
Apuntes FYTI(2009)
154
Introducción a la Inmunología Documento2 01/03/2007 Pag.1.4 FUNDAMENTOS Y TÉCNICAS INMUNOLÓGICAS Tema 1. Introducción a la Inmunología Inmunidad natural y adquirida Concepto del Sistema inmunitario El organismo opone a la penetración y establecimiento de los organismos patógenos exteriores (antígenos), una serie de barreras cuyo fin es el mantenimiento de la salud del individuo. La inmunología trata sobre las respuestas del hospedador a los diversos agentes infecciosos. En ella se estudia cuando como y porque se produce la respuesta, así como todas aquellas células y moléculas involucradas en la misma. La primera de esas barreras es la denominada INMUNIDAD NATURAL, INNATA O NATIVA, cuyo primer nivel de protección esta constituido por una serie de barreras tanto físicas, (epitelio y mucosas que recubren las estructuras tubulares del organismo), como químicas, pH de la piel y estómago, que forman una barrera entre el medio interno y el externo. El segundo nivel lo constituye la denominada INMUNIDAD INNATA INDUCIDA. En esta fase entran en juego ya ciertas moléculas (Receptores, citoquinas y células (Macrófagos, células NK, neutrófilos) que mediante un proceso inflamatorio, logran en la mayoría de los casos, controlar, de forma inespecífica, a los agentes patógenos Todos estos factores de protección de los que hemos hablado, se caracterizan por dos propiedades fundamentales. Una es la INESPECIFICIDAD,o NO DISCRIMINACION, es decir son medidas de tipo general independientes del tipo de organismo invasor; la otra, podríamos denominarla como la INTEMPORALIDAD, ya que son medidas que existen siempre, incluso antes de que tenga lugar la infección. Por esta razón este tipo de inmunidad, es la que actúa en las primeras horas de la infección. Cuando el organismo extraño logra superar esta Inmunidad innata, entra en acción un segundo grupo de medidas agrupadas todas ellas bajo el nombre de INMUNIDAD ADQUIRIDA o ADAPTATIVA, cuyos mecanismos de activación empiezan a sucederse a partir de las 24 horas de la entrada del microorganismo. La principal característica de este tipo de inmunidad es que es ESPECÍFICA, lo que quiere decir que va dirigida directamente de forma particular, y exclusiva contra el agente invasor. Este tipo de defensa, esta mediada básicamente por un tipo de células: los linfocitos, y un tipo de sustancias solubles: los anticuerpos. Los anticuerpos se producen solo en respuesta a infecciones específicas, por lo cual los anticuerpos presentes en un individuo dado reflejan de forma directa las infecciones que ha sufrido. Ambos sistemas defensivos, no son independientes ni actúan de forma descoordinada, sino que ambos, aunque solapándose, actúan de forma secuencial, comunicándose entre sí a través de una serie de sustancias solubles conocidas genéricamente como citoquinas. De estas, las secretadas por los macrófagos serian constituyentes de la Inmunidad natural, mientras que las secretadas por los linfocitos lo serán de la Inmunidad específica. formulandoalgoqueaportar.blogspot.com
-
Upload
cristina-coronilla-batres -
Category
Documents
-
view
135 -
download
5
Transcript of Apuntes FYTI(2009)

Introducción a la Inmunología
Documento2 01/03/2007 Pag.1.4
FUNDAMENTOS Y TÉCNICAS INMUNOLÓGICAS
Tema 1. Introducción a la Inmunología Inmunidad natural y adquirida Concepto del Sistema inmunitario
El organismo opone a la penetración y establecimiento de los organismos patógenos exteriores (antígenos), una serie de barreras cuyo fin es el mantenimiento de la salud del individuo.
La inmunología trata sobre las respuestas del hospedador a los diversos agentes infecciosos. En ella se estudia cuando como y porque se produce la respuesta, así como todas aquellas células y moléculas involucradas en la misma.
La primera de esas barreras es la denominada INMUNIDAD NATURAL, INNATA O NATIVA, cuyo primer nivel de protección esta constituido por una serie de barreras tanto físicas, (epitelio y mucosas que recubren las estructuras tubulares del organismo), como químicas, pH de la piel y estómago, que forman una barrera entre el medio interno y el externo. El segundo nivel lo constituye la denominada INMUNIDAD INNATA INDUCIDA. En esta fase entran en juego ya ciertas moléculas (Receptores, citoquinas y células (Macrófagos, células NK, neutrófilos) que mediante un proceso inflamatorio, logran en la mayoría de los casos, controlar, de forma inespecífica, a los agentes patógenos
Todos estos factores de protección de los que hemos hablado, se caracterizan por dos propiedades fundamentales. Una es la INESPECIFICIDAD, o NO DISCRIMINACION, es decir son medidas de tipo general independientes del tipo de organismo invasor; la otra, podríamos denominarla como la INTEMPORALIDAD, ya que son medidas que existen siempre, incluso antes de que tenga lugar la infección. Por esta razón este tipo de inmunidad, es la que actúa en las primeras horas de la infección.
Cuando el organismo extraño logra superar esta Inmunidad innata, entra en acción un segundo grupo de medidas agrupadas todas ellas bajo el nombre de INMUNIDAD ADQUIRIDA o ADAPTATIVA, cuyos mecanismos de activación empiezan a sucederse a partir de las 24 horas de la entrada del microorganismo. La principal característica de este tipo de inmunidad es que es ESPECÍFICA, lo que quiere decir que va dirigida directamente de forma particular, y exclusiva contra el agente invasor. Este tipo de defensa, esta mediada básicamente por un tipo de células: los linfocitos, y un tipo de sustancias solubles: los anticuerpos. Los anticuerpos se producen solo en respuesta a infecciones específicas, por lo cual los anticuerpos presentes en un individuo dado reflejan de forma directa las infecciones que ha sufrido.
Ambos sistemas defensivos, no son independientes ni actúan de forma descoordinada, sino que ambos, aunque solapándose, actúan de forma secuencial, comunicándose entre sí a través de una serie de sustancias solubles conocidas genéricamente como citoquinas. De estas, las secretadas por los macrófagos serian constituyentes de la Inmunidad natural, mientras que las secretadas por los linfocitos lo serán de la Inmunidad específica.
formulandoalgoqueaportar.blogspot.com

Introducción a la Inmunología
Documento2 01/03/2007 Pag.2.4
Filogenéticamente hablando, la inmunidad específica es más tardía, pues aparece con el desarrollo de los vertebrados. En su transcurso evolutivo la Inmunidad específica ha retenido muchos de los mecanismos de la Inmunidad natural, a la vez que ha ido añadiendo nuevas cualidades. Entre ellas cabe destacar:
Memoria: El Sistema inmunitario es ahora capaz de Recordar cada encuentro con el agente invasor, de modo que los sucesivos encuentros con aquel aumentan la respuesta.
Amplificación: De los mecanismos propios de la Inmunidad Innata.
Centrándonos en la Inmunidad adquirida y basándonos en los componentes que median la respuesta, la INMUNIDAD ADQUIRIDA, se clasifica en dos tipos:
Inmunidad humoral: Es aquella inmunidad mediada por anticuerpos, moléculas secretadas por Linfocitos B, que están mayoritariamente en suero y plasma y que son capaces de reconocer a los antígenos, ayudando a su eliminación.
inmunidad celular: Aquella mediada por células en particular por linfocitos T. Sus principales dianas son los organismos intracelulares, destruyéndolos bien directamente, o bien en aquellos casos en que así se exija destruyendo también la célula hospedadora.
En función de la forma de inducción a una persona, se puede hablar de tres tipos de inmunidad:
Activa: Cuando es el propio sistema inmunitario del individuo el que responde a la invasión externa; por ejemplo a través de las vacunas se induce, a un individuo, inmunidad frente a un determinado organismo.
Pasiva: Cuando la protección a un individuo se consigue transfiriéndole las sustancias propias de la inmunidad humoral.
Adaptativa: Cuando lo que se transfieren son los elementos de la inmunidad celular, es decir los linfocitos.
Sistema Inmunitario
La principal característica del sistema inmunitario, es que es un sistema disperso, sin continuidad anatómica y cuyos elementos se hallan coordinados entre sí, a través de una serie de sustancias denominadas citoquinas, (ya mencionadas). Asimismo, tampoco es un sistema aislado, sino que se halla relacionado con otros sistemas como el Nervioso o el Endocrino.
Las características principales de este sistema, se pueden resumir en cinco puntos
formulandoalgoqueaportar.blogspot.com

Introducción a la Inmunología
Documento2 01/03/2007 Pag.3.4
fundamentales:
1. Especificidad. 2. Capacidad de discriminación de lo NO PROPIO 3. Diversidad 4. Memoria 5. Autolimitación.
1. Especificidad: La respuesta es específica para cada antígeno. En realidad la respuesta se dirige contra ciertos componentes estructurales (proteínas, polisacáridos), reconociéndose pequeñas porciones de los mismos denominadas DETERMINANTES ANTIGÉNICOS Ó EPITOPOS. Esto tiene lugar gracias a que los linfocitos presentan en su membrana unas moléculas que actúan de receptores y que son capaces de unirse a moléculas complementarias, que serian los antígenos. 2. Diversidad: Las células del sistema inmunitario tienen unos receptores en su superficie que van a reconocer al agente extraño. Dichos receptores se forman al azar, calculándose que hay 10 9 receptores distintos, denominándose a dicho número Repertorio linfocitario. Así los linfocitos difieren en la estructura de sus receptores y por lo tanto en su especificidad 3. Capacidad de discriminación entre LO PROPIO y LO NO PROPIO: El sistema inmunitario, en su correcto funcionamiento, ha de distinguir entre los antígenos extraños, a los que debe ser capaz de reconocer y eliminar, y los antígenos propios frente a los que no debe reaccionar de forma perjudicial, es decir estableciéndose un estado de no respuesta frente a lo propio que se denominan este caso un estado Tolerancia. El fallo en este proceso conduce al desarrollo de enfermedades autoinmunes. Esto es lo que se denomina un proceso de Selección negativa. 4. Memoria inmunitaria: Es una característica, por la cual ante un segundo contacto (Respuesta secundaria) con el mismo antígeno, la respuesta será más rápida, más intensa, y por lo tanto más eficaz, que en el primer contacto (Respuesta primaria). Esto es debido a la formación de unas células de vida muy larga (células de memoria) preparadas para responder con mayor rapidez a una nueva estimulación. 5. Autolimitación: La respuesta inmune disminuye y cesa una vez que el estimulo antigénico ha sido controlado
Inmunidad y patología
En el caso de la respuesta, nos encontramos con dos situaciones:
1. Respuesta protectora: Aquella que elimina al organismo patógeno sin dañar a las estructuras propias.
2. Respuesta patológica: Cuando la respuesta daña al propio individuo. Esta última respuesta se produce por dos causas fundamentales:
a) Por ser la respuesta demasiado larga o demasiado intensa, dando lugar a las denominadas reacciones de hipersensibilidad. Puede estar mediada tanto por los mecanismos de inmunidad humoral como celular.
b) Por fallos en el estado de tolerancia, es decir por la existencia de
formulandoalgoqueaportar.blogspot.com

Introducción a la Inmunología
Documento2 01/03/2007 Pag.4.4
errores en el sistema de reconocimiento de lo NO PROPIO, que lleva a provocar una respuesta frente a antígenos propios. Este proceso dará lugar a fenómenos de Autoinmunidad y a la aparición consiguiente de enfermedades autoinmunes.
c) Por debilidad o formación defectuosa del Sistema Inmunitario, se producen la denominada Inmunodeficiencia, por las cuales el organismo “permite”, el establecimiento de infecciones repetidas u oportunistas. Esto puede deberse a causas genéticas, en cuyo caso se denominan primarias, o a daños provocados en el Sistema inmune (SIDA, terapia anticancerosa, terapia de transplantados...etc.), en cuyo caso se denominaran secundarias.
formulandoalgoqueaportar.blogspot.com

Bases Morfológicas del Sistema Inmunitario
Documento5 01/03/2007 Pag..1.9
Tema 2: Bases Morfológicas del Sistema inmunitario
Células del sistema inmunitario: Origen, Morfología y función. Órganos linfoides primarios y secundarios. Breve reseña Ontogénica Recirculación linfocitaria Moléculas de Adhesión
Las células del sistema inmunitario se pueden dividir en tres grandes grupos: 1. Las células presentadoras de antígenos, Son las células que capturan y
exponen los antígenos. 2. Las células que reconocen y responden de forma específica a antígenos
extraños 3. Las células efectoras, que serian las encargadas de la destrucción final de los
agentes patógenos La pertenencia a uno de los grupos no excluye la posibilidad de poder ser
incluidas en otro. Todas las células del S.I. y el resto de las células sanguíneas, como glóbulos rojos
y plaquetas, proceden de un único tipo celular existente en la médula ósea denominada CELULA PLURIPOTENCIAL (STEM CELL).
La división de estas tiene lugar de una forma poco corriente, pues en lugar de dar lugar a una progenie idéntica, se dividen de manera funcionalmente asimétrica, de forma que una de las células hijas es y seguirá siendo igual que la célula madre manteniéndose el número de células pluripotenciales. Pero, la segunda célula hija, se diferencia, primero en la denominada “Unidad formadora de Colonias” que a su vez dará lugar a las células precursoras (Eritroide, Mieloide y Linfoide), de las que derivaran los diferentes tipos de células hematopoyéticas.
El progenitor mieloide, es el precursor de los macrófagos, mastocitos y la serie granulocitica del sistema inmune. El progenitor linfoide dará paso a las diferentes líneas de linfocitos
Macrófagos
Están distribuidos por todos los tejidos y juegan un papel primordial en la Inmunidad innata, a través de la fagocitosis.
Proceden de los Monocitos. Estos se forman en la médula ósea y entran en la circulación periférica (sangre), aún sin diferenciar. La maduración de estos tiene lugar cuando pasan a los tejidos convirtiéndose en Macrófagos. Durante el proceso de activación experimentan un gran aumento del citoplasma, (células epiteloides). Incluso, en algunos casos, se fusionan varios de ellos para dar lugar a células gigantes multinucleadas.
Una vez en los tejidos, los macrófagos adoptan diferentes nombres en función del tejido donde se asienten.
Entre sus funciones principales destacan: . Citocida
formulandoalgoqueaportar.blogspot.com

Bases Morfológicas del Sistema Inmunitario
Documento5 01/03/2007 Pag..2.9
. Presentación de antígenos
. Eliminación de Inmunocomplejos
. Liberación de citoquinas
. Fagocitosis
Mastocitos
Proceden igualmente de la línea mieloide. Residen fundamentalmente en las mucosas y tejidos epiteliales, cerca de los vasos, estando presentes en todos los tejidos vascularizados, excepto SNC y retina. Son uno de los principales mediadores en la respuesta alérgica, al expresar receptores de alta afinidad para IgE.
Granulocitos
Los granulocitos, o polimorfonucleares, se denominan así por presentar granulaciones en su citoplasma y núcleos lobulados de formas diversas. Hay tres tipos;
Neutrófilos. Son los más abundantes, pero tienen una vida media muy corta. Su función principal es la fagocitaria y son los componentes más importantes de la respuesta inflamatoria aguda al responder muy rápidamente a los estímulos quimiotácticos. Sus gránulos citoplasmáticos contienen numerosas sustancias microbicidas.
La principal diferencia, en cuanto a la función fagocitica entre los neutrofilos y los macrófagos, es que los primeros mueren tras la fagocitosis, siendo los componentes principales del pus; en cambio los macrófagos son células de vida larga
Eosinófilos. Son abundantes en los infiltrados celulares de la inflamación tardia. Expresan en su superficie receptores para la IgE, y por ello son particularmente eficaces contra infecciones que estimulan la producción de este anticuerpo como los helmintos. De hecho, estos parásitos son mas eficazmente destruidos por las proteínas especializadas de los eosinófilos (proteína básica principal y proteína cationica eosinofilica) que por las lisoenzimas de neutrófilos y macrófagos. El crecimiento y diferenciación de los eosinófilos es estimulado por una citoquina (IL5).
Basófilos, tienen una acción muy similar a la presentada por los mastocitos, y como estos expresan receptores de alta afinidad para la IgE.
Linfocitos
Provienen de los progenitores linfoides. Son las únicas células del organismo capaces de reconocer específicamente diferentes determinantes antigénicos y responsables de las dos principales características del sistema inmunitario, como son la especificidady la memoria.
Hay dos clases fundamentales de Linfocitos: Los linfocitos B y los linfocitos T. Los linfocitos B, una vez activados se transformaran en plasmocitos productores de anticuerpos, siendo las únicas células capaces de producirlos. Los linfocitos T, se subdividen en tres poblaciones Th, Tc y Tr). Se diferencian por que las tres presentan
formulandoalgoqueaportar.blogspot.com

Bases Morfológicas del Sistema Inmunitario
Documento5 01/03/2007 Pag..3.9
diferentes moléculas en su superficie. Los linfocitos que presentan en su superficie el complejo CD4, se denominan CD4+ o linfocitos T helper (LTh) y son esenciales para la activación de otras células como linfocitos B y macrófagos. Los CD8+ o linfocitos T citotóxicos (LTc) son los encargados de la destrucción de células alteradas o infectadas por organismos intracelulares. Los Linfocitos Tr, median la regulación de la respuesta inmunitaria. Existen varias subpoblaciones de linfocitos supresores.
El receptor del linfocito B (BCR), es un anticuerpo anclado a la membrana. Una vez activado ese linfocito B secretara un anticuerpo con la misma especificidad.
Tanto los linfocitos B como los T se originan en la médula ósea, pero solo maduran en ella los linfocitos B. Los linfocitos T, migran al Timo, de ahí su nombre, para madurar.
Otro tipo de células linfoides que carecen de receptor específico son las denominadas células NK, y que serán objeto de un tema específico mas adelante.
Órganos linfoides
Pueden dividirse en: 1. Primarios o centrales que son aquellos en los que se producen y maduran los linfocitos, es decir la médula ósea y el timo. . Médula Ósea : La médula ósea es un tipo de tejido que se encuentra en el interior de los grandes huesos, sobre todo de los centrales del cuerpo como cráneo, vértebras (hueso irregular), costillas, esternón, cintura escapular y pelvis.
Es donde se producen la generación, (Hematopoyesis), de todas las células sanguíneas y dentro de ellas, como es lógico, los linfocitos a partir de la célula Madre pluripotencial y donde maduran, en los mamíferos las células B. En la pubertad la hematopoyesis tiene lugar principalmente en el esternón, las vértebras y las costillas. La proliferación y maduración en la médula ósea de las células precursoras es estimulada por citoquinas, que reciben, en general el nombre de factores estimuladores de colonias (CSF). . Timo:
Es un órgano bilobulado situado en el mediastino anterior. Es el lugar donde maduran los linfocitos T. Esquemáticamente presenta, la corteza, rica en Linfocitos T y la médula escasamente poblada de linfocitos. Dispersas por el timo existen diversos tipos de células epiteliales así como células dendríticas y macrófagos. Los linfocitos del Timo, llamados timocitos, se encuentran en diversos estados de maduración. En el proceso de maduración, solo son seleccionados para sobrevivir aquellos que no reconocen a los antígenos propios, por tanto solo lo harán frente a antígenos extraños (Selección negatriva). En su proceso de maduración los precursores se pueden distinguir por la expresión de diferentes moléculas en su superficie como son la CD44, CD25, CD3, CD4 y CD8. La mayoria de los timocitos (98%) mueren durante este proceso de maduración. Los macrofagos del timo eliminan continuamente las células muertas que no han completado su maduración. El timo presenta su máxima actividad en la juventud, produciéndose posteriormente su involución que se completa, aproximadamente, a los 30 años. 2. Secundarios o periféricos. Donde comienza la respuesta inmune adaptativa. Los
formulandoalgoqueaportar.blogspot.com

Bases Morfológicas del Sistema Inmunitario
Documento5 01/03/2007 Pag..4.9
agentes patógenos penetran en el organismo por muy diversas vías, pero al final van a ser dirigidos hacia los órganos linfoides secundarios donde se van a encontrar con los linfocitos. Los principales órganos linfoides secundarios serían:
Ganglios linfáticos: El sistema inmunitario no inicia una respuesta inmunitaria adaptativa en los innumerables sitios en los que se puede establecer una infección, sino que captura parte de ese patógeno y lo conduce a los tejidos linfoides secundarios organizados Ganglios, Bazo y tejido linfoide. Los ganglios, son estructuras linfoides muy especializadas, donde, convergen los líquidos extracelulares. De esta manera, los antígenos que penetran por la piel u otras vías, terminan en los vasos linfáticos, siendo así, transportados a los ganglios. De esta manera, el Sistema linfático proporciona el mecanismo básico de recolección de antígenos.
El ganglio, internamente esta constituido por una corteza externa, en las que hay agregados de células constituyendo los folículos. Dentro de estos existen los folículos primarios, que son aquellos en los que se encuentran los linfocitos B en reposo. Cuando entran en contacto con el antígeno se transforman en los folículos secundarios donde se desarrolla el Centro Germinal. En este Centro germinal se produce la maduración y diferenciación de los linfocitos B hasta células plasmáticas, o o células de memoria que quedan en su mayoría retenidas en el ganglio o migrando, en su gran mayoría hacia la médula osea.
La parte mas interna del ganglio contiene la médula con linfocitos y fagocitos mononucleares. La linfa que penetra en el ganglio, se filtra a través de la corteza y la médula y sale a través de un único vaso eferente. Mientras que los folículos son ricos en células B, la médula lo es en linfocitos T.
El bazo, recolecta antígenos procedentes de la sangre, así como glóbulos rojos viejos. La mayoría del bazo, esta constituido por la pulpa roja, donde se encuentran los eritrocitos, rodeando a de la pulpa blanca. Esta está constituida por una acumulación de linfocitos T rodeados de una corona de células B. En estas zonas, se encuentran folículos linfoides, algunos de ellos con centro germinal.
La funcionalidad del bazo y los ganglios es muy similar. La principal diferencia es que en el bazo, se produce la respuesta a antígenos procedentes de la sangre, mientras que en los ganglios se produce frente a antígenos procedentes de tejidos,. Recolectados por medio de la linfa.
El Tejido linfoide asociado a intestino (GALT), incluye amígdalas, adenoide, apéndice y unas estructuras especializadas del intestino delgado denominadas Placas de Péyer. Se encargan de recolectar los antígenos procedentes del epitelio gastrointestinal. En las placas de Péyer, el antígeno se captura por unas células epiteliales especializadas denominadas células M. Igualmente hay Tejido linfoide asociado a bronquios (BALT) a mucosas (MALT) y a dermis y epidermis. Estos están constituidos por una gran masa de linfocitos B rodeados de unos pocos linfocitos T.
Por último, en ciertos lugares, de intensa respuesta inmunitaria, se pueden formar tejidos linfoides ectópicos. Parece ser que la producción local de citoquinas, particularmente linfotoxina provoca la formación de tejido linfoide ectópico.
formulandoalgoqueaportar.blogspot.com

Bases Morfológicas del Sistema Inmunitario
Documento5 01/03/2007 Pag..5.9
Recirculación de los linfocitos
Los linfocitos, que han madurado en médula y timo, y que todavía no se han encontrado con el antígeno se denominan linfocitos vírgenes. Estas células circulan continuamente entre la sangre y los tejidos linfoides periféricos. Cuando existe una infección los linfocitos se detienen en los órganos linfoides donde proliferan y se convierten en células efectoras, capaces de combatir la infección. Así pues y como vemos los linfocitos están en continua circulación, y los que están en la sangre (Hemograma), son solo una pequeña porción de los que están circulando por los distintos tejidos.
Existen cuatro patrones principales de migración de linfocitos:
Ø Migración de los precursores entre los tejidos linfoides primarios Ø Migración de los linfocitos vírgenes a los tejidos linfoides secundarios Ø Migración de los linfocitos activados de los tejidos linfoides a los focos de inflamación. Ø Migración de células entre los distintos tejidos linfoides secundarios.
Migración de los precursores entre los tejidos linfoides primarios
Los precursores de las células T originadas en la médula ósea, abandonan la misma, circulan por la sangre y penetran en la corteza tímica. Estas células denominadas ahora Timocitos, son estimuladas para proliferar. Estas células supervivientes, migran desde la corteza del timo a la médula y finalmente son liberadas a la periferia como células T maduras y vírgenes.
En el caso de los linfocitos B al producirse la formación y maduración en la médula no se produce circulación a este nivel.
Migración de los linfocitos vírgenes a los tejidos linfoides secundarios
La extravasación de los linfocitos desde la sangre al interior del ganglio linfático, se produce de forma selectiva por la existencia de vénulas especializadas en el interior del ganglio. Estas venas, se hallan revestidas de células endoteliales y que le dan un aspecto morfológico distinto al resto del sistema venoso y que por ello se denominan vénulas endoteliales altas (HEV). Estas HEV se encuentran también en otros tejidos linfoides pero faltan en el bazo.
La forma en que se produce la extravasación, es por la mayor adhesividad de este tipo de endotelio. En condiciones normales los linfocitos colisionan al azar, y o bien rebotan o se adhieren al endotelio durante una fracción de segundo antes de ser arrastrados por la corriente sanguínea. En cambio en las HEV, la unión perdura durante mas tiempo, lo que permite que se refuerce la unión. El linfocito, así comienza a “rodar” por la superficie de la vena, para finalmente y una vez fijado mas firmemente pasar a través de los espacios intercelulares al interior del tejido linfoide. La unión de los linfocitos al endotelio esta mediado a unos ligandos expresados en las células endoteliales, que de modo general se denominan moléculas de adhesión. En este caso se conocen particularmente como diriginas. Estas diriginas se unen específicamente a otra molécula situada sobre la superficie de los linfocitos T vírgenes que se denomina SelectinaL. Si estos linfocitos no interactuan con su antígeno , salen del ganglio y regresan al sistema circulatorio, vía conducto torácico.
formulandoalgoqueaportar.blogspot.com

Bases Morfológicas del Sistema Inmunitario
Documento5 01/03/2007 Pag..6.9
Migración de los linfocitos activados de los tejidos linfoides a los focos de inflamación.
Una vez activados, los linfocitos T ya no se asientan eficazmente en los ganglios linfáticos por que expresan niveles inferiores de SelectinaL. En cambio si lo hacen con eficacia en los focos inflamatorios, que son, a menudo, los lugares de entrada y persistencia del antígeno. La razón de esto es que los estímulos producidos en el proceso inflamatorio, aumentan la expresión de otras moléculas de adhesión en las células T activadas, en este caso denominadas Integrinas, y a su vez aumentan la expresión de sus correspondientes ligandos en las células endoteliales del foco inflamatorio.
Migración de células entre los distintos tejidos linfoides secundarios. Los linfocitos que recirculan son células de vida larga, que pueden penetrar en los ganglios linfáticos, bien a través de los canales aferentes, o bien a través del endotelio vascular por los mecanismos citados anteriormente.
Moléculas de adhesión
al disociar mecánicamente las células de dos especies diferentes de esponjas marinas, y colocar en medio líquido una mezcla de ellas, éstas se reúnen nuevamente formando las mismas esponjas originales. Se demostró así, que las células de un organismo multicelular se reconocen entre sí y se adhieren específicamente. Este proceso está basado en la presencia de moléculas específicas, denominadas "Moléculas de Adhesión Celular" (CAMS, Las CAMs son glicoproteínas mediante las cuales se efectúan las interacciones específicas célula célula y célulamatriz. Estas glicoproteínas tienen en un extremo un grupo carboxilo. El otro extremo, extracelular, es un grupo amino, que da la especificidad a la molécula para unirse a otras CAMS.
Todas las funciones biológicas parecen requerir, o son influenciadas, por estas interacciones, especialmente la embriogénesis, la forma celular, el desarrollo tisular, la adhesión celular, la migración de células, los procesos inflamatorios e inmunológicos y muchos otros.
La expresión de las distintas moléculas sobre la superficie de los leucocitos depende de la línea celular y del estado de activación y diferenciación. Según sea el agente inductor variara el tipo de molécula expresada. Así distintos grupos de citoquinas inducen la expresión de grupos diferentes de moléculas de adhesión endoteliales. Asimismo los mediadores del proceso inflamatorio también inducen la expresión de un determinado tipo de moléculas de adhesión.
Las principales familias de moléculas de adhesión estructuralmente emparentadas serian las siguientes:
v Selectinas v Integrinas v Inmunoglobulinas v Carbohidratos
formulandoalgoqueaportar.blogspot.com

Bases Morfológicas del Sistema Inmunitario
Documento5 01/03/2007 Pag..7.9
Selectinas
Son una grupo de tres moléculas que se denominan genéricamente Moléculas de adhesión tipo lectina (LECAM), que median la adhesión de los leucocitos a las células endoteliales. Estructuralmente presentan un glucoproteína transmembrana, una serie de carbohidratos denominados lectinas de tipo C, un dominio homologo al factor de crecimiento epidérmico, otros dominios, relacionados con las proteínas de control el Complemento y un extremo amino terminal externo. La unión de la selectina con su correspondiente ligando es dependiente de Ca++.
La SelectinaL o CD62L, es, como ya hemos visto el factor que actúa como asentamiento de los linfocitos en las HEV. Pero no sólo se expresa en los linfocitos, sino que también lo hace en otras células como neutrófilos para facilitar su persistencia en los focos inflamatorios. Una propiedad de esta selectina es que la unión de la misma a su es de baja afinidad, aunque lo suficiente como para permitir las rodadura de la célula (rolling).
La SelectinaE conocida también como ELAM1 o CD62E, es expresada únicamente por las células endoteliales activadas por citoquinas. El ligando de la selectinaE en los linfocitos se conoce como CLA1, (Cutaneus Lymphocytantigen 1), y se le ha involucrado con el asentamiento de linfocitos T en la piel. En general la expresión de selectinaE en las células endoteliales es un distintivo de la inflamación aguda.
La SelectinaP o CD62P, se identifico por primera vez en las plaquetas, de ahí su nombre., aunque como la E, se expresa también en células endoteliales. Media la unión de neutrófilos y monocitos. Su expresión, aumenta en el foco inflamatorio.
La falta la expresión de los ligandos para aquellas se produce el Síndrome de deficiencia de adhesión leucocitaria de tipo II
Integrinas
Son proteínas heterodiméricas, (cadenas α y β) que actúan fundamentalmente como moléculas de adhesión, aunque también pueden hacerlo como transmisoras de señales. Existen varias subfamilias denominadas en función de la cadena común que comparten (α3 β1, α4 β1, α1 β2 etc.). Una característica de las integrinas es que a partir de las señales intracelulares generadas en la célula a partir de otros receptores, por ejemplo TCR o citoquinas, se produce una señal, denominada DentroFuera, por el cual aumenta la afinidad de las integrinas por su ligando. Una vez unido al ligando, las integrinas proporcionan una señal, ahora FueraDentro, a la célula, que por ejemplo pueden favorecer la expresión de los genes de citoquinas. En general se van a expresar en linfocitos T, linfocitos B y monocitos
La subfamilia β2, representado por LFA1, o también CD11aCD18¸, comparten la cadena CD18, es expresado por mas del 90% de los timocitos y las células T maduras. Media en la adhesión de los linfocitos al endotelio y en la adhesión de los linfocitos T a las CPA (Células presentadoras de antígeno). Sus ligandos correspondientes son ICAM
formulandoalgoqueaportar.blogspot.com

Bases Morfológicas del Sistema Inmunitario
Documento5 01/03/2007 Pag..8.9
1 (Molécula de adhesión intercelular), ICAM2 e ICAM3, y su avidez por ellos aumenta después de la estimulación de los Linfocitos T a través de su TCR. Se ha demostrado que el ligando de LFA1, ICAM1 es un receptor específico para los rinovirus, agentes causantes del resfriado común. Ciertas citoquinas pueden aumentar la avidez de esta integrina por los ligandos que se expresan en las células endoteliales. Esto hace que se favorezca la migración de los linfocitos T a los focos inflamatorios.
Así las principales moléculas de esta subfamilia serian: Molécula Cadenas Ligandos Expresión LFA1 αLβ2 CD11aCD18 ICAM1,2,3 Leucocitos CR3 αMβ2 CD11bCD18 ICAM1,2; iC3b, iC4b Fagoc y Neutrófilos CR4 αXβ2 CD11cCD18 Macrófagos
La deficiencia de estas integrinas lleva a la deficiencia de adhesión leucocitaria de tipo I, caracterizada sobre todo por infecciones bacterianas y micoticas recurrentes y severas.
Otra de las subfamilias es la β1. Comparten la cadena CD29, y se denominan en general, VLA (Very Late Antigens), ya que aparecen de 2 a 4 semanas después de la activación de los linfocitos T. Tres de los miembros de esta familia, los VLA4, 5 y 6, se expresan sobre los linfocitos en reposo. El ligando de VLA4, denominado VCAM1, se expresa en células endoteliales activadas por citoquinas. De este modo esta pareja, al igual que LFA1 con ICAM1, vista con anterioridad pueden controlar la salida de los linfocitos de los vasos sanguíneos hacia los focos inflamatorios.
Las principales moléculas de esta familia serian: Molécula Cadenas Ligandos Expresión VLA1 Α1β1 CD49aCD29 Colágeno y Laminina LT, Fibroblastos VLA2 Α1β1 CD49bCD29 Colágeno y Laminina LT y plaquetas VLA3 Α1β1 CD49cCD29 Colágeno, Laminina y
Fibronectina Riñón y Tiroides
VLA4 Α1β1 CD49dCD29 VCAM1 y Fibronectina LT, LB, Monocitos VLA5 Α1β1 CD49eCD29 Fibronectina Leucocitos, Plaquetas VLA6 Α1β1 CD49fCD29 Laminina Leucocitos, Plaquetas
Inmunoglobulinas
Son conocidas primordialmente por su función de anticuerpos, aunque algunas de ellas se expresan o inducen sobre el endotelio y actúan como moléculas de adhesión. Así ICAM1, como vimos el ligando de LFA1, se expresa también sobre la superficie de linfocitos, contribuyendo a la adhesión entre los linfocitos T y entre estos y las CPAs, teniendo por ello un importante papel en la presentación antigénica. Por otra parte ICAM2, perteneciente también a esta subfamilia, solo se expresa en la superficie de células endoteliales.
Otra importante molécula de adhesión perteneciente a esta subfamilia es la VCAM1 o CD106. Es como se ha visto receptor de la integrina VLA4, y es expresada sobre todo por
formulandoalgoqueaportar.blogspot.com

Bases Morfológicas del Sistema Inmunitario
Documento5 01/03/2007 Pag..9.9
células endoteliales activadas. También es expresada en células dendríticas foliculares, participando ene l adhesión de estas a los linfocitos B.
Por último la molécula PECAM1. Es importante, al encontrarse en los espacios intercelulares, para la extravasación leucocitaria. Se ha postulado que su interacción con los linfocitos es una señal de activación de las integrinas, actuando mas como una señal que como molécula de adhesión. Asimismo parece ser que tiene una función limitante de la permeabilidad vascular.
PAPEL DE LAS MOLECULAS DE ADHESIÓN EN LA INFLAMACIÓN
La primera etapa en el fenómeno inflamatorio es el rodamiento de los leucocitos sobre la pared endotelial, tal y como hemos visto con anterioridad. Esto da lugar a la adhesión de las células a la pared, gracias a la expresión de moléculas de adhesión tanto en los linfocitos y otras células como en el endotelio como consecuencia de la liberación de ciertas citoquinas proinflamatorias. Como consecuencia de esta adhesión del leucocito, se produce la extravasación y correspondiente salida de estos del torrente circulatorio hacia el foco inflamatorio.
PAPEL DE LAS MOLECULAS DE ADHESIÓN EN LAS METASTASIS
Muchas Células tumorales expresan receptores de adhesión idénticos a los expresados por los leucocitos. Como consecuencia de ello, las células cancerosas desprendidas del tumor primario van a ser capaces de fijarse a determinados endotelios y con ello colonizar y provocar un tumor secundario. La importancia de esto radica es que interfiriendo por medio de determinadas sustancias la expresión de estas moléculas se puede, en teoría disminuir la capacidad mestatásica de un tumor.
formulandoalgoqueaportar.blogspot.com

TEMA 3. MOLECULAS DE ADHESION. RECIRCULACION LINFOCITARIA Los leucocitos vigilan constantemente nuestro organismo utilizando el sistema circulatorio; cuando se detecta una infección las células atraviesan la pared de los vasos y se dirigen al sitio de infección. Todo ello es posible gracias a la existencia de las llamadas moléculas de adhesión, que detienen a los leucocitos en la pared de los vasos locales (del tejido infectado) y los inmovilizan hasta que la acción de otros mediadores (las quimiocinas) los dirigen hacia el foco de infección. MOLÉCULAS DE ADHESION CELULAR Las células de nuestros tejidos se mantienen unidas gracias a las interacciones moleculares que hay entre ellas. En cambio, las células del SI pueden atravesar de una a otra parte del organismo. Lo curioso es que las mismas moléculas que se utilizan para mantener juntas a las células de los tejidos (Moléculas de adhesión celular = CAMs) se usan por os leucocitos para interaccionar con las células de los tejidos. El endotelio vascular actúa de portero, regulando el paso de moléculas y células desde la sangre a los tejidos; el proceso, en lo que se refiere a las células, se denomina extravasación. Para que los linfocitos alcancen los órganos linfoides periféricos o los sitios de inflamación Las células endoteliales expresan CAMs que son específicas para los leucocitos, aunque algunas de ellas se expresan constitutivamente y otras lo hacen solo en respuesta a la concentración local de citocinas inflamatorias que se producen durante una respuesta inflamatoria. Aquellas células circulantes con un receptor que se une a las CAMs que hay sobre las células del epitelio vascular son las que pueden extravasarse a los tejidos. Los linfocitos recirculantes tienen receptores que se fijan a las CAM del endotelio vascular, lo que les permite extravasarse hacia los tejidos. Además de su papel en la unión de los leucocitos a las células endoteliales las CAMs que hay sobre los leucocitos aumentan la fuerza de las interacciones funcionales entre las células del SI. Hay varias moléculas de adhesión que contribuyen a las interacciones entre: Células Th y CPAs Th y células B Células Tc y células blanco La mayoría de las CAMs pertenecen a cuatro familias de proteínas, que son:
• Selectinas • Tipo mucina • Integrinas • Inmunoglobulinas (Ig)
Selectinas Son glucoproteínas de membrana que tienen un dominio extracelular tipo lectina, lo que les permite unirse de forma específica a grupo de hidratos de carbono (azúcares). La interacción primaria de las selectinas es con una porción hidrocarbonada que tiene ácido siálico (sialilada) y que se denomina sialil-Lewisx y que se encuentra unida con frecuencia a las moléculas tipo mucina. La familia incluye tres moléculas, llamadas:
formulandoalgoqueaportar.blogspot.com

• L-Selectina (CD62L) que se expresa sobre la mayoría de los leucocitos circulantes
• E-Selectina (CD62E) que se expresa sobre las células endoteliales durante una respuesta inflamatoria, tras la estimulación de su síntesis por citocinas pro-inflamatorias
• P-Selectina (CD62P), se almacena en unos gránulos de las células endoteliales (llamados de Weibel-Palade) que, tras la estimulación de la célula endotelial, se funden con la membrana plasmática quedando las P-Selectinas sobre la superficie celular.
La mayoría de los leucocitos circulantes expresan L-Selectina, mientras que las otras dos se expresan sobre las células endoteliales durante la respuesta inflamatoria. Estas selectinas son las responsables de la adhesión inicial de los leucocitos al endotelio vascular. Mucinas Constituyen un grupo de proteínas ricas en serina y treonina y muy glucosiladas. Su estructura extendida les permite presentar porciones como sialil-Lewis y otras estructuras hidrocarbonadas para que se capturen por las selectinas. Integrinas Son proteínas hetero-diméricas con una cadena α y otra β, asociadas de forma no covalente, que se expresan sobre la superficie celular. La mayoría interacciona con las moléculas de la matriz extracelular y mantienen la estructura de los tejidos en todo el organismo. Algunas subfamilias (con una subunidad común) se unen a las moléculas de adhesión de la superficie celular e intervienen en las interacciones célula-célula. Los leucocitos expresan una subfamilia específica de integrinas (conocida como β2), así como otras varias, que también se expresan sobre otros tipos celulares. Las β2 se unen a las proteínas de la SPF de las Ig, y a proteínas asociadas a la respuesta inflamatoria. Algunas integrinas necesitan activarse para unirse con alta afinidad a sus ligandos y su agrupamiento sobre la membrana celular aumenta la probabilidad de unión efectiva. Su importancia se refleja en la deficiencia de adhesión de los leucocitos (LAD) tipo 1, unas enfermedades autosómica recesiva que se caracteriza por la presencia de infecciones bacterianas recurrentes y dificultad para curar las heridas. Una deficiencia similar se ve en individuos que tienen alterada la expresión de selectinas y se denomina LAD tipo 2. La combinación de integrinas que expresa un tipo dado de célula es lo que la permite unirse a las diferentes CAMs que hay sobre la superficie del endotelio vascular. Superfamilia de las Ig (ICAMs) Esta SPF incluye moléculas de adhesión con dominios tipo Ig, como ICAM-1 (CD54), ICAM-2 (CD102), ICAM-3 (CD50) y VCAM (CD106), que se expresan sobre las células del endotelio vascular y se unen a varias moléculas de integrinas. Hay una molécula de adhesión (llamada MadCAM-1) que tiene dominios tipo Ig y tipo mucina, que se expresa sobre el endotelio de la mucosa y dirige la penetración de los linfocitos en la misma; se une a una integrina vía dominio tipo Ig y a L-Selectina vía dominio tipo mucina. También hay adhesiones homotípicas entre miembros de la SPF..
formulandoalgoqueaportar.blogspot.com

La molécula PECAM-1 ([molécula 1 de adhesión células endoteliales-plaquetas] CD31) está en la superficie de leucocitos (neutrófilos, monocitos y un subtipo de linfocitos T) y en complejos que unen a las células endoteliales. Presenta unión homotípica, donde el PECAM-1 de una célula se une al PECAM-1 de otra. La molécula de adhesión JAM-1 (CD321) también se encuentra en las uniones endoteliales y puede interaccionar consigo misma y con Integrinas, lo que le otorga un papel en la migración trans-endotelial. También puede encontrarse unión homotípica entre miembros de la SPF Ig en otros tipos celulares, como ocurre con L1 y NCAM sobre las células neurales.
Ejemplos de moléculas de adhesión celular (CAM) de distintas familias Tipo mucina Selectinas SPF Ig Integrinas
GliCAM-1 Selectina L ICAM-1, 2 y 3 VLA-4 (α4β1) CD34 Selectina P VCAM-1 VLA-6 (α6β1) PSGL-1 Selectina E LFA-2 (CD2) LFA-1 (α1β2) MadCAM-1 LFA-3 (CD58) MaC-1 (αMβ2) MadCAM-1 RECIRCULACION DE LOS LINFOCITOS Al contrario del resto de las células, los linfocitos recirculan constantemente a través de la sangre, la linfa y los órganos linfoides secundarios. Tras un breve tránsito por la sangre (unos 30 m.) casi el 45% de ellos la abandona directamente a través del bazo, donde residen aproximadamente unas 5 horas. Una cantidad casi igual de linfocitos (42%) sale de la sangre a través de los ganglios linfáticos, donde residen unas 12 horas. Un número pequeño de linfocitos (10%) migra a los tejidos extra-linfoides terciarios atravesando a través de las células endoteliales de los capilares. Estos tejidos normalmente albergan pocos (o ninguno) linfocitos a menos que se “reclamen” por la presencia de una respuesta inflamatoria. Los tejidos terciarios extra-linfoide más activos son lo que forman la interfase con el exterior, tales como piel y epitelios mucosos gastro-intestinales, pulmonares y genito-urinarios. La recirculación mantiene una dotación óptima de linfocitos específicos distintos, que pueden “encontrar” a cualquier antígeno. Un linfocito individual hace el circuito sangre – tejidos – linfa – sangre entre una y dos veces diarias. Como solo uno entre 100.000 linfocitos reconoce un antígeno particular parece que este mecanismo permite a un ayor número de células T y B reconocer al antígeno (sobre células presentadoras o en forma soluble) y desencadenar la respuesta adaptativa en menos tiempo. Los contactos entre células presentadoras y células T se aumentan mucho gracias a los factores que regulan, organizan y dirigen la circulación de linfocitos y células presentadoras de antígenos. PASO DE LOS LINFOCITOS A TRAVÉS DEL ENDOTELIO VASCULAR 1.- Por las vénulas endoteliales altas Aunque algunos linfocitos abandonan la sangre a través de vénulas no especializadas, la ruta de salida principal en los mamíferos es a través de una sección especializada de las vénulas post-capilares que se conocen como vénulas endoteliales altas (HEV). Estas HEV abundan en la zona para-cortical de los GL, son menos abundantes en la zona cortical y faltan en la zona medular. Además de en los GL, las HEV también se encuentran en el MALT (Tejido linfoide asociado a las mucosas) y en el timo. Algunos linfocitos, principalmente los T, llegan a través de los vasos linfáticos aferentes (en lugar de a través de las HEV), que es la ruta principal de llegada de los antígenos al GL.
formulandoalgoqueaportar.blogspot.com

Las HEV son estructuras permanentes de los tejidos linfoides secundarios, pero también pueden desarrollarse a partir del epitelio normal en los sitios de reacciones inflamatorias crónicas; esto puede significar que las células T específicas tengan una vía directa de acceso (a través de las HEV recién formadas) a la zona inflamatoria. El movimiento de los linfocitos a través del endotelio vascular se controla mediante moléculas de adhesión y quimiocinas, como p.e.:
Mad-CAM-1 que se expresa en las células endoteliales de los tejidos intestinales VCAM-1 que está presente en las células endoteliales de pulmones y piel
En los linfocitos hay otras moléculas que, interaccionan directamente con estas moléculas de adhesión, les llevan directamente a órganos concretos. En el caso del intestino son las integrinas alfa4beta7 (a4b7) las que median la adhesión de los linfocitos en las HEV de las Placas de Peyer (que expresan Mad-CAM-1). Extravasación de los linfocitos Varios subtipos de linfocitos presentan una extravasación dirigida a los sitios inflamatorios y órganos linfoides secundarios. La recirculación está cuidadosamente controlada para asegurar el reclutamiento apropiado de poblaciones T y B hacia los diferentes tejidos. Como ocurre con los neutrófilos, la extravasación de linfocitos incluye una serie de interacciones entre moléculas de adhesión y las cuatro fases de rodamiento, activación, detención y adhesión y, finalmente, migración trans-endotelial. Las vénulas con endotelios altos (vénulas endoteliales altas o HEV) son los sitios de extravasación de los linfocitos Algunas regiones del endotelio vascular de las vénulas post-capilares de distintos órganos linfoides están compuestas por células especializadas de formas cuboides y rechoncha (son “altas”), denominándose a estas regiones “vénulas endoteliales altas o HEV”. El aspecto de sus células contrasta mucho con el aplanado de las células endoteliales que recubren el resto del capilar. Todos los órganos linfoides secundarios, con excepción del bazo, contienen HEV. Cuando se incuban con linfocitos cortes de ganglios linfáticos, placas de Peyer o amígdalas y se lavan luego para eliminar a las células no fijadas, cerca del 85% de las células fijadas están adheridas a las HEV, aunque estas vénulas representan solo el 1-2% del área total del corte. Se estima que cada segundo se extravasan, a través de las HEV, unos 140.000 linfocitos en un solo ganglio linfático. Las citocinas producidas en respuesta a la captación de antígenos está comprobado que intervienen en el desarrollo y conservación de las HEV en los órganos linfoides; por ejemplo, los animales criados en condiciones axénicas (sin gérmenes) no desarrollan HEVs, mientras que si se bloquea quirúrgicamente el linfático aferente del ganglio (con lo que bloquea la entrada de los antígenos) al cabo de poco tiempo las HEVs presentan pérdida de funcionalidad y pueden volver a una morfología más aplanada. Las HEV expresan una variedad de moléculas de adhesión. Como cualquier otra células del endotelio vascular expresan: E y P-Selectinas, moléculas tipo mucina (GlyCAM-1 y CD34) y moléculas de la superfamilia de las Ig(ICAm-1, ICAM-2, ICAM-3, VCAM-1 y MadCAM-1). A algunas moléculas de adhesión que se distribuyen específicamente en un tejido se les ha llamado “adresinas vasculares” pues dirigen la extravasación de diferentes poblaciones de linfocitos re-circulantes a órganos linfoides particulares. La figura 13-8 presenta las interacciones típicas en la extravasación de las células T vírgenes a través de las HEV en los ganglios linfáticos. El primer paso es, normalmente,
formulandoalgoqueaportar.blogspot.com

una interacción selectina-carbohidrato similar a la de la adhesión de los neutrófilos. Los linfocitos T vírgenes se unen inicialmente a las HEVs a través de L-Selectina, que sirve como un receptor de albergue que dirige los linfocitos a tejidos que expresan las correspondientes adresinas vasculares tipo mucina, tales como CD34 o GlyCAM-1. El rodamiento de los linfocitos es menos acusado que el de l os neutrófilos. Aunque la interacción inicial selectina-carbohidrato es muy débil, el flujo tan lento de la sangre en las vénulas post-capilares, especialmente en la región de HEV, reduce la probabilidad de que el flujo arrastre al linfocito que se “arrastra”. En el segundo paso, el estímulo de activación de las integrinas está mediado por quimiocinas que bien se sitúan sobre la membrana endotelial o se secretan localmente. El grueso glicocalix que recubre las HEV puede retener estos factores quimio-atractantes in situ. La fijación de quimiocinas a los receptores que hay sobre los linfocitos, acoplados a G-proteína, lleva ala activación de las moléculas de integrina que sobre la membrana (como ocurre en la extravasación de los neutrófilos). Una vez activada la molécula de integrina interacciona con moléculas de la superfamilia de las Ig (p.e. ICAM-1), lo que permite a los linfocitos adherirse firmemente al endotelio. Los mecanismos moleculares que rigen el paso final, la migración trans-endotelial, parece que comprenden a las moléculas de adhesión PECAM-1 (CD31) y JAM-1 (CD321). El albergue de los linfocitos está dirigido por perfiles de receptores y señalesLa extravasación de los linfocitos es similar, en general, a la extravasación de los neutrófilos. Un carácter importante que distingue a ambos procesos es que los distintos subtipos de linfocitos migran diferencialmente a tejidos diferentes. Esta migración específica depende de combinaciones únicas de moléculas de adhesión y quimiocinas; los receptores que dirigen la migración de varias poblaciones de lilnfocitos a tejidos linfoides concretos y tejidos inflamatorios reciben el nombre de “receptores de albergue” o “receptores guías”. Los linfocitos vírgenes recirculan a los tejidos linfoides secundariosLos linfocitos vírgenes no son capaces de montar una respuesta inmune hasta que se han activado para pasar a células efectoras. La activación de las células vírgenes tiene lugar en un microambiente especializado en los tejidos linfoides secundarios (ganglios linfáticos periféricos, placas de Peyer, amígdalas y bazo). En este microambiente las células dendríticas capturan antígenos y los presentan a los linfocitos vírgenes; el resultado es que los linfocitos vírgenes se activan. Las células vírgenes no presentan preferencia por un tipo particular de tejido linfoide secundario, sino que circulan indiscriminadamente por todos los tejidos linfoides secundarios del cuerpo. La unión inicial de los linfocitos vírgenes a las HEV está generalmente mediada por la unión de L-Selectina del linfocito a moléculas de adhesión tales como GlyCAM-1 y CD34 sobre las HEV. Recientemente se ha encontrado que LFA-1 y VLA-4 también median el rodamiento en su conformación de baja afinidad (antes de la activación). Son las quimiocinas CCL21, CCL19 y CXCL12 las que inducen el cambio conformacional a la alta afinidad de LFA-1 y VLA-4, que lleva a la adhesión fuerte. Además de estas quimiocinas CXCL13 activa a las integrinas sobre las células B vírgenes. Otras quimiocinas atraen a las subpoblaciones de linfocitos a sus zonas en los ganglios linfáticos. El patrón de tráfico de las células T vírgenes está diseñado para tener a estas células recirculando constantemente a través de los tejidos linfoides secundarios, cuya función principal es atrapar a los antígenos de la sangre y los tejidos. Una vez que los linfocitos vírgenes encuentran a su antígeno atrapado en un tejido linfoide secundario se activan y aumentan su tamaño pasando a linfoblastos. La
formulandoalgoqueaportar.blogspot.com

activación lleva alrededor de 48 horas y, durante este tiempo, los blastos están retenidos en la región paracortical de los tejidos linfoides secundarios. En esta fase, también llamada “de cierre”, los linfocitos específicos del antígeno no pueden detectarse en la circulación. Es durante la fase de cierre cuando se produce la rápida proliferación y diferenciación de las células vírgenes. Las células efectoras y de memoria que se forman dejan el tejido linfoide y comienzan su propia recirculación. Las células efectoras y de memoria se mueven de forma distinta que las T vírgenesLos patrones migratorios de ambos tipos de células difieren del patrón de los linfocitos T vírgenes. Las células efectoras tienden a albergarse en las zonas de infección pues reconocen el endotelio inflamado así como a las moléculas quimiotácticas que se producen durante la respuesta inflamatoria. Los linfocitos de memoria tienden a albergarse selectivamente en el tipo de tejido en el que encontraron por primera vez al antígeno. Al parecer esto asegura que una célula de memoria particular retorne al tejido donde hay mayor probabilidad de que pueda enfrenarse a la amenaza que supone su antígeno. Las células efectoras y de memoria expresan cantidades elevadas de ciertas moléculas de adhesión, como LFA-1, que interaccionan con ligandos presentes en los tejidos extra-linfoides terciarios (como piel y epitelios mucosos) y en los sitios de inflamación, lo que permite que las células efectoras y de memoria los alcancen. Las células inmaduras carecen de las moléculas de adhesión correspondientes y no alcanzan ni se albergan en estos sitios. El endotelio inflamado expresa las Selectinas E y P y las moléculas de la superfamilia de las Ig VCAM-1 e ICAM-1, que se unen a los receptores correspondientes expresados en gran cantidad por células efectoras y de memoria. El resultado de todo ello es que estas células presentan una clara vocación tisular, mientras que evitan a los tejidos linfoides secundarios como consecuencia de su poca expresión de L-Selectina. Un segundo subgrupo de células efectoras y de memoria tienen una clara vocación dérmica y su dotación se caracteriza por la expresión baja de L-Selectina y alta de LCA (antígeno linfocítico cutáneo) y LFA-1, moléculas que se fijan a la E-Selectina e ICAMs que hay dentro de las vénulas dérmicas de la piel. Aunque las células efectoras y de memoria, que expresan cantidades reducidas de L-Selectina, no tienden a dirigirse a los tejidos linfoides secundarios a través de las HEV si pueden alcanzar los ganglios linfáticos secundarios a través de la linfa aferente.
formulandoalgoqueaportar.blogspot.com

