







Idiomas
Páginas
Jurídico


UNIVERSIDAD CENTRAL DEL ECUADOR
FACULTAD DE CIENCIAS BIOLÓGICAS
CARRERA DE CIENCIAS BIOLÓGICAS Y AMBIENTALES
Análisis de la variación morfológica de Hoplias
malabaricus (Bloch 1794), asociada al tipo de
hábitat utilizando morfometría geométrica.
Trabajo de titulación presentado como requisito previo a la
obtención del Título de Licenciado en Ciencias Biológicas y
Ambientales
AUTOR: Cocha Alulema Andrés Patricio
TUTOR: MSc. Juan Francisco Rivadeneira Romero
Quito, Enero 2018

ii
DERECHOS DE AUTOR
Yo Andrés Patricio Cocha Alulema, en calidad de autor y titular de los
derechos morales y patrimoniales del trabajo de titulación: Análisis de la
variación morfológica de Hoplias malabaricus (Bloch 1794), asociada al
tipo de hábitat utilizando morfometría geométrica, de conformidad con el
Art. 114 del CÓDIGO ORGÁNICO DE LA ECONOMÍA SOCIAL DE LOS
CONOCIMIENTOS, CREATIVIDAD E INNOVACIÓN, concedo a favor
de la Universidad Central del Ecuador una licencia gratuita, intransferible y
no exclusiva para el uso no comercial de la obra, con fines estrictamente
académicos. Conservo a mi favor todos los derechos de autor sobre la obra,
establecidos en la normativa citada.
Así mismo, autorizo a la Universidad Central del Ecuador para que realice
la digitalización y publicación de este trabajo de titulación en el repositorio
virtual, de conformidad a lo dispuesto en el Art. 144 de la Ley Orgánica de
Educación Superior.
El autor declara que la obra objeto de la presente autorización es original en
su forma de expresión y no infringe el derecho de autor de terceros,
asumiendo la responsabilidad por cualquier reclamación que pudiera
presentarse por esta causa y liberando a la Universidad de toda
responsabilidad.

iii
APROBACIÓN DEL TUTOR
DEL TRABAJO DE TITULACIÓN
Yo Juan Francisco Rivadeneira Romero en mi calidad de tutor del trabajo de
titulación, modalidad Proyecto de investigación, elaborado por el estudiante
Andrés Cocha, cuyo título es: Análisis de la variación morfológica de
Hoplias malabaricus (Bloch 1794), asociada al tipo de hábitat utilizando
morfometría geométrica, previo a la obtención del Grado de Licenciada en
Ciencias Biológicas y Ambientales; considero que el mismo reúne los
requisitos y méritos necesarios en el campo metodológico y epistemológico,
para ser sometido a la evaluación por parte del tribunal examinador que se
designe, por lo que lo APRUEBO, a fin de que el trabajo sea habilitado para
continuar con el proceso de titulación determinado por la Universidad
Central del Ecuador.
En la ciudad de Quito, a los 11 días del mes de enero de 2018.

iv
APROBACIÓN DE LA PRESENTACIÓN ORAL/TRIBUNAL
El tribunal constituido por: la Dra. María Mercedes Gavilánez Endara PhD, Dra. Ana
Soto-Vivas PhD y el Dr. Iván Vinicio Jácome Negrete MSc.
Luego de receptar la presentación oral del trabajo de titulación previo a la obtención del
título de Licenciado en Ciencias Biológicas y Ambientales presentado por el señor Cocha
Alulema Andrés Patricio.
Con el título:
Análisis de la variación morfológica de Hoplias malabaricus (Bloch 1794), asociada al
tipo de hábitat utilizando morfometría geométrica
Emite el siguiente veredicto: APROBADO
Fecha: lunes, 9 de abril de 2018
Para constancia de lo actuado firman:

v
DEDICATORIA
A mi madre por haberme apoyado en todo momento, por sus consejos, sus
valores, por la motivación constante que me ha permitido ser una persona de
bien, pero más que nada, por su amor.
A mis amigos: Henry Robayo, Manuel Jiménez, Pablo Sannt, Deysi Gualavisi,
Carolina Cachiguango, Ibeth Morales por el apoyo mutuo en nuestra formación
profesional y por haberme ayudado a realizar este trabajo.
Finalmente a los maestros, aquellos que marcaron cada etapa de mi camino
universitario, y que me ayudaron en asesorías y dudas presentadas en
la elaboración de la tesis

vi
ÍNDICE DE CONTENIDOS
LISTA DE ANEXOS…………………………………………………………………………………….vii
LISTA DE TABLAS ...................................................................................... viii
LISTA DE GRÁFICOS .................................................................................... ix
RESUMEN ................................................................................................... x
ABSTRACT................................................................................................... xi
INTRODUCCIÓN .......................................................................................... 1
METODOLOGÍA .......................................................................................... 8
Área de estudio ....................................................................................... 8
Diseño metodológico ............................................................................ 10
Población y muestra .............................................................................. 10
Métodos ................................................................................................ 10
Procesamiento de las muestras ............................................................. 11
CONSIDERACIONES ÉTICAS ....................................................................... 12
ANÁLISIS ESTADÍSTICO ............................................................................. 13
RESULTADOS ............................................................................................ 14
DISCUSIÓN ............................................................................................... 28
CONCLUSIONES ........................................................................................ 34
RECOMENDACIONES ................................................................................ 35
REFERENCIAS BIBLIOGRÁFICAS ................................................................. 36
ANEXOS .................................................................................................... 37

vii
LISTA DE TABLAS
Tabla 1. Resultados del test MANOVA para la variable canónicas 1 en las
cuatro localidades analizadas……………………………………………...20
Tabla 2. Matriz de clasificación para las cuatro localidades analizadas. Las
filas presentan los grupos asignados a priori y las columnas los grupos a los
que fueron asignados por el programa……………………………………22
Tabla 3. Resultados del test MANOVA para la variable canónica 1 de la
subcuenca río Aguarico y subcuenca río San Miguel……………………...27
Tabla 4. Matriz de clasificación para las subcuencas analizadas…………..27
Tabla 5. Ficha de registro de datos en campo…………………………...…50
Tabla 6. Valores propios de los componentes y porcentaje de varianza…...51
Tabla 7. Matriz de valores de significación de las comparaciones pareadas
de acuerdo con la prueba de Hotelling para las cuatro localidades……….52

viii
LISTA DE FIGURAS
Figura 1. Mapa político del cantón Cáscales –
Sucumbíos……………….............................................................................8
Figura 2 Mapa de ubicación de los puntos de muestreo en el cantón
Cáscales………………………………………………………………….....9
Figura 3. Imagen del costado izquierdo de un individuo de Hoplias
malabaricus con los 12 landmarks que indican la posición de las estructuras
biológicas comparadas…………………………………………………….15
Figura 4. Dispersión de puntos que indica las variaciones en la localización
de cada uno de los landmarks en las distintas configuraciones de los
especímenes de H. malabaricus………………………………………...…15
Figura 5. Conformación consenso generada a partir de los 12 landmarks en
los 96 individuos de H. malabaricus……………………………………...15
Figura 6. Morfoespacio definido por análisis de componentes
principales……………………………………….………………………..16
Figura 7. Espécimen colectado en un ecosistema léntico y ecosistema lótico
...…………………………………………………………………………..18
Figura 8. Mallas de deformación con vectores del primer eje del análisis de
componentes principales…………………………………………………..18
Figura 9. Los cambios en la forma se muestran mediante rejillas de
deformación con una gradiente de color de azul a rojo……………………18
Figura 10. Morfoespacio entre los ejes 1 y 2 resultantes del análisis de
variables canónicas aplicado a las 4 poblaciones de H. malabaricus……...21
Figura 11. Conformación consenso a partir de los 12 landmarks en los 96
individuos de H. malabaricus generada con el análisis Thin Plate
Spline...……...…………………………………………………………….23
Figura 12. Forma consenso generada con el análisis Thin Plate Spline de H.
malabaricus en cuatro localidades………………………………………...24
Figura 13. Morfoespacio definido por análisis de componentes principales
subcuenca río San Miguel y subcuenca río Aguarico……………………..26

ix
LISTA DE ANEXOS
Anexo 1. Permiso de investigación otorgado por el Ministerio del
Ambiente de Sucumbíos………..………………………..........................53
Anexo 2. Localidades de estudio................................................................54
Anexo 3. Trabajo de campo………………………………………………55
Anexo 4. Trabajo de laboratorio.................................................................57

x
TEMA: “Análisis de la variación morfológica de Hoplias malabaricus (Bloch 1794), asociada al tipo de hábitat utilizando morfometría geométrica”.
RESUMEN
Los peces de agua dulce a menudo presentan patrones de variación en la forma
del cuerpo al desarrollarse en hábitats divergentes, que en la mayoría de los
casos, estas diferencias observadas se han atribuido a las presiones
ambientales de vivir en hábitats lénticos o lóticos. Hoplias malabaricus presenta
una de las distribuciones más amplias de las especies de agua dulce de América
Latina y se encuentra desde Panamá hasta Argentina. En este estudio
empleamos herramientas de morfometría geométrica como: análisis
generalizado de Procrustes, análisis componentes principales y análisis de
variables canónicas, para describir la variabilidad morfológica de 96 individuos
de H. malabaricus, colectados en cuatro localidades pertenecientes a
ecosistemas lénticos y lóticos del cantón Cáscales. Se encontró que existen
diferencias morfológicas significativas entre poblaciones en diferentes tipos de
hábitats (lénticos y lóticos). La altura del cuerpo y el alargamiento y
ensanchamiento de la aleta caudal fueron los cambios morfológicos de mayor
peso en la diferenciación de estas poblaciones. Los especímenes colectados en
la subcuenca río Aguarico y subcuenca río San Miguel, no presentan diferencias
significativas en la morfología corporal. Este trabajo aporta nueva información
sobre la variación morfológica dentro de poblaciones de H. malabaricus, lo cual
puede explicar en parte la diversidad poblacional de esta especie en Ecuador.
Términos Descriptivos: Hoplias malabaricus, morfología, morfometría geométrica,
puntos anatómicos.

xi
TITLE: “Analysis of the morphological variation of Hoplias malabaricus
(Bloch 1794), associated at type of habitat using geometric morphometry”
ABSTRACT
Freshwater fishes often show large amounts of body shape variation across
divergent habitats and, in most cases, the observed differences have been
attributed to the environmental pressures of living in lentic or lotic habitats. H.
malabaricus is a neotropical fish and has one of the widest distributions of
freshwater species in Latin America, occurring from Panama to Argentina. The
objective of the present study was to describe and analyze with geometric
morphometric tools such as: generalized analysis of Procrustes, principal
component analysis and analysis of canonical variables, to describe the
morphological variability of 96 individuals of H. malabaricus, collected in four
locations belonging to the lentic and lotic ecosystems. The shape showed
significant differences among the studied populations. It was evidenced that there
are significant morphological differences between populations in different types
of habitats (lentic and lotic). The body height and the widening of the tail fin are
the most important variables in this differentiation. The sub-basins in which the
specimens were collected, there were no significant differences in body
morphology. This work provides new information on the intra-population
morphological variation of H. malabaricus, which may explain in part the
diversification of this specie in Ecuador.
Keywords: Hoplias malabaricus, morphotypes, geometric morphometrics,
landmarks.
I CERTIFY that the above and foregoing is a true and correct translation of the original
document in Spanish
Firma Certified Translator: Galo David Sierra ID: 1710905579

1
INTRODUCCIÓN
Los teleósteos son el grupo de peces más numeroso (Froese y Pauly 2017), este
grupo es el más antiguo y amplio entre los vertebrados y han evolucionado en
una gran diversidad de formas que les ha permitido colonizar todos los hábitats
acuáticos (Galvis et al. 2006). Este grupo de peces se originó en el agua dulce y
posteriormente fueron colonizando el mar (Audesirk et al. 2004, Albuja y
Merizalde 2012). En el transcurso de la adaptación y evolución de estos
organismos, las estructuras de sus sistemas y órganos, también han evidenciado
cambios funcionales en sus formas y tamaños, presentando diversas
adaptaciones (Vizcaíno et al. 2013, Wagner 2001a). Estos peces presentan
variabilidad morfológica, posiblemente producto de factores bióticos y abióticos
que interactúan entre sí; presentando especies con nuevos rasgos y formas
propias que afectan de manera diferencial en la biología y ecología de estos
organismos (Hass et al. 2010). Esta diversificación ha provocado el interés de
científicos, ocupados en identificar patrones morfológicos en estos organismos y
en estudios comparativos sobre la forma de especies de peces (Sepúlveda et al.
2012).
Por lo tanto, la morfometría se presenta como una herramienta robusta para
estudiar la forma de los organismos y se define como “el estudio cuantitativo de
la variación de la forma biológica” y su covariación con otras variables,
identificando así patrones morfológicos inter-específicos e intra-específicos
(Salas et al. 2012). En sus orígenes, la morfometría utilizaba variables lineales
tales como medidas de longitud, ancho y ángulos de un organismo; a partir de
estas medidas, se obtenía un conjunto de datos analizados posteriormente por
métodos estadísticos multivariados (Rohlf y Marcus 1993, Larraín et al. 2012). A

