orlando Mora (Bogotá, Colombia), Jorge Oliva (Trujillo, Perú) y Leo Walter González (Isla Margarita, Venezuela). También un especial reconocimiento a Pedro Trillo por su valiosa e incansable asistencia en las diversas fases de la producción de este volumen y al apoyo de Sara Bravo en la corrección ortográfica, Gaby Matsumoto en el cuidado de las figuras y Giancarlo Pinasco en algunas traducciones. Por último, queremos agradecer a la Universidad Nacional Mayor de San Marcos por habernos acogido como editores y por todo el apoyo brindado en la producción de este número especial.

5
El Jurel Trachurus murphy en el Perú
Rev. peru. biol. special issue 20(1): 005 - 008 (September 2013)Ecology, fisheries and conservation of Jack mackerel (Trachurus murphyi) in Peru. Editors: Csirke J., R. Guevara-Carrasco & M. Espino
Rev. peru. biol. número especial 20(1): 005 - 008 (Septiembre 2013)Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú. Editores: Csirke J., R. Guevara-Carrasco & M. Espino
Facultad de ciencias Biológicas unMsMISSN-L 1561-0837
El Jurel Trachurus murphy en el Perú
Jorge Csirke
The Jack mackerel Trachurus murphy in Peru
Instituto del Mar del Perú, esquina Gamarra y General Valle S/N Chucuito, Callao, Perú.
Email Jorge Csirke: [email protected]
introducción
© Los autores. Publicado por la Revista Peruana de Biología de la Facultad de Ciencias Biológicas, Universidad Nacional Mayor de San Marcos. Este es un artículo de acceso abierto, distribuido bajo los términos de la Licencia de Atribución Reconocimiento-NoComercial-SinObraDerivada 3.0 de Creative Commons (http://creativecommons.org/licenses/by-nc-nd/3.0/deed.es_ES), que permite el uso no comercial, distribución y reproducción en cualquier medio, siempre que la obra original sea debidamente citadas. Para uso comercial, por favor póngase en contacto con [email protected].
Citación:Csirke J. 2013. El Jurel Trachurus murphy en el Perú. En: Csirke J., R. Guevara-Carrasco & M. Espino (Eds.). Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú. Rev. peru. biol. número especial 20(1): 005 - 008 (Septiembre 2013)
En el Perú se ha capturado y consumido jurel Trachurus murphyi desde épocas re-motas, y merecidamente se le considera como uno de los principales recursos pesqueros del país. Sin embargo, hay evidencias de que tanto el interés en esta especie como su abundancia y disponibilidad han experimentado grandes cambios y fluctuaciones a través del tiempo.
El interés que hay en la actualidad en el jurel T. murphyi es evidente y está ple-namente justificado. Lo confirman el volumen de sus capturas, su disponibilidad y precio accesible en los mercados, su buena aceptación y amplio consumo popular, además de la dimensión internacional que tiene su explotación y la ordenación de sus pesquerías al tratarse del recurso pesquero transzonal más importante del momento en el Pacífico Sur. Dentro de este contexto, la creación y entrada en funciones de la Organización Regional de Ordenación Pesquera del Pacífico Sur (SPRFMO, del inglés South Pacific Regional Fisheries Management Organisation) está contribuyendo a ordenar la pesca en alta mar, donde antes no estaba regulada, y le ha dado una nueva dimensión internacional al interés que ya había a nivel nacional y regional en la pesca y conservación de esta especie.
Es el reconocimiento de este gran interés nacional e internacional lo que ha mo-tivado esta obra. Pero son los grandes cambios y fluctuaciones en la abundancia y disponibilidad de esta especie lo que ha determinado la trama principal de este número especial de la Revista Peruana de Biología dedicado al jurel en el Perú, donde se da cuenta de los resultados de las más recientes investigaciones del Instituto del Mar del Perú (IMARPE) sobre los diversos aspectos de la historia, el ambiente, la biología y la pesquería de esta importante especie.
Al indagar sobre la historia del jurel en el Perú, encontramos que el interés por esta especie no es sólo un hecho reciente. Hay evidencias de que los antiguos pobladores de lo que hoy conocemos como el Perú ya lo pescaban y consumían en cantidades significativas. En efecto, excavaciones recientes en la Quebrada de los Burros en el sur del país han puesto al descubierto evidencias de que entre 10000 y 6000 años a.p., en el Holoceno temprano y medio (periodo pre cerámico temprano), ya había pescadores y recolectores de moluscos que se dedicaban habitualmente a la pesca de jurel T. murphyi (Lavallée et al. 2011).
Los estudios de Lavallée et al. (2011) dan cuenta que hace 10000 años los antiguos peruanos que ocuparon por milenios la Quebrada de los Burros ya dominaban el arte de la pesca y se alimentaban mayormente de moluscos y, sobre todo, de peces marinos. Entre los peces capturados y consumidos regularmente por estos primeros pescadores del litoral sur peruano destacan la lorna Sciaena deliciosa y en segundo lugar el jurel T. murphyi, seguidos de machete Ethmidium maculatum, cabinza Isacia conceptionis, bonito Sarda chiliensis, pintadilla Cheilodactylus variegatus, y al menos otras 25 especies de peces marinos.
Otras excavaciones en las zonas arqueológicas de Chivateros en el litoral central del Perú y en Paiján, Cupisnique, Moche, Casma y otras en el litoral norte confirman que pescadores primitivos también ocuparon esas zonas desde hace aproximadamente 10000 años, en el Holoceno temprano o pre cerámico temprano (Lumbreras et al.
Publicado online: 11/10/2013Publicado impreso: 15/10/2013

6
Csirke
Rev. peru. biol. número especial 20(1): 005 - 008 (Septiembre 2013)Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú.
Editores: Csirke J., R. Guevara-Carrasco & M. Espino
2008). Aunque no hay indicación sobre las especies de peces encontradas en los restos dejados por los ocupantes de estas zonas en el Holoceno temprano, es muy probable que entre las especies que capturaban y consumían se encontrara también el jurel T. murphyi. Excavaciones de épocas más recientes corres-pondientes al Holoceno medio y tardío (pre cerámico medio y tardío) confirman su presencia en los restos dejados por anti-guas comunidades que ocuparon algunos de estos y otros sitios arqueológicos a lo largo del litoral peruano 8000 a 3800 años a.p., como en Áspero y Caral (Shady & Cáceda 2008), Cardal (Lumbreras et al. 2008), Hauynuná (Vásquez et al. 2012) y Bandurria (Chu 2008).
Luego, con el inicio del periodo cerámico 3800 años a.p. se encuentran numerosas evidencias del desarrollo de aldeas de pescadores de diversas culturas prehispánicas que ocuparon el litoral peruano, como las culturas Ancón, Chancay, Chimú, Cupisnique, Lima, Moche, Nazca, Pachacámac, Paita, Paracas, Sicán, Wari y otras que precedieron a la cultura Inca (Lumbreras et al. 2008, Pérez et al. 1976). Entre estas evidencias, cabe des-tacar que en los restos dejados por los Wari entre los años 600 y 1000 d.C. en la zona de Cerro Baúl, a 65 km en línea de aire del litoral entre Mollendo e Ilo en el sur del Perú, se encontró gran cantidad de huesos de peces entre los cuales predomina el jurel T. murphyi, con indicaciones de que esta especie se consumía regularmente, y en particular cantidad en banquetes ceremoniales (Moseley et al. 2005).
Por otro lado, los numerosos huacos, vasijas y otros ceramios escultóricos con escenas de navegación y pesca dejados por los antiguos peruanos (Pérez et al. 1976) no dejan dudas de que los pescadores del periodo cerámico ya dominaban la navegación y la pesca costera. Estos ceramios muestran que se usaban embar-caciones como los “caballitos de totora”, en la actualidad aún en uso en el norte del Perú, y también otras un poco más grandes a juzgar por algunos ceramios que muestran embarcaciones con dos y tres tripulantes. También existen indicios de que en el periodo pre cerámico ya se usaban embarcaciones para la pesca, que podrían haber sido semejantes a las del periodo cerámico aunque no hay evidencias sobre ello. Como señalan Quilter y Stocker (1983), cualquier tipo de embarcación y parte de los equipos de pesca usados en el pre cerámico habrían sido dejados a la intemperie en las playas en lugar de ser transportados a los campamentos o aldeas donde podrían haber sido encontrados entre restos arqueológicos. Sin embargo, en muchos sitios pre cerámicos se han encontrado anzuelos, redes, flotadores y pesos de redes, y se aprecia que la pesca con redes flotantes recibió progresivamente más énfasis a través del tiempo durante dicho periodo (Quilter & Stocker 1983). Todo esto indica que ya desde aquel periodo y hasta el incaico los antiguos peruanos empleaban embarcaciones pequeñas de tipo rudimentario para la pesca.
Con la llegada de los españoles, en el siglo XVI se inicia el em-pleo de embarcaciones y materiales europeos más evolucionados para la pesca y se desarrolla más como actividad de subsistencia, pero además como una actividad comercial local. Crónicas de la época virreinal e inicios de la época republicana dan cuenta de los pregoneros que pasaban por las calles de Lima y de otras ciudades del país ofreciendo sus mercancías y productos. Entre ellos destacan los pescadores, que solían pasar antes del mediodía o al final de la tarde después de haber terminado sus faenas de pesca. Sin embargo, el interés que hubo en esos años en el jurel parece haber sido bajo, y estaría reflejado por lo que podemos observar documentadamente ya a principios del siglo XX.
A principios del siglo pasado, se describía al jurel como una especie abundante a lo largo de toda la costa peruana (Coker 1907, 1910). Sin embargo, los registros oficiales de desembar-
ques nos indican que, antes y durante gran parte del siglo XX, estuvo relegado a los últimos lugares en las preferencias de los pescadores y del poblador peruano en general. Por lo tanto, la importancia de esta especie en esta época algo más reciente resulta discutible.
Según los registros oficiales, hasta 1963 solamente se captu-raban pocos cientos de toneladas de jurel por año a lo largo del litoral peruano. Hasta 1963 ocupó los últimos lugares en las estadísticas de desembarque nacionales, siendo superado amplia-mente por especies como anchoveta Engraulis ringens, bonito S. chiliensis, listado o barrilete Katsuwonus pelamis, rabíl Thunnus albacares, caballa Scomber japonicus, machete E. maculatum, corvina Cilus gilbert, cabrilla Paralabrax humeralis, cachema Cynoscion analis, sardina Sardinops sagax, coco Paralonchurus peruanus y cabinza I. conceptionis, además de cojinovas Seriolella spp., sucos Micropogonias spp, tollos Mustelus spp., meros Epine-phelus spp., lisas (Mugilidae), pejerreyes (Atherinidae) y otros.
Es recién a fines de la década de los sesenta que el jurel pasa a ocupar lugares intermedios en las estadísticas de desembarques, con capturas anuales crecientes que pasan rápidamente de los cientos a los miles de toneladas por año, hasta que en 1974 se llegó a capturar 129 mil toneladas de esta especie. Con ello el jurel T. murphyi pasó a ocupar el segundo lugar en las estadísticas de desembarque nacionales, después de la anchoveta E. ringens. Desde entonces, se ha mantenido entre el segundo y cuarto lugar en las estadísticas de volumen desembarcado, siendo precedido alternadamente, durante uno o más años, por especies como anchoveta E. ringens, sardina S. sagax, calamar o jibia gigante Dosidicus gigas y caballa S. japonicus. Con estas especies, ha venido compartiendo los méritos de ser uno de los recursos pesqueros de mayor importancia para el país, aunque mantiene el primado como el recurso pesquero para el consumo humano directo de más importancia a nivel nacional.
Es así que llegamos a la época actual y a la producción de este número especial, donde se analiza la vida y la pesquería del jurel desde diversos ángulos y en varias escalas de tiempo. La mayor parte de la información y datos que se analizan en los 13 artículos que siguen, provienen de los muestreos y programas de investigación instituidos por el IMARPE desde su creación hace casi 50 años. Pero en otros casos se ha podido extender los periodos de observación a años y hasta decenios anteriores, o hasta milenios como se ha intentado hacer en esta breve nota introductoria.
Los 13 artículos de este volumen están agrupados en cuatro grandes temas: el medio ambiente, la biología, la ecología, y la pesca y la conservación del jurel T. murphyi en el Perú que, como se intenta establecer en los artículos que siguen, son los aspectos principales que rigen los cambios y fluctuaciones en la abundancia y disponibilidad de esta especie en aguas peruanas. Además, el orden en que se presentan estos 13 artículos no es casual. Como se establecerá más adelante, va de la descripción del ambiente marino, que se considera el principal factor ge-nerador de los cambios y fluctuaciones del jurel en el Perú, a los principales aspectos de su biología como la reproducción y el crecimiento que parecen no haber cambiado mucho en el tiempo, para seguir con aspectos de su interacción con el ecosistema donde sí se observan cambios notables, y se termina con la pesca, que está supeditada a los cambios y fluctuaciones que le impone la naturaleza y que trata de imbuir una cierta estabilidad con regulaciones pesqueras, en un contexto donde la naturaleza impone sus propias reglas.
Los dos primeros artículos de este volumen (Espino 2013 y Flores et al. 2013) describen y analizan diversas variables

7
El Jurel Trachurus murphy en el Perú
Rev. peru. biol. special issue 20(1): 005 - 008 (September 2013)Ecology, fisheries and conservation of Jack mackerel (Trachurus murphyi) in Peru. Editors: Csirke J., R. Guevara-Carrasco & M. Espino
ambientales que determinan el clima y sus fluctuaciones en el mar peruano y la alta mar adyacente. Describen las complejas interacciones de las diversas masas de agua e identifican los principales patrones de variabilidad, que al compararlos con información de la pesca, la abundancia y el comportamiento y biología general del jurel permiten explicar fases de cambio en el comportamiento y la distribución espacial y temporal de la especie, así como sus fluctuaciones en diversas escalas de tiempo. Espino (2013) y Flores et al. (2013) presentan claras evidencias de que es el estrés ambiental el principal desencadenante de los grandes cambios observados en la abundancia y la disponibilidad del jurel T. murphyi en las aguas jurisdiccionales peruanas y en el alta mar adyacente, y explican los bajos niveles de biomasa y disponibilidad de los últimos 15 años como consecuencia de un cambio de régimen ambiental.
Siguen cinco artículos sobre la biología del jurel T. murphyi en el Perú, donde primero se describen y examinan sus aspectos reproductivos en los artículos de Perea et al. (2013) y Sánchez et al. (2013), seguidos de tres estudios sobre la edad y el crecimiento de Dioses (2013a), Goicochea et al. (2013) y Díaz (2013).
Al analizar los aspectos reproductivos, Perea et al. (2013) encuentran que la talla a la primera madurez del jurel en el Perú no ha cambiado en 45 años, manteniéndose entre los 25-27 cm de longitud total, esto a pesar de los grandes cambios ambientales y en su distribución y abundancia observados. También se observa que, en el Perú, el jurel tiene un máximo de desove en noviembre, desfasado en un mes con respecto al jurel que se encuentra frente a Chile, y el ciclo de desove es más amplio y muestra variabilidad mucho mayor. Ello constituiría una adaptación de la estrategia reproductiva del jurel peruano a las condiciones más cambiantes e inciertas del ambiente marino en el Perú. A ello Sánchez (2013) agrega la confirmación de la tipificación como desovante parcial del jurel y da a conocer la escala de madurez gonadal macroscópica ya en uso en el Perú, validada con estudios microscópicos.
En cuanto a la edad y crecimiento, este volumen presenta los resultados de una combinación de varios enfoques que incluyen la interpretación convencional de marcas anuales en otolitos de Dioses (2013a) y Goicochea et al. (2013); la validación del pri-mer anillo anual mediante lectura de microincrementos diarios y el análisis del crecimiento del radio del otolito entre el segundo y tercer anillo, siguiendo una cohorte, de Goicochea et al. (2013); y el análisis de frecuencias de longitud mensual para estimar los parámetros de crecimiento (Díaz 2013). Los resultados de todos estos estudios fueron fuertemente congruentes y confirman las estimaciones de los parámetros de la función de crecimiento de von Bertalanffy para el jurel peruano en uso desde la década de los 1990 donde: L∞ = 80.77 cm de longitud total, k = 0.1553 año-1 y to =-0.3562 de Dioses (1995, 2013a).
Se destaca que tanto Dioses (2013a) como Goicochea et al. (2013) y Díaz (2013) concluyen que tanto las estimaciones de longitud (y peso) a la edad como los parámetros de crecimiento del jurel peruano son marcadamente diferentes, con longitudes (y peso) a la edad y tasa de crecimiento (k) mayores que las esti-madas para el jurel chileno, aun en los casos en que la longitud asintótica (L∞) estimada sea muy semejante. Se especula que estas marcadas diferencias podrían ser de origen ontológico o metodológico, o ser una combinación de ambos.
Se hace notar que el valor de longitud asintótica L∞ indicado líneas arriba guarda relación con la talla máxima encontrada en más de 40 años de muestreos de longitudes del IMARPE a lo largo de todo el litoral peruano, que es Lmax = 79 cm (en febre-ro de 1974). Sobre este particular, cabe destacar que el récord
del jurel T. murphy más grande (Lmax?) capturado en el litoral peruano les correspondería a antiguos pescadores peruanos que ocuparon los sitios arqueológicos de Huaynuná en el valle de Casma, en la costa norte del Perú. Precisamente en dicho lugar se encontraron restos óseos de esta especie asociados cronológica-mente al pre cerámico tardío (aproximadamente unos 4200 años a.p.) que corresponderían a un ejemplar de 150 cm de longitud total (Vásquez et al. 2012). Es probable que esta información no estuviera a disposición de Dioses (2013a), Goicochea et al. (2013) y Díaz (2013) al escribir sus contribuciones en este volumen, pero, aunque así hubiera sido, es seguro que no la habrían podido incorporar en sus análisis por no proceder de un muestreo regular de las capturas y por caer fuera de sus periodos de observación declarados.
Los siguientes cuatro artículos analizan cuatro aspectos rele-vantes de la ecología del jurel T. murphyi desde diferentes ángulos y usando diferentes fuentes de información. Dioses (2013b) examina sus patrones de abundancia y distribución espacial y temporal, Alegre et al. (2013) examinan sus cambios en la ecología trófica, Ayón y Correa (2013) la variabilidad espacial y temporal de la presencia de sus larvas, y Segura y Aliaga (2013) presentan y discuten los estimados acústicos de biomasa y de distribución de los últimos años.
Cruzando información oceanográfica y de la pesca comercial, Dioses (2013b) observa tres patrones de distribución vertical y profundidad de las capturas de jurel, y llega a determinar que: cuando ocurren fenómenos El Niño fuertes, los cardúmenes se distribuyen entre 100 y 200 m de profundidad y las capturas son altas y varían poco entre día y noche; cuando hay fuertes afloramientos los cardúmenes se distribuyen entre 0 y 75 m de profundidad y la pesca es más oceánica y superficial; y, cuando se refuerza la extensión sur de la Corriente de Cromwell, los cardúmenes están entre 0 y 300 m de profundidad, la pesca es más costera y a mayor profundidad de día y es más superficial de noche.
Al analizar los cambios en la dieta del jurel T. murphyi, Alegre et al. (2013) identifican dos escenarios claramente diferenciados: el periodo 1977 – 2000 que corresponde a condiciones cálidas y donde su dieta estuvo fuertemente dominada por Euphausiidae, y otro del 2000 a la fecha que corresponde a condiciones frías y donde consumió mayor diversidad de presas, principalmente Euphausiidae, pero también camaroncito rojo Pleuroncodes monodon y Zoeas. Este cambio en su dieta hacia el año 2000 guarda buena correspondencia con los cambios decadales en el ambiente que describen Espino (2013) y Flores et al. (2013). Alegre et al. (2013) destacan también que T. murphyi es un de-predador facultativo oportunista que en el ecosistema pelágico peruano se alimenta tanto en el ambiente nerítico como en el ambiente oceánico epipelágico, pudiéndose adaptar fácilmente a situaciones cambiantes.
En el artículo siguiente, Ayón y Correa (2013) confirman la presencia constante de larvas de T. murphyi en el mar peruano y la alta mar adyacente durante todo el periodo de 40 años estudiado y a lo largo de todo el litoral, con algunas variaciones a escala decadal relacionadas con factores ambientales. La coincidencia entre estas observaciones de Ayón y Correa (2013) y las del ciclo de desove más aplanado, prolongado y variable de Perea et al. (2013) es de particular interés teniendo en cuenta la alta variabilidad y alta productividad de las aguas peruanas, con una heterogeneidad espacial apropiada para el éxito del desove por la presencia de puntos de generación de eddies (Chaigneau et al. 2013). Se resalta que además de ser centros de fertilización, estos eddies o remolinos actúan como centros de retención y transporte de una variedad de organismos marinos, incluyendo

8
Csirke
Rev. peru. biol. número especial 20(1): 005 - 008 (Septiembre 2013)Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú.
Editores: Csirke J., R. Guevara-Carrasco & M. Espino
los productos del desove, reduciendo el transporte hacia fuera de la costa y aumentando la retención de las larvas y, posiblemente, también sus probabilidades de sobrevivencia.
Los resultados de las evaluaciones hidroacústicas del jurel son analizados por Segura y Aliaga (2013), quienes destacan el gran cambio en la distribución y abundancia de esta especie en las áreas observadas alrededor de 1998, con altos estimados de biomasa acústica entre 1982-1998, con máximos de 8,5 millones de toneladas en 1983 y 1993, y un periodo de bajos estimados de 1998 a la fecha, con estimados inferiores al millón de toneladas. Este cambio a fines de los años 1990 guarda buena relación con los cambios ambientales decadales descritos por otros autores y, en particular, se resalta el buen paralelismo entre la brusca disminución de las áreas de las isotermas comprendidas entre 22 y 25 °C (típicas de Aguas Subtropicales Superficiales) mostradas en la Figura 11 de Espino (2013) y las biomasas acústicas en la Figura 1 de Segura y Aliaga (2013). Se destaca también que estos cambios en la abundancia han ocurrido en ausencia de una presión de pesca importante, con capturas muy bajas en las décadas de los ochenta y noventa.
Los dos últimos artículos de este número especial describen y analizan diversos aspectos de la pesquería de jurel en el Perú: Ñiquen et al. (2013) realizan una detallada descripción de la pesquería y Zuzunaga (2013) presenta una revisión de las regu-laciones pesqueras que han normado su pesca en el Perú en los últimos 40 años. Ñiquen et al. (2013) describen los cambios en las capturas en los últimos 74 años y en el tamaño y estructura de la flota dedicada a la pesca de jurel en los últimos 43 años, y muestra que los cambios mensuales en sus capturas han tenido una buena relación con la variación de parámetros ambientales descritos por otros autores en este volumen y, en particular, con las anomalías mensuales de la temperatura superficial del mar en Chicama. Por su lado, Zuzunaga (2013) da cuenta de las diversas medidas de ordenación de la pesquería de jurel que fueron aplicadas y que están vigentes en el Perú, destacando la obligación que rige desde 2002 de destinar la pesca de sardina S. sagax, jurel T. murphyi y caballa S. japonicus exclusivamente para el consumo humano directo.
Para terminar, se hace una breve referencia a los cambios en la distribución y marcada reducción en la disponibilidad de jurel T. murphyi en el Perú en el curso del año 2013, mientras se edita y publica este volumen. Como explican Espino (2013), Flores et al. (2013), Dioses (2013b) y otros autores en este volumen, estos cambios en la distribución y la disponibilidad están asociados con las condiciones de La Niña, que se han desarrollado en el curso del año 2013 y están causando que el jurel se encuentre más disperso y más alejado de la costa. Este aumento en la distancia de la costa y la menor concentración de los cardúmenes hace que en las condiciones actuales su pesca sea más difícil y menos atractiva para la flota. Esto no hace más que corroborar en el corto plazo la fuerte interacción entre la abundancia, distribución y la disponibilidad del jurel y la dinámica ambiental del mar peruano descritas ampliamente en este volumen.
Literatura citadaAlegre A., P. Espinoza & M. Espino. 2013. Ecología trófica del jurel Trachurus
murphyi en el Perú entre 1977-2011. Perú. Rev. peru. biol. número especial 20(1): 075- 082.
Ayón P. & J. Correa. 2013. Variabilidad espacial y temporal de larvas de jurel Trachurus murphyi en el Perú entre 1966-2010. Rev. peru. biol. número especial 20(1): 083- 086.
Chaigneau A., N. Domínguez, G. Eldin et al. 2013. Near-coastal circulation in the Northern Humboldt Current System from shipboard ADCP data. Journal of Geophysical Research: Oceans. DOI: 10.1002/jgrc.20328.
Chu A. 2008. Bandurria: arena, mar y humedad en el seguimiento de la civi-lización andina. Proyecto Arqueológico Bandurria, Perú: 185pp.
Coker R.E. 1907. Condición en que se encuentra la pesca marina desde Paita hasta bahía de la Independencia. Boletín del Ministerio de Fomento, Año VI, N.o 2, 30 de noviembre de 1907, 89-117.
Coker R.E. 1910. Condiciones de la pesquería de Mollendo. Boletín del Minis-terio de Fomento, Año VIII, N.o 8, 31 de octubre de 1910, 64-114.
Díaz E. 2013. Estimación del crecimiento del jurel Trachurus murphyi cap-turado en el Perú, a partir del análisis de las frecuencias por tallas. Rev. peru. biol. número especial 20(1): 061- 066.
Dioses T. 2013a. Edad y crecimiento del jurel Trachurus murphyi en el Perú. Rev. peru. biol. número especial 20(1): 045- 052.
Dioses T. 2013b. Patrones de distribución y abundancia del jurel Trachurus mur-phyi en el Perú. Rev. peru. biol. número especial 20(1): 067- 074.
Espino M. 2013. El jurel Trachurus murphyi y las variables ambientales de macroescala. Rev. peru. biol. número especial 20(1): 009- 020.
Flores R., M. Espino, G. Luque & J. Quispe. 2013. Patrones de variabilidad ambiental en el mar peruano. Rev. peru. biol. número especial 20(1): 021-028 .
Goicochea C., J. Mostacero, P. Moquillaza et al. 2013. Validación del ritmo de formación de los anillos de crecimiento en otolitos del jurel Trachurus murphyi Nichols 1920. Rev. peru. biol. número especial 20(1): 053- 060.
Lavallée D., M. Julien, P. Béarez et al. 2011. Quebrada de los Burros: Los primeros pescadores del litoral Pacífico en el extremo sur peruano. Chungara, Arica, 43 (Num. Especial 1): 333-351
Lumbreras L.G., P. Kaulicke, J. I. Santillana & W. Espinoza-Soriano. 2008. Compendio de historia económica del Perú I: Economía prehispá-nica. BCRP, IEP, Lima: 444 pp.
Moseley M.E., D.J. Nash, P.R. Williams et al. 2005 Burning down the brewery: Establishing and evacuating an ancient imperial colony at Cerro Baul, Peru. PNAS 102 (48): 17264-17271
Ñiquen M., M. Bouchon, D. Ulloa & A. Medina. 2013. Análisis de la pesquería del jurel Trachurus murphyi en el Perú. Rev. peru. biol. número especial 20(1): 097-106 .
Perea A., J. Mori, B. Buitrón & J. Sánchez. 2013. Aspectos reproductivos del jurel Trachurus murphyi en el Perú. Rev. peru. biol. número especial 20(1): 029- 034.
Pérez A., H. Buse, E. Sarmiento et al. 1976. La pesca en el Perú prehispánico. Empresa Pública de Producción de Harina y Aceite de Pescado. Editoriales Unidas S.A., Lima: 253 pp.
Quilter J. & T. Stocker. 1983. Subsistence economies and the origins of An-dean complex societies. American Anthropologist 85 (3): 545-562
Sánchez J., A. Perea, B. Buitrón & L. Romero. 2013. Escala de madurez gonadal del jurel Trachurus murphyi Nichols 1920. Rev. peru. biol. número especial 20(1): 035 - 044.
Segura M. & A. Aliaga. 2013. Biomasa acústica y distribución del jurel Trachurus murphyi en el Perú. Rev. peru. biol. número especial 20(1): 087- 096.
Shady R. & D. Cáceda. 2008. Áspero, la ciudad pesquera de la civilización Caral: recuperamos su historia para vincularla con nuestro presente. Proyecto Especial Arqueológico Caral-Supe (Perú): 24 pp.
Vásquez V.F., E. Tsurumi, T. Pozorski, S. Pozorski, T. Rosales-Tham. 2012. Evidencias zooarqueológicas y escultóricas de pesca oceánica en la costa norte. Archaeobios 6 (1): 92-96
Zuzunaga J. 2013. Medidas de conservación y ordenación pesquera del jurel Trachurus murphyi (Nichols 1920) en el Perú. Rev. peru. biol. número especial 20(1): 107-113 .

9
El jurel Trachurus murphyi y las variables ambientales de macroescala
Rev. peru. biol. special issue 20(1): 009- 020 (September 2013)Ecology, fisheries and conservation of Jack mackerel (Trachurus murphyi) in Peru. Editors: Csirke J., R. Guevara-Carrasco & M. Espino
0Rev. peru. biol. número especial 20(1): 009- 020 (Septiembre 2013)Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú. Editores: Csirke J., R. Guevara-Carrasco & M. Espino
Facultad de ciencias Biológicas unMsMISSN-L 1561-0837
El jurel Trachurus murphyi y las variables ambientales de macroescala
Marco Espino
The Jack mackerel Trachurus murphyi and the environmental macro-scale variables
Instituto del Mar del Perú, esquina Gamarra y General Valle S/N Chucuito, Callao, Perú.
Email Marco Espino: [email protected]
el medioambiente del jurel (Trachurus murphyi) en el Perú
© Los autores. Publicado por la Revista Peruana de Biología de la Facultad de Ciencias Biológicas, Universidad Nacional Mayor de San Marcos. Este es un artículo de acceso abierto, distribuido bajo los términos de la Licencia de Atribución Reconocimiento-NoComercial-SinObraDerivada 3.0 de Creative Commons (http://creativecommons.org/licenses/by-nc-nd/3.0/deed.es_ES), que permite el uso no comercial, distribución y reproducción en cualquier medio, siempre que la obra original sea debidamente citadas. Para uso comercial, por favor póngase en contacto con [email protected].
Citación:Espino M. 2013. El jurel Trachurus murphyi y las variables ambientales de macroescala. En: Csirke J., R. Guevara-Carrasco & M. Espino (Eds.). Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú. Rev. peru. biol. número especial 20(1): 009- 020 (Sep-tiembre 2013)
ResumenSe analiza información de diversas variables ambientales de macroescala disponibles desde 1876 para el Pacífico Sudoriental y datos más recientes de desembarques y biomasas de jurel Trachurus murphyi Nichols 1920 en el mar peruano en relación con áreas probables de masas de agua equivalentes a Aguas Costeras Frías (ACF) y Aguas Subtropicales Superficiales (ASS). Se concluye que el Índice de Oscilación Decadal del Pacífico (PDO) presenta expresiones de variabilidad coherentes con las encontradas para el Índice de Oscilación del Sur (SOI) y que los cambios observados en las biomasas de T. murphyi en el mar peruano reflejan cambios en la disponibilidad del recurso asociado a patrones de variabilidad seculares (SOI) y decadales (PDO). Tales fluctuaciones en la disponibilidad del recurso afectan las pesquerías de Ecuador, Perú y norte de Chile, las que muestran variaciones importantes en sus desembarques y habrían dado una imagen sesgada de la abundancia conduciendo a diagnósticos errados de la real situación del recurso. Estos patrones de variabilidad afectarían también las manifestaciones de El Niño, haciendo que se inicien en otoños o primaveras australes dependiendo de si el PDO imperante es negativo o positivo. Por otro lado, en función a la distancia euclidiana de las varianzas del SOI se identifican periodos de alta (1876 – 1925 y 1976 – 2012) y baja (1926 – 1975) variabilidad; y con el PDO se diferencian periodos cálidos (1925 – 1944 y 1975 – 1994), frio (1945 – 1974) y templados o interfaces (1895 – 1924 y 1995 – 2012), siendo estos últimos producto de la interacción con los momentos de alta variabilidad.
Palabras clave: Jurel, variabilidad ambiental, PDO, SOI, distancia euclidiana
AbstractThis paper analyses information on various macro environmental variables available since 1876 for the Southeast Pacific and more recent data on Jack mackerel Trachurus murphyi (Nichols, 1920) landings and biomass in the Peruvian sea, relating them to probable areas of water masses equivalent to Cold Coastal Waters (CCW) and Subtropical Surface Waters (SSW). It is concluded that the index of the Pacific Decadal Oscillation (PDO) presents expressions of variability that are consistent with those found for the Southern Oscillation Index (SOI) and that the detected changes in biomass of Jack mackerel T. murphyi in the Peruvian sea reflect changes in the availability of the fish stock associated with secular (SOI) and decadal (PDO) variability patterns. These fluctuations in stock availability impact fisheries in Ecuador, Peru and northern Chile, which show significant variations in their landings and would have given a biased picture of the state of abundance, leading to wrong diagnoses of the real situation of the exploited stocks. These patterns of variability would also affect the appearance of El Niño, making them start in the southern hemisphere autumn or spring depending on whether the current PDO is positive or negative. Periods of high (1876 – 1925 and 1976 – 2012) and low (1926 – 1975) variability are also identified in relation to the Euclidean distance of the variances of the SOI; and in relation to the PDO a distinction is made between warm (1925 – 1944 and 1975 – 1994), cold (1945 – 1974) and tempered or interface periods (1895 – 1924 and 1995 – 2012), the latter being explained by the interaction between periods of high variability.
Keywords: Jack mackerel, environmental variability, PDO, SOI, Euclidian distance.
Publicado online: 11/10/2013Publicado impreso: 15/10/2013

10
Espino
Rev. peru. biol. número especial 20(1): 009- 020 (Septiembre 2013)Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú.
Editores: Csirke J., R. Guevara-Carrasco & M. Espino
Introducción El Pacífico Sudoriental está sometido permanentemente a
procesos de variabilidad ambiental que afectan los principales aspectos de la dinámica poblacional de los recursos pesque-ros, como el reclutamiento, crecimiento, mortalidad natural, mortalidad por pesca (capturabilidad) y los patrones de com-portamiento. Estos efectos se manifiestan a escala, intensidad y frecuencia diferentes, condicionando respuestas que finalmente constituyen estados de la misma población frente a la combina-ción de los factores de variabilidad correspondientes a cada uno de los momentos o periodos ambientales prevalentes, y que como ya se ha observado pueden ser de corto (estacionales), mediano (interanuales-ENSO), largo plazo (decadales) y muy largo plazo (seculares) (Espino 2003, Espino & Yamashiro 2012).
En este trabajo se presenta un análisis de algunos factores de la variabilidad ambiental a macroescala, en búsqueda de regularidades o patrones normativos ambientales de variabi-lidad (Espino 2003). Los que se plantean a través de modelos conceptuales que permiten caracterizar e identificar escenarios ambientales que explican cambios observados en las variables biológico-pesqueras del jurel Trachurus murphyi Nichols 1920 y su entorno ambiental, propios del mar peruano.
De lograrse, ello contribuiría a plantear acciones de ma-nejo acordes con cada momento o escenario ambiental y el probable estado poblacional coherente con dicho momento. En este contexto, la planificación de la producción pesquera en el corto, mediano y largo plazo podría contribuir a evitar cambios socioeconómicos y financieros traumáticos, y tam-bién favorecería el mejor aprovechamiento del recurso en cada estado y en particular en momentos de bonanza causados por el advenimiento de condiciones poblacionales favorables, bajo escenarios ambientales propicios para esta especie.
Material y métodos Los datos mensuales del Índice de Oscilación del Sur (SOI,
Southern Oscillation Index) de 1876 a 2012 fueron obtenidos del Bureau of Meteorology, National Climate Center of Australia. Basándose en la oscilación armónica definida de acuerdo a la siguiente expresión:
Var IOS = 7.5 – 1.5 [sen(2 * 3.1416 Ti – 2050 + 5)/ 100] (Espino, 2003)
Mediante la cual se plantea un modelo conceptual en el que se diferencian periodos o momentos de alta y baja variabilidad para el Pacífico Sur, en función a las desviaciones estándar del SOI. También se promediaron las desviaciones estándar anuales del SOI cada 50 años (1876 – 1925, 1926 – 1975 y 1976 – 2012) con lo cual se obtuvieron agrupamientos por distancia euclidiana.
Los datos mensuales del Índice de Oscilación Decadal del Pacífico (PDO, del inglés Pacific Decadal Oscillation) de 1900 a 2012 fueron obtenidos del Joint Institute for the Study of the Atmosphere and Ocean (JISAO), con los cuales Espino (2003) determinó una oscilación armónica asimétrica que responde a un modelo conceptual de acuerdo a la siguiente expresión:
PDO = – 0.25 – 1.5 [sen(2 *3.1416 Ti – 1880 + 5)/ 50 (Espino 2003)
Siguiendo los arreglos: 1895 – 1924, 1925 – 1944, 1945 – 1974, 1975 – 1994 y 1995 – 2012, se promediaron los datos
anuales de PDO, para luego definir agrupamientos térmicos diferenciados por distancia euclidiana.
Entre 1950 y 2012, los datos mensuales del Índice Multi-variado ENSO (MEI, Multivariate ENSO Index) se obtuvieron del Earth System Research Laboratory de la NOAA y los datos mensuales de temperatura superficial del mar (TSM) de Chicama fueron obtenidos de la base de datos institucional del IMARPE.
De acuerdo a la serie disponible de las variables ambientales MEI y TSM-Chicama, se promediaron por cuarto de siglo (1925 – 1949, 1950 – 1974, 1974 – 1999 y 2000 – 2012), los cuales se agruparon por distancia euclidiana para definir escenarios ambientales térmicos.
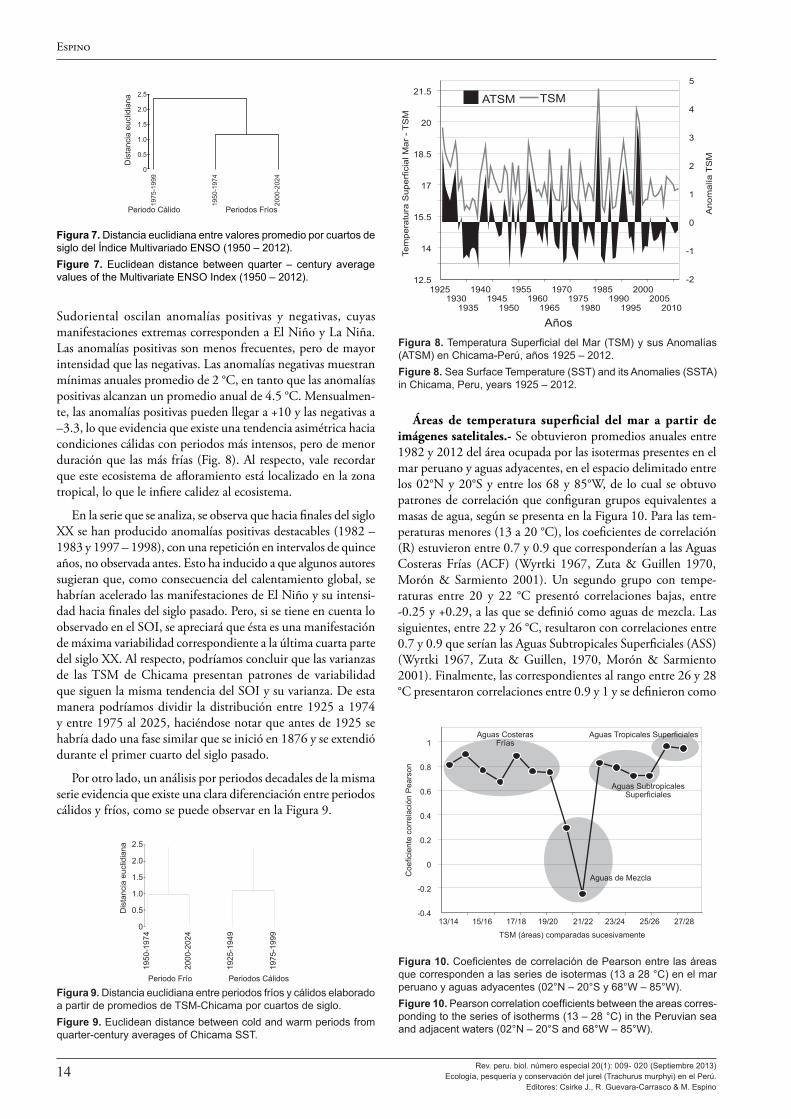
Las áreas de las isotermas (13 a 28 °C) fueron proporcionadas por la Unidad de Percepción Remota y Sistemas de Informa-ción Geográfica (UPRSIG) del IMARPE y correspondieron a promedios anuales del espacio geográfico comprendido entre los 02°N – 20°S y los 85 – 68°W. En el periodo 1982 – 2012, los tamaños promedio anuales de las áreas de cada isoterma (13 a 28 °C) se correlacionaron una a una (13/14, 14/15, 15/16, 16/17, 17/18, 18/19, 19/20, 20/21, 21/22, 22/23, 23/24, 24/25, 25/26, 26/27 y 27/28), obteniéndose coeficientes de correlación de Pearson (r), a partir de los cuales se planteo una propuesta operacional de identificación de masas de aguas superficiales, mediante el levantamiento de cartas satelitales de TSM. Además, se calcularon valores de TSM Medias Ponderadas (TSMP) al área promedio anual de las isotermas correspondientes, de acuerdo a la siguiente formulación:
TSMP=Σ(Tº isoterma*Área (km2) isoterma)/(ΣÁreas de las isotermas)
De ésta se derivaron las anomalías de la TSMP (ATSM) según la siguiente ecuación:
ATSM=TSMP(i) – Promedio de la TSMP(1982 – 2012)
Se analizaron los desembarques anuales de 1960 a 2012, obtenidos del Ministerio de la Producción del Perú, de doce especies de peces que representan en promedio el 96.5% de los desembarques nacionales, los cuales fueron estandarizados para cada año de acuerdo a lo propuesto por Noakes y Beamish (2009).
Las especies seleccionadas fueron el jurel T. murphyi, ancho-veta Engraulis ringens, sardina Sardinops sagax, caballa Scomber japonicus, bonito Sarda chiliensis chiliensis, merluza Merluccius gayi peruanus, cojinova Seriolella violacea, pejerrey Odontesthes regia, cavinza Isacia conceptionis, machete Ethmidium maculatum, sierra Scomberomorus sierra y la pota o jibia Dosidicus gigas.
Se establecieron relaciones entre los desembarques de T. murphyi con los de las otras especies, para lo cual se empleó la correlación de Pearson (r) para arreglos decadales (décadas de los sesenta, setenta, ochenta, noventa, y el periodo comprendido entre el 2000 y el 2010). Por otro lado, las biomasas T. murphyi fueron relacionadas con las variables ambientales también bajo los mismos arreglos decadales.
Los análisis de agrupamiento fueron realizados por distancia euclidiana y análisis de similitud (similaridad) de Bray Curtis, empleando el programa PRIMER 6.0.

11
El jurel Trachurus murphyi y las variables ambientales de macroescala
Rev. peru. biol. special issue 20(1): 009- 020 (September 2013)Ecology, fisheries and conservation of Jack mackerel (Trachurus murphyi) in Peru. Editors: Csirke J., R. Guevara-Carrasco & M. Espino
Figura 2. Distancia euclidiana aplicada a la desviación estándar del SOI, que agrupa los periodos de alta variabilidad (1876 – 1925 y 1976 – 2012) diferenciándolos de los de baja variabilidad (1926 – 1975).Figure 2. Euclidean distance applied to the standard deviation of the SOI, which groups high variability periods (1876 – 1925 and 1976 – 2011) distinguishing them from those of low variability (1926 – 1975).
Figura 1. Índice de Oscilación del Sur (SOI) y su varianza expresada como Índice de Variabilidad Secular del Pacífico (PSV) desde 1876 al 2012.Figure 1. Southern Oscillation Index (SOI) and its variance expressed as Pacific Secular Variability Index (PSV) from 1876 to 2012.
Resultados Variables ambientales de macroescala espacial.- El aná-
lisis de mediano y largo plazo de las variables ambientales de macroescala permitiría explicar los patrones de distribución de T. murphyi en el mar peruano y en el Pacífico Sudoriental en general. El efecto de estas variables en la dinámica poblacional podría apreciarse a partir del mejor conocimiento de las mismas.
El Índice de Oscilación del Sur (SOI).- Desde 1876 la serie disponible del SOI refleja una alta variabilidad en pro-cesos estacionales (verano-invierno), interanuales (El Niño-La Niña), decadales (décadas cálidas-décadas frías) y seculares (alta y baja variabilidad). Es así que entre 1876 y 1925 se presentan valores más extremos, negativos en los años 1877, 1888, 1896, 1905 y positivos en 1879, 1910, 1917, reflejando una mayor variabilidad o relativa inestabilidad. Entre 1925 y 1975, las manifestaciones extremas negativas (1940, 1941) y positivas (1950) fueron menos frecuentes y menos intensas, lo que expresó un patrón de menor variabilidad o relativa estabilidad. Luego, desde 1976 a la actualidad se repite una tendencia similar a la observada entre 1876 y 1925 (Figs. 1 y 2).
Un análisis de agrupamiento utilizando la distancia euclidiana evidencia la separación de esos periodos (Fig. 2). De acuerdo a ello se identifican momentos de alta y baja variabilidad en el Pacífico Sudoriental, expresados en manifestaciones más y menos intensas de El Niño y La Niña, respectivamente.
Los momentos de alta variabilidad con expresiones máximas o muy fuertes de El Niño se produjeron en los primeros y últimos cuartos de cada siglo coincidentemente con lo que planteó la serie de Quinn et al. (1987) donde las expresiones “muy fuertes” (VS: Very Strong) de El Niño se observaron en los años 1578, 1728, 1791, 1828, 1877-78, 1891, 1925-26, 1982-83 y 1997-98. Obsérvese que tanto en el siglo XIX como en el siglo XX se presentaron tres manifestaciones extremas de El Niño, dos de las cuales se observaron en el último cuarto de los referidos siglos y una en el comienzo (1925).
Probablemente, en los siglos anteriores ocurrieron manifes-taciones similares a las acaecidas en el siglo XX, pero no pueden ser definidas claramente, porque la información utilizada esta
basada en reportes de cronistas e historiadores sobre los impac-tos del evento en el ambiente, en la pesquería y en la vida de las aves y mamíferos marinos, que podrían estar distorsionados por la apreciación subjetiva de los autores de tales registros. Sin embargo, a pesar de ello no dejan de ser señales útiles para el análisis de las manifestaciones extremas de El Niño.
Quinn et al. (1987) reportaron manifestaciones de El Niño desde 1525, observándose que eventos muy fuertes no han sido registrados más de tres en cada siglo. Podemos destacar el siglo XVII en que no se produjo ninguna manifestación de tal mag-nitud (Tablas 1 y 2), esto es coincidente con la pequeña edad glacial del medioevo, cuando se produjo el Mínimo de Maunder (MM), caracterizado por extremas temperaturas bajas que afecta-ron a grandes áreas del planeta (Maenza & Compagnucci 2010).
Por su parte, en los cuartos de siglo intermedios (1925 – 1975), de menor variabilidad, las manifestaciones de El Niño son menos frecuentes y de menor intensidad, no mostrando manifestaciones fuertes. Tal es el caso de los años 1940 y 1941, en los que la magnitud (SOI) del fenómeno fue significativa (fuerte), pero como expresión de variabilidad no lo fue, pues su desviación estándar no fue importante, con valores medios anuales de -13.8 y -12.9, pero con una baja desviación estándar.
En conclusión, los datos expresan que en este periodo, en intervalos alternados de aproximadamente 50 años, el SOI y su varianza definen momentos de alta y baja variabilidad, con manifestaciones fuertes y muy fuertes de El Niño y La Niña, en los últimos y primeros cuartos de cada siglo. En los cuartos de siglo intermedios, son menos probables las manifestaciones severas de este fenómeno. Estos momentos con dominancia de SOI positivos aumentarían la probabilidad de mayor disponibili-dad del stock norte (far north) del jurel del Pacífico Sudoriental.
El patrón de variabilidad expresado por los valores anuales altos y bajos de la desviación estándar del SOI se ha denomi-nado Variabilidad Secular del Pacífico (PSV) (Espino 2003), por corresponder a ciclos armónicos simétricos completos de 100 años, que significan una expresión de intensidad de varia-bilidad en ambos extremos: positivo y negativo. Sin embargo, este patrón no constituye manifestación térmica de variabilidad como el PDO.
La Oscilación Decadal del Pacífico (PDO).- La Oscilación Decadal del Pacífico (PDO) (Hare 1996, Mantua et al. 1997) es un patrón de variabilidad climática espacial persistente (20 a
Índi
ce d
e O
scila
ción
Sur
(SO
I)
Des
viac
ión
Está
ndar
del
SO
I - P
SV
25
15
5
-5
-15
-25
Años
20
15
10
5
0
1884 1900 1916 1932 1948 1964 1980 1996 2012
79
77 8896
5
10
17
4041
50 75
778287
811
298
919792
93
1876 1892 1908 1924 1940 1956 1972 1988 2004
Dis
tanc
ia e
uclid
iana
Alta variabilidad Baja variabilidad
10
8
6
4
2
0
1876
-192
5
1976
-202
5
1926
-197
5

12
Espino
Rev. peru. biol. número especial 20(1): 009- 020 (Septiembre 2013)Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú.
Editores: Csirke J., R. Guevara-Carrasco & M. Espino
Año Int Año Int Año Int
1525 – 1526 S 1687 – 1688 S+ 1871 S1531 – 1532 S 1696 S 1877 – 1878 VS1539 – 1541 S 1701 S+ 1884 S+
1552 S 1707 – 1708 S 1891 VS1567 – 1568 S+ 1714 – 1715 S 1899 – 1900 S
1574 S 1720 S+ 1911 – 1912 S1578 VS 1728 VS 1917 S
1591 – 1592 S 1747 S 1925 – 1926 VS1607 S 1761 S 1932 S1614 S 1775 S 1940 – 1941 S
1618 – 1619 S 1785 – 1786 S 1957 – 1958 S1624 S 1791 VS 1972 – 1973 S1634 S 1803 – 1804 S+ 1982 – 1983 VS1652 S+ 1814 S 1997 – 1998 VS1660 S 1828 VS 2009 – 2010 S1671 S 1844 – 1845 S+1681 S 1864 S
Int = Intensidad; S = Fuerte (Strong); VS = Muy fuerte (Very Strong)
Tabla 1. Eventos El Niño fuertes y muy fuertes acaecidos en el Perú desde 1525 a la actualidad (actualizado después de Quinn et al. 1987)Table 1. Strong and very strong El Niño events occurred in Peru since 1525 to date (updated after Quinn et al. 1987)
Figura 3. Oscilación Decadal del Pacífico (1900 – 2012).Figure 3. Pacific Decadal Oscillation (1900 – 2012).
30 años), que abarca todo el Océano Pacífico (Fig. 3) con ma-nifestaciones muy claras en el Pacífico Norte (Norteamérica) y efectos secundarios en los trópicos. Todo lo contrario de lo que sucede con el proceso ENSO, cuyo escenario es la zona inter-tropical. Mantua (1999a y b) describe manifestaciones de este evento entre 1890 y 1924 (fase fría), entre 1925 y 1946 (fase cálida), entre 1947 y 1976 (fase fría) y, finalmente, de 1977 hasta mediados de la década de los noventa (fase cálida). Observando la Figura 3 se puede pronosticar que en la segunda mitad de la década de los noventa se habría iniciado una fase fría (Espino & Yamashiro 2012).
En esa medida, para la zona ecuatorial, también se definen fases positivas y negativas del PDO, que se alternan entre sí condicionando escenarios fríos y cálidos para la zona norte del Pacífico Sudoriental, es decir, en el mar peruano y ecuatoriano. Durante el primer cuarto del siglo XX, se alternan años cálidos y fríos, lo cual coincide con un periodo de alta variabilidad ex-presada en la varianza del SOI (Fig. 1). Luego, se observa una fase positiva que se extiende hasta 1946. Entre 1946 y 1975 se observan valores negativos del PDO, con 5 manifestaciones positivas (1948, 1958, 1960, 1961 y 1970), lo que representa sólo el 17% de la serie. Es decir, entre 1926 y 1975, se puede observar que está claramente definido que a una fase cálida le si-gue una fría, lo cual coincide con un periodo de baja variabilidad del SOI. De 1976 a 1995 predominaron condiciones positivas con sólo 4 desviaciones negativas (1989, 1990, 1991 y 1995) que representan el 20% de la serie. Luego, entre 1996 y 2012 se alternan desviaciones positivas y negativas, con comportamiento similar al observado previamente a 1925.
El análisis de agrupamiento por distancia euclidiana entre periodos diferenciados del PDO determina tres grupos de años que corresponden a periodos cálidos (predominio de anomalías positivas), periodos fríos (predominio de anomalías negativas) y periodos intermedios o de interfase que, en adelante, se denominarán periodos templados porque en tales momentos se intercalan desviaciones positivas y negativas con la misma
frecuencia (9) hasta el 2012 (Fig. 4). Esto sugiere que en las fases de SOI de alta variabilidad se producen recurrentemente desviaciones en ambos sentidos, lo cual imposibilita la definición clara de un estado cálido o frío, según corresponda, mientras que las manifestaciones del PDO son más evidentes en su estado positivo o negativo en las fases de baja variabilidad.
Se puede concluir que el PDO presenta expresiones de varia-bilidad coherentes con las encontradas para el SOI. Entre estas manifestaciones, en la fase intermedia (1926 – 1975) de baja variabilidad del SOI, se destaca la definición clara de un periodo cálido (entre 1925 y 1944) y un periodo frío (entre 1945 y 1974), mientras que los denominados periodos templados se diferencian
Año Int Año Int Año Int
1803-1804 S+ 1867-1868 M 1925-1926 VS
1806-1807 M 1871 S+ 1930-1931 W/M
1812 M 1874 M 1932 S
1814 S 1877-1878 VS 1939 M+
1817 M+ 1880 M 1940-1941 S
1819 M+ 1884 S+ 1943 M+
1821 M 1887-1889 W/M 1951 W/M
1824 M 1891 VS 1953 M+
1828 VS 1896-1897 M+ 1957-1958 S
1832 M 1899-1900 S 1965 M+
1837 M 1902 M+ 1972-1973 S
1844-1845 S+ 1905 W/M 1976 M
1850 M 1907 M 1982-1983 VS
1854 W/M 1911-1912 S 1987 M
1857-1858 M+ 1914 M+ 1991-1994 M
1860 M 1917 S 1997-1998 VS
1864 S 1918-1919 W/M 2002 M
1866 M 1923 M 2009-2010 S
Int = Intensidad; W/S = Casi moderado (Near Moderate); M = Moderado (Moderate) S = Fuerte (Strong); VS = Muy fuerte (Very Strong)
Tabla 2. Eventos El Niño casi moderados, moderados, fuertes y muy fuertes acaecidos en el Perú desde 1803 a la actualidad (Actualizado de Quinn et al. 1987).Table 2. Almost moderate, moderate, strong and very strong El Niño events in Peru occurred since 1803 to date (Updated from Quinn et al. 1987)
Osc
ilaci
ón D
ecad
al d
el P
acífi
co
1900 1914 1928 1942 1956 1970 1984 1998 2012
2
1
0
-1
-2
-3
Años
1925-1926
1945-19461975-1976
1995-1996
2025-2026

13
El jurel Trachurus murphyi y las variables ambientales de macroescala
Rev. peru. biol. special issue 20(1): 009- 020 (September 2013)Ecology, fisheries and conservation of Jack mackerel (Trachurus murphyi) in Peru. Editors: Csirke J., R. Guevara-Carrasco & M. Espino
Figura 4. Distancia euclidiana aplicada al PDO diferenciando tres agrupamientos térmicos correspondientes a periodos cálidos (1925 – 1944 y 1975 – 1994), templados o interfases (1895 – 1924 y 1995 – 2024) y frío (1945 – 1974).Figure 4. Euclidean distance applied to the PDO distinguishing three thermal groupings, corresponding to warm (1925 – 1944 and 1975 – 1994), temperate or interface (1895 – 1924 y 1995 – 2024) and cold (1945 – 1974) periods.
Indi
ce M
ultiv
aria
do E
NS
O
1950 1960 1970 1980 1990 2000 2010 2020 2030
2
1.5
1
0.5
0
-0.5
-1
-1.5
2
Años
Figura 5. Índice multivariado ENSO – El Niño Southern Oscillation (1950 – 2012).Figure 5. Multivariate ENSO Index (1950 – 2012).
Indi
ce M
ultiv
aria
do E
NS
O
1950 1960 1970 1980 1990 2000 2010 2020 2030
0.6
0.4
0.2
0
-0.2
-0.4
-0.6
Décadas
Figura 6. Índice Multivariado ENSO por décadas (1950 – 2010).Figure 6. Multivariate ENSO Index by decades (1950 – 2010).
claramente en los momentos de alta variabilidad, debido a que en ellos se alternan desviaciones positivas y negativas, que responden al patrón de alta variabilidad imperante. Al respecto, de 1995 a 2012, hemos tenido 9 desviaciones positivas y 9 negativas, lo que coincide con lo observado el siglo pasado. Es de esperar que esta dinámica persista en tanto dure el periodo de alta variabilidad que atravesamos (1976 – 2025) y que, luego de concluido, se den condiciones similares a las obtenidas a mediados del siglo XX, repitiéndose una fase de baja variabilidad (2026 – 2075) (Figs. 2, 3 y 4).
Respecto al jurel, esta especie estaría más disponible a la pes-quería de Perú y Ecuador cuando predominen valores positivos del PDO, tal como lo que se observó entre finales de la década de los setenta e inicios de los noventa, así como recientemente cuando han predominado condiciones cálidas (2003 – 2006). Se puede pronosticar que en los próximos años (al 2025) habrá desviaciones alternadas, negativas y positivas del PDO, las cuales alejarán o acercarán a T. murphyi de las costas del norte de Chile, Perú y Ecuador, lo que dependerá del periodo del SOI que atra-vesemos. Luego, tendremos el advenimiento de un persistente PDO positivo (+) cuando las condiciones sean manifiestamente favorables para aumentar la disponibilidad del stock norte (far north) del jurel T. murphyi del Pacífico Sudoriental.
Índice Multivariado ENSO (El Niño Southern Oscilla-tion).- El Índice Multivariado ENSO (MEI) (Wolter 1987, Wolter & Timlin 1993, 1998) es el mejor indicador para el monitoreo de El Niño debido a que integra seis variables que reflejan el acoplamiento entre el océano y la atmósfera, siendo menos sensible a las variaciones ocasionales, por efectos locales de algunas variables. El MEI integra la presión a nivel del mar (P), componentes zonales (U) y meridionales (V) del viento superficial, temperatura superficial del mar (S), temperatura superficial del aire (A) y cobertura total del cielo (C).
En la serie disponible entre 1950 y el 2011, se revelan dos escenarios ambientales: uno con dominancia de valores negativos, entre 1950 y 1976; y un segundo, entre 1977 y 2006, con domi-nancia de valores positivos. Luego, son recurrentes las desviaciones negativas (Fig. 5) en los últimos cinco años. Se podría concluir que éstos corresponderían a escenarios ambientales fríos y cálidos,
respectivamente. El primero de ellos coincide con la declinación de una fase de baja variabilidad y con una fase fría del PDO, mientras que el segundo coincide con el inicio de una fase de alta variabilidad y con una fase cálida del PDO. Asimismo, debemos considerar que además de ser muy útil para monitorear El Niño, el MEI también es de utilidad para definir escenarios ambientales como sucede con el SOI y TSM de Chicama.
Ahora bien, si se analiza la serie del Índice Multivariado ENSO por cuartos de siglo, por distancia euclidiana se obtiene dos agrupamientos que reflejan dos periodos fríos (1950 – 1974 y 2000 – 2012) y uno cálido (1975 – 1999) (Figs.6 y 7).
Temperatura superficial del mar (TSM) y sus anomalías (ATSM).- En el Perú y el Pacífico Sudoriental, la serie más am-plia de información (1925 – 2012) de Temperatura Superficial del Mar (TSM) y sus anomalías (ATSM) es la que corresponde a la estación costera de Chicama (07°41’41”S – 79°26’03”W). Esta serie revela que la variabilidad es una característica en el Ecosistema de Afloramiento Peruano (EAP). Es decir, en el mar peruano la “normalidad” es la variabilidad (Espino & Yamashiro 2012). En esa medida, debemos considerar que en el Pacífico
5
4
3
2
1
0
Dist
ancia
euc
lidia
na
1945
-197
4
1925
-194
4
1975
-199
4
1995
-202
4
1895
-192
4
Periodo FríoPeriodos Cálidos
Periodos Templados
14
Espino
Rev. peru. biol. número especial 20(1): 009- 020 (Septiembre 2013)Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú.
Editores: Csirke J., R. Guevara-Carrasco & M. Espino
Figura 7. Distancia euclidiana entre valores promedio por cuartos de siglo del Índice Multivariado ENSO (1950 – 2012).Figure 7. Euclidean distance between quarter – century average values of the Multivariate ENSO Index (1950 – 2012).
Tem
pera
tura
Sup
erfic
ial M
ar -
TS
M
1925 1940 1955 1970 1985 20001930 1945 1960 1975 1990 2005
1935 1950 1965 1980 1995 2010
21.5
20
18.5
17
15.5
14
12.5
Años
5
4
3
2
1
0
-1
-2
Ano
mal
ía T
SM
ATSM TSM
Figura 8. Temperatura Superficial del Mar (TSM) y sus Anomalías (ATSM) en Chicama-Perú, años 1925 – 2012.Figure 8. Sea Surface Temperature (SST) and its Anomalies (SSTA) in Chicama, Peru, years 1925 – 2012.
Figura 9. Distancia euclidiana entre periodos fríos y cálidos elaborado a partir de promedios de TSM-Chicama por cuartos de siglo.Figure 9. Euclidean distance between cold and warm periods from quarter-century averages of Chicama SST.
Coe
ficie
nte
corr
elac
ión
Pea
rson
1
0.8
0.6
0.4
0.2
0
-0.2
-0.413/14 15/16 17/18 19/20 21/22 23/24
TSM (áreas) comparadas sucesivamente
25/26 27/28
Aguas CosterasFrías
Aguas Tropicales Superficiales
Aguas SubtropicalesSuperficiales
Aguas de Mezcla
Figura 10. Coeficientes de correlación de Pearson entre las áreas que corresponden a las series de isotermas (13 a 28 °C) en el mar peruano y aguas adyacentes (02°N – 20°S y 68°W – 85°W).Figure 10. Pearson correlation coefficients between the areas corres-ponding to the series of isotherms (13 – 28 °C) in the Peruvian sea and adjacent waters (02°N – 20°S and 68°W – 85°W).
Sudoriental oscilan anomalías positivas y negativas, cuyas manifestaciones extremas corresponden a El Niño y La Niña. Las anomalías positivas son menos frecuentes, pero de mayor intensidad que las negativas. Las anomalías negativas muestran mínimas anuales promedio de 2 °C, en tanto que las anomalías positivas alcanzan un promedio anual de 4.5 °C. Mensualmen-te, las anomalías positivas pueden llegar a +10 y las negativas a –3.3, lo que evidencia que existe una tendencia asimétrica hacia condiciones cálidas con periodos más intensos, pero de menor duración que las más frías (Fig. 8). Al respecto, vale recordar que este ecosistema de afloramiento está localizado en la zona tropical, lo que le infiere calidez al ecosistema.
En la serie que se analiza, se observa que hacia finales del siglo XX se han producido anomalías positivas destacables (1982 – 1983 y 1997 – 1998), con una repetición en intervalos de quince años, no observada antes. Esto ha inducido a que algunos autores sugieran que, como consecuencia del calentamiento global, se habrían acelerado las manifestaciones de El Niño y su intensi-dad hacia finales del siglo pasado. Pero, si se tiene en cuenta lo observado en el SOI, se apreciará que ésta es una manifestación de máxima variabilidad correspondiente a la última cuarta parte del siglo XX. Al respecto, podríamos concluir que las varianzas de las TSM de Chicama presentan patrones de variabilidad que siguen la misma tendencia del SOI y su varianza. De esta manera podríamos dividir la distribución entre 1925 a 1974 y entre 1975 al 2025, haciéndose notar que antes de 1925 se habría dado una fase similar que se inició en 1876 y se extendió durante el primer cuarto del siglo pasado.
Por otro lado, un análisis por periodos decadales de la misma serie evidencia que existe una clara diferenciación entre periodos cálidos y fríos, como se puede observar en la Figura 9.
Áreas de temperatura superficial del mar a partir de imágenes satelitales.- Se obtuvieron promedios anuales entre 1982 y 2012 del área ocupada por las isotermas presentes en el mar peruano y aguas adyacentes, en el espacio delimitado entre los 02°N y 20°S y entre los 68 y 85°W, de lo cual se obtuvo patrones de correlación que configuran grupos equivalentes a masas de agua, según se presenta en la Figura 10. Para las tem-peraturas menores (13 a 20 °C), los coeficientes de correlación (R) estuvieron entre 0.7 y 0.9 que corresponderían a las Aguas Costeras Frías (ACF) (Wyrtki 1967, Zuta & Guillen 1970, Morón & Sarmiento 2001). Un segundo grupo con tempe-raturas entre 20 y 22 °C presentó correlaciones bajas, entre -0.25 y +0.29, a las que se definió como aguas de mezcla. Las siguientes, entre 22 y 26 °C, resultaron con correlaciones entre 0.7 y 0.9 que serían las Aguas Subtropicales Superficiales (ASS) (Wyrtki 1967, Zuta & Guillen, 1970, Morón & Sarmiento 2001). Finalmente, las correspondientes al rango entre 26 y 28 °C presentaron correlaciones entre 0.9 y 1 y se definieron como
2.5
2.0
1.5
1.0
0.5
0
Dis
tanc
ia e
uclid
iana
1950
-197
4
2000
-202
4
1975
-199
9
Periodo Frío Periodos Cálidos
1925
-194
9
2.5
2.0
1.5
1.0
0.5
0
Dis
tanc
ia e
uclid
iana
1975
-199
9
1950
-197
4
2000
-202
4
Periodos FríosPeriodo Cálido

15
El jurel Trachurus murphyi y las variables ambientales de macroescala
Rev. peru. biol. special issue 20(1): 009- 020 (September 2013)Ecology, fisheries and conservation of Jack mackerel (Trachurus murphyi) in Peru. Editors: Csirke J., R. Guevara-Carrasco & M. Espino
Áre
as (k
m2 )
Mile
s
500
400
300
200
100
0
TSM
- P
onde
rada
23
22.5
22
21.5
21
20.5
20
19.51980 1985 1990 1995 2000 2005 2010
Años
Isotermas 13 - 20ºC Isotermas 22 - 25ºC TSM ponderada
Figura 11. Distribución de áreas de conjunto de isotermas (13 – 20 °C y 22 – 25 °C) en el mar peruano y áreas adyacentes (02°N – 20°S y 68 – 85°W), curvas de tendencia de las ASS (líneas negra) y ACF (línea gris), y Temperatura Superficial Media del Mar Ponderada (TSMP).Figure 11. Distribution of the areas two sets of isotherms (13 – 20 °C and 22 – 25 °C) in the Peruvian sea and adjacent waters (02°N – 20°S and 68 – 85°W), trend curves of the SSW (black line) and CCW (grey line), and Weighted Average Sea Surface Temperature (WASST).
Biom
asas
tota
les
mill
ones
(t)
ATSM
P &
BE
Años
2
1
0
-1
-2
1982 1987 1992 1997 2002 2007 2012
ATSMP (+)Biomasa estandarizada (BE)
ATSMP (-)
Biomasa total
Biomasa media
periodo frío
periodo cálido
0
2
4
8
10
6
Figura 12. Biomasas acústicas totales y biomasas estandarizadas de jurel T. murphyi y ATSMP, evidenciando periodos cálidos con dominan-cia de ASS y desviaciones positivas de las biomasas, y periodos fríos con predominio de ACF y desviaciones negativas de las biomasas.Figure 12. Total acoustic biomass and standardized biomass of Jack mackerel T. murphyi, and Weighted Surfase See Temperature Ano-maly (WSSTA) showing warm periods with dominance of SSW and positive deviations of biomass and cold periods with CCW prevalence and negative biomass deviations.
Aguas Tropicales Superficiales (ATS) (Wyrtki 1967, Morón & Sarmiento 2001). Entre 1982 y 2012, con estas consideraciones se definen escenarios térmicos que se diferencian claramente a partir del año 2000 (Fig. 11) y si se comparan con la tendencia de la Temperatura Superficial Media Ponderada (TSMP), la tendencia es muy similar a la que corresponde a las áreas de las isotermas comprendidas entre 22 y 25 °C (ASS).
De esta manera, las condiciones térmicas del mar peruano estarían regidas por la interacción acercamiento-alejamiento de las ASS y las ACF, lo cual define escenarios que constituyen estados ambientales fríos y cálidos. En esa medida, a partir de las áreas de TSM determinadas a través de imágenes satelitales, debemos concluir que durante las décadas de los ochenta y de los noventa han prevalecido condiciones cálidas en el mar peruano y a partir del 2000 se ha iniciado un periodo con dominancia de condiciones frías. Al respecto, subrayamos que en los últimos 12 años se ha reducido la TSMP en 1.4 °C debido a un incremento significativo de las áreas de las isotermas comprendidas entre 13 y 20 °C correspondiente a las ACF.
Por otro lado, si se relacionan las denominadas ACF y ASS con las variables ambientales de macroescala (Tabla 3) como la TSM-Chicama, TSMP, SOI, PDO y MEI, las correlaciones son significativas, evidenciando que lo obtenido a partir de imágenes satelitales explica parte del comportamiento del ecosistema de esta zona del Pacífico Sudoriental.
La pesquería del jurel y las variables de macroescala en el mar peruano.- El jurel T. murphyi es un recurso pelágico que cuenta con registros históricos de desembarques desde la década de los treinta. Ñiquen et al. (2013) describen detalladamente esta pesquería, documentando registros de desembarque y otros datos pesqueros desde comienzos del siglo XX. Los primeros años muestran valores bajos porque la pesquería dirigida a esta especie era muy reducida; los registros de desembarques recién aumentaron en la segunda mitad de la década de los setenta, llegando a 504992 y 386793 toneladas en 1977 y 1978 respecti-vamente. En la década de los ochenta, los desembarques fueron más bajos, pero en 1985 llegaron a 189000 toneladas y hacia finales de la década superaban las 100000 toneladas. La década de los noventa mostro un incremento de los desembarques, man-teniéndose en promedio por encima de las 200000 toneladas. En el 2001, los desembarques superaron las 700000 toneladas, y después descendieron paulatinamente hasta alcanzar su mínima expresión en el 2010, para luego volver a recuperarse por encima de las 200000 toneladas el 2011. En general, los desembarques no mostraron una tendencia determinada, predominando una alta variabilidad.
Para el caso del jurel T. murphyi en el mar peruano, por la elevada variabilidad que condiciona un alto desplazamiento del recurso, las biomasas acústicas sólo expresan su disponibilidad y muestran dos momentos claramente diferenciados (Fig. 12). El primero corresponde a las décadas de los ochenta y los noventa, que coinciden con ATSMP positivas y altas biomasas, las cuales promedian los 4.9 millones de toneladas; y el segundo, que corresponde a los años 2000 con ATSMP negativas y con bajas biomasas que en promedio no superan las 500000 toneladas anuales. Es decir, en los últimos años la biomasa ha mostrado una reducción del 90%. Sin embargo, las capturas han mostrado un incremento promedio del 30%, lo que se explica en que, si bien las biomasas son mayores durante la dominancia de ATSMP
positivas, su distribución en el mar peruano es más amplia, siendo menos accesible a la flota pesquera de cerco dedicada a la pesca de este recurso. Asimismo, durante los periodos cálidos, las mínimas de oxígeno (Bertrand et al. 2011) y la isoterma de 15 °C (Flores et al. 2013) se ubican a mayor profundidad, lo cual favorece la dispersión vertical del jurel T. murphyi, haciendolo menos vulnerable a la pesquería de cerco. A esto se debe agregar que durante los últimos 50 años las principales especies objetivo de la pesquería peruana de cerco han sido la anchoveta E. ringens en la década de los sesenta, la sardina S. sagax sagax de mediados de la década de los setenta hasta inicios de los noventa, y luego nuevamente la anchoveta, persistiendo hasta la actualidad (2013).
En general, hasta hace algunos años la flota peruana se ha dedicado a la pesca de jurel T. murphyi más como una alternativa para los periodos en que las otras pesquerías eran poco abundantes o no se ejercían.
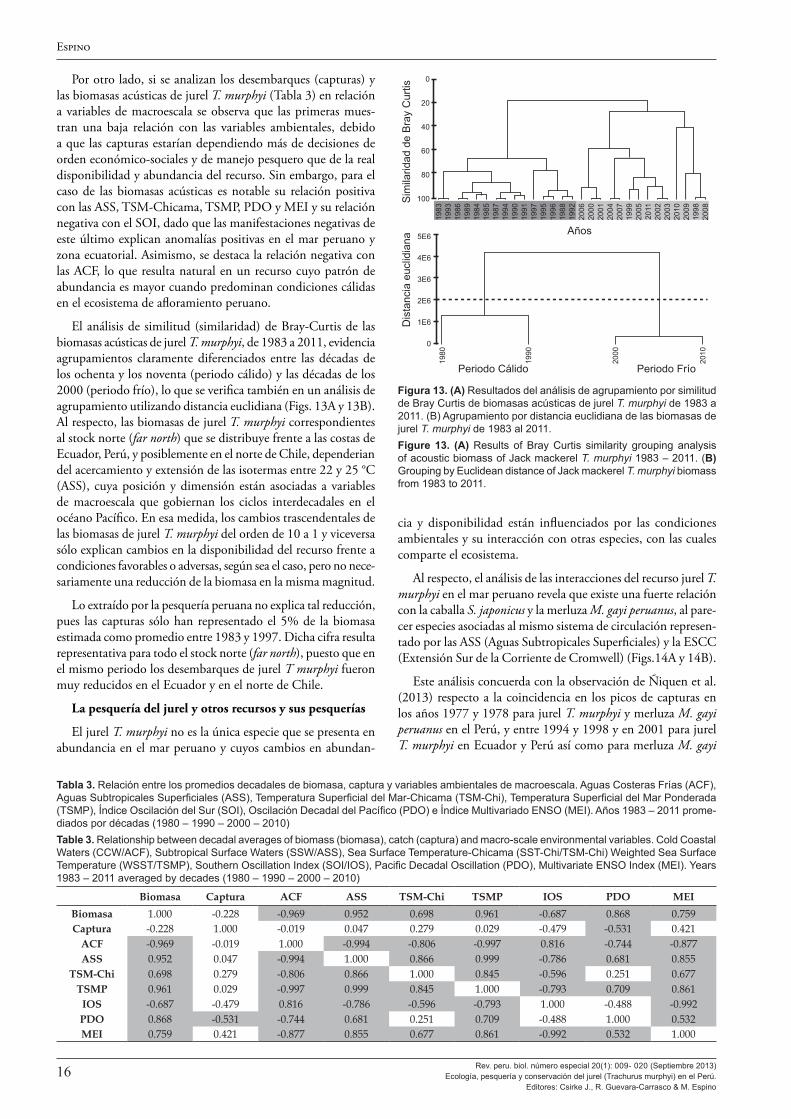
16
Espino
Rev. peru. biol. número especial 20(1): 009- 020 (Septiembre 2013)Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú.
Editores: Csirke J., R. Guevara-Carrasco & M. Espino
Tabla 3. Relación entre los promedios decadales de biomasa, captura y variables ambientales de macroescala. Aguas Costeras Frías (ACF), Aguas Subtropicales Superficiales (ASS), Temperatura Superficial del Mar-Chicama (TSM-Chi), Temperatura Superficial del Mar Ponderada (TSMP), Índice Oscilación del Sur (SOI), Oscilación Decadal del Pacífico (PDO) e Índice Multivariado ENSO (MEI). Años 1983 – 2011 prome-diados por décadas (1980 – 1990 – 2000 – 2010)Table 3. Relationship between decadal averages of biomass (biomasa), catch (captura) and macro-scale environmental variables. Cold Coastal Waters (CCW/ACF), Subtropical Surface Waters (SSW/ASS), Sea Surface Temperature-Chicama (SST-Chi/TSM-Chi) Weighted Sea Surface Temperature (WSST/TSMP), Southern Oscillation Index (SOI/IOS), Pacific Decadal Oscillation (PDO), Multivariate ENSO Index (MEI). Years 1983 – 2011 averaged by decades (1980 – 1990 – 2000 – 2010)
Figura 13. (A) Resultados del análisis de agrupamiento por similitud de Bray Curtis de biomasas acústicas de jurel T. murphyi de 1983 a 2011. (B) Agrupamiento por distancia euclidiana de las biomasas de jurel T. murphyi de 1983 al 2011. Figure 13. (A) Results of Bray Curtis similarity grouping analysis of acoustic biomass of Jack mackerel T. murphyi 1983 – 2011. (B) Grouping by Euclidean distance of Jack mackerel T. murphyi biomass from 1983 to 2011.
Por otro lado, si se analizan los desembarques (capturas) y las biomasas acústicas de jurel T. murphyi (Tabla 3) en relación a variables de macroescala se observa que las primeras mues-tran una baja relación con las variables ambientales, debido a que las capturas estarían dependiendo más de decisiones de orden económico-sociales y de manejo pesquero que de la real disponibilidad y abundancia del recurso. Sin embargo, para el caso de las biomasas acústicas es notable su relación positiva con las ASS, TSM-Chicama, TSMP, PDO y MEI y su relación negativa con el SOI, dado que las manifestaciones negativas de este último explican anomalías positivas en el mar peruano y zona ecuatorial. Asimismo, se destaca la relación negativa con las ACF, lo que resulta natural en un recurso cuyo patrón de abundancia es mayor cuando predominan condiciones cálidas en el ecosistema de afloramiento peruano.
El análisis de similitud (similaridad) de Bray-Curtis de las biomasas acústicas de jurel T. murphyi, de 1983 a 2011, evidencia agrupamientos claramente diferenciados entre las décadas de los ochenta y los noventa (periodo cálido) y las décadas de los 2000 (periodo frío), lo que se verifica también en un análisis de agrupamiento utilizando distancia euclidiana (Figs. 13A y 13B). Al respecto, las biomasas de jurel T. murphyi correspondientes al stock norte (far north) que se distribuye frente a las costas de Ecuador, Perú, y posiblemente en el norte de Chile, dependerian del acercamiento y extensión de las isotermas entre 22 y 25 °C (ASS), cuya posición y dimensión están asociadas a variables de macroescala que gobiernan los ciclos interdecadales en el océano Pacífico. En esa medida, los cambios trascendentales de las biomasas de jurel T. murphyi del orden de 10 a 1 y viceversa sólo explican cambios en la disponibilidad del recurso frente a condiciones favorables o adversas, según sea el caso, pero no nece-sariamente una reducción de la biomasa en la misma magnitud.
Lo extraído por la pesquería peruana no explica tal reducción, pues las capturas sólo han representado el 5% de la biomasa estimada como promedio entre 1983 y 1997. Dicha cifra resulta representativa para todo el stock norte (far north), puesto que en el mismo periodo los desembarques de jurel T murphyi fueron muy reducidos en el Ecuador y en el norte de Chile.
La pesquería del jurel y otros recursos y sus pesquerías
El jurel T. murphyi no es la única especie que se presenta en abundancia en el mar peruano y cuyos cambios en abundan-
cia y disponibilidad están influenciados por las condiciones ambientales y su interacción con otras especies, con las cuales comparte el ecosistema.
Al respecto, el análisis de las interacciones del recurso jurel T. murphyi en el mar peruano revela que existe una fuerte relación con la caballa S. japonicus y la merluza M. gayi peruanus, al pare-cer especies asociadas al mismo sistema de circulación represen-tado por las ASS (Aguas Subtropicales Superficiales) y la ESCC (Extensión Sur de la Corriente de Cromwell) (Figs.14A y 14B).
Este análisis concuerda con la observación de Ñiquen et al. (2013) respecto a la coincidencia en los picos de capturas en los años 1977 y 1978 para jurel T. murphyi y merluza M. gayi peruanus en el Perú, y entre 1994 y 1998 y en 2001 para jurel T. murphyi en Ecuador y Perú así como para merluza M. gayi
Biomasa Captura ACF ASS TSM-Chi TSMP IOS PDO MEIBiomasa 1.000 -0.228 -0.969 0.952 0.698 0.961 -0.687 0.868 0.759Captura -0.228 1.000 -0.019 0.047 0.279 0.029 -0.479 -0.531 0.421
ACF -0.969 -0.019 1.000 -0.994 -0.806 -0.997 0.816 -0.744 -0.877ASS 0.952 0.047 -0.994 1.000 0.866 0.999 -0.786 0.681 0.855
TSM-Chi 0.698 0.279 -0.806 0.866 1.000 0.845 -0.596 0.251 0.677TSMP 0.961 0.029 -0.997 0.999 0.845 1.000 -0.793 0.709 0.861
IOS -0.687 -0.479 0.816 -0.786 -0.596 -0.793 1.000 -0.488 -0.992PDO 0.868 -0.531 -0.744 0.681 0.251 0.709 -0.488 1.000 0.532MEI 0.759 0.421 -0.877 0.855 0.677 0.861 -0.992 0.532 1.000
Sim
ilarid
ad d
e Br
ay C
urtis
1983
1993
1986
1989
1984
1985
1987
1994
1990
1991
1997
1995
1996
1988
1992
2006
2000
2001
2004
2007
1999
2005
2011
2002
2003
2010
2009
1998
2008
0
20
40
60
80
100
Años5E6
4E6
3E6
2E6
1E6
0
Dis
tanc
ia e
uclid
iana
2010
2000
1990
1980
Periodo Cálido Periodo Frío