TEMA 3. MOLECULAS DE ADHESION. RECIRCULACION LINFOCITARIA Los leucocitos vigilan constantemente nuestro organismo utilizando el sistema circulatorio; cuando se detecta una infección las células atraviesan la pared de los vasos y se dirigen al sitio de infección. Todo ello es posible gracias a la existencia de las llamadas moléculas de adhesión, que detienen a los leucocitos en la pared de los vasos locales (del tejido infectado) y los inmovilizan hasta que la acción de otros mediadores (las quimiocinas) los dirigen hacia el foco de infección. MOLÉCULAS DE ADHESION CELULAR Las células de nuestros tejidos se mantienen unidas gracias a las interacciones moleculares que hay entre ellas. En cambio, las células del SI pueden atravesar de una a otra parte del organismo. Lo curioso es que las mismas moléculas que se utilizan para mantener juntas a las células de los tejidos (Moléculas de adhesión celular = CAMs) se usan por os leucocitos para interaccionar con las células de los tejidos. El endotelio vascular actúa de portero, regulando el paso de moléculas y células desde la sangre a los tejidos; el proceso, en lo que se refiere a las células, se denomina extravasación. Para que los linfocitos alcancen los órganos linfoides periféricos o los sitios de inflamación Las células endoteliales expresan CAMs que son específicas para los leucocitos, aunque algunas de ellas se expresan constitutivamente y otras lo hacen solo en respuesta a la concentración local de citocinas inflamatorias que se producen durante una respuesta inflamatoria. Aquellas células circulantes con un receptor que se une a las CAMs que hay sobre las células del epitelio vascular son las que pueden extravasarse a los tejidos. Los linfocitos recirculantes tienen receptores que se fijan a las CAM del endotelio vascular, lo que les permite extravasarse hacia los tejidos. Además de su papel en la unión de los leucocitos a las células endoteliales las CAMs que hay sobre los leucocitos aumentan la fuerza de las interacciones funcionales entre las células del SI. Hay varias moléculas de adhesión que contribuyen a las interacciones entre: Células Th y CPAs Th y células B Células Tc y células blanco La mayoría de las CAMs pertenecen a cuatro familias de proteínas, que son:
• Selectinas • Tipo mucina • Integrinas • Inmunoglobulinas (Ig)
Selectinas Son glucoproteínas de membrana que tienen un dominio extracelular tipo lectina, lo que les permite unirse de forma específica a grupo de hidratos de carbono (azúcares). La interacción primaria de las selectinas es con una porción hidrocarbonada que tiene ácido siálico (sialilada) y que se denomina sialil-Lewisx y que se encuentra unida con frecuencia a las moléculas tipo mucina. La familia incluye tres moléculas, llamadas:
formulandoalgoqueaportar.blogspot.com

• L-Selectina (CD62L) que se expresa sobre la mayoría de los leucocitos circulantes
• E-Selectina (CD62E) que se expresa sobre las células endoteliales durante una respuesta inflamatoria, tras la estimulación de su síntesis por citocinas pro-inflamatorias
• P-Selectina (CD62P), se almacena en unos gránulos de las células endoteliales (llamados de Weibel-Palade) que, tras la estimulación de la célula endotelial, se funden con la membrana plasmática quedando las P-Selectinas sobre la superficie celular.
La mayoría de los leucocitos circulantes expresan L-Selectina, mientras que las otras dos se expresan sobre las células endoteliales durante la respuesta inflamatoria. Estas selectinas son las responsables de la adhesión inicial de los leucocitos al endotelio vascular. Mucinas Constituyen un grupo de proteínas ricas en serina y treonina y muy glucosiladas. Su estructura extendida les permite presentar porciones como sialil-Lewis y otras estructuras hidrocarbonadas para que se capturen por las selectinas. Integrinas Son proteínas hetero-diméricas con una cadena α y otra β, asociadas de forma no covalente, que se expresan sobre la superficie celular. La mayoría interacciona con las moléculas de la matriz extracelular y mantienen la estructura de los tejidos en todo el organismo. Algunas subfamilias (con una subunidad común) se unen a las moléculas de adhesión de la superficie celular e intervienen en las interacciones célula-célula. Los leucocitos expresan una subfamilia específica de integrinas (conocida como β2), así como otras varias, que también se expresan sobre otros tipos celulares. Las β2 se unen a las proteínas de la SPF de las Ig, y a proteínas asociadas a la respuesta inflamatoria. Algunas integrinas necesitan activarse para unirse con alta afinidad a sus ligandos y su agrupamiento sobre la membrana celular aumenta la probabilidad de unión efectiva. Su importancia se refleja en la deficiencia de adhesión de los leucocitos (LAD) tipo 1, una enfermedad autosómica recesiva que se caracteriza por la presencia de infecciones bacterianas recurrentes y dificultad para curar las heridas. Una deficiencia similar se ve en individuos que tienen alterada la expresión de selectinas y se denomina LAD tipo 2. La combinación de integrinas que expresa un tipo dado de célula es lo que la permite unirse a las diferentes CAMs que hay sobre la superficie del endotelio vascular. Superfamilia de las Ig (ICAMs) Esta SPF incluye moléculas de adhesión con dominios tipo Ig, como ICAM-1 (CD54), ICAM-2 (CD102), ICAM-3 (CD50) y VCAM (CD106), que se expresan sobre las células del endotelio vascular y se unen a varias moléculas de integrinas. Hay una molécula de adhesión (llamada MadCAM-1) que tiene dominios tipo Ig y tipo mucina, que se expresa sobre el endotelio de la mucosa y dirige la penetración de los linfocitos en la misma; se une a una integrina vía dominio tipo Ig y a L-Selectina vía dominio tipo mucina. También hay adhesiones homotípicas entre miembros de la SPF.. La molécula PECAM-1 ([molécula 1 de adhesión células endoteliales-plaquetas] CD31) está en la superficie de leucocitos (neutrófilos, monocitos y un subtipo de linfocitos T) y
formulandoalgoqueaportar.blogspot.com

en complejos que unen a las células endoteliales. Presenta unión homotípica, donde el PECAM-1 de una célula se une al PECAM-1 de otra. La molécula de adhesión JAM-1 (CD321) también se encuentra en las uniones endoteliales y puede interaccionar consigo misma y con Integrinas, lo que le otorga un papel en la migración trans-endotelial. También puede encontrarse unión homotípica entre miembros de la SPF Ig en otros tipos celulares, como ocurre con L1 y NCAM sobre las células neurales.
Ejemplos de moléculas de adhesión celular (CAM) de distintas familias Tipo mucina Selectinas SPF Ig Integrinas
GliCAM-1 Selectina L ICAM-1, 2 y 3 VLA-4 (α β ) 4 1CD34 Selectina P VCAM-1 VLA-6 (α β ) 6 1PSGL-1 Selectina E LFA-2 (CD2) LFA-1 (α1β ) 2MadCAM-1 LFA-3 (CD58) MaC-1 (αMβ2) MadCAM-1
Familias de moléculas de
adhesión
Tipo MUCINA INTEGRINAS
SELECTINAS SPF Ig
formulandoalgoqueaportar.blogspot.com

TEMA 3. MOLECULAS DE ADHESIÓ. CIRCULACIÓN LINFOCITARIA (2) RECIRCULACION DE LOS LINFOCITOS Al contrario del resto de las células, los linfocitos recirculan constantemente a través de la sangre, la linfa y los órganos linfoides secundarios. Tras un breve tránsito por la sangre (unos 30 m.) casi el 45% de ellos la abandona directamente a través del bazo, donde residen aproximadamente unas 5 horas. Una cantidad casi igual de linfocitos (42%) sale de la sangre a través de los ganglios linfáticos, donde residen unas 12 horas. Un número pequeño de linfocitos (10%) migra a los tejidos extra-linfoides terciarios atravesando a través de las células endoteliales de los capilares. Estos tejidos normalmente albergan pocos (o ninguno) linfocitos a menos que se “reclamen” por la presencia de una respuesta inflamatoria. Los tejidos terciarios extra-linfoide más activos son lo que forman la interfase con el exterior, tales como piel y epitelios mucosos gastro-intestinales, pulmonares y genito-urinarios. La recirculación mantiene una dotación óptima de linfocitos específicos distintos, que pueden “encontrar” a cualquier antígeno. Un linfocito individual hace el circuito sangre – tejidos – linfa – sangre entre una y dos veces diarias. Como solo uno entre 100.000 linfocitos reconoce un antígeno particular parece que este mecanismo permite a un mayor número de células T y B reconocer al antígeno (sobre células presentadoras o en forma soluble) y desencadenar la respuesta adaptativa en menos tiempo. Los contactos entre células presentadoras y células T se aumentan mucho gracias a los factores que regulan, organizan y dirigen la circulación de linfocitos y células presentadoras de antígenos.
BAZO (5 h)
GANGLIOS LINFATICOS
(12 h)
Pool de linfocitos
sanguíneos (30 m)
Células no recirculantes
Médula ósea Superficies epitelios
Peritoneo
Tejidos extralinfoidesterciarios:
Epitelios mucosos de intestino, pulmones y tracto genito-urinario
Hígado, Cerebro, Piel
?
45% Linfocitos vírgenes
10%
Linfocitos activados
42% Linfáticos aferentes10%
Linfáticos eferentes
(52%)
Recirculación linfocitos
PASO DE LOS LINFOCITOS A TRAVÉS DEL ENDOTELIO VASCULAR 1.- Por las vénulas endoteliales altas Aunque algunos linfocitos abandonan la sangre a través de vénulas no especializadas, la ruta de salida principal en los mamíferos es a través de una sección especializada de las vénulas post-capilares que se conocen como vénulas endoteliales altas (HEV). Estas
formulandoalgoqueaportar.blogspot.com

HEV abundan en la zona para-cortical de los GL, son menos abundantes en la zona cortical y faltan en la zona medular. Además de en los GL, las HEV también se encuentran en el MALT (Tejido linfoide asociado a las mucosas) y en el timo. Algunos linfocitos, principalmente los T, llegan a través de los vasos linfáticos aferentes (en lugar de a través de las HEV), que es la ruta principal de llegada de los antígenos al GL. Las HEV son estructuras permanentes de los tejidos linfoides secundarios, pero también pueden desarrollarse a partir del epitelio normal en los sitios de reacciones inflamatorias crónicas; esto puede significar que las células T específicas tengan una vía directa de acceso (a través de las HEV recién formadas) a la zona inflamatoria. El movimiento de los linfocitos a través del endotelio vascular se controla mediante moléculas de adhesión y quimiocinas, como p.e.:
Mad-CAM-1 que se expresa en las células endoteliales de los tejidos intestinales VCAM-1 que está presente en las células endoteliales de pulmones y piel
En los linfocitos hay otras moléculas que, interaccionan directamente con estas moléculas de adhesión, les llevan directamente a órganos concretos. En el caso del intestino son las integrinas alfa4beta7 (a4b7) las que median la adhesión de los linfocitos en las HEV de las Placas de Peyer (que expresan Mad-CAM-1).
Extravasación en las vénulas de endotelio alto
Extravasación de los linfocitos Varios subtipos de linfocitos presentan una extravasación dirigida a los sitios inflamatorios y órganos linfoides secundarios. La recirculación está cuidadosamente controlada para asegurar el reclutamiento apropiado de poblaciones T y B hacia los diferentes tejidos. Como ocurre con los neutrófilos, la extravasación de linfocitos incluye una serie de interacciones entre moléculas de adhesión y las cuatro fases de rodamiento, activación, detención y adhesión y, finalmente, migración trans-endotelial. Las vénulas con endotelios altos (vénulas endoteliales altas o HEV) son los sitios de extravasación de los linfocitos
formulandoalgoqueaportar.blogspot.com

Algunas regiones del endotelio vascular de las vénulas post-capilares de distintos órganos linfoides están compuestas por células especializadas de formas cuboides y rechoncha (son “altas”), denominándose a estas regiones “vénulas endoteliales altas o HEV”. El aspecto de sus células contrasta mucho con el aplanado de las células endoteliales que recubren el resto del capilar. Todos los órganos linfoides secundarios, con excepción del bazo, contienen HEV. Cuando se incuban con linfocitos cortes de ganglios linfáticos, placas de Peyer o amígdalas y se lavan luego para eliminar a las células no fijadas, cerca del 85% de las células fijadas están adheridas a las HEV, aunque estas vénulas representan solo el 1-2% del área total del corte. Se estima que cada segundo se extravasan, a través de las HEV, unos 140.000 linfocitos en un solo ganglio linfático. Las citocinas producidas en respuesta a la captación de antígenos está comprobado que intervienen en el desarrollo y conservación de las HEV en los órganos linfoides; por ejemplo, los animales criados en condiciones axénicas (sin gérmenes) no desarrollan HEVs, mientras que si se bloquea quirúrgicamente el linfático aferente del ganglio (con lo que bloquea la entrada de los antígenos) al cabo de poco tiempo las HEVs presentan pérdida de funcionalidad y pueden volver a una morfología más aplanada. Las HEV expresan una variedad de moléculas de adhesión. Como cualquier otra células del endotelio vascular expresan: E y P-Selectinas, moléculas tipo mucina (GlyCAM-1 y CD34) y moléculas de la superfamilia de las Ig(ICAm-1, ICAM-2, ICAM-3, VCAM-1 y MadCAM-1). A algunas moléculas de adhesión que se distribuyen específicamente en un tejido se les ha llamado “adresinas vasculares” pues dirigen la extravasación de diferentes poblaciones de linfocitos re-circulantes a órganos linfoides particulares. La figura 13-8 presenta las interacciones típicas en la extravasación de las células T vírgenes a través de las HEV en los ganglios linfáticos. El primer paso es, normalmente, una interacción selectina-carbohidrato similar a la de la adhesión de los neutrófilos. Los linfocitos T vírgenes se unen inicialmente a las HEVs a través de L-Selectina, que sirve como un receptor de albergue que dirige los linfocitos a tejidos que expresan las correspondientes adresinas vasculares tipo mucina, tales como CD34 o GlyCAM-1. El rodamiento de los linfocitos es menos acusado que el de l os neutrófilos. Aunque la interacción inicial selectina-carbohidrato es muy débil, el flujo tan lento de la sangre en las vénulas post-capilares, especialmente en la región de HEV, reduce la probabilidad de que el flujo arrastre al linfocito que se “arrastra”. En el segundo paso, el estímulo de activación de las integrinas está mediado por quimiocinas que bien se sitúan sobre la membrana endotelial o se secretan localmente. El grueso glicocalix que recubre las HEV puede retener estos factores quimio-atractantes in situ. La fijación de quimiocinas a los receptores que hay sobre los linfocitos, acoplados a G-proteína, lleva ala activación de las moléculas de integrina que sobre la membrana (como ocurre en la extravasación de los neutrófilos). Una vez activada la molécula de integrina interacciona con moléculas de la superfamilia de las Ig (p.e. ICAM-1), lo que permite a los linfocitos adherirse firmemente al endotelio. Los mecanismos moleculares que rigen el paso final, la migración trans-endotelial, parece que comprenden a las moléculas de adhesión PECAM-1 (CD31) y JAM-1 (CD321). El albergue de los linfocitos está dirigido por perfiles de receptores y señalesLa extravasación de los linfocitos es similar, en general, a la extravasación de los neutrófilos. Un carácter importante que distingue a ambos procesos es que los distintos subtipos de linfocitos migran diferencialmente a tejidos diferentes. Esta migración
formulandoalgoqueaportar.blogspot.com

específica depende de combinaciones únicas de moléculas de adhesión y quimiocinas; los receptores que dirigen la migración de varias poblaciones de lilnfocitos a tejidos linfoides concretos y tejidos inflamatorios reciben el nombre de “receptores de albergue” o “receptores guías”. Los linfocitos vírgenes recirculan a los tejidos linfoides secundariosLos linfocitos vírgenes no son capaces de montar una respuesta inmune hasta que se han activado para pasar a células efectoras. La activación de las células vírgenes tiene lugar en un microambiente especializado en los tejidos linfoides secundarios (ganglios linfáticos periféricos, placas de Peyer, amígdalas y bazo). En este microambiente las células dendríticas capturan antígenos y los presentan a los linfocitos vírgenes; el resultado es que los linfocitos vírgenes se activan. Las células vírgenes no presentan preferencia por un tipo particular de tejido linfoide secundario, sino que circulan indiscriminadamente por todos los tejidos linfoides secundarios del cuerpo. La unión inicial de los linfocitos vírgenes a las HEV está generalmente mediada por la unión de L-Selectina del linfocito a moléculas de adhesión tales como GlyCAM-1 y CD34 sobre las HEV. Recientemente se ha encontrado que LFA-1 y VLA-4 también median el rodamiento en su conformación de baja afinidad (antes de la activación). Son las quimiocinas CCL21, CCL19 y CXCL12 las que inducen el cambio conformacional a la alta afinidad de LFA-1 y VLA-4, que lleva a la adhesión fuerte. Además de estas quimiocinas CXCL13 activa a las integrinas sobre las células B vírgenes. Otras quimiocinas atraen a las subpoblaciones de linfocitos a sus zonas en los ganglios linfáticos. El patrón de tráfico de las células T vírgenes está diseñado para tener a estas células recirculando constantemente a través de los tejidos linfoides secundarios, cuya función principal es atrapar a los antígenos de la sangre y los tejidos.
CD34 GlyCAM-1
L-Selectina
Célula T virgen: adhesión
Célula endotelial alta
Una vez que los linfocitos vírgenes encuentran a su antígeno atrapado en un tejido linfoide secundario se activan y aumentan su tamaño pasando a linfoblastos. La activación lleva alrededor de 48 horas y, durante este tiempo, los blastos están retenidos en la región paracortical de los tejidos linfoides secundarios. En esta fase, también llamada “de cierre”, los linfocitos específicos del antígeno no pueden detectarse en la circulación. Es durante la fase de cierre cuando se produce la rápida proliferación y diferenciación de las células vírgenes. Las células efectoras y de memoria que se forman dejan el tejido linfoide y comienzan su propia recirculación.
formulandoalgoqueaportar.blogspot.com

Las células efectoras y de memoria se mueven de forma distinta que las T vírgenesLos patrones migratorios de ambos tipos de células difieren del patrón de los linfocitos T vírgenes. Las células efectoras tienden a albergarse en las zonas de infección pues reconocen el endotelio inflamado así como a las moléculas quimiotácticas que se producen durante la respuesta inflamatoria. Los linfocitos de memoria tienden a albergarse selectivamente en el tipo de tejido en el que encontraron por primera vez al antígeno. Al parecer esto asegura que una célula de memoria particular retorne al tejido donde hay mayor probabilidad de que pueda enfrenarse a la amenaza que supone su antígeno. Las células efectoras y de memoria expresan cantidades elevadas de ciertas moléculas de adhesión, como LFA-1, que interaccionan con ligandos presentes en los tejidos extra-linfoides terciarios (como piel y epitelios mucosos) y en los sitios de inflamación, lo que permite que las células efectoras y de memoria los alcancen. Las células inmaduras carecen de las moléculas de adhesión correspondientes y no alcanzan ni se albergan en estos sitios. El endotelio inflamado expresa las Selectinas E y P y las moléculas de la superfamilia de las Ig VCAM-1 e ICAM-1, que se unen a los receptores correspondientes expresados en gran cantidad por células efectoras y de memoria. El resultado de todo ello es que estas células presentan una clara vocación tisular, mientras que evitan a los tejidos linfoides secundarios como consecuencia de su poca expresión de L-Selectina. Un segundo subgrupo de células efectoras y de memoria tienen una clara vocación dérmica y su dotación se caracteriza por la expresión baja de L-Selectina y alta de LCA (antígeno linfocítico cutáneo) y LFA-1, moléculas que se fijan a la E-Selectina e ICAMs que hay dentro de las vénulas dérmicas de la piel. Aunque las células efectoras y de memoria, que expresan cantidades reducidas de L-Selectina, no tienden a dirigirse a los tejidos linfoides secundarios a través de las HEV si pueden alcanzar los ganglios linfáticos secundarios a través de la linfa aferente.
Células T efectoras: adhesión
formulandoalgoqueaportar.blogspot.com

INMUNIDAD INNATA Introducción Los vertebrados están protegidos por dos sistemas de inmunidad: la natural y la adaptativa. La inmunidad innata consiste en una serie de defensas frente a la infección que están listas para activarse antes del ataque de un patógeno. Este sistema innato incluye barreras físicas, químicas y celulares. Las barreras físicas principales son la piel y las mucosas. Entre las barreras químicas están la acidez del estómago y unas moléculas solubles con acción antimicrobiana (péptidos antimicrobianos). La línea celular de la defensa innata incluye una serie de células con receptores especiales para detectar productos bacterianos y desencadenar un contraataque. La respuesta a la invasión de un agente microbiano que sobrepasa las barreras iniciales (piel y mucosas) es muy rápida y típicamente se desarrolla a los pocos minutos de la invasión. A pesar de las múltiples capas del sistema innato algunos patógenos consiguen evadir las defensas innatas y es el momento de llamar a un segundo sistema, denominado inmunidad adaptativa (o adquirida) que se induce por la exposición al microbio y responde a la infección con una respuesta específica “a la medida” en forma de grandes poblaciones de linfocitos que reconocen específicamente al invasor. Pero producir una respuesta adaptativa lleva tiempo (una semana o más) antes de que sea totalmente efectiva. La inmunidad adaptativa tiene la propiedad de la memoria inmune que consiste en que una vez estimulada por un patógeno en particular, las futuras exposiciones al mismo patógeno desencadenan respuestas más rápidas y potentes. El reconocimiento de los invasores se realiza por medio de los anticuerpos y los receptores T, que son los “sensores” de la inmunidad adaptativa. Estas moléculas son el producto de genes con una característica extraordinaria: experimentan modificaciones y diversificación (recombinación genética) en el hospedador para generar una población gigantesca de centinelas para vigilar a los invasores. La inmunidad innata es el mecanismo de defensa más antiguo de los vertebrados frente a los microbios y también se han encontrado formas de la misma en todas las plantas y animales pluricelulares. La inmunidad adaptativa ha evolucionado en los vertebrados y es de aparición mucho más reciente. En los vertebrados, la inmunidad adaptativa complementa a un sistema innato bien desarrollado. Está ahora claro que ambas inmunidades han co-evolucionado en los vertebrados y que entre ambos sistemas se han desarrollado muchas interacciones e interdependencias. De hecho, si un patógeno evade completamente a las primeras líneas de defensa (el sistema innato) la respuesta adaptativa es muy débil; el reconocimiento por el sistema innato pone las bases para una respuesta inmune efectiva del sistema adaptativo. Barreras anatómicas Lo primero que destaca en la inmunidad innata son las barreras externas que se oponen a la invasión microbiana (piel y membranas mucosas) e incluyen a los epitelios mucosos que recubren el tracto respiratorio, el gastrointestinal y el urogenital, aislando así el interior del cuerpo de los patógenos del mundo exterior. La piel consta de dos capas distintas: una externa (epidermis) y otra situada debajo y mucho más gruesa (dermis). La epidermis consta de varias capas de células epiteliales, muy apretadas entre si, y se divide en una capa externa formada básicamente por células
formulandoalgoqueaportar.blogspot.com

muertas llenas de una proteína impermeabilizante denominada queratina. La dermis está formada por tejido conectivo y contiene vasos sanguíneos, folículos pilosos, glándulas sebáceas y glándulas sudoríparas. La piel y los epitelios aportan una especie de “lámina adhesiva” que recubre y protege las cavidades internas del cuerpo del mundo exterior. Pero las barreras anatómicas son algo más que un envoltorio pasivo y tienen también defensas bioquímicas activas que producen y secretan péptidos y proteínas con actividad antimicrobiana. Entre las muchas sustancias que produce la piel humana se ha identificado recientemente la psoriasina, una pequeña proteína que tiene una actividad antimicrobiana potente frente a Escherichia coli. Su existencia aclara el por qué la piel humana resiste a la colonización por E. coli a pesar de estar expuesta constantemente a la bacteria (otras bacterias son menos sensibles a la la psoriasina). La capacidad de piel y epitelios para producir varios agentes antimicrobianos es importante ya que las heridas y erosiones cutáneas son rutas de infección que se explotarían inmediatamente por los microbios patógenos si no existieran estas defensas bioquímicas. En lugar de por la piel, los tractos alimentario, respiratorio, urogenital y los ojos están revestidos por membranas mucosas, formadas por una capa epitelial externa y otra subyacente de tejido conectivo. Muchos microbios penetran en el cuerpo atravesando esas membranas y, para evitarlo, hay muchos sistemas de defensa inespecíficos; p.e., saliva, lágrimas y secreción mucosa arrastran (lavan) a los invasores potenciales además de contener sustancias antibacterianas y antivirales. El fluido viscoso llamado mucus, secretado por las células epiteliales de las membranas mucosas, atrapa a los organismo foráneos. En el tracto respiratorio inferior la membrana mucosa está recubierta por un epitelio ciliado y el movimiento sincronizado de los cilios arrastra al mucus, y a los microorganismos atrapados, hacia la garganta para su deglución y destrucción por la barrera ácida gástrica. Con cualquier comida ingerimos un gran número de microorganismos, pero estos tienen que enfrentarse a los elementos antimicrobianos de saliva y de los epitelios de la boca y al ambiente hostil que supone el conjunto de acidez y enzimas digestivas del estómago. Además de las barreras bioquímicas y anatómicas los microbios deben competir por los recursos con numerosos microorganismo no patogénicos que colonizan las superficies mucosas. La flora normal está muy adaptada al ambiente interno que impide la unión de los patógenos a la superficie mucosa y compite favorablemente por los nutrientes necesarios. Algunos microorganismos han desarrollado métodos para evadir las defensas que hay en las superficies mucosas como, p.e., el virus de la gripe tiene una molécula superficial que le permite fijarse a las células de la membrana mucosa del tracto respiratorio, lo que impide su arrastre por la corriente ciliar. El organismo que produce la gonorrea (Neisseria gonorreae) tiene unas proyecciones superficiales que se unen a las células epiteliales de la membrana mucosa del tracto urogenital. LA adherencia de las bacterias a las membranas mucosas está generalmente mediada por unas proyecciones parecidas a pequeños flagelos (llamadas fimbrias o pili) que interaccionan con ciertas glicoproteínas o glicolípidos que solo se expresan por las células epiteliales de la membrana mucosa de tejidos particulares. Por esta razón, algunos tejidos son susceptibles a la invasión por patógenos particulares, a pesar de la eficacia protectora general de las barreras epiteliales. Cuando esto ocurre, son los receptores de la inmunidad innata los que se encargan de detectar la infección y poner en marcha una defensa efectiva frente a ella. LAS CELULAS DE LA INMUNIDAD INNATA
formulandoalgoqueaportar.blogspot.com

En la respuesta innata intervienen muchos tipos celulares distintos, pero los actores claves (las estrellas) son: neutrófilos, monocitos/macrófagos, Células Nk y células dendríticas Los neutrófilos fagocitan y matan Son las primeras células que emigran desde la sangre a los sitios de infección y tienen una serie de armas para oponerse a los microbios invasores. Son esenciales para la defensa innata frente a bacterias y hongos. Su arma principal es la fagocitosis, pero también tienen otras armas como la presencia de varios receptores que les permiten detectar los peptidoglicanos de las bacterias Gram+ y el lipopolisacárido (LPS) de las Gram-. Aunque los neutrófilos pueden reconocer directamente a los patógenos, la fijación y fagocitosis mejoran drásticamente cuando los microbios están opsonizados (en la respuesta innata inicial por fragmentos del complemento que se depositan sobre ellos). Los macrófagos tienen varios mecanismos antimicrobianos Los macrófagos en reposo se activan por una serie de estímulos. Los receptores TLR de su superficie reconocen componentes microbianos tales como LPS, peptidoglicanos y flagelina, mientras que sus receptores de citocinas detectan los mensajes solubles (citocinas) producidos por otras células que intervienen en la respuesta inflamatoria. La activación de los macrófagos supone que adquieren una mayor capacidad fagocitaria y que aumenta su capacidad para destruir a los microbios fagocitados, además de producir y secretar mediadores inflamatorios. También expresan niveles altos de moléculas CMH Clase II esenciales para la presentación de antígenos a los linfocitos T (que es otro punto importante en la colaboración entre la inmunidad innata y la adaptativa. Las células NK son la primera línea defensiva frente a los virus Estas células (procedentes de la línea linfoide) son capaces de discriminar entre células infectadas y no infectadas por virus; respetan a las no infectadas y destruyen a las infectadas mientras se pone en marcha la respuesta adaptativa que aportará las células T citotóxicas específicas y los anticuerpos, aunque hay algunas infecciones que se eliminan totalmente por la inmunidad innata sin la ayuda de la adaptativa. Las NK activadas son grandes productoras de citocinas que regulan a otras células del sistema inmune, sobre todo interferón gamma (IFN-g) y el factor de necrosis tumoral alfa (TNF-a), dos citocinas inmuno-reguladoras potentes y versátiles. Estas dos citocinas pueden estimular la maduración de las células dendríticas (DCs), que son las coordinadoras clave entre la inmunidad innata y la adaptativa. Las células dendríticas capturan patógenos e inducen la respuesta adaptativa a través de las células T Las células dendríticas maduras son capaces de activar a las células T, tanto a las Th (CD4+) como a las Tc (CD8+) ya que, además de tener moléculas CMH Clase I y Clase Il, presentan una fuerte actividad coestimuladora. Como agentes de la inmunidad innata las DCs inmaduras presentan una gran variedad de PRRs, especialmente TLRs, para reconocer patógenos. El reconocimiento provoca la activación de las DCs y su entrada en eñ proceso de maduración, durante el que aumentan la síntesis de moléculas Clase II
formulandoalgoqueaportar.blogspot.com

y moléculas coestimuladoras destinadas a activar a las células T. Durante la maduración migran a los órganos linfoides secundarios para presentar los antígenos a los linfocitos T. La respuesta de las DCs no solo interviene en la comunicación entre los dos niveles del sistema inmune, sino que también ataca directamente a los patógenos que detecta, ya que producen ROI, NO y sintetizan péptidos antimicrobianos. Es decir las DCs usan mecanismos comunes con los macrófagos para destruir a los microbios fagocitados. Además, hay un subtipo de DCs (las células dendríticas plasmacitoides) que produce grandes cantidades de interferones antivirales (IFN-a e IFN-b). Una característica de los virus es la expresión de su genoma en la célula hospedadora y el reconocimiento de ácidos nucleicos extraños por parte de los TLRs de las DCs plasmacitoides induce la producción de estos interferones, que bloquean la replicación viral. Otros subtipos de DCs producen IL-12, TNF-a e IL-6, que son potentes inductores de inflamación. IL-12 determina el tipo de respuesta T de la inmunidad adaptativa. CONEXIONES ENTRE LA INMUNIDAD INNATA Y LA ADAPTATIVA. Una vez que un patógeno sobrepasa las barreras anatómicas y fisiológicas puede producir la infección y la enfermedad. El sistema inmune responde a la invasión detectándola mediante dos tipos de sensores y elaborando sistemas de defensa que atacan al invasor. La primera detección tiene lugar cuando el invasor interacciona con receptores solubles o unidos a la membrana de las células del hospedador, capaces de discriminar entre lo propio (del hospedador) y lo no propio (del patógeno). Estos sensores reconocen moléculas (o patrones moleculares) que se han conservado en grupos de microbios (normalmente porque son imprescindibles para su supervivencia) y que normalmente no existen en el hospedador. Como reconocen conjuntos de moléculas concretas (patrones moleculares), tales receptores se denominan Receptores de Reconocimiento de Patrones (PRRs). Como estos conjuntos moleculares se encuentran sobre o en los patógenos se llaman Patrones Moleculares Asociados a Patógenos (PAMPs). Los PAMPs reconocidos por PRRs incluyen combinaciones de azúcares, ciertas proteínas, moléculas particulares portadoras de lípidos y algunos motivos de ácidos nucleicos. Esta restricción del reconocimiento innato a los patrones moleculares que se encuentran en los microbios permite al sistema innato centrarse sobre las entidades productoras de infección y no sobre sustancias que solo son no propias, tales como las prótesis artificiales de cadera. Los receptores de la inmunidad adaptativa (anticuerpos y receptores de los linfocitos T), por el contrario, reconocen detalles más finos de las estructuras moleculares y pueden discriminar con una especificidad exquisita entre antígenos con diferencias estructurales mínimas. Típicamente, la capacidad de los PRRs para distinguir entre lo propio y lo no propio es perfecta, ya que los patrones moleculares detectados por el receptor solo se producen por los patógenos y nunca por el hospedador. Esto contrasta claramente con el reconocimiento ocasional de antígenos propios por los receptores de la inmunidad adaptativa, una disfunción potencialmente peligrosa que puede convertirse en una enfermedad autoinmune. Para la detección de los PAMPs por los receptores de la inmunidad innata tienen que intervenir múltiples componentes de la inmunidad. Los mediadores solubles incluyen a iniciadores del sistema del complemento, tales como la Lectina fijadora de manosa (MBL) y la Proteína C reactivan (CRP). Si el patógeno porta PAMPs reconocibles por estos mediadores se activa el sistema del complemento. Una parte del sistema del
formulandoalgoqueaportar.blogspot.com

complemento consiste en una serie de proteínas que cuando se activan forman poros en la membrana de los microbios blanco (Lisis celular). El sistema del complemento también incluye varias glicoproteínas que, cuando se activan, promocionan la fagocitosis de los microorganismos por los fagocitos (Opsonización). El sistema del complemento está a caballo sobre los sistemas inmunes innato y adaptativo: la activación de la cascada del complemento lleva a la opsonización o lisis de los invasores y puede activarse tanto por moléculas que reconocen PAMPs (I innata) o por anticuerpos (I adaptativa) que reconocen a antígenos específicos. Además, algunos subproductos de la activación del complemento promueven la Inflamación y, de este modo, conducen a los leucocitos a los sitios de infección y promueven otra de las capas de la respuesta. Las células dendríticas inmaduras (iDCs) de los tejidos y los macrófagos tienen una amplia serie de receptores, incluyendo los TLRs, que detectan los productos microbianos. Hasta ahora se han descrito 12 TLRs para los ratones y 11 para los humanos; cada TLR reacciona con un producto microbiano específico y esto permite a DCs y macrófagos detectar a un amplio espectro de patógenos. Las señales que se hincan en los TLrs de macrófagos estimulan la actividad fagocitaria y y la producción de mediadores químicos que son tóxicos para los microbios fagocitados. Los macrófagos activados también secretan una serie de moléculas conocidas colectivamente como citocinas y que son hormonas o factores de crecimiento que actúan vía receptores para inducir respuestas celulares específicas. Las iDCs del sitio de infección interiorizan y procesan al antígeno, maduran y migran a los tejidos linfoides, donde su presentación de antígenos a las células T es el paso clave para que se inicie la respuesta adaptativa frente al patógeno invasor. Esta función es el puente que se establece entre la inmunidad innata y la adaptativa. Las DCs también secretan una serie de citocinas que promueven la inflamación y ayudan directamente a la respuesta adaptativa del hospedador. Todas las funciones innatas se realizan al comienzo de la infección, antes de que se forme una población significativa de células T específicas del patógeno y de anticuerpos específicos para el patógeno por parte de las células B. Además, las citocinas liberadas por las células que intervienen en la respuesta innata afectan a la naturaleza de la subsiguiente respuesta adaptativa a la infección. En muchos casos, el sistema innato puede controlar y eliminar a una infección por si mismo y, cuando no puede, el patógeno se encuentra frente a un ataque coordinado por parte del sistema adaptativo. Durante la respuesta adaptativa, las células T citotóxicas detectan y destruyen a los patógenos que se esconden dentro de las células del hospedador y los anticuerpos neutralizan la capacidad del invasor para invadir o infectar a otras células mientras aumentan la probabilidad e fagocitosis de los invasores por parte de neutrófilos y macrófagos (opsonización). Los anticuerpos también colaboran con el sistema del complemento para inducir la lisis de los patógenos. Cuando se elimina (aclara) la infección algunas de las células B y T generadas durante la fase adaptativa de la respuesta persisten en el hospedador durante tiempos largos en forma de células de memoria. Una nueva infección por el patógeno se encontrará con una reserva de linfocitos específicos lista para detectar al patógeno y montar una respuesta rápida y eficaz. MOLÉCULAS SOLUBLES Y RECEPTORES ASOCIADOS A LA MEMBRANA El sistema innato es polifacético, usando moléculas solubles y receptores de membrana como sus efectores. Ciertas moléculas solubles se producen en el sitio de la infección o
formulandoalgoqueaportar.blogspot.com

daño y actúan localmente. Estas moléculas incluyen péptidos antimicrobianos como defensinas y catelicinas así como a los interferones, que son un grupo importante de citocinas con acción antiviral. Otros efectores solubles se producen en sitios alejados y llegan al tejido blanco a través de la sangre; dentro de este grupo están las proteínas del complemento y las proteínas de fase aguda. Péptidos antimicrobianos Se han aislado de fuentes tan diversas como humanos, ranas, moscas, nematodos y varias especies de plantas. Su pronta aparición en la evolución y la conservación de esta estrategia defensiva, unida a la identificación de más de 800 péptidos antimicrobianos diferentes, testifica su efectividad. Su tamaño varía entre 6 y 59 aminoácidos y la mayoría presentan son catiónicos (con carga positiva), como las magaininas de la piel de las ranas y las defensinas de humanos y otras especies. Las defensinas humanas son péptidos catiónicos con 29-35 residuos de aminoácidos y 6 cisteínas constantes que forman 3 puentes disulfuro y estabilizan una estructura tridimensional relativamente rígida. Matan a una serie de bacterias, incluyendo Staphylococcus aureus, Streptococcus pneumoniae, E. coli, Pseudomonas aeruginosa y Hemophilus influenzae. Los neutrófilos son una fuente abundante de estos péptidos, pero también se producen por las células de Paneth del intestino y las células epiteliales de páncreas y riñón a producen y secretan rápidamente al suero. Las defensinas matan a las bacterias con rapidez (pocos minutos y, las más lentas, en unos 90). El mecanismo de acción más frecuente es alterar las membranas bacterianas pero también producen una serie de efectos intracelulares, tales como inhibición de la síntesis de DNA, RNA y proteínas o activando enzimas antimicrobianas líticas. Los péptidos antimicrobianos no solo atacan a bacterias y hongos sino que se ha visto que se unen a las lipoproteínas de la envoltura de algunos virus (influenza, herpesvirus). La amplitud de su espectro, su probada eficacia antimicrobiana y el aumento de las resistencias a los antibióticos son factores que favorecen el planteamiento de su uso terapéutico, aunque todavía faltan muchos datos para su aprobación por las autoridades sanitarias. Las proteínas de fase aguda contribuyen a la inmunidad innata Durante las infecciones por neumococos (antes de la aparición de los antibióticos) se vio que cambiaban las concentraciones de varias proteínas del suero durante la fase aguda de la enfermedad, fase que precedía a la curación o la muerte. Estos cambios del suero se llamaron colectivamente Respuesta de fase aguda y las proteínas que suben o bajan durante la fase aguda se laman todavía Proteínas de fase aguda. El significado fisiológico de muchas de estas proteínas todavía se desconoce, pero ahora sabemos que algunas, como las proteínas del complemento o la proteína C reactiva (CRP), son parte de la respuesta inmune innata a la infección. La respuesta de fase aguda se induce por señales que viajan a través de la sangre desde el sitio de daño o infección. El sitio principal de producción es el hígado y las señales responsable de ella son las citocinas IL-1, IL-6 y TNF-a; estos mensajeros se producen en la fase inicial de la respuesta por los fagocitos. La proteína C reactiva (CRP) pertenece a una familia de proteínas pentaméricas llamadas pentraxinas, que se unen a sus ligandos (polisacáridos del neumococo y, sobre todo a la fosforil-colina que abunda en la superficie de la mayoría de los microbios) en forma Ca-dependiente. La unión de la CRP a estos ligandos sobre las
formulandoalgoqueaportar.blogspot.com

superficies microbianas promueve su fagocitosis y activa un ataque mediado por el complemento contra los microbios. Otra proteína de fase aguda es la Lectina fijadora de manosa (MBL), que reconoce patrones moleculares con manosa sobre los microbios, pero no sobre las células de vertebrados. . También dirige la acción del complemento contra los microbios a los que se fija. La inmunidad innata usa una serie de receptores para detectar la infección Presentes en sangre y líquidos tisulares como proteínas solubles o unidas a la membrana de neutrófilos, macrófagos y células dendríticas. Como receptores solubles están MBL y CRP que se unen a las membranas microbianas y promueven la fagocitosis o hacen del invasor un blanco probable para la lisis mediada por complemento. La proteína fijadora de polisacárido (LBP) es una parte importante del sistema que reconoce y pone en marcha la cascada de señales en respuesta al lipopolisacárido (LPS), un componente de la capa externa de la pared de las bacterias Gram -. Las proteínas NOD (de nucleotide binding oligomerization domain) son de localización citosólica. NOD-1 reconoce un tripéptido procedente de la degradación del peptidoglicano, mientras que NOD-2 reconoce al muramil-dipéptido procedente de la degradación el peptidoglicano de las bacterias Gram -. Los receptores basureros de la membrana (Scavenger receptors o SRs) está en los macrófagos y varios tipos de células dendríticas e intervienen en la fijación e internalización de las bacterias tanto Gram + como Gram -, así como en la fagocitosis de las células apoptóticas del hospedador. RECEPTORES TOLL-LIKE Una historia muy reciente La proteína Toll empezó a intrigar a los investigadores alemanes en la década de los 1980s cuando vieron que las moscas en desarrollo no desarrollaban un eje dorso-ventral adecuado en ausencia de tal proteína. (Toll, referido a estas moscas mutantes de anatomía casi imposible procede de “extraño” en alemán). Se trata de una proteína transmembrana receptora de señales y las moléculas relacionadas con papeles en la inmunidad innata se conocen como TLRs. Tres descubrimientos han llevado al conocimiento del papel central de los TLRs en la inmunidad innata: La primera observación viene de la mosca de la fruta. En 1996, Jules Hoffman y Bruno Lemaitre encontraron que las mutaciones en Toll (conocido previamente su papel en el desarrollo embrionario de la mosca) hacían a las moscas muy susceptibles a las infecciones letales por Aspergillus fumigatus, un hongo al que son inmunes las moscas de vida libre. La segunda observación viene de un año después (1997), cuando Ruslan Medzhitov y Charles Janeway cuando descubrieron que cierta proteína humana (identificada por la homología de su dominio citoplásmico con el de Toll) activaba la expresión de genes de respuesta inmune cuando se transfectaba a una línea celular humana. Esta proteína humana se denominó TLR4 y es la primera evidencia de una vía de respuesta inmune conservada y compartida por moscas de la fruta y humanos La tercera evidencia es de 1998 y prueba que los TLRs forman parte de la fisiología inmune normal de los mamíferos. Los ratones mutantes del Laboratorio de de Bruce Beutler, homozigóticos para el local lps, resisten al lipopolisac-arido (endotoxina) de las
formulandoalgoqueaportar.blogspot.com

bacteria Gram -. Las infecciones severas en humanos pueden producir el shock séptico, en que pueden fallar órganos vitales tales como cerebro, corazón, riñón e hígado. Cada año mueren unas 20.000 de shock séptico producido por infecciones con Gram -, por lo que resultaba raro que algunas cepas mutantes de ratones resistieran dosis letales de LPS. La secuenciación del DNA reveló que el gen murino lps codificaba una forma mutante de TLR4, que se diferencia de la forma normal en un solo aminoácido. Este trabajó fue la demostración inequívoca de que TLR4 es indispensable para el reconocimiento de LPS. En los pocos años siguiente múltiples investigadores han determinado la existencia de muchos TLRs; hasta ahora, 11 en humanos y 12 en ratones. Estructura Los TLRs son proteínas que “salen” de la membrana y tienen elementos estructurales comunes en la región extracelular; son las repeticiones ricas en Leucina (LRRs) (segmentos repetidos de 24 a 29 aa que contienen la secuencia xLxxLxLxx, donde x es cualquier aminoácido y L es Leucina). Todos los TLRs contienen varios LRRs y es una porción de LRR la que conforma el sitio de fijación para el ligando. La parte (dominio) intracelular del TLR se denomina TIR (de TLR e IL-1R) por la similitud de los dominios citoplásmicos de TLRs y receptor de IL-1. Los dominios TIR tienen tres regiones muy conservadas que son las que sirven de muelle de anclaje para las proteínas intracelulares que participan en las vías de señales mediadas por los TLRs. Funciones Se han determinado las funciones de 9 de los 11 TLRs humanos. Curiosamente, cada TLR detecta un repertorio distinto de moléculas de patógenos muy conservadas. La dotación de TLRs permite detectar virus, bacterias, hongos e, incluso, algunos protozoos. Hay que destacar que los ligandos que fijan los TLRs son componentes indispensables para los patógenos: un virus no funciona sin sus ácidos nucleicos, una bacteria Gram- es inviable sin su LPS. Los TLRs que econocen ligandos extracelulares están en la superficie de la célula, mientras que los que reconocen ligandos intracelulares (RNA, viral, fragmentos de DNA de bacterias, etc) se localizan en compartimentos intracelulares. El emparejamiento de TLRs afecta a la especificidad. La unión TLR2-TLR6 detecta una amplia variedad de moléculas de microbios (peptidoglicanos, zymosanes, lipopéptidos bacterianos, etc.); pero cuando TLR2 se empareja con TLR1 reconoce lipoproteínas bacterianas y y algunas proteínas características de la superficie de parásitos. TLR4 es el receptor clave para la mayoría de los lipopolisacáridos bacterianos TLR5 reconoce la flagelina (componente estructural de los flagelos bacterianos) TLR3 reconoce el RNA de doble hélice (dsRNA) que aparece en las células tras la infección por virus RNA TLR7 y TLR8 reconocen al RNA viral de hélice simple (ssRNA). TLR9 reconoce y responde a la secuencia de DNA CpG (Citosina no metilada unida a Guanidina). Estas secuencias no metiladas abundan en el DNA bacteriano y son muy raras en el DNA de vertebrados. TLRs Ligandos Microbios blanco TLR1 Triacil lipopéptidos Mycobacterias TLR2 Peptidoglicanos, Bacterias G+
formulandoalgoqueaportar.blogspot.com

Proteínas con anclaje GPI Lipoproteínas Zymosan
Tripanosomas Mycobacterias Levaduras y otros hongos
TLR3 RNA de doble hélice (dsRNA) Virus TLR4 LPS
Proteína F Bacterias Gram – Virus respiratorio sincitial (RSV)
TLR5 Flagelina Bacterias TLR6 Diacil lipopéptidos
Zymosan Mycobacterias Levaduras y hongos
TLR7 RNA de hélilce simple (ssRNA) Virus TLR8 RNA de hélilce simple (ssRNA) Virus TLR9 Dinucleótidos CpG no metilados
Dinucleótidos en infección por herpesvirus
DNA bacteriano Algunos herpesvirus
TLR10, 11 Desconocido Desconocido VIAS DE TRANSDUCCION DE SEÑALES Los receptores de la superficie celular reciben las señales iniciales. El paso siguiente, la transmisión de esas señales al interior de la célula (Transducción de señales), es un mecanismo universal de los sistemas biológicos. La respuesta a la señal requiere tres elementos:
• La propia señal • El receptor • La vía de transducción de señales, que conecta la detección con la respuesta
efectora
Señal Receptor Transducción de señales Mecanismo efector
En el caso de la inmunidad innata, la señal será un producto microbiano, el receptor será un PRR y la señal se transduce por la interacción de moléculas intracelulares específicas. El mecanismo efector (la acción que se produce como consecuencia de la señal) tiende a la eliminación del organismo invasor. Las señales a través de TLRs ponen en marcha vías de transducción típicas La vía empieza por la interacción del receptor con la señal.Los productos microbianos se unen a la porción extracelular del TLR. En el lado citoplásmico del receptor está la estructura (muy conservada) TIR, que ofrece sitios de unión para otros componentes de la vía. La señal induce el ensamblaje de los componentes de la vía, en el que interviene una molécula adaptadoraLas propias moléculas adaptadoras, que también contienen dominios TIR, interaccionan con el dominio TIR del TLR. La proteína adaptadora más habitual en las vías TLR es MyD88, que promueve la asociación de dos protein-kinasas y la formación del complejo IRAK1 - IRAK4
formulandoalgoqueaportar.blogspot.com

Fosforilación mediada por protein-kinasasEl componente 4 del complejo anterior fosforila al componente 1 y este nuevo fosfato supone un punto de atraque sobre IRAK1 para TRAF6, que se fija y luego se disocia en compañía de IRAK1 formando el complejo intermediario IRAK1-TRAF6. Ahora, otra protein-kinasa (TAK1) se une al complejo junto con otras proteínas y el resultado es la activación de la actividad kinasa TAK1. Comienza una cascada enzimáticaLa actividad de TAK1 permite la activación, mediada por fosforilación, de otros dos módulos de transducción: Uno de ellos va a la vía de la protein-kinasa activada por mitógeno (MAP-kinasa) que pone en marcha una cascada enzimática (conservada en todas las células eucariotas) cuyo producto final penetra en el núcleo y promueve la fosforilación de uno o más factores de transcripción. El otro va a la vía NF-kB. TAK1 también fosforila a la protein-kinasa IKK, que es la enzima clave para activar la vía NF-kB. El factor NF-kB está inhibido por la unión a una proteína plasmática no fosforilada, IkB. La activación de IKK fosforila a IkB y lo separa del complejo, con lo que NF-kB, ya libre, puede migrar al núcleo e inducir la transcripción de muchos genes implicados en las funciones efectoras de la inmunidad innata, tales como citocinas inflamatorias, moléculas de adhesión y otros efectores. NF-kB también tiene un papel importante en algunas vías de transducción de las células T y B y es, por tanto, también importante en la inmunidad adaptativa. La activación de las vías de señales TLR tiene muchos efectos: Promueve la expresión de genes que contribuyen a la inflamación Induce cambios en las CPAs, que las hace más eficientes como células presentadoras Induce la síntesis de moléculas de señales intercelulares que afectan al comportamiento de leucocitos y otras células La intervención de los TLRs puede aumentar la capacidad fagocitaria de macrófagos y neutrófilos y cambiar su fisiología a vías que aumenten su capacidad para matar y eliminar a los patógenos.
formulandoalgoqueaportar.blogspot.com

TEMA 05. INMUNIDAD INNATA (II)
Respuesta Innata. - Complemento Activación Vía clásica Activación vía alternativa Activación vía de las lectinas (manosa) Regulación Receptores del Complemento Papel biológico del Complemento Sustancias solubles de la Inmunidad innata inducida: Complemento Es uno de los factores solubles, integrante de la Respuesta Innata inducida y quizás uno de los mas importantes en la defensa frente a bacterias. Bordet, a finales del siglo XiX ya comprobó la perdida de capacidad lítica expe-rimentada por los antisueros cuando se calentaban a 56 ºC, capacidad que recuperaban si se añadía suero normal. Dado que los anticuerpos son termoestables, este fenómeno no era debido a los ellos sino a otras sustancias termolábiles que complementaban la acción de los anticuerpos, dándoseles el nombre de Sistema del Complemento. La relación entre ambos componentes se parece a la existente entre la llave de contacto y el motor de un vehículo: la molécula de anticuerpo sirve para poner en marcha el sistema del complemento, que como el motor del coche, hace el verdadero trabajo. El sistema del complemento consta de mas 30 proteínas que actúan de forma secuencial, de modo similar a como lo hace el sistema de la coagulación, y contribuye a muchas de las funciones efectoras de la Inmunidad. El proceso de activación se basa en que cada una molécula, inicialmente inactiva, se activa por proteolisis y, a su vez, activa al siguiente componente y así sucesivamente. El carácter amplificador del proceso viene dado porque la molécula activada en un paso puede activar a muchas moléculas en el paso siguiente. Las proteínas del sistema del complemento, así como las que se producen durante su activación, cumplen una serie de funciones entre las que podríamos destacar las siguientes:
Opsonizacion. Esta mediada por moléculas que se unen covalentemente a la superficie de los microbios y facilitan su fagocitosis por las células fagocitarías.. Estas tienen en su membrana receptores específicos para estos fragmentos del complemento.
Quimiotaxis.. Los fragmentos pequeños, que se producen por la proteolisis de algunas de las proteínas del complemento, atraen a los fagocitos hacia el lugar de activación del complemento. Además, también activan a dichos fagocitos.
Inflamación: Alguno de los fragmentos anteriores también activa a los mastocitos para que liberen mediadores inflamatoriois.
Acción Lítica: Los componentes terminales de la cascada dañan a los microorganismos introduciéndose en las membranas bacterianas y formando poros en las mismas (Lisis celular).
Aclaramiento de inmunocomplejos: Los receptores y fragmentos del complemento contribuyen a la eliminación de los complejos antígeno/anticuerpos que se forman en el organismo como consecuencia de la respuesta adaptativa.
TEMA 05. INMUNIDAD INNATA (II) Pág. 1 de 10
formulandoalgoqueaportar.blogspot.com

