
Anatomía y Función del Nicho de Células Madre
19
Anatomía y función del nicho de células madre Las células madre son responsables para el crecimiento, la homeostasis y la reparación de muchos tejidos. El mantenimiento y la supervivencia de las células madre está regulada por las aportaciones de su microambiente local a que se refiere como el "nicho de células madre" a menudo. El nicho de células madre hipótesis fue desarrollada en 1978 por Schofield, quien propuso que las células madre residen dentro de los compartimentos fijos, o nichos, que son propicias para el mantenimiento de la definitivos propiedades de células madre 1. Por lo tanto, el nicho representa un compartimiento anatómica definida que proporciona señales a las células madre en la forma de secretada y las moléculas de la superficie celular para controlar la tasa de proliferación de células madre, determinar el destino de las hijas de células madre, y proteger a las células madre a partir de agotamiento o la muerte. Experimentos elegantes en organismos modelo como los gusanos y las moscas siempre la primera visualización de nichos de células madre in vivo y experimentos genéticos posteriores han confirmado la importancia del nicho en la regulación del comportamiento de células madre. Recientemente, nuevas herramientas para etiquetar las células madre in situ también han facilitado la localización y caracterización de los nichos de células madre en tejidos de mamíferos. Además de proporcionar evidencia concreta que los nichos son esenciales para la función adecuada de células madre, estos estudios han revelado que los nichos de células madre son tan variadas como las células madre que apoyan. Por otra parte, el trabajo reciente indica la existencia de clases funcionales diferentes de nicho, cada uno especializado para sostener las funciones únicas de tejidos particulares. Por último, cada vez más pruebas implica la desregulación de la nicho de células madre como una causa proximal de muchas patologías asociadas a la degeneración de los tejidos, el envejecimiento y la tumorigénesis. Como se discute más adelante, los estudios en organismos modelo tales como Drosophila melanogaster y Caenorhabditis elegans han revelado varias características de nichos de células madre que son importantes para controlar el comportamiento de células madre. En primer lugar, las señales que emanan de la regulan nicho de células madre de auto-renovación, la supervivencia y el mantenimiento. En segundo lugar, la relación espacial particular entre las células madre y las células de apoyo puede polarizar las células madre en el nicho para promover divisiones de células madre asimétricos. En tercer lugar, la adhesión entre las células madre y las células del estroma de apoyo y / o los anclajes de la matriz extracelular (ECM) las células madre dentro del nicho en estrecha proximidad a las señales de supervivencia de auto-renovación y. Debido a que los acontecimientos recientes han facilitado la localización y visualización de las células madre dentro de los tejidos demamífero in vivo, está quedando claro que estas características clave de nichos de células madre también se utilizan en sistemas de células madre más complejas. Por lo tanto, el nicho de células madre proporciona un soporte estructural, soporte trófico,
-
Upload
usmp-fn-archivos -
Category
Documents
-
view
442 -
download
3
Transcript of Anatomía y Función del Nicho de Células Madre

Anatomía y función del nicho de células madreLas células madre son responsables para el crecimiento, la homeostasis y la reparación de muchos tejidos. El mantenimiento y la supervivencia de las células madre está regulada por las aportaciones de su microambiente local a que se refiere como el "nicho de células madre" a menudo. El nicho de células madre hipótesis fue desarrollada en 1978 por Schofield, quien propuso que las células madre residen dentro de los compartimentos fijos, o nichos, que son propicias para el mantenimiento de la definitivos propiedades de células madre 1. Por lo tanto, el nicho representa un compartimiento anatómica definida que proporciona señales a las células madre en la forma de secretada y las moléculas de la superficie celular para controlar la tasa de proliferación de células madre, determinar el destino de las hijas de células madre, y proteger a las células madre a partir de agotamiento o la muerte.
Experimentos elegantes en organismos modelo como los gusanos y las moscas siempre la primera visualización de nichos de células madre in vivo y experimentos genéticos posteriores han confirmado la importancia del nicho en la regulación del comportamiento de células madre. Recientemente, nuevas herramientas para etiquetar las células madre in situ también han facilitado la localización y caracterización de los nichos de células madre en tejidos de mamíferos. Además de proporcionar evidencia concreta que los nichos son esenciales para la función adecuada de células madre, estos estudios han revelado que los nichos de células madre son tan variadas como las células madre que apoyan. Por otra parte, el trabajo reciente indica la existencia de clases funcionales diferentes de nicho, cada uno especializado para sostener las funciones únicas de tejidos particulares. Por último, cada vez más pruebas implica la desregulación de la nicho de células madre como una causa proximal de muchas patologías asociadas a la degeneración de los tejidos, el envejecimiento y la tumorigénesis.
Como se discute más adelante, los estudios en organismos modelo tales como Drosophila melanogaster y Caenorhabditis elegans han revelado varias características de nichos de células madre que son importantes para controlar el comportamiento de células madre. En primer lugar, las señales que emanan de la regulan nicho de células madre de auto-renovación, la supervivencia y el mantenimiento. En segundo lugar, la relación espacial particular entre las células madre y las células de apoyo puede polarizar las células madre en el nicho para promover divisiones de células madre asimétricos. En tercer lugar, la adhesión entre las células madre y las células del estroma de apoyo y / o los anclajes de la matriz extracelular (ECM) las células madre dentro del nicho en estrecha proximidad a las señales de supervivencia de auto-renovación y. Debido a que los acontecimientos recientes han facilitado la localización y visualización de las células madre dentro de los tejidos demamífero in vivo, está quedando claro que estas características clave de nichos de células madre también se utilizan en sistemas de células madre más complejas. Por lo tanto, el nicho de células madre proporciona un soporte estructural, soporte trófico, información topográfica y las señales fisiológicas apropiadas para regular la función de las células madre en ambos organismos invertebrados y vertebrados (Figura 1).
En esta revisión, se discuten los conceptos y las cuestiones que rodean nichos de células madre y su papel en la regulación de mantenimiento y reparación de los tejidos actuales. Las células madre tienen un enorme potencial para revelar los mecanismos fundamentales de la especificación del destino celular y el crecimiento del tejido, así como para estimular nuevos enfoques para la reparación y reemplazo de tejidos. Sin embargo, uno de los mayores obstáculos para la mejor comprensión de estas células y su uso en medicina regenerativa es el establecimiento de sistemas ex vivo que apoyan la función de las células madre normales - incluyendo la auto-renovación y diferenciación apropiada linaje específico.

Sólo al descubrir la íntima relación entre las células madre y sus alrededores podemos aspirar a alcanzar los conocimientos necesarios que permitan el desarrollo y la utilización de dichos sistemas.
