Cardio Arritmia Vol I Libro - tienda.grupocto.estienda.grupocto.es/pdf/ME_CardioA_CapM.pdf · A...
15
1 01 Capítulo 1 . 1 . Introducción El entendimiento de los mecanismos electrofisiológicos ce- lulares y tisulares cardíacos resulta esencial para el correcto diagnóstico de las arritmias, así como su tratamiento, tanto far- macológico como invasivo. Este capítulo tiene como objetivo principal revisar los conceptos más generales de bioelectrici- dad de las membranas y electrofisiología celular para estable- cer las bases del entendimiento de los mecanismos básicos de las arritmias y su tratamiento a nivel clínico. 1 . 2 . Bioelectricidad A nivel teórico son muchas las similitudes entre el flujo de iones que atraviesa la membrana plasmática y la corriente que generan, con el comportamiento de la corriente en un circuito eléctrico. Algunos conceptos básicos de bioelectricidad permiten com- prender los mecanismos que dirigen la excitabilidad cardíaca, el movimiento de iones y las corrientes a través de la membrana plasmática de la célula (véase la Figura 1.1 en página siguiente). Carga Los elementos químicos tienden a conservar el mismo número de protones (carga positiva) que de electrones (carga negativa), manteniendo una carga neta cero. Sin embargo, dentro de la es- tabilidad química que confiere completar eléctricamente la últi- ma capa de electrones de los átomos, algunos elementos pue- den perder o ganar electrones, dando lugar a moléculas en las que el número de electrones y protones difieren. Estas partículas cargadas eléctricamente se denominan iones, y son la base de la excitabilidad cardíaca, puesto que su movimiento a través de los canales iónicos de la membrana plasmática genera la corriente responsable del potencial de acción (PA) cardíaco. Los iones car- gados positivamente (que han perdido electrones) se conocen como cationes (K + , Na + o Ca 2+ ) y los iones cargados negativamen- te (que han ganado electrones) son los aniones (Cl - o I - ). El movi- miento de atracción entre partículas cargadas de diferente signo es lo que genera la corriente eléctrica. Voltaje La perturbación espacial que genera una partícula cargada en el espacio es lo que se conoce como campo eléctrico. Toda partí- cula cargada genera una fuerza de atracción hacia las cargas de diferente signo, y de repulsión hacia las del mismo, de manera que las cargas se moverán en el espacio de acuerdo a estas fuer- zas para conseguir un entorno eléctrico neutro. El trabajo ejercido por el campo eléctrico para mover una carga desde un punto A hasta un punto B se conoce como diferencia de potencial (o diferencia de voltaje), se puede medir con un vol- tímetro y su unidad es el voltio (V ). Desde un punto de vista más práctico, una diferencia de potencial se genera cuando existe una distribución diferencial de cargas entre dos puntos aislados, que equivaldría a una acumulación asimétrica de cargas en un condensador. A nivel electrofisiológico, existe una diferencia de potencial entre ambos lados de la membrana plasmática de la Electrofisiología celular José Manuel Alfonso Almazán * Manuel Marina Breysse *, ** Conrado J. Calvo Sáiz *, *** David Filgueiras Rama *, **** * Fundación Centro Nacional de Investigaciones Cardiovasculares Carlos III (CNIC). Madrid. España ** Hospital Universitario de Móstoles. Madrid. España *** Electrofisiología Cardíaca y Bioingeniería (GEB). BioITACA-IIS Investigación Biomédica INCLIVA. Universidad Politécnica de Valencia. Valencia. España **** Hospital Universitario Clínico San Carlos. Madrid. España
Transcript of Cardio Arritmia Vol I Libro - tienda.grupocto.estienda.grupocto.es/pdf/ME_CardioA_CapM.pdf · A...

1
01C a p í t u l o
1.1. Introducción
El entendimiento de los mecanismos electrofi siológicos ce-lulares y tisulares cardíacos resulta esencial para el correcto diagnóstico de las arritmias, así como su tratamiento, tanto far-macológico como invasivo. Este capítulo tiene como objetivo principal revisar los conceptos más generales de bioelectrici-dad de las membranas y electrofi siología celular para estable-cer las bases del entendimiento de los mecanismos básicos de las arritmias y su tratamiento a nivel clínico.
1.2. Bioelectricidad
A nivel teórico son muchas las similitudes entre el flujo de iones que atraviesa la membrana plasmática y la corriente que generan, con el comportamiento de la corriente en un circuito eléctrico.
Algunos conceptos básicos de bioelectricidad permiten com-prender los mecanismos que dirigen la excitabilidad cardíaca, el movimiento de iones y las corrientes a través de la membrana plasmática de la célula (véase la Figura 1.1 en página siguiente).
Carga
Los elementos químicos tienden a conservar el mismo número de protones (carga positiva) que de electrones (carga negativa), manteniendo una carga neta cero. Sin embargo, dentro de la es-
tabilidad química que confi ere completar eléctricamente la últi-ma capa de electrones de los átomos, algunos elementos pue-den perder o ganar electrones, dando lugar a moléculas en las que el número de electrones y protones difi eren. Estas partículas cargadas eléctricamente se denominan iones, y son la base de la excitabilidad cardíaca, puesto que su movimiento a través de los canales iónicos de la membrana plasmática genera la corriente responsable del potencial de acción (PA) cardíaco. Los iones car-gados positivamente (que han perdido electrones) se conocen como cationes (K+, Na+ o Ca2+) y los iones cargados negativamen-te (que han ganado electrones) son los aniones (Cl- o I-). El movi-miento de atracción entre partículas cargadas de diferente signo es lo que genera la corriente eléctrica.
Voltaje
La perturbación espacial que genera una partícula cargada en el espacio es lo que se conoce como campo eléctrico. Toda partí-cula cargada genera una fuerza de atracción hacia las cargas de diferente signo, y de repulsión hacia las del mismo, de manera que las cargas se moverán en el espacio de acuerdo a estas fuer-zas para conseguir un entorno eléctrico neutro.
El trabajo ejercido por el campo eléctrico para mover una carga desde un punto A hasta un punto B se conoce como diferencia de potencial (o diferencia de voltaje), se puede medir con un vol-tímetro y su unidad es el voltio (V). Desde un punto de vista más práctico, una diferencia de potencial se genera cuando existe una distribución diferencial de cargas entre dos puntos aislados, que equivaldría a una acumulación asimétrica de cargas en un condensador. A nivel electrofi siológico, existe una diferencia de potencial entre ambos lados de la membrana plasmática de la
Electrofisiología celular
José Manuel Alfonso Almazán*
Manuel Marina Breysse*, **
Conrado J. Calvo Sáiz*, ***
David Filgueiras Rama*, ****
* Fundación Centro Nacional de Investigaciones Cardiovasculares Carlos III (CNIC). Madrid. España** Hospital Universitario de Móstoles. Madrid. España*** Electrofi siología Cardíaca y Bioingeniería (GEB). BioITACA-IIS Investigación Biomédica INCLIVA. Universidad Politécnica de Valencia. Valencia. España **** Hospital Universitario Clínico San Carlos. Madrid. España

2
Cardio Arr i tmias
célula cardíaca (que actuaría a modo de condensador eléctrico) debido a la distribución asimétrica de iones: en el líquido intra-celular se encuentran más cargas negativas (aniones) que en el extracelular, por lo que el interior celular es más electro-negativo que el exterior.
Corriente
Siempre que los dos puntos sobre los que existe una diferencia de potencial permanezcan aislados, dicha diferencia de voltaje se mantendrá constante. Sin embargo, si el punto A y el punto B se conectan a través de un elemento conductor, las cargas se mo-verán de un punto al otro por la fuerza de atracción entre cargas de distinto signo. Este movimiento de cargas por un elemento conductor genera corriente eléctrica, cuya unidad es el amperio (A) y puede medirse con un amperímetro.
Dentro del contexto de un cardiomiocito (o cualquier célula del organismo) los iones se encuentran en constante movi-miento entre el interior y el exterior celular a través de los
canales iónicos. Estos canales son proteínas integrales de membrana que, cuando se encuentran en estado abierto, comunican el interior y el exterior de la célula permitiendo el paso de iones de manera masiva a favor de la diferencia de potencial. Este flujo de iones genera una corriente eléc-trica como si del movimiento de electrones por el cable de un circuito se tratara. La corriente que generan los distintos canales iónicos cuando están activados se puede medir me-diante experimentos de patch clamp, siendo del orden de pA (10-12 A).
