
Funcionalidad de la interacción simbiótica entre ... · Funcionalidad de la interacción...
187
Funcionalidad de la interacción simbiótica entre variedades de yuca y genotipos de Rhizophagus irregularis en la Orinoquía Colombiana Isabel Cristina Ceballos Rojas Universidad Nacional de Colombia Facultad de Ciencias, Instituto de Biotecnología Bogotá, Colombia 2016
Transcript of Funcionalidad de la interacción simbiótica entre ... · Funcionalidad de la interacción...

Funcionalidad de la interacción simbiótica entre
variedades de yuca y genotipos de Rhizophagus
irregularis en la Orinoquía Colombiana
Isabel Cristina Ceballos Rojas
Universidad Nacional de Colombia
Facultad de Ciencias, Instituto de Biotecnología
Bogotá, Colombia
2016

Funcionalidad de la interacción
simbiótica entre variedades de yuca y
genotipos de Rhizophagus irregularis en
la Orinoquía Colombiana
Isabel Cristina Ceballos Rojas
Tesis presentada como requisito parcial para optar al título de:
Doctora en Biotecnología
Directora:
Ph.D. Alia Rodríguez Villate
Codirector:
Ph.D. Ian Robert Sanders
Línea de Investigación:
Biotecnología Agrícola
Grupo de Investigación:
Biotecnología de Hongos Formadores de Micorrizas Arbusculares
Universidad Nacional de Colombia
Facultad de Ciencias, IBUN
Bogotá, Colombia
2016

Para todos los seres vivos de este planeta…
especialmente para mis seres queridos.

Agradecimientos
A la Fundación Suiza para la Ciencia y a Colciencias por el apoyo económico para el
desarrollo de este proyecto. Al personal del Instituto de Biotecnología de la Universidad
Nacional de Colombia (Unal) por la gestión para mi formación como investigadora. A la
Universidad de la Salle (Unisalle) por la participación dentro de este proyecto. A Alia
Rodríguez (Unal) e Ian Sanders (Universidad de Lausanne - Unil), directora y codirector,
por permitirme hacer parte de esta investigación, por su incondicional asesoría, su
excelente gestión administrativa, y por todas las enseñanzas brindadas en el campo
investigativo, docente y personal. Al grupo de investigación de “Biotecnología de Hongos
Formadores de Micorrizas” (Unal) por la retro-alimentación constante. Al grupo de
investigación “Ecology and Evolution of Symbiotic Organisms” (Unil) por enseñarme
tantas cosas durante la pasantía. A Daniela León y Marcela Ordoñez por su amistad e
incomparable compañía desde el inicio de este trabajo. A Cristian Fernández, por sus
gestiones y buenos aportes. A los estudiantes de Utopía (Unisalle) que hicieron parte de
este trabajo porque su participación fue esencial en el establecimiento de los
experimentos, y en la recolección y análisis de los datos. A Ricardo Peña y Ricardo
Bueno, directores de Utopía (Unisalle), por todas las gestiones y aportes realizados para
el desarrollo de esta investigación. A Michael Ruiz y su bonita familia, por permitir
establecer los experimentos en sus terrenos, por su hospitalidad, valiosa ayuda y cálida
compañía. A Luis F. Cadavid, por compartirme su valiosa experiencia y conocimiento
sobre el cultivo de la yuca y, por sus cálculos de fertilizaciones para los experimentos que
se establecieron. Al personal de Clayuca (CIAT), por la formación que nos brindaron. A
Fernando Calle y Frankling Beltrán (CIAT), por la semilla donada para los experimentos y
su excelente disposición para el envío. A Julián García, por toda la asesoría que me
brindó para realizar la evaluación económica en el proyecto. Y finalmente a mis queridos
padres, Luz Helena Rojas y Fabio Ceballos, mi hermana Luisa Ceballos y a Andrés
Peláez, por su apoyo incondicional, la fortaleza constante y el amor que me brindaron
para poder realizar este proyecto.

Resumen y Abstract
Resumen
En este trabajo se encontró que la inoculación con un aislado de Rhizophagus irregularis
producido in vitro aumentó significativamente las producciones y la eficiencia en la
aplicación de fertilizantes fosfatados en un cultivo de la variedad de yuca MCOL2737
sembrado en Yopal (Casanare). Luego, un experimento en campo fue establecido para
evaluar el efecto de líneas genéticamente diferentes de esta especie de hongo en la
producción, calidad y colonización de raíces y en el crecimiento y supervivencia de las
plantas de tres variedades diferentes de yuca (MCOL2737, CM4574 y CM6438). Este
experimento fue establecido en Yopal en dos años consecutivos. La producción, calidad
y colonización de las raíces y el crecimiento de las plantas fueron diferentes según la
línea inoculada y, las respuestas inducidas por cada línea genética dependieron de la
variedad de yuca. Además, los efectos generados en la producción por las líneas
fúngicas fueron reproducibles en las dos repeticiones de los experimentos. Algunas de
estas líneas, producidas en el laboratorio a partir del cultivo in vitro de una espora
tomada de líneas iniciales, produjeron efectos diferentes que los de sus parentales. El
experimento también se estableció en Santana, donde el clima y el suelo son diferentes
a los de Yopal. Se concluyó que: 1) La diversidad funcional causada por la variabilidad
genética de R. irregularis fue suficiente para ser detectada en campo en Yopal; 2)
Algunas líneas de R. irregularis obtenidas en el laboratorio en cultivos in vitro, generaron
efectos diferentes a los de sus parentales, en la producción y la calidad de la yuca en
Yopal; 3) El efecto de esas líneas dependió de la variedad de yuca sembrada y del
ambiente donde ocurrió la interacción. Con este trabajo se demostró que la selección de
líneas genéticas de R. irregularis puede ser utilizada para obtener líneas fúngicas que
produzcan un efecto deseado en un determinado cultivo como el de la yuca y en un
ambiente particular como el que ofrece Yopal. Palabras clave: HFMA, cultivos de yuca,
variabilidad intra-específica, funcionalidad de la simbiosis, beneficio de la inoculación,
micorrizas.

Resumen y Abstract VI
Abstract
In this work, it was found that the in vitro mass-produced Rhizophagus irregularis
significantly increased the root production and the phosphate fertilizers efficiency in a
commercial crop with MCOL2737 cassava variety located in Yopal (Eastern Plains of
Colombia). Then, a field experiment was established to study if R. irregularis genetic lines
lead to differences on root production, quality and colonization and on plant growth and
survival for three cassava varieties (MCOL2737, CM4574 and CM6438). This experiment
was established in Yopal in two different years. Root production, quality and colonization
and plant growth varied according to the inoculated AMF line and, the responses induced
by each fungal line were also different between cassava varieties. Further, fungal lines
effects on root production were reproducible in the two years. Some fungal lines were
obtained in the lab by culturing one AMF spore isolated from initial AMF lines, and some
of them, did not produce the same effect than their parental lines in cassava plants. The
same experiment was also established in Santana, where the climate and soil are
different than in Yopal. We conclude that: 1) Functional diversity within R. irregularis was
enough to be detected in field conditions in Yopal; 2) Inoculation with R. irregularis lines
obtained from culturing in the lab altered production and quality cassava crops in Yopal in
a different way than their parental lines; 3) AMF lines effect depended on the cassava
variety and the environment. We demonstrated that selection of R. irregularis genetic
lines produced in vitro could be used to find lines that have a desired effect on a given
crop such as cassava in a particular environment like Yopal.
Keywords: AMF, cassava crops, intra-specific variability, AMF symbiosis functionality,
AMF symbiosis benefit, mycorrhizae.

Contenido VII
Contenido
Pág.
Agradecimientos ....................................................................................... IV Resumen ..................................................................................................... V Abstract ...................................................................................................... VI Lista de figuras ........................................................................................... X Lista de tablas ......................................................................................... XIV Lista de símbolos y abreviaturas ........................................................... XV Introducción ................................................................................................ 1
Objetivo general ......................................................................................... 4
Objetivos específicos ................................................................................. 4
Marco Teórico ............................................................................................. 5
La yuca ........................................................................................................ 5
La yuca como cultivo clave de la Orinoquía Colombiana ........................ 6
Micorrizas arbusculares ............................................................................. 7
La yuca y la aplicación agronómica de los hongos formadores de
micorrizas arbusculares............................................................................. 9
La diversidad funcional de la simbiosis entre plantas y hongos
formadores de micorrizas arbusculares ................................................. 10
La variabilidad intra-específica de Rhizophagus irregularis ................. 11
La variabilidad genética intra-específica de Rhizophagus irregularis
y su potencial para mejorar la productividad de los cultivos ................ 12
1 Capítulo 1: Efecto de un inóculo comercial de Rhizophagus
irregularis en el cultivo de la yuca en Yopal (Casanare) ........................ 14
1.1 Introducción ....................................................................................... 14 1.2 Metodología ....................................................................................... 17 1.3 Resultados ......................................................................................... 22 1.4 Discusión ........................................................................................... 32

VIII Funcionalidad de la simbiósis entre variedades de yuca y genotipos de
Rhizophagus irregularis en la Orinoquía Colombiana
1.5 Conclusión ......................................................................................... 36
2 Capítulo 2: Funcionalidad de la simbiosis entre genotipos de
Rhizophagus irregularis y variedades de yuca ...................................... 39
2.1 Introducción ....................................................................................... 39 2.2 Metodología ....................................................................................... 40 2.3. Resultados ........................................................................................ 50 2.4. Discusión .......................................................................................... 62 2.5. Conclusiones .................................................................................... 67
3 Capítulo 3: Influencia del ambiente sobre la funcionalidad de
la simbiosis entre genotipos de Rhizophagus irregularis y
cultivares de yuca ..................................................................................... 69
3.1 Introducción ....................................................................................... 69 3.2 Metodología ....................................................................................... 71 3.3 Resultados ......................................................................................... 74 3.4 Discusión ........................................................................................... 80 3.5 Conclusiones ..................................................................................... 84
4 Conclusiones y recomendaciones ............................................... 87
4.1 Conclusiones ..................................................................................... 87 4.2 Perspectivas y recomendaciones ....................................................... 88
5 Productos generados .................................................................... 89
6 Consideraciones éticas ................................................................ 93
7 Anexo A : Permisos, trámites y/o licencias necesarias para
desarrollar esta investigación ...................................................................... 95
8 Anexo B : Análisis fisicoquímicos de los suelos antes de
establecer los experimentos ....................................................................... 99
9 Anexo C: Soporte estadístico del experimento con inóculo
comercial en Yopal ................................................................................... 104
Modelos de regresión y gráficas complementarias para los
análisis del BI a lo largo del ciclo del cultivo .............................................. 105
10 Anexo D: Soporte estadístico del experimento de dosis en
Yopal. .................................................................................................... 109
11 Anexo E: Soporte estadístico de las dos repeticiones del
experimento de líneas de Rhizophagus irregularis en Yopal ..................... 114
12 Anexo F: Condiciones del clima, topografía y fertilidad del suelo
para Santana y Yopal. .............................................................................. 150

Contenido IX
13 Anexo G: Estadísticas del efecto del ambiente sobre la
funcionalidad de la simbiosis entre las líneas de Rhizophagus irregularis
y la variedad de yuca MCOL2737. ............................................................ 151
14 Anexo H: Soporte estadístico del experimento de las líneas de
Rhizophagus irregularis y el beneficio de la inoculación sobre la
producción de yuca COL2215 en Santana ............................................... 155
Bibliografía .............................................................................................. 159

Contenido X
Lista de figuras
Pág.
Figura 1. Distribución de cinco bloques perpendiculares a la pendiente y efecto borde en
el experimento de dosis. ................................................................................................. 20
Figura 2. Biomasa seca total, de la raíz y de la parte aérea de la planta a lo largo del
ciclo del cultivo de yuca en Yopal ................................................................................... 24
Figura 3. Variables de crecimiento y producción para plantas de yuca con inóculo y sin
inóculo bajo tres niveles de fertilización fosfatada. ......................................................... 26
Figura 4. Beneficio de la inoculación en términos de biomasa seca total de la planta a lo
largo del ciclo del cultivo. ................................................................................................ 27
Figura 5. Beneficio de la inoculación en términos de la biomasa seca total de las plantas
para cada uno de los muestreos a lo largo del ciclo fenológico de la planta, con diferentes
niveles de fertilización fosfatada. .................................................................................... 29
Figura 6. Efecto de la concentración de inóculo sobre la biomasa fresca de las raíces y
de toda la planta………………………………………………………………………………….30
Figura 7. Retorno de la inversión obtenido por la venta de yuca fresca del cultivo
establecido en Yopal por Ceballos et al, 2013 para la concentración de inóculo real
utilizada; la concentración de inóculo recomendada por la casa comercial; y el 25% de la
concentración de inóculo recomendada por la casa comercial. ....................................... 32
Figura 8. Cultivo simultáneo de dos esporas (líneas) y cultivo monospórico in vitro para
producir líneas genéticas de R. irregularis en cultivos in vitro en el laboratorio. .............. 43

Contenido XI
Figura 9. Distribución de tratamientos y efecto borde para los experimentos establecidos
en Yopal con las líneas genéticas de Rhizophagus irregularis y las tres variedades de
yuca. .............................................................................................................................. 46
Figura 10. Efecto de la inoculación de 15 líneas de Rhizophagus irregularis sobre la
producción y colonización de plantas de yuca en dos variedades diferentes en el
experimento de Yopal (primera repetición). .................................................................... 51
Figura 11. Beneficio en términos de las raíces frescas y secas producido por la
inoculación con líneas de R. irregularis para cada variedad de yuca sembrada en Yopal
(consolidado de ambos experimentos). .......................................................................... 52
Figura 12. Resultados del efecto de las líneas genéticas de Rhizophagus irregularis
sobre la producción de raíces secas en dos variedades de yuca en Yopal para los
experimentos sembrados en dos años diferentes. .......................................................... 53
Figura 13. Efecto de la interacción entre líneas genéticas de Rhizophagus irregularis y la
variedad de yuca sobre el contenido de almidón extractable y el porcentaje de
colonización para cada tratamiento en el momento de la cosecha ................................. 54
Figura 14. Mosaico que representa la frecuencia de plantas muertas inoculadas con las
líneas genéticas de Rhizophagus irregularis para cada variedad de yuca en ambas
repeticiones. ................................................................................................................... 56
Figura 15. Producción de yuca en plantas de la variedad CM4574 y MCOL2737 que
fueron inoculadas con las líneas parentales D1, C2 y C3. .............................................. 58
Figura 16. Producción de yuca de las variedades MCOL2737 y CM4574 que fueron
inoculadas con la línea parental C2 y sus líneas descendientes. ................................... 59
Figura 17. Producción de plantas de yuca de la variedad CM4574 que fueron inoculadas
con la línea parental C3 y sus líneas descendientes. ..................................................... 60
Figura 18. Análisis de correspondencia entre las variedades de yuca y la supervivencia
de las plantas para las dos repeticiones de los experimentos. ...................................... 61

XII Funcionalidad de la simbiósis entre variedades de yuca y genotipos de
Rhizophagus irregularis en la Orinoquía Colombiana
Figura 19. Beneficio de la inoculación en términos de peso seco de raíces producidas
por plantas que fueron inoculadas con líneas genéticamente diferentes de Rhizophagus
irregularis en dos ambientes diferentes. .......................................................................... 75
Figura 20. Diferencias en el beneficio de la inoculación que produjeron líneas de
Rhizophagus irregularis en términos del peso seco de raíces producidas por plantas de
yuca de la variedad MCOL2737 y que fueron sembradas en diferentes ambientes. ....... 77
Figura 21. Colonización Total de HFMA en raíces de yuca de plantas que fueron
inoculadas con diferentes líneas de Rhizophagus irregularis en dos ambientes diferentes.
....................................................................................................................................... 78
Figura 22. Beneficio de la inoculación en términos de la producción de biomasa seca y
fresca de las plantas inoculadas con las diferentes líneas de Rhizophagus irregularis para
las dos variedades de yuca sembradas en Santana. ...................................................... 79
Figura 23. Producción de plantas de yuca de las variedades MCOL2737 y CM4574 que
fueron inoculadas con la línea parental C2 y sus líneas descendientes. ....................... 133
Figura 24. Producción de plantas de yuca de la variedad MCOL2737 que fueron
inoculadas con la línea parental C2 y sus líneas descendientes. .................................. 134
Figura 25. Producción de plantas de yuca de la variedad CM4574 que fueron inoculadas
con la línea parental C2 y sus líneas descendientes. .................................................... 135
Figura 26. Producción de plantas de yuca de las variedades MCOL2737 y CM4574 que
fueron inoculadas con la línea parental C3 y sus líneas descendientes. ....................... 137
Figura 27. Producción de plantas de yuca de la variedad MCOL2737 que fueron
inoculadas con la línea parental C3 y sus líneas descendientes. .................................. 138
Figura 28. Producción de plantas de yuca de la variedad CM4574 que fueron inoculadas
con la línea parental C3 y sus líneas descendientes. .................................................... 139

Contenido XIII
Figura 29. Producción de plantas de yuca de las variedades MCOL2737 y CM4574 que
fueron inoculadas con la línea parental C3 y sus líneas descendientes. .......................140
Figura 30. Producción de plantas de yuca de la variedad MCOL2737 que fueron
inoculadas con la línea parental C3 y sus líneas descendientes. ..................................141
Figura 31. Producción de plantas de yuca de la variedad CM4574 que fueron inoculadas
con la línea parental C3 y sus líneas descendientes. ....................................................142
Figura 32. Producción de plantas de yuca de las variedades MCOL2737 y CM4574 que
fueron inoculadas con la línea parental C3 y sus líneas descendientes. .......................143
Figura 33. Producción de plantas de yuca de la variedad MCOL2737 que fueron
inoculadas con la línea parental C3 y sus líneas descendientes. ..................................144
Figura 34. Producción de plantas de yuca de la variedad CM4574 que fueron inoculadas
con la línea parental C3 y sus líneas descendientes. ....................................................145
Figura 35. Beneficio de la inoculación en términos de peso fresco de raíces producidas
por plantas que fueron inoculadas con líneas genéticamente diferentes de Rhizophagus
irregularis en dos ambientes diferentes. ........................................................................152
Figura 36. Efecto de líneas de R. irregularis sobre el peso fresco de raíces por planta en
Santana. ........................................................................................................................157

Contenido XIV
Lista de tablas
Pág.
Tabla 1. Fases fenológicas del cultivo de yuca de la variedad MCOL2737 en Yopal
(Casanare) para plantas no inoculadas.. ..........................................................................23
Tabla 2. Información sobre las 15 líneas genéticas de Rhizophagus irregularis que
fueron evaluadas en el experimento de Yopal .................................................................44
Tabla 3. Análisis de varianza para las variables de calidad y los porcentajes de
colonización total de los hongos formadores de micorrizas arbusculares. ........................55
Tabla 4. Análisis de varianza con el efecto de las líneas de Rhizophagus irregularis y
las variedades sobre el crecimiento de la parte aérea de la planta para los datos de la
segunda repetición del experimento. ................................................................................55
Tabla 5. Condiciones del clima, topografía y fertilidad del suelo para los sitios de
Santana y Yopal ............................................................................................................. 150

Contenido XV
Lista de símbolos y abreviaturas
En esta sección se incluyen símbolos generales (con letras latinas y griegas), subíndices,
superíndices y abreviaturas utilizadas en este documento.
Símbolos con letras latinas
Símbolo Término
cm Centímetros cm2 Centímetros cuadrados ºC Grados Celsius CO2 Dióxido de carbono g Gramos h Horas ha Hectárea K Potasio HCl Acido clorhídrico KCl Cloruro de potasio Kg Kilogramo m Metro mm Milímetros ml Mililitros N Nitrógeno, concentración normal de una solución NaOH Hidróxido de Sodio P Fósforo R2 Coeficiente de determinación t Tiempo T Toneladas valor p o p Probabilidad que mide evidencia en contra de la hipótesis nula v/v Concentración porcentual en volumen de una especie química
Símbolos con letras griegas
Símbolo Término
α Nivel de significancia estadística
Χ2 Test de Chi-cuadrado
Subíndices
Subíndice Término
+AMF Plantas inoculadas con HFMA
-AMF Plantas no inoculadas con HFMA

XVI Funcionalidad de la simbiósis entre variedades de yuca y genotipos de
Rhizophagus irregularis en la Orinoquía Colombiana
Abreviaturas
Abreviatura Término
AFLP Polimorfismos en la Longitud de Fragmentos Amplificados ANOVA Análisis de varianza BI Beneficio de la inoculación CIAT Centro de Investigación de Agricultura Tropical Col Colonización total por HFMA DAP Fosfato di-amónico dds Días después de siembra FIL Tratamiento con gel sin propágulos GLO Tratamiento con producto comercial Glomygel® HFMA Hongos Formadores de Micorrizas Arbusculares FMA Formadores de Micorrizas Arbusculares msnm Metros sobre el nivel del mar prop. Propágulos REML Restricted Maximum Likelihood ROI Retorno de la inversión variable+AMF Variable en plantas inoculadas con HFMA variable+AMF Variable en plantas inoculadas con HFMA var. variedad

Introducción
Actualmente, uno de los mayores retos de la humanidad se centra en buscar estrategias
para alimentar una población en constante crecimiento. Según las estimaciones de las
Naciones Unidas, para el 2050 el planeta superará los 9 billones de personas (FAO
2013). Sin embargo, para aumentar la productividad de los cultivos, la opción no es
extender la frontera agrícola, pues los recursos naturales cada vez son más escasos y el
componente de sostenibilidad debe estar obligatoriamente ligado a cualquier estrategia
que busque superar este reto.
La yuca (Manihot esculenta Crantz) es un cultivo muy importante para la seguridad
alimentaria y la subsistencia de agricultores de regiones del trópico y sub-trópico. Este
arbusto leñoso produce raíces almidonadas que son esenciales para la dieta básica de
más de un millón de personas en el mundo, y además, sus productos son la tercera
fuente de calorías más importante para los países del trópico (FAO 2013).
Las condiciones típicas de los suelos ácidos del trópico, limitan la producción de la
mayoría de sus cultivos, demandando la aplicación de grandes cantidades de fertilizantes
y generando problemas económicos y ambientales. Por esta razón, es necesario el
desarrollo y la aplicación de tecnologías sostenibles que permitan elevar la
competitividad de los cultivos a través de una mejor productividad, menores costos de
procesamiento y mayor eficiencia en el aprovechamiento de los recursos biológicos de la
región.
Los hongos formadores de micorrizas arbusculares (HFMA) forman naturalmente una
simbiosis mutualista con las raíces de la mayoría de las plantas terrestres, otorgando
beneficios a sus hospederos. Esta simbiosis puede incrementar la biomasa de las plantas
y mejorar su producción debido a que el/los hongo(s) ayuda(n) a la planta a obtener agua
y nutrientes del suelo (Smith & Read 2008).

2 Introducción
Varios experimentos en Colombia y África, realizados con plantas in vitro (Carretero et al.
2009), en invernadero (Sieverding 1991) e inclusive en campo en suelos no estériles
(Howeler & Sieverding 1983; Oyetunji & Osonubi 2007; Liasu et al. 2006), demostraron
que la inoculación con los HFMA tiene efectos sobre el crecimiento de la yuca. Con la
aplicación de estos hongos en Colombia se obtuvieron incrementos en el rendimiento del
cultivo de yuca de más de 5 T/ha y efectos en los requerimientos de fertilizantes
fosfatados (Howeler & Sieverding 1983). Sin embargo, a pesar del gran potencial que
tiene la yuca para ser beneficiada por estos hongos, en los experimentos establecidos en
Colombia, se encontraron respuestas muy variables de acuerdo a los tipos de suelos y al
manejo de los cultivos (Howeler & Sieverding 1983). Esto demuestra la necesidad de
profundizar en la respuesta de la simbiosis del sistema yuca-HFMA en condiciones de
campo, considerando la interacción con el ambiente y con la microbiota local presente.
Adicionalmente, la producción de estos hongos hasta hace poco, demandaba técnicas
tradicionales de propagación en grandes cantidades de suelo, mediante la utilización de
cultivos trampa. Esto no permitía una producción apropiada con calidad, es decir, un
inóculo con concentraciones de propágulos conocidas y en bajos volúmenes, que pudiera
aplicarse en condiciones de campo. Con esto se afectaba la viabilidad de aplicar esta
tecnología, debido a los altos costos en el transporte, la presencia de potenciales
patógenos que podían ser introducidos al suelo, entre otros aspectos (Sanders 2010).
Actualmente, es posible producir inóculos puros y concentrados en sistemas estériles a
partir de cultivos in vitro de los HFMA, en un medio artificial que utiliza raíces
transformadas con Agrobacterium rhizogenes (Declerck et al. 2005). Con esto se abre
inmediatamente la posibilidad de utilizar los HFMA producidos in vitro para incrementar el
rendimiento de los cultivos de yuca en el trópico (Rodriguez & Sanders 2016). Sin
embargo, no todas las especies de HFMA pueden crecer en este medio artificial.
Entre las pocas especies que pueden crecer en este sistema in vitro, se encuentra
Rhizophagus irregularis, anteriormente conocido como Glomus intraradices (Stockinger
et al. 2009). Este hongo es una especie clave dentro de los HFMA ya que ha sido
ampliamente estudiado, y además, ha sido encontrado en casi todos los tipos de suelos
del mundo (Smith & Read 2008). En Colombia, por ejemplo, R. irregularis se encontró

Introducción 3
como una de las especies mas abundantes en los ecosistemas y en los suelos agrícolas
del amazonas (León 2015; León 2006).
Las poblaciones de R. irregularis exhiben una alta variabilidad genética (Croll et al. 2008;
Börstler et al. 2008; Gamper et al. 2008), la cual es importante en términos de la
funcionalidad de la simbiosis. Estas variaciones genéticas intra-específicas se han
encontrado asociadas con variaciones fenotípicas (Koch et al. 2004; Croll et al. 2009), las
cuales a su vez pueden presentar diferencias en la forma como el HFMA afecta el
crecimiento y la producción de las plantas (Angelard et al. 2010; Croll et al. 2009).
Recientemente, experimentos con R. irregularis mostraron que a partir de hongos
aislados de campo, se pueden obtener líneas genéticamente diferentes a sus parentales
y/o entre ellas mismas por medio de procesos realizados en laboratorio (Croll & Sanders
2009; Angelard et al. 2010). Esto permitió obtener, en condiciones in vitro, nuevas líneas
genéticas de R. irregularis (Ehinger et al. 2012). Adicionalmente, en estudios recientes
realizados en condiciones de invernadero, se demostró que la diversidad de las líneas de
R. irregularis, afectó de forma diferencial el crecimiento de algunas plantas y la
colonización de sus raíces (Angelard et al. 2010; Angelard & Sanders 2011; Croll et al.
2009; Koch et al. 2004). Con esto, se abre una posibilidad muy interesante para un
desarrollo biotecnológico con los HFMA, donde a través de procesos naturales que
incrementan la variabilidad genética de estos hongos en el laboratorio se pueden
encontrar líneas de HFMA que mejoren la productividad de un determinado cultivo
(Sanders 2010).
La posibilidad de obtener líneas genéticas de R. irregularis en condiciones in vitro que
pueden provocar efectos diferenciales en la funcionalidad de la simbiosis y, la facilidad de
producirlas rápidamente como inóculo en cantidades comerciales, permitieron por
primera vez evaluar el efecto de la variabilidad genética intra-específica de esta especie
de HFMA en campo durante el desarrollo de esta investigación, cuyos objetivos fueron:

4 Introducción
Objetivo general
Evaluar el efecto de la simbiosis entre diferentes genotipos de Rhizophagus irregularis y
variedades de yuca (Manihot esculenta) sobre el rendimiento de los cultivos en la llanura
mal drenada de la Orinoquía Colombiana.
Objetivos específicos
Evaluar el efecto de un inóculo comercial de R. irregularis en el cultivo de la yuca para la
zona de estudio.
Comparar la funcionalidad de la simbiosis entre genotipos de R. irregularis y variedades
de yuca en términos de la producción de las plantas y la colonización de los HFMA.
Analizar la influencia del ambiente sobre la funcionalidad de la simbiosis entre genotipos
de R. irregularis y cultivares de yuca.

Marco Teórico
Este capítulo contiene el marco teórico general que soporta el planteamiento de esta
investigación. Inicialmente se hace una descripción del cultivo de la yuca, el cual fue
utilizado como modelo para evaluar la funcionalidad de la simbiosis en los experimentos
de campo establecidos. Luego, se enuncian las características del ambiente donde se
desarrolló la investigación. Y finalmente, se exponen las bases teóricas en las áreas de
biología, ecología y genética de los hongos formadores de micorrizas arbusculares
(HFMA) que soporta la hipótesis planteada.
La yuca
La yuca (Manihot esculenta Crantz) es una planta dicotiledónea perteneciente a la familia
Euphorbiaceae (Ceballos & De la Cruz 2002). Es un arbusto leñoso, perenne, con
variaciones en la altura de la planta que oscila entre 1 y 5 m. La principal característica
de las raíces de yuca es su capacidad de almacenamiento de almidones, razón por la
cual este órgano tiene un gran valor económico (Ceballos & De la Cruz 2002).
La yuca es un cultivo de amplia adaptación ya que se siembra desde el nivel del mar
hasta los 1.800 msnm, en temperaturas entre 20 y 30ºC, en humedades relativas entre
50 y 90 % y en zonas con precipitaciones anuales entre 600 y 3000 mm (Cock & Rosas
1975).
Los cultivos de yuca tienen un ciclo desde la siembra hasta la cosecha entre 7 y 24
meses dependiendo de las condiciones ambientales y de la variedad sembrada (Ospina
& Ceballos 2002). Las plantas se pueden propagar vegetativamente. Al sembrar sus
estacas, brotan las yemas de la parte superior y salen las raíces en la base de la estaca,
de los nudos inferiores. Durante las primeras semanas, la planta forma raíces fibrosas;
dos o tres meses después de la siembra, algunas de esta raíces, comienzan a acumular

6 Marco teórico
almidón continuamente hasta la cosecha (Cock & Rosas 1975). Durante los tres primeros
meses, la formación de hojas tiene prioridad sobre la formación de raíces de
almacenamiento. Luego del tercer mes comienza una etapa de engrosamiento de la raíz,
hasta el sexto mes, dependiendo del cultivar. Finalmente, ocurre una acumulación de
almidón hasta el final del ciclo del cultivo (Cock & Rosas 1975).
Adicionalmente, esta planta tiene características morfológicas y fisiológicas que
favorecen su supervivencia y producción en condiciones climáticas y edáficas adversas,
como las que se encuentran en la Orinoquía Colombiana (Edwards & Kang 1978; Connor
et al. 1981).
La yuca como cultivo clave de la Orinoquía Colombiana
Durante los últimos años, a pesar del potencial agrícola de la Región Oriental de
Colombia, gran parte de sus tierras han sido utilizadas para ganadería, afectando la
fertilidad del suelo (Corpoorinoquía 2011). Los sistemas ganaderos deterioran la
diversidad, modifican el balance de nutrientes, aumentan la compactación y propician la
erosión del suelo (Sadeghian 2009). Por esta razón, convertir tierras utilizadas para
ganadería en modelos productivos agrícolas sostenibles puede constituirse en una
opción para mejorar la calidad del suelo de la región.
La yuca es un cultivo que tiene: tolerancia a la sequía, capacidad de producirse en suelos
degradados, resistencia a plagas y enfermedades, tolerancia a los suelos ácidos
(predominantes en las sabanas tropicales del mundo) y flexibilidad en cuanto al momento
de la plantación y cosecha (Ceballos & De la Cruz 2002). Estas ventajas lo convierten en
un cultivo importante para la Orinoquía (Ospina & Ceballos 2002), la cual se caracteriza
por su baja fertilidad química (Sanchez & Salinas 1981); erosión; deficiencias de fósforo
(P), nitrógeno (N) y potasio (K); la acidez (pH< 5.3); toxicidad por aluminio y la alta
capacidad de fijación del fósforo (Rivas et al. 2004).
En Colombia, se producen un poco más de 2 millones de toneladas de yuca por año en
un área de 190.000 ha, con un rendimiento promedio de 10,6 T/ha (FAO 2013; Agronet

Marco teórico 7
2013). De la producción anual del país, la Orinoquía tiene una participación del 20%
(Sipsa et al. 2014) presentando rendimientos promedio en la región de 11,4 T/ha
(Agronet 2013). Las limitaciones de la producción pueden reducir considerablemente los
rendimientos, haciendo que los cultivos sean menos rentables en el mercado competitivo
de los carbohidratos. Es por eso, que en este país, donde el cultivo de la yuca es
tradicional y constituye una importante fuente de alimentación y de ingresos de los
agricultores, es clave mejorar la competitividad de este cultivo y convertirlo en una fuente
de recursos atractiva.
Dentro de las estrategias que se pueden utilizar para solucionar el problema de baja
productividad de los suelos ácidos del trópico, se encuentra el uso de microorganismos
que incrementen la dinámica de la obtención de nutrientes por las plantas o que mejoren
las propiedades de los suelos. En el caso de la yuca, una de las principales razones, que
podría explicar el éxito de este cultivo en suelos con baja fertilidad química, es la
simbiosis mutualista que las raíces de estas plantas establecen con hongos formadores
de micorrizas arbusculares (HFMA), pues se ha demostrado que la yuca es una planta
que depende casi completamente de esta asociación simbiótica para poder crecer
(Howeler & Sieverding 1983) y para la toma de fósforo en suelos con bajos contenidos
disponibles de este nutriente (Howeler & Asher 1982; Yost & Fox 1979). Por esta razón,
esta simbiosis se convierte en un punto de interés en la búsqueda del manejo integrado,
sostenible y eficiente de estos cultivos para la zona.
Micorrizas arbusculares
Las micorrizas arbusculares son una de la simbiosis mutualistas mas abundantes en
ecosistemas terrestres del planeta. Esta simbiosis ocurre entre las raíces del 80% de las
plantas terrestres (Wang & Qiu 2006) y los hongos del filo Glomeromycota (Schüβler et
al. 2001), conocidos como hongos formadores de micorrizas arbusculares (HFMA). Los
HFMA han co-evolucionado con las plantas terrestres por más de 400 millones de años
(Remy et al. 1994) y a cambio de carbono, proveen de beneficios a las plantas,
incrementando la adquisición de nutrientes (Marschner & Dell 1994; Smith & Smith 2011),
mejorando el uso eficiente del agua (Augé 2001), protegiendo a la planta de patógenos

8 Marco teórico
(Borowicz 2001) e incrementando la tolerancia a estreses de tipo biótico y abiótico (Feng
et al. 2002).
Los HFMA son biótrofos obligados que crecen y se proliferan dentro de la raíz de una
planta hospedera y afuera, en el suelo circundante. Antes de la colonización, el hongo
reconoce los potenciales hospederos y estimula el crecimiento de sus hifas (Requena et
al. 2007), luego ocurren una serie de eventos regulados por ambos simbiontes que
finalmente llevan a la colonización. El reconocimiento para que se inicien los eventos de
la simbiosis se puede definir como una compatibilidad, la cual esta genéticamente pre-
determinada. Existen iniciadores secretados por las plantas e identificados como
estrigolactonas (López-Ráez et al. 2008) que estimulan la actividad metabólica del hongo
(Tamasloukht et al. 2003) produciendo ramificación en su micelio (Buee et al. 2000). El
hongo a través del micelio secreta señales difusibles hacia las raíces de la planta y se
induce la activación de la simbiosis en las raíces que se encuentran en contacto con el
hongo, incluyendo la expresión de genes relacionados con la simbiosis (Kosuta et al.
2003). Posteriormente, algunas células forman un apresorium y un aparato de
penetración (Genre et al. 2005) que permite que el hongo ingrese a la raíz a través de la
epidermis. Dentro de la raíz, la hifa del hongo crece intercelularmente hasta que penetra
las paredes de las células corticales y forma estructuras intercelulares (Genre et al.
2005). Para completar su ciclo de vida, el hongo sale de la raíz de la planta y comienza a
tener un crecimiento extensor de la hifa extra-radical, para finalmente, formar esporas en
las terminaciones de algunas de estas hifas. A través de todo el ciclo se mantiene un
intercambio de señales entre los simbiontes (Genre et al. 2005) que induce el desarrollo
de patrones de expresión génica tanto en la raíz de la planta como en el hongo.
Los beneficios que pueden otorgar los HFMA a las plantas, incluso con las de cultivos de
importancia mundial como el de la yuca, convierten a los HFMA en organismos con
potencial de aplicación en agricultura y en el desarrollo de aplicaciones biotecnológicas.

Marco teórico 9
La yuca y la aplicación agronómica de los hongos formadores de micorrizas
arbusculares
Aunque se han encontrado efectos significativos de la aplicación de los HFMA en cultivos
de yuca, existían grandes limitaciones para evaluar la eficiencia real de estos hongos.
Una limitación era que muchos estudios realizaron los experimentos en suelos estériles,
donde se eliminaba completamente la interacción de los organismos introducidos con los
que se encontraban presentes en el suelo. Howeler y Sieverding (1983), demostraron
que al inocular HFMAs, las diferencias en el rendimiento de los cultivos de yuca entre las
plantas inoculadas y no inoculadas eran mayores en suelos estériles que en suelos no
estériles. Esto, demostró que la microbiota presente en el suelo altera los efectos de la
inoculación. Los resultados de experimentos con suelos estériles, aunque garantizaban
que el efecto era debido a un organismo determinado, se alejaron de las condiciones
reales que se encuentran en campo, y la interpretación de sus resultados debe ser
cuidadosa cuando el objetivo real de un trabajo apunta al desarrollo de herramientas
biotecnológicas para aplicar en agricultura.
Otra limitación era que la mayoría de las especies de HFMA utilizadas en la inoculación
habían sido multiplicados en condiciones no estériles a partir de labores intensivas y
costosas de propagación que, finalmente, no garantizaban inóculos puros, concentrados
y con posibilidad de seguimiento (Sanders 2010). Sin este control era complicado atribuir
un efecto a un determinado organismo. Se hacía entonces necesario hacer evaluaciones
en campo del efecto de los HFMA con inóculos controlados y puros teniendo en cuenta
las interacciones con las comunidades locales.
Otra limitación, es que ciertos HFMA pueden mejorar el crecimiento de unas plantas pero
no de otras, lo que elimina la posibilidad de tener un inóculo universal (Sanders 2010) ya
que se da una diversidad en los efectos de la simbiosis dependiendo de quienes están
involucrados en la relación simbiótica.