17
El jurel Trachurus murphyi y las variables ambientales de macroescala
Rev. peru. biol. special issue 20(1): 009- 020 (September 2013)Ecology, fisheries and conservation of Jack mackerel (Trachurus murphyi) in Peru. Editors: Csirke J., R. Guevara-Carrasco & M. Espino
Coe
ficie
nte
Cor
rela
ción
1
0.8
0.6
0.4
0.2
0
-0.2
-0.4
-0.6
-0.8
Especies1 2 3 4 5 6 7 8 9 10 11
Bonito
Sierra Cojinoba
Caballa
MerluzaPejerrey
Cavinza Pota
SardinaAnchoveta
Machete
0
20
40
60
80
100
Sim
ilarid
ad d
e Br
ay C
urtis
Anc
hove
ta
Sar
dina
Pot
a
Jure
l
Cab
alla
Mer
luza
Cav
inza
Sie
rra
Bon
ito
Coj
inob
a
Pej
erre
y
Mac
hete
Figura 14. (A) Correlación de Pearson entre los desembarques de jurel T. murphyiy y los de otras especies seleccionadas (1960 – 2012). (B) Agrupamiento por similitud de Bray-Curtis de desembarques (1960 – 2012) de las principales especies del ecosistema de aflo-ramiento peruano.Figure 14. (A) Pearson's correlation between the landings of Jack mackerel T. murphyi and that of other selected species (1960 – 2012). (B) Results of Bray Curtis similarity grouping analysis of landings (1960 – 2012) of the main species of the Peruvian upwelling ecosystem.
peruanus en el Perú, coincidentes con desviaciones positivas del PDO. Esto sugiere la posibilidad de pronosticar periodos favorables para la pesca de jurel T. murphyi y merluza M. gayi peruanus en Perú y Ecuador a partir de expresiones positivas del PDO. Sin embargo, las desviaciones positivas del PDO estarán seguidas de desviaciones negativas, y viceversa, por el periodo de alta variabilidad ambiental por el que atravesamos. En ese senti-do, dado que en los últimos años han predominado condiciones frías (en 4 años de 5), es de esperar que en los próximos años se mantenga la predominancia de manifestaciones positivas del PDO, lo cual favorecería la pesquería de jurel T. murphyi para Ecuador, Perú y el norte de Chile, según se ha venido dando en los últimos años.
Por otro lado, son menos significantes las relaciones con pejerrey O. regia regia, cabinza I. conceptionis y pota D. gigas. Los desembarques de esta última especie han mostrado un in-cremento, en contraste con lo acaecido con jurel T. murphyi en los últimos años, lo que explica la diferencia con lo obtenido por Espino y Yamashiro (2012), con información hasta el 2009.
Asimismo, es notable la relación inversa con bonito S. chi-liensis chiliensis, seguido de sierra S. sierra y cojinova S. violacea, pero en mucha menor magnitud. Un análisis de similitud de Bray-Curtis entre las principales especies de la pesquería peruana revela una asociación entre jurel T. murphyi, caballa S. japonicus, merluza M. gayi peruanus y, en menor grado, con la pota D. gigas, lo cual indicaría tendencias similares en su disponibilidad a las pesquerías. La asociación jurel-caballa se explica por la re-lación con las ASS, y la asociación jurel-merluza por la relación con la ESCC. En tanto que en la relación jurel-pota, debido a que los frentes entre ASS y ACF son más amplios en periodos
de abundancia de jurel T. murphyi, el desarrollo de la pota D. gigas se posibilita (Arguelles & Tafur 2010, Ñiquen et al. 2004, Swartzman et al. 2008).
DiscusiónEl Sistema de la Corriente Peruana está sometido a patrones
de variabilidad decadales y multidecadales que afectan la di-námica de circulación oceánica y los patrones de distribución, abundancia y disponibilidad de los recursos pesqueros, en la medida que las principales masas de aguas ACF y ASS alternan su dominancia.
En esa medida, se encuentran evidencias de que las variaciones seculares estarían vinculadas a fluctuaciones de amplia escala del SOI, como se reflejan en oscilaciones detectadas para el Pacífico Norte en anillos de árboles de Baja California, lo cual sugiere la existencia de éstas en el clima de esta región entre 1706 y 1977, con grandes oscilaciones decadales en 1750, 1905 y 1947 (Biondi et al. 2001), con una intermitencia de casi 50 años en el último siglo. Asimismo, en un rango de escalas de tiempo que se remontan por lo menos a 1600 años, reconstrucciones climáticas a partir de anillos de árboles y de corales del Pacífico sugieren variaciones de PDO, existiendo variaciones importantes en diferentes periodos. Sin embargo, para el siglo XX las fluctuaciones PDO fueron más enérgicas con dos periodicidades generales, una de 15 a 25 años, y otra de 50 a 70 años (Mantua & Hare 2002).
En otras latitudes, también existen evidencias del efecto de los regímenes ambientales en los recursos pesqueros. Como ejemplos más saltantes, están los casos de la influencia de re-gímenes ambientales en la dinámica poblacional del lenguado del Pacífico Norte “halibut” Hippoglossus stenolepis, el colín de Alaska o “Alaska pollock”, Theragra chalcogramma, la sardina de California Sardinops caeruleus (=Sardinops sagax), la sardina japonesa S. melanostictus (=Sardinops sagax) y los lenguados del mar de Bering: Hippoglossoides elassodon, Lepidopsetta polyxystra y Atheresthes stomias, entre los más importantes, y en cuyos casos se han documentado efectos sobre los reclutamientos, procesos de denso-dependencia y patrones de crecimiento individual y poblacionales (Quinn & Niebauer 1995, Jacobson & MacCall 1995, Wada & Jacobson 1998, Francis et al. 1998, Walters & Wilderbuer 2000, Clark & Hare 2002).
Con mayor frecuencia, los elementos determinantes que expli-can la variabilidad ambiental en el Pacífico Norte son asociados al PDO (Hare & Mantua 2001, Lluch-Cota et al. 2003) y en el hemisferio sur al fenómeno ENSO (Mantua et al. 1997). Aunque en el hemisferio sur se hace principalmente referencia al fenóme-no ENSO, existe la posibilidad de que se produzcan procesos de oscilación interdecadales que pueden influenciar la dinámica poblacional de los recursos en formas y a niveles diferentes. Estos procesos han sido descritos por Chávez et al. (2003), quienes plantean la existencia de ciclos ambientales de alrededor de 50 años, en los que se alternan fases frías y cálidas. Las fases frías en los periodos de 1900 – 1925 y de 1950 – 1975 y las fases cálidas en los periodos de 1925 – 1950 y de 1975 a finales del siglo pasado indican que en el momento actual estamos atravesando por una fase fría con dominancia de anchoveta E. ringens y ausencia de sardina S. sagax y, asimismo, por el alejamiento y disminución ostensible de la disponibilidad de jurel T. murphyi.
Por otro lado, la variabilidad decadal también puede modular la frecuencia, intensidad y duración del ciclo ENSO (Wang &

18
Espino
Rev. peru. biol. número especial 20(1): 009- 020 (Septiembre 2013)Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú.
Editores: Csirke J., R. Guevara-Carrasco & M. Espino
Talla
med
ia (c
m)
50
45
40
35
30
25
20
15
Años
Jurel Merluza
1970 1785 1980 1985 1990 1995 2000 2005 2010 2015
Figura 15. Tallas medias anuales de jurel T. murphyi (gris) y merluza M. gayi peruanus (negro) en las capturas comerciales, Perú, 1972 a 2012Figure 15. Annual average sizes of Jack mackerel T. murphyI (grey) and hake M. gayi peruanus (black) in commercial catches, Peru, 1972 – 2012
Picaut 2004, Wang et al. 2004, Wang & Fiedler 2006, Cobb et al. 2003) así como el momento de inicio de El Niño (Espino 2003). Estos cambios de baja frecuencia en el entorno físico pueden resultar en "cambios de régimen" en la estructura del ecosistema marino en la cuenca del Pacífico (Chávez et al. 2003, Mantua & Hare 2002, Schwartzlose et al. 1999, Hsieh et al. 2005, Di Lorenzo & Ohman 2013).
Dentro del Ecosistema de la Corriente Costera Peruana se identifican dos "regímenes de anchoveta E. ringens" (desde el inicio de la pesca en la década de los cincuenta hasta inicios de la década de los setenta, y desde principios de la década de los noventa hasta la actualidad), separados por un "régimen de sar-dina S. sagax " (a partir de mediados de la década de los setenta hasta principios de la década de los noventa) (Csirke at al. 1996, Chávez et al. 2003, Alheit & Ñiquen 2004, Ayón et al. 2011).
Por otro lado, Ballón et al. (2008) describen cambios en las tallas de merluza M. gayi peruanus, diferenciándolas entre las décadas de los setenta y ochenta con respecto a la de los noventa y la década del 2000 al 2010, de un rango de 35 a 50 cm a otro de 20 a 35 cm, respectivamente, lo cual evidencia una madura-ción más temprana y desaparición de adultos por presión de la pesca (Wosnitza-Mendo et al. 2004, Guevara-Carrasco 2004, Guevara-Carrasco & Lleonart 2008).
Sin embargo, anteriormente se ha definido (Ñiquen et al. 2013) una relación entre jurel T. murphyi y merluza M. gayi peruanus, basándose en que ambas especies están asociadas a una dominancia de las ASS que favorece la extensión de la ESCC. Por lo tanto, tal reducción de tallas estaría también asociada a cambios en los patrones de distribución por tallas del recurso, que se afectan por la retracción de la ESCC, lo que explicaría la presencia de impor-tantes biomasas de merluza M. gayi peruanus en el mar ecuatoriano (328000 toneladas) el 2010 (González Troncoso 2010).
En el Perú, las merluzas M. gayi peruanus de mayor tamaño se distribuyen hacia el norte y las más pequeñas hacia el sur (Armstrong, 1981; Samamé, 1981; Espino et al. 1995). Con la retracción de ESCC las más pequeñas se ubican en la principal área de pesca de merluza M. gayi peruanus del Perú (04 – 05 ºS) y las más grandes en el Ecuador, e incluso más al norte.
Al respecto, se subraya que en la Figura 15, en el caso del jurel T. murphyi y la merluza M. gayi peruanus, se aprecia que ambas curvas muestran la misma tendencia, con una relativa estabilidad hasta 1990, excepto en 1987 en que la talla media del primero disminuye drásticamente por debajo de 25 cm y se recupera hacia 1990. A partir de este año muestra una tendencia declinante que persiste los veinte años siguientes, hasta 2011, con un aumento notorio en 2012. Al estar sometidas al mismo régimen de circulación oceánica y a los mismos patrones de variabilidad de macroescala, ambas especies siguen el compor-tamiento de las condiciones que les son propias. La merluza M. gayi peruanus está vinculada a la ESCC que al retraerse hacia el norte retrae consigo a los individuos de mayor tamaño, dejando a disponibilidad de la pesquería los más pequeños, cuya ubicación era más al sur en la década de los setenta y ochenta.
Por su parte, el jurel T. murphyi se aleja en la medida que las ASS se alejan, portando entre sí a los individuos de mayor tama-ño y dejando disponibles los más pequeños en la zona costera.
Gutierrez et al. (2008) caracterizan este periodo (mediados de
los noventa al año 2000) como de condiciones más frías, debido al dominio de aguas frías costeras a lo largo del Perú, basándose en las relaciones existentes entre munida Pleuroncodes monodon y anchoveta E. ringens. Esta característica favorece el incremento de la productividad primaria y la producción de zooplancton (Swartzman et al. 2008, Ayón et al. 2008), lo cual diferencia este periodo de uno anterior menos frío o más cálido.
Por su parte Barange et al. (2009) indican que en general las poblaciones de anchoveta E. ringens y sardina S. sagax expanden sus áreas de distribución con aumentos en su densidad y concen-tración, en consistencia con el modelo de la cuenca que habitan. La relación entre el área de distribución y biomasa de la población es alométrica, lo que implica la existencia de un límite a la expansión en función de las nuevas áreas. Los patrones de ocupación de los espacios difieren entre la sardina S. sagax y la anchoveta E. ringens en algunas regiones. En el sur de África y Japón, la anchoveta ocupa un área mayor por unidad de biomasa que la sardina, de acuerdo con los requerimientos de hábitat determinados por su ecología de alimentación. En California y Perú, los resultados confirman que en general, estas especies amplían sus áreas de distribución con el tamaño de la población, pero los modelos específicos son menos claros. Se puede inferir que las poblaciones se acercarán o alejarán de las áreas de pesca en la medida que las condiciones les sean favorables, dando imágenes sesgadas de cambios en la abundancia. Estas imágenes serían más bien reflejos de cambios en la disponibilidad, en función a los cambios en las condiciones del ambiente (Vert- pre et al. 2013), pero no necesariamente cambios en la abundancia o biomasa total.
Finalmente, se reconocen patrones decadales con cambio de régimen o salto climático alrededor de 1925/1926 y 1976/1977 hacia un periodo cálido, con manifestaciones en el ambiente y en el comportamiento de recursos pesqueros (Noakes et al. 2000, McFarlane & Beamish 2001, Wilderbuer 2002, Lavaniegos & Ohman 2003, Peterson & Schwing 2003, Gray et al. 2003, Miller & Sydeman 2004, Hoff 2006, Saitoh 2006, Chhak & Di Lorenzo 2007, d'Orgeville & Peltier 2007, Zhuguo Ma 2007, Noakes & Beamish 2009, Jacques-Coper 2009, Ceballos et al. 2009).

19
El jurel Trachurus murphyi y las variables ambientales de macroescala
Rev. peru. biol. special issue 20(1): 009- 020 (September 2013)Ecology, fisheries and conservation of Jack mackerel (Trachurus murphyi) in Peru. Editors: Csirke J., R. Guevara-Carrasco & M. Espino
En conclusión, los cambios en magnitud detectados en las biomasas observadas de jurel T. murphyi en el mar peruano están reflejando cambios en la disponibilidad del recurso asociados a patrones de variabilidad de macroescala seculares y multidecada-les. A su vez, estos patrones afectan también las manifestaciones de El Niño haciendo que sean otoñales o primaverales si el PDO imperante es negativo o positivo, respectivamente (Espi-no 2003). Asimismo, las fluctuaciones en la disponibilidad del recurso están afectando las pesquerías de Ecuador, Perú y norte de Chile, las mismas que han mostrado variaciones importantes en sus desembarques y habrían dado una imagen sesgada de su abundancia, lo que conduce a diagnósticos errados de la real situación del recurso. Por ello, es necesario que se promuevan programas de investigación mancomunados más activos entre los países costeros de la región que participan en la pesca del stock norte (far north) de jurel T. murphyi, con el objeto de co-nocer su real dimensión y dinámica, en función de las variables ambientales que gobiernan la parte norte del Ecosistema de la Corriente Costera Peruana.
AgradecimientosDeseo expresar mi agradecimiento al ingeniero Luis Escudero
por haber proporcionado las áreas satelitales de las diferentes iso-termas (13 – 28°C) en la serie 1982 – 2011. A Carmen Yamashiro por la revisión del trabajo y sus valiosas sugerencias para mejorarlo.
Literatura citadaArmstrong, D. 1981. Investigación de la merluza en IMARPE. Informe Inst.
Mar. Perú, Callao, 79 I parte. 47p.Alheit J. & M. Ñiquen. 2004. Regime shifts in the Humboldt Current ecosys-
tem. Progress in Oceanography): 201 – 222. DOI: 10.1016/j.pocean.2004.02.006
Argüelles J. & R. Tafur. 2010. New insights on the biology of the jumbo squid Dosidicus gigas in the Northern Humboldt Current System: Size at maturity, somatic and reproductive investment. Fisheries Research 106: 185 – 192. Doi 10.1016/j.fishres.2010.06.005
Ayón P., G. Swartzman, P. Espinoza & A. Bertrand. 2011. Longterm changes in zooplankton size distribution in the Peruvian Humboldt Current System: conditions favouring sardine or anchovy. Mar Ecol. Prog. Ser. 422: 211 – 222. DOI: 10.3354/meps08918
Ayón P., G. Swartzman, A. Bertrand, M. Gutiérrez & S. Bertrand. 2008. Zooplankton and forage fish species off Peru: Large – scale bottom – up forcing and local – scale depletion. Progress in Oceanography 79: 208 – 214.
Ballón M., C. Wosnitza – Mendo, R. Guevara – Carrasco & A. Bertrand. 2008. The impact of overfishing and El Niño on the condition factor and reproductive success of Peruvian hake, Merluccius gayi peruanus. Progress in Oceanography 79: 300 – 307. DOI: 10.1016/j.po-cean.2008.10.016
Barange M., J. Coetzee, A. Takasuka, K. Hill, M. Gutiérrez, Y. Oozeki, C. van der Lingen & V. Agostini. 2009. Habitat expansion and contraction in anchovy and sardine populations. Progress in Oceanography 83(1 – 4): 251 – 260.
Bertrand A., A. Chaigneau, S. Peraltilla, J. Ledesma, M. Graco, F. Monetti & Francisco P. Chávez. 2011. Oxygen: A Fundamental Property Regulating Pelagic Ecosystem Structure in the Coastal Southeastern Tropical Pacific. PLoS ONE 6(12): e29558. doi:10.1371/journal.pone.0029558.
Biondi F., A. Gershunov & D.R. Cayan, 2001: North Pacific Decadal Climate Variability since 1661. Journal of Climate, 14: 5 – 10. doi: http://dx.doi.org/10.1175/1520-0442(2001)014<0005:NPDCVS>2.0.CO;2
Ceballos L., E. Di Lorenzo, N. Schneider & B. Taguchi. 2009. North Pa-cific Gyre Oscillation synchronizes climate fluctuations in the eastern and western North Pacific. Journal of Climate, DOI: 10.1175/2009JCLI2848.1.
Chhak K.& E. Di Lorenzo. 2007. Decadal variations in the California Current upwelling cells, Geophys. Res. Lett. 34, L14604, doi:10.1029/2007GL030203.
Chávez F.P., J. Ryan, S.E. Lluch-Cota & M. Ñiquen. 2003. From anchovies to sardines and back: Multidecadal change in the Pacific Ocean. Review: Climate. Science 299: 217 – 221. DOI: 10.1126/scien-ce.1075880
Clark W.G. & S.R. Hare. 2002. Effects of Climate and Stock Size on Re-cruitment and Growth of Pacific Halibut. N. Am. J. Fish. Mgmt. 22: 852 – 862.
Cobb K.M., C.D. Charles, H. Cheng & R.L. Edwards. 2003. El Niño – Southern Oscillation and tropical Pacific climate during the last millennium. Nature 424: 271 – 276. doi:10.1038/nature01779
Csirke J., R. Guevara – Carrasco, G. Cárdenas, M. Ñiquen & A. Chipollini. 1996. Situación de los recursos anchoveta (Engraulis ringens) y sardina (Sardinops sagax sagax) a principios de 1994 y perspectivas para la pesca en el Perú de 1994, conparticular referencia a las regiones norte y centro de la costa peruana. Bol. Inst. Mar Perú – Callao 15(1): 1 – 23.
Di Lorenzo E. & M. D. Ohman. 2013. A double – integration hypothesis to explain ocean ecosystem response to climate forcing. PNAS 110 (7): 2496–2499. DOI: 10.1073/pnas.1218022110
d’Orgeville M. & W.R. Peltier. 2007. On the Pacific Decadal Oscillation and the Atlantic Multidecadal Oscillation: Might they be related? Geophys. Res. Lett. 34, L23705, doi:10.1029/2007GL031584.
Espino, M., R. Castillo & F. Fernández. 1995. Biology and Fisheries of Peruvian Hake (M. gayi peruanus). P. 339 – 359. In J. Alheit and T. Pitcher (eds). HAKE, fisheries, ecology and markets. Chapman & Hall, Fish and fisheries Series 15. First edition.
Espino M. 2003. Estrategia de gestión oriental para el Pacífico Oriental con especial mención a la pesquería peruana. Tesis para optar el grado de Magíster en Geografía con mención en Ordenamiento y Gestión Ambiental. Unidad de Post Grado, Facultad de Ingeniería Geológica, Minera, Metalúrgica y Geográfica, Universidad Nacional Mayor de San Marcos, Lima. 102 pp.
Espino M. & C. Yamashiro. 2012. La variabilidad climática y las pesquerías en el Pacífico Sudoriental. En: P.M. Arana (Guest Editor). International Conference: “Environment and Resources of the South Pacific”. Latin American Journal of Aquatic Research 40(3): 705 – 721. DOI: 103856/vol40 – issue3 – fulltext – 18.
Flores R., M. Espino, G. Luque & J. Quispe. 2013. Patrones de variabilidad ambiental en el mar peruano. En: Csirke J., R. Guevara-Carrasco & M. Espino (Eds.). Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú. Rev. peru. biol. número especial 20(1): 021 – 028
Francis R.C., S.R. Hare, A.B. Hollowed & W.S. Wooster. 1998. Effects of interdecadal climate variability on the oceanic ecosystems of the Northeast Pacific Ocean. Fish. Oceanogr. 7: 1 – 21.
González Troncoso D.M. 2010. Informe de la campaña de investigación pes-quera. Ecuador 2010. B/O Miguel oliver, 16/10/2010 ‐05/11/2010. Campañas Oceanográficas de Coorperación España – Ecuador 2008 – 2010. Gobierno de España, Ministerio del Medio Ambiente, Me-dio Rural y Marino, Instituto Español de Oceanografía. República del Ecuador, Instituto Nacional de Pesca. 130 pp.
Gray S.T., J.L. Betancourt, C.L. Fastie & S.T. Jackson. 2003. Patterns and sources of multidecadal oscillations in drought – sensitive tree – ring records from the central and southern Rocky Mountains. Geophys.Res. Lett. 30(6), 1316, doi:10.1029/2002GL016154.
Guevara-Carrasco R. 2004. Sobrepesca de la merluza peruana: lecciones mal entendidas. Boletín 21(1-2):27-32
Guevara-Carrasco R., & J. Lleonart. 2008. Dynamics and fishery of the Peruvian hake: Between nature and man. Journal of Marine Systems 71 (3–4): 249-259. doi:10.1016/j.jmarsys.2007.02.030.
Gutiérrez M., A. Ramirez, S. Bertrand, O. Móron & A. Bertrand. 2008. Ecological niches and areas of overlap of the squat lobster ‘munida’ (Pleuroncodes monodon) and anchoveta (Engraulis ringens) off Peru. Progress in Oceanography 79: 256–263.
Hare, S.R. 1996. Low frequency climate variability and salmon production. Ph.D. Dissertation. Univ. Washington, School of Fisheries. 306 pp.
Hare S.R. & N.J. Mantua. 2001. An historical narrative on the Pacific Decadal Oscillation, interdecadal climate variability and ecosystem impacts. Report of a talk presented at the 20th NE Pacific Pink and Chum workshop, Seattle, WA, 22 March 2001.
Hare S.R. & N.J. Mantua. 2002. The Pacific Decadal Oscillation. Journal of Oceanography 1: 35 – 44, DOI: 10.1023/A: 1015820616384.
Hoff G.R. 2006. Biodiversity as an index of regime shift in the eastern Bering Sea Fish. Bull. 104:226 – 237.

20
Espino
Rev. peru. biol. número especial 20(1): 009- 020 (Septiembre 2013)Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú.
Editores: Csirke J., R. Guevara-Carrasco & M. Espino
Hsieh C.H., S.M. Glaser, A.J. Lucas & G. Sugihara. 2005. Distinguishing random environmental fluctuations from ecological catastrophes for the North Pacific Ocean, Nature 435:336 – 340, doi:10.1038/nature02553.
Jacobson L.D. & A.D. MacCall. 1995. Stock – recruitment models for pacific sardine (Sardinops sagax). Canadian Journal of Fisheries and Aquatic Sciences 52: 566 – 577.
Jacques-Coper R. 2009. Caracterización del salto climático de mediados de los setenta en Sudamérica. Primer Congreso de Oceanografía Física, Meteorología y Clima. Universidad de Concepción, Concepción, Chile. 30 septiembre – 02 de octubre de 2009. Sesión: Variabilidad Climática y Cambio Global / Paleoclima.
Lavaniegos B.E. & M.D. Ohman. 2003. Long – term changes in pelagic tunicates of the California Current. Deep – Sea Research II, 50: 2473 – 2498.
Lluch-Cota D.B., W.S. Wooster, S.R. Hare, D. Lluch – Belda & A. Parés – Sierra. 2003. Principal Modes and Related Frequencies of Sea Surface Temperature Variability in the Pacific Coast of North America. Journal of Oceanography 59 (4): 477 – 488 (12). DOI: 10.1023/A:1025592616562
Maenza R. A. & R. H. Compagnucci. 2010. Simulación de la pequeña edad de hielo usando el modelo EdGCM. GEOACTA 35: 78 – 91.
Mantua N.J., S.R. Hare, Y. Zhang, J.M. Wallace & R.C. Francis. 1997. A Pacific interdecadal climate oscillation with impacts on salmon production. Bull. Amer. Meteor. Soc. 78: 1069 – 1079. DOI: http://dx.doi.org/10.1175/1520-0477(1997)078<1069:APICOW>2.0.CO;2
Mantua, N.J. 1999 (a). The Pacific Decadal Oscillation. A brief overview for non – specialists, to appear in the Encyclopedia of Environmental Change. <http://www.atmos.washington.edu/~mantua/REPORTS/PDO/PDO_egec.htm> [accedido: 09/07/2013]
Mantua, N.J. 1999 (b). The Pacific Decadal Oscillation and Climate Forecasting for North America. To appear in premiere issue of “Climate Risk So-lutions” newsletter. <http://www.atmos.washington.edu/~mantua/REPORTS/PDO/PDO_cs.htm>[accedido: 09/07/2013]
Mantua N. J. & S. R. Hare. 2002. The Pacific Decadal Oscillation. Journal of Oceanography (58): 35 – 44. DOI 10.1023/A:1015820616384
McFarlane G.A. & R.J. Beamish. 2001. The re-occurrence of sardines off British Columbia characterises the dynamic nature of regimes. Progress in Oceanography, 49 (1 – 4): 151 – 165. DOI: 10.1016/S0079-6611(01)00020-9
Miller A.K. & W.J. Sydeman. 2004. Rockfish response to low – frequency ocean climate change as revealed by the diet of a marine bird over multiple time scales. Mar Ecol. Prog. Ser. 281: 207 – 216.
Morón O. & M. Sarmiento. 2001. Aspectos oceanográficos de El Niño 1997 – 98 y su relación con los recursos pelágicos. En: J. Tarazona, W.E. Arntz y E. Castillo de Maruenda (eds.), El Niño en América Latina: Impactos Biológicos y Sociales. Consejo Nacional de Ciencia y Tecnología. Lima, pp. 5 – 27.
Noakes D.J., R.J. Beamish & M.L. Kent. 2000. On the decline of Pacific salmon and speculative links to salmon farming in British Co-lumbia. Aquaculture 183(3–4): 363 – 386. doi:10.1016/S0044-8486(99)00294-X.
Noakes D.J. & R.J. Beamish. 2009. Synchrony of Marine Fish Catches and Climate and Ocean Regime Shifts in the North Pacific Ocean. Ma-rine and Coastal Fisheries: Dynamics, Management and Ecosystem Science1:155 – 168, 2009. DOI: 10.1577/C08 – 001.1.
Ñiquen M., A. Chipollini, A. Echevarría, J. Mori & B. Buitrón. 2004. Cambios biológicos en los principales recursos marinos peruanos en el verano 2001. Inf. Inst. Mar. Perú – Callao 32 (1): 57 – 62.
Ñiquen M., M. Bouchon, D. Ulloa & A. Medina. 2013. Análisis de la pesquería del jurel Trachurus murphyi en el Perú. En: Csirke J., R. Guevara-Carrasco, M. Espino (Eds.). Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú. Rev. peru. biol. número especial 20(1): 097- 106
Peterson W.T. & F.B. Schwing. 2003. A new climate regime in nor-theast pacific ecosystems, Geophys. Res. Lett. 30(17), 1896, doi:10.1029/2003GL017528, 2003.
Quinn W.H., V.T. Neal & S.E. Antunez de Mayolo. 1987. El Niño Occurrences Over the Past Four and Half Centuries. Journal of Geophysical Re-search, 92 (C13):14449 – 14461. DOI 10.1029/JC092iC13p14449
QuinnT.J.,II. & H.J. Niebauer.1995. Relation of eastern Bering Sea wallege pollack (Theragra chalcogramma) recruitment to environmental and oceanographic variables. Pp. 497 – 507. In R.J.Beamish, ed. Climate change and northern fish populations. Canadian Special Publications in Fisheries and Aquatic Sciences 121.
Samamé M., 1981. Las Pesquerías Demersales en el Perú. CPPS, Seminario Regional sobre los Recursos Demersales y su Medio Ambiente en el Pacífico Oriental. Lima, Perú 24–28 de noviembre de 1980. CPPS, Serie Seminarios y Estudios, 3, pp. 213–247.
Saitoh T., B. Cazelles, J. Olav Vik, H. Viljugrein & N.Chr. Stenseth. 2006. Effects of regime shifts on the population dynamics of the grey – sided vole in Hokkaido, Japan. Clim. Res.32:109 – 118 (2006), doi:10.3354/cr032109.
Schwartzlose R.A., J. Alheit, A. Bakun, T. R. Baumgartner, R. Cloete, R.J.M. Crawford, W.J.Fletcher, Y. Green – Ruiz, E. Hagen, T. Kawasaki, D. Lluch – Belda, S.E. Lluch – Cota, A.D. MacCall, Y. Matsuura, M.O. Nevarez – Martínez, R.H. Parrish, C. Roy, R. Serra, K.V. Shust, M.N. Ward & J.Z. Zuzunaga 1999. Worldwide large – scale fluctuations of sardina and anchovy populations. South African Journal of Marine Science 21:289 – 347. DOI: 10.2989/025776199784125962
Swartzman G., A. Bertrand, M. Gutiérrez, S. Bertrand & L. Vásquez. 2008. The relationship of anchovy and sardine to water masses in the Peruvian Humboldt Current System from 1983 – 2005. Prog. Oceanogr. 79: 228 – 237.
Vert- pre K. A., R. O. Amoroso, O. P. Jensenc & R. Hilborn. 2013. Frequency and intensity of productivity regime shifts in marine fish stocks. PNAS. January 29, 2013. 110. (5): 1779–1784
Wada T. & L.D.Jacobson. 1998. Regimes and stock-recruitment relationships in Japanese sardine (Sardinops melanostictus), 1951 – 1995. Ca-nadian Journal of Fisheries and Aquatic Sciences 55: 2455 – 2463.
Walters G.E. & T.K.Wilderbuer. 2000. Decreasing length at age in a rapidly expanding population of northern rock sole in the Bearing Sea and its effect on managment advice. Netherlands Journal of Sea Research 44: 17 – 26.
Wang C. & J. Picaut. 2004. Understanding ENSO physics – A review, in: Earth’s Climate: The Ocean-Atmosphere Interaction, edited by: C. Wang, S.P. Xie andJ.A. Carton., AGU Geophysical Monograph Series 147: 21 – 48.
Wang C., S.P. Xie & J.A. Carton.2004. A global survey of oceanatmosphere interaction and climate variability, in: Earth’s Climate: The Ocean – Atmosphere Interaction, edited by: C. Wang, S.P. Xie and J.A. Carton, AGU Geophysical Monograph Series 147: 1 – 19.
Wang C. & P. C. Fiedler. 2006. ENSO variability and the eastern tropical Pacific: A review. Progress in Oceanography 69: 239 – 266.
Wilderbuer T.K., A.B. Hollowed, W.J. Ingraham, Jr., P.D. Spencer, M.E. Conners, N.A. Bond & G.E. Walters. 2002. Flatfish recruitment response to decadal climatic variability and ocean conditions in the eastern Bering Sea. Progress in Oceanography 55: 235 – 247. DOI: 10.1016/S0079-6611(02)00081-2
Wolter, K., 1987: The Southern Oscillation in surface circulation and climate over the tropical Atlantic, Eastern Pacific, and Indian Oceans as captured by cluster analysis. Journal of Climate and Applied Meteorology 26, 540 – 558. doi: http://dx.doi.org/10.1175/1520-0450(1987)026<0540:TSOISC>2.0.CO;2
Wolter, K., & M.S. Timlin, 1993: Monitoring ENSO in COADS with a seasonally adjusted principal component index. Proc. of the 17th Climate Diagnostics Workshop, Norman, OK, NOAA/N MC/CAC, NSSL, Oklahoma Clim. Survey, CIMMS and the School of Meteor., Univ. of Oklahoma, 52 – 57.
Wolter, K., & M.S. Timlin, 1998: Measuring the strength of ENSO – how does 1997/98 rank? Weather, 53, 315 – 324. DOI: 10.1002/j.1477-8696.1998.tb06408.x
Wosnitza-Mendo C., R. Guevara-Carrasco & M. Ballón. 2004. Possible causes of the drastic decline in mean length of Peruvian hake in 1992, vol. 21. Boletín Instituto del Mar del Perú. pp. 1 – 26.
Wyrtki K. 1967. Circulation and water masses in the Eastern Equatorial Pacific Ocean. Int. Jour. Oceanol. and Limnol. 1(2):117 – 147.
Zhuguo Ma. 2007. The interdecadal trend and shift of dry/wet over the central part of North China and their relationship to the Pacific Decadal Oscillation (PDO). Chinese Science Bulletin 52 (15): 2130 – 2139
Zuta S. & O. Guillén. 1970. Oceanografía de las Aguas Costeras del Perú. Bol. Inst. Mar del Perú – Callao, 2 (5): 161 – 223.

21
Patrones de variabilidad ambiental en el mar peruano
Rev. peru. biol. special issue 20(1): 021 – 028 (September 2013)Ecology, fisheries and conservation of Jack mackerel (Trachurus murphyi) in Peru. Editors: Csirke J., R. Guevara-Carrasco & M. Espino
Rev. peru. biol. número especial 20(1): 021 – 028 (Septiembre 2013)Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú. Editores: Csirke J., R. Guevara-Carrasco & M. Espino
Facultad de ciencias Biológicas unMsMISSN – L 1561 – 0837
Patrones de variabilidad ambiental en el mar peruano
Roberto Flores, Marco Espino, Gertrudes Luque y Jorge Quispe
Environmental variability patterns in the Peruvian waters
Instituto del Mar del Perú, esquina Gamarra y General Valle S/N Chucuito, Callao, Perú.
Email Roberto Flores: [email protected]
el medioambiente del jurel (Trachurus murphyi) en el Perú
© Los autores. Publicado por la Revista Peruana de Biología de la Facultad de Ciencias Biológicas, Universidad Nacional Mayor de San Marcos. Este es un artículo de acceso abierto, distribuido bajo los términos de la Licencia de Atribución Reconocimiento – NoComercial – SinObraDerivada 3.0 de Creative Commons (http://creativecommons.org/licenses/by – nc – nd/3.0/deed.es_ES), que permite el uso no comercial, distribución y reproducción en cualquier medio, siempre que la obra original sea debidamente citadas. Para uso comercial, por favor póngase en contacto con [email protected].
Citación:Flores R., M. Espino, G. Luque & J. Quispe. 2013. Patrones de variabilidad ambiental en el mar peruano. En: Csirke J., R. Guevara-Carrasco & M. Espino (Eds.). Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú. Rev. peru. biol. número especial 20(1): 021 – 028 (Septiembre 2013)
Resumen Se describen los escenarios ambientales y los patrones de variabilidad observados en el mar peruano a partir del análisis de información obtenida por el Instituto del Mar del Perú entre los años 1960 – 2011. Se examinan datos de cruceros de investigación oceanográfica, de evaluación de recursos pesqueros y bío-oceanográficos, además de información de diversas publicaciones sobre el mar peruano y sus recursos. Los datos de temperatura, salinidad y contenido de oxígeno, fueron interpolados para obtener promedios mensuales y construir series espaciales y temporales. En la superficie, las Aguas Costeras Frías (ACF) y Aguas Subtropicales Superficiales (ASS) crean condiciones ambientales diferentes a las de la parte subsuperficial donde predomina la intromisión de las aguas ecuatoriales a través de la Corriente Subsuperficial Perú-Chile (CSPC). En el norte, durante el verano y otoño, predominan en la capa superficial aguas cálidas de origen ecuatorial. En el centro, aguas de origen ecuatorial interactúan con las ACF del afloramiento costero. El sur está influenciado por las ASS y el afloramiento costero con mayor intensidad, indicando una mayor actividad en el Sistema de la Corriente del Perú (SCP).
Palabras clave: Escenarios ambientales, Perú, ACF, ASS, CSPC
AbstractEnvironmental scenarios and variability patterns observed in the Peruvian waters are de-scribed based on the analysis of information obtained by the Instituto del Mar del Peru be-tween 1960 and 2011. This information includes data from oceanographic, stock assessment and bio-oceanographic scientific research surveys, published information was also used to complete this study. Sea temperature, salinity and oxygen content data was interpolated to obtain monthly averages and build temporal and spatial series. Surface Cold Coastal Waters (ACF) and Subtropical Surface Water (SSW) create environmental conditions that are differ-ent from those in the subsurface, where the intrusion of equatorial waters dominates through the Peru-Chile Subsurface Current (CSPC). In the north, during the summer and fall, there is predominance in the surface layer of waters from the equatorial system. In the central zone the equatorial waters interact with ACF from coastal upwelling. The south is more strongly influenced by the coastal upwelling and the ASS, which is an indicator of increased activity in the Current System of Peru (SCP).
Keywords: Environmental scenarios, Peru, ACF, ASS, CSPC
IntroducciónFrente a la costa occidental de Sudamérica se encuentra una de las áreas de mayor
productividad biológica, que influencia todos los niveles tróficos del ecosistema marino. El área marítima frente a la costa del Perú representa 0.1% del área oceánica del planeta y produce más del 10% del total de capturas de peces a nivel mundial (Chávez et al. 2008). Esta alta productividad biológica se debe principalmente a las condiciones físicas muy especiales existentes en el Sistema de la Corriente del Perú (SCP) y en particular a la presencia de celdas de afloramiento costero, y a la dinámica oceánica (Zuta & Guillén 1970, Chávez et al. 1989, Bakun & Sweeks 2008, Graco et al. 2007).
Cerca de la costa peruana, y a lo largo de ésta, los vientos dominantes hacia el ecuador y el rotacional del esfuerzo del viento ciclónico dan lugar a intensas celdas de
Publicado online: 11/10/2013Publicado impreso: 15/10/2013

22
Flores et al.
Rev. peru. biol. número especial 20(1): 021 – 028 (Septiembre 2013)Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú.
Editores: Csirke J., R. Guevara-Carrasco & M. Espino
afloramiento caracterizadas por una franja de aguas frías (Fig. 1a) y de alta productividad biológica. Este régimen de viento costero permanente conduce a un sistema típico de corriente de afloramiento compuesto por flujos superficiales hacia el ecuador y flujos subsuperficiales hacia el polo. La circulación superfi-cial hacia el ecuador está compuesta por la Corriente Costera Peruana (CCP) y la Corriente Oceánica Peruana (COP) (Fig. 1b). Estas corrientes cerca a la superficie están principalmente impulsadas por el viento. Sin embargo, la CCP es reforzada a través del ajuste geostrófico asociado con fuertes gradientes de temperatura y densidad (Fig. 1a). La circulación subsuperficial hacia el Polo está principalmente compuesta por la Corriente Subsuperficial Perú-Chile (CSPC) a lo largo de la pendiente y plataforma continental. En la parte norte es reforzada con la Corriente Subsuperficial Ecuatorial (CSE) y la Contracorriente Subsuperficial primaria (CCSSp). Éstas corrientes subsuperficia-les son capaces de alcanzar la costa peruana, mientras que más al sur la CSPC recibe el aporte adicional de otro flujo ecuatorial subsuperficial conocido como la Contracorriente Subsuperficial secundaria (CCSSs) o Jet Tsuchiya secundaria, (Fig. 1a) (Montes et al 2010, Chaigneau et al 2013).
En el presente trabajo, se describen los escenarios ambienta-les existentes en el mar peruano y los patrones de variabilidad imperantes, a partir del análisis de información obtenida in situ en los últimos 50 años por el Instituto del Mar del Perú (IMAR-PE) en sus cruceros de investigación oceanográfica, cruceros de evaluación de recursos pesqueros, cruceros de investigación internacionales, y otras publicaciones orientadas al conocimiento del mar peruano y sus recursos.
Material y métodosSe recopilaron datos de temperatura, salinidad y contenido
de oxígeno, del periodo 1960 – 2011, de 256 cruceros de in-vestigación nacional e internacional, totalizando 8959 perfiles verticales distribuidos entre los 04 y 18°S. Así mismo se incluye información de prospecciones biológicas, oceanográficas y pes-queras realizadas en el mismo periodo. En la capa superficial de 0 – 30 m de profundidad entre las 50 – 100 mn de distancia se obtuvo la climatología mensual y estacional de la temperatura.
Para la determinación de la profundidad de la isoterma de 15 °C (1961 – 2011) se trabajó información obtenida de perfiles hasta los 300 m de profundidad entre los 05 y 18°S, que se separaron en 3 zonas: norte (05 – 08°S), centro (08 – 13°S) y sur (13 – 18°S), entre las 50 – 100 mn de la costa. Usando el método de estimador geoestadístico kriging se completaron los datos faltantes y se interpolaron por cada metro hasta los 300 m de profundidad (Bardossy et al. 1988, Bourgault & Marcotte 1991, Journel & Huijbrenchts 1978), y luego se extrajeron los valores de la profundidad de la isoterma de 15 °C (Z15 °C) con un error menor a un metro. También se calcularon promedios mensuales y promedios simples de todas las estaciones del año, desde 1961 hasta 2011.
Todas las series de la Z15 °C se filtraron con un promedio móvil de 5 meses centrado en 2 meses. Es decir, para cada mes se calculó el promedio incluyendo los 2 meses antes y 2 meses después. Las series obtenidas se promediaron de manera simple y se obtuvo una serie única de la Z15°C.
Para evaluar las condiciones de corrientes marinas a lo largo de la costa peruana se aplicó la metodología de procesamiento
empleada por Chaigneau et al. (2013). Los perfiles verticales y distribución de las corrientes horizontales fueron obtenidos en los cruceros realizados por el IMARPE utilizando el BIC José Olaya, entre los años 2008 y 2012, con un Perfilador de Corrientes Acústico Doppler (ADCP) marca RD Instrumentos 75 kHz de banda ancha, montado en el casco del barco. Para evitar la interferencia entre el ADCP y la Ecosonda Simrad de doble-frecuencia EK60, se utilizó un sistema sincronizador.
Los datos del ADCP fueron procesados y calibrados utilizan-do el sistema estándar Common Ocean Data Access (CODAS) desarrollado por la Universidad de Hawaii. Se hicieron pruebas a los datos para obtener señales de retornos con errores razonables. Las velocidades relativas fueron rotadas desde el transductor respecto a la referencia de la Tierra usando el Girocompás del barco. Para determinar la velocidad absoluta de la corriente, se utilizó el Girocompás a bordo y el Sistema de Posicionamiento Global (GPS). Para el ancho de celda de 30 minutos se utilizó un filtro de paso bajo para las velocidades en la capa de referencia de 50 a 180 m. Finalmente los resultados corresponden a pro-medios interpolados cada 10 m entre 25 a 600 m (Chaigneau et al. 2013).
Resultados y discusiónSobre la base de datos e información descritos por muchos
autores y con información procesada para este artículo, es posible caracterizar los siguientes escenarios ambientales y los siguientes patrones de variabilidad en el mar peruano.
Escenarios ambientales en el Sistema de la Corriente del Perú.- El mar peruano presenta las siguientes características (Fig. 1):
• Una extensa área de afloramiento costero, que normal-mente puede abarcar de 0 a 30 millas de la costa en pro-medio (Morón 2000), pudiendo extenderse a más o me-nos, en función de la estación del año (estacionalidad), de los procesos derivados del ENSO (interanual), de la oscilación decadal (periódicos) y la secularidad (Espino 2003, Gaxiola et al. 2008, Espino & Yamashiro 2012).
• Una reducida área influenciada por las Aguas Tropicales Superficiales al norte de los 04ºS, donde encontramos una alta biodiversidad y bajas biomasas.
• Esto deja una zona de mezcla entre los 04 y los 06°S, con influencia alternada del sur y el norte, dependiendo de la estación imperante del año. En el invierno-primavera prevalecen condiciones del sur y en el verano-otoño condiciones del norte. Esta zona también se caracteriza por una alta biodiversidad, pero menor a la anterior y con bajas biomasas.
• Por fuera de las zonas de afloramiento costero, se localizan las áreas oceánicas dominadas por las Aguas Subtropicales Superficiales (ASS) de condiciones cálidas y que se acercan a la zona costera con mayor intensidad en los veranos y otoños, pero que son influenciadas e inclusive desplazadas por las aguas costeras de afloramiento, durante los inviernos y primaveras. En estas áreas, en la columna de agua que va desde la superficie hasta profundidades mayores a los 1000 metros, domina la Vinciguerria lucetia que constituye alimento de atunes Thunnus spp., dorados Coryphaena hippurus y calamares Dosidicus gigas (Rosas Luis 2007) y

23
Patrones de variabilidad ambiental en el mar peruano
Rev. peru. biol. special issue 20(1): 021 – 028 (September 2013)Ecology, fisheries and conservation of Jack mackerel (Trachurus murphyi) in Peru. Editors: Csirke J., R. Guevara-Carrasco & M. Espino
Figura 1. Sistema de la Corriente del Perú (SCP), a) corrientes subsuperficiales, (b) corrientes superficiales. (Chaigneau et al. 2013)Figure 1. Peruvian Current System (PCS), a) subsurface currents, b) surface currents. (Chaigneau et al. 2013)
también del jurel T. murphy. Al respecto, Alegre et al. (2013) plantean que fuera de las 130 mn de la costa la presencia de peces mesopelágicos se incrementa ligeramente en la dieta de T. murphy; sin embargo, existe poca información disponible de pesquería de T. murphy a distancias mayores a 150 mn donde la abundancia de peces mesopelágicos como la Vinciguerria lucetia es mayor. Cabe destacar asimismo que durante la última década, la mayor abundancia de V. lucetia se localizó al sur de los 16ºS, siendo en dicha zona donde se han realizado las mayores capturas de T. murphy (Gutiérrez Torero 2002).
Subsuperficialmente, por el norte se identifican los siguientes subsistemas:
• La zona comprendida entre los 3°30’S hasta los 6°S está influenciada por la presencia de la CSE y la CCSSp, estas dos corrientes ecuatoriales alcanzan la costa peruana y contribuyen con la parte norte de la CSPC.
• Al sur de los 6°S la CSPC recibe aporte adicional de otro flujo ecuatorial subsuperficial (CCSSs) o Jet Tsuchiya secundario, que se desplaza hasta más allá de los 40°S.
Con información de cruceros del verano del 2011 y prima-vera del 2012, se determinó que los flujos superficiales están modulados por la presencia de giros anticiclónicos (Fig. 2a), formando frentes asociados a vorticidades del orden del Radio de Deformación de Rossby (Cushman-Roisin 1986), los cuales predominan en el centro y sur del Sistema de la Corriente del Perú (SCP). Estos frentes son procesos de convergencia de dos masas de agua, y producen una zona de transición de régimen dinámico intenso y de turbulencia geostrófica. Estas caracterís-ticas condicionan vórtices o giros anticiclónicos de movimientos de masas de agua y, por la dinámica de Ekman, se estimaría que la acción de los vientos podría afectar hasta una profundidad promedio de 100m (Cushman & Beckers 2007). Estas caracte-
rísticas podrían mantener la productividad marina constante, lo cual favoreceria la presencia de recursos pesqueros.
En la zona centro, la configuración de la plataforma con un ancho mayor a las 60 mn frente a Chimbote jugaría un papel importante, permitiendo que los procesos fotosintéticos se desarrollen plenamente en presencia de las aguas afloradas que llevan gran cantidad de nutrientes y por lo tanto fortalecerían la cadena alimenticia y como consecuencia la concentración de recursos pelágicos. Esta plataforma también se encuentra influenciada por flujos subsuperficiales de origen ecuatorial que transporta agua de la zona ecuatorial (CSPC). Asimismo, por el sur tenemos el subsistema constituido por las Aguas Templadas de la Subantártica (ATSA), al que está asociado el bacalao de profundidad Dissostichus eleginoides y su fauna acompañante.
A estos escenarios se agregan los ambientes litorales en los cuales predominan moluscos, crustáceos y equinodermos de alto valor económico y que sustentan importantes pesquerías artesanales, sobre todo en el sur del Perú.
Variabilidad regional a mediano y largo plazo.- A pesar de encontrarse en la zona tropical, en el mar peruano predominan condiciones templadas a frías que eventualmente son interrum-pidas por el fenómeno de El Niño que causa la prevalencia temporal de condiciones tropicales (Fig. 3).
a) Temperatura Superficial del Mar (TSM) .- Las propie-dades hidrográficas como la temperatura aportan evidencias de los procesos físicos que modulan su distribución espacial. Las máximas temperaturas ocurren en verano entre las 50 y 100 mn de la costa desde Punta Sal hasta el Callao, y en otoño, desde Punta Negra hasta Supe, mientras que las mínimas se observan en invierno y primavera en las primeras 50 mn. La distribución superficial de temperatura (Fig.3) muestra valores mínimos cerca de la costa (0 – 50 mn) y con una orientación perpendicular a la costa (E-W). Este patrón refleja la importancia del afloramiento

24
Flores et al.
Rev. peru. biol. número especial 20(1): 021 – 028 (Septiembre 2013)Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú.
Editores: Csirke J., R. Guevara-Carrasco & M. Espino
costero. Los mínimos relativos de temperatura cercanos a la costa se observan a lo largo del año y reflejan aguas subsuperficiales afloradas principalmente en la región sur.
b) Profundidad de la isoterma de 15 °C.- Para el análisis de la variabilidad termal en la vertical, se analizó la profundi-dad media de la isoterma de 15 °C (Z15°C) como la base de la termoclina, donde en la capa por encima de la termoclina se observaron las variaciones temporales.
La profundidad media de la isoterma de 15 °C (Z15°C) es mayor durante el otoño (abril-junio) en la zona norte y centro,
lo cual coincide con la mayor intensificación de la CSPC en la parte norte, que transporta agua cálida con mayor cantidad de oxígeno disuelto de la zona ecuatorial. La menor profundización se presenta en octubre (primavera) cuando se intensifican los afloramientos costeros procedentes del sur. En el sur, en donde los afloramientos costeros son más intensos, la profundidad de la Z15°C es menor (Flores et al. 2009).
En el norte, la Z15°C varía de 108 m en primavera a 123 m en otoño, variación significativa debido al incremento de las velocidades de las corrientes subsuperficiales que vienen de la zona ecuatorial. En el centro varía, de 86 m en invierno a 94 m en otoño. En el sur, de 60 m en invierno a 66 m en verano, presentándose más estable (Fig. 4).
El análisis de la serie de tiempo (1961 – 2011) de la profun-didad de la isoterma de 15 °C (Z15°C) para la zonas norte (05 – 08°S), centro (08 – 13°S) y sur (13 – 18°S) en el mar peruano, y entre las 50 – 100 mn (Fig. 5a), revela una alta variabilidad, principalmente en la zona norte, mientras que para toda el área las mayores fluctuaciones se presentan durante los eventos ENSO. En los eventos extraordinarios del fenómeno El Niño de los años 1982 – 1983 y 1997 – 1998, los valores promedio de la Z15°C estuvieron entre 140 – 200 m, mientras que durante los eventos fríos La Niña fue mucho más superficial y se localizó en alrededor de 30 m. Es decir, durante El Niño, la Z15°C se profundiza en función a la naturaleza y duración del evento, y se hace más superficial durante La Niña.
La Z15°C presentó una tendencia hacia la variabilidad decadal que revela una profundización de dicha isoterma en las déca-das de los ochenta y noventa, definiendo un escenario a nivel subsuperficial favorable a los recursos demersales asociados a corrientes de origen ecuatorial como la CSPC (Flores et al.
Figura 2. Corrientes marinas obtenidas con ADCP (Acoustic Doppler Current Profiler): (a) Capa vertical integrada entre 12-20m de profun-didad (crucero 2012-0910), (b) Secciones verticales (crucero 2012-0506).Figure 2. Marine currents obtained with ADCP (Acoustic Doppler Current Profiler): (a) Integrated vertical layer from 12-20m depth (cruise 2012-0910), (b) Vertical sections (cruise 2012-0506).
(a)
(b)

25
Patrones de variabilidad ambiental en el mar peruano
Rev. peru. biol. special issue 20(1): 021 – 028 (September 2013)Ecology, fisheries and conservation of Jack mackerel (Trachurus murphyi) in Peru. Editors: Csirke J., R. Guevara-Carrasco & M. Espino
Figura 3. Climatología de la temperatura en la capa de 0-30 m de profundidad entre las 0-100 mn de la costa.Figure 3. Climatology of temperature in the layer of depth 0-30 m between the 0-100 mn from the coast.
Figura 4. Climatología estacional de la Z15°C
Figure 4. Seasonal climatology of Z15°C
E F M A M J J A S O N D
0
50
100
150
200
250
300
(m)
E F M A M J J A S O N D
Zona surZona centroZona norte
MesesE F M A M J J A S O N D

26
Flores et al.
Rev. peru. biol. número especial 20(1): 021 – 028 (Septiembre 2013)Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú.
Editores: Csirke J., R. Guevara-Carrasco & M. Espino
Figura 6. Salinidad superficial del mar entre los 03 y 17°S y hasta las 60 mn de la costa en el periodo 1961-2008 (Morón 2011).Figure 6. Sea surface salinity between 03 and 17°S and up to 60 nm offshore in the period 1961-2008 (Morón 2011).
Figura 5. a) Profundidad de la isoterma de 15 °C de las zonas norte, centro y sur. b) Promedio (norte, centro y sur) de la profundidad de la isoterma de 15 ºC (Z15°C) entre las 50-100 mn de la costa en el periodo comprendido entre 1961 y 2011.Figure 5. a) Depth of the 15 °C isotherm of the northern-center, and southern. b) Average (north, center and south) of 15 °C isotherm (Z15°C) depth, between the 50-100 mn from the coast in the period between 1961 and 2011.
1961 1965 1969 1973 1977 1981 1985 1989 1993 1997 2001 2005 2009
0
50
100
150
200
250
Área Norte
Área Centro
1961 1965 1969 1973 1977 1981 1985 1989 1993 1997 2001 2005 2009
0
20
40
60
80
100
120
140
160
180
Promedio de las 3 zonas
(a)
(b)
Área Sur
Pro
fund
idad
(m)
Pro
fund
idad
(m)
2009, Bertrand et al. 2011) (Fig. 5b). Para la zona comprendida entre las 50 – 100 mn de la costa, la variabilidad de la Z15°C tiene el mismo comportamiento para la zona comprendida entre 0 – 60 mn de la costa, con ligeras variaciones en las profundidades.
c) Salinidad Superficial del Mar.- Por otro lado, para esta-blecer los patrones de variabilidad de las masas de agua frente al Perú se analizó la serie de la Salinidad Superficial del Mar (SSM) dentro de las 60 mn de la costa para el periodo com-prendido entre 1960 y 2008 (Morón 2011). Por lo general, las Aguas Tropicales Superficiales (ATS) y las Aguas Ecuatoriales Superficiales (AES) se mantienen al norte de los 04 y de los 06ºS
respectivamente, proyectándose en cada caso más al sur, hasta aproximadamente los 06 y 08°S. Especialmente en relación a El Niño (EN) o al arribo de ondas Kelvin, que en muchos casos no generan un evento EN frente a la costa peruana.
En la decada de los sesenta predominaron las salinidades relativamente bajas correspondientes a ACF. En la decada de los setenta hasta mediados de los ochenta se presentó un predominio de ASS, mientras que desde mediados de los ochenta hasta antes de El Niño 1997 – 1998 se presentaron fluctuaciones de ASS y de ACF. En la década del 2000 predominaron ACF (Fig. 6) lo que coincide con lo planteado por Espino (2013).
Las principales manifestaciones de los eventos EN con catego-
36
35.5
35
34.5
34
33.5
33
32.5
32
31.5
31
30.5
3º
5
7
9
11
13
15
17º
AE
SA
SS
ACF
AT
S
Latit
ud S
ur
1960 1972 1976 1980 19841964 1968 1988 1992 1996 2000 2004 2008

27
Patrones de variabilidad ambiental en el mar peruano
Rev. peru. biol. special issue 20(1): 021 – 028 (September 2013)Ecology, fisheries and conservation of Jack mackerel (Trachurus murphyi) in Peru. Editors: Csirke J., R. Guevara-Carrasco & M. Espino
ría de fuertes o extraordinarios aproximaron a lo largo de la costa ASS de elevada salinidad (35.3 – 35.6). Posteriormente, son las AES y ATS las que se proyectan hacia el sur con baja salinidad, incluso menor a 29.9, lo que da lugar al repliegue hacia el sur de las ASS registradas al inicio del evento, hasta que los vientos del sureste fortalecen los afloramientos y repliegan estas aguas (Zuta et al. 1983, Morón 2011).
d) Profundidad de la oxiclina (Z2ML/L).- De acuerdo a Ber-trand et al. (2011), la profundidad de la oxiclina (Z2ml/L) presentó valores bajos en la década de los sesenta, seguidos de un incremento en la década de los setenta, alcanzando un valor máximo en los años 80. Estos autores destacan que las mayores profundizaciones se presentaron durante los años El Niño (Fig.7). El valor óptimo de contenido de oxígeno para la presencia de jurel T. murphyi (Dioses 2013) estaría entre 1 y 3 ml/L, por encima de la oxiclina. En las dos últimas décadas (de mil novecientos noventa y de los años dos mil) hubo dominancia de las ACF y la pesca fue más superficial, lo que explicaría las mayores capturas obtenidas en este periodo.
De acuerdo al patrón promedio de captura por áreas de pesca del jurel T. murphyi, las mayores concentraciones en el mar pe-ruano se localizan entre las 50 – 100 mn de la costa (Espinoza et al. 2008). Esta zona se caracteriza por la alta dinámica de los afloramientos costeros y las manifestaciones de los patrones de variabilidad interdecadal (periodos cálidos y fríos), las cuales definen un escenario favorable para el T. murphyi, como sucedió en las décadas de los setenta, ochenta y parte de los noventa. Asimismo, estudios realizados por Dioses (1995) y Grechina et al. (1998) indican que los frentes oceánicos formados por la convergencia de las aguas costeras frías y las aguas subtropicales superficiales serían el hábitat preferente del recurso T. murphyi, lo que se evidencia por las capturas de este recurso. Así mismo Bertrand et al. (2004) refieren que el T. murphyi se ubica en aguas oxigenadas (encima de la oxiclina).
Literatura citadaAlegre A., P. Espinoza & M. Espino. 2013. Ecología trófica del jurel Trachurus
murphyi en el Perú entre 1977-2011. En: Csirke J., R. Guevara-Carrasco & M. Espino (Eds.). Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú. Rev. peru. biol. número especial 20(1): 075 – 082
Bakun A., & S. J. Weeks. 2008. The marine ecosystem off Peru: what are the secrets of its fishery productivity and what might its future hold? Progress In Oceanography 79:290 – 299. DOI: 10.1016/j.pocean.2008.10.027
Bárdossy A., 1988. Notes on the robustness of kriging systems. Math. Geology, v. 20, no. 3, pp. 189 – 203. DOI: 10.1007/BF00890253
Bertrand A., M. Segura, M. Gutiérrez & L. Vásquez. 2004. From small-scale habitat loopholes to decadal cycles: a habitat-based hypothesis explaining fluctuation in pelagic fish populations off Peru. Fish and Fisheries 5: 296 – 316. DOI: 10.1111/j.1467-2679.2004.00165.x
Bertrand A., A. Chaigneau, S. Peraltilla, J. Ledesma & M. Graco. 2011. Oxygen: A Fundamental Property Regulating Pelagic Ecosystem Structure in the Coastal Southeastern Tropical Pacific. PLoS ONE6(12): e29558. doi:10.1371/ journal.pone. 0029558.
Bougault, G., & D. Marcotte. 1991. The multivariable variogram and its appli-cation to the linear coregionalization model. Journal Mathematical Geology. 23 (10):899 – 928. DOI: 10.1007/BF02066732
Chaigneau, A., Domínguez, N., Eldin, G., Vásquez, L., Flores, R., Grados, C., & Echevin, V. 2013. Near – coastal circulation in the Northern Humboldt Current System from shipboard ADCP data. Journal of Geophysical Research: Oceans. DOI: 10.1002/jgrc.20328
Chávez F.P., R.T. Barber & M.P. Sanderson. 1989. The potential Primary Production of the Peruvian Upwelling System. Instituto del Mar del Peru (IMARPE), Deutsche Gesellschaft fur Technische Zu-sammenarbeit (GTZ) and International Center for Living Aquatic Resources Management (ICLARM).
Chávez F.P., A. Bertrand, R. Guevara-Carrasco, P. Soler & J. Csirke. 2008. The northern Humboldt Current System: Brief history, present status and a view towards the future. En A. Bertrand, R. Guevara-Carrasco, P. Soler, J. Csirke & F. Chavez (eds.). Progress in Oceanography 79(2 – 4): 95 – 105. DOI: 10.1016/j.pocean.2008.10.012
Cushman-Roisin B. 1986. Frontal geostrophic dynamics. Jou-na l o f Phys ica l Oceanography 16:132-143. DOI: 10.1175/1520-0485(1986)016<0132:FGD>2.0.CO;2
Cushman R.B. & J.M. Beckers. 2007. Introduction to Geophysical Fluid Dynamics, Academic Press. Pp851
Dioses T. 1995. Análisis de la distribución y abundancia de los recursos jurel y caballa frente a la costa peruana. Informe Progresivo del Instituto del Mar del Perú 3, 55 pp.
Dioses T. 2013. Patrones de distribución y abundancia del jurel Trachurus mur-phyi en el Perú. En: Csirke J., R. Guevara-Carrasco & M. Espino (Eds.). Ecología, pesquería y conservación del jurel (Trachurus mur-phyi) en el Perú. Rev. peru. biol. número especial 20(1): 067 – 074
Espino M. 2003. Estrategia de gestión oriental para el Pacífico Oriental con especial mención a la pesquería peruana. Tesis para optar el grado de Magíster en Geografía con mención en Ordenamiento y Gestión Ambiental. Unidad de Post Grado, Facultad de Ingeniería Geológica, Minera, Metalúrgica y Geográfica, Universidad Nacional Mayor de San Marcos. 102 pp.
Espino M. 2013. El jurel Trachurus murphyi (Nichols 1920) y las variables ambientales de macroescala. En: Csirke J., R. Guevara-Carrasco & M. Espino (Eds.). Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú. Rev. peru. biol. número especial 20(1): 009 – 020
Espino M. y C. Yamashiro. 2012. La variabilidad climática y las pesquerías en el Pacífico suroriental. Lat. Am. J. Aquat. Res., 40(3): 705 – 721 International Conference: “Environment and Resources of the
0
10
20
30
40
50
60
70
80
90
100
1960 1975 1980 1985 19901965 1970 1995 2000 2005
5
4.5
4
3.5
3
2.5
2
1.5
1
0.5
Figura 7. Patrón de variabilidad de la profundidad de la oxiclina (Z2ML/L) (A. Bertrand, pers. com.).Figure 7. Variability pattern of the depth of oxycline (Z2ML/L) (A. Bertrand, pers. com.).

28
Flores et al.
Rev. peru. biol. número especial 20(1): 021 – 028 (Septiembre 2013)Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú.
Editores: Csirke J., R. Guevara-Carrasco & M. Espino
South Pacific” P.M. Arana (Guest Editor) DOI: 103856/vol40 – issue3 – fulltext – 18.
Espinoza E., M. Ñiquen & Flores R. 2008. Changes in the latitudinal and longitudinal distribution of the aggregations of Jack mackerel (Tra-churus murphyi) in the Peruvian sea during 1996 – 2007. Chilean Jack Mackerel Workshop 30 June – 4 July 2008.
Flores R., J. Tenorio & N. Domínguez. 2009. Variaciones de la Extensión Sur de la Corriente Cromwell frente al Perú entre 3 y 14°S. Bol. Inst. Mar Perú – Callao 24 (1 y 2).
Gaxiola-Castro G, R. Durazo, B. Lavaniegos, M.E. De la Cruz-Orozco, E. Millan-Nunez, L. Soto-Mardones, J. Cepeda-Morales. 2008. Pelagic ecosystem response to interanual variability off Baja California. Cienc. Mar. 34: 263: 26.
Graco, M, J. Ledesma, G. Flores & M. Girón. 2007. Nutrientes, oxígeno y procesos biogeoquímicos en el sistema de surgencias de la corriente de Humboldt frente a Perú. Revista Peruana de Biología 14 (1): 117 – 128
Grechina A., S. Núñez & D. Arcos. 1998. El desove del recurso jurel, Trachurus symmetricus Murphy (Nichols), en el océano Pacífico Sur. En D. Arcos (Ed.). Biología y ecología del jurel en aguas chilenas. Instituto de Investigación Pesquera, Chile: 117 – 140.
Gutiérrez Torero M. 2002. An approach based in acoustic data to the study of the variability in distribution and abundance of small pelagics in the Humboldt Ecosystem. Ices Symposium on Acoustics in Fisheries and Aquatic Ecology, Montpellier, France 10 – 14 June 2002: 12 p.
Journel A. G. & C. J. Huijbregts. 1978. Mining Geostatistics. Academic Press. London, pp 600
Montes I., F. Colas, X. Capet & W. Schneider. 2010. On the pathways of the equatorial subsurface currents in the eastern equatorial Pacific and their contributions to the Peru‐Chile Undercu-rrent. Journal of Geophysical Research: Oceans, 115, C09003, doi:10:1029/2009JC005710.
Morón O. 2000. Características del ambiente marino frente a la costa peruana. Bol. Inst. Mar Perú 19(1 – 2): 179 – 204.
Morón O. 2011. Climatología de la salinidad superficial frente a la costa peruana (1960 – 2008), IMARPE 38 (1): 7 – 39.
Rosas Luis R. 2007. Descripción de la alimentación de calamar gigante Dosidi-cus gigas (D`Orbigny, 1835) en la costa occidental de la península de Baja California. Tesis para obtener el grado de Maestro en Ciencias. Uso, manejo y preservación de los recursos naturales. Orientación: Biología Marina. Centro de Investigaciones Biológicas del Noroeste, S.C. La Paz, Baja California Sur. Septiembre 2007.
Zuta S. & O.G. Guillén. 1970. Oceanografía de las aguas costeras del Perú. Bol. Inst. Mar Perú (2): 157-324
Zuta S., I. Tsukayama & R. Villanueva. 1983. El ambiente marino y las fluctua-ciones de las principales poblaciones pelágicas de la costa peruana. FAO, informe de pesca 291(2): 179 – 253.