Resistencia
Todo conductor opone cierta resistencia al paso de la corriente. La unidad de resistencia es el ohmio (), aunque generalmen-te se recurre a la unidad inversa, la conductancia (g = 1/R). Las unidades de la conductancia son los siemens (S). Los resistores más simples presentan una relación voltaje-corriente lineal que sigue la Ley de Ohm, por lo que reciben el nombre de resistores óhmicos:
Figura 1.1. Canal iónico sensible a voltaje y modelo simplifi cado de circuito eléctrico asemejando el funcionamiento de la membrana celular en las células excitables cardíacas. A: canal iónico sensible a voltaje (panel superior) y modelo eléctrico equivalente (panel inferior); B: representación simplifi cada de la célula cardíaca en equilibrio electroquímico y la disposición de los canales sensibles a voltaje más relevantes; C: circuito equivalente de la célula cardíaca (miocito). La membrana lipídica está representada por un condensador Cm. El voltaje transmembrana, Vm, representa la diferencia de potencial entre los medios extracelular e intracelular (CSV: canales iónicos sensibles a voltaje)

01 Electrof is io logía celu lar
3
V = I x R
Donde V es la diferencia de potencial, I la intensidad de corrien-te y R la resistencia eléctrica. La Ley de Ohm indica que, ante un aumento en la diferencia de potencial entre ambos lados de la membrana, se producirá un aumento directamente proporcio-nal en la corriente eléctrica. Los canales iónicos, como conduc-tores de la corriente, se pueden representar como resistencias eléctricas, que en estado cerrado se oponen al paso de iones, y que al activarse aumentan su conductancia permitiendo el fl ujo de los iones entre el medio intracelular y extracelular.
1.3. Bases de la electrofisiología cardíaca normal: la membrana plasmática
Estructura de doble capa y propiedades físicas
La membrana plasmática de los cardiomiocitos, denominada sarcolema, está formada por una bicapa lipídica de naturaleza principalmente hidrofóbica. La función del sarcolema es aislar el medio intracelular del extracelular, así como regular el tráfi co de moléculas entre la célula y su entorno para mantener una correc-ta homeostasis. El comportamiento hidrofóbico de la membrana se debe a que está compuesta por fosfolípidos, que se disponen formando una bicapa, de manera que las regiones hidrófi las se encuentren en la parte exterior de la membrana y las regiones hi-drofóbicas en la parte interna. Esta diferencia de composición ori-gina un gradiente de concentración que induce a la difusión de las moléculas (a favor de gradiente), desde el espacio en que se encuentran en mayor concentración hacia el espacio en el que la concentración es menor. A su vez, y debido a la distribución asi-métrica de los distintos iones entre ambos lados de la membrana (Tabla 1.1)(1), existe un gradiente eléctrico y una diferencia de potencial eléctrico, denominado potencial transmembrana (Vm). El Vm viene determinado por la concentración relativa de los dis-tintos iones en el medio intracelular y extracelular, así como por
la permeabilidad de la membrana plasmática a cada uno de los iones (Na+, K+, Ca2+ y Cl- principalmente).
Movimiento de iones a través del sarcolema
Imagínese una situación ideal en la que se separan dos medios de diferente concentración iónica por una membrana semiper-meable; los iones se moverían a favor de gradiente de concentra-ción desde el medio de mayor concentración al de menor con-centración, lo que originaría un gradiente eléctrico entre ambos medios. Este gradiente eléctrico se opondría al gradiente quí-mico. Los iones continuarían moviéndose de un espacio al otro hasta alcanzar una situación de equilibrio en la que el gradiente químico y el gradiente eléctrico se igualarían. Como consecuen-cia de estas dos fuerzas opuestas, en la situación de equilibrio existe una diferencia de concentración entre ambos espacios y al mismo tiempo una diferencia de potencial.
Aplicando el ejemplo anterior a los distintos iones celulares por separado, si el sarcolema fuera totalmente permeable al Na+ (e impermeable al resto de los iones), el Na+ se introduciría en la célula hasta alcanzar el equilibrio electroquímico en el que el fl u-jo de iones Na+ a través de la membrana fuese nulo. El potencial transmembrana alcanzaría en ese momento lo que se conoce como potencial de equilibrio, que para el Na+ tiene un valor de +65 mV. El potencial de equilibrio para el K+, por el contrario, ten-dría un valor negativo de –96 mV, mientras que para el Ca2+ sería de +125 mV (véase la Tabla 1.1).
Teniendo en cuenta todos los iones en su conjunto y que la per-meabilidad de la membrana para cada ion es diferente, se puede calcular el potencial transmembrana de reposo. El potencial de re-poso para las células musculares auriculares y ventriculares, y para las células del sistema de conducción His-Purkinje oscila entre –85 y –90 mV, mientras que para las células del nodo sinusal y del nodo auriculoventricular (NAV) oscila entre –55 y –60 mV. Por tanto, el potencial de reposo depende del equilibrio entre la capacidad de los distintos iones para atravesar el sarcolema a favor de su gra-diente electroquímico, así como del transporte activo de dichos iones en contra de su gradiente, mediado principalmente por la bomba Na+/K+ ATPasa o el cotransportador Na+/Ca2+.
La diferencia entre el valor del potencial de equilibrio y el poten-cial transmembrana de reposo es la razón por la que los iones se moverán siempre en la misma dirección a favor de gradiente electroquímico: los iones Na+ y Ca2+ tenderán a entrar en la célula, siendo los responsables de la despolarización durante la gene-ración y propagación del PA cardíaco; mientras que los iones K+ tenderán a salir al medio extracelular para controlar el potencial de reposo (véase la Figura 1.1). De hecho, es fácil observar que el potencial de equilibrio del K+ difi ere muy poco del potencial de reposo (–96 mV frente a –85 mV, respectivamente), debido principalmente a la gran densidad celular de canales de potasio.
Ion Concentración extracelular (mM)
Concentración intracelular (mM)
Potencial de equilibrio (mV)
Sodio 145 10 +67
Potasio 4 150 -96
Calcio 1,5 10-4 +125
Cloro 120 4 -90
Tabla 1.1. Concentración intracelular y extracelular de los iones principales responsables de la generación del potencial de acción en condiciones fi siológicas (estimación matemática del valor del potencial de equilibrio para una temperatura de 37 ºC)

4
Cardio Arr i tmias
Canales iónicos y estructura
El movimiento de los iones se puede producir a favor de gra-diente electroquímico (transporte pasivo) o en contra del mismo (transporte activo), mediante la traslocación de iones de un lado al otro de la membrana plasmática. Este movimiento coordinado y organizado de iones entre el medio intracelular y extracelular es la base molecular de la excitabilidad celular y de la genera-ción y propagación del PA cardíaco. Funcionalmente se pueden clasifi car en: • Canales iónicos. • Bombas iónicas. • Cotransportadores.
Los canales iónicos representan la familia más numerosa y son responsables de originar el PA mediante su activación e inacti-vación secuencial y coordinada. Son proteínas transmembrana que conforman un poro hidrofílico selectivo a través del cual los distintos iones pueden atravesar el sarcolema a favor de gra-diente electroquímico de manera masiva (del orden de hasta 108 iones/s), generando corriente eléctrica. Los canales iónicos presentan dos propiedades principales que los hacen tan impor-tantes: • Selectividad iónica. Esta selectividad se consigue por impedi-
mento estérico y termodinámico, que se basa tanto en el diá-metro de apertura del canal, que sólo permite el paso de su ion en su forma hidratada, así como en la carga del ion, de modo que ciertos residuos del canal se alinean en el poro e interac-cionan con los iones formando barreras termodinámicas que favorecen el paso de unos iones frente a otros.
• Gating. Los canales se activan y abren, o cierran ante deter-minados estímulos, propiedad que se denomina en inglés gating. Principalmente los canales pueden activarse por vol-
taje, ante cambios en el potencial transmembrana (canales dependientes de voltaje), o por ligando, ante la unión de una molécula específi ca al canal (canales dependientes de ligan-do), o por estiramiento, ante un estímulo mecánico en la célula (canales dependientes de estiramiento). Los principales canales de Na+ y Ca2+ responsables del PA cardíaco y la gran mayoría de canales de K+ son dependientes de voltaje. Los canales iónicos dependientes de ligando (también denomi-nados receptores ionotrópicos) generalmente responden ante distintas hormonas, neurotransmisores o moléculas se-ñalizadoras para producir una respuesta celular determinada ante un estímulo externo (Tabla 1.2).
