
TEMA%9% …aulaoncologia.com/AulaOncologia/Downloads/curso_onco_molecular/... ·...
17
1 TEMA 9 CARACTERÍSTICAS INMUNOLÓGICAS DE CÁNCER El sistema inmunitario se encarga de defender al individuo de sustancias extrañas, a través de la protección por la inmunidad innata y la inmunidad adaptativa. En el caso del cáncer, una cuestión que no se ha resuelto todavía es si el sistema inmune favorece o impide el desarrollo de tumores. Por una parte, se conoce que los cánceres asociados a infecciones están aumentando a una velocidad alarmante y que, los tumores con capaces de crear un ambiente inflamatorio que les ayuda a progresar a estados más malignos. Sin embargo, también se sabe que los pacientes cuyos tumores poseen infiltraciones de células inmunes tienen un mejor pronóstico y que las personas inmunodeprimidas son más propensas a padecer cáncer, que las que no lo son. Así, parece que el sistema inmune vigila la formación de tumores para evitar que estos progresen. No obstante, las células cancerígenas son capaces de adquirir la capacidad de evadir la vigilancia del sistema inmune y limitar el ataque de éste, lo que las posibilita a formar tumores. En este tema se estudia la relación entre el sistema inmune y las células cancerígenas, analizando el papel que tiene la inflamación en la carcinogénesis, así como los mecanismos que adquieren las células cancerígenas para evitar la vigilancia y el ataque del sistema inmune. Además se analizan las nuevas estrategias de inmunoterapia para combatir el cáncer. ÍNDICE CARACTERÍSTICAS INMUNOLOGÍCAS DEL CÁNCER INFLAMACIÓN PROMOVIDA POR TUMORES Células inmunes inflamatorias INMUNOEDICIÓN DEL CÁNCER CAPACIDADES INMUNOLÓGICAS ADQUIRIDAS POR LAS CÉLULAS TUMORALES Capacidad para prosperar en un microambiente de inflamación crónica Capacidad para evadir el reconocimiento inmune Capacidad para suprimir la reactividad inmunitaria ESTRATEGIAS PARA CONTRARRESTAR LAS CARACTERÍSTICAS INMUNOLÓGICAS DE CÁNCER Procedimientos de activación de la reactividad inmune Caracterización de oncoantígenos no eliminables Métodos para neutralizar la supresión inmune CONCLUSIONES Y PERSPECTIVAS
Transcript of TEMA%9% …aulaoncologia.com/AulaOncologia/Downloads/curso_onco_molecular/... ·...

1
TEMA 9
CARACTERÍSTICAS INMUNOLÓGICAS DE CÁNCER El sistema inmunitario se encarga de defender al individuo de sustancias extrañas, a través de la protección por la inmunidad innata y la inmunidad adaptativa. En el caso del cáncer, una cuestión que no se ha resuelto todavía es si el sistema inmune favorece o impide el desarrollo de tumores. Por una parte, se conoce que los cánceres asociados a infecciones están aumentando a una velocidad alarmante y que, los tumores con capaces de crear un ambiente inflamatorio que les ayuda a progresar a estados más malignos. Sin embargo, también se sabe que los pacientes cuyos tumores poseen infiltraciones de células inmunes tienen un mejor pronóstico y que las personas inmunodeprimidas son más propensas a padecer cáncer, que las que no lo son. Así, parece que el sistema inmune vigila la formación de tumores para evitar que estos progresen. No obstante, las células cancerígenas son capaces de adquirir la capacidad de evadir la vigilancia del sistema inmune y limitar el ataque de éste, lo que las posibilita a formar tumores. En este tema se estudia la relación entre el sistema inmune y las células cancerígenas, analizando el papel que tiene la inflamación en la carcinogénesis, así como los mecanismos que adquieren las células cancerígenas para evitar la vigilancia y el ataque del sistema inmune. Además se analizan las nuevas estrategias de inmunoterapia para combatir el cáncer. ÍNDICE CARACTERÍSTICAS INMUNOLOGÍCAS DEL CÁNCER INFLAMACIÓN PROMOVIDA POR TUMORES
Células inmunes inflamatorias INMUNOEDICIÓN DEL CÁNCER CAPACIDADES INMUNOLÓGICAS ADQUIRIDAS POR LAS CÉLULAS TUMORALES
Capacidad para prosperar en un microambiente de inflamación crónica Capacidad para evadir el reconocimiento inmune Capacidad para suprimir la reactividad inmunitaria
ESTRATEGIAS PARA CONTRARRESTAR LAS CARACTERÍSTICAS INMUNOLÓGICAS DE CÁNCER
Procedimientos de activación de la reactividad inmune Caracterización de onco-‐antígenos no eliminables Métodos para neutralizar la supresión inmune
CONCLUSIONES Y PERSPECTIVAS

