
Fisiología médica II
178
APUNTES DEL SEMESTRE - 2º de MEDICINA Fisiología médica II Cristóbal Muñoz Pérez UNIVERSITAT de BARCELONA TEMARIO SEMINARIOS CAMPUS de BELLVITGE 1. Eje hipotálamo-hipósis-gónadas (2) 13. Control de la frecuencia cardíaca (85) 2. A. F. del ap. reproductor masculino (4) 14. Circulación coronaria (88) 3. A. F. del ap. reproductor femenino (12) 15. Sist. circulatorio y microcirculación (94) 4. Sangre (28) 16. Control de la circ. periférica (102) 5. Eritrocitos y hemoglobina (34) 17. Sistema linfático y venoso (111) 6. Metabolismo del hierro (44) 18. Sistema renal (115) 7. Grupos sanguíneos (46) 19. Procesos de transporte renal (120) 8. Hemostasia (49) 20. Concentración y dilución de la orina (127) 9. Sistema cardiovascular (59) 21. Regulación de la función renal (130) 10. Ciclo cardíaco (61) 22. Transporte de potasio (132) 11. Electrosiología cardíaca (68) 23. Transporte de ácidos y bases (136) 12. Gasto cardíaco (80) 24. Aparato respiratorio (139) S1. Seminarios I y II de cardiovascular (159) S3. Seminarios de respiratorio (I-II) (173) S2. Casos de renal (1-4) (162) k
-
Upload
cristobal-munoz-perez -
Category
Documents
-
view
236 -
download
16
description
Apuntes de Fisiología II (2º de medicina). Escritos por Cristóbal Muñoz en el curso 2011-12, alumno de Medicina en la Universitat de Barcelona (Campus de Bellvitge). Profesores de la asignatura: Dr. C. Javierre, Dr. J. R. Barbany.
Transcript of Fisiología médica II

APUNTES DEL SEMESTRE - 2º de MEDICINA
Fisiología médica II
Cristóbal Muñoz Pérez
UNIVERSITAT de BARCELONA
TEMARIO
SEMINARIOS
CAMPUS de BELLVITGE
1. Eje hipotálamo-hipó!sis-gónadas (2) 13. Control de la frecuencia cardíaca (85)
2. A. F. del ap. reproductor masculino (4) 14. Circulación coronaria (88)
3. A. F. del ap. reproductor femenino (12) 15. Sist. circulatorio y microcirculación (94)
4. Sangre (28) 16. Control de la circ. periférica (102)
5. Eritrocitos y hemoglobina (34) 17. Sistema linfático y venoso (111)
6. Metabolismo del hierro (44) 18. Sistema renal (115)
7. Grupos sanguíneos (46) 19. Procesos de transporte renal (120)
8. Hemostasia (49) 20. Concentración y dilución de la orina (127)
9. Sistema cardiovascular (59) 21. Regulación de la función renal (130)
10. Ciclo cardíaco (61) 22. Transporte de potasio (132)
11. Electro!siología cardíaca (68) 23. Transporte de ácidos y bases (136)
12. Gasto cardíaco (80) 24. Aparato respiratorio (139)
S1. Seminarios I y II de cardiovascular (159) S3. Seminarios de respiratorio (I-II) (173)
S2. Casos de renal (1-4) (162)
k

FISIOLOGÍA MÉDICA II
1. EJE HIPOTÁLAMO-HIPOFISARIO-GÓNADAS
La función reproductiva está muy regulada por el hipotálamo. Como ya sabemos por Fisiolo-gía I y otras materias, el hipotálamo es el centro rector de la respuesta tanto vegetativa como endocrina. Justo por debajo del hipotálamo está la hipófisis (o glándula pituitaria), que consta de dos lóbulos: uno anterior (adenohipófisis), y otro posterior (neurohipófisis). La hipófisis posterior está unida al hipotálamo mediante el tallo hipofisario, que con-tiene axones originados en los núcleos hipotalámicos (núcleos paraventricular y supraóptico). La hipófisis posterior almacena y segrega dos hormonas: la antidiurética (ADH o vasopresina) y la oxitocina (llegan por transporte axonal). La hipófisis anterior no se encuentra conectada directamente con el hipotálamo, sino que se conecta a él mediante los vasos sanguíneos de la circulación portal hipofisaria. El hipo-tálamo sintetiza hormonas (factores) hipotalámicas liberadoras e inhibidoras que llegan a la ade-nohipófisis y que inciden sobre distintas poblaciones celulares, provocando que se estimule o inhiba la liberación de las hormonas formadas por estas células. Existen cinco grandes po-blaciones celulares (en clase se habló de céls. neurosecretoras):
Las células gonadotropas representan entre el 10 y el 15% del total que hallamos en la ade-nohipófisis. A grosso modo, sabemos que las dos hormonas liberadas por estas células tienen efectos sobre las gónadas tanto masculinas como femeninas. Más concretamente, la FSH fa-
vorece la espermatogénesis o la foliculogénesis, mientras que la LH actúa sobre las céls. de Leydig (que forman testosterona) o sobre las céls. de la teca (que forman estrógenos y progesterona).
Hormona liberadora de gonatropina (GnRH)
La GnRH es una decapéptido que tiene una vida media muy corta (un par de minutos). Las neuronas hipotalámicas encargadas de secretar GnRH tienen un origen común con las neu-ronas olfatorias, por lo que hay una evidente relación entre el sistema reproductor y el olfatorio. Es por eso que mujeres que conviven durante mucho tiempo, llegan a sincronizar sus ciclos menstruales (feromonas). Se sabe que si se aplican feromonas de una mujer en la fase folicu-lar a otra, ésta última verá como sufre un pico de LH y su menstruación se acorta.
1
Poblaciones celulares de la adenohipófisisPoblaciones celulares de la adenohipófisisCéls. gonadotropas Son las encargadas de formar la hormona luteinizante (LH) y la foliculoestimulan-
te (FSH), ante la recepción de GnRH (o LHRH)Céls. lactotropas Forman prolactina (PRL). Tienen dos factores hipotalámicos que inciden sobre su
secreción: PRH (↑secreción) y PIH (↓secreción; es dopamina).Céls. adrenocorticotropas Forman la hormona adrenocorticotropa (ACTH). La CRH estimula su liberación.Céls. somatotropas Forman la hormona del crecimiento (GH). Tienen dos factores hipotalámicos que
inciden sobre su secreción: GHRH (↑secreción) y GHIH (↓secreción; es somatos-tatina).
Céls. tirotropas Forman la tirotropina (TSH). Tienen dos factores hipotalámicos que inciden sobre su secreción (véase Fisiología I): TRH (↑secreción) y GHIH (↓secreción).

La secreción de GnRH, y por ende la liberación de FSH y LH, es pulsátil. Con ello se quiere evitar que las céls. gonadotropas se desensibilicen. Las pulsaciones lentas de GnRH tienen una mayor incidencia sobre la liberación de FSH mientras que las rápidas afectan más a la LH.
Regulación de la secreción de GnRH
Existen varios elementos que modifican el ritmo de secreción de GnRH por parte del hipotá-lamo. A continuación se describen los más importantes:
Diferenciación sexual
Los cromosomas sexuales son los que determinan el sexo de una persona (XX: mujer; XY: hombre). Hasta la séptima u octava semana del desarrollo, las gónadas son bipotenciales y están formadas por dos conductos accesorios (de Wolff y de Müller). Cuando la gónada se dife-rencie únicamente quedará uno de los dos conductos. El gen SRY (Eng: sex-determining region Y) es el que determinará que la gónada se vuelva masculina (y su ausencia que sea femenina). Este gen da lugar a la proteína SRY (factor
determinante del testículo), que hace que la gónada pase a ser un testículo. Los testículos dan lugar a tres hormonas: (a) hormona antimulleriana (o AMH, formada por las céls. de Sertoli); (b) testosterona (formada por las céls. de Leydig); y (c) dihidrotestosterona (o DHT, formada a partir de la testosterona gracias al enzima 5α-reductasa). La AMH hace que los conductos de Müller degeneren, dejando a los de Wolff. Los conductos de Wolff dan lugar al epidídimo, el conducto deferente y la vesícula seminal. Más adelante, la testosterona controla la bajada de los testículos desde el abdomen al escroto. La DHT es la encargada de la diferenciación de los genitales masculinos externos. Los embriones femeninos verán como sus conductos de Wolff degeneran (al no haber testosterona) y los de Müller permanecen intactos, dando lugar a la vagina, el útero y las
2
Factores reguladores de la liberación de GnRHFactores reguladores de la liberación de GnRHSistema catecolaminérgico La noradrenalina aumenta la secreción mientras que la serotonina (5-HT) y la
dopamina la inhiben.Opiáceos endógenos Las endorfinas, encefalinas y dinorfinas inhiben.Neuropéptidos Cuando los niveles de estrógenos son normales, los neuropéptidos (como el NP-
Y) estimulan la secreción de GnRH. Si no son normales entonces inhiben.CRH La hormona liberadora de corticotropina y los glucocorticoides inhiben la libera-
ción de GnRH.GABA En función de la clase de receptor con la que interacciones, estimula o inhibe.PRL La prolactina inhibe la liberación de GnRH.Feedback Feedback largo: los esteroides sexuales formados por las gónadas (estrógenos y
progestágenos) inhiben la GnRH. No obstante, las céls. hipotalámicas responsa-bles de la liberación de GnRH son poco sensibles a dichos esteroides. Para resol-verlo, en su membrana tienen unos enzimas con actividad hidroxilasa que con-vierten los estrógenos en catecolestrógenos (y éstos sí que inciden más).
Feedback
Feedback corto: las gonadotropinas (FSH y LH) actúan sobre la liberación de GnRH. Por definición, un feedback corto es: “una hormona de la hipófisis anterior inhibe la liberación de su hormona hipotalámica” (Netter pág. 312)
Feedback
Feedback ultracorto: la secreción de GnRH inhibe su propia liberación.

trompas de Falopio. La ausencia de DHT hace que no se desarrollen genitales externos mas-culinos.
3

2. ANATOMÍA FUNCIONAL DEL APARATO REPRODUCTOR MASCULINO
Introducción - conductos
El aparato reproductor masculino está forma-do por unos elementos internos y otros exter-nos. Los internos son: (a) testículos, (b) vías ge-nitales extratesticulares (epidídimos, conductos deferentes), y (c) glándulas anexas (vesículas seminales, próstata y glándulas bulbouretrales o de Coowper). Los elementos externos son el escroto y el pene. Los testículos se encuentran fuera de la cavidad abdominal (temperatura 2-3 ºC por debajo de la corporal) y se forman a partir de unas estructuras cercanas a los riñones. El pa-rénquima testicular consiste en numerosos túbulos seminíferos donde se produce la esper-
matogénesis. Desde éstos, los espermatozoides pasarán por toda una serie de conductos: los túbulos rectos, la rete testis, los conductos efe-
rentes (hasta aquí dentro del testículo), el epi-dídimo, el conducto deferente, y finalmente la uretra.
Glándulas anexas
Como ya hemos dicho, existen toda una serie de glándulas accesorias en el aparato repro-ductor masculino:
(a) Vesículas seminales. Las vesículas seminales forman un fluido alcalino rico en fructosa (soporte energético para los espermatozoides), prostaglandinas (favorecen contraccio-nes uterinas, por lo que se facilita el transporte de espermatozoides), y fibrinógeno (se forma un coágulo)1. Aquí se forma el 60% del semen.
(b) Próstata. La próstata forma un líquido blanquecino con ácido cítrico y enzimas que
hacen coagular al semen. Aquí se forma el 30% del semen y sin próstata los esper-matozoides serían inviables.
(c) Gl. de Coowper. Secretan un líquido lubricante alcalino que protege a los esperma-tozoides y lubrica el glande durante el acto sexual. Se forma el 10% del semen.
4
1 En clase se comentó que el coágulo de semen que se forma tras la eyaculación retiene cerca del 90% de los espermatozoides y sirve a modo de selección. Según el Guyton (pág. 976) la función de este coágulo es mante-ner el semen en las regiones profundas de la vagina, donde está situado el cuello uterino.
Aparato reproductor masculino. (Guyton pág. 973). En clase se detalló que el conducto deferente se ensancha y da lugar a la ampolla. Recordemos que todo esto se detalla más organografía (tema 19).

La alcalinidad de las secreciones de las glándulas anexas es importante para así neutralizar el pH ácido de la vagina con el que se encontrarán los espermatozoides tras ser eyaculados.
Túbulos seminíferos
Los túbulos seminíferos están constituidos por una luz central revestida de un epitelio seminí-fero. El epitelio seminífero podría clasificarse como estratificado y está formado por dos po-blaciones celulares: (a) las células de Sertoli, y (b) las células espermatogénicas. Por la parte ex-terna del epitelio hay una membrana basal y una pared formada por fibras de colágeno, fi-broblastos y células mioides contráctiles. La mayor parte de las células del epitelio germinal son las células espermatogénicas, que están dispuestas de manera que las más indiferenciadas están en la periferia del túbulo seminífero (espermatogonias) y las más diferenciadas están casi en contacto con la luz o di-rectamente en la misma (espermatozoides). El proceso por el cual las espermatogonias pasan a ser espermatozoides es lo que se conoce con el nombre de espermatogénesis.
Espermatogénesis. El proceso se describe con más detalle en organografía, pero a grosso modo es el que se comenta a continuación. Espermatogonias (2n) hay de dos clases: A y B. Las A son las que proliferan
por mitosis (céls. madre) y las B están diferenciadas (pasan a ser espermatocitos primarios 2n). Los es-permatocitos 1º entran en meiosis y dan: dos espermatocitos 2º (primera división) y despúes al proseguir
la meiosis cuatro espermátidas (segunda división). Se observan puentes citoplasmáticos.
El paso desde espermátida (n) a espermatozoide es lo que se conoce como espermiogénesis, proceso que se caracteriza por el aplanamiento del núcleo y la condensación de la cromati-na, la formación del acrosoma a partir del aparato de Golgi, la migración de los centríolos para formar un flagelo, la migración de las mitocondrias, y el desprendimiento de parte del citoplasma (los cuerpos residuales son captados por las céls. de Sertoli). La espermatogénesis al completo dura una 74 días aproximadamente. Atención, en un inicio los espermatozoides no son móviles. Se pasarán unas dos se-manas en el epidídimo donde adquirirán una motilidad parcial (insuficiente para fecundar al óvulo). La motilidad completa la obtienen en el tracto reproductor femenino.
Células de Sertoli
Las células de Sertoli se proyectan desde la lámina basal hasta la luz del túbulo. Emiten unas prolongaciones laterales que se conectan entre sí (las de las distintas células) por medio de uniones oclusivas (tight). Estas uniones, debido a su alta impermeabilidad, forman la barrera
hematotesticular, que separa la sangre de la serie espermatogénica. En un principio, se creía que las células de Sertoli únicamente ejercían una función de soporte. Conforme se han ido estudiando, se han descubierto diferentes funciones: (a)
control de la espermatogénesis, (b) producción de sustancias que regulan los niveles de FSH; (c) regulación del metabolismo de las céls. germinales (serie espermatogénica); (d) transporte de sus-
tancias dentro del túbulo seminífero; (e) formar la barrera hematotesticular; y (f) fagocitar los
cuerpos residuales.
5
Las céls. de Sertoli son sensibles a la FSH (tienen receptores), formando toda una serie de sus-tancias nada más recibirla:
(1) Lactato. Intermediario metabólico(2) Transferrina y ceruloplasmina. Metabolitos empleados por las céls. germinales.(3) ABP. La ABP (Eng: androgen-binding protein) se une a la testosterona para que ésta
pase a ser dihidrotestosterona. (4) Otros. Como factores de crecimiento (FGF), somatomedina C (IGF-1), clusterina,
factor de crecimiento seminífero (SGF: favorece el crecimiento de las céls. de Serto-li), factor de las céls. madre (estimula la división de las espermatogonias antes de entrar en meiosis), inhibina y activina (menor cantidad).
Células de Leydig
Las céls. de Leydig derivan de los fibro-blastos y constituyen un 20% de las cé-lulas que encontramos en el testículo (están en el tejido conectivo intersticial). Se encargan de formar testosterona y pe-queñas cantidades de estradiol 17β. La testosterona se forma a partir del colesterol, el cual se forma en los microsomas de la célula de Leydig a par-tir del acetato. Una vez tenemos coleste-rol en el interior de la cél. de Leydig, és-te pasa a la mitocondria para dar lugar a los precursores de la testosterona, que pa-sarán otra vez al microsoma para dar la testosterona (y el estradiol)2. Las céls. de Leydig no las vamos a encontrar en niños, por lo que no tendrán testosterona en su organismo. No obstante, se sa-be que durante el primer año de vida, el neonato tiene un pico de testosterona por causa desconocida. Además, durante el desarrollo embrionario es obvio que el feto debe tener tes-tosterona: para la diferenciación sexual (evitar que los conductos de Wolff degeneren).
Relación entre las diferentes células
Existe una relación entre las diferentes células del testículo:- Las FSH actúa sobre las céls. de Sertoli, favoreciendo la espermatogénesis y haciendo
que las céls. de Leydig expresen más receptores para la LH.- La LH actúa sobre las céls. de Leydig, haciendo que formen testosterona (que tiene
varios efectos, entre ellos incidir sobre las céls. de Sertoli).
6
2 La testosterona pasa a estradiol por acción de la FSH y a DHT por acción del enzima 5α-reductasa.
Concentraciones plasmática de testosterona (rojo) y pro-ducción de espermatozoides (azul). (Guyton pág. 981).

- Las céls. mioides tienen receptores para la testosterona y con sus contracciones se favorece que las células de la serie espermatogénica vayan avanzando hacia la luz. Además, activan a las céls. de Sertoli.
Formación y transporte de la testosterona
La testosterona se forma a partir del colesterol en las céls. de Leydig. Cuando la LH interacciona con los receptores para la LH de las céls. de Leydig, se da una cascada de señalización intracelular que cul-mina con un aumento del cAMP. El aumento de es-te segundo mensajero provoca: (a) activación de las
proteínas StAR, (b) aumento de la producción de otras
proteínas, y (c) promueve la renovación celular. El punto que nos interesa es la activación de las pro-teínas StAR, ya que permite la entrada del colesterol a la mitocondria, y que éste pueda transformarse pregnenolona. Esta pregnenolona saldrá de la mitocondria para volver al microsoma y así formar dehidroepiandrosterona y androstenedio-
na. Finalmente, a partir de la androstenediona se forma la testosterona. En función de la etapa vital en la que se encuentra el hombre, la producción de tes-tosterona es una u otra. A continuación se describen los principales picos:
- Desarrollo fetal. Hacia los dos primeros meses de vida hay un pico de testosterona inducido por la gonadotropina coriónica humana (hCG). Esto permite que los conductos de Wolff no degeneren y que se formen las gónadas masculinas. Después del primer trimestre de embarazo, la hCG decae y con ella los niveles de testosterona.
- Infancia. A excepción del primer año vida (por causas desconocidas), no hay forma-ción de testosterona. Una teoría que explica la aparición de testosterona en el primer año de vida es por inmadurez del eje hipotálamo-hipofisario (no hay mecanismo de feedback negativo).
- Pubertad. En la pubertad hay una secreción de GnRH en forma de picos que se dan a lo largo de la noche (y por tanto, si hay picos de secreción de GnRH, lo mismo ocurre con la FSH y la LH). A medida que la pubertad avanza, los picos de GnRH también se observan durante el día.
- Ritmo circadiano. Una vez los picos aparecen durante la noche y el día, ya hablamos de un ritmo circadiano. El máximo de dicho pico está a primera hora de la mañana (6.00 h - 8.00h) y el mínimo hacia la noche (20.00h - 22.00h).
Al pasar a la sangre, la testosterona puede ir en tres formas distintas: (a) conjugada una globu-
lina sexual (SHBG; sex hormone binding globulin); (b) conjugada con la albúmina (aunque la testosterona tiene menos afinidad por ésta); o (c) libre. Para hacernos una idea, el 60% viaja en las globulinas sexuales, entre el 20 y el 40% en albúmina y únicamente entre el 1 y el 3% viaja libre. Dura en sangre unos 30 minutos y actúa sobre diferentes órganos diana.
7

Atención, las mujeres también forman testosterona: producen un poco en la zona reticular de la corteza suprarrenal. Para hacernos una idea de lo poco que se forma, en un hombre úni-camente representa el 5%3 del total de testosterona formada. En la sangre, la testosterona puede llegar a una célula diana (y producir toda una serie de efectos), puede llegar a una célula que no sea diana y ser degradada, o puede ser trans-formada en androsterona y dehidroepiandrosterona en el hígado. Estos dos últimos metabolitos son conjugados con glucorónicos y son excretados junto a la bilis o la orina.
Mecanismo molecular de la testosterona
La testosterona es un esteroide, por lo que puede pasar por difusión a través de las membra-nas biológicas. Si pasa al interior de una célula diana, automáticamente será transformada en dehidrotestosterona gracias al enzima 5α-reductasa. Una vez tenemos DHT, éste se une al re-
ceptor de andrógenos (AR)4, que en realidad es un factor de transcripción que será fosforilado y dimerizará para irse hacia el núcleo. Una vez en el núcleo contactará los elementos de res-
puesta a andrógenos (ARE) y se incrementará la expresión del PSA, se incrementa el crecimiento y se aumenta la supervivencia. Atención, para aquella testosterona que no viaja libre por la sangre (ya sea en albúmi-na o una globulina5 sexual), precisará de un receptor de membrana para ser interiorizada.
Regulación testicular
Tanto la FSH como la LH, producidas en las céls. gonadotropas de la hipófisis anterior, son glucoproteínas6 formadas por dos cadenas (alfa y beta) codificadas en genes ubicados en cromosomas distintos. Los receptores para la FSH y la LH están acoplados a proteínas G, im-plicando un aumento del cAMP intracelular y la activación de la fosfolipasa C y A2. A conti-nuación se describen los diferentes efectos de la FSH:
- En función de la madurez. En función de si la gónada está más o menos madura, la FSH tendrá un efecto u otro. En gónadas inmaduras aumenta el número de céls. de Sertoli así como la longitud de los túbulos seminíferos. En gónadas maduras aumenta la pro-
ducción de la ABP, que se encargará de transportar la testosterona por el túbulo seminí-fero.
- Estimulación de la espermatogénesis. La FSH estimula la espermatogénesis incrementan-do la producción, por parte de la cél. de Sertoli, de transferrina, ceruloplasmina, lac-tato, inhibina, etc.
- Espermatogonia → espermátida. En presencia de testosterona, la FSH estimula este paso.- Favorece la unión entre la cél. germinal y Sertoli. Gracias a que se incrementa la produc-
ción de F-actina y vinculina.
8
3 Es insuficiente para desarrollar los órganos sexuales masculinos.
4 El AR también puede unirse a la testosterona, pero con menor afinidad.
5 Las globulinas también transportan el 17β-estradiol. La ABP es familia de la SHBG.
6 Se comentó en clase que guardan similitud estructural con la hCG y la TSH.
- Disminución de mRNA. La FSH disminuye los mRNA que, traducidos, dan lugar a los receptores para la DHT. Con esto logramos una desensibilización que dura unas 16h.
- Producción de SCF. La FSH hace que las céls. de Sertoli formen el ligando SCF, que se une al protooncogen c-kit ubicado en las células germinales. La interacción entre es-tos dos elementos estimula el crecimiento y la diferenciación de las céls. germinales.
- Inhibina. Después de la fagocitosis de los cuerpos residuales, la FSH estimula produc-ción de inhibina (y en menor cantidad de activina) para así regular la producción de FSH.
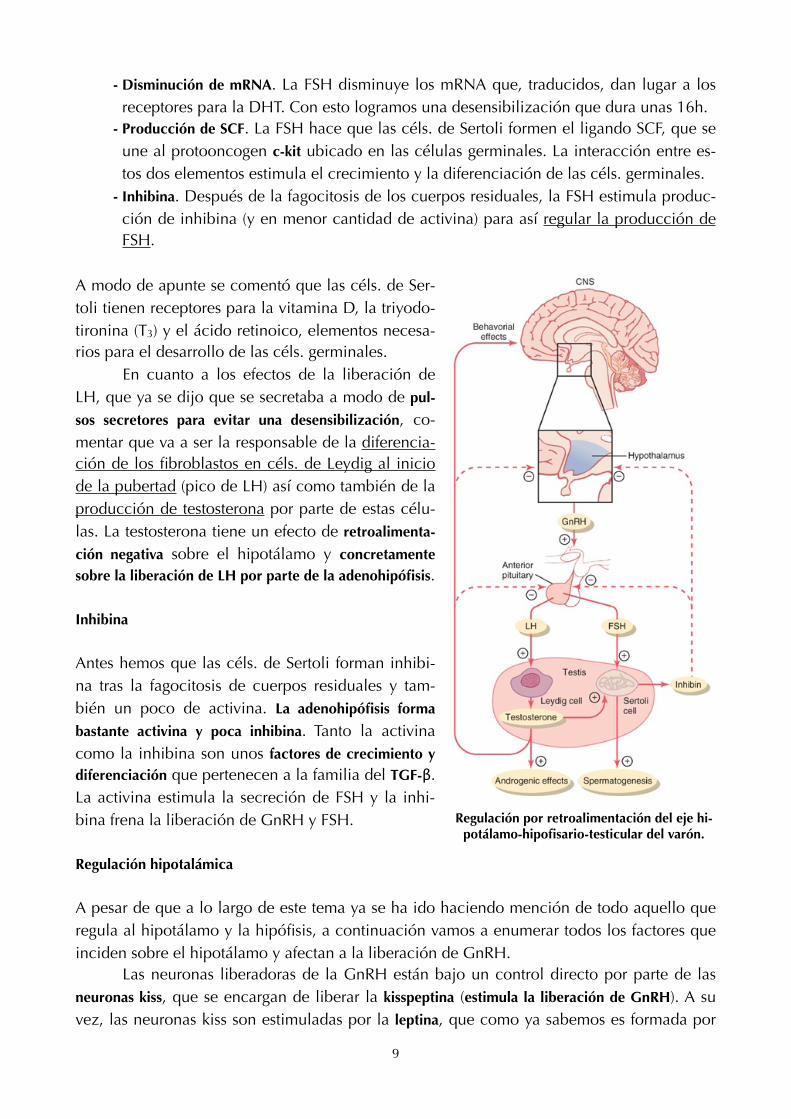
A modo de apunte se comentó que las céls. de Ser-toli tienen receptores para la vitamina D, la triyodo-tironina (T3) y el ácido retinoico, elementos necesa-rios para el desarrollo de las céls. germinales. En cuanto a los efectos de la liberación de LH, que ya se dijo que se secretaba a modo de pul-
sos secretores para evitar una desensibilización, co-mentar que va a ser la responsable de la diferencia-ción de los fibroblastos en céls. de Leydig al inicio de la pubertad (pico de LH) así como también de la producción de testosterona por parte de estas célu-las. La testosterona tiene un efecto de retroalimenta-
ción negativa sobre el hipotálamo y concretamente sobre la liberación de LH por parte de la adenohipófisis.
Inhibina
Antes hemos que las céls. de Sertoli forman inhibi-na tras la fagocitosis de cuerpos residuales y tam-bién un poco de activina. La adenohipófisis forma
bastante activina y poca inhibina. Tanto la activina como la inhibina son unos factores de crecimiento y diferenciación que pertenecen a la familia del TGF-β. La activina estimula la secreción de FSH y la inhi-bina frena la liberación de GnRH y FSH.
Regulación hipotalámica
A pesar de que a lo largo de este tema ya se ha ido haciendo mención de todo aquello que regula al hipotálamo y la hipófisis, a continuación vamos a enumerar todos los factores que inciden sobre el hipotálamo y afectan a la liberación de GnRH. Las neuronas liberadoras de la GnRH están bajo un control directo por parte de las neuronas kiss, que se encargan de liberar la kisspeptina (estimula la liberación de GnRH). A su vez, las neuronas kiss son estimuladas por la leptina, que como ya sabemos es formada por
9
Regulación por retroalimentación del eje hi-potálamo-hipofisario-testicular del varón.

los adipocitos. La liberación de GnRH se produce durante unos minutos cada 1 a 3h. Si la frecuencia de secreción aumenta, la acción de las hormona gonadotrópicas disminuye (de-sensibilización). Además del control directo por parte de las neuronas kiss, existen mecanis-mos de feedback que sería interesante conocer:
- Feedback ultracorto. Es aquel en que se controla la liberación de GnRH por parte de un metabolito que se forma en el mismo hipotálamo (dopamina, noradrenalina, 5-HT, melatonina, prostaglandinas, endorfinas, óxido nítrico, etc) o la propia GnRH. La GnRH inhibe su propia liberación.
- Noradrenalina y óxido nítrico: activan a las neuronas kiss y por tanto aumenta la
liberación de GnRH.- Dopamina y endorfinas: inhiben a las neuronas kiss y por tanto disminuye la libe-
ración de GnRH.- Feedback corto. Es aquel en que las dos hormonas gonadotropas (FSH + LH) controlan
la formación de GnRH. Cuanta mayor sea la cantidad de estas dos hormonas, menor
será la liberación de GnRH (en el ciclo sexual femenino esto cambia). La secreción de FSH sólo aumenta y disminuye ligeramente con las fluctuaciones de GnRH; sin em-bargo, varía de una forma más lenta a lo largo de periodos de muchas horas en res-puesta a las variaciones a largo plazo de la GnRH. Esto se debe a que la liberación de FSH depende de la GnRH, de la activina y de la inhibina. Contrariamente la relación entre la secreción de GnRH y de LH es mucho más estrecha, por lo que la GnRH también se conoce como hormona liberadora de LH (LHRH).
- Feedback largo. Las hormonas hipotalámicas e hipofisarias forman un eje endocrino que se inhibe por la hormona de la glándula diana (testosterona).
Funciones generales de los andrógenos
A continuación se van a detallar las funciones que tienen los andrógenos a lo largo de la vi-da del individuo y en los diferentes órganos diana. Empezamos con las funciones biológicas de la testosterona (desarrollo fetal).
- Diferenciación sexual. En el primer trimestre del embarazo, el pico de hCG hace que haya un pico de testosterona que favorece la diferenciación sexual (hacia el sexo masculino). Se forma un pene y un escroto en lugar de un clítoris y una vagina. Tam-bién induce la formación de la glándula prostática, las vesículas seminales y los con-ductos genitales masculinos, así como suprime la formación de órganos sexuales fe-meninos.
- Cerebro. La testosterona tiene un efecto indirecto sobre el cerebro; ésta es transforma-da en estradiol, que será el encargado de masculinizar el cerebro.
- Descenso testicular. Como norma general, los testículos desciende al escroto durante los últimos dos o tres meses de gestación, cuando empiezan a secretar cantidades su-ficientes de testosterona.
Tras la pubertad, el aumento de la secreción de testosterona hace que el pene, el escroto y los testículos aumenten unas ocho veces de tamaño antes de los 20 años de edad. Además, la testosterona induce también el desarrollo simultáneo de los carácteres sexuales secunda-
10

rios del varón, comenzando en la pubertad y terminando en la madurez. Estos carácteres sexuales secundarios, además de los propios órganos sexuales, distinguen al varón de la mu-jer en los siguientes aspectos.
- Efecto sobre la distribución del vello corporal. La testosterona hace crecer el pelo: (1) so-bre el pubis; (2) hacia arriba a lo largo de la línea alba; (3) en la cara; (4) habitualmen-te en el tórax; (5) con menor frecuencia en la espalda. También hace que el vello de otras regiones del cuerpo prolifere más.
- Calvicie. La testosterona reduce el crecimiento del pelo en la parte superior de la ca-beza. Sin embargo, muchos hombres viriles no se quedan calvos; esto se debe a que puede ser que la calvicie depende de dos factores: (a) genéticos, y la (b) propia testos-
terona (hormonas androgénicas). - Efectos sobre la voz. La testosterona produce una hipertrofia de la mucosa laríngea y un
aumento del tamaño de la laringe. Los efectos originan primero una voz relativamente discorde, “cascada”, que poco a poco se acaba conviertiendo en la típica voz grave del varón adulto.
- Efectos sobre la piel. La testosterona aumenta el grosor de la piel de todo el cuerpo y la dureza de los tejidos subcutáneos. También incrementa la secreción de algunas, y quizá de todas, las glándulas sebáceas (relación con el acné7).
- Efectos sobre la formación de proteínas y el desarrollo muscular. La testosterona aumenta la masa corporal hasta un 50% más en el hombre que en la mujer. Aumenta la estatu-ra.
- Efectos sobre el hueso. Se favorece la absorción de Ca2+ y se retiene más en los huesos (aumentan su densidad). Aumenta el crecimiento de los huesos largos y cierra las epí-fisis óseas.
- Metabolismo basal. Se incrementa en un 15%.- Riñón. La testosterona sobre el riñón estimula la liberación de eritropoyetina (EPO) y así
regula la síntesis de hemoglobina (Guyton pág. 982: “A pesar de la estrecha relación entre tes-
tosterona y aumento del hematocrito, la testosterona no parece aumentar directamente los niveles de
EPO”). Aumenta el número de eritrocitos.
La próstata, las vesículas seminales, las glándulas sebaceas, los riñones, la piel y el cerebro disponen del enzima 5α-reductasa, por lo que transforman la testosterona en dihidrotestoste-
rona. La DHT tiene los siguientes efectos:- Aumenta la vascularización de los tejidos sexuales.- Determina el desarrollo y el aumento de la secreción de la próstata y las v. seminales.- Incrementa el tamaño del pene y hace que la piel del escroto sea rugosa y pigmenta-
da.- Favorece el desarrollo de los túbulos seminíferos.- Posibilita la acción de la FSH sobre el epitelio germinal.- Junto con los andrógenos, estimula la libido y la potencia sexual.
11
7 El acné es un rasgo común de la adolescencia del varón cuando el organismo experimenta por primera vez el aumento de testosterona. Tras varios años de secreción de testosterona, la piel suele adaptarse de alguna mane-ra a ella, lo que favorece la desaparición del trastorno.

3. ANATOMÍA FUNCIONAL DEL APARATO REPRODUCTOR FEMENINO
Introducción
La estructura macroscópica del aparato reproductor fue discutida en Anatomía. A continua-ción haremos un breve repaso anatómico y funcional de las partes que nos interesa para así después hablar de su funcionamiento. Empezaremos hablando de los ovarios, unas estructu-ras ovoides de unos dos a cinco cm de longitud. En cada uno de los dos ovarios distinguimos una parte externa (epitelio germinativo), que es donde están los folículos; una parte interna (médula) rica en tej. conjuntivo, céls. contráctiles e intersticiales; y un hilio, el lugar de entra-da y de salida de vasos sanguíneos y linfáticos además de nervios. Los ovarios tienen dos funciones principales: (a) formación de los gámetos; y (b) función
endocrina (producción de hormonas esteroideas). Las hormonas liberadas por los ovarios son necesarias tanto para la formación de los óvulos como para la preparación del endometrio en el csao que haya una fecundación (y por tanto, que se pueda dar la implantación). A diferencia del hombre, que desde la pubertad hasta que muere forma espermato-zoides, la mujer ovula unos 400 óvulos a lo largo de su vida. El número máximo de ovogonias de las que dispone una mujer es a los 20 meses de gestación (entre 6 y 7·106). A partir de este punto, el número disminuye hasta alcanzar la cifra de unas 400.000 al inicio de la pu-bertad. El resto de ovocitos no ovulados entrarán en atresia.
Ovogénesis
Las ovogonias pasan a ser ovocitos primarios en el útero (se da en el feto), los cuales iniciarán
la meiosis y se detendrán en la profase de la 1a división meiótica. Una vez alojados en el ova-rio, el ovocito primario es rodeado por una sóla capa de capa de células de la granulosa, dan-do lugar al folículo primordial. Las mujeres disponen de un pool de folículos primordiales que se encuentra en la corteza del ovario. Cada mes, entre 6 y 12 folículos primarios empiezan a madurar. De éstos, sólo uno llegará a ser folículo de De Graaf. La ovulación se dará por un pico de LH. Acto seguido, el ovocito (que aún seguía siendo primario) pasa a ser un folículo secundario detenido en la metafase de la 2a división meió-
tica. El paso de ovocito secundario a óvulo (la continuación de la división meiótica) única-mente se da si hay fecundación. Huelga decir que con cada división meiótica aparece un corpúsculo polar8. Por normal general, únicamente se ovula un óvulo por ovario.
Crecimiento del folículo ovárico [fase folicular del ciclo ovárico]
Una niña al nacer presenta folículos primordiales; es decir, ovocitos primarios rodeados por una capa de céls. de la granulosa. Se cree que hasta que no llega la pubertad, las céls. de la granulosa nutren al ovocito y secretan un factor inhibidor de la maduración del ovocito (OMI),
12
8 La existencia de corpúsculos polares se explica fácilmente. En lugar de hacer una división simétrica del cito-plasma, hacemos que una de las dos células hija se quede con la mayor parte del mismo (y por tanto, más nu-trientes).

manteniendo su estado de secuestro en la profase de la primera división meiótica. La llegada de la pubertad es acompañada por el inicio de las secre-
ciones de GnRH, por lo que también empieza a aparecer la FSH y la LH en el sistema. A partir de este momento, algunos folículos primordiales podrán empezar su crecimiento. En primer lugar, se observa que el ovocito primario aumenta su tamaño unas dos o tres veces. Además, se de-sarrollan nuevas capas de céls. de la
granulosa. En este momento ya ha-blamos de folículos primarios. Durante unos pocos días al inicio de cada ciclo sexual mensual femenino, las concentraciones de FSH y LH experimentan un aumento ligero o moderado; el ascenso de FSH es algo mayor y precede en unos días al de LH. Estas hormonas, sobre todo la
LH, inducen el crecimiento acelerado de 6 a 12 folículos primarios cada mes. El efecto inicial es la proliferación rápida de las céls. de la granulosa, con lo que las capas de dichas céls. se multiplican. Las céls. de la granulosa empizan a formar y liberar mucopolisacáridos que dan lugar a una capa translúcida que rodea al ovocito primario, la zona pelúcida (ZP). Las células fusiformes, derivadas del intersticio ovárico, se agrupan formando varias capas por fuera de las céls. de la granulosa, dando origen a una segunda masa de células denominada teca. Esta se divide en dos capas. En la teca interna encontramos células que forman hormonas sexuales esteroideas (estrógenos + progesterona). La teca externa consiste en tejido conectivo muy vascularizado que reviste al folículo en desarrollo. Tras la fase proliferativa inicial, que dura unos días, la masa de céls. de la granulosa secreta un líquido folicular9 que contiene una elevada concentración de estrógenos. En un inicio este líquido se acumula en unas pequeñas vesículas antrales (folículo secundario), que se fusionarán para formar la cavidad antral que observamos ya en el folículo de De Graaf. El folículo de De Graaf mide unas 2.000 µm y el ovocito que contiene está rodeado por una capa de céls. de la granulosa denominada cumulus oophurus (a posteriori será la co-rona radiata). Es el único folículo que puede dar un óvulo maduro. El resto de folículos que iniciaron el ciclo entrarán en atresia.
Papel de las hormonas
El crecimiento inicial del folículo hasta la etapa antral depende sobre todo de la FSH sola. Después se da un crecimiento muy acelerado, que forma folículos mucho más grandes de-
13
9 Según la Dra. Jiménez, el líquido antral también es formado por las céls. de la teca interna.
Etapas del crecimiento folicular en el ovario, incluyendo la formación del cuerpo lúteo. (Guyton pág. 989)

nominados folículos antrales. Este crecimiento acelerado se da por las razones que se van a exponer a continuación:
- Céls. de la granulosa. Las céls. de la granulosa, ante la la llegada de FSH, forman estró-
genos. Estos estrógenos afectan a las mismas céls. de la granulosa e inducen la expre-
sión de receptores para la FSH, con lo que se logra una mayor sensibilización a la FSH (feedback positivo).
- LH + estrógenos. La combinación de la hormona luteinizante y estrógenos actúa sobre las céls. de la granulosa para aumentar la expresión de receptores para la LH. Así hace-mos que las céls. de la granulosa sean más sensibles a la FSH y a la LH. Esto incre-menta la secreción folicular (se forman más estrógenos).
- Céls. de la teca. Las células proliferan y secretan mucho más gracias al aumento de la LH hipofisaria y de la cantidad creciente de estrógenos en el folículo. En clase se co-mentó que se incrementan los receptores para la LH.
Ovulación
Poco antes de la ovulación, la pared externa del folículo, que hace relieve, se hincha con rapidez y una pequeña zona del centro de la cápsula folicular, denominada estigma, forma una protuberancia similar a un pezón. El líquido folicular empezará a rezumar a través del estigma y poco tiempo después se producirá una gran rotura y un líquido más viscoso, que ha ocupado la porción central del folículo, se vierte hacia fuera. Este líquido viscoso lleva consigo al óvulo rodeado por una masa de pequeñas céls. de la granulosa denominadas co-rona radiada.
Líquido folicular/antral. Este líquido está formado por mucopolisacáridos, proteínas plasmáticas, enzi-mas para la síntesis de esteroides, estrógenos, progesterona, andrógenos, FSH, LH, inhibina, activina,
citocinas diversas, OMI, factores de crecimiento (IGF, EGF), oxitocina, prostaglandinas, etc.
Para que se dé la ovulación, debe producirse un pico de LH. Unas 16h antes de la ovulación vemos un pico una LH que ha ido viendo como su concentración iba aumentando (hasta multiplicarse entre 6 y 10 veces). Junto con la FSH, la LH logra que el folículo se agrande. La LH además convierte a las células de la granulosa y de la teca en céls. secretoras de progesterona, por lo que los niveles de estrógenos empiezan a disminuir y aumentan los de progesterona. La gran cantidad de LH liberada provoca que se secreten hormonas esteroideas folicula-
res (progesterona). Para que se dé la ovulación se producen dos hechos importantísimos:(1) Teca externa. La teca externa empieza a liberar enzimsa proteolíticas (colagenasa) que
debilita la pared del folículo, incrementando la hinchazón del mismo y degeneran-do el estigma.
(2) Vasos sanguíneos. Al mismo tiempo se produce un crecimiento rápido de nuevos va-sos sanguíneos en el interior de la pared del folículo y comienzan a secretarse pros-taglandinas (que provocan una vasodilatación) en los tejidos foliculares. Estos dos efectos producen trasudación de plasma al interior del folículo, que también contribu-ye a que este se hinche.
14

La combinación del hinchazón del folículo con el debilitamiento del estigma hace que el folículo se rompa y expulse el óvulo.
Ciclo ovárico
El primer día del ciclo es el primer día de la menstruación. A partir de aquí dividimos el ciclo en tres etapas: (1) fase folicular, (2) ovulación y (3) fase lútea. Algunas de estas fases ya se han ido medio explicando a lo largo del tema, sobre todo las dos primeras.
Fase folicular
En los primeros días del ciclo menstrual las concentraciones FSH y de LH empiezan a au-mentar, sobre todo la de FSH. La FSH induce el crecimiento de entre seis y doce folículos. Es-tos folículos en crecimiento ven aumentar el número de sus células de la granulosa y apare-cen las tecas interna y externa. Recordemos que las céls. de la teca interna forman esteoroi-des (estrógenos y progesterona)
- Céls. de la teca. Estas células disponen de receptores de la LH. Ante la llegada de la LH forman androstenediona (un andrógeno) a partir del colesterol. Esta androstenediona se va a la circulación y después acabará en las céls. de la granulosa.
- Céls. de la granulosa. Las células de la granulosa disponen de receptores para la FSH. Ante la llegada de la FSH, y gracias a la acción del enzima citocromo P450 aromatasa, transforma la androstenediona en estrógenos (estradiol). Este estradiol pasará hacia el líquido antral.
Únicamente uno de los folículos antrales verá sus vesículas con líquido antral fusionadas y será ya considerado como folículo de De Graaf.
15
Interacción de las céls. foliculares de la teca y la granulosa para la producción de estrógenos. (Guyton pág. 993). Las céls. de la teca, bajo el control de la LH, producen andrógenos que se difunden hacia las céls. de la granulosa. En los folículos maduros, la FSH actúa sobre las céls. de la granulosa para estimular la actividad aromatasa, que convierte los andrógenos en estrógenos.

A medida que el folículo de De Graaf va incrementando su tamaño, las céls. de la granulosa experimentan una mayor respuesta a la FSH, con lo que se forma más estradiol. La acción conjunta de la FSH y el estradiol hace que las céls. de la granulosa empiecen a expresar re-
ceptores de la LH, por lo que ahora la célula de la granulosa también podrá formar progeste-rona a partir del colesterol (en forma de LDL). La progesterona formada por esta nueva fuente ejerce un mecanismo de feedback po-sitivo sobre la adenohipófisis, estimulando la liberación de LH. Este aumento de la concentra-ción de LH se traduce en un incremento de la producción de androstenediona por parte de las céls. de la teca y por ende también un aumento de la formación de estradiol gracias a la acción sinérgica existente entre las céls. de la teca y de la granulosa. Durante la fase folicular temprana, las céls. de la granulosa forman inhibina, que ejer-ce un feedback positivo (sí, está bien) sobre la adenohipófisis que provoca una disminución
de los niveles de FSH. Esto refuerza la atresia (véase más adelante) de los folículos que no se han desarrollado del todo. A medida que avanza la foliculogénesis, los niveles de FSH van disminuyendo debido a la acción de la inhibina y el estradiol (estrógenos y estradiol son si-nónimos). El 95% de los estrógenos en circulación proceden del folículo maduro que ovula-rá. En cuanto a la atresia, es lo que le sucederá al resto de folículos que iniciaron el ciclo. Se cree que se da porque la gran cantidad de estrógenos formada por el folículo de De Graaf hace que la adenohipófisis disminuya la formación de FSH (sobre todo) y de LH. Aquellos folículos que no son lo suficientemente grandes, degenerarán. El folículo más avanzado (en términos de madurez), se hace un feedback sobre si mismo que culmina con la expresión de
más receptores para la FSH, por lo que aunque los niveles de la hormona bajen, apenas notará los efectos de esta caída.
Ovulación
De 34 a 36h antes de la ovulación, las concentraciones de FSH y sobre todo de LH aumen-tan. La máxima liberación de LH se da unas 12h antes de ovular. Este pico de LH reactiva la meiosis del ovocito (que se volverá a detener pero en la metafase de la segunda división meiótica). El pico también es el responsable de la expulsión del óvulo.
Fase lútea
Durante las primeras horas después de la expulsión del óvulo, lo que resta del folículo son las células de la granulosa y de la teca interna, que se transforman en células luteinicas. Estas céls. aumentan su tamaño y se llenan de inclusiones lipídicas que les confieren un aspecto amarillento. Este proceso recibe el nombre de luteinización y el conjunto de la masa de célu-las se denomina cuerpo lúteo. Las células de la granulosa forman estrógenos y sobre todo progesterona. Las céls. de la teca del cuerpo lúteo forman dos andrógenos: androstenediona y testosterona (que difundi-rán hacia las céls. de la granulosa). La progesterona es importantísima para que se dé la im-plantación del óvulo en el endometrio (si no, se produce un aborto).
16

Si a los doce días no ha habido una fecundacion del óvulo, el cuerpo lúteo regresiona y pasa a ser corpus albicans. Esta involución se produce por la disminución hasta niveles muy bajos de la FSH y LH: recordemos que la progesterona y los estrógenos ejercen un efecto inhibidor sobre la liberación de estas dos hormonas y, además, el cuerpo lúteo forma inhibina, que frena también la producción de FSH. Cuando el cuerpo lúteo degenera, la producción de FSH y LH de-
ja de estar inhibida por estos tres factores (estrógenos + progesterona + inhibina), por lo que el ciclo se restaura (la FSH y LH vuelven a adquirir valores normales).
Si se produce la fecundación del óvulo, el cuerpo lúteo no regresiona ya que el trofoblasto forma la gonadotropina coriónica humana (hCG), que tiene casi las mismas propiedades que la LH y permite que el cuerpo lúteo se mantenga hasta más o menos el cuarto mes. Por tanto, hasta ese cuarto mes, el cuerpo lúteo se convierte en una glándula endocrina que forma hormonas. Al llegar el cuarto mes, los niveles de hCG disminuyen y el cuerpo lúteo pasa a albicans. La barrera se coloca en el cuarto mes ya que es el momento en que la placenta puede formar hormonas (y relega al cuerpo lúteo).
Ciclo endometrial
Las fluctuaciones cíclicas de estrógenos y progesterona por parte de los ovarios es lo que ha-rá que se produzcan unos cambios en el aparato reproductor y en el endometrio. Estos cam-bios que se dan en el endometrio es lo que conocemos como ciclo endometrial mensual, que se da unas 400 veces a lo largo de la vida de las mujeres (igual que ovulaciones). Se divide en tres etapas: (a) proliferativa, (b) secretora, y (c) menstrual.
Fase proliferativa
La fase proliferativa también se puede denominar fase estrogénica o preovulatoria. Al inicio de esta fase el endometrio está descamado por la menstruación previa. Como ya sabemos, el
17
Concentraciones plasmáticas aproximadas de gonadotropinas (FSH y LH) y hormonas ováricas (estradiol y progesterona) durante el ciclo sexual femenino normal. (Guyton pág. 988).
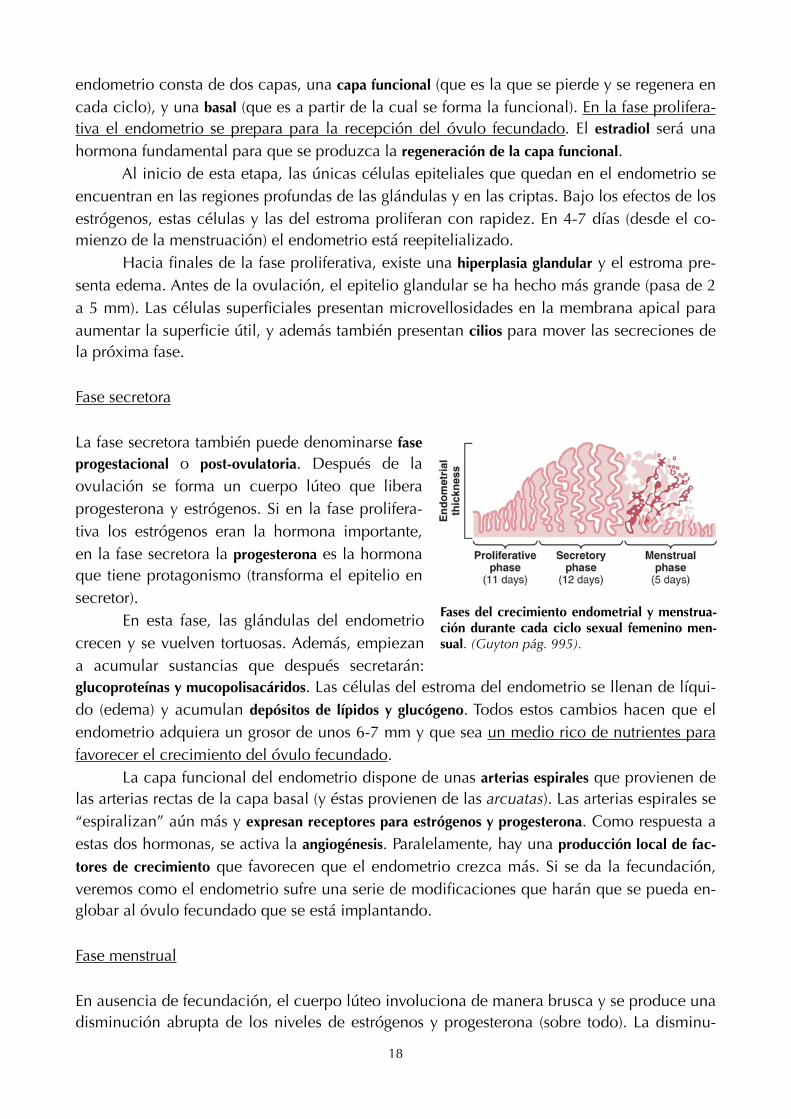
endometrio consta de dos capas, una capa funcional (que es la que se pierde y se regenera en cada ciclo), y una basal (que es a partir de la cual se forma la funcional). En la fase prolifera-tiva el endometrio se prepara para la recepción del óvulo fecundado. El estradiol será una hormona fundamental para que se produzca la regeneración de la capa funcional. Al inicio de esta etapa, las únicas células epiteliales que quedan en el endometrio se encuentran en las regiones profundas de las glándulas y en las criptas. Bajo los efectos de los estrógenos, estas células y las del estroma proliferan con rapidez. En 4-7 días (desde el co-mienzo de la menstruación) el endometrio está reepitelializado. Hacia finales de la fase proliferativa, existe una hiperplasia glandular y el estroma pre-senta edema. Antes de la ovulación, el epitelio glandular se ha hecho más grande (pasa de 2 a 5 mm). Las células superficiales presentan microvellosidades en la membrana apical para aumentar la superficie útil, y además también presentan cilios para mover las secreciones de la próxima fase.
Fase secretora
La fase secretora también puede denominarse fase progestacional o post-ovulatoria. Después de la ovulación se forma un cuerpo lúteo que libera progesterona y estrógenos. Si en la fase prolifera-tiva los estrógenos eran la hormona importante, en la fase secretora la progesterona es la hormona que tiene protagonismo (transforma el epitelio en secretor). En esta fase, las glándulas del endometrio crecen y se vuelven tortuosas. Además, empiezan a acumular sustancias que después secretarán: glucoproteínas y mucopolisacáridos. Las células del estroma del endometrio se llenan de líqui-do (edema) y acumulan depósitos de lípidos y glucógeno. Todos estos cambios hacen que el endometrio adquiera un grosor de unos 6-7 mm y que sea un medio rico de nutrientes para favorecer el crecimiento del óvulo fecundado. La capa funcional del endometrio dispone de unas arterias espirales que provienen de las arterias rectas de la capa basal (y éstas provienen de las arcuatas). Las arterias espirales se “espiralizan” aún más y expresan receptores para estrógenos y progesterona. Como respuesta a estas dos hormonas, se activa la angiogénesis. Paralelamente, hay una producción local de fac-
tores de crecimiento que favorecen que el endometrio crezca más. Si se da la fecundación, veremos como el endometrio sufre una serie de modificaciones que harán que se pueda en-globar al óvulo fecundado que se está implantando.
Fase menstrual
En ausencia de fecundación, el cuerpo lúteo involuciona de manera brusca y se produce una disminución abrupta de los niveles de estrógenos y progesterona (sobre todo). La disminu-
18
Fases del crecimiento endometrial y menstrua-ción durante cada ciclo sexual femenino men-sual. (Guyton pág. 995).

ción de progesterona provoca la descamación del endometrio (capa funcional) y la menstrua-ción. El primer efecto (de la bajada de estas dos hormonas) es la disminución de la estimula-
ción de las céls. endometriales (que era realizada por estas dos hormonas), seguida de una rá-pida involución del propio endometrio. A continuación se produce una infiltración de neutrófi-
los que hacen que el estroma presente un aspecto pseudoinflamado. La infiltración de los leucocitos es esencial para que el endometrio se empiece a fragmentar (gracias a la libera-ción de metaloproteasas que degradan la matriz). A esto hay que sumarle que las propias cé-lulas del estroma también forman y liberan proteasas que favorecen que la matriz se siga fragmentando. Las arterias espirales presentan una mayor “espiralización”, de tal manera que se pro-duce una resistencia al flujo sanguíneo que se traduce en una hipoxia. Finalmente, estas arte-rias espirales sufrirán una vasoconstricción inducida por las prostaglandina vasoconstrictora F2α (PGF2α), sintetizada por las céls. del estroma ante la bajada de progesterona. Otras prosta-glandinas liberadas inician unas contracciones del miometrio que favorecen el desprendimien-to de la capa funcional. Las prostaglandinas además tienen un efecto proinflamatorio. La vasoconstricción de las arterias espirales, junto con la bajada de nutrientes y la pérdida de la estimulación hormonal (sobre todo de la progesterona), produce la necrosis del
endometrio y de los vasos sanguíneos. Lo primero que se pierde es el estrato vascular (arterias espirales) y después las sucesivas capas necróticas hasta llegar a la capa basal. La masa de tejido descamado junto con la sangre que se ha generado, sale gracias a la acción del mio-metrio (prostaglandinas). Con la regla se pierden 40 ml de sangre y 35 ml de líquido seroso. Junto con el material necrótico se libera fibrinolisina, para así evitar la coagulación de la sangre. No obstante, si se forma demasiada sangre, es posible que se observen algunos coágulos. Aunque el epitelio ahora se encuentre descamado, hay una producción masiva de leucocitos que evita las infecciones (y parte se van con la regla; leucorrea).
Funciones de las hormonas ováricas
El ovario forma dos clases de hormonas: (a) estrógenos, y (b) gestágenos (progesterona). La principal función de los estrógenos es preparar el aparato reproductor femenino y en general el cuerpo para promover los carácteres sexuales secundarios de la mujer. La principal fun-ción de los gestágenos es preparar el útero para la gestación y a las mamas para la lactancia.
Estrógenos
Los estrógenos se forman sobre todo en los ovarios. Recordemos, no obstante, que se forman una pequeña cantidad en la zona reticular de las glándulas suprarrenales. Cuando una mujer está embarazada, también puede formarlos en la placenta. Existen tres estrógenos: el 17β-estradiol (el más importante y abundante), la estrona y el estriol. La estrona se sintetiza en tejidos periféricos a partir de los andrógenos (androstene-diona) formados por las céls. de la teca y las glándulas suprarrenales. El estriol es un estróge-no débil, un producto oxidativo formado en el hígado derivado tanto de la estrona como del
19

estradiol. La potencia estrogénica del estradiol es 12 veces mayor comparada con la estrona y 80 veces mayor si se compara con el estriol.
A continuación se exponen las funciones de los estrógenos. Su principal función, no obstan-te, consiste en estimular la proliferación celular y el crecimiento de los tejidos de los órganos sexuales y de otros tejidos relacionados con la reproducción.
- Efectos sobre el útero y los órganos sexuales externos femeninos. Durante la niñez apenas se secretan estrógenos, será con la llegada de la pubertad que su secreción se incre-mentará notablemente (efecto de las gonadotropinas hipofisarias). En general, se in-
crementará el tamaño de los órganos sexuales femeninos internos (trompas de Falopio, útero y vagina) y de los elementos externos del mismo (monte de Venus, labios mayores y menores). Se incrementa la resistencia del epitelio vaginal (cúbico → estratificado). Re-cordemos que los estrógenos son importantes para la regeneración de la capa funcio-nal del endometrio en el marco del ciclo endometrial mensual.
- Efectos sobre las trompas de Falopio. Tiene un efecto similar al que tiene en el endome-trio (proliferación de tejido glandular e incremento de céls. epiteliales ciliadas). In-
crementa la actividad de los cilios (ayudan a propulsar al óvulo).- Sobre las mamas. En un inicio, las mamas primordiales de la mujer y del varón son
exactamente iguales. Los estrógenos favorecen: (a) el desarrollo de los tejidos del estro-
ma mamario; (b) el crecimiento del sistema de conductos; (c) el depósito de grasa en las mamas. Los efectos de los estrógenos en el desarrollo de los lobulillos y los alvéolos mamarios es escasa (esto lo hace más la progesterona y la prolactina). En resumen, los estrógenos inician el crecimiento de las mamas y del aparato productor de leche. Son
20
Síntesis de las principales hormonas femeninas. (Guyton pág. 992).

también responsables del crecimiento y del aspecto externo característicos de la ma-ma madura. Sin embargo, no completan la tarea de convertir las mamas en órganos productores de leche.
- Sobre el esqueleto. Estimula el crecimiento óseo (inhibe la actividad osteoclástica) y fomenta la fusión temprana de las epífisis con las diáfisis de los huesos largos (efecto mayor que el de la testosterona en los hombres, por lo que las mujeres dejan de cre-cer antes). Con la menopausia, el cese de producción de estrógenos hace que se incre-mente la actividad osteoclástica, se pierda matriz ósea y que haya un descenso de los depósitos de Ca2+ y fosfato. En algunas mujeres este efectos es muy pronunciado y se da la osteoporosis.
- Sobre los depósitos de proteína. Se incrementa ligeramente la producción proteica (ba-
lance nitrogenado positivo). Este efecto es menor al que produce la testosterona en los varones.
- Sobre el metabolismo corporal y los depósitos de grasa. Se incrementa el metabolismo corporal (aunque menos que en el hombre por efecto de la testosterona). Se producen depósitos de grasa adicionales en las nalgas y los muslos.
- Sobre la distribución del pelo. Los estrógenos apenas tienen efectos sobre la distribución del pelo (aunque se dearrolla vello en el pubis y las axilas, principalmente por los an-
drógenos formados en la corteza suprarrenal).- Sobre la piel. Los estrógenos vuelven la piel fina y blanda. Se incrementa su vasculari-
zación.- Sobre el equilibrio electrolítico. Hay una similitud química entre las hormonas estrogé-
nicas y las hormonas corticosuprarrenales. Se produce una retención de Na+ y agua en los túbulos renales (retención de líquidos en la gestación).
Progesterona
En cuanto a la progesterona, su función más importante es la promoción de la capacidad secre-
tora del endometrio uterino, preparando así el útero para la implantación. Además de este efecto sobre el endometrio, reduce la frecuencia e intensidad de las contracciones uterinas (evitando la expulsión del óvulo implantado).
- Efectos sobre las trompas de Falopio. Promueve la secreción en las mismas (nutrición del óvulo).
- Efectos sobre el desarrollo de las mamas. Estimula el desarrollo de los lobulillos y los al-veolos mamarios. Sin embargo, no hace que los alveolos secreten leche, ello depende de la prolactina. Incrementa el tamaño de las mamas (en parte por el desarrollo de las estructuras productoras de leche y también por el aumento del líquido presente en el tejido).
Ciclo del cérvix
El cérvix también se modificará a lo largo del ciclo ovárico y aceptará mejor los espermato-zoides cuando haya ovulación. El cérvix presenta dos orificios (o esfínteres): uno interno que
21

mira hacia el útero y otro externo que está orientado hacia la vagina. También presenta unas glándulas secretoras y está revestido por una mucosa especial que genera una secreción densa. Las cantidad de glándulas está modulada por hormonas (estrógenos y progestágenos). Los cambios que sufre el cérvix son los siguientes:
- A medida que avanza la foliculogénesis, los dos esfínteres de los que antes hemos ha-blado se van relajando, llegando al máximo diámetro de obertura en la ovulación.
- Por defecto, las secreción del cérvix son densas (así se protege el útero). No obstante, en el momento de la ovulación las secreciones se vuelven más fluidas, facilitando el paso de posibles espermatozoides.
- El pH de las secreciones cervicales es, por naturaleza, ácido. Al llegar la ovulación este pH se hace alcalino. Recordemos que la vagina presenta un pH ácido (los eosinó-filos generan lactato). Además de ver modificado el pH, la secreción contiene más agua y la disposición de las glucoproteínas cambia10 para así facilitar el paso de los es-permatozoides (en la ovulación).
Depósito de espermatozoides en la vagina
Tras el coito se depositan en la vagina entre 100 y 200 millones de espermatozoides. Estos espermatozoides, gracias a las sustancias de la próstata y de las vesículas seminales (p. ej., fibrinógeno), se forma un coágulo de espermatozoides. De este coágulo únicamente partirán algunos espermatozoides (que siguen siendo muchos), el resto serán fagocitados por los ma-crófagos. A este coágulo además debemos sumarle el pH ácido de la vagina, por lo que de cada 1.000 espermatozoides, únicamente 1 atraviesa el cuello del útero. Los espermatozoides se moverán gracias a las contracciones del miometrio inducidas por
las prostaglandinas del semen. El útero presenta toda una serie de criptas en las cuales parte de los espermatozoides se perderán. Sólamente llegarán a las trompas entre 100 y 200 esperma-tozoides11.
Capacitación de los espermatozoides
El espermatozoide debe sufrir toda una serie de cambios para fecundar el óvulo. Estos cam-bios son una continuación de los que tuvo en el epidídimo y se van a dar en el útero.
- Pérdida de los factores inhibidores. Cuando los espermatozoides están en el hombre o cuando han sido eyaculados están completamente rodeados por factores inhibidores. A medida que avanzan por el tracto reproductor femenino, los van perdiendo.
- Vesículas de colesterol. El núcleo del espermatozoide está recubierto por vesículas de colesterol, que también tapan al acrosoma (lo protegen). De camino a las trompas el espermatozoide pierde las vesículas de colesterol y la membrana del acrosoma que-dará descubierta (irán saliendo los enzimas hidrolíticos).
22
10 En condiciones normales (cuando no hay la ovulación), las glucoproteínas están dispuestas de tal manera que se impide el paso de los espermatozoides.
11 A todos estos datos hay que sumar el hecho de que, en la especie humana, el 20% de los espermatozoides son anómalos.