Molecularmente, la transición entre los estados abierto y cerrado del canal se debe a cambios conformacionales y secuenciales en la proteína. Los canales iónicos se pueden encontrar en estado cerrado (o de reposo), activado (o abierto) e inactivado. Solamen-te en el estado abierto se permite el paso de los iones para gene-rar corriente eléctrica.
En el caso de los canales dependientes de voltaje, la gran mayoría de los canales se activan cuando la membrana se despolariza. Generalmente la apertura del canal es muy breve y rápidamente pasa al estado inactivo, aunque la membrana permanezca des-polarizada. En estado inactivo el poro del canal se encuentra ce-rrado (no conductor), y no responderá a nuevos estímulos duran-te el periodo de tiempo en que se encuentre en este estado. Este fenómeno de inactivación es muy importante para el correcto funcionamiento de los canales iónicos, puesto que impide que el canal esté activo constantemente a valores de voltaje superiores a su umbral. La correcta coordinación de activación e inactiva-ción de todos los canales de manera secuencial permite que las corrientes eléctricas sean de la duración necesaria para originar
Receptores Acción
Receptores betaadrenérgicos
· Modulan: Los canales I
Ca-L: pueden inducir pospotenciales y actividad desencadenada.
La corriente If: aumentan la frecuencia de descarga de las células automáticas.
Varias corrientes de potasio (IK1
e Ito
). La corriente de cloro y bajo ciertas circunstancias las de Na+.
· Incrementan los niveles de calcio intracelular. · Incrementan la acción de la bomba Na+/K+ (hiperpolarizan las células).
Comentarios:Tanto los efectos sobre el calcio como el potasio producen una aceleración de la conducción auriculoventricular y un acortamiento de su periodo refractario.Son los responsables de la taquicardización y acortamiento del intervalo QT.
Receptores alfaadrenérgicos
Dos o tres subtipos de receptores 1 adrenérgicos interaccionan con: · La bomba Na+/K+: (disminuyen la fase 4 de las células automáticas). · Las corrientes I
K1 e I
to, prolongando la repolarización.
· Pueden inducir arritmias por pospotenciales o automatismo anormal.
Receptores muscarínicos
· El receptor muscarínico cardíaco dominante es el receptor M2. · Su densidad es entre 2 y 5 veces mayor en aurícula que en ventrículo. · Se relacionan directamente a la corriente I
K,ACh para el control vagal del ritmo cardíaco.
Tabla 1.2. Principales receptores de las células cardíacas y su acción

01 Electrof is io logía celu lar
5
el PA cardíaco. Además, la cinética de inactivación de los canales da lugar a un periodo de refractariedad, durante el que la célula no será capaz de responder a nuevos estímulos para generar un nuevo PA, puesto que sus canales se encuentran inactivos.
El número de estados que presenta cada canal, así como el tiem-po en que se encuentra en cada estado y la velocidad a la que pasa de uno a otro es lo que se conoce como cinética de activa-ción. Por ejemplo, el canal de Na+ se caracteriza por una rápida activación y casi inmediata inactivación, que dura hasta casi el fi nal del PA. Razón por la que la entrada de Na+ en la célula es la responsable principal de la fase de despolarización rápida del PA. Los canales de K+ no presentan estado inactivo, únicamente se pueden encontrar abiertos o cerrados.
Canales de sodio dependientes de voltaje
Los canales de Na+ dependientes de voltaje son fundamentales en la génesis y propagación de la señal eléctrica en tejidos ex-citables como el corazón, el músculo esquelético o el sistema nervioso.
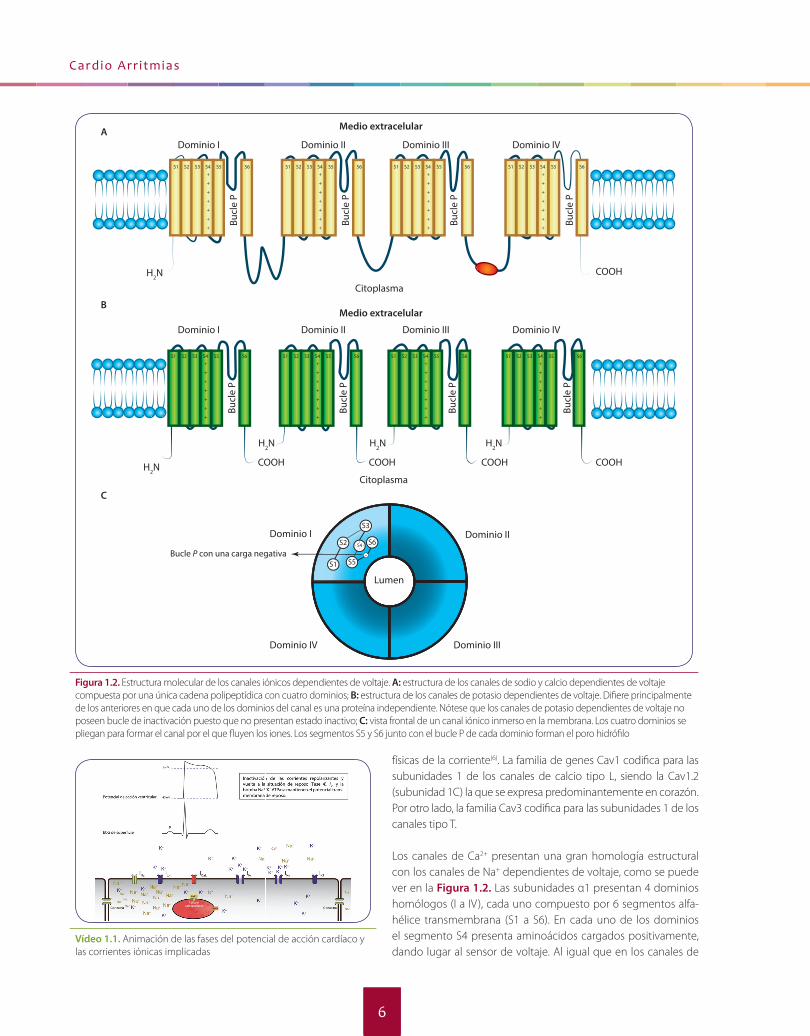
El canal de sodio está constituido por una subunidad alfa prin-cipal y una o varias subunidades accesorias beta, dependiendo del tejido donde se exprese. La subunidad alfa es la subunidad formadora del canal y está constituida por una única proteína de 260 kDa formada por cuatro dominios homólogos (I-IV), con seis segmentos alfa-hélice transmembrana cada uno de ellos (S1-S6). Los segmentos de cada dominio se conectan por secuencias pep-tídicas sin estructura secundaria (loops) no conservadas. Entre el segmento S5 y S6 de cada dominio se encuentra el bucle P (o P loop), que está incrustado en la región transmembrana del canal y forma un estrecho fi ltro selectivo de iones en el extremo extrace-lular del poro. Los extremos amino terminal y carboxilo terminal, así como las secuencias que conectan los cuatro dominios entre sí son intracelulares(2). Toda la proteína se pliega de manera perfec-ta para que en la membrana celular los segmentos S5 y S6, junto con el bucle P de cada dominio conformen el lumen del canal (el poro). Los segmentos S1, S2 y S3 quedan apartados del centro, po-sicionados en la periferia del canal y rodeados de los fosfolípidos de la membrana. Justo entre estos tres segmentos se encuentra el segmento S4, que presenta un aminoácido cargado positivamen-te (arginina o lisina) cada tres aminoácidos y actúa como sensor de voltaje del potencial transmembrana(3). La secuencia peptídica que une el dominio III con el dominio IV forma el bucle de inactivación, mecanismo por el que se inactiva el canal. Por otro lado, cuatro aminoácidos presentes en los bucles P forman un anillo denomi-nado locus DEKA: ácido aspártico en el dominio I, ácido glutámico en dominio II, lisina en el dominio III y alanina en el dominio IV. Esta estructura determina la conductancia y la selectividad iónica del canal al Na+ y puede ser diana de algunos fármacos y toxinas(4). En la Figura 1.2 (véase en página siguiente) se muestra con detalle la arquitectura molecular del canal de Na+.