2
CARACTERÍSTICAS INMUNOLOGÍCAS DEL CÁNCER
En la última década se ha puesto de manifiesto que las células cancerígenas deben adquirir la habilidad de aprovechar los mecanismos del sistema inmune, así como evadir la respuesta inmune, con el fin de dar lugar a un tumor. Se ha demostrado que la modificación de los mecanismos inmunes es una capacidad adquirida por la mayoría, o incluso, todos los tumores. Por ello, cada vez se están teniendo más en cuenta las características inmunológicas distintivas de los tumores malignos, como una buena diana para el diseño de nuevas estrategias antitumorales. El conocimiento a nivel molecular y genético de las evidencias obtenidas hasta ahora han identificado tres características inmunológicas que las células tumorales deben adquirir:
• Capacidad para prosperar en un microambiente de inflamación crónica. • Capacidad para evadir el reconocimiento inmune. • Capacidad para suprimir la reactividad inmunitaria.
La adquisición de los cambios genéticos necesarios para adquirir estas tres capacidades, así
como para ponerlas en práctica, es una característica esencial en cánceres naturales y experimentales. Sin embargo, su intensidad puede variar de un tipo de tumor a otro, siendo aún más variables los mecanismos mediante los cuales los diversos tipos de cáncer llevan a cabo estas actividades. Por tanto, mientras la adquisición de un cambio específico en el genoma puede no ser importante, la adquisición de estas capacidades es crucial, independientemente de los mecanismos implicados.
La existencia de células inmunes que tienen la capacidad tanto de promover tumores como de antagonizarlos, se explica atendiendo a las diversas funciones que desempeña el sistema inmune. Por una parte, el sistema inmune detecta y se dirige específicamente contra agentes infecciosos a través de la respuesta inmune adaptativa, que se lleva a cabo por las células del sistema inmune innato. Por otra parte, el sistema inmune innato está implicado en la cicatrización de heridas y en la limpieza las células muertas y los desechos celulares. Estas tareas especializadas son realizadas por diferentes subclases de células inflamatorias, los macrófagos y los neutrófilos convencionales, encargados de la inmunidad adaptativa, así como diversas subclases de macrófagos, neutrófilos y progenitores mieloides “alternativamente activados”. Estos últimos subtipos de células inmunes son una de las principales fuentes de factores de crecimiento angiogénicos, epiteliales y estromales y de enzimas implicadas en la remodelación de la matriz, que son necesarias para la curación de heridas, favoreciéndose, de ese modo, la progresión neoplásica. Del mismo modo, las subclases de linfocitos B y T pueden facilitar el reclutamiento, la activación y la persistencia de macrófagos y neutrófilos implicados, tanto en la curación de heridas, como en la promoción de tumores. Por supuesto, otras subclases de linfocitos B y T y de tipos celulares de la respuesta inmune innata también pueden ejercer acciones antitumorales (figura 1). El equilibrio entre ambas respuestas inflamatorias contradictorias en el pronóstico tumoral y, muy posiblemente, en las terapias diseñadas para redirigir estas células hacia la destrucción del tumor resulta fundamental.

3
Figura 1. Respuesta inmunitaria antitumoral mediada por los linfocitos T. Las células tumorales producen antigénos que son reconocidos por los linfocitos, activándolos. Esto da lugar a la liberación de citoquinas que atraen a otras células del sistema inmune que rompen las células tumorales, por lo que producen la regresión tumoral.
4
INFLAMACIÓN PROMOVIDA POR TUMORES Los patólogos han reconocido desde hace tiempo que algunos tumores presentan una alta
densidad de células responsables de las respuestas innata y adaptativa del sistema inmune y, por lo tanto, son un reflejo de la influencia de las condiciones inflamatorias en tejidos no neoplásicos. Gracias a la utilización de mejores marcadores para la identificación precisa de los distintos tipos de células del sistema inmune, ahora está claro que prácticamente todas las lesiones neoplásicas contienen células inmunes en cantidades que van, desde infiltraciones detectables sólo con anticuerpos específicos, hasta inflamaciones graves que son evidentes incluso mediante técnicas de tinción histoquímica.
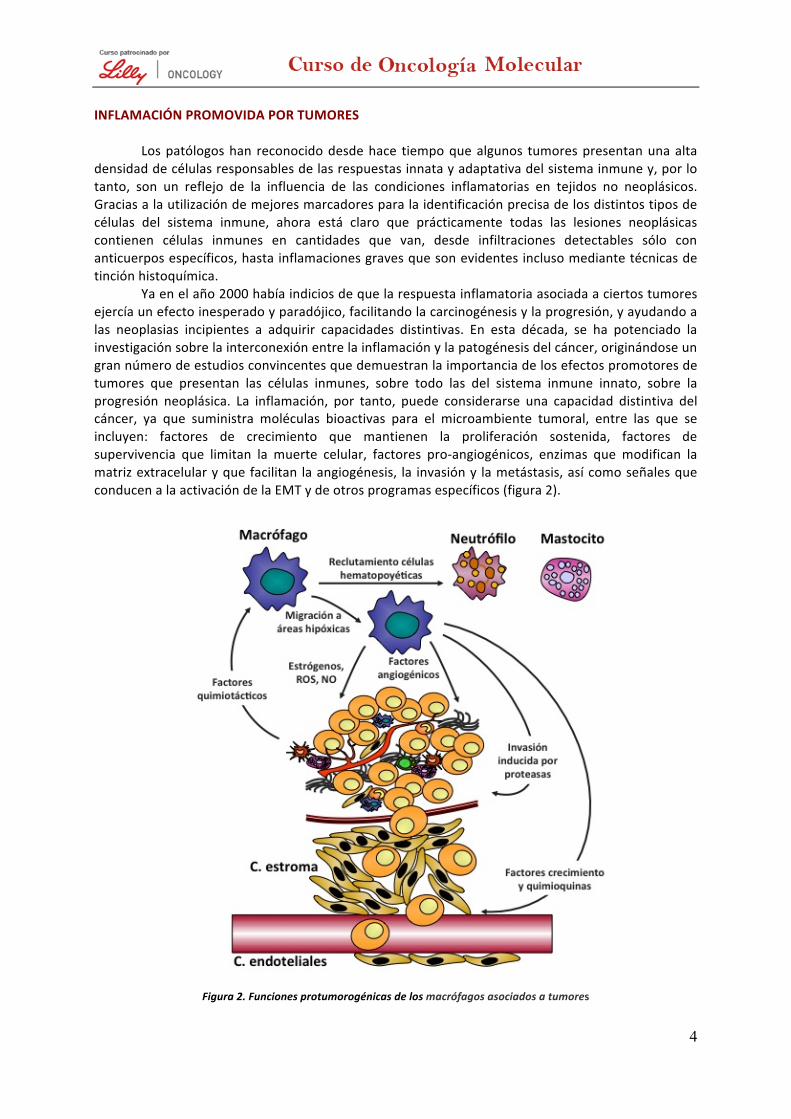
Ya en el año 2000 había indicios de que la respuesta inflamatoria asociada a ciertos tumores ejercía un efecto inesperado y paradójico, facilitando la carcinogénesis y la progresión, y ayudando a las neoplasias incipientes a adquirir capacidades distintivas. En esta década, se ha potenciado la investigación sobre la interconexión entre la inflamación y la patogénesis del cáncer, originándose un gran número de estudios convincentes que demuestran la importancia de los efectos promotores de tumores que presentan las células inmunes, sobre todo las del sistema inmune innato, sobre la progresión neoplásica. La inflamación, por tanto, puede considerarse una capacidad distintiva del cáncer, ya que suministra moléculas bioactivas para el microambiente tumoral, entre las que se incluyen: factores de crecimiento que mantienen la proliferación sostenida, factores de supervivencia que limitan la muerte celular, factores pro-‐angiogénicos, enzimas que modifican la matriz extracelular y que facilitan la angiogénesis, la invasión y la metástasis, así como señales que conducen a la activación de la EMT y de otros programas específicos (figura 2).
Figura 2. Funciones protumorogénicas de los macrófagos asociados a tumores

