






Idiomas
Páginas
Jurídico


~ ~
CICY
Centro de Investigación Científica de Yucatán, A.C.
Posgrado en Ciencias Biológicas
AGRESIVIDAD DE CEPAS DE Mycosphaerella fijiensis AISLADAS DE FINCAS BANANERAS CON
DIFERENTE MANEJO DE FUNGICIDAS
Tesis que presenta
EDUARDO ENRIQUE ZAPIEN DE LA ROSA
En opción al título de
MAESTRO EN CIENCIAS (Ciencias Biológicas: Opción Biotecnología)
Mérida, Yucatán, México
Febrero 2014


~ :S
CICY RECONOCIMIENTO
POSGRADO EN
CIENCIAS BIOLÓGICAS
/
Por medio de la presente, bago constar que el trabajo de tesis titulado
Agresividad de cepas de Mycosphaerella fijiensis aisladas de fincas
bananeras con diferente manejo de fungicidas, fue realizado en los
laboratorios de la Unidad de Biotecnología del Centro de Investigación
Científica de Yucatán, A.C. bajo la dirección de la (los) Dra. Cecilia
Mónica Rodríguez García, dentro de la Opción Maestría en
Biotecnología de Plantas, perteneciente al Programa de Posgrado en
Ciencias Biológicas de este Centro.
Atentamente,
Coordinador de Docencia
Centro de Investigación Científica de Yucatán, AC.

Mérida, Yucatán , México; a 06 de Febrero de 2014
DECLARACIÓN DE PROPIEDAD
Declaro que la información contenida en la sección de Materiales y Métodos Experimentales, los Resultados y Discusión de este documento proviene de las actividades de experimentación realizadas durante el período que se me asignó para desarrollar mi trabajo de tesis, en las Unidades y Laboratorios del Centro de Investigación Científica de Yucatán, A.C. , y que a razón de lo anterior y en contraprestación de los servicios educa.tivos o de apoyo que me fueron brindados, dicha información, en términos de la Ley Federal del Derecho de Autor y la Ley de la Propiedad Industrial , le pertenece patrimonialmente a dicho Centro de Investigación. Por otra parte, en virtud de lo ya manifestado, reconozco que de igual manera los productos intelectuales o desarrollos tecnológicos que deriven o pudieran derivar de lo correspondiente a dicha información, le pertenecen patrimonialmente al Centro de Investigación Científica de Yucatán, A.C., y en el mismo tenor, reconozco que si derivaren de este trabajo productos intelectuales o desarrollos tecnológicos, en lo especial , estos se regirán en todo caso por lo dispuesto por la Ley Federal del Derecho de Autor y la Ley de la Propiedad Industrial, en el tenor de lo expuesto en la presente Declaración.
N ornbre: Eduardo Enrique Zapien De la Rosa

AGRADECIMIENTOS
Al Consejo Nacional de Ciencia y Teología (CONACYT) por la beca otorgada número 254671 , la cual cubrió los gastos de manutención y servicio médico, indispensables en la realización de este trabajo.
Al proyecto número 116886 del Fondo Institucional de Fomento Regional para el Desarrollo Científico, Tecnológico y de Innovación (FORDECYT), otorgado por CONACYT, mediante el cual se adquirió el equipo, reactivos y material necesarios para realizar el presente trabajo de investigación.
Al Centro de Investigación Científica de Yucatán (CICY), A.C., por abrirme sus puertas y formar parte de sus alumnos de posgrado, proporcionándome las instalaciones, reactivos y equipo requeridos para realizar este trabajo de investigación.
A mi directora de tesis la Doctora Cecilia Mónica Rodríguez García, por sus enseñanzas, consejos y recomendaciones en el ámbito académico y personal, por ser un ejemplo de ética profesional y organización, así como por su amistad.
Al personal técnico que fue de gran apoyo para llevar a cabo las actividades experimentales en el laboratorio, así también por su amistad y compañerismo. En concreto:
• M. C. Leticia Peraza Echeverría, tu tutelaje fue vital para hacer posible este trabajo
• M.C. Miguel Alonso Tzéc Simá • M.C. Bartolomé Chi Manzanero • Q. B. B. Rosa Grijalva Arango • M.C. Eduardo Castillo Castro • I.Q.I. Silvia B. Andrade Canto • M.C. Andrés Quijano Ramayo
Al personal administrativo de posgrado:
• Lic. Landy Rodríguez Salís • Lic. Nery Alejandra Arcea García
• Lic. Gilma Michell
Al personal de biblioteca, por su ayuda para proporcionar la literatura científica requerida para respaldar los resultados de este trabajo:
• Lic. Sergio de Jesús Pérez • Lic. Miriam Juan Qui Valencia • Lic. Narcedalia Gamboa Angula

AGRADECIMIENTOS
A Dios (El Gran Arquitecto del Universo) , por concederme la vida y permitirme ver realizados mis esfuerzos.
A mis padres Eduardo y Fanny, por apoyarme en todo momento y ser un ejemplo a seguir. Con todo mi cariño y mi amor para ustedes que hicieron todo en la vida para que yo pudiera lograr mis sueños, por motivarme y darme la mano cuando sentra que el camino se terminaba, a ustedes por siempre mi corazón y mi agradecimiento.
A mi hermano Carlos, por ser parte de mi vida y representar la unidad familiar, por brindarme apoyo y ánimo cuando más los necesité. A mi tía Mago, gracias por todo tu apoyo, porque nunca te olvidaste de mí y por ser fuente de consejos cuando más los necesité.
A mis maestros, quienes influyeron con sus lecciones y experiencias en formarme como un profesionista de bien y preparado para los retos que impone la vida, especialmente los Ores. Blondy Canto y Andrew James, por orientarme durante la experimentación y discusión de los resultados, así también por proporcionarme literatura científica de apoyo.
Gracias a todos mis amigos y compañeros que siempre estuvieron listos para brindarme toda su ayuda, por todo el apoyo que recibí de ustedes en el laboratorio, en campo y en el salón de clases:
Abril Canché, José Cruz, Miguel Canseco, Muhilan Mahendhiran, Frank Bailete, Erika Chan , Yamili Burgos, Blanca Lara, Xenia Mena, Gabriela Medina, Efrén Pech , Yoreni Hernández, Inés Arana, Nuvia Kantún , Roberto Vásquez, Yeni Couoh, Mayra Díaz, Yasmín Sánchez y Carmela Alcacer .. .
Y a quienes involuntariamente omití, mi más sincero agradecimiento.

LISTADO DE ABREVIATURAS
ÍNDICE DE FIGURAS
ÍNDICE DE CUADROS
RESUMEN
ABSTRACT
CAPITULO l. INTRODUCCIÓN
Introducción
Antecedentes
Aspectos generales del cultivo de banano
Enfermedades que afectan al banano
Distribución geográfica de Mycosphaerella fijiensis
Biología y ciclo de vida de Mycosphaerella fijiensis
Etapas de la enfermedad
Interacción M. fijiensis-Musa spp.
Interacción compatible: Cultivar susceptible
Interacción incompatible: Cultivar resistente
Diversidad genética de M. fijiensis
Control químico de la Sigatoka negra
El cultivo de banano en México y la Sigatoka negra
Resistencia y agresividad de M. fijiensis
Cuantificación de la severidad de la enfermedad
Justificación
Hipótesis
fNDICE
Número de página
V
VIl
XII
1
3
5
5
7
7
8
8
8
11
12
12
12
13
14
15
15
17
18
21

fNDICE
Objetivo general 21
Objetivos específicos 21
Estrategia experimental 21
Bibliografía 22
CAPITULO 11. INFECCIÓN in vitro DE FRAGMENTOS FOLIARES DE Musa spp. CON
CEPAS DE Mycosphaerel/a fijiensis PROVENIENTES DE FINCAS CON DIFERENTE
MANEJO CON FUNGICIDAS 33
Introducción 33
Fragmentos foliares en la interacción Musa spp.-M. fijiensis 35
Materiales y métodos 36
Material fúngico 36
Etapa 1 37
Crecimiento de las cepas en medio V8C 37
Producción de conidios en medio V8E 37
Cosecha de esporas 38
Conteo y ajuste de la suspensión de esporas utilizando la cámara
Sedgwick-Rafter 39
Etapa 11 39
Preparación de medio AB (Agua-Bencimidazol) 39
Preparación de fragmentos foliares 40
Inoculación y muestro de fragmentos foliares 40
11

[N DICE
Resultados 42
Reactivación de las cepas fúngicas 46
Recuperación de cepas de M. fijiensis contaminadas 47
Experimentos utilizando las cepas reactivadas 49
Experimento Gran enano-C1233 50
Experimentos Gran enano-CR4b 51
Discusión 54
Bibliografía 56
CAPITULO 111. CUANTIFICACIÓN DEL DAÑO INDUCIDO POR CEPAS DE M. fijiensis
AISLADAS DE FINCAS CON DIFERENTE MANEJO DE FUNGICIDAS EN
FRAGMENTOS FOLIARES DE Musa spp 61
Introducción 61
Características del programa Assess 2.0 63
Características del programa lmageJ 1.46r 64
Materiales y métodos 68
Material vegetal 69
Captura y procesamiento de imágenes digitales 69
Formación de imágenes binarias 70
Cuantificación de la severidad de la enfermedad 70
Análisis estadístico 71
Resultados 72
Cuantificación esporas producidas por aislados de M. fijiensis 72
Cuantificación del área dañada y número de lesiones en fragmentos foliares de
Musa spp. 74
111

fNDICE
Daño inducido por la cepa C1233 74
Daño inducido por la cepa CR4b 76
Daño inducido por la cepa Oz1 b 79
Daño inducido por la cepa Jaguar C3 82
Severidad foliar inducida por cepas de M. fijiensis en el cultivar Gran enano
84
Severidad foliar en Calcuta IV 86
Severidad foliar en Fougamou 88
Severidad foliar inducida a los 30 ddi en Gran enano, Fougamou y Calcuta IV 90
Discusión 93
Bibliografía 99
CAPITULO IV. DISCUSIÓN GENERAL 105
Cuantificación de la enfermedad con los programas Assess 2.0 e lmage J 1.46r.
11 o
Conclusiones 112
Perspectivas 112
Bibliografía 113
IV

Abreviatura
ddi
M. fijiensis
P. fijiensis
M. musicola
M. eumusae
BBTV
BSV
CMV
R. so/anacearum
mm
F
IJL
GIRAD
mL
L
g
pH
AB
%
ppm
cv.
LISTADO DE ABREVIATURAS
Significado
días después de inoculación
Mycosphaerella fijiensis
Pseudocercospora fijiensis
Mycosphaerel/a musicola
Mycosphaerella eumusae
Banana Bunchy Top Virus
Banana Streak Virus
Cucumber Mosaic Virus
Ralstonia solanacearum
Milímetros cúbicos
Milímetros cuadrados
Milímetros
Número de campos contados
Micro litro
Centré de Coopération lnternationalé en Recherche Agronomic pour le Développement
Mililitro
Litro
Gramo
Potencial de Hidrógeno
Agar Bencimidazol
Tanto por ciento
Grados centígrados
Partes por millón
Cultivar
V

rpm
dpi
SIAP
LISTADO DE ABREVIATURAS
Revoluciones por minuto
Puntos por pulgada (del inglés dots per
inch)
Servicio de Información
Agroalimentaria y Pesquera
VI

rNDICE DE FIGURAS
Número de página
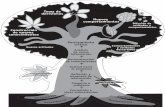
Figura 1. Ciclo de vida de Mycosphaerella fijiensis y su interacción con Musa spp. 11
Figura 2. Estrategia experimental para cuantificar la agresividad de aislados de M. fijiensis 21
Figura 3. Fotografías y micrografías de fragmentos foliares del cultivar Gran enano a O ddi con las
cepas C1233 (A y B) , CR4b (C y D) y Oz1 b (E y F). 42
Figura 4. Fotografías y micrografías de fragmentos foliares del cultivar Gran enano a 5 ddi con las
cepas C1233 (A y B), CR4b (C y D) y Oz1b (E y F) . 43
Figura 5. Fotografías y micrografías de fragmentos foliares del cultivar Gran enano a 15 ddi con
las cepas C1233 (A y B), CR4b (C y D) y Oz1b (E y F) . 44
Figura 6. Fotografías y micrografías de fragmentos foliares del cultivar Gran enano a 30 ddi con
las cepas C 1233 (A y B), CR4b (C y D) y Oz1 b (E y F). 45
Figura 7. Fragmentos foliares inoculados con M. fijiensis mostrando contaminación en medio V8C.
A) Fragmentos inoculados con C 1233, B) Fragmentos inoculados con Oz1 b, C) Fragmentos
inoculados con CR4b. 47
Figura 8. Cepa Oz1 b contaminada. A) vista macroscópica de la contaminación creciendo en medio
V8E. B) vista microscópica de la contaminación observada en el medio V8E. 48
Figura 9. M. fijiensis en medio V8C después de 48 horas de agitación continua en medio PDB
adicionado con A) Ampicilina+Kanamicina+Cefotaxima, B) Kanamicina+Cefotaxima, C)
Kanamicina; no se observa presencia de contaminación sobre el medio de cultivo. 49
VIl

rNDICE DE FIGURAS
Figura 1 O. Lesiones en fragmentos foliares del cultivar Gran enano causadas por la cepa e 1233 a
30 ddi. A) Cepa sin reactivar, 8) Cepa reactivada. 51
Figura 11. Micrografía de fragmentos foliares de cultivar Gran enano a 30 ddi. A) Fragmento
control. 8) Fragmento inocu lado con C1233 reactivada. 51
Figura 12. Lesiones en fragmentos foliares del cultivar Gran enano causadas por la cepa CR4b a
30 ddi. A) Cepa sin reactivar, 8) Cepa reactivada. 52
Figura 13. Micrografía de fragmentos foliares del cultivar Gran enano a 30 ddi. A) Fragmento
control. 8) Fragmento inoculado con CR4b reactivada. P: penetración . 52
Figura 14. Presencia de contaminación sobre la superficie del fragmento foliar de Gran enano A)
fragmento control y 8) Fragmento inoculado con CR4b . 53
Figura 15. Diagrama general de flujo de la cuantificación de la enfermedad empleando los programas Assess 2.0 e lmageJ 1.46r. 66
Figura 16. Fotografía digital de un fragmento foliar del cultivar Gran enano a 30 dlas después de ser inoculado con la cepa CR4b. 69
Figura 17. Procedimientos de ajuste de las escalas de medida y espacios de color en los programas Assess 2.0 e lmage J 1.46r. 71
Figura 18. Sobre estimación del área foliar dañada a 30 ddi por M. fijiensis empleando el programa Assess 2.0. 73
Figura 19. Imagen digital binaria y cuantificación del daño en un fragmento foliar a 30 ddi inducido por M. fijiensis. 73
Figura 20. Área foliar afectada por C1233 (1) en Gran enano (G), Calcuta IV (C) y Fougamou (F) a 5, 15 y 30 ddi , cuantificado con Assess 2.0 (A) e lmage J 1.46r (8) . 74
VIII

ÍNDICE DE FIGURAS
Figura 21 . Número de lesiones inducidas por la cepa C1233 (1) en los cultivares Gran enano (G) , Calcuta IV (C) y Fougamou (F) a 5, 15 y 30 días después de inoculación, cuantificados con el programa Assess 2.0 (A) e lmage J 1.46r (8) . 75
Figura 22. Daño inducido por la cepa C1233 en fragmentos foliares de los cultivares A) Gran enano, 8) Fougamou y C) Calcuta IV a 30 ddi. 76
Figura 23. Área foliar dañada por la cepa CR4b (2) en Gran enano (G) y Calcuta IV (C) a 5, 15 y 30 ddi , cuantificado con Assess 2.0 (A) e lmage J 1.46r (8) . 77
Figura 24. Número de lesiones inducidas por la cepa CR4b (2) en los cultivares Gran enano (G) y Calcuta IV (C) a 5, 15 y 30 días después de inoculación, cuantificados con los programas Assess 2.0 (A) e lmage J 1.46r (8). 78
Figura 25. Daño inducido por la cepa CR4b en fragmentos foliares de los cultivares A) Gran enano y 8) Calcuta IV a 30 ddi. 79
Figura 26. Área foliar dañada por Oz1 b (3) en Gran enano (G), Calcuta IV (C) y Fougamou (F) a 5, 15 y 30 ddi, cuantificado con Assess 2.0 (A) e lmageJ 1.46r (8) . 80
Figura 27. Número de lesiones inducidas por la cepa Oz1 b (3) en los cultivares Gran enano (G) , Calcuta IV (C) y Fougamou (F) a 5, 15 y 30 días después de inoculación, cuantificados con los programas Assess 2.0 (A) e lmage J 1.46r (8) . 81
Figura 28 . Daño inducido por la cepa Oz1 b en fragmentos foliares de los cultivares A) Gran enano, 8) Fougamou y C) Calcuta IV a 30 ddi. 81
Figura 29. Área foliar dañada por Jaguar C3(4) en Gran enano (G) , Calcuta IV (C) y Fougamou (F) a 5, 15 y 30 ddi, cuantificado con Assess 2.0 (A) e lmage J 1.46r (8). 82
Figura 30. Número de lesiones inducidas por la cepa Jaguar C3 (4) en los cultivares Gran enano (G), Calcuta IV (C) y Fougamou (F) a 5, 15 y 30 dlas después de inoculación, cuantificados con los programas Assess 2.0 (A) e lmage J 1.46r (8) . 84
IX

rNDICE DE FIGURAS
Figura 31 . Daño inducido por la cepa Jaguar C3 en fragmentos foliares de los cultivares A) Gran enano, 8) Fougamou y C) Calcuta IV a 30 ddi. 84
Figura 32. Área foliar afectada por las cepas C1233 (1), CR4b (2), Oz1b (3) y Jaguar (4) a 5, 15 y 30 ddi en fragmentos foliares del cultivar Gran enano, cuantificada con los programas Assess 2.0 (A) e lmage J 1.46r (8). 85
Figura 33. Número de lesiones inducidas por C1233 (1), CR4b (2) , Oz1b (3) y Jaguar (4) en el cultivar Gran enano (G) a 5, 15 y 30 días después de inoculación, cuantificadas con los programas Assess 2.0 (A) e lmage J 1.46r (8) . 86
Figura 34. Área foliar afectada por las cepas C 1233 ( 1 ), CR4b (2), Oz1 b (3) y Jaguar (4) a 5, 15 y 30 ddi en fragmentos foliares del cultivar Calcuta IV, cuantificada con los programas Assess 2.0 (A) e lmage J 1.46r (8) . 87
Figura 35. Comparación entre lesiones en fragmentos foliares de Calcuta IV en A) Fragmento control y 8) fragmento inoculado con C1233 a 30 ddi. 87
Figura 36 . Número de lesiones inducidas por C1233 (1) , CR4b (2) , Oz1 b (3) y Jaguar (4) en el cultivar Calcuta IV (C) a 5, 15 y 30 dias después de inoculación, cuantificadas con los programas Assess 2.0 (A) e lmage J 1.46r (8). 88
Figura 37. Área foliar dañada por las cepas C1233 (1 ), Oz1 b (3) y Jaguar (4) a 5, 15 y 30 ddi, en fragmentos foliares del cultivar Fougamou, cuantificada con los programas Assess 2.0 (A) e lmage J 1.46r (8). 89
Figura 38. Número de lesiones inducidas por las cepas C1233 (1) , Oz1b (3) y Jaguar (4) en el cultivar Fougamou (F) a 5, 15 y 30 dias después de inoculación , cuantificadas con los programas Assess 2.0 (A) e lmage J 1.46r (8). 90
Figura 39. Área foliar afectada por C1233 (1) , CR4b (2), Oz1b (3) y Jaguar (4) a 30 ddi, en fragmentos foliares de los cultivares Gran enano (G) , Calcuta IV (C) y Fougamou (F) cuantificada con los programas Assess 2.0 (A) e lmage J 1.46r (8). 91
X

ÍNDICE DE FIGURAS
Figura 40 . Número de lesiones inducidas por C1233 (1), CR4b (2), Oz1b (3) y Jaguar (4) en los cultivares Gran enano (G), Calcuta IV (C) y Fougamou (F) a 30 días después de inoculación, cuantificadas con los programas Assess 2.0 (A) e lmage J 1.46r. 92
XI

rNDICE DE CUADROS
Número de página
Cuadro 1. Aislados de M. fijiensis provenientes de fincas con diferente manejo de fungicidas
36
Cuadro 2. Composición del medio de crecimiento va. 37
Cuadro 3. Composición del medio de esporulación Va. 37
Cuadro 4. Arreglo factorial 23 empleando tres factores (Ampicilina, Kanamicina y Cefotaxima)
a dos niveles cada uno (presencia o ausencia del antibiótico en el medio). Presencia (+) o
ausencia (-) de contaminación en medio PDB; crecimiento (+) o inhibición (-) de P. fijiensis.
49
Cuadro 5. Experimentos empleando cepas reactivadas de M. fijiensis . 50
Cuadro 6. Nomenclatura de los experimentos realizados por la M.C. Leticia Peraza Echeverria.
6a
Cuadro 7. Recuento de esporas producidas por cepa de M. fijiensis 72
XII


RESUMEN
El banano (Musa spp.) es el cuarto cultivo de importancia mundial después del
arroz, el trigo y el maíz, su comercialización genera ganancias anuales multibillonarias.
Su cultivo tiene factores limitantes para su producción, entre estos está la Sigatoka
negra, enfermedad foliar más destructiva del banano de etiología fúngica, es causada por
el ascomiceto Mycosphaerel/a fijiensis en su fase sexual en donde se producen
ascosporas y por Pseudocercospora fijiensis en la asexual , en donde se producen
conidios; en este trabajo se utilizaron conidios de Pseudocercospora fijiensis para realizar
los experimentos, sin embargo, para ser congruentes con toda la literatura, y a falta de
una acuerdo internacional que permita uniformizar el uso de este término, en adelante se
hará referencia a la fase sexual del hongo, Mycosphaerella fijiensis.
Diferentes tácticas se han implementado para su control , siendo hasta el momento
la aplicación de fungicidas sintéticos la forma más eficiente y recurrente en los sistemas
de producción. Sin embargo, debido a la alta variabilidad genética del fitopatógeno, se
han reportado cepas resistentes a determinados fungicidas.
Poco se sabe sobre el efecto que los fungicidas sintéticos, aplicados en el campo,
podrían tener en la agresividad de las cepas del fitopatógeno, por lo cual este trabajo de
investigación se enfocó al estudio de la interacción in vitro de cepas del fitopatógeno
aisladas de fincas con y sin aplicaciones de fungicidas, en fragmentos foliares de
cultivares de Musa spp. con diferente grado de susceptibilidad al hongo. El daño inducido
por el patógeno en los fragmentos foliares se cuantificó utilizando los programas
informáticos Assess 2.0 e lmage J 1.46r con imágenes mejoradas en el programa
Photoshop 3.0
Los resultados de esta investigación muestran que la cepa Oz1 b (aislada de una
finca sin manejo de fungicidas) fue la que indujo mayor superficie dañada en fragmentos
foliares del cultivar susceptible Gran enano a 30 días después de inoculación (ddi) ,
mientras que los cultivares parcialmente resistente (Fougamou) y resistente (Calcuta IV)
mostraron menor daño, por otra parte la cepa CR4b (aislada de una finca con manejo de
fungicidas) indujo menor severidad en los tres cultivares al igual que la cepa control
(C1233). En general, la cepa Jaguar C3 (aislada de una finca con manejo de fungicidas)
fue la que indujo menor severidad en los tres cultivares de Musa spp. utilizados.
1


ABSTRACT
Bananas (Musa spp.) are grown in many tropical and subtropical areas worldwide,
irs the fourth most important crop just behind rice, wheat and maize. lt's trading generates
multibillionaire incomes every year.
There are factors limiting its cultivation though , among these factors, the Black Leaf
Streak Disease is the most important foliar disease caused by the ascomycete
Mycosphaerella fijiensis, in its sexual phase, where ascospores are produced , and by
Pseudocercospora fijiensis which is the asexual phase and produces conidia . In this work,
conidia from Pseudocercospora fijiensis were used to perform the experiments, however,
given the lack of an international agreement that allows to standardize the use of this term
and in order to agree with the literature, we ' ll refer to the sexual phase of the fungus
known as Mycosphaerella fijiensis from now on.
Different control techniques have been employed , being synthetic fungicides the
most efficient strategy to control the pathogen so far. Nonetheless, given the pathogen 's
high genetic variability, resistant populations to certain fungicides have arisen.
Little is known regarding the effect that synthetic fungicides, applied in field , might
cause on the aggressiveness of pathogenic strains, therefore this work focused on the in
vitre interaction of fungal pathogenic strains, isolated from fields with and without
applications of fungicides, on foliar fragments of Musa spp. cultivars, with different degree
of susceptibility to the pathogen. The damage induced by the fungus in the foliar fragments
was measured using the Assess 2.0 and lmage J 1.46r programs, using improved images
by the Photoshop 3.0 program.
The results from this work show that the Oz1 b strain (isolated from a banana
plantation with no fungicide application) induced the biggest damaged area in foliar
fragments of the susceptible cultivar Great dwarf at 30 dpi, while the CR4b strain (isolated
form a banana plantation with fungicide application) induced less damage and there was
no statistical difference compared with the severity induced by the control strain (C1233).
In general, the Jaguar C3 strain (isolated form a banana plantation with fungicide
application) was the less aggressive strain of all , inducing the smallest sev rity in the foliar
fragments of the three cultivars used.
3


CAPITULO 1
CAPITULO l. INTRODUCCIÓN
El banano (Musa spp.) se cultiva en las regiones tropicales y subtropicales en 150
países de acuerdo a la Conferencia de las Naciones Unidas para el Comercio y el
Desarrollo en 2012 (UNCTAD por sus siglas en inglés), su producción varía entre 105 y
120 millones de toneladas anuales (UNCTAD, 2012), siendo La India el primer país
productor de este cultivo (FAOSTAT, 2011) . De acuerdo con datos oficiales de la
Organización Mundial de las Naciones Unidas para la alimentación y agricultura 2011
(FAOSTAT por sus siglas en inglés) , México ocupa el décimo lugar en la lista de los veinte
principales países productores con 2 138 690 toneladas.
Esta fruta es un alimento importante en los países en desarrollo y una fuente de
ingresos cuya comercialización a nivel mundial alcanza los 7.9 billones de dólares anuales
de acuerdo con cifras reportadas por la UNCTAD (2012).
En cuanto a exportaciones se refiere, Ecuador encabezó el primer lugar con 5,
156, 477 toneladas (FAOSTAT, 2011), seguido por Costa Rica y Colombia; en tanto
México ocupó el décimo quinto lugar entre los 20 principales exportadores de banano, con
un registro promedio anual de 161 , 028 toneladas exportadas (FOSTAT, 2011).
La producción de banano es seriamente afectada, como el resto de cultivos de
importancia alimentaria, por diversas enfermedades entre las cuales destaca la Sigatoka
negra, por ser la más devastadora a nivel foliar y por ocasionar grandes pérdidas de
producción (Carlier et al., 2003). Esta enfermedad es causada por el hongo ascomiceto
Mycosphaerel/a fijiensis (anamorfo Pseudocercospora fijiensis), uno de los tres miembros
del complejo Sigatoka, el cual también está constituido por Mycosphaerelfa musicola
(anamorfo Pseudocercospora musae) que causa la enfermedad de la Sigatoka amarilla y
Mycosphaerel/a eumusae (anamorfo Pseudocercospora eumusae) que causa la
enfermedad de la mancha eumusae de la hoja (Jones, 2003; Crous y Mourichon 2002,
mencionado por Arzanlou , 2008).
La defoliación que provoca M. fijiensis en el banano es tan severa que en
ausencia de fungicidas, la calidad y la cantidad de la fruta se afectan, generando pérdidas
de la producción hasta en un 80%. Asimismo induce la maduración precoz del fruto
(Graenen y Ortiz, 2003, citado por Hidalgo et al., 2006).
S