2
este enfoque se le conoce actualmente como morfometría tradicional
(Mitteroecker et al. 2013). Sin embargo estas variables de tipo lineal no recrean
en esencia la forma biológica y son difíciles de interpretar, presentando
incongruencias en las medidas y la significancia biológica que implica cada una
de ellas (Klingenberg 2010). Para superar estas limitaciones, a finales de los
años 80 surgió un método más sofisticado llamado morfometría geométrica
(Bookstein 1996, Muñoz et al. 2011).
La morfometría geométrica es la caracterización cuantitativa de la forma
biológica en un espacio bi o tridimensional, cuyo objetivo es analizar y comparar
patrones de variación (Bookstein 1991, Bechara et al. 2012); se diferencia de la
morfometría tradicional en que el estudio de la forma se realiza mediante
coordenadas, representadas en un plano cartesiano (Ríos et al. 2013). Esto
significa que la forma del organismo en estudio no se describirá en términos de
sus dimensiones, sino a partir de desplazamientos en el plano o espacio de un
conjunto de puntos de referencia llamados landmarks (Rodríguez 2015). Los
landmarks, también llamados puntos anatómicos de referencia (PAR) son
caracteres discretos, localizados de acuerdo algún criterios de homologia sobre
una imagen del organismo a estudiar (Ibáñez 2014); estos puntos se
corresponden en su localización con otros especímenes del mismo tipo,
presentando una equivalencia morfológica o funcional (Charlin et al. 2016). Se
traducen en coordenadas cartesianas x, y, en un espacio bidimensional; x, y, z,
en un espacio tridimensional, permitiendo así su análisis cuantitativo y haciendo
posible la visualización de los cambios morfológicos a través de
representaciones gráficas (Gonzales et al. 2012).

3
Los landmarks deben ser seleccionados por el investigador bajo los siguientes
criterios propuestos por Bookstein (1991); Toro et al. (2010); Benítez et al.
(2014):
Homología: Los landmarks deben estar ubicados sobre el mismo rasgo
morfológico a lo largo de toda la muestra, por lo que las estructuras
estudiadas deben ser homólogas, es decir deben tener similitud en su
ubicación dentro de la topografía del espécimen y en su origen ya que son
derivadas a partir de una estructura ancestral común.
Cobertura adecuada: Hace referencia al número de landmarks
marcados en las imágenes. Cantidades insuficientes de landmarks
suprimen valiosa información de los organismos estudiados; por el
contrario, una cantidad exagerada en relación del número de muestras
puede generar información errónea.
Repetibilidad: Considera que cada landmarks debe ser fácil de localizar
y estar claramente definido en todas las imágenes.
Coplanariedad: Otro criterio para seleccionar las marcas es la dimensión,
al no tomar en cuenta este criterio la interpretación puede ser errónea. Al
trabajar con imágenes que presentan una profundidad muy pronunciada
se debe evaluar la información presente en el plano posterior de la imagen
para validar la forma pura del organismo. En estos casos se emplean
imágenes tridimensionales.
Además Bookstein (1991) define los distintos tipos de landmarks, basados en la
información que pueden entregar y su localización:

4
Tipo 1: Son fácilmente reconocibles en cada una de las estructuras
analizadas y se encuentran en la intersección de dos o más tejidos.
Tipo 2: Están ubicados en zonas con curvaturas muy pronunciadas
Tipo 3: Corresponden a landmarks localizados en estructuras muy
distantes y están asociados a datos del tamaño como diámetros, longitud
y radio, por lo que la información sobre la forma es bastante escasa.
Una vez obtenido el mapa de coordenadas a partir de los diferentes tipos de
landmarks, se requiere de un proceso de normalización de las variables para
poder hacer comparaciones entre las muestras (Gonzales et al. 2012). Para esto
se aplica el método de superposición de puntos propuesto por Rohlf y Silice
(1990), denominado Análisis Procrustes Generalizado (APG). Este método
permite hacer un ajuste por mínimos cuadrados mediante el escalado, rotación
y traslación de cada una de las configuraciones de coordenadas, con la finalidad
de remover la información referente a la orientación, el tamaño y la posición, de
esta manera, la única variable a analizar es la forma (Torcida et al. 2012, Charlin
et al. 2016). Finalmente, estos datos se analizan con métodos estadísticos
multivariados para cuantificar la formación de grupos entre o dentro de las
poblaciones con base en las deformaciones que muestren los organismos
(Zelditch et al. 2004, López 2015).
A pesar de que la morfometría geométrica es una técnica relativamente nueva,
su uso en las ciencias naturales está extendido a nivel mundial y América Latina
no es la excepción. Por ejemplo en un trabajo hecho en Colombia, denominado:
“Estudio la variabilidad morfológica en Neoleucinodes elegantalis (Guenée
1854)” conocido como gusano perforador, se demuestra la relación entre la
forma y factores como ecorregión, altitud y hospedero (Obando 2011). Calle et

5
al. (2008), con ayuda de la morfometría geométrica diferenció once especies de
Anopheles, con la finalidad de identificar los principales vectores de malaria en
Colombia. En Venezuela se encontró diferencias en la estructura alar de la tribu
Rhodniini, permitiendo clasificar e identificar especies, muchas de ellas vectores
de enfermedades (Soto et al. 2011). Berchara y Liria (2012), estudiaron la
variabilidad morfológica de cinco especies de las familias Buthidae y
Scorpionidae en Venezuela, generando información para resolver los problemas
taxonómicos de estos grupos. Bissaro et al. (2013) analizaron la forma del
cefalotórax de Xyphopeneaus kroyeri (Heller 1862) en cuatro pesquerías,
revelando diferencias morfológicas entre poblaciones de las zonas de pesca en
el este de Brasil. En Ecuador Malato et al. (2017) estudio la divergencia de la
forma del cuerpo en el género de peces Rhoadsia, a lo largo de los gradientes
altitudinales de las laderas occidentales de los Andes ecuatorianos en el que
encontró que la forma corporal varía continuamente entre las poblaciones dentro
de los drenajes en función de la elevación encontrando que la forma del cuerpo
varia significativamente entre los individuos en función de la elevación aportando
al conocimiento de la adaptación y especiación de este género. Además de los
trabajos mencionados, la morfometría geométrica se puede emplear en una
amplia variedad de estudios, como cambios en la forma, enfoques
ontogenéticos, cambios de alometría, diferencias en la forma entre sexos,
estudios morfométricos con enfoques filogenéticos, cambios morfométricos entre
grupos con diferente distribución geográfica, entre muchos otros (Bonilla 2016).
Un modelo adecuado para estos tipos de estudios lo ofrecen los peces del Orden
Characiformes, ya que poseen la mayor riqueza de especies entre los peces de
agua dulce en Ecuador y presentan una diversidad extraordinaria de formas y

6
adaptaciones, así como preferencias de hábitat, alimentación y comportamiento
(Barriga 2012, Vanegas et al. 2013). Hoplias malabaricus (Bloch 1794), también
conocido como guanchiche, es una especie de la familia Erythrinidae, puede
llegar a alcanzar tallas de hasta 46 cm de longitud y peso de hasta 595 gr
(Valdiviezo et al. 2012). De acuerdo a estudios recientes, presenta una de las
distribuciones más amplias de las especies de agua dulce de América Latina y
se encuentra desde Panamá hasta Argentina (Zambrano et al. 2015). Su cuerpo
es alargado y semicilíndrico con escamas cicloideas, la cabeza está formada por
placas óseas, con los ojos ubicados ligeramente hacia arriba y unas poderosas
mandíbulas cubiertas de dientes cónicos muy filosos; su coloración varia de gris
verdoso a verde oscuro, aletas de color marrón y bandas irregulares de color
negro (Gómez 2016). Son depredadores solitarios y agresivos, de hábitos
nocturnos y con una gran tolerancia a ambientes con bajas concentraciones de
oxigeno ya que tiene la capacidad de respirar aire atmosférico (Galvis et al.
2006). Esta especie no es de importancia comercial, pero es importante en la
pesca de subsistencia local (Gómez 2005).
Existen estudios sobre el género Hoplias en Ecuador: Aguirre et al. (2013)
analizó la divergencia fenotípica y genética de las poblaciones de Hoplias
microlepis en ríos y embalses artificiales del drenaje del río Guayas en el oeste
de Ecuador; los resultaron evidenciaron que los peces de embalses artificiales
tienden a tener ojos, aletas dorsal y aleta caudal más largas y cuerpos más
delgados que los peces de río. Montero y Granda (2015), emplearon morfometría
geométrica para determinar si existen diferencias morfológicas entre poblaciones
de Hoplias microlepis (Günther 1864) y Hoplias malabaricus (Bloch 1794), en
relación a su distribución geográfica, hábitat, dieta y sexo; evidenciando que

7
existen diferencias morfológicas entre poblaciones de Hoplias geográficamente
aisladas, aunque no se identificaron diferencias en cuanto a la dieta y el sexo.
El objetivo del presente trabajo consiste en comparar la posible variación
morfológica de Hoplias malabaricus en hábitats lénticos y lóticos, mediante el
uso de la morfometría geométrica.

8
METODOLOGÍA
Área de estudio
El cantón Cáscales, se encuentra ubicado al nororiente del territorio ecuatoriano
y al noroccidente de la provincia de Sucumbíos (ver Figura 1), se encuentra
dividido en tres parroquias: El Dorado de Cáscales (cabecera cantonal), Sevilla
y Santa Rosa. Sus límites son: al Norte con la República de Colombia, al sur
con las provincias de Napo y Orellana, al este con la República del Perú y al
oeste con las provincias de Carchi, Imbabura y Pichincha (Sasintuña et al. 2015).
Posee una extensión total de terreno de 1244.82 𝐾𝑚², con temperaturas
promedio que oscilan entre los 25 °C. Se identifican cinco tipos de ecosistemas
segun Holdridge y Tosi (1978): Bosque húmedo Tropical, Bosque muy húmedo
Tropical, Bosque muy húmedo Pre-Montano, Bosque pluvial Pre-Montano y
Bosque muy húmedo Montano Bajo.
Figura 1. Mapa político del cantón Cáscales – Sucumbíos.
9
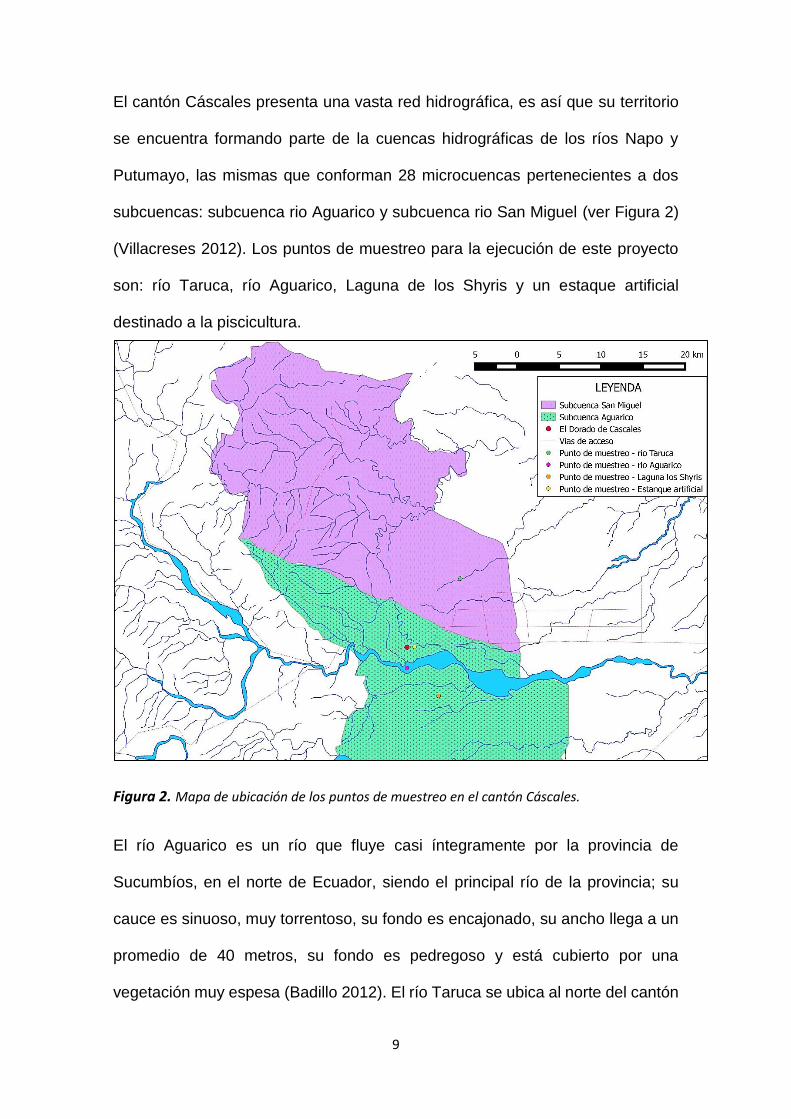
El cantón Cáscales presenta una vasta red hidrográfica, es así que su territorio
se encuentra formando parte de la cuencas hidrográficas de los ríos Napo y
Putumayo, las mismas que conforman 28 microcuencas pertenecientes a dos
subcuencas: subcuenca rio Aguarico y subcuenca rio San Miguel (ver Figura 2)
(Villacreses 2012). Los puntos de muestreo para la ejecución de este proyecto
son: río Taruca, río Aguarico, Laguna de los Shyris y un estaque artificial
destinado a la piscicultura.
Figura 2. Mapa de ubicación de los puntos de muestreo en el cantón Cáscales.
El río Aguarico es un río que fluye casi íntegramente por la provincia de
Sucumbíos, en el norte de Ecuador, siendo el principal río de la provincia; su
cauce es sinuoso, muy torrentoso, su fondo es encajonado, su ancho llega a un
promedio de 40 metros, su fondo es pedregoso y está cubierto por una
vegetación muy espesa (Badillo 2012). El río Taruca se ubica al norte del cantón