Nichos de células madrePara entender cómo el microambiente local puede proteger a las células madre e influir en su comportamiento, es necesario en primer lugar determinar dónde residen las células madre. La identificación y caracterización de nichos de células madre se ha complicado por el hecho de que las células madre son extremadamente raros y, en muchos casos, los marcadores específicos que permiten la identificación definitiva de las células madre in vivo se carece. Sin embargo, recientemente se ha hecho un gran progreso en la identificación de nichos de células madre, especialmente en los tejidos de mamíferos. Por ejemplo, muchas células madre hematopoyéticas (HSC) residen a lo largo de la superficie interna del hueso
trabecular en estrecha proximidad a ambos osteoblastos formadores de hueso y las células endoteliales que recubren los vasos sanguíneos. HSC puede dejar este lugar, entrar en la circulación y volver al nicho, y su proximidad a las células endoteliales puede facilitar la movilización de la médula ósea hacia la circulación. Las células madre neurales (NSC) se pueden encontrar en dos lugares diferentes en el cerebro: dentro de la zona subventricular del hipocampo y en el bulbo olfatorio. En ambos nichos, NSC se encuentran adyacentes a las células endoteliales, similares a las CMH. Esta estrecha asociación de las células madre con la vasculatura del tejido podría ser importante para exponer estas células a los factores sistémicos que pueden promover su supervivencia, regular la auto-renovación y potencial de diferenciación, y / o señales de 'daños' se comunican para activar su proliferación.
Las células madre epiteliales residen dentro de una región especializada de la vaina radicular externa del folículo piloso, conocida como la protuberancia folicular. Estas células madre multipotentes pueden contribuir a la regeneración del folículo piloso y las glándulas sebáceas, así como la epidermis interfolicular, aunque no parecen ser necesarias para el normal, la sustitución homeostático de las células epidérmicas. Las células madre que repueblan la epidermis interfolicular, conocido como queratinocitos basales, se encuentran en la base de la epidermis, inmediatamente por encima de una membrana basal que los separa de la dermis subyacente. Dentro del intestino delgado de mamíferos, gut células madre residen en la base de las criptas intestinales y se dividen para producir células hijas que se diferencian a medida que migran hacia arriba, hacia las vellosidades que se extienden en el lumen intestinal. Aunque las células madre intestinales (ISC) se pensaron inicialmente a residir inmediatamente por encima células de Paneth de las criptas (en la posición 4), análisis reciente rastreo de linaje en cambio, ha puesto de manifiesto que su actividad sigue a una nueva población de células columnares base de la cripta (CBC) que se caracterizan por la expresión de LGR5 (Leu-rico-repetir que contienen la proteína G-receptor acoplado a-5) y se entrelazan entre las células de Paneth. Las células madre de espermatogonias (SSC) mantienen la espermatogénesis durante toda la vida de los hombres adultos. SSC se encuentran adyacentes a la membrana basal a lo largo de la periferia de los túbulos seminíferos de los testículos. Estudios recientes han demostrado, sin embargo, que la distribución de indiferenciado espermatogonias, que probablemente incluye SSC, no es al azar. Estas células parecen localizar preferentemente cerca de la red vascular y las células intersticiales que existen entre los túbulos adyacentes. Células madre de músculo esquelético, un subconjunto de las células satélite-fibra-musculares asociados, se encuentran a lo largo de la longitud de la myofibre, en estrecho contacto con la membrana plasmática myofibre y por debajo de su membrana basal. Curiosamente, parece que las células madre de tejido a menudo se encuentran en ubicaciones donde están relativamente protegidas de daños (tales como toxinas ambientales o radiación ultravioleta) en comparación con las células más diferenciadas que producen. Las características específicas de cada uno de estos nichos se discuten con más detalle a continuación (Tabla1).

Componentes de nichos de células madreNichos de células madre prototípicos, incluidos los que la sangre de apoyo, las células madre foliculares bulto línea germinal y epitelial, han puesto de manifiesto varias características físicas y funcionales que parecen ser características de un nicho de células madre (figura 1). Al sintetizar los datos de numerosos sistemas, podemos generar "lista de partes" de un hipotético para nichos de células madre, incluyendo: la propia de células madre, las células del estroma de apoyo que interactúan directamente con las células madre y con los demás a través de receptores de la superficie celular, uniones y los factores solubles, proteínas ECM que proporcionan estructura, organización y señales mecánicas en el lugar; vasos sanguíneos que transportan las señales sistémicas y proporcionan un conducto para la contratación de inflamación y otras células que circulan en el lugar, y las entradas neuronales que podrían igualmente comunicar señales fisiológicas distantes al microambiente de células madre (figura 1). Aunque no todos los nichos necesariamente incorporan todos estos componentes distintos, es evidente a partir de esta suma que el nicho representa una entidad compleja y dinámica en la que la integración de múltiples entradas lleva a cabo un control exquisito del número de células madre y la función.
Factores secretados. Comunicación dentro del nicho es esencial para el mantenimiento de la función adecuada de células madre y para la determinación del tipo de células madre de auto-renovación. Factores secretados pueden actuar localmente (dentro de 1-2 diámetros celulares) o pueden difundirse por todo el lugar a las decisiones del destino de células madre directos. Estudios en moscas y gusanos indicaron que las células de apoyo, que se encuentran adyacentes a las células madre, factores secretan que se requieren para el mantenimiento de la identidad de células madre y para la especificación de células madre de auto-renovación. Factores de crecimiento secretados también se han demostrado para regular el comportamiento de células madre en sistemas de células madre de mamíferos (ver más abajo). A medida que nuestra comprensión de las interacciones celulares en el nicho sigue siendo bastante rudimentario, es probable que el trabajo futuro revelará células madre de tejidos específicos de las vías de señalización que pueden ser diferentes para cada tipo de células madre y dentro de cada nicho de células madre.
Un estudio de las vías de señalización que se han identificado en sistemas de células madre caracterizadas revela notable conservación de las cascadas de señalización utilizados, pero las consecuencias de la activación de estas vías puede ser diferente entre los diversos tejidos (TABLA 1). Un ejemplo, que consideramos específicamente en la siguiente discusión para ilustrar este punto, es la vía de transducción de señales Wnt. A pesar de la señalización de Wnt se ha implicado en muchos sistemas de células madre, parece ser explotada para fines distintos en cada uno (figura 2). En el epitelio intestinal, la señalización de Wnt parece estar implicado en el apoyo a la proliferación de las células madre y las células de amplificación
de tránsito, así como los que dirigen la diferenciación de un subconjunto específico de células, las células de Paneth, en la base de las criptas dentro del intestino delgado. Aunque las células específicas que secretan las proteínas Wnt no se han identificado en el intestino, células mesenquimales rodean las criptas y podrían servir como una fuente local.
