
[SEPARATA DE LA EDICIÓN DE JUNIO-2006 (NO · importantes (por ejemplo con los tiburones, las...
49
0
Transcript of [SEPARATA DE LA EDICIÓN DE JUNIO-2006 (NO · importantes (por ejemplo con los tiburones, las...

0

1
diciembre 2009 - Nº 38 - ISSN 1409-2158
Revista semestral de la
Escuela de Ciencias Ambientales,
Universidad Nacional
Teléfono: 2277-3688 Fax: 2277-3289
Apartado postal: 86-3000
Heredia, Costa Rica
[email protected] www.ambientico.una.ac.cr
Consejo editor
Marielos Alfaro, Universidad Nacional
Gerardo Budowski, Universidad para la Paz Enrique Lahmann, UICN
Enrique Leff, UNAM (México)
Olman Segura, Universidad Nacional
Rodrigo Zeledón, Universidad Nacional
Director y editor
Eduardo Mora
Fotografía www.galeriaambientalista.una.ac.cr
Asistencia, administración y diagramación Rebeca Bolaños
Foto de portada Gregory Basco
C O N T E N I D O
Editorial
Estimación de poblaciones de jaguar (Panthera onca) en el
Área de Conservación Guanacaste, Costa Rica Ronit Amit, Luis Diego Alfaro y Eduardo Carrillo
Relación entre la presencia de jaguar (Panthera onca) y de
sus presas en el Área de Conservación Guanacaste, Costa
Rica Ronit Amit, Luis Diego Alfaro y Eduardo Carrillo
Riqueza, diversidad y estado de conservación de murciélagos
en el Corredor Biológico Chorotega Elías José Gordillo, Jeannette Úrdiales y Joel C. Sáenz
Condiciones socio-económicas en dos sectores del Corredor
Biológico Chorotega: Percepciones de la comunidad Yusselfy Alarcón, Guillaume Gama, Marcela Rodríguez y Joel C. Sáenz
Gremios de aves en agroecosistemas del Pacífico Central de
Costa Rica y su importancia para la conservación
Martha Lucía Enríquez, Joel C. Sáenz y Muhammad Ibrahim
Hábitat y población de mono tití (Cebidae: Saimiri oerstedii oerstedii) en la península de Osa, Costa Rica Daniela Solano y Grace Wong
Normas mínimas para la presentación de artículos a la re-
vista AMBIENTALES con vistas a su publicación
2
3
6
11
20
26
33
47
Ambientales, revista semestral de la Escuela de Cien-
cias Ambientales de la Universidad Nacional, fue creada
en 1980. Su objetivo es estimular y difundir la investiga-ción y la discusión científica en el campo de lo ambien-
tal. Publica resultados de investigación referentes a situaciones ambientales y, secundariamente, ensayos
teóricamente consistentes con algún enfoque científico
atinentes a problemas de ese mismo ámbito temático. Se dirige principalmente, a académicos, a estudiantes
de nivel superior y a funcionarios e integrantes de organizaciones gubernamentales y no gubernamentales
con competencias en lo ambiental.
Ambientales/ Escuela de Ciencias Ambienta-
les.— No. 38 (2009) – Heredia, C.R.
Semestral
1. Ecología Publicaciones periódicas. I. Universidad Nacional.
Escuela de Ciencias Ambientales.
634.9
C569C

2
Fauna Silvestre en Costa Rica hoy
n un espacio que representa el 0,03% de la superficie mundial, Costa Rica posee el 3,6% de la biodiversi-dad del planeta, colocándolo entre los 20 países más ricos en diversidad de especies; pero si se considera el número de éstas por km2, podría ser el país con mayor diversidad del planeta. Sin embargo, el proceso de
desarrollo ha sometido a mucha especies de fauna silvestre a un escenario de amenazas múltiples y complejas. A pesar de las innovadoras políticas en conservación de la biodiversidad que han puesto a Costa Rica entre los paí-ses más avanzados en esta materia en Latinoamérica, la tendencia actual es a un aumento de las especies de fau-na silvestre colocadas en alguna categoría de amenaza (véase: Obando, V. 2002. Biodiversidad en Costa Rica y,
también, Obando, V. 2008. Biodiversidad en cifras. Inbio. Costa Rica). De las 1.050 especies de fauna silvestre
(aves, mamíferos, anfibios y reptiles), el 12% de las aves poseen algún grado de amenaza (amenazadas y/o en peligro de extinción), con una especie extinta (no ha sido registrada por más de 25 años) y cuatro probablemente extintas. El 33% y el 16% de los mamíferos y reptiles, respectivamente, están en la misma situación. Los anfibios son probablemente el grupo más amenazado, con un 47% de sus especies; a lo que hay que sumar que los exper-tos consideran que ya se han extinguido unas 18 especies de ellos, aunque la lista oficial solo registra una. No está claro si esto se debe a que se dispone de mayor información, a una mejora en los procesos de consulta, o a un aumento real de las amenazas.
En los últimos años se han dado algunos avances en políticas de conservación en el país. Así, por ejemplo, se ha legislado a favor de la protección de la lapa verde, especie muy amenazada, protegiendo su hábitat y creando áreas silvestres para tal fin; y se ha hecho esfuerzos en la península de Osa para conservar mamíferos en peligro de extinción. También se ha legislado a favor de la conservación de tortugas, protegiendo sus áreas de anidación y regulando la pesca, que es una de los causas de mortalidad de ellas; de igual manera, leyes que regulan el apro-vechamiento del tiburón han sido publicadas. Sin embargo, a la par de estos logros hay retrocesos debidos a polí-ticas gubernamentales de desarrollo, sobre todo en el campo turístico, que están amenazando la fauna silvestre, como por ejemplo la autorización de desarrollos urbanísticos en playas de anidación de tortugas, en ecosistemas críticos (e.g. humedales), en bosques con remanentes importantes de fauna silvestre, y -casos extremos- autori-zando explotaciones mineras en ecosistemas con fauna amenazada. Esto es paradójico, porque más de la mitad de los turistas vienen al país a ver fauna silvestre.
Queda todavía mucho por hacer para garantizar la conservación de la fauna silvestre, por lo que la investiga-ción académica tiene un reto por delante. Tradicionalmente, las investigaciones se han realizado en áreas silves-tres protegidas haciendo avances significativos, y muchas han dado paso a decisiones políticas de conservación importantes (por ejemplo con los tiburones, las tortugas, las lapas verdes, etc.). Sin embargo, se ha olvidado que las áreas silvestres no son islas, y el solo hecho de conservarlas no garantiza la preservación de su fauna; o sea, es equivocado relegar a un segundo plano las áreas circundantes a las áreas protegidas, porque en el contexto de la dinámica de las poblaciones silvestres ellas son fundamentales para la sobrevivencia a largo plazo de las especies de un paisaje. Muchos estudios han demostrado que fuera de las áreas protegidas (agropaisajes) todavía existe una cantidad importante de fauna silvestre que debería ser incluida en políticas nacionales de conservación; caso contrario, perderemos especies, incluso dentro de las áreas silvestres protegidas. Además, diversos biólogos de la conservación han sugerido que muchas de la áreas protegidas del país no tienen el tamaño adecuado para mante-ner algunas poblaciones de fauna silvestre, siendo ésta, probablemente, una de las causas que mantienen la ten-dencia al aumento de las especies amenazadas en el país.
El escenario de amenazas múltiples al que se ve sometida la fauna silvestre en la actualidad trasciende en al-gunos casos las fronteras nacionales. A las amenazas de los procesos de deforestación y cambio de uso del suelo hoy se agrega la del cambio climático global, cuyos efectos se asocian ya a la extinción de algunas especies. El desafío que tenemos ante nosotros es complejo, urgente y requiere de información pertinente para la toma de decisiones que puedan garantizar a largo plazo el mantenimiento de la mayor parte de la fauna silvestre del país.
Esta edición de Ambientales, hecha conjuntamente con la dirección del Instituto para la Conservación y Mane-
jo de Vida Silvestre, de la Universidad Nacional, presenta resultados de varias investigaciones que muestran la importancia y la problemática de la fauna silvestre costarricense tanto en áreas silvestres protegidas como en zo-nas de amortiguamiento y agropaisajes.
E

3
Inicio – Siguiente
Amit, R., L. Alfaro y E. Carrillo. “Estimación de poblaciones de jaguar (Panthera onca) en el Área de Conservación Guanacaste, Costa Rica”, Ambientales, No. 38, diciembre 2009, Costa Rica. Págs. 3-5.
Estimación de poblaciones de jaguar (Panthera onca)
en el Área de Conservación Guanacaste, Costa Rica
RONIT AMIT, LUIS DIEGO ALFARO y EDUARDO CARRILLO
RESUMEN/ABSTRACT
Se estimó la población de jaguares del Área de Conservación Guanacaste (ACG), al noroeste de Costa Rica, mediante el
uso de cámaras trampa. Se aplicó el método de captura-recaptura para obtener la abundancia y se calculó la densidad según el área efectiva de muestreo. Para el Sector San Cristóbal el área efectiva de muestreo fue 164,81 km2. La abundan-
cia se estimó en 9,50 ± 7,7 jaguares y la densidad fue 0,057 ± 0,046 ind/km2. El área efectiva de muestreo para los parques nacionales Santa Rosa y Guanacaste fue 103,92 km2. La abundancia encontrada fue 3,00 ± 2,46 jaguares para una densi-
dad de 0,028 ± 0,023 ind/km2. Los resultados resaltan que aún no es posible estimar el número total de jaguares por en-contrarse valores tan diferentes entre los dos sectores muestreados. Con base en este estudio se rediseñó una Unidad de
Conservación del Jaguar (UCJ) propuesta por expertos para el noroeste de Costa Rica, ya que la cordillera de Guanacaste
probablemente presenta una alta densidad de jaguares.
We estimated the jaguar population in two sites in Guanacaste Conservation Area (GCA), northwestern Costa Rica, using camera traps.
We applied a capture-recapture method to obtain abundance and density was calculated for the effective sample area. For Sector San Cris-tobal the effective area was 164,81 km2, abundance was 9,50 ± 7,7 jaguars and density was 0,057 ± 0,046 ind/km2. For Santa Rosa and Guanacaste National Parks the effective area was 103,92 km2, abundance was 3,00 ± 2,46 jaguars for a density of 0,028 ± 0,023 in-
d/km2. For the GCA, our results suggest that it is not possible to estimate the total number of jaguars because of the different population densities we found in two close areas. After the results of this study, we redesign a Jaguar Conservation Unit (JCU) proposed for northwes-
tern Costa Rica, because of the probability that the Guanacaste Mountain Range presents a high jaguar density.
Palabras clave: jaguar, Área de Conservación Guanacaste, cámaras trampa, captura-recaptura, estimación poblacional. Key words: jaguar, Guanacaste Conservation Area, camera traps, capture-recapture, population estimation.
pesar de que el jaguar (Panthera onca) presen-
ta una amplia distribución en el continente americano, la pérdida y la fragmentación de
su hábitat son las principales amenazas a su subsis-tencia, sumado a la matanza en represalia por los ataques a animales domésticos. Los jaguares como depredadores tienen un papel significativo en el mantenimiento del ecosistema y si esta especie des-aparece el balance natural de las poblaciones de sus presas se verá afectado (Aranda 2002, Renata y Galvao 2002).
En los últimos años, la investigación sobre las poblaciones de jaguares se ha incrementado en va-rios países (Harmsen et al. 2009). En Costa Rica se
ha venido generando información localizada, prin-
cipalmente en la península de Osa, que muestra la urgencia de aumentar el conocimiento sobre el esta-do de conservación de este felino. El presente estu-dio presenta la estimación de abundancia y densidad de jaguar en dos sectores del Área de Conservación Guanacaste, al noroeste de Costa Rica.
Área de estudio El estudio se realizó en dos sectores del Área de
Conservación Guanacaste (ACG), el primero in-cluyó los parques nacionales Santa Rosa (PNSR; 10°51’55” N, 85°39’22” W) y Guanacaste (PNG; 10°55’ 01” N, 85°31’02” W), y el segundo el Sector San Cristóbal (SCS; 10°52’06” N, 85°22’34” W). El área combinada es de 76.300 ha con elevaciones desde el nivel del mar hasta los 1.300 msnm. Entre los hábitats presentes está el bosque lluvioso en las
A
Los autores son especialistas en conservación y manejo de vida silvestre y profesores e investigadores de la Universidad Nacional. [Fecha de recepción: enero, 2010. Fecha de aceptación: marzo, 2010.]

4
mayores elevaciones y el bosque seco en las zonas bajas, lo que resalta la heterogeneidad espacial de la disponibilidad de agua. La precipitación varía entre 900 y 4.000 mm anuales y la temperatura ambiental entre 16 y 38° C, con una estación seca y una lluvio-sa (Janzen 1986, Pacheco 1994).
El área se encuentra en un proceso de regenera-ción pasiva y está rodeada principalmente por fincas agrícolas y ganaderas. Existen algunos asentamien-tos humanos alrededor pero la población está dis-persa. El ACG alberga poblaciones de mamíferos que son presas importantes para el jaguar, como el chancho de monte (Tayassu pecari), el saíno (Pecari
tajacu), el tepezcuintle (Cuniculus paca), la guatusa
(Dasyprocta punctata), el armadillo (Dasyprocta puncta-
ta) y la danta (Tapirus bairdii), entre otros (Alfaro
2006, Amit 2006, Ramírez 2003).
Métodos En el SSC se muestreó de octubre de 2005 a ene-
ro de 2006 durante 43 días, mientras que en el PNSR-PNG se muestreó entre febrero y mayo de 2006 durante 70 días. Para registrar la presencia de jaguares se usó cámaras-trampa con sensores infra-rrojos activados por calor y movimiento (Stealth-CamTM, LLC, P.O.Box 211662, Bedford, TX 76095, U.S.A. y CamTrakker 1050 Industrial Drive, Watkinsville, Georgia, U.S.A.). Se aplicó el método de captura-recaptura (Otis et al. 1978) para estimar la
abundancia de jaguar mediante sus patrones indivi-duales de manchas (Silver et al. 2004, Karanth &
Nichols 1998). Se instaló seis estaciones de cámaras-trampa en
el PNSR-PNG y 15 en el SSC. Cada estación consis-tió en dos cámaras, una frente a otra; se colocó sobre
senderos existentes donde se halló previamente sig-nos de la presencia de jaguar o de sus presas (como excretas o huellas) y se geo-referenció mediante un receptor de GPS. La distancia entre estaciones fue de 2,5-3,6 km. La identificación individual de jagua-res se llevó a cabo comparando sus patrones únicos de manchas. Para lograr la independencia de los datos se consideró como una captura exitosa una fotografía por día por individuo; cada periodo de muestreo abarcó menos de tres meses para cumplir con el supuesto de población cerrada (Silver 2004).
La densidad poblacional se estimó con el método de captura-recaptura descrito por Williams et al.
(2002), siguiendo los estudios sobre tigres Panthera
tigris (Karanth & Nichols 1998). La densidad está
dada por la relación entre la abundancia estimada
y el área efectiva de muestreo Ä(Ẅ). Para
obtener el valor de en cada sitio de
estudio, se creó una matriz de capturas para su análisis mediante el programa Capture (Otis et al. 1978); éste ajusta los
datos al mejor modelo de estimación (Rexstad & Burmham 1991). El valor de Ä(Ẅ) se estimó usando el polígono míni-
mo compuesto (Worton 1989) generado desde la localización geográfica de las estaciones de cámaras-trampa, al que se suma un área de amortiguamiento (área buffer) calculada como la mitad de la dis-
tancia media máxima (1/2 DMM) reco-rrida por un individuo (Soisailo & Caval-canti 2006).
Resultados Para el SSC el área efectiva de mues-
treo fue 164,81 km2. La abundancia se estimó en 9,50 ± 7,7 jaguares y la densi-
dad fue 0,057 ± 0,046 individuos por km2. El esfuer-zo de muestreo fue de 645 días/trampa. Las foto-grafías correspondieron a cuatro jaguares adultos, tres hembras y uno de sexo no identificado. De siete foto-capturas, una correspondió a una hembra activa durante el día, mientras que el resto fueron capturas nocturnas.
El área efectiva de muestreo para el PNSR-PNG fue 103,92 km2. La abundancia encontrada fue 3,00 ± 2,46 jaguares para una densidad de 0,028 ± 0,023 individuos por km2. El esfuerzo de muestreo fue de 420 días/trampa. Se identificaron cuatro individuos, dos machos adultos y dos hembras adultas.
Discusión La densidad de jaguares encontrada en el SSC es
relativamente alta comparada con las encontradas
Jaguar Programa Jaguar, W.C. S.

5
en la cordillera de Talamanca (0,049 ind/km2 du-rante la estación seca y 0,033 ind/km2 en la estación lluviosa, Gonzalez-Maya et al. 2009), mientras que
es menor que la estimada para el Parque Nacional Corcovado al sur del país (0,07 ind/km2, Salom-Pérez et al. 2007). En el PNSR-PNG es similar a los
estimados de El Chaco Boliviano (0,04 ind/km2, Maffei et al. 2004).
La variación entre sectores, aun los separados por cortas distancias como el SSC y el PNSR-PNG (distancia), dificulta hacer una estimación para grandes extensiones territoriales pues depende de la heterogeneidad espacial. En el caso del ACG, los resultados resaltan que aún no es posible estimar el número total de jaguares por encontrarse valores tan diferentes entre los dos sectores muestreados. Aun-que las estimaciones del presente estudio son la línea base para comprender el estado de conservación de los jaguares en el ACG, se requiere un plan de se-guimiento a largo plazo que contemple la heteroge-neidad de hábitats.
La disponibilidad de agua podría explicar la me-nor densidad de jaguares en el PNSR-PNG que en el SSC dado que éste presenta condiciones menos es-tresantes para los mamíferos de la región. Por otro lado, las montañas probablemente estén desempe-ñando un papel como fuente de la población de jaguares porque aún conservan una cobertura bosco-sa y disponibilidad de presas, mientras que en las tierras bajas, que no ofrecen refugio y alimento óptimo, se da un efecto de sumidero poblacional.
Con base en este estudio se ha rediseñado la Unidad de Conservación del Jaguar (UCJ) propues-ta para el noroeste de Costa Rica por los expertos en el taller El Jaguar en el Nuevo Milenio (Sanderson et
al. 2002). Al considerar que la cadena de montañas
llamada cordillera de Guanacaste probablemente presenta una alta densidad de jaguares como se en-contró en el SSC, la nueva UCJ se extendió desde la costa del ACG hasta el Parque Nacional Volcán Tenorio que es parte del Área de Conservación Are-nal-Tempisque (ACAT) (Zeller 2007).
Referencias biblográficas Alfaro, L. D. 2006. Estado de la población de jaguar (Panthera onca) y sus
presas en el Área de Conservación Guanacaste, Costa Rica. Tesis de Ma-
estría. Universidad Nacional. Costa Rica. Amit, R. 2006. El jaguar (Panthera onca) en el sector San Cristóbal del Área de Conservación Guanacaste, Costa Rica: Densidad, abundancia de
presas y depredación de ganado. Tesis de Maestría. Universidad Nacio-
nal. Costa Rica. Aranda, M. “Importancia de los pecaríes para la conservación del jaguar en México”, en Medellín, R. et al. (eds.). 2002. El jaguar en el
nuevo milenio. Universidad Nacional Autónoma de México y Wildlife
Conservation Society. México D.F. Gonzalez-Maya, J. et al. 2009. Final Technical Report: Jaguar (Panthera onca) density estimation in Cordillera Talamanca, Costa Rica. Proyecto de Conservación del Área Talamanca, ProCAT.
Harmsen, B. et al. “Spatial and temporal interactions of two sympa-
tric cats in a neotropical forest: the jaguar (Panthera onca) and the
puma (Puma concolor)”, en Journal of Mammalogy 90(3), 2009.
Janzen, D. H. 1986. Parque Nacional Guanacaste, restauración ecológica
y cultural en el trópico. Mimeografiado. San José.
Karanth, K. y J. Nichols. “Estimation of tiger densities in India using photographic captures and recaptures”, en Ecology 79, 1998.
Maffei, L., E Cuéllar y A. Noss. “One thousand jaguars (Panthera
onca) in Bolivia’s Chaco? Camera trapping in the Kaa-Iya National
Park”, en J. Zool. Lond. 262, 2004.
Otis, D. L. et al. “Statistical inference from capture data on closed
populations”, en Wildlife Monographs 62, 1978.
Pacheco, C. 1994. Hábitos alimentarios y uso estacional de hábitat de la pava crestada (Penelope purpurascens) en el Bosque Tropical Seco, Parque
Nacional Santa Rosa, Costa Rica. Programa Regional de Manejo de
Vida Silvestre, Universidad Nacional. Costa Rica. Ramírez, S. 2003. Abundancia relativa y tipo de hábitats preferidos por el jaguar (Panthera onca) y el puma (Puma concolor) en el Parque Nacional
Santa Rosa, Guanacaste, Costa Rica. Programa Regional de Manejo de
Vida Silvestre, Universidad Nacional. Costa Rica. Renata, M. y F. Galvao. “El jaguar, el puma y el hombre en tres áreas protegidas del bosque atlántico costero de Paraná, Brasil”, en Medellín, R. et al. (eds.). 2002. El jaguar en el nuevo milenio. Universi-
dad Nacional Autónoma de México y Wildlife Conservation Socie-ty. México D.F. Rexstad, E. y K. Burnham. User’s guide for interactive program CAP-
TURE. Abundance estimation of closed animal populations. Fort Collins,
Colorado State University. U.S.A. Salom-Pérez, R. et al. “Critical condition of the jaguar Panthera
onca population in Corcovado National Park, Costa Rica”, en Oryx
41 (1), 2007. Sanderson, E. et al. “Planning to save a species: The jaguar as a
model”, en Conserv. Biol. 16, 2002.
Silver, S. et al. “The use of camera traps for estimating jaguar Pant-
hera onca abundance and density using capture/recapture analysis”,
en Oryx 38, 2004.
Silver, S. 2004. Estimando la abundancia de jaguares mediante trampas-
cámara. Wildlife Conservation Society. En: http://www.panthera.
org/documents/SilverJaguarCamera-TrappingProtocol_spanish.pdf. Soisailo, M. y S. Cavalcanti. “Estimating the density of jaguar popu-lation in the Brazilian Pantanal using camera-traps and capture-recapture sampling in combination with GPS radio-telemetry”, en Biological Conservation 129, 2006.
Williams, B., J. Nichols y M. Conroy. 2002. Analysis and Management
of Animal Populations. Academic Press.
Worton, B. J. “Kernel Methods for Estimating the Utilization Dis-tribution in Home-Range Studies”, en Ecology 70, 1989.
Zeller, K. 2007. Jaguars in the New Millennium Data Set Update: The State of the Jaguar in 2006. Report to the Wildlife Conservation Society’s
Jaguar Conservation Program. Wildlife Conservation Society. Takoma
Park, MD, U.S.A.
Agradecimientos
Este estudio fue financiado por el Servicio de Pesca y Vida Silvestre de Estados Unidos (USFWS), la Sociedad para la Conservación de la Vida Silvestre (WCS), el Cleveland Metroparks Zoo y el Fondo de Incentivos del Micit-Conici.