10
Cáscales y pertenece a la subcuenca rio San Miguel; su lecho es de piedra y
arena, su curso es muy tortuoso (Moncayo 2011). La Laguna Los Shyris se
caracteriza por sus aguas oscuras muy común en estos sistemas lacustre, se
encuentra ubicada en la comunidad Pastaza, al sur de la parroquia el Dorado de
Cáscales, presenta una gran biodiversidad, de flora y fauna, aguas de color
oscuro donde se desarrollan diversidad de especies acuáticas; (Sasintuña et al.
2015).
Diseño metodológico
Este tipo de investigación es descriptiva de conjunto, dado que se describe la
morfología corporal de poblaciones de Hoplias malabaricus que habitan en
ecosistemas lénticos y lóticos en el cantón Cáscales, provincia de Sucumbíos.
Población y muestra
Para este estudio se trabajó con 24 ejemplares de cuatro localidades,
considerando que se tomaron 12 landmarks, de acuerdo a lo sugerido por
Bookstein (1991) y Toro et al. (2010).
Métodos
Para obtener ejemplares de H. malabaricus se realizaron salidas de campo en
los meses de agosto, septiembre, octubre y noviembre, bajo el permiso de
investigación: Oficio Nro. 01-IC-FAU/-DPS/MAFAUNAX, otorgado por el
Ministerio del Ambiente de Sucumbíos (Anexo 1). Se tomaron muestras de
cuatro localidades, dos correspondientes a ecosistemas lénticos (Laguna Shyris,
Estanques artificiales) y dos a ecosistemas lóticos (Río Aguarico, Río Taruka)
obteniendo un total de 24 individuos por cada localidad. El tamaño de la muestra
se justifica según lo propuesto por Bookstein (1991) y Toro et al. (2010), que

11
indica que el número de individuos debe ser el doble, respecto al número de
landmarks; se requiere entonces muestras n= 24, por cada unidad de muestreo
para un esquema de doce landmarks. Solo los individuos adultos con tallas a
partir de los quince centímetros fueron colectados, para evitar las diferencias
ontogénicas en la morfología de esta especie (Bohórquez 2010).
Se combinaron diferentes artes de pesca, así durante una misma faena se
utilizaron atarrayas con diámetros de 2 a 4 m y distancia entre nudos de 0.5 a 2
cm; redes de arrastre con ancho de 1 a 2.5 y 3 a 12 m de largo y ojo de malla
0.1 a 0.5 cm; red agallera con longitud de 15 m y 2 m de alto, con distancias
entre nudos de 1 a 3 cm.
Procesamiento de las muestras
Para el análisis de las muestras se tomaron fotografías de cada espécimen por
el lado izquierdo con una cámara digital Nikon D200. Las fotografías se
realizaron en una mesa de trabajo con soporte para cámara fotográfica y
conexiones para luminarias. Los peces se colocaron sobre una bandeja y en el
borde inferior se colocó un ictiómetro para poder hacer la escala de tamaño
respectiva.
Con el programa tpsUtil (Rohlf 2008), se escogieron y agruparon en un archivo
96 fotografías de los peces correspondiendo cada una a un individuo con el fin
de transformar estas imágenes a formato tps.
Luego se establecieron un total de doce marcas de puntos homólogos
(landmark): (1) punta de mandíbula superior, (2) parte superior de la región
supraoccipital, (3) inicio de la aleta dorsal, (4) final de la aleta dorsal, (5) punto
de inserción superior de la aleta caudal, (6) final de la línea lateral, (7) punto de

12
inserción inferior de la aleta caudal, (8) punto de inserción posterior de la aleta
anal, (9) punto de inserción anterior de la aleta anal, (10) origen de la aleta
pélvica, (11) origen de la aleta pectoral y (12) extremo posterior de la mandíbula
inferior; el proceso de digitalización de estos puntos se realizó con el programa
tpsDig (Rohlf 2010). Para facilitar la ubicación de los puntos anatómicos de
referencia se colocaron agujas en el cuerpo de los especímenes, marcando los
landmarks y haciéndolos más visibles en las fotografías.
Posteriormente, se realizó el método de superimposición de análisis
generalizado de Procrustes (Rohlf y sílice 1990, Bohórquez 2010) con ayuda del
programa tps Relative Warps (Rolhf 2015), este método permite eliminar los
efectos de translación, rotación y escala de las imágenes con el fin de minimizar
las diferencias entre las configuraciones de landmarks para poder compararlos
y realizar inferencias de dichas comparaciones.
CONSIDERACIONES ÉTICAS
Todo el procedimiento fue llevado a cabo según la guía del Canadian Council on
Animal Care (2005), para el manejo ético de peces en investigación. Se indujo
de manera humanitaria, la muerte de los animales (eutanasia) empleados en la
investigación, con el propósito de eliminar o disminuir al mínimo el dolor y el
estrés previo y durante el procedimiento (Martínez y Osorio 2015). Para esto, los
individuos fueron anestesiados con Benzocaína (0,1gL-1), sumergiéndolos en un
baño de esta solución (Gómez et al. 2011). Una vez que los individuos perdieron
el eje de nado y los movimientos bucal y opercular, se procedió a realizar un
corte transversal en la médula espinal para sacrificarlos.

13
ANÁLISIS ESTADÍSTICOS
Se realizó un Análisis de Componentes Principales (ACP) de los datos
morfométricos, sobre una matriz de covarianza, para la separación entre grupos;
este método describe las variaciones de los individuos en un conjunto de datos,
reduciendo la dimensionalidad de las variables y mostrando las principales
tendencias de cambio (Miralles 2015). Para describir las diferencias en la figura
de cada población se obtuvieron las mallas de deformación con los vectores
derivados de este análisis.
Para evaluar las diferencias estadísticas en cada tipo de hábitat y subcuencas,
se realizó un análisis de variación canónica (CVA), que permite encontrar
diferencias significativas entre grupos conocidos, mediante una prueba de
análisis de varianza multivariado (MANOVA). El CVA es un análisis discriminante
para más de dos muestras, cuyo fin es determinar si los grupos previamente
establecidos difieren entre sí y en qué forma (Toro et al. 2010).
Posteriormente se realizaron comparaciones múltiples entre pares de
localidades mediante la prueba de Hotelling, la cual es una generalización
multivariante de la distribución t de Student (Cuadras 2007).
Todos estos análisis fueron procesados en el programa Paleontological Statistics
Past 1.81 (Hammer et al. 2001).
Por último, se realizara un análisis de Thin Plate Spline con el programa tpsPLS
versión 1.23 (Rohlf 2015). Este análisis genera un mapa de hitos de acuerdo a
la forma promedio de todos los puntos, con el fin de visualizar el grado de
deformación de cada grupo, mediante rejillas de deformación (López 2015).

14
RESULTADOS
Se analizaron un total de 96 peces colectados de cuatro localidades: río Taruca,
río Aguarico, laguna “los Shyris” y un estanque artificial, que corresponden a dos
subcuencas: río San Miguel y río Aguarico.
Análisis de componentes principales
Las coordenadas generadas luego de la superimposición de Procrustes, fueron
analizadas mediante un análisis de componentes principales (PCA). Los
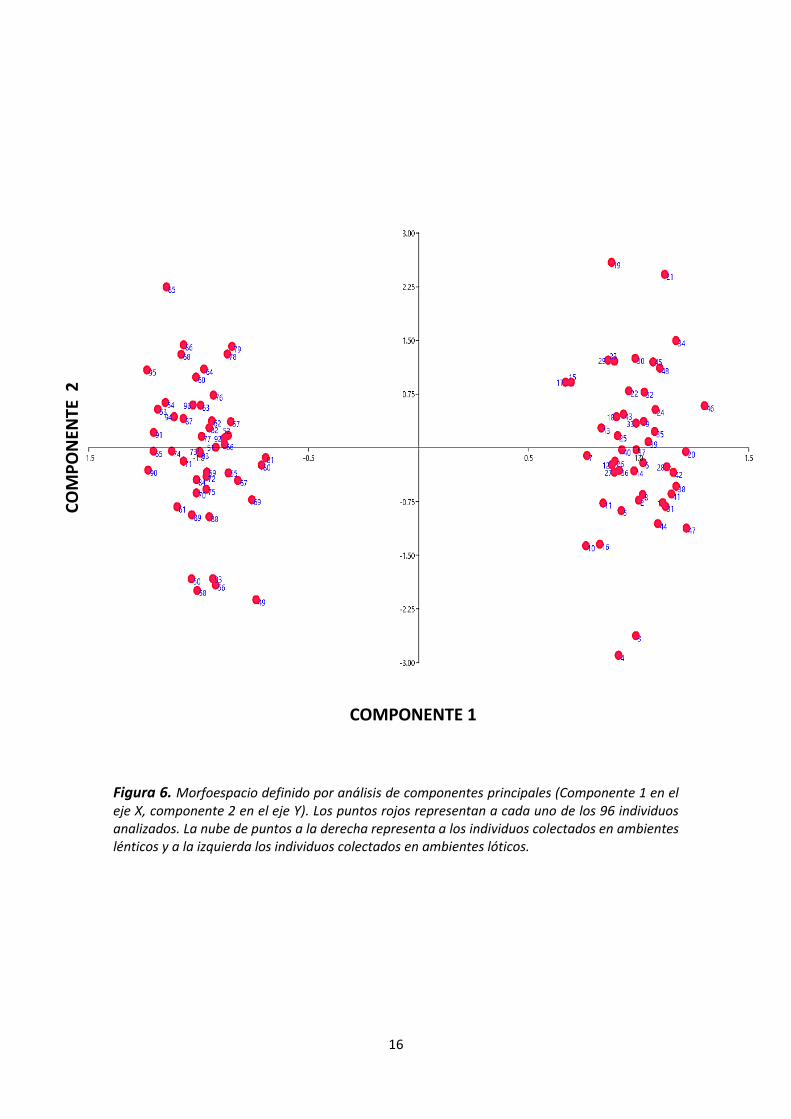
resultados del análisis de componentes principales muestran que los dos
primeros componentes explican el 85,26 % de la varianza total, siendo el primer
componente (83,18%) el que más aporta a la discriminación entre formas (ver
figura 6) (ver tabla 6), lo que indica que hay una alta diferenciación en la forma
de Hoplias malabaricus con respecto al hábitat en el que fueron colectados.
La Figura 6 muestra el grafico de componentes principales con 96 puntos rojos
distribuidos a lo largo del morfoespacio que representan el total de los individuos
colectados 24 por cada zona de captura. Se observa una clara tendencia de los
especímenes pertenecientes al río Taruca y río Aguarico a ubicarse en los
cuadrantes inferior y superior izquierdo mientras que los especímenes de la
laguna de los Shyris y el estanque artificial se ubicaron en el cuadrante inferior y
superior derecho. Esta gradiente de izquierda a derecha corresponde al primer
componente y nos permitió separar los individuos colectados en ambientes
lénticos de los colectados en ambientes lóticos, correspondiendo los valores
negativos (lóticos) a cuerpos más delgados y alargados y los valores positivos
(lénticos) a cuerpos más gruesos.
15
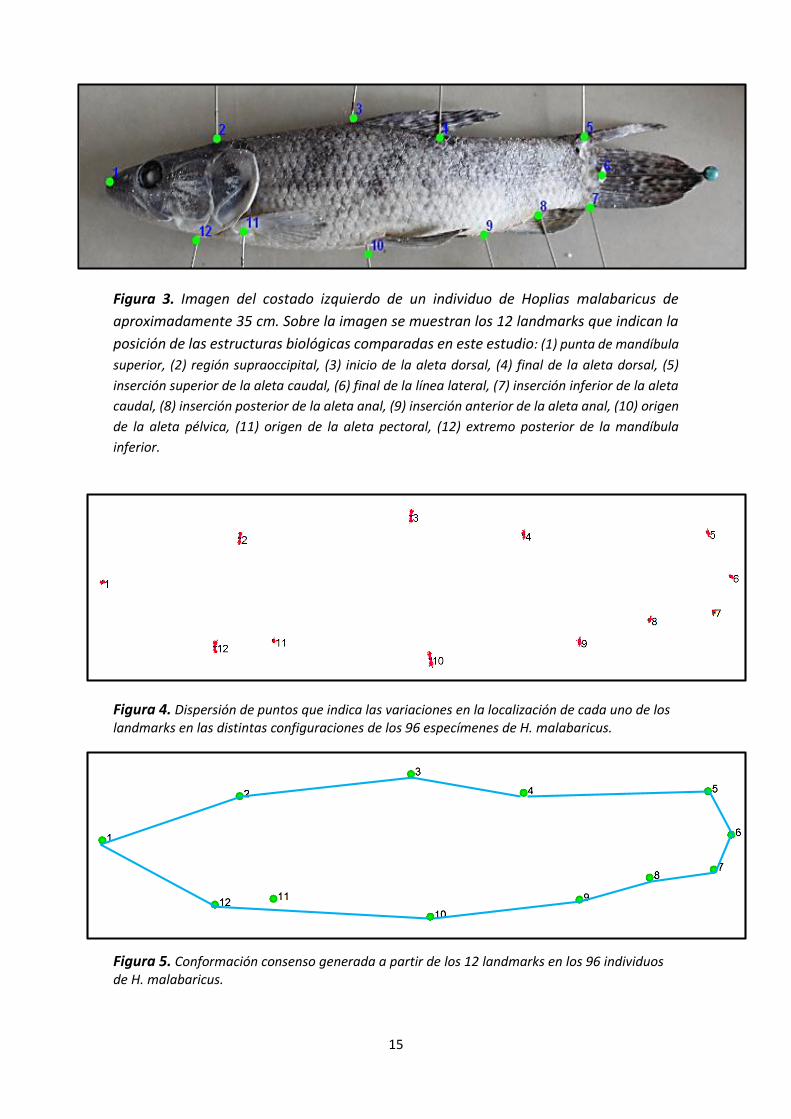
Figura 3. Imagen del costado izquierdo de un individuo de Hoplias malabaricus de
aproximadamente 35 cm. Sobre la imagen se muestran los 12 landmarks que indican la
posición de las estructuras biológicas comparadas en este estudio: (1) punta de mandíbula
superior, (2) región supraoccipital, (3) inicio de la aleta dorsal, (4) final de la aleta dorsal, (5)
inserción superior de la aleta caudal, (6) final de la línea lateral, (7) inserción inferior de la aleta
caudal, (8) inserción posterior de la aleta anal, (9) inserción anterior de la aleta anal, (10) origen
de la aleta pélvica, (11) origen de la aleta pectoral, (12) extremo posterior de la mandíbula
inferior.
Figura 4. Dispersión de puntos que indica las variaciones en la localización de cada uno de los landmarks en las distintas configuraciones de los 96 especímenes de H. malabaricus.
Figura 5. Conformación consenso generada a partir de los 12 landmarks en los 96 individuos de H. malabaricus.
16
Figura 6. Morfoespacio definido por análisis de componentes principales (Componente 1 en el eje X, componente 2 en el eje Y). Los puntos rojos representan a cada uno de los 96 individuos analizados. La nube de puntos a la derecha representa a los individuos colectados en ambientes lénticos y a la izquierda los individuos colectados en ambientes lóticos.
COMPONENTE 1
CO
MP
ON
ENTE
2