- Permeabilidad al Ca2+. El hecho de perder los factores inhibidores y las vesículas de colesterol hacen que la membrana sea más permeable al calcio (y así el espermato-zoide se vuelve hiperactivo).
Fertilización
El óvulo tiene una corona radiada y una zona pelúcida que lo rodean. Las céls. de la granu-losa están unidas por ácido hialurónico. La ZP está formada por tres glucoproteínas: ZP1, ZP2 y ZP3, la más importante de las cuales es la última (ZP3) ya que es un receptor de especie (y será reconocida por los espermatozoides). En el acrosoma de los espermatozoides está la hialuronidasa, que se encargará de de-gradar la matriz que une a todas las céls. de la granulosa entre sí (corona radiada). Como el espermatozoide está hiperactivo, irá haciéndose paso a través de la corona radiada hasta lle-gar a la ZP. Una vez en la ZP, se producirá la interacción entre el espermatozoide y la gluco-proteína ZP3 (reconocimiento); esta reacción induce la reacción acrosomal. En este punto, la membrana del espermatozoide se fusionará con la del óvulo, haciendo que el óvulo englobe el núcleo, el cuello y una parte de la cola del espermatozoide. Las mitocondrias del cuello y el flagelo se desprenderán, dejando únicamente el núcleo del espermatozoide en el interior del óvulo. En este momento se reactivará la división meiótica (finalizará la 2º división) que culmina con la formación del segúndo corpúsculos polar y la generación de un pronúcleo mas-
culino y otro femenino. Una vez el espermatozoide ha reconocido la ZP3, el óvulo, para evitar la polispermia, realiza una reacción cortical que se divide en dos tiempos:
- Primera reacción. Una primera reacción rápida (a los 5 min.) que consiste en que la membrana del óvulo modifica sus proteínas para evitar la entrada de otros espermato-zoides.
- Segunda reacción. La entrada de Ca2+ en el ovocito provoca la liberación de unos grá-nulos corticales que contienen enzimas que hidrolizan las ZP y hacen que la zona pelúcida no pueda ser reconocida por los espermatozoides (explicación bastante sim-ple, mejor explicado en Biología celular del desarrollo).
Implantación
Una vez el óvulo ha sido fertilizado pasa a denominarse cigoto. Este cigoto experimentará toda una serie de divisiones (se formarán blastómeros) que hacen que el nº de céls. aumente pero no el volumen total del cigoto (así evitamos que se quedé atrapado en las trompas). El nº de céls. tampoco incrementa mucho porque la energía de la que dispone el cigoto en sus reservas es limitada. En este punto la corona radiada y las céls. de la granulosa ya se han perdido (no son lo mismo?), pero aún queda la ZP. La presencia de la zona pelúcida impide que el sistema in-munitario reconozca el cuerpo extraño. A medida que avanzan las divisiones, el cigoto pasa a denominarse morula y empieza a haber una oganización interna: unas células se ubican en la periferia ((cito)trofoblasto) y
23

otras dentro (embrioblasto). Progresivamente se va generando una cavidad en el interior (blastocele) y cuando esto ocurre ya habla-mos de blástula o blastocisto. Tras unas 90 a 150h flotando, la blástula pierde la zona pelúcida y ya puede implantarse. Recordemos que esta implan-tación se producía por el polo embrionario, que es la zona de la blástula donde se en-cuentra el embrioblasto. El (cito)trofoblasto es el responsable, incluso antes de que se produzca la implantación, de la formación de la hCG necesaria para el mantenimiento del cuerpo lúteo. La progesterona formada por el cuerpo lúteo hace que las células del endometrio acumulen líquido, lípidos y glucógeno (↑nutrientes). El citotrofoblasto se fusionará con las células del endometrio y se formará un sincitiotrofoblasto que irá rodeando progresivamente al blastocisto (el embrioblasto ha empezado a diferenciarse). Entre la sexta y la duodécima semana hay un pico de hCG que se da para mantener el cuerpo luteo y que así el endometrio siga siendo rico en nutrientes. La pastilla abortiva Ru486 (que puede ser usada hasta los 49 días) inhibe los receptores de progesterona de las céls.
endometriales, produciéndose el desprendimiento del blastocisto.
Placenta
Las células del endometrio que acumulan nutrien-tes bajo los efectos de la progesterona se denomi-nan células deciduales. La fusión entre las céls. de-ciduales y las del trofoblasto dan lugar al sincitio-
trofoblasto. A partir de éste se forman la placenta y las membranas extraembrionarias, y dentro de las membranas extraembrionarias distinguimos las ve-
llosidades coriónicas. La placenta tiene varias funciones: de intes-
tino (aporte de nutrientes), respiratoria (aporte de O2 y captación del CO2), excretora, y endocrina. Hacia el tercer mes de vida ya está formada, mo-mento en el cual actuará sobre el cuerpo lúteo y hará que éste ya no responda a la hCG. En un inicio, la nutrición es trofoblástica y
24
(A) Ovulación, fecundación del óvulo e implantación del blastocisto en el útero. (B) Intervención de las céls. trofoblásticas en la implantación del blastocisto en el endometrio. (Guyton pág. 1004)
Nutrición del feto. En un principio, la nutrición procede de la digestión trofoblástica y absor-ción de nutrientes a partir de la decidua endo-metrial. A posteriori, toda la nutrición se debe a la difusión a través de la membrana placenta-ria.

después pasa a ser placentaria. Un primer paso para la formación de la placenta es que las arterias espirales pasen a tener forma de em-budo (hacia las 16 semanas). Estos embudos cada vez se hacen más anchos y dan lugar a las arterias uteroplacentales. Para que nos ha-gamos una idea: el flujo de sangre de una ar-teria espiral es de unos pocos ml/min, mien-tras que el de estas arterias es de entre 600 y 1.000 ml/min. En las vellosidades coriónicas es donde se da el intercambio entre la madre y el feto (nutrientes, productos del metabolismo, gases, etc). Este intercambio se realiza por di-ferentes mecanismos de transporte: difusión
simple (agua, electrólitos), difusión facilitada
(oxígeno, glucosa), pinocitosis (anticuerpos), etc. Antes hemos comentado que en la pla-centa se forman hormonas. En este órgano se da la esteroidogénesis: se forman estrógenos y progesterona. Para que la placenta realice esta función, precisa tanto de la madre como del feto. La madre tiene los precursorse de los es-trógenos y el feto los de la progesterona. En la placenta, el estrógeno más importante es el es-
triol (no el estradiol).
Gemelos (apunte): Si los blastómeros se separan y dan lugar a dos morulas, la mujer gestará a gemelos mono-cigóticos (cada uno en su placenta). Si el embrioblasto se separa cada uno dará un embrión (gemelos monoco-riónicos mono/diamnióticos). Finalmente, si el embrioblasto se separa en dos partes y después se vuelve unir,
hablaremos de siameses.
Hormonas durante el embarazo
A lo largo del embarazo12, los niveles de estrógenos siempre se mantienen por debajo de los de progesterona. Hacia el final del embarazo, los estrógenos superan a la progesterona. Ade-más de estas dos hormonas, la placenta forma el lactógeno placentario humano (hLP) y prolac-
tina (cuya acción lactogénica queda inhibida durante el embarazo por acción de la progeste-rona). La hLP hace que la madre obtenga energía a partir de los ácidos grasos en lugar de a partir de la glucosa (que la cede al feto)13.
25
12 Un embarazo dura unas 40 semanas (±2).
13 Es por esta razón que las mujeres embarazadas presentan una glucemia un poco más elevada de lo normal, pudiendo llegar a padecer una diabetes gestacional.
Parte superior: Organización de la placenta madura. Parte inferior: relación entre la sangre fetal presente en los capilares de las vellosidades y la sangre ma-terna contenida en los espacios intervellosos.
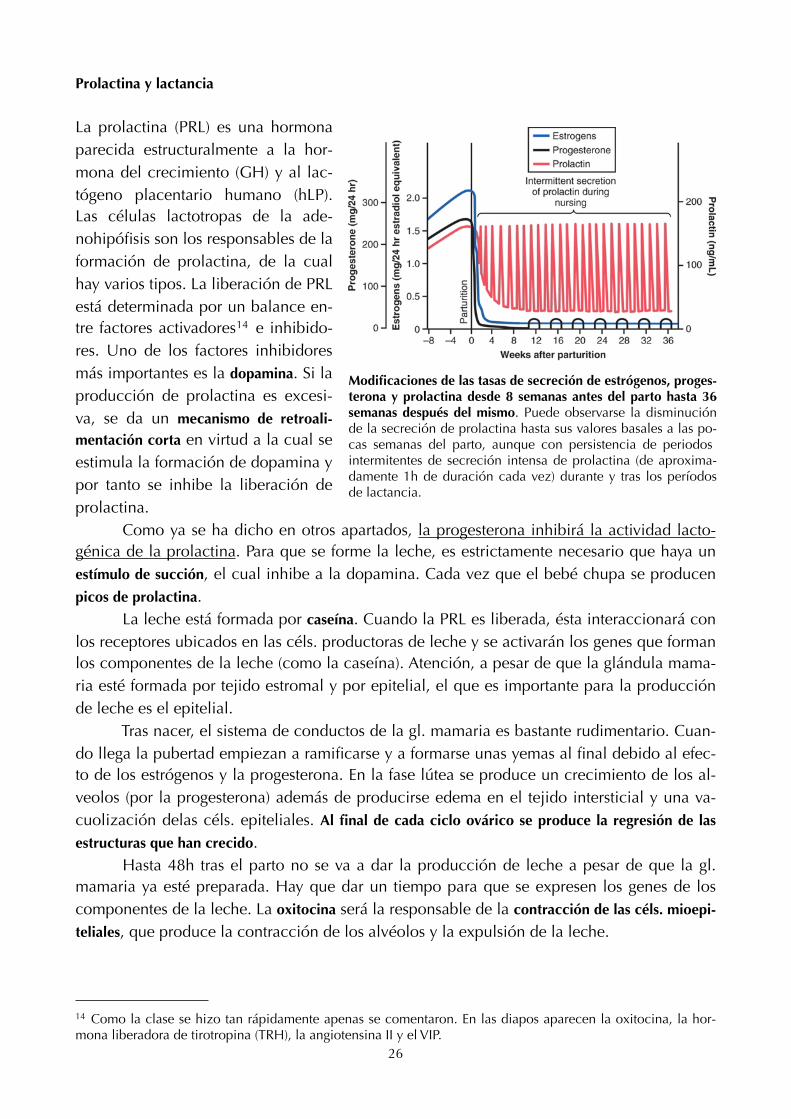
Prolactina y lactancia
La prolactina (PRL) es una hormona parecida estructuralmente a la hor-mona del crecimiento (GH) y al lac-tógeno placentario humano (hLP). Las células lactotropas de la ade-nohipófisis son los responsables de la formación de prolactina, de la cual hay varios tipos. La liberación de PRL está determinada por un balance en-tre factores activadores14 e inhibido-res. Uno de los factores inhibidores más importantes es la dopamina. Si la producción de prolactina es excesi-va, se da un mecanismo de retroali-mentación corta en virtud a la cual se estimula la formación de dopamina y por tanto se inhibe la liberación de prolactina. Como ya se ha dicho en otros apartados, la progesterona inhibirá la actividad lacto-génica de la prolactina. Para que se forme la leche, es estrictamente necesario que haya un estímulo de succión, el cual inhibe a la dopamina. Cada vez que el bebé chupa se producen picos de prolactina. La leche está formada por caseína. Cuando la PRL es liberada, ésta interaccionará con los receptores ubicados en las céls. productoras de leche y se activarán los genes que forman los componentes de la leche (como la caseína). Atención, a pesar de que la glándula mama-ria esté formada por tejido estromal y por epitelial, el que es importante para la producción de leche es el epitelial. Tras nacer, el sistema de conductos de la gl. mamaria es bastante rudimentario. Cuan-do llega la pubertad empiezan a ramificarse y a formarse unas yemas al final debido al efec-to de los estrógenos y la progesterona. En la fase lútea se produce un crecimiento de los al-veolos (por la progesterona) además de producirse edema en el tejido intersticial y una va-cuolización delas céls. epiteliales. Al final de cada ciclo ovárico se produce la regresión de las
estructuras que han crecido. Hasta 48h tras el parto no se va a dar la producción de leche a pesar de que la gl. mamaria ya esté preparada. Hay que dar un tiempo para que se expresen los genes de los componentes de la leche. La oxitocina será la responsable de la contracción de las céls. mioepi-
teliales, que produce la contracción de los alvéolos y la expulsión de la leche.
26
14 Como la clase se hizo tan rápidamente apenas se comentaron. En las diapos aparecen la oxitocina, la hor-mona liberadora de tirotropina (TRH), la angiotensina II y el VIP.
Modificaciones de las tasas de secreción de estrógenos, proges-terona y prolactina desde 8 semanas antes del parto hasta 36 semanas después del mismo. Puede observarse la disminución de la secreción de prolactina hasta sus valores basales a las po-cas semanas del parto, aunque con persistencia de periodos intermitentes de secreción intensa de prolactina (de aproxima-damente 1h de duración cada vez) durante y tras los períodos de lactancia.

Correo de la profesora para acabar la clase
A la classe de lactància m’ha quedat per explicar-vos que quan una dona embarassada desenvo-lupa la glàndula mamària cap a una glàndula productora de llet, quan deixa d’alletar, les estructu-res involucionen i la mama esdevé una glàndula amb estructura de glàndula mamària puberal (època pubertat), però tot i que és igual que la mama d’una dóna nulípara (que no ha tingut mai fills), és molt més diferenciada. Es a dir, totes dues, dóna nulípara i mare que ja no alleta tenen la estructura de la glàndula mamària similar (amb els conductes i les gemes terminals) però la glàn-dula de la mare que ja no alleta és molt més diferenciada que la de la noia, ja que les seves cèl·lu-les s’han diferenciat cap a cèl·lules productores de llet (la màxima diferenciació de la glàndula mamària) i li queden cèl·lules amb memòria que es tornaran a transformar en glàndules producto-res de llet al proper embaràs. Totes dues glàndules, mare i noia, experimenten els canvis cíclics deguts al cicle ovàric, però la glàndula mamària de la noia és més indiferenciada que la de la ma-re. D’aquí a que les dones que han estat mares tinguin una glàndula més diferenciada i això supo-sa un menor risc a tenir càncer de mama. Un altre aspecte que tampoc us vaig dir és que “en teoria” mentres una mare està alletant, no li vé la regla. Fisiològicament parlant, mentres s’alleta, es produeix Prolactina, i aquesta inhibeix l’eix hipotàlem-hipòfisi de manera que no hi ha producció de LH ni FSH per tant no hi ha ovulació. Quan una mare deixa d’alletar, es deixa de produir prolactina i per tant l’eix es torna a activar. Però us dic “en teoria” perquè l’alletament no es pot pendre com a “mètode anticonceptiu” ja que ha hagut casos de dones que s’han quedat embarassades mentres alletaven als seus fills. Depèn de la dona, el cicle ovàric es pot tornar a re-establir més aviat o més tard, i ho farà més aviat si ets dona treballadora, on deixes d’alletar tant sovint al fill, llavors els nivells de prolactina no estan tant alts i l’eix es re-estableix.
Així doncs, per una banda el reflex de la succió fa que es produeixi oxitocina que contrau a les cèl·lules mioepitelials dels alvèols i fa que es secreti la llet a la llum i d’aquí als ductes i conductes galactòfors, aquest reflex fa que hi hagi producció de prolactina (que activa la síntesi de llet), i per altra banda manté l’eix inhibit.
Espero que aquesta explicació acabi d’arrodonir la classe de lactància.
Pepita 1-02-12
27

4. SANGRE
Fluidos corporales
El líquido corporal total se distribuye sobre todo en dos compartimentos: el líquido extracelu-
lar y el líquido intracelular. A su vez, el líquido extracelular puede dividirse en el líquido in-
tersticial y el plasma sanguíneo. En el varón adulto medio de 70kg, el agua corporal total es alrededor del 60% del peso corporal o unos 42 l. A medida que envejecemos, este porcenta-je se va reduciendo. En las mujeres, el porcentaje es de un 50% y en bebés prematuros o neonatos la cantidad relativa de agua puede alcanzar valores de hasta el 75% del peso cor-poral. En una persona media, 2/3 partes del agua total corporal corresponden al líquido intracelu-lar, mientras que el 1/3 restante corresponde al fluido extracelular. De este fluido extracelular, 3/4 corresponden al líquido intersticial y 1/4 parte al plasma sanguíneo. En este contexto, el plasma, que son unos 3 l, es el medio que sufre directamen-
te las pérdidas y ganancias de líquidos. Después estos cambios se transmiten al fluido intersticial y de ahí al medio intracelular. El plasma nos permite mantener constantes una serie de características del
medio intracelular. La sangre es un estrato de paso a nuestro medio interno. Ingresamos agua en nuestro organismo por medio del agua que bebemos directamente, la que se encuentra en los alimentos y la que ge-neramos por nuestro propio metabolismo. Las pérdidas se dan por las heces, la orina, la sudo-ración y la pérdida insensible a través de la piel y cuando exhalamos. Para hacernos una idea, en condiciones normales ganamos y perdemos
entre 2.500 ml diarios de agua.
Adición de soluciones
Si añado agua al medio interno (a la sangre), ésta pasará al medio extracelular y verá su os-molaridad reducida (dilución). Pasará un flujo osmótico desde el espacio extracelular al in-tracelular para equilibrar las presiones osmóticas. El resultado es una reducción de la osmo-laridad de los dos compartimentos y un aumento del volumen en ambos. Si añado una solución isotónica únicamente incrementará el volumen del líquido extra-celular. Esto es ideal cuando ha habido una pérdida de fluidos (hemorragia, diarrea).
28
Principales compartimentos de líquidos del cuerpo y membranas que los separan. (Guyton pág. 286). Los valores son para una persona me-dia con un peso de 70kg.

Finalmente, si añado una solución hipertónica veremos como el volumen del compartimento intracelular disminuye para que así se equilibre la osmolaridad de los dos compartimentos. Esto puede ser útil para problemas a nivel del SNC.
Sangre
La sangre es un tejido formado por células inmersas en una matriz extracelular líquida. Por tanto, es correcto afirmar que es un tejido conectivo fluida. Consta del plasma y de los elemen-
tos formes.- Plasma. El plasma15 es un 90% agua y un 10% de otros elementos. Dentro de ese 10%
encontramos proteínas (70%), moléculas de bajo peso molecular (20%) y electrólitos (10%). El plasma tiene una densidad de 1,8 respecto al agua y una osmolaridad de 290 mOsm. El plasma contiene entre 65 y 80 g de proteínas por litro.
- Elementos formes. El 99% de los elmentos formes son eritrocitos, que son los que le dan el color rojo a la sangre. El 1% restante está formado por leucocitos y plaquetas. El color rojo de la sangre es variable: rojo intenso en la sangre arterial y oscuro en la sangre venosa.
Disponemos de unos 5·106 hematíes/mm3 (4,5·106 en mujeres). De leucocitos tenemos entre 4.000 y 10.000 células/ml (67% granulocitos, 27% linfocitos y 6% monocitos). Finalmente, de plaquetas tenemos entre 150.000 y 300.000/ml. Si extraemos sangre y la dejamos reposar con algunos anticoagulantes (ya que si no se formaría el coágulo y en lugar de plasma tendríamos suero) obtendremos tres fases bien dife-renciadas: (a) plasma (55%); (b) hematíes (por debajo del plasma; 45%); (c) una película blan-ca entre el plasma y los hematíes (plaquetas + leucocitos; < 1%). Las funciones de la sangre pueden clasificarse en función a distintos criterios, pero en clase se optó por dividirlas en funciones de transporte y de regulación.
La sangre representa entre el 7 y el 8% del perso corporal (unos 5 l aprox.). El mantenimiento de este volumen es imprescindible y además es determinante para la presión venosa central.
29
15 La diferencia entre plasma y suero es que el segundo ya no tiene fibrinógeno (se ha formado el coágulo).
Funciones de transporteFunciones de transporteRespiratoria Transporte de O2 (hematíes) y de CO2 (plasma).Nutritiva Lleva elementos que deben ser transformados y/o almacenados.Depuradora Lleva los productos del catabolismo hacia el riñón, pulmón, intestino y piel.Endocrina Es el medio de transporte de las hormonas.Funciones de regulación (homeostasis corporal)Funciones de regulación (homeostasis corporal)Temperatura corporal La sangre se comporta como un sistema conductor del calor y del frío. Permite enviar
calor o frío de una región del organismo a otra. En reposo liberamos unas 120 kcal/h (y si hacemos ejercicio esta cantidad se multiplica).
Presión osmótica Cantidad y tamaño de las proteínas contenidas en el plasma (hay algunas con carga negativa e índice de reflexión de 1; es decir, que se quedan siempre en la sangre).
Presión arterial La presión arterial puede modificarse por el volumen sanguíneoOtras Eq. ácido-base, eq. hidro-salino, hemostasia.

Por presión venosa central entendemos la presión de la sangre en la vena cava inferior. Esta medida nos indica la cantidad de sangre que regresa al corazón y la capacidad del mismo para bombear la sangre hacia el sistema arterial. Disponemos de unos 61 a 66 ml de sangre por kg (hematíes = 24-30 ml/kg; plasma = 33-35 ml/kg). También podemos expresarlo de otro modo: tenemos unos 2,89 l de sangre por metro cuadrado (2,44 l/m2 en mujeres). Los jóvenes y los niños pueden tener valores más al-tos y durante el embarazo se da una hemodilución (aumenta el volumen sanguíneo para ga-nar plasma). La sangre tiene una presión osmótica de unos 5.016 mmHg. De esta presión, aproxi-madamente 25 mmHg corresponden a la presión oncótica, que es la presión dependiente de las proteínas. El pH de la sangre oscila entre 7,35 y 7,40. Está condicionado por los ácidos y bases presentes en la sangre, proteínas y el CO2 (y esto hace que el pH venoso sea menor que el arterial).
Conducción de la sangre
La sangre es un fluido con un peso específico de unos 1.060 kg/m3. Cuando sale del cora-zón, es un fluido viscoso que parte a gran presión y velocidad, comportándose como un fluido
turbulento en que sus componentes celulares van agrediendo el endotelio por donde pasan. Esto explica que la gran mayoría de aneurismas se den en la aorta (y concretamente en la abdominal). A medida que la sangre se aleja del corazón, ésta se comporta como un flujo laminar. Un flujo laminar consiste en que el fluido forma una serie de láminas que se man-tienen a una cierta distancia de la pared y que van a velocidades distintas. Las láminas más cercanas a la pared van más lentas que las que ocupan una posición central.
La viscosidad de la sangre depende de la viscosidad del plasma, de la cantidad de eritrocitos y de la agregabilidad o capacidad de deformación por parte de los elementos formes. A ma-yor viscosidad, mayor resistencia periférica, mayor trabajo cardíaco, menor velocidad de cir-culación y peor oxigenación de los tejidos.
30
Flujo laminar y flujo turbulento. (Netter pág. 111). Normalmente se produce un flujo laminar o aerodinámico en la mayor parte del sistema vascular. En situaciones patológicas, como la estenosis (estrechamiento del vaso), las alteraciones valvulares y la baja viscosidad de la sangre (como en la anemia) se producen turbulencias que se asocian a soplos en el corazón o en los grandes vasos, o soplos vasculares que pueden escucharse mediante la auscultación. La turbulencia aumenta el gradiente de presión necesario para producir el flujo.

Eritrocitos
Los eritrocitos son los elementos formes transportadores de la hemoglobina. Esta hemogobina es sintetizada en los precurso-res medulares del eritrocito (el 80% de la síntesis se da en el eritroblasto policromatófi-
lo). En hombres, hay unos 14 g/dl y en mu-jeres 13 g/dl. Debido a la destrucción de los hematíes, hay una producción media diaria de hemoglobina de 6 g. El hematocrito es el porcentaje ocupado por los eritrocitos del total del volumen sanguíneo. En condicio-nes normales, un adulto varón tiene un he-matocrito del 45% (y es menor en las muje-res). Podemos conocer rápidamente el he-matocrito si centrifugamos una muestra de sangre y analizamos la porción de hema-tíes. A continuación se comentan toda una serie de factores de los que depende el he-matocrito.
- Volumen del hematíe. Si tenemos hematíes hinchados (exposición a soluciones hipotó-nicas), con una forma distinta (anisocitosis/poiquilocitosis) o con modificaciones en su deformabilidad (anemia falciforme o talasemia).
- Hipoxemia. Si una persona está expuesta a un ambiente hipóxico (p. ej., al fumar) du-rante un tiempo prolongado, podemos encontrarnos con una eritrocitosis secundaria.
- Patología. Como la policitemia Rubra Vera16 o la anemia.- Otras causas. Como la edad o la hidratación.
Existen toda una serie de valores relacionados con los eritrocitos que son bastante inetere-santes. La velocidad de sedimentación globular (VSG) es el tiempo que tarda una muestra de sangre en ver sedimentar a sus hematíes (nos describe la velocidad con que se produce este proceso). La VSG normal es de 10 mm/h. Los procesos inflamatorios son los responsables del aumento de la VSG. Los índices eritrocíticos nos indican cómo son los hematíes. Existen tres índices eritro-cíticos (no tenemos que saber los valores): HCM (hemoglobina corpuscular media), CHCM (concentración de hemoglobina corpuscular media)17 y VCM (volumen corpuscular medio). La hemoglobina corpuscular media (HCM) nos indica la cantidad de hemoglobina que hay en cada uno de los hematíes.
31
16 Se trata de un cáncer en la serie eritropoyética que hace que los valores de eritrocitos se disparen (hasta 11 millones/mm3) y den lugar a un hematocrito de hasta el 80%. La policitemia secundaria se da cuando la con-centración de eritrocitos se incrementa pero por una deshidratación o por hipoxemia.
17 De ésta si que habló de un dato concreto: el valor normal es del 30% aprox.
Saladin: Anatomy & Physiology: The Unity of Form and Function, Third Edition
18. The Circulatory System: Blood
Text © The McGraw!Hill Companies, 2003
Cha
pter 18
remove their wastes, substances must pass between thebloodstream and tissue fluid through the capillary walls.This transfer of fluids depends on a balance between thefiltration of fluid from the capillary and its reabsorption byosmosis (see fig. 3.15, p. 108). The rate of reabsorption isgoverned by the relative osmolarity of the blood versus thetissue fluid. If the osmolarity of the blood is too high, thebloodstream absorbs too much fluid, which results in highblood pressure and a potentially dangerous strain on theheart and arteries. If its osmolarity drops too low, toomuch fluid remains in the tissues. They become edema-tous (swollen) and the blood pressure may drop to dan-gerously low levels because of the amount of fluid lostfrom the bloodstream.
It is therefore important that the blood maintain anoptimal osmolarity. The osmolarity of the blood is a productmainly of its sodium ions, protein, and erythrocytes. Thecontribution of protein to blood osmotic pressure—calledthe colloid osmotic pressure (COP)—is especially impor-tant, as we see from the effects of extremely low-proteindiets (see insight 18.1).
Before You Go OnAnswer the following questions to test your understanding of thepreceding section:
1. From your body weight in kilograms, predict how manykilograms and how many liters of blood you have.
2. What are the two principal components of the blood?3. What percentage of the blood is composed of erythrocytes?
What is the term for this percentage?4. Why is blood viscosity important? What are the main factors
that contribute to blood viscosity?5. Why is blood osmolarity important? What are the main factors
that contribute to blood osmolarity?
Insight 18.1 Clinical Application
Starvation and Plasma Protein DeficiencySeveral conditions can lead to hypoproteinemia, a deficiency of plasmaprotein: extreme starvation or dietary protein deficiency, liver diseasesthat interfere with protein synthesis, kidney diseases that result in pro-tein loss through the urine, and severe burns that result in protein lossthrough the body surface. As the protein content of the blood plasmadrops, so does its osmolarity. The bloodstream loses more fluid to thetissues than it reabsorbs by osmosis. Thus, the tissues become edema-tous and a pool of fluid may accumulate in the abdominal cavity—acondition called ascites (ah-SY-teez).
Children who suffer severe dietary protein deficiencies often exhibita condition called kwashiorkor (KWASH-ee-OR-cor) (fig. 18.3). Thearms and legs are emaciated for lack of muscle, the skin is shiny andtight with edema, and the abdomen is swollen by ascites. Kwashiorkoris an African word for a “deposed” or “displaced” child who is no longerbreast-fed. Symptoms appear when a child is weaned and placed on adiet consisting mainly of rice or other cereals. Children with kwashior-kor often die of diarrhea and dehydration.
682 Part Four Regulation and Maintenance
Formedelements
Plasma(55% of whole blood)
Buffy coat: leukocytes and platelets(<1% of whole blood)
Erythrocytes(45% of whole blood)
Centrifuge
Withdrawblood
Figure 18.2 The Hematocrit. A small sample of blood is taken in aglass tube and spun in a centrifuge to separate the cells from the plasma.The percent volume of red cells (hematocrit) is then measured. In thisexample, the hematocrit is 45%.
Table 18.2 General Properties of Blood*
Mean Fraction of Body Weight 8%
Volume in Adult Body Female: 4–5 L; male: 5–6 L
Volume/Body Weight 80–85 mL/kg
Mean Temperature 38°C (100.4°F)
pH 7.35–7.45
Viscosity (relative to water) Whole blood: 4.5–5.5; plasma: 2.0
Osmolarity 280–296 mOsm/L
Mean Salinity (mainly NaCl) 0.9%
Hematocrit (packed cell Female: 37%–48% volume) male: 45%–52%
Hemoglobin Female: 12–16 g/dL male: 13–18 g/dL
Mean RBC Count Female: 4.2–5.4 million/!Lmale: 4.6–6.2 million/!L
Platelet Count 130,000–360,000/!L
Total WBC Count 5,000–10,000/!L
*Values vary slightly depending on the testing methods used.Hematocrito. Por medio de este sencillo procedimiento (que se puede realizar en la clínica) podemos compro-bar el hematocrito. En hombres tiene un valor del 45% aprox.

Plasma
El plasma es un fluido coloidal amarillo claro (por la bilirrubina) y opalescente (por las gra-sas) con múltiples componentes que pueden ser clasificados en elementos inorgánicos y en elementos orgánicos.
- Elementos inorgánicos. En esta categoría encontramos principalmente el agua (son 3,5 l y representan el 90% del total), electrólitos y minerales.
- Elementos orgánicos. En esta categoría encontramos multitud de elementos que serán discutidos más adelante: proteínas plasmáticas, lípidos, carbohidratos, hormonas, ga-ses, productos del metabolismo (bilirrubina), enzimas y proteínas de transporte.
Proteínas
Disponemos de unos 7 g de proteínas por dl de sangre. Mayormente se forman en el hígado aunque hay otras, como las inmunoglobulinas, que se forman en otros compartimentos cor-porales. Con un proteinograma las podemos separar en función a su tamaño, pudiendo dis-tinguir cinco bandas principales de proteínas; la más importante de éstas por cantidad es la albúmina.
Albúmina
La albúmina es una proteína de síntesis hepática y gran tamaño. Es la más importante desde el punto de vista cuantitativo (banda principal del proteinograma; 53-66%) y disponemos de 40 g/l aprox. Además de su función transportadora, que se discute a continuación, la albúmina es responsable de 3/4 partes de la presión oncótica. La albúmina es una proteína transportado-ra que puede llevar consigo una gran cantidad de elementos:
- Elementos propios. Como por ejemplo ácidos grasos, colesterol, lipoproteínas, fosfogli-céridos, bilirrubina, Ca2+ y hormonas esteroideas (estradiol y progesterona entre otras).
- Medicamentos. Como por ejemplo antibióticos como la penicilina y otros fármacos (digoxina, salicilatos). Si ocupo la albúmina con un medicamento, aquello que está siendo transportado queda liberado.
α1 globulinas
Las α1 globulinas representan aproximadamente el 2% de las proteínas plasmáticas. Se ven aumentadas en procesos inflamatorios agudos, neoplasias, infartos y necrosis. En clase se destacaron las siguientes:
- α1 glicoproteína ácida. Es un buen marcador, aunque inespecífico, de procesos infla-matorios tanto agudos como crónicos.
- α1 antitripsina. Es inhibidor de algunas proteasas inflamatorias (sobre todo a la tripsi-na).
- α1 fetoproteína. Puede ser un marcador tumoral de hepatocarcinoma.
32

α2 globulinas
Representan el 10% aprox. de las proteínas plasmáticas. En clase se destacaron dos: la ceru-
loplasmina (encargada de transportar cobre); y la haptoglobina, que se une a la hemoglobina impidiendo que ésta pase al riñón y pueda bloquear los túbulos renales (importante cuando se den hemólisis intravasculares).
β globulinas
Las β globulina representan también un 10% aproximado del total de proteínas plasmáticas (diapositivas: 8-13%). En clase se destacaron aquellas que pertenecen al sistema del comple-
mento (aumenta en procesos inflamatorios), la transferrina (que transpora hierro) y la transco-balamina (transporta la vit. B12).
γ-globulinas
Las γ globulinas representan entre el 9 y el 17% de las proteínas plasmáticas y son formadas por los linfocitos B y las células plasmáticas. También se pueden denominar inmunoglobulinas y participan en la inmunidad específica. Existen cinco isotipos de γ globulinas:
- IgG. Es la más abundante (80%) y la que tiene más hemivida (20 días). Puede atravesar la placenta y entrar en el feto (lo que es bueno y puede ser contraproducente a la vez).
- IgA. Es la secretada por las mucosas y la piel. Tiene 4 días de hemivida, representa el 13% del total y la encontramos en el plasma, el tracto digestivo, el aparato genitouri-nario, la leche, el calostro, etc.
- IgM. Tiene seis días de hemivida y son el 6% del total.- IgE. Su número aumenta en reacciones alérgicas.- IgD. No se explicó.
Además de las que acabamos de explicar, hay una serie de proteínas cuyo aumento o dismi-nución está relacionado con patología. Por ejemplo, un aumento de la amilasa es indicador de problema pancreático, y lo mismo con el hígado en el caso de aparición de transaminasas. Las creatinfosfoquinasas son indicador de contracción muscular.
33
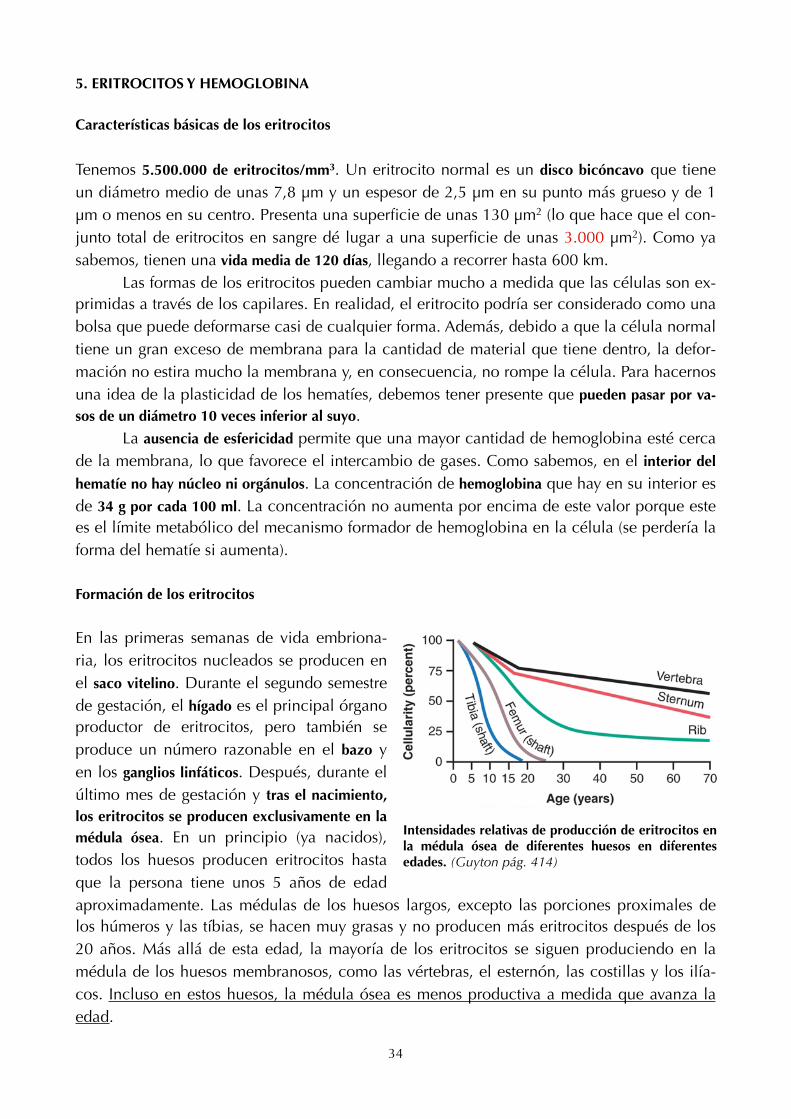
5. ERITROCITOS Y HEMOGLOBINA
Características básicas de los eritrocitos
Tenemos 5.500.000 de eritrocitos/mm3. Un eritrocito normal es un disco bicóncavo que tiene un diámetro medio de unas 7,8 µm y un espesor de 2,5 µm en su punto más grueso y de 1 µm o menos en su centro. Presenta una superficie de unas 130 µm2 (lo que hace que el con-junto total de eritrocitos en sangre dé lugar a una superficie de unas 3.000 µm2). Como ya sabemos, tienen una vida media de 120 días, llegando a recorrer hasta 600 km. Las formas de los eritrocitos pueden cambiar mucho a medida que las células son ex-primidas a través de los capilares. En realidad, el eritrocito podría ser considerado como una bolsa que puede deformarse casi de cualquier forma. Además, debido a que la célula normal tiene un gran exceso de membrana para la cantidad de material que tiene dentro, la defor-mación no estira mucho la membrana y, en consecuencia, no rompe la célula. Para hacernos una idea de la plasticidad de los hematíes, debemos tener presente que pueden pasar por va-sos de un diámetro 10 veces inferior al suyo. La ausencia de esfericidad permite que una mayor cantidad de hemoglobina esté cerca de la membrana, lo que favorece el intercambio de gases. Como sabemos, en el interior del
hematíe no hay núcleo ni orgánulos. La concentración de hemoglobina que hay en su interior es de 34 g por cada 100 ml. La concentración no aumenta por encima de este valor porque este es el límite metabólico del mecanismo formador de hemoglobina en la célula (se perdería la forma del hematíe si aumenta).
Formación de los eritrocitos
En las primeras semanas de vida embriona-ria, los eritrocitos nucleados se producen en el saco vitelino. Durante el segundo semestre de gestación, el hígado es el principal órgano productor de eritrocitos, pero también se produce un número razonable en el bazo y en los ganglios linfáticos. Después, durante el último mes de gestación y tras el nacimiento, los eritrocitos se producen exclusivamente en la médula ósea. En un principio (ya nacidos), todos los huesos producen eritrocitos hasta que la persona tiene unos 5 años de edad aproximadamente. Las médulas de los huesos largos, excepto las porciones proximales de los húmeros y las tíbias, se hacen muy grasas y no producen más eritrocitos después de los 20 años. Más allá de esta edad, la mayoría de los eritrocitos se siguen produciendo en la médula de los huesos membranosos, como las vértebras, el esternón, las costillas y los ilía-cos. Incluso en estos huesos, la médula ósea es menos productiva a medida que avanza la edad.
34
Intensidades relativas de producción de eritrocitos en la médula ósea de diferentes huesos en diferentes edades. (Guyton pág. 414)

Eritropoyesis
Por eritropoyesis entendemos el proceso de formación de eritrocitos en la médula ósea. Todas las células sanguíneas comienzan sus vidas en la médula ósea a partir de un solo tipo de célula llamado célula precursora hematopoyética pluripotencial (PHSC), de la cual derivan todas las células de la sangre. Esta célula, al dividirse, dará lugar a dos tipos celulares: (a) una célula
exactamente igual a la original: para así mantener un pool de células madre pluripotenciales (el nº se va reduciendo con la edad); y (b) una célula precursora/colonia formadora18, las cuales presentan una menor capacidad de diferenciación (están comprometidas) y están destinadas a formar uno o varios tipos celulares únicamente.
Los hematíes tardan unos 5 días en madurar. Se renuevan un 0,8% cada día, lo que viene a ser renovar 160·106 eritrocitos/min. La regulación de la eritropoyesis está determinada por toda una serie de moléculas que serán discutidas más adelante, ahora hablaremos de los pa-sos desde el precursor del eritrocito hasta el mismo eritrocito. La primera célula que puede identificarse como perteneciente a la serie eritrocítica es el proeritroblasto. Bajo el estímulo adecuado, se forman grandes número de estas células a partir de las células precursoras CFU-E.
35
18 Por colonia formadora en realidad nos estamos refiriendo a unidad formadora de colonias (CFU). Si nosotros hacemos crecer estas CFUs en un cultivo, darán lugar a un tipo celular que le dará el apellido a la CFU; p. ej., unidad formadora de colonias de eritrocitos (CFU-E).
Formación de las diferentes células sanguíneas a partir de la PHSC. (Guyton pág. 414). Estrictamente, las PHSC pueden dar lugar a tres tipos celulares: (a) uno igual al original (otra PHSC para mantener el pool); (b) el precursor linfoide (que da lugar a los linfocitos B y T); (c) la CFU-S (unidad formadora de colonias esplénicas). Será la CFU-S la que pasará por un proceso de diferenciación para llegar a CFU-B (de blastos) y de ahí a la CFU-E (de eritrocitos).

Una vez se ha formado el proeritroblasto, se divide múltiples ve-ces formando finalmente muchos eritrocitos maduros. Las células de primera generación se llaman eritroblastos basófilos porque se tiñen con colorantes básicos; la célula ha acumulado en este momento muy poca hemoglobina. En las generaciones siguientes, las células se llenan de hemoglobina hasta una concentración del
34%, el núcleo se condensa hasta un tamaño pequeño y su resto final se absorbe o expulsa de la célula. Al mismo tiempo se reab-sorbe el RER. La célula en este estadio se llama reticulocito porque todavía contiene una pequeña cantidad de material basófilo (mitocondrias, restos del Golgi y algunos orgánulos citoplasmáti-cos). Durante el estadio de reticulocito, la célula pasa de la médula ósea a los capilares sanguíneos mediante diapédesis. El material basófilo restante en el reticulocito desaparece normalmente en 1-2 días, y la célula es después un eritrocito ma-
duro. Debido a la corta vida de los reticulocitos, su concentración entre los eritrocitos sanguíneos es normalmente algo menor del 1%. Si la concentración de reticulocitos es mayor, eso indica que se ha acelerado la formación de hematíes (por ejemplo, tras una anemia). En clase se hizo algún apunte sobre las diferentes anemias: (a) anemia hipocrómica microcítica (hematíes más pequeños y páli-dos porque contienen una menor cantidad de hemoglobina); (b) falciforme; (c) megaloblástica; y (d) eritoblastosis fetal (se comentará más adelante).
Regulación de la eritropoyesis
La eritropoyesis está regulada principalmente por la eritropoyetina (EPO). Además, hay toda una serie de moléculas que también regulan: IL-3, andrógenos, insulina y hormonas tiroi-deas. Es importante que se regule la cantidad de eritrocitos que se forman para que: (a) haya el número adecuado de eritrocitos que transporten el suficiente O2 desde los pulmones hasta los tejidos; (b) no sean tan numerosos como para impedir el flujo sanguíneo. La oxigenación tisular es el regulador más importante de la producción de eritrocitos. Hay toda una serie de factores que disminuyen esta oxigenación: volumen sanguíneo bajo, anemia, hemoglobina baja, mal flujo sanguíneo y enfermedades pulmonares. La hipoxia del tejido renal19 conduce a la formación del factor 1 inducible por hipoxia (HIF), el cual actúa como un factor de transcripción de genes inducibles por hipoxia, entre ellos el gen de la eri-
tropoyetina. Hasta el 90% de la EPO se forma en el riñón, el 10% restante se forma en el hí-gado. El efecto de la EPO es estimular la producción de proeritroblastos a partir de las céls. pre-
cursoras hematopoyéticas de la méd. ósea. Además, una vez se forman los proeritroblastos, la
36
19 La hipoxia de otras partes del cuerpo también puede estimular la liberación de EPO en el riñón.
Eritropoyesis. En la etapa del eritroblasto policromatófilo se produce hasta el 80% de la hemoglobina. En el si-guiente paso (ortocromático) se reabsorbe el núcleo y las mitocondrias.

EPO hace que estas células pasen con mayor rapidez de lo normal a través de los diferentes estadios eritro-blásticos, lo que acelera la producción de nuevos eri-trocitos.
Factores necesarios para la eritropoyesis
Existen una serie de elementos que son necesarios pa-ra que se formen los eritrocitos: (a) hierro; (b) vitamina
B12; y (c) ácido fólico.(a) Hierro. Sin él no se puede formar la hemo-
globina.(b) Vitamina B12 y ácido fólico. Debido a la necesidad continua de reponer los eritrocitos,
las células eritropoyéticas de la médula ósea se encuentran entre las céls. de todo el organismo que más rápidamente crecen y se reproducen. Su maduración y su pro-ducción están influidas mucho por el estado nutricional de la persona. Tanto la vita-
mina B12 como el ácido fólico son esenciales para la síntesis del DNA, ya que cada una de estas dos vitaminas es necesaria para la formación de trifosfato de timidina. La falta de una de estas dos vitaminas da lugar a un DNA anormal o reducido y, en consecuencia, que no se produzcan la maduración y la división celular. Además, las céls. eritroblásticas de la médula ósea, además de no proliferar con rapidez, produ-cen sobre todo eritrocitos mayores de lo normal llamados macrocitos, y la propia cé-lula tienen una membrana frágil, a menudo irregular, grande y oval en lugar del disco bicóncavo habitual. Estas céls. mal formadas, tras entrar en la circulación, son capa-ces de transportar oxígeno normalmente, pero su fragilidad les acorta la vida a la mitad o a un tercio de lo normal.
Ciclo vital de los hematíes (véase: metabolismo de los hematíes)
Como ya hemos dicho antes, los hematíes, desde el momento en que salen de la médula ósea, viven unos 120 días antes de ser destriuidos. Pese a que los eritrocitos maduros no dis-pongan de un núcleo, ni mitocondrias, ni ribosomas, ni RER, dispone de enzimas citoplasmá-ticos capaces de metabolizar la glucosa y formar pequeñas cantidades de ATP. Estos enzimas, además, son capaces de:
- Mantener la flexibilidad de la membrana.- Mantener el transporte de los iones en la membrana.- Mentener el hierro de la hemoglobina en la forma ferrosa en lugar de la férrica.- Impide la oxidación de las proteínas en los eritrocitos.
37
Función del mecanismo eritropoyético para aumentar la pro-ducción de eritrocitos cuando se reduce la oxigenación tisular. (Guyton pág. 416) En el esquema falta el factor 1 inducible por hipoxia (HIF), que se expresaría en el riñón para incrementar la formación de EPO.

Una vez la membrana del eritrocito se hace frágil, la célula se rompe durante el paso a través de puntos rígidos de la circulación. Muchos de los eritrocitos se destruyen en el bazo. El bazo es una víscera protegida (aunque frágil ante golpes directos)20 que se caracteriza por ser un re-
servorio de eritrocitos y por su función de limpieza de la sangre. El bazo dispone de dos áreas independientes para almacenar la sangre: los senos venosos y la pul-
pa. Los senos pueden ingurgitarse igual que cual-quier otra parte del sistema venoso y almacenar la sangre total. En la pulpa los capilares son tan per-meables que la sangre total, incluidos los eritrocitos, rezuma a través de las paredes de los capilares hacia la malla trabecular, formando la pulpa roja. Los eri-trocitos quedan atrapados en esta malla (reservorio
de eritrocitos) y el plasma fluye hacia la circulación general. Si el SN simpático excita el bazo y hace que el bazo y los vasos se contraigan, se puede libe-rar semejante cantidad de eritrocitos que hagan aumentar el hematocrito hasta en un 2%. En cuanto a la función del bazo relacionada con la limpieza de la sangre, ésta se basa en dos elementos:
- Eliminación de céls. viejas. Las céls. sanguíneas que atraviesan la pulpa esplénica son cuidadosamente exprimidas, por lo que se puede esperar que los eritrocitos frágiles no superen este traumatismo. Por tal motivo, muchos de los eritrocitos destruidos en el organismo encuentran su destino final en el bazo. Después de la rotura de las célu-las la hemoglobina liberadas y el estroma celular son digeridos por las células reticu-
loendoteliales del bazo y los productos de la digestión son reutilizados en su mayor parte en en el organismo como nutrientes, a menudo para elaborar céls. sanguíneas nuevas.
- Células reticuloendoteliales en el bazo [macrófagos tisulares]. Estas céls., que están en los senos venosos, funcionan dentro del sistema de limpieza de la sangre, actuando en concierto con un sistema similar de céls. reticuloendoteliales de los senos venosos del
hígado. Cuando la sangre está invadida por microorganismos infecciosos las céls. de este sistema eliminan rápidamente los restos, las bacterias, parásistos, etc. Además, el bazo incrementa su tamaño en muchos procesos infecciosos crónicos, de la misma forma en que los ganglios linfáticos aumentan de tamaño.
Hemocateresis
La destrucción de los hematíes es un proceso que se conoce con el nombre de hemocateresis. Este proceso puede producirse de manera extravascular (90%) o intravascular (10%). Empeza-remos hablando de la hemocateresis mayoritaria: la extravascular o celular.
38
20 Una pequeña fisura puede derivar en una hemorragia interna y se deberá retirar el bazo entero (esplenecto-mía). La retirada del bazo hace que en la sangre haya una mayor presencia de hematíes de mala calidad (p. ej., con cuerpos de Heinz, siderocitos, falciformes, etc.
Estructuras funcionales del bazo.

La destrucción extravascular consiste en la fagocitosis de los hematíes viejos por parte de los macrófagos, ubicados por fuera del vaso sanguíneo. Existen toda una serie de órganos donde la presencia de los macrófagos es moderada: médula ósea, bazo e hígado. A menudo el bazo recibe el calificativo de cementerio de los hematíes. Recordemos que el bazo dispone de determinados puntos en los que se “prueba” la elasticidad del hematíe, de tal manera que un eritrocito que haya alcanzado su vida útil se fragmentará. Una vez el macrófago ha incorpo-rado al hematíe, éste quedará incluido en una vacuola en la que actuarán diferentes enzimas que degradarán una parte importante de los elementos que conforman al hematíe. En refe-rencia a la hemoglobina, ésta se descompondrá en las globinas, que darán lugar a aas que pueden ser reutilizados, y en el grupo hemo. A su vez, el grupo hemo sufre las siguientes transformaciones:
- Hierro. El hierro será liberado a la sangre (a través de la ferroportina) y se unirá a la transferrina para ser transportado. Otra opción es que se quede en el macrófago.
- Cadena recta de cuatro anillos pirrólicos. La parte no férrica del grupo hemo (es decir, la mayoría) será transformada a biliverdina por parte de un enzima que tiene el macrófa-go, la hemo-oxigenasa (A). A continuación, también en el macrófago, la biliverdina pa-sa a bilirrubina indirecta (o no conjugada) gracias a la acción de la biliverdina-reductasa (B). Finalmente, la bilirrubina no conjugada formada será liberada y se conjugará con la albúmina.
- CO2. Hay una cierta producción endógena de dióxido de carbono.
Destrucción intravascular
La destrucción intravascular es aquella que se produce en el seno del hematíe. La lisis de la membrana del mismo hace que todo su interior pase al plasma, por lo que la hemoglobina quedará expuesta en el plasma. A esta hemoglobina le pueden suceder dos cosas:
- Haptoglobina. Si hay la suficiente haptoglobina, toda la hemoglobina formada quedará unida a ésta y así será transportada hacia el hígado. El complejo Hb-haptoglobina de-saparece del plasma en unos 9-30 min. La hemoglobina pasará por un proceso idénti-
co al que se observa en los macrófagos. Como la haptoglobina empleada no será de-vuelta al plasma, un nivel elevado de destrucción intravascular implica una disminu-ción de la cantidad de haptoglobina en plasma.
- Circulación libre. Si no hay suficiente haptoglobina, la hemoglobina tendrá que circu-lar por el plasma sin ser acompañada por una proteína de transporte. A partir de aquí hay tres posibilidades:
- Hígado. La hemoglobina libre llega sin ningún problema al hígado y allí es degradada tal y como hemos visto en la hemocateresis extravascular.
- Oxidación. La hemoglobina puede oxidarse a metahemoglobina (Fe3+) y enton-ces disociarse en el poco soluble (a pH fisiológico) grupo hemo y en globina. El grupo hemo se unirá a la hemopexina o a la albúmina para así poder ser trans-portado hasta el hígado (donde se metabolizará).
39
Anillo pirrólico A⎯ →⎯ Biliverdina B⎯ →⎯ Bilirrubina indirecta

- Disociación. La hemoglobina se disocia en un dímero α y β otro. Estos dos dí-meros pueden atravesar la cápsula de Bowmann y ser filtrados para después ser reabsorbidos por los propios glomérulos renales. Las céls. del epitelio de los túbulos renales almacernarán estos dímeros en forma de ferritina y hemo-
siderina.
Si la ferritina y la hemosiderina21 almacenadas en las céls. del epitelio de los túbulos renales no es movilizada, tarde o temprano aparecerán en la orina por la propia renovación del epi-telio (descamación de las células antiguas). Si la capacidad de absorción de hemoglobina queda limitada por un exceso de ésta, aparecerá en el orina.
Hemoglobina
A lo largo de la explicación acerca de la hemo-cateresis hemos estado hablando de la hemoglo-bina. Puede que lo recordemos del año pasado, pero no está de más repasarla un poco. Cada eri-trocito contiene unos 280 millones de moléculas de hemoglobina. La hemoglobina consiste en cuatro cadenas proteicas llamadas globinas (2 ca-denas α de 141 aas y 2 cadenas β de 146 aas) conjugadas cada una de ellas con un grupo hemo. El grupo hemo está formado por un anillo orgá-nico complejo, la protoporfirina, a la que se le une un átomo de hierro en estado ferroso (Fe2+). El átomo de hierro puede unirse al O2. Si cada grupo hemo se puede unir a un O2 y cada molé-cula de hemoglobina contiene cuatro grupos hemo (uno por cada cadena de globina), una hemoglobina puede transportar 4 O2. Además, aproximadamente un 5% del CO2 que va por el torrente sanguíneo puede unirse a la hemoglobi-na, aunque acoplándose a la cadena proteica de globina y no al grupo hemo. La hemoglobina tiene diferentes formas. La que acabamos de describir es la forma adulta (hemoglobina adulta o HbA)22. El feto forma una forma específica denominada hemoglobina fetal
40
21 La hemosiderina es un derivado de la ferritina que se encuentra en el hígado, el bazo y la médula ósea, es insoluble en disoluciones acuosas y forma agregados que liberan lentamente hierro cuando hay una deficien-cia. Se considera que el hierro almacenado en la hemosiderina está menos disponible respecto a la ferritina.
22 Existe una variación: el 2,5% de la hemoglobina de un adulto es de la forma HbA2, en que las cadenas β pasan a ser δ.
Saladin: Anatomy & Physiology: The Unity of Form and Function, Third Edition
18. The Circulatory System: Blood
Text © The McGraw!Hill Companies, 2003
Cha
pter 18
called globins (fig. 18.10a). Two of these, the alpha (!)chains, are 141 amino acids long, and the other two, thebeta (") chains, are 146 amino acids long. Each chain isconjugated with a nonprotein moiety called the hemegroup (fig. 18.10b), which binds oxygen to a ferrous ion(Fe2#) at its center. Each heme can carry one molecule ofO2; thus, the hemoglobin molecule as a whole can trans-port up to 4 O2. About 5% of the CO2 in the bloodstreamis also transported by hemoglobin but is bound to the glo-bin moiety rather than to the heme. Gas transport byhemoglobin is discussed in detail in chapter 22.
Hemoglobin exists in several forms with slight dif-ferences in the globin chains. The form we have justdescribed is called adult hemoglobin (HbA). About 2.5%of an adult’s hemoglobin, however, is of a form calledHbA2, which has two delta ($) chains in place of the "chains. The fetus produces a form called fetal hemoglobin(HbF), which has two gamma (%) chains in place of the "chains. The $ and % chains are the same length as the "chains but differ in amino acid sequence. HbF binds oxy-gen more tightly than HbA does; thus it enables the fetusto extract oxygen from the mother’s bloodstream.
Insight 18.2 Evolutionary Medicine
The Packaging of HemoglobinThe gas-transport pigments of earthworms, snails, and many other ani-mals are dissolved in the plasma rather than contained in blood cells.
690 Part Four Regulation and Maintenance
Nucleus
Lysosomes
(b)
Figure 18.9 Comparison of RBCs and WBCs as Seen with theTransmission Electron Microscope. (a) RBCs in a small bloodvessel, showing their strongly biconcave shape and lack of organelles.(b) An eosinophil, a representative leukocyte, showing a nucleus, manylysosomes, and other organelles.What is the approximate width of this blood vessel?
Bloodvessel
Erythrocytes
(a)
!
!
Hemegroups
(a)
(b)
"
"
C
CH3 CH2
C
C
C
CC
C
C
CC
CC
N
N
NN
CH
CH
CH
CH
CH2
COOH
CH3 CH3
CH2
CH2
CH2
COOH
CH2 CH3
HC
C
C C
CHC
Fe2+
Figure 18.10 The Structure of Adult Hemoglobin (HbA).(a) The hemoglobin molecule consists of two ! proteins and two "proteins, each conjugated to a nonprotein heme group. (b) Structure ofthe heme group. Oxygen binds to Fe2# at the center of the heme.
HbA y grupo hemo. La protoporfirina está forma-da por cuatro anillos de cinco átomos (pirrol), por lo que se denomina también tetrapirrol. Los anillos están unidos por puentes metileno.