29
Reproducción del jurel Trachurus murphyi en el Perú
Rev. peru. biol. special issue 20(1): 029 - 034 (September 2013)Ecology, fisheries and conservation of Jack mackerel (Trachurus murphyi) in Peru. Editors: Csirke J., R. Guevara-Carrasco & M. Espino
Rev. peru. biol. número especial 20(1): 029 - 034 (Septiembre 2013)Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú. Editores: Csirke J., R. Guevara-Carrasco & M. Espino
Facultad de ciencias Biológicas unMsMISSN-L 1561-0837
Aspectos reproductivos del jurel Trachurus murphyi
Ángel Perea, Julio Mori, Betsy Buitrón y Javier Sánchez
Reproductive aspects of Jack mackerel Trachurus murphy
Instituto del Mar del Perú, esquina Gamarra y General Valle S/N Chucuito, Callao, Perú.
Email Ángel Perea: [email protected]
biología del jurel (Trachurus murphyi) en el Perú
© Los autores. Publicado por la Revista Peruana de Biología de la Facultad de Ciencias Biológicas, Universidad Nacional Mayor de San Marcos. Este es un artículo de acceso abierto, distribuido bajo los términos de la Licencia de Atribución Reconocimiento-NoComercial-SinObraDerivada 3.0 de Creative Commons (http://creativecommons.org/licenses/by-nc-nd/3.0/deed.es_ES), que permite el uso no comercial, distribución y reproducción en cualquier medio, siempre que la obra original sea debidamente citadas. Para uso comercial, por favor póngase en contacto con [email protected].
Citación:Perea A., J. Mori, B. Buitrón & J. Sánchez. 2013. Aspectos reproductivos del jurel Trachurus murphyi en el Perú. En: Csirke J., R. Guevara-Carrasco & M. Espino (Eds.). Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú. Rev. peru. biol. número especial 20(1): 029- 034 (Septiembre 2013)
Resumen Se analiza y discute la serie temporal del Índice Gonadosomático (IGS) y la talla de primera madurez del jurel Trachurus murphyi Nichols 1920 en el Perú calculada mensualmente entre los años 1967 – 2012. Sobre la base de estos análisis se determinó la variación anual e in-teranual del ciclo reproductivo, además se demostró que frente a la costa del Perú T. murphyi tiene un solo periodo de desove relativamente amplio y que ocurre con un máximo en el mes de noviembre de cada año. Asimismo, se evidenció que durante más de cuatro décadas T. murphyi ha desovado regularmente cada año en el mar peruano. La actividad reproductiva del T. murphyi tiene una mayor variabilidad en el Perú y el periodo de desove tiene picos de menor magnitud pero de mayor amplitud con respecto al desove que ocurre frente a la costa de Chile. El análisis de las tallas de primera madurez no presentó cambios significativos a lo largo de toda la serie en el mar peruano.
Palabras clave: Reproducción, jurel, Trachurus murphyi, ciclo reproductivo, desove.
Abstract The time series of monthly gonadosomatic index (GSI) and biannual size at first maturity of Jack mackerel Trachurus murphyi Nichols 1920 in Peru between 1967 and 2012 is analyzed and discussed. The annual and interannual variation of the reproductive cycle was determined. It is shown that in Peru T. murphyi has a single relatively extended spawning period with a maximum in November each year. It is also shown that for more than four decades T. murphyi has spawned regularly every year in Peruvian waters. The reproductive activity of T. murphyi has a greater variability off Peru and the spawning period has peaks of lesser magnitude but extend longer than observed in the spawning occurring off Chile. The analyses of the sizes at first maturity of Jack mackerel in Peruvian waters did not show significant changes throughout the entire period observed.
Keywords: Reproduction, Jack mackerel, Trachurus murphyi, reproductive cycle, spawning.
Introducción Trachurus murphyi Nichols 1920 es una especie pelágica de amplia distribución en
el Pacífico Sudoriental que incluye zonas de jurisdicción de Ecuador, Perú y Chile, como en alta mar. Desde el punto de vista reproductivo, se trata de un recurso con características gonocóricas (sexos separados) e iteróparos (la descendencia se produce en tandas sucesivas de desove), sin dimorfismo sexual visible. Estudios histológicos demostraron que su desarrollo ovocitario es de tipo asincrónico (Dioses et al. 1988, Gordo et al. 2008) y se determinó que esta especie tiene un patrón de desove parcial, ajustándose a la descripción de desovantes parciales de Hunter y Macewicz (1984) y de Murua y Saborido-Rey (2003).
En el Perú, Dioses et al. (1988) determinaron la talla media de madurez gonadal en 21 cm de longitud total (LT), verificando que T. murphyi desova con mayor in-tensidad entre setiembre y diciembre. Además, precisaron que durante los últimos años la principal área de desove en el Perú se ubicó entre los 14°00’S y 18°30’S, en el frente oceánico limitado por las aguas costeras frías de intenso afloramiento y las Publicado online: 11/10/2013
Publicado impreso: 15/10/2013

30
Perea et al.
Rev. peru. biol. número especial 20(1): 029 - 034 (Septiembre 2013)Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú.
Editores: Csirke J., R. Guevara-Carrasco & M. Espino
0
0,5
1
1,5
2
2,5
3
3,5
4
E F M A M J J A S O N D
IGS
CICLO REPRODUCTIVO DEL JUREL 2001 - 2012NORTE - PERU
0
0,5
1
1,5
2
2,5
3
3,5
4
E F M A M J J A S O N D
IGS
CICLO REPRODUCTIVO DEL JUREL 2001 - 2012NORTE - CHILE
0
0,5
1
1,5
2
2,5
3
3,5
4
E F M A M J J A S O N D
IGS
MESES
CICLO REPRODUCTIVO DEL JUREL 2001 - 2012
CENTRO - SUR - CHILE
Figura 1. Ciclo reproductivo de jurel T. murphy: a) en Perú, b) al norte de Chile; y, c) al centro-sur de Chile.Figure 1. Reproductive cycle of Jack mackerel T. murphyi: a) in Peru with information from 1962 to 2012; b) in northern Chile; and c) in southern-central Chile.
aguas subtropicales superficiales, generalmente entre 100 y 150 mn de la costa.
El potencial reproductivo del jurel T. murphyi en el Perú, medido a través de la fecundidad parcial, muestra que esta espe-cie es capaz de desovar en promedio 78789 ovocitos por tanda de desove, pudiendo alcanzar en las tallas mayores a 35 cm de longitud total, fecundidades parciales de hasta 162590 ovocitos por desove (Dioses et al. 1988).
En este trabajo se presentan resultados del análisis de infor-mación biológica proveniente del seguimiento de la pesquería desde el año 1967 al 2012, lo cual ha permitido conocer la variabilidad anual e interanual del ciclo reproductivo del T. murphyi en la costa peruana, sus cambios a lo largo del tiempo, así como estimar la talla de primera madurez con resolución bianual y de toda la serie.
Material y métodos Fueron analizados un total de 145466 individuos colectados
entre 1967 y 2012, por los Laboratorios Costeros del IMARPE de Paita (5°S), Chimbote (9°S), Huacho (11°S), Callao (12°S), Pisco (14°S) e Ilo (17°S), y que forman parte del programa de seguimiento de la pesquería pelágica. Para estos análisis se siguió la metodología descrita por Bouchon et al. (2002) que tomaron como referencia lo descrito por Holden y Raitt (1975), y se consideraron los siguientes datos: longitud total (LT), peso total, peso eviscerado y peso de la gónada. Para conocer los periodos de máxima madurez y desove se utilizó el valor promedio men-sual del Índice Gonadosomático (IGS), el que se calculó como:
IGS=100*(Wg/We)
Donde:
Wg = Peso de la gónada y We = Peso eviscerado.
Para los cálculos de IGS se consideraron sólo hembras adultas iguales o mayores a 26 cm LT, en concordancia con la talla de primera madurez histórica estimada en este trabajo.
Con la finalidad de conocer los cambios anuales del IGS, se calcularon los valores medios por año, considerando solamente los de setiembre a diciembre (periodos de mayor actividad repro-ductiva), a los cuales se denominó “valor crítico”. Se determinó el intervalo de confianza, integrando los valores de la desviación estándar, el número de muestra y la probabilidad al 95%.
Para comparar el ciclo reproductivo de T. murphyi en Perú y Chile, se analizaron datos del IGS del jurel de Chile desde 2001 hasta 2012. Estos datos están disponibles en boletines publicados periódicamente por el IFOP en su página web (ésta página web se actualiza periódicamente y sólo mantiene los boletines más re-cientes). Con esos datos se calcularon el promedio y la desviación estándar mensuales, para construir los patrones que se presentan en la Figura 1. Al momento de imprimir esta publicación, el patrón para la región centro-sur se puede observar en IFOP (2013a), y el patrón para la zona norte en IFOP (2013b). Dada la amplia distribución de T. murphyi, para la comparación del ciclo reproductivo se consideraron tres zonas, dos para la parte chilena y una para la peruana, de la siguiente manera:
Norte de Chile (Arica – Antofagasta – Coquimbo)
Centro – sur de Chile (San Antonio – Guateca)
Perú (Paita – Ilo)
Para comparar los ciclos reproductivos entre zonas, los valo-res mensuales de IGS fueron estandarizados tomándose como máximo valor 1 y recalculándose los valores mensuales para cada serie. Con los nuevos valores calculados se obtuvieron los ciclos reproductivos para cada zona.
La talla de primera madurez fue estimada para toda la serie histórica (1967 – 2012) agrupando los datos cada dos años para tener un tamaño de muestra representativo debido a la intermitencia de la actividad pesquera. Para construir las ojivas de madurez se aseguró que las muestras utilizadas provengan de periodos de desove y correspondan a un amplio rango de tallas e incluyan individuos virginales (Vazzoler 1982, Buitrón et al. 2011). Las proporciones fueron ajustadas a una curva logística de la forma:
y=a/(1+b*exp( – cx))
Donde:
x= Longitud total; a, b y c =constantes, y= L50 o longitud de proporción 50% o 0.5.
Resultados y discusión Ciclo reproductivo.- La serie histórica completa del IGS
muestra que en el Perú T. murphyi tiene un periodo de madu-ración y desove relativamente amplio, que se extiende entre sep-

31
Reproducción del jurel Trachurus murphyi en el Perú
Rev. peru. biol. special issue 20(1): 029 - 034 (September 2013)Ecology, fisheries and conservation of Jack mackerel (Trachurus murphyi) in Peru. Editors: Csirke J., R. Guevara-Carrasco & M. Espino
1.0
0.8
0.6
0.4
2 4 6 8 10 12
Meses
IGS
est
anda
rizad
o
Perú
Chile
Figura 2. Ciclo reproductivo de jurel T. murphyi en Perú y sur de Chile con IGS estandarizado, años 2001-2010.Figure 2. Reproductive cycle of Jack mackerel T. murphyi in Peru and southern Chile with standardized GSI, years 2001-2010.
Características del ciclo reproductivo
Amplitud de la actividad
reproductiva (meses)
Mes de máxima actividad
reproductiva (Máximo valor
de IGS)
Variabilidad de la actividad reproductiva
(SD)
Perú 4 Noviembre Amplia (± 1.25)
Chile (norte) 3 Noviembre Baja (± 0.84)
Chile (centro-sur) 3 Diciembre Baja (±0.54)
Tabla 1. Principales características del ciclo reproductivo del jurel T. murphyi de acuerdo a la variación mensual del IGS.Table 1. Main characteristics of the reproductive cycle of Jack macke-rel T. murphyi according the monthly variability in the gonadosomatic index (GSI).
tiembre y diciembre con máximos valores de IGS en noviembre. El patrón anual del IGS de T. murphy en Perú es claramente diferente al que se observa para el T. murphyi en el centro-sur de Chile, donde según Oyarzún et al. (1998) y Aracena et al. (1998) los valores máximos se producen en diciembre (Fig. 1).
Por otro lado, Grechina et al. (1998) afirmaron que en la subdivisión norte del Pacífico Sudoriental, T. murphyi presen-taría un desove intenso entre agosto y septiembre. Sin embargo nuestros resultados muestran claramente que los máximos valores de desove de T. murphyi frente a la costa peruana se producen en noviembre.
Las diferencias temporales entre los máximos desoves que se observan T. murphyi entre la jurisdicción de Perú y Chile, también han sido observadas en otras especies pelágicas de la región, tal como la anchoveta Engraulis ringens, la cual presenta un máximo de actividad reproductiva de julio a febrero en Chile (Claramunt 2012) y de setiembre a marzo en el norte-centro de Perú (Chirinos & Alegre 1969 , Buitron & Perea 2000).
En peces la actividad de desove está fuertemente influenciada por las condiciones ambientales y las diferentes poblaciones y especies adaptan su calendario de desove para aprovechar ven-tanas de tiempo o periodos de menor incertidumbre ambiental (Wooton 1995) y una mayor probabilidad de supervivencia para sus productos de desove. Por lo tanto, la asincronía en el plazo máximo de actividad reproductiva entre el T. murphyi en Perú y el de la zona centro-sur de Chile puede ser el resultado de una respuesta diferente de estas poblaciones a los diferentes regímenes ambientales.
En la Figura 2 se muestran los valores de IGS estandarizados de T. murphyi de Perú y de la parte sur de Chile, donde se apre-cian las diferencias en la amplitud y los meses que registran la máxima actividad reproductiva. Aunque no se cuenta con una serie de tiempo basada en estudios microscópicos que permita estimar la fracción desovante de T. murphyi, la característica de tener un periodo de desove más amplio con valores de IGS más bajos podría interpretarse como una estrategia reproductiva inducida por la alta variabilidad ambiental en el mar peruano, y que llevaría a T. murphyi a producir desoves menos intensos pero de mayor duración que permitirían aumentar la probabilidad de que sus estadios tempranos de vida encuentren condiciones
adecuadas para su sobrevivencia en una situación de mayor incertidumbre.
Hay tres diferencias interesantes en los ciclos reproductivos que se muestran en la Tabla 1 y que podrían ser los indicadores de diferentes adaptaciones a largo plazo en la estrategia repro-ductiva: (1) La amplitud del periodo máximo de desove en el Perú es de 4 meses, mientras que en el norte y en el centro-sur de Chile es de 3 meses; (2) los valores medios mensuales del IGS de T. murphyi frente a la costa peruana son inferiores a los de T. murphyi frente a la costa de Chile; y, (3) la desviación estándar del IGS en la actividad reproductiva máxima es mayor en Perú que en Chile. La mayor amplitud del periodo de desove y la desviación estándar más alta de IGS son señales claras de una respuesta biológica diferencial a un entorno más variable respecto al sur de Chile.
En general, los máximos valores de IGS son inferiores en el Perú comparados con los de T. murphyi en Chile, lo cual podría interpretarse como una menor actividad reproductiva en el mar peruano, que coincide con lo reportado por Cubillos et al. (2008). Sin embargo, en algunos años los mayores valores del IGS superan al patrón establecido, como lo registrado en los años 2001, 2003 y 2005 (Fig. 3).
Por otro lado, las estimaciones de abundancia de larvas de T. murphyi obtenidas con los cruceros de evaluación desde 1966 al 2010 (Ayón & Correa 2013) confirman que efectivamente T. murphyi desova y recluta en el mar peruano. Además, en algunos años dichas estimaciones muestran altas densidades de larvas, las cuales alcanzaron valores superiores a 500 larvas/m2 indicando desoves abundantes. Por lo tanto, y considerando que el IGS es un índice que representa el crecimiento en peso del ovario debido a los procesos de maduración y desove, el registro de valores más bajos de IGS en Perú describe un ciclo reproductivo con características distintas, que lo distingue del ciclo reproductivo del T. murphyi en Chile.
Variación interanual del IGS.- Desde el año 1967 a la fecha, la variación interanual del IGS describe tres periodos en función de valores críticos (Fig. 3): el primero, desde 1967 hasta 1985, con valor medio crítico de 1.09 (DE = 1.18); el segundo, desde 1986 hasta 1998 con valor crítico de 1.54 (DE = 1.36); y el último desde 1999 hasta la actualidad, con valor crítico de 0.94 (DE = 1.06). En general, el IGS ha presentado valores más altos entre 1967 y 1999, periodo en el cual la pesca de T. murphyi para uso industrial fue más alta frente a Perú
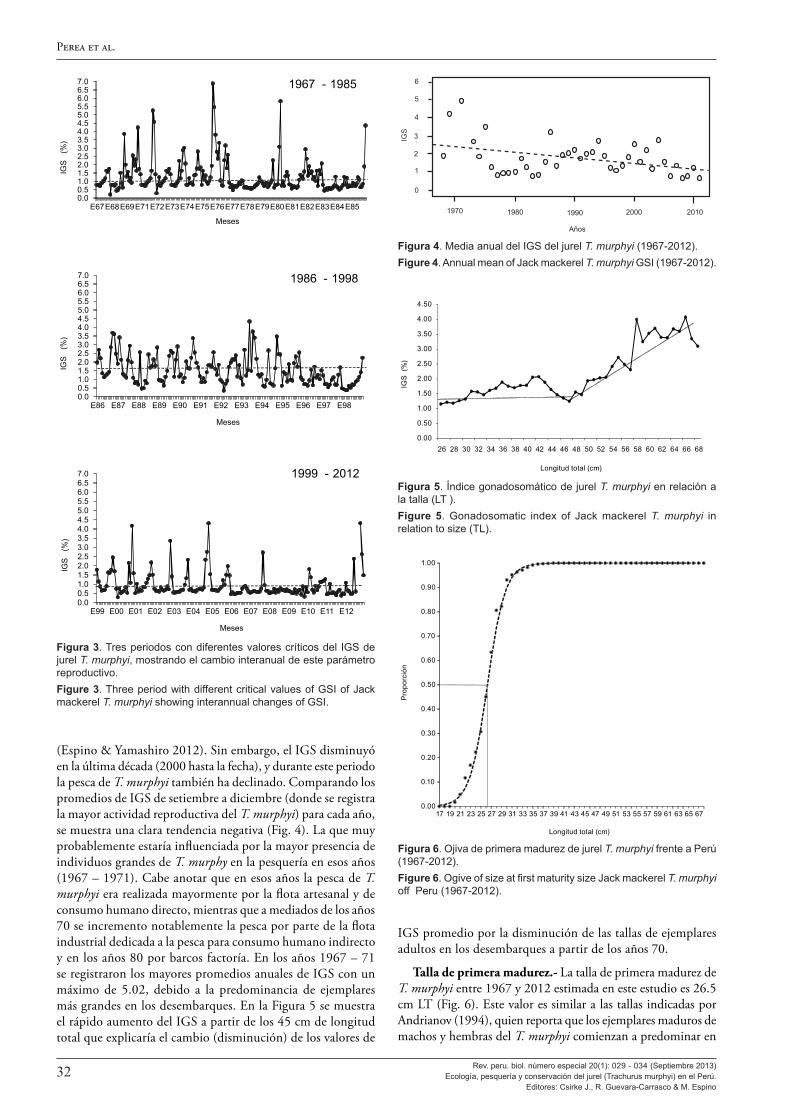
32
Perea et al.
Rev. peru. biol. número especial 20(1): 029 - 034 (Septiembre 2013)Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú.
Editores: Csirke J., R. Guevara-Carrasco & M. Espino
Figura 3. Tres periodos con diferentes valores críticos del IGS de jurel T. murphyi, mostrando el cambio interanual de este parámetro reproductivo.Figure 3. Three period with different critical values of GSI of Jack mackerel T. murphyi showing interannual changes of GSI.
1970 1980 1990 2000 2010
6
5
4
3
2
1
0
IGS
Años
Figura 4. Media anual del IGS del jurel T. murphyi (1967-2012).Figure 4. Annual mean of Jack mackerel T. murphyi GSI (1967-2012).
0.00
0.50
1.00
1.50
2.00
2.50
3.00
3.50
4.00
4.50
26 28 30 32 34 36 38 40 42 44 46 48 50 52 54 56 58 60 62 64 66 68
IGS
(%
)
Longitud total (cm)
Figura 5. Índice gonadosomático de jurel T. murphyi en relación a la talla (LT ).Figure 5. Gonadosomatic index of Jack mackerel T. murphyi in relation to size (TL).
0.00
0.10
0.20
0.30
0.40
0.50
0.60
0.70
0.80
0.90
1.00
17 19 21 23 25 27 29 31 33 35 37 39 41 43 45 47 49 51 53 55 57 59 61 63 65 67
Longitud total (cm)
Pro
porc
ión
Figura 6. Ojiva de primera madurez de jurel T. murphyi frente a Perú (1967-2012). Figure 6. Ogive of size at first maturity size Jack mackerel T. murphyi off Peru (1967-2012).
(Espino & Yamashiro 2012). Sin embargo, el IGS disminuyó en la última década (2000 hasta la fecha), y durante este periodo la pesca de T. murphyi también ha declinado. Comparando los promedios de IGS de setiembre a diciembre (donde se registra la mayor actividad reproductiva del T. murphyi) para cada año, se muestra una clara tendencia negativa (Fig. 4). La que muy probablemente estaría influenciada por la mayor presencia de individuos grandes de T. murphy en la pesquería en esos años (1967 – 1971). Cabe anotar que en esos años la pesca de T. murphyi era realizada mayormente por la flota artesanal y de consumo humano directo, mientras que a mediados de los años 70 se incremento notablemente la pesca por parte de la flota industrial dedicada a la pesca para consumo humano indirecto y en los años 80 por barcos factoría. En los años 1967 – 71 se registraron los mayores promedios anuales de IGS con un máximo de 5.02, debido a la predominancia de ejemplares más grandes en los desembarques. En la Figura 5 se muestra el rápido aumento del IGS a partir de los 45 cm de longitud total que explicaría el cambio (disminución) de los valores de
IGS promedio por la disminución de las tallas de ejemplares adultos en los desembarques a partir de los años 70.
Talla de primera madurez.- La talla de primera madurez de T. murphyi entre 1967 y 2012 estimada en este estudio es 26.5 cm LT (Fig. 6). Este valor es similar a las tallas indicadas por Andrianov (1994), quien reporta que los ejemplares maduros de machos y hembras del T. murphyi comienzan a predominar en
0.00.51.01.52.02.53.03.54.04.55.05.56.06.57.0
E67E68E69E71E72E73E74E75E76E77E78E79E80E81E82E83E84E85
IGS
(%
)
Meses
1967 - 1985
0.00.51.01.52.02.53.03.54.04.55.05.56.06.57.0
E86 E87 E88 E89 E90 E91 E92 E93 E94 E95 E96 E97 E98
IGS
(%
)
Meses
1986 - 1998
0.00.51.01.52.02.53.03.54.04.55.05.56.06.57.0
E99 E00 E01 E02 E03 E04 E05 E06 E07 E08 E09 E10 E11 E12
IGS
(%
)
Meses
1999 - 2012

33
Reproducción del jurel Trachurus murphyi en el Perú
Rev. peru. biol. special issue 20(1): 029 - 034 (September 2013)Ecology, fisheries and conservation of Jack mackerel (Trachurus murphyi) in Peru. Editors: Csirke J., R. Guevara-Carrasco & M. Espino
24.5 2527
24.325.6 25.3
21.1
27
21.9
25.824.7
25.727.2
25.126
26.927.8
25.6 25.6 25.1 25
15
20
25
30
35
40
1963
- 64
1967
- 69
1971
- 72
1973
- 74
1975
- 76
1977
- 78
1979
- 80
1981
- 82
1983
- 84
1985
- 86
1987
- 88
1989
- 90
1991
- 92
1993
- 94
1995
- 96
1997
- 98
1999
- 20
00
2001
- 20
02
2003
- 20
04
2005
- 20
06
2007
- 20
08
2009
- 20
10
2011
- 20
12
L 50
Long
itud
tota
l
Años
Figura 7. Tallas de primera madurez (L50) de jurel T. murphyi para el Perú, valores bianuales de 1967 a 2012.Figure 7. Size at first maturity (L50) of T. murphyi for Peru, biannual values from 1967 to 2012.
39,7
34,8
27,2
24
2625,2
15
20
25
30
35
40
1963
- 64
1967
- 69
1971
- 72
1973
- 74
1975
- 76
1977
- 78
1979
- 80
1981
- 82
1983
- 84
1985
- 86
1987
- 88
1989
- 90
1991
- 92
1993
- 94
1995
- 96
1997
- 98
1999
- 20
00
2001
- 20
02
2003
- 20
04
2005
- 20
06
2007
- 20
08
2009
- 20
10
2011
- 20
12
L 50
Long
itud
tota
l
AñosFigura 8. Tallas de primera madurez (L50) de jurel T. murphyi para Chile, valores bianuales de 1964 a 2012.Figure 8. Size at maturity of Jack mackerel T. murphyi for Chile, biannual values from 1964 to 2012.
tallas de 25 a 27 cm LT en el océano abierto de la subdivisión norte del Pacífico Sudoriental.
Las serie de estimaciones de primera madurez de T. murphyi en Perú muestran que no existe una tendencia en los valores de L50 en el periodo analizado (Fig. 7). A este respecto se destaca que en otras pesquerías se ha demostrado que la presión de pesca intensa y sostenida induce la maduración temprana de los recursos explotados (Trippel 1995). No han sido encontradas evidencias de un proceso de este tipo en la información analizada y, mas bien, los estimados de tallas de primera madurez de este estudio indican que no se ha producido maduración temprana alguna en el periodo observado. Se destaca que tampoco se han observado efectos sobre la talla de primera madurez relacionados con eventos tipo El Niño o con cambios de régimen.
Se hace notar que para el caso de Chile, para el mismo perio-do analizado la talla de primera madurez gonadal bianual de T. murphyi ha venido disminuyendo de 39.7 cm LT en el periodo 1963 – 1964 (Kaiser 1973) a 26.0 cm LT en el periodo 1999 – 2000 (Cubillos & Alarcón 2010), y a 25.2 cm LT en el periodo 2011 – 2012 (Fig. 8).
Literatura citadaAndrianov D. 1994. Sobre la biología reproductiva del jurel peruano. En:
Biología y pesca comercial del jurel peruano en el Pacífico Sur. Dagoberto Arcos y Alexanders Grechina (Eds). Instituto de Inves-tigación Pesquera Talcahuano, Chile: 25 – 35.
Aracena O., R. Alarcón, S. Collado, I. Lépez & D. Arriagada. 1998. Aspectos reproductivos del jurel (Trachurus symmetricus murphyi) de la pesquería de cerco de la Octava región, Chile, 1994 – 1995. En: Biología y ecología del jurel en aguas chilenas. Instituto de Investi-gación Pesquera, Talcahuano-Chile, 216 pp.
Ayón P. & J. Correa. 2013. Variabilidad espacial y temporal de larvas de jurel Trachurus murphyi en el Perú entre 1966-2010. En: Csirke J., R. Guevara-Carrasco & M. Espino (Eds.). Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú. Rev. peru. biol. número especial 20(1): 009 – 020
Bouchon M., M. Ñiquen, J. Mori, A. Echevarría & S. Cahuín. 2002. Manual de muestreo de la pesquería pelágica. Inf. Prog. Inst. Mar Perú – Callao. 157: 38pp.
Buitrón B., A. Perea, J. Mori, J. Sánchez & C. Roque. 2011. Protocolo para estudios de peces pelágicos y demersales. Inf. Inst. Mar Perú – Callao 38 (4): 373 – 384.
Buitron B. & A. Perea. 2000. Aspectos reproductivos de la anchoveta peruana durante el periodo 1992 - 2000. Bol Inst. Mar Perú. Bol 19 (1-2) : 45-53.

34
Perea et al.
Rev. peru. biol. número especial 20(1): 029 - 034 (Septiembre 2013)Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú.
Editores: Csirke J., R. Guevara-Carrasco & M. Espino
Chirinos A. & B. Alegre.1969. Madurez sexual de la anchoveta (Engraulis ringens J) en los periodos reproductivos 1961 – 1968. Bol. Inst. Mar Perú 2 (3).
Claramunt, G .2012. Inter-annual reproductive trait variation and spawning habitat preferences of Engraulis ringens off northern Chile. Revista de Biología Marina y Oceanografía, 47 (2): 227-243
Cubillos L.A., J. Páramo, P. Ruiz, S. Núñez & A. Sepúlveda. 2008. The spatial structure of the oceanic spawning of Jack mackerel (Trachurus murphyi) off central Chile (1998–2001). Fisheries Research 90: 261–270. DOI: 10.1016/j.fishres.2007.10.016
Cubillos L.A. & C. Alarcón. 2010. Estimación de la talla media de madurez sexual en Trachurus murphyi mediante parámetros del consumo relativo de oxígeno. Lat. Am. J. Aquat. Res. 38(2): 178 – 187.
Dioses T., V.H. Alarcón, M.E. Nakama & A. Echevarría. 1988. Desarrollo ovocitario, fecundidad parcial y distribución vertical de los cardú-menes en desove del jurel Trachurus murphyi (N). En: Memorias del Simposio Internacional de los Recursos Vivos y las Pesquerías en el Pacífico Sudeste. Pp. 287 – 294.
Espino M. & C. Yamashiro. 2012. La variabilidad climática y las pesquerías en el Pacífico Suroriental. Lat. Am. J. Aquat. Res. 40(3): 705 – 721, 2012. Latin American Journal of Aquatic Research 705. International Con-ference: “Environment and Resources of the South Pacific”, P.M. Arana (Guest Editor). DOI: 103856/vol40 – issue3 – fulltext – 18.
Grechina A., S. Núñez & D. Arcos. 1998. Biología reproductiva del jurel (Trachurus symmetricus murphyi) en el Pacífico Sur. En: Biología y ecología del jurel en aguas chilenas. Instituto de Investigación Pesquera, Talcahuano-Chile, 216 pp.
Gordo L., A. Costa, P. Abaunza, P. Lucio, A. Eltink & I. Figueiredo. 2008. Determinate versus indeterminate fecundity of horse mackerel. Fisheries Research 89: 181 – 185
Holden M.J. & D.F.S. Raitt. 1975. Manual de ciencia pesquera. Parte 2: Mé-todos para investigar los recursos y su aplicación. Doc. Téc. FAO Pesca (115) Rev. 1: 211 pp.
Hunter J.R. & B.J. Macewicz. 1984. Measurement of spawning frequency in multiple spawning fishes. En: R. Lasker (ed). An egg production method for estimating spawning biomass of pelagic fish: Application to the northern anchovy (Engraulis mordax): 330 pp.
IFOP, 2013a. Informe Quincenal N°11 (01 al 15 Junio, 2013). Asesoría Integral para la toma de decisiones en Pesca y Acuicultura, 2013. Programa de Seguimiento de las Pesquerías Pelágicas, Zona Centro - Sur 2013. Subsecretaría de Economía / Julio 2013. 08p. (www.ifop.cl)
IFOP, 2013b. Boletín Semanal N°23 (XV, I, II, III, IV Regiones) (03 al 09 Junio, 2013). Convenio I: Asesoría Integral para la Pesca y Acuicultura, 2013. Proyecto 1.10. Programa de Seguimiento de las Pesquerías Pelágicas Zona Norte 2013. Subsecretaría de Economía / Junio 2013. 15p. (www.ifop.cl)
Kaiser C. 1973 Gonadal Maturation and Fecundity of Horse Mackerel Tra-churus murphyi (Nichols) off the Coast of Chile. Transactions of the American Fisheries Society 102(1): 101 – 108.
Murua H. & F. Saborido-Rey. 2003. Female reproductive strategies of marine fish species of the North Atlantic. J. Northw. Atl. Fish. Sci. 33: 23 – 31.
Oyarzun C., J. Chong & M. Malagueño. 1998. Fenología reproductiva en el jurel Trachurus symetricus (Ayres 1855) (Perciformes, Carangidae) en el área Talcahuano-Chile: 1982 – 10984. En: Biología y ecología del jurel en aguas chilenas. Instituto de Investigación Pesquera, Talcahuano-Chile, 216 pp.
Trippel E. 1995. Age at maturity as a stress indicator in fisheries. BioScience 45 (11): 759 – 771.
Vazzoler A. 1982. Manual de métodos para estudos biológicos de populações de peixes. Reprodução e crescimento. CNPq, Programa Nacional de Zoologia, Brasília, 2: 108p.
Wooton R.J. 1995. Ecology of teleost fishes. Published by Chapman & Hall, 2 – 6 Boundary Row, London SE1 8HN 404 pp.

35
Escala de madurez gonadal del jurel Trachurus murphyi
Rev. peru. biol. special issue 20(1): 035 - 044 (September 2013)Ecology, fisheries and conservation of Jack mackerel (Trachurus murphyi) in Peru. Editors: Csirke J., R. Guevara-Carrasco & M. Espino
Rev. peru. biol. número especial 20(1): 035 - 044 (Septiembre 2013)Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú. Editores: Csirke J., R. Guevara-Carrasco & M. Espino
Facultad de ciencias Biológicas unMsMISSN-L 1561-0837
Escala de madurez gonadal del jurel Trachurus murphyi Nichols 1920
Javier Sánchez, Ángel Perea, Betsy Buitrón y Liz Romero
Scale of gonad maturity stages of Jack mackerel Trachurus murphyi Nichols 1920
Instituto del Mar del Perú, esquina Gamarra y General Valle S/N Chucuito, Callao, Perú.
Email Javier Sánchez: [email protected]
biología del jurel (Trachurus murphyi) en el Perú
© Los autores. Publicado por la Revista Peruana de Biología de la Facultad de Ciencias Biológicas, Universidad Nacional Mayor de San Marcos. Este es un artículo de acceso abierto, distribuido bajo los términos de la Licencia de Atribución Reconocimiento-NoComercial-SinObraDerivada 3.0 de Creative Commons (http://creativecommons.org/licenses/by-nc-nd/3.0/deed.es_ES), que permite el uso no comercial, distribución y reproducción en cualquier medio, siempre que la obra original sea debidamente citadas. Para uso comercial, por favor póngase en contacto con [email protected].
Citación:Sánchez J., A. Perea, B. Buitrón & L. Romero. 2013. Escala de madurez gonadal del jurel Trachurus murphyi Nichols 1920. En: Eds. Csirke J., R. Guevara-Carrasco & M. Espino. Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú. Rev. peru. biol. número especial 20(1): 035 - 044 (Septiembre 2013)
ResumenEl presente trabajo describe la escala de madurez gonadal macroscópica del jurel Trachurus murphyi Nichols 1920, validada con estudios microscópicos y en uso en Perú. La escala consta de seis estadios de maduración para hembras y machos, que van desde el estadio 0 (virginal) al estadio V (recuperación en hembras, posexpulsante en machos). Para el estudio, se contó con 350 ejemplares provenientes del plan de seguimiento de la pesquería pelágica y de cruceros de evaluación de recursos pelágicos; dichos ejemplares fueron colectados desde el 2006 al 2009. La escala cataloga a los individuos virginales en el denominado estadio 0, lo cual permite la separación práctica entre juveniles (estadio 0) y adultos (estadios I a V). Se discute las bondades de la escala validada para el jurel T. murphyi, la cual se compara con otras escalas de madurez propuestas para especies de la misma familia. El análisis de las características microscópicas de los ovarios confirman la tipificación de desove parcial ya descrita por otros autores.
Palabras claves: jurel, Trachurus murphyi, madurez gonadal, reproducción, estadios de madurez.
Abstract This paper describes the macroscopic scale of gonad maturity stages of Jack mackerel Tra-churus murphy Nichols 1920, validated by microscopic studies, and being used in Peru,. A scale of six maturity stages are identified for females and males, ranging from stage 0 (virgin) to stage V (recovery in females, post-spermiation in males). The gonads of 350 specimens were analyzed. These specimens were collected as part of a pelagic fishery monitoring programme and during pelagic stock assessment surveys between 2006 and 2009. Virgin individuals are classified as stage 0 (virgin), which draws the distinction between (non-reproductive) juveniles and (reproductive) adults in stages I to V. The advantages of using the validated stages of gonad development of Jack mackerel T. murphyi are discussed and are compared to other scales of maturity stages proposed for other species in the same family. The analysis of the microscopic characteristics of the ovaries confirms that Jack mackerel is a batch spawner, which has already been described by other authors.
Keywords: Jack mackerel, Trachurus murphyi, gonadal maturity, reproduction, maturity stages.
Introducción El estudio de la reproducción de los stocks de peces es un elemento importante en
el desarrollo de modelos de evaluación y el manejo de los recursos, proporcionando variables como las tallas de primera madurez y el potencial reproductivo.
En la mayoría de los casos, la determinación de la condición reproductiva de la población a través de una escala de madurez basada en la morfología gonadal externa emplea criterios que no permiten identificar a todas las hembras en desove (Gluyas-Millán 1994). Sin embargo, el uso de este método permite analizar un gran número de individuos con relativamente poco esfuerzo.
Por otro lado, la elaboración de escalas deben basarse en criterios que permitan la identificación de los diferentes estados de maduración para proporcionar el mínimo error posible (Gonçalves et al. 2004), lo cual puede obtenerse por el análisis histológico, que en la actualidad es fácil y rápido de aplicar, resultando en la creación de escalas
Publicado online: 11/10/2013Publicado impreso: 15/10/2013

36
Sánchez et al.
Rev. peru. biol. número especial 20(1): 035 - 044 (Septiembre 2013)Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú.
Editores: Csirke J., R. Guevara-Carrasco & M. Espino
basadas en criterios microscópicos (Pérez & Figueiredo 1992, Costa & Borges 1996, Costa 2001).
Como señalan Ferreri et al. (2009), es conveniente considerar las causas de discrepancias entre las observaciones macroscópicas e histológicas, incluidas la subjetividad del operador en la cata-logación, distinción clara entre las descripciones de los estadios de madurez, muestreo durante el día y el tiempo transcurrido desde la captura hasta la catalogación de las gónadas. Si con-tamos con una escala de madurez bien elaborada, con estadios bien definidos (Hunter & Macewicz 2003, Gonçalves et al. 2004, Gerritsen & McGrath 2006), esta podría ser utilizada en el trabajo de rutina y dejar el trabajo histológico solamente para los casos en que sea estrictamente necesario.
Desde que se iniciaron los estudios sobre los aspectos repro-ductivos de jurel Trachurus murphyi en el Perú hace más de 40 años, se ha venido utilizando la escala adaptada de madurez gonadal, descrita por Johansen (1924) que consta de 8 estadios y que fue descrita para el arenque (Clupea harengus). También ha sido adaptada y aplicada al jurel T. murphyi la escala general de 5 estadios para reproductores parciales sugerida por Holden & Raitt (1975) (Aracena et al. 1998, Cubillos 2003). Poste-riormente, Andrianov (1987) propuso una escala de madurez gonadal para el jurel, de 7 estadios, donde el estadio maduro está subdividido en tres etapas de desarrollo. Dioses et al. (1989) describieron las características citológicas del desarrollo ovocita-rio del jurel, pero no incluyeron la descripción macroscópica de las gónadas, y no elaboraron una escala de madurez.
La manera y los objetivos de describir escalas macroscópi-cas validadas en peces quedaron documentados en la reunión mundial realizada en Bergen, Noruega, en septiembre de 2001, con participación de investigadores de varios países que incluyó sudamericanos como Argentina, Chile y Perú. En esta reunión se elaboró un documento (Kjesbu et al. 2003) que entre otras recomendaciones, sugiere la elaboración de escalas macroscópicas que deberían de contar con sustento histológico y criterios que simplifiquen la identificación de los estadios de madurez.
En el presente trabajo describimos las características de una
escala de madurez gonadal de seis estadios para el jurel T. mur-phyi, tanto para hembras como para machos. Cada estadio es descrito a nivel microscópico y macroscópico, lo cual facilita la identificación macroscópica de cada estadio de madurez gonadal.
Material y métodos Se analizaron en total 350 individuos de jurel T. murphyi,
204 gónadas (76 testículos y 128 ovarios) colectados entre enero 2006 y diciembre 2009 de los muestreos del programa de seguimiento de la pesquería pelágica; y 146 individuos (81 testículos y 65 ovarios) de los cruceros hidroacústicos realizados entre el 2008 y 2011 (Tabla 1).
Las gónadas frescas fueron rotuladas, registrando sus carac-terísticas macroscópicas, tomando fotografías digitales de cada una. Adicionalmente, como en trabajos similares (Gonçalves et al. 2004, Ferreri et al. 2009), fueron anotados aspectos comple-mentarios como grado de vascularización, turgencia, textura y en algunos casos tipos de ovocitos visibles a simple vista. En el caso particular de los machos, se anotó la turgencia testicular y el semen fluyente.
Las gónadas fueron procesadas histológicamente usando la técnica de infiltración con parafina (Humason 1979). Con el uso del micrótomo de rotación, se realizaron cortes de 7 μm para ovarios y 5 μm para testículos. La diferencia en el espesor de los cortes de machos y hembras toma en cuenta las diferencias en la estructura ovocitaria y espermatogénica de las gónadas. Las células sexuales masculinas son de menor tamaño que las células sexuales femeninas, y por lo tanto necesitan un corte más fino para evitar su aglomeración y poder observarlas mejor.
Las secciones histológicas obtenidas fueron coloreadas con Hematoxilina-Eosina contrastante y analizadas en un micros-copio compuesto. Las secciones histológicas fueron observadas en detalle y los distintos tipos de ovocitos fueron clasificados en función del número y distribución de nucléolos, grado de vitelogénesis y gotas lipídicas y ; además se determinó la presencia o no de folículos post-ovulatorios, así como ovocitos en estado de atresia. Las descripciones se basaron en las realizadas por Hunter y Goldberg (1980) y Hunter y Macewicz (1985). En el caso de los machos se identificó la histoarquitectura testicular y el desarrollo espermatogénico en los túbulos seminíferos (Abascal et al. 2004, Dadzie & Abou-Seedo 2004), lo cual nos permitió diferenciar los estadios de maduración gonadal, teniendo en cuenta que el proceso de espermatogénesis es dinámico y algunos estadios de maduración podrían confundirse con otros.
Procedencia Sexo Rango de talla (cm)
N°. ejemplares
Seguimiento 2006 Hembra 23 – 49 46
Seguimiento 2006 Macho 23 – 52 28
Seguimiento 2008 Hembra 22 – 54 73
Seguimiento 2008 Macho 26 – 43 46
Seguimiento 2009 Hembra 33 – 43 9
Seguimiento 2009 Macho 40 – 41 2
Crucero 2008 (Nov-Dic) Hembra 22 4
Crucero 2008 (Nov-Dic) Macho 22 – 23 6
Crucero 2011 (Oct-Dic) Hembra 27 – 37 61
Crucero 2011 (Oct-Dic) Macho 28 - 38 75
Total 22 - 54 350
Tabla 1. Muestras de gónadas de jurel Trachurus murphyi utilizadas para la elaboración de la escala de madurez gonadal validada.Table 1. Gonad samples of Jack mackerel Trachurus murphyi used for preparing the scale of gonad maturity stages being validated.
Estadio Hembra Macho
Estadio 0 Virginal Virginal
Estadio I Reposo Reposo
Estadio II En maduración En maduración
Estadio III Maduro Maduro
Estadio IV Desovante Expulsante
Estadio V Recuperación Posexpulsante
Tabla 2. Estadios de madurez gonadal de hembras y machos de jurel Trachurus murphyi.Table 2. Gonadal maturity stages of females and males of Jack mackerel Trachurus murphyi.
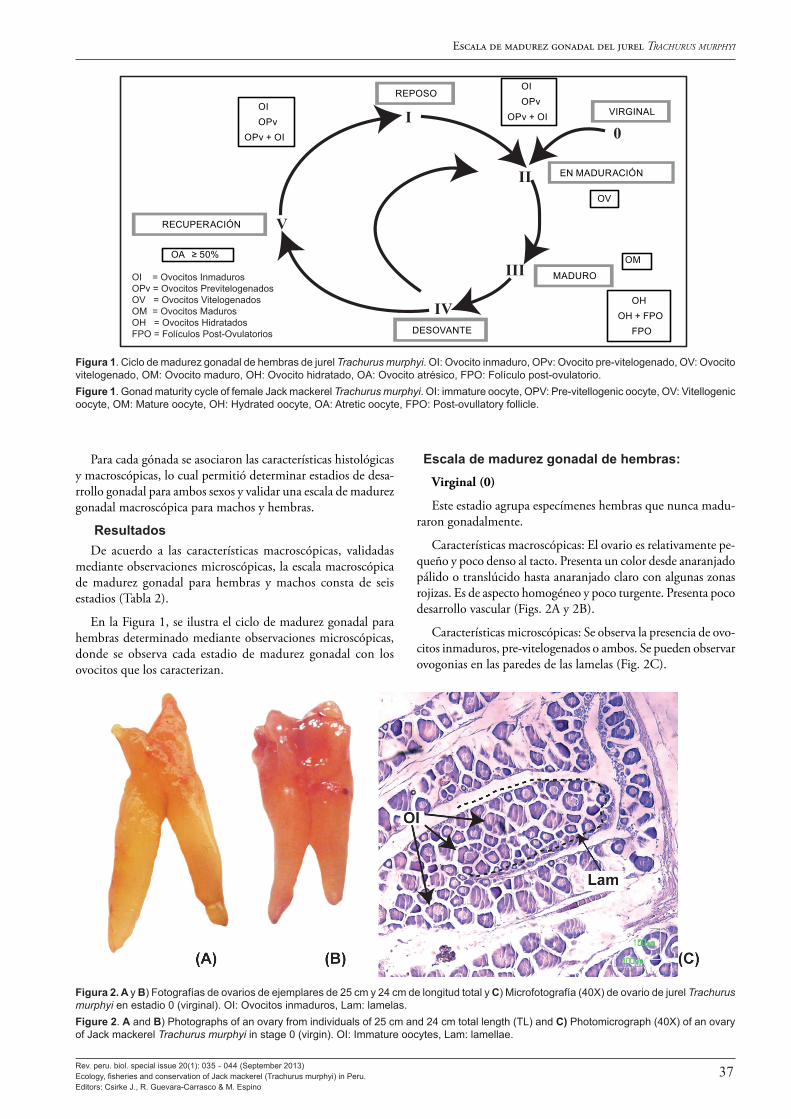
37
Escala de madurez gonadal del jurel Trachurus murphyi
Rev. peru. biol. special issue 20(1): 035 - 044 (September 2013)Ecology, fisheries and conservation of Jack mackerel (Trachurus murphyi) in Peru. Editors: Csirke J., R. Guevara-Carrasco & M. Espino
Para cada gónada se asociaron las características histológicas y macroscópicas, lo cual permitió determinar estadios de desa-rrollo gonadal para ambos sexos y validar una escala de madurez gonadal macroscópica para machos y hembras.
ResultadosDe acuerdo a las características macroscópicas, validadas
mediante observaciones microscópicas, la escala macroscópica de madurez gonadal para hembras y machos consta de seis estadios (Tabla 2).
En la Figura 1, se ilustra el ciclo de madurez gonadal para hembras determinado mediante observaciones microscópicas, donde se observa cada estadio de madurez gonadal con los ovocitos que los caracterizan.
Escala de madurez gonadal de hembras:Virginal (0)
Este estadio agrupa especímenes hembras que nunca madu-raron gonadalmente.
Características macroscópicas: El ovario es relativamente pe-queño y poco denso al tacto. Presenta un color desde anaranjado pálido o translúcido hasta anaranjado claro con algunas zonas rojizas. Es de aspecto homogéneo y poco turgente. Presenta poco desarrollo vascular (Figs. 2A y 2B).
Características microscópicas: Se observa la presencia de ovo-citos inmaduros, pre-vitelogenados o ambos. Se pueden observar ovogonias en las paredes de las lamelas (Fig. 2C).
Figura 1. Ciclo de madurez gonadal de hembras de jurel Trachurus murphyi. OI: Ovocito inmaduro, OPv: Ovocito pre-vitelogenado, OV: Ovocito vitelogenado, OM: Ovocito maduro, OH: Ovocito hidratado, OA: Ovocito atrésico, FPO: Folículo post-ovulatorio.Figure 1. Gonad maturity cycle of female Jack mackerel Trachurus murphyi. OI: immature oocyte, OPV: Pre-vitellogenic oocyte, OV: Vitellogenic oocyte, OM: Mature oocyte, OH: Hydrated oocyte, OA: Atretic oocyte, FPO: Post-ovullatory follicle.
Figura 2. A y B) Fotografías de ovarios de ejemplares de 25 cm y 24 cm de longitud total y C) Microfotografía (40X) de ovario de jurel Trachurus murphyi en estadio 0 (virginal). OI: Ovocitos inmaduros, Lam: lamelas.Figure 2. A and B) Photographs of an ovary from individuals of 25 cm and 24 cm total length (TL) and C) Photomicrograph (40X) of an ovary of Jack mackerel Trachurus murphyi in stage 0 (virgin). OI: Immature oocytes, Lam: lamellae.
OIOPv
OPv + OI
OIOPv
OPv + OI
OV
OM
OHOH + FPO
FPO
OA
VIRGINAL
EN MADURACIÓN
MADURO
DESOVANTE
RECUPERACIÓN
REPOSO
≥ 50%
I
II
III
IV
V
0
OI = Ovocitos InmadurosOPv = Ovocitos PrevitelogenadosOV = Ovocitos Vitelogenados OM = Ovocitos MadurosOH = Ovocitos HidratadosFPO = Folículos Post-Ovulatorios
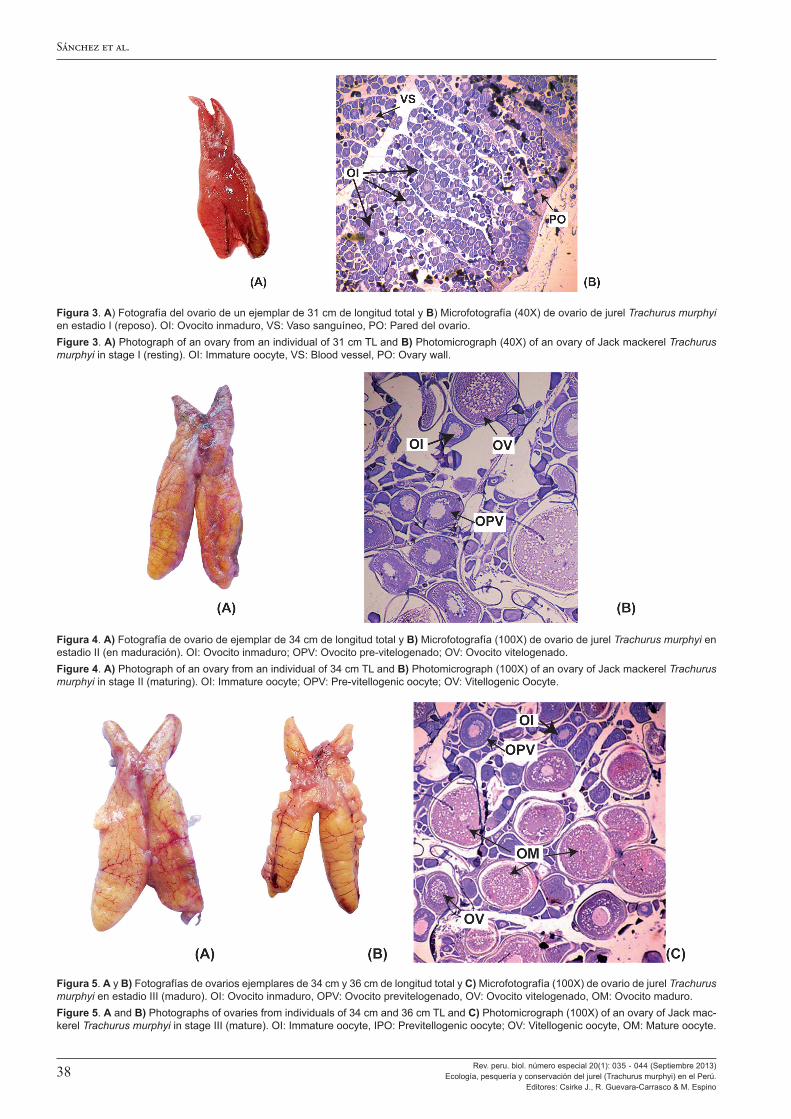
38
Sánchez et al.
Rev. peru. biol. número especial 20(1): 035 - 044 (Septiembre 2013)Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú.
Editores: Csirke J., R. Guevara-Carrasco & M. Espino
Figura 3. A) Fotografía del ovario de un ejemplar de 31 cm de longitud total y B) Microfotografía (40X) de ovario de jurel Trachurus murphyi en estadio I (reposo). OI: Ovocito inmaduro, VS: Vaso sanguíneo, PO: Pared del ovario.Figure 3. A) Photograph of an ovary from an individual of 31 cm TL and B) Photomicrograph (40X) of an ovary of Jack mackerel Trachurus murphyi in stage I (resting). OI: Immature oocyte, VS: Blood vessel, PO: Ovary wall.
Figura 4. A) Fotografía de ovario de ejemplar de 34 cm de longitud total y B) Microfotografía (100X) de ovario de jurel Trachurus murphyi en estadio II (en maduración). OI: Ovocito inmaduro; OPV: Ovocito pre-vitelogenado; OV: Ovocito vitelogenado.Figure 4. A) Photograph of an ovary from an individual of 34 cm TL and B) Photomicrograph (100X) of an ovary of Jack mackerel Trachurus murphyi in stage II (maturing). OI: Immature oocyte; OPV: Pre-vitellogenic oocyte; OV: Vitellogenic Oocyte.
Figura 5. A y B) Fotografías de ovarios ejemplares de 34 cm y 36 cm de longitud total y C) Microfotografía (100X) de ovario de jurel Trachurus murphyi en estadio III (maduro). OI: Ovocito inmaduro, OPV: Ovocito previtelogenado, OV: Ovocito vitelogenado, OM: Ovocito maduro.Figure 5. A and B) Photographs of ovaries from individuals of 34 cm and 36 cm TL and C) Photomicrograph (100X) of an ovary of Jack mac-kerel Trachurus murphyi in stage III (mature). OI: Immature oocyte, IPO: Previtellogenic oocyte; OV: Vitellogenic oocyte, OM: Mature oocyte.

39
Escala de madurez gonadal del jurel Trachurus murphyi
Rev. peru. biol. special issue 20(1): 035 - 044 (September 2013)Ecology, fisheries and conservation of Jack mackerel (Trachurus murphyi) in Peru. Editors: Csirke J., R. Guevara-Carrasco & M. Espino
Reposo (I)
La identificación de este estadio es sumamente importante porque se trata de individuos considerados como adultos, pero son reproductivamente inactivos.
Características macroscópicas: Los ovarios son lisos de color naranja y medianamente turgentes, no son traslúcidos como los virginales. Los ovocitos no son visibles a simple vista, la pared ovárica es engrosada y los vasos sanguíneos están desarrollados (Figura 3A).
Características microscópicas: Desde el punto de vista ovoci-tario, este estadio es igual al virginal. Sin embargo, lo diferencia, principalmente, la pared gruesa del ovario y la mayor cantidad de vasos sanguíneos. En algunos casos, se observan algunos ovocitos atrésicos (Fig. 3B).
En maduración (II)
Características macroscópicas: Los ovarios son de color naran-ja claro. Pueden observarse zonas rosadas o rojizas dependiendo
del grado de maduración en el que se encuentren (dichas zonas contienen gran cantidad de ovocitos inmaduros y previteloge-nados). Son ovarios poco densos al tacto. Se pueden observar ovocitos pequeños a simple vista. Se identifica con claridad el vaso sanguíneo principal en la cara ventral del ovario (Fig. 4A).
Características microscópicas: En este estadio se observan ovocitos inmaduros, previtelogenados y ovocitos vitelogenados. (Fig. 4B).
Maduro (III)
Características macroscópicas: Los ovarios presentan diversas tonalidades de naranja, en algunos casos con zonas sanguinolen-tas producto de la marcada presencia de vasos sanguíneos. Son ovarios turgentes. Como en el caso del estadio en maduración, se pueden observar ovocitos a simple vista, siendo éstos de mayor tamaño (Figs. 5A y 5B).
Características microscópicas: En este estadio se observan ovocitos inmaduros, previtelogenados, vitelogenados y ovocitos
Figura 6. A y B) Fotografías de ovarios de ejemplares de 35cm y de 36cm de longitud total y C) Microfotografía (100X) de ovario de jurel Trachurus murphyi en estadio IV (desovante), OM: Ovocito maduro, FPO: Folículo post-ovulatorio.Figure 6. A and B) Photographs of ovaries from individuals of 35 cm and 36 cm TL and C) Photomicrograph (100X) of an ovary of Jack mac-kerel Trachurus murphyi in stage IV (spawning). OM: Mature oocyte; FPO: Post-ovulatory follicle.
Figura 7. A) Fotografía de ovario de ejemplar de 35 cm de longitud total y B) Microfotografía (40X) de ovario de jurel Trachurus murphyi en estadio V (recuperación). OA: Ovocito atrésico.Figure 7. A) Photograph of an ovary of an individual of 35 cm TL and B) Photomicrograph (40X) of an ovary of Jack mackerel Trachurus murphyi stage V (recovery). OA: Atresic oocyte.

40
Sánchez et al.
Rev. peru. biol. número especial 20(1): 035 - 044 (Septiembre 2013)Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú.
Editores: Csirke J., R. Guevara-Carrasco & M. Espino
maduros. En algunos casos, es posible identificar ovocitos con núcleo migratorio. Igualmente, es posible identificar ovocitos vitelogenados o sin vitelo en proceso de atresia, pero con pro-porción no mayoritaria o frecuencia menor al 50% (Fig. 5C).
Desovante (IV)
Son considerados como desovantes aquellos individuos que están en pleno desove o han desovado recientemente.
Características macroscópicas: Ovarios de gran tamaño y turgentes cuando están en pleno desove o previos a éste. Los ovocitos hidratados se pueden observar a simple vista. Los que ya han desovado presentan una coloración que va desde el verdoso hasta el ámbar. Cuando están parcialmente desova-dos son un tanto flácidos. En este estadio, es común apreciar una sectorización de los ovocitos hidratados en la gónada, en relación al resto de ellos de distribución más homogénea (Fig. 6A y 6B).
Características microscópicas: En el caso de las hembras que se encuentran en pleno desove, la característica más resaltante es la presencia de los ovocitos hidratados y/o folículos post-ovu-latorios. Si tienen folículos post-ovulatorios, éstos se visualizan de diferentes formas dependiendo del grado de degeneración en el que se encuentren. Estas características microscópicas que tipifican a ovarios desovantes, afortunadamente, para esta especie son de fácil identificación (Fig. 6C).
Recuperación (V)
Características macroscópicas: Este tipo de ovarios son poco frecuentes y se caracterizan por ser flácidos o no turgentes. Los ovocitos atrésicos visibles se observan a manera de grumos ama-rillos claros o puntos rojos u oscuros en la superficie ovárica. El ovario suele adquirir también este color (Fig. 7A).
Características microscópicas: El tejido ovárico presenta ovo-citos atrésicos (atresia α) en frecuencia igual o mayor al 50%. También puede observarse restos de ovocitos vitelogenados o no vitelogenados en proceso de atresia en diferentes grados de dege-neración (α, β, o γ). Los ovocitos inmaduros y/o previtelogenados son los que prevalecen. Es perfectamente visible gran vasculariza-ción que le da el aspecto sanguinolento a este estadio (Fig. 7B).
Escala de madurez gonadal de machos:Virginal (0)
Este estadio agrupa a los individuos machos que nunca han madurado gonadalmente.
Características macroscópicas: El testículo es pequeño, delgado y poco consistente, es de tonalidad rosada o rojiza, de apariencia translúcida, y se puede observar poco o nulo desarrollo vascular (Fig. 8A y 8B).
Características microscópicas: Pueden observarse zonas de crecimiento con la presencia de espermatogonias, donde igual-mente se pueden encontrar algunos espermatocitos. (Fig. 8C).
Reposo (I)
Este estadio corresponde a individuos que han madurado gonadalmente alguna vez a lo largo de su vida, pero que se en-cuentran por el momento en una etapa de inactividad gonadal.
Características macroscópicas: Testículo de tamaño mediano, poco denso al tacto, de tonalidad anaranjada rojiza con algunas
zonas claras y se puede observar un desarrollo vascular moderado (Fig. 9A y 9B).
Características microscópicas: Se observan los túbulos semi-níferos con la presencia de espermatogonias y pueden observarse además algunos espermatocitos, con presencia de tejido conjun-tivo que ocupan el espacio entre los túbulos seminíferos. Pared del testículo gruesa (Fig. 9C).
En maduración (II)
Estadio que agrupa a los individuos que dan inicio a la ma-duración testicular.
Características macroscópicas: Testículo de tamaño mediano a grande, que consiste en algunas zonas de color, desde blanco humo con presencia o no de algunas zonas rojizas hasta blanco cremoso y se puede observar desarrollo vascular moderado (Fig. 10A).
Características microscópicas: Túbulos seminíferos con presen-cia de espermatogonias, espermatocitos y espermátides, los cuales se observan de tamaño homogéneo. En ciertos sectores pueden o no observarse la presencia de algunos espermatozoides (Fig. 10-B).
Maduro (III)
Características macroscópicas: Gónada de gran tamaño, color blanquecino a blanco lechoso, turgente y con desarrollo vascular evidente (Fig. 11A y 11B).
Características microscópicas: Túbulos seminíferos con pre-sencia de espermatogonias, espermatocitos y espermátides. En el lumen de los túbulos se observa la presencia de espermatozoides. El tubo colector común se presenta completamente lleno y de aspecto turgente (Fig. 11C).
Expulsante (IV)
Características macroscópicas: En su etapa inicial, los tes-tículos tienen un tamaño máximo, muy turgentes, de color blanco lechoso y el líquido espermático se expele fácilmente con una suave presión. En su etapa avanzada, van disminuyendo de tamaño y disminuye la turgencia. De color blanquecino, y pueden presentar algunos sectores con tonalidades rosadas o rojizas (Fig. 12A).
Características microscópicas: Túbulos seminíferos con poca presencia de espermatogonias y espermatocitos, dominados por espermátides y espermatozoides que rellenan los túbulos. Es posible observar otras zonas vacías producto de la expulsión (Fig. 12B).
Post-explusante (V)
Características macroscópicas: Los testículos disminuyen de tamaño y su coloración varía a tonos más rojizos, debido al incremento de la irrigación sanguínea por efecto del evento de evacuación del líquido espermático. Son de consistencia flácida, debido al término del proceso de evacuación (Figs. 13A y 13B).
Características microscópicas: Se puede observar esperma-togonias, espermatocitos y restos de espermatozoides en los túbulos seminíferos. En una etapa avanzada se pueden observar túbulos casi vacíos con material residual (Fig. 13C).
DiscusiónLa escala de madurez propuesta en este trabajo contempla
las recomendaciones sugeridas por el “Grupo de trabajo sobre

41
Escala de madurez gonadal del jurel Trachurus murphyi
Rev. peru. biol. special issue 20(1): 035 - 044 (September 2013)Ecology, fisheries and conservation of Jack mackerel (Trachurus murphyi) in Peru. Editors: Csirke J., R. Guevara-Carrasco & M. Espino
Figura 8. A y B) Fotografías de testículos de ejemplares de 25 cm y 26 cm de longitud total y C) Microfotografía (100X) de testículo de jurel Trachurus murphyi en estadio 0 (virginal). Eg: Espermatogonia.Figure 8. A and B) Photographs of testicles of individuals of 25cm and 26cm TL and C) Photomicrograph (100X) of a testicle of Jack mackerel Trachurus murphyi in stage 0 (virgin). Eg: Spermatogonia.
Figura 9. A y B) Fotografías de testículos de ejemplares 40 cm y 42 cm de longitud total y C) Microfotografía (40X) de testículo de jurel Tra-churus murphyi en estadio I (reposo). TS: Túbulo seminífero, TC: Tejido conjuntivo, PT: Pared testicular.Figure 9. A and B) Photographs of testicles of individuals of 40cm and 42cm TL and C) Photomicrograph (40X) testicles of Jack mackerel Trachurus murphyi in stage I (resting). TS: Seminiferous tubule, TC: Connective tissue, PT: Testicular Wall.
Figura 10. A) Fotografía de testículo de ejemplar de 29 cm de longitud total y B) Microfotografía (100X) de testículo de jurel Trachurus murphyi en estadio II (en maduración). Eg: Espermatogonia, Ec: Espermatocito, Et: Espermátide, Ez: Espermatozoide.Figure 10. A) Photograph of a testicle of an individual of 29 cm total length and B) Photomicrograph (100X) of a testicle of Jack mackerel Trachurus murphyi in stage II (maturing). Eg: Spermatogonia, Ec: Spermatocytes, Et: Spermatid; Ez: Sperm.
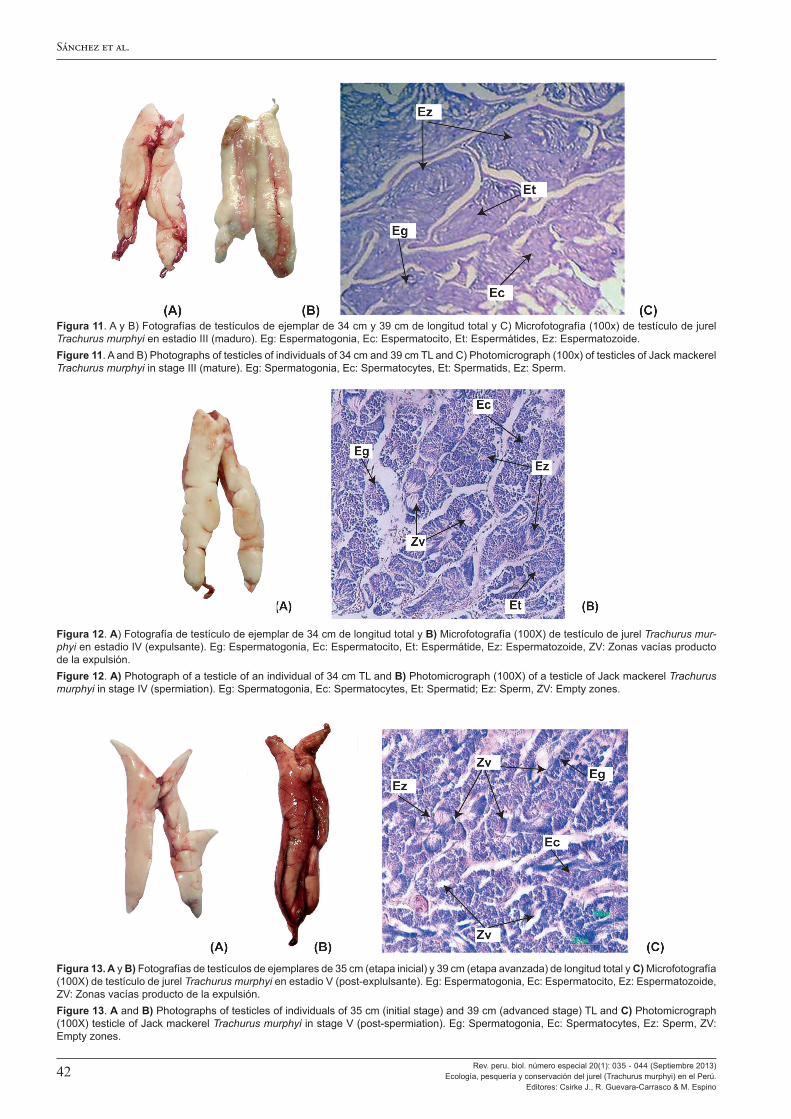
42
Sánchez et al.
Rev. peru. biol. número especial 20(1): 035 - 044 (Septiembre 2013)Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú.
Editores: Csirke J., R. Guevara-Carrasco & M. Espino
Figura 11. A y B) Fotografías de testículos de ejemplar de 34 cm y 39 cm de longitud total y C) Microfotografía (100x) de testículo de jurel Trachurus murphyi en estadio III (maduro). Eg: Espermatogonia, Ec: Espermatocito, Et: Espermátides, Ez: Espermatozoide.Figure 11. A and B) Photographs of testicles of individuals of 34 cm and 39 cm TL and C) Photomicrograph (100x) of testicles of Jack mackerel Trachurus murphyi in stage III (mature). Eg: Spermatogonia, Ec: Spermatocytes, Et: Spermatids, Ez: Sperm.
Figura 12. A) Fotografía de testículo de ejemplar de 34 cm de longitud total y B) Microfotografía (100X) de testículo de jurel Trachurus mur-phyi en estadio IV (expulsante). Eg: Espermatogonia, Ec: Espermatocito, Et: Espermátide, Ez: Espermatozoide, ZV: Zonas vacías producto de la expulsión.Figure 12. A) Photograph of a testicle of an individual of 34 cm TL and B) Photomicrograph (100X) of a testicle of Jack mackerel Trachurus murphyi in stage IV (spermiation). Eg: Spermatogonia, Ec: Spermatocytes, Et: Spermatid; Ez: Sperm, ZV: Empty zones.
Figura 13. A y B) Fotografías de testículos de ejemplares de 35 cm (etapa inicial) y 39 cm (etapa avanzada) de longitud total y C) Microfotografía (100X) de testículo de jurel Trachurus murphyi en estadio V (post-explulsante). Eg: Espermatogonia, Ec: Espermatocito, Ez: Espermatozoide, ZV: Zonas vacías producto de la expulsión.Figure 13. A and B) Photographs of testicles of individuals of 35 cm (initial stage) and 39 cm (advanced stage) TL and C) Photomicrograph (100X) testicle of Jack mackerel Trachurus murphyi in stage V (post-spermiation). Eg: Spermatogonia, Ec: Spermatocytes, Ez: Sperm, ZV: Empty zones.

43
Escala de madurez gonadal del jurel Trachurus murphyi
Rev. peru. biol. special issue 20(1): 035 - 044 (September 2013)Ecology, fisheries and conservation of Jack mackerel (Trachurus murphyi) in Peru. Editors: Csirke J., R. Guevara-Carrasco & M. Espino
enfoques modernos para evaluar la madurez y la fecundidad de los peces y calamares de aguas cálidas y frias” (Kjesbu et al. 2003), y tiene base en observaciones microscópicas que permiten plantear una escala de seis estadios, incluyendo a los individuos virginales catalogados como estadio 0. El estadio 0 permite discriminar con facilidad a los individuos virginales y estimar los estadios de madurez sólo con la fracción adulta. Al mismo tiempo, hace posible obtener estructuras de madurez gonadal de la población más claras, con las que se pueden estimar proporciones de estadios de madurez por tallas, que son usadas para construir las ojivas de madurez.
Las características microscópicas de los ovarios denotan la tipificación de desove parcial que ya fue observado por Dio-ses (1988) y confirmado por Perea et al. (2003). Este hecho también ha sido observado en otras especies de jurel como Trachurus trachurus (Arruda 1984, Karlou-Riga & Economidis 1997). En nuestro trabajo, las características de los tipos de ovocitos de jurel T. murphyi observados también coinciden con lo reportado por Dioses (1998), así como para otras especies del mismo género (Karlou-Riga & Economidis 1997).
Cabe mencionar que para muchas especies no es posible distinguir a simple vista, las hembras inmaduras de las que se encuentran en “reposo” (Trippel & Morgan 1997, Saborido-Rey & Junquera 1998, Domínguez-Petit 2007). Sin embargo, las gónadas del jurel T. murphyi tienen un mayor volumen, y por ello es mucho más fácil discriminar ambos estadios sin la necesidad de hacer cortes histológicos como lo sugieren Fe-rreri et al. (2009), que elaboraron la escala de madurez de la anchoveta Engraulis encrasicolus y recomendaron realizar cortes histológicos de las gónadas de los individuos durante los picos de desove, para poder diferenciar las hembras virginales de las que ya habían tenido desoves, y que se encontraban inactivas. Esta recomendación se realizaba para evitar que se confundan y clasifiquen erróneamente los estadios virginal e inactivo, lo que introduciría errores al estimar la proporción de individuos maduros de la población, sobre todo si parte de las hembras en reposo que ya habían contribuido a la biomasa reproductora son consideradas inmaduras.
El estadio en recuperación puede ser considerado como un estadio “confuso” por algunos autores (Hunter & Macewicz 2003), pero en el caso del jurel T. murphyi este estadio es reconocible a simple vista por el color, pérdida de turgencia y disminución del volumen del ovario. Por otro lado, al igual que lo observado en los ovarios de merluza, debido a su dimensión, los ovocitos atrésicos de jurel T. murphyi se reconocen a simple vista al hacer un corte transversal del ovario, observándose como grumos que van desde el color amarillo claro hasta el rojizo. Se destaca que los niveles de atresia observados en jurel T. murphyi, al igual que en T. trachurus (Gordo et al. 2008) y en merluza Merluccius gayi peruanus, son mayores que los niveles de atresia observados en ovarios de anchoveta Engraulis ringens.
En conclusión, en el presente trabajo han sido descritas las fases de madurez gonadal del jurel T. murphyi basadas en ca-racterísticas que pueden diferenciarse a simple vista y validadas con exámenes histológicos. Esta escala permitirá la estimación rápida de la fase de madurez de la población. Por otro lado, es recomendable que el uso de esta escala sea intercalibrada entre los usuarios de diferentes zonas, así como otros países, para estandarizar su uso, lo que conllevaría a disminuir la subjeti-
vidad en la catalogación y el sesgo en los resultados (Gerritsen & McGrath 2006, Ferreri et al.2009).
Literatura citada Abascal F., C. Megina & A. Medina. 2004. Testicular development in migrant
and spawning bluefin tuna Thunnus thynnus (L.) from the eastern Atlantic and Mediterraneam. Fish. Bull. 102: 407-417.
Andrianov D.D. 1987. Sobre la biología reproductiva del jurel peruano. En: Bio-logía y pesca del jurel peruano. VNIRO, Moscú: 20-34. (En ruso).
Aracena O., R. Alarcón, S. Collado, I. Lépez, & D. Arriagada. 1998. Aspectos reproductivos del jurel (Trachurus symmetricus murphyi) de la pesquería de cerco de la Octava Región, Chile, 1994-1995. In: Arcos, D. (ed.), Biología y ecología del jurel en aguas chilenas, Instituto de Investigación Pesquera, Talcahuano, Chile, p. 101-114.
Arruda L.M. 1984. Sexual maturation and growth of Trachurus trachurus (L.) along the Portuguese coast. Inv. Pesq. 48(3): 419-430.
Costa A.M. 2001. Contribuição para o estudo da fecundidade temporal e espa-cial do carapau (Trachurus trachurus L.) na costa Portuguesa. Tese para acesso à categoria de Investigador Auxiliar – IPIMAR, 180 pp.
Costa A.M. & M.F. Borges. 1996. Total fecundity and atresia of horse mackerel (Trachurus trachurus L.). ICES, C. M.1996/ H: 26.
Cubillos L. (Ed.) 2003. Informe final, proyecto FIP 2001-12, condición bio-lógica del jurel en alta mar, año 2001. Fondo de Investigación Pes-quera - Instituto de Investigación Pesquera, Chile, Octava Región: 212 pp. <http://www2.udec.cl/~coyarzun/inffinalFIP2001-12.pdf > [acceso 16/08/2013]
Dadzie S. & F. Abou-Seedo. 2004. Testicular structure and spawning cycle in the silvery croaker, Otolithes ruber (Perciformes: Scianidae) in the Kuwaiti waters of the Arabian Gulf. Ichthyol. Res. 51:263-268 .
Dioses T., V. Alarcón, M.H. Nakama & A. Echeverría. 1989. Desarrollo ovo-citario, fecundidad parcial y distribución vertical de los cardúmenes en desove del jurel Trachurus murphyi (N). En: R. Jordán, R. Kelly, O. Mora, A. de Vildoso & N. Henríquez (eds.). Memorias del Sim-posio internacional sobre recursos vivos y pesquerías en el Pacífico sudeste. Rev. Com. Perm. Pacífico Sur. Número especial: 287-294.
Domínguez-Petit R. 2007. Study of Reproductive Potential of Merluccius Merluccius in the Galician Shelf. Doctoral Thesis. University of Vigo, Vigo, Spain: 253 pp.
Ferreri R., G. Basilone, M. D’Elia, A. Traina, F. Saborido-Rey & S. Mazzola. 2009. Validation of macroscopic maturity stages according to mi-croscopic histological examination for european anchovy. Marine Ecology 30: 181–187. doi:10.1111/j.1439-0485.2009.00312.x.
Gerritsen H.D. & D. McGrath. 2006. Variability in the assignment of maturity stages of plaice (Pleuronectes platessa L.) and whiting (Merlangius merlangus L.) using macroscopic maturity criteria. Fisheries Research 77:72-77.
Gonçalves P., E. Cunha & A. Costa. 2004. Escala de maturação microscópica e macroscópica das gónadas femininas de pescada (Merluccius merluccius). Relat. Cient. Téc. IPIMAR, Série digital (http://ipimariniap.ipimar.pt) 21. 27 pp.
Gordo L.S., A. Costa, P. Abaunza et al.2008. Determinate versus indeterminate fecundity in horse mackerel. Fisheries Research 89:181-185.
Gluyas-Millán M.G. 1994. Reproducción, edad y talla de primera madurez de la macarela Scomber japonicus (Houttuyn 1872) en Bahía Vizcaíno. Ciencias Marinas 20(3): 409-419.
Holden M.J. & D.F.S. Raitt. 1975. Manual de ciencia pesquera. Parte 2: Mé-todos para investigar los recursos y su aplicación. Doc. Téc. FAO Pesca (115). Rev 1: 211 p.
Humason G.L. 1979. Animal tissue techniques. Fourth edition. W. H. Freeman Co. San Francisco. 661 pp.
Hunter J. & S. Goldberg. 1980. Spawning incidence and batch fecundity in northern anchovy, Engraulis mordax. Fish. Bull. 77: 641-652.
Hunter J.R. & B.J. Macewicz. 1985. Measurement of spawning frequency in multiple spawning fish. In R. Lasker (Ed.). An Egg Production Method for Estimating Spawning Biomass of Pelagic Fish: Applica-tion to the Northern Anchovy, Engraulis mordax. NOAA Technical Report NMFS 36. Pp. 79-94
Hunter J.R. & B.J. Macewicz. 2003. Improving the accuracy and precision of reproductive information used in fisheries. In: O.S. Kjesbu , J.R. Hunter and P. Witthames (Eds.), Report of the Working Group on Modern Approaches to Assess Maturity and Fecundity of Warm- and Coldwater Fish and Squids. Fisken og 12.

44
Sánchez et al.
Rev. peru. biol. número especial 20(1): 035 - 044 (Septiembre 2013)Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú.
Editores: Csirke J., R. Guevara-Carrasco & M. Espino
Johansen A.C. 1924. On the summer and autumn spawning herring of the North Sea. Meddelelser fra Kommissionen for Havundersøgelser. Serie Fiskeri, Bind VII, Nr.5, 119 pp., 15 figs. København.
Karlou-Riga C. & P.S. Economidis. 1997. Spawning frequency and batch fe-cundity of horse mackerel, Trachurus trachurus (L.), in the Saronikos Gulf (Greece). J. Appl. Ichthyol. 13:97-104.
Kjesbu O., R. Hunter & P. Witthames (Ed.). 2003. Plenary document. In: Report of the working group on modern approaches to assess maturity and fecundity of warm- and cold-water fish and squids. Fisken og havet 2003-12. http://brage.bibsys.no/imr/handle/URN:NBN:nobibsys_brage_3594. [acceso: 18/07/2013]
Pérez N. & I. Figueiredo. 1992. First approach to the study of atresia in the ovary of sardine Sardine pilchardus (Walb.). Bol. Inst. Esp. Ocea-nogr. 8(1):191-199.
Perea A., J. Mori, B. Buitrón & J. Sánchez. 2013. Aspectos reproductivos del jurel Trachurus murphyi (Nichols 1920) en el Perú. En: Csirke J., R. Guevara-Carrasco, M. Espino (Eds.). Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú. Rev. peru. biol. número especial 20(1): 029 - 034
Saborido-Rey F. & S. Junquera. 1998. Histological assessment of variations in sexual maturity of cod (Gadus morhua L.) at the Flemish Cap (north-west Atlantic). ICES Journal of Marine Science 55:515-521.
Trippel E.A. & M.J. Morgan. 1997. Skewed sex ratios in spawning shoals of Atlantic cod (Gadus morhua). Oceanographic Literature Review 44, 511 pp.

45
Edad y crecimiento del jurel Trachurus murphyi en el Perú
Rev. peru. biol. special issue 20(1): 045- 052 (September 2013)Ecology, fisheries and conservation of Jack mackerel (Trachurus murphyi) in Peru. Editors: Csirke J., R. Guevara-Carrasco & M. Espino
Rev. peru. biol. número especial 20(1): 045- 052 (Septiembre 2013)Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú. Editores: Csirke J., R. Guevara-Carrasco & M. Espino
Facultad de ciencias Biológicas unMsMISSN-L 1561-0837
Edad y crecimiento del jurel Trachurus murphyi en el Perú
Teobaldo Dioses
Age and growth of Jack mackerel Trachurus murphyi in Peru
Instituto del Mar del Perú, esquina Gamarra y General Valle S/N Chucuito, Callao, Perú.
Email Teobaldo Dioses: [email protected]
biología del jurel (Trachurus murphyi) en el Perú
© Los autores. Publicado por la Revista Peruana de Biología de la Facultad de Ciencias Biológicas, Universidad Nacional Mayor de San Marcos. Este es un artículo de acceso abierto, distribuido bajo los términos de la Licencia de Atribución Reconocimiento-NoComercial-SinObraDerivada 3.0 de Creative Commons (http://creativecommons.org/licenses/by-nc-nd/3.0/deed.es_ES), que permite el uso no comercial, distribución y reproducción en cualquier medio, siempre que la obra original sea debidamente citadas. Para uso comercial, por favor póngase en contacto con [email protected].
Citación:Dioses T. Edad y crecimiento del jurel Trachurus murphyi en el Perú. En: Csirke J., R. Guevara-Carrasco & M. Espino (Eds.). Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú. Rev. peru. biol. número especial 20(1): 045- 052 (Septiembre 2013)
ResumenSe determinó la edad y se estimó el crecimiento del jurel Trachurus murphyi Nichols 1920, mediante la lectura y medición de anillos anuales de crecimiento en 977 pares de otolitos de especímenes entre 3 y 71 cm de longitud total colectados dentro del dominio marítimo peruano durante el año 1978 y parte de 1977 y 1979. Se comprobó la validez de las marcas anuales en 50 otolitos con 5 marcas o zonas anuales, y también con el análisis del incremento marginal mensual en 240 otolitos de especímenes entre 35 y 39 cm obtenidos entre julio 1977 y junio 1979. A partir de las medidas de los anillos anuales y sus respectivas tallas retrocalculadas se estimaron los parámetros de crecimiento en longitud y peso de la ecuación de von Bertalanffy: longitud infinita (L∞) =80.77 cm, peso infinito (W∞) = 3744.10 g, coeficiente de crecimiento (k) = 0.155 y edad teórica a la longitud 0 (t0) = –0.356. Se discuten las diferencias y semejanzas con las curvas y parámetros de crecimiento calculados por otros autores, haciendo énfasis en aquéllos cuyos resultados contribuyen a validar los estimados en el presente trabajo. Según los parámetros estimados, el T. murphyi peruano tendría una velocidad de crecimiento mayor que el T. murphyi de más al sur.
Palabras clave: Edad, crecimiento, otolitos, jurel, Trachurus murphyi.
AbstractThe age and growth of Jack mackerel Trachurus murphyi Nichols 1920 was determined by the reading and measuring annual growth rings from 977 pairs of otoliths from specimens between 3 and 71 cm total length collected within the Peruvian maritime domain during 1978 and part of 1977 and 1979. The validity of the annual marks was confirmed with 50 otolith with 5 marks or annual zones, and also with the analysis of the monthly marginal increment in 240 otoliths of specimens between 35 and 39 cm obtained from July 1977 to June 1979. From the annual rings measurements and their respective retroestimated sizes, the following parameters of the von Bertalanfy ecuation for growth in length and weight were estimated: infinite length (L∞) = 80.77 cm, infinite weight (W∞) = 3744.10 g, growth coefficient (k) = 0.155, and theoretical age at length 0 (t0) = –0.356. The estimated parameters suggest that the Peruvian T. murphyi has a faster growth rate than the more southern T. murphyi.
Keywords: Age, growth, otoliths, Jack mackerel, Trachurus murphyi.
IntroducciónEl jurel Trachurus murphyi Nichols 1920 es una de las especies pelágicas comerciales
más importantes del Pacífico Sudoriental. En Chile alcanzó una captura máxima de más de 4 millones de toneladas en 1994 y en Perú de alrededor de 800 mil toneladas en 2001. Para analizar la dinámica poblacional y evaluar los cambios en la abundancia de este importante recurso pesquero, es de gran utilidad el poder contar con parámetros confiables de edad y crecimiento de la especie a fin de evaluar el estado del stock en el Perú.
Entre los métodos comúnmente aceptados y más empleados en la determinación de la edad y estimación del crecimiento individual, figuran aquellos basados en la lec-tura de los anillos de crecimiento en otolitos, aplicando diversas metodologías según la especie. En el caso del T. murphyi, en el Perú se han aplicado la lectura tradicional (lectura de anillos de crecimiento en la cara externa de los otolitos enteros), lectura de secciones transversales de otolitos y lectura de anillos diarios. En este trabajo se presen-tan los resultados de las determinaciones de su edad y crecimiento mediante la lectura tradicional y se comparan con los obtenidos por otros autores con métodos similares.
Publicado online: 11/10/2013Publicado impreso: 15/10/2013

46
Dioses
Rev. peru. biol. número especial 20(1): 045- 052 (Septiembre 2013)Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú.
Editores: Csirke J., R. Guevara-Carrasco & M. Espino
Material y métodos El material y los respectivos datos analizados en este estudio
fueron tomados de los muestreos biológicos efectuados tanto en el laboratorio central del Callao como en los laboratorios costeros de Paita, Chimbote, Pisco e Ilo del Instituto del Mar del Perú (IMARPE). Se trabajó con muestras del año 1978, complementándose para las longitudes extremas con muestras de los años 1977 y 1979. Los muestreos biológicos que realiza el IMARPE corresponden a muestreos sistemáticos estratifi-cados. En el caso de T. murphyi se analizaron mensualmente 10 ejemplares en detalle, por cada centímetro de longitud. De estos ejemplares se obtuvo la siguiente información: longitud total, peso total, longitud y peso de las gónadas, sexo y grado de madurez sexual, contenido de grasa, contenido estomacal. Además se colectan otolitos para la determinación de la edad y gónadas para el seguimiento histológico del desarrollo ovocitario.
En la determinación de la edad de T. murphyi se utilizó el otolito sacular o sagitta. Este otolito cambia gradualmente su apariencia y se observa que toma una forma regular después que ha completado su primer año de vida. Para la lectura de anillos se utilizaron muestras de otolitos provenientes de todo el litoral peruano, tratándose de cubrir el mayor rango posible de tallas presentes en la pesquería. En total se hizo la lectura tradicional en 977 otolitos de otros tantos especímenes machos y hembras de T. murphyi. Para cada otolito se tuvo la medida de longitud total del ejemplar y se agruparon las muestras en rangos de longitud con intervalos de un centímetro. Se contó con ejemplares desde los 3 hasta los 71 cm de longitud total y, en lo posible, se tomó 15 especímenes por intervalo de longitud. Además, en el análisis del borde de los otolitos y en la determinación del incremento marginal se utilizaron 240 otolitos; mensualmente se tomaron 10 otolitos de especímenes de 35 a 39 cm de longitud, desde julio de 1977 a junio de 1979.
La metodología de lectura de los anillos de crecimiento es la que utilizaron Misu y Hamasaki (1971) en la merluza pe-ruana (Merluccius gayi peruanus), Samamé (1977) en la sardina (Sardinops sagax), Geldenhuys (1973) en el jurel de Sudáfrica (Trachurus trachurus) y Dioses (1985) en la merluza M. gayi pe-ruanus. Para visualizar los anillos se probaron diferentes técnicas. La que dio mejores resultados fue la de quemar ligeramente la cara interna de los otolitos con un mechero de alcohol, hasta que tomen un color marrón claro. Esto facilitó la observación de las zonas hialinas oscuras así como las opacas un poco blanquecinas.
Los otolitos fueron leídos en una cámara oscura utilizando un proyector de contornos Nikon 6CT-2 a 10 aumentos, utilizando luz reflejada. Las mediciones se efectuaron en milímetros, del núcleo hacia el borde posterior de cada anillo hialino (rn), así como del núcleo hacia el borde posterior o radio del otolito (R), como se puede observar en la Figura 1A. Para ver con mayor nitidez los anillos, el otolito fue colocado en una placa Petri de fondo negro y pincelado con aceite de cedro. Para las lecturas, generalmente se utilizó el otolito derecho, empleándose el iz-quierdo cuando el primero se encontraba en malas condiciones.
El incremento marginal fue calculado mediante la ecuación:
IM=(R–rn)/(rn–rn–1)
Donde :
R = radio del otolito
rn = radio del último anillo formado
Mediante el método del retrocálculo a edades pretéritas se de-terminó la longitud que tendría el pez al completar la formación de cada anillo hialino anual. Con este fin se utilizó la ecuación R = a’ L2 + b’ L + c’ que relaciona la longitud (L) del pez con la medida del radio (R) del otolito.
Los parámetros de longitud infinita (L∞) y del coeficiente de crecimiento se estimaron por el método gráfico de Ford-Walford (Ford 1933, Walford 1946), para lo cual se plotearon las longi-tudes medias (L) del T. murphyi a la edad t contra la longitud media a la edad t+1, de ello se obtuvo una recta interpolada que corta la bisectriz en un punto correspondiente a la estimación de L∞. La recta ajustada mediante el método de mínimos cuadrados corresponde a la ecuación de Ford- Walford:
Lt+1=a + bLt
donde: a = L∞(1 – e-k)
b = e-k
y de donde: L∞= a/(1–b)
k=–Lnb
La edad teórica (t0) en la cual el T. murphyi tendría longitud cero se estimó mediante la ecuación de Gulland (1977), que es una derivación de la expresión de crecimiento de von Bertalanffy, según la ecuación:
t0= t + (1/k)*Ln((L∞– Lt)/L∞)
De donde el t0 final se calcula como el promedio de los t0 obtenidos para las diferentes edades observadas.
ResultadosDeterminación de la edad
Descripción y medidas de los otolitos.- Esta descripción se hace tomando el otolito de un espécimen de cinco años (Fig. 1a y b). El otolito tiene una forma un poco lanceolada, mode-radamente redondeada en el borde posterior o cauda, mientras que el borde anterior o rostro es alargado y puntiagudo. En el borde dorsal, en dirección al rostro se encuentra el antirrostro, un poco corto y terminado en punta, por encima de una pequeña hendidura. La parte que va del antirrostro hacia el borde ventral es ligeramente lisa y curvada. La cara externa es cóncava, mientras que la interna es convexa y en ella se encuentra el surco acústico, canal que corre en la parte media horizontal y en sentido antero-posterior, terminando en un punto ciego cerca de la cauda.
Figura 1. Otolito derecho: a) Cara externa, b) Cara internaFigura 1. Right otolith: a) External surface, b) Internal surface
(a)
(b)

47
Edad y crecimiento del jurel Trachurus murphyi en el Perú
Rev. peru. biol. special issue 20(1): 045- 052 (September 2013)Ecology, fisheries and conservation of Jack mackerel (Trachurus murphyi) in Peru. Editors: Csirke J., R. Guevara-Carrasco & M. Espino
En la cara externa del otolito, usando luz reflejada, se ob-serva una disposición concéntrica de anillos o zonas opacas (blanquecinas) y hialinas (traslúcidas) que se forman alrededor de un núcleo central o foco. Para la interpretación de la edad se utilizaron los anillos hialinos, por ser éstos más nítidos, con-siderándose en estos estudios a un anillo opaco más un anillo hialino como una zona anual.
Se observa que durante la formación del primer anillo anual hialino (Fig. 2) se desarrolla una mayor continuidad en el en-torno del otolito a medida que alcanza un determinado grosor. Un criterio para determinar los siguientes anillos anuales y ad-judicarlos como tales fue que éstos debían tener continuidad en la mayor parte del otolito, además de tener un grosor similar o equivalente al grosor observado en el primer anillo anual (Figs. 3, 4, 5 y 6). Por ejemplo, en la Figura 3, entre el primer y segundo anillo anual se puede observar la presencia de dos anillos que se descartaron al ser considerados como anillos falsos, por no satisfacer los criterios indicados.
Se comprobó la validez de esta metodología a través de la comparación con dos aspectos del crecimiento de los otolitos como son la similitud de crecimiento entre la medida de las marcas (cuyo crecimiento decrece con la formación de una nueva marca) y la variación mensual del incremento marginal.
Similitud entre las medidas de las marcas de los otolitos.- Se usaron 50 otolitos con 5 marcas o zonas anuales (anillos hiali-nos) y se relacionó la medida de cada marca (rn) con la longitud o radio (R) del otolito. En la Figura 7 se pueden apreciar las líneas de regresión para cada marca en relación a la longitud del otolito. Se hace notar la evidente similitud de formación entre las marcas de este grupo de otolitos, cuyo crecimiento decrece con la formación de una nueva marca.
Variación del incremento marginal.- Para analizar la pe-riodicidad de variación de los anillos anuales de los otolitos se midió el incremento marginal de especímenes cuyas longitudes oscilaron entre 35 y 39 cm, para cada mes entre julio de 1977 y junio de 1979. En la Figura 8 se puede apreciar que a partir del mes de junio hasta setiembre de 1978 (periodo invernal) se
Figura 2. Otolitos mostrando la secuencia de formación del primer anillo anual en jureles T. murphyi de 3 a 17 cm de longitud total (LT): a) 3 cm, b) 4 cm, c) 5 cm, d) 6 cm, e) 7 cm, f) 8 cm, g) 9 cm, h) 13 cm, i) 14 cm, j) 15 cm y k) 17 cm.Figura 2. Otoliths showing the evolution in the formation of the first annual ring in Jack mackerels T. murphyi from 3 to 17 total length (TL): a) 3 cm, b) 4 cm, c) 5 cm, d) 6 cm, e) 7 cm, f) 8 cm, g) 9 cm, h) 13 cm, i) 14 cm, j) 15 cm and k) 17 cm.
Figura 3. Otolito de edad 3+ años de un jurel T. murphyi de 33 cm LT.Figure 3. Age 3+ years otolith of Jack mackerel T.murphyi of 33 cm TL
Figura 4. Otolito de edad 4+ años de un jurel T. murphyi de 42 cm LT.Figure 4. Age 4+ years otolith of Jack mackerel T. murphyi of 42 cm TL.
Figura 5. Otolito de edad 5+ años de un jurel T. murphyi de 54 cm LT.Figure 5. Age 5+ years otolith of Jack mackerel T. murphyi of 54 cm TL.
Figura 6. Otolito de edad 7+ de un jurel T. murphyi de 63 cm LT.Figure 6. Age 7+ years otolith of Jack mackerel T. murphyi of 63 cm TL.
6 mm
3
0
a b c d
e f g
h i j
k
0 3 6mm
presentan los valores mínimos, lo que estaría indicando el final de la formación de un anillo y el inicio de la formación de un nuevo anillo hialino. De esta observación podemos concluir que la formación de una nueva zona de crecimiento se inicia en los meses de primavera, lo que se evidencia por el aumento del incremento marginal. Igualmente, para el año 1977 los menores valores se observan en los meses de invierno, especialmente en agosto y setiembre. Para el año 1979, en el mes de junio ya se observó cierta tendencia a la disminución de este incremento lue-go de haber llegado al máximo valor en abril de ese mismo año.