17
Los gráficos de las rejillas de deformación confirman los resultados obtenidos en
el gráfico de dispersión del análisis de componentes principales, confirmando
que las poblaciones provenientes de ambientes lénticos y lóticos difieren
principalmente en la profundidad del cuerpo de H. malabaricus. Los
desplazamientos de landmarks más significativos entre las configuraciones de
consenso involucraron a aquellos asociados con la parte superior de la región
supraoccipital (2), inicio de la aleta dorsal (3), final de la aleta dorsal (4), cuyas
variaciones se desplazan ventralmente, y los landmarks correspondientes a el
punto de inserción anterior de la aleta anal (9), origen de la aleta pélvica (10),
extremo posterior de la mandíbula inferior (12) los cuales se desplazan
dorsalmente (ver Figura 8); estos desplazamientos indicaron una compresión
dorsal-ventral en individuos de ambientes lóticos en relación con individuos de
ambientes lénticos que presentaron un cuerpo más pronunciado.
Otros puntos de referencia que exhibieron desplazamientos conspicuos incluyen
aquellos en el punto de inserción superior de la aleta caudal (5), final de la línea
lateral (6), punto de inserción inferior de la aleta caudal (7) que se mueven
posteriormente y hacia afuera en los individuos de ambientes lóticos, lo que
resulta en un ligero alargamiento y ensanchamiento de la aleta caudal con
respecto a individuos de ambientes lénticos (ver Figura 8). Así, las mallas de
deformación derivadas muestran que de acuerdo aún gradiente de variación de
negativo a positivo para el PCA 1, los organismos capturados en ecosistemas
lóticos presentan menor altura, con las aletas caudales más alargadas y anchas,
mientras que los organismos de ecosistemas lénticos presentan mayor altura,
con las aletas caudales más pequeñas y angostas. El resto de características no
parecen ser distintas.

18
Figura 7. Espécimen colectado en un ecosistema léntico (derecha) y ecosistema lótico
(izquierda).
A B
Figura 8. Mallas de deformación con vectores del primer eje del análisis de componentes
principales. Las líneas apuntan en la dirección del cambio de forma para cada hito, donde A indica
especímenes colectados en hábitats lóticos y B representa especímenes de hábitats lénticos.
A B
Figura 9. Los cambios en la forma se muestran mediante rejillas de deformación con una
gradiente de color de azul a rojo, donde; los colores celeste y azul indican contracción y el
amarillo y rojo indican expansión en los especímenes de ambientes lénticos (A) y lóticos (B).
CP1 (+) CP1 (-)
CP1 (-) CP1 (+)

19
Análisis de variables canónicas
El diagrama de dispersión del análisis de variables canónicas indica la
separación de dos grupos: el primero está conformado por las poblaciones del
río Taruca y río Aguarico; en el segundo se encuentran las poblaciones de
Laguna de los Shyris y el estanque artificial (ver figura 10). Dentro de cada grupo
se evidenció un solapamiento de sus poblaciones, lo que indica que las
diferencias morfológicas son más grandes entre poblaciones de distintos hábitats
que dentro de ellas. La variación de la forma del cuerpo asociada con la
divergencia a lo largo de CV1 representó el 99.01 % de la variación de la forma
corporal entre los grupos, y fue atribuible en gran medida a las diferencias en
profundidad corporal consistentes con las descritas anteriormente por el análisis
de componentes principales. Los valores positivos a lo largo de este eje indican
cuerpos más profundos, mientras que los valores negativos indican cuerpos más
hidrodinámicos.
Probablemente los peces al movilizarse en hábitats más turbulentos o
torrentosos (ecosistemas lóticos), implican que aumente la fricción con el medio
acuático; de esta manera podría decir que estos peces han modificado su
morfología para reducir la superficie de contacto con el agua, y así se reduce
dicha fricción. A diferencia de los ecosistemas lenticos en donde el sedentarismo
es potencialmente mayor. Además considerando que los ecosistemas lénticos
por lo general presentan mayores recursos alimenticios es posible que las
poblaciones de H. malabaricus presentes en estos hábitats se alimenten mejor
que en los ríos y se vuelvan más robustos.

20
En general el análisis de MANOVA mostró que existen diferencias significativas
(p= 2.22045e-16) entre las poblaciones colectadas en ecosistemas lóticos y
lénticos a lo largo del eje de variable canónica 1 (ver Tabla 1). El eje de la
variable canónica 2 no presentó diferencias significativas (p= 0.023).
Posteriormente se realizó un análisis de comparación por pares usando la
prueba de hotelling; se encontraron diferencias significativas entre las
poblaciones de: río Taruca y laguna de los Shyris (p: 4.4916E-41), río Taruca y
Estanque artificial (p: 5.2589E-41), río Aguarico y Laguna de los Shyris (p:
5.7631E-41), río Aguarico y Estanque artificial (p: 4.0108E-41), mientras que la
comparación entre río Taruca y río Aguarico (p: 0.14778) y Laguna de los Shyris
y Estanque artificial (0.068988) no presentan valores significativos (ver Tabla 7).
Tabla 1. Resultados del test MANOVA para la variable canónicas 1 en las cuatro localidades analizadas.
Wilks´ lambda: 2.163E-92
df1: 20
df2: 75
F: 1.734E92
p : 2.22045e-16

21
Figura 10. Morfoespacio entre los ejes 1 y 2 resultantes del análisis de variables canónicas
aplicado a las 4 poblaciones de H. malabaricus: río Taruca, río Aguarico, laguna de los Shyris y
estanque artificial.
VARIABLE CANONICA 1
VA
RIA
BLE
CA
NO
NIC
A 2
RIO TARUCA
RIO AGUARICO
LAGUNA SHYRIS
ESTANQUE ARTIFICIAL

22
Matriz de clasificación
La clasificación de las 96 muestras en los grupos a priori fue basada según la
localidad donde fueron colectadas. La matriz de confusión clasificó
correctamente el 76,04% de los individuos (ver Tabla 2). Las muestras
reclasificadas correctamente al 100% fueron las pertenecientes a los
ecosistemas lóticos.
Tabla 2. Matriz de clasificación para las cuatro localidades analizadas. Las filas presentan
los grupos asignados a priori y las columnas los grupos a los que fueron asignados por
el programa.
LENTICOS
LOTICOS
SHYRIS
ESTANQUE
TARUCA
AGUARICO
TOTAL
SHYRIS
66.6%
33.3% - -
(100%) 24
ESTANQUE
12.5
87.5% - -
(100%) 24
TARUCA - -
75%
25%
(100%) 24
AGUARICO - -
25%
75%
(100%) 24
TOTAL
79.5% (19)
120.8%
(29)
100% (24)
100% (24)
96 individuos
CLASIFICADOS CORRECTAMENTE : 76,04%

23
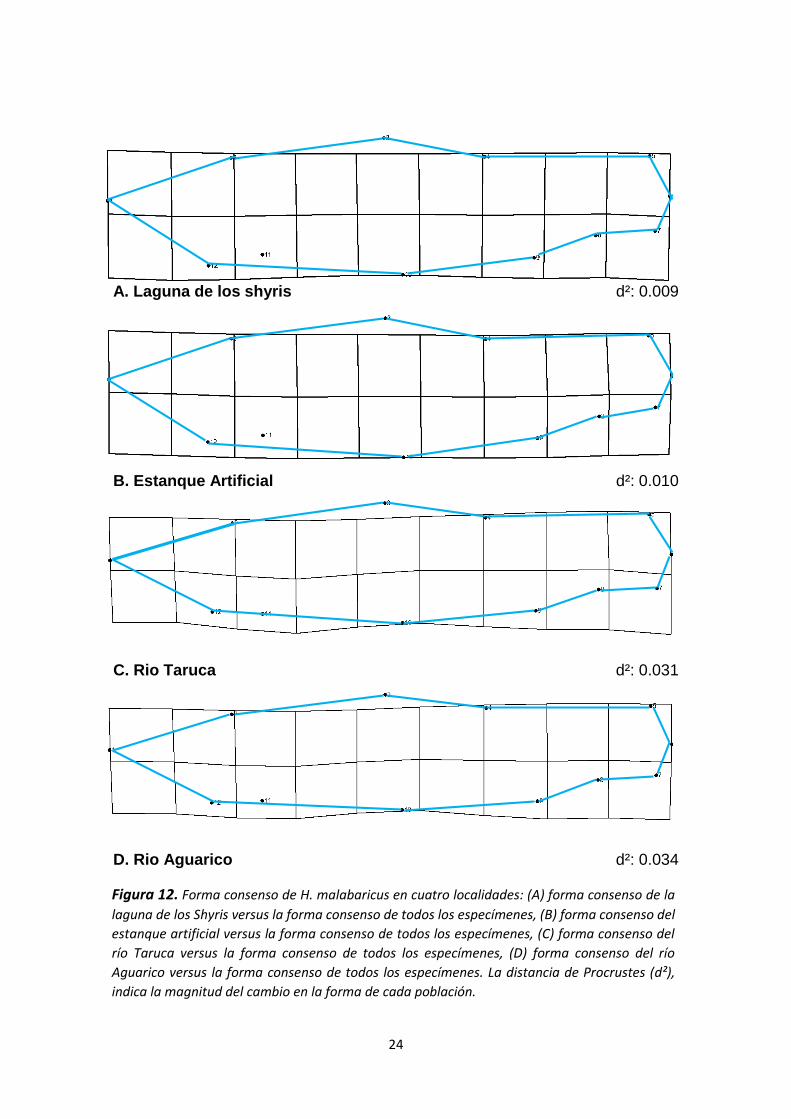
Thin Plate Spline
Se generaron mapas de hitos con su respectiva deformación en una gradilla de
acuerdo a la forma promedio de todos los puntos con el fin de visualizar
diferencias en la forma del cuerpo entre las figuras consenso de las muestras
(ver Figura 11). Los especímenes se clasificaron según el sitio de captura y luego
fueron contrastados versus la forma consenso de todos los especímenes (ver
Figura 12).
La distancia Procrustes, d², indica la magnitud del cambio en la forma de los
especímenes; mientras mayor es este valor, mayores son las diferencias entre
la forma promedio y la estudiada (ver figura 10). Los peces de la laguna Shyris y
del Estanque artificial (lénticos) tienen características muy similares, se
distinguen por tener la zona abdominal prominente y el valor de “d²” para ambos
casos confirma que las imágenes son muy parecidas. En el caso de los
especímenes del río Aguarico y río Taruca, se observa que son un poco más
delgados y alargados y también presentan valores “d²” similares.
Figura 11. Conformación consenso a partir de los 12 landmarks en los 96 individuos de H.
malabaricus generada con el análisis Thin Plate Spline.
24
A. Laguna de los shyris d²: 0.009
B. Estanque Artificial d²: 0.010
C. Rio Taruca d²: 0.031
D. Rio Aguarico d²: 0.034
Figura 12. Forma consenso de H. malabaricus en cuatro localidades: (A) forma consenso de la
laguna de los Shyris versus la forma consenso de todos los especímenes, (B) forma consenso del
estanque artificial versus la forma consenso de todos los especímenes, (C) forma consenso del
río Taruca versus la forma consenso de todos los especímenes, (D) forma consenso del río
Aguarico versus la forma consenso de todos los especímenes. La distancia de Procrustes (d²),
indica la magnitud del cambio en la forma de cada población.