6
Inicio – Siguiente
Amit, R., L. Alfaro y E. Carrillo. “Relación entre la presencia del jaguar (Panthera onca) y de sus presas en el Área de Conservación Guanacastes, Costa Rica”, Ambientales, No. 38, diciembre 2009,
Costa Rica. Págs. 6-10.
Relación entre la presencia de jaguar (Panthera onca) y
de sus presas en el Área de Conservación Guanacaste,
Costa Rica
RONIT AMIT, LUIS DIEGO ALFARO y EDUARDO CARRILLO
RESUMEN/ABSTRACT
Mediante la búsqueda de rastros se estudió la relación entre la presencia del jaguar y la de mamíferos medianos y grandes
que son sus presas potenciales en el Área de Conservación Guanacaste. El área de estudio fue dividida en 34 cuadrículas
de 1 km2 donde se registró cada pista de rastros de mamíferos grandes (>12kg), mamíferos medianos (1,5 – 12 kg) y pre-sencia (1) – ausencia (0) para jaguar. Para cada categoría fue calculado un índice de abundancia relativa y un índice de ri-
queza en cada cuadrícula que fueron analizados mediante modelos lineales generalizados. Se detectaron 16 especies de mamíferos silvestres y se registró la presencia de jaguar en ocho ocasiones. Nuestros resultados indican que la presencia del
felino aumenta entre mayor sea la abundancia relativa para mamíferos grandes. Entre éstos se destacan la danta (Tapirus
bairdii) y el chancho de monte (Tayassu pecari) reportados en nuestro estudio y cuyas poblaciones están en peligro de des-
aparecer.
We studied the relationship between the presence of jaguar and that of medium and big sized mammals which are its potential prey. The study area was divided in 34 blocks of 1km2 where we record track series (signs of presence of an individual) of big mammals (>12kg), me-dium sized mammals (1.5 – 12 kg) and presence (1) – absence (0) of jaguar. We calculated a Relative Abundance Index (RAI) and a
richness index for each block, and we analyzed that with generalized linear models. We detected 16 species of wild mammals and recorded the presence of jaguar 8 times. Our results suggest that jaguar presence increases at the more relative abundance of big sized mammals.
Among those, the tapir (Tapirus bairdii) and the white-lipped peccaries (Tayassu pecari) are important and endangered as the jaguar it-self.
Palabras clave: jaguar, Área de Conservación Guanacaste, rastreo, índice de abundancia relativa, relación predador-presa. Key words: jaguar, Guanacaste Conservation Area, tracking, relative abundance index, prey-predator relationship.
l rastreo, una técnica económica y efectiva, pone en evidencia la importancia de la canti-dad de presas para un carnívoro en la cúspide
de la cadena alimenticia como lo es el jaguar (Pant-
hera onca). Un carnívoro mantiene en equilibrio po-
blaciones de presas y a su vez reduce el impacto de poblaciones de herbívoros sobre comunidades de plantas (Lindstrom et al. 1994). El equilibrio existen-
te entre la relación predador-presa se desarrolla me-diante ciclos estables (Krebs et al. 2001) que si se
rompen pueden originar diversos problemas ecológi-
cos como disminución de capacidad de carga, altas tasas de herbivoría y hasta conflictos con el ser humano por competencia de recursos, como ha su-cedido con la depredación de animales domésticos por jaguares (Azevedo y Murray 2007).
Algunos estudios sugieren clasificar las especies de carnívoros según el efecto sobre sus presas y vice-versa, siendo una de estas categorías la de carnívoro residente especialista (Hanski et al. 1991, Korpimaki
y Krebs 1996). El jaguar puede ser considerado de-ntro de esta categoría, ya que está fuertemente liga-do a sus presas, donde cambios en el tamaño de las poblaciones de ellas provocan una respuesta retar-dada en la población del jaguar. Estudiar poblacio-nes de jaguares y sus presas resulta costoso dadas
E
Los autores son especialistas en conservación y manejo de vida silvestre y profesores e investigadores de la Universidad Nacional. [Fecha de recepción: enero, 2010. Fecha de aceptación: marzo,
2010.]

7
estas características, de manera que técnicas confia-bles y financieramente viables, como la búsqueda sistemática de rastros de las especies, deben ser in-corporadas al estudio de estas poblaciones de mamí-feros.
Área de estudio El área de estudio se ubica entre los volcanes Ca-
cao y Rincón de la Vieja, que corresponden a los parques nacionales Guanacaste y Rincón de la Vieja (10°50.8’–10°57.9’ N, 85°21.5’–85°25.8’ W). La topografía es irregular con elevaciones desde los 400 hasta los 1.300 msnm. La precipitación reportada durante el periodo de estudio fue de 4.340 mm. Las formaciones vegetales corresponden a las zonas de vida de bosque húmedo tropical, bosque lluvioso montano bajo y bosque lluvioso premontano (según Holdridge 1967). Los bosques son primarios hacia el interior de los parques nacionales y en los alrededo-res se presenta un gradiente hacia bosques secunda-rios y áreas fragmentadas, encontrando zonas de charrales y pastizales.
Fincas privadas, dedicadas a ganadería (lechera principalmente), agricultura (predominan los cítricos Citrus spp.), palmito (Bactris gasipaes) y plátanos
(Mussa acuminata)) y un moderado turismo del tipo
agroecológico y rural, rodean las áreas protegidas. La población humana está dispersa y los pueblos
principales son Quebrada Grande de Liberia y Dos Ríos de Upala. La gente local tiene bajos ingresos y pocas oportunidades de trabajo; usualmente entran en las áreas protegidas para cazar y pastorear su
ganado ilegalmente.
Métodos El área de estudio fue dividida en 34 cuadrículas
de 1 km2. Entre los meses de agosto y septiembre se realizaron cuatro recorridos dentro de las cuadrícu-las utilizando senderos existentes y/o recientemente establecidos. En cada cuadrícula se realizó una búsqueda de rastros (huellas, escarbaderos, marcas, etc.) de mamíferos medianos y grandes, incluyendo las presas potenciales del jaguar y se registraron observaciones directas. Al encontrar un conjunto de huellas o rastros correspondientes a un individuo, definido como una pista (Aranda 2000), se tomó el registro de presencia de la especie. Para animales que se desplazan en grupos se registró una pista de presencia del grupo. Se caminó a una velocidad media de 1 km/h, con un equipo de 2-3 personas. Los rastros fueron identificados in situ con la ayuda de guías de rastros de mamíferos (Aranda 2000, Reid 1997), en caso de alguna duda se tomó mues-tras de yeso y fotografías digitales para posterior identificación con ayuda de expertos.
Jaguar Programa Jaguar, W.C. S.

8
Análisis Las pistas de rastros fueron clasificadas en mamí-
feros grandes (>12 kg), mamíferos medianos (1,5 – 12 kg) y presencia (1) – ausencia (0) para jaguar. Para cada categoría fue calculado un índice de abundancia relativa (IAR) en cada cuadricula. El IAR se calculó dividiendo la cantidad de pistas por categoría entre la distancia recorrida en kilómetro durante la colecta de datos. Además se calculó un índice de riqueza mediante la relación entre número de especies y la distancia recorrida en km. Se ajusta-ron modelos lineales generalizados para relacionar la presencia – ausencia de jaguar con el IAR de mamíferos grandes y menores, IAR para mamíferos en general y la riqueza de especies. Se utilizó el cri-terio de información de Akaike (CIA) para definir el modelo con mejor ajuste. Los análisis se realizaron mediante el programa R versión 2.10.0 (2009).
Resultados Durante agosto y septiembre de 2005 se recorrió
una distancia acumulada de 94,8 km, se detectaron 16 especies de mamíferos silvestres y se registró la presencia de jaguar en ocho ocasiones (cuadro 1). La presencia de jaguar se relacionó positivamente con el índice de abundancia de mamíferos grandes, medianos y para la combinación de ambos según el valor de CIA (figura 1), esto con relación al modelo nulo que supone la no relación entre presencia de jaguar y sus presas. Sin embargo, el índice de rique-za de presas no se relacionó con la presencia de ja-guar (cuadro 2).
Discusión El aumento o disminución en las poblaciones de
predadores se relaciona con la densidad de sus pre-sas (O’Donoghue et al. 1997). En el caso del jaguar,
nuestros datos indican que la presencia del felino aumenta cuanto mayor sea la abundancia relativa de mamíferos grandes. Entre éstos se destacan la danta y el chancho de monte reportados en nuestro estudio y cuyas poblaciones corren peligro de extinción, al igual que las del jaguar. Otros estudios sugieren la importancia de mamíferos de mediano y grande porte para mantener poblaciones de jaguares (Wec-kel et al. 2006, DeOliveira 2002), siendo el chancho
de monte una de sus principales presas (Carrillo et
al. 2009).
Según nuestros datos y lo reportado por Alfaro (2006), las poblaciones de jaguar y sus mamíferos presa tienen importante representación en el sector montañoso del ACG, con respecto a las partes bajas hacia el litoral del Pacífico. La complejidad de hábi-tats y la disponibilidad de agua en la región monta-ñosa permiten una mayor abundancia de recursos tanto de abrigo como de alimento para comunidades de herbívoros de mediano y grande porte que consti-tuye la dieta del jaguar.
En el Área de Conservación Guanacaste el área de estudio presenta cacería ilegal y pastoreo de ga-nado dentro del área protegida, mientras que en los alrededores se da la pérdida de bosques para cambio en el uso del suelo a ganadería y agricultura, lo cual pone en peligro la biodiversidad de esta área (Amit 2006).
Cuadro 1. Especies de mamíferos encontradas en agosto y septiembre de 2005 en el Área de Conservación Guanacaste.
Especies N° pistas IAR
Alouatta palliata 9 0,09
Agouti paca 32 0,34
Ateles geoffroyi 3 0,03
Cebus capuccinus 4 0,04
Conepatus semiastriatus 3 0,03
Dasyprocta punctata 231 2,44
Dasypus novemcinctus 63 0,66
Leopardus pardalis 9 0,09
Leopardus wiedii 2 0,02
Nasua narica 2 0,02
Odocoileus virginianus 11 0,12
Potos flavus 3 0,03
Tapirus bairdii 42 0,44
Tayassu pecari 1 0,01
Tayassu tajacu 44 0,46
Urocyon cinereoargenteus 1 0,01 Nota: IAR: índice de abundancia relativa en el área de estudio = n° de pistas/km recorrido

9
Cuadro 2. Parámetros para los modelos ajustados de índice de abundancia de mamíferos (IA general), índice de abundancia de mamíferos
grandes (IA .grandes), índice de abundancia de mamíferos medianos (IA medianos), índice de riqueza de mamíferos (IR presas) y presencia de jaguar (Panthera onca) en comparación con el modelo nulo. Área de Conservación Guanacaste. 2010.
Modelo CIAc Gl dCIAc Peso
IA.general 33,5 2 0 0,292 IA.grandes 33,6 2 0,1 0,274 IA.medianos 34 2 0,5 0,226 Modelo nulo 35,3 2 1,8 0,12 IR.presas 35,9 2 2,4 0,088
Nota: CIAc= criterio de información de Akaike corregido, gl= grados de libertad, dCIA= delta del CIA.
Figura 1. Relación entre presencia de jaguar (Panthera onca) e índice de abundancia de mamíferos grandes (valor de CIA = 34,0, valor de CIA
para el modelo nulo = 35,3, p = 0,0145, modelo lineal generalizado de tipo binomial ajustado). Área de Conservación Guanacaste. 2010.
Técnicas simples, económicas y de fácil aplicación, como uso de rastros e índices de abundancia, permi-ten el seguimiento efectivo de las especies dentro y fuera de nuestras áreas protegidas, sabiendo que ellas cuentan con pocos recursos financieros para este tipo de actividades. Algunas variables que podr-ían incorporarse a nuestros modelos son los datos climáticos y los tipos de cobertura vegetal, lo que mejoraría la relación explicada por el modelo. Otras variables que tienen efecto directo sobre poblaciones de fauna silvestre, como la tasa de cacería ilegal, presentan mayor complejidad de medición, mas la cooperación privada unida a iniciativas de investiga-ción universitaria ayuda a suplir estos vacios de información.
Se recomienda la aplicación de estas herramien-tas de muestreo con repetidas mediciones a diferen-tes escalas espaciales y temporales, ya que nos podr-ía conducir a describir patrones causados por impac-tos antropogénicos en nuestras áreas protegidas. Estos datos indicarían zonas críticas para priorizar
acciones de manejo de los tipos compensatorias y preventivas.
Referencias bibliográficas Alfaro, L. D. 2006. Ocupación y patrones de actividad de especies presa potenciales para el Jaguar (Panthera onca) en el Área de Conservación
Guanacaste, Costa Rica. Tesis de Maestría. Universidad Nacional.
Costa Rica. Amit, R. 2006. Estado de manejo del Sector San Cristóbal, Área de Conser-
vación Guanacaste. Informe Técnico. Icomvis-Una. Costa Rica.
Aranda, M. 2000. Huellas y otros rastros de los mamíferos grandes y
medianos de México. Instituto de Ecología, A.C. México.
Azevedo, F. y D. Murray. “Evaluation of potential factors predispo-sing livestock to predation by jaguars”, en Journal of Wildlife Manage-
ment 71, 2007.
Carrillo, E., T. Fuller y J. Sáenz. “Jaguar (Panthera onca) hunting
activity: effects of prey distribution and availability”, en Journal of
Tropical Ecology 25, 2009.
De Oliveira, T. “Ecología comparativa de la alimentación del jaguar y del puma en el geotrópico”, en Medellín, R. et al. (eds.). 2002. El
jaguar en el nuevo milenio. Universidad Nacional Autónoma de Méxi-
co y Wildlife Conservation Society. México D.F. Hanski, I., H. Henttonen y L. Korpimaki. “Small-rodent dynamics and predation”, en Journal of Ecology 60, 1991.
Holdridge, L. 1967. Life Zone Ecology. Tropical Science Center: S.L.

10
Korpimaki, E. y C. J. Krebs. “Predation and population cycles of small mammals”, en BioScience 46, 1996.
Krebs, C. J. et al. “What drives the 10-year Cycle of Snowshoe
hares?”, en BioScience 51, 2001.
Lindstrom, E. R. et al. “Disease reveals the predator: Sarcoptic
Mange, red fox predation, and prey populations”, en Ecology 75,
1994. O'Donoghue, M. et al. “Numerical responses of coyotes and lynx to
the snowshoe hare cycle”, en Oikos 80, 1997.
Reid, F. A. 1997. A field guide to the mammals of Central America and
Southeast Mexico. Universidad de Oxford. Nueva York.
Weckel, M., W. Giuliano y S. Silver. “Jaguar (Panthera onca) feeding
ecology: distribution of predator and prey through time and space”, en Journal of Zoology 270, 2006.
Agradecimientos Este estudio fue financiado por el Servicio de Pesca y Vida Silvestre de Estados Unidos (USFWS), la Sociedad para la Conservación de la Vida Silvestre (WCS), el Cleveland Metroparks Zoo y el Fondo de Incentivos del MICIP-Conicit.
Jaguar Programa Jaguar, W.C. S.

11
Inicio – Siguiente
Gordillo, E., J. Úrdiales y J. C.. Sáenz. “Riqueza, diversidad y estado de conservación de murciélagos en el Corredor Biológico Chorotega”, Ambientales, No. 38, diciembre 2009, Costa Rica. Págs. 11-19.
Riqueza, diversidad y estado de conservación de
murciélagos en el Corredor Biológico Chorotega
ELÍAS JOSÉ GORDILLO, JEANNETTE ÚRDIALES y JOEL C. SÁENZ
RESUMEN/ABSTRACT
La fragmentación de la cobertura de bosque primario ha ocasionado que muchas especies y grupos taxonómicos se vean
afectados por la pérdida de conectividad, el efecto de borde y la disminución de recursos. Por tal razón, han surgido los co-rredores biológicos como una estrategia de conservación. Considerando la necesidad de evaluar la viabilidad tanto social
como biológica del Corredor Biológico Chorotega (sector Diriá-Ostional), seleccionamos el grupo de murciélagos como un
elemento de importancia para la evaluación biológica de dicho corredor. En el presente estudio realizamos una evaluación rápida de la riqueza y diversidad de murciélagos a lo largo del Corredor con el objetivo de determinar la viabilidad de él
para este grupo así como para obtener información base que permita la toma de decisiones. Durante el presente estudio re-gistramos 25 especies de murciélagos por medio de un esfuerzo de muestreo de 54 horas red en cuatro puntos a lo largo del
Corredor. La mayoría de las especies registradas corresponden a especies generalistas pero de gran importancia para la re-generación de los bosques por su amplia capacidad de dispersión de semillas.
The fragmentation of primary forest cover has afected many especies and taxonomic groups due the lost of conectivity, the border effect and the lack of resources. For that reason biological corridors have been created as a conservation estrategy in many different countries. Consi-
dering the need for evaluating the social and biological viability of the Chorotega Biological Corridor (Diriá-Ostional), we selected the bat assembly as an important element in the biological evaluation. For that reason, in the present study we evaluated the diversity and assem-blage of the bat comunity within the biological corridor. The main goal of the study was to evaluate the biological viability of the corridor
for the bat assembly and to obtain information that will allow in the future the decition making in the area. During the presented study we registred 25 especies with a sampling effort of 54 hrs/net in 4 sampling points along the corridor. The majority of the especies registred
are considered to be generalistic but important for the regeneration of the forest within the corridor given their wide seed dispersal capabili-ty.
Palabras claves: murciélagos, diversidad, corredor biológico, Parque Nacional Diriá. Key words: bat, diversity, biological corridor, Diriá National Park.
a fragmentación de la cobertura de bosque primario ha ocasionado que muchas especies y grupos taxonómicos se vean afectados por
la pérdida de conectividad, el efecto de borde y la disminución de recursos (Harvey et al. 2006, Hermes
2007), lo que ha ocasionado que en los últimos años muchas especies hayan desaparecido local o global-mente. Por esta razón, es importante conocer la biodiversidad presente dentro y fuera de las áreas protegidas, para tomar mejores decisiones de mane-jo y conservación (Gascon et al. 1999, Newell 1999,
Baguette et al. 2000).
Ante los severos efectos de la fragmentación sur-gen los corredores biológicos como una estrategia de conservación, con el objetivo de buscar la conectivi-dad estructural y funcional de los fragmentos de cobertura vegetal a lo largo de matrices inhóspitas para las especies (Bennett 2004). Sin embargo, mu-chos de estos corredores han sido resultado de agen-das políticas, alejadas de los verdaderos objetivos de la conservación (Andelman y Willig 2002). En otros casos se han creado a partir de datos de consulta de expertos apoyados en sistemas de información ge-ográfica, careciendo de datos que indiquen si en realidad esos corredores son viables desde el punto de vista biológico, lo que hace necesario contar con
L
Los autores, especialistas en manejo y conservación de vida silves-tre, son investigadores del Instituto de Conservación y Manejo de Vida Silvestre de la Universidad Nacional. [Fecha de recepción: enero, 2010. Fecha de aceptación: marzo, 2010.]

12
herramientas que permitan evaluar la viabilidad e impacto que los corredores biológicos presentan sobre la biodiversidad presente en una región.
Dado que la fragmentación del hábitat tiene efec-tos desiguales sobre el ensamble de especies presen-tes en una comunidad, su evaluación puede ser un indicador de las condiciones del hábitat (Hermes 2007). Aspectos como la tolerancia de las especies a las perturbaciones, la habilidad de ellas para cruzar a través de áreas abiertas permitiendo la coloniza-ción de nuevos fragmentos, y las interacciones de-ntro de la matriz formada por la modificación del hábitat, son claves en el impacto que la fragmenta-ción tiene sobre las especies existentes en una región (Bennett 2004).
Se ha considerado que la evaluación del ensam-ble de murciélagos en un ecosistema puede ser de importancia como indicador del estado general del hábitat, dadas las características propias del grupo. Aspectos como la alta sensibilidad que muestran a la fragmentación y el impacto que ésta tiene sobre los procesos ecológicos que los murciélagos realizan, y que a su vez comprometen la dinámica y regenera-ción de los bosques, hacen que utilizarlos como indicadores sea de gran relevancia (Schulze et al.
2000, Bernard y Fenton 2002, Aguirre et al. 2003,
Gorresen y Willig 2004). Asimismo, la gran varie-dad de roles tróficos que desempeñan en los ecosis-temas, su diversidad, su abundancia, su movilidad, su interacción con un amplio espectro de organis-mos y, en el caso de algunas especies, su fácil captu-ra (Bonaccorso y Humphrey 1984, Fleming 1988, Findley 1993, Altringham 1996), son aspectos que permiten que la evaluación de este grupo sea de utilidad para determinar la viabilidad de los corredo-res biológicos desde el punto de vista biológico.
Considerando la necesidad de evaluar la viabili-dad tanto social como biológica del Corredor Bio-lógico Chorotega (Diriá-Ostional), se seleccionó el grupo de murciélagos como un elemento de impor-tancia para la evaluación biológica del corredor. Por tal razón, en el presente estudio realizamos una evaluación rápida de la riqueza y diversidad de mur-ciélagos a lo largo del Corredor con el objetivo de determinar la viabilidad suya para este grupo, así como para obtener información base que permita la toma de decisiones en el futuro.
Área de estudio El corredor biológico Diriá se ubica al suroeste
de la provincia de Guanacaste y se extiende hasta el Parque Marino Las Baulas, pasando por las áreas protegidas de Parque Nacional Diriá, Cerro La Palma, Cerro de Jesús, Fila Reyes, Cerro La Cruz y Humedales de Corral de la Piedra y Mata Redonda (figura 1).
Las principales asociaciones vegetales que están presentes en el Corredor son bosques semi-deciduos, bosques deciduos de bajura, semi-deciduos de baju-ra, bosque siempre-verde, pastizales arbolados y manglares. Con respecto a la flora y la fauna del Corredor, de acuerdo con información de la Organi-zación de Estudios Tropicales, de Parques Naciona-les y de Fundación Neotrópica, la diversidad del área está compuesta por 26 especies de anfibios, 43 de reptiles, 306 de aves, 109 de mamíferos (61 de murciélagos), 139 de flora y 144 de plantas acuáti-cas.
Debido a la extensión del Corredor y el tiempo disponible para evaluar la diversidad del área, en este estudio únicamente se evaluó el sector Chorote-ga, el cual se extiende desde el Parque Nacional Diriá hasta el Refugio de Vida Silvestre Ostional, abarcando las comunidades Juan Díaz, Cola de Gallo, Arado, Vista al Mar, La Esperanza, El Pro-greso, Jazminal, Ostional, El Rayo y Playa Pelada. Por razones de accesibilidad a los sitios de muestreo, la evaluación de la diversidad de este Corredor se centró en cuatro localidades del sector Chorotega (Parque Nacional Diriá, Vista al Mar, Jazminal y El Rayo), procurando incluir el área protegida, las áre-as que limitan con el Parque y las partes media baja y baja del Corredor. En este estudio no fue posible muestrear el sector medio del Corredor por razones de accesibilidad.
La vegetación presente en los fragmentos mues-treados dentro de las comunidades constituye bos-ques maduros y en regeneración conectados por una matriz de potreros arbolados y remanentes boscosos que han sido dejados por los pobladores debido a las fuertes pendientes que tienen. La altura promedio de los sitios de muestreo es de 120 m.s.n.m en el Par-que Nacional Diriá, 960 m.s.n.m. en Vista al Mar, 200 m.s.n.m. en la parte media baja (Jazminal) y 23 en El Rayo.
Metodología
Selección de sitios de muestreo Seleccionamos cuatro puntos de muestreo a lo
largo del Corredor Biológico para lo cual conside-ramos la altura promedio de sitio y la accesibilidad a él. El primero de ellos ubicado dentro del Parque Nacional Diriá (PND) y los tres restantes fuera del Parque, en las localidades Vista al Mar, Jazminal y El Rayo. Los puntos de muestro se ubicaron sobre parches de bosque o charrales maduros. Debido a que los cuerpos de agua son utilizados por los ani-males para trasladarse de un sitio a otro, le dimos preferencia a aquellos parches asociados a ríos o arroyos.

13
Captura de murciélagos Para la captura de los murciélagos utilizamos
cuatro redes de niebla de 12 m de largo x 2,5 m de ancho. Colocamos las redes dentro de fragmentos de bosque, en sitios potenciales para la captura de mur-ciélagos (árboles con flores, frutos o túneles y cerca de cuerpos de agua). Las redes estuvieron abiertas seis horas (de 17:30 a 23:30) durante dos días, con excepción de la localidad Jazminal, donde mues-treamos tres días consecutivos. Identificamos las especies con la ayuda de la guía de campo de Me-dellín (1997) y LaVal y Rodríguez (2002).
Análisis de datos
Diversidad alfa Estimamos la diversidad alfa de cada sitio por
medio del índice de Brillouin, el cual es recomenda-do para datos que no son tomados con un diseño de muestreo riguroso. Este índice expresa la uniformi-dad de los valores de importancia a través de todas las especies de la muestra, midiendo de esta forma la equidad de las especies en una comunidad (Magu-rran 1988). Para ello utilizamos el programa Espe-cies Diversity and Richness v.4.0.
Comparación de riqueza y diversidad Para comparar la riqueza de especies entre sitios
de muestreo utilizamos la prueba de rangos de
Kruskal-Wallis, la cual se basa en la comparación de las medianas de cada muestra y en la que se asume como hipótesis nula que la riqueza de los diferentes sitios de muestreo es igual (Sokal y Rohlf 1991). Para ello utilizamos el programa R v. 2.0 8 (R Deve-lopment Core Team 2008).
Para comparar la diversidad de especies presen-tes en los cuatro puntos de muestreo del Corredor Biológico, contrastamos el índice de diversidad de cada sitio mediante una prueba de Randomization con un nivel de confianza del 5%, realizada con el
programa Species Diversity and Richness v.4.0.
Estimación de la riqueza esperada Para conocer cuál es el número de especies espe-
radas para cada sitio de muestreo de acuerdo al es-fuerzo realizado y la riqueza observada, utilizamos curvas de rarefacción a través del número de indivi-duos capturados (Moreno 2001). Las curvas de rare-facción fueron calculadas por medio del programa Species Diversity and Richness v.4.0.
Descripción del ensamble de murciélagos Para describir el ensamble de las especies de
murciélagos registrados en el Corredor, identifica-mos la familia taxonómica, el gremio trófico (in-sectívoro; frugívoro; omnívoro; nectívoro, hemató-fago), el estado de conservación y la categoría de
Figura 1.