48
Dioses
Rev. peru. biol. número especial 20(1): 045- 052 (Septiembre 2013)Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú.
Editores: Csirke J., R. Guevara-Carrasco & M. Espino
Fluctuación de las zonas hialinas y opacas en el borde de los otolitos.- Se realizó un análisis de la formación periódica de los bordes del otolito a través de la observación de la distribución de material hialino y opaco acumulado mensualmente en el borde de los otolitos entre julio 1977 y junio 1979. Se determinó que el aumento de la frecuencia de bordes opacos se produjo durante el verano, que coincide con el periodo de mayor crecimiento del pez y que es seguido por el aumento de la frecuencia de bordes hialinos durante el invierno, cuando el crecimiento del pez es menor. Estas fluctuaciones confirman lo observado en el incremento marginal. El mayor porcentaje de bordes hialinos se presenta en los meses de invierno, en agosto del año 1977 y en los meses de julio y setiembre en 1978. El porcentaje de bordes hialinos disminuye a medida que los bordes opacos aumentan. Estos bordes opacos alcanzan su máxima frecuencia en los meses de febrero, marzo y abril en el año 1977 y en enero, febrero y marzo en el año 1979.
De lo observado se deduce que existe una formación cíclica de los bordes hialinos y opacos. Esta periodicidad corresponde
a la formación de un anillo anual durante los meses de mayor intensidad del desove de esta especie, lo que nos permite sugerir el 1° de enero como fecha de nacimiento arbitraria biológica, para la elaboración de las claves edad-longitud.
Determinación del crecimiento
Relación entre la longitud del pez y el radio total (R) del oto-lito.- La medida promedio de la distancia núcleo-borde posterior de los 977 otolitos (R) se relacionó con las respectivas longitudes totales (L) de los peces, según los valores de la Tabla 1. Al correlacionar estas variables, la regresión que mejor se ajustó a las observaciones es la de tipo cuadrática, que responde a la expresión:
R=-0.0008798 L2+0.141083 L+0.283965
El ajuste y datos observados están representados gráficamente en la Figura 9.
Cálculo de los parámetros de crecimiento.- En el material observado se encontró otolitos hasta con 11 marcas. En la Tabla 2 se dan las medidas promedios de los radios de los otolitos por cada grupo de marca. Las longitudes promedio del pez a cada edad se determinaron por el método del retro-cálculo. La Tabla 3 presenta las longitudes resultantes de la sustitución de rn, por R de la ecuación anterior. Las longitudes promedio (Ln) obtenidas y presentadas en la Tabla 3 para cada marca anual serán utilizadas más adelante en el cálculo de la longitud asintótica (L∞) y el parámetro de crecimiento k..
Ploteo de Ford-Walford.- El ploteo de Ford-Walford se puede observar en la Figura 10 y la ecuación resultante de este ploteo es la siguiente:
L (t+1)=11.6208+0.8561Lt
Figura 7. Líneas de regresión entre la medida total o radio del otolito (R) y la medida de los radios de las marcas anuales (rn) para especímenes con 5 anillos.Figura 7. Regression lines of the relationship between otoliths radius (R) and the radius of annual rings (rn) for individuals with 5 rings.
Figura 8. Variación mensual media del incremento marginal de los otolitos de jurel T. murphyi periodo julio 1977 - junio 1979.Figure 8. Monthly variation of the marginal increment of Jack mackerel T. murphyi otoliths, for the period July 1977 – June 1979.
Long. total (cm)
f R (mm)
Long total (cm)
f R (mm)
Long. total (cm)
f R (mm)
3 3 0.53 26 15 3.45 49 15 5.014 15 0.71 27 15 3.47 50 15 4.985 15 0.82 28 15 3.55 51 15 5.086 15 0.97 29 15 3.61 52 15 5.147 15 1.11 30 15 3.74 53 15 5.208 15 1.22 31 15 3.93 54 15 5.269 15 1.38 32 15 3.98 55 15 5.2810 15 1.58 33 15 4.07 56 15 5.3811 15 1.68 34 15 4.14 57 15 5.3812 15 1.81 35 15 4.15 58 15 5.4713 15 1.97 36 15 4.28 59 15 5.5214 15 2.16 37 15 4.26 60 15 5.5215 15 2.32 38 15 4.43 61 15 5.5916 15 2.42 39 15 4.45 62 15 5.7517 15 2.57 40 15 4.51 63 15 5.7218 15 2.72 41 15 4.49 64 15 5.7319 15 2.82 42 15 4.58 65 15 5.9520 15 2.85 43 15 4.63 66 15 5.9421 15 2.97 44 15 4.68 67 15 5.9822 15 3.04 45 15 4.72 68 7 5.9623 14 3.19 46 15 4.78 69 8 6.0224 10 3.25 47 15 4.90 70 3 6.0325 15 3.36 48 15 4.96 71 2 6.10
Tabla 1. Valores de longitud total, radio del otolito (R) y frecuencias (f) utilizados en la determinación de la edad del jurel T. murphyi.Table 1. Total length, otolith radius (R) and frequency (f) of specimens used to determine the age of Jack mackerel T. murphyi.
Long
itud
de m
arca
(mm
)
Longitud de otolito (mm)
5.00
4.00
3.00
2.00
1.00
4.00 4.50 5.00 5.50 R
rn
Incr
emen
to m
argi
nal (
mm
)
Años (meses)
0.6
0.5
0.4
0.3
0.2
0.1
0
1977 1978 1979
Jul
Ago Set
Oct
Nov Dic
Ene Feb
Mar
Abr
May Jun
Jul
Ago Set
Oct
Nov Dic
Ene Feb
Mar
Abr
May Jun

49
Edad y crecimiento del jurel Trachurus murphyi en el Perú
Rev. peru. biol. special issue 20(1): 045- 052 (September 2013)Ecology, fisheries and conservation of Jack mackerel (Trachurus murphyi) in Peru. Editors: Csirke J., R. Guevara-Carrasco & M. Espino
Parámetros de crecimiento en longitud.- Como resultado de estos análisis y el ajuste de las ecuaciones indicadas se esti-maron los siguientes parámetros de crecimiento para el jurel: L∞=80.77 cm, k= 0.155 y to= –0.356. La curva de crecimiento teórico resultante (línea continua) y los valores observados de longitud media a la edad (puntos) aparecen representados gráfi-camente en la Figura 11, donde es de destacar la correspondencia entre los puntos de las longitudes promedio observadas y los valores de la curva de crecimiento teórico calculada.
Parámetros de crecimiento en peso.- Primero se determinó la relación longitud-peso del T. murphyi con datos de 1654 es-pecímenes, cuyas longitudes variaron entre 3.0 y 71.0 cm, que incluyeron 27 ejemplares por rango de longitud (Tabla 4). Con estos valores se ajustó con un r = 0,9984 la ecuación longitud-peso W = 0.0137Lt
2.8 que se presenta gráficamente en la Figura 12 junto a los valores observados. En esta ecuación se destaca el exponente (2.85) que al ser diferente de 3 está indicando que el T. murphyi tiene un crecimiento alométrico. Según estos estimados, la ecuación de crecimiento en peso según el modelo de von Bertalanffy fue la siguiente:
Figura 9. Regresión cuadrática entre la longitud total y el radio del otolito de jurel T. murphyi.Figure 9. Quadratic regression between total length and otolith radius of Jack mackerel T. murphyi.
Grupo/Edad F r1 r2 r3 r4 r5 r6 r7 r8 r9 r10 r11
0 155 1 123 2.22 119 2.18 3.143 143 2.16 3.13 3.964 127 2.16 3.12 3.88 4.435 111 2.2 3.15 3.95 4.44 4.96 70 2.18 3.16 3.94 4.45 4.87 5.167 55 2.17 3.14 3.95 4.42 4.88 5.14 5.448 38 2.2 3.15 3.93 4.43 4.87 5.15 5.4 5.579 21 2.18 3.15 3.94 4.42 4.86 5.14 5.42 5.55 5.6710 8 2.18 3.13 3.95 4.44 4.85 5.16 5.44 5.54 5.65 5.7311 7 2.2 3.14 3.92 4.45 4.9 5.14 5.4 5.56 5.65 5.74 5.78
Frecuencia total 977 822 699 580 437 310 199 129 74 36 15 7Media de los radios (r) 2.18 3.14 3.94 4.44 4.88 5.15 5.42 5.56 5.66 5.74 5.78
Tabla 2. Frecuencia (f) y medidas promedio de los radios (r, en mm) de las marcas de crecimiento en otolitos de jurel T. murphyi.Table 2. Frequency (f) and mean radius (r, in mm) of the growth rings in Jack mackerel T. murphy otoliths.
Figura 10 . Ploteo de Ford – Walford.Figure 10. Ford-Walford Plot.
Long
itud
tota
l (cm
)
Edad (años)
80
70
60
50
40
30
20
10
105 15
L∞ = 80.77 cm
Lt = 80.77[1 – e–0.155347(t+0.356215)]
Figura 11. Crecimiento en longitud del jurel según von Bertalanffy.Figure 11. von Bertalanffy growth in length of Jack mackerel.
10
20
30
40
50
60
70
80
90
Lt+
1
10 20 30 40 50 60 70 80 90
L t
L∞ = 80.77 cm
Lt+1= 11.6208 + 0.8561 Lt
Rad
io o
tolit
o (m
m)
Longitud Total (cm)
7.00
6.00
5.00
4.00
3.00
2.00
1.00
0 10 20 30 40 50 7060
media
Rango de valoresobservados
R = -0.0008798L2 + 0.141083L + 0.283965

50
Dioses
Rev. peru. biol. número especial 20(1): 045- 052 (Septiembre 2013)Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú.
Editores: Csirke J., R. Guevara-Carrasco & M. Espino
Wt=3744.10[1 – e(–0.1553*(t + 0.3562))]2.84919
Discusión En los estudios de la edad y el crecimiento del jurel T. murphyi
en el Pacífico Suroriental se han venido aplicando tres técnicas de preparación e interpretación de los anillos de crecimiento presentes en los otolitos sagita. Dichas técnicas son las siguientes: (a) la lectura tradicional (rutinaria) de anillos anuales en otolitos enteros, (b) la lectura de anillos anuales en cortes transversales de los otolitos y (c) la lectura de microincrementos o anillos diarios.
Al analizar los resultados de la aplicación de estas técnicas en la lectura de otolitos de jurel T. murphyi en la región se observa que
Grupo/Edad f r1 r2 r3 r4 r5 r6 r7 r8 r9 r10 r111 123 14.99 2 119 14.81 23.773 143 14.64 23.67 32.744 127 14.64 23.57 31.79 38.755 111 14.99 23.87 32.62 38.88 45.786 70 14.81 23.97 32.5 39.02 45.29 50.387 55 14.73 23.77 32.62 38.61 45.46 50 56.38 38 14.99 23.87 32.38 38.75 45.29 50.19 55.37 59.629 21 14.9 23.87 32.5 38.61 45.13 50 55.83 59.07 62.59
10 8 14.81 23.67 32.62 38.88 44.97 50.38 56.3 58.81 61.95 64.6411 7 14.99 23.77 32.26 39.02 45.78 50 55.37 59.34 61.95 65.01 66.58
Frecuencia total 822 822 699 580 437 310 199 129 74 36 15 7Longitudes medias 14.85 23.78 32.45 38.82 45.39 50.16 55.83 59.21 62.16 64.83 66.58
Tabla 3. Frecuencia (f) y longitudes medias (L) de jurel T. murphyi calculadas de las medias de los radios de los otolitos.Table 3. Frequency (f) and mean length (L) of Jack mackerel T. murpyi calculated from average otoliths radius.
Long. total (cm)
f W (g)Long. total (cm)
f W (g)Long. total (cm)
f W (g)
3 4 0.31 26 27 142.85 49 27 901.594 6 0.77 27 27 159.67 50 27 966.445 20 1.45 28 27 173.48 51 27 985.406 20 2.17 29 27 196.00 52 27 1065.597 21 3.52 30 27 225.33 53 27 1106.378 25 5.02 31 27 244.96 54 27 1161.859 27 7.07 32 27 251.37 55 27 1276.0010 27 10.27 33 27 283.00 56 27 1385.6211 27 13.14 34 27 311.60 57 27 1413.9312 27 16.97 35 27 332.85 58 27 1465.2213 27 21.37 36 27 348.11 59 27 1568.8114 27 26.67 37 27 377.93 60 27 1685.4415 27 34.60 38 27 398.44 61 27 1677.7416 27 40.66 39 27 432.15 62 27 1780.9617 27 49.27 40 27 479.96 63 27 1933.8518 27 59.29 41 27 514.89 64 27 2081.5019 23 57.17 42 27 585.48 65 27 2162.1620 27 66.22 43 27 589.52 66 27 2245.0421 27 79.81 44 27 662.59 67 27 2465.3822 14 93.21 45 27 688.78 68 9 2485.1123 9 104.67 46 27 743.44 69 12 2550.9224 10 107.50 47 27 821.93 70 4 2644.5025 27 125.92 48 27 855.07 71 2 2785.00
Tabla 4. Longitud total, peso (W) y frecuencias (f) de los especíme-nes de jurel T. murphyi utilizados en la determinación de la relación longitud–peso.Table 4. Total length, weight (W) and frequency (f) of the specimens of Jack mackerel T. murphyi used in the length-weigh relationship.
Figura 12. Relación longitud total – peso del jurel T. murphyi.Figure 12. Weight – length relationship for Jack mackerel T. murphyi.
existen diferencias en las determinaciones de la edad y en los estima-dos de los parámetros de crecimiento. Algunas de estas diferencias podrían deberse a las diversas técnicas de preparación y dificultades en la aplicación de criterios de interpretación y separación de los anillos anuales o a distinto instrumental óptico empleado. Sin embargo, es evidente que estas diferencias reflejan diversidad de velocidades de crecimiento expresadas en sus correspondientes tasas o coeficientes (k) y, asimismo, en las tallas máximas de crecimiento (L∞) del T. murphyi proveniente de zonas y latitudes distintas o de stocks o substocks más o menos independientes.
Para fines comparativos, en la Tabla 5 se presentan los pará-metros de crecimiento de T. murphyi proveniente de distintas zonas del Pacífico Sudoriental que han sido estimados por diversos autores siguiendo alguna de las tres técnicas señaladas anteriormente para la lectura de anillos en los otolitos. En la Figura 13 se representan gráficamente las longitudes teóricas calculadas para cada edad con estos parámetros. Tanto los pará-metros como las curvas teóricas de crecimiento que se presentan están expresados en longitud a la horquilla, y se puede observar claramente las diferencias entre los tres parámetros y las curvas de crecimiento teórico resultantes tanto entre autores, como entre zonas y técnicas de preparación y lectura.
Pes
o (g
)
Longitud Total (cm)
3000
2000
1000
0 10 20 30 40 50 7060
W = 0.013744Lt2.85
r = 0.9984 n = 1654
media
Rango de valoresobservados
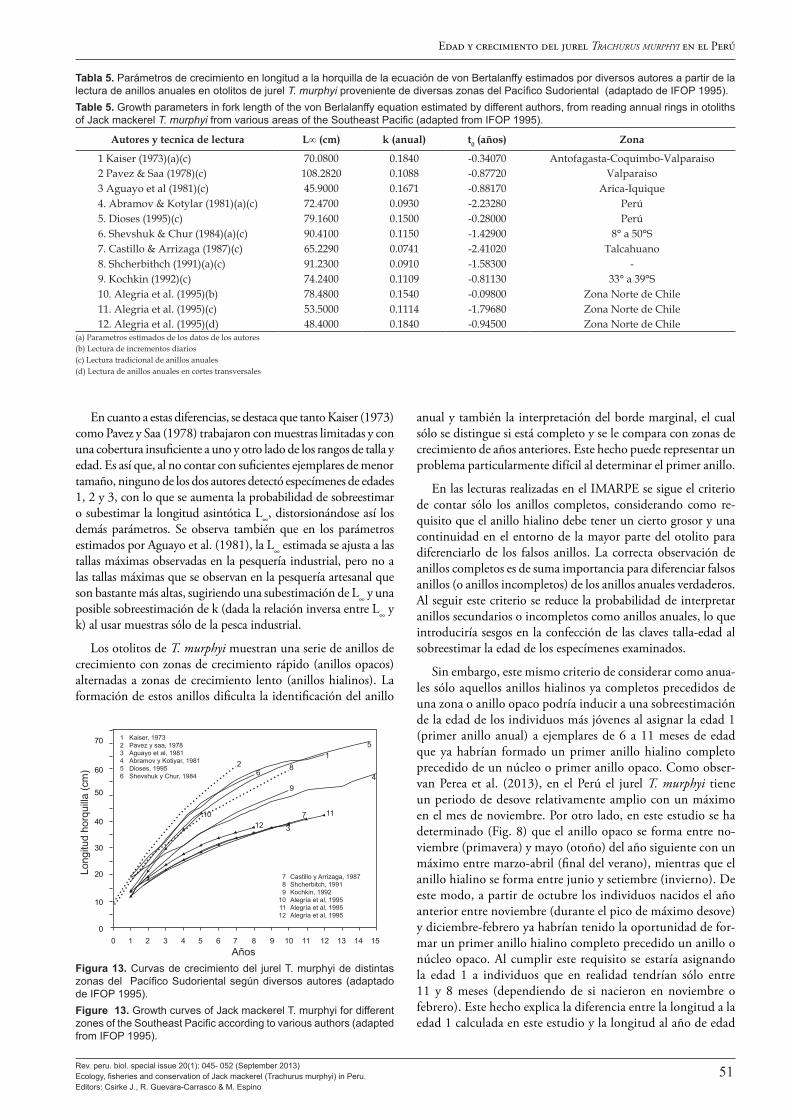
51
Edad y crecimiento del jurel Trachurus murphyi en el Perú
Rev. peru. biol. special issue 20(1): 045- 052 (September 2013)Ecology, fisheries and conservation of Jack mackerel (Trachurus murphyi) in Peru. Editors: Csirke J., R. Guevara-Carrasco & M. Espino
En cuanto a estas diferencias, se destaca que tanto Kaiser (1973) como Pavez y Saa (1978) trabajaron con muestras limitadas y con una cobertura insuficiente a uno y otro lado de los rangos de talla y edad. Es así que, al no contar con suficientes ejemplares de menor tamaño, ninguno de los dos autores detectó especímenes de edades 1, 2 y 3, con lo que se aumenta la probabilidad de sobreestimar o subestimar la longitud asintótica L∞, distorsionándose así los demás parámetros. Se observa también que en los parámetros estimados por Aguayo et al. (1981), la L∞ estimada se ajusta a las tallas máximas observadas en la pesquería industrial, pero no a las tallas máximas que se observan en la pesquería artesanal que son bastante más altas, sugiriendo una subestimación de L∞ y una posible sobreestimación de k (dada la relación inversa entre L∞ y k) al usar muestras sólo de la pesca industrial.
Los otolitos de T. murphyi muestran una serie de anillos de crecimiento con zonas de crecimiento rápido (anillos opacos) alternadas a zonas de crecimiento lento (anillos hialinos). La formación de estos anillos dificulta la identificación del anillo
anual y también la interpretación del borde marginal, el cual sólo se distingue si está completo y se le compara con zonas de crecimiento de años anteriores. Este hecho puede representar un problema particularmente difícil al determinar el primer anillo.
En las lecturas realizadas en el IMARPE se sigue el criterio de contar sólo los anillos completos, considerando como re-quisito que el anillo hialino debe tener un cierto grosor y una continuidad en el entorno de la mayor parte del otolito para diferenciarlo de los falsos anillos. La correcta observación de anillos completos es de suma importancia para diferenciar falsos anillos (o anillos incompletos) de los anillos anuales verdaderos. Al seguir este criterio se reduce la probabilidad de interpretar anillos secundarios o incompletos como anillos anuales, lo que introduciría sesgos en la confección de las claves talla-edad al sobreestimar la edad de los especímenes examinados.
Sin embargo, este mismo criterio de considerar como anua-les sólo aquellos anillos hialinos ya completos precedidos de una zona o anillo opaco podría inducir a una sobreestimación de la edad de los individuos más jóvenes al asignar la edad 1 (primer anillo anual) a ejemplares de 6 a 11 meses de edad que ya habrían formado un primer anillo hialino completo precedido de un núcleo o primer anillo opaco. Como obser-van Perea et al. (2013), en el Perú el jurel T. murphyi tiene un periodo de desove relativamente amplio con un máximo en el mes de noviembre. Por otro lado, en este estudio se ha determinado (Fig. 8) que el anillo opaco se forma entre no-viembre (primavera) y mayo (otoño) del año siguiente con un máximo entre marzo-abril (final del verano), mientras que el anillo hialino se forma entre junio y setiembre (invierno). De este modo, a partir de octubre los individuos nacidos el año anterior entre noviembre (durante el pico de máximo desove) y diciembre-febrero ya habrían tenido la oportunidad de for-mar un primer anillo hialino completo precedido un anillo o núcleo opaco. Al cumplir este requisito se estaría asignando la edad 1 a individuos que en realidad tendrían sólo entre 11 y 8 meses (dependiendo de si nacieron en noviembre o febrero). Este hecho explica la diferencia entre la longitud a la edad 1 calculada en este estudio y la longitud al año de edad
Autores y tecnica de lectura L∞ (cm) k (anual) t0 (años) Zona
1 Kaiser (1973)(a)(c) 70.0800 0.1840 -0.34070 Antofagasta-Coquimbo-Valparaiso2 Pavez & Saa (1978)(c) 108.2820 0.1088 -0.87720 Valparaiso3 Aguayo et al (1981)(c) 45.9000 0.1671 -0.88170 Arica-Iquique4. Abramov & Kotylar (1981)(a)(c) 72.4700 0.0930 -2.23280 Perú5. Dioses (1995)(c) 79.1600 0.1500 -0.28000 Perú6. Shevshuk & Chur (1984)(a)(c) 90.4100 0.1150 -1.42900 8° a 50°S7. Castillo & Arrizaga (1987)(c) 65.2290 0.0741 -2.41020 Talcahuano8. Shcherbithch (1991)(a)(c) 91.2300 0.0910 -1.58300 -9. Kochkin (1992)(c) 74.2400 0.1109 -0.81130 33° a 39°S10. Alegria et al. (1995)(b) 78.4800 0.1540 -0.09800 Zona Norte de Chile11. Alegria et al. (1995)(c) 53.5000 0.1114 -1.79680 Zona Norte de Chile12. Alegria et al. (1995)(d) 48.4000 0.1840 -0.94500 Zona Norte de Chile
(a) Parametros estimados de los datos de los autores(b) Lectura de incrementos diarios(c) Lectura tradicional de anillos anuales(d) Lectura de anillos anuales en cortes transversales
Tabla 5. Parámetros de crecimiento en longitud a la horquilla de la ecuación de von Bertalanffy estimados por diversos autores a partir de la lectura de anillos anuales en otolitos de jurel T. murphyi proveniente de diversas zonas del Pacífico Sudoriental (adaptado de IFOP 1995).Table 5. Growth parameters in fork length of the von Berlalanffy equation estimated by different authors, from reading annual rings in otoliths of Jack mackerel T. murphyi from various areas of the Southeast Pacific (adapted from IFOP 1995).
Long
itud
horq
uilla
(cm
)
Años0 1 2 3 4 5 76 8 9 10 11 12 13 1514
Kaiser, 1973Pavez y saa, 1978Aguayo et al, 1981Abramov y Kotiyar, 1981Dioses, 1995Shevshuk y Chur, 1984
123456
Castillo y Arrizaga, 1987Shcherbitch, 1991Kochkin, 1992Alegría et al, 1995Alegría et al, 1995Alegría et al, 1995
789
101112
12
3
4
5
6
7
8
9
10 1112
0
10
20
30
40
50
60
70
Figura 13. Curvas de crecimiento del jurel T. murphyi de distintas zonas del Pacífico Sudoriental según diversos autores (adaptado de IFOP 1995).Figure 13. Growth curves of Jack mackerel T. murphyi for different zones of the Southeast Pacific according to various authors (adapted from IFOP 1995).

52
Dioses
Rev. peru. biol. número especial 20(1): 045- 052 (Septiembre 2013)Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú.
Editores: Csirke J., R. Guevara-Carrasco & M. Espino
(365 días) calculada por análisis de incrementos diarios por Goicochea et al. (2013).
El método basado en el análisis de la formación de los in-crementos de crecimiento diarios permite validar los resultados obtenidos por el método tradicional de lectura de anillos anuales en los otolitos. Al respecto, Araya et al. (2003) demostraron que los microincrementos en los otolitos de T. murphyi corresponden a formaciones diarias.
En la Figura 13 se observa que las longitudes teóricas de la curva 10 de Alegría et al. (1995) calculadas en base a lectura de microincrementos para las edades 1 a 5 se aproximan a los valo-res de la curva 5, que corresponde a los resultados de la lectura directa de otolitos enteros de Dioses (1995) confirmados en este estudio. Estos resultados contribuirían a validar los parámetros estimados en el presente trabajo, por lo menos hasta la edad 5. Siempre con respecto a la Figura 13, es probable que en las lecturas y los cálculos de los parámetros de Aguayo et al. (1981) (curva 3), de Castillo y Arrizaga (1987) (curva 7), y de Alegría et al. (1985) por lectura rutinaria (curva 11) y por corte (curva 12), se estarían sobreestimando las edades. Como se puede apreciar, en las curvas 3, 7, 11 y 12 las longitudes teóricas corresponden a edades que son casi el doble que las mismas longitudes en las curvas 10 de Alegría et al. (1995), por incrementos diarios, y 5 de Dioses (1995), por lectura tradicional.
En Perú, Goicochea et al. (2013) han estudiado el periodo de formación del primer anillo de crecimiento en los otolitos, aplican-do la misma técnica de análisis de microincrementos o lectura de anillos diarios aplicada por Alegría et al. (1995) para el T. murphyi de la zona norte de Chile. Con las lecturas de esta técnica y el ajuste del modelo de crecimiento de Laird-Gompertz se podría suponer que un individuo de un año de edad (365 días) tendría 195,3 mm de longitud total (17,7 cm de longitud a la horquilla).
Otro método que se emplea para validar las edades y las tasas de crecimiento estimadas por lecturas de otolitos es el que se basa en el seguimiento de una clase anual a través del tiempo mediante la progresión modal de frecuencia de tallas. Aplicando este método, Goicochea et al. (2013) encontraron que en 12 meses, entre enero 2011 y enero 2012, el crecimiento de los otolitos entre el segundo y el tercer anillo fue de 0,62 mm en promedio. En el presente trabajo se encontró que el incremento del segundo al tercer radio fue de 0,80 mm. Sin embargo, a pesar de esta diferencia, los parámetros de crecimiento estimados por Goicochea et al. (2013) son similares a los estimados en este estudio.
Paralelamente, Díaz (2013) hace un análisis de las frecuencias de tallas del T. murphyi peruano y estima parámetros de creci-miento similares a los de este estudio.
El T. murphyi que habita en el mar peruano y aguas adyacentes presenta un crecimiento alométrico y diferente (mayor) a otros stocks o substocks de T. murphyi presentes en el Pacífico Sudorien-tal y que ocupan espacios más australes y en algunos casos muchos más alejados de la costa sudamericana. Según los parámetros estimados, el jurel T. murphyi peruano tendría una velocidad de crecimiento mayor que el jurel T. murphyi de zonas más australes.
Literatura citadaAbramov A.A & A.N. Kotlyar . 1980. Some Biological Features of the Peru-
vian Jack Mackerel, Trachurus symmetricus murphyi. J. Ichthyol. 20(1) 25-31.
Aguayo M., A. Estay & V. Ojeda 1981. Determinación de edad y crecimiento del jurel (Trachurus murphyi) en las zonas Arica-Coquimbo y Tal-cahuano. Informe Subsecretaría de Pesca. Inst. Fom. Pesq. Santiago, Chile, 88p.
Alegría V., J. Oliva, H. Robotham et al. 1995. Estudio biológico pesquero sobre el recurso jurel en la zona norte de Chile (Regiones I y II). Informe Final, Proyecto FIP 93-18. Fondo de Investigación Pesquera. Instituto de Fomento Pesquero, Chile: 221 pp.
Araya M., M. Medina & H. Arancibia . 2003. Preliminary results of the empirical validation of daily increments in otoliths of Jack mackerel Trachurus symetricus (Aires, 1855) marked with oxytetracycline. Scientia marina 67 (4): 471-475.
Castillo G. & A. Arizaga. 1987. Edad y crecimiento del jurel Trachurus symme-tricus murphyi (Nichols, 1920) en aguas de Chile Central. Biología Pesquera. Pont. Univ. Católica de Chile (Talcahuano) 16: 19-33.
Díaz E. 2013. Estimación del crecimiento del jurel (Trachurus murphyi) cap-turado en el Perú a partir del análisis de las frecuencias por tallas. En: Csirke J., R. Guevara-Carrasco & M. Espino (Eds.). Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú. Rev. peru. biol. número especial 20(1): 061 – 066
Dioses T. 1985. Edad y crecimiento de la merluza Merluccius gayi peruanus (G). Anales I Congreso Nacional de Biología Pesquera. Alvaro E. Tresierra Aguilar (editor). Colegio de Biólogos Regional del Norte. Trujillo, Perú . pp. 65-69.
Dioses T. 1995. Análisis de la distribución y abundancia de los recursos jurel y caballa frente a la costa peruana. Inf. Prog. Inst. Mar Perú-Callao (3): 65-69 (mimeo).
Ford, E. 1933. An account of the herring investigation conducted at Plymouth during the years from 1924-1933. J. Mar. Biol. Assoc., 19: 305-384.
Geldenhuys N.D. 1973. Growth of the South African Maasbanker Trachurus trachurus L. and age composition of the catches, 1950-1971. Rep. of S. African Dept. Ind., Sea Fish. Branch Invest. Rep. 101, 1–24
Goicochea C., J. Mostacero, P. Moquillaza, T. Dioses, Y. Topiño & Renato Guevara-Carrasco. 2013. Validación de los anillos de crecimiento de otolitos del jurel Trachurus murphyi. En: Csirke J., R. Guevara-Carrasco & M. Espino (Eds.). Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú. Rev. peru. biol. número especial 20(1): 053 – 060
Gulland J.A. (Ed.) 1977. Fish population dynamics. London. J.Wiley and Sons pp. 372
IFOP. 1995. Estudio biológico pesquero sobre el recurso jurel en la zona norte (Regiones I y II). Informes Técnicos FIP, Informe Final Proyecto FIP-IT/93-17. IFOP, enero 1995: 374 pp.
Kaiser C.E. 1973. Age and growth of Horse Mackerel Trachurus murphyi of Chile. Trans. Ame. Fish. Soc. 102 (3): 591-595 .
Kochkin P.N. 1994. Age Determination and Estimate of Growth Rate for the Peruvian Jack Jack Mackerel, Trachurus symmetricus murphyi. J. of Ichthyology 34 (3):39-50 .
Misu H. & S. Hamasaki. 1971. Age and growth of Peruvian hake Merluccius gayi (Guichenot). Bull. Saikai Reg. Fish. Res. Lab. (41):93-106 .
Pavez C.P. & C.E. Saa. 1978. Determinación de edad y crecimiento del jurel Trachurus murphyi Nichols, 1920. Inv. Mar. Valparaíso , Chile. Vol. 6 (6): 89-100 .
Perea A., J. Mori, B. Buitrón & J. Sánchez. 2013. Aspectos reproductivos del jurel Trachurus murphyien el Perú. En: Csirke J., R. Guevara-Carrasco & M. Espino (Eds.). Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú. Rev. peru. biol. número especial 20(1): 029 – 034
Samamé M. 1977. Determinación de la edad y crecimiento de la sardina Sardi-nops sagax sagax (J.). Bol. Inst. Mar Perú – Callao, 3 (3): 95-112 .
Shcherbitch, L. V. 1991. Fishery-ecological researches in the South Pacific Ocean. Trudy AtlantNIRO:35-53 (en ruso)
Shevchuk L.V. & V.N. Chur . 1984. Métodos de determinación de la edad del jurel (Trachurus murphyi N.) del Pacífico Suroriental. En: Investiga-ciones pesqueras d las regiones abiertas del Océano Mundial. VNIRO, Moscú : 97-105 (en ruso).
Walford, L.A. 1946. A New graphic method of describing the growth of animals. Bio. Bull. 90(2): 141-147.

53
formación de los anillos de crecimiento en otolitos del jurel
Rev. peru. biol. special issue 20(1): 053- 060 (September 2013)Ecology, fisheries and conservation of Jack mackerel (Trachurus murphyi) in Peru. Editors: Csirke J., R. Guevara-Carrasco & M. Espino
Rev. peru. biol. número especial 20(1): 053- 060 (Septiembre 2013)Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú. Editores: Csirke J., R. Guevara-Carrasco & M. Espino
Facultad de ciencias Biológicas unMsMISSN-L 1561-0837
Validación del ritmo de formación de los anillos de crecimiento en otolitos del jurel Trachurus murphyi Nichols 1920
Carlos Goicochea, Jorge Mostacero, Patricia Moquillaza, Teobaldo Dioses, Yeny Topiño y Renato Guevara-Carrasco
Validation of the formation rate of growth rings in otoliths of Jack mackerel Trachurus murphyi Nichols 1920
Instituto del Mar del Perú, esquina Gamarra y General Valle S/N Chucuito, Callao, Perú.
Email Carlos Goicochea: [email protected]
biología del jurel (Trachurus murphyi) en el Perú
© Los autores. Publicado por la Revista Peruana de Biología de la Facultad de Ciencias Biológicas, Universidad Nacional Mayor de San Marcos. Este es un artículo de acceso abierto, distribuido bajo los términos de la Licencia de Atribución Reconocimiento-NoComercial-SinObraDerivada 3.0 de Creative Commons (http://creativecommons.org/licenses/by-nc-nd/3.0/deed.es_ES), que permite el uso no comercial, distribución y reproducción en cualquier medio, siempre que la obra original sea debidamente citadas. Para uso comercial, por favor póngase en contacto con [email protected].
Citación:Goicochea C., J. Mostacero, P. Moquillaza, T. Dioses, Y. Topiño & R. Guevara-Carrasco. 2013. Validación del ritmo de formación de los anillos de crecimiento en otolitos del jurel Trachurus murphyi Nichols 1920. En: Csirke J., R. Guevara-Carrasco & M. Espino (Eds.). Ecología, pes-quería y conservación del jurel (Trachurus murphyi) en el Perú. Rev. peru. biol. número especial 20(1): 053- 060 (Septiembre 2013)
ResumenSe presentan resultados de la validación del crecimiento del jurel Trachurus murphyi por tres técnicas: 1) validación del primer anillo anual mediante lectura de microincrementos diarios; 2) análisis del crecimiento del radio del otolito entre el segundo y tercer anillo, siguiendo una cohorte; y, 3) lectura convencional de anillos anuales. Los resultados demostraron que la edad promedio de jureles T. murphyi con el primer anillo completamente formado fue de 364.8 días, con una talla media de 19.5 cm, y que el incremento promedio del segundo al tercer anillo fue de 0.62 mm. Se estimaron los parámetros de crecimiento Loo = 75.17 cm, k = 0.165/año, to = -0.817año y se discuten estos resultados destacándose que estos parámetros indican que el jurel T. murphyi peruano crece más rápido que el jurel T. murphyi chileno. Las mediciones del radio del primer anillo anual alcanzaron valores hasta 18% más altos que los reportados por otros autores para el jurel chileno, y las medidas del radio de otolitos de individuos de dos y tres años fueron hasta 20% más altas. La longitud promedio de ejemplares de jurel peruano de un año de edad es aproximadamente 35% mayor que la del jurel chileno obtenida por otros autores. El índice de crecimiento phi prima (Ø’) de jurel peruano obtenido en este estudio y en otros, evidencia patrones de crecimiento similares, por el bajo valor del coeficiente de variación (1%), y que es menor al 7% obtenido por otros autores para el jurel T. murphyi de más al sur.
Palabras claves: Crecimiento, anillos diarios, radio, Trachurus murphyi.
AbstractWe present the results of the validation of Jack mackerel Trachurus murphy growth with three techniques: 1) validation of the first annual ring by reading daily microincrements; 2) analysis of the growth of the otolith radius between the second and third ring, following-up one cohort; and, 3) conventional interpretation of annual rings. The results show that the average age of Jack mackerels T. murphyi with the first fully formed ring was 364.8 days, with a mean total length of 19.5 cm, and that the increment from the second to the third ring is on average 0.62 mm. The estimated growth parameters were Loo = 75.17 cm, k = 0.165 /year, to = -0.817 year. These results are discussed, highlighting that the Peruvian Jack mackerel T. murphyi appears to grow faster than Chilean Jack mackerel T. murphyi. The radius of the first annual ring is up to 18% larger than reported by other authors for the Chilean Jack mackerel, while the radius of the otoliths of two and three year-olds was up to 20% higher. Also the average length of a one year old Peruvian Jack mackerel is approximately 35% larger than that of the Chilean Jack mackerel, obtained by other authors. The growth index phi prime (Ø') estimated for the Peruvian Jack mackerel in this study and others show similar growth patterns, given the low value of the coefficient of variation (1%), lower than the (7%) reported by others authors for the more southern Jack mackerel T. murphyi.
Keywords: Growth, daily rings, radio, Trachurus murphyi.
IntroducciónKaiser (1973) fue unos de los primeros investigadores que realizaron estudios sobre
edad y crecimiento de jurel T. murphyi utilizando otolitos (sagitta), en especímenes capturados en aguas frente a la costa de Chile. Posteriormente, se sumaron otros in-vestigadores que estudiaron este tema (Pavez & Collantes 1978, Abramov & Kotlyar 1980, Aguayo et al. 1981, Nekrasov 1994, Kochkin 1994, Cubillos & Arancibia 1995, Gili et al. 1995), con resultados muy variables. Asimismo, Dioses (1995, 2013) realizó investigaciones sobre edad y crecimiento del jurel T. murphyi y su trabajo se constitu-
Publicado online: 11/10/2013Publicado impreso: 15/10/2013

54
Goicochea et al.
Rev. peru. biol. número especial 20(1): 053- 060 (Septiembre 2013)Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú.
Editores: Csirke J., R. Guevara-Carrasco & M. Espino
yó en el primer estudio del crecimiento del jurel con muestras provenientes de capturas realizadas en el mar jurisdiccional del Perú. Por otro lado, otros investigadores utilizaron el método de frecuencias de tallas para la determinación de grupos de edad y crecimiento como Cubillos y Grechina (1998) para el jurel T. murphyi chileno, y Díaz (2013) para el jurel T. murphyi peruano.
La investigación de Panella (1971 y 1974) y posteriores traba-jos como el de Campana y Neilson (1985), ayudaron a entender que el incremento diario de las microestructuras en los otolitos de peces está regulado por un ritmo endógeno circadiano. Este ritmo circadiano de depósito de carbonato de calcio está bajo un control endocrino (Simkiss 1974), y con este concepto, Araya et al. (2003) validaron la formación de los anillos diarios en otolitos de jureles T. murphyi en cautiverio, capturados frente a Chile, confirmando preliminarmente este comportamiento.
Con estos antecedentes, y en la necesidad de validar el ritmo del crecimiento del jurel T. murphyi peruano, en el presente trabajo estudiamos el periodo de formación del primer anillo de crecimiento en los otolitos, mediante la identificación y análisis de microincrementos diarios, así como la tasa de crecimiento mensual, a través del monitoreo de sus dimensiones durante un año consecutivo.
Material y métodosLa lectura de otolitos para la determinación de la edad y
estimación del crecimiento del jurel T. murphyi consistió en tres pasos: (1) validación del primer anillo anual, (2) análisis del crecimiento del radio del otolito entre el segundo y tercer anillo, siguiendo una cohorte y (3) conteo convencional del número de anillos anuales.
1) Validación del primer anillo anual
Para la validación del primer anillo anual, se hizo un análisis de microincrementos, asumiendo la validación concebida por Araya et al. (2003), quienes comprobaron la formación diaria de los microincrementos en el jurel chileno. Para ello, se utilizaron los otolitos de 31 individuos juveniles, de los cuales 9 tenían formado el primer anillo de crecimiento, mientras que los 22 otolitos restantes fueron de tallas menores sin formación del primer anillo. Las muestras provinieron de capturas realizadas durante el verano del 2011 en la región sur dentro del mar jurisdiccional peruano.
Se asumió que en el primer anillo anual, desde el centro al borde final del primer anillo hialino completamente for-mado (identificado en la microestructura como una zona translúcida antes del borde del otolito), existen alrededor de 365 microincrementos o anillos diarios (Victor & Brothers 1982). La preparación de los otolitos se realizó siguiendo la metodología utilizada por Buratti (2003). La fijación de los otolitos a la lámina porta-objeto fue realizada utilizando cristal bond, en el pulido final, se utilizó alúmina de 0.3 μm; esta última técnica se realizó sólo para los otolitos que todavía no tenían el primer anillo formado. Debido al cambio del plano del otolito, para este tamaño y para evitar la eliminación de los anillos diarios de los bordes, se aplicó la técnica de corte transversal (Morales–Nin 1991). La observación y conteo de los anillos de crecimiento se realizaron por tres especialistas en edad y crecimiento, utilizando un microscopio compuesto a 400X de aumento.
Obtenidos los promedios de las edades en días y las longitu-des de los individuos y los otolitos en milímetros, se procedió a ajustar el modelo de crecimiento de Laird-Gompertz (Gamito, 1998, Santos & Renzi 1999):
Lt=L∞*(e−e(−k*(T−t0))
) (1)
Donde:
Lt = la longitud total del pez en el tiempo t,
L∞ = Longitud asintótica,
k = la velocidad diaria a la cual Lt se acerca a su asíntota, y
t0 = es la edad teórica en la cual el pez tendría una longitud
igual a zero.
Se hizo la conversión de la longitud total a longitud de la horquilla (Lh) (Serra 1976) con la ecuación siguiente:
Lh= 0,3873 + 0,8893 (Lt),
para los individuos de un año de edad, con la finalidad de comparar los resultados obtenidos en este estudio con los re-sultados de investigaciones de crecimiento del jurel T. murphyi que se distribuye frente a Chile.
2) Análisis del crecimiento del radio del otolito entre el segundo y tercer anillo, siguiendo una cohorte
Las muestras de otolitos de jurel T. murphyi peruano se obtu-vieron de los muestreos biológicos realizados por los laboratorios costeros del IMARPE de las regiones centro y sur, entre enero y octubre del 2011 y enero del 2012; y se constituyeron en 11 muestras y 448 otolitos de jurel adulto, entre hembras y machos. Durante este periodo de tiempo, las capturas estuvieron consti-tuidas por individuos que formaron parte de un mismo grupo de edad (o cohorte) que pudo ser seguido a través del tiempo, con tallas modales de 28 hasta 36 cm Lt. Se decidió utilizar esta información para comprobar el incremento en el radio de los otolitos de un año a otro, a través de su monitoreo mensual, y de ese modo validar el criterio de que la formación de los anillos de crecimiento es anual.
Los otolitos fueron remojados y limpiados previamente, luego fueron introducidos dentro de una placa Petri con fondo negro y alcohol (medio aclarante). Posteriormente, utilizando un estereoscopio de discusión a 1X de aumento y una regla micrométrica, se midieron el radio total (del núcleo al borde de la zona posrostral) y los anillos de crecimiento, según el criterio tomado por Dioses (2013). Finalmente, las medidas fueron convertidas a milímetros. Las observaciones fueron realizadas por tres especialistas en edad y crecimiento.
Por tratarse solamente de una cohorte, la muestra del primer mes fue tomada como inicio de la cronología para el año en estudio. Para precisar el origen y el fin del crecimiento del oto-lito durante el año, se establecieron promedios de tamaños de los anillos de crecimiento de los otolitos de la misma muestra.
3) Lectura convencional de anillos anuales
Una vez validados el primer anillo anual mediante la lectura de microincrementos y el segundo y tercer anillo mediante la medición de los radios de los otolitos según indicado líneas arriba, se procedió a contar los anillos de crecimiento en otolitos de jurel hembras y machos, de una muestra de 1179 otolitos

55
formación de los anillos de crecimiento en otolitos del jurel
Rev. peru. biol. special issue 20(1): 053- 060 (September 2013)Ecology, fisheries and conservation of Jack mackerel (Trachurus murphyi) in Peru. Editors: Csirke J., R. Guevara-Carrasco & M. Espino
colectados en los laboratorios costeros durante el año 1995 frente a las costas peruanas, con rango de tallas de 6 a 60 cm Lt.
Para las estimaciones de la edad se usó la metodología usada por Dioses (1985, 2013). De las edades estimadas se elaboró una clave talla-edad y a la vez se obtuvieron los parámetros de crecimiento de la ecuación de von Bertalanffy mediante un ajuste con la rutina SOLVER de MS Excel. Haciendo:
Lt=L∞*(1−e(−k*(t−t0))
) (2)
Lt = la longitud total del pez en el tiempo t,
L∞ = Longitud asintótica,
k = es la tasa de crecimiento anual; y,
t0 = es la edad hipotética a la longitud 0.
Para fines comparativos con otros autores que estudiaron la edad y crecimiento del jurel peruano, se utilizó el índice de crecimiento phi prima (Ø') (Pauly & Munro, 1984), donde:
Ø' = log k + 2 * log L∞ (3)
Resultados y discusión Validación del primer anillo anual
Los radios de los otolitos que tenían el primer anillo formado oscilaron entre 2.36 y 2.85 mm y los días contabilizados fueron de 369 a 437. Asimismo, el radio para el primer anillo anual estuvo entre los 2.26 a 2.78 mm, con un promedio de 2.49 mm y 364.8 días respectivamente (Tabla 1).
Por otro lado, como resultado del ajuste de los datos observa-dos de los jureles juveniles a la ecuación de Laird-Gompertz, las edades de los individuos fluctuaron entre 65 a 437 días, mientras que la longitud total varió entre 35 y 210 mm (Tabla 2).
El ajuste del modelo de Laird-Gompertz a los datos obser-vados (relación entre la longitud y la edad) fue significativo (p<0.001). De acuerdo a este ajuste, un individuo de 1 año de edad tendría una longitud total de 195.3 mm (Fig. 1).
Esta longitud total convertida a longitud a la horquilla resultó en 17.7 cm, valor mayor en 35% al del jurel T. murphyi chileno reportado por Nekrasov (1982), Araya et al. (1993) y Shcherbich (1991). Asimismo, las estimaciones de longitud para un año de
Edad en días
Lt (mm) RO (mm) días R1 (mm) Días R1 Lector 1 Lector 2 Lector 3 CV%190 2.36 415 2.26 374 380 369 374 1.47190 2.58 369 2.54 356 340 360 369 4.17200 2.47 396 2.40 366 376 362 360 2.38200 2.50 375 2.46 358 363 360 352 1.59200 2.52 437 2.36 372 376 363 377 2.10200 2.52 391 2.45 365 369 355 370 2.30210 2.63 380 2.60 367 361 372 369 1.55210 2.85 390 2.78 361 355 369 358 2.04210 2.63 405 2.53 364 353 372 368 2.75
Promedio 2.49 364.8 363.7 364.7 366.3 2.26DE 0.151 5.932 13.019 6.041 8.047
Lt= Longitud total, RO= Radio del otolito, R1= radio al primer anillo
Tabla 1. Medidas del radio total y primer anillo de crecimiento de los otolitos de jurel T. murphyi y lectura de sus edades en días por tres lectores.Tabla 1. Measurements of the total radio and first growth ring of Jack mackerel T. murphyi otoliths and their ages (in days) estimated by three readers.
Edad en díasN° ind
Long. (mm)
Edad (días) Lec-1 Lec-2 Lec-3 CV %
1 35 65 67 65 64 2.3382 38 59 58 59 59 0.9843 57 96 98 94 95 2.1764 58 99 100 95 101 3.2585 67 123 120 127 122 2.9316 71 115 113 118 115 2.1827 96 139 135 139 142 2.5338 84 139 137 144 137 2.9019 89 144 142 150 140 3.67510 81 145 152 142 141 4.19511 79 111 107 111 115 3.60412 78 141 135 145 142 3.64813 98 156 162 150 156 3.84614 162 303 315 299 295 3.49315 155 240 233 242 244 2.44516 190 369 355 377 375 3.29717 160 225 220 232 222 2.86218 156 224 218 231 223 2.92719 159 274 263 281 278 3.52020 161 270 280 272 259 3.92121 163 278 287 282 265 4.14822 160 245 255 235 246 4.08323 200 335 322 346 337 3.61924 190 415 430 410 404 3.28325 200 396 404 382 401 3.01526 200 375 387 372 365 3.00027 200 437 422 441 448 3.07928 200 391 404 389 380 3.10129 210 380 381 384 375 1.20630 210 390 370 397 402 4.41831 210 405 390 418 408 3.501
Tabla 2. Longitud total (mm) y número de incrementos diarios leídos en 31 individuos de jurel T. murphyi capturado en aguas peruanas durante el verano del 2011.Tabla 2. Total length (mm) and number of daily increments in 31 individuals of Jack mackerel T. murphyi caught in Peruvian waters during the summer of 2011.
edad del jurel T. murphyi peruano que más se aproximan a nues-tros resultados fueron reportados por Abramov y Kotlyar (1980) con 19.1 cm Lh y Nekrasov (1987) con 18.2 cm Lh (Tabla 3).

56
Goicochea et al.
Rev. peru. biol. número especial 20(1): 053- 060 (Septiembre 2013)Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú.
Editores: Csirke J., R. Guevara-Carrasco & M. Espino
Análisis del crecimiento del radio del otolito entre el segundo y tercer anillo
El principio por el que se rige la asignación de la edad a partir de la lectura de otolitos se basa en la acertada identificación de los anillos de crecimiento y en la adecuada discriminación de los falsos anillos, para lo cual se establece un patrón definido. Así tenemos que, por ejemplo, el radio del primer anillo anual suele medir en promedio 2.48 mm, el segundo anillo mide en promedio 3.36 mm, mientras que el radio del tercer anillo mide en promedio 3.97 mm. En este estudio, el radio del primer anillo anual encontrado es mayor en un 18% y esta diferencia se incrementa al analizar las medidas del radio de otolitos de individuos de dos y tres años, pudiendo llegar a ser 20% mayores a los reportados para el jurel T. murphyi chileno por Aguayo et al. (1981), Gili et al. (1995) y Alegría et al. (1995). Kochkin (1994) realizó estudios de edad en jurel peruano y reportó el valor más cercano con 2.32 mm (Tabla 4).
En la Tabla 4 podemos apreciar la diferencia que existe en las medidas de los anillos de crecimiento de los otolitos dadas a conocer por varios autores. Sólo en el caso de Kochkin (1994) existe una aproximación con la medida del primer anillo obte-nida en este estudio.
Una manera de corroborar la asignación de la edad a partir de los anillos claramente definidos, es mediante el monitoreo mensual del incremento del radio total de los otolitos durante al menos un año a partir de la presencia de una cohorte o clase
anual claramente identificable en la pesquería durante ese pe-riodo (Fig. 2).
Es decir, se debería estar en la capacidad de verificar si efec-tivamente el radio de los anillos crece en promedio desde 2.48 hasta 3.36 mm, al pasar del primer al segundo año, y desde 3.36 hasta 3.97 mm, al pasar del segundo al tercer año.
Al respecto, en la lectura de las muestras de enero 2011, los otolitos contaron en su mayoría con dos anillos y tuvieron en promedio 3.32 mm de radio total, mientras que, en las muestras de enero 2012, algunos de los otolitos comenzaron a evidenciar un tercer anillo y midieron en promedio 3.94 mm. En doce meses, los otolitos crecieron en promedio 0.62 mm (Fig. 3).
Figura 1. Curva de crecimiento de jurel juvenil peruano T. murphyi, estimada mediante el modelo de Laird-Gompertz. Figure 1. Laird-Gompertz growth curve for juveniles of Peruvian Jack mackerel T. murphyi.
Tabla 3. Longitudes medias a la horquilla (cm) de jurel T. murphyi con 1 año de edad obtenida por varios autores en Perú y Chile y el presente estudio.Tabla 3. Average fork lengths (cm) of 1 year old Jack mackerel T. murphyi, obtained by several authors in Peru and Chile, and the present study.
Autores Paises LhAbramov & Kotlyar (1980) (Perú) 19.1Nekrasov (1982) (Chile) 17.4Nekrasov (1987) (Perú) 18.2Nosov et al. (1989) (Perú) 14.1Kochkin (1994) (Perú) 13.4Shcherbich (1991) (Chile) 8.2Araya et al. (1993) (Chile) 8.6Presente estudio (Perú) 17.7
Edad (años)1 2 3
Aguayo et al. (1981) (Chile) 2125 2781 3218Gili et al. (1995) (Chile) 1950 2540 3060
Alegría et al. (1995) (Norte-Chile) 1978 2625 3118Kochkin (1994) (Perú) 2325 2910 3400
Shcherbich (1991) (Chile) 2070 2800 3265Este estudio (Perú) 2485 3360 3970
Tabla 4. Medidas de anillos de crecimiento de otolitos de jurel T. mur-phyi (µm) por diferentes autores en Perú y Chile y el presente estudio. Tabla 4. Measurements of Jack mackerel T. murphyi growth rings (µm) by different authors in Peru and Chile and the present study.
Figura 2. Frecuencias por talla mensual del jurel T. murphyi desde enero 2011 hasta enero 2012.Figure 2. Monthly size frequencies of Jack mackerel T. murphyi from January 2011 to January 2012.
4 7 10 13 16 19 22 25 28 31 34 37 40 43 46 49 52
Frec
uenc
ia (%
)
Longitud total (cm)
ene-11
feb-11
mar-11
abr-11
may-11
jun-11
jul-11
ago-11
set-11
oct-11
nov-11
dic-11 ene-12
0
50
100
150
200
250
0 100 200 300 400 500
edad (días)
modeloobservado
R2=0.97
Long
itud
tota
l (m
m)

57
formación de los anillos de crecimiento en otolitos del jurel
Rev. peru. biol. special issue 20(1): 053- 060 (September 2013)Ecology, fisheries and conservation of Jack mackerel (Trachurus murphyi) in Peru. Editors: Csirke J., R. Guevara-Carrasco & M. Espino
Estos resultados corroboran la asignación de edades a partir de la lectura de anillos, pues se ha demostrado que los otolitos, de los individuos pertenecientes a un mismo grupo de edad, crecieron desde un radio previamente asignado para individuos de dos años hasta un radio previamente asignado para individuos de tres años, durante un lapso de doce meses.
La información completa sobre la longitud de los individuos, el número de anillos y el radio de los otolitos se presentan en la Tabla 5.
Lectura convencional de anillos anuales
El rango de tallas observadas fluctuó entre 6 y 60 cm de longitud total. El máximo número de anillos observados fue de siete. Por cada grupo de edad, la longitud media fluctuó entre 9.5 y 53.7 cm. La separación de los grupos de edad fue clara, a excepción de los dos últimos (Tabla 6).
Dioses (1995, 2013) utilizó una muestra de 977 otolitos de jurel colectados en los principales puertos de desembarque durante los años 1977 – 1979. Se trató de la muestra con el rango de tallas más amplio con el que se haya contado (3 a 71 cm de longitud total) y el máximo número de anillos observados fueron once. Este autor encontró más anillos de crecimiento que los encontrados en el presente estudio debido a que las muestras analizadas contenían individuos de mayor tamaño (hasta 71 cm de longitud total, contra 60 cm en el presente estudio).
El ajuste del modelo de crecimiento de von Bertalanffy a los datos observados (Tabla 6) fue significativo (p <0.001). Los parámetros de crecimiento resultantes de dicho ajuste fueron L∞= 75.17 cm; k = 0.165 anual; y, t0 = –0.817 años.
En la Figura 4 se presenta la curva de crecimiento resultante, junto a las curvas de crecimiento según los parámetros de creci-miento estimados por otros autores mediante lectura de otolitos (Dioses 1995, 2013) y por análisis de frecuencias de tallas (Diaz 2013), y en la Tabla 7 se presentan los parámeros de crecimiento según estos autores.
3.2
3.3
3.4
3.5
3.6
3.7
3.8
3.9
4.0
4.1
0 100 200 300 400
Rad
io o
tolit
o (m
m)
Cronología - 2011 (días)
r3
r2
Figura 3. Radio medio de los otolitos de jurel T. murphyi medidos desde enero 2011 hasta enero 2012 y radio medio del segundo (r2) y tercer (r3) anillo de crecimiento.Figure 3. Average radius of Jack mackerel T. murphyi otoliths mea-sured from January 2011 to January 2012 and average radius of the second (r2) and third (r3) growth ring.
Fech
a m
uest
reo
Día
s tr
ansc
urri
dos
Núm
ero
ejem
plar
es
Long
itud
med
ia p
ez (c
m)
Rad
io m
edio
(mm
)
Des
viac
ión
esta
ndar
(mm
)
19/01/2011 0 50 27.8 3.32 0.21707/02/2011 18 62 28.9 3.42 0.20714/03/2011 51 46 29.7 3.53 0.15918/04/2011 86 42 31.1 3.54 0.15711/05/2011 119 34 31.1 3.60 0.15021/06/2011 159 34 32.2 3.70 0.14322/07/2011 190 31 32.8 3.68 0.20917/08/2011 215 44 32.7 3.75 0.19314/09/2011 242 33 32.8 3.77 0.13827/10/2011 285 35 32.8 3.81 0.18217/01/2012 365 37 35.5 3.94 0.183
Tabla 5. Crecimiento del radio del otolito (mm) de jurel T. murphyi desde enero 2011 hasta enero 2012. Tabla 5. Growth of otolith radius (mm) of Jack mackerel T. murphyi from January 2011 to January 2012.
Como se puede observar, tanto los parámetros de crecimiento como las curvas de crecimiento resultantes de este estudio son muy semejantes a los estimados por Dioses (1995, 2013) y Díaz (2013). Del análisis comparativo de estos parámetros (Tabla 7) resulta un valor bajo, de 1% del coheficiente de variacion (CV) del indice de crecimiento phi prima (Ø'), lo que evidencia pa-trones de crecimiento similares del jurel T. murphyi peruano. Se destaca además que este coeficiente de variación es menor que el 7% obtenido por Cubillos y Arancibia (1995) para el jurel T. murphyi de Chile.
0
10
20
30
40
50
60
70
80
0 1 2 3 4 5 6 7 8 9 10
Long
itud
tota
l (cm
)
Edad (años)
Este estudio
Diaz 2013
Dioses 1995
Figura 4. Curvas de crecimiento de jurel T. murphyi peruano esti-madas mediante el modelo de von Bertalanffy en este estudio y por otros autores.Figure 4. Von Bertalanffy Growth Curves of Peruvian Jack mackerel T. murphyi, estimated in this study and by other authors.

58
Goicochea et al.
Rev. peru. biol. número especial 20(1): 053- 060 (Septiembre 2013)Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú.
Editores: Csirke J., R. Guevara-Carrasco & M. Espino
Edad (años)
Lt (cm) 0 1 2 3 4 5 6 7 Total6 1 17 1 18 2 29 2 2
10 3 311 2 1 212 1 5 413 1 10 1114 16 1615 23 2316 27 2717 30 3018 30 3019 30 3020 29 1 3021 27 3 3022 22 8 3023 15 15 3024 11 19 3025 7 23 3026 4 26 3027 2 28 3028 1 28 1 3029 27 3 3030 25 5 3031 23 7 3032 20 10 3033 17 13 3034 13 17 3035 8 21 1 3036 5 21 4 3037 2 19 9 3038 12 18 3039 9 21 3040 7 22 1 3041 5 22 3 3042 4 20 6 3043 2 16 12 3044 1 12 16 1 3045 8 20 2 3046 6 20 4 3047 5 20 6 3148 3 18 7 1 2949 1 14 7 2 2450 1 9 6 3 1951 7 6 4 1752 5 4 4 1353 3 5 7 1554 1 5 6 1255 4 6 1056 3 4 757 2 3 558 1 1 259 2 260 1 1
Total 13 290 291 157 169 155 63 44 1179Lt media (cm) 9.5 18.7 28.4 35.6 41.2 46.6 50.6 53.7
Tabla 6. Clave talla - edad de jurel T. murphyi peruano, año 1995Tabla 6. Age-length key for Peruvian Jack mackerel T. murphyi, year 1995
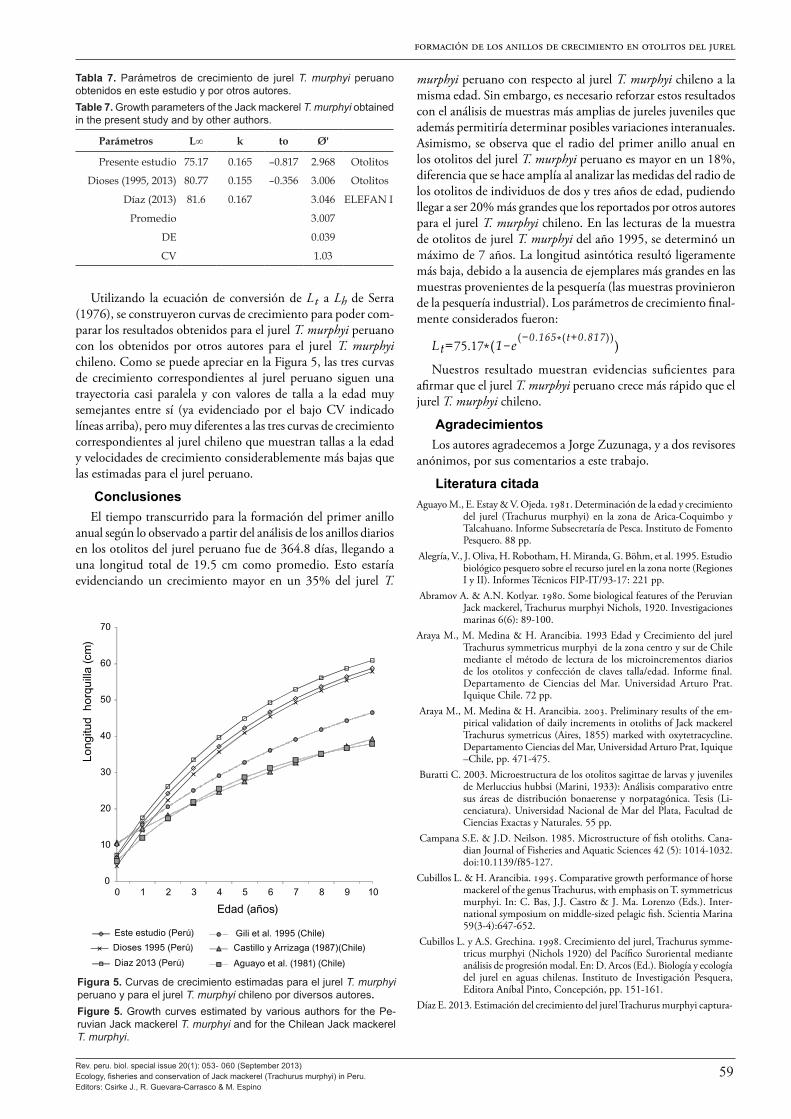
59
formación de los anillos de crecimiento en otolitos del jurel
Rev. peru. biol. special issue 20(1): 053- 060 (September 2013)Ecology, fisheries and conservation of Jack mackerel (Trachurus murphyi) in Peru. Editors: Csirke J., R. Guevara-Carrasco & M. Espino
Utilizando la ecuación de conversión de Lt a Lh de Serra (1976), se construyeron curvas de crecimiento para poder com-parar los resultados obtenidos para el jurel T. murphyi peruano con los obtenidos por otros autores para el jurel T. murphyi chileno. Como se puede apreciar en la Figura 5, las tres curvas de crecimiento correspondientes al jurel peruano siguen una trayectoria casi paralela y con valores de talla a la edad muy semejantes entre sí (ya evidenciado por el bajo CV indicado líneas arriba), pero muy diferentes a las tres curvas de crecimiento correspondientes al jurel chileno que muestran tallas a la edad y velocidades de crecimiento considerablemente más bajas que las estimadas para el jurel peruano.
Conclusiones El tiempo transcurrido para la formación del primer anillo
anual según lo observado a partir del análisis de los anillos diarios en los otolitos del jurel peruano fue de 364.8 días, llegando a una longitud total de 19.5 cm como promedio. Esto estaría evidenciando un crecimiento mayor en un 35% del jurel T.
murphyi peruano con respecto al jurel T. murphyi chileno a la misma edad. Sin embargo, es necesario reforzar estos resultados con el análisis de muestras más amplias de jureles juveniles que además permitiría determinar posibles variaciones interanuales. Asimismo, se observa que el radio del primer anillo anual en los otolitos del jurel T. murphyi peruano es mayor en un 18%, diferencia que se hace amplía al analizar las medidas del radio de los otolitos de individuos de dos y tres años de edad, pudiendo llegar a ser 20% más grandes que los reportados por otros autores para el jurel T. murphyi chileno. En las lecturas de la muestra de otolitos de jurel T. murphyi del año 1995, se determinó un máximo de 7 años. La longitud asintótica resultó ligeramente más baja, debido a la ausencia de ejemplares más grandes en las muestras provenientes de la pesquería (las muestras provinieron de la pesquería industrial). Los parámetros de crecimiento final-mente considerados fueron:
Lt=75.17*(1−e(−0.165*(t+0.817))
)
Nuestros resultado muestran evidencias suficientes para afirmar que el jurel T. murphyi peruano crece más rápido que el jurel T. murphyi chileno.
AgradecimientosLos autores agradecemos a Jorge Zuzunaga, y a dos revisores
anónimos, por sus comentarios a este trabajo.
Literatura citadaAguayo M., E. Estay & V. Ojeda. 1981. Determinación de la edad y crecimiento
del jurel (Trachurus murphyi) en la zona de Arica-Coquimbo y Talcahuano. Informe Subsecretaría de Pesca. Instituto de Fomento Pesquero. 88 pp.
Alegría, V., J. Oliva, H. Robotham, H. Miranda, G. Böhm, et al. 1995. Estudio biológico pesquero sobre el recurso jurel en la zona norte (Regiones I y II). Informes Técnicos FIP-IT/93-17: 221 pp.
Abramov A. & A.N. Kotlyar. 1980. Some biological features of the Peruvian Jack mackerel, Trachurus murphyi Nichols, 1920. Investigaciones marinas 6(6): 89-100.
Araya M., M. Medina & H. Arancibia. 1993 Edad y Crecimiento del jurel Trachurus symmetricus murphyi de la zona centro y sur de Chile mediante el método de lectura de los microincrementos diarios de los otolitos y confección de claves talla/edad. Informe final. Departamento de Ciencias del Mar. Universidad Arturo Prat. Iquique Chile. 72 pp.
Araya M., M. Medina & H. Arancibia. 2003. Preliminary results of the em-pirical validation of daily increments in otoliths of Jack mackerel Trachurus symetricus (Aires, 1855) marked with oxytetracycline. Departamento Ciencias del Mar, Universidad Arturo Prat, Iquique –Chile, pp. 471-475.
Buratti C. 2003. Microestructura de los otolitos sagittae de larvas y juveniles de Merluccius hubbsi (Marini, 1933): Análisis comparativo entre sus áreas de distribución bonaerense y norpatagónica. Tesis (Li-cenciatura). Universidad Nacional de Mar del Plata, Facultad de Ciencias Exactas y Naturales. 55 pp.
Campana S.E. & J.D. Neilson. 1985. Microstructure of fish otoliths. Cana-dian Journal of Fisheries and Aquatic Sciences 42 (5): 1014-1032. doi:10.1139/f85-127.
Cubillos L. & H. Arancibia. 1995. Comparative growth performance of horse mackerel of the genus Trachurus, with emphasis on T. symmetricus murphyi. In: C. Bas, J.J. Castro & J. Ma. Lorenzo (Eds.). Inter-national symposium on middle-sized pelagic fish. Scientia Marina 59(3-4):647-652.
Cubillos L. y A.S. Grechina. 1998. Crecimiento del jurel, Trachurus symme-tricus murphyi (Nichols 1920) del Pacífico Suroriental mediante análisis de progresión modal. En: D. Arcos (Ed.). Biología y ecología del jurel en aguas chilenas. Instituto de Investigación Pesquera, Editora Aníbal Pinto, Concepción, pp. 151-161.
Díaz E. 2013. Estimación del crecimiento del jurel Trachurus murphyi captura-
Parámetros L∞ k to Ø'
Presente estudio 75.17 0.165 –0.817 2.968 Otolitos
Dioses (1995, 2013) 80.77 0.155 –0.356 3.006 Otolitos
Díaz (2013) 81.6 0.167 3.046 ELEFAN I
Promedio 3.007
DE 0.039
CV 1.03
Tabla 7. Parámetros de crecimiento de jurel T. murphyi peruano obtenidos en este estudio y por otros autores. Table 7. Growth parameters of the Jack mackerel T. murphyi obtained in the present study and by other authors.
Figura 5. Curvas de crecimiento estimadas para el jurel T. murphyi peruano y para el jurel T. murphyi chileno por diversos autores.Figure 5. Growth curves estimated by various authors for the Pe-ruvian Jack mackerel T. murphyi and for the Chilean Jack mackerel T. murphyi.
0
10
20
30
40
50
60
70
0 1 2 3 4 5 6 7 8 9 10
Long
itud
hor
quill
a (c
m)
Edad (años)
Este estudio (Perú)
Diaz 2013 (Perú)
Dioses 1995 (Perú) Castillo y Arrizaga (1987)(Chile)Gili et al. 1995 (Chile)
Aguayo et al. (1981) (Chile)

60
Goicochea et al.
Rev. peru. biol. número especial 20(1): 053- 060 (Septiembre 2013)Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú.
Editores: Csirke J., R. Guevara-Carrasco & M. Espino
do en el mar peruano a partir del análisis de las frecuencia por tallas. En: Csirke J., R. Guevara-Carrasco, M. Espino (Eds.). Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú. Rev. peru. biol. número especial 20(1): 061 - 066
Dioses T. 1985. Edad y crecimiento de merluza. Anales: I Congreso Nacional de Biología Pesquera. Ed. A, Tresierra. 65-68.
Dioses T. 1995. Análisis de la distribución y abundancia de los recursos jurel y caballa frente a la costa peruana. Informe Progresivo del Instituto del Mar del Perú (3). 55 pp.
Dioses T. 2013. Edad y crecimiento del jurel Trachurus murphyi, (Nichols 1920) en el Perú. En: Csirke J., R. Guevara-Carrasco, M. Espino (Eds.). Ecología, pesquería y conservación del jurel (Trachurus mur-phyi) en el Perú. Rev. peru. biol. número especial 20(1): 045- 052
Gamito S. 1998. Growth models and their use in ecological modelling: an application to a fish population. Ecological Modelling 113 (1–3): 83-94. doi:10.1016/S0304-3800(98)00136-7.
Gili L.,V. Bocic & R. Serra. 1995. Análisis de la periodicidad de la formación del anillo hialino en otolitos de jurel de la costa de Chile. Informe interno. IFOP.
Kochkin P.1994. Age determination and estimate of growth rate for the Pe-ruvian Jack mackerel trachurus symmetricus murphyi. J. Ichthyol. 34(3):39-50.
Kaiser C. 1973. Age and Growth of Horse Mackerel Trachurus murphyi off Chi-le. Transactions of the American Fisheries Society 102(3): 591-595.
Morales-Nin B. 1991. Determinación del crecimiento de peces óseos en base a la microestructura de los otolitos. FAO. Documento técnico de pesca Nº 322. Roma, FAO. 58 pp.
Nekrasov V. 1982. Distribution, biology and fisheries potential of horse macke-rel (Trachurus simmetricus murphyi) in the south – eastern Pacific. VNIRO. Internal report. Moscow. 43 pp. (In Russian).
Nekrasov V. 1987. Age and growth of horse mackerel. Pp. 44-46. In collected papers on the biology and fisheries of Peruvian horse mackerel. VNIRO. Moscow. (In Russian).
Nekrasov V. 1994. Edad y ritmo de crecimiento. En: D. Arcos & A.S. Gre-china (Eds.). Biología y pesca comercial del jurel en el Pacífico Sur. Instituto de Investigación Pesquera. Talcahuano, Chile. Edición especial, Aníbal Pinto, Concepción, Chile. pp. 43-45.
Nosov E., N. Marquina & P. Kachugin. 1989, Methods for evaluating espaw-
ning stock dynamics, age and growth of Peruvian horse mackerel. P 3-63. In: Arch TINRO. Vladi Vostock (in Russian).
Pannella G. 1971. Fish otoliths: daily growth layers and periodical patterns. Science (Wsh., D.C.). 173:1124-1127.
Pannella G. 1974. Otolith growth patterns: an aid in age determination in temperate and tropical fishes, p. 28-39. In: TB Bagenal (ed.), Ageing of fishes. Unwin Bros. Ltd. London. 234pp.
Pauly, D. and J.L. Munro, 1984. Once more on the comparison of growth in fishes and invertebrates. Fishbyte. Newsletter of the Network of Tropical Fisheries Scientists 2(1): 21.
Pavez C. & S. Collantes. 1978. Determinación de edad y crecimiento del jurel Trachurus murphyi (Nichols 1920). Investigaciones marinas 6(6): 89-100.
Santos B. & M. Renzi. 1999. Estudios de edad en el grupo 0 de Merluza (Merluccius hubbsi). Seminario final proyecto UNIDEP – JICA. Pp. 111-113.
Serra R. 1976. Relación longitud total – longitud a la horquilla del jurel, Trachurus murphyi Nichols, de la zona de Arica-Talcahuano. Rev. Com. Per. Pacífico Sur 5: 173-174.
Shcherbich L. V. 1991. Aging methods for Pacific horse mackerel in the Subarea of Chile. En: informe final Proyecto FIP N.o 95-10. Validación de los métodos aplicados en la determinacion de la edad y crecimiento, y determinación de la mortalidad de jurel de la zona centro-sur. Instituto de investigación pesquera VIII región S.A. Talcahuano 1998. 170 pp.
Simkiss K. 1974. Calcium metabolism of fish in relation to ageing. Pp. 1-12. En:. Bagenal T.B (Ed.). Ageing of fishes. Unwin Bros. Ltd. London. 234pp.
Victor B. & Brothers E. 1982. Age and growth of the fallfish Semotilus corporalis with daily otolith increments as a method of annulus verification. Canadian Journal of Zoology 60 (11): 2543-2550. doi:10.1139/z82-326.