El canal de Na+ presenta al menos tres estados conformacionales (reposo, activo e inactivo). Durante la diástole eléctrica, el canal se encuentra en estado de reposo. La despolarización de la mem-brana por encima del umbral de activación del canal de sodio, en torno a los –70 mV(5), produce un cambio conformacional en la estructura del canal que causa su apertura durante tan solo 0,5-2 ms, generándose una corriente rápida de entrada de Na+, deno-minada I
Na, responsable de la fase 0 de despolarización rápida del
PA. A continuación, el canal se inactiva rápidamente, lo que pro-duce el cese de la entrada de Na+. La inactivación no sólo cierra el canal, sino que impide la reapertura del mismo hasta que haya pasado el tiempo sufi ciente para su recuperación, determinando así la frecuencia máxima de excitación celular y preservando el gradiente iónico intracelular, lo que impide la muerte celular. El Vídeo 1.1 (véase en página siguiente) muestra las distintas fases del PA y las corrientes iónicas implicadas.
Canales de calcio dependientes de voltaje
Estos canales permiten la entrada de Ca2+ en respuesta a la despo-larización de la membrana y son esenciales para acoplar las señales eléctricas con la respuesta fi siológica a nivel celular. En las células del miocardio se diferencian principalmente dos tipos de canales de Ca2+ dependientes de voltaje(5): • Canales de Ca2+ tipo T. Se activan rápidamente a un poten-
cial transmembrana de –50 mV y se inactivan también rápi-damente.
• Canales de Ca2+ tipo L. Se activan cuando se despolariza la membrana hasta aproximadamente los –30 mV y su inactiva-ción es mucho más lenta, pudiendo prolongarse hasta 100 ms. Bajo la mismas condiciones, la conductancia del canal de Ca2+ tipo L es aproximadamente el doble que la del canal T.
Los canales tipo L se localizan principalmente en los túbulos T del sarcolema, próximos y enfrentados a los receptores de rianodina (RyR) del retículo sarcoplá smico, y tienen dos papeles fundamen-tales: • Median la corriente I
Ca-L, que prolonga la despolarización del
PA dando lugar a la fase 2 o meseta del PA cardíaco. • Son los responsables del acoplamiento electromecánico.
Por su parte, los canales tipo T se localizan principalmente en las células auriculares, en las células del nodo sinusal y en las fi bras del His-Purkinje, mientras que su presencia en las células ventri-culares es mínima.
Los canales de Ca2+ dependientes de voltaje son heterotetrá meros compuestos por las subunidades α1, β y α2/δ, y en algunos tejidos, la subunidad γ. Dicha subunidad γ no está presente en los cardio-miocitos. Los diferentes tipos de corriente de entrada de Ca2+ (I
Ca)
se defi nen en función de la subunidad α1, subunidad principal que forma el canal, mientras que las subunidades reguladoras α2/δ, β y γ modulan su tráfi co en la membrana y/o las propiedades bio-
6
Cardio Arr i tmias
físicas de la corriente(6). La familia de genes Cav1 codifi ca para las subunidades 1 de los canales de calcio tipo L, siendo la Cav1.2 (subunidad 1C) la que se expresa predominantemente en corazón. Por otro lado, la familia Cav3 codifi ca para las subunidades 1 de los canales tipo T.
Los canales de Ca2+ presentan una gran homología estructural con los canales de Na+ dependientes de voltaje, como se puede ver en la Figura 1.2. Las subunidades α1 presentan 4 dominios homó logos (I a IV), cada uno compuesto por 6 segmentos alfa-hé lice transmembrana (S1 a S6). En cada uno de los dominios el segmento S4 presenta aminoácidos cargados positivamente, dando lugar al sensor de voltaje. Al igual que en los canales de
Figura 1.2. Estructura molecular de los canales iónicos dependientes de voltaje. A: estructura de los canales de sodio y calcio dependientes de voltaje compuesta por una única cadena polipeptídica con cuatro dominios; B: estructura de los canales de potasio dependientes de voltaje. Difi ere principalmente de los anteriores en que cada uno de los dominios del canal es una proteína independiente. Nótese que los canales de potasio dependientes de voltaje no poseen bucle de inactivación puesto que no presentan estado inactivo; C: vista frontal de un canal iónico inmerso en la membrana. Los cuatro dominios se pliegan para formar el canal por el que fl uyen los iones. Los segmentos S5 y S6 junto con el bucle P de cada dominio forman el poro hidrófi lo
Vídeo 1.1. Animación de las fases del potencial de acción cardíaco y las corrientes iónicas implicadas

01 Electrof is io logía celu lar
7
Na+, el bucle P une los S5 y S6 de los cuatro dominios y contri-buye a formar el poro del canal. Cada uno de los bucles P de los cuatro dominios contiene un residuo de ácido glutá mico (“locus EEEE”). Estos residuos forman una zona de alta afi nidad por el Ca2+ y conforman el fi ltro de selectividad del canal.
Canales de potasio dependientes de voltaje
Los canales de K+ conforman la familia más diversa, numerosa y compleja de canales iónicos. Se encuentran en todas las células del organismo y desempeñan múltiples funciones fi siológicas, como la repolarización y mantenimiento del potencial trans-membrana de reposo, la duración del PA, el control de la frecuen-cia de disparo de las células automáticas, la secreción de insulina, el transporte de electrolitos, la regulación de la contracción del músculo liso, etc.
La estructura de los canales de K+, al igual que la de los canales de Na+ y de Ca2+, también está compuesta por cuatro dominios (I-IV) que al plegarse conforman la subunidad alfa con el poro hidrófi -lo, pero difi ere principalmente en que los canales de K+ cada uno de estos dominios es una cadena polipeptídica independiente, al contrario que los canales de Na+ y Ca2+, en los que toda la proteína era un único gran péptido (véase la Figura 1.2). Los canales de K+ se clasifi can principalmente de acuerdo al número de segmentos transmembrana en sus dominios y al número de poros(7): • Canales formados por 2 segmentos transmembrana y 1 poro
(2TM/1P). También llamados canales de K+ rectifi cadores in-ternos (canales Kir).
• Canales formados por 4 segmentos transmembrana y 2 po-ros (4TM/2P). Denominados canales K2P.
• Canales formados por 6 segmentos transmembrana y 1 poro (6TM/1P). Incluye a los canales de K+ dependientes de voltaje (canales Kv) y a los canales de K+ activados por Ca2+ (canales KCa), que presentan un segmento TM adicional (S0) en el ex-tremo N-terminal.
Las cinéticas de activación e inactivación de cada canal deter-minan su participación en el control de la repolarización. Las corrientes que se activan e inactivan rápidamente (corriente transitoria de salida de K+; I
to y la corriente ultrarrápida rectifi ca-
dora tardía de K+; IKur
) intervienen en la repolarización durante las primeras fases del PA, encontrándose en mayor medida en la aurícula. Por otro lado, los canales que generan corrientes rectifi -cadoras tardí as (I
Kr e I
Ks, con rápida y lenta cinética de activación,
respectivamente) se activan cuando el potencial transmembrana alcanza –40 mV, son de activación mucho más lenta y participan fundamentalmente en la repolarización durante la fase 3 del PA mediante una corriente de salida de iones K+ de la célula.
La corriente rectifi cadora de entrada de K+ (Ik1
) es la corriente prin-cipal del mantenimiento del valor de potencial transmembrana de reposo. Esta corriente es abundante en las células musculares au-
riculares y ventriculares, y en las fi bras de conducción His-Purkinje, pero está prácticamente ausente en las células nodales. Esta distri-bución diferencial de la corriente en el tejido podría ser la explica-ción de los distintos valores de potencial de reposo en las células.
En cuanto a las corrientes de potasio activadas por distintas molécu-las, cabe destacar tanto la corriente rectifi cadora de K+ dependiente de acetilcolina (I
K,ACh) y la dependiente de ATP (I
K,ATP). I
K,ACh se activa por
la unión de acetilcolina a los receptores muscarínicos del sarcolema de los cardiomiocitos, produciendo una corriente de salida que hi-perpolariza la célula. Esta corriente es muy importante en el control vagal del ritmo cardíaco(8). En condiciones fi siológicas, a concentra-ciones normales de ATP, I
K,ATP se encuentra inactivada. Sin embargo,
ante situaciones de depleción de ATP (como durante la isquemia) la corriente se activa produciendo una salida de K+ de la célula y una hiperpolarización de la membrana.