La vía de transducción de señales Wnt también tiene un papel en la especificación de células madre de auto-renovación de las CMH. Sin embargo, en lugar de ser expresado por las células del estroma circundante, Wnt puede ser secretado a partir de las HSC sí mismos y podrían actuar en un bucle autocrino para controlar la proliferación de células madre (Figura 2). La señalización de Wnt puede ser particularmente importante para la mediación de la supervivencia de las HSC fetal y neonatal, porque inducida experimentalmente deleción del marcador de células madre fetalesSOX17 induce la expresión del antagonista Wnt Dickkopf-1 (DKK1) por HSC y posteriormente causa la muerte HSC. Es interesante considerar que la producción de factores inhibidores del crecimiento por las propias células madre podría proporcionar un mecanismo simple para determinar el número de células madre dentro de un nicho o tejido dado. Las células madre que están presentes

más, cuanto mayor sea la concentración de factores inhibidores del crecimiento, hasta que se alcanza un umbral que provoca una detención o retraso en la proliferación de células madre.Por el contrario, como el nicho de células madre se agota de células madre endógenas, la concentración de factores inhibitorios se reduciría, dando lugar a la activación de la proliferación de células madre (o la atracción de la migración de las células madre) para repoblar el nicho. Este modelo bien especulativo pone de relieve la importancia de las células madre en sí como un componente activo del nicho, y el papel del nicho como una unidad estructural que se concentra factores moduladores de células madre.
Considerando que la vía de señalización de Wnt es importante para células madre de auto-renovación en el intestino y la sangre, que se utiliza para la diferenciación específica de tejido directa en otros sistemas de células madre. Por ejemplo, en la epidermis de mamífero, la señalización de Wnt parece tener un papel complejo en la diferenciación de los precursores del folículo piloso, en lugar de la auto-renovación de las células madre multipotentes en la protuberancia folicular (Figura 2). En el folículo del pelo, las células madre bulto expresan altos niveles de inhibidores de la señalización de Wnt. Además, la activación excesiva de la señalización de Wnt puede acelerar el ciclo del pelo y promover el crecimiento de nuevos folículos, llevando eventualmente a los tumores de piel. Del mismo modo, en el músculo esquelético, la señalización de Wnt promueve la diferenciación terminal y la fusión de mioblastos proliferantes, y también tiene una función dependiente de la edad recién apreciado en la determinación del equilibrio entre la miogénesis y la fibrosis en los músculos lesionados.
Además de los factores de proteínas secretadas, pequeñas moléculas e iones pueden proporcionar señales importantes en nichos de células madre. En la médula ósea, las concentraciones locales altas de Ca 2 + aparecen para facilitar la localización de las HSC adyacentes a los osteoblastos en el endostio. Del mismo modo, el microambiente hipóxico de nichos de HSC en el hueso puede ser importante para limitar la exposición de las HSC a las especies reactivas de oxígeno, que aparecen para inducir una respuesta de estrés oxidativo que conduce a la disfunción de HSC. Por lo tanto, múltiples factores solubles y moléculas pequeñas que inciden sobre la función de células madre se pueden concentrar en el nicho de tal manera que sus actividades pueden ser integrados con las entradas adicionales para determinar las respuestas de células madre apropiadas a los estímulos fisiológicos.
La adhesión celular. Unión física de las células del estroma de soporte o a una lámina basal también es importante para la regulación del comportamiento de células madre y ayuda a mantener las células madre en el nicho, en estrecha proximidad a las señales de auto-renovación. Uniones adherentes son contactos célula-célula que están formado por las interacciones entre las proteínas transmembrana homotípicos llamadas cadherinas. La pérdida de la función de las cadherinas específicamente dentro de las células madre de línea germinal (GSCS) en el ovario o testículo mosca interrumpe uniones adherentes entre las células madre y las células de apoyo y hace que la pérdida posterior de células madre, lo que indica que la adhesión célula-célula se requiere para el mantenimiento de células madre en la D. melanogaster gónadas (J. Voog y DLJ, observaciones no publicadas). Basado en estudios de expresión, la adhesión celular mediada por cadherina También se ha sugerido para facilitar la asociación de HSC con osteoblastos (a través de N-cadherina), y se ha implicado en la determinación de la correcta colocación de las células satélites musculares a lo largo de la fibra muscular (a través de M-cadherina) . Sin embargo, los ratones que carecen M-cadherina no muestran defectos en el desarrollo del músculo esquelético o de la regeneración, y estudios recientes argumentan en contra de la participación de N-cadherina en la regulación de las HSC; esta proteína no se expresa de manera detectable por CMH de médula ósea identificadas fenotípicamente, y todo el hematopoyética la actividad de la médula ósea normal reconstituir parece residir en el antagonista Dickkopf-1 (DKK1) por HSC y posteriormente causa la muerte HSC. Es interesante considerar que la producción de factores inhibidores del crecimiento por las propias células madre podría proporcionar un mecanismo

simple para determinar el número de células madre dentro de un nicho o tejido dado. Las células madre que están presentes más, cuanto mayor sea la concentración de factores inhibidores del crecimiento, hasta que se alcanza un umbral que provoca una detención o retraso en la proliferación de células madre. Por el contrario, como el nicho de células madre se agota de células madre endógenas, la concentración de factores inhibitorios se reduciría, dando lugar a la activación de la proliferación de células madre (o la atracción de la migración de las células madre) para repoblar el nicho. Este modelo bien especulativo pone de relieve la importancia de las células madre en sí como un componente activo del nicho, y el papel del nicho como una unidad estructural que se concentra factores moduladores de células madre.
Considerando que la vía de señalización de Wnt es importante para células madre de auto-renovación en el intestino y la sangre, que se utiliza para la diferenciación específica de tejido directa en otros sistemas de células madre. Por ejemplo, en la epidermis de mamífero, la señalización de Wnt parece tener un papel complejo en la diferenciación de los precursores del folículo piloso, en lugar de la auto-renovación de las células madre multipotentes en la protuberancia folicular (Figura 2). En el folículo del pelo, las células madre bulto expresan altos niveles de inhibidores de la señalización de Wnt. Además, la activación excesiva de la señalización de Wnt puede acelerar el ciclo del pelo y promover el crecimiento de nuevos folículos, llevando eventualmente a los tumores de piel. Del mismo modo, en el músculo esquelético, la señalización de Wnt promueve la diferenciación terminal y la fusión de mioblastos proliferantes, y también tiene una función dependiente de la edad recién apreciado en la determinación del equilibrio entre la miogénesis y la fibrosis en los músculos lesionados.
Además de los factores de proteínas secretadas, pequeñas moléculas e iones pueden proporcionar señales importantes en nichos de células madre. En la médula ósea, las concentraciones locales altas de Ca 2 + aparecen para facilitar la localización de las HSC adyacentes a los osteoblastos en el endostio. Del mismo modo, el microambiente hipóxico de nichos de HSC en el hueso puede ser importante para limitar la exposición de las HSC a las especies reactivas de oxígeno, que aparecen para inducir una respuesta de estrés oxidativo que conduce a la disfunción de HSC. Por lo tanto, múltiples factores solubles y moléculas pequeñas que inciden sobre la función de células madre se pueden concentrar en el nicho de tal manera que sus actividades pueden ser integrados con las entradas adicionales para determinar las respuestas de células madre apropiadas a los estímulos fisiológicos.