25
Comparación entre subcuencas
Se realizó un análisis de componentes principales para identificar las principales
variaciones en la forma de los especímenes colectados en la subcuenca río San
Miguel y subcuenca río Aguarico (ver figura 13). El primer componente explicó
apenas el 14,86 % de la variación, mientras que el segundo el 11,36 %. Así los
10 primeros componentes representan el 80,47 % de la variación morfológica, lo
que significa que este análisis resulto poco efectivo para separar los individuos.
En la gráfica de dispersión a lo largo de los dos primeros componentes, se
observa dos nubes de puntos dispersas en color verde para la subcuenca río
San Miguel y rojo para la subcuenca río Aguarico. Estas poblaciones presentaron
un gran solapamiento entre sí; por lo que no se pudo discriminar ningún patrón
de diferenciación entre los morfotipos, indicando la imposibilidad de reconocer
distintas formas entre los individuos colectados en la subcuenca río San Miguel
y subcuenca río Aguarico. Esto significa que existe una gran similitud en forma
del cuerpo entre estas poblaciones.
Estas poblaciones posiblemente presentan algunas zonas de contacto ya que no
existen barreras geográficas evidentes que las aíslen, además gracias a la
capacidad de Hoplias malabaricus de utilizar el oxígeno atmosférico, puede
atravesar caminos de tierra para cruzar de un cuerpo de agua a otro, por lo que
puede existir flujo génico entre ellas

26
Figura 13. Morfoespacio definido por análisis de componentes principales (Componente 1 en el eje X, componente 2 en el eje Y). Subcuenca río San Miguel (color verde) y subcuenca río Aguarico (color rojo).
Subcuenca rio San Miguel
Subcuenca rio Aguarico
COMPONENTE 1
CO
MP
ON
ENTE
2
27
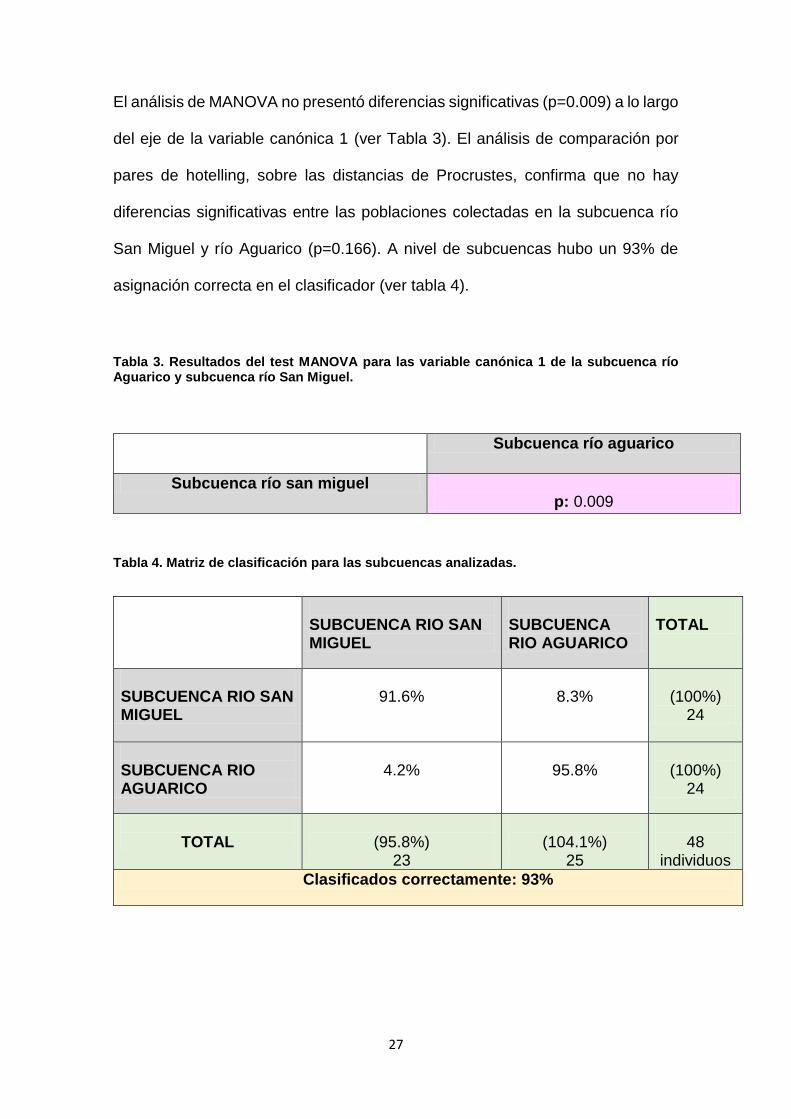
El análisis de MANOVA no presentó diferencias significativas (p=0.009) a lo largo
del eje de la variable canónica 1 (ver Tabla 3). El análisis de comparación por
pares de hotelling, sobre las distancias de Procrustes, confirma que no hay
diferencias significativas entre las poblaciones colectadas en la subcuenca río
San Miguel y río Aguarico (p=0.166). A nivel de subcuencas hubo un 93% de
asignación correcta en el clasificador (ver tabla 4).
Tabla 3. Resultados del test MANOVA para las variable canónica 1 de la subcuenca río Aguarico y subcuenca río San Miguel.
Tabla 4. Matriz de clasificación para las subcuencas analizadas.
SUBCUENCA RIO SAN MIGUEL
SUBCUENCA RIO AGUARICO
TOTAL
SUBCUENCA RIO SAN MIGUEL
91.6%
8.3%
(100%)
24
SUBCUENCA RIO AGUARICO
4.2%
95.8%
(100%)
24
TOTAL
(95.8%)
23
(104.1%)
25
48
individuos
Clasificados correctamente: 93%
Subcuenca río aguarico
Subcuenca río san miguel p: 0.009

28
DISCUSIÓN
Las poblaciones de una misma especie que habitan en diferentes ambientes
exhiben una considerable diversidad en sus formas (Ruzzante et al. 1998, Soria
et al. 2011, Foster et al. 2015). Esto es particularmente cierto en el entorno
acuático, que es muy variable tanto desde el punto de vista espacial como
temporal. La capacidad de un pez para moverse eficientemente a través del agua
depende en gran medida de su forma corporal, limitando así las especies a
ciertos hábitats o gradientes ambientales. En el caso particular de Hoplias
malabaricus se han realizado varios trabajos sobre la biología básica de la
especie (Menin 2015, Rodríguez et al. 2017), composición de la dieta
(Llamazares et al. 2011, Granada y Valencia 2015, Vegh 2015), análisis
genéticos (Reiss et al. 2015, Grassi et al. 2017), sobre su edad, tasas de
crecimiento y alimentación (Corrêa et al. 2014) y sobre el cariotipo (Perrin et al.
2016). Pero existe una carencia desde el punto de vista poblacional, ya que muy
poco se conoce de Hoplias malabaricus a lo largo de su distribución geográfica.
Los cambios morfológicos son el resultado de la plasticidad fenotípica y permiten
tener respuestas rápidas a los cambios ambientales, al generar diferentes
fenotipos en una especie, sin realizar cambios en el genoma, facilitando así su
adaptabilidad (Faster et al. 2015). Numerosos factores bióticos y abióticos
(profundidad, distribución, edad, ambientes contrastantes), han demostrado
contribuir en esta diversidad morfológica en muchos peces (Haas et al. 2010,
Hankison et al. 2006, Krabbenhoft et al. 2009). Es tal la variación y la cantidad
de factores que inciden en la forma de los organismos que se puede decir que el
fenotipo es producto de un modelo explicativo llamado “efecto - área”; esta teoría
asume que las micro condiciones del hábitat presentes en pequeñas escalas

29
espaciales y/o temporales son las que determinan el fenotipo (Cameron y Dillon
1984, Urbano 2011). La plasticidad fenotípica entonces explica los cambios en
la forma de poblaciones de ecosistemas lénticos y lóticos, ya que al ser
ambientes diferentes en ciertos aspectos como flujo de agua, temperatura,
disponibilidad de alimento, profundidad, nutrientes, influencias antropogénicas y
variación de oxígeno disuelto, pueden dar origen a cambios morfológicos
significativos (Montero y Granda 2015). La divergencia morfológica asociada al
hábitat es común en los peces (Krabbenhoft et al. 2009, Schaefer et al. 2011,
Webster et al. 2011). Foster et al. (2015) realizo un estudio en Goodea atripinnis
(Goodeidae) y Chirostoma jordani (Atherinopsidae), demostrando que los peces
de una misma especie que habitan en ambientes lénticos y lóticos sufren
modificaciones, no solo genéticas, también morfológicas; en sus resultados se
observó que el alto del cuerpo variaba mucho entre las poblaciones dependiendo
del hábitat en el que fueron capturados. Esta característica también fue descrita
por Franssen (2011) quien realizó un estudio de la divergencia morfológica de
Cyprinella lutrensis en ríos y embalses artificiales de EE.UU. En Ecuador Aguirre
et al. (2013) y Montero y Granda (2015) realizaron estudios de morfología en
Hoplias microlepis colectados en ríos, embalses y represas; con resultados
similares.
Los resultados del presente estudio concuerdan con los anteriormente descritos
demostrando la divergencia morfológica de Hoplias malabaricus en dos hábitats
contrastantes. En los gráficos de dispersión del CPA, se observa una clara
separación de la nube de puntos, discriminando las cuatro localidades en dos
grandes agrupaciones (lénticos y lóticos). El grafico del CVA presenta una
distribución de puntos muy parecida, pero en esta se evidencia un solapamiento

30
de las poblaciones dentro de cada hábitat. El MANOVA mostró que existen
diferencias significativas entre los organismos de las cuatro poblaciones
analizadas a lo largo de la variables canónica 1. Sin embargo, de acuerdo con
las comparaciones pareadas al usar la prueba de Hotelling, se evidencia que las
diferencias son significativas solo entre los especímenes de hábitats diferentes.
Las diferencias más notorias que se observó mediante las mallas de
deformación, fue que las poblaciones que habitan los ríos tienden a ser más
delgadas y alargadas, mientras que las que habitan en lagunas y estanques son
más grandes, especialmente en el abdomen presentando un cuerpo más
robusto. El largo y ancho de la aleta caudal también tuvo cierta variación,
encontrándose que en los ecosistemas lóticos son más largas y anchas que en
los peces de ecosistemas lénticos. Tales diferencias morfológicas pueden haber
surgido como resultado de presiones de selección divergentes en ambos
hábitats, como las diferencias de flujo de agua, disponibilidad de alimento y
depredación.
Prácticamente todos los peces se mueven constantemente a través de su medio
acuático para realizar tareas críticas para la supervivencia y la reproducción; en
consecuencia, las capacidades locomotoras de los peces están relacionadas con
el régimen del flujo de agua (Rojas et al. 2015). Los ambientes lóticos presentan
una corriente continua de agua, por lo que tienden a seleccionar formas
corporales fusiformes que reducen la resistencia en ambientes acuáticos,
permitiendo la propulsión efectiva y el mantenimiento de la velocidad a un menor
costo de energía, mientras que los peces de ambientes lénticos potencialmente
requieren de menos movilidad, ya que se desenvuelven en aguas estancadas
sin ningún flujo de corriente (Foster et al. 2015). Otra de las adaptaciones