CAPITULO 1
En el control de la Sigatoka negra se incluyen diversas estrategias, tales como
prácticas culturales y la utilización de fungicidas sintéticos de contacto y sistémicos; los
fungicidas de contacto y sistémicos se utilizan alternadamente de manera estratégica para
manejar el riesgo de resistencia de las poblaciones del fitopatógeno a los fungicidas (Chin
et al., 2001) .
La estrategia más eficiente en el control de la Sigatoka negra ha sido la utilización
de fungicidas sintéticos, sin embargo, en diversos estudios (Cañas-Gutiérrez et al., 2009,
2006; Chin et al., 2001; Gisi et al. , 2000) se tienen reportes sobre la generación de
resistencia a dichos fungicidas en algunas poblaciones del fitopatógeno. También, la
agresividad del fitopatógeno ha sido poco evaluada (Romero y Sutton , 1998) la cual es un
componente cuantificable de las cepas sometidas a diferente manejo agronómico (con y
sin aplicaciones de fungicidas) .
Con base en lo anterior mencionado, en este trabajo se evaluó la agresividad de
cepas de Mycosphaerella fijiensis aisladas de fincas de banano con aplicación de
fungicidas y de cepas provenientes de fincas sin aplicación de fungicidas en fragmentos
foliares de Gran enano, genotipo AAA (susceptible) , Calcuta IV, genotipo AA (resistente) y
Fougamou genotipo ABB, (tolerante) . Con esto se determinó que las cepas de M. fijiensis
aisladas de fincas con aplicaciones de fungicidas son menos agresivas que las cepas del
fitopatógeno aisladas de fincas sin aplicaciones de fungicidas.
6

CAPITULO 1
Antecedentes
Aspectos generales del cultivo de banano
El banano (Musa spp.) cuyo lugar de origen se encuentra en el sudeste asiático,
probablemente Malasia, Borneo, Indonesia, Sumatra y China Meridional (AI-Busaidi,
2013) , se cultiva en más de 150 países (UNCTAD, 2012) y anualmente se producen
alrededor de 120 millones de toneladas, de las cuales el 15% se produce en América
Latina, de donde Ecuador aporta aproximadamente un tercio de la exportación mundial.
Esta fruta es un alimento importante en los países en desarrollo y una fuente de ingresos
cuya comercialización a nivel mundial alcanza los 8 billones de dólares anuales
(UNCTAD, 2012).
En México, la producción anual de plátano representa 1.4% del valor de la
producción agrícola nacional, lo que equivale aproximadamente a 5, 780, 121 millones de
pesos. En esta actividad participan más de 5,000 productores de los estados de
Campeche, Chiapas, Colima, Guerrero, Jalisco, Estado de México, Michoacán, Morelos,
Nayarit, Oaxaca, Puebla, Quintana Roo, Tabasco, Veracruz y Yucatán, en este último
estado se produjeron 1, 046 toneladas en 2012, lo que representa un 0.05% de la
producción nacional total (SIAP, 2011).
Enfermedades que afectan al banano
Existen diversas enfermedades que afectan el cultivo de banano como las
causadas por virus, por ejemplo Banana Bunchy Top Virus (BBTV), Banana Streak Virus
(BSV) y Cucumber Mosaic Virus (CMV) (Lockhart y Jones 2002; Caruana 1999; Reichal et
al. , 1997). Por bacterias e.g . Ralstonia. solanacearum la cual produce la enfermedad
conocida como Moko del banano.
Entre los hongos que causan enfermedades se encuentran Fusarium oxysporum,
causante del mal de Panamá (Pioetz et al., 2003), Mycosphaerella musicola agente
causal de la Sigatoka amarilla y Mycosphaerella fijiensis (Pseudocercospora fijiensis en su
fase asexuaf) responsable de la Sigatoka negra (Stover, 1987). Así mismo, un nuevo tipo
de enfermedad fúngica con síntomas muy parecidos a los causados por la Sigatoka negra
fue descubierta en el sur de La India, Sri Lanka, Tailandia, Malasia, Vietnam, islas
Mauricios y Nigeria. Esta enfermedad es causada por el hongo Mycosphaerella eumusae
7

CAPITULO 1
(fase asexual Pseudocercospora eumusae) con ciclos muy parecidos a los de Sigatoka
negra. Sin embargo, M. fijiensis que es el hongo causante de la Sigatoka negra, presenta
una alta competitividad por nichos, siendo la forma más virulenta, debido a esto llega a
desplazar a Mycosphaerel/a eumusae, (Crous y Mourichon, 2002).
Distribución geográfica de M. fijiensis
El ascomiceto fitopatógeno se encuentra distribuido ampliamente en países
tropicales y subtropicales de todo el mundo (Romero y Sutton, 1997).
La enfermedad fue identificada por primera vez en 1963 en las islas Fiji en la costa
sudeste del valle de Viti Levu, (Rhodes, 1964). En África, el primer reporte de la
enfermedad ocurrió en Zambia en 1973 (Reamakers, 1975), posteriormente en Gabón en
1978 (Frossard, 1980). La enfermedad se siguió diseminando en la parte central y este de
África, reportándose en Nigeria (1986), Ghana (1986), Uganda y Malawi en 1990 y en
Madagascar en el 2000.
La primera aparición de Sigatoka negra en América Latina ocurrió en Honduras en
1972 (Mourichon y Fullerton, 1990). Entre 1973 y 1980 ocurrieron epidemias en Belice,
Guatemala, El Salvador, Nicaragua y Costa Rica. Posteriormente en 1981 se reportó la
enfermedad en México, Panamá y Colombia. En 1986 se presentó en Ecuador (Martínez
et al., 1999; Fullerton, 1990) y durante la década siguiente se reportó en Cuba (1990) y
Venezuela (1991) . En Perú se reportó su presencia en 1994 y en Bolivia dos años
después (Tejerina et al., 1998). En los años siguientes se presentaron reportes en
Jamaica y República Dominicana (1995-1996). La primera detección de Sigatoka negra
en Estados Unidos ocurrió en 1997, en el estado de Florida (Pioetz y Mourichon, 1999). El
último registro en Suramérica se reportó en Brasil en 1998 (Jones, 2000, citado por
Rivas-Piatero, 2002).
Biología y ciclo de vida de M. fijiensis
M. fijiensis se encuentra agrupado taxonómicamente de la siguiente forma: Reino:
Fungi , Subreino: Dikarya, Phylum: Ascomicota, Subphylum: Pezizomycotina, Clase:
Dothideomycetes, Subclase: Dothideomycetidae, Orden: Capnodiales, Familia:
Mycospharellaceae, Género: Mycosphaerella y Especie: Mycosphaerella fijiensis (Schoch
et al. 2006).
8

CAPITULO 1
Independientemente de su genoma (AA, AAA, AAB, ABB), los cultivares de
banano son afectados por la Sigatoka negra en diferentes grados al causar una
defoliación severa, la cual en ausencia de fungicidas se traduce en pérdidas (20-1 00%)
en la producción del fruto, así como en su calidad . Paralelamente la Sigatoka negra
induce la maduración precoz del fruto (Graenen y Ortiz, 2003, citado por Hidalgo et al.
2006) que también se traduce en grandes pérdidas económicas.
La Sigatoka negra o enfermedad de la raya negra del banano es causada por el
hongo heterotálico (requiere la unión de gametos posicionados en talos complementarios
para poder reproducirse) (Mourichon y Zapater, 1990) Mycosphaerella fijiensis (anamorfo
Pseudocercospora fijiensis) , que se caracteriza por ser más infectivo y tener mayor
plasticidad genética debido a la recombinación (Pérez et al. , 2004). Los estados
anamórficos y teleomórficos están presentes en las hojas infectadas y las ascosporas
producidas en la fase sexual son determinantes en el progreso de la enfermedad
(Mourichon y Zapater, 1990).
Durante la enfermedad se observa la presencia de 1 - 4 conidióforos que emergen
de los estomas en el envés de las hojas. Los conidióforos producen conidios que son
desprendidos por el viento, sin embargo, la principal fuente de inóculo son las ascosporas,
que pueden dispersarse a grandes distancias (Mülder y Holliday, 1974). En los
conidióforos se encuentran grupos de 4 - 8 conidios, estos son cilíndricos y pueden estar
formados por 1 - 1 O segmentos, aparecen extendidos o curvados, con forma obtusa en el
ápice y truncados o redondeados en la base. Su longitud varía entre 30 y 132 ¡Jm (Stover
y Simmonds, 1987), sin embargo, se han reportado diversas medidas que varían en
longitud desde 17.4 hasta 114.2 ¡Jm (Rodríguez-Garcfa et al. 2007).
Los peritecios, son estructuras esféricas que contienen las ascosporas de la fase
sexual del hongo, varían en cuanto a sus características, pero en su mayoría son
globulares con un diámetro entre 47 - 85 ¡Jm, se observan incrustados en el tejido de la
hoja y se pueden visualizar en ambas superficies, aunque son más abundantes en el haz.
Las paredes del peritecio son de color marrón oscuro y están formados por tres o cuatro
capas de células con forma poligonal. Dentro del peritecio se forman muchas aseas, las
cuales son bitunicadas y obclavadas. Las ascosporas alcanzan dimensiones de 12.5-16.5
¡Jm x 2.5-3.8 ¡Jm y son biceldadas con la célula más predominante unida al asea (Mülder y
Holiday, 1974).
9

CAPITULO 1
El ciclo de vida de M. fijiensis (Figura 1) se inicia con la germinación de las
esporas, seguida de la penetración. Ambos procesos requieren de alta humedad que
forme una película sobre la superficie de las hojas. La germinación de las esporas inicia
en aproximadamente 2 horas, se observan tubos germinativos los cuales se alargan, se
ramifican y posteriormente penetran si las condiciones de humedad (92-100%) y
temperatura (25-28 oc) son favorables (Jacome y Schuh, 1992); en su fase biotrófica el
patógeno coloniza exclusivamente los espacios intercelulares entre las células del
mesófilo (Beveraggi, 1992) y obtiene nutrientes del apoplasto. El hongo permanece en
esta fase de 3 - 4 semanas antes de que aparezca alguna lesión necrótica (Portal et a/. ,
2011 ). Los conidióforos se desarrollan principalmente en el envés de la hoja durante las
primeras lesiones (Stover, 1987}, aunque también se han observado en etapas tardías de
la enfermedad (Rodríguez-García, 2013; comunicación personal); los conidióforos se
forman a partir de las hitas presentes en las cavidades subestomáticas y producen
múltiples conidios; los conidios son producidos continuamente entre el segundo estado de
raya y segundo estado de mancha (Meredith y Lawrence, 1969). A los 28 días de la
infección aparecen los primeros conidios y a los 49 días finaliza el ciclo cuando se inicia
con la liberación de las primeras ascosporas (Merchán, 1998).
En campo, el período entre la penetración y la aparición de los primeros síntomas
de la enfermedad se conoce como el tiempo de incubación (Brun, 1963 y Klein , 1960;
citados por Henderson et a/., 2006). Gauhl et al. (2000) definieron el tiempo de progreso
de la enfermedad como el período entre la infección y la formación de manchas maduras.
Al igual que en el período de incubación, el periodo de progreso de la enfermedad está
influenciado por las condiciones climáticas, susceptibilidad del hospedero y la cantidad de
inóculo. El período de transición es el tiempo requerido para el desarrollo de los síntomas
de rayas a manchas (Henderson et a/., 2006). Durante la época húmeda se presenta una
alta descarga de ascosporas y un rápido progreso de la infección (Marín et a/. , 2003).
10

CAPITULO 1
Fas.e asexu.at Esporod.o u os sobre las ho s
(
1P netr2Ciótt a 1rav~s de los 2$lomas
Figura 1. Ciclo de vida de Mycosphaerel/a fijiensis y su interacción con Musa spp. (Imagen
tomada de http:
www. ba yercropscience. com. mx/baye r/cropscience/bcsmexico. nsf/files/LifeCycle/$fi 1 e/ E 1 Osigatoka N
egra.gif).
Etapas de la enfermedad
Según Fouré (1985), el progreso de la enfermedad conocida como Sigatoka negra
presenta las seis etapas siguientes:
Etapa 1. Se presenta una pequeña decoloración que solo se observa en el envés de la
hoja. Incluye una pequeña "pizca" de color café rojizo dentro del área decolorada.
Etapa 2. La mancha o estría inicial de color café rojizo se alarga y continua visible solo en
el envés de las hojas.
Etapa 3. La estría aumenta de grosor y longitud; se mantiene de color café rojizo y se
hace visible tanto en el haz como en el envés de las hojas.
11

CAPITUlO 1
Etapa 4. La estría continua aumentando su grosor y longitud, ocurre un cambio de color a
café oscuro y negro. Este signo se considera como mancha y se hace claramente visible
en el haz de la hoja.
Etapa 5. La mancha negra está rodeada por un halo amarillo y se presenta una ligera
depresión en la hoja, producto de la coalescencia de las manchas.
Etapa 6. La mancha se seca en el centro de la lesión y adquiere un color gris claro y la
rodea un anillo bien definido de color negro, rodeado a su vez por un halo de color
amarillo brillante. Estas manchas, se podrán observar aún después de que la hoja se ha
secado ya que el anillo persiste.
Interacción M. fijiensis-Musa spp.
Interacción compatible: cultivar susceptible
La mayor parte de la información sobre esta interacción proviene de estudios de
los cultivares susceptibles Gran enano y Williams los cuales pertenecen al subgrupo
Cavendish de genotipo AAA. En estos cultivares durante las primeras etapas de infección
ocurre una extensa colonización del tejido laminar (en el espacio apoplástico) del
hospedero por parte del micelio del hongo, sin evidenciar células necrosadas en etapas
tempranas de infección (Beveraggi et al., 1995; Riveros, 1995; Sallé et al., 1989). Los
patosistemas Gran enano - M. fijiensis y Williams - M. fijiensis muestran un período largo
de compatibilidad, 28 días aproximadamente según reportado por Sallé et al., (1989),
tiempo durante el cual ocurre una intensa colonización del hospedero antes de la
aparición del primer estado de estría. Durante este período ocurren modificaciones
citológicas de las células mesófilas, caracterizadas por acumulaciones intravacuolares de
glóbulos esféricos y finalmente plasmólisis (Sallé et al. , 1989).
Interacción incompatible: cultivar resistente
La reacción de hipersensibilidad a M. fijiensis, representa el primer paso hacia la
caracterización de esta interacción. Esta reacción de resistencia vertical, a menudo opera
dentro de una relación gen por gen y podría convertir la resistencia en inestable y poco
duradera en el tiempo (Riveros, 2002). En los cultivares resistentes las reacciones de
hipersensibilidad aparecen más temprano que en los cultivares susceptibles como Gran
12

CAPITULO 1
enano Sallé et al., (1989) reportaron la presencia de sustancias osmofílicas
intravacuolares en células de banano infectadas con el fitopatógeno; la presencia de estas
sustancias es mayor en los tejidos de los cultivares resistentes (Fougamou y Yangambi
Km 5). La afinidad de estas inclusiones intravacuolares por colorantes catiónicos sugirió
que son de naturaleza polifenólica. La resistencia está ligada, en parte, a la acumulación
de compuestos fenólicos que se almacenan en las células del parénquima (Torres et al.
2009). Los genotipos resistentes muestran una rápida inducción de los mecanismos de
defensa, que inducen una muerte rápida de las células hospedantes en los sitios de la
infección (Beveraggi et al. , 1993)
Diversidad genética de M. fijiensis
Carlier et al., (1996) realizaron estudios para determinar la diversidad genética del
patógeno en cinco poblaciones geográficas representativas de las áreas productoras de
banano: sureste Asiático, que incluye a Filipinas y Papúa Nueva Guinea, África, América
Latina y las islas del Pacífico. En este trabajo se demostró que los mayores niveles de
diversidad genética se encontraron en las poblaciones del sureste Asiático , coincidiendo
con el centro de origen del hospedero y proponiéndose por lo tanto también como centro
de origen del fitopatógeno. La mayoría de los aleles detectados en las poblaciones de
África, América Latina y las islas del Pacífico también fueron detectados en las
poblaciones del sureste Asiático. Basados en la frecuencia de a lelos de 19 RFLP,
estimaron el nivel de diversidad genética y diferenciación genética entre las cinco
poblaciones geográficas. Carlier et al., (1996) citan el trabajo de Weir (1990) en el cual el
análisis filogenético evidencia que las poblaciones de América Latina y África fueron
diferentes lo que sugiere introducciones independientes en estos continentes.
En América Latina, se encontró un alto nivel de diferencia genética entre las
poblaciones, las más diversas correspondieron a Honduras y Costa Rica, mientras que se
presentó una marcada diferenciación genética entre las islas del Caribe. Las poblaciones
genéticamente más distantes fueron encontradas en República Dominicana-Panamá y
Cuba-Panamá (Rivas-Piatero et al., 2004). Robert et al., (2012), confirmaron que la
diseminación del patógeno en el continente Americano se debió a múltiples eventos de
introducción, en tanto que en África todas las poblaciones del patógeno descienden de un
único evento fundador exitoso.
13

CAPITULO 1
El conocimiento de la magnitud y distribución de la variación genética de M.
fijiensis es importante para el mejoramiento y manejo de la resistencia a la Sigatoka negra
y además, permite identificar el potencial de resistencia de los cultivares en áreas donde
exista diversidad patogénica. Así mismo, identificar zonas de mayor diversidad genética
del fitopatógeno donde se pudieran realizar pruebas de resistencia de los materiales
vegetales resistentes al patógeno (Rivas-Piatero et al., 2004).
Control químico de la Sigatoka negra
Dada la susceptibilidad de los bananos comerciales a esta enfermedad, la
Sigatoka negra es controlada principalmente por medio de la aplicación continua y a
frecuencias relativamente cortas de fungicidas químicos ya sean de contacto (e.g.
Mancozeb® y Clorotalonil®) o sistémicos (e.g . Propiconazol® y Benomil®) aplicados en
emulsiones con adherentes, ya que su control por medio de otras alternativas menos
agresivas (como la técnica cultural de deshoje) aún resulta ineficaz (Romero y Sutton
1997, 1998). En la medida en la que se ha incrementado la superficie de cultivo del
banano así como el uso de fungicidas, se han detectado aislados resistentes a los
agentes químicos (Chin et al. , 2001 ; Romero y Sutton, 1998, 1997a).
A partir de 1991 se detectó resistencia del hongo hacia el fungicida sistémico
Benomil (Romero y Sutton , 1997a), aunque en un trabajo posterior (Romero y Sutton,
1998) hacen referencia a reportes de resistencia del hongo en Honduras desde 1977. La
aplicación intensiva de algunos fungicidas así como la naturaleza cíclica de esta
enfermedad, resulta en la exposición de grandes poblaciones del patógeno a una presión
de selección (Chin et al., 2001), lo cual puede generar poblaciones que no sean lo
suficientemente sensibles para ser controladas de forma adecuada (Marín et al., 2003;
Guzmán, 2002; Guzmán y Romero, 1997).
En México, los fungicidas más comúnmente utilizados en el control de la Sigatoka
negra son Mancozeb® y Clorotalonil® (contacto) y Benzimidazoles, Triazoles,
Estrobirulinas y Anilopirimidinas (sistémicos) (Martínez-Bolaños et al. , 2012). Según lo
reportado por el FRAC (2012), en México anualmente se tienen de 48 a 52 aplicaciones
de Mancozeb®, Propiconazol® y Tridemorf® para controlar la Sigatoka negra.
14

CAPITULO 1
El cultivo de banano en México y la Sigatoka negra
En México la Sigatoka negra se identificó por primera vez en 1981 , en los estados
de Chiapas y Tabasco (Contreras, 1983) actualmente se encuentra distribuida en todos
donde se cultiva el banano.
La región de mayor producción de banano es la región Sur-Sureste que aporta el
78.6% de la producción nacional, siendo Chiapas el estado de donde se obtiene la mayor
parte. La región que ocupa el segundo lugar en producción de banano es la Centro
Occidente con el 20.9% del volumen total producido. Colima es el estado que obtiene el
volumen más alto de esta región .
Los principales genotipos cultivados en México son AAA ("Gran Enano" y "Valery",
subgrupo Cavendish) , ("Macho" o "Falso Cuerno" y "Dominico", subgrupo Plantain) , AAB
("Manzano" o "Silk"), ABB ("Pera" o "Cuadrado") y AA ("Dátil") (Orozco-Santos et al.,
1998).
La presencia de la Sigatoka negra en México ha ocasionado pérdidas en todas las
regiones productoras de banano debido a que modificó el manejo de las fincas , en
especial los programas de aspersión de fungicidas, lo cual trajo como consecuencia un
incremento en los costos de producción (Manzo-Sánchez et al., 2001).
Resistencia y agresividad de Mycosphaerella fijiensis
Con el propósito de estudiar la resistencia de genotipos de banano al hongo
fitopatógeno se han abordado diferentes enfoques en cuanto a la forma de inducir la
infección en condiciones in vitro , Twizeyimana et al. (2007) inocularon fragmentos foliares
de cultivares susceptibles y resistentes, con micelio y suspensiones de esporas. No hubo
diferencia significativa en el área foliar dañada cuando se comparó entre ambos métodos
y ambos se correlacionaron . Los cultivares resistentes Calcuta IV y PITA-17 mostraron
menor área foliar afectada que el cultivar susceptible Agbagba y el tolerante FHIA-23, de
esta forma se estimó el nivel de tolerancia de los cultivares a los aislados del hongo y
paralelamente se evaluó la agresividad de cada aislado. Ambos métodos son rápidos y
requieren de poco espacio para realizarse, los autores recomiendan utilizar suspensiones
conidiales en lugar de fragmentos miceliales del hongo debido a la dificultad que implica
homogenizar la cantidad de fragmentos a ser inoculados en la planta.
15

CAPITULO 1
En contraste, Donzelli y Churchill en 2007 reportaron que es preferible utilizar
fragmentos miceliales del hongo en lugar de suspensiones de esporas para inocular
plántulas, debido a que la esporulación puede ser inconsistente en un mismo aislado o
entre diferentes aislados y, en algunos casos son significativamente deficientes en su
producción in vitre de esporas. Desarrollaron un método para evaluar la agresividad de
diferentes aislados inoculando fragmentos de micelio del hongo en diferentes hojas de
una misma planta, y cuantificaron la severidad foliar empleando el programa de análisis
de imágenes Assess versión 1.0. Empleando este método pudieron aumentar el número
de cepas que se pueden evaluar, lo cual es crítico para comparar simultáneamente la
agresividad de diferentes cepas.
Estos bioensayos son importantes debido a que permiten evaluar la agresividad de
diferentes aislados que han estado en contacto con fungicidas sintéticos en campo, los
cuales pueden tener un efecto diferencial en la agresividad
El control químico de la Sigatoka en la exportación del banano utiliza tanto
fungicidas de contacto como sistémicos, los cuales se emplean de forma alternada como
estrategia para retrasar o manejar la resistencia a los fungicidas tal como mencionan Chin
et al. (2001) quienes comprobaron la resistencia cruzada de aislados del fitopatógeno a
los fungicidas Trifloxistrobin, Azoxistrobin , Famoxadona y Fenamidona.
En la actualidad , los ocho principales fungicidas de uso son los que inhiben la
desmetilación (DMis), aminas, inhibidores de la quinona (Qols; estrobirulinas),
anilinopirimidinas (Aps), bencimidazoles (BCMs), inhibidores de la Succinato
deshidrogenasa (SDHis), N-Fenilcarbamatos y guanidinas (FRAC, 2012). Las principales
estrategias de control, tal como lo remarca el Comité de Acción de Resistencia a
Fungicidas (FRAC por sus siglas en inglés) del Grupo de Trabajo Global para el Banano,
han sido aplicar mezclas de fungicidas con modos diferentes de acción para minimizar el
riesgo de selección de cepas resistentes, así como reducir el número de aplicaciones
anuales de cada fungicida y emplear manejos integrados de la enfermedad (FRAC, 201 O;
citado por Churchill , 2011 ).
El FRAC en 201 O (citado por Churchill 2011 ), indicó que el desarrollo de
resistencia a los Qols (estrobilurinas) se ha convertido en una problemática en Colombia,
Costa Rica, Guatemala y Panamá. La resistencia adquirida por M. fijiensis a las
estrobirulinas ha sido ampliamente atribuida ya sea a una mutación específica o a una vía
16

CAPITULO 1
alterna de oxidación (Chin et a/., 2001). En relación a la pérdida de sensibilidad del
fitopatógeno a ciertos fungicidas (e.g . Benomil®), Cañas-Gutiérrez et al. (2006) estudiaron
un fragmento del gen de la f3-tubulina (sitio de unión del Benomil®) de diez aislados del
patógeno con diferente nivel de resistencia . El análisis de las secuencias mostró un
cambio de adenina en lugar de citosina en el codón 198 de todos los fragmentos del gen
de la f3-tubulina obtenidos de aislados con niveles medio y alto de resistencia al Benomil®.
Por otro lado, Romero y Sutton (1998) reportaron un incremento en la agresividad del
patógeno, reflejado en el número de lesiones con respecto al tiempo que indujeron los
aislados resistentes a Benomil® en hojas del cultivar susceptible Gran Enano.
Contrastando con este comportamiento Jacome y Schuh (1992) reportaron que la cepa
sin manejo de fungicidas de M. fijiensis se comportó más agresiva que las cepas
expuestas a manejo con fungicida. Por el contrario, en otro patosistema, patosistema
So/anum tuberosum-Phytophtora infestans se observó que los aislados resistentes al
fungicida Metalaxil® fueron menos agresivos que las cepas sensibles (Day y Shattock,
1997; Dowley, 1987). Hermanto et al. (201 0) , reportaron que las cepas fúngicas de M.
fijiensis aisladas de plantaciones con y sin aplicaciones de fungicidas, fueron igualmente
agresivas; esto demuestra que la agresividad del patógeno en función de los fungicidas
sintéticos es muy heterogénea.
La experiencia en manejo ha evidenciado que la aplicación de fungicidas no ha
sido una solución consistente, debido a la naturaleza compleja del fitopatógeno de la
Sigatoka negra (alta tasa reproductiva y de recombinación , patogenicidad, diseminación,
entre otros) y a las características del hospedero (uniformidad genética, plantaciones
extensas y tejido susceptible disponible todo el año) esto ha permitido una estrecha
relación entre hospedero y parásito.
Cuantificación de la severidad de la enfermedad
Para estudiar la interacción del patosistema M. fijiensis-Musa spp. se han
empleado diferentes bioensayos en los cuales se han empleado tanto plántulas como
fragmentos foliares. Como mencionaron Twizeyimana et al. (2007) y Donzelli y Churchill
(2007), estos bioensayos permiten realizar estudios evaluando una gran número de cepas
en diferentes cultivares del Musa spp. con diferentes grados de susceptibilidad, con el
objetivo de tener cultivares tolerantes o cuantificar la severidad por el fitopatógeno.
17