10 Marco teórico
La diversidad funcional de la simbiosis entre plantas y hongos formadores de micorrizas
arbusculares
La funcionalidad de la simbiosis se define como el efecto que produce la relación
simbiótica de una determinada combinación entre planta y hongo/s (Feddermann et al.
2010), pues una planta puede estar asociada con diferentes especies o individuos de
HFMAs (Magrow 1936; Gerdemann 1955). La funcionalidad de la simbiosis puede ser
medida en la planta o en el hongo de acuerdo con los intereses del investigador. Por
ejemplo, Munkvold et al. (2004), midieron la funcionalidad de la simbiosis en términos del
crecimiento y la toma de fósforo en plantas de pepino y también, en términos del
crecimiento del hongo dentro de la raíz. De esta forma, es fundamental que cuando se
desee analizar la funcionalidad de la simbiosis se aclare que variables se están
evaluando y se conozca de antemano como se evaluará la eficiencia de la interacción.
El efecto de la inoculación sobre las plantas depende de varios factores como la
identidad del hongo FMA aplicado (Sieverding 1991; Munkvold et al. 2004; Jones & Smith
2004; Helgason et al. 2002) y de la planta hospedera (Helgason et al. 2002; Baon et al.
1993; Howeler & Sieverding 1983); las condiciones ambientales (Mohammad et al. 2003);
el tipo de suelo y las comunidades de organismos allí presentes (Howeler & Sieverding
1983).
En experimentos de invernadero, se encontró que diferentes combinaciones entre planta
hospedera – hongos, exhiben una alta diversidad funcional no sólo cuando se varían las
especies sino también los genotipos de las plantas y/o los aislados de los hongos (Baon
et al. 1993; van der Heijden et al. 1998; Munkvold et al. 2004; Avio et al. 2006; Jansa et
al. 2008). Los HFMA se consideran organismos asexuales y hasta el momento se han
identificado unas 230 especies (www.mycobank.org).
La mayoría de los estudios que han evaluado las divergencias funcionales de la
simbiosis, se han enfocado en las diferencias que pueden generar las especies de los
HFMA o de las plantas (Klironomos 2003). Sin embargo, parece ser que la diversidad
genética y fenotípica intra-específica de estos hongos es extremadamente amplia

Marco teórico 11
(Sanders & Croll 2010; Sanders & Rodriguez 2016) y estas diferencias pueden afectar
directamente la funcionalidad de la simbiosis (Munkvold et al. 2004; Oliveira et al. 2010).
La variabilidad intra-específica de Rhizophagus irregularis
Aunque R. irregularis ha sido la especie de HFMA mas estudiada en términos de
variabilidad genética intra-específica, existen también estudios que han demostrado esto
en otras especies de hongos FMA, como Glomus geosporum (Oliveira et al. 2010). R.
irregularis se ha convertido en el modelo para investigaciones del filo Glomeromycota,
debido a que es el primero en ser cultivado bajo el sistema de raíces transformadas de
zanahoria in vitro (Stockinger et al. 2009). Como simbionte es altamente efectivo en la
movilización, la toma y la transferencia de diferentes nutrientes minerales (Govindarajulu
et al. 2005) y coloniza rápidamente las plantas hospederas, dentro de las cuales se
encuentran el arroz, la alfalfa y la yuca (Martin et al. 2008). Además, se ha estudiado el
genoma y transcriptoma de esta especie (Tisserant et al. 2013), lo cual permite un mejor
entendimiento de procesos relacionados con la biología y genética de este hongo (Martin
et al. 2008).
Para el caso de R. irregularis, Kock et al., (2004), utilizando la técnica de Polimorfismos
en la Longitud de Fragmentos Amplificados (AFLP), encontraron una gran cantidad de
polimorfismos entre 16 diferentes aislados de esta especie provenientes de un mismo
campo en Suiza. En el estudio de Koch et al. (2004), un análisis AMOVA (Analysis of
Molecular Variance) reveló diferencias genéticas amplias entre los aislados de la
población. Croll et al. (2008) desarrollaron marcadores de secuencias simples repetidas
(SSR), de intrones de genes nucleares y mitocondriales con el fin de caracterizar los
aislados recolectados por Koch et al. (2004). Los marcadores mostraron una fuerte
diferenciación a nivel nuclear y mitocondrial entre los aislados.
Estas variaciones genéticas intra-específicas pueden promover una diversidad funcional
en la planta. Las diferencias genéticas pueden, por ejemplo, producir diferencias en el
tamaño externo de la hifa, incrementando el área de contacto entre el hongo y el suelo, y
alterando la cantidad de fósforo que el hongo le entrega a la planta. Esta hipótesis fue

12 Marco teórico
probada por Munkvold et al. (2004), quienes encontraron una alta diversidad funcional
dentro de aislados de la misma especie para Glomus mosseae, G. claroideum, G.
caledonium y G. geosporum sobre plantas de pepino al evaluar la colonización de estos
hongos en las raíces y el crecimiento de las plantas en invernadero.
La variabilidad genética intra-específica de Rhizophagus irregularis y su potencial para
mejorar la productividad de los cultivos
El nivel de variabilidad genética en R. irregularis, una de las especies de HFMA que
puede ser cultivada en un sistema in vitro, podría entonces ser utilizado para buscar
efectos deseados en las plantas. Los HFMA cuentan con un micelio cenocítico donde
hospedan muchos núcleos en un citoplasma común (Smith & Read 2008) y hace poco,
se comprobó que la anastomosis y la segregación generaron variabilidad genética en
líneas cultivables de R. irregularis. En estos dos procesos que ocurren naturalmente en
los HFMA (Giovannetti et al., 1999; Giovannetti et al., 2001; Angelard et al., 2001), la
progenie resultante presentó genotipos y fenotipos diferentes a sus parentales (Angelard
et al., 2010; Croll et al., 2009) y adicionalmente, afectó el crecimiento de plantas de arroz
a nivel de invernadero de una forma diferente a la de sus parentales (Angelard et al.,
2010).
La posibilidad de producir y seleccionar líneas genéticas de R. irregularis en laboratorio
que afecten de una forma deseable el crecimiento de las plantas hospederas, abre
entonces una opción de utilizar este desarrollo biotecnológico para mejorar la producción
de los cultivos. Pero para ésto, se hace necesario hacer evaluaciones en campo del
efecto de esa variabilidad intra-específica de los HFMA con inóculos controlados en
condiciones de campo, y teniendo en cuenta, las interacciones con las comunidades del
suelo.
Este trabajo es importante y único, porque por primera vez, se evaluó el efecto en
campo de la diversidad intra-específica de una especie importante de HFMA sobre el
crecimiento de una planta altamente mico-trófica y de gran interés comercial como la

Marco teórico 13
yuca. Los experimentos en campo correctamente diseñados se convierten en
herramientas ideales para entender procesos, donde las variables de estudio se miden
teniendo en cuenta todas las interacciones reales con el ambiente (Harrison & List 2004).
De esta forma, la evidencia en campo del efecto de la variabilidad intra-específicas de los
HFMA, sobre la respuesta funcional de la simbiosis, proporciona el racional requerido
para la selección de líneas del hongo FMA en el contexto de aumentar la productividad
de plantas de cultivos de interés agronómico, bajo parámetros de sostenibilidad y
racionalidad en el uso de los recursos naturales.

1 Capítulo 1: Efecto de un inóculo comercial
de Rhizophagus irregularis en el cultivo de
la yuca en Yopal (Casanare)
1.1 Introducción
La yuca es un cultivo extremadamente adaptado a los suelos del trópico ácido (Sanchez
& Salinas 1981), pero en este tipo de suelos, las deficiencias de fósforo son las mayores
limitantes para su producción. La fertilización inorgánica ha sido empleada para
incrementar la producción en los cultivos de yuca en esta región (Howeler 1981) ya que
este cultivo responde fuertemente a la fertilización fosfatada. Se han encontrado
respuestas en el rendimiento hasta con aplicaciones de 400 Kg de fosfato di-amónico
(DAP) por hectárea aunque los niveles usualmente recomendados se encuentran entre
100 y 150 Kg DAP/ha (Cock 1985).
Las condiciones de estos suelos exigen grandes cantidades de agroquímicos y, en el
caso del fósforo, mientras las reservas mundiales de este nutriente cada vez van
disminuyendo, las demandas de su uso van incrementando para poder alimentar una
población global creciente (Gilbert 2009). Los altos costos de los fertilizantes y los
problemas ambientales asociados a su aplicación hacen que sea necesario buscar
alternativas como el uso de hongos formadores de micorrizas Arbusculares (HFMA) para
mejorar la producción del cultivo de la yuca e incrementar la toma de fósforo en este tipo
de suelos (Howeler & Asher 1982; Yost & Fox 1979).
En la aplicación de los HFMA se han encontrado respuestas negativas y positivas en los
rendimientos de los cultivos, dependiendo de la especie fúngica inoculada (van der
Heijden et al. 1998; Bever et al. 2001; Vogelsang et al. 2013). Esto demuestra la

Capítulo 1 15
necesidad de considerar la especie de hongo FMA que se está aplicando. En esta
investigación se realizaron inoculaciones con Rhizophagus irregularis, debido a que esta
especie de HFMA: 1) Ha sido encontrada en la mayoría de los ecosistemas alrededor del
mundo (León 2006; Smith & Read 2008); 2) Fue encontrada como una de las mas
abundantes en las raíces de plantas silvestres de yuca en la Amazonía Colombiana
(León 2015); 3) Puede ser producida como inóculo en grandes cantidades, convirtiéndola
en una candidata potencial para la elaboración de inóculos que se pueden utilizar para
mejorar la producción de cultivos comerciales como el de la yuca; 4) Se ha convertido en
el hongo modelo de los HFMA para los investigadores debido a que puede ser crecido
fácilmente en el laboratorio en un sistema in vitro, similar al utilizado para producirlo a
escala comercial y 5) su variabilidad intra-específica es importante en términos de la
funcionalidad de la simbiosis. Por estas razones, en este capítulo, se evaluó el efecto que
esta especie particular de hongo modelo produjo sobre los rendimientos y las
producciones de un cultivo comercial de yuca en los Llanos Orientales de Colombia.
El uso de inoculantes de HFMA en sistemas agrícolas también se puede abordar desde
la perspectiva de aislar los hongos FMA que se encuentren en los suelos de interés,
preseleccionarlos de acuerdo a atributos deseables, llevarlos a sistemas in vitro para su
producción y evaluar su efecto en campo. Sin embargo, esta opción es dispendiosa en
tiempo y recursos, pues la mayoría de especies de HFMA no logran crecer fácilmente en
los cultivos in vitro. Se calcula que este abordaje requiere como mínimo 6 años de trabajo
continuo, y se puede convertir en una alternativa costosa (Sanders 2010).
En este capítulo no sólo se midió el efecto de la simbiosis en términos del crecimiento y
la producción en la planta y sus órganos, sino que también se midió en términos del
porcentaje de la raíz que estaba colonizado por los hongos FMA. La colonización es una
variable que se mide en casi todos los estudios que involucran este tipo de hongos. Es
importante ya que representa el porcentaje de raíz que se encuentra colonizada por
estos simbiontes. Sin embargo, la metodología de tinción utilizada (Vierheilig et al., 1998)
no permite diferenciar entre las especies o aislados de HFMA que se encuentran
presentes en esa raíz, y por lo tanto, no distingue los hongos inoculados de los que
estaban localmente presentes. Además, esta variable no siempre presenta correlación
con el efecto observado en las plantas (Isobe et al. 2014).

16 Efecto de un inóculo comercial
En los sistemas agrícolas, los efectos de la inoculación con HFMA en plantas de cultivo
están influenciados por factores abióticos como la disponibilidad de agua y nutrientes del
suelo, la precipitación y otros, y factores bióticos, como la composición de las comunidad
de microorganismos presentes en el suelo (Johnson et al. 2013; Hoeksema et al. 2010;
Feddermann et al. 2010) entre otros. Estos factores del ambiente donde ocurre la
interacción pueden variar constantemente en el tiempo y en el espacio (Feddermann et
al. 2010), y por esta razón, las investigaciones recientes se han enfocado en evaluar
desde una perspectiva dinámica el efecto de la micorrización (Finlay 2008). En este
capítulo se realizaron diferentes muestreos con el fin de comparar la respuesta de la
simbiosis a lo largo del ciclo del cultivo.
El efecto de la inoculación con los HFMA generalmente se ha evaluado en términos del
crecimiento de las plantas (Smith & Read 2008) y las variables más comunes para medir
este efecto han sido el rendimiento y la biomasa seca y fresca de las plantas y/o de sus
producciones. Sin embargo, hacer comparaciones entre plantas inoculadas y no
inoculadas a lo largo del tiempo es difícil, debido a que existe un efecto particular en cada
muestreo. Por esta razón, el beneficio de la inoculación (BI) es un parámetro muy útil
para describir la dinámica del efecto que genera la aplicación de cierta especie de HFMA
a lo largo de un ciclo, por ejemplo un ciclo de cultivo. En el BI se comparan los resultados
de las plantas inoculadas y las no inoculadas en términos de una variable de interés
(Raju et al. 1990). Los cálculos de BI para un sistema agrícola definido y sembrado en un
ambiente determinado permiten: 1) Evaluar si la inoculación esta aumentando o
disminuyendo la variable medida en una condición ambiental o en un estado de
desarrollo de la planta; 2) Reconocer los tiempos en que es necesario realizar las
mediciones para evaluar la funcionalidad de la simbiosis; 3) Conocer en que momento se
maximiza o minimiza el efecto de la simbiosis sobre la variable de interés; y 4) Identificar
cuáles factores bióticos y abióticos se asocian con la respuesta de la simbiosis. La
aplicación de HFMA puede estar limitada cuando no se conoce la dinámica del sistema y,
por esta razón, en este capítulo se muestra la dinámica del beneficio de la inoculación de
R. irregularis en un cultivo de yuca. Por primera vez, se presenta un reporte en campo
del beneficio de la inoculación de un hongo FMA sobre las plantas de un cultivo a lo largo
de todo su ciclo.

Capítulo 1 17
Adicionalmente, teniendo en cuenta que los progresos de la biotecnología agrícola
buscan incrementar la productividad de los cultivos, especialmente mediante la reducción
de los costos de producción, en este capítulo se presenta una evaluación económica de
la aplicación de HFMA para mejorar el rendimiento de los cultivos de yuca en la zona.
El objetivo principal de este capítulo fue evaluar el efecto de un inóculo comercial de R.
irregularis producido masivamente en condiciones in vitro sobre el cultivo de la yuca para
la zona de estudio. Pero también se abordaron los siguientes objetivos: 1) Caracterizar el
ciclo fenológico del cultivo de yuca en la zona de estudio; 2) Evaluar el efecto de un
inóculo comercial de R. irregularis sobre el crecimiento de las plantas de yuca var.
MCOL2737 y sobre el rendimiento del cultivo bajo tres niveles de fertilización fosfatada;
3) Comparar el beneficio de la inoculación en los muestreos realizados a lo largo del ciclo
del cultivo; 4) Establecer los tiempos de medición, los niveles de fertilización fosfatada y
las variables agronómicas relevantes para evaluar el efecto de R. irregularis en un cultivo
de yuca para la zona de interés; y 5) Determinar la viabilidad económica de aplicar el
inóculo comercial de R. irregularis para la producción de yuca en la zona.
1.2 Metodología
Para responder al objetivo principal de este capítulo, se estableció un experimento en
campo con la variedad de yuca más sembrada en la zona de estudio para consumo en
fresco (var. MCOL2737) y un inóculo comercial de Rhizophagus irregularis producido in
vitro. Además, se estableció otro experimento en campo con el fin de evaluar el efecto de
la dosis del mismo inóculo comercial de R. irregularis sobre la producción de yuca en las
plantas del cultivo.
1.2.1 Metodología del experimento en campo con inóculo
comercial de Rhizophagus irregularis y plantas de yuca var.
MCOL2737
La descripción del sitio de estudio del experimento principal, el material vegetal y fúngico
utilizado, el diseño y la forma como se estableció el experimento en campo, la manera
cómo se midieron las variables de crecimiento en la planta y los hongos, y los análisis

18 Efecto de un inóculo comercial
estadísticos y económicos, se encuentran detallados en Ceballos et al., (2013). La
metodología utilizada para el establecimiento del experimento de dosis y para otros
ensayos que no fueron publicados en Ceballos et al., (2013) son explicados a
continuación.
Ciclo fenológico del cultivo de yuca var. MCOL2737 en la zona de estudio
La caracterización del ciclo fenológico de la variedad de yuca MCOL2737 en Yopal se
realizó con los resultados del crecimiento de las plantas del experimento establecido por
Ceballos et al., (2013). La frecuencia de los muestreos (45 días) permitió siete
mediciones durante todo el ciclo del cultivo. Para esta caracterización se utilizaron los
datos de las plantas no inoculadas que fueron fertilizadas con 201 Kg*ha-1 de fosfato di-
amónico. Este nivel de fertilización fue seleccionado ya que representaba las condiciones
que tendría el cultivo con la cantidad de fertilizante que normalmente aplican los
agricultores de la zona. También se compararon los resultados de variables de
crecimiento de plantas inoculadas y no inoculadas durante el ciclo del cultivo.
Beneficio de la inoculación (BI) a lo largo del ciclo del cultivo de yuca
El beneficio de la inoculación (BI) de R. irregularis en plantas de yuca fue calculado en
cada uno de los tratamientos para cada uno de los muestreos realizados en el
experimento de campo establecido en Yopal por Ceballos et al., (2013). Los muestreos
se hicieron cada 45 días hasta la cosecha final a los 320 días después de siembra (dds).
El BI fue calculado como un porcentaje según la fórmula (Variable +AMF – Variable -AMF) /
Variable-AMF * 100, propuesta por Raju et al., (1990). Donde: BI: Beneficio de la
inoculación; Variable+AMF: Valor de la variable en plantas inoculadas con HFMA; Variable-
AMF: Valor de la variable para las plantas no inoculadas. Cuatro réplicas fueron tomadas
por tratamiento en cada muestreo. Cada réplica correspondió al promedio de tres plantas
por parcela. Las variables utilizadas para calcular el BI fueron la biomasa seca total de la
planta y la de sus diferentes órganos (pecíolos, tallos, raíces y hojas).

Capítulo 1 19
Las condiciones ambientales durante el experimento fueron medidas con la estación
meteorológica localizada en el campus de la Universidad de la Salle en Yopal, Colombia
(ver: http://www.weatherlink.com/user/utopia/).
1.2.2 Experimento de dosis del inóculo comercial de
Rhizophagus irregularis
Un experimento en campo fue establecido en Yopal para evaluar el efecto de diferentes
dosis del inóculo comercial de R. irregularis sobre la producción de las plantas de yuca
var. MCOL2737. La descripción del lugar se encuentra en Ceballos et al., (2013). Las
propiedades físico-químicas del suelo del lote donde se estableció el experimento se
presentan en el Anexo B: numeral 8.1. El material vegetal y fúngico para este
experimento fueron los mismos utilizados en Ceballos et al., (2013) en el experimento de
Yopal.
Diseño y establecimiento del experimento en campo
Para este experimento la dosis se definió como la combinación entre la cantidad de
aplicaciones y las concentraciones del inóculo. Un diseño en bloques completamente
aleatorio fue empleado para evaluar el efecto de los factores: 1) Cantidad de aplicaciones
(doble o simple) y 2) Concentración, definida como el porcentaje de inóculo utilizado con
respecto a lo recomendado por la casa comercial. Así el 100% correspondió a la
concentración recomendada de 250 propágulos (prop.) de hongo por planta (0,125 ml de
producto comercial sin diluir). Los niveles de concentraciones aplicadas a las plantas de
yuca fueron de 25% (62 prop.), 50% (125 prop.), 75%(187 prop.), 100% (250 prop.),
125% (312 prop.), 150% (375 prop.), 175% (437 prop.) y 200% (500 prop.). Se
establecieron cinco bloques con una réplica de cada uno de los 16 tratamientos y los
controles. Las plantas sin tratamiento de inoculación (0%) recibieron la misma cantidad
de agua sin hongo. Los bloques fueron ubicados en línea a lo largo de la pendiente del
terreno (Figura 1). Dos filas de plantas de yuca var. MCOL2737 fueron sembradas
alrededor del área que contenía las plantas tratamiento, para reducir el efecto borde. La

20 Efecto de un inóculo comercial
siembra de la yuca y el manejo del cultivo se realizaron tal y como se encuentra descrito
en Ceballos et al., (2013) con una densidad de siembra de 10.000 plantas*ha-1.
La fertilización se aplicó a los 45 y 90 dds según los requerimientos del cultivo, los
contenidos iniciales de nutrientes en el suelo y la eficiencia de la fertilización en la zona.
Las plantas recibieron 8,4 g/planta de fosfato di-amónico (DAP), 4,1 g/planta de Kieserita
(fertilizante con 3% de potasio soluble, 24% de magnesio y 19% de sulfuro), 5,4 g/planta
de cloruro de potasio (KCl) y 2,2 g/planta de Vicor (fertilizante granular con 15% de
nitrógeno, 5% de calcio, 3% de magnesio, 2% de sulfuro, 0,02% de boro, 0,02% de
cobre, 0,02% de molibdeno y 2,5% de zinc).
Figura 1. Distribución de cinco bloques perpendiculares a la pendiente (Blanco) y efecto borde (Amarillo) en el
experimento de dosis. X: Plantas de yuca var. MCOL2737 para disminuir el efecto borde. O: Plantas tratamiento
var. MCOL2737 que fueron inoculadas una o dos veces con ocho concentraciones diferentes.
La inoculación con todas las concentraciones se realizó a los 20 dds para las plantas con
una sola aplicación y, adicionalmente, se realizó otra a los 40 dds para aquellas con
doble inoculación. Se tomaron diferentes volúmenes del producto comercial y se
diluyeron en 10 ml de agua destilada con el fin de obtener las concentraciones requeridas
para cada planta. Las plantas no inoculadas recibieron la misma cantidad de agua. Los
10 ml del inóculo fueron aplicados sobre la raicillas de la planta luego de levantar
levemente el cangre sembrado.

Capítulo 1 21
Medición del crecimiento en plantas y hongos
Para obtener la biomasa de la parte aérea, todas las plantas fueron cortadas 20 cm por
encima de la superficie del suelo a los 340 dds. La biomasa fresca aérea fue pesada
directamente en campo. El material vegetal fue recolectado en costales y secado en
horno a 70ºC hasta peso constante (Aprox. 49 h.). A los 360 dds, las raíces fueron
cosechadas y pesadas en campo para medir la producción en peso fresco por planta. El
peso seco de las raíces fue calculado utilizando el método de gravedad específica
propuesto por Toro & Cañas, (1983) y validado por el CIAT (Aristizábal et al. 2007).
La colonización total de HFMA fue medida en raíces de menos de 2 mm de diámetro a
los 12 meses después de siembra. Las estructuras fúngicas fueron visualizadas con tinta
negra Scheffer (Vierheilig et al. 1998). El porcentaje de colonización en las raíces fue
calculado con el método de intersección de la grilla (Giovannetti & Mosse 1980) para
cada uno de los tratamientos.
1.2.3 Análisis estadísticos
Los datos de todos los experimentos fueron analizados con el programa estadístico
JMP® (Statistical Analysis Systems Institute, version 10). Para evaluar diferencias
significativas entre los tratamientos en el beneficio de la inoculación (BI), la cantidad de
raíces producidas, el diámetro ecuatorial, la biomasa seca y fresca y la colonización, se
realizaron análisis de varianza (ANOVA) de una y dos vías. Los supuestos de normalidad
y homoceasticidad en los datos fueron verificados antes del análisis. Luego de encontrar
diferencias significativas en los ANOVAS, se realizaron pruebas de Tukey o de t de
Student para determinar las diferencias entre los tratamientos con un nivel de
significancia de α = 0,05.
Cuando se cumplieron los supuestos de normalidad pero no los de homoceasticidad, se
corrieron ANOVAS sólo en los casos en que el número de individuos para cada grupo
fueran similares (es decir cuando el numero de muestras del grupo mayor no fueron 1 ½
más que el número de muestras del grupo menor) y cuando las desviaciones estándar
entre los grupos de tratamientos no fueron mayores a un 20% (Dai 2009). En algunas

22 Efecto de un inóculo comercial
variables, como en el caso de la colonización, se realizó una tranformación Arco seno de
los datos.
El grado de relación entre dos variables se determinó con el coeficiente de correlación de
Pearson. Se buscaron modelos de ajuste para entender la relación entre algunas
variables. La selección de los modelos se realizó teniendo en cuenta la medición del
mejor ajuste utilizando el programa JMP®.
1.3 Resultados
1.3.1 Ciclo fenológico del cultivo de yuca en la zona de estudio
En la caracterización del ciclo del cultivo de la yuca variedad MCOL2737 (Tabla 1) se
identificaron tres fases fenológicas: 1) La fase de formación de raíces tuberosas,
caracterizada por el incremento en el número de raíces, la cual finalizó antes de 112 días
después de siembra (dds). 2) La fase de endurecimiento de raíces, caracterizada por la
tasa máxima en el incremento del diámetro ecuatorial de la raíz luego de su formación.
Esta fase comenzó a los 112 dds y finalizó a los 151 dds. 3) La tasa de acumulación de
biomasa seca en raíces, caracterizada por la tasa máxima en el incremento de la
biomasa seca en raíces y la alta pérdida de área foliar. Comenzó a los 151 dds y terminó
en la cosecha.
La biomasa seca total de la planta fue incrementando con el tiempo (Figura 2). Hubo una
estabilización de la biomasa aérea luego de los 190 dds, mientras que la biomasa de
raíces siguió incrementando. Estos valores y los cambios en la relación biomasa parte
aérea/raíces a lo largo del ciclo del cultivo, muestra una dinámica en la distribución de la
biomasa de la planta con el tiempo (Tabla 1).

Capítulo 1 23
Tabla 1. Fases fenológicas del cultivo de yuca var. MCOL2737 en Yopal (Casanare) para plantas no inoculadas. Fertilización como es normalmente
aplicada en la zona.
Tiempo de
muestreo
Días
después de
siembra
Cantidad de
raíces
Perímetro
ecuatorial (cm)
Peso seco
de raíces (g)
Relación
Biomasa parte
aérea/Biomasa
raíces
Área Foliar (cm2) Fases fenológicas
de la raíces
T1 70 7,0 ± 1,15 b 6,7 ± 0,5 f 10,9 ± 2,5 c 25,20 ± 0,02 a 5.847,7 ± 776,3 b Formación de raíz
T2 112 9,2 ± 1,3 a 17,5 ± 0,7 e 293,2 ± 42,8 bc 1,22 ± 0,07 b 20.177,8 ± 2300,4 b Endurecimiento de
raíces
T3 151 8,0 ± 1,1 a 27,7 ± 0,8 d 438,8 ± 58,2 b 1,59 ± 0,05 b 41.701,8 ± 6863,7 a
T4 190 9,2 ± 0,9 a 33,4 ± 0,9 c 1085,8 ± 92,34
a
0,76 ± 0,12 b 38.114,0 ± 4104,7 a Acumulación
máxima de biomasa
seca de raíces
T5 237 9,41 ± 0,6 a 42,0 ± 0,9 b 1066,3 ± 156,3
a
1,19 ± 0,10 b 9.772,4 ± 923,1 b
T6 277 10,9 ± 1,16 a 39,9 ± 1,0b 1223,4 ± 106,5
a
0,87 ± 0,07 b 7.924,9 ± 1202,8 b
T7 320 9,4 ± 1,0 a 40,8 ± 1,2 b 1434,1 ± 72, 8 a 0,76 ± 0, 13 b 19.161,2 ± 3919, 5 b
a,b,c,d,e,f Los intervalos representan los errores estándar de la media. Letras diferentes representan diferencias significativas en los diferentes muestreos para un p
< 0,05

24 Efecto de un inóculo comercial
Figura 2. Biomasa seca total (verde), de la raíz (azul) y de la parte aérea de la planta (rojo) a lo largo del ciclo del
cultivo de yuca en Yopal. Cada barra de error corresponde a un error estándar de la media. T1: 70 dds; T2: 112
dds; T3: 151 dds; T4: 190 dds; T5: 237 dds; T6: 277 dds; T7: 320 dds.
1.3.2 Efecto de un producto comercial de Rhizophagus
irregularis sobre el crecimiento de las plantas de yuca var.
MCOL2737 y sobre el rendimiento del cultivo bajo tres
niveles de fertilización fosfatada
Los resultados de esta sección se encuentran publicados en Ceballos et al., 2013. Se
encontró que la inoculación con un aislado de Rhizophagus irregularis producido in vitro
aumentó significativamente el rendimiento del cultivo de yuca var. MCOL2737 en Yopal,
produciendo rendimientos mayores a los esperados para la región. El aumento en la
cosecha fue de un 18% más con respecto a las plantas que no fueron inoculadas. De
todas las variables evaluadas en el momento de la cosecha sólo el peso de las raíces fue
afectado por la inoculación (Ceballos et al., 2013). En Yopal, los mayores rendimientos
se obtuvieron en plantas inoculadas independientemente del nivel de fósforo aplicado
(P). Sorprendentemente, las plantas inoculadas y fertilizadas con 0% P generaron
rendimientos estadísticamente iguales a aquellos generadas por plantas que no fueron

Capítulo 1 25
inoculadas pero que fueron fertilizadas con el 50 % y el 100 % de la fertilización fosfatada
que es utilizada en la región.
Analizando los datos obtenidos para todos los muestreos, se encontró que la producción
de raíces y la biomasa total de la planta se afectaron por la inoculación, y este efecto fue
dependiente del tiempo de muestreo (ver Anexo C: numerales 9.1, 9.2; Figura 3).
Cuando se analizó muestreo por muestreo se estableció de forma detallada la dinámica
del crecimiento de la planta a través del tiempo para los diferentes niveles de fertilización
fosfatada. La Figura 3, muestra que los comportamientos del crecimiento de los
diferentes órganos son similares en plantas inoculadas y no inoculadas. La inoculación
generó efectos significativos en la producción de biomasa de algunas partes de la planta
dentro de cada uno de los muestreos y este efecto dependió del nivel de fertilización que
la planta recibió. Los efectos significativos de la inoculación que generaron disminución
de biomasa se observaron al inicio del ciclo del cultivo sobre variables relacionadas con
el crecimiento y la capacidad fotosintética de la planta. Los efectos significativos de la
inoculación que generaron aumento, por el contrario, se observaron sólo al final del
cultivo y se asociaron con la producción de raíces (Figura 3).
1.3.3 Beneficio de la inoculación a lo largo del ciclo del cultivo de
yuca var. MCOL2737 en Yopal
El beneficio de la inoculación (BI) fue calculado en términos de la biomasa seca total de
la planta y de sus órganos para todos los tratamientos y en todos los muestreos del
experimento establecido en Yopal por Ceballos et al., (2013) . En la Figura 4 se muestran
los resultados de la dinámica del BI a lo largo del ciclo del cultivo para las plantas que
fueron inoculadas y fertilizadas con 100 Kg*ha-1 de fosfato di-amónico (50% de la
cantidad de fertilizante que normalmente aplican los agricultores de la zona).

26 Efecto de un inóculo comercial
Figura 3. Variables de crecimiento y producción para plantas de yuca con inóculo (azul) y sin inóculo (rojo) bajo tres niveles de fertilización fosfatada.
Las barras de error representan el error estándar de la media.*: diferencias significativas entre plantas inoculadas y no inoculadas para un nivel de
significancia de 0,05 (negro) 0,1 (negro y gris).

Capítulo 1 27
Figura 4. Beneficio de la inoculación (BI: barras de color naranja) en términos de biomasa seca total de la planta a
lo largo del ciclo del cultivo. Línea azul: precipitación acumulada (mm). Línea verde punteada: Colonización total
de HFMA en raíces de plantas inoculadas. Eje X: Días después de siembra (dds) y fases fenológicas de las raíces
de yuca (Tabla 1). Las barras de error representan ± 1 error estándar de la media. Letras diferentes encima de las
barras representan diferencias significativas para un p < 0,05. Diagrama: Cristhian Fernández.
El BI cambio a lo largo del tiempo (Figura 4). Su valor fue negativo durante el período de
0 a 151 dds, definiéndose así, una fase de depresión de crecimiento por la inoculación
como respuesta en la simbiosis. Luego, el valor de BI se volvió positivo hasta el momento
de la cosecha (320 dds) definiéndose así, la fase de promoción de crecimiento en la
simbiosis debido a la inoculación del producto comercial.
La dinámica del BI para las plantas que recibieron el 50% de la fertilización fosfatada en
el tiempo se ajustó a un modelo polinomial de grado tres: BI= -0,581626 + 0,150756*t -
0,0219199*(t - 4)^2 - 0,0074729*(t - 4)^3 (Anexo C: numeral 9.3). Donde BI: Beneficio de
la inoculación y t: Tiempo de muestreo. Para este modelo, el valor en donde el BI es cero

28 Efecto de un inóculo comercial
y a partir del cual el efecto de la simbiosis es positivo (t = 3,86) fue un punto cercano al
cuarto muestreo. Se obtuvo un punto máximo en la función (t = 5,79) que corresponde al
máximo beneficio de la inoculación cerca del muestreo 6 (Anexo C: numeral 9.3).
De acuerdo con el ciclo fenológico del cultivo (Tabla 1), la disminución en la biomasa seca
total de la planta y la de los órganos no fotosintéticos (biomasa seca de tallo y raíces -
datos no mostrados) se presentó a lo largo de la etapa de formación de raíces y durante
la mayor parte de la etapa de endurecimiento. La fase de promoción de crecimiento
comenzó simultáneamente con la acumulación de biomasa seca de raíces (Figura 4).
Adicionalmente, se encontró una correlación significativa entre el BI y dos variables
ambientales: La precipitación acumulada (Correlación = -0,8064; valor p = 0,04999*) y la
presión barométrica del aire (Correlación = - 0,9329; valor p= 0,0066*). En la Figura 4, se
puede observar que la fase de promoción de la simbiosis con la aplicación del inóculo
coincide con el inicio del período de sequía en la región.
En los otros niveles de fertilización fosfatada también se encontraron cambios del BI a
través del tiempo, pero los cambios (depresión y promoción) en el patrón de crecimiento
a lo largo del ciclo del cultivo, fueron diferentes dependiendo del nivel de fósforo aplicado
(Anexo C: numeral 9.5). Los niveles de fertilización afectaron el beneficio de la
inoculación cuando se analizaron las plantas de todos los muestreos en conjunto
(ANOVA F ratio = 7,4811, p ≤ 0,0012). Sin embargo, se encontró que sólo en los dos
primeros muestreos, hubo diferencias significativas en el beneficio de la inoculación entre
las plantas con diferentes niveles de fertilización. Luego, este beneficio fue el mismo
independientemente del nivel de fósforo aplicado (Figura 5).
Los porcentajes de raíz colonizada por HFMA en la mayoría de los muestreos fueron los
mismos para las plantas inoculadas y las control. La colonización total de HFMA en las
raíces de las plantas inoculadas y no inoculadas fue dinámica en el tiempo (Figura 4).
Durante el ciclo del cultivo, esta variable cambió del 40% al 80 % siguiendo un patrón
similar al del beneficio de la inoculación (Anexo C: numeral 9.4). El modelo de mejor
ajuste para la colonización en el tiempo también fue un polinomio de tercer grado: Col =
49,765873 - 0,1438492*t + 1,421131*(t - 4)^2 + 0,7673611*(t- 4)^3. Donde Col:

Capítulo 1 29
Colonización total de HFMA en raíces de yuca y t: Tiempo de muestreo. No se encontró
una correlación entre la colonización y el BI.
Figura 5. Beneficio de la inoculación (BI) en términos de la biomasa seca total de las plantas para cada uno de los
muestreos a lo largo del ciclo fenológico de la planta, con diferentes niveles de fertilización fosfatada. Las barras
de error representan ± 1 error estándar de la media. Letras diferentes encima en las barras representan diferencias
significativas entre los tres niveles de fósforo para un p < 0,05

30 Efecto de un inóculo comercial
1.3.4 Efecto de la dosis del inóculo comercial de Rhizophagus
irregularis sobre la producción de las plantas de yuca var.
MCOL2737 en Yopal.
Las concentraciones del inóculo alteraron la producción de las raíces en la variedad de
yuca MCOL2737 (ANOVA F ratio = 6,4997, p ≤ 0,0001) y la producción de biomasa seca
total de la planta (ANOVA F ratio = 2,5407, p ≤ 0,0233) en el momento de la cosecha
(Figura 6). Pero la cantidad de aplicaciones del inóculo no tuvo un efecto sobre estas
variables (Anexo D: numerales 10.2, 10.3). La biomasa aérea de la planta no se vio
afectada por la dosis del inóculo (Anexo D: numeral 10.1). De forma interesante, las
concentraciones de inóculo menores (25%, 50% y 75%) que la recomendada por la casa
comercial para Europa (100%) generaron las mayores producciones de raíces (Figura 6).
La aplicación con el 50% de lo recomendado incrementó en promedio en casi dos veces
la producción de raíces.
Figura 6. Efecto de la concentración (%) de inóculo sobre la biomasa fresca de las raíces (A) y de toda la planta
(B). Las barras representan ± 1 error estándar de la media. Letras diferentes encima de las barras representan
diferencias significativas para un p < 0,05.
La producción de raíces y de biomasa en plantas de yuca de la variedad MCOL2737 se
ajustaron a modelos polinomiales de grado cuatro cuando se varía la concentración del
inóculo comercial (Anexo D: numerales 10.4, 10.5). Esto mostró que no hubo una

Capítulo 1 31
correlación lineal entre la concentración de inóculo y la producción de raíces o de
biomasa en la planta. Los modelos de ajuste para estas dos variables presentaron puntos
máximos en las concentraciones cercanas al 50% para ambos casos (Anexo D:
numerales 10.4, 10.5; Figura 6).
La colonización total de HFMA en las raíces de la yuca MCOL2737 evaluada en el
momento de la cosecha, por el contrario, tuvo un efecto debido a la dosis (ANOVA F ratio
= 8,9395, p ≤ 0,0043) pero no a la concentración (Anexo D: numeral 10.6). Las plantas
que tuvieron doble inoculación presentaron mayores porcentajes de colonización (74,15 ±
3,14 %) que las que recibieron sólo una dosis (59,58 ± 3,26 %). Sin embargo, esta
variable no tuvo correlación con el peso fresco ni de las raíces producidas ni de la planta.
1.3.5 Estudio económico teniendo en cuenta el efecto de las
dosis de Rhizophagus irregularis en la producción de yuca
var. MCOL2737 en Yopal
La concentración de inóculo utilizada en el experimento de Ceballos et al., (2013)
correspondió al 200% de la recomendada por la casa comercial en Europa. Teniendo en
cuenta los precios europeos del inóculo, la concentración utilizada generó costos de 1.3
millones de pesos Colombianos por hectárea (Ceballos et al., 2013). El retorno de la
inversión (ROI) obtenido para esta concentración fue de 117% y esta representado por la
letra A en la Figura 7. Este ROI es menor que el obtenido cuando se aplicó el 50% del
fertilizante fosfatado sin HFMA (ROI=126,8%, línea roja en Figura 7). Con la
concentración del inóculo recomendada por la casa comercial (concentración = 100%) se
produjo la misma cantidad de yuca fresca (Figura 6) y su ROI (135%) fue mayor que el
obtenido para el tratamiento sin HFMA (Figura 7). Sin embargo, al aplicar el 25% de la
concentración recomendada, se obtuvo la misma producción que con la dosis
recomendada (Figura 6) y el ROI para este caso (150%) fue más del 20% que el del
tratamiento sin inoculación.

32 Efecto de un inóculo comercial
Figura 7. Retorno de la inversión (ROI) obtenido por la venta de yuca fresca del cultivo establecido en Yopal por
Ceballos et al, 2013 para la concentración de inóculo real utilizada (A); la concentración de inóculo recomendada
por la casa comercial (B); y el 25% de la concentración de inóculo recomendada por la casa comercial (C). La línea
verde diagonal representa el ROI para el tratamiento con HFMA mas rentable y depende de la cantidad de inóculo
aplicado. La línea roja horizontal representa el ROI para el tratamiento mas rentable sin HFMA.
1.4 Discusión
Existen estudios que muestran resultados de la inoculación con HFMA sobre cultivos en
campo, medidos en diferentes tiempos (Quiñones-Aguilar et al. 2014; Shamshiri et al.
2012; Solaiman & Hirata 1997; Wiseman & Wells 2009). Sin embargo, la cantidad de
muestreos y las variables evaluadas en este estudio permitieron caracterizar la dinámica
del efecto de la inoculación sobre el crecimiento de las plantas de yuca y su
productividad en campo a lo largo de todo el ciclo del cultivo. Investigaciones como ésta,
en sistemas complejos y a través del tiempo, son fundamentales para mejorar la
comprensión sobre el funcionamiento de estos sistemas y para que los factores
ambientales puedan ser integrados en el análisis de la funcionalidad de la simbiosis
micorrícica (Johnson et al. 1997).