14
vulnerabilidad para Costa Rica (amenazado, peligro de extinción [Minae 2007], y alta, moderada y baja respectivamente [Daily et al. 2003]) a la cual perte-
necen los individuos capturados.
Resultados
Riqueza específica Con un esfuerzo de muestreo de 54 horas red pa-
ra todo el Corredor registramos 152 individuos de 25 especies de murciélagos, pertenecientes a cinco fami-lias, siendo la familia Phillostomidae la más abun-dante, con 21 especies. Las especies más abundantes fueron Artibeus jamaicensis, Desmodus rotundus, Nocti-
lio leporinus, Carollia brevicauda, Carollia perpicillata,
Sturnira lillium (cuadro 1).
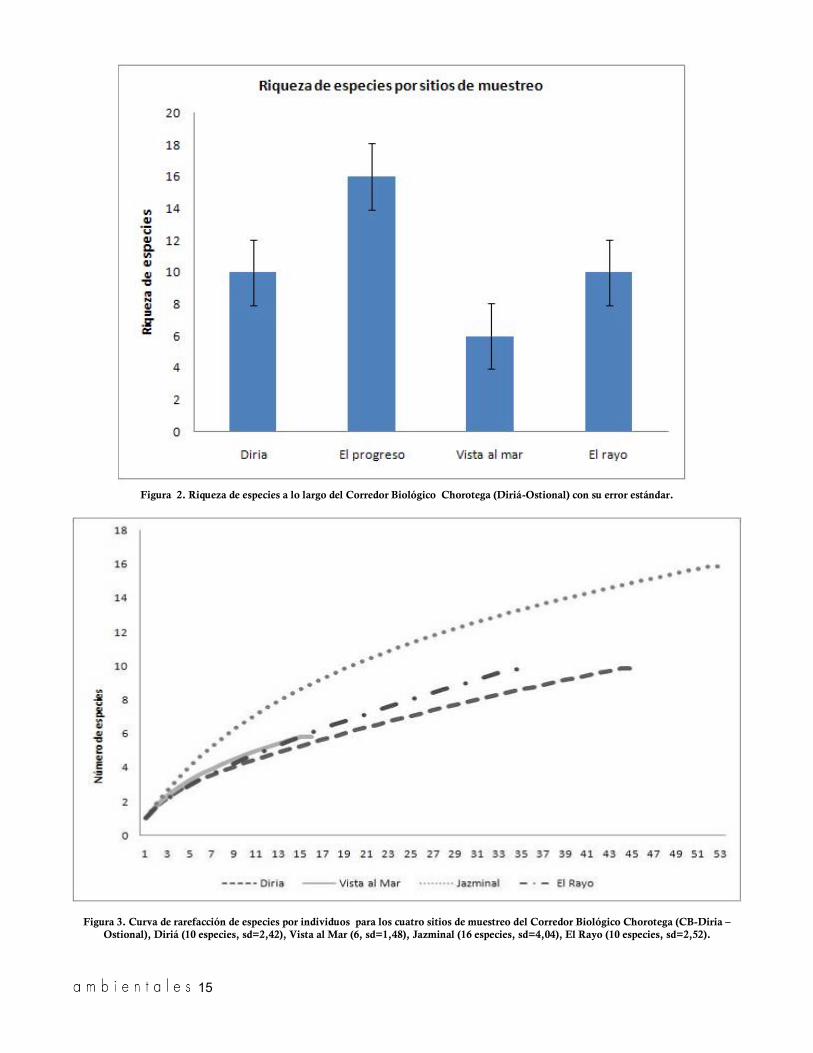
El sitio con mayor riqueza específica fue Jazmi-nal, con 17 especies, y el sitio con menor riqueza fue Vista al Mar, con seis (figura 2). Al comparar la diversidad de especies entre sitios mediante la prue-ba de rangos de Kruskal-Wallis no se encontraron diferencias significativas entre sitios (H=2,98 p=0,39). Con respecto al estado de conservación, únicamente el vampiro espectrum es considerado una
especie con amenaza de extinción, de acuerdo a los datos reportados por Minae 2007.
Riqueza estimada El análisis de rarefacción para los sitios de mues-
treo estimó el mismo número de especies registrado (figura 3). Sin embargo, al comparar el conjunto de valores estimados mediante un gráfico de medias con un intervalo de confianza del 95%, encontramos
que los sitios Jazminal y Vista al Mar difieren en el número de especies estimadas por las curvas de rare-facción (figura 4).
Diversidad alfa De acuerdo al índice de diversidad de Brillouin,
el sitio con mayor diversidad de especies es Jazmi-nal, con un índice de 2.024 (IC=1,7-2,1), seguido por Diriá (1,31 IC=0,95-1,5), El Rayo (1,241, IC=0,79-1,48) y Vista al Mar (1.142, IC=0,70-1,28), que reporta la menor diversidad de especies. Al comparar los índices de diversidad mediante una prueba de ramdomización encontramos que la di-versidad encontrada en el Parque Nacional Diriá no difiere de los sitios muestreados fuera del Parque, con excepción de Jazminal, donde la diversidad es mayor a la registrada en los sitios restantes.
Ensamble de murciélagos Respecto del ensamble de murciélagos de las 25
especies registradas, el gremio mejor representado es el frugívoro con un 64% del total de individuos, seguido del insectívoro con un 24%, y con menor riqueza los gremios nectívoro, hematófago y piscí-voro con un 4%. Es importante mencionar que del total de individuos capturados el 57% (87 indivi-duos) fueron machos. En cuanto a las especies indi-cadoras únicamente el Vampiro espectrum y el Micro-
nycteris sylvestris son catalogados como especies con
requerimientos específicos de hábitat, sin embargo este último en algunos estudios se ha encontrado en ambientes perturbados.
Murciélago fantasma (Ectophyla Alba) Juan José Pucci
15
Figura 2. Riqueza de especies a lo largo del Corredor Biológico Chorotega (Diriá-Ostional) con su error estándar.
Figura 3. Curva de rarefacción de especies por individuos para los cuatro sitios de muestreo del Corredor Biológico Chorotega (CB-Diria –
Ostional), Diriá (10 especies, sd=2,42), Vista al Mar (6, sd=1,48), Jazminal (16 especies, sd=4,04), El Rayo (10 especies, sd=2,52).

16
Aunque el grupo de mamíferos terrestres no fue el objetivo de esta investigación, durante los recorri-dos realizados en los sitios de muestreo registramos 11 especies de mamíferos terrestres de forma opor-tunista. De éstos, únicamente el mono congo y el cacomixtle son catalogados por el Minae y Cites con algún grado de amenaza (cuadro 2).
Discusión La riqueza de especies registradas en el Corredor
Biológico Chorotega (Diriá-Ostional) fue de 25 es-pecies de murciélagos y 11 especies de mamíferos terrestres. Esto representa el 41% y 20%, respecti-vamente, de las especies reportadas para dicho Co-rredor por otros autores (Maldonado 1995). Sin embargo, esta riqueza con un mayor esfuerzo de muestreo podría incrementarse.
Es importante mencionar que el bajo número de mamíferos terrestres reportados para el Corredor se debe a que no realizamos un muestreo sistematizado para este grupo, sin embargo se registraron especies como el armadillo y el venado, las cuales son de importancia socioeconómica por ser utilizadas como fuentes de alimento a lo largo de su rango de distri-bución geográfica (Ojasti 2002), y, en el caso del venado, es altamente cazado en la zona (Rodríguez et al. 2008), lo que demuestra la importancia del
PND como refugio para estas especies. Al comparar la riqueza y los índices de diversi-
dad obtenidos para el Corredor, encontramos que las mayores diversidad y riqueza se registraron fuera del Parque, lo cual es contrario a lo esperado. Esto
puede deberse a que el Parque es un área en regene-ración sujeta a una fuerte presión antropogénica. Por otro lado, el muestreo dentro del Parque fue menor, y durante noches claras, lo que pudo afectar la captura dentro del sitio (Weller y Lee 2007, Lang et al. 2006).
Las pruebas de randomización del índice de di-versidad de Brillouin muestran que únicamente el sitio Jazminal tiene una diversidad diferente con respecto al Parque y a los otros sitios. La razón por la cual Jazminal registró una mayor diversidad pro-bablemente sea el mayor esfuerzo de muestreo des-plegado en este sitio y que ésa es zona de poca in-fluencia antropogénica; sin embargo, esta última inferencia carece de datos que la respalden.
La abundancia y la diversidad de especies regis-tradas muestran que el ensamblaje de murciélagos está fuertemente dominado por algunas especies
como Artibeus jamaicensis, Desmodus rotundus, Noctilio
leporinus, Carollia brevicauda, Carollia perpicillata y
Sturnira lillium, las cuales reflejan la perturbación
antropogénica que existe a lo largo del Corrredor (Medellín et al. 2000), siendo esto concordante con
otros estudios en ambientes fragmentados (Estrada et al. 1993, Moreno y Halfter 2000, Montiel y Estra-
da 2006). Aunque el ensamblaje de especies en el Corredor
esté dominado por algunas especies, la presencia de las de los géneros Carollia, Artibeus y Sturnira son muy importantes para el Corredor por tratarse de especies que favorecen la regeneración natural de los bosques, al ser buenas dispersoras de semillas y al
Figura 4. Comparación de media de la riqueza estimada por las curvas de rarefacción, para el Corredor Biológico Diriá,
sector Chorotega (CB-Diriá – Ostional).

17
Cuadro 1. Murciélagos registrados en el Corredor Biológico Diriá sector Chorotega (Diriá-Ostional).
Familia Especie Gremio Diriá Vista al Mar Jazminal El Rayo Ind.
Phyllostomidae Artibeus intermedius Frugívoro 5 5
Phyllostomidae Artibeus jamaicensis Frugívoro 20 7 5 4 36
Phyllostomidae Artibeus lituratus Frugívoro 1 1
Phyllostomidae Carollia brevicauda Frugívoro 2 9 1 12
Phyllostomidae Carollia perpicillata Frugívoro 1 4 1 4 10
Phyllostomidae Carollia sp Frugívoro 2 2
Phyllostomidae Centurio senex Frugívoro 1 1
Phyllostomidae Chiroderma salvini Frugívoro 1 1
Phyllostomidae Chiroderma villosusm Frugívoro 1 1 2
Phyllostomidae Choeromiscus godmani Nectívoro 1 3 4
Phyllostomidae Dermanura phaeotis Frugívoro 3 1 4
Phyllostomidae Dermanura watsoni Frugívoro 2 2
Phyllostomidae Desmodus rotundus Hematófago 14 13 27
Molossidae Eumops glaucinus Insectívoro 2 2
Phyllostomidae Micronycteris sylvestris Insectívoro 1 1
Vespertilionidae Myotis keasy Insectívoro 1 1 2
Noctilionidae Noctilio leporinus Piscivoro 20 20
Phyllostomidae Phyllostomus discolor Frugívoro 1 1
Phyllostomidae Platyrrhinus helleri Frugívoro 1 1 2
Emballonuridae Rhychonycteris naso Insectívoro 1 1
Emballonuridae Saccopteryx bileneata Insectívoro 1 1
Phyllostomidae Sturnira lillium Frugívoro 3 2 4 9
Phyllostomidae Sturnira ludovici Frugívoro 1 1
Phyllostomidae Tonatia brasilensis Insectívoro 1 1
Phyllostomidae Uroderma bilobatum Frugívoro 1 1 2
Phyllostomidae Vampyrum spectrum Carnívoro 1 1
Cuadro 2. Mamíferos terrestres registrados en el Corredor Biológico Chorotega (Diriá-Ostional).
Especie Nombre común Minae Cites
Peromiscus sp Ratón
Spilogale poutorius Zorrillo
Didelphis virginiana Tlacuache o zorro pelón
Potos favus Martilla
procyon lotor Mapache
Dasypus novemcinctus Armadillo
Alouatta palliatta Mono congo PE I
Philander opossum Tlacuache cuatro ojos
Caluromys derbianus Zorro platanero o tlacuachillo dorado
Odocoileus virginianus Venado cola blanca
Bassariscus sumichrasti Cacomixtle AE III PE=peligro de extinción, AE=con amenaza de extinción.

18
aumentar su éxito de germinación (Janzen 1983, Schulze et al. 2000,
López y Vaughan 2004). Este rol ecológico resulta indispensable para mantener la conexión entre las áreas protegidas, al contribuir con la am-pliación del rango de dispersión de semillas y al incremento en la po-blación de plantas (Fleming 1986). En este sentido, esta función tam-bién es complementada por otros géneros presentes en el Corredor como son Platyrrihnus, Dermanuda, Uroderma y Chiroderma, los cuales son buenos dispersores de pequeñas semillas (Emmons 1997).
Con respecto a las especies indi-cadoras reportadas por la literatura y que fueron registradas dentro del Corredor, únicamente el Vampiro
espectrum es catalogado como una
especie con algún grado de amenaza y con restricción a hábitats poco alterados y de gran extensión (LaVal y Rodriguez 2002, Emmons 1997). La captura de esta especie contrasta con la ausencia de este murciélago fuera de ambientes perturbados (Schulze et al. 2000), lo cual es indi-
cador de que el PND tiene capacidad de albergar especies con requeri-mientos estrictos de hábitat conser-vados. Por otro lado, la rareza de este murciélago y la presencia del Chrotopterus auritus, capturado por
Pineda (2003) dentro del PND, su-gieren que estas especies pueden persistir en algunas partes del Corre-dor, resaltando de esta manera la importancia de éste para la conser-vación de aquéllas, debido a que por sus requerimientos de hábitat son altamente sensibles a la fragmenta-ción (Estrada y Coates-Estrada 2002).
Aunque la mayoría de las especies registradas a lo largo del Corredor son generalistas, destaca el importante número de frugívoras, las cuales son importantes para el Corredor dado el rol que desem-peñan en la regeneración de los bosques a través de la dispersión de semillas (López y Vaughan 2004, Schulze et al. 2000). Este rol ecológico resulta indis-
pensable para mantener la conexión entre las áreas protegidas, al contribuir a la ampliación del rango de dispersión de semillas y al incremento en la po-blación de plantas (Fleming 1986). En este sentido, de acuerdo a las especies registradas en el presente
estudio, la dispersión de semillas grandes está siendo llevada a cabo por el género Artibeus sp., y las semi-
llas pequeñas están siendo dispersadas por los géne-ros Carollia, Platyrrihnus, Dermanuda, Uroderma y Chiroderma (Emmons 1997).
Referencias bibliográficas Aguirre, L. F. et al. “Consistency and variation in the bat assemblage
inhabiting two forest islands within a neotropical savanna in Boli-via”, en Journal of Tropical Ecology, Julio-2003.
Andelman, S. J. y M. R. Willig. “Alternative Configurations of Conservation Reserves for Paraguayan Bats: Considerations of Spatial Scale”, en Conservation Biology, Octubre-2003.
Altringham, J. D. 1996. Bats – Biology and Behaviour. Oxford Univer-
sity Press. Oxford.
Murciélago Gregory Basco

19
Baguette, M., S. Petit y F. Queva. “Population spatial structure and migration of three butterfly species within the same habitat network: consequences for conservation”, en Journal of Applied Ecology, Fe-
brero-2000. Bennett, A. F. 2004. Enlazando el Paisaje: El papel de los corredores y la
conectividad en la conservación de la vida silvestre. UICN. San José.
Bernard, E. y M. B. Fenton. “Species diversity of bats (Mammalia: Chiroptera) in forest fragments, primary forests, and savannas in Central Amazonia, Brazil”, en Canadian Journal of Zoology, Junio-
2002. Bonaccorso, F. J. y S. R. Humphrey. “Fruit bat niche dynamics: their role in maintaining tropical forest diversity”, en Chadwick, A. C. y S. L. Sutton (eds.). Tropical Rain Forests: the Leeds Symposium.
Leeds Philosophical y Literary Society. Leeds. Brosset, A. et al. “Bat communities and deforestation in French
Guiana”, en Canadian Journal of Zoology, Noviembre-1996.
Daily, G. et al. “Countryside biogeography of neotropical mammals:
conservation oportunities in agricultural landscapes of Costa Rica”, en Conservation biology, 2003.
Emmons, L. H. 1997. Neotropical rainforest mammals, a field guide. The
University of Chicago Press. Chicago - London. Estrada, A., R. Coates-Estrada y D. Meritt. “Bat species richness and abundance in tropical rain forest fragments and in agricultural hábi-
tat at Los Tuxtlas, México”, en Ecography, Octubre-1993.
Estrada, A. y R. Coates-Estrada. 2002. “Bats in continuous forest, forest fragments and in anagricultural mosaic habitat-island at Los Tuxtlas, México”, en Biological Conservation, Febrero-2002.
Fleming, T. H. 1986. “The structure of Neotropical bat communities: a preliminary analysis”, en Revista Chilena de Historia Natural, Octu-
bre-1986. Fleming, T. H. 1988. The Short-tailed Fruit Bat. The University of
Chicago Press. Chicago. Findley, J. S. 1993. Bats: a Community Perspective. University Press.
Cambridge. Gascon, C. et al. “Matrix habitat and species richness in tropical
forest remnants”, en Biological Conservation, Diciembre-1999.
Gorresen, P. M. y M. R. Willig. “Landscape responses of bats to habitat fragmentation in Atlantic Forest of Paraguay”, en Journal of
Mammalogy, 2004.
Janzen, D. H. “Dispersal of seeds by vertebrate guts”, en Futuyama, D. J. y M. Slatkin (eds.). 1983. Coevolution. Sinauer Associates Inc.
Sunderland. LaVal, R. K. y B. Rodríguez. 2002. Murciélagos de Costa Rica. Insti-
tuto Nacional de Biodiversidad. Costa Rica. Lang, A. et al. 2006. “Activity levels of bats and katydids in relation
to the lunar cicle”, en Oecologia, Enero-2006.
López, J. E. y C. Vaughan. “Observations on the role of frugivorous bats as seed dispersers in Costa Rican secondary humid forests”, en Acta chiropterologica, 2004.
Magurran, A. E. 1988. Ecological Diversity and its Measurement. Prince-
ton University Press. New Jersy, U.S.A. Medellín., R. A, H. T. W. Arita y O. Sánchez. 1997. Identificación de
los murciélagos de México: clave de campo. Asociación Mexicana de
Mastozoología, A. C. México D.F. Minae. 2007. Lista oficial de las especies de flora y fauna silvestre con
poblaciones reducidas y en peligro de extinción. En:
http://www.sinaccr.net/vidasilvestre.php. Moreno, C. y G. Halffter. “Assesing the completeness of bat biodi-versity inventories using species acumulation curves”, en Journal of
Applied Ecology, Febrero-2000.
Moreno, C. E. 2001. Métodos para medir la biodiversidad. M & T-
Manuales y Tesis SEA. México. Montiel, S., A. Estrada y P. León. “Bats assemblages in naturally fragmented ecosystem in the Yucatan Peninsula, México: species richness, diversity and spatio-temporal dynamics”., en Journal of
Tropical Ecology, Mayo-2006.
Newell, G. R. “Response of Lumholtz’s tree-kangaroo (Dendrolagus
lumholtzi) to loss of habitat within a tropical rainforest fragment”, en
Biological Conservation, Diciembre-1999.
Ojasti, J. 2000. Manejo de Fauna Silvestre Neotropical. Smithsonian
Institute. Washington D.C.
Pineda, L. W. “Mamíferos”, en Villarreal, Johnny (ed.). 2003. Eva-luación ecológica rápida en el Refugio de Vida Silvestre Bosque Nacional
Diriá. Costa Rica.
R Development Core Team (2008). R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. ISBN 3-900051-07-0, URL http://www.R-project.org. Sokal, R. R. y F. J. Rohlf. 1991. Biometry. W. H. Freeman and
Company. San Francisco. Schulze, M. D, N. E. Seavy y D. F. Whitacre. “A comparison of the phyllostomid bat assemblages in undisturbed Neotropical forest and forest fragments of a slash-and-burn farming mosaic in Petén, Gua-temala”, en Biotropica, Marzo-2006.
Weller, T. y D. Lee. “Mist net effort required to inventory a forest bat species assemblage”, en Journal of Wildlife Management, Febrero-
2007.

20
Inicio – Siguiente
Alarcón, Y., G. Gama, M. Rodríguez y J. C. Sáenz “Condiciones socio-económicas en dos sectores del Corredor Biológico Chorotega: Percepciones de la comunidad”, Ambientales,
No. 38, diciembre 2009, Costa Rica. Págs. 20-24.
Condiciones socio-económicas en dos sectores
del Corredor Biológico Chorotega:
Percepciones de la comunidad
YUSSELFY ALARCÓN, GUILLAUME GAMA, MARCELA RODRÍGUEZ y JOEL C. SÁENZ
RESUMEN/ABSTRACT
Identificamos los aspectos socio-económicos involucrados en la conservación del Corredor Biológico Chorotega en los sec-
tores Diriá-Ostional y Cerros del Rosario para noviembre de 2008. Empleamos metodologías de investigación social cuali-
tativa y cuantitativa para recolectar información sobre la percepción de la gente e instituciones involucradas en el área de estudio. El enfoque principal utilizado para el diseño de las herramientas fue identificar los usos, el manejo, las amenazas y
el conocimiento que tienen los actores involucrados sobre el concepto de corredor biológico. La información obtenida permite afirmar que el conocimiento respecto a los corredores biológicos es pobre. La falta de conocimientos sobre temas
ambientales, el escaso desarrollo de programas de capacitación por parte de las instituciones ambientales y la ausencia de coordinación entre instituciones y municipalidad, limitan las iniciativas para la conservación de los recursos naturales.
Socio-economics aspects involved in the conservation of Chorotega Biological Corridor, sectors Diriá-Ostional and Cerros
del Rosario were identified in November 2008. We used qualitative and quantitative methodologies for social research to collect information about perception by people and institutions involved in the study area. Design of tools was focused on
identify use, management, threat and knowledge about biological corridor from communities involved. Information ob-tained to suggest that knowledge about biological corridors is poor. Absent of knowledge on environmental issues, sparse
development of training programs by institutions related and lack of coordination between these and municipality, limit ef-forts to conserve natural resources.
Palabras clave: amenaza, Corredor Biológico Chorotega, investigación social, percepción, Guanacaste, Costa Rica.
Keywords: threat, Chorotega biological corridor, social research, perception, Guanacaste, Costa Rica.
os corredores biológicos en Costa Rica han sido planteados como escenarios de partici-pación activa de las comunidades que en ellos
se encuentran; la comunicación y la coordinación entre comunidad e instituciones que gestionan el manejo de los recursos naturales en estos escenarios es fundamental, como también lo son los estudios que se realicen para conocer el estado en que se encuentran estos aspectos para generar opciones de manejo que permitan equilibrar esas participaciones a través del conocimiento de los diferentes actores.
En este estudio identificamos los aspectos socio-económicos involucrados en la conservación de los sectores del Corredor Biológico Chorotega conoci-dos como Diriá-Ostional y Cerros del Rosario a través de actores claves. El objetivo fue recolectar información sobre la percepción de la gente e insti-tuciones involucradas en el área de estudio, a través del empleo de metodologías de investigación social cualitativa y cuantitativa. El enfoque principal utili-zado para el diseño de las herramientas fue el de identificar los usos, el manejo, las amenazas y el conocimiento que tienen los actores involucrados del concepto de corredor biológico.
Para la consolidación del corredor biológico se deben vincular los esfuerzos a nivel regional, nacio-
L
Los autores, especialistas en conservación y manejo de vida silvestre, están adscritos al Instituto de Conservación y Manejo de Vida Silvestre de la Universidad Nacional. [Fecha de recep-ción: enero, 2010. Fecha de aceptación: marzo, 2010.]