CAPITULO 1
Además estos bioensayos requieren de poco espacio y la respuesta de las plántulas o los
fragmentos se correlaciona con la respuesta observada en campo.
No obstante las ventajas que conlleva utilizar ensayos in vitro, uno de los
problemas que enfrentan estos ensayos son la rápida senescencia del tejido vegetal, por
esta razón se han empleado diferentes agentes para retrasar la senescencia de los
fragmentos foliares, algunos de estos incluyen al Bencimidazol en arroz (Mishra y Samal,
1972), en trigo (Pearson et al. 1957, Wang et al. 1961), Elodea (Yoshida, 1970),
cacahuate y arroz (Misra y Mishra, 1968). El Bencimidazol se difunde a través de los
tejidos vegetales, evitando la degradación de la clorofila, proteínas y ácidos ribonucleicos
(ARN), y a bajas concentraciones no resulta tóxico para el hongo (Arraino et al., 2001 ).
Otros agentes incluyen sales de níquel y cobalto , extracto de levadura y leche de coco
(Bushnell , 1966); sin embargo, el níquel llega a ser tóxico para el tejido a concentraciones
de 25 - 50 ppm y además induce manchas cloróticas en la superficie foliar (Malavolta y
Moraes, 2007), por su parte el cobalto es absorbido por vía foliar y es prácticamente
inmóvil , tendiendo a formar quelatos. El extracto de levadura y la leche de coco pueden
favorecer la contaminación de los tejidos vegetales.
Teniendo en cuenta todos los factores anteriormente mencionados, los bioensayos
son de gran utilidad para permitir inducir la infección del tejido vegetal por el hongo y de
esta forma evaluar la severidad del daño. La cuantificación precisa de la severidad de la
enfermedad es esencial para predecir cuantitativamente el progreso de la enfermedad o la
estimación de pérdidas de producción, así como para elucidar variedades resistentes a
una determinada enfermedad (Sherwood et al., 1982). Los procedimientos más sencillos
consisten en evaluar el progreso de la enfermedad, asignándole a la lesión un valor
comprendido en una escala, de acuerdo con el tamaño observado por el evaluador en
campo. Estas escalas se basan en progresiones logarítmicas diagramáticas las cuales se
hacen muy imprecisas cuando el tamaño de la lesión excede el 50% del área de la hoja,
por lo tanto la evaluación visual resulta subjetiva y tiene limitaciones en cuanto a la
precisión (Wijekoon et al., 2008). La severidad de la enfermedad también ha sido
estimada empleando cuantificadores de área (Sutton, 1985), o en porcentaje de luz solar
reflejada (Nutter et a/. , 1993).
Debido a la limitada precisión en estos métodos de evaluación de las
enfermedades, se han desarrollado métodos computacionales tendientes a determinar
18

CAPITULO 1
con mayor precisión el daño al área foliar; los avances en el desarrollo de equipos y
programas ha permitido que muchos sistemas de cómputo estén disponibles en una gran
cantidad de laboratorios de forma rutinaria para la evaluación de enfermedades en
plantas. Esto permite procesar un gran número de muestras y por lo tanto se les conoce
como sistemas de alto rendimiento.
Uno de los programas de análisis de imágenes más versátiles y sencillos de
operar es el programa Assess: programa para el análisis de imágenes y cuantificación de
enfermedades vegetales, que fue desarrollado por Lakhdar Lamari (2002), un experto en
el campo de la fitopatología . Sin embargo, pese a su gran versatilidad, una de las
limitantes de este programa es que no es de dominio público; únicamente se puede
adquirir cubriendo su costo, lo cual limita a muchos laboratorios a adquirir esta
herramienta.
Afortunadamente existen otros programas disponibles gratuitamente en internet,
que pueden ser descargados en los equipos personales sin tener que cubrir su licencia de
uso. Uno de estos programas es el lmage J (Abramoff et al., 2004). Este programa se
desarrolló en el National lnstitutes of Health (NIH por sus siglas en inglés), por Wayne
Rasband . Puede ejecutarse en Windows®, MacOS®, Linux®; además es capaz de leer
distintitos tipos de formato : TIFF, JPGE, PNG, etc. Tiene compatibilidad con funciones
estándar de procesamiento de imágenes tales como manipulación de contraste, nitidez,
suavizado, detección de bordes entre otros; limitado únicamente por la memoria
disponible del equipo empleado (Abramoff et al., 2004).
Los programas Assess 2.0 e lmage J 1.46r pueden ser utilizados para cuantificar
el daño inducido por el hongo fitopatógeno, brindar información sobre aspectos de
patogenicidad en la interacción M. fijiensis-Musa spp. y contribuir al escrutinio de
variedades resistentes. El programa Assess 2.0 se adquirió debido a su especificidad para
cuantificar el daño en sistemas vegetales. Por otro lado, se utilizó el programa lmage J
1.46r el cual es un programa universal de análisis de imágenes para comparar los
resultados obtenidos con el Assess 2.0 y determinar si puede aplicarse al estudio de la
agresividad en Musa spp.
19

CAPITULO 1
Justificación
El cultivo del banano es una de las principales actividades agrícolas a nivel
mundial ; en México este cultivo aporta el 1.4% de la producción agrícola total, sin
embargo, su producción es afectada severamente por la Sigatoka negra.
En la actualidad , la táctica más efectiva para combatir la Sigatoka negra es el
control químico empleando fungicidas con diferentes mecanismos de acción, aplicados de
forma individual o en mezclas.
Por otro lado, poco se conoce con respecto al grado en que la agresividad de M.
fijiensis es influenciada por efecto de los fungicidas que se emplean para su control en
campo.
Por lo que es necesario establecer un método utilizando fragmentos foliares in
vitro para evaluar la interacción de aislados de M. fijiensis provenientes de fincas
nacionales con diferente manejo agronómico (con y sin aplicación de fungicidas) en
fragmentos foliares de cultivares de Musa spp. (Gran enano, Fougamou y Calcuta IV) que
presentan diferente grado de susceptibilidad para cuantificar el daño foliar inducido por
cada aislado del fitopatógeno y determinar de esta forma el grado de agresividad de las
cepas y si la agresividad mantiene relación con el manejo agronómico que recibe la finca
de origen del patógeno
20

CAPITULO 1
Hipótesis
1. Las cepas de M. fijiensis aisladas de fincas con manejo intensivo de fungicidas,
son más agresivas que las cepas aisladas de fincas sin manejo de fungicidas.
Objetivo general
Evaluar la agresividad de cepas de M. fijiensis aisladas de fincas con y sin
aplicaciones de fungicidas, en fragmentos foliares de cultivares de banano (cultivar
susceptible, tolerante o resistente).
Objetivos específicos
• Inducir in vitro la Sigatoka negra en fragmentos foliares de tres cultivares de
banano para reproducir la interacción M. fijiensis-Musa spp.
• Cuantificar el número de lesiones y área foliar dañada en fragmentos foliares de
tres cultivares de banano infectados con diferentes cepas de M. fijiensis.
Estrategia experimental
Material fúnglco : C1233 CR4b Ozlb
Jaguar C3
Crecimiento de las cepas en medioVSC
Esporulación de las cepas y cosecha de conldlos
Preparación de medio de soporte: Agar-Bencimldazol
Corte y desinfección de fragmentos foliares
Inmovilización de fragmentos foliares en medio de soporte
Inoculación de fragmentos foliares
Captura de Imágenes digitales
Muestreos: Oddi Sddi 15 ddl 30 ddi
Cuantificación del da~o foliar
1 Análisis estadlstlco 1
Observación microscópica
Figura 2. Estrategia experimental para cuantificar la agresividad de aislados de M. fijiensis
21

CAPITULO 1
Bibliografía
Abramoff, M. Magalhaes, P. Ram, S. (2004). lmage processing with lmageJ.
Biophotonics international. 11, 36-42.
AI-Busaid i, K. (2013). Banana domestication on the Arabian Peninsula: A review of their
domestication history. Journal of Horticulture and Forestry. 5, 194-203.
Arraino, L. S. Brading , P.A. Brown J.K. M. (2001). A detached seedling leaf technique
to study resistance to Mycosphaerella graminicola (anamorph Septoria tritici) in
wheat. Plant Pathology. 50, 339-346.
Arzanlou, M. Groenewald, J.Z. Fullerton, R. A. Abeln , E.C.A. Carlier, J. Zapater,
M.F. Buddenhagen, I.W. Viljoen , A. Crous, P.W. (2008). Multiple gene genealogies
and phenotypic characters differentiate several novel species of Mycosphaerella
and related anamorphs on banana. Persoonia. 20, 19-37.
Bayer Crop Science México. [Online] (Actualizado 09 de Marzo del 2011) Disponible
en:
http://www.bayercropscience.com.mx/bayer/cropscience/bcsmexico.nsflid/Sigatoka
Diseases_BCS [Acceso 31de Octubre del2012].
Beveragg i, A. (1992). Etude des interactions hote-parasite chez des bananier sensibles
et résistance inoculés par Cercosporafijiensisresponsable de la maladie des raíces
noires. Thése de 3éme cycle , Université de Montpellier 11. USTL. 7 4.
Beveraggi, A. Mourichon, X. Salle, G. (1993). Study of host-parasite interactions in
susceptible and resistant cultivated bananas inoculated with Cercospora fijiensis,
pathogen of black leaf streak disease. Breeding banana and plantain for resistance
to diseases and pests. CIRAD, INIBAP: Montpellier, Francia. 213-220.
Beveraggi, A; Mourichon, X; Salle, G. (1995). Etude comparée des premieresétapes de
L'infeccion chez les bananiers sensibles et résistants infectés per le
Cercosporafijiensis agent responsable de la maladie des raices noire. Canadian
Journal of Botany. 73, (1328-1337).
22

CAPITULO 1
Bushnell, W.R. Smillie, W.R. (1966). Delay of senescence in wheat leaves by
cytokinins, nickel and other substanes. Can. J. Bot. (44) , 1485-1493.
Cañas-Gutiérrez, G.P. Angarita-Velásquez, M.J. Restrepo-Fiores, J.M. Rodríguez, P.
Moreno, C.X. Arango, R. (2009). Analysis of the CYP51 gene and encoded protein
in Propiconazole-resistan isolates of Mycosphaerella fijiensis. Journal of
Phytopathology. Volumen 65. Número 8. pp. 892-899.
Cañas-Gutiérrez, G.P. Patiño L.F. Rodríguez-Arango, E. Arango, R. (2006). Molecular
characterization of Benomyl-resistant isolates of Mycosphaerella fijiensis, collected
in Colombia. Journal of Phytopathology. 154(7-8), 403-409.
Carlier, J; Leburn, M.H; Zapater, M.F. Dubois,C; mourichon, X. (1996). Genetic
structure of the global population of banana black leaf streak fungus,
Mycosphaerel/a fijiensis. Molecular Ecology. 5, 499-51 O.
Carlier, J. Hayden H. Rivas, G. Zapater, M. F. Abadie, C. Aitken , E. (2003). Genetic
differentiation in Mycosphaerella leaf spot pathogens. Mycosphaerella Leaf Spot
Diseases of Banana: Present Statuts and Outlook. 123-129.
Caruana, M.L.1999. Banana bunchy top virus the most serious virus disease of banana.
Australasian Plant Pathology 71 p.
Chin , K.M. Wiz, M. Laird, D. (2001). Sensitivity of Mycosphaerella fijiensis from banana
to trifloxystrobin. Plant disease. 85, 1264-1270.
Churchill , A. (2011). Mycosphaerella fijiensis, the black leaf streak pathogen of banana:
progress towards understanding pathogen biology and detection, disease
development, and the challenges of control. Molecular Plant Pathology. 12(4), 307-
328.
Contreras, M. de E.M. (1983). El chamusco negro (Sigatoka) una nueva enfermedad de
la hoja de los plátanos. Universidad Autónoma de Chapingo. México. Revista de
Geografía Agrícola , 4, 61-102.
23

CAPITULO 1
Crous, P.W. Mourichon. (2002). Mycosphaerella eumusae and its anamoph
Pseudocercospora emusae spp. Nov. Causal agent of emusae leaf spot disease.
Sydonia . 54(1), 35-43.
Day, J.P. Shattock, R.C. (1997). Aggressiveness and other factors relating to
displacement of populations of Phytophtora infestans in England and Wales.
European Journal of Plant Pathology. 103, 379-391.
Donzelli, B. Churchill, A. (2007), a quantitative assay using mycelial fragments to
assess virulence of Mycosphaerella fijiensis. Phytopathology. (97), 916-929.
Dowley, L.J . (1987). Factors affecting the survival of metalaxil resistant strains of
Phytophtora infestans (Mont.) de Bary in lreland. Potato Research. 30, 473-475.
Food and Agriculture Organization of The United Nations (FAOSTAT), 2011 [Online] .
(actualizado 07 de agosto de 2012) Disponible en :
http://faostat.fao.org/site/339/default.aspx [consulta 3 Agosto, 2013].
Fouré, E. 1985. Black leaf streak disease of Banana and plantains (Mycosphaerella
fijiensis Morelet), study of the symptoms and stages of the disease in Gabon IRFA
París.
Frossard, P. (1980). Apparition d'une nouvelle et grave maladie foliaire des bananiers
et plantains au Gabon: la maladie des reices noires, Mycosphaerella fijiensis
Morelet. Fruits 35, 443-453.
Fullerton, RA. (1990). Studies on Mycosphaerella fijiensis Morelet in the Pacific lslands.
In : Fullerton RA, Stover RH (eds) Sigatoka Leaf Spot Diseases of Banana INIBAP:
Montpellier, 29-37.
Fungicide Resistance Action Committee, 201 O. Summary of FRAC guidelines for
banana in 2012. [Online] (Actualizado 22-23 marzo 2012) . Disponible en:
http :/ /www.frac.info/frac/index.htm [Consulta 02 noviembre 2012].
24

CAPITULO 1
Gauhl , F. Pasberg-Gauhl, C. Jones, D.R. 2000. Black leaf streak: Disease cycle and
epidemiology. Diseases of Banana, Abacá y Enset. pp. 56-62.
Gisi, U. Chin , K.M. Knapova, G. Küng , R. Farber. Mohr U. Parisi , S. Sierotzki, H.
Steinfeld, U. (2000). Recent devolpments in elucidating modes of resistance to
Phenylamide, DMI and Strobilurin fungicides. Novartis Crop Protection. Research
Biology. 19, (8-1 0) , 863-872.
Guzmán, M. (2002) . Situación de la Sigatoka Negra en Costa Rica y opciones para el
manejo de la enfermedad. Memorias ACORBAT. Cartagena de Indias, Colombia.
18-191 .
Guzmán, M. Romero, R. (1997). Comparación de los funguicidas azoxistrobina,
propiconalzole y difeconazole en el control de la Sigatoka negra (Mycosphaerella
fijiensis Morelet) en Banano (Musa AAA). Corbana, Costa Rica. 22(47), 49-59.
Henderson, J. Pattemore, J.A. Porchun , S.C. Hayden, H.L. Van Brunschot, S. Grice,
K.RE. Peterson, R.A. Thomas-Hall , S.R. Aitken, E.A.B. (2006) . Black Sigatoka
Disease: new technologies to strengthen eradication strategies in Austral ia.
Australasian Plant Pathology. 35, 181-193.
Hermanto, C. Opina, O. Natural , M. (201 0). Assessment of a fungicide resistance in a
population of Mycosphaerella Spp. on Señorita Banana Variety (Sucrier Group).
Tree and Forestry Science and Biotechnology. 4 (2) , 85-90.
Hidalgo, M. Tapia , A. Rodríguez, W. Serrano, E. (2006). Efecto de la Sigatoka negra
(Mycosphaerella fijiensis) sobre la fotosíntesis y transpiración fol iar del banano
(Musa spp. AAA, cv. Valery) . Agronomía costarricense. 30(1), 35-41 .
Jacome, L.H . Schuh, W . (1992). Effect of temperature on growth and conidial
production in vitro, and comparison of infection and aggressiveness in vivo among
isolates of Mycosphaerella fijiensis var. difformis. Tropical Agriculture. 70(1 ), 51-
59.
25

CAPITULO 1
Jones, D.R. (2003). The distribution and importance of the Mycosphaerella leaf spot
diseases of banana. En: Mycosphaerella Leaf Spot Diseases of Bananas: Present
Status and Outlook. Proceedings of the Workshop on Mycosphaerella Leaf Spot
Diseases, San José, Costa Rica, 20-23 May 2002 (Jacome, L., Lepoivre, P.,
Marin , D., Oriz, R. , Romero, R. and Escalant, J.V., eds) , 25-41. Montpellier: The
lnternational Network for the lmprovement of Banana and Plantain.
Lakhdar, L. (2002). [Online] (actualizado Noviembre 2002). Disponible en:
http://www.amazon .com/s/ref=ntt_athr _dp_sr _1? _encoding=UTF8&field
author=Lakhdar%20Lamari&search-alias=books&sort=relevancerank. [Consulta 16
de enero de 2014]
Lockhart, B.E. Jones, D.R. 2002 . Bacilliform Virus Related Serologically to Banana
Streak Virus. Plant Disease 72, 230-233.
Malavolta, E. Moraes, M. F. (2007). Nickel, from toxic to essential nutrient. Better Crops
with Plant Food. 91 (3) , 26-27.
Manzo-Sánchez, G. Orozco-Santos, M. Guzmán-Gonzales, S. (2001 ). Caracterización
morfológica de Mycosphaerella fijiensis Morelet de la región del pacífico-centro de
México y su desarrollo en medios líquidos. Revista Mexicana de Fitopatolog ía.
Volumen 19. Pp. 66-71.
Marín,D; Romero,R; Guzmán,M; Sutton, T. (2003). Black Sigatoka and increasing
threat to banana cultivation. Plant disease. 87(3) , 208-222.
Martines-Bolaños, L. Téliz-Ortíz, D. Rodríguez-Maciel, J. Mora-Aguilera, A. Nieto
Angel , D. Cortés-Flores, l. Méjia-Sánchez, D. Nava-Díaz, C. Silva-Aguayo, G.
(2012). Resistencia a Fungicidas en Poblaciones de Mycosphaerella fijiensis del
Suereste Mexicano. Agrociencia. 46, 707-717.
Martínez, G. Pargas, R. Muñoz, D. (1999). La Sigatoka negra y su avance en el
territorio Venezolano. Implicaciones Socioeconómicas. CENIAP.
26

CAPITULO 1
Merchán V, VM. (1998). Manejo de problemas fitosanitarios del cultivo del plátano en la
zona central cafetera. Seminario internacional sobre la producción de plátano,
Armenia , Colombia. 177-191.
Meredith , D.S. Lawrence, J.S. (1969). Black leaf streak disease of bananas
(Mycosphaere/la fijiensis) : symptoms of disease in Hawaii , and notes on the
conidial state of the causal fungus. Transactions of the British Mycological Society.
52(3) , 459-476.
Mishra, D. Samal, B. (1972). Retardation of senesce in rice (Oryza sativa L.) leaves by
Bencimidazol and nickel chloride. Bacteria! and Fungallnfection. (40), 100-106.
Mishra, D. Misra, B. (1968). Effect of growth regulating chemicals on degradation of
chlorophyll and starch in detached leaves of crop plants. Pflanzenphysiol. (58) ,
207-211 .
Mourichon, X. Fullerton , R.A. (1990). Geographical distribution of two species
Mycosphaerella musico/a Leach . (Cercospora musae) and M. fijiensis Morelet (C.
fijiensis) , respectively agents of Sigatoka disease and Black Leaf Streak Disease in
bananas and plantains.Fruits. 45, 213-218.
Mourichon, X. Zapater, M. F. 1990. Obtention in vitro du stade Mycosphaerella fijiensis ,
forme parfaite de Cercospora fijiensis . Fruits 45(6) , 553-557.
Mülder, J.L. Holliday, P. 1974. Mycosphaerella fijiensis. CMI. Descriptions of
Pathogenic Fungi and bacteria. No. 413.
Nutter, J.R. Gleason, M.L. Jenco, J.H. Christians, N.C. (1993). Assessing the accuracy,
intra-rater repeatability and intra-rater reliability of disease assessment systems.
Phytopathology. 83, 806-812.
Orozco-Santos, M. Farías-Larios, J. Manzo-Sánchez, G. Guzmán-Gonzales, S. (1998).
La Sigatoka negra (Mycosphaerella fijiensis Morelet) en México. INFOMUSA.
10(1), 33-37.
27

CAPITULO 1
Pearson , C. Samborski , D. J. Forsyth , F.R. (1957). Effect of Benzimidazole in detached
wheat leaves. Nature. (180}, 1294-1295.
Pérez, M. Rebullído, R. Pérez, L. (2004) . Lineas base de sensibilidad de las
poblaciones de Mycosphaerella fijiensis Morelet de Cuba, a los fungicidas
azoxystrobin y trifloxystrobin . Fitosanidad. 8, 41-47.
Ploetz, R. C. Mourichon. (1999). First report of Black Sigatoka in Florida. Plant Disease.
83, 300.
Ploetz, RC. Thomas, J. E. Slabaug, W.R. (2003). Diseases of banana and plantain.
Diseases of Tropical Fruit Crop. 73-134.
Portal, O. Izquierdo, Y. De Vleesschauwer, D. Sánchez-Rodríguez, A Rodríguez
Mendoza, M. Acosta-Suárez, M. Ocaña. B. Jiménez, E. Hofte, M. (2011). Analysis
of expressed sequence tags derived from a compatible Mycosphaerella fijiensis
banana interaction. Plant Cell Reports. 30, 913-928.
Reamakers, R. (1975). Black leaf spot disease in Zambia. PANS. 21, 396-400.
Reichel, H. Belalcázar, S. Múnera, G. Perez, R. Avévalo, E. 1997. The presence of
banana streak virus infecting sugarcane and has been confined in plantain (musa
AAB commonds) Sugar cane (Sacharumofficinarum) and edible cane
(cannaedrelis) in Colombia. INFOMUSA. 6 (1), 9-12.
Rivas-Piatero, G.G. 2002. Genética de poblaciones de Mycosphaerella fijiensis en el
trópico americano. Memorias XV reuinión. Realizada en Cartagena de Indias,
Colombia. 27 de octubre al 02 de noviembre de 2002. Medellín (COL) : asociación
de bananeros de Colombia AUGURA, 2002
Rhodes, P.L. (1964) . A new banana disease in Fiji. Common. Phytopathology News.
10, 38-41 .
28

CAPITULO 1
Rivas-Piatero, G.G. Zapater, M. Abadie, C. Carlier, J. (2004). Founder effects and
stochastic dispersa! at the continental scale of the fungal pathogen of bananas
Mycosphaerella fijiensis. Molecular Ecology. 13(2), 471-482.
Rivas-Piatero, G.G. (2002). Genética de poblaciones de Mycosphaerella fijiensis en el
trópico americano. Memorias XV reuinión . Realizada en Cartagena de Indias,
Colombia. 27 de octubre al 02 de noviembre de 2002. Medellín (COL) : asociación
de bananeros de Colombia AUGURA, 2002.
Riveros, A.S. (1995). Etude D elicitoursasocies a la resistance du cultivar de bananier
Yangambi Km.S a Mycosphaerella fijiensis Morellet. Faculté de Sciences
Agronomiques de Gembloux. 170p.
Riveros, A.S. (2002). Preparación de inductores exógenos a partir de suspensión
conidial de Mycosphaerella fijiensis en germinación. En : Rosales FE y LE
Pocasangre (eds) oferta tecnológica de banano y plátano para America Latina y
el Caribe Turrialba; Costa Rica. 12-13p.
Robert, S. Ravigne, V. Zapater, M.F. Abadie, C. Carlier, J . (2012). Contrasting
introduction scenarios among continents in the worldwide invasion of the banana
fungal pathogen Mycosphaerella fijiensis. Molecular Ecology. 21 , 1098-1114.
Rodríguez-García, C.M. Navarrete-Mapen, R.Z. Peraza-Echeverría, L. (2007).
Characterisation of the in vitro conidial morphology of Mycosphaerella fijiensis, a
major pathogen of banana. Australasian Plant Pathology. 36, 369-372.
Romero, R. A. Sutton, T. B. (1997a). Sensitivity of Mycosphaerella fijiensis, Causal
Agent of Black Sigatoka of Banana, to Propiconazole. Phytopathology. 87, 96-1 OO.
Romero, R. A. Sutton, T. B. (1998). Characterization of benomyl resistance in
Mycosphaerella fijiensis, cause of black Sigatoka of banana, in Costa Rica. Plant
Disease. 82, 931-934.
29

CAPITULO 1
Sallé, G. Pichard, V. Mourichon, X. (1989). Cytological study of the interaction between
Mycosphaere/la fijiensis Morelet and three cultivars of Musa presenting different
levels of resistance. In INIBAP Workshop of Sigatoka Leaf Spots Diseases
(Mycosphaerella spp.), San José, Costa Rica. 88.
Schoch, C.L. Shoemaker, R.A. Seifert, K.A. Hambleton, S. Spatafora, J.W. Crous, P.W.
(2006). A multigene phylogeny of the Dothideomycetes using four nuclear loci.
Mycologia. 98, 1042-1052.
Servicio de Información Agroalimentaria y Pesquera (SIAP), 2011 [consultado en línea].
Disponible en:
http://www.siap.gob.mx/index.php?option=com_wrapper&view=wrapper<emid=35
O [consulta: 3 Agosto 2013].
Sherwood, R.T. Berg, C.C. Hoover, M.R. Zeiders, K.E. (1982). lllusions in visual
assessment of Stagonospora Leaf Spot of Orchardgrass. Phytopathology. 73, 173-
177.
Stover, R.H. 1987. Producción de plátano en presencia de Sigatoka negra. In:
Pinochet, J. Plagas y Enfermedades de carácter epidémico en cultivos frutales de
la zona centroamericana. Informe Técnico No. 10. Centro agronómico tropical de
investigación y enseñanza, CATIE. Panamá. 27-36.
Stover, R.H . Simmonds, N. (1987). Bananas. Longman scientific and tecnichal. Tercera
edición. Inglaterra. 467p.
Sutton, J.C. (1985). Effectiveness of fungicides for managing foliar diseases and
promoting yields of Ontario winter wheat. Phytoprotection . 66, 141-152.
Tejerina, J.C. (1998). First report of Black Sigatoka in Bolivia. Plant Diseases. 81 (11 ),
1332.
30

CAPITULO 1
Torres, J. Rodríguez H.A. Rodríguez E. Arango, R. 2009. Aspectos bioquímicos de la
resistencia del banano (Musa acuminata) al ataque del hongo Mycosphaerella
fijiensis Morelet. Tumbaga. Número 4. pp. 85-96.
Twizeyimana, M. Ojiambo, P.S. Tenkouano, A. lkotun, T. Bandyopadhyay, R. (2007).
Rapid Screening of Musa species for resistance to Black Streak Disease using in
vitro plantlets intubes and detached leaves. Plant Disease. (91), 308-314.
United Nations Conference on Trade and Development (UNCTAD) , 2012. [Consultado
en línea]. (última actualización: 05/09/2012) Disponible en:
http://www.unctad .info/en/lnfocomm/AACP-Products/COMMODITY-PROFILE---Banana/
[consulta: 3 Agosto 2013].
Wang , D. Hao, M.S.H. Waygood , E.R. (1961). Effect of Benzimidazole analogues on
steam rust and chlorophyll metabolism. Can. J. Bot. (39), 1029-1036.
Wijekoon, C.P. Goodwin, P.H. Hsiang, T. (2008). Quantifying fungal infection of plant
leaves by digital image analysis using Scion lmage software. Journal of
Microbiological Methods. 74, 94-101 .
Yoshida, Y. (1970). Control of chloroplast senescence of detached Elodea leaves. Bot.
Mag. (83) , 137-143.
31