Capítulo 1 33
Las principales discusiones de este capítulo se presentan en Ceballos et al., (2013), pero
a continuación se complementa la discusión.
Las mediciones de las variables de crecimiento de las plantas de yuca variedad
MCOL2737, permitieron precisar las etapas del ciclo fenológico con respecto a las
presentadas por Cadavid (2005), teniendo en cuenta el genotipo de la planta y los
factores del clima de la región (Tabla 1). Las etapas del ciclo, presentadas en la Tabla 1, se
pueden generalizar para plantas con y sin inoculación y con diferentes niveles de
fertilización, debido a que las fases fenológicas encontradas y los tiempos en los cuales
éstas ocurrieron fueron iguales para todos los casos (datos no mostrados). La
descripción del ciclo del cultivo de la yuca, var. MCOL2737, no sólo es importante para
establecer estrategias de manejo efectivas, sino también para conocer los momentos en
los cuales ocurren cambios fenológicos en las plantas de yuca bajo estas condiciones
ambientales.
La aplicación de Rhizophagus irregularis producido in vitro conlleva a una mayor
producción de yuca y a disminuir la aplicación de fertilizantes fosfatados en Yopal
La aplicación de un aislado de Rhizophagus irregularis en cultivos de yuca en Yopal
mostró un gran potencial para aumentar la producción y disminuir la aplicación de
fertilizantes fosfatados en este lugar. Con estos resultados se mejora el aprovechamiento
de los recursos en los suelos del trópico como, por ejemplo, el fósforo. Los resultados
mostraron que la inoculación con HFMA permitió que con sólo la mitad de los fertilizantes
fosfatados aplicados en la región para los requerimientos del cultivo se obtuvieron la
misma producción que aplicando el 100% de los fertilizantes. Esto es importante,
especialmente en el trópico, ya que permite incrementar la productividad de los cultivos a
menores costos y mejora la competitividad de los cultivos de una forma sostenible.
Adicionalmente, como R. irregularis se puede cultivar in vitro, esto permite que se puedan
producir inóculos puros y concentrados con este HFMA para hacer aplicaciones en
cualquier sistema agrícola.

34 Efecto de un inóculo comercial
El beneficio de la inoculación es dinámico a lo largo del ciclo del cultivo de yuca en
Yopal
El beneficio de la inoculación sobre la biomasa total de las plantas fue diferente a lo largo
del ciclo del cultivo de la yuca sembrado en Yopal, y también, para las plantas que
recibieron diferentes niveles de fertilización fosfatada (Figura 4, Figura 5). Estas
diferencias se pueden explicar porque la funcionalidad de la simbiosis esta mediada por
efectos directos e indirectos de factores bióticos y abióticos en los alrededores de la
rizósfera de la planta, del cultivo e inclusive del ecosistema (Johnson et al. 1997;
Johnson & Graham 2013; Feddermann et al. 2010).
La depresión en el crecimiento de las plantas de yuca meses después de la inoculación y
el efecto retardado de la promoción en su crecimiento ya ha sido reportado para otras
plantas (Bethlenfalvay et al. 1982; Koide 1985). El costo energético que tiene la planta
por el carbono que entrega al hongo, puede ser mayor que el costo resultante por
producir mayor biomasa (representada en mayor área foliar o mayor cantidad de raíces)
en ese momento o para esa condición específica. Pero esas pérdidas en biomasa a corto
plazo por el establecimiento de la simbiosis le puede representar a la planta beneficios a
largo plazo. Los resultados encontrados corroboran lo afirmado por Johnson et al. (1997),
donde se plantea que el balance entre los costos y los beneficios netos es dinámico en el
establecimiento de una asociación micorrícica y depende de la interacción con el
ambiente.
Es común encontrar que los niveles de fertilización fosfatada tengan un efecto sobre el
beneficio de la simbiosis, y esto, ya ha sido reportado para cultivos de yuca en Colombia
(Howeler & Sieverding 1983). Cuando las plantas crecen en suelos con una mayor
cantidad de fósforo, los costos energéticos para establecer la simbiosis pueden ser
mayores que los beneficios nutricionales que recibe la planta por la misma simbiosis
(Smith et al. 2003; Grace et al. 2009; Johnson et al. 1997). Sin embargo el estudio del
sistema de una forma dinámica permitió evidenciar que el efecto de la fertilización sobre
el beneficio de la simbiosis no es constante durante todo el ciclo del cultivo de la yuca
(Figura 5). Para el caso del experimento establecido por Ceballos et al., (2013), esos
costos sólo se reflejaron en los dos primeros muestreos, cuando probablemente se

Capítulo 1 35
estaba dando el establecimiento de la simbiosis entre la planta y el hongo inoculado. En
los últimos cinco muestreos, el nivel de fósforo aplicado no generó efecto alguno sobre el
beneficio de la simbiosis (Figura 5). Estos resultados invitan a reflexionar sobre la
importancia en la selección del tiempo de muestreo, para evaluar el efecto de una
simbiosis, teniendo en cuenta el objetivo de la investigación. De la misma forma, se debe
tener cuidado al hacer conclusiones sobre efectos encontrados en una etapa
determinada sin tener en cuenta la dinámica del sistema.
Los porcentajes de raíz colonizada por HFMA iguales en plantas inoculadas y control,
demuestran que la inoculación no tuvo un efecto sobre esta variable en casi ningún
muestreo. Sin embargo, como la metodología de tinción utilizada no permite identificar
los hongos que están involucrados en la colonización, explicar los patrones que siguen
los HFMA involucrados a lo largo del ciclo del cultivo no fue posible. Incluso, las
herramientas moleculares existentes no tienen el alcance aún para explicar esa dinámica
en un sistema establecido en campo. Puede que la colonización total en la raíz no se
afecte, pero en realidad la estructura de la comunidad si tenga cambios significativos que
estén asociados con la respuesta del crecimiento de la planta.
La aplicación de Rhizophagus irregularis para la producción de yuca es viable en la
región
El estudio de dosis permitió identificar que para obtener los mejores rendimientos de la
variedad de yuca MCOL2737 en la zona se puede hacer una sola aplicación del producto
en una concentración de 25 propágulos por planta. Aunque la casa comercial que
produce R. irregularis in vitro, recomienda aplicar este producto en una concentración de
250 propágulos por planta para hortalizas en Europa, se encontró que aplicando sólo el
25% de lo recomendado se obtienen las mismas producciones. Los resultados
encontrados reiteran la necesidad de ajustar las dosis de inoculantes dependiendo del
sistema en donde serán aplicados, pues en muchas ocasiones lo recomendado no va a
representar la condición ideal. La selección adecuada de la dosis permite tener una
mayor eficiencia en el uso de insumos y en los costos de producción como se explica a
continuación.

36 Efecto de un inóculo comercial
Para hacer el análisis económico presentado en Ceballos et al., (2013), el costo del
inóculo se calculó para el doble de la cantidad recomendada por la casa comercial, ya
que esto fue lo que se aplicó en los experimentos. El costo del producto aplicado, el cual
representó entre un 12 y 18% de los costos totales, fue determinante en la viabilidad
económica de esta aplicación (Ceballos et al., 2013). Por esta razón, y teniendo en
cuenta que el costo esta directamente relacionado con la cantidad de inóculo aplicada, se
estableció un experimento en campo para evaluar el efecto de las dosis del inóculo
comercial sobre la producción de plantas de yuca var. MCOL2737.
Los resultados del experimento de dosis mostraron que la aplicación de R. irregularis
para la producción de yuca en la zona es viable. En términos económicos, el retorno de
la inversión de un agricultor que aplique el 25% de la dosis recomendada, se incrementa
en un 10% comparado con uno que aplique el 100%. Pero además, cuando un agricultor
aplica el 25% de la dosis recomendada, es decir 62 propágulos por planta, su retorno de
la inversión es mayor que cuando no aplica inóculo.
Con estos resultados se muestra el potencial que tiene la aplicación de este aislado de R.
irregularis en los suelos de la zona de estudio, no sólo para incrementar los rendimientos
de yuca y para disminuir la aplicación de fertilizantes en estos cultivos, sino también para
mejorar el ingreso de los agricultores de este producto en Yopal.
1.5 Conclusión
La inoculación de un aislado de Rhizophagus irregularis producido en condiciones in vitro
aumentó significativamente los rendimientos y disminuyó la aplicación de los fertilizantes
fosfatados en un cultivo de yuca de la variedad MCOL2737 sembrado en Yopal (Ceballos
et al. 2013). Estos resultados implican mejoras en la productividad y confirman el
potencial que tienen algunos aislados de R. irregularis para mejorar la producción de
cultivos de interés agronómico en suelos ácidos del trópico como el de Yopal, donde se
limita la producción de la mayoría de las plantas y donde la aplicación de grandes
cantidades de fertilizantes generan problemas económicos y ambientales. Esto además,
puede ayudar a mejorar significativamente la economía de los agricultores en la zona
teniendo en cuenta que pueden obtener incrementos significativos en la producción de

Capítulo 1 37
yuca utilizando ciertos aislados de R. irregularis producidos in vitro incluso cuando los
fertilizantes escaseen o tengan fluctuaciones fuertes de su precio en el futuro.
Adicionalmente, la aplicación de este aislado de R. irregularis para producir yuca de la
variedad MCOL2737, en la zona, es económicamente viable cuando se utilizan
concentraciones del inóculo menores o iguales a 250 propágulos por planta. Con el uso
de este tipo de tecnología, se puede mejorar entonces la rentabilidad del sistema del
cultivo de la yuca en la región. En Colombia, el cultivo de yuca tradicionalmente es una
fuente importante de alimentación y de ingresos de los agricultores, y es indispensable,
mejorar la competitividad de este cultivo para convertirla en una fuente de recursos
atractiva en el mercado de los carbohidratos.
Por otra parte, se encontró que el beneficio de la inoculación con este aislado sobre la
biomasa total de las plantas de yuca fue diferente a lo largo del ciclo del cultivo. Hubo
una disminución en la biomasa de la planta meses después de la inoculación, la cual se
observó en las raíces y la parte aérea de las plantas, afectando directamente el
crecimiento y el área foliar. Al finalizar el ciclo, hubo una promoción en el crecimiento que
se observó únicamente en la biomasa de las raíces beneficiando directamente la
producción del cultivo. Este beneficio también dependió de los niveles de fertilización
fosfatada que recibieron las plantas, pero sólo en los primeros muestreos. Con esto se
demuestra la importancia de considerar la simbiosis entre esta especie de HFMA y esta
variedad de yuca como un sistema dinámico que responde a las condiciones del
ambiente.
Finalmente, el efecto que genera un aislado de R. irregularis en la producción de yuca y
en la aplicación de fertilizantes en este tipo de suelos, lo convierten en un candidato
potencial para un programa de selección de genotipos de esta especie de HFMA en la
búsqueda de líneas mejoradas del hongo que tengan un efecto deseado en la planta.


2 Capítulo 2: Funcionalidad de la simbiosis
entre genotipos de Rhizophagus
irregularis y variedades de yuca
2.1 Introducción
La funcionalidad de la simbiosis se define como el efecto que genera una determinada
combinación entre planta y hongo/s dentro de una relación simbiótica (Feddermann et al.
2010). Este efecto puede ser medido en la planta o en el hongo de acuerdo a los
intereses del investigador.
Las variaciones en la funcionalidad de la simbiosis entre hongos formadores de
micorrizas arbusculares (HFMAs) y plantas puede depender de diferencias genotípicas
entre los simbiontes que se expresan en un ambiente determinado (Koch et al. 2006). Se
ha reportado que distintas especies de HFMA alteran el crecimiento de las plantas de
una forma diferente (Feddermann et al. 2010; Munkvold et al. 2004; Smith et al. 2004),
pero también, que las especies vegetales responden de manera desigual cuando son
inoculadas por la misma especie de hongo FMA (Angelard et al., 2010), incluso al nivel
de variedad (Howeler & Sieverding 1983). Esto demuestra que no existe un inóculo
universal de HFMA que pueda ser efectivo para todas las especies vegetales, y
consecuentemente, para todos los cultivos (Sanders 2010; Sanders & Rodriguez 2016).
Munkvold et al., (2004) encontraron que la variabilidad intra-específica de los HFMA era
muy importante en términos de la funcionalidad de la simbiosis. Cuando inocularon
aislados de HFMA de una misma especie, observaron que ellos producían diferencias en
el crecimiento del pepino y en la toma de nutrientes por la planta, y en algunos casos,
esas diferencias fueron mayores que las obtenidas entre las especies de los hongos
evaluados. También se encontró que aislados de una población de R. irregularis y

40 Diversidad intra-específica y funcionalidad de la simbiosis vares de yuca
provenientes de una misma localización geográfica no sólo presentaron diferencias
genéticas y fenotípicas entre ellos (Koch et al. 2004), sino que también produjeron
diferencias en el crecimiento de Plantago lanceolata en invernadero. Estos estudios
demuestran que la amplia diversidad funcional intra-específica de los HFMA, es un rasgo
característico de estos hongos y de gran relevancia para la aplicación práctica de esta
simbiosis mutualista en la agricultura.
Adicionalmente, en estudios recientes, se encontró que líneas de Rhizophagus
irregularis, alteraron significativamente el crecimiento de las plantas cultivadas en
sistema axénico (Koch et al. 2004) y en invernadero (Angelard et al. 2010). Estas líneas
genéticas se produjeron por medio del cultivo en laboratorio de aislados del hongo.
Teniendo entonces la posibilidad de crear líneas mejoradas del hongo en laboratorio,
gracias a la alta variabilidad genética que presenta R. irregularis, se abren preguntas
como: ¿Es esta variabilidad genética intra-específica suficiente para promover diferencias
en campo? Esto, evidencia la importancia de estudiar estas variables en campo a un
nivel ecológico para obtener resultados en cultivos de interés dentro de un sistema y
ambiente definidos.
El objetivo de este capítulo fue comparar la funcionalidad de la simbiosis entre genotipos
de R. irregularis y variedades de yuca en términos de la producción de las plantas y la
colonización de los HFMA. Un experimento en campo fue establecido en Yopal en dos
años diferentes para cumplir con el objetivo general y además para: 1) Comparar el
efecto entre las líneas genéticas de R. irregularis o entre grupos de éstas y 2) Evaluar el
efecto de las líneas del hongo en la calidad de las raíces, en los niveles de supervivencia
y en el porcentaje de colonización total radical de HFMA en las plantas de cada variedad.
2.2 Metodología
Los resultados del experimento utilizando inóculo comercial (Capítulo Uno), permitieron
establecer los tiempos de medición, los niveles de fertilización fosfatada y las variables
agronómicas relevantes, para evaluar el efecto del inóculo con R. irregularis en un cultivo
de yuca en la zona de estudio. Teniendo en cuenta estos resultados, los experimentos de
este capítulo se diseñaron bajo estas condiciones: 1) Se realizó la aplicación del 50% de
la fertilización fosfatada, ya que este nivel de fósforo produjo los mismos rendimientos

Capítulo 2 41
que los obtenidos con la aplicación del 100% de los fertilizantes cuando se inoculó el
HFMA (Ceballos et al. 2013); 2) El producto comercial de R. irregularis se empleó como
un control positivo para la variedad de yuca MCOL2737 y 3) Las mediciones para evaluar
la funcionalidad de la simbiosis se realizaron únicamente en el momento de la cosecha.
2.2.1 Lugar del experimento
Los experimentos fueron establecidos en el campus de Utopía de la Universidad de la
Salle (72° 179 4899 W, 5° 199 3199 N) en los Llanos Orientales de Colombia (Yopal,
Casanare) en dos años consecutivos (2 repeticiones). La descripción del lugar se
encuentra en Ceballos et al., (2013). Las propiedades físico-químicas del suelo del lote
donde se establecieron los experimentos para ambos años se presentan en el Anexo CB:
numerales 8.2 y 8.3. Cada montaje tuvo una duración de un año, el cual corresponde al
ciclo completo del cultivo de yuca para estas variedades en la zona.
2.2.2 Material vegetal y fúngico
En las dos repeticiones de los experimentos se sembraron tres variedades de yuca, dos
de las cuales hacen parte de la colección de recursos genéticos del Centro Internacional
de Agricultura Tropical (CIAT 2016). Las variedades fueron la MCOL2737, conocida
como Brasilera, utilizada para consumo en fresco; la CM4574, conocida como La
Cubana, utilizada para doble propósito (industrial y consumo) y la CM6438, conocida
como Corpoica Vergara, utilizada para doble propósito (industrial y consumo). Las tres
variedades fueron mejoradas para la zona de los Llanos Orientales, donde los suelos son
ácidos, hay una alta humedad relativa y una precipitación entre 1500 y 4000 mm por año.
Los clones de la variedad MCOL2737 se obtuvieron de cultivos previamente establecidos
en la Universidad de la Salle. Los clones de las otras dos variedades se obtuvieron de
cultivos de yuca de fincas experimentales del CIAT. Estas tres variedades, fueron
seleccionadas porque su crecimiento responde a la fertilización con fósforo (Calle, F,
comunicación personal. 2012, Junio 12) y a la inoculación con HFMA (Cadavid, L.F.
comunicación personal. 2011, Diciembre 8). Las semillas de estacas clonales fueron
obtenidas de la misma forma que en Ceballos et al., (2013).

42 Diversidad intra-específica y funcionalidad de la simbiosis vares de yuca
Las líneas genéticas de R. irregularis evaluadas en estos experimentos fueron cultivadas
en condiciones in vitro bajo el sistema de raíces transformadas de zanahoria con
Agrobacterium rhizogenes (Bécard & Fortin 1988). Las 15 líneas del hongo que se
inocularon en los experimentos fueron obtenidas por el grupo de investigación
colaborador “Ecología y Evolución de Organismos Simbióticos” de la Universidad de
Lausana en Suiza, en condiciones de laboratorio (Koch et al. 2004; Croll, Wille, Hannes
A. Gamper, et al. 2008; Croll et al. 2009; Angelard et al. 2010). En la Tabla 2 se muestran
algunas características de estas líneas. El material fúngico comprendió: 1) Tres líneas de
cultivos que iniciaron a partir de una sola espora de tres aislados diferentes de R.
irregularis provenientes de un campo en Suiza (Koch et al. 2004). Estas son las líneas
parentales D1, C2 y C3; y 2) Doce líneas no clonales obtenidas en el laboratorio a partir
de cultivos in vitro (Croll et al. 2009; Angelard et al. 2010) que son: S4, S4a, S4b, Sc1,
Sc1a, Sc1d, Sc1e, Sc2, Sc2a, Sc2b, Sc2c y Sc2d.
Durante el montaje de los experimentos se creía que las líneas S4, Sc1 y Sc2 eran líneas
cruzadas, pues fueron obtenidas en el laboratorio (ver Croll et al., 2009) a partir de
esporas que crecían luego de procesos de anastomosis entre líneas parentales en cultivo
in vitro (Figura 8 A). Del cultivo simultáneo de C2 y C3 se obtuvo la línea S4 y del cultivo
simultáneo de C3 y D1 se obtuvieron las líneas Sc1 y Sc2. Sin embargo, en la medida en
que las metodologías moleculares mejoraron, se encontró en las caracterizaciones
genotípicas que no había suficiente evidencia para decir que ocurría un cruce entre las
líneas parentales. Una caracterización genotípica reciente de las líneas de R. irregularis
con RadSeq realizada por el grupo de Ian Sanders, mostró que las líneas resultantes de
un cruce contenían una baja frecuencia de alelos diferentes con respecto a un parental y
alta con respecto al otro (Sanders, I. R. comunicación personal. 2015, Octubre 12).
Además, Croll et al., (2009), utilizaron marcadores genéticos nucleares diferentes con el
fin de evaluar el ADN transmitido por las líneas parentales a sus descendientes. Ellos
encontraron que la línea S4 proveniente del cruce C2 y C3, sólo mostró alelos del aislado
parental C2. Y que las líneas Sc1 y Sc2 provenientes del cruce entre C3 y D1, sólo
mostraron alelos del aislado C3. Por esta razón, en esta investigación, se definió que las
líneas S4, Sc1 y Sc2, resultantes de un cultivo simultáneo de dos líneas diferentes,
deberían ser consideradas como líneas de primera generación provenientes de la línea
parental C3 (Sanders I.R. Comunicación personal. 2015, Octubre 12).

Capítulo 2 43
Las otras líneas producidas en laboratorio se obtuvieron luego de cultivar S4, Sc1 y Sc2
(Figura 8 B). De S4 se obtuvo una espora para iniciar el cultivo de la línea S4a y otra
para el cultivo de la línea S4b; de Sc1 se tomaron otras esporas para el cultivo
monospórico de las líneas Sc1a, Sc1d y Sc1e y de Sc2 otras para el cultivo de las líneas
Sc2a, Sc2b, Sc2c y Sc2d. Estas nuevas líneas son líneas de segunda generación
provenientes de C3.
Todas las líneas empleadas en el experimento han sido caracterizadas como líneas no
clonales y sus diferencias genéticas han sido reportadas en los artículos de Angelard et
al. (2010); Croll et al. (2008) y Wyss et al. (2016).
Todas las líneas fueron crecidas en condiciones in vitro utilizando como hospedero raíces
transformadas de zanahoria con el plásmido T-DNA para obtener propágulos suficientes
en las inoculaciones (Bécard & Fortin 1988). Luego, con los propágulos de estas líneas
se prepararon formulaciones de inóculos con el mismo vehículo que el empleado para el
producto comercial evaluado en el Capítulo Uno, el cual fue uno de los controles
positivos del experimento para la variedad MCOL2737 (Cano & Vago, 2007. Patente
CSIC WO/2007/014974: 343425).
Figura 8. Cultivo simultáneo de dos esporas (líneas) (A) y cultivo monospórico in vitro (B) para producir líneas
genéticas de R. irregularis en cultivos in vitro en el laboratorio.
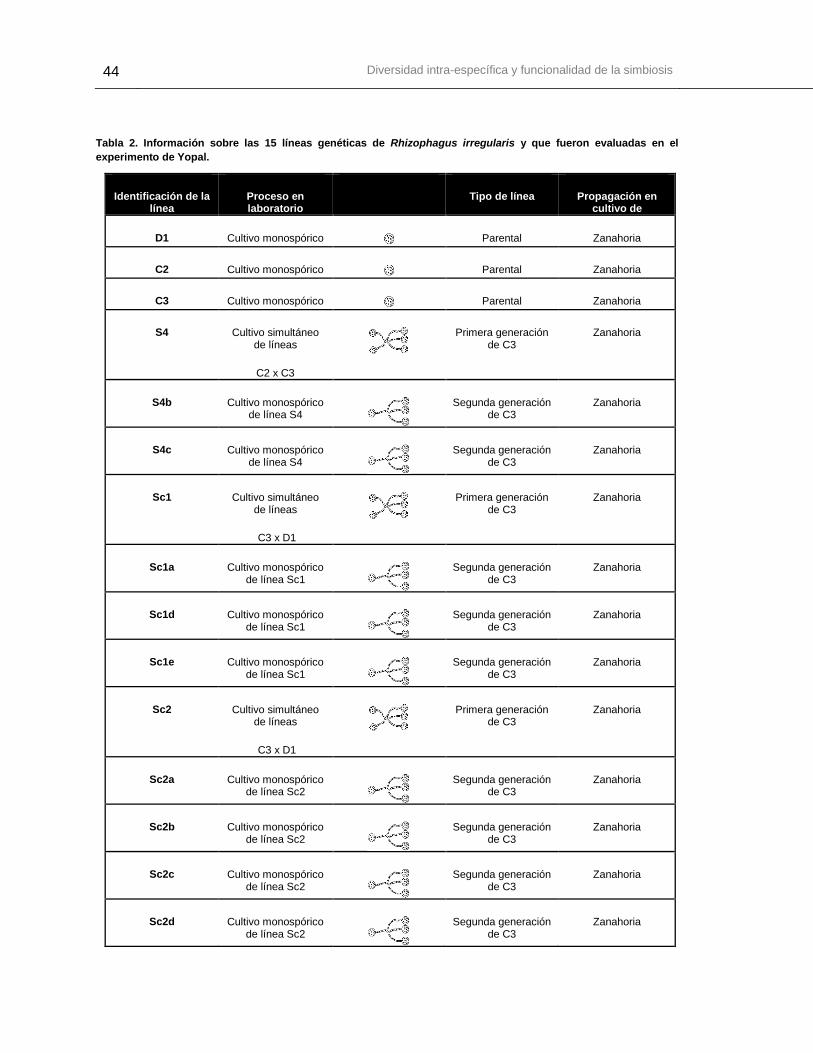
44 Diversidad intra-específica y funcionalidad de la simbiosis vares de yuca
Tabla 2. Información sobre las 15 líneas genéticas de Rhizophagus irregularis y que fueron evaluadas en el
experimento de Yopal.
Identificación de la línea
Proceso en laboratorio
Tipo de línea Propagación en cultivo de
D1 Cultivo monospórico Parental Zanahoria
C2 Cultivo monospórico Parental Zanahoria
C3 Cultivo monospórico Parental Zanahoria
S4 Cultivo simultáneo de líneas
C2 x C3
Primera generación de C3
Zanahoria
S4b Cultivo monospórico de línea S4
Segunda generación de C3
Zanahoria
S4c Cultivo monospórico de línea S4
Segunda generación de C3
Zanahoria
Sc1 Cultivo simultáneo de líneas
C3 x D1
Primera generación de C3
Zanahoria
Sc1a Cultivo monospórico de línea Sc1
Segunda generación de C3
Zanahoria
Sc1d Cultivo monospórico de línea Sc1
Segunda generación de C3
Zanahoria
Sc1e Cultivo monospórico de línea Sc1
Segunda generación de C3
Zanahoria
Sc2 Cultivo simultáneo de líneas
C3 x D1
Primera generación de C3
Zanahoria
Sc2a Cultivo monospórico de línea Sc2
Segunda generación de C3
Zanahoria
Sc2b Cultivo monospórico de línea Sc2
Segunda generación de C3
Zanahoria
Sc2c Cultivo monospórico de línea Sc2
Segunda generación de C3
Zanahoria
Sc2d Cultivo monospórico de línea Sc2
Segunda generación de C3
Zanahoria

Capítulo 2 45
2.2.3 Diseño y establecimiento del experimento en campo
El diseño experimental utilizado fue en bloques completos al azar. El efecto de la línea de
R. irregularis y la variedad de yuca fueron evaluados sobre variables agronómicas y
biológicas de las plantas. Los tratamientos incluyeron la combinación de 15 líneas de R.
irregularis (C2, C3, D1, S4, Sc1, Sc2, S4b, S4c, Sc1a, Sc1d, Sc1e, Sc2a, Sc2b, Sc2c,
Sc2d) con las tres variedades de yuca (MCOL2737, CM4574 y CM6438). Se emplearon
tres controles: 1) Un control positivo del producto comercial Glomygel®; 2) Un control
negativo de agua y 3) Un control del vehículo del gel sin propágulos. Se establecieron
nueve bloques perpendiculares a la pendiente que contenían una réplica de todos los
tratamientos y controles.
Las plantas fueron sembradas en caballones. Dos filas de plantas de la variedad
MCOL2737 fueron sembradas alrededor de cada tratamiento (efecto borde) para evitar
cruce de líneas del hongo durante el ciclo del cultivo (Figura 9). Las estacas de la yuca
fueron sembradas siguiendo la metodología empleada en Ceballos et al., (2013). En
ninguna de las repeticiones de los experimentos se aplicó irrigación artificial y el manejo
agronómico en cada caso, se realizó de acuerdo a las plagas, enfermedades e incidencia
de malezas en el cultivo.
La siembra de las plantas de la primera repetición del experimento se realizó en Octubre
del 2012. En este caso, las plantas a los 3 meses estuvieron sometidas a la sequía típica
de Diciembre - Enero en la zona y a una alta incidencia de insectos. La siembra de las
plantas para la segunda repetición se realizó en Mayo del 2013. En este caso, por el
contrario, las plantas a los 3 meses tuvieron disponibilidad de agua por las lluvias
constantes típicas de ésta época del año. De hecho, en esta segunda repetición hubo
una inundación en el cultivo a los 70 dds que afectó la supervivencia de algunas plantas
empleadas como efecto borde.
La mitad de la fertilización se aplicó a los 45 dds (días después de siembra) y la otra
mitad a los 90 dds. La cantidad de fertilizantes fue determinada de acuerdo al contenido
nutricional del suelo donde se estableció el cultivo, los requerimientos nutricionales de la
yuca y la eficiencia de la fertilización en la zona. Se fertilizó con el 50% de la cantidad de

46 Diversidad intra-específica y funcionalidad de la simbiosis vares de yuca
fosfatos requeridos por el cultivo, como resultado de lo reportado por Ceballos et al
(2013). Las plantas de la primera repetición del experimento recibieron 233 Kg ha-1 de
urea, 125 Kg ha-1 de fosfato di-amónico y 100 Kg ha-1 de cloruro de potasio (KCl). Las
plantas del experimento de la segunda repetición recibieron 84 Kg ha-1 de fosfato di-
amónico (DAP), 54 Kg ha-1 de cloruro de potasio (KCl), 41 Kg ha-1 de Kieserita
(fertilizante con 3% de potasio soluble, 24% de magnesio y 19% de sulfuro), y 22 Kg ha-1
de Vicor (fertilizante granular con 15% de nitrógeno, 5% de calcio, 3% de magnesio, 2%
de sulfuro, 0,02% de boro, 0,02% de cobre, 0,02% de molibdeno y 2,5% de zinc).
Figura 9. Distribución de tratamientos y efecto borde para los experimentos establecidos en Yopal con las líneas
genéticas de Rhizophagus irregularis y las tres variedades de yuca. Se presentan los nueve bloques (B1 – B9) con
un acercamiento que muestra la distribución de las plantas en el experimento. Las plantas encerradas en un
cuadro rosado representan las plantas tratamiento y los controles. Las plantas no encerradas representan las del
efecto borde.
Las plantas fueron inoculadas 20 dds con 500 propágulos de hongo por planta. La
aplicación se realizó cerca de las primeras raíces emergentes del cangre. Las plantas no
inoculadas recibieron la misma cantidad de agua utilizada para la dilución de las
formulaciones (10 ml).

Capítulo 2 47
2.2.4 Mediciones del crecimiento de las plantas y del hongo
Las mediciones del crecimiento de las plantas y del hongo se realizaron con las mismas
metodologías implementadas para el experimento de dosis, explicado en la página 21 del
Capítulo Uno.
Luego de evaluar todas las variables de crecimiento medidas, se realizó una
comparación entre los efectos de los grupos de líneas genéticas de R. irregularis sobre la
producción de plantas de diferentes variedades de yuca en Yopal. La variable respuesta
que se seleccionó para correr esta prueba fue el peso seco de las raíces, donde se
encontró una mayor diferencia entre los efectos de los tratamientos. De igual forma, es
importante anotar que esta variable tiene importancia en campo y en términos de
producción, ya que expresa la biomasa de raíces producida por la planta,
independientemente de la cantidad de agua presente en el ambiente durante la cosecha.
Los contrastes se hicieron entre parejas de tratamientos o sus grupos (líneas parentales
y/o líneas de primera y segunda generación producidas en laboratorio).
2.2.5 Mediciones de la calidad del almidón extraído de las raíces
de yuca
En este estudio también se evaluó el efecto de las líneas genéticas del hongo en la
calidad del almidón de dos variedades de yuca en Yopal (MCOL2737 y CM4574). Las
muestras se tomaron de la primera repetición del experimento de las líneas de R.
irregularis. Las variables medidas fueron: producción de almidón, porcentaje de pulpa en
el almidón, y acidez titulable, reportadas por el CIAT como importantes para la yuca de
consumo humano y de uso industrial. Los protocolos utilizados fueron publicados por el
CIAT en Aristizábal, Sánchez & Mejía (2007).
Las raíces de yuca fueron cosechadas al final del ciclo del cultivo y llevadas al laboratorio
para su conservación. Allí, las raíces fueron lavadas y cortadas en piezas de
aproximadamente 2 cm de grosor para luego conservar a -20ºC en bolsas de polietileno
(Sánchez & Alonso 2002).

48 Diversidad intra-específica y funcionalidad de la simbiosis vares de yuca
La extracción del almidón se realizó siguiendo la metodología presentada en Aristizábal
et al., (2007). Antes de iniciar el proceso, los pedazos de raíz fueron descongelados y
lavados nuevamente. Se removió la cáscara de las raíces y se realizó un pesaje para
obtener la biomasa fresca de las raíces de donde se obtuvo el almidón. Las raíces se
trituraron con un procesador de alimentos (Black and Decker Food Processor 110 V)
para liberar los gránulos de almidón. El triturado obtenido se lavó en un cedazo de 1,7
mm con agua, hasta que el agua saliera cristalina de nuevo. El líquido lavado de cada
muestra fue recolectado en un recipiente y se decantó por 24 horas. El agua fue retirada
y la pasta blanca decantada (almidón) fue secada por 48 horas a temperatura ambiente.
El almidón obtenido fue secado a 50ºC hasta peso constante antes de cada medición.
Contenido de almidón seco
El contenido de almidón fue calculado por el método experimental luego de la extracción.
Este valor correspondió al almidón seco extraído de una cantidad determinada de
biomasa de raíz de yuca (Aristizábal et al. 2007). Sólo se evaluaron los tratamientos que
contaron con al menos tres réplicas y las plantas que no fueron inoculadas se
consideraron como un control negativo.
Contenido de pulpa
La pulpa es el grupo de partículas de fibra (pared celular) que se encuentra en el
almidón. Para determinar el contenido de pulpa se siguió la metodología presentada en
Aristizábal et al. (2007). Dos gramos de almidón fueron disueltos en 100 ml de HCL al
0,4% v/v. Esta dilución se llevó a ebullición por una hora y se pasó por un filtro Whatman
Nº1. Para calcular la cantidad de pulpa extraída, la muestra en el filtro fue pesada y
secada a 80ºC hasta peso constante.
Acidez titulable
La acidez titulable fue calculada por la presencia de ácido láctico y fue determinada por
titulación con hidróxido de sodio (Aristizábal et al. 2007). Quince gramos de almidón se

Capítulo 2 49
mezclaron con 100 ml de agua fría (previamente hervida para eliminar CO2) y esta
solución se pasó por un filtro Whatman Nº 1. Luego, 50 ml del filtrado fueron titulados con
NaOH 0,01N usando fenolftaleína como indicador. La acidez titulable se midió en las
variedades MCOL2737 y CM4574, pero sólo en las plantas inoculadas con las líneas
parentales C2, C3 y D1.
2.2.6 Análisis estadísticos
Los datos de todos los experimentos fueron analizados con el programa estadístico
JMP® (Statistical Analysis Systems Institute, version 10). Para evaluar diferencias
significativas entre los tratamientos en el beneficio de la inoculación (BI), la cantidad de
raíces producidas, la biomasa de la planta, la calidad y cantidad de almidón y la
colonización en raíces se realizaron análisis de varianza (ANOVA) de una y dos vías. El
modelo mixto incluyó dos factores fijos (variedad de yuca y líneas de HFMA) y dos
factores aleatorios (repetición del experimento y bloques). El método utilizado para el
análisis del modelo con factores aleatorios fue el REML (Restricted Maximum Likelihood).
Cuando la variable fue evaluada en las dos repeticiones, se ejecutó el mismo modelo sin
incluir el factor tiempo dentro de ese modelo. Este valor se presenta en los resultados
como RSquare without "Time". Con esto se pudo comprobar la cantidad de variabilidad
explicada por el factor Repetición.
El efecto generado por las líneas del hongo sobre los porcentajes de supervivencia de las
plantas se realizó con un análisis de Chi-cuadrado. Se utilizaron las pruebas estadísticas
de Likelihood Ratio y Pearson para evaluar la hipótesis relacionada con las proporciones
de la población de plantas que sobrevivieron.
La comparación entre los efectos de los grupos de líneas genéticas de R. irregularis
sobre la producción de plantas de diferentes variedades de yuca en Yopal se realizó con
ANOVAS entre los grupos de tratamientos.

50 Diversidad intra-específica y funcionalidad de la simbiosis vares de yuca
2.3 Resultados
Un experimento en campo fue establecido en Yopal en dos años consecutivos para
comparar la funcionalidad de la simbiosis entre líneas genéticas de Rhizophagus
irregularis y variedades de yuca, en dos épocas diferentes. Considerando la disminución
de las réplicas en algunos tratamientos de la variedad CM6438, se decidió eliminar esta
variedad para correr los análisis del efecto de la inoculación con las líneas del HFMA
sobre las variables de crecimiento, producción y calidad de las raíces de las plantas. Los
datos obtenidos para la variedad CM6438 sólo fueron utilizados en los análisis de
supervivencia.
2.3.1 Funcionalidad de la simbiosis entre genotipos de
Rhizophagus irregularis y variedades de yuca en Yopal
(Casanare)
La funcionalidad de la simbiosis se midió en términos de producción, calidad y
colonización total de HFMA de raíces y en el crecimiento y supervivencia de las plantas.
Producción de raíces
El análisis de varianza realizado con los resultados de las dos repeticiones de este
experimento mostró que hubo un efecto significativo de la interacción entre las diferentes
líneas de R. irregularis y las variedades de yuca para el peso seco (ANOVA F ratio =
3,4128; p ≤ 0,0001) y fresco (ANOVA F ratio = 2,9382; p ≤ 0,0001) de la raíces
producidas por las plantas (Anexo E: numerales 11.5, 11.6). En la Figura 10 se presentan
los resultados del peso seco de las raíces producidas por planta para cada uno de los
tratamientos en la primera repetición de este experimento. Los resultados del peso fresco
se muestran en el numeral 11.5 del anexo E.
Para ambas variedades, algunas líneas tuvieron mayor producción que otras en términos
de biomasa seca y fresca de raíces. Además, estas respuestas en la producción fueron
diferentes dependiendo de la variedad (Anexo E: numeral 11.9, 11.10). Por ejemplo, la
línea S4b en la variedad CM4574, produce la misma cantidad de raíces secas (0,63

Capítulo 2 51
0,10 Kg/planta) que las plantas inoculadas con el producto comercial - GLO (0,39 0,10
Kg/planta), mientras que en la variedad MCOL2737 produce menos cantidad de raíces
secas (0,15 0,06 Kg/planta) que las plantas inoculadas con el producto comercial (0,37
0,06 Kg/planta).
Figura 10. Efecto de la inoculación de 15 líneas de Rhizophagus irregularis sobre la producción y
colonización de plantas de yuca en dos variedades diferentes en el experimento de Yopal (primera
repetición). H2O: Tratamiento con agua sin propágulos (Control negativo); FIL: Tratamiento con “vehículo”
sin propágulos (Control de vehículo) y GLO: Tratamiento con producto comercial de un aislado de R.
irregularis (Control positivo para variedad MCOL2737). D1, C2 y C3: Líneas parentales. S4, Sc1, Sc2 S4b,
S4c, Sc1a, Sc1d, Sc1e, Sc2a, Sc2b, Sc2c, Sc2d: Líneas producidas en laboratorio. Las barras representan la
producción por planta y la línea punteada roja los porcentajes de colonización total de HFMA. Las barras de
error representan un error estándar de la media. La línea horizontal punteada de color negro corresponde
al promedio del tratamiento sin inoculación.