21
nal y local integrando el progreso integrado en los ámbitos social y ambiental, ya que ambos son base para el crecimiento económico y mejora de la cali-dad de vida de los habitantes de estas áreas. El pre-sente trabajo es un diagnóstico social, económico y ambiental de las comunidades que se encuentran dentro del Corredor Biológico Cerros del Rosario (CBCR) y Corredor Biológico Diriá (CBD), y de su relación con éstos, el conocimiento que las comuni-dades tienen de ellos y de las amenazas tanto socia-les como ambientales que rodean dichos corredores. El diagnóstico constituye solo un acercamiento a la dinámica de estas comunidades ya que el tiempo y la logística limitan los alcances del estudio en cuanto a área y número de comunidades cubiertas, sin em-bargo se espera sea una herramienta que brinde pau-tas y recomendaciones para la gestión de estos co-rredores.
Aspectos socioeconómicos
Metodología
Para la identificación de los aspectos socio-económicos involucrados en la conservación del Corredor Biológico en los sectores Diriá-Ostional (CB-DO) y Cerros del Rosario (CB-CR) acudimos a actores claves relacionados con el corredor, quienes nos facilitaron la documentación relacionada al desarrollo del Corredor Biológico Chorotega (CBCh).
Con el objetivo de recolectar información sobre la percepción de la gente e instituciones involucra-das en el área de estudio, empleamos metodologías de investigación social cualitativa y cuantitativa, como son la entrevista semi-estructurada y el cues-tionario estructurado, respectivamente. Combina-mos estas metodologías con el objetivo de lograr la triangulación metodológica (Taylor y Bogdan 1996, Vargas 2002).
El enfoque principal utilizado para el diseño de las herramientas fue el identificar los usos y el mane-jo de los recursos naturales por parte de los habitan-tes de la zona, el identificar las posibles amenazas para estos recursos y el conocer cuál es el concepto de corredor biológico que tienen los actores involu-crados. Aplicación de las herramientas
Corredor Biológico Cerros del Rosario (CB-CR): Iden-
tificamos y trabajamos en 16 comunidades aledañas al Parque Nacional Barra Honda (PNBH): Rosario, Puerto Humo, Pozo de Agua, Cañal, Monte Galán, Moracia, Corralillo, Caballito, Corral de Piedra, Roblar, Sonzapote, Quebrada Honda, Barra Honda, Santa Ana, El Flor de Corralillo y San Antonio.
Corredor Biológico Diriá-Ostional (CB-DO): En este
Corredor entrevistamos a funcionarios de algunas de las instituciones gubernamentales y no gubernamen-tales involucradas en el diseño del Corredor Biológi-co: el director del Área de Conservación Tempisque (ACT) Sinac-Minaet; la fundadora y presidenta de la Comisión del Corredor Biológico CB-DO y el presi-dente de la Asociación de Desarrollo de Arado.
Por otro lado, el trabajo se dividió en dos zonas. Un grupo aplicó las herramientas metodológicas a los funcionarios del PND y pobladores de las comu-nidades vecinas: Juan Díaz (PND), Arado, Cola de Gallo, La Esperanza, Los Ángeles, Hojancha y San-ta Cruz. El otro grupo aplicó las herramientas a los funcionarios del Refugio Nacional de Vida Silvestre Ostional (RNVSO), el director de la Asociación de Desarrollo Integral de Ostional, guías locales y fun-cionarios de la Policía Turística, así como las princi-pales comunidades vecinas: El Progreso, Nosara y Playa Pelada. En total se realizaron 26 cuestionarios y nueve entrevistas.
Modelo conceptual, pautas y recomendaciones
Para poner en evidencia una representación de la realidad de la zona de estudio, se estructuró un mo-delo conceptual de amenazas, basado en los datos generados durante el trabajo de campo, la metodo-logía de triangulación y tomando como referencia la metodología para el diseño de modelos conceptuales desarrollada por Margoluis y Salafsky (1998). Con-sideramos que dicho modelo es la base para el plan-teamiento de las actividades dirigidas a revertir las principales amenazas que enfrenta el CBCh.
Realizamos el modelo teniendo como escenario deseado que el Corredor Biológico Diriá-Ostional y el Corredor Biológico Cerros de Rosario se gestio-nan de manera integrada como parte del Corredor Biológico Chorotega.
Resultados
Corredor Biológico Chorotega sector Diriá-
Ostional Los recursos naturales identificados por los en-
trevistados comprenden la fauna, la flora, cuerpos de agua, montañas y semillas. En las comunidades cercanas al Refugio Natural de Vida Silvestre Ostio-nal (RNVSO) los venados, tepezcuintles, saínos y urracas fueron los animales terrestres más mencio-nados. Además, la especie más identificada en Os-tional fue la tortuga lora (Lepidochelys olivacea), debi-
do a que la comunidad está asentada a orillas de la playa de Ostional, que desde hace más de seis déca-das recibe estas arribadas. Otros recursos identifica-dos son las plantas, como el espavel, cedro, pochote, guanacaste, cocobolo, entre otras

22
La principal actividad productiva para las comunidades aledañas al Parque Nacional Diriá es la agricultura. La caída del precio de la carne en los últimos años ha producido una disminución en la actividad ganadera. Sin embargo el turismo y las actividades asociadas, principalmente en la zona costera, y la inscripción de terrenos en el régimen de pago por servicios ambientales son las actividades productivas más importantes. En la región costera asociada al RNVSO se identificó como actividad productiva relevante la extracción y venta de huevos de tortuga lora.
Los problemas identificados por los entrevistados son: la cacería ilegal de tepezcuintles y venados, la deforestación, las quemas para siembra y ganadería, el comercio de especies como orquídeas, loros y pericos. La destrucción de nidos de tortuga (por animales silvestres, domésticos y por el hombre) es identificada por los habitantes de las comunidades aledañas al RNVSO como el principal problema de los recursos naturales de la zona. En cuanto a la deforestación, los habitantes precisaron que ésta ocurre fuera de las áreas protegidas para el cultivo de maíz y fríjol.
Por otro lado, los pobladores mencionaron que el mal manejo de la basura es un problema, ya que no se cuenta con una planta de selección ni rellenos sanitarios. Además, se menciona la contaminación por agroquímicos que afecta a las cuencas. Finalmente, el turismo es percibido como una amenaza debido al desarrollo no planificado.
Las posibles soluciones identificadas por los pobladores son: incrementar la presencia de la autoridad (gubernamental) y seguimiento (más funcionarios y más puestos de control) por parte de una entidad encargada exclusivamente de esta actividad. Se exige también una capacitación, por parte de entidades del Gobierno, en actividades de producción alternativas como el cultivo de orquídeas, para evitar su extracción y venta ilegal. En el sector del RNVSO se identificó la generación de viveros para la reforestación de orillas de ríos como una actividad alternativa a la extracción y comercialización de huevos de tortuga lora. Los pobladores propusieron también la creación y el desarrollo de un programa de educación ambiental con el fin de concientizar a la gente sobre la importancia de conservar y proteger los recursos.
Corredor Biológico Chorotega sector Cerros
del Rosario Los entrevistados identificaron como recursos
naturales relevantes la laguna Mata Redonda y las cavernas del Parque Nacional Barra Honda (PNBH)
así como plantas y animales del bosque seco. Tam-
bién identificaron algunas plantas como cedro, po-chote, guanacaste, canelo, etc.
La agricultura y la ganadería fueron señaladas como las principales actividades, aunque la ganader-ía ha disminuido en los últimos años. El sector del CB-CR no presenta ninguna fuente de turismo direc-to. Algunos pobladores trabajan como guías en el Parque Nacional Palo Verde (PNPV). Aunque no hay muchas fuentes de empleo, los pobladores no han migrado y viven de la producción agrícola pro-pia.
Los principales problemas de los recursos natura-les en los alrededores del PNBH son la tala de árbo-les, la cacería (para autoconsumo), las quemas (es-pecialmente en época seca) y la contaminación del agua por la ganadería y el uso de agroquímicos. Para revertir las amenazas sobre los recursos natura-les, la mayoría de los entrevistados son conscientes de la necesidad de capacitación, hoy insuficiente. Además se considera que el trabajo del Ministerio de Ambiente (Minaet) y del Sistema Nacional de Áreas de Conservación (Sinac) en la implicación de las comunidades es ineficaz. Como solución, los entrevistados propusieron la integración de las co-munidades a las actividades de estas instituciones en la zona y que se ponga en marcha programas de capacitación. Sin embargo, gracias a la labor del actual administrador del PNBH, la relación entre Sinac y comunidades aledañas (dueños de las fincas y demás miembros) ha mejorado (Campos y Cam-pos 2008). También se planteó la sensibilización de la comunidad sobre la importancia del manejo de los desechos y de elementos de descomposición, como las hojas secas. Además, se propone desarro-llar iniciativas locales con el Minaet para reforestar principalmente las áreas aledañas a los ríos.
Discusión
Problemas y modelo conceptual El siguiente análisis se basa en los problemas
identificados por los entrevistados. A partir de estos problemas elaboramos un modelo conceptual para relacionar el efecto de éstos sobre la viabilidad de los corredores (figura 1). El escenario deseado es la gestión integrada de los corredores CBCR y CBDO como parte del CBCh. Las amenazas sociales y bio-lógicas corresponden a la deficiente viabilidad social de los corredores y a la degradación de los ambien-tes naturales.
Un origen de este problema es la desvinculación entre instituciones ambientales (Sinac, Minaet) y municipalidades, que se hace evidente en la defi-ciencia de planificación, pues no existe un plan de desarrollo urbano establecido y funcionando que regule la compra de tierras por parte de extranjeros y

23
defina las pautas normativas para la construcción de hoteles o megaproyectos turísticos. La compra de tierra y la inversión extranjera genera una inflación en su valor y un incremento en los impuestos debido a la construcción de carreteras y vías que facilitan el acceso a estos predios.
Además, existe una falta de recursos para el pago de funcionarios que ocasiona un déficit en la contra-tación de personal. Esto impide que las acciones de control y protección en las áreas protegidas sean eficientes y facilita que actividades como la cacería se practiquen sin control.
Por otra parte, la deficiente planificación del de-sarrollo urbano ha hecho que se construyan carrete-ras en sitios que son paso de fauna silvestre. Otra presión son las prácticas agrícolas tradicionales no sostenibles, que ocasionan la deforestación para el cultivo de maíz, fríjol y para la ganadería. Estas actividades producen, además, la contaminación de las aguas superficiales y subterráneas por el uso de agroquímicos para el control de plagas.
La deficiente gestión del Minaet conduce al des-conocimiento de los pobladores del funcionamiento de la institución. La percepción de la ineficiencia de los funcionarios provoca un debilitamiento en los puentes de comunicación entre miembros de la co-munidad y miembros de la institución. De la misma manera, los problemas de gestión se traducen en la débil apropiación del concepto corredor biológico.
Otra amenaza son las escasas fuentes de empleo y los bajos ingresos económicos de los pobladores. Esto les obliga a trabajar como jornaleros en sitios alejados o a migrar a las ciudades, generando una pérdida de la cultura local. Esta situación provoca una alta deserción escolar y un bajo nivel educativo que tiene un impacto sobre la calidad de vida de las comunidades. Además, amenazas como los conflic-tos y la inadecuada comunicación generan una defi-ciente organización comunal.
Puesta en perspectiva de los problemas
identificados La información obtenida permite afirmar que el
conocimiento respecto a los corredores biológicos es pobre. Esto debido a la falta de conocimientos sobre temas ambientales, lo que limita las iniciativas para la conservación de los recursos naturales. Sinac y Minaet no han desarrollado programas de capacita-ción al respecto.
Es remarcable la percepción de disminución de la biodiversidad en el área. Muchos de los entrevista-dos concordaron en que las especies de flora y fauna han disminuido drásticamente en las últimas déca-das. Identifican la tala, los incendios, la quema de los bosques y la agricultura (café, maíz y arroz) co-mo las principales causas del descenso de poblacio-
nes silvestres como la del venado, la del chancho de monte y la del tepezcuintle. Las especies vegetales identificadas en esa condición son el cedro, el gua-nacaste, el pochote y el cenízaro.
Es importante destacar que la tala, los incendios y la quema provocados en los años anteriores gene-raron una conciencia ambiental en los pobladores mayores, aun cuando éstos no han participado en programas de educación ambiental. Ellos señalan la importancia de las acciones para revertir este efecto. Una muestra es las actividades de la Universidad Estatal a Distancia, que ha desarrollado programas de educación ambiental y capacitación para la refo-restación de áreas pertenecientes al CB-DO, en es-pecial del distrito de Santa Cruz (Rodríguez 2008).
También se reconocieron prácticas tradicionales como la cacería para consumo y la cacería deportiva permitidas legalmente bajo ciertas condiciones. Sin embargo, también se identificó la cacería ilegal, tanto en fincas privadas como en áreas protegidas. Pobladores y miembros del Minaet reconocen que la falta de personal en las áreas protegidas limita el control de estas actividades.
Una concientización por parte de miembros de instituciones y municipalidades podría evitar el par-celamiento de los terrenos y permitir su conserva-ción. En efecto, la demanda de terrenos por parte de extranjeros y los bajos ingresos que perciben los pobladores rurales hacen que ellos estén dispuestos a vender sus tierras. Éstas se destinan a la construc-ción de estructuras para el turismo. La ausencia de incentivos para la conservación y la invisibilidad de otras alternativas de producción prolongan este es-cenario.
Los pobladores pasan de ser propietarios a ser empleados en cargos inferiores dentro de los com-plejos turísticos, debido a que no cuentan con un nivel educativo alto. De la misma forma, la genera-ción de fuentes de empleo, especialmente en las áreas costeras, ha hecho que muchos pobladores migren hacia las zonas turísticas, como en el caso de la zona de Diriá y Ostional, ocasionando el abando-no y el escaso desarrollo de las comunidades.
En el caso particular de Ostional, los entrevista-dos identificaron la extracción de huevos de tortuga lora como la principal actividad económica, seguida por la creciente industria del turismo. El desarrollo de esta actividad económica ha llevado a que en los últimos años turistas extranjeros compren terrenos para la construcción de casas de veraneo. Este pro-ceso ha alarmado a las comunidades aledañas al RNVSO porque perciben este comportamiento co-mo una amenaza para el desarrollo de sus activida-des, y consideran que los inversionistas solo están interesados en promover sus proyectos turísticos con el fin de generar ingresos, mientras que la comuni-

24
dad dejaría de percibir los ingresos asociados al alo-jamiento y guía de turistas (Rojas 2008, Morera 2008).
Finalmente, percibimos una ausencia de coordinación entre las instituciones ambientales y las municipalidades. Por ejemplo, los entrevistados mencionan que el Minaet no otorga permisos para la tala de árboles, sin embargo la Municipalidad sí autoriza la tala. Dentro de las actividades de ambas instituciones no se identifican acciones que promuevan el conocimiento de los corredores biológicos CB-D y CB-CR.
Referencias bibliográficas Margoluis, R. y N. Salafsky. 1998. Medidas de Éxito: Diseño, manejo y
monitoreo de proyectos de conservación y desarrollo. Island Press.
Taylor, S. J. y R. Bogdan.1996. Introducción a los Métodos Cualitativos
de Investigación Social. La Búsqueda de Significados. Paidós. Barcelona.
Vargas, E. “Las comunidades rurales y la conservación: Bases conceptuales y metodológicas”, en Curso EPS001: Sociología Rural (Ciencias Sociales y Vida Silvestre) 2007. Programa Regional en Manejo de
Vida Silvestre, Universidad Nacional de Costa Rica.
Entrevistas Campos, Stella de y Luis Campos (miembros comunidad Santa Ana, aledaña al PNBH). 2008. Costa Rica. Rodríguez, Rosa. Presidenta de la Comisión del Corredor Biológico Diriá. 2008. Costa Rica. Rojas, Gilbert. Presidente Asociación de Desarrollo Integral Ostional. 2008. Costa Rica. Morera, Rodrigo. Biólogo regente de la Asociación de Desarrollo Integral Ostional. 2008. Costa Rica.
Agradecimientos A The Nature Conservancy por el apoyo financiero aportado para el desarrollo del trabajo de campo. Al Instituto para la Conservación y el manejo de la Vida Silvestre, de la Una, por el apoyo logístico y financiero aportado. Al U.S. Fish & Wildlife Service por el finan-ciamiento del curso y por las becas de estudiantes de maestría del Instituto. Al personal del Parque Nacional Barra Honda y del Parque Nacional Diriá y el Refugio Nacional de Vida Silvestre Ostional por facilitarnos sus instalaciones. A Eduardo Carrillo, Jorge Fallas, Luis Diego Alfaro y Joel C. Sáenz por su apoyo en el campo. A los miembros de las comunidades que accedieron a brindar información para el presente estudio y a los propietarios de fincas que permitieron realizar los muestreos biológicos en sus propiedades. A Carlos Espi-
noza y Gerardo Barrantes por el apoyo en el desarrollo del análisis de los aspectos socio-económicos.
Costa Rica Julio Díaz Orias
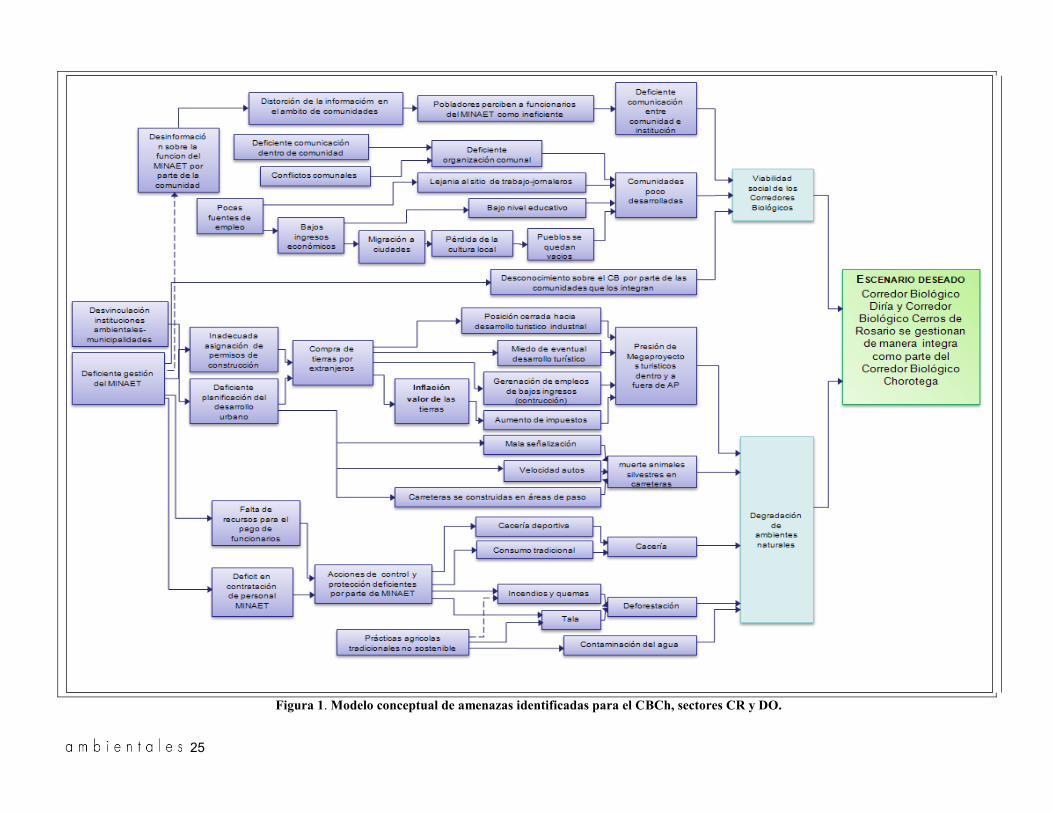
25
Figura 1. Modelo conceptual de amenazas identificadas para el CBCh, sectores CR y DO.

26
Inicio – Siguiente
Enríquez, M., J. C. Sáenz, y M. Ibrahim. “Gremios de aves en agroecosistemas del Pacífico Central de Costa Rica y su importancia para la conservación”, Ambientales, No. 38,
diciembre 2009, Costa Rica. Págs. 26-32.
Gremios de aves en agroecosistemas del Pacífico
Central de Costa Rica y su importancia
para la conservación
MARTHA LUCÍA ENRÍQUEZ, JOEL C. SÁENZ y MUHAMMAD IBRAHIM
RESUMEN/ABSTRACT
Entre julio y diciembre de 2004, se realizaron muestreos de aves en 12 fincas ganaderas del Pacífico Central de Costa Ri-ca, para evaluar la riqueza y la abundancia de gremios de aves y la presencia de especies de importancia para la conser-
vación. Para el muestreo de aves se establecieron parcelas en todos los usos de la tierra existentes dentro de cada finca, y en el centro de éstas se ubicaron puntos de conteo de aves con un radio fijo de 25 m, haciendo registros durante 10 minu-
tos. Se encontró que el gremio más abundante es el de las insectívoras y se registraron 23 especies de importancia para la
conservación. Se concluye que para la conservación de estas especies, resulta fundamental la conservación de los relictos de bosque dentro de este tipo de sistemas productivos.
Between July and December 2004, avian guilds were evaluated within twelve livestock farms, in the area of Esparza, Province of Punta-renas, Costa Rica. In each farm, one agricultural plot of each available land use type was randomly selected, and birds were recorded
from the centre of the plots during 10 minutes using 25 m fixed-radius point counts. According to the results, insectivorous birds were the most abundant, and at landscape scale, 23 species of particular conservation value were recorded. In conclusion, the presence of forest
fragments clearly influence the composition of bird guilds in the agro-ecosystems studied, which underlines the importance of the conser-vation of remnant forest patches inside of this kind of productive systems.
Palabras clave: aves, gremios, fragmentos de bosque, sistemas silvopastoriles. Keywords: birds, guilds, forest fragments, silvopastoral systems.
urante las últimas décadas, gran parte del área boscosa original fue deforestada para promover la ganadería extensiva en Latino-
américa (Kaimowitz 1996, Serrao y Toledo 1990). En muchas regiones, especialmente en zonas bajas, grandes áreas de bosque fueron convertidas a pastu-ras para la producción ganadera, dando como resul-tado un paisaje de pequeños fragmentos de bosque dentro de una matriz agropecuaria (Lang et al.
2003). En América Central, el área de pasturas re-presenta un 46% del total (18,4 millones de hectáre-as), siendo uno de los más importantes usos de la
tierra (Murgueitio y Ibrahim 2000). La degradación de hábitats, la fragmentación y el
aislamiento de bosques disminuyen las poblaciones de aves y afectan de manera distinta los diferentes gremios (Keyser et al. 1998). Por ejemplo, las rapa-
ces de interior de bosque, las insectívoras terrestres y las frugívoras grandes son altamente propensas a la extinción, mientras que las nectarívoras, granívoras, frugívoras pequeñas e insectívoras aéreas son más flexibles a los cambios en el paisaje (Naranjo y Chacón de Ulloa 1997, Renjifo 1999, Wilson et al.
1999, Crooks et al. 2001). Las aves de gran tamaño o
de interior de bosque han sido reportadas como propensas a la extinción debido a que son renuentes a cruzar áreas abiertas, mientras que las especies de borde con alta movilidad han sido reportadas como resistentes a ésta (Karr 1982, Stouffer y Bierregaard
D
Martha Enríquez y Joel Sáenz son biólogos especialistas en manejo de vida silvestre, y él además es director del Instituto para la Con-servación y el Manejo de Vida Silvestre de la Universidad Nacional. M. Ibrahim, agrónomo, es jefe del grupo de Ganadería y Manejo del Medio Ambiente del Catie, en Costa Rica. [Fecha de recepción: enero, 2010. Fecha de aceptación: marzo, 2010.]