5
Los macrófagos son reclutados a los tumores por factores quimiotácticos y proporcionan muchas funciones tróficas que promueven la progresión de los tumores y la metástasis. Estos macrófagos asociados a tumores (TAM) migran a las zonas hipóxicas dentro del tumor, donde estimulan la angiogénesis por activación de la expresión de factores proangiogénicos como el factor de crecimiento vascular endotelial (VEGF), las angiopoyetinas 1 y 2 (Ang-‐1 y Ang-‐2) y reclutan otras células hematopoyéticas, como los mastocitos y neutrófilos, que pueden realizar tareas similares. Los TAM también promueven la invasión tumoral por la producción de proteasas del tipo uroquinasa del plasminógeno (uPA), la metaloproteasa de la matriz 9 (MMP-‐9) y catepsinas, que rompen la membrana basal y remodelan la matriz estromal. La MMP-‐9 también contribuye a la angiogénesis. Varios factores de crecimiento y quimioquinas, como el factor de crecimiento epidérmico (EGF), el factor de crecimiento transformante-‐β (TGF-‐β), la interleuquina-‐8 (IL-‐8) y el factor de necrosis tumoral-‐α (TNF-‐α), contribuyen a la migración de las células tumorales hacia los vasos y proporcionan señales de proliferación y antiapoptóticas a estas células. Los macrófagos atraídos por los sitios de inflamación o ruptura de tejidos también pueden iniciar o promover la carcinogénesis a través de la síntesis de los estrógenos y la generación de mutágenos como un subproducto de la producción de especies reactivas de oxígeno y los radicales del óxido de nitrógeno.
Es importante destacar que, en algunos casos, la inflamación es evidente en las primeras
etapas neoplásicas, habiéndose demostrado su capacidad de promover la progresión de neoplasias incipientes hacia auténticos cánceres. Además, las células inflamatorias pueden liberar sustancias químicas, en particular especies reactivas de oxígeno, que son mutagénicas para las células de la vecindad del tumor, acelerando su evolución genética hacia estados de mayor malignidad. Por ello, la inflamación puede considerarse una característica que contribuye a la adquisición de capacidades distintivas básicas por parte del tumor.
Células inmunes inflamatorias
Como se ha mencionado anteriormente, cada vez está más aceptado que la infiltración de
células del sistema inmune es un constituyente genérico de tumores. Estas células inflamatorias funcionan de dos formas contradictorias, ya que en la mayoría de las lesiones neoplásicas, si no en todas, se han encontrado leucocitos que pueden actuar, bien como antagonistas tumorales, o bien como promotores tumorales. Aunque la presencia de los antagonistas tumorales linfocitos T citotóxicos (CTLs) y de las células “natural killers” (NK) pueden inhibir el tumor, también se ha propuesto que las células inmunes mejoran las capacidades tumorales de las células cancerígenas. En la década de los años 90 comenzaron a acumularse evidencias de que la infiltración de los tejidos neoplásicos por las células del sistema inmune servía para promover la progresión tumoral. Los primeros trabajos que pusieron de manifiesto la existencia de una asociación de los sitios de inflamación crónica con la formación de tumores sugerían que los tumores podrían ser tratados como “heridas que nunca sanan”. En el curso de curación normal de las heridas, las células inmunes inflamatorias aparecen transitoriamente y después desaparecen. Por el contrario, éstas persisten en los sitios de inflamación crónica, donde su presencia se ha asociado con varias patologías tisulares, incluyendo fibrosis, angiogénesis aberrante, y neoplasia.
Durante la última década, la manipulación de genes implicados en funciones efectoras de células inmunes de diversos tipos, junto con el estudio de inhibidores farmacológicos de tales células o de sus funciones, ha demostrado que dichas células desempeñan diversos roles que son críticos en el fomento de la carcinogénesis. En la actualidad, la lista de células inflamatorias que actúan como promotores tumorales incluye los subtipos de macrófagos, mastocitos y neutrófilos, así como linfocitos T y B. Estos estudios, también, están aumentando la lista de moléculas de señalización liberadas por las células inflamatorias que sirven como efectores en sus acciones como promotores tumorales. Entre dichas moléculas se incluyen el factor de crecimiento epidermico EGF, el factor de crecimiento angiogénico VEGF, otros factores proangiogénicos, tales como FGF2, quimioquinas y citocinas que amplifican el estado inflamatorio. Además, estas células pueden producir enzimas proangiogénicas y/o proinvasivas, que degradan la matriz, como las metaloproteasas de la matriz, entre las que se incluye MMP-‐9 (figura 2). En concordancia con la expresión de todos estos efectores, se ha demostrado que las células inflamatorias infiltrantes de un tumor inducen y ayudan a

6
mantener la angiogénesis tumoral, con el fin de estimular la proliferación de las células cancerígenas, de facilitar la invasión de tejidos a través de su presencia en los márgenes de los tumores y de apoyar la diseminación metastática y el asentamiento de las células cancerígenas.
Además de las células inmunes totalmente diferenciadas presentes en el estroma del tumor, también se han identificado una variedad de progenitores mieloides parcialmente diferenciados en ciertos tumores. Estas células actúan como intermediarios entre las células circulantes de la médula ósea y las células inmunes diferenciadas que se encuentran típicamente en tejidos normales e inflamados. Es importante destacar que estos progenitores, al igual que sus derivados más diferenciados, muestran una actividad promotora de tumores (figura 3). De particular interés es una clase de células mieloides infiltrantes de tumores, que coexpresan el marcador de macrófagos CD11b y el marcador de neutrófilos Gr1, y que se identificaron de forma independiente como las células supresoras derivadas de mieloides (MDSCs), las cuales se ha demostrado que suprimen la actividad de células CTL y NK. Esta capacidad plantea la posibilidad de que el reclutamiento de ciertas células mieloides pueda ser doblemente beneficioso para el desarrollo del tumor; por una parte, mediante la promoción directa de la angiogénesis y de la progresión tumoral, y por otra parte, proporcionando al mismo tiempo un medio para evadir la destrucción inmunitaria (figura 3).
Figura 3. Células inmunes del microambiente tumoral Las células tumorales reclutan macrófagos (ver figura 2), activan a los linfocitos T, por la presencia de antígenos tumorales, y atraen células mieloides, que, a su vez, activan a los linfocitos. El conjunto de todas interacciones puede provocar efectos beneficiosos o perjudiciales para el tumor, dependiendo de la inmunogenicidad de éste.