52 Diversidad intra-específica y funcionalidad de la simbiosis vares de yuca
Teniendo en cuenta los resultados de los controles en ambas repeticiones, se encontró
que tanto para la variedad MCOL2737 como para la CM4574, las producciones de las
plantas tratadas con FIL fueron las mismas que las de las plantas control sin inoculación.
Adicionalmente, los resultados muestran que el producto comercial (GLO), en la variedad
MCOL2737 funcionó como control positivo en este experimento. De la misma forma
como se demostró en Ceballos et al., (2013), su efecto fue mayor que el de las plantas
control no inoculadas (Anexo E: numeral 11.9).
El beneficio de la inoculación en términos de la producción de biomasa fresca (ANOVA F
ratio = 2,6535, p ≤ 0,0011) y seca (ANOVA F ratio = 2,1056, p ≤ 0,0112) de raíces
también se vio afectado por la combinación entre las líneas de R. irregularis inoculadas y
la variedad de yuca para los resultados de ambas repeticiones (Anexo E: numerales
11.7, 11.8).
Figura 11. Beneficio en términos de las raíces frescas y secas producido por la inoculación con líneas de R.
irregularis para cada variedad de yuca sembrada en Yopal (consolidado de ambos experimentos). Las barras de
error se construyen con 1 error estándar de la media. Letras diferentes encima de las barras representan diferencias
significativas para un p < 0,05. Las letras minúsculas representan diferencias dentro de la variedad MCOL2737 y las
mayúsculas las diferencias dentro de la variedad CM4574

Capítulo 2 53
En la Figura 11 se observa que dentro de cada variedad la inoculación con algunas líneas
aumentó la producción seca y fresca de las raíces de las plantas, mientras la inoculación
con otras la disminuyó. Pero además, este efecto dependió de la variedad de yuca
sembrada. La inoculación de la línea S4, por ejemplo, incrementó la producción de yuca
seca en la variedad MCOL2737, pero la disminuyó en plantas de la variedad CM4574.
El método de análisis del modelo mixto REML, mostró que la repetición o “Time” explica
en un porcentaje muy bajo la variabilidad del peso seco (-0,264 %) y fresco (-0,309 %)
de las raíces producidas por las plantas de yuca en los experimentos y de los beneficios
de las inoculación en términos de peso seco (2,993 %) y fresco (1,742 %) (Anexo E:
numerales 11.5, 11.6, 11.7 y 11.8; ver RSquare without “Time” as a factor in the model).
Adicionalmente, al correr los modelos sin el factor “Time” no se observaron cambios
fuertes en los RSquare. Es decir la variabilidad explicada por el modelo incluyendo el
factor aleatorio de repetición o “Time” fue similar a la del modelo sin incluir este factor
(Anexo E: numerales 11.5, 11.6, 11.7 y 11.8). Estos resultados demuestran que no hubo
un efecto significativo de la repetición o “Time” sobre la producción de yuca (Figura 12)
ni sobre el beneficio de la inoculación.
Figura 12. Resultados del efecto de las líneas genéticas de Rhizophagus irregularis sobre la producción de raíces
secas en dos variedades de yuca en Yopal para los experimentos sembrados en dos años diferentes. La primera
repetición fue sembrada en Octubre de 2012 y la segunda en Mayo de 2013. En rosado se encuentran subrayadas las
líneas parentales o iniciales del hongo. En azul, las líneas de primera generación provenientes de C3, obtenidas en
laboratorio. En amarillo, las líneas de segunda generación provenientes de C3, obtenidas en laboratorio.
0,00
0,20
0,40
0,60
0,80
1,00
0,00
0,20
0,40
0,60
0,80
1,00
K
F
G
S4
S4
b
S4
c
Sc1
Sc1
a
Sc1
d
Sc1
e
Sc2
Sc2
a
Sc2
b
Sc2
c
Sc2
d
H2
0
FIL
GLO
K
F
G
S4
S4
b
S4
c
Sc1
Sc1
a
Sc1
d
Sc1
e
Sc2
Sc2
a
Sc2
b
Sc2
c
Sc2
d
H2
0
FIL
GLO
MCOL 2737 CM 4574
Tratamientos
Exp. en tiempo 2
Exp. en tiempo 1
Pe
so
se
co
po
r p
lan
ta (
Kg
)
D1
C2
C3
D1
C2
C3

54 Diversidad intra-específica y funcionalidad de la simbiosis vares de yuca
Calidad de raíces de yuca
La calidad en la raíces fue evaluada debido a que este parámetro es afectado por
muchos factores fisiológicos de la planta y por factores ambientales que finalmente son
directa o indirectamente alterados por los HFMA. La funcionalidad de la simbiosis en
términos de calidad fue medida para la inoculación de genotipos diferentes de R.
irregularis en dos variedades de yuca (MCOL2737 y CM4574) en la primera repetición de
este experimento. Las variables de calidad medidas fueron la producción, el porcentaje
de pulpa y la acidez titulable del almidón.
En estos experimentos se encontró que las proporciones de almidón extraído por gramo
de biomasa de yuca estuvieron entre el 17,5% y el 39,4%. Los genotipos inoculados de
R. irregularis también tuvieron un efecto significativo sobre el porcentaje de almidón
producido dependiendo de la variedad de yuca (Anexo E: numeral 11.16; Figura 13), pero
en las otras variables de calidad no se encontró este tipo de efecto combinado (Tabla 3).
Figura 13. Efecto de la interacción entre líneas genéticas de Rhizophagus irregularis y la variedad de yuca sobre el
contenido de almidón extractable (barras) y el porcentaje de colonización para cada tratamiento en el momento de
la cosecha (líneas punteadas de color naranja). H2O: Tratamiento control: plantas sin inoculación. Las barras de error
representan mas o menos un error estándar de la media. Las letras mayúsculas representan diferencias relacionadas con
la colonización y las minúsculas diferencias relacionadas con el contenido de almidón.
Las líneas tuvieron efectos diferentes en la cantidad de almidón producido dentro de
cada variedad y, además, el efecto de cada línea dependió de la variedad inoculada
(Figura 13). Sin embargo, las líneas que produjeron mayor cantidad de biomasa no

Capítulo 2 55
fueron las mismas que produjeron mayor cantidad de almidón por gramo de yuca.
Ninguna línea de R. irregularis produjo un efecto sobre la producción de almidón
diferente al de las plantas control.
Tabla 3. Análisis de varianza para las variables de calidad evaluadas y los porcentajes de colonización total de
HFMA.
Fuente de variación
Contenido de almidón / gramo
de biomasa
Acidez titulable
Porcentaje de pulpa en el almidón
Colonización Total por HFMA
Probabilidad de efecto
Variedad de yuca < 0,0001* 0,2563 0,0033* 0,4938
Línea de HFMA 0,0033* 0,7543 0,2312 0,5923
Variedad de yuca X Línea de HFMA
0,005* 0,3330 0,1764 0,0041*
Para el grupo de muestras a las cuales se les evaluó la calidad de almidón producido, se
encontró que la colonización fue afectada por la interacción entre las Líneas genéticas
del hongo inoculado y la variedad de yuca (Figura 13; Anexo E: numeral 11.17). Además,
se encontró una correlación entre la colonización y la producción de almidón (Correlación
= 0,4614; Prob. de significancia = 0,0079*) para este grupo de muestras.
Supervivencia y crecimiento de las plantas de yuca
El crecimiento de la parte aérea de las plantas fue medida solamente en la segunda
repetición de este experimento. Para esta variable se encontró que también hubo un
efecto de la interacción entre las diferentes líneas inoculadas y la variedad de yuca
(Tabla 4; Anexo E: numeral 11.18).
Tabla 4. Análisis de varianza con el efecto de las líneas de R. irregularis y las variedades sobre el crecimiento de la
parte aérea de la planta para los datos de la segunda repetición del experimento.
Source DF Sum of Squares F Ratio Prob > F
Variedad de yuca 1 204,8 7,2664 0,0076*
Línea de R. irregularis 17 205 1,0714 0,3839
Variedad de yuca X Línea de R. irregularis
17 205,2 1,7487 0,0368*
Nota: Source: Nombre de los efectos dentro del modelo. DF: Grados de libertad para la prueba de los efectos. Sum of squares: Suma de los
cuadrados para la hipótesis para la cual la lista de los efectos es cero; F Ratio: Es el estadístico F para probar que no hay efecto.
Probabilidad de significancia para la prueba F. Prob>F: Es la probabilidad de significancia para F; es la probabilidad de que la hipótesis nula
sea falsa.

56 Diversidad intra-específica y funcionalidad de la simbiosis vares de yuca
Las líneas de R. irregularis no tuvieron un efecto sobre las proporciones de las plantas
que sobrevivieron en el momento de la cosecha para ninguna variedad (Anexo E:
numerales 11.2, 11.3, 11.4). Es decir, no hubo diferencias en la supervivencia de las
plantas luego de ser inoculadas con las diferentes líneas del hongo (Figura 14).
Figura 14. Mosaico que representa la frecuencia de plantas muertas inoculadas con las líneas genéticas de
Rhizophagus irregularis para cada variedad de yuca en ambas repeticiones.
Un análisis de Chi-cuadrado (Anexo E: numeral 11.1) mostró que la supervivencia de las
plantas fue diferente para las dos repeticiones de este experimento. Los niveles de
supervivencia de las plantas de la primera repetición del experimento (56,27 3,25 %)
fueron menores que los de la segunda repetición (72,57 2,67 %).
Colonización total de HFMA en raíces
La funcionalidad de la simbiosis en términos de la colonización total de HFMA en raíces
fue medida en ambas repeticiones del experimento. Para cumplir con los supuestos
necesarios de los análisis estadísticos, fue necesario realizar una transformación arco
seno en los datos de colonización. Para esta variable transformada se encontró un efecto
de la interacción entre Líneas genéticas de R irregularis y la variedad en los datos de
ambas repeticiones en Yopal (ANOVA F = 1,7245; valor p < 0,0395; Anexo E: numeral
11.11). Sin embargo, para esta variable, los resultados de la primera repetición fueron
diferentes que los de la segunda. En la segunda repetición no hubo efectos de los
factores evaluados, mientras que en la primera el arco seno de la colonización se vio
afectada por la interacción Líneas genéticas de R. irregularis y la variedad (Anexo E:
numeral 11.12). Sólo en la segunda repetición, las líneas inoculadas afectaron el arco
seno de la colonización de los HFMA en las raíces de las plantas. Adicionalmente, no

Capítulo 2 57
hubo una correlación entre la colonización y la producción de raíces en la planta, ni entre
la colonización y el beneficio de la inoculación (Correlación de Pearson = 0,0491; valor p
= 0,4292).
2.3.2 Comparación entre los efectos de los grupos de líneas de
Rhizophagus irregularis sobre la producción de raíces de
yuca en Yopal (Casanare)
Con el fin de comparar los efectos de la inoculación de las diferentes líneas de R.
irregularis sobre la producción de las plantas de yuca, se realizaron ANOVAS entre los
grupos de líneas (iniciales o parentales, de primera y de segunda producción) dentro de
cada una de las variedades evaluadas. A continuación se presentan los resultados de la
comparación entre los efectos de:
1) Líneas parentales genéticamente diferentes
Las líneas parentales D1,C2 y C3 fueron clasificadas como genotipos diferentes de R.
irregularis a partir de marcadores de secuencias simples repetidas (SSR), de intrones de
genes nucleares y de intrones de genes mitocondriales (Croll et al., 2008). Al comparar
los resultados para estas líneas se encontró que tuvieron un efecto diferente sobre la
producción de raíces secas al ser inoculadas en la variedad CM4574, pero su producción
fue la misma en la variedad MCOL2737 (Figura 15 ; Anexo E: numerales 11.13, 11.14).

58 Diversidad intra-específica y funcionalidad de la simbiosis vares de yuca
Figura 15. Producción de yuca en plantas de la variedad CM4574 y MCOL2737 que fueron inoculadas con las
líneas parentales D1, C2 y C3. Las barras de error se construyen con 1 error estándar de la media. Letras diferentes
encima de las barras representan diferencias significativas para un p < 0,05.
2) Líneas producidas en laboratorio a partir de las líneas iniciales C2 y C3.
Dentro de un programa de selección de genotipos del hongo se deben buscar líneas
producidas en el laboratorio que generen un efecto deseado en las plantas. Por esta
razón, se hizo una comparación entre los resultados obtenidos con la inoculación de las
líneas obtenidas en el laboratorio y sus parentales (Anexo E: numeral 11.15).
En todos los grupos evaluados se encontró un efecto de la interacción Línea de HFMA x
Variedad de yuca (Anexo E, numeral 11.15: Figura 23, Figura 26, Figura 29, Figura 32).
Es decir hubo un efecto de las líneas, pero ese efecto dependió de la variedad de yuca.
Aunque los resultados en los grupos de las líneas del hongo evaluadas fueron diferentes
para la dos variedades de yuca, en ambas variedades se observó que líneas
provenientes de un mismo parental produjeron efectos diferentes en la producción de
yuca de las plantas (Figura 16; Anexo E, numeral 11.15: Figura 24, Figura 25, Figura 28 y
Figura 34).
Un ejemplo de este resultado se muestra en la Figura 16, en donde se presentan los
resultados del grupo de las líneas provenientes de la línea C2. En ambas variedades,
todas las líneas producidas en el laboratorio (S4, S4b, S4c) presentaron los mismos
rendimientos que su línea parental (C2), pero no tuvieron los mismos efectos en las

Capítulo 2 59
producciones al compararlas entre ellas. En la variedad MCOL2737, la línea segregada
S4, generó mayor producción de yuca que la línea S4b. En la variedad CM4574, la
inoculación de la línea S4, por el contrario causó una menor producción de raíces que la
S4b.
En otros casos, las líneas obtenidas en el laboratorio presentaron diferentes
producciones que las de sus parentales (Figura 17; Anexo E, numeral 11.15: Figura 28,
Figura 31 y Figura 34). La Figura 17 muestra los resultados del efecto del grupo de las
líneas Sc1, Sc1a, Sc1d y Sc1e provenientes de la línea C3 sobre la variedad CM4574. La
línea Sc1 y Sc1e tuvieron una mayor producción de raíces que lo que generó su parental
C3, mientras que Sc1a y Sc1d tuvieron el mismo efecto que su parental para esta
variable. Para este grupo de líneas (C3, Sc1, Sc1a, Sc1d y Sc1e) no se encontró un
efecto de la inoculación de las líneas en la producción de la variedad MCOL2737 (Anexo
E, numeral 11.15: Figura 30).
Figura 16. Producción de yuca de las variedades MCOL2737 y CM4574 que fueron inoculadas con la línea parental
C2 y sus líneas descendientes. C2: Línea parental; S4, S4b y S4c: Líneas obtenidas en el laboratorio. Las barras de
error se construyen con 1 error estándar de la media. Letras diferentes encima de las barras representan diferencias
significativas para un p < 0,05.

60 Diversidad intra-específica y funcionalidad de la simbiosis vares de yuca
Figura 17. Producción de plantas de yuca de la variedad CM4574 que fueron inoculadas con la línea parental C3 y
sus líneas descendientes. C3: Línea parental; Sc1, Sc1a, Sc1d y Sc1e Líneas generadas en el laboratorio. Las barras de
error se construyen con 1 error estándar de la media. Letras diferentes encima de las barras representan diferencias
significativas para un p < 0,05.
2.3.3 Efecto de la variedad sobre las variables agronómicas de
los cultivos de yuca en Yopal
Para los resultados de las dos repeticiones del experimento, la variedad tuvo un efecto
sobre la supervivencia (Anexo E: numeral 11.1). La mayoría de las plantas de la variedad
MCOL2737 sobrevivieron (81,0 ± 17,5%), las de la variedad CM4574 tuvieron una
supervivencia de 68,4 ± 15,9 % y las de la variedad CM6438 tuvieron una supervivencia
muy baja (43,5 ± 15,9%). La Figura 18 muestra un análisis de correspondencia entre la
supervivencia y las variedades para los resultados de las dos repeticiones del
experimento.
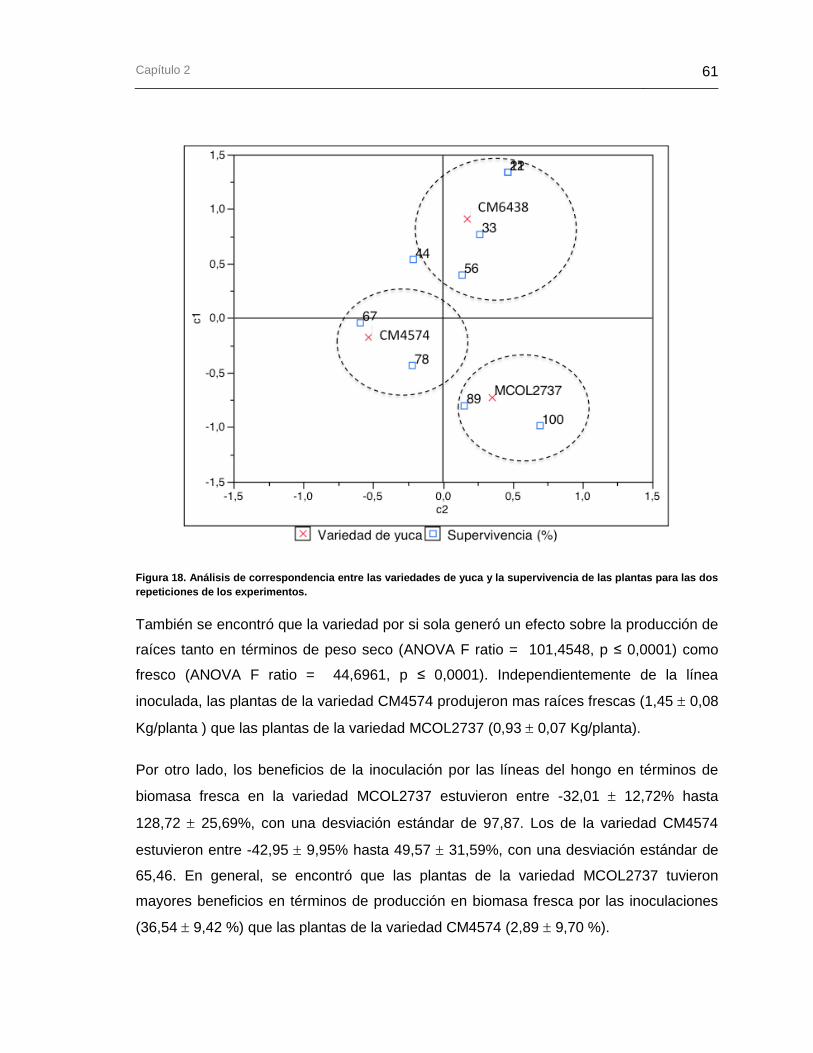
Capítulo 2 61
Figura 18. Análisis de correspondencia entre las variedades de yuca y la supervivencia de las plantas para las dos
repeticiones de los experimentos.
También se encontró que la variedad por si sola generó un efecto sobre la producción de
raíces tanto en términos de peso seco (ANOVA F ratio = 101,4548, p ≤ 0,0001) como
fresco (ANOVA F ratio = 44,6961, p ≤ 0,0001). Independientemente de la línea
inoculada, las plantas de la variedad CM4574 produjeron mas raíces frescas (1,45 0,08
Kg/planta ) que las plantas de la variedad MCOL2737 (0,93 0,07 Kg/planta).
Por otro lado, los beneficios de la inoculación por las líneas del hongo en términos de
biomasa fresca en la variedad MCOL2737 estuvieron entre -32,01 12,72% hasta
128,72 25,69%, con una desviación estándar de 97,87. Los de la variedad CM4574
estuvieron entre -42,95 9,95% hasta 49,57 31,59%, con una desviación estándar de
65,46. En general, se encontró que las plantas de la variedad MCOL2737 tuvieron
mayores beneficios en términos de producción en biomasa fresca por las inoculaciones
(36,54 9,42 %) que las plantas de la variedad CM4574 (2,89 9,70 %).

62 Diversidad intra-específica y funcionalidad de la simbiosis vares de yuca
2.4 Discusión
Los resultados de este capítulo mostraron que existe, en condiciones de campo, una
diversidad funcional entre líneas de R. irregularis cuando son inoculadas en variedades
de yuca. Las líneas del hongo presentaron un efecto sobre: la producción (Figura 10,
Figura 15 - 17), la calidad (Figura 13; Tabla 3) y la colonización de las raíces (Figura 10)
y sobre el crecimiento de las plantas (Anexo E: numeral 11.18) en cultivos de diferentes
variedades de yuca en los Llanos Orientales de Colombia. El efecto significativo de la
interacción entre genotipos del HFMA y de la planta sobre las variables evaluadas
demostró que la diversidad genética intra-específica de ambos simbiontes afectó la
funcionalidad de la simbiosis en condiciones de campo
El cambio del efecto de las líneas sobre la funcionalidad de la simbiosis según el
hospedero fue reportado para R. irregularis en plantas de diferentes especies por
Angelard et al., (2010), pero no había sido reportado hasta el momento, para variedades
de plantas de una misma especie como se encontró en este trabajo. El efecto que tiene
la variabilidad genética intra-específica de R. irregularis sobre la funcionalidad de la
simbiosis ya se había demostrado en cultivos axénicos (Koch et al. 2006; Croll et al.
2009) y en condiciones de invernadero (Koch et al. 2006; Angelard et al. 2010). En esta
investigación se demuestra que el efecto de la diversidad intra-específica de los HFMA
fue suficiente para ser detectada en condiciones de campo en Yopal, y que existe una
interacción entre las líneas del hongo FMA y su hospedante, inclusive al nivel de
variedad. Los estudios en campo representan condiciones reales y tienen un gran valor
ya que evalúan las variables teniendo en cuenta la interacción con los factores que hacen
parte del ambiente.
Diversidad funcional intra-específica de Rhizophagus irregularis en condiciones de
campo
Los efectos generados por la inoculación de las tres líneas parentales de R. irregularis
sobre la producción de las plantas de yuca fueron diferentes en la variedad CM4574
(Figura 15). Como estas tres líneas corresponden a tres genotipos diferentes según lo
encontrado por Croll et al., (2008), se sugiere que las diferencias en el genotipo del
hongo son las que producen cambios en la funcionalidad de la simbiosis. La falta de

Capítulo 2 63
efecto en el resultado obtenido con estas tres líneas para la variedad MCOL2737 (Figura
15), indica que esas diferencias genéticas no se expresan de la misma forma para cada
variedad de yuca. Y que el hospedero responde de una forma diferente. Estos resultados
demuestran que los efectos de las líneas dependieron del hospedero. Además, la
producción de raíces secas varió significativamente cuando las plantas fueron inoculadas
con líneas provenientes de un mismo parental (Figura 16). Un análisis de marcadores
específicos asociados con fenotipos particulares podría ayudar a elucidar las bases
genéticas de la funcionalidad de esta simbiosis.
Los procesos de cultivo de líneas de Rhizophagus irregularis en el laboratorio
promueven diversidad funcional en condiciones de campo
La mayoría de las líneas de R. irregularis evaluadas en estos experimentos (menos las
parentales o iniciales) fueron producidas en el laboratorio a partir de esporas únicas, las
cuales se sometieron a procesos de cultivo para su crecimiento. Al igual que en los
resultados encontrados por Angelard et al., (2010), en esta investigación, la respuesta
de las plantas de yuca en condiciones de campo fue diferente luego de ser inoculadas
con algunas de esas líneas producidas en el laboratorio. Hubo líneas que causaron
efectos en la producción de yuca diferentes, e incluso mayores, a los generados por sus
parentales (Figura 17). Estos resultados y los rangos de variación encontrados indican
que hay un gran potencial para que la variabilidad genética intra-específica de R.
irregularis se utilice en la búsqueda de líneas genéticas del hongo que promuevan la
producción y calidad de los cultivos de yuca en suelos ácidos del trópico.
El beneficio de la inoculación es una herramienta importante en el programa de
selección de genotipos de R. irregularis
Las evaluaciones de la producción de raíces teniendo en cuenta la interacción Líneas del
hongo X Variedades de yuca, permitieron indagar sobre la combinación de los simbiontes
que generan una mayor o menor producción de raíces. Pero para medir el efecto de las
líneas eliminando el efecto que tiene la variedad por si sola sobre la producción, fue
necesario tener en cuenta el beneficio de la inoculación. Esta medida, es de gran
importancia dentro de cualquier programa de selección de líneas de R. irregularis, porque
permite evidenciar el efecto real debido a la inoculación por cada línea a partir de una

64 Diversidad intra-específica y funcionalidad de la simbiosis vares de yuca
comparación entre las plantas que fueron y no inoculadas. Desde 1981, los
investigadores se han interesado en cuantificar los costos y beneficios de la asociación
con HFMA (Yocum 1981); y luego, este beneficio se ha utilizado para normalizar efectos
y poder comparar como afectan los hongos diferentes especies de plantas (Klironomos
2000).
El beneficio de la inoculación también dependió de la línea de R. irregularis inoculada y
de la variedad de yuca sembrada. Los resultados para ambas variedades fueron
positivos para algunas líneas y negativos para otras, indicando que algunas líneas
incrementaron la producción mientras que otras la disminuyeron con respecto a los
resultados encontrados, por ejemplo, con el inóculo comercial. Esto demostró, que dentro
de los procesos utilizados en el laboratorio para generar variabilidad genética, se pueden
obtener también líneas con efectos no deseados en las plantas. Ya se han reportado
resultados en otros estudios donde los HFMA no siempre son benéficos en un sistema
vegetal determinado (Johnson et al. 1997; Koch et al. 2006; van der Heijden et al. 1998).
Por esta razón, es necesario que este tipo de programa de selección de líneas de HFMA
sea enriquecido con información molecular sobre la función que cumplen y los efectos
que promueven diferentes genes del hongo sobre la planta. Esto haría mas eficiente la
búsqueda de líneas con efectos deseables para los cultivos.
El efecto de las líneas de Rhizophagus irregularis sobre la producción de raíces
fue reproducible en los dos experimentos
Las dos repeticiones de este experimento presentaron resultados similares en términos
de los efectos que mostraron las líneas sobre las variables evaluadas más importantes:
la producción de raíces y el beneficio de la inoculación. Se encontró que el factor tiempo
no explicó significativamente la variabilidad de los datos, indicando que el efecto de las
líneas para las variedades evaluadas no dependió del año en que se estableció el
experimento. Es decir, el efecto de las líneas sobre las plantas de yuca en dos
variedades fueron reproducibles de un año para otro. Los experimentos realizados
mostraron resultados consistentes (no iguales) para estas variables al ser repetidos en
dos años diferentes.

Capítulo 2 65
Adicionalmente, los resultados obtenidos para los controles mostraron que también hubo
reproducibilidad en el control positivo (GLO) pues al inocularlo en los experimentos en
plantas de la variedad MCOL2737 se obtuvieron incrementos en la producción de raíces
comparados con las plantas no inoculadas. La reproducibilidad de los experimentos es
fundamental en la ciencia, sobre todo cuando se realiza desarrollo tecnológico aplicado,
incluso reconociendo las dificultades que se tienen para lograr previsibilidad absoluta en
el mundo natural (Fang 2010).
La colonización total de los hongos formadores de micorrizas arbusculares en las
raíces de las plantas inoculadas
En los experimentos establecidos en este capítulo se encontraron diferencias en la
colonización total de las raíces de las plantas, debidas a la interacción entre las líneas del
hongo inoculadas y la variedad de yuca sembrada. Angelard y Sanders, (2011)
encontraron que las líneas del hongo R. irregularis, producidas en el laboratorio, tuvieron
diferentes patrones de desarrollo y crecimiento dentro de las raíces de plantas crecidas
en invernadero y que además, el desarrollo del hongo también fue afectado por el
hospedero. Esta condición no se pudo verificar en estos experimentos debido a la
existencia de una comunidad de HFMA local presente en el suelo. Sin embargo, el
resultado obtenido en estos experimentos es muy interesante ya que esos niveles de
colonización podrían estar representando una diferencia en la habilidad que tiene cada
una de las líneas para coexistir con los HFMA de la comunidad establecida en ese suelo.
Los resultados del efecto de las líneas sobre la colonización, a diferencia de lo
encontrado para la producción de raíces, no fueron reproducibles de un año para otro.
La variabilidad genética intra-específica de estos hongos también es importante en
términos de calidad
Los valores encontrados en el porcentaje de almidón para todas las muestras se
encontraron en el rango normal reportado por el CIAT (Aristizábal et al. 2007). Fue
interesante también encontrar que la variable de calidad evaluada más importante, la
producción de almidón por gramo de biomasa seca, también se vio afectada por la
interacción de la inoculación con las líneas del HFMA y las variedades de yuca. Con esto
se demuestra que la variabilidad genética intra-específica de estos hongos, no sólo es

66 Diversidad intra-específica y funcionalidad de la simbiosis vares de yuca
importante en la producción de yuca sino también en su calidad. Estos resultados pueden
explicarse porque la producción de almidón esta asociado con la nutrición y la
disponibilidad de agua que tenga la planta durante las primeras etapas de crecimiento
(Defloor et al. 1986; Prammanee et al. 2010; Santisopasri 2001). Y estos dos parámetros
son afectados directa o indirectamente por los HFMA (Smith & Smith 2011; Smith & Read
2008). Los hongos al hacer una extensión de las raíces de la planta, tiene más área de
exploración de agua y nutrientes y una mayor área de reserva. Esto ayuda a que la
planta con HFMA tenga una mejor nutrición y una mayor tolerancia al estrés hídrico
(Smith & Read 2008). Teniendo en cuenta esto, los resultados sugieren que algunas
líneas pueden tener fenotipos que presenten micelios mas extensos y con mayor
capacidad de exploración de agua y nutrientes en el suelo, representando una ventaja
para mejorar la producción de almidón. Estos resultados son importantes teniendo en
cuenta que la yuca es considerada como un cultivo de seguridad alimentaria, en donde
es clave mejorar la producción de carbohidratos.
Efecto de la variedad sobre las variables agronómicas
Las líneas de R. irregularis en promedio generaron mayores beneficios en la variedad
MCOL2737 que en la variedad CM4574. También hubo mayor cantidad de líneas que
incrementaron la producción de raíces en la variedad MCOL2737 que en la CM4574
(Figura 11). Esto puede interpretarse como si la variedad MCOL2737 tuviera mayor
dependencia a la inoculación que la variedad CM4574.
Adicionalmente, las desviaciones estándar obtenidas en el beneficio de la inoculación
evidencian que hubo mayor variabilidad en la diversidad funcional por la inoculación de
las líneas en la variedad MCOL2737 que en la CM4574. Este rango de variación en la
funcionalidad de la simbiosis no necesariamente sugiere que la variedades MCOL2737
es mas sensible que la CM4574 para responder a las variaciones genéticas de las líneas
evaluadas, debido a que las variaciones entre las repeticiones de un mismo tratamiento
pueden ser diferentes para las dos variedades. Y, esto afectaría directamente la
comparación entre las medias de los tratamientos en un análisis de varianza. Sin
embargo, dentro de un programa de selección de líneas de R. irregularis es importante

Capítulo 2 67
identificar las variedades que presentan respuestas sensibles a los genotipos y que
permitan evidenciar diferencias funcionales entre las diferentes líneas del hongo.
Los resultados encontrados mostraron, por primera vez que, la selección de líneas de R.
irregularis producidas en el laboratorio podría ser utilizada para encontrar líneas de esta
especie de HFMA que tengan un efecto deseado en ciertos cultivos como el de la yuca
para determinados suelos y ambientes como los que ofrece Yopal. Las diferencias en
producción generadas por la diversidad genética de R. irregularis, encontradas en estos
experimentos, muestran el gran potencial que tiene un programa de selección de líneas
de R. irregularis y cuyas bases pueden presentar una alternativa viable frente al
mejoramiento de la producción y la calidad de cultivos agrícolas de importancia mundial.
Igualmente, es importante realizar experimentos con mayor cantidad de réplicas,
diferentes hospederos y en mayor cantidad de ambientes para poder comprobar su
potencial en diversos sistemas agrícolas reales.
2.5 Conclusiones
La variabilidad genética intra-específica de una población de R. irregularis afectó la
producción, la calidad y la colonización total de las raíces y el crecimiento de plantas de
yuca de la variedad MCOL2737 y CM4574 en condiciones de campo en Yopal
(Casanare). La variación en la funcionalidad de la simbiosis dependió de la línea de R.
irregularis inoculada y la respuesta inducida por su inoculación fue diferente para cada
variedad de yuca. Los efectos generados para las líneas de R. irregularis fueron
reproducibles en las dos repeticiones de este experimento realizadas en dos épocas
diferentes del año.
En Yopal, las líneas de R. irregularis producidas en el laboratorio no necesariamente
tuvieron el mismo efecto sobre la producción de raíces de las plantas de yuca en las
variedades sembradas que el generado por sus parentales. Esto indica que los procesos
realizados en el laboratorio para incentivar la variabilidad genética en individuos o líneas
de R. irregularis pueden promover diversidad funcional en plantas de yuca de ciertas
variedades (var. MCOL2737 y CM4574) que son sembradas en campo en ambientes
como el de Yopal.

68 Diversidad intra-específica y funcionalidad de la simbiosis vares de yuca
Las diferencias obtenidas en estas variables agronómicas para los tratamientos
evaluados demuestran el gran potencial que tiene un programa de selección de líneas de
de esta especie de hongo en la búsqueda de mejores producciones en los cultivos de
este lugar. Sin embargo, también se encontraron individuos del hongo que produjeron
características no deseadas, y por esta razón, es necesario que se trabaje bajo un
esquema de programa de selección de líneas que sea asistido molecularmente.
La variabilidad genética intra-específica de R. irregularis también generó un efecto sobre
la producción de almidón en las variedades MCOL2737 y CM4574. Esta es una de las
características de calidad mas importante para cualquier cultivo de seguridad alimentaria.
Esto sugiere que la variabilidad intra-específica de los HFMA puede utilizarse dentro de
un programa de selección de líneas de R. irregularis en la búsqueda de diversos efectos
deseados en la planta.

3 Capítulo 3: Influencia del ambiente sobre la
funcionalidad de la simbiosis entre
genotipos de Rhizophagus irregularis y
cultivares de yuca
3.1 Introducción
En los capítulos anteriores se demostró que la yuca no sólo responde positivamente a la
inoculación con hongos formadores de micorrizas arbusculares (HFMA) para mejorar su
producción sino también que algunas de sus variedades alteran su crecimiento y
producción al ser inoculadas con genotipos diferentes de Rhizophagus irregularis.
También se encontró que la simbiosis micorrícica entre las variedades de yuca y los
genotipos de R. irregularis fueron altamente sensitivas al estrés ambiental, como por
ejemplo, el producido por las deficiencias de fósforo. Entender los cambios generados
por el ambiente en las producciones de los cultivos establecidos en esta investigación es
fundamental para comprender la respuesta del sistema durante el desarrollo de las
simbiosis.
Cuando se habla de mejoramiento genético de plantas dentro del cual se busca hacer
una selección de genotipos, el ambiente es definido como: “Todos los factores que
afectan las plantas que no son de origen genético”. Incluye factores ambientales
predecibles (ej: radiación solar; tipo y fertilidad de suelo; fecha, densidad y método de
siembra) y factores ambientales impredecibles (ej: cantidad y distribución de lluvias;
temperatura y humedad relativa; presiones repentinas de insectos o enfermedades, etc.)
(Vallejo & Estrada 2002).

70 Diversidad funcional en ambientes
Variedades de plantas cultivadas en diferentes ambientes pueden tener desempeños
diferentes. La alteración en el desempeño relativo de los genotipos, en virtud de
diferencias de ambiente, se denomina interacción genotipo x ambiente (G x A) (Fox &
Cameron 1997). Las interacciones G x A han sido definidas también como la
imposibilidad de los genotipos para obtener los mismos desempeños en los diferentes
ambientes (Baker, 1988). Por esta razón, dentro de cualquier programa de mejoramiento
genético es clave: 1) Identificar las características del ambiente que contribuyen a
mejorar los rendimientos; 2) Conocer el efecto de las interacciones G x A; y 3) Evaluar la
estabilidad de los genotipos en los diferentes ambientes.
La yuca es reconocida por su alta capacidad para crecer y producir competitivamente
bajo condiciones limitantes (Cock & Rosas 1975). A pesar de su fácil adaptación a
diferentes ecosistemas, la yuca también se caracteriza por su acentuada interacción G x
A. Una misma variedad difícilmente se comporta de forma semejante en todas las
localidades (Damba 2008).
Se conoce también que las respuestas de las asociaciones micorrícicas no son
homogéneas (Smith, Jakobsen, & Smith, 2004). Cada asociación genera una respuesta
fenotípica que depende fuertemente de los genotipos de las plantas y hongos
involucrados y además, del ambiente donde ellos se encuentran (Feddermann et al.
2010). Así, cuando se piensa en un programa de selección de genotipos de HFMA para
buscar líneas de R. irregularis que produzcan efectos deseados en las plantas de yuca,
es importante tener en cuenta las respuestas de las plantas a la simbiosis en diferentes
ambientes.
En paralelo, en un programa de mejoramiento genético de plantas, las respuestas de
éstas varían dependiendo de los factores ambientales de cada lugar y son una función de
las interacciones que existen entre el ambiente y el genotipo vegetal. En el caso del
mejoramiento genético o selección de genotipos de HFMA, el sistema se vuelve más
complejo, pues en él se deberá evaluar el efecto que produce el genotipo de un hongo
FMA sobre una planta con un genotipo determinado en diferentes ambientes. El principio
se mantiene, pero en este caso, es necesario separar el efecto que genera el ambiente
sobre la planta per se, del efecto que genera el ambiente sobre la respuesta de la

Capítulo 3 71
simbiosis. Por esta razón, en este trabajo, los efectos del ambiente sobre la simbiosis se
obtuvieron a partir de comparaciones entre los beneficios de las inoculaciones (BI). Esta
variable BI, la cual ya fue descrita en el Capítulo Dos, permitió comparar los beneficios
obtenidos por la inoculación teniendo en cuenta los resultados de las plantas control en
cada uno de los ambientes.
El objetivo principal de este capítulo fue analizar la influencia del ambiente sobre la
funcionalidad de la simbiosis entre genotipos de R. irregularis y cultivares de yuca. En
Santana (Boyacá) se estableció un experimento en campo para comparar, con los
resultados obtenidos en Yopal (Casanare), la influencia del ambiente, sobre la
funcionalidad de la simbiosis, entre genotipos de R. irregularis y cultivares de yuca de la
variedad MCOL2737. Las condiciones del clima, la topografía y la fertilidad del suelo en
Santana son diferentes a las encontradas en Yopal (Anexo F). Con este experimento se
buscó: 1) Identificar el efecto del ambiente sobre el beneficio de la inoculación de líneas
genéticamente diferentes de R. irregularis sobre la producción de raíces de las plantas de
yuca de la variedad MCOL2737 en la cosecha final; 2) Comparar la colonización total de
HFMA en raíces de plantas de yuca de la variedad MCOL2737 en los dos ambientes; y 3)
Evaluar el efecto de las diferentes líneas genéticas de R. irregularis, sobre la producción
y la colonización total de sus raíces, para diferentes variedades de yuca en un ambiente
diferente al de Yopal.
3.2 Metodología
La funcionalidad de la simbiosis se midió en términos de la producción de raíces y su
colonización en el momento de la cosecha, teniendo en cuenta que en los resultados
obtenidos en Ceballos et al., (2013), el efecto de la inoculación sobre la producción de
yuca en campo, se observó en esta etapa del cultivo. El material fúngico utilizado, el
diseño y la forma como se estableció el experimento en campo, la manera cómo se
midieron las variables de producción en la planta y la colonización total de los hongos
fueron similares a lo presentado para el experimento de líneas de R. irregularis en la
página 40 del Capítulo Dos. Los complementos y diferencias en la metodología para este
experimento, en la localidad de Santana (Boyacá) se explican a continuación:

72 Diversidad funcional en ambientes
3.2.1 Sitio de estudio
El experimento se estableció en la finca comercial de yuca “El Salto” de la vereda San
Roque en el Municipio de Santana, Boyacá. La descripción del sitio donde se estableció
el experimento se presenta en Ceballos et al., (2013). Las características físico-químicas
del suelo se presentan en el Anexo B: numeral 8.4.
3.2.2 Material vegetal
Las variedades sembradas en este experimento fueron la variedad MCOL2737, también
sembrada en los experimentos de Yopal, y la variedad COL2215, como material
referencia para la zona, pues es una de las variedades más utilizadas en Boyacá para
consumo en fresco (Ruiz, M; Comunicación personal, Octubre 12, 2011). La semilla
(estacas del tallo con mínimo tres gemas) de la variedad MCOL2737 fue donada por la
Universidad de la Salle y la de COL2215 se obtuvo de cultivos anteriormente
establecidos en la finca “El Salto”. Las estacas tuvieron los mismos manejos reportados
en Ceballos et al., (2013).
3.2.3 Establecimiento del experimento
El manejo del cultivo y la siembra de la yuca se realizó de la misma forma que en
Ceballos et al., (2013). La fertilización se aplicó a los 50 días después de siembra (dds)
según los requerimientos del cultivo, los contenidos iniciales de nutrientes en el suelo y la
eficiencia de la fertilización en la zona. La cantidad de los fertilizantes fosfatados
correspondió al 50% de los fertilizantes normalmente aplicados a los cultivos en la región.
Las plantas recibieron 270 Kg/ha de Urea, 150 Kg/ha de fosfato di-amónico (DAP), 110
Kg/ha de Kieserita (fertilizante con 3% de potasio soluble, 24% de magnesio y 19% de
sulfuro) y 240 Kg/ha de cloruro de potasio (KCl).