27
1995). Por otra parte, las especies que dependen de recursos variables espacial y temporalmente, como frutos o néctar, tienen la necesidad de moverse sobre grandes áreas, por lo que son también altamente susceptibles a la fragmentación (Loiselle y Blake 1992). Sekercioğlu et al. (2002) encontró que las aves
insectívoras de sotobosque son particularmente sen-sibles a la perturbación de hábitat y a la fragmenta-ción, debido a que están más confinadas al interior del bosque, y a su poca habilidad para dispersarse a través de lugares deforestados.
Para la conservación de la biodiversidad en pai-sajes fragmentados o áreas productivas son de espe-cial importancia los elementos arbóreos que éstas poseen, ya que estos elementos crean una variedad de hábitats que no están presentes en lugares mane-jados intensivamente por el hombre (Naranjo 1992, Greenberg et al. 1997, Griffith 1999). Los sistemas
silvopastoriles combinan el manejo de árboles y pasturas de varios tipos con la producción ganadera, y al integrar árboles dentro de potreros estos siste-mas pueden cumplir diferentes funcio-nes y proveer múltiples servicios. Entre otros, ayudan a incrementar la producti-vidad de la finca mientras protegen los recursos naturales, favorecen la diversi-dad y almacenan buena cantidad de carbono, mitigando los efectos de inver-nadero (Harvey 2001, Ibrahim y Mora-Delgado 2001, Cordero y Boshier 2003).
Este estudio evaluó la composición de los gremios de aves presentes en sis-temas silvopastoriles, e identificó espe-cies de importancia para la conserva-ción.
Metodología
Área de estudio Se estudiaron 12 fincas ubicadas en-
tre los cantones de Montes de Oro, San-tiago y Esparza y el distrito de Barranca, provincia de Puntarenas, al noroeste de Costa Rica. Dentro de cada finca se muestrearon parcelas al azar de cada tipo de uso de suelo presente, teniendo en promedio ocho usos del suelo por finca. Los usos muestreados fueron bos-que secundario (BSEC), bosque ripario (BR), frutales (FRU), tacotales (TAC), potrero con alta densidad de árboles (PAD), potrero con baja densidad de árboles (PBD), cercas vivas (CV), made-rables (MAD), bancos forrajeros (BF), pasturas sin árboles (PSA) y granos básicos (GB), según la clasificación de
usos de suelo propuesta por Murgueitio et al. (2003).
Muestreo de aves Se realizaron cuatro conteos de aves entre julio y
diciembre de 2004 mediante el método de puntos de conteo (Reynolds et al. 1980). Los puntos fueron
ubicados con la ayuda de un GPS en el centro de cada parcela (uso de suelo) y tuvieron un radio fijo de 25 m. En cada punto de conteo se registraron las aves que estaban haciendo uso del área (perchadas, en el suelo, forrajeando, etc.) y se clasificaron de acuerdo al gremio al que pertenecen, distinguiendo entre carnívoras (incluyendo las carroñeras), piscí-voras, frugívoras, nectarívoras, granívoras y omní-voras, siguiendo la clasificación de la guía de aves de Costa Rica (Stiles y Skutch 2003). Algunas aves fueron clasificadas de acuerdo a lo que comen en mayor proporción, aunque su dieta incluyera en menor cantidad otro tipo de alimentos.
Yigüirro (Turdus grayi) Gino Biamonte

28
Análisis de datos La abundancia relativa de aves se calculó como
el cociente entre la abundancia de la especie i y el
número de parcelas muestreadas en cada uso de suelo (pi =ni/n). El dato de abundancia de cada una
de estas parcelas se obtuvo previamente promedian-do los valores de abundancia de cada especie.
Especies de importancia para
la conservación El análisis de amenazas e importancia para la
conservación de las especies registradas en la zona, se evaluó con base en las siguientes referencias:
1. Especies consideradas por la Convención sobre el Comercio Internacional de Especies Amenaza-
das de Fauna y Flora Silvestre (Cites), listadas
en tres diferentes apéndices de acuerdo al ti-po de protección que necesiten por sobre-explotación. El Apéndice I incluye las espe-cies que están en peligro de extinción, el Apéndice II incluye especies que no están necesariamente en peligro de extinción, pero podrían llegar a estarlo si no se controla es-trictamente su comercio, y el Apéndice III incluye las especies que están siendo regula-das por algún país en particular y que nece-sita la cooperación internacional para pre-venir su explotación (Cites 2005).
2. Especies de interés para la conservación, ca-tegorizadas en la Lista de Fauna de Importan-cia para la Conservación en Centroamérica y
México: Listas rojas, listas oficiales y especies
en Apéndices Cites (UICN 1999). Están in-cluidas las especies con poblaciones reduci-das o amenazadas en Costa Rica.
3. Amenazas a nivel mundial: Especies consi-deradas amenazadas por la Comisión de Supervivencia de Especies de la Unión Mundial para la Naturaleza (UICN), citadas en la Lista Roja. Incluye las siguientes cate-
gorías: EX (extinto), EW (extinto en estado silvestre), CR (críticamente en peligro), EN (en peligro), VU (vulnerable, LR (bajo ries-go), DI (datos insuficientes) y NE (no eva-luado) (UICN 2004).
Resultados y discusión
Estructura de los gremios tróficos
Descripción general
Se registraron 1.901 individuos de 113 especies, pertenecientes a 31 familias y 90 géneros, que co-rresponden a siete diferentes gremios tróficos (Stiles y Skutch 2003). El gremio de las insectívoras fue el que dominó la composición de la comunidad de
aves, lo cual ha sido un patrón general en varios estudios realizados en agroecosistemas (Cárdenas 2003, Cárdenas et al. 2003, Estrada et al. 1997, Lang
et al. 2003, Menacho y Sáenz 2004). El gremio de las
omnívoras, siguiente en abundancia, es muy versátil en el uso de diferentes recursos y puede acoplarse muy bien a los ambientes antrópicos, como en el caso de Turdus grayi o Crotophaga sulcirostris, las cua-
les son especies extremadamente abundantes en la región (Stiles y Skutch 2003). Aunque la mayoría de aves encontradas son típicas de agroecosistemas, algunas especies fueron exclusivas de usos con alta cobertura arbórea, debido a sus requerimientos ali-menticios y de hábitats boscosos o arbolados. Algu-nos ejemplos son Geranospiza caerulescens, Glaucis
aenea e Hylocinchla mustelina, especies que según
Stiles y Skutch (2003) son altamente dependientes de bosque.
Insectívoras
Fue el gremio con mayor abundancia y riqueza, con 691 individuos observados de 53 especies. El 27% de las especies estuvo presente en los bosques, 19% en pastos con alta densidad de árboles y el me-nor porcentaje se dio en los usos con cobertura arbó-rea muy baja o nula. Posee una amplia gama de especies, que van desde las muy dependientes de bosque hasta las que prefieren habitar sitios abiertos. La mayor parte de las especies insectívoras son muy flexibles a la perturbación de hábitat y a la fragmen-tación, como en el caso de Pitangus sulfuratus y Cam-
pylorhynchys rufinuca (Stiles y Skutch 2003). En cuan-
to a las insectívoras dependientes de bosque, se en-contró a la especie Thamnophilus doliatus, la cual
estuvo presente únicamente en el bosque secundario debido a su alta especificidad de habitar el sotobos-que. Según Sekercioğlu et al. (2002), las aves insectí-
voras de sotobosque son particularmente sensibles a las perturbaciones y están más confinadas al interior del bosque.
Omnívoras
Presentaron 19 especies y 515 individuos. La mayor abundancia relativa de este gremio se dio en los frutales, seguido de los pastos con baja cobertura arbórea, mientras que tuvieron una presencia muy baja en los bancos forrajeros. Las especies Crotop-
haga sulcirostris, Turdus grayi y Aimophila ruficauda
fueron las especies más abundantes a escala de pai-saje. En cuanto a la riqueza, el 16% de las especies se encontró en el bosque ripario y, en segundo lugar (con un 14% cada uso), se reportaron en los frutales, pastos con alta y pastos con baja densidad de árbo-les.

29
Granívoras
Se avistaron 313 individuos de nueve especies, siendo más abundantes en los pastos con alta densi-dad, en granos básicos (cultivo de maíz) y pastos con baja densidad arbórea. En cuanto a la riqueza, presentaron un 14% de las especies en los pastos con baja densidad y cercas vivas, contrastando con un 4% en bosque seco, en donde presentaron también una abundancia muy baja. Columbina passerina y
Leptotila verreauxi fueron las especies más abundantes
de este gremio (con 80 y 69 individuos respectiva-mente) como también algunas de las especies más abundantes a escala de paisaje.
Debido a sus hábitos alimenticios, las especies omnívoras y granívoras fueron más abundantes en usos de suelo abiertos y con poca cobertura vegetal. La mayoría de las especies de estos gremios son muy abundantes en hábitats rurales y urbanos, ya que son capaces de explotar los recursos que ofrece la matriz antropogénica (flores, frutos, sitios de des-canso y percha, refugio, etc) (Laurence et al. 2002),
por lo que en general no han sido catalogadas como especies de importancia para la conservación. La única especie típica de bosque registrada fue Hilo-
cinchla mustelina (omnívora), que es una especie mi-
gratoria.
Frugívoras
Se observaron 296 individuos de 13 especies dife-rentes. Presentaron su mayor abundancia en los pastos con alta densidad, seguido de los frutales. El 18% de las especies se presentó tanto en los bosques riparios como en pasturas con alta densidad. Un 14% estuvo en pastos de baja densidad y un 12% en frutales y cercas vivas. Dos de las especies frugívoras más abundantes a escala de gremio y paisaje fueron Amazona albifrons y Brotogerys jugularis. La gran
abundancia de especies frugívoras en pasturas con alta cobertura arbórea, se puede deber a que en la zona es muy común la siembra de árboles frutales dentro de los potreros, pues esto les permite a los finqueros tener un mejor aprovechamiento de estos usos, aunque generalmente solo utilizan las frutas para autoconsumo. Terborgh y Winter (1980) han identificado a las aves frugívoras como un grupo propenso a la extinción, lo cual puede estar relacio-nado a su dependencia de recursos claves o tempora-les (Howe 1984, Terborgh 1986). En concordancia con estos autores, durante el transcurso de esta in-vestigación se encontró que la mayor abundancia y riqueza de frugívoras se dio cuando había disponibi-lidad de frutos (principalmente papaya, mango y maíz) e, inclusive, una de las especies de este gremio (Ramphastos sulfuratus) se registró solo una vez, pre-
Tucán pico iris (Ramphastos sulfuratus) Gregory Basco

30
cisamente durante la cosecha de un frutal mixto de papaya, banano, guayaba y cítricos.
Nectarívoras
Con 42 individuos de siete especies presentaron el mayor porcentaje de especies en los tacotales y en las cercas vivas (18% en cada uno), siguiendo los frutales (17%) y bosques secundarios (14%). Las especies más abundantes fueron Amazilia tzacatl y,
en segundo lugar, Amazilia rutila. Este grupo se vio
favorecido en usos tales como cercas vivas y tacota-les, por su mayor oferta de plantas con néctar. Es una práctica común entre los finqueros sembrar árboles o arbustos que tengan flores, sobre todo cer-ca de sus casas, como una manera de embellecer su finca, lo que es aprovechado por los colibríes, sobre todo por especies poco sensibles a la perturbación como las Amazilia spp.
Carnívoras y piscívoras
Las carnívoras estuvieron representadas por 41 individuos de 11 especies. Más del 60% de las espe-cies estuvieron presentes en las pasturas (ocho espe-cies en los pastos con baja densidad y seis en los pastos con alta densidad). La especie más abundante de este gremio fue Coragips atratus (con 14 indivi-
duos), seguida de Milvago chimachima (con nueve
individuos). Las piscívoras solo contaron con una especie (Cloroceryle amazona) de la cual se realizaron
tres avistamientos en bosques riparios. Estos dos gremios se caracterizan por tener poblaciones redu-cidas, por lo que su observación se hace poco proba-ble, siendo las dos subfamilias con menor número de registros. Todas las especies de carnívoras obser-vadas en este estudio están incluidas en el Apéndice II de Cites (Cites 2005), siendo por tanto de impor-tancia para la conservación. Las piscívoras, por su parte, son aves especialistas, lo que las hace más vulnerables a los cambios en el paisaje (Hansson et
al. 1995, With et al. 1997), sobre todo en la época
seca, cuando las fuentes de agua dentro de las fincas se secan o quedan reducidas a pequeñas corrientes de agua.
Especies de importancia para
la conservación De acuerdo a la Convención sobre el Comercio Inter-
nacional de Especies Amenazadas de Fauna y Flora Sil-
vestre (Cites 2005), en la zona de estudio hay 22 es-
pecies que están citadas en el Apéndice II, de las cuales 20 -17,7%- están amenazadas por sobre-explotación o comercio ilegal. Dentro de éstas se encuentran todas las especies de los órdenes Falco-niformes (gavilanes, águilas, elanios y caracaras) y Psittaciformes (loras y pericos), todas las especies de la familia Trochilidae (colibríes) y una especie de la
familia Ramphastidae (tucán pico iris, Ramphastos
sulfuratus). Según la Lista de Fauna de Importancia
para la Conservación en Centroamérica y México, hay
siete especies en el área de estudio con poblaciones reducidas o amenazadas en Costa Rica, que inclu-yen a una rapaz (Geranospiza caerulescens), cinco es-
pecies de loras y pericos (Amazona albifrons, Amazona
autumnalis, Aratinga canicularis, Aratinga finschi y Bro-
togeris jugularis) y el pato real (Cairina moschata) que
fue registrado en la zona de estudio pero fuera del tiempo de muestreo. A nivel mundial, la especie Dendroica cerulea se encuentra citada como vulnera-
ble en la Lista Roja de la UICN, debido a que sus
poblaciones están disminuyendo por causa de la fragmentación de bosques y pérdida de hábitat (UICN 2004). (Todas las especies con importancia para la conservación y su estatus a nivel nacional y mundial, se pueden apreciar en la cuadro 1.)
La mayor abundancia de las especies de impor-tancia para la conservación fue encontrada por este estudio en los pastos con alta densidad de árboles, cercas vivas, bosques riparios, frutales y bosques secundarios, que son usos de suelo con alta cobertu-ra arbórea. Esto concuerda con los estudios realiza-dos por Cárdenas et al. (2003), Naranjo (1992) y
Petit y Petit (2003), que encontraron que los bosques secundarios, bosques riparios y charrales, son los que poseen la mayor parte de aves de interés para la conservación, por lo que concluyen que los esfuer-zos de conservación deben enfocarse en los fragmen-tos de bosque secundario y bosque ripario que aún persisten en este paisaje, como también en aumentar la cobertura arbórea en los potreros activos.
Para poder mantener las poblaciones actuales de aves es necesario, entonces, que exista una gran variedad de especies de plantas dentro de los usos de suelo, y que éstos a su vez tengan diferente cantidad de cobertura arbórea para favorecer tanto a las espe-cies de áreas abiertas como a las de áreas boscosas, siendo una buena opción la implementación de una gama de sistemas silvopastoriles (cercas vivas, ban-cos de forraje, árboles en potreros y cultivos made-rables entre otros).
Dentro de los planes de manejo futuros de las fincas se debe incluir, por lo tanto, la siembra de cercas vivas diversificadas, potreros arbolados y recomendar la siembra de árboles frutales, ya sea como un uso de suelo o dentro de otros usos de la finca. Es necesario, también, incentivar políticas de conservación de los relictos de bosque secundario y riparios, que son fundamentales para la superviven-cia de las aves con algún tipo de dependencia al bosque.

31
Cuadro 1. Especies de importancia para la conservación a escala mundial según la Lista Roja de UICN, Apéndices de Cites y, a escala nacional,
la Lista de Fauna de Importancia para la Conservación en Centroamérica y México.
Taxón Nombre común Situación nacional
Situación mun-dial
Orden Anseriformes
Familia Anatidae Patos Cairina moschata Pato real PR
Orden Falconiformes
Familia Accipitridae Gavilanes, águilas y elanios Buteo magnirostris Gavilán chapulinero Cites II Buteo nitidus Gavilán pollero Cites II Buteo platypterus Gavilán aludo Cites II Elanus leucurus Gavilán bailarín Cites II Geranospiza caerulescens Gavilán ranero PR Cites II
Familia Falconidae Halcones y caracaras Caracara plancus Caracara cargahuesos Cites II Herpetotheres cachinnans Guaco Cites II Milvago chimachima Caracara cabecigualdo Cites II
Orden Psittaciformes
Familia Psittacidae Loros Amazona albifrons Loro frentiblanco PR Cites II Amazona autumnalis Loro frentirrojo PR Cites II Aratinga canicularis Perico frentinaranja PR Cites II Aratinga finschi Perico frentirrojo PR Cites II Brotogeris jugularis Periquito barbinaranja PR Cites II
Orden Apodiformes
Familia Trochilidae Amazilia rutila Amazilia canela Cites II Amazilia saucerrottei Amazilia culiazul Cites II Amazilia tzacatl Amazilia colirrufa Cites II Anthracothorax prevostii Manguito pechiverde Cites II Archilochus colubris Colibrí garganta de rubí Cites II Glaucis aenea Ermitaño bronceado Cites II Phaethornis guy Ermitaño verde Cites II
Orden Coraciformes
Familia Ramphastidae Ramphastos sulfuratus Tucán pico iris Cites II
Orden Passeriformes
Familia Parulidae Dendroica cerulean Reinita cerúlea Vu Situación nacional: PR= poblaciones reducidas. Situación mundial: Vu= vulnerable.
Referencias bibliográficas Cárdenas, G. “Composición y estructura de la avifauna en diferentes sistemas de producció”, en Sánchez, M. D. y M. Rosales (eds.). 2003. Agroforestería para la producción animal en América Latina. Memo-
rias Segunda conferencia electrónica Agosto 2000-Marzo 2001. Fao-Cipav.
Cárdenas, G. et al. “Diversidad y riqueza de aves en diferentes hábi-
tats en un paisaje fragmentado en Cañas, Costa Rica”, en Agrofores-
tería en las Américas 10(39-40), 2003.
Cites. 2005. Convención sobre el comercio internacional de especies amena-
zadas de Fauna y Flora silvestres. Apéndices I, II y III en vigor a partir
de 17-2-05. En: www.cites.org/esp/app/index.shtml. Cordero, J. y D. H. Boshier (eds.). 2003. Árboles de Centroamérica: Un
manual para extensionistas [CD-ROM] Oxford Forestry Institute,
Catie. Oxford, UK - Costa Rica. Crooks, K. R. et al. “Extinction and colonization of birds on habitats
islands”, en Conservation Biology 15(1), 2001.
Estrada, A., R.. Coates-Estrada y D. A. Meritt Jr. “Anthropogenic
landscape changes and avian diversity at Los Tuxtlas, México”, en Biodiversity and Conservation 6, 1997.
Greenberg, R., P. Bichier y J. Sterling. “Acacia cattle and migratory birds in southeastern México”, en Biological Conservation 80, 1997.
Griffith, D. M. “Agroforestry: a refuge for tropical biodiversity after FIRE”, en Conservation Biology 14(1), 1999.
Harvey, C. A., “The conservation of biodiversity in silvopastoral systems”, en Ibrahim, M. et al. (eds.). 2001. International Symposium on silvopastoral systems and second Congress on agroforestry and livestock production in Latin America, silvopastoral systems for restoration of degraded
tropical pasture ecosystems. 3-7th April, San José, Costa Rica.
Howe, H. F. “Implications of seed dispersal by animals for tropical reserve management”, en Biological Conservation 30, 1984.
Ibrahim, M. y J. Mora-Delgado. 2001. Conferencia electrónica: Potencia-lidades de los sistemas silvopastoriles para la generación de servicios ambien-
tales (EAD-PFI-ECONF-L), Octubre-Noviembre 2001. En: http://lead-
es.virtualcentre.org/es/ele/conferencia3/articulo1.htm. Kaimowitz, D. “Will livestock intensification help save Latin Americas’s tropical forest?”, en Angelsen, A. y D. Kaimowitz (eds.). 2001. Agricultural technologies and tropical deforestation. Walling-ford,
UK. Karr, J. R. “Avian extinction on Barro Colorado Island, Panama: a reassessment”, en American Naturalist 119, 1982.
Keyser, A. J., G. E. Hill y E. C. Soehren. “Effects of forest fragment size, nest density and proximity to edge on the risk of predation to ground-nesting passerine birds”, en Conservation Biology 16(5), 1998.
Lang, I. et al. “Composición de la comunidad de aves en cercas vivas
de Río Frío, Costa Rica”, en Agroforestería en las Américas 10(39-40),
2003.
Loiselle, B. A. y J. G. Blake. “Population variation in a tropical bird community: implications for conservation”, en BioScience 42, 1992.
Menacho, R. y J. Sáenz. 2004. Informe de Biodiversidad de primer año. Proyecto Enfoques Silvopastoriles Integrados para el Manejo de Ecosistemas.
Una. Costa Rica.
Murgueitio, E. y M. Ibrahim. 2000. Agroforestería pecuaria para la reconversión de la ganadería en Latinoamérica. Ponencia en XVII Congreso Panamericano de Ciencias Veterinarias, Panamá, Septiembre del 2000.
Murgueitio, E. et al. 2003. Usos de la tierra en fincas ganaderas: Guía para el pago de servicios ambientales en el proyecto Enfoques Silvopastoriles
Integrados para el Manejo de Ecosistemas. Cipav, Catie, Uca-Nitlapan.
Apotema Ltda. Colombia.