(HbF) que se caracteriza por tener una mayor afinidad por el oxígeno respecto a la HbA (ca-denas γ en lugar de β), lo que le permite extraer oxígeno a partir de la sangre materna.
Degradación de la hemoglobina
Una vez el hematíe es demasiado frágil, su membrana se rompe y la hemoglobina libe-rada es fagocitada por los macrófagos tisu-lares. La hemoglobina se escinde primero en globina y hemo, y el anillo hemo sirve para dar: (1) hierro libre que la transferrina transporta en la sangre; y (2) una cadena rec-
ta de cuatro núcleos pirrólicos, que constituye el sustrato final a partir del cual se forma la bilirrubina. La primera sustancia que se forma es la biliverdina (gracias a la hemo oxigenasa), aunque enseguida se reduce ha-cia la bilirrubina libre, también denominada bilirrubina no conjugada (o indirecta), que va liberándose poco a poco de los macrófagos en el plasma. Esta forma de bilirrubina se une de manera inmediata e intensa a la al-
búmina del plasma23, que la transporta por la sangre y los líquidos intersticiales. En muy pocas horas, la bilirrubina no conjugada se absorbe por la membrana del hepatocito. Al entrar se desliga de la albú-mina plasmática y muy pronto se conjuga, en un 80%, con el ácido glucorónico para formar glucoronato de bilirrubina, en un 10% con el ácido sulfúrico para formar sulfato de bilirrubina, y en un 10% final con muchas otras sustan-cias. De esta manera, la bilirrubina (ya conjugada; liposoluble) sale de los hepatocitos a tra-vés de un mecanismo de transporte activo y se excreta a los canalículos biliares y, desde aquí, hacia el intestino. Casi la mitad de la bilirrubina conjugada se transforma, una vez en el intestino y por el efecto bacteriano, en urobilinógeno (muy soluble). Parte del urobilinógeno se reabsorbe por la mucosa intestinal hacia la sangre, pero la mayoría de veces vuelve a eliminarse por el hígado hacia el intestino; cerca del 5% se elimina por los riñones con la orina. Después de la expo-sición de la orina al aire, el urobilinógeno se oxida a urobilina; si se trata de las heces, el urobilinógeno se modifica y oxida para dar la estercobilina (pasando previamente por el es-tercobilinógeno).
41
23 Dos moléculas de bilirrubina por cada albúmina.
Formación y eliminación de la bilirrubina. (Guyton pág. 841)

Metabolismo del hematíe
Durante la eritropoyesis los hematíes pierden sus mitocondrias, y con ellas la capacidad de realizar un metabolismo dependiente del oxígeno (ciclo de Krebs, fosforilación oxidativa). La única manera que tiene el hematíe para formar ATP es por medio de la glucólisis que se da en el citoplasma. El balance de 2 moles de ATP por cada mol de glucosa oxidada ya es suficiente para cubrir la actividad metabólica del hematíe (bombas Na+/K+, fosfolípidos de membrana, etc). La vía metabólica principal del hematíe será, por tanto, el ciclo de Embden-Meyerhof (glucólisis), el cual dispone de tres cortocircuitos: vía de Rapaport-Luebering, vía de la hexosa
monofosfato y vía de la metahemoglobina reductasa.
Ciclo de Embden-Meyerhof
El 90% de la energía del hematíe se obtiene a partir de este ciclo. Los cortocircuitos que se le añaden permitirán el buen funcionamiento del hematíe.
- Vía de Rapaport-Luebering. Consiste en el paso de 1,3-bifosfoglicerato a 2,3-bifosfoglice-
rato (BPG) por medio de una mutasa. Este metabolito reduce la afinidad de la hemo-globina por el oxígeno, por lo que facilitará la liberación del oxígeno hacia el tejido por el que está pasando el hematíe.
Las personas que se han adaptado a grandes altitudes (↓pO2) o son fumadoras expe-rimentan un incremento de los niveles de BPG. Este ajuste del nivel de BPG tiene un efecto pequeño sobre la unión de O2 en los pulmones pero afecta de manera impor-tante a la liberación de O2 en los tejidos. La hemoglobina fetal tiene poca afinidad por el BPG, por lo que tiene una mayor afinidad por el O2 (mejora la transferencia de O2 por la placenta).
- Vía de la hexosa monofosfato. Es la también conocida como vía de las pentosas. Ofrece el 10% de la energía restante y es fundamental porque forma dos moléculas de NADPH reducido. Este NADPH reducido sirve para reducir el glutation, que es la prin-cipal defensa frente a la oxidación de proteínas, fosfolípidos de membrana, etc. Si este sistema antioxidante no funciona correctamente, los hematíes envejecerán con una mayor rapidez.
- Vía de la metahemoglobin-reductasa. El paso de gliceraldehído 3-fosfato a 1,3-bifosfo-glicerato genera NADH, que permite la transformación de la metahemoglobina (Fe3+; forma oxidada que no puede unirse al O2) a hemoglobina (Fe2+).
El glutation también permite que la metahemoglobina se reduzca a la hemoglobina. Si hay un fallo en el mecanismo de reducción de la metahemoglobina aparecerán los corpúsculos
de Heinz (agregados de metahemoglobina). Los hematíes con estos agregados son más pro-pensos a la hemólisis (riesgo de anemia).
42
HbBPG+ O2HbO2 +BPG

43
Metabolismo del hematíe. Vemos como la glucólisis llega hasta la formación del lactato (fermentación láctica) para así recuperar las moléculas de NAD oxidado que se precisan para la glucólisis.
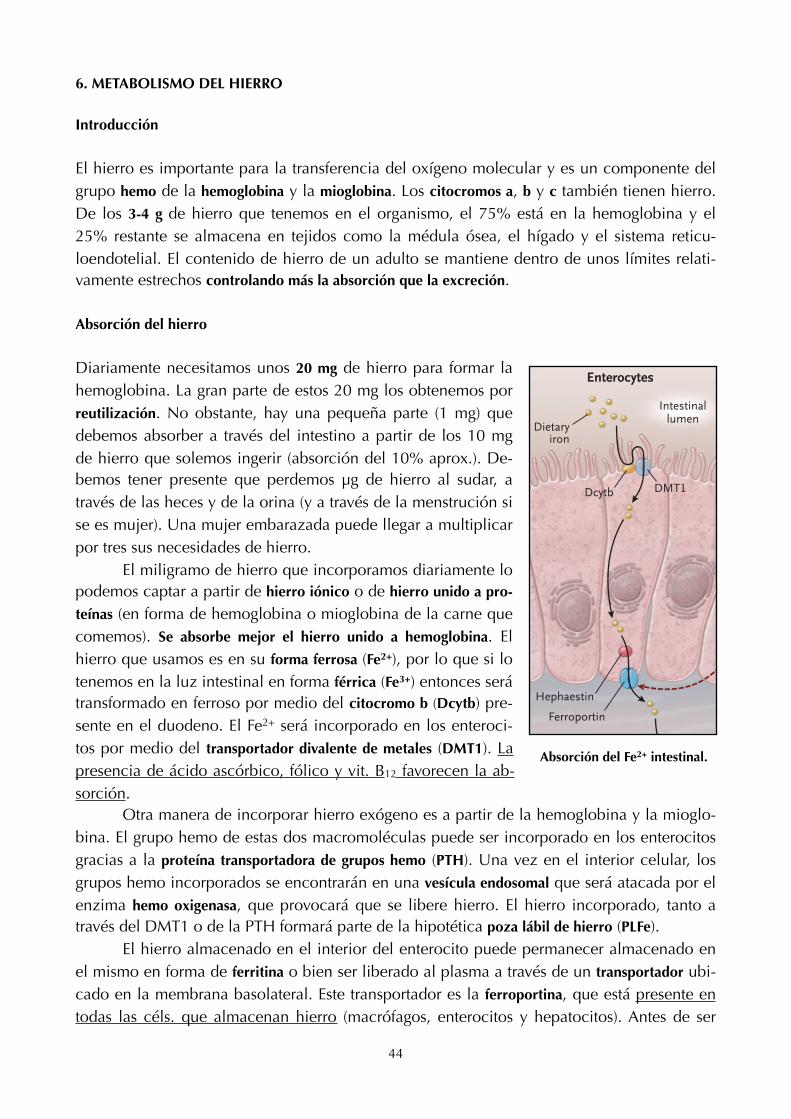
6. METABOLISMO DEL HIERRO
Introducción
El hierro es importante para la transferencia del oxígeno molecular y es un componente del grupo hemo de la hemoglobina y la mioglobina. Los citocromos a, b y c también tienen hierro. De los 3-4 g de hierro que tenemos en el organismo, el 75% está en la hemoglobina y el 25% restante se almacena en tejidos como la médula ósea, el hígado y el sistema reticu-loendotelial. El contenido de hierro de un adulto se mantiene dentro de unos límites relati-vamente estrechos controlando más la absorción que la excreción.
Absorción del hierro
Diariamente necesitamos unos 20 mg de hierro para formar la hemoglobina. La gran parte de estos 20 mg los obtenemos por reutilización. No obstante, hay una pequeña parte (1 mg) que debemos absorber a través del intestino a partir de los 10 mg de hierro que solemos ingerir (absorción del 10% aprox.). De-bemos tener presente que perdemos µg de hierro al sudar, a través de las heces y de la orina (y a través de la menstrución si se es mujer). Una mujer embarazada puede llegar a multiplicar por tres sus necesidades de hierro. El miligramo de hierro que incorporamos diariamente lo podemos captar a partir de hierro iónico o de hierro unido a pro-
teínas (en forma de hemoglobina o mioglobina de la carne que comemos). Se absorbe mejor el hierro unido a hemoglobina. El hierro que usamos es en su forma ferrosa (Fe2+), por lo que si lo tenemos en la luz intestinal en forma férrica (Fe3+) entonces será transformado en ferroso por medio del citocromo b (Dcytb) pre-sente en el duodeno. El Fe2+ será incorporado en los enteroci-tos por medio del transportador divalente de metales (DMT1). La presencia de ácido ascórbico, fólico y vit. B12 favorecen la ab-sorción. Otra manera de incorporar hierro exógeno es a partir de la hemoglobina y la mioglo-bina. El grupo hemo de estas dos macromoléculas puede ser incorporado en los enterocitos gracias a la proteína transportadora de grupos hemo (PTH). Una vez en el interior celular, los grupos hemo incorporados se encontrarán en una vesícula endosomal que será atacada por el enzima hemo oxigenasa, que provocará que se libere hierro. El hierro incorporado, tanto a través del DMT1 o de la PTH formará parte de la hipotética poza lábil de hierro (PLFe). El hierro almacenado en el interior del enterocito puede permanecer almacenado en el mismo en forma de ferritina o bien ser liberado al plasma a través de un transportador ubi-cado en la membrana basolateral. Este transportador es la ferroportina, que está presente en todas las céls. que almacenan hierro (macrófagos, enterocitos y hepatocitos). Antes de ser
44
n engl j med
352;17
www.nejm.org april
28, 2005
1742
P E R S P E C T I V E
brush border by means of a duodenal ferric reduc-tase (Dcytb). Ferrous iron is then transported acrossthe apical plasma membrane of the enterocyte bydivalent metal transporter 1 (DMT1). Iron taken upby the enterocyte may be stored intracellularly asferritin (and excreted in the feces when the senes-cent enterocyte is sloughed) or transferred acrossthe basolateral membrane to the plasma. This ironis transferred out of the enterocyte by the basolater-al transporter ferroportin — a process that is facil-itated by the ferroxidase activity of the ceruloplas-min homologue hephaestin. The expression of eachof the genes involved in these steps is subject to
regulation. Changes in the expression of each ofthe identified molecules that participate in iron ab-sorption have been examined in various conditions.The best characterized of these conditions is ane-mia caused by dietary iron deficiency, in which theup-regulation of duodenal DMT1, Dcytb, and fer-roportin messenger RNA and protein is observed.
Iron released into the circulation binds to trans-ferrin and is transported to sites of use and storage.Production of hemoglobin by the erythron accountsfor most iron use. High-level expression of trans-ferrin receptor 1 in erythroid precursors ensuresthe uptake of iron into this compartment. Hemo-
Interplay of Key Proteins in Iron Homeostasis.
In the duodenal enterocyte, dietary iron is reduced to the ferrous state by duodenal ferric reductase (Dcytb), transported into the cell by divalent metal transporter 1 (DMT1), and released by way of ferroportin into the circulation. Hephaestin facilitates enterocyte iron release. Hepatocytes take up iron from the circulation either as free iron or transferrin-bound iron (through transferrin receptor 1 and transferrin receptor 2). Transferrin receptor 2 may serve as a sensor of circulat-ing transferrin-bound iron, thereby influencing expression of the iron regulatory hormone hepcidin. The hepcidin re-sponse is also modulated by HFE and hemojuvelin. Hepcidin is secreted into the circulation, where it down-regulates the ferroportin-mediated release of iron from enterocytes, macrophages, and hepatocytes (dashed red lines).
Hemojuvelin
HFE
Dietaryiron
Transferrin
DMT1Dcytb
Hephaestin
Ferroportin
Transferrinreceptor 2
Transferrinreceptor 1
Ferroportin Ferroportin
Transferrin-boundiron
Erythrocyte
Macrophage
Hepatocyte
Enterocytes
Hepcidin
Hepcidin
Intestinallumen
Hepcidin
Orchestration of Iron Homeostasis
The New England Journal of Medicine Downloaded from nejm.org at CRAI UNIVERSITAT DE BARCELONA on March 4, 2012. For personal use only. No other uses without permission.
Copyright © 2005 Massachusetts Medical Society. All rights reserved.
Absorción del Fe2+ intestinal.

transportado al plasma, el hierro ferroso volveráa su for-ma férrica (Fe3+) por medio de la hefastina. En el plasma, el hierro férrico será tranportado por la transferrina24, que lo llevará o bien a tejidos que lo precisen o donde se pueda almacenar. La transferrina se satura con un 30% del metal y cuando toda la transferrina plasmática está ocupada, el hierro no tendrá más remedio que permane-cer en el enterocito (con el riesgo de que se pierda cuan-do éste se descame). Para que el hierro que transporta la transferrina pueda ser liberado, tendrá que haber una in-teracción entre la transferrina y su receptor (receptor de
transferrina). Estos receptores los podemos encontrar en las céls. precursoras de los eritrocitos (eritroblasto basófi-
lo)25 o en los hepatocitos.
Hierro libre. El hierro libre es tóxico (puede formar radicales libres),
por lo que debe ser siempre transportado por una proteína. Las cifras extremas de hierro pueden ocasionar trastornos en el miocardio, y
aunque existen sistemas celulares antioxidantes, puede ser que las propiedades altamente oxidativas del hierro lleguen a generar lesio-nes.
Hepcidina
El hepatocito dispone de unos receptores de transferrina que le sirven, además de que para la transferrina le dé el hierro, para conocer la cantidad de transferrina plasmática ocupada por
hierro. Si se detecta que hay una gran cantidad de transferrina ocupada, los hepatocitos libe-ran un péptido hormonal denominado hepcidina. Una vez liberada al torrente sanguíneo, la hepcidina tendrá los siguientes efectos:
- Ferroportina. Degrada la ferroportina por lo que evita que el hierro sea liberado a par-tir de los hepatocitos, los enterocitos y los macrófagos.
- DMT1. Disminuye la actividad del transportador divalente de metales.- DCytb. Impide que el Fe3+ pase a Fe2+ en el duodeno.
En suma, la hepcidina lo que impide es una absorción del hierro. La eritropoyesis tiene el efecto contrario: impide que se forme hepcidina en los hepatocitos y por tanto se favorece la absorción de hierro. Otros factores, como por ejemplo una hipoxia tisular por anemia, pue-den también disminuir la producción de hepcidina.
45
24 La transferrina es considerada como el compartimento más activo, ya que su hierro es por lo general reem-plazado o recambiado al menos 10 veces cada 24h.
25 Recordemos que en esta etapa se forma la hemoglobina.
Ferritina. La ferritina es la principal pro-teína de almacenamiento de hierro que se encuentra en la mayoría de céls. del organismo. Actúa como la reserva de hierro en el hígado, bazo y en la médu-la ósea. La concentración de ferritina en plasma es proporcional a la cantidad de hierro almacenado; por ello la medi-ción de ferritina en plasma es uno de los mejores indicadores de deficiencia de hierro.

7. GRUPOS SANGUÍNEOS
Introducción
Cuando se intentaba por primera vez realizar una transfusión de sangre de una persona a otra, ocurría a menudo la aglutinación inmediata o tardía y la hemólisis de los eritrocitos de la sangre, lo que daba como resultado reacciones transfusionales típicas que llevan frecuen-temente a la muerte. Pronto se descubrió que la sangre de personas diferentes tiene antíge-nos y propiedades inmunitarias diferentes, por lo que los anticuerpos del plasma de un tipo de sangre reaccionarán con los antígenos que hay en las superficie de los eritrocitos de otro tipo sanguíneo. Existen más de 30 familias de antígenos en la membrana de los hematíes. Desde el punto de vista clínico, los que más se usan son el grupo ABO y el grupo Rh.
Grupos sanguíneos ABO
En la superficie de los hematíes encontramos dos antígenos (tipo A y tipo B), los cuales tam-bién pueden recibir el nombre de aglutinógenos (puesto que son los responsables de la mayo-ría de las reacciones transfusionales sanguíneas). Debido a la forma en que se heredan estos aglutinógenos, es posible que las personas no tengan ninguno de ellos en sus células, tengan uno o tengan ambos a la vez. Existen seis genotipos que dan lugar a cuatro fenotipos distin-tos.
Los grupos sanguíneos también pueden ser representados de la siguiente manera: O = OAB (no expresa antígeno y tiene anticuerpos anti-A y anti-B); A = AB; B = BA; AB = AB. A modo de ejemplo, si yo añado anticuerpos (aglutininas) anti-A a una muestra de sangre del tipo A, como tiene aglutinógenos A (o antígeno A), la muestra se aglutinará. Inmediatamente después edel nacimiento, la cantidad de aglutininas en el plasma es casi nula. De 2 a 8 meses después del nacimiento, el niño empieza a formar aglutininas anti-X (donde X es el aglutinógeno que no presenta). La concentración máxima de aglutininas se al-canza a los 10 años de edad (a partir de ese momento empieza a declinar). La formación de estas aglutininas se realiza gracias a las pequeñas cantidades de antígenos de los tipos A y B que entran en el cuerpo a través de la comida, las bacterias y otras formas. Las aglutininas pueden ser del tipo IgG, con dos puntos de anclaje, o del tipo IgM, con hasta 10 puntos de anclaje (la aglutinación será mayor con las IgM).
46
Genotipos Tipos sanguíneos Aglutinógenos Aglutininas
OO O - Anti-A y anti-B
OA o AA A A Anti-B
OB o BB B B Anti-A
AB AB A y B -

Nota. Si alguien del grupo O le da sangre a una persona del grupo A, el plasma del donante contiene una pe-
queña cantidad de aglutininas anti-A que producen una hemólisis reducida (podría apreciarse un pequeño au-mento de la bilirrubina).
Tipos sanguíneos Rh
Existen seis tipos frecuentes de antígenos Rh, cada uno llamado factor Rh. Estos tipos se de-signan C, D, E, c, d, y e. El que tiene una mayor capacidad para desencadenar una reacción antigénica es el D, que se transmite de manera dominante (D > d). Por tanto, únicamente si presentas los dos “d” (dd), serás Rh negativo (Rh-). Si presenta el antígeno D entonces habla-mos de Rh positivo (Rh+). En la raza europea, el 85% de las personas son Rh+, mientras que en los africanos de raza negra esta cantidad relativa asciende hasta el 100%. Mientras que el sistema ABO hay una serie de aglutininas que se forman de manera espontánea, en el sistema Rh únicamente se forman tras exponer a la persona de forma muy intensa a un antígeno Rh (por ejemplo a través de un transfusión). En otras palabras, por na-turaleza, una persona Rh- no dispondrá de anticuerpos anti-Rh. Una vez se produce una ex-posición intensa, se da una sensibilización por la cual el individuo adquiere los anticuerpos anti-Rh.
Eritroblastosis fetal
La eritoblastosis fetal es una enfermedad del feto y de los niños recién nacidos caracterizada por la aglutinación y la fagocitosis de los eritrocitos del feto. En la mayoría de los casos de eritroblastosis fetal, la madre es Rh- y el padre Rh+. El bebé hereda el antígeno Rh+ del pa-dre y la madre produce aglutininas anti-Rh por la exposición al antígeno Rh del feto. Estas agluti-ninas son IgG, por lo que pueden difundir (usan un receptor) a través de la placenta hasta el feto y aglutinar los eritrocitos. Con el primer hijo no suele haber problema debido a que no se forma el número sufi-ciente de aglutininas. El problema vendría cuando viene el segundo (o un tercero). En el caso de que se dé la enfermedad, la destrucción masiva de eritrocitos produce una gran cantidad de bilirrubina26 que intenta ser procesada por el hígado (se da ictericia). Además, la hemólisis
intravascular que se está dando hará que el feto produzca un mayor número de hematíes, in-crementándose el tamaño del hígado y el bazo (hepato-/esplenomegalia). La formación ma-siva de hematíes hace que éstos se liberen en forma de eritroblastos (aún con orgánulos). La única manera que hay para tratar esta enfermedad es reemplazar la sangre del re-cién nacido con sangre Rh- (esto se puede hacer en el mismo útero). Con esto se intenta evi-tar la anemia y, sobre todo, los niveles altos de bilirrubina (riesgo de daño neurológico). Un modo de prevenir la eritroblastosis fetal es la administración, a la madre Rh-, de un anticuerpo anti-D (el antígeno D es el principal responsable de la inmuninización de una madre Rh- a un feto Rh+). Estos anticuerpos inhiben la producción del antígeno inductor del anticuerpo linfocito B en la madre embarazada. Además, también se une a los sitios antigé-nicos D de los eritrocitos fetales Rh+ que pueden atravesar la placenta.
47
26 Previamente se forma una gran cantidad de hemoglobina que intenta ser compensada con la haptoglobina.

Grupo ABO [molecularmente]
El sistema de grupos sanguíneos humano (ABO) viene determinado por unos gan-gliósidos y, más concretamente, por los azúcares que tienen acoplados. Estos gan-gliósidos (glucoesfingolípidos) se encuentran en la membrana (parte externa) de nuestras células sanguíneas, de modo que los glú-cidos dan al exterior. Por tanto, el sistema inmunitario puede reconocer si dicha célu-la pertenece o no a nuestro organismo. Si nos fijamos el grupo O presenta la estruc-tura básica a partir de la cual se formará A y B; es por ello que se dice que O es el donante universal. El grupo AB presenta una mezcla de ambas formas. A nivel ge-nético una persona de grupo A lo que tiene es una enzima que le permite acoplar una GalNAc al gangliósido básico (el caracte-rístico del grupo O). El grupo O lo único que tiene son las enzimas necesarias para acoplar a la ceramida el resto de glúcidos y así conformar el gangliósido característico.
48

8. HEMOSTASIA
Introducción
La hemostasia es el mecanismo mediante el cual se mantiene la sangre fluida. En el caso de que exista un daño en un vaso sangúneo, se producirá una reparación rápida, potente y que se circunscriba al territorio lesionado. En la hemostasia van a participar:
- Sangre. Con sus diferentes elementos celulares y aquellos elementos disueltos.- Endotelio. El endotelio del vaso será el principal factor (cuando no está lesionado) que
mantiene fluida la sangre. Al igual que será uno de los principales desencadenantes de los procesos de coagulación cuando se daña.
- Flujo sanguíneo.
Por un lado tenemos la formación del coágulo y por el otro la fibrinólisis (términos antagóni-cos). En cuanto a la formación del cóagulo, ésta se separa en dos grandes etapas: (a) hemos-
tasia primaria (o vascular-plaquetaria), y la (b) hemostasia secundaria (o plasmática). Para que se forme un coágulo habrá una adhesión, activación y agregación plaquetarias, y después la formación de la trombina, la fibrina y el coágulo.
Hemostasia primaria
En la hemostasia primaria participan dos elementos: el propio vaso sanguíneo y las plaque-tas. A continuación se van a tratar cada uno de ellos con cierto detalle.
Vaso sanguíneo
Si se produce una lesión importante en un vaso, éste responderá con una vasoconstricción. Generalmente, este mecanismo va a ser poco relevante a excepción de algunas situaciones concretas y especialmente graves; p. ej., la pérdida del miembro superior producirá la vaso-constricción de la arteria braquial que permitirá que el individuo desmembrado viva unas horas (sin este mecanismo se desangraría en minutos). La vasoconstricción será más efectiva
cuanto menos homogéneo (o limpio) haya sido el corte. Pensemos que los restos de adventicia (y demás capas del vaso) que quedan tras romper un vaso violentamente pueden ayudar a taponar el vaso. Un fenómeno curioso es la compresión hística: si un vaso se rompe y la sangre que se pierde no va a la superficie, la sangre que se ha extravasado produce la suficiente presión como para que se detenga el sangrado.
Plaquetas
Excepto en determinadas circunstancias, el elemento más importante en la hemostasia pri-maria son las plaquetas. Las plaquetas (o trombocitos) son unos elementos formes anucleados que provienen del megacariocito y que tienen forma de disco (ø = 1-4 µm). En condiciones
49

normales hay 300.000/mm3 y de éstas unas 2/3 partes están circulando y el 1/3 restante está secuestrado en el bazo. La estructura de la plaqueta va a ser importante para comprender su funcionamiento, por lo que a continuación se describen algunos de sus elementos:
- Membrana plasmática. Como en las otras células, la membrana plasmática de las pla-quetas es una bicapa lipídica en la que hallamos colesterol, glucolípidos, glucoproteí-nas y fosfolípidos. Algunos fosfolípidos. como la fosfatidilserina, tienen una importante función procoagulante y en condiciones de reposo estarán en la monocapa interna. Las glucoproteínas tienen una función de receptor (se evita la adherencia al endotelio sano y la adhesión a las zonas dañadas).
- Glucocáliz. El glucocáliz está más allá de la membrana plasmática y está constituido por glucoproteínas, mucopolisacáridos y una carga negativa conferida por el ácido siá-
lico (Neu5Ac). Se comentó que habían proteínas que quedaban absorbidas por el plasma.
- Sistema canalicular abierto. La membrana de la plaqueta se invagina para formar una serie de vesículas y canales que se aproximan a los gránulos y los comunican con el medio extracelular.
- Sistema canalicular cerrado. Se trata del sistema reticuloendotelial (el RE) en el cual habrá una alta concentración de Ca2+ que será liberado cuando la plaqueta se active en el marco del proceso de coagulación.
- Citoesqueleto. Está constituido por pequeños fragmentos de actina que, al fosforilarse, modificarán la conformación espacial de la plaqueta. Además de actina, el citoesque-leto del trombocito contiene microtúbulos que mantienen su forma cuando la plaque-ta está en reposo.
- Gránulos alfa (α). Son los más numerosos y los que tienen una mayor variedad de sus-tancias: fibrinógeno, fibrinonectina, factor de von Willebrand y factores plaquetarios como el XII y el XIII.
- Gránulos densos. Presentan una gran cantidad de Ca2+ (importante para el proceso de activación de la plaqueta), ATP (mitocondrias), ADP y serotonina. Tanto el ADP como la serotonina son los responsables de amplificar el proceso de activación plaquetaria.
Mecanismo de acción de las plaquetas
Cuando se produce una discontinuidad en la pared endotelial de un vaso sanguíneo, ésta expone colágeno o factor de von Willebrand (vWF), que contacta con el complejo GP Ib/VIX presente en la membrana de las plaquetas. En este momento ya se ha producido una unión entre elementos subendoteliales con las plaquetas. La unión entre el complejo y el colágeno induce la expresión de la glucoproteína VI, que a su vez activa a la integrina α2β1, que da es-tabilidad a la unión que se acaba de producir.
Por tanto, acabamos de ver la primera etapa de la acción plaquetaria: la adhesión. A conti-nuación tocan la secreción y la activación (no obstante, mientras que se va dando la adhesión, simultáneamente ya se está dando la secreción y la activación). Hasta el momento, debemos tener presente que no se ha dado ningún cambio en la estructura de la plaqueta. La activa-
50
Colágeno/vWF → complejo GP Ib/VIX → GpVI → Integrina α2β1 → adhesión plaquetaria

ción de la plaqueta está inducida por varios factores: colágeno (unido a la GpVI), trombina (no necesita de daño en el vaso para que se dé la activación), ADP (esencial para la amplifica-ción), y otros. El ADP es tan importante para llamar a otras plaquetas que inhibidores del mismo (de su receptor en todo caso) son usados como anticoagulantes. La activación de la plaqueta produce:
- Activación metabólica. Se activan la fosfolipasa C (PLC) y la fosfolipasa A2. La PLC fosfo-rila proteinas de la membrana y favorece la movilización del Ca2+. La fosfolipasa A2 es la responsable de la producción del tromboxano A2, un importante vasoconstrictor e inductor de la agregación plaquetaria (papel de la aspirina como anticoagulante).
- Cambios en la morfología. La activación metabólica favorece que se produzcan toda una serie de cambios morfológicos: las plaquetas se vuelven esferas y emiten pseudó-
podos, lo cual facilita que unas plaquetas se unan con otras (metamorfosis viscosa). Además, se incrementa la concentración de Ca2+ intraplaquetario y se produce la se-creción de los gránulos α y de los densos.
Una vez la plaqueta ha sido activada y ha secretado el contenido de los gránulos, se dará la agregación plaquetaria para completar el tapón plaquetario. Las plaquetas comienzan a expre-sar la integrina αIIbβ3 que sirve para que el fibrinógeno se ancle y mantenga unidas a todas las plaquetas. En la activación de la plaqueta se observan además los siguientes acontecimien-tos:
- Reorientación de los fosfolípidos de la membrana. Aquellos fosfolípidos (p. ej., fosfatidil-serina) que presentan una carga negativa y que estaban en la monocapa interna en condiciones de reposo, pasarán a la monocapa externa donde ejercerán un efecto procoagulante (relacionado con la hemostasia plasmática).
- Expresión de genes. Se expresan toda una serie de genes importantes para el anclaje de
factores de coagulación como el VIII y el V. Estos factores permitirán la activación del factor IX y del factor X. Esto hará que tengamos una gran cantidad de factor X y de trombina.
- Formación de micropartículas. Las micropartículas son fracciones de las propias plaque-tas que expresan glucoproteínas que favorecen la exposición de los fosfolípidos anióni-
cos. Además, en las microparticulas hay una gran cantidad de factor tisular, que ya ve-remos que es un fundamental para la hemostasia secundaria.
- Expresión de P-selectina. La P-selectina es una proteína de adhesión que facilita que los factores de coagulación estén adheridos al trombo plaquetario además de proteger a los mismos frente a factores fibrinolíticos o anticoagulantes.
Hemostasia secundaria
Mientras que la hemostasia primaria es aquella en la que las plaquetas forman un tapón pla-quetario, la hemostasia secundaria (o plasmática) tiene como objetivo formar la trombina res-ponsable de la transformación del fibrinógeno en la fibrina, que forma una red que da consis-tencia al trombo plaquetario. En la hemostasia secundaria participan toda una serie de facto-res.
51

Cascada de coagulación
El elemento inicial de la coagulación plasmática es el denominado factor tisular (FT). El FT es formado por diferentes estructuras: (a) endotelio lesionado y activado; (b) plaquetas activadas; (c)
pericitos (en la adventicia de los vasos); (d) micropartículas (en las que la P-selectina ya se ha-brá encargado de activarlas para que formen el FT). El FT activa al factor VII. Atención, de manera basal existen pequeñas cantidades de f. VIIa27 circulantes por el plasma. El complejo VIIa + FT activa al f. X. El f. Xa es fundamental ya que es el responsable de activar la protrom-
bina (f. II) y transformarla en trombina (f. IIa).
Como vemos, el f. Xa va a ser importantísimo, ya que hace que tengamos trombina. Existe, no obstante, otro mecanismo o vía para activar el f. X. Esta vía consiste en el paso del f. XII a XIIa (se desconoce como), el XIIa hace que el f. XI se active, y el XIa hace que el IX se active. El f. IXa junto con el VIIIa, los fosfolípidos aniónicos y el Ca2+, activan al f. X.
Por lo tanto, vemos dos vías de activación del f. X: la del complejo TF + VIIa y la del complejo
IXa + VIIIa + FLP + Ca2+. La actuación simultánea de estas dos vías multiplica la capacidad de formación de trombina enormemente. El factor Xa es capaz de transformar la protrombina (II) en trombina (IIa), aunque lenta-
mente. Podemos incrementar la velocidad si esta reacción se da junto al f. Va, un ambiente
rico en Ca2+ y sobre una superficie de fosfolípidos aniónicos.
Una vez la trombina ha sido activada, ésta realiza tres acciones fundamentales:- Sobre el XIIa. Estimula la acción del f. XIIa sobre el f. XI, por lo que la vía del f. IX se
acelera (y es mucho más importante que la del FT).
52
27 Nomenclatura: cuando al número del factor se le añade una “a” quiere decir que está activado.
Factores que participan en la hemostasia secundariaFactores que participan en la hemostasia secundaria
Zimógenos Los factores de coagulación XII, XI, X, VI y II circulan por el plasma a modo de proenzimas, por lo que precisan de una activación. La activación consiste en una proteólisis limitada a un residuo de serina. Una parte importante de los f. de coagulación están formados por el híga-do (hepatopatía = posible coagulopatía). Los factores II, VII, IX y X son vitamina K-depen-dientes.
Cofactores Tienen la función de acelerar, fijar o limitar las reacciones de la coagulación plasmática.
Fosfolípidos aniónicos
Ca2+ Casi todas las reacciones precisan de calcio.
Fibrinógeno Es la proteína estructural diana de la trombina.
FT → paso de VII a VIIa* → Complejo FT + VIIa → paso de X a Xa → paso de II a IIa (trombina) (* + cantidad circulante por el plasma)
Paso de XII a XIIa → paso de XI a XIa → paso de IX a IXa → IXa + VIIIa + FLP + Ca2+ → paso de X a Xa
Paso de X a Xa → Xa + Va + FLP + Ca2+ → paso de II a IIa [300.000 veces mayor]

- Sobre el f. V. Favorece el paso de f. V a f. Va. Favorece una mayor formación de trom-bina.
- Sobre el f. VIII. Favorece el paso de f. VIII a f. VIIIa.
La vía del FT y del VIIa también recibe el nombre de vía extrínseca, debido a que precisa de una lesión (FT). La otra vía, la del IXa, se denomina también vía intravascular. La vía extrínse-ca es la inicial y es poco potente, su cometido es formar un poco de trombina (IIa) para que así, por los circuitos de retroalimentación positiva que hemos observado, se incremente aún más la formación de trombina. Hemos visto como algunas de las reacciones precisan de cal-cio. Existen agentes quelantes, como el EDTA, que secuestran el calcio para ejercer una fun-ción anticoagulatoria. La trombina no es la única que tiene mecanismos de retroalimentación, el mismo fac-
tor Xa estimula el paso de IX a IXa y del VII a VIIa. Como vemos, la cascada de factores es bas-tante explosiva, pasamos de un poco de trombina generada por la vía extrínseca a montones de trombina formados por la acción conjunta de las dos vías (sobre todo de la intravascular). Atención, el f. Xa genera un inhibidor (IVFT) que actúa sobre el f. VIIa y el mismo f. Xa. Con esto logramos que la coagulación se atenúe si la vía intravascular no se activa.
La coagulación únicamente debe producirse en los lugares donde hay lesión y no más allá. En condiciones normales existe un balance entre elementos coagulantes y anticoagulantes, don-de los anticoagulantes se encuentran en una mayor medida. La coagulación únicamente se dará si hay una lesión, puesto que dicha lesión implicará la activación (por trombina, colá-geno, ADP) de unas plaquetas que ofrecerán el sustrato para que se den todas las reacciones que acabamos de ver (fosfolípidos aniónicos, Ca2+).
53
Xa
V
X
TF
EndotelioPlaquetasPericitosP-selectina
VIIa + TF
VII
VIIa
IIa
II
VaCa2+ FLP
IVFT
VIIIaCa2+ FLPIXa
VIII
IX
XIaXI
XII XIIa
Cascada de coagulación. Con un triángulo naranja ( ) se marcan los procesos favorecidos por el factor Xa. Con un triángulo verde ( ) se marcan los procesos favorecidos por el factor IIa.

Nota. En la arteriosclerosis lo que hay es una lesión del endotelio, por lo que se van formando tapones.
Recordemos que en las lesiones se forma un tapón y además hay vasoconstricción.
Formación de la fibrina
Como hemos visto, tanto la vía extravascular como intrínseca tienen como objetivo pasar la protrombina (f. II) a trombina (f. IIa), pues esta última se encargará de formar la fibrina a partir del fibrinógeno (por proteólisis limitada). El fibrinógeno es la proteína más abundante del plasma (de entre las de coagulación) y está formado por tres cadenas (α, β y γ). Se precisa de un mínimo de fibrinógeno para se mantenga nuestra capacidad de coagulación. Si consumi-mos todo el fibrinógeno o éste es anómalo, tendremos tendencia a no coagular [coagulación intravascular diseminada = se consume el fibrinógeno y aparecen hemorragias]. La formación de la fibrina se divide en tres etapas: (a) ruptura del fibrinógeno; (b) con-
seguir fibrina ensamblada; y (c) estabilización de esa fibrina. En la primera fase, la trombina rea-liza una proteólisis limitada formando dos fibrinopéptidos: uno que es la parte terminal de la cadena α (FBPA), y otro que es la parte final de la cadena β (FBPB). Estos dos monómeros tie-nen tendencia a ensamblarse entre ellos, formándose unos aglomerados no del todo estables y aún solubles. El paso final (estabilización) consiste en la formación de enlaces covalentes entre las ca-denas α y γ de las diferentes fibrinas. Estos puentes se formarán gracias al f. XIIIa, que será ac-tivado por la propia trombina. El f. XIIIa además sirve como fijador de la α-antiplasmina (inhi-bidor de la plasmina).
Todo este proceso que acabamos de describir se da desde el principio, nada más haber un poco de trombina, ésta empieza a formar fibrina, pero como el trombo no es lo suficiente-mente estable necesitaremos una mayor cantidad de trombina. En otras palabras: en cuanto la primera plaqueta se adhiere, ya empieza a formarse una red de fibrina que se irá estabili-zando a medida que los otros componentes de la coagulación se van incorporando. Una vez el tejido se ha regenerado (proceso que inicia simultáneamente al de coagulación), el cóagu-lo ya no nos sirve y necesitaremos de un proceso de fibrinolisis.
Fibronolisis (plasminógeno → plasmina)
La ruptura del coágulo dependerá principalmente de la acción de la plasmina, un enzima que hidroliza las uniones Arg-Lys. La plasmina proviene del plasminógeno, una proteína que exis-te en dos formas: glu-plasminógeno y lys-plasminógeno. La forma lys- es bastante más activa y se forma a partir de la actuación de la plasmina sobre el glu-plasminógeno.
¿Cómo se produce la conversión de plasminógeno a plasmina? Pues existen varias vías, la primera que se explicó (y la más potente) es la del factor tisular de activación del plasminógeno (t-PA). Este t-Pa es formado por las céls. endoteliales activas en el proceso de coagulación.
54
Fibrinógeno → FBPA + FBPB → fibrina estable (por el XIIIa)
Glu-plasminógeno → Plasmina → Transformación de glu- en lys-plasminógeno

Hay una serie de factores que hacen que el endotelio exprese el t-PA: epinefrina, trombina, f. Xa, éstasis (estancamiento de la sangre) e hipoxia. Atención, el paso de plasminógeno a plasmina por acción del t-PA está además favorecido por la fibrina (cofactor), así aseguramos que el proceso trombolítico quede relegado allá donde hata un coágulo. Una segunda vía es la de la urocinasa. La urocinasa es liberada por las céls. endotelia-les en forma de prourocinasa. El t-PA activa a la urocinasa y ésta, sobre todo en presencia de
fibrina, potencia la reacción de la t-PA sobre el plasminógeno: el paso de éste a plasmina. Tanto la primera como la segunda vía precisan de unas céls. endoteliales dañadas o acti-
vadas, es por eso que estas dos vías se agrupan bajo la categoría de vía extrínseca. La vía an-tagónica a ésta es la intravascular, que se caracteriza por la activación del plasminógeno gra-cias al f. XII, al f. XI y a la calicreína, siendo la calicreína el elemento más importante. Para que ésta vía se active no precisamos de lesión.
Fibronolisis (ruptura de la fibrina)
La plasmina producirá una fibrinolisis progresiva sobre la fibrina. Paralelamente, la plasmina logra los siguientes efectos:
- Incrementa la producción de urocinasa.- Activa a los f. V y VIII, por lo que aumentamos la producción de trombina, y con ello in-
crementamos la fibrinolisis.
El hecho de que el f. XIIIa esté lo suficientemente activo hace que los trombos sean de buena calidad e insolubles. La presencia de uniones covalentes entre las fibrinas hace que, cuando la plasmina realiza una fibrinolisis progresiva, se mantienen toda una serie de polímeros que no serán proteolizados por las plasmina ya que están unidos al f. XIIIa.
Existe toda una serie de factores que inhiben la fibrinolisis:- Factor inhibidor de la activación del plasminógeno (PAI-1). Actúa a dos niveles: (a) sobre
el t-PA; y (b) sobre el paso de prourocinasa a urocinasa. La fibrina, a su vez, impide la producción del PAI-1.
- α2-antiplasmina. Se trata de una glucoproteína que, como su nombre indica, impide la acción de la plasmina. Está unida a la fibrina a través del f. XIIIa, por lo que actúa a nivel del trombo.
- TAFI. El f. inhibidor de la fibrinolisis activable por la protrombina impide que el t-PA y el plasminógeno se unan a la fibrina.
Pequeña aclaración. Debemos comprender que la fibrina es un excelente cofactor de la fibrinolisis. A la fibrina
se le va a unir el t-PA para que acelere la reacción de plasminógeno a plasmina. Es la propia fibrina del trombo la que favorece la fibrinolisis del trombo.
Elementos en la fase plaquetaria
El Dr. Javierre habló de tres elementos que están relacionados con la fase plaquetaria: el óxi-do nítrico, las prostaciclinas y la adenosina.
55

- Óxido nítrico. El óxido nítrico es un vasodilatador y un inhibidor de la adhesión plaqueta-
ria. Su producción depende del cGMP sintetizado por las plaquetas activadas. El óxi-do nítrico se formará en una mayor cantidad ante un aumento de ATP, de trombina, de serotonina o de bradicinina.
- Prostaciclinas. Son formadas por la oxidación del ácido araquidónico en el endotelio (céls. endoteliales activadas). Disminuye la agregación plaquetaria y es un vasodilatador. Su acción está vinculada con un aumento del cAMP.
- Adenosina. Es un inhibidor de la función plaquetaria y un potente vasodilatador. Cuan-do una célula no recibe el suficiente oxígeno, tendrá dificultades para formar ATP. Po-drá formarlo a partir de la reacción que se ilustra a continuación.
Esta reacción, no obstante, implicará que salga adenosina. Resumiendo: si falta oxíge-no encontraremos adenosina en el medio y, gracias a su función vasodilatadora, in-tentaremos solventar esa hipoxia.
Elementos en la fase plasmática
En el territorio plasmático nos vamos a encontrar con toda una serie de factores que favore-cen la inactivación de la cascada.
- Antitrombina (antitrombina III según algunos manuales). Inhibe la acción de la trombi-
na y del f. Xa. Esta antritrombina tendrá receptores endoteliales para moléculas que formará el propio endotelio, como el heparán sulfato. Este heparán, combinado con la antitrombina, incrementa la actividad de ésta última unas 10.000 veces. La antitrom-bina también tiene un efecto inhibitorio sobre los factores IXa, XIa y XIIa. La antitrom-bina es muy potente, y más cuando está fijada al endotelio.
- Trombomodulina (TM). La trombomodulina se une a la trombina (f. IIa) disminuyendo su eficacia. El complejo TM-IIa se unirá a la proteína C, activando a ésta última y permi-tiendo que también ésta última inhiba a los factores Va y VIIIa. La proteína C es de origen hepático y vit. K-dependiente (precisará de Ca2+).
- Proteína S. La proteína S se forma en diferentes lugares: el endotelio, el hígado y me-gacariocitos. Favorece la acción de la proteína C. Además, activa al factor inhibidor del
factor tisular (IVFT; regulador de la vía extravascular).- Proteína Z. Inactiva el f. Xa.
Aplicaciones prácticas
Vitamina K
Una parte de la vit. K es formada por las bacterias que tenemos en nuestro intestino. En los primeros días de vida, al no tener flora bacteriana, se nos suelen administrar vitamina K. Hay algunos alimentos que tienen vit. K, como las hojas verdes. La vit. K permite que en el hígado se dé la carboxilación de algunos residuos glutámicos de la proteína Gla (residuos Gla), una im-
56
2 ADP→ ATP + AMP

portante familia de proteínas hepáticas. Estos residuos Gla se podrán unir al Ca2+ y facilitarán la acción de la proteína. Un déficit de vit. K provocará una disminución de todos aquellos factores dependien-tes de la vit. K: II, VII, IX y X, quedando afectada la vía plasmática de la coagulación. Las de-ficiencias de vitamina K pueden darse o bien por un problema hepático (no tiene la capaci-dad de realizar la carboxilación de la proteína Gla) o bien por una litiasis biliar (dificultad pa-ra absorber grasas y entre ellas la vit. K, que es liposoluble). Siempre que haya una mínima función hepática, podemos administrar vit. K antes de una intervención para que el paciente coagule correctamente28. Una deficiencia grave de vit. K puede hacer que hayan hemorra-gias ante una agresión.
Disminución del número de plaquetas
La disminución del número de plaquetas se denomina trombocitopenia. Podemos tener me-nos plaquetas de lo normal y preservar nuestra capacidad de coagulación; ahora bien, hay límites: mínimo de 50.000 plaquetas/mm3. Cuando faltan plaquetas, con el tiempo van apare-ciendo pequeñas lesiones que no son taponadas por un trombo plaquetario, por lo que em-piezan a aparecer pequeñas manchas de sangre: púrpura. La gran mayoría de púrpuras trom-
bocitopénicas son idiopáticas, aunque se sospecha que puede haber un trasfondo autoinmune. Estos cuadros suelen responder bien a una esplenectomía (ya que es en el bazo donde se des-truyen las plaquetas).
Hemofilia
La hemofilia es una enfermedad genética recesiva ligada al sexo (Chr. X). La mujer es la por-tadora y el varón es la que la sufre. Hay dos tipos de hemofilia, aquella en la que se ve afec-tada la producción de f. VIII (mayoritaria) y aquella en la que se afecta la producción del f. IX (menos frecuente).
Tromboembolismo
Un coágulo anormal que aparece en un vaso sanguíneo se llama trombo. Una vez el coágulo ha aparecido, es probable que el flujo continuo de sangre que pasa por el coágulo lo des-prenda y haga que el coágulo fluya con la sangre, lo que se conoce como émbolo. Los émbo-los que se originan en las arterias grandes o en el lado izquierdo del corazón pueden fluir hacia la periferia y taponar arterias o arteriolas en el cerebro, los riñones o cualquier otro lu-gar. Los émbolos que se originan en el sistema venoso o en la parte derecha del corazón van generalmente a los vasos pulmonares, donde pueden causar una embolia arterial pulmonar. En condiciones normales se nos forman una gran cantidad de pequeños trombos en diferentes partes del cuerpo. Por ejemplo, al estar sentado mucho rato puedes favorecer la formación de estos trombos en el sistema venoso de las piernas. Ahora bien, el propio flujo
57
28 Si no hay esa función hepática mínima, siempre se puede recurrir a la administración de un concentrado de factores de coagulación.

así como la dilución del ambiente procoagulante hará que este trombo se disuelva. Debemos tener presente que hay agente anticoagulante en la sangre además de cúmulos de heparina en diferentes partes del cuerpo. Uno de los primeros cúmulos de la heparina con los que se en-contraría un trombo formado en el sistema venoso sería el que está a nivel de los pulmones. Pocos tejidos forman heparina, uno de ellos son los pulmones. La heparina actúa co-mo cofactor uniéndose a la antitrombina y así inhibiendo la consolidación del coágulo. Al inhibir la trombina disminuimos la actividad de los factores X y IX. Podemos tratar un tromboembolismo por medio de la administración de heparina29 (con sus riesgos) o con la administración local de t-PA.
Cumarinas
Las cumarinas bloquean a la vit. K, por lo que todos aquellos factores que dependen de ésta (trombina, VII, IX y X) verán sus cantidades reducidas. Esto hace que progresivamente vaya disminuyendo la capacidad coagulatoria del paciente al que se le administra la cumarina (p. ej., sintrom), su capacidad coagulatoria no se inhibe de golpe. Esta disminución progresiva permite que podamos adaptar la dosis en función de las necesidades del paciente (y periódi-camente se van realizando pruebas de coagulación para ajustar la dosis). En altas dosis el sintrom puede ser usado como veneno. La vit. K es formada por las bacterias intestinales y también la podemos incorporar a través de la dieta. Eso quiere decir que podemos alterar nuestra coagulabilidad por medio de modificaciones en la dieta.
58
29 Cuando alguien está hospitalizado (y en cama), una medida profiláctica es la administración periódica de heparina para disminuir el riesgo de padecer una trombosis. Hay casos en los que no se puede poner heparina: traumatismo craneoencefálico en el que hay riesgo de que se produzca una hemorragia a nivel del SNC.

9. SISTEMA CARDIOVASCULAR
Introducción
En este bloque el sistema cardiovascular será explicado como algo constante y estable, cuando en realidad es un sistema en el cambiarán toda una serie de elementos para que to-do se mantenga igual (p. ej., sistema capilar de las personas que hacen ejercicio en compa-ración con las que no). Este sistema tiene un núcleo muy claro, el corazón, que se trata de un músculo extremadamente especial que cuando se contrae lo hace de manera unitaria y al máximo de su capacidad. El corazón es una bomba pulsátil que introduce presión en un sistema cerrado para así mover un gran volumen a grandes distancias. La sangre contenida en los vasos, por tanto, fluirá en virtud a un gradiente de presión. La bomba cardíaca, las arterias y las venas envían la sangre de manera desordenada, mientras que en los capilares se producirá una reordenación para que haya una difusión de elementos entre la sangre y el tejido (bidireccional). La difusión sigue un movimiento browniano (aleatorio), por lo que nos interesa que la distancia que separa el origen y el destino de una molécula que difunde tiene que ser lo más pequeña posible para reducir el tiempo del trayecto.
La capacidad de difusión de una molécula depende de su coeficiente de difusión, de la dis-tancia que debe recorrer, del área que hay para poder difundir, del contenido hídrico y del gradiente de presión.
Circulación mayor y menor
La circulación mayor es aquella que se encarga de llevar todo tipo de elementos hacia la peri-feria. Este circuito empieza en el ventrículo izquierdo y acaba en el atrio derecho. En este cir-cuito será muy importante la velocidad, por lo que tendrá una elevada presión (al menos en el territorio arterial). La circulación menor, en cambio, es aquella que se encarga de oxigenar la sangre (y librarse del CO2) en los pulmones. Comienza en el ventrículo derecho y acaba en el atrio iz-
quierdo. Tendrá una presión baja para facilitar el intercambio gaseoso. Nuestro corazón dispone de cuatro cavidades para diferenciar estos dos sistemas, para aislar aquella sangre oxigenada de la que es pobre en oxígeno. Además, pensemos que el retorno venoso se basa en un gradiente de presión que disminuye conforme nos vamos acer-
59
Movimiento dirigido (izquierda) vs movimiento browniano (derecha). Supongamos que la distancia que sepa-ran los dos puntos es de 10 cm. Si la partícula A se mueve a una velocidad de 1 cm/s, tardará 10 s en recorrer la distancia que separa los dos puntos. En el movimiento browniano (aleatorio), la partícula B tardará el cuadrado de la distancia en llegar a su destino (102 = 100 s). Como las moléculas que van por difusión u ósmosis siguen este tipo de movimiento, nos interesa acortar las distancias.
A B

cando al corazón, y eso no sería compatible con el gradiente de presión que generamos en el ventrículo izquierdo para bombear la sangre a grandes distancias. La sangre enviada por el corazón irá a los diferentes tejidos en función de la cantidad de sangre que necesiten en ese momento (el flujo de sangre para cada tejido será determinado por el SNV o bien por la acti-vidad de ese territorio). A pesar de ser una bomba pulsátil, el corazón debe generar un flujo
continuo. El circuito siempre estará lleno de sangre. Si perdemos sangre del sistema venoso, no pasará nada (a no ser que perdamos mucho); en cambio, si perdemos sangre del territorio arterial, probablemente entraremos en shock. ¿Por qué? Pues porque en el sistema arterial hay menos sangre que en el venoso y ésta será importante para mantener el fundamental gradiente de presión (la pérdida de volumen, por poca que sea, ya descompensa y produce una abrupta caída de la presión).
60

10. CICLO CARDÍACO
Introducción
Nuestro corazón es una bomba pulsátil que se llena de sangre de una manera más o menos constante y que expulsa la sangre de una manera más o menos discontinua. Sin embargo, esto no quiere decir que en algún momento los vasos estén vacíos; al contrario, los vasos san-
guíneos siempre estarán llenos de sangre.
Corazón izquierdo
El atrio izquierdo está comunicado con el ventrículo izquierdo a través de la válvula mitral (bicúspide o atrioventricular izquierda). La sangre llegará al atrio izquierdo desde el territorio pulmonar cuando el corazón haya finalizado la sístole ventricular. Recordemos que sístole es contracción y diástole es relajación (y casi siempre nos estaremos refiriendo al ventrículo, aunque también hay sístole y diástole auricular). En este momento (finalización de la sístole ventricular), la válvula mitral y la válvula sigmoidea (aórtica) estarán cerradas. Recordemos también que la válvula mitral se abre hacia abajo y se cierra hacia arriba, mientras que la sigmoidea se abre hacia arriba y se cierra hacia abajo. A medida que el atrio izquierdo se vaya llenando, la presión en el mismo se irá in-crementando. La presión del atrio izquierdo acabará superando a la del ventrículo izquierdo, por lo que la válvula mitral cederá y permitirá el paso de sangre hacia un ventrículo que aún
tenía algo de sangre. Una vez el ventrículo se ha llenado con parte de la sangre de la aurícula, comienza la contracción auricular. Esta contracción acaba de vaciar la aurícula (y por tanto acaba de rellenar al ventrículo). Cuando el ventrículo izquierdo está repleto de sangre procedente de la aurícula, de-bemos imaginar como las valvas de la válvula mitral reposan sobre el volumen de sangre ventricular. Además, contrariamente a lo que podríamos pensar, la presión del ventrículo iz-quierdo será, en este momento, extremadamente baja, tanto que la presión del territorio pulmo-nar (3-4 mmHg) es suficiente para empujar la sangre desde la aurícula al ventrículo. La baja presión en el ventrículo izquierdo hace éste reciba el apelativo, en ese momento, de cámara
de baja presión. Una vez el estímulo que ha producido que el atrio se contraiga llega al ventrículo, éste se transformará en una cámara de alta presión: la presión se incrementará, fugaz y brus-camente, a una velocidad de 1.400 mmHg/s. Este incremento de presión tan brusco hace que la válvula mitral se cierre pasivamente [1º tono cardíaco: lub, grave; inicio de la sístole]. La sangre no saldrá del ventrículo hasta que la presión en el mismo sea superior a la
presión que hay en la aorta, momento en el que se podrá abrir la válvula sigmoidea. Una vez superada la presión aórtica, inyectaremos sangre en la aorta y la contracción proseguirá has-ta que la presión de la aorta venza a la ventricular (ya que la aorta tiene un calibre determi-nado y un volumen sanguíneo preexistente). Cuando la presión aórtica supera a la ventricu-lar, se produce el cierre brusco de la válvula sigmoidea [2º tono cardíaco: dub, corto y agudo;
final de la sístole].
61
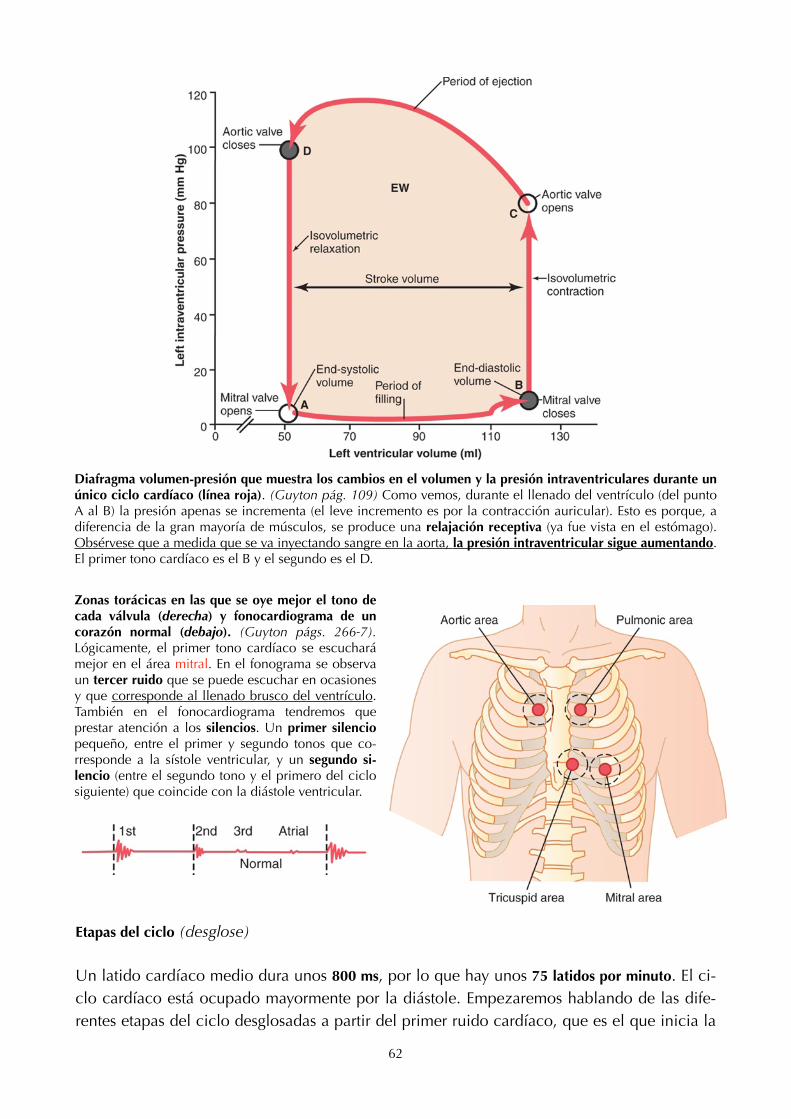
Etapas del ciclo (desglose)
Un latido cardíaco medio dura unos 800 ms, por lo que hay unos 75 latidos por minuto. El ci-clo cardíaco está ocupado mayormente por la diástole. Empezaremos hablando de las dife-rentes etapas del ciclo desglosadas a partir del primer ruido cardíaco, que es el que inicia la
62
Diafragma volumen-presión que muestra los cambios en el volumen y la presión intraventriculares durante un único ciclo cardíaco (línea roja). (Guyton pág. 109) Como vemos, durante el llenado del ventrículo (del punto A al B) la presión apenas se incrementa (el leve incremento es por la contracción auricular). Esto es porque, a diferencia de la gran mayoría de músculos, se produce una relajación receptiva (ya fue vista en el estómago). Obsérvese que a medida que se va inyectando sangre en la aorta, la presión intraventricular sigue aumentando. El primer tono cardíaco es el B y el segundo es el D.
Zonas torácicas en las que se oye mejor el tono de cada válvula (derecha) y fonocardiograma de un corazón normal (debajo). (Guyton págs. 266-7). Lógicamente, el primer tono cardíaco se escuchará mejor en el área mitral. En el fonograma se observa un tercer ruido que se puede escuchar en ocasiones y que corresponde al llenado brusco del ventrículo. También en el fonocardiograma tendremos que prestar atención a los silencios. Un primer silencio pequeño, entre el primer y segundo tonos que co-rresponde a la sístole ventricular, y un segundo si-lencio (entre el segundo tono y el primero del ciclo siguiente) que coincide con la diástole ventricular.
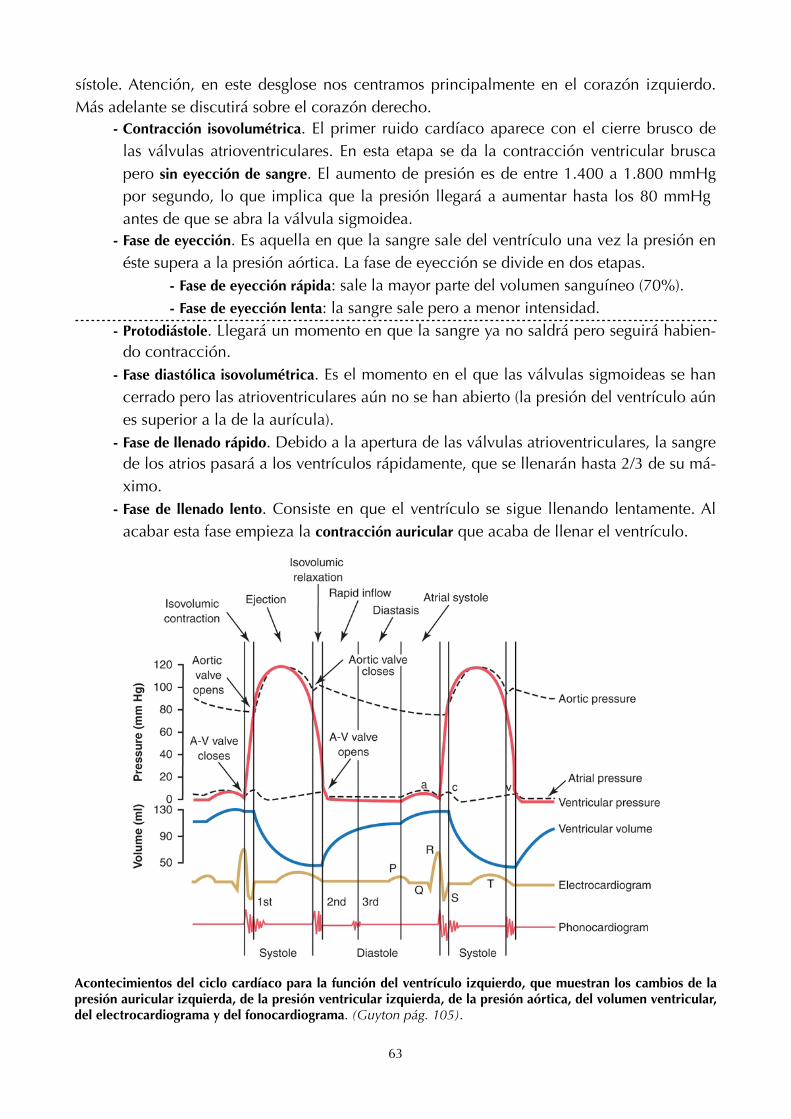
sístole. Atención, en este desglose nos centramos principalmente en el corazón izquierdo. Más adelante se discutirá sobre el corazón derecho.
- Contracción isovolumétrica. El primer ruido cardíaco aparece con el cierre brusco de las válvulas atrioventriculares. En esta etapa se da la contracción ventricular brusca pero sin eyección de sangre. El aumento de presión es de entre 1.400 a 1.800 mmHg por segundo, lo que implica que la presión llegará a aumentar hasta los 80 mmHg antes de que se abra la válvula sigmoidea.
- Fase de eyección. Es aquella en que la sangre sale del ventrículo una vez la presión en éste supera a la presión aórtica. La fase de eyección se divide en dos etapas.
- Fase de eyección rápida: sale la mayor parte del volumen sanguíneo (70%).- Fase de eyección lenta: la sangre sale pero a menor intensidad.
- Protodiástole. Llegará un momento en que la sangre ya no saldrá pero seguirá habien-do contracción.
- Fase diastólica isovolumétrica. Es el momento en el que las válvulas sigmoideas se han cerrado pero las atrioventriculares aún no se han abierto (la presión del ventrículo aún es superior a la de la aurícula).
- Fase de llenado rápido. Debido a la apertura de las válvulas atrioventriculares, la sangre de los atrios pasará a los ventrículos rápidamente, que se llenarán hasta 2/3 de su má-ximo.
- Fase de llenado lento. Consiste en que el ventrículo se sigue llenando lentamente. Al acabar esta fase empieza la contracción auricular que acaba de llenar el ventrículo.
63
Acontecimientos del ciclo cardíaco para la función del ventrículo izquierdo, que muestran los cambios de la presión auricular izquierda, de la presión ventricular izquierda, de la presión aórtica, del volumen ventricular, del electrocardiograma y del fonocardiograma. (Guyton pág. 105).
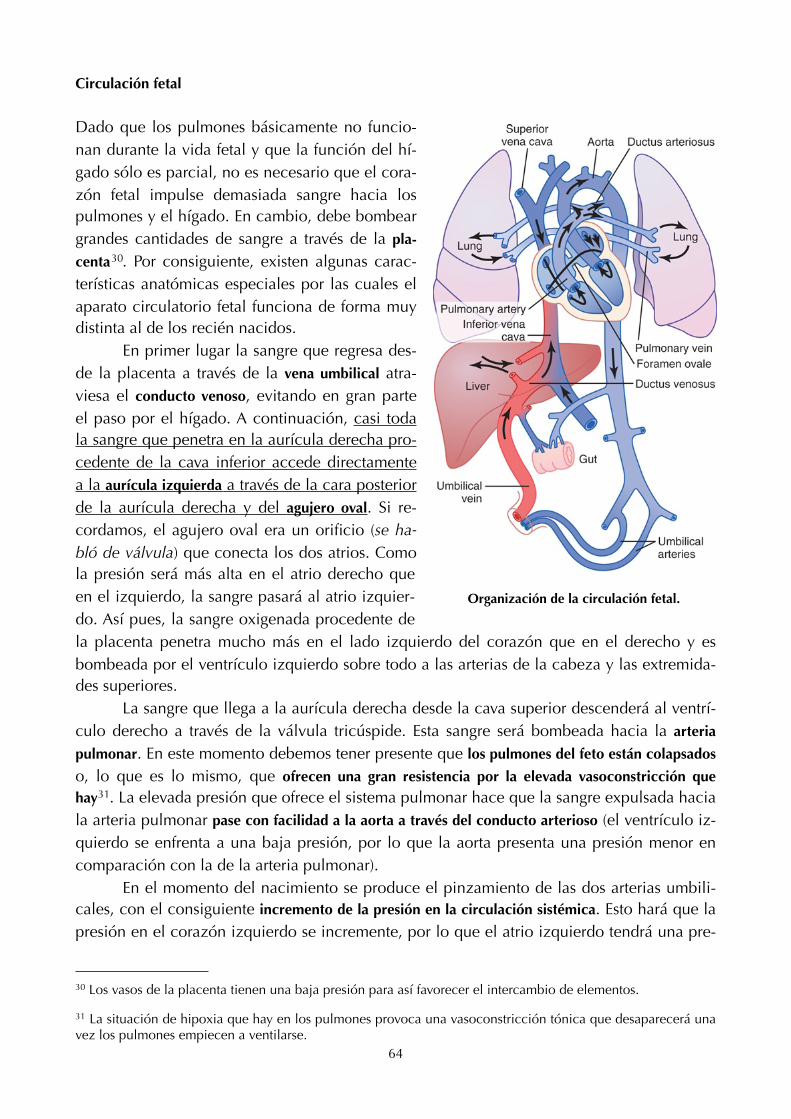
Circulación fetal
Dado que los pulmones básicamente no funcio-nan durante la vida fetal y que la función del hí-gado sólo es parcial, no es necesario que el cora-zón fetal impulse demasiada sangre hacia los pulmones y el hígado. En cambio, debe bombear grandes cantidades de sangre a través de la pla-
centa30. Por consiguiente, existen algunas carac-terísticas anatómicas especiales por las cuales el aparato circulatorio fetal funciona de forma muy distinta al de los recién nacidos. En primer lugar la sangre que regresa des-de la placenta a través de la vena umbilical atra-viesa el conducto venoso, evitando en gran parte el paso por el hígado. A continuación, casi toda la sangre que penetra en la aurícula derecha pro-cedente de la cava inferior accede directamente a la aurícula izquierda a través de la cara posterior de la aurícula derecha y del agujero oval. Si re-cordamos, el agujero oval era un orificio (se ha-bló de válvula) que conecta los dos atrios. Como la presión será más alta en el atrio derecho que en el izquierdo, la sangre pasará al atrio izquier-do. Así pues, la sangre oxigenada procedente de la placenta penetra mucho más en el lado izquierdo del corazón que en el derecho y es bombeada por el ventrículo izquierdo sobre todo a las arterias de la cabeza y las extremida-des superiores. La sangre que llega a la aurícula derecha desde la cava superior descenderá al ventrí-culo derecho a través de la válvula tricúspide. Esta sangre será bombeada hacia la arteria
pulmonar. En este momento debemos tener presente que los pulmones del feto están colapsados o, lo que es lo mismo, que ofrecen una gran resistencia por la elevada vasoconstricción que hay31. La elevada presión que ofrece el sistema pulmonar hace que la sangre expulsada hacia la arteria pulmonar pase con facilidad a la aorta a través del conducto arterioso (el ventrículo iz-quierdo se enfrenta a una baja presión, por lo que la aorta presenta una presión menor en comparación con la de la arteria pulmonar). En el momento del nacimiento se produce el pinzamiento de las dos arterias umbili-cales, con el consiguiente incremento de la presión en la circulación sistémica. Esto hará que la presión en el corazón izquierdo se incremente, por lo que el atrio izquierdo tendrá una pre-
64
30 Los vasos de la placenta tienen una baja presión para así favorecer el intercambio de elementos.
31 La situación de hipoxia que hay en los pulmones provoca una vasoconstricción tónica que desaparecerá una vez los pulmones empiecen a ventilarse.
Organización de la circulación fetal.

sión mayor respecto al derecho y el foramen oval se bloqueará (como si fuera una válvula) para después acabar de cerrarse. Cuando el recién nacido empiece a respirar, ventilará y la resis-tencia ofrecida por el sistema pulmonar descenderá, por lo que la sangre que antes iba desde la arteria pulmonar a la aorta, ahora irá en sentido contrario. La sangre que ahora va desde la aorta a la pulmonar se caracteriza por estar muy oxigenada (de 15-20 mmHg de PO2 a unos 100 mmHg de PO2), lo que producirá una vasoconstricción potente. Esta vasoconstricción se-rá un cierre funcional que con el tiempo pasará a ser un cierre anatómico (únicamente queda-rá un vestigio fibroso). Si este conducto no se cerrase completamente nos encontraríamos ante un soplo: un ruido que ocuparía toda la sístole.
Nota: conceptos importantes. El Dr. Javierre introdujo cuatro conceptos al final de la clase que describen diver-
sas capacidades del corazón: (a) inotropismo/contractilidad: capacidad del músculo cardíaco para convertir la energía química en mecánica; (b) cronotropismo/automatismo: capacidad para, independientemente de estí-
mulos externos, producir la contracción; (c) excitabilidad/batmotropismo: capacidad para responder a un es-tímulo; (d) conductibilidad/dromotropismo: capacidad para conducir un estímulo.
Presión auricular (PAI) [ mejor que pases y ya lo darás más adelante; esto confunde]
La forma de la curva de presión en la aurícula izquierda refleja tanto los fenómenos cardíacos como el retorno venoso (el flujo de sangre que vuelve hacia la aurícula). Está formada por las ondas a, c y v. La contracción auricular provoca la onda a. Durante la contracción isovolu-métrica se observa otro aumento ascendente en la curva de la PAI, la onda c, causada por la protusión de la válvula mitral de nuevo hacia la aurícula izquierda, cuando el ventrículo in-tenta contraerse contra un volumen fijo. Durante el periodo de eyección del corazón iz-quierdo, la PAI aumenta lentamente, mientras que el retorno venoso desde la circulación pulmonar va llenado la aurícula (la válvula mitral se cierra durante este periodo), producien-do la onda v. Cuando la presión ventricular izquierda (PVI) cae por debajo de la PAI, la vál-vula mitral se abre y la PAI disminuye cuando la sangre que fluye va entrando en la aurícula izquierda. Durante el resto de la diástole, la PAI se mantiene por encima de la PVI cuando se llena el corazón. Como no hay válvulas entre las venas cava y la aurícula derecha (o vena pulmonar y aurícula izquierda), las curvas del pulso venoso y de la presión auricular tienen una forma similar.
65
Curva de presión auricular. (Guyton pág. 105). En Netter pág. 114 también sale una buena figura. Nótese co-mo el equilibrio entre PAI y PVI determina la apertura o no de la válvula mitral. La curva del pulso venoso tiene una forma bastante similar a la que forma la PAI.