Canales HCN y corriente de células marcapasos
La corriente funny, If, es una de las corrientes responsables del au-
tomatismo normal de las células. Sólo se expresa en células con actividad de marcapasos: nodo sinusal, NAV y fi bras de His-Pur-kinje). I
f se genera en los canales HCN, y se caracteriza por tratarse
de una corriente despolarizante de entrada lenta de Na+, activa-da por la hiperpolarización, cuando el potencial transmembrana está entre –45 mV y –65 mV, alcanzando su máxima amplitud para un potencial de –100 mV.
En las células marcapasos, la fase 4 o periodo entre dos poten-ciales no es isoeléctrico, sino que gracias a la corriente I
f permite
la existencia de una ligera pendiente de despolarización positiva que lleva el valor de potencial transmembrana desde el potencial de reposo diastólico hasta el valor de potencial umbral, para el que se inicia un nuevo PA en las células automáticas.
Corriente a través de las uniones estrechas o gap junctions
Los cardiomiocitos están unidos entre sí mediante discos inter-calares, que permiten el acoplamiento eléctrico y la propagación del PA. Los discos intercalares están compuestos por grupos de conexones, proteínas transmembrana que conectan los citoplas-mas de dos células adyacentes. Los conexones de cada célula se encuentran dispersos por todo el sarcolema y se unen con los co-nexones de las células vecinas para formar un canal por el que fl ui-rán los iones. A esta estructura formada por dos conexones enfren-tados que conectan dos células vecinas se le conoce como unión estrecha o gap junction. Las gap junctions siempre están abiertas y permiten el paso continuo de iones en todo momento sin presen-tar prácticamente resistencia al paso de la corriente.
Cada conexón está constituido por seis subunidades denomina-das conexinas formando una estructura hexagonal con un poro central. Se han identifi cado diferentes tipos de conexinas y se ha

8
Cardio Arr i tmias
demostrado que en cada región del corazón se expresa preferen-temente una variante: la conexina 43 (Cx43) es la predominante en el tejido muscular ventricular y la conexina 40 (Cx40) en las fi bras de Purkinje(5, 8).
Bombas iónicas y cotransportadores
Existen otras proteínas transportadoras integrales de membrana que traslocan iones de un lado al otro del sarcolema en contra de gradiente electroquímico, mediante un proceso denominado transporte activo. Estas proteínas son las bombas iónicas y los cotransportadores.
Las bombas iónicas dependen de la energía metabólica, utilizan la energía liberada en la hidrólisis de trifosfato de adenosina (ATP) para transportar iones en contra de gradiente. Debido a su me-canismo se les denomina también bombas ATPasas. Destaca la bomba Na+/K+ ATPasa, que actúa de manera prácticamente con-tinua a lo largo de todo el ciclo cardíaco, y es la responsable de que exista la gran diferencia de concentración de Na+ y K+ entre el interior y el exterior celular: expulsa 3 Na+ e introduce 2 K+ en cada ciclo de acción.
Los cotransportadores de iones no dependen directamente de la energía metabólica, sino que utilizan el gradiente electroquímico favorable de un ion para mover otros iones a través del sarcole-ma en contra de gradiente, de manera que transportan un ion a favor de su gradiente y otro en contra. De entre ellos destaca el cotransportador Na+/Ca2+, que expulsa un ion Ca2+ por cada 3 iones Na+ que introduce en la célula.
1.4. Potencial de acción
El PA es esencialmente una transición eléctrica que se puede pro-pagar a través de largas distancias conservando su amplitud. Esta característica principal del PA se logra al basar su iniciación en los fenómenos autorregenerativos comentados previamente.
Fases del potencial de acción transmembrana
El PA cardíaco es el resultado de una serie de cambios secuencia-les en la permeabilidad de la membrana a distintos iones, que va-
BA
t = 0 s t = 0,04 s
60-100 lpm
40-50 lpm
Nodo sinoauricular
células auricularesNodo AV
His
Ramas del His
Purkinje
Células ventriculares
Tiempo sg
30-40 lpm
30-40 lpm30-40 lpm
15-30 lpm
15-30 lpm
HRA
HPPRI
I aVR V1 V4
II aVL V2 V5
III aFV V3 V6
RD
Nodo AV
Nodo sinusal
t = 0,08 s t = 0,1 s t = 0,16 s t = 0,20 s t = 0,24 s t = 0,28 s
Figura 1.3. Sistema de conducción cardíaco y representación electrocardiográfi ca en la superfi cie del torso. A: la activación cardíaca iniciada en el nodo sinusal se transmite a través del nodo auriculoventricular a los ventrículos. Se muestran los potenciales de acción característicos de las células del sistema de conducción y del músculo miocárdico que contribuyen a la formación del ECG de superfi cie; B: representación de la actividad eléctrica cardíaca sobre la superfi cie corporal de un paciente durante ritmo sinusal. Se muestran electrodos y ECG promedio para las derivaciones convencionales superpuestas sobre la rejilla de papel del ECG (25 mm/s, 0,5 mV/div, 0,2 s/div)

01 Electrof is io logía celu lar
9
rían en función de dos variables: el Vm y el tiempo. La Figura 1.3 muestra el PA cardíaco en las células musculares auriculares, ven-triculares y sistema His-Purkinje. La Figura 1.4 muestra las distintas corrientes iónicas que participan en la generación del PA auricular y ventricular(5, 8, 9-11).
Fase 0 de despolarización rápida dependiente de sodio
Ante un estímulo despolarizante, si éste es lo suficiente-mente intenso como para elevar el Vm hasta el valor umbral que activa los canales de sodio (–70 mV), se desencadena-
INA
IKI
* Presente en situaciones de isquemia
ICaL
ICaL
ICa-L
INCX
IKur
Ito
Ito
IK1
IKs
IKs
INaK
IKACh
IKATP
IKr
IKr
IKsIKr
Ito
IKl IKl
IKlIKATP*
IK,ATP*IKur
IK,ACh CA-IK,ACh
INa
INa
100 ms
10 mV
100 ms
10 mV-83 -86
0-0-
AD
A B
AI VD VI
Figura 1.4. Potencial de acción cardíaco y contribución de las principales corrientes a su formación. A: panel superior, anatomía detallada de aurícula humana coloreada para representar la aurícula derecha (‘AD’, en marrón claro) y aurícula izquierda (‘AI’, en rojo). Panel inferior, potencial de acción auricular y principales corrientes que contribuyen en las distintas fases; B: panel superior, anatomía detallada de un ventrículo humano coloreado representando las cavidades ventricular derecha (VD, verde claro) y ventricular izquierda (VI, rojo claro). Panel inferior, representación del potencial de acción ventricular y la contribución de las distintas corrientes en cada fase

10
Cardio Arr i tmias
rá el PA. Al abrirse los canales de Na+, se produce la entra-da masiva de iones Na+ a favor de gradiente electroquímico (cargas positivas en el medio extracelular son atraídas por cargas negativas del medio intracelular) y gradiente de con-centración (145 mM en el medio extracelular frente a 10 mM en el miocito). Este flujo brusco de iones genera la denomi-nada corriente rápida de entrada de sodio (I
Na), invirtiendo la
polaridad del Vm, que se vuelve positivo hasta +20/+30 mV. La activación de I
Na es un proceso muy rápido (0,5-2 ms) y la
magnitud de INa
determina la amplitud, la velocidad máxima de despolarización del PA y, por tanto, la velocidad de con-ducción intracardíaca.
Los estímulos más pequeños que no llevan el potencial de repo-so hasta el potencial umbral provocan PA pequeños, no propaga-dos, denominados potenciales electrotónicos. Estos potenciales son importantes porque pueden afectar a la conducción de PA subsiguientes.
Fase 1 de rápida repolarización
Es la fase inicial de la repolarización al producirse una salida de K+ por gradiente de concentración (4 mM en el medio extracelular frente a 150 mM en el interior del miocito), que compensa la in-versión de polaridad que ha sufrido la célula en la fase 0. La fase 1 se produce por la inactivación de I
Na y la activación de canales
de potasio dependientes de voltaje, que originan dos corrientes de salida de iones K+: • I
to, que se activa e inactiva rápidamente.
• IKur
, de rápida activación y lenta inactivación, con especifi ci-dad auricular.