La adhesión celular. Unión física de las células del estroma de soporte o a una lámina basal también es importante para la regulación del comportamiento de células madre y ayuda a mantener células madre dentro del nicho, en estrecha proximidad a las señales de auto-renovación. Uniones adherentes son contactos célula-célula que están formado por las interacciones entre las proteínas transmembrana homotípicos llamadas cadherinas. La pérdida de la función de las cadherinas específicamente dentro de las células madre de línea germinal (GSCS) en el ovario o testículo mosca interrumpe uniones adherentes entre las células madre y las células de apoyo y hace que la pérdida posterior de células madre, lo que indica que la adhesión célula-célula se requiere para el mantenimiento de células madre en la D. melanogaster gónadas (J. Voog y DLJ, observaciones no publicadas). Sobre la base de estudios de expresión, la adhesión celular mediada por cadherina También se ha sugerido para facilitar la asociación de HSC con osteoblastos (a través de N-cadherina), y se ha implicado en la determinación de la correcta colocación de las células satélite de músculo a lo largo de la fibra muscular (a través de M-cadherina) . Sin embargo, los ratones que carecen M-cadherina no muestran defectos en el desarrollo del músculo esquelético o de la regeneración, y estudios recientes argumentan en contra de la participación de N-cadherina en la regulación de las HSC; esta proteína no se expresa de manera detectable por CMH de médula ósea identificadas fenotípicamente, y todo el hematopoyética la actividad de la médula ósea normal reconstituir parece residir en la membrana dentro de los grupos o nidos de 2-3 células diploides basales localizados que se intercalan entre los enterocitos

poliploides. Análisis de la orientación del husillo mitótico en ISC divisorias indica que estas células madre se dividen de forma no aleatoria, de tal manera que la hija de CAI que queda adyacente a la membrana basal sigue siendo un ISC, mientras que la célula hija que se desplaza se diferencia para formar un enteroblast. División asimétrica de ISC está mediada por la vía de señalización de Notch. Aunque todas las células en los nidos que contienen las células madre son Notch +, sólo los ISC en contacto directamente con la membrana basal y las manchas positivas para el Delta del ligando Notch. Curiosamente, la señalización de Notch se activa exclusivamente en el enteroblast hija, en lugar de en el ISC. Por lo tanto, parece que la señal de ISC a través de Delta para activar genes diana de Notch en enteroblasts. Descubrir cómo la señalización Notch está bloqueada dentro del ISC para facilitar esta división asimétrica es probable que revele nuevos paradigmas de cómo se regulan las células madre auto-renovación y mantenimiento.
Establecimiento y facturación de los nichosDado el número de componentes y la complejidad de las interacciones dentro del nicho de células madre, es evidente que la formación y la actividad de nichos deben ser regulados cuidadosamente para controlar apropiadamente el número de células madre y el comportamiento. En muchos sistemas, nichos parecen formar en tiempos de desarrollo discretas, y su aspecto a menudo permite el establecimiento o reclutamiento de células madre en localizaciones anatómicas particulares. Es significativo que, una vez formados, estos nichos responden dinámicamente a las señales homeostáticas y regenerativa y puede exhibir importantes alteraciones fisiológicas que afectan la forma en que interactúan con las células madre que soportan.
Establecimiento. El establecimiento de nichos de células madre puede proceder por al menos dos mecanismos distintos. En primer lugar, los nichos pueden surgir durante el desarrollo de tipos de células heterólogas, y pueden existir de manera estable si o no las células madre están presentes para ocupar ellos. En la D. melanogaster ovario y testículo, el componente somático de la gónada (incluyendo las células que en última instancia contribuir al nicho) se forma en la ausencia de GSCS. En los testículos de mamíferos, células de Sertoli apoyan muchos aspectos de la espermatogénesis, incluyendo SSC auto-renovación. Similar a la situación que se produce en el D. melanogaster gónadas, las células de Sertoli están presentes y mantenido incluso en los testículos aspermic (tales como las de c-kit mutante (W / W v) ratones). Sin embargo, a pesar de la ausencia de células germinales en los testículos, las células de Sertoli son plenamente competentes para apoyar la espermatogénesis, como lo demuestra la fertilidad restaurada siguiendo SSC trasplante. Curiosamente, en los ratones que carecen del ligando c-kit SLF (que son infértiles debido a un defecto en el apoyo de la diferenciación de células de Sertoli SSC), el trasplante de las células normales de Sertoli restaura la función del lugar y de la fertilidad, lo que sugiere que las SSC en realidad pueden ser mantenidas en ausencia de estas células nicho.
En segundo lugar, las células madre y las células de apoyo que interactúan con ellos pueden desarrollar de forma conjunta, con la aparición de cada ser dependiente de especificación correcta, la localización y las interacciones con la otra. Este escenario se demuestra dramáticamente en los estudios de trasplante de células madre epidérmicas de mamíferos de la región protuberancia folicular. Cuando las células madre foliculares bulto y queratinocitos diferenciados purificada se trasplantaron juntos sobre el dorso de ratones desnudos heridos sin pelo, las células trasplantadas generado por todos los linajes epidérmicas, incluyendo grandes mechones de pelo tallo-derivado de células y nuevos nichos de células madre dentro de la región protuberancia de donante derivados de folículos pilosos de la piel de acogida. Del mismo modo, en el músculo esquelético, los mismos precursores embrionarios que delaminate de los somitas y de mediar en la formación de los músculos esqueléticos en la extremidad también dan lugar a células musculares satélite, que se convierten encerrado debajo de la lámina basal de las miofibras maduras y se mantienen como miogénica músculo-residente las células madre. Por otra parte, la regeneración muscular, mediado por las células

musculares satélite adultos, implica tanto la reforma de miofibras maduras y de re-siembra de las células madre de músculo en el nicho myofibre regenerada. Por lo tanto, en tanto embriogénesis y regeneración adulto, el músculo nicho de células madre se forma a partir de los mismos precursores que generan células madre de músculo a sí mismos.Por último, varias poblaciones de células madre de tejidos, tales como las HSC y células germinales
primordiales (PGC), encuentran múltiples nichos en todo el desarrollo que podrían ser personalizados para dar soporte a divisiones simétricas frente asimétrica; para facilitar la rápida proliferación o imponer quiescencia de células madre, o a la diferenciación de sesgo las células progenitoras hacia un linaje particular, (discutido a continuación). En el desarrollo de los ratones, las HSC aparecen secuencialmente en el saco vitelino, aorta dorsal, placenta, hígado fetal, bazo y en última instancia, la médula ósea, que sirve como el sitio predominante de la hematopoyesis en la edad adulta. Curiosamente, en esta progresión, parece que el desarrollo de cada nicho generalmente precede a la siembra con células madre. Debido a que las CMH están constantemente presentes en la circulación fetal, esto podría indicar que la formación de nichos adecuados es el paso limitante de la velocidad en la siembra de las HSC en estos tejidos. Por el contrario, la desaparición de desarrollo temporizada de las HSC a partir de tejidos tales como el hígado puede reflejar una pérdida de nichos en estos sitios. Alternativamente, la siembra desarrollo programado de los órganos hematopoyéticos por las HSC puede ser resultado de la adquisición secuencial inducida por la exposición con el nicho "corriente" de los receptores de homing o de adhesión que son necesarios para la localización en el "siguiente" sitio. Esta hipótesis sería consistente con los datos que sugieren que la transición a o secuencial a través de cada uno de estos compartimentos anatómicos distintos puede proporcionar señales importantes que son necesarios para la maduración completa de las CMH.