Fonocardiograma
A pesar de que se describe en la práctica, como no se explicó mucho en clase comento al-gunos de los aspectos más interesantes del fonocardiograma (extraído de Netter pág. 115). Podemos llegar a escuchar hasta cuatro sonidos cardíacos, representados como S1, S2, S3 y S4.
- S1. Es el primer ruido cardíaco. Se genera por el cierre de las válvulas mitral y tricús-pide (la válvula mitral se cierra ligeramente antes que la triscúspide).
- S2. Se asocia al cierre de las válvulas aórtica y pulmonar. De nuevo, la válvula iz-quierda se cierra ligeramente antes que la válvula derecha. Durante la inspiración, el cierre de la válvula pulmonar está ligeramente retrasado, dando lugar normalmente a un desdoblamiento fisiológico audible de S2. Este retraso se debe al aumento del llena-do del ventrículo derecho, debido al descenso de la presión intratorácica durante la inspi-
ración.- S3. Es normal en niños y se asocia a un llenado ventricular rápido. No se escucha en
adultos normales y puede ser signo de sobrecarga de volumen, como en la insuficien-
cia cardíaca congestiva.- S4. Se debe al llenado ventricular activo, tampoco es audible en los adultos normales,
pero puede estar presente en algunas patologías cardíacas.
Asincronía entre corazones
Hay una ligera asincronía entre el corazón izquierdo y el corazón derecho, en términos de abertura y cierre de válvulas, aunque las secuencias son las mismas. La secuencia conjunta es la siguiente: (1) cierre mitral, (2) cierre triscúspide, (3) abertura pulmonar, (4) abertura aór-tica, (5) cierre aórtico, (6) cierre pulmonar, (7) abertura tricúspide, y (8) abertura mitral.
Corazón derecho
En el corazón derecho se producen las aberturas y cierre de las válvulas bajo presiones unas cinco veces por debajo de las que encontramos en el corazón derecho. Mientras que las dos aurículas más o menos se contraen a la vez, con los ventrículos eso no ocurre: la válvula
pulmonar se abrirá antes que la aórtica ya que para abrir esta última habrá que superar una mayor presión.
Trabajo cardíaco (Netter págs. 121-2)
Cuando decimos que el corazón expulsa unos 5 L/min, en realidad nos estamos refiriendo a que el corazón expulsa 5 l por la aorta. En realidad se expulsan cinco por la aorta y cinco por
la pulmonar, dando un total de 10 L/min. El ventrículo izquierdo tendrá menos tiempo para expulsar el volumen que contiene. Cuando hablamos de gasto cardíaco (volumen cardíaco minuto o VCM) en realidad nos estamos refiriendo a la cantidad de sangre que expulsa el corazón por el ventrículo izquier-do. El VCM es igual al producto de la frecuencia cardíaca y el volumen sistólico. El gasto car-
66

díaco está determinado por nuestras necesidades. Si nuestras necesidades son unas y el gasto cardíaco está por debajo, entramos en un estado de insuficiencia cardíaca. El gasto cardíaco también puede calcularse con la frecuencia cardíaca multiplicada por la diferencia entre el
volumen telediastólico (VTD) y el volumen telesistólico (VTS). El VTD es el máximo volumen que puede alojar el ventrículo izquierdo (volumen diastólico final) y el VTS es la cantidad de san-gre que permanece en el ventrículo tras la eyección de parte del mismo y el posterior cierre de la válvula aórtica. El VTD está relacionado con la precarga.
Se ha demostrado que el gasto cardíaco aumenta32 en proporción con la edad. En conse-cuencia, el gasto cardíaco se expresa en términos de índice cardíaco, que es el resultado del cociente entre el gasto cardíaco y la superficie corporal.
Finalmente, la fracción de eyección es el porcentaje de corazón que se vacía. La normalidad está entre el 60 y el 70%.
Presión arterial
Para una frecuencia cardíaca (FC) de unos 70 latidos/min, tengo unos 8 mmHg de presión en la aurícula izquierda y 4,5 mmHg en la aurícula derecha. Como ya se ha comentado antes, el corazón derecho estará bajo presiones notablemente menores respecto al corazón iz-quierdo. El cambio de presión que sufre el ventrículo izquierdo desde la telediástole (máxi-mo volumen) y la sístole es de 9,5 a hasta 125 mmHg. Esta última presión (125 mmHg) es la máxima que consigo inyectar en la aorta: es la presión arterial sistólica (máxima) y en reposo está sobre los 120 mmHg. Por el otro lado, cuando el corazón está relajado y llenándose el sis-tema alcanzará las presiones mínimas: la presión arterial diastólica (mínima), que en reposo suele ser 80 mmHg. Estas presiones son, claro está, del corazón izquierdo; en el corazón de-recho la presión máxima y mínima serían 25/10 mmHg.
67
32 Corrección (3 de mayo): en el seminario se comentó que con la edad disminuye la secreción tiroidea, por lo que baja el metabolismo y por tanto nuestras necesidades (bajará el gasto cardíaco).
Gasto cardíaco = VCM = FC ⋅VS
Gasto cardíaco = VCM = FC ⋅ VTD− VTS( )
Índice cardíaco =Gasto cardíaco L / min( )Superficie corporal m2( )
Fracción eyección =
VTD− VTSVTD
⋅100

11. ELECTROFISIOLOGÍA CARDÍACA
Células del corazón
En el corazón disponemos de tres grandes tipos celulares:- Céls. con capacidad autoexcitatoria. Representan aproximadamente el 1% del total de
células que hay en el corazón. Son pequeñas, no generan tensión y se encuentran principalmente en los nódulos sinusal y auriculoventricular (AV). Como sabemos, el co-razón tiene la capacidad de autoexcitarse; no dispone de placas motoras33, si no que dispone de estas céls. con capacidad autoexcitatoria que se despolarizarán y enviarán estímulos al resto de las células.
- Céls. que facilitan la conducción. Se trata de unas células que facilitan la transmisión de los potenciales de acción. En su conjunto forman el tejido de conducción.
- Céls. musculares. Son similares a las céls. musculares esqueléticas. Son pequeñas, ge-neralmente disponen de un sólo núcleo y presentan ramificaciones que les permite conectarse con otras células. Las uniones intercelulares (discos intercalares) se caracte-rizan por presentar desmosomas (transmiten la fuerza) y unas uniones comunicantes que aceleran la conducción cardíaca. En cuanto a su arquitectura intracelular: túbulos T más gruesos y ramificados, RE pequeño y hasta 1/3 de su espacio dedicado a las mi-tocondrias (aprovechamiento del O2).
Pese a que el corazón pueda contraerse de manera autónoma, y que por ello si lo extraemos de un cuerpo seguirá bombeando durante un tiempo, precisa de un medio rico en calcio. Si bañamos un corazón en un medio con una gran cantidad de calcio daremos lugar a contrac-ciones potentes.
Conducción del impulso eléctrico
El nódulo sinusal o nódulo sinoauricular (SA) se encuentra en la base de la vena cava superior. Se despolarizará, en reposo, a una frecuencia aproximada de unas 70 veces/min. Este nódu-lo, gracias a que está conectado con toda una serie de células, podrá transmitir el potencial de acción por las dos aurículas. La velocidad de transmisión es de 0,3 m/s, aunque en algunos puntos alcanza valores de 1 m/s. Estos puntos son tres bandas interauriculares (anterior, media y posterior) que se encargan de favorecer que el impulso vaya más rápido a la aurícula iz-quierda y así lograr una contracción casi al unísono de los dos atrios. En reposo la contracción auricular no tiene una gran trascendencia hemodinámica, es más importante la contracción ventricular. El sistema de conducción auricular está organiza-do de modo que el impulso no viaja desde las aurículas hacia los ventrículos demasiado rá-pidamente; se deja un tiempo para que las aurículas se vacíen antes de que comience la contracción ventricular. Este retraso se da gracias a un nódulo auriculoventricular (AV), que está en el rodete fibroso que separa aurícula de ventrículo.
68
33 Con esto decimos que no habrá acetilcolina para iniciar una contracción, como mucho para regular la fre-cuencia de contracción.
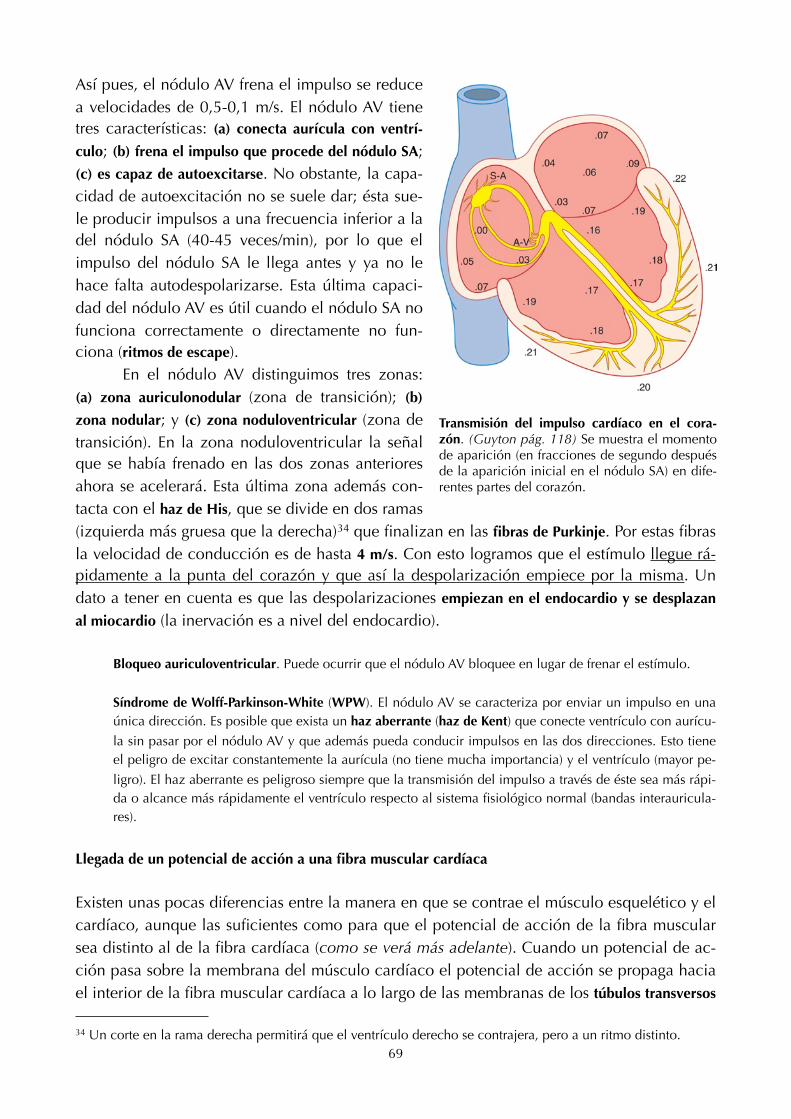
Así pues, el nódulo AV frena el impulso se reduce a velocidades de 0,5-0,1 m/s. El nódulo AV tiene tres características: (a) conecta aurícula con ventrí-
culo; (b) frena el impulso que procede del nódulo SA; (c) es capaz de autoexcitarse. No obstante, la capa-cidad de autoexcitación no se suele dar; ésta sue-le producir impulsos a una frecuencia inferior a la del nódulo SA (40-45 veces/min), por lo que el impulso del nódulo SA le llega antes y ya no le hace falta autodespolarizarse. Esta última capaci-dad del nódulo AV es útil cuando el nódulo SA no funciona correctamente o directamente no fun-ciona (ritmos de escape). En el nódulo AV distinguimos tres zonas: (a) zona auriculonodular (zona de transición); (b)
zona nodular; y (c) zona noduloventricular (zona de transición). En la zona noduloventricular la señal que se había frenado en las dos zonas anteriores ahora se acelerará. Esta última zona además con-tacta con el haz de His, que se divide en dos ramas (izquierda más gruesa que la derecha)34 que finalizan en las fibras de Purkinje. Por estas fibras la velocidad de conducción es de hasta 4 m/s. Con esto logramos que el estímulo llegue rá-pidamente a la punta del corazón y que así la despolarización empiece por la misma. Un dato a tener en cuenta es que las despolarizaciones empiezan en el endocardio y se desplazan
al miocardio (la inervación es a nivel del endocardio).
Bloqueo auriculoventricular. Puede ocurrir que el nódulo AV bloquee en lugar de frenar el estímulo.
Síndrome de Wolff-Parkinson-White (WPW). El nódulo AV se caracteriza por enviar un impulso en una única dirección. Es posible que exista un haz aberrante (haz de Kent) que conecte ventrículo con aurícu-
la sin pasar por el nódulo AV y que además pueda conducir impulsos en las dos direcciones. Esto tiene el peligro de excitar constantemente la aurícula (no tiene mucha importancia) y el ventrículo (mayor pe-
ligro). El haz aberrante es peligroso siempre que la transmisión del impulso a través de éste sea más rápi-da o alcance más rápidamente el ventrículo respecto al sistema fisiológico normal (bandas interauricula-res).
Llegada de un potencial de acción a una fibra muscular cardíaca
Existen unas pocas diferencias entre la manera en que se contrae el músculo esquelético y el cardíaco, aunque las suficientes como para que el potencial de acción de la fibra muscular sea distinto al de la fibra cardíaca (como se verá más adelante). Cuando un potencial de ac-ción pasa sobre la membrana del músculo cardíaco el potencial de acción se propaga hacia el interior de la fibra muscular cardíaca a lo largo de las membranas de los túbulos transversos
69
34 Un corte en la rama derecha permitirá que el ventrículo derecho se contrajera, pero a un ritmo distinto.
Transmisión del impulso cardíaco en el cora-zón. (Guyton pág. 118) Se muestra el momento de aparición (en fracciones de segundo después de la aparición inicial en el nódulo SA) en dife-rentes partes del corazón.

(o túbulos T). Este cambio de potencial en los túbulos T produce, como en el músculo cardía-co, que el retículo sarcoplasmático (RS) abra sus canales de Ca2+, liberándolo al sarcoplas-ma. No obstante, este Ca2+ no será suficiente para que se produzca una contracción fuerte (el RS de la fibra cardíaca no está tan desarrollado como el de la fibra esquelética). El Ca2+
que nos falta se introducirá en el interior de la fibra gracias a la apertura de unos canales de
Ca2+ voltaje dependientes presentes en la membrana de los túbulos T. Huelga decir que es de suma importancia que el medio extracelular sea rico en calcio. Como en el músculo esquelético, el Ca2+ produce la contracción de los sarcómeros. Ya en el marco de la relajación, parte del Ca2+ volverá al RS y otra parte saldrá al espacio ex-tracelular siendo intercambiado por Na+ (un catión calcio por tres de sodio)35. El sodio inte-riorizado será intercambiado por potasio gracias a la ATPasa Na+/K+.
Potencial de acción en el músculo cardíaco
En la fibra esquelética el potencial de acción es bastante fugaz. Partimos de un potencial de membrana moderadamente negativo y, tras una pequeña despolarización inicial que alcanza un potencial de membrana umbral, se produce una despolarización abrupta desencadenada por un circuito de retroalimentación positiva en el que participan unos canales de Na+ Vdep rá-pidos que se van abriendo a medida que la membrana se va despolarizando. Los canales de
70
35 Esta bomba de membrana es bastante importante. Hay un amplio arsenal terapéutico focalizado en la activi-dad de esta bomba.
Mecanismos para el acoplamiento de excitación-contracción y relajación en el músculo cardíaco. (Guyton pág. 118) Parece ser que el potencial de acción desencadena la apertura de los canales de Ca2+ en los túbulos T y será éste calcio el que permita que salga el calcio del retículo sarcoplasmático (al menos así lo dijo el Dr. Ja-vierre). Nótese la salida del calcio del retículo sarcoplasmático en forma de chispas o pulsos.

K+ Vdep, que se estaban empezando a abrir desde un primer momento (pero tienen una ciné-tica más lenta), serán los responsables de la repolarización. El potencial de acción en el músculo cardíaco dura más en el tiempo; presenta una meseta tras la espiga. La espiga se da por la apertura de los mismos canales de Na+ presentes en el músculo esquelético. La me-seta, en cambio, es fruto de la apertura de unos canales calcio-sodio36 lentos en su mem-brana que siguen introduciendo cationes en el sarcoplasma de la fibra (la membrana se-guirá despolarizada)37. Además de esta des-polarización mantenida en el tiempo, la per-
meabilidad de la membrana al K+ disminuye tras el inicio del potencial de acción. Esto impide la salida de cationes K+ durante la meseta y, por tanto, impide el regreso rápido del voltaje del potencial de acción a su nivel de reposo. Cuando los canales lentos de calcio-sodio se cierran después de 0,2 a 0,3 s y se interrumpe el flujo de entrada de iones calcio y sodio, también aumenta rápidamente la permeabilidad al
K+ de la membrana, devolviendo el potencial de membrana a su nivel de reposo.
Hasta ahora hemos visto el potencial de acción en el músculo cardíaco, ahora veremos lo que sucede con el potencial de acción que se da en el nódulo SA. Recordemos que los mio-
71
36 Parece ser que también son los responsables de favorecer la contracción.
37 El Dr. Javierre hizo una aclaración que ya se comentó el año pasado en Biofísica: a nivel de cargas, los cam-bios se dan en zonas adyacentes en la membrana. Recordemos que el medio intracelular es conductor, por lo que los excesos de cargas se irán hacia la interfase entre este conductor (citoplasma) y el medio extracelular.
Fuerza de contracción del músculo cardíaco ventri-cular, que muestra también la duración del periodo refractario y del periodo refractario relativo. (Guyton pág. 103) El tiempo refractario es mayor que en el músculo esquelético, dando tiempo a que se produz-ca la fase mecánica.
Potencial de acción (en mV) de una fibra de Purkinje del corazón, que muestra una meseta (plateau). (Guyton pág. 66) (0) En la espiga inicial hay un incremento de la permeabilidad al Na+; (1) la permeabilidad al Na+ disminuye e incrementa la de K+; (2) disminuye la permeabilidad al K+ y se incrementa la del Ca2+; (3) incre-menta la permeabilidad al K+ y disminuye la permeabilidad al Ca2+; (4) potencial en reposo.
(0)
(1)(2)
(3)
(4)

cardiocitos especializados que forman el nó-dulo SA no son contráctiles. El potencial en membrana en reposo38 de estas células es de -60 mV (compárese con los -85 a -90 mV del músculo cardíaco). La causa de esta menor ne-gatividad es que las membranas celulares de las fibras sinusales son permeables naturalmente a los iones sodio y calcio. La despolarización que se observa en el complejo lento (así se denomina esta clase de potencial de acción) se debe principalmente a la entrada de sodio y calcio a través de los ca-nales calcio-sodio lentos (iCa2+ > if [Na+]). Esta despolarización más acelerada se dará gracias a que se llega a un potencial umbral (threshold en la imagen). La repolarización que viene a continuación se debe a un incremento de la permeabilidad de la membrana al potasio y la disminución de la permeabilidad al calcio. El complejo lento que observamos tanto en el nódulo SA como en el AV presenta tres parámetros que pueden ser modificados para que así la frecuencia cardíaca sea distinta:
- Potencial de reposo. Si el potencial de reposo se hace más negativo, se tardará más en llegar al potencial umbral, por lo que la frecuencia cardíaca será menor. Este es el efec-to que tiene el parasimpático. Por contra, el simpático hace que el potencial en reposo sea menos negativo, por lo que la frecuencia cardíaca será mayor.
- Pendiente. Nos estamos refiriendo a la pendiente de la recta que une un punto ubica-do a nivel del potencial de reposo y otro ubicado en el potencial umbral. Cuando mayor sea la pendiente, mayor será frecuencia cardíaca puesto que se tardará menos en alcanzar el potencial umbral. El parasimpático disminuye la pendiente y el simpático la aumenta.
- Potencial umbral. El potencial umbral es la diferencia de potencial que permite la apertura progresiva de los canales de calcio que aceleran la despolarización. Si este parámetro es alto, la frecuencia cardíaca será menor (y viceversa).
Despolarizaciones vistas como vectores
Las células con capacidad para autodespolarizarse (p. ej. nódulo SA) transmitirán el poten-cial de acción que generan a lo largo de todo el músculo cardíaco. La despolarización de una fibra cardíaca es un fenómeno de membrana; durante ésta el medio extracelular inmediato a la fibra presenta un exceso de carga negativa y el medio intracelular inmediato a la mem-brana presenta un exceso de carga negativa (al contrario que normalmente). En la figura de la siguiente página (apartado A) vemos como esta despolarización se representa en forma de un vector que va de izquierda a derecha. Además, esta fibra dispone de dos electrodos: el
72
38 Habitualmente se habla de un potencial de reposo inestable.
Descarga rítmica de una fibra del nódulo sinusal. (Guyton pág. 116) El complejo lento experimenta una despolarización progresiva ya que en ella el sodio no juega un papel decisivo. Observamos po-tenciales de esta clase en neuronas que participan en automatismos (p. ej., neuronas del SNE).

izquierdo está en una zona de negatividad y el derecho en una de positividad, por lo que el medidor registra un valor positivo. Cuando la onda despolarizante ha pasado por los dos electrodos, el registro muestra un valor de 0 (los dos electrodos están en zonas de igual nega-
tividad). Con la repolarización ocurrirá todo lo contrario: se registrarán valores negativos (ya que el electrodo izquierdo está en una zona más positiva que el derecho) que pro-gresivamente que alcanzarán un máximo para después llegar al 0.
Si hemos asumido que la despolarización de una única fibra cardíaca da lugar a un vector, la suma de todas estas fibras cardíacas darán lugar a un vector producto de la suma de todas las
despolarizaciones. Si conozco dónde está el órgano (corazón) y dónde he colocado los elec-trodos, mirando el registro que obtenga podré averiguar hacia donde se está produciendo la despolarización. Si un frente positivo se está acercando a un electrodo éste detectará valores positivos, si se aleja detectará valores negativos y si avanza perpendicular a éste obtendré valores neutros (registro isoeléctrico).
Electrocardiograma normal
El electrocardiograma (ECG) normal nos permite registrar la actividad eléctrica del corazón. Estrictamente el ECG es un registro de los cambios de potencial de la superficie del cuerpo producidos por la despolarización y repolarización del corazón. Se forma a partir de la suma de los potenciales de acción de todas las células que forman el músculo cardíaco (véase imagen de la pág. siguiente). En el ECG distinguimos varias ondas:
- Onda P. Corresponde al nódulo SA y a la despolarización siguiente de las aurículas.- Complejo QRS. Nos informa de la despolarización del ventrículo. La repolarización
auricular se da aquí también, pero no se observa porque es muy pequeña en compa-ración con la despolarización ventricular.
- Onda T. Representa la repolarización ventricular. Atención, si la despolarización ven-tricular genera una onda tan abrupta, lo lógico es que la repolarización generara una onda en sentido contrario a la del QRS. Esto no es así (de ahí que la onda T sea positi-va) porque la repolarización, al contrario que la despolarización, se da desde el mio-
cardio al endocardio.
73
Registro de la onda de despolarización (A y B) y de la onda de repolarización (C y D) de una fibra muscular cardíaca. (Guyton pág. 122) Para comprender el re-gistro de la derecha es necesario saber que restamos el potencial detectado por el electrodo (-) al detectado por el electrodo (+) [electrodo derecho - electrodo izquierdo].

Derivaciones electrocardiográficas
Los electrodos que nos permiten registrar los fenómenos eléctricos del corazón deben estar colocados en puntos específicos para que podamos sacar conclusiones. Existen tres clases de derivaciones:
- Derivaciones estándar (I, II y III). También pueden denominarse derivaciones bipolares. Por bipolar entendemos que el ECG se registra a partir de dos electrodos que están localizados en lados diferentes del corazón. Por tanto, una de estas derivaciones no es un único cable que procede del cuerpo, sino una combinación de dos cables y sus electrodos para formar un circuito completo entre el cuerpo y el electrocardiógrafo.
- Derivaciones unipolares ampliadas (aVR, aVL y aVF). aVR (Eng: augmented vector right) = brazo derecho; aVL = brazo izquierdo; aVF (Eng: augmented vector foot) = pie.
- Derivaciones torácicas (precordiales; V1-V6). Si bien las derivaciones estándar y las uni-polares ampliadas nos informaban sobre la actividad eléctrica en el plano coronal las torácicas nos informan en un plano axial.
Triángulo de Einthoven
El triángulo de Einthoven es un triángulo equilátero formado por la unión de estos tres pun-tos: la raíz de los dos miembros inferiores y el final del tronco. En el centro de este triángulo se encuentra el corazón. Sobre este triángulo se estudian las tres derivaciones estándar:
- Derivación I. Mide la diferencia de potencial existente entre el brazo derecho y el bra-zo izquierdo. La negatividad está en el brazo derecho y la positividad en el izquierdo.
74
Actividad eléctrica del corazón. (Netter pág. 103) Los potenciales de acción varían a medida que la onda de despolarización avanza a través de las vías de conducción del corazón, comenzando en el nodo SA y termi-nando en el músculo ventricular.
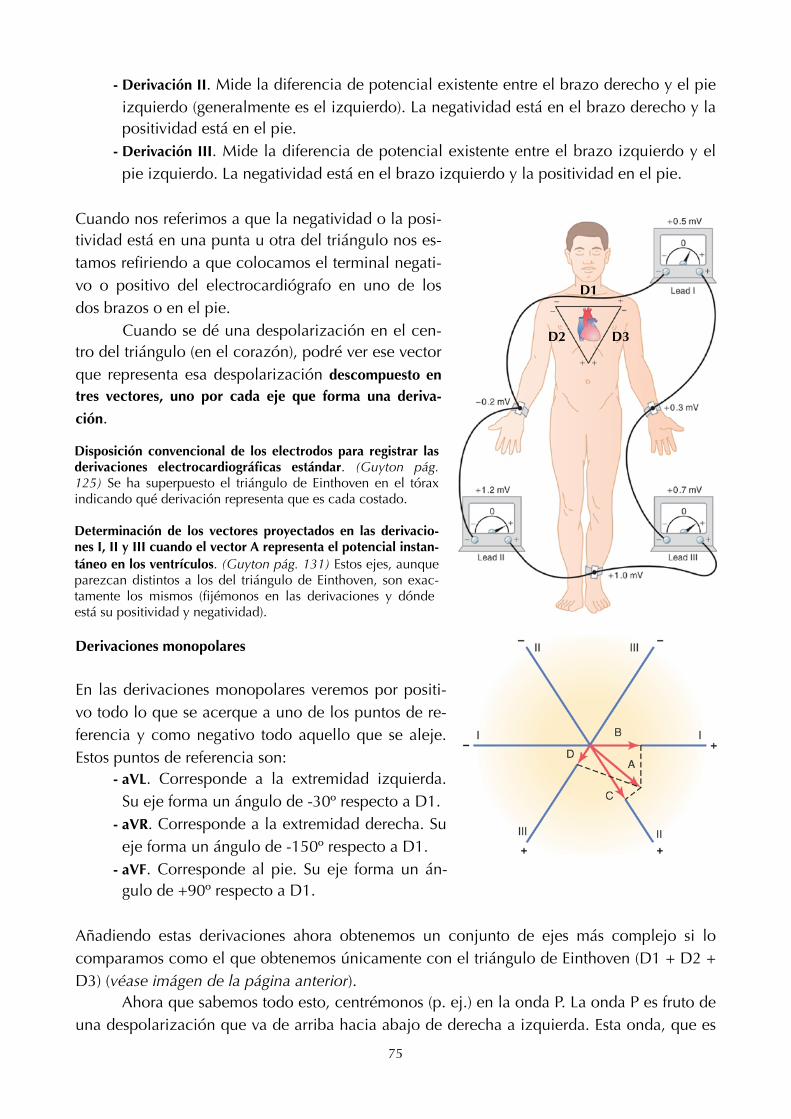
- Derivación II. Mide la diferencia de potencial existente entre el brazo derecho y el pie izquierdo (generalmente es el izquierdo). La negatividad está en el brazo derecho y la positividad está en el pie.
- Derivación III. Mide la diferencia de potencial existente entre el brazo izquierdo y el pie izquierdo. La negatividad está en el brazo izquierdo y la positividad en el pie.
Cuando nos referimos a que la negatividad o la posi-tividad está en una punta u otra del triángulo nos es-tamos refiriendo a que colocamos el terminal negati-vo o positivo del electrocardiógrafo en uno de los dos brazos o en el pie. Cuando se dé una despolarización en el cen-tro del triángulo (en el corazón), podré ver ese vector que representa esa despolarización descompuesto en tres vectores, uno por cada eje que forma una deriva-
ción.
Derivaciones monopolares
En las derivaciones monopolares veremos por positi-vo todo lo que se acerque a uno de los puntos de re-ferencia y como negativo todo aquello que se aleje. Estos puntos de referencia son:
- aVL. Corresponde a la extremidad izquierda. Su eje forma un ángulo de -30º respecto a D1.
- aVR. Corresponde a la extremidad derecha. Su eje forma un ángulo de -150º respecto a D1.
- aVF. Corresponde al pie. Su eje forma un án-gulo de +90º respecto a D1.
Añadiendo estas derivaciones ahora obtenemos un conjunto de ejes más complejo si lo comparamos como el que obtenemos únicamente con el triángulo de Einthoven (D1 + D2 + D3) (véase imágen de la página anterior). Ahora que sabemos todo esto, centrémonos (p. ej.) en la onda P. La onda P es fruto de una despolarización que va de arriba hacia abajo de derecha a izquierda. Esta onda, que es
75
Disposición convencional de los electrodos para registrar las derivaciones electrocardiográficas estándar. (Guyton pág. 125) Se ha superpuesto el triángulo de Einthoven en el tórax indicando qué derivación representa que es cada costado.
Determinación de los vectores proyectados en las derivacio-nes I, II y III cuando el vector A representa el potencial instan-táneo en los ventrículos. (Guyton pág. 131) Estos ejes, aunque parezcan distintos a los del triángulo de Einthoven, son exac-tamente los mismos (fijémonos en las derivaciones y dónde está su positividad y negatividad).
D1
D3D2

pequeña, se observará bien en D2, mientras que en aVR la veremos negativa (ya que va justa-mente en el sentido opuesto). El espacio que sigue a la onda P (nódulo AV) es isoeléctrico.
Igual que hemos hecho con la aurícula, conociendo los ejes y estudiando el registro que ob-tenemos podremos averiguar hacia donde se está produciendo la despolarización y si ésta es normal o no.
Tiempo y voltaje en electrocardiograma
Si observamos un ECG estándar veremos como éste se imprime en un papel cuadriculado. Cada milímetro (pequeño cuadrado) representa 0,04 s en el eje horizontal y 0,1 mV en el eje vertical. En otras palabras: 25 mm (x) es igual a 1 s y 10 mm (y) es igual a 10 mV. Si, por la razón que sea, queremos ver mejor el tiempo, podemos hacer pasar más rápidamente el papel (p. ej., de 25 mm/s a 50 mm/s).
El intervalo P-R o P-Q es el tiempo que transcurre entre el inicio de la onda P y el comienzo del complejo QRS, representa que es el intervalo existente entre la excitación eléctrica de las
76
Ejes de las derivaciones estándar y monopolares (izquierda) y despolarización auricular (derecha). (Guyton pág. 133) En cuanto a la imagen de la derecha, observamos como se observa la onda P en las tres derivaciones estándar. Nótese que en las tres aparece un registro positivo.
Electrocardiograma normal. (Guyton pág. 121)

aurículas y el inicio de la excitación de los ventrículos (nos informa sobre la transmisión del impulso a través de la aurícula). Su duración aproximada es de unos 0,16 s. El intervalo Q-T nos informa sobre la contracción ventricular y dura aproximadamente unos 0,35 s. Finalmente, el intervalo RR nos indica cual puede ser la frecuencia cardíaca, ya que su valor es el tiempo que hay entre dos latidos cardíacos. El RR casi nunca se mantiene a lo largo del tiempo, varía ligeramente aunque la frecuencia cardíaca por minuto sea la mis-ma (variabilidad cardíaca). El Dr. Javierre nos dijo que en un ECG debíamos identificar los siguientes elementos: (a) ritmo cardíaco (midiendo el RR); (b) ritmicidad (si hay arritmia o no); (c) ritmo sinusal; (d)
nódulo AV (midiendo el PR, que debe oscilar entre 0,12 y 0,20 s); (e) despolarización ventricu-
lar adecuada (complejo QRS); (f) sístole eléctrica ventricular (intervalo QT de entre 0,3 y 0,4 s). El ritmo cardíaco en reposo es de 60 a 80 lpm. Por debajo de 60 lpm se habla de una bradi-cardia moderada y por debajo de 40 lpm ya se habla de una bradicardia severa. Un ritmo en reposo por encima de 80 lpm es propio de una persona con taquicardia. El ritmo sinusal es aquel cumple las siguientes condiciones: (a) ritmo cardíaco de 60 a 100 lpm; (b) ritmicidad; (c) onda P que precede al complejo QRS; (d) onda P positiva en aVF y D2 y negativa en aVR; (e)
intervalo PR (0,12-0,20 s); (f) QRS menor a 0,12 s.
Bloqueo AV. Cuando hay un problema en el sistema de conducción cardíaco hablamos de arritmias. El término bloqueo auriculoventricular (bloqueo cardíaco) se refiere a un grupo de arritmias cardíacas que
se asocian a alteraciones de la conducción a través del nódulo AV. El bloqueo AV de tercer grado (hay de primero y de segundo también) se produce cuando la onda de despolarización no es conducida a través del nodo AV. En esta situación puede darse un ritmo de escape producido por la autoexcitación del nodo
AV (40-50 lpm).
Derivaciones torácicas o precordiales
Es común registrar seis derivaciones estándar del tórax, que van desde la pared anterior del tórax hasta la parte lateral izquierda/posterior. Las seis derivaciones son: V1, V2, V3, V4, V5 y V6. Como las superficies del corazón están próximas a la pared torácica, cada una de las derivaciones precordiales registra principalmente el potencial eléctrico de la muscula-tura cardíaca que está inmediatamente debajo del electrodo. Esto es especialmente útil para detectar anomalías en la pared
ventricular anterior.
77
Bloqueo AV de tercer grado. (Netter pág. 102). Se observan intervalos P-P y R-R regulares, pero las ondas P están disociadas de los complejos QRS.
18
D. torácicas. Derivaciones según Wilson V1, V2, V3, V4, V5, V6
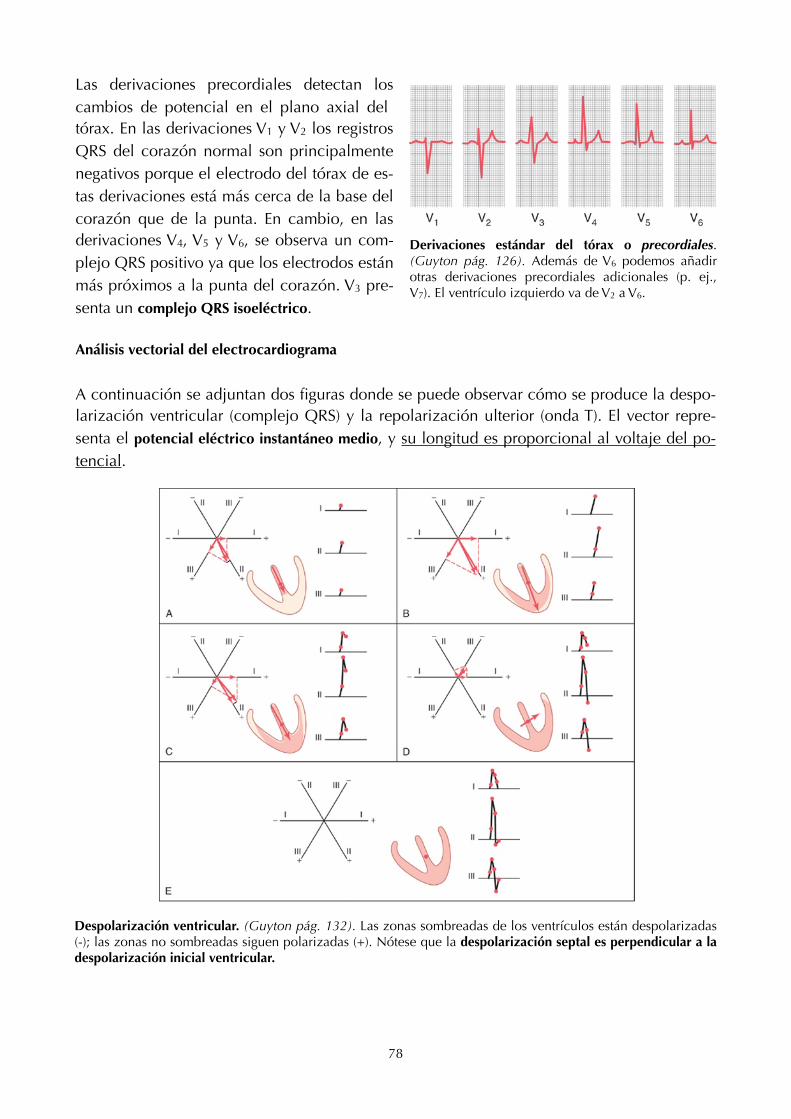
Las derivaciones precordiales detectan los cambios de potencial en el plano axial del tórax. En las derivaciones V1 y V2 los registros QRS del corazón normal son principalmente negativos porque el electrodo del tórax de es-tas derivaciones está más cerca de la base del corazón que de la punta. En cambio, en las derivaciones V4, V5 y V6, se observa un com-plejo QRS positivo ya que los electrodos están más próximos a la punta del corazón. V3 pre-senta un complejo QRS isoeléctrico.
Análisis vectorial del electrocardiograma
A continuación se adjuntan dos figuras donde se puede observar cómo se produce la despo-larización ventricular (complejo QRS) y la repolarización ulterior (onda T). El vector repre-senta el potencial eléctrico instantáneo medio, y su longitud es proporcional al voltaje del po-tencial.
78
Despolarización ventricular. (Guyton pág. 132). Las zonas sombreadas de los ventrículos están despolarizadas (-); las zonas no sombreadas siguen polarizadas (+). Nótese que la despolarización septal es perpendicular a la despolarización inicial ventricular.
Derivaciones estándar del tórax o precordiales. (Guyton pág. 126). Además de V6 podemos añadir otras derivaciones precordiales adicionales (p. ej., V7). El ventrículo izquierdo va de V2 a V6.

79
Generación de onda T durante la repolarización de los ventrículos (Guyton pág. 133). El tiempo total desde el inicio de la onda T hasta su final es de aproximadamente 0,15 s.

12. REGULACIÓN DEL GASTO CARDÍACO
Introducción
Como ya se ha dicho en temas anteriores, el gasto cardíaco es igual al producto de la fre-cuencia cardíaca por el volumen sistólico. Éste puede ser considerado como un índice indi-
recto de las necesidades de oxígeno39 de nuestro organismo, tal y como refleja la siguiente fór-mula:
El gasto cardíaco también está relacionado con la presión arterial y con las resistencias peri-féricas totales, siendo éste igual al cociente entre estos dos últimos factores citados:
Al igual que ocurría con la frecuencia cardíaca, que a pesar de decir un valor por minuto ésta variaba a cada latido (variabilidad cardíaca), el gasto cardíaco también cambia a cada la-tido, puesto que el corazón no se llena exactamente igual en cada ciclo cardíaco. Por tanto, cuando hablamos de gasto cardíaco en realidad nos referimos al volumen de sangre medio que
expulsa el corazón por minuto (l/min).
Ejemplo diapositivas. Se observa como el corazón, ante una bajada de la fracción de eyección del 60%
al 55%, mantenga el volumen sistólico (incrementaremos el volumen telediastólico). La disminución de la fracción de eyección es lo mismo que decir que ha disminuido la contractibilidad del corazón.
Para modificar el rendimiento cardíaco tenemos dos grandes clases de factores: (a) factores homeométricos y (b) factores heterométricos. Los primeros modifican el rendimiento sin alterar la longitud de la fibra cardíaca (con la misma distensión cardíaca), mientras que los segun-dos sí que modifican la distensión.
Ley de Frank-Starling
La ley de Frank Starling dice que “la energía liberada al pasar del estado de reposo al estado de contracción depende de la longitud de la fibra muscular, esto es, de la superficie quími-camente activa”. En otras palabras, cuando la precarga se incremente (VTD), también lo hará el
80
39 Y otras como por ejemplo la necesidad de repartir el calor corporal (termorregulación). No obstante, bajo condiciones estables, lo que nos modificará el gasto cardíaco son las necesidades de oxígeno. Las necesidades de oxígeno dependerán en parte de la capacidad de nuestros tejidos para aprovechar este oxígeno, para poder extraerlo de la sangre. Si tras un infarto estamos mucho tiempo en cama, empeoramos esta capacidad de apro-vechamiento y por tanto, cuando volvemos a la actividad, ponemos de nuevo a prueba nuestro corazón porque tendrá que realizar un mayor gasto cardíaco. Estar en cama también disminuye el volumen sanguíneo.
GC =
presión arterialresistencias totales periféricas
Consumo O2 = GC ⋅diferencia arteriovenosa para el O2
El gasto cardíaco es directamente proporcional a la presión arterial e inversamente proporcional a las resis-tencias periféricas. Si hay una vasodilatación periférica, para mantener la presión arterial hay que incrementar el gasto cardíaco.
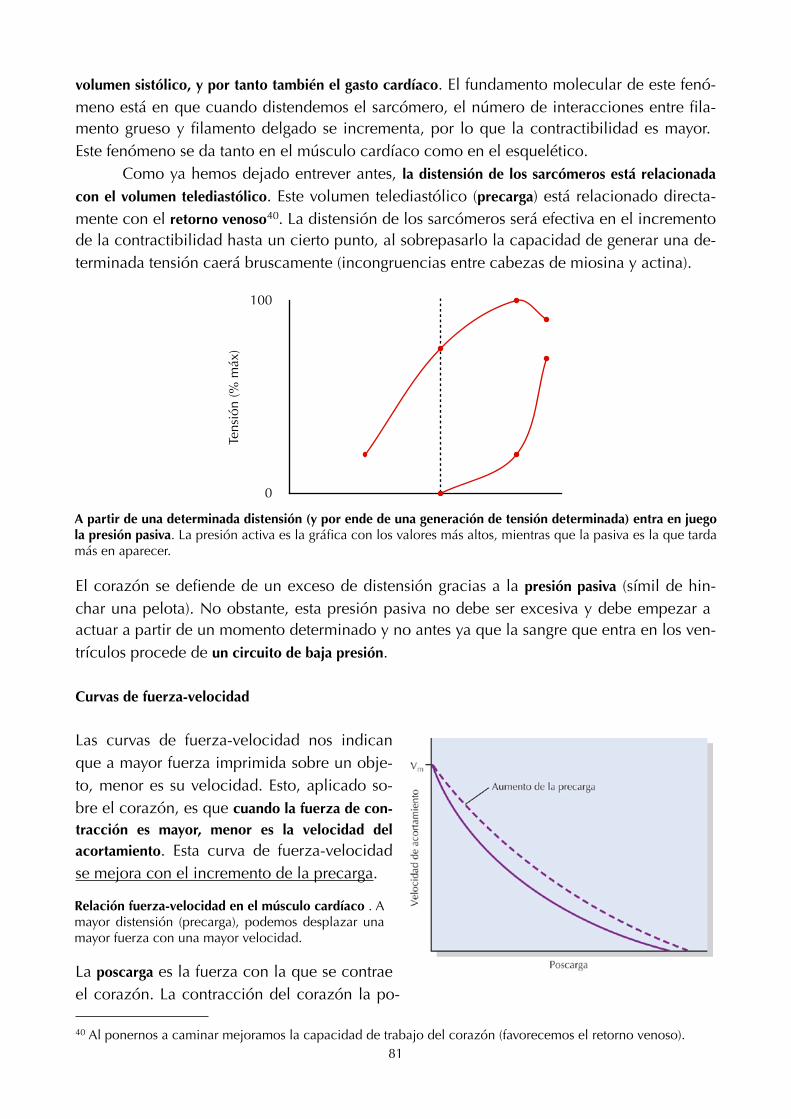
volumen sistólico, y por tanto también el gasto cardíaco. El fundamento molecular de este fenó-meno está en que cuando distendemos el sarcómero, el número de interacciones entre fila-mento grueso y filamento delgado se incrementa, por lo que la contractibilidad es mayor. Este fenómeno se da tanto en el músculo cardíaco como en el esquelético. Como ya hemos dejado entrever antes, la distensión de los sarcómeros está relacionada
con el volumen telediastólico. Este volumen telediastólico (precarga) está relacionado directa-mente con el retorno venoso40. La distensión de los sarcómeros será efectiva en el incremento de la contractibilidad hasta un cierto punto, al sobrepasarlo la capacidad de generar una de-terminada tensión caerá bruscamente (incongruencias entre cabezas de miosina y actina).
El corazón se defiende de un exceso de distensión gracias a la presión pasiva (símil de hin-char una pelota). No obstante, esta presión pasiva no debe ser excesiva y debe empezar a actuar a partir de un momento determinado y no antes ya que la sangre que entra en los ven-trículos procede de un circuito de baja presión.
Curvas de fuerza-velocidad
Las curvas de fuerza-velocidad nos indican que a mayor fuerza imprimida sobre un obje-to, menor es su velocidad. Esto, aplicado so-bre el corazón, es que cuando la fuerza de con-tracción es mayor, menor es la velocidad del acortamiento. Esta curva de fuerza-velocidad se mejora con el incremento de la precarga.
La poscarga es la fuerza con la que se contrae el corazón. La contracción del corazón la po-
81
40 Al ponernos a caminar mejoramos la capacidad de trabajo del corazón (favorecemos el retorno venoso).
0
100
Tens
ión
(% m
áx)
A partir de una determinada distensión (y por ende de una generación de tensión determinada) entra en juego la presión pasiva. La presión activa es la gráfica con los valores más altos, mientras que la pasiva es la que tarda más en aparecer.
Relación fuerza-velocidad en el músculo cardíaco . A mayor distensión (precarga), podemos desplazar una mayor fuerza con una mayor velocidad.

dremos ver manifestada con la presión aórtica, que recordemos que ésta debe ser superada para que podamos expulsar el contenido del ventrículo. Una persona con hipertensión pre-sentará un ventrículo hipertrofiado precisamente para compensar ese exceso de presión en la aorta. Con el tiempo el corazón entra en insuficiencia y, con el fin de preservar la cantidad de sangre que se eyecta en cada sístole, ve su tamaño incrementado (au-
mento de su circunferencia). La gran mayo-ría de insuficiencias van apareciendo con la edad y no suelen dar una gran patolo-gía. Este corazón agrandado se puede ob-servar con facilidad en una radiografía.
Factores heterométricos
Para regular el gasto o rendimiento cardíaco tenemos dos grandes clases de factores como ya es ha dicho previamente. A continuación vamos a discutir los denominados heterométricos, que son aquellos que tienen que ver con modificaciones en la longitud del sarcómero.
- Volemia. La volemia es uno de los principales factores heterométricos. Reducciones de la volemia de hasta el 15-20% podrán ser bien toleradas41 y compensadas gracias a la gran cantidad de sangre que hay en el territorio venoso. Si la pérdida de volemia es superior (el doble de esos valores de tolerancia) ya se pone en compromiso el mante-nimiento del gasto cardíaco y se tendrán que adoptar medidas para restaurar la vole-mia (p. ej., transfusión sanguínea).
- Posición corporal. Cuando estamos en decúbito supino las diferencias de presión a nivel del retorno venoso son bajas, pudiendo encontrar presiones de unos 12 mmHg en los pies y en el corazón derecho de 5 mmHg. En cambio, en bipedestación nos encon-tramos con que se acumula sangre en las venas que están por debajo del corazón (grave-dad). Esto puede suponer una reducción del retorno venoso del 20%, por lo que tam-bién se reduce el volumen sistólico (e incrementaremos la frecuencia cardíaca para mantener el gasto). Cuando nos acostamos, por tanto, la frecuencia cardíaca no dis-minuye únicamente por la ausencia de actividad, si no por una causa hemodinámica.
- Contracción muscular. La contracción muscular colabora en el retorno venoso, ya que al acortar el músculo se incrementa el diámetro del mismo y se presiona a la vena. Recordemos que las venas tienen un sistema de válvulas que impide el flujo retrógra-do.
- Tono venoso. Por tono venoso entendemos el tono de contracción de la musculatura lisa
de las venas, que está fuertemente regulado por el simpático. La acción del simpático reduce el calibre de las venas y dificulta el retorno venoso.
82
41 A lo sumo habrá una disminución de la presión arterial hasta los 70-80 mmHg.
Corazón normal Insuf. cardíacaCircunferencia (cm)Final diástole 35 48Final sístole 26 45Acortamiento 25,71% 6,25%Volumen eyectado 60 ml 60 mlFracción de eyección 60% 25%
Diferencia entre un corazón sano y otro insuficiente.

- Barrera toraco-abdominal. Al inspirar incrementamos la presión negativa en el territorio pulmonar, favoreciendo la llegada de sangre al mismo. Atención, inspirando se difi-culta el retorno de la sangre a la aurícula izquierda.
- Pericardio. El pericardio consta de dos capas entre las cuales se encuentra el líquido
pericárdico. Un incremento de este líquido (p. ej., pericarditis) deja con menos espa-cio al corazón y dificulta la distensión miocárdica.
- Distensibilidad miocárdica. Es la capacidad del corazón para distenderse. Esta capaci-dad disminuye con la edad, incrementándose el riesgo de padecer infartos.
Factores homeométricos
Los factores homeométricos regulan el gasto cardíaco independientemente de la longitud del sarcómero. Dentro de los factores homeométricos distinguimos los (a) factores tónicos, que actúan en todo momento; y los (b) factores fásicos, que actúan en situaciones concretas. Dentro de los factores tónicos encontramos la edad (disminuye la actividad ATPásica de las cabezas de misiona) y factores de tipo endocrino como el hiper- y el hipotiroidismo (in-crementa y disminuye la contractibilidad respectivamente). En cuanto a los factores fásicos, quizá el más conocido sea el efecto Treppe (o efecto
escalera), en virtud al cual la contractibilidad irá aumentando de manera proporcional al in-cremento de la frecuencia cardíaca (y en sentido contrario también). La explicación molecu-lar de este hecho es porque el Ca2+ citoplasmático será mayor, favoreciéndose la contrac-ción42.
En la gráfica de la derecha vemos como la presión a la que se debe abrir la válvula sigmoidea es la mis-ma con o sin aplicación del factor homeométrico positivo. En otras palabras: se modifica la contracti-bilidad, que no es lo mismo que decir que aumenta la
poscarga. La poscarga es la tensión que debe generar la pared cardíaca. En la siguiente página veremos lo que ocurre cuando se incrementa la poscarga por, p. ej., un incremento de la presión arterial. En este caso lo que ocurrirá es que el corazón deberá generar una mayor presión puesto que debe vencer una oposición (en la aorta) supe-rior. Como la presiones en las que me moveré serán mayores, las válvulas sigmoideas tardarán
83
42 No hay que confundir esto con una tetanización. La tetanización es un proceso que se da en la musculatura y que consiste en que ante una frecuencia de estímulos elevada, se va acumulando la contracción. También se habló del opistótonos, que es cuando un paciente tiene tensa toda su musculatura, adquiriendo la posición de arco tenso (paciente sostenido por talones y coronilla).
1
Efecto de los factores homeométricos. En azul vemos la gráfica volumen-presión normal y en rojo la gráfica tras aplicar un fac-tor homeométrico positivo. Sin modificar la poscarga se produ-ce: (1) incremento de la fracción de eyección, y (2) disminu-ción del VSF.

más en abrirse y se cerrarán antes, resultando en una bajada del volumen sistólico y un incremento del volumen residual (volumen telesistólico). Finalmente, si se observa un incremento en la
precarga, eso quiere decir que aumentamos el vo-lumen telediastólico pero que, como eyectamos más sangre del corazón (aumento del volumen sistóli-co), el volumen telesistólico se mantiene.
Síntesis
El gasto cardíaco se verá modificado por mis nece-sidades, aunque siempre teniendo en mente lo que sucede en la periferia (p. ej., un aumento de la ex-tracción de O2 puede hacer que el GC sea menor). Como ya sabemos el GC es el producto de la FC y el VS, y el VS depende tanto de la precarga como de la contractibilidad. El GC además va a ser dependiente de: (1)
resistencias periféricas, entendiendo éstas como la suma de la contracción de las arteriolas de mi or-ganismo; y (2) la presión arterial, que a su vez tam-bién depende de las resistencias periféricas y de como funciona el corazón. La presión arterial será la poscarga, ya que la tensión que debe realizar la pared cardíaca está relacionada con la presión que se debe solventar en la base de la aorta (siempre en condiciones de normalidad). Todo esto será así si no tenemos en cuenta el diámetro del corazón. Si el diámetro aumenta, la posibilidad de incrementar el GC se limita.
84
2
Control de la
función cardíaca
Medicina. Bellvitge, 2012.
1

13. CONTROL DE LA FRECUENCIA CARDÍACA
Introducción: sistema vegetativo
Antes hemos visto factores que regulan el gasto cardíaco, ahora veremos los que regulan la frecuencia cardíaca (FC). Eminentemente, la frecuencia cardíaca depende de un control vegeta-
tivo (simpático + parasimpático). Las fibras simpáticas que inervan a nuestro corazón parten de los cinco primeros segmentos torácicos y de los dos últimos cervicales. A partir de estas fibras se generará el plexo cardíaco. El control de la frecuencia cardíaca desde el punto de vista simpático es asimétrico: el lado derecho sí que actúa fundamentalmente sobre la FC, mientras que el lado izquierdo tiene un mayor efecto sobre la contractibilidad. Esto se observa con facilidad si estimulamos una parte u otra del simpático: al estimular el ganglio estrellado izquierdo se incrementa la
presión sistólica (modificación de la contractibilidad), mientras que si estimulamos el ganglio estrellado derecho incrementamos la FC43. Mientras que el simpático es taquicardizante, el parasimpático disminuye la FC. Se ori-gina en el núcleo ambiguo y el núcleo motor dorsal del bulbo. De estos dos núcleos parten unas fibras que serán conducidas por el nervio vago (X). Formará parte del plexo cardíaco e inerva-rá directamente al nódulo SA (vago derecho) y al nódulo AV (vago izquierdo). Debido a la ele-vada cantidad de acetilcolinesterasa presente en el músculo cardíaco, el efecto del parasimpá-tico disminuirá rápidamente. El equilibrio entre la acción del parasimpático y la del simpático determinará la fre-cuencia cardíaca resultante. Si administramos un inhibidor del parasimpático (atropina; inhi-ve a los receptores muscarínicos) y después un inhibidor del simpático (propanolol; β-blo-queante), obtendremos la frecuencia cardíaca intrínseca, aquella que se observa sin control vegetativo. Esta frecuencia cardíaca intrínseca es superior a la basal que solemos tener: 110 latidos/min. Esto nos indica que en el control vegetativo, predomina el parasimpático.
Reflejos
Hemos visto como el sistema nervioso vegetativo va a ser el principal responsable del con-trol de la frecuencia cardíaca. Ahora bien, ¿qué es lo que activa a cada una de las divisiones del vegetativo? Principalmente los reflejos.
Reflejo barorreceptor
El reflejo barorreceptor depende de unos receptores sensibles al estiramiento que encontramos en el seno carotideo y en el cayado aórtico. Los receptores ubicados en el seno carotídeo emi-ten sus fibras hacia el nervio glosofaríngeo (IX), mientras que los receptores distribuidos en el cayado aórtico emitirán sus fibras hacia el nervio vago (X). Las fibras procedentes de ambas ubicaciones llegarán, vía tracto solitario, hasta el centro bulbar.
85
43 La presión sistólica también experimenta un leve aumento, nada que ver con el observado al estimular el ganglio estrellado izquierdo.