32
Naranjo, L. G. “Estructura de la avifauna en un área ganadera en el valle del Cauca, Colombia”, en Caldasia 17, 1992.
Naranjo, L. G. y P. Chacón de Ulloa. “Diversidad de insectos y aves insectívoras de sotobosque en hábitats perturbados de selva lluviosa tropical”, en Caldasia 19(3), 1997.
Petit, L. J. y D. R. Petit. “Evaluating the importance of human-modified lands for neotropical bird conservation”, en Conservation
Biology 17(3), 2003.
Renjifo, L. M. “Composition changes in a subandean avifauna after long-term forest fragmentation”, en Conservation Biology 13, 1999.
Reynolds, R. T., J. M. Scott y R. A. Nussbaum. “A variable circu-lar-plot method for estimating bird numbers”, en Condor 82, 1980.
Sekercioğlu, C. et al. “Disappearance of insectivorous birds from
tropical forest fragments”, en PNAS 99(1), 2002.
Serrao, E. A. S. y J. M. Toledo. “The search for sustainability in Amazonian pastures”, en Anderson, A. B. (ed.). 1990. Columbia University Press. NY. Stiles , G. y A. Skutch. 2003 . Guía de las aves de Costa Rica. Inbio.
Stouffer, P. C. y R. O. Bierregaard Jr. “Use of Amazonian forest fragments by understory insectivorous birds”, en Ecology 76, 1995.
Terborgh, J. “Keystone plant resources in the tropical forest”, en Soulé, M. E. (ed.). 1986. Conservation Biology: an evolutionary-
ecological perspective. Sinauer Associates. Massachussets.
Terborgh, J. y B. Winter. “Some causes of extinction”, en Soulé, M. E. y B. A. Wilcox (eds.). 1980. Conservation Biology: an evolutionary-
ecological perspective. Sinauer Associates. Massachussets.
UICN – Unión Mundial para la Naturaleza. 1999. Listas de fauna de importancia para la conservación en Centroamérica y México: Listas rojas,
listas oficiales y apéndices CITES. UICN-ORMA/WWF Centroaméri-
ca. San José. UICN 2004. 2004 IUCN Red List of Threatened Species. En:
www.redlist.org/search/search-basic.html. Wilson, J. D. et al. “A review of the abundance and diversity of
invertebrate and plant foods of granivorous birds in Northern Europe in relation to agricultural change”, en Agriculture, Ecosys-
tems and Environment 75, 1999. Gavilán aludo (Buteo platypterus) Gregory Basco

33
Inicio – Siguiente
Solano, D. y G. Wong. “Hábitat y población de mono tití (Cebidae: Saimiri oerstedii oerstedii)”, Ambientales, No. 38, diciembre 2009, Costa Rica. Págs. 33-46.
Hábitat y población de mono tití (Cebidae: Saimiri
oerstedii oerstedii) en la península de Osa, Costa Rica
DANIELA SOLANO y GRACE WONG
RESUMEN/ABSTRACT
Se evaluó el estado del hábitat del mono tití fuera del Parque Nacional Corcovado en la Península de Osa, entre septiem-bre de 2005 y mayo de 2006. Se evaluó 22 sitios boscosos entre el río Rincón de Osa y el río Carate con presencia de mo-
nos tití, para determinar su composición florística y estructura horizontal, y las características demográficas (tamaño de tropa y composición por sexos y edades) de las tropas de mono tití. Se establecieron 63 parcelas, distribuidas entre los sitios
boscosos, con un tamaño de 20 x 50 m. Se encontró un total de 4.416 árboles de 354 especies diferentes. Los sitios bosco-sos ubicados cerca del poblado de La Palma fueron los más diversos y compartieron similaridad en su composición florís-
tica. Las tropas tuvieron en promedio 19 individuos. Es necesario implementar herramientas de conservación voluntaria de
bosques secundarios fuera de áreas protegidas y educar a los pobladores de las comunidades cercanas a los sitios boscosos utilizados por el mono tití.
The squirrel monkey’ habitat status was evaluated between September 2005 and May 2006, outside of Corcovado National Park in Osa Peninsula. Twenty two forested sites were evaluated between Rincon River and Carate River, to determine the floral composition and ho-
rizontal composition of the forest and the demographic characteristics of squirrel monkey troops (size and sex composition). Sixty three 20 by 50 meters plots were established and a total of 4416 trees were found belonging to 354 different species. Forested sites near La Palma
town were more diverse and shared a similarity in floral composition. It is urgent to implement voluntary conservation tools among pri-vate landowners outside of protected areas and to educate the community members in the surroundings of forested sites used by squirrel
monkey troops.
Palabras clave: paisaje, hábitat, demografía, índices de diversidad, península de Osa, Saimiri oerstedii oerstedii
Keywords: landscape, habitat, demography, Diversity Index, Osa Peninsula, Saimiri oerstedii oerstedii
a península de Osa, ubicada en el Pacífico Sur de Costa Rica, ha tenido una fuerte historia social y política, la cual influenció
el uso del suelo que actualmente existe: una mez-cla de bosques y tierras utilizadas para la agricul-tura y la ganadería.
Este agropaisaje resultante de la fragmentación del hábitat con áreas boscosas remanentes utilizadas por muchos animales (Kapelle et al. 2002, Daily et al.
2003, Marsh 2003b, Rodríguez-Toledo et al. 2003,
Chapman et al. 2003) es el hogar de la subespecie de
mono tití centroamericano, S. o. oerstedii, el cual se
distribuye del río Sierpe hasta la frontera sureste con
Panamá en la península de Burica y de Osa, y se encuentra protegida dentro del Parque Nacional Corcovado (PNC), en parte.
Dentro del ámbito altitudinal donde se desplaza, el mono tití utiliza diferentes tipos de bosque, como el bosque primario, el secundario, los bosques de galería, los manglares y los cultivos de palma aceite-ra (Elais oleifera), siendo el bosque secundario joven
y maduro el hábitat más utilizado, sobre todo en el estrato bajo y medio del bosque (Boinski 1987, Wong 1990a y 1990b, Arauz 1993, Rodríguez-Vargas 2003).
El uso del hábitat y las especies de árboles y otros componentes de dieta de los monos tití de Costa Rica han sido estudiados con anterioridad (Boinski 1987, Wong 1990a y 1990b). Su dieta compuesta de artrópodos fitófagos, pequeñas bayas y frutas culti-
L
Las autoras son especialistas en manejo de vida silvestre y conser-vación. D. Solano es cofundadora de la Coalición Sur y G. Wong es profesora en la Universidad Nacional. [Fecha de recepción: enero, 2010. Fecha de aceptación: marzo, 2010.]

34
vadas cuando están disponibles, explican la prefe-rencia clara por hábitat en sucesión temprana y hábitat perturbados en comparación con el bosque primario (Janzen y Schoener 1968, Wong 1990a y 1990b).
Dentro del Parque Nacional Corcovado, a pesar de su extensión aproximada de 41.788 has. (42 km2), los monos tití se encuentran principalmente en los bosques secundarios cercanos a la estación Sirena o en sitios donde existen diferentes etapas sucesionales (Boinski 1987). Allí mismo, tropas de monos tití no utilizaron más la meseta de La Llorona, debido a la maduración del bosque secundario (Boinski et al.
1997). Estas características intrínsecas de la historia natural de los monos tití centroamericanos se han considerado también como amenazas para la espe-cie (Boinski et al 1997), debido a que este tipo de
hábitat preferido por ellos por lo general no se en-cuentra protegido bajo alguna categoría de manejo establecida por el Estado.
Se han descrito también como amenazas para la especie el declive de la disponibilidad de alimento durante la estación lluviosa, e incluso se ha determi-nado que en poblaciones aisladas existe una alta vulnerabilidad cuando el tamaño del grupo es menor de 15 individuos y el hábitat disponible es menor a 30 ha (Boinski y Sirot 1997), sin conexión con otros sitios de bosque.
Actualmente, el mono tití, S. o. oerstedii, está en la
lista de especies en peligro de extinción según el grupo de especialistas en primates de la UICN (UICN 1996), y se han señalado como amenazas también la distribución geográfica restringida; el desarrollo de un mosaico de cultivos agrícolas, la actividad ganadera, los asentamientos humanos y fragmentos de bosque en lo que eran principalmente áreas boscosas; el futuro incierto en propiedades privadas de poblaciones del mono tití, en su gran mayoría casi aisladas y sin comunicación entre ellas. También la merma en las poblaciones de titís en los años 60, por una epidemia de fiebre amarilla y por prácticas agrícolas como la fumigación de banana-les; el tráfico de mascotas (en menor medida) y el desarrollo turístico en las últimas décadas, que ha impactado sobre todo a la subespecie del Pacífico Central, el S. o. citrinellus (Wong 2002), el cual co-
mienza a impactar el Pacifico Sur también. Por otro lado, las estimaciones de la población de
monos tití, Saimiri o. oerstedii, han sugerido la exis-
tencia de 3.000 individuos en Costa Rica (Boinski y Sirot 1997), y Rodríguez-Vargas (2003) estima 4.755 individuos en Panamá. En los estudios poblaciona-les se enfatiza que algunos grupos de Saimiri o. oers-
tedii no están conectados unos con otros dentro del
paisaje, otros están aislados y son más vulnerables (Rodríguez-Vargas 2003), al menos en Panamá.
Adicionalmente, existe gran controversia sobre cuál es el tamaño mínimo que una población de animales necesita para asegurar su supervivencia en un largo plazo, ya que esto dependerá de cada organismo (Chiarello 2000).
Por ejemplo, Franklin y Frankham (1998) argu-mentan que una población efectiva (Ne) de 500–1.000 individuos mantiene la diversidad genética de una población. Algunos estudios enfatizan la ten-dencia de que a mayores tamaños de fragmentos boscosos existe mayor densidad de primates, y en general de mamíferos (Chiarello 2000, Chiarello y de Melo 2001), por lo que la supervivencia a largo plazo de una población efectiva dependerá también del tamaño del hábitat disponible.
A medida que los parches de hábitat se vuelven más pequeños y más separados, hay un incremento de las tasas de extinción y una disminución de la tasa de recolonización (Hanski 1997), lo que tam-bién afecta la dinámica de supervivencia de una población, sobre todo para especies en peligro de extinción, cuya respuesta a los cambios producidos por la fragmentación del hábitat (Saunders et al.
1991, Turner et al. 2001) y a la complejidad del pai-
saje es percibida de manera diferencial (Turner et al.
2001, Morrison 2002), por lo que habría que deter-minar estos parámetros para cada subpoblación de monos tití para asegurar su conservación y poder determinar su respuesta a estos cambios
Lo anterior es de vital importancia, sobre todo cuando diversos autores, en estudios previos sobre el tamaño y la composición de las tropas de las dife-rentes especies de Saimiri spp., han determinado que
en lugares donde la calidad del hábitat es menor el número de individuos por tropa también es menor, al igual que las tropas de hábitat fragmentados, en contraste con tropas en bosques tropicales más ex-tensos, que pueden llegar hasta a 300 individuos por tropa, por ejemplo en Brasil, Perú y Colombia (Baldwin y Baldwin 1972 y 1986, Wong 1990a, Wong 1990b, Boinski 1987a, Vaughan et al. 1980,
Thorington 1968, Terborgh 1983, Klein y Klein 1973, Klein [en: Robinson y Janson 1987], Freese et
al. 1982, Neville 1976, Baldwin y Baldwin 1971,
Altricher et al. 1996, Sierra et al. 2003, Rodríguez
1999, Cáceres et al. 2002).
La fragmentación, por lo tanto, afecta a los pri-mates en los remanentes boscosos, con el potencial de afectarlos en diferentes aspectos, como los de-mográficos, de comportamiento, genéticos, distribu-ción, diversidad (Chiarello 2003, Gonçalvez et al.
2003, Marsh 2003a, Rodríguez-Toledo 2003). Además, el tipo y la calidad del hábitat que utilicen los grupos de tití están asociados con la variación en el número de individuos por tropa (Baldwin y Bald-win 1981, Wong 1990b). Por lo que, a pesar de que

35
los monos tití y otras especies, como el mono cara-blanca y el aullador, son especies que se han adap-tado a ambientes fragmentados (Sáenz y Sáenz 2007, Bicca-Marquez 2003, Rodríguez-Vargas 2003), las tropas pueden ser más susceptibles a en-fermedades o simplemente no poseer un tamaño de tropa adecuado para la supervivencia a largo plazo. Al encontrarse las poblaciones en áreas cada vez más pequeñas se favorece la oportunidad para la transmisión de parásitos e incremento de enferme-dades, factores influyentes en la distribución y den-sidad de especies, e identificados como componen-tes críticos a considerar en la biología de la conser-vación de las especies (Anderson 1979, May 1998, Scott 1998, Laboratorio de Primatología de los Tux-tlas 2004).
Por otro lado, el hábitat óptimo para especies como el mono tití (aunque no para otros tipos de animales silvestres) en la península de Osa son tie-rras donde existe mezcla de diferentes etapas suce-sionales de bosque. Este mosaico de diferentes tipos de vegetación y bajuras utilizados por el mono tití en mayor proporción se encuentra precisamente en esta área vulnerable a la fragmentación, fuera de áreas protegidas, en las tierras privadas de la franja coste-ra, tierras que no poseen protección estatal formal y que pertenecen principalmente a campesinos y ex-tranjeros, lo que los hace aun más vulnerables, debi-do a que la conservación del hábitat depende del uso que sus propietarios le den y el cual puede resultar en impactos ecológicos para la especie y el mismo hábitat (Chapman et al. 2003).
La desaparición o aislamiento de primates puede tener consecuencias extremas: su función ecológica como dispersores de semillas y, por lo tanto, su in-fluencia en la distribución y composición de árboles (March 2003b) podría generar la extinción local de algunos árboles al perderse estos primates (Chap-man y Chapman 1995, Chapman y Onderdonk 1998, Dominy y Duncan 2005). Es por esta razón que hay necesidad de entender y analizar los frag-mentos en los que habitan primates como un poten-cial importante para la población total, aun si el fragmento es muy pequeño para la supervivencia de los primates que allí se encuentran (Saunders et al.
1991, March 2003b). Finalmente, al ser el Parque Nacional Corcovado
el área protegida más grande en la zona y el área donde -en teoría- los monos tití se encuentran prote-gidos, y debido a las amenazas existentes sobre los fragmentos de bosque fuera de las áreas protegidas donde existen monos tití, existe una creciente nece-sidad de diseñar redes de conservación entre las áreas fragmentadas y las áreas con alguna categoría de conservación. Para ello se necesita primero eva-luar la situación de las tropas en estos remanentes de
bosque y entender claramente los problemas en el paisaje creados por la fragmentación, para así poder dar una solución para la conservación a largo plazo de las poblaciones de mono tití.
Materiales y métodos Este estudio se realizó del 1 de septiembre de
2005 hasta el 17 de mayo de 2006 en remanentes de bosque ubicados en la franja costera de la boca del río Rincón hasta la boca del río Carate, colindando con la Reserva Forestal Golfo Dulce y el Refugio Nacional Mixto Platanares, en la península de Osa, cantón de Golfito, provincia de Puntarenas en el Área de Conservación Osa, sobre el Pacífico Sur de Costa Rica (figura 1).
Para evaluar las tropas de monos tití y su hábitat se escogieron sitios con cobertura boscosa. Estos sitios se definieron como los sitios utilizados por los monos tití para realizar actividades como forrajeo, desplazamiento o descanso y que no necesariamente cumple con los requisitos para clasificarse como un parche de bosque, ya que no se encuentran delimi-tados en su totalidad por coberturas incompatibles con los monos tití. Algunos de estos sitios fueron bosques secundarios contiguos a bosques primarios, pero utilizados por los monos tití principalmente en el área evaluada (bosque secundario); otros fueron parches o fragmentos de bosque aislados.
El número de sitios a evaluar se escogió según la probabilidad de hallar al menos una tropa de mono tití. Para este fin se realizaron previamente entrevis-tas informales con informantes claves (Ander-Egg 1987), a quienes se les preguntó sobre la localización de las tropas. Del total de sitios visitados y mencio-nados en las entrevistas informales se escogieron 22 sitios de bosque cuando en la primera visita se loca-lizó a un grupo de monos tití.
Estos 22 sitios se visitaron al menos tres veces, cada uno durante la totalidad del período de estudio (aproximadamente nueve meses). La primera visita se realizó durante la época lluviosa (septiembre, octubre y noviembre) y se recorrieron 16 sitios con presencia de monos. Se recorrió cada sitio en su totalidad o hasta detectar una tropa de monos tití. Al detectar una tropa se siguió hasta establecer la dirección de sus movimientos. Cuando la tropa salió del área de bosque (maduro, tacotal, charral) y uti-lizó otro tipo de cobertura diferente a bosque en sus estados sucesionales (plantaciones, frutales, etc.), se registró pero no se siguió sus movimientos, debido a que estas coberturas no serían evaluadas estructu-ralmente.
En la primera visita a cada sitio de muestreo se determinaron las características demográficas (número de individuos y clase de edad) a través de conteos directos (Rudran et al. 1996) cuando fue

36
posible. Posteriormente, en la segunda visita a cada sitio, se establecieron las parcelas al azar para carac-terizar la vegetación (composición florística y estruc-tura horizontal) dentro de cada sitio de bosque y en la dirección de los movimientos de la primera tropa detectada (determinados en la primera visita). Las parcelas de muestreo de vegetación se realizaron durante la transición de la época lluviosa a la seca (diciembre, enero y febrero). Cuando fue posible se verificaron los conteos de avistamientos directos de monos tití durante la evaluación de la vegetación. En este mismo período (transición época lluviosa-seca) se visitaron seis sitios nuevos (Agujas, Bambú, Quebrada Tigre, Ojo de Agua, Osa Vida y Pitosa) y se siguió la misma metodología. La última visita a los sitios de muestreo se realizó durante la época seca (marzo, abril, mayo) con el propósito de com-pletar la evaluación de la vegetación (parcelas de muestreo) en los sitios de bosque nuevos y se realizó observaciones ad libitum de algunas tropas, ya que
durante esta época se dan cambios estacionales en el uso de los sitios de bosque.
Vegetación y relación con las tropas
de monos tití En cada sitio de muestreo delimitado por la tra-
yectoria de los movimientos de la tropa de monos tití y los diferentes tipos de cobertura se establecie-
ron entre 2 y 11 parcelas de 20 x 50 m (0,1 ha) para el muestreo de la vegetación. Las parcelas fueron establecidas dentro de la trayectoria de los movi-mientos de la tropa de monos tití. El número de parcelas dentro de cada sitio se estableció depen-diendo del largo de la trayectoria utilizada por los monos tití. En áreas donde el bosque cubría menos de 50 ha, y la ruta o trayectoria era menor a 1 km, se establecieron como mínimo dos parcelas de mues-treo (cuadro 1). Cuatro parcelas del primer sitio (Puerto Escondido) se eliminaron de los análisis por estar en un sitio de bosque inundable donde los mo-nos tití no fueron vistos.
Cada una de estas parcelas se estableció a partir de un carril central, dejando 10 m a cada lado del carril, cubriendo una franja hacia la derecha y hacia la izquierda del carril. La línea central del carril se colocó dentro de la trayectoria aproximada de los movimientos de la tropa de monos tití determinada en la primera visita al sitio de muestreo. Las parcelas se establecieron de esta manera para tratar de eva-luar la mayor cantidad de área de vegetación utili-zada por los monos tití para sus actividades diarias. Se estimó el número de parcelas que cabrían a lo largo de la ruta y se realizó el mayor número de parcelas dejando al menos entre 50 y 100 m entre cada parcela.
Dentro de cada parcela se contabilizó el número
Figura 1. Área de estudio: de boca de río Rincón a boca de río Carate.

37
de árboles mayores a 5 cm de diámetro a la altura del pecho (DAP). El DAP fue medido con una cinta diamétrica (± 0,50 cm) y la altura fue estimada vi-sualmente en metros (± 1 m). Los árboles y arbustos se identificaron hasta especie con las guías disponi-bles para Costa Rica y la península de Osa (Quesada et al. 1997, Zamora y Pennington 2001). Aquellos
árboles que no pudieron ser identificados in situ se colectaron y se herborizaron, algunos especímenes fueron identificados por el botánico Reinaldo Agui-lar y otros en el Herbario del Instituto Nacional de Biodiversidad. Los árboles que por su altura u otro factor no pudieron ser colectados y los colectados que no fue posible identificar se catalogaron como no identificados. Para cada sitio se determinó el número de árboles y se calculó la riqueza de especies (S) en 0,2 ha, la diversidad de la altura del follaje (FHD), el área basal por hectárea, la diversidad de
DAP (DH), así como la diversidad alfa () de árbo-
les por parcela.
Determinación del tamaño y composición de
las tropas Para determinar el tamaño y la composición de
cada una de las tropas de monos observadas, se contó la totalidad de los miembros de la tropa (cuando fue posible). Los conteos totales por avis-tamientos directos (Rudran et al. 1996) fueron reali-
zados durante los recorridos dentro de los 22 sitios de muestreo escogidos. Se georreferenció la ubica-ción de las tropas, para lo cual se utilizó un receptor Garmin Plus III y un receptor Garmin ETrex. Du-rante el establecimiento de las parcelas temporales de vegetación, también fue posible confirmar los conteos directos efectuados previamente, al igual que se realizó un conteo final en el último trimestre de la toma de datos. Sin embargo, debido al uso estacional del hábitat por las tropas de monos tití, se vuelve menos probable detectarlos en esa época (marzo-abril-mayo), y en algunos de los sitios eva-luados no fue posible efectuar este conteo final.
El sexo y la edad de los individuos por tropa se estimaron de acuerdo con la visibilidad de los geni-tales, en el caso de los machos. Las hembras, juveni-les y crías se estimaron con base en el comporta-miento y el tamaño, aunque se puede hacer uso de otras características morfológicas como depósito de grasa, color del pelaje para la identificación (Boinski 1987, Wong 1991a y b, Kunz et al. 1996).
Análisis de datos La diversidad florística (PH) por sitio se calculó
por parcela usando el índice de Shannon-Wiener, el cual a su vez se promedió por cada sitio, debido a que existieron sitios con más parcelas que otros
(cuadro 1). Se utilizó el programa Species Diversity and Richness 3.02 (Henderson y Seaby 2002) para el cálculo de la diversidad. El número de individuos y especies de árboles por sitio se estandarizó a 0,2 ha para poder realizar comparaciones y por ser ésta el área mínima evaluada en cada sitio de bosque.
Para la estimación de la diversidad de la altura del follaje (FDH) y de la diversidad de clases dia-métricas (DH) en cada sitio de estudio se utilizó el índice de Shannon-Wiener con el programa Species Diversity and Richness 3.02 (Henderson y Seaby 2002). Las alturas estimadas de cada árbol por sitio de bosque se agruparon en siete clases de la siguien-te manera: Clases: 1 = 0 a 5 m; 2 = 5,1 a 10 m; 3 = 10,1 a 15; 4 = 15,1 a 20; 5 = 20,1 a 25 m; 6 = 25,1 a 30 m; 7 = >30 m. Los DAP de cada individuo por sitio se agruparon en seis clases de la siguiente ma-nera: Clases: 1 : 5-14,9; 2 : 15-24,9; 3 : 25-34,9; 4 : 35-44,9; 5 : 45-54,9; 6 : > 55.
Se realizó un escalonamiento multidimensional no métrico (NMS) con PC-ORD 4.0 (McCune & Mefford 1999) para determinar cuáles de los 22 si-tios de muestreo fueron más similares según la abundancia de las 354 especies encontradas en la totalidad de los sitios. Se utilizó la distancia de Bray-Curtis (Sorensen) para el cálculo de la similitud en-tre matrices.
Para el análisis de los datos demográficos, se rea-lizaron solamente estadísticas básicas como prome-
dio de los conteos de avistamiento y se obtuvo la proporción de sexos.
Resultados
Vegetación y relación con las tropas
de mono tití Se estableció un total de 63 parcelas distribuidas
en los 22 sitios evaluados. Dentro de ellas, un total de 354 especies de árboles fueron identificadas y 69 árboles (19%) fueron clasificados como no identifi-cados. Los sitios de bosque ubicados cerca del po-blado La Palma fueron los más diversos dentro del total de 22 sitios. El denominado Cabinas Tití fue el sitio con mayor número de especies (S=69) y con una diversidad florística (HP) más alta H=3,46.
Se midieron un total de 4.416 árboles y sus fustes en los 22 sitios de bosque. Se encontró que la estruc-tura de la vegetación de los sitios fue muy similar. Todos los sitios evaluados poseen una estructura de “jota” invertida (estructura discetánea), donde la mayoría de los individuos medidos dentro de la parcela se encuentran distribuidos en las primeras clases diamétricas (91,64% ± 4,08 DE), o sea, tuvie-ron un DAP menor a 35 cm. Lo mismo ocurre con la altura de los árboles. Para el total de sitios mues-treados, el mayor porcentaje de individuos se en-

38
contró entre los 5 a 10 metros de altura (43,14 % ± 9,96 DE).
Se encontró un promedio de 144 árboles (DE=34) por 0,2 ha para todos los sitios muestrea-dos (n=22). El sitio de bosque con mayor número de individuos fue Bosque del Cabo (234), de los cuales la mayoría de árboles fueron botarrama (Vochysia
guatemalensis, 33). Por otro lado, el sitio con mayor
área basal (G) fue el El Bambú, cerca del poblado Puerto Jiménez (G=46,8) (cuadro 1).
Las especies arbóreas más abundantes dentro de los sitios de bosque muestreados fueron el peine’e mico (Apeiba tibourbou, 161), el Croton shiedeanus
(100), el guácimo (Guazuma ulmifolia, 107), la guaba
de mono o guabilla (Inga thibaudiana, 137), el guá-
cimo colorado (Luehea seemani, 148) y el jobo (Spon-
dias mombin, 129). Sin embargo, algunas de estas
especies fueron más abundantes en unos sitios que otros. En algunos sitios, árboles frutales no nativos como la guayaba (Psidium guayaba) fueron también
abundantes y utilizados por los monos tití como alimento cuando sus frutos están disponibles.
Los sitios ubicados cerca de La Palma son simi-lares en su composición florística, con la excepción de playa Colibrí (NMS: estrés final=13,20859; ines-tabilidad final=0,00001; figura 2). En playa Colibrí existen nueve especies de árboles (20%) que no se encontraron en otros sitios, pero son característicos de sitios modificados por el ser humano, por ejem-
plo, el marañón (Anacardium occidentale), el achiote
(Bixa orellana) y el mamón chino (Nephelium lappa-
eum). Este sitio, además, posee el menor número de
árboles por 0,2 ha (83) y un número menor de árbo-les de la clase diamétrica de 15 a 24,9 cm que los demás sitios.
Se encontró una especie que no ha sido reporta-da para la península de Osa con anterioridad, el árbol Phyllocarpus reidelly. Esta especie se localizó en
cabo Matapalo, en el sitio Osa Vida, el cual es di-símil en composición florística con respecto a los demás sitios de bosque (NMS: estrés fi-nal=13,20859; inestabilidad final=0,00001) (figura 2) y, además, posee uno de los mayores porcentajes de especies de bosque que no se encontraron en ningún otro sitio (18%).
Tamaño y composición de las tropas Se registraron 20 tropas dentro del área de estu-
dio (cuadro 2). La densidad y el total de individuos en el área de estudio no fueron calculados debido a que el registro de las tropas estudiadas no fue ex-haustivo ni excluyente. Se sabe de otras tropas exis-tentes en el área de estudio, pero debido a que se pudo comprobar su existencia apenas al final de la investigación, no se incluyeron en el estudio. Estas tropas se localizaron en: barrio Bonito (Agujas), Platanares y Agua Buena.
Figura 2. Similitud entre los 22 sitios de bosque evaluados según la abundancia de especies de plantas (NMS: estrés final=13,20859,
inestabilidad final=0,00001). ∆= Sitios de bosque; += especies arbóreas.