TEMA 05. INMUNIDAD INNATA (II)
Los componentes séricos, están normalmente inactivos o con un nivel espontáneo de activación muy bajo y hay varias formas (vías) de poner en marcha (activar) al sistema del complemento Estas vías difieren en las moléculas iniciadoras pero luego convergen para producir el mismo grupo de moléculas efectoras. VIAS DE ACTIVACIÓN DEL COMPLEMENTO En la respuesta innata
• Vía de las lectinas: Se pone en marcha cuando una lectina soluble (Lectina fijadora de manosa o MBL) detecta a este azúcar en la superficie de un microorganismo. Para que se produzca la activación MBL tiene que asociarse a dos proteasas (MASP1 y MASP2).
• Vía alternativa: Es filogenéticamente anterior a la vía clásica. Su activación se produce de forma espontánea a un nivel muy bajo y los daños al tejido (traumas, estrés, infección) aumentan a nivel funcional la activación.
• Vía de la proteína C reactiva (RCP): Esta proteína se une a la fosforilcolina (abundantemente expuesta en las membranas de los microorganismos e inaccesible en las células animales) y activa el complemento a partir del primer componente de la vía clásica. Solo funciona hasta el nivel de C3.
En la respuesta adaptativa • Vía clásica: Se activa por los complejos antígeno/anticuerpos, aunque solo son
capaces de activarla los anticuerpos de las clases IgG e IgM. NOMENCLATURA Las proteínas del complemento se encuadran en tres categorías:
• Funcionales: Las que intervienen en las cascadas de activación. Los componentes de la vía clásica se denominan con la letra C seguida de los subíndices 1-9 (C1... C9). Para la vía alternativa existen 5 proteínas denominadas como Factores B,H,I,D y properdina.
• Reguladoras: Reciben nombres específicos (p.e. Factor I, DAF, C1q – inhibidor, etc.)
• Receptores: Se encuentran en la superficie de diversa células y se denominan CR, seguido de un número romano y, a veces, de letras minúsculas (p.e. CRIIIb)
CASCADAS DEL COMPLEMENTO
1. - Vía Clásica.
Es uno de los mecanismos efectores de la inmunidad Humoral, que se activa por la unión del primer componente C1 a las regiones Fc (constantes) de ciertas clases de anticuerpos, cuando están unidos a su antígeno. En humanos sólo fijan el complemento IgM, IgG1, IgG2 e IgG3, mientras que IgG4 no lo hace. Esto ocurre porque , tras la unión al antígeno, hay cambios conformacionales en la estructura del anticuerpo, cambios que hacen accesible al componente C1 del complemento el sitio e fijación, situado en el fragmento Fc del anticuerpo. Otra de las condiciones que debe cumplirse para la activación del complemento es que el fragmento C1 debe unirse, al menos, a dos fragmentos Fc.
Secuencia de activación del Complemento:
TEMA 05. INMUNIDAD INNATA (II) Pág. 2 de 10
formulandoalgoqueaportar.blogspot.com

TEMA 05. INMUNIDAD INNATA (II)
C1: Es una molécula formada por una subunidad C1q asociada a dos C1r y dos C1s. La C1q, es quien se fija al Fc de los anticuerpos.. Es una molécula (410 KD) que se asemeja a un ramo de flores, siendo la porción N-terminal la que se abre a modo de cabeza globular. Las C1r y C1s, son péptidos sencillos (85 kD), que dependientes de Ca++, se combinan para formar la molécula C1r-C1s-C1s-C1r, que a su vez se unen a la C1q. La unión de C1q al antígeno provoca un cambio conformacional en C1r, que a su vez activa a C1s que asimismo actúa sobre los siguientes componentes de la cascada C2 y C4.
C4: Es un heterotrímero (α, β, γ) de 210 kD, que cuando se activa se separa en dos fragmentos C4a de 8,6(kD) y el resto denominado C4b. La mayoría de las moléculas C4b reaccionan con el agua formando un intermediario de vida muy corta C4bi, pero otras forman enlaces covalentes con la superficie celular, tanto proteínas como carbohidratos, asegurándose así la activación en la superficie celular en la que se unió el anticuerpo. Este fragmento C4b no tiene propiedad enzimática sobre el C2, y lo único que hace es permitir que la siguiente molécula en la cascada del Complemento “C2” se fije a él en presencia de iones Magnesio.
C2: Es el tercer componente sérico de la vía clásica. Tiene 110 kD y, como ya se ha mencionado, se une al fragmento C4b sobre las superficies celulares en presencia de Mg++. Así unido es asequible a la fracción C1s que ahora si es capaz de escindir el C2 en dos fragmentos C2a y C2b. El fragmento C2b, de 35 kD, y el fragmento C2a que permanece unido al C4b para formar el C4b2a, que es la C3 Convertasa de la Vía clásica.
C3: Es el más abundante (0,5 a 1,2 mg/ml) de todos los componentes del Complemento, ya que es la piedra angular donde convergen todos las vías de activación. Es un heterodímero (cadenas α y β). Por la acción de la convertasa formada en la fase anterior, se escinde en dos fragmentos. Uno pequeño de 9 kD (C3a), y el resto C3b. La mayor parte del C3b, y al igual que ocurría con el C4b reacciona con el agua dando como resultado una molécula inactiva. Una pequeña porción, alrededor del 10%, se une covalentemente a la superficie de las células formándose un nuevo complejo C4b2a3b, que es la C5 Convertasa.
Vía alternativa (pag 347 janeway)
En el caso de la vía alternativa la C3 convertasa se genera sin necesidad de la presencia de anticuerpos.
En la vía alternativa, el C3b se produce constantemente en pequeñas cantidades por la C3 convertasa de la fase fluida que se genera a expensas de la hidrólisis espontánea del enlace tioester de C3, pero no tiene efectos perniciosos debido a que se hidroliza a una forma inactiva en fracciones de segundo. El proceso sería como sigue: En condiciones normales el factor C3 se combina con una molécula de agua para formar el C3(H20) o C3i. Este complejo se une en la fase fluida al factor B (similar al factor C2 de la vía clásica), para formar el C3(H20)B. Unido al anterior el factor B es susceptible a la acción del factor D, que lo rompe en Ba y Bb, factor, este último que queda unido al complejo C3(H20) para formar el C3(H20)Bb, que es un factor muy lábil, y que para estabilizarse necesita unirse al factor P (Properdina). De esta forma el factor C3Bb, unido a la properdina (C3BbP) conforma la C3 convertasa de la vía alternativa factor y que al igual que la C3 convertasa de la vía clásica, puede escindir el C3 en C3a
TEMA 05. INMUNIDAD INNATA (II) Pág. 3 de 10
formulandoalgoqueaportar.blogspot.com

TEMA 05. INMUNIDAD INNATA (II)
y C3b. El factor C3b se puede unir a las membranas celulares, incluso las propias, captando más factor B y amplificando el proceso. Cuando no existe infección, este proceso se produce de forma constante y las pocas moléculas de C3b no inactivadas por la segunda hidrólisis, se fijan a la superficie de las células propias.
El mecanismo de control, para que esta vía no se dispare y se produzca la lisis de
esas células, es que sobre ellas existen unas proteínas reguladores de inhibición, que impide el acoplamiento del factor B, con lo que se frena la vía, quedando así protegidas las células. En el caso de que exista infección al no existir sobre la superficie de los microorganismos esas proteínas reguladoras, si se produce la unión del factor B, con la subsiguiente amplificación de la vía1
La siguiente fase de esta vía es similar a la mencionada anteriormente para la vía clásica. La C3 convertasa actúa sobre el C3, escindiéndolo y dando lugar al complejo C3Bb3b, que es la C5 convertasa de la vía alternativa. En este factor confluyen ambas vías y con su formación tiene lugar el inicio de la siguiente fase de actuación del Complemento que es la fase lítica. Finalmente hay que destacar, que además de su forma de activación particular, la vía alternativa supone un mecanismo de amplificación del sistema del complemento iniciado a través de la vía clásica ya que las moléculas generadas por la vía clásica se activan, a partir de ahí tanto por la vía clásica como por la vía alternativa. Fase lítica
Durante esta fase se va a formar el denominado “Complejo de ataque a la membrana” (CAM), que culminara con la lisis celular.
C5: Es un heterodímero de 190 kD similar a C3 y C4. Tras sufrir el ataque proteolítico de la C5 convertasa, desprende un pequeño fragmento de 11 kD (C5a), con ciertas propiedades biológicas, la principal quimiotactica, y otro mayor, con el resto de la molécula (C5b). Este C5b mantiene de forma transitoria una conformación capaz de unir a C6, formando un complejo estable C5b6 que fija, a su vez, a C7. El complejo C5b67 es muy lipofílico y se inserta en la bicapa lipídica de las membranas celulares y virales, donde se convierte en un receptor de membrana de alta afinidad para C8. Este C8 es un trímero formado por tres cadenas de las que dos estan unidas covalentemente entre si. Este C8 se inserta en la bicapa y el complejo C5b678 queda asi asegurado de forma estable a la superficie celular. Este complejo inicia ya la lisis de algunos microorganismos y células eucariotas y se le considera como una forma del CAM. Sin embargo, la verdadera actividad lítica se produce con la fijación del factor C9. Este, es una proteína monomérica que se polimeriza en el sitio donde está C5b-8 y forma poros en la membrana plasmática (similares a los provocados por las proteínas formadoras de poros llamadas perforinas o citolisinas de los LTc y de las células NK) que provocan la lisis de muchos microorganismos y células eucariotas. Estos poros permiten el intercambio pasivo de pequeñas moléculas, iones y agua, aunque son demasiado pequeños para permitir el paso de moléculas grandes tales como las proteínas; como resultado hay un flujo de agua hacia el interior de la célula que lleva a la lisis osmótica.
1 En el caso del veneno de cobra, la muerte sobreviene porque el veneno posee un factor B, similar al humano, pero que no es inhibido por el factor H, por lo que permite que se dispare la vía.
TEMA 05. INMUNIDAD INNATA (II) Pág. 4 de 10
formulandoalgoqueaportar.blogspot.com

TEMA 05. INMUNIDAD INNATA (II)
Además de este mecanismo letal la muerte celular puede producirse por el paso de Ca++ al interior de la célula. 3.- Vía de las Proteínas de respuesta de fase aguda. Estas son proteínas , que en situaciones de infección son secretadas por el hígado bajo la acción de ciertas citoquinas secretadas por macrófagos y neutrófilos. Su importancia reside en que mimetizan ciertas moléculas inmunitarias, como Anticuerpos, pero al contrario de estos poseen amplia especificidad por las moléculas patógenas. a.- La Proteína C reactiva La PCR es una proteína pentamérica capaz de unirse a los lipopolisacaridos de ciertas paredes bacterianas. Es miembro de una familia de proteínas que se conoce como pentaxinas debido a estar formadas por cinco subunidades idénticas. En muchas paredes de bacterias y hongos hay lipopolisacaridos con porciones fosforilcolina y es en estas porciones donde se fija la proteína C reactiva (aunque también hay fosforilcolina en la membrana de las células de mamíferos la CRP no se une a ella por estar en una conformación distinta a la que no reconoce). Cuando la CRP se une a la pared bacteriana no solo opsoniza a la bacteria sino que también puede activar a la cascada del complemento ya que fija a C1q, el primer componente de la vía clásica del complemento. La interacción con C1q se realiza a otro nivel de donde lo hacen los anticuerpos2, pero se pone en marcha la misma cascada de reacciones.
b.- La lectina fijadora de manano (MBL). Es una proteína, como ya se ha dicho con capacidad de unirse a residuos hidrocarbonatos, preferentemente a los extremos de manosa, fucosa y glucosamina de polisacáridos o glucoproteínas de membrana de gran variedad de bacterias3. A diferencia de la anterior lo que mimetiza este componente es el componente C1 del Complemento. Es un componente parecido estructuralmente al C1q, constituido por hexámeros con 18 cadenas polipeptídicas idénticas enrolladas de tres en tres en α-helice. La unión a los restos carbohidratados, provoca un efecto similar a lo que ocurre con el complejo C1. Es decir, sufre un cambio conformacional, que se traduce en una activación de unas serin proteasas asociadas (MASP-1 y MASP-2)homologas con C1r o C1s, que lleva finalmente a la actuación sobre los fragmentos C2 y C4 con la consiguiente formación de la C3 Convertasa.
De este modo, en un par de días, la respuesta de fase aguda proporciona al hospedador dos proteínas que actúan con propiedades funcionales de anticuerpos pero que tienen un rango de fijación mucho más amplio.
Hasta aquí hemos visto, como se produce la activación, y las condiciones que tienen que ocurrir, formación de IC, para la vía clásica penetración de microorganismos para la vía alternativa y la vía de las proteínas de fase aguda y el proceso final que da lugar a la destrucción final de la célula. Sin embargo, es necesario que existan ciertos mecanismos reguladores que frenen la cascada del complemento una vez que esta no es necesaria. En caso contrario la activación incontrolada del complemento pudiera llevar a la formación del CAM sobre los tejidos propios y a la formación exagerada de mediadores inflamatorios, que serían altamente perjudiciales. Mecanismos reguladores
2 En las porciones tipo colágeno y no en las zonas globulares como hacen con los anticuerpos 3 En los mamíferos estos residuos se hallan cubiertos por otros restos de azucares
TEMA 05. INMUNIDAD INNATA (II) Pág. 5 de 10
formulandoalgoqueaportar.blogspot.com

TEMA 05. INMUNIDAD INNATA (II)
La eficacia de la vía alternativa, depende en gran medida de su capacidad de distinguir lo propio de lo no propio, es decir de permitir la unión del C3b sobre la superficie de las células extrañas y no sobre las propias. La vía clásica. No tiene este problema, toda vez que al activarse a través de los anticuerpos son estas las moléculas encargadas de tal diferenciación. De este modo, la regulación de la vía alternativa esta mediada por una serie de moléculas reguladoras que son:
Factor I. Factor proteolítico H: proteína sérica. Une C3b y bloquea la formación de C3 convertasa de la vía
alternativa. Permite la escisión por el Factor I MCP: Se une a C3b y C4b, haciéndolos susceptibles al Factor I DAF: Se une a C3b y C4b, inhibe la interacción del primero con el factor B y
del segundo con el C2 CR1: Se une a C3b y C4b. inhibe la formación de la C3 convertasa. Permite la
acción del factor I De todas ellas, las tres últimas se expresan en la superficie de las células propias
que de este modo quedan protegidas. Por el contrario, al no expresarse sobre la superficie de los microorganismos, estos no cuentan con esta protección, pudiendo el complemento activarse sobre su superficie.
El factor H, es una proteína plasmática podría actuar en ambos casos. Para evitar la activación de este factor, el factor clave es el ácido siálico. Para poder actuar el factor H se debe unir a la superficie celular. Esta unión esta mediada por ácido siálico. Las células de mamíferos presentan un alto contenido de ácido siálico, mientras que la mayoría de los organismos o carecen de él o tienen un contenido poco uniforme. De esta manera, en estos últimos, como el factor H no se une a la membrana del patógeno, no puede efectuar su acción protectora.
Todos los factores anteriores funcionan como cofactores bien desplazando o impidiendo la unión de los siguientes componentes de la cadena del Complemento. En esas situaciones, esos factores aislados son fraccionados por el Factor I, que es el que verdaderamente tiene la acción proteolítica
Factor I: Es una serina proteasa que escinde tanto el C3b como el C4b, por ello se agrupa dentro del grupo de “cofactores de la degradación proteolítica” y es el que tiene de verdad la acción proteolítica, impidiéndose la formación de las convertasas C3 y C5. Sin embargo, para la actividad de factor I, es necesaria la intervención de otros cofactores.
REGULACION DE LA VIA CLASICA. A pesar de lo mencionado anteriormente de que la vía clásica, de por si es capaz de reconocer entre lo propio y lo no propio, por su propia esencia de activación, también son necesarios mecanismos que regulen esta vía una vez que se ha puesto en marcha, para desactivarla una vez que ya no es necesaria. Esta regulación se realiza a nivel de la activación de C1, de la formación y actividad de las C3 y C5 convertasas y de la formación del Complejo de ataque a la membrana Las moléculas reguladoras serian C1-NIH: Se une a C1r y C1s, impidiendo la actividad proteolítica de C1s C4BP: Se une al C4b y bloquea la formación de la C3 convertasa. Permite la acción del Factor I. La diferencia con los factores Cr1 y MCP es que no se une a C3b. Vitronectina: Se une al complejo C5b67, impidiendo su inserción en la membrana Agrupina: Igual que la anterior. Presenta en la superficie de los espermatozoides
TEMA 05. INMUNIDAD INNATA (II) Pág. 6 de 10
formulandoalgoqueaportar.blogspot.com

TEMA 05. INMUNIDAD INNATA (II)
Evolución del proceso El control del inicio de la vía clásica se produce por la acción de C1 inhibidor (C1 INH) quien interfiere en la acción proteolítica de C1r y C1s. Se une a la parte activa y la disocia del C1q sobre sus sustratos respectivos.. A esto hay que añadir que el C1INH, puede inactivar a cualquier C1r2-C1s2 activo que pueda formarse en los inmunocomplejos circulantes. Los factores C4BP y DAF4, compiten con C2b por la unión a C4b. cuando el C4BP se une al C4b, este se vuelve susceptible al ataque del factor I. En el caso del factor H o del receptor CR15, estos se unen a C3b, desplazando a C2b o a Bb y hacen a C3b susceptible de rotura por el factor I. Otro de los factores de regulación es el cofactor MCP, que se encuentra unido a la membrana de todos los tipos celulares excepto eritrocitos, faltando en la membrana de los microorganismos por lo que contribuye a la activación selectiva del complemento sobre la superficie de estos últimos. Es decir es la forma por la que el sistema del complemento distingue entre LO PROPIO y LO NO PROPIO. El ácido siálico, también interviene en que el complemento se active preferentemente en las superficies bacterianas. Se ha comprobado que cuando existe abundancia de ácido siálico se favorece la unión del factor H a la C3 convertasa, mientras que una deficiencia favorece la unión del factor B. Muchas bacterias contienen cantidades menores de ácido siálico en su superficie en comparación con las células de mamíferos, lo que favorece la activación del complemento en estas últimas. La actividad de los componentes finales del componente esta también regulada por proteínas de superficie de las que la mas importante es la CD59, que evita el ensamblaje final del CAM en la etapa C8-C9. En cuanto a los inhibidores plasmáticos, se encuentran la proteína S o vitronectina y la agrupina. Ambas ejercen su función uniéndose al Complejo C5b-7 soluble, e impidiendo su inserción en la superficie de las células, aunque no impide la unión del C8 y C9. La agrupina, se encuentra en gran cantidad en tracto reproductor masculino, seguramente con la función de proteger a los espermatozoides cuando penetran en el aparato reproductor femenino. Patologías derivadas de deficiencias en componentes del Complemento Las deficiencias en los factores C1, C2 y C4, curiosamente están asociadas mas a problemas de autoinmunidad, debido fundamentalmente a la acumulación de IC circulantes que a una mayor susceptibilidad a las infecciones. Las personas con estas deficiencias sufren una enfermedad parecida al “Lupus eritomatoso sistémico”. Esto indica que la vía alternativa es suficiente para al defensa del hospedador en la mayoría de los casos.
4 Se expresa en la mayoría de las células sanguíneas, endoteliales y epiteliales. Este factor se une a la C3 convertasa, liberando bien el factor C2a o el Bb. Así pues la función del DAF es bloquear la activación del complemento en la misma célula en la que se expresa, siendo por ello junto al MCP, responsable de la distinción entre lo propio y lo no propio, ya que solo esta presente en las células propias 5 Es una proteína que se distribuye casi exclusivamente entre las células sanguíneas, por lo que su papel se limita a bloquear el complemento en aquellas células que han expresado el receptor CR1
TEMA 05. INMUNIDAD INNATA (II) Pág. 7 de 10
formulandoalgoqueaportar.blogspot.com

TEMA 05. INMUNIDAD INNATA (II)
En cambio la deficiencia de C3 si se asocia a una mayor susceptibilidad a infecciones, que incluso pueden llegar a ser mortales. Asimismo también cursan con una mayor susceptibilidad a las infecciones, las deficiencias en componentes de la vía alternativa como properdina o factor D. Por ultimo las deficiencias de los componentes del CAM es una propensión a infecciones diseminadas por determinados tipos de bacterias como Neisseria, lo que indica que la lisis bacteriana es fundamental en estos casos. Receptores del Complemento
Muchas de las acciones biológicas del Complemento dependen de la unión de fragmentos de los componentes a receptores de membrana específicos, expresados sobre varios tipos celulares. CR1 (CD35). Glicoproteína integral de membrana de una sola cadena que une con alta afinidad a C3b y C4b; se encuentra en la mayoría de las células, pero sobre todo en fagocitos y eritrocitos. Entre sus funciones están: a) Favorece la fagocitosis, al actuar como opsonizante al fijar los componentes C3b y C4b. b) Aclaramiento de IC circulantes gracias a su presencia en los hematíes. c) Regulador de la activación del Complemento. Al unirse a los Componentes C3b y C4b, compite con los sustratos activadores Bb y C2a, e impidiendo así la formación de la C3 convertasa CR2 (CD21). Glicoproteína integral de membrana de una sola cadena que estimula la respuesta inmunitaria humoral. Fija a C3bi (C3b escindido por el Factor I); se encuentra sobre LB, células dendríticas foliculares y células epiteliales. En las células dendríticas foliculares el CR26 sirve para atrapar los inmunocomplejos, recubiertos por el componente C3bi, con lo que facilita la presentación del antígeno a los linfocitos B activados y provocando así la formación de LB de memoria. En el caso de los linfocitos B la presencia de CR2 en su membrana facilita una segunda señal necesaria para la activación de los mismos. La primera seria el reconocimiento antigénico directo y una segunda, la unión del CR2 al C3bi unido a la superficie del microorganismo. De este modo el CR2 es parte integrante del complejo correceptor del linfocito B CR3(Mac-1) y CR4 ( CD11bCD18) (CD11cCD18). 6 CR2 fija también específicamente al virus de Epstein-Barr (EBV), causante de la mononucleosis infecciosa. Es un herpesvirus que tras la infección se mantiene latente en las células B o en las del epitelio faríngeo durante toda la vida. Además EBV está ligado a varios procesos malignos, tales como el linfoma de Burkitt en África (un tumor maligno de las células B), linfomas B asociados a quimioterapia o VIH y carcinoma nasofaríngeo; estos tumores derivan de las únicas células normales que expresan CR2. EBV es un potente activador policlonal de las células B e, in vitro, puede transformar linfocitos B normales en líneas celulares linfoblastoides inmortales. Todos los efectos del virus dependen de la expresión de CR2 sobre las células B.
TEMA 05. INMUNIDAD INNATA (II) Pág. 8 de 10
formulandoalgoqueaportar.blogspot.com

TEMA 05. INMUNIDAD INNATA (II)
Ambas integrinas, se unen a C3bi y actúan como opsoninas. El Mac-1, se encuentra sobre Neutrofilos y Monocitos y puede reconocer directamente ciertas moléculas microbianas Papel Biológico del Complemento 1. - CITOLISIS MEDIADA POR EL COMPLEMENTO. Sería su función primordial y ya se ha explicado como donde y porque se produce. 2. - OPSONIZACION Y FAGOCITOSIS.
Como hemos visto la activación del Complemento lleva a la generación de C3b y C3bi unidos covalentemente a las superficies celulares (opsonización) y a su unión específica a los receptores CR expresados por macrófagos y neutrófilos. La fagocitosis dependiente de C3b y C3bi es posiblemente el mecanismo defensivo principal frente a las infecciones por bacterias y hongos. Los receptores del complemento y los receptores para el fragmento Fc de la IgG (Fc R) cooperan en la adherencia de las partículas opsonizadas y en la transducción de señales que estimulan la capacidad fagocitaria del leucocito. 3. - ANAFILATOXINAS E INFLAMACION.
C3a, C4a y C5a se denominan anafilatoxinas debido a que inducen la liberación
desde los mastocitos de sustancias (histamina) que originan el aumento de la permeabilidad vascular y la contracción de la musculatura lisa, característico de la anafilaxis. Los receptores para C3a y C4a se expresan sobre mastocitos, basófilos, células de músculo liso y linfocitos, mientras que el receptor para C5a lo hace sobre mastocitos, basófilos, neutrófilos, monocitos/MCFs y células endoteliales. C5a es el más potente como anafilatoxina (20 veces más que C3a y 2500 veces más que C4a) y también estimula la liberación de TNF por los mastocitos. Sobre los neutrófilos C5a tiene acción quimiotáctica, potencia la expresión de moléculas de adhesión y estimula la explosión respiratoria con producción de intermediarios reactivos del oxígeno. La acción de C5a sobre mastocitos, neutrófilos y células endoteliales contribuye a la inflamación en los sitios de activación del complemento. 4. - SOLUBILIZACION Y ACLARAMIENTO DE INMUNOCOMPLEJOS.
En la circulación se están formando constantemente pequeñas cantidades de IC y
la concentración de los mismos aumenta de modo brutal cuando se produce una respuesta inmune frente a un antígeno circulante presente en gran cantidad. Estos IC son potencialmente peligrosos debido a que pueden depositarse en las paredes de los vasos, activar al complemento y provocar las reacciones inflamatorias subsecuentes que llevan al daño del tejido local. Además de lo anterior el C3b formado y unido covalentemente a las Ig del IC se une específicamente a CR1 sobre la superficie de los eritrocitos, quienes los transportan hasta el hígado y bazo donde son eliminados por las células fagocitarias cuando los eritrocitos atraviesan los sinusoides de estos órganos7.
7 La formación de IC grandes (potencialmente peligrosos) no solo requiere la fijación de los fragmentos
Fab de los anticuerpos específicos al antígeno, sino que precisa también de las interacciones no covalentes entre los fragmentos Fc de las moléculas de anticuerpo yuxtapuestas; la fijación del complemento a las Ig bloquea estéricamente estas interacciones Fc-Fc solubilizando a los IC y desestabilizándolos.
TEMA 05. INMUNIDAD INNATA (II) Pág. 9 de 10
formulandoalgoqueaportar.blogspot.com

TEMA 05. INMUNIDAD INNATA (II)
TEMA 05. INMUNIDAD INNATA (II) Pág. 10 de 10
formulandoalgoqueaportar.blogspot.com

TEMA 06 CELULAS NK 07/08
CELULAS NK Introducción Las células NK son linfocitos grandes que circulan en la sangre (representan el 10- 15% de los linfocitos circulantes) y que tienen un citoplasma bien desarrollado que contiene gránulos citotóxicos. En la primera descripción las células NK recibieron el nombre de “Grandes Linfocitos Granulares (GLG)” Estas células aportan la inmunidad innata frente a infecciones intracelulares para lo que migran desde la sangre hasta los tejidos infectados, en respuesta a las citocinas inflamatorias. Las células NK, a diferencia de lo que ocurre con los linfocitos de la inmunidad adaptativa (que circulan en estado de reposo y necesitan varios días para activarse y expandirse) circulan en un estado de activación parcial, listas para penetrar y actuar en los tejidos infectados en cuanto los macrófagos den la alarma (mediante la producción de IL-12). La actividad citotóxica de las células NK no precisa una exposición previa o sensibilización al patógeno • Las células NK (asesinas naturales) constituyen una primera línea defensiva de la inmunidad innata frente a los agentes infecciosos intracelulares y frente a las células tumorales • Las NK se desarrollan en la médula ósea a expensas de una célula progenitora hematopoyética División Las células NK se caracterizan por la expresión de los marcadores: CD56 (moléculas de adhesión de las células nerviosas [N-CAM]) y CD16 (Receptor de baja afinidad para el fragmento Fc de anticuerpos IgG [FcRgIII] De acuerdo con la densidad de la expresión de la molécula CD56 sobre la superficie se distinguen dos subpoblaciones, que participan en la inmunidad mediante dos mecanismos: CITOTÓXICAS CD56 dim (↓) CD16 bright (↑): Median citotoxicidad natural y ADCC SECRETORAS CD56 bright (↑) CD16 dim (↓):Secretan citocinas y quimiocinas Las flechas indican la intensidad de la fluorescencia (débil o brillante). • Las citotóxicas predominan en la circulación, mientras que las secretoras predominan en los órganos linfoides secundarios. • La célula progenitora expresa el marcador CD34; se diferencia a una célula precursora intermedia que expresa el receptor para IL-15 quien, bajo la acción de la IL-15 secretada por las células estromales, genera la célula CD56 bright (secretora). Parece que estas células, finalmente, maduran a CD56 dim (↓) (citotóxica) en los ganglios linfáticos, inducidas por las señales procedentes de las DCs. • La mayoría de las células NK expresan el receptor de afinidad intermedia para IL-2 (que carece de la cadena alfa [CD25]), pero las NK secretoras (CD56 bright) expresan de forma constitutiva el receptor de alta afinidad, lo que quiere decir que cantidades mínimas de IL-2 inducen su proliferación, mientras que son necesarias concentraciones alta de IL-2 para la proliferación de las NK citotóxicas (CD56 dim).
Página 1 de 5
formulandoalgoqueaportar.blogspot.com

TEMA 06 CELULAS NK 07/08
Citocinas activadoras de las NK • IFNs Tipo 1: Aumentan la actividad citotóxica y la proliferación • IL-12 e IL-18: Aumentan la producción de citocinas • IL-15: Activa la maduración, crecimiento y actividad citolítica
Las NK recién aisladas de la sangre pueden destruir ciertos tipos de células diana; este nivel básico de citotoxicidad aumenta entre 20 y 100 veces después de la exposición a los interferones (IFN) alfa y beta producidos por la célula afectada como respuesta a la infección viral. Además, también se activan por la IL-12 y el TNF-a, ambas producidas por los macrófagos al comienzo de muchas infecciones. La acción de estas cuatro citocinas produce una oleada de células NK activadas durante la primera fase de una infección viral que puede eliminarla o
En las primeras etapas de una infección, las células NK son las principales productoras de IFN-g, citocina que luego activa a los macrófagos para que estos secreten las citocinas que inician la respuesta adaptativa de las células T. Cuando se forman los linfocitos T efectores y llegan al sitio infectado se convierten en los productores principales de IFN-g y en los mediadores de la citotoxicidad. La llegada de los linfocitos T efectores inactiva a las células NK (el inactivador es la IL-10, una citocina producida por los linfocitos T citotóxicos). Mecanismos de citotoxicidad Las NK desencadenan la citotoxicidad mediante dos procesos distintos: la lisis natural y la dependiente de anticuerpos (ADCC). Según el mecanismo bioquímico la lisis natural puede depender de: • La exocitosis del contenido de las granulaciones (mecanismo secretor); es el más importante • La activación de receptores letales en la célula blanco (mecanismo no secretor). Mecanismo secretor El reconocimiento de la célula blanco y la activación NK llevan a la polimerización de los microtúbulos del citoesqueleto que movilizan los gránulos NK hacia el lugar del contacto. Los gránulos contienen principalmente Granzima B (una serin-proteasa activadoras de caspasas) y perforina (que altera la membrana de forma similar a C9). Ambas proteínas están formando un complejo con una tercera, denominada serglicina, que actúa como proteína transportadora. El trímero se libera en la zona de contacto y se endocita por la célula blanco mediante un denominado Receptor de Manosa 3-fosfato (MPR); El pH ácido de la vacuola endocítica activa a las perforinas, que forman poros en la membrana de la vacuola permitiendo el paso al citosol del granzima-B que activa la vía de las caspasas, lo que lleva a la apoptosis de la célula blanco. Mecanismo no secretor Las NK en reposo almacenan FasL (Fas ligando) en los lisosomas y, tras la activación, este se trasloca a la membrana y desde allí puede reconocer al Fas (CD95), que se expresa constitutivamente por distintos tipos celulares. La interacción induce la
Página 2 de 5
formulandoalgoqueaportar.blogspot.com

TEMA 06 CELULAS NK 07/08
apoptosis de la célula blanco. La ADCC (Citotoxicidad dependiente de anticuerpos) indica la capacidad de las NK para destruir células recubiertas con anticuerpos IgG específicos frente a epitopos del patógeno. El mecanismo se basa en la unión a estos anticuerpos de la moléculas CD16 (FcgRIIIR) y liberación posterior del contenido de las granulaciones de la células NK sobre la célula blanco. Interacciones de las NK con otras células Interacciones con macrófagos La citocina principal que liberan las NK es el IFN-g, que activa al macrófago. Y produce la liberación por parte de este de IL-12. La secreción de IL-12 por el macrófago y de IFN-g por la NK crea un sistema de retroalimentación positiva que aumenta el grado de activación de ambos tipos de células, dentro de un tejido infectado. Interacciones con DCs (“diálogo continuo”) Ambos células interaccionan en los tejidos periféricos y se modulan continuamente. Las DCs, activan a las NK en reposo (mediante IL-12 e IL-15) e inducen su proliferación, producción de citocinas y aumento de actividad citotóxica. Pero la activación NK tiene la contrapartida de modular la funcionalidad y supervivencia de las DCs inmaduras (iDC), bien positivamente (induciendo su maduración mediante IFN-g y TNF-a) o negativamente (reconociendo a las iDCs mediante el receptor NKp30 y El concepto de la vigilancia inmune Se considera que las células NK son las vigilantes permanentes de la población celular de nuestro organismo. Como todas las células de un organismo portan las mismas moléculas de histocompatibilidad de Clase I, se las puede considerar como el “carné de identidad” que acredita que son propias y sanas. Las células NK son las encargadas de verificar que la célula examinada tiene carné, que este es adecuado y que no está alterado (cosa que ocurre con algunos parásitos intracelulares). Si no se cumplen las condiciones anteriores, la célula NK se activa y destruye a la célula blanco. Receptores de las células NK Para mantener a las NK circulantes en un estado de activación parcial y disponibilidad inmediata frente a la infección y controlar su potencial de atacar a los tejidos sanos hay una serie de receptores de superficie; algunos generan señales activadoras y otros emiten señales inhibidoras. Los receptores inhibidores producen señales intracelulares que mantienen a las NK en reposo, mientras que los receptores activadores producen señales intracelulares que activan a las células NK y ponen en marcha la maquinaria que lleva a la destrucción de la célula que porta el ligando. El que la NK permanezca en reposo o se active dependerá del equilibrio de señales. El equilibrio puede alterarse durante la infección intracelular, ya que un patógeno puede: • Estimular la expresión, en la célula infectada, de ligandos para los receptores activadores de las NK • Inhibir la expresión, en la célula infectada, de ligandos para los receptores inhibidores NK Ligandos para los receptores NK Los principales son las moléculas CMH de Clase I; su reconocimiento por los receptores inhibidores impide la activación NK. Pero las células infectadas o neoplásicas suelen expresar menos moléculas Clase I, provocando señales inhibidoras
Página 3 de 5
formulandoalgoqueaportar.blogspot.com

TEMA 06 CELULAS NK 07/08
de menor intensidad y predominando entonces las de activación a través de receptores activadores. Esta ausencia de señales inhibidores (mediadas por las moléculas Clase I) que llevan a la activación de las células NK se conoce como “Hipótesis de la pérdida de lo propio”. Dicho de otro modo, las NK vigilan constantemente la expresión de moléculas Clase I en las células del organismo y se activan si los niveles disminuyen. Los receptores NK pertenecen a dos tipos estructurales: En uno de ellos el sitio de unión al ligando es un dominio similar al de las Inmunoglobulinas (Ig), mientras que el otro tiene un dominio de unión al ligando del tipo lectina C (similar al de MBL) Receptores KIR Un carácter de los 14 genes que codifican para los receptores KIR es que, en un mismo individuo, distintas células NK pueden expresar combinaciones diferentes de receptores KIR. Son los receptores de mayor diversidad y es muy raro que 2 personas no relacionadas (<1% de los casos) presenten el mismo juego de alelos KIR. Hay tres factores que se combinan para crear esta diversidad: 1) Las personas tienen números diferentes de genes KIR (7 a 15) 2) Los haplotipos de KIR tienen distinto número de genes que codifican para receptores activadores e inhibidores 3) Los genes KIR son polimórficos Algunas personas tienen genotipos con solo genes que codifican para KIR inhibidores, mientras que otras tienen un número equivalente que codifica para KIR activadores e inhibidores. Las diferencias en el tipo de KIR parecen correlacionar con distintos parámetros de enfermedades infecciosas, autoinmunes y en trasplantes. Solo hay un KIR (KIR2DL4) inhibidor que se expresa por todas las células NK humanas. Los otros se expresan en subpoblaciones NK, con una distribución casi al azar del número y la combinación de genes KIR que se expresan por cualquier célula determinada.. Una vez establecido el patrón de expresión de KIR durante el desarrollo de la célula NK, este permanece estable en el clon posterior. Ligandos de los genes KIR La mayoría son productos codificados por los genes de Clase I del CMH (A, B y C) propio. Al expresar, al menos, un receptor inhibidor que se une al alotipo del HLA Clase I propio, cada célula NK tolera a las células sanas de su propio cuerpo (células autólogas) y no las ataca. No ocurre lo mismo con las células sanas de otro individuo (células alogénicas); si la molécula de Clase I es similar la célula sobrevivirá, pero si es distinta la matará. Esto se puede aprovechar para eliminar a las células leucémicas residuales y prevenir las recidivas tras el trasplante de médula para tratar a la leucemia mieloide aguda. Se dice que las células NK “ven lo propio que falta”. Receptores LIR Son un grupo de receptores inhibidores que se expresan en NK y otras células Ligandos de los genes LIR Moléculas Clase I del CMH Receptores NKp (NKR) Se conocen como “Receptores de citotoxicidad natural”. Se expresan exclusivamente sobre células NK y estimulan (activadores) la citotoxicidad NK contra células con expresión deficiente o ausente de moléculas Clase I. En los humanos estos receptores incluyen NKp80, NKp46, NKp44 y NKp30. Los
Página 4 de 5
formulandoalgoqueaportar.blogspot.com

TEMA 06 CELULAS NK 07/08
subrayados se expresan constitutivamente en todas las NK presentes en sangre periférica, mientras que NKp44 solo se expresa tras la estimulación por IL-2. NKp30 es el responsable de las interacciones entre las células NK y las DC. Ligandos de los genes NKR Parecen ser productos virales en la superficie de las células infectadas. Receptores de la familia de las lectinas Tipo-C (NKC): NKG2 y CD94 La molécula CD94 (que no tiene tallo citoplásmico para trasmitir las señales) se asocia con diferentes miembros (hay 5) de la familia NKG2, excepto NKG2D Funcionalmente, los heterodímeros pueden ser inhibidores o activadores, en función de los dominios citoplásmicos de la molécula NKG2: Funcionalmente, los heterodímeros pueden ser inhibidores o activadores, en función de los dominios citoplásmicos de la molécula NKG2: • El heterodímeros CD94/NKG2A tiene tallo citoplásmico largo (con dominios ITIM), por lo que es inhibidor • Los heterodímeros CD94/NKG2C y CD94/NKG2F tienen tallo corto, pero se asocian a moléculas transductoras de señales, con lo que median señales de activación Ligandos para CD94/NKG2 CD94/NKG2A y CD94+/NKG2C reconocen a la molécula Clase I no clásica HLA-E, que es un péptido derivado de las secuencias líder de las moléculas clásicas de Clase I; así, controlando a HLA-E, las NK “saben” como va la expresión de las moléculas Clase I clásicas y actúan si un proceso patológico interfiere con la expresión correcta de este indicador sobre una célula propia. La actitud y distribución de estos dos receptores es radicalmente distinta (es muy raro que una NK exprese estos dos receptores) : CD94/NKG2A es un receptor inhibidor que se expresa con mucha más frecuencia e impide que las NK ataquen a células humanas sanas que expresan un conjunto normal de moléculas HLA Clase I. CD94/NKG2C se asocia a un adaptador y emite señales activadoras Ligandos para NKG2D NKG2D no se asocia con CD94 y se expresa como un homo-dímero. En humanos, reconoce las proteínas transmembrana MIC-A y MIC-B (codificadas en el CMH), que son cadenas pesadas similares a las de Clase I, pero que no se asocian con alfa-2- microglobulina. Son proteínas que se inducen por el estrés y solo existen de forma constitutiva (y en pequeña cantidad) en el epitelio intestinal. Sin embargo, cuando alguna célula intestinal se infecta, envejece o sufre un proceso canceroso, se aumenta la expresión de estas proteínas en el epitelio intestinal. En este momento, las células intestinales se convierten en el blanco del ataque de las NK (a través de su receptor NKG2D). A la porción citoplásmica del receptor se acoplan dos pequeñas moléculas adaptadoras que ponen en marcha la cascada de señales que llevan a la liberación de gránulos citotóxicos y citocinas. Normalmente estas moléculas tienen una expresión mínima y se inducen por células malignas o infectadas. Son un verdadero semáforo de la alteración celular.
Página 5 de 5
formulandoalgoqueaportar.blogspot.com

Tema 7. (I) INFLAMACION
Tema 7: Focalización de la respuesta: Inflamación. Comunicación celular: Citoquinas - Inflamación aguda y crónica
o Fase vascular o Fase celular o Fase de regeneración
- Citoquinas o Principales citoquinas o Subpoblaciones Th1 y Th2
La inflamación.
La Respuesta inflamatoria es un proceso complejo que comienza
cuando hay daño tisular (en los tejidos) debido a factores, endógenos (necrosis de tejidos o fracturas óseas) o exógenos (mecánicos, químicos, infecciosos, inmunológicos [hipersensibilidad]).
Este proceso se produce tanto por elementos celulares como solubles y es un proceso normal de la inmunidad innata y adaptativa. Se pone en marcha a expensas de los macrófagos residentes y básicamente consiste en la atracción de leucocitos y extravasación de proteínas plasmáticas hacia las zonas de infección. La inflamación aguda se corresponde con la inmunidad innata. Si esta no logra resolver la infección, se pone en marcha la inmunidad adaptativa, que puede o bien resolver la infección, o bien si no lo consigue intentar aislarlo, provocando una inflación crónica, con la formación final de un granuloma.
El proceso inflamatorio juega tres papeles fundamentales:
Suministrar moléculas efectoras adicionales y células a los sitios de infección para potenciar la acción de los macrófagos de la primera línea.
Formar una barrera física que impida la diseminación de la infección.
Reparar el tejido dañado Se pueden distinguir tres fases: - Fase vascular - Fase Celular - Fase de restauración
Fase Vascular Los principales inductores de esta fase son las anafilotoxinas C3a, C4a y C5a, Estos componentes estimulan la producción de componentes lípídicos de la inflamación(leucotrieno, PAF) e histamina por las células
TEMA 07 (I). INFLAMACION 06/05/2008 Pág.1 de 5
formulandoalgoqueaportar.blogspot.com

Tema 7. (I) INFLAMACION
endoteliales y mastocitos respectivamente. Efectos provocados por estos mediadores inflamatorios: Vasodilatación aumento de la viscosidad sanguínea aumento de la permeabilidad vascular Accion quimiotáctica Fase celular
Da comienzo con la migración de los neutrofilos hacia el foco infeccioso. Los mediadores inflamatorios, junto a una serie de citoquinas proinflamatorias (IL-1, IL-6, TNF, IL-8 e IL-12) producidas por los macrófagos y neutrofilos, comienzan a inducir la expresión de moléculas de adhesión para permitir la migración de estas células desde la sangre hasta los tejidos circundantes. La primera en aparecer es la selectina E, inducida por TNF. Poco después es reforzada por las ICAM-1 y VCAM-1. Estas moléculas se reorganizan a las 24 horas agrupándose en las uniones intercelulares, donde se refuerza por otra molécula de adhesión perteneciente, como las dos anteriores, al grupo de las inmunoglobulinas, denominada PECAM-1, favoreciendo así la extravasación y diapédesis de los leucocitos hacia el foco inflamatorio
Después de la extravasación, el proceso continúa con la migración de los leucocitos hasta el lugar dañado. Esta fase esta regulada por medio de un subgrupo de citoquinas denominadas quimioquinas (IL-8). El papel de las quimioquinas es triple: estabilizan a los leucocitos que van “rodando” sobre el endotelio, dirigen su migración a los largo de un gradiente de concentración hasta el sitio inflamatorio y en algunos casos los activan (activan la explosion respiratoria). El gradiente se consigue por la fijación de pequeñas quimioquinas solubles a los proteoglicanos de la matriz extracelular y de la superficie de las células endoteliales, con lo que se forma un sustrato sólido de quimioquinas a lo largo del cual se desplazan los leucocitos. Al comienzo de la inflamación la población celular predominante son los neutrófilos. Así, en estos, que en condiciones normales solo viven unas pocas horas, la acción de las citoquinas IL-8 y TNF, activa la “explosión respiratoria” que produce radicales libres de oxígeno y de nitrógeno y la degranulación; Asimismo los neutrófilos fagocitan a los patógenos opsonizados. Si todavía no se ha producido la respuesta adaptativa utilizan como opsoninas a los fragmentos C3b y C3bi generados en la vía alternativa del complemento. Además, los neutrófilos reconocen directamente, a través de sus receptores, distintos componentes de las paredes bacterianas1. Así pues y como vemos todo 1 La importancia de los neutrófilos en la defensa se ilustra por las consecuencias de los defectos genéticos en la maduración o funciones antibacterianas; los pacientes con estas deficiencias sufren infecciones recurrentes, frecuentemente de bacterias y hongos que forman parte de la flora normal y que
TEMA 07 (I). INFLAMACION 06/05/2008 Pág.2 de 5
formulandoalgoqueaportar.blogspot.com

Tema 7. (I) INFLAMACION
ello contribuye a la defensa y al daño tisular local en los sitios de infección por bacterias piogénicas (formadoras de pus).
Además de su acción local las citoquinas producidas por
neutrófilos y macrófagos tienen efectos generales que contribuyen a la defensa del hospedador. Una de ellas es la elevación de la temperatura corporal producida por los “pirógenos endógenos” IL-1, IL-6 y TNF. La fiebre es, en principio, beneficiosa para el hospedador: la mayoría de los patógenos crece mejor a menos temperatura, la respuesta adaptativa es más intensa a temperaturas altas y las células propias también quedan protegidas de los efectos nocivos del TNF a temperaturas altas.
Un efecto final de las citoquinas producidas por los fagocitos es inducir la neutrofilia mediante el aumento de producción en la médula ósea y la liberación de los que están unidos en forma débil al endotelio en los órganos internos. Finalmente, el TNF induce la migración de las células dendríticas desde los tejidos periféricos hasta los ganglios linfáticos, donde maduran y se convierten en CPAs. Estas células son cruciales para la iniciación de la respuesta adaptativa. Por tanto, los efectos de las citoquinas producidas como respuesta a la infección contribuyen a su control mientras se está desarrollando la respuesta innata Fase de resolución. Durante esta fase de resolución, se producen mediadores anti-inflamatorios como TGF-∃ e IL-10, que inhiben la producción de las citocinas proinflamatorias. Asimismo cesa la expresión de las selectinas sobre las células endoteliales y linfocitos. Una vez que el proceso ha terminado, si ha tenido éxito, da comienzo la fase final de cicatrización. Los productos secretados por los macrófagos llevan a la activación y proliferación de los fibroblastos, que comenzaran su proceso particular de "fabricación" de fibras de colágeno para entretejer una red sobre la que proliferan las células epiteliales (desde el borde hacia el interior), en un intento de restaurar el tejido2.
En este proceso de reparación juegan un papel importante las proteasas neutras producidas por los MCFs (tales como elastasa y colagenasa) que intervienen actuando sobre la matriz extracelular durante el proceso de curación. escapan del sitio inflamatorio y pasan a la sangre (septicemia). Si el defecto está en los mecanismos antibacterianos los patógenos fagocitados pueden escapar de los neutrófilos muertos (el neutrófilo es una célula terminal y muere tras la activación) y proseguir la colonización.
2En este sentido la cicatriz, es una parte de la red que no ha podido ser colonizada por las células epiteliales
TEMA 07 (I). INFLAMACION 06/05/2008 Pág.3 de 5
formulandoalgoqueaportar.blogspot.com

Tema 7. (I) INFLAMACION
Además de todos los procesos anteriores, cuando se produce una
herida, el daño de los vasos sanguíneos pone en marcha inmediatamente otras dos cascadas enzimáticas protectoras (que también pueden comenzar, en ausencia de herida o daño, por las células endoteliales activadas):
El sistema de las kininas, que produce varios mediadores inflamatorios incluyendo la Bradikinina que aumentan la permeabilidad vascular. El sistema de la coagulación, con formación de un coágulo que
impide la penetración de cualquier microorganismo en la circulación.
Todos los acontecimientos anteriormente expuestos lleva a que en el sitio de infección aparezcan la tétrada de signos descrita por Galeno: Calor, Rubor, Dolor y Tumor que caracterizan a la respuesta inflamatoria.
Vasodilatación (aumento del diámetro de los vasos), que lleva a un aumento del flujo de sangre local (Calor y Rubor) y a la disminución de la velocidad del flujo sanguíneo, sobre todo en los vasos pequeños.
Activación de las células endoteliales (que forman la pared interior de estos vasos) que ahora expresan moléculas de adhesión que retienen a los leucocitos circulantes como paso previo a su migración al tejido infectado. Estos dos primeros cambios se inician a expensas de las citoquinas producidas por los macrófagos residentes activados. Las primeras células que llegan son los neutrófilos, seguidos por los monocitos (que, en el tejido, maduran a macrófagos).
Aumento de la permeabilidad vascular (separación de las células endoteliales que limitan la pared de los vasos locales), que permite el paso de líquidos y proteínas desde la sangre y su acumulación en el tejido (Edema o Tumor y Dolor)
Inflamación crónica Cuando el proceso infeccioso no logra ser eliminado, se acumulan macrófago activados en la zona, que siguen secretando sustancias que en su función “curativa” no discriminan entre los microorganismos y lo tejidos propios. En esta situación la inflamación pasa a ser un proceso crónico, que en la mayoría de los casos resulta “perjudicial” para el hospedador. Esto es debido a que los factores liberados van modificando el ambiente tisular local, que conlleva a la sustitución del tejido normal por tejido conjuntivo que acarrea la consiguiente fibrosis y la consiguiente pérdida de funcionalidad. Los macrófagos de la zona aumentan el citoplasma recordando a las células epiteliales por lo que se les conoce como células epitelioides. Asimismo los macrófago se pueden fusionar entre si, dando
TEMA 07 (I). INFLAMACION 06/05/2008 Pág.4 de 5
formulandoalgoqueaportar.blogspot.com

Tema 7. (I) INFLAMACION
lugar a las denominadas células gigantes multinucleadas. Esto junto a la acumulación en la zona de Linfocitos T da lugar a la formación de un infiltrado celular que origina el granuloma. Esta reacción es una reacción de curación que trata de aislar al germen patógeno, pero en ese empeño interfiere en la funcionalidad del tejido normal que pasa a ser afuncional. En definitiva, en estas ocasiones y como suele decirse “Es peor el remedio que la enfermedad”
TEMA 07 (I). INFLAMACION 06/05/2008 Pág.5 de 5
formulandoalgoqueaportar.blogspot.com

Tema 7 (II). Citocinas
Tema 7: Focalización de la respuesta: Inflamación. Comunicación celular: Citoquinas - Inflamación aguda y crónica
o Fase vascular o Fase celular o Fase de regeneración
- Citoquinas o Principales citoquinas o Subpoblaciones Th1 y Th2
CITOQUINAS
La respuesta inmunitaria se basa principalmente en interacciones celulares. Estas interacciones se producen gracias a las citoquinas, que son hormonas peptídicas producidas por unas células y captadas por otras que tienen los receptores adecuados. Por lo tanto, el mundo de las citoquinas está constituido tanto por los factores solubles como por los receptores de membrana.
A las citoquinas se les han atribuido multitud de nombres,
relacionados con la célula productora. De ellos el más extendido, aunque no el más exacto desde ese punto de vista, es el de Interleucina, debido a que se producen y actúan sobre leucocitos. No obstante es el término utilizado en la actualidad para clasificar a las distintas citoquinas.
Propiedades de las citoquinas. 1.- Autolimitación: La secreción es breve, cesa casi inmediatamente que desaparece el estímulo (RNAm de vida corta). 2.- Afinidad: Se necesitan cantidades muy pequeñas para producir el efecto biológico, 3.- Redundancia:, la misma acción es ejercida por varias citoquinas. 4.- Multisecreción: La misma citoquina, puede ser sintetizada por tipos celulares distintos 5.- Antagonismo y sinergismo: las citoquinas pueden actuar de modo antagónico, en unas ocasiones, y de forma sinérgica en otras. 6.- Pleitropismo: Las citoquinas pueden actuar sobre distintos tipos celulares. 7.- Secuencialidad: Las citoquinas actúan con frecuencia en cascada influyendo una en la síntesis de otra...etc. 8.- Multiacción:Las citoquinas pueden dar lugar a efectos diferentes sobre la misma célula diana. 9.- Regulación:La expresión de muchos receptores de las citoquinas esta regulada por señales específicas, que puede ser por una señal externa, por otra citoquina o por la misma citoquina que provoca una retroalimentación
TEMA 07 (II). CITOCINAS 06/05/2008 Pág.1 de 8
formulandoalgoqueaportar.blogspot.com