Fase 2 de meseta
Es la fase de repolarización lenta. En esta fase persisten corrien-tes de salida de K+ que se compensan principalmente por una corriente de entrada lento de Ca2+ al interior de la célula. En esta fase, el Vm se mantiene constante y cercano a 0 mV gracias al equilibrio entre dos corrientes de entrada (I
Na [su componente de
inactivación lenta] e ICa-L
), y tres corrientes rectifi cadoras tardías de salida de K+ (de activación ultrarrápida, rápida y lenta [I
Kur, I
Kr e
IKs
, respectivamente]).
ICa-L
se produce por gradiente de concentración (1,5 mM en el medio extracelular frente a 10-4 mM en el interior del miocito). ICa-L
se activa entre –30 y –20 mV, y alcanza su máxima ampli-tud entre 0 y +20 mV. Sin embargo, su cinética de activación es muy lenta, por lo que se empieza a activar durante la fase 0 del PA, pero alcanza su máxima amplitud durante la fase 2. I
Ca-L
es responsable del acoplamiento electromecánico del músculo miocárdico. La entrada de Ca2+ al interior celular activa los re-ceptores de rianodina del retículo sarcoplásmico, cuya función es liberar el contenido de Ca2+ del retículo sarcoplásmico al ci-
tosol y activar los miofi lamentos del citoesqueleto, provocando la contracción.
Fase 3 de repolarización
Es la fase terminal de la repolarización. Persisten las corrientes re-polarizantes de salida de K+ (I
Kr e I
Ks) mediadas por gradiente de
concentración, pero se cierran los canales de Ca2+. Al fi nal de la fase 3, la célula alcanza de nuevo el valor del potencial de reposo de –85 mV, pero eléctricamente sus cargas han cambiado por lo que la célula es aún inexcitable.
En esta fase, otras corrientes repolarizantes de potasio se pueden activar bajo determinadas circunstancias, como durante la sobre-carga de calcio (I
K-Ca), la sobrecarga de sodio (I
K-Na), la estimulación
colinérgica (IK,ACh
) o la depleción de ATP que ocurre durante la isquemia (I
K,ATP).
Fase 4 o periodo entre dos potenciales
Describe el Vm durante la diástole donde, aunque sea isoeléctri-co, al inicio de esta fase la bomba Na+/K+ ATPasa expulsa Na+ del interior celular e introduce K+. La célula vuelve a la situación de equilibrio, con el K+ de nuevo en su interior, el Na+ en el exterior y el interior celular cargado negativamente.
En los miocitos auriculares y ventriculares, que expresan en gran medida I
K1, el Vm se estabiliza en un valor próximo a la inversión
de corriente y se requiere una fuente de corriente signifi cativa para volver a excitar la célula. El mantenimiento del Vm en ni-veles constantes se debe principalmente a I
K1, aunque también
participan la bomba Na+/K+ ATPasa y el intercambiador Na+/Ca2+, para mantener las diferencias de concentraciones de iones entre el medio intracelular y extracelular. Si la expresión de I
K1
es pobre (miocitos del Purkinje) o ausente (células nodales), la fase 4 se hace más positiva e inestable. En tales condiciones, pequeñas corrientes hacia el interior pueden causar despolari-zación progresiva, llevando a la reexcitación (comportamiento automático).
La fase 4 del PA se inicia una vez que el potencial de la célula alcanza de nuevo su valor de reposo y fi naliza al comienzo del siguiente PA.
Morfología del potencial de acción en los tejidos cardíacos especializados
La morfología del PA varía en las distintas células cardíacas se-gún su localización. Las dos variedades más diferenciadas de PA son el de las células musculares o células de respuesta rápida (véase la Figura 1.3) y el PA de las células marcapasos o PA de las células de respuesta lenta (principalmente células nodales) (Figura 1.5).

01 Electrof is io logía celu lar
11
Una característica general de todas ellas es su larga duración. A diferencia del músculo esquelético y de las neuronas, cuya du-ración es de 1 a 5 ms (al no tener fase de meseta), el potencial de las fi bras cardíacas oscila entre 140 y 300 ms en condiciones normales. La duración del PA cardíaco tiene importantes conse-cuencias funcionales: • La superposición en el tiempo del PA (sístole eléctrica) con la
contracción del miocito (sístole mecánica). • La prolongación de los periodos refractarios, garantizando
que el músculo no pueda reexcitarse en ningún momento (periodo refractario absoluto), excepto muy al fi nal de la con-tracción (periodo refractario relativo).
Células del nodo sinusal
Las células nodales presentan un Vm en la fase 4 menos nega-tivo que las células de trabajo (–60 mV), que además presentan una pendiente ascendente debido a la despolarización diastólica espontánea mediada por I
f y regulada por I
K,ACh (frecuencia de dis-
paro entre 60-100 lpm). Cuando se alcanza el potencial umbral (–40 mV) en estos grupos celulares, se inicia el PA, con una fase 0 más lenta, mediada por la corriente de entrada de calcio (I
Ca-L)(12).
Células del nodo auriculoventricular
El PA se propaga a través de las células musculares auriculares a 0,3-1 m/s y sus haces preferenciales. Al llegar al NAV la veloci-dad de propagación del impulso disminuye hasta 0,01-0,5 m/s. Al igual que en el nodo sinusal, I
Ca-L es también la corriente lenta
responsable de la fase 0 a nivel de las células del NAV. Este retraso en la conducción permite el llenado telediastólico ventricular tras la contracción auricular (acoplamiento electromecánico). Las cé-lulas del NAV también presentan una pendiente ascendente en fase 4 (despolarización diastólica espontánea) aunque de menor inclinación, por lo que la frecuencia de disparo de su automatis-mo oscila entre 40-50 lpm, observándose en la clínica cuando el nodo sinusal falla (ritmo de escape nodal).
Sistema His
El sistema His tiene una función predominante de propagación del PA a gran velocidad (1-4 m/s), mediada por I
Na. Las células del siste-
ma His también presentan en fase 4 una despolarización diastólica espontánea aunque de menor grado que las nodales, por lo que la frecuencia de disparo de su automatismo oscila entre 30-40 lpm.
Longitud ciclo cardíaco
k+ k+
k+
k+ k+
k+
k+
Na+Na+
Na+
Na+
k+
k+k+
k+
Ca2+
Ca2+
Ca2+
Ca2+Ca2+
Ca2+
Ca2+
ICaT
ICaL
IKs
IKACh
ICaL
ICaT
If
IKr
IKs
IKACh
If
IKrPotencial umbral
Fase 0lenta (Ca2+)
Potencial diastólico máximo
20 mV
100 ms
Figura 1.5. Activación de los marcapasos cardíacos, células autoexcitables del sistema de conducción. Representación del potencial de acción de las células automáticas del nodo sinusal. Se representan los niveles de potencial diastólico máximo, donde comienza la despolarización diastólica y el nivel umbral que da lugar a la generación del nuevo potencial de acción. Se representan los principales canales que participan en la fase de hiperpolarización, despolarización diastólica, despolarización lenta y repolarización

12
Cardio Arr i tmias
Fibras de Purkinje
Las fi bras de Purkinje son el claro ejemplo de un tejido cardíaco especializado cuya función principal es también la de propagar los PA a gran velocidad para despolarizar de forma sincrónica las células contráctiles ventriculares. Este proceso está mediado por INa
durante la fase 0 (velocidad de conducción de hasta 10 m/s). Las células de Purkinje también presentan una despolarización diastólica espontánea, aún más lenta que las células hisianas, por lo que la frecuencia de disparo de su automatismo será menor (entre 15-30 lpm).
Una propiedad del automatismo normal que resulta clave para comprender gran parte de los mecanismos electrofi siológicos cardíacos es el fenómeno llamado supresión por sobreestimu-lación (overdrive supression), que consiste en una depresión transitoria de la frecuencia de descarga de un determinado foco automático inmediatamente después de que éste se haya despolarizado a una frecuencia más rápida que su frecuencia in-trínseca durante un cierto tiempo. Este fenómeno viene deter-minado por una acumulación de Na+ en el interior de las células sobreestimuladas, lo que lleva a un aumento de la actividad de la bomba Na+/K+ATPasa, y se traduce en un potencial de reposo más negativo y más alejado del potencial umbral, disminuyen-do así la frecuencia de descarga. La frecuencia espontánea de las fi bras suprimidas permanece baja hasta que la concentración de Na+ vuelve a sus niveles previos. Las fi bras de Purkinje, más dependientes del fl ujo de Na+, son más fácilmente suprimibles. A nivel clínico, las terapias de estimulación eléctrica cardíaca (mar-capasos, etc.) producen la supresión de todos los automatismos cardíacos, afectando a todas las células con automatismo alcan-zadas por el PA derivado de esa estimulación eléctrica.