7
INMUNOEDICIÓN DEL CÁNCER
El conocimiento de que el sistema inmune no sólo protege al huésped contra la formación de tumor, sino que también existen tumores inmunogénicos, ha permitido desarrollar la hipótesis de la inmunoedición del cáncer.
Se ha postulado que el proceso de inmunoedición del cáncer, en su realización más compleja, se da secuencialmente a través de tres fases diferenciadas que se han denominado "eliminación", "equilibrio" y "escape" (figura 4). Sin embargo, en algunos casos, las células tumorales pueden entrar directamente en la fase de equilibrio o de escape sin pasar a través de una fase anterior. Además, factores externos pueden influir en la direccionalidad de este flujo. Esta última consideración puede ayudar a explicar las influencias del estrés ambiental, el deterioro del sistema inmune que acompaña el envejecimiento e, incluso, la intervención inmunoterapéutica en la eliminación de tumores en pacientes humanos con cáncer.
Figura 4. El concepto de inmunoedición del cáncer La inmunoedición del cáncer es un mecanismo supresor tumoral extrínseco que se activa sólo cuando ha ocurrido la transformación celular y cuando el mecanismo intrínseco de la supresión tumoral ha fallado. En su forma más compleja, la inmunoedición del cáncer se desarrolla en tres fases secuenciales: eliminación, equilibrio y escape. En la fase de

8
eliminación, la inmunidad innata y adaptativa trabajan juntas para destruir tumores incipientes antes de que sean aparentes desde el punto de vista clínico. Se han identificado muchas de las moléculas inmunes y de las células que participan en la fase de eliminación, aunque se necesita profundizar más para determinar la secuencia exacta de su acción. Si esta fase se completa, entonces el huésped se mantiene libre de cáncer, representando la eliminación la totalidad del proceso. Sin embargo, si existe una variante celular rara de cáncer que no se destruye en la fase de eliminación comienza la fase de equilibrio, en la que se previene el crecimiento de dicho tumor a través de mecanismos inmunológicos. Las células T, la IL-‐12 y el IFN-‐γ se requieren para mantener las células tumorales en un estado latente, mientras que no se requieren ni las células NK ni moléculas, que participan en el reconocimiento o en la función efectora de la inmunidad innata. Esto indica que el equilibrio sólo es una función de la inmunidad adaptativa. La edición de la inmunogenicidad tumoral ocurre en la fase de equilibrio. El equilibrio también puede representar el último paso del proceso de inmunoedición y puede restringir el crecimiento de cánceres ocultos durante toda la vida del huésped. Sin embargo, como consecuencia de la presión de selección inmune que tiene lugar en células tumorales genéticamente inestables, algunas variantes de células tumorales pueden escapar porque, (I) no son reconocidas por la inmunidad adaptativa (variantes con pérdida de antígenos o células tumorales que desarrollan defectos en el procesamiento o en la presentación de antígenos), (II) se convierten en insensibles a los mecanismos efectores de la inmunidad o (III) inducen un estado inmunosupresivo dentro del microambiente tumoral, dando lugar a la fase de escape. Estas células tumorales que emergen causan la enfermedad aparente clínicamente. Eliminación
La fase de eliminación se describe como la etapa de la inmunovigilancia del cáncer, en la que los sistemas de inmunidad innata y adaptativa trabajan conjuntamente para detectar la presencia de un tumor en desarrollo y destruirlo antes de que sea clínicamente evidente.
La fase de eliminación no se ha observado aún directamente in vivo, pero su existencia se ha deducido por la aparición más temprana y la mayor penetrancia de neoplasias en ratones que carecen de ciertos tipos de células inmunes, de moléculas de reconocimiento, de vías efectoras o de citoquinas, así como de los estudios que comparan la iniciación del tumor, el crecimiento y metástasis en individuos normales frente a ratones inmunodeficientes. Estos estudios han revelado que los componentes específicos del sistema inmune son necesarios para la eliminación efectiva de cualquier tumor.
Equilibrio
Algunas variantes raras de células tumorales pueden sobrevivir a la fase de eliminación y entrar en la fase de equilibrio, en la que el sistema adaptativo inmune evita el reconocimiento de las células tumorales y se forma la inmunogenicidad de las células cancerígenas. Se piensa que el equilibrio es la etapa más larga del proceso de inmunoedición del cáncer y que, tal vez, se extienda a lo largo de la vida del anfitrión. En el equilibrio, el sistema inmunitario mantiene a las células tumorales residuales en un estado funcional de latencia, término usado para describir a las células tumorales latentes que pueden residir en pacientes durante décadas antes de reanudar de nuevo el crecimiento, ya sea como tumores recurrentes primarios o metástasis distales. En resumen, la fase de equilibrio representa una fase en la cual el tumor permanece oculto para el sistema inmune, pero que, a su vez, está controlado específicamente por inmunidad. Escape
En la fase de escape las células tumorales, que han adquirido la capacidad de eludir el reconocimiento inmune y/o la destrucción, pueden reanudar progresivamente el crecimiento, formando tumores visibles. La progresión de la fase de equilibrio a la de escape se puede producir debido a cambios en la población de células tumorales de población en respuesta a las funciones de edición del sistema inmunológico, a cambios del sistema inmune del huésped en respuesta a un aumento de la inmunosupresión inducida por el cáncer o al deterioro del sistema inmunológico. El escape de las células tumorales puede ocurrir a través de muchos diferentes mecanismos. A nivel de la célula tumoral, pueden ser alteraciones que disminuyan el reconocimiento por parte del sistema inmune, como una pérdida de antígenos, o el aumento de la resistencia a los efectos citotóxicos de la inmunidad, por ejemplo, a través de la inducción de anticuerpos antiapoptóticos, mecanismos que

9
implican la activación persistente de factores de transcripción pro-‐oncogénicos, tales como STAT3, (ver tema 4) o expresión de moléculas antiapoptóticas, como Bcl-‐2. De todos ellos, la pérdida de expresión de antígenos tumorales es uno de los mejor estudiados, y puede ocurrir de al menos tres formas: (i) a través de aparición de las células tumorales que carecen de expresión de antígenos de rechazo, (ii) a través de la pérdida del complejo mayor de histocompatibilidad (MHC) clase I, proteínas que presentan los antígenos específicos del tumor a las células T, o (iii) a través de la pérdida del tratamiento del antígeno. Todas estas alteraciones se deben, probablemente, a una combinación de inestabilidad genética inherente a todas las células tumorales y al proceso de inmunoselección de las células cancerígenas.