Tema 7 (II). Citocinas
positiva. 10.- la acción de las citoquinas puede ser autocrina, paracrina o en ciertos casos endocrina Estas propiedades de las citoquinas dan lugar a una serie de funciones que se pueden resumir en los siguientes puntos: a) Mediadores de la inmunidad innata y adaptativa b) Reguladores de activación, crecimiento y diferenciación de los linfocitos en la respuesta específica frente a los antígenos. c) Estimuladores de la hematopoyesis.
Dada la diversidad de propiedades y funciones, se ha tratado de
establecer numerosas clasificaciones de las citoquinas. De estas, la más amplia es las que las agrupa en función de las acciones biológicas principales que ejercen.
Así se pueden distinguir tres grupos principales: 1.- Citoquinas Proinflamatorias 2.- Reguladoras de la Respuesta Inmune
De la inmunidad innata De la inmunidad adaptativa Inmunosupresoras
3.- Citoquinas estimuladoras de la hematopoyesis 4.- Quimiocinas A su vez los factores de membrana serían los receptores encargados de captar estas moléculas. Existen varios tipos: Receptor Tipo Ig: IL-1, M-CSF Tipo I (hematopoyetina): La mayoría de las IL y CSF. Tienen la cadena de señales (proteína g) común Tipo II: IFNs e IL-10 Familia TNFR: TNF, CD40, Fas, NGF Familia quimiocinas R (7 pasos y acoplados a Proteína G) Como en el caso de los factores solubles, la expresión de los receptores esta regulada por el estado en que se encuentra la célula. Las células en reposo o no expresan el receptor o presentan una versión del mismo de afinidad baja o media. Esto asegura, que solo las células “preparadas” responderán a la citoquina. Una vez unida la citoquina a su receptor, este debe producir señales intracelulares que llevan a la producción de factores de trascripción activos y, finalmente, a la expresión de genes Vamos a ver ahora brevemente las principales citoquinas agrupadas por su principal función biológica
TEMA 07 (II). CITOCINAS 06/05/2008 Pág.2 de 8
formulandoalgoqueaportar.blogspot.com

Tema 7 (II). Citocinas
1.- Proinflamatorias: En general, estas citoquinas, promueven la migración de los fagocitos hacia el foco infeccioso, aumentan la expresión de moléculas de adhesión, inducen una elevación de la temperatura (fiebre), así como la síntesis de las proteínas de fase aguda y estimulan la migración de las células dendríticas
Factor de Necrosis Tumoral (TNF) (proinflamatoria) Los lípidos bacterianos, conocidos en general como endotoxinas o lipopolisacaridos (LPS), a concentraciones bajas estimulan a los fagocitos. Estos fagocitos estimulados por el LPS son la principal fuente de TNF, Entre las acciones que provoca se encuentran las siguientes:
De modo general la principal acción del TNF es atraer y activar a los neutrófilos y monocitos hacia las zonas de infección. 1.- Provoca la expresión de moléculas de adhesión en las células endoteliales, especialmente selectinas y los ligandos de las integrinas. 2.- Provoca la activación de los neutrófilos y monocitos 3.-Estimula a los macrófagos para que produzcan quimiocinas (citocinas quimiotácticas) y otras citoquinas como IL-1 4.- Induce la formación de nuevos vasos (angiogénesis) y es un factor de crecimiento para los fibroblastos. 5.- Induce la apoptosis. El defecto de TNF, puede llevar a un fracaso en la resolución de la infección, pero el exceso nos conduce a una serie de alteraciones patológicas, entre las que destacamos: 1.- El TNF es un pirógeno endógeno 2.- El TNF activa el sistema de la coagulación. Si la secreción es masiva el sujeto muere por colapso circulatorio y coagulación intravascular diseminada. 3.- Suprime la división de la célula madre en la médula ósea. Por ello la administración crónica de TNF puede dar lugar a linfopenia e inmunodepresión. 4.- Induce una perdida de apetito que puede llevar a la caquexia. En ciertos canceres se liberan cantidades anormales de TNF que explica el estado caquéctico que presentan estos enfermos. 5.- Por último destacar que la infusión de dosis elevadas de TNF es por si mismo letal, dando lugar a un Shock y a la muerte. IL-1 La principal fuente de esta citoquina, al igual que la de TNF es el macrófago activado. Aunque en esta caso también puede ser producido por otros tipos celulares como neutrófilos, células epiteliales y endoteliales.
TEMA 07 (II). CITOCINAS 06/05/2008 Pág.3 de 8
formulandoalgoqueaportar.blogspot.com

Tema 7 (II). Citocinas
A concentraciones bajas la IL-1 es un mediador de la inflamación local, aumentando la expresión de moléculas de adhesión. A dosis elevadas comparte con el TNF, su capacidad para producir fiebre, inducir la síntesis de proteínas hepáticas de fase aguda e iniciar el desgaste metabólico que conduce a la caquexia. Sin embargo presentan ciertas diferencias con el TNF como son: a) no llega a ser tan perjudicial dosis elevada como es el TNF. A concentraciones sistémicas elevadas no es letal. b) No es capaz de inducir apoptosis celular. c) No tiene capacidad para aumentar la expresión de moléculas del CMH d) Al contrario que el TNF potencia la división, no la suprime de las células de la médula ósea. IL-6 Parece que es secretada como respuesta al TNF y a la IL-1. Sus acciones fundamentales son estimular a los hepatocitos para la síntesis de fibrinógeno y de proteínas de fase aguda. También actúa como factor de crecimiento para los linfocitos B activados. El problema es que también actúa como factor de crecimiento para células plasmáticas neoplásicas. 2. Reguladoras de la respuesta inmunitaria IL-12 Es secretada por macrófagos y células dendríticas y es la única que funciona como un heterodímero (p35) y (p40). Es un mediador de la respuesta innata a organismos intracelulares y un inductor de la respuesta adaptativa frente a estos organismos. 1.- Es un potente inductor de IFN-γ tanto en las células NK como en las T 2.- Aumenta la actividad lítica de las células NK y células T CD8+ 3.- Estimula la diferenciación de los linfocitos CD4 hacia la población Th1, es decir actúa como potenciador de la inmunidad celular.
Todo ello proporciona a esta citoquina, un papel importante de conexión entre la inmunidad innata y la específica. Interferones (IFN) tipo 1 Dentro de ellos se encuadran los interferones αy β. La El primero es producido, principalmente por fagocitos, mientras que el beta es fundamentalmente secretado por fibroblastos. El estimulo, parece ser el ARN bicatenario producido durante al replicación de los virus.
Entre las funciones principales de estos interferones se encuentran: 1.- Inhiben la replicación viral, a través de las síntesis de algunos
TEMA 07 (II). CITOCINAS 06/05/2008 Pág.4 de 8
formulandoalgoqueaportar.blogspot.com

Tema 7 (II). Citocinas
enzimas que interfieren la replicación viral. La acción es sobre todo paracrina. Las células infectadas secretan IFN para proteger a las células vecinas sanas. 2.- Estimula el desarrollo de los linfocitos Th1, principalmente por inducir a los linfocitos T a expresar receptores para la IL-12, que como veíamos es la principal inductora de esta población. 3.- Aumentan el potencial lítico de las células NK 4.- Aumentan la expresión de moléculas MHC de clase I e inhiben las de clase II. 5.- Inhiben la proliferación celular (por los mismos mecanismos que lo hacen para el virus. Por ello se han utilizado como terapia antitumoral. Como se ve todas las acciones van encaminadas a la lucha antiviral. 2. b. Reguladoras de la inmunidad adaptativa En general, este grupo de citoquinas, van a activar la proliferación celular y a inducir la formación de órganos linfoides secundarios. Asimismo son la responsables del cambio de isotipo de los anticuerpos y van a orientar la respuesta hacia el tipo humoral Th2) o celular (Th1) IL-2 Es una glucoproteína, producida por los linfocitos T, especialmente por los CD4+. La secreción dura un periodo máximo de 12 horas y es la principal citoquina que actúa activando el ciclo celular de los linfocitos T. Es por tanto, la responsable de la expansión clonal de los linfocitos T. Tiene una acción autocrina y paracrina. Otras acciones serían 1.- Estimula la producción de otras citoquinas como el IFN-( y la linfotoxina 2.- Estimula a las células NK, que también tienen receptores para la IL-2. Aunque en este caso al poseer estas un receptor de menos afinidad se necesitan cantidades elevadas para provocar el estímulo 3.- Actúa como factor de crecimiento de Linfocitos B 4.- Puede actuar como factor de apoptosis para las células T activadas por el antígeno IL-4 Las principales fuente de la IL-4 es la subpoblación Th2 de los linfocitos CD4+, así como mastocitos y basófilos. Su acción primordial es promover el cambio de clase en los linfocitos B hacia la IgE, así como estimular la diferenciación de los linfocitos T a Th2. Además es un factor de crecimiento para la subpoblación Th2 diferenciados. Asimismo estimula la expresión de ciertas moléculas de adhesión como la VCAM-1, con lo que aumenta la adhesión de diversos tipos celulares, especialmente eosinófilos. IL-5
TEMA 07 (II). CITOCINAS 06/05/2008 Pág.5 de 8
formulandoalgoqueaportar.blogspot.com

Tema 7 (II). Citocinas
Esta producida por la subpoblación Th2 y mastocitos activados. Su principal actividad se centra en estimular el crecimiento y diferenciación de eosinófilos. Interferón de tipo II (IFN-() Esta producido por los linfocitos T (Th1), tanto CD4 como CD8 activados y por las células NK. El principio de la secreción comienza con la activación por el antígeno y aumenta por la acción de la IL-2 e IL-12. Actúa sobre todo como inmurregulador, característica esta que lo diferencia de los interferones de tipo I. Es la principal citoquina activadora de macrófagos Entre sus acciones cabe destacar: 1.- Es un potente activador de los fagocitos mononucleares. 2.- Aumenta la expresión de moléculas del CMH tanto de clase 1 como las de tipo II. 3.- favorece la diferenciación de los linfocitos T CD4+ hacia la subpoblación Th1, e inhibe la proliferación de la Th2. Induce la respuesta de anticuerpos opsonizantes. 4.- Activa a los neutrofilos y a las células NK, de forma mas potente que lo hacen los de tipo 1. 5.- Es un activador de la expresión de moléculas de adhesión en las células endoteliales, favoreciendo la adhesión de los linfocitos CD4+. Linfotoxina (LT) Es homologa en un 30% al TNF. Por eso se le llama también TNF-β. Es producida exclusivamente por los linfocitos T activados. Su acción mas diferenciada es su participación en el desarrollo de órganos linfoides secundarios. Así se ha visto que ratones con inhibición génica de esta citoquina son incapaces de formar ganglios linfáticos, y al contrario un exceso de la misma provoca la formación de tejido linfoide ectópico. IL-13 Es una citoquina con estructura similar a IL-4, producida por linfocitos T Th2 y células epiteliales. 3. Estimuladoras de la hematopoyesis:
En general se llaman Factores estimuladores de colonias. A nivel de las células progenitoras pluripotenciales, también existen
citoquinas estimuladoras necesarias para llevar a cabo las respuestas innatas y adaptativa. No olvidemos que las respuestas consumen leucocitos, que por tanto deben ser reemplazados. GM-CSF, (factor estimulador de colonias de granulocitos monocitos). Esta producido por linfocitos T activados, macrófagos y
TEMA 07 (II). CITOCINAS 06/05/2008 Pág.6 de 8
formulandoalgoqueaportar.blogspot.com

Tema 7 (II). Citocinas
células del la médula ósea. El GM-CSF tiene un amplio impacto sobre la hematopoyesis, estimulando el desarrollo y la maduración de las células madres que van a dar origen a los neutrófilos (PMN), eosinófilos, monocitos y megacariocitos. G-CSF (factor estimulador de granulocitos): Tiene una actividad más específica, estimula la proliferación, diferenciación y función de la línea de los granulocitos. IL-3, que es un factor estimulante de colonias de líneas múltiples, siendo secretada por los CD4+, y actúa sobre los progenitores más inmaduros de la médula ósea, favoreciendo la expansión de todos los tipos celulares. IL-7, Es secretada por las células del estroma de la médula ósea. Parece que actúa sobre la proliferación de los linfocitos. 4. Citoquinas supresoras Factor transformador de crecimiento (TGF) TGF-β es secretado por diversas células del sistema inmunitario, como células T activadas y macrófagos. Su principal acción es inhibir la proliferación y activación de linfocitos y macrófagos. En cierto modo actúa para contrarrestar los efectos de las citoquinas proinflamatorias. También estimula ciertas acciones que conducen a al reparación del tejido, como la angiogénesis Otra de sus acciones es provocar en los linfocitos B el cambio de clase hacia la síntesis de IgA. IL-10 Es producida por macrófagos activados y algunos linfocitos. Su principal acción es inhibitoria de los macrófagos y células dendríticas. También inhibe la expresión de de moléculas del CMH de clase II. La deficiencia en esta citoquina provoca una aumento del proceso inflamatorio y una elevacion de la liberación de oxido nítrico, con un mayor infiltrado celular. Por el contrario su exceso provoca shock séptico y un aumento de citoquinas proinflamatorias. Esta citoquina es, por tanto clave en la regulación del proceso inflamatorio. 4 Quimioquinas
Son pequeños polipéptidos estrechamente relacionados. Se producen por las células fagocitarías y otros muchos tipos celulares.
Estructuralmente, las quimioquinas tienen una secuencia de aminoácidos similar y se dividen estructuralmente en dos grandes grupos, según que tengan dos cisteínas contiguas (grupo CC) o separadas por otro aminoácido (grupo CxC). Ambos grupos actúan sobre juegos distintos de receptores y distintos tipos celulares. En general, las
TEMA 07 (II). CITOCINAS 06/05/2008 Pág.7 de 8
formulandoalgoqueaportar.blogspot.com

Tema 7 (II). Citocinas
CC promueven la migración de monocitos y otros tipos celulares, mientras que las CxC promueven la migración de neutrófilos. Se conoce una quimioquina con solo una cisteína (C): es la linfotactina que parece que es la que atrae a los precursores T hacia el timo.
Se secretan inducidas por ciertos microorganismos y citoquinas inflamatorias, especialmente TNF e IL-1.
Su acción es fundamentalmente quimiotáctica de monocitos, macrófagos e incluso actúan de atrayentes de los precursores de los linfocitos T hacia el Timo. En otras palabras son citoquinas que estimulan el movimiento de los leucocitos y regulan su migración desde la sangre a los tejidos así como intervienen en la migración de los linfocitos entre los órganos linfáticos periféricos. Además de las funciones relacionadas con el proceso inmunitario, se ha descubierto que ciertas quimiocinas intervienen en el desarrollo de ciertos órganos (morfogenia)1
Ya se han identificado 17 receptores diferentes para las quimioquinas, que se expresan en los leucocitos, la mayor parte en linfocitos T. De ellos, hay uno que actúa como correceptor para el virus de la Inmunodeficiencia humana (VIH). De esta forma, algunos linfocitos T activados secretan quimiocinas que se unen a este receptor y bloquean la infección por VIH, o al menos compiten con el virus.
1 El déficit de un receptor (CxCR4) en los ratones se ha visto que conlleva defectos mortales de formación en el desarrollo del corazón y cerebelo
TEMA 07 (II). CITOCINAS 06/05/2008 Pág.8 de 8
formulandoalgoqueaportar.blogspot.com

FYTI. Tema 07. ANTIGENOS Curso 07-08
Tema 7
• Inmunógenos, antígenos y haptenos. • Adyuvantes
La respuesta inmune adaptativa se produce como consecuencia de la exposición a moléculas extrañas. Estas sustancias extrañas se definen del siguiente modo: Inmunógeno, sustancia que induce la respuesta inmune adaptativa. Antígeno, cualquier agente capaz de unirse específicamente a los componentes de la respuesta adaptativa (receptor del linfocito B [BCR] y anticuerpos solubles, así como al receptor del LT [TCR]) Hay compuestos que no son capaces de inducir una respuesta adaptativa pero, en cambio, reaccionan con los componentes de la respuesta elaborada frente a ellos. Es decir, todos los inmunógenos son antígenos, pero no todos los antígenos son inmunógenos. Hapteno. Esta diferencia se hace obvia en el caso de las sustancias de bajo peso molecular (grupo en el que se incluyen muchos antibióticos y drogas), que no son capaces, por si mismos, de inducir una respuesta inmune pero que cuando se unen a moléculas de peso molecular alto (tales como las proteínas) forman conjugados capaces de inducir una respuesta inmune. Estas moléculas, de peso molecular bajo, se denominan Haptenos y la molécula a la que se acoplan Portadores (“carrier”). Por tanto, un Hapteno es un compuesto incapaz, por si mismo, de inducir una respuesta inmune pero que reacciona con los componentes de la respuesta que se induce cuando se conjuga con una molécula portadora. Se ha comprobado que se puede producir respuesta inmune frente a todas las familias de compuestos bioquímicos conocidas, incluyendo carbohidratos, lípidos, proteínas y ácidos nucleicos. También se pueden inducir la respuesta a drogas, antibióticos, aditivos alimentarios, cosméticos y pequeños péptidos sintéticos, pero solo cuando se acoplan a un portador. Características de los inmunógenos Para que una sustancia sea inmunogénica debe tener las siguientes características:
• Ser extraña (1) • Peso molecular alto (2) • Complejidad química (3) • Estructura (4) • Poderse degradar a péptidos que interaccionen con las proteínas del CMH (5)
(1) El organismo normalmente no responden frente a lo propio; esta es la es la primera condición para que un compuesto sea inmunogénico. Cuanto más extraña sea la sustancia (más distancia génica) mayor será su inmunogenicidad. (2) Las sustancias necesitan un peso molecular mínimo para ser inmunogénicas; 5-6 KDa se consideran el peso mínimo para la inmunogenicidad y esta aumenta en función del peso molecular.
Página 1 de 5
formulandoalgoqueaportar.blogspot.com

FYTI. Tema 07. ANTIGENOS Curso 07-08
(3) Un inmunógeno debe tener una complejidad química mínima. Homopolímeros de un solo aminoácido con peso molecular alto (p.e. poli-lisina de Pm 30 KDa) es muy raro que sean inmunogénicos. Pero si se les unen moléculas pequeñas o se introducen otros aminoácidos (sobre todo si son aromáticos), la molécula se torna inmunogénica. En general, el aumento de la complejidad química es directamente proporcional al aumento de la inmunogenicidad. (4) La mayoría de los inmunógenos son proteínas y la respuesta adaptativa reconoce muchos caracteres estructurales de estos compuestos. Por ejemplo, los anticuerpos pueden reconocer características estructurales de una proteína, tales como las estructuras: Primaria (secuencia de los aminoácidos en la cadena) Secundaria (plegamientos de la cadena primaria, que pueden ser en hélices alfa o en estructuras planas beta) Terciaria (formada por los plegamientos de la estructura secundaria mantenidos por puentes disulfuro, puentes de hidrógeno, interacciones hidrofóbicas, etc.) Cuaternaria (formada por la yuxtaposición de varias subunidades proteicas) (5) Para que los antígenos interaccionen con las células T (que son las que ponen en marcha y coordinan la respuesta adaptativa) tienen que ser capaces de unirse a las moléculas CMH que se expresan sobre las CPAs. Estas células primero degradan mediante enzimas a los antígenos (procesamiento) y luego unen los fragmentos producidos (epitopos) a las moléculas CMH para enseñar el conjunto sobre la superficie (presentación). Otros requerimientos Hay, además, otros factores que influyen en la inmunogenicidad, tales como la base genética (genotipo) del hospedador, la dosis, vía de administración, etc. Las dosis insuficientes y las demasiado abundantes no provocan la puesta en marcha de la respuesta adaptativa, ya que inducen la tolerancia de los linfocitos. Pero además de la dosis adecuada también influye en la respuesta el número de dosis administradas (pauta de inoculación). En cuanto a la vía de administración, la subcutánea da normalmente muy buenos resultados, ya que el antígeno se deposita en la vecindad de las células de Langerhans (DCs inmaduras de la piel), que son las CPAs más potentes del organismo. Estas células emigran a los ganglios linfáticos próximos, donde se elabora la respuesta adaptativa. Los antígenos administrados por vía intravenosa llegan al bazo, donde pueden inducir tolerancia o, si se presentan por CPAs, inducir respuesta. La vía oral induce respuestas locales de anticuerpos en la lámina propia del intestino y, con frecuencia, provoca tolerancia sistémica a tal antígeno. La vía respiratoria (intranasal) es frecuente que provoque respuestas alérgicas. Como la respuesta adaptativa depende de las interacciones celulares, el tipo y la intensidad de la respuesta refleja la población celular presente en el órgano al que llega finalmente el antígeno. Respuestas primaria y secundaria El primer contacto con un inmunógeno provoca una respuesta primaria, tras una serie de interacciones entre células T, CPAs y células B. La curva de anticuerpos que se produce es característica.
Página 2 de 5
formulandoalgoqueaportar.blogspot.com

FYTI. Tema 07. ANTIGENOS Curso 07-08
El siguiente contacto con el mismo inmunógeno induce una respuesta secundaria que difiere de la primaria (más rápida, más amplia, más eficaz), como si el organismo “recordara” que ya había estado expuesto a ese inmunógeno antes. De hecho, la respuesta secundaria y las subsecuentes explotan el aumento del número de linfocitos específicos para el inmunógeno producidos durante la respuesta primaria. Esta respuesta también se llama “respuesta de memoria o anamnésica” y los linfocitos que participan en ella se denominan células de memoria. Antigenicidad y sitio de unión al antígeno La respuesta inmune genera anticuerpos y/o linfocitos que reaccionan específicamente con alguna porción del antígeno, a la que se denomina epitopo (también se le llama determinante antigénico). El sitio de unión de un anticuerpo o del receptor de un linfocito al epitopo presenta una estructura única que es complementaria del mismo. La porción del anticuerpo (Ig) que se une específicamente al epitopo está situada en una zona hipervariable de la molécula, que forma la llamada región determinante de la complementariedad (CDR). El tamaño del epitopo del antígeno que se combina con el CDR de un anticuerpo es de 5-7 aminoácidos. Lo que reconoce el receptor del linfocito T son secuencias de 8-12 aminoácidos asociados en forma no covalente a proteínas del Complejo Principal de Histocompatibilidad (CMH) en la superficie de las CPAs (células presentadoras de antígenos).
Reconocimiento antigénico por parte de los linfocitos B y T Características Linfocitos B Linfocitos T Interacción con el Ag Naturaleza del Ag Fijación Ags solubles Epitopos reconocidos
El receptor B (Ig de membrana) fija al Ag Proteínas, hidratos de carbono, lípidos SI Accesibles (secuenciales o conformacionales)
El receptor T fija al conjunto Ag-CMH Péptidos NO Secuenciales, procedentes del procesamiento
Los epitopos que reconocen las células B y T. Como se ve en la tabla, los epitopos reconocidos por las células B difieren de los que reconocen las células T. En general, los receptores de los linfocitos B reconocen y fijan antígenos libres, en solución. Por tanto, los epitopos tienen que estar en la superficie de la molécula antigénica donde son accesibles al receptor B. La parte terminal de las cadenas laterales de polisacáridos y las porciones hidrofílicas de las moléculas de proteína generalmente forman los epitopos B, aunque también los hay procedentes de los plegamientos (epitopos conformacionales). Las células T no son capaces de fijar a los antígenos solubles. La interacción de un epitopo con el receptor T requiere que una CPA procese enzimáticamente al antígeno (lo degrade a péptidos pequeños) para que las porciones se asocien a las moléculas CMH. Por tanto, los receptores T solo pueden reconocer epitopos lineales (secuenciales), pues los conformacionales se destruyen durante el procesamiento de la proteína. Normalmente, estos epitopos lineales son porciones hidrofóbicas internas de la proteína. El resultado de todo esto es que los polisacáridos solo contienen epitopos reconocibles por los linfocitos B, mientras que las proteínas contienen epitopos reconocible tanto por las células B como por las células T.
Página 3 de 5
formulandoalgoqueaportar.blogspot.com

FYTI. Tema 07. ANTIGENOS Curso 07-08
El número de epitopos de una molécula puede ser solo uno (hapteno) o múltiple y en una proteína normalmente hay muchos y distintos, mientras que en los azúcares suelen ser iguales (repetitivos). Clases de antígenos Polisacáridos. Son inmunógenos cuando se asocian a portadores proteicos, como ocurre con los que forman parte de moléculas más complejas (glicoproteínas), que inducen una respuesta inmune parte de la cual es específica para la parte hidrocarbonada de la molécula. Pero también se puede inducir una respuesta inmune de anticuerpos frente a muchos tipos de polisacáridos, como los que forman parte de microorganismos y células eucariotas. El ejemplo clásico de antigenicidad de los polisacáridos es la respuesta a los antígenos sanguíneos del sistema ABO, que son polisacáridos de la superficie de los glóbulos rojos. Lípidos. Son normalmente poco inmunogénicos, pero pueden inducir respuesta si se unen a proteínas portadoras. Los lípidos de las mycobacterias se reconocen por un subtipo especial de células T; los Tgd, que forman parte de la respuesta ancestral. Ácidos nucleicos. No son muy inmunogénicos, salvo que se asocien con proteínas portadoras. Los anticuerpos anti-DNA son característicos de varios procesos autoinmunes. Proteínas. Virtualmente, todas las proteínas son inmunogénicas y la gran mayoría de los inmunógenos son proteínas. Cuanto mayor es la complejidad de la proteína más fuerte es la respuesta. Normalmente contienen muchos epitopos. Reacciones cruzadas Se producen por la presencia de epitopos comunes en moléculas distintas. Un ejemplo es la toxina diftérica que puede manipularse para que pierda su toxicidad conservando la capacidad inmunogénica; se ha transformado en toxoide. La inmunización con el toxoide (inocuo) da lugar a la formación de anticuerpos que neutralizan a la toxina diftérica cuando se produce la infección. Estamos usando determinantes comunes para vacunar, sin usar la toxina. Hay otros casos de reactividad cruzada donde las sustancias que reaccionan no están relacionadas, excepto en que tienen una o más áreas con características tridimensionales similares. A estas sustancias se las llama antígenos heterófilos. Por ejemplo el antígeno sanguíneo A reacciona con los anticuerpos formados frente al antígeno capsular (polisacárido) de un tipo de neumococo. Adyuvantes Son los “aditivos” que se usan para aumentar la respuesta a un inmunógeno dado. El nombre viene del latín adjuvare (ayudar). Aunque su uso viene de antiguo, el interés actual se debe a que los inmunógenos que se están desarrollando para hacer nuevas vacunas (sobre todo las basadas en péptidos sintéticos) presentan una inmunogenicidad muy pobre. El mecanismo de los adyuvantes incluye:
• Aumento de la vida media biológica o inmunológica de los antígenos de las vacunas
• Aumento de la producción local de citocinas inflamatorias • Mejora en la liberación del antígeno y en su procesamiento y presentación por
las CPAs, especialmente por las células dendríticas.
Página 4 de 5
formulandoalgoqueaportar.blogspot.com

FYTI. Tema 07. ANTIGENOS Curso 07-08
Se sabe de modo empírico que los mejores adyuvantes son los que contienen componentes microbianos (p.e. extractos de mycobacterias). Estos componentes de patógenos inducen la expresión de moléculas coestimuladoras en las CPAs (macrófagos y DCs) así como la producción de citocinas. Ahora se ha comprobado que tal inducción por los componentes microbianos se debe a que contienen patrones moleculares (PAMPs); su unión a los TLRs de las CPAs genera las señales que llevan a la expresión de moléculas coestimuladoras y a la síntesis de citocinas. Los adyuvantes autorizados hasta ahora para uso humano son los que se basan en hidróxido de aluminio o fosfato de aluminio (alúmina). La alúmina se fija a las proteínas y las hace precipitar, induciendo así una respuesta inflamatoria que potencia de forma inespecífica la inmunogenicidad del antígeno. El antígeno precipitado se libera más despacio en el sitio de inyección, lo que unido al aumento de tamaño que se genera en la precipitación propicia una mayor probabilidad de que el antígeno se fagocite por una célula presentadora. En los animales de experimentación se han usado muchos adyuvantes, siendo uno de los más corrientes el Adyuvante Completo de Freund (FCA), consistente en mycobacterias muertas suspendidas en aceite mineral. Al añadir la solución acuosa del antígeno se genera una emulsión que, inyectada, libera lenta y continuamente al antígeno con lo que mantiene constante la estimulación durante mucho más tiempo. Otros microorganismos usados como adyuvantes son el bacilo de Calmette-Guerin (BCG), que es una micobacteria atenuada, Corynebacterium parvum y Bordetella pertussis. En realidad, todos utilizan las propiedades estimuladoras de las moléculas asociadas al microbio cuando se fijan sobre los TLRs de las células presentadoras (especialmente las DCs). Los ISCOM (complejos inmuno-estimuladores) son unas mezclas de colesterol y saponinas que forman unas estructuras similares a jaulas en las que queda atrapada la solución acuosa del antígeno. Esta “jaulas” se funden con los lípidos de las membranas celulares y depositan así su contenido en el citosol. El resultado es una vacuna que induce una respuesta celular mediada por linfocitos T citotóxicos, en lugar de los anticuerpos que inducen las restantes vacunas.
Página 5 de 5
formulandoalgoqueaportar.blogspot.com

EL SITIO DE UNION AL ANTIGENO Las moléculas de anticuerpos tienen dos papeles: la unión al antígeno y mediar las funciones efectoras. La unión al antígeno se realiza por el extremo amino-terminal y las funciones efectoras por el extremo carboxi-terminal. Las regiones variables La variabilidad viene dada por el número de aa. diferentes en una posición dada / frecuencia de los aa. más comunes en una posición dada. P.e., la comparación de secuencias de 100 cadenas H revela que la serina se encuentra en posición 7 en 51 de las secuencias (es el aa. más frecuente, con frecuencia 0,51); el examen de las otras 49 secuencias muestra que la posición 7 puede estar ocupada por glutamina, histidina, prolina o triptófano (5 aa. distintos, si sumamos la serina), por lo que la variabilidad de esta posición debe ser 9,8 (5:0,51). Las representaciones gráficas de los dominios VH y VL muestran que la máxima variación se ve que corresponde a las porciones de secuencias que unen las hebras β. Esas regiones se llamaron originalmente hipervariables. Como el sitio de unión al antígeno es complementario a la estructura del epitopo, ahora se llaman regiones determinantes de la complementariedad (CDRs). El resto de secuencias de los dominios V presentan mucha menos variabilidad y se las conoce como regiones estructurales (FRs; Framework Regions) El gran número de especificidades que presentan los anticuerpos se debe a las variaciones en longitud y secuencia de aa.s de los 6 CDRs que configuran el sitio de fijación. La región estructural actúa como un andamiaje que soporta esas seis regiones. La estructura tridimensional de la región estructural de cualquier anticuerpo es superponible entre los distintos anticuerpos, mientras que las regiones hipervariables (CDRs) son únicas para cada anticuerpo. Los dominios constantes Contribuyen a diferentes funciones de los anticuerpos que vienen determinadas por la secuencia de aa. de cada dominio. Dominios C1H y CLSirven para extender los brazos Fab de la moléculas de anticuerpos, facilitando la interacción con los antígenos y aumentando la rotación máxima de los brazos Fab. La región bisagra Las cadenas γ, δ y α presentan una secuencia extendida de péptidos entre los dominios CH1 y CH2. Esta región (llamada región bisagra) es ricas en restos de prolina y es flexible, lo que permite la deformación de las IgG, IgD a IgA. Como resultado los dos brazos Fab pueden adoptar varios ángulos cuando se unen al antígeno. Esta es la región más susceptible a la degradación enzimática y la que, de hecho, se rompe por la pepsina y la papaina. Los dos aminoácidos prominentes en la región bisagra son prolina y cisteína; La prolina es responsable de la flexibilidad de la molécula y las cisteínas establecen los puentes disulfuro entre las dos cadenas H. El número de puentes disulfuro intra-catenarios en la región bisagra varía considerablemente entre las distintas clases de anticuerpos y entre las especies. Aunque las cadenas µ y ε carecen de región bisagra, tienen en cambio un dominio adicional (CH2) de 110 aa. con caracteres “tipo bisagra”.
formulandoalgoqueaportar.blogspot.com

Otros dominios constantes Aunque los dominios CH2 de IgM e IgE ocupan la misma posición que la cadena polipeptídica de la región bisagra en las otras clases de Igs, todavía no ha sido posible asignarle ninguna función. Los dominios CH2 de IgG, IgD e IgA (y los CH3 de IgE e IgM) están aislados por cadenas laterales de polisacáridos lo que les permite ser mucho más accesibles que los otros al ambiente acuático. Esta accesibilidad es uno de los elementos que contribuye a la actividad biológica de estos dominios en la activación de los componentes del complemento por IgG e IgM. El dominio carboxi-terminal es CH3 (en IgG, IgD e IgA) y CH4 (en IgM e IgE). Todas las moléculas pueden expresarse como inmunoglobulinas secretadas (sIg) o como inmunoglobulinas unidas a la membrana (mIg). El dominio carboxi-terminal de sIg difiere tanto en estructura como en función del de mIg. Las sIg tienen una secuencia, de diferentes longitudes, de aa hidrofílicos en el extremo carboxi-terminal, mientras que en las mIg hay tres regiones:
• Una secuencia “espaciadora” extracelular formada por 26 aa • Una secuencia hidrofóbica transmembrana • Un tallo citoplásmico corto.
La longitud de la región transmembrana es constante entre todos los isotipos de Ig, mientras que las secuencias espaciadoras y los tallos citoplásmicos son de longitud variable.
FUNCIONES EFECTORAS MEDIADAS POR ANTICUERPOS Además de fijar a los antígenos, los anticuerpos participan en una gran variedad de acciones biológicas efectoras. La simple fijación generalmente no mata ni elimina a ningún patógeno. Son las regiones constantes de las cadenas pesadas las que interaccionan con otras proteínas, células y tejidos y el resultado lo conocemos como funciones efectoras de la respuesta humoral. Como las funciones efectoras son el resultado de las interacciones de las cadenas H con otras proteínas séricas o receptores de membrana, no todas las clases de Ig tienen las mismas propiedades funcionales. 1.- Anticuerpos y opsonización En la superficie de macrófagos y neutrófilos (y también de otras células que no intervienen en la fagocitosis) hay moléculas proteicas llamadas receptores Fc (FcRs) que pueden unirse a la región constante de las moléculas de Ig. Si varias moléculas de anticuerpo están unidas al mismo antígeno (p,e,, a una bacteria) su unión a los receptores Fc provoca una interacción que fija al patógeno sobre la membrana del fagocito y pone en marcha una vía de transducción de señales que lleva a la fagocitosis del complejo antígeno-anticuerpos. Dentro del fagocito, el complejo es el blanco de varios procesos destructores (digestión enzimática, mecanismos oxidativos, péptidos antibacterianos, etc.) 2.- Los anticuerpos activan el complemento IgM y, en humanos, la mayoría de las subclases de IgG pueden activar el sistema del complemento por la vía clásica (dependiente de anticuerpos). Un producto importante derivado de la cascada de activación es C3b que se fija de horma no específica a las células y a los complejos Ag-Ac próximos al sitio de activación.
formulandoalgoqueaportar.blogspot.com

Muchos tipos celulares tienen receptores para C3b y también fijan células o complejos a los que se ha asociado C3b. La fijación del C3b adherente por los macrófagos lleva a la fagocitosis de la célula o complejo unidos a C3b. La fijación de l os complejos Ag-Ac por los C3bR de los glóbulos rojos permite llevarlos al hígado o bazo, donde se capturan por los macrófagos (sin dañar al GR), promoviendo asi la eliminación (aclaramiento) de los IC. La colaboración entre los Ac y el sistema del complemento es importante para la inactivación y eliminación de antígenos y para la muerte de patógenos. 3.- La citotoxicidad dependiente de anticuerpos (ADCC) mata a células La unión del anticuerpo sobre la célula blanco (p.e., células del hospedador infectadas por virus) y de su fragmento Fc a receptores existentes en otras células (p.e., células NK) puede dirigir la actividad citotóxica de la célula efectora hacia la célula blanco. 4.- Algunos anticuerpos pueden cruzar las barreras epiteliales por transcitosis. La distribución de anticuerpos por las superficies mucosas de los sistemas pulmonar, gastrointestinal y urogenital, así como su exportación a través de la leche materna, requiere que las Igs se muevan a través de las capas epiteliales, en un proceso denominado transcitosis. La capacidad de un anticuerpo para ser transportado depende de las propiedades de la región constante. En humanos y ratones IgA es el anticuerpo principal que sufre la transcitosis, aunque también puede transportarse la IgM hasta las superficies mucosas. Algunas especies de mamíferos, como humanos y ratones, también transfieren cantidades significativas de la mayoría de subclases de IgG de la madre al feto. Como los sistemas circulatorios materno y fetal están separados, los anticuerpos tienen que ser transportados a través de la placenta, que separa a madre de feto. En los humanos el transporte se realiza en el tercer trimestre de la gestación. La consecuencia principal es que el feto recibe una muestra del repertorio de anticuerpos de la madre a modo de regalo que le protege de los patógenos. Como ocurre con otras funciones efectoras, la capacidad de transporte depende de las propiedades de la región constante de la molécula de anticuerpo. La transferencia de anticuerpos de la madre al feto es una forma de inmunización pasiva, sistema por el que se adquiere inmunidad al recibir anticuerpos ya preformados, en lugar de producirlos tras la exposición al antígeno. Esta posibilidad de transferir inmunidad de un individuo a otro mediante la transferencia de anticuerpos es la base de la terapia con anticuerpos, un tratamiento importante en la medicina actual. También las subclases de IgG se transfieren de la madre al recién nacido en los humanos, mediante los receptores que hay en la mucosa intestinal del recién nacido y que perduran hasta que se desarrolla la flora intestinal (aprox. 1 año). 5.- Los anticuerpos de la clase IgE activan a mastocitos, eosinófilos y basófilos Se verá posteriormente en las hipersensibilidades.
CLASES DE ANTICUERPOS Y ACTIVIDADES BIOLOGICAS Cada clase se distingue por una secuencia única de aa. en las regiones constantes de la cadena H que le confiere propiedades estructurales y funcionales específicas. Inmunoglobulina G
formulandoalgoqueaportar.blogspot.com

La más abundante en el suero (representa, aproximadamente, el 80%). Es monomérica tiene tres dominios constantes en la cadena H. Hay, en humanos, 4 subclases que se distinguen por diferencias en la secuencia de las regiones constantes de la cadena γ y se numeran según disminuye la media de la concentración sérica desde 1 hasta 4. Las secuencias de aa. que distinguen las 4 subclases están codificadas por genes CH diferentes de la línea germinal, con una homología de secuencias del 90-95%. Estructuralmente se distinguen por el tamaño de la región bisagra y el número y posición de los puentes disulfuro intercatenarios. Estos sutiles cambios en la estructura afectan a las propiedades biológicas de las moléculas:
• IgG1, 3 y 4 cruzan fácilmente la placenta y tienen un papel importante en la protección del feto en desarrollo.
• IgG3 es la más efectiva en la activación del complemento, seguida en eficiencia por IgG1 e IgG2. La subclase IgG4 no es capaz de activar al complemento
• IgG1 e IgG3 se fijan con alta afinidad a los receptores Fc que hay sobre las células fagocitaria y, por tanto, median la opsonización. IgG4 tiene una afinidad intermedia para los receptores Fc e IgG2 tiene una afinidad extremadamente baja
Inmunoglobulina M Supone el 5-10% de las Ig del suero, con una concentración media de 1,5 mg/ml. La IgM monomérica se expresa como mIg sobre las células B. LA sIg se secreta por las células plasmáticas en forma de pentámero; cada pentámero contiene un péptido adicional (cadena J) que se une, formando puentes disulfuro, a los residuos de cisteína carboxi-terminales de dos cadenas µ. Parece que se requiere a la cadena J para la polimerización de los monómeros; se añade en el momento anterior a la secreción del pentámero. Es la primera Ig que se forma en la respuesta primaria a un antígeno, la primera sintetizada por el neonato y la primera que aparece en la evolución. Aunque tiene 10 sitios de fijación solo se une simultáneamente, por problemas de configuración estérica, a cinco o menos moléculas de los grandes antígenos. Debido a su valencia es más eficiente que otros isotipos para fijar antígenos con epitopos repetitivos La activación del complemento requiere al menos dos regiones Fc muy próximas y la estructura pentamérica de IgM cumple este requisito. IgM no difunde bien debido a su tamaño y, por ello, está en concentraciones muy bajas en los líquidos tisulares. La presencia de la cadena J permite a la IgM fijarse a receptores sobre células secretoras, que la transportan a través de las capas mucosas hasta formar parte de la secreción que baña las superficies mucosas. Aunque el isotipo principal que se encuentra en estas secreciones es IgA, IgM también tiene un papel accesorio importante como Ig secretora. Inmunoglobulina A Aunque solo representa el 10-15% de las Ig séricas es la que predomina en las secreciones, tales como leche materna, saliva, lágrimas y mucus de las vías bronquiales, genitourinarias y digestivas. En el suero está, sobre todo, como monómero. También hay formas poliméricas (di, tri y penta), y todas incluyen un polipéptido (cadena J). La IgA de las secreciones externas se conoce como IgA secretora (IgAs), formada por un dímero (o tetrámero), una cadena J y una pieza secretora. La síntesis diaria de IgAs es mayor que la de cualquier otro tipo de Ig y las células plasmáticas encargadas de su producción están concentradas a lo largo de las superficies mucosas. Un ejemplo: en toda la extensión del yeyuno hay 2,5 x 1010 células
formulandoalgoqueaportar.blogspot.com

plasmáticas que secretan IgA (¡una cifra superior al total de células plasmáticas que hay entre médula ósea, linfa y bazo!). Un humano envía diariamente entre 5 y 15 gr de IgA hacia las excreciones mucosas. Las células plasmáticas productoras de IgA migran preferentemente hacia los tejidos sub-epiteliales, y se alojan en ellos; allí es donde la IgA secretada se une a un receptor específico para moléculas de Ig poliméricas (el Poli-Ig), que se expresa en la superficie basolateral de la mayoría de las células de los epitelios mucosos y epitelios glandulares (de las mamas, glándulas salivares y lagrimales). Una vez unida la Ig el complejo se transporta por el interior de la célula hasta la membrana luminal y la luz; una vez allí el receptor sufre una degradación enzimática parcial que deja a parte del mismo envolviendo a la Ig (pieza secretora). Este pieza secretora “tapa” sitios susceptibles de degradación enzimática, tales como las regiones bisagra, y permite una vida media que no sería posible sin tal protección en un medio mucoso, rico en proteasas. Este mecanismo también transporta IgM pentamérica a las secreciones mucosas, aunque en una proporción muy inferior a la de la IgA. El receptor Poli-Ig interacciona con la cadena J de ambos tipos de anticuerpos. Las superficies mucosas son los sitios de entrada de la mayor parte de microorganismos patógenos y allí es donde desempeña la IgAs su función principal, capturando antígenos solubles y, sobre todo, antígenos de la superficie d virus y bacterias que así no pueden establecer contacto directo con las células mucosas. Los complejos inmunes quedan atrapados en el mucus y se eliminan bien por la ciliatura (bronquios), bien por los movimientos peristálticos (intestino) o se eliminan con la orina. Está comprobada la acción protectora de la IgA frente a bacterias como Salmonella, Vibrio cholerae, Neisseria gonorrhoeae y virus como los de la poliomielitis, la gripe y también el reovirus. La leche materna tiene IgAs y otras muchas moléculas que ayudan a proteger al recién nacido contra las infecciones durante el primer mes de vida. Como el sistema inmune de los lactantes todavía no funciona plenamente, la lactancia materna tiene un papel esencial en la conservación de la salud de los recién nacidos y durante la primera infancia. Los anticuerpos pasan del intestino del lactante hacia su interior por medio de unos receptores “tipo IgN” que hay en el epitelio intestinal y que desaparecen cuando se establece la flora intestinal y se completa la dotación de enzimas proteolíticos. Inmunoglobulina E La potente actividad biológica de IgE permitió su identificación en el suero a pesar de su baja concentración en el mismo (0,3 ug/ml). Los Acs IgE median las reacciones de hipersensibilidad inmediata, responsables de los síntomas de la fiebre del heno, asma, urticaria y shock anafiláctico. La presencia de un factor del suero responsable de la fiebre alérgica se demostró en 1921 por K. Prausnitz y H. Krustner. Inyectaron por vía subcutánea suero de un alérgico a otro individuo no alérgico y, después, el antígeno responsable de fiebre del heno en el mismo sitio; el resultado fue una reacción local con formación de una pápula con picor (“wheal and flare). Esta reacción, llamada P-K, fue la base para el primer ensayo biológico de actividad IgE. La identificación de la IgE se realizó por K. y T. Ishizaka en 1966. El nuevo anticuerpo se llamó IgE (en recuerdo del antígeno del polen de ambrosía, que es el inductor más potente de este tipo de Ac). La IgE se une a receptores de membrana sobre los basófilos de la sangre y mastocitos de los tejidos. Cuando la IgE ligada sobre la célula se une a su antígeno correspondiente (alergeno) se produce el entrecruzamiento de los receptores, lo que induce a la célula a trasladar sus granulaciones a la membrana celular y liberar su contenido
formulandoalgoqueaportar.blogspot.com

(degranulación). El resultado es que se liberan una serie de mediadores farmacológicos activos y se producen las manifestaciones alérgicas. La degranulación localizada de los mastocitos, inducida por IgE, también puede liberar mediadores que facilitan la concentración de varias células (eosinófilos) necesarias para una defensa ADCC frente a parásitos. Inmunoglobulina D Descubierta a partir del mieloma múltiple de un paciente, cuya proteína no reaccionaba frente a los sueros anti-sotipo frente a los anticuerpos conocidos hasta entonces (IgG, IgA, IgM). Tiene una concentración sérica de 30 ug/ml y representa el 2% de los anticuerpos del suero. IgD, junto con IgM monomérica, son los receptores de membrana principales que se expresan sobre las células B maduras. No se le conocen efectos biológicos como anticuerpo.
DETERMINANTES ANTIGENICOS DE LAS Ig Cuando se inyectan moléculas de Ig de un animal en otro de especie diferente se comportan como inmunógenos potentes que inducen la formación de anticuerpos. Estos Ac anti-Ig se usan para el estudio del desarrollo de los linfocitos B y de la respuesta humoral. Los determinantes antigénicos (epitopos) de las moléculas de Ig pertenecen a una de tres categorías:
• Isotópicos • Alotípicos • Idiotípicos
que se localizan en partes determinadas de la molécula. Isotipos Los determinantes idiotípicos forman parte de las regiones constantes y definen, colectivamente, las clases y subclases de las cadenas H y los tipos y subtipos de cadenas L de una especie. Cada isotipo está codificado por un gene de región constante y todos los miembros de la especie tienen la misma colección de genes de regiones constantes (que pueden incluir múltiples alelos). En una especie, cada individuo normal debe expresar todos los isotipos en el suero. Las diferentes especies heredan genes de regiones constantes distintos y, por tanto, expresan isotipos diferentes. Así, al inyectar anticuerpos de una especie en otra, los determinantes isotópicos se clasifican como extraños (no se reconocen como propios) e inducen una respuesta inmune humoral (de anticuerpos) frente a ellos. Los anticuerpos anti-isotipo sirven para determinar las clases y subclases de anticuerpos que se producen en una respuesta inmune de la primera especie. También para caracterizar la clase de los receptores de membrana que hay en un LB. Alotipos Aunque todos los miembros de la misma especie heredan el mismo juego de genes de isotipos, algunos de ellos tienen múltiples alelos. Esos alelos codifican diferencias sutiles de aminoácidos, que conforman los llamados determinantes alotípicos y que solo se presentan en algunos (pero no en todos ) los individuos de la especie. La suma de los determinantes alotípicos individuales en una molécula de anticuerpo determina su alotipo.
formulandoalgoqueaportar.blogspot.com

En los humanos se han encontrado alotipos en las cuatro subclases de IgG, en una subclase de IgA y en la región constante de la cadena ligera kappa. Los alotipos de la cadena gamma se refieren como marcadores Gm.. Se han identificado por lo menos 25 alotipos Gm, que se designan por la clase y subclase, seguidas por un número de orden [p.e., G1m (1), G2m (23), G3m (11), G4m (4ª)]. De las dos subclases de IgA solo IgA2 tiene alotipos [À2m (1) y A2m (2)]. Los alotipos de la cadena kappa son 3 [ km (1), km(2) y km(3)]. Cada uno de estos determinantes alotípicos presenta diferencias de 1 a 4 aminoácidos con los alelos diferentes del mismo gene. También pueden obtenerse anticuerpos frente a los determinantes alotípicos, inyectando anticuerpos de un miembro de la especie en otro con alotipos diferentes. Esos anticuerpos se producen a veces por una madre gestante en respuesta a los determinantes alotípicos paternos en las Ig fetales. También pueden producirse anticuerpos anti-alotipo por transfusiones sanguíneas. Idiotipo La secuencia de aminoácidos de los dominios VH y VL es única, dado que actúa o solo como sitio de fijación al antígeno sino también como un conjunto de determinantes antigénicos. Cada determinate individual en estas regiones se denomina idiotopo y cada anticuerpos presenta múltiples idiotopos ; algunos de ellos están en el sitio de fijación para el antígeno y otros en secuencias de la región variable que no forman parte del sitio de fijación. La suma de los idiotopos individuales es lo que se denomina idiotipo del anticuerpo (o del receptor). Como los anticuerpos producidos por células B individuales derivadas del mismo clon tienen secuencias idénticas en la región variable, todos ellos tienen el mismo idiotipo. Los anticuerpos anti-idiotipo se producen cuando se inyectan Ac que tienen variaciones mínimas tanto en su isotipo como en su alotipo, de odo que puedan reconocerse sus diferencias idiotípicas. Con frecuencia se usan para ello proteínas homogéneas (como las de mieloma o anticuerpos monoclonales) inyectadas a un receptor genéticamente idéntico al donante, lo que provoca la producción de anticuerpos anti-idiotipo.
EL RECEPTOR B Durante mucho tiempo la pregunta del millón ha sido el como la mIg (inmunoglobulina de membrana) de las células B puede inducir una señal de activación tras el contacto con el antígeno. El dilema se basa en que los isotipos de mIg tienen tallos citoplásmicos muy cortos:
• mIgM y mIgD tienen solo 3 aa. • mIgA……………………14 aa. • mIgG y mIgE …………..28 aa.
En todos los casos estos tallos son demasiado cortos para asociarse con moléculas de señales intracelulares (p.e., protein-kinasas y Proteínas G). La respuesta a este puzzle es que la mIg no constituye la totalidad del receptor del LB. El complejo receptor B (BCR) es un complejo proteico transmembrana formado por mIg y heterodímeros denominados Igα/Igβ. Las moléculas del heterodímero se asocian con mIg para formar el BCR y, como tienen tallos citoplásmicos largos (65 aa. en la cadena α y 48 aa. en la cadena β) su longitud les permite interaccionar con las moléculas de señales intracelulares. El descubrimiento del heterodímero Igα/Igβ por Michael Reth y col. al comienzo de los 90 hizo posible responder a la pregunta del millón
formulandoalgoqueaportar.blogspot.com

LOS FcR SE UNEN A LOS FRAGMENTOS Fc DE LAS Ig
Son glucoproteínas de membrana presentes en muchas células que tienen afinidad por el fragmento Fc de las moléculas de anticuerpo. Tales receptores son esenciales para muchas de las funciones biológicas de los anticuerpos. Los FcR son los responsables de:
• El movimiento de los anticuerpos a través de las membranas celulares • La transferencia de anticuerpos de la Clase IgG desde la madre al feto a través
de la placenta • La adquisición pasiva de anticuerpos por muchos tipos celulares, lo que da lugar
a la opsonización y a la citotoxicidad dependiente de anticuerpos (ADCC) • La producción de señales inmuno-moduladoras (sobre todo en los LB), que
afectan a la activación celular, a la diferenciación y, en algunos casos, la regulación negativa de las respuestas celulares.
Hay muchos receptores Fc, entre los que se pueden destacar: Poli-Ig; esencial para el transporte de Ig poliméricas (IgA dimérica y, en menor extensión IgM pentamérica) a través de las superficies epiteliales.
• FcNR; el receptor Fc neonatal transfiere IgGs desde la madre hasta el feto, a través de la placenta. También interviene en la regulación de los niveles séricos de Ig.
• FcαR; Fija a la IgA • FcεR; Fija a la IgA • FcγR; Capaces de fijar a IgG y sus subclases. Entre ellos están:
FcγRI FcγRII FcγRIIB1 FcγRIIB2 FcγRIII En muchos casos, el entrecruzamiento de estos receptores tras la unión de los complejos antígeno-anticuerpos (Ag-Acs) provoca la puesta en marcha de las cascadas de señales, que finalizan en procesos tales como la fagocitosis o la ADCC. Los FcR forman parte, con frecuencia, de complejos de transducción de señales en lo que también intervienen otras cadenas accesorias. En el caso de FcαR las cadenas accesorias están representadas por un dímero transmembrana formado por dos cadenas γ y en el caso de PCER las cadenas accesorias son dos γ una β. Esta asociación de un receptor extracelular con una o más unidades transductoras de señales existe en el BCR y también es característica fundamental del TCR.
LAS SUPERFAMILIA (SPF) DE LAS Ig.
Las estructuras tipo Ig comparten varias características, lo que sugiere que tienen un origen evolutivo común. Una propiedad que se presenta sistemáticamente es el plegamiento en dominios; la presencia de estos dominios en las cadenas H y L indica que los genes que los codifican proceden de un gen ancestral común que codificaba un péptido de unos 110 aa. La duplicación génica y divergencias posteriores parecen haber generado los varios genes de cadenas H y L. Se ha comprobado que un gran número de proteínas de membrana tiene una o más regiones homólogas a los dominios de Ig y a estas proteínas de membrana se las
formulandoalgoqueaportar.blogspot.com

clasifica dentro de la SPF de las Ig. Entre las proteínas que forman parte de la SPF están, además de las Ig,: Heterodímeros Igα / Igβ del BCR Receptores poli-Ig que contribuyen con la pieza secretora a la formación de las formas secretadas de IgA e IgM. TCR Proteínas accesorias de células T, que incluyen CD2 CD4 CD8 CD28 Cadenas γ, δ y ε de CD3 Moléculas CMH Clases I y II Β2-microglobulina que forma parte de las moléculas Clase I Moléculas de adhesión celular, tales como: VCAM-1 ICAM-1 ICAM-2 LFA-3 Factor de crecimiento derivado de plaquetas
formulandoalgoqueaportar.blogspot.com