31
biomecánicas para aumentar el rendimiento de la natación en los peces es la
presencia de aletas caudales robustas. Esta característica reduce los costos de
natación en comparación con las aletas caudales más pequeñas (Franssen
2011), y presumiblemente explica los cambios que presentaron en este estudio
las poblaciones de Hoplias malabaricus en la forma de la aleta caudal. De hecho,
la necesidad de aumentar la capacidad de natación en ambientes con mayor
intensidad de flujo de agua puede ser un fenómeno generalizado, impulsando
variaciones fenotípicas repetibles y predecibles en diferentes grupos de peces
(Morales 2017). A pesar de la evidencia de que la velocidad del agua puede
impulsar la variación fenotípica en los peces, pocos estudios han investigado
directamente el papel de los regímenes de flujo divergentes en la especiación
(Bhagat et al. 2006, Langerhans 2008, Langerhans et al. 2010). Claramente, se
requiere más trabajo para comprender el papel del flujo de agua en la generación
de diferencias fenotípicas.
Otro factor que puede influenciar en la divergencia morfológica de Hoplias
malabaricus puede ser disponibilidad de alimento; los ecosistemas lénticos
presentan mayores recursos alimenticios y por consecuencia los peces en estos
hábitats se alimentan mejor que en los ríos y se vuelven más robustos. Martins
(2009) relata que Hoplias malabaricus presenta diferentes nichos alimentarios,
siendo piscívora en la fase adulta e insectívora en la fase juvenil. Mello et al.
(2006) cita otro nicho alimentario para esta especie, afirmando que en la fase
larval se comporta como herbívora. Este cambio en el régimen alimentario junto
con el crecimiento, es una adaptación de esta especie para aprovechar la mayor
gama de ítems alimentarios posibles (Martins 2009). En el caso de las
poblaciones de H. malabaricus que se desarrollan en ambientes lénticos,

32
acceden fácilmente y en gran cantidad a estos recursos (Perreira et al. 2004,
Granitto et al. 2016). Algunos estudios sobre el contenido estomacal en esta
especie revelan una alta incidencia de estómagos vacíos en poblaciones de
diferentes ríos (Fernandez et al. 2012, Vegh 2016, Saigo 2016). Resultados
contrastantes con los anteriores fueron descritos por Piraquive et al. (2015) y
Cardoso (2015) para H. malabaricus en lagos de la Amazonia, evidenciando
poblaciones en mejores condiciones alimenticias. Es posible entonces afirmar
que para esta especie los ambientes lénticos presentan condiciones más
favorables para su desarrollo y establecimiento, evidenciadas por su alta
actividad reproductiva y su flexibilidad alimentaria.
Lo cambios en la profundidad del cuerpo y la aleta caudal pueden estar también
relacionados con los hábitos depredadores de esta especie, así por ejemplo, es
posible que las poblaciones que habitan en ríos, necesiten desplazarse
constantemente y más rápido para capturar a sus presas las cuales se
encuentran distribuidas irregularmente en aguas abiertas (Martins et al. 2012),
resultando una gran ventaja en ambientes lóticos el aumento de la natación en
ráfaga, gracias al aumento de la musculatura en la región caudal y un cuerpo
más pequeño y fusiforme. De esta manera resultan más eficientes las
actividades inestables de natación, como mayor velocidad y aceleración durante
los ataques y mayor estabilidad durante los giros rápidos (Langerhans y Reznick
2010). Tomando en cuenta la actividad depredadora y la adaptación locomotora
a través de las gradientes de flujo de agua, se puede predecir la divergencia en
la forma del cuerpo y la forma de la aleta caudal en esta especie. Sin embargo
se necesita más investigación si queremos obtener una comprensión más clara

33
de la posible relación entre la energía de la natación, la estructura del hábitat,
alimentación, régimen de depredación y la variación morfológica.
Finalmente, las poblaciones de río Taruca y río Aguarico que corresponden a las
subcuencas río San Miguel y río Aguarico respectivamente, no mostraron
diferencias significativas al realizar el análisis de Manova. Estas poblaciones
posiblemente presentan algunas zonas de contacto, ya que no existen barreras
geográficas evidentes que las aíslen, por lo que puede existir flujo génico entre
ellas y no se encuentran aisladas completamente (Chávez 2016).
Los estudios de morfometría geométrica en peces que habitan en ecosistemas
contrastantes coinciden siempre en la separación de las muestras en dos grupos,
donde siempre se ven separadas las muestras de ecosistemas lóticos de las
muestras de ecosistemas lénticos. Estos resultados indican que los hábitats
divergentes impulsan la diversificación fenotípica intraespecífica, y son
importantes para predecir las respuestas de adaptación de las especies de peces
de agua dulce. Las diferencias morfológicas encontradas en el alto del cuerpo y
la aleta caudal en el presente estudio sugieren que existe variabilidad
intraespecífica y la presencia de dos fenotipos de H. malabaricus en las zonas
estudiadas.

34
CONCLUSIONES
Los análisis indican diferencias morfológicas entre los peces de
ecosistemas lénticos y lóticos, y se presentan como dos grupos que
difieren significativamente en su morfología.
Las diferencias en la variación de la forma encontradas a nivel de
subcuencas no fueron significativas para las poblaciones de río Taruca y
río Aguarico, lo cual indica que no existen diferencias en la forma corporal
de estas poblaciones.
La disparidad entre los individuos de H. malabaricus analizados en este
estudio se debe principalmente a diferencias de forma relacionadas con
la altura del cuerpo y en la región caudal.

35
RECOMENDACIONES
Se recomienda realizar análisis moleculares de los especímenes
colectados, con el objetivo de esclarecer si las diferencias observadas en
los diferentes grupos se deben a un caso de plasticidad fenotípica o de
evolución contemporánea.
Se podría cuantificar la variación de la forma de H. malabaricus con otros
métodos y compararlos con los resultados obtenidos en este estudio con
el fin de elucidar si efectivamente la cuantificación morfológica fue
cubierta en su totalidad. En este contexto el estudio de contornos por
medio de análisis elípticos de Fourier es una técnica de morfometría
geométrica que podría ser considerada.
Es importante comparar la variación morfológica con otros parámetros no
incluidos en este análisis, por ejemplo el sexo, diferentes tallas y
diferentes gradientes altitudinales.

36
REFERENCIAS BIBLIOGRÁFICAS
Abrahão V, Pupo F. 2014. Técnica de dissecção do neurocrânio de Siluriformes
para estudo do encéfalo. Boletim Sociedade Brasileira de Ictiología 112 21-26.
Aguirre W, Shervette V, Calle S, Navarrete R. 2013. "Morphological and Genetic
Divergence of Hoplias microlepis (Characiformes, Erythrinidae) in Rivers and
Artificial Impoundments of Western Ecuador.
Albuja L, Merizalde C. 2012. Zoologia: Vertebrados, Instituto de Ciencias
Biologicas, Escuela Politecnica Nacional, Quito, Ecuador, 8, 60-61.
Alvarez S, Barraza D, Mendivil M. 2013. Optimización de la deshidratación
osmótica de filete de tilapia roja (oreochromis ssp) para el mejoramiento de su
vida útil (Doctoral dissertation, Universidad de Cartagena).
Audesirk T, Audesirk G, Byers B. 2004. Biología: La vida en la Tierra. Sexta
edición. Prentice Hall – Pearson Educación Latinoamericana, México. 980 págs.
Badillo, J. 2012. Implementación de Buenas Prácticas de Turismo Sostenible en
la comunidad Shayari, parroquia El Dorado de Cáscales, cantón Cáscales de la
provincia de Sucumbíos (Programa Rainforest Alliance) (Bachelor's thesis,
Escuela Superior Politécnica de Chimborazo).
Barriga, R. 2012. Lista de peces de agua dulce e intermareales del Ecuador.
Politécnica; 30 (3): 83-86.
Bechara W, Liria J. 2012. Morfometría geométrica en cinco especies de Buthidae
y Scorpionidae de Venezuela. Revista mexicana de biodiversidad, 83(2), 421-
431.
Benítez H, Püschel T. 2014. Modelando la varianza de la forma: morfometría
geométrica aplicaciones en biología evolutiva. International Journal of
Morphology, 32(3), 998-1008.

37
Bhagat Y, Fox M, Ferreira, M. 2006. Diferenciación morfológica en Lepomis
gibbosus de semilla de calabaza introducida que ocupa diferentes zonas de
hábitat en reservorios portugueses. Journal of Fish Biology, 69 (sc), 79-94.
Bissaro F, Gómez J, Madeira A. 2013. "Morphometric variation in the shape of
cephalothorax of shrimp Xyphopeneaus kroyeri on the east coast of Brazil.,"
Journal of the Marine Biological Association of the United Kingdom., vol. 93, pp.
683-691.
Bohorquez J. 2010. Ecomorfología alimentaria de algunas especies de peces
asociadas a fondos blandos (Tesis doctoral, Instituto Politécnico Nacional,
Bolivia).
Bonilla Rivero A. 2016. Morfometría geométrica comparativa entre las especies
del género Gephyrocharax de Venezuela y Colombia, vol. 2, pág. 15 -23.
Bookstein F. 1991. Morphometric tools for landmark data: Geometry and Biology.
Advances in morphometrics. Pp. 101-120. Plenum Press, New York. Cambridge
University Press, Cambridge.
Bookstein F. 1996. Combining the tools of geometric morphometrics. En Marcus,
L.F., Corti, M., Loy, A., Naylor, G. J. P. & D. Slice (eds.): Advances in
morphometrics. Pp. 131-151. Plenum Press, New York. Cambridge University
Press, Cambridge.
Calle D, Quiñones M, Erazo H, Jaramillo N. 2008. Discriminación por
morfometría geométrica de once especies de Anopheles (Nyssorhynchus)
presentes en Colombia. Biomédica 28(3).
Cameron R, Dillon J. 1984. Habitat stability, population histories and patterns of
variation in Cepaea. Malacologia 25(2):271-290.

38
Canadian Council on Animal Care (CCAC). 2005. Guidelines on: the care and
use of fish in research, teaching and testing. Canadian Council on Animal Care,
Ottawa, 85p.
Cardoso D. 2015. Alimentação e ecomorfologia de peixes predadores em lagos
de várzea amazônica. Instituto de Biologia Programa de Pós-Graduação em
Ecologia e Conservação de Recursos Naturais, pág. 45-89.
Carrillo M. 2016. Preferencia alimentaria de tres especies de peces marinos
comerciales según su estructura dental en la provincia de esmeraldas,
ecuador (Bachelor's thesis, QUITO/UIDE/2016).
Charlin J, Llosas M. 2016. Morfometría geométrica y representaciones rupestres:
explorando las aplicaciones de los métodos basados en landmarks. Arqueología
22(1) 103-125.
Chávez L. 2016. El manejo de las cuencas hidrográficas compartidas. Revista
Verbum. Vol 11(11).
Corrêa L, Souza G, Takemoto R, Ceccarelli P, Adriano E. 2014. Behavioral
changes caused by Austrodiplostomum spp. in Hoplias malabaricus from the São
Francisco River, Brazil. Parasitology research 113(2), 499-503.
Cuadras C. 2007. Nuevos métodos de análisis multivariante. CMC Editions, pág
10-17.
Dujardin J, Costa J, Bustamante D, Jaramillo N, Catalá S. 2009. Deciphering
morphology in Triatominae: The evolutionary signals. Acta Tropica. 110:101-111.
Escobar D. 2013. Diversidad Críptica y Relaciones Filogenéticas de la Familia
Characidae en el Yasuní, Ecuador (Bachelor's thesis, QUITO/PUCE/2013).