CAPITUlO 11
CAPITULO 11. INFECCIÓN in vitro DE FRAGMENTOS FOLIARES DE Musa
spp. CON CEPAS DE Mycosphaerella fijiensis PROVENIENTES DE FINCAS
CON DIFERENTE MANEJO DE FUNGICIDAS
Introducción
Debido a la importancia económica que representa el impacto de la Sigatoka negra
en el cultivo del banano, es básico estudiar este patosistema desde diferentes
perspectivas, para lo cual es necesario inducir el proceso de infección que se lleva a cabo
cuando estos dos organismos interactúan; con base en lo anterior, el método debe ser lo
más parecido posible a lo que ocurre en condiciones de campo.
En general, se ha recurrido a pruebas de campo cuando se trabaja en el escrutinio
de cultivares resistentes a una determinada enfermedad (Eyal et al., 1985; Van Ginkel y
Scharen, 1988; Kema y Van Silfhout, 1997) sin embargo, algunas de las desventajas de
estas pruebas, requieren de una gran cantidad de tiempo y espacio. Otras desventajas de
realizar experimentos en campo son la incidencia natural de otros microorganismos en las
plantas evaluadas, lo cual dificulta o enmascara la interacción entre el hospedero y el
fitopatógeno en estudio; y la influencia de factores ambientales en los experimentos.
Una opción para realizar un escrutinio de cultivares tolerantes a la Sigatoka negra
es la utilización de plantas completas que han sido propagadas in vitro. Sin embargo,
algunas de las desventajas que supone el uso de plántulas es que se requiere emplear
cámaras de crecimiento con temperatura, luz y humedad controladas lo cual puede
resultar costoso aun cuando las condiciones sean favorables para que la interacción
resulte en compatibilidad.
Los bioensayos con hojas desprendidas también han sido reportados en estudios
de interacción planta-fitopatógeno; Farlow (1885), fue el primero en usar hojas
desprendidas para pruebas de inoculación de plantas del género Pomaceae
inoculándolas con hongos parasíticos del género Gymnosporangium. Brown y Wolfe
(1990) utilizaron hojas desprendidas de trigo (Triticum aestivum) y cebada (Hordeum
vulgare) inoculadas con moho velloso (Erysiphe graminis) para comprobar la alta
correlación con las pruebas de campo. Una desventaja de estas pruebas es que el
material foliar se encuentra en un medio artificial , tal como lo mencionó Karjalainen
33

CAPITULO 11
(1984), al estudiar el patosistema Septoria nodorum-Triticum aestivum. Yarwood (1946)
definió el cultivo de hojas desprendidas como el mantener las hojas en condiciones de
vida durante un largo período de tiempo después de ser desprendidas de la planta de la
cual formaron parte alguna vez. En el cultivo de hojas desprendidas, las principales
preocupaciones son el desarrollo y función de la hoja misma, debido a algunos endófitos
habitando en el interior de la hoja (Zinniel et al., 2002), los cuales podrían dañar el tejido
escindido de la planta cuando se encuentre en condiciones in vitro.
Una de las principales desventajas en la utilización de fragmentos foliares y de
hojas desprendidas es la muerte temprana de las mismas ocasionada por deshidratación.
En general, las hojas desprendidas tardan más tiempo en deshidratarse que los
fragmentos foliares si se les proporcionan las condiciones de humedad requeridas (Mishra
y Pradhan, 1973). Las hojas desprendidas de las monocotiledóneas (e.g. Musa spp.)
sobreviven menos tiempo que las hojas de las plantas dicotiledóneas, esto se puede
deber a que las hojas de las monocotiledóneas generalmente guardan sus carbohidratos
lábiles como azúcares solubles, en tanto que en las hojas de la mayoría de las
dicotiledóneas el almidón prevalece. Además ocurre mayor lesión y superficie dañada
cuando las hojas de las monocotiledóneas son desprendidas, con respecto al daño que
ocurre en las dicotiledóneas (Arraino et al., 2001).
Con respecto al uso de fragmentos foliares, un problema adicional que se presenta
además de la deshidratación es el catabolismo de proteínas hidrosolubles asociadas a la
clorofila (WSCP por sus siglas en inglés) (Damaraju, 2010). Por ejemplo, la enzima
RuBisCo (ribulosa-1 ,5-bisfosfato carboxilasa oxigenasa) que se encarga de fijar C02 a
una forma orgánica disminuye, con lo cual disminuye la actividad fotosintética necesaria
para mantener el fragmento en buenas condiciones (Hildbrand et al., 1994).Por otro lado,
una ventaja de utilizar fragmentos foliares es que el espacio requerido para manipular y
almacenar los fragmentos es menor al espacio que se necesitaría para las hojas
desprendidas, lo cual representa una ventaja importante; además, los problemas de
deshidratación y senescencia se pueden minimizar al mantener los fragmentos en un
medio saturado con agua y que contenga un agente que retrase el deterioro del tejido
vegetal.
34

CAPITULO 11
Fragmentos foliares en la interacción Musa spp.-Mycosphaerella fijiensis
Twizeyimana et al., (2007) emplearon fragmentos foliares de 12 cm2 de superficie
para el escrutinio de diferentes genotipos tolerantes a la Sigatoka negra, utilizando
fitohormonas (ácido giberélico, citoquininas) y Bencimidazol para retardar la senescencia
de los fragmentos; de las tres hormonas evaluadas, el ácido giberélico fue el agente más
eficiente, retrasando la senescencia de los fragmentos hasta por 52 días, en tanto que
para Arraino et al., (2001 ), el mejor agente para retrasar la senescencia de las hojas de
trigo fue el Bencimidazol en el rango de 50 - 200 ppm. Debido a que uno de los efectos
fisiológicos que el ácido giberélico tiene en las plantas es la expansión de las hojas
(Falkowska et al., 201 0) , en este trabajo se utilizó Bencimidazol para evitar que los
fragmentos foliares se expandieran fuera del molde.
El uso de fragmentos foliares es una herramienta valiosa para el estudio de la
interacción Musa spp.-Mycosphaerella fijiensis ya sea en la búsqueda de genotipos
tolerantes a este fitopatógeno o para evaluar la severidad inducida por cepas del
fitopatógeno aisladas de fincas tratadas con diferente manejo agronómico (uso de
fungicidas).
Por lo anterior, es necesario evaluar ambos sistemas biológicos (hongo
fitopatógeno y banano) en condiciones in vitro para estudiar su interacción libre de
cualquier otro componente que pudiera afectar el proceso de infección.
35

CAPITULO 11
Materiales y métodos
Material fúngico
Los materiales de banano conteniendo al fitopatógeno fueron colectadas de los
estados productores de banano de Chiapas y Tabasco a partir de fincas productoras con
diferente manejo agronómico de fungicidas. Las cepas del fitopatógeno fueron aisladas a
partir de dichos materiales por los Doctores Ignacio Islas Flores y Andrew James Kay;
posteriormente la selección de las cepas fue realizada por la Doctora Cecilia Rodríguez
García con base en la ubicación geográfica y el tipo de manejo de fungicidas de las fincas
de dónde provinieron las cepas (Cuadro 1). La M. C. Leticia Peraza Echeverría evaluó la
producción masiva de esporas asexuales de dichas cepas y después se seleccionaron
dos cepas por estado, cada cepa proveniente de una finca con diferente ubicación y
manejo de fungicidas.
Cuadro 1. Aislados de M. fijiensis provenientes de fincas con diferente manejo de fungicidas .
Cepa Ubicación Manejo
C1233 Yucatán Sin fungicidas
CR4b Chiapas Mancozeb lB> , Calixin® y
Bravo® 720
Ozlb Chiapas
Jaguar C3 Tabasco Mancozeb iB>,
Estrobilurinasl!> y Triazoles®
Cruse C3 Tabasco Sin fungicidas
La cepa CIRAD C1233 con número de acceso IMI 392976, del lnternational
Mycological lnstitute, fue aislada de Musa acuminata, de la península de Yucatán, y en
este estudio se usó como cepa control ya que se tiene caracterizada su producción de
conidios (Rodríguez-García et al., 2007).
La infección in vitre de fragmentos foliares se realizó en dos etapas.
36

CAPITULO 11
Etapa 1
Crecimiento de las cepas en medio VSC
Las cepas de M. fijiensis se cultivaron en medio de cultivo denominado V8C. Este
medio fue reportado por Mourichon et al., (1987) y modificado por Puch-Ceh, (2001),
Peraza-Echeverría et al., (2008); Su composición se detalla en el Cuadro 2.
Cuadro 2. Composición del medio de crecimiento va
Componente composición
CaC03
Jugo de verduras V8 Hérdez®
Agar microbiológico
2g/L
200 mL/L
20 g/L
El medio V8C se esterilizó en autoclave a 120 oc en un período de 20 minutos. Para la
siembra en el medio V8C se cortaron fragmentos de micelio de una o varias colonias en
crecimiento de las cepas anteriormente mencionadas (procedentes de la colección de
cepas del proyecto FORDECYT). Las cajas se colocaron en un cuarto de cultivo con
temperatura de 25-30 oc y con fotoperiodo 12h luz/12h oscuridad.
Producción de conidios en medio VSE
Para la obtención de conidios, las cepas empleadas en este estudio se sembraron
en medio de esporulación V8E, reportado por Mourichon et al., (1987) que tiene la
siguiente composición:
Cuadro 3. Composición del medio de esporulación Va
Componente composición
CaC03
Jugo de verduras V8 Hérdez®
Agar microbiológico
pH final
0.2 g/L
100 ml/L
20 g/L
6.0
El medio de esporulación se esterilizó en autoclave a 120°C por 20 minutos.
37

CAPITULO 11
La siembra en el medio de esporulación se llevó a cabo de la siguiente manera:
con un bisturí se cortaron fragmentos de aproximadamente 1 cm2 de superficie de la parte
con crecimiento más joven del hongo, posteriormente el fragmento de hongo se depositó
en un mortero estéril y se procedió a macerar durante 5 minutos agregando 2 mL de agua
estéril, posteriormente se agregó 1 mL más de agua estéril y se continuó macerando hasta
observar una consistencia no tan pastosa, de tal forma que fuera fácilmente distribuido
sobre el medio de cultivo.
El macerado se homogenizó empleando una micropipeta de 1 mL con punta
cortada y con filtro y posteriormente se tomaron 2,4 mL (tres alícuotas de 800 ¡.¡.L) los
cuales se depositaron sobre cajas Petri conteniendo el medio V8E.
Posteriormente el macerado se distribuyó dentro de la caja Petri mediante
movimientos circulares; todas las cajas sembradas se incubaron a temperatura de 20 ± 2
oc con luz blanca y luz negra permanente, durante 6 días (Peraza-Echeverría et al.,
2008).
Cosecha de esporas
Para cosechar las esporas del hongo a los seis días después de inoculación del
medio V8E, se depositó una alícuota de 2.4 mL de grenetina al1% (en tres alícuotas de
800 ¡.¡L) en cada caja Petri para cosechar las esporas, las cuales se removieron utilizando
un pincel de cerdas de camello , haciendo un barrido suave sobre toda la superficie de la
caja Petri. Todo el procedimiento se realizó en condiciones estériles en campana de flujo
laminar. Después de la primera cosecha de esporas, las cajas se sellaron y se regresaron
a la misma condición de cultivo (luz continua blanca y negra, 20 oc ± 2°C), para realizar la
siguiente cosecha al día 6.
Las esporas se recuperaron inclinando la caja Petri y haciendo un lavado a toda la
caja con la misma suspensión, en las partes superior y media, finalmente la suspensión
de esporas se depositó en un tubo Falcón de 50 mL. Se filtró la suspensión de esporas
para eliminar restos de micelio, haciéndola pasar a través de cuatro capas de gasa y
recuperándola en otro tubo Falcón estéril. Después de cosechadas las esporas de todas
las cajas, se obtuvo una suspensión turbia clara en la primera cosecha y turbia oscura en
la segunda cosecha debido a una mayor cantidad de esporas.
38

CAPITULO 11
Conteo y ajuste de la suspensión de esporas en cámara Sedgwick-Rafter
Para la cuantificación de esporas se preparó una dilución 1:1 O de la suspensión
madre en un volumen final de 12001-JI, utilizando una solución de grenetina comercial al
1% p/v y homogenizando la suspensión . Asimismo se utilizó una cámara Sedgwick-Rafter
en la cual se colocaron cuidadosamente, sin burbujas, 1 mL de la suspensión de esporas
diluida para proceder al conteo (5 campos por cuadrante y parte central) utilizando un
microscopio óptico (marca Axioplan® Zeiss 20X, programa Action visión 3.1 ©). Este
procedimiento se repitió dos veces consecutivas utilizando dos alfcuotas diferentes, se
obtuvo la media y para calcular la concentración se aplicó la fórmula propuesta por Hotzel
y Croome (1999):
. {Células) Concentración ml =
Dónde:
N: Número de células contadas
A: Área del campo (mm2)
D: Profundidad del campo (mm)
F: Número de campos contados
Etapa 2
¿N* 1000 mm3
A*D * F
Preparación de medio AB (Agua-Bencimidazol)
* Factor de dilución
Se preparó medio de soporte para colocar los fragmentos foliares, utilizando agar
agua al 1% (p/v de agar microbiológico) y Bencimidazol a 200 ppm, para retrasar la
senescencia
Se procedió de la siguiente manera: se prepararon dos soluciones, el medio de
soporte utilizando agar-agua al 1% y una solución retardadora de la senescencia:
Bencimidazol a 200 ppm. Esta última se diluyó en agua utilizando un agitador magnético
durante 2h y se esterilizó por filtración (membrana Nalgene® con 0.22 1Jm de poro). Las
dos soluciones se mezclaron y la mezcla se vertió en cajas Petri, las cuales se sellaron
empleando una película plástica comercial (Kieen pack®). Como control para evaluar el
39

CAPITULO 11
efecto del Bencimidazol se emplearon cajas Petri conteniendo únicamente medio M
(agar-agua).
Preparación de fragmentos foliares
Para esta etapa se siguió el protocolo de EI-Hadrami et al. , (2000) modificado por
el equipo de la Dra. Rodríguez (comunicación personal). Se emplearon fragmentos de la
lámina foliar de plantas de banano (aproximadamente de 5 a 6 meses de edad) crecidas
en condiciones protegidas, por cada planta se tomaron las dos primeras hojas cortándolas
desde el peciolo y se removió la nervadura principal, posteriormente se lavaron con agua
corriente y se desinfestaron en condiciones estériles en una dilución (0.6% v/v) a partir de
hipoclorito de sodio comercial (6% v/v) durante 15 minutos, agitando de forma circular el
vaso para que la solución de cloro penetrara todos los espacios de la hoja.
Posteriormente se cortaron fragmentos foliares de 25 cm2 de superficie.
Inoculación y muestreo de fragmentos foliares
Los fragmentos foliares se inmovilizaron en medio agar-agua al 1% p/v
conteniendo una solución de Bencimidazol (200 ppm). Se tomó una alícuota de 125 IJL de
la suspensión de esporas previamente ajustada a una concentración de 200 esporas IJL-1
y se depositaron sobre la superficie de cada fragmento foliar; la suspensión de esporas se
esparció sobre la superficie del fragmento empleando un pincel de cerdas de camello
estéril.
Posterior a la inoculación , las cajas Petri conteniendo los fragmentos inoculados se
colocaron en anaqueles en un cuarto de incubación a temperatura controlada y
fotoperiodo 12h/12h. Para cuantificar la severidad causada por el hongo a los fragmentos
foliares, se realizaron muestreos a los O, 5, 15 y 30 días después de inoculación (ddi) ,
fotografiando 3 fragmentos/tiempo de muestreo/cepa. Al igual, en cada muestreo se tomó
una sección del fragmento foliar de aproximadamente 7 x 4 mm de superficie, se tiñó con
azul de anilina y se almacenó a 4°C durante un día para permitir que el tinte se fijara en la
muestra. Posteriormente las secciones teñidas se observaron en microscopio óptico
(Axioplan® Zeiss 20X, programa Action visión 3.1©). Se realizó el mismo procedimiento
40

CAPITULO 11
con los fragmentos control tanto del patosistema M. fijiensis-Musa spp. (Agar + Agua -
Hongo) como del Bencimidazol (medio Agar-Agua).
41

CAPITULO 11
Resultados
Se inocularon fragmentos foliares del cultivar Gran enano con conidios
cosechados de las cepas C1233, CR4b y Oz1 b. Únicamente las cajas conteniendo los
fragmentos foliares correspondientes a O ddi se mantuvieron dentro de la campana de
flujo laminar. El resto de las cajas fueron selladas y almacenadas en un cuarto de cultivo
para su posterior muestreo. Se cortaron secciones de 7x4 mm aproximadamente y se
tiñeron con azul de anilina para observar los fragmentos al microscopio. En la Figura 3 se
observan las fotografías y micrografías de los fragmentos foliares del cultivar Gran enano
inoculados con la suspensión de esporas de tres cepas de M. fijiensis a tiempo O.
Figura 3. Fotografías y micrografías de fragmentos foliares del cu ltivar Gran enano a O ddi
con las cepas C1233 (A y 8), CR4b (C y O) y Oz1b (E y F).
42

CAPITULO 11
A los cinco d ías posterior,es a ll a ,inoculación de los fragmentos foliares se observó
creaimiiento d,e las hitas de llas tr~es cepas sobne ia superficie de los !fragmentos sin que se
pudiier.a o'b.serv.atr penetr-;adón ,en ning1una de las muestras {Fig.ura 4).
Figura 4. Fotografías y micrografías de fragmentos foliares del cultivar Gran enano a 5 ddi con las cepas C1233 (A y B) , CR4b (C y D) y Oz1b (E y F).
A los quince días posteriores a la inoculación únicamente en uno de los
fragmentos inoculados con C 1233 se pudo observar la aparición de pequeñas lesiones
características de la enfermedad (Figura 5).
43

CAPITULO 11
Figura 5. Fotografías y micrografías de fragmentos foliares del cultivar Gran enano a 15
ddi con las cepas C1233 {A y B), CR4b {C y D) y Oz1b {E y F).
A 30 ddi se observaron lesiones sobre la superficie de los fragmentos foliares del
cultivar Gran enano inoculados con las cepas C1233 y CR4b, sin embargo no hubo
presencia de lesiones en los fragmentos inoculados con Oz1 b, a pesar de que en todas
las micrografías se observó la presencia de las hitas del hongo (Figura 6).
44

CAPITULO 11
Figura 6. Fotografías y micrografías de fragmentos foliares del cultivar Gran enano a 30 ddi con las cepas C1233 (A y B), CR4b (C y O) y Oz1b (E y F).
Los resultados de estos experimentos contrastaron con los de Peraza Echeverría
(comunicación personal), quien a 30 ddi observó lesiones en etapa 5 de la escala de Fouré
(1985) en los fragmentos foliares de Gran enano inoculados con la cepa Oz1b.
45

CAPITULO 11
En general, se observaron bajos niveles de severidad en todos los fragmentos
foliares a 30 ddi con las tres cepas de M. fijiensis (C1233, CR4b y Oz1b), de hecho, como
lo muestra la figura 6E, no se observaron lesiones visibles en los fragmentos inoculados
con la cepa Oz1b.
Las posibles causas de que los fragmentos foliares no presentaron los daños
esperados a nivel 5 de la escala de Fouré (1985) pusieron ser:
1) La pérdida de la capacidad infectiva de las cepas, ocasionada por el mantenimiento
continuo de éstas en condiciones in vitro. S un y Heitman (2011) proponen que las cepas
fúngicas mantenidas en laboratorio tienden a perder su capacidad de reproducción
sexual, debido en parte a que los genes de virulencia se encuentran físicamente cerca de
los genes MAT, al perderse los genes MAT se pueden perder también los de viru lencia.
11 ) La inhibición de la infección fúngica ocasionada por contaminación bacteriana. Las
pruebas con medio LB no mostraron turbidez luego de 48 h, pero estas pruebas sólo
sirvieron para verif icar que la cepa no estuviera contaminada al momento de inocularla en
el fragmento. No descarta la posibilidad de alguna bacteria endófita en el material vegetal.
111 ) La germinación debilitada de los conidios, ocasionada por fluctuaciones de la
temperatura (25-21 oc registrados en el termómetro del cuarto) en el cuarto de cultivo que
pudo favorecer la deshidratación del medio, así como la no disponibilidad de los
nutrientes, como lo reportaron Faria et al., (201 O) al estudiar la germinación de Beauveria
bassiana y Metarhizium anisopliae.
IV) La concentración de Bencimidazol (200 mg/L) en el medio de soporte AB para los
fragmentos, pudo inhibir el crecimiento del hongo. Diversos autores reportan haber
utilizado concentraciones menores a 120 mg/L de Bencimidazol como agente para
retrasar la senescencia de las hojas de banano {Twizeyimana et al., 2007; Pérez-Vicente
et al., 2006; Abadie et a/., 2001) y de arroz (Singh y Mishra, 1975).
Reactivación de las cepas fúngicas
Debido a los resultados obtenidos anteriormente se decidió seguir con el postulado
de Koch para reactivar las cepas C1233, Oz1b, CR4b, Jaguar y Cruse C3 inoculándolas
en su hospedero. Posterior a esto se utilizaron fragmentos foliares (1 5 mm2) infectados
46

CAPITULO 11
que mostraron una o dos manchas negras. Siete de estos fragmentos se colocaron sobre
mediio V8 de crec'imiento por caja Petr¡; las cajas se guardaron en un cuarto de cultivo con
fotoper.iodo de 12/12h, durante un período de tres semanas para permitir el crecimiento
de llas cepas en coloniias y post.e niormente se cortaron fragmentos de las colonias en
crecimiento y se transfirieron a medio V8C fresco en donde se cultivaron por un período
de tres semanas hasta alcanzar la !biomasa requerida para cont·inuar con los
·experimentos.
Recuperación de ,cepas de M~cosphaerella :fijiensis .contaminadas
Durante .el proceso de reactivación de los aislados de M. fijiensis después de
haber sido inocu'lados 'en fragmentos de su hospedero natural, los fragmentos infectados
presentaron contaminación sobre el medio V8C (Figura 7). Dicha contam inación (cuyo
.origen fue determinado con la colaboración del M.C. Andrés Quijano) fue causada por
bacterias Gram +; Figura 8) las cuales inhibieron el crecimiento del hongo en el medio de
cultivo .. No se :determinó si 1la contaminación se debió a la manipulación de los aislados
durante la inoculación de los fragmentos o por la presencia de microorgan ismos endófitos
de la !lámina fo:liar del banano, los cuales emergieron de los tej idos vegetales cuando se
realizó un daño mecánico al cortar los fragmentos con el bisturí (Zinniel et al., 2002).
Figura 7. Fragmentos foliares inoculados con M. fíjiensis mostrando contaminación en medio VBC.
A) Fragmentos inoculados con C1233, B) Fragmentos inoculados con Oz1b, C) Fragmentos
inoculados con CR4b
47

CAPITULO 11
Figura 8. Cepa Oz1 b contaminada. A) vista macroscópica de la contaminación creciendo en medio
V8E. B) vista microscópica de la contaminación observada en el medio V8E.
Se cortaron fracciones de aproximadamente 1 cm2 de los fragmentos foliares
inoculados con las cepas de M. fíjiensis y fueron transferidos a matraces conteniendo 20
ml de medio líquido papa-dextrosa (PDB) con Tetraciclina (10 mg ml-1) para obtener
cultivos axénicos. Los matraces se colocaron en agitación continua y después de 48
horas se observó turbidez en todos los matraces los cuales fueron descartados.
Se procedió a repetir los experimentos en matraces utilizando esta vez los
antibióticos Ampicilina, Kanamicina y Cefotaxima empleando un diseño factorial 23, en
donde los tres factores a evaluar fueron los antibióticos y los dos niveles fueron presencia
(+)y ausencia(-) de los mismos, dando un total de 8 tratamientos (Cuadro 4). Después de
48 horas en agitación constante las cepas en los tratamientos en donde no se observaron
turbidez (Ampicilina+Kanamicina+Cefotaxima; Kanamicina+Cefotaxima; Kanamicina) se
depositaron sobre la superficie de medio de crecimiento V8C (Figura 9), en el cual no se
observaron agentes contaminantes, aunque también se inhibió el crecimiento de las
cepas fúngicas luego de 72 horas en estudio (Cuadro 4 ).
48

CAPITULO 11
Cuadr.o 4. Arreglo factorial 23 utilizando tres factores (Ampicilina, Kanamicina y Cefotaxima) a dos
niveles cada uno (presencia o ausencia del antibiótico en el medio). Presencia (+) o ausencia (-)
de contaminación en medio PDB; crecimiento(+) o inhibición(-) de M. fijiensis
Tratamiento Contaminación Crecimiento del hongo
Control + +
Ampicilina.Cefotaxima + +
Ampicilina + +
Ampicilin.Kanamicina .Cefo taxima
Cefotaxima.Kanamicina
Cefotaxima + +
Kanamicina
Ampicil ina.Kanamicina +
Figura 9. M. fijiensis en medio V8C después de 48 horas de agitación continua en medio POS
adicionado con A) Ampicilina+Kanamicina+Cefotaxima, 8) Kanamicina+Cefotaxima, C)
Kanamicina; no se observa presencia de contaminación sobre el medio de cultivo.
Finalmente, utilizando la técnica de punta de hifa se recuperaron las cepas de M.
fijiensis en forma axénica sin recurrir al uso de antibióticos. Chi-Manzanero (comunicación
personal) proporcionó las cepas limpias para continuar los experimentos.
Experimentos utilizando las cepas reactivadas
Teniendo las cepas libres de contaminación y luego de ser reactivadas en su
hospedero natural, se iniciaron los experimentos enlistados en el cuadro 5. Debido a la
49

CAPITUlO 11
baja producción de esporas ocasionada aparentemente por una baja intensidad de luz en
el cuarto de esporulación, se abortó el experimento con la cepa Oz1b no siendo posible
evaluar la interacción entre esta cepa y el cultivar Gran enano.
El desarrollo de la infección se evaluó durante los 30 días que duraron los
experimentos con las cepas C1233 y CR4b en Gran enano.
Cuadro 5. Experimentos utilizando cepas reactivadas de M. fljiensis.
Experimento Fecha Estado Justificación
1: C1233-Gran 19/03/12
enano
2: C1233-Gran 25/03/12
enano
3: CR4b-Gran enano 13/03/12
4: CR4b-Gran enano 20/03/12
5: Oz1b-Gran enano 21/04/12
6: Oz1b-Gran enano 27/04/12
7: C1233-Gran 05/06/12
enano
8: CR4b-Gran enano 06/06/12
Experimento Gran enano-C1233
Concluido
Abortado
Concluido
Abortado
Abortado
Abortado
Concluido
Abortado
Baja producción de conidios
Baja producción de conidios
Baja producción de conidios
Contaminación
Baja producción de conid ios
En contraste con el experimento utilizando la cepa C1233 sin reactivar, los
fragmentos foliares inoculados con la cepa C1233 reactivada (inoculada y aislada de Gran
enano) mostraron mayor número y tamaño de lesiones (Figura 10). Lo cual aporta
evidencias para inferir que la cepa no perdió su agresividad en cierta medida, si bien no
se observó el mismo daño reportado por Peraza-Echeverría (comunicación personal).
so

CAPITULO 11
f 1igura 10 .. Lesiones en fragmentos foliares del cultivar Gran enano causadas por la cepa C1233 a
30 .ddii. A) Cepa sin r-eactivar, 8 ) Cepa reactivada
La observación microscóp.ica del tejido vegetal permitió apreciar el crecimiento
mice!ial del hongo sobre la superficie de los fragmentos foliares (Figura 11 ), sin embargo,
no se observó penetración de los estomas por las hifas en ninguna de las muestras,
aunque esto no significó que ·la penetración no haya ocurrido, debido a que solo se
observó una sección de:l fragmento fo ~li ar .
Figura 11 . Micrografía de fragmentos foliares de cultivar Gran enano a 30 ddi. A) Fragmento
control. B) Fragmento inoculado con C1233 reactivada.
Experimento Gran enano-CR4b
A treinta días posteriores a la inoculación en fragmentos foliares de Gran enano
con la cepa CR4b reactivada, se observaron mayor número de lesiones en comparación
al experimento anterior en donde se empleó la cepa sin haber sido reactivada (Figura 12).
51

CAPITULO 11
Figura 12. Lesiones en fragmentos foliares del cultivar Gran enano causadas por la cepa CR4b a
30 dd i. A} Cepa sin reactivar, B} Cepa reactivada.
Se observó crecimiento epifílico de las hifas sobre el fragmento inoculado con la
cepa CR4b reactivada y penetración en uno de los estomas (Figura 13).
Figura 13. Micrografía de fragmentos foliares del cultivar Gran enano a 30 ddi. A} Fragmento
control. B} Fragmento inoculado con CR4b reactivada. P: penetración.
Se observó contaminación por bacterias Gram+ distribuidas sobre toda la
superficie de los fragmentos tanto control como inoculados con CR4b (Figura 14), podría
tratarse de una bacteria endófita (Lodewyckx et al., 2002) o de un error operativo durante
la manipulación de los fragmentos foliares.
52