61
Estimación del crecimiento de Trachurus murphyi por del análisis de las frecuencias por tallas
Rev. peru. biol. special issue 20(1): 061- 066 (September 2013)Ecology, fisheries and conservation of Jack mackerel (Trachurus murphyi) in Peru. Editors: Csirke J., R. Guevara-Carrasco & M. Espino
Rev. peru. biol. número especial 20(1): 061- 066 (Septiembre 2013)Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú. Editores: Csirke J., R. Guevara-Carrasco & M. Espino
Facultad de ciencias Biológicas unMsMISSN-L 1561-0837
Estimación del crecimiento del jurel Trachurus murphyi capturado en el Perú, a partir del análisis de las frecuencias por tallas
Erich Díaz
Estimation of growth parameters of Jack mackerel Trachurus murphyi caught in Peru, from length frequency analysis
Instituto del Mar del Perú, esquina Gamarra y General Valle S/N Chucuito, Callao, Perú.
Email Erich Díaz: [email protected]
biología del jurel (Trachurus murphyi) en el Perú
© Los autores. Publicado por la Revista Peruana de Biología de la Facultad de Ciencias Biológicas, Universidad Nacional Mayor de San Marcos. Este es un artículo de acceso abierto, distribuido bajo los términos de la Licencia de Atribución Reconocimiento-NoComercial-SinObraDerivada 3.0 de Creative Commons (http://creativecommons.org/licenses/by-nc-nd/3.0/deed.es_ES), que permite el uso no comercial, distribución y reproducción en cualquier medio, siempre que la obra original sea debidamente citadas. Para uso comercial, por favor póngase en contacto con [email protected].
Citación:Díaz E. 2013. Estimación del crecimiento del jurel Trachur-us murphyi capturado en el Perú, a partir del análisis de las frecuencias por tallas. En: Csirke J., R. Guevara-Carrasco & M. Espino (Eds.). Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú. Rev. peru. biol. número especial 20(1): 061- 066 (Septiembre 2013)
Resumen Se presentan las estimaciones de los parámetros de crecimiento del modelo de von Berta-lanffy para el jurel Trachurus murphyi Nichols 1920 capturado en el mar peruano entre 1972 y 2012, a partir de análisis de frecuencias por tallas. La L∞ fue estimada a partir de la máxima talla observada a lo largo de la historia de su pesquería y se le mantuvo como parámetro fijo en la estimación del parámetro de crecimiento k a partir del ajuste directo de una curva de crecimiento sobre la progresión mensual de modas. Los valores estimados fueron: L∞ = 81.6 cm y k = 0.167 año-1. Se destaca que este valor de k es similar al estimado por otros autores y con otros métodos para el Perú, y es a su vez más alto que el estimado para el T. murphyi capturado frente a las costas de Chile. Estos resultados sugieren que el T. murphyi en el Perú tiene una velocidad de crecimiento más alta que el T. murphyi de Chile.
Palabras clave: Jurel, Trachurus murphyi, crecimiento, frecuencias por tallas, parámetros de crecimiento.
Abstract This paper presents estimates of the von Bertalanffy growth parameters for Jack mackerel T. murphyi caught in Peruvian waters from 1972 to 2012, based on length frequency analysis. L∞ was estimated from the maximum length observed throughout the history of the fishery, and was kept as a fixed parameter in the estimation of k obtained by fitting a growth curve to the monthly progression of modes. The estimated values were: L∞ = 81.6 cm and k = 0.167y-1. It is highlighted that this k values is similar to those estimated by other authors for Peru, and is higher than those estimated for the T. murphyi caught off in Chile. These results suggest that T. murphyi caught in Peru has a faster growth rate than the T. murphyi from Chile.
Keywords: Jack mackerel, Trachurus murphyi, growth, length frequencies, growth parameters.
IntroducciónHistóricamente, la estimación de los parámetros de crecimiento del T. murphyi
capturado en aguas peruanas se ha basado en la lectura de estructuras duras como los otolitos, y las lecturas y estimados han sido muy similares y las pequeñas diferencias pueden ser atribuidas principalmente a los periodo de estudio y las características de las muestras (Abramov & Kotlyar 1980, Dioses 1995, Goicochea et al. 2013). Sin em-bargo, estas estimaciones para el jurel de aguas peruanas muestran marcadas diferencias con las hechas para el T. murphyi capturado frente a Chile (Kochkin 1994, Cubillos & Arancibia 1995, Gili et al. 1995 en Serra & Canales, 2009). La mayoría de estas diferencias son atribuibles a las diferentes interpretaciones que se da a los anillos de crecimiento de los otolitos (Dioses 2013a).
Los métodos basados en el Análisis de las Frecuencias por Tallas (AFT) se presentan como una alternativa metodológica aceptable para comparar y validar los parámetros estimados por la lectura de otolitos. Además de ofrecer una fuente de análisis indepen-diente, el empleo del AFT permite el uso de información abundante y fácil de obtener, como son las frecuencias por tallas, y el menor tiempo requerido para las estimaciones ahora que se cuenta con mejores facilidades computacionales. Una desventaja de los métodos basados en el AFT son su alta sensibilidad a la baja representatividad de las
Publicado online: 11/10/2013Publicado impreso: 15/10/2013
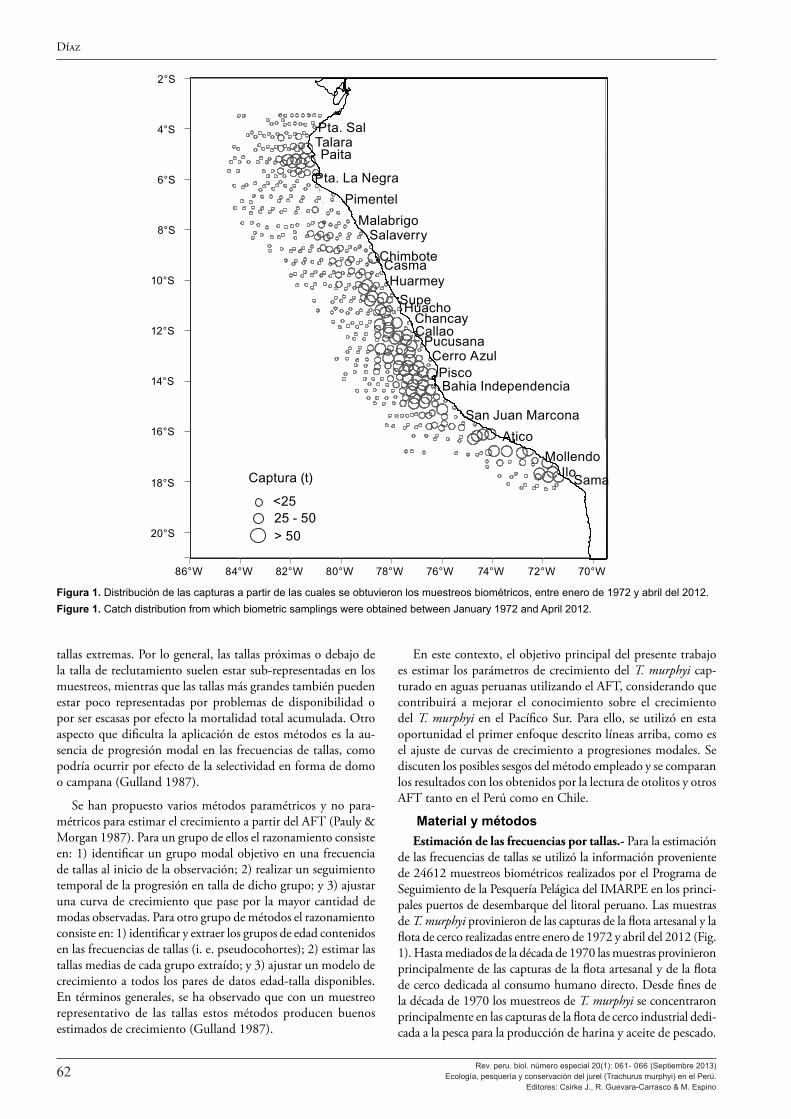
62
Díaz
Rev. peru. biol. número especial 20(1): 061- 066 (Septiembre 2013)Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú.
Editores: Csirke J., R. Guevara-Carrasco & M. Espino
Figura 1. Distribución de las capturas a partir de las cuales se obtuvieron los muestreos biométricos, entre enero de 1972 y abril del 2012. Figure 1. Catch distribution from which biometric samplings were obtained between January 1972 and April 2012.
tallas extremas. Por lo general, las tallas próximas o debajo de la talla de reclutamiento suelen estar sub-representadas en los muestreos, mientras que las tallas más grandes también pueden estar poco representadas por problemas de disponibilidad o por ser escasas por efecto la mortalidad total acumulada. Otro aspecto que dificulta la aplicación de estos métodos es la au-sencia de progresión modal en las frecuencias de tallas, como podría ocurrir por efecto de la selectividad en forma de domo o campana (Gulland 1987).
Se han propuesto varios métodos paramétricos y no para-métricos para estimar el crecimiento a partir del AFT (Pauly & Morgan 1987). Para un grupo de ellos el razonamiento consiste en: 1) identificar un grupo modal objetivo en una frecuencia de tallas al inicio de la observación; 2) realizar un seguimiento temporal de la progresión en talla de dicho grupo; y 3) ajustar una curva de crecimiento que pase por la mayor cantidad de modas observadas. Para otro grupo de métodos el razonamiento consiste en: 1) identificar y extraer los grupos de edad contenidos en las frecuencias de tallas (i. e. pseudocohortes); 2) estimar las tallas medias de cada grupo extraído; y 3) ajustar un modelo de crecimiento a todos los pares de datos edad-talla disponibles. En términos generales, se ha observado que con un muestreo representativo de las tallas estos métodos producen buenos estimados de crecimiento (Gulland 1987).
En este contexto, el objetivo principal del presente trabajo es estimar los parámetros de crecimiento del T. murphyi cap-turado en aguas peruanas utilizando el AFT, considerando que contribuirá a mejorar el conocimiento sobre el crecimiento del T. murphyi en el Pacífico Sur. Para ello, se utilizó en esta oportunidad el primer enfoque descrito líneas arriba, como es el ajuste de curvas de crecimiento a progresiones modales. Se discuten los posibles sesgos del método empleado y se comparan los resultados con los obtenidos por la lectura de otolitos y otros AFT tanto en el Perú como en Chile.
Material y métodos Estimación de las frecuencias por tallas.- Para la estimación
de las frecuencias de tallas se utilizó la información proveniente de 24612 muestreos biométricos realizados por el Programa de Seguimiento de la Pesquería Pelágica del IMARPE en los princi-pales puertos de desembarque del litoral peruano. Las muestras de T. murphyi provinieron de las capturas de la flota artesanal y la flota de cerco realizadas entre enero de 1972 y abril del 2012 (Fig. 1). Hasta mediados de la década de 1970 las muestras provinieron principalmente de las capturas de la flota artesanal y de la flota de cerco dedicada al consumo humano directo. Desde fines de la década de 1970 los muestreos de T. murphyi se concentraron principalmente en las capturas de la flota de cerco industrial dedi-cada a la pesca para la producción de harina y aceite de pescado.
86°W 84°W 82°W 80°W 78°W 76°W 74°W 72°W 70°W
20°S
18°S
16°S
14°S
12°S
10°S
8°S
6°S
4°S
2°S
Pta. Sal Talara Paita
Pta. La NegraPimentel
MalabrigoSalaverry
ChimboteCasmaHuarmey
SupeHuachoChancayCallao
PiscoBahia Independencia
San Juan MarconaAtico
MollendoIloSama
PucusanaCerro Azul
Captura (t)
<2525 - 50> 50
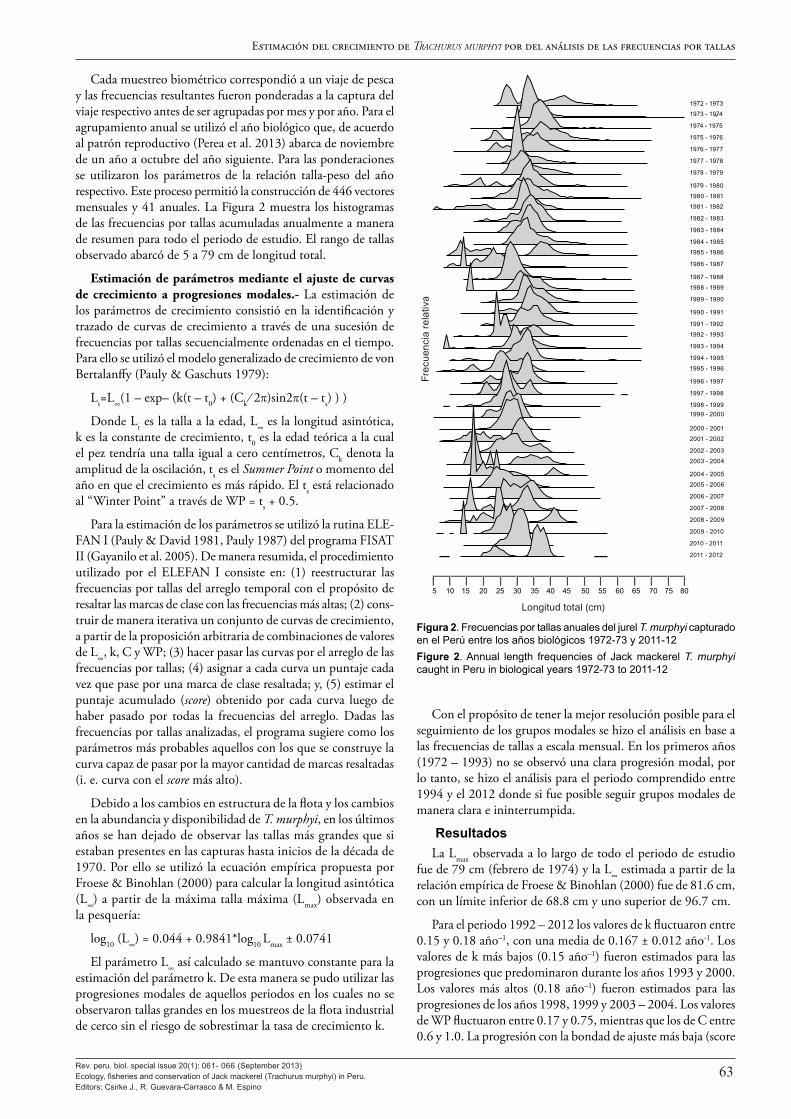
63
Estimación del crecimiento de Trachurus murphyi por del análisis de las frecuencias por tallas
Rev. peru. biol. special issue 20(1): 061- 066 (September 2013)Ecology, fisheries and conservation of Jack mackerel (Trachurus murphyi) in Peru. Editors: Csirke J., R. Guevara-Carrasco & M. Espino
Figura 2. Frecuencias por tallas anuales del jurel T. murphyi capturado en el Perú entre los años biológicos 1972-73 y 2011-12Figure 2. Annual length frequencies of Jack mackerel T. murphyi caught in Peru in biological years 1972-73 to 2011-12
Cada muestreo biométrico correspondió a un viaje de pesca y las frecuencias resultantes fueron ponderadas a la captura del viaje respectivo antes de ser agrupadas por mes y por año. Para el agrupamiento anual se utilizó el año biológico que, de acuerdo al patrón reproductivo (Perea et al. 2013) abarca de noviembre de un año a octubre del año siguiente. Para las ponderaciones se utilizaron los parámetros de la relación talla-peso del año respectivo. Este proceso permitió la construcción de 446 vectores mensuales y 41 anuales. La Figura 2 muestra los histogramas de las frecuencias por tallas acumuladas anualmente a manera de resumen para todo el periodo de estudio. El rango de tallas observado abarcó de 5 a 79 cm de longitud total.
Estimación de parámetros mediante el ajuste de curvas de crecimiento a progresiones modales.- La estimación de los parámetros de crecimiento consistió en la identificación y trazado de curvas de crecimiento a través de una sucesión de frecuencias por tallas secuencialmente ordenadas en el tiempo. Para ello se utilizó el modelo generalizado de crecimiento de von Bertalanffy (Pauly & Gaschuts 1979):
Lt=L∞(1 – exp– (k(t – t0) + (Ck ⁄ 2π)sin2π(t – ts) ) )
Donde Lt es la talla a la edad, L∞ es la longitud asintótica, k es la constante de crecimiento, t0 es la edad teórica a la cual el pez tendría una talla igual a cero centímetros, Ck denota la amplitud de la oscilación, ts es el Summer Point o momento del año en que el crecimiento es más rápido. El ts está relacionado al “Winter Point” a través de WP = ts + 0.5.
Para la estimación de los parámetros se utilizó la rutina ELE-FAN I (Pauly & David 1981, Pauly 1987) del programa FISAT II (Gayanilo et al. 2005). De manera resumida, el procedimiento utilizado por el ELEFAN I consiste en: (1) reestructurar las frecuencias por tallas del arreglo temporal con el propósito de resaltar las marcas de clase con las frecuencias más altas; (2) cons-truir de manera iterativa un conjunto de curvas de crecimiento, a partir de la proposición arbitraria de combinaciones de valores de L∞, k, C y WP; (3) hacer pasar las curvas por el arreglo de las frecuencias por tallas; (4) asignar a cada curva un puntaje cada vez que pase por una marca de clase resaltada; y, (5) estimar el puntaje acumulado (score) obtenido por cada curva luego de haber pasado por todas la frecuencias del arreglo. Dadas las frecuencias por tallas analizadas, el programa sugiere como los parámetros más probables aquellos con los que se construye la curva capaz de pasar por la mayor cantidad de marcas resaltadas (i. e. curva con el score más alto).
Debido a los cambios en estructura de la flota y los cambios en la abundancia y disponibilidad de T. murphyi, en los últimos años se han dejado de observar las tallas más grandes que si estaban presentes en las capturas hasta inicios de la década de 1970. Por ello se utilizó la ecuación empírica propuesta por Froese & Binohlan (2000) para calcular la longitud asintótica (L∞) a partir de la máxima talla máxima (Lmax) observada en la pesquería:
log10 (L∞) = 0.044 + 0.9841*log10 Lmax ± 0.0741
El parámetro L∞ así calculado se mantuvo constante para la estimación del parámetro k. De esta manera se pudo utilizar las progresiones modales de aquellos periodos en los cuales no se observaron tallas grandes en los muestreos de la flota industrial de cerco sin el riesgo de sobrestimar la tasa de crecimiento k.
1972 - 1973
1973 - 1974
1974 - 1975
1975 - 1976
1976 - 1977
1977 - 1978
1978 - 1979
1979 - 1980
1980 - 1981
1981 - 1982
1982 - 1983
1983 - 1984
1984 - 1985
1985 - 1986
1986 - 1987
1987 - 1988
1988 - 1989
1989 - 1990
1990 - 1991
1991 - 1992
1992 - 1993
1993 - 1994
1994 - 1995
1995 - 1996
1996 - 1997
1997 - 1998
1998 - 19991999 - 2000
2000 - 2001
2001 - 2002
2002 - 2003
2003 - 2004
2004 - 2005
2005 - 2006
2006 - 2007
2007 - 2008
2008 - 2009
2009 - 2010
2010 - 2011
2011 - 2012
Longitud total (cm)
5 10 15 20 25 30 35 40 45 50 55 8065 70 7560
Frec
uenc
ia re
lativ
a
Con el propósito de tener la mejor resolución posible para el seguimiento de los grupos modales se hizo el análisis en base a las frecuencias de tallas a escala mensual. En los primeros años (1972 – 1993) no se observó una clara progresión modal, por lo tanto, se hizo el análisis para el periodo comprendido entre 1994 y el 2012 donde si fue posible seguir grupos modales de manera clara e ininterrumpida.
Resultados La Lmax observada a lo largo de todo el periodo de estudio
fue de 79 cm (febrero de 1974) y la L∞ estimada a partir de la relación empírica de Froese & Binohlan (2000) fue de 81.6 cm, con un límite inferior de 68.8 cm y uno superior de 96.7 cm.
Para el periodo 1992 – 2012 los valores de k fluctuaron entre 0.15 y 0.18 año–1, con una media de 0.167 ± 0.012 año-1. Los valores de k más bajos (0.15 año–1) fueron estimados para las progresiones que predominaron durante los años 1993 y 2000. Los valores más altos (0.18 año–1) fueron estimados para las progresiones de los años 1998, 1999 y 2003 – 2004. Los valores de WP fluctuaron entre 0.17 y 0.75, mientras que los de C entre 0.6 y 1.0. La progresión con la bondad de ajuste más baja (score

64
Díaz
Rev. peru. biol. número especial 20(1): 061- 066 (Septiembre 2013)Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú.
Editores: Csirke J., R. Guevara-Carrasco & M. Espino
Grupo de edadseguido durante
Periodo de pre-dominancia
k(año-1)
WP C Score
1992-1994 1993 0.15 0.50 0.9 0.150
1994-1995 1995 0.17 0.75 0.8 0.135
1994-1997 1996 0.16 0.75 0.7 0.126
1998-1999 1998 0.18 0.25 0.6 0.152
1998-2000 1999 0.18 0.58 0.9 0.136
1999-2001 2000 0.15 0.58 0.6 0.128
2000-2003 2001 0.16 0.17 0.9 0.130
2002-2005 2003-2004 0.18 0.08 0.9 0.131
2005-2008 2006-2007 0.17 0.58 1.0 0.193
2010-2012 2011-2012 0.17 0.58 0.6 0.220
Promedio 0.167
Tabla 1. Estimados de k, WP, C y score de la bondad de ajuste para los grupos de edad seguidos durante el periodo 1993 – 2012. Table 1. Estimated k, WP, C and goodness of fit scores for the age groups followed during the period 1993 – 2012.
= 0.126) fue la que predominó durante 1996, mientras que la que presentó la bondad de ajuste más alta (score = 0.220) fue la predominante durante el periodo 2011 – 2012 (Fig. 3, Tabla 1).
Discusión
En este trabajo se prefirió usar la ecuación de Froese & Bi-nohlan (2000) para calcular L∞ dado que esta relación empírica basada en el análisis de más de 550 observaciones sobre diferentes especies, diferentes stocks de una misma especie y diferentes sexos de un mismo stock permite calcular límites de confianza, aunque la conocida aproximación Lmax/L∞ (Pauly 1983, después de Taylor 1958) da valores semejantes.
Para la estimación del parámetro de crecimiento k se estimó primero L∞ y se le mantuvo fijo para todo el periodo de estudio, esto con la finalidad de utilizar las progresiones modales de los últimos años, en los cuales no se observaron tallas grandes en los muestreos de la flota industrial de cerco, sin el riesgo de
sobrestimar la tasa de crecimiento k. Al respecto, cabe destacar que independientemente del método empleado, las estimacio-nes de L∞ a partir de frecuencias de talla sin presencia de tallas mayores suele resultar en la subestimación de éste parámetro y la sobreestimación de k dada la relación inversa entre estos parámetros (Quinn & Deriso 1999). Además, existe alguna evidencia que sugiere que el parámetro L∞ sería más estable que el parámetro k (Isaac 1990).
La ausencia de individuos de mayor tamaño en las muestras recientes puede ser explicada por diferentes factores (Chen & Watanabe 1989, Lorenzen 1996, Pauly & Morgan eds. 1987), entre los cuales cabe destacar los cambios ambientales de corto, mediano y largo plazo, los cambios en la abundancia, distribu-ción y disponibilidad, y los cambios en las regulaciones pesqueras y en la estructura y operación de la flota (Flores et al. 2013, Espino 2013, Dioses 2013b, Segura & Aliaga 2013, Zuzunaga 2013, Ñiquen et al. 2013).
En cuanto al ajuste de curvas de crecimiento a las frecuencias por tallas, se consideró apropiado iniciar el proceso a partir del periodo de tiempo en que se evidenciaron las progresio-nes modales (1993). Una de las reconocidas limitaciones de los AFT y en especial del ELEFAN I es la imposibilidad de estimar el crecimiento a partir de frecuencias por tallas que no evidencian una sucesión temporal de modas (Pauly & David 1981). Dichas progresiones no fueron evidentes en el periodo 1972 – 1992, por lo que forzar el ajuste de curvas de crecimiento a estas frecuencias con modas casi constantes hubiera dado resultados contradictorios, poco consistentes y con estimaciones de k poco realistas.
Es importante destacar la similitud existente entre los pará-metros de crecimiento estimados para el T. murphyi capturado en el Perú (métodos del M2 al M4, Tabla 2), sobre todo si tenemos en cuenta que las estimaciones han sido realizadas utilizando metodologías independientes tanto en lo referente a las fuentes de información como a los procedimientos. Los valores de k son muy próximos entre sí, pero consistentemente más altos que los estimados para el T. murphyi capturado frente
Figura 3. Ajuste de una curva de crecimiento a la progresión de modas que pudo ser seguida desde enero del 2002 (flecha izquierda-derecha, moda en 12 cm) hasta marzo del 2005 (flecha derecha-izquierda, moda en 40 cm).Figure 3. Fitting of a growth curve to the modal progression followed from January 2012 (left-right arrow, mode in 12 cm) to March 2005 (right-left mode, mode in 40 cm).
50
40
30
20
10
0
Leng
th (c
m)
2002 2003 2004 2005J F M A M J J A S O N D J F M A M J J A S O N D J F M A M J J A S O N D J F M A M J J A S O N D

65
Estimación del crecimiento de Trachurus murphyi por del análisis de las frecuencias por tallas
Rev. peru. biol. special issue 20(1): 061- 066 (September 2013)Ecology, fisheries and conservation of Jack mackerel (Trachurus murphyi) in Peru. Editors: Csirke J., R. Guevara-Carrasco & M. Espino
Método Autor Lugar Base del Método L∞ (cm) K t0
M1 Díaz (2013) PerúRelación empírica (Froese
& Binohlan. 2000)81.6
M2 Díaz (2013) Perú AFT (ELEFAN I) 0.167
M3 Goicochea et al. (2013) Perú Lectura de otolitos 75.17 0.165 -0.817
M4 Dioses (1995) Perú Lectura de otolitos 80.8 0.155 -0.356
M5 Cubillos & Arancibia (1995) Chile (Centro–Sur) AFT (ELEFAN) 78.6* 0.138
M6 Gili et al. (1995) en Serra & Canales 2007 Chile (Centro–Sur) Lectura de otolitos 77.8* 0.094 -0.896
* Convertidas de L∞ a la horquilla (Lh∞) a L∞ total (Lt∞) a partir de la relación Lt=0,514+1,091LH (Cubillos & Arancibia 1995)
Tabla 2. Comparación de los parámetros de crecimiento estimados por diferentes autores para el jurel T. murphyi capturado frente al Perú y Chile.Table 2. Comparison of the growth parameters estimated by different authors and methods for the Jack mackerel T. murphyi caught off Peru and Chile.
a Chile (métodos M5 y M6, Tabla 2). Esto sugiere que frente al Perú el T. murphyi tendría una tasa de crecimiento más alta que frente a Chile.
Existen múltiples ejemplos de especies donde sus diversos stocks tienen parámetros de crecimiento diferentes y que se ex-plicarían por estar expuestas a distintas condiciones ambientales (Jennings & Beverton 1991, Ormseth & Norcross 2009), niveles de explotación (Law 2000) o estructuras comunitarias diferentes (Gislason et al. 2008).
Por las facilidades para su aplicación, consideramos que el método empleado en este estudio podría ser utilizado de forma rutinaria para futuras estimaciones del crecimiento del T. murphyi en todo el ámbito de distribución de la especie en el Pacífico Sur. Sin embargo, también es recomendable utilizar otros métodos y sugerimos que en futuros estudios de la edad y crecimiento del T. murphyi en el Pacífico Sudeste se incluyan: 1) estimaciones a partir de uno o más de los diferentes metodos de AFT disponibles, tanto paramétricos (e.g. MULTIFAN, NORMSEP) como no paramétricos (e.g. SLCA, PROJMAT, Powell-Wetherall); 2) análisis de sensibilidad de los resultados a los datos de entrada; 3) análisis de las potencialidades y de-bilidades de cada método, medidos a través de indicadores de desempeño; y, 4) una comparación final de todos los resultados. Otro aspecto que no se debe dejar de lado es procurar que el muestreo siempre sea representativo tanto en términos de espacio como de tiempo.
AgradecimientosEl autor desea expresar su más sincero agradecimiento a la
Dirección General de Investigaciones en Recursos Pelágicos del IMARPE por las facilidades otorgadas para la preparación de este trabajo, a Ricardo Oliveros-Ramos y al revisor anónimo por los aportes al trabajo y a Giancarlo Morón por el apoyo computacional y de revisión bibliográfica.
Literatura citada Abramov A. & A. Kotlyar. 1980. Some biological features of the Peruvian
Jack mackerel, Trachurus murphyi Nichols, 1920. Invest. Mar. 6(6):89-100.
Cubillos L. & H. Arancibia. 1995. Comparative growth performance of horse mackerel of the genus Trachurus, with emphasis on T. symmetricus murphyi in Chile. Sci. Mar. 59(3-4):647-652.
Chen S. & S. Watanabe. 1989. Age dependence of natural mortality coefficient in fish population dynamics. Bulletin of the Japanese Society of Scientific Fisheries 55(2): 205-208.
Dioses T. 1995. Análisis de la distribución y abundancia de los recursos jurel y caballa frente a la costa peruana. Informe Progresivo del Instituto del Mar del Perú (IMARPE), Callao, Perú, 1995, Nº 3, 55 p.
Dioses T. 2013a. Edad y crecimiento del jurel Trachurus murphyi en el Perú. En: Csirke J., R. Guevara-Carrasco & M. Espino (Eds.). Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú. Rev. peru. biol. número especial 20(1): 045 – 052
Dioses T. 2013b. Patrones de distribución y abundancia del jurel Trachurus murphyien el Perú. En: Csirke J., R. Guevara-Carrasco & M. Espino (Eds.). Ecología, pesquería y conservación del jurel (Tra-churus murphyi) en el Perú. Rev. peru. biol. número especial 20(1): 067 – 074
Espino M. 2013. El jurel Trachurus murphyi y las variables ambientales de macroescala. En: Csirke J., R. Guevara -Carrasco & M. Espino (Eds.). Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú. Rev. peru. biol. número especial 20(1): 009 – 020
Flores R., M. Espino, G. Luque & J. Quispe. 2013. Patrones de variabilidad ambiental en el mar peruano. En: Csirke J., R. Guevara-Carrasco & M. Espino (Eds.). Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú. Rev. peru. biol. número especial 20(1): 021 – 028
Froese R. & C. Binohlan. 2000. Empirical relationships to estimate asymp-totic length, length at first maturity and length at maximum yield per recruit in fishes, with a simple method to evaluate length frequency data. Journal of Fish Biology 56(4): 758-773. DOI: 10.1111/j.1095-8649.2000.tb00870.x
Gayanilo Jr. F., P. Sparre & D. Pauly. 2005. The FAO-ICLARM Stock As-sessment Tools II Windows Version (FISAT II) user’s guide (Re-vision I). FAO Computerized Information Series Fisheries, Nº 8.
Gislason H., J. Pope, J. Rice, & N. Daan. 2008. Coexistence in North Sea fish communities: implications for growth and natural mortality. – ICES Journal of Marine Science, 65: 514–530. DOI: 10.1093/icesjms/fsn035
Goicochea C., J. Mostacero P. Moquillaza, T. Dioses, Y. Topiño & Renato Guevara-Carrasco. 2013. Validación de los anillos de crecimiento de otolitos del jurel Trachurus murphyi. En: Csirke J., R. Guevara-Carrasco & M. Espino (Eds.). Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú. Rev. peru. biol. número especial 20(1): 053 – 060

66
Díaz
Rev. peru. biol. número especial 20(1): 061- 066 (Septiembre 2013)Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú.
Editores: Csirke J., R. Guevara-Carrasco & M. Espino
Gulland J. 1987. Length-based methods in fisheries research: from theory to application, p. 335-342. In D. Pauly and G.R. Morgan (eds.) Length-based methods in fisheries research. ICLARM Conference Proceedings 13, 468 p. International Center for Living Aquatic Resources Management, manila, Philippines, and Kuwait Institute for Scientific Research, Safat, Kuwait.
Isaac V. 1990. The accuracy of some length-based methods for fish population studies. ICLARM Tech. Rep. 27.81 p.
Jennings S. & R. Beverton. 1991. Intraspecific variation in the life history tactics of Atlantic herring (Clupea harengus L.) stocks. ICES Journal of Marine Science, 48: 117 –125. DOI: 10.1093/icesjms/48.1.117
Kochkin P. 1994. Age determination and estimate of growth rate for the Pe-ruvian Jack mackerel Trachurus symmetricus murphyi. J. Ichthyol. 34(3):39-50.
Law R. 2000. Fishing, selection, and phenotypic evolution. – ICES Journal of Marine Science, 57: 659–669. DOI: 10.1006/jmsc.2000.0731
Lorenzen K. 1996. The relationship between body weight and natural morta-lity in juvenile and adult fish: a comparison of natural ecosystems and aquaculture. Journal of Fish Biology 49:627–647. DOI: 10.1111/j.1095-8649.1996.tb00060.x
Ñiquen M., M. Bouchon, D. Ulloa & A. Medina. 2013. Análisis de la pesquería del jurel Trachurus murphyi en el Perú. En: Csirke J., R. Guevara-Carrasco, M. Espino (Eds.). Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú. Rev. peru. biol. número especial 20(1): 097- 106
Ormseth O. & B. Norcross. 2009. Causes and consequences of life-history variation in North American stocks of Pacific cod. – ICES Journal of Marine Science, 66: 349–357. DOI: 10.1093/icesjms/fsn156
Pauly D. 1983. Some simple methods for the 1983 assessment of tropical fish stocks. FAO Fish.Tech.Pap., 234:52 p.
Pauly D. 1987. A review of the ELEFAN system for analysis of length-frequency data in fish and aquatic invertebrates, p. 7-34. In D. Pauly and G.R. Morgan (eds.) Length-based methods in fisheries research. ICLARM Conference Proceedings 13, 468 p. International Center for Living Aquatic Resources Management, manila, Philippines, and Kuwait Institute for Scientific Research, Safat, Kuwait.
Pauly D. & N. David. 1981. ELEFAN I, a BASIC program for the objective extraction of growth parameters from length-frequency data. Berichte der Deutschen Wissensschaftlichen Kommission für Meeresforschung 28(4): 205-211.
Pauly, D. & G. Gaschuts. 1979. A simple method for fitting oscillating growth data, with a program for pocket calculators. ICES C. M. 1979/G: 24. 26 pp.
Pauly D. & G. Morgan (Eds.) 1987. Length-based methods in fisheries research. ICLARM Conference Proceedings 13, 468 p. International Center for Living Aquatic Resources Management, manila, Philippines, and Kuwait Institute for Scientific Research, Safat, Kuwait.
Perea A., J. Mori, B. Buitrón & J. Sánchez. 2013. Aspectos reproductivos del jurel Trachurus murphyien el Perú. En: Csirke J., R. Guevara -Carrasco & M. Espino (Eds.). Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú. Rev. peru. biol. número especial 20(1): 029 – 034
Quinn T. & R. Deriso. 1999. Quantitative fish dynamics. Oxford University Press, New York, N.Y. pp: 560.
R. 2012. The R Foundation for Statistical Computing. R version 2.15.1 Segura M. & A. Aliaga M. 2013. Biomasa acústica y distribución del jurel
Trachurus murphyien el Perú. En: Csirke J., R. Guevara -Carrasco & M. Espino (Eds.). Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú. Rev. peru. biol. número especial 20(1): 087 – 096
Serra R. & C. Canales. 2009. Short review of some biological aspects of the Chi-lean Jack mackerel, Trachurus murphyi. SP-07-SWG-JM-SA-05, 7p.
Taylor C.C. 1958. Cod growth and temperature. J. Cons. perm. Int. Explor. Mer., 23(3): 365-370
Zuzunaga J. 2013. Medidas de conservación y ordenación pesquera del jurel Trachurus murphyi en el Perú. En: Csirke J., R. Guevara -Carras-co, M. Espino (Eds.). Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú. Rev. peru. biol. número especial 20(1): 097- 106

67
Patrones de distribución y abundancia deTrachurus murphyi en el Perú
Rev. peru. biol. special issue 20(1): 067- 074 (Septembre 2013)Ecology, fisheries and conservation of Jack mackerel (Trachurus murphyi) in Peru. Editors: Csirke J., R. Guevara-Carrasco & M. Espino
Rev. peru. biol. número especial 20(1): 067- 074 (Septiembre 2013)Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú. Editores: Csirke J., R. Guevara-Carrasco & M. Espino
Facultad de ciencias Biológicas unMsMISSN-L 1561-0837
Patrones de distribución y abundancia del jurel Trachurus murphyi en el Perú
Teobaldo Dioses
Abundance and distribution patterns of Jack mackerel Trachurus murphyi in Peru
Instituto del Mar del Perú, esquina Gamarra y General Valle S/N Chucuito, Callao, Perú.
Email Teobaldo Dioses: [email protected]
ecología del jurel (Trachurus murphyi) en el Perú
© Los autores. Publicado por la Revista Peruana de Biología de la Facultad de Ciencias Biológicas, Universidad Nacional Mayor de San Marcos. Este es un artículo de acceso abierto, distribuido bajo los términos de la Licencia de Atribución Reconocimiento-NoComercial-SinObraDerivada 3.0 de Creative Commons (http://creativecommons.org/licenses/by-nc-nd/3.0/deed.es_ES), que permite el uso no comercial, distribución y reproducción en cualquier medio, siempre que la obra original sea debidamente citadas. Para uso comercial, por favor póngase en contacto con [email protected].
Citación:Dioses T. 2013. Patrones de distribución y abundancia del jurel Trachurus murphyi en el Perú. En: Csirke J., R. Guevara-Carrasco & M. Espino (Eds.). Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú. Rev. peru. biol. número especial 20(1): 067- 074 (Septiembre 2013)
ResumenSe analiza información de cruceros bio-oceanográficos del IMARPE con estaciones hasta 100 mn de la costa y entre 0 y 180 m de profundidad entre 1961 y 2011; información de observa-dores científicos a bordo de buques arrastrero-factoría de media agua nacionales, soviéticos y cubanos que operaron entre 1983 y 1998; y series de tiempo de anomalías de temperatura superficial del mar, profundidad de la isoterma de 15 °C y salinidad de 1950 a la fecha. Se identifican y describen tres patrones de distribución vertical y profundidad de las capturas de jurel Trachurus murphyi en relación a cambios en condiciones ambientales: (1) cuando ocurren fenómenos El Niño (EN) fuertes los cardúmenes se distribuyen entre 100 y 200 m de profundidad, las capturas son altas y varían poco entre día y noche; (2) cuando hay fuertes afloramientos los cardúmenes se distribuyen entre 0 y 75 m de profundidad, la pesca es más oceánica y superficial y (3) cuando se refuerza la Extensión Sur de la Corriente de Cromwell (ESCC), los cardúmenes están entre 0 y 300 m de profundidad, la pesca es más costera, a mayor profundidad de día y más superficial de noche. T. murphyi es menos abundante en períodos fríos, cuando dominan Aguas Costeras Frías (ACF) y se alejan de la costa las Aguas Subtropicales Superficiales (ASS), y más abundante en periodos cálidos, asociados a eventos EN y dominancia de la ESCC. Se observa segregación latitudinal por tamaños en relación a la Oscilación Decadal del Pacífico.
Palabras claves: Jurel, distribución espacial, comportamiento, El Niño
Abstract
This paper analyzes information from IMARPE bio-oceanographic surveys conducted between 1961 and 2011 covering an area from 0 to 100 nm from the coast with observations at depths from 0 to 180 m; information from scientific observers on board Peruvian, Soviet and Cuban factory trawlers operating between 1983 – 1998; as well as time series of sea surface tempera-ture anomalies, depth of the 15 °C isotherm and sea surface salinity from 1950 to date. Three patterns in the vertical distribution and depth of catches of Jack mackerel Trachurus murphyi are identified and described in relation to changing environmental conditions: (1) During strong El Niño (EN) events shoals are at depths between 100 – 200 m, catches are higher and don’t change much between day and night; (2) During strong upwellings shoals are shallower, at depths between 0 – 75 m and catches are more oceanic (offshore) and closer to the surface; and, (3) When there is a strengthen Southern Extension of the Cromwell Current (SECC) shoals are at depths between 0 and 300 m, fishing takes place closer to the coast being deeper during the day and shallower during the night. During cold periods there is lower abundance of T. murphyi due to the prevalence of Cold Coastal Waters (CCW) and the offshore shift of the Subtropical Surface Waters (SSW), while abundance is higher during warmer periods as-sociated with EN events and the strengthening of the SECC. A certain latitudinal segregation by sizes in relation to the Pacific Decadal Oscillation is also observed.
Keywords: Jack mackerel, spatial distribution, behavior, El Niño
Publicado online: 11/10/2013Publicado impreso: 15/10/2013

68
Dioses
Rev. peru. biol. número especial 20(1): 067- 074 (Septiembre 2013)Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú.
Editores: Csirke J., R. Guevara-Carrasco & M. Espino
IntroducciónEl ambiente marino entre los 04°S y el extremo sur del
dominio marítimo peruano está dominado por el Sistema de la Corriente del Perú (SCP), que sustenta una alta producción pesquera y que recurrentemente se ve afectada por fuertes va-riaciones del clima y de las condiciones oceanográficas en ciclos o estadios superpuestos de amplitud diversa. Estas variaciones pueden ser de corta duración o estacionales, de mediano plazo como los eventos ENSO (El Niño-Oscilación del Sur) con sus fases cálidas (El Niño) y frías (La Niña), los de largo plazo con periodos decadales cálidos y fríos de 25 años a más (Chávez, et al. 2003, Chávez et al. 2008); y por ultimo otros seculares (mayores a 50 años) (Espino 2003, Espino & Yamashiro 2012).
La distribución y concentración de Trachurus murphyi Ni-chols 1920 se relacionan con la variación e interacción de las masas de agua que existen frente a la costa peruana, según se trate de años normales, o anormales con calentamiento (El Niño) o enfriamiento (La Niña). El hábitat preferido de T. murphyi es el frente oceánico formado por las Aguas Costeras Frías (ACF) y las Aguas Subtropicales Superficiales (ASS). Los frentes y remo-linos que se forman entre estas masas de agua tendrían un efecto sobre algunas características biológicas y de comportamiento de T. murphyi (áreas y épocas de reproducción, sobrevivencia de larvas, cambios en distribución y disponibilidad, entre otros). La abundancia y la disponibilidad de este recurso se explican por las variaciones de este frente.
En este trabajo se describe y analiza las concentraciones de los cardúmenes del jurel T. murphyi con los cambios ambienta-les de corto y mediano plazo que se han presentado durante el desarrollo de esta pesquería.
Material y métodosLos datos analizados fueron obtenidos de los cruceros reali-
zados por el IMARPE frente a la costa del Perú, en una malla de estaciones bio-oceanográficas distribuidas hasta las 100 mn de la costa y secciones oceanográficas verticales hasta los 180 m de profundidad estándar, en el periodo comprendido entre 1961 – 2011. También se analizó la información obtenida por los Técnicos Científicos de Investigación (TCI) a bordo de bu-ques arrastrero-factoría (BAF) de media agua nacionales (1996 – 1998), soviéticos (1983 – 1987) y cubanos (1988 – 1991) que han operado en la pesca de T. murphyi en aguas peruanas. Además se analizaron series de tiempo de 1950 a la fecha de las anomalías de la temperatura superficial del mar (ATSM), de la profundización de la isoterma de 15 °C y de la salinidad.
En función de los datos tomados por los TCI, tales como fecha, área de pesca, según los cuadrados Marsden (1° Lat. x 1° Long.) e información de los arrastres (hora, posición, rumbo, profundidad de la red, captura estimada, temperatura del agua y del aire, etc.), Icochea et al. (1989) analizaron el periodo febrero 1983 a marzo 1987 y Chirinos (1992) analizó el periodo mayo 1988 a abril 1991. Estos investigadores estudiaron las operaciones de pesca en que se capturaron especies pelágicas, con especial énfasis en el jurel. El área de estudio estuvo comprendida entre los 05°S y los 09°S. El límite norte de operación de la flota arrastrera determinada por el Gobierno Peruano fue los 06°S, para el periodo febrero 1983 – marzo 1985, y a partir de abril de 1985 fue los 05°S. Dioses et al. (1989) explican la distribución batimétrica de los cardúmenes de T. murphyi en desove entre octubre y diciembre de 1986.
El periodo 1996 – 2007 fue analizado por Espinoza, Ñiquen y Flores (2008) y el periodo 2008 – 2012 por Espinoza (com. pers.) en base a la información del Programa Bitácoras de Pesca. Este Programa registra las operaciones a bordo de la flota de cerco autorizada para la pesca de T. murphyi a nivel industrial, que es una flota equipada con sistema de enfriamiento por circulación de agua de mar (RSW).
Se establecieron patrones de comportamiento migratorio vertical y horizontal del recurso mediante la revisión y análisis de los ecogramas más representativos, de la distribución horizontal y batimétrica de los cardúmenes en relación al ambiente.
Para el análisis de la relación recurso-ambiente se presenta la distribución vertical de los cardúmenes de T. murphyi asociada con el ambiente (temperatura, salinidad, oxígeno), en lances de pesca seleccionados.
Resultados y discusiónEn el mar peruano, el hábitat preferido por el T. murphyi es el
frente oceánico formado por las Aguas Costeras Frías (ACF) con las Aguas Subtropicales Superficiales (ASS). En tales condiciones, los cardúmenes se ubican en la columna de agua con rangos de temperatura entre 14 y 24 °C, Ganoza (1998). En relación a la profundidad, conforme más cerca de la costa, las isotermas adoptan una configuración de divergencia vertical, unas por encima de la termoclina y que se dirigen hacia la superficie (afloramiento costero) y otro grupo de isotermas por debajo de la termoclina que se profundizan y que están asociadas a la ESCC. La profundidad de la isoterma de 15 °C constituye el valor inmediato por debajo de la termoclina y es la que marca la profundidad donde se produce la divergencia de las isotermas (Flores et al. 2009).
De la información anterior y de los trabajos de Icochea et al. (1989), Chirinos (1992), Ganoza (1998) y Dioses et al. (1989) se describen los siguientes patrones de comportamiento del T. murphyi frente a su ambiente:
El primer patrón se distingue durante la ocurrencia del fe-nómeno El Niño, cuando las especies pelágicas se concentran debajo de una marcada y profunda termoclina. Las capturas se realizan a profundidades mayores de 100 m, en temperaturas de 15 a 20 °C, salinidades entre 35.0 – 35.1 ups y niveles de oxígeno mayores a 1 mL/L. Bajo estas condiciones las capturas tienden a ser altas y no varían significativamente durante el día debido a la reducción del rango de distribución de los cardú-menes. La presencia de elevadas temperaturas superficiales, por encima de los 29 °C, cinco grados más sobre lo normal y de una marcada termoclina que llega hasta los 200 m, determinan que los cardúmenes de T. murphyi se presenten a profundidades mayores a 100 m, entre 15 y 20 °C. Las capas superficiales del mar son ocupadas por las Aguas Ecuatoriales Superficiales (AES) y las Aguas Tropicales Superficiales (ATS) de altas temperaturas y bajas salinidades. En estas condiciones el recurso es más ac-cesible y vulnerable a la flota de arrastre que a la cerquera (Fig. 1a y Fig. 2a y b).
El segundo patrón se suele observar posterior a los eventos El Niño, con dominio de las ACF. Las isotermas mayores a 15 °C se encuentran más superficiales conforme más cerca de la costa, con salinidades mayores a 34.95 ups, y concentraciones de oxígeno entre 1 y 3 mL/L. (Fig. 1b y Fig. 3a, b y c). La captura es más superficial y oceánica, fluctuando con la extensión del

69
Patrones de distribución y abundancia deTrachurus murphyi en el Perú
Rev. peru. biol. special issue 20(1): 067- 074 (Septembre 2013)Ecology, fisheries and conservation of Jack mackerel (Trachurus murphyi) in Peru. Editors: Csirke J., R. Guevara-Carrasco & M. Espino
5°
10°
15°
a) Febrero - Julio 1983
PAITA
PTA AGUJA
CHICAMA
CHIMBOTE
SUPE
CALLAO
PISCO
SAN JUAN
ATICO
ILO
050
100150200250300350
PAITA
PTA AGUJA
CHICAMA
CHIMBOTE
SUPE
CALLAO
PISCO
SAN JUAN
ATICO
ILO
F M A M J J1983
Sur
TM TM/Hr
1491 1151 1014 1296 1593 25876.9 8.0 9.8 6.2 6.8 6.1
b) Noviembre 1984 - Marzo 1985
050
100150200250300350
D E F M19851984
N
TM TM/Hr
157854.7
180186.9
121935.0
127796.4
113246.1
Figura. 1 Principales zonas de pesca y distribución vertical de las capturas de jurel T. murphyi: a) Periodo El Niño febrero – julio 1983, b) Periodo noviembre 1984 – marzo 1985. (Modificada de Icochea et al. 1989).Figure. 1 Major fishing areas and vertical distribution of Jack mac-kerel T. murphyi: a) El Niño Period February to July, 1983, b) Period November 1984 - March 1985. (Modified after Icochea et al. 1989).
Figura 2.- Distribución vertical de a) Temperatura y b) Salinidad frente a Punta Aguja en febrero – marzo 1983. (de Icochea et al. 1989)Figure 2. - Vertical distribution of a) temperature and b) salinity, off Punta Aguja in February-March 1983. (from Icochea et al. 1989)
T (°C)
Est 231 230 229 228
50
20
25
20
15
0m
50
100
200
30013
Millas
S (ups)
Est 231 230 229 228
0m
50
100
200
50 Millas
32.033.034.0 34.8
35.035.1
35.1
35.0
34.9
a) b)
Profundidad
Profundidad
T (°C) S (ups)
80 60 40 20 0
Est 60 59 58 57 56
Millas Millas Millas80 60 40 20 0 80 60 40 20 0
Est 60 59 58 57 56 Est 60 59 58 57 56
a) b) c)
50 50 50
2019
18
17
16
15
14
34.95
34.9 1
2
3
0,5
O2 (mL/L)
Figura 3.- Distribución vertical de a) Temperatura, b) Salinidad y c) Oxígeno frente a Punta Aguja en diciembre 1984. (de Icochea et al. 1989)Figure 3. - Vertical distribution of a) temperature, b) salinity and c) off Punta Aguja Oxygen in December 1984. (from Icochea et al. 1989)
5°
10°
15°
PAITA
PTA AGUJA
CHICAMA
CHIMBOTE
SUPE
CALLAO
PAITA
PTA AGUJA
CHICAMA
CHIMBOTE
SUPE
CALLAO
Sur
a) Enero - Mayo 1986
050
100150200250300
F M A M1986
TM TM/Hr
3295 1965 3161 1891 281611.6 15.9 10.9 8.2 7.7
b) Enero - Marzo 1987
050
100150200250300350
E F M
1987
TM TM/Hr
250714.3
167011.3
21.4319.6
E
Figura 4. Principales zonas de pesca y distribución vertical de las capturas de jurel T. murphyi a) Periodo enero – mayo 1986, b) Periodo enero – marzo 1987. (Modificada de Icochea et al. 1987)Figure 4. Main fishing areas and vertical distribution of Jack mackerel T. murphyi: a) Period January to May 1986, b) Period January to March 1987. (Modified from Icochea et al. 1987)
0m
50
100
150
200
250
300
35050
Millas
100 0
26 34.75 34.80 25 2134.9 24
35.020
3.0
192.0
18
17
16
15
35.05
35.05
35.01.0
14
1312
Cr. R/V D. MENDELEVChicama 2/13 Feb 1987T (°C)S (ups)O2M/P RIO CAONAO
Pro
fund
idad
(m)
Figura 5.- Distribución vertical de los cardúmenes de jurel T. murphyi, en relación con los parámetros oceanográficos de temperatura, sali-nidad y oxígeno frente a Chicama (2/13 Febrero 1987).Figure 5.- Vertical distribution of Jack mackerel T. murphyi schools in relation to oceanographic parameters of temperature, salinity and oxygen off Chicama (February 2/13, 1987).
frente oceánico. En este segundo patrón, el valor de 1.0 mL/L de oxígeno se ubica aproximadamente a los 50 m de profundidad, lo que explica la presencia superficial de los cardúmenes de T. murphyi y la ausencia de este recurso a mayores profundidades.
El tercer patrón se observa cuando se refuerza la ESCC, detec-tada por la profundización a niveles cercanos a los 300 m de las isotermas de 14 °C y 15 °C cerca de la costa. Trachurus murphyi es capturado a profundidades mayores a 100 m en el día y cerca de la superficie durante la noche. Las capturas alcanzan valores más altos en el día, debido a que en la noche se encuentra más disperso en la superficie debido a la dispersión de su alimento (Fig. 4). Además se observan migraciones verticales diurnas hasta 300 m de profundidad favorecidas por la ausencia de una marcada termoclina y la presencia de aguas con alto contenido de oxígeno disuelto a mayores profundidades. El valor de oxí-geno de 1,0 mL/L alcanza profundidades cercanas a los 300 m y ocurre lo mismo con la isoterma de 14 °C. En este caso, los cardúmenes prefieren rangos de temperatura entre 15 y 20 °C, contenido de oxígeno entre 1.0 y 5.0 mL/L y de salinidad de 35.0 ups (Fig. 5).
Mediante la comparación de la distribución batimétrica de los cardúmenes en desove (con ovocitos hidratados), con los parámetros oceanográficos (temperatura, salinidad y oxígeno) de

70
Dioses
Rev. peru. biol. número especial 20(1): 067- 074 (Septiembre 2013)Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú.
Editores: Csirke J., R. Guevara-Carrasco & M. Espino
Millas
0m
50
100
150
200 150 100 50 0
LANCES L-23 L-21 L-20 L-18 L-22 L-19 L-43 L-42 L-41
BIC HUMBOLDT BIC SNP-1
SECCIÓN SAN JUAN
CRUCERO 8610-12
BIC HUMBOLDT
BIC SNP-1
T°C
S %
O2
J - JURELC - CABALLA
M - MEDUSAMy - MYCTOPHIDOS 9 - 10 NOV 86
J 84%C 10S 1M 4
J 76%C 9.5S 2.1M 12.54
J 51%C 7My 9M 33
35.1 35.0 5.0M 100% M 100%
J 94%C 6
J 19%S 4M 79
J 30%M 70
34.9
A 87%J 2My 1M 10
34.8
35.3
50
40
3020
10
18.0°
17.0°16.0°
15.0°
14.0°
35.32
34.8
S - SARDINAA - ANCHOVETA O - OTROS
Figura 6.- Distribución vertical de los cardúmenes en desove de jurel T. murphyi, en relación con los parámetros oceanográficos de tempe-ratura, salinidad y oxígeno frente a San Juan (9/10 noviembre 1986) (de Dioses et al. 1989).Figure 6.- Vertical distribution of Jack mackerel T. murphyi spawning schools in relation to oceanographic parameters of temperature, salinity and oxygen off San Juan (November, 9/10, 1986) (from Dioses et al. 1989).
0
50
100
150
200
250
300
1 2 3
J J
J
J
Pro
fund
idad
(m)
021 mL/L
021 mL/L
021 mL/L
1. El Niño 1982-19832. Post El Niño3. Presencia de la Extensión Sur de la Corriente de Cromwell
32
1
Figura 7. Patrones de distribución vertical de jurel T. murphyi en relación a la isooxígena de 1 mL/L (Modificada de Chirinos.1992)Figure 7. Vertical distribution patterns Jack mackerel T. murphyi in relation to oxygen isoline of 1.0 mL/ L (Modified from Chirinos. 1992)
la sección San Juan (Cr. 8610-12), Dioses et al. (1989) determi-naron que los cardúmenes de T. murphyi en desove se localizan entre las 100 y 150 millas de la costa, en el frente delimitado por las ACF, producto del intenso afloramiento de Punta San Juan (salinidades menores de 35.0 ups) y las ASS (salinidades mayores a 35.1 ups), con temperaturas y concentraciones de oxígeno mayores a 18 °C y 5,0 mL/L respectivamente. (Fig. 6)
La característica del desove de T. murphyi, de efectuarse en la capa superficial, sobre los 50 m y de requerir altos valores de oxigeno determinan que los cardúmenes en desove de T. murphyi sean pequeños y estén dispersos entre el dominio marítimo sur de Perú y los 14°S. Por ser esta especie desovadora de primavera no tiene mayor interacción con los desoves de anchoveta Engraulis ringens, sardina Sardinops sagax y caballa Scomber japonicus, por ocupar diferentes áreas de desove.
El desove en esta área dentro de la zona eufótica y por encima de los 50 m permite que las larvas de T. murphyi sean desplazadas hacia la zona del frente de las ACF con las ASS por efecto del intenso transporte de Ekman (fenómeno de deriva larvaria). Este frente se despliega por fuera de las 90 mn de la costa en julio, agosto y setiembre principalmente (Santander & Flores 1983). Esto también explica la disponibilidad estacional de este recurso cuando el frente oceánico se acerca a la costa, que lo hace más accesible a la pesquería.
Por otro lado, se ha observado que entre la isoterma de 15 °C y el contenido de 1.0 mL/L de oxígeno existe una importante relación que sirve como indicador de la presencia de esta especie para la flota pesquera. Sobre la base de este valor de contenido de oxígeno, cercano al mínimo (0.5 mL/L), se han esquematizado los tres patrones de comportamiento de la distribución vertical de los cardúmenes de T. murphyi (Fig. 7). Para el patrón 1, se observa que el tenor de 1.0 mL/L de oxígeno alcanza los 200 m de profundidad y los cardúmenes se ubican entre esta profun-didad y los 100 m, como ocurre durante los eventos El Niño
fuertes como en 1983. En el patrón 2, de fuertes afloramientos, el valor de 1.0 mL/L se halla sobre los 100 m de profundidad y los cardúmenes se presentan sobre los 75 m. En el patrón 3, característica de una marcada presencia de la ESCC (en ausencia de un evento El Niño o en presencia de eventos suaves o mode-rados), el valor de 1.0 mL/L alcanza profundidades cercanas a los 300 m y permite una amplia migración de los cardúmenes entre esta profundidad y la superficie.
La variabilidad ambiental de largo plazo afecta la distribu-ción y abundancia del T. murphyi a lo largo del mar peruano, tal como se puede observar en la distribución latitudinal de norte (Paita) a sur (Ilo) de las anomalías de la TSM entre 1950 y 2010 (Fig. 8). Además, el periodo cálido (1972 – 1998) es el más apto para la presencia de altas biomasas de T. murphyi, en

71
Patrones de distribución y abundancia deTrachurus murphyi en el Perú
Rev. peru. biol. special issue 20(1): 067- 074 (Septembre 2013)Ecology, fisheries and conservation of Jack mackerel (Trachurus murphyi) in Peru. Editors: Csirke J., R. Guevara-Carrasco & M. Espino
Figura 8. Anomalías de la temperatura superficial del mar de Paita a Ilo (1950 – 2010) y biomasas acústicas de jurel T. murphyi (1983 – 2010).Figure 8. Sea surface temperature anomalies from Paita to Ilo (1950 - 2010), and Jack mackerel T. murphyi acoustic biomass (1983 – 2010).
1996 1997 1998 1999 2000 2001 2002 2003 2004 2005 2006 2007 2008 2009 2010 2011 2012
-4°-6°-8°
-10°-12°-14°-16°-18°-20°
Figura 9. Distribución de las posiciones geográficas de las capturas de jurel T. murphyi registradas a bordo, durante las actividades de pesca de la flota de cerco entre 1996 – 2007. (Espinoza et al. 2008)Figure 9. Geographic distribution of recorded catches of Jack mackerel T. murphyi, on board the purse seine fleet, between 1996 and 2007. (Espinoza et al. 2008)
el cual destacan las mayores abundancias en años de El Niño, con notables disminuciones en años fríos.
Durante el periodo cálido (1972 – 1998) se presentaron los tres patrones de distribución, en tanto que en el actual periodo frío, se ha observado principalmente el patrón 2.
En cuanto a la variabilidad de la profundización de la isoterma de 15 °C, como indicador de flujos dirigidos hacia el sur, Flores et al. (2013) mencionan que las mayores profundizaciones se han producido durante los eventos El Niño 1982-83 y 1991-92, tanto al norte, centro y sur de la costa peruana. Esto nos indica una presencia fuerte de la ESCC, la cual favorece la disponibi-lidad y abundancia del jurel durante estas etapas. Después de estos eventos, los flujos dirigidos hacia el sur se repliegan hacia el norte. En este proceso se produce una disminución en la abundancia y disponibilidad de este recurso.
El acercamiento a la costa de las ASS (Flores et al. 2013) durante periodos cálidos de largo plazo (Espino & Yamashiro 2012, Chávez et al. 2003) favorece la presencia, disponibilidad y abundancia del T. murphyi. Eso se aprecia en las altas biomasas de la década de los 80 y mediados de los 90. A partir del año 2001, entre los 08 y 14°S, se produce una mayor disponibilidad de jurel, probablemente debido al acercamiento de las ASS a esta área y a la presencia de las AES al norte de los 08°S. (Fig. 9)
El análisis de la estacionalidad de las biomasas acústicas de T. murphyi por grados de latitud durante el periodo cálido 1972 – 1998 muestra un desplazamiento norte-sur desde el verano hasta la primavera (Fig. 10). Durante el verano, las mayores biomasas se ubican en el norte (05 – 11°S) mientras que en otoño se extienden tanto al norte, centro y sur. En invierno se observa una tendencia a ser más abundante en el sur y en pri-mavera se aprecia una mayor abundancia en la zona centro-sur, que coincide con la época principal del desove. Se destaca que al sur de los 14°S se encuentra una de las principales áreas de desove de T. murphyi en el mar peruano.
Después de El Niño 1997-98 a la fecha se viene experi-mentando a lo largo del mar peruano lo que se considera un periodo frío, durante el cual se observa un desplazamiento de las concentraciones de biomasa por latitud y estacionalmente hacia el sur de los 06°S. Los mayores valores de biomasa latitudinales se presentan en la zona centro-sur, especialmente al sur de los 11°S, con los valores más bajos en la estación de otoño (Fig. 11).
Dioses y Ñiquen (1988) observaron el aumento latitudinal, de sur a norte, más o menos gradual en la estructura por tamaños del T. murphyi en las capturas de muestreo realizadas durante un crucero científico en agosto-setiembre 1987 (Fig. 12B). Poste-riormente Dioses (1995) observó algo semejante en las capturas comerciales realizadas en enero-mayo 1990 (Fig. 12A). En ambos
1950 1972 1982
Periodo Frío
1997 2010
Periodo Cálido Periodo Frío
Paita
Chicama
Chimbote
Callao
Pisco
Ilo
4.00 − 6.00 2.00 – 4.00 0.00 – 2.00 -2.00 – 0.00
Anomalías Positivas de TSM en °C: Anomalías Negativas de TSM en °C:
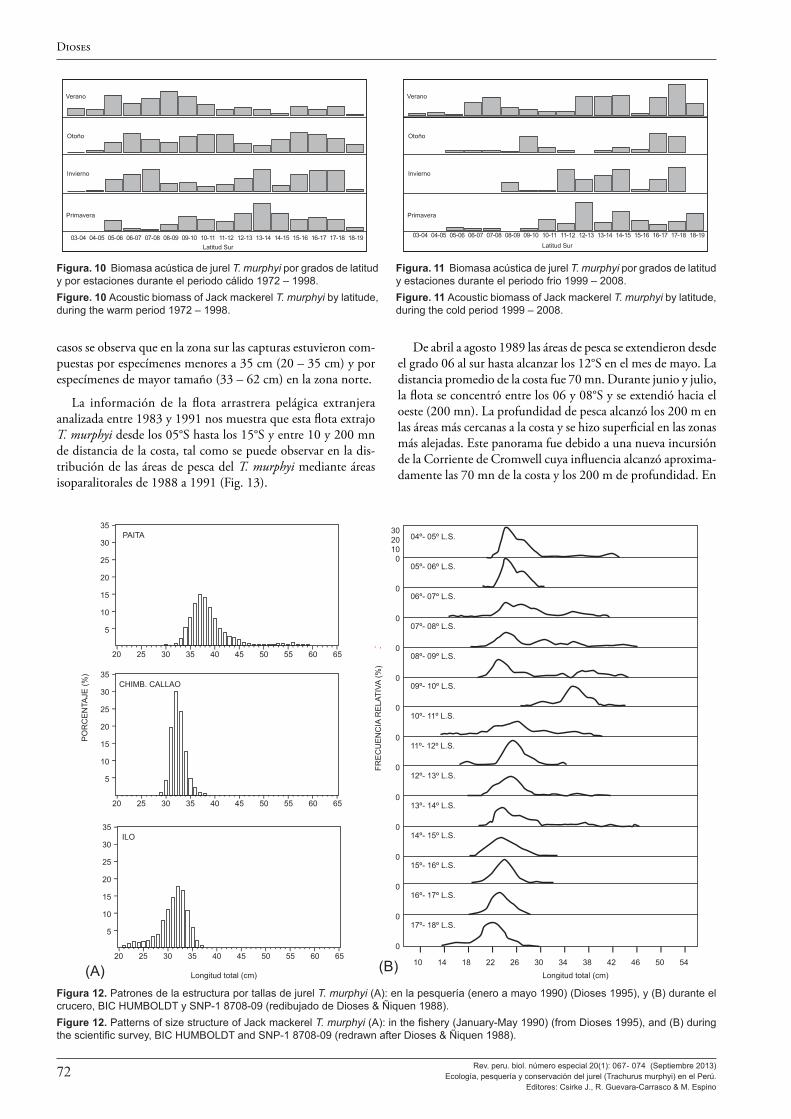
72
Dioses
Rev. peru. biol. número especial 20(1): 067- 074 (Septiembre 2013)Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú.
Editores: Csirke J., R. Guevara-Carrasco & M. Espino
Figura. 10 Biomasa acústica de jurel T. murphyi por grados de latitud y por estaciones durante el periodo cálido 1972 – 1998.Figure. 10 Acoustic biomass of Jack mackerel T. murphyi by latitude, during the warm period 1972 – 1998.
Verano
Otoño
Invierno
Primavera
03-04 04-05 05-06 06-07 07-08 08-09 09-10 10-11 11-12 12-13 13-14 14-15 15-16 16-17 17-18 18-19Latitud Sur
Verano
Otoño
Invierno
Primavera
03-04 04-05 05-06 06-07 07-08 08-09 09-10 10-11 11-12 12-13 13-14 14-15 15-16 16-17 17-18 18-19Latitud Sur
Figura. 11 Biomasa acústica de jurel T. murphyi por grados de latitud y estaciones durante el periodo frio 1999 – 2008.Figure. 11 Acoustic biomass of Jack mackerel T. murphyi by latitude, during the cold period 1999 – 2008.
PO
RC
EN
TAJE
(%)
Longitud total (cm) Longitud total (cm)
35
30
25
20
15
10
5
20 25 30 35 40 5045 55 60 65
PAITA
35
30
25
20
15
10
5
20 25 30 35 40 5045 55 60 65
CHIMB. CALLAO
35
30
25
20
15
10
5
20 25 30 35 40 5045 55 60 65
ILO
3020100
0
0
0
0
0
0
0
0
0
0
0
0
0
FRE
CU
EN
CIA
RE
LATI
VA (%
)
04º- 05º L.S.
05º- 06º L.S.
06º- 07º L.S.
07º- 08º L.S.
08º- 09º L.S.
09º- 10º L.S.
10º- 11º L.S.
11º- 12º L.S.
12º- 13º L.S.
13º- 14º L.S.
14º- 15º L.S.
15º- 16º L.S.
16º- 17º L.S.
17º- 18º L.S.
10 14 18 22 26 30 34 38 42 46 50 54(A) (B)
Figura 12. Patrones de la estructura por tallas de jurel T. murphyi (A): en la pesquería (enero a mayo 1990) (Dioses 1995), y (B) durante el crucero, BIC HUMBOLDT y SNP-1 8708-09 (redibujado de Dioses & Ñiquen 1988).Figure 12. Patterns of size structure of Jack mackerel T. murphyi (A): in the fishery (January-May 1990) (from Dioses 1995), and (B) during the scientific survey, BIC HUMBOLDT and SNP-1 8708-09 (redrawn after Dioses & Ñiquen 1988).
casos se observa que en la zona sur las capturas estuvieron com-puestas por especímenes menores a 35 cm (20 – 35 cm) y por especímenes de mayor tamaño (33 – 62 cm) en la zona norte.
La información de la flota arrastrera pelágica extranjera analizada entre 1983 y 1991 nos muestra que esta flota extrajo T. murphyi desde los 05°S hasta los 15°S y entre 10 y 200 mn de distancia de la costa, tal como se puede observar en la dis-tribución de las áreas de pesca del T. murphyi mediante áreas isoparalitorales de 1988 a 1991 (Fig. 13).
De abril a agosto 1989 las áreas de pesca se extendieron desde el grado 06 al sur hasta alcanzar los 12°S en el mes de mayo. La distancia promedio de la costa fue 70 mn. Durante junio y julio, la flota se concentró entre los 06 y 08°S y se extendió hacia el oeste (200 mn). La profundidad de pesca alcanzó los 200 m en las áreas más cercanas a la costa y se hizo superficial en las zonas más alejadas. Este panorama fue debido a una nueva incursión de la Corriente de Cromwell cuya influencia alcanzó aproxima-damente las 70 mn de la costa y los 200 m de profundidad. En

73
Patrones de distribución y abundancia deTrachurus murphyi en el Perú
Rev. peru. biol. special issue 20(1): 067- 074 (Septembre 2013)Ecology, fisheries and conservation of Jack mackerel (Trachurus murphyi) in Peru. Editors: Csirke J., R. Guevara-Carrasco & M. Espino
Pto. Pizarro
Paita
Pta. Sal
Pta. La Negra
Áreas de Pesca de Jurel - Pesca Arrastre1988 - 1991
Pimentel
ChicamaSalaverry
Chimbote
HuarmeySupeHuacho
ChancayCallao
PucusanaCerro Azul
PiscoBahía Independencia
Punta CaballasSan Juan
TicoQuilca
MollendoIlo
Sama
4º
5º
6º
7º
8º
9º
10º
11º
12º
13º
14º
15º
16º
17º
18º
19º
20º
21º
85º 80º 75º 70º
0 - 500 g500 - 1000t1000 - 5000t
Figura 13. Áreas de pesca de jurel T. murphyi de la flota de arrastre pelágico (abril 1988 – mayo1991).Figure 13. Jack mackerel T. murphyi fishing areas of the midwater trawl fleet (April 1988 – May1991).
2ºS
4ºS
6ºS
8ºS
10ºS
12ºS
14ºS
16ºS
18ºS
20ºS
Pta. SalTalaraPaita
Pta. La Negra
86ºW 84ºW 82ºW 80ºW 78ºW 76ºW 74ºW 72ºW 70ºW 68ºW
Crucero de investigaciónde calamar gigante
Kaiyo Maru, Nov-Dic 2007
Operaciones de pesca concaptura de jurel
Longitudes de losejemplares (cm)
Sin muestreo8 5 5
3 6 4 4
4
4
4
5
3 36
7
Pimentel
MalabrigoSalaverry
ChimboteCasmaHuarmey
SupeHuachoChancayCallao
PucusanaCerro AzulPiscoBahía Independencia
San Juan Marcona
AticoMollendo
IloSama
Figura 14.- Distribución de juveniles de jurel T. murphyi obtenidos con Red de media agua Nichimo Larva Catcher. Fuente: IMARPE 2008.Figure 14. – Distribution of juveniles of Jack mackerel T. murphyi obtai-ned with Nichimo Midwater Net Larva Catcher. Fuente:IMARPE 2008.
setiembre-octubre de 1989 la pesca fue mayormente superficial, favorecida por un intenso afloramiento.
Entre julio 1990 y febrero de 1991 ocurrió la distribución más amplia de todo el período analizado, del grado 06°S al 15°S. Trachurus murphyi estuvo bastante disperso entre las 30 y 200 mn de distancia a la costa. La ESCC se presentó entre Punta Falsa y Paita debilitada por el proceso de enfriamiento del invierno.
Al sur de los 15°S esta flota no operó, porque esta área es la zona habitual de desove y de cría de esta especie, siendo por lo tanto los jureles de esta zona pequeños y no rentables para este tipo de barcos factoría.
Es importante destacar que estas observaciones (estructura por tamaños y zona de desove) fueron realizadas entre los años 1972 y 1997, periodo decadal cálido (PDO+) y que muestran cierta segregación espacial por tamaños, con los especímenes más pequeños en el sur, lo que sugiere una migración hacia el norte durante el ciclo de vida de esta especie.
Actualmente, después de El Niño 1997-98, estamos viviendo un período decadal frío (PDO-), período en el cual venimos observando fuertes afloramientos con valores de oxigeno de 1 mL/L desde 80 – 100 m de profundidad a las 90 mn de distancia a la costa hasta 20 – 30 m de profundidad cerca a la costa. Con frentes apropiados para la captura de T. murphyi entre las 40 – 60 mn. Esta es una de las razones por la cual el T. murphyi se ubica más superficial en la actualidad, estando más disponible y accesible a la flota cerquera. Esta flota con sistema de refrigeración RSW y dedicada a su captura viene operando entre los 05°S y los 17°S y desde las 10 a 110 mn de la costa.
Entre junio y diciembre de 1998 debido a un fuerte afloramiento el recurso se ubicó entre 05 y 08°S, concentrándose en el grado 06°S. En junio – julio la flota operó dentro de las 70 mn, para luego desplazarse hacia áreas más alejadas alcanzando las 150 mn. La profundidad de pesca estuvo sobre los 100 m en este período.
En la última década se viene presentando un amplio domi-nio de las ACF retrayendo la ESCC, situación que ha causado cambios en las áreas de pesca, en la estructura latitudinal por tamaños y en las áreas de desove de T. murphyi. Las áreas de pesca, a partir del año 2000 se desplazaron de la zona norte hacia los 17°S hasta el año 2007, a partir del cual se ubican en la zona central, tal como observamos en la Figura 9. Comportamiento similar observamos con las biomasas acústicas para este mismo periodo frio. En cuanto al desove durante este periodo frio, observamos un desplazamiento del área principal del desove hacia el norte, tal como lo demuestra la captura de juveniles de T. murphyi (3 – 8 cm LT) mediante una red de media agua Nichimo Larva Catcher durante el Crucero Kaiyo Maru, de noviembre a diciembre 2007 (Fig. 14). De acuerdo con la serie de tiempo disponible del IGS de T. murphyi, este desove fue exitoso, el cual no se ha repetido en años posteriores al 2007, dando lugar a una cohorte de T. murphyi que ha soportado las capturas de los últimos años.
En cuanto a la distribución latitudinal de los tamaños, para el periodo enero-mayo 2011, la Figura 15, nos permite apreciar que en enero, febrero y marzo la pesquería se inició al norte entre los grados 06 y 08°S y las capturas tuvieron tallas entre 28 y 30 cm. En tanto que de abril a mayo, la pesquería se realizó entre los 11°S y 14°S con tallas entre 31 y 32 cm, evidenciando una progresión de tallas de norte a sur lo que indicaría que en este último periodo los reclutamientos se estarían produciendo en este sentido; es decir de norte a sur.
La importancia de la dinámica de las masas de agua frente a la costa peruana y su relación con la disponibilidad de T. murphyi se corrobora una vez más en la distribución de las áreas de pesca y la dominancia de esta especie en las capturas industriales del año 2008, especialmente en marzo. Esas capturas están relacionadas con la aproximación de los frentes subtropical y/o ecuatorial hacia la costa peruana (Fig. 16).