El efecto de este reflejo es inhibir la actividad simpática, por lo que disminuirá tanto la frecuencia cardíaca como la contractibilidad. Un discreto aumento de presión po-drá ser detectado por estos barorreceptores y producir una disminución del tono simpático y un ligero aumen-to del tono vagal. Si el incremento de presión es eleva-do (mayor de 25 mmHg), el reflejo barorreceptor provo-cará la supresión del simpático. Una disminución de la presión arterial provocará efectos contrarios: disminución del tono vagal e incre-mento del simpático. Existe una bajada umbral de pre-sión, de entre 20 y 25 mmHg, que cuando ocurre se da un importante incremento del tono simpático. Si el volumen sistólico es modificado, p. ej. se incrementa, este aumento provocará una distensión en el cayado aórtico que se traducirá en la inducción del reflejo barorreceptor y la disminución de la actividad simpática. Con esto controlamos rápidamente la respuesta
cardíaca ante modificaciones de presión.
Nitroprusiato y fenilefrina. El nitroprusiato es un vasodilatador, hará que la presión arterial baje y que la
frecuencia cardíaca se incremente (estímulo simpático). La fenilefrina incrementa la presión arterial y baja el ritmo cardíaco (disminución de la actividad simpática).
Reflejo de Bainbridge
También se puede denominar reflejo de los receptores auriculares. Si añadimos un cierto vo-lumen al retorno venoso observaremos un incremento de la frecuencia cardíaca (a FC bajas). Este aumento se da gracias a unos receptores presentes en la unión entre las grandes venas y las aurículas, que experimentarán una distensión cuando haya un incremento del volumen. A través del vago, estos receptores enviarán sus impulsos al SNC, respondiendo éste con una modulación de los sistemas simpático y parasimpático44. Si el volumen de retorno aumenta demasiado, también ascenderá el volumen sistólico, por lo que el reflejo barorreceptor sustituirá al de Bainbridge (es lo más probable). El SNC puede regular la volemia incrementando el volumen urinario por medio de dos vías: (a) vasopresina y (b) péptido natriurético auricular, que estimula la pérdida de sodio.
Arritmia sinusal respiratoria
Cuando hay una activación del nervio frénico se incrementa la actividad simpática, y viceversa cuando el nervio frénico es inhibido. Por tanto, por el simple hecho de estar respirando, ten-dré una arritmia respiratoria (al espirar no hay actividad simpática y sí parasimpática). Cuan-
86
44 Nunca es uno u otro, si no la acción combinada de los dos.

do hay espiración la longitud del ciclo aumenta (disminuye la FC), y cuando hay inspiración esta longitud se acorta (incrementa la FC). Cuando se da la inspiración, la distensión de bronquios y bronquiolos genera también una respuesta simpática. Por tanto, al hecho de modi-ficar la accción del vegetativo en función de la actividad frénica, hay que sumarle este efecto ventilatorio. El centro respiratorio se encarga de inducir una determinada ventilación y también va a tener un efecto directo sobre el centro cardíaco del bulbo (que modifica la FC). La propia respiración tendrá efectos indirectos sobre la FC:
- Cambios en la presión intratorácica. Los cambios en el volumen pulmonar excitan unos receptores de estiramiento que afectan al centro cardíaco.
- Cambios en el retorno venoso. La inspiración favorece el retorno venoso, por lo que se dará el reflejo de Bainbridge.
- Cambios en la tensión arterial. Los posibles cambios en la tensión arterial que puedan darse modificarán la FC gracias al reflejo barorreceptor.
Quimiorreceptores
Los quimiorreceptores pueden ser centrales (sensibles a H+ y al CO2) o periféricos (sensibles a modificaciones del O2). Los quimiorreceptores periféricos estimulan al centro vagal bulbar, disminuyendo así la frecuencia cardíaca. Simultáneamente, estos mismos quimiorreceptores incrementan la actividad respiratoria, lo que se traduce en un aumento del estiramiento pulmo-
nar y en una hipocapnia (disminución de la presión parcial de CO2). Estos dos efectos de la actividad respiratoria inhiben al centro vagal bulbar, por lo que la frecuencia cardíaca se mo-dificará en función al equilibrio entre el efecto primario de los quimiorreceptores periféricos (acción directa sobre el centro vagal) y el efecto secundario (actividad respiratoria inducida por los mismos quimiorreceptores).
Mirar última diapositiva del tema (los dos últimos esquemas)
Reflejo ventricular
Es similar al reflejo auricular (de Bainbridge). Se basa en unos receptores de presión ubica-dos en el ventrículo, tanto en el endocardio como en el epicardio. Su estimulación produce una disminución de la frecuencia cardíaca y las resistencias periféricas.
87

14. CIRCULACIÓN CORONARIA
Introducción
La circulación coronaria es especial e importante. Especial por las condiciones en que debe aportar sangre, e importante porque su defecto es una de las principales causas de morbili-dad o muerte en nuestro medio. Como ya dijimos en el tema introductorio al sistema cardiocirculatorio, la circulación pulmonar o menor está conectada en serie con la sistémica o mayor (todo lo que sale por el corazón izquierdo llegará al derecho). Por contra, los órganos que reciben la sangre están conectados en paralelo, y cada órgano/tejido recibirá un % de sangre distinto, que no tiene nada que ver con su extensión (p. ej., la piel recibe un 5% y el cerebro un 15%). Los tejidos que reciben más sangre serán aquellos que ofrezcan una menor resistencia a ella (mirar tablas de las diapositivas; comparación riñón vs musculatura esquelética). Nuestro sistema cardiocirculatorio tiende a preservar el volumen que destina a cada órgano. Por ejemplo, aunque el gasto cardíaco se duplique, a nuestro cerebro45 le seguirá llegando la misma cantidad de sangre por minuto (unos 750 mL/min). De igual manera en el caso contrario (disminución del volumen; asegurar el suministro a cerebro y corazón). El músculo esquelético en reposo consume poca sangre, mientras que el corazón, un músculo activo constantemente, le llegan unos 250 mL/min (84 ml · 100 g · min-1), gracias a su baja resistencia al paso de sangre.
Diferencia arterio-venosa de O2
Hay tejidos, como el renal, que reciben una gran cantidad de sangre pero que apenas la uti-lizan para uso propio. Esto se traduce en una gran llegada de sangre (25% del gasto cardía-co) y un bajo consumo de O2 (ligeramente superior al 7%). Si se consume poco O2, querrá decir que no habrá gran diferencia, en cuanto a oxigenación, entre la sangre arterial que en-tra en el órgano y la venosa que sale. Esta diferencia es la diferencia arterio-venosa de O2. El corazón, en cambio, recibe el 5% del gasto cardíaco y consume el doble de O2 (11%), por lo que aprovecha mucho el aporte de O2 que le llega (diferencia arterio-venosa = 114 vs 14 que observamos en el riñón). Es interesante conocer esto para tener presente que si bien es cierto que si queremos hacer trabajar al doble el corazón, tendremos que duplicarle el aporte de sangre puesto que la que recibe ya la aprovecha al máximo, con otros tejidos eso no sucede así. El músculo es-quelético, por ejemplo, cuando se activa aprovechará mejor la sangre que recibe en lugar de solicitar recibir más (y por lo tanto no sobrecarga al corazón).
Sistema coronario
Como ya se vio en Esplacnología, el árbol coronario parte de dos arterias coronarias que na-cen de la base de la aorta: las coronarias derecha e izquierda. De éstas ya surgirán otras que se
88
45 También se comentó que nuestro cerebro debe estar aislado de los cambios en la presión arterial.

encargarán de proporcionar nutrientes a otras partes del corazón, como las descendentes, la circunfleja, la marginal, etc. Podemos dividir la circulación coronaria en tres partes.
- Porción epicárdica (o subepicárdica). Cumple una función conductora. Aquí encontra-mos arterias musculares (ø = 0,3 mm) que ofrecen una muy baja resistencia al flujo (no se modificará significativamente la presión a pesar de que modifiquemos el flujo), manteniendo así la resistencia periférica. Podrán asumir una gran cantidad de flujo.
- Ramas musculares intramiocárdicas. Éstas sí que ofrecen una mayor resistencia al flujo sanguíneo. Producirán una disminución en la presión del mismo y están influenciadas por la presión que genera la pared muscular. Durante la sístole se comprometerá el aporte hacia las mismas.
- Capilares. Aquí es donde se produce el intercambio de elementos. Hay una gran ri-queza capilar (hasta 4.000/mm2). La densidad capilar está relacionada con la cantidad de músculo y su actividad (por tanto, en caso de hipertrofia, habrá también una hiper-plasia capilar).
La circulación coronaria se caracteriza también por ser funcionalmente terminal, esto quiere decir que tiene una gran cantidad de anastomosis. Nótese que se dice que es “funcionalmen-te” terminal y no sobre la práctica. Esto es así porque estas redes arteriales son poco eficien-tes y, por tanto, si se da un bloqueo en una de las arterias, estas anastomosis no serán lo sufi-cientemente efectivas como para paliar ese taponamiento. Ahora bien, si el bloqueo es pro-gresivo (p. ej., arteriosclerosis), esta circulación lateral sí que se puede estimular (e incluso pueden aparecer flujos retrógrados).
Metabolismo cardíaco
La riqueza vascular que encontramos en el corazón es extraordinaria. Podemos observar has-ta cuatro capilares flanqueando a una única fibra muscular. En estas fibras observamos una gran cantidad de mitocondrias, lo cual significa que: (1) al haber menor tejido contráctil la ca-
pacidad de tensión es menor (en comparación con la musculatura esquelética); y (2) serán fi-
bras muy poco fatigables (metabolismo oxidativo). Los miocitos cardíacos son células con una gran cantidad de mitocondrias (metabolis-mo aeróbico), por lo que pueden oxidar prácticamente cualquier sustrato: los productos de la β-oxidación (68% del total de la energía), el ácido láctico46 (15%) y la glucosa (16%). En
reposo el consumo de O2 es de 8 mL · min-1 · 100 g-1. Atención, en reposo no quiere decir en asistolia, si no que no estemos realizando una gran actividad física (70 lpm). Cuando reali-zamos una actividad física, éste consumo se incrementa hasta los 70 mL O2 · min-1 · 100 g-1. Este incremento del consumo de oxígeno se hará para incrementar el metabolismo aeróbico de las fibras musculares, no el anaeróbico. En condiciones de asistolia, el consume de oxíge-no será menor al que encontramos en reposo: 1,3 - 1,5 mL O2 · min-1 · 100 g-1. La asistolia puede darse en una parada cardíaca (RCP para introducir algo de sangre en las coronarias y
89
46 En el caso de estar realizando una actividad física, el lactato producido será aprovechado por el corazón (se convertirá en una fuente importante de energía). El corazón es considerado como un gran consumidor de lacta-to.
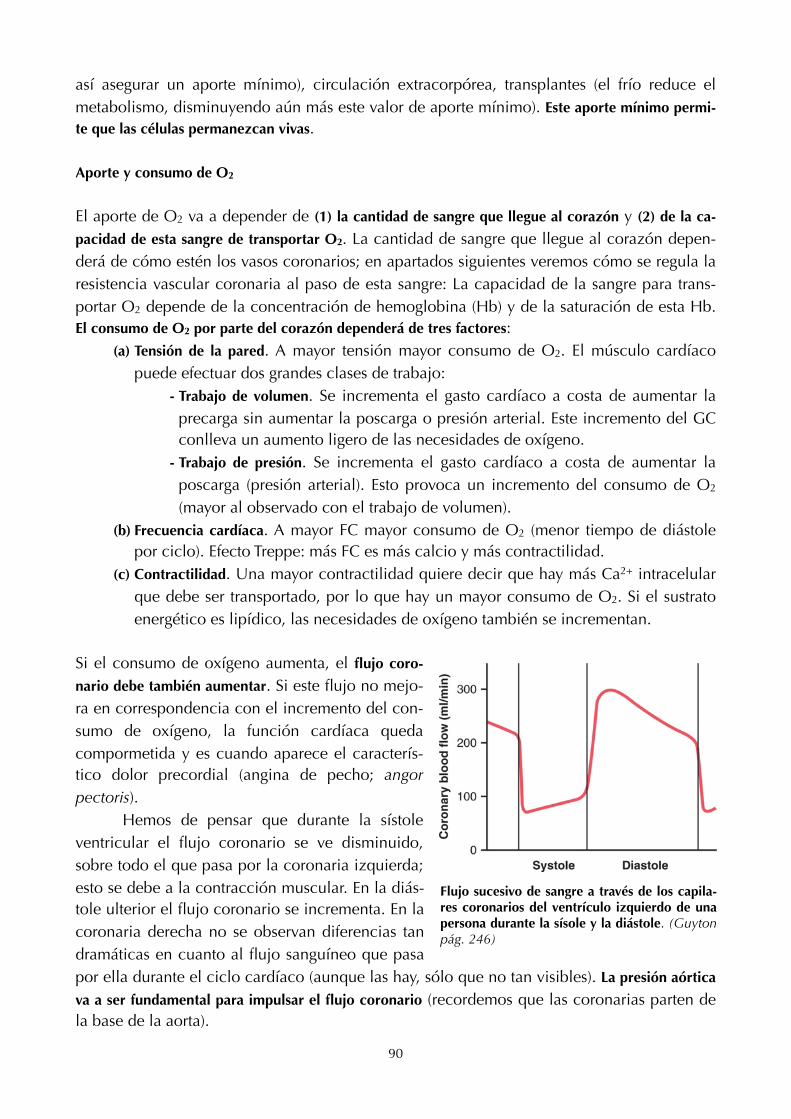
así asegurar un aporte mínimo), circulación extracorpórea, transplantes (el frío reduce el metabolismo, disminuyendo aún más este valor de aporte mínimo). Este aporte mínimo permi-te que las células permanezcan vivas.
Aporte y consumo de O2
El aporte de O2 va a depender de (1) la cantidad de sangre que llegue al corazón y (2) de la ca-
pacidad de esta sangre de transportar O2. La cantidad de sangre que llegue al corazón depen-derá de cómo estén los vasos coronarios; en apartados siguientes veremos cómo se regula la resistencia vascular coronaria al paso de esta sangre: La capacidad de la sangre para trans-portar O2 depende de la concentración de hemoglobina (Hb) y de la saturación de esta Hb. El consumo de O2 por parte del corazón dependerá de tres factores:
(a) Tensión de la pared. A mayor tensión mayor consumo de O2. El músculo cardíaco puede efectuar dos grandes clases de trabajo:
- Trabajo de volumen. Se incrementa el gasto cardíaco a costa de aumentar la precarga sin aumentar la poscarga o presión arterial. Este incremento del GC conlleva un aumento ligero de las necesidades de oxígeno.
- Trabajo de presión. Se incrementa el gasto cardíaco a costa de aumentar la poscarga (presión arterial). Esto provoca un incremento del consumo de O2 (mayor al observado con el trabajo de volumen).
(b) Frecuencia cardíaca. A mayor FC mayor consumo de O2 (menor tiempo de diástole por ciclo). Efecto Treppe: más FC es más calcio y más contractilidad.
(c) Contractilidad. Una mayor contractilidad quiere decir que hay más Ca2+ intracelular que debe ser transportado, por lo que hay un mayor consumo de O2. Si el sustrato energético es lipídico, las necesidades de oxígeno también se incrementan.
Si el consumo de oxígeno aumenta, el flujo coro-
nario debe también aumentar. Si este flujo no mejo-ra en correspondencia con el incremento del con-sumo de oxígeno, la función cardíaca queda compormetida y es cuando aparece el caracterís-tico dolor precordial (angina de pecho; angor pectoris). Hemos de pensar que durante la sístole ventricular el flujo coronario se ve disminuido, sobre todo el que pasa por la coronaria izquierda; esto se debe a la contracción muscular. En la diás-tole ulterior el flujo coronario se incrementa. En la coronaria derecha no se observan diferencias tan dramáticas en cuanto al flujo sanguíneo que pasa por ella durante el ciclo cardíaco (aunque las hay, sólo que no tan visibles). La presión aórtica
va a ser fundamental para impulsar el flujo coronario (recordemos que las coronarias parten de la base de la aorta).
90
Flujo sucesivo de sangre a través de los capila-res coronarios del ventrículo izquierdo de una persona durante la sísole y la diástole. (Guyton pág. 246)

Balón de contrapulsación intraaórtico (BIAC). Este balón consiste en un globo situado en la arteria aorta des-
cendente (desde la salida de la subclavia izquierda hasta las arterias renales) conectado a una consola que su-ministra gas (normalmente helio) para su inflado y desinflado. El BIAC se hincha durante la diástole para incre-
mentar la presión en la base de la aorta y así favorecer la perfusión coronaria. Durante la sístole se deshincha para permitir la salida de sangre por el ventrículo. Limitaciones: arritmia y que las válvulas cardíacas estén co-rrectamente.
Regulación del flujo coronario
Existen diferentes factores que pueden regular el flujo coronario (la resistencia que se ofrece al flujo): éstos pueden ser: (a) factores metabólicos; (b) factores endoteliales; (c) autorregulación; (d) factores humorales; (e) factores neurales y (f) fuerzas de compresión extravascular. Podemos afirmar, que el flujo coronario depende de la fase diastólica y de la resistencia vascular.
Factores metabólicos
El flujo coronario tiene una relación directa con la actividad metabólica. Si no hay un co-rrecto aporte de O2, el ATP de las fibras musculares se irá gastando, por lo que se recurrirá a la formación de un mol de ATP a costa de dos moles de ADP. Esta reacción tiene el inconve-niente de generar también AMP, que ya debe ser metabolizado y dará adenosina. La adenosi-na es un potentísimo vasodilatador, por lo que el flujo sanguíneo se incrementará, se mejora-rá el aporte de O2, se volverá a un equilibrio entre aporte y demanda de O2 y por tanto deja-rá de formarse adenosina.
Si el aporte es insuficiente por un problema estructural (p. ej., placa de ateroma), ya se puede estar formando adenosina y que se esté produciendo la vasodilatación consecuente que el flujo no mejorará. Pasado un tiempo, las células cardíacas habrán perdido tanta adenosina que aunque el flujo se restaure no podrán volver a formar ATP.
Mecanismos endoteliales
A mayor flujo se producirá una mayor vasodilatación. El flujo sanguíneo activa a un mecano-rreceptor ubicado en la membrana apical de las células endoteliales que, tras una cascada de señalización intracelular, produce la activación de la óxido nítrico sintasa. Este enzima for-ma óxido nítrico (NO) a partir de la L-arginina. El NO difundirá hasta la musculatura lisa del vaso sanguíneo, que quedará relajada y se dará la vasodilatación. Éste es un mecanismo im-portante para mantener los vasos relajados así como también para recuperarnos tras una is-quema transitoria. El aumento del CO2, la disminución de la tensión de O2, el aumento de hidrogeniones, y un aumento del K+ también producen una vasodilatación.
91
2 ADP→ ATP + AMP

Autorregulación
Consiste en que el músculo liso vascular se contraerá cuando haya un incremento de la pre-sión, para así mantener el flujo en las mismas condiciones.
Factores neurales
La estimulación de los nervios autónomos que van hacia el corazón afectan al flujo sanguí-neo coronario, tanto directa como indirectamente. Los efectos directos son consecuencia de los neurotransmisores (ACh, adrenalina y noradrenalina) sobre los propios vasos coronarios. Los efectos indirectos son consecuencia de los cambios asociados del flujo sanguíneo corona-rio provocado por el aumento o descenso de la actividad cardíaca. Los efectos indirectos son contrarios a los directos y tienen un papel más importante en el control del flujo sanguíneo coronario.
(1) Estimulación simpática. El efecto directo del simpático es provocar una vasoconstric-ción (receptores α: vasoconstricción; receptores β: vasodilatación). El efecto indirec-to incrementa la FC, la contractibilidad y la actividad metabólica, por lo que se da una vasodilatación secundaria.
(2) Estimulación parasimpática. El efecto directo del parasimpáticoes provocar una vaso-dilatación. El efecto indirecto es disminuir la FC, la contractibilidad y la actividad metabólica, por lo que se da una vasoconstricción secundaria.
Vemos como el corazón es un ejemplo claro de predominio de los sistemas de control local
frente a los centrales (escapa al control neural).
Factores humorales
Son aquellos vehiculizados por sangre. Algunos ejemplos son:- Adrenalina: incrementa el flujo coronario de manera indirecta. Efecto α-adrenérgico.- Angiotensina II: es un potente vasoconstrictor que producirá una incremento de la ten-
sión arterial, por lo que también producirá un aumento de la tensión del músculo ventricular y de ahí la vasodilatación.
- Hormonas tiroideas: incrementan el metabolismo, por lo que se da una vasodiatación.
Sistema de control del flujo coronario a medio/largo plazo
Como ya se ha dicho, la circulación coronaria es funcionalmente terminal. Si se le da el sufi-ciente tiempo, la obturación de un pequeño vaso puede ser paliada por medio de la forma-ción de una ruta colateral (angiogénesis). Si la obturación es abrupta no habrá esta posibilidad y se dará una isquemia que puede afectar a un área mayor o menor. La isquemia generará: (a)
angina (dolor); (b) liberación de enzimas (por muerte celular); (c) alteración en el ECG (proble-mas en la despolarización); (d) disfunción muscular; y (e) arritmias.
92

Clínicamente la isquemia se da progresiva-mente. En un primer momento tenemos una isquemia reversible: si se restaura el aporte las células seguirán funcionales. En el mar-co de esta isquemia reversible observamos como se incrementa la glucólisis anaeróbi-ca, la producción de ácido láctico, el in-cremento de hidrogeniones, la disminución del ATP y el incremento también del calcio intracelular (ya que éste no se reintroduce). Si esta isquemia reversible pasa de los 20-30
minutos (23 min según la diapositiva), pasará a ser irreversible. En la isquemia irreversible las células han perdido mucho ATP y ade-nosina, y los niveles de calcio intracelular y mitocondrial estarán muy elevados. En este ca-so, si restauramos el aporte, tendremos tan poca adenosina (liberada para inducir la vasodila-tación) en las células que no podremos formar ATP de nuevo. El área necrótica progresivamente se va haciendo más pequeña, por lo que tras un proceso isquémico se da una ligera recuperación.
93
Etapas de la recuperación de un infarto de miocardio. (Guyton pág. 251)

15. SISTEMA CIRCULATORIO (Arterias)
Introducción
La circulación sanguínea nos permite una transferencia rápida de diferentes elementos desde la bomba cardíaca hacia la periferia. Dentro del sistema circulatorio distinguimos tres gran-des compartimentos: (a) arterial; (b) capilar y (c) venoso. Las arterias se encuentran en un territorio de alta presión, por lo que tendrán que ser distensibles (para que la presión no aumente excesivamente) a la vez que elásticas (para que la presión no disminuya mucho durante la diástole). Conforme menor sea el diámetro de la arteria menor será esta capacidad de adaptación (capacidad que también se va perdiendo con la edad). Distinguimos tres segmentos funcionales dentro de las arterias:
- Vasos elásticos (vasos windkessel): son los grandes vasos, encargados de absorber la energía del corazón y adaptarla durante la diásole para asegurar un flujo continuo en los territorios de intercambio (en contraposición al flujo pulsátil emitido por el corazón). Nuestros vasos siempre están llenos de sangre.
- Vasos de resistencia. Son las arterias terminales y las arteriolas. Tienen una gran pared, predominantemente formada por músculo. No tienen una gran capacidad de defor-mación pasiva, pero pueden regular su diámetro hasta llegar a cerrar completamente la luz. Regulan el flujo sanguíneo.
- Vasos esfínter (esfínteres precapilares). Determinan el paso de sangre a un territorio ca-pilar.
Tras el sistema arterial vemos los capilares, que no tienen capacidad contráctil. Finalmente, tras los capilares vienen las venas, que también se pueden dividir en diferentes segmentos funcionales:
- Vénulas. Tienen una gran capacidad contráctil, por lo que pueden modificar de mane-ra activa su diámetro y por tanto también la resistencia al flujo.
- Resto de venas. Serán vasos de capacidad, con una gran distensibilidad.
Si nos fijamos en la presión en cada uno de los compartimentos veremos como en el com-partimento arterial la presión se mantiene constante dentro de un marco (se modifica de ma-
nera pulsátil pero dentro de unos márgenes; 80-120 mmHg). En las arteriolas estos cambios de presión desaparecen y se produce una progresiva caída de la presión (cambios también en la velocidad). Los vasos se irán haciendo más pequeños, pero la sección será mayor (entiéndase la suma de la sección de todos los vasos, no individualmente). La mayor sección transversal la encontramos en los capilares, y esto sumado a la baja presión da una muy baja velocidad
de circulación. En las vénulas y las venas es donde encontraremos un mayor volumen sanguí-neo (reservorio de sangre). En Organografía se discutieron los cambios en la histología de los diferentes vasos. Todos estos datos que acabamos de comentar (presión, velocidad) se verán incremen-tados cuando aumente la actividad cardíaca (ejercicio físico).
94

Arterias
Los grandes vasos tienen como objetivo transformar el flujo de sangre discontinuo que sale del corazón en un flujo continuo (que no quiere decir regular). Esto se logrará por medio de dos características de estos vasos: (a) distensibilidad y (b) elasticidad.
(a) Distensibilidad. La distensibilidad relaciona cómo se modifica la presión en depen-dencia a un pulso determinado de volumen. Cuanto más distensible sea el vaso, menos incrementa la presión a pesar de que se aumente el volumen. La distensibili-dad del vaso, propiedad que se va perdiendo con la edad (igual que la elasticidad), es importante para que las presiones sistólicas no sean excesivas.47
(b) Elasticidad. La elasticidad relaciona los cambios de diámetro con los cambios en la presión. Cuanto más elástico sea el vaso mayor mantenimiento de la presión (pro-mueve el mantenimiento de la presión durante la diástole). La elasticidad está impli-cada en la presión de empuje.
Cuando el corazón expulsa la sangre de su ventrículo, las paredes de la aorta la reciben y se deforman, almacenando una determinada cantidad de energia potencial. Cuando la presión que ofrece el ventrículo empieza a caer esa energía potencial se transformará en energía ciné-
tica y permitirá el avance de la sangre. De manera continua el flujo irá avanzando y la de-formación de la pared se irá transmitiendo hacia segmentos siguientes de la arteria (onda de
pulso). La onda de pulso va más rápida que el flujo sanguíneo (2 m/s según el punto; versus 33 cm/s el flujo sanguíneo); para hacer nos una idea, cuando observamos esta onda en la arteria pedia, la sangre va por la aorta descendente. La velocidad de propagación de la onda de pulso depende del grosor de la pared y del diámetro del vaso.
95
47 No sé exactamente a qué vino, pero el Dr. Javierre comentó que la aorta se va dilatando con la edad, lo cual podría ser problemático si nuestras válvulas sigmoideas se ajustaran al diámetro de la base de la aorta que en-contramos cuando acabamos de desarrollarnos. Al contrario, las válvulas sigmoideas podrían llegar a cubrir una superficie de casi el doble de la sección de la base de la aorta. Si así no fuera se daría una insuficiencia de la válvula sigmoidea por la distensión excesiva de la aorta.
Presiones sanguíneas normales en las distintas porciones del aparato circulatorio cuando una persona está en decúbito. (Guyton pág. 159)

Las curvas de pulso nos informan sobre lo que está ocurriendo en el corazón. En el pulso po-demos ver los siguientes elementos:
(1) Existencia de pulso. Hay algunas agenesias de troncos braquiocefálicos que cursan sin pulso.
(2) Frecuencia. Por debajo de 60 pulsaciones hablamos de bradicardia y por debajo de 40 de bradicardia severa. Por encima de 80 pulsaciones hablamos de taquicardia. Con la edad la frecuencia va disminuyendo y ésta también puede ser alterada por la temperatura (alta = aumenta; baja = disminuye)48.
(3) Ritmo. Que puede ser regular o irregular. Si es irregular no quiere decir que estemos ante una arritmia cardíaca.
(4) Tamaño. Es la relación entre el momento de menor y mayor tono; se relaciona con el volumen sistólico. Un mayor tamaño se observa en una insuficiencia sigmoidea.
(5) Pendiente. La velocidad con la que aumenta la presión. Si fuéramos capaces de de-terminar la pendiente podríamos saber como están las resistencias periféricas.
(6) Tensión. Es la fuerza o presión que tengo que aplicar para deprimir ese pulso. Nos indica la presión arterial media.
El pulso debe ser bilateral. Además de poderlo tomar en los puntos de referencia normales, puede aparecer en otros lugares en caso de patología (p. ej., arterias intercostales en el caso de una estenosis aórtica). El pulso es el resultado de la transmisión de la onda de presión. Den-tro de la presión arterial podemos distinguir entre:
- Presión sistólica (Ps). Es el máximo de la curva del pulso de presión durante la sístole.- Presión diastólica (Pd). Es el mínimo de la curva del pulso de presión durante la diás-
tole. La amplitud es la diferencia entre estas dos presiones (Ps - Pd)- Presión arterial media (PAM). No es el resultado de sumas Ps y Pd y dividir entre dos, al
menos no en todo el cuerpo. Depende de la zona del cuerpo en que nos hallemos. En las arterias centrales la PAM es igual a la Pd más la mitad de la amplitud. En las arterias periféricas la PAM es igual a la Pd más un tercio de la amplitud.
Transmisión del pulso de presión
Cuando el corazón expulsa la sangre hacia la aorta durante la sístole, primero se distiende sólo la porción proximal de la aorta porque la inercia de la sangre impide el movimiento brusco de la sangre hacia la periferia. No obstante, el aumento de la presión en la aorta pro-
96
48 Hay patologías que cursan con cruces en este comportamiento. La fiebre tifoidea cursa con temperaturas altas y bradicardia. La fiebre reumática cursa con poca fiebre y taquicardia. Un aforismo de la fiebre reumática es que lame las articulaciones y muerde al corazón (sintomatología en las articulaciones pero afecta bastante al corazón).
Cambios de perfil de la presión aórtica. En la arte-riosclerosis vemos unas presiones sistólicas más ele-vadas y en la estenosis aórtica vemos unas presiones diastólicas similares a la normalidad y una menor presión sistólica (menor volmen sistólico).

ximal supera rápidamente esta inercia y el frente de onda de distensión se va extendiendo a lo largo de la aorta. Esto es lo que se conoce como transmisión del pulso de la presión en las arterias. La velocidad de transmisión del pulso de la presión en la aorta normal es de 3 a 5 m/s, de 7 a 10 m/s en las ramas arteriales grandes y de 15 a 35 m/s en las pequeñas arterias. Cuanto mayor sera la compliancia de cada segmento vascular, más lenta será la velocidad, lo que explica la transmisión lenta en la aorta y mucho más rápida en las arterias distales pe-queñas, mucho menos distensibles.
Gráfica del pulso de presión
En la aorta partimos de una presión mínima en la diástole de unos 80 mmHg. Esta presión se irá incrementando progresivamente a me-dida que el ventrículo izquierdo vaya expul-sando sangre, llegando hasta los 120 mmHg en la base de la aorta. Al finalizar la contracción ventricular, esta presión va disminuyendo y se producirá una incisura fruto de la deformación de las válvulas sigmoideas hacia el interior del ventrículo (al cerrarse). Esta incisura tam-bién está relacionada con un flujo retrógrado momentáneo (véase gráfica de flujo). El incre-mento de la presión que se observa durante la sístole va a estar muy relacionado con la dis-tensibilidad de la aorta. En el caso de una ar-teriosclerosis, como se observa en la figura de la página anterior, la presión sistólica alcanza valores elevados porque las arterias se han en-durecido. A medida que se va progresando por el árbol arterial, las presiones sistólicas tien-den a aumentar y las diastólicas a disminuir. La velocidad máxima del pulso de flujo se irá haciendo menor a medida que avanza-mos en el árbol arterial; ahora bien, el flujo durará más en el tiempo. La compliancia del árbol arterial va a reducir las pulsaciones de la presión hasta que prácticamente desaparecen en el momento en que la sangre alcanza los capilares, por lo que el flujo sanguíneo tisular es prácticamente contínuo con un escaso carácter pulsátil.
Este pulso de flujo sera progresivamente menor en cuanto a la velocidad máxima que alcance pero mayor res-
pecto a la duracion. En condiciones normales, a nivel de la aorta abdominal e incluso en las femorales observa-remos un flujo retrogrado, que dependerá del aumento de presión generado por el pulso de presión que esta-mos transfiriendo.
Hipertensión arterial (HTA). La HTA es el asesino silencioso, ya que no produce daño a corto plazo pero puede
provocar incluso la muerte a medio/largo plazo. Se incrementa la poscarga, se da una hipertrofia ventricular izquierda y finalmente se da una insuficiencia cardíaca. Habrán problemas isquémicos cardíacos y la dilata-ción de la aorta será menor. A nivel del ECG veremos ondas T negativas (signo de isquemia). Hay problemas
97
Perfil de pulso de presión registrado en la aorta as-cendente. (Guyton pág. 169)

neurológicos, como mareos, vértigos, cefalalgia occipital (matutina), infarto cerebral y hemorragia cerebral. Se
afectan las arterias retinianas, escotomas (puntos negros sin visión). Además de todo este cuadro: hematemesis, metrorragia, epistaxis y hemoptisis.
Método oscilatorio de la toma de presión. Revisar la práctica de toma de presión.
98
4
Pulso de Flujo (I)
Vc de flujo/Sección transversal
Vel
ocid
ad (c
m/s
)
Secc
ión
trans
vers
al (c
m)
arterias arteriolas capilares
Vc
�

15(2). MICROCIRCULACIÓN
Introducción
Hasta que llegamos a las arteriolas lo que nos interesa es aportar rápidamente un volumen de sangre. Una vez alcanzadas las arteriolas la velocidad del flujo disminuirá para así favore-
cer el intercambio de elementos entre el medio intersticial y la sangre. El territorio en el que se dará este intercambio comprende tres elementos: (a) arteriolas terminales; (b) capilares; y (c)
vénulas.(a) Arteriolas terminales. Son vasos en los que encontramos células endoteliales con
agregados de células musculares, capaces de modificar el diámetro de manera acti-va. Un tipo de arteriola terminal son las metaarteriolas, un circuito de conducción preferencial (permite el paso rápido del flujo). Encontraremos derivaciones arteriove-
nosas que nos permitirán evitar el territorio capilar. (b) Capilares. Hay tres grandes clases: continuos, fenestrados y sinusoides (de menor a
mayor permeabilidad). La riqueza de capilares está directamente relacionada con la ratio metabólica del tejido (↓capilares: cartílago, tejido subcutáneo; ↑capilares: muco-sa gástrica, glándulas salivales).
(c) Vénulas. Existen dos grandes tipos: pos-capilares y colectoras. Las primeras (pos-capi-lares) recogen la sangre procedente de los capilares y se producirán procesos de in-tercambio similares a los que se ven en el territorio capilar. Las segundas (colectoras) disponen de musculatura y pueden regular el diámetro de manera activa, pudiendo llegar a incrementar la presión de un segmento anterior (se favorece el paso de agua al espacio intersticial).
Intercambio de elementos
El intercambio de elementos se puede realizar por medio de tres métodos. El primero de to-dos es el mecanismo de los solutos y es dependiente de los poros y de las hendiduras interce-lulares del endotelio. Por medio de este mecanismo podemos pasar moléculas de un calibre
menor a esos espacios. Aquí nos encontraremos con moléculas liposolubles (ácidos grasos, O2 y
CO2) que pueden difundir a través de la membrana. Estas moléculas se comportan según las leyes de difusión (pasan de un medio en el que están más concentrados a otro en el que me-nos). La difusión se rige por la siguiente fórmula:
Al territorio capilar llega sangre rica en O2 que pasará hacia un espacio intersticial pobre en O2 (aunque la diferencia de concentración debe de ser elevada). Con el CO2 sucede lo con-trario, llega sangre pobre en CO2 que recibirá el CO2 del medio intersticial. El coeficiente de
permeabilidad del CO2 es mucho mayor que el del O2 (no precisa de diferencias de concentra-
99
D = P ⋅A ⋅ ΔCDifusión. P: coeficiente de permeabilidad de la sustancia; A: área de intercambio; ΔC: diferencia de concentra-ciones entre los dos medios.
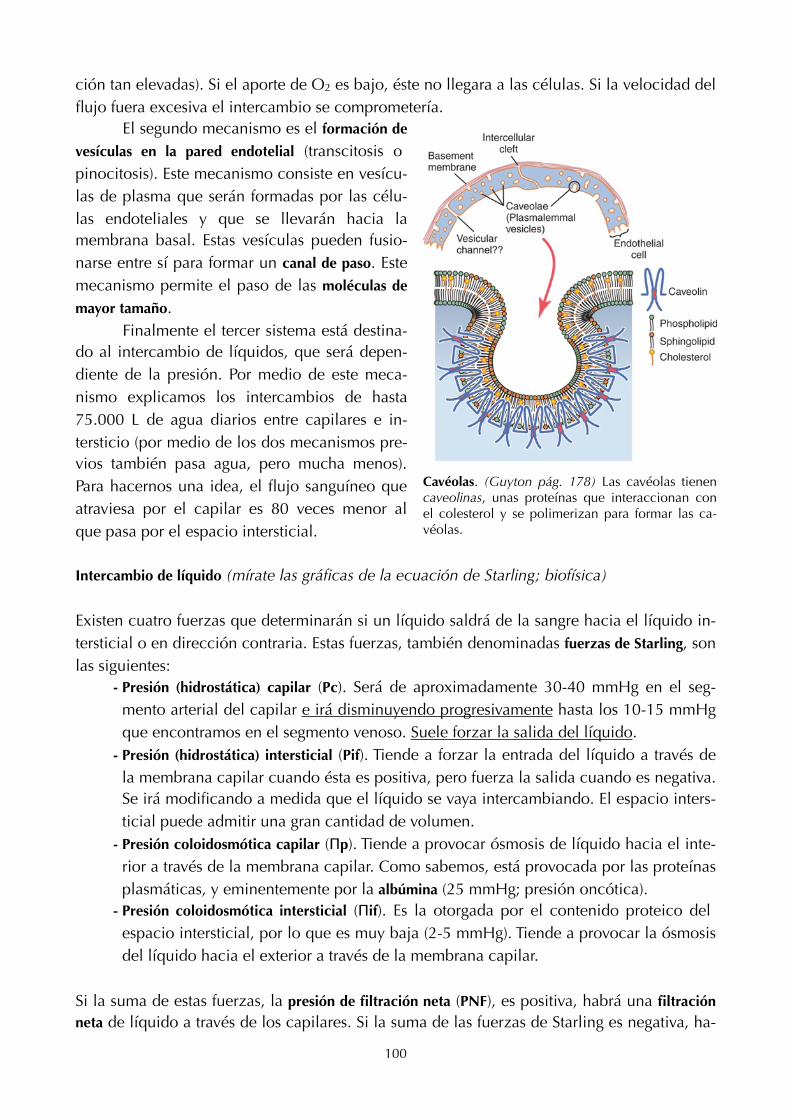
ción tan elevadas). Si el aporte de O2 es bajo, éste no llegara a las células. Si la velocidad del flujo fuera excesiva el intercambio se comprometería. El segundo mecanismo es el formación de
vesículas en la pared endotelial (transcitosis o pinocitosis). Este mecanismo consiste en vesícu-las de plasma que serán formadas por las célu-las endoteliales y que se llevarán hacia la membrana basal. Estas vesículas pueden fusio-narse entre sí para formar un canal de paso. Este mecanismo permite el paso de las moléculas de
mayor tamaño. Finalmente el tercer sistema está destina-do al intercambio de líquidos, que será depen-diente de la presión. Por medio de este meca-nismo explicamos los intercambios de hasta 75.000 L de agua diarios entre capilares e in-tersticio (por medio de los dos mecanismos pre-vios también pasa agua, pero mucha menos). Para hacernos una idea, el flujo sanguíneo que atraviesa por el capilar es 80 veces menor al que pasa por el espacio intersticial.
Intercambio de líquido (mírate las gráficas de la ecuación de Starling; biofísica)
Existen cuatro fuerzas que determinarán si un líquido saldrá de la sangre hacia el líquido in-tersticial o en dirección contraria. Estas fuerzas, también denominadas fuerzas de Starling, son las siguientes:
- Presión (hidrostática) capilar (Pc). Será de aproximadamente 30-40 mmHg en el seg-mento arterial del capilar e irá disminuyendo progresivamente hasta los 10-15 mmHg que encontramos en el segmento venoso. Suele forzar la salida del líquido.
- Presión (hidrostática) intersticial (Pif). Tiende a forzar la entrada del líquido a través de la membrana capilar cuando ésta es positiva, pero fuerza la salida cuando es negativa. Se irá modificando a medida que el líquido se vaya intercambiando. El espacio inters-ticial puede admitir una gran cantidad de volumen.
- Presión coloidosmótica capilar (Πp). Tiende a provocar ósmosis de líquido hacia el inte-rior a través de la membrana capilar. Como sabemos, está provocada por las proteínas plasmáticas, y eminentemente por la albúmina (25 mmHg; presión oncótica).
- Presión coloidosmótica intersticial (Πif). Es la otorgada por el contenido proteico del espacio intersticial, por lo que es muy baja (2-5 mmHg). Tiende a provocar la ósmosis del líquido hacia el exterior a través de la membrana capilar.
Si la suma de estas fuerzas, la presión de filtración neta (PNF), es positiva, habrá una filtración neta de líquido a través de los capilares. Si la suma de las fuerzas de Starling es negativa, ha-
100
Cavéolas. (Guyton pág. 178) Las cavéolas tienen caveolinas, unas proteínas que interaccionan con el colesterol y se polimerizan para formar las ca-véolas.

brá una absorción neta de líquido desde los espacios intersticiales hacia los capilares. La pre-sión neta de filtración se calcula como:
La presión hidrostática capilar tiende a disminuir, la coloidosmótica capilar es estable, la presión hidrostática intersticial es ligeramente negativa, y la presión coloidosmótica intersti-cial es despreciable. Por tanto, la única fuerza que trata de hacer que el líquido entre en el capilar es la presión coloidosmótica capilar (insistimos que la Πif no la tenemos en cuenta). En la primera parte del capilar, la zona más arterial, ocurrirá lo siguiente: Pc - Pif + Πif > Πp, por lo que se producirá la filtración. Ahora bien, el principal motor de dicha filtración será la presión hidrostática capilar (Pc), por lo que a medida que vayamos avanzando a tra-vés del capilar y ésta vaya disminuyendo, la filtración también irá descendiendo. Finalmente la presión oncótica capilar será superior a la suma de las otras fuerzas, dándose la absorción (ya hacia el final del capilar). Para hacernos una idea, cuando (Pc - Pif + Πif) < 25 mmHg, se producirá la absorción.
El compartimento intersticial tiene un intervalo de volúmenes en los que mantiene una pre-sión más o menos constante. Si el volumen sobrepasa ese intervalo de seguridad entonces se dará un edema y si baja demasiado entonces podría darse una deshidratación. A continuación veremos algunas variaciones que se pueden dar en el intercambio de fluidos:
- Presión hidrostática incrementada (↑Pc). Si en los capilares la Pc está incrementada, se favorecerá la filtración e incluso podría no darse la absorción (si la Pc es muy alta, su caida a medida que se avanza en el sistema capilar puede no ser suficiente; la Πp po-dría no superarla). Atención, que la presión arterial sistémica esté elevada no quiere decir que la capilar, que es la que nos interesa, también lo esté.
- Vasodilatación. Habrá un mayor flujo de sangre que pase a los capilares, incrementán-dose la Pc. Hay una tendencia al edema.
- Vasoconstricción. La Pc será menor. Se favorece la absorción. La vasoconstricción se puede inducir (por medio de frío) para paliar una vasodilatación.
- Presión venosa incrementada. Si hay problemas con el retorno venoso entonces la caída de la presión hidrostática capilar será menor, por lo que la absorción apenas se dará. Tendencia al edema.
- Deshidratación. Hay una mayor absorción por el incremento de la Πp.
101
PNF = Pc −Pif − Πp +Πif
Fuerzas de Starling. (Guyton pág. 178) La diferencia de presión hidrostática entre el segmento arterial del capi-lar y el segmento venoso (ΔPc) es de unos 7 mmHg aproximadamente (32-25).

16. CONTROL DE LA CIRCULACIÓN PERIFÉRICA
Introducción
La circulación periférica es controlada por dos grandes mecanismos: uno central o extrínseco, que es dependiente del sistema nervioso, y otro local y dependiente de la actividad de los ór-ganos.
Control extrínseco del flujo sanguíneo periférico
El control extrínseco del flujo sanguíneo periférico puede dividirse en cuatro elementos: (a)
simpático; (b) parasimpático; (c) factores humorales; y (d) reflejos vasculares.
SN simpático
La división simpática del vegetativo ejercerá su función sobre los vasos sanguíneos a partir del centro vasomotor, ubicado en el tercio inferior de la protuberancia y los dos tercios supe-riores del bulbo. Este centro transmite los impulsos parasimpáticos a través de los nervios va-gos hacia el corazón y transmite los impulsos simpáticos a través de la médula espinal y los nervios simpáticos periféricos prácticamente hacia todas las arterias, arteriolas y venas del organismo. La anatomía del centro vasomotor aún no se tiene clara, pero se sabe que hay una región vasoconstrictora (región presora)49 que envía señales continuamente hacia las fibras vasoconstrictoras simpáticas a una frecuencia de entre 0,5 y 2 impulsos/segundo (tono vaso-constrictor simpático). Estos impulsos mantienen un estado parcial de contracción de los va-sos sanguíneos, el tono vasomotor.
102
49 En el centro vasomotor hay una región antagónica al centro vasoconstrictor, el centro vasodilatador, que en-vía señales que inhiben la actividad vasoconstrictora del primer centro.
Efecto de la anestesia espinal total sobre la presión arterial. Se observa como la presión disminuye como con-secuencia de la pérdida del tono vasomotor. Una inyección de noradrenalina, el neurotransmisor empleado por el SN simpático para generar dicho tono, restaura la presión hasta que ésta es eliminada.

La noradrenalina usada por el simpático también incrementa la frecuencia cardíaca y la con-tractilidad, provocando un aumento de la presión arterial. En pocos segundos, gracias a la estimulación cardíaca, la contracción arteriolar y la contracción de grandes vasos (recorde-mos que las venas contienen la mayor parte del volumen sanguíneo), el centro vasomotor podrá alterar la presión arterial en pocos segundos (multiplicarla por dos o reducirla a la mitad).
Región sensorial. Una parte del centro vasomotor es la región sensorial o zona sensitiva. Las neuronas de
esta zona reciben señales nerviosas sensitivas desde el sistema circulatorio, principalmente a través de los pares IX y X, y emite señales eferentes desde esta zona sensitiva que facilitan las actividades de con-trol de las zonas tanto vasoconstrictoras como vasodilatadoras, con lo que se consigue el control reflejo
de muchas funciones circulatorias.
SN parasimpático
El control parasimpático es, por lo general, poco importante. Ni la piel ni el músculo esquelé-tico son inervados por esta división. Afectará a la cabeza, las vísceras y la región sacra (geni-tales, vejiga e intestino grueso). Como vemos, la regulación de la presión, al menos a nivel vegetativo, depende más del simpático.
Factores humorales
A continuación se discutirán toda una serie de factores humorales:- Adrenalina y noradrenalina. La adrenalina a bajas concentraciones tiene un efecto β-a-
drenérgico a nivel muscular (vasodilatación), mientras que a altas concentraciones produce justo el efecto contrario (α-adrenérgico). La producción más importante de estas dos hormonas se da en el sistema nervioso vegetativo. La médula suprarrenal también las produce, sobre todo noradrenalina. La noradrenalina está más destinada a potenciar el tropismo cardíaco.
- Angiotensina. Se trata de un potente vasoconstrictor que en determinados territorios puede incluso producir isquemia.
- Vasopresina (u hormona antidiurética). Es el agente biológico con mayor potencial vaso-constrictor. Estará siempre a unas bajas concentraciones. En situaciones de shock hi-povolémico permite mantener la presión arterial.
- Serotonina. En la piel tiene un efecto vasodilatador.- Histamina. Producida por mastocitos y basófilos, tiene un efecto proinflamatorio.- Prostaglandinas. Pueden ser vasoconstrictoras o vasodilatadoras en función del tipo.
Además de todos estos metabolitos, modificaciones en la concentración de determinados iones pueden producir vasodilatación (Mg2+, K+) o vasoconstricción (Ca2+). No obstante los cambios en el calibre del vaso se producirán por los cambios en la presión osmótica fruto de los cambios en la concentración de estos iones. Un metabolito al que se le dio bastante importancia es la bradicinina. La α2-globulina pasa a calidina por el efecto de la calicreína. La calicreína se encuentra inactiva en condicio-nes normales, pero se activará ante señales inflamatorias. La calidina, por efecto de enzimas
103

tisulares, pasará a ser bradicinina (por tanto, la secuencia es: α2-globulina → calidina → bra-dicinina). La bradicinina tiene dos efectos principales: (a) dilatar las arteriolas e (b) incrementar la permeabilidad. Sobre un territorio inflamado estos dos efectos permiten que la inflamación prosiga. La bradicinina durará unos minutos en el sistema porque será rápidamente degrada-da por una carboxipeptidasa o enzima convertidora (la responsable de la activación de la an-giotensina). La calicreína podrá ser destruida por el inhibidor de la calicreína. La bradicinina es importante para la regulación del flujo sanguíneo en la piel, en las glándulas salivares y gastrointestinales.
Reflejos neurales
Como ya se comentó en temas anteriores, encontramos unos barorreceptores ubicados en las carótidas y en el arco aórtico. Los del seno carotídeo son sensibles a presiones que oscilan entre los 60 y los 180 mmHg, mientras que los aórticos detectan cambios que van entre los 90 y 210 mmHg. Ambos barorreceptores envían información hacia el centro vasomotor, uno usando el noveno par (carotídeos) y otro el décimo (aórtico). La estimulación de los barorre-ceptorse provoca una inhibición simpática. Los barorreceptores no responden a los cambios de presión de manera lineal, lo ha-cen siguiendo una función sigmoide. De esta manera logramos que para cambios de presión acaecidos en la zona de los 80 mmHg se produzca una gran variación en la respuesta. Los barorreceptores van a ser importantes en el mantenimiento de la presión ante cambios agudos (p. ej., cambios de posición corporal). Los barorreceptores pueden llegar a adaptarse a pre-siones altas (individuos con hipertensión). En caso de una denervación, la presión arterial os-cilará mucho. Además de estos receptores encontramos otros ubicados en el territorio cardiopulmo-nar que pueden alterar la función cardíaca además de controlar la presión. Para modificaciones de, por ejemplo, 300 mL en el retorno venoso, se generan modificaciones de presión del or-den de 10-15 mmHg. Para este pequeño incremento del retorno venoso, si bloqueamos los receptores aórticos y carotídeos, la presión se incrementaría hasta 50 mmHg. Si además blo-queamos los receptores de retorno, el incremento ascendería hasta los 120 mmHg. Los re-ceptores de retorno están unidos a la función renal gracias al péptido natriurético auricular. Finalmente, a parte de los receptores de presión encontramos los quimiorreceptores. Los quimiorreceptores detectan modificaciones de algunos parámetros químicos de la sangre, como el pH, el CO2 y el O2. Los receptores de O2, que son los únicos que comentaremos, se encuentran en la carótida y tienen una rica irrigación arteriolar. A partir de presiones parciales
de O2 (PaO2) menores de 80 mmHg (PaO2 normal = 100 mmHg) se empiezan a activar. Para va-lores de <60 mmHg de PaO2 los quimiorreceptores cogerían las riendas y controlarán la respi-
ración. Como vemos, el margen de activación de estos receptores es grande (no empiezan a activarse hasta que la PaO2 baja bastante). Con la PaCO2 ocurre todo lo contrario, cambios de 1 mmHg ya producen una respuesta. La respuesta que inducen estos quimiorreceptores es un incremento del tono vascular y la capacitancia. En el corazón disponemos de unos qui-miorreceptores con fibras aferentes simpáticas activados por isquemia que transmiten dolor. Este dolor es el clásico precordial, la angina de pecho (aporte inadecuado de sangre).
104

Mecanismos locales y agudos de control del flujo sanguíneo
Los mecanismos que veremos a continuación afectarán al flujo en un determinado territorio. El control local depende de elementos estructurales (endotelio y músculo) y otros dinámicos (flujo y presión) Los tres mecanismos son: (a) autorregulación; (b) endotelio y (c) factores meta-
bólicos.(a) Autorregulación (o mecanismo miogénico). Depende de la actividad del músculo liso,
que responde a un estiramiento con una contracción. Un incremento en la perfusión a un determinado territorio produce la distensión inicial del vaso, lo que provoca que el músculo se contraiga y se retorne al nivel de perfusión previo.
Ejemplo. Se puso como ejemplo el hecho de pasar de decúbito a bipedestación. Este cambio
de posición incrementa la presión hidrostática en las zonas bajas del cuerpo, aumentando la presión intramural (la que tiende a deformar el vaso). Esta deformación produce una vasocons-tricción que reduce la filtración capilar y por tanto modifica la presión oncótica y la presión
intersticial (...).
(b) Endotelio. Se trata quizá del mecanismo más importante. Hasta el 80% de las resis-tencias periféricas totales dependen de la actividad endotelial. A mayor velocidad de flujo, mayor vasodilatación. Si el endotelio no está sano favorecerá la activación de la cascada de coagulación. El endotelio tendrá la vasodilatación que comentamos por tres factores:
- Hiperpolarización. Cambios en la permeabilidad del K+ dificultará que las cé-lulas se contraigan.
- Prostaglandina I2. Favorece el incremento del cAMP intracelular y por ende la vasodilatación.
- Óxido nítrico (NO). El óxido nítrico lo generan las células endoteliales ante las fuerzas de cizallamiento (flujo sanguíneo). Se forma a partir de la arginina y oxígeno y difunde hacia las células musculares lisas. En éstas activa la guanila-
to ciclasa, incrementándose el cGMP. El cGMP disminuye la sensibilidad de los canales de Ca2+ y disminuye la respuesta mitocondrial (relajación muscu-
lar). El NO será importante para producir una vasodilatación ante una isque-
mia aguda (hiperemia reactiva)50. En el corazón, durante la contracción oclusiva en la sístole, se va a observar este mecanismo.
(c) Regulación metabólica. El aporte sanguíneo a un tejido es porporcional a su tasa me-tabólica. Existen dos explicaciones (probablemente participen los dos) que justifican esta premisa:
- Explicación A. Si la tasa metabólica aumenta, se forma más ácido láctico, CO2, K+, H+, PO4
-, y sobre todo adenosina. Todos estos metabolitos son vasodilata-dores (aumenta el flujo). Como antes de que se dé esta vasodilatación habrá una pequeña isquemia, se expresará el factor hipóxico tisular que dará lugar a la formación de NO. Una vez incrementado el flujo, todos estos metabolitos serán arrastrados y se diluirá este cóctel vasodilatador.
105
50 Por hiperemia entendemos la “abundancia extraordinaria de sangre en una parte del cuerpo”. En la respueta hiperémica el NO es imporante, pero no es el único metabolito que se forma.
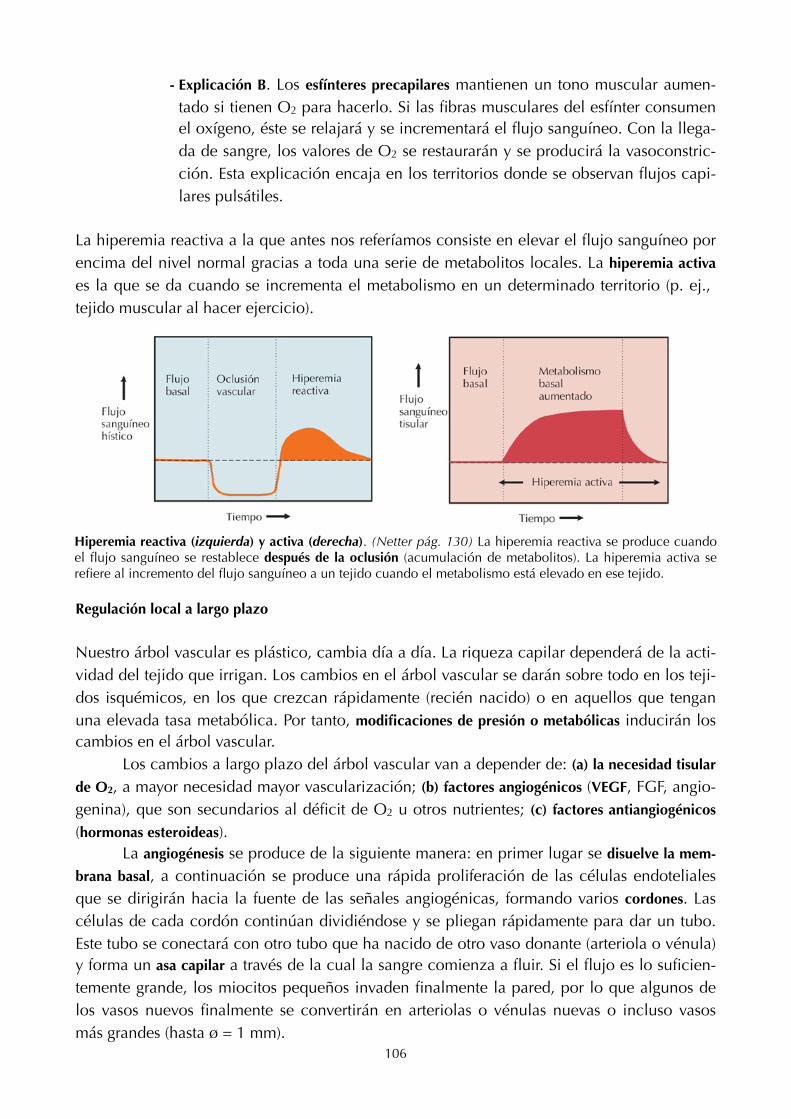
- Explicación B. Los esfínteres precapilares mantienen un tono muscular aumen-tado si tienen O2 para hacerlo. Si las fibras musculares del esfínter consumen el oxígeno, éste se relajará y se incrementará el flujo sanguíneo. Con la llega-da de sangre, los valores de O2 se restaurarán y se producirá la vasoconstric-ción. Esta explicación encaja en los territorios donde se observan flujos capi-lares pulsátiles.
La hiperemia reactiva a la que antes nos referíamos consiste en elevar el flujo sanguíneo por encima del nivel normal gracias a toda una serie de metabolitos locales. La hiperemia activa es la que se da cuando se incrementa el metabolismo en un determinado territorio (p. ej., tejido muscular al hacer ejercicio).
Regulación local a largo plazo
Nuestro árbol vascular es plástico, cambia día a día. La riqueza capilar dependerá de la acti-vidad del tejido que irrigan. Los cambios en el árbol vascular se darán sobre todo en los teji-dos isquémicos, en los que crezcan rápidamente (recién nacido) o en aquellos que tengan una elevada tasa metabólica. Por tanto, modificaciones de presión o metabólicas inducirán los cambios en el árbol vascular. Los cambios a largo plazo del árbol vascular van a depender de: (a) la necesidad tisular
de O2, a mayor necesidad mayor vascularización; (b) factores angiogénicos (VEGF, FGF, angio-genina), que son secundarios al déficit de O2 u otros nutrientes; (c) factores antiangiogénicos (hormonas esteroideas). La angiogénesis se produce de la siguiente manera: en primer lugar se disuelve la mem-
brana basal, a continuación se produce una rápida proliferación de las células endoteliales que se dirigirán hacia la fuente de las señales angiogénicas, formando varios cordones. Las células de cada cordón continúan dividiéndose y se pliegan rápidamente para dar un tubo. Este tubo se conectará con otro tubo que ha nacido de otro vaso donante (arteriola o vénula) y forma un asa capilar a través de la cual la sangre comienza a fluir. Si el flujo es lo suficien-temente grande, los miocitos pequeños invaden finalmente la pared, por lo que algunos de los vasos nuevos finalmente se convertirán en arteriolas o vénulas nuevas o incluso vasos más grandes (hasta ø = 1 mm).
106
Hiperemia reactiva (izquierda) y activa (derecha). (Netter pág. 130) La hiperemia reactiva se produce cuando el flujo sanguíneo se restablece después de la oclusión (acumulación de metabolitos). La hiperemia activa se refiere al incremento del flujo sanguíneo a un tejido cuando el metabolismo está elevado en ese tejido.