CAPITULO 11
Figura 14 . Presencia de contaminación sobre la superficie del fragmento fo liar de Gran
enano A) fragmento control y B) Fragmento 1i noculado con CR4b .
53

CAPITULO 11
Discusión
En los experimentos realizados con las cepas reactivadas (inoculadas en su
hospedero natural y re aisladas a partir de lesiones necróticas de tejido foliar) se observó
un aumento en la agresividad de las cepas (Figuras 1 O y 12) con respecto a los
experimentos anteriores en los cuales se emplearon estas mismas cepas después de
haber sido subcultivadas seis o siete veces en medio V8C. Como mencionan García et
al., (2007), quienes observaron disminución tanto en el número de esporas como en la
agresividad del hongo entomopatógeno Beavuveria bassiana. Los subcultivos pueden
ejercer un efecto indeseable en las características macroscópicas, de germinación,
velocidad de crecimiento y actividad del hongo. Cuevas-Rangel (1979), reportó que los
subcultivos indujeron una baja actividad enzimática del hongo micorrízico Pisolithus
tinctorius por lo cual no tuvo un efecto favorable en Pinus montezumae. Cruz-Martín et al.
(2011) reportaron que tanto la producción de esporas como la agresividad de aislados de
M. fijiensis se redujeron drásticamente después de ocho subcultivos.
En nuestro estudio, los subcultivos de las cepas de M. fijiensis pudieron disminuir
la agresividad del fitopatógeno, agresividad que se evidenció nuevamente al inocular las
cepas en su hospedero natural y reaislarlas.
Sin embargo, contrastando con los resultados de García et al., (2007), Peteira et
al., (2007) no observaron cambios en la agresividad de Pochonia chlamydosporia
después de haber sido conservada in vitro mediante subcultivos, aunque estos autores
mencionan que en algunos casos el subcultivo puede resultar en pérdidas de ciertas
características originales de la cepa, como cambios morfológicos, alteraciones en los
componentes de la pared celular, y pérdida o atenuación de la agresividad. Con respecto
a esto, Hall (1980), mencionó que el subcultivo repetido de Verticillium lecanii en
diferentes medios de cultivo indujo cambios fisiológicos y morfológicos en la cepa, pero no
atenuó la agresividad del hongo.
Otro aspecto importante que se debe considerar en relación al problema de
esporulación de los aislados de M. fijiensis , es que el cuarto de esporulación que se
empleó en este trabajo, tiene problemas para conservar las condiciones de humedad,
temperatura e intensidad luminosa, parámetros que son críticos para inducir la óptima
esporulación de las cepas.
54

CAPITULO 11
En diferentes ocasiones se observó una fluctuación de temperatura registrada en
el termómetro que varió entre 21 a 25 oc, esto puede afectar la cantidad de agua
disponible en el medio de cultivo para el hongo, y por lo mismo estar afectando su
esporulación. Como mencionan Aguirre et al., (2009), se necesitan altos valores de
actividad de agua (cantidad de agua disponible para el microorganismo) en el rango de
95-98% para inducir la esporulación en Nomuraea rileyi (hongo entomopatógeno).
En general, se logró inducir la infección de los fragmentos foliares en los
experimentos en donde se emplearon tanto las cepas subcultivadas como las cepas
reactivadas, sin embargo no se lograron obtener los mismos niveles de daño foliar
observados anteriormente en los experimentos realizados por la M.C. Leticia Peraza.
Debido a esta variación en el daño, los resultados obtenidos en estos experimentos se
usaron únicamente como material para establecer el método de cuantificación de la
enfermedad, empleando los programas Assess 2.0 e lmage J 1.46r, los cuales se abordan
en el siguiente capítulo.
55

CAPITULO 11
Bibliografía
Abadie, C. El Hadrami. Rivas, G. Zapater, M.F. Carlier, J. (2001). Studies of
Mycosphaerella fijiensis Population Structure and Partial Resistance of Bananas.
lnfomusa. 10. 14-15
Aguirre, N. Villamizar, L. Espine! , C. Cotes, A. (2009). Efecto del pH y de la actividad de
agua sobre el desarrollo de Nomuraea rileyi (Hyphomycetes) . Revista colombiana
de entomología. 35(2) , 138-149.
Arraino, L.S. Brading, P.A. Brown, J.K.M. (2001) . A detached seedling leaf technique to
study resistance to Mycosphaerella gramíneo/a (anamorph Septoria tritic1) in wheat.
Plant pathology. 50, 339-346.
Brown, J.K.M. Wolfe, M.S. (1990). Structure and evolution of a population of Erysiphe
graminis f. sp. Hordei. Plant Pathology. 39, 376-390.
Cruz-Martín , M. Alvarado-Capó, Y. Acosta-Suárez, M. Leiva-Mora, M. Sánchez-García, C.
Roque, B. (2011). Effect of the in vitro subculture in conidial production and
aggressiveness of Mycosphaerella fijiensis Morelet. Centro Agrícola. 38 (3) , 11-16.
Cuevas-Rangel , R. (1979). Prueba de inoculación con el hongo micorrízico Pisolithus
tinctorius (Pers.) Coker and Couch, en plántulas de Pinus montezumae Lamb. en
suelos de vivero. Revista Mexicana de Ciencias Forestales. 4(19) , 46-62.
Damajaru, S. (201 O). Analysis of Proteins lnvolved in Chlorophyll Catabolism: The impact
of Chlorophyllase and Water Soluble Chlorophyll Protein on the reduction of
undesirable pigments in crop plants. Tesis de Doctorado. Mathematisch
Naturwissenschaftlichen Fakultat 1 der Humboldt-Universitat zu Berlin. Alemania .
26 p.
El Hadrami , A. Abadie, C. Carlier, J. (2000) . Evaluation de la résistance partielle des
bananiers a Mycosphaerella fijiensis (maladie des raies noires) en conditions
56

CAPITULO 11
contrólées et au champ. Journées Jean Chevaugeon : Rencontres de mycologie
phytopathologie. Aussois (FRA), 2000/03/05-09.
Eyal , Z. Scharen, A L. Huffman, M.O. Prescott, J.M. (1985). Global insights into virulence
frequencies of Mycosphaerella graminico/a. Phytopathology. 75, 1456-1462.
Falkoswka, M. Pietryczuk, A Piotrowska, A Bajguz, A Grygoruk, A Czerpak, R. (201 O).
The Effect of Gibberellic Acid (GA3) on Growth, Metal Biosorption and Metabolism
of the Green Algae Chlorella vulgaris (Chlorophyceae) Beijerinck Exposed to
Cadmium and Lead Stress. Polish Journal Of Environment Studies. 20, 53-59.
Faria M. Hotchkiss J. H. Hajek AE. Wraight S. P. (201 O) . Debilitation in conida of the
entomopathogenic fungi Beauveria bassiana and Metharhizium anisopliae and
impilication with respect to viability determinations and mycopesticide quality
assessments. Journal of invertebrade pathology, 105, 7 4-83.
Farlow, W .G. (1885). Notes on sorne species of Gymnosporangium and Chrysomyxa of
the united states. Arts and Science. 20, 311-323.
Fouré, E. 1985. Black leaf streak disease of Banana and plantains (Mycosphaerella
fijiensis Morelet) , study of the symptoms and stages of the disease in Gabon IRFA
Paris.
Garcfa, M. Villamizar, L. Torres, L. Cotes, A (2007). Efecto de subcultivos sucesivos de
Beauveria bassiana sobre sus caracterfsticas y actividad contra Premnotrypes
vorax. Manejo Integrado de Plagas y Agroecologfa. 77, 50-56.
Hall , R. A (1980). Effect of repeting subculturing on agar and passing through an insect
host on pathogenicity, morphology and growth rate of Vertilillium lecanii. Journal of
lnvertebrate Pathology. 36(2) , 216-222.
Hildbrand, M. Fischer, A Feller, U. (1994). Protein Catabolism in Sean Leaf Discs:
Accumulation of a Soluble Fragment of Ribulose 1,5 bisphosphate/oxygenase
Under Oxygen Deficiency. Journal of Experimental Bothany. 45 (9) , 1197-1204.
57

CAPITULO 11
Hotzel, G. y Croome R. (1999). A phytoplankton methods manual for Australian
freshwaters. Land and Water Resources Research Development Corporation.
Canberra. 58.
Karjalainen, M. (1984). Evaluation of detached seedling leaves for use in spring wheat
cultivars to Septoria nodorum Berk. Acta agricultura Scandinavia. 34, 386-390.
Kema, G.H.J. Van Silfhout, C.H. (1997). Genetic variation for virulence and resistance in
the wheat-Mycosphaerella graminico/a pathosystem. Comparative seedling and
adult plant and adult plant experiments. Phytopathology. 87, 266-272.
Lodewyckx C. Vangronsveld J. Porteous F. R.B. E. Taghavi S. Mezgeay M. van der Lelie
D. (2002). Endophytic Bacteria and Their Potential Applications. Critica! Reviews in
Plant Sciences. 21, 583-606.
Mishra, D. Pradhan, P. (1973). Regulation of senesce in detached rice leaves by light,
Benzimidazole and kinetin. Experimental gerontology. 8, 153-155.
Mourichon X., Peter , D. Zapater, M.F. (1987). lnoculation expérimentale de
Mycosphaerella fijiensis Morelet sur de jeunes plantules de bananiers issues de
culture in vitre. Fruits. 42(4), 195-198.
Peraza-Echeverría, L., Rodríguez-García, C.M. and Zapata-Salazar, D.M. (2008) A rapid ,
effective method for profuse in vitro conidial production of Mycosphaerella fijiensis.
Australas. Plant Pathol. 37, 460-463.
Pérez-vicente, L. Pérez-Miranda, M. Jiménez, M. Jama, M. (2006). Ensayo en Fragmentos
de Hojas de Bananos y Plátanos (Musa Spp.) para Estudio a Nivel Monocíclico de
la Evolución de los Síntomas de la Sigatoka Negra Causada por Mycosphaerel/a
fijiensis. Fitosanidad . 1 O, 3-9.
Peteira, B. Esteves, l. Montes de Oca, N. Hidalgo-Díaz, L. (2007). Estabilidad de la cepa
IMI SD 187 de Pochonia chlamydosporia val. catenulata en medio sólido. Revista
de Protección Vegetal. 22(2), 124-127.
58

CAPITULO 11
Puch-Ceh, M.A. (2001). Evaluación de la actividad fitotóxica en cultivos de
Mycosphaerella fijiensis Morelet mediante el uso de diferentes técnicas de
bioensayo. Centro de Investigación Científica de Yucatán, A.C. Mérida, Yucatán,
México.
Rodríguez-García, C.M. Navarrete-Mapen, R.Z. Peraza-Echeverría, L. (2007).
Characterization of the in vitro conidial morphology of Mycosphaerella fijiensis, a
majar pathogen of banana. Australian Plant Pathology. 36, 369-372 .
Singh , M. Mishra, D. (1975). The Effect of Benzimidazole and Red Light on the
Senescense of Detached Leaves of Oryza sativa cv. Ratna. Plant Phisiology. 34.
67-74.
Sun, S. Heitman, J. 2011 . ls sex necessary? BMC Biology. lncorporating Journal of
Biology 9(56), 1-4.
Twizeyimana, M. Ojiambo, P.S. Tenkouano, A. lkotun, T. Bandyopadhyay, R. (2007).
Rapid Screening of Musa Species for Resistance to Black Leaf Streak Using In
Vitro Plantlets in Tu bes and Detached Leaves. Plant disease. 91, 308-314.
Van Ginkel, M. Scharen, AL. 1988. Host-pathogen relationships of wheat and Septoria
tritici. Phytopathology. 78, 72-766.
Yarwood , C. E. (1946). Detached leaf culture. The botanical review. 7, 1-56.
Zinn iel , D. Lambrecht, P. Harris, N. Feng , Z. Kuczmarski , D. Higley, P. lshimaru , C.
Arunakumari , A. Barletta, R. Vidaver, A. (2002). lsolation and Characterization of
Endophytic Colonizing Bacteria from Agronomic Crops and Prairie Plants. Applied
and Environmental Microbiology. 2198-2208.
59


CAPITULO 111
CAPITULO 111. CUANTIFICACIÓN DEL DAÑO INDUCIDO POR CEPAS DE M.
fijiensis AISLADAS DE FINCAS CON DIFERENTE MANEJO DE FUNGJCJDAS
EN FRAGMENTOS FOLIARES DE Musa spp.
Introducción
En la agricultura actual se emplean cultivos genéticamente uniformes que ofrecen
características agronómicas deseables como alta productividad, elevada densidad de
siembra y fácil manejo cultural ; tal es el caso del cultivo de banano. Sin embargo las
enfermedades fúngicas limitan en gran medida el rendimiento de los cultivos y su
incidencia e importancia se ha incrementado debido a la uniformidad genética de los
cultivos (Baum et al., 2011 ).
La cuantificación del daño inducido durante la enfermedad es de gran importancia
en la evaluación de la interacción entre el patógeno y su hospedero, dado que permite
vislumbrar si el manejo intensivo con fungicidas en las fincas ha incrementado la
agresividad de las cepas de los fitopatógenos. Para cumplir con este propósito, los
métodos utilizados en la cuantificación de la severidad de la enfermedad deben ser
simples, baratos, fáciles , flexibles , y precisos (Wijekoon et al., 2008). Muchas de las
técnicas utilizadas hasta hace 15 años dependieron del criterio visual de un evaluador.
Aunque estas técnicas han sido aplicadas exitosamente en la evaluación del control de la
enfermedad y la identificación de variedades vegetales resistentes, muchos estudios han
demostrado la limitada confiabilidad del ojo humano para la determinación objetiva de los
síntomas (Sherwood et al., 1982).
Cuando los síntomas se muestran en la superficie de la hoja, la severidad de la
enfermedad se puede medir como un porcentaje o proporción de daño foliar en un
diagrama hecho a mano (Filho et al., 1997), empleando una escala semi-cuantitativa
(Siopek, 1989) o pesando réplicas en papel de fotografías de hojas enfermas (Sherwood
et al., 1982). Las escalas de cuantificación, basadas usualmente en la progresión
logarítmica del valor de la media han resultado ser un intento por suplir la falta de
precisión y reproducibilidad en las mediciones.
El uso de diagramas de área estandarizados, usualmente incorporando escalas
logarítmicas, es una forma común de valorar la severidad de las enfermedades en
plantas. Sin embargo, el empleo de tales diagramas está sujeto al operador en cuestión y
61

CAPITULO 111
se requiere un juego diferente de diagramas para cada patosistema. Sherwood et al.
(1982), reportaron que aun usando diagramas estandarizados de área, tanto el número de
lesiones como el área dañada pueden diferir sustancialmente del número real de lesiones
y área dañada.
Estos métodos se han empleado frecuentemente para cuantificar la enfermedad
en hojas infectadas, asumiendo que el daño inducido por el crecimiento del patógeno se
correlaciona con el grado de susceptibilidad de la planta (Baum et al., 2011). Sin
embargo, algunos estudios (Lim y Gaunt, 1981; Sherwood et al. , 1982) han demostrado
que los datos basados en la evaluación visual de los síntomas son imprecisos cuando el
porcentaje de lesión es cercano al 50% y contiene sesgos correlacionados con el número,
área y forma de las lesiones de la enfermedad (Camargo y Smith, 2009).
La limitada precisión en los métodos de evaluación de las enfermedades de
plantas, aún en conjunto con herramientas tales como los diagramas de áreas, ha llevado
al desarrollo de métodos de evaluación basados en programas computacionales. Los
avances en el desarrollo de equipos y programas han hecho que muchos sistemas de
cómputo empleados rutinariamente en la evaluación de enfermedades fúngicas de plantas
estén disponibles para muchos laboratorios actualmente (Baum et al. 2011).
Casi desde su comienzo, las computadoras han sido empleadas en la evaluación
de las enfermedades de plantas, ya que a diferencia de la visión de un evaluador, la
respuesta de una cámara a la luz incidente no se ve afectada por la severidad de las
lesiones y la cuantificación computacional objetiva de las áreas dañadas no es propensa a
los sesgos (e.g. sobreestimación del área dañada) relacionados con el número o forma de
las lesiones (Dammer et al., 2011 ).
Muchos programas comerciales de análisis de imágenes (HyphArea, Support
Vector Machine, Neo lmage Analysis Software, LAMINA, GROWSCREEN, PHENOPSIS,
PHENODYN, lmage J , Assess, etc}, usualmente respaldados por un software robusto,
están disponibles y son utilizados intensivamente en la evaluación de las enfermedades
de plantas (Kolukisaoglu y Thurow, 201 0).
Uno de los programas de análisis de imágenes más versátiles y sencillos de
manejar es el Assess 2.0, empleado previamente en el estudio del patosistema M.
fijiensis-Musa spp. (Donzelli y Churchill 2007, 2009}, desarrollado por el Dr. Lakhdar
62

CAPITULO 111
Lamari en 2002. Asses 2.0 permite la cuantificación rápida y sencilla del número de
lesiones así como el porcentaje de área dañada sobre la hoja o cualquier otra parte de la
planta, además se puede utilizar como una herramienta interactiva al adaptarse a
cualquier tipo de imagen ya sea que provenga de un escáner, un microscopio o una
cámara digital. Lo anterior ha facilitado a los fitopatólogos la evaluación rápida y precisa
del progreso de la enfermedad, gracias a la interface intuitiva basada en Windows®,
permitiendo que la investigación de la fitopatología alcance un nuevo nivel.
El programa es independiente del equipo y funciona en una computadora con
sistema operativo de 32 bits (capacidad del procesador para trabajar con imágenes de 32
bits de ancho, o de "color verdadero" cuando se habla de la profundidad de color, es decir,
la cantidad de bits necesarios para representar el color de un píxel en una imagen digital).
Características del programa Assess 2.0
• Cuantifica automáticamente diferentes lesiones en enfermedades de
plantas: independiente del usuario, el resultado será siempre el mismo.
• Reconoce automáticamente y mide el porcentaje de enfermedad de hojas
individuales en la misma imagen: esta característica permite ahorrar
tiempo. Se pueden escanear muchas hojas en la misma pantalla y obtener
una medición de la enfermedad para cada hoja con un solo movimiento del
"ratón".
• No requiere calibración cuando las imágenes se escanean en el Assess
2.0. Si se conoce la resolución de la imagen (dpi) , el usuario puede
simplemente introducir los valores en Assess 2.0. entonces se pueden
convertir las unidades físicas de medida en el programa, se puede optar
por cualquiera de las 1 O unidades sin tener que calibrar el programa.
• Soporta todos los formatos estándar de archivos y muchos de los no
estándar de lectura y escritura: Assess 2.0 puede manejar más de 60 tipos
de formatos, incluyendo varios formatos RAW (tipo de formato que contiene
la totalidad de los datos de la imagen tal y como ha sido captada por el
sensor digital de la cámara fotográfica) para cámaras digitales. No hay
necesidad de convertir las imágenes empleando otros programas.
• Cuenta con interfaz para escáner.
63

CAPITULO 111
Se debe considerar que aunque Assess 2.0 posee una gran versatilidad , continúa
siendo un programa de uso privado, solamente se puede acceder a esta herramienta
mediante el pago de licencias (costo en el año 2011: $11 890.79 pesos mexicanos, de
acuerdo al precio de compra en nuestro grupo de trabajo para cinco licencias). Aunque
este programa es bueno y utilizado en estudios agrícolas para medir área de daño foliar,
evidentemente esto limita su uso a únicamente grupos de trabajo o laboratorios que
cuentan con la capacidad económica para adquirirlo. Afortunadamente, existen otras
herramientas informáticas de libre acceso que permiten la manipulación y análisis de
imágenes digitales, tales como el programa lmage J 1.46r (Abramoff et al. 2004).
Características del programa lmage J 1.46r
Este programa es la creación de Wayne Rasband, desarrollador de programas de
procesamiento de imágenes en el National lnstitutes of Health (NIH por sus siglas en
inglés), quien también desarrolló el programa Scion lmage el cual fue precursor del lmage
J.
lmage J (versión 1.46r) está programado en Java (lenguaje de programación de
alto nivel orientado a objetos). Fue diseñado con una arquitectura abierta que proporciona
extensibilidad mediante complementos de Java y macros (macroinstrucciones), lo cual
hace posible resolver problemas de procesado y análisis de imágenes, desde imágenes
en vivo de las células en tres dimensiones, procesado de
imágenes radiológicas, comparaciones de múltiples datos de sistema de imagen, hasta
sistemas automáticos de hematología.
lmage J 1.46r puede ejecutarse en Microsoft Windows®, Mac OS®, Mac OS
X®, Linux®, y Sharp Zaurus PDA®, permite mostrar, editar, analizar, procesar, guardar, e
imprimir imágenes de 8, 16 y 32 bits. Puede leer varios formatos de imagen incluyendo
TIFF, PNG, GIF, JPEG, BMP, DICOM, FITS, así como formatos RAW (formato de Imagen
sin modificaciones).
Es compatible con las funciones estándar de procesamiento de imágenes tales
como operaciones lógicas y aritméticas entre imágenes, manipulación de contraste,
nitidez, suavizado, detección de bordes, etc. El programa es compatible con cualquier
número de imágenes al mismo tiempo, limitado solamente por la memoria disponible
(Abramoff et al., 2004)
64

CAPITULO 111
El algoritmo con el cual operan los programas Assess 2.0 e lmage J 1.46r, es un
algoritmo Watershed, o de línea divisoria. Es una técnica de segmentación basada en
morfología, que permite encontrar regiones de una imagen; es una técnica sencilla y
eficiente, ampliamente utilizada para el análisis de manchas (La Serna y Pró, 201 0). La
transformación empleando el algoritmo de línea divisoria es una poderosa herramienta
para la segmentación de imágenes. Sin embargo, la efectividad de los métodos de
segmentación de imágenes basados en transformaciones de línea divisoria está limitados
por la calidad del gradiente de la imagen (Wang, 1998).
Muchos modelos de color han sido desarrollados para la estandarización de los
sistemas de medición. Un modelo de color es la determinación de un sistema coordinado
de tres dimensiones y es un sub-espacio de este sistema donde cada color puede ser
representado por un punto determinado. Los modelos de color más utilizados para
equipos son los RGB (Red , Green, Blue), los cuales se emplean en pantallas y video
cámaras (lsik etal., 2010).
Como primer paso, la imagen se captura en el programa y el usuario determina los
valores de referencia. Este valor constituye la unidad de medición de la imagen el cual
puede estar dado en micras, mil ímetros, centímetros, metros o kilómetros. En el segundo
paso, el color de la región de referencia es determinado por el usuario. En el tercer paso,
se escoge el algoritmo y se calcula el área; el usuario puede evaluar los resultados
(Figura 15).
65

Capturar imagen
Determinar el área de interés
Determinar umbral de color
Cuantificación de las regiones seleccionadas
Análisis y reporte de resultados
CAPITULO 111
Figura 15. Diagrama general de flujo de la cuantificación de la enfermedad empleando los programas Assess 2.0 e lmageJ 1.46r.
El algoritmo para cuantificar el área dañada determina el número de regiones
similares en la imagen. Las regiones por debajo del umbral seleccionado por el usuario
son ignoradas.
La segmentación de la imagen consiste en asignar etiquetas lógicas a cada pixel
de la imagen. A la discriminación (es decir, distinción) de una entidad del resto de la
imagen, típicamente el fondo, se le llama segmentación binaria (Baum et al., 2011 ). La
segmentación se puede realizar ya sea por enfoques no supervisados, basados por
ejemplo en la categorización de similitudes/diferencias de la textura de la imagen y la
detección del borde, o por métodos supervisados, los cuales consisten en separar la
imagen en términos de las texturas, cuyas funciones de distribución fueron
completamente especificadas (Camargo y Smith, 2009).
En general, la segmentación separa la lesión causada por el patógeno del tejido
vegetal en una imagen, para extraer solamente las características de la lesión en sí. Por
lo mismo se necesita de una representación matemática para separar ambas imágenes.
66

CAPITULO 111
Estas imágenes binarias son la base para un análisis posterior, tales como calcular el
área dañada o el número de lesiones. Estos métodos son adecuados para imágenes con
alta resolución y con amplia información del espacio de color, es decir, del sistema de
color en el que los colores son inequívocos, esto es, donde las interpretaciones de los
colores en el espacio están definidos colorimétricamente sin referencias a factores
externos (Tastl y Purgathofer, 1994). En el caso de imágenes que contienen alta
información de color, se puede aplicar un procesamiento adicional para mejorar el
contraste entre la lesión y el fondo antes de realizar la cuantificación (Baum et al. , 2011).
Dadas las características de las imágenes, debe haber un juego de rasgos más
informativos sobre el dominio de la imagen. Los rasgos relacionados con la textura
pueden ser utilizados como factores para distinguir cuando una imagen no sigue un
patrón de forma o color definido. Una vez que la imagen es capturada digitalmente,
pueden utilizarse un conjunto de algoritmos de procesamiento de imagen para extraer
rasgos de la imagen; la utilidad de cada uno de estos dependerá de los patrones
particulares a ser resaltados en la imagen. Los patrones son rasgos particulares de una
imagen y deberían ser invariables con el traslado de imagen, su rotación y cambio de la
escala. La ventaja en la clasificación de las imágenes por evaluación de rasgos es que los
patrones permanecen idénticos si las condiciones preliminares son cambiadas (Camargo
y Smith, 2009).
Una práctica común para solucionar el problema de tener múltiples rasgos, es
iniciar con tantos de ellos como sea posible y luego implementar un filtro para descartar
aquellos que aportan poca o ninguna información al sistema (McNitt-Gray et al., 1995).
Otro enfoque consiste en usar un pequeño número de rasgos que son los más
representativos de la imagen, y confiar en este criterio de clasificación (Sena et al., 2003;
Guo et al., 2001; Tang et al., 2000; King et al., 2000) . Para el caso específico del
patosistema M. fijiensis-Musa spp., se empleó la escala de Fouré (1985) para seleccionar
las lesiones que a criterio del evaluador, con base en el color y la forma, fueron
ocasionadas por el ascomiceto durante la interacción. El éxito de cada sistema de
reconocimiento de patrones depende no solo del clasificador, sino también de la calidad
de la información de cada uno de los patrones.
67

CAPITULO 111
Materiales y métodos
Para la estandarización del método de cuantificación de la severidad foliar, causada
por la Sigatoka negra, se utilizaron las imágenes digitales obtenidas en ·los experimentos
realizados por la M.C. Leticia Peraza Echeverría (cuadro 6); en los cuales se cuantificó el
área foliar dañada y número de lesiones.
Cuadro 6. Nomenclatura asignada a los experimentos realizados por la M.C. Leticia Peraza
Echeverría
Nomenclatura Aislados Cultivar
Exp1F-2011 C1233, CR4b y Oz1b Gran enano
Exp2F-2011 C1233, CR4b y Oz1b Calcuta IV
Exp3F-2011 C1233 y Oz1b Fougamou
Exp4F-2011 C1233 y Jaguar C3 Gran enano
ExpSF-2011 C1233 y Jaguar C3 Fougamou
ExpGF-2011 C1233 y Jaguar C3 Calcuta IV
Para realizar los experimentos amiba mencionados, se caracterizó la producción
de esporas de las cepas C1233, CR4b, Oz1b y Jaguar C3, empleando el conteo de
conidios en cámara Sedgwick-Rafter tal como se describió en el capítulo anterior. La
producción de esporas se evaluó en la primera y segunda cosecha. El número de esporas
producidas en cada cosecha se muestra en el Cuadro 7.
Debido a que no se lograron mantener constantes las condiciones operativas del
cuarto de esporulación, no fue posible realizar el experimento de inoculación del cultivar
tolerante Fougamou con la cepa CR4b.
68