10
CAPACIDADES INMUNOLÓGICAS ADQUIRIDAS POR LAS CÉLULAS TUMORALES Capacidad para prosperar en un microambiente de inflamación crónica
Los modelos de ratón genéticamente modificados han puesto de manifiesto el potencial cancerígeno intrínseco de la inflamación y han permitido la definición molecular de algunos de los mecanismos mediante los cuales un tumor incipiente se aprovecha del microambiente inflamatorio. Así, la activación continua de poblaciones leucocitarias desencadenada por infecciones, el daño tisular o el comportamiento anómalo de las células mutadas contribuye a la progresión tumoral, a través de múltiples mecanismos que involucran desde especies reactivas de oxígeno hasta factores de crecimiento o angiogénicos. El equilibrio entre inmunidad e inflamación se altera a favor del segundo durante el envejecimiento, periodo en el que la incidencia de tumores está muy relacionada con la inflamación.
La importancia de un microambiente inflamatorio es tan grande que, incluso una población celular “adicta” a un oncogén, que da lugar a un tumor en ratones transgénicos, es modulada por el estroma reactivo que rodea la lesión cancerígena. La inflamación y la carcinogénesis están relacionadas, incluso en ausencia de estímulos inflamatorios externos. Las señales promovidas por oncogenes activan vías proinflamatorias que afectan a la aparición y al progreso de un carcinoma. Mediante análisis de expresión diferencial de genes en micromatrices o “microarrays” del genoma de animales transgénicos, se ha identificado que las células cancerígenas aumentan la expresión de citoquinas en el microambiente tumoral, que son necesarias para la progresión del tumor. Las citoquinas inflamatorias producidas pueden, por una parte, estar involucradas en bucles autocrinos que promueven directamente la proliferación de las células tumorales y, por otra parte, pueden ser liberadas por células inmunes/inflamatorias que son reclutadas al microambiente tumoral. Estos datos moleculares están directamente respaldados por muchos estudios epidemiológicos en humanos, que vinculan las vías inflamatorias extrínseca e intrínseca con un aumento del riesgo de cáncer. Por ejemplo, el aumento de riesgo de cáncer gástrico en el contexto de infecciones bacterianas está vinculado a los polimorfismos en genes que codifican para citoquinas proinflamatorias. Estos hallazgos muestran cómo las vías inflamatorias extrínseca e intrínseca están interconectadas para dirigir el cáncer. Por todo esto, la definición de los mecanismos moleculares por los que la inflamación crónica contribuye a la carcinogénesis en humanos está abriendo nuevas perspectivas en el campo de la inmuno-‐prevención. Capacidad para evadir el reconocimiento inmune
La teoría de la vigilancia inmunológica fue postulada en la década de 1960. En ella se destacaba la capacidad de identificar y destruir tumores nacientes por parte del sistema inmunitario, pero más tarde esta idea se desechó al no observar un aumento de la incidencia tumoral en ratones atímicos. Sin embargo, el trabajo de los últimos diez años ha demostrado que dichos ratones no constituían un modelo apropiado para la investigación de la vigilancia inmunológica, mientras que el empleo de ratones genéticamente modificados, que asemejan defectos inmunitarios estables, ha demostrado plenamente esta teoría. Los ratones con alteraciones genéticas que conducen a deficiencias de células T y B son más propensos a la carcinogénesis espontánea y química que los ratones de tipo salvaje. Además, los defectos genéticos adicionales que afectan a las respuestas inmunológicas naturales aumentan el riesgo de producir tumores más agresivos y precoces. Por otra parte, los mecanismos inmunes mantienen en un estado latente el cáncer durante períodos equivalentes a la duración de la vida natural del ratón, mientras que la inmunodepresión temporal permite que el cáncer progrese.
Los mecanismos de vigilancia inmunológica limitan el desarrollo del cáncer, pero no son totalmente eficientes. Los tumores que surgen eventualmente son aquellos que presentan poca o nula inmunogenicidad. Una característica fundamental que distingue las lesiones neoplásicas