1.5. Estudio de las corrientes iónicas a nivel celular
La técnica de patch clamp (Figura 1.6) permite registrar la acti-vidad eléctrica de los canales iónicos en forma de cambios del potencial transmembrana, que refl ejan el fl ujo de iones hacia el interior o exterior de la célula. Los avances en esta técnica, desde su desarrollo en la década de 1980 por Erwin Neher y Berk Sak-mann, junto con las técnicas actuales para aislar cardiomiocitos individuales, hacen posible que se pueda medir con total preci-sión la corriente generada por una célula, e incluso por un único canal iónico. Se pueden llevar a cabo dos abordajes principales: • Fijar el potencial transmembrana y registrar las corrientes
generadas ante el estímulo de dicho potencial (modalidad denominada voltage-clamp).
• Fijar una corriente determinada para registrar el potencial transmembrana (modalidad denominada current-clamp).
La modalidad más utilizada y clásica es la de voltage-clamp, puesto que permite determinar la cinética de activación de los distintos canales, registrar la corriente iónica en función del po-tencial transmembrana, así como los umbrales de apertura de los canales iónicos. • Experimentalmente, se utiliza un capilar de vidrio al que se le
aplica una fuente de calor en uno de los extremos para for-mar una punta de 1-4 m de diámetro, dando lugar a lo que se conoce como micropipeta. Esta micropipeta se llena con solución salina (que constituirá la solución interna) y se co-necta a un sistema de registro eléctrico y amplifi cador para servir tanto como electrodo para estimular la célula como para registrar las corrientes iónicas generadas. La micropipe-ta se aproxima a la superfi cie de la membrana celular y con una ligera succión se consigue formar un sello de alta resis-tencia (del orden de gigaohmios) entre la luz de la micropi-peta y la membrana celular, que se invagina hacia el interior de la pipeta. Este sello de alta resistencia consigue aislar eléctricamente a los canales iónicos presentes en la región invaginada de la membrana, evitando el contacto de iones del medio extracelular. En función del experimento que se vaya a llevar a cabo se puede variar la solución interna de la pipeta para controlar en todo momento qué compuestos e iones van a interaccionar con los canales iónicos. A través de la micropipeta se puede estimular la célula a distintos vol-tajes con secuencias de pulsos despolarizantes o hiperpo-larizantes y registrar la corriente eléctrica generada por los
Figura 1.6. Esquema de las modalidades de patch clamp

01 Electrof is io logía celu lar
13
canales iónicos. A partir de este momento se pueden llevar a cabo distintas confi guraciones, cada una de ellas con el objetivo de estudiar los canales iónicos desde distintos pun-tos de vista(13):
• Confi guración de parche sobre célula entera (cell-attached patch mode).
• Confi guración de célula entera (whole-cell patch). • Confi guración inside-out patch. • Confi guración outside-out patch.
Confi guración de parche sobre célula entera
La confi guración de parche sobre célula entera (cell-attached patch mode) es la confi guración más simple a nivel experimen-tal. Consiste directamente en registrar las corrientes presentes en la porción de la membrana invaginada en la micropipeta tras la realización del sello de alta resistencia. Al tratarse de una región sumamente pequeña de la membrana, esta técnica permite el registro de la corriente iónica unitaria o microscópica, genera-da por la activación de un único canal presente en el parche de membrana.
Esta confi guración presenta la ventaja de ser la más fi siológica de las cuatro mencionadas porque no modifi ca el medio intra-celular y es la más adecuada para estudios que requieran la par-ticipación de todos los sistemas celulares moduladores del canal iónico. Esta confi guración permite además estudiar la acción de distintas moléculas o fármacos sobre el canal iónico añadiendo dichas moléculas a la solución interna de la micropipeta.
Confi guración de célula entera
A partir de la confi guración anterior, y aplicando una ligera suc-ción adicional, se puede romper la porción de membrana invagi-nada dentro de la pipeta y ganar acceso al citoplasma. Por consi-guiente, la solución interna de la micropipeta entra en contacto con el citoplasma y se produce un recambio casi inmediato de la composición del mismo (lo que se denomina diálisis intracelular). En estas condiciones es posible fi jar totalmente el voltaje trans-membrana al valor deseado y, tras la aplicación de los protocolos experimentales adecuados, registrar la corriente iónica resultante de la actividad de todos los canales presentes en la membrana celular.
Esta confi guración permite controlar y modifi car a voluntad el medio intracelular, pero también diluye los mediadores intrace-lulares responsables de la regulación de los canales iónicos.
Confi guración inside-out patch y confi guración outside-out patch
En la confi guración inside-out patch, partiendo de la confi gura-ción de parche sobre célula entera y elevando la célula del fon-
do del baño, se puede separar el parche de membrana incluido en la luz de la micropipeta del resto de la célula, quedando así el lado citosólico de la membrana expuesto al medio externo y el lado extracelular en contacto con la solución interna de la micropipeta.
En la confi guración outside-out patch, si a partir de la confi gura-ción de célula entera se eleva la célula ligeramente, la membra-na se romperá, y gracias a las propiedades de los fosfolípidos se plegará sobre sí misma para formar un sello cubriendo la pipeta. En esta confi guración, la cara extracelular de la membrana es la que queda en contacto con la solución externa. En esta ocasión, la solución interna de la pipeta debe mimetizar el ambiente in-tracelular, puesto que es la cara citosólica de la membrana la que queda en contacto con ella.
Ambas confi guraciones se denominan también “de parche es-cindido” y permiten el registro de corrientes unitarias. Son ade-cuadas para el estudio del mecanismo de acción de fármacos o mediadores que actúen selectivamente sobre las superfi cies ex-tracelular o intracelular de la membrana. Sin embargo, presentan el inconveniente de que desligan los canales iónicos del conteni-do citosólico que modula su actividad.
1.6. Propagación del potencial de acción
Mecanismo general de propagación
Los cardiomiocitos están unidos entre sí por los previamente co-mentados discos intercalares, que permiten el acoplamiento eléc-trico, y por los desmosomas, que facilitan el acoplamiento contrác-til. De esta forma, el miocardio funciona como un sincitio funcional (estructura formada por la fusión de varias células)(14).
La velocidad a la que se desplaza el frente de activación depende esencialmente de cómo muchas células a lo largo de la trayectoria de propagación pueden excitarse simultáneamente por la carga suministrada por la “fuente”. Esto, a su vez, depende de la cantidad de la carga de la fuente (densidad de I
Na en el miocardio de trabajo
y la densidad de ICa-L
en los nodos), y de la distancia a lo largo del eje de la célula, a la que puede propagarse la carga sin disiparse al espacio extracelular.
Hay mecanismos patológicos que pueden hacer que una zona tenga una velocidad de conducción disminuida, como ocurre ante potenciales de reposo menos negativos, lo que se asocia a menos canales de Na+ disponibles para su activación. Este fenó-meno se conoce como depresión del potencial de reposo trans-

14
Cardio Arr i tmias
membrana y suele ser secundario a la isquemia. Otro mecanismo que puede disminuir la velocidad de conducción es la sobrecar-ga de Ca2+ intracelular a través de un aumento de la resistencia de las gap junctions(15).
Refractariedad: concepto de safety factor de la propagación
El periodo refractario se defi ne como el intervalo de tiempo poste-rior a la despolarización, durante el que la célula no es excitable(10). En las células que generan PA dependientes de Na+, el periodo refractario viene determinado por la cinética de activación de I
Na.
Los canales de sodio se encuentran en estado cerrado (reposo) durante la diástole eléctrica (fase 4), se abren en la fase 0 y, a conti-nuación, pasan a estado inactivo, en el que permanecen hasta que la repolarización alcanza valores negativos por debajo de –50 mV. Dado que el estado inactivo no permite la entrada de iones Na+, la aplicación de un estímulo durante las fases 1, 2 y el comienzo de la fase 3 es incapaz de generar una respuesta propagada. A este periodo de tiempo se le denomina periodo refractario absoluto, y se corresponde con el inicio del complejo QRS hasta el pico de la onda T del electrocardiograma de superfi cie (aproximadamente 200 ms), ocupando dos tercios de la refractariedad total.