74
Dioses
Rev. peru. biol. número especial 20(1): 067- 074 (Septiembre 2013)Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú.
Editores: Csirke J., R. Guevara-Carrasco & M. Espino
33
32
31
30
29
28
Grados de Latitud
Mod
as (c
m)
-14 -13 -12 -11 -10 -9 -8 -7 -6
5
4
2
3
1
Figura 15.- Modas por grados de latitud de jurel T. murphyi en el mar peruano (enero – mayo 2011, números 1 a 5) (Espino, com. pers.).Figure 15. - Mode by degrees latitude of Jack mackerel T. murphyi in the Peruvian sea (January – May 2011, numners 1 to 5) (Espino, pers. com.).
0º
1º
2º
3º
4º
5º
6º
7º
8º
9º
10º
11º
12º
13º
14º
15º
16º
17º
18º
19º
20º
Puerto PizarroMáncora
TalaraPaitaParachique
Santa RosaChicama
SalaverryCoishcoChimboteCasmaCulebras
SupeChancayCallaoPucusana
Cerro AzulT. De MoraBahía IndependenciaInfiernillos
San JuanChala
TicoCamaná
MataraniIloSama
Zonas de pescadel jurelATS
AES
ASS
Acf
Figura 16. Distribución de las zonas de pesca de jurel T. murphyi y su relación con las masas de agua frente a Perú en marzo 2008. ASS (>35.1 ups) en gris punteado, AES (33.8 a 34.8 ups) en negro entero y ATS (>33.8 ups) en gris entero.Figure 16. Distribution of Jack mackerel T. murphyi fishing areas and their relationship with the water masses of Peru in March 2008. ASS (> 35.1 psu) in gray dotted line , AES (from 33.8 to 34.8 psu) in black line, and ATS (> 33.8 psu) in gray line.
En síntesis, de este estudio podemos confirmar que el hábi-tat preferido por T. murphyi es el frente oceanográfico de alta productividad (Dioses et al. 1989), formado por las ASS con las ACF. Este frente determina la abundancia, cercanía o profundi-zación del recurso T. murphyi, y probablemente de otras especies pelágicas de importancia comercial. Al sur de los 6°S, las aguas con 35.0 a 35.1 ups tienen mayor relación con las ACF típicas y las ASS, mientras que al norte de los 6°S, la mezcla se produce entre las ACF típicas, AES y ASS (Morón 2011). Estas mezclas presentan variaciones espacio-temporales durante el año y sobre ellas ejerce un importante rol la intensidad de los vientos alisios que influyen directamente en el afloramiento costero (Morón 2011), así como en la dinámica del ENSO. Estos y otros aspectos deberán ser profundizados para mejorar nuestro entendimiento del comportamiento de especies como el T. murphyi.
Literatura citadaChávez F.P., J. Ryan,S.E. Lluch-Cota & M. Ñiquen. 2003. From anchovies to
sardines and back: Multidecadal change in the Pacific Ocean. Review: Climate. Science 299: 217-221. DOI: 10.1126/science.1075880
Chávez, F. P., A. Bertrand, R. Guevara-Carrasco, P. Soler, & J. Csirke. 2008. The northern Humboldt Current System: Brief history, present status and a view towards the future. Progress in Oceanography 79 (2008) 95-105. DOI: 10.1016/j.pocean.2008.10.012
Chirinos J. G. 1992. La pesca del jurel (Trachurus murphyi) y la caballa (Scomber japonicus peruanus) por la Flota Arrastrera Cubana, en el periodo enero 1986-marzo 1987. Tesis para optar el título de ingeniero pesquero. Universidad Nacional Agraria La Molina. Facultad de Pesquería.
Dioses T., V. H. Alarcón, M. E. Nakama & A. Echevarría. 1989. Desarrollo ovocitario, fecundidad parcial y distribución vertical de los cardú-menes en desove de jurel, Trachurus murphyi (N). En: Memorias del Simposio Internacional de los Recursos Vivos y las Pesquerías en el Pacífico Sudeste. Viña del Mar, 9 – 13 Mayo 1988. Rev. Pacífico Sur (Número Especial), pp. 287 – 294.
Dioses T. 1995. Análisis de la distribución y abundancia de los recursos jurel y caballa frente a la costa peruana. Inst. Mar Perú. Inf. Progresivo 3. pp. 55
Dioses T. & M. Ñiquen, 1988. Aspectos generales sobre la biología y pesquería de recurso jurel (Trachurus murphyi) en Perú. En FORUM: Ca-racterísticas en el diseño de una embarcación ideal para la captura de especies pelágicas7. PESCA PERU. 36 pp.
Espino M. 2003. Estrategia de gestión oriental para el Pacífico Oriental con especial mención a la pesquería peruana. Tesis para optar el grado de Magíster en Geografía con mención en Ordenamiento y Gestión Ambiental. Unidad de Post Grado, Facultad de Ingeniería Geológica, Minera, Metalúrgica y Geográfica, Universidad Nacional Mayor de San Marcos, Lima. 102 pp.
Espino M. & C. Yamashiro. 2012. La variabilidad climática y las pesquerías en el Pacífico Suroriental. Lat. Am. J. Aquat. Res., 40(3): 705-721, 2012 Latin American Journal of Aquatic Research 705. International Conference: “Environment and Resources of the South Pacific”. P.M. Arana (Guest Editor) DOI: 103856/vol40-issue3-fulltext-18.
Espinoza E., M. Ñiquen & Flores R. 2008. Changes in the latitudinal and longitudinal distribution of the aggregations of Jack mackerel (Trachurus murphyi) in the Peruvian sea during 1996 – 2007. SPRFMO Chilean Jack Mackerel Workshop, Santiago, Chile, 30 June 4 - July 2008. SPRFMO/CHJMWS Paper 4: 11 pp (http://www.southpacificrfmo.org/chilean-Jack-mackerel-workshop-Meeting-Documents/)
Flores R., J. Tenorio & N. Domínguez. 2009. Variaciones en la Extensión Sur de la Corriente de Cromwell frente al Perú entre los 3 y 14° S. Bol. Inst. Mar Perú. 24(1–2):45-58.
Flores R., M. Espino, G. Luque & J. Quispe. 2013. Patrones de variabilidad ambiental en el mar peruano. En: Csirke J., R. Guevara & M. Espino (Eds.). Ecología, pesquería y conservación del jurel (Trachurus mur-phyi) en el Perú. Rev. peru. biol. número especial 20(1): 021 – 028
Ganoza F., A. 1998. Abundancia y comportamiento del jurel (Trachurus picturatus murphyi) a través de proyecciones acústicas de 1983 a 1994. Inf. Inst. Mar Perú 136: 23-47.
Icochea L., A. Chipollini & M. Ñiquen. 1989. Análisis de la pesquería de arrastre pelágica en la costa peruana durante 1983-1987 y su relación con el medio ambiente. En: Memorias del Simposio Internacional de Recursos Vivos y las Pesquerías en el Pacífico Sudeste. Viña del Mar, 9 – 13 Mayo 1988. Rev. Pacífico Sur (Número especial), pp. 455-465.
IMARPE. 2008. Informe ejecutivo del crucero de investigación del calamar gigante BIC Kaiyo Maru 0711-12, noviembre-diciembre 2007. Informe Ejecutivo IMARPE: 14 pp.
Morón O. 2011. Climatología de la salinidad superficial del mar frente a la costa peruana. 1960 – 2008. Inf. Inst Mar Perú Vol. 38 N° 1: 07 - 39. Enero-marzo 2011.
Santander H. & R. Flores. 1983. Los desoves y distribución larval de ciertas especies pelágicas y sus relaciones con las variaciones del ambiente marino frente al Perú. FAO Fish Report / FAO Inf. Pesca (291), 3: 835 – 867.

75
Ecología trófica del jurel Trachurus murphyi en el Perú
Rev. peru. biol. special issue 20(1): 075- 082 (September 2013)Ecology, fisheries and conservation of Jack mackerel (Trachurus murphyi) in Peru. Editors: Csirke J., R. Guevara-Carrasco & M. Espino
Rev. peru. biol. número especial 20(1): 075- 082 (Septiembre 2013)Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú. Editores: Csirke J., R. Guevara-Carrasco & M. Espino
Facultad de ciencias Biológicas unMsMISSN-L 1561-0837
Ecología trófica del jurel Trachurus murphyi en el Perú entre 1977 – 2011
Ana Alegre, Pepe Espinoza y Marco Espino
Trophic ecology of Jack mackerel Trachurus murphyi in Peru, 1977 – 2011
Instituto del Mar del Perú, esquina Gamarra y General Valle S/N Chucuito, Callao, Perú.
Email Ana Alegre: [email protected]
ecología del jurel (Trachurus murphyi) en el Perú
© Los autores. Publicado por la Revista Peruana de Biología de la Facultad de Ciencias Biológicas, Universidad Nacional Mayor de San Marcos. Este es un artículo de acceso abierto, distribuido bajo los términos de la Licencia de Atribución Reconocimiento-NoComercial-SinObraDerivada 3.0 de Creative Commons (http://creativecommons.org/licenses/by-nc-nd/3.0/deed.es_ES), que permite el uso no comercial, distribución y reproducción en cualquier medio, siempre que la obra original sea debidamente citadas. Para uso comercial, por favor póngase en contacto con [email protected].
Citación:Alegre A., P. Espinoza & M. Espino. 2013. Ecología trófica del jurel Trachurus murphyi en el Perú entre 1977-2011. En: Csirke J., R. Guevara-Carrasco & M. Espino (Eds.). Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú. Rev. peru. biol. número especial 20(1): 075- 082 (Septiembre 2013)
ResumenSe analizó la variabilidad espacio-temporal de la dieta de jurel Trachurus murphyi frente a Perú de 1977 a 2011. Un total de 21058 estómagos fueron analizados utilizando métodos bivariados y el Modelo de Árbol de Regresión Multivariado (ARM). Se utilizó como variable respuesta la composición gravimétrica de la dieta, con respecto a diferentes variables pre-dictivas como años, profundidad de la isoterma de 15 °C, Índice de Oscilación del Sur (IOS), Índice de Oscilación Decadal del Pacífico (PDO), anomalía de la temperatura superficial del mar, zona (norte, centro y sur) y distancia de la costa. Nuestros resultados nos permitieron identificar dos escenarios diferentes, uno desde el año 1977 al 2000, donde la dieta del jurel T. murphyi estuvo fuertemente dominada por Euphausiidae, y otro escenario después del 2000, donde el jurel T. murphyi consumió mayor diversidad de presas, principalmente Euphausiidae, camaroncito rojo Pleuroncodes monodon y Zoeas. Se destaca que el primer escenario cor-responde a condiciones cálidas y el segundo a condiciones frías, ambas coincidentes con los periodos multidecadales. A diferencia de estudios anteriores que muestran que la ancho-veta Engraulis ringens es una presa abundante, en este estudio no fue significativamente importante. Espacialmente, los Euphausiidae dominaron la dieta en el norte-centro de Perú, mientras que al sur, la composición de las presas fue más diversa. La dieta también varió significativamente según la distancia a la costa, con mayor proporción de P. monodon cerca a la costa y con respecto a su etapas de desarrollo ontogenético.
Palabras clave: Jurel, alimentación, Perú.
AbstractWe analyzed the spatiotemporal variability of Jack mackerel Trachurus murphyi diet off Peru from 1977 to 2011. A total of 21058 stomachs were analyzed using bivariate methods and the Multivariate Regression Tree Model (MRT). The gravimetric composition of the diet was used as response variable with respect to different predictor variables such as years, depth of the 15°C isotherm, Southern Oscillation Index (SOI), Index of the Pacific Decadal Oscillation (PDO), sea surface temperature anomaly, zone (north, center and south) and distance to the coast. Our results allowed us to identify two different scenarios. From 1977 to 2000 the Jack mackerel T. murphy diet was strongly dominated by Euphausiidae, and since 2000 when T. murphyi consumed a greater diversity of prey, mainly Euphausiidae, squat lobster Pleuron-codes monodon and Zoea. It is noted that the first scenario corresponds to warm conditions and the second to cold conditions, both coincident with observed multidecadal periods. In contrast to previous studies showing that the anchovy Engraulis ringens is an abundant prey, in this study anchovy was not significantly important. Spatially, the Euphausiidae dominated the diet in central-northern Peru, while further south the prey composition was more diverse. The diet also varied significantly according to the distance from the coast (higher proportion of P. monodon closer to the coast) and the ontogenic stages.
Keywords: Jack mackerel, food, Peru.
IntroducciónEl jurel Trachurus murphyi Nichols 1920 es uno de los componentes de la red trófica
marina peruana que se asocia principalmente con el subsistema pelágico oceánico, y junto a la anchoveta Engraulis ringens, sardina Sardinops sagax sagax y caballa Scomber japonicus han posibilitado el desarrollo de una de las principales actividades pesqueras del ámbito mundial, localizada frente al Perú.
Cada vez con mayor frecuencia se recomienda que los administradores pesqueros deban tener en cuenta las más amplias consideraciones ecológicas en el manejo de los
Publicado online: 11/10/2013Publicado impreso: 15/10/2013

76
Alegre et al.
Rev. peru. biol. número especial 20(1): 075- 082 (Septiembre 2013)Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú.
Editores: Csirke J., R. Guevara-Carrasco & M. Espino
stocks de peces explotados (Botsford et al. 1997). Dentro de las consideraciones ecológicas, se menciona que las relaciones tróficas de las especies de importancia comercial representan uno de los principales aspectos a estudiar.
Un método común para estudiar las relaciones tróficas es el que se basa en los análisis de los contenidos estomacales (Caddy & Sharp 1988). Mediante este método, se puede conseguir información de utilidad que nos aproxime a cuantificar el flujo de energía en la red trófica marina, identificándose las especies de mayor relevancia en dicha estructura, sea como predadores o presas.
Trachurus murphyi ha sido definido como un depredador del zooplancton y de peces pelágicos tales como la anchoveta E. rin-gens (Sánchez de Benites et al. 1985). También, se le ha definido como un predador facultativo en el ecosistema pelágico de Perú (Konchina 1980 y 1983) debido a su naturaleza migratoria, que lo lleva a acceder a ambientes neríticos y oceánicos epipelágicos para alimentarse (Medina & Arancibia 2002). En los últimos años se han observado cambios notables en las capturas, así como en la abundancia, distribución y disponibilidad espacial del jurel, lo que ha llevado a interrogarnos sobre las posibles consecuencias en su rol trófico. En el presente estudio se postula que el jurel T. murphyi es una especie altamente plástica en su dinámica de alimentación y para entender su rol trófico se analiza la información colectada para el mar peruano desde 1977 a 2011.
Material y métodos Muestreo.- La muestra está conformada por datos de 21058
estómagos de jurel T. murphyi de los cuales en 8693 (41,3%) se
encontró contenido. Las muestras de estómagos fueron colec-tadas de los cruceros de investigación de IMARPE, operaciones Eureka y la flota industrial, cuyas capturas se realizaron a lo largo de todo el litoral peruano (zonas de Paita, San José, Chimbote, Huacho, Callao, Pisco e Ilo), desde 1,5 a 205 mn de distancia a la costa, desde 1977 a 2011 (Fig. 1).
La colecta de estómagos se efectuó de manera estratificada, para lo cual se consideraron todos los rangos de talla registrados en el momento de la captura, en las principales zonas de pesca. Las muestras colectadas entre 1977 y 1992 provienen de opera-ciones Eureka y cruceros de investigación de recursos pelágicos y las muestras colectadas de 1994 al 2011 son procedentes de la flota industrial.
Los estómagos colectados en la zona del Callao fueron fijados en alcohol al 96% y los estómagos procedentes de los laborato-rios costeros (San José, Chimbote, Huacho, Callao, Pisco e Ilo), cruceros de investigación y operaciones Eureka fueron fijadas en formol al 10%.
Análisis de las muestras.- Los contenidos estomacales fueron pesados con precisión al 0.01 g y las presas fueron diferenciadas con microscopio estereoscópico al 10X de aumento, llegando al mínimo taxón posible mediante el empleo de listas y claves taxonómicas especializadas para teleósteos (Fitch & Brownell 1968, García-Godos 2001, Chirichigno 1998), cefalópodos (Wolf 1984), crustáceos (Newel 1963, Méndez 1981), zoo-plancton (Trégouboff & Rosse 1957) y moluscos (Álamo & Valdivieso 1987).
Tratamiento de los datos
Composición de la dieta.- El índice gravimétrico fue calcula-do como el porcentaje en peso de las presas (Berg 1979, Hyslop 1980) y agrupadas según años (de 1977 al 2011) por intervalos de talla (cada 10 cm), estadios de madurez (8 estadios), latitud (por estratos de 1°S) y distancia a la costa (desde la línea de costa, por estratos de cada 10 mn):
%P = pp * pt –1*100
Donde:
%P = índice gravimétrico (%)pp = peso de la presa (g)pt = peso total del depredador (g)
Índice de llenura estomacal.- Con respecto al peso evis-cerado para cada individuo, el índice de llenura estomacal se calculó mediante la fracción del peso de contenido estomacal, de acuerdo a la relación de Hureau (1970), modificado por Rasero et al. (1996):
F= (pce * 100) / (pt – pce)
Donde:
F = índice de llenura estomacal (%)pce = peso de contenido estomacal (g)pt = peso total del individuo (g)
En función a este índice se agrupó la información disponible calculando la media aritmética (sin considerar los estómagos vacíos), según latitud, distancia a la costa y talla. Con esta in-formación se realizaron comparaciones múltiples entre el índice de llenura estomacal en función a talla (cada 10 cm), estadios
Figura 1. Distribución de las muestras de estómagos de jurel T. murphyi frente a Perú de 1977 al 2011.Figure 1. Distribution of stomach samples of Jack mackerel T. mur-phyioff Peru, 1977 to 2011.

77
Ecología trófica del jurel Trachurus murphyi en el Perú
Rev. peru. biol. special issue 20(1): 075- 082 (September 2013)Ecology, fisheries and conservation of Jack mackerel (Trachurus murphyi) in Peru. Editors: Csirke J., R. Guevara-Carrasco & M. Espino
de madurez, latitud (por estratos de 1°S) y distancia a la costa (desde la línea de costa por estratos de 10 mn), para validar los resultados de estas comparaciones se utilizó la prueba estadística no paramétrica de Kruskal-Wallis (Iman et al. 1975).
Árbol de Regresión Multivariado.- Este método forma gru-pos homogéneos respecto al comportamiento de variables pre-dictivas con respecto a la variable respuesta (De’ath & Fabricius 2000). La ventaja de usar este método es que no es necesario que las variables tengan una distribución normal (Zuur et al. 2009) lo que se ajusta muy bien a los datos de contenido estomacal. El proceso de determinar el mejor tamaño de árbol es llamado “poda” (Breiman et al., 1984). La poda del árbol se puede llevar a cabo mediante varios criterios. Un criterio primario es utilizar el parámetro de complejidad (cp). El cp es un parámetro de aviso y está especificado de acuerdo a la fórmula:
RSScp=RSS+CP * size of tree
RSS es la suma de los cuadrados de los residuos y size of tree es el tamaño del árbol (número de divisiones más 1). Un valor de cp=1 resultará en un árbol sin divisiones. El valor de cp=0.01 es un valor razonable para una poda inicial del árbol, pero algunas veces se sobreestima la poda sobre todo para conjuntos grandes de datos. Sin embargo, se puede obtener podas más confiables optimizando el valor de cp mediante validación cruzada (CV). Para el presente trabajo, se usó como variable respuesta la composición de grupos dietarios en proporción gravimétrica denominada “Grupo” y como variables predictivas el año, pro-fundidad de la isoterma de 15 °C, Índice de Oscilación Decadal
del Pacífico (PDO), zonas (norte, centro y sur) y distancia a la costa en millas náuticas(mn). Donde:
Grupo ≅ Año+Isot 15°C+PDO+Zona+Distancia (mn)
ResultadosCambios en la dieta
1. Temporales.- En términos de porcentaje en peso húmedo, la composición de la dieta varió con respecto al tiempo. En la Figura 2 se observan dos escenarios en la composición de la dieta en términos de porcentaje en peso húmedo: el primero que co-menzó en 1977 hasta el año 2000, siendo la especie dominante los Euphausiidae (%W≤ 96.0), después del periodo interdecadal cálido y el ENSO 1997-98 (Chávez et al. 2008). A partir del 2000 aumentó la proporción de zoeas (%W≤ 45.1) y P. monodon (%W≤ 61.8), indicador de Aguas Costeras Frías (ACF), mientras que la presencia de Euphausiidae disminuyó (%W≤ 55.8) sin dejar de ser representativa. El índice de llenura fue variable en todo el periodo de estudio (p-valor < 0.0001; H15= 408.79) (Fig. 2).
2. Según la talla.- Respecto de la talla, en los individuos menores de 21 cm de LT, la dieta estuvo compuesta mayormente por Copepoda (%W= 34.0), la proporción de Euphausiidae fue aumentando con la talla hasta los 50 cm de LT (%W= 27.0 – 49.5). A partir de 51 cm de LT, aumentó la ingesta de peces mesopelágicos (%W= 15.8), otros Teleosteii (%W= 16.9) y Mollusca (%W= 10.8). El índice de llenura estomacal disminuyó en relación al incremento del tamaño (p-valor < 0.0001; H4= 217.14) (Fig. 3).
Figura 2. PDO, Índice de llenura estomacal y proporción de presas en estómagos de jurel T. murphyi frente a Perú de 1977 al 2011.Figure 2. PDO, Index of fullness and proportion of prey in stomachs of Jack mackerel T. murphyi off Peru from 1977 to 2011.
100%
80%
60%
40%
20%
0%
1977
1978
1979
1980
1981
1982
1983
1984
1985
1986
1987
1988
1989
1990
1991
1992
1993
1994
1995
1996
1997
1998
1999
2000
2001
2002
2003
2004
2005
2006
2007
2008
2009
2010
2011
Años
% P
eso
Mollusca Copepoda Euphaussidae Pleuroncodes monodon
Engraulis ringens
Otros
Zoea MesopelágicosCrustacea
Teleosteii Tamaño de muestra
4.0
3.0
2.0
1.0
0.0
% L
lenu
ra3.0
2.0
1.0
0.0
-1.0
-2.0
PD
O
Escenario 1 Escenario 2
6477 20127 144 173
351 282180
432 386232
21 18100
268169 170
93 54
573
296
806
1009
348
944
418314
185
236 203

78
Alegre et al.
Rev. peru. biol. número especial 20(1): 075- 082 (Septiembre 2013)Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú.
Editores: Csirke J., R. Guevara-Carrasco & M. Espino
3. Según el sexo y madurez gonadal.- No se presentan diferencias según el sexo en la composición de la dieta. Sin em-bargo, se observan ligeros cambios respecto al índice de llenura estomacal, que fue mayor en los machos (p-valor< 0.0001; H1= 34.37). La dieta respecto a la madurez tuvo un comportamiento similar a lo observado con la talla, pero se puede apreciar que en los estadios del 1 al 3, la proporción de zoeas (%W= 18.1 – 22.2) y P. monodon (%W=11.4 – 14.2) son más altas que en los siguientes estadios de madurez. La proporción de Euphausiidae fue alta en los estadios del 4 al 6 (%W= 55.0 – 59.9). El índice de llenura disminuyó desde el estadio 1 al estadio 6, a partir del cual se aprecia un ligero aumento (p-valor < 0.0001; H7= 131.46), lo que corresponde al periodo de desove y recuperación (Fig. 4).
4. Según la distribución espacial.- En la Figura 5, con relación a la latitud, se observó que los Copepoda fueron las presas más destacadas (%W= 77.9) en los 03°S. Esta zona está fuertemente influenciada por las Aguas Ecuatoriales Superficiales (AES) y las Aguas Superficiales Tropicales (AST) (Morón et al. 2001). A partir de la ampliación de la plataforma continental (de 04 a 09°S), los Euphausiidae constituyeron las presas do-minantes (%W= 59.1 – 86.8). En la zona centro-sur, la dieta se tornó más variada, las zoeas (%W= 7.5 – 25.7) y P. monodon (%W= 4.6 – 20.0) fueron muy importantes. En el sur, entre los 17 y 18°S, se observó un ligero aumento de peces mesopelágicos (%W= 10.7 – 12.8); asimismo, el índice de llenura estomacal a lo largo del litoral fue variable (p-valor < 0.0001; H15= 408.79).
Hasta las 120 mn, los Euphausiidae (%W= 35,1 – 60.1) fueron las presas más representativas en la dieta de T. murphyi; sin embargo, P. monodon fue importante en términos de propor-ción en peso hasta 40 mn (%W= 17 9 – 11.7). A partir de 91 mn, la proporción de las presas fue más variada y destacaron los Copepoda (%W= 12.1 – 22.8), las zoeas (%W= 9.0 – 46.5) y los Teleosteii en la dieta (%W= 10.3 – 24.7). Asimismo, se observó después de las 130 mn un ligero incremento en la proporción de mesopelágicos en la dieta (%W= 12.9 – 13.4) (Fig. 6). El índice de llenura estomacal disminuyó respecto a la distancia a la costa, excepto a partir de las 131 mn (p-valor < 0.0001; H12= 352.31). Cabe destacar que en estómagos del jurel, P. monodon estuvo presente hasta las 80 mn entre el 2004 y 2008.
Relación recurso-ambienteEn el Árbol de Regresión Multivariado (Fig. 7), los resulta-
dos del árbol de regresión podado (cp= 0.012) muestran que la variable predictiva más importante fue el tiempo, con un nivel de importancia de 39.59%. Esta variable discrimina 2 periodos en el comportamiento trófico de T. murphyi: el primero, desde 1977 al 2000, en el que se tiene a Euphausiidae como la presa más representativa; y, el segundo, del 2000 hacia adelante, carac-terizado principalmente por tener a Euphausiidae, P. monodon y zoea, como presas muy importantes. En este segundo periodo, surge nuevamente el tiempo como variable discriminante y clasifica el comportamiento alimentario de T. murphyi para un corto periodo entre 2000 y 2004, en el que la profundidad de la isoterma de 15 °C es la variable predictora de importancia (16.61%). Por esta variable, se distinguen dos agrupaciones: una cuando la isoterma de 15°C se encuentra a más de 88 m de profundidad y es caracterizada por la dominancia de P. monodon en la dieta, y la otra agrupación caracterizada por la ingesta de Copepoda, Euphausiidae y Teleosteii.
Figura 3. Índice de llenura estomacal y proporción de presas en estó-magos de jurel T. murphyicon respecto a la talla (cm) de 1977 al 2011.Figure 3. Stomach fullness index and proportion of prey in stomachs of Jack mackerel T. murphyi in relation to size (cm) from 1977 to 2011.
Figura 4. Índice de llenura estomacal y proporción de presas en estómagos de jurel T. murphyicon respecto al estadio de madurez de 1977 al 2011.Figure 4. Stomach fullness index and proportion of prey in stomachs of Jack mackerel T. murphyi in relation to maturity stage from 1977 to 2011.
100%
80%
60%
40%
20%
0%
Talla (cm)
% P
eso
Mollusca
Copepoda
Euphaussidae
Pleuroncodes monodon
Engraulis ringens
Otros
Zoea
Mesopelágicos
Crustacea
Otros Teleosteii
Tamaño de muestra
4.00
3.00
2.00
1.00
0.00
% L
lenu
ra
5000
4000
3000
2000
1000
0
Nº d
e es
tóm
agos
219
<21] [21-30] [31-40] [41-50] >51]
3273
4486
643
72
100%
80%
60%
40%
20%
0%
Estadío de madurez
% P
eso
Mollusca
Copepoda
Euphaussidae
Pleuroncodes monodon
Engraulis ringens
Otros
Zoea
Mesopelágicos
Crustacea
Otros Teleosteii
Tamaño de muestra
3.00
2.00
1.00
0.00
% L
lenu
ra
4000
3000
2000
1000
0
Nº d
e es
tóm
agos
174
1 2 3 4 5
1853
3526
821513
852
389189
6 7 8
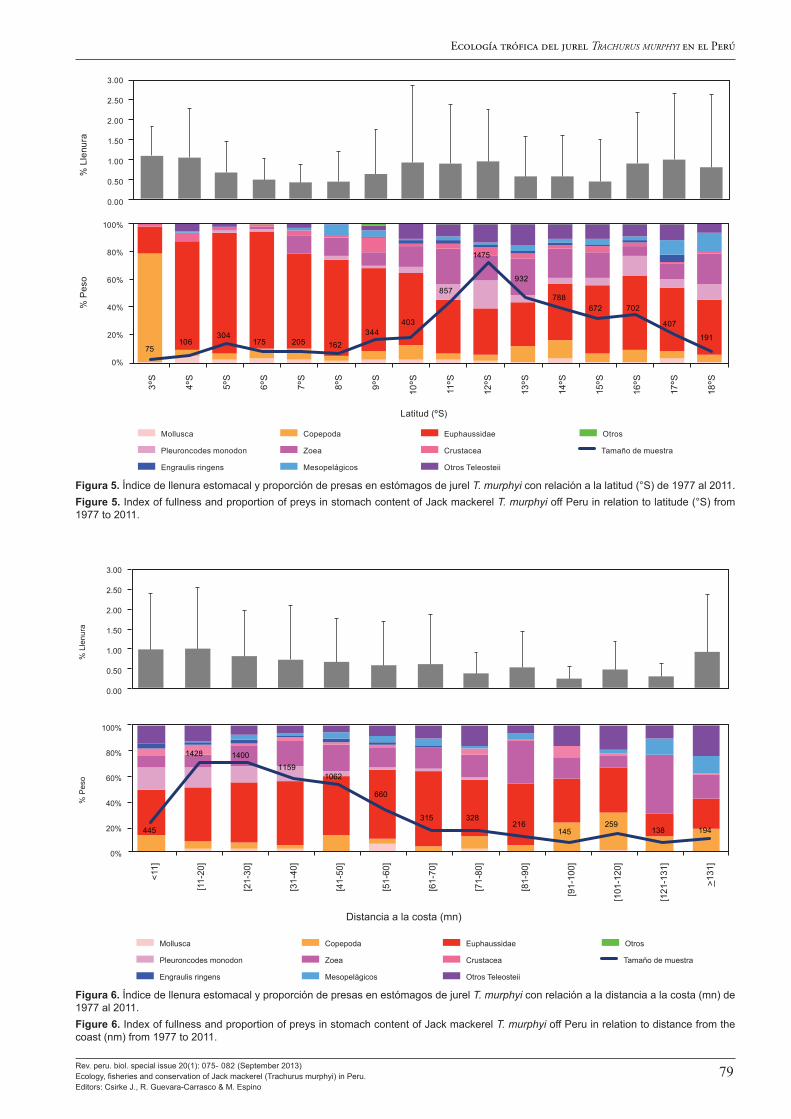
79
Ecología trófica del jurel Trachurus murphyi en el Perú
Rev. peru. biol. special issue 20(1): 075- 082 (September 2013)Ecology, fisheries and conservation of Jack mackerel (Trachurus murphyi) in Peru. Editors: Csirke J., R. Guevara-Carrasco & M. Espino
Figura 5. Índice de llenura estomacal y proporción de presas en estómagos de jurel T. murphyi con relación a la latitud (°S) de 1977 al 2011.Figure 5. Index of fullness and proportion of preys in stomach content of Jack mackerel T. murphyi off Peru in relation to latitude (°S) from 1977 to 2011.
Figura 6. Índice de llenura estomacal y proporción de presas en estómagos de jurel T. murphyi con relación a la distancia a la costa (mn) de 1977 al 2011.Figure 6. Index of fullness and proportion of preys in stomach content of Jack mackerel T. murphyi off Peru in relation to distance from the coast (nm) from 1977 to 2011.
100%
80%
60%
40%
20%
0%
3ºS
4ºS
5ºS
6ºS
7ºS
8ºS
9ºS
10ºS
11ºS
12ºS
13ºS
14ºS
15ºS
16ºS
17ºS
18ºS
Latitud (ºS)
% P
eso
Mollusca Copepoda Euphaussidae
Pleuroncodes monodon
Engraulis ringens
Otros
Zoea
Mesopelágicos
Crustacea
Otros Teleosteii
Tamaño de muestra
3.00
2.50
2.00
1.50
1.00
0.50
0.00
% L
lenu
ra
75106
304175 205 162
344403
857
1475
932
788672 702
407
191
445
1428 14001159
1062
660
315 328216
145259
138 194
100%
80%
60%
40%
20%
0%
<11]
[11-
20]
[21-
30]
[31-
40]
[41-
50]
[51-
60]
[61-
70]
[71-
80]
[81-
90]
[91-
100]
[101
-120
]
[121
-131
]
>131
]
Distancia a la costa (mn)
% P
eso
Mollusca Copepoda Euphaussidae
Pleuroncodes monodon
Engraulis ringens
Otros
Zoea
Mesopelágicos
Crustacea
Otros Teleosteii
Tamaño de muestra
3.00
2.50
2.00
1.50
1.00
0.50
0.00
% L
lenu
ra
80
Alegre et al.
Rev. peru. biol. número especial 20(1): 075- 082 (Septiembre 2013)Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú.
Editores: Csirke J., R. Guevara-Carrasco & M. Espino
En el caso de los individuos agrupados después del 2004, la otra variable predictora que discrimina las muestras es la dis-tancia a la costa (15.12%), que permite clasificar las muestras en aquellas que están a más de 53.3 mn y que se alimentaron mayormente de zoea, mientras que aquellos individuos colec-tados a menor distancia de 41.59 mn, estuvieron influenciados por el PDO (16.55% de importancia). De este modo, a valores ≥ -1.06, la dieta estuvo dominada por Euphausiidae, y a valores menores < -1.06, la dieta estuvo compuesta mayormente de zoea, Euphausiidae y P. monodon.
Debido a que el árbol de regresión no incluye las variaciones térmicas como una variable predictora, hemos realizado una comparación entre las anomalías de la temperatura superficial del mar en Chicama, que son consideradas como un buen in-dicador de condiciones El Niño (Espino 2013) y la proporción de Euphausiidae en la dieta del jurel T. murphyi (Fig. 8). Como se puede apreciar, no se aprecia una relación clara entre la pro-porción dietaria de Euphausiidae con la anomalía térmica de la estación de Chicama, lo que estaría indicando que los cambios en la proporción de esta presa en la dieta de T. murphyi no son influenciados por la ocurrencia o no de eventos El Niño. También, comparamos la extensión del área de la isoterma de 22 – 25 °C, la cual corresponde a las masas de Aguas Subtropi-cales Superficiales (ASS) frente al Perú (Wyrtki 1967, Morón & Sarmiento 2001) con la proporción dietaria de Euphausiidae y nuevamente no encontramos una relación clara entre estas variables. Trachurus murphyi se alimenta de Euphausiidae en las mismas proporciones durante eventos El Niño y eventos que no tienen relación con El Niño (Fig. 9).
DiscusiónEl análisis de la dieta de T. murphyi entre 1977 y el 2011
refleja que en el Perú han habido por lo menos dos escenarios temporales: el primero, desde 1977 hasta alrededor del 2000, y el segundo, posterior al año 2000. Estos escenarios coinciden con los periodos multidecadales cálidos y fríos propuestos por
Figura 7. Árbol de Regresión Multivariado de la proporción de presas en estómagos de jurel T. murphyi, con relación a 5 variables predictivas: año (de 1977 a 2011); Isot15 (profundidad de la isoterma de 15°C); PDO (Pacific Decadal Oscillation); zona (zona 1: norte, zona 2: centro, zona 3: sur); y, distancia (distancia a la costa en millas náuticas). Figure 7. Multivariate Regression Tree of preys proportion in stomach content of Jack mackerel T. murphyi off Peru in relation to 5 predictor variables: year (from 1977 to 2011); Isot15 (depth of the 15°C isotherm); PDO (Pacific Decadal Oscillation); zone (zone 1: north, zone 2: centre, zone 3: south; and, distancia (distance to the coast in nautical miles).
año<2000
año>=2004Nodo 1
distancia<41.59 Isot15>=88
PDO>=-1.055 zona=1
distancia<61.92
año>=2002
Nodo 2 Nodo 3
Nodo 4 Nodo 5
Nodo 6
Nodo 7 Nodo 8
Nodo 9
OtroTele
MesoAnchCrusZoeaPleurEuphCope
Moll0 0.5 1
OtroTele
MesoAnchCrusZoeaPleurEuphCope
Moll0 0.5 1
OtroTele
MesoAnchCrusZoeaPleurEuphCope
Moll0 0.5 1
OtroTele
MesoAnchCrusZoeaPleurEuphCope
Moll0 0.5 1 0 0.5 1
OtroTele
MesoAnchCrusZoeaPleurEuphCope
MollOtroTele
MesoAnchCrusZoeaPleurEuphCope
Moll
0 0.5 1
OtroTele
MesoAnchCrusZoeaPleurEuphCope
Moll
0 0.5 1
OtroTele
MesoAnchCrusZoeaPleurEuphCope
Moll
0 0.5 1
OtroTele
MesoAnchCrusZoeaPleurEuphCope
Moll
0 0.5 1
Moll: MolluscaCope: CopepodaEuph: EuphausiidaePleur: Pleuroncodes monodonZoea: ZoeaCrus: CrustaceaAnch: Engraulis ringensMeso: MesopelágicosTele: TeleosteiiOtro: Otros
100
70
40
10-1.5 -0.5 0.5 1.5 2.5 3.5 4.5
Con
teni
do E
upha
úsiid
ae (%
)
Anomalía TSM-Chicama
1989
19941990
1988
19841995
1991 1992
1997
1987
1998
1983
19791999
19862005
19961978
2000
2003 20061977
20112001
19822010
20022008
2009
20042007
Figura 8. Relación de la anomalía de la temperatura super-ficial del mar de Chicama (°C) con la proporción en la dieta de Euphausiidae en la dieta del jurel T. Murphyi frente a Perú de 1977 al 2011.Figure 8. Relationship of Chicama sea surface temperature anomaly (°C) with the proportion in the diet of Euphausiidae in the diet of Jack mackerel T. murphyi off Peru, 1977 to 2011.

81
Ecología trófica del jurel Trachurus murphyi en el Perú
Rev. peru. biol. special issue 20(1): 075- 082 (September 2013)Ecology, fisheries and conservation of Jack mackerel (Trachurus murphyi) in Peru. Editors: Csirke J., R. Guevara-Carrasco & M. Espino
Chávez et al. (2003), Espino (2003) y Chávez et al. (2008). El primer escenario corresponde a condiciones cálidas y el segundo a condiciones frías.
En cuanto al efecto de la talla en la alimentación de T. murphyi, se observa que, a medida que crece y madura, el peso relativo del contenido estomacal disminuye. Al respecto, Konchina et al. (1996) señalan que el jurel T. murphyi juvenil está relacionado con la zona epipelágica marina y que en etapas adultas se traslada a la zona nerítica; siendo los nichos ecoló-gicos en ambas zonas diferentes, la alimentación más activa la observó en los individuos juveniles de T. murphyi en las aguas epipelágicas oceánicas fuera de los límites de la plataforma. Sin embargo, nuestros resultados muestran que la proporción peso del cuerpo y peso del estómago disminuye con la edad, lo cual podría sostenerse en lo que afirma Pauly (1984), de que los peces requieren tanto de una adecuada provisión de alimento como de oxígeno para crecer, cumpliendo con la regla general de que los individuos juveniles tienen mayor consumo por sus altos requerimientos metabólicos.
Según nuestros resultados, T. murphyi se alimenta de zoo-plancton en una proporción semejante al 80% en términos de peso, desde tallas menores a mayores, con una mayor contri-bución de mesozooplancton en los individuos menores de 21 cm y de macrozooplancton en los de tallas mayores de 21 cm. Estos resultados entran en clara controversia con la afirmación de que T. murphyi cambia de estrategia zooplanctófaga a ictiófaga en la talla de 30 cm (Muck & Sánchez 1987). Según nuestros resultados, T. murphyi, desde tallas menores ingiere peces en una proporción semejante a 20% en peso, con algunas particu-laridades, como es el incremento en la proporción de E. ringens en la dieta de los individuos con tallas entre 31 y 50 cm y que fluctúan entre 2.2 y 5.7%. Nuevamente, esta última apreciación contradice la afirmación de que E. ringens es la presa principal de T. murphyi (Paredes 1972, Sánchez de Benites & Muck 1987, Muck & Sánchez 1987, Dioses 1995).
En cuanto a los estadios de madurez gonadal, no hemos encontrado un patrón definido que diferencie la dieta de los individuos de estadios de madurez inicial con los de estadio
tardío. Los individuos de estadios iniciales pueden comer la misma proporción de E. ringens que los individuos de estadios de madurez gonadal avanzados, sólo observamos cambios referidos a la composición del zooplancton ingerido por T. murphyi. Al respecto, según Konchina (1979) y Konchina et al. (1996), T. murphyi cuando es juvenil se asocia más con la zona epipelágica marina y cuando es sexualmente maduro se concentra en la zona nerítica. La mayor ingesta de Euphausiidae por parte de indivi-duos con estadio de madurez avanzados parece ser suficiente para proveerles de energía y continuar con su proceso reproductivo.
Según nuestros resultados, el patrón latitudinal de la dieta de T. murphyi rechaza la afirmación de que T. murphyi es ictiófago de norte a sur, en condiciones normales, y que ante un evento El Niño se torna mayormente zooplanctófago y puede depredar sobre mictófidos y juveniles de E. ringens (Sánchez de Benites et al. 1985, Sánchez de Benites & Muck 1987, Dioses 1995). Estos mismos autores indican que hay cambios en la ictiofagia, con predominio de Anchoa sp. en la zona norte, E. ringens en la zona centro y peces mesopelágicos en el sur. Nuestros resultados muestran que la ictiofagia es de menor importancia de norte a sur y que se incrementa de 5 a 30%, sin mostrar predominio de E. ringens ni de peces mesopelágicos como se ha mencionado en la literatura mencionada.
Según la distancia a la costa, la tendencia del índice de llenura estomacal es decreciente. Esta tendencia es contradictoria con la hipótesis de segregación por tallas postulada por Konchina (1979) y Konchina et al. (1996). Sin embargo, a partir de las 130 mn, fuera del límite de la plataforma continental, se observó un incremento del índice de llenura estomacal y según lo observado, estaría asociándose con la mayor ingesta de peces a esa distancia, en especial los peces mesopelágicos. En toda el área estudiada, el predominio de Euphausiidae guarda relación con su amplia distribución en el mar peruano (Aronés et al. 2009, Ballón et al. 2011). Pleuroncodes monodon, que está principalmente aso-ciado con las Aguas Costeras Frías (Gutiérrez et al. 2008), fue ingerido con singular importancia por T. murphyi hasta 40 mn de distancia a la costa.
En cuanto a la relación recurso-ambiente, el Árbol de Regre-sión Multivariado muestra la gran importancia que tienen en la dieta las variables explicativas tiempo (definiendo dos periodos, antes y después de 2000), la distancia a la costa (fuertemente ligada con el PDO), y la profundidad de la isoterma de 15°C (ligada fuertemente a la ingesta del camaroncito rojo P. monodon.
ConclusionesSe observan dos escenarios en la dieta del jurel T. murphyi
frente al Perú: el primero dentro de un periodo cálido desde 1977 hasta el 2000, que se caracterizó por la presencia de Euphausiidae, y el último dentro de un periodo frío a partir del 2001, en el que la dieta fue más diversa y las presas dominantes fueron el camaroncito rojo P. monodon y los Euphausiidae. Los escenarios descritos en el tiempo estarían obedeciendo a cambios ambientales observados en los últimos 30 años.
Este trabajo rebate el paradigma de la dominancia de la anchoveta E. ringens en la dieta del jurel T. murphyi y queda comprobado que los Euphausiidae son el componente que sustenta su dieta a lo largo del tiempo.
Se confirma la gran plasticidad del comportamiento alimenta-rio del jurel T. murphyi. Esta especie puede adaptar su régimen de
100
80
60
40
20
00 100000 20000 300000 400000
Con
teni
do E
upha
úsiid
ae (%
)
Área (Km2) Isoterma de 22 a 25 °C
1982
1983
19981998
1997
1995
19921994
1984
1990
19891991
1997
1988
1986
1996
19992000
2005
2007
2010
2003
2004 2006
2002
200920082001
2011
Figura 9. Relación del área de la isoterma de 22-25 °C (ASS), con la proporción de Euphausiidae en la dieta de jurel T. murphyi frente a Perú de 1977 al 2011.Figure 9. Relationship of the area of isotherm of 22-25 °C (SSW) with the proportion of Euphausiidae in the diet of Jack mackerel T. murphyi off Peru, 1977 to 2011.

82
Alegre et al.
Rev. peru. biol. número especial 20(1): 075- 082 (Septiembre 2013)Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú.
Editores: Csirke J., R. Guevara-Carrasco & M. Espino
alimentación para hacer frente a los cambios ambientales espacio-temporales que impactan la disponibilidad y composición de las presas, siendo capaz de modificar la amplitud de su hábitat.
Literatura citadaÁlamo V.& V. Valdivieso. 1987. Lista sistemática de moluscos marinos del
Perú. Boletín Instituto del Mar del Perú. Vol.extraordinario: 205 pp.Aronés K., P. Ayón, H.J. Hirche et al. 2009. Hydrographic structure and zo-
oplankton abundance and diversity off Paita, northern Peru (1994 to 2004) – ENSO effects, trends and changes. Journal of Marine Systems 78:582-598. DOI:10.1016/j.jmarsys.2009.01.002.
Ballón M., A. Bertrand, A. Lebourges-Dhaussy et al. 2011. Is there enough zooplankton to feed forage fish population off Peru? An acoustic (positive) answer. Progress in Oceanography 91: 360-381. DOI: 10.1016/j.pocean.2011.03.001.
Berg J. 1979. Discussion of methods of investigating the food of fishes, with reference to a preliminary study of the prey of Gobiusculus flavescens (Gobiidae). Mar. Biol. 50: 263-273. DOI: 10.1007/BF00394208
Botsford L., J. Castilla & C. Peterson. 1997. The management of fisheries and marine ecosystems. Science 277: 509-515. DOI: 10.1126/science.277.5325.509
Breiman L., J. Friedman, R. Olshen &C. Stone. 1984. Classification and RegressionTrees. Wadsworth & Brooks/Cole Advanced Books & Software, Pacific Grove, CA.
Caddy J. & G. Sharp. 1988. Un marco ecológico para la investigación pesquera. FAO Doc. Tec. Pesca 283:155 pp.
Chávez F., J.Ryan,S. Lluch-Cota & M. Ñiquen. 2003. From anchovies to sar-dines and back: multidecadal change in the Pacifc Ocean. Science 299: 217-221. DOI: 10.1126/science.1075880
Chávez F., A. Bertrand, R. Guevara-Carrasco, P. Soler & J. Csirke. 2008. The northern Humboldt Current System: Brief history, present status and a view towards the future. Progress in Oceanography 79: 95-1051. DOI: 10.1016/j.pocean.2008.10.012.
Chirichigno N. 1998. Clave para identificar los peces marinos del Perú, 2ª ed. Instituto del Mar de Perú. Publicación Especial. Callao: 496 pp.
De’ath G. & K. Fabricius. 2000. Classification and regression trees: a powerful yet simple technique for ecological data analysis. Ecology 81(11): 3178-3192. DOI: 10.1890/0012-9658(2000)081[3178:CARTAP]2.0.CO;2
Dioses T. 1995. Análisis de la distribución y abundancia de los recursos jurel y caballa frente a la costa peruana. Inf. Prog. Inst. Mar Perú. 03:55 pp.
Espino M. 2003. Estrategia de gestión oriental para el Pacífico Oriental con especial mención a la pesquería peruana. Tesis para optar el grado de Magíster en Geografía con mención en Ordenamiento y Gestión Ambiental. Unidad de Post Grado, Facultad de Ingeniería Geológica, Minera, Metalúrgica y Geográfica, Universidad Nacional Mayor de San Marcos, Lima. 102 pp.
Espino M. 2013. El jurel Trachurus murphyi y las variables ambientales de ma-croescala. En: Csirke J., R. Guevara-Carrasco & M. Espino (Eds.). Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú. Rev. peru. biol. número especial 20(1): 009- 020
Fitch J. & R. Brownell. 1968. Fish Otoliths in Cetacean Stomachs and Their Importance in Interpreting Feeding Habits. Journal of Fisheries Re-search Board of Canada 25(12):2561-2574. DOI: 10.1139/f68-227
García-Godos I. 2001. Patrones morfológicos del otolito sagitta de algunos peces óseos del mar peruano. Boletín Instituto del Mar del Perú 20 (1-2): 83 pp.
Gutiérrez M., A. Ramírez, S. Bertrand et al. 2008. Ecological niches and areas of overlap of the squat lobster ‘munida’ (Pleuroncodes monodon) and anchoveta (Engraulis ringens) off Peru. Progress in Oceanography 79:256-263. DOI : 10.1016/j.pocean.2008.10.019
Hureau J. 1970. Biologie compare de quelques poissons antarctiques (No-totheniidae). Bulletin de l’Institut Oceanographique Monaco 68(1391): 244 pp.
Hyslop E. 1980. Stomach contents analysis-a review of methods and their applications. Journal of Fish Biology 17 (4): 411-429. DOI: 10.1111/j.1095-8649.1980.tb02775.x
Iman R., D. Quade & Alexander D. 1975. Exact Probability Levels for Kruskal-Wallis Test. In:H. Harter and D. Owen (Eds.). Select Tables in Mathematical Statistics (Vol 3). American Mathematical Society, Providence, Rhode Island.
Konchina Y. 1979. The feeding of the Peruvian Jack mackerel, Trachurus sym-metricus murphyi. Journal of Ichthyology 19: 52-61.
Konchina Y. 1980. The Peruvian Jack mackerel, Trachurus symmetricus mur-phyi, a facultative predator in the coastal upwelling ecosystem. Journal of Ichthyology 20(5): 46-59.
Konchina Y. 1983. The feeding niche of the hake Merluccius gayi(Merlucciidae), and the Jack mackerel, Trachurus symmetricus(Carangidae) in the trophic system of the Peruvian coastal upwelling. Journal of Ichthyology 23(2): 87-98.
Konchina Y., A.V. Nesin, N.A. Onishchik et al. 1996. On the migration and feeding of the Jack mackerel Trachurus symmetricus murphyi in the Eastern Pacific. Journal of Ichthyology 36(9):753-766.
Medina M. & H. Arancibia. 2002. Dinámica trófica del jurel (Trachurus sym-metricus murphyi) en el norte de Chile. Invest. Mar., Valparaíso, 30(1): 45-55.
Méndez M. 1981. Claves de identificación y distribución de los langostinos y camarones (Crustacea: Decapoda) del mar y ríos de la costa del Perú. Boletín Instituto del Mar del Perú 5:170 pp.
Morón O. & M. Sarmiento. 2001. Aspectos oceanográficos de El Niño 1997-98 y su relación con los recursos pelágicos. En:J. Tarazona, W.E. Arntz y E. Castillo de Maruenda(Eds.). El Niño en América Latina: Impactos Biológicos y Sociales. Consejo Nacional de Ciencia y Tecnología. Lima: 5-27.
Muck P. & G. Sánchez . 1987. The importance of mackerel and horse mackerel predation for the Peruvian anchoveta stock (a population and fee-ding model). In: D. Pauly and I. Tsukayama (Eds.). The Peruvian Anchoveta and Its Upwelling Ecosystem: Three Decades of Change. ICLARM Studies and Reviews 15: 351 pp.
Newell G. 1963. Marine plankton a practical guide. Hutchison Biological Monographs. 207 pp.
Paredes V. 1972. Estudio sobre contenido estomacal de jurel Trachurus murphyi Nichols del área de pesca del Callao. Tesis para optar el grado de Bachiller en Ciencias Biológicas. Universidad Nacional Mayor de San Marcos. Lima, Perú.
Pauly D. 1984. A mechanism for the juvenile-to-adult transition in fishes. Jo-urnal Du Conseil 41 (3):280-284. DOI:10.1093/icesjms/41.3.280.
Rasero M., A. González, B. Castro et al. 1996. Predatory relationships of two sympatric squid, Todaropsis eblanae and Illex coindetii (Cephalo-poda: Ommastrephidae) in Galician waters. Journal of the Marine Biological Association of the United Kingdom 76: 73-87. DOI: 10.1017/S0025315400029027
Sánchez de Benites G., A. Álamo & H. Fuentes. 1985. Alteraciones en la dieta alimentaria de algunos peces comerciales por efecto del fenómeno “El Niño”. En W. Arntz, A. Landa y J. Tarazona (Eds.). El Niño: Su impacto en la fauna marina. Boletín Instituto Mar Perú. Vol. Extraordinario: 135-142.
Sánchez de Benites G. & P. Muck. 1987. Situación alimentaria del jurel Tra-churus murphyi Nichols en un año normal (1979) y en El Niño 1982-1983. Boletín Instituto Mar Perú 11(4):152-171.
Trégouboff G. & M. Rosse. 1957. Manuel de plactonologie mediterraneénne. Centre National de la Recherche Scientifique. Paris. Tome I, 587 pp; Tome II, 207 planches.
Wolff G.1984. Identification and estimation of size from the beaks of 18 species of cephalopods from the Pacific Ocean. NOAA Technical Report NMFS, 17: 1-50.
Wyrtki K. 1967. Circulation and water masses in the eastern Equatorial Pacific Ocean. Int. Jour Oceanol and Limnol 1(2):117-147.
Zuur F., E. Ieno, N. Walker et al. 2009. Mixed effects models and extensions, in Ecology with R. Springer. 530pp.

83
Variabilidad espacial y temporal de las larvas de jurel Trachurus murphyi
Rev. peru. biol. special issue 20(1): 083- 086 (September 2013)Ecology, fisheries and conservation of Jack mackerel (Trachurus murphyi) in Peru. Editors: Csirke J., R. Guevara-Carrasco & M. Espino
Rev. peru. biol. número especial 20(1): 083- 086 (Septiembre 2013)Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú. Editores: Csirke J., R. Guevara-Carrasco & M. Espino
Facultad de ciencias Biológicas unMsMISSN-L 1561-0837
Variabilidad espacial y temporal de larvas de jurel Trachurus murphyi en el Perú entre 1966 – 2010
Patricia Ayón y Jonathan Correa
Spatial and temporal variability of Jack mackerel Trachurus murphyi larvae in Peru between 1966 – 2010
Instituto del Mar del Perú, esquina Gamarra y General Valle S/N Chucuito, Callao, Perú.
Email Patricia Ayón: [email protected]
ecología del jurel (Trachurus murphyi) en el Perú
© Los autores. Publicado por la Revista Peruana de Biología de la Facultad de Ciencias Biológicas, Universidad Nacional Mayor de San Marcos. Este es un artículo de acceso abierto, distribuido bajo los términos de la Licencia de Atribución Reconocimiento-NoComercial-SinObraDerivada 3.0 de Creative Commons (http://creativecommons.org/licenses/by-nc-nd/3.0/deed.es_ES), que permite el uso no comercial, distribución y reproducción en cualquier medio, siempre que la obra original sea debidamente citadas. Para uso comercial, por favor póngase en contacto con [email protected].
Citación:Ayón P. & J. Correa. 2013. Variabilidad espacial y temporal de larvas de jurel Trachurus murphyi en el Perú entre 1966-2010. En: Csirke J., R. Guevara-Carrasco & M. Espino (Eds.). Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú. Rev. peru. biol. número especial 20(1): 083- 086 (Septiembre 2013)
ResumenEl análisis espacial y temporal de larvas de jurel Trachurus murphyi en el mar peruano entre los años 1966 y 2010 confirma su presencia constante durante todo el periodo estudiado. En general, en este periodo la abundancia de larvas de jurel T. murphyi muestra una tendencia ligeramente ascendente destacándose cambios en la distribución espacial y una alta variabi-lidad interanual, con cortos periodos de alta abundancia promedio, intercalados con periodos más prolongados y de baja abundancia. El promedio anual estimado varió entre 3 y 79 larvas/m2. La distribución espacial presentó tres periodos importantes: uno con mayores densidades de larvas al sur de los 14°S, entre 1970 y 1979; cambió hacia el norte de esta latitud, entre 1980 y 1999; y otro de mayor densidad al sur de los 18°S, en el último periodo 2000 – 2010. Los centros de gravedad de la distribución de larvas presentaron tres periodos muy notorios: fueron más costeros hacia el sur de los 14°S, entre 1966 y1978; más oceánicos y al norte de los 14°S, entre 1979 y 1994; y con una posición intermedia entre 1995 – 2010. Se discute la relación entre la distribución de las larvas con las variables oceanográficas y la distribución de T. murphyi adultos y juveniles, además se destaca que el mar del Perú es un centro principal y permanente de actividad reproductiva de esta especie.
Palabras clave: larvas, jurel, centro de gravedad de larvas, mar peruano.
AbstractConstant presence of Jack mackerel Trachurus murphyi larvae in the Peruvian sea between 1966 and 2010 was confirmed by an analysis of their spatial and temporal variability. In general, there is a slight positive trend in abundance of Jack mackerel T. murphyi larvae, with notice-able changes in their spatial distribution and high interannual variability. High abundance of larvae is observed in some years, which are usually followed by a longer sequence of poor abundance years. Yearly averages varied between 3 and 79 larvae/m2. The spatial distribution of average larvae density showed three notorious periods along time: the first between 1970 and 1979 with higher larvae densities south of 14°S; the second between 1980 and 1999 with the higher densities to the north of 14°S; and the third between 2000 and 2010 with higher densities south of 18°S. The displacement of the centers of gravity of the spatial distribution marked three noticeable periods: between 1966 and 1978 were more coastal, south of 14°S; between 1979 and 1994 more oceanic and north of 14°S; and between 1995 and 2010 which were at an intermediate position. The relationship of larvae density with oceanographic variables as well as with the distribution of adult and juvenile Jack mackerel T. murphyi are discussed, stressing the observation that there is a main center of permanent spawning activity of this species in Peruvian waters.
Keywords: larvae, Jack mackerel, larval gravity center, Peruvian sea
IntroducciónEl jurel Trachurus murphyi Nichols 1920 se encuentra asociado a la interacción
de las masas de agua del Sistema de las Corrientes del Perú, hábitat formado por las Aguas Subtropicales Superficiales (ASS) con Aguas Costeras Frías (ACF) de intensos afloramientos y las Aguas Ecuatoriales Superficiales (AES) (Dioses et al. 1989).
Esta especie es un desovador parcial. Se ha definido la ocurrencia de huevos y larvas en el mar peruano desde fines del invierno hasta la primavera, con mayor intensidad en los meses de octubre y noviembre, ubicando su principal zona de desove entre los
Publicado online: 11/10/2013Publicado impreso: 15/10/2013

84
Ayón & Correa
Rev. peru. biol. número especial 20(1): 083- 086 (Septiembre 2013)Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú.
Editores: Csirke J., R. Guevara-Carrasco & M. Espino
Figura 1. Estaciones de muestreo entre 1966 y 2010.Figure 1. Sampling stations from 1966 to 2010
Figura 2. Abundancia anual promedio de larvas de jurel Trachurus murphyi (1966 – 2010).Figure 2. Annual average abundance of Jack mackerel Trachurus murphyi larvae (1966 – 2010)
14°00’S y los 18°30’S. En el caso del jurel, el desove está asocia-do a concentraciones elevadas de oxígeno mayores a 5.0 ml/L (Santander & Flores 1983, Dioses et al. 1989).
El presente trabajo muestra los resultados de la distribución espacial y temporal de larvas de jurel T. murphyi y se discuten algunas relaciones entre las condiciones ambientales y la distri-bución de los adultos.
Material y métodosPara el presente estudio se ha trabajado con información
proveniente de 65 cruceros de investigación ejecutados por el Instituto del Mar del Perú entre los años 1966 y 2010. El área de trabajo se encuentra ubicada a lo largo del dominio marítimo peruano y las longitudes comprendidas entre 70°49.80’W – 87°12’W (Fig. 1). Las colectas se realizaron con una red Hensen de 0.60 m de diámetro de boca, provista con malla nytal de 330 μm y operada en jales verticales desde 50 m de profundidad hasta la superficie. El conteo de las larvas se hizo en el total de
la muestra y todos los resultados fueron estandarizados, expre-sando la abundancia como número de larvas/m2. Se calculó la abundancia promedio considerando solamente las estaciones positivas con larvas de jurel.
Se analizó la dinámica espacial mediante la posición geo-gráfica de los centros de gravedad de larvas para cada crucero. Los centros de gravedad se calcularon para la longitud (X) y la latitud (Y) mediante las siguientes ecuaciones:
Xt= (∑Ct,iXt,i) /∑Ct,i
Yt= (∑Ct,iYt,i) /∑Ct,i
En donde: Ct,i = densidad de larvas de jurel
Xt = longitud
Yt = latitud
Adicionalmente se determinó la mediana de los centros de gravedad. Los mapas de distribución y abundancia de larvas de jurel T. murphyi se elaboraron con el programa SURFER, mientras que el análisis de los datos así como los gráficos se realizaron en R (http://www.r-project.org/).
ResultadosLa abundancia de las larvas de jurel T. murphyi por estación
de muestreo fluctuó entre 3 y 561 larvas/m², con una mediana de 21 larvas/m². En general, se observó una baja frecuencia y abundancia, especialmente en los cruceros de verano. La abun-dancia promedio anual varió entre 3 y 79 larvas/m². Las series de tiempo de la abundancia total y la abundancia promedio mostraron una tendencia ligeramente ascendente, con una alta variabilidad interanual con breves periodos de alta abundancia promedio intercaladas con periodos más largos de baja abun-dancia (Fig. 2).
La presencia constante de larvas de jurel T. murphyi durante las épocas de desove en todos los años entre 1966 y 2010 de-muestra que frente a las costas del Perú se ubica un área perma-nente de actividad reproductiva, con la presencia de un stock desovante estable. Además, observamos en varias ocasiones la presencia de larvas en áreas donde ni las flotas pesqueras ni los cruceros científicos habían detectado la presencia de cardúmenes de T. murphyi, este hecho manifestaba la presencia previa de ejemplares desovantes en la zona, aun cuando estos no hayan sido detectados. Además, la presencia de larvas sería la evidencia
0ºS
2ºS
4ºS
6ºS
8ºS
10ºS
12ºS
14ºS
16ºS
18ºS
20ºS
22ºS
Pta. SalTalaraPaita
Pta. La Negra
PimentelMalabrigo
Salaverry
ChimboteCasmaHuarmey
SupeHuacho
ChancayCallao
PucusanaCerro AzulPiscoBahia Independencia
San Juan MarconaAtico
MollendoIlo
Sama
Área evaluada1966 - 2010
84ºW 80ºW 76ºW 72ºW 68ºW88ºW
1970 1980 1990 2000 2010
Años
0
20
40
60
80
Larv
as/m
2

85
Variabilidad espacial y temporal de las larvas de jurel Trachurus murphyi
Rev. peru. biol. special issue 20(1): 083- 086 (September 2013)Ecology, fisheries and conservation of Jack mackerel (Trachurus murphyi) in Peru. Editors: Csirke J., R. Guevara-Carrasco & M. Espino
Figura 3. Patrones de distribución de larvas/m2 de jurel Trachurus murphyi (1970 – 2010).Figure 3. Distribution patterns of Jack mackerel Trachurus murphyi larvae/m2 (1970 – 2010).
Figura 4. Ubicación anual de los centros de gravedad de la abun-dancia de larvas de jurel Trachurus murphyi en el mar peruano. Los puntos indican los valores anuales y las líneas punteadas indican el valor de la mediana del centro de gravedad para cada periodo.Figure 4. Annual location of the centers of gravity of Jack mackerel Trachurus murphyi larvae in Peruvian waters. The dots indicate the annual values and the broken lines indicate the median value of the centers of gravity for each period.
de un cambio en el comportamiento gregario de T. murphyi durante la fase de reproducción (Francois Gerlotto, com. pers.).
La distribución espacial de las larvas de jurel T. murphyi presentan cambios importantes en el tiempo (Fig. 3). Así, entre 1970 y 1979, las larvas estuvieron distribuidas principalmente en la zona sur de Perú, mientras que entre 1980 y 1989 se observa una distribución amplia frente a toda la costa peruana, con una mayor densidad al norte de los 14°S, la misma que se mantuvo en el periodo 1990 – 1999 con una expansión hacia el sur, en-tre el 2000 y 2010. Estos patrones de distribución observados modificaron también la ubicación de los centros de gravedad de las larvas calculados para toda la serie (Fig. 4). Se identifican tres periodos de esta modificación: el primero, entre 1966 y 1978, con el centro de gravedad situado principalmente entre los 14°S y 18°S, cercano a la costa; el segundo, entre 1979 y 1994, con el centro ubicado algo más al norte, entre los 4°S y 14°S, y algo más oceánico; y, finalmente, el tercero, entre 1995 y 2010, con una posición intermedia entre los dos periodos antes mencionados.
Discusión El presente trabajo muestra la continuada presencia de larvas
de jurel T. murphyi en el mar peruano, indicando que siempre ha desovado en esta zona, poniendo en evidencia que el mar del Perú es una de las principales zonas de permanente actividad reproductiva de T. murphyi. Parte de la variabilidad observada en la presencia de larvas podría ser explicada por un defecto de muestreo al no cubrirse toda el área longitudinal de su dis-tribución, que en algunos años es más costera y en otros años estarían más alejadas de la costa. En efecto, en varios cruceros las larvas se encontraban hacia el borde exterior de las estaciones más oceánicas, lo que sugiere la existencia de una distribución más oceánica, en zonas más alejadas de la costa y en parte fuera de las zonas muestreadas.
La presencia constante de larvas del jurel T. murphyi en el mar peruano a lo largo del tiempo refuerza dos planteamientos sobre su distribución en el en el Pacífico Sudoriental. Una es la hipótesis de trabajo sobre la existencia de dos stocks o substocks, en gran parte independientes (Serra 1991, SPRFMO 2008) siendo considerado en el ámbito de la Organización Regional de Ordenamiento Pesquero del Pacífico Sur y otra es la planteada por
Gerlotto et al. (2010) sobre la existencia de una subpoblación peruana o stock norte separado del stock chileno más al sur por la Zona de Convergencia Subtropical (Wyrtki 1967 y Fariña et al. 2008), cuya dinámica e intensidad condicionaría periodos de aislamiento y de migración entre estas subpoblaciones.
La variabilidad que se presenta en escala interanual, que co-rresponde a la dinámica de El Niño Oscilación Sur (ENOS), y en escala interdecadal, correspondiente a la Oscilación Decadal del Pacífico (PDO) (Chávez et al. 2003, Espino 2003), estarían influyendo en la disponibilidad de las larvas de jurel T. murphyi a lo largo del mar peruano en distintas épocas. Al respecto, la amplia distribución de larvas observada durante el Niño 97/98 y la que hubo durante el año 2006 (Fig. 5) estuvieron asocia-das a un acercamiento de las Aguas Subtropicales Superficiales que habrían generado un intenso proceso de mezcla en la zona
Latit
ud S
Años
Long
itud
W
72°
76°
80°
4°
8°
12°
16°
1970 1980 1990 2000 2010
1970 1980 1990 2000 2010

86
Ayón & Correa
Rev. peru. biol. número especial 20(1): 083- 086 (Septiembre 2013)Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú.
Editores: Csirke J., R. Guevara-Carrasco & M. Espino
costera. Por otro lado, los cambios en el centro de gravedad de la distribución espacial de las larvas indicarían la existencia de cambios a escalas de décadas, en variables abióticas que estarían determinando su distribución. En ese sentido, la profundización de la isoterma de 15 °C, la distribución espacial de la salinidad y la profundización de la mínima de oxígeno durante las décadas de los años 1980 y 1990 estarían favoreciendo altas abundancias de larvas de jurel al norte de los 10°S, asociadas a una intromisión de Corrientes Ecuatoriales y Aguas Subtropicales Superficiales (Bertrand 2004, Flores el at. 2013).
De otro lado, al comparar la distribución de las larvas captu-radas durante los cruceros de investigación con la de juveniles y adultos de jurel T. murphyi, provenientes de la captura comercial entre 1996 al 2012, no encontramos una correspondencia total entre todos los años comparados, solo en algunos años se observa una superposición entre ellos, como el año 2006 que muestra una cobertura espacial similar. Otros años como 1999, 2000 y 2007 coinciden en algunos puntos, pero al analizar la información sobre si provienen del mismo periodo de muestreo, solamente el año 1999 son de periodos similares, mientras que los otros dos años tienen un desfase temporal de más de seis meses. En los años 2001 y 2003 se presentan larvas al centro y al centro-sur de similar distribución latitudinal, pero en posición más oceánica que la de los adultos, sólo el 2001 tiene correspondencia tem-poral. Asimismo, la distribución espacial entre juveniles y larvas tampoco mostró una relación directa en los años analizados (Fig. 5), pero se observó una superposición clara en 1998.
Es evidente que la distribución espacial de adultos detectada por la flota comercial no guarda relación con la proveniente de las larvas de los cruceros, en algunos casos ello se explicaría porque los muestreos se hicieron en momentos diferentes. Sin embargo, la distribución de las larvas tampoco coincidió con la de adultos en los años 1996, 2000 y 2006, cuando los datos provenían de un mismo periodo. La discrepancia podría deberse a que la flota pesquera normalmente se concentra en las zonas de mayor densidad o biomasa del jurel T. murphyi, mientras que los muestreos de larvas se hacen en lugares y segun trayectos prefijados. Sin embargo, las observaciones de las larvas en zonas donde no se han registrados adultos por medios convencionales es relevante, porque estarían indicando la presencia previa de ejemplares desovantes en esas zonas.
Las observaciones presentadas este trabajo, mostrarían que la alta variabilidad entre décadas en la distribución y abundancia de larvas de jurel T. murphyi tiene coherencia con la variabilidad de largo plazo de las condiciones ambientales del mar peruano, por lo cual esta variabilidad de largo plazo debe ser considerada necesariamente en el proceso de evaluación de los niveles po-blacionales de los adultos.
Literatura citadaBertrand A., M. Segura, M. Gutiérrez & L. Vásquez. 2004. From small-scale
habitat loopholes to decadal cycles: a habitat-based hypothesis ex-plaining fluctuation in pelagic fish populations off Peru. En: Fish and fisheries, 5: 296–316. DOI: 10.1111/j.1467-2679.2004.00165.x
Chávez F.P., J. Ryan, S. E. Lluch-Cota & M. Ñiquen. 2003. From anchovies to sardines and back: multidecadal change in the Pacific ocean. Review: Climate. ScienceVol. 299: 217-221. DOI: 10.1126/science.1075880
Dioses T., V. H. Alarcón, M.E. Nakama & A. Echevarría. 1989. Desarrollo ovocitario, fecundidad parcial y distribución vertical de los cardú-menes en desove de jurel, Trachurus murphyi (N). En: Memorias del Simposio Internacional de los Recursos Vivos y las Pesquerías en el Pacífico Sudeste. Viña del Mar, 9-13 de mayo de 1988. Rev. Pacífico Sur (Número especial) 287-294.
Espino M. 2003. Estrategia de gestión para el Pacífico Oriental con especial mención a la pesquería peruana. Tesis para optar el grado de Magíster en Geografía con mención en Ordenamiento y Gestión Ambiental. Unidad de Post Grado, Facultad de Ingeniería Geológica, Minera, Metalúrgica y Geográfica, Universidad Nacional Mayor de San Marcos. 102 p.
Espinoza E., M. Ñiquen & Flores R. 2008. Changes in the latitudinal and longitudinal distribution of the aggregations of Jack mackerel (Tra-churus murphyi) in the Peruvian sea during 1996 – 2007. Chilean Jack Mackerel Workshop 30 June - 4 July. 2008.
Fariña J.M., P.G. Ossa & J.C Castilla. 2008. Diversidad de los ecosistemas marinos. En: CONAMA (Coordinador). Biodiversidad de Chile, patrimonio y desafíos. Ocho Libros editores. Santiago de Chile. pp: 96-104.
Flores R., M. Espino, G. Luque & J. Quispe. 2013. Patrones de variabilidad ambiental en el mar peruano. En: Csirke J., R. Guevara-Carrasco & M. Espino (Eds.). Ecologia, pesqueria y conservacion del jurel (Trachurus murphyi) en el Peru. Rev. peru. biol. numero especial 20(1): 021 – 028
Gerlotto F., M. Gutiérrez & A. Bertrand. 2010. The distribution of Jack macke-rel Trachurus murphyi in the South Pacific Ocean: a metapopulation hypothesis. FAST-ICES-WG annual meeting. SWNFS, NOAA, San Diego, April 2010.
Santander H. & R. Flores. 1983. Spawning and larval distribution of four pe-lagic species and their relations to marine environmental variations off Peru. En: G.D. Sharp and J. Csirke (Eds.), Expert Consultation to Examine Changes in Abundance and Species Composition of Neritic Fish Resources, San Jose (Costa Rica), 18 Apr 1983. Procee-dings: FAO Fisheries Report, FAO, Rome, Italy, 291(2-3): 835-870.
Serra, J.R. 1991. Important life history aspects of the Chilean Jack mackerel, Trachurus symmetricus murphyi. Invest. Pesq. (Chile)36:67-83.
SPRFMO. 2008. Chilean Jack mackerel workshop. Report of the South Pacific Fisheries Management Organisation (SPRFMO), Chilean Jack Mackerel Workshop, Santiago, Chile, 30 June-4July 2008:71pp
Wyrtki K. 1967. Circulation and Water Masses in the Eastern Equatorial Pacific Ocean. Int. J. Oceanology & Limnology, 1(2):117-147.
Figura 5. Distribución anual de jurel en estado larva, juvenil y adulto (1996 a 2012) (adaptado de Espinoza et al. 2008)Figure 5. Annual distribution of Jack mackerel in larvae, juvenile and adult stages (1996 to 2012) (adapted from Espinoza et al. 2008)

87
Biomasa acústica y distribución del jurel Trachurus murphyi en el Perú
Rev. peru. biol. special issue 20(1): 087- 096 (September 2013)Ecology, fisheries and conservation of Jack mackerel (Trachurus murphyi) in Peru. Editors: Csirke J., R. Guevara-Carrasco & M. Espino
Rev. peru. biol. número especial 20(1): 087- 096 (Septiembre 2013)Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú. Editores: Csirke J., R. Guevara-Carrasco & M. Espino
Facultad de ciencias Biológicas unMsMISSN-L 1561-0837
Biomasa acústica y distribución del jurel Trachurus murphyi en el Perú
Marceliano Segura y Aníbal Aliaga
Acoustic biomass and distribution of Jack mackerel Trachurus murphyi in Peru
Instituto del Mar del Perú, esquina Gamarra y General Valle S/N Chucuito, Callao, Perú.
Email Marceliano Segura: [email protected]
ecología del jurel (Trachurus murphyi) en el Perú
© Los autores. Publicado por la Revista Peruana de Biología de la Facultad de Ciencias Biológicas, Universidad Nacional Mayor de San Marcos. Este es un artículo de acceso abierto, distribuido bajo los términos de la Licencia de Atribución Reconocimiento-NoComercial-SinObraDerivada 3.0 de Creative Commons (http://creativecommons.org/licenses/by-nc-nd/3.0/deed.es_ES), que permite el uso no comercial, distribución y reproducción en cualquier medio, siempre que la obra original sea debidamente citadas. Para uso comercial, por favor póngase en contacto con [email protected].
Citación:Segura M. & A. Aliaga. 2013. Biomasa acústica y distri-bución del jurel Trachurus murphyi en el Perú. En: Csirke J., R. Guevara-Carrasco & M. Espino (Eds.). Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú. Rev. peru. biol. número especial 20(1): 087- 096 (Septiembre 2013)
Resumen Se analizan los resultados de las evaluaciones hidroacústicas del recurso jurel Trachurus mur-phyi Nichols 1920 realizadas en aguas peruanas entre 1983 – 2012. Desde 1983 se incluyó al T. murphyi como especie de estudio durante los cruceros de evaluación de recursos pelágicos ejecutados por el Instituto del Mar del Perú. Debido al énfasis en la estimación de biomasa de la anchoveta Engraulis ringens y de la sardina Sardinops sagax cuando esta última es más abundante, los cruceros se llevan a cabo durante el verano austral y las áreas de evaluación están circunscritas a las zonas más costeras hasta 100 mn, con sólo algunas exploraciones en otras estaciones y hasta 200 millas. El máximo valor de biomasa de 8.51 millones de toneladas de T. murphyi en aguas peruanas estimado con las evaluaciones hidroacústicas fue encontrado durante el crucero realizado en otoño (marzo-mayo) de 1983. En los años siguientes los estimados de biomasa acústica fluctuaron entre 180 mil toneladas en 1985 y otro máximo de 8.47 millones de toneladas en 1993, para luego disminuir gradualmente hasta un mínimo de 1239 t en 2010, con una ligera recuperación en los años 2011 y 2012. El área de distribución de T. murphyi fue muy fluctuante en todo el periodo observado.
Palabras clave: Jurel, biomasa acústica, recursos pelágicos
AbstractThis paper analyzes the results of acoustic stock assessment surveys for Jack mackerel Tra-churus murphy Nichols 1920 conducted in Peruvian waters between 1983 and 2012. Since 1983 Jack mackerel T. murphyi was included as a target species to be assessed during the pelagic stock assessment surveys executed by the Peruvian Institute of Marine Research. Due to the emphasis on the assessment of anchovy Engraulis ringens and sardine Sardinops sagax when this species was abundant, these surveys usually took place during the austral summer and only covered the first 100 nautical miles from the coast, and only occasional surveys were conducted in other seasons or surveyed as far as 200 nm offshore. A maximum biomass of 8.51 million t of T. murphyi in Peruvian waters estimated through acoustics during a survey carried out in autumn (March-May) 1983. In the following years acoustic biomass estimates ranged between 180 thousand t in 1985 and another maximum of 8.47 million t in 1993, to then decreased gradually to a minimum of 1239 t in 2010, with a slight recovery in 2011 and 2012. The areas of distribution of T. murphyi have been fluctuating markedly throughout the observed period.
Keywords: Jack mackerel, acoustic biomass, pelagic resources.
IntroducciónTrachurus murphy Nichols 1920 es una especie pelágica transzonal que presenta una
amplia distribución asociada a las fluctuaciones de distinta escala en el ambiente del Pacífico Sudoriental, las que han dado lugar también a cambios en el largo plazo en la abundancia y distribución de los recursos pesqueros pelágicos y en sus pesquerías (Jordán 1983, Zuta et al. 1983, Serra 1983 y 1991, Csirke 1995). En el Perú, en algunas áreas y bajo ciertas condiciones oceanográficas T. murphyi se encuentra compartiendo las mismas áreas con otras especies como la caballa Scomber japonicus, anchoveta E. ringens y la sardina S. sagax.
En este trabajo se presentan los resultados de las estimaciones de biomasa y áreas de distribución de T. murphyi con el método hidroacústico obtenidos de los cruceros de
Publicado online: 11/10/2013Publicado impreso: 15/10/2013

88
Segura & Aliaga
Rev. peru. biol. número especial 20(1): 087- 096 (Septiembre 2013)Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú.
Editores: Csirke J., R. Guevara-Carrasco & M. Espino
evaluación de los principales recursos pelágicos realizados entre 1983 – 2012 por el Instituto del Mar del Perú. La evaluación directa del T. murphyi por hidroacúastica se efectúa en el Perú como parte de la evaluación del principal recurso pesquero na-cional que es la anchoveta E. ringens. Por ello estos cruceros de evaluación se realizan por lo general en verano y sólo abarcaron la franja costera hasta 100 millas náuticas (mn) de distancia de la costa. Ocasionalmente se han ejecutado este tipo de cruceros en otras estaciones del año y en algunos casos se llegó hasta las 200 mn de la costa, como los cruceros hechos en la primavera de 1986 y verano de 1990 (Ganoza 1998). Por ello se considera que los estimados de las biomasas de T. murphyi por el método hidroacústico representan sólo una fracción de la biomasa total disponible en toda el área potencial de su distribución frente a la costa peruana.
Material y métodos Durante el periodo 1983 – 2012, los cruceros para evaluar la
biomasa acústica y la distribución de recursos pelágicos ejecuta-dos por el Instituto del Mar del Perú han mantenido en general la misma cobertura espacial desde el extremo norte al extremo sur del dominio marítimo peruano, siguiendo transectos siste-máticos paralelos (Johannesson & Robles 1977, MacLennan & Simmonds 1992) perpendiculares a la costa, con una separación de 12 a 17 mn entre transectos. Estos cruceros se realizaron a bordo de los buques de investigación científica (BIC) SNP-1, Humboldt, Olaya y SNP-2. Además, en 1990 se realizó una evaluación conjunta con el R/V Nansen (Ex–URSS).
La identificación de cada crucero se hace mediante un código compuesto de dos letras seguidas de cuatro o seis números. Las dos primeras letras “Cr” indican que se trata de un crucero, los dos números siguientes indican el año en que se realizó el crucero y los dos o cuatro números siguientes indican el mes o meses en que se realizó el mismo. Es así que, por ejemplo “Cr.1102-04” identifica el crucero realizado el año 2011 entre los meses de febrero y abril.
A lo largo del periodo de estudio se utilizaron equipos cien-tíficos vigentes en su época, los que siempre se mantuvieron actualizados. En los primeros años se emplearon equipos analó-gicos tales como las ecosondas científicas SIMRAD de modelos EK y ecointegradores Hewlett Packard, y en los últimos años se vienen empleando equipos digitales SIMRAD modelo EK60 en frecuencia de trabajo 38, 120 y 200 kHz, y con software de procesamiento y ecointegración Echoview.
Al inicio de cada exploración se efectúa la calibración del sistema de evaluación hidroacústico para determinar la constante del instrumento (Ci). A partir de 1992 se empezó a calibrar con blanco estándar (Foote 1987, SIMRAD 1992) utilizando esferas de cobre de 23 mm (38 kHz), 60mm (120 kHz) y 13.7mm (200 kHz). Asimismo, cuando la evaluación se realiza con más de una embarcación y con el objeto de estandarizar el rendimiento del sistema acústico, se efectúa una intercalibración entre las embarcaciones participantes, navegando en forma paralela muy cerca una de la otra y ejecutando ecointegración simultánea.
Teniendo en cuenta la fuerza del eco del blanco (TS) es-pecífica para cada especie, el procedimiento de cuantificación durante el muestreo acústico se hace a través de un proceso de ecointegración por Unidades Básicas de Muestreo (UBM). Se integra la señal de los ecos por rangos de distancia de 1 mn, a
velocidad aproximada de 10 nudos. La ecointegración consiste en una integración del area de retrodispersión transversal de los ecos recibidos, también conocida como back-scattering strength area. Los resultados de la ecointegración se dan clásicamente en m2 por mn (Simmonds & MacLennan 2005),
Durante las exploraciones acústicas se realizan operaciones de pesca para la comprobación de los ecotrazos y para realizar muestreos biológicos. Las operaciones de pesca se realizan con red de arrastre pelágica 124/1800 mm de poliamida (PA), con una longitud total de 73 m y una abertura vertical promedio de 12 m. Esta red tiene un copo interno de tamaño de malla de 12 mm y los lances tienen una duración efectiva entre 15 y 30 minutos, dependiendo de la densidad del número de cardú-menes. El comportamiento de la red de pesca es monitoreado con una netsonda.
La separación de los valores integrados por especies se hace por reconocimiento directo del cardumen y/o asignación de valores, para lo cual se toman en cuenta los resultados de las operaciones de pesca (Simmonds & MacLennan 2005). Luego de hacer la separación de los valores integrados (m2/mn) por especie, se determinan las áreas de distribución y concentración con el software SURFER mediante el método de interpolación kriging y contorneo (Gutiérrez 1997). Se han establecido cua-tro categorías de concentración: muy disperso (0 – 10m2/mn), disperso (> 10 – 100m2/mn), denso (> 100 – 1000m2/mn) y muy denso (>1000 m2/mn).
La biomasa es calculada por áreas isoparalitorales que tienen una dimensión de 0.5° de latitud por 10 mn de ancho siguiendo un trazado paralelo a la línea de costa. Este es un sistema propio desarrollado por el IMARPE en los años 70 y luego fue definido en formato digital (Gutiérrez & Peraltilla 1999). Previamente los datos son tratados con el software MapInfo y los cálculos se hacen con el software R (http://www.r-project.org/).
A partir de 1992 se siguió un sistema de procesamiento para los estimados de la biomasa algo diferente al seguido en años anteriores. Entre 1983 y 1991 la biomasa se obtenía del producto del área del estrato, los valores integrados de la especie en el estrato y una constante de calibración estimada a partir de mediciones electrónicas, acústicas y con peces vivos en jaula (Johannesson & Vílchez 1981). De 1992 a la fecha se sigue el mismo procedimiento sólo que la constante de calibración se estima a partir de la calibración con blanco estándar y la relación fuerza de blanco (TS)/longitud (L) del jurel (TS/L).
Entre 1992 y 1997, se trabajó con ecosonda de 38 kHz y para calcular la fuerza de blanco (TS) se utilizó la ecuación (Foote 1987):
TS = 20 log L – 71.9 dB
A partir de 1998 se trabajó con ecosonda de 120 kHz y se uso la siguiente ecuación para calcular la fuerza de blanco (Gutiérrez et al. 1999, Castillo et al. 2009):
TS = 20 log L – 70.8 dB
De este modo, la biomasa por área isoparalitoral, que esta referida a distancia de la costa, se calcula de acuerdo a la siguien-te fórmula, donde el área de retrodispersión transversal (σ), la densidad de peces (ρ), el número de individuos (n) y la biomasa en el estrato es calculada de:

89
Biomasa acústica y distribución del jurel Trachurus murphyi en el Perú
Rev. peru. biol. special issue 20(1): 087- 096 (September 2013)Ecology, fisheries and conservation of Jack mackerel (Trachurus murphyi) in Peru. Editors: Csirke J., R. Guevara-Carrasco & M. Espino
Figura 1. Biomasa estimada por hidroacústica y desembarques de jurel T. murphyi en toneladas, años 1983-2012.Figure 1. Jack mackerel T. murphyi acoustic biomass estimates and landings in tons, years 1983-2012.
σ = 4 (π) 10TS/10
ρ = 10000 Ci Sa / σ
n = ρ A
Biomasa = n*W / 1000000
donde,
Ci : Constante del instrumentoρ : Densidad peces/m2
A : Área isoparalitoral (mn2)Sa : valor ecointegrado promedio (m2/mn2)W : peso promedio del jurel (g)n : número de peces
Los límites de confianza fueron determinados según lo pro-puesto por Bazigos (1975):
Donde:
B: biomasa o abundancia total de la evaluación (toneladas)Sj
2: varianza del estrato jnj: número de UBMs en el j-ésimo estrato.Aj: área del j-ésimo estrato (mn2)NASCj: Coeficiente Náutico de Dispersión Acústica prome-
dio en el j-ésimo estrato
Resultados y discusión Biomasa.- Durante el periodo 1983 – 2012 se registraron dos
valores máximos en los estimados de biomasa hidroacústica: en el otoño de 1983 (Cr.8303-05) con 8512674 t; y en el verano de 1993 (Cr.9301-03) con 8471096 t (Figura 1, Tabla 1). Después de El Niño 1982-83, en el invierno de 1984 (Cr.8406-09), la biomasa estimada disminuyó a 5001913 t. El proceso de en-friamiento de 1985 originó el desplazamiento longitudinal del frente oceánico, lo cual produjo el alejamiento de T. murphyi a
distancias mayores de la costa, fuera del área prospectada (Ga-noza 1998), lo que explicaría los bajos estimados de biomasa ese año. En los años siguientes se notó una recuperación en los estimados, hasta alcanzar otro máximo valor en 1993. Los eventos cálidos de 1987-88 y 1992 afectaron la distribución y biomasa de este recurso, dando lugar a desplazamientos hacia el sur. Luego, a partir de 1994, se produjo una disminución paulatina de la biomasa estimada, con ligeras recuperaciones en el 2000-2001, 2006 y 2011.
Con relación a las densidades promedio en función a las áreas de distribución, los mayores valores de áreas de distribución de T. murphyi en la franja costera evaluada se dieron en la década de los 80 y principios de los 90, mientras que los valores más altos de densidad se dieron en los primeros años de la década de los 90 (Tabla 1 y Fig. 2). En general se observo una tendencia a encontrar densidades medias más altas cuando las áreas de T. murphyi eran menores y más pegadas a la costa.
Luego de El Niño de 1992 se observo un incremento de la densidad, que alcanzo sus máximos valores en 1993 y 1994 con 336 y 368 t/mn2 respectivamente, para disminuir a 185 t/mn2 en 1996 en áreas de evaluación semejantes. En años más recientes las densidades y las biomasas acústicas se han mantenido bajas y los valores más bajos de los últimos años se encontraron en 2010, con un ligero repunte en 2011 donde la biomasa estimada por acústica fue de 363 mil toneladas con densidades medio-altas pero reducidas para descender nuevamente en 2012 (Tabla 1).
Las áreas exploradas durante los cruceros científicos de evaluación acústica han cubierto todos los años la franja más costera del mar peruano y es relevante destacar que la extensión de estas áreas se mantuvo más o menos constante durante el período estudiado (Fig. 3). En la Figura 3, también se puede observar que hasta principios de los años 90 (Crucero 9301-03) se encontró T. murphyi en buena parte (casi el 50%) de las áreas exploradas. En los años siguientes la porción ocupada por las áreas de distribución del T. murphyi disminuyen significativa-mente, y disminuyen con mayor notoriedad a partir de 2001.
0
1
2
3
4
5
6
7
8
9
1983 1985 1987 1989 1991 1993 1995 1997 1999 2001 2003 2005 2007 2009 2011
Bio
mas
a m
illon
es (t
onel
adas
)
Años
Biomasa Acústica (t)
Captura (t)
Despues de 1998 la biomasa
acústica no superó el millón de
toneladas.
Antes de 1998 la máxima
biomasa acústica fue 8.5
millones de toneladas.