Similar a las HSC, las CGP (los precursores embrionarios de la línea germinal) se forman en un sitio anatómico que es distinta de su lugar de descanso final en el organismo completamente desarrollado. En el ratón, las CGP se especifican a partir de células del epiblasto proximal y deben cumplir una poligonal complicado y desarrollo temporizada a través del organismo en desarrollo, que pasa secuencialmente a través de la línea primitiva, endodermo definitivo y visceral, alantoides, intestino grueso y el mesenterio dorsal para llegar a los genitales crestas. Curiosamente, esta cuidadosamente orquestada migración parece estar regulada en parte por proteínas quimiotácticas como CXCR4 SDF1α y en parte por la localización anatómica de los factores de supervivencia PGC, como c-kit-SLF, ya que la inhibición de la muerte es el resultado PGC apoptóticas en la acumulación ectópica de PGC en lugares inapropiados.
Volumen de negocios del lugar. Teniendo en cuenta que la disponibilidad de nichos en muchos sistemas puede controlar el número de células madre, los mecanismos que determinan el número de nichos es probable que tengan un impacto directo en la actividad de las células madre en un determinado tejido. En general, hay poca información disponible sobre cómo se determina el número de nichos, ¿cómo se mantiene el mismo lugar después de que se establezca, ni la frecuencia puede ser reemplazado los componentes celulares de nichos. Significativamente, la dinámica de la renovación de las células nicho pueden variar de tejido a tejido o en diferentes etapas de desarrollo. Sin embargo, en varios sistemas, una disminución de la función o nicho en el número total de nichos puede conducir a la pérdida posterior o desregulación de las células madre de tejidos (Cuadro 1). La importancia potencial de la disfunción nicho se destacó en particular por los datos recientes que sugieren que hay una entrada de microambientales significativa en las deficiencias asociadas con la edad de la función de células madre. Mantenimiento y la regeneración de tejidos tales como la piel, el hígado, la sangre y el músculo disminuyen enormemente con la edad. En algunos casos, se han propuesto cambios autónomos de células que tienen un papel en la disminución observada en la función de células madre de tejido, sin embargo, los cambios locales y sistémicos de células extrínsecos también contribuyen a la disminución de la capacidad de las células madre de edad para reparar adecuadamente los tejidos dañados.

El trabajo reciente ha demostrado directamente que los cambios relacionados con la edad se producen dentro del nicho de células madre que podría contribuir a la deficiente número de células madre y / o actividad en los tejidos envejecidos. Por ejemplo, el trasplante de las ESC de jóvenes, ratones machos fértiles en los testículos atrofiados de edad, los hombres no para producir la espermatogénesis robusta o el aumento de peso de los testículos, lo que indica una disminución relacionada con la edad en la capacidad del nicho de células madre para apoyar la colonización y / o auto renovación de las ESC trasplantados. Los estudios complementarios mostraron una disminución de 73% en la expresión de la citocina factor de célula
glial-derivado de la línea neurotrófico (GDNF) en el envejecimiento (15-19 meses de edad), los varones infértiles relativos a los jóvenes (2-4 meses de edad), fértil control de los hombres. Debido a que el GDNF, que es secretada por las células de Sertoli, se ha demostrado que se requiere para SSC auto-renovación y mantenimiento (TABLA 1), una disminución significativa en los niveles de GDNF en los testículos de los machos viejos podría proporcionar un mecanismo por el cual el envejecimiento de el nicho conduce a la disminución de la actividad SSC y la espermatogénesis en machos viejos. Estos datos son similares a la disminución relacionada con la edad en la expresión de las señales de auto-renovación de células de apoyo clave en el D. melanogaster macho nicho de SGC, y por lo tanto puede sugerir un mecanismo molecular conservada que contribuye al envejecimiento de la nicho de células madre.
Consistente con esta hipótesis, los cambios dentro del nicho también parecen afectar a la función de células madre en el músculo esquelético de mamíferos de edad. Trasplante de pequeñas cantidades de tejido de músculo entero o picada, procedentes de donantes jóvenes viejos en camas receptores musculares o, a la inversa, a partir de donantes jóvenes en camas musculares de edad, indica que la actividad regenerativa del músculo está determinada en gran parte por la edad de la microambiente de acogida. Sorprendentemente, la "edad" del músculo del nicho de células madre parece estar determinada principalmente por la circulación de factores sistémicos. La exposición de las células satélite de músculo viejos a la sangre transmitidas por factores rejuvenece su actividad regenerativa, tanto in vivo como in vitro, mientras que por el contrario, la exposición de las células satélite jóvenes de edad suero inhibe su potencial miogénica, favoreciendo en cambio la fibrosis y la formación de cicatrices.
La flexibilidad y la funciónAl igual que las células madre en diferentes tejidos se regulan de manera diferente en respuesta a diversas demandas para el reemplazo de células, los nichos que apoyan estas células exhiben una diversidad de funciones para proporcionar señales de desarrollo y homeostático que sean apropiadas a cada tejido. Por ejemplo, en muchos sistemas, tales como el intestino y epidermis, las células madre deben funcionar continuamente para reponer las células maduras que muestran normalmente una duración de vida limitada. En estos sistemas, el nicho debe suministrar señales apropiadas para equilibrar de células madre de auto-renovación y diferenciación para mantener la producción en curso de células especializadas sin agotamiento catastrófico de la piscina de células madre. Por el contrario, en otros tejidos tales como músculo esquelético, la progenie de las células madre madura (fibras musculares) son de larga vida, y por lo general requieren el reemplazo sólo cuando dañado por una lesión o enfermedad. En este caso, el nicho debe proporcionar estas células madre facultativos con señales inhibitorias que promueven su supervivencia y prevenir su diferenciación bajo condiciones de estado estacionario de mantenimiento y, al mismo tiempo mantener la capacidad de respuesta a las señales regenerativas de manera que estas células madre pueden ser movilizados en acción cuando sea necesario . Por último, en algunos sistemas, tales como la sangre, se necesitan ambas funciones debido a las HSC no sólo deben mantener la producción diaria de células sanguíneas, sino que también deben responder robusta para expandir células precursoras primitivas después de insulto hematopoyética.