FYTI. Apoyo teórico al Tema 10 (I) El receptor del linfocito B La inmunidad adaptativa depende de los linfocitos B y T. Hay muchos miles de millones de clones distintos y cada uno tiene el receptor para el antígeno de una especificidad única. Este repertorio tan amplio es el que nos permite poner en marcha una respuesta adaptativa adecuada contra cualquiera de los patógenos con los que nos encontremos a lo largo de la vida. Las Inmunoglobulinas (Ig) o anticuerpos (Ac) están formadas por cuatro cadenas peptídicas, iguales dos a dos. Un par es de peso molecular (Pm) alto y se denomina “Cadena pesada (H)” y el otro par tiene menor Pm y se denomina “Cadena ligera (L)” Las cadenas se asocian entre si mediante puentes disulfuro. Cada cadena tiene un fragmento terminal que es distinto en cada molécula y se llama “Variable (V)”, seguido por fragmentos conservados a los que denominamos “Constantes (C)”. Los fragmentos están plegados formando unos bucles a expensas de puentes disulfuro intracatenarios que se establecen entre aminoácidos azufrados (cisteinas) que están separados en la estructura primaria; estas estructuras reciben el nombre de “Dominios”. Por tanto, las Ig están formadas por 2 cadenas H y 2 cadenas L que presentan un dominio terminal V y uno o más dominios C. Las cadenas ligeras pueden ser de dos tipos: κ (kappa) y λ (lambda), pero son funcionalmente iguales. Las cadenas pesadas tienen 3 o 4 dominios C y, según el patrón estructural, se distinguen cinco tipos principales, que son la base de las cinco clases de anticuerpos que presentan los humanos: IgG, IgA, IgM, IgE e IgD. Los dominios variables de la cadena L y la cadena C configuran el sitio de unión para el antígeno. Estos dominios V concentran la variabilidad en tres lugares diferentes (tanto en H como en L), denominados “Regiones determinantes de complementariedad (CDR)” o hipervariables. En cada cadena H y L los CDRs se disponen muy próximos entre si y el conjunto define el sitio de unión para el antígeno o “paratopo”, que es distinto para los receptores de cada linfocito. Las células B y T reconocen al antígeno de forma diferente. Las B, a través de las Ig de membrana, reconocen directamente a los epitopos del antígeno en su configuración nativa y siempre que están en la superficie (accesibles). Por el contrario, los linfocitos T no pueden reconocer alLa exclusión alélica. Los genes delta se localizan en el complejo génico alfa y se eliminan durante el reordenamiento alfa. Esto supone una exclusión irrevocable de los genes δ, localizados en el mismo cromosoma que los α que se están reordenando. La organización de los segmentos génicos J y C de la cadena β en dos grupos duplicados permite que, si se realiza un reordenamiento no productivo, el timocito pueda intentar un segundo reordenamiento con lo que aumenta la probabilidad de reordenamiento productivo de la cadena β. Si el primer reordenamiento es productivo se inhibe el segundo. Las excepciones a la regla de la exclusión alélica son más numerosas en l0s genes de la cadena α. Como la exclusión alélica no completa para la cadena α del TCR hay veces en las que se expresa más de una cadena alfa sobre la membrana de una célula T dada. La pregunta obligada es: ¿Cómo estas células T tan taras, que expresan dos receptores αβ, mantienen la especificidad para unirse a un solo antígeno? Una posible respuesta es que cuando una célula T expresa dos receptores Tαβ diferentes solo uno esté CMH-restringido y sea, por tanto, el único funcional
Página 1 de 2
formulandoalgoqueaportar.blogspot.com

antígeno de forma directa. El TCR solo puede reconocer péptidos derivados de proteínas antigénicas cuando se les “enseña” de una forma especial: esta forma es unidos a moléculas CMH sobre la superficie de otra célula a la que se denomina “Célula presentadora de antígeno (CPA)”. Dicho de otro modo, las células T necesitan a otras células para que les preparen (procesen) y presenten el antígeno. El receptor B (BCR) consiste en una Ig de superficie asociada a un heterodímero formado por las cadenas Igα (CD79a) e Igβ (CD79b). La Ig de superficie es la responsable del reconocimiento antigénico y el heterodímero lo es de la transducción de la señal al interior de la célula. El resultado del reconocimiento y activación es la producción de millones de copias de la Ig de membrana en forma soluble, que se vierten al medio y son los que llamamos “anticuerpos”. El receptor T se llama TCR y está formado por dos cadenas distintas entre si, denominadas α y β, responsables del reconocimiento del péptido antigénico asociado a la molécula CMH. El complejo asociado CD3 es el responsable de translucir la señal de activación al interior de la célula T. El heterodímero αβ se parece a las cadenas H y L de las Ig en que también tienen dominios V (cuyas regiones CDR conforman el sitio de fijación para el antígeno) y C. La generación de los receptores antigénicos se produce antes del ingreso del antígeno durante la maduración de los linfocitos (en la médula ósea para los LB y en timo para los LT). Los mecanismos que generan la gran diversidad de las Ig del BCR son similares a los que intervienen en la formación de la diversidad del TCR. La generación de la diversidad del BCR se debe al reordenamiento del DNA que codifica para los dominios V de las Ig durante el desarrollo de las células B. El dominio V de la cadena L está codificado por dos genes diferentes; el primer gen, denominado V codifica la mayor parte de la porción variable (95-110 aa), mientras que el segundo gen (denominado J) codifica para los restantes 13 aa del dominio V. La asociación de un gen V con uno J produce un exón continuo que codifica todo el dominio V de la cadena L. La unión a los genes de l dominio constante se realiza por procesos de corte y empalme a lo largo de la cadena de DNA.. En el caso de las cadenas H el dominio variable está codificado por tres genes distintos, denominados V, D y J. Igual que en el caso anterior, forman un exón continuo que se asocia a los genes de los dominios constantes mediante procesos de corte y empalme a lo largo de la cadena de DNA.. La generación de la diversidad del repertorio de las Ig se produce mediante los siguientes mecanismos:
• El uso de genes diferentes para el dominio variable por parte de los distintos LB • La asociación de cadenas H y L, cada una de las cuales se reordena de forma
independiente • La unión poco precisa de los segmentos génicos, lo que ocasiona más diversidad
por la adición y sustracción de nucleótidos • La hipermutación somática, que aumenta el repertorio B después del
reconocimiento antigénico. Una vez que se ha reconocido al antígeno, los linfocitos que consiguen activarse proliferan para diferenciarse después a células efectoras. Hay tal diversidad de receptores antigénicos en los linfocitos que cada sujeto es capaz de montar una respuesta adaptativa frente a cualquier microorganismo a lo largo de la vida.
Página 2 de 2
formulandoalgoqueaportar.blogspot.com

TEMA 10 (II) El receptor T (TCR) El receptor T es un hetero-dímero con una estructura de dominios (V y C) muy similar a la del receptor B y también estabilizado mediante un puente disulfuro. Las regiones transmembrana de ambas cadenas son poco corrientes, pues contienen residuos de aminoácidos (aa.) con carga positiva, lo que les permite interaccionar con las cadenas del complejo receptor que se encargan de la transducción de las señales. La mayor parte de las células T humanas expresan receptores codificados por los genes alfa y beta, que reconocen antígenos peptídicos previamente procesados y presentados en condiciones especiales por células especializadas, llamadas Células Presentadoras de Antígenos (CPAs). Siempre hay excepciones Pero datos iniciales indicaban que ciertas células T reaccionaban con antígenos no peptídicos, lo que no empezó a aclarase hasta que se comprobó que las moléculas CD1 (distintas de las CMH) podían presentar antígenos hidrocarbonados y lipídicos a la subpoblación T gamma-delta. Dicho de otro modo: el dogma “las células T solo reconocen a los antígenos cuando se les presentan procesados y unidos a moléculas del complejo principal e histocompatibilidad (CMH)” es válido para las células T αβ pero no parece serlo para otros tipos de células T. Trabajos estructurales más recientes indican que el receptor γδ puede fijar directamente moléculas proteicas (sin restricción CMH), lo que sugiere que este receptor se parece mucho, desde el punto de vista funcional, a los PRRs de la inmunidad innata. El reconocimiento de antígenos comunes a grupos de microorganismos y su capacidad para unirse a moléculas CMH no clásicas sugiere una respuesta rápida, que es más característica de la inmunidad innata que de la adaptativa. El número de células T γδ circulantes es pequeño, comparado con el de células T αβ, y sus segmentos génicos V tienen una diversidad limitada. Muchas de estas células carecen de CD4 y CD8 y la mayoría expresan la misma pareja de cadenas γδ en su receptor. En los humanos, el receptor predominante que se expresa en las células γδ circulantes reconoce un antígeno, que es un fosfolípido microbiano (3-formil-1-butil pirofosfato), propio de mycobacterium tuberculosis y otras bacterias y parásitos. Esta especificidad para antígenos de patógenos frecuentes apoya la hipótesis de que las γδ actúan como una rama de la inmunidad innata, presentando una respuesta rápida a ciertos antígenos sin necesidad de una fase de procesamiento. Es interesante notar que la especificidad de las células γδ circulantes de ratones, y otras especies estudiadas, no es paralela a la humana, lo que indica que esta respuesta debe dirigirse hacia los patógenos comunes a cada una de las distintas especies animales. EL COMPLEJO RECEPTOR T (TCR) El receptor T, de forma similar al B, se asocia con un grupo de proteínas (denominado aquí CD3). El receptor se encarga del reconocimiento del antígeno, mientras que CD3 se encarga de la transducción de las señales que se producen como consecuencia del reconocimiento.
formulandoalgoqueaportar.blogspot.com

CD3 es un complejo de cinco polipéptidos invariantes que se asocian para formar tres dímeros: γε (gamma-epsilon), δε (delta-epsilon) y ζζ (zeta-zeta) o ζη (zeta-teta). Zeta y teta están codificados en el mismo gene, pero difieren en sus extremos C-terminales debido a diferencias en la maduración del trascrito primario de RNA. Aproximadamente el 90% de los CD3 incorporan el homodímero ζζ y el resto el ζη. Se puede concebir un TCR formado por cuatro dímeros: El heterodímero αβ (o el γδ) del receptor, que determina la especificidad para el ligando, y los dímeros que componen CD3 (γε, δε y ζζ o ζη), que son necesarios para que se exprese el receptor en la membrana durante la maduración de la célula T y para la transducción de señales. Las cadenas γ, δ y ε pertenecen a la SPF de las Ig, con un dominio extracelular seguido por una región transmembrana y un dominio citoplásmico de unos 40 aminoácidos. La cadena ζ tiene una estructura diferente, con una región extracelular muy pequeña (solo 9 aa.), una transmembrana y un tallo citoplásmico largo (113 aa.). Las regiones transmembrana de todas las cadenas de CD3 tienen aa. con carga negativa (aspártico o glutámico) que interaccionan con 1-2 aa. con carga positiva del dominio transmembrana del TCR. Los tallos citoplásmicos de CD3 tienen un motivo ITAM (Immunoreceptor tyrosine-based activation motif) , que también se encuentra en otros receptores (p.e. las cadenas asociadas Igα/Igβ en el BCR). Los ITAM interaccionan con tirosin-kinasas y juegan un papel importante en la transducción de señales. En CD3, las cadenas γ, δ y ε tienen un ITAM, mientras que ζ y η contienen tres copias. Las moléculas accesorias de las células T Son moléculas de superficie Algunas de ellas refuerzan la interacción entre las células T y las células presentadoras (CPAs) o las células blanco, mientras que otras actúan en la transducción de señales y otras, finalmente, en ambos sentidos. Los co-receptores CD4 y CD8 se unen a regiones conservadas de las moléculas de histocompatibilidad (CMH) de Clase II y Clase I, respectivamente. Las células T se pueden dividir en dos subpoblaciones en función de que expresen CD4 o CD8. CD4 y CD8 se clasifican como co-receptores porque reconocen a las moléculas CMH que forman parte del complejo de presentación antigénico y por su papel transductor de señales. Sus dominios extracelulares se unen a regiones conservadas de las moléculas CMH sobre las CPAs o sobe las células blanco. CD4 es un glicopéptido de membrana con 4 dominios extracelulares tipo Ig (D1-D4), una región transmembrana hidrofóbica y un largo tallo citoplásmico que contiene tres residuos de serina (que pueden fosforilarse). CD4 interacciona con las moléculas CMH de Clase II a través del dominio distal que se une a una depresión hidrofílica formada a expensas de residuos de los dominios α2 y β2 de la moléculas Clase II. CD4 facilita la transducción de señales y la activación de las células T que reconocen complejos Péptido-Clase II. CD8 se presenta como hetero-dímero (αβ) o como homo-dímero (αα), estabilizados mediante un puente disulfuro. Las cadenas son pequeñas y cada una tiene un dominio
formulandoalgoqueaportar.blogspot.com

tipo Ig extracelular, una región transmembrana hidrofóbica y un tallo citoplásmico de 27-29 residuos, entre los que hay varios capaces de fosforilarse. CD8 contacta con los dominios α2 y α3 de la molécula Clase I (también tiene algún contacto con β2-microglobulina) La afinidad del TCR por los complejos péptido-CMH aumenta gracias a los co-receptores La afinidad entre TCR y el complejo péptido-CMH es de afinidad baja a media (Kd 10-4
a 10-7 M) si se compara con la de la unión antígeno-anticuerpo (Kd 10-6 a 10-10 M). Sin embargo, las interacciones no dependen solo del TCR, pues las moléculas de adhesión aumentan la fuerza de la unión. Las células T tienen varias moléculas de adhesión (CD2, LFA-1, CD28 y CD45R) que se fijan, con independencia de otros ligandos, sobre las moléculas complementarias de las CPAs o de las células blanco. Una vez establecido el contacto célula-célula, a través de las moléculas de adhesión, el receptor T puede explorar la membrana de la otra célula en busca de complejos péptido-CMH. Durante la activación de la célula T, por la detección de un complejo particular péptido-CMH, hay un aumento transitorio de su expresión de moléculas de adhesión que permite un contacto más estrecho entre las células y la transferencia efectiva de citocinas o sustancias tóxicas. Al poco tiempo de la activación, disminuye el nivel de moléculas de adhesión y la célula T se desprende de la CPA o de la célula blanco correspondiente. Como ocurre con CD4 y CD8, algunas moléculas de adhesión también actúan como co-estimuladoras y transductoras de señales.
formulandoalgoqueaportar.blogspot.com

EL COMPLEJO PRINCIPAL DE HISTOCOMPATIBILIDAD (CMH) Al contrario de lo que ocurre con los anticuerpos, o receptores B, que pueden reconocer antígenos directamente, el receptor T solo reconoce antígenos tras su procesamiento (degradación) y presentación en conjunto con moléculas codificadas en el Complejo Principal de Histocompatibilidad (CMH). Este complejo se estudió inicialmente como conjunto de genes que influía en la capacidad de un organismo para aceptar o rechazar los trasplantes de tejidos entre individuos de la misma especie. Los estudios pioneros de P. Doherty, B. Benacerraf y otros, dejaron claro que las moléculas codificadas por el CMH son básicas para la inducción de la respuesta inmune adaptativa así como que las moléculas CMH particulares expresadas por un individuo influyen en el repertorio de antígenos al que pueden responder los linfocitos T. Como el CMH determina como responde un organismo a los agentes infecciosos, está implicado en la susceptibilidad a las enfermedades, así como en el desarrollo de autoinmunidad. El hallazgo reciente de que las células NK expresan receptores para las moléculas Clase I del CMH y el hecho de que la interacción receptor-Clase I puede llevar a la inhibición o activación de estas células expande el papel conocido de esta familia de genes.
ORGANIZACIÓN GENERAL Y HERENCIA DEL CMH
A mediados de los años treinta, tras el trabajo de Peter Gorer, surge el concepto de que el rechazo de tejido extraño es el resultado de la respuesta inmune a moléculas de la superficie celular del tejido extraño, moléculas denominadas ahora antígenos de histocompatibilidad. Gorer uso cepas de ratones endogámicos para identificar antígenos de grupos sanguíneos e identificó cuatro grupos de genes, designados del I a IV, que codificaban los antígenos de células sanguíneas. Los trabajos posteriores de Gorer y George Snell en los cuarenta y los cincuenta establecieron que los antígenos codificados por los genes del grupo II participaban en el rechazo de tejidos trasplantados y tumores.. Snell denominó a estos genes “de histocompatibilidad II”; su designación actual (H-2) hace referencia a los antígenos del grupo sanguíneo II de Gorer. Aunque Gorer murió antes de que estos trabajos se reconocieran plenamente, Snell recibió el Premio Nóbel en 1980 por ellos.. El MCH codifica tres clases principales de moléculas Este complejo es un conjunto de genes dispuestos a lo largo de un segmento continuo y largo del DNA en el cromosoma 6 humano y en el 17 murino. En humanos se llama HLA y en ratones H-2. Aunque la disposición de los genes es algo distinta, en ambos casos se organizan en regiones que codifican tres clases de moléculas:
• Genes de clase I; codifican glucoproteínas que se expresan en la superficie de casi todas las células nucleadas. La función principal de estas proteínas es la presentación de antígenos peptídicos a los linfocitos Tc
• Genes clase II; Codifican glucoproteínas que se expresan sobre todo en células presentadoras e antígenos (macrófagos, células dendríticas y linfocitos B), donde presentan péptidos procesados a los linfocitos Th.
• Genes clase III; Codifican, además de otros productos, varias proteínas con funciones inmunitarias (moléculas del complemento y moléculas relacionadas con la inflamación).
formulandoalgoqueaportar.blogspot.com

Las moléculas clase I, codificadas por los loci A, B y C en humanos, se descubrieron las primeras y se expresan sobre un rango celular amplio; se las llama moléculas Clase I clásicas. En el complejo HLA hay grupos adicionales de genes que también codifican para moléculas clase I; a estos genes se les denomina genes Clase I no clásicos. La expresión de los productos no clásicos se limita a ciertos tipos específicos de células. Aunque no se conocen las funciones de todos estos productos génicos, algunos pueden tener papeles muy específicos en la inmunidad. Un ejemplo es la expresión de las moléculas HLA-G sobre los citotrofoblastos de la interfase maternal-fetal, donde se las ha implicado en la protección del feto para que no se reconozca como extraño (esto puede ocurrir cuando los antígenos paternos empiezan a aparecer) y se rechace por las células T citotóxicas de la madre. Las dos cadenas de las moléculas clase II están codificadas por las regiones DP, DQ y DR en humanos. Como en los loci Clase I, los de Clase II también codifican moléculas adicionales, con funciones específicas en el proceso inmune. Las moléculas Clase I y Clase II tienen caracteres estructurales comunes y ambas participan en el procesamiento y presentación antigénicas. En cambio, la región Clase III, situada en los humanos entre Clase I y Clase II, codifican moléculas críticas para la función inmune, pero que no tienen nada en común con las I ni con las II. Los productos clase III incluyen los componentes del complemento C4, C2 y Factor B y varias citocina pro-inflamatorias, incluyendo TNF. Las formas alélicas de los genes CMH se heredan como grupos de genes unidos, llamados haplotipos Los locus CMH son muy polimórficos, lo que quiere decir que en la población hay muchas copias del gen (alelos). Los genes de los locus CMH se encuentra muy cerca entre si, por lo que la frecuencia de fenómenos de cruzamiento cromosómicos durante la mitosis es bajísima (apenas un 0,5%). Esto hace que casi todas las personas hereden los alelos codificados por estos locus como dos grupos, uno de cada padre. En las poblaciones exogámicas la descendencia suele ser heterocigoto en muchos locus y expresará alelos del CMH tanto maternos como paternos (que se expresan de forma co-dominante, es decir, en la misma célula se expresan productos paternos y maternos). En las cepas endogámicas (alelos idénticos en todos los locus) cada loci es homocigoto, ya que los haplotipos materno y paterno son idénticos, y toda la descendencia expresará haplotipos idénticos. (Las cepas congénicas son las que tienen alelos idénticos en todos los locus, excepto en el del CMH) En una población exogámica cada individuo suele ser heterocigoto en cada locus. El sistema HLA humano es muy polimórfico y hay muchos alelos de cada gen de clase I y II. Sin embargo, los locus están estrechamente unidos y suelen heredarse como haplotipo. Si el padre y la madre tienen haplotipos distintos hay una posibilidad entre cuatro de que la descendencia herede los mismos haplotipos paternos y maternos y, en consecuencia, sea histocompatible con cada uno entre si. Ninguno de la descendencia será histocompatible con los padres. Aunque el índice de recombinación por cruzamiento es bajo, todavía contribuye de forma significativa a la diversidad de los locus en las poblaciones. La recombinación génica genera nuevas combinaciones alélicas y el número de generaciones humanas intermedias desde la aparición de la especie permite una recombinación extensa, por lo que es excepcional que dos personas no relacionadas tengan grupos idénticos de genes HLA.
formulandoalgoqueaportar.blogspot.com

MOLECULAS CMH Y GENES
Estas moléculas (I y II) son glucoproteínas unidas a la membrana (similares en estructura y función) que actúan como moléculas presentadoras de antígenos altamente especializadas, que forman complejos inusualmente estables con ligandos peptídicos, presentándolos sobre la superficie de la célula para que se reconozcan por los linfocitos T . Por el contrario, las moléculas clase III son un grupo de moléculas no relacionadas sin semejanzas estructurales ni funcionales con las moléculas clase I y II. Estructura de las moléculas clase I y II Las moléculas de Clase I constan de una cadena α (45 kDa) que se asocia de forma no covalente a una molécula de β2-microgobulina (12 kDa). La naturaleza de la cadena α es de glucoproteína transmembrana codificada por genes polimórficos en las regiones A. B y C de locus Clase I del HLA. β2-microgobulina es una proteína codificada por un gen muy conservado localizado en un cromosoma diferente. Es necesario que la cadena α se asocie con la β2-microgobulina para que las moléculas Clase I se expresen sobre la membrana. La cadena α se ancla mediante su segmento transmembrana hidrofóbico y su tallo citoplásmico hidrófilo. La cadena α está estructurada en tres dominios externos (α1, α2 y α3) de unos 90 aa, la fracción transmembrana de unos 25 aa hidrofóbicos y el tallo citoplásmico de 30 aa. La β2-microgobulina es similar en tamaño y organización al dominio α3. Los dominios α1y α2 interaccionan para formar una plataforma de ocho plegamientos β anti-paralelos bordeada por dos largas regiones de plegamientos α helicoidales. La estructura forma un surco profundo denominada sitio de unión con el péptido y su longitud le permite unir péptidos de hasta 10 aminoácidos. α3 y β2-microgobulina tienen la estructura de dominio de las Ig. El dominio α3 parece estar muy conservado entre las moléculas Clase I y contiene una secuencia que interacciona con la molécula CD9 presente sobre al superficie de las células Tc. β2-microgobulina interacciona fuertemente con α3 y también con aa de los dominios α1 y α2.; esta interacción es necesaria para el plegamiento correcto de la molécula Clase I. Pero esta unión es poco estable y se estabiliza por la fijación de un péptido, formando la estructura trimérica nativa. Este complejo es el que finalmente se transporta a la superficie celular. Las moléculas Clase II están formadas por dos cadenas (α, de 33kDa y β, de 28 kDa) que se asocian e forma no covalente. Como las moléculas Clase I las clase II con glucoproteínas transmembrana que contienen dominios externos, transmembrana y citoplásmicos; los dominios distales de la membrana (α1 y β1) forman el surco de unión con el péptido. La cristalografía de rayos X muestra una gran similitud de este surco con el de clase I, excepto en que es un surco abierto. Polimorfismo de los sitios de unión con el péptido Polimorfismo de los surcos de fijación En humanos se han identificado varios cientos de variantes alélicas de moléculas Clase I y II. Cualquier individuo, sin embargo solo expresa un pequeño número de ellas (hasta 6 diferentes de clase I y hasta12 diferentes de clase II). Pero este número limitado de moléculas debe ser capaz de presentar una cantidad enorme de péptidos antigénicos a la células T, y permitir al sistema inmune responder de modo específico a una enorme variedad estímulos antigénicos. Esto quiere decir que los péptidos que se unen a los surcos de las moléculas CMH no presentan la especificidad estricta que encontramos en
formulandoalgoqueaportar.blogspot.com

la unión de los antígenos a los anticuerpos o al TCR. En lugar de ello, una moléculas CMH concreta puede fijar a numerosos péptidos diferentes y, además, algunos péptidos pueden unirse a varias moléculas CMH distintas. Debido a esta amplia especificidad la unión entre un péptido y una molécula CMH se denomina, con frecuencia, “promiscua”. Dada la similitud de los surcos en clase I y II no es sorprendente que presenten algunas de las características de fijación de péptidos comunes. En ambos casos las moléculas de péptido ligadas están en una conformación extendida a lo largo del surco. El surco de la moléculas clase I está bloqueado en ambos extremos, mientras que el de la clase II está abierto; el resultado es que las moléculas clase I solo pueden fijar péptidos de 8-10 residuos de aminoácidos, mientras que las de clase II acomodan péptidos de mayor longitud (12-18 aa.). Otra diferencia es que la fijación por clase I requiere que el péptido tenga residuos de aminoácidos específicos cerca de los extremos carboxi y amino-terminales, mientras que este requerimiento no se da para la unión a moléculas clase II. La unión péptido-CMH es muy estable en condiciones fisiológicas (Kd ≈10-6 – 10-10) y, por lo tanto, la mayoría de la moléculas CMH expresadas sobre la membrana de una célula estarán asociadas con un péptido, sea endógeno o exógeno. Interacción Clase I-péptido Las moléculas clase I fijan péptidos y los presentan a las células T CD8+. En general, estos péptidos derivan de proteínas endógenas intracelulares que se degradan en el citosol. Los péptidos se transportan luego desde el citosol hasta las cisternas del retículo endoplásmico (RE), donde interaccionan con las moléculas Clase I. Cada tipo de moléculas Clase I (A, B, C) fija a un único grupo de péptidos. Además, cada variante alélica de de una molécula clase I fija a un grupo distinto de péptidos. Como una célula nucleada expresa unos 105 copias de cada variante alélica de clase I se deben expresar muchos péptidos diferentes simultáneamente sobre la superficie de una célula nucleada por las moléculas Clase I. Los péptidos eluídos de moléculas clase I tienen dos caracteres distintivos: longitud de 8 a 10 residuos de aminoácidos (lo más común es 9) y aa específicos que parecen esenciales para la fijación a una moléculas CMH particulares. La capacidad de una molécula clase I para fijar un grupo diverso de péptidos se debe a la presencia de los mismos o similares aa. en varias posiciones definidas a lo largo del péptido. Como estos residuos son los que anclan el péptido al surco, se llaman “residuos de anclaje”. Las cadenas laterales de los residuos de anclaje son complementarias con los caracteres estructurales de la superficie del surco de fijación de la molécula Clase I. Los residuos de aa que bordean los sitios de fijación varían entre distintas variantes alélicas de clase I y determinan la identidad de los residuos de anclaje que pueden interaccionar con una molécula dada de clase I. Todos los péptidos que se unen a clase I parecen tener un anclaje carboxi-terminal, formado generalmente por residuos hidrofóbicos. También se encuentran otros anclajes en la posición 2 o en la 2 y 3 del extremo amino-terminal. Como norma, cualquier péptido de la longitud adecuada y que tenga los mismo (o similares) residuos de anclaje debe unirse a la misma molécula CMH clase I. (figuras). Interacción Clase II-péptidoLas moléculas clase II fijan péptidos y los presentan a las células T CD4+. Como las moléculas clase I, las clase II también fijan a una variedad de péptidos que, en general, derivan de proteínas exógenas (tanto propias como no propias) que se degradan en la vía endocitica de procesamiento. La mayoría de los péptidos asociados a moléculas clase II
formulandoalgoqueaportar.blogspot.com

derivan de proteínas propias ancladas a la membrana o de proteínas extrañas interiorizadas por fagocitosis o por endocitosis mediada por receptor y luego procesadas a través de la vía endocítica. Por ejemplo, los péptidos derivados de la digestión de las moléculas clase I ancladas a la membrana se encuentran con frecuencia unidos a moléculas clase II. Los péptidos aislados de las moléculas clase II van de 13 a 18 aminoácidos (más largos que los de unión a clase I) pero como el surco de fijación está abierto por ambos lados los péptidos pueden sobresalir. Los péptidos unidos al surco de las moléculas clase II mantienen una elevación más o menos constante sobre el suelo del surco, lo que también distingue la fijación entre clase I (elevación brusca, mayor o menor en función de la longitud dl péptido) y clase II (elevación suave constante). Es el núcleo central de 13 aa. es el que determina si el péptido se une o no a la moléculas clase II. Los péptidos que se unen a la misma molécula tienen una secuencia interna conservada (motivo) y, a diferencia de lo que ocurre con los péptidos que se unen a las moléculas clase I, no tiene residuos de anclaje conservados sino que se establecen puentes de hidrógeno entre la “columna vertebral” del péptido y los extremos del surco. Los péptidos que se unen presentan una secuencia interna de 7-10 aa que es la que aporta la mayoría de puntos de contacto. Normalmente esta secuencia tiene un aa aromático o residuos hidrófobos en su extremo amino-terminal y tres residuos hidrofóbicos adicionales en las porciones media y carboxi-terminal del péptido. Además, aproximadamente el 30% de los péptidos eluídos de moléculas clase II tienen un residuo de prolina en posición 2 y un grupo de prolinas en la porción carboxi-terminal Diversidad del HLA en la especie y en los individuos Hay una diversidad enorme en ambos casos, que recuerda la de anticuerpos y TCRs, aunque la causa no es la misma. La diversidad de anticuerpos y receptores T se forma por procesos somáticos (recombinación génica y mutaciones somáticas), por lo que la formación de estos receptores es dinámica y varía con el tiempo en el individuo. Por el contrario, las moléculas CMH que expresa un individuo está fijas en sus genes y no hay cambios con el tiempo. La diversidad del CMH en una especie brota del polimorfismo (la presencia de múltiples alelos en un locus dado, en la especie). En cuanto a la diversidad del CMH en un individuo es el resultado no solo de tener alelos diferentes de cada gen, sino también de la presencia de genes duplicados (somos diploides) con funciones similares o que se solapan. El CMH contiene un gran número de alelos diferentes en cada locus y es uno de los complejos genéticos más polimórficos de los vertebrados superiores. Los alelos difieren en la secuencia de DNA del alelo, de un individuo a otro, entre el 5 y el 10%. La diferencia a nivel de aminoácidos puede ser muy significativa, ya que hay hasta 20 aa que contribuyen a la naturaleza estructural única de cada alelo. (Cuadro de alelos). Desequilibrio de uniónLos cálculos astronómicos anteriores del número de haplotipos asumen combinaciones de alelos totalmente al azar. Pero se sabe que la diversidad actual es menor, ya que hay combinaciones de alelos en los haplotipos son más frecuentes que la frecuencia que predice la combinación al azar, fenómeno que se conoce como desequilibrio de unión. Este desequilibrio es la diferencia entre la frecuencia observada de una combinación de alelos determinada y de la frecuencia esperada de los alelos particulares. La frecuencia esperada. La frecuencia esperada de la combinación puede calcularse multiplicando la
formulandoalgoqueaportar.blogspot.com

frecuencia de los dos alelos; p.e., si HLAA1 está presente en el 16% en la población (frecuencia = 0,16) y HLAB8 en un 9% (frecuencia = 0,09) se espera que alrededor del 1,4% del grupo tengan ambos alelos (0,16 x 0,09 = 0,014). Sin embargo, los datos muestran que esta asociación está presente en el 8,8% de los individuos estudiados. Esta cifra es la mediad del desequilibrio de unión entre estos dos alelos de Clase I. La causa puede ser que todavía han trascurrido pocas generaciones como para que los cruzamientos permitan la distribución homogénea de todos los alelos e la población fundadora, Otra teoría es evolutiva; se basa en que ciertas combinaciones sobre-representadas en la población podrían aportar resistencia a ciertas enfermedades por lo que se seleccionarían y distribuirían entre la descendencia. Por el contrario, si la asociación induce susceptibilidad a enfermedades autoinmunes irá despareciendo y estará sub-representada en la población actual. Otra hipótesis indica que las fusiones (crossovers) son más frecuentes en ciertas regiones de la secuencia del DNA y la presencia o ausencia de regiones propicias a la fusión (“hot points”) entre alelos puede dictar a frecuencia de la asociación alélica. Hay datos experimentales en apoyo de esta última hipótesis y parece que deben existir “puntos calientes” de recombinación que influyan en le desequilibrio de unión en las poblaciones. A pesar del desequilibrio de unión, todavía existe u enorme polimorfismo en el CMH humano. Este se ha originado por procesos de recombinación, mutaciones puntuales y conversión génica, todos los cuales contribuyen a la diversidad de los genes CMH en la población. Esta diversidad entre locus CMH de los diferentes individuos (haplotipos) es el principal obstáculo para los trasplantes.
MAPA GENÓMICO DETALLADO DEL HLA El CMH ocupa unas 4.000 kb del DNA humano. El proyecto genoma muestra que la región rebosa de genes, muchos de ellos de función conocida (figura). La clase I se extiende unas 2000 kb en el extremo telomérico del complejo HLA Contiene unos 20 genes, entre los que se incluyen los de las moléculas clásicas (A, B y C); también hay muchos genes no clásicos, incluyendo HLA-E, HLA-F, HLA-G, HFE, HLA-J y HLA-X, además de una familia de genes de reciente descubrimiento llamada MIC, que incluye desde MICA hasta MICE. Algunos de los genes no clásicos son pseudogenes (no codifican productos proteicos) pero otros, como HLA-G y HFE, codifican productos tipo clase I con funciones muy especializadas. La familia MIC tiene solo un 15-30% de identidad de secuencia con las Clase I clásicas y MICA es muy polimórfica. Los productos de los genes MIC se expresan a niveles bajos en las células epiteliales y se inducen por calor u otros estímulos que influyen en la producción de proteínas de choque térmico. La función de las moléculas clase I no clásicas no se conoce mucho, aunque hay trabajos que sugieren que algunas de estas moléculas, tal como hacen las clásicas, pueden presentar péptidos a los LT. Hay datos que apuntan a una posible función presentadora de péptidos, procedentes de procariotas intracelulares. Hay una molécula no clásica murina capaz de presentar péptidos con formal-metionina en el extremo amino-terminal. Ste tipo de péptido es raro en eucariotas y frecuente en procariotas, por lo que es posible que la tal molécula no clásica se dedique a presentar péptidos de microorganismos intracelulares. Lysteria monocytogene es uno de esos organismos y hay moléculas de clase I murinas que presentan péptidos con las características anteriores procedentes de este organismo.
formulandoalgoqueaportar.blogspot.com

La clase II está en el extremo centromérico del HLA Contiene genes que codifican cadenas alfa y cadenas beta de las moléculas Clase II. La región está dividida en tres subregiones: DP, DQ y DR. Hay múltiples genes de cadena beta en varias regiones; todos los productos beta pueden expresarse junto con los productos de cadena alfa en una célula dada, lo que aumenta el número de moléculas presentadoras de antígenos diferentes sobre la célula
Nº de genes
Subregión
Región
3-4*13222
DRBDRADQBDQADPBDPA
DRDQDP
GENES HLA Clase II
* Hay más genes, pero solo son funcionales 3-4
También en esta región se han identificado genes no clásicos, como DM y DO, así como LMP, LMP7, TAP1 y TAP2. DM codifica un producto tipo clase II (αβ) que facilita la carga de péptidos antigénicos en las moléculas II clásicas. DO (también αβ) solo se expresa en el timo y en los LB maduros y actúa como un regulador del procesamiento (parece inhibir a DM). La clase III está entre la Clase I y la Clase II Estos genes codifican para:
• Componentes del complemento, como C2, C4a, Cab de la vía clásica y el Factor B de la vía alternativa.
• Esteroide 21-hidrolasas. Hay dos genes (CYP21 y CYP21P) • Proteínas de choque térmico. 2 genes (HSP) • Citocinas: 2 genes (TNF-α y TNF-β)
Algunos d los productos de estos genes juegan un papel en ciertas enfermedades como, por ejemplo, las mutaciones en los genes CYP (que codifican esteroide 21-hidrolasas) parece que están ligados a la hiperplasia adrenal congénita. La presencia de este grupo de genes unidos se conserva en todas las especies que tienen región CMH.
EXPRESION CELULAR DE LAS MOLECULAS HLA En general, las moléculas Clase I clásicas se expresan sobre todas las células nucleadas, pero el nivel de expresión difiere entre los distintos tipos celulares. Los niveles más altos los expresan los linfocitos (representan el 1% de las proteínas de membrana, lo que equivale a 4 x105 moléculas por célula). El ejemplo contrario son los fibroblastos, hepatocitos y células neurales que expresan niveles muy bajos de estas moléculas. Este bajo nivel de moléculas clase I en las células del hígado contribuye de forma considerable al éxito de los trasplantes de hígado, reduciendo la probabilidad de rechazo
formulandoalgoqueaportar.blogspot.com

por las células Tc del receptor. Unas pocas células (neuronas y células del esperma en ciertos estados de diferenciación) carecen totalmente de moléculas Clase I. Como ya se expuso, cualquier molécula HLA individual puede fijar muchos péptidos diferentes. Como los alelos se expresan de forma codominante, los heterocigóticos expresan en sus células los productos génicos codificados por ambos alelos de cada locus HLA. Se expresan 6 alelos de Clase I (2 A, 2B y 2C procedentes de cada uno de los progenitores) sobre la membrana de las células nucleadas. La expresión de tantos alelos clase I permite a cada célula individual presentar una gran cantidad de péptidos. En las células sanas, normales, las moléculas clase I deben presentar auto-péptidos, procedentes del metabolismo normal de las proteínas. En las células infectadas por virus, los péptidos virales (así como los auto-péptidos) se presentan sobre la membrana. Una simple célula infectada por virus hay que imaginarla con varios tipos de moléculas Clase I sobre su membrana, cada una conteniendo y presentado distintos grupos de péptidos virales. Como la diferencias alélicas están en los surco de fijación, los distintos individuos de una especie deben tener la capacidad de presentar distintos grupos de péptidos procedentes de cualquier virus. Por el contrario, las moléculas Clase II se expresan constitutivamente solo sobre las células presentadoras de antígenos (CPAs) como macrófagos, células dendríticas maduras y linfocitos B; las células epiteliales tímicas y algunos otros tipos celulares pueden, en ciertas condiciones y estimuladas por citocinas, expresar moléculas clase II y funcionar como células presentadoras. En algunos casos, la expresión de clase II depende del estado de diferenciación de las células; p.e., no pueden detectarse moléculas clase II sobre las células pre-B, pero se expresan constitutivamente sobre la membrana de las células B maduras. También monocitos y macrófagos expresan niveles bajos de moléculas clase II hasta que se activan tras interaccionar con un antígeno, tras lo que aumenta significativamente el nivel de expresión. Como cada moléculas cae II clásica está formada por dos cadenas diferentes, codificada en un loci distinto, un individuo heterocigótico expresa no solo las moléculas clase II parentales, sino también cadenas alfa y beta procedentes de distintos cromosomas; no hay ninguna restricción sobre el origen genético de las parejas de cadenas α y β, por lo que pueden expresarse conjuntas. Como el HLA contiene tres genes Clase II (DP, DQ y DR) un individuo heterocigótico expresa seis moléculas parentales de clase II y otras seis que contienen combinaciones de cadenas α y β de cada progenitor. El número de moléculas de clase II expresadas por un individuo se aumenta, además, por la presencia de de múltiples cadenas β y también α. La diversidad que aportan estos mecanismos presumiblemente aumenta el número de péptidos antigénicos distintos que se pueden presentar, lo que es ventajoso para el organismo
REGULACION DE LA EXPRESION DEL CMH El estudio de los mecanismos que controlan la expresión del CMH en las distintas células todavía está en la fase inicial. Tanto los genes de clase I como los de clase II están flaqueados por secuencias 5promotoras, que fijan secuencias específicas de los factores de trascripción. La regulación transcripcional está controlada por elementos activadores e inhibidores. La expresión también está regulada por varias citocinas: Los IFNs (α, β y γ) y el TNF-α aumentan la expresión de clase I; el IFN-γ, por ejemplo, parece inducir la formación de un factor de transmisión específico que se une a las secuencias promotoras que flaquean a los genes de Clase I. Como consecuencia de esta
formulandoalgoqueaportar.blogspot.com

unión parece ser la expresión coordinada de los genes que codifican la cadena α de la molécula clase I, el de la β2-microgobulina y los de otras proteínas que intervienen en el procesamiento y presentación de los antígenos. IFN-γ también induce la expresión del factor trascripcional de las molécula Clase II, CIITA (“Class II Transcription Activation”) por lo que, indirectamente, aumenta la expresión de las moléculas clase I sobre una serie de células (CPAs, queratinocitos de la piel, células epiteliales intestinales, células del epitelio vascular, células de la placenta y células beta del páncreas) Otras citocinas influyen en la expresión de las moléculas CMH solo en ciertas células como, por ejemplo, IL-4 activa la expresión de moléculas clase II sobre los linfocitos B en reposo; en cambio, el IFN-γ, corticoides y prostaglandinas regulan negativamente tal expresión. La expresión de moléculas CMH sobre las superficies celulares disminuye por la infección con ciertos virus, como CMV (citomegalovirus), VHB (Virus de la hepatitis B) y adenovirus. En algunos casos la expresión reducida de clase I se debe a la disminución del nivel de los componentes que se necesitan para el transporte de péptidos o moléculas HLA, más que a una disminución de la trascripción. En la infección por CMV, por ejemplo, una proteína viral fija a β2-microglobulina, impidiendo el ensamblaje de las moléculas de clase I y su transporte a la membrana plasmática. En la infección por adenovirus 12 hay una disminución de la transcripción de los genes de transporte TAP1 y TAP2. El bloqueo de la expresión de TAP impide el paso de los péptidos al retículo endoplásmico; el resultado es que la moléculas Clase I no pueden unir al péptido ni la cadena alfa queda ensamblada con β2-microglobulina, con lo que las moléculas clase I no se expresan sobre la membrana. La disminución de la expresión de clase I, sea por el mecanismo que sea, debe ayudar a los virus a evadir la respuesta inmune disminuyendo la probabilidad de que la célula infectada pueda presentar complejos péptido viral-Clase I, que la hacen blanco adecuado para los LTc.
CMH Y SUSCEPTIBILIDAD A LA ENFERMEDAD Algunos Alelos se presentan con más frecuencia en los individuos que sufre una enfermedad que en la población general. Estas enfermedades asociadas al HLA incluyen procesos autoinmunes, ciertas enfermedades virales, enfermedades por alteraciones del sistema del complemento, algunas enfermedades neurológicas y varias alergias diferentes. La asociación enfermedad-alelo puede cuantificarse determinando la frecuencia con la que aparece el alelo en los enfermos de esa patología y la frecuencia con la que aparece dicho alelo en la población general. Esta comparación permite calcular el riesgo relativo (RR): RR = [ (Ag+/Ag-) en el grupo de enfermos] / [(Ag+/Ag-) en el grupo control] El RR = 1 implica que el alelo se expresa con la misma frecuencia en los pacientes que en la población general, lo que supone que tener el alelo no aumenta el riesgo de padecer la enfermedad. Un valor sustancialmente >1 indica una asociación entre alelo HLA y enfermedad. Por ejemplo, los individuos HLAB-27+ son 90 veces más susceptibles a la espondilitis anquilosante que los que carecen de este alelo; esta enfermedad se trata de un proceso inflamatorio de las articulaciones vertebrales que lleva a la destrucción el cartílago. Otra asociación con RR importante es HLA-DR2-narcolepsia
formulandoalgoqueaportar.blogspot.com

La existencia de una asociación alelo-enfermedad no implica que el alelo sea el causante de la enfermedad. En el caso de espondilitis anquilosante, la proximidad de los genes de TNF con el locus HLA-B hace probable que la expresión de tales citocinas intervenga en la destrucción del cartílago. Cuando la asociación es débil (RR bajo) es probable que sean varios genes los que influyan en la susceptibilidad, uno de los cuales pertenece al CMH. Se ha estudiado en profundidad el origen genético de procesos autoinmunes tales como esclerosis múltiple (asociada a DR2, con RR = 5) y artritis reumatoide (asociada a DR4, con RR = 10). Que estas enfermedades no se heredan por simple segregación Mendeliana de los alelos HLA puede verse en los gemelos: ambos heredan el factor de riesgo, pero no es normal que ambos desarrollen la enfermedad. Todo lo anterior indica que múltiples genes y factores ambientales intervienen en el desarrollo de la enfermedad, especialmente si es autoinmune, donde el papel de los genes HLA es importante, pero no exclusivo. Una dificultad adicional para asociar un gen HLA a una enfermedad es el fenómeno genético del desequilibrio de unión y el hecho de que algunos alelos de clase I estén en desequilibrio de unión con alelos clase II hace que su contribución a la susceptibilidad a la enfermedad parezca mayor de lo que es en realidad. Por ejemplo, si DR4 contribuye al RR de una enfermedad y está asociado con frecuencia a A3 (por desequilibrio de unión) puede asociarse incorrectamente a A3 como con la enfermedad. ¿Cómo interviene el HLA en la susceptibilidad a la enfermedad? Entre las hipótesis que se barajan están:
• la distinta habilidad de los diferentes alelos para presentar antígenos procesados a las células T.
• que los alelos codifiquen moléculas que se reconozcan como receptores por virus o toxinas bacterianas.
Hay datos que indican que la reducción el polimorfismo CMH en una especie la predispone a las enfermedades infecciosas; guepardos y otros felinos de vida libre, tales como las panteras de Florida, son muy susceptibles a las enfermedades por virus debido a una capacidad reproductora baja que se traduce en una pérdida de diversidad CMH. El aumento de la susceptibilidad de los guepardos a ciertos procesos virales puede ser el resultado de una reducción en el número de distintas moléculas CMH disponible por la especie y la correspondiente limitación en el rango de antígenos procesados con los que pueden interaccionar esas moléculas. Por tanto, el amplio polimorfismo que se observa en varias especies les aporta la ventaja de tener una gran reserva de moléculas presentadoras de antígenos. Aunque un simple individuo de la especie probablemente no sea capaz de desarrollar la respuesta inmune frente a un patógeno dado (por consiguiente es susceptible a la infección), el gran polimorfismo asegura que al menos algunos miembros de esa especie será capaz de responder y será resistente. De esta forma, la diversidad CMH parece proteger a una especie frente a un gran número de enfermedades infecciosas.
formulandoalgoqueaportar.blogspot.com

CMH Y RESPUESTA INMUNE La capacidad de un animal para montar una respuesta inmune (valorada mediante la producción de anticuerpos) depende de su haplotipo CMH y, más concretamente, de la pesentación antigénica controlada por los genes de clase II. Hay dos posibles explicaciones para la diferencia de respuesta entre distinto haplotipos. De acuerdo con el modelo de selección de determinantes las distintas moléculas de clase II difieren en su capacidad para fijar antígenos procesados. El modelo alternativo de “agujeros en el repertorio” defiende que aquellas células T con receptores que reconocen antígeno extraños muy parecidos a autoantígenos se eliminan durante la selección tímica. Como en la respuesta T a un antígeno intervienen TCR, péptido antigénico y moléculas Clase II, ambos modelos pueden ser correctos.
RESTRICCION CMH
Una vez establecida la relación entre CMH y respuesta inmune se comprobó que las células T CD4+ y CD8+ solo pueden reconocer a un antígeno cuando se le presenta con una molécula propia del CMH, una cualidad conocida como restricción CMH. Experiencias posteriores mostraron que la restricción se ejerce por las moléculas Clase II frente a las células T CD4+ y por las moléculas Clase II frente a las células T CD8+. Los descubridores de estas restricciones (Doherty y Zinkernagel) recibieron el Premio Nóbel en 1996 por su “contribución al conocimiento del papel del CMH en la inmunidad mediada por células”.
PAPEL DE LAS CELULAS PRESENTADORAS DE ANTIGENOS (CPAs) Ya en 1959 los inmunólogos tuvieron que enfrentarse con la dura realidad: los datos experimentales indicaban que células T y células B reconocían a los antígenos mediante mecanismos diferentes. El dogma en aquellos tiempos (que persistió hasta los 80) era que las células del sistema inmune reconocían a la proteína entera y en su conformación nativa. El procesamiento es imprescindible para el reconocimiento por las células T El dogma “el reconocimiento antigénico por las células b y T es básicamente similar” se viene abajo con los trabajos de K. Ziegler y E.R. Unanue, que comprobaron que la activación de las células Th (CD4+) por antígenos proteicos bacterianos no se produce si se trata a las CPAs con p-formaldehído antes de la exposición al antígeno. Pero si se fijaban con p-formaldehido entre 1 y 3 horas después de añadir el antígeno ya lo habían procesado y presentado sobre la membrana en una forma capaz de activar a las células T. Otros trabajos indican que no hacen falta proteínas nativas, y que la activación puede conseguirse con péptidos procedentes de las proteínas nativas, finalmente se concluye que el procesamiento es un proceso metabólico que degrada las proteínas a péptidos que entonces se pueden presentar sobre la membrana celular junto con moléculas CMH La mayoría de las células pueden presentar antígenos con clase I, pero la presentación con clase II está restringida a las CPAs Como todas las células que expresan moléculas CMH pueden presentar péptidos a las células T por lo que, en un sentido estricto, todas son células presentadoras. Sin embargo, se denominan CPAs a las que expresan de forma constitutiva moléculas clase
formulandoalgoqueaportar.blogspot.com

II (que presentan péptidos y activan a las células T CD4+), mientras que las que los presentan con clase I a las T CD8+ (Tc) se denominan “células diana o células blanco”. Hay una serie de células que pueden actuar como CPAs; el carácter que las distingue es su capacidad de expresar moléculas clase II y producir señales coestimuladoras. Células dendríticas (DCs), macrófagos (MCFs) y células B (LB) actúan como presentadoras, aunque difieren entre ellas en los mecanismos de ingestión de antígenos, en la expresión constitutiva de las moléculas clase II y en la actividad coestimuladora.
• Las DCs son las más efectivas como CPAs. Expresan constitutivamente niveles altos de clase II y tienen acción coestimuladora, por lo que pueden activar a los linfocitos Th vírgenes.
• Los MCFs tienen que activarse fagocitando antigenos antes de expresar las moléculas clase II y las moléculas coestimuladoras de membrana.
• Los LB expresan constitutivamente moléculas clase II, pero deben activarse para expresar las moléculas coestimuladoras.
Hay otras células, clasificadas como “CPAs no profesionales” en las que se puede inducir la expresión de clase II o de señales coestimuladoras. La mayoría solo presentan antígenos por periodos cortos, durante una respuesta inflamatoria mantenida.
CELULAS PRESENTADORAS DE ANTIGENOS PROFESIONALES NO PROFESIONALES
DCs MCFs
LB
Fibroblastos (piel) Células gliales (SNC) Células β del páncreas
Células epiteliales (timo) Células epiteliales (tiroides) Células endoteliales (vasos)
Como las células nucleadas expresan moléculas clase I (que presentan antígenos endógenos) suelen ser dianas apropiadas para las células Tc (citotóxicas). La mayoría son células infectadas por virus u otros patógenos intracelulares, pero también pueden serlo células cancerosas, de viejos y procedentes de trasplantes.
EVIDENCIAS DE QUE HAY DOS VIAS DE PROCESAMIENTO El sistema inmune usa vías diferentes para eliminar a los antígenos intra y extracelulares Como norma general, los antígenos endógenos (producidos por la célula) se procesan por la vía citosólica y se presentan sobre la membrana con moléculas clase II. En cambio, los exógenos (capturados del exterior por endocitosis) se procesan por la vía endocítica (también conocida como vía vesicular) y se presentan sobre la membrana con moléculas clase II. Esto se pudo comprobar experimentalmente mediante el uso alternativo de inhibidores de la síntesis proteica (inhiben la presentación con clase I) y de cloroquina (sustancia anti-palúdica que inhibe la presentación con clase II). Estos y otros datos llevan a la conclusión de que los péptidos presentados por ambas clases de moléculas CMH discurren por compartimentos celulares distintos y separados.
ANTIGENOS ENDOGENOS: LA VIA CITOSOLICA Los niveles de proteínas están estrechamente regulados en las células eucariotas y la velocidad de degradación se expresa como vida media (Vm); algunas proteínas normales (p.e. ciclinas, etc.,) y las anormales (desnaturalizadas, mal plegadas, etc.,) se
formulandoalgoqueaportar.blogspot.com

degradan con rapidez. Los péptidos mal sintetizados (o derivados defectuosos de ribosomas [DRiPs]) son el núcleo principal de los productos que se degradan con rapidez. La Vm de las proteínas celulares es de unos dos días, pero muchas se degradan en 10 minutos y la consecuencia de esta renovación constante (tanto de proteínas normales como defectuosas) es una avalancha de productos de degradación en las células. La mayoría se degradan hasta el nivel de aminoácidos, peo otros persisten en el citosol como péptidos a los que el sistema inmune chequea para determinar si se deben presentar sobre la membrana, asociados a moléculas clase I, o no. Los péptidos destinados a la presentación se generan en complejos de proteasas llamados PROTEASOMAS. Las proteínas intracelulares se degradan a pequeños péptidos mediante un sistema citosólico, llamado proteasoma, presente en todas las células, formado por 14 subunidades (algunas con actividad proteasa y otras no) agrupadas en forma de cilindro de anillos simétricos, con un tamaño 20S; en cada extremo hay unos canales por donde entran las proteínas. Muchas de las proteínas destinadas a la proteolisis tienen acoplada a un extremo otra pequeña proteína llamada ubiquitina. Estos conjugados U-P pueden degradarse en otro tipo de proteasoma, formado por la unidad 20S más otra unidad reguladora 19S. La estructura resultante (26S) rompe los enlaces peptídicos en forma ATP-dependiente. Además del proteasoma estándar, presente en todas las células, hay otro del mismo tamaño, pero con algunas subunidades diferentes, inducido por IFN-γ y TNF-α en las células con actividad inmune y denominado inmunoproteasoma. Su presencia en las células infectadas por virus indica un papel en el procesamiento de las proteínas virales para su presentación junto con moléculas clase I. El inmunoproteasoma degrada más rápido que el proteasoma normal y es posible que este mayor nivel de degradación tenga otras consecuencias que el “marcado” de las células infectadas por virus. Es posible que la autoinmunidad sea consecuencia del aumento del nivel de procesamiento de proteínas propias, en células con niveles altos de inmunoproteasomas. Los péptidos pasan desde el citosol al retículo endoplásmico rugoso (RER) Hay una línea celular (RMA-S) que solo expresa sobre la membrana el 5% de la tasa normal de moléculas Clase I, a pesar de sintetizar cantidades normales de la cadena α.Towsend y col. Comprobaron que “alimentando” a estas células con péptidos se restauraba el nivel de expresión sobre la membrana, e interpretaron que los péptidos son necesarios para estabilizar la unión entre cadenas α y β2-microglobulina. La restauración del número de moléculas clase I sobre la membrana indica que esta línea celular debe tener un defecto en el transporte de péptidos. Experiencias posteriores indican que el defecto está en las proteínas que transportan los péptidos desde el citosol hasta el RER, donde se sintetizan las moléculas clase I. Ciando estas células se transfectan con los genes que codifican las proteínas transportadoras expresan cantidades normales de moléculas clase I sobre la membrana. La proteína transportadora, denominada TAP (Transporters Associated with antigen Processing) , es un heterodímero formado por dos proteínas (TAP-1 y TAP-2), cuyo fragmento transmembrana atraviesa esta repetidamente: Además de sus múltiples segmentos transmembrana, las proteínas TAP tienen un dominio que se proyecta en el lumen del RER y un dominio fijador que se proyecta en el citosol.. TAP 1 y 2 pertenecen a la familia de proteínas “casette” fijadoras de ATP que se encuentran en la membrana de
formulandoalgoqueaportar.blogspot.com

muchas células m incluidas bacterias. Estas proteínas median el transporte de aminoácidos, azúcares, iones y péptidos. Los péptidos generados en el citosol por el proteasoma se transportan por las proteínas TAP al RER, en un proceso que requiere la hidrólisis de ATP. TAP fija preferentemente péptidos de 8-16 aminoácidos, que se acercan al tamaño ideal para la fijación a las moléculas clase I. Pregunta ¿Cómo se consigue este tamaño? Respuesta: gracias a las amino-peptidasas presentes en el RER, que recortan los péptidos trasportados. Además, las moléculas TAP muestran preferencia por los péptidos con aminoácidos carboxi-terminales hidrófobos o básicos; Es decir, TAP está optimizada para transportar los péptidos que pueden interaccionar con las moléculas clase I. Los genes de TAP 1-2 están en el locus clase II del CMH (adyacentes a los que codifican para subunidades del proteasoma) y tienen distintos alelos distribuidos en la población. La deficiencia en TAp puede llevar a síndromes patológicos con aspectos de inmunodeficiencia y autoinmunidad. Los péptidos se asocian a las moléculas clase I ayudados por chaperones. Como otras proteínas, la cadena α y la β2-microglobulina se sintetizan en los polisomas del RER. La asociación de estos componentes en un complejo estable (que puede salir del RER) necesita que el péptido se una al surco de fijación. Esta unión puede tener varios pasos e incluye la participación de chaperones moleculares que facilitan el plegamiento de los polipéptidos. El primer chaperón que interviene se denomina calnexina (proteína residente en la membrana del RER) que se asocia a la cadena α y promueve su plegamiento y se separa tras la unión de β2-microglobulina a la cadena α. Ahora la molécula clase I se asocia con los chaperones calreticulina y tapasina. La tapasina arrastra al transportador TAP a las proximidades de la molécula recién formada facilitando la unión al péptido antigénico. Hay una proteína adicional con función enzimática, ERp57 que se une al complejo anterior, lo estabiliza y permite la liberación del complejo trimolecular (cadena α, β2-microglobulina-péptido).que pasa a la superficie celular vía complejo de Golgi En el RER las endopeptidasas actúan sobre los péptidos que no se han asociado y los degradan
ANTIGENOS EXOGENOS: LA VIA ENDOCITICA DCs y MCFs pueden fagocitar y endocitar, mientras que el resto de las CPAs no fagocitan y solo internalizan los antígenos por endocitosis (tanto mediada por receptor como por pinocitosis). LA células B internalizan a los antígenos muy eficientemente por endocitosis mediada por receptor, usando el BCR para capturar específicamente a los antígenos. Los péptidos se generan en las vesículas endocíticas Una vez internalizado el antígeno se degrada a péptidos en los compartimentos de la vía endocítica de procesamiento. El Ag tarda 1-3 horas en atravesar el compartimento y aparecer sobre la membrana en forma de complejo péptido-Clase II. Parece que hay tres compartimentos que intervienen en la vía endocítica:
• Endosomas tempranos, con un pH 6,0-6,5 • Endosomas tardíos (también llamados endolisosomas) con un pH 5,0-6,0
formulandoalgoqueaportar.blogspot.com