11
latentes de los tumores en desarrollo es su susceptibilidad al control inmunitario y, por tanto, la capacidad para evadir la vigilancia inmunológica es otra característica del cáncer. En este sentido, los datos de los pacientes con deficiencias inmunológicas adquiridas, tales como el SIDA y la supresión inmunológica post-‐trasplante, muestran un aumento dramático en la incidencia de diferentes tipos de tumores, incluyendo el cáncer de pulmón, linfomas y tumores relacionados con infecciones virales, como el sarcoma de Kaposi o los carcinomas ano-‐genitales inducidos por el virus del Papiloma humano (VPH).
En los últimos 10 años, se ha hecho evidente que un tumor es capaz de reconocer su susceptibilidad al ataque inmunológico y elabora muchas defensas contra ella. En cuanto a los mecanismos celulares y moleculares, se ha observado que el aumento de la inestabilidad del genoma de células precancerígenas favorece la aparición de clones de diferente inmunogenicidad. Los clones que son inmunogénicamente débiles son los que escapan a la inmunovigilancia. Así, la clandestinidad de tumores clínicos puede verse como uno de los resultados de una inmunovigilancia eficaz. A nivel molecular, la pérdida de la expresión de las glicoproteínas del complejo mayor de histocompatibilidad (MHC) en la membrana celular es uno de los mecanismos por los cuales se adquiere la clandestinidad tumoral. Además, ésta puede ser una consecuencia de sobrexpresión de proteínas codificadas por oncogenes o a la alteración de la maquinaria de procesamiento de péptidos antigénicos. La disminución de la expresión de glicoproteínas del MHC y de péptidos antigénicos en la superficie de la célula tumoral impide el reconocimiento directo de los antígenos tumorales por las células T y bloquea la respuesta inmune sobre el tumor, ya que anula la fase efectora de la reacción de las células T contra los antígenos asociados a tumores. El bloqueo de estas dos funciones es un aspecto crucial en el desarrollo del tumor dado que la citotoxicidad mediada por las células T constituye un mecanismo de inhibición tumoral muy eficaz. Capacidad para suprimir la reactividad inmunitaria
Dado que el crecimiento de las células tumorales que se produce como consecuencia de su evasión al reconocimiento inmune no es suficiente para “combatir” contra los ataques del sistema inmune, la gran mayoría de los tumores adquieren la capacidad de liberar una serie de factores y citoquinas para contrarrestar los mecanismos de defensa normales. Cuando un tumor adquiere la capacidad de liberar cantidades significativas del factor estimulante de colonias (CSF) o del factor de crecimiento endotelial vascular (VEGF), provoca la expansión de una población de células mieloides inmaduras que, no sólo pueden ayudar a los tumores a suprimir la reacción inmune, sino que también ayudan a la construcción de nuevos vasos sanguíneos para el crecimiento del tumor (ver tema 8). Aún más, un tumor puede convertir células T naive en células T reguladoras (TReg), a través de la liberación directa del factor de crecimiento transformante β (TGFβ), IL-‐10 e indoleamina 2,3-‐dioxigenasa (IDO) o, a través de la activación de estas secreciones por las células supresoras mieloides, macrófagos asociados a tumores (TAM) y células dendríticas. Así, la expansión de estas células es otra manera por la cual un tumor detiene la reactividad inmunológica del huésped (ver figura 12 del tema 1).
Los tumores también aprovechan el papel fisiológico de las células TReg naturales para bloquear las reacciones inmunes. Estas células reconocen con alta afinidad los antígenos propios y bloquean la inducción de la autoinmunidad. Al igual que ocurre con la expansión de las células tumorales que sobrexpresan productos oncogénicos, la sobrexpresión de unos pocos autoantígenos tolerados conduce a la activación de las células TReg naturales. En consecuencia, un tumor en crecimiento puede derivar la respuesta inmune hacia la inmunosupresión, tanto a través del empleo de un mecanismo de salvaguardia fisiológica, para controlar la autoinmunidad, como gracias a su capacidad de convertir células T naive en una población supresora. La activación de las células TReg naturales y adaptativas se dispara por la activación específica de sus receptores de las células T. Los mecanismos supresores de las TReg están mediados por diferentes funciones:

12
1. La exposición en la membrana celular de moléculas portadoras de señales negativas para las células dendríticas (CTLA4 y LAG3). Estas señales inhiben la maduración de las células dendríticas, bloquean la expresión de MHC y de las moléculas co-‐estimuladoras (CD80 y CD86), activan su capacidad para producir IDO, que conduce a la generación del mediador inmunosupresor kynurenina e indirectamente suprimen genes que codifican para IL-‐6 y TNF.
2. La liberación de adenosina y la secreción de TGFβ, IL-‐10, e IL 35, que interfieren con la activación y las funciones efectoras de las células T.
3. La secreción de granzimas y perforina, que podrían tener efectos citolíticos en células T diana, así como en células dendríticas.
El mismo grupo de señales desencadena la activación y el mantenimiento de las funciones anómalas de las células dendríticas tolerogénicas y de los macrófagos asociados al tumor. De esta manera, un tumor en crecimiento organiza una red de distintas, pero a la vez integradas, actividades represivas.

13
ESTRATEGIAS PARA CONTRARRESTAR LAS CARACTERÍSTICAS INMUNOLÓGICAS DE CÁNCER
El conocimiento adquirido en la última década ofrece la oportunidad de aprender cómo implementar medidas específicas para invertir la situación en favor del sistema inmune y, finalmente, del paciente. Esta nueva información podría ser canalizada para hacer frente a lo que parecen ser las tres principales características para el control inmunológico de la progresión del cáncer: • Procedimientos eficaces para activar la reactividad inmune. • Caracterización de onco-‐antígenos no eliminables. • Métodos para neutralizar la supresión inmune. Procedimientos de activación de la reactividad inmune
La inflamación crónica puede ser paliada con fármacos antinflamatorios, como el Sulindac y la aspirina, que, en algunos casos, reducen el riesgo de cáncer. Sin embargo, una estrategia más adecuada consiste en reorientar la inflamación, de tal modo que sea un factor de prevención tumoral, en lugar de actuar como un promotor tumoral. Tanto la inmunización pasiva (anticuerpos) como la activa (vacunas) protegen eficientemente al huésped de la aparición del tumor, aunque hay muchas más evidencias a favor de la inmunización activa (figura 5).
La alta eficacia de las vacunas en la prevención de infecciones por virus carcinogénicos y otros agentes infecciosos causantes de cáncer está teniendo un extraordinario impacto social. Las vacunas dirigidas a la eliminación de factores de riesgo infecciosos han comenzado a ser de uso común (ver distintas estrategias terapéuticas enfocadas a la modificación inmunológica en los diferentes temas).
El carcinoma hepatocelular es el responsable de más de un 4% de todos los cánceres humanos y el 80% de los casos se asocian con infecciones virales. La vacunación contra la hepatitis B (VHB) reduce notablemente la incidencia de carcinoma hepatocelular post-‐hepatitis. Dado que la inflamación crónica tiene un papel importante en la aparición del cáncer de hígado que aparece tras la infección por VHB, esta vacuna puede ser considerada como una forma de prevención primaria de una inflamación carcinogénica crónica.
El VPH causa trastornos neoplásicos, que van desde verrugas benignas hasta carcinomas cervicales y ano-‐genitales malignos. La implementación de programas de vacunación en todo el mundo contra el VPH comenzó sólo hace unos pocos años y su eficacia a largo plazo en la prevención del carcinoma cervical todavía no se ha evaluado completamente. Los resultados iniciales son extremadamente favorables, y se prevé que se alcance una casi completa prevención de dicha carcinogénesis. Las vacunas actuales son eficaces en la prevención del cáncer, pero carecen de eficacia terapéutica, por lo que en la actualidad están siendo estudiadas vacunas capaces de curar carcinomas cervicales.
El virus de Epstein-‐Barr (EBV) está implicado en una variedad de enfermedades en todo el mundo: la mononucleosis infecciosa en países occidentales, el carcinoma nasofaríngeo en Asia, el linfoma de Burkitt en África, y las enfermedades linfoproliferativas en pacientes inmunodeficientes. Por ello, uno de los objetivos que se está persiguiendo en este campo es el uso de algunas vacunas prometedoras.
El uso de las vacunas para prevenir tumores que están relacionados con algún agente infeccioso está convirtiéndose en una realidad médica, pero, además, un gran número de estudios realizados en ratones genéticamente modificados sugieren que las vacunas también pueden ser una forma de prevención de tumores que no están relacionados con agentes infecciosos. Existen muchos datos de estudios realizados en ratones inmunocompetentes portadores de oncogenes, que les predestinan a sufrir cáncer letal y que muestran que las vacunas frente a ciertos productos oncogénicos bloquean la aparición de las lesiones neoplásicas. Además, la repetición de la administración de la vacuna proporciona una protección permanente que puede durar toda la vida