90
Segura & Aliaga
Rev. peru. biol. número especial 20(1): 087- 096 (Septiembre 2013)Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú.
Editores: Csirke J., R. Guevara-Carrasco & M. Espino
Áre
as d
e di
strib
ució
n (m
n2)
Densidad (t/m
n2)
0
100
200
300
400
500
600
0
10000
20000
30000
40000
50000
60000
70000
80000
1981 1985 1989 1993 1997 2001 2005 2009 2013
área de distribución del jureldensidad media
Años
Figura 2. Áreas de distribución (mn2) y densidades medias (t/mn2) de jurel T. murphyi y sus medias móviles (lineas continua y punteadas) por años, 1983 – 2012. Figure 2. Distribution areas (mn2) and mean densities (t/mn2) of Jack mackerel T. murphyi with their moving averages (solid and broken lines), by year, 1983 – 2012.
Tabla 1. Información general de los cruceros de evaluación acústica ejecutados entre 1983 y 2012, incluyendo información sobre el área evaluada y las áreas de distribución, biomasa acústica y densidad promedio de jurel T. murphyi estimadas en cada crucero. Table 1. General information of the acoustic surveys conducted between 1983 and 2012, including information on the surveyed area and the areas of distribution, acoustic biomass and mean densities of Jack mackerel T. murphyi estimated on each survey.
Año Crucero Área distribución del jurel (mn2)
Área total evaluada (mn2) Biomasa acustica Densidad Área jurel Densidad media
1983 8309-12 28368 66711 8512674 300.080161 10.253017 5.70404964
1984 8406-09 50922 126627 5001913 98.226955 10.8380503 4.58728067
1985 8503-05 4592 76750 180259 39.2550087 8.43207094 3.67007905
1986 8604-05 51480 85973 4329874 84.1078866 10.8489487 4.43210034
1987 8708-09 66898 100227 6471955 96.7436246 11.1109244 4.57206443
1988 8805-07 52701 115813 2811590 53.3498416 10.8723897 3.97687101
1989 8907-09 71470 120712 4365164 61.0768714 11.1770331 4.11213326
1990 9002-04 43522 94323 5971710 137.211295 10.6810218 4.92152204
1991 9106-08 53748 104269 5597492 104.143261 10.8920617 4.64576746
1992 9201-03 11893 74952 2957578 248.68225 9.38370527 5.51617598
1993 9301-03 25173 100496 8471096 336.515155 10.1335273 5.81864319
1994 9401-03 17430 107606 6413607 367.963683 9.76594814 5.90798425
1995 9502-04 11077 69444 3251207 293.509705 9.31262617 5.6819107
1996 9602-04 16633 103683 3375866 202.961943 9.71914395 5.31301849
1997 9709-10 3452 50927 1892286 548.170915 8.14670905 6.30658713
1998 9803-05 3789 50948 107236 28.3019266 8.23985741 3.34292988
1999 9902-03 6136 89618 661714 107.846557 8.72187945 4.68070945
2000 0001-02 20841 110375 1070788 51.3779012 9.94469711 3.93920814
2001 0103-04 16492 114736 1066544 64.6699094 9.71063821 4.16929601
2002 0202-03 8869 82029 446782 50.3773522 9.0902836 3.91954171
2003 0302-03 4067 98898 453597 111.543714 8.31054888 4.71441657
2004 0402-03 3155 104461 239356 75.8700393 8.05668472 4.32902187
2005 0502-04 2511 104387 561535 223.607457 7.82853715 5.40989209
2006 0602-04 3736 88189 879266 235.350091 8.22576881 5.46107416
2007 0702-04 2494 108580 236235 94.7294407 7.8215585 4.55102484
2008 0802-04 2245 140171 109643 48.8463125 7.71630411 3.88867889
2009 0902-03 1204 89618 70074 58.1791896 7.09377692 4.06352772
2010 1002-04 69 72690 23429 339.544435 4.2341065 5.82760482
2011 1102-04 2798 89114 362777 129.6558 7.93666016 4.86488325
2012 1202-04 2101 97636 44934 21.3848845 7.65026568 3.06268434

91
Biomasa acústica y distribución del jurel Trachurus murphyi en el Perú
Rev. peru. biol. special issue 20(1): 087- 096 (September 2013)Ecology, fisheries and conservation of Jack mackerel (Trachurus murphyi) in Peru. Editors: Csirke J., R. Guevara-Carrasco & M. Espino
Figura 3. Áreas exploradas durante los cruceros de evaluación de recursos pelágicos (línea negra) y áreas de distribución de jurel T. murphyi (línea gris) entre 1983 y 2012. Figure 3. Explored areas during the pelagic stock assessment acoustic surveys (black line) and areas of distribution of Jack mackerel T. murphyi (gray line). 1983 – 2012.
0
50000
100000
150000
200000
250000
04°
Tone
lada
s
06° 08° 10° 12° 14° 16° 18°
Latitud S
-0
200000
400000
600000
800000
1000000
1200000
1400000
1600000
1800000
2000000
2200000
2400000
2600000
2800000
3000000
Cruceros
Gra
dos
de la
titud
S
Mag
nitu
d de
la b
iom
asa
de ju
rel (
tone
lada
s)
18°
16°
14°
12°
10°
8°
6°
4°
18°
16°
14°
12°
10°
8°
6°
4°
1983
1984
1985
1986
1987
1988
1989
1990
1991
1992
1993
1994
1995
1996
1997
1998
1999
2000
2005
2006
2007
2008
2009
Figura 4. Biomasas promedio de jurel T. murphyi por grados de latitud de la serie 1983 – 2012.Figure 4. Jack mackerel T. murphyi average biomass per latitude degrees for the period 1983 – 2012.
Figura 5. Distribución latitudinal de la biomasa de jurel T. murphyi estimada acústicamente entre 1983 y 2009.Figure 5. Distribution by latitude of the estimated Jack mackerel T. murphyi acoustic biomass between 1983 and 2009.
Según Barbieri et al. (2004, 2005) el T. murphyi tendría una cierta capacidad de adaptación a las características cambiantes del ecosistema, y en particular a los cambios relacionados con eventos como El Niño y La Niña, y esto podría explicar en parte los cambios en las áreas de distribución y densidad obser-vados. Desde 2000 los cardúmenes de T. murphyi que fueron detectados durante las exploraciones acústicas se encontraban en forma dispersa e individual (no agregados), razón por la cual las densidades fueron por lo general muy bajas.
Distribución.- Entre 1983 y 2012 (Fig. 4), la distribución promedio de la biomasa de T. murphyi por grados de latitud en el mar peruano entre 1983 y 2012 confirma la presencia de esta especie a lo largo de todo el litoral peruano y, especialmente, de los 5°S hacia el sur. Esta observación general de la distribución de T. murphyi detectado acústicamente guarda buena relación con la presencia de huevos y larvas de la misma especie en toda la costa del Perú (Ayón & Correa 2013).
La imagen sinóptica de la variación latitudinal de la biomasa acústica con el tiempo (Fig. 5) muestra como se han dispuesto los principales focos de concentración a lo largo de la costa
0
1
2
3
4
5
1983 1985 1987 1989 1991 1993 1995 1997 1999 2001 2003 2005 2007 2009 2011
Log(
UB
M)
Años
UBM total
UBM con Jurel
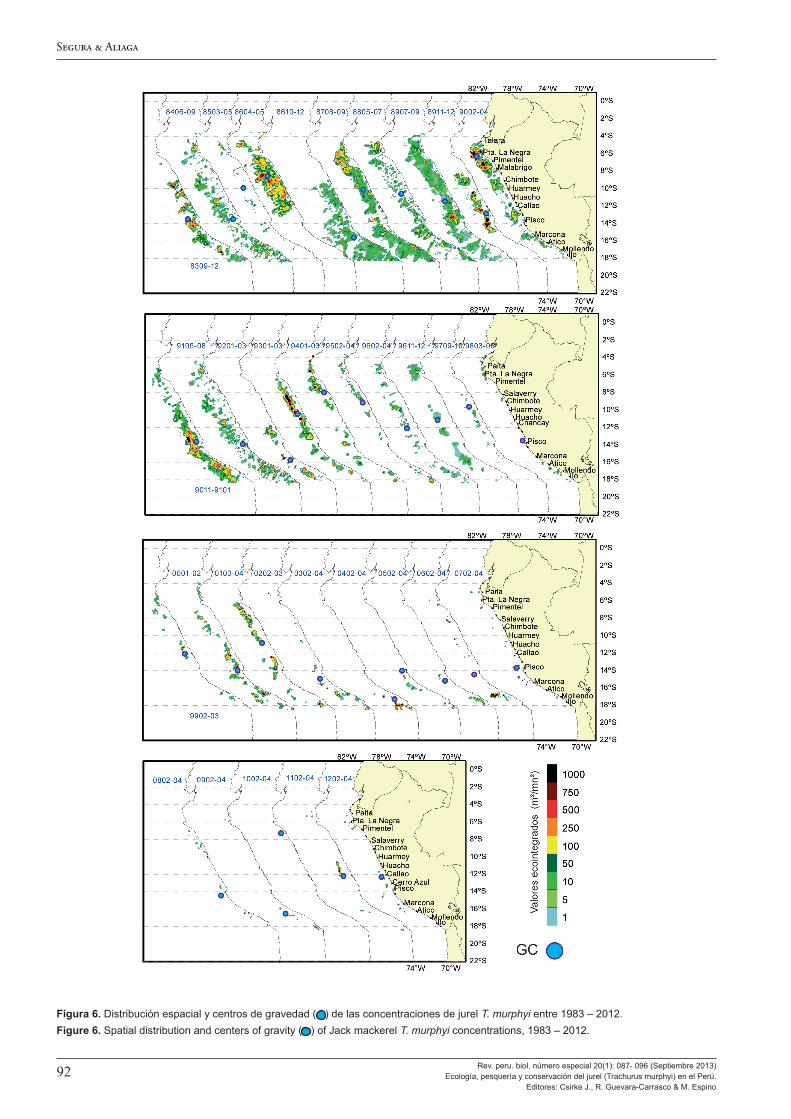
92
Segura & Aliaga
Rev. peru. biol. número especial 20(1): 087- 096 (Septiembre 2013)Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú.
Editores: Csirke J., R. Guevara-Carrasco & M. Espino
Figura 6. Distribución espacial y centros de gravedad ( ) de las concentraciones de jurel T. murphyi entre 1983 – 2012.Figure 6. Spatial distribution and centers of gravity ( ) of Jack mackerel T. murphyi concentrations, 1983 – 2012.

93
Biomasa acústica y distribución del jurel Trachurus murphyi en el Perú
Rev. peru. biol. special issue 20(1): 087- 096 (September 2013)Ecology, fisheries and conservation of Jack mackerel (Trachurus murphyi) in Peru. Editors: Csirke J., R. Guevara-Carrasco & M. Espino
0
20
40
60
80
100
2002Años
Prof
undi
dad
med
ia d
el c
ardu
men
(m)
2003 2004 2005 2006 2007 2008 2009 2010 2011 2012
Figura 7. Profundidades medias y distribución vertical de cardúmenes de jurel T. murphyi, 2002 – 2012.Figure 7. Average depths and vertical distribution of Jack mackerel T. murphyi shoals, 2002 – 2012.
Figura 8. Distribución de la salinidad superficial y las principales masas de agua, 2006 – 2009. ATS (Aguas Tropicales Superficiales); AES (Aguas Ecuatoriales Superficiales); ACF (Aguas Costeras Frías); ASS (Aguas Subtropicales Superficiales).Figure 8. Surface salinity and main water masses distribution, 2006 – 2009. ATS (superficial tropical waters or STW); AES (superficial equatorial waters or SEW); ACF (Cold coastal waters or CCW); ASS (superficial subtropical waters or SSW)
entre 1983 y 2009. Se destaca que estos focos de concentración guardan buena correspondencia con la variabilidad de la con-figuración de las masas de agua en el mar peruano descrita por Flores et al. (2013). En esta figura se puede observar el efecto de un evento El Niño extraordinario como el de 1997-98 en la disponibilidad de jurel T. murphyi en el área evaluada, que causó la notable reducción de la biomasa acústica luego de ese evento. Es interesante anotar la concordancia de estas observaciones con los resultados de los análisis de ecología trófica presentados por Alegre et al. (2013).
Entre 1983 y 1990 se observaron concentraciones de T. mur-phyi en forma continua en casi todo el área evaluada (Fig. 6),
con concentraciones densas al norte de Pisco (Pisco-Chimbote en 1983, Talara-Huarmey en 1986, Supe-Pisco en 1989, y Paita-Malabrigo en 1990) y dispersas en las áreas restantes. En la primavera de 1983 (Cr. 8309-12), T. murphyi se distribuyó hasta las 120 mn de la costa, con mayores concentraciones en los 10°S y 14°S. En 1988 y en 1989 alcanzó las 130 mn, y en 1990 se le detectó hasta las 140 mn.
Entre 1990 y 1998 T. murphyi ocupó pequeñas áreas de distribución en las áreas exploradas. Sólo se detectaron con-centraciones densas hasta 1994, entre Pisco y Malabrigo. La distribución fue tornándose más cercana la costa, desde 120 mn (1990) hasta 70 mn de distancia de costa (1994). De 1995 a 1998 el jurel se observó en pequeñas concentraciones dispersas y aisladas cerca de la costa.
Entre el 2000 y 2007, T. murphyi se encontró muy disperso, con pequeñas áreas de concentración densa al sur de Pisco. Del 2008 al 2012, sus áreas de concentración continuaron siendo muy dispersas y replegadas hacia la costa, mientras que las áreas con densidades altas siguieron ausentes en el área de evaluación.
Los cambios en la distribución de T. murphyi se reflejaron también en la localización de los centros de gravedad de las con-centraciones de jurel (Fig. 6), con un desplazamiento latitudinal que demuestra el dinamismo en la distribución de esta especie en el mar peruano.
En general, se observa que entre 1983 y 2012 el área ocupada por el T. murphyi fluctuó considerablemente dentro de un área total de evaluación cuya extensión no varió significativamente. A partir de 1992 se presento un cambio considerable en la ex-tensión de las áreas de distribución de T. murphyi. Analizando
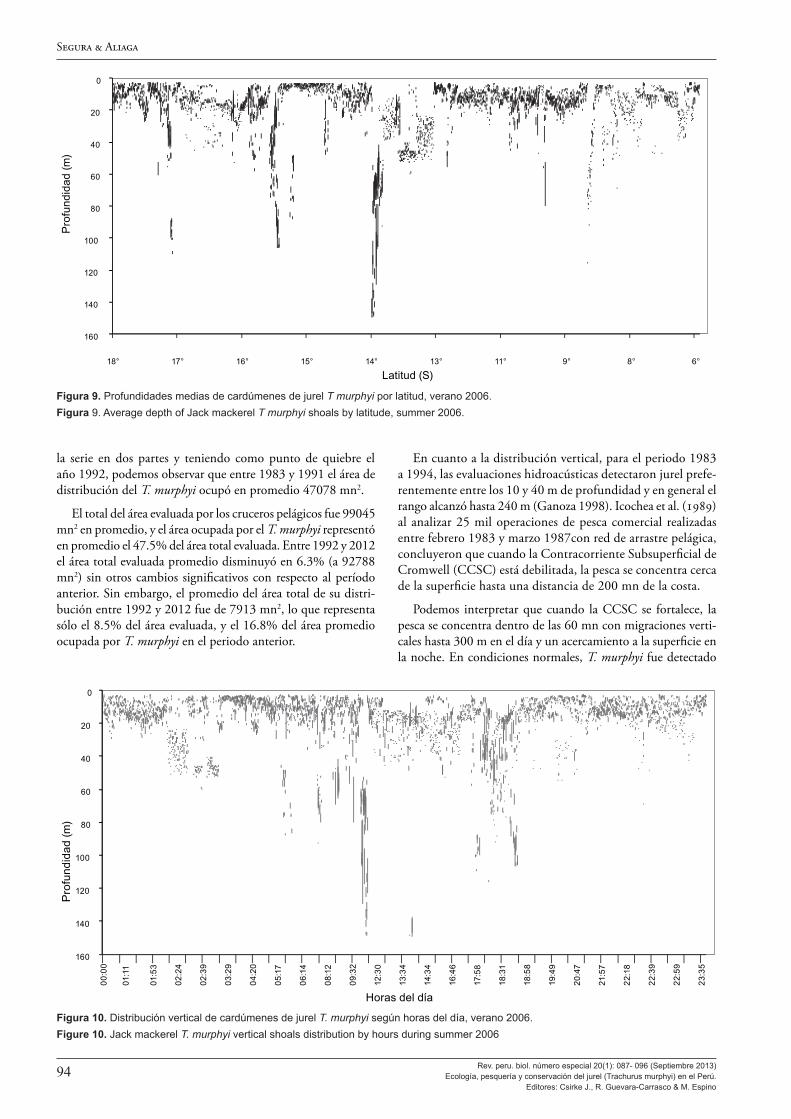
94
Segura & Aliaga
Rev. peru. biol. número especial 20(1): 087- 096 (Septiembre 2013)Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú.
Editores: Csirke J., R. Guevara-Carrasco & M. Espino
Figura 9. Profundidades medias de cardúmenes de jurel T murphyi por latitud, verano 2006.Figura 9. Average depth of Jack mackerel T murphyi shoals by latitude, summer 2006.
Figura 10. Distribución vertical de cardúmenes de jurel T. murphyi según horas del día, verano 2006.Figure 10. Jack mackerel T. murphyi vertical shoals distribution by hours during summer 2006
la serie en dos partes y teniendo como punto de quiebre el año 1992, podemos observar que entre 1983 y 1991 el área de distribución del T. murphyi ocupó en promedio 47078 mn2.
El total del área evaluada por los cruceros pelágicos fue 99045 mn2 en promedio, y el área ocupada por el T. murphyi representó en promedio el 47.5% del área total evaluada. Entre 1992 y 2012 el área total evaluada promedio disminuyó en 6.3% (a 92788 mn2) sin otros cambios significativos con respecto al período anterior. Sin embargo, el promedio del área total de su distri-bución entre 1992 y 2012 fue de 7913 mn2, lo que representa sólo el 8.5% del área evaluada, y el 16.8% del área promedio ocupada por T. murphyi en el periodo anterior.
En cuanto a la distribución vertical, para el periodo 1983 a 1994, las evaluaciones hidroacústicas detectaron jurel prefe-rentemente entre los 10 y 40 m de profundidad y en general el rango alcanzó hasta 240 m (Ganoza 1998). Icochea et al. (1989) al analizar 25 mil operaciones de pesca comercial realizadas entre febrero 1983 y marzo 1987con red de arrastre pelágica, concluyeron que cuando la Contracorriente Subsuperficial de Cromwell (CCSC) está debilitada, la pesca se concentra cerca de la superficie hasta una distancia de 200 mn de la costa.
Podemos interpretar que cuando la CCSC se fortalece, la pesca se concentra dentro de las 60 mn con migraciones verti-cales hasta 300 m en el día y un acercamiento a la superficie en la noche. En condiciones normales, T. murphyi fue detectado
0
20
40
60
80
100
120
140
160
18° 17° 16° 15° 14° 13° 11° 9° 8° 6°
Pro
fund
idad
(m)
Latitud (S)
0
20
40
60
80
100
120
140
160
00:0
0
01:1
1
01:5
3
02:2
4
02:3
9
03:2
9
04:2
0
05:1
7
06:1
4
08:1
2
09:3
2
12:3
0
13:3
4
14:3
4
16:4
6
17:5
8
18:3
1
18:5
8
19:4
9
20:4
7
21:5
7
22:1
8
22:3
9
22:5
9
23:3
5
Pro
fund
idad
(m)
Horas del día

95
Biomasa acústica y distribución del jurel Trachurus murphyi en el Perú
Rev. peru. biol. special issue 20(1): 087- 096 (September 2013)Ecology, fisheries and conservation of Jack mackerel (Trachurus murphyi) in Peru. Editors: Csirke J., R. Guevara-Carrasco & M. Espino
con mayor frecuencia por encima de los 50 m de profundidad. En eventos El Niño, T. murphyi se profundiza hasta 200 m con desplazamientos verticales desde la superficie hasta cerca a los 350 m de profundidad (Dioses & Ñiquen, 1988).
En general, durante el periodo 2002 al 2012, la profun-didad media de los cardúmenes de jurel se ubicó alrededor de los 15 m, sin embargo durante este periodo observamos que T. murphyi disminuyó su rango de distribución vertical (Fig. 7). Entre 2002 y 2006 los cardúmenes se encontraron principalmente a profundidades que alcanzaron los 70 m y en algunos casos llegaron hasta 150 m. Pero entre 2007 y 2012 los cardúmenes se encontraron preferentemente en los primeros 40 m profundidad.
En el mar peruano, la distribución de T. murphyi muestra una estrecha relación con variables ambientales como la temperatura y la salinidad (Fig. 8). Varios estudios y análisis han sustentado el rol de la Zona Mínima de Oxigeno (ZMO) como regulador de los procesos claves de la productividad y la distribución de los recursos pelágicos (Bertrand et al 2010). Esta relación es notoria, particularmente durante eventos El Niño o La Niña, cuando se observa gran fluctuación de estas variables y T. murphyi muestra gran capacidad de respuesta modificando tanto su distribución horizontal como vertical en la época de estos eventos (Figs. 6, 7, 9 y 10).
Se ha observado que la distribución vertical de las concen-traciones de T. murphyi cambia según la latitud y horas del día. En la Figura 9 se puede apreciar que en el verano de 2006 los cardúmenes alcanzan mayor profundidad en la zona centro-sur, especialmente en el paralelo 14°S. Respecto a las horas, existe una tendencia a que los cardúmenes vayan más hacia la superficie durante la noche y las primeras horas del día debido a su ciclo alimenticio y la agregación de macro-zooplancton en forma de cordón en las noches (Fig. 10).
Conclusiones.- La biomasa de T. murphyi estimada por el método de evaluación hidroacústica tuvo dos valores máximos: el primero en 1983, con 8512674 t; y el otro en 1993, con 8471096 t. Luego se observa una tendencia decreciente, con un estimado de biomasa de solamente 1239 t para el año 2010.
Los estimados de biomasa del T. murphyi dentro del área evaluada disminuyeron drásticamente a partir de 1992, año en el cual se da un punto de quiebre en su distribución y concen-tración en aguas peruanas. La misma tendencia se observo en la extensión de sus áreas de concentración.
Trachurus murphyi muestra una marcada variabilidad en su abundancia y sus áreas de distribución entre 1983 – 2012. Hasta antes de 1994, las concentraciones densas se encontraron al norte de Pisco, mientras que de 1995 al 2012 las concen-traciones se reportaron al sur en concentraciones aisladas y muy reducidas.
Frente a las costas de Perú, la distribución del T. murphyi muestra una estrecha interrelación con parámetros ambientales como la temperatura, la salinidad y el oxígeno.
Tanto en su distribución horizontal como vertical, la especie muestra una gran capacidad de adaptación a las condiciones de los eventos ENSO que afectan los tipos, formas, dinámica de cardumen y dinámica nictemeral.
Las evaluaciones de T. murphyi han estado condicionadas a la distribución de la anchoveta E. ringens que tiene una distribución más costera y es muy probable que subestimen las verdaderas existencias de T. murphyi frente a las costas peruanas en la medida que parte de la población se distribuye a mayor distancia de la costa en aguas peruanas o también en alta mar.
Literatura citadaAlegre A., P. Espinoza & M. Espino. 2013. Ecología trófica del jurel Trachurus
murphyi en el Perú entre 1977-2011. En: Csirke J., R. Guevara-Carrasco & M. Espino (Eds.). Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú. Rev. peru. biol. número especial 20(1): 075 – 082
Ayón P. & J. Correa. 2013. E Variabilidad espacial y temporal de larvas de jurel Trachurus murphyi en el Perú entre 1966-2010. En: Csirke J., R. Guevara-Carrasco & M. Espino (Eds.). Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú. Rev. peru. biol. número especial 20(1): 083 – 086
Barbieri M.A., V. Correa & J. Córdova. 2005. Pesca de investigación: “De-terminación de la distribución espacial de jurel entre la V y X Regiones”. Otoño 2005. Informe final - Subpesca. Instituto de Fomento Pesquero. Chile.
Barbieri M.A , V. Correa & J. Córdova. 2004. Pesca de investigación “Determi-nación de la distribución espacial de jurel entre la V y X Regiones. Invierno 2004. Informe final -Subpesca. Instituto de Fomento Pesquero. Chile. 20 pp. (más tablas y figuras).
Bazigos G.P. 1975. The statistical efficiency of echo surveys with special reference to Lake Tanganyika. FAO Fisheries Technical Paper 139. 43 pp.
Bertrand, A., Ballón, M & A. Chaigneau. 2010 Acoustic Observation of Living Organisms Reveals the Upper Limit of the Oxygen Minimum Zone. PLoS ONE S(4): e10330. doi:10.1371/journal.pone.0010330.
Csirke J. 1995. Fluctuations in abundance of small and mid-sized pelagic. Science Marine 59(3-4): 481-490.
Castillo, P.R., S. Peraltilla, A. Aliaga, M. Flores, M. Ballón, J. Calderón & M. Gutiérrez, 2009. Protocolo técnico para la evaluación acústica de las áreas de distribución y abundancia de recursos pelágicos en el mar peruano. Versión 2009. Informe Inst. Mar Perú, 36(1-2): 7-28.
Dioses T. & M. Ñiquen, 1988. Aspectos generales sobre la biología y pesquería de recurso jurel (Trachurus murphyi) en Perú. En FORUM: Ca-racterísticas en el diseño de una embarcación ideal para la captura de especies pelágicas 7. PESCA PERU. 36 pp.
Flores R., M. Espino, G. Luque & J. Quispe. 2013. Patrones de variabilidad ambiental en el mar peruano. En: Csirke J., R. Guevara & M. Espino (Eds.). Ecología, pesquería y conservación del jurel (Trachurus mur-phyi) en el Perú. Rev. peru. biol. número especial 20(1): 021 – 028
Foote K. 1987. Fish target strengths for use in echo integrator surveys. Journal of the Acoustical Society of America 82:981-987. DOI: 10.1121/1.395298
Ganoza F. 1998. Abundancia y comportamiento del jurel (Trachurus picturatus murphyi) a través de prospecciones acústicas de 1983 a 1994. Inf. Inst. Mar Perú. 136: 23-47.
Gutiérrez M. & S. Peraltilla. 1999. Aplicación de un sistema de información geográfica y de la carta electrónica isoparalitoral en las evaluaciones hidroacústicas de las biomasas de recursos pesqueros en el litoral peruano. Inf. Inst. Mar Perú 146: 25-29.
Gutiérrez M. 1997. Aplicación de software de interpolación en las evaluaciones hidroacústicas de la biomasa y distribución de los recursos pelágicos. Inf. Prog. Inst. Mar Perú 67: 21-30.
Gutiérrez M., N. Herrera & D. Marín. 1999. Distribución y abundancia de anchoveta y otras especies pelágicas entre los eventos El Niño de 1982-83 y 1997-98. Bol. IMARPE, 18(1-2): 77-88.
Icochea L., A. Chipollini & M. Ñiquen. 1989. Análisis de pesquería de arrastre pelágica en la costa peruana durante 1983-1987 y su relación con el medio ambiente. Memorias del Simposio Internacional de los Recursos Vivos y las Pesquerías en el Pacífico Sudeste. Viña del mar, 9-13 mayo, 1988.
Johannesson K. & A. Robles. 1977. Echo surveys of Peruvian anchoveta. Rapp. P.-V. Réu. Cons. Int. Explor. Mer. 170: 237-244.
Johannesson K. & R. Vilchez. 1981. Application and some results of echo-integration methods of monitoring Peruvian anchovy resources. In: J. Suomala (Ed.). Meeting on hydroacoustical methods for the esti-mation of marine fish populations, 25-19 June, 1979. Charles Stark Draper Laboratory Inc. Cambridge, MA. Vol. 2b: pp. 756-816.

96
Segura & Aliaga
Rev. peru. biol. número especial 20(1): 087- 096 (Septiembre 2013)Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú.
Editores: Csirke J., R. Guevara-Carrasco & M. Espino
Jordán J. 1983. Variabilidad de los recursos pelágicos en el Pacífico Sudeste. En G.D. Sharp y J. Csirke (Eds.). Actas de la Consulta de Expertos para examinar los cambios en la abundancia y composición por especies de recursos de peces neríticos. San José, Costa Rica, 18-29 abril 1983. Una reunión preparatoria para la Conferencia Mundial de la FAO sobre Ordenación y Desarrollo Pesqueros. FAO Fish.Rep./FAO Inf. Pesca (291), Vol. 2: pp. 113-129.
MacLennan D. & J. Simmonds. 1992. Fisheries Acoustics. London, Chapman and Hall Eds. 325pp.
Serra J.R. 1991. Important life history aspects of the chilean Jack mackerel,Trachurus symmetricus murphyi. Invest. Pesq. (Chile) 36:67-83.
Serra J. R. 1983. Changes in the abundance of pelagic resources along the Chi-lean coast. In Proceedings of the Expert Consultation to Examine Changes in Abundance and Species Composition of Neritic Fish Resources. San Jose, Costa Rica, April 1983. Sharp, G.D. and J. Csirke (Eds). FAO Fish. Rep. 291 (2): 255-284.
Simmonds E.J. & D.N. MacLennan. 2005. Fisheries acoustics, theory and practice. Blackwell Science, 472 pp.
SIMRAD. 1992. Instruction Manual, Simrad EK 500 scientific echo sounder. P217E. Calibration of the EK 500 P2260E. Simrad Norge AS. Norway: 15-16
Zuta S., I. Tsukayama & R. Villanueva. 1983. El ambiente marino y las fluctua-ciones de las principales poblaciones pelágicas de la costa peruana. En G.D. Sharp & J. Csirke (Eds.). Actas de la Consulta de Expertos para examinar los cambios en la abundancia y composición por especies de recursos de peces neríticos. San José, Costa Rica, 18-29 abril 1983. Una reunión preparatoria para la Conferencia Mundial de la FAO sobre ordenación y desarrollo pesqueros. FAO Fish. Rep./FAO Inf. Pesca (291), Vol. 2: pp. 179-253.

97
Análisis de la pesquería del jurel Trachurus murphyi en el Perú
Rev. peru. biol. special issue 20(1): 097 - 106 (September 2013)Ecology, fisheries and conservation of Jack mackerel (Trachurus murphyi) in Peru. Editors: Csirke J., R. Guevara-Carrasco & M. Espino
Rev. peru. biol. número especial 20(1): 097 - 106 (Septiembre 2013)Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú. Editores: Csirke J., R. Guevara-Carrasco & M. Espino
Facultad de ciencias Biológicas unMsMISSN-L 1561-0837
Análisis de la pesquería del jurel Trachurus murphyi en el Perú
Miguel Ñiquen, Marilú Bouchon, Danny Ulloa y Ana Medina
Analysis of the Jack mackerel Trachurus murphyi fishery in Peru
Instituto del Mar del Perú, esquina Gamarra y General Valle S/N Chucuito, Callao, Perú.
Email Miguel Ñiquen: [email protected]
Email Marilú Bouchon: [email protected]
Pesquería, manejo y conservación del jurel (Trachurus murphyi) en el Perú
© Los autores. Publicado por la Revista Peruana de Biología de la Facultad de Ciencias Biológicas, Universidad Nacional Mayor de San Marcos. Este es un artículo de acceso abierto, distribuido bajo los términos de la Licencia de Atribución Reconocimiento-NoComercial-SinObraDerivada 3.0 de Creative Commons (http://creativecommons.org/licenses/by-nc-nd/3.0/deed.es_ES), que permite el uso no comercial, distribución y reproducción en cualquier medio, siempre que la obra original sea debidamente citadas. Para uso comercial, por favor póngase en contacto con [email protected].
Citación:Ñiquen M., M. Bouchon, D. Ulloa y A. Medina. 2013. Análisis de la pesquería del jurel Trachurus murphyi en el Perú. En: Csirke J., R. Guevara-Carrasco & M. Es-pino (Eds.). Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú. Rev. peru. biol. número especial 20(1): 097 - 106 (Septiembre 2013)
ResumenLa presencia del jurel Trachurus murphyi ha sido registrado en los desembarques de la pesca artesanal en Perú desde 1907. A partir del año 2000, las capturas de T. murphyi se ubicaron en el tercer lugar después de la anchoveta Engraulis ringens y el calamar gigante Dosidicus gigas. Las capturas de T. murphyi son realizadas por la flota artesanal, industrial cerquera y barcos arrastreros factoría. En el año 2011, la flota industrial cerquera con permiso de pesca para T. murphyi en la costa peruana estaba constituida por 62 embarcaciones industriales de acero (30177 m3) y 42 industriales de madera (3082 m3). Las capturas de T. murphyi tuvieron un notable incremento a partir de 1972, alcanzando sus máximos valores en 1977, 1996-97 y 2001, a los que siguieron años de muy baja disponibilidad que llevó al mínimo registro en las capturas en 2010. Sin embargo en el 2011 se revirtió esta baja disponibilidad y las capturas alcanzaron casi las 260000 toneladas. En el periodo 1972 – 2012 se observó una relación inversa entre los estimados de biomasa acústica del jurel T. murphyi y los desembarques anuales de anchoveta E. ringens. Las capturas mensuales de T. murphyi por la flota indus-trial y artesanal antes del 2002 fueron altas en la zona norte de Perú (Mancora-Chimbote), mientras que después del 2002 las capturas fueron altas en la zona centro-sur (Huacho-Ilo).
Palabras clave: Jurel, recursos pelágicos, pesquería peruana, esfuerzo de pesca.
AbstractThere are records of the presence of Jack mackerel Trachurus murphyi in the artisanal fishery landings in Peru since 1907. Since 2000 Jack mackerel T. murphyi catches are ranked third after anchovy Engraulis ringens and jumbo squid Dosidicus gigas. Artisanal, industrial purse seine and offshore (factory trawlers) fleets take part in the Peruvian catches of T. murphyi. In 2011 the industrial purse-seine fleet with license to fish for T. murphyi along the Peruvian coast comprised of 62 industrial steel vessels (30177 m3) and 42 industrial wood vessels (3082 m3). Catches of T. murphyi had a significant increase after 1972, with maximum catches in 1977, 1996-97 and 2001, after which several years of low availability followed, with the lowest catches in 2010. However, this low availability trend was reversed in 2011, and catches reached almost 260 thousand tons that year. A general inverse relationship has been observed between the T. murphyi estimated acoustic biomass and the annual landings of anchovy E. ringens during the the period 1972 – 2012. The monthly catches of T. murphyi by the industrial and artisanal fleets before 2002 were higher in the northern part of Peru (Mancora-Chimbote) while after 2002 the catches were higher in the south-central zone (Huacho-Ilo).
Keywords: Jack mackerel, pelagic, Peruvian fishery, fishing effort.
IntroducciónEl jurel, Trachurus murphyi Nichols 1920 es una especie transzonal de amplia dis-
tribución en el mar peruano. Es un recurso importante de la pesquería pelágica del Perú, que se caracteriza por ubicarse en diferentes condiciones oceanográficas y realiza desplazamientos a áreas favorables ante alteraciones climáticas (Tsukayama 1983, Zuta 1983, Bertrand et al. 2004).
Desde sus inicios, la extracción de T. murphyi ha sido realizada por diferentes tipos de flota, tanto en tamaño como en estrategias de pesca. Coker en el año 1907 reporta capturas de T. murphyi en el mar peruano. Sin embargo, los registros de desembarques empezaron en 1939 con cantidades que no superaban las 10 toneladas anuales. Esta
Publicado online: 11/10/2013Publicado impreso: 15/10/2013

98
Ñiquen et al.
Rev. peru. biol. número especial 20(1): 097 - 106 (Septiembre 2013)Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú.
Editores: Csirke J., R. Guevara-Carrasco & M. Espino
70 72 74 76 78 80 82 84 86 88 90 92 94 96 98 20 02 04 06 08 10 12
Años
Arrastre
Anchovetera
Sardinera
RSW
Artesanal
Figura 1. Periodos de operación de las diversas flotas pesqueras dirigidas a la captura de jurel T. murphyi en el mar peruano durante los años 1970-2012 (RSW = con sistema de refrigeración por agua de mar refrigerada, del inglés Refrigerated Sea Water)Figure. 1. Periods of operation of the various fishing fleets targeting on Jack mackerel T. murphyi in Peruvian waters during 1970-2012 (RSW = Refrigerated Sea Water)
situación se mantuvo hasta 1962, año en el que se superaron las 1000 toneladas. Desde los años sesenta, las estadísticas ubican a T. murphyi entre las tres primeras especies de peces pelágicos en la pesquería e, inclusive a partir del año 2000 se ubicó en segundo lugar después de la anchoveta.
Antes de 2002, cerca de 500 embarcaciones de un total aproximado de 1200 que conformaban la flota de cerco in-dustrial dirigieron su pesca al jurel T. murphyi, principalmente en periodos de veda de la anchoveta Engraulis ringens. En su mayoría, estas embarcaciones contaban con aparejos de pesca y sistemas de conservación que les permitían alternar entre una y otra espacie pelágica.
El año 2002 se promulgó el DS N°. 001-2002-PRODUCE, que estableció el uso exclusivo para consumo humano directo del jurel T. murphyi junto con la sardina Sardinops sagax y la caballa Scomber japonicus. Ese decreto limitó las capturas y permitió el desarrollo de una flota industrial con sistema de refrigeración RSW (Refrigerated Sea Water), con características especiales para consumo humano directo. Posteriormente, el 12 de abril del 2007 se estableció el Reglamento de Ordenamiento Pesquero de Jurel y Caballa, aprobado por DS N°. 011-2007-PRODUCE, que norma y promueve la explotación racional de los recursos jurel T. murphyi y caballa S. japonicus en un contexto de con-servación y sostenibilidad de estos recursos.
El presente trabajo describe el comportamiento del esfuerzo de pesca y las capturas de T. murphyi efectuadas en Perú, desde 1939 a 2012, y de esta manera proporciona la información básica para la evaluación del stock existente en el mar peruano.
Material y métodos La zona de estudio comprende todo el mar peruano, desde
el extremo norte hasta el extremo sur del dominio marítimo y desde la línea de costa hasta las 200 millas, considerándose tres regiones: Norte (3°24’ – 09°59’S), Centro (10° – 15°59’S) y Sur (16°S – Extremo Sur).
El análisis está basado en las capturas mensuales (toneladas) de toda la serie histórica de la pesquería de T. murphyi, tanto de la flota industrial como de la artesanal. Para el periodo comprendido entre 1939 – 1962 se consideró la serie obtenida por Tilic (1963), y a partir de 1963 hasta el 2012 los datos fueron obtenidos de los Anuarios Pesqueros del Ministerio de Producción, comple-mentados con informaciones del IMARPE basadas en datos del Seguimiento de la Pesquería Pelágica para la pesca industrial y la Unidad de Estadística para la pesca artesanal. En el caso del esfuer-zo pesquero se utilizó el número de embarcaciones, reconstruido en base a informaciones obtenidas por Bouchon et al. (2000).
Los datos de condiciones ambientales en el mar peruano provienen de cruceros de evaluación hidroacústica de recursos pelágicos ejecutados durante el periodo 1983 – 2012 con los BIC Olaya y SNP-2, cubriendo el área comprendida entre la costa y 100 mn del dominio marítimo peruano. Los datos mensuales de temperatura superficial del mar (TSM) provienen de los registros efectuados en el puerto de Chicama por el IMARPE.
Resultados y discusiónFlota: estructura y distribución (cerco y arrastre).- En el
periodo comprendido entre los años 1970 a 2012 de las diferentes flotas (artesanal, industrial de cerco y de arrastre factoría) que actuaron sobre los recursos jurel T. murphyi y caballa S. japonicus,
destacó la actividad permanente de la flota artesanal y la flota cerquera, que presento variaciones en su estructura pasando de ser una flota anchovetera en sus inicios hacia una flota sardinera que impulsó la captura de T. murphyi para el consumo humano directo, situación que actualmente se realiza mediante las actividades de la flota con sistema de refrigeración RSW (Fig. 1).
La flota artesanal emplea embarcaciones de pequeño calado, que cuentan hasta con 30 m3 de capacidad de bodega y operan en la zona costera cerca a los puertos y caletas de origen. La flota industrial está conformada por todas aquellas embarcaciones ma-yores o iguales a 32.6 m3 de capacidad de bodega, comúnmente conocidas como “bolicheras”, que usan redes de cerco, algunas de ellas con sistema de refrigeración a bordo. Estas embarcaciones capturan incidentalmente el jurel T. murphyi, pues normalmente se dedican a la pesquería de sardina S. sagax o anchoveta E. ringens. Esta flota estuvo constituida por 500 embarcaciones, cuyas capacidades de bodega fluctuaron entre 32.6 y 800 m3, con un promedio de 199 m3 y una capacidad total acumulada de aproximadamente 99600 m3. Aquéllas que tienen permiso para pescar anchoveta E. ringens (flota anchovetera) emplean redes con abertura de malla de 13 mm (½ pulgada), mientras las que tienen permiso múltiple tanto para anchoveta E. ringens como para sardina S. sagax, jurel T. murphyi y caballa S. japonicus (flota sardinera), cuando se dirigen a pescar T. murphyi emplean redes con abertura de malla de 38 mm (1 y ½ pulgada).
Por otra parte, durante los años 1984 – 1991, mediante convenios especiales con países como la ex-URSS y Cuba, se permitió la operación de buques arrastreros factoría (BAF). Esta flota arrastrera de media agua operó generalmente fuera de las 20 millas de la costa en la región norte, entre Paita y Chimbote, contando con 6 a 14 barcos pertenecientes a la flota cubana y entre 22 a 26 barcos pertenecientes a la flota de la ex-URSS (Ta-bla 1). Al inicio de esta pesquería esta flota presentó sus mejores rendimientos con capturas de 561 t/barco/mes.
En la década de los setenta, la flota cerquera pelágica presentó un comportamiento irregular debido principalmente a la ines-tabilidad y reducción de la población de anchoveta E. ringens, que trajo como consecuencia la disminución considerable del número de embarcaciones. Esta situación empezó a revertirse a partir de 1985 y en la década de los noventa la flota cerquera pelágica aumentó en número y capacidad de bodega. Además se observó un importante desarrollo tecnológico de la flota, registrándose variaciones en su composición, renovación de los equipos de pesca, modificaciones en las redes, inclusión de

99
Análisis de la pesquería del jurel Trachurus murphyi en el Perú
Rev. peru. biol. special issue 20(1): 097 - 106 (September 2013)Ecology, fisheries and conservation of Jack mackerel (Trachurus murphyi) in Peru. Editors: Csirke J., R. Guevara-Carrasco & M. Espino
PeriodoNacionalidad de la
flotaNúmero de meses
operativosNúmero de
barcosCaptura jurel
(toneladas)Toneladas de jurel/
barco/mesMayo 1984 – agosto 1985 ex-URSS 16 22 197 621 561Enero 1986 – marzo 1987 Cuba 15 06 37 774 420Enero – diciembre 1988 Cuba 12 14 45 656 272Enero 1989 – mayo 1991 ex-URSS 29 26 227 191 301
Tabla 1. Periodos de operación de barcos arrastreros factoría y sus capturas de jurel T. Murphyi en Perú, 1984 – 1991Table 1. Operative periods of factory trawlers and their catches of Jack mackerel T. murphyi in Peru, 1984 – 1991
Características Barco Cerquero (flota Perú)
Artes de pesca (método) Activa (Niveles superficiales)Tipo de pesca Embolsamiento
Materiales redes Tenaz, alta densidad
Cardúmenes Plumas o concentraciones mayores, superficiales
Volúmenes más de 500 t por lanceCobertura Costera
Días de operación efectiva 4 – 5Número de lances por día 01 – 04
Procesos de la captura Fresco en agua refrigerada
Sistema de frío Agua de mar refrigerada (RSW). Agua de mar enfriada (CSW)
Tabla 2. Características de la flota cerquera dedicada a la pesca de jurel T. murphyi.Table 2. Features of the purse seine fleet devoted to fish for Jack mackerel T. murphyi.
Figura 2. Número de embarcaciones operativas de la flota de cerco en la pesquería de jurel T. murphyi frente a Perú durante 1970 – 2011 (la línea punteada indica fecha en que el jurel comienza a ser usado exclusivamente para consumo humano directo)Figure 2. Number of operational vessels of the purse seine fleet in the Peruvian Jack mackerel T. murphyi fishery during 1970 – 2011 (the dotted line indicates the date that the Jack mackerel begin to be used exclusively for direct human consumption)
Flota industrial de madera (usando sistema de cajas/hielo)
Rango de capacidad de bodega
(m3)
Número barcos
Total de la capacidad de bodega (m3)
Promedio de la capacidad de bodega (m3)
31 - 60 14 556 39.761 – 90 13 991 76.291 - 110 15 1535 102.3
Total 42 3082 73.4
Flota industrial de acero (usando sistema RSW)
111 - 300 2 348 174.0301 – 400 6 2167 361.2401 – 500 29 12 913 445.3501 – 600 19 10 537 554.6601 – 700 4 2477 619.3
> 700 2 1735 867.5Total 62 30 177 486.7
Tabla 3. Número y capacidad de bodega (m3) de la flota operativa dirigida al jurel T. murphyi usando red de cerco durante el 2011.Table 3. Number of vessels and hold capacity (m3) of the operational fleet targeting on Jack mackerel T. murphyi with purse seine during 2011.
100
200
300
400
500
1970
1971
1972
1973
1974
1975
1976
1977
1978
1979
1980
1981
1982
1983
1984
1985
1986
1987
1988
1989
1990
1991
1992
1993
1994
1995
1996
1997
1998
1999
2000
2001
2002
2003
2004
2005
2006
2007
2008
2009
2010
2011
Núm
ero
de e
mba
rcac
ione
s
Años
algún sistema de refrigeración y mayor autonomía y capacidad de desplazamiento hacia las áreas y núcleos de concentración, lo cual significó un incremento en el poder de pesca de la flota (Bouchon et al. 2000, Ñiquen et al. 2000).
Características de la flota cerquera.- La flota cerquera que opera en el mar peruano está conformada por embarcaciones con un amplio rango de capacidad de bodega que va desde 100 hasta las 868 m3, con un promedio de 500 m3. La cobertura de acción de la flota peruana dirigida a la pesca de T. murphyi se caracteriza por ser costera, pues en promedio las actividades de extracción se realizan en la franja de las 100 primeras millas náuticas. En la Tabla 2 se presentan algunas características de la flota cerquera dedicada a la pesca de T. murphyi.
En el año 2006 la estructura de la flota con permiso de pesca de T. murphyi para consumo humano directo en la costa peruana contó con la participación de embarcaciones de la flota industrial
de acero y de la flota industrial de madera. El número total de embarcaciones fue de 542, de las cuales 84 fueron industriales de acero y 458 industriales de madera. La capacidad de bodega acumulada de toda la flota con permiso para jurel fue de 50587 m3, de las cuales 36488 m3 corresponden a la flota industrial de acero y 28892 m3 a la flota industrial de madera.
A partir de 2011 se redujo la flota con permiso de pesca para T. murphyi en la costa peruana a 104 embarcaciones, de las cuales 62 son industriales de acero y 42 industriales de ma-dera. La capacidad de bodega acumulada de toda la flota con permiso para T. murphyi es de 33 359 m3, de las cuales 30 177 m3 corresponden a la flota industrial de acero y 3082 m3 a la flota industrial de madera (Tabla 3).

100
Ñiquen et al.
Rev. peru. biol. número especial 20(1): 097 - 106 (Septiembre 2013)Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú.
Editores: Csirke J., R. Guevara-Carrasco & M. Espino
Figura 3. Desembarques anuales de jurel T. murphyi en Perú durante 1972 – 2012.Figure 3. Annual landings of Jack mackerel T. murphyi in Peru during 1972 – 2012.
Localidad Comentarios sobre presencia de jurel
Paita Siempre abundante
Bahía de Sechura Presencia de jureles
Lobos de TierraAbundan por épocas. Los pescadores cogieron ejemplares grandes que tenían cerca de un metro de largo.
GuañapeSe observó un cardumen constituido por un escaso número de peces, que eran visiblemente jureles.
CallaoSe les considera regulares. Se presentan todo el año y abundan en verano. Se les saca con redes, chichorro y cordel.
Chilca Abundantes, pescados con cordeles y redes.
Cerro Azul El jurel es uno de los principales pescados de esta región.
Pisco Anteriormente eran más abundantes que hoy. Se les considera buenos.
Mollendo Es abundante.
Tabla 4. Comentarios sobre presencia de jurel en Perú (de Coker 1907, 1910).Table 4. Comments on the presence of Jack mackerel in Peru (from Coker 1907, 1910).
Tabla 5. Desembarques anuales de jurel Trachurus murphyi (tonela-das) según flotas 1939-2012.Table 5. Annual landings of Jack mackerel Trachurus murphyi (tonnes) according to fleets 1939-2012
Año Cerco Arrastre Año Cerco Arrastre
1939 10 1976 54 1541940 34 1977 504 992 1941 46 1978 386 793 1942 15 1979 151 591 1943 13 1980 123 380 1944 23 1981 37 875 1945 17 1982 50 013 1946 43 1983 43 433 33 3921947 59 1984 54 607 129 7261948 44 1985 40 717 46 7491949 48 1986 37 006 31 1561950 30 1987 31 588 14 7161951 89 1988 72 420 45 6561952 81 1989 57 516 83 2041953 69 1990 90 544 100 5951954 62 1991 92 212 44 1251955 138 1992 93 065 1956 673 1993 121 309 1957 366 1994 213 220 1958 173 1995 386 748 1959 448 1996 362 197 1960 281 1997 371 485 1961 174 1998 314 123 1962 666 1999 82 541 1963 1954 2000 296 581 1964 1718 2001 774 603 1965 2561 2002 154 218 1966 4271 2003 217 735 1967 3071 2004 187 368 1968 2790 2005 80 663 1969 4176 2006 277 567 1970 4711 2007 254 427 1971 9191 2008 169 537 1972 18 782 2009 74 694 1973 42 781 2010 17 559 1974 129 211 2011 257 241 1975 37 899 2012 168 779
El número de embarcaciones operativas de la flota industrial cerquera que participaron en la pesquería de T. murphyi durante el periodo 1970 – 2011 se presenta en la Figura 2. Este número alcanza un máximo cercano a las 500 embarcaciones en la década de los años noventa y se destaca que ha sido mucho menor y se ha mantenido casi constante desde el 2002 (Fig. 2).
Capturas de jurel T. murphyi: análisis de largo plazo y tendencia.- La extracción de T. murphyi se ha desarrollado en el Perú de manera intermitente, tanto como pesca objetivo o como pesca incidental de la pesquería de E. ringens.
En Coker (1907) se describió el estado de la pesquería en Perú durante 1907, explicando que “no hay en el mundo otras aguas que estén más copiosamente provistas de pequeños pes-cados que las del Perú, favorecidas por las enormes bandadas de anchovetas” (p. 95). Coker (1907, 1910) realizó un estudio sobre la pesca marina desde Paita (05°S) hasta Mollendo (17°S) y presentó un listado de los peces y mariscos existentes, donde destaca la importante presencia de jurel T. murphyi, según lo descrito en la Tabla 4.
En 1972 se observó un notable incremento con capturas superiores a las 10000 t, que llegaron a alcanzar en 1977 uno de
sus mayores registros con 505000 t (Fig. 3). Posteriormente, las capturas se mantuvieron entre 100 a 200 mil toneladas anuales, destacando los pulsos de 1984 y 1990, seguidos de un fuerte incremento en 1997 con 650000 t, hasta alcanzar su mayor captura en el 2001 con 723000 t (Fig. 3).
0
100
200
300
400
500
600
700
800
1972 1974 1976 1978 1980 1982 1984 1986 1988 1990 1992 1994 1996 1998 2000 2002 2004 2006 2008 2010 2012
Mile
s de
tone
lada
s
Año

101
Análisis de la pesquería del jurel Trachurus murphyi en el Perú
Rev. peru. biol. special issue 20(1): 097 - 106 (September 2013)Ecology, fisheries and conservation of Jack mackerel (Trachurus murphyi) in Peru. Editors: Csirke J., R. Guevara-Carrasco & M. Espino
Figura 5. Captura mensual de jurel T. murphyi por: a) flota industrial y b) flota artesanal en el periodo 1997 – 2011.Figure 5. Monthly catch of Jack mackerel T. murphyi by: a) industrial fleet and b) artisanal fleet in the period 1997 – 2011
Figura 6. Capturas de jurel T. murphyi por la flota artesanal en el periodo 1997 – 2011, según zonas.Figure 6. Jack mackerel T. murphyi catches by the artisanal fleet in the period 1997 – 2011, by zones.
Figura 4. Estacionalidad de capturas de jurel T. murphyi en Perú, basada en promedios mensuales, periodos 1970-2002 y 2003-2011.Figure 4. Catch seasonality of the Jack mackerel T. murphyi in Peru, based on monthly averages for two periods, 1970-2002 and 2003-2011.
0
0.5
1
1.5
2
2.5
3
3.5
4
4.5
5
ene
may
sep
ene
may
sep
ene
may
sep
ene
may
sep
ene
may
sep
ene
may
sep
ene
may
sep
ene
may
sep
ene
may
sep
ene
may
sep
ene
may
sep
ene
may
sep
ene
may
sep
ene
may
sep
ene
may
sep
1997 1998 1999 2000 2001 2002 2003 2004 2005 2006 2007 2008 2009 2010 2011
artesanal
0
20
40
60
80
100
120
140
160
180
ene
may se
p
ene
may se
p
ene
may se
p
ene
may se
p
ene
may se
p
ene
may se
p
ene
may se
p
ene
may se
p
ene
may se
p
ene
may se
p
ene
may se
p
ene
may se
p
ene
may se
p
ene
may se
p
ene
may se
p
1997 1998 1999 2000 2001 2002 2003 2004 2005 2006 2007 2008 2009 2010 2011
industrialM
iles
de to
nela
das
Mile
s de
tone
lada
sTo
nela
das
Tone
lada
s
0
500
1000
1500
2000
2500
3000
3500
4000
ene
may
sep
ene
may
sep
ene
may
sep
ene
may
sep
ene
may
sep
ene
may
sep
ene
may
sep
ene
may
sep
ene
may
sep
ene
may
sep
ene
may
sep
ene
may
sep
ene
may
sep
ene
may
sep
ene
may
sep
1997 1998 1999 2000 2001 2002 2003 2004 2005 2006 2007 2008 2009 2010 2011
Mancora - Pimentel 04° - 06° S
0
500
1000
1500
2000
2500
3000
ene
may se
p
ene
may se
p
ene
may se
p
ene
may se
p
ene
may se
p
ene
may se
p
ene
may se
p
ene
may se
p
ene
may se
p
ene
may se
p
ene
may se
p
ene
may se
p
ene
may se
p
ene
may se
p
ene
may se
p
1997 1998 1999 2000 2001 2002 2003 2004 2005 2006 2007 2008 2009 2010 2011
San Juan - Ilo 15° - 18° S
En el periodo 2002 – 2011 los reportes sobre desembarques anuales de T. murphyi en el litoral peruano presentaron una clara tendencia a disminuir desde el 2007 y tuvieron un regis-tro mínimo en el 2010, debido a una baja disponibilidad de dicha especie (Fig. 3). Sin embargo, en el 2011 incremento su disponibilidad, lo cual representa un cambio en la tendencia decreciente de su captura, pues alcanzó una cifra cercana a las 260000 t, superior a lo obtenido en los años 2007 y 2008 (Tabla 5, Fig. 3).
En el periodo 1970 – 2002 se observó una marcada esta-cionalidad en las capturas de T. murphyi (Fig. 4), con mejores rendimientos en verano y primavera. Debemos destacar que en los últimos años (2003 – 2011) las mayores capturas han ocurrido con más frecuencia en verano.
50001000015000200002500030000
Cap
tura
s (t) 1970 - 2002
05000
1000015000200002500030000
E F M A M J J A S O N D
Cap
tura
s (t)
Mes
2003 - 2011

102
Ñiquen et al.
Rev. peru. biol. número especial 20(1): 097 - 106 (Septiembre 2013)Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú.
Editores: Csirke J., R. Guevara-Carrasco & M. Espino
Tone
lada
sTo
nela
das
20 000
40 000
60 000
80 000
100 000
120 000
140 000
ene
may se
p
ene
may se
p
ene
may se
p
ene
may se
p
ene
may se
p
ene
may se
p
ene
may se
p
ene
may se
p
ene
may se
p
ene
may se
p
ene
may se
p
ene
may se
p
ene
may se
p
ene
may se
p
ene
may se
p
1997 1998 1999 2000 2001 2002 2003 2004 2005 2006 2007 2008 2009 2010 2011
Mancora - Chimbote 04° - 09° S
20 000
40 000
60 000
80 000
100 000
120 000
140 000
160 000
ene
may se
p
ene
may se
p
ene
may se
p
ene
may se
p
ene
may se
p
ene
may se
p
ene
may se
p
ene
may se
p
ene
may se
p
ene
may se
p
ene
may se
p
ene
may se
p
ene
may se
p
ene
may se
p
ene
may se
p
1997 1998 1999 2000 2001 2002 2003 2004 2005 2006 2007 2008 2009 2010 2011
Huacho - Ilo 11° - 18° S
Figura 7. Capturas de jurel T. murphyi por la flota industrial en el periodo 1997 – 2011, según zonas.Figure 7. Jack mackerel T. murphyi captures by the industrial fleet in the period 1997 – 2011 by zones.
0
10
20
30
40
50
60
70
80
90
100
1970
1972
1974
1976
1978
1980
1982
1984
1986
1988
1990
1992
1994
1996
1998
2000
2002
2004
2006
2008
2010
2012
Pro
porc
ión
(%)
JUREL
CABALLA
AñosFigura 8. Composición porcentual de las capturas de jurel T. murphyi y caballa S. japonicus por años, periodo 1970 – 2012.Figure 8. Percentage composition of the catches of Jack mackerel T. murphyi and mackerel S. japonicus by year, period 1970 – 2012.
La información mensual sobre capturas de T. murphyi efec-tuadas por la flota industrial y la flota artesanal en el periodo 1997 – 2011 (Fig. 5a y 5b) mostró una tendencia similar, con periodos de mayor disponibilidad en el 2001 y entre los años 2006 a 2008. Los principales puntos de desembarque para la flota artesanal fueron Ilo, Pisco, Huacho, Matarani, Paita, Talara, Callao y Chimbote, mientras que en el caso de la flota industrial fueron Chimbote, Callao, Paita y Pisco.
Esta información también permitió observar variaciones en las principales zonas y niveles de pesca de T. murphyi de las flotas artesanal (Fig. 6) e industrial (Fig. 7). En ambos casos se observa que las capturas de T. murphyi en la parte norte (Máncora-Pimentel y Máncora-Chimbote) fueron mayores antes del 2002. Lo inverso sucedió en la parte centro sur (San Juan-Ilo y Huacho-Ilo), donde las capturas fueron mayores despues del 2001.
La información biológica provenientes de las capturas efec-tuadas por la flota cerquera dirigida al recurso jurel muestra una constante presencia del jurel T. murphyi y la caballa S. japonicus, lo que significaría que ambas comparten el mismo hábitat en
rangos de tamaños similares. Durante el periodo 1970 – 2012, la información de los desembarques anuales efectuados por la flota de cerco industrial sobre composición por especies de jurel T. murphyi y caballa S. japonicus (Fig. 8) muestra que hubo pre-dominancia de T. murphyi, en un promedio de 72% respecto al 28% de la caballa S. japonicus.
Cambios en la disponibilidad de jurel T. murphyi en función a variables ambientales.- Desde los primeros registros sobre la disponibilidad de T. murphyi en el Perú hasta el año 1996, tuvo una amplia distribución en todo el mar peruano, con estimaciones de biomasa acústica promedio anual de 4.8 millones de toneladas y promedio anual de desembarques de 142000 t. Sin embargo, la información posterior a 1996 mues-tra una distribución restringida a la parte centro-sur del mar peruano, con estimaciones de biomasa acústica promedio anual de 420000 t y promedio anual de desembarques de 256000 t.
Una característica ambiental común observada en los años de mejor pesca de T. murphyi (1977, 1996-97 y 2001) fue la incidencia de anomalías negativas de la temperatura superficial del mar (Fig. 9). Esta incidencia de bajas temperaturas no fue

103
Análisis de la pesquería del jurel Trachurus murphyi en el Perú
Rev. peru. biol. special issue 20(1): 097 - 106 (September 2013)Ecology, fisheries and conservation of Jack mackerel (Trachurus murphyi) in Peru. Editors: Csirke J., R. Guevara-Carrasco & M. Espino
0
20
40
60
80
100
120
140
160
180
-4
-3
-2
-1
0
1
2
3
4
1972
1973
1974
1975
1976
1977
1978
1979
1980
1981
1982
1983
1984
1985
1986
1987
1988
1989
1990
1991
1992
1993
1994
1995
1996
1997
1998
1999
2000
2001
2002
2003
2004
2005
2006
2007
2008
2009
2010
2011
2012
2013
Ano
mal
ias
térm
icas
(°C
)
Años
Capturas jurel (m
iles toneladas)
Capturas de Jurel
Anomalias Termicas
Figura 9. Capturas mensuales de jurel T. murphyi en relación a las anomalías mensuales de la Temperatura Superficial del Mar (TSM) en Chicama durante 1972 – 2012. El área sombreada resalta la asociación entre la anomalía negativa de la TSM y la mayor disponibilidad de jurel.Figure 9. Monthly catches of Jack mackerel T. murphyi in relation to the monthly anomalies of the SuperfaceSea Temperature (SST) in Chi-cama during 1972 – 2012. The shaded area highlights the association between negative anomaly of the SST and the greater availability of Jack mackerel.
0.0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.0
2.0
4.0
6.0
8.0
10.0
1972
1974
1976
1978
1980
1982
1984
1986
1988
1990
1992
1994
1996
1998
2000
2002
2004
2006
2008
2010
2012
Jurel (milones toneladas)
Anc
hove
ta (m
illon
es to
nela
das)
AnchovetaJurel
Figura 10. Desembarques anuales de jurel T. murphyi en relación con los desembarques de anchoveta durante 1972 – 2012.Figure 10. Annual ladings of Jack mackerel T. murphyi in relation with the landings of anchovy during 1972 – 2012.
de corto plazo, sino que se observo en varios años seguidos (1976 a 1978, de 1994 a mediados de 1997, y de 1999 al 2001). Estas condiciones favorables para la captura de T. murphyi podemos observarla en particular en el año 2001. En este año predominaron anomalías térmicas negativas al sur del Callao, asociadas a la presencia de las Aguas Templadas de la Subantártica y durante este año se observó la ocurrencia de intensos afloramientos y la intromisión de Aguas Subtropicales Superficiales, características ambientales que favorecieron la disponibilidad de T. murphyi. La persistencia del enfriamien-to en estos periodos favoreció la presencia de masas de agua homogéneas que permitieron un mayor desplazamiento de los cardúmenes de T. murphyi, tanto de sur a norte como de oeste a este, haciéndolos más disponibles a la acción de la flota cerquera pelágica (Gutierrez et al. 2004).
Simultáneamente a la mayor disponibilidad de T. murphyi en los años 1977, 1996-97 y 2001, se observó una menor dispo-nibilidad de E. ringens (Fig. 10). Esta relación inversa se explica por la tendencia de la anchoveta E. ringens a concentrarse hacia
la región norte, mientras que el jurel T. murphyi se concentra en las regiones centro-sur en estos periodos, haciéndose más disponible a la acción de la flota cerquera pelágica. Esta relación inversa entre las capturas de T. murphyi y E. ringens también fue registrada en los estimados de biomasa acústica en el mar peruano durante el periodo 1983 – 2012, observándose además que después de 1995 dominó claramente E. ringens y dismi-nuyó las biomasas de jurel T. murphyi (Fig. 11).
El jurel T. murphyi y la caballa S. japonicus son recursos que habitan distintos espacios de altamar y zonas costeras, en particularmente en los frentes oceánicos, que son zonas productivas donde se encuentran dos tipos de masas de agua (Grechina 1998). En particular, T. murphyi es una especie cuya distribución frente al Perú, en la columna de agua, está asociada a la profundidad de la capa de mínimo oxígeno y a las Aguas Subtropicales Superficiales (ASS) (Dioses 2013). Los indicadores ambientales de largo plazo señalan que la profundidad de la capa de mínimo oxígeno (Fig. 12) ha su-frido cambios decadales frente a la costa peruana (Bertrand

104
Ñiquen et al.
Rev. peru. biol. número especial 20(1): 097 - 106 (Septiembre 2013)Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú.
Editores: Csirke J., R. Guevara-Carrasco & M. Espino
0
2000
4000
6000
8000
10000
12000
14000
1983
1984
1985
1986
1987
1988
1989
1990
1991
1992
1993
1994
1995
1996
1997
1998
1999
2000
2001
2002
2003
2004
2005
2006
2007
2008
2009
2010
2011
2012
Bio
mas
a (m
iles
tone
lada
s)
Anchoveta Jurel
Figura 11. Estimados de biomasa acústica de jurel T. murphyi y anchoveta E. ringens en el mar peruano durante el periodo 1983 – 2012.Figure 11. Acoustic biomass estimates of Jack mackerel T. murphyi and anchovy E. ringens in Peruvian waters during the period 1983 – 2012
Figura 12. Profundidad de la capa de mínimo oxígeno en Perú durante 1960 – 2005, (Bertrand et al. 2011, Flores et al. 2013). Figure 12. Depth of the layer of minimum oxygen in Peru during 1960 – 2005, (Bertrand et al. 2011, Flores et al. 2013).
et al. 2011, Flores et al. 2013), y también probablemente en una parte importante del Pacífico Sudoriental (Espino 2013). Estos cambios en la profundidad de la capa de mínimo oxígeno asociadas a la temperatura superficial del mar y a la salinidad superficial podrían explicar los cambios en la disponibilidad de T. murphyi y S. japonicus observados frente a la costa peruana, especialmente desde fines de los años noventa.
Variaciones en la disponibilidad espacial de jurel y otros recursos.- Se observa que las variaciones en la distribución de T. murphyi hacia las regiones centro y norte de Perú se presen-tan asociadas a la mayor velocidad y dirección sur-norte de las corrientes, que se producen durante e inmediatamente después de un evento La Niña. En estas circunstancias, que correspon-dería al patrón 2 descrito por Dioses (2013), la disponibilidad de T. murphyi es mayor y consecuentemente se incrementan los niveles de desembarque. En este sentido se destacan los altos desembarques alcanzados en el año 2001, y que también se registraron en Ecuador (Fig. 13). Algo similar se observó en los desembarques alcanzados durante eventos La Niña (Trasmonte & Silva 2008) como en 1995-96 (Niña moderada, con una duración de 9 meses), 2001 (Niña débil, con una duración de 7 meses) y 2006-2007 (Niña fuerte, con una duración 9 meses). Un evento frío similar ocurrió en el 2010 (ENFEN, 2010) que
fue seguido por una recuperación de los desembarques en el mar peruano durante el 2011 y que también tuvo repercusiones positivas en Ecuador.
SumarioLa documentación existente revela la presencia de jurel T.
murphyi en los desembarques artesanales en Perú desde 1907. Desde los años sesenta, las estadísticas ubicaron al jurel entre las tres primeras especies de peces pelágicos en la pesquería, y partir del año 2000 se ubicó en segundo lugar después de la anchoveta E. ringens.
Las capturas de T. murphyi son realizadas por la flota artesanal, industrial cerquera y de altura (barcos arrastreros). Históricamen-te, la flota dedicada a la pesquería de anchoveta y sardina paso a capturar alternativamente T. murphyi durante sus periodos de mayor disponibilidad y en épocas de veda de E. ringens.
En el 2011, la estructura de la flota con permiso de pesca para T. murphyi estuvo constituida por 62 embarcaciones industriales de acero y 42 industriales de madera. La capacidad de bodega acumulada de toda la flota es de 33259 m3, de las cuales 30177 m3 corresponden a la flota industrial de acero y 3082 a la flota industrial de madera.
30
35
40
45
50
55
60
65
1960
1962
1964
1966
1968
1970
1972
1974
1976
1978
1980
1982
1984
1986
1988
1990
1992
1994
1996
1998
2000
2002
2004
2006
2008
Pro
fund
idad
Oxi
clin
a (m
)
Años
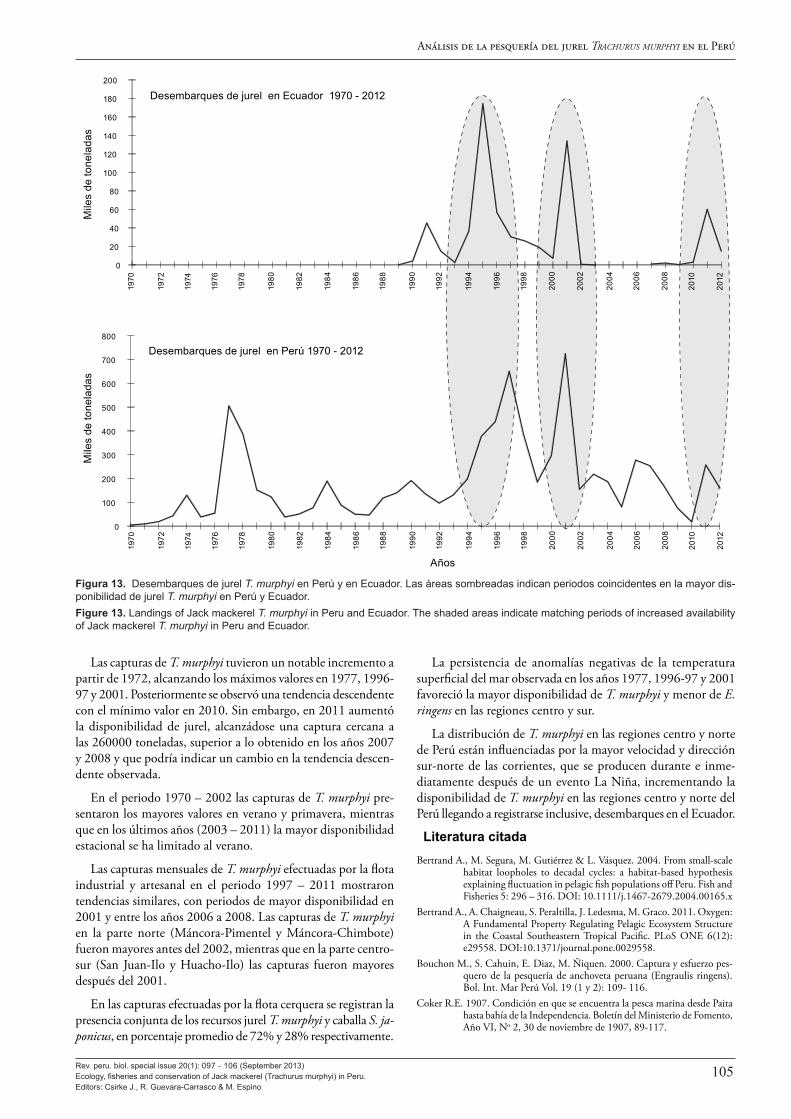
105
Análisis de la pesquería del jurel Trachurus murphyi en el Perú
Rev. peru. biol. special issue 20(1): 097 - 106 (September 2013)Ecology, fisheries and conservation of Jack mackerel (Trachurus murphyi) in Peru. Editors: Csirke J., R. Guevara-Carrasco & M. Espino
Figura 13. Desembarques de jurel T. murphyi en Perú y en Ecuador. Las áreas sombreadas indican periodos coincidentes en la mayor dis-ponibilidad de jurel T. murphyi en Perú y Ecuador.Figure 13. Landings of Jack mackerel T. murphyi in Peru and Ecuador. The shaded areas indicate matching periods of increased availability of Jack mackerel T. murphyi in Peru and Ecuador.
Desembarques de jurel en Ecuador 1970 - 2012
Desembarques de jurel en Perú 1970 - 2012
0
20
40
60
80
100
120
140
160
180
200
0
100
200
300
400
500
600
700
800
Mile
s de
tone
lada
s
Años
Mile
s de
tone
lada
s
1972
1974
1976
1978
1980
1982
1984
1986
1988
1990
1992
1994
1996
1998
2000
2002
2004
2006
2008
2010
2012
1970
1972
1974
1976
1978
1980
1982
1984
1986
1988
1990
1992
1994
1996
1998
2000
2002
2004
2006
2008
2010
2012
1970
Las capturas de T. murphyi tuvieron un notable incremento a partir de 1972, alcanzando los máximos valores en 1977, 1996-97 y 2001. Posteriormente se observó una tendencia descendente con el mínimo valor en 2010. Sin embargo, en 2011 aumentó la disponibilidad de jurel, alcanzádose una captura cercana a las 260000 toneladas, superior a lo obtenido en los años 2007 y 2008 y que podría indicar un cambio en la tendencia descen-dente observada.
En el periodo 1970 – 2002 las capturas de T. murphyi pre-sentaron los mayores valores en verano y primavera, mientras que en los últimos años (2003 – 2011) la mayor disponibilidad estacional se ha limitado al verano.
Las capturas mensuales de T. murphyi efectuadas por la flota industrial y artesanal en el periodo 1997 – 2011 mostraron tendencias similares, con periodos de mayor disponibilidad en 2001 y entre los años 2006 a 2008. Las capturas de T. murphyi en la parte norte (Máncora-Pimentel y Máncora-Chimbote) fueron mayores antes del 2002, mientras que en la parte centro-sur (San Juan-Ilo y Huacho-Ilo) las capturas fueron mayores después del 2001.
En las capturas efectuadas por la flota cerquera se registran la presencia conjunta de los recursos jurel T. murphyi y caballa S. ja-ponicus, en porcentaje promedio de 72% y 28% respectivamente.
La persistencia de anomalías negativas de la temperatura superficial del mar observada en los años 1977, 1996-97 y 2001 favoreció la mayor disponibilidad de T. murphyi y menor de E. ringens en las regiones centro y sur.
La distribución de T. murphyi en las regiones centro y norte de Perú están influenciadas por la mayor velocidad y dirección sur-norte de las corrientes, que se producen durante e inme-diatamente después de un evento La Niña, incrementando la disponibilidad de T. murphyi en las regiones centro y norte del Perú llegando a registrarse inclusive, desembarques en el Ecuador.
Literatura citadaBertrand A., M. Segura, M. Gutiérrez & L. Vásquez. 2004. From small-scale
habitat loopholes to decadal cycles: a habitat-based hypothesis explaining fluctuation in pelagic fish populations off Peru. Fish and Fisheries 5: 296 – 316. DOI: 10.1111/j.1467-2679.2004.00165.x
Bertrand A., A. Chaigneau, S. Peraltilla, J. Ledesma, M. Graco. 2011. Oxygen: A Fundamental Property Regulating Pelagic Ecosystem Structure in the Coastal Southeastern Tropical Pacific. PLoS ONE 6(12): e29558. DOI:10.1371/journal.pone.0029558.
Bouchon M., S. Cahuin, E. Diaz, M. Ñiquen. 2000. Captura y esfuerzo pes-quero de la pesquería de anchoveta peruana (Engraulis ringens). Bol. Int. Mar Perú Vol. 19 (1 y 2): 109- 116.
Coker R.E. 1907. Condición en que se encuentra la pesca marina desde Paita hasta bahía de la Independencia. Boletín del Ministerio de Fomento, Año VI, No 2, 30 de noviembre de 1907, 89-117.