• Lisosomas, con un pH de 4,5-5,0 Los antígenos se mueven de los endosomas tempranos a los tardíos y, finalmente, a los lisosomas; durante el trayecto encuentran en cada compartimento enzimas hidrolíticas y menor pH. Los lisosomas, p. e., tienen más de 40 hidrolasas ácido-dependientes. En estos compartimentos los antígenos se degradan a oligo-péptidos de 13-18 aminoácidos que se unen a las moléculas de clase II y así quedan protegidos de una degradación hidrolítica posterior. Como las enzimas solo son activas a pH acido puede inhibirse el procesamiento mediante sustancias que eleven el ph de los compartimentos (p.e., con el antimalárico cloroquina) así como por inhibidores de proteasas. El mecanismo mediante el que los antígenos atraviesan el compartimento endocítico no está plenamente demostrado;
• puede que los endosomas tempranos de la periferia se muevan hacia el interior para volverse endosomas tardíos y, finalmente, lisosomas
• otra posibilidad es que los antígenos pasen de una zona del compartimento a la siguiente vehiculados en pequeñas vesículas de transporte.
En un momento dado, los compartimentos endocíticos (o parte de ellos) vuelven a la superficie, donde se fusionan con la membrana plasmática. Este es el modo en el que se reciclan los receptores de superficie. La cadena invariante dirige el transporte de la molécula clase II a las vesículas endocíticas Como las CPAs expresan clase I y clase II tiene que haber algún mecanismo que impida a las moléculas clase II unirse a los péptidos que se unen a las moléculas clase I. Cuando las cadenas de la molécula clase II se sintetizan y caen al RER se asocian con una proteína llamada cadena invariante (CD74) que interacciona con el surco de fijación e impide que los péptidos de la vía endógena se unan mientras permanece en el retículo. La cadena invariante también interviene en el plegamiento de las cadenas de la molécula clase II, en su salida del RER y en la elección de la ruta que lleva a la vía endocítica desde el complejo de Golgi. Las experiencias han demostrado el papel de la cadena invariante en la dirección del movimiento de las moléculas clase II. Esta cadena contiene “señales de salida” en su tallo citoplásmico que dirigen el transporte del complejo desde la red trans-Golgi hasta el compartimento endocítico. Los péptidos desplazan a los resto de la cadena invariante y se unen a la molécula Clase II Los complejos cadena invariante-Clase II se mueven desde los endosomas tempranos, a través de los tardíos, hasta los lisosomas. Como la actividad proteolítica aumenta en cada compartimento sucesivo, la cadena invariante se degrada de modo progresivo hasta que solo queda un fragmento de la cadena, llamado CLIP (“Class II associated Invariant chaín Peptide) bloqueando el surco de fijación. Hace falta el concurso de una moléculas clase II no clásica (HLA-DM) para catalizar el intercambio de CLIP por péptidos antigénicos. DM es una molécula monomórfica que no se encuentra sobre la membrana celular, sino que se encuentra preferentemente en el compartimento endosomal. DM se expresa solo en aquellas células que expresan las moléculas clásicas clase II. La acción de HLA-DM se altera en presencia de HLA-DO, ya que esta molécula se une a HLA-DM y disminuye su eficiencia en la catálisis del intercambio CLIP-péptido. DO difiere de DM en que solo se expresa por las células B y en el timo y es posible que
formulandoalgoqueaportar.blogspot.com

intervenga también en la selección de los péptidos que se unen a las moléculas clase II en las células B. Como en el caso de las moléculas clase I, se requiere la unión el péptido para mantener la estructura y estabilidad de la molécula clase II. Tras la unión, el complejo clase II-péptido se transporta a la membrana celular, donde el pH neutro permite al complejo asumir una forma compacta estable. El que un péptido antigénnico se asocie con clase I o clase II viene dictado por el mode de penetrar en la célula y por el sitio de procesamiento.
PRESENTACION CRUZADA DE ANTIGENOS EXOGENOS
Hay ciertos casos en los que las CPAs pueden presentar antígenos exógenos a las células Tc (citotóxicas) en el contexto de moléculas de clase I. El primero en comunicar tal hecho fue Michael Bevan y, más recientemente, se ha descrito con un cierto detalle por Meter Cresswell. El fenómeno de la presentación cruzada requiere que el antígeno internalizado, que debería manipularse normalmente por la vía exógena y presentarse en el contexto de moléculas de clase II, se cruce con la vía endógena y se una a moléculas clase I. Estos péptidos se presentan ahora como si fuesen endógenos. Entre los mecanismos propuestos para explicar la presentación cruzada está el cambio desde el compartimento endosomal al compartimento donde se realiza la unión de los péptidos a las moléculas clase I. También es posible que los péptidos exógenos tengan acceso al retículo endoplásmico para entrar en la secuencia de procesamiento. No se sabe si las CPAs capaces de presentación cruzada la usan como un mecanismo alternativo a la presentación normal de antígenos endógenos o si este mecanismo es la vía exclusiva de presentación en el contexto de las moléculas Clase I en estas células. Los ejemplos más claros de presentación cruzada están en las DCs, sin que esté claro si otras CPAs son capaces de ello. La presentación cruzada por las DCs supone una gran ventaja para el hospedador, pues permite a las DCs capturar virus, procesar los antígenos virales y generar linfocitos T citotóxicos que pueden atacar a los virus y a las células infectadas por virus antes de que se disemine la infección viral. Aunque las DCs carecen de los receptores celulares específicos que explotan los virus para su penetración en las células, pueden captar muchos tipos de virus diferentes para su procesamiento y presentación. La mayor incógnita que persiste respecto a la presentación cruzada es como y donde coinciden dos vías de presentación, que están relativamente bien definidas, para permitir que un antígeno exógeno se presente como un antígeno endógeno.
PRESENTACION DE ANTIGENOS NO PEPTIDICOS
Además del procesamiento y presentación de antígenos peptídicos CMH restringida el sistema inmunitario también puede reconocer y responder a antígenos no proteicos. A mediados de la década de los 1980s comienzan los informes de proliferación de células T en presencia de antígenos no proteicos derivados de agentes infecciosos. Informes más recientes indican que las células T con receptor γδ reaccionan con glucolípidos derivados de bacterias tales como Mycobacterium tuberculosis y que la presentación de
formulandoalgoqueaportar.blogspot.com

estos antígenos corre a cargo de moléculas de la familia CD1, asimilables a moléculas clase I no clásicas. Las proteínas CD1 se asocian con β2-microgobulina y, en humanos, la familia está representada por cinco genes (CD1A-E), que se localizan fuera del CMH, en el cromosoma 1. En función de sus homologías se clasifican en dos grupos: grupo 1 (CD1.A, B, C, E) y grupo 2 (CD1.D). Todos los mamíferos estudiados hasta la fecha tienen genes CD1, aunque su número varía. Por ejemplo, en los roedores solo existen genes del grupo 2. El surco de unión al antígeno de las moléculas CD1 es más profundo y voluminosos que el de las moléculas clase I clásicas. La distribución de las moléculas CD1 individuales varía entre las distintas células y su expresión se puede inducir por la exposición a ciertas citocinas, tales como GM-CSF e IL-3. En cuanto al patrón migratorio intracelular difiere entre las moléculas de la familia:
• CD1a se encuentra, sobre todo, en endosomas tempranos o en la superficie celular
• CD1b y CD1d se encuentran en endosomas tardíos • CD1c se encuentra presente en todo el sistema endocítico
Ciertas moléculas CD1 se reconocen por las células T en ausencia de antígenos extraños y, en esas reacciones, puede demostrarse la auto-restricción. El examen de los antígenos presentados por las moléculas CD1 revela que son lípidos (ac. micólico) que forman parte de la pared celular de Mycobacterium tuberculosis. Estudios posteriores de presentación de lípidos indican que estas moléculas también pueden presentar unglucolípido (lipoarabinomanano) de M. leprae. Los datos concernientes a la presentación por CD1 de antígenos indican la existencia de una tercera vía para el procesamiento; una vía con distintos pasos intracelulares en la que no intervienen las moléculas que facilitan el procesamiento con Clase I. Por ejemplo, las moléculas CD1 son capaces de procesar antígenos en células TAP-deficientes. Datos más recientes también indican que hay diferencias en el tráfico de las moléculas de la familia CD1, con CD1a en la superficie o en los compartimentos endocíticos de reciclaje y CD1b, y CD1c en los compartimentos lisosomales. El como la vía CD1 interacciona o se cruza con las mejor conocidas vías Clase I y Clase II está en discusión. En cuanto a los tipos de células T que reaccionan con CD1 encontramos primero a los que tienen: TCRγδ y carecen de CD4 y CD8 o TCR con cadenas αα específicos de CD1 NKT (células T asesinas naturales) reconocen moléculas CD1 que presentan antígenos autólogos, así como esfingolípidos bacterianos, lo que sugiere un papel en las respuestas inmunes de tipo innato frente aciertas bacterias. Las células Tγδ El receptor T es un hetero-dímero con una estructura de dominios (V y C) muy similar a la del receptor B y también estabilizado mediante un puente disulfuro. Las regiones transmembrana de ambas cadenas son poco corrientes, pues contienen residuos de aminoácidos (aa.) con carga positiva, lo que les permite interaccionar con las cadenas del complejo receptor encargadas de la transducción de las señales.
formulandoalgoqueaportar.blogspot.com

La mayor parte de las células T humanas expresan receptores codificados por los genes alfa y beta, que reconocen antígenos peptídicos previamente procesados y presentados en condiciones especiales por células especializadas, llamadas Células Presentadoras de Antígenos (CPAs). Pero datos iniciales indicaban que ciertas células T reaccionaban con antígenos no peptídicos, lo que no empezó a aclarase hasta que se comprobó que las moléculas CD1 (distintas de las del complejo de histocompatibilidad encargadas de presentar los antígenos peptídicos) podían presentar antígenos hidrocarbonados y lipídicos a la subpoblación T gamma-delta. Más recientemente, se ha visto que ciertas células T γδ reconocen antígenos que ni se procesan ni se presentan en el contexto del CMH. Trabajos estructurales más recientes indican que el receptor γδ puede fijar directamente moléculas proteicas (sin restricción CMH), lo que sugiere que este receptor se parece mucho, desde el punto de vista funcional, a los PRRs de la inmunidad innata. Mientras que el receptor αβ reconoce a los antígenos que se le presentan en el contexto de moléculas CMH, los datos acumulados indican que las células T γδ no precisan ni procesamiento ni presentación en el contexto CMH para reconocer a los antígenos. La función precisa de estas células todavía no se conoce y hay que aprender cual es su papel en la inmunidad frente a patógenos y, posiblemente, en la autoinmunidad. El reconocimiento de antígenos comunes a grupos de microorganismos y su capacidad para ligar a moléculas CMH no clásicas sugiere una respuesta rápida, que es más característica de la inmunidad innata que de la adaptativa. El número de células T γδ circulantes es pequeño, comparado con el de células T αβ, y sus segmentos génicos V tienen una diversidad limitada. Muchas de estas células carecen de CD4 y CD8 y la mayoría expresan la misma pareja γδ en su receptor. En los humanos, el receptor predominante que se expresa en las células γδ circulantes reconoce un antígeno que es un fosfolípido microbiano (3-formil-1-butil pirofosfato), propio de mycobacterium tuberculosis y otras bacterias y parásitos. Esta especificidad para antígenos de patógenos frecuentes apoya la hipótesis de que las γδ actúan como una rama de la inmunidad innata, presentando una respuesta rápida a ciertos antígenos sin necesidad de una fase de procesamiento. Es interesante notar que la especificidad de las células γδ circulantes de ratones, y otras especies estudiadas, no es paralela a la humana, lo que indica que esta respuesta debe dirigirse hacia los patógenos comunes de las distintas especies animales. En ciertos tipos de infección el número de células γδ circulantes o en ciertos órganos aumenta de forma dramática. Infecciones con una serie de bacterias, incluyendo mycobacterias y Haemophilus influenzae, así como por parásitos, como los de la malaria o la leishmaniosis, lleva al aumento del número de células T γδ. Estas células pueden matar a células infectadas u organismos por los mismos mecanismos que usan los linfocitos T citotóxicos y las células NK. Datos adicionales sobre las células T γδ las implican en procesos autoinmunes crónicos, incluyendo lupus, miositis y esclerosis múltiple. Además, los datos indican que las T γδ pueden secretar un espectro de citocinas y quimiocinas que sugieren lo que puede ser un papel regulador en el reclutamiento de células T αβ al sitio de la invasión por los patógenos. Estas células reclutadas deben presentar, presumiblemente, un amplio espectro de receptores entre los que se activarán y expandirán selectivamente los de mayor afinidad por el patógeno. Mecanismo y funciones de las T γδ son un campo activo de investigación actual, especialmente en lo que concierne a los antígenos que reconocen y a si pueden reconocer antígenos en el contexto de las moléculas CMH no clásicas. Estudios más
formulandoalgoqueaportar.blogspot.com

recientes también sugieren que las T γδ pueden servir como células presentadoras de antígenos (CPAs), lo que expandiría enormemente su potencial funcional.
formulandoalgoqueaportar.blogspot.com
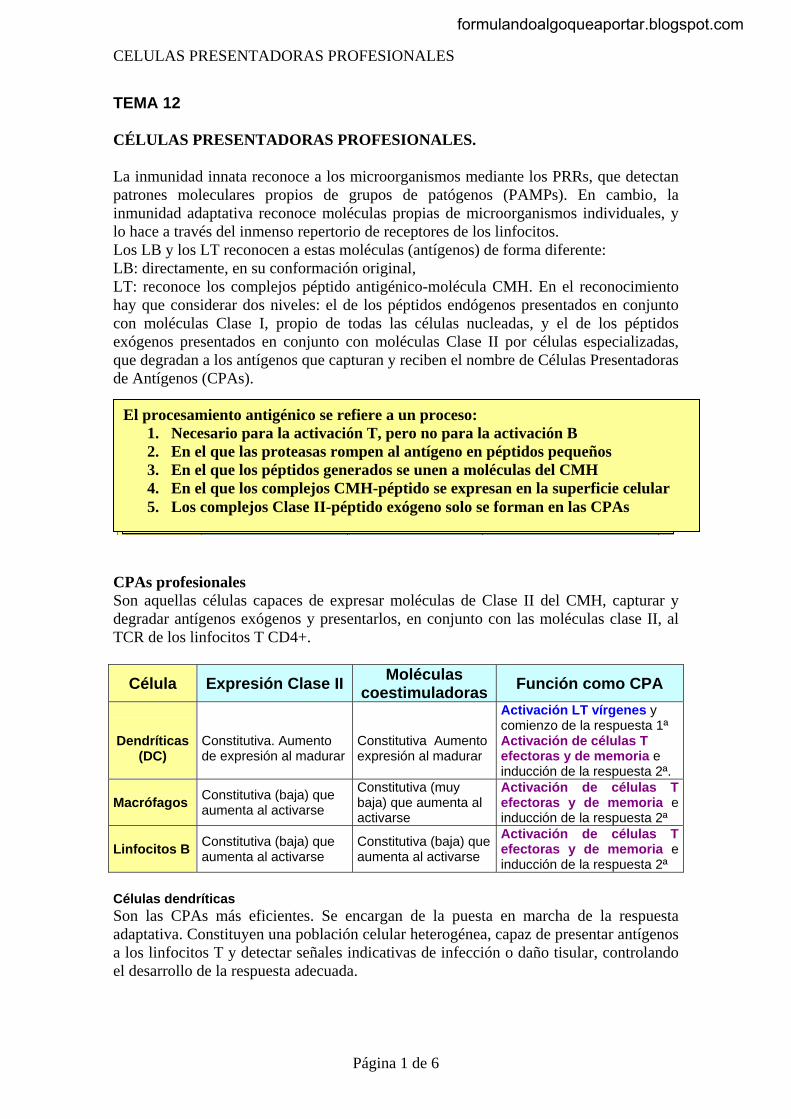
CELULAS PRESENTADORAS PROFESIONALES
TEMA 12 CÉLULAS PRESENTADORAS PROFESIONALES. La inmunidad innata reconoce a los microorganismos mediante los PRRs, que detectan patrones moleculares propios de grupos de patógenos (PAMPs). En cambio, la inmunidad adaptativa reconoce moléculas propias de microorganismos individuales, y lo hace a través del inmenso repertorio de receptores de los linfocitos. Los LB y los LT reconocen a estas moléculas (antígenos) de forma diferente: LB: directamente, en su conformación original, LT: reconoce los complejos péptido antigénico-molécula CMH. En el reconocimiento hay que considerar dos niveles: el de los péptidos endógenos presentados en conjunto con moléculas Clase I, propio de todas las células nucleadas, y el de los péptidos exógenos presentados en conjunto con moléculas Clase II por células especializadas, que degradan a los antígenos que capturan y reciben el nombre de Células Presentadoras de Antígenos (CPAs).
El procesamiento antigénico se refiere a un proceso: 1. Necesario para la activación T, pero no para la activación B 2. En el que las proteasas rompen al antígeno en péptidos pequeños 3. En el que los péptidos generados se unen a moléculas del CMH 4. En el que los complejos CMH-péptido se expresan en la superficie celular 5. Los complejos Clase II-péptido exógeno solo se forman en las CPAs
CPAs profesionales Son aquellas células capaces de expresar moléculas de Clase II del CMH, capturar y degradar antígenos exógenos y presentarlos, en conjunto con las moléculas clase II, al TCR de los linfocitos T CD4+.
Célula Expresión Clase II Moléculas coestimuladoras Función como CPA
Dendríticas
(DC)
Constitutiva. Aumento de expresión al madurar
Constitutiva Aumento expresión al madurar
Activación LT vírgenes y comienzo de la respuesta 1ª Activación de células T efectoras y de memoria e inducción de la respuesta 2ª.
Macrófagos Constitutiva (baja) que
aumenta al activarse
Constitutiva (muy baja) que aumenta al activarse
Activación de células T efectoras y de memoria e inducción de la respuesta 2ª
Linfocitos B Constitutiva (baja) que
aumenta al activarse Constitutiva (baja) que aumenta al activarse
Activación de células T efectoras y de memoria e inducción de la respuesta 2ª
Células dendríticas Son las CPAs más eficientes. Se encargan de la puesta en marcha de la respuesta adaptativa. Constituyen una población celular heterogénea, capaz de presentar antígenos a los linfocitos T y detectar señales indicativas de infección o daño tisular, controlando el desarrollo de la respuesta adecuada.
Página 1 de 6
formulandoalgoqueaportar.blogspot.com

CELULAS PRESENTADORAS PROFESIONALES
Origen Se forman en la médula ósea a partir de progenitores linfoides y mieloides. Constituyen menos del 1% de los linfocitos circulantes. Las mieloides son las presentadoras, mientras que las linfoides producen grandes cantidades de interferones de Tipo I. DCs mieloides Tipos funcionales Tienen dos estadios diferentes, con distintas capacidades funcionales:
DCs inmaduras DCs maduras
Las DCs inmaduras (DCi) Especializadas en capturar y procesar antígenos en tejidos periféricos. Las características de los procesos inflamatorios / infecciosos en los tejidos
condiciona su perfil de maduración. Estos perfiles se diferencian entre si por el patrón de citocinas y quimiocinas que
van a producir Estos perfiles condicionarán el curso de la respuesta inmune adaptativa Las DCi se localizan en los tejidos periféricos, sobre todo en piel y mucosas,
donde patrullan vigilando la presencia de patógenos potenciales. Las DCs maduras
Proceden de las DCi que, tras endocitar el antígeno en la periferia, emigran a través de la circulación a los órganos linfoides secundarios.
La maduración se caracteriza por: - El aumento de moléculas Clase II del CMH - El aumento de moléculas coestimuladoras - La capacidad para unir los péptidos antigénicos a las moléculas Clase II - La expresión en la superficie de los complejos péptido-Clase II.
Son las únicas capaces de activar a los linfocitos T vírgenes DCs inmaduras DCs maduras Localización Tejidos periféricos Órganos linfoides 2ºs Capacidad endocítica Alta Baja Procesamiento Alta Baja Moléculas coestimuladoras, Expresión baja Expresión alta Moléculas Clase I y Clase II Expresión baja Expresión alta Presentación a LT vírgenes Baja Alta Expresión de CCR7 Baja Alta DCs inmaduras Están en la periferia, sobre todo bajo la piel y las mucosas, ejerciendo funciones de vigilancia. Su función es la de capturar patógenos/antígenos, detectando la aparición de procesos infecciosos o inflamatorios. Los fenómenos inflamatorios locales inducen la expresión de moléculas de adhesión por el endotelio y la producción de quimiocinas, circunstancias en las que se reclutan con rapidez precursores y DCs inmaduras hacia el tejido lesionado. Las DCi capturan y procesan antígenos de forma eficaz, pero son malas activadoras de las T vírgenes, ya que expresan concentraciones bajas de moléculas coestimuladoras y moléculas Clase II (las moléculas Clase II están “secuestradas” en los endosomas).
Página 2 de 6
formulandoalgoqueaportar.blogspot.com

CELULAS PRESENTADORAS PROFESIONALES
Una DCi tipo es la célula de Langerhans de la epidermis que, en número escaso (< 1%), es capaz de contactar con el 25% de la epidermis gracias a sus largos procesos citoplásmicos. Ante la llegada de un antígeno y el comienzo de una respuesta inflamatoria local estas DCs
Detectan señales de alarma Capturan y procesan antígenos Inician la migración hacia los ganglios linfáticos regionales y, durante ella,
completan el proceso de maduración. La captura del antígeno por parte de las DCi Estas células presentan una capacidad endocítica extraordinaria, mediante dos procesos:
Endocitosis mediada por receptores (vía clatrinas) Macropinocitosis
Los FcR intervienen en el reconocimiento y endocitosis de los antígenos que han interaccionado con anticuerpos específicos. La internalización a través de FcgR lleva a la liberación del antígeno en el citosol y a su presentación en conjunto con moléculas de Clase I (PRESENTACIÓN CRUZADA) Los CR intervienen en el reconocimiento y endocitosis de microorganismos opsonizados y, también, en la internalización de
células apoptóticas.
Receptores para el fragmento Fc de las Ig (IgG e IgE)
Receptores para el Complemento (CR3 y CR4)
Receptores lectina Tipo C (CLRs)
Receptores para proteínas de choque térmico (HSP)
Receptores basurero (SR)
Los CLR son los responsables del reconocimiento de las estructuras hidrocarbonadas presentes en los antígenos. La internalización mediada por LCR lleva a la presentación a través de Clase II, pero también a la Presentación cruzada con moléculas de Clase I. El HSPR es, en realidad, el receptor para a2-macroglobulina, que es capaz de reconocer e internalizar péptidos “marcados” con las proteínas de choque térmico hsp70 y/o hsp96 procedentes de células necróticas o apoptóticas. Los SR pueden reconocer e internalizar componentes presentes en parásitos y bacterias. Las DCi tienen, además, una capacidad fagocítica alta; la de microorganismos opsonizados está mediada por los receptores Fc y los CR, mientras que la de microorganismos no opsonizados depende de algunos receptores CLR (DC-SIGN) y MR. La Macropinocitosis es un mecanismo en el que no intervienen receptores y permite internalizar antígenos mediante la proyección de seudópodos que forman vesículas endocíticas de gran tamaño. Esta actividad endocítica es constitutiva en las DC, mientras que en macrófagos y células epiteliales solo se induce tras la activación celular. De este modo, las DCs muestrean su entorno de forma permanente, sin necesidad de receptores que reconozcan los distintos componentes microbianos.
Página 3 de 6
formulandoalgoqueaportar.blogspot.com

CELULAS PRESENTADORAS PROFESIONALES
La migración y maduración de las DCs Durante la maduración:
Aumentan la expresión del receptor de quimiocinas CCR7 y se dirigen a los órganos linfoides secundarios
Disminuye la capacidad endocítica, con lo que los antígenos que presenta son los adquiridos en la periferia
Aumentan la expresión de las moléculas coestimuladoras CD40, CD80 y CD86 (B27-1 y B27-2).
Aumentan la expresión de los complejos antigénicos péptido-Clase II (por aumento de la síntesis pero, sobre todo, a expensas de las moléculas Clase II secuestradas en el compartimiento endosómico).
En realidad, las DCi periféricas migran, en tasas muy bajas, de modo constante hacia los ganglios regionales, sin experimentar la maduración. Esta migración basal (sin estímulos inflamatorios) seguramente interviene en la inducción de la tolerancia periférica frente a antígenos propios. Por el contrario, la migración inducida por estímulos inflamatorios aporta un gran número de DCs maduras a los ganglios linfáticos que, después, inducirán una respuesta adaptativa adecuada. Cambios en el patrón de expresión de los receptores de quimiocinas Las DCi expresan muchos receptores de quimiocinas (CCR1, 2, 4, 5, 6, CXCR1, 2) En las DC, mientras maduran, disminuye mucho su expresión y aparece CCR7. Los ligandos de CCR7 se expresan en las células endoteliales linfáticas y en las HEV (vénulas endoteliales altas) de las zonas T-dependientes de los ganglios, hacia donde se dirigirán ahora, obligatoriamente, las DCs que han entrado en la maduración. Estímulos que inducen la maduración de las DCs El principal es el reconocimiento de PAMPs por los TLRs de las DCi. Las DC mieloides expresan TLR2 y TLR4, lo que les permite reconocer componentes de las bacterias G+ y G--. El reconocimiento induce la producción de quimiocinas y citocinas inflamatorias, entre ellas IL-12 (que dirige la diferenciación de las células T vírgenes hacia Th1). También expresan TLR3 intracelular que detecta el RNA de doble hélice de ciertos virus. Otro estímulo lo representan las citocinas y otros mediadores inflamatorios procedentes de las células del foco infeccioso. (TNF-a, IFN Tipo I, NO y PGE2) También la interacción con células CD4+ y CD8+. Los mecanismos incluyen: Interacción entre el CD40 de la DC y CD40L de las CD4+* Interacción entre el Fas de la DC y el FasL de la CD4+* La acción mediada por IFN-g y TNF-a procedentes de las CD8+*
Página 4 de 6
formulandoalgoqueaportar.blogspot.com

CELULAS PRESENTADORAS PROFESIONALES
Factores que influyen en la gran capacidad como CPA de las DCm La gran carga antigénica que acumulan en la fase DCi Su capacidad migratoria La expresión muy alta de complejos péptido-Clase II y su estabilidad en la
superficie celular La expresión muy alta de moléculas coestimuladoras Las citocinas que producen
La maduración puede llevar a distintos perfiles funcionales de la DC; según los estímulos activadores de la periferia, la maduración puede llevarlas a orientar de distinto modo la maduración de las células T vírgenes. Células dendríticas plasmocitoides Cuando se habla de DCs presentadoras la referencia es, en general, a las DCs mieloides Hay una segunda población de DCs, denominadas DCs Plasmocitoides, que difieren en las características fenotípicas (CD11c- y CD123+alto) y funcionales. Morfológicamente, las mieloides se asemejan a monocitos y las plasmocitoides a células plasmáticas (plasmocitos). También difieren en la localización: las mieloides inmaduras están en piel y mucosas y las plasmocitoides inmaduras están en la circulación y en los órganos linfoides secundarios. Diferencias fundamentales entre ambas poblaciones son la poca capacidad endocítica de las plasmocitoides inmaduras y la aparente incapacidad de las maduras para activar a los linfocitos T vírgenes. Las DCs plasmocitoides representan el 0,2-0,8% de las células mononucleares de sangre periférica. Expresan selectivamente TLR7 y TLR9 (reconocen RNA viral de cadena simple y DNA viral) y producen grandes cantidades de IFNs de tipo I (con niveles entre 100 y 1000 veces + que el resto de las células nucleadas). Son la fuente principal de estos IFN en la infección viral aguda. También producen algo de TNF-a e IL-6 Generación y migración de las DC plasmocitoides La citocina FLT3L es su factor principal de crecimiento y diferenciación. Tras abandonar la m.o. pasan a la circulación para dirigirse, a través de las HEV, a las áreas T-dependientes de los órganos linfoides secundarios Activación y maduración de las DC plasmocitoides Tras activarse y producir IFNs Tipo I maduran, reduciéndose de forma llamativa su capacidad de producción de IFNs Tipo I. Esta maduración puede ser:
A expensas de los IFNs Tipo I y TNF-a producidos por las propias DCs plasmocitoides, activadas por la infección viral, lo que lleva a la diferenciación de las CD4+ a Th1.
A expensas de la IL-3 producida por eosinófilos, basófilos y mastocitos, en respuesta a los estímulos del tipo que se producen en las infecciones parasitarias. Esta segunda vía induce la producción de IL-4, 5 y 10 y, por tanto, el perfil Th2.
Es decir, las DC plasmocitoides también tienen la plasticidad funcional que les lleva, en función del tipo de maduración, a orientar la diferenciación T en uno u otro sentido.
Página 5 de 6
formulandoalgoqueaportar.blogspot.com
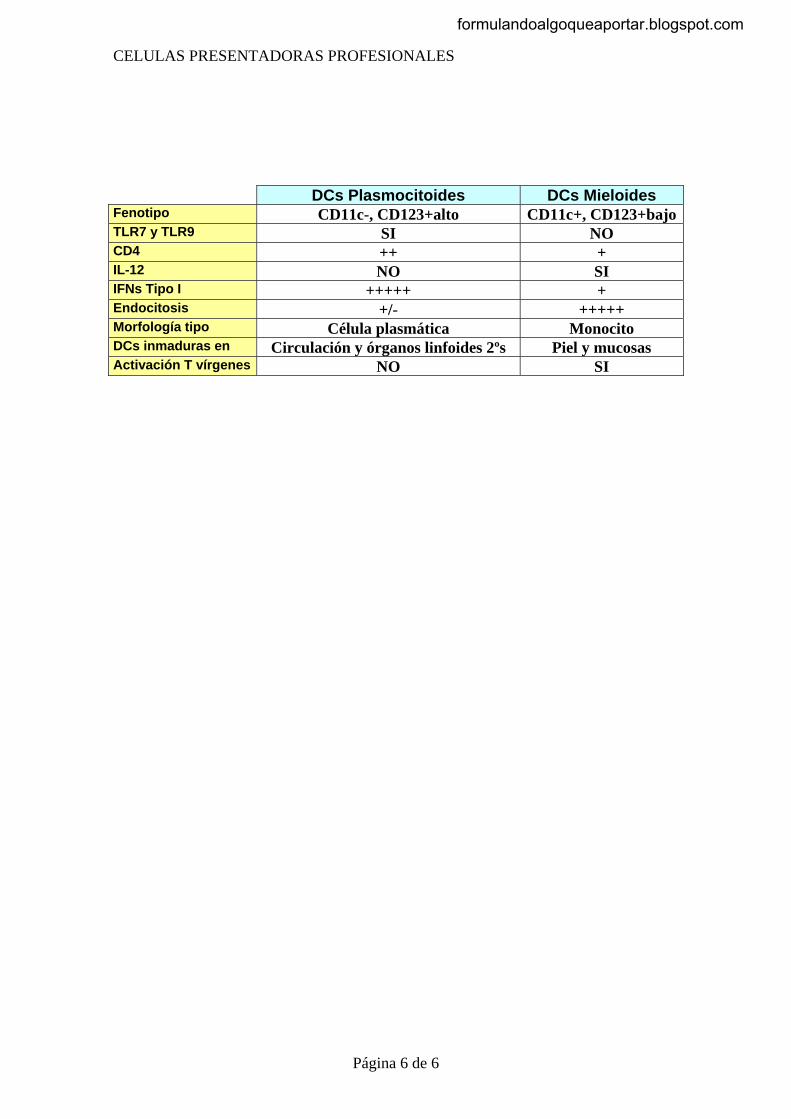
CELULAS PRESENTADORAS PROFESIONALES
DCs Plasmocitoides DCs Mieloides Fenotipo CD11c-, CD123+alto CD11c+, CD123+bajoTLR7 y TLR9 SI NO CD4 ++ + IL-12 NO SI IFNs Tipo I +++++ + Endocitosis +/- +++++ Morfología tipo Célula plasmática Monocito DCs inmaduras en Circulación y órganos linfoides 2ºs Piel y mucosas Activación T vírgenes NO SI
Página 6 de 6
formulandoalgoqueaportar.blogspot.com

MADURACION DE LOS LINFOCITOS T La característica que distingue el reconocimiento del antigeno por los linfocitos B y T es la restricción CMH. Tanto la maduración de las células progenitoras en el timo como la activación de los LT vírgenes en la periferia están condicionadas por la intervención de las moléculas CMH. La diversidad de reconocimiento antigénico de la población T se reduce durante la maduración tímica, mediante un proceso de selección que solo permite madurar a las células que reconocen al CMH y reaccionan frente a lo no propio. Los estados finales de la maduración discurren a lo largo de dos vías, que generan dos subpoblaciones (CD4+ y CD8+) funcionalmente distintas y que presentan restricción por moléculas CMH de Clase II y Clase I, respectivamente. MADURACION T EN EL TIMO Los progenitores T empiezan pronto a emigrar al timo desde los sitios de hematopoyesis iniciales (11 días de gestación en el ratón y 8-9 semana en humanos). La maduración supone el reordenamiento de los genes para el TCR de la línea germinal y la expresión de una serie de marcadores de membrana. Las formas de desarrollo en el timo se llaman timocitos, que proliferan y se diferencian a lo largo de vías de desarrollo que producen distintas subpoblaciones T. El desarrollo T comienza cuando arriba al timo un pequeño número de Precursores linfoides desde la sangre. Hasta ahora se pensaba que la arquitectura tímica era necesaria para el desarrollo T, pero se ha comprobado por experiencias in vitro que lo necesario es que las células estromales de la médula ósea expresen el ligando para el receptor Notch; las células que portan este receptor se comprometen desde este momento con la línea de desarrollo T. Tras la llegadla timo las células precursoras, que todavía no han reordenado sus genes, penetran en el cortex y proliferan lentamente. Durante unas tres semanas pasan por una serie de estados que se caracterizan por cambios fenotípicos en su superficie. Al comienzo del desarrollo carecen de CD4 y CD8, por lo que se denominan células doble negativas (DN). Estas DN pueden dividirse un cuatro subpoblaciones (DN1-4), caracterizadas por la presencia o ausencia de ciertas moléculas como c-Kit (el receptor para el factor de crecimiento de células madre o CSF), CD44 (una molécula de adhesión) y CD25 (cadena alfa del receptor para IL-2). Durante la fase DN2 empieza el reordenamiento génico de las cadenas, γ, δ y ε; el locus α no se reordena, posiblemente porque el DNA está muy compactado y no es accesible a la maquinaria de recombinación. Cuando las células progresan hacia DN3, prosigue el reordenamiento de TCRγ, TCRδ y TCRε. Las células destinadas a ser Tγδ suponen un pequeño porcentaje (< 5% de los timocitos maduros) divergen en la transición entre DN2 y DN3 y maduran con pocos cambios superficiales. La mayor parte de las células están destinadas a ser Tαβ, adquieren el fenotipo DN3 (c-Kit-, CD44-, CD25+) y ya expresan la cadena β del receptor. Esta cadena β se combina con una glucoproteína de 33 kDa, conocida como cadena pre-T, y se asocia con CD3 para formar el complejo receptor pre-T (las proteínas Notch tienen un papel crítico en esta fase del desarrollo T; las que no expresan Notch no siguen la maduración). La formación del pre-TCR activa una vía de transducción de señales, con varias consecuencias:
• Indica que una célula ha conseguido un reordenamiento productivo de la cadena β del TCR y la señala para la posterior proliferación y maduración
• Suprime reordenamientos posteriores de gen de cadenas β (exclusión alélica)
formulandoalgoqueaportar.blogspot.com

• Vuelve a la célula susceptible al reordenamiento de la cadena α del TCR • Induce la progresión del desarrollo hacia el estado doble positivo (CD4+CD8+)
Tras el reordenamiento de la cadena β la célula progresa rápidamente por DN3 y DN4 y da lugar a la población doble positiva (DP) (se expresan los co-receptores CD4 y CD8), que es una fase de proliferación rápida. Sin embargo, en este punto todavía no se ha reordenado el locus α, que solo comienza cuando termina la proliferación de las DP y aumentan los niveles de RAG-2. Esta estrategia aumenta la diversidad del TCR, pues la multiplicación de una población con el gen β ya reordenado permite que esta cadena se asocie con una cadena α diferente en cada célula particular del clon. El desarrollo T es un proceso derrochador para el hospedador, pues el 98% de todos los timocitos no llegan a madurar. Los DP que expresan el complejo FCR-CD3 y sobreviven a la selección positiva tímica se desarrollan hasta células positiva simple (SP) CD4+ o CD8+. Estas células PS sufren una selección negativa adicional y migran desde el cortex hasta la médula para abandonar el timo con la circulación. SELECCIÓN TIMICA DEL REPERTORIO T Selección positiva Tiene lugar en la zona cortical del timo, donde los timocitos inmaduros interaccionan con las células epiteliales del cortex. Los TCRs de los timocitos se unen a grupos de moléculas CMH sobre la superficie de las células epiteliales, lo que permite a los timocitos recibir señales de supervivencia. La selección negativa asegura la auto-tolerancia Los timocitos que sobreviven a la selección positiva tienen afinidades muy distintas para los auto-antígenos presentados por las moléculas CMH. Los que muestran afinidad muy alta se eliminan durantes esta fase de selección. Las células que interaccionan con los timocitos en esta fase son macrófagos y células dendríticas de la zona cortico-medular, quienes presentan complejos Clase I-péptido y clase II-péptido., Así se consigue la tolerancia en el timo frente a los antígenos propios.
formulandoalgoqueaportar.blogspot.com

TEMA 13. Maduración de linfocitos Papel de la médula ósea y el timo Tolerancia central Introducción a la maduración de los linfocitos B. El proceso de maduración culmina con la producción de plasmocitos y células de memoria y se puede dividir en tres fases:
PRODUCCION → ACTIVACION → DIFERENCIACION 1. Producción de células B maduras, inmunocompetentes 2. Activación de las células B maduras por interacción con el antígeno 3. Diferenciación de las células B activadas en plasmocitos y células de memoria
Fase antígeno independiente En muchos vertebrados, incluyendo ratón y hombre, es la médula ósea la productora de linfocitos B, mediante una sucesión ordenada de reordenamientos génicos, en ausencia de antígeno. Fase antígeno dependiente Las células B inmaduras (con IgM de membrana) abandonan la médula ósea y evolucionan hasta expresar simultáneamente IgM e IgD de membrana, con la misma especificidad antigénica. Esta Célula B virgen circula por sangre y linfa hasta los órganos linfoides secundarios (OL2º). Si reconoce a un antígeno se activa, lo que se traduce en proliferación (expansión clonal) y diferenciación en una población células productoras de anticuerpos y otra de células de memoria. Durante la vida se producen muchas células B en la médula ósea, pero solo maduran unas pocas de ellas. En el ratón, el tamaño del pool circulante es de 2x109; la mayoría son células vírgenes con vida media corta (entre 3 días y algunas semanas) que mueren si no encuentran a su antígeno o no compiten con éxito por el. Esto quiere decir que, como el sistema inmune tiene una diversidad total de anticuerpos superior a 1x1010, solo circula en un momento dado una pequeña fracción (aproximadamente un 10%) del repertorio total. Maduración de los linfocitos B Comienza en el estado embrionario y prosigue durante toda la vida. Durante el periodo fetal (en ratones y humanos) la maduración tiene lugar en el saco embrionario, hígado fetal y médula ósea fetal. Tras el nacimiento las células B se forman y maduran en la médula ósea. Las células progenitoras (Pro-B) maduran en la médula óseaLas células Pro-B proceden del progenitor linfoide común y se distinguen por tener en su membrana una tirosin-fosfatasa denominada CD45R. Para proliferar y diferenciarse a células Pre-B (precursoras) estas células necesitan el microambiente que aportan las células estromales de la médula ósea. Las células estromales interaccionan directamente con las células Pro y Pre-B y secretan varias citocinas, sobre todo IL-7, que promueven el desarrollo. Al comienzo del desarrollo, las Pro-B necesitan contactar con las células estromales por medio de moléculas de adhesión; tras este contacto anterior el receptor c-Kit de la célula Pro-B interacciona con el factor de células madre o SCF que hay en la membrana de la célula estromal.(ver diapositiva). La interacción dispara la actividad tirosin-kinasa de c-Kit que lleva a la célula a proliferar y diferenciarse a Pre-B.
formulandoalgoqueaportar.blogspot.com

La IL-7 secretada por las células estromales se une a su receptor sobre las células Pre-B y dirige el proceso posterior de maduración, sub-regulando las moléculas de adhesión para que las células proliferantes puedan separarse de las estromales. Esta Pre-B ya no necesitan el contacto directo con las estromales, pero siguen precisando la presencia de IL-7. El reordenamiento génico produce células B inmadurasLa progresión a células B inmaduras depende de los reordenamientos del DNA de las Ig en las células progenitoras y precursoras.
• Pro-B. En los genes de las cadenas H de las células Pro-B se produce el reordenamiento D-J seguido del reordenamiento V-DJ. Si este primer reordenamiento no es productivo sigue en el otro cromosoma.
• Pre-B. Terminado el reordenamiento de la cadena H tienen que reordenarse los genes de la cadena L. Debido a la exclusión alélica que se produce en las cadenas L solo se expresa un isotipo sobre la membrana de una célula B.
• B-inmadura. Ya expresa mIgM junto con Igα/Igβ, lo que supone expresar un BCR capaz de trasmitir señales tras la unión al antígeno.
• B-madura. Expresa IgM e IgD sobre la membrana. Para ello hay un cambio previo en el procesamiento del trascrito primario de RNA que permite la producción de dos mRNA (mensajeros), uno de los cuales codifica para la forma de membrana de la cadena µ y el otro para la cadena δ.
Como era de esperar, las recombinasas RAG-1/2, necesarias para el reordenamiento de los genes de las cadenas H y L, se expresan durante las fases Pro y Pre-B. La enzima desoxinucleotidil-transferasa-terminal (TdT) que cataliza la inserción de nucleótidos en las uniones D-J y V-DJ de las cadenas H, está activa durante la fase Pro-B y desaparece durante la subfase Pre-B en la que se reordenan las cadenas L, por lo que normalmente no hay nucleótidos en las uniones V-J. La fase B inmadura termina con la formación de un BCR que no es totalmente funcional y la unión al antígeno induce la muerte o anergia, en lugar de la activación. El receptor Pre-B es esencial para el desarrollo.En las células Pre-B la cadena µ de membrana se asocia con una cadena L “sustituta”, que es un complejo formado por dos pequeñas proteínas que se asocian en forma no covalente; una de ellas es similar al dominio V y la otra al dominio C de las cadenas ligeras. El complejo cadena µ- cadena sustituta aparece sobre las células Pre-B asociado a Igα/Igβ. Solo las células Pre-B que presentan este seudo-BCR pueden progresar hacia la maduración. Tras el establecimiento de este receptor las células Pre-B experimentan 6-8 divisiones que producen unos 256 descendientes. Cada una de las células de esta progenie tiene que reordenar ahora sus cadenas L. con lo que aumentan la diversidad total del repertorio de anticuerpos. Marcadores de superficie y estado de desarrollo.
• Pro-B. CD45R, Igα/Igβ, CD19 (parte del co-receptor B), CD43 (leucosialina), así como c-Kit (receptor para el SCF de la superficie de las células estromales) • Pre-B. Pierde C-Kit y CD43 y expresan CD25 (cadena α del receptor de IL-2). Expresan el Pre-BCR
formulandoalgoqueaportar.blogspot.com

• B inmaduras. Pierden el Pre-BCR y CD25 y expresan el BCR con IgM de superficie.
Células B-1 B. Un subgrupo auto-renovable. En humanos y ratón estas células suponen aproximadamente un 5% del total de células B, mientras que en otras especies, como conejo y vaca, son la población predominante. Proceden de las células madre hematopoyéticas durante la vida fetal y expresan Ig de superficie, aunque la inmensa mayoría tienen IgM (en lugar de IgG o IgA) y poca o ninguna IgD. A diferencia de las B vírgenes clásicas (B-2) las B-1 B son auto-renovables y pueden producir más B-1 B vírgenes. También se distinguen por presentar el marcador CD5 (una molécula normalmente asociada a los linfocitos T), aunque no se trata de una molécula indispensable para la línea B-1 B y, de hecho, las B-1 de ratas careeccen de ella. En humanos y ratones, donde B-2 es la población principal, las B-1 B representan una población menor en los órganos linfoides secundarios. Pero, a pesar de su escasez en los tejidos linfoides, son la población principal de las cavidades pleural y peritoneal. Es en estas cavidades donde encuentran los antígenos (autoantígenos y ambientales) que dirigen su expansión y mantenimiento, así como otras señales, incluida IL-10, que promueve su supervivencia. Algunos de los antígenos que dirigen la expansión B-1, tales como fosforilcolina, se encuentran en la superficie de las bacterias que colonizan el intestino Además de la auto-renovación y expresar CD5, también se diferencian de las B-2 en algunas otras características, como:
• El repertorio de las regiones V en B-1 es mucho más restringido que el de la población B-2
• Los anticuerpos producidos por B-1 son de menor afinidad que los producidos por B-2
• La activación de B-1 es más por hidratos de carbono que por proteínas • No requieren ayuda T y raramente se diferencian a células de memoria • No presentan cambio de clase, por lo que la mayoría tienen IgM superficial • Tienen poca o nula hipermutación somática, por lo que no presentan maduración
de la afinidad. • Muchos de los anticuerpos producidos son multi-específicos; pueden unirse a
varios antígenos diferentes, incluyendo algunos que están presentes sobre patógenos.
Se considera a estos anticuerpos multi-específicos como parte de la inmunidad ancestral del hospedador, más próxima a la inmunidad innata que a la adaptativa. Anticuerpos naturales Son componentes de la inmunidad innata. Esta “IgM natural” está codificada por genes reordenados de anticuerpos sin diversificación posterior por hipermutación somática. Constituyen una parte importante de la IgM circulante en los humanos y no parecen ser el resultado de una respuesta adaptativa a la infección. Tienen afinidad baja para muchos patógenos microbianos y muchas reacciones cruzadas, incluso con moléculas propias. Además, no se sabe si se producen en respuesta a la flora normal de las superficies epiteliales del cuerpo o en respuesta a lo propio. Sin embargo, puede que intervengan en la defensa frente a Streptococcus pneumoniae, fijándose a la fosforilcolina de la pared bacteriana y contribuyendo a su eliminación antes de que se vuelva peligroso.
formulandoalgoqueaportar.blogspot.com

Linfocitos de la zona marginal (BMZ) Residen en los senos marginales de la pulpa blanca y parecen ser células B maduras en reposo, pues todavía tienen un juego de proteínas de superficie distintas a las que porta la población mayoritaria situada en los folículos. Tienen una especificidad antigénica restringida, dirigida hacia antígenos ambientales comunes e incluso autoantígenos y puede que estén adaptados para responder rápidamente si tales antígenos alcanzan la circulación. No parecen necesitar ayuda T para activarse. Fenotípica y funcionalmente se parecen a las células B-1 y experiencias recientes sugieren una selección positiva por ciertos autoantígenos positivamente (tal como ocurre con las células B-1). Las funciones de las células B-1 y la de los BMZ se dirigen a la defensa de las cavidades corporales y frente a los patógenos que han penetrado en la sangre. El repertorio de receptores tan restringido en ambos grupos los equipa para actuar en la respuesta innata inicial. Además, los segmentos de genes V que usan para codificar el receptor de B-1 y BMZ parecen haber evolucionado mediante selección natural para reconocer antígenos bacterianos comunes, por lo que podrían también participar en la fase precoz de la respuesta adaptativa. Pero, en la práctica, está claro que las células B-1 contribuyen poco a la respuesta adaptativa frente a los antígenos proteicos, pero mucho a la respuesta frente a algunos antígenos hidrocarbonados.
formulandoalgoqueaportar.blogspot.com

TEMA 14. Activación LT vírgenes
Tema 14. ACTIVACION DE LINFOCITOS VIRGENES Células T vírgenes Tanto las CD4+ como las CD8+ dejan el timo a través de la circulación como células en la fase de reposo del ciclo celular (Go). Estas células se caracterizan por:
• Tener cromatina muy condensada • Muy poco citoplasma • Poca actividad trascripcional
Recirculan y pasan por los tejidos linfoides cada 12-24 h. Como solo 1 entre cada 105
células vírgenes es específica para un antígeno determinado, esta recirculación a gran escala aumenta las probabilidades de que una célula T virgen encuentre al antígeno adecuado. Las células T activadas producen células T efectoras (Te) y células T de memoria (Tm). Cuando una célula T virgen reconoce a su antígeno se activa y comienza una respuesta primaria. A las 48 h. de la activación la célula se transforma en un blasto de gran tamaño que experimenta varias rondas sucesivas de división celular. Las señales 1+2 inducen la trascripción del gen de IL-2 y del de la cadena α (CD25) para el receptor de alta afinidad de IL-2 (IL-2R). La señal 2 estabiliza el mRNA de IL-2 con lo que aumenta la producción del IL-2 unas 100 veces. La unión de IL-2 a IL-2R induce a la célula activada a proliferar y diferenciarse. Estas células activadas se dividen 2-3 veces al día durante 4-5 días, generando una descendencia clonal que se diferencia a poblaciones de memoria y efectoras. Células T efectorasRealizan funciones especializadas, como secretar citocinas y ayudar a otras células inmunes, tales como células B, células T CD8+ (citotóxicas) y macrófagos. Derivan tanto de las células T vírgenes como de las de memoria. Las células Te son células de vida corta (unos días a pocas semanas) y con un patrón circulatorio diferente del de las T vírgenes. A esta población pertenecen las células:
• Th1, que secretan IL-2, IFN-γ y TNF. Colaboran con la inmunidad celular • Th2, que secretan IL-4, IL-5, IL-6 e IL-10. Colaboran con la inmunidad humoral
Células T de memoria Derivan tanto de las T vírgenes como de los linfocitos T efectores tras su activación por el antígeno. Son células quiescentes, de vida larga, que responden con gran intensidad al mismo antígeno original, desencadenando una respuesta secundaria. Cuando va finalizando esta respuesta la población efectora declina, pero permanece una población abundante de células de memoria. Como las T vírgenes, las T de memoria son células en fase de reposo (Go), pero mucho menos exigentes en lo que respecta a sus requisitos para la activación, por lo que las pueden activar todas las CPAs; es la abundancia en su superficie de moléculas de adhesión lo que las permite unirse a cualquier CPA. Sus patrones circulatorios son distintos de los de las T vírgenes y las T efectoras.
Página 1 de 4
formulandoalgoqueaportar.blogspot.com