14
del animal. Sorprendentemente, la respuesta citotóxica mediada por las células T juega un papel menor en la protección otorgada por varias de estas vacunas. Teniendo en cuenta que, en estos casos, los productos oncogénicos son moléculas propias, éstos provocan una especie de tolerancia que ocasiona, principalmente, la desaparición de células T CD8+ de alta afinidad. Además, esta respuesta es inhibida por la expansión de las células TReg naturales, que reconocen el antígeno diana como una proteína endógena. Por lo tanto, la mayor parte de la acción antitumoral provocada por las vacunas preventivas se basa en la actividad antitumoral directa e indirecta de los anticuerpos.
Figura 5. Células de sistema inmune que deben ser activadas para producir vacunas antitumorales eficaces Caracterización de onco-‐antígenos no eliminables
Las vacunas que deben mantener una respuesta inmunitaria sostenida a lo largo de toda la vida, conllevan el riesgo de la pérdida de modulación o la desactivación del antígeno diana por parte de las células neoplásicas. Un antígeno diana apropiado que previene la pérdida de reconocimiento inmune debería cumplir los siguientes requisitos:
• Desempeñar un papel esencial en el crecimiento o la progresión del tumor. • Ser una diana de las células citotóxicas y anticuerpos.
Se ha adoptado el término “onco-‐antígeno” para los antígenos tumorales que cumplen
ambos requisitos. En la carcinogénesis provocada por oncogenes, pueden existir variantes celulares que pierdan el antígeno, lo que haría que su potencial tumorogénico estuviera notablemente afectado. En las últimas etapas de la progresión del tumor, el papel del oncogén podría ser asumido por diferentes genes, cuyos productos, a su vez, producirían onco-‐antígenos diana adicionales.
Los tumores evaden el reconocimiento por las células T a través de la desactivación de la maquinaria de procesamiento del antígeno y de la expresión de la glicoproteína del MHC. Sin embargo, el reconocimiento de moléculas accesibles no se ve afectado, y los anticuerpos todavía aseguran una inhibición funcional del onco-‐antígeno diana, así como una activación de la citotoxicidad mediada por el complemento y la ADCC. Los onco-‐antígenos de clase I expresados en la superficie de la célula pueden ser atacados por los anticuerpos, así como por la respuesta inmunitaria dependiente de las células T, y son probablemente las mejores dianas para una vacuna preventiva. Los onco-‐antígenos de clase II son moléculas que se secretan por los tumores o que se encuentran en el microambiente tumoral y que juegan papeles esenciales en la expansión del tumor. Estos pueden ser el blanco de anticuerpos, pero no de la respuesta inmunitaria dependiente de las

15
células T. Los onco-‐antígenos de clase III son moléculas tumorales que no pueden ser atacadas por los anticuerpos debido a su localización intracelular y, por lo tanto, sólo pueden ser dianas de las células T.
Se podría pensar que, en un futuro, las vacunas para prevenir el cáncer se podrían administrar a la población en general, tal y como está haciéndose en la actualidad para prevenir tumores infecciosos. Desde una perspectiva más realista, determinados grupos de riesgo de cáncer podrían beneficiarse de vacunas específicas, especialmente en el caso de riesgo genético, de síndromes pre-‐neoplásicos, de grupos de individuos previamente expuestos a agentes carcínogenos ambientales y de supervivientes de cáncer con mayor riesgo de padecer un nuevo tumor primario. Es de particular interés el hallazgo de que la vacuna contra ErbB2 (Trastuzumab, Herceptin), un onco-‐antígeno arquetípico de clase I que, además de actuar sobre tumores de mama ya formados, impide la carcinogénesis química en hámsters, lo que abre un nuevo camino para el tratamiento de personas sanas con un riesgo específico de padecer cáncer inducido químicamente, para quienes no existe una opción terapéutica en la actualidad. Métodos para neutralizar la supresión inmune
La eficacia de las vacunas está disminuida por la expansión de las células inmunosupresoras mediada por el tumor, incluyendo las células TReg y las células supresoras mieloides (MDSCs), lo que provoca una disminución significativa de la respuesta inmune y una supresión de su mecanismo efector. Las estrategias que contrarrestan la supresión durante la vacunación pueden marcar la diferencia entre una vacuna poco efectiva y una efectiva. Las células TReg se acumulan en tumores humanos y en tumores de ratón, así como en los órganos linfoides secundarios, y son reclutadas y expandidas a partir de la proliferación de células TReg pre-‐existentes o de la conversión de células T CD25-‐negativas. La expansión de las células TReg promovida por el tumor también cambia el repertorio de células T específicas de tumores e inhibe la reacción de las células T de baja avidez contra los antígenos tumorales.
Cuando la vacunación está asociada a la disminución de células TReg, ocasionada por la administración de anticuerpos monoclonales contra CD25, se induce una inmunidad tumoral de larga duración y la respuesta al anticuerpo mejora. Además, la baja avidez de la respuesta de los linfocitos T citotóxicos (CTLs) contra el péptido inmuno-‐dominante se restaura, debido a la liberación de células T CD8+ desde células TReg. Estos efectos de la disminución de células TReg hacen eficaz la vacunación en etapas tumorales en las que la vacunación sola no lo era. Del mismo modo, la inhibición de las células TReg funcionales, por medio de la activación de OX40, protege a los ratones del desarrollo tumoral e induce un rechazo total de los nódulos ya establecidos.
Las TReg no son las únicas células supresoras que pueden ser atacadas para contrarrestar la supresión inmune. De creciente interés son las MDSCs, que suponen una diana no desarrollada todavía. Aunque se ha demostrado que hay vacunas potentes que inhiben las MDSCs, una estrategia más directa puede ser más eficaz. En un estudio reciente se han indicado cuatro líneas de actuación: la inducción de la maduración de las MDSC, así como la inhibición de la generación de MDSC, de su acumulación y de su función supresora.
Muchos pacientes de cáncer se podrán beneficiar del uso clínico de los abordajes anti-‐supresivos, en particular aquellos que presenten estadíos más avanzados, que frecuentemente muestran niveles más altos de supresión inmune y de células supresoras. Esta línea de nuevas inmuno-‐terapias se ha intentado por primera vez en pacientes avanzados, y se piensa que la tasa de éxito de tales ensayos clínicos se vería significativamente mejorada por la aplicación simultánea de estrategias anti-‐supresoras del sistema inmune.