Capítulo 3 73
3.2.4 Efecto del ambientes sobre el beneficio de la inoculación
(BI) medida en términos de la producción de raíces
Para determinar la influencia que tuvo el ambiente sobre el efecto de las líneas de R.
irregularis en la producción de la variedad de yuca MCOL2737, se hicieron
comparaciones con los resultados obtenidos en ambos sitios: Yopal y Santana. Sin
embargo, se sabe, que independientemente de la inoculación, el ambiente por sí solo
genera un efecto sobre el crecimiento de las plantas. Para hacer la comparación fue
necesario entonces normalizar los datos con la fórmula propuesta por Raju et al., (1990)
y siguiendo la metodología aplicada para el experimento con inóculo comercial descrito
en el Capítulo Uno (pág. 18). Así, se compararon los beneficios para los ambientes
evaluados. El beneficio de la inoculación (BI) fue calculado en términos de la biomasa
seca y fresca de las raíces producidas en el momento de la cosecha.
3.2.5 Análisis estadístico
Los datos de todos los experimentos fueron analizados con el programa estadístico
JMP® (Statistical Analysis Systems Institute, version 10) siguiendo la misma metodología
explicada en la página 21 del Capítulo Uno. Cuando se cumplieron los supuestos de
normalidad pero no los de homoceasticidad, se corrieron ANOVAS sólo en los casos en
que el número de individuos para cada grupo fueran similares (es decir cuando el numero
de muestras del grupo mayor no fuera 1 ½ más que el número de muestras del grupo
menor) y cuando las desviaciones estándar entre los grupos de tratamientos no fueron
mayores a un 20% (Dai 2009). Para los otros casos se aplicó el test de Welch que no
exige igualdad en las varianzas. Para los casos en que no hubo normalidad, ni
homoceasticidad ni posibilidad de transformar los datos, se utilizó el test de Kruskal-
Wallis para pruebas no paramétricas, con el fin de determinar diferencias significativas
entre los tratamientos. Y para hacer las comparaciones múltiples se utilizó el test de
Wilcoxon para pruebas no paramétricas.

74 Diversidad funcional en ambientes
Para evaluar el efecto del ambiente se realizó un análisis de varianza del diseño
completamente aleatorio con el siguiente modelo: Yijk = + Gi + Ej + GEij + Bjk + ijk.
Donde es la media del beneficio de la inoculación, Gi es el efecto de la i-ésima línea
genéticamente diferente de R. irregularis, Ej es el efecto de j-ésimo ambiente, GEij es la
interacción i-ésima línea de R. irregularis con el j-ésimo ambiente, Bjk es el efecto de la k-
ésima réplica en el j-ésimo ambiente, y ijk es error aleatorio del modelo. Para analizar los
datos del modelo se utilizó el método de análisis de varianza REML (Restricted Maximum
Likelihood).
3.3 Resultados
En el momento de la cosecha en Santana todas las plantas se observaron saludables. El
porcentaje de supervivencia de la variedad MCOL2737, la cual fue sembrada en ambos
ambientes, fue mayor en Santana (94%) que en Yopal (60%). En esta variedad, las
plantas que no fueron inoculadas tuvieron una producción de raíces en biomasa fresca
significativamente mayor en Santana (6,35 1,80 Kg/planta) que en Yopal (0,87 0,34
Kg/planta) (ANOVA F ratio = 70,8535, p < 0,0001).
Para determinar la influencia que tuvo el ambiente sobre el efecto de las líneas de R.
irregularis en la producción de la variedad de yuca MCOL2737, se compararon los
beneficios de la inoculación de las líneas en términos de las producciones de biomasa
seca y fresca de raíces en ambos sitios.
3.3.1 Efecto del ambiente sobre el beneficio de la inoculación en
términos de la producción de raíces en plantas inoculadas
con líneas genéticamente diferentes de Rhizophagus
irregularis para la variedad de yuca MCOL2737
Los análisis de varianza del beneficio que generó la inoculación de las líneas muestran
respuestas diferentes para cada uno de los ambientes. En Yopal, se encontró que las
líneas afectaron la producción de yuca MCOL2737 en términos de biomasa seca
(ANOVA F ratio = 2,28, p ≤ 0,0078, Anexo E: numeral 11.8), mientras que en Santana no

Capítulo 3 75
(Anexo H: numeral 14.3). Adicionalmente, se encontró lo mismo en la producción de
biomasa fresca (Anexo E: numeral 11,7; Anexo H:numeral 14,2).
Un análisis de varianza combinado del beneficio de la inoculación para los dos ambientes
permitió identificar diferencias significativas en la interacción Líneas del hongo X
Ambiente (Anexo G: numerales 13.1, 13.2). La significancia de la interacción se observó
tanto en el beneficio sobre la producción de raíces frescas (Welch´s ANOVA F ratio =
1,7125, p ≤ 0,0294; Figura 35 del Anexo G: numeral 13.1) como en el de las raíces secas
(Welch´s ANOVA F ratio = 1,7014, p ≤ 0,0344; Figura 19).
Figura 19. Beneficio de la inoculación (BI) en términos de peso seco de raíces producidas por plantas que fueron
inoculadas con líneas genéticamente diferentes de Rhizophagus irregularis en dos ambientes diferentes. Las
barras de error se construyen con 1 error estándar de la media. Letras diferentes encima de las barras representan
diferencias significativas para un p < 0,05.
El beneficio en términos de peso fresco y seco de raíces recibido por la inoculación de
las diferentes líneas de R. irregularis dependió del ambiente donde se encontraba el
cultivo. En general, las plantas en Yopal (33,31 10,33 %) tuvieron beneficios por la
inoculación con los HFMA en términos de peso seco que las plantas en Santana (-0,55
abc
abc abc
ab
abc abc
c
abc
c
abc abc
c
abc
c
c
a
bc
c c
c c
bc
c
abc
bc
bc bc
c
bc
bc
c
bc
-90
-40
10
60
110
160
210
D1
C2
C3
S4
S4
b
S4c
Sc1
Sc
1a
Sc1
d
Sc
1e
Sc2
Sc
2a
Sc2
b
Sc
2c
Sc2
d
GL
O
D1
C2
C3
S4
S4
b
S4c
Sc1
Sc
1a
Sc1
d
Sc
1e
Sc2
Sc
2a
Sc2
b
Sc
2c
Sc2
d
GL
O
Yopal Santana
IB (
Pro
du
cció
n d
e r
aíc
es e
n p
eso
se
co
po
r p
lan
ta)
(%)

76 Diversidad funcional en ambientes
3,34 % ). En Santana, el beneficio de la inoculación en términos de la producción de
biomasa seca de raíces para la variedad MCOL2737 varió entre 28,3 % hasta -18,3 %
mientras que en Yopal varió desde 157,1 % hasta -63,3 % (Figura 19). El valor de la
desviación estándar de esta variable para todos los tratamientos en Santana fue de 39,94
y en Yopal de 109,41. Para la variedad MCOL2737, los coeficientes de determinación de
los ANOVA utilizados para evaluar las diferencias en el beneficio de la inoculación por las
líneas genéticas de R. irregularis fueron mayores en Yopal (39%) que en Santana (11%)
(Anexo G, numerales: 13.4, 13.5). En Yopal, el 39% de la variabilidad generada en el
beneficio de la inoculación fue explicado por las línea inoculada, mientras que en
Santana este factor sólo explicó el 11% de la variabilidad.
La Figura 20 muestra que el ambiente tuvo una mayor influencia sobre el efecto de
algunos genotipos del hongo (Ej: S4 y GLO), y una menor influencia sobre el efecto de
otros genotipos (Ej: Sc2b y D1). Las líneas del hongo D1 y Sc2d (pendientes de 0,07 y
1,06 respectivamente) presentaron resultados similares de producción de biomasa seca
de raíces en ambos ambientes. Pero las líneas S4 y el producto comercial GLO
(pendientes de -167,5 y -146,7 respectivamente) tuvieron un efecto completamente
diferente. La línea S4 en Yopal generó una promoción en el crecimiento de las raíces de
yuca de la variedad evaluada, y en Santana por el contrario, generó una depresión.
La mayoría de las líneas del hongo mostraron un mayor beneficio (medido en términos
de producción de raíces secas) en Yopal que en Santana, pero las líneas D1, Sc1, Sc1d,
Sc2a, Sc2c, Sc2d produjeron los mismo beneficios de la inoculación tanto en Yopal como
en Santana. Ninguna línea generó mayores beneficios estadísticamente significativos en
Santana que en Yopal.

Capítulo 3 77
Figura 20. Diferencias en el beneficio de la inoculación que produjeron líneas de Rhizophagus irregularis en
términos del peso seco de raíces producidas por plantas de yuca de la variedad MCOL2737 y que fueron
sembradas en diferentes ambientes. Las ecuaciones de la derecha representan una línea que une las medias del
beneficio de la inoculación para una misma línea de R. irregularis en cada ambiente.
3.3.2 Efecto del ambiente sobre la colonización total por HFMA
en raíces de plantas inoculadas con líneas genéticamente
diferentes de Rhizophagus irregularis
En el momento de la cosecha, el efecto que generó la inoculación con las diferentes
líneas de R. irregularis sobre la colonización total de HFMA en las raíces de yuca
(MCOL2737) dependió del lugar donde la yuca fue cultivada (ANOVA F ratio = 2,1088 p <
0,0130; Anexo G: numeral 13.3). En Santana, no hubo efecto por la inoculación de las
diferentes líneas de R. irregularis sobre la colonización. En Yopal, por el contrario, las
líneas D1 y Sc1d tuvieron porcentajes de colonización mayores que los de la línea Sc2b.
El ambiente además, generó también un efecto sobre la colonización en las plantas

78 Diversidad funcional en ambientes
inoculadas con todas las líneas de R. irregularis (ANOVA F ratio = 14,1410 p < 0,0004;
Anexo G: numeral 13.3). En Santana los porcentajes de colonización en promedio para
todas las líneas fueron mayores (75,20 % 2,00) que en Yopal (65,51 1,79 ).
Figura 21. Colonización Total de HFMA en raíces de yuca de plantas que fueron inoculadas con diferentes líneas
de Rhizophagus irregularis en dos ambientes diferentes. Las barras de error se construyen con 1 error estándar de la
media. Letras diferentes encima de las barras representan diferencias significativas para un valor p < 0,05.
En el momento de la cosecha no se encontró correlación entre la colonización total de
HFMA y la producción de raíces secas ni frescas en ninguno de los ambientes evaluados
para la variedad MCOL2737.
Luego de abordar el objetivo principal de este capítulo, se presenta a continuación los
resultados del experimento establecido en Santana con la inoculación de las líneas
genéticas de R. irregularis sobre plantas de dos variedades de yuca.

Capítulo 3 79
3.3.3 Efecto de la inoculación de diferentes líneas de
Rhizophagus irregularis en la producción de raíces de dos
variedades de yuca cultivadas en Santana.
La inoculación con líneas genéticamente diferentes de R. irregularis no alteró
significativamente la producción de la biomasa seca o fresca de yuca para ninguna de las
variedades evaluadas en Santana (Anexo H: numerales 14.2, 14.3, 14.4, 14.5 y Figura
366). Los valores de R2 en los ajustes para los diferentes modelos utilizados indicaron
una baja proporción de la variación en las producciones de yuca alrededor de la media
que fue explicada por la inoculación de las líneas.
De la misma forma, el beneficio de la inoculación en términos de la producción de raíces
tampoco presentó cambios significativos dependiendo de la línea de R. irregularis
inoculada para ninguna de las dos variedades en Santana. Sin embargo, el beneficio en
términos de producción de yuca seca que recibió cada una de las variedades por la
inoculación en promedio de todas las líneas fue significativamente diferente (Anexo H:
numeral 14.67). Las plantas de la variedad COL2215 tuvieron beneficios por las
inoculaciones mayores que las plantas de la variedad MCOL2737 (Figura 22).
Figura 22. Beneficio de la inoculación en términos de la producción de biomasa seca y fresca de las plantas
inoculadas con las diferentes líneas de Rhizophagus irregularis para las dos variedades de yuca sembradas en
Santana. Las barras representan ± 1 error estándar de la media. Letras diferentes encima de las barras representan
diferencias significativas para un p < 0,05.

80 Diversidad funcional en ambientes
Los porcentajes de colonización total por HFMA de las raíces de las plantas de yuca no
fueron afectados por ninguno de los factores evaluados. Tampoco se encontró
correlación entre la colonización y las producciones para los resultados de Santana.
3.3.4 Efecto de las variedades de yuca en la producción de raíces
para los cultivos en Santana.
Cuando se compararon los promedios de todas las plantas (inoculadas y no) no se
observaron diferencias en la producción de yuca en biomasa fresca entre las variedades,
pero si en la producción de yuca en biomasa seca (ANOVA F ratio = 13,4508, p ≤
0,0003) (Anexo H: numeral 14.1). La variedad COL2215 produjo 1,75 0,08 Kg de raíces
secas por planta, mientras que la variedad MCOL2737 produjo 1,47 0,08 Kg de raíces
secas por planta.
Bajo las condiciones de este experimento en Santana, el promedio de la producción de
yuca fresca de las plantas de la variedad COL2215 inoculadas con el producto comercial
(1,56 0,23 Kg/planta) fue 13% mayor que el promedio de las plantas control (1,48
0,25 Kg/planta), pero en este caso no se encontraron diferencias significativas entre los
dos tratamientos.
3.4 Discusión
En Santana (Boyacá) se estableció un experimento en campo con la misma metodología
y el mismo diseño experimental del experimento explicado en el Capítulo Dos. El objetivo
fue comparar la influencia del ambiente sobre la funcionalidad de la simbiosis entre
genotipos de R. irregularis y plantas de yuca. Primero, se compararon los resultados de
la variedad MCOL2737 con los obtenidos en Yopal para de evaluar el efecto del
ambiente sobre el beneficio de la inoculación durante la cosecha final. Luego se evaluó el
efecto de las diferentes líneas de HFMA sobre la producción y la colonización total en las
raíces para diferentes variedades de yuca en este lugar de la misma forma como se hizo
en Yopal.

Capítulo 3 81
El ambiente tuvo un efecto sobre el beneficio de la simbiosis y sobre la
colonización en raíces de plantas inoculadas con diferentes genotipos de
Rhizophagus irregularis.
En este capítulo se comparó el beneficio en la producción de yuca y la colonización que
tuvieron plantas de la variedad MCOL2737 al ser inoculadas con líneas genéticamente
diferentes de R. irregularis en dos ambientes: Yopal y Santana. Estos ambientes
presentan características climáticas, topográficas y edáficas diferentes (Anexo F: Tabla
5).
Las diferencias entre los niveles de supervivencia y las producciones obtenidas en cada
lugar para las plantas no inoculadas de la variedad MCOL2737 mostraron un fuerte
efecto del ambiente sobre las plantas de yuca de esa variedad independientemente de la
inoculación. Esto indica que la variedad de yuca MCOL2737 tiene un mejor desempeño
en Santana que en Yopal, ya que en este ambiente sobreviven mayor cantidad de
plantas y se incrementa en más de seis veces la producción de este cultivo comparada
con la producción en Yopal. Estos resultados demuestran que el ambiente por si sólo
tuvo un efecto en la producción de yuca de la variedad estudiada. Así, para poder
identificar el efecto que generó el ambiente sobre la simbiosis, fue indispensable hacer
las comparaciones de los lugares en términos del beneficio de la inoculación (BI). Esta
variable propuesta por Raju et al., 1990, realiza una comparación entre las plantas
inoculadas y no inoculadas en un mismo sistema. Y esto, permitió eliminar el efecto que
por si genera el ambiente sobre las plantas y evaluar el que produce sobre la simbiosis
en términos de la variable de interés.
La significancia encontrada para la interacción Líneas genéticas del hongo x Ambiente,
es un indicativo de la respuesta diferencial en el beneficio de la simbiosis que
presentaron las líneas de R. irregularis a las variaciones ambientales en las dos
localidades evaluadas. También se encontró un efecto del ambiente sobre el beneficio de
la inoculación de todas las líneas en promedio sobre la producción de raíces. Las plantas
inoculadas con las líneas tuvieron en promedio un mayor beneficio en Yopal (33,31%)
que en Santana (-0,55%). La explicación mas plausible es que Yopal ofreció un ambiente
de menor calidad que Santana para las plantas de esta variedad de yuca teniendo en

82 Diversidad funcional en ambientes
cuenta los resultados de producción y supervivencia encontrados. Con esto, se corrobora
entonces que las interacciones simbióticas positivas suelen ocurrir y mantenerse en
ambientes con menores calidades (Hochberg et al. 2000; Thrall et al. 2007). Este
resultado, se vuelve mucho mas interesante cuando se tiene en cuenta que el sistema
que se esta evaluando es agrícola. La mayoría de la producción mundial de alimentos se
realiza en sistemas con limitaciones edáficas y climatológicas, como la acidez del suelo,
la presencia de Aluminio, la compactación del suelo, la falta de agua, etc. Teniendo en
cuenta esto, el rol que tiene la simbiosis en este tipo de sistemas cada vez mas
degradados es clave para el mejoramiento de la producción y calidad de los cultivos.
Para la variedad MCOL2737, la inoculación con las líneas explicó una mayor proporción
de la variabilidad en la respuesta del beneficio de la inoculación en Yopal (39%) que en
Santana (11%)(Anexo G: numerales 13.4,13.5). Esto sugiere que Yopal permite una
mejor discriminación del efecto de los genotipos de las líneas comparado con Santana.
Adicionalmente, la comparación entre los beneficios de la inoculación que generó cada
una de las líneas en los dos ambientes evaluados (Figura 20) permitió evaluar la
estabilidad del efecto que se obtienen en la variedad MCOL2737 por la inoculación con
esas líneas genéticas en los dos lugares evaluados. Un estudio con mayor cantidad de
ambientes: 1) aumentaría la información con respecto a la estabilidad general que tienen
las líneas para promover o no un beneficio en términos de producción de raíces en la
variedad de yuca MCOL2737; 2) permitiría detectar los genotipos del hongo que generen
altos beneficios en las plantas y que sean estables para diferentes ambientes y 3)
identificaría grupos de ambientes que presenten un mismo patrón de respuesta a la
inoculación de las líneas.
Parte de la variación encontrada en el beneficio que generó la inoculación de las líneas
en ambos lugares podría ser explicada por la interacción entre la comunidad de
organismos del suelo presentes en cada ambiente. A pesar de las limitaciones
metodológicas que presenta la evaluación de la colonización micorrícica, se encontró que
este parámetro cambió según la línea genética del hongo inoculada y esos cambios,
dependieron del ambientes donde creció la planta. Las variaciones en los niveles de
colonización de un genotipo que es inoculado en dos ambientes diferentes no sólo

Capítulo 3 83
podrían estar asociadas con la forma como cada línea de HFMA puede co-existir con las
comunidades de HFMA presente en el suelo de cada uno de los lugares, sino también
con diferentes características de cada ambiente que pueden estar regulando la forma
como cierto aislado coloniza la raíz, como por ejemplo: la forma como la planta altera el
microambiente del suelo (Wu et al. 2009); las características biológicas de la rizósfera, la
diversidad y composición de las comunidades de HFMA (Gange et al. 1990), las
demandas nutricionales del hospedero (Muthukumar et al. 2003) y, las variaciones de las
condiciones del ambiente durante el año (Jakobsen et al. 2002).
Efecto de la inoculación de las diferentes líneas de Rhizophagus irregularis en
términos de la producción de raíces para dos variedades de yuca cultivadas en
Santana
En Santana, no se evidenció un efecto de las líneas de R. irregularis ni sobre la
producción de las plantas, ni sobre el beneficio de la inoculación, ni sobre la colonización
de las raíces en ninguna de las variedades de yuca evaluadas. Estos resultados fueron
diferentes a los encontrados en Yopal en donde si se evidenció efecto de la inoculación
de las líneas fúngicas sobre estas variables. La diversidad en la funcionalidad de esta
simbiosis ya ha sido reportada en investigaciones y sugiere que las diferencias en las
condiciones que ofrece cada ambiente tienen una influencia sobre el efecto de la
inoculación (Avio et al. 2006; Feddermann et al. 2010; Munkvold 2004). Las interacciones
de las condiciones bióticas y abióticas (ej: nutrientes del suelo, condiciones
climatológicas, microorganismos presentes en el suelo, etc.) de Santana podrían ofrecer
un mayor grado de beneficios para el hospedero que las que ofrece Yopal (Jones &
Smith 2004).
En una investigación previa, se evaluó el efecto del mismo inóculo comercial de R.
irregularis sobre la variedad de yuca COL2215 en Santana y bajo las mismas
condiciones de fertilización fosfatada de este experimento. En los resultados previos, los
cuales se encuentran publicados en Ceballos et al., 2013, la inoculación con este
producto generó un incremento del 14% sobre la producción de yuca fresca en plantas
inoculadas. Para este experimento se encontró un incremento similar del 13% en
promedio, pero en este caso, no se encontraron diferencias significativas al comparar

84 Diversidad funcional en ambientes
este tratamiento con las plantas control. La menor cantidad de réplicas utilizadas en este
experimento con respecto a las utilizadas en el reportado por Ceballos et al, (2013),
podría explicar la falta de significancia.
A pesar de que en Santana no se encontró un efecto de las líneas del hongo sobre las
variables evaluadas, si se encontró un efecto de la variedad de yuca sobre el beneficio
de la inoculación en general. Es ya bien reconocido que existen variaciones en la
respuesta a la micorrización dependiendo del hospedero (Klironomos 2000; Husband et
al. 2002; Croll, Wille, Gamper, et al. 2008; Bever 2002). La variedad de yuca COL2215
utilizada en la zona presentó una mayor respuesta a la inoculación que la variedad
MCOL2737 (Figura 22). La variabilidad intra-específica de plantas de yuca afectó la
respuesta en la funcionalidad de la simbiosis, y por lo tanto, existen variedades de yuca
que tienen una mayor capacidad de responder a la micorrización que otras. Este
resultado muestra la importancia de tener en cuenta los simbiontes involucrados a nivel
intra-específico en un ambiente determinado cuando se esta pensando en utilizar este
tipo de asociación para mejorar la productividad de los cultivos. Así, las variedades de
yuca recomendadas para un ambiente determinado deberían llegar a ser re-
seleccionadas cuando se considere una aplicación conjunta con HFMA en busca de
mayores producciones. Y el proceso de selección debe tener en cuenta la respuesta de
cada variedad a la asociación simbiótica con líneas que potencialicen la producción del
sistema agrícola.
3.5 Conclusiones
Los resultados encontrados indican que hubo un efecto del ambiente sobre la producción
de raíces y la supervivencia de las plantas de la variedad MCOL2737 independiente de la
inoculación. Esto justificó el uso del beneficio de la simbiosis, como parámetro, para
eliminar el efecto que genera el ambiente por si, sobre las plantas y poder evaluar el
efecto que produce sobre la simbiosis.
El beneficio de la inoculación con líneas de R. irregularis y la colonización total de HFMA
en las raíces de yuca de la variedad MCOL2737 dependieron del ambiente en donde se
encontraba el cultivo. Ninguna línea fúngica provocó mayores beneficios en términos de
producción de yuca en Santana que en Yopal. Sin embargo, es necesario un mejor

Capítulo 3 85
entendimiento del papel que juegan los diferentes factores ambientales de tipo biótico y
abiótico para establecer estrategias de aplicación y manejo de los genotipos de R.
irregularis en los sistemas productivos de yuca.
Este análisis mostró como las condiciones de dos ambientes evaluados alteraron el
desempeño de los diferentes genotipos de R. irregularis sobre las plantas de yuca
dependiendo del lugar donde se encontraba el cultivo. Esto reitera la importancia del
efecto del ambiente sobre el desempeño de las líneas fúngicas producidas, lo cual es
clave en cualquier programa de selección de genotipos de esta especie de hongo con
miras a su aplicación en la producción agrícola. Sin embargo se recomienda ampliar la
cantidad de ambientes evaluados con el fin de poder tener conclusiones mas generales
sobre la estabilidad del efecto que las líneas pueden tener sobre las plantas de una
variedad.


4 Conclusiones y recomendaciones
generales
4.1 Conclusiones
En este trabajo se encontró que un inóculo comercial de un aislado de R.
irregularis producido en condiciones in vitro aumentó significativamente los rendimientos
y disminuyó la aplicación de los fertilizantes fosfatados en un cultivo de yuca de la
variedad MCOL2737 sembrado en Yopal (Casanare). El sistema de producción in vitro
del inóculo garantiza una calidad controlada y facilita su transporte incluso a zonas
remotas (Sanders 2010). Por esta razón, esta aplicación biológica podría ser viable para
incrementar las producciones de yuca en otras zonas del trópico donde la productividad
de los cultivos también es limitada (Rodriguez & Sanders 2016).
Adicionalmente, se encontró que la diversidad funcional generada por la variabilidad
genética de R. irregularis fue suficiente para ser detectada en condiciones de campo en
Yopal y, que algunas de las líneas fúngicas que se produjeron en el laboratorio por
procesos de cultivo in vitro, no tuvieron el mismo efecto que sus parentales. Aunque se
demostró que un aislado de R. irregularis producido in vitro se puede utilizar para
aumentar significativamente los rendimientos de la yuca, el hecho de que los diferentes
genotipos producidos a nivel in vitro generen efectos diferentes en el crecimiento y
producción de las plantas en determinados lugares, hace que esta especie de hongo sea
un fuerte candidato para un programa de selección de genotipos con el fin de buscar
líneas que incrementen aún más los rendimientos de yuca en el futuro.
En Yopal, la producción, calidad y colonización de las raíces y el crecimiento de las
plantas fueron diferentes según el genotipo de R. irregularis inoculado. En algunas
ocasiones se encontraron respuestas mayores en estas variables que las generadas por
las plantas no inoculadas, pero en otras ocasiones se encontraron respuestas menores.

88 Conclusiones y recomendaciones
Además, las respuestas inducidas por cada genotipo cambiaron dependiendo de la
variedad de yuca y del ambiente donde ocurrió la interacción entre los simbiontes. Esto
muestra que en los procesos de variabilidad genética se pueden generar gran cantidad
de líneas con funcionalidades diferentes en la planta sin tener conocimiento de todas las
funciones de los genes de los simbiontes, pero de la misma forma, esto es una
desventaja ya que algunas de las líneas producidas podrían promover características
indeseables en la funcionalidad de la simbiosis (Rodriguez & Sanders 2016).
4.2 Perspectivas y recomendaciones
Los resultados de estos experimentos son un tamizado preliminar de los efectos que
tienen líneas genéticas de R. irregularis en condiciones de campo en diferentes
variedades de yuca y en diferentes ambientes. Sin embargo, es necesario probar algunas
de estas líneas en condiciones de un cultivo con mayor cantidad de réplicas, para así,
poder confirmar su potencial en sistemas agrícolas reales (Rodriguez & Sanders 2016).
El tamizaje de genotipos en diferentes variedades de yuca y en diferentes ambientes
puede convertirse en un proceso lento, tedioso y costoso. Por esta razón es
indispensable identificar las bases genéticas en el hongo que puedan explicar las
diferencias encontradas en el crecimiento de la yuca. Esto permitiría trabajar bajo un
esquema de programa de selección de genotipos asistido con marcadores moleculares
(Rodriguez & Sanders 2016).
De la misma manera es importante entender cómo ocurre el proceso de variabilidad intra-
específica en estos hongos. Con esto se podrían definir estrategias para obtener una
mayor variabilidad y eficiencia en la obtención de líneas mejoradas del hongo. Sin
embargo otro reto fundamental en este proceso es comprender la estabilidad de esas
líneas a nivel de ambiente, variedad de yuca y a lo largo del tiempo. Pues este tema
garantizaría un mayor éxito en los efectos de la inoculación. Con la combinación de
estudios como éste y lo obtenido del mejoramiento genético asistido molecularmente se
podría en un futuro predecir cuál hongo va a mejorar las producciones de las plantas de
yuca para una variedad específica en un suelo y en un ambiente determinado.

5 Productos generados
Los productos generados a partir de esta investigación son presentados a
continuación por categorías
5.1 Artículos Científicos Publicados
En el 2013, se publicó un artículo científico en una revista internacional con los
resultados del primer objetivo de este trabajo:
1. Ceballos I., Ruiz M., Fernández C., Peña R., Rodríguez A., Sanders I.R., 2013. The In
Vitro Mass-Produced Model Mycorrhizal Fungus, Rhizophagus irregularis,
Significantly Increases Yields of the Globally Important Food Security Crop Cassava.
PLoS One 8(8) pp. e70633.
5.2 Artículos Científicos en Preparación
Actualmente, se encuentran en elaboración cuatro artículos derivados de este trabajo:
2. Artículo en preparación sobre el efecto de la inoculación de las diferentes líneas de R.
irregularis sobre el crecimiento de plantas para diferentes cultivares de yuca en
campo.
3. Artículo en preparación sobre el beneficio de la inoculación con un producto
comercial de R. irregularis a lo largo del ciclo del cultivo
4. Artículo en preparación sobre el efecto de la diversidad genética intra-específica de
R. irregularis sobre la calidad de la yuca.
5. Artículo en preparación sobre el efecto de las dosis de un inóculo comercial sobre la
producción de las plantas de yuca en campo

90 Productos esis o trabajo de investigación
Se buscará someter al menos dos artículos en revistas internacionales.
5.3 Formación de Estudiantes
Durante el desarrollo de este trabajo, se llevó un proceso de formación en investigación
con dos estudiantes de pregrado, quienes participaron en cada una de las etapas de
investigación para cada proyecto en el que participaron.
6. Formación de la estudiante de pregrado de Ing. Química, Camila Andrea Delgadillo.
Proyecto: Efecto de la diversidad genética intra-específica de R. irregularis sobre la
calidad de la yuca. Con los resultados de este trabajo Camila participó en la 8va
Conferencia Internacional de Micorrizas realizada en Flagstaff (USA, 2105).
7. Formación del estudiante de pregrado Michael Ruiz, en el desarrollo de su trabajo de
grado para optar por el título de Ingeniero Agrónomo. Título del Proyecto: “Efecto de
las Micorrizas en la Absorción de Fósforo por el Cultivo de Yuca (Manihot esculenta
Krants) cv COL2215”. Parte de los resultados de su trabajo fueron publicados en
Ceballos et al., 2013.
5.4 Participación en Conferencias Internacionales
Los trabajos presentados en Conferencias Internacionales con los resultados derivados
de esta investigación son:
New Phytologist Symposium (14 - 16 de Mayo de 2014) en Zurich (Suiza)
8. Poster: Ceballos, I., Fernández, C., Rodríguez, A., & Sanders, I. R. (2014). Within-
species genetic differences in Rhizophagus irregularis induce a wide range of growth
responses of cassava in the field and in the presence of a native AMF community. Zurich:
ICOM Committee.
9. Poster: Fernández, C., Ceballos, I., Rodríguez, A., & Sanders, I. R. (2014). Benefit of
the in vitro mass-produced Rhizophagus irregularis on the globally important food security
crop cassava. Zurich: ICOM Committee.

Productos 91
8th International Conference on Mycorrhiza (ICOM8) Flagstaff Arizona (USA) (2- 7 de
Agosto del 2015)
10. Poster: Delgadillo, C., Ceballos, I., Rodríguez, A., & Sanders, I. R. (2015). Intra-
specific Genetic Variability in Arbuscular Mycorrhizal Fungi had an Effect on Starch
Production in Field but not on Starch Quality. In 8th International Conference on
Mycorrhizal. Flagstaff (USA): Northern Arizona University.
11. Charla corta: Ceballos, I., Rodríguez, A., & Sanders, I. R. (2015). Mass-Produced
Genetically Modified Rhizophagus irregularis Alter Cassava Production in Field. In 8th
International Conference on Mycorrhizal. Flagstaff (USA): Northern Arizona University.
Esta charla fue premiada con el Biozynterra Prize, como la mejor presentación oral con
énfasis en investigación aplicada de HFMA.
5.5 Pasantía de Investigación
12. Pasantía de investigación en el semestre 2015-II. Grupo de investigación del profesor
Ian Sanders, el cual estudia la ecología y evolución de organismos simbióticos.
Universidad de Lausanne, Suiza.
5.6 Divulgación del proyecto
Las divulgaciones que se realizaron sobre el proyecto y algunos de sus resultados en
medios no científicos fueron:
Nacionales
13. Divulgación en Periódico Nacional: Castaño, L. (2013, September). 20% más de yuca
con 50% menos agroquímicos. UN Periódico, p. 9. Bogotá.
14. Divulgación en periódico Nacional: Graber, P. C., & Rodríguez, A. (2014). Microbios ,
el secreto para evitar una hambruna, El Espectador, p 1–5.

92 Productos esis o trabajo de investigación
Internacional
15. Video Internacional : Condayan, C. (2015). The Power of Fungal Genetics. USA:
American Sociaty of Microbiology. Retrieved from https://vimeo.com/119007230
16. Divulgación en Revista Internacional: Conniff, R. (2013). Enlisting bacteria and fungi
from the soil to support crop plants is a promising alternative to the heavy use of fertilizer
and pesticides. Scientific American, 76–79.
17. Divulgación en Revista Internacional: Graber, C. (2014). The Next Green Revolution
May Rely on Microbes. PBS online, pp. 1–5.

6 Consideraciones éticas
Esta investigación fue realizada bajo la normatividad colombiana y teniendo en cuenta la
asesoría del Ministerio de Medio Ambiente sobre los trámites y/o licencias necesarias
para el establecimiento de los experimentos (Ver Anexo A)
De acuerdo con la normatividad colombiana, la Autoridad Nacional de Licencias
Ambientales de la Subdirección de Instrumentos, Permisos y Trámites Ambientales
determinó que para este proyecto no era necesario un permiso especial para el uso de
las diferentes líneas de HFMA en esta investigación científica debido a que los inóculos
fueron utilizados en actividades de investigación en agricultura sin involucrar
especímenes de fauna o flora en peligro de extinción.
El permiso CITIES firmado por Colombia para la exportación e importación del inóculo no
es requerido debido a que las especies involucradas en el estudio no pertenecen a la
lista de especies en peligro de extinción.
Todos los experimentos fueron establecidos en tierras privadas, con el respectivo
permiso de sus dueños.
No fueron necesarios permisos adicionales bajo lo exigido por la ley colombiana, debido
a que los organismos de este estudio han sido también encontrados en casi todos los
suelos y ecosistemas colombianos.


7 Anexo A : Permisos, Trámites y/o Licencias
Necesarias Para Desarrollar Esta
Investigación

96 Anexo A: Permisos, trámites y/o licencias necesarias para esta investigación Título de la tesis o trabajo de investigación

Anexo A: Permisos, trámites y/o licencias necesarias para esta investigación 97

98 Anexo A: Permisos, trámites y/o licencias necesarias para esta investigación Título de la tesis o trabajo de investigación

8 Anexo B : Análisis Fisicoquímicos de los
suelos antes de establecer los
experimentos en Yopal y en Santana

100 Anexo B: Análisis fisico-químicos Título de la tesis o trabajo de investigación
8.1 Experimento con inóculo comercial en Yopal

Anexo B: Análisis fisico-químicos 101
8.2 Experimento con líneas de Rhizophagus irregularis en Yopal (Primera repetición).
NOTA: Los parámetros, métodos de análisis, las valoraciones y los niveles generales de
referencia son los empleados por el laboratorio de suelos de la Facultad de Agronomía
de la Universidad Nacional de Colombia, presentados en numeral 8.1 del Anexo B de
este trabajo.

102 Anexo B: Análisis fisico-químicos Título de la tesis o trabajo de investigación
8.3 Experimento con líneas de Rhizophagus irregularis en Yopal (Segunda repetición).
NOTA: Los parámetros, métodos de análisis, las valoraciones y los niveles generales de
referencia son los empleados por el laboratorio de suelos de la Facultad de Agronomía
de la Universidad Nacional de Colombia, presentados en numeral 8.1 del Anexo B de
este trabajo.