TEMA 14. Activación LT vírgenes
La activación La piedra central de la respuesta inmune, tanto humoral como celular, es la activación y expansión clonal de los linfocitos T. La activación comienza por la unión del complejo receptor (TCR-CD3) al complejo péptido antigénico/molécula CMH. En la interacción y activación intervienen además otras moléculas de superficie de la CPA y de la célula T. Tras las interacciones comienza una cascada de señales que inducen a la célula T a salir de la fase de reposo (Go) y entrar en el ciclo celular para proliferar y posteriormente diferenciarse a células T efectoras y células T de memoria. Los genes que intervienen en el proceso pueden clasificarse en tres categorías:
• Genes inmediatos. Se expresan entre ½ y 1 tras el reconocimiento del antígeno y codifican para una serie de factores de transcripción entre los que están c-Myc, c-Fos, NFAT y NF-kB.
• Genes tempranos. Se expresan a la 1-2 h. del reconocimiento del antígeno y codifican para citocinas y receptores de citocinas, tales como IL-2, IL-2R, IL-3, IL-6, IFN-γ y otras muchas proteínas
• Genes tardíos. Se expresan a partir de los 2 días del estímulo antigénico y codifican para moléculas de adhesión.
Estos cambios tan profundos son el resultado de las vías de transducción de señales que se activan tras la interacción TCR-péptido-CMH La unión al TCR dispara varias vías de señales La detección e interpretación de las señales del entorno es una propiedad indispensable de todas las células, incluidas las inmunitarias. Estas vías, a pesar de ser muy diferentes, tienen una serie de caracteres comunes, tales como:
• La transducción de la señal comienza tras la interacción con el receptor • La recepción de la señal lleva, con frecuencia, a la producción de “segundos
mensajeros”, que son moléculas o iones que difunden a otros sitios dentro de la célula e inducen cambios metabólicos.
• Se produce la activación o inhibición de protein-kinasas y protein-fosfatasas • Las señales se amplían mediante cascadas enzimáticas
Lo que pasa en el periodo que transcurre entre el reconocimiento del antígeno y la expresión génica incluye muchos de los procesos anteriores. El reconocimiento cataliza una serie de procesos intracelulares que comienzan en la cara interna de la membrana plasmática y culminan a nivel nuclear mediante la activación de genes que dirigen el ciclo celular y/o la diferenciación de la célula T. Como ya sabemos, el TCR consiste en una unidad de reconocimiento extracelular y un sistema de traducción de señales, intracelular. En la inducción de la respuesta T se distinguen dos fases: Fase de iniciación. Las señales que recibe la célula T tienen que estar tan bien reguladas que aseguren que solo se produce la activación como resultado del reconocimiento específico del antígeno. En una célula T en reposo, p56Lck (una protein-tirosin-kinasa esencial para que se ponga en marcha la vía de señales dependiente del TCR) está en las “balsas lipídicas” de la membrana (macrodominios ricos en esfingomielina, glicoesfingolípidos y colesterol) y el complejo TCR está excluido de estos dominios. Una vez que se une al antígeno, el complejo TCR se asocia a estas “balsas lipídicas” y p56Lck se une a los tallos citoplásmicos del complejo, donde fosforila ITAMs (Inmuno-receptores con motivos de activación basados en tirosina). Las tirosinas fosforiladas de los ITAMs de las cadenas zeta constituyen el punto de amarre de otra protein-tirosin-kinasa (llamadea ZAP-70) que cataliza la fosforilación de una serie de moléculas adaptadoras asociadas a la
Página 2 de 4
formulandoalgoqueaportar.blogspot.com

TEMA 14. Activación LT vírgenes
membrana y que son el andamiaje para el reclutamiento de moléculas que participan en distintas vías intracelulares de transducción de señales. Producción de muchas señales intracelularesComo consecuencia de la fase anterior se activan muchas vías de señales (ver diapositiva) Sinapsis inmunológica (SI) Una característica de la fase de iniciación de las señales procedentes del TCR es la formación de una estructura supra-molecular conocida como “sinapsis inmunológica”. Se trata de una estructura altamente organizada que se forma cuando las células T contactan con las CPAs. Parece que la entrada de los TCRs en las “balsas lipídicas” es un paso crítico para la formación de la SI; en la región central de la sinapsis se sitúan los TCRs y otras proteínas individuales asociadas al complejo, mientras que en la periferia se sitúan las moléculas de adhesión. Aunque la formación de la SI no es imprescindible para la fase de iniciación de las señales procedentes del TCR, su presencia hace que el mantenimiento de la activación sea más efectivo. ¿Cuántos TCRs tienen que unirse al antígeno para activar a una célula T? Los resultados de experiencias concluyentes se publicaron en 2004 por un equipo de Stanford (M. Davis et al.) basadas en la detección de la liberación de Ca iónico (2+) (este Ca2+ es un indicador d activación celular) y la conclusión es que la unión de un solo TCR es suficiente para detectar liberación de Ca2+ y que el máximo se alcanza con un número de TCRs tan limitado como 10. Las señales coestimuladoras La activación T requiere la intervención dinámica de muchas moléculas asociadas a la membrana (para poner en marcha las vías de señales), pero para activar totalmente a una célula T virgen esto no es bastante, y se requiere más de una señal:
• Señal 1. Es la que se produce como consecuencia de la unión del antígeno al TCR
• Señal 2. Coestimuladora, no antígeno-específica y debida a la interacción entre CD28 (célula T) y B7 (célula dendrítica).
Hay dos isoformas de B7: ambas son muy similares en la porción extracelular (tipo Ig), pero difieren profundamente en la citosólica. Ambas se expresan constitutivamente sobre las DCs y se inducen tras la activación en macrófagos y linfocitos B. Los ligandos para estas moléculas son CD28 y CTLA-4, que se expresan como homodímeros S-S sobre la membrana de las células T. Ambas moléculas son estructuralmente semejantes, pero su acción es antagonista: B7 --- CD28 ---- Activación B7 --- CTLA-4 - Inhibición En las células en reposos no se detecta CTLA-4 y es la unión del TCR lo que induce su formación. El máximo se detecta a los 2-3 días después de la estimulación y aunque su cantidad es menor que la de CD28, compite favorablemente por B7 debido a su mayor afinidad. Es significativo que el nivel de CTLA-4 se “dispare” por las señales coestimuladoras de la unión CD28-B7, lo que indica una regulación negativa. Efectivamente, los ratones CTLA-4 KO presentan una respuesta T masiva, con linfoadenopatías (agrandamiento ganglios linfáticos) y esplenomegalia (agrandamiento del bazo), con la muerte del
Página 3 de 4
formulandoalgoqueaportar.blogspot.com

TEMA 14. Activación LT vírgenes
animal a las 3-4 semanas del nacimiento. Por tanto, está claro el papel de las señales inhibidoras producidas por CTLA-4 en la homeostasis. Anergia clonal A veces, el reconocimiento del complejo P-CMH no lleva a la activación de la célula sino a un estado de falta de respuesta denominado anergia clonal. Que se produzca respuesta o anergia depende de la presencia o ausencia de señales coestimuladoras, tales como las que se producen por la interacción CD28/B7. Hay evidencias de que se pueden anergizar tanto las células T CD4+ como las CD8+. Una célula T CD4+ puede energizarse por otras vías, además de la generada por la falta de señal coestimuladora. La aparición de la molécula CTLA-4 sobre células T activadas puede bloquear la coestimulación, resultando una anergia funcional (CTLA-4 presenta mayor afinidad por B7 que CD28, por lo que interfiere con ella; el resultado de la unión CTLA-4/B7 es una señal de inhibición). Si bien no conocemos las vías de la anergia, recientemente se han identificado varios componentes de las mismas en las células T anérgicas, incluyendo varias ubiquitin-ligasas que parecen capturar a componentes claves de la vía de señales del TCR y llevarlos a la vía proteasómica de degradación.
Página 4 de 4
formulandoalgoqueaportar.blogspot.com

TEMA 15 POBLACIONES DE LINFOCITOS. DIFERENCIACION LINFOCITARIA Th1 Y Th2. CELULAS T EFECTORAS. RESPUESTA Th1. La célula progenitora linfoide T, en la médula ósea, da lugar a precursores inmaduros que emigran al timo y se desarrollan hasta alcanzar la madurez. Los timocitos DN dan lugar, mayoritariamente, a células Tαβ, pero también pueden formar Tγδ. La decisión sobre el tipo de receptor se toma en el estado DN3, ya que el reordenamiento de los genes de las cadenas β, γ y δ comienza de modo simultáneo. Si gamma y delta se reordenan con éxito se detiene el reordenamiento de la cadena β y el TCRγδ (TCR1) se puede ya expresar sobre la membrana. Pero la mayoría de los timocitos reordenan con éxito la cadena beta, antes que la gamma y delta, con lo que el desarrollo prosigue hacia la expresión del TCRαβ (TCR2). Linfocitos T γδ. Son una subpoblación menor de los linfocitos T, también conocida como “linfocitos intraepiteliales” (IEL). Una de sus características más notables es la división en dos grupos:
• Uno se encuentra en los tejidos linfoides de todos los vertebrados y tienen el receptor diversificado (como los Tαβ)
• El otro grupo lo componen los intraepiteliales que se presentan de forma variable en distintos vertebrados y tienen un receptor de muy poca diversidad. Como tampoco recirculan se ha propuesto que reconocen ligandos derivados del epitelio en el que residen, pero que solo se expresan cuando una célula se ha infectado.
El reconocimiento no lo hacen de forma CMH-restringida, sino que parecen reconocer a sus ligandos de forma directa, por lo que podrían reconocer y responder rápidamente a moléculas expresadas por tipos celulares diferentes. La capacidad de reconocer moléculas que se expresan como consecuencia de la infección y no antígenos específicos del mismo patógeno los distingue de los restantes linfocitos y sitúa a los Tγδ intraepiteliales en la inmunidad ancestral, que separa las respuestas innata y adaptativa. Como consecuencia de la activación producen rápidamente citocinas, pero no parecen desarrollar ningún tipo de memoria. Linfocitos Tαβ Dentro de la subpoblación alfa/beta se distinguen los linfocitos T CD4+ y CD8+, según expresen una u otra de estas moléculas accesorias en su superficie. Los CD4+ son los linfocitos T cooperadores (Th), que prestan su ayuda a las células encargadas de la defensa frente a los patógenos intracelulares (Inmunidad celular) o frente a los patógenos extracelulares (Inmunidad humoral), mientras que los CD8+ son las células T citotóxicas encargadas de destruir a las células infectadas. Células NKT También hay un pequeño número de células T que expresan un receptor T αβ especial sobre su superficie; entre sus características, destacan:
• Reconocen antígenos lipídicos presentados en conjunto con la molécula CMH no clásica CD1d.
• Secretan grandes cantidades de citocinas • Hay 2 subpoblaciones en humanos: CD4+ y DN.
formulandoalgoqueaportar.blogspot.com

• Las CD4+ secretan IL-4 e IL-13 y dirigen el perfil de diferenciación hacia Th2
• Las DN secretan IF-γ y dirigen al perfil Th1 • Escasas, < 0,1-1% de leucocitos periféricos
Se encuentran tanto en el timo como en los órganos linfoides periféricos. Expresan, como parte de su receptor, una cadena alfa invariable que se empareja con una de las tres cadenas beta que es capaz de sintetizar, por lo que su receptor es poco variable (“cuasi invariable”). La respuesta principal es la producción de grandes cantidades de citocinas, incluyendo IL-4, IL-10 e IFN-g, por lo que parece que su función debe ser reguladora El reconocimiento se hace en conjunto con moléculas CD1, que no están codificadas en el CMH. Aunque no se sabe la misión exacta de las moléculas CD1, se sabe que CD1b presenta el ácido micólico (lípido de micobacterias) a células T αβ, mientras que otras moléculas CD1 se reconocen por los T γδ. Tras la activación estas células secretan grandes cantidades de IL-4 e IFN-g. El déficit en NKT se asocia con enfermedades autoinmunes, tales como diabetes, lo que las implica como células reguladoras. DIFERENCIACION LINFOCITARIA Th1-Th2 Las células T que emigran del timo son células maduras CD4+ o CD8+ en estado de reposo, denominados “vírgenes”, ya que todavía no han encontrado a su antígeno específico. Estas células pasan a los órganos linfoides secundarios (OLS) a la búsqueda del antígeno. Una vez en el área T correspondiente interaccionan con las DCs (que han llegado desde la periferia) “examinando” los complejos antigénicos que estas presentan en su superficie. Si tiene lugar el reconocimiento los linfocitos T se activan, se expanden y se diferencian a linfocitos T efectores. La activación de las células CD4+ vírgenes puede llevar a dos perfiles diferentes de células efectoras: Th1 y Th2. Los primeros median en la respuesta frente a patógenos intravesiculares (intracelulares) o endocitados al compartimiento vacuolar; para ello inducen la activación del macrófago, que es la principal célula efectora de la respuesta Th1. Las células Th2 conducen el enfrentamiento con los patógenos extracelulares; para ello colaboran con los linfocitos B para que estos desarrollen una respuesta humoral efectiva. Producción de células T efectoras Interacción T virgen-DC. Sinapsis inmunológica La unión temporal T virgen-DC, necesaria para el “examen”, se produce mediante interacciones inespecíficas de pares de moléculas de adhesión, sobre todo las que se establecen entre LFA-1 del linfocito T y sus ligandos (ICAM-1, ICAM-2) sobre las DCs. En esta interacción las moléculas actuantes se disponen en círculos concéntricos, donde los TCR que reconocen y los complejos péptido-Clase II reconocidos ocupan el área central y las moléculas de adhesión están en la periferia; a estos grupos se les denomina clusters supramoleculares de activación centrales (c-SMAC) y periféricos (p-SMAC), respectivamente. Los clusters (grupos) se forman después del comienzo de las señales generadas por el complejo TCR-CD3. El complejo supramolecular recibe el nombre de Sinapsis inmunológica y su formación potencia las señales inducidas a través del TCR. También impone un límite temporal, ya que la agregación extensiva del c-SMAC
formulandoalgoqueaportar.blogspot.com

favorece la regulación negativa (disminución) de los TCRs expresados sobre la superficie celular. La activación y la expansión clonal de la célula T dependen de dos señales diferentes
• 1: depende del péptido antigénico presentado junto al CMH a través del TCR • 2: depende de las moléculas coestimuladoras expresadas por las DCs
En ausencia de la señal 2 la célula T entra en un estado de anergia La interacción CD40L-CD40 mejora la activación T Al activarse, las células T expresan la molécula CD40L, que interacciona con la molécula CD40 expresada por las CPAs de forma constitutiva. Esto produce señales estimuladoras en el LT e induce en la CPA el aumento de la expresión de B7, lo que se traduce en una mayor activación T. Estas moléculas pertenecen a la gran familia TNF. Control de la expansión clonal CTLA-4 (CD152) controla el resultado de la expansión clonal Su expresión se induce en las células T después de la activación. Tiene cierta homología con CD28 por lo que no es extraño que comparta sus ligandos (B7). Pero la interacción CTLA-4-B7 no lleva a la activación T sino que induce una señal inhibidora fuerte. Su expresión (36-72 h. tras la activación) desplaza a CD28 de la unión gracias a la mayor afinidad (unas 20 veces más) por los ligandos. Los ratones deficientes en CTLA-4 mueren a las 3-4 semanas por el infiltrado masivo de sus órganos por LT expandidos. El sistema Fas-FasL en el control de la expansión clonalLos linfocitos T efectores (y no los vírgenes) expresan la proteína FasL (de la familia TNF), que interacciona con el Fas (CD95, de la familia TNFR) que se expresa de modo constitutivo sobre los linfocitos. La interacción lleva a la apoptosis. Los pacientes defectivos en Fas o FasL presentan procesos linfo-proliferativos normalmente mortales. Presencia del antígenoLa expansión requiere la presencia de antígeno, por lo que la eliminación del mismo, como consecuencia de la respuesta, redunda en el cese de la expansión clonal. CÉLULAS T EFECTORAS: Th1 y Th2 La estimulación T y la polarización Th1 / Th2 requiere tres señales derivadas de las DCs La señal 1 es la específica del antígeno, obtenido de los péptidos procedentes del procesamiento de las moléculas del patógeno tras la internalización por PRRs especializados, y que se libera presentándolo al TCR en conjunto con la molécula CMH II. La señal 2 es la co-estimuladora, mediada principalmente por B7.1/2 (CD80/86) y CD28. B7.1/2 se expresa por las DCs tras la unión de PRRs (TLRs) especializados en detectar la infección mediante el reconocimiento de PAMPs o TFs La señal 3 es la polarizadora, mediada por varios factores solubles o ligados a la membrana, tales como IL-12 y la quimiocina CCL2, que promueven el desarrollo respectivamente de células Th1 o Th2. La naturaleza de la señal 3 depende de la activación de PRRs particulares o de los factores de transcripción (TFs).
formulandoalgoqueaportar.blogspot.com

Los PAMPs Tipo 1 y Tipo 2 y los TFs pueden definirse como aquellos que marcan a las DCs para que produzcan altos niveles de factores polarizantes 1 o 2. Mientras que el perfil polarizante T viene condicionado por el reconocimiento de PAMPs, su expresión óptima requiere normalmente la retroalimentación por el CD40L expresado por las células T tras la activación por las señales 1 y 2. Características y funciones La respuesta inmune debe inducir una serie adecuada de funciones inmunes que puedan eliminar del hospedador al agente tóxico o a sus toxinas. Un ejemplo de función adecuada es la neutralización de toxinas bacterianas solubles, que requiere anticuerpos, mientras que para enfrentarse a un patógeno intracelular hace falta una respuesta celular (citotóxica o de hipersensibilidad retardada). Para ello hacen falta que se activen distintas subpoblaciones Th (CD4+), productoras de patrones de citocinas diferentes. La mayoría de las funciones de las células T CD4+ se ejercen mediante la secreción de citocinas, que pueden actuar sobre las propias células que las producen (acción autocrina) o modular la respuesta de otras células a través de la vía paracrina. Las subpoblaciones Th1 y Th2 producen GM-CSF e IL-3, pero difieren en el resto de la citocinas que producen. Ambas subpoblaciones se distinguen por las siguientes funciones que desempeñan:
• La subpoblación Th1 es responsable de muchas funciones mediadas por células, como la hipersensibilidad retardada y la activación de las células T citotóxicas (CD8+), así como de la producción de anticuerpos IgG opsonizantes (que se fijan con alta afinidad a los receptores Fc de los fagocitos e interaccionan con el sistema del complemento). Esta subpoblación también se asocia con la producción excesiva de inflamación y con el daño tisular
• La subpoblación Tg2 induce la activación y diferenciación de los eosinófilos, ayuda a las células B y promueve la producción relativamente abundante de IgM, IgE y de de isotipos de IgG que no activan (o poco?) el complemento. Las células Th2 también son las responsables de las reacciones alérgicas.
Subpoblaciones Th1 y Th2. Patrón de citocinas y funciones de
las mismas Citocina Th1 Th2 Función Th1 Th2 IL-2 + - Ayuda para la producción total de Ac + ++ IFN-γ ++ - Ayuda para la producción de IgE - ++ TNF-β ++ - Ayuda para la producción de IgG2 ++ + GM-CSF ++ + Producción de eosinófilos y mastocitos - ++ IL-3 ++ ++ Activación de macrófagos ++ - IL-4 - ++ Hipersensibilidad retardada ++ - IL-5 - ++ Activación Tc ++ - IL-10 - ++ IL-13 - ++
Las distintas citocinas que producen las células Th1 y Th2 determinan la diferencia en las funciones biológicas: Subpoblación Th1 Las Th1 median una respuesta inflamatoria a nivel de los tejidos periféricos, donde la célula efectora principal es el macrófago activado por el IFN-g. En los tejidos infectados
formulandoalgoqueaportar.blogspot.com

este macrófago recibe, además, señales a través de los PRRs, lo que da como resultado un macrófago con alta potencialidad microbicida e inflamatoria. IFN-γ, además de activar a los macrófagos en su capacidad microbicida, hace que aumente la expresión de moléculas Clase II (actividad presentadora) y la secreción de citocinas que, como IL-12, llevan a las células T vírgenes a diferenciarse a Th1. El IFN-γ producido por las células Th1 también induce el cambio de clase de anticuerpos a subclases de IgG que promueven la fijación del complemento (opsonizantes) y la fagocitosis. La secreción de TNF e IFN-γ es parte de la contribución de las células Th1 a procesos inflamatorios, tales como la hipersensibilidad retardada. IL-2 e IFN-γ promueven la diferenciación de los precursores T citotóxicos (CD8+ vírgenes) a Tc funcionales. Además, y como colofón, IFN-γ inhibe la expansión de la subpoblación Th2. Subpoblación Th2 La secreción de IL-4 e IL-5 induce la producción de IgE y es el soporte para el ataque a los gusanos por parte de los eosinófilos. IL-4 promueve el cambio de clase a las IgG que no fijan el complemento y también el cambio de IgM a IgE. La producción de IgE se coordina con la diferenciación y activación de eosinófilos, que presentan gran cantidad de receptores para el fragmento Fc de la IgE (FcεR). Los anticuerpos IgE reaccionan con los helmintos e interaccionan después con los receptores fc de los eosinófilos, formando así un puente antígeno-específico entre gusano y eosinófilo. La unión a la IgE promueve el entrecruzamiento de los receptores, lo que desencadena el ataque de los eosinófilos a los gusanos. Además de esta función defensiva, la IgE es también la responsable de la alergia. Finalmente, IL-4 e IL-10 suprimen la expansión de las poblaciones Th1 Th1 y Th2 tienen pautas migratorias diferentesAl activarse las T vírgenes pierden la L-Selectina por lo que, una vez que emigran, ya no regresan a los OLS. Las T vírgenes expresan CCR7, que reconoce a las quimiocinas producidas en el área T de los órganos linfoides secundarios, por lo que se acumulan en los mismos. Después de la activación pierden este receptor de quimiocinas y expresan nuevos receptores. Las células Th1 expresan los receptores de quimiocinas CCR5 y CXCR3, que capturan las quimiocinas inflamatorias producidas por la activación de las células endoteliales, epiteliales y leucocitos del foco inflamatorio. Además, el endotelio de los vasos del foco inflamatorio aumenta enseguida la expresión de E y P- selectina, y las Th1 se caracterizan por expresar altas concentraciones de los ligandos para estas selectinas. Estas 2 características (expresión de receptores de quimiocinas y expresión de ligandos para E y P-selectinas) garantizan que las Th1 van al foco inflamatorio naciente. Las células Th2 aumentan la expresión de CCR3, que es el receptor para una quimiocina (eotaxina) que también participa en el reclutamiento de eosinófilos en las mucosas digestiva y pulmonar y se asocia con procesos antiparasitarios y alérgicos. Las citocinas Th2 estimulan la producción de eotaxina en los tejidos periféricos. CCR3 se expresa también en eosinófilos y mastocitos, lo que permitiría la localización de los tres tipos celulares en los sitios donde se produce la quimiocina.
formulandoalgoqueaportar.blogspot.com

Th1 y Th2 son células efectoras que intervienen en la protección frente a patógenos diferentes y, también, en la etiología de procesos autoinmunes distintos. Diferenciación a Th1 Las DCs que llegan a los OLS tienen distintas propiedades funcionales según las interacciones establecidas por sus PRRs con los microbios (o sus productos) en el tejido infectado (Teoría de la tercera señal). Esto condiciona las citocinas y quimiocinas que expresan en el OLS y que refleja, en definitiva, su historia en el tejido infectado. Las citocinas IL-12, 18, 23 y 27, producidas por las DCs, orientan la diferenciación de las CD4 vírgenes hacia Th1. La citocina principal parece ser IL-12. IL-12Heterodímero formado por dos subunidades (p35 y p40) que ejerce sus efectos a través de un receptor de alta afinidad también formado por dos cadenas (IL-12Rbeta1 e IL-12Rbeta2) y que no se expresa sobre las T vírgenes pero si en las activadas. Las que pasan a Th1 mantienen la expresión del receptor pero las que pasan a Th2 la pierden. Por tanto, además de dirigir al perfil Th1 IL-12 también estimula su proliferación. También actúa como factor de crecimiento para las células NK, que expresan el receptor de alta afinidad. IL-18Favorece la diferenciación a Th1 inducida por IL-12 y aumenta, en estas células, la producción de IFN-g IL-23Estimula la producción de IFN-g por las Th1 y su expansión clonal IL-27Recientemente descrita, comparte propiedades con IL-12 (diferenciación de CD4+ a perfil Th1 y estimulación de expansión clonal) y con IL-18 e IL-23 (potencia la producción de IFN-g por las Th1) Diferenciación a Th2 Parece requerir la presencia de IL-4. Esta citocina se une a su receptor y al de IL-13, lo que explica el que ambas citocinas medien funciones similares y contribuyan a la diferenciación de las células T CD4+ a un perfil Th2. Las células productoras de IL-4 han sido motivo de polémica. Los datos recientes indican que son las NKT las que desempeñan un papel central.
formulandoalgoqueaportar.blogspot.com

Tema 16. Respuesta efectora Th2 e inmunidad humoral
TEMA 16. RESPUESTA EFECTORA Th2: COOPERACION CON LOS LINFOCITOS B EN LA RESPUESTA HUMORAL LA RESPUESTA HUMORAL La respuesta primaria y la secundaria difieren significativamente Activación de Linfocitos B vírgenes →→→→ Respuesta 1ª Activación de Linfocitos B de memoria →→→ Respuesta 2ª El primer contacto con un antígeno exógeno lleva a la respuesta primaria, caracterizada por la formación de células secretoras de anticuerpos (plasmocitos) y células B de memoria. Tras la penetración del antígeno hay una fase de latencia que se corresponde a:
• La célula B virgen endocita al antígeno, lo procesa y lo presenta unido a moléculas Clase II a las células T específicas
• Expansión de la población de células B • Diferenciación a células plasmáticas productoras de anticuerpos
Seguida por una fase logarítmica de producción de anticuerpos que se estabiliza en un máximo y comienza a descender paulatinamente. Para los antígenos proteicos la fase de latencia es de aproximadamente una semana y el pico máximo de producción de anticuerpos se alcanza en unos 14 días. Los anticuerpos secretados durante una respuesta primaria son inicialmente IgM que luego cambian a IgG. Las células de memoria que se forman durante la respuesta primaria cesan de dividirse y entran en la fase de reposo (Go) del ciclo celular. La capacidad para desarrollar una respuesta secundaria depende de la existencia de esta población de células B de memoria (Bm), así como de las Tm. La activación por el antígeno de las células de memoria lleva a una respuesta con menor tiempo de latencia, mayor magnitud de producción de anticuerpos y más duradera. También se caracteriza por la producción de anticuerpos con mayor afinidad para el antígeno. Cuadro comparativo Respuesta 1ª Respuesta 2ª Tipo de LB que interviene B virgen B de memoria Tiempo de latencia 4-7 días 1-3 días Máximo de respuesta de Ac. 7-10 días 3-5 días Magnitud de la respuesta Según el Ag. 100 a 1000 veces + Isotipo predominante IgM IgG
T-dependiente y T-independiente Tipo de antígeno T-dependiente
Afinidad de los Acs Baja Alta La menor duración de la fase de latencia en la respuesta 2ª se debe a que la población de células B de memoria, específicas para el antígeno, es considerablemente mayor que la población correspondiente de células B vírgenes específicas. Además, las células de memoria se activan con más facilidad que las vírgenes. Sitios donde tiene lugar la respuesta inmune La activación y diferenciación de las células B se produce en sitios anatómicamente definidos, cuya estructura permite las interacciones celulares necesarias. Cando un
Página 1 de 4
formulandoalgoqueaportar.blogspot.com

Tema 16. Respuesta efectora Th2 e inmunidad humoral
antígeno penetra en el cuerpo se concentra en varios órganos linfoides periféricos; los que van por la sangre se filtran en el bazo, mientras que los tisulares llegan con la linfa a los ganglios linfáticos (GL). Un GL es un filtro eficiente, capaz de atrapar >90% e cualquier antígeno que llegue por los linfáticos aferentes. Los antígenos (o los complejos Ag-Ac) entran los GL solos o transportados por células presentadoras (DCs) y, mientras percolan por la arquitectura celular del ganglio tienen que encontrar alguna CPA profesional que se encargará d presentarlo a las células T (recordar que las células B son células presentadoras). Las células T y B específicas para el antígeno se aproximan unas a otras con rapidez (<1m.) hasta formar un conjugado en el para-cortex (interfase de las zonas T y B dependientes). Una vez que el antígeno ha activado a las células B se forman pequeños focos de proliferación en el borde de la zona T-dependiente y estas células luego se diferencia a plasmocitos y células de memoria. Estos plasmocitos crucen principalmente IgM e IgG y la mayoría de los anticuerpos producidos durante una respuesta 1ª proceden de las células plasmáticas de esos focos de proliferación. En el bazo hay una secuencia similar, produciéndose los focos de proliferación de las células B en la vaina linfoide peri-arteriolar (T-dependiente). Pocos días después de la formación de los focos de proliferación hay una emigración de células T y B desde ellos hasta los folículos linfoides primarios, que con la llegada de estas células se transforman en secundarios y aportan un entorno favorable para la interacción entre LB, LT y CDFs (células dendríticas foliculares, que no proceden de la médula ósea, que no expresan moléculas Clase II, ni presentan antígenos a las T CD4+). Las CDFs tienen muchas ramificaciones, en las que se acumulan los receptores para el fragmento Fc de los anticuerpos y para fragmentos del complemento, que les permiten capturar y retener los complejos Ag-Ac durante tiempos prolongados. Las células B activadas, junto con algunas células T activadas, pueden emigrar hacia el centro del folículo 2ª formando un centro germinal (CG). Centros germinales y diferenciación B inducida por el Ag Los CG aparecen a los 7-10 días de la exposición a un antígeno timo-dependiente. En el CG se producen tres acontecimientos principales:
• La maduración de la afinidad de los anticuerpos • El cambio de clase de los anticuerpos • La formación de células plasmáticas y células de memoria
Pero en lugares fuera del CG también se pueden producir algunos cambios de clase y formarse un número significativo de células plasmáticas productoras de anticuerpos. Durante la primera fase de formación del CG las células B activadas experimentan una intensa proliferación; durante este periodo se las conoce como Centroblastos y están en la zona oscura del CG. Los centroblastos se caracterizan por mayor tamaño, citoplasma expandido, cromatina difusa y ausencia (o casi ausencia) de Ig de superficie. Los centroblastos dan lugar a Centrocitos, que son células pequeñas que ya expresan Ig sobre la membrana. Estos centrocitos se mueven desde la zona oscura hasta otra zona clara, caracterizada por la presencia de CDFs que “enseñan” en la superficie de sus ramificaciones los complejos Ag-Ac retenidos por los receptores para el fragmento Fc y para fracciones del complemento.
Página 2 de 4
formulandoalgoqueaportar.blogspot.com

Tema 16. Respuesta efectora Th2 e inmunidad humoral
La maduración de la afinidad es el resultado de mutación y selección.La afinidad media de los anticuerpos que se producen durante la respuesta humoral aumenta notablemente, hasta el punto de hacerlo unas 140 veces entre la semana 2 y la 8 post-inoculación. La maduración de la afinidad de los anticuerpos es el resultado del proceso de hipermutación somática. La hipermutación somáticaSe produce sobre todo en las células B del centro germinal, aunque también las hay en células B activadas por el antígeno en otros focos. Estas hipermutaciones se producen en los genes V(D)J de los dominios variables y, concretamente, a nivel de las CDRs (regiones determinantes de la complementariedad o hipervariables). Esta tasa de mutación se ha calculado en 10-3 /pares de bases/ división, lo que supone una tasa de mutación un millón de veces superior a la que se presenta en otros genes humanos. Como entre los genes variables de las cadenas H y L suman unos 700 pares de bases, cada centroblasto debe sufrir una mutación cada dos divisiones (en una u otra cadena). Como la hipermutación somática es al azar habrá unas pocas células con receptores de alta afinidad, mientras que el resto o no ha cambiado su afinidad o la presentan menor. Aquí es donde entra la selección, que permitirá el desarrollo de una población de alta afinidad, mientras que las células de media o baja afinidad no pueden competir con éxito en la captura del antígeno y mueren en el centro germinal. El papel de la selecciónLa hipermutación ocurre cuando los centroblastos proliferan en la zona oscura del CG y la selección se realiza sobre los centrocitos en la zona clara. La clave es la capacidad de la Ig de superficie de los centrocitos para reconocer y unirse a los antígenos que les “enseñan” las CDFs; estos centrocitos deben competir por la pequeña cantidad de antígeno retenida sobre las CDFs y si consiguen unirse reciben, a través del receptor, una señal imprescindible para la supervivencia. Los que no reciben la señal mueren. Aunque esta señal es necesaria para los centrocitos no es suficiente; debe recibir, además, otras señales en la interacción con células Th CD4+ para sobrevivir. Para esta interacción es indispensable la unión de CD40 del centrocito con la CD40L de la célula T. Como es lógico, también es necesario que el antígeno capturado mediante el receptor, endocitado y procesado, se presente a la célula Th unido a moléculas Clase II. El cambio de clase de las IgEl reconocimiento en los anticuerpos depende de las regiones variables H y L, mientras que las propiedades biológicas dependen del isotipo de regiones constantes de las cadenas H. El cambio de clase es el proceso mediante el cual un dominio V se puede asociar con los dominios constantes de cualquier isotipo. La decisión depende de las citocinas que aporta la interacción con las células Th. Las células B de memoria y las plasmáticas se forman en el centro germinal Tras la selección en el centro germinal, algunas células B se diferencian a células plasmáticas, mientras que otras se transforman en células de memoria. Aunque los centros germinales son sitios importantes para la generación de células plasmáticas, también pueden formarse células productoras de anticuerpos en otros lugares.
Página 3 de 4
formulandoalgoqueaportar.blogspot.com

Tema 16. Respuesta efectora Th2 e inmunidad humoral
BASES DE LA HIPERMUTACION SOMATICA Y EL CAMBIO DE CLASE AID media la hipermutación y el cambio de clase En las Ig humanas y murinas hay tres tipos de modificaciones génicas:
1. El reordenamiento y unión de segmentos génicos de la línea germinal, que durante la recombinación V(D)J forma un gen variable funcional
2. La hipermutación somática introduce cambios en las regiones variables de los genes de las Ig, lo que puede afectar a las propiedades de fijación al antigeno de los anticuerpos correspondientes
3. La recombinación durante el cambio de clase lleva a un gen V(D)J a asoiarse con regiones constantes diferentes, lo que modifica las funciones efectoras de la molécula de Ig.
Las recombinasas RAG-1 y RAG-2, son las responsables de la recombinación V(D)J, mientras que la enzima AID es la mediadora clave de la hipermutación y de la recombinación que lleva al cambio de clase. AID pertenece a una familia de “enzimas editoras de RNA” y actúa desaminando a citosinas seleccionadas en ciertas cadenas de RNA mensajero, con lo que estas pasan a uracilos, con lo que se alteran (se editan) las instrucciones codificadoras de proteínas de ese mRNA, Parece que esta enzima también es capaz de modificar directamente al DNA mediante el mismo procedimiento, que lleva a la formación de uracilo; pero este nucleótido no es de los habituales del DNA y el sitio de deaminación tiene que repararse para formar un par A-T en lugar del C-G original o, alternativamente, eliminando directamente al uracilo y sustituyéndolo por cualquiera de las 4 bases del DNA
Las células plasmáticas generalmente carecen de Ig de membrana y, en su lugar, sintetizan grandes cantidades de anticuerpos. La diferenciación de las células B maduras a plasmocitos requiere un cambio en el procesamiento del RNA, de modo que se sintetice la forma soluble de la cadena H. Además, la velocidad de trascripción es mucho mayor en las células plasmáticas que en las células B maduras, llegando
a producir un millón de moléculas de anticuerpo/segundo/célula plasmática.
Nucleósido
Nucleótido
AID
Acción de AID
En cuánto a las células B de memoria, excepto por las Ig de membrana, casi no se diferencian de las células B vírgenes.
Página 4 de 4
formulandoalgoqueaportar.blogspot.com

Tema 17. REGULACION: TOLERANCIA. MEMORIA INMUNE TOLERANCIA A comienzos del S XX Paul Ehrlich indicó que el sistema inmune también podía equivocarse y centrar su ataque, además de en los antígenos extraños, en los antígenos propios. Esta situación, que Ehrlich describió como horror autotoxicus, puede provocar una serie de enfermedades. En definitiva, estas enfermedades son la consecuencia de no distinguir (o hacerlo mal) entre lo propio y lo no propio. Hay unos mecanismos que protegen frente a los linfocitos potencialmente autoinmunes y el conjunto de tales mecanismos recibe el nombre de Tolerancia. El mecanismo inicial es la Tolerancia Central, que elimina a clones B y T, antes de su maduración, si portan receptores que reconocen a los antígenos propios con mayor afinidad de la adecuada. Esta tolerancia central se realiza en los órganos linfoides primarios (OLP). Como la tolerancia central no es perfecta siempre habrá linfocitos auto-reactivos capaces de alcanzar los órganos linfoides periféricos, por lo que se pone en marcha a este nivel una segunda línea de control destinada a limitar su actividad; entre los mecanismos de este segundo nivel destaca la Tolerancia Periférica que inactiva o vuelve anérgicos a los linfocitos auto-reactivos. La posibilidad de que los linfocitos auto-reactivos produzcan daño también se limita por la expectativa de vida de los linfocitos activados, que ponen en marcha su programa de destrucción (apoptosis) cuando reciben la señal. Pero, a pesar del control ejercido a distintos niveles, hay clones de linfocitos auto-reactivos B y T que se activan y responden frente a los antígenos propios. Tal respuesta recibe el nombre de Autoinmunidad. Establecimiento y mantenimiento de la tolerancia La tolerancia se define como el estado en el que no se responde a un antígeno, ya que normalmente el contacto con el antígeno lleva a la respuesta. Esta tolerancia puede deberse a un modo imperfecto de presentación o a la alteración del propio antígeno, El que un antígeno se comporte como Tolerogénico o Inmunogénico depende de factores tales como la vía de penetración, la dosis, etc.
Señales en la presentación.
Linfocito maduro
Ag Tolerogénico Ag Inmunogénico
ProliferaciónApoptosis Anergia
Linfocito maduro
Ag Tolerogénico Ag Inmunogénico
ProliferaciónApoptosis Anergia
La falta de señal coestimuladora durante la presentación lleva a la anergia, mientras que si la presentación se realiza con las dos señales preceptivas se produce una respuesta potente. Estado de maduración de la DC. La presentación por DCs inmaduras lleva a la tolerancia, mientras que la presentación por
DCs maduras provoca la respuesta.
formulandoalgoqueaportar.blogspot.com

Factores que promueven la tolerancia • Dosis altas de antígeno • Persistencia del antígeno • Administración oral o subcutánea • Niveles coestimuladores bajos
Tolerancia oral Cuando un antígeno, inmunogénico por vía subcutánea o intradérmica, se administra por vía oral puede que no se produzca la respuesta frente a el. Se provoca una tolerancia antígeno-específica frente al antígeno tolerogénico sin que haya una inmunosupresión general. La tolerancia puede inducirse En 1953 se publicó en Nature el artículo de Medawar et al. “Actively Adquired Tolerance of Foreign Cells” que sentó las bases de un dogma central de la inmunología: se puede instruir al sistema inmune para que no rechace a un antígeno particular. El fenómeno se denominó Tolerancia inmunitaria. La inoculación sistemática de un antígeno extraño (p.e. albúmina de otra especie) a hembras preñadas induce la tolerancia a ese antígeno en la descendencia, ya que los fetos han estado en contacto constante con dicho antígeno durante el periodo de selección clonal y lo consideran como propio. La recirculación de células maduras auto-reactivas no lleva obligatoriamente a la autoinmunidad. Está comprobado que de los órganos linfoides primarios salen células auto-reactivas maduras, pero que esto no lleva obligatoriamente a la autoinmunidad pues existen otros mecanismos reguladores, sobre todo la limitación a la activación celular que imponen las células T reguladoras. TOLERANCIA CENTRAL Limita el número de células auto-reactivas mediante la delección clonal, durante la maduración en los órganos linfoides primarios, de las que reaccionan con Ag propios. Parte de la diversidad de las células B y T dependen del reordenamiento génico al azar de V(D)J (diversidad combinatoria), lo que hace previsible el que algunos receptores reaccionen con lo propio. La tolerancia central elimina a estos clones mediante (entre otros mecanismos) la Selección Negativa. Antes de esta eliminación las células auto-reactivas tienen la oportunidad de editar un nuevo receptor, que tendrá que pasar por la selección pertinente. Delección clonal y edición del receptor son los mecanismos básicos de esta Tolerancia. TOLERANCIA PERIFERICA Controla a las células auto-reactivas que pasan a la circulación, pues la Tolerancia Central no elimina a todos los linfocitos auto-reactivos, ya que:
1. Los Órganos Linfoides Primarios no expresan todos los auto-antígenos y es en ellos en los que se realiza la selección
2. La afinidad de la unión condiciona la selección, lo que permite a algunos clones débilmente auto-reactivos sobrevivir a la selección.
La Tolerancia Periférica se encarga de inactivar a estas células auto-reactivas para que no puedan reaccionar contra lo propio.
formulandoalgoqueaportar.blogspot.com

El mecanismo básico de inactivación T es el reconocimiento a través del receptor (señal 1) sin las señales coestimuladoras procedentes de la unión B7-CD28 (señal 2). CTLA-4 Tras la activación celular se induce la expresión por el linfocito de otra molécula de superficie con mayor avidez para B7 que CD28. La nueva unión genera una señal inhibidora que impide la activación celular. La expresión de CTLA-4 asegura el control y la regulación de la respuesta. De hecho, los animales CTLA-4 KO presentan una proliferación masiva de linfocitos y enfermedades autoinmunes lo que nos da una idea de la importancia de esta molécula en el mantenimiento de la tolerancia periférica. La muerte celular Tiene un papel importante en el mantenimiento de la tolerancia, tanto central como periférica. La unión de la pareja Fas-FasL induce una apoptosis rápida, conocida como AICD (muerte celular inducida por activación). Hay animales que presentan mutaciones espontáneas de estos receptores, por lo que no entran en la vía AICD tras la activación y desarrollan con rapidez enfermedades autoinmunes. Las células T reguladoras forman parte de la tolerancia periférica. La tolerancia periférica se mantiene también mediante células T reguladoras, que se activan en los órganos linfoides 2º (o en los sitios de inflamación) y controlan los procesos autoinmunes. Estas células T reguladoras (Treg) son las únicas células CD4+ que expresan niveles altos de la cadena α (CD25) del receptor para IL-2 y proceden de una subpoblación tímica que expresa receptores con afinidad media para auto-antígenos; algunas de ellas regulan de modo positivo la expresión del factor represor de trascripción Foxp-3, lo que las dirige hacia Treg capaces de inhibir la respuesta frente a los auto-antígenos. Células Treg naturales (Sakaguchi, 1995) Representan, aproximadamente, el 10% de las células T circulantes. El marcador CD25 Además de CD4 y CD25, estas células expresan también moléculas como CTLA-4, el receptor TNF inducido por glucocorticoides (GITR) y el receptor de quimiocinas CCR4 El marcador Foxp-3 Los ratones Scurfy desarrollan una enfermedad linfo-proliferativa muy parecida a la enfermedad humana IPEX. Ambas, ligadas al cromosoma X, son consecuencia de la mutación o falta del gen Foxp-3. El gen codifica para la proteína escurfina, que actúa como represor de la transcripción y regula la activación de las células T. Migración y tráfico de las T reguladoras naturales.
formulandoalgoqueaportar.blogspot.com

Emigran desde el timo como una población T madura. Sus moléculas de adhesión y receptores de quimiocinas les permiten alcanzar los OLS y los tejidos inflamados. Mecanismo de acción Tras su estimulación, a través del TCR (por antígenos propios o extraños), inhiben la activación, expansión y producción de citocinas por las células Th1, Th2 y Tc. Detectan concentraciones mínimas de antígenos (entre 10 y 100 veces menos que las necesarias para activar a las T CD4+ CD25- de la misma especificidad).
Las CD4+ CD25+ se activan de forma específica por su antígeno pero, una vez activadas, la acción supresora sobre las Th1, Th2 y Tc es inespecífica.
Entre los mecanismos supresores están: Secreción de citocinas supresoras (IL-10 y TGF-b) Inducción de señales inhibidoras en las CD4+ y CD8+ por contacto directo
célula-célula (cada vez hay más evidencias de que es el mecanismo básico) Inhibición de la capacidad presentadora de las DCs
Células Treg inducibles Células Tr-1 Proceden de células T vírgenes que adquieren el perfil regulador tras su activación por las DCs semi-maduras presentes en los OLS, caracterizadas por la baja expresión de CD40, baja producción de IL-12 y alta producción de IL-10. Estas DCs semi-maduras son células que emigran desde la periferia sin haber recibido señales indicativas de un proceso inflamatorio (LPS o TNF-a como 3ª señal) y arriban a los OLS sin alcanzar la madurez, actuando como Tolerogénicas. En cambio, las que reciben señales de inflamación en la periferia (a través de sus TLRs o de receptores de citocinas) maduran y se convierten en DCs activadoras de las CD4+ vírgenes. Un punto crucial es que estas DCs maduras producen IL-6, que es capaz de bloquear la acción inhibidora mediada por las células T reguladoras. Producen, fundamentalmente IL-10, y también TGF-b, e inhiben la activación, expansión y producción de citocinas por las Th1, Th2 y Tc. Su papel parece ser el silenciar clones auto-reactivos de la periferia, así como el control de la respuesta inmune frente a microbios y tumores. CELULAS Th3 La administración de un antígeno por vía oral o la estimulación in vitro de células T CD4+CD25- en presencia de TGF-b induce la formación de células T CD4+CD25+ productoras de TGF-b. Las células Th3 controlan, mediante TGF-b, la respuesta T frente a los componentes de la flora intestinal. La acción supresora se basa en inhibir la diferenciación de las células T vírgenes, tanto CD4+ como CD8+, a células efectoras o en suprimir la actividad de tales células cuando ya se han diferenciado. TGF-β actúa a nivel molecular inhibiendo la expresión de los factores de trascripción Gata-3 y T-bet, críticos para la diferenciación de las células T vírgenes a Th2 y Th1,
formulandoalgoqueaportar.blogspot.com

respectivamente. También puede ejercer un efecto supresor indirecto por su capacidad inhibitoria de la funcionalidad de las DCs. El secuestro antigénico protege de los auto-antígenos (sitios de privilegio inmune) Además de los mecanismos de tolerancia inmune anteriores, un sistema muy eficaz para evitar la auto-reactividad es el secuestro o compartimentación de los auto-antígenos. De este modo, en circunstancias normales, tales antigenos no pueden entrar en contacto con los linfocitos reactivos y es imposible la reactividad. Estos sitios de privilegio inmune, caracterizados por aceptar trasplantes alogénicos. son únicos por dos motivos:
1. La comunicación entre el sitio y el resto del cuerpo es atípica, ya que sus fluidos extracelulares no pasan por los linfáticos convencionales.
2. En ellos se producen citocinas que afectan al microambiente de la respuesta inmune. En este aspecto TGF-β parece ser muy importante: los antígenos mezclados con TGF-β parecen inducir respuestas que no dañan los tejidos, tales como la Th2.
Ciertas partes del cuerpo, como ojo, cerebro, sistema gastro-intestinal y reproductor, suprimen la respuesta inmune frente a patógenos. Si se respondiese, los delicados tejidos del ojo se destruirían por la inflamación y el tracto gastro-intestinal no toleraría la ingestión de alimento. El mecanismo supresor consiste en la llegada de los antígenos a los OLS, donde su presentación a los LT, en lugar de inducir su activación, induce su falta de respuesta (tolerancia).
El caso del ojo. Hay células presentadoras que contienen una proteína (F4/80) y que trasportan los antígenos desde el ojo al bazo, donde estimulan la producción de células T reguladoras que detienen la respuesta inmune, tanto a nivel general como en el sitio de invasión (en este caso el ojo). En una respuesta efectora se estimulan las células T que desencadenan el ataque inmune, con inflamación y destrucción de tejidos. En los animales sin F4/80 no hay inmuno-supresión cuando los Ag extraños se presentan en el bazo por estas CPAs, lo que indica un papel directo para F4/80 en la supresión de la respuesta inmune. Otra consecuencia del secuestro antigénico es que la falta de contacto con el antígeno durante el desarrollo de los linfocitos no induce tolerancia frente a el. Si se rompen las barreras entre antígeno y linfocitos (por inoculación, trauma, etc.) el sistema inmune responderá. Tras la ruptura de la barrera hemato-encefálica, por ejemplo, puede haber una reacción inmune frente a componentes del SNC. EL EJE INMUNO-NEURO-ENDOCRINO Cuando se profundiza en el estudio del sistema inmune se corre el riesgo de mirar al organismo como una serie de células linfoides y mieloides que se desplazan dentro de un gran saco, olvidando que la fisiología de cualquier organismo es un sistema integral. Influencia neuro-endocrina sobre los procesos inmunes Las células inmunitarias tienen receptores para una serie de hormonas: corticosteroides, insulina, hormona del crecimiento, estradiol, testosterona, acetilcolina, endorfinas y encefalinas, etc. En general:
• glucocorticoides y andrógenos disminuyen la respuesta, • estrógenos, hormona del crecimiento, tiroxina e insulina la potencian.
formulandoalgoqueaportar.blogspot.com

La producción de glucocorticoides es la principal respuesta al estrés, aunque también se liberan durante la respuesta inmune, actuando como factor limitante a través de un eje neuro-endocrino. Estos glucocorticoides suprimen la respuesta Th1 y aumentan la Th2, por lo que las personas con predisposición genética a producir grandes cantidades de glucocorticoides en situaciones de estrés presentan susceptibilidad alta a la infección por patógenos intracelulares (pues estos se eliminan por la respuesta Th1). Se considera a los estrógenos como los responsables principales de que la respuesta sea mayor en la mujer que en el hombre. Ellas tienen mayores niveles de Ig séricas y de IgA secretora, más respuesta a antígenos T-independientes, más resistencia a las infecciones y una resistencia relativa a desarrollar tolerancia T. Son, en cambio, más susceptibles a los procesos autoinmunes. Influencia inmune sobre los mecanismos neuroendocrinos. IL-1 estimula la síntesis de glucocorticoides. También actúa sobre el SNC, donde puede llegar vehiculada en la sangre o bien producirse localmente; su acción se manifiesta en: 1.- Fiebre. Actúa sobre el centro de regulación térmica del hipotálamo; una temperatura superior a 38,5ºC: Aumenta:
• La capacidad microbicida de los PMN, • La acción antiviral de los IFNs, • La proliferación linfocitaria, • La respuesta frente a los antígenos T-dependientes, • La acción citolítica de los LTc, • La activación de la vía alternativa del complemento
No afecta: • A la actividad de los macrófagos (fuente principal de IL-1)
2.- Sueño. Hay una asociación entre la actividad de IL-1 y la inducción de la fase lenta del sueño. 3.- Regulación. IL-1 actúa sobre el hipotálamo e induce la liberación del Factor Liberador de Corticotropina (CRF) quien, a su vez, actúa sobre la hipófisis para que libere la hormona corticotrófica (ACTH); también puede inducir la producción de la hormona del crecimiento, la luteinizante y la estimuladora del tiroides (TSH). La acción de IL-1 a nivel del SNC tiene tendencia a disminuir la reactividad del sistema inmune:
• Menos actividad NK, • Menos proliferación • Menos producción de IL-2;
La acción del ACTH sobre las suprarrenales provoca la liberación de corticosteroides quienes suprimen la producción de IL-1 por los MCFs (regulación negativa). Además IL-1 induce la producción del factor de crecimiento nerviosa (NGF) por las células de Schwann y los fibroblastos; bajo la acción de NGF las neuronas producen la sustancia P, que potencia la producción de IL-1 por los MCFs (regulación positiva). MEMORIA INMUNE Memoria T.
formulandoalgoqueaportar.blogspot.com