16
CONCLUSIONES Y PERSPECTIVAS
Existen evidencias recientes que muestran que la actuación del sistema inmune puede controlar el cáncer. Aunque los datos no son concluyentes todavía, hay una serie de informes recientes que sugieren que la inmunoterapia puede ser una opción real en el tratamiento de pacientes con cáncer. De hecho, las vacunas de diversos tipos han dado resultados iguales o mejores que la mayoría de los tratamientos convencionales que se usan en la actualidad en una gama de enfermedades neoplásicas, tales como linfomas, melanomas, cáncer de próstata y cáncer de pulmón. Estos resultados, junto con la aprobación por la FDA de EE.UU. de una vacuna para la terapia del cáncer de próstata, impulsarán nuevas estrategias más racionales basadas en el uso de vacunas en la terapia del cáncer. Por tanto, es previsible que las nuevas vacunas basadas en tecnologías innovadoras lleguen a alcanzar progresivamente la eficacia de los tratamientos contra el cáncer más convencionales, minimizando los devastadores efectos secundarios de la quimioterapia. En algunos casos, el uso de vacunas podrá ser aún más eficiente, hablando en términos de curación del paciente, que la quimioterapia.
Es digno de mención que los resultados de estudios preclínicos y clínicos convergen en la importancia de los anticuerpos en las respuestas inmunes antitumorales. En la última década, Herceptin y otros anticuerpos monoclonales se han convertido en nuevos medicamentos altamente eficaces, que se están utilizando comúnmente con gran éxito. El mecanismo de la actividad antitumoral de los anticuerpos monoclonales es complejo y depende del antígeno diana. Los anticuerpos contra los onco-‐antígenos de clase I y clase II no sólo inhiben funcionalmente la actividad de sus dianas e inducen mecanismos de muerte dependientes de anticuerpos y complemento, sino que, además, son capaces de activar la respuesta adaptativa del huésped, actuando en cierta medida, como una vacuna. A pesar de estos importantes resultados clínicos, la mayoría de las vacunas antitumorales se diseñan para activar sólo la inmunidad celular. Es de reseñar que el ensayo clínico de la primera vacuna aprobada puso de manifiesto una correlación significativa de la supervivencia del paciente con el título del anticuerpo específico, pero no con la respuesta mediada por células T. Probablemente, ha llegado el momento de integrar todos los conceptos adquiridos desde entonces en el diseño de nuevas vacunas terapéuticas contra el cáncer.
En conclusión, la evaluación de las características inmunológicas del cáncer, así como de su posible control, abre las puertas no sólo a la generalización de la inmuno-‐prevención contra el cáncer, sino también a inmunoterapias innovadoras y más eficaces contra el cáncer.

17
BIBLIOGRAFÍA Bindea G., Mlecnik B., Fridman WH., Pagés F. y Galon, J. (2010). Natural immunity to cancer in humans. Curr. Opin. Immunol. 22:215–222. Colotta F., Allavena P., Sica A., Garlanda C. y Mantovani, A. (2009). Cancer-‐related inflammation, the seventh hallmark of cancer: links to genetic instability. Carcinogenesis 30:1073–1081. de Visser KE., Eichten A. y Coussens LM. (2006). Paradoxical roles of the immune system during cancer development. Nat. Rev. Cancer 6:24–37. Grivennikov SI., Greten FR. y Karin M. (2010). Immunity, inflammation and cancer. Cell 140:883–899. Johansson M., Denardo DG. y Coussens LM. (2008). Polarized immune responses differentially regulate cancer development. Immunol. Rev. 222:145–154. Kim R., Emi M. y Tanabe K. (2007). Cancer immunoediting from immune surveillance to immune escape. Immunology 121:1–14. Mantovani A. (2010). Molecular pathways linking inflammation and cancer. Curr. Mol. Med. 10:369–373. Mantovani A., Allavena P., Sica A. y Balkwill F. (2008). Cancer-‐related inflammation. Nature 454:436–444. Mougiakakos D., Choudhury A., Lladser A., Kiessling R. y Johansson CC. (2010). Regulatory T cells in cancer. Adv. Cancer Res. 107:57–117. Pagès F., Galon J., Dieu-‐Nosjean MC., Tartour E., Sautès-‐Fridman C. y Fridman WH. (2010). Immune infiltration in human tumors: a prognostic factor that should not be ignored. Oncogene 29:1093–1102. Qian BZ. y Pollard JW. (2010). Macrophage diversity enhances tumor progression and metastasis. Cell 141:39–51. Smyth MJ., Dunn GP. y Schreiber RD. (2006). Cancer immunosurveillance and immunoediting: the roles of immunity in suppressing tumor development and shaping tumor immunogenicity. Adv. Immunol. 90:1–50.