39
El tamaño promedio de las tropas fue de 19,3 in-dividuos (DE= ± 4,7; n=20). Se observaron tropas de 9 a 28 individuos. La determinación de la com-posición de cada una de las tropas observadas por edad y sexo de los individuos fue difícil debido a la movilidad de los individuos que componen las tro-pas (cuadro 2). Los juveniles, debido a su compor-tamiento, fueron más fáciles de observar. Los pro-medios de individuos adultos, machos adultos, hembras adultas, juveniles y crías fueron 11,8 (DE= ± 4,7), 4,0 (DE= ± 1,6), 7,0 (DE= ± 3,4), 5,5 (DE= ± 2,9), 4,2 (DE= ± 2,6), respectivamente. La pro-porción de sexos y edad encontrada fue de 1M: 2H: 2J:1C.
Discusión
Vegetación y relación con las tropas
de monos tití La mayor diversidad florística encontrada en si-
tios cercanos a la cuenca del río Rincón, cerca del poblado La Palma, puede deberse a que esta cuenca es uno de los sitios donde existe un mayor ende-mismo y diversidad de especies de árboles en la península de Osa (Quesada 1997, Maldonado 1997, Aguilar 2006), a pesar de que esta zona es también la más vulnerable a la fragmentación por su cercanía a actividades humanas como agricultura, ganadería e infraestructura, y el cambio del uso del suelo que estas actividades implican (Rosero-Bixby et al. 2002,
Barrantes y Lobo 2005). La presencia de relictos de bosque primario y
bosques riparios que han permanecido a pesar de la deforestación de la década de los 40, y que se en-cuentran en los sitios de bosque muestreados utili-zados por el mono tití, puede ser fuente de especies arbóreas menos comunes en bosques en regenera-ción y aumentar la diversidad encontrada en algu-nos sitios, como es el caso de Cabinas Tití, sitio de bosque con la mayor riqueza de árboles y que colin-da con bosques primarios de la Reserva Forestal Golfo Dulce.
La estructura horizontal de los 22 sitios de bos-que evaluados, descrita mediante la distribución del número de árboles por clase diamétrica, es una es-tructura discetánea, donde los individuos arbóreos del bosque se encuentran distribuidos en las prime-ras clases diamétricas, que se representa en la distri-bución de “jota” invertida (Louman et al. 2001). Esta
estructura implica que los 22 sitios de bosque eva-luados han sido modificados y se encuentran en proceso de regeneración, o sea, son bosques secun-darios jóvenes. Este tipo de bosque es preferido por los monos tití (Boinski 1987, Wong 1990a y 1990b, Arauz 1993).
Playa Colibrí fue el sitio de bosque más modifi-cado o alterado, con mayor influencia humana. Lo evidencia la presencia de especies únicas caracterís-ticas de sitios usados por el ser humano como el achiote (B. orellana), la palma africana (E. guianensis)
y el marañón (A. occidentale), entre otros. Además, el
bajo número de individuos en la clase diamétrica de 15 a 24,9 cm indica que se ha dado una explotación forestal mayor que en todos los demás sitios. En este sitio se detectó la tropa de monos tití más pequeña, producto de la baja calidad de este hábitat (Baldwin y Baldwin 1981, Wong 1990b, Rodríguez 1999).
Por otro lado, el sitio de bosque playa Colibrí se conectaba por medio de 186 ha de melina (Gmelina
arborea) con el sitio de bosque Puerto Escondido y
con la tropa de monos tití de ese sitio. La tropa de Puerto Escondido fue detectada en playa Colibrí en algunas ocasiones. Además, la plantación fue utili-zada por ambas tropas de mono tití para desplazarse de un sitio a otro, ocasionalmente para alimentarse de los insectos existentes entre las hojas de melina y los frutos de los árboles que forman pequeños rema-nentes de bosque ripario en medio de la plantación. Asimismo, las 186 ha que constituyen la plantación de melina proveen conectividad con otros sitios de bosques. Estas características podrían contribuir con la supervivencia de la tropa de monos tití de playa Colibrí, a pesar de la baja calidad del hábitat y, por lo tanto, esta conexión es crítica en el sitio.
Otros autores (Sierra et al. 2003) han documenta-
do también monos tití usando plantaciones foresta-les como conexión entre fragmentos de bosque; sin embargo, su importancia dentro del paisaje se ve amenazada por la susceptibilidad a los efectos an-tropogénicos y debido a que la conservación de este hábitat depende del uso que el propietario le quiera dar, por ser establecida desde su inicio para el apro-vechamiento maderable.
Hábitat en regeneración, con un gran número de árboles medianos en altura y área basal, como los 22 sitios de bosque estudiados, es preferido por los mo-nos tití en comparación con el bosque primario, debido a la mayor abundancia de alimento, uno de los principales factores que influencian la preferencia de primates por un determinado hábitat (Colishaw 1997, Ostro et al. 2000, Li 2004, Wong et al. 2006).
Los bosques en regeneración, tienen una mayor abundancia de artrópodos debido a que existe ma-yor proporción de biomasa foliar comparados con bosques primarios (Janzen y Schoener 1968, Wong 1990a y 1990b) y a que la diversidad en la altura del follaje (F.H.D; capas de herbáceas, troncos leñosos, sotobosque, entre otros) (MacArthur y MacArthur 1961 [en MacArthur 1972]) que caracteriza a los bosques secundarios, ofrece mayor cantidad de sus-

40
Cuadro 1. Número de individuos y riqueza de especies arbóreas en 0,2 hectáreas,
área basal y número de parcelas realizada por sitio de bosque.
Parche Individuos
(0,2 ha) Especie (0,2 ha)
Área basal (m2/ha)
# Parcelas
Pto. Escondido 133 30 21,4 11 (7*)
Playa Colibrí 83 35 26,1 2
Osa Palma 111 47 42,8 3
Danta Lodge 145 54 29,4 2
Fela 154 63 34,5 2
Köbö 147 57 42,5 2
Monterrey 137 54 20,7 2
Cabinas Tití 179 69 33,9 2
Agujas 122 46 32,8 2
Río Tigre 125 35 19 2
El Bambú 166 36 46,8 4
Río Nuevo 117 32 11,6 2
Quebrada Tigre 154 24 23,3 2
Ojo de Agua 135 59 43,7 2
Lapa Ríos 160 32 25,4 2
Osa Vida 87 33 37,6 2
Bosque del Cabo 234 36 43,7 3
Pitosa 194 27 13,6 2
Piro 174 32 20,1 3
Piro-FOO 157 29 35,8 3
Laguna 120 20 20,4 4
Jungla Camp 127 21 15,3 4
Promedio
(± =D.E.) 143,68 (34,22) 39,59 (14,18) 29,11 (10,85) 2,68 (1,21)
*Número de parcelas utilizadas para el análisis de datos
trato para insectos y frutos, parte de la alimentación de los monos tití.
El bosque primario es utilizado por los monos tití como conexión entre parches de bosques en regene-ración, o estacionalmente, cuando la disponibilidad de alimento como artrópodos, frutos y flores es muy baja en los bosques secundarios (Boinski 1986) y los monos aprovechan frutos de especies que se desarro-llan en el bosque primario (Wong 1990a). Diferen-cias en el uso de un hábitat, dependiendo de la esta-cionalidad de la disponibilidad de alimento, se han encontrado en estudios anteriores con otros primates neotropicales también (Ostro et al. 1999). Esto podr-
ía explicar la baja probabilidad de hallar a los monos tití cuando se realizó el tercer conteo en la mayoría de los sitios de bosque, probablemente por su des-plazamiento en busca de especies fructificando en el bosque primario (Wong 1990b). Esto sugiere que el hábitat de bosque primario debe ser considerado también en futuros estudios.
El número de especies comestibles en cada sitio podría ser una variable importante que influye en su uso por parte de las tropas de monos tití. Los árboles
de las familias Moraceae, Lauraceae y Myristicaceae son fuente de frutos comestibles principalmente durante la época seca, de enero a mayo. Especies arbóreas de estas familias fueron las más abundantes en los sitios de estudio, sin embargo, el tamaño de las tropas de mono tití en los sitios de bosque no puede explicarse solo por la presencia de estas espe-cies comestibles, como se ha encontrado también en otros estudios (Wong et al. 2006). Es necesario con-
siderar, además, todo el espectro de la dieta de los monos tití (insectos) y variables biofísicas como la distancia a otros bosques y la conexión entre sitios de bosques, las cuales aumentan la probabilidad de encontrar tropas de monos tití en un sitio determi-nado (Sáenz y Sáenz 2007), variables que no fueron medidas cuantitativamente en este estudio.
Todos los sitios de bosque evaluados pueden ser clasificados por su estructura como bosques secun-darios, sin embargo son diferentes en su riqueza y composición florística, variables que no se relacio-naron con el tamaño de las tropas de mono tití (So-lano-Rojas 2007).

41
Cuadro 2. Observaciones directas de mono tití en península de Osa: de cuenca baja de río Rincón a cuenca baja de río Carate.
Localidad Coordenadas Altitud Tamaño del
grupo Hábitat Comentarios
1 Lat 8.6694 Lon 83.4606
0-10 m.s.n.m. 22 Bosque secundario Inundable-ripario
2 Lat 8.6542 Lon 83.4349
0-10 m.s.n.m. 9 Bosque secundario Muy intervenido
3 Lat 8.6288 Lon 83.4593
40-90 m.s.n.m. 21 Bosque secundario y prima-rio
Ripario
4 Lat 8.6148 Lon 83.4735
40-60 m.s.n.m. 20 Bosque secundario y prima-rio
Ripario
5 Lat 6.8428 Lon 59.7880
10 m.s.n.m. 23 Bosque secundario y prima-rio
Ripario
6 Lat 8.6081 Lon 83.4465
20 m.s.n.m. 16 Bosque secundario y prima-rio
Ripario
7 Lat 8.5732 Lon 83.3956
12 m.s.n.m. 16* Bosque secundario y prima-rio
Ripario
8 Lat 6.4278 Lon 59.9052
20-50 m.s.n.m. 28 Bosque secundario y prima-rio
Ripario
9 Lat 6.2039 Lon 60.3750
20 m.s.n.m. 16* Bosque secundario Ripario
10 Lat 8.5271 Lon 83.4091
115 m.s.n.m. 20 Bosque secundario Ripario
11 Lat 8.5225 Lon 83.3263
28-70 m.s.n.m. 18 Bosque secundario y prima-rio
Ripario
12 Lat 8.4977 Lon 83.3817
60-170 m.s.n.m. 25 Bosque secundario Ripario
13 Lat 5.1052 Lon 61.6115
0-10 m.s.n.m. 17 Bosque secundario Ripario-muy joven
14 Lat 4.6669 Lon 61.3878
100-150 m.s.n.m. 20 Bosque secundario y prima-rio
Ripario
15 Lat 4.4264 Lon 61.5444
40- 120 m.s.n.m. 20 Bosque secundario Ripario
16 Lat 4.2261 Lon 61.5892
100 m.s.n.m. 18 Bosque secundario Bajura
17 Lat 8.3851 Lon 83.2988
170-230 m.s.n.m. 19 Bosque secundario Ripario-pendiente pronun-ciada
18 Lat 8.3939 Lon 83.3097
135-175 m.s.n.m. 11 Bosque secundario Muy joven
19 Lat 8.4076 Lon 83.3418
20-30 m.s.n.m. 15 Bosque secundario Ripario
20 Lat 8.4018 Lon 83.3395
20-30 m.s.n.m. 20 Bosque secundario Ripario
21 Lat 8.4397 Lon 83.4406
20-40 m.s.n.m. 28** Bosque secundario Inundable - joven
22 Lat 8.4395 Lon 83.4497
0-10 m.s.n.m. 28** Bosque secundario Ripario
*-** En este sitio las tropas son probablemente las mismas
Sitios= 1 Pto. Escondido, 2 Playa Colibrí, 3 Osa Palma, 4 Danta Lodge, 5 Fela, 6 Kobo, 7 Monterrey , 8 Cabinas Tití, 9 Agujas, 10 Río Tigre, 11 El Bambu, 12 Río Nuevo, 13 Queb. Tigre, 14 Ojo de Agua, 15 Lapa Ríos, 16 Osa Vida, 17 Bosque Cabo, 18 Pitosa, 19 Piro, 20 Piro FOO, 21 Laguna, 22 Jungle Camp.
La abundancia de especies arbóreas comestibles
fue más importante. Chapman (2003) también men-ciona la abundancia de plantas de bosque secunda-rio y el aumento de insectos en los fragmentos de bosque como una variable que podría permitir la supervivencia de primates en hábitat fragmentado como es el bosque atlántico brasileño.
Otras investigaciones con primates apoyan tam-bién estos resultados; por ejemplo, Ostro et al. (2000)
encontraron que sitios con árboles más grandes y mayor cobertura relativa de las principales especies consumidas son utilizados cinco veces más por mo-nos congo (A. pigra), incluso cuando estos sitios
tienen una diversidad y riqueza total menor en com-paración con otros sitios con presencia de estos pri-
mates. Oates (1977) también encontró que la com-posición entre sitios donde habita Colobus guereza era
diferente, pero especies de bosque secundario y es-pecies de borde eran comunes, lo que permitía a estos primates explotar incluso hábitat de bosque primario. Wong et al. (2006) también encontraron
que C. vellerosus es capaz de ajustarse a ciertas espe-
cies arbóreas en su dieta para acomodarse a las dife-rencias en composición de especies de flora en los diferentes fragmentos de bosque que utiliza.
En el sitio de bosque La Laguna, donde se en-contró la tropa con mayor número de monos tití y se determinó la menor riqueza de especies arbóreas, se obtuvo un alto porcentaje de especies consumidas por los monos tití como la guabilla (Inga vera) y el
sotobosque fue dominado por un arbusto (Alchornea

42
costarricensis) descrito como hospedero de larvas de
mariposas (DeVries 1987), que son utilizadas como alimento por estos primates (Boinski 1986, Wong 1990b). Se debe tener en cuenta que este estudio no consideró especies de insectos consumidas, ítem importante en la dieta del mono tití en todos los sitios de bosque (observación personal) y que podría estar influenciando también el tamaño de tropas, y en los resultados de este estudio.
Además, Boinski et al. (2003) sugieren que el
riesgo de depredación al cual están expuestas las tropas de mono tití influye en el tamaño de sus tro-pas y que la estructura de la vegetación de un deter-minado tipo de hábitat a su vez influye sobre el ries-go de depredación. Todos los sitios de bosque fue-ron utilizados en el estrato medio y bajo y sus árbo-les poseyeron alturas bajas entre los 5 y 15 mts. El uso de estos estratos podría estar relacionado con la protección contra aves rapaces y otros depredadores
naturales existentes en la zona, por existir mayor cantidad de obstáculos para el vuelo de las aves y el paso de depredadores. También, en los estratos me-dios y bajos de los bosques secundarios existe gran cantidad de árboles medianos y hay mayor disponi-bilidad de alimento (insectos, murciélagos y lagarti-
jas, entre otros) (MacArthur y MacArthur 1961 [en MacArthur, 1972]).
Árboles más altos (mayores a 15 m) fueron usa-dos ocasionalmente, cuando las tropas se desplaza-ban de un sitio de forrajeo hacia otro, o debido a la presencia de una amenaza terrestre. La influencia de potenciales amenazas hacia los monos tití sobre el tamaño de la tropa se debe considerar en próximos estudios también.
Tamaño y composición de las tropas En la península de Osa, en el área de estudio, los
tamaños de tropa variaron de 9 a 28 individuos; en estudios recientes (Morera 2007), en el sector del área de amortiguamiento del Parque Nacional La Amistad-Pacifico (21.307 ha) se encontró grupos de entre 7 y 68 individuos por tropa. Además, se en-contró un mayor porcentaje de hembras, seguido por los juveniles, semejante a lo encontrado en este
estudio. Sin embargo, existen diferencias marcadas en la composición de tropas en las diferentes investi-gaciones realizadas en el área de distribución del género Saimiri (Rodríguez 1999). Algunos autores
sugieren que existen errores de apreciación por parte de los investigadores en la distinción de la edad y el sexo y que para evitar estos sesgos es recomendable
Mono tití Eduardo Carrillo

43
estudios a largo plazo (Baldwin y Baldwin 1981, Rodríguez 1999). Boinski y Sirot (1997) sugieren que una tropa debe estar compuesta por 40% de hembras adultas, 10% de machos adultos, 25% de juveniles y 25% de crías.
El promedio de individuos por tropa en los sitios de bosque evaluados en este estudio fue de 19,3, promedio menor al reportado para una sola tropa dentro del área protegida PNC, que fue de 41 indi-viduos (Boinski 1986). Para S. o. oerstedii se ha repor-
tado en Panamá 18,5 individuos por tropa (Rodrí-guez 1999), con un mínimo de cuatro y un máximo de 48 individuos. En áreas fragmentadas del Pacífi-co Central de Costa Rica se han detectado prome-dios de tropas de 32 individuos para S. o. citrinellus
con un rango de 15 a 80 individuos por tropa y un promedio de tres crías por tropa (9% a 25% crías por tropa) (Sierra et al. 2003) y de 14 a 80 individuos
por tropa en estudios más recientes (Sáenz y Sáenz 2007).
En estos agropaisajes del Pacifico Central se ha reportado tamaños promedio de tropas de 19 indivi-duos en plantación de frutales y de 23 en plantacio-nes de palma africana, estando estos promedios siempre dentro del rango encontrado en parches de bosque (29,14 ±13,9 individuos) (Estrada et al.
2005). Sáenz y Sáenz (2007) en el Pacifico Central concluyen que los tamaños de tropa no fueron dife-rentes de los reportados para áreas continuas o bos-ques protegidos, lo cual hace viable la conservación de los primates en áreas fragmentadas, siempre y cuando existan bosques continuos cercanos y co-nexión entre los diferentes sitios de bosque utiliza-dos por el mono tití.
Sin embargo, en estos sitios fragmentados y sin protección estatal, donde además los animales son atrapados o cazados, el tamaño de las tropas dismi-nuye debido a la relación que existe entre el tamaño de ellas y la calidad y el tipo de hábitat usado (Baldwin y Baldwin 1981, Wong 1990b, Rodríguez 1999). Por ejemplo, en playa Colibrí, sitio de bosque que resultó ser el sitio más degradado y con mayor influencia humana (composición florística y menor número de árboles que otros sitios), se detectó la tropa de monos tití más pequeña. En este sitio pro-bablemente la disponibilidad de alimento es menor, sumado a la cercanía de las actividades humanas como siembra de cultivos no arbolados de subsisten-cia (frijoles, maíz, etc.).
En el Pacifico Central, Wong (1990b) detectó 14 tropas que variaron entre 22 y 66 individuos, con un valor promedio de 41 individuos por tropa. Las tro-pas menos numerosas y la menor densidad se encon-traron en las zonas aledañas al Parque Nacional Manuel Antonio (PNMA), fuera de las áreas prote-gidas, las dos tropas más cercanas a la ciudad de
Quepos. Sin embargo, Wong (1990b) no encontró diferencias entre los tamaños de grupos dentro del Parque Manuel Antonio y los grupos encontrados afuera, que van de 28 a 66 y de 22 a 63 respectiva-mente. Para el periodo en que se efectuó el estudio, el bosque dentro del PNMA estaba en proceso de regeneración, lo cual seguramente influyó en este resultado. Además, existieron ocho tropas utilizan-do bosques fuera del Parque mientras que dentro se localizaron seis grupos.
En el área que comprende este estudio, los sitios donde existen tropas de mayor tamaño y mayor número de tropas de mono tití y de las otras tres especies de monos simpátricas, es en aquellos donde el bosque continuo es más cercano y donde existe un menor porcentaje de potreros y cultivos, es decir, en los sitios de bosque de cabo Matapalo hacia Carate. Resultados similares han sido encontrados en Brasil, en un paisaje fragmentado, donde todos los frag-mentos de bosque con presencia de monos congo (Alouatta guariba clamitans) se localizaron a una dis-
tancia no mayor de un kilómetro de otros fragmen-tos de bosque de galería (Ribeiro y Bicca-Marques 2005).
En Panamá, Rodríguez-Vargas (2003) ha men-cionado la vulnerabilidad de los grupos de monos tití en ese país, debido a que algunos grupos no están conectados con otros dentro del paisaje, otros están aislados y esto los torna más vulnerables. Adicio-nalmente, existe gran controversia sobre cuál es el tamaño mínimo que una población de animales necesita para asegurar su supervivencia en un largo plazo, ya que esto dependerá de cada especie (Chia-rello 2000).
Chiarello (2003) remarca la importancia de to-mar en cuenta, para cada especie, la movilidad entre sitios de bosque en la matriz circundante. Si las tro-pas son capaces de utilizar pastos y cultivos para pasar de un sitio de bosque a otro, las subpoblacio-nes de una metapoblación se mantendrían interco-nectadas y en ese escenario subpoblaciones de déci-mas de individuos podrían sobrevivir al estar de-mográfica y genéticamente en contacto una con otra (Chiarello 2003). Los monos tití, en el área de estu-dio, fueron observados cruzando por cercas de alambre y cultivos (observación personal); habitan-tes de las comunidades mencionaron verlos cruzar calles de cinco metros de ancho que dividen sitios de bosque, siempre y cuando no exista tránsito frecuen-te. Sin embargo, son más vulnerables a ataques por perros o a otras amenazas cuando realizan este tipo de actividad.
Se debe recalcar, además, que a pesar de que to-das las especies del género Saimiri son ecológica-
mente similares con respecto a sus preferencias de hábitat y alimento, existen diferencias en la distribu-

44
ción de los recursos alimenticios a través de su rango de distribución geográfica, lo que ha influenciado la divergencia de su organización social y la forma en que compiten por los recursos (Terborgh 1983, Boinski 1987b, Boinski 1989, Wong 1990 a y b, Cropp et al. 2002). Esto a su vez puede influenciar
las diferencias en los tamaños de tropas de Centro-américa con respecto a sus congéneres de Suraméri-ca, además de considerar el tamaño de los bosques utilizados y la interconexión entre ellos; y las enfer-medades, componentes críticos a considerar en la biología de la conservación de las especies (Baldwin y Baldwin 1971, Anderson 1979, May 1998, Scott 1998, Laboratorio de Primatología de los Tuxtlas 2004).
Por otro lado, el riesgo de depredación es otro factor que puede estar influyendo en el tamaño de las tropas de mono tití (Colishaw 1997, Boinski et al.
2003). En la zona, uno de los principales depre-dadores son las águilas grandes, sobre todo las blan-cas y/o crestadas (Boinski et al. 2003), las cuales
fueron más comunes en las zonas colindantes con la Reserva Forestal Golfo Dulce y hacia cabo Matapalo y Carate (observación personal). En estos sitios se encontraron tropas más grandes, lo cual se ha relacionado con una reducción del riesgo de depredación. Sin embargo, tamaños de tropa más grandes aumenta la competencia por recursos alimenticios, entre otras cosas (Boinski et al. 2003).
Estudios acerca de las amenazas potenciales y los depredadores son necesarios para llegar a conclusiones más determinantes en este sentido.
Implicaciones para el manejo y conservación
del mono tití Los monos tití están habitando sitios de bosque
con una composición florística diferente entre sí. Esta diversidad de bosques secundarios, compuesta por el 50% de los árboles descritos en la península de Osa, debe ser protegida. El uso de herramientas de conservación privada (pago de servicios ambien-tales, servidumbres ecológicas) debe ser incentivado en la zona por organizaciones no gubernamentales locales e instituciones gubernamentales.
Se debe conservar al menos un sitio boscoso ca de cada poblado para preservar la diversidad florística de la península. Sitios como Cabinas Tití, Danta Logde, Osa Palma, Fela, cabo Matapalo y El Bambú, son prioritarios para la conservación de la diversidad florística y, además, contienen tropas con importante número de individuos. Algunos de estos sitios se encuentran en régimen de conservación voluntaria, lo que facilita su conservación. Los sitios
de bosque El Bambú y Fela son sitios donde aún falta incentivar y/o fortalecer herramientas de con-servación voluntaria para asegurar la existencia de estos bosques a largo plazo.
Es necesario manejar los bosques secundarios y agropaisajes fuera de áreas protegidas con el fin de mantenerlos en regeneración, debido a su importan-cia como hábitat para los monos tití y otros prima-tes, así como otros mamíferos. Las plantaciones forestales de melina, teca, amarillón, entre otras, así como las plantaciones de palma aceitera, cumplen un rol muy importante para la conservación del mono tití: proveen conexión entre diferentes sitios boscosos, son hábitat de insectos parte de la dieta de los monos tití y brindan protección a las tropas mientras se desplazan de un sitio a otro, por lo que su aprovechamiento debe ser regulado estrictamen-te, asegurando no perder la conectividad entre los sitios boscosos utilizados por el mono tití.
Estudios futuros deben enfocarse en el enrique-cimiento y mejoramiento de la calidad de hábitat de bosques secundarios jóvenes y mejorar conexiones entre sitios boscosos, así como el estudio del uso de los bosques primarios como parte del hábitat necesa-rio para el buen desarrollo de las tropas de monos tití encontradas durante este estudio.
Referencias bibliográficas Ander-Egg, E. 1987. Técnicas de investigación social. El Ateneo.
México. Anderson, R. “Parasite pathogenicity and the depression of host population equilibria”, en Nature 279, 1979.
Arauz, J. 1993. Estado de conservación del mono tití (Saimiri oerstedii
citrinellus) en su área de distribución original. Tesis de Maestría en Vida
Silvestre. Programa Regional de Manejo de Vida Silvestre. Universi-dad Nacional. Costa Rica. Baldwin, J. y J. Baldwin. “Squirrel Monkeys (Saimiri) in natural
habitats in Panamá, Colombia, Brasil and Perú”, en Primates. 12 (1),
1971. Baldwin, J. y J. Baldwin. “The ecology and behavior of squirrel monkeys (Saimiri oerstedii) in natural forest in western Panamá”, en
Folia Primatologica 18, 1972.
Baldwin, J. y J. Baldwin. “The squirrel monkey, genus Saimiri”, en
Coimbra-Filho, A. y R. Mittermeier (eds.). 1981. Ecology and behavior
of neotropical primates. Vol. 1. Academia Brasileira de Ciencias.
Barrantes, G. y J. Lobo. “Conservación y deforestación”, en Lobo, J. y F. Bolaños (eds.). 2005. Historia Natural de Golfito. Editorial Inbio.
Costa Rica. Bicca-Marques, J. C. “How do howling monkeys cope with fragmen-tation?”, en Marsh, L. (ed). 2003. Primates in Fragments. Ecology and
conservation. Kluwer Academic/Plenum Publishers. USA.
Boinski, S. 1986. The Ecology of Squirrel Monkey in Costa Rica. Disserta-tion. Presented to the Faculty of the Graduate School of The University of
Texas at Austin. United States.
Boinski, S. “Status of the squirrel monkey (Saimiri oerstedii citrinellus)
in Costa Rica”, en Primate Conservation 8, 1987a.
Boinski, S. “Habitat use by squirrel monkeys (Saimiri oerstedii) in
Costa Rica”, en Folia Primatologica 49, 1987b.