Exactamente cómo afecta el nicho de la conversión de regulada homeostática a los modos de regeneración de células madre de mantenimiento y auto-renovación todavía no está

claro. Sin embargo, esto podría implicar la inducción de reclutamiento o de nichos alternativos, alteración programada de nichos existentes, o ambos. Por lo tanto, se derivan nichos de células deben ser dinámico en ough para proporcionar señales de desarrollo y homeostático adecuados para regular el comportamiento de células madre en una manera específica de tejido, y en respuesta a diversos estímulos fisiológicos y condiciones patológicas. Curiosamente, el número de células somáticas de apoyo se expande en la ausencia de GSCS en el D. melanogaster gónadas, lo que resulta en una aparente expansión del nicho de células madre. Además, trabajar en el D. melanogaster ovario ha demostrado que los nichos de células madre que se agotan de células madre endógenas mantienen la capacidad de señal para soportar la proliferación y ectópica de células entrantes, lo que indica que los nichos vacíos pueden ser capaces de promover la auto-renovación, la proliferación y / o supervivencia de las células que normalmente no residen dentro de ese nicho. Los aumentos en el número de células de apoyo han sido inducida experimentalmente en el sistema de la sangre de mamíferos, utilizando enfoques genéticos o farmacológicos para expandir los osteoblastos de hueso-revestimiento. Los osteoblastos proporcionan por lo menos uno de los nichos que son importantes para la función HSC (Tabla 1) , y la expansión inducida de los osteoblastos in vivo provoca la expansión concomitante de las HSC (BOX 2,. Figura
3) . Estos datos apoyan la noción de que los nichos de células madre son dinámicos, debido a que el número de células de apoyo y nichos disponibles parece responder a las señales fisiológicas complejas, así como la presencia o ausencia de la piscina de células madre endógenas.
Lecciones del sistema hematopoyético. Como se ha indicado anteriormente, muchas células madre presentan cambios en el desarrollo o contextuales en su función y actividad, y estos pueden reflejarse en diferencias en los nichos que ocupan. Este paradigma está particularmente ejemplificado por el sistema hematopoyético, en la que el sitio primario de la producción de células sanguíneas cambia secuencialmente por la transición a múltiples localizaciones anatómicas antes de establecerse en última instancia, en la médula ósea justo antes del nacimiento. Curiosamente, además de su localización, el potencial destino de la célula de las HSC también cambia durante el desarrollo, con varios linajes de células sanguíneas que surjan durante la hematopoyesis prenatales que no pueden ser generados por las HSC adultas (Vγ5 + células T, por ejemplo).
HSC fetales y adultas también presentan notables diferencias en su superficie celular marcador expresión y la actividad proliferativa. En el ratón, la transición de las HSC de fetal a las propiedades adultos se produce dentro de las primeras semanas después del nacimiento, y parece implicar un desarrollo intrínsecamente cronometrada reprogramación que se asocia con la pérdida de la expresión del ADN SOX17 factor de unión dentro de las CMH. SOX17 es necesario para la supervivencia de las HSC fetales y neonatales, pero es prescindible para la supervivencia HSC adulto. Precisamente cómo la transición de SOX17 + CMH de tipo fetal al SOX17 - es orquestada CMH de tipo adulto sigue siendo poco clara, pero es posible que esta maduración se inicia o se ven favorecidos por la exposición a las señales de las CMH microambientales diferentes que son inducidos poco después del nacimiento. Esta hipótesis, que aún no se ha probado experimentalmente, sugiere que las propiedades funcionales distintas de diferentes nichos de células madre en diferentes momentos del desarrollo propiedades HSC el grado de desarrollo específicos directos y especificación de distintos linajes de células sanguíneas.
Del mismo modo, incluso en el sistema de la sangre de adultos, pueden existir múltiples nichos para las CMH (incluyendo otros osteoblásticas, vasculares y tal vez) y pueden proporcionar funciones de regulación distintos para equilibrar la auto-renovación y diferenciación, lo que lleva a la producción de células sanguíneas maduras. La dificultad hasta la fecha en el desciframiento de la importancia fisiológica relativa de distintos nichos HSC surge en parte del hecho de que los estudios de nichos HSC in vivo , incluso en los animales adultos, son particularmente desafiados por la naturaleza migratoria inusual de la sangre de formación de las CMH. Utilizando ratones parabiotic, nos habíamos hecho la sorprendente observación de que HSC adultas constitutivamente recirculan en condiciones fisiológicas normales. Estas células pasan claramente de la médula ósea al torrente sanguíneo, y pueden

volver a la médula ósea para volver a injertar y semillas hematopoyesis en curso (Figura
1) . Curiosamente, las HSC también aparecen al tráfico de la circulación sanguínea en los tejidos no hematopoyéticos periféricos, en los que pueden participar en la respuesta inmune o inflamatoria locales, o pueden pasar de nuevo a la circulación a través del sistema linfático. Las estimaciones del flujo normal de las HSC en el cuerpo sugieren una sorprendentemente alta magnitud del tráfico de HSC en animales normales, lo que hace difícil para inferir la función primaria de un nicho (es decir, ¿promueve HSC auto-renovación, quiescencia, diferenciación o migración?) únicamente por la visualización directa de su interacción con las CMH in situ. Futuros estudios de acoplamiento de alta resolución en vivo serán necesarios microscopía, aislamiento de células nicho inequívoca, y los indicadores funcionales o genéticos directos de las decisiones de destino celular HSC para aclarar estas cuestiones importantes.
ConclusionesNichos de células madre son estructuras complejas e interactivas que integran señales locales y sistémicas para la regulación positiva y negativa de las actividades de células madre de una manera definida espacial y temporalmente. 'Lista de piezas' El componente de nichos es extensa, incluyendo entidades celulares y acelulares, solubles y moléculas de señalización unido a la membrana, entradas mecánicas y químicas, y de dirección, así como de control de retroalimentación. Recientemente, grandes avances se han hecho en la identificación de los circuitos de control que median las interacciones de células madre con el nicho a nivel molecular. Estos estudios han puesto de manifiesto una notable conservación de la función y, en algunos casos de los efectores moleculares de estas funciones, entre los nichos de células madre. Además, los avances en la tecnología de formación de imágenes y la identificación de marcadores de células madre han mejorado sustancialmente la capacidad de visualizar las células madre in situ dentro del nicho, revelar nuevos conocimientos sobre la organización anatómica y estructural de estos compartimentos. Con estas herramientas en la mano, el campo está preparado para responder a varias preguntas que son de importancia fundamental para frenar la biología celular. Por ejemplo, ¿cuál es la tasa de rotación de los nichos, y cómo afecta esto a la función de las células madre? ¿Cómo son diferentes señales equilibradas e integradas en el nicho? Es el movimiento del nicho necesario para la diferenciación de células madre en todos los sistemas de células madre? Cómo (y por qué) no nichos de células madre cambian en el contexto de la enfermedad y el envejecimiento? Las respuestas a estas y otras preguntas nos movernos más cerca de alcanzar la compleja comprensión multidimensional de las células madre y sus nichos que sean necesarias para que la recapitulación de estos microambientes nativos fuera del cuerpo, así como para dirigir in vivo la manipulación del nicho para modular la función de las células madre endógenas. Tales habilidades se dió un conocimiento más sofisticado de la función del tejido y facilitarán terapias basadas en células madre nuevas y mejoradas.