39
Fernández E, Ferriz R, Bentos C, López G. 2012. Dieta y ecomorfología de la
ictiofauna del arroyo Manantiales, provincia de Buenos Aires, Argentina. Revista
del Museo Argentino de Ciencias Naturales 14(1) 1-13.
Flores A., Brown A. 2010. Peces nativos de agua dulce de América del Sur de
interés para la acuicultura: Una síntesis del estado de desarrollo tecnológico de
su cultivo. FAO.
Foster K, Bower L, Piller K. 2015. Getting in shape: habitat-based morp-hological
divergence for two sympatric fishes. Biological Journal of the Linnean Society
114 152–162.
Froese R, Pauly D. Editors. 2017. FishBase. World Wide Web electronic
publication. www.fishbase.org, versión (02/2017).
Galvis G, Mojica J, Duque S, Castellanos C, Sánchez-Duarte P, Arce M,
Arbeláez F. 2006. Peces del Medio Amazonas: Región de Leticia. Conservatión
Internacional.
García V, Zaragoza C, Ramirez J, Guerrero A, Ruiz C. 2015. Caracterización
rápida de la biodiversidad usando morfometría geométrica: Caso de estudio con
abejas sin aguijón (Apidae) del sur de Ecuador. ACI Avances en Ciencias e
Ingenierías, 7(1).
Gómez E, Obando M, Tovar M, Caldas M, Hurtado H. 2011. Estudio histológico
del tracto digestivo del Neón Cardenal Paracheirodon axelrodi (Characidae).
International Journal of Morphology 29 (3):782-786.
Gómez S. 2005. Representación y economía ecológica de la cacería de fauna
mayor y la pesca en la Reserva Biológica de Limoncocha (Tesis de grado previa
a la obtención del título de Ingeniería Ambiental). Universidad Internacional SEK,
Quito, Ecuador.

40
Gómez S. 2016. Comunidad de peces de la cuenca del Rio Salado del Sur y
notas para su conservación (Argentina, Sud América). Bioikos 29(1) 21-28.
González Salazar R. 2017. Modelado dinámico y control de un sistema de
locomoción biomimético. Tesis maestría en Ciencias en Ingeniería Mecánica,
Instituto Politécnico Nacional, pág. 146.
González C, De las Nieves M. 2012. ¿Podemos establecer un conjunto mínimo
de landmarks que defina el sexo de un coxal?: morfometría geométrica aplicada
al conocimiento de las poblaciones del pasado. Bachelor's thesis, Universidad
Autonoma de Madrid.
Granada M, Valencia A. 2015. Contenido estomacal de Hoplias malabaricus en
un ecosistema léntico del piedemonte Amazónico. Momentos de Ciencia, 3(2).
Granitto M, Rosso J, Boveri M, Rennella, A. 2016. Impacto del uso del suelo
sobre la condición de ribera en arroyos pampeanos y su relación con la
estructura de la comunidad de peces. Biología Acuática.
Grassi D, Swarça A, Dergam J, Pastori M, Fenocchio A. 2017. Cytogenetic
characterization of Hoplias malabaricus (Bloch 1794) from the Ctalamochita
River (Córdoba, Argentina): first evidence for southernmost populations of this
species complex and comments on its biogeography. Comparative Cytogenetics
11 (1): 15–28.
Haas T, Blum M, Heins D. 2010. Morphological responses of a stream fish to
water impoundment. Proceedings of the Royal Society Biological Letters 6: 803–
806.
Hammer Ø, Harper D, Ryan P. 2001. PAST: Paleontological Statistics Software
Package for Education and Data Analysis. Paleontología Electrónica 4: 9.

41
Hankison S, Childress M, Schmitter J, Ptacek M. 2006. Morphological divergence
within and between the Mexican sailfin mollies, Poecilia velifera and Poecilia
petenensis. Journal of Fish Biology 68: 1610–1630.
Ibáñez M. 2014. Diferencias de forma entre Microlophus quadrivittatus y
Microlophus atacamensis (Tropiduridae) Una aproximación desde la
morfometría geométrica (Doctoral dissertation, Universidad de Concepción.
Facultad de Ciencias Naturales y Oceanográficas. Departamento de Zoología).
José C, Riofrío Q. 2013. Aspectos biométricos y reproductivos de Boquichico
Prochilodus nigricans Agassiz 1829 (Prochilodontidae) en Ucayali, Perú.
Klingenberg C. 2010. Evolution and development of shape: integrating
quantitative approaches. Nature Reviews. Vol. 11. 623-635.
Klingenberg C. 2011. MorphoJ: an integrated software package for geometric
morphometrics. Molecular Ecology Resources 11:353-357.
Krabbenhoft T, Collyer M, Quattro J. 2009. Differing evolutionary patterns
underlie convergence on elongate morphology in endemic fishes of Lake
Waccamaw, North Carolina. Biological Journal of the Linnean Society 98: 636–
645.
Krabbenhoft T, Collyer M, Quattro J. 2009. Patrones evolutivos diferentes
subyacen a la convergencia en la morfología alargada en peces endémicos del
lago Waccamaw, Carolina del Norte. Revista Biológica de la Sociedad Linneana
98: 636-645.
Langerhans R, Reznick D. 2010. Ecology and evolution of swimming
performance in fishes: predicting evolution with biomechanics. Fish locomotion:
an eco-ethological perspective 200-248.

42
Larraín M, Díaz N, Lamas C, Vargas C, Araneda C. 2012. Genetic composition
of Mytilus species in mussel populations from southern Chile. Latin American
Journal of Aquatic Research., 40 1077-1084.
Londoño C, Hurtado Giraldo H. 2010. Preliminary morphologic and morphometric
study of the encephalon of the tiburoncito Ariopsis seemanni
(Ariidae). Universitas Scientiarum 15(2) 101-109.
Lopez A. 2015. Morfometría geométrica: el estudio de la forma y su aplicación
en biología. Temas de Ciencia y Tecnología. Vol. (19), 53-59.
López H, Ponte Gómez J. 2015. Serie Arte y Sociedad, Comentarios y
digresiones sobre las viejas de agua. ProBiota: Serie Arte y Sociedad en la
Ictiología Nacional.
Malato G, Shervette V, Amaya R, Rivera J, Salazar F, Delgado P, Aguirre W.
2017. Parallel body shape divergence in the Neotropical fish genus Rhoadsia
(Characidae) along elevational gradients of the western slopes of the Ecuadorian
Andes. PLoS One 12(6), e0179432.
Martínez C, Osorio A. 2015. Ética en investigación con animales: una actitud
responsable y respetuosa del investigador con rigor y calidad científica. Revista
Latinoamericana de Bioética, 8(15), 46-71.
Martins J. 2009. Biología de Hoplias malabaricus (Bloch 1794) (Erythrinidae) na
Represa de Capim Branco I, Rio Araguari, MG.
Menin E, Mimura O. 2015. Anatomía funcional da cavidad e bucofaringeana de
Hoplias malabaricus(Bloch,1794)(Erythrinidae). Ceres, 38(217).
Miralles H. 2015. Estudio de la variabilidad del patron cromático plastral en
Testudo hermanni hermanni Gleim 1789 mediante elípticas de
Fourier,Bachelor's thesis, Universistat de Lleida.

43
Mitteroecker P, Gunz P, Windhager S, Schaefer K. 2013. A brief review of shape,
form, and allometry in geometric morphometrics, with applications to human
facial morphology. Hystrix, the Italian Journal of Mammalogy.
Mojica J, Castellanos C, Sánchez-Duarte P, Díaz C. 2006. Peces de la cuenca
del río Ranchería, La Guajira, Colombia. Biota Colombiana, 7(1) 129-142.
Mojica J, Ortega H, Hidalgo M, Alonso J. 2017. Listado de los peces de la cuenca
del río Putumayo en su sector colombo-peruano, 3(2), 9-10.
Moncayo C. 2011. Investigación de las actividades turísticas sostenibles
manejadas por comunidades indígenas en tres Provincias Amazónicas del
Ecuador: Orellana, Sucumbíos y Zamora Chinchipe (Bachelor's thesis, Quito:
USFQ 2011).
Montero C, Granda J. 2015. Aplicación de morfometría geométrica para la
comparación de distintas poblaciones de guanchiche (hoplias spp) en
ecosistemas lenticos y lóticos del ecuador (Bachelor's thesis).
Morales Flores G. 2017. Variabilidad morfólogica de poecilia sphenops y poecilia
nelsoni (Poeciliidae) en aguas dulces y salobres en el sur de México (Doctoral
dissertation, Instituto en Ciencias Biológicas-Licenciatura en Biología-
UNICACH).
Muñoz F, Ventura J, López M. 2011. Estudio de la variación morfológica en una
zona de polimorfismo Robertsoniano de ratón doméstico, Mus musculus
domesticus (Schwarz y Schwarz 1943). Universitat Autònoma de Barcelona.
Navarro B, Pisco G. 2015. Identificación de recursos ecoturísticos de una
comunidad ashéninka de la amazonía peruana. Investigación Universitaria.
Noboa M. 2015. Caracterización morfométrica de la biodiversidad del barrenador
del fruto Neoleucinodes elegantalis (Gueneé) en el cultivo de naranjilla Solanum
quitoense en diez localidades (Bachelor's thesis, Quito: UCE).

44
Obando V. 2011. Variabilidad morfometrica de Neoleusinodes elegantalis
(Guanee): perforador de frutos de solanácidas de importancia económicas.
Medellín, Colombia: Universidad Nacional de Colombia.
Perin D, Palhano A, Bianchin S, Pietricoski L. 2016. An á lise cromossômica em
hoplias malabaricus (erythrinidae) no rio sarandi, em realeza paraná. anais da
jic-jornada de iniciação científica e tecnológica 1(6).
Piraquive E, Duque S, González A. 2015. Estructura trófica del ensamblaje íctico
de los lagos de Yahuarcaca, Amazonas, Colombia. Mundo Amazónico, 6(2).
Reis D. 2015. Hematological And Histopathological Changes in Hoplias
Malabaricus from the Sao Francisco River, Brazil Caused by Larvae of
Contracaecum Sp (nematoda, Anisakidae). Helminthologia.
Revelo W. 2010. "Aspectos biológicos y pesqueros de las principales peces del
sistema hídrico de la provincia de Los Ríos.," Instituto Nacional de Pesca., Los
Ríos., Boletin científico y técnico, 4-5.
Rios L, Rodríguez F, Velázquez A, Hernández A. 2013. Morfometría geométrica
del corazón de Hyla plicata a través de un gradiente altitudinal en el eje
neovolcánico mexicano. International Journal of Morphology, 31(3), 905-910.
Rivera J. 2014. Ampliación de la distribución de Brycon dentex (Characidae) y
Hamuelopsis elongatus (Heamulidae) en Ecuador. ACI Avances en Ciencias e
Ingenierías, 6(2).
Rodrigues L, Santos A, Ferreira E, Teófilo T, Pereira D, Costa F. 2017.
Parasitologic aspects of traíra (Hoplias malabaricus) from the São Bento
city, Arquivo Brasileiro de Medicina Veterinária e Zootecnia, 69(1) 264-268.
Rodríguez, A. 2015. Comparación de la ubicación interna y externa de puntos
morfológicos clave para describir la variación morfológica en conchas de Cerion
(Cerionidae)/Comparison of internal and external location of landmarks to

45
describe the morphological... Revista Cubana de Ciencias Biológicas, 4(1), 63-
68.
Rohlf F. 2008. tpsUtil. Version 1.41. departament of Ecology and Evolution, State
Unv. Of New York at Stony Brook, Stony Brook. Disponible en linea
(http://life.bio.sunysb.edu/morph/index.html).
Rohlf F. 2010. tpsDig, digitize landmarks and outlines. Version 2.16. Department
of Ecology and Evolution. State University of New York at Stony Brook.
Rohlf F. 2015. The tps series of software. Hystrix, the Italian Journal of
Mammalogy 26(1), 9-12.
Rohlf F, Marcus L. 1993. A revolution in morphometrics. Trends in Ecology and
Evolution, 8 129-132.
Rohlf F, Silice D. 1990. Extensions of the Procrustes method for the optimal
superimposition of Landmarks. Systematic zool.
Rojas M, Pellón M, Del Sol M. 2015. Desarrollo de la Aleta Caudal del Salmón
(Salmo salar). International Journal of Morphology, 33(2), 514-521.
Ruzzante D, Walde S, Cussac V, Macchi P. 1998. Trophic polymorphism, habitat
and diet segregation in Percichthys trucha (Pisces: Percichthyidae) in the Andes.
Biological Journal of the Linnean Society 65: 191– 214.
Saigo M. 2016. Estructura trófica del bentos de sistemas río-laguna de la llanura
aluvial del río Paraná Medio a través de análisis de dieta e isótopos estables del
carbono y nitrógeno. Tesis doctoral, Universidad Nacional del Litoral.
Salas D, Véliz D, Scott S. 2012. Diferenciación morfológica en especies del
género Cheirodon (Characidae) mediante morfometría tradicional y geométrica.
Gayana (Concepción). 76 142-152.