45
Boinski, S. “The positional behavior and substrate use of squirrel monkeys: Ecological Implications”, en J. Hum Evol 18, 1989.
Boinski, S. y L. Sirot. “Uncertain conservation status of squirrel
monkeys in Costa Rica, Saimiri oerstedii oerstedii and Saimiri oerstedii
citrinellus”, en Folia Primatol. 68, 1997.
Boinski, S. et al. “Squirrel monkeys in Costa Rica: drifting to extinc-
tion”, en Oryx 32, 1998.
Boinski, S. et al. “Are vigilance, risk from avian predators and group
size consequences of habitat structure? A comparison of three species of Squirrel monkey (Saimiri oerstedii, S. boliviensis, and S. sciureus)”, en
Behaviour 140, 2003.
Chapman, C. y L. Chapman. “Survival without dispersers? Seedling recruitment under Parents”, en Cons. Bio. 9, 1995.
Chapman, C. A. y D. A. Onderdonk. “Forests without primates: Primate/plant Codependency”, en Am. J. Primatol. 45, 1998.
Chapman, C. et al. “Primate survival in commu-
nity-owned forest fragments: Are metapopulation models useful amidst intensive use?”, en Marsh, L. (ed). 2003. Primates in Fragments. Ecology and
conservation. Kluwer Academic/Plenum Publish-
ers. USA. Chiarello, A. G. “Density and Population Size of Mammals in Remnants of Brazilian Atlantic Forest”, en Conservation Biology 14(6), 2000.
Chiarello, A. G. y F. R. de Melo. “Primate Population Densities and sizes in Atlantic Forest. Remnants of Northen Espirito Santo, Brazil”, en International Journal of Primatology 22, 2001.
Chiarello, A. G. 2003. “Primates of the Brazilian Atlantic forest: the influence of forest fragmenta-tion on survival”, en Marsh, L. K. (ed.) 2003. Primates in Fragments: Ecology and Conservation.
Kluwer/Plenum Press. New York. Cropp, S., S. Boinski y W. Li. “Allelic Variation in the Squirrel Monkey X-Linked Color Vision Gene: Biogeographical and Behavioral Corre-lates”, en J Mol Evol 54, 2002.
Dominy, N. J. y B.W. Duncan. “Seed-spitting
Primates and the Conservation and Dispersion of Large-seeded Trees”, en International Journal of
Primatology 26(3), 2005.
DeVries, P. J. 1987. The butterflies of Costa Rica and
their natural history. Princeton University Press.
Princeton NJ. Estrada, A. et al. 2005. “Primates in agroecosys-
tems: conservation value of some agricultural practices in Mesoamerican Landscape”, en Estrada, A. et al. (eds.). New perspectives in the study of mesoamerican primates: Distribution, ecology,
behavior and conservation. Kluwer/Springer Press.
Hanski, I. “Metapopulation dynamics: from concepts and observations to predictive models”, en Hanski, I. y M. E. Gilpin (eds.). 1997. Metapo-
pulation biology: ecology, genetics and evolution.
London Academic. Franklin, I. R. y R. Frankham. “How large must population be to retain evolutionary potential?”, en Animal Conservation 1, 1998.
Janzen, D. y T. Schoener. “Differences in insect abundance and diversity between wetter and drier sites during a tropical dry season”, en Ecology 49,
1968. Kunz, T., C. Wemmer y V. Hayssen. “Sex, Age, and Reproductive Condition of Mammals” (Appendix 5), en Wilson, D. et al. (eds). 1996. Measuring and Monitoring Biological Diversity.
Standard Methods for Mammals. Smithsonian
Institution Press. Washington. Laboratorio de Primatología de los Tuxtlas. 2004. www.primatesmx.com/inicio.html. Li, Y. “The effect of forest clear-cutting on habi-tat use in Sichuan snub-nosed monkey (Rhinopi-
thecus roxellana) in Shennongjia Nature Reserve”,
en China Primates 45, 2004.
Louman, B., A. Mejía y L. Núñez. “Inventarios Especiales”, en
Orozco, L. y C. Brumer. 2002. Inventarios Forestales para Bosques
Latifoliados en América Central. CATIE. Costa Rica.
Maldonado, T. 1997. Uso de la Tierra y Fragmentación de Bosques.
Algunas Áreas Críticas en el Área de Conservación Osa, Costa Rica. Centro
de Estudios Ambientales y Políticas. Fundación Neotrópica. San José. Marsh, L. K. “Wild Zoos: Conservation of Primates in situ”, en
Marsh, L. K. (ed.) 2003a. Primates in Fragments:Ecology and Conserva-
tion. Kluwer/Plenum Press. New York.
Marsh, L. K. “The Nature of Fragmentation”, en Marsh, L. K. (ed.) 2003b. Primates in Fragments: Ecology and Conservation. Kluw-
er/Plenum Press. New York.
Mono tití Luis Diego Alfaro

46
MacArthur, R. 1972. Geographical Ecology. Patterns in the Distribution
of Species. Princeton University Press. New Jersey.
McCune, B. y J. Mefford. 1999. PC-ORD. Multivariate Analyses of
Ecological Data. Version 4.34. Oregon.
Morrison, M. L. 2002. Wildlife Restoration. Techniques for Habitat
Analysis and Animal Monitoring. Island Press. Washington.
Morera, R. 2007. Análisis de la situación del mono tití en el área de amor-tiguamiento del Parque Internacional La Amistad. II Simposio de Primates en Costa Rica: Estrategia Nacional para su Conservación. Libro de resúme-
nes. San José.
Ostro, L. E. T. et al. “Ranging behavior of translocated and estab-
lished groups of black howler monkeys Alouatta pigra in Belize,
Central America”, en Biological Conservation 87, 1999.
Ostro, L. E. T. et al. “Habitat selection by translocated black howler
monkeys in Belize”, en Animal Conservation 3, 2000.
Quesada, F. J. et al. 1997. Árboles de la península de Osa. Inbio-Sida.
Costa Rica. Ribeiro, S. y J. C. Bicca-Marques. “Landscape characteristics and their influence on the occurrence of Brown howling monkeys (Aloua-
tta guariba clamitans Cabrera) in forest fragments in the Vale do Ta-
quari, RS – Brazil”, en Naturaza & Conservacao 3(2), 2005.
Rodríguez, A. 1999. Estatus de la Población y Hábitat del Mono Tití,
Saimiri oerstedii, en Panamá. Tesis de Maestría en Vida Silvestre.
Programa Regional de Manejo de Vida Silvestre. Universidad Na-cional. Costa Rica. Rodríguez-Toledo, E., S. Mandujano y F. García-Orduña. “Rela-tionships between forests fragments and howler monkeys (Alouatta
palliata mexicana) in southern Veracruz, México”, en Marsh, L. K.
(ed.) 2003. Primates in Fragments: Ecology and Conservation. Kluw-
er/Plenum Press. New York. Rodríguez-Vargas, A. “Analysis of the hypothetical population structure of the Squirrel Monkey (Saimiri oerstedii) in Panamá”, en
Marsh, L. K. (ed.) 2003. Primates in Fragments: Ecology and Conserva-
tion. Kluwer/Plenum Press. New York.
Rosero-Bixby, L., T. Maldonado-Ulloa y R. Bonilla-Carrión. 2002. “Bosque y población en la Península de Osa, Costa Rica”, en Rev.
Biol. Trop. 50(2), 2002.
Rundran, R. et al. “Observational Techniques for Nonvolant Mam-
mals” (Chapter 6), en Wilson, D. et al. (eds). 1996. Measuring and
Monitoring Biological Diversity. Standard Methods for Mammals. Smith-
sonian Institution Press. Washington. Sáenz, J. y P. Sáenz. “Influencia de las variables de hábitat y paisaje sobre la presencia-ausencia del mono tití y mono carablanca en un área fragmentada del Pacífico Central de Costa Rica”, en Harvey, C. y J. Sáenz, (eds). 2007. Evaluación y conservación de biodiversidad en
paisajes fragmentados de Mesoamérica. Inbio. Costa Rica.
Saunders, D., R. Hobbs y C. Margules. “Biological consequences of Ecosystem Fragmentation: A Review”, en Conservation Biology 5(1),
1991. Sierra, C. et al. “New Data on the Distribution and Abundance of
Saimiri oerstedii citrinellus”, en Primate Conservation 19, 2003.
Solano-Rojas, D. 2007. Evaluación del hábitat, paisaje y la población del mono tití (Cebidae, Plathyrrini: Saimiri oerstedii oerstedii) en la Península
de Osa, Costa Rica. Tesis de Maestría. Instituto Internacional de
Conservación y Manejo de Vida Silvestre. Universidad Nacional. Costa Rica. Terborgh, J. 1983. Five New World primates: A study in comparative
ecology. Princeton University Press. Princeton NJ.
Turner, M., R. Gardner y R. O´Neill. 2001. Landscape ecology in
theory and practice. Springer Science-Business Media Inc. USA.
Wong, G. 1990a. Ecología del mono tití (Saimiri oerstedii citrinellus) en el
Parque Nacional Manuel Antonio, Costa Rica. Tesis de Licenciatura.
Universidad Nacional. Costa Rica. Wong, G. 1990b. Uso de hábitat, estimación de la composición y densidad poblacional del mono tití (Saimiri oerstedii citrinellus) en la zona de Manuel
Antonio, Quepos, Costa Rica. Tesis de Maestría. Programa Regional en
Manejo de Vida Silvestre. Universidad Nacional. Costa Rica. Wong, G. “Mono tití en extinción”, en Ambientico 107, 2000.
Wong, S., T. Saj y P. Sicotte. “Comparison of habitat quality and diet of Colobus vellerosus in forest fragments in Ghana”, en Primates
47, 2006. Zamora, N. y T. D. Pennington. 2001. Guabas y cuajiniquiles de Costa
Rica (Inga spp.). Instituto Nacional de Biodiversidad. Costa Rica.
Entrevistas Aguilar, R. 2006. Comunicación Personal.

47
Normas mínimas para la presentación de artículos a la revista
Ambientales con vistas a su publicación
Formato de presentación
El artículo ha de ser presentado impreso en papel -a doble
espacio- y en soporte informático –a espacio sencillo- en lenguaje
Word.
Resumen o abstract El artículo debe estar precedido por un resumen (en el que,
¡atención!, debe evitarse dar información adicional al artículo, ni
de presentación de él) de unos 10 renglones con su traducción al
inglés.
Palabras claves Debajo del resumen habrá de consignarse palabras clave -con
su traducción en inglés- que indiquen al lector los temas princi-
pales del documento.
Tamaño y características tipográficas El artículo no debiera exceder las 9.000 palabras, o sea: unas
12 páginas tamaño carta (8 ½ x 11 pulgadas), a espacio sencillo y
escritas en letra Times New Roman tamaño nº 11.
Apoyos gráficos Las figuras, cuadros e ilustraciones que el artículo contuviera
o llevara como anexos -que se justifican solo si hacen al artículo verdaderamente más comprensible- deberán caber en aquella
docena de páginas, disminuyendo así el número de palabras del
estricto texto y habrán de ser entregadas dos versiones de la
misma imagen, figura o gráfico, ambas en formato JPG. La
primera en alta resolución y en blanco y negro y, la segunda, en mediana resolución y a color (ésta última para la edición digital
de la revista). Respecto de tales gráficos, figuras e ilustraciones,
es importante que en el texto venga señalado, entre corchetes, los
lugares en que preferiblemente deben aparecer (los cuadros sí pueden venir en el mismo archivo del texto del artículo en len-
guaje Word).
Las figuras, cuadros e ilustraciones que se incluya, además
de tener soporte informático, deberán ser acompañados de una muy nítida impresión en papel (recuérdese que la revista no usa
colores en su interior). A cada uno de esos apoyos debe corres-
ponder una o varias referencia(s) en el texto. Tales figuras e
ilustraciones deben ser adjuntadas en formato jpg, a color, además de la versión blanco y negro para la edición impresa. Los
cuadros que vengan como imágenes y no como tablas, deberán
ser también enviados adjuntamente en formato jpg.
Los títulos de figuras e ilustraciones van situados al pie de ellas. Los de los cuadros van situados encimas de éstos.
Estilo de los subtítulos En el artículo puede haber subtítulos de tres rangos distintos
que deben distinguirse claramente.
Los subtítulos de primer rango deberán escri-
birse en letra tamaño 14 sin negrita;
los de segundo rango en letra tamaño 12 con ne-
grita,
y los de tercer rango en letra tamaño 11 en cursivas sin
negrita.
Citas textuales Citas textuales no han de ponerse en cursivas, ni usando
sangría ni en párrafo aparte, sino entrecomillando.
Comunicaciones personales y entrevistas
La mención en el texto de comunicaciones personales o en-
trevistas se hará así: luego de una apertura de paréntesis se
consigna el apellido del entrevistado y, después, y sin ningún
signo de puntuación, por el año en que se efectuó la comunica-
ción (sin hacer la tradicional aclaración de Com. pers.). Ejemplo:
(Obama 1999.)
Las entrevistas referenciadas se consignarán en una subsec-ción de la sección Referencias bibliográficas (en letra 14), llama-
da Entrevistas (subtítulo, éste, escrito con negrita y letra tamaño
nº 12).
Notas al pie de página Aparte de las referencias bibliográficas, y solo en casos impres-
cindibles, podrá hacerse notas a pie de página con el fin de ir
aclarando o ampliando al lector (se aconseja insertar el conteni-do de las potenciales notas-al-pie en el texto, entre paréntesis).
Uso de cursivas y uso (excepcional) de comillas (nunca negri-
tas, ni subrayado) Se usará cursivas para enfatizar conceptos, pero no negritas,
ni subrayado ni mayúsculas. Vocablos en otras lenguas no acep-
tados por la Real Academia Española de la Lengua, y neologis-mos, han de escribirse también en cursivas. Asimismo, irán en
cursivas nombres de obras de teatro y cinematográficas, de li-
bros, de folletos, de periódicos, de revistas y de documentos, y
también de capítulos de libros y de artículos de publicaciones periódicas cuando a la par no se esté mencionando el nombre del
libro o la publicación que los contiene –en caso de hacerse esto
último el capítulo o artículo se escribirá entre comillas-.
Uso de números Cuando las cantidades sean escritas numéricamente ha de
usarse la coma solo antes de los decimales (no usar puntos); y el
punto para señalar miles y millones (no usar comas). Las unidades de medida, en caso de consignarse abreviadamen-
te, habrán de escribirse en singular y en minúsculas. Ejemplo:
“... la bestia, de 100 k de peso, recorrió 90 m antes de caer exan-
güe...”.
Originalidad Los artículos publicados en la revista deben de ser estricta-
mente originales, es decir, no pueden haber sido publicados con anterioridad en ninguna otra revista o medio de comunicación
escrito.
Información del autor Dado que la revista, en la primera página de cada artículo
hace una muy breve presentación del autor, en dicha página,
abajo del nombre del autor, habrá de consignarse la especializa-
ción científica y los cargos -académicos, gubernamentales o de otro tipo- de él. Ejemplo: El autor, biólogo especialista en mane-
jo de vida silvestre, es director del Instituto de Investigaciones
sobre Animales Hipotéticos.
Inicio

48
Referencias bibliográficas (dentro del texto) Las referencias bibliográficas deben hacerse intercaladas en
el texto, entre paréntesis, con los siguientes datos:
Libros:
El apellido del autor al que se hace referencia, el año de pu-
blicación de la obra y, en caso necesario, la(s) página(s). Por
ejemplo: (Pérez 1999: 83) Artículos de revistas y periódicos:
Tratándose de un artículo de revista se procederá igual. Tratándose de un artículo aparecido en un periódico habrá de
consignarse la fecha de manera abreviada y apretada. Ejemplo:
(Núñez 16-3-09: 22).
Si el artículo o pieza periodística careciera de autor explícito habría que consignar, en vez de éste, el nombre del periódico en
letras cursivas. Ejemplo: (La Nación 15-3-99: 1)
Presentación de la bibliografía Al final del artículo, debajo del subtítulo Referencias bi-
bliográficas (de primer rango), habrá de consignarse todas las obras referenciadas:
1. Libros:
Primero se anotará el apellido del autor, luego, precedido de una coma, su nombre; después, e inmediatamente luego de un
punto, el año de publicación de la obra; seguidamente, y en
cursivas, el título de la obra; posteriormente, y después de un
punto, el nombre de la editorial, y, finalmente, antecedido por un punto, el lugar de publicación de la obra (si la editorial tiene
como parte de su nombre la palabra “editorial”, ésta debe poner-
se, pero, si no, no; si la ciudad es internacionalmente conocida
no hace falta señalar el país, pero, si no es reconocida fácilmente, solo el país). Ejemplo: Pérez, Juan. 1999. La ficción de las áreas
silvestres. Anagrama. Barcelona.
2. Artículo contenido en un libro o en una revista:
Si lo referenciado es un artículo contenido en un libro o en
una revista, el artículo deberá consignarse entre comillas, des-
pués de lo cual, seguido de una coma, habrá de ponerse la prepo-
sición “en” y luego el nombre del libro o revista –éste sí con cursivas-. Para indicar la fecha de publicación procédase según el
caso:
2.1. Artículo contenido en un libro de varios autores:
La fecha de publicación se anotará inmediatamente des-
pués del nombre del autor (o compilador, o editor) al que se
le atribuye principalmente el volumen, y no del autor del artículo. Ejemplo: Fernández, Pedro. “Las huellas de los di-
nosaurios en las áreas silvestres protegidas”, en Ordóñez,
Jaime. 1950. Aunque usted no lo crea. Editorial Aguaclara.
Managua.
2.2. Artículo contenido en una revista:
La fecha iría después del nombre de la revista (que se es-
cribiría en cursivas) y del número de edición de la misma.
Ejemplo: Fernández, Pedro. “Las huellas de los dinosaurios
en áreas silvestres protegidas”, en Fauna prehistórica 15, enero
de 1999. Costa Rica.
3. Periódicos:
Si la referencia fuera a un diario o semanario habría de pro-
cederse igual que si se tratara de una revista, con la diferencia de
que la fecha de publicación se consignará abreviada y apretada-mente. Ejemplo: Núñez, Alicia. “Descubren vida inteligente en
Marte”, en La Nación 16-3-99.
Y si el artículo o pieza periodística referenciada careciera de
autor explícito habría que anotar como autor el nombre del
órgano de prensa, en letras cursivas (porque es a la vez autor y
nombre de la obra). Ejemplo: La Nación. “¡Vida en Marte!”. 15-
3-09.
4. Documentos en línea:
Si se trata de un autor personal se anotará el apellido de éste, luego, precedido de una coma, su nombre; en el caso de un
autor corporativo, se pone la sigla oficial, seguida entre parénte-
sis del nombre completo de la institución, y el país donde está
localizada en forma abreviada. Ejemplo: C.R.; en el caso de que carezca de siglas, se introduce directamente el nombre de la
institución. Si el documento no tiene la indicación del autor
personal o corporativo, y carece de editor (es) responsable (s), la
referencia bibliográfica se detalla por el título de la publicación. Posteriormente, se introduce el año de publicación, salvo en
aquellas obras anónimas en las cuales se indica el año a conti-
nuación del título, separado por punto y un espacio. Si no ha
sido posible identificar el año de publicación, se registra en la
referencia bibliográfica la del derecho de autor (copyright). Si este dato se desconoce pero es posible estimar el año aproxima-
do de publicación, se indica esta información con signo de
interrogación. Ejemplo: ¿1998?. En los casos en que no se ha
podido determinar la fecha de publicación de un documento, esta condición se señala con la anotación s.f. (sin fecha).
Inmediatamente, luego de un punto, se incorpora el título
de la obra o artículo, la referencia bibliográfica, entonces, se
sigue elaborando tal y como se especifica en los puntos 1, 2 y 3, sea un libro, un artículo de revista, o de periódico, respectiva-
mente; si el material carece de título, redáctese uno de acuerdo
al tema que se describe y anóteselo encerrado entre paréntesis.
Posteriormente, se incorpora la fecha en que se consultó el material; este dato se indica después de la ciudad o el país de la
publicación, separado por un punto y anotando el término “con-
sultado”; para la mención de la fecha, se registra primero el día,
luego el mes abreviado y por último el aña. Ejemplo: 10-6-2001.
Finalmente, e inmediatamente luego de un punto, se señala la dirección electrónica en la que el documento está disponible
anotando el término “en” seguido de dos puntos y el enlace
electrónico.
5. Números de páginas:
No habrá de consignarse número de páginas de libros ni de
artículos referenciados, ni tampoco el dato referente a en qué página comienza y en cuál termina un artículo.
6. Autores múltiples:
Cuando el texto referenciado tenga dos o tres autores, el primero será consignado empezando por el apellido, seguido de
la inicial de su nombre, y el segundo y el tercero por la inicial de
sus nombres, seguido por el primer apellido, y entre el segundo
autor y el tercero (o entre el primero y el segundo si solo fueran dos) se pondrá la conjunción “y”. Ejemplo: Fernández, J., A.
Otárola y M. Sáenz. 1985. La enfermedad principal de las vacas.
Euned. San José.
Tratándose de más de tres autores, después de la consigna-
ción del primer autor, con apellido y nombre completo, se escri-
birá et al. (en cursivas), y se prescindirá de consignar los nombres
de los otros.


