CAPITULO 111
Material vegetal
P,ara rea lizar los experimentos anteriormente enlistados se emplearon plantas de
Musa spp. cv. Gran enano (genotipo susceptible), Calcuta IV (genotipo resistente) y
Fougamou (genotipo tolerante), provenientes de cultivo in vitro (material propagado por la
Q.B.B. Rosa Grijalva Arango) y mantenidas en condiciones de invernadero hasta 6
semanas al momento de ser utilizadas.
Captura y procesamiento de las imágenes digitales
Se tomaron fotografías dig itales de los fragmentos fol iares inoculados empleando
una cámara digital SONY ® Cyber-shot full HD 1080. Los momentos registrados
fotográficamente fueron los tiempos de O, 5, 15 y 30 días después de la inoculación con
cada una de las cepas (Figura 16). Las imágenes digitales se guardaron con formato JPG
y se cargaron en los programas Assess 2.0 e lmage J 1.46r para cuantificar el porcentaje
de área dañada, así como el número de lesiones por fragmento, en cada tiempo de
muestreo. En ambos programas se ajustaron los umbrales de color con el sistema RGB
(Red, Green, Black) para determinar tanto el fondo (hoja) de la imagen, como el color de
las lesiones que debería detectar el programa. Se seleccionó el algoritmo correspondiente
para cada operación y se procedió a la cuantificación de las variables área afectada y
número de lesiones.
Figura 16. Fotografía digital de un fragmento foliar del cultivar Gran enano a 30 días después de ser inoculado con la cepa CR4b
69

CAPITULO 111
Formación de imágenes binarias
Para formar imágenes binarias, es decir, imágenes que solo contienen dos
elementos (fondo verde y lesión) se empleó el programa de diseño gráfico Photoshop
Cs3®. Las imágenes se procesaron de la siguiente manera:
1) Se cargó el archivo de imagen y se abrió en el programa Photoshop, 2) Se eliminó el
resto de la imagen con la herramienta de corte y únicamente se trabajó con el fragmento
foliar, ajustando a un tamaño de 25 cm2 para uniformizar todos los fragmentos, 3) Se
mejoró la calidad de la fotografía aplicando filtros de enfoque, aumentando el bri llo y el
contraste, según lo requiriera cada imagen, 4) Se amplificó la imagen hasta 300% de su
tamaño original y se dividió en cuadrantes, 5) Se dibujaron las lesiones de cada cuadrante
empleando la herramienta "pincel" con un color café que simula el color marrón de las
lesiones causadas por el hongo, 6) el fondo de la imagen se re llenó de color verde
empleando la herramienta "bote de pintura", 7) f inalmente todas las capas de la imagen se
acoplaron para formar una sola y se procedió de la misma forma con todas las fotografías
digitales de los fragmentos.
El tiempo requerido para formar cada imagen binaria comprendió entre 3 y 4
horas, para fragmentos con 500 lesiones; en ocasiones el tiempo fue mayor cuando se
trató de fragmentos con más de 1000 lesiones. De esta manera se formó un sistema
binario con contornos de color uniforme y definido; cada imagen binaria pudo ser
reconocida y cuantificada tanto con el programa Assess 2.0 como por ellmage J 1.46r.
Cuantificación de la severidad de la enfermedad
Las imágenes binarias se cargaron en los programas Assess 2.0 e lmage J 1.46r,
en ambos programas se seleccionó la unidad de medida en centímetros y el espectro de
color se ajustó empleando el sistema RGB. Los rangos de color tanto de las lesiones
como del fondo se ajustaron para que fueran los mismos en cada programa y las
imágenes se pudieran cuantificar en condiciones homogéneas. En la Figura 17 se detallan
los procedimientos que se siguieron para ajustar los parámetros de medida en ambos
programas.
70

lmage J 1.46r
Menú: Analyse
Set Scale
Distance in pixles: 118.2 Known distance: 1.0
Pixel aspect ratio: 1.0 Unit of length: cm
Menú: lmage
Adjust: Treshold color
Color space: RGB
Red: 31-110 Green: 0-199
CAPITULO 111
Assess 2.0
Manual panel
Calibrate: cm
Video and digital cameras: • Real size of object
• Area: 25.00 • Horizontal: 5.00
• Vertical: 5.00
Calibrate: si
Apply
OK
Lession: 31-1 10 Leaf: 0-199
Figura 17. Procedimientos de ajuste de las escalas de medida y espacios de color en los
programas Assess 2.0 e lmage J 1.46r
Análisis estadístico
El conjunto de datos se analizó apl icando una prueba de varianzas para
determinar si los datos seguían una distribución normal; en los casos en donde los datos
no mostraron diferencia significativa en sus varianzas, se procedió a realizar un anál isis
de varianza (ANOVA) y se graficaron las medias con sus respectivas desviaciones
estándar. En los casos en donde los datos siguieron una distribución no paramétrica, se
aplicó la prueba de Kruskai-Wallis, la cual es adecuada cuando las desviaciones estándar
de los diferentes grupos no son iguales entre sí y se realizaron gráficos de caja y bigotes.
71

CAPITULO 111
Resultados
Cuantificación de esporas producidas por aislados de M. fijiensis.
El número de esporas producidas por las cepas del hongo fitopatógeno en la
primera y segunda cosecha (6 y 12 ddi. respectivamente) se muestra en el cuadro 7.
Cuadro 7. Recuento de esporas producidas por cepa de M. fijiensis.
A' 1
d No. de esporas.ml 1 No. de esporas.mLI IS a 0 lra . cosecha 2ra. cosecha
C1233 105, 550 838, 000
CR4b 819, 200 587, 200
Ozlb 333, 400 229, 200
Jaguar C3 152, 800 38, 100
Los fragmentos infectados presentaron superficies cloróticas, las cua les no fueron
ocasionadas por el ataque del hongo sino por la manipulación durante el proceso de
preparación de los fragmentos. Debido a esto, ambos programas sobrestimaron tanto el
número de lesiones como el área dañada, representando en varias ocasiones un grupo de
lesiones necróticas dentro de un halo clorótico como una sola lesión . (Figura 18). Los
programas no pudieron discriminar entre las lesiones causadas por M. fijiensis y las
lesiones mecánicas debido a la manipulación del tejido vegetal durante el corte ya que
ambos tipos de lesiones presentaron la misma coloración, por lo que fue necesario
realizar una calca digital de las lesiones sobre la fotografía del fragmento (Figura 19),
incluyendo únicamente las lesiones inducidas por el hongo, a criterio del evaluador y
basándose en la escala de Fouré (1985).
72

CAPITULO 111
Figura 18. Sobreestimación del área foliar dañada por M. fijiensis a 30 ddi utilizando el programa Assess 2.0
Figura 19. Imagen digital binaria y cuantificación del daño inducido por M. fijiensis en un fragmento foliar del cultivar Gran enano a 30 ddi
73

CAPITULO 111
Cuantificación del área dañada y número de lesiones en fragmentos foliares
de Musa spp.
Daño inducido por la cepa C1233
El aislado control (cepa C1233) indujo mayor daño en la superficie foliar del
cultivar susceptible a partir del día 15 posterior a la inoculación, en tanto que en el cultivar
Fougamou indujo en su superficie foliar el daño equivalente, pero en el doble de tiempo, a
los 30 ddi, lo cual muestra la capacidad del cultivar para tolerar al patógeno (Figura 20).
Además, en la misma figura se aprecia que la cepa no logró inducir una severidad
significativa en el cultivar resistente (Calcuta IV), a pesar del tiempo de interacción con el
tej ido vegetal, cuya área afectada siempre fue menor a la de los otros dos cultivares; esto
pone de manifiesto la resistencia del cultivar Calcuta IV.
A pesar de las diferencias entre los programas (Assess 2.0 e lmage J 1.46r), la
superficie dañada de los cultivares fue muy similar tal como se puede apreciar en la
Figura 20.
8 A a
6
ro 4 -o b ro b IC ro 2 -o e e e e e .... e ro- o - -= ·-N
:2 E TSG TSF TSC TlSG TlSF T15C T30G T30F T30C u
(!)- e B "ü ;.;::; .... (!) e a o. ::1
Vl
4
2 b b e e e e e e
o --TSG TSF TSC T15G TlSF TlSe T30G T30F T30C
Tratamientos
Figura 20. Área foliar afectada por la cepa C1233 en fragmentos foliares de los cultivares Gran
enano {G), Fougamou (F) y Calcuta IV (C) a 5, 15 y 30 ddi, cuantificados con !os programas
Assess 2.0 (A) e lmage J 1.46r (B). Los datos se analizaron empleando la prueba de Kruskai-Wallis
a un nivel de P~0.05 .
74



CAPITULO 111
En cuanto .al número de .lesiones inducidas por la cepa control, el mayor número
se observó en el cu:ltivar susceptible (Gran enano) a partir del día 15 y no varió incluso
hasta e.l final del experimento (día 30), Jo cual no significa que la severidad haya
permanecido igua·l, ya .que aunque el número de lesiones no aumentó, se fusionaron entre
s í, causando mayor superficie afectada. En ambos programas se estimó un
comportamiento similar del número de lesiones con respecto al tiempo, sin embargo, el
programa 'lmage J 1.46r cuantificó mayor número de lesiones en los fragmentos foliares
d.e ~ os tres ·genot1ipos (Figura 21 ). En los cu ltivares Fougamou (tolerante) y Calcuta IV
(resistente) el número de lesiones cuant ificadas durante todo el experimento fue
consid:erab.lemente menor a las observadas en el cultivar susceptible . En la Figura 22 se
observa cualitativamente el daño foliar inducido por C1233 en los tres cult ivares.
540 A a
I a 340 I
tn (1) b e 140 b b b b o I b b I I I tn
I I I (1)
<lJ -60 '"O
o TSG TSF TSC TlSG TlSF TlSC T30G T30F T30C ~ 1200 B (1)
E a a -:::;:¡
I I z 700
b
I b b b b b b
I I I I 200
I I -300
TSG TSF TSC TlSG TlSF TlSC T30G T30F T30C
Tratamiento
Figura 21 . Número de lesiones inducidas por la cepa C1233 en fragmentos foliares de los cultivares Gran enano (G), Fougamou (F) y Calcuta IV (C) a 5, 15 y 30 ddi, cuantificadas con los programas Assess 2.0 (A) e lmage J 1.46r (8) . Los datos se analizaron empleando la prueba de la diferencia mínima significativa de Fisher (LSD por sus siglas en inglés) a un nivel de p::; 0.05.
75

CAPITULO 111
Figura 22. Daño inducido por la cepa C1233 en fragmentos foliares de los cultivares A) Gran enano, B) Fougamou y C) Calcula IV a 30 ddi.
Daño inducido por la cepa CR4b
Se cuantificó mayor superficie dañada por la cepa CR4b en los fragmentos foliares
del cultivar susceptible a partir de 15 ddi y se incrementó hasta el final del experimento.
Como era de esperarse, los fragmentos foliares del cultivar resistente fueron poco
afectados y estadísticamente el área dañada no se incrementó a medida que transcurrió
el tiempo de infección, siendo el área dañada muy inferior con respecto al área dañada de
los fragmentos foliares del cultivar susceptible. Debido a que no fue posible realizar el
experimento para evaluar a la cepa CR4b en el cultivar tolerante Fougamou, este cultivar
no se incluye en la Figura 23.
76

CAPITULO 111
8 A a
6
ro 4 -o ro
lC ro
2 -o lo....
ro - e N
b e e e
~ E o u
Q) TSG TSC TlSG TlSC T30G T30C u ~ 8 r- B lo.... Q) c.. a ::::::J
V) 6 1- T
4 r-
2 1-j_
b e e e e
o 1-
TSG TSC TlSG T15C T30G T30C
Tratamientos
Figura 23. Área foliar afectada por la cepa CR4b en fragmentos foliares de los cu ltivares Gran
enano (G) y Calcuta IV (C) a 5, 15 y 30 ddi, cuantificada con los programas Assess 2.0 (A) e lmage
J 1.46r (8). Los datos se anal izaron empleando la prueba de Kruskai-Wallis a un nivel de Ps 0.05.
Tanto con el programa Assess 2.0 como con el lmage J 1.46r, se pudo evidenciar
que la cepa CR4b, la cual proviene de manejo intensivo en campo, indujo mayor número
de lesiones en el cultivar susceptible Gran enano a partir del día 15 y que las lesiones
aumentaron hasta el día 30, sin embargo, es notoriamente mayor el número de lesiones
(5 veces aproximadamente) detectado por lmage J 1.46r con respecto al número
detectado por Assess 2.0 (Figura 24).
77

CAPITULO 111
380 A
a
I b
180 I V)
(]) e: e o
I ed
V) d ed I (])
I I (]) -20 -e o TSG TSC TlSG TlSC T30G T30C l.... 1500 B a (])
E á ':::J z 1000
500
b e e e e
o TSG TSC TlSG TlSC T30G T30C
Tratamiento
Figura 24. Número de lesiones inducidas por la cepa CR4b en fragmentos foliares de los cultivares Gran enano (G) y Calcuta IV (C) a 5, 15 y 30 ddi, cuantificadas con tos programas Assess 2.0 (A) e tmage J 1.46r (B). Los datos se analizaron empleando la prueba de la diferencia mínima significativa de Fisher (LSD por sus siglas en inglés) y la prueba de Kruskai-Watlís a un nivel de PS 0.05.
En la Figura 25 se puede observar cualitativamente que la cepa CR4b indujo
mayor número de lesiones así como un mayor daño en la superficie foliar del cultivar
susceptible Gran enano que en el cultivar resistente Calcuta IV a los 30 ddi.
78

CAPITULO 111
Figura 25. Daño inducido por la cepa CR4b en fragmentos fol iares de los cu ltivares A) Gran enano y B) Ca lcuta IV a 30 dd i.
Daño inducido ¡por la cepa Oz1b
El área foliar con mayor daño inducido por la cepa Oz1b (manejo sin fungicidas) se
observó en Gran enano en comparación con los demás cultivares; asimismo se observó
que :la cepa Oz1 b fue la que ocasionó mayor daño fol iar (aprox. 2 veces más que las
cepas C1233 y CR4b) que las cepas C1233 y CR4b. La cuantificación con el programa
Assess 2.0 (Figura 26A) mostró que a partir del día 15 la cepa indujo mayor daño sobre la
superficie de los fragmentos con respecto a los demás tratamientos, sin embargo cuando
se cuantificó el área dañada con el programa lmage J 1.46 r (Figura 26B), el daño fue el
mismo en todos los cu ltivares y tiempos, a excepción de Gran enano a 30 días, en donde
la superficie dañada superó a todos los demás tratamientos; esto sugiere que el programa
Assess 2.0 tiene mayor sensibilidad que el programa lmage J 1.46r, con lo que consigue
detectar diferencias más suti les en la superficie dañada del material vegetal.
79

CAPITULO 111
a 11 A I
7
ro
"'' ro zc ro 3 b "'' I .... e e e e e e e ro-
I I I I I I · - N I .E E
u ·1
(1) - TSG · TSF TSC TlSG TlSF TlSC T30G T30F T30C ·o '+= a .... 9 B (1)
I o. ::J
V') 6
3
b
I b
I b
I b
I b
I b
I b
I b
I o
-3 L----------------------------------------------------------TSG TSF TSC T15G TlSF TlSC T30G T30F T30C
Tratamientos
Figura 26. Área foliar afectada por la cepa Oz1 b en fragmentos foliares de los cultivares Gran
enano (G), Fougamou (F) y Calcuta IV (C) a 5, 15 y 30 ddi, cuantificada con los programas Assess
2.0 (A) e lmageJ 1.46r (8 ). Los datos se analizaron empleando la prueba de la diferencia mínima
significativa de Fisher (LSD por sus siglas en inglés) a un nivel de P!> 0.05.
Como se observa en la Figura 27, el cultivar susceptible fue el que mostró mayor
número de lesiones a los 15 días de haber iniciado el experimento. A los 30 días, que es
cuando se esperaría un incremento en el número de lesiones, se observó lo contrario,
debido a que todas las lesiones aumentaron de tamaño y se fusionaron, pareciendo que
el número de lesiones disminuía; además, a 30 ddi tanto el cultivar susceptible como el
cultivar resistente (Calcuta IV) mostraron el mismo número de lesiones, aunque en
Calcuta IV las lesiones permanecieron sin cambio durante el tiempo de muestreo (30
días). Cabe mencionar que si bien el número de lesiones es una variable importante a
considerar cuando se evalúa el progreso de la enfermedad, esta variab~e no refleja la
severidad de la enfermedad, como lo hace el área dañada. Resulta interesante mencionar
que el cultivar Fougamou mantuvo el mismo número de lesiones y área foliar dañada
durante el tiempo de evaluación (5, 15 y 30 ddi) (Figuras 26 y 27).
80

CAPITULO 111
350 A a
I 150
b b V) I I be (]) e I e e e e e o I I I I I V) (])
-50 (])
TSG TSF TSC TlSG TlSF T15C T30G T30F T30C '"O o
530 l... B (])
E · ::J z a
I b 230 b
I I be e e e I e
I e
I I I I -70
TSG TSF TSC TlSG T15F TlSC T30G T30F T30C
Tratamiento
F1igura 27. Número de lesiones inducidas por la cepa Oz1 b en fragmentos foliares de los cultivares Gran enano (G), Fougamou (F) y Calcuta IV (C) a 5, 15 y 30 ddi, cuantificadas con los programas Assess 2.0 (A) e lmage J 1.46r (8). Los datos se analizaron empleando la prueba de la diferencia mínima significativa de Fisher (LSD por sus siglas en inglés) a un nivel de Ps 0.05.
En la Figura 28 se puede apreciar el daño que Oz1 b indujo en los tres cultivares;
evidentemente el cultivar susceptible fue el que mostró mayor grado de daño.
Figura 28. Daño inducido por la cepa Oz1 b en fragmentos foliares de los cultivares A) Gran enano, 8 ) Fougamou y C) Calcuta IV a 30 ddi.
81

CAPITUlO 111
Daño inducido por la cepa Jaguar C3
De las cuatro cepas evaluadas, Jaguar C3 (manejo intensivo) indujo menor daño
en los fragmentos fol iares en comparación con las otras cepas (C1233, CR4b y Oz1b), ya
que el área foliar dañada no superó los 0.8 cm2 de superficie (Figura 29); Los fragmentos
foliares de los cultivares Gran enano (susceptible) y Fougamou (tolerante) presentaron
mayor daño a 30 ddi , mientras que Calcuta IV presentó la misma área dañada que
Fougamou. Esto indica que para llegar a determinar de manera absoluta cuán agresiva
es una cepa, es necesario evaluarla sobre varios genotipos, utilizando la mayor cantidad
posible de réplicas y repeticiones.
12 A a
0.8
ro 0.4 -e ro
lC ro o -e .... ro -= N -0.4 .E E
u (]) -
b I ab b b I b
1 b b
I I I I I TSF TSC TlSG TlSF TlSC T30G T30F T30C
·o 12 B ~ .... (]) a.
0.8 :J V')
a 0.4
o
b I ab b b b b I b
I I I I I I -0.4
TSF TSC TlSG TlSF TlSC T30G T30F T30C
Tratamientos
Figura 29. Área foliar afectada por la cepa Jaguar C3 en fragmentos foliare.s de los cultivares Fougamou (F), Calcuta IV (C) y Gran enano (G) a 5, 15 y 30 ddi, cuantificada con los programas Assess 2.0 (A) e lmage J 1.46r (8 ). Los datos se analizaron empleando la prueba de la diferencia mínima significativa de Fisher (LSD por sus siglas en inglés) a un nivel de p:s; 0.05.
En general con el programa lmage J 1.46r se cuantificó mayor número de lesiones
con respecto al programa Assess 2.0 (Figura 30). Una desventaja del programa Assess
es su falta de sensibilidad para detectar y cuantificar lesiones pequeñas (Wijekoon et al.,
82

CAPITULO 111
2008) aunque sí logró detectar diferencias significativas cuando se cuantificó el área
dañada por la cepa Oz1 b, como se mencionó en los párrafos anteriores. Además, como
se observa en la Figura 30, con el programa lmage J 1.46r no se detectó diferencia
significativa en el número de lesiones, independientemente del tiempo que duró la
interacción del patógeno con el cultivar evaluado. En la Figura 31 se observa que la
variable que cuantifica mejor el daño es el área foliar dañada y no el número de lesiones.
58 A a a
ab I I ab
I ab
I ab ab
I b
I 18
1 V) Q) e o V) Q)
·22 Q) TSF TSC T15G T15F T15C T30G T30F T30C -o o lo..
130 Q)
E a •::J z a
I I a
70
I a
a a I I a
I I 10
-50
TSF TSC T15G TlSF T15C T30G T30F T30C
Tratamiento
Figura 30. Número de lesiones inducidas por la cepa Jaguar C3 en fragmentos foliares de los Fougamou (F), Calcuta IV (C) y Gran enano (G) a 5, 15 y 30 ddi , cuantificadas con los programas Assess 2.0 (A) e lmage J 1.46r (B). Los datos se analizaron empleando la prueba de la diferencia mínima significativa de Fisher (LSD por sus siglas en inglés) a un nivel de P~ 0.05.
83

CAPITULO 111
Figura 31. Daño inducido por la cepa Jaguar en fragmentos foliares de los cultivares A) Gran enano, B) Fougamou y C) Calcuta IV a 30 ddi.
Severidad foliar inducida por cepas de M. fijiensis en el cultivar Gran enano
Como se observa en la Figura 32, al comparar el área foliar afectada por las cuatro
cepas en los tres tiempos de muestreo, la cepa Oz1b (finca sin manejo de fungicidas) fue
la que indujo mayor severidad en Gran enano a 30 ddi. En este mismo tiempo, las cepas
C1233 (control, finca sin manejo de fungicidas) y CR4b (finca con manejo de fungicidas)
mostraron haber inducido igual severidad en el área foliar de Gran enano; la severidad fue
mayor que en cualquier otro tiempo de muestreo. Independientemente del programa
empleado, el comportamiento de las cepas fue el mismo, sin embargo, el programa
Assess 2.0, detectó mayor área foliar afectada que el programa lmage J 1.46.
84

CAPITULO 111
1.2 A a
8 b b
ro
"' 4 rtl !C e e rtl
"' e
.... d d d d c::=:J rtl - =--·-N o = o E '+- u TS(l) T5(2) T5(3) TlS(l) T15(2) T15(3) T15(4) T30(1) T30(2) T30(3) T30(4) .Q)-
·o 10 .8 B ¡_¡::: ,_ a Q)
I 0.. :::::¡ 6.8 V) b b
I I 2.8
1 e
e e e e
I e e
I I I I I I -1 .2
TS(l) T5(2) T5(3) T15(1) T15(2) T15(3) T15(4) T30(1) T30(2) T30(3) T30(4)
cv. Gran enano
Figura 32. Área foliar afectada por las cepas C1233 (1 ), CR4b (2), Oz1 b (3) y Jaguar (4) a 5, 15 y 30 ddi en fragmentos foliares del cultivar Gran enano, cuantificada con los programas Assess 2.0 (A) e lmage J 1.46r (8). Los datos se analizaron empleando la prueba de la diferencia mínima significativa de Fisher (LSD por sus siglas en inglés) y la prueba de Kruskai-Wallis a un nivel de P~ 0.05.
En la Figura 33A se observa que la cepa C1233 indujo el mayor número de
lesiones en el cultivar susceptible a partir del décimo quinto día del experimento, y el
número de lesiones se mantuvo hasta llegar al día treinta , en donde tuvo igual número de
lesiones que las inducidas por la cepa CR4b; sin embargo, el área foliar dañada por
ambas cepas fue menor en comparación al área dañada por Oz1 b (Figura 32), la cual
desde el día 15 ddi mantuvo el mismo número de lesiones hasta el final del experimento,
aunque estas aumentaron el tamaño del área dañada al fundirse unas con otras. Con
ambos programas se logró detectar que la cepa CR4b se retrasó el doble de tiempo en
inducir el mismo número de lesiones que la cepa control (C1233). El número de lesiones
inducidas por la cepa Jaguar C3 (finca con manejo de fungicidas) permaneció constante,
la cepa no logró inducir más lesiones en el tejido vegetal ni siquiera en los últimos días de
la interacción con su hospedero.
85

CAPITULO 111
560 A
a
I ab 360 b I b
I I e 1/l I cd Q) 160 cd cd I e
I d d o I d
I 1/l I I Q)
Q) -40 ""O TS (1) TS (2) T5(3) TlS (1) TlS (2) TlS (3) TlS (4) T30(1) T30(2) T30 (3) T30 (4) o l... Q) 1300 B
a E I '::::1 a a z
I I 800
b b
I b b I b
300 b I b I b
I I I I -200
TS (1) TS {2) TS (3) TlS (1) TlS (2) TlS (3) TlS (4) T30(1) T30{2) T30{3) T30(4)
cv. Gran enano
Figura 33. Número de lesiones inducidas por las cepas C1233 (1), CR4b (2), Oz1b (3) y Jaguar (4) en el cultivar Gran enano a 5, 15 y 30 ddi, cuantificadas con los programas Assess 2.0 (A) e lmage J 1.46r (B). Los datos se analizaron empleando la prueba de la diferencia mlnima significativa de Fisher (LSD por sus siglas en inglés) a un nivel de Ps 0.05.
Severidad foliar en Calcuta IV
El resultado de la cuantificación del área foliar dañada en el cultivar Calcuta IV fue
muy similar con ambos programas (Figura 34). En general, el daño causado por las cepas
no superó los 0.18 cm2 (0.72%) de superficie foliar, que fue mucho menor al área dañada
que se observó en el cultivar Gran enano, lo cual es congruente con la resistencia del
cultivar Calcuta IV a M. fijiensis. Como se observa en la Figura 34, con el programa
Assess 2.0 no se logró observar diferencia significativa en el daño foliar inducido por las
cepas a ningún tiempo, esto solo fue posible detectarse con el programa lmage J 1.46r.
Sin embargo, se debe tomar en cuenta cierto margen de error durante la selección de las
lesiones ocasionadas por el hongo con respecto a las lesiones causadas por daño
mecánico con el pincel y la solución de grenetina. Tanto el pincel como la grenetina
86

CAPITULO 111
indu3eron reacciones de hipersensibilidad en Calcuta IV, muy similares a las inducidas por
M. fijiensis (Figura 35), es decir, presentan la m isma coloración marrón y se encuentran
para1e'las a los conductos vasculares de los fragmentos foliares.
ab
ab I ~- ,,. I I ! l = NE -0.06 ------------------------------.E u TS(l) TS(2) T5(3) T5(4) TlS(l) T15(2) T15(3) T15(4) T30(1) T30(2) T30(3) T30(4)
-~- 0.18
'E <lJ a. :::1
1/)
0 .08
B
e
I e
a
I e e e
I I
ab
I abe be
I e be
e
I I I I I I -0.02 e:::_ __________________________ _
TS( l ) T5(2) T5(3) 15(4) T15(1) T15(2) T15(3) 115(4) T30(1) T30(2) T30(3) 130(4)
cv. Calcuta IV
Figura 34. Área foliar afectada por las cepas C1233 (1), CR4b (2), Oz1b (3) y Jaguar (4) a 5, 15 y 30 ddi en fragmentos foliares del cultivar Calcuta IV, cuantificada con los programas Assess 2.0 (A) e lmage J 1.46r (8). Los datos se analizaron empleando la prueba de la diferencia mínima significativa de Fisher (LSD por sus siglas en inglés) a un nivel de P~ 0.05.
Figura 35. Comparación entre lesiones en fragmentos foliares de Calcuta IV en A) Fragmento control y 8) fragmento inoculado con C1233 a 30 ddi.
87