Un mecanismo con una finalidad similar es el de la circulación colateral. En el caso de que un vaso se bloquee, se estimula la dilatación prácticamente al instante de los vasos colatera-les. No obstante, esta primera dilatación es insuficiente (25% de las necesidades) y no será hasta el final del día en que el aporte empezará a ser más notable (50%). Al cabo de uno meses y con factores angiogénicos la recuperación puede ser del 100%. ¿Qué factores pro-mueven la proliferación celular y la optimización de la circulación colateral? El aumento del flujo (fuerzas de cizallamiento), aumento del NO, de citocinas proinflamatorias, de moléculas que favorecen la adhesión, etc (microambiente inflamatorio). La inflamación está asociada con enzimas que degradan la pared vascular y con factores de crecimiento que inducen la proliferación de las céls. endoteliales.
VEGF como abordaje a los tumores. Los tumores suelen crecer rápidamente, lo cual generalmente no
suele estar asociado con un crecimiento paralelo de su vascularización. Esto por un lado es bueno, ya que disminuye el riesgo de metástasis, pero por el otro lado impide que la quimioterapia sea del todo
efectiva. A pesar de que pueda parecer arriesgado, actualmente se contempla el uso del VEGF para pro-ducir una angiogénesis local y así poder potenciar el efecto de la quimioterapia.
107

16(2). CIRCULACIONES ESPECIALES
Circulación muscular
El conjunto de músculo de una persona alcanza fácilmente entre el 30-40% del peso corpo-ral. No obstante, a pesar de ser un importante porcentaje del peso, recibe, en reposo, muy poco del gasto cardíaco (15-20%). Ahora bien, cuando se está realizando un esfuerzo, prác-ticamente recibe la totalidad del mismo (80-90%), en función de la intensidad del ejercicio. El flujo que irá a parar a los músculos estará regulado por diferentes factores:
- Regulación neural. El simpático puede actuar a nivel de α y β adrenorreceptores (α: va-soconstricción; β: vasodilatación). La noradrenalina es agonista de los α y la adrenali-na de ambos, aunque con preferencia por los β. El músculo también dispone de re-
ceptores colinérgicos (parasimpático) que producen vasodilatación, aunque la vasodila-tación inducida neuralmente va a ser pobre (depende más de factores metabólicos).
- Factores metabólicos locales. La hipoxia, el incremento de la pCO2 y la liberación de metabolitos como H+, adenosina, K+, ácido láctico, bradicinina, histamina y ACh van a tener un efecto vasodilatador (↑metabolismo = ↑vasodilatación). Estos metabolitos se oponen al efecto α-adrenérgico del simpático.
- Factores mecánicos. La contracción muscular disminuye la perfusión. Al relajarse el músculo se dará una hiperemia reactiva.
- Regulación del intercambio. Durante el ejercicio se abre el 100% de los capilares.
En el músculo el control neural será importante, pero cuando el músculo esté activo el con-trol por factores metabólicos será el predominante. Cuando hacemos ejercicio tendremos un tono simpático que generará una vasoconstricción periférica a excepción de en aquellos músculos activos, ya que localmente se liberarán metabolitos vasodilatadores que se opon-drán al efecto vasoconstrictor del simpático.
Circulación esplácnica
La circulación esplácnica recibe un porcentaje importante del gasto cardíaco (25-30%). Tie-ne dos funciones principales: (a) transportar los nutrientes absorbidos en la digestión; (b) reser-
vorio de volumen sanguíneo (mejora el retorno venoso). Veamos los factores que modulan esta circulación:
- Factores metabólicos. Un incremento en el metabolismo o una disminución de la pO2 produce un incremento del flujo.
- Motilidad. Las contracciones peristálticas compromente el flujo sanguíneo. Cuando haya una relajación se dará una hiperemia reactiva.
- Control nervioso. El simpático produce una vasoconstricción que desaparecerá si se mantiene durante mucho tiempo gracias a un escape autorregulador inducido por la formación de metabolitos vasodilatadores.
108

Circulación hepática
El hígado recibe el 29% del gasto cardíaco, aunque de una manera peculiar. Recibe unos 1.000 mL a través de la vena porta (que le da el 25-30% del O2) y unos 500 mL de la arteria
hepática. Si hubiese una disminución del aporte a través de la hepática, seríamos capaces de extraer oxígeno de la sangre de la porta (tiene riqueza suficiente).
Circulación esplénica
En la mayoría de los animales el bazo actúa como un gran reservorio sanguíneo, capaz in-cluso de generar una autotransfusión. En los humanos esta capacidad está muy mermada, aunque si se el bazo se contrae podemos incrementos hasta en un 2% el hematocrito.
Circulación cerebral
El cerebro apenas representa el 2% del peso corporal pero consume el 15% del GC. Como sabemos, es muy dependiente del aporte de O2 y glucosa que le proporciona la sangre. Además, debemos tener en cuenta que el cerebro está en un armazón óseo, por lo que las
condiciones de intercambio (filtración/absorción) deben permanecer estables. El flujo sanguíneo cerebral será constante bajo diversas circunstancias (p. ej., incremento de la presión arterial sistémica). Centrándonos en la circulación cerebral, comentar que apenas va a haber mezcla en-tre los dos hemsferios y que el polígono de Willis dificulta la existencia de grandes anastomo-sis que faciliten el intercambio de volúmenes importantes de sangre. La circulación va a te-ner un marcado carácter terminal, por lo lesiones en un determinado vaso generan isquemias en el territorio dependiente de ese vaso. El drenaje es a través de las yugulares. A continua-ción se detallan algunas características diferenciales de esta circulación:
- Revestimiento endotelial continuo. Por lo que apenas se dejan espacios intercelulares.- Muchas zonas ocludens. Se impide la filtración.- Membrana basal continua. Rodeada además por pericitos y por glía. Vemos como todo
esto dificulta la filtración, por lo que los elementos hidrosolubles o de gran tamaño lo de tendrán complicado para pasar.
- Pobre transporte vesicular. No habrá mucha pinocitosis.- Densidad capilar variable. Dependiente de la actividad metabólica.
La barrera hematoencefálica (BBB; Eng: blood-brain barrier) mantiene constante el ambiente del SNC, nos protege de toxinas (tanto exógenas como endógenas) y previene de la salida de los neurotransmisores. La madurez de la barrera es determinante para evitar infecciones (ni-ños y meningitis). La BBB determina el paso de fármacos (dar levodopa en lugar de dopami-na, o triptófano en lugar de serotonina51). La circulación cerebral es regulada por los siguientes factores:
- Factores metabólicos. Dependientes del O2 y el CO2.
109
51 Apenas tenemos serotonina circulante: únicamente en plaquetas y en basófilos.

- Autorregulación cerebral. Se mantiene la presión constante si está entre valores de 8 a 20 kPa52 (se dará una vasoconstricción). Por debajo de 8 kPa la presión disminuye muy pronunciadamente. Por encima de 20 kPa la presión subirá rápidamente y habrá una tendencia al edema.
- Vegetativo. El simpático únicamente será importante en condiciones extremas en las que el sistema autorregulador fracase. Si en el SNC se produce un edema, se dificulta el aporte de sangre, por lo que habrá una vasodilatación dependiente del descenso de la PaO2, lo cual nos lleva de nuevo al edema (círculo vicioso).
110
52 1 mmHg = 133,32 Pa.

17. SISTEMA LINFÁTICO Y VENOSO
Introducción
El sistema linfático se encarga de drenar el exceso de líquido que pueda haber en el espacio
intersticial fruto de una excesiva filtración. Cuantitativamente estaríamos hablando de unos 3 L de líquido con un bajo contenido proteico que tendrá que retornar al territorio vascular. El sistema vascular linfático se inicia en unos fondos de saco (capilares ciegos) y se caracteriza por contener un flujo que va a una baja velocidad y que tiene una presión muy baja (1 mmHg). En los ganglios linfáticos se producirá parte de la reabsorción y el resto irá a las venas sub-clavias. Ni en el cartílago, ni el en el hueso, ni en los epitelios ni en el SNC hay sistema lin-fático alguno.
Características de los vasos linfáticos
Los vasos linfáticos se caracterizan por, en primer lugar, tener unas células endoteliales unidas entre sí por uniones muy laxas. Esto permite una gran permeabilidad y el reingreso de moléculas gran-des como las proteínas. Al igual que en el sistema venoso, los vasos linfáticos disponen de un siste-
ma pseudovalvular que impide la circulación retró-grada. A pesar de ser unos vasos finos, disponen de músculo liso con actividad miogénica espontá-
nea (ante un incremento de presión se produce la contracción). Finalmente comentar que los vasos linfáticos están unidos entre sí por medio de una red anastomósica, mezclándose la linfa de varios territorios (p. ej., ganglios de la axila que drenan la linfa de la mama y el brazo).
Composición lipoproteica
La composición lipoproteica de la linfa dependerá de la zona donde se haya formado. Re-cordemos que gran parte de las grasas de la dieta irán por ésta en forma de quilomicrones. Hasta el 50% del fibrinógeno plasmático estará en la linfa (la linfa se podrá coagular)53.
Bomba linfática
La linfa avanza por los vasos linfáticos gracias a la actividad contráctil de la musculatura lisa y a las válvulas. Las válvulas dividen un vaso en diferentes segmentos o compartimentos. El
111
53 Un edema linfático se endurece y es duro de revertir, mientras que un edema vascular se deprime si lo apre-tamos.
Estructura de un vaso linfático. Vemos como el vaso está unido al tejido conectivo circundante por medio de unos filamentos de anclaje. Si la presión en el espacio intersticial aumenta mu-cho se permitirá el paso de líquido hacia el capi-lar ciego.

llenado de un segmento provoca su contracción, con lo que el líquido se bombea a través de la válvula siguiente hacia el siguiente segmento. A diferencia de lo que habíamos visto hasta ahora, el flujo pasa de una zona de menor presión a otro de mayor. El retorno linfático está favorecido por toda una serie de elementos adyacentes a los propios vasos linfáticos: músculos, paquetes vasculonerviosos y modificaciones de las pre-siones intratorácica e intraabdominal.
Papel fisiológico de la linfa
La linfa es importante por los motivos siguientes:- Equilibrio entre el espacio intravascular e intersticial. Se encarga de drenar el exceso de
líquido filtrado respecto al reabsorbido. Previene los edemas.- Proteínas. Permite el reingreso de proteínas a la circulación sistémica.- Sistema inmunitario. Mantiene a los linfocitos y las inmunoglobulinas en circulación
tras su paso por el espacio intersticial y favorece la extracción de microorganismos.- Absorción. En los periodos post-prandiales es importante para facilitar el aclaramiento
de los lípidos absorbidos durante la digestión. Favorece y participa en la reabsorción de los líquidos en terceros espacios.
- Presión negativa. Está implicada en la generación de una presión negativa en el espa-cio intersticial.
Sistema venoso
El sistema venoso es el que se encarga de transportar la sangre desde la periferia hacia el co-razón. Contiene la mayor parte de la sangre del cuerpo y se caracteriza por dos rasgos: (a) presión y velocidad bajas, y (b) paredes delgadas y distensibles (vasos de capacitancia). Las venas tienen una morfología ovalada que se vuelve redondeada cuando el flujo que pasa por el vaso aumenta (y así no se modifica la presión).
Diferencias entre el sistema arterial y el venoso
En primer lugar, el sistema venoso dispone de toda una serie de plexos, formando una exten-
sa red venosa. Para hacernos una idea, el árbol venoso duplica al arterial. En segundo lugar, y tal como hemos visto en el sistema linfático, las venas disponen de unas válvulas que se abren en sentido proximal para impedir el flujo retrógrado y facilitar el retorno venoso. En tercer lugar, las venas disponen de una cantidad más pequeña de musculatura lisa si las comparamos con las arterias. Esta musculatura, que no es tan necesaria porque la presión en el sistema venoso es baja, estará inervada, pero tendrá muy poca actividad espontánea (a excep-
ción de las venas cutáneas, que serán muy sensibles a modificaciones térmicas). En cuarto lugar tenemos el asunto de las presiones, todas menores en comparación con el sistema arterial: en las venas intratorácicas encontramos 5 mmHg y en la aurícula derecha 3-4 mmHg. En quinto lugar está el almacenamiento de sangre, mucho mayor en las venas que en las arterias (2/3 partes del total de sangre). El flujo venoso será viscoso, lento y turbulento (válvulas).
112

Control del sistema venoso
El control del sistema venoso puede ser nervioso o humoral:- Control nervioso. El simpático actúa a nivel de los receptores α-adrenérgicos y produce
una venoconstricción.- Control humoral. La angiotensina II y la vasopresina producen venoconstricción. Los
mismos factores que trataban de subsanar una disminución de la volemia (vasocons-tricción para mantener la presión) ahora sirven para favorecer el retorno venoso y así mantener la presión arterial.
Pulso venoso
Por pulso venoso entendemos las modificaciones de presión que se dan en las venas adyacen-tes al corazón o en la propia aurícula (izquierda y derecha). En la onda del pulso venoso en primer lugar se observa un incremento brusco dependiente de la contracción auricular (onda a). Tras dicha contracción la presión desciende hasta que se produce un pico (onda c) que tiene que ver con la protusión de las válvulas auriculo-ventriculares al inicio de la contrac-
ción ventricular. Durante la contracción ventricular el ventrículo hace descender el plano valvular (se incrementa el espacio que hay en la aurícula, produciendo un gran descenso en la onda de presión venosa que observamos en forma de un gran valle (onda x). Finalmente, durante el llenado de la aurícula la presión va ascendiendo (onda v) y despues baja al abrirse las válvulas auriculo-ventriculares (onda y)
Retorno venoso
La bomba cardíaca determina el retorno venoso. Cuando empieza el llenado ventricular o cuando se produce la eyección ventricular se va a producir una caída en el pulso venoso que favorecerá el retorno de la sangre. De igual manera, con el llenado auricular o cuando
113
Onda de presión venosa.

la aurícula se contrae se va a impedir el retorno venoso. La bomba respiratoria también influen-ciará en el retorno venoso. La inspiración tiene un efecto de succión que favorecerá dicho re-torno. Finalmente, un tercer elemento que mo-dula el retorno venoso es la bomba músculo-es-
quelética. Ésta consiste que cuando se contrae la musculatura, ésta comprime al plexo profundo, que discurre entre la musculatura, y, si las vál-vulas son competentes, entonces se hace pro-gresar el volumen sanguíneo en sentido proxi-mal. Cuando este volumen avanza, el segmento venoso que se acaba de vaciar queda con una presión negativa que servirá para que los plexos superficiales envian la sangre hacia ese segmen-to. La bomba músculo-esquelética va a ser im-portante para evitar el edema.
114
Efecto de la presión gravitacional en las presiones veno-sas en todo el cuerpo en una persona en bipedestación. (Guyton pág. 173). Al caminar la presión de retorno dis-minuye, tan solo con hacer unos pocos pasos. El paseo con que sea a ritmo lento es efectivo (y de hecho es el más recomendable). Muy interesante para mujeres emba-razadas (retorno venoso dificultado = se les hinchan las piernas).
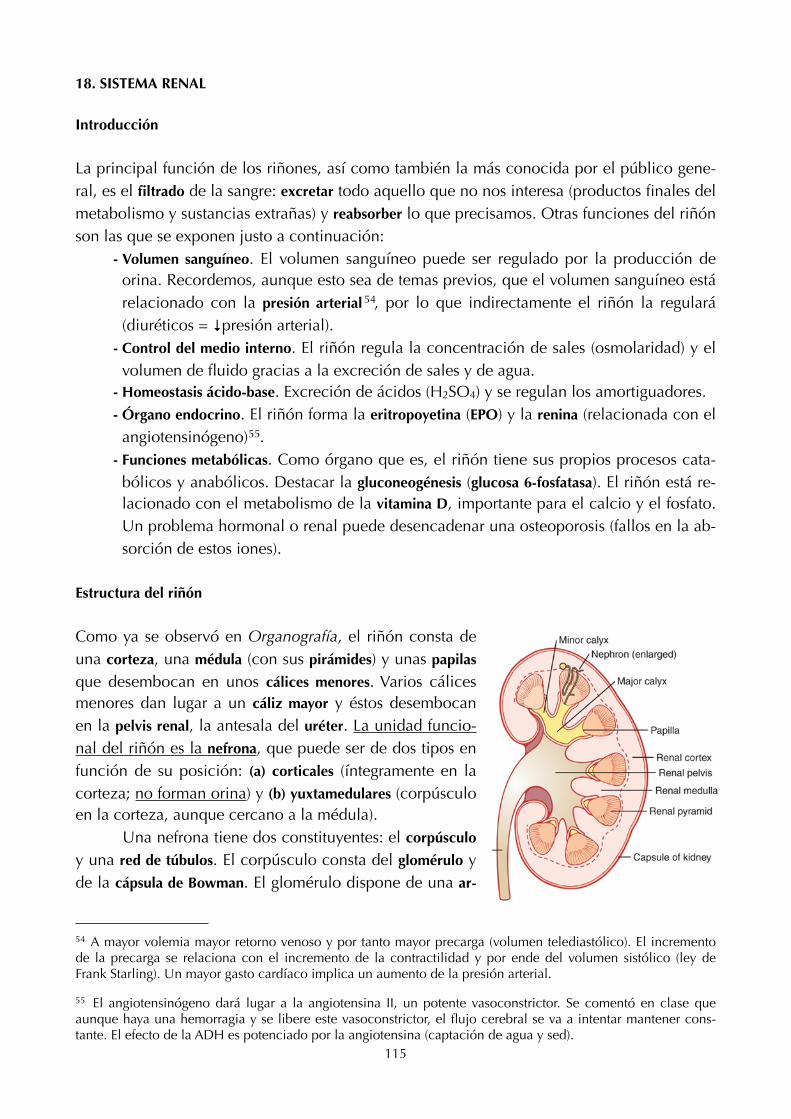
18. SISTEMA RENAL
Introducción
La principal función de los riñones, así como también la más conocida por el público gene-ral, es el filtrado de la sangre: excretar todo aquello que no nos interesa (productos finales del metabolismo y sustancias extrañas) y reabsorber lo que precisamos. Otras funciones del riñón son las que se exponen justo a continuación:
- Volumen sanguíneo. El volumen sanguíneo puede ser regulado por la producción de orina. Recordemos, aunque esto sea de temas previos, que el volumen sanguíneo está relacionado con la presión arterial 54, por lo que indirectamente el riñón la regulará (diuréticos = ↓presión arterial).
- Control del medio interno. El riñón regula la concentración de sales (osmolaridad) y el volumen de fluido gracias a la excreción de sales y de agua.
- Homeostasis ácido-base. Excreción de ácidos (H2SO4) y se regulan los amortiguadores.- Órgano endocrino. El riñón forma la eritropoyetina (EPO) y la renina (relacionada con el
angiotensinógeno)55.- Funciones metabólicas. Como órgano que es, el riñón tiene sus propios procesos cata-
bólicos y anabólicos. Destacar la gluconeogénesis (glucosa 6-fosfatasa). El riñón está re-lacionado con el metabolismo de la vitamina D, importante para el calcio y el fosfato. Un problema hormonal o renal puede desencadenar una osteoporosis (fallos en la ab-sorción de estos iones).
Estructura del riñón
Como ya se observó en Organografía, el riñón consta de una corteza, una médula (con sus pirámides) y unas papilas que desembocan en unos cálices menores. Varios cálices menores dan lugar a un cáliz mayor y éstos desembocan en la pelvis renal, la antesala del uréter. La unidad funcio-nal del riñón es la nefrona, que puede ser de dos tipos en función de su posición: (a) corticales (íntegramente en la corteza; no forman orina) y (b) yuxtamedulares (corpúsculo en la corteza, aunque cercano a la médula). Una nefrona tiene dos constituyentes: el corpúsculo y una red de túbulos. El corpúsculo consta del glomérulo y de la cápsula de Bowman. El glomérulo dispone de una ar-
115
54 A mayor volemia mayor retorno venoso y por tanto mayor precarga (volumen telediastólico). El incremento de la precarga se relaciona con el incremento de la contractilidad y por ende del volumen sistólico (ley de Frank Starling). Un mayor gasto cardíaco implica un aumento de la presión arterial.
55 El angiotensinógeno dará lugar a la angiotensina II, un potente vasoconstrictor. Se comentó en clase que aunque haya una hemorragia y se libere este vasoconstrictor, el flujo cerebral se va a intentar mantener cons-tante. El efecto de la ADH es potenciado por la angiotensina (captación de agua y sed).

teriola aferente y de una arteriola eferente (particularidad: entra y sale una arteria). La nefrona en sí va estar rodeada por dos redes capilares: (1) glomerular y (2) peritubular (al-rededor de los túbulos y desembocará en la vena renal). El segundo sistema de capilares (peritubular) es el que tiene una presión más baja y además tiene la misma forma que la parte de la nefrona que rodea (vasa recta re-
nis), que suele ser el asa de Henle. En el glomérulo nos vamos a encon-trar con tres capas: (a) céls. endoteliales; (b) membrana basal; y (c) podocitos. ¿De qué va a depender la filtración de elementos de la sangre? De su tamaño y de su carga (mem-brana basal cargada negativamente). La red de túbulos está formada por: (1) túbulo proximal; (2) asa de Henle (que puede ser dividida en una rama delgada descendente, una rama delgada ascendente y una rama gruesa ascendente); (3) túbulo
contorneado distal; y (4) túbulo colector. En el túbulo contorneado distal encontramos el apa-
rato yuxtaglomerular o mácula densa, en contacto con el glomérulo.
Filtración glomerular
Las mismas fuerzas de Starling que vimos que intervenían en el intercambio de líquidos en el capilar (filtración/absorción) también estarán presentes en el paso de fluidos desde los capi-lares glomerulares hacia el espacio de Bowman (véase imagen superior). Las presiones que determinan la dinámica de la filtración glomerular son la presión hidrostática capilar glomeru-
lar (PGC), que fuerza el líquido fuera de los capilares, la presión oncótica capilar glomerular (πGC), que atrae el líquido hacia el capilar glomerular, la presión hidrostática del espacio de
Bowman (PBS) que se opone a la PGC, y la presión oncótica del espacio de Bowman (πBS), que atrae el líquido hacia ese espacio. Normalmente la cantidad de proteínas presentes en el es-pacio de Bowman es irrisoria (tal y como ocurría con la Πif), por lo que se asume que πBS es 0. La presión neta de filtración (PNF) es la siguiente:
A diferencia del resto de capilares, en los que la presión hidrostática capilar caía bastante a medida que se avanzaba en el vaso, los capilares glomerulares no ven su PGC muy disminui-da en el extremo distal debido a que la arteriola eferente se puede contraer y mantener la pre-
sión. Con esto se logra que la PBS y la πGC juntas nunca superen a la PGC y por tanto única-mente se dé la filtración a lo largo de todo el capilar glomerular (siempre PNF > 0).
116
PNF = PGC − PBS( ) − πGC
(A) Ultraestructura básica de los capilares glomerula-res. (B) Sección transversal de la membrana capilar glomerular y sus principales componentes.

Velocidad de filtración glomerular
La velocidad de filtración glomerular (VFG o FG o FGR; Eng: filtration glomerular rate;) se con-sidera el parámetro de referencia de la función renal. La VFG es la cantidad de plasma (sin proteínas ni células) filtrada por todos los glomérulos de los riñones por unidad de tiempo. En un adulto sano, la VFG es de ∼100-125 mL/min, y la cifra es mayor en los hombres que en las mujeres. La VFG se puede mantener en un nivel bastante constante a pesar de las fluctua-ciones de la presión arterial media entre 80 y 180 mmHg. Este mantenimiento no se tiene muy claro como se logra, el Dr. Estévez mencionó dos hipótesis:
- Mecanismo miogénico. Las céls. endoteliales del riñón se contraen y la favorecen.- Regulación del calibre de las arteriolas aferente y eferente. Si el aparato yuxtaglomerular
detecta un incremento de los solutos en la nefrona entonces se produce una vaso-constricción en la arteriola aferente y se reduce la presión (y con ello la filtración).
La VFG está determinada por la PNF y por factores físicos relacionados con los glomérulos o Kf (coeficiente de filtración y área total de la superficie, que refleja el número de nefronas y su tamaño). La ecuación es la siguiente:
La VFG debe mantenerse para eliminar el exceso de líquidos y electrólitos de la sangre y mantener la hemostasis global. El cambio abrupto de uno de los parámetros de la ecuación alterará significativamente la VFG. Por ejemplo, una hemorragia puede hacer caer la presión arterial por debajo de 80 mmHg e impedir que haya filtración glomerular porque la PGC ha-brá descendido demasiado.
Aclaramiento renal
Puesto que la VFG es una medida importante de la función renal, se mide de forma habitual. La medida se puede hacer de varias formas. Experimentalmente se pueden medir los factores físicos y las presiones, pero no resulta práctico hacerlo en humanos. En su lugar, se utiliza el principio del aclaramiento renal, que es el volumen de plasma depurado de una sustancia por unidad de tiempo56. La ecuación de aclaramiento incorpora las concentraciones urinaria y plasdmática de la sustancia y el flujo urinario, y se suele expresar en mL/día o L/día:
117
56 Guyton pág. 340: el aclaramiento de una sustancia es el volumen de plasma que queda completamente des-provisto de la sustancia por unidad de tiempo.
VFG = Kf ⋅ PGC − PBS( ) − πGC⎡⎣ ⎤⎦
Cx =
Ux ⋅Vi⎛
⎝⎞⎠
PxAclaramiento de la sustancia x (Cx). (Netter pág. 204) Ux es la concentración en la orina de x; Ṿ (el punto va arriba, pero no hay dicho símbolo en el editor, en el cuerpo del texto se usará este símbolo) es el volumen de orina producido por unidad de tiempo; Px es la concentración plasmática de la sustancia.

El aclaramiento de una sustancia es igual a la VFG si la sustancia se filtra libremente pero no
se reabsorbe ni se secreta57. Para complicarlo un poco más: si la carga tubular de una sustancia es igual a la excreción urinaria de dicha sustancia, la velocidad de filtración glomerular será igual al aclaramiento de esa sustancia. Retrocedamos un poco y definamos lo que es carga tubular y excreción urinaria, que así lo entenderemos mejor: La carga tubular de una sustancia (Ctx) es la cantidad de una sustancia específica filtrada por unidad de tiempo. En inglés es “filtered load of x”.
En cuanto a la excreción urinaria de una sustancia (Exx), es la concentración de la sustancia en orina (Ux) multiplicada por el volumen de orina producido por unidad de tiempo (Ṿ):
Si estableces la equivalencia Ctx = Ux, llegarás a la conclusión de que Cx = VFG (los cálculos te los haces en tu libreta). Las sustancias que cumplen estas condiciones (Ctx = Exx) son los denominados indicadores. La creatinina o la inulina (un polisacárido de fructosa) son dos indi-cadores que suelen usarse para evalular la función renal. Para medir el aclaramiento de la inulina (y determinar así la VFG), se infunde inulina intravenosa y cuando alcanza una con-centración plasmática estable, se hacen recogidas de orina cronometradas.
Habitualmente no se usa la infusión de inulina para determinar el aclaramiento debido a la naturaleza agresiva del procedimiento. En su lugar se usa el aclaramiento de un indicador endógeno, la creatinina (producto de degradación del metabolismo muscular). La creatinina no es un perfecto indicador, ya que si bien no se absorbe, sí que se secreta (∼10%). Cuando se
usa la creatinina como indicador se hace una pequeña corrección. Es posible calcular el aclaramiento de cualquier sustancia filtrada y cuando el aclara-miento se compara con la VFG, proporciona una idea general de si existe reabsorción neta o secreción neta de esa sustancia. Si Cx > VFG quiere decir que hay una secreción neta, puesto que la cantidad de plasma limpiado es mayor que la atribuible a la VFG sola. En cambio, si Cx < VFG, existe reabsorción neta.
Sustancias que se reabsorben o se secretan
A modo de introducción, porque más adelante se hablará de cada molécula en detalle, ha-brán sustancias que una vez filtradas se reabsorberán (como la glucosa), otras que se secreta-
118
57 Esto se verá cuando hablemos de lo que le ocurre a cada molécula en su paso por la nefrona. No obstante, y creo que ya se comentó en Organografía, comentar que una molécula puede ser captada por los vasos peritu-bulares o bien excretada desde los mismos hacia la luz del túbulo.
Ctx = Px ⋅VFG
Exx =Ux ⋅Vi
Cinulina = VFGLey de la conservación de masa de Fick. El Dr. Estévez insistió que en el caso de un indicador (como la inulina) la ley de la conservación de masa se cumplía: toda masa filtrada será después excretada (al no haber fenóme-nos de secreción y absorción).
Pinulina ⋅VFG =Uinulina ⋅Vi

rán (como el PAH o ácido paraaminohipúrico) y otras que se se reabsorberán y secretarán (co-mo la urea). Si nos centramos en la glucosa, cuyo aclaramiento tiene que ser 0 mL/min (el plasma no se limpia de glucosa), veremos que su filtrado (o CtGlc, que es lo mismo) será me-nor a la cantidad que se excreta por la orina (EGlc). Si hiciéramos el cociente entre el aclara-miento de la glucosa con el de la inulina, nos saldría < 1. En cambio, si esa misma opera-ción la hiciéramos con un xenobiótico (como el PAH), nos saldría > 1 (porque hay secreción activa).
La velocidad de reabsorción de una sustancia (Rx) es igual a la carga filtrada de la sustancia (Ctx) menos su excreción urinaria (Ex).
La velocidad de secreción de una sustancia (Sx) es equivalente a la velocidad de excreción me-nor la carga filtrada de la sustancia.
119
CtGlc = PGlc ⋅VFG
Rx = Ctx − Exx
Sx = Exx − Ctx

19. PROCESOS DE TRANSPORTE RENAL
Introducción
Una vez el plasma se ha filtrado al espacio de Bowman, nos vamos a encontrar con toda una serie de procesos de reabsorción y de secreción a lo largo de los diferentes segmentos en los que se divide el sistema de túbulos de la nefrona. El túbulo contorneado proximal (TCP o TP, en clase se omitió el término contorneado) se divide en tres segmentos (S1, S2 y S3) y es donde se reabsorberá de manera masiva los nutrientes, el NaCl, el HCO3
- y el agua. El asa de Henle tendrá un papel muy importante en la captación de agua. Tanto el túbulo contorneado
distal (TCD) como el túbulo colector (TC) se van a encargar de ajustar finamente la cantidad de agua y sal que tendrá la orina final. En el túbulo colector encontramos tres tipos celulares: (a)
céls. intercalares α, implicadas en la reabsorción de K+ y la secreción de H+; (b) céls. intercala-
res β, implicadas en la secreción de bicarbonato y la reabsorción de Cl-; (c) céls. principales, encargadas de reabsorber NaCl y secretar K+.
Mecanismos generales de transporte
En los túbulos renales vamos a ver toda una serie de mecanismos de transporte tanto en un sentido (reabsorción) como en otro (secreción). A continuación vamos a clasificarlos. En primer lugar podemos distinguir si la molécula pasará o no a tra-vés de la célula. El transporte transcelular es aquel en el que la sustancia debe atra-vesar la célula (empleando el medio que sea) para alcanzar su destino. Por el otro lado tenemos el transporte paracelular, en el que la molécula podrá atravesar una superficie epitelial a través de las uniones estrechas que unen las células del epitelio (zonula occludens). Estas dos clases de transporte, así como los que veremos a continuación, se rigen por el equilibrio electroquímico (para simplificar: gradiente de concentración). Centrándonos en el transporte transcelular, la molécula que debe ir a un destino de-terminado tendrá diferentes maneras para acceder a la célula. Si la interiorización no implica un gasto energético, puesto que es a favor de gradiente, entonces hablamos de un transporte
pasivo. Si, por contra, precisara de energía, hablaríamos de un transporte activo. Dentro del transporte pasivo encontramos la difusión simple (moléculas liposolubles) y la difusión facilita-da (interviene una proteína; p. ej., iones). Dentro del transporte activo podemos hablar de un transporte activo primario, otro secundario (dependiente del primario) y otro terciario (depen-diente del secundario y por ende también del primario). El transportador activo primario típi-co es el ATPasa Na+/K+: hidrolizando una molécula de ATP lleva tres cationes Na+ al medio
120
Vías transcelulares y paracelulares de transporte transe-pitelial. En el transporte paracelular, las moléculas se mueven extracelularmente a través de las uniones estre-chas, cuya permeabilidad ante moléculas pequeñas e io-nes depende de la composición de los componentes de la unión y del estado fisiológico de las céls. epiteliales.

extracelular y dos cationes K+ al medio in-tracelular. Un transportador activo secunda-rio es dependiente del primario, me explico: el cotransportador glucosa/Na+ (SGLT1) que encontramos en diferentes lugares del cuer-po (riñón, intestino) aprovecha la entrada a favor de gradiente del sodio para interiorizar glucosa en contra de gradiente. ¿Por qué decimos que es un transporte activo si apa-rentemente no hay gasto de ATP? Porque el sodio interiorizado debe ser extraido de nuevo para mantener la concentración in-tracelular, y quien lo extrae es la ya men-cionada ATPasa Na+/K+, que sí que gasta ATP. Finalmente, el transportador activo terciario lo veremos más adelante, pero para hacernos una idea podemos decir que sería aquel que de-pende de la actividad de un secundario (p. ej., un transportador que intercambie la sustancia X por glucosa intracelular; precisaría por narices un transportador del estilo SGLT1).
Transporte de urea
La urea es una carbodiamida que se forma en el hígado (ciclo de la urea) para detoxificar el amonio que se forma al oxidar las proteínas (glutamato deshidrogenasa). La urea es elimina-da del cuerpo a través de las heces, el sudor y la orina. En el TP, el 50% de la urea filtrada es reabsorbida, por lo que aún resta un 50% al final del TP. En el asa de Henle, hay una secre-
ción, haciendo que al final del asa tengamos un 110% (un 10% más que la filtrada inicialmente). Finalmente, en el túbulo colector se reabsorbe un 70%, por lo que excretaremos un 40% de la orina filtrada inicialmente. La urea es fundamental en el mecanismo de concentración de la orina. La urea es secretada o reabsor-bida por medio de diferentes meca-nismos. En el TP se hará pasar al inters-ticio por mecanismo trans- y paracelu-lar (solvatada o por difusión facilitada). En Henle se observa una difusión faci-litada para la secreción. En el TC tam-bién habrá una difusión facilitada, pero en sentido contrario a la observada en Henle y regulada hormonalmente (por vasopresina; a mayor reabsorción de urea, mayor reabsorción de agua).
121
Cotransportador de glucosa/sodio. (Guyton pág. 55). En este caso, estamos hablando de un transportador activo secundario simporter (ambas moléculas van en la mismo sentido). El antiporter es lo contrario.
Relación entre flujo de orina y urea excretada. Para un flujo de orina diario normal (1,5 L/día) tenemos una excreción de urea de poco menos de 0,3 mmoles/min. Si la cantidad de orina formada es mayor, la urea excre-tada también lo es (y viceversa).
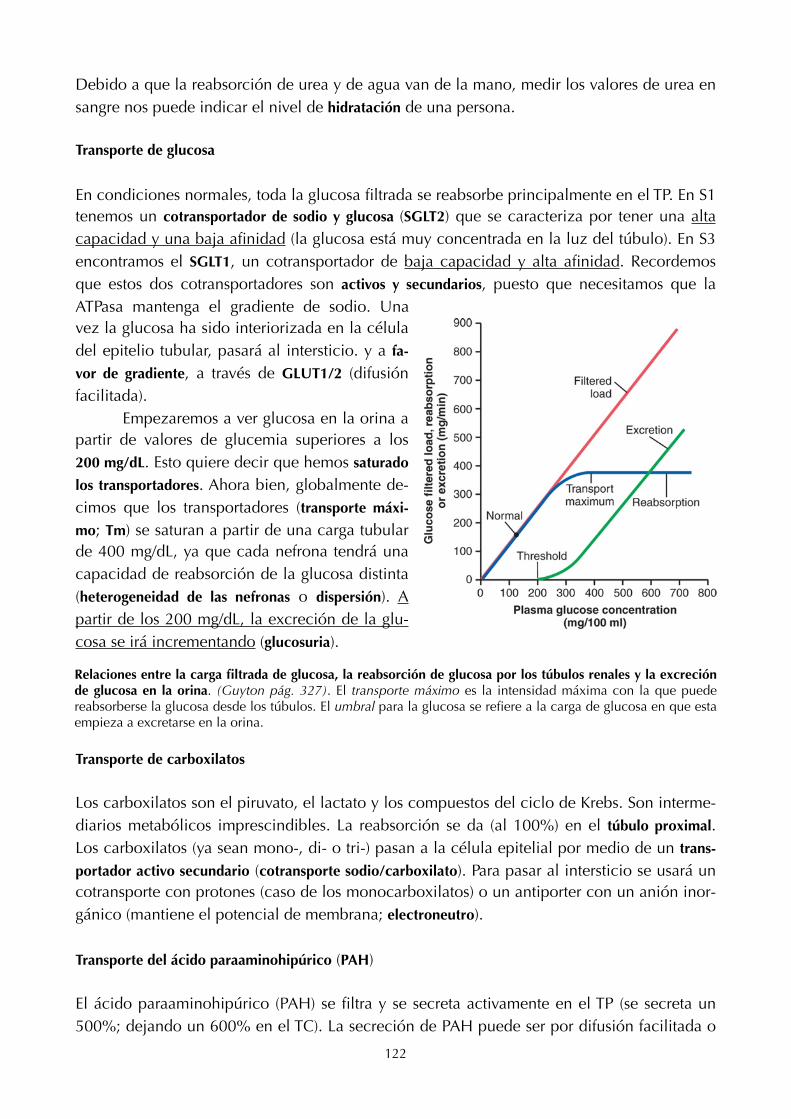
Debido a que la reabsorción de urea y de agua van de la mano, medir los valores de urea en sangre nos puede indicar el nivel de hidratación de una persona.
Transporte de glucosa
En condiciones normales, toda la glucosa filtrada se reabsorbe principalmente en el TP. En S1 tenemos un cotransportador de sodio y glucosa (SGLT2) que se caracteriza por tener una alta capacidad y una baja afinidad (la glucosa está muy concentrada en la luz del túbulo). En S3 encontramos el SGLT1, un cotransportador de baja capacidad y alta afinidad. Recordemos que estos dos cotransportadores son activos y secundarios, puesto que necesitamos que la ATPasa mantenga el gradiente de sodio. Una vez la glucosa ha sido interiorizada en la célula del epitelio tubular, pasará al intersticio. y a fa-
vor de gradiente, a través de GLUT1/2 (difusión facilitada). Empezaremos a ver glucosa en la orina a partir de valores de glucemia superiores a los 200 mg/dL. Esto quiere decir que hemos saturado
los transportadores. Ahora bien, globalmente de-cimos que los transportadores (transporte máxi-
mo; Tm) se saturan a partir de una carga tubular de 400 mg/dL, ya que cada nefrona tendrá una capacidad de reabsorción de la glucosa distinta (heterogeneidad de las nefronas o dispersión). A partir de los 200 mg/dL, la excreción de la glu-cosa se irá incrementando (glucosuria).
Transporte de carboxilatos
Los carboxilatos son el piruvato, el lactato y los compuestos del ciclo de Krebs. Son interme-diarios metabólicos imprescindibles. La reabsorción se da (al 100%) en el túbulo proximal. Los carboxilatos (ya sean mono-, di- o tri-) pasan a la célula epitelial por medio de un trans-
portador activo secundario (cotransporte sodio/carboxilato). Para pasar al intersticio se usará un cotransporte con protones (caso de los monocarboxilatos) o un antiporter con un anión inor-gánico (mantiene el potencial de membrana; electroneutro).
Transporte del ácido paraaminohipúrico (PAH)
El ácido paraaminohipúrico (PAH) se filtra y se secreta activamente en el TP (se secreta un 500%; dejando un 600% en el TC). La secreción de PAH puede ser por difusión facilitada o
122
Relaciones entre la carga filtrada de glucosa, la reabsorción de glucosa por los túbulos renales y la excreción de glucosa en la orina. (Guyton pág. 327). El transporte máximo es la intensidad máxima con la que puede reabsorberse la glucosa desde los túbulos. El umbral para la glucosa se refiere a la carga de glucosa en que esta empieza a excretarse en la orina.

intercambiándolo por un anión. Podríamos plantear-nos cómo ha llegado el PAH a la célula epitelial del túbulo proximal. El mecanismo es el siguiente: el PAH entra en la célula a cambio de un anión dicarboxilato (transporte activo terciario). Este mismo dicarboxilato se reintroduce en la célula por medio de un simporter
que introduce también tres cationes de sodio (transporte activo secundario). El gradiente de sodio se mantiene por la ATPasa (transporte activo primario). Si la concentración plasmática de PAH se in-crementa, llegará un momento que superaremos la Tm de los transportadores de PAH, por lo que la excre-ción en orina se mantendrá pero el aclaramiento irá
descendiendo (recordemos que el aclaramiento se re-fiere al volumen de plasma depurado de esa molécu-la).
Transporte de cationes orgánicos
Los cationes orgánicos se secretan (a un % variable) en el TP. El transporte desde el intersticio hacia la célula epitelial se hace por difusión facilitada (recordemos que el medio intracelular presenta una carga neta negativa por la acción de la ATPasa). El paso del catión al túbulo se hace por intercambio con un protón (antiporter; transporte activo terciario)58.
Transporte de ácidos y bases débiles
El transporte de ácidos y bases débiles depende del pH de la orina primaria59. Debemos par-tir de la base que una forma neutra (sin carga) pasará mejor que una forma cargada. Las formas neutras pasan por difusión. Por tanto, una orina primaria con un pH bajo implicará que ten-drá una gran cantidad de protones y por ende el equilibrio de la reacción de disociación de un ácido débil se desplazará hacia la formación de la forma neutra. En otras palabras, a pH ácido, un ácido débil se reabsorberá mejor. Con una base débil sucederá lo contrario.
El porcentaje de absorción de una molécula cargada depende del pH del fluido tubular. Su-pongamos que una persona se intoxica con aspirina, que es un ácido débil (ácido acetilsali-cílico). Lo que tendríamos que hacer es darle una base (p. ej., bicarbonato en vena) para así aumentar el pH de la orina y evitar la reabsorción del ácido (incrementar el aclaramiento). Otra opción es dar un diurético: diluir la orina y así facilitar el transporte reverso (únicamen-te funciona en el caso de los metabolitos que pasan por difusión facilitada).
123
58 El secundario del que depende es una bomba Na+/H+ (introduce sodio en el interior celular y saca protones).
59 Decimos orina primaria debido a que aún estamos en una parte inicial de los túbulos.
Secreción del ácido paraaminohipúrico. Nos interesa que la concentración de dicar-boxilato sea elevada para que así el trans-portador OAT1 lo capte con facilidad y lo intercambie por el PAH.
AH A− + H + BH
+ B + H +

Transporte de fostato
Los fosfatos son necesiarios para la formación de la matriz ósea - que está en forma de Ca3(PO4)2 - y para los mecanismos de alta energía intracelulares (formación de ATP). El fosfa-to plasmático puede estar asociado a proteínas, a iones o bien ir libre; únicamente la forma
libre se filtrará. El 80% del fosfato filtrado se reabsorbe en el TP, el 10% en el TCD y el 10% se pierde por la orina. Por tanto, a concentraciones fisiológicas de fosfato en plasma, ya per-demos una parte a través de la orina. Las células que tapizan el túbulo proximal disponen de unos transportadores activos
secundarios de Na+/Pi (simporter), tanto para la forma monoprotonada (3 cationes Na+) como para la forma biprotonada (2 cationes Na+). Se desconoce cómo se transfiere este fosfato al intersticio. La paratohormona (PTH) es secretada- por las glándulas paratiroides en respuesta a al-tas concentraciones plasmáticas de fosfato o bajas de calcio. La hormona disminuye los co-transportadores Na+-Pi luminales (por endocitosis), lo que reduce la reabsorción y aumenta la excreción urinaria de Pi. El pH también regula la reabsorción de Pi, la forma monoproto-nada se reabsorbe mejor.
Transporte del calcio
El calcio plasmático puede estar, como el fosfato, unido a proteínas (albúmina), a iones o libre. El libre es el que se filtrará y un 65% se reabsorberá en el TP. En el asa gruesa ascendente de Henle se reabsorbe el 25%, en el TCD un 8% y en el TC el 1%. En el TP el calcio puede pasar al intersticio empleando la vía paracelular (claudinas) o la vía
transcelular (difusión facilitada para entrar en la célula y diferentes tipos de transporte para salir; ATPasa de Ca2+). La regulación del transporte de calcio va a afectar sobre todo a la reabsorción del mismo en el asa de Henle y en el TCD. Las glándulas parati-roides liberan PTH cuando la concentración de Ca2+ plasmática es baja (o la de Pi es elevada)60; ésta favorece la reabsorción del Ca2+. La vitamina D también regula el transporte de calcio por medio de: (a) incrementar la expresión de la calcio-ATPasa; e (b) incrementar la expresión de la calcium-
124
60 Si con la PTH alta hubiera mucho Pi intracelular, éste podría precipitar junto al Ca2+ reabsorbido.
H 3PO4 H2PO4
− + H + HPO32− + 2H + PO4
3− + 3H +
Equilibrios del ácido fosfórico. En rosado se marca la forma monoprotonada, que se transporta mejor en el túbulo proximal a través de los canales Na+/Pi.
Mecanismos de reabsorción de calcio por las rutas paracelular y transcelular en las células tubulares proximales. (Guyton pág. 368). Nó-tese la calcio-ATPasa presente en la membrana basolateral.

binding protein (CBP), que secuestra el calcio en el interior celular y por tanto deja de contar para el gradiente. Un exceso de calcio en sangre, y por ende en el intersticio, activa un sensor de calcio ubicado en la membrana basolateral de la célula que tapiza el asa ascendente gruesa de Henle. Ente sensor da lugar a una cascada de señalización intracelular que culmina con la inhibición de un canal de K+, produciendo una despolarización y por tanto disminuyendo el gradiente electroquímico del calcio.
Transporte del magnesio
El magnesio (Mg2+) es un componente esencial de los huesos y del músculo esquelético. Su patrón de reabsorción se parece mucho al del potasio. La mayor parte del transporte se da en la porción gruesa ascendente del asa de Henle (70%) aunque el 15% que se reabsorbe en el TP se hace por vía paracelular.
Transporte de sodio (y cloro)
Los riñones mantienen el volumen de líquido extra-celular, regulando de pasada la presión arterial (volumen telediastólico; Frank-Starling). El principal componente de dicho fluido es el sodio. En el transporte de sodio a lo largo de los túbulos de la nefrona es complejo y por ello se discutirá como se transporta en cada uno de los segmentos. En el túbulo proximal se reabsorbe un 67%
del sodio filtrado (VFG: 180 L/día; PNa: 142 mmol/L; CtNa: 25.500 mmol/día). Entra una gran cantidad de sodio gracias a los famosos transportadores activos
secundarios (de glucosa, de fosfato, de aas y de áci-dos orgánicos). También están los antiporte Na+/H+ (como vemos, todo es transcelular). La entrada de sodio va también de la mano de la entrada de agua, por lo que el líquido tubular (orina) que abandona el túbulo proximal tiene la misma osmo-
laridad que el filtrado inicialmente. El asa de Henle es conocida como el ele-mento de la nefrona que diluye. Esto entra en con-tradicción con lo que hemos dicho antes: donde vaya el sodio, el agua le sigue. ¿Cómo se logrará esta dilución? Porque la nefrona dispone de distinas
permeabilidades para la sal y para el agua (el riñón debe ser capaz de separar el agua de la sal). En el asa gruesa ascendente el 25% del sodio pasa al
125
Mecanismos del transporte de sodio, el coloro y el potasio en el asa ascendente gruesa de Henle. (Guyton pág. 331). La bomba ATPasa Na+/K+ en la porción basolateral de la membra-na celular mantiene una concentración intrace-lular de sodio baja y un potencial eléctrico ne-gativo en la célula. El cotransportador Na+/K+/2Cl- en la membrana luminal transporta estos tres iones desde la luz tubular hacia las células usando la energía potencial liberada por difu-sión del sodio siguiente un gradiente electro-químico dentro de las células. El sodio también se transporta al interior celular mediante un cotransporte sodio-hidrógeno. La carga positiva (+8 mV) de la luz tubular respecto al líquido intersticial fuerza a los cationes a difundir des-de la luz al líquido intersticial a través de la vía paracelular.

intersticio por la vía paracelular y el resto a través del antiporte Na+/H+ y del simporte electroneutro Na+/K+/2Cl-. El sodio introducido sale al intersti-cio por medio de la ATPasa y el cloro por medio de un canal. El potasio interiorizado por este simporte electroneutro sale de nuevo al túbulo o al intersticio por un canal. La furosamida es un diurético (diurético del
asa) que bloquea el cotransportador Na+/K+/2Cl-, perdiéndose parte de la diferencia de potencial que hacía que cationes como el Na+, el Mg2+ y el Ca2+ pasaran al intersticio por la vía transcelu-lar (véase imagen pág. anterior). En el túbulo contorneado distal hay un co-transporte Na+/Cl- sensible a diuréticos tiacídicos. El cloruro sale al intersticio por medio de un ca-nal y el sodio por la ATPasa. Finalmente, en las células principales del túbulo colector encontramos unos canales de sodio cuya presencia estará modulada por la aldosterona y la hormona antidiu-
rética. El TC representa la reabsorción de un 3% de sodio, pero es el que es regulado hormonal-mente. El diurético amilorida bloquea ese canal. Del transporte de cloro podemos afirmar que es más o menos similar al de sodio (no se hizo hincapié en los mecanismos).
126
Cambios en la concentración de diferentes sus-tancias en el líquido tubular a lo largo del túbulo contorneado proximal respecto a las concentra-ciones de estas sustancias en el plasma y en el fliltrado glomerular. (Guyton pág. 330). Un valor de 1 indica que la concentración en el líquido tubular es la misma que la plasmática. Por debajo de 1 implica que se reabsorbe con más avidez que el agua, mientras que los valores superiores a 1 indican que o bien la sustancia se reabsorbe en menor grado que el agua o se secreta. Nótese como la osmolaridad no cambia. El indicador creatinina aumenta por la disminución de agua (lo mismo sucedería con la inulina).

20. MECANISMOS DE CONCENTRACIÓN Y DILUCIÓN DE LA ORINA
Introducción
Si observamos una gráfica de la osmolaridad del líquido tubular a medida que pasa a través de los diferentes segmentos tubulares, veremos como la tendencia neta es que ésta descienda, a no ser que haya una restricción de la diuresis por el motivo que sea (p. ej., deshidratación). No obstante, este descenso no es lineal: en el TP la osmolaridad no varía y en el asa de Henle
sufre un incremento y después un descenso. A partir del asa, la osmolaridad irá descendiendo de una manera más o menos pronunciada en función de nuestras necesidades.
Asa de Henle
En el asa de Henle se produce un cambio tan atípico de la osmolaridad debido a que su par-te descendente y su parte ascendente tienen propiedades distintas. Las ramas descedentes se
caracterizan por ser impermeables a la reabsorción de solutos y permeables al agua (por tanto, concentran). Por el otro lado, las ramas ascendentes son permeables a los solutos e impermeables al agua. Esto quiere decir que:
- Asa delgada descendente. La osmolaridad se irá incrementando progresivamente y el agua podrá salir al intersticio por flujo osmótico usando canales específicos (acuapori-
nas). Esto quiere decir que el intersticio medular debe ser hiperosmótico, para que así el agua pase del fluido tubular al intersticio. La hiperosmolaridad gracias a la alta con-centración de urea.
- Asa delgada ascendente. La osmolaridad irá disminuyendo ya que el sodio, muy con-centrado, pasará por difusión facilitada hacia el intersticio.
- Asa gruesa ascendente. La osmolaridad seguirá disminuyendo gracias a la acción del cotransportador Na+/K+/2Cl-.
127
Cambios en la osmolaridad del líquido tubular a través de los segmentos de los túbulos de la nefrona. (Guyton pág. 352). Nótese como en presencia de la hormona antidiurética (ADH) la osmolaridad final de la orina se incrementa enormemente.

Mecanismo multiplicador de contracorriente medular
El intersticio renal tiene un gradiente osmolar que es menor en la corteza y va aumentando hasta llegar a la médula interna. Esta hiperosmolaridad del intersticio es la que permitirá el flujo osmótico de agua hacia el intersticio. ¿Cómo se logra dicha hiperosmolaridad? Por me-dio de la propia geometría de la nefrona y del ciclo de reabsorción/secreción de la urea. Veamos lo que sucede por partes:
(1) Cotransportadores Na+/K+/2Cl-. Estos cotransportadores presentes en el asa gruesa ascendente de Henle permiten el paso de iones desde la luz del túbulo hacia el in-tersticio. Con esto estamos logrando incrementar la osmolaridad del intersticio y disminuir la del líquido tubular.
(2) Flujo osmótico de agua. La osmolaridad del intersticio ha sido incrementada en virtud al punto anterior, lo que favorecerá el paso de agua en el asa descendente delgada. Esto también nos lleva a incrementar la osmolaridad del líquido tubular.
(3) Avance del flujo tubular. Una vez este flujo tubular, cuya osmolaridad está aumenta-da por el punto anterior, pasa a la rama gruesa ascendente y, en virtud al primer punto, hara incrementar más la osmolaridad del intersticio.
Este ciclo de tres puntos se repite hasta establecer el gradiente intersticial completo y es lo que se conoce como multiplicador a contracorriente. Precisa de que la geometría de la nefro-na sea la que es. La concentración osmolar del intersticio de la profundidad de la médula depende de la longitud de las asas de Henle: cuanto más largas sean las asas, mayor será la ca-
pacidad de concentración (1.200 mOsm/L en humanos, lo que implica que el líquido tubular también podrá alcanzar esa osmolaridad en lo más profundo del asa de Henle).
El reciclado de la urea (ciclo fútil) contribuye al desarrollo y mantenimiento del gradiente os-molar intersticial, puesto que:
- La urea permanece en el líquido tubular mientras que el agua es reabsorbida en la rama descendente, lo que contribuye a la osmolaridad del líquido tubular.
128
Sistema multiplicador por contracorriente en el asa de Henle para la producción de una médula renal hipe-rosmótica. (Guyton pág. 349). Los valores numéricos corresponden a mOsm/L.

- La ADH aumenta la reabsorción de agua y urea en los TC medulares, pero no en los corticales; la urea se recicla en la médula interna, con lo que contribuye al gradiente de concentración intersticial.
Vasos rectos del riñón
Los vasos rectos del riñón (lat: vasa recta renis) se caracterizan por tener la misma morfología que la nefrona y por alojar en su interior un flujo que va muy lentamente. Este flujo tendrá la misma osmolaridad que la existente en el intersticio. La perfusión sanguínea se mantiene pe-se a la elevada osmolaridad gracias a estos tres puntos: geometría, velocidad y equilibrio.
129

21. REGULACIÓN DE LA FUNCIÓN RENAL
Introducción
A lo largo de los temas previos en los que se hablaba de diferentes aspectos del sistema renal hemos dejado entrever como algunos parámetros de su función pueden ser modificados por factores ajenos al riñón o del mismo.
Vasopresina
La hormona antidiurética (ADH o AVP por sus siglas en inglés: arginine vasopressin) hace que se capte agua en la parte distal de la nefrona (TC) incrementando la expresión de acuaporinas
en la membrana apical. La ADH la libera la hipófisis posterior en el caso en que se note un in-cremento de la osmolaridad del plasma en el hipotálamo. La ADH también está asociada con la captación de urea y de sodio en el túbulo colector, por lo que incrementa la osmola-ridad del intersticio.
Péptido natriurético auricular
El péptido natriurético auricular (ANP) es liberado si se nota un incremento de la presión en la
aurícula derecha. Para regular la VFG, el ANP dilata la arteriola aferente y constriñe la eferen-te, incrementado la PCG y por ende la VFG. El flujo aumentado eleva la excreción de sodio (na-triuresis) y agua con lo que reduce el volumen sanguíneo.
Simpático
Los nervios simpáticos liberan catecolaminas en respuesta a la reducción de la presión arte-rial sistémica, produciendo una vasoconstricción de las arterias y arteriolas renales. A niveles tónicos de actividad nerviosa simpática, los sistemas intrínsecos contrarestan ese efecto para asegurar que los vasos renales permanecen dilatados y se conserva la VFG. En caso de acti-vidad nerviosa simpática elevada (hemorragia, ejercicio extenuante), el simpático anula los mecanismos reguladores intrínsecos y reduce el flujo sanguíneo renal y la VFG.
Sistema renina-angiotensina-aldosterona (SRAA)
Ante un flujo vascular renal bajo61 se activa el SRAA, que libera renina (céls. yuxtaglomerula-res). La renina escinde el angiotensinógeno (proteína plasmática secretada por el hígado) en angiotensina I (A-I), que es convertida A-II por la enzima conversora de la angiotensina (ECA) presente en el pulmón y otros tejidos. La A-II, además de ser un poderoso vasoconstrictor
130
61 En el caso de una estenosis renal (que afecta a un riñón), el SRAA se activaría y la aldosterona formada afec-taría a ambos riñones.