A medida que el Vm continúa descendiendo en la fase 3 hasta el valor del potencial transmembrana de reposo (–90 mV), una porción de los canales de Na+ pasan del estado inactivo al estado de reposo. Al periodo en que la célula es excitable, pero en el que aún no ha recuperado la excitabilidad total, se le denomi-na periodo refractario efectivo, donde un estímulo supraumbral (mayor al umbral basal de excitación) es capaz de provocar una respuesta local en el cardiomiocito, pero no un PA propagado.
A este periodo refractario efectivo le sigue el periodo refractario relativo, en el que el número de canales de Na+ disponibles es mayor, y en el que un estímulo supraumbral sí es capaz de provo-car un PA propagado, pero de menor amplitud y duración.
Finalmente, durante un breve intervalo de la fase 3, justo antes de que la célula recupere su potencial de reposo, un estímulo más débil de lo normal podría despolarizar la célula. A este periodo se le conoce como periodo supranormal.
Propagación anisotrópica
La distribución de las gap junctions genera una diferencia fi sio-lógicamente relevante en la velocidad de propagación entre el sentido paralelo a la orientación de las células (propagación longitudinal) y el sentido perpendicular a la misma (propagación transversal)(12). Esta diferencia se denomina anisotropía eléctrica.
Los miocitos cardíacos son alargados (70-100 μm de largo x 10-20 μm de ancho). Cuando estas unidades celulares (miocitos) se
organizan para formar el miocardio, se establecen conexiones de extremo a extremo y de lado a lado. Por tanto, la corriente axial viajará en ambas direcciones. Sin embargo, para recorrer una misma distancia de tejido, la corriente tiene mayor resistencia en la dirección transversal que en la dirección longitudinal. La conducción lenta (anisotrópica) se produce por el aumento de la resistencia a la conducción en el eje transversal. La propagación del impulso será más rápida en el eje longitudinal de los miocitos que en el eje transversal, lo que supone la base de las arritmias por reentrada anisotrópica.
Propagación tridimensional miocárdica
A excepción de las fi bras de Purkinje y otros delgados haces subendocárdicos y trabéculas, la propagación del PA en el cora-zón difícilmente puede considerarse unidimensional. Cuando la propagación del impulso se produce en dos o tres dimensiones, entran en juego factores adicionales. Por ejemplo, en dos dimen-siones (2D) la curvatura del frente de onda es un determinante importante del éxito o fracaso de la propagación; cuanto más convexo es un frente de onda, menor es su velocidad de propa-gación. Más allá de una cierta curvatura crítica, la propagación del frente de onda no podría continuar.
El miocardio es tridimensional (3D), por lo que una comprensión completa de la propagación requiere el conocimiento de lo que ocurre en el espesor de la pared auricular o ventricular, ya que el impulso eléctrico se mueve de forma transmural. Algunos estu-dios han demostrado que, para un impulso iniciado por un pun-to de estimulación de esa superfi cie, el frente de activación cerca del sitio de estimulación epicárdico es elíptico, pero se vuelve asimétrico y aparecen pliegues y ondulaciones a medida que penetra la pared. Además, estas elipses en cada plano giran en sentido horario hacia el endocardio(8). Este complejo patrón de propagación 3D se explica, en parte, por la estructura anatómi-ca de los ventrículos, incluyendo la anisotropía de rotación. Los ejes de fi bras musculares giran transmuralmente hasta 120º en algunas áreas. Por otra parte, los patrones de excitación también se ven afectados por la red de fi bras de Purkinje en el subendo-cardio y por las discontinuidades macroscópicas y tabiques de tejido conjuntivo que separan los haces musculares.
Ideas para recordar
El potencial transmembrana de reposo de las células muscula-res auriculares, ventriculares y de las células del sistema de con-ducción His-Purkinje oscila entre –85 y –90 mV, mientras que el de las células del nodo sinusal y el NAV oscila entre –55 y –60 mV.
Los iones se encuentran en constante movimiento entre el interior y exterior celular a través de proteínas transmembra-

01 Electrof is io logía celu lar
15
na denominadas canales iónicos, generando una corriente eléctrica. Estos canales tienen dos propiedades fundamen-tales: selectividad iónica y gating (canales dependientes de voltaje, canales dependientes de ligando o canales depen-dientes de estiramiento).
A nivel molecular, la transición entre los estados cerrado, abierto e inactivado de los canales se deben a cambios con-formacionales y secuenciales de sus proteínas.
La cinética de inactivación de los canales dependientes de voltaje da lugar a un periodo de refractariedad, mecanismo de gran importancia para la comprensión de los mecanismos electrofi siológicos.
El gradiente electroquímico es el responsable de que los iones Na+ y Ca2+ tiendan a entrar en los cardiomiocitos (despolariza-ción de la membrana celular), mientras que los iones K+ tiendan a salir al medio extracelular (repolarización de la membrana).
El potencial de acción es una transición eléctrica que se pro-paga a través de largas distancias conservando su amplitud. Su morfología varía en las distintas células cardíacas según su localización: células musculares (respuesta rápida) y células marcapasos (respuesta lenta), dependiendo del canal iónico responsable de la fase 0 de dicho potencial de acción.
Los cardiomiocitos están unidos entre sí mediante discos in-tercalares compuestos por grupos de conexones, permitien-do el acoplamiento eléctrico y la propagación del potencial de acción a nivel tisular. Las gap junctions son estructuras formadas por dos conexones enfrentados que siempre están abiertas y permiten el paso continuo de iones.
La técnica de patch clamp permite registrar la actividad eléc-trica de los canales iónicos en forma de cambios del poten-cial transmembrana, que refl ejan el fl ujo de iones hacia el interior o exterior de la célula.
Bibliografía
1. Alberts, B, Johnson A, Lewis J, Raff M, Rober ts K, Walter P. Bio-logía molecular de la célula. Barcelona. Omega. 2007.
2. Yu FH, Yarov-Yarovoy V, Gutman GA, Catterall WA. Overview of molecular relationships in the voltage-gated ion channel superfamily. Pharmacol Rev 2005; 57: 387-395.
3. Catterall WA. Cellular and molecular biology of voltage-gated sodium channels. Physiol Rev 1992; 72: S15-S48.
4. Noda M, Suzuki H, Numa S, Stuhmer W. A single point mu-tation confers tetrodotoxin and saxitoxin insensitivity on the sodium channel II. FEBS letters 1989; 259: 213-216.
5. Dubin D. Ion adventure in the heartland. Tampa. Cover Publish-ing Company. 2003.
6. Nerbonne JM, Kass RS. Molecular physiology of cardiac repo-larization. Physiol Rev 2005; 85: 1.205-1.253.
7. Tamargo J, Caballero R, Gomez R, Valenzuela C, Delpon E. Pharmacology of cardiac potassium channels. Cardiovasc Res 2004; 62: 9-33.
8. Jalife J, Delmar M, Anumonwo J, Berenfeld O, Kalifa J. Basic cardiac electrophysiology for the clinician. Chichester. John Wi-ley & Sons Ltd. 2009.
9. Tamargo J. Electrofi siología cardíaca. En: Moro C, Hernández Madrid A, García Cosío F (ed.). Electrocardiografía clínica. Ma-drid. McGraw-Hill Interamericana. 2001; 1-22.
10. Zaza A, Wilders R, Opthof T. Cellular Electrophysiology. En: Macfarlane PW, van Oosterom A, Pahjm O, Janse M, Camm J (ed.). Comprehensive Electrocardiology. Londres. Springer. 2011; 105-144.
11. Antzelevitch C, Burashnikov A. Overview of basic mecha-nisms of cardiac arrhythmia. Card Electrophysiol Clin 2011; 3: 23-45.
12. Gaztañaga L, Marchlinski FE, Betensky BP. Mechanisms of car-diac arrhythmias. Rev Esp Cardiol 2012; 65: 174-185.
13. Molleman A. Patch Clamping An introductory Guide to Patch Clamp Electrophysiology. Chichester. John Wiley & Sons Ltd. 2003.
14. Rubart M, Zipes DP. Génesis de las arritmias cardíacas: aspectos electrofi siológicos. En: Braunwald E, Zipes DP, Libby P, Bonow R (ed.). Tratado de cardiología. Barcelona. Elsevier Saunders. 2009; 727-762.
15. Waldo A, Witt AL. Mechanisms of cardiac arrhythmias and con-duction disturbances. En: Fuster V (ed.). Hurst’s the heart. New York. McGraw-Hill. 2001; 751-796.