CAPITUlO 111
Cuando se cuantificó el número de lesiones inducidas en el cultivar resistente
(Figura 36), únicamente la cepa Oz1 b mostró inducir un número significativamente mayor
de lesiones en comparación con las demás cepas, a 15 ddi.
130 A a
I b 80 b be bcd I bcd I bcd I bcd
Vl I d d d cd I I I Q) 30
I e
I I I .Q Vl
..9::! Q) -20
"' TS(l) T5(2) TS(3) TS(4) T15(1) T15(2) T15(3) T15(4) T30(1) T30(2) T30(3) T30(4) e 210 B a Q)
I E ' ::::1 2 130
bcd be b bcd bcd
I bcd I I I I I bcd
50 d cd cd I I I I -30
TS(l) T5(2) T5(3) TS(4) T1S(1) T15(2) Tl5(3) T15(4) T30(1) T30(2) T30(3) T30(4)
cv. Calcuta IV
Figura 36. Número de lesiones inducidas por las cepas C1233 (1), CR4b (2), Oz1b (3) y Jaguar (4) en fragmentos foliares del cultivar Calcuta IV a 5, 15 y 30 ddi, cuantificadas con los programas Assess 2.0 (A) e lmage J 1.46r (B). Los datos se analizaron empleando la prueba de la diferencia mínima significativa de Fisher (LSD por sus siglas en inglés) a un nivel de P::> 0.05.
Severidad foliar en Fougamou
La mayor superficie afectada en los fragmentos foliares se observó a 30 ddi y fue
causada por la cepa control (C1233). Ninguna otra cepa indujo severidad significativa en
la superficie de los fragmentos, pese al tiempo de interacción entre el fitopatógeno y su
hospedero. Tanto con el programa Assess 2.0 como con el programa lmage J 1.46r se
observó el mismo comportamiento, aunque el primer programa permitió cuantificar mayor
área dañada que el segundo (Figura 37).
88

CAPITULO 111
1.5 A a
I b b b
ro 0 .5 b b
I b
I b b I -o
ro
I I I I I IC ro
-o ~
ro-· - N
.0.5 .E E TS{l) T5{3) T5(4) TlS{l) Tl5(3) Tl5{4) T30(1) T30{3) T30(4) u OJ-
1.7 'ü B 'E
OJ o. ::J a V)
0.7 I b b b
b b
I b
I b b I I I I I I
-0.3 TS{l) T5{3) TS(4) TlS(l) T15(3) T15{4) T30{1) T30{3) T30{4)
cv. Fougamou
Figura 37. Área foliar afectada por las cepas C1233 (1), Oz1b (3) y Jaguar (4) a 5, 15 y 30 ddi, en fragmentos foliares del cultivar Fougamou, cuantificada con los programas Assess 2.0 (A) e lmage J 1.46r (8 ). Los datos se analizaron empleando la prueba de la diferencia mínima significativa de Fisher (LSD por sus siglas en inglés) a un nivel de p::; 0.05.
Las cepas C1233 y Jaguar indujeron mayor número de lesiones en Fougamou al
tiempo final de los experimentos (30 días); durante los tiempos 5 y 15 el número de
lesiones fue el mismo con respecto a las demás cepas (Figura 38A).
En tanto, en la Figura 388, aun cuando se aprecia un comportamiento similar de
los tratamientos con respecto a la Figura 38A, en general, el programa lmage J 1.46r
cuantificó más lesiones que Assess 2.0.
89

Vl Q)
50
5 10
Vl Q)
Q)
! I be be
I be
I I
CAPITULO 111
a
I ab
I e
I e
I \J -30 ~-----------------------------------------------------------2 TS(l) T5(3) T5(4) TlS(l)
E 130 B ' :::J z
70
r r b
b
I 10
T15(3) T15(4)
b b
I
T30(1)
a
I T30(3) T30(4)
ab
b I I
-50 ~---------------------------------------------------------TS(l) T5(3) T5(4) TlS(l) T15(3) T15(4) T30(1) T30(3) T30(4)
cv. Fougamou
Figura 38. Número de lesiones inducidas por las cepas C1233 (1), Oz1b (3) y Jaguar (4) en
fragmentos foliares del cultivar Fougamou a 5, 15 y 30 ddi, cuantificadas con los programas Assess
2.0 (A) e lmage J 1.46r (8 ). Los datos se analizaron empleando la prueba de la diferencia mínima
significativa de Fisher (LSD por sus siglas en inglés) a un nivel de Ps 0.05.
Severidad foliar inducida a 30 ddi en Gran enano, Fougamou y Calcuta IV
Se comparó el efecto de las cuatro cepas evaluadas (C1233, CR4b, Oz1b y
Jaguar C3) al final de los experimentos con los tres cultivares: Gran enano, Fougamou y
Calcuta IV, para observar cual cepa causó mayor severidad tanto en área como en
número de lesiones en los fragmentos foliares de los tres cultivares.
De las cuatro cepas evaluadas, Oz1b (finca sin manejo de fungicidas) indujo
mayor daño en la superficie foliar de los fragmentos del cultivar susceptible Gran enano a
los treinta días después de haber sido inoculada sobre los fragmentos. Las cepas C1233
y CR4b le siguieron en orden de magnitud, dañando por igual al cultivar susceptible. Los
90

CAPITULO 111
cu'ltivares Fougamou y Calcuta IIV mostraron menor severidad que Gran enano,
independientemente de lla cepa que se les inoculó y fueron muy s.imilares entre sí (Figura
39), esto era de esperarse ya que tanto Fougamou como Calcuta IV son genotipos más
nesiistentes a M fijiensis que Gran enano.
ro
"' ro u: ro
"'
12 a
8
4
e
G(l) G(2) G(3) G(4) F(l) F(3) F(4) C(l) C(2) C(3) C(4)
a
b
I I
6 b
I 2 e
I e
I e e e e e e
I I I I I I -2 ~----------------------------------------------------
G(l) G(2) G(3) G(4) F(l) F(3) F(4) C(l) C(2) C(3) C(4)
Tratamientos
Figura 39. Área fo liar afectada por C1233 (1), CR4b (2), Oz1b (3) y Jaguar (4) a 30 ddi, en fragmentos fol iares de los cultivares Gran enano (G), Calcuta IV (C) y Fougamou (F) cuantificada con los programas Assess 2.0 (A) e lmage J 1.46r (8). Los datos se analizaron empleando la prueba de la diferencia mínima significativa de Fisher (LSD por sus siglas en inglés) y la prueba de Kruskai-Wallis a un nivel de p:s; 0.05.
Como se observa en la Figura 40A, las cepas C1233 y CR4b, indujeron mayor
número de lesiones sobre la superficie de los fragmentos de Gran enano a treinta días
después de haber sido inoculadas, la cepa CR4b provino de manejo intensivo con
fungicidas.
En tanto que la cuantificación realizada con el programa lmage J 1.46r (Figura
408) mostró que la cepa CR4b fue la que indujo mayor número de lesiones en Gran
enano a tiempo treinta, seguido por la C1233 para este mismo tiempo. Todas las demás
cepas en cualquier otro tiempo indujeron el mismo número de lesiones y se mantuvieron
por debajo del daño ocasionado por CR4b y C1233 en Gran enano, lo cual pone de
91

CAPITULO 111
manifiesto la resistencia o tolerancia de los cultivares Calcuta IV y Fougamou a M.
fijiensis.
600
400
\1) 200 Q) b e o e e \1) d ~ Q)
o Q)
G(l) G(2) G(3) G(4) F(l) F(3) F(4) C(l) C{2) C{3) C(4)
"'' e 1600
Q) a E
I -:::J b z 1000
I 400 e
I e e e e e e e e
I I I I I I I I -200
G(l) G(2) G(3) G(4) F(l) F(3) F(4) C(l) C(2) C(3) C(4)
Tratamiento
Figura 40. Número de lesiones inducidas por las cepas C1233 (1), CR4b (2), Oz1b (3) y Jaguar (4) en fragmentos foliares de los cultivares Gran enano (G), Calcuta IV (C) y Fougamou (F) a 30 ddi, cuantificadas con los programas Assess 2.0 (A) e lmage J 1.46r. Los datos se analizaron empleando la prueba de la diferencia mínima significativa de Fisher (LSD por sus siglas en inglés) y la prueba de Kruskai-Wallis a un nivel de Ps 0.05
92

CAPITULO 111
l a agresividad de las cepas de P. fijiensis, definida como la cantidad de daño
iinduddo en .eJ hospedero en un período de tiempo (Romero y Sutton, 1998; Jacome y
Schuh, 1'992) ha s ido correlacionada con la apl icación de fungicidas en las fincas
plataneras, es decir, ~las cepas que han sido aisladas de fincas con aplicaciones de
fung1icidas han sido más agresivas que las cepas aisladas de fincas sin aplicaciones de
fungicidas. Contrario a esto, en este estudio se encontró que los aislados CR4b y Jaguar
G3 de M fijiensis provenientes de fincas con manejo de fungicidas fueron menos
agresivos que la cepa Oz1 b aislada de una finca sin aplicación de fungicidas.
Jacome y Schuh (1992) reportaron que su cepa control (aislada de una finca sin
aplicaciones de fungicidas) fue más agresiva que las cepas que fueron aisladas de fincas
con aplicaciones de fung icidas, lo cual coincide con los resultados de este estudio debido
a que la cepa control de referencia indujo mayor severidad en los fragmentos foliares del
cultivar susceptible que la cepa Jaguar C3, pero similar severidad que la cepa CR4b a 30
dd i. La cepa Oz1 b que también se aisló de una plantación sin manejo de fungicidas,
indujo mayor severidad que las demás cepas. Sin embargo, Jacome y Schuh (1992)
utilizaron plantas completas y estas fueron humidificadas constantemente en un
invernadero durante los experimentos, a diferencia de este estudio en donde se utilizaron
fragmentos foliares los cuales no se mantuvieron a humedad constante.
Independientemente del tipo de material vegetal, el resultado fue el mismo.
Aunque la respuesta a la enfermedad es similar en ambos bioensayos (plantas
completas y fragmentos fo liares) , cada modelo es diferente y puede mostrar resultados
diferentes. Twizeyimana et al. (2007) reportaron que un problema de utilizar plantas
completas es que se producen resultados variables, aunque como se observó en las
imágenes de los fragmentos fol iares, éstos también presentaron respuestas
heterogéneas.
La diferencia en la severidad foliar inducida por los aislados de M. fijiensis y
traducido como agresividad puede estar asociada, en parte, con un mayor número de
tubos gérminativos por esporas producidas en alta humedad relativa que a baja humedad
relativa. A su vez, el número de tubos germinativos puede estar correlacionado con
múltiples penetraciones estomáticas por conidio como mencionaron Jacome y Schuh
(1992), quienes evaluaron la germinación, crecimiento y producción de esporas de M.
93

CAPITULO 111
fijiensis. Por otro lado, Zandjanakou et al., (2013) mencionaron que la alta humedad
relativa favorece a la Sigatoka negra, debido a esto, la falta de humedad constante en
este estudio pudo influir una expresión diferencial de la enfermedad en los fragmentos
foliares inoculados con una misma cepa, lo cual se evidenció durante los muestreos, en
donde se observaron fragmentos con diferente número y tamaño de lesiones. En este
trabajo el único factor ambiental controlado fue la temperatura (27 °C), por lo tanto se
observaron fragmentos con abundante humedad y otros con poca humedad sobre la
lámina foliar. Esto pudo influenciar la capacidad infectiva de las cepas, por lo que en
futuros trabajos de investigación es deseable utilizar una cámara de cultivo para controlar
la humedad, la temperatura y la intensidad luminosa. A pesar de que hubo variación entre
réplicas, la tendencia de la respuesta del material vegetal en términos generales coincide
con lo que se sabe ocurre en la naturaleza, el fragmento del genotipo susceptible se
comportó como tal y el fragmento del genotipo resistente por lo consiguiente, esto cuando
se inoculó con una cepa agresiva como la Oz1 b.
A diferencia de Jacome y Schuh (1992) quienes utilizaron diagramas a escala,
basados en la evaluación visual del daño, los cuales son subjetivos como se explicó
previamente, en este trabajo de investigación el área dañada se cuantificó a partir de
imágenes digitales, las cuales reproducen fielmente el área dañada en los fragmentos
foliares. Por lo tanto, la cuantificación de la agresividad de las cepas del patógeno fue
más precisa.
Los fragmentos foliares de Gran enano, Fougamou y Calcuta IV inoculados con las
cepas aisladas de fincas con diferente manejo de fungicidas presentaron un daño
diferencial , lo cual era de esperarse ya que dichos genotipos (GE, F y C) se han
caracterizado por ser susceptible (Cavalcante et al. 2011 ), tolerante (Lepoivre et al. 2003)
y resistente (Twizeyimana et al., 2007), respectivamente. Las cepas C1233, CR4b y Oz1 b
a partir del día 15 lograron inducir mayor número de lesiones en el cultivar susceptible
(Gran enano) en comparación con los otros dos cultivares. La cepa Oz1 b dañó mayor
superficie de los fragmentos foliares a los 30 días (Figura 39), aunque indujo menor
número de lesiones que C1233 y CR4b (Figura 40). Esto se debe a que en el transcurso
de la infección, las lesiones aumentaron de tamaño, uniéndose unas con otras, lo cual
redujo el número de lesiones y aumentó el tamaño del área foliar afectada (Figura 28).
Con base en estos resultados se sugiere que el área foliar afectada es la variable más
adecuada para evaluar la agresividad de cepas de M. fijiensis.
94

CAPITULO 111
Los fragmentos foliares de Calcuta IV fueron resistentes a las cepas evaluadas,
independientemente de su procedencia, lo cual es consistente con los reportes en donde
se evalúa su resistencia a Sigatoka negra (Twizeyimana et al., 2007; Cavalcante et al.,
2011 ). Considerando el daño diferencial inducido en el material vegetal, esto parece
sugerir una relación entre la agresividad de las cepas y su lugar de procedencia, salvo
para las cepas C1233 (aislada de una plantación sin manejo de fungicidas) y CR4b
(aislada de una plantación con manejo de fungicidas) (Yucateca y Chiapaneca
respectivamente) las cuales fueron igual de agresivas.
En el cultivar susceptible se pudo observar una agresividad diferencial de todos los
aislados y comparando el área foliar dañada resulta evidente que aislados de diferentes
áreas geográficas tienen diferente agresividad. Esto concuerda con los resultados de
Zandjanakou et a/., (2013) quienes observaron que las cepas de M. fijiensis aisladas de
diferente área geográfica, dentro de un mismo país, tienen diferente agresividad , la cual
está influenciada por factores climáticos como la temperatura y la humedad relativa .
Los resultados aquí reportados y los de Zandjanakou et al. , (2013) son contrarios a
los resultados de Romero y Sutton (1997b) quienes no observaron diferencia significativa
en la agresividad de los aislados de una misma región ; solamente observaron diferencia
significativa en la agresividad entre aislados de diferentes países. Como mencionaron
Fullerton et al. , (1995), las cepas individuales del fitopatógeno pueden mostrar patrones
consistentes pero diferentes de agresividad .
En este trabajo y los de Zandjanakou et al. , (2013), se utilizaron fragmentos
foliares y los resultados fueron similares en cuanto a la agresividad diferencial de los
aislados en relación al área geográfica de procedencia. Esto fue muy evidente en el
cultivar Gran enano en donde se observaron diferencias muy marcadas en el área foliar
dañada.
En el cultivar resistente, Oz1 b indujo mayor número de lesiones únicamente a los
quince días (Figura 36) pero posteriormente no hubo diferencia significativa en el número
de lesiones que indujo con respecto a las demás cepas; al final del experimento (30 ddi)
todas las cepas indujeron igual área foliar dañada y número de lesiones.
La cepa control aislada de una finca de Yucatán sin manejo de fungicidas, indujo
un dar'lo significativamente mayor a los 30 ddi en comparación con las otras cepas. En el
95

CAPITULO 111
cultivar Fougamou (tolerante), el daño fue mucho menor al ocasionado en el cultivar Gran
enano (susceptible) , lo cual era de esperarse (Figura 37). Probablemente la cepa C1233
que fue aislada de la parcela experimental deiiNIFAP en donde se encuentran plantas del
cultivar Fougamou, pudo estar en contacto con este material y desarrollar estrategias
infectivas más eficientes para atacar a Fougamou, como por ejemplo, mejor
reconocimiento de la topografía de las hojas; Allen et al., (1991) mencionaron que las
características físicas y químicas de la hoja pueden jugar un papel importante en
determinar la compatibilidad o incompatibilidad de la interacción.
Considerando el comportamiento de la cepa control C1233 en el cultivar tolerante,
era de esperarse que afectara de manera similar al cultivar Calcuta IV que también estuvo
presente en la parcela experimental del INIFAP, sin embargo, se debe tomar en cuenta
que este material vegetal es altamente resistente a la Sigatoka negra (Twizeyimana et al.,
2007), lo que significa que responde más rápidamente al ataque del patógeno y detiene el
desarrollo de la infección (Cavalcante et al., 2011).
La cepa Jaguar C3 indujo un daño mínimo en los fragmentos foliares de los tres
genotipos, sin embargo, fue detectable y significativo (PsO.OS) al utilizar los programas
Assess 2.0 e lmage J 1.46r; fragmentos de Gran enano y Fougamou presentaron mayor
daño a los 30 ddi (Figura 29). El programa Assess detectó diferencias significativas
(PsO.OS) en el número de lesiones no así el programa lmage J 1.46r, lo cual sugiere que
Assess 2.0 es más sensible que el otro (Figura 30). Esto lleva a pensar que cuando la
agresividad de la cepa es baja no permite inducir un número significativo de lesiones que
puedan ser detectadas por el programa lmage J 1.46r, independientemente de la
susceptibilidad del cultivar en donde es inoculada.
La diferencia en el daño (superficie dañada y número de lesiones) inducido en los
fragmentos foliares, pone de manifiesto la susceptibilidad, tolerancia o resistencia que
posee cada cultivar a la enfermedad ocasionada por aislados de M. fijiensis.
La heterogeneidad en el daño que causan las cepas de M. fijiensis no depende
exclusivamente de la susceptibilidad o resistencia del material vegetal en el cual fueron
inoculados, se debe tomar en cuenta la capacidad de recombinación genética y de
selección de poblaciones resistentes (Pérez et al. , 2004) de este fitopatógeno. Se podría
esperar que en una población diversa, se encuentren aislados altamente agresivos,
contrario a esto, los resultados obtenidos por Zandjanakou et al., (2013) muestran que los
96

CAPITULO 111
aislados menos agresivos del fitopatógeno provinieron de la población geográfica con
mayor diversidad. Cabe mencionar, que los autores no realizaron ningún estudio
molecular, únicamente determinaron la variabilidad de los aislados con base en el índice
de Shannon, el cual solo considera el número de especies, número de individuos de cada
especie, y la proporción que existe entre ellos. Por lo tanto, la agresividad de los aislados
está en función no solamente de las características ambientales del área geográfica de
donde provienen, sino además de factores artificiales (fungicidas) que ejercen una presión
de selección la cual induce un cambio en la agresividad del fitopatógeno (Chin et al.,
2001; Romero y Sutton, 1998; Jacome y Shuh, 1992).
La aplicación de fungicidas en campo ejerce una fuerza de selección, la cual
influye en la agresividad de los aislados. Romero y Sutton (1998) pusieron de manifiesto
que las cepas resistentes a Benomil (fungicida sistémico del grupo de los bencimidazoles)
mostraron mayor agresividad en hojas del cultivar Gran enano procedentes de cultivo in
vitro; esto contrasta con los resultados de este estudio ya que se observó que la cepa
Oz1 b, la cual se aisló de una finca sin manejo de fungicidas, mostró ser más agresiva en
el cultivar susceptible que las dos cepas (CR4b y Jaguar C3) aisladas de fincas con
manejo de fungicidas. Además la cepa CR4b fue igual de agresiva que la cepa control en
Gran enano; y la cepa Jaguar C3 que también se aisló de una finca con aplicaciones de
fungicidas fue la cepa menos agresiva, logrando causar mayor daño solo en los cultivares
Gran enano y Fougamou únicamente a 30 días del experimento.
Los resultados reportados en la literatura son contradictorios en referencia al
efecto que los fungicidas ejercen sobre la agresividad de las cepas del fitopatógeno.
Romero y Sutton (1998) reportaron mayor agresividad de las cepas que fueron aisladas
de una plantación con aplicación de fungicidas, esto es congruente con los resultados
reportados por Yang et al. , (2013) y Cowger y Mundt (2002), quienes encontraron una
correlación positiva entre la aplicación de fungicidas y mayor agresividad de las cepas de
M. graminicola. Contrastando con este comportamiento Jacome y Schuh (1992)
reportaron que la cepa silvestre de M. fijiensis resultó ser más agresiva que las cepas
expuestas a manejo con fungicida. En otro patosistema como Solanum tuberosum
Phytophtora infestans se observó que los aislados resistentes al fungicida Metalaxil®
fueron menos agresivos que las cepas sensibles (Day y Shattock, 1997; Dowley, 1987).
97

CAPITULO 111
Se debe considerar que el tipo de fungicida utilizado en cada finca de donde se
obtuvieron los aislados pudo ejercer un efecto diferente tanto en la agresividad como en la
capacidad de esporulación, ya que CR4b fue más agresiva y produjo casi 6 veces más
cantidad de esporas que la cepa Jaguar C3 (ambas cepas aisladas de una finca con
manejo de fungicidas); la cepa C1233 (control) produjo más esporas que Jaguar C3 y fue
igual de agresiva que CR4b. Finalmente, Oz1 b que fue aislada de una finca sin manejo de
fungicidas, fue la cepa más agresiva y su producción de esporas fue inferior a las de
C1233 y CR4b, (cuadro 7). Esto sugiere que el número de esporas producidas no
determina la agresividad de una cepa.
Se logró cuantificar la agresividad de los aislados de M. fijiensis inoculados en
fragmentos foliares de tres cultivares con diferente grado de susceptibilidad, utilizando los
programas Assess 2.0 e lmage J 1.46r; en general, el programa Assess 2.0 detectó mayor
superficie foliar afectada que el programa lmage J 1.46r, en tanto que éste último detectó
mayor número de lesiones que el programa Assess 2.0. Resultó interesante combinar
estas dos herramientas informáticas para comparar los resultados de cada una, observar
la agresividad de las cepas y determinar cuál programa es más sensible para detectar una
determinada variable de interés. Si bien existen diversos estudios en donde se han
empleado los programas Assess (Carretero et al. , 201 O; Serrago et al., 2009; Donzelli y
Churchill, 2007) e lmage J (Kathiria et al., 2010; Ushada et al., 2007; Horiguchi et al. ,
2006) para evaluar sistemas biológicos vegetales y sus interacciones con agentes
fitopatógenos, es la primera vez que se conjuntan ambas herramientas para estudiar
robustamente al patosistema M. fijiensis-Musa spp.
Con base en los resultados de este estudio, es evidente que la aplicación o no
aplicación de fungicidas en las fincas plataneras tiene un efecto diferencial en los aislados
del fitopatógeno, pero no fue posible determinar si la aplicación de fungicidas en las fincas
plataneras tiene un efecto positivo en la agresividad de los aislados provenientes de cada
una de ellas. Para poder determinar esto último sería necesario ampliar el número de
aislados de cada finca con los diferentes manejos.
98

CAPITULO 111
Bibliografía
Abramoff, M. Magalhaes, P. Ram, S. (2004). lmage processing with lmageJ. Biophotonics
international. 11 , 36-42.
Allen, E. Hoch, H. Steadman, J. Stavely, R. (1991). lnfluence of Leaf Surface Features on
Spore Deposition and the Epiphytic Growth of Phytopathogenic Fungi.
Contemporary Bioscience. 87-11 O.
Baum, T. Navarro-Quesada, A. Knogge, W. Douchkov, D. Schweizer, P. Seiffert, U.
(2011). HyphArea-Automated analysis of spatiotemporal fungal patterns. Journal of
Plant Physiology. 168, 72-78.
Camargo, A. Smith, J.S. (2009). lmage pattern classification for the identification of
disease causing agents in plants. Computers and Electronics in Agriculture. 66,
121-125.
Carretero, R. Serrago, R. Bancal, M. Perelló, A. Miralles, D. (201 O). Absorbed radiation
and radiation use efficiency as affected by foliar diseases in relation to their vertical
composition into de canopy in wheat. Field Crops Research . (116), 184-195.
Cavalcante, M. Escoute, J. Madeira J.P. Romero,R. Nicole, M. Oliveira, L. Hamelin, C.
Lartaud, M. Verdeil, J.L. 2011 . Reactive Oxygen Species and Cellular lnteractions
Between Mycosphaerella fijiensis and Banana. Tropical Plant Biology. 4. pp. 134-
143.
Chin, K.M. Wirz, M. Laird , D. (2001). Sensitivity of Mycosphaerella fijiensis from banana to
trifloxystrobin . Plant disease. 85, 1264-1270.
Cowger, C. Mundt, C. (2002). Aggressiveness of Mycosphaerella graminicola lsolates
from Susceptible and Partially Resistant Wheat Cultivars. Phytopathology. 92, 624-
630.
99

CAPITULO 111
Dammer, K. H. Moller, B. Rodemann, B. Hepnner, D. (2011 ). Detection of head blight
(Fusarium ssp.) in winter wheat by color and multispectral image analysis. Crop
Protection. 30, 420-428.
Day, J.P. Shattock, R.C. (1997). Aggressiveness and other factors relating to
displacement of populations of Phytophtora infestans in England and Wales.
European Journal of Plant Pathology. 103, 379-391.
Donzelli , B.Churchill, A. (2009). A dose-response approach differentiating virulence of
Mycosphaerella fijiensis strains on banana leaves uses either spores or mycelia as
inocula. Acta Horticulture. 828, 153-160.
Donzelli, B. Churchill, A. (2007). A quantitative assay using micelial fragments to assess
virulence of Mycosphaerella fijiensis. Phytopathology. 97, 916-929.
Dowley, L.J. (1987). Factors affecting the survival of metalaxil resistant strains of
Phytophtora infestans (Mont.) de Bary in lreland. Potato Research. 30, 473-475.
Fillho, A.B. Carneiro, S.M.T.P.G. Godoy, C.V. Amorim, L. Berger, R.O. Hau, B. (1997).
Angular leaf spot of Phaseolus beans: Relationships between disease, healthy leaf
area and yield. Phytopathology. 87, 506-515.
Fouré, E. 1985. Black leaf streak disease of Banana and plantains (Mycosphaerella
fijiensis Morelet), study of the symptoms and stages of the disease in Gabon IRFA
Paris.
Fullerton, R.A. Olsen, T.L. (1995). Pathogenic variability in Mycospaerella fijiensis Morelet,
Cause of Black Sigatoka in Banana and Plantain. New Zealand Journal of Crop
and Horticultura! Science. 23. 39-48.
Guo, G. Li, S.Z. Chan, K.L. (2001). Support vector machines for face recognition . lmage
and Vision Computing. 19, 631-638.
100

CAPITULO 111
Horiguchi, G. Fujikura, U. Ferjani, A. lshikawa, N. Tsukaya, H. (2006). Large-scale
histological analysis of leaf mutants using two simple leaf observation methods:
identification of novel genetic pathways governing the size and shape of leaves. The
Plant Journal. (48) , 638-644.
lsik, H. Sezgin , E. Avunduk, M.C. (201 O). A new software program for pathological data
analysis. Computers in biology and medicine. 40, 715-722.
Jacome, L. H. Schuh, W. (1992). Effect of temperature on growth and conidial production in
vitro , and comparison of infection and aggressiveness in vivo among isolates of
Mycosphaerella fijiensis var. difformis. Tropical Agriculture. 70(1 ), 51-59.
Kathiria , P. Sidler, C. Golubov, A. Kalischuk, M. Kawchuk, L. Kovalchuk, l. (201 0).
Tobacco mosaic virus results in an increase in recombination frequency and resistance
to viral, bacteria! and fungal pathogens in the progeny of infected tobacco plants. Plant
Physiology. (153) , 1859-1870.
King, S.L. Bennett, K.P. List, S. (2000). Modelling noncatastrophic individual tree mortality
using logistic regression, neural networks, and support vector methods. Computers
and Electronics in agriculture. 27, 401-406.
Kolukisaoglu , O. Thurow, K. (201 O) . Future and frontiers of automated screening in plant
sciences. Plant Science. 178, 476-484.
La Serna-Palomino, N. Pró-Concepción, L. (2010). Watershed : un algoritmo eficiente y
flexible para segmentación de imágenes en geles 2-DE. Revista de Investigación de
Sistemas e Informática. 7, 35-41 .
Lepoivre, P. Busorogo, J. Etame, j . El Hadrami, A. Carlier, J. Harelimana, G. Mourichon,
X. Panis, B. Stella-Rivero, A. Sallé, G. Strosse, H. Swennen, R. 2003. Banana
Mycosphaerella fijiensis lnteractions. 2nd lnternational workshop on Mycosphaerella
leaf spot diseases, San José. 151-159.
101