Anexo B: Análisis fisico-químicos 103
8.4 Experimento de líneas de Rhizophagus irregularis en Santana
NOTA: Los parámetros, métodos de análisis, las valoraciones y los niveles generales de
referencia son los empleados por el laboratorio de suelos de la Facultad de Agronomía
de la Universidad Nacional de Colombia, presentados en numeral 8.1 del Anexo B de
este trabajo.
ElSaltoMichaelRuizSantanaBoyacá

9 Anexo C: Soporte Estadístico del
Experimento Con Inóculo Comercial en
Yopal
9.1 Response Root Dry Weight (g) Summary of Fit
RSquare 0,754791 RSquare Adj 0,73303 Root Mean Square Error 306,8411 Mean of Response 767,558 Observations (or Sum Wgts) 504 Fixed Effect Tests Source Nparm DF DFDen F Ratio Prob > F
AMF 1 1 6 0,1652 0,6985 P level 2 2 456 0,9419 0,3906 Time 6 6 456 222,8870 <,0001* AMF*P level 2 2 456 0,3006 0,7405 AMF*Time 6 6 456 3,9603 0,0007* P level*Time 12 12 456 1,3641 0,1797 AMF*P level*Time 12 12 456 0,2903 0,9908
9.2 Response Plant Dry Weight (g) Summary of Fit
RSquare 0,782504 RSquare Adj 0,763202 Root Mean Square Error 523,1633 Mean of Response 1506,507 Observations (or Sum Wgts) 504 Fixed Effect Tests Source Nparm DF DFDen F Ratio Prob > F
AMF 1 1 6 0,0053 0,9443 P level 2 2 456 1,5327 0,2171 Time 6 6 456 255,0628 <,0001* AMF*P level 2 2 456 0,5863 0,5568 AMF*Time 6 6 456 2,7584 0,0121* P level*Time 12 12 456 1,6839 0,0673 AMF*P level*Time 12 12 456 0,5596 0,8744

Anexo C: Soporte Estadístico del Experimento Con Inóculo Comercial en Yopal 105
Modelos de regresión y gráficas complementarias para los análisis del BI a lo largo del ciclo del cultivo
9.3 Modelo de regresión para el BI en el tiempo (nivel de P = 50%)
Summary of Fit
RSquare 0,345105 RSquare Adj 0,263243 Root Mean Square Error 0,32001 Mean of Response -0,06628 Observations (or Sum Wgts) 28
Polynomial Fit Degree=3
BI= -0,581626 + 0,150756*Sample time - 0,0219199*(Sample time-4)^2 - 0,0074729*(Sample time-4)^3 When BI=0, the equation has three real roots: x1 = 7.0395881, x2 = -2.8085451 and x3 = 3.8616386, where x: sample time After first derivation f´(x) = -(224187*x^2-1355098*x+325840)/10000000 When f´(x) = 0 ; x = 5.793631830598907. There is a maximum After second derivation f´´(x) = -(224187*x-677549)/5000000 When f´´(x) = 0; x = 677549/224187. There is a inflexion point in x = 3,022 Analysis of Variance Source DF Sum of Squares Mean Square F Ratio
Model 3 1,2951428 0,431714 4,2157 Error 24 2,4577493 0,102406 Prob > F
C. Total 27 3,7528921 0,0157*
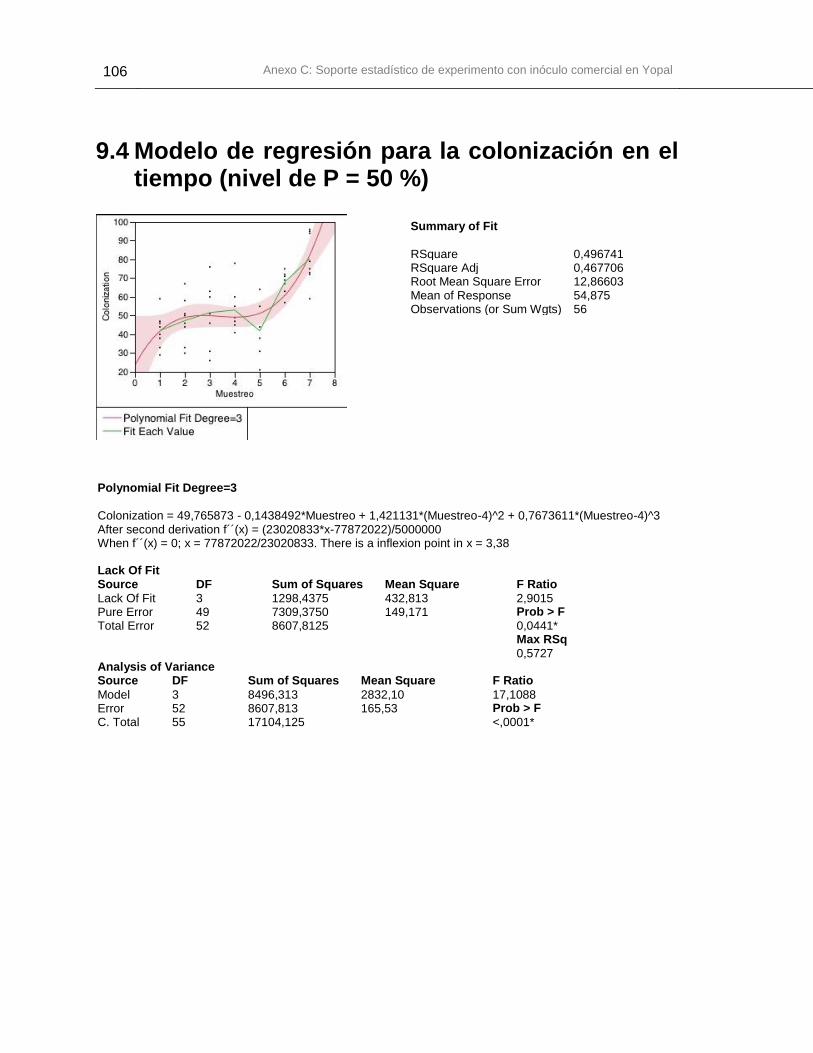
106 Anexo C: Soporte estadístico de experimento con inóculo comercial en Yopal Título de la tesis o trabajo de investigación
9.4 Modelo de regresión para la colonización en el tiempo (nivel de P = 50 %)
Summary of Fit
RSquare 0,496741 RSquare Adj 0,467706 Root Mean Square Error 12,86603 Mean of Response 54,875 Observations (or Sum Wgts) 56
Polynomial Fit Degree=3
Colonization = 49,765873 - 0,1438492*Muestreo + 1,421131*(Muestreo-4)^2 + 0,7673611*(Muestreo-4)^3 After second derivation f´´(x) = (23020833*x-77872022)/5000000 When f´´(x) = 0; x = 77872022/23020833. There is a inflexion point in x = 3,38 Lack Of Fit Source DF Sum of Squares Mean Square F Ratio
Lack Of Fit 3 1298,4375 432,813 2,9015 Pure Error 49 7309,3750 149,171 Prob > F
Total Error 52 8607,8125 0,0441* Max RSq
0,5727 Analysis of Variance Source DF Sum of Squares Mean Square F Ratio
Model 3 8496,313 2832,10 17,1088 Error 52 8607,813 165,53 Prob > F
C. Total 55 17104,125 <,0001*

Anexo C: Soporte Estadístico del Experimento Con Inóculo Comercial en Yopal 107
9.5 Beneficio de la inoculación a lo largo del ciclo del cultivo para las plantas fertilizadas con tres niveles de fertilización fosfatada
Each error bar is constructed using 1 standard error from the mean. Response IB Summary of Fit
RSquare 0,408505 RSquare Adj 0,220729 Root Mean Square Error 0,304463 Mean of Response -0,04401 Observations (or Sum Wgts) 84 Analysis of Variance Source DF Sum of Squares Mean Square F Ratio
Model 20 4,0332589 0,201663 2,1755 Error 63 5,8399596 0,092698 Prob > F C. Total 83 9,8732184 0,0102* Effect Tests Source Nparm DF Sum of Squares F Ratio Prob > F
Sample time 6 6 1,9122920 3,4382 0,0053* P level 2 2 1,3869548 7,4811 0,0012* P level*Sample time 12 12 1,5853637 1,4252 0,1784
108 Anexo C: Soporte estadístico de experimento con inóculo comercial en Yopal Título de la tesis o trabajo de investigación
LS Means Plot
LSMeans Differences Tukey HSD (α=0,050)
Level Least Sq Mean 7 A 0,1646583 6 A B 0,0783500 5 A B 0,0702000 4 A B -0,0378750 3 A B -0,0816333 1 B -0,2316750 2 B -0,2701250 Levels not connected by same letter are significantly different. P level LSMeans Differences Tukey HSD (α=0,050)
Level Least Sq Mean 0% A 0,2323250 50% B -0,3546000 100% B -0,5727500 Levels not connected by same letter are significantly differente

Anexo : Soporte estadístico del experimento de dosis en Yopal 109
10 Anexo D: Soporte Estadístico del Experimento
de Dosis en Yopal
10.1 Response Fresh_Above Ground Biomass(Kg) Summary of Fit
RSquare 0,378612 RSquare Adj 0,189976 Root Mean Square Error 0,30667 Mean of Response 0,640851 Observations (or Sum Wgts) 74 Fixed Effect Tests Source Nparm DF DFDen F Ratio Prob > F
Concentration 8 8 52,61 0,4862 0,8605 Dosis 1 1 52,98 1,2326 0,2719 Concentration*Dosis 8 8 52,79 2,1150 0,0505
10.2 Response Fresh_Root_Biomass Summary of Fit
RSquare 0,546781 RSquare Adj 0,392687 Root Mean Square Error 0,137234 Mean of Response 0,305904 Observations (or Sum Wgts) 68 Fixed Effect Tests Source Nparm DF DFDen F Ratio Prob > F
Concentration 8 8 47,93 6,4997 <,0001* Dosis 1 1 47,74 0,0538 0,8175 Concentration*Dosis 8 8 48,09 1,2087 0,3142

110 Anexo D: Soporte estadístico del experimento de dosis en Yopal Título de la tesis o trabajo de investigación
Effect Details Concentration LSMeans Differences Tukey HSD α=0,050 Level Least Sq Mean
50 A 0,51118971 75 A B 0,44950965 25 A B C 0,40171675 175 A B C D 0,32418212 100 B C D 0,27997185 150 B C D 0,24762541 200 B C D 0,20557711 125 C D 0,18388983 0 D 0,14910000 Levels not connected by same letter are significantly different.
10.3 Response Fresh_Total_Plant_Biomas(Kg) Summary of Fit
RSquare 0,547897 RSquare Adj 0,38437 Root Mean Square Error 0,352377 Mean of Response 0,976577 Observations (or Sum Wgts) 65 Fixed Effect Tests Source Nparm DF DFDen F Ratio Prob > F
Concentration 8 8 42,78 2,5407 0,0233* Dosis 1 1 42,46 0,7114 0,4037 Concentration*Dosis 8 8 43,01 1,8250 0,0985 Effect Details Concentration LSMeans Differences Tukey HSD α=0,050 Level Least Sq Mean
50 A 1,3614671 75 A B 1,1988378 25 A B 1,1171668 175 A B 1,0546926 125 A B 1,0309257 150 A B 0,8839313 100 A B 0,8415840 200 A B 0,7581101 0 B 0,7381000 Levels not connected by same letter are significantly different.

Anexo : Soporte estadístico del experimento de dosis en Yopal 111
10.4 Bivariate Fit of Fresh_Root_Biomass By Concentration
Polynomial Fit Degree=4
Fresh_Root_Biomass = 0,6832315 - 0,0039777*Concentration + 0,000039*(Concentration-93,3824)^2 + 5,4574e-7*(Concentration-93,3824)^3 - 5,8617e-9*(Concentration-93,3824)^4 Summary of Fit
RSquare 0,427853 RSquare Adj 0,391526 Root Mean Square Error 0,138358 Mean of Response 0,305904 Observations (or Sum Wgts) 68 Lack Of Fit Source DF Sum of Squares Mean Square F Ratio
Lack Of Fit 4 0,0936509 0,023413 1,2418 Pure Error 59 1,1123487 0,018853 Prob > F Total Error 63 1,2059996 0,3032 Max RSq Analysis of Variance
Source DF Sum of Squares Mean Square F Ratio Model 4 0,9018501 0,225463 11,7779 Error 63 1,2059996 0,019143 Prob > F C. Total 67 2,1078496 <,0001*

112 Anexo D: Soporte estadístico del experimento de dosis en Yopal Título de la tesis o trabajo de investigación
10.5 Bivariate Fit of Fresh Total Plant Biomass (Kg) By Concentration
Polynomial Fit Degree=4
Fresh_Total_Plant_Biomas(Kg) = 1,4466134 - 0,0052848*Concentration + 9,4454e-5*(Concentration-92,6923)^2 + 7,7788e-7*(Concentration-92,6923)^3 - 1,2588e-8*(Concentration-92,6923)^4
Summary of Fit
RSquare 0,177343 RSquare Adj 0,122499 Root Mean Square Error 0,407461 Mean of Response 0,976577 Observations (or Sum Wgts) 65 Lack Of Fit Source DF Sum of Squares Mean Square F Ratio
Lack Of Fit 4 0,5717445 0,142936 0,8525 Pure Error 56 9,3897315 0,167674 Prob > F Total Error 60 9,9614759 0,4982 Max RSq Analysis of Variance Source DF Sum of Squares Mean Square F Ratio Model 4 2,147433 0,536858 3,2336 Error 60 9,961476 0,166025 Prob > F C. Total 64 12,108909 0,0181*

Anexo : Soporte estadístico del experimento de dosis en Yopal 113
10.6 Response AMF Total Colonization (%) Response Arc sin Colonization Summary of Fit RSquare 0,147651 RSquare Adj 0,099405 Root Mean Square Error 0,267311 Mean of Response 0,771774 Observations (or Sum Wgts) 57 REML Variance Component Estimates Random Effect Var Ratio Var Component Std Error 95% Lower 95% Upper Pct of Total Random Block -0,032553 -0,002326 0,0029829 -0,008172 0,0035202 -3,365 Residual 0,071455 0,0143576 0,0499488 0,1106642 103,365 Total 0,0691289 100,000 -2 LogLikelihood = 41,540615174 Fixed Effect Tests Source Nparm DF DFDen F Ratio Prob > F Concentration 1 1 49,48 3,3112 0,0749 Dosis 1 1 52,07 8,9395 0,0043* Concentration*Dosis 1 1 52,73 1,0608 0,3077 Effect Details Dosis LSMeans Differences Student's t α= 0,050 Level Least Sq Mean 2 A 0,87785652 1 B 0,66394512 Levels not connected by same letter are significantly different.

11 Anexo E: Soporte Estadístico de las Dos
Repeticiones del Experimento de Líneas de
Rhizophagus irregularis en Yopal
11.1 Contingency Analysis of Supervivencia By Time and Variety
Contingency Analysis of Supervivencia By Replica Mosaic Plot
Tests
N DF -LogLike RSquare (U) 972 1 6,8205258 0,0105
Test ChiSquare Prob>ChiSq Likelihood Ratio 13,641 0,0002* Pearson 13,602 0,0002* Fisher's Exact Test
Prob Alternative Hypothesis
Left 0,9999 Prob(Supervivencia=viva) is greater for Replica=Rep 1 than Rep 2 Right 0,0001* Prob(Supervivencia=viva) is greater for Replica=Rep 2 than Rep 1 2-Tail 0,0003* Prob(Supervivencia=viva) is different across Replica

Anexo E : Soporte estadístico del experimento de líneas en Yopal 115
Analysis of Means for Proportions
Contingency Analysis of Supervivencia By Cassava variety Mosaic Plot
Tests
N DF -LogLike RSquare (U) 972 2 59,439531 0,0916
Test ChiSquare Prob>ChiSq Likelihood Ratio 118,879 <,0001* Pearson 116,833 <,0001* Analysis of Means for Proportions

116 Anexo E: Soporte estadístico del experimento de líneas en Yopal Título de la tesis o trabajo de investigación
11.2 Contingency Analysis of Superviencia By Linea de HFMA Variedad de yuca=MCOL2737
Tests
N DF -LogLike RSquare (U) 324 17 10,072931 0,0615
Test ChiSquare Prob>ChiSq Likelihood Ratio 20,146 0,2668 Pearson 19,180 0,3183
Analysis of Means for Proportions
11.3 Contingency Analysis of Superviencia By Linea de HFMA Variedad de yuca=CM4574
Tests
N DF -LogLike RSquare (U) 324 17 8,3007464 0,0397
Test ChiSquare Prob>ChiSq Likelihood Ratio 16,601 0,4817 Pearson 16,484 0,4898

Anexo E : Soporte estadístico del experimento de líneas en Yopal 117
Analysis of Means for Proportions
11.4 Contingency Analysis of Superviencia By Linea de HFMA Variedad de yuca=CM6438
Tests
N DF -LogLike RSquare (U) 324 17 9,8276428 0,0454
Test ChiSquare Prob>ChiSq Likelihood Ratio 19,655 0,2922 Pearson 19,169 0,3189 Analysis of Means for Proportions

118 Anexo E: Soporte estadístico del experimento de líneas en Yopal Título de la tesis o trabajo de investigación
11.5 Response Root fresh weight per plant (kg) _ 2 repeticiones en Yopal
Growth variable: Root fresh weight per plant (Kg per plant)
Variable measured in both years
Statistical analysis for both years Random factors and levels
Time: Time 1, Time 2 Blocks: 1,2, 3, 4, 5, 6, 7, 8, 9.
Fixed factors and levels
AMF line (D1, C2, C3, S4, Sc1, Sc2, S4a, S4b, Sc1a, Sc1d, Sc1e, Sc2a, Sc2b, Sc2c, Sc2d, GLO, FIL, H2O). Cassava variety (MCOL2737, CM4574)
Residuals data is from the Normal distribution. Residuals variances are equal. REML recomeded method
Summary of Fit
RSquare 0,286859 RSquare Adj 0,229348 Root Mean Square Error 0,728118 Mean of Response 1,156704 Observations (or Sum Wgts) 470 RSquare without “Time” as a factor in the model: 0,292853 REML Variance Component Estimates Random Effect
Var Ratio Var Component Std Error 95% Lower 95% Upper Pct of Total
Block 0,0761953 0,0403954 0,0251768 -0,00895 0,089741 7,102 Time -0,003311 -0,001755 0,0008386 -0,003399 -0,000112 -0,309 Residual 0,5301565 0,0363483 0,4655505 0,6092854 93,207 Total 0,5687967 100,000 -2 LogLikelihood = 1095,9518911 Fixed Effect Tests Source Nparm DF DFDen F Ratio Prob > F
Cassava variety 1 1 426,9 44,6961 <,0001* AMF line 17 17 425,3 2,5375 0,0007* AMF line*Cassava variety 17 17 423,1 2,9382 <,0001* Effect Details Block LSMeans Differences Tukey HSD α= 0,050 Level Least Sq Mean
5 A 1,3978163 6 A 1,3927407 7 A B 1,3208232 1 A B C 1,2388578 2 A B C 1,2176000 4 A B C 1,1994239 8 A B C 1,1539952 3 B C 0,9383569 9 C 0,8933038
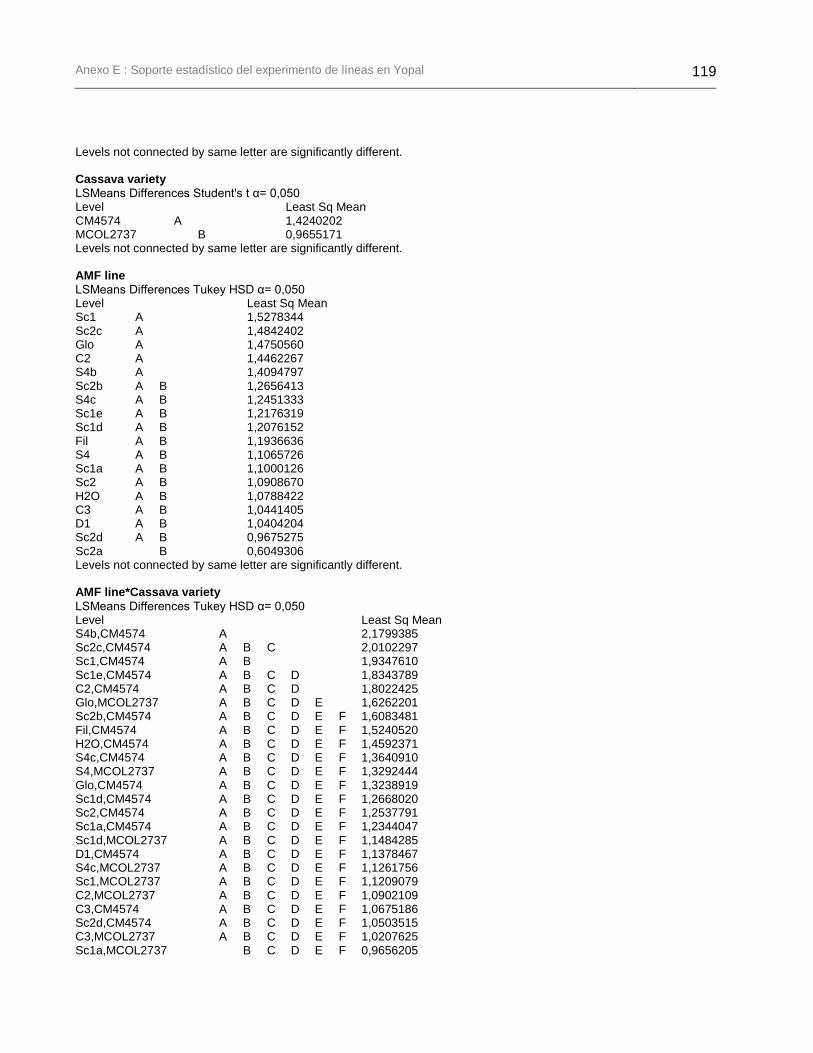
Anexo E : Soporte estadístico del experimento de líneas en Yopal 119
Levels not connected by same letter are significantly different. Cassava variety
LSMeans Differences Student's t α= 0,050 Level Least Sq Mean CM4574 A 1,4240202 MCOL2737 B 0,9655171 Levels not connected by same letter are significantly different. AMF line
LSMeans Differences Tukey HSD α= 0,050 Level Least Sq Mean Sc1 A 1,5278344 Sc2c A 1,4842402 Glo A 1,4750560 C2 A 1,4462267 S4b A 1,4094797 Sc2b A B 1,2656413 S4c A B 1,2451333 Sc1e A B 1,2176319 Sc1d A B 1,2076152 Fil A B 1,1936636 S4 A B 1,1065726 Sc1a A B 1,1000126 Sc2 A B 1,0908670 H2O A B 1,0788422 C3 A B 1,0441405 D1 A B 1,0404204 Sc2d A B 0,9675275 Sc2a B 0,6049306 Levels not connected by same letter are significantly different. AMF line*Cassava variety
LSMeans Differences Tukey HSD α= 0,050 Level Least Sq Mean S4b,CM4574 A 2,1799385 Sc2c,CM4574 A B C 2,0102297 Sc1,CM4574 A B 1,9347610 Sc1e,CM4574 A B C D 1,8343789 C2,CM4574 A B C D 1,8022425 Glo,MCOL2737 A B C D E 1,6262201 Sc2b,CM4574 A B C D E F 1,6083481 Fil,CM4574 A B C D E F 1,5240520 H2O,CM4574 A B C D E F 1,4592371 S4c,CM4574 A B C D E F 1,3640910 S4,MCOL2737 A B C D E F 1,3292444 Glo,CM4574 A B C D E F 1,3238919 Sc1d,CM4574 A B C D E F 1,2668020 Sc2,CM4574 A B C D E F 1,2537791 Sc1a,CM4574 A B C D E F 1,2344047 Sc1d,MCOL2737 A B C D E F 1,1484285 D1,CM4574 A B C D E F 1,1378467 S4c,MCOL2737 A B C D E F 1,1261756 Sc1,MCOL2737 A B C D E F 1,1209079 C2,MCOL2737 A B C D E F 1,0902109 C3,CM4574 A B C D E F 1,0675186 Sc2d,CM4574 A B C D E F 1,0503515 C3,MCOL2737 A B C D E F 1,0207625 Sc1a,MCOL2737 B C D E F 0,9656205

120 Anexo E: Soporte estadístico del experimento de líneas en Yopal Título de la tesis o trabajo de investigación
Level Least Sq Mean Sc2c,MCOL2737 B C D E F 0,9582508 D1,MCOL2737 B C D E F 0,9429941 Sc2,MCOL2737 B C D E F 0,9279549 Sc2b,MCOL2737 B C D E F 0,9229346 Sc2d,MCOL2737 B C D E F 0,8847035 S4,CM4574 A B C D E F 0,8839007 Fil,MCOL2737 C D E F 0,8632751 H2O,MCOL2737 E F 0,6984473 Sc2a,CM4574 D E F 0,6965894 S4b,MCOL2737 E F 0,6390209 Sc1e,MCOL2737 E F 0,6008849 Sc2a,MCOL2737 F 0,5132719 Levels not connected by same letter are significantly different.
11.6 Response Root dry weight per plant (Kg)_ 2 repeticiones en Yopal
Growth variable: Root dry weight per plant (Kg per plant)
Variable measured in both years
Statistical analysis for both years Random factors and levels
Time: Time 1, Time 2 Blocks: 1,2, 3, 4, 5, 6, 7, 8, 9.
Fixed factors and levels
AMF line (D1, C2, C3, S4, Sc1, Sc2, S4a, S4b, Sc1a, Sc1d, Sc1e, Sc2a, Sc2b, Sc2c, Sc2d, GLO, FIL, H2O). Cassava variety (MCOL2737, CM4574)
Residuals data is from the Normal distribution. Residuals variances are equal. REML (Recommended method) Response Root dry weight per plant (Kg) Summary of Fit
RSquare 0,360457 RSquare Adj 0,30888 Root Mean Square Error 0,201305 Mean of Response 0,310423 Observations (or Sum Wgts) 470 RSquare without “Time” as a factor in the model: 0,364234 REML Variance Component Estimates Random Effect
Var Ratio Var Component Std Error 95% Lower 95% Upper Pct of Total
Block 0,0741035 0,003003 0,00188 -0,000682 0,0066877 6,917 Time -0,002828 -0,000115 9,1571e-5 -0,000294 0,0000649 -0,264 Residual 0,0405238 0,0027802 0,0355825 0,0465767 93,347 Total 0,0434122 100,000 -2 LogLikelihood = -19,79654191 Fixed Effect Tests Source Nparm DF DFDen F Ratio Prob > F

Anexo E : Soporte estadístico del experimento de líneas en Yopal 121
Source Nparm DF DFDen F Ratio Prob > F
Cassava variety 1 1 426,4 101,4548 <,0001* AMF line 17 17 425,8 2,6706 0,0004* AMF line*Cassava variety 17 17 425,1 3,4128 <,0001* Effect Details Block LSMeans Differences Tukey HSD α=0,050 Level Least Sq Mean
6 A 0,37882563 5 A 0,37812329 7 A B 0,35437428 2 A B C 0,33373678 1 A B C 0,32673164 4 A B C 0,32659137 8 A B C 0,30911724 3 B C 0,25649838 9 C 0,23873875 Levels not connected by same letter are significantly different. Cassava variety LSMeans Differences Student's t α= 0,050 Level Least Sq Mean
CM4574 A 0,41801666 MCOL2737 B 0,22703608 Levels not connected by same letter are significantly different. AMF line LSMeans Differences Tukey HSD α= 0,050
Level Least Sq Mean
Sc2 A 0,43497341 C2 A 0,40224603 Sc2c A 0,39699760 Glo A 0,38625300 S4b A 0,38014870 Sc1 A 0,37792140 Sc2b A B 0,33665558 Fil A B 0,32589004 S4c A B 0,31714465 Sc1e A B 0,30503990 Sc1a A B 0,29456738 Sc1d A B 0,29380985 S4 A B 0,29129311 H2O A B 0,28267694 D1 A B 0,28246980 Sc2d A B 0,26435040 C3 A B 0,26267067 Sc2a B 0,17036628 Levels not connected by same letter are significantly different. AMF line*Cassava variety
LSMeans Differences Tukey HSD α= 0,050 Level Least Sq Mean
Sc2,CM4574 A 0,65118098 S4b,CM4574 A B 0,60853154

122 Anexo E: Soporte estadístico del experimento de líneas en Yopal Título de la tesis o trabajo de investigación
Level Least Sq Mean
Sc2c,CM4574 A B C 0,57894980 C2,CM4574 A B C D 0,55204261 Sc1,CM4574 A B C D E 0,50620727 Sc1e,CM4574 A B C D E F 0,46952013 Sc2b,CM4574 A B C D E F G 0,46253906 Fil,CM4574 A B C D E F G H 0,44034088 H2O,CM4574 A B C D E F G H 0,39988491 Glo,CM4574 A B C D E F G H 0,38890719 Glo,MCOL2737 A B C D E F G H 0,38359880 S4c,CM4574 A B C D E F G H 0,38069589 D1,CM4574 A B C D E F G H 0,34986542 Sc1a,CM4574 A B C D E F G H 0,34877981 S4,MCOL2737 A B C D E F G H 0,34368388 Sc1d,CM4574 A B C D E F G H 0,33272834 Sc2d,CM4574 B C D E F G H 0,32782125 C3,MCOL2737 C D E F G H 0,26342326 C3,CM4574 C D E F G H 0,26191809 Sc1d,MCOL2737 C D E F G H 0,25489136 S4c,MCOL2737 D E F G H 0,25359340 C2,MCOL2737 C D E F G H 0,25244945 Sc1,MCOL2737 D E F G H 0,24963553 Sc1a,MCOL2737 D E F G H 0,24035495 S4,CM4574 C D E F G H 0,23890234 Sc2a,CM4574 E F G H 0,22548442 Sc2,MCOL2737 E F G H 0,21876583 D1,MCOL2737 E F G H 0,21507418 Sc2c,MCOL2737 E F G H 0,21504540 Fil,MCOL2737 E F G H 0,21143919 Sc2b,MCOL2737 E F G H 0,21077209 Sc2d,MCOL2737 E F G H 0,20087955 H2O,MCOL2737 F G H 0,16546897 S4b,MCOL2737 G H 0,15176587 Sc1e,MCOL2737 G H 0,14055966 Sc2a,MCOL2737 H 0,11524814 Levels not connected by same letter are significantly different.
11.7 Response IB_ Root fresh weight per plant (kg) - 2 years
Summary of Fit
RSquare 0,270392 RSquare Adj 0,211124 Root Mean Square Error 75,43687 Mean of Response 15,94504 Observations (or Sum Wgts) 387 RSquare without “Time” as a factor in the model: 0,265337 REML Variance Component Estimates Random Effect Var Ratio Var Component Std Error 95% Lower 95% Upper Pct of Total
Random Block 0,1112152 632,89476 384,10651 -119,9402 1385,7297 9,926 Replica 0,0092414 52,59005 117,60224 -177,9061 283,0862 0,825 Residual 5690,7208 431,42747 4931,487 6640,8489 89,249 Total 6376,2056 100,000

Anexo E : Soporte estadístico del experimento de líneas en Yopal 123
-2 LogLikelihood = 4222,7819793 Fixed Effect Tests Source Nparm DF DFDen F Ratio Prob > F
Cassava variety 1 1 349,4 14,0454 0,0002* AMF line 14 14 348,8 1,7986 0,0374* AMF line*Cassava variety 14 14 348,9 2,6535 0,0011* Effect Details
Cassava variety LSMeans Differences Student's t α= 0,050 Level Least Sq Mean
MCOL2737 A 31,010819 COL4574 B 1,729836 AMF line LSMeans Differences Tukey HSD α= 0,050 Level Least Sq Mean
C2 A 43,06103 Sc1 A 39,81493 S4 A B 32,15339 Sc2c A B 31,05385 S4c A B 23,90405 S4b A B 22,79831 Sc1a A B 21,33597 Sc2b A B 19,08143 Sc1d A B 17,07642 Sc2 A B 14,87186 C3 A B 11,55357 D1 A B 7,66594 Sc1e A B -0,37561 Sc2d A B -1,30050 Sc2a B -37,13971 AMF line*Cassava variety LSMeans Differences Tukey HSD α= 0,050 Level Least Sq Mean
S4,MCOL2737 A 104,3478 C3,MCOL2737 A B 54,4771 S4b,COL4574 A B 54,2399 Sc1a,MCOL2737 A B 53,5392 S4c,MCOL2737 A B 50,8923 Sc1,MCOL2737 A B 49,4980 Sc1d,MCOL2737 A B 49,4618 C2,MCOL2737 A B 47,3799 Sc2c,COL4574 A B 45,4331 C2,COL4574 A B 38,7422 Sc2,MCOL2737 A B 32,6036 Sc1,COL4574 A B 30,1318 D1,MCOL2737 A B 25,9831 Sc2b,MCOL2737 A B 22,0024 Sc1e,COL4574 A B 17,9586 Sc2c,MCOL2737 A B 16,6746 Sc2b,COL4574 A B 16,1604 Sc2d,MCOL2737 A B 14,9465 Sc2,COL4574 A B -2,8599 S4c,COL4574 A B -3,0842 S4b,MCOL2737 B -8,6432

124 Anexo E: Soporte estadístico del experimento de líneas en Yopal Título de la tesis o trabajo de investigación
Level Least Sq Mean
D1,COL4574 B -10,6513 Sc1a,COL4574 B -10,8673 Sc1d,COL4574 B -15,3090 Sc2d,COL4574 B -17,5475 Sc1e,MCOL2737 B -18,7098 Sc2a,MCOL2737 B -29,2909 C3,COL4574 B -31,3700 S4,COL4574 B -40,0410 Sc2a,COL4574 B -44,9885 Levels not connected by same letter are significantly different.
11.8 Response IB_ Root dry weight per plant (kg). (2 years)
Summary of Fit
RSquare 0,257273 RSquare Adj 0,19694 Root Mean Square Error 74,86156 Mean of Response 14,09855 Observations (or Sum Wgts) 387 RSquare without “Time” as a factor in the model: 0,256053 REML Variance Component Estimates Random Effect Var Ratio Var Component Std Error 95% Lower 95% Upper Pct of Total
Random Block 0,1042659 584,3327 358,74442 -118,7934 1287,4589 9,428 Replica 0,0016906 9,4743403 55,995219 -100,2743 119,22295 0,153 Residual 5604,2537 424,89719 4856,5166 6540,0069 90,419 Total 6198,0607 100,000 -2 LogLikelihood = 4216,1663722 Fixed Effect Tests Source Nparm DF DFDen F Ratio Prob > F
Cassava variety 1 1 349,4 16,8490 <,0001* AMF line 14 14 348,9 1,8612 0,0294* AMF line*Cassava variety 14 14 349 2,1056 0,0112* Effect Details
Cassava variety LSMeans Differences Student's t α= 0,050 Level Least Sq Mean
MCOL2737 A 29,76830 COL4574 B -2,05752 AMF line LSMeans Differences Tukey HSD α=0,050 Level Least Sq Mean
Sc1 A 45,54890 C2 A 36,60400 Sc2c A B 27,76713 S4c A B 26,21211 Sc1d A B 22,99561

Anexo E : Soporte estadístico del experimento de líneas en Yopal 125
Level Least Sq Mean
S4b A B 19,36476 Sc2b A B 18,40442 S4 A B 17,26487 Sc1a A B 15,81626 Sc2 A B 8,01668 D1 A B 4,63704 C3 A B 4,32463 Sc1e A B 3,45151 Sc2d A B -3,75637 Sc2a B -38,82073 AMF line*Cassava variety LSMeans Differences Tukey HSD α=0,050 Level Least Sq Mean
S4,MCOL2737 A 74,82800 S4c,MCOL2737 A 58,42800 Sc1d,MCOL2737 A 58,15902 Sc1,MCOL2737 A 57,36802 C2,MCOL2737 A B 49,34883 S4b,COL4574 A B 48,96437 Sc1a,MCOL2737 A B 46,24171 Sc2c,COL4574 A B 37,38085 Sc1,COL4574 A B 33,72978 C3,MCOL2737 A B 32,85756 D1,MCOL2737 A B 30,52064 Sc2,MCOL2737 A B 27,14645 Sc2b,MCOL2737 A B 26,32558 Sc1e,COL4574 A B 24,32862 C2,COL4574 A B 23,85917 Sc2d,MCOL2737 A B 18,98174 Sc2c,MCOL2737 A B 18,15341 Sc2b,COL4574 A B 10,48326 S4c,COL4574 A B -6,00378 S4b,MCOL2737 A B -10,23486 Sc2,COL4574 A B -11,11309 Sc1d,COL4574 A B -12,16780 Sc1a,COL4574 A B -14,60919 Sc1e,MCOL2737 A B -17,42560 D1,COL4574 A B -21,24657 Sc2a,MCOL2737 A B -24,17400 C3,COL4574 A B -24,20830 Sc2d,COL4574 A B -26,49448 S4,COL4574 A B -40,29826 Sc2a,COL4574 B -53,46745 Levels not connected by same letter are significantly different.
11.9 Response: Root dry and fresh weight per plant (Kg) Cassava variety=MCOL2737 (two years)

126 Anexo E: Soporte estadístico del experimento de líneas en Yopal Título de la tesis o trabajo de investigación
Response Root fresh weight per plant (kg) Cassava variety=MCOL2737 Summary of Fit RSquare 0,334885 RSquare Adj 0,287773 Root Mean Square Error 0,590964 Mean of Response 0,954275 Observations (or Sum Wgts) 258 REML Variance Component Estimates Random Effect Var Ratio Var Component Std Error 95% Lower 95% Upper Pct of Total Block 0,1293831 0,0451856 0,0289224 -0,011501 0,1018725 9,487 Time 0,2343508 0,0818444 0,1196903 -0,152744 0,3164331 17,184 Residual 0,3492387 0,0324967 0,2933713 0,4228282 73,328 Total 0,4762686 100,000 -2 LogLikelihood = 497,86022292 Fixed Effect Tests Source Nparm DF DFDen F Ratio Prob > F AMF line 17 17 231,7 2,6598 0,0005* Effect Details AMF line LSMeans Differences Tukey HSD α= 0,050 Level Least Sq Mean Glo A 1,6021595 S4 A B 1,2581737 S4c A B 1,1113959 Sc1d A B 1,1032559 Sc1 A B 1,0838001 C2 A B 1,0702351 Sc1a A B 1,0030243 C3 A B 0,9970981 D1 A B 0,9485437 Sc2b A B 0,9021820 Sc2c A B 0,8827673 Sc2 A B 0,8726910 Fil B 0,8550737 Sc2d A B 0,8386058 H2O B 0,7202672 S4b B 0,6451313 Sc1e B 0,6049820 Sc2a B 0,5062296 Levels not connected by same letter are significantly different. Response Root dry weight per plant (Kg) Cassava variety=MCOL2737 Summary of Fit RSquare 0,351506 RSquare Adj 0,305571 Root Mean Square Error 0,13853 Mean of Response 0,225713 Observations (or Sum Wgts) 258 REML Variance Component Estimates Random Effect Var Ratio Var Component Std Error 95% Lower 95% Upper Pct of Total Block 0,1382876 0,0026538 0,0016765 -0,000632 0,0059397 10,023 Time 0,2413509 0,0046317 0,006767 -0,008631 0,0178947 17,494 Residual 0,0191907 0,0017858 0,0161207 0,0232346 72,483 Total 0,0264762 100,000 -2 LogLikelihood = -198,0105705 Fixed Effect Tests Source Nparm DF DFDen F Ratio Prob > F AMF line 17 17 231,6 2,9771 0,0001*

Anexo E : Soporte estadístico del experimento de líneas en Yopal 127
Effect Details AMF line LSMeans Differences Tukey HSD α= 0,050 Level Least Sq Mean Glo A 0,37800232 S4 A B 0,33237185 C3 A B C 0,26125483 S4c A B C 0,25203888 C2 A B C 0,24858919 Sc1d A B C 0,24786424 Sc1a A B C 0,24519995 Sc1 A B C 0,24451126 D1 A B C 0,21821640 Fil A B C 0,21080845 Sc2 A B C 0,21029602 Sc2b A B C 0,20780992 Sc2c A B C 0,20616742 Sc2d B C 0,19220897 H2O B C 0,16938460 S4b B C 0,15375310 Sc1e B C 0,14087597 Sc2a C 0,11265703 Levels not connected by same letter are significantly different.
11.10 Response: Root dry and fresh weight per plant (Kg) Cassava variety=CM4574 (2 years)
Response Root fresh weight per plant (kg) Cassava variety=CM4574 Summary of Fit RSquare 0,315045 RSquare Adj 0,255024 Root Mean Square Error 0,797827 Mean of Response 1,403057 Observations (or Sum Wgts) 212 REML Variance Component Estimates Random Effect Var Ratio Var Component Std Error 95% Lower 95% Upper Pct of Total Block 0,0873303 0,0555881 0,0422958 -0,02731 0,1384864 7,152 Time 0,1338147 0,0851767 0,1293299 -0,168305 0,3386586 10,958 Residual 0,6365274 0,0662122 0,5243549 0,7891917 81,890 Total 0,7772923 100,000 -2 LogLikelihood = 524,24868387 Fixed Effect Tests Source Nparm DF DFDen F Ratio Prob > F AMF line 17 17 186,4 3,1249 <,0001* Effect Details AMF line LSMeans Differences Tukey HSD α= 0,050 Level Least Sq Mean S4b A 2,2253311 Sc2c A B 2,0154661 Sc1 A B 1,9806421 Sc1e A B 1,8741194 C2 A B C 1,7894486
128 Anexo E: Soporte estadístico del experimento de líneas en Yopal Título de la tesis o trabajo de investigación
Level Least Sq Mean Sc2b A B C 1,6209562 Fil A B C 1,5736261 H2O A B C 1,4584996 S4c A B C 1,4034989 Glo A B C 1,3285089 Sc1d A B C 1,2821859 Sc2 A B C 1,2679960 Sc1a A B C 1,2670414 D1 A B C 1,1482542 C3 B C 1,0829995 Sc2d B C 1,0237494 S4 B C 0,8354563 Sc2a C 0,7190418 Levels not connected by same letter are significantly different. Response Root dry weight per plant (Kg) Cassava variety=CM4574 Summary of Fit RSquare 0,336145 RSquare Adj 0,277972 Root Mean Square Error 0,239536 Mean of Response 0,413514 Observations (or Sum Wgts) 212 REML Variance Component Estimates Random Effect Var Ratio Var Component Std Error 95% Lower 95% Upper Pct of Total Block 0,0903694 0,0051852 0,0038759 -0,002411 0,0127818 7,149 Time 0,1736618 0,0099643 0,0148915 -0,019223 0,0391512 13,739 Residual 0,0573776 0,0059668 0,0472687 0,0711344 79,112 Total 0,0725271 100,000 -2 LogLikelihood = 57,839471854 Fixed Effect Tests Source Nparm DF DFDen F Ratio Prob > F AMF line 17 17 186,4 3,2948 <,0001* Effect Details AMF line LSMeans Differences Tukey HSD α= 0,050 Level Least Sq Mean Sc2 A 0,65739779 S4b A 0,63428811 Sc2c A B 0,58273961 C2 A B 0,54486249 Sc1 A B 0,53125555 Sc1e A B 0,49049425 Sc2b A B 0,46261957 Fil A B 0,46176677 H2O A B 0,40358950 S4c A B 0,39745580 Glo A B 0,39057710 Sc1a A B 0,36470833 D1 A B 0,35944837 Sc1d A B 0,33752019 Sc2d A B 0,30965588 C3 B 0,27060966 Sc2a B 0,23088786 S4 B 0,22721453 Levels not connected by same letter are significantly different.