Figura 1 | Componentes y funciones de nichos de células madre. El nicho es una estructura compleja y dinámica que transmite y recibe señales a través de mediadores celulares y acelulares. Este esquema representa un compuesto nicho hipotética, que resume los componentes conocidos de los descritos anteriormente nichos de mamíferos y no mamíferos: la célula madre en sí, las células del estroma, factores solubles, matriz extracelular, entradas neurales, red vascular y los componentes de adhesión celular. Es importante tener en cuenta que, aunque muchos componentes de nicho se conservan, es poco probable que todos los nichos incluye necesariamente todos los componentes enumerados. En cambio, los nichos son propensos a incorporar una selección de estas posibles vías de comunicación, adaptados específicamente a las funciones particulares de ese nicho, lo que podría ser la de proporcionar un soporte estructural, soporte trófico, información topográfica y / o señales fisiológicas.
Cuadro 1 | Un nicho de las células madre del cáncer?
Descripciones clásicas de transformación celular y tumorigénesis citan a menudo sobreproliferación apropiado y la pérdida de inhibición de contacto como las propiedades característica de las células malignas. Dado que las células de nicho normalmente limitan o controlan la división de células madre, estos datos argumentan que la pérdida de entrada desde el nicho puede permitir la proliferación excesiva de células madre, que podrían predisponer a la transformación. Por lo tanto, a la luz del hecho de que los cánceres son ahora cada vez más reconocidos como enfermedades que son mantenidos por auto-renovación de células madre del cáncer (CSC), este modelo sugiere que la independencia de los requisitos de nicho podría diferenciar entre un tumor-la propagación de células madre cancerosas de sus homólogos normales.

Por el contrario, la evidencia adicional apoya un modelo alternativo mediante el cual el apoyo directo de nichos especializados CSC puede estar implicada en la iniciación del tumor y / o es esencial para el mantenimiento del tumor. En un ejemplo convincente, Parada y colegas hicieron la sorprendente observación de que, en un modelo de ratón de neurofibroma inducida por la inactivación de la NF1 supresor de tumores en las células gliales, haploinsuficiencia de NF1 en tipos de células no neuronales del microambiente del tumor en realidad promueve el desarrollo de tumores, en parte, por la contratación de alteración funcional mastocitos , que proporcionan factores tróficos para apoyar el crecimiento del tumor. Además, los estudios que utilizan modelos de ratón indican que las células de apoyo no neoplásicas especializados pueden actuar como balizas para el reclutamiento de las células metastásicas en el establecimiento de tumores secundarios en sitios distales. Además, los perfiles de expresión de las células del estroma que están asociados con carcinomas de células basales humanos demostró que el tumor-estroma asociado, pero no estroma asociado con la piel no tumoral, expresa los factores secretados que bloquean la diferenciación de las células epiteliales dentro del tumor. Por lo tanto, una fuerte evidencia soporta múltiples funciones del microambiente tumoral en la promoción y el mantenimiento de un crecimiento anormal de células en el contexto de la malignidad, y alienta además la novela investigación del nicho de CSC como un nuevo objetivo potencial para la terapia del cáncer (véase también el Recuadro 2 ).
Figura 2 | funciones múltiples para la señalización de Wnt dentro de nichos de células madre. señalización Wnt puede promover la proliferación celular, tales como la auto-renovación de células madre hematopoyéticas (HSC) ( a ) o la proliferación de las células de amplificación de tránsito dentro de las criptas intestinales (no se muestra). Dentro de algunos tejidos, sin embargo, la señalización de Wnt dirige la diferenciación de linajes celulares específicos, tales como precursores de folículos pilosos ( b ), en lugar de promover la autorrenovación de las células madre multipotentes de la protuberancia folicular. Ver el texto principal y Tabla 1 para más detalles. ANG1, angiopoyetina-1; BMP4, la proteína morfogenética ósea 4; EPHR, ephrin receptor; GSK3, glucógeno sintasa quinasa-3β; TIE2, angiopoyetina-1 receptor; LEF, el factor potenciador linfoide; OPN, osteopontina; TCF, factor de células T .
Cuadro 2 | modulante el nicho para la terapia
Varios estudios intrigantes ahora apoyan la noción de que la actividad de células madre puede ser modulada indirectamente por dirigidas específicamente a las células de nicho. Por ejemplo, en una elegante serie de experimentos de prueba-en-concepto, Adams ha demostrado recientemente que los tratamientos hormonales que estimulan la activación de la nicho osteoblástica podrían aumentar el número de células madre hematopoyéticas (HSC) in vivo. Por lo tanto, tales tratamientos pueden ser utilizados terapéuticamente para aumentar el número de las HSC que pueden ser recogidos para el trasplante, para mejorar la expansión de estas células inmediatamente después del trasplante, y para proteger a los animales de haematotoxins. Asimismo, estudios recientes con ratones parabiotic sugieren que los cambios en el microambiente del músculo esquelético que se acumulan con la edad están

regulados por señales sistémicas que inclinan la balanza regenerativa en el músculo viejo para favorecer la fibrosis durante la regeneración muscular. Cabe destacar que el tratamiento con agonistas o antagonistas de Wnt Notch parece revertir estas alteraciones en el lugar y rejuvenece la función de reparación muscular. Por último, la asociación frecuente de un microambiente del estroma alterado con tumores primarios y metastásicos en realidad puede colaborar con los cambios genéticos y epigenéticos intrínsecas en las células madre tumorales para promover la tumorigénesis. Por lo tanto, dirigirse directamente al lugar de la célula del tumor, o prevenir el acceso de las células tumorales a su nicho, podría revertir o retrasar la progresión del tumor y por lo tanto proporcionar una terapia de cáncer de novela.
En conjunto, estos resultados tienen importantes implicaciones para el diseño de nuevos enfoques para manipular la actividad de células madre para la terapia, y sugieren que proceda en vivo la modificación de células de nicho puede ser útil. Por ejemplo, la modificación de las células de nicho aumenta la proliferación y función de las células madre endógenas, facilita el injerto y la expansión de las células madre trasplantadas, influye en las decisiones de destino celular de diferenciación de las células madre, y / o inhibe la proliferación o promueve la apoptosis de cáncer de tumor de multiplicación células madre (figura
3) . Estas posibilidades son propensos a abrir una amplia gama de nuevas posibilidades terapéuticas en medicina regenerativa, y puede ser de especial importancia para detener o revertir los déficits relacionados con la edad en la función del tejido, que aparecen en varios tejidos como resultado del deterioro de la actividad regulada extrínsecamente células madre.