(constricción de la arteriola aferente y eferente, por lo que la VFG desciende), provoca la libe-
ración del mineralocorticoide aldosterona, que retiene el sodio62 y agua.
Autorregulación
Ya se comentó que aunque la presión arterial vaya variando, la VFG se mantendrá constante. Si la presión arterial disminuye mucho, el riñón libera prostaglandinas (PGE2 y prostaciclina
PGI2), que son vasodilatadoras y contrarrestan de forma primaria la vasoconstricción mediada por la angiotensina II. Los AINEs (como la aspirina) bloquean la síntesis de prostaglandinas y restringen la vasodilatación renal compensadora (riesgo de insuficiencia renal aguda; VFG < 25 mL/min).
131
62 Se dijo que la aldosterona atraviesa la membrana e incrementa los enzimas mitocondriales, por lo que au-menta el ATP, la ATPasa funcionará mejor y por tanto el gradiente electroquímico mejora.
Cuadro resumen de las hormonas que afectan al riñónCuadro resumen de las hormonas que afectan al riñónCuadro resumen de las hormonas que afectan al riñónHormona Producida por y como respuesta a: Efectos:Aldosterona Producida por las células de la zona glomeru-
lar de la corteza suprarrenal como respuesta a la angiotensina II (SRAA) o a un incremento de la concentración plasmática de potasio.
Estimula la secreción de potasio en los túbulos renales (céls. ppales). Favorece la actividad de la ATPasa Na+/K+ e indu-ce la expresión de canales de K+ lumina-les
Aldosterona Producida por las células de la zona glomeru-lar de la corteza suprarrenal como respuesta a la angiotensina II (SRAA) o a un incremento de la concentración plasmática de potasio.
Estimula la reabsorción de Na+ en las células ppales.
Aldosterona Producida por las células de la zona glomeru-lar de la corteza suprarrenal como respuesta a la angiotensina II (SRAA) o a un incremento de la concentración plasmática de potasio.
Captando sodio, de pasada capta agua.
Aldosterona Producida por las células de la zona glomeru-lar de la corteza suprarrenal como respuesta a la angiotensina II (SRAA) o a un incremento de la concentración plasmática de potasio.
Pérdida de protones en las células inter-calares del TC (alcalosis metabólica).
Vasopresina Producida por la hipófisis posterior como res-puesta a un incremento de la osmolaridad del plasma detectada en el hipotálamo.
Estimula la captación de urea en los túbulos corticales medulares.
Vasopresina Producida por la hipófisis posterior como res-puesta a un incremento de la osmolaridad del plasma detectada en el hipotálamo. Estimula la captación de Na+ en las céls.
ppales.
Vasopresina Producida por la hipófisis posterior como res-puesta a un incremento de la osmolaridad del plasma detectada en el hipotálamo.
Estimula la captación de agua gracias al incremento de la expresión de acuapo-rinas en el túbulo colector (medular).
Péptido natriurético auricular
Producido por la aurícula derecha como res-puesta a un incremento del volumen sanguí-neo.
Dilata la arteriola aferente y contrae la eferente, incrementando la VFG por el aumento de la PCG. Favorece la excre-ción de agua y de sodio (natriuresis)

22. TRANSPORTE DEL POTASIO
Introducción
La concentración de potasio en el líquido extracelular está regulada normalmente en unos 4,2 mEq/L, y raramente aumenta o disminuye en ± 0,3 mEq/L. Este control preciso es necesa-rio porque muchas funciones celulares son muy sensibles a los cambios en la concentración de potasio en el líquido extracelular. Por ejemplo, el mantenimiento de un buen gradiente electroquímico de potasio es esencial para mantener el potencial de membrana de todas las células, sean excitables o no. También interviene en el control del pH63. Uno de los principales problemas para regular la cantidad de potasio que tenemos en el cuerpo es que la mayoría está en el interior de las células (98%). La ingesta de alimentos ricos en potasio, como los plátanos, pueden incrementar los niveles de potasio a cotas de-masiado elevadas (hiperpotasemia64), incluso mortales. La vuelta a la normalidad vendrá de la mano de la excreción fecal y, sobre todo, de la excreción renal.
Interiorización del potasio en las células
Ante el incremento de la concentración de potasio extracelular, lo primero que se hará será pasar ese exceso al interior de las células hasta que los riñones puedan hacerse cargo del ex-ceso. A continuación se discuten algunos factores que modifican la distribución del K+:
- Insulina. La insulina incrementa la captación de K+. Riesgo de los diabéticos de tener hiperpotasemias tras las comidas. Se comentó que la insulina incrementa la actividad de la ATPasa Na+/K+ y permite también la entrada de glucosa, por lo que podremos generar una mayor cantidad de ATP y así interiorizar potasio del medio extracelular.
- Aldosterona. La aldosterona favorece la captación de potasio por las células.- Estímulo β-adrenérgico. La secreción de adrenalina puede provocar el movimiento del
potasio desde el líquido extracelular al intracelular. - Alteraciones ácido-básicas. Una concentración de hidrogeniones elevada (pH ácido)
reduce la actividad de la ATPasa Na+/K+, lo que implica que se captará menos potasio del medio extracelular (y éste estará incrementado). Por tanto, la acidosis metabólica está asociada con un incremento del K+ extracelular, mientras que la alcalosis metabó-
lica se asocia con una disminución del K+ extracelular. - Lisis celular. La lisis celular permite que el potasio intracelular se libere.
Visión general de la excreción renal de potasio
La excreción renal de potasio está determinada por la suma de tres procesos renales: (1) fil-
tración; (2) reabsorción; y (3) excreción. La filtración de potasio es relativamente constante en personas sanas. La filtración (mencionada en temas anteriores como carga tubular) se puede
132
63 El potasio también está implicado en el control del volumen celular. Hay enzimas, como la succinato deshi-drogenasa (ciclo de Krebs) que precisan de potasio.
64 También podemos referirnos a ésta como hiperkalemia (lat: hiper - alto, kalium - potasio).
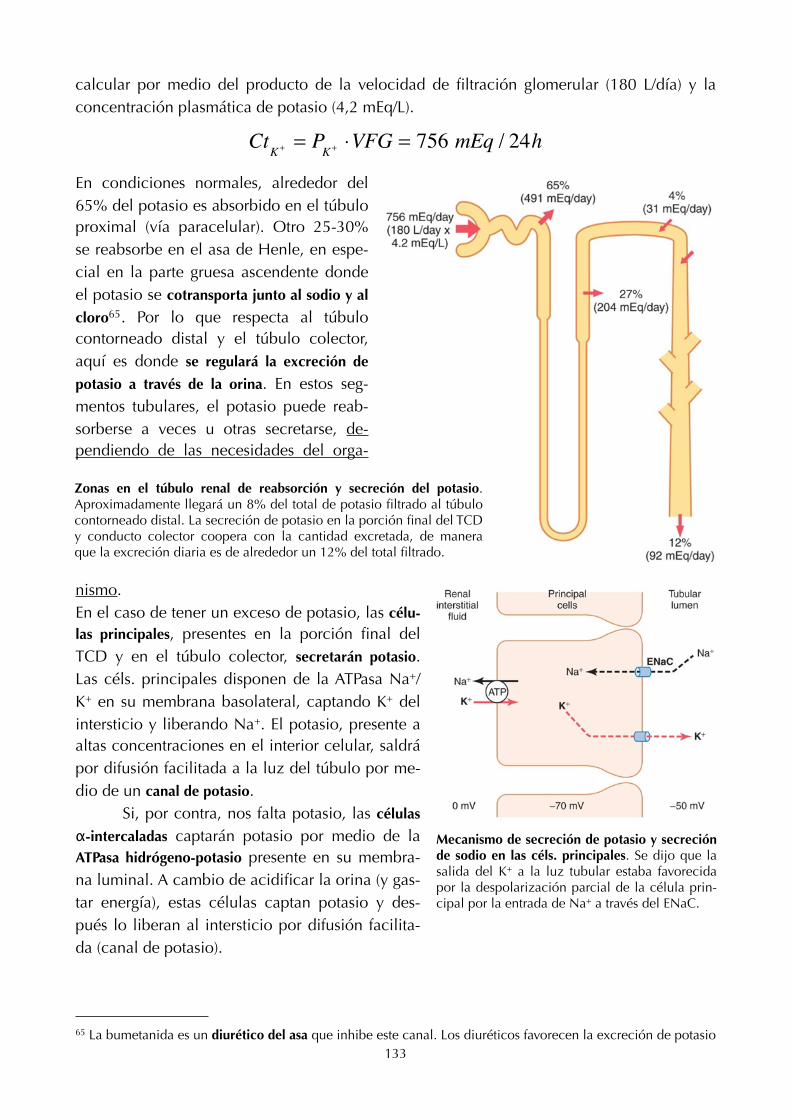
calcular por medio del producto de la velocidad de filtración glomerular (180 L/día) y la concentración plasmática de potasio (4,2 mEq/L).
En condiciones normales, alrededor del 65% del potasio es absorbido en el túbulo proximal (vía paracelular). Otro 25-30% se reabsorbe en el asa de Henle, en espe-cial en la parte gruesa ascendente donde el potasio se cotransporta junto al sodio y al
cloro65. Por lo que respecta al túbulo contorneado distal y el túbulo colector, aquí es donde se regulará la excreción de
potasio a través de la orina. En estos seg-mentos tubulares, el potasio puede reab-sorberse a veces u otras secretarse, de-pendiendo de las necesidades del orga-
nismo. En el caso de tener un exceso de potasio, las célu-las principales, presentes en la porción final del TCD y en el túbulo colector, secretarán potasio. Las céls. principales disponen de la ATPasa Na+/K+ en su membrana basolateral, captando K+ del intersticio y liberando Na+. El potasio, presente a altas concentraciones en el interior celular, saldrá por difusión facilitada a la luz del túbulo por me-dio de un canal de potasio. Si, por contra, nos falta potasio, las células
α-intercaladas captarán potasio por medio de la ATPasa hidrógeno-potasio presente en su membra-na luminal. A cambio de acidificar la orina (y gas-tar energía), estas células captan potasio y des-pués lo liberan al intersticio por difusión facilita-da (canal de potasio).
133
65 La bumetanida es un diurético del asa que inhibe este canal. Los diuréticos favorecen la excreción de potasio
CtK + = PK + ⋅VFG = 756 mEq / 24h
Zonas en el túbulo renal de reabsorción y secreción del potasio. Aproximadamente llegará un 8% del total de potasio filtrado al túbulo contorneado distal. La secreción de potasio en la porción final del TCD y conducto colector coopera con la cantidad excretada, de manera que la excreción diaria es de alrededor un 12% del total filtrado.
Mecanismo de secreción de potasio y secreción de sodio en las céls. principales. Se dijo que la salida del K+ a la luz tubular estaba favorecida por la despolarización parcial de la célula prin-cipal por la entrada de Na+ a través del ENaC.
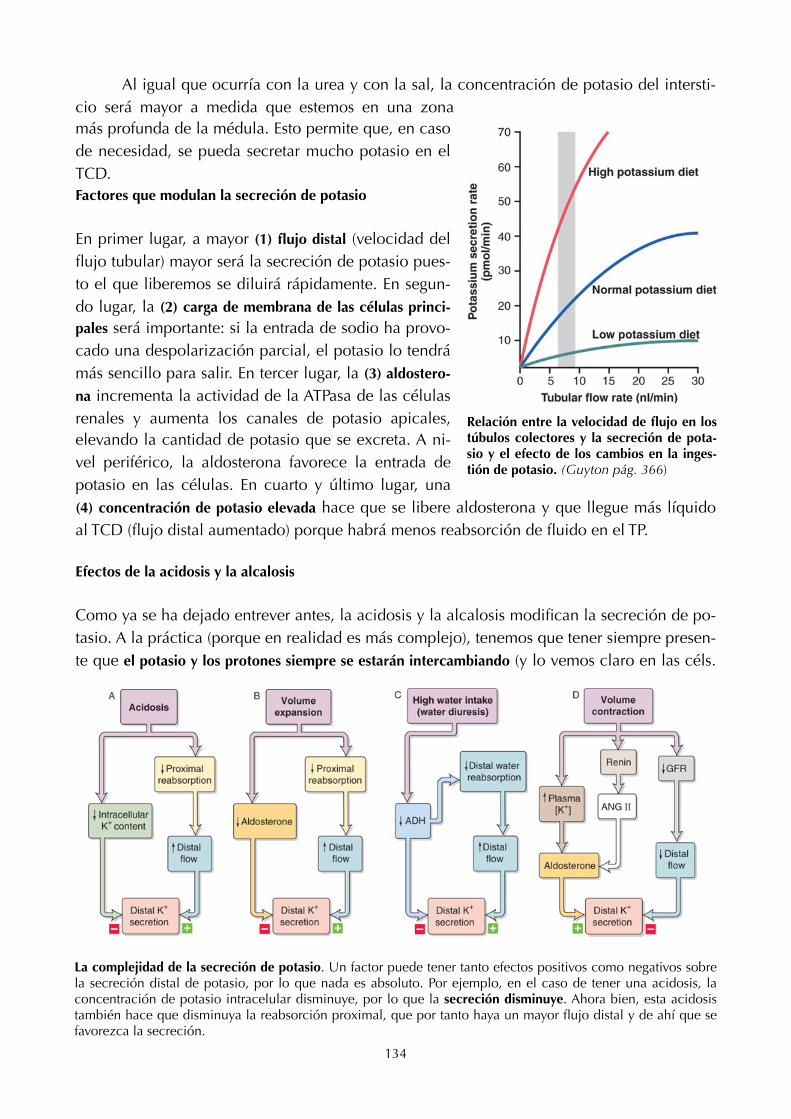
Al igual que ocurría con la urea y con la sal, la concentración de potasio del intersti-cio será mayor a medida que estemos en una zona más profunda de la médula. Esto permite que, en caso de necesidad, se pueda secretar mucho potasio en el TCD.Factores que modulan la secreción de potasio
En primer lugar, a mayor (1) flujo distal (velocidad del flujo tubular) mayor será la secreción de potasio pues-to el que liberemos se diluirá rápidamente. En segun-do lugar, la (2) carga de membrana de las células princi-pales será importante: si la entrada de sodio ha provo-cado una despolarización parcial, el potasio lo tendrá más sencillo para salir. En tercer lugar, la (3) aldostero-
na incrementa la actividad de la ATPasa de las células renales y aumenta los canales de potasio apicales, elevando la cantidad de potasio que se excreta. A ni-vel periférico, la aldosterona favorece la entrada de potasio en las células. En cuarto y último lugar, una (4) concentración de potasio elevada hace que se libere aldosterona y que llegue más líquido al TCD (flujo distal aumentado) porque habrá menos reabsorción de fluido en el TP.
Efectos de la acidosis y la alcalosis
Como ya se ha dejado entrever antes, la acidosis y la alcalosis modifican la secreción de po-tasio. A la práctica (porque en realidad es más complejo), tenemos que tener siempre presen-te que el potasio y los protones siempre se estarán intercambiando (y lo vemos claro en las céls.
134
Relación entre la velocidad de flujo en los túbulos colectores y la secreción de pota-sio y el efecto de los cambios en la inges-tión de potasio. (Guyton pág. 366)
La complejidad de la secreción de potasio. Un factor puede tener tanto efectos positivos como negativos sobre la secreción distal de potasio, por lo que nada es absoluto. Por ejemplo, en el caso de tener una acidosis, la concentración de potasio intracelular disminuye, por lo que la secreción disminuye. Ahora bien, esta acidosis también hace que disminuya la reabsorción proximal, que por tanto haya un mayor flujo distal y de ahí que se favorezca la secreción.

α-intercalares). Por tanto, en el caso de una acidosis (incremento de protones), vamos a se-
cretar protones y vamos a reabsorber potasio (hiperkalemia). Si sucede lo contrario, una al-calosis, excretaremos potasio a cambio de mantener los protones (hipokalemia).
135

23. TRANSPORTE DE ÁCIDOS Y BASES
Introducción
Es importante que el pH de nuestro medio interno se mantenga estable. Nuestro metabolis-mo, por medio de sus productos, modifica dicho pH. Los pulmones, los riñones y el intesti-no66 son estructuras que usa el organismo para intentar revertir esos cambios. A continuación vamos a estudiar principalmente lo que sucede cuando el pH del medio interno disminuye, lo que se conoce como acidosis. Esta acidosis puede darse por la aparición de ácidos volátiles, fácilmente excretables por medio de los pulmones incrementando la frecuencia respiratoria (exhalando más CO2), o de ácidos no-volátiles, la eliminación de los cuales dependerá emi-nentemente del riñón.
Papel del riñón en la regulación ácido-base
Como hemos dicho, el riñón es el encargado de neutralizar los ácidos no-volátiles, puesto que el pulmón no puede hacerlo. El rol que tiene el riñón en la regulación ácido-base se puede desgranar en dos grandes puntos: (1) compensar el exceso de protones que se produce
diariamente por el propio metabolismo; (2) evitar la pérdida del bicarbonato filtrado. Para hacernos una idea, excretamos vía orina unos 70 mmol de hidrogeniones por día. No obstante, esta carga de protones no puede ser liberada sin más, puesto que el pH de
la orina disminuiría demasiado, más allá de un mínimo tolerable (que está en torno al 4,4). En su lugar, los protones se liberan combinados con otras moléculas: con amoníaco para formar amonio y con otros ácidos titulables (fosfatos) [Eng: titrable acids]. Atención, los protones que van hacia la orina en forma de estas moléculas no tan “agresivas” no son los mismos que se forman en la sangre y que son los causantes de la acido-sis. El origen de estos protones es la producción de
novo de HCO3- para compensar aquel que ha consu-
mido en sangre para tamponar el exceso de proto-
nes67. Las células α-intercaladas del túbulo colec-tor pueden secretar protones. Éstos, una vez en la luz del TC, pueden tener tres destinos: (a) ayudar a
la reabsorción de bicarbonato; (b) formar ácidos ti-
tulables; (c) formar amonio. El bicarbonato, por la acción de la anhidrasa carbónica (que en este caso la hallamos en la membrana apical) puede ser transformado en dióxido de carbono y un hidroxilo. El CO2 difunde a través de la membrana apical sin
136
66 Se comentó que a través de las heces podemos perder base (10 mmol OH-/día).
67 En sangre el exceso de protones puede compensarse: (a) con bicarbonato; (b) con una base, formando una sal.
Anhidrasa carbónica. En el interior de la célula, una molécula de agua se escinde en un hidroxi-lo y en un protón. El protón sale al exterior y el hidroxilo ayuda a la formación del bicarbonato. Vemos como se forma un ciclo.

dificultad (difusión pasiva) y ahí una anhidrasa carbónica citosólica hace la reacción en sen-tido contrario, formando bicarbonato, que saldrá al medio intersticial.
Con los fosfatos y el amonio ocurre algo similar. El protón formado por la escisión de una molécula de agua puede salir al túbulo y combinarse con una forma protonada del ácido fosfórico para dar lugar a otra (p. ej., de mono- a biprotonada). En el interior celular el amo-nio puede escindirse en amoníaco y un protón para salir al lumen del túbulo y después vol-ver a formar amonio en él. La formación de los diferentes compuestos protonados depende del pH que haya en el túbulo, la concentración de la molécula aceptora de protones y de su pKa (captará más protones si el pH es cercano a su pKa). Tanto la exteriorización de protones al lumen como la liberación de bicarbonato se realiza de diferentes maneras en función de la parte de la nefrona en que estemos. Por tanto, la liberación de protones no se limita a las células α-intercaladas como se ha podido sospe-char antes. Los diferentes mecanismos se resumen en la imagen inferior.
137
CO2 + H2O HCO3− + H +
Reacción “oficial” de la anhidrasa carbónica. Como producto también a veces se pone que se forma ácido carbónico (H2CO3). Lo de la imagen de la página anterior, el tema de la hidrólisis del agua y tal, es lo mismo. La reacción puede darse sin el enzima, pero ésta acelera el proceso sustancialmente.
Mecanismos de liberación de bicarbonato y secreción de protones. El bicarbonato tiene un cotransportador con sodio o con cloro ubicado en la membrana basolateral. Los protones pueden ser intercambiados con sodio, potasio o bombeados directamente con un gasto de ATP.

El riñón en sus mitocondrias, es capaz de tranformar la glutamina en amonio y glutamato y desaminar oxidativamente este último por medio de la glutamato deshidrogenasa, formando amo-nio. Esta es una manera que tenemos de deshacernos del nitrógeno de las proteínas. Otra está en el hígado (ciclo de la urea). El amonio es muy tóxico (sustituye al potasio), puede bloquear la cadena respiratoria. Para evitar que éste llegue a la circulación sanguínea, se re-cicla entre diferentes puntos de la nefrona distantes a la circulación (zonas medulares). El transporte de amonio, por tanto, no se da en segmentos corticales de la nefrona. Tanto la urea, como el potasio y el amonio favorecerán el incremento la osmolaridad del intersticio medular.
Detección de una caída de pH
Ante una caida de pH (a través de unos receptores que no se comentaron) van a haber unas respuestas celulares: (a) se incrementa la expresión del enzima NHE3 (intercambiador Na+/H+); (b) aumenta la NBC (extrae bicarbonato); (c) mayor cantidad de transportador Na+/citrato (lo pa-só de largo); (d) mayor expresión de los enzimas sintetizadores de amonio. En caso de una alcalosis metabólica, las células intercaladas tenderán a diferenciarse en el tipo β. Este tipo se caracteriza por tener la pendrina, un intercambiador que secreta bi-carbonato (HCO3-/Cl- antiporter).
138

24. APARATO RESPIRATORIO
Introducción
Los componentes del aparato respiratorio son los pulmones, las vías aéreas y los músculos respiratorios. La principal función de este sistema es facilitar el intercambio de O2 y CO2 entre el cuerpo y el medio ambiente. El aparato respiratorio también participa en importantes pro-cesos fisiológicos adicionales que incluyen entre otros:
- Equilibrio ácido-base. El riñón también participa en esta función, ejerciéndola de una manera mucho más precisa, aunque más lenta, que los pulmones.
- Termorregulación. Cuando tenemos calor jadeamos, por lo que hay una mayor ventila-ción del espacio muerto anatómico68 y perdemos una mayor cantidad de vapor de agua. Evaporar agua nos supone una gran pérdida calórica.
- Función inmunitaria.- Funciones metabólicas. En el pulmón se forma el enzima conversor (sistema de la renina-
angiotensina-aldosterona), el surfactante pulmonar y prostaglandinas. Se destruyen trombos, catecolaminas y bradicinina.
- Reserva de volemia.- Otras. Como por ejemplo el habla (y el canto). El sistema respiratorio favorece funcio-
nes fisiológicas como la defecación, la émesis y el parto.
Paso del aire a las vías respiratorias
Antes de llegar a los alvéolos pulmonares, el aire que inspiramos debe ser condicionado. Esto consiste, principalmente, en: (a) calentarlo, a la temperatura corporal (37 ºC); (b) humidificarlo (saturarlo con agua); y (c) limpiarlo. El acondicionamiento del aire se da principalemente en las fosas nasales. Hemos de pensar que los cornetes que hay en su interior ofrecen una gran resistencia y hacen que el flujo de aire, ahora turbulento, se caliente, se humidifique y se limpie (esto último en parte). Las partículas de mayor tamaño (> 6 µm) serán eliminadas en la nariz. Esto se logra por los pelos de las narinas (vibrisas) y por la precipitación turbulenta (debido al choque de las partículas con los cornetes, el tabique y la pared faríngea). Las partículas de menor tamaño (< 6 µm) accederán a zonas más allá de las fosas na-sales y la faringe. Las partículas de entre 1 y 5 µm se depositarán en los bronquíolos más pe-queños (o antes) por precipitación gravitacional (se adhieren al moco producido por las céls. caliciformes). Las de menor tamaño aún llegarán a los alvéolos y podrán adherirse a las pa-redes o bien quedar suspendidas. En ambos casos, los macrófagos del polvo (o alveolares) po-drán deshacerse de éstas en la gran mayoría de casos. Las partículas atrapadas por el moco serán transportadas hacia arriba (fuera de los pulmones) gracias al movimiento de los cilios (ascensor mucociliar69).
139
68 Se trata de la zona de conducción (vías respiratorias) donde no hay intercambio de gases.
69 Permiten mover partículas a una velocidad de hasta 1,5 mm/min. El moco que ha sido ascendido puede ser o bien deglutido o bien exteriorizado (vía tos).

El moco liberado a la luz respiratoria humidifica y cumple una función defensiva. Entre sus compuestos encontramos proteínas que ya habíamos visto en la saliva, como la lactoferrina70 y la lisozima. La mucosa respiratoria, además, tiene la capacidad para liberar IgA71.
Circulación pulmonar
En primer lugar, convendría decir que el pulmón tie-ne dos circulaciones, una pulmonar cuyo cometido principal es hacer pasar la sangre por los alvéolos para el intercambio gaseoso, y otra bronquial cuyo objetivo es nutrir a los pulmones. La primera circu-lación se caracteriza por tener un gran flujo y una baja presión, mientras que la segunda es todo lo contrario, bajo flujo y una elevada presión (ligera-mente inferior a la aórtica). La circulación pulmonar empieza en la arteria pulmonar, que surge del ventrículo derecho. Tras 5 cm de recorrido, se divide en una rama derecha y otra izquierda. Todas las arterias de la circulación pul-monar tienen un diámetro mayor que sus correspon-dientes sistémicas y unas paredes finas. Estos dos fac-tores combinados hacen que estas arterias tengan una elevada distensibilidad, lo que les permite alojar el volumen sistólico del ventrículo derecho (5 L/min en reposo). La presión sistólica del ventrículo derecho es de 25 mmHg, que se transmite hacia la arteria pulmonar. Al acabar la sístole, la presión ventricular disminuye abruptamente, mien-tras que la presión arterial pulmonar disminuye más lentamente a medida que la sangre fluye a través de los capilares de los pulmones. La presión arterial pulmonar sistólica es de 25 mmHg, la diastólica es de unos 8 mmHg y la media es de unos 15 mmHg. La presión capilar pulmonar es de unos 7 mmHg. Como veremos más adelante, es im-portante que esta presión se mantenga así de baja (prevención de edemas). El sistema circu-latorio pulmonar tendrá mecanismos para mantener esa presión (bajas las resistencias en el caso de que se incremente el flujo; ley de Poiseuille). Si seguimos avanzando en el árbol arterial pulmonar, al final llegaremos a los capila-res. La longitud de los vasos es tan pequeña y el entramado es tan denso que se forma, lite-ralmente, una película de sangre alrededor del alvéolo. El espacio entre el espacio alveolar y la sangre es el más pequeño posible para así facilitar el intercambio de gases.
140
70 Quelante del hierro. Priva de hierro a las bacterias.
71 Los deportistas de élite, paradójicamente, tienden a padecer infecciones respiratorias. Esto se debe a que el ejercicio físico de gran intensidad disminuye la secreción de IgA en las mucosas. Por el otro lado, el ejericicio físico moderado (el que no es de competición) intensifica la liberación de IgA. Una actividad física moderada podría ser considerada aquella que hace que nuestra FC sea el 75% de la máxima (FCmáx = 220 - edad).
Presión en el ventrículo derecho, la aorta y la arteria pulmonar. (Guyton pág. 478) Nó-tese como la presión ventricular derecha disminuye súbitamente tras la sístole).

Debido a la gran cantidad de capilares que hay en la circulación pulmonar, la sección total es
elevada, lo que implica que la velocidad será muy baja. Un hematíe, estando nosotros en repo-so, puede tardar unos 0,75 s en recorrer la red capilar perialveolar. Únicamente necesitará 0,25 s de ese tiempo para que se dé el intercambio gaseoso. ¿Cómo es que sobra tanto tiem-po? Para así tener margen cuando esta velocidad se incrementa (al hacer ejercicio por ejem-plo) o bien cuando, por patología, se necesita más tiempo para el intercambio de gases (p. ej., edema).
Respecto a las presiones de la circulación pulmonar, comentar que éstas son más bajas que la circulación sistémica porque el ventrículo derecho tiene una menor masa muscular que el izquierdo, por lo que puede transmitir una menor presión a la columna de sangre. La presión
diferencial, que es la diferencia entre la máxima y la mínima, es mayor (relativamente) en la circulación pulmonar que en la sistémica.
Intercambio capilar en los pulmones
Tal y como habíamos visto en la circulación sistémica y también en el sistema renal, hay to-da una serie de fuerzas que influyen en el intercambio capilar (fuerzas de Starling). La diná-mica del intercambio de líquido a través de las membranas capilares pulmonares es cualita-tivamente la misma que en los tejidos periféricos. Sin embargo, cuantitativamente hay algunas diferencias. La presión hidrostática capilar pulmonar (Pcp) es de unos 7 mmHg, bastante inferior que la que habíamos visto en la circulación sistémica. La Pcp favorece la salida de líquido hacia el medio intersticial (filtración). La presión hidrostática tisular pulmonar (Ptp) se opone teórica-mente a la Pcp. Ahora bien, como la Ptp es negativa, en realidad favorece la filtración. La pre-
sión coloidosmótica (u oncótica) capilar (πcp) favorece la reabsorción mientras que la presión
coloidosmóstica intersticial (tisular) (πtp) favorece la filtración. Tras medir todas estas fuerzas, se observa como hay una pequeña tendencia hacia la filtración. Esto implicaría que progresivamente el medio intersticial se iría llenando de fluido y, por tanto, la distancia entre espacio alveolar y sangre se incrementaría (edema). Además, también podría suceder que parte del líquido del espacio intersticial pasara a los alvéolos, inundándolos. ¿Cómo se evita esto? Gracias a que el intersticio es drenado constantemente por el linfático. La presión hidrostática tisular pulmonar estaría actuando aquí como un meca-nismo protector frente al edema: si aumenta demasiado, el linfático drenará más72.
141
72 Si el linfático no drena bien entonces probablemente tendremos un edema pulmonar. Una causa de que el linfático no drene correctamente es el cáncer de pulmón. Generalmente esta neoplasia debuta con edema pulmonar.
Q = S ⋅ v Q =ΔPz
Ley de continuidad (izquierda) y ley de Poiseuille (derecha). La primera ley nos dice que, a flujo constante, si la sección es muy elevada la velocidad tendrá que ser muy baja. La segunda relaciona el flujo con la presión y las resistencias (z). Si la presión es muy baja en la circulación pulmonar, y el flujo es constante, las resistencias serán muy bajas.

Surfactante pulmonar
El pulmón puede ser considerado como un conjunto de 300 millones de burbujas, cada una de 0,3 mm de diámetro. Tal estructura es inherentemente inestable. Como consecuencia de la tensión superficial73 del líquido que cubre el alvéolo, se desarrollan fuerzas relativamente importantes que tienden a colapsar los alvéolos. La tensión superficial debe ser vencida para poder realizar la inspiración, por lo que nos interesa disminuirla para que así el trabajo respi-ratorio cueste lo menos posible. Las células de Clara (células epiteliales alveolares de tipo II) secretan el surfactante pulmonar, un tensioactivo (reduce la tensión superficial). El surfactante se sintetiza y se libera hacia el final del desarrollo fetal (antes no se necesita). Éste consiste en una mezcla de lípidos (90%) y proteínas (10%). La dipalmitoil fosfatatidilcolina (o dipalmitoil
lecitina) es el lípido mayoritario (80%). Después también encontramos palmítico y colesterol. En cuanto a las proteínas, tenemos dos grandes tipos: (a) con PM elevado e hidrofílicas, y (b)
con PM bajo e hidrofóbicas. Las primeras tienen una función defensiva (son proteínas tipo in-munoglobulina, IgA?) y las segundas ayudan en la función tensioactiva.
El surfactante está diluido en el líquido pulmonar. En virtud a la ley de Laplace (véase arriba), para mantener la máxima superficie alveolar debemos disminuir la tensión superficial. Las funciones del surfactante son: (1) disminuir el trabajo respiratorio; (2) favorecer la inspiración;
142
73 Esto se discute en el tema de biofísica de la respiración (Bioquímica y biofísica de sistemas). Para hacernos una idea, cuando el agua forma una superficie con el aire, las moléculas de agua de la superficie del agua tie-nen una atracción especialmente intensa entre sí. En consecuencia, la superficie del agua está intentando con-traerse. En las paredes alveolares tenemos el líquido alveolar que, por este mismo principio, intenta contraerse y así hacer evacuar el aire que hay en el interior de los alvéolos para finalmente acabar colapsando el alvéolo. Para evitar esto tenemos el surfactante.
Algunos factores que influyen en el intercambio capilarAlgunos factores que influyen en el intercambio capilarIncremento de la Pcp Supongamos que estamos en una altura lo suficientemente elevada como para que la
PO2 atmosférica sea más baja. Habrán zonas del pulmón que no ventilarán, por lo que habrá una vasoconstricción (ya veremos por qué). Las que ventilan recibirán un exceso de flujo, lo que se traduce en un aumento de la presión ya que la sección es más baja (Q = ΔP · z). La hipertensión pulmonar (de altura) consiste en que la Pcp se eleva demasiado, favoreciendo la filtración y finalmente dando lugar al edema pul-monar agudo de altura.
Inflamación En la inflamación se incrementa la permeabilidad de los vasos (bradicinina), favore-ciendo la filtración.
Desnutrición La desnutrición implica una disminución de la concentración de las proteínas plasmá-ticas o, lo que es lo mismo, la bajada de la presión oncótica capilar. Esto favorece la filtración.
Surfactante La falta de surfactante favorece la filtración.
P =2Τr
Ley de Laplace. La ley de Laplace relaciona la presión (P) con la tensión superficial (T) y con el radio del alvéo-lo (r). El surfactante pulmonar hace disminuir esta tensión. Hemos de pensar que, tal y como veremos en la figu-ra de la página siguiente, una gota de agua de menor radio sufrirá una presión mucho mayor. También es cierto que conforme más pequeño es el radio, el surfactante estará más concentrado, por lo que la tensión superficial disminuirá.

(3) mantener la estabilidad alveolar; (4) prevenir el edema pulmonar y la atelectasia. La atelectasia es el colapso de los alvéolos (que puede darse tanto por la falta de surfactante como por la obstrucción total de las vías aéreas).
Algunos recién nacidos, especialmente los prematuros, no han formado el suficiente surfac-tante pulmonar, por lo que los pulmones del niño tienen tendencia a colapsarse o a llenarse de líquido. Esta enfermedad es el síndrome de distrés respiratorio del recién nacido (antaño: en-
fermedad de las membranas hialinas, ya que antes se creía que había un velo que evitaba que los pulmones se inflaran con facilidad). Uno de los principales reguladores de la síntesis de surfactante es la distensión alveolar, por lo que un tratamiento con respiración asistida a pre-sión positiva podría favorece la síntesis de surfactante y resolver el problema. También po-demos usar corticoides (que estimulan la producción de surfactante) o introducir directamen-te el surfactante vía aerosol.
Correlaciones ventilación-perfusión
Las zonas que reciben una buena ventilación deben recibir una gran perfusión74. La perfusión sanguínea para los dos pulmones es de unos 5,5 L/min, mientras que la ventilación alveolar es de 4,5 L/min. En virtud a esto, la relación ventilación-perfusión, también denominada índice
VA/QA, está en un valor de 0,8 o 1. Este índice no es homogéneo en todo el pulmón, todo lo contrario. Desequilibrios en este índice pueden tener una causa fisiológica (normal) o pato-lógica. La causa fisiológica principal es la gravedad: la ventilación y la perfusión es mayor en la base del pulmón debido a la fuerza que ejerce la gravedad. ¿Por qué las bases ventilan más que los vértices? Bien, para entenderlo en primer lu-gar debemos asumir que la presión pleural es menos negativa en las bases que en los vértices. Pensemos que la pleura de la base recibe todo el peso de los pulmones, por lo que la presión es más positiva. En los vértices ocurre justo lo contrario: el pulmón tiende a alejarse de la pleura más apical por su propio peso, haciendo que la presión sea más negativa (hay más
143
74 Es preferible hablar de perfusión más que de irrigación. No obstante, ambos términos son correctos.
Efectos de la tensión superficial. Para acabar de ilustrar el concepto de tensión superficial, vemos a continua-ción el comportamiento de dos gotas de agua unidas entre sí. La gota de menor tamaño (menor radio), como tiene una mayor presión (ley de Laplace) se vaciará en la gota de mayor radio.

espacio del que se necesita). Recordemos que, sin tener en cuenta el efecto de la gravedad, la presión pleural es negativa ya que hay una aspiración continua del líquido pleural75. Sabien-do todo esto, nos vamos al gráfico inferior. Veremos como las presiones pleurales menos ne-gativas se asocian con cambios en el volumen pulmonar más grandes, mientras que las más negativas se asocian con cambios en el volumen pulmonar más pequeños. En otras palabras: una presión pleural más positiva (dentro de la negatividad) favorece la ventilación = las bases ven-
tilan más.
Que las bases ventilen más que los vértices no debe confundirnos: a pesar de esto, los vérti-
ces tienen un mayor volumen pulmonar que las bases (fíjate en la gráfica). Es por eso que el ápex pulmonar se ve más oscuro en los RX. Aprovechando que hemos introducido una gráfica volumen/presión pleural, podemos hacer cuatro pinceladas del trabajo respiratorio. El traba-jo respiratorio se divide en inspiratorio y espiratorio (que suele ser gratuito). Al inspirar nos enfrenta-mos a la elasticidad del pulmón y a la tensión super-
ficial.
Hasta ahora hemos hablado de la ventilación pulmonar y las diferencias regionales de la misma. ¿Qué sucede con la perfusión? Pues también por el efecto de la gravedad, la perfu-sión de las bases es mayor que la de los vértices. En primer lugar, la presión hidrostática capi-
144
75 Y también porque el pulmón tiende a irse por un lado y la caja torácica al otro. Los cambios en la presión pleural los podemos medir gracias a un globo que colocamos en la faringe.
Relación entre los cambios en el volumen pulmonar y la presión pleural. Vemos como las presiones pleurales menos negativas (base) se asocian con cambios de volumen mayores (ΔV1 < ΔV2)
Vol
umen
pul
mon
ar (
L)
Pr. pleural (cm H2O)-2 -14-9-7
ΔV1
ΔV2
Trabajo respiratorio. El área sombreada en gris representa el trabajo inspiratorio. Si sombreáramos el área que representa el trabajo espiratorio, este área estaría incluida dentro del área del trabajo inspiratorio, es por eso que la espiración nos sale gratis. La morfología de la gráfica se puede ver mo-dificada por patología.
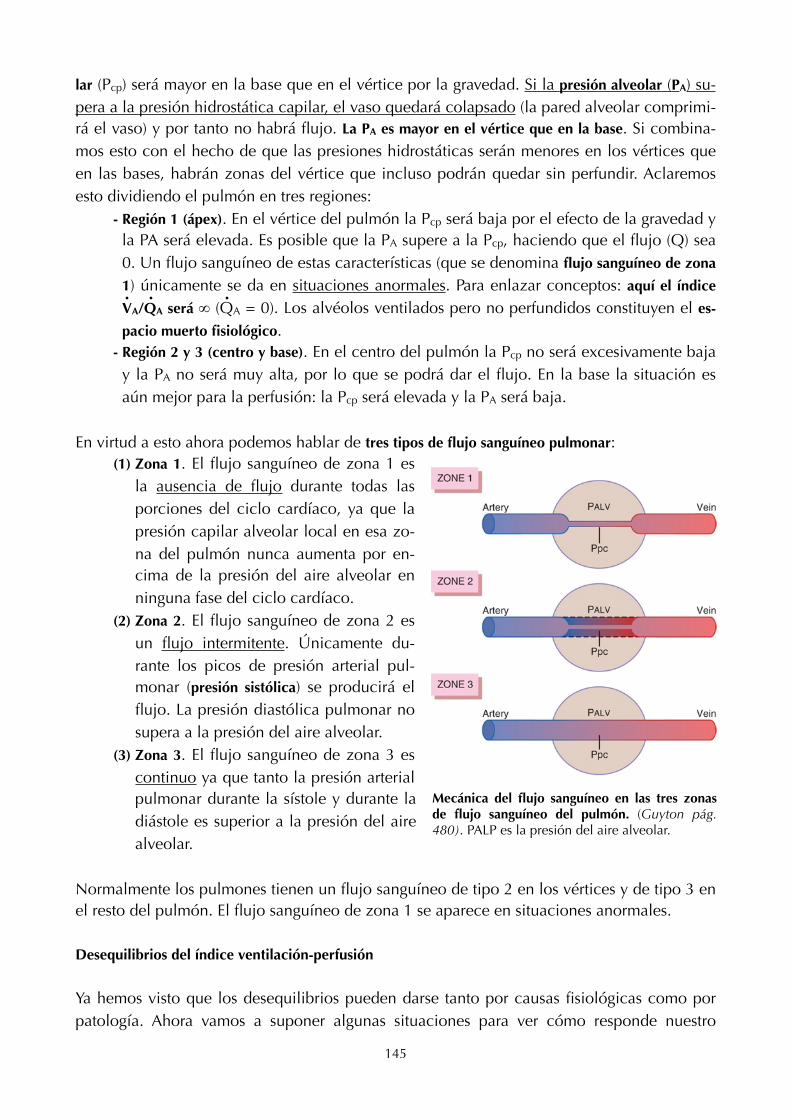
lar (Pcp) será mayor en la base que en el vértice por la gravedad. Si la presión alveolar (PA) su-pera a la presión hidrostática capilar, el vaso quedará colapsado (la pared alveolar comprimi-rá el vaso) y por tanto no habrá flujo. La PA es mayor en el vértice que en la base. Si combina-mos esto con el hecho de que las presiones hidrostáticas serán menores en los vértices que en las bases, habrán zonas del vértice que incluso podrán quedar sin perfundir. Aclaremos esto dividiendo el pulmón en tres regiones:
- Región 1 (ápex). En el vértice del pulmón la Pcp será baja por el efecto de la gravedad y la PA será elevada. Es posible que la PA supere a la Pcp, haciendo que el flujo (Q) sea 0. Un flujo sanguíneo de estas características (que se denomina flujo sanguíneo de zona
1) únicamente se da en situaciones anormales. Para enlazar conceptos: aquí el índice
VA/QA será ∞ (QA = 0). Los alvéolos ventilados pero no perfundidos constituyen el es-
pacio muerto fisiológico.- Región 2 y 3 (centro y base). En el centro del pulmón la Pcp no será excesivamente baja
y la PA no será muy alta, por lo que se podrá dar el flujo. En la base la situación es aún mejor para la perfusión: la Pcp será elevada y la PA será baja.
En virtud a esto ahora podemos hablar de tres tipos de flujo sanguíneo pulmonar:(1) Zona 1. El flujo sanguíneo de zona 1 es
la ausencia de flujo durante todas las porciones del ciclo cardíaco, ya que la presión capilar alveolar local en esa zo-na del pulmón nunca aumenta por en-cima de la presión del aire alveolar en ninguna fase del ciclo cardíaco.
(2) Zona 2. El flujo sanguíneo de zona 2 es un flujo intermitente. Únicamente du-rante los picos de presión arterial pul-monar (presión sistólica) se producirá el flujo. La presión diastólica pulmonar no supera a la presión del aire alveolar.
(3) Zona 3. El flujo sanguíneo de zona 3 es continuo ya que tanto la presión arterial pulmonar durante la sístole y durante la diástole es superior a la presión del aire alveolar.
Normalmente los pulmones tienen un flujo sanguíneo de tipo 2 en los vértices y de tipo 3 en el resto del pulmón. El flujo sanguíneo de zona 1 se aparece en situaciones anormales.
Desequilibrios del índice ventilación-perfusión
Ya hemos visto que los desequilibrios pueden darse tanto por causas fisiológicas como por patología. Ahora vamos a suponer algunas situaciones para ver cómo responde nuestro
145
Mecánica del flujo sanguíneo en las tres zonas de flujo sanguíneo del pulmón. (Guyton pág. 480). PALP es la presión del aire alveolar.
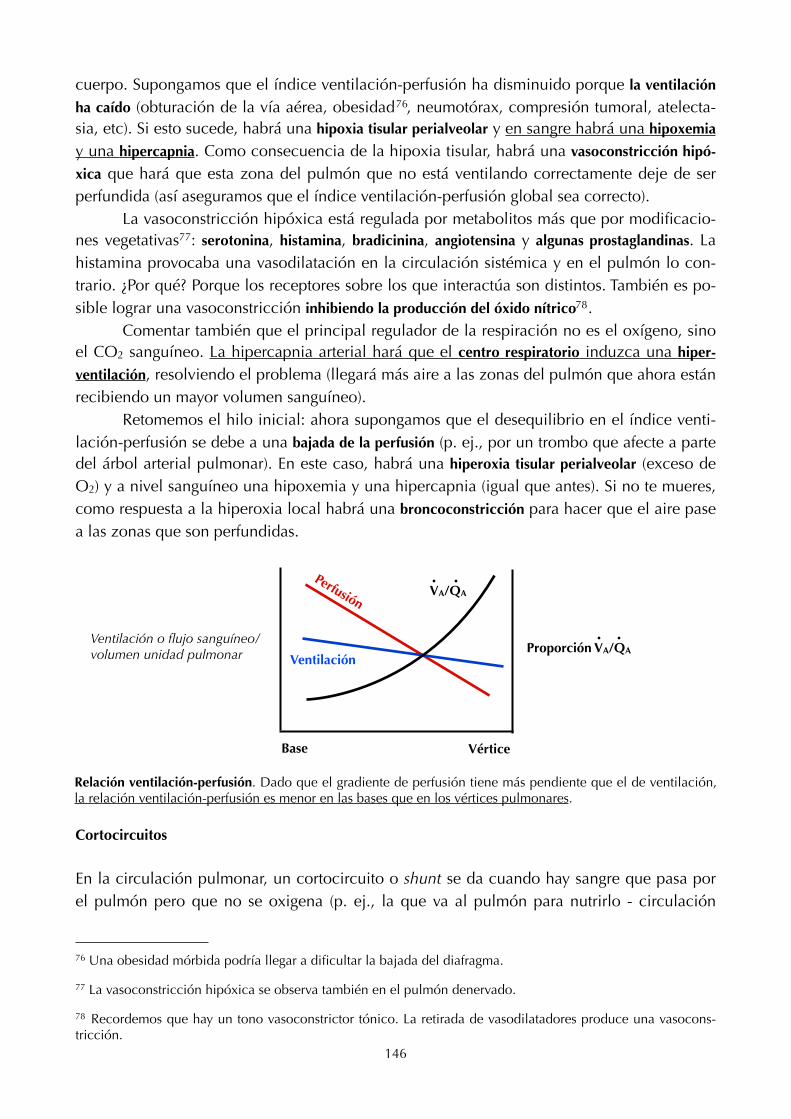
cuerpo. Supongamos que el índice ventilación-perfusión ha disminuido porque la ventilación
ha caído (obturación de la vía aérea, obesidad76, neumotórax, compresión tumoral, atelecta-sia, etc). Si esto sucede, habrá una hipoxia tisular perialveolar y en sangre habrá una hipoxemia y una hipercapnia. Como consecuencia de la hipoxia tisular, habrá una vasoconstricción hipó-
xica que hará que esta zona del pulmón que no está ventilando correctamente deje de ser perfundida (así aseguramos que el índice ventilación-perfusión global sea correcto). La vasoconstricción hipóxica está regulada por metabolitos más que por modificacio-nes vegetativas77: serotonina, histamina, bradicinina, angiotensina y algunas prostaglandinas. La histamina provocaba una vasodilatación en la circulación sistémica y en el pulmón lo con-trario. ¿Por qué? Porque los receptores sobre los que interactúa son distintos. También es po-sible lograr una vasoconstricción inhibiendo la producción del óxido nítrico78. Comentar también que el principal regulador de la respiración no es el oxígeno, sino el CO2 sanguíneo. La hipercapnia arterial hará que el centro respiratorio induzca una hiper-
ventilación, resolviendo el problema (llegará más aire a las zonas del pulmón que ahora están recibiendo un mayor volumen sanguíneo). Retomemos el hilo inicial: ahora supongamos que el desequilibrio en el índice venti-lación-perfusión se debe a una bajada de la perfusión (p. ej., por un trombo que afecte a parte del árbol arterial pulmonar). En este caso, habrá una hiperoxia tisular perialveolar (exceso de O2) y a nivel sanguíneo una hipoxemia y una hipercapnia (igual que antes). Si no te mueres, como respuesta a la hiperoxia local habrá una broncoconstricción para hacer que el aire pase a las zonas que son perfundidas.
Cortocircuitos
En la circulación pulmonar, un cortocircuito o shunt se da cuando hay sangre que pasa por el pulmón pero que no se oxigena (p. ej., la que va al pulmón para nutrirlo - circulación
146
76 Una obesidad mórbida podría llegar a dificultar la bajada del diafragma.
77 La vasoconstricción hipóxica se observa también en el pulmón denervado.
78 Recordemos que hay un tono vasoconstrictor tónico. La retirada de vasodilatadores produce una vasocons-tricción.
Ventilación o flujo sanguíneo/volumen unidad pulmonar
Base Vértice
Perfusión
Proporción VA/QAVentilación
VA/QA
Relación ventilación-perfusión. Dado que el gradiente de perfusión tiene más pendiente que el de ventilación, la relación ventilación-perfusión es menor en las bases que en los vértices pulmonares.

bronquial-). Esto hace que la PO2 arterial será un poco menor de lo que nos esperábamos (ya veremos más adelante) porque mezclaremos en la vena pulmonar sangre muy oxigenada con sangre que no ha sido oxigenada. En condiciones normales, el 98% de la sangre que llega a la aurícula izquierda proviene directamente de los capilares alveolares. El 2% restan-te procede de la circulación bronquial. La diferencia de PO2 entre la que realmente esperá-bamos y la real es lo que se conoce como gradiente alvéolo-arterial de PO2 (o gradiente A-a
PO2). En personas normales este gradiente oscila entre los 6 y los 9 mmHg. Este gradiente puede aumentar en caso de patología (p. ej., edema o engrosamiento de la membrana alvéo-lo-capilar).
Composición del aire atmosférico y del aire alveolar
Podemos hablar de cuatro tipos distintos de aire: (a) atmosférico; (b) inspirado; (c) alveolar y (d)
espirado. Los tres últimos dependerán en gran medida de las características del primero. Su-pongamos que inspiramos aire seco a nivel del mar. La presión atmosférica a nivel del mar es de 760 mmHg. En virtud a la ley de Dalton, la suma de las presiones parciales de los gases (ideales) que forman una mezcla deberá ser igual a la presión total (PA + PB + PC = PT). Las presiones parciales de los gases atmosféricos son (en mmHg):
Una vez hemos inspirado el aire, éste será acondicionado. Que sea calentado y limpiado no tiene ninguna influencia sobre las presiones parciales. Ahora bien, al humidificarlo estamos incrementando la PH2O:
Al llegar a los alvéolos, la composición del aire inspirado (ahora llamado aire alveolar) cam-biará porque habrá un intercambio de CO2 y de O2 con la sangre. Finalmente, el aire espira-do será una mezcla del alveolar con el del espacio muerto anatómico (que será idéntico al inspirado). A continuación se muestran las presiones parciales de ambos aires:
(*) El aire alveolar es el aire alveolar promedio entre el que acaba de llegar al alvéolo (sin apenas CO2) y el que
está a punto de abandonarlo (rico en CO2). Nótese como el aire espirado sigue teniendo bastante oxígeno y tampoco demasiado CO2. Es útil saber este dato por si nos preguntan para qué narices sirve hacer el boca a
boca (además, el CO2 estimula el centro respiratorio).
Intercambio de gases
La sangre que llega al capilar alveolar tiene una PO2 de 40 mmHg. La PO2 del aire alveolar es de 104 mmHg, por lo que en el inicio, la ΔPO2 es de 64 mmHg. No obstante, recalcamos
147
PN2 PO2 PCO2 PH2O Ptotal
Aire atmosférico 597 159 0,3 (trazas) 3,7 760
PN2 PO2 PCO2 PH2O Ptotal
Aire inspirado 563,4 149,3 0,3 (trazas) 47 (↑) 760
PN2 PO2 PCO2 PH2O Ptotal
Aire alveolar* 569 104 40 47 760Aire espirado 566 120 27 47 760

que esto es en un inicio, puesto que a medida que la PO2 sanguínea aumenta y la alveolar va disminuyendo, esta diferencia se va haciendo más pequeña. Al eritrocito le basta 1/3 del reco-rrido total a través del capilar para oxigenarse. La PCO2 de la sangre que llega al capilar alveolar es de 47 mmHg. Al abandonar el capi-lar, la PCO2 ha descendido hasta 40 mmHg. A pesar de que la ΔPO2 es más grande que la ΔPCO2, captamos prácticamente el mismo oxígeno
que dióxido de carbono que liberamos. Hemos de pensar que la difusión está regida por la ley de Fick. Según ésta la diferencia de presiones par-ciales (ΔP) determina el grado de difusión de un gas. Ahora bien, no es el único factor: la cons-
tante de difusión (D) también determina esta di-fusión. A pesar de que la ΔPO2 es mayor que la ΔPCO2, la constante de difusión del CO2 es mayor que la del O2 porque el primer gas es
veinte veces más soluble que el segundo.
La presiones parciales de oxígeno y de anhídrido carbónico en sangre arterial y en sangre venosa son las siguientes:
(*) Podríamos esperar que la PO2 en la sangre arterial fuera de 105 mmHg. En realidad es de 100 (o de 95)
mmHg por el shunt (gradiente alvéolo-arterial de PO2).
El coeficiente de difusión (DL) es un parámetro que nos permite evaluar la permeabilidad de la membrana a los gases79. Se define como los mililitros de gas que atraviesan una membrana por minuto y por cada mmHg de gradiente de presión. El DLO2 normal es de 20-25 mL · min -1
· mmHg-1. La difusión del pulmón puede analizarse mediante un estudio funcional conocido como la capacidad de difusión pulmonar para CO (DLCO). El monóxido de carbono nos sirve
148
79 Por lo tanto, en caso de edema o de atelectasia, en que el gas de debe de atravesar un mayor espacio o hay una menor superficie útil, este coeficiente ve disminuido su valor. Al hacer ejercicio el coeficiente aumenta y también lo hace cuando estamos en altura (hipoxia crónica) y nos aclimatamos (disminución del grosor de la membrana).
Vi
=A ⋅D ⋅ P1 − P2( )
TD =
SolPM
Ley de Fick. (A) área de difusión; (D) constante de difusión; (P1-P2) diferencia de presión parcial; (T) espesor de la lámina; (Sol) solubilidad del gas; (PM) peso molecular. A pesar de que el CO2 y el O2 tengan un peso mole-cular similar, su solubilidad es muy distinta, lo cual hace que sus constantes de difusión sean diferentes.
Captación de oxígeno por la sangre capilar pul-monar. (Guyton pág. 496)
Sangre arterial Sangre venosaPO2 100* 40 (variable)PCO2 40 47

porque su transferencia está limitada por la difusión (explicación más chula en Netter págs. 159-60). Una vez sabemos la DLCO, podemos calcular la DLO2 sabiendo su constante de difu-
sión (D) La constante de difusión es una característica del gas e indica la facilidad con que puede pasar a través de una membrana. Depende del peso molecular y de la solublidad. La constante de difusión disminuye con el edema y aumenta al hacer ejercicio. La constante de difusión del oxígeno es el producto de la DLO2 por la diferencia de presiones parciales de oxígeno (ΔPO2). Esta diferencia en un principio es 64 mmHg (104 - 40) pero después va disminuyendo a medida que la sangre pasa por el capilar alveolar. La diferencia de presión media es 15 mmHg aproximadamente. La capacidad de difusión del oxígeno
es el producto de su coeficiente de difusión por la diferencia de presiones parciales:
Entre 300 y 350 mL de O2 difunden por minuto. Esto puede ser de interés para calcular el volu-
men de oxígeno consumido por unidad de tiempo (VO2) o bien el volumen de oxígeno máximo consumido por unidad de tiempo (VO2 máx)80.
Transporte de oxígeno a través de la sangre
El oxígeno es poco soluble en agua. Por tanto, la mayor parte del oxígeno viajará combinado con el catión hierro ferroso (Fe2+) del grupo hemo de la hemoglobina. Hasta un 97% del O2 es transportado por la hemoglobina; el 3% está disuelto en plasma. Cada gramo de hemo-globina permite el transporte de 1,34 mL de O2. Si tenemos 15 g de Hb/dL, la capacidad de
unión de O2 es:
1,34 mL O2/g Hb · 15 g Hb/100 mL de sangre = 20,1 mL O2/100 mL sangre
No obstante, esta cantidad no representa la cantidad de oxígeno que transporte nuestra san-gre. Debemos tener presente también aquel que va disuelto en el plasma. Según la ley de Hen-ry, la concentración de un gas disuelto en un líquido es directamente proporcional a la pre-sión parcial del gas en la atmósfera a la que el líquido se haya expuesto y a su solubilidad en el disolvente. Por cada mmHg de PO2, únicamente se disuelven 0,003 mL O2 en 100 mL de sangre a temperatura corporal. Debido a que la PO2 arterial en los pulmones alcanza los 100 mmHg, la cantidad de oxígeno disuelto en la sangre arterial es de 0,3 mL O2/100 mL sangre. El contenido de oxígeno en sangre es, por tanto:
Contenido de O2 = % saturación (de 0 a 1) · capacidad de unión de O2 + O2 disueltoContenido de O2 = 1 · 20,1 + 0,3 = 20,4 mL O2/100 mL sangre
149
80 Estos valores son importantes para los deportistas. En una prueba no gana aquel que tenga una capacidad aeróbica máxima, sino aquel que puede estar el máximo tiempo posible haciendo un ejercicio a la velocidad media empleando los menores recursos posibles (menor gasto metabólico). Esto es lo que se conoce como efi-ciencia energética o economía de carrera. El consumo de oxígeno está relacionado con los capilares y las mito-condrias (hiperplasia capilar en el caso que nos acomodemos y un mayor nº de mitocondrias).
CdO2 = DLO2 ⋅ ΔPO2 = 20 ⋅15 = 300
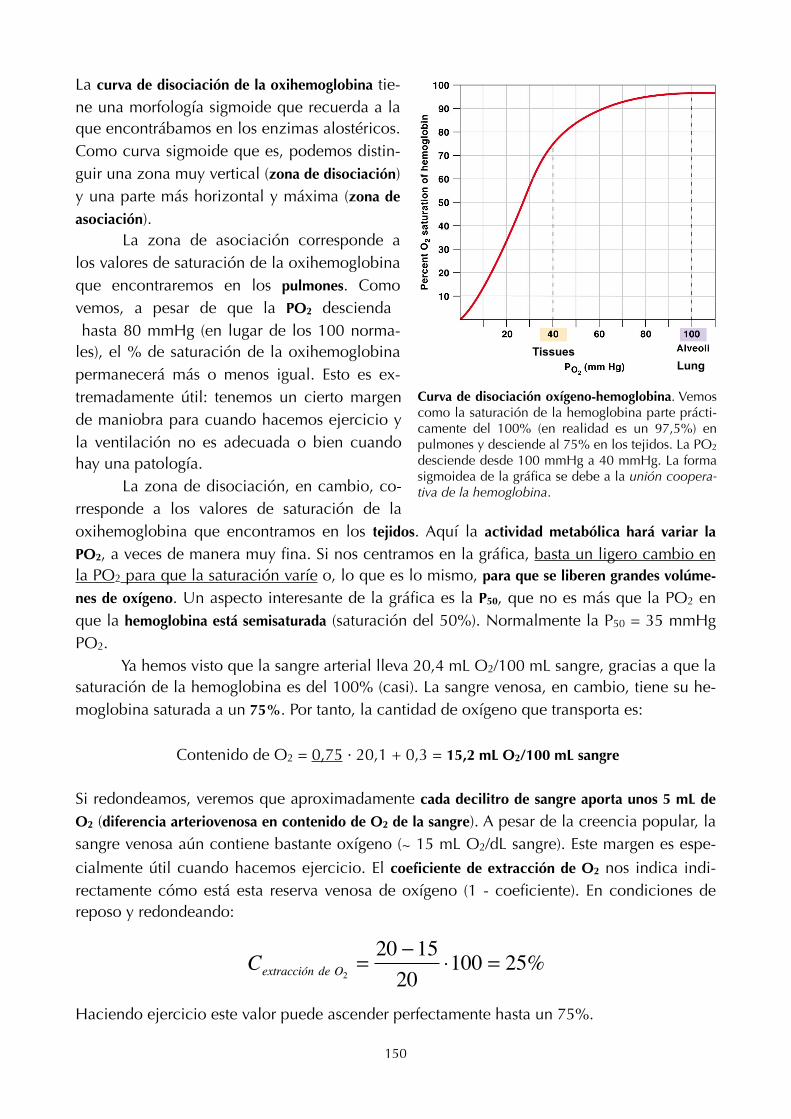
La curva de disociación de la oxihemoglobina tie-ne una morfología sigmoide que recuerda a la que encontrábamos en los enzimas alostéricos. Como curva sigmoide que es, podemos distin-guir una zona muy vertical (zona de disociación) y una parte más horizontal y máxima (zona de
asociación). La zona de asociación corresponde a los valores de saturación de la oxihemoglobina que encontraremos en los pulmones. Como vemos, a pesar de que la PO2 descienda hasta 80 mmHg (en lugar de los 100 norma-les), el % de saturación de la oxihemoglobina permanecerá más o menos igual. Esto es ex-tremadamente útil: tenemos un cierto margen de maniobra para cuando hacemos ejercicio y la ventilación no es adecuada o bien cuando hay una patología. La zona de disociación, en cambio, co-rresponde a los valores de saturación de la oxihemoglobina que encontramos en los tejidos. Aquí la actividad metabólica hará variar la
PO2, a veces de manera muy fina. Si nos centramos en la gráfica, basta un ligero cambio en la PO2 para que la saturación varíe o, lo que es lo mismo, para que se liberen grandes volúme-
nes de oxígeno. Un aspecto interesante de la gráfica es la P50, que no es más que la PO2 en que la hemoglobina está semisaturada (saturación del 50%). Normalmente la P50 = 35 mmHg PO2. Ya hemos visto que la sangre arterial lleva 20,4 mL O2/100 mL sangre, gracias a que la saturación de la hemoglobina es del 100% (casi). La sangre venosa, en cambio, tiene su he-moglobina saturada a un 75%. Por tanto, la cantidad de oxígeno que transporta es:
Contenido de O2 = 0,75 · 20,1 + 0,3 = 15,2 mL O2/100 mL sangre
Si redondeamos, veremos que aproximadamente cada decilitro de sangre aporta unos 5 mL de
O2 (diferencia arteriovenosa en contenido de O2 de la sangre). A pesar de la creencia popular, la sangre venosa aún contiene bastante oxígeno (∼ 15 mL O2/dL sangre). Este margen es espe-
cialmente útil cuando hacemos ejercicio. El coeficiente de extracción de O2 nos indica indi-rectamente cómo está esta reserva venosa de oxígeno (1 - coeficiente). En condiciones de reposo y redondeando:
Haciendo ejercicio este valor puede ascender perfectamente hasta un 75%.
150
TissuesLung
Normal Hb-O2 dissociation curveCurva de disociación oxígeno-hemoglobina. Vemos como la saturación de la hemoglobina parte prácti-camente del 100% (en realidad es un 97,5%) en pulmones y desciende al 75% en los tejidos. La PO2 desciende desde 100 mmHg a 40 mmHg. La forma sigmoidea de la gráfica se debe a la unión coopera-tiva de la hemoglobina.
Cextracción de O2=
20 −1520
⋅100 = 25%

Existen varios factores que pueden modificar la morfología de la curva de disociación oxígeno-hemoglobina: (a) pH; (b) temperatura; (c) PCO2; y (d) 2,3-bifosfoglicerato (BPG). Un descenso del pH o un aumento de los otros tres factores pro-voca que la gráfica se desplace hacia la dere-cha (y si sucede lo contrario, que se desplace hacia la izquierda). Los desplazamientos afec-tan mayormente a la zona de disociación más que a la de asociación, por lo que la mayoría de los efectos por cambios en la morfología de la gráfica afectarán a la periferia que a la cap-tación de oxígeno en los pulmones. El desplazamiento de la curva de diso-ciación hacia la derecha en respuesta a los aumentos de CO2 y de los iones H+ (↓pH) de la sangre tiene un efecto significativo porque aumenta la liberación de O2 desde la sangre ha-cia los tejidos y mejora la oxigenación en los pulmones. Esto se denomina efecto Bohr, y se puede explicar como sigue: cuando la sangre atraviesa los tejidos, el CO2 difunde desde las céls. tisulares hacia la sangre. Esto aumenta la PCO2 sanguínea, lo que a su vez eleva la con-centración sanguínea de ácido carbónico (H2CO3) y de H+. Estos efectos desplazan la curva de disociación de oxígeno-hemoglobina hacia la derecha y hacia abajo, haciendo que el oxígeno se disocie de la hemoglobina y liberando de esta manera mayores cantidades de O2 a los tejidos (aumenta la P50).
En los pulmones ocurre justo lo contrario: el CO2 difunde desde la sangre hacia los alvéolos, reduciendo la PCO2 sanguínea y desplazando la curva hacia la izquierda y hacia arriba. Esto hace que la cantidad de O2 que se une a la Hb a cualquier PAO2 dada aumenta considerable-
mente, permitiendo de esta manera un mayor transporte de oxígeno hacia los tejidos.
El efecto Bohr es el aumento de la liberación de oxígeno hacia los tejidos cuando el dióxido de carbono y los iones hidrógeno desplazan la curva de disociación oxígeno-hemoglobina.
El 2,3-bifosfoglicerato (BPG) es un metabolito que se produce en uno de los cortocircuitos del ciclo de Embden-Meyerhof en los eritrocitos (glucólisis). Éste, al unirse a la hemoglobina, disminuye su afinidad por el oxígeno. Por tanto, un incremento de este metabolito (p. ej., por una hipoxia) tendrá el mismo efecto que un incremento de la PCO2 o una disminución del pH. El Dr. Barbany señaló que la vía de formación del BPG implica un gasto adicional de ATPs, por lo que no tiene mucho sentido que en una situación de hipoxia se induzca la for-mación de BPG (hay otros mecanismos mejores). La síntesis adicional de 2,3-bifosfoglicerato cobra especial relevancia cuando nos aclimatamos a la altura (hipoxia crónica).
151
Desplazamiento de la curva de disociación oxíge-no-hemoglobina. Se enumeran los factores que ha-cen que la gráfica vaya hacia la izquierda o hacia la derecha.
CO2(d ) + H2O H2CO3 H + + HCO3−