46
Santos J. 2011. Identificación de nemátodos parásitos en peces dulceacuícolas
colectados en los ríos: San Pablo, Caracol y Babahoyo. Guayaquil: Universidad
Estatal de Guayaquil., Tesis de pregrado.
Sasintuña G, Elizabeth J. 2015. Plan de difusión turística del cantón Cáscales
ubicado en la provincia de sucumbíos, con la finalidad de lograr un desarrollo
turístico del sector (Bachelor's thesis).
Schaefer J, Duvernell R, Kreiser S. 2011. Variabilidad de la forma en topminnows
(complejo de especies de Fundulus notatus) a lo largo del continuo del río.
Revista Biológica de la Sociedad Linneana 103: 612-621.
Sepúlveda R, Ibáñez C, Slice D. 2012. Clinal variation in the shell morphology of
intertidal snail Acanthina monodon in the Southeastern Pacific Ocean. Marine
Biology Research., 8, 363- 372.
Sheets H. 2001. Software IMP © (Integrated Morphometric Package) Department
of Physics, Canisius College, Buffalo, NY 1408, Department of Geology State
University of New York atn Buffalo, NY 14260. Disponible en:
http://www.canisius.edu/~sheets/ morphsoft.html.
Sheets H. 2011b. Software IMP © (Integrated Morphometric Package) PCAGen.
Department of Physics, Canisius College, Buffalo, NY 1408, Department of
Geology State University of New York atn Buffalo, NY 14260. Disponible en:
http://www.canisius.edu/~sheets/ morphsoft.html.
Sheets H. 2011c. Software IMP © (Integrated Morphometric Package) CVAGen.
Department of Physics, Canisius College, Buffalo, NY 1408, Department of
Geology State University of New York atn Buffalo, NY 14260. Disponible en:
http://www.canisius.edu/~sheets/ morphsoft.html.
Soria M, Rodiles R, González-Díaz A. 2011. Morfometría de las especies de
Vieja (Cichlidae) en ríos de la cuenca del Usumacinta, Chiapas, México. Revista
mexicana de biodiversidad, 82(2), 569-579.

47
Soto A, Liria J, De Luna E. 2011. "Morfometría Geométrica y Filogenia en
Rhodniini (Hemiptera, Reduviidae) de Venezuela.," Acta zoológica Mexicana,
vol. 27, pp. 87-102.
Torcida S, Perez S. 2012. Análisis de procrustes y el estudio de la variación
morfológica. Revista argentina de antropología biológica 14(1) 131-141.
Toro M, Manriquez G, Suazo I. 2010. Morfometría geométrica y el estudio de las
formas biológicas: de la morfología descriptiva a la morfología
cuantitativa. International Journal of Morphology 28(4), 977-990.
Valdivieso J, Carrillo C, Madera R, Abarracin M. 2012. Guía de peces de
Limoncocha. Universidad Internacional SEK. Quito, Ecuador.
Vanegas J, Meza A, Azpelicueta M. 2013. Extensión de la distribución geográfica
de Chrysobrycon hesperus y C. myersi (Characidae, Stevardiinae) para varios
drenajes fluyendo hacia la cuenca del Amazonas. Revista mexicana de
biodiversidad, 84(1), 384-387.
Vegh S. 2015. Análisis de Contenidos Estomacales de las Especies de Interés
Deportivo y Comercial. Dirección de Pesca Continental, Subsecretaría de Pesca
y Acuicultura, MAGyP. Bs. As. Informe Técnico 29:1-19.
Vegh S, Lozano I, Dománico A. 2011. composición de la dieta de hoplias
malabaricus y salminus brasiliensis en la localidad de victoria (entre ríos,
argentina) durante la primavera. Comisión de Investigaciones Científicas de la
Provincia de Buenos Aires.
Villacreses X. 2012. Propuesta de un modelo de gestión institucional para el plan
de desarrollo y ordenamiento territorial del cantón Cáscales, provincia de
Sucumbíos 2011-2030.

48
Vizcaino L, Rincón L, Bulla M, Ramírez E, Giraldo H. 2013. Estudio Morfológico
y Morfométrico del Cerebro del Bagre Tigrito Pimelodus Pictus (Pimelodidae)
Steindachner (1876). Revista Facultad de Ciencias Básicas, 9(1), 94-107.
Wagner H. 2001a. Brain Areas in Abyssal Demersal Fishes. Brain, Behavior and
Evolution, 57 (6): 301-316.
Webster M, Atton N, Ciervo P, Sala A. 2011. Variación morfológica específica
del hábitat entre los espinosos triespinos (Gasterosteus aculeatus) dentro de una
cuenca de drenaje.
Zambrano B, Stalin L, Cucalón R. 2015. Filogeografìa molecular del guanchiche
hoplias spp. (ERYTHRINIDAE) de la costa ecuatoriana. Tesis de grado, Escuela
Superior Politécnica Del Litoral.
Zelditch M, Swiderski H, Sheets W. 2004. Geometric morphometric for biologists:
a primer, San Diego, CA, Elsevier Academic Press.

49

50
Tabla 5. Ficha de registro de datos en campo
Fecha de colecta: _______________ Hora: ______________ Número de campo: _____ Localidad:______________________________________________________________ Subcuenca:_____________________________________________________________Coordenadas:_________________________________Elevacion:_________________________________________________________________________________________Colector:_______________________________________________________________ Hábitat (ambiente): _____________Tipo de agua: ____________ Profundidad: ______ Ancho: ________________ Corriente: ___________ _______ Color: ________________ Vegetación:_________________________________________Sustrato:__________________________________________________________________________________ Sustrato:______________________________________________________________________________________________________________________________________Observaciones:________________________________________________________________________________________________________________________________________________________________________________________________________
N° CODIGO OBSERVACIONES
Responsable: Fecha:
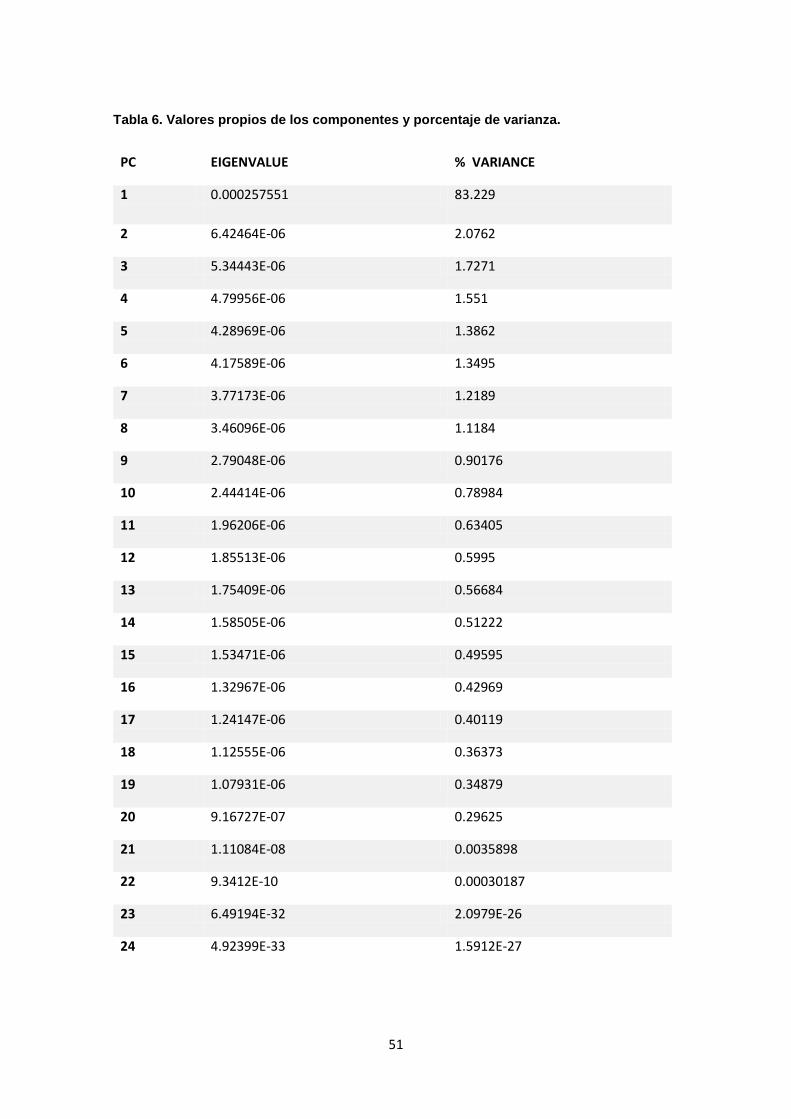
51
Tabla 6. Valores propios de los componentes y porcentaje de varianza.
PC EIGENVALUE % VARIANCE
1 0.000257551 83.229
2 6.42464E-06 2.0762
3 5.34443E-06 1.7271
4 4.79956E-06 1.551
5 4.28969E-06 1.3862
6 4.17589E-06 1.3495
7 3.77173E-06 1.2189
8 3.46096E-06 1.1184
9 2.79048E-06 0.90176
10 2.44414E-06 0.78984
11 1.96206E-06 0.63405
12 1.85513E-06 0.5995
13 1.75409E-06 0.56684
14 1.58505E-06 0.51222
15 1.53471E-06 0.49595
16 1.32967E-06 0.42969
17 1.24147E-06 0.40119
18 1.12555E-06 0.36373
19 1.07931E-06 0.34879
20 9.16727E-07 0.29625
21 1.11084E-08 0.0035898
22 9.3412E-10 0.00030187
23 6.49194E-32 2.0979E-26
24 4.92399E-33
1.5912E-27
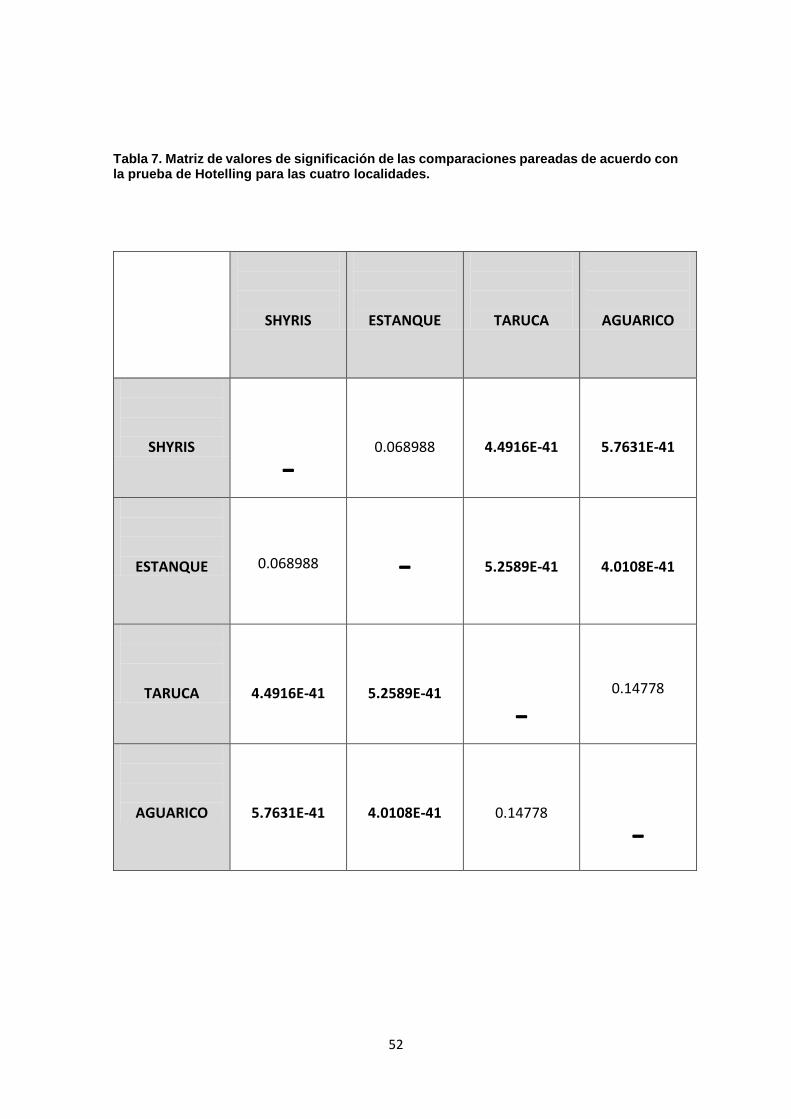
52
Tabla 7. Matriz de valores de significación de las comparaciones pareadas de acuerdo con la prueba de Hotelling para las cuatro localidades.
SHYRIS
ESTANQUE
TARUCA
AGUARICO
SHYRIS
-
0.068988
4.4916E-41
5.7631E-41
ESTANQUE
0.068988
-
5.2589E-41
4.0108E-41
TARUCA
4.4916E-41
5.2589E-41
-
0.14778
AGUARICO
5.7631E-41
4.0108E-41
0.14778
-

53
Anexo 1. Permiso de Investigación otorgado por el Ministerio del Ambiente de
Sucumbíos.

54
Anexo 2. Localidades de estudio
a) Localidad Laguna de los Shyris 0°01'41.6"N / 77°10'41.1"W
b) Localidad Estanque Artificial 0°05'00.2"N / 77°12'17.4"W
c) Localidad río Aguarico 0°03'35.8"N / 77°12'57.0"W
d) Localidad río Taruca 0°07'09.9"N / 77°11'54.4"W

55
Anexo 3. Trabajo de campo.

56

57
Anexo 4. Trabajo de laboratorio.
Top Related