Figura 3 | Orientación del nicho para la terapia de la deficiencia de células madre o la desregulación contribuye a varias patologías humanas, y la evidencia acumulada sugiere que la orientación de la terapéutica nicho de células madre puede proporcionar una nueva y eficaz estrategia para mejorar el tratamiento de estos trastornos. a | Por ejemplo , la corrección de envejecimiento-o enfermedad asociada a alteraciones en el nicho podría ser utilizado para impulsar el número de células madre endógenas o función, y de este modo mejorar la función del tejido. b | Del mismo modo, mejorar la función nicho de apoyo durante el trasplante podría mejorar la eficiencia de injerto o acelerar vástago reconstitución de células, tal vez la reducción del número de células madre necesarias para la reconstitución de tejido eficaz. c | Además, debido a que el nicho puede tener un papel importante para influir en las decisiones del destino de células madre, así como la promoción de células madre de auto-renovación, modificación apropiada de las señales desde el nicho podría ser utilizado para alterar los resultados de la diferenciación de células madre para favorecer la producción de un tipo de célula sea necesario o inhibir la producción de un efecto perjudicial uno. d | Por último, a la luz de la acumulación de evidencia que sugiere que las células madre de cáncer de tumor de propagación dependen de señales de su nicho de mercado, al igual que sus homólogos no malignas, la ablación terapéutica de los componentes del nicho de células madre del cáncer podría proporcionar una nueva estrategia para eliminar los factores de apoyo del tumor, y así lograr la remisión del cáncer.
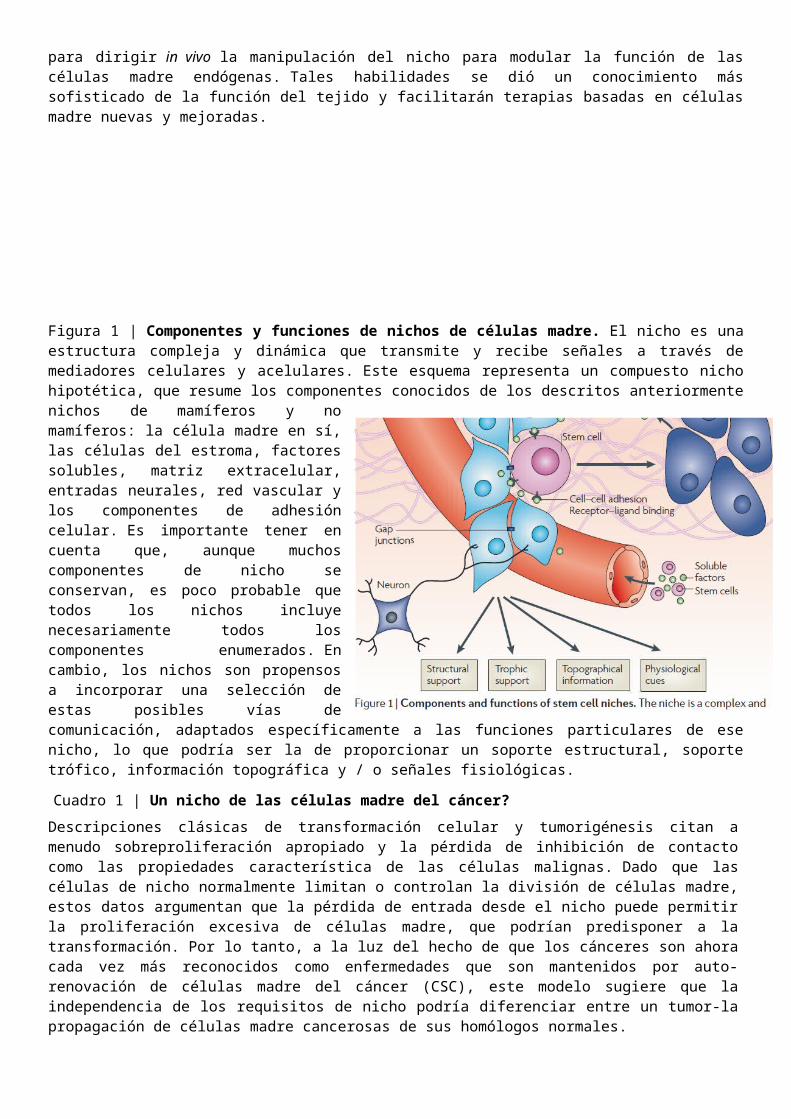
Tabla 1 | Ejemplos de células madre de tejido y sus nichos
Tejido Stem Cell Las células de apoyo
Las vías de señalización
Adhesión
C. elegans gónadas GSC La punta distal celular *
Muesca NI
D. melanogastertestículo
GSC Células Hub * JAK-STAT DE-cadherina, β-catenina
D. melanogasterovario GSC Células Cap * y CES DPP-BMP DE-cadherina, β-catenina
D. melanogastertestículo
CPC Células Hub * JAK-STAT DE-cadherina, β-catenina
D. melanogasterovario ESC NI JAK-STAT NI
D. melanogasterovario FSC NI Erizo DE-cadherina, β-catenina
D. melanogasterintestino medio
ISC NI Muesca Posiblemente para ECM
Ratón músculo esquelético
Células satélite NI Muesca β1 integrina
Ratón de la médula ósea
HSC * Los osteoblastos, células vasculares
SLF, Wnt, Notch, ANG1, OPN
β1 integrina
Intestino delgado Ratón CBC Cripta fibroblastos, células de Paneth
Wnt, BMP β-catenina
Piel de ratón Queratinocitos interfolicular
NI Wnt, Shh, Notch E-cadherina, β-catenina, β1 integrina
Piel de ratón Células madre protuberancia folicular
Fibroblastos dérmicos Wnt, BMP β-catenina, β1 integrina
Cerebro de ratón (ventrículo lateral)
Células madre ZVS
Células vasculares, astrocitos
Shh, BMP N-cadherina, β-catenina
Cerebro de rata (hipocampo)
Células madre SGZ
Células vasculares, astrocitos
Shh, Wnt N-cadherina, β-catenina
Ratón testículo SSC * Células de Sertoli, las células intersticiales vasculatura,
GDNF, SLF α6 integrina, β1 integrina
ANG1, angiopoyetina-1, BMP, proteína morfogenética ósea, CBC, cripta base de células columnares; C. elegans , Caenorhabditis elegans , CPC, células progenitoras quiste (células madre somáticas), DPP, Decapentaplegic; D. melanogaster , Drosophila melanogaster , ECM, matriz extracelular, ESC, células madre escolta, FSC, células madre del folículo; GDNF, factor neurotrófico línea celular derivada de células gliales, GSC, células madre de línea germinal;, células madre hematopoyéticas HSC; ISC, células madre intestinales ; JAK, Janus quinasa; NI, ninguno identificado, OPN, osteopontina, SGZ, zona subgranular; Shh, sonic hedgehog, SLF, factor de acero; SSC, células madre espermatogonias, STAT, transductor de señal y activador de la transcripción; SVZ, zona subventricular; UO, J. Voog y DLJ, observaciones no publicadas. * Indica las células de apoyo que se han demostrado para regular directamente el comportamiento de las células madre que se encuentran dentro de ese nicho.


