Que la temperatura afecte a la gráfica tiene una especial importancia. Pensemos que cuando hacemos ejercicio, hay una hipertermia81 local que calienta la sangre, incrementando la diso-ciación del oxígeno a los tejidos. Finalmente, también las hormonas tiroideas y la hormona del
crecimiento desplazan la gráfica hacia la derecha (incrementan el metabolismo).
Hemoglobina
La hemoglobina está formada por cuatro cadenas
proteicas llamadas globinas (2 α y 2 β) conjugadas cada una de ellas con un grupo hemo. Este grupo tiene un átomo de hierro en estado ferroso (Fe2+) que le permite unirse a una molécula de O2. Si la hemoglobina interacciona con el oxígeno pasa a llamarse oxihemoglobina. La afinidad del Fe2+ por el oxígeno es 200 veces menor que la que tiene por
el monóxido de carbono. En otras palabras: una vez el CO se une a la hemoglobina, no la suelta (carboxihemoglobina). El CO2 también puede unir-se a la hemoglobina, pero únicamente a radica-les amino de las cadenas proteicas (globinas). Si esto sucede la hemoglobina pasa a denominarse carbaminohemoglobina (le da un tinte azulado a la sangre). Supuestamente la carbaminohemoglobi-na podría unirse al oxígeno y dar lugar a la car-
baminooxihemoglobina, pero es muy complicado. Si el hierro ferroso del grupo hemo pasa a ser férrico (Fe3+), la hemoglobina pasa a denomi-narse metahemoglobina, y no tiene capacidad pa-ra unirse al oxígeno. El cianuro oxida el catión ferroso y además se une a la hemoglobina, blo-queándola y dando lugar a la cianmetahemoglobi-na. Hasta ahora hemos estado hablando de la hemoglobina adulta o HbA. La hemoglobina fetal (HbF) es similar a la adulta, aunque tiene una mayor afinidad por el oxígeno. Si observáramos su curva de disociación veríamos que es exactamente igual que la de la adulta, únicamente que muy desplazada hacia la izquierda. Cuando el niño nace, debe deshacerse en poco tiempo de la HbF. Esto implica una hemoca-teresis impresionante, con la consiguiente producción de pigmentos biliares. No es por tanto extraño que el niño tenga una ictericia fisiológica neonatal. La hemoglobina S (HbS) es una variante anormal (defecto en la globina β) de la hemo-globina (patológica). La HbS hace que el eritrocito tenga forma de hoz, haciéndolo más pro-
152
81 Decimos hipertermia y no fiebre ya que esta última se asocia con un proceso infeccioso.
Saladin: Anatomy & Physiology: The Unity of Form and Function, Third Edition
18. The Circulatory System: Blood
Text © The McGraw!Hill Companies, 2003
Cha
pter 18
called globins (fig. 18.10a). Two of these, the alpha (!)chains, are 141 amino acids long, and the other two, thebeta (") chains, are 146 amino acids long. Each chain isconjugated with a nonprotein moiety called the hemegroup (fig. 18.10b), which binds oxygen to a ferrous ion(Fe2#) at its center. Each heme can carry one molecule ofO2; thus, the hemoglobin molecule as a whole can trans-port up to 4 O2. About 5% of the CO2 in the bloodstreamis also transported by hemoglobin but is bound to the glo-bin moiety rather than to the heme. Gas transport byhemoglobin is discussed in detail in chapter 22.
Hemoglobin exists in several forms with slight dif-ferences in the globin chains. The form we have justdescribed is called adult hemoglobin (HbA). About 2.5%of an adult’s hemoglobin, however, is of a form calledHbA2, which has two delta ($) chains in place of the "chains. The fetus produces a form called fetal hemoglobin(HbF), which has two gamma (%) chains in place of the "chains. The $ and % chains are the same length as the "chains but differ in amino acid sequence. HbF binds oxy-gen more tightly than HbA does; thus it enables the fetusto extract oxygen from the mother’s bloodstream.
Insight 18.2 Evolutionary Medicine
The Packaging of HemoglobinThe gas-transport pigments of earthworms, snails, and many other ani-mals are dissolved in the plasma rather than contained in blood cells.
690 Part Four Regulation and Maintenance
Nucleus
Lysosomes
(b)
Figure 18.9 Comparison of RBCs and WBCs as Seen with theTransmission Electron Microscope. (a) RBCs in a small bloodvessel, showing their strongly biconcave shape and lack of organelles.(b) An eosinophil, a representative leukocyte, showing a nucleus, manylysosomes, and other organelles.What is the approximate width of this blood vessel?
Bloodvessel
Erythrocytes
(a)
!
!
Hemegroups
(a)
(b)
"
"
C
CH3 CH2
C
C
C
CC
C
C
CC
CC
N
N
NN
CH
CH
CH
CH
CH2
COOH
CH3 CH3
CH2
CH2
CH2
COOH
CH2 CH3
HC
C
C C
CHC
Fe2+
Figure 18.10 The Structure of Adult Hemoglobin (HbA).(a) The hemoglobin molecule consists of two ! proteins and two "proteins, each conjugated to a nonprotein heme group. (b) Structure ofthe heme group. Oxygen binds to Fe2# at the center of the heme.
HbA y grupo hemo. La protoporfirina está forma-da por cuatro anillos de cinco átomos (pirrol), por lo que se denomina también tetrapirrol. Los anillos están unidos por puentes metileno.

clive a romperse por su falta de deformabilidad. Si la hemólisis es masiva, tendremos la de-nominada anemia falciforme. En regiones con malaria (Plasmodium falciparum) es habitual que los individuos tengan esta hemoglobina defectuosa, puesto que les otorga un beneficio. Finalmente, para acabar con este apartado, comentar que la mioglobina es una proteí-na muscular muy similar, tanto estructural como funcionalmente, a la hemoglobina. Su prin-cipal función es ejercer a modo de reserva de oxígeno en el músculo82. La mioglobina tiene más afinidad por el oxígeno que la hemoglobina (su curva de disociación es una hipérbole).
Transporte de anhídrico carbónico a través de la sangre
El anhídrico carbónico puede viajar de distintas formas a través de la sangre. En primer lugar, debido a que es veinte veces más soluble que el oxígeno, una parte considerable de éste podrá ir disuelto en el plasma (7%). En segundo lugar, hasta un 23% viaja unido a radicales amino de di-ferentes proteínas plasmáticas, principalmente a la hemoglobina (formando la ya vista carbami-nohemoglobina). Finalmente, el 70% restante viaje en forma de anión bicarbonato. El dióxido de carbono se une al agua del eritrocito y, gracias a la anhidrasa carbónica, se transforma en ácido carbónico. Este ácido, pese a ser débil, se disocia rápidamente en hidrogeniones y bicarbonato, y este bicarbonato saldrá al medio extracelular (la sangre) intercambiándose por cloruro para mantener el equilibrio electroquímico (fenómeno conocido como desplazamiento del cloruro). Los hidrogeniones formados se asociarán con la hemoglobina, sustituyendo al potasio que estaba combinado con ella. El potasio, ahora libre en el citoplasma del eritrocito, se asociará con el cloruro en-trante para formar cloruro potásico (KCl). Ahora bien, la concentración de KCl de los eritroci-tos venosos no será mayor que la de los eritrocitos arteriales puesto que la entrada de agua por ósmosis hace que la concentración sea idéntica. Lo que sí que ocurrirá es que los hema-
tíes venosos estarán más hinchados, por lo que el hematocrito venoso será mayor que el arterial.
Efecto Haldane
El efecto Haldane explica las modificaciones del transporte de dióxido de carbono que se dan en presencia del oxígeno. Hemos de pensar, que la hemoglobina es importantísima para el transporte de CO2, ya que o bien lo lleva consigo (carbaminohemoglobina) o bien actúa como aceptor de los hidrogeniones que se forman al combinar CO2 y agua. La oxihemoglo-bina es más ácida que la hemoglobina normal, por lo que: (1) no acepta H+ con tanta facilidad; y (2) no es tan carbamizable. Esto hace que el CO2 tenga que difundir hacia otro lado: hacia el
153
82 Los humanos no tenemos mucha mioglobina en los músculos de las extremidades inferiores porque la llega-da de oxígeno está favorecida (por la gravedad).
Transporte del dióxido de carbono en la sangre.

espacio alveolar. En otras palabras: la unión del oxígeno a la hemoglobina tiende a desplazar el dióxido de carbono desde la sangre. En la periferia sucede lo contrario: el au-mento de la desoxihemoglobina (más básica) favorece la captación de CO2.
Coeficiente respiratorio
Ya hemos visto que por cada 100 mL de sangre, transportamos unos 20 mL de O2 (redon-deando). Gracias a que el transporte de CO2 no es tan problemático, podemos transportar más del doble de CO2 que de O2: 47 mL de CO2 por 100 mL de sangre. El transporte normal de O2 desde los pulmones a los tejidos por cada 100 mL de sangre (diferencia arterio-venosa de O2) es de unos 5 mL, mientras que el mismo parámetro para el CO2 es de 4 mL. En condi-ciones de reposo, por tanto, únicamente se elimina a través de los pulmones una cantidad de CO2 que es aproximadamente el 82% de la cantidad de oxígeno que captamos. El cociente de la producción de CO2 respecto a la captación de O2 se denomina coeficiente respiratorio (QR):
El QR depende del sustrato que estemos oxidando83. Si oxidamos glucosa (1 mol de glucosa implica el consumo de 6 moles de O2 y la producción 6 moles de CO2), el QR = 1. Si oxi-damos un lípido, consumiremos más O2 que CO2 producido (QR < 0,8). Para una persona que hace una dieta normal y que consume cantidades medias de carbohidratos, grasas y pro-teínas, se considera que el valor medio de QR es 0,825.
Control de la respiración
Si bien la respiración puede controlarse voluntariamente, básicamente es un proceso invo-luntario que controla muy de cerca la PaO2 y la PaCO2. Los cambios tanto en la intensidad co-mo en la frecuencia de la respiración participan en este proceso. Los tres componentes esen-ciales del sistema de control involuntario son los siguientes: (1) centros respiratorios del tronco
encefálico; (2) quimiorreceptores centrales y periféricos; (3) otros receptores*.
154
83 Y también del equilibrio ácido-base, pero no se dijo nada más.
Porciones de la curva de disociación del dióxido de car-bono cuando la PO2 es de 100 mmHg y de 40 mmHg. La flecha representa el efecto Haldane sobre el transporte de dióxido de carbono. Como vemos, este efecto permite que la cantidad de volumen de CO2 que se puede trans-portar sea mayor (paso de 52 a 48 volúmenes de CO2 por ciento, en lugar de sólo de 52 a 50; mira el eje verti-cal).
QR =
CO2 liberadoO2 captado
0,8

(*) Ya lo veremos más adelante pero la regulación del pulmón puede basarse en el análisis de la composición
química de la sangre o de otros fluidos como el LCR (regulación humoral) gracias a los quimiorreceptores, o bien basarse en factores mecánicos (regulación no humoral) gracias a otros receptores.
Las señales que intervienen en el control de la respiración son integradas por el centro respi-
ratorio bulbar, lo que resulta en la regulación de la actividad de los músculos respiratorios, afectando al volumen corriente (VT), a la frecuencia y patrón respiratorios. En el bulbo en-contramos dos núcleos o grupos neuronales:
(a) Grupo respiratorio dorsal (GRD). Inerva a los músculos inspiratorios (en este caso, diafragma84). Este grupo neuronal cobra protagonismo cuando estamos realizando una inspiración tranquila, en la que únicamente invertimos trabajo en inspirar y la espiración es un acto pasivo. El patrón activatorio de estas neuronas es disparar po-tenciales de acción a alta frecuencia cuando se está inspirando y que después la fre-cuencia caiga prácticamente a 0 cuando se espira. Se desconoce cómo se logra esta descarga cíclica.
Las neuronas del GRD reciben información del nervio vago (X). El vago capta, entre otras cosas, información de unos receptores de distensión pulmonares. Cuando se da la inspiración, y ésta es suficiente (el volumen pulmonar ha aumentado hasta un cierto grado), el vago inhibirá a las neuronas del GRD.
(b) Grupo respiratorio ventral (GRV). Inerva tanto a los músculos inspiratorios como a los espiratorios. Este grupo neuronal cobra protagonismo en la respiración activa. El pa-trón de activación de las neuronas espiratorias es justo el contrario que las inspirato-rias. Este centro se activará cuando la señal aferente de regulación sea notable85.
El centro respiratorio bulbar recibe entradas de dos áreas importantes de la protuberancia: (1)
centro neumotáxico y (2) centro apnéustico. El primero regula la frecuencia y la profundidad
155
84 También podríamos hablar de los intercostales externos, pero hemos de tener presente que estamos en una respiración tranquila.
85 El Dr. Barbany dijo que esto era bastante complejo y que aún se estaba estudiando, pero que para hacernos una idea podíamos pensar en que el GRD es la primera barrera y el GRV es la segunda barrera. Si la señal de regulación es de poca intensidad (respiración tranquila), dicha señal únicamente llegará al GRD. Si la señal, en cambio, es de mayor intensidad (respiración forzada), la señal atravesará el GRD y activará al GRV.
Patrón activatorio de las neuronas del grupo respiratorio dorsal (GRD). Vemos como la frecuencia de descarga disminuye abruptamente tras la inspiración.

de la respiración por inhibición cíclica de la inspiración. Este centro recibe entradas de la corteza cerebral. El segundo estimula la inspiración (es el antagonista al centro neumotáxico). Una lesión en la protuberancia o del bulbo raquídeo superior puede causar apneusia86. Expe-rimentalmente, la apneusia se origina por la ablación del centro neumotáxico y sección del nervio vago.
Regulación no humoral de la respiración
La regulación no humoral se basa en el análisis de parámetros mecánicos, como el grado de distensión pulmonar. Podemos hablar de unos (a) mecanismos pulmonares y de unos (b) meca-
nismos no pulmonares.
Mecanismos pulmonares
- Mecanorreceptores pulmonares. Responden a la distensión excesiva si se hincha mucho el pulmón y dan lugar a la finalización de la inspiración. Las señales aferentes de es-tos receptores situados en la musculatura lisa de las paredes de las vías aéreas se transmiten a través del nervio vago a la médula, donde inhiben al centro apnéustico, finalizando así la inspiración. Esta respuesta a la distensión pulmonar se conoce como el reflejo de Hering-Breuer. El cometido de este reflejo es que no inspires más allá de los necesario (ahorrar energía)87.
- Paradójico Heath. Se trata de un reflejo en el que, con el fin de renovar el aire del es-pacio muerto anatómico, inspiras y sigues inspirando para hacer una espiración final (suspiro).
- Receptores yuxtacapilares (receptores J). Estos receptores están ubicados en los alvéolos y son estimulados por el hinchado de los pulmones y por varios estímulos químicos. Detectan cambios en el flujo alveolar capilar (correlación ventilación-perfusión)88 y responden produciendo un aumento de la frecuencia respiratoria.
- Receptores irritantes. Presentes en las vías aéreas superiores y responden a gases noci-vos y a partículas. La activación de estos receptores da lugar a señales aferentes hacia el SNC, principalmente por el nervio vago, causando la broncoconstricción refleja y la tos.
Mecanismos no pulmonares
Aquí ya entrarían todos los receptores que no miden parámetros pulmonares. Encontramos los nociceptores (el dolor modifica la actividad respiratoria), los propiorreceptores en las extre-
156
86 Respiración caracterizada por esfuerzos inspiratorios prolongados y exhalaciones breves e intermitentes.
87 Peligro con el enfermo enfisematoso: como tiene un mayor volumen de aire en los pulmones, los mecanorre-ceptores pulmonares están permanentemente activos y por tanto se corre el riesgo de que la actividad respirato-ria disminuya demasiado.
88 Responden también al edema pulmonar provocando una hiperventilación

midades (durante el movimiento aumenta la actividad respiratoria), y la termorregulación (res-piración jadeante cuando aumenta la temperatura).
Regulación humoral de la respiración
La regulación humoral de la respiración depende de la detección de una serie de parámetros químicos de la sangre, tales como la PaCO2, la PaO2 y el pH. Los quimiorrreceptores detectan estos cambios, y pueden ser centrales o periféricos. Los quimiorreceptores centrales se encuentran en la superficie ventrolateral del bulbo -están en contacto directo con el líquido cefalorraquídeo (LCR)- y responden indirectamente a cambios de la PaCO2. La barrera hematoencefálica es prácticamente impermeable al bicarbona-to89 y a los hidrogeniones, pero el dióxido de carbono puede atravesarla fácilmente, modifi-
cando el pH del LCR (equilibrio inferior).
Si nos fijamos en el equilibrio superior, si incrementa la PaCO290, una parte del exceso de
CO2 difundirá hacia el LCR y se incrementará la concentración de protones y de bicarbona-to, disminuyendo el pH91. Este aumento de la acidez estimula a los quimiorreceptores cen-trales y se da un incremento de la frecuencia respiratoria. Si sucediera lo contrario (caída de la PaCO2), el pH aumentaría y los quimiorreceptores centrales harán que disminuya la frecuen-cia respiratoria. La regulación de la respiración por parte de los quimiorreceptores centrales es bastan-te lenta (hemos de esperar a que el pH del LCR se modifique). Esto en reposo y estando sanos no es problema. En cuanto a los quimiorreceptores periféricos, éstos se encuentran en la bifurcación ca-
rotidea y en el cayado aórtico. A diferencia de los quimiorreceptores centrales, detectan cam-bios directamente de la PaCO2 y de la PaO2 (del oxígeno y dióxido de carbono disuelto), así como también del pH:
- Cambios de la PaO2. Los quimiorreceptores periféricos no son demasiado sensibles a los cambios de la PaO2. Apenas se darán cambios en la ventilación si la PaO2 está por en-cima de los 60 mmHg. Por debajo de esa presión, los quimiorreceptores se volverán muy sensibles y los cambios en la ventilación serán mayores. Hemos de pensar tam-bién que una disminución del oxígeno en sangre hace que los tejidos tengan que re-currir a un metabolismo anaeróbico, por lo que se formará lactato e hidrogeniones (dis-minución del pH).
157
89 Si la concentración de bicarbonato sanguíneo aumenta mucho, éste podría llegar a pasar a través de la barre-ra hematoencefálica y neutralizar la caída de pH necesaria para estimular la respiración.
90 Si aumenta mucho, al final “saturamos” al centro respiratorio y se anula. Riesgo de fallo respiratorio en per-sonas que producen mucho CO2 y que no pueden evacuarlo correctamente.
91 Si por un defecto de la barrera hematoencefálica (que ésta no esté madura, cosa normal si eres niño) o si tie-nes una meningitis (incrementa la permeabilidad de la barrera) han entrado proteínas al LCR, éstas pueden lle-gar a neutralizar a los protones que se forman a partir de la difusión de CO2 y por tanto hacer que éstos no ten-gan ningún efecto sobre el pH. Riesgo de alteración respiratoria.
CO2(d ) + H2O H2CO3 H + + HCO3−
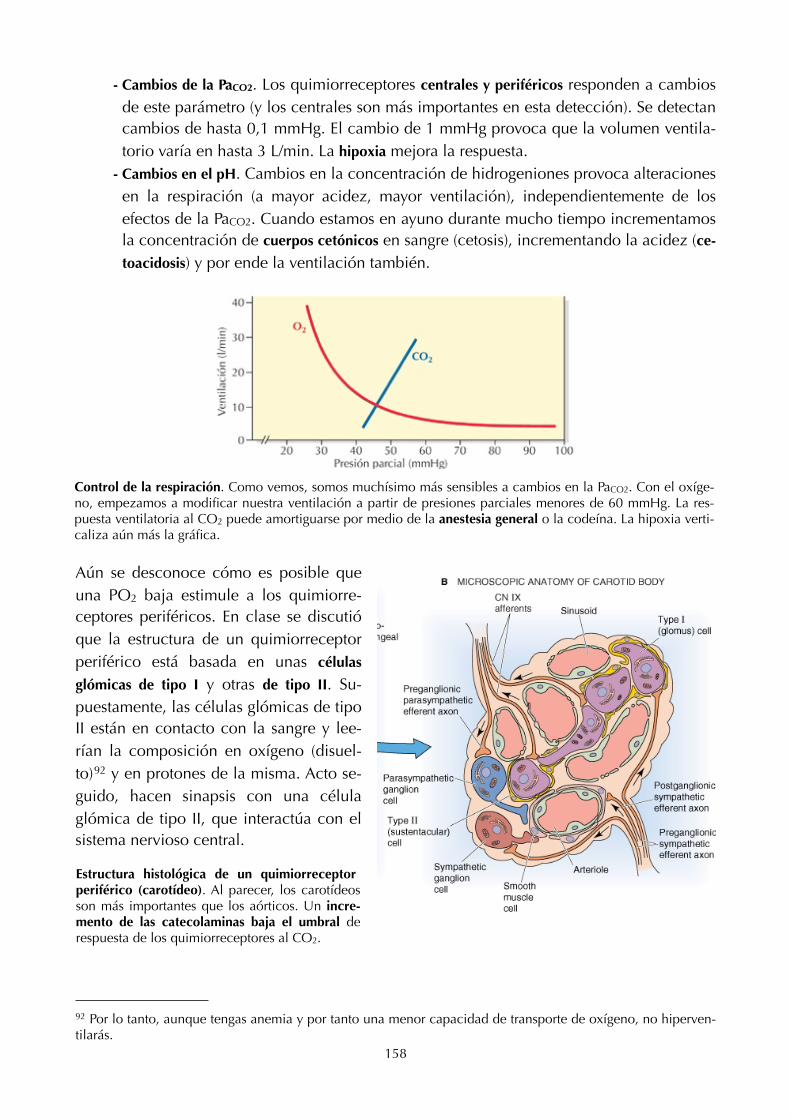
- Cambios de la PaCO2. Los quimiorreceptores centrales y periféricos responden a cambios de este parámetro (y los centrales son más importantes en esta detección). Se detectan cambios de hasta 0,1 mmHg. El cambio de 1 mmHg provoca que la volumen ventila-torio varía en hasta 3 L/min. La hipoxia mejora la respuesta.
- Cambios en el pH. Cambios en la concentración de hidrogeniones provoca alteraciones en la respiración (a mayor acidez, mayor ventilación), independientemente de los efectos de la PaCO2. Cuando estamos en ayuno durante mucho tiempo incrementamos la concentración de cuerpos cetónicos en sangre (cetosis), incrementando la acidez (ce-
toacidosis) y por ende la ventilación también.
Aún se desconoce cómo es posible que una PO2 baja estimule a los quimiorre-ceptores periféricos. En clase se discutió que la estructura de un quimiorreceptor periférico está basada en unas células
glómicas de tipo I y otras de tipo II. Su-puestamente, las células glómicas de tipo II están en contacto con la sangre y lee-rían la composición en oxígeno (disuel-to)92 y en protones de la misma. Acto se-guido, hacen sinapsis con una célula glómica de tipo II, que interactúa con el sistema nervioso central.
158
92 Por lo tanto, aunque tengas anemia y por tanto una menor capacidad de transporte de oxígeno, no hiperven-tilarás.
Control de la respiración. Como vemos, somos muchísimo más sensibles a cambios en la PaCO2. Con el oxíge-no, empezamos a modificar nuestra ventilación a partir de presiones parciales menores de 60 mmHg. La res-puesta ventilatoria al CO2 puede amortiguarse por medio de la anestesia general o la codeína. La hipoxia verti-caliza aún más la gráfica.
Estructura histológica de un quimiorreceptor periférico (carotídeo). Al parecer, los carotídeos son más importantes que los aórticos. Un incre-mento de las catecolaminas baja el umbral de respuesta de los quimiorreceptores al CO2.

COMENTARIOS SEMINARIO I FISIOLOGÍA
Caso 1
Paciente de 85 años, delgada y frágil. Internada para hacerle un diagnóstico y tratamiento a partir de una anemia descubierta en un examen rutinario. Mostraba palidez y refería cansancio tras un pequeño esfuerzo. En la consulta se le dedición realizar un hematocrito que dio bastante ajo, de manera que se le decidión hacerle análisis más completos. Los resultados se aparecen en el semina-rio original.
La mujer únicamente presenta 1,6 · 106 hematíes/mm3, valor que se aleja bastante de los 4-4,5 · 106 hematíes/mm3 que se establece como normal para las mujeres. No obstante, para esa edad es probable que ese valor de referencia no sea válido. Se vea por donde se vea, el
número de hematíes es bastante bajo. Los hematíes además presentan diferente forma (anisoci-tosis), poca hemoglobina (hipocromía) y pequeño tamaño (microcitosis). En cuanto al hematocrito, está al 11% y la hemoglobina está en 3 g/dl (normal en mu-jeres: 13 g/dl). Ambos valores son bastante bajos, parece difícil pensar que la mujer esté viva a no ser que haya pasado a este estado de manera progresiva. Los leucocitos son normales, por lo que se descarta infección o respuesta a una.
Teorías expuestas en clase
- Insuficiencia de eritropoyetina: una insuficiencia de EPO justificaría el número anormal de hematíes y que el número de plaquetas y de leucocitos se mantengan normales. No obstan-te, la EPO es formada en un 90% por los riñones, y un defecto en su producción indica un fallo renal, que daría otra clínica.
- Insuficiencia hepática: no podría ser, la mujer presenta ictericia (acumulación de bilirrubina en los tejidos grasos).
- Aplasia medular: explicaría el número bajo de hematíes, pero no explica que las plaquetas y los leucocitos estén normales.
- Déficit de B12: a pesar de que la atrofia de la mucosa gástrica sea típica en personas mayo-res, una anemia por déficit de B12 se caracteriza por ser megaloblástica (y no microcítica, como la que tiene esta paciente).
- Cáncer de colon ascendente: el cáncer de colon ascendente no presenta mucha sintomatolo-gía y produce pérdida de sangre en heces. Esto provocaría una anemia progresiva, dejando normales los niveles de leucocitos y plaquetas.
Caso 2
Se trata de un hombre de 50 años que consulta al médico quejándose de fatiga. Cuando sube esca-leras se detiene frecuentemente y respira rápida e intensamente. Esta dificultad ha empeorado en los últimos meses. El paciente no hace ejercicio regularmente. Trabaja en un ambiente estresante y fuma, bebe unas seis tazas de café al día y toma diariamente dos o tres copas de licor tras el trabajo. Tiene do-
159

lor en el estómago y ha ido tomando aspirinas para el dolor durante los seis últimos meses. En los tres últimos meses a intentado bajar el aporte de calorías con tal de perder peso, pero con poco éxi-to. Los valores de sus pruebas aparecen en el seminario original.
Vemos que la temperatura es normal pero que la respiración (FR: 24 respiraciones/min) y la frecuencia cardíaca (FC: 105 lpm) están aumentadas. En estas condiciones una persona se ma-
rearía, el paciente respira así porque lo necesita. Observamos un hematocrito bajo (30%) y una transferrina poco saturada (13% res-pecto al 30% máximo). A diferencia del caso anterior, éste es un caso crítico: el dolor de es-tómago y el consumo de aspirinas pueden referirnos a una úlcera péptica por la que el pa-ciente pierde sangre. El menor número de hematíes justifica el incremento de la FC (deben circular más rápido)
COMENTARIOS SEMINARIO II
Caso 1
Una paciente de 25 años acude al médico porque sangra repetidamente por la boca. A esta persona le habían diagnosticado hace dos años una embolia pulmonar, y desde entonces es tratada con he-parina. Datos originales en el seminario.
Temperatura, FC y FR normales. Aparecen púrpuras y petequias signos claros de una tromboci-
topenia. Además han aparecido anticuerpos inducidos por la heparina: en ocasiones la hepa-rina puede hacer que se generen anticuerpos anti-plaquetas. Como posibles tratamientos podemos usar: (a) una concentrado de plaquetas; (b) TPO (trombopoyetina)93; (c) inmunosupresores; (d) esplenectomía.
Caso 2
Un niño de tres años es llevado al pediatra por sus padres, los cuales han notado que su hijo sangre excesivamente alrededor de las rodillas y los codos. También han notado que cualquier corte en la piel le provoca un sangrado excesivo. En la historia de la familia de la madre hay antecedentes de hemofilia A. Datos originales en el seminario
La frecuencia cardíaca es baja para un niño (60 lpm), el hematocrito es algo bajo pero las plaquetas son normales. Que te digan que hay antecedentes de hemofilia A ya te dice que el niño sufre hemofilia A, un déficit en la formación del f. VIII. Hay algunas pruebas que mere-ce la pena que se comenten:- Tiempo de protrombina: se explora la vía extrínseca. Consiste en coger plasma al que se le
añade un reactivo (Ca2+, fosfolípidos, FT) y se espera cuanto tiempo tarda en coagular.- Tiembo de tromboplastina parcial activada: se explora la vía intrínseca. Se coge sangre, se in-
cuba, se centrifuga y se le extrae el plasma. A este plasma se le inactivan unos factores y se
160
93 La trombopoyetina se forma principalmente en el hígado, y también en el rión y en el músculo. La TPO se parece a la EPO, por lo que también se puede estimular la eritropoyesis.

le añade f. IX, fosfolípidos y calcio. Si la persona está sana, se formará un coágulo porque presenta factor VIII en su plasma. En el caso de los hemofílicos A, el tiempo de protrombina parcial activada está aumentado.
161

CASO 1 RENAL (Cálculos esenciales en fisiología renal)
Emplea los siguientes datos (PAH: ácido paraaminohipúrico; Px: concentración plasmática de x; Ux: concentración en la orina de x; RAx: concentración en la arteria renal de x; VAx: con-centración en la vena renal de x;
V (velocidad flujo orina): 1 mL/minPinulina: 100 mg/mLUinulina: 12 g/mLRAPAH: 1,2 mg/mLRVPAH: 0,1 mg/mLUPAH: 650 mg/mLPA: 10 mg/mLUA: 2 g/mLPB: 10 mg/mLUB: 10 mg/mLHematocrito: 0,45
1) ¿Cuál es el valor de la velocidad de filtración glomerular?
La velocidad de filtración glomerular (VFG) es igual al aclaramiento de un indicador (en este caso, la inulina). Por tanto, VFG = Cinulina.
La VFG es 120 mL/min.
2) ¿Cuál es el valor del “verdadero” flujo renal de plasma? ¿Cuál es el valor para el “verdadero” flujo sanguíneo? ¿Cuál es el valor del flujo renal de plasma “efectivo”? ¿Por qué es diferente el valor efectivo del verdadero?
El flujo plasmático renal (FPR) es la cantidad de plasma que pasa por el riñón. Éste se podría calcular a partir de la sustancia teórica x que se aclarara al 100% del plasma, y que por tanto todo volumen de plasma que entre en el riñón por unidad de tiempo quedaría desprovisto de esa sustancia. Como esa sustancia no existe, se realiza una aproximación con el PAH (cuyo cociente de extracción es de un 90% aproximadamente).
La RAPAH es de 1,2 mg/mL y la RVPAH es de 0,1 mg/mL, lo que quiere decir que se ha elimi-nado el 91,67% del PAH plasmático (cociente de extracción del PAH).
162
Cinulina =
Uinulina ⋅Vi
Pinulina=
12.000 mg / mL ⋅1 mL / min100 mg / mL
= 120 mL / min
CPAH =
UPAH ⋅Vi
PPAH=
650 mg / mL ⋅1 mL / min1,2 mg / mL
= 541,67 mL / min

Para facilitar próximos cálculos, comentar que el coeficiente de extracción del PAH (EPAH) se calcula dividiendo la diferencia entre la concentración de PAH en arteria y vena renal por la concentración de PAH en la arteria renal (RAPAH = PPAH).
Cuando hablamos de valor efectivo o verdadero quizá nos referimos a que generalmente de-cimos que el valor de RVPAH es 0 (valor efectivo) mientras que en este problema nos revelan ese dato (0,1 mg/dL). El flujo renal plasmático efectivo (FPRE) es, por tanto, el aclaramiento del PAH. En cuanto al flujo sanguíneo total (FSR), éste se calcula tomando en cuenta el he-matocrito:
3) ¿Cuál es el valor de la fracción de filtración, y cuál es el significado de este valor?
La fracción de filtración (FF) es el % del flujo plasmático renal que se filtra en el glomérulo. El resto del plasma irá por la red peritubular.
4) Asumiento que la sustancia A se filtra de forma libre (no unida a proteínas plasmáticas), ¿cuál es la carga filtrada de la sustancia A? ¿Se reabsorbe o se secreta? ¿Cuál es la velocidad de reabsor-ción o secreción?
La carga filtrada es en lo que los apuntes hablábamos de carga tubular (Ct). Por tanto:
Para saber si la sustancia A se reabsorbe o se secreta es tan sencillo como comprar esta carga tubular con lo que se secreta (ExA):
Claramente hay un fenómeno de secreción (CtA < ExA). Podríamos calcular también el co-ciente de aclaramiento (CC), que no es más que el cociente entre el aclaramiento de una sustancia y el de la inulina (saldrá superior a 1, por lo que hay secreción). La velocidad de secreción me imagino que será de 800 mg/min.
163
EPAH =PPAH − RVPAH
PPAH
FSR =FPR
1− hematocrito= 1.074,29 mL / min
FF =VFGFPR
⋅100 = 20,31%
CtA = PA ⋅VFG = 1.200 mg / min
ExA =UA ⋅Vi
= 2.000 mg / min
FPR =CPAH
EPAH
=541,67 mL / min
0,9167= 590,89 mL / min

5) ¿Cuál es la fracción de excreción de la sustancia A?
La fracción de excreción se calcula por medio del cociente entre el aclaramiento de una sus-tancia (la que nos interese) y el aclaramiento de un indicador. Se expresa en %. Lo hemos visto en el apartado previo: es un 167%.
6) ¿Cuál es el aclaramiento de la sustancia A? Está de acuerdo este valor con la conclusión obtenida en la pregunta 4 sobre si A se reabsorbe o se secreta?
Sí, nos sale un aclaramiento superior al del indicador (lo que implica que la sustancia se se-creta).
7) La sustancia B está unida en un 30% a proteínas. ¿Se reabsorbe o se secreta B? ¿Cuál es la velo-cidad de reabsorción o secreción?
Si el 30% de la sustancia B está unida a proteínas, únicamente un 70% se podrá filtrar. Para saber si se reabsorbe o si se secreta lo que haremos será calcular el cociente de aclaramien-to.
Se reabsorbe, además claramente. La velocidad de reabsorción se puede calcular haciendo la diferencia entre la cantidad filtrada por minuto (carga tubular) y la excreción por minuto.
La velocidad de reabsorción es de 830 mg/min. No nos olvidemos que la concentración plasmática de B (PB) debe ser multiplicada por 0,7 (porque el 30% de la sustancia está unida a proteínas).
164
CA =
UA ⋅Vi
PA=
2000 mg / mL ⋅1 mL / min10 mg / mL
= 200 mL / min
CC =CA
Cinulina
=200120
= 1,67
CB =
UB ⋅Vi
PB ⋅0,7=
10 mg / mL ⋅1 mL / min10 mg / mL ⋅0,7
= 1,43 mL / min
CC =CB
Cinulina
=1,43120
= 0,01
CtB = PB ⋅VFG = 840 mg / min
ExB =UB ⋅Vi
= 10 mg / min

CASO 2 RENAL (diabetes mellitus)
Un chico joven padece sed constantemente (bebe entre 4 y 5 L de agua diariamente) y orina cada 30-40 min. Los análisis indicaban que:
TA: 90/55 mmHg (estirado supino); 70/45 mmHg (alzado)Glucemia: 320 mg/dL (o 320 mg/100 mL)[Na+]plasmático: 143 mEq/LGlucosa en orina: +++Cetonuria: ++Sodio en orina: incrementado
En la exploración física, el chico mostraba turgencia disminuida en la piel, ojos hundidos y boca seca.
a) Cómo es tratada la glucosa en la nefrona (discutid la filtración, el transporte y la excreción de glucosa)? Qué transportadores están implicados en el proceso de reabsorción de la glucosa?
La mayor parte de glucosa se reabsorbe en el túbulo proximal a través de los transportadores SGLT1/2, que serán de baja afinidad y alta capacidad en S1, y de alta afinidad y baja capa-cidad en S2. En condiciones normales (glucemia normal), la excreción de glucosa es 0. Aho-ra bien, en el caso de que se supere un determinado transporte máximo (teórico de 200 mg/dL y práctico de 400 mg/dL por la heterogeneidad de las nefronas) empiezan a aparecer tra-zas de orina en la orina (glucosuria).
b) ¿Por qué el paciente excreta glucosa por la orina? Por qué hay cetonuria? La glucosuria del pa-ciente desapareció al recibir inyecciones de insulina. ¿Por qué?
Aparece glucosa en la orina porque se sobrepasa el límite de reabsorción renal (200 mg/dL). La diabetes se caracteriza por falta de insulina (al menos la de tipo I) y una gran cantidad de
glucagón. Además de que los tejidos periféricos no podrán recaptar la glucosa sanguínea, el glucagón induce la gluconeogénesis hepática, dejando el ciclo de Krebs sin metabolitos. En este contexto metabólico, habrá una lipólisis: los acetil-CoA formados por β-oxidación no podrán interiorizarse en Krebs (falta de oxalacetato) y el hígado empezará a formar cuerpos
cetónicos (y se incrementará su concentración en sangre: cetonuria). La insulina revierte este cuadro.
c) ¿Por qué razones se produce la poliuria? Por qué encontramos una excreción incrementada de sodio en orina?
Hay poliuria porque hay una gran cantidad de glucosa en el líquido tubular, llevándose con-sigo agua (es un osmolito). Al haber una mayor cantidad de agua, habrá más volumen en que diluir el sodio, por lo que ésta estará más diluido, lo que favorecerá la excreción de so-dio.
165

d) La osmolaridad plasmática puede ser estimada por la fórmula que se expone a continuación. Cal-culad la osmolaridad plasmática del paciente y comparadla con el valor fisiológico normal (290 mOsm/L).
Sale una osmolaridad más elevada de lo normal, sobre todo debido al exceso de glucosa.
e) Discutid por qué el paciente tiene sensación de sed.
La osmolaridad elevada de la sangre es detectada por unos osmorreceptores ubicados en el hipotálamo, lo que dará lugar a una sensación de sed.
f) Por qué la presión arterial es menor que en condiciones normales? Por qué disminuye cuando el paciente se alza?
La elevada diuresis (poliuria) nos indica que el paciente ha perdido volumen sanguíneo, por lo que el retorno venoso será menor, el volumen telediastólico también (VTD) y, por la ley de Frank Starling, la contractibilidad cardíaca también. El volumen sistólico (VS) se verá resenti-co, lo que implica una disminución del gasto cardíaco (GC). El gasto y la presión arterial es-tán directamente relacionadas (GC = TA/RPT), de ahí que ésta sea baja. Cuando el paciente se alza, dificulta aún más el retorno venoso, por lo que la presión aún bajará más.
g) ¿Por qué presenta turgencia disminuida de la piel?
Por la falta de líquidos.
h) ¿Cómo crees que será la frecuencia cardíaca (FC) tumbado respecto a la normalidad? ¿Y al alzar-se?
Al tumbarnos la FC será menor respecto a la normalidad porque mejoramos el retorno veno-so. Al alzarnos, para preservar el GC, habrá un incremento de la FC gracias a los barorrecep-tores carotídeos y aórticos (cambio de presión agudo).
166
Osmpl =2 ⋅ Na+⎡⎣ ⎤⎦ pl + Glc[ ]pl
18+BUN2,8

CASO 3 RENAL (sistema renal y fisiología ácido-base)
Simon Papel es un profesor de física en la Universidad de 54 años de edad y con un estilo de vida saludable. Hace ejercicio de forma regular, no fuma ni bebe alcohol, y mantiene su pe-so dentro de un peso normal. Recientemente, sin embargo, ha tenido debilidad muscular y dolores de cabeza. Él atribuyó los dolores de cabeza al estrés causado por la renovación de su beca de investigación. La medicación para quitar el dolor no funcionaba. Su mujer, que estaba preocupada, pidió una cita con el doctor. Al realizar un examen físico, el aparecía sano. Sin embargo, su presión sanguínea es-taba elevada a 180/100, ya sea estando sentado o estando de pie. Su médico le pidió distin-tos tests en el laboratorio de sangre y de orina que le dieron el siguiente resultado:
SangrepH 7.5 (normal 7.4)PCO2 48 mm Hg (normal 40 mm Hg)Na+: 142 mEq/L (normal 140 mEq/L)K+: 2,0 mEq/L (normal 4.5 mEq/L)Total CO2 (HCO3-): 36 mEq/L (normal 24 mEq/L)Cl-: 98 mEq/L (normal 105 mEq/L)Creatinina: 1,1 mg/dL (normal 1.2 mg/dL)
OrinaExcreción de Na+: 200 mEq/24 hr (normal)Excreción de K+: 1.350 mEq/24 hr (elevada)Excreción de creatinina: 1.980 mg/24 hrCatecolaminas de 24 hr: normal
(1) La presión arterial de Simon estaba elevada sentado o de pie. Considera los factores que regulan la presión arterial y sugiere posibles causas para esta hipertensión. ¿Qué enfermedad está des-cartada por el valor normal de la excreción urinaria catecolaminas durante 24 hr?
La presión arterial se puede calcular como el producto del gasto cardíaco y las resistencias periféricas totales (RPT). Podríamos pensar que éstas últimas están elevadas, pero sus hábitos saludables nos hacen descartar la arteriosclerosis que incrementaría las RPT. Por el otro lado, si nos centramos en el gasto cardíaco, éste depende de la frecuencia cardíaca (FC) y del vo-lumen sistólico (VS). Una posible causa para la hipertensión es un incremento del volumen sanguíneo que, por extensión, incrementa el volumen telediastólico (VTD) y por ende el VS. La excreción normal de catecolaminas nos indica que el incremento de presión no está aso-ciado a un problema con el simpático.
(2) El médico pensó que Simon tendría un problema en el sistema renina-angiotensina II-aldostero-na. Pidió más test, incluyendo la actividad de renina en el plasma, la aldosterona en suero y el cortisol en suero, con los siguientes resultados:
167

Actividad renina en plasma: DisminuidaAldosterona en suero: IncrementadaCortisol en suero: normal
Indica una posible explicación para esta patología.
La aldosterona incrementada en suero debería estar asociada con un incremento de la reni-na. Que no lo esté nos indica que la secreción de aldosterona es independiente a dicha re-nina y que por tanto no se debe a la activación del SRAA. Los niveles normales de cortisol nos hacen descartar la enfermedad de Cushing. Probablemente será un tumor de las células de la zona glomerular de la corteza suprarrenal.
(3) El médico sospecho que Simon tenía hiperaldosteronismo, lo que significa que el principal pro-blema es que su glándula adrenal secretaba demasiada aldosterona. ¿Por qué un incremento en los niveles de aldosterona provoca un incremento en la presión arterial?
La aldosterona provoca un incremento de la reabsorción de sodio, lo cual favorece la reab-sorción de agua y por tanto incrementa la volemia. Este aumento de la volemia es el causan-te de la elevada presión arterial.
(4) ¿Qué efecto esperáis que tenga el hiperaldosteronismo primario en la excreción de sodio urina-rio? De acuerdo con tu predicción, explica como es que la secreción urinaria de sodio era nor-mal.
El hiperaldosteronismo primario debería provocar niveles bajos de sodio en la orina, pero observamos que los niveles son normales (200 mEq/24h). Como este hiperaldosteronismo está ahí desde hace tiempo (el paciente ha tardado en ir al médico), habrán habido adapta-ciones. Ante el incremento de volumen sanguíneo, los barorreceptores auriculares se habrán distendido y habrán inducido la liberación del péptido natriurético auricular (ANP), el cual favorece la natriuresis. Esta secreción de sodio compensar la reabsorción favorecida por la aldosterona (hay otra causa: natriuresis y diuresis por presión; escape de la aldosterona; Guyton p. 925).
(5) ¿Qué explicación puedes dar para la hipokalemia de Simon? Si el médico le hubiera dado una inyección de KCl, ¿habría la inyección corregido esta hipokalemia?
La aldosterona incrementa la actividad de la ATPasa Na+/K+. La entrada de sodio provoca una despolarización que revierte el gradiente electroquímico y favorece la secreción de po-tasio. A menudo se dice que la aldosterona tiene dos caras: permite la recuperación de la presión a costa de perder potasio. Una inyección de KCl solucionaria la hipokalemia mo-mentáneamente, pero como el riñón seguiría secretando potasio, a largo plazo no serviría.
(6) Explica la debilidad muscular de Simon basándote en esta hipokalemia.
168

El gradiente electroquímico de potasio es fundamental para preservar el potencial de mem-brana. En reposo, la célula tiene canales de K+ abiertos. Si la concentración de potasio ex-tracelular disminuye, el potencial de membrana se hace más negativo (compruébalo con la fórmula del potencial de Nernst), por lo que es más difícil llegar al potencial umbral (se pro-duce una hiperpolarización; - 91 mV aprox).
(7) ¿Qué anormalidad ácido-base tiene Simon? ¿Cuál era su causa? ¿Cuál es la compensación apro-piada para esta enfermedad? ¿Ocurrió esta compensación?
Tiene una alcalosis metabólica producida por la aldosterona (que estimula la pérdida de hidro-geniones a través de la orina). El principal órgano que reacciona para compensar los cambios del pH sanguíneo es el pulmón, que compensa modulando la frecuencia respiratoria (FR). Los riñones y el hígado participan también en la regulación del pH. Ante una alcalosis, el pulmón disminuye la FR para incrementar el CO2 sanguíneo en virtud al equilibrio siguiente:
La compensación se da puesto que vemos como la pCO2 aumenta de 40 a 48 mmHg. Un dato: la pKa del ácido carbónico es de 3,57 a 37ºC, por lo que sobre el papel no es un buen tampón a pH fisiológico (recordemos que tampona a pH ±1 de su pKa. La explicación (Lehninger págs. 62-63) se basa en la gran capacidad de reserva de CO2(g) en el espacio aé-reo de los pulmones.
(8) ¿Cuál era la velocidad de filtración glomerular?
En lugar de darte la producción de orina y la Ucreatinina de manera independiente, te dan ya el producto en los datos (excreción de creatinina). Vemos que la VFG es de 180 L/24h.
(9) ¿Cuál era su excreción fraccional de sodio?
La excreción fraccional de sodio se calcula haciendo el cociente de aclaramientos (Csodio/Ccreatinina):
169
VNernstK +
= −R ⋅Tz ⋅F
⋅ lnK +⎡⎣ ⎤⎦iK +⎡⎣ ⎤⎦e
⎛
⎝⎜
⎞
⎠⎟
CO2(d ) + H2O H2CO3 H + + HCO3−
VFG = Ccreatinina =
Ucreatinina ⋅Vi
Pcreatinina=
1.980 mg / 24h1,1 mg / dL
= 1.800 dL / 24h

(10) Un scanner tomográfico computerizado mostró la presencia de un adenoma simple en la glán-dula adrenal izquierda. Se le llevó a un cirujano, el cuál quiso operar inmediatamente para qui-tar el adenoma. Simon pidió dos semanas de retraso para poder acabar la renovación de la be-ca. El cirujano estuvo de acuerdo, pero siempre que Simon tomara un diurético específico. ¿Qué diurético habrías tu prescrito y con que mecanismo? ¿Qué anormalidades serían corregi-das por este diurético?
Habría recetado o bien amilorida, que bloquea los canales de Na+, o espironolactona, que es un antagonista de la aldosterona. Corregirá sobre todo la presión arterial (no habrá tanta cap-tación de sodio y por ende de líquido).
170
CC =CNa+
Ccreatinina
=1,41180
= 0,0078
CNa+
=U
Na+⋅V
i
PNa+
=200 mEq / 24h142 mEq / L
= 1,41 L / 24h

CASO 4 RENAL (diabetes insípida)
Paciente de 19 años. Desde hace un tiempo presenta poliuria (cada hora tiene que ir a orinar y polidipsia (beve constantemente, unos 5 L al día, siempre lleva una botella de agua). Su médico contempla dos alternativas: o bien padece un desorden psiquiátrico que implica be-ber compulsivamente (polidipsia primaria) o bien padece diabetes insípida. La exploración física muestra una persona normal. Su presión arterial es de 105/70, la frecuencia cardíaca es de 85 latidos/min y los campos visuales son normales. Las analíticas de sangre y orina dan los siguientes valores:Na+ plasmático: 147 mEq/LOsmolaridad plasmática: 301 mOsm/L (normal: 290 mOsm/L)Osmolaridad urinaria: 70 mOsm/LGlucemia: 90 mg/dL (normal: 70-100 mg/dL)Glucosa en orina: 0
A partir de estos datos, el médico hizo una privación de agua durante dos horas. Al final de este periodo la osmolaridad de la orina permaneció a 70 mOsm/L y la osmolaridad plasmá-tica se incrementó a 325 mOsm/L. En un test siguiente se le administró ADH subcutánea. A partir de aquí la osmolaridad urinaria se incrementó a 500 mOsm/L y la osmolaridad plas-mática bajó hasta 290 mOsm/L. Una RMN excluyó un tumor cerebral. En consecuencia se le diagnosticó diabetes in-sípida central. Probablemente se desarrolló por anticuerpos contra las neuronas que secretan ADH. La paciente inició un tratamiento con un análogo de ADH en spray nasal con buenos resultados.
(1) ¿Cuál es el valor normal de la osmolaridad urinaria? Describir los mecanismos que regulan la osmolaridad urinaria.
El rango de valores normales de osmolaridad urinaria van desde los 50 a los 1.200 mOsm/L. Lo que modula la osmolaridad urinaria es la osmolaridad plasmática detectada en el hipota-lamo. A partir de aquí, la osmolaridad urinaria será una u otra en función de la cantidad de agua reabsorbida en el túbulo colector.
(2) Los valores iniciales de los parámetros en sangre y orina sugieren que la causa de la poliuria no es primariamente la polidipsia. ¿Por qué? El test de privación de agua durante dos horas también apoya que la poliuria no es debida primariamente a la polidipsia. Discutidlo.
Esencialmente lo que sucede es que la persona, beba agua o no, presenta una orina muy di-luida (con una muy baja osmolaridad).
(3) No hay glucosa en la orina, discutidlo.
171

No tiene por qué haber glucosa en la orina. La glucemia es normal y toda la glucosa filtrada es reabsorbida en el túbulo proximal.
(4) Después de los análisis iniciales de sangre y orina, los médicos sospecharonque se trataba o bien de una diabetes insípida central o bien de una diabetes insípida nefrogénica. Comentad por qué cada una de ellas podría explicar los valores de osmolaridad plasmática y urinaria encontradas.
La diabetes insípida central se basa en que hay un problema con la secreción de hormona antidiurética, que se forma en la neurohipófisis. Este problema puede ser o bien que las célu-las no están, o un tumor o bien una causa autoinmune. Por el otro lado, la nefrogénica se basa en que pese a que la liberación de ADH es normal, ésta no puede realizar su efecto puesto que hay un problema con el receptor, la adenilato ciclasa u otras causas. En ambos casos, la ADH no ejerce su efecto, por lo que la orina no se concentra, se pierde fluido y se incrementa la osmolaridad plasmática.
(5) Discutir de qué manera confirmó el médico que el problema era una diabetes insípida central y no nefrogénica.
Pues o bien se podrían estudiar los niveles en sangre de ADH o bien introducir ADH direc-tamente y ver si la orina se concentra. Como se logró que la orina se concentrara, se asume que la diabetes insípida es central.
(6) ¿Cómo encontraríamos la ADH plasmática en una diabetes insípida central? ¿Y en una nefrogé-nica?
En la central estarían bajos. En la nefrogénica estarían altos.
(7) Cuando el médico administró ADH en la paciente, se esperaba que la osmolaridad urinaria fuera la máxima (1.200 mOsm/L), pero únicamente alcanzó 500 mOsm/L. ¿Por qué? Qué pasará al cabo de un tiempo de tratamiento?
El incremento en la osmolaridad urinaria no es el esperado porque el intersticio renal del pa-ciente no es demasiado hiperosmolar (el transporte de agua precisa de ello). La paciente ne-cesitará un tiempo para generar esa hiperosmolaridad.
(8) Discutir íntegramente el caso, comenzando a partir de que el hipotálamo no forma ADH (o muy
poca).
(Cuando estudies renal lo redactas). La persona no podrá controlar la osmolaridad de la ori-na puesto que se aplica ADH de manera “constante”. Tendrá que beber poca agua.
La tiazida es un diurético que bloquea la absorción de sal en el túbulo contorneado distal. Esto afectará al aparato yuxtaglomerular, que provocará una bajada de la filtración para compensar la pérdida de líquido (?).
172
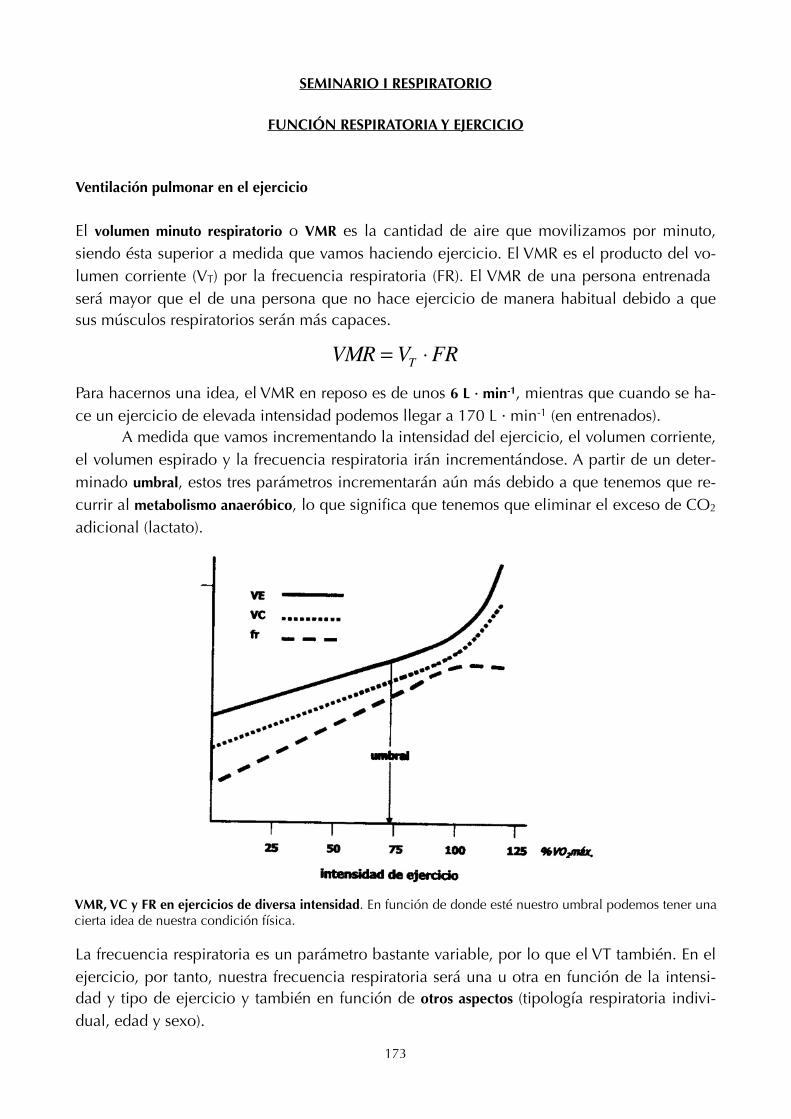
SEMINARIO I RESPIRATORIO
FUNCIÓN RESPIRATORIA Y EJERCICIO
Ventilación pulmonar en el ejercicio
El volumen minuto respiratorio o VMR es la cantidad de aire que movilizamos por minuto, siendo ésta superior a medida que vamos haciendo ejercicio. El VMR es el producto del vo-lumen corriente (VT) por la frecuencia respiratoria (FR). El VMR de una persona entrenada será mayor que el de una persona que no hace ejercicio de manera habitual debido a que sus músculos respiratorios serán más capaces.
Para hacernos una idea, el VMR en reposo es de unos 6 L · min-1, mientras que cuando se ha-ce un ejercicio de elevada intensidad podemos llegar a 170 L · min-1 (en entrenados). A medida que vamos incrementando la intensidad del ejercicio, el volumen corriente, el volumen espirado y la frecuencia respiratoria irán incrementándose. A partir de un deter-minado umbral, estos tres parámetros incrementarán aún más debido a que tenemos que re-currir al metabolismo anaeróbico, lo que significa que tenemos que eliminar el exceso de CO2 adicional (lactato).
La frecuencia respiratoria es un parámetro bastante variable, por lo que el VT también. En el ejercicio, por tanto, nuestra frecuencia respiratoria será una u otra en función de la intensi-dad y tipo de ejercicio y también en función de otros aspectos (tipología respiratoria indivi-dual, edad y sexo).
173
VMR = VT ⋅FR
VMR, VC y FR en ejercicios de diversa intensidad. En función de donde esté nuestro umbral podemos tener una cierta idea de nuestra condición física.

Circulación pulmonar en el ejercicio
Es importante que la circulación pulmonar sea un circuito de baja presión y un alto flujo. Para evitar incrementos de la presión, que como ya sabemos se asocian con edemas, lo que ha-remos será disminuir la resistencia del circuito (ley de Poiseuille). Las resistencias vasculares ya de por sí serán bajas por la morfología de los vasos pulmonares (paredes finas94 y diámetro elevado = muy distensibles). En el caso de que la resistencia deba disminuir aún más, se re-curre al reclutamiento de más capilares. Los vasos pulmonares apenas responden a las cateco-
laminas. La baja presión se traduce en una baja velocidad, lo que nos da un margen para que se dé el intercambio de gases y hace que la circulación pulmonar constituya un reservorio de sangre (hasta 1 L de sangre en reposo). En el caso de que estemos haciendo ejercicio (o que tengamos una hemorragia), la movilización de esta reserva de sangre incrementa la volemia
efectiva.
Trabajo respiratorio
Respirar no nos supone un elevado gasto energético. En reposo, si bien para inspirar debe-mos superar unas resistencias elásticas y de tensión superficial, espirar es un proceso pasivo y gratuito. Un parámetro interesante es el equivalente respiratorio (VMR/VO2), que la cantidad de aire que necesitas ventilar para incorporar 1 L de oxígeno. Conforme menor sea este equivalente, mejor (mayor eficiencia respiratoria).
Aumento del intercambio de gases
Tanto la capacidad como el coeficiente de difusión se incrementan cuando hacemos ejercicio. El coeficiente de difusión es mayor debido a que tenemos una mayor superficie de intercam-bio (mayor ventilación y mayor perfusión). La capacidad de difusión es mayor porque el coeficiente de difusión se ha incrementado y porque el gradiente de presión es mejor (más PCO2 y menos PO2 en sangre venosa; menos PACO2 y más PAO2
95).
Principales factores de la regulación de la respiración
Como ya se vio previamente, la respiración se puede regular por medio de factores humora-les y factores no humorales. Veamos algunos de estos parámetros y cómo se relacionan con el ejercicio:
- Composición plasmática. La PaCO2, la PaO2 y el pH afecta a los quimiorreceptores cen-trales y periféricos. Ahora bien, durante el ejercicio no suele haber hipoxemia ni hiper-capnia. Si bien el pH puede verse modificado por el lactato (acidosis), la ventilación ya habrá aumentado bastante antes.
174
94 Apenas tienen musculatura lisa, por lo que se distienden fácilmente ante un incremento de la presión arterial.
95 Discutible (lo decimos entre comillas).

- Regulación neurogénica. Vía corteza motora y propiorreceptores musculares y tendino-sos. Se supone que ésta tiene una especial importancia en la regulación de la respira-ción durante el ejercicio físico.
- Termorregulación. Sí, aunque ésta suele darse al final del ejercicio.- Catecolaminas. Recordemos que las catecolaminas pueden bajar el umbral de sensibi-
lidad al CO2 en los quimiorreceptores. - Reflejos de estiramiento de Hering y Breuer.- Otros factores. Se discute si en el ejercicio la respiración puede ser modulada por re-
ceptores tisulares o por un incremento del potasio plasmático (incremento de la per-meabilidad de la membrana de la fibra muscular).
175

SEMINARIO II RESPIRATORIO
ADAPTACIONES A LA ALTURA
Introducción
En el siguiente seminario hablaremos sobre cómo nos adaptamos a las grandes alturas (nada de Tibidabo). La altitud tendrá un efecto en nosotros que dependerá de, en primer lugar y como es lógico, la altura alcanzada (no es lo mismo 5.000 msnm que 8.000 msnm; msnm = metros sobre el nivel del mar). Después también entran en juego otros factores, como la ve-locidad de ascenso, la sensibilidad individual, las posibles patologías previas que tengamos, los días de permanencia, nuestro grado de adaptación y la aclimatación. Hay una cosa que está clara y que tenemos que tener muy en cuenta: a más altura,
menor PO2 atmosférica. Además, la curva de disociación de la oxi-hemoglobina quedará afecta-da.
Adaptación inmediata a la altura (> 1.500 msnm)
En primer lugar habrá una vasoconstricción hipóxica96 (riesgo de hipertensión pulmonar hipó-xica) y una hiperventilación (hipocapnia, hipoxemia y alcalosis). La hiperventilación nos pro-ducirá una disnea por esfuerzo y además nos deshidrataremos un poco (el aire que inspira-mos es muy seco y perdemos vapor de agua al espirar). A nivel sanguíneo, nuestro hematocrito aumentará, aunque no por poliglobulia (será por pérdida de líquidos = hemoconcentración)97. Ante la hipoxia crónica que sufriremos, nuestros eritrocitos formarán una mayor cantidad de 2,3-bifosfoglicerato para desplazar hacia la derecha la curva de disociación de la oxi-hemoglobina. A nivel cardíaco, nuestro gasto cardíaco aumenta a costa de incrementar la frecuencia cardíaca (el volumen sistólico se mantiene o incluso disminuye porque la contractibilidad es peor)98. El GC aumenta para así incrementar la perfusión pulmonar. Las resistencias periféricas aumentan por el frio, por lo que la presión arterial aumenta99. La sangre estará más viscosa, por lo que la presión será también mayor por esta razón. Otros efectos a nivel periférico es el incremento de la diuresis, insomnio, cefalea100 o hemicránea (dolor en la mitad de la cabeza), y una disminución de la espermatogénesis.
176
96 A modo de curiosidad: ésta también se da en el feto.
97 Una vez nos aclimatamos, tendremos una mayor volemia, por lo que nuestro hematocrito puede ser que no se incremente por el hecho de estar mucho tiempo en altura (la producción mayor de eritrocitos se compensa con un mayor volumen de plasma).
98 Si bien el GC aumenta, las cotas máximas de VS y FC se reducen.
99 Se discute si esto implica un mayor riesgo cardiovascular.
100 Por incremento de la perfusión cerebral (aumenta la presión intracraneal).

Nuestra capacidad de trabajo se verá mermada por causas neurológicas, por la alcalosis res-piratoria y porque podremos transportar una menor cantidad de O2 a los músculos.
Edema cerebral agudo o mal de altura
Se piensa que se debe a la vasodilatación local de los vasos sanguíneos cerebrales producida por la hipoxia. Esto hace que la presión capilar se incremente, favoreciendo la filtración y en última instancia el edema. La fuga de líquido hacia los tejidos cerebrales se asocia con la ce-falea, la anorexia, una disminución en las funciones cognitivas, desorientación e insomnio con somnolencia.
Edema pulmonar agudo de altura
La hipoxia grave hace que algunas arteriolas pulmonares se constriñan de manera intensa, derivando el flujo hacia otros vasos de la circulación pulmonar. Esto hace que las regiones con un exceso de perfusión vean su presión capilar pulmonar incrementada, favoreciendo la filtración y el edema. Este edema provoca una disfunción pulmonar que puede llegar a ser mortal. El mecanismo por el cual se logra la vasoconstricción es gracias a la eliminación del óxido nitroso, un potente vasodilatador (hipótesis actual). El edema pulmonar agudo de altura se manifiesta en forma de cianosis, taquicardia, taquipnea, fiebre… El tratamiento más efec-tivo es bajar a la persona. Si esto no es posible, hacerle respirar oxígeno puro o colcarlo en una cámara hiperbárica portátil podría salvarlo.
Adaptación a largo plazo
La adaptación a largo plazo es lo que se conoce como aclimatación. En clase se comentaron los siguientes cambios corporales:
- Poliglobulia. Supuestamente el hematocrito tendría que incrementar, pero hemos de pensar que también retenemos más líquido (mayor volemia).
- Aumenta el 2,3-bifosfoglicerato.- Aumenta la capacidad de difusión de los gases. Ya sea porque la membrana alveolar se
hace más fina, porque incrementa la perfusión o porque incrementa la ventilación.- Compensación urinaria a la alcalosis.- Angiogénesis. Mayor cantidad de capilares (hiperplasia capilar).- Modificaciones musculares. Ya sean de tipo metabólico o enzimático. Se discute si el
contenido el mioglobina varía.
177


