CAPITULO 111
Lim, L.G. Gaunt, R.E. (1981). Leaf area as a factor in disease assessment. Journal of
Agricultura! Science. 97,481-483.
McNitt-Gray, M. Huang, H.K. Sayre, W.J. (1995). Feature selection in the patter
classification problem of digital chest radiograph segmentation. IEEE Transaction
Medicallmaging. 14, 537-547.
Pérez, M. Rebullído, R. Pérez, L. (2004). Líneas base de sensibilidad de las poblaciones
de Mycosphaerella fijiensis Morelet de Cuba, a los fungicidas azoxystrobin y
trifloxystrobin . Fitosanidad. 8, 41-47.
Romero, R. A. Sutton , T. B. (1998). Characterization of benomyl resistance in
Mycosphaerella fijiensis, cause of black Sigatoka of banana, in Costa Rica. North
Carolina State University, Raleigh 27695-7616. Plant Dis. 82, 931-934.
Romero, R. A. Sutton, T. B. (1997b). Reaction of four Musa genotypes at three
temperatures to isolates of Mycosphaerella fijiensis from different geographical
regions. Plant Disease. 81. 1139-1142.
Sena, D.G. Pinto, F.A.C. Queiroz, D.M. Viana, P.A. (2003). Fall armyworm damaged
maize plant identification using digital images. Biosystems Engineering. 85, 449-454.
Serrago, R. Carretero, R. Bancal, M. Miralles, D. (2009). Foliar diseases affect the eco
physiological attributes linked with yield and biomass in wheat (Triticum aestivum L.).
European Journal of Agronomy. (31 ), 195-203.
Sherwood, R.T. Berg , C.C. Hoover, M.R. Zeiders, K.E. (1982). lllusions in visual
assessment of Stagonospora Leaf Spot of Orchardgrass. Phytopathology. 73, 173-
177.
Slopek, S.W. (1989). An improved method for estimating percent leaf area diseased using
a 1 to 5 disease assessment scale. Canadian Journal of Plant Pathology. 11 , 381-387.
102

CAPITULO 111
Tang , Y.Y. Tao, Y. Lam, E.C.M. (2000). New method for feature extraction based on
fractal behavior. Pattern Recognition. 35, 1071-1081.
Taslt, l. Purgathofer, W. (1994). Color Spaces and Human Color Perception. Photorealistic
Rendering in Computer Graphics. 219-226.
Twizeyimana, M. Ojiambo, P.S. Tenkouano, A. lkotun, T. Bandyopadhyay, R. (2007).
Rapid Screening of Musa Species for Resistance to Black Leaf Streak Using In Vitro
Plantlets inTubes and Detached Leaves. Plant disease. 91, 308-314.
Ushada, M. Murase, H. Fukuda, H. (2007). Non-destructive sensing and it's inverse model
for canopy parameters using texture analysis and artificial neural network.
Computers and Electronics in Agriculture. (57) , 149-165.
Wang , D. (1998). Unsupervised image segmentation based on watersheds and temporal
tracking. IEEE Transactions on Circuits and Systems for Video Technology. 8, 539-
546.
Wijekoon , C.P. Goodwin , P.H. Hsiang, T. (2008). Quantifying fungal infection of plant
leaves by digital image analysis using Scion lmage software. Journal of Microbiological
Methods. 74, 94-101.
Zandjanakou , M. Ojiambo, P.S. Vroh-bi , l. Tenkouano, A. Gumedzoe, Y.M.
Bandyopadhyay, R. (2013) . Pathogenic variation of Mycosphaerella species infecting
banana and plantain in Nigeria. Plant Pathology. 62, 298-308.
103


CAPITULO IV
CAPITULO IV. DISCUSIÓN GENERAL
En muchos países en desarrollo el cultivo de banano constituye un alimento básico
y una fuente significativa de ingresos económicos. Es cultivado en las áreas tropicales y
subtropicales de diversos países (Marín et al., 2003), siendo la India el principal productor
de esta fruta en tanto que el mayor exportador es Ecuador, su comercialización genera
anualmente alrededor de 5 billones de dólares (Arias et al., 2003).
Sin embargo, así como otros cultivos, el banano es atacado por diversas plagas y
enfermedades, siendo la Sigatoka negra la principal enfermedad foliar, ya que llega a
causar hasta el 100% de pérdidas en la producción. Esta enfermedad es causada por el
hongo fitopatógeno Mycosphaerella fijiensis (Jones, 2003).
En las fincas plataneras la forma más eficiente de control de la enfermedad es
mediante la aplicación alternada de fungicidas sintéticos con diferente modo de acción
(sistémicos o de contacto) para prevenir o retrasar el desarrollo de resistencia hacia algún
fungicida específico (Chin et al., 2001 ). Sin embargo, en diversos estudios (Cañas
Gutiérrez et al., 2009; 2006; Chin et al., 2001; Romero y Sutton 1998) se ha evidenciado
que existen poblaciones del fitopatógeno las cuales han desarrollado resistencia a
determinados tipos de fungicida.
La influencia de los fungicidas puede inducir cambios no solo en el nivel de
sensibilidad (resistencia) hacia estas moléculas, sino que también pueden estar
modificando la agresividad de las cepas de M. fijiensis, tal como Romero y Sutton (1998)
observaron en su estudio, en el cual las cepas que fueron resistentes al fungicida Benomil
fueron más agresivas que las cepas sensibles al fungicida, esto se evidenció en un mayor
daño en el área foliar del cultivar Gran Enano en menor tiempo.
Debido a la importancia de la agresividad en la capacidad patogénica del hongo y
a las desventajas que plantea realizar estudios de interacción con su hospedero a nivel de
campo (se requiere mucho tiempo, condiciones climáticas cambiantes, se requiere de
mucho espacio y por tanto la capacidad para evaluar diferentes cultivares se reduce), se
han utilizado otros bioensayos que puedan ofrecer una respuesta similar a la observada
en campo, sin que presenten las desventajas de estos. Para ello se han utilizado
fragmentos foliares (Twizeyimana et al., 2007) y plántulas de banano (Donzelli y Churchill ,
lOS

CAPITULO IV
2007) y estos ensayos han mostrado una correlación con la infección que ocurre de forma
natural en campo. Utilizando estos bioensayos, se ha podido realizar el escrutinio de
cultivares resistentes a la enfermedad, así como evaluar el desarrollo de la enfermedad
empleando diferentes cepas y cantidades de inóculo.
En lo que se refiere a fragmentos foliares, debido al problema de la rápida
senescencia de los tejidos vegetales cuando son desprendidos del resto de la planta, se
han tenido que utilizar diferentes agentes para evitar la degradación de la clorofila así
como de otros componentes celulares importantes. Entre los diferentes agentes
empleados se encuentran el Bencimidazol, sales de níquel, cobalto, fitohormonas,
extracto de levadura, leche de coco, entre otros. Sin embargo, debido a las características
de cada compuesto, en este estudio se optó por emplear el Bencimidazol para retrasar la
senescencia de los fragmentos foliares de los cultivares Gran enano, Fougamou y Calcuta
IV.
Paralelamente a la preparación de los fragmentos foliares de los tres cultivares, se
indujo la esporulación de las cepas C1233, CR4b, Oz1 b y Jaguar C3 en medio de
esporulación reportado por Mourichon et al., (1987) y las cepas se mantuvieron en un
cuarto acondicionado para mantener temperaturas de 20 ± 2 oc y luz blanca y ultravioleta
constantes. Sin embargo, como se reportó en el capítulo 11 , muchos de los experimentos
programados se tuvieron que abortar debido a que las cepas no produjeron una cantidad
masiva de esporas, lo cual no permitió tener la cantidad requerida de inóculo.
Entre los posibles factores que pudieron afectar la producción de esporas se
encuentra el que el cuarto de esporulación no fue el adecuado para mantener constantes
las condiciones de temperatura e intensidad luminosa, ya que durante los períodos de
esporulación la temperatura fluctuó de 21 a 25 oc, lo cual se sale del rango óptimo de
temperatura para inducir la esporulación del patógeno (20°C}. Como se ha reportado en
los modelos fúngicos Beauveria bassiana , Metharhizum anisopliae y Nomuraca rileyi
(Faria et al., 201 O; Aguirre et al. , 2009) la actividad de agua en el medio de cultivo es
esencial tanto para la esporulación de las cepas como para prevenir el debilitamiento de
los con idios, y para esto la temperatura juega un papel crucial para disminuir la pérdida de
agua en el medio, por lo cual resulta de suma importancia que el cuarto de esporulación
tenga la capacidad de mantener temperatura constante a 20 oc.
106

CAPITULO IV
Sin embargo, se debe mencionar que en este trabajo no se midió pérdida de agua
en el medio de cultivo, por lo cual no se cuenta con evidencia experimental para
argumentar que una posible baja actividad de agua causó debilitamiento de las esporas
de M. fijiensis, a consecuencia de una elevación de la temperatura en el cuarto de
esporulación. Únicamente se considera este factor como una posible causa, con base en
la literatura consultada.
La intensidad de las lámparas de luz ultravioleta en el cuarto de esporulación
también varió considerablemente, ya que en un período de 53 días la intensidad
descendió en un 71%, lo cual indudablemente afectó la esporulación en todas las cepas.
La exposición a luz continua es necesaria para inducir la esporulación de M. fijiensis
(Romero y Sutton 1997a). Con respecto a otros modelos fúngicos, Leach (1967)
menciona que la esporulación abundante de Fusarium nivale, Helmintosporium
catenarium y Cercosporella herpotricoides se ve favorecida cuando las cepas son
sometidas a luz ultravioleta constante. Por esta razón es crucial que las lámparas
ultravioletas mantengan constante la intensidad luminosa, por lo que se recomienda
cuantificar constantemente la intensidad.
Las micrografías revelaron la presencia de las hifas de las cepas sobre los
fragmentos foliares inoculados, sin embargo, el daño inducido por las cepas fue mucho
menor al obtenido en los experimentos realizados por la M.C. Leticia Peraza Echeverría,
esto se puede deber a que las condiciones fluctuantes de temperatura e intensidad
luminosa en el cuarto de esporulación debilitaron las esporas y con esto las cepas no
lograron inducir los niveles anteriormente observados de daño foliar.
Otro factor que debe ser considerado es el número de subcultivos de las cepas ya
que inicialmente se observó baja esporulación en cepas que tenían 6-7 subcultivos. En
cuanto a este factor, los resultados son muy variados y contrastantes en diferentes
modelos fúngicos, García et al., (2007) observaron que conforme aumentó el número de
subcultivos de Beauveria bassiana el número de esporas producidas fue menor y
disminuyó la agresividad del agente de control biológico. La influencia de los subcultivos
indujo menor actividad enzimática del hongo micorrízico Pisolithus tinctorium, según lo
reportado por Cuevas-Rangel (1979). Sin embargo, contrario a estos resultados, Peteira
et al., 2007 no observaron cambios en la estabilidad de Pocho nía chlamydosporia y Hall
107

CAPITULO IV
(1980) únicamente observaron cambios en la morfología de Verticil/ium lecanii pero su
agresividad no varió.
Debido a las diferentes respuestas de los modelos fúngicos al número de
subcultivos, es necesario evaluar el efecto que este factor puede tener en la esporulación
y debilitamiento de las cepas de M. fijiensis.
Debido a la influencia de los diferentes factores anteriormente mencionados sobre
la esporulación, y las respuestas de los diferentes modelos fúngicos ante cada uno, es
necesario inducir la esporulación de M. fijiensis en una cámara de crecimiento, la cual
tenga la capacidad de mantener cond iciones constantes de temperatura, intensidad
luminosa y humedad, al mismo tiempo que se pone especial cuidado en el número de
subcultivos realizados en las cepas.
Cuando las cepas se reactivaron inoculándolas en su hospedero natural, en
general se observó mayor daño en los fragmentos foliares del cultivar susceptible Gran
enano, sin embargo, el daño siguió siendo inferior al obtenido en los experimentos de la
M.C. Leticia Peraza. Debido a esto, los fragmentos foliares dañados por las cepas
reactivadas, únicamente se emplearon para establecer el método de cuantificación con
los programas Assess 2.0 e lmage J 1.46r.
Para la cuantificación del daño en los fragmentos foliares, inicialmente se formaron
sistemas binarios realizando calcas digitales sobre cada fragmento. Esta técnica ha sido
empleada con anterioridad por Wijekoon et al., (2008) quienes emplearon un programa de
diseño gráfico para remarcar los bordes de las lesiones, así como para mejorar el
contraste y el brillo de las imágenes, sin embargo, esta técnica no se ha empleado para el
estudio del patosistema M. fijiensis-Musa spp. Donzelli y Churchill (2007) desarrollaron un
método para cuantificar el progreso de la enfermedad en plántulas empleando el
programa Assess 1.0, sin embargo, debido las características de los bioensayos en donde
se emplearon fragmentos foliares, los síntomas de la enfermedad se mostraron diferentes
a los obtenidos por Donzelli y Churchill , lo cual no permitió evaluar directamente las
imágenes con los programas Assess 2.0 e lmage J 1.46r.
En algunos casos, se observaron áreas oscuras sobre la superficie de los
fragmentos lo cual puede deberse a ciertas modificaciones del tejido después de ser
108

CAPITULO IV
desprendido de la lámina foliar completa (Karjalainen, 1984), el cual no conserva las
características y la funcionalidad de una planta completa. En otras ocasiones los halos
cloróticos de las lesiones se fusionaron y la cuantificación del área dañada se
sobreestimó, o aparecieron lesiones en la superficie de los fragmentos causados por la
manipulación mas no inducidos por el fitopatógeno, debido a esto fue necesario utilizar el
programa Photoshop® CS3, para crear calcas digitales únicamente de las lesiones
inducidas por M. fijiensis con base en la escala de Fouré (1985) para discriminar del resto
de las lesiones. Como mencionaron Wijekoon et al., (2008) cada imagen debe ser
modificada según lo requieran sus características.
Posterior a establecer el método de cuantificación y convertir todas las imágenes
digitales a sistemas binarios, se realizó la cuantificación del daño foliar y el número de
lesiones. Así, el cultivar Gran Enano (susceptible) mostró mayor severidad foliar, siendo la
cepa Oz1 b (aislada de una finca sin aplicaciones de fungicidas) más agresiva y las cepas
CR4b (aislada de una finca con aplicaciones de fungicidas) y C1233 (control), las cuales
mostraron ser igual de agresivas en el cultivar susceptible.
La cepa Jaguar C3 (aislada de una finca con aplicación de fungicidas) fue la que
indujo menor severidad foliar en el cultivar a 30 ddi, siendo comparable este daño al
inducido en el cultivar tolerante Fougamou por la misma cepa y la C1233. Gran enano fue
el cultivar en donde se pudo observar mejor la diferencia de agresividad de las cuatro
cepas, en tanto que en los cultivares Fougamou y Calcuta IV se observó poca diferencia
en el área foliar y en el número de lesiones inducidas entre las 4 cepas a 30 ddi, lo cual
indicó que independientemente de la agresividad de las cepas, los cultivares resistente y
tolerante mostraron una resistencia consistentemente mayor a la de Gran enano; esto
sugirió que el tejido susceptible es el mejor candidato para posteriores estudios de
interacción en donde se desee estudiar la agresividad diferencial de aislados del hongo,
esto se puede deber a que el tejido susceptible ejerce menor presión de selección en las
cepas de M. fijiensis (Abadie et al., 2003).
Con los resultados de este estudio se observó que las dos cepas aisladas de
fincas con manejo de fungicidas (CR4b y Jaguar C3) fueron menos agresivas que la cepa
Oz1 b la cual se aisló de una finca sin manejo de fungicidas , pero aún entre estas cepas la
diferencia en la agresividad fue muy grande, lo cual podrfa indicar un efecto diferencial del
109

CAPITULO IV
tipo de fungicida al que cada cepa estuvo expuesta en las fincas de donde se aislaron. No
se observó una relación entre el número de conidios producidos por la cepa y su manejo
en campo, ya que CR4b y Jaguar C3 produjeron mayor número de conidios que la cepa
control, sin embargo, Jaguar C3 fue la cepa que indujo menor daño, CR4b causó el
mismo daño que C1233, y la cepa Oz1b aislada de manejo sin fungicidas, produjo mejor
cantidad de conidios que CR4b sin embargo fue la más agresiva.
Los resultados de este estudio contrastan con los obtenidos por Romero y Sutton
(1998), quienes observaron que las cepas aisladas de fincas con aplicaciones de Benomil
causaron mayor severidad que las cepas no resistentes. Sin embargo debe considerarse
en primer lugar que, las cepas evaluadas en este estudio no fueron aisladas de fincas en
las cuales se haya utilizado el Benomil, sino que provienen de fincas en donde se
utilizaron fungicidas de diferente grupo químico tal como se mencionó previamente en el
capítulo 11 , por lo tanto, la influencia de los fungicidas en la agresividad y producción de
esporas no es la misma. En este estudio, tampoco se determinó si las cepas aisladas de
fincas con aplicaciones de fungicidas son sensibles o resistentes a los fungicidas a los
que fueron sometidas, en este rubro la información generada por el grupo del Dr. Beltrán
(participante del proyecto FORDECYT) puede complementar los resultados de este
estudio y así tener un panorama más claro sobre la influencia de los fungicidas en la
producción de esporas y la agresividad de las cepas.
Cuantificación de la enfermedad con los programas Assess 2.0 e lmage J
1.46r
En general, con el programa Assess 2.0 se cuantificó mayor área foliar dañada así
como número de lesiones detectables que con el programa lmage J 1.46r, esto se puede
deber a que el programa Assess 2.0 está diseñado exclusivamente para cuantificar
enfermedades de plantas, en tanto que el programa lmage J 1.46r es un programa
universal de análisis de imágenes. Sin embargo, esto no le resta valor al programa lmage
J, ya que se podría recurrir a él cuando se desee verificar los datos cuantificados con el
programa Assess, además de ser una herramienta gratuita y de fácil acceso por internet.
Debido a esto se recomienda utilizar ambas herramientas informáticas para cuantificar la
severidad de la enfermedad, teniendo en cuenta que el número de lesiones no reflejó el
110

CAPITULO IV
nivel de daño inducido por las cepas, ya que como se observó en el capítulo 111, Oz1 b fue
la cepa que indujo mayor superficie foliar dañada en Gran enano, sin embargo, el número
de lesiones que indujo estuvo por debajo de las causadas por C1233 y CR4b, pero esto
se debió a que las lesiones p.or Oz1 b fueron más grandes y coalescieron.
Al realizar la regresión lineal de los datos analizados, se observó una fuerte
correlación positiva (¡2= 0.93) entre ambos programas en el área dañada por la cepa
C1233 en el cultivar Gran enano; en el cultivar Fougamou (tolerante) se observó una
correlación menos fuerte (¡2= 0.81); finalmente no se observó correlación en el área foliar
dañada entre ambos programas en el cultivar resistente Calcuta IV (r2=0.09). Cuando se
evaluó el área foliar dañada por la cepa CR4b en los cultivares susceptible y resistente
(no se logró evaluar la interacción con el cultivar tolerante), se observó una fuerte
correlación entre ambos programas (¡2=0.96) cuando se cuantificó el área dañada en
Calcuta IV, en tanto que la correlación fue menos fuerte (¡2=0.82) en la interacción con el
cultivar susceptible.
Se detectó una correlación fuerte (r2=0.87) entre ambos programas en el área
dañada por la cepa Oz1 b en el cultivar Gran enano, sin embargo no se logró observar una
correlación fuerte entre ambos programas cuando se cuantificaron las áreas dañadas en
los cultivares Calcuta (¡2=0.34) y Fougamou (¡2=0.03).
Cuando se cuantificó el área dañada por la cepa Jaguar C3, nuevamente solo se
observó una fuerte correlación entre los programas en el cultivar susceptible (¡2=0.86), en
los cultivares resistente y tolerante las correlaciones fueron más débiles (r2=0.48 y 0.79,
respectivamente).
Una desventaja considerable en el uso del programa lmage J. 1.46r, es que los
parámetros de umbral de color y unidades de medida deben ser ajustados para cada
imagen que se necesita cuantificar, lo cual puede requerir de mucho tiempo y esfuerzo
dependiendo del número de imágenes; en tanto que en el programa Assess 2.0 los
parámetros se ajustan una sola vez, esto facilita manejar un número grande de imágenes.
111

CAPITULO IV
CONCLUSIONES
1. Los resultados no confirman la hipótesis de que las cepas de M. fijiensis aisladas
de fincas con aplicaciones de fungicidas son más agresivas que las cepas que
provienen de fincas sin aplicaciones de fungicidas.
2. Se determinó que el programa Assess 2.0 se adecúa mejor para cuantificar la
agresividad de cepas de M. fijiensis.
3. Se cuantificó la agresividad de las cepas C1233, CR4b, Oz1 b y Jaguar C3 en los
fragmentos foliares de Gran enano, Fougamou y Calcuta IV, siendo Oz1 b la cepa
más agresiva y Jaguar C3 la cepa menos agresiva.
PERSPECTIVAS
1. Inducir la esporulación de cepas de M. fijiensis en una cámara de crecimiento, que
mantenga condiciones constantes de temperatura, humedad e intensidad luminosa
para asegurar una producción constante y masiva de esporas.
2. Evaluar la severidad inducida por las cepas reactivadas C1233, CR4b, Oz1 b y
Jaguar C3.
3. Evaluar otras cepas de M. fijiensis aisladas de fincas con diferente manejo de
fungicidas para confirmar o descartar la hipótesis nula.
112

CAPITULO IV
BIBLIOGRAFÍA
Abadie, C. El Hadrami, A. Fouré, E. Carlier, J. (2003). Efficiency and durability of partial
resistance against black leaf streak disease. Mycosphaerella leaf spot diseases of
banana: present status and outlook. 161-168.
Aguirre, N. Villamizar, L. Espinel , C. Cotes, A. (2009). Efecto del pH y de la actividad de
agua sobre el desarrollo de Nomuraea rileyi (Hyphomycetes). Revista colombiana
de entomología. 35(2), 138-149.
Arias, P., Dankers, C., Liu , P. and Pilkauskas, P. (2003). The World Banana Economy,
1985-2002. Reme: Food and Agriculture Organization of the United Nations.
Cañas-Gutiérrez, G.P. Angarita-Velásquez, M.J. Restrepo-Fiores, J.M. Rodríguez, P.
Moreno, C.X. Arango, R. (2009). Analysis of the CYP51 gene and encoded protein
in Propiconazole-resistan isolates of Mycosphaerella fijiensis. Journal of
Phytopathology. 65 (8) . 892-899.
Cañas-Gutiérrez, G.P. Patiño L.F. Rodríguez-Arango, E. Arango, R. (2006). Molecular
characterization of Benomyl-resistant isolates of Mycosphaerella fijiensis, collected
in Colombia. Journal of Phytopathology. 154(7 -8) , 403-409.
Chin, K.M. Wirz, M. Laird , D. (2001). Sensitivity of Mycosphaerella fijiensis from banana to
trifloxystrobin . Plant Disease. 85, 1264-1270.
Cuevas-Rangel , R. (1979). Prueba de inoculación con el hongo micorrízico Piso/ithus
tinctorius (Pers.) Coker and Couch, en plántulas de Pinus montezumae Lamb. en
suelos de vivero. Revista Mexicana de Ciencias Forestales. 4(19) . 46-62.
Donzell i, B. Churchill , A. (2007), A quantitative assay using mycelial fragments to assess
virulence of Mycosphaerella fijiensis. Phytopathology. (97). 916-929.
113

CAPITULO IV
Faria M. Hotchkiss J. H. Hajek A. E. Wraight S. P. (201 0) . Debilitation in conida of the
entomopathogenic fungi Beauveria bassiana and Metharhizium anisopliae and
impilication with respect to viability determinations and mycopesticide quality
assessments. Journal of invertebrade pathology, 105, 7 4-83.
Fouré, E. 1985. Black leaf streak disease of Banana and plantains (Mycosphaerella
fijiensis Morelet) , study of the symptoms and stages of the disease in Gabon IRFA
Paris.
García, M. Villamizar, L. Torres, L. Cotes, A. (2007). Efecto de subcultivos sucesivos de
Beauveria bassiana sobre sus características y actividad contra Premnotrypes
vorax. Manejo Integrado de Plagas y Agroecología. 77, 50-56.
Hall , R. A. (1980). Effect of repeting subculturing on agar and passing through an insect
host on pathogenicity, morphology and growth rate of Vertilillium lecanii. Journal of
lnvertebrate Pathology. 36(2) , 216-222.
Jones, D.R. (2003). The distribution and importance of the Mycosphaerella leaf spot
diseases of banana. En: Mycosphaerella Leaf Spot Diseases of Bananas: Present
Status and Outlook. Proceedings of the Workshop on Mycosphaerella Leaf Spot
Diseases, San José, Costa Rica, 20-23 May 2002 (Jacome, L., Lepoivre, P.,
Marin, D., Oriz, R. , Romero, R. and Escalant, J.V., eds) , 25-41 . Montpellier: The
lnternational Network for the lmprovement of Banana and Plantain.
Karjalainen , M. (1984). Evaluation of detached seedling leaves for use in spring wheat
cultivars to Septoria nodorum Berk. Acta Agriculture Scandinavia. 34, 386-390.
Leach, C. (1967). lnteraction of near-ultraviolet light and temperature on sporulation of the
fung i Alternaría, Cercosporel/a , Fusarium, Helminthosporum, and Stemphylium.
Canadian Journal of Bothany. 45(11 ), 1999-2016.
Marin,D; Romero,R; Guzmán,M; Sutton, T. (2003). Black Sigatoka and increasing threat to
banana cultivation. Plant Disease. 87(3). 208-222.
114

CAPITULO IV
Mourichon X., Peter , D. Zapater, M.F. (1987). lnoculation expérimentale de
Mycosphaerel/a fijiensis Morelet sur de jeunes plantules de bananiers issues de
culture in vitro. Fruits. 42(4), 195-198.
Peteira, B. Esteves, l. Montes de Oca, N. Hidalgo-Díaz, L. (2007). Estabilidad de la cepa
IMI SO 187 de Pochonia ch/amydosporia val. catenulata en medio sólido. Revista
de Protección Vegetal. 22(2), 124-127.
Romero, R. A. Sutton, T. B. (1997a). Sensitivity of Mycosphaerella fijiensis, Causal Agent
of Black Sigatoka of Banana, to Propiconazole. Phytopathology. 87, 96-1 OO.
Romero, R. A. Sutton, T. B. (1998). Characterization of benomyl resistance in
Mycosphaerel/a fijiensis, cause of black Sigatoka of banana, in Costa Rica. Plant
Disease. 82, 931-934.
Twizeyimana, M. Ojiambo, P.S. Tenkouano, A. lkotun, T. Bandyopadhyay, R. (2007).
Rapid Screening of Musa species for resistance to Black Streak Disease using in
vitro plantlets intubes and detached leaves. Plant Disease. (91), 308-314.
Wijekoon, C.P. Goodwin, P.H. Hsiang, T. (2008). Quantifying fungal infection of plant
leaves by digital image analysis using Scion lmage software. Journal of
Microbiological Methods. 74, 94-101 .
115
Top Related