Anexo E : Soporte estadístico del experimento de líneas en Yopal 129
11.11 Response Arcsin AMF Total Colonization (2 years)
Growth variable: AMF Total Colonization (%)
Variable measured in both years.
Statistical analysis for both years Random factors and levels Time: Time 1, Time 2 Blocks: 1,2, 3, 4, 5, 6, 7, 8, 9. Fixed factors and levels
AMF line (D1, C2, C3, S4, Sc1, Sc2, S4a, S4b, Sc1a, Sc1d, Sc1e, Sc2a, Sc2b, Sc2c, Sc2d, GLO, FIL, H2O). Cassava variety (MCOL2737, CM4574)
Data were Arcsin transformed Response ArcSin AMF Total Colonization Summary of Fit RSquare 0,202267 RSquare Adj 0,083456 Root Mean Square Error 0,245972 Mean of Response 0,775985 Observations (or Sum Wgts) 271 RSquare without “Time” as a factor in the model: 0,169485
REML Variance Component Estimates Random Effect Var Ratio Var Component Std Error 95% Lower 95% Upper Pct of Total Time 0,0674212 0,0040791 0,0064302 -0,008524 0,0166821 6,316 Residual 0,0605021 0,0055934 0,0508783 0,0731562 93,684 Total 0,0645813 100,000 -2 LogLikelihood = 118,78875474 Fixed Effect Tests Source Nparm DF DFDen F Ratio Prob > F Cassava variety 1 1 234 0,3903 0,5328 AMF line 17 17 234,1 0,9142 0,5578 Cassava variety*AMF line 17 17 234 1,7245 0,0395* Effect Details Time LSMeans Differences Student's t α=0,050 Level Least Sq Mean Time 2 A 0,81531284 Time 1 B 0,72976052 Levels not connected by same letter are significantly different. Cassava variety*AMF line LSMeans Differences Student's t α= 0,050 Level Least Sq Mean CM4574,Sc1 A 0,98984260 MCOL2737,S4c A B 0,93811875 MCOL2737,C3 A B C 0,90230837 MCOL2737,Sc1d A B C 0,89580585 CM4574,C2 A B C D 0,87807473 CM4574,Sc2a A B C D 0,85927174 MCOL2737,Sc1e A B C D 0,85690458 MCOL2737,D1 A B C D 0,85322754

130 Anexo E: Soporte estadístico del experimento de líneas en Yopal Título de la tesis o trabajo de investigación
Level Least Sq Mean CM4574,S4 A B C D 0,84624361 MCOL2737,H2O A B C D 0,84508445 CM4574,Sc1e A B C D 0,84434449 CM4574,S4b A B C D 0,82780678 MCOL2737,Sc2c A B C D 0,82621691 MCOL2737,S4 A B C D E 0,81778157 MCOL2737,Glo A B C D E 0,80118257 CM4574,Sc2b A B C D E 0,79986575 CM4574,Sc2 A B C D E 0,79853184 MCOL2737,Sc1 A B C D E 0,79535884 MCOL2737,S4b A B C D E 0,79280972 MCOL2737,Fil A B C D E 0,78624745 CM4574,S4c A B C D E 0,76597293 CM4574,Glo A B C D E 0,74609640 CM4574,Sc1a B C D E 0,73816311 MCOL2737,Sc1a B C D E 0,73542793 MCOL2737,Sc2a B C D E 0,72511864 CM4574,H2O B C D E 0,72393771 CM4574,Sc2c B C D E 0,71567311 CM4574,Sc2d B C D E 0,69337321 MCOL2737,C2 C D E 0,66885083 CM4574,Fil C D E 0,66356465 MCOL2737,Sc2d C D E 0,65384891 CM4574,C3 C D E 0,64260913 MCOL2737,Sc2 D E 0,63227103 CM4574,Sc1d D E 0,61543385 CM4574,D1 E 0,58702706 MCOL2737,Sc2b E 0,54892387 Levels not connected by same letter are significantly different.
11.12 Response Arcsin Colonization (By year) Response ArcSin AMF Total Colonization Time=Time 1 (first year) Summary of Fit RSquare 0,619483 RSquare Adj 0,469841 Root Mean Square Error 0,192002 Mean of Response 0,714446 Observations (or Sum Wgts) 125 REML Variance Component Estimates Random Effect Var Ratio Var Component Std Error 95% Lower 95% Upper Pct of Total Block 0,0307948 0,0011352 0,00211 -0,003 0,0052707 2,987 Residual 0,0368647 0,0057284 0,0277859 0,0512782 97,013 Total 0,038 100,000 -2 LogLikelihood = 41,762373492 Fixed Effect Tests Source Nparm DF DFDen F Ratio Prob > F Cassava variety 1 1 85,41 0,0264 0,8713 AMF line 17 17 85,83 2,2165 0,0088* Cassava variety*AMF line 17 17 85,58 5,5611 <,0001* Effect Details Cassava variety*AMF line LSMeans Differences Tukey HSD α= 0,050 Level Least Sq Mean CM4574,Sc2a A 1,1149588 CM4574,C2 A B 1,0847542 MCOL2737,D1 A B C 1,0360806 CM4574,Sc1 A B C 1,0299290 MCOL2737,Sc1d A B C D 0,9536218 MCOL2737,S4c A B C D 0,9217790

Anexo E : Soporte estadístico del experimento de líneas en Yopal 131
Level Least Sq Mean MCOL2737,Glo A B C D 0,9130905 CM4574,S4b A B C D 0,9066609 CM4574,Sc2 A B C D 0,9065976 CM4574,Sc1e A B C D 0,8980363 MCOL2737,S4b A B C D 0,8578706 CM4574,S4c A B C D E 0,8427915 MCOL2737,Sc2c A B C D 0,8347442 CM4574,Glo A B C D E 0,8195410 MCOL2737,C3 A B C D E 0,8140249 CM4574,S4 A B C D E 0,7757262 CM4574,Sc2b A B C D E 0,7364519 MCOL2737,Sc1a A B C D E 0,7342317 MCOL2737,S4 A B C D E 0,7293556 MCOL2737,H2O A B C D E 0,7253601 MCOL2737,Fil A B C D E 0,6769178 CM4574,Sc1a A B C D E 0,6589761 MCOL2737,Sc2 A B C D E 0,6493634 CM4574,Sc2c A B C D E 0,6440505 MCOL2737,Sc2a A B C D E 0,6390051 MCOL2737,Sc1e A B C D E 0,6379488 MCOL2737,Sc1 A B C D E 0,6017219 MCOL2737,Sc2d A B C D E 0,5929765 CM4574,Sc1d A B C D E 0,5572518 CM4574,Fil A B C D E 0,5450481 CM4574,Sc2d B C D E 0,5202246 MCOL2737,C2 C D E 0,5064917 CM4574,H2O C D E 0,4424420 CM4574,C3 D E 0,4201173 MCOL2737,Sc2b D E 0,3998592 CM4574,D1 E 0,2177772 Levels not connected by same letter are significantly different. Response ArcSin AMF Total Colonization Time=Time 2 (second year) Summary of Fit RSquare 0,050624 RSquare Adj -0,25145 Root Mean Square Error 0,249246 Mean of Response 0,828673 Observations (or Sum Wgts) 146 REML Variance Component Estimates Random Effect Var Ratio Var Component Std Error 95% Lower 95% Upper Pct of Total Block -0,041269 -0,002564 0,0004829 -0,00351 -0,001617 -4,305 Residual 0,0621236 0,0084626 0,0483787 0,0827218 104,305 Total 0,0595598 100,000 -2 LogLikelihood = 83,88939155 Fixed Effect Tests Source Nparm DF DFDen F Ratio Prob > F Cassava variety 1 1 109,7 0,0839 0,7726 AMF line 17 17 62,86 1,1565 0,3254 Cassava variety*AMF line 17 17 61,21 0,7974 0,6892
11.13 Response Root dry weight per plant (Kg) Cassava variety=MCOL2737 for lines C2, C3 and D1

132 Anexo E: Soporte estadístico del experimento de líneas en Yopal Título de la tesis o trabajo de investigación
Response Root dry weight per plant (Kg) Cassava variety=MCOL2737 Summary of Fit RSquare 0,720295 RSquare Adj 0,704312 Root Mean Square Error 0,111266 Mean of Response 0,238921 Observations (or Sum Wgts) 38
RSquare without “Time” as a factor in the model: 0,631556 REML Variance Component Estimates Random Effect Var Ratio Var Component Std Error 95% Lower 95% Upper Pct of Total Block 1,427477 0,0176723 0,0105127 -0,002932 0,0382769 49,073 Time 0,4814142 0,00596 0,009434 -0,01253 0,0244502 16,550 Residual 0,0123801 0,003414 0,0076964 0,023154 34,377 Total 0,0360124 100,000 -2 LogLikelihood = -27,27838726 Fixed Effect Tests Source Nparm DF DFDen F Ratio Prob > F AMF line 2 2 27,24 0,5660 0,5743
11.14 Response Root dry weight per plant (Kg) Cassava variety=CM4574. Lines = C2, C3 and D1
Response Root dry weight per plant (Kg) Cassava variety=CM4574 Summary of Fit RSquare 0,543969 RSquare Adj 0,51791 Root Mean Square Error 0,227679 Mean of Response 0,378105 Observations (or Sum Wgts) 38
RSquare without “Time” as a factor in the model: 0,504733 REML Variance Component Estimates Random Effect Var Ratio Var Component Std Error 95% Lower 95% Upper Pct of Total Block 0,5834641 0,0302456 0,0236925 -0,016191 0,076682 35,046 Time 0,0813847 0,0042188 0,010288 -0,015945 0,024383 4,888 Residual 0,0518379 0,0147188 0,0318308 0,0990663 60,066 Total 0,0863023 100,000 -2 LogLikelihood = 16,114651767 Fixed Effect Tests Source Nparm DF DFDen F Ratio Prob > F AMF line 2 2 25,94 6,1907 0,0063* Effect Details AMF line Least Squares Means Table Level Least Sq Mean Std Error C2 0,56516035 0,10173762 C3 0,22984403 0,09935272 D1 0,34118482 0,09716052

Anexo E : Soporte estadístico del experimento de líneas en Yopal 133
LSMeans Differences Tukey HSD α=0,050 Level Least Sq Mean C2 A 0,56516035 D1 A B 0,34118482 C3 B 0,22984403 Levels not connected by same letter are significantly different.
11.15 Contrates entre grupos de líneas de Rhizophagus irregularis: ¿El efecto de las líneas obtenidas del cultivo de un mismo parental es diferente entre ellas o al de su parental?
Grupo línea parental C2
Parental Descendencia Proceso en laboratorio
C2 S4 – 1era generación Cruce de líneas C2 x C3
S4b – 2nda generación Cultivo monospórico de línea S4
S4c – 2nda generación Cultivo monospórico de línea S4
Analysis for both cassava varieties
Figura 23. Producción de plantas de yuca de las variedades MCOL2737 y CM4574 que fueron inoculadas con la línea parental
C2 y sus líneas descendientes. C2: Línea parental; S4, S4b y S4c: Líneas obtenidas en el laboratorio a partir de C2. Las barras
de error se construyen con 1 error estándar de la media.
Response Root dry weight per plant (Kg) Summary of Fit RSquare 0,323096 RSquare Adj 0,273739 Root Mean Square Error 0,224353 Mean of Response 0,346221 Observations (or Sum Wgts) 104 RSquare without “Time” as a factor in the model: 0,339736
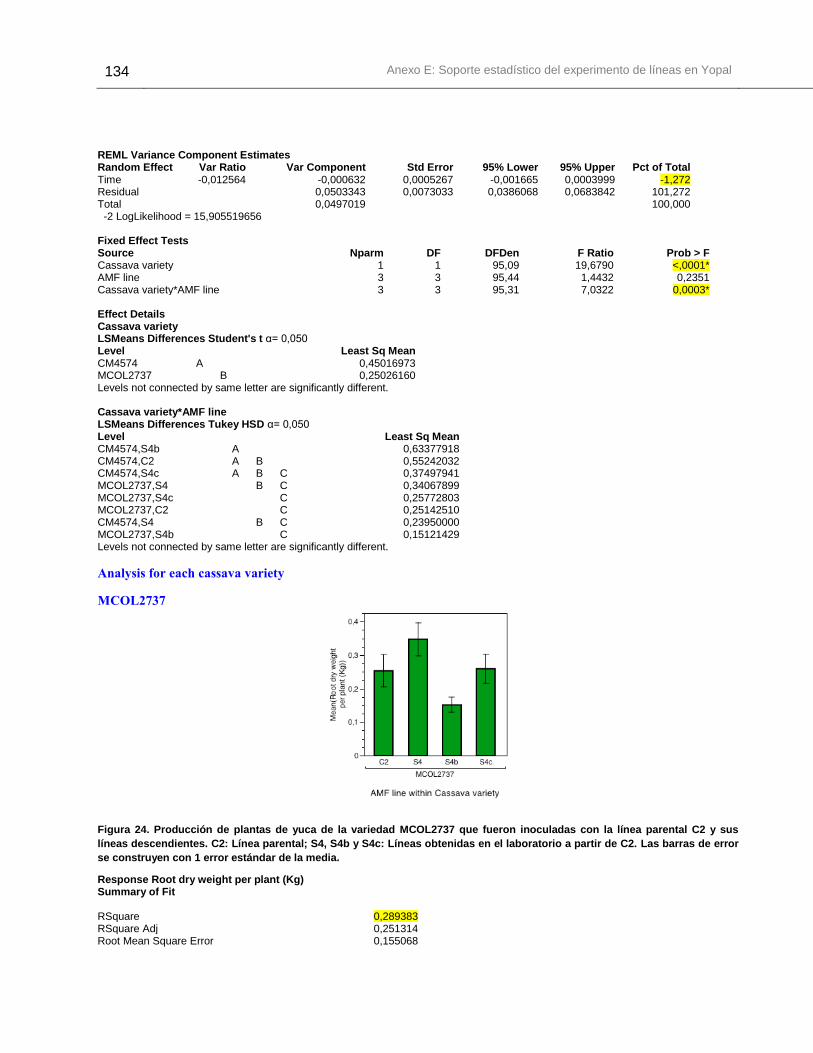
134 Anexo E: Soporte estadístico del experimento de líneas en Yopal Título de la tesis o trabajo de investigación
REML Variance Component Estimates Random Effect Var Ratio Var Component Std Error 95% Lower 95% Upper Pct of Total Time -0,012564 -0,000632 0,0005267 -0,001665 0,0003999 -1,272 Residual 0,0503343 0,0073033 0,0386068 0,0683842 101,272 Total 0,0497019 100,000 -2 LogLikelihood = 15,905519656 Fixed Effect Tests Source Nparm DF DFDen F Ratio Prob > F Cassava variety 1 1 95,09 19,6790 <,0001* AMF line 3 3 95,44 1,4432 0,2351 Cassava variety*AMF line 3 3 95,31 7,0322 0,0003* Effect Details Cassava variety LSMeans Differences Student's t α= 0,050 Level Least Sq Mean CM4574 A 0,45016973 MCOL2737 B 0,25026160 Levels not connected by same letter are significantly different. Cassava variety*AMF line LSMeans Differences Tukey HSD α= 0,050 Level Least Sq Mean CM4574,S4b A 0,63377918 CM4574,C2 A B 0,55242032 CM4574,S4c A B C 0,37497941 MCOL2737,S4 B C 0,34067899 MCOL2737,S4c C 0,25772803 MCOL2737,C2 C 0,25142510 CM4574,S4 B C 0,23950000 MCOL2737,S4b C 0,15121429 Levels not connected by same letter are significantly different.
Analysis for each cassava variety
MCOL2737
Figura 24. Producción de plantas de yuca de la variedad MCOL2737 que fueron inoculadas con la línea parental C2 y sus
líneas descendientes. C2: Línea parental; S4, S4b y S4c: Líneas obtenidas en el laboratorio a partir de C2. Las barras de error
se construyen con 1 error estándar de la media.
Response Root dry weight per plant (Kg) Summary of Fit RSquare 0,289383 RSquare Adj 0,251314 Root Mean Square Error 0,155068

Anexo E : Soporte estadístico del experimento de líneas en Yopal 135
Mean of Response 0,253017 Observations (or Sum Wgts) 60 RSquare without “Time” as a factor in the model: 0,134246 REML Variance Component Estimates Random Effect Var Ratio Var Component Std Error 95% Lower 95% Upper Pct of Total Block 0,0279236 0,0006715 0,0023051 -0,003846 0,0051894 2,192 Time 0,2462403 0,0059211 0,0095466 -0,01279 0,0246321 19,326 Residual 0,0240461 0,0049737 0,0166491 0,0377784 78,483 Total 0,0306386 100,000 -2 LogLikelihood = -32,7851609 Fixed Effect Tests Source Nparm DF DFDen F Ratio Prob > F AMF line 3 3 48,55 3,0679 0,0365* Effect Details Time LSMeans Differences Student's t α= 0,050 Level Least Sq Mean Time 2 A 0,29796448 Time 1 B 0,19598984 Levels not connected by same letter are significantly different. AMF line LSMeans Differences Tukey HSD α= 0,050 Level Least Sq Mean S4 A 0,33225124 S4c A B 0,25531100 C2 A B 0,24873973 S4b B 0,15160668 Levels not connected by same letter are significantly different.
CM4574
Figura 25. Producción de plantas de yuca de la variedad CM4574 que fueron inoculadas con la línea parental C2 y sus líneas
descendientes. C2: Línea parental; S4, S4b y S4c: Líneas obtenidas en el laboratorio a partir de C2. Las barras de error se
construyen con 1 error estándar de la media.
Response Root dry weight per plant (Kg) Cassava variety=CM4574 Summary of Fit RSquare 0,368601 RSquare Adj 0,321247 Root Mean Square Error 0,262466
136 Anexo E: Soporte estadístico del experimento de líneas en Yopal Título de la tesis o trabajo de investigación
Mean of Response 0,473318 Observations (or Sum Wgts) 44 RSquare without “Time” as a factor in the model: 0,207449 REML Variance Component Estimates Random Effect Var Ratio Var Component Std Error 95% Lower 95% Upper Pct of Total Block 0,0143332 0,0009874 0,0089938 -0,01664 0,018615 1,085 Time 0,3071263 0,0211574 0,0344846 -0,046431 0,0887459 23,241 Residual 0,0688881 0,0176073 0,0441676 0,1222595 75,674 Total 0,0910329 100,000 -2 LogLikelihood = 21,351014681 Fixed Effect Tests Source Nparm DF DFDen F Ratio Prob > F AMF line 3 3 34,6 4,7613 0,0070* Effect Details Time LSMeans Differences Student's t α= 0,050 Level Least Sq Mean Time 1 A 0,55521949 Time 2 B 0,36358561 Levels not connected by same letter are significantly different. AMF line LSMeans Differences Tukey HSD α= 0,050 Level Least Sq Mean S4b A 0,66115625 C2 A B 0,54121179 S4c A B 0,39552870 S4 B 0,23971344 Levels not connected by same letter are significantly different.
Grupo línea parental C3 completo
Parental Descendencia Procedimiento en el laboratorio
C3 Sc1 – 1era generación Cruce de líneas D1 x C3
Sc1a – 2nda generación Cultivo monospórico de línea Sc1
Sc1d – 2nda generación Cultivo monospórico de línea Sc1
Sc1e – 2nda generación Cultivo monospórico de línea Sc1
Sc2 – 1era generación Cruce de líneas D1 x C3
Sc2a – 2nda generación Cultivo monospórico de línea Sc2
Sc2b – 2nda generación Cultivo monospórico de línea Sc2
Sc2c – 2nda generación Cultivo monospórico de línea Sc2
Sc2d – 2nda generación Cultivo monospórico de línea Sc2

Anexo E : Soporte estadístico del experimento de líneas en Yopal 137
Analysis for both cassava varieties
Figura 26. Producción de plantas de yuca de las variedades MCOL2737 y CM4574 que fueron inoculadas con la línea parental
C3 y sus líneas descendientes. C3: Línea parental; Sc1, Sc1a, Sc1d, Sc1e, Sc2, Sc2a, Sc2b, Sc2c, Sc2d: Líneas obtenidas en el
laboratorio a partir de C3. Las barras de error se construyen con 1 error estándar de la media.
Response Root dry weight per plant (Kg) Summary of Fit RSquare 0,372256 RSquare Ad 0,321717 Root Mean Square Error 0,203841 Mean of Response 0,300418 Observations (or Sum Wgts) 256 RSquare without “Time” as a factor in the model: 0,389161 REML Variance Component Estimates Random Effect Var Ratio Var Component Std Error 95% Lower 95% Upper Pct of Total Block 0,0906485 0,0037666 0,0025992 -0,001328 0,008861 8,364 Time -0,006884 -0,000286 8,4919e-5 -0,000452 -0,00012 -0,635 Residual 0,0415513 0,0039028 0,0348497 0,0504018 92,271 Total 0,0450318 100,000 -2 LogLikelihood = 1,1178034345 Fixed Effect Test Source Nparm DF DFDen F Ratio Prob > F Cassava variety 1 1 230 62,3832 <,0001* AMF line 9 9 224,2 3,2921 0,0009* Cassava variety*AMF line 9 9 218,7 2,9414 0,0026* Effect Details Cassava variety LSMeans Differences Student's t α= 0,050 Level Least Sq Mean CM4574 A 0,41588572 MCOL2737 B 0,21038286 Levels not connected by same letter are significantly different. Tukey-Kramer applicable only if more than 2 levels AMF line LSMeans Differences Tukey HSD α= 0,050 Level Least Sq Mean Sc2 A 0,43508413 Sc2c A 0,39589625 Sc1 A 0,37384777 Sc2b A B 0,33648139 Sc1e A B 0,30055852

138 Anexo E: Soporte estadístico del experimento de líneas en Yopal Título de la tesis o trabajo de investigación
Level Least Sq Mean Sc1a A B 0,29493652 Sc1d A B 0,29206553 C3 A B 0,26635134 Sc2d A B 0,26537120 Sc2a B 0,17075025 Levels not connected by same letter are significantly different. Cassava variety*AMF line LSMeans Differences Tukey HSD α= 0,050 Level Least Sq Mean CM4574,Sc2 A 0,65336221 CM4574,Sc2c A B 0,58047600 CM4574,Sc1 A B C 0,50049208 CM4574,Sc2b A B C D 0,46515377 CM4574,Sc1e A B C D 0,46057454 CM4574,Sc1a B C D E 0,34504177 CM4574,Sc1d B C D E 0,33378707 CM4574,Sc2d B C D E 0,33207411 MCOL2737,C3 B C D E 0,26751924 CM4574,C3 B C D E 0,26518345 MCOL2737,Sc1d C D E 0,25034399 MCOL2737,Sc1 C D E 0,24720346 MCOL2737,Sc1a C D E 0,24483127 CM4574,Sc2a C D E 0,22271219 MCOL2737,Sc2 C D E 0,21680605 MCOL2737,Sc2c C D E 0,21131650 MCOL2737,Sc2b C D E 0,20780902 MCOL2737,Sc2d D E 0,19866830 MCOL2737,Sc1e E 0,14054250 MCOL2737,Sc2a E 0,11878831 Levels not connected by same letter are significantly different.
Analysis for each cassava variety
MCOL2737
Figura 27. Producción de plantas de yuca de la variedad MCOL2737 que fueron inoculadas con la línea parental C3 y sus
líneas descendientes. C3: Línea parental; Sc1, Sc1a, Sc1d, Sc1e, Sc2, Sc2a, Sc2b, Sc2c, Sc2d: Líneas obtenidas en el
laboratorio a partir de C3. Las barras de error se construyen con 1 error estándar de la media.
Response Root dry weight per plant (Kg) Cassava variety=MCOL2737
Summary of Fit RSquare 0,330613 RSquare Adj 0,282799 Root Mean Square Error 0,138431 Mean of Response 0,208287 Observations (or Sum Wgts) 136

Anexo E : Soporte estadístico del experimento de líneas en Yopal 139
RSquare without “Time” as a factor in the model: 0,242428 REML Variance Component Estimates Random Effect Var Ratio Var Component Std Error 95% Lower 95% Upper Pct of Total Block 0,1664265 0,0031893 0,0022561 -0,001233 0,0076112 11,892 Time 0,2330409 0,0044658 0,0067371 -0,008739 0,0176703 16,652 Residual 0,0191633 0,0025044 0,0150665 0,0252016 71,456 Total 0,0268184 100,000 -2 LogLikelihood = -97,43308346 Fixed Effect Tests Source Nparm DF DFDen F Ratio Prob > F AMF line 9 9 118,2 1,5994 0,1231
CM4574
Figura 28. Producción de plantas de yuca de la variedad CM4574 que fueron inoculadas con la línea parental C3 y sus líneas
descendientes. C3: Línea parental; Sc1, Sc1a, Sc1d, Sc1e, Sc2, Sc2a, Sc2b, Sc2c, Sc2d: Líneas obtenidas en el laboratorio a
partir de C3. Las barras de error se construyen con 1 error estándar de la media.
Response Root dry weight per plant (Kg) Cassava variety=CM4574 Summary of Fit RSquare 0,369492 RSquare Adj 0,317905 Root Mean Square Error 0,24218 Mean of Response 0,404833 Observations (or Sum Wgts) 120 RSquare without “Time” as a factor in the model: 0,307084 REML Variance Component Estimates Random Effect Var Ratio Var Component Std Error 95% Lower 95% Upper Pct of Total Block 0,1188857 0,0069728 0,0061655 -0,005111 0,019057 9,458 Time 0,1381028 0,0080999 0,0129125 -0,017208 0,033408 10,987 Residual 0,0586512 0,0082925 0,0452718 0,0790152 79,555 Total 0,0737239 100,000 -2 LogLikelihood = 39,13122821 Fixed Effect Tests Source Nparm DF DFDen F Ratio Prob > F AMF line 9 9 102 3,9917 0,0002* Effect Details AMF line LSMeans Differences Tukey HSD α= 0,050 Level Least Sq Mean Sc2 A 0,66808831 Sc2c A B 0,58865740 Sc1 A B C 0,52736053
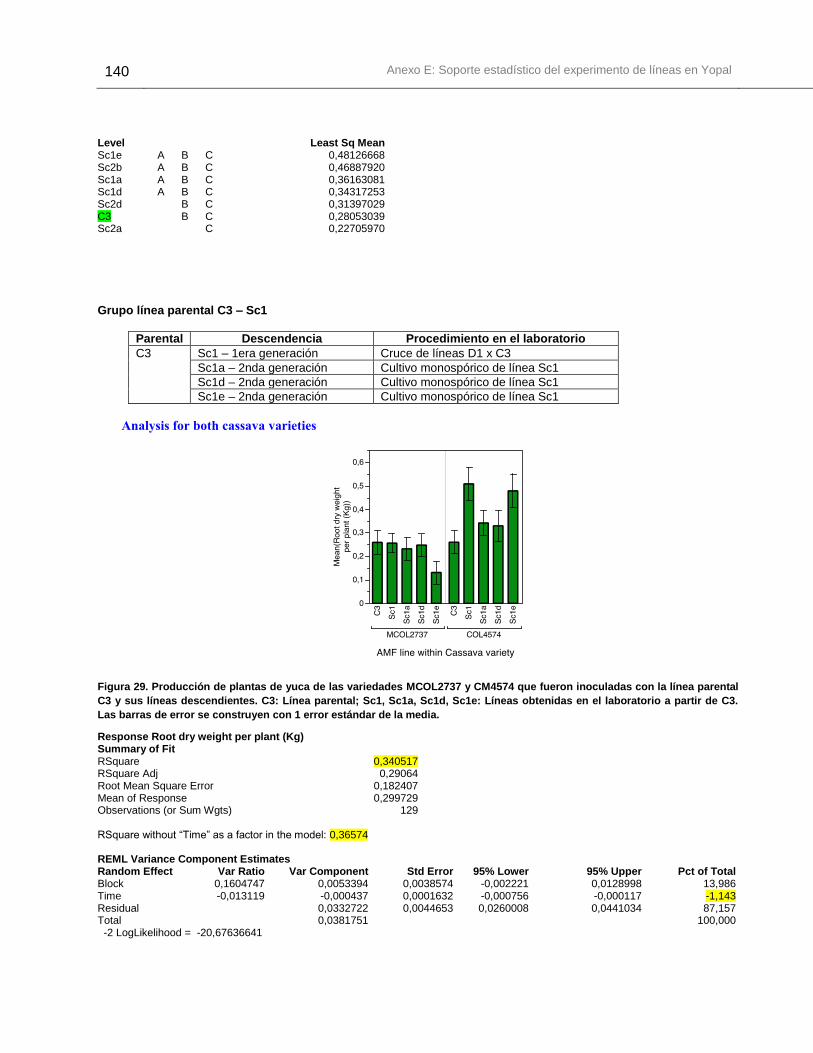
140 Anexo E: Soporte estadístico del experimento de líneas en Yopal Título de la tesis o trabajo de investigación
Level Least Sq Mean Sc1e A B C 0,48126668 Sc2b A B C 0,46887920 Sc1a A B C 0,36163081 Sc1d A B C 0,34317253 Sc2d B C 0,31397029 C3 B C 0,28053039 Sc2a C 0,22705970
Grupo línea parental C3 – Sc1
Parental Descendencia Procedimiento en el laboratorio
C3 Sc1 – 1era generación Cruce de líneas D1 x C3
Sc1a – 2nda generación Cultivo monospórico de línea Sc1
Sc1d – 2nda generación Cultivo monospórico de línea Sc1
Sc1e – 2nda generación Cultivo monospórico de línea Sc1
Analysis for both cassava varieties
Figura 29. Producción de plantas de yuca de las variedades MCOL2737 y CM4574 que fueron inoculadas con la línea parental
C3 y sus líneas descendientes. C3: Línea parental; Sc1, Sc1a, Sc1d, Sc1e: Líneas obtenidas en el laboratorio a partir de C3.
Las barras de error se construyen con 1 error estándar de la media.
Response Root dry weight per plant (Kg) Summary of Fit RSquare 0,340517 RSquare Adj 0,29064 Root Mean Square Error 0,182407 Mean of Response 0,299729 Observations (or Sum Wgts) 129 RSquare without “Time” as a factor in the model: 0,36574 REML Variance Component Estimates Random Effect Var Ratio Var Component Std Error 95% Lower 95% Upper Pct of Total Block 0,1604747 0,0053394 0,0038574 -0,002221 0,0128998 13,986 Time -0,013119 -0,000437 0,0001632 -0,000756 -0,000117 -1,143 Residual 0,0332722 0,0044653 0,0260008 0,0441034 87,157 Total 0,0381751 100,000 -2 LogLikelihood = -20,67636641

Anexo E : Soporte estadístico del experimento de líneas en Yopal 141
Fixed Effect Tests Source Nparm DF DFDen F Ratio Prob > F Cassava variety 1 1 112 23,5712 <,0001* AMF line 4 4 111,7 1,4340 0,2274 Cassava variety*AMF line 4 4 108 3,2510 0,0147* Effect Details Cassava variety LSMeans Differences Student's t α= 0,050 Level Least Sq Mean CM4574 A 0,39353691 MCOL2737 B 0,23114015 Levels not connected by same letter are significantly different. Cassava variety*AMF line LSMeans Differences Tukey HSD α= 0,050 Level Least Sq Mean CM4574,Sc1 A 0,51033865 CM4574,Sc1e A B 0,49241274 CM4574,Sc1a A B C 0,36045353 CM4574,Sc1d A B C 0,33608677 CM4574,C3 A B C 0,26839285 MCOL2737,C3 A B C 0,26605708 MCOL2737,Sc1d B C 0,26325771 MCOL2737,Sc1 B C 0,25964809 MCOL2737,Sc1a B C 0,23360375 MCOL2737,Sc1e C 0,13313414
Analysis for each cassava variety
MCOL2737
Figura 30. Producción de plantas de yuca de la variedad MCOL2737 que fueron inoculadas con la línea parental C3 y sus
líneas descendientes. C3: Línea parental; Sc1, Sc1a, Sc1d, Sc1e: Líneas obtenidas en el laboratorio a partir de C3. Las barras
de error se construyen con 1 error estándar de la media.
Response Root dry weight per plant (Kg) Cassava variety=MCOL2737 Summary of Fit RSquare 0,330001 RSquare Adj 0,286775 Root Mean Square Error 0,1522 Mean of Response 0,226955 Observations (or Sum Wgts) 67 RSquare without “Time” as a factor in the model: 0,266885 REML Variance Component Estimates Random Effect Var Ratio Var Component Std Error 95% Lower 95% Upper Pct of Total Block 0,1801609 0,0041734 0,0036148 -0,002911 0,0112583 13,531 Time 0,1512623 0,003504 0,0059844 -0,008225 0,0152332 11,361

142 Anexo E: Soporte estadístico del experimento de líneas en Yopal Título de la tesis o trabajo de investigación
Random Effect Var Ratio Var Component Std Error 95% Lower 95% Upper Pct of Total Residual 0,0231649 0,0044654 0,0164094 0,0351778 75,108 Total 0,0308423 100,000 -2 LogLikelihood = -32,95515848 Fixed Effect Tests Source Nparm DF DFDen F Ratio Prob > F AMF line 4 4 55,66 1,1412 0,3467
CM4574
Figura 31. Producción de plantas de yuca de la variedad CM4574 que fueron inoculadas con la línea parental C3 y sus líneas
descendientes. C3: Línea parental; Sc1, Sc1a, Sc1d, Sc1e: Líneas obtenidas en el laboratorio a partir de C3. Las barras de
error se construyen con 1 error estándar de la media.
Response Root dry weight per plant (Kg) Cassava variety=CM4574 Summary of Fit RSquare 0,486243 RSquare Adj 0,45019 Root Mean Square Error 0,181889 Mean of Response 0,378371 Observations (or Sum Wgts) 62 RSquare without “Time” as a factor in the model: 0,466557 REML Variance Component Estimates Random Effect Var Ratio Var Component Std Error 95% Lower 95% Upper Pct of Total Block 0,5252643 0,0173776 0,0121649 -0,006465 0,0412205 33,589 Time 0,038524 0,0012745 0,0034201 -0,005429 0,0079778 2,464 Residual 0,0330836 0,0068524 0,0228961 0,0520128 63,947 Total 0,0517357 100,000 -2 LogLikelihood = -4,015626907 Fixed Effect Tests Source Nparm DF DFDen F Ratio Prob > F AMF line 4 4 48,07 2,8372 0,0343* Effect Details AMF line LSMeans Differences Student's t α= 0,050 Level Least Sq Mean Sc1e A 0,49827451 Sc1 A 0,49365635 Sc1a A B 0,36756544 Sc1d A B 0,36325886 C3 B 0,28641170 Levels not connected by same letter are significantly different.
Grupo línea parental C3 – Sc2

Anexo E : Soporte estadístico del experimento de líneas en Yopal 143
Parental Descendencia Procedimiento en el laboratorio
C3 Sc2 – 1era generación Cruce de líneas D1 x C3
Sc2a – 2nda generación Cultivo monospórico de línea Sc2
Sc2b – 2nda generación Cultivo monospórico de línea Sc2
Sc2c – 2nda generación Cultivo monospórico de línea Sc2
Sc2d – 2nda generación Cultivo monospórico de línea Sc2
Analysis for both cassava varieties
Figura 32. Producción de plantas de yuca de las variedades MCOL2737 y CM4574 que fueron inoculadas con la línea parental
C3 y sus líneas descendientes. C3: Línea parental; Sc2, Sc2a, Sc2b, Sc2c, Sc2d: Líneas obtenidas en el laboratorio a partir de
C3. Las barras de error se construyen con 1 error estándar de la media.
Response Root dry weight per plant (Kg) Summary of Fit RSquare 0,408743 RSquare Adj 0,361614 Root Mean Square Error 0,21254 Mean of Response 0,294713 Observations (or Sum Wgts) 150 RSquare without “Time” as a factor in the model: 0,419095 REML Variance Component Estimates Random Effect Var Ratio Var Component Std Error 95% Lower 95% Upper Pct of Total Block 0,0916299 0,0041392 0,0033815 -0,002488 0,0107669 8,461 Time -0,008703 -0,000393 0,0003535 -0,001086 0,0002997 -0,804 Residual 0,045173 0,0056171 0,0359082 0,0585717 92,342 Total 0,0489191 100,000 Fixed Effect Tests Source Nparm DF DFDen F Ratio Prob > F Cassava variety 1 1 130,6 34,2386 <,0001* AMF line 5 5 130,9 5,2325 0,0002* Cassava variety*AMF line 5 5 131,1 3,7847 0,0031* Effect Details Cassava variety LSMeans Differences Student's t α= 0,050 Level Least Sq Mean CM4574 A 0,41886026 MCOL2737 B 0,20464813 Levels not connected by same letter are significantly different.

144 Anexo E: Soporte estadístico del experimento de líneas en Yopal Título de la tesis o trabajo de investigación
AMF line LSMeans Differences Tukey HSD α= 0,050 Level Least Sq Mean Sc2 A 0,43608250 Sc2c A 0,40268943 Sc2b A B 0,33153906 C3 A B 0,26924477 Sc2d A B 0,26301401 Sc2a B 0,16795540 Levels not connected by same letter are significantly different. Cassava variety*AMF line LSMeans Differences Tukey HSD α= 0,050 Level Least Sq Mean CM4574,Sc2 A 0,65422282 CM4574,Sc2c A B 0,58834268 CM4574,Sc2b A B C 0,45942552 CM4574,Sc2d B C D 0,32945504 MCOL2737,C3 B C D 0,27449803 CM4574,C3 C D 0,26399151 MCOL2737,Sc2 C D 0,21794219 CM4574,Sc2a C D 0,21772399 MCOL2737,Sc2c C D 0,21703619 MCOL2737,Sc2b C D 0,20365260 MCOL2737,Sc2d C D 0,19657298 MCOL2737,Sc2a D 0,11818680 Levels not connected by same letter are significantly different.
Analysis for each cassava variety
MCOL2737
Figura 33. Producción de plantas de yuca de la variedad MCOL2737 que fueron inoculadas con la línea parental C3 y sus
líneas descendientes. C3: Línea parental; Sc2, Sc2a, Sc2b, Sc2c, Sc2d: Líneas obtenidas en el laboratorio a partir de C3. Las
barras de error se construyen con 1 error estándar de la media.
Response Root dry weight per plant (Kg) Cassava variety=MCOL2737 Summary of Fit RSquare 0,441914 RSquare Adj 0,403689 Root Mean Square Error 0,118898 Mean of Response 0,198848 Observations (or Sum Wgts) 79 RSquare without “Time” as a factor in the model: 0,31408 REML Variance Component Estimates Random Effect Var Ratio Var Component Std Error 95% Lower 95% Upper Pct of Total Block 0,2953816 0,0041758 0,0030099 -0,001724 0,010075 17,654 Time 0,377764 0,0053404 0,0080983 -0,010532 0,0212128 22,578 Residual 0,0141368 0,0025035 0,0102755 0,020683 59,768

Anexo E : Soporte estadístico del experimento de líneas en Yopal 145
Random Effect Var Ratio Var Component Std Error 95% Lower 95% Upper Pct of Total Total 0,0236529 100,000 -2 LogLikelihood = -72,02124303 Fixed Effect Tests Source Nparm DF DFDen F Ratio Prob > F AMF line 5 5 64,8 2,1057 0,0759
CM4574
Figura 34. Producción de plantas de yuca de la variedad CM4574 que fueron inoculadas con la línea parental C3 y sus líneas
descendientes. C3: Línea parental; Sc2, Sc2a, Sc2b, Sc2c, Sc2d: Líneas obtenidas en el laboratorio a partir de C3. Las barras
de error se construyen con 1 error estándar de la media.
Response Root dry weight per plant (Kg) Cassava variety=CM4574 Summary of Fit RSquare 0,4042 RSquare Adj 0,35837 Root Mean Square Error 0,263086 Mean of Response 0,40138 Observations (or Sum Wgts) 71 RSquare without “Time” as a factor in the model: 0,340248 REML Variance Component Estimates Random Effect Var Ratio Var Component Std Error 95% Lower 95% Upper Pct of Total Block 0,0885846 0,0061313 0,0073663 -0,008306 0,020569 7,128 Time 0,1541813 0,0106715 0,0179591 -0,024528 0,0458707 12,406 Residual 0,0692142 0,0129476 0,0494888 0,1036863 80,466 Total 0,0860171 100,000 -2 LogLikelihood = 35,130970347 Fixed Effect Tests Source Nparm DF DFDen F Ratio Prob > F AMF line 5 5 59,12 5,2412 0,0005* Effect Details AMF line LSMeans Differences Tukey HSD α=0,050 Level Least Sq Mean Sc2 A 0,65866942 Sc2c A B 0,59770748 Sc2b A B C 0,45317405 Sc2d B C 0,30955232 C3 B C 0,27098877 Sc2a C 0,22007591 Levels not connected by same letter are significantly different.