106
Ñiquen et al.
Rev. peru. biol. número especial 20(1): 097 - 106 (Septiembre 2013)Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú.
Editores: Csirke J., R. Guevara-Carrasco & M. Espino
Coker R.E. 1910. Condiciones de la pesquería de Mollendo. Boletín del Minis-terio de Fomento, Año VIII, No 8, 31 de octubre de 1910, 64-114.
DS N°. 001-2002-PRODUCE. 2002. Establecen que los recursos sardina, jurel y caballa serán destinados al consumo humano directo. 06 de setiembre 2002. El Peruano. Normas Legales: 229522-229523.
DS N°. 011-2007-PRODUCE. 2007. Aprueban Reglamento de Ordenamiento Pesquero de Jurel y Caballa. 13de abril de 2007. El Peruano Normal Legales: 343399-343402.
Dioses T. 2013. Patrones de distribución y abundancia del jurel Trachurus mur-phyi en el Perú. En: Csirke J., R. Guevara-Carrasco & M. Espino (Eds.). Ecología, pesquería y conservación del jurel (Trachurus mur-phyi) en el Perú. Rev. peru. biol. número especial 20(1): 067- 074
Flores R., M. Espino, G. Luque & J. Quispe. 2013. Patrones de variabilidad ambiental en el mar peruano. En: Csirke J., R. Guevara-Carrasco & M. Espino (Eds.). Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú. Rev. peru. biol. número especial 20(1): 021 – 028
Espino M. 2013. El jurel Trachurus murphyi y las variables ambientales de ma-croescala. En: Csirke J., R. Guevara-Carrasco & M. Espino (Eds.). Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú. Rev. peru. biol. número especial 20(1): 009 – 020
ENFEN. 2010. Nota de Prensa No 08 del Comité Multisectorial encargado del Estudio Nacional del Fenómeno El Niño (ENFEN). 03 de agosto de 2010.
Gutiérrez M., P.R. Castillo, F. Ganoza, N. Herrera, S. Peraltilla & A. Aliaga. 2004. Análisis de la distribución y abundancia de algunos recursos pelágicos peruanos acústicamente evaluados en la primavera 2001. Inf. Inst. Mar Perú, 32(2): 155-165.
Grechina A. 1998. Historia de investigación y aspectos básicos de la ecología del jurel Trachurus symmentricus murphyi (Nichols) en alta mar del Pacífico Sur. En: D. Arcos (Ed.). Biología y ecología del jurel en aguas chilenas , Instituto de Investigaciones Pesqueras S.A. Talcahuano, Chile: 11-34.
Jordán R. 1983. Variabilidad de los recursos pelágicos en el Pacífico Sudeste. En G.D. Sharp y J. Csirke (Eds.), Actas de la Consulta de Expertos para examinar los cambios en la abundancia y composición por especies de recursos de peces neríticos. San José, Costa Rica, 18-29 abril de 1983. Una reunión preparatoria para la Conferencia Mundial de la FAO sobre ordenación y desarrollo pesqueros. FAO Fish.Rep./FAO, Inf. Pesca (291) Vol. 2:113-129.
Ñiquen M., M. Bouchon, S. Cahuín & E. Díaz. 2000. Pesquería de anchoveta en el mar peruano. Bol. Inst. Mar Perú Vol. 19(1 y 2): 117- 124.
Tilic I. 1963. Material estadístico sobre la industria peruana de harina de pes-cado. Inf. Inst. Inv. Rec. Marinos No 14, 51p.
Trasmonte G., & Y. Silva. 2008. Evento La Niña: Propuesta de definición y clasificación según las Anomalías de Temperatura Superficial del Mar en el Área Niño 1+2. Inf. Inst. Mar Perú 35(3): 199-207.
Tsukayama I. 1983. Recursos pelágicos y sus pesquerías en Perú. Rev. Com. Perm. Pacífico Sur 13: 25-63.
Zuta S. 1983. El ambiente marino y las fluctuaciones de las principales pobla-ciones pelágicas de la costa peruana. En: G.D. Sharp y J. Csirke (Eds.), Actas de la Consulta de Expertos para examinar los cambios en la abundancia y composición por especies de recursos de peces neríticos. San José, Costa Rica, 18-29 abril 1983. Una reunión pre-paratoria para la Conferencia Mundial de la FAO sobre ordenación y desarrollo pesqueros. San José, Costa Rica, 18 - 29 April 1983. FAO Fish. Rep. 291(2): 179-254.

107
conservación y ordenación pesquera del jurel Trachurus murphyi
Rev. peru. biol. special issue 20(1): 107 - 113 (September 2013)Ecology, fisheries and conservation of Jack mackerel (Trachurus murphyi) in Peru. Editors: Csirke J., R. Guevara-Carrasco & M. Espino
Rev. peru. biol. número especial 20(1): 107 - 113 (Septiembre 2013)Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú. Editores: Csirke J., R. Guevara-Carrasco & M. Espino
Facultad de ciencias Biológicas unMsMISSN-L 1561-0837
Medidas de conservación y ordenación pesquera del jurel Trachurus murphyi en el Perú
Jorge Zuzunaga
Conservation and fishery management regulations of Jack mackerel Trachurus murphyi in Peru
Instituto del Mar del Perú, esquina Gamarra y General Valle S/N Chucuito, Callao, Perú.
Email Jorge Zuzunaga: [email protected]
Pesquería, manejo y conservación del jurel (Trachurus murphyi) en el Perú
© Los autores. Publicado por la Revista Peruana de Biología de la Facultad de Ciencias Biológicas, Universidad Nacional Mayor de San Marcos. Este es un artículo de acceso abierto, distribuido bajo los términos de la Licencia de Atribución Reconocimiento-NoComercial-SinObraDerivada 3.0 de Creative Commons (http://creativecommons.org/licenses/by-nc-nd/3.0/deed.es_ES), que permite el uso no comercial, distribución y reproducción en cualquier medio, siempre que la obra original sea debidamente citadas. Para uso comercial, por favor póngase en contacto con [email protected].
Citación:Zuzunaga J. 2013. Medidas de conservación y ordenación pesquera del jurel Trachurus murphyi en el Perú. En: Csirke J., R. Guevara-Carrasco & M. Espino (Eds.). Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú. Rev. peru. biol. número especial 20(1): 107 - 113 (Septiembre 2013)
Resumen Las pesquerías peruanas como actividades realizadas sobre recursos naturales renovables son reguladas por un marco jurídico, cuyo objetivo es asegurar su conservación y uso sos-tenible. La Ley General de Pesca (DL N°. 25977) señala que para cada tipo de pesquería y según la situación del recurso explotado debe establecerse un sistema de ordenamiento que concilie el principio de sostenibilidad de los recursos pesqueros o conservación en el largo plazo, con la obtención de los mayores beneficios económicos y sociales. En el ordenamiento pesquero del jurel Trachurus murphyi, las medidas incluyen entre otras, el régimen de acceso constituido por las autorizaciones de incremento de flota y permisos de pesca, que se otor-gan solo a los buques cuyas capturas son destinadas únicamente para el consumo humano directo, de acuerdo a lo dispuesto en el Reglamento de Ordenamiento Pesquero (DS N.° 011-2007-PRODUCE). La obligación de destinar los recursos sardina Sardinops sagax, jurel T. murphyi y caballa Scomber japonicus al consumo humano directo rige desde septiembre 2002 (DS N°. 001-2002-PRODUCE). Otras medidas vigentes restringen el tamaño mínimo de malla de las redes, prohíben la captura de ejemplares menores de 31 cm de longitud to-tal, determinan el inicio y cierre de las temporadas de pesca, los límites de captura o cuotas anuales. Estas disposiciones son complementadas por medidas de seguimiento, control y vigilancia para la efectiva conservación y uso sostenido del recurso.
Palabras clave: Conservación, ordenación, jurel, pesquerías
AbstractThe Peruvian fisheries exploit renewable natural resources and as such are regulated by a legal framework aiming at ensuring their conservation and sustainable use. The General Fisheries Act, DL N°. 25977, points out that fisheries management systems shall be established for each type of fishery taking into account to the state of exploitation of the fishery resources, reconciling the principles of sustainability and long-term conservation with those aiming at the attainment of the maximum possible economic and social benefits. According to the provi-sions of the Fisheries Management Regulations approved by SD N°. 011-2007-PRODUCE, the management of the Jack mackerel Trachurus murphyi fisheries includes among others, a fishing access regime that regulates the fleet size and the allocation of fishing licenses, which can only be granted to vessels fishing only for direct human consumption. The obligation to allocate the resources sardine Sardinops sagax, Jack mackerel T. murphyi, and mackerel Scomber japonicus, for direct human consumption has been in force since September 2002, following the ruling established by SD N°. 001-2002-PRODUCE. Other measures currently in force include those establish a minimum mesh size, the ban to catch or land specimens under 31 cm total length limit, the opening and closing of the fishing seasons, and catch limits or annual quotas. These provisions are supplemented by measures for monitoring, control and surveillance, and for the effective conservation and sustainable use of the fishery resources.
Keywords: Conservation, management, Jack mackerel, fisheries
Publicado online: 11/10/2013Publicado impreso: 15/10/2013

108
Zuzunaga
Rev. peru. biol. número especial 20(1): 107 - 113 (Septiembre 2013)Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú.
Editores: Csirke J., R. Guevara-Carrasco & M. Espino
IntroducciónEl Perú ha adoptado un marco normativo jurídico e insti-
tucional que establece medidas para la conservación y el uso sostenible a largo plazo de los recursos pesqueros. Este marco normativo se inspira y está plenamente en concordancia con los principios rectores del Código de Conducta para la Pesca Responsable (FAO 1995). Es así que en la legislación peruana se establecen medidas de conservación y ordenación, aplicables al aprovechamiento de los recursos naturales en general (Ley N°. 26821) y pesqueros en particular (DL N°. 25977, Ley General de Pesca).
En principio, la Ley General de Pesca norma esta actividad con el fin de promover su desarrollo sostenido como fuente de alimentación, empleo e ingresos y de asegurar un aprovecha-miento responsable de los recursos hidrobiológicos, optimizando los beneficios económicos, en armonía con la preservación del medio ambiente y la conservación de la biodiversidad. Pero a su vez, las medidas de ordenamiento deben basarse en los datos científicos más rigurosos disponibles y deben estar diseñadas para garantizar la sostenibilidad a largo plazo de los recursos pesqueros.
En el presente trabajo ofrecemos una breve descripción de las medidas de gestión aplicables a los stocks peruanos del jurel Trachurus murphyi Nichols 1920 y de la caballa Scomber japo-nicus Linnaeus 1758, que como se describe más adelante, para los fines de ordenamiento fueron considerados como partes de una misma pesquería, en razón de encontrarse frecuentemente distribuidos en las mismas áreas y por lo tanto, ser accesibles a las mismas flotas y artes de pesca.
Marco legal e institucional La Constitución Política del Perú establece los principios que
rigen la propiedad y el acceso a la explotación de los recursos naturales. Así, los recursos naturales renovables y no renovables son patrimonio de la Nación y el Estado es soberano en su uso (Artículo 66°). También dispone que las condiciones sean establecidas por ley orgánica, para su uso por particulares. Asimismo, el Artículo 67° declara que el Estado determina la política nacional del ambiente y promueve el uso sostenible de los recursos naturales.
Sobre la base de este mandato constitucional la Ley sobre el Uso Sostenible de los Recursos Naturales Renovables (Ley N.° 26821) establece el marco general para promover y regular la sostenibilidad, persiguiendo un equilibrio dinámico entre el crecimiento económico, la conservación de los recursos naturales y el medio ambiente. De acuerdo con este principio, las condi-ciones de acceso para cada recurso deben ser reguladas por leyes especiales. En el caso de los recursos pesqueros, la misma norma dispone que la Ley General de Pesca (DL N°. 25977) conserve su plena vigencia, a pesar de que fue dictada con anterioridad a la Ley sobre el Uso Sostenible de los Recursos Naturales.
En este orden de ideas, las disposiciones contenidas en la Ley General de Pesca constituyen el marco jurídico adecua-do para regular la actividad pesquera y la promoción de su desarrollo sostenible, mientras que el Reglamento de la Ley General de Pesca (DS N°. 012-2001-PE) establece normas para la determinación de principios y medidas reguladoras que han de aplicarse a las actividades de pesca o extracción de los recursos acuáticos vivos. De acuerdo al principio establecido en
la ley y en su reglamento, cada pesquería es tratada como una unidad diferenciada (Artículo 12° DL N°. 25977), teniendo presente que para definir el nivel de explotación apropiado a los objetivos del ordenamiento se debe delimitar la unidad de población. Tal es el caso del stock de jurel que se distribuye la mayor parte de las aguas jurisdiccionales del Perú y del cual se ocupa la ordenación pesquera como una unidad de población independiente a la cual se administra de manera sostenible, en el presente caso mediante las disposiciones del Reglamento de Ordenamiento Pesquero de Jurel y Caballa (DS N°. 011-2007). En dicho reglamento se fija como principio el objetivo de promover el aprovechamiento racional de las poblaciones del jurel T. murphyi y la caballa S. japonicus, proteger el eco-sistema marino y salvaguardar la biodiversidad, contribuyendo a la diversificación y desarrollo de la pesca como fuente de alimento, empleo e ingresos económicos.
Como se observa para el caso de la pesquería de jurel y ca-balla, el ordenamiento específico de cada pesquería se efectúa mediante reglamentos de ordenamiento pesquero, los cuales en su elaboración son producto de un proceso que toma en cuenta lo que la FAO define como:
“(…) proceso integrado de recolección de información, análisis, planificación, consulta, adopción de decisiones, asignación de recursos y formulación y ejecución, así como imposición cuando sea necesario, de reglamentos o normas que rijan las actividades pesqueras para asegurar la produc-tividad de los recursos y la consecución de otros objetivos” (FAO 1999, p. 7).
Para el efecto, la Ley General de Pesca considera que para establecer el ordenamiento de cada pesquería concurren el conocimiento del recurso y las condiciones de la actividad, señalando que:
“(…) sobre la base de evidencias científicas disponibles y de factores socioeconómicos, se determinará, según el tipo de pesquerías, los sistemas de ordenamiento pesquero, las cuotas de captura permisible, las temporadas y zonas de pesca, la regulación del esfuerzo pesquero, los métodos de pesca, las tallas mínimas de captura y demás normas que requieran la preservación y explotación racional de los recursos hidrobio-lógicos” (DL N°. 25977, Art. 9).
Por lo tanto, el Reglamento de Ordenamiento Pesquero de Jurel y Caballa (DS N°. 011-2007-PRODUCE) contiene las medidas de ordenamiento pesquero para acceder a la actividad extractiva de estas especies, aplicables a las autorizaciones de incremento de flota y permisos de pesca que se otorgan a arma-dores de embarcaciones de bandera nacional, de cerco, arrastre de media agua, multipropósito (cerco/arrastre de media agua) y de pesca con anzuelo.
Medidas de ordenamiento pesquero del jurel En lo que se refiere a la pesca de cerco del jurel, desde 1974 en
adelante se establecieron medidas de regulación pesquera como el tamaño de malla, fijándola en 38 mm, y la talla mínima de captura de ejemplares. Desde 1992 (Ley General de Pesca, art. 33; Reglamento de la Ley General de Pesca, DS Nº. 017-92-PE), con relación a la protección de las áreas de pesca se dispuso la reserva de las primeras 5 millas náuticas (mn) a partir de la línea de costa para la pesca artesanal, por lo que la pesca de jurel por la flota industrial, en cualquiera de sus formas, debía realizarse

109
conservación y ordenación pesquera del jurel Trachurus murphyi
Rev. peru. biol. special issue 20(1): 107 - 113 (September 2013)Ecology, fisheries and conservation of Jack mackerel (Trachurus murphyi) in Peru. Editors: Csirke J., R. Guevara-Carrasco & M. Espino
fuera de esta zona.
En la década de los ochenta, la pesca de jurel se compartió entre la flota de cerco y la flota de arrastre factoría, normada por el Reglamento para la Operación de Buques Pesqueros de Arrastre (DS N°. 012-84-PE), que establecía que el Ministerio de Pesquería lleve el control de los permisos de pesca conciliando el esfuerzo pesquero con los volúmenes de captura recomendados por el IMARPE. Con el mismo dispositivo, se reservaba para el Estado el 25% del total de la captura permisible anual de los recursos de jurel, caballa y merluza Merluccius gayi peruanus u otras especies, para cuya explotación podría celebrar convenios con armadores nacionales o extranjeros. Además, normaba el tamaño de malla en 76 mm, restringiendo las áreas de pesca a aquéllas con profundidades no menores a 100 m y distancia de la costa no menor a 20 mn, sin desmedro de las profundidades; es decir, cuando las profundidades eran mayores a 100 m a una distancia menor de 20 mn, la prohibición era aplicada.
En el caso del jurel, el Reglamento de Ordenamiento Pesquero de Jurel y Caballa (DS N°. 011-2007-PRODUCE) considera un régimen de acceso restringido sólo a las embarcaciones que cuentan con sistemas operativos de preservación de las capturas a bordo. En relación a esta medida, y como se ha mencionado anteriormente, ya mediante el DS N°. 001-2002-PRODUCE se estableció que las capturas del jurel deben ser destinadas ex-clusivamente al consumo directo. Esta norma está vigente desde hace once años y viene siendo cumplida a cabalidad.
En el caso de las embarcaciones pesqueras de cerco, el acceso sólo está autorizado cuando sustituye (reemplaza) igual capaci-dad de bodega de la flota operando con permisos para la extrac-ción de jurel y caballa. Los armadores de las embarcaciones que son autorizadas para realizar actividades pesqueras están sujetos al pago de derechos de pesca y la flota autorizada para extraer jurel y caballa sólo puede operar fuera de las 10 millas de costa. Las embarcaciones de arrastre no pueden operar en áreas con menos de 200 m de profundidad (DS N°. 011-2007-PRODUCE).
Está determinado un tamaño mínimo de malla para las redes. Para la red de cerco, el tamaño mínimo es de 38 mm (1½ pulgadas), y para la red de arrastre de media agua, es de 76 mm (3 pulgadas) en el copo. También se prohíbe la extracción, procesamiento y comercialización de ejemplares de jurel con tallas inferiores a 31 cm de longitud total y caballa con tallas inferiores a 29 cm de longitud a la horquilla (equivalente a 32 cm de longitud total).
Complementariamente al reglamento de ordenamiento pesquero, mediante normas específicas, como resoluciones ministeriales, son establecidas las vedas y temporadas de pesca y los correspondientes límites (cuotas) de captura, así como las acciones de monitoreo, control y vigilancia.
El marco actual de ordenamiento de la pesquería de jurel incluye las siguientes medidas y restricciones:
• Tamaño de malla 38 mm o 1½ pulgada (red de cerco).
• Tamaño de malla 76 mm o 3 pulgadas en el copo (red de arrastre).
• Tamaño mínimo de captura: 31 cm de longitud total.
• Tolerancia de jureles juveniles en la captura: 30% del número total.
• Reglamento de Ordenamiento Pesquero de Jurel y Ca-balla (DS N°. 011-2007-PRODUCE. 2007).
• El uso exclusivo del jurel para consumo humano directo (DS N°. 001-2002-PRODUCE).
• Implementación del monitoreo satelital para la flota industrial de acero y de madera (DS N°. 031-2003-PRO-DUCE)
• Establecimiento de medidas para la explotación de recursos como el jurel y la caballa, por embarcacio-nes nacionales de gran escala en la alta mar (DS N°. 022-2009-PRODUCE)
• Desde el 2003, la aplicación de cuotas de captura para el jurel por determinados periodos. En este informe se incluye una relación de las normas emitidas en los dos últimos años.
• Principales medidas de ordenamiento y conservación de jurel y caballa en el periodo 2010 – 2012
A continuación se presenta una relación de las normas de ordenamiento y conservación para los stocks de jurel y caballa, para el periodo más reciente 2010 – 2012. Estas normas han sido dirigidas a establecer los regímenes de pesca y regular las actividades de extracción de estos dos stocks en el marco general del ordenamiento pesquero peruano:
-La RM N°. 026-2010-PRODUCE, vigente desde el 3 de febrero del 2010, estableció un régimen provisional para la extracción de jurel y caballa en todo el litoral peruano hasta el 31 de marzo de 2010, de alcance a las capturas provenientes de la actividad pesquera artesanal y de las embarcaciones pesqueras de mayor escala, con permiso vigente para la extracción de dichos recursos.
-La RM N°. 027-2010-PRODUCE, vigente desde el 3 de febrero del 2010, estableció un régimen de pesca exploratoria de recursos pelágicos asociados al evento El Niño en todo el litoral peruano, que autoriza la extracción de los recursos bonito (Sarda chilensis), barrilete (Katsuwonus pelamis), melva (Auxis rochei), jurel (T. murphyi) y caballa (S. japonicus) hasta el 31 de marzo de 2010.
-La RM N°. 096-2010-PRODUCE, vigente desde el 17 de abril del 2010, estableció un régimen provisional para la extracción de jurel y caballa en todo el litoral peruano, hasta el 31 de mayo de 2010, de alcance a las capturas provenientes de la actividad pesquera artesanal y de las embarcaciones pesqueras de mayor escala con permiso de pesca vigente. Establece un límite de captura mensual de 15000 t. Una vez cumplido el límite de captura para cada recurso, el Ministerio de la Producción suspendía las actividades extractivas por los días que resten del mes correspondiente. Las actividades extractivas se sujetaron al cumplimiento de las disposiciones del reglamento de Ordenamiento Pesquero de Jurel y Caballa (DS N°. 011-2007-PRODUCE).
-La RM N.° 097-2010-PRODUCE, vigente del 18 de abril del 2010, estableció un régimen de pesca exploratoria de recursos pelágicos de oportunidad en todo el litoral peruano, asociados al evento El Niño, y que autoriza la extracción de los recursos bonito (S. chilensis), barrilete (K. pelamis), melva (A. rochei), jurel (T. murphyi), caballa (S. japonicus) y

110
Zuzunaga
Rev. peru. biol. número especial 20(1): 107 - 113 (Septiembre 2013)Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú.
Editores: Csirke J., R. Guevara-Carrasco & M. Espino
otros, hasta el 31 de mayo de 2010. Prohibió la extracción, recepción y procesamiento de la anchoveta Engraulis rin-gens y merluza Merluccius gayi peruanus, incluida su fauna acompañante.
-La RM N.° 202-2010-PRODUCE, vigente desde el 18 de agosto del 2010, estableció un régimen provisional para la extracción del jurel T. murphyi y caballa S. japonicus en todo el litoral peruano por un plazo de sesenta (60) días, de alcance a las capturas provenientes de la actividad pesquera artesanal y de las embarcaciones pesqueras de mayor escala, con permiso de pesca vigente para dichos recursos.
-La RM N°. 035-2011-PRODUCE, vigente del 6 de febrero del 2011, suspendió las actividades de extracción y recepción del jurel, a partir de las 00:00 horas del 7 de febrero hasta las 24:00 horas del 28 de febrero de 2011, en el marco del régimen provisional de extracción establecido mediante RM N.° 025-2011-PRODUCE.
-La RM N°. 039-2011-PRODUCE, vigente el 9 de febrero del 2011, modifico los Artículos 1º y 2º de la RM N°. 035-2011-PRODUCE, del 6 de febrero del 2011 con el siguiente texto: "Artículo 1°.- Suspender a partir de las 00:00 horas del 7 de febrero hasta las 24:00 horas del 28 de febrero de 2011, las actividades de extracción del recurso jurel, en el marco del Régimen Provisional de extracción establecido mediante RM N.° 025-2011-PRODUCE. Artículo 2°.- La actividad de procesamiento del recurso jurel podrá desarrollarse hasta las 24:00 horas del 9 de febrero de 2011".
-La RM N°. 058-2011-PRODUCE, vigente desde el 26 de febrero del 2011, modifico el primer párrafo del Artículo 2° de la RM N°. 025-2011-PRODUCE, determinándose que el límite de captura establecido es de aplicación para las capturas provenientes de las embarcaciones pesqueras de mayor escala y será hasta la conclusión del régimen provisional de extracción del jurel y la caballa; se amplió el límite de captura a 45000 toneladas. Preciso que del límite de captura determinado en el presente régimen provisional, aplicable a las embarcaciones de mayor escala, queda por extraer 25236 t de jurel.
-La RM N°. 099-2011-PRODUCE, vigente desde el 20 de marzo del 2011, suspendió las actividades de extracción del recurso jurel, a partir de las 00:00 horas del día siguiente de su publicación, en el marco del régimen provisional de extracción dispuesto por la RM N°. 058-2011-PRODUCE.
-La RM N°. 107-2011-PRODUCE, vigente desde el 25 de marzo del 2011, prorrogo la vigencia del régimen provi-sional de pesca del jurel y la caballa, establecido por la RM N°. 025-2011-PRODUCE, hasta el 30 de abril de 2011, en todo el litoral peruano, de acuerdo con las razones expuestas en la parte considerativa de la resolución. Amplía el límite de captura aplicable a las capturas de las embarcaciones de mayor escala, en aplicación de la primera disposición final, complementaria y transitoria del Reglamento de Ordena-miento Pesquero (DS N°. 011-2007-PRODUCE) hasta 55000 t de jurel y caballa, y mantiene la vigencia de las demás disposiciones contenidas en la RM N°. 025-2011, modificada por la RM N°. 058-2011-PRODUCE.
-La RM N°. 124-2011-PRODUCE, vigente del 10 de abril del 2011, suspendió las actividades de extracción del jurel a partir de las 00:00 horas del día siguiente de su publicación, en el marco del régimen provisional de extracción dispuesto por la RM N°. 025-2011-PRODUCE, modificado por la RM N°. 058-2011-PRODUCE y prorrogado por la RM N°. 107-2011-PRODUCE.
-La RM N°. 130-2011-PRODUCE, vigente desde 13 de abril del 2011, amplio el límite de las capturas provenientes de las embarcaciones pesqueras de mayor escala, hasta 62000 t para el jurel, en el marco de lo dispuesto en el régimen provisional establecido por la RM N.° 025-2011-PRODUCE.
-La RM N°. 146-2011-PRODUCE, vigente desde el 28 de abril del 2011, prorrogo la vigencia del régimen provisional de extracción del jurel y la caballa, hasta el 30 de junio de 2011 en todo el litoral peruano, establecido por la RM N°. 025-2011-PRODUCE.
-La RM N.° 198-2011-PRODUCE, vigente del 29 de junio del 2011, estableció un límite de captura de 45000 t de jurel y caballa, a partir de las 00:00 horas del 1 de julio hasta las 24:00 horas del 30 de setiembre de 2011, en todo el litoral peruano.
-La RM N°. 277-2011-PRODUCE, vigente desde el 01 de octubre del 2011, amplio el límite de captura del jurel y la caballa, establecido mediante la RM N.° 198-2011-PRO-DUCE, a 75000 t, desde el día siguiente de su publicación hasta las 24:00 horas del 31 de diciembre de 2011, en todo el litoral peruano.
-La RM N°. 034-2012-PRODUCE, vigente desde el 24 de enero del 2012, estableció el límite de captura del jurel y la caballa en 40000 y 20000 t respectivamente, correspondiente al primer trimestre del año 2012. Una vez cumplido el límite de captura para cada recurso, el Ministerio de la Producción suspendió las actividades extractivas por los días que restaron del periodo autorizado.
-La RM N°. 055-2012-PRODUCE, vigente desde el 03 de febrero del 2012, suspendió las actividades extractivas del jurel en todo el litoral peruano a partir de las 00:00 horas del día siguiente de su publicación. Los armadores que se encontraban pescando en el litoral peruano tuvieron un plazo de dos días para el desembarque. La RM N°. 133-2012-PRO-DUCE la dejo sin efecto.
-La RM N°. 132-2012-PRODUCE, vigente del 22 de marzo del 2012, preciso que los alcances de la RM N°. 034-2012-PRODUCE en el límite de captura del jurel y caballa correspondiente al primer trimestre del año 2012, es para las actividades extractivas realizadas por la flota industrial pesquera. Lo cual indicaba que la suspensión de las actividades extractivas del recurso jurel, regulada en la RM N°. 055-2012-PRODUCE, no era de aplicación a la flota artesanal.
-La RM N°. 133-2012-PRODUCE, vigente desde el 23 de marzo del 2012, estableció el límite de captura del jurel y caballa en 30 mil y 20 mil t respectivamente para la flota industrial, lo que correspondió al segundo trimestre del año 2012, (1 de abril al 30 de junio de 2012). Una vez cumplido el límite de captura para cada recurso, el Ministerio de la

111
conservación y ordenación pesquera del jurel Trachurus murphyi
Rev. peru. biol. special issue 20(1): 107 - 113 (September 2013)Ecology, fisheries and conservation of Jack mackerel (Trachurus murphyi) in Peru. Editors: Csirke J., R. Guevara-Carrasco & M. Espino
Producción suspendió las actividades extractivas por los días que restaban al periodo autorizado. Dejo sin efecto las dis-posiciones contenidas en la RM N°. 055-2012-PRODUCE.
-La RM N°. 142-2012-PRODUCE, vigente desde el 24 de marzo del 2012, modifico el Artículo 1° de la RM N°. 133-2012-PRODUCE 23/03/2012, que estableció el límite de captura de jurel y caballa.
-La RM N°. 173-2012-PRODUCE, vigente desde 15 de abril del 2012, estableció la cuota de captura total permisible para el jurel en 120000 t correspondiente al año 2012. La cuota establecida comprendía las capturas efectuadas al amparo de la RM N°. 034-2012-PRODUCE, precisada por la RM N°. 132-2012-PRODUCE y la RM N°. 133-2012-PRODUCE, modificada por la RM N°. 142-2012_PRODUCE. En caso de cumplirse la cuota de captura establecida, el Ministerio de la Producción suspen-dería las actividades extractivas.
-La RM N°. 191-2012-PRODUCE, del 19 de abril del 2012, suspendió las actividades extractivas del jurel a partir de las 00:00 horas del día 21 de abril de 2012, en todo el litoral peruano. En consecuencia, se prohibió transportar y comercializar jurel a partir del 4º día desde la entrada en vigencia de la suspensión de sus actividades extractivas. Los armadores de embarcaciones pesqueras industriales que se encontraban capturando jurel en el litoral peruano tuvieron un plazo de dos días para efectuar el desembarque del vo-lumen capturado.
Medidas de seguimiento, control y vigilancia Un marco jurídico e institucional eficaz de ordenación pes-
quera exige que se apliquen medidas de seguimiento, control y vigilancia que garanticen el cumplimiento de las normas, principalmente en lo que se refiere a las actividades de pesca por parte de los administrados, incluyendo cuando procedan, programas de observadores, mecanismos de inspección y sistemas de vigilancia de buques (FAO 1995) que cumplan con evitar la pesca ilegal y la depredación de los recursos y, en general, que se asegure obtener los objetivos del ordenamiento pesquero establecidos.
En el caso de Perú, institucionalmente el Ministerio de Producción, a través del Viceministro de Pesquería, tiene la función de diseñar, establecer, ejecutar y supervisar, la política nacional y sectorial aplicable al sector de pesquería, la de dictar normas y lineamientos técnicos para la adecuada gestión de los recursos, así como para el otorgamiento, reconocimiento de derechos, la sanción, fiscalización y ejecución coactiva. Estas funciones se traducen a la labor del Estado en cuanto a regular el manejo integral y la explotación racional de los recursos hidrobiológicos; asimismo entre sus labores están el ser responsable del seguimiento y controlar las actividades pesqueras. Además, estas funciones incluyen el control de las embarcaciones de pesca mediante los sistemas de vigilancia sa-telital. Dichas acciones perfeccionan los tradicionales sistemas de control de las capturas que existen en los puertos donde los desembarcos permiten conocer y fiscalizar el cumplimiento de las cuotas de captura.
Si como efecto de la fiscalización de las actividades pesque-ras se constata faltas a la normatividad vigente y se realiza un previo informe técnico sustentatorio, se apertura el procedi-
miento administrativo sancionador a cargo de la Dirección General de Sanciones del Ministerio de la Producción. Este organismo deberá determinar si corresponde o no aplicar la sanción administrativa correspondiente, decisión que de ser impugnada por el armador y/o titular pesquero es sometida a revisión por parte del Consejo de Apelación de Sanciones, quien en su calidad de superior jerárquico hará una revisión de la legalidad del procedimiento seguido por la Dirección General de Sanciones.
Concluido el procedimiento sancionador en vía adminis-trativa, el infractor deberá cumplir con las sanciones impuestas por el órgano sancionador, caso contrario se procederá a la ejecución coactiva de las mismas. Cabe señalar que, agotada la vía administrativa, el armador tiene la posibilidad de ejercer su derecho de defensa en la vía judicial, la cual hará una revisión del procedimiento administrativo a fin de que se hayan respetado las garantías del debido procedimiento.
Complementariamente a las responsabilidades del Ministerio de la Producción para formular, aprobar y supervisar las polí-ticas de alcance nacional aplicables a las actividades extractivas y productivas del sector pesquería, y dictar normas de alcance nacional y supervisar su cumplimiento, el Ministerio de Defensa a través de la Dirección General de Capitanías y Guardacostas de la Marina de Guerra del Perú ejerce la autoridad marítima, fluvial y lacustre. Es decir, esta dirección es responsable de nor-mar y velar por la seguridad de la vida humana, la protección del medio ambiente y sus recursos naturales, así como reprimir todo acto ilícito, ejerciendo el control y vigilancia de todas las actividades que se realizan en el medio acuático, de esta manera contribuye al desarrollo nacional.
Hacia la aplicación del enfoque ecosistémicoTambién se acepta que el concepto tradicional del orde-
namiento pesquero basado en un enfoque dirigido a una sola especie ha de cambiar hacia un concepto ecosistémico, recono-ciendo que, en general, la actividad extractiva no sólo afecta a la población objetivo, sino a todos los elementos del ecosistema.
Por lo tanto, se requiere un enfoque diferente, cuyo objetivo fundamental sea el aprovechamiento sostenible del ecosiste-ma. Es conveniente que este planteamiento que dio origen a la aplicación del enfoque ecosistémico a la pesca en general, sea incorporado al ordenamiento del jurel y caballa. Sobre la base del enfoque ecosistémico a la pesca se podrá comprender la planificación, desarrollo y ordenación de la pesca de estas especies, de un modo que la actividad satisfaga las múltiples necesidades de nuestra sociedad actual, sin poner en riesgo la posibilidad de que las generaciones futuras se beneficien también de los bienes y servicios que pueden obtenerse de la integridad del ecosistema marino.
En este enfoque, la estrategia para incluir la pesquería del jurel y la caballa demanda que el sistema de ordenamiento actual incorpore algunas necesidades o requerimientos, como obtener mejor información para formular las políticas y los planes de ordenamiento. Estos planes deben considerarse como la base técnica de los reglamentos de ordenamiento pesquero en la perspectiva jurídica, y los planes de ordenación en la descripción de la biología de las especies que son objeto de nuestra atención en el presente caso, y en su interrelación con el ecosistema. Es imperativo el establecimiento de objetivos e indicadores ope-

112
Zuzunaga
Rev. peru. biol. número especial 20(1): 107 - 113 (Septiembre 2013)Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú.
Editores: Csirke J., R. Guevara-Carrasco & M. Espino
racionales para la pesquería de jurel y caballa, en tanto estas especies continúen tratándose como integrantes de una actividad.
Una vez establecidos los objetivos operacionales, será necesa-rio determinar las medidas de ordenamiento, las cuales podrán darse sobre la base de las medidas vigentes, pero orientadas a mantener o restablecer la estructura y funcionamiento del ecosistema.
Finalmente, ratificamos que una buena base técnica para avanzar hacia un ordenamiento pesquero basado en el enfoque ecosistémico a la pesca lo viene proporcionando el sistema de observación del ambiente, los recursos vivos y sus pesquerías, que el IMARPE desarrolla desde hace casi 50 años y se evidencia en la presente publicación, que comprende muchos de los aspectos biológicos y pesqueros del jurel, así como del ambiente en el cual se desarrolla.
Conclusiones La actividad pesquera es una fuente importante de suminis-
tros para la seguridad alimentaria de la población peruana y pro-porciona beneficios económicos y sociales para la nación. Uno de los principales recursos hidrobiológicos de la pesca marina es el jurel que ha representado en los últimos años cerca del 30% de la captura total de especies marinas con destino al consumo humano directo. Estas consideraciones obligan a que la gestión pesquera del recurso sea efectiva y exitosa. Se entiende como gestión exitosa el mantener la sostenibilidad de la pesquería, la conservación del recurso y el mantenimiento de su aporte a la producción de consumo humano directo. Correspondiendo a este interés nacional, se ha establecido un sistema de ordena-miento y regulación de la pesca de jurel que se fundamenta en los principios de las leyes de Aprovechamiento Sostenible de los Recursos Naturales y General de Pesca, y se hace efectivo con el Reglamento de Ordenamiento Pesquero de los recursos jurel y caballa vigente.
En el presente caso sobre el ordenamiento pesquero del jurel, se demuestra que la legislación nacional es adecuada. Para ello se tiene en consideración que abarca un conjunto de principios normativos y reglamentos nacionales. Estos principios normativos y reglamentos definen con claridad los objetivos del ordenamiento y las medidas de gestión de la pesca del jurel peruano T. murphyi, como también la consiguiente protección del ambiente asociado a los recursos en explotación. El reglamento respectivo incluye medidas como la fijación de cuotas de capturas permisibles con un único destino para el consumo humano, las temporadas de pesca, el esfuerzo pesquero, los métodos de pesca, tamaño mínimo de malla de las redes, el tamaño mínimo de captura de los ejemplares de jurel. Asimismo, entre las medidas incluidas está el estable-cimiento de otras normas que son en el presente necesarias para la conservación y explotación racional de los recursos en general y de esta especie en particular. Además, hace posible la definición de más medidas, de acuerdo a la condición o estado del recurso hidrobiológico. Muchas de las normas que se aplican actualmente en la pesquería de la especie han sido descritas en el presente trabajo, cuya finalidad ha sido mostrar que el esquema de gestión del jurel cumple con el propósito de mantener al recurso con la capacidad de proporcionar cap-turas sostenidas destinadas al consumo humano, así como la viabilidad económica y social de su desarrollo.
Más aún, se puede afirmar que mediante el establecimiento y aplicación de este sistema de ordenamiento se cumple con el derecho internacional, el cual está reflejado en las disposiciones pertinentes de la Convención de las Naciones Unidas sobre el Derecho del Mar de 1982. Para los estados ribereños que tienen aguas bajo jurisdicción nacional, en ese documento se establece el privilegio de ejercer sus derechos de soberanía para los fines de exploración, explotación, conservación y gestión de la pesca de los recursos y la conservación de los recursos marinos vivos sobre los que la pesca tiene un impacto.
Si bien con este sistema de gestión de la pesca se demuestra el éxito de su implementación a nivel nacional, ha existido y existe permanentemente el riesgo potencial del desarrollo indiscrimi-nado de la pesca sobre esta unidad de stock de jurel en alta mar, lo cual puede desestabilizar su equilibrio poblacional y, por lo tanto, afectar la pesquería peruana. Por esta razón, el Perú ha participado activamente en la conformación de una entidad de ordenamiento pesquero para el alta mar, la OROP del Pacífico Sur (Organización Regional de Ordenación Pesquera del Pacífico Sur), la cual es una organización intergubernamental que debe estar permanentemente comprometida con la conservación a lar-go plazo, el uso sostenible de los recursos pesqueros en el océano Pacífico Sur y la protección de los ecosistemas marinos en los que dichos recursos se producen. Estos principios son reconocidos por la convención al igual que la existencia de consideraciones económicas y geográficas, además de las necesidades especiales de los estados y sus comunidades costeras, en relación con el de-sarrollo de la conservación y la gestión sostenible de los recursos pesqueros, así como es reconocida la explotación equitativa de esos recursos, por la cual se acordó establecer dicha organización.
Literatura citadaConstitución Política del Perú. 1993. Congreso de la República del Perú. 1993.
<http://www.tc.gob.pe/legconperu/constitucion.html>. Acceso 20 de junio del 2013.
DL N°. 25977. 1992. Ley General de Pesca. Congreso de la República del Perú. 21 de de diciembre de 1992. <http://www2.produce.gob.pe/dispositivos/publicaciones/2013/rm077-2013-produce.pdf >. Acceso 16 de junio del 2013.
DS N°. 012-84-PE. 1984. Reglamento para la Operación de Buques Pesque-ros de Arrastre. LEX-FAOC018459. 19/09/1984. Acceso 24 de agosto 2013.
DS N°. 017-92-PE. 1992. Ministerio de Pesquería. Prohíben la pesca con el uso de métodos que modifiquen las condiciones bioecológicas del medio marino en la zona adyacente a la costa, comprendida entre las cero y cinco millas marinas. Revista de Legislación y Jurisprudencia. Normas Legales, Tomo 196, setiembre 1992. pagina 212.
DS N°. 012-2001-PE. 2001. Aprueban el Reglamento de la Ley General de Pesca, Ministerio de la Producción. 13 de marzo de 2001. El Peruano Normal Legales: 199905-199921
DS N°. 001-2002-PRODUCE. 2002. Establecen que los recursos sardina, jurel y caballa serán destinados al consumo humano directo. 06 de setiembre 2002. El Peruano. Normas Legales: 229522-229523.
DS N°. 031-2003-PRODUCE. 2003. Establecen Marco Provisional para la Implementación del Sistema de Seguimiento Satelital. 04 de noviembre 2003. El Peruano Normas Legales: 254393-254394.
DS N°. 011-2007-PRODUCE. 2007. Aprueban Reglamento de Ordenamiento Pesquero de Jurel y Caballa. 13de abril de 2007. El Peruano Normal Legales: 343399-343402
DS N°. 022-2009-PRODUCE. 2009. Decreto Supremo que establece medidas de ordenamiento para la extracción comercial a mayor escala de recursos hidrobiológicos tranzonales (jurel y caballa) en la alta mar. 05 de julio 2009. El Peruano Normas Legales: 398635-398638.
FAO. 1995. Código de Conducta para la Pesca Responsable. 1995. Roma. FAO. 46 p.
FAO. 1999. La ordenación pesquera. Orientaciones Técnicas para la Pesca Responsable N.° 4. Roma. FAO. 81 p.

113
conservación y ordenación pesquera del jurel Trachurus murphyi
Rev. peru. biol. special issue 20(1): 107 - 113 (September 2013)Ecology, fisheries and conservation of Jack mackerel (Trachurus murphyi) in Peru. Editors: Csirke J., R. Guevara-Carrasco & M. Espino
Ley N°. 26821. 1997. Aprueba Ley Orgánica para el aprovechamiento. Soste-nible de los recursos naturales. Congreso de la República del Perú. 26 de junio de 1997. <http://www2.produce.gob.pe/Repositorio-APS/1/jer/PROPESCA_OTRO/marco-legal/1.3.%20Ley26821.pdf>. Acceso 15 de junio del 2013.
RM N°. 0171-74-PE. 1974. Ministerio de Pesquería. 1974. Prohíben extracción de especies hidrobiológicas cuya longitud sea inferior a lo autorizado. Revista de Legislación y Jurisprudencia Normas Legales, Tomo 70, enero- marzo 1974: pagina 145.
RM N°. 026-2010-PRODUCE. 2010. Establecen régimen provisional para la extracción de los recursos jurel y caballa en todo el litoral peruano. 03 de febrero 010. El Peruano Normas Legales: 412689-412690.
RM N°. 027-2010-PRODUCE. 2010. Establecen régimen de pesca exploratoria de recursos pelágicos de oportunidad asociados al evento El Niño en todo el litoral. 03 de febrero 010. El Peruano Normas Legales: 412690-412692.
RM N°. 096-2010-PRODUCE. 2010. Establecen Régimen Provisional para la extracción de los recursos jurel y caballa en todo el litoral peruano. 18 de abril 2010. El Peruano Normas Legales: 417356-417357.
RM N°. 097-2010-PRODUCE. 2010. Establecen Régimen de Pesca Explo-ratoria de recursos pelágicos de oportunidad asociados al evento El Niño en todo el litoral peruano. 18 de abril 2010. El Peruano Normas Legales: 417357-417359.
RM N°. 202-2010-PRODUCE. 2010. Establecen Régimen Provisional para la extracción de los recursos jurel y caballa en todo el litoral peruano. 18 de agosto de 2010. El Peruano Normas Legales: 423977-423978
RM N°. 025-2011-Produce. 2011. Establecen Régimen Provisional para la extracción de los recursos jurel y caballa en todo el litoral peruano. 26 de enero 2011. El Peruano Normas Legales: 434925-434927.
RM N°. 035-2011-PRODUCE. 2011. Suspenden actividades de extracción y recepción del recurso jurel en el marco establecido mediante R.M. N.° 025-2011-PRODUCE. 06 de febrero 2011. El Peruano Normas Legales: 435774-435775.
RM N°. 039-2011-PRODUCE. 2011. Modifican la R.M. Nº 035-2011-PRO-DUCE mediante la cual se dispuso suspender las actividades de extracción y recepción del recurso jurel. 09 de febrero de 2011. El Peruano Normas Legales: 435847-435848.
RM N°. 058-2011-PRODUCE. 2011. Modifican R.M. N.° 025-2011-PRO-DUCE en lo referente al límite de captura establecido en el Régimen Provisional para la extracción de los recursos jurel y caballa. 26 de febrero 2011. El Peruano Normas Legales: 437661-437662.
RM N°. 099-2011-PRODUCE. 2011. Suspenden actividades de extracción de los recursos jurel, anchoveta y anchoveta blanca. 20 de marzo 2011. El Peruano Normas Legales: 439295-439296.
RM N°. 107-2011-PRODUCE. 2011. Prorrogan vigencia del Régimen Provi-sional de pesca de los recursos jurel y caballa establecida mediante RM N°. 025-2011-PRODUCE. 25 de marzo 2011. El Peruano Normas Legales: 439597-439598.
RM N°. 124-2011-PRODUCE. 2011. Suspenden actividades de extracción del jurel. 10 de abril de 2011. El Peruano Normas Legales: 440684-440686
RM N°. 130-2011-PRODUCE. 2011. Amplían límite de capturas provenientes de las embarcaciones pesqueras de mayor escala, en el marco estable-cido mediante R.M. N.° 025-2011-PRODUCE. 13 de abril 2011. El Peruano Normas Legales: 440876-440877.
RM N°. 146-2011-PRODUCE. 2011. Prorrogan vigencia del Régimen Pro-visional para la extracción de los recursos jurel y caballa establecido por R.M. N.° 025-2011-PRODUCE. 28 de abril 2011. El Peruano Normas Legales: 441665-441666.
RM N°. 198-2011-PRODUCE. 2011. Establecen límite de captura de recur-sos jurel y caballa. 29 de junio 2011. El Peruano Normas Legales: 445535-445536.
RM N°. 034-2012-PRODUCE. 2012. Establecen límite de captura de recursos jurel y caballa, correspondiente al primer trimestre del año 2012. 24 de enero 2012. El Peruano Normas Legales: 459721-459722.
RM N°. 055-2012-PRODUCE. 2012. Suspenden actividades extractivas del recurso jurel en todo el litoral. 03 de febrero 2012. El Peruano Normas Legales: 460291.
RM N°. 132-2012-PRODUCE. 2012. Precisan alcances de la R.M. N.° 034-2012-PRODUCE en el extremo referente al límite de captura de los recursos jurel y caballa correspondientes al primer trimestre del año 2012. 22 de marzo 2012. El Peruano Normas Legales: 462911.
RM N°. 133-2012-PRODUCE. 2012. Establecen límite de captura de los recursos jurel y caballa para la flota industrial pesquera, correspon-diente al segundo trimestre del año 2012. 23 de marzo 2012. El Peruano Normas Legales: 462958.
RM N°. 142-2012-PRODUCE. 2012. Modifican la R.M. N.° 133-2012-PRO-DUCE, que estableció el límite de captura de los recursos jurel y caballa para el segundo trimestre de 2012. 24 de marzo 2012. El Peruano Normas Legales: 463015-463016.
RM N°. 173-2012-PRODUCE. 2012. Establecen cuota de captura total permisible para el recurso jurel correspondiente al año 2012. 15 de abril 2012. El Peruano Normas Legales: 464166-464167.
RM N°. 191-2012-PRODUCE. 2012. Suspenden actividades extractivas del recurso jurel efectuadas por la flota industrial, en todo el litoral peruano, a partir del 21 de abril de 2012. 20 de abril 2012. El Peruano Normas Legales: 464560-464561.
RM N°. 277-2011-PRODUCE. 2011. Amplían límite de captura de los recursos jurel y caballa establecido mediante RM N°. 198-2011-PRODUCE. 01 de octubre 2011. El Peruano Normas Legales: 450954-450955.

114
Zuzunaga
Rev. peru. biol. número especial 20(1): 107 - 113 (Septiembre 2013)Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú.
Editores: Csirke J., R. Guevara-Carrasco & M. Espino

115
Colofón

116
Suscripciones y CanjeSubscriptions and Exchange programs
La Revista Peruana de Biología es publicada en abril. agosto y diciembre y esta dedicada a la publicación de los resultados de investigaciones originales e inéditas en las áreas de Biodiversidad, Biotecnología, Manejo ambiental, Ecología y Biomédicas. Los trabajos recibidos son evaluados por árbitros según criterios internacionales de calidad, creatividad, originalidad y contribución al conocimiento.
The Revista Peruana de Biología is published in April, August and December, the aims is to disseminate the results of original research in Biodiversity, Biotechnology, Environmental management, Ecology and Biomedical areas. Papers in Spanish or English are peer-reviewed using international criteria of quality, creativity, originality and the knowledge contribution.
Revista Peruana de Biología
Suscripción anual, costo incluye envío.Annual subscription, mailing is included.Perú 300 nuevos solesOther countries US$ 250
Nombre/name:……………………………………………………………..
Dirección postal/Full postal address: ……………………………………………
…………………………………………………………………………………
…………………………………………………………………………………
…………………………………………………………………………………
…………………………………………………………………………………
Favor de extender Cheque certificado u orden de pago a nombre de la Facultad de Ciencias Biológicas-UNMSM.
Please make check or money order payable to Facultad de Ciencias Biológicas-UNMSM.
Enviar a:Send to:
Leonardo Romero Editor, Revista Peruana de Biología Facultad de Ciencias Biológicas-UNMSM Casilla Postal: 11-0058 Lima-11, Perú
Mayor información dirigirse a:For futher information contact:Editor Jefe, Leonardo RomeroTeléfono (511) 619-7000-1502/ Telefax (511) 619-7000-1509email: [email protected]

117
IdentIdad y propósIto
La Revista Peruana De Biología es una publicación científica arbitrada producida por el Instituto de Investigaciones de Ciencias Biológicas Antonio Raimondi, Facultad de Ciencias Biológicas, Universidad Nacional Mayor de San Marcos, Lima, Perú; es publicada tres veces al año y los números aparecen en abril, agosto y diciembre, tanto en su versión impresa como Online.
La Revista Peruana De Biología publica artículos completos, originales e inéditos que contribuyan al conocimiento científico en los temas de biodiversidad, biotecnología, ecología, manejo ambiental y biomedicina elaborados según las normas indicadas en las presentes pautas.
evaluacIón de los trabajos
Los trabajos que cumplan con las pautas solicitadas serán incluidos en el la lista para evaluación. Un editor será encargado de conducir el proceso enviando el trabajo a árbitros. El trabajo será evaluado según criterios internacionales de calidad, creatividad, originalidad y contribución al conocimiento. El artículo será aceptado luego del proceso de revisión por árbitros y de realizadas las modificaciones indicadas. El artículo aceptado será editado y una prueba enviada al autor para la aceptación y consentimiento de publicación.
sumIsIón para edIcIón y lIcencIa de uso
1. Consideraciones sobre la edición El envío de trabajo mencionado a la Revista Peruana de Biología, implica el consentimiento de los autores para la revisión
por pares, la corrección, la edición y posteriormente el consentImIento de la publIcacIón del trabajo tal como se presentara después de la prueba final del trabajo.
Los autores reconocen que la Revista Peruana de Biología se adhiere a la política de acceso abIerto dando inmediato acceso libre a su contenido bajo el principio de: hacer disponible gratuitamente la investigación al público fomenta un mayor intercambio de conocimiento global.
Los autores garantizan la calidad del trabajo mediante una carta de presentacIón, la carta será considerada una declaración jurada y debe afirmar que:
a. Garantizan que han leído las pautas de presentación y entendido los requerimientos legales y éticos que se mencionan en las PAUTAS PARA PRESENTACION DE TRABAJOS.
b. Garantizan que todos los datos contenidos son exactos y todos los hechos declarados proceden de observaciones o investigaciones realizadas por los autores.
c. Garantizan la autoría de todos los nominados en el trabajo, y su participación de manera relevante en el diseño, ejecución y análisis y se responsabilizan por el trabajo publicado.
d. Garantizan que los el trabajo presentado no presenta problemas de competencia en los resultados o conclusiones vertidas.e. Garantizan que el manuscrito enviado, nunca ha sido publicado en su totalidad o en parte, en el idioma presentado o en cu-
alquier otro idioma, y no se encuentra sometido otra revista. f. Los autores garantizan que no infringieron leyes ni normas éticas o bioéticas nacionales ni internacionales en la elaboración del
artículo. g. Esta carta debe ser enviada por cada uno de los autores, por email debidamente firmada.2. Entendimientos sobre la licencia de uso Entrando en vigencia en el momento de su consentImIento para publIcacIón, los autores convienen que el artículo será
publicado por la Revista Peruana de Biología, en medio impreso y digital, además de ser adaptado a sistemas informáticos que amplíen su visibilidad y difusión, incluyendo su reproducción o publicación en formatos aptos para ser leídos por máquinas y su incorporación en sistemas de recuperación y bases de datos bibliográficos.
Los autores, en conocimiento que la Revista Peruana de Biología tiene una política de acceso abierto (Open Access) aceptan las condiciones de la lIcencIa creatIve commons atrIbucIón – No Comercial – Sin obra derivada 3.0 (CC BY-NC-ND 3.0) [http://creativecommons.org/licenses/by-nc-nd/3.0/deed.es_ES] que permite copiar, distribuir y comunicar públicamente la obra. Esta Licencia será colocada en toda forma y medio en que el artículo se publique por la Revista Peruana de Biología.
Todas las copias de los artículos de la Revista Peruana de Biología, en papel o digitales, o cualquier otro uso de la in-formación deberán incluir una cita de la fuentes de publicación en su totalidad.
Una vez terminado el proceso de edición, el Editor Jefe de la Revista Peruana de Biología enviará a los autores una prueba del trabajo en formato PDF, para su revisión final; deberán efectuar los cambios pertinentes, corregirán los errores de impresión y una vez subsanados los errores el autor enviara la carta de consentImIento para publicación del artículo, en la cual: acepta la publicación del trabajo en la Revista Peruana de Biología y está de acuerdo con la lIcencIa de uso.
3. Los autores retienen sus derechos como: a. Los derechos de marca y patente y derechos sobre cualquier proceso o procedimiento descrito en el artículo. b. El derecho de compartir, copiar, distribuir, ejecutar y comunicar públicamente la obra.c. El derecho posterior a la publicación, de utilizar el artículo o cualquier parte de aquel, en una compilación impresa de sus
propios trabajos, como ser una selección de escritos o notas para conferencias, en una tesis, o para ampliar el artículo a formato de libro para publicación, siempre que se haga cita de las fuentes de la publicación en su totalidad.
PAUTAS PARA LA PRESENTACIóN DE LOS ARTÍCULOS EN LA REVISTA PERUANA DE BIOLOGÍA
Julio 2013

118
cartas de presentacIón
Las coordinaciones se llevaran entre el Editor Jefe y el responsable del trabajo. El trabajo será acompañado de una carta del responsable y dirigido al Editor Jefe, indicando haber leído las presentes Pautas para Presentación de Trabajos; asegurando la originalidad, carácter inédito y completo del trabajo presentado y su disposición para que sea revisado y editado. Esta carta tendrá carácter de declaración jurada según lo estipulan las directivas del Vicerrectorado de Investigación de la Universidad Nacional Mayor de San Marcos referidos a las autorías de trabajos publicados. Cada autor del trabajo deberá de enviar por correo electrónico una Carta de Presentación del trabajo dando las garantías mencionadas anteriormente.
presentacIón de los trabajos
Los archivos del trabajo serán enviados por correo electrónico al editor: [email protected] Los trabajos pueden ser presentados en idioma inglés o castellano. El trabajo debe tener tres partes básicas: (a) identificación, (b) cuerpo y (c) literatura citada(a) La identificación debe contener: Título (en inglés y castellano), nombre y apellido del autor o los autores, afiliación institucional
de los autores, correo electrónico de cada uno de los autores, y dirección postal del autor para correspondencia. Resumen no mayor de 250 palabras (en inglés y castellano), 5 palabras clave (en inglés y castellano).
Información adicional. Debe incluirse información sobre la contribución de cada uno de los autores y de no incurrir en conflicto de intereses.
Fuente de financiamiento. Debe indicarse la fuente de financiamiento del trabajo, e incluir el código o numero de referencia.(b) El cuerpo del trabajo. variará según la sección de la Revista:trabajos orIgInales. Son artículos primarios, inéditos que exponen los resultados de trabajos de investigaciones completas y finales,
y que constituyen aportes al conocimiento. Aquí también se incluyen las descripciones de especies nuevas, que cumplan con las Normas y características de la información en los trabajos (ver abajo). El cuerpo está organizado en: Introducción, Material y métodos, Resultados, Discusión y Agradecimientos. Debe abarcar un texto promedio de 30 páginas, las ilustraciones (Tablas y Figuras) deben ser sólo las necesarias para una mejor exposición de los resultados.
notas cIentífIcas. Son artículos primarios, se incluyen aquí: (a) Trabajos de interés taxonómico como reportes de distribución, inventarios taxonómicos, notas taxonómicas, con la información necesaria para cubrir los estándares de Darwin core. También se incluyen resultados de ensayos de laboratorio, cuya información es de interés para la comunidad científica. La extensión del texto no será mayor de 15 páginas. El cuerpo de la Nota puede estar organizada: Introducción, Material y métodos, Resultados y Discusión y Agradecimientos.
comentarIos. Son artículos donde se discute y exponen temas o conceptos de interés para la comunidad científica. Se incluyen aquí ensayos de opinión y monografías. Deben contar con las siguientes partes: cuerpo del comentario y Agradecimientos. Todo el artículo debe tener un texto promedio de 10 páginas.
comentarIos de lIbros. Son artículos que comentan recientes publicaciones de interés para la comunidad científica. Puede solicitarse al Comité Editor la elaboración de un comentario enviando dos copias del libro a la dirección postal de la Revista Peruana de Biología.
los agradecImIentos. Deben dedicarse solamente a las personas e instituciones que colaboraron directamente en la realización del trabajo.
cItacIón. Las citas en el texto deben incluir el apellido del autor y año sin comas que los separen (ejemplo: (Carrillo 1988) o «... de acuerdo a Sánchez (1976)…»
Cuando dos autores son citados en el texto se usa la conjunción “y”: …la concentración de nitrógeno fue medida según Chávez y Castro (1998)…. Cuando son citados entre paréntesis se usa el símbolo “&”: …de nitrógeno se midió por el método colorimétrico (Chávez & Castro 1998)…. Citas de varias referencias entre paréntesis, van separadas por comas, en secuencia de importancia: … la diversidad de especies es considerada elevada (Simoniz 1968, Perez 2012, Aquino 1988). Si hay varios trabajos de un autor en un mismo año, se citará con una letra en secuencia adosada al año (ejemplo: Castro 1952a). Citas de leyes y normas: "tal como se indica en la Ley N.° 26821..." Cuando hay más de dos autores se citará al primer autor y se colocará “et al.” sin itálicas: (e.g. Smith et al. 1981) o “… según
Smith et al. (1981)”). Citas directas o literales se deben hacer entre comillas dentro del texto cuando son menos de 40 palabras, idiomas diferentes al
texto en itálicas, indicar página del extracto: … Finalmente, después de revisar la distribución de Lu. felina, se estableció un área entre el Estrecho de Magallanes, Patagonia,
Chile y Perú, pero incurrió en un error al extender los limites hasta el Ecuador mencionando “It has been found, in Perú, and in Ecuador it has been recorded from San Lorenzo” (Thomas 1889, p: 198). En realidad esta era la localidad tipo de L. peruviensis que Gervais (1841) indico como “… procuré, à San-Lorenzo du Pérou, une portion de crâne trouvée sur la plage, et qui provient incontestablement d'une Loutre” (p: 15).
Citas directas o literales de más de 40 palabras, deben separarse del texto en un párrafo sangrado, la cita como en el ejemplo anterior.
(c) La lIteratura cItada incluirá todas las referencias citadas en el texto dispuestas solamente en orden alfabético y sin numeración. La cita se inicia con el apellido del primer autor a continuación, sin coma, las iniciales del nombre separadas con puntos y sin

119
espacio. El segundo y tercer autor deben de tener las iniciales de los nombre y a continuación el apellido. El último autor se diferenciara por que le antecede el símbolo &. Si hubiesen más de tres autores pueden ser indicados con la abreviatura et al. Los nombres de las publicaciones periódicas (revistas) pueden ir en la abreviatura oficial considerada según su código ISSN. El código DOI debe ser colocado al final de la referencia. En la literatura citada solamente se usa letra tipo normal, no itálica, no versalita. Ejemplos:
• Andrén H., & H. Andren. 1994. Effects of Habitat Fragmentation on Birds and Mammals in Landscapes with Different Proportions of Suitable Habitat: A Review. Oikos 71(3):355-366. doi:10.2307/3545823.
• Dormann C., J.M. McPherson, M.B. Araújo, et al. 2007. Methods to Account for Spatial Autocorrelation in the Analysis of Species Distributional Data: a Review. Ecography 30(5):609–628. doi:10.1111/j.2007.0906-7590.05171.x.
• Buhrnheim C.M. & L.R. Malabarba. 2006. Redescription of the type species of Odontostilbe Cope, 1870 (Teleostei: Chara-cidae: Cheirodontinae), and description of three new species from the Amazon basin. Neotrop. ichthyol. 4 (2): 167-196.
• Laub B.G. & Palmer M.A., 2009. Restoration Ecology of Rivers, in: G.E. Likens (Ed.), Encyclopedia of Inland Waters. Aca-demic Press, Oxford, pp. 332–341. dx.doi.org/10.1016/B978-012370626-3.00247-7
• Rohde, K., 2002. Ecology and biogeography of marine parasites, in: Advances in Marine Biology. Academic Press, pp. 1–83. dx.doi.org/10.1016/S0065-2881(02)43002-7,
• McLachlan A. & A.C Brown. 2006. The Ecology of Sandy Shores. Elsevier Science & Technology Books. 373pp.• Crawford D.J. 1983. Phylogenetic and systematic inferences from electrophoretic studies. In: S.D. Tanksley and T.J. Orton,
eds. Isozymes in Plant Genetics and Breeding, Part A. Elsevier, Amsterdam. Pp. 257-287.• Pianka E.R. 1978. Evolutionary ecology. 2nd edn. New York: Harper & Row.• Carroll S.B. 2005. Evolution at Two Levels: On Genes and Form. PLoS Biol 3(7): e245. <http://biology. plosjournals.org/
archive/1545-7885/3/7/pdf /10.1371_journal.pbio.0030245-S.pdf >. Acceso 31/07/2005.• Food and Drug Administrations (FDA). 2001. Fish and Fishery Products Hazards and Controls Guidance. Third Edition
June 2001. <http://www.cfsan.fda.gov/~comm/haccp4.html> (acceso 24/12/07).• CONAM. 2005. (en línea). Informe nacional del estado del ambiente 2001. <http://www.conam.gob.pe /sinia/INEA2001.
shtml>. Acceso 31/07/2005.• IMARPE. 2002. (en línea). Segundo informe del BIC José Olaya Balandra. Paita – Salaverry. 24 febrero- 05 Marzo 2002.
<http://www.imarpe.gob.pe /imarpe/informeolaya02-032002.php>. Acceso 01/07/2005.• Solari S.A. 2002. Sistemática de Thylamys (mammalia: didelphimorphia: marmosidae). Un estudio de las poblaciones asignadas
a Thylamys elegans en Perú. Tesis, Magíster en Zoología, mención Sistemática y Evolución. Facultad de Ciencias Biológicas Universidad Nacional Mayor de San Marcos. <http://www.cybertesis.edu.pe/sisbib/2002/solari_ts /html/index-frames.html>. Acceso 31/07/2005
• Ley N°. 26821. 1997. Ley orgánica para el aprovechamiento sostenible de los recursos naturales. Compendio de la legislación ambiental peruana. Volumen I. Marco normativo general. Ministerio del Ambiente –MINAM, Editado por la Dirección General de Políticas, Normas e Instrumentos de Gestión Ambiental del Ministerio del Ambiente. Actualizado al 30 de junio de 2010. pp: 85-91
• DS N°. 012-2001-PE. 2001. Aprueban el Reglamento de la Ley General de Pesca, Ministerio de la Producción. 13 de marzo de 2001. El Peruano Normal Legales: 199905-199921
• DS N° .179-2004-EF. 2004. Texto único ordenado de la ley del impuesto a la renta. 08 de diciembre de 2004. El Peruano Normas Legales: 281912-281940
Las citas de artículos en prensa deben incluir el volumen, el año y el nombre de la revista donde saldrán publicados; de lo contrario deberán ser omitidos.
No se aceptan las citas a resúmenes de eventos académicos (congresos y otros).normas y característIcas de la InformacIón en los trabajos
La Revista Peruana de Biología se guía de los conceptos éticos de autoría mencionados en los Uniform Requirements for Manu-scripts Submitted to Biomedical Journals (http://www.icmje.org/). La revista asume la veracidad de la carta de presentación y de la carta de consentimiento de publicación.
Cuando el artículo exponga sobre experimentos con humanos y animales, los procedimientos deben de ceñirse a la Declara-ción de Helsinki de 1975 y a las leyes peruanas vigentes (Ley 27265). Deben ser presentadas las declaraciones pertinentes y mencionadas en el texto.
Cuando el artículo exponga sobre nuevas especies, nuevos registros, ampliaciones biogeográficas o inventarios taxonómicos debe tomarse en cuenta la información requerida en el Darwin Core. También debe indicarse el depósito de los ejemplares en una institución taxonómica reconocida, comprometida en la preservación del material, que permita el libre acceso al mate-rial, publique constantemente la relación del material que posee en custodia y que brinde la información necesaria sobre los ejemplares cuando sea requerido. No son consideradas las colecciones particulares.
Los nombres científicos del género y especie irán en cursivas. La primera vez que se cita un organismo deberá hacerse con su nombre científico completo (género, especie y autor); posteriormente podrá citarse solamente la inicial del nombre genérico y el nombre específico completo. Para el caso de las abreviaturas de autores en nombres botánicos pueden referirse a la base de datos TROPICOS (http://www.tropicos.org/) y AlgaeBase (http://www.algaebase.org/).
Las localidades y puntos de colecta deben ser referenciadas geográficamente, usando coordenadas cartesianas sexagesimales, tanto en el texto como en las ilustraciones.

120
Deben usarse los símbolos de las unidades del Sistema Internacional de Medidas. Si fuera necesario agregar medidas en otros sistemas, las abreviaturas correspondientes deben ser definidas en el texto.
Debe usarse el punto decimal (ejemplo correcto: 0.5; incorrecto: 0,5). Las cifras mantenerse juntas (ejemplo correcto: 1994, 30302; incorrecto: 1,994, 30 302).
IlustracIones
Las Figuras (mapas, esquemas, diagramas, dibujos, gráficos, fotos, etc.) serán numeradas correlativamente con números arábigos; de igual manera las Tablas. Las leyendas de las Figuras y Tablas deben presentarse a continuación del texto y ser suficientemente explicativas.
Las figuras escaneadas deben guardarse en un archivo TIFF, tamaño del original, 300 dpi. Las gráficas deben de enviarse en formato nativo editable (achivo.xls, archivo.wmf, archivo.svg, archivo.eps), no como imágenes (JPGE, TIFF, PNG). Los mapas pueden enviarse en formatos SHP.
Fotos de cámaras digitales en formato JPGE mayor a 3 Mpixel. Otros archivos de imágenes en TIFF, BMP, JPGE de alta reso-lución y tamaño, y figuras vectoriales en Ai, PSD.
Formatos procedentes de software como R, en formatos editables de PDF, EPS (Postscript). Costos por ilustraciones a color serán asumidos por el autor.
Los archivos deben presentarse por separado, esto es, un archivo con el texto y leyendas en formato MS-Word. Otro archivo para las tablas en MS-Excel o como tablas en MS-Word. Otros archivos en formatos nativos, no como imágenes insertadas en otros archivos (por ejemplo no enviar imágenes pegadas en una hoja de MS-Word o Excel).
consentImIento de publIcacIón El autor responsable recibirá una prueba del trabajo en formato PDF, el cual deberá revisarlo cuidadosamente, y consultarlo con los
otros autores. Una vez solucionados errores de edición, el autor responsable enviara una carta o email confirmando que el trabajo fue revisado por todos los autores y están conformes, y mencionar explícitamente su consentimiento para la publicación del trabajo como artículos de la Revista Peruana de Biología y estan de acuerdo con la lIcencIa de uso.
Este es un requisito para la publicación del trabajo. Con la prueba, el autor responsable recibirá el costo por publIcacIón en los casos que corresponda.
costo por publIcacIón. El costo por publicación se refiere a gastos extras por manipulación, edición extraordinaria (v.g. elaboración de ilustraciones, traduccio-
nes, etc.) y por impresión de ilustraciones a color. El pago por publicación se realiza en las oficinas de la Facultad de Ciencias Biológicas de la Universidad Nacional Mayor de San Marcos, o en coordinación con el Editor Jefe.
El autor principal podrá solicitar cuatro ejemplares de la revista en las oficinas de la Facultad de Ciencias Biológicas de la Universidad Nacional Mayor de San Marcos.
Comité EditorEmail: [email protected] postal: Leonardo Romero (Editor Jefe)Revista Peruana de BiologíaUNMSM-FCBApartado 11-0058Lima 11Perú

83 Variabilidad espacial y temporal de larvas de jurel Trachurus murphyi en el Perú entre 1966 – 2010 Spatial and temporal variability of Jack mackerel Trachurus murphyi larvae in Peru between 1966 – 2010 Patricia Ayón y Jonathan Correa
87 Biomasa acústica y distribución del jurel Trachurus murphyi en el Perú Acoustic biomass and distribution of Jack mackerel Trachurus murphyi in Peru Marceliano Segura y Aníbal Aliaga
Pesquería, manejo y conservación del jurel (Trachurus murPhyi) en el Perú
97 Análisis de la pesquería del jurel Trachurus murphyi en el Perú Analysis of the Jack mackerel Trachurus murphyi fishery in Peru Miguel Ñiquen, Marilú Bouchon, Danny Ulloa y Ana Medina
107 Medidas de conservación y ordenación pesquera del jurel Trachurus murphyi en el Perú Conservation and fishery management regulations of Jack mackerel Trachurus murphyi in Peru Jorge Zuzunaga

Revista PeRuana de Biología
Volumen 20 Septiembre, 2013 Número 1Rev. peru. biol. ISSN-L 1561-0837
Número especial: Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú
Special issue: Ecology, fisheries and conservation of Jack mackerel (Trachurus murphyi) in PeruCONTENIDO
PresenTación
3 El IMARPE y el número especial Ecología, pesquería y conservación del jurel (Trachurus murphyi) en el Perú The IMARPE and the special issue Ecology, fisheries and conservation of jack mackerel (Trachurus murphyi) in Peru Germán Vásquez Solís Talavera
4 Agradecimientos
inTroducción
5 El Jurel Trachurus murphy en el Perú The Jack mackerel Trachurus murphy in Peru Jorge Csirke
el medioambienTe del jurel (Trachurus murPhyi) en el Perú 9 El jurel Trachurus murphyi y las variables ambientales de macroescala The Jack mackerel Trachurus murphyi and the environmental macro-scale variables Marco Espino
21 Patrones de variabilidad ambiental en el mar peruano Environmental variability patterns in the Peruvian waters Roberto Flores, Marco Espino, Gertrudes Luque y Jorge Quispe
biología del jurel (Trachurus murPhyi) en el Perú
29 Aspectos reproductivos del jurel Trachurus murphyi Reproductive aspects of Jack mackerel Trachurus murphy Ángel Perea, Julio Mori, Betsy Buitrón y Javier Sánchez
35 Escala de madurez gonadal del jurel Trachurus murphyi Nichols 1920 Scale of gonad maturity stages of Jack mackerel Trachurus murphyi Nichols 1920 Javier Sánchez, Ángel Perea, Betsy Buitrón y Liz Romero
45 Edad y crecimiento del jurel Trachurus murphyi en el Perú Age and growth of Jack mackerel Trachurus murphyi in Peru Teobaldo Dioses
53 Validación del ritmo de formación de los anillos de crecimiento en otolitos del jurel Trachurus murphyi Nichols 1920 Validation of the formation rate of growth rings in otoliths of Jack mackerel Trachurus murphyi Nichols 1920 Carlos Goicochea, Jorge Mostacero, Patricia Moquillaza, Teobaldo Dioses, Yeny Topiño y Renato Guevara-Carrasco
61 Estimación del crecimiento del jurel Trachurus murphyi capturado en el Perú, a partir del análisis de las frecuencias por tallas Estimation of growth parameters of Jack mackerel Trachurus murphyi caught in Peru, from length frequency analysis Erich Díaz
ecología del jurel (Trachurus murPhyi) en el Perú
67 Patrones de distribución y abundancia del jurel Trachurus murphyi en el Perú Abundance and distribution patterns of Jack mackerel Trachurus murphyi in Peru Teobaldo Dioses
75 Ecología trófica del jurel Trachurus murphyi en el Perú entre 1977 – 2011 Trophic ecology of Jack mackerel Trachurus murphyi in Peru, 1977 – 2011 Ana Alegre, Pepe Espinoza y Marco Espino
(Continúa...)