Es consecuencia de la existencia de poblaciones, pequeñas pero persistentes, de células específicas para los antígenos con los que ha entrado en contacto el sistema inmune del individuo. Aunque la mayoría de las células de memoria están en estado de reposo, hay una pequeña proporción de las mismas que están dividiéndose de modo constante. Las células T vírgenes requieren el contacto con los complejos CMH-péptido propio para su supervivencia a largo plazo en la periferia, pero las T memoria sobreviven gracias a IL-7 e IL-15, aunque para un funcionalismo óptimo también parecen necesitar el contacto con los complejos CMH-péptido propio. Tras el contacto de una T virgen con su antígeno se produce un gran número de células de memoria, que luego decrecen y se estabilizan en una cantidad 100-1000 veces mayor que se mantiene durante toda la vida. Las células T de memoria en reposos se caracterizan por presentar la proteína de superficie CD45RO y, tras su activación, dan lugar a dos tipos celulares: a) Las que maduran con rapidez a células T efectoras y secretan grandes cantidades de IFN-γ, IL-4 e IL-5; expresan los receptores para quimiocinas inflamatorias CCR3 y CCR5, por lo que migran a los tejidos inflamados b) Las que expresan el receptor de quimiocinas CCR7 y permanecen recirculando por los órganos linfoides secundarios Memoria B Las células B de memoria abundan 100 veces más que las B vírgenes específicas para el mismo antígeno y los anticuerpos que producen son de mayor afinidad. Estas células de memoria ya han sufrido el cambio de clase y expresan una concentración mayor de moléculas Clase II y coestimuladoras, por lo que el procesamiento antigénico es más rápido y eficiente. A diferencia de lo que ocurre con las células T de memoria (que pueden emigrar a los tejidos inflamados debido a que han cambiado los receptores de quimiocinas), las B de memoria siguen circulando a través de los órganos linfoides secundarios, especialmente por los folículos de los ganglios linfáticos, bazo y placas de Peyer. Como ocurría en la respuesta primaria, hay una proliferación abundante de células T y B en la interfase entre las zonas T y B-dependientes, con una amplia producción de células plasmáticas productoras de IgG. Algunas de estas células B no completan la diferenciación terminal y migran al folículo, donde constituyen las células B del centro germinal. Allí entran en una segunda fase de proliferación antes de dar lugar a la formación de células de memoria y nuevas células productoras de anticuerpos. El pecado antigénico original Una persona que se infecte varias veces con el virus del sarampión solo produce anticuerpos frente a los epitopos de la variante original. Los anticuerpos existentes inhiben la respuesta de las células B vírgenes específicas para los nuevos epitopos. Este supone una ventaja, pues el hospedador solo usa las células B que pueden responder más rápida y eficazmente al virus. La única forma de romper esta dinámica es que la persona se exponga a una variante del virus que carezca de todos los epitopos de la cepa original, pues no hay anticuerpos que puedan neutralizar al virus que quedará libre para que las células B vírgenes específicas puedan responder frente a el.
formulandoalgoqueaportar.blogspot.com

FYTI TEMA 18. SISTEMA INMUNE REGIONAL
INMUNIDAD ASOCIADA A LAS MUCOSAS Aunque las IgA solo representan entre el 10 y 20% de las Igs del suero, suponen el 60-70% de todas las Igs producidas cada día por las personas normales y sanas. La mayoría de las IgA se secreta, atravesando células epiteliales especializadas, a las superficies mucosas. En los grandes epitelios mucosos de
• Tubo gastrointestinal • Aparato respiratorio • Aparato lacrimal • Aparato genitourinario
así como en las secreciones:
• Lágrimas • Saliva • Leche materna • Algunas secreciones urogenitales
hay grandes cantidades de IgA. La parte del sistema inmune asociado a las superficies mucosas se define con frecuencia como una parte separada e independiente del resto del sistema inmunitario y se conoce como MALT (mucosa-associated lymphoid tissue), mientras que el sistema inmune que actúa en los tejidos no mucosos se denomina sistema periférico. Los MALT tienen estructuras linfoides secundarias que son folículos linfáticos similares a los que hay en los ganglios linfáticos o en el bazo del sistema periférico; estas estructuras se encuentran en las amígdalas de la faringe y las Placas de Peyer del intestino delgado. Aunque ambos sistemas tienen grandes diferencias no están totalmente aislados y pueden interactuar e influirse mutuamente. Para explicar las características del sistema inmune asociado a las mucosas (MALT) se describe a continuación, como ejemplo, la parte que está asociada de forma permanente al tracto gastro intestinal (GI) El sistema inmune del tracto GI contiene dos zonas diferentes: epitelio intestinal: además de las células epiteliales (con sus funciones características) hay Células M, (especializadas en capturar antígenos de la luz intestinal) y linfocitos intra-epiteliales (IEL) lámina propia: Está debajo del epitelio y en ella se sitúan las Placas de séller y una amplia representación de linfocitos B y T, así como células dendríticas, macrófagos y otros leucocitos. Tejido epitelial Contiene las células que realizan la mayor parte de la toma de contacto con los antígenos procedentes de la luz intestinal. Las células epiteliales no solo expresan moléculas de clase I, sino también de clase II y Ib y pueden ingerir, procesar y presentar las moléculas procedentes de la luz intestinal a las células T IEL (intraepiteliales) que están diseminados entre ellas. Además de esta presentación antigénica las células epiteliales secretan citocinas como:
Página 1 de 4
formulandoalgoqueaportar.blogspot.com

FYTI TEMA 18. SISTEMA INMUNE REGIONAL
• IL-7, que favorecen el desarrollo de los IEL • TGF-β e IL-10, que inhiben las respuestas celulares inflamatorias
Los IEL tienen una variedad limitada de TCRs y 2/3 partes de ellos expresan la molécula CD8. Alrededor del 10% de estas células son Tγδ mientras que el resto es Tαβ que, además, presentan un fenotipo poco corriente. Entre los IEL Tαβ solo una pequeña proporción es “típica” mientras que la mayoría presentan características atípicas o poco corrientes y parecen tener propiedades distintas:
• IEL Tαβ cuya molécula de CD8 está compuesta por dos cadenas α en lugar de por una α y otra β (fenotipo TCRαβ; CD8αα)
• IEL citolíticos naturales (células NKT), que expresan tanto el TCRαβ como receptores NK (p.e. NKG2D)
La labor conjunta de estos IEL contribuye a la eliminación de células infectadas y al comienzo de la cicatrización.
• Algunos linfocitos T (tanto Tαβ como Tγδ) con CD8αα reconocen una molécula indicadora de estrés (TL en el ratón y por identificar en humanos) que se expresa sobre células infectadas o dañadas. El reconocimiento del ligando lleva al aumento de producción de citocinas por parte de la célula T pero no aumenta su actividad citolítica.
• Los linfocitos Tαβ CD8αα pueden reconocer a unas moléculas de estrés (en humanos HLA-G) que se expresan sobre células infectadas o dañadas. El resultado de la unión es la destrucción de la célula alterada.
• Los linfocitos Tγδ;CD8αβ parecen reconocer glucolípidos o lipo-polisacáridos presentados por CD1c (una molécula similar a las de clase I e implicada en la presentación de fragmentos de moléculas no proteicas por parte de un gran numero de células, entre ellas las del epitelio intestinal).
• Los linfocitos NKT utilizan a el receptor NKG2D para reconocer a las moléculas de estrés MICA y MICB y destruir a células con estrés o alteradas y que expresan de forma simultánea niveles muy bajos de moléculas Clase I. Tras la activación, las NKT infiltradas entre los IEL comienzan a secretar IL-4 y otras citocinas.
Los IEL actúan en la frontera del epitelio para eliminar las células infectadas o dañadas, con lo que contribuyen a la reparación del epitelio intestinal. Las IL-4, IL-10 y TNFα producidas por las NKT y las células del epitelio intestinal crean un microambiente en la superficie epitelial que inhibe el desarrollo de las respuestas inmunes inflamatorias.
• Las células M del epitelio intestinal proceden de células que migran desde las criptas del intestino delgado. Estas células, que se localizan sobre las Placas de Peyer, presentan formas irregulares, con recovecos e invaginaciones que permiten a las DCs y linfocitos que están por encima de la lámina propia se aproximen a la superficie luminar de la célula M. Estas células M endocitan productos procedentes de la luz intestinal (cara apical) y los transportan a la cara baso-lateral de la
Página 2 de 4
formulandoalgoqueaportar.blogspot.com

FYTI TEMA 18. SISTEMA INMUNE REGIONAL
célula, donde los pueden capturar las DCs y linfocitos que estaba “esperando”. Hay cierta controversia respecto a si las células M pueden procesar y presentar los antígenos que transportan pero las DCs que captan a estos antígenos “transportados” realizan esta función con gran eficiencia.
Lámina propia Esta lámina propia, comparada con el tejido epitelial, es un oasis de “normalidad” ya que contiene linfocitos Tαβ convencionales (la mayoría CD4+), DCs, LB, células plasmáticas y fagocitos. Las DCs ingieren los productos transportados por las células M que, tras su procesamiento, se presentan a los linfocitos T. Pero las DCs de la lámina propia también pueden extender proyecciones de su citoplasma entre las células epiteliales y capturar directamente los productos de la luz intestinal. Además, en la lámina propia y por debajo de las células M se encuentran las Placas de Peyer, donde CPAs, LT y LB están expuestas al antígeno e interaccionan unas con otras. Las DCs migran desde la lámina propia a los ganglios linfáticos (normalmente los mesentéricos) donde activan a los linfocitos T y les “informan” de su procedencia (por medios todavía desconocidos) para que los LT efectores se dirijan hacia la lámina propia. Los LT y las CPAs pueden participar en las Placas de Peyer y GL mesentéricos en la activación de los linfocitos B. Igual que ocurría con los LT, los LB y las células plasmáticas alcanzan la lámina propia (generalmente a través de la circulación periférica). Los linfocitos B que pasan por las placas de Peyer son “inducidos” a producir IgA y las células plasmáticas productoras de IgA se trasladan a las criptas, entre las micro-vellosidades, donde secretan IgA dimérica que se transporta a través de células epiteliales especializadas para pasar al glucocaliz que recubre el epitelio intestinal. Mecanismos de la inmunidad de las mucosas. Igual que ocurre con la inmunidad periférica, la inmunidad de las mucosas usan moléculas de adhesión y quimiotácticas para dirigir el movimiento de las células hacia la lámina propia y el epitelio intestinal. Por ejemplo: los linfocitos T migran a los tejidos mucosos usando a L-selectina y a la integrina α4β7 para detectar a MadCam-1 en las células del endotelio vascular de los MALT. La unión con MadCam-1 facilita la extravasación de los LT a los tejidos mucosos. Una vez en el tejido mucoso del tacto GI los LT usan los receptores de quimiocinas CCR9 y CCR10 para detectar a las quimiocinas CCL25 y CCL28, producidas por los epitelios de los intestinos grueso y delgado, respectivamente. Finalmente, utilizan la integrina αEβ7 para detectar y unirse a la E-cadherina del epitelio vascular del intestino delgado. Los linfocitos T y B activados en el entorno de las mucosas, aunque puedan volver a circular por el cuerpo, tienden a volver a los tejidos mucosos. El medio mucoso suele tener carácter no inflamatorio y ha desarrollado mecanismos que previenen la aparición de respuestas inflamatorias, especialmente las celulares. La respuesta inflamatoria podría ser contraproducente en el medio intestinal, donde el sistema inmune asociado a
Página 3 de 4
formulandoalgoqueaportar.blogspot.com

FYTI TEMA 18. SISTEMA INMUNE REGIONAL
las mucosas está continuamente expuesto a grandes cantidades de antígenos extraños procedentes de la alimentación. Para responder de forma adecuada y constante habría que producir una inflamación crónica (permanente e intensa) que dañaría y destruiría el epitelio intestinal. Por lo tanto, la presencia de un medio “Th2” no inflamatorio (como demuestra el predominio de IgA sobre igG y la presencia de citocinas tales como IL-4, TGF-β e IL-10) contrasta con lo que ocurre en la respuesta periférica. De esta forma se crea un entorno donde la tolerancia hacia los antígenos externos es la norma y no la excepción. Aunque podría parecer que los sistemas periférico y mucoso trabajan de forma independiente, en realizad no está aislados el uno del otro y las células circulan por uno y otro, o viceversa. De hecho, un antígeno que debería de ser inmunogénico en los tejidos parenterales podría dejar de serlo si se administra por vía oral, de forma que su primer contacto con el sistema inmune fuera a través de las mucosas; es lo que se conoce como Tolerancia oral.
Página 4 de 4
formulandoalgoqueaportar.blogspot.com

TEMA 18-19 RESPUESTA INMUNE REGIONAL
EVOLUCION DE LOS ANTICUERPOS SEGÚN LA EDAD
LA INMUNOSENESCENCIA RESPUESTA INMUNE EN LAS MUCOSAS
El sistema inmune de las mucosas, es el más complejo del organismo, desde el punto de vista de las células que lo componen El de la mucosa intestinal tiene que responder frente a los patógenos que penetran (o intentan penetrar) y, a la vez, no responder (tolerar) frente a los antígenos alimentarios y a los de las bacterias comensales.
Los animales criados en ausencia de gérmenes (axénicos) tienen intestinos con estructuras linfoides poco desarrolladas y tasas de linfocitos en mucosas disminuidas. Hace falta el estímulo antigénico de la flora para el desarrollo inmune de la mucosa.
BARRERA INTESTINAL E INMUNIDAD INNATA
El epitelio colabora de forma crítica en la inmunidad innata de las mucosas, mediante: Una función de barrera, que reduce el ingreso de antígenos desde la luz intestinal Una función secretora de sustancias que aumentan la resistencia a la invasión de la mucosa por parte de los patógenos: estas sustancias pueden ser constitutivas o inducidas por el reconocimiento de PAMPs microbianos por los PRRs que hay en las células del epitelio. Entre ellas hay citocinas, quimiocinas, glucoproteínas, defensinas, lisozima (Células de Paneth), lactoferrina, etc. La gruesa capa de mucus impide el contacto directo de los microorganismos con las células epiteliales. Además, el borde apical del enterocito tiene microvellosidades (borde en cepillo) recubiertas por un glucocaliz que actúa como barrera física y biológica.
INMUNIDAD ADAPTATIVA EN LAS MUCOSAS
El tejido linfoide asociado a las mucosas
Las mucosas que recubren digestivo, respiratorio y urogenital tienen una superficie combinada de unos 400 m
2
y son el sitio de entrada de la mayoría de los patógenos. Esta gran superficie basa su defensa en el MALT (Tejido Linfoide Asociado a Mucosas). Estos tejidos varían mucho en su estructura, que va desde grupos laxos y apenas organizados de células linfoides en la lámina propia de las vellosidades intestinales hasta estructuras muy organizadas, como amígdalas , apéndice y las Placas de Peyer que se encuentran en la capa submucosa del revestimiento intestinal.
La importancia del MALT en la defensa del cuerpo se comprueba por su población considerable de células plasmáticas productoras de anticuerpos, cuya cifra excede en mucho al sumatorio de plasmocitos de bazo, ganglios linfáticos y médula ósea.
A pesar de todas las barreras hay un paso continuo de antígenos a través del epitelio, lo que supone un flujo de información permanente para el sistema inmune desde el medio externo. En las mucosas hay que diferenciar: Los sitios inductores de la respuesta adaptativa. Entre ellos están las Placas de Peyer (estructura con 30- 40 folículos linfoides, situados en submucosa y lámina propia, coronada por
formulandoalgoqueaportar.blogspot.com

una célula M) y los ganglios linfáticos mesentéricos (que recogen la linfa del tejido intestinal) donde se generan las células efectoras. Los sitios efectores de la respuesta adaptativa, son aquellos a los que se dirigen las células efectoras para participar en respuestas producidas en sitios distantes, tales como la lámina propia del intestino, el tracto respiratorio, lágrimas, saliva y glándulas mamarias. Es decir, que las estructuras del tejido linfoide asociado a las mucosas (MALT) están vinculadas entre si merced a la recirculación celular entre los distintos sitios y funcionan como una unidad integrada.
Localización de las células linfoides en la mucosa
Las células linfoides están en distintas localizaciones del tejido: La capa epitelial mucosal externa contiene, entre las células epiteliales especializadas (enterocitos) a los llamados linfocitos intraepiteliales (IEL), cuya mayoría expresa TCRγδ; la situación de estos IEL es óptima, pero presentan poca diversidad de receptor y todavía se desconoce en parte su función real. La lámina propia, que está debajo de la capa epitelial, contiene gran cantidad de células B, células plasmáticas, macrófagos y células Th activadas en grupos laxos. Los cortes histológicos revelan más de 15.000 folículos linfoides en la lámina propia intestinal de un niño sano. La capa submucosa, situada debajo de la lámina propia contiene Placas de Peyer, que son nódulos de 30-40 folículos linfoides, que pueden convertirse en folículos secundarios con centros germinales. El tejido linfoide en las placas de Peyer está muy relacionado con el epitelio que lo separa de la luz intestinal y forma una unidad integrada llamada FAE (Epitelio Asociado al Folículo) en la que el elemento distintivo es la célula M. El comienzo de la respuesta adaptativa
La respuesta efectora La mucosa mejor estudiada es la que recubre el tubo digestivo. Tiene la capacidad (como también ocurre en las mucosas respiratoria y urogenital) de capturar antígenos de la cavidad (luz intestinal) por endocitosis. La respuesta inmune a estos antígenos lleva a la formación de anticuerpos que pasan a la luz intestinal para combatir in situ a los patógenos invasores.
La respuesta de anticuerpos Las células M se sitúan en los llamados sitios inductivos (sitios pequeños en la membrana mucosa, situados sobre los folículos linfoides organizados) y los antígenos transportados por las células M pueden activar a células B dentro de esos folículos linfoides. Las células B activadas se diferencian a células plasmáticas que abandonan los folículos y secretan anticuerpos de la clase IgA. Estos se movilizan seguidamente a través de las células epiteliales y pasan a la zona luminal en forma de IgA-secretora, donde pueden interaccionar con el antígeno. La IgA es la mayoritaria en las secreciones y alcanza un 80% en lágrimas, saliva, calostro, leche y secreciones nasal e intestinal. Más del 90% de esta igA se presenta en forma de dímero, asociado a la cadena J y a un componente secretor procedente del receptor Poli-Ig. Esta IgA secretora tiene una función neutralizante.
Células de los sitios efectores En los sitios efectores hay una colección de plasmocitos productores de IgA, células T efectoras (Th1 y Th2), células T CD8+ citotóxicas, macrófagos y NK, situadas de forma difusa en la lámina propia del intestino.
formulandoalgoqueaportar.blogspot.com

Las células T y B efectoras usan moléculas de adhesión y quimiocinas para volver a las mucosas Piel y mucosa intestinal están expuestas de forma continua al entorno y son los sitios ideales para la penetración de patógenos. Los linfocitos efectores producidos en respuesta a infecciones en estos sitios expresan patrones de moléculas de adhesión y receptores de quimiocinas que garantizan su tropismo hacia ellos. Por ejemplo, los linfocitos T efectores con tropismo por la mucosa intestinal expresan la Integrina α4/β7, que reconocerá al MadCAM-1 expresado sobre las células del endotelio vascular. Otras combinaciones de moléculas de adhesión y receptores de quimiocinas son las responsables de regular el tropismo de los linfocitos efectores a otros epitelios mucosos.
El sitio en el que se realiza la activación linfocitaria determina el patrón de expresión de los receptores de albergue y, como consecuencia, dirige a las células efectoras al lugar donde encontrar a su antígeno específico en los tejidos periféricos
La activación de los linfocitos T en los Ganglios Linfáticos Mesentéricos y en las Placas de Peyer induce la expresión de receptores de albergue intestinal en las células efectoras (pero no se expresa las mismas moléculas cuando las células T efectoras se activan en ganglios cutáneos o en el bazo). Estas células efectoras pasan a la circulación y vuelven, en función de las moléculas de albergue que expresan y los receptores de quimiocinas al tejido intestinal
Los linfocitos B. El reclutamiento de los LB involucra a los mismos receptores de albergue que los de los LT. Los LB vírgenes que contactan con el antígeno aumentan la expresión del receptor CCR7, receptor que los dirige hacia el área T del ganglio (que es donde se están produciendo sus ligandos) y contactan con los LTh2 específicos para ese antígeno. Durante la respuesta se producen en el centro germinal la hipermutación somática y el Cambio de Clase. Los plasmoblastos no tienen receptores para las quimiocinas propias de los OLS, por lo que se facilita su salida de los mismos y la llegada a la circulación. Una fracción importante emigra a la médula ósea, atraídos por la quimiocina CXCL 12, que producen las células estromales, y se transforman en plasmocitos productores de anticuerpos Pero una proporción alta de los LB activados en los tejidos linfoides asociados a las mucosas sufren el cambio de isotipo a IgA y los plasmoblastos generados migran preferentemente a los tejidos mucosos. La localización del tejido linfoide en el que se produjo la inducción es la que condiciona el reclutamiento hacia una mucosa determinada; p.e., la inmunización oral conduce a la producción de IgA específica tanto en la mucosa intestinal como en las glándulas salivares, glándulas mamarias y mucosa pulmonar, mientras que la inmunización intranasal lleva a una producción de IgA restringida a las mucosas del tracto respiratorio, digestivo superior y tracto urogenital.
La diferencia migratoria hay que buscarla en la expresión diferencial por los plasmoblastos de ciertos receptores de quimiocinas y de integrinas que reconocerán a sus ligandos en unos tejidos, pero no en otros. Participación de los linfocitos B1
Gran parte de la IgAs que hay en las mucosas es el resultado de la activación de las células peritoneales B1 por las bacterias comensales. Estas B1 migran al ganglio linfático regional donde proliferan y se diferencian a plasmocitos que se dirigen al tejido linfoide asociado a las mucosas (p.e. lámina propia del intestinos) donde secretan grandes cantidades de IgAs. En caso de infección por las bacterias comensales no aparece IgA sérica específica sino IgG específica producida por las células B2. La conclusión es que la IgAs de las mucosas, producida por las B1, tiene un papel relevante en la defensa de las mismas impidiendo la penetración de bacterias comensales.
formulandoalgoqueaportar.blogspot.com

Algunos patógenos penetran a través de la mucosa
Las células epiteliales de la mucosa están adheridas entre si por unas uniones apretadas, que dificultan el paso de los agentes patógenos. Hay algunas bacterias y virus que han aprovechado las células M como vía de entrada a través de barrera mucosa: En algunos casos la célula M internaliza al patógeno y lo transporta a la bolsa En otros casos el patógeno se une a la célula M y la altera, lo que hace posible su
penetración. Entre los patógenos que usan estas vías están las cepas invasoras de Salmonella, Shigella, Vibrio cholerae y el virus de la poliomielitis.
Inducción de tolerancia El sistema inmune del intestino no solo tiene que prevenir la entrada de patógenos y comensales, sino que está en contacto permanente con antígenos extraños procedentes de los alimentos. Pero estos antígenos no provocan la respuesta del sistema sino que, por el contrario, inducen una respuesta dirigida a la tolerancia. En el proceso intervienen las células Treg y, especialmente, las Th3, que producen el inmuno-modulador TGF-b que impide la estimulación y respuesta de las células T convencionales.
A modo de resumen Las mucosas tienen sitios inductivos, en los que se produce la estimulación de los linfocitos por los antígenos locales. Estos linfocitos, así como las DCs, van a los folículos linfoides asociados a la mucosa o a los ganglios linfáticos mesentéricos, donde interaccionan y se producen las células efectoras. Las células efectoras T pasan por la linfa a la circulación sanguínea y, en función de las moléculas de adhesión que expresan, se reclutan para los distintos tejidos mucosos. Algo similar ocurre con las células B, pero estas se desplazan en la forma de plasmoblastos y plasmocitos productores de anticuerpos antes de llegar a los sitios efectores de las distintas mucosas. La inducción se produce en una mucosa y la función efectora se manifiesta a nivel de muchas mucosas. ANTICUERPOS Y EDAD
La tasa de las distintas clases de anticuerpos se manifiesta, sobre todo, durante el periodo fetal y los primeros años de la vida, alcanzándose la madurez del sistema inmunitario hacia el final de la segunda infancia (8-10 años).
Durante el comienzo de la vida fetal el sistema inmunitario del feto está formándose, por lo que su defensa inmune humoral depende de los anticuerpos que le transfiere la madre. El feto está aislado del resto del organismo materno por la placenta y las células de la misma presentan una serie de características especiales, orientadas muchas de ellas a evitar el rechazo del feto (a fin de cuentas, la “mitad” del mismo es un aloinjerto con moléculas CMH distintas), Una de las características es la de presentar un receptor especial llamado IgnR, que captura la IgG en la superficie de las células placentarias y, mediante un sistema de transporte activo, la deposita en el interior. La mayoría, si no toda, la IgG fetal procede de la madre. Hacia la semana 20 de la gestación el sistema inmune del feto comienza a producir IgM en pequeñas cantidades que representan, en el momento del nacimiento, menos del 20% de la cifra del adulto. La producción de IgG también comienza hacia la semana 24, pero las cantidades de este anticuerpo son mínimas, representando, en el momento del nacimiento, menos del 5% del valor normal del adulto. En cuanto a la IgA comienza a producirse después del nacimiento. El resultado es que el niño, en el momento del nacimiento, depende prácticamente de los anticuerpos transferidos por la madre a través de la placenta y a partir de ese momento, de los pocos que produce y d los aportados nuevamente por la madre a través de la lactancia. El intestino del recién nacido tiene unos receptores similares a los IgnR placentarios, encargados de capturar a los anticuerpos IgG y llevarlos a través de la mucosa hasta la circulación. La
formulandoalgoqueaportar.blogspot.com

protección materna cubre al recién nacido durante los primeros meses de su vida, mientras su sistema inmune va madurando rápidamente. Al año de vida las tasas de producción de anticuerpos IgM, IgG e IgA se sitúa entre el 60 y el 80% de la del adulto.
INMUNOSENESCENCIA
Si el problema del recién nacido era la falta de elementos defensivos (como los anticuerpos) el del anciano es el cambio o remodelación de sus sistema inmunitario. La inmunidad innata se mantiene muy bien con el avance de la edad, siendo la adaptativa la que presenta los cambios.
Inmunidad adaptativa celular El primer cambio que se observa es la involución del timo, que comienza muy pronto en la vida (segunda década) y parece completarse a finales de la quinta década. Como consecuencia hay una disminución gradual de su peso, lo que se debe a distintas causas, como: Menor migración de precursores desde la médula ósea Menor diferenciación de estos precursores en la glándula Disminución progresiva de reordenamiento génico a nivel de la cadena beta del TCR Disminución de la producción de citocinas estimuladoras de la proliferación, como IL-7 A pesar de todos estos cambios la cifra de linfocitos T periféricos no disminuye con la edad, lo que puede deberse a la existencia de clones de vida larga y la auto-perpetuación en la periferia a causa de los estímulos antigénicos locales. De todos modos, la respuesta a antígenos nuevos depende de la producción de células vírgenes por parte del timo y es, precisamente, esta característica la que va disminuyendo. Con la edad hay un aumento de células de memoria y una disminución paralela de células vírgenes, así como una disminución del número de células Tgd circulantes. Los cambios también se reflejan en el perfil de citocinas, ya que las células vírgenes producen un perfil tipo 1 (IL-2, IFN-g) mientras que las células de memoria producen más bien un perfil tipo 2 (IL-4 e IL-6). También se afectan por el envejecimiento las moléculas coestimuladoras, tal como ocurre con LFA-1 que no es capaz de realizar el cambio conformacional que la llevaría a la forma de alta avidez durante la cooperación T/B. La disminución de la respuesta a los antígenos comunes en las pruebas cutáneas de hipersensibilidad retardada (DTH) es un indicador de la disminución funcional de la rama celular de la inmunidad y la falta de respuesta (anergia) es un indicador de mortandad próxima.
Inmunidad adaptativa humoral
Durante la vejez también disminuye la producción de linfocitos B, debido a una disminución en los linfocitos Pre-B de la médula ósea, que presentan una alta susceptibilidad a la apoptosis. La diversidad del repertorio depende de la producción de células vírgenes en la médula ósea, por lo que la disminución de l a producción se refleja en la diversidad. El número de células B periféricas se mantiene, lo que se debe seguramente al aumento de células de memoria de vida larga y a la auto-renovación local antígeno-dependiente. Lo que aumenta es el número relativo de linfocitos B1 (CD5+), que son los encargados de producir los anticuerpos naturales y el resultado es el aumento de los autoanticuerpos que se produce en el anciano. Como la respuesta T a nuevos antígenos está disminuida la respuesta d anticuerpos T-dependiente también lo estará; la importancia de esto se manifiesta en la respuesta a las vacunas durante la tercera edad. Aunque los niveles de Ig no disminuyen si lo hace la afinidad de estos anticuerpos, junto con los diferentes procesos de los centros germinales (expansión clonal, hipermutación somática, cambio de clase), lo que se refleja en una marcada reducción del tamaño de los centros germinales.
formulandoalgoqueaportar.blogspot.com

Inmunodisfunciones
TEMA 20. INMUNODISFUNCIONES AUTOINMUNIDAD HIPERSENSIBILIDAD El sistema inmunitario tiene que mantener un difícil equilibrio entre una respuesta potente y eficaz frente a los patógenos y la tolerancia frente a los antígenos propios y a los antígenos ambientales no perjudiciales. Si se rompe el equilibrio en contra de la tolerancia la consecuencia es la aparición de patología. La respuesta frente a los antígenos propios da lugar a las patologías conocidas como Enfermedades Autoinmunes, mientras que las respuestas (por falta de regulación) frente a antígenos foráneos provoca las patologías conocidas como Hipersensibilidades. Las reacciones de hipersensibilidad provocan el daño al tejido usando los mismos mecanismos efectores que se usan en la respuesta frente a los patógenos. Las Hipersensibilidades se clasificaron se clasificaron, teniendo en cuenta el mecanismo inmunitario y su etiopatogenia, por Gell y Coombs en: LA HIPERSENSIBILIDAD TIPO I Está mediada por IgE, que se produce como consecuencia de la producción de células efectoras Th2 en respuesta a antígenos que, con la debida tolerancia, son inofensivos. Los sujetos Atópicos son aquellos con predisposición a producir IgE en respuesta a antígenos ambientales. El término viene del griego “atopos”, que quiere decir “fuera de lugar” y estos individuos presentan, con frecuencia, una o más enfermedades de esta naturaleza. Los antígenos ambientales que selectivamente evocan una respuesta efectora de tipo Th2, con producción de IgE, se denominan Alergenos y presentan, con frecuencia, las siguientes propiedades:
• Proteínas de Pm bajo, solubilidad y estabilidad alta, capaces de desencadenar reacciones de hipersensibilidad Tipo I, incluso a concentraciones bajas.
Todos estamos expuestos permanentemente a estos alergenos y la gran mayoría respondemos produciendo cantidades bajas de anticuerpos IgG1 e IgG4 (respuesta efectora Th1). Por el contrario, los atópicos responden produciendo niveles altos de anticuerpos específicos de la Clase IgE. Estos sujetos suelen reaccionar positivamente frente a ciertos extractos de alergenos usados en las pruebas diagnósticas. Los procesos alérgicos son procesos inflamatorios La llegada del alergeno pone en marcha la respuesta con la colaboración B/T a expensas de las Th2 en un microambiente dominado por IL-4 e IL-5, que dirigen el cambio de clase a IgE y la eosinofilia respectivamente. La IgE se une por el fragmento Fc a los receptores específicos (FceR) que hay sobre mastocitos y basófilos. Una vez que empieza, la síntesis de IgE persiste meses (incluso años), por lo que mastocitos y basófilos están preparados (sensibilizados) para reaccionar rápidamente cuando se produzca el próximo contacto con el alergeno.
Página 1 de 3
formulandoalgoqueaportar.blogspot.com

Inmunodisfunciones
En la siguiente penetración el alergeno se une a la IgE ligada sobre los mastocitos, lo que provoca el entrecruzamiento de los receptores, la activación y la degranulación, con liberación de los mediadores preformados que hay en las granulaciones (sobre todo Histamina) y los de nueva síntesis, entre los que están: factor de agregación plaquetaria (PAF), prostaglandinas (PG) y leucotrienos (L). Es la liberación brusca y masiva de mediadores la que provoca la inflamación responsable de la reacción alérgica aguda. La prevalencia de respuesta Th2 en los atópicos se asocia a defectos en la actividad supresora de las células T reguladoras (Treg). Los pacientes que carecen de Treg naturales, debido a mutaciones en Foxp-3, desarrollan un eczema severo, niveles muy altos de IgE, eosinofilia y alergias alimentarias. Las células Th3 (productoras de TGF-b), y también las Tr1, (IL-10), reducen eficazmente la hiper-reactividad bronquial. HIPERSENSIBILIDAD TIPO II Están mediada fundamentalmente por IgG (aunque también intervienen Acs IgM), que reconocen antígenos sobre la superficie celular o en la matriz extracelular. La unión de los anticuerpos específicos “marcan” a estas células como blancos de la activación del complemento. Dentro de este grupo de reacciones están las:
• Reacciones trasfusionales • Anemia hemolítica del recién nacido (incompatibilidad Rh) • Anemias hemolíticas autoinmunes
Sistema ABO de grupos sanguíneos Los hematíes tienen antígenos hidrocarbonados en su superficie, que proceden de una estructura básica común, llamada Sustancia H (propia del grupo O) a la que se puede añadir una N-acetil-galactosamina (grupo A) o una Galactosa (grupo B), o ambos azúcares (grupo AB). El hecho de que los sujetos del grupo A tengan anticuerpos anti-B, los del grupo B tengan anticuerpos anti-A, los del grupo O tengan ambos tipos de anticuerpos y los del Grupo AB no tengan ningún anticuerpo, depende de un proceso de reacción cruzada entre antígenos compartidos por las bacteria Gram - de la flora intestinal y los GR. Un individuo del grupo A se hace tolerante, durante la educación tímica, a los antígenos A pero no a los B y estos estimularán la formación de anticuerpos (Isohemaglutininas, de la clase IgM). Incompatibilidad Rh (Rhesus) Depende fundamentalmente del llamado antígeno D. Una madre RhD- puede fácilmente sensibilizarse por los GR de su niño RhD+ durante el parto, pues las hemorragias de la placenta pueden transferir muchos GR del niño a la madre. Los anticuerpos que se forman (preferentemente IgG) son capaces de atravesar la placenta en otro embarazo subsiguiente y unirse a los GR fetales, opsonizándolos y llevándolos a la destrucción, lo que provoca la enfermedad hemolítica del recién nacido. Las madres RhD- se tratan profilácticamente con IgG-anti-D inmediatamente después del primer parto, lo que reduce mucho los riesgos de sensibilización. HIPERSENSIBILIDAD TIPO III La razón del comportamiento de los IC in vivo no depende solo de las cantidades absolutas de antígeno y anticuerpos sino, sobre todo, de sus proporciones relativas: entre exceso de anticuerpos y exceso débil de antígeno los IC precipitan rápidamente y tienden a localizarse en el sitio de penetración del antígeno, mientras que entre exceso
Página 2 de 3
formulandoalgoqueaportar.blogspot.com

Inmunodisfunciones
medio y alto de antígeno se forman IC solubles. La lesión tisular se caracteriza por la aparición de necrosis con infiltrado celular de neutrófilos La fijación del complemento inhibe la precipitación de IC por unión covalente de C3b a los mismos, lo que impide las interacciones Fc-Fc que se requieren para la formación de grandes agregados insolubles. Estos pequeños IC que contienen C3b se aclaran a través de los glóbulos rojos (Ver “propiedades del Complemento” en el Tema correspondiente) Entre las enfermedades por IC destacan la glomérulonefritis post-estreptocócica, el Lupus eritematoso sistémico, las vasculitis agudas, etc. El Lupus eritematosos sistémico es una enfermedad autoinmune en la que se producen auto-anticuerpos frente al DNA o nucleoproteínas propias y su depósito como IC provoca glomérulonefritis y artritis. HIPERSENSIBILIDAD TIPO IV Esta hipersensibilidad se encuentra en muchas reacciones alérgicas a bacterias, virus, hongos y parásitos, en las dermatitis de contacto frente a ciertas sustancias químicas, y en el rechazo de tejidos transplantados. El ejemplo más conocido es la reacción de Mantoux que se produce cuando se inyecta tuberculina en la piel de un sujeto en el que la infección previa con el bacilo tuberculoso ha inducido un estado de inmunidad mediada por células (IMC). La reacción se caracteriza por eritema e induración que aparecen al cabo de varias horas (retardada) y alcanza el máximo a las 24-48 horas. Las lesiones resultan de la interacción “exagerada” entre el antígeno y el mecanismo normal de la IMC. Tras los primeros contactos, las células T de memoria reconocen al antígeno cuando entra otra vez y, si son Th1, liberan citocinas que atraen y activan a los macrófagos y también ayudan a los precursores T CD8+ citotóxicos para transformarse en células asesinas y colaborar al daño tisular. La continuidad de la hipersensibilidad retardada por antígenos persistentes da lugar a la formación de granulomas. La hipersensibilidad de contacto: dermatitis por contacto Es un ejemplo clásico de hipersensibilidad Tipo IV. La sensibilización empieza cuando un hapteno o el antígeno penetran en la piel la primera vez. Se capturan por las células de Langerhans y se presentan a las T vírgenes en el ganglio regional, lo que produce linfocitos T efectores. En un 2º contacto los linfocitos T de memoria van rápidamente hacia el sitio de contacto en la piel, reconocen al antígeno y desencadenan una respuesta inflamatoria con participación de los macrófagos. Se llaman dermatitis por contacto por desencadenarse al entra la piel (por contacto) las sustancias desencadenantes. Entre ellas están: componentes de tintes de pelo, joyas, cosméticos, calzados, venenos y vegetales. A menudo se trata de haptenos que penetran con facilidad por la piel. El penta-deca-catecol, responsable el rash cutáneo, es el hapteno de la hiedra venenosa. El níquel de las aleaciones en joyas también produce estas reacciones en el sitio de contacto con la piel.
Página 3 de 3
formulandoalgoqueaportar.blogspot.com

TÉCNICAS PARA LA CARACTERIZACIÓN DE LINFOCITOS Y RESPUESTA EFECTORA La caracterización de los linfocitos y otras células implicadas en respuesta inmunitaria requiere de la separación e identificación de las subpoblaciones celulares separadas. -Los linfocitos pueden aislarse a partir de sangre, de médula, órganos linfoides, epitelios o sitios donde se haya producido una reacción inflamatoria:
Los linfocitos se pueden separar de sangre periférica tratada con heparina medinte un gradiente de Ficoll (mezcla de polímeros de carbohidratos) y metrizamida. (Figura 2.22).
La centrifugación de la sangre diluida depositada sobre el “Ficoll-Hypaque” permite la separación de los eritrocitos, polimorfonucleares y granulocitos (que van al fondo) de las células mononucleares de sangre periférica (PBLs) que incluyen linfocitos y algunos monocitos que quedan en la interfase. En animales de experimentación y ocasionalmente en humanos se pueden separar linfocitos a partir de otras locacalizaciones (bazo, timo, médula osea, ganglios linfáticos, tejido linfoide asociado a mucosas). Sin embargo, normalmente el estudio de las poblaciones de linfocitos se hace en sangre periférica. -Obtención de poblaciones de linfocitos con especificidades homogeneas: El estudio de la función efectora de células T depende en buena medida de contar con poblaciones monoclonales de células T (especificidad única). Esto se puede alcanzar de varias formas: 1-Como en el caso de los hibridomas B, células T específicas para un ag pueden fusionarse con líneas celulares derivadas de linfomas de células T para generar híbridos T. Estos híbridos son excelentes herramientas para el análisis de la especificidad, sin embargo no se pueden utilizar para el estudio de la proliferación de células T en respuesta a un ag específico puesto que están continuamente dividiendo. Por esta razón, el estudio de las funciones efectoras de células T requiere de la utilización de clones de células T. Son clones de células T derivadas de una sola célula con especificidad única, a partir de cultivos heterogéneos denominados líneas de células T, cuyo crecimiento depende de la reestimulación por ag periodica así como de la adición de factores de crecimiento (Figura 2.31). La obtención de los clones de células T se obtiene mediante clonación por el método de dilución límite. Cuando se lleva a cabo esta dilución límite de una mezcla heterogénea de células T, algunos pocillos no recibirán ninguna célula T específica de un ag determinado, otras una otras 2 etc. Las células T en los pocillos son activadas con APCs, ag y factores de crecimiento. Después de varios días en cultivo (necesarios para su crecimiento y diferenciación) las células de cada pocillo son testadas en cuanto a su respuesta frente al ag (producción de citoquinas , función efectora etc). -Marcadores de diferenciación: Marcador: cualquier característica funcional o estructural que permite definir un tipo o población de células e incluso separarlas físicamente de las demás. El desarrollo de anticuerpos monoclonales y tecnologías, como la citometría de flujo, que permite una detección rápida de las células reconocidas por esos anticuerpos monoclonales, ha permitido el conocimiento de un gran número de ags. de superficie celular que son útiles como marcadores de diferenciación. Algunos de estos antígenos son exclusivos de
1
formulandoalgoqueaportar.blogspot.com

un determinado linaje celular y otros se expresan en un determinado estadio de diferenciación y sirven para saber el estado de madurez de un determinada célula. Estas moléculas han sido incluidas en un sistema de nomenclatura conocido como “CD” (Se conocen más de 120 CDs diferentes). La distribución de las subpoblaciones linfocitarias en sangre períferica humana en base a la expresión de determinados marcadores puede caracterizarse de esta forma (Figura 2.23). La herramienta más poderosa para la caracterización de poblaciones celulares heterogéneas es sin duda la citometría de flujo -Citometría de Flujo: Es una técnica analítica que permite la caracterización multiparamétrica e individualizada de una población heterogénea de células. Permite no solo la cuantificación de diversos parámetros de la célula, sino que lo hace en poco tiempo (en minutos se pueden analizar miles de células) e incluso se pueden separar fisicamente las poblaciones elegidas por el operario. Componentes básicos de un citómmetro: (Figura) -Sistema hidraúlico: rodea la suspensión celular en flujo, con una vaina externa, formada por un fluido libre de partículas, que mueve la muestra a velocidad constante y controlada. Produce el enfoque hidrodinámico de la muestra: la suspensión de células se inyecta en un embudo en el que sea su vez se inyecta una corriente de tampón a diferente presión. La diferencia de presioenes crea un efecto hidrodinámico que hace que el tampón envuelva la suspensión de células y consiga que las células de la suspensión se alineen una a una verticalmente.
-Sistema de iluminación: La fuente de luz es una haz de laser (normalmente de argon) que incide sobre la corriente de células alineadas.
-Sistema óptico: Enfoca la iluminación de las partículas de la muestra de tal forma que cuando el laser no incide sobre una célula no se registra ninguna señal por el aprato. Sin embargo, cuando el laser incide sobre una célula, está prodece una dispersión de luz en todos los ángulos.
-Sistema electrónico: La luz dispersada es recogida y amplificada por fotodetectores (fotomultiplicadores) situados en la misma dirección de incidencia del laser y en 90º con respecto al ángulo de incidencia. Estas señales, recogidas en forma de pulsos son registradas y almacenadas por un ordenador.
¿Qué parámetros se pueden medir? La cantidad de luz dispersada en la misma dirección del laser y con la misma longitud de onda del haz incidente es proporcional al volumen de la célula y por tanto esta medida nos permite cuantificar el tamaño de las células. (Figura) La luz dispersada por las células en un angulo de 90º (Figura) con una longitud de onda igual a la del laser de excitación también es proporcional al volumen de la célula pero también es afectada por la topografía de la célula (rugosidades, relación nucleo:citoplasma, homogeneidad del citoplasma-presencia y tamaño de vacuolas etc). Este parámetro se denomina complejidad. Por ejemplo: linfoctios y neutrófilos con tamaño similar tendrían diferentes señales de “light scatter” (complejidad) (90º). Pero además de estos parámetros, si las células van marcadas con un fluorocromo (por ejemplo un anticuerpo conjugado con un fluorocromo específico para un marcador de superficie CD de la célula), el haz de laser excita el fluocromo y se produce una emisión de fluorescencia (diferente longitud de onda a la luz incidente y dependiente del fluorocromo utilizado) que es proporcional a la cantidad de fluocromo y por tanto a la cantidad de marcador que se esté investigando. Por tanto, la luz recogida
2
formulandoalgoqueaportar.blogspot.com

en 90º con una longitud de onda superior a la de excitación permite la cuantificación de las señales procedentes de los fluorocromos. Teniendo en cuenta que existe un elevado número de fluorocromos que se excitan en un rango de longitudes de onda parecido y que emiten en un rango muy diferente de longitudes de onda (Tabla), es posible llevar a cabo el marcaje simultaneo de diferentes marcadores celulares diferentes mediante la utilización de anticuerpos conjugados a diferentes fluorocromos. El citómetro, mediante una combinación de espejos y filtros (Figura) es capaz de separar las señales de emisión de cada uno de los fluorocromos y registrar y cuantificar por tanto la presencia del marcador celular respectivo. Análisis de los resultados: El software del equipo permite llevar a cabo un análisis posterior de las señales registradas. Se puede llevar a cabo un análisis mono o multiparamétrico, puesto que conocemos todos los parámetros que hemos analizado (volumen, complejidad y cada una de las fluorescencias analizadas de todas y cada una de las células de la población). Se obtienen así diagramas mono o multiparamétricos (Enseñar gráficas) y en cualquier caso el equipo nos informa de la la cuantificación de cada una de las señales así como del porcentaje exacto de la población heterogenénea con determinadas carcterísticas (si es + o – para un determinado parámetro). Pero además, determinados tipos de citómetros de flujo nos permiten llevar a cabo la separación física de las poblaciones analizadas (FACS-citómetros separadores-lo vemos ahora). -Separación de poblaciones celulares:
1-FACS: Determinados citómetros con capacidad de separación son capaces de cargar positiva o negativamente las gotas que contienen células individuales generadas en el flujo hidrodinámico con determinadas características (presencia de CD4 por ejemplo o presencia simultanea de varios parametros CD3, CD44, tamaño). Estás gotas, al pasar por un condensador son separadas en función de la carga (hacia el polo + o -) y recogidas en placas multipocillo o frascos de cultivo.
Aunque el FACS es la técnica de elección para una separación con elevado grado de pureza, cuando se trata de separar linfocitos de forma rápida y una pureza elevada se pueden utilizar métodos mecanicos de separación. En muchas ocasiones se utilizan estos métodos como pasos previos al FACS.
2-Separación magnética: Consiste en la utilización de bolas magnéticas conjugadas con anticuerpos monoclonales que reconocen CDs espec´ficos de las células a separar. Las células se incuban con estas bolas y se hacen pasar por una columna que contiene material que atrae las bolas magéticas en presencia de un campo magnético. Las células que unieron el Ac (las que se quieren separar) son retenidas en la columna cuando se amplica este campo magnético mientras que aquellas que no expresan el CD no se retienen. La retirada posterior del campo magnético permite la elución y recogida de las células de interés. (Figura)
3-Panning: unión de Acs específcos de CD a superficies plásticas; incubación de las células en las placas; lavado de las placas; Las células que expresan el CD que quiero detectar son retenidas en las placas.
4-Eliminación de las células no deseadas con anticuerpo más complemento.
3
formulandoalgoqueaportar.blogspot.com

Medida de la activación de linfocitos: Medida de proliferación celular:
Los linfocitos T cuando reconocen péptidos antigénicos específicos se activan y proliferan antes de diferenciarse en células efectoras para dar lugar a un clón de células con la misma especificidad. Esta respuesta mitógena es acompañada de cambios morfológicos hacia blastos. El grado de activación o proliferación de linfocitos in “vitro” en respuesta a un estímulo se puede evaluar determinando el pocentaje de blastos en cultivo o mediante la medida de incorporación de un análogo de nucleótido radioactivo al DNA que se sintetiza de novo. De cualquier forma el análisis de la proliferación de linfocitos resulta esencial para el estudio de la respuesta celular. Activación por mitógenos policlonales: Ciertas substancias estimulan la proliferación de linfocitos in vitro de forma similar al antígeno pero independientemente de la especicidad de su receptor. A estas substancias se les denomina mitógenos mitógenos policlonales. Al contrario que el ag, que solo induce in vitro la proliferación de aquellos clones específicos, estos mitógenos inducen la proliferación de una mayoría de la población de linfocitos. Sin embargo, existe una adecuada correlación entre la respuesta de linfocitos de un individuo a mitógenos (medida in vitro) y su status inmunológico. Es por ello que estos mitógenos (Tabla 2.26 Janeway) se utilizan in vitro para evaluar la capacidad de proliferación de linfocitos de sangre periférica. Este ensayo se utiliza en clínica para testar la capacidad de los linfocitos de un paciente con sospecha de inmunodeficiencia. Los pacientes con deficiencias inmunitarias se detectan clínicamente por historias clínicas de infecciones recurrentes. Para determinar la competencia del sistema inmune de estos individuos, se somete al paciente a una serie de pruebas (Tabla 2.34 Janeway) entre las que se incluyen la medida in vitro de proliferación celular en respuesta a mitógenos. Activación de linfocitos dependiente de antígeno: Además de medir in vitro la proliferación de linfocitos en respuesta a mitógenos, se puede evaluar de la misma forma la respuesta de linfocitos a antígenoa específicos. Este test se puede utilizar para medir respuestas mediadas por células T después de una inmunización (Figura 2.27 de Janeway). ¿Cómo medir la proliferación celular?
-Ensayos de incorporación de timidina tritiada. Las células que proliferan sintetizan DNA. Se incorpora al medio de cultivo un análogo de nucleótido como la timidina tritiada. Este análogo solo se incorpora en células que estén dividiendo puesto que se une a DNA sintetizao de novo. Midiendo la radioactividad en células en presencia y ausencia de señal estimuladora se puede evaluar la proliferación en función de las cuentas radioactivas incorporadas (Figura). Medida de respuesta efectora: La medida de la proliferación celular nos da una idea de la capacidad de las poblaciones de linfocitos de activarse pero no informa sobre la capacidad funciónal de las mismas. Existen sin embargo modos de medir las funciones efectoras: 1-Lisis de la células diana: mediada por células citotóxicas y NKs. Células citotóxicas CD8 son capaces de lisar células que presenten el péptido específcio asociado a su molécula de MHC de clase I. Se puede medir la actividad
4
formulandoalgoqueaportar.blogspot.com

citotóxica de una población de linfocitos T. Se incuban las células diana con Na251CrO4.
Las células vivas lo toman pero no lo liberan espontaneamente. Se lavan bien las células después del marcaje y se incuban con las células citotóxicas y al cabo de un tiempo se evalua la radioactividad en el sobrenadante, que será directamente proporcional a la actividad citotóxica de las células efectoras. Necesidad de establecer unos ratios de efectoras:dianas: (3:1/6:1/12:1) que dependen de la capacidad citotóxica de la scélulas. (Figura). 2-La funcionalidad de linfocitos T CD4 implica la activación por parte de estas células de macrófagos (en el caso de TH1) o de células B (en el caso de células TH2). Los efectos de las células T sobre macrófagos y LB están mediados, en buena parte, por citoquinas producidas por LT cuando se reconoce especificamente el complejo MHC-péptido. Estas citoquinas son diferentes y se producen en diferente cantidad dependiendo del tipo de célula efectora. Por esta razón, la funcionalidad de células TH se mide normalmente identificando y cuantificando las citoquinas producidas por estas poblaciones (Poner tabla de citoquinas producidas por distintas poblaciones Ts. -Detección y cuantificación de citoquinas: Se puede realizar mediante ELISA o mediante RT-PCR. ELISA: se suele utilizar un ELISA en sandwich en el cual la citoquina se caracteriza por su capacidad de formar un puente entre dos anticuerpos monoclonales que reconocen diferente epítopos de la citoquina. Figura 2.29 Janeway: Se recubre la placa donde se hace el ensayo con anticuerpo anti IL-2 (se incuba la placa con el anticuerpo durante unas horas), se lava bien el exceso de Ac y se añade el fluido sobre el que se quiere cuantificar la citoquina; después de un tiempo de incubación se lava la placa y se añade el 2º anticuerpo acoplado a un sistema enzimático de identificación (métodos colorimétricos) o acoplado a un fluorocromo (se detecta la fluorescencia emitida). RT-PCR: Una segunda opción para cuantificar citoquinas es la valoración de la cantidad de RNA mensajero específico para la citoquina en cuestión. Se utiliza el método de RT-PCR. Este método consiste en la obtención de RNAm total a partir de las células que se quieren cuantificar y obtención de cDNA total utilizando transcriptasa reversa. A partir de este cDNA total, se lleva a cabo una reacción de PCR utilizando oligonucleótidos específicos del gen de la citoquina que se quiere cuantificar. La cantidad de DNA amplificado será proporcional a la cantidad de transcrito para esa citoquina. Explicar la reacción de PCR (Figura) 3-Fagocitosis: Se puede evaluar la capacidad fagocítica de macrófagos o neutrófilos “in vitro” de varias formas. En cualquier caso, el ensayo se suele hacer en placas multipocillo, las células fagocíticas se ponen en contacto con el microorganismo (bacterias, hongos..) durante unas ) durante un tiempo, en el medio de cultivo adecuado (p ejemplo RPMI 1640) y al cabo de ese tiempo se evalúa la fagocitosis:
1-Mediante tinción de Giemsa y observación microscópica de los macrófagos. 2-Mediante citometría de flujo: se cuantificar la fagocitosis mediante la
cuantificación del “burst oxidativo”-aumento de la concentración de peróxido de hidrógenio y otros radicales de oxígeno. Las células fagocíticas se tiñen con diacetato de diclofluoresceína, que cuando entra en la célula sufre la acción de esterasas celulares. La diclorofluoresceína, en presencia de peróxido de hidrógeno (consecuencia del burst
5
formulandoalgoqueaportar.blogspot.com

oxidativo) sufre un proceso de oxidación que lo transforma en un compuesto fluorescente. La fluorescencisa es proporcioanl al burst oxidaticvo y por tanto a la fagocitosis. Otra forma de cuantificar fagocitosis es marcar los microorganismos con compuestos fluorescentes y luego cuantificar señal de fluorescencia de los macrófagos en el citómetro.
6
formulandoalgoqueaportar.blogspot.com










![[APUNTES] Shambhala - Sistema de Sanacion Multidimensional - Manual Verdadero 2009](https://static.fdocuments.es/doc/165x107/55cf9d19550346d033ac3be1/apuntes-shambhala-sistema-de-sanacion-multidimensional-manual-verdadero.jpg)