146 Anexo E: Soporte estadístico del experimento de líneas en Yopal Título de la tesis o trabajo de investigación
11.16 Response: Starch %. Para tratamientos que tuvieron evaluación del efecto de la líneas sobre la calidad del almidón en las raíces. Datos de la primera repetición del experimento en Yopal.
Response Arco seno Strach Summary of Fit RSquare 0,86408 RSquare Adj 0,765915 Root Mean Square Error 0,038567 Mean of Response 0,253942 Observations (or Sum Wgts) 32 Analysis of Variance Source DF Sum of Squares Mean Square F Ratio Model 13 0,17020279 0,013093 8,8023 Error 18 0,02677305 0,001487 Prob > F C. Total 31 0,19697585 <,0001* Effect Tests Source Nparm DF Sum of Squares F Ratio Prob > F Variety 1 1 0,05589599 37,5799 <,0001* Line 6 6 0,04534413 5,0809 0,0033* Line*Variety 6 6 0,06412793 7,1857 0,0005* Variety LSMeans Differences Student's t α= 0,050 t= 2,10092 Level Least Sq Mean MCOL 2737 A 0,29333426 CM4574 B 0,20833630 Levels not connected by same letter are significantly different. Line LSMeans Differences Tukey HSD α=0,050 Q=3,3044 Level Least Sq Mean 11 A 0,32016998 14 A B 0,28594694 7 A B 0,25821190 5 A B 0,24099803 h2o B 0,22355241 10 B 0,22238170 1 B 0,20458600 Levels not connected by same letter are significantly different. Line*Variety LSMeans Differences Tukey HSD α=0,050 Q=3,83873 Level Least Sq Mean Sc2,MCOL 2737 A 0,40124871 Sc2c,MCOL 2737 A 0,38194930 Sc1,MCOL 2737 A B 0,29178119 C3,MCOL 2737 A B 0,28610158 Sc1e,CM4574 A B 0,26726271 h2o,MCOL 2737 A B C 0,25935149 S4b,MCOL 2737 A B C 0,25540684 Sc2,CM4574 B C 0,23909126

Anexo E : Soporte estadístico del experimento de líneas en Yopal 147
Level Least Sq Mean S4b,CM4574 B C 0,22658921 Sc1,CM4574 B C 0,22464262 Sc2c,CM4574 B C 0,18994458 h2o,CM4574 B C 0,18775333 Sc1e,MCOL 2737 B C 0,17750070 C3,CM4574 C 0,12307042 Levels not connected by same letter are significantly different.
11.17 Response Colonization %. Para tratamientos que tuvieron evaluación del efecto de la líneas sobre la calidad del almidón en las raíces. Datos de la primera repetición del experimento en Yopal
Response Arco seno Col Summary of Fit RSquare 0,518904 RSquare Adj 0,295538 Root Mean Square Error 0,223516 Mean of Response 0,816558 Observations (or Sum Wgts) 42 Analysis of Variance Source DF Sum of Squares Mean Square F Ratio Model 13 1,5087932 0,116061 2,3231 Error 28 1,3988599 0,049959 Prob > F C. Total 41 2,9076532 0,0301* Effect Tests Source Nparm DF Sum of Squares F Ratio Prob > F Variety 1 1 0,0240176 0,4807 0,4938 Line 6 6 0,2339174 0,7804 0,5923 Line*Variety 6 6 1,2508582 4,1729 0,0041* Line*Variety LSMeans Differences Tukey HSD α= 0,050 Q= 3,66041 Level Least Sq Mean C3,MCOL 2737 A 1,0996806 Sc1,CM4574 A 1,0367039 Sc1e,MCOL 2737 A B 0,9785123 h2o,MCOL 2737 A B 0,9273118 Sc2,CM4574 A B 0,9109546 S4b,CM4574 A B 0,9022285 Sc2c,CM4574 A B 0,8979520 Sc2c,MCOL 2737 A B 0,8276072 h2o,CM4574 A B 0,8209560 Sc1e,MCOL 2737 A B 0,7113194 Sc1,MCOL 2737 A B 0,6778234 Sc2,MCOL 2737 A B 0,6610467 S4b,CM4574 A B 0,6395705 C3,CM4574 B 0,3401491 Levels not connected by same letter are significantly different.

148 Anexo E: Soporte estadístico del experimento de líneas en Yopal Título de la tesis o trabajo de investigación
11.18 Response: Above ground fresh weight per plant (kg). Segunda repetición del experimento
Summary of Fit RSquare 0,302907 RSquare Adj 0,187821 Root Mean Square Error 0,738134 Mean of Response 1,285387 Observations (or Sum Wgts) 248 Fixed Effect Tests Source Nparm DF DFDen F Ratio Prob > F Cassava variety 1 1 204,8 7,2664 0,0076* AMF line 17 17 205 1,0714 0,3839 Cassava variety*AMF line 17 17 205,2 1,7487 0,0368* Effect Details Cassava variety Least Squares Means Table Level Least Sq Mean Std Error MCOL2737 1,4170157 0,10486182 CM4574 1,1588037 0,10837217 LSMeans Differences Student's t, α=0,050 Level Least Sq Mean MCOL2737 A 1,4170157 CM4574 B 1,1588037 Levels not connected by same letter are significantly different. Cassava variety*AMF line LSMeans Differences Student's t, α=0,050 Level Least Sq Mean MCOL2737,Sc1e A 1,9129321 MCOL2737,Sc2b A 1,8901192 MCOL2737,S4 A 1,8072222 CM4574,Sc2 A B C 1,7954112 MCOL2737,GLO A C 1,7809924 MCOL2737,Sc1a A B C D 1,7596638 MCOL2737,Sc1 A B C D 1,7053674 MCOL2737,S4c A B C D E 1,6505556 CM4574,Sc1d A B C D E F 1,5985417 MCOL2737,Sc2a A B C D E F G 1,4956900 MCOL2737,F A B C D E F G 1,4863906 CM4574,Sc2c A B C D E F G H 1,4491440 MCOL2737,Sc2c A B C D E F G H 1,3969458 CM4574,H2O A B C D E F G H 1,3923797 CM4574,S4c A B C D E F G H 1,3751180 CM4574,Sc1e A B C D E F G H 1,3592984 MCOL2737,H2O A B C D E F G H 1,3583614 CM4574,Sc2d A B C D E F G H 1,3371114 MCOL2737,G A B C D E F G H 1,2601180 MCOL2737,Sc2d A B C D E F G H 1,2559924 MCOL2737,K A B C D E F G H 1,2476695 CM4574,GLO A B C D E F G H 1,2026362 CM4574,Sc1 A B C D E F G H 1,1857280 CM4574,FIL A B C D E F G H 1,0789449 MCOL2737,Sc1d B D E F G H 1,0716667 CM4574,Sc1a B C D E F G H 1,0476906 MCOL2737,FIL D E F G H 1,0249864 CM4574,Sc2b B C D E F G H 0,9878893 CM4574,Sc2a D E F G H 0,9866441

Anexo E : Soporte estadístico del experimento de líneas en Yopal 149
Level Least Sq Mean CM4574,K E F G H 0,9652244 CM4574,S4b F G H 0,8756742 CM4574,G F G H 0,7853674 MCOL2737,Sc2 G H 0,7493494 CM4574,F F G H 0,7208352 CM4574,S4 F G H 0,7148278 MCOL2737,S4b H 0,6522589 Levels not connected by same letter are significantly different.

12 Anexo F: Condiciones del clima,
topografía y fertilidad del suelo para
Santana y Yopal.
Tabla 5. Condiciones del clima, topografía y fertilidad del suelo para Santana y Yopal

13 Anexo G: Estadísticas del Efecto del
Ambiente Sobre la Funcionalidad de la
Simbiosis entre las Líneas de R. irregularis
y la Variedad de Yuca MCOL2737.
13.1 Analysis of IB root fresh weight per plant (%) By Treatment
Level Count Std Dev MeanAbsDif to Mean MeanAbsDif to
Median SantanaC2 9 25,7372 19,79197 19,65004 SantanaC3 9 29,1554 21,09070 20,06999 SantanaD1 9 50,9123 35,57889 35,62555 SantanaGLO 9 59,5591 43,89618 43,74453 SantanaS4 8 34,0380 22,99213 22,79528 SantanaS4b 8 45,1516 35,52165 28,95669 SantanaS4c 9 47,7429 40,99543 40,94488 SantanaSc1 9 35,8027 28,21036 29,43132 SantanaSc1a 9 37,5594 28,71974 27,62905 SantanaSc1d 8 35,0283 27,66240 26,39764 SantanaSc1e 7 38,2156 29,40061 31,51856 SantanaSc2 8 47,9965 39,65059 38,44488 SantanaSc2a 7 41,1809 26,80379 26,38920 SantanaSc2b 7 24,7569 18,49912 18,96513 SantanaSc2c 9 33,6964 27,68154 25,17935 SantanaSc2d 9 35,9481 25,69457 25,86177 YopalC2 8 101,7737 78,29316 78,27522 YopalC3 6 86,5464 68,24509 68,24509 YopalD1 7 113,9213 74,65364 75,12453 YopalGLO 8 72,2837 60,14493 59,09026 YopalS4 9 82,7752 60,20312 60,07557 YopalS4b 8 54,7090 41,33305 40,75190 YopalS4c 9 64,2486 50,72765 51,54255 YopalSc1 9 74,8176 55,70489 56,00676 YopalSc1a 6 112,9379 79,06762 75,01794 YopalSc1d 9 103,2153 79,08037 78,49365 YopalSc1e 6 113,2748 87,34555 67,49892 YopalSc2 9 62,0260 56,65443 53,49405 YopalSc2a 7 48,8863 41,37273 40,34192 YopalSc2b 7 96,3791 70,55854 73,18943

152 Anexo G: Estadísticas del efecto del ambiente sobre la funcionalidad de la simbiosis Título de la tesis o trabajo de investigación
Level Count Std Dev MeanAbsDif to Mean MeanAbsDif to Median
YopalSc2c 9 94,6923 78,79693 78,64671 YopalSc2d 8 53,0658 41,15368 41,15368 Test F Ratio DFNum DFDen Prob > F O'Brien[.5] 1,5123 31 227 0,0472* Brown-Forsythe 1,7291 31 227 0,0129* Levene 2,4353 31 227 <,0001* Bartlett 2,6737 31 . <,0001* Welch's Test Welch Anova testing Means Equal, allowing Std Devs Not Equal F Ratio DFNum DFDen Prob > F 1,7125 31 78,41 0,0294*
Figura 35. Beneficio de la inoculación (BI) en términos de peso fresco de raíces producidas por plantas que fueron
inoculadas con líneas genéticamente diferentes de Rhizophagus irregularis en dos ambientes diferentes. Las
barras de error se construyen con 1 error estándar de la media. Letras diferentes encima de las barras representan
diferencias significativas para un p < 0,05.
13.2 Analysis of IB root dry weight per plant (%) By Treatment
Level Count Std Dev MeanAbsDif to Mean MeanAbsDif to Median
SantanaC2 9 25,7372 19,7920 19,6500 SantanaC3 9 29,1554 21,0907 20,0700

Anexo G: Estadísticas del efecto del ambiente sobre la funcionalidad de la simbiosis 153
Level Count Std Dev MeanAbsDif to Mean MeanAbsDif to Median
SantanaD1 9 50,9123 35,5789 35,6255 SantanaGLO 9 59,5591 43,8962 43,7445 SantanaS4 8 34,0380 22,9921 22,7953 SantanaS4b 9 35,8027 28,2104 29,4313 SantanaS4c 9 37,5594 28,7197 27,6290 SantanaSc1 8 45,1516 35,5217 28,9567 SantanaSc1a 8 35,0283 27,6624 26,3976 SantanaSc1d 7 38,2156 29,4006 31,5186 SantanaSc1e 8 47,9965 39,6506 38,4449 SantanaSc2 9 47,7429 40,9954 40,9449 SantanaSc2a 7 41,1809 26,8038 26,3892 SantanaSc2b 8 33,5777 26,2156 24,8622 SantanaSc2c 9 26,7795 19,4926 18,4952 SantanaSc2d 9 36,5378 27,5221 27,1391 YopalC2 7 91,2345 73,7974 76,9133 YopalC3 4 58,6846 48,2143 48,2143 YopalD1 6 94,7283 81,2500 62,9464 YopalGLO 7 126,8620 96,7566 102,1684 YopalS4 5 183,5066 138,2143 144,6429 YopalS4b 7 127,9721 96,1006 76,9133 YopalS4c 8 120,2244 95,4241 95,4241 YopalSc1 7 28,9201 21,3192 21,8112 YopalSc1a 6 87,7329 66,9643 66,9643 YopalSc1d 6 20,9546 16,0714 16,0714 YopalSc1e 6 133,3461 94,6429 79,0179 YopalSc2 8 164,3432 129,0737 111,4955 YopalSc2a 7 51,2441 41,3265 33,2908 YopalSc2b 6 80,1560 65,6250 65,6250 YopalSc2c 7 41,7180 35,0948 36,7347 YopalSc2d 7 63,0542 50,8382 50,5102 Test F Ratio DFNum DFDen Prob > F O'Brien[.5] 2,5353 31 207 <,0001* Brown-Forsythe 2,5290 31 207 <,0001* Levene 4,6991 31 207 <,0001* Bartlett 4,4415 31 . <,0001* Welch's Test Welch Anova testing Means Equal, allowing Std Devs Not Equal F Ratio DFNum DFDen Prob > F 1,7014 31 69,283 0,0344*
13.3 Response AMF Total Colonization (%) Summary of Fit RSquare 0,515557 RSquare Adj 0,327162 Root Mean Square Error 13,2654 Mean of Response 69,38357 Observations (or Sum Wgts) 126 Fixed Effect Tests

154 Anexo G: Estadísticas del efecto del ambiente sobre la funcionalidad de la simbiosis Título de la tesis o trabajo de investigación
Source Nparm DF DFDen F Ratio Prob > F AMF line 17 17 87,18 1,8298 0,0365* Place 1 1 59,22 14,1410 0,0004* AMF line*Place 17 17 87,28 2,1088 0,0130*
13.4 Response IB Root dry weight per plant (%) Environment=Santana Var=MCOL2737
Summary of Fit RSquare 0,111407 RSquare Adj -0,00143 Root Mean Square Error 40,26498 Mean of Response -0,55173 Observations (or Sum Wgts) 143 Fixed Effect Tests Source Nparm DF DFDen F Ratio Prob > F AMF line 16 16 117,9 0,7142 0,7749
13.5 Response IB root dry weight per plant (%) Environment=Yopal Var=MCOL2737
Summary of Fit RSquare 0,390768 RSquare Adj 0,28816 Root Mean Square Error 94,76555 Mean of Response 33,30676 Observations (or Sum Wgts) 112 Fixed Effect Tests Source Nparm DF DFDen F Ratio Prob > F AMF line 16 16 88,38 2,2784 0,0078*

14 Anexo H: Soporte Estadístico del
Experimento de las líneas de R. irregularis
y el beneficio de la inoculación sobre la
producción de yuca COL2215 en Santana
14.1 Response Root dry weight per plant (Kg) - Santana
Summary of Fit RSquare 0,248612 RSquare Adj 0,150115 Root Mean Square Error 0,648755 Mean of Response 1,621322 Observations (or Sum Wgts) 303 Fixed Effect Tests Source Nparm DF DFDen F Ratio Prob > F Cassava variety 1 1 259,2 13,4508 0,0003* AMF line 17 17 259,2 1,3474 0,1635 Cassava variety*AMF line 17 17 259,2 1,0667 0,3870 Effect Details Cassava variety Least Squares Means Table Level Least Sq Mean Std Error MCOL2737 1,4762233 0,08656237 COL2215 1,7510679 0,08642876 LSMeans Differences Student's t α=0,050 Level Least Sq Mean COL2215 A 1,7510679 MCOL2737 B 1,4762233 Levels not connected by same letter are significantly different.

156 Anexo H: Soporte estadístico del experimento en Santana
14.2 Oneway Analysis of Root fresh weight per plant (kg) By AMF line Cassava variety=MCOL2737 - Santana
Summary of Fit Rsquare 0,084336 Adj Rsquare -0,03359 Root Mean Square Error 2,548324 Mean of Response 6,313667 Observations (or Sum Wgts) 150 Analysis of Variance Source DF Sum of Squares Mean Square F Ratio Prob > F AMF line 17 78,95175 4,64422 0,7152 0,7830 Error 132 857,20233 6,49396 C. Total 149 936,15408
14.3 Oneway Analysis of Root dry weight per plant (Kg) By AMF line Cassava variety=MCOL2737 - Santana
Summary of Fit Rsquare 0,079345 Adj Rsquare -0,03833 Root Mean Square Error 0,598296 Mean of Response 1,484453 Observations (or Sum Wgts) 151 Analysis of Variance Source DF Sum of Squares Mean Square F Ratio Prob > F AMF line 17 4,103054 0,241356 0,6743 0,8235 Error 133 47,608503 0,357959 C. Total 150 51,711556
14.4 Oneway Analysis of Root fresh weight per plant (kg) By AMF line Cassava variety=COL2215 - Santana
Summary of Fit Rsquare 0,131522 Adj Rsquare 0,021342 Root Mean Square Error 2,853492 Mean of Response 6,611349

Anexo H: Soporte estadístico del experimento en Santana 157
Observations (or Sum Wgts) 152 Analysis of Variance Source DF Sum of Squares Mean Square F Ratio Prob > F AMF line 17 165,2338 9,71964 1,1937 0,2779 Error 134 1091,0840 8,14242 C. Total 151 1256,3178
14.5 Oneway Analysis of Root dry weight per plant (Kg) By AMF line Cassava variety=COL2215 - Santana
Summary of Fit Rsquare 0,146561 Adj Rsquare 0,038289 Root Mean Square Error 0,751909 Mean of Response 1,75729 Observations (or Sum Wgts) 152 Analysis of Variance Source DF Sum of Squares Mean Square F Ratio Prob > F AMF line 17 13,010101 0,765300 1,3536 0,1700 Error 134 75,759163 0,565367 C. Total 151 88,769265
Figura 36. Efecto de líneas de R. irregularis sobre el peso fresco de raíces por planta (barras). Las barras representan ± 1 error estándar de la media.

158 Anexo H: Soporte estadístico del experimento en Santana
14.6 Response BI Root fresh weight per plant (%)- Santana
Summary of Fit RSquare 0,210933 RSquare Adj 0,108416 Root Mean Square Error 42,4075 Mean of Response 2,760069 Observations (or Sum Wgts) 288 Fixed Effect Tests Source Nparm DF DFDen F Ratio Prob > F Cassava variety 1 1 246,5 3,5494 0,0607 AMF line 16 16 246,3 1,4223 0,1314 Cassava variety*AMF line 16 16 246,3 0,8996 0,5703
14.7 Response BI Root dry weight per plant (%) -
Santana Summary of Fit RSquare 0,264125 RSquare Adj 0,167761 Root Mean Square Error 45,78077 Mean of Response 12,78566 Observations (or Sum Wgts) 286 Fixed Effect Tests Source Nparm DF DFDen F Ratio Prob > F Cassava variety 1 1 244,5 24,0052 <,0001* AMF line 16 16 244,4 1,2984 0,1985 Cassava variety*AMF line 16 16 244,3 1,0498 0,4050 Effect Details Random Block Cassava variety Least Squares Means Table Level Least Sq Mean Std Error MCOL2737 -0,95654 5,8668122 COL2215 25,72946 5,8663056 LSMeans Differences Student's t, α=0,050 Level Least Sq Mean COL2215 A 25,72946 MCOL2737 B -0,95654 Levels not connected by same letter are significantly different.

Bibliografía
Agronet: Ministerio de Agricultura y Desarrollo, 2013. Producción de Yuca en Colombia, 2012-
2013. Area cosechada y producción. Disponible en: http://www.agronet.gov.co [Consulatdo
May 15, 2015].
Angelard, C. et al., 2010. Segregation in a mycorrhizal fungus alters rice growth and symbiosis-
specific gene transcription. Current biology : CB, 20(13), pp.1216–21. Disponible en:
http://www.ncbi.nlm.nih.gov/pubmed/20541408 [Consultado Marzo 12, 2012].
Angelard, C. & Sanders, I.R., 2011. Effect of segregation and genetic exchange on arbuscular
mycorrhizal fungi in colonization of roots. New Phytologist, 189(3), pp.652–657.
Aristizábal, J., Sánchez, T. & Mejía, D., 2007. Guía técnica para producción y análisis de almidón
de yuca CIAT., Roma. 163 páginas.
Augé, R., 2001. Water relations, drought and vesicular-arbuscular mycorrhizal symbiosis.
Mycorrhiza, 11, pp.3–42.
Avio, L. et al., 2006. Functional diversity of arbuscular mycorrhizal fungal isolates in relation to
extraradical mycelial networks. The New phytologist, 172(2), pp.347–57. Disponible en:
http://www.ncbi.nlm.nih.gov/pubmed/16995921 [Consultado Marzo 12, 2012].
Baon, J.B., Smith, S.E. & Alston, A. M., 1993. Mycorrhizal responses of barley cultivars differing in
P efficiency. Plant and Soil, 157(1), pp.97–105.
Bécard, G. & Fortin, a. J., 1988. Early events of vesicular-arbuscular mycorrhiza formation on Ri
T-DNA transformed roots. New Phytologist, 108(2), pp.211–218.
Bethlenfalvay, G.J., Pacovsky, R.S. & Brown, M.S., 1982. Parasitic and Mutualistic Association
Between a Mycorrhizal Fungus and Soybean: Development of the Endophyte. Ecology and
Epidemiology, 72(7), pp.894–897.
Bever, J.D. et al., 2001. Arbuscular Mycorrhizal Fungi: More Diverse than Meets the Eye, and the

160 Funcionalidad de la simbiósis entre variedades de yuca y genotipos de
Rhizophagus irregularis en la Orinoquía Colombiana
Ecological Tale of Why. BioScience, 51(11), pp.923–932. Disponible en:
http://www.bioone.org/doi/abs/10.1641/0006-3568(2001)051[0363:PSIUEN]2.0.CO;2.
Bever, J.D., 2002. Host-specificity of AM fungal population growth rates can generate feedback on
plant growth. Plant and Soil, 244, pp.281–290.
Borowicz, V.A., 2001. Do Arbuscular Mycorrhizal Fungi Alter Plant-Pathogen Relations? Ecology,
82(11), pp.3057–3068. Disponible en: http://www.jstor.org/stable/2679834.
Börstler, B. et al., 2008. Genetic diversity of the arbuscular mycorrhizal fungus Glomus intraradices
as determined by mitochondrial large subunit rRNA gene sequences is considerably higher
than previously expected. New Phytologist, 180(2), pp.452–465.
Buee, M. et al., 2000. The pre-symbiotic growth of arbuscular mycorrhizal fungi is induced by a
branching factor partially purified from plant root exudates. Molecular plant-microbe
interactions : MPMI, 13(6), pp.693–698.
Cadavid, L.F., 2005. Manual de producción de yuca Clayuca, Ed. Caramba, Cali, Colombia.
Carretero, C.L. et al., 2009. Growth Responses of Micropropagated Cassava Clones as Affected
by Glomus Intraradices Colonization. Journal of plant nutrition, 32(2), pp.261–273.
Ceballos, H. & De la Cruz, G.A., 2002. Taxonomía y Morfología de la Yuca. En La Yuca en el
Tercer Mileno:Sistemas Modernos de Producción, Procesamiento, Utilización y
Comercialización. Cali: Publicaciones CIAT, pp. 16–33.
Ceballos, I. et al., 2013. The in vitro mass-produced model mycorrhizal fungus, Rhizophagus
irregularis, significantly increases yields of the globally important food security crop cassava.
PloS one, 8(8), p.e70633. Disponible en:
http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=3737348&tool=pmcentrez&rendert
ype=abstract.
CIAT, 2016. Programa de Recursos Genéticos - CIAT. Colección de yuca. Disponible en:
http://isa.ciat.cgiar.org/urg/cassavacollection.do [Consultado Junio 7, 2016].

Bibliografía 161
Cock, J.H., 1985. Cassava: New potential for a neglected Crop, Boulder, Colorado: Westview
Press. Pág 5.
Cock, J.H. & Rosas, S.C., 1975. Ecophysiology of cassava. Ed CIAT. En Internal Symposium on
Ecophysiology of Tropical Crops.
Connor, D.J., Cock, J.H. & Parra, G.E., 1981. Response of cassava to water shortage. Field Crops
Research, 4(3), pp.285–296.
Corpoorinoquía, 2011. Foro Nacional Ambiental. En La Mejor Orinoquía que Podemos Construir.
Bogotá: Corpoorinoquía, p. 20 p.
Croll, D., Wille, L., Gamper, H.A., et al., 2008. Genetic diversity and host plant preferences
revealed by simple sequence repeat and mitochondrial markers in a population of the
arbuscular mycorrhizal fungus Glomus intraradices. New Phytologist, 178(3), pp.672–687.
Croll, D. et al., 2009. Non-self vegetative fusion and genetic exchange in the arbuscular
mycorrhizal fungus Glomus intraradices. The New phytologist, 181(4), pp.924–37. Disponible
en: http://www.ncbi.nlm.nih.gov/pubmed/19140939 [Consultado Marzo 12, 2012].
Croll, D. & Sanders, I.R., 2009. Recombination in Glomus intraradices, a supposed ancient
asexual arbuscular mycorrhizal fungus. BMC evolutionary biology, 9, p.13. Disponible en:
http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=2630297&tool=pmcentrez&rendert
ype=abstract [Consultado Noviembre 20, 2014].
Dai, M., 2009. Undersatnading the One-way ANOVA. En Diseño estadístico para experimentos de
campo. p. 236 p.
Damba, G.P., 2008. Evaluación de métodos para análisis de estabilidad en diferentes ambientes
en genotipos de yuca (Manihot esculenta Crantz). Universidad Nacional de Colombia - Sede
Palmira. pág. 56-60
Declerck, S., Strulle, D.G. & Fortin, A.J., 2005. In Vitro Culture of Mycorrhizas. Springer, ed.,
Heidelberg. p. 53 p.
Defloor, I., Dehing, I., & Delcour, J. A. (1998). Physico-Chemical Properties of Cassava Starch.

162 Funcionalidad de la simbiósis entre variedades de yuca y genotipos de
Rhizophagus irregularis en la Orinoquía Colombiana
Starch - Stärke, 50(2-3), 58–64. doi:10.1002/(sici)1521-379x(199803)50:2/3<58::aid-
star58>3.0.co;2-n
Edwards, D.J. & Kang, B.T., 1978. Tolerance of cassava (Manihot esculenta Crantz) to high soil
acidity. Field Crops Research, 1, pp.337–346.
Ehinger, M.O. et al., 2012. Significant genetic and phenotypic changes arising from clonal growth
of a single spore of an arbuscular mycorrhizal fungus over multiple generations. The New
phytologist, 196(3), pp.853–61. Disponible en:
http://www.ncbi.nlm.nih.gov/pubmed/22931497.
Fang, F.C., 2010. Reproducible Science. American Society for Microbiology, Editorial. Disponible
en: http://iai.asm.org/content/78/12/4972.full#xref-fn-1-1.
FAO, 2013. FAOSTAT. Rome, Italy: FAO publications. Disponible en:
http://faostat3.fao.org/home/index.html. [Consultado Junio 17, 2013].
Feddermann, N. et al., 2010. Functional diversity in arbuscular mycorrhiza – the role of gene
expression, phosphorous nutrition and symbiotic efficiency. Fungal Ecology, 3(1), pp.1–8.
Disponible en: http://linkinghub.elsevier.com/retrieve/pii/S1754504809000877 [Consultado
Febrero 5, 2013].
Feng, G. et al., 2002. Improved tolerance of maize plants to salt stress by arbuscular mycorrhiza is
related to higher accumulation of soluble sugars in roots. Mycorrhiza, 12(4), pp.185–190.
Finlay, R.D., 2008. Ecological aspects of mycorrhizal symbiosis: with special emphasis on the
functional diversity of interactions involving the extraradical mycelium. Journal of
experimental botany, 59(5), pp.1115–26. Disponible en:
http://www.ncbi.nlm.nih.gov/pubmed/18349054 [Consultado Julio 9, 2014].
Fox, B. & Cameron, A.G., 1997. Ciencia De los alimentos, nutrición y salud Primera Ed., México:
Editorial Limusa.
Gamper, H. a et al., 2008. Real-time PCR and microscopy: are the two methods measuring the
same unit of arbuscular mycorrhizal fungal abundance? Fungal genetics and biology : FG &

Bibliografía 163
B, 45(5), pp.581–96. Disponible en: http://www.ncbi.nlm.nih.gov/pubmed/17964831
[Consultado Septiembre 25, 2014].
Gange, A.C., Brown, V.K. & Farmer, L.M., 1990. A test of mycorrhizal benefit in an early
successional plant community. New Phytologist, 115. pp. 85-91.
Genre, A. et al., 2005. Arbuscular mycorrhizal fungi elicit a novel intracellular apparatus in
Medicago truncatula root epidermal cells before infection. The Plant cell, 17(12), pp.3489–
3499.
Gerdemann, J.W., 1955. Relation of a large soil-borne spore to phycomycetous mycorrhizal
infections. Mycologia, 47, pp.619–632.
Gilbert, N., 2009. Environment: The disappearing nutrient. Nature, 461(7265), pp.716–718.
Giovannetti, M. et al., 2001. The occurrence of anastomosis formation and nuclear exchange in
intact arbuscular mycorrhizal networks. New Phytologist, 151(3), pp.717–724. Disponible en:
http://doi.wiley.com/10.1046/j.0028-646x.2001.00216.x.
Giovannetti, M., Azzolini, D. & Citernesi, a S., 1999. Anastomosis formation and nuclear and
protoplasmic exchange in arbuscular mycorrhizal fungi. Applied and environmental
microbiology, 65(12), pp.5571–5. Disponible en: http://www.pubmedcentral.nih.gov/
Giovannetti, M. & Mosse, B., 1980. An evaluation of techniques for measuring vesicular
arbuscular mycorrhizal infection in roots. New Phytologist, 84, pp.489–500.
Govindarajulu, M. et al., 2005. Nitrogen transfer in the arbuscular mycorrhizal symbiosis. Nature,
435(7043), pp.819–823.
Grace, E.J. et al., 2009. Arbuscular mycorrhizal inhibition of growth in barley cannot be attributed
to extent of colonization , fungal phosphorus uptake or effects on expression of plant
phosphate transporter genes. , pp.938–949.
Harrison, G.W. & List, J.A., 2004. Field Experiments. Journal of Economic Literature, 42(4),
pp.1009–1055.

164 Funcionalidad de la simbiósis entre variedades de yuca y genotipos de
Rhizophagus irregularis en la Orinoquía Colombiana
van der Heijden, M.G.A. et al., 1998. Different arbuscular mycorrhizal fungal species are potential
determinants of plant community structure. Ecology, 79(6), pp.2082–2091.
van der Heijden, M.G.A. et al., 1998. Mycorrhizal fungal diversity determines plant biodiversity,
ecosystem variability and productivity. Nature, 396(6706), pp.69–72. Disponible en:
http://dx.doi.org/10.1038/23932.
Helgason, T. et al., 2002. Selectivity and Functional Diversity in Arbuscular Mycorrhizas of Co-
Occurring Fungi and Plants from a Temperate Deciduous Woodland. Journal of Ecology,
90(2), pp.371–384. Disponible en: http://www.jstor.org/stable/3072140.
Hochberg, M. et al., 2000. Weak sinks could cradle mutualistic symbioses – strong sources should
harbour parasitic symbioses. Journal of Evolutionary biology, 13, pp.213–222.
Hoeksema, J.D. et al., 2010. A meta-analysis of context-dependency in plant response to
inoculation with mycorrhizal fungi. Ecology letters, 13(3), pp.394–407.
Howeler, R., 1981. Mineral Nutrition and fertilization of Cassava. Centro Internacional de
Agricultura Tropical, p.52.
Howeler, R. & Asher, C., 1982. Establishment of an effective endomycorrhizal association in
cassava in flowing solution culture and its effect on phosphorus nutrition. New Phytologist,
90, pp.229–238.
Howeler, R. & Sieverding, E., 1983. Potentials and limitations of mycorrhizal inoculation by
experiments with field-grown cassava. Plant and Soil, 75, pp.245–261.
Howeler, R.H. & Sieverding, E., 1983. Potentials and limitations of mycorrhizal inoculations
ilustrated by experiments with field-grown cassava. Plant and Soil, 75, pp.245–261.
Husband, R. et al., 2002. Molecular diversity of arbuscular mycorrhizal fungi and patterns of host
association over time and space in a tropical forest. Molecular ecology, 11(12), pp.2669–78.
Disponible en: http://www.ncbi.nlm.nih.gov/pubmed/12453249.
Isobe, K. et al., 2014. Effect of Winter Crop Species on Arbuscular Mycorrhizal Fungal

Bibliografía 165
Colonization and Subsequent Soybean Yields. Plant Production Science, 17(3), pp.260–267.
Disponible en: https://www.jstage.jst.go.jp/article/pps/17/3/17_260/_article.
Jakobsen, I., Smith, S.E. & Smith, F.A., 2002. Function and diversity of arbuscular mycorrhizae in
carbon and mineral nutrition. In M. G. A. Van Der Heijden & I. R. Sanders, eds. Mycorrhizal
ecology. Berlín: Springer, pp. 75–88.
Jansa, J., Smith, F. a & Smith, S.E., 2008. Are there benefits of simultaneous root colonization by
diferent arbuscular mycorrhizal fungi? New Phytologist, 177, pp.779–789.
Johnson, N.C. et al., 2013. Predicting community and ecosystem outcomes of mycorrhizal
responses to global change. Ecology letters, 16 Suppl 1, pp.140–53.
Johnson, N.C. & Graham, J.H., 2013. The continuum concept remains a useful framework for
studying mycorrhizal functioning. Plant and Soil, 363(1-2), pp.411–419.
Johnson, N.C., Graham, J.H. & Smith, F.A., 1997. Functioning of mycorrhizal associations along
the mutualism – parasitism continuum *. New Phytologist, (135), pp.575–585.
Jones, M.D. & Smith, S.E., 2004. Exploring functional definitions of mycorrhizas: Are mycorrhizas
always mutualisms? Canadian Journal of Botany, 82(8), pp.1089–1109.
Klironomos, J.J., 2000. Host-specificity and functional diversity among arbuscular mycorrhizal
fungi. Microbial biosystems: New frontiers, pp.845–851. Disponible en:
http://plato.acadiau.ca/isme/Symposium26/klironomos.PDF.
Klironomos, J.N., 2003. Variation in Plant Response to Native and Exotic Arbuscular Mycorrhizal
Fungi. Ecology, 84(9), pp.2292–2301.
Koch, A.M. et al., 2004. High genetic variability and low local diversity in a population of arbuscular
mycorrhizal fungi. Proceedings of the National Academy of Sciences of the United States of
America, 101(8), pp.2369–74. Disponible en:
http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=356957&tool=pmcentrez&renderty
pe=abstract.
Koch, A.M., Croll, D. & Sanders, I.R., 2006. Genetic variability in a population of arbuscular

166 Funcionalidad de la simbiósis entre variedades de yuca y genotipos de
Rhizophagus irregularis en la Orinoquía Colombiana
mycorrhizal fungi causes variation in plant growth. Ecology letters, 9(2), pp.103–10.
Disponible en: http://www.ncbi.nlm.nih.gov/pubmed/16958874 [Consultado Noviembree 18,
2014].
Koide, R., 1985. The Nature of Growth Depression in Sunflower Caused by Vesicular-Arbuscular
Mycorrhizal Infection. New Phytology, 99, pp.449–462.
Kosuta, S. et al., 2003. A diffusible factor from arbuscular mycorrhizal fungi induces symbiosis-
specific MtENOD11 expression in roots of Medicago truncatula. Plant physiology, 131(3),
pp.952–962.
León, D., 2015. Comunidades Nativas de Hongos Formadores de Micorrizas Arbusculares
Asociadas a Yuca Silvestre en la Amazonía Colombiana en Época Leca y lluviosa Bajo Dos
Tipos de Paisaje (Denudación y Llanura Aluvial. Universidad Potificia Javeriana.
León, D., 2006. Evaluación y Caracterización de Micorrizas Arbusculares Asociadas a Yuca
(Manihot esculenta sp.) en dos regiones de la amazonía colombiana. Universidad Pontificia
Javeriana.
Liasu, M.O., Atayese, M.O. & Osonubi, O., 2006. Effect of mycorrhiza and pruning regimes on
seasonality of hedgerow tree mulch contribution to alley-cropped cassava in Ibadan , Nigeria.
Journal of Biotechnology, 5(July), pp.1341–1349.
López-Ráez, J.A. et al., 2008. Tomato strigolactones are derived from carotenoids and their
biosynthesis is promoted by phosphate starvation. New Phytologist, 178(4), pp.863–874.
Magrow, M.J., 1936. Culture et inoculation du champignon symbiotique d’Arum maculatum.
Comptes Rendues hebd Seanc Acad. Sci, 203, pp.887–888.
Marschner, H. & Dell, B., 1994. Nutrient uptake in mycorrhizal symbiosis. Plant and Soil, 159(1),
pp.89–102. Disponible en: http://link.springer.com/article/10.1007/BF00000098.
Martin, F. et al., 2008. The long hard road to a completed Glomus intraradices genome. New
Phytologist, 180(4), pp.735–738.

Bibliografía 167
Mohammad, M.J., Malawi, H.I. & Shilbi, R., 2003. Effects of arbuscular mycorrhizal fungi and
phosphorus fertilization on growth and nutrient uptake of barley grown on soils with different
levels of salts. Journal of plant nutrition, 26(1), pp.125–137. Disponible en:
http://cat.inist.fr/?aModele=afficheN&cpsidt=14520863 [Consultado Septiembre 20, 2012].
Munkvold, L. et al., 2004. High functional diversity within species of arbuscular mycorrhizal fungi.
New Phytologist, 164(2), pp.357–364. Disponible en: http://doi.wiley.com/10.1111/j.1469-
8137.2004.01169.x [Consultado Noviembre 18, 2014].
Muthukumar, T. et al., 2003. Distribution of roots and arbuscular mycorrhizal associations in
tropical forest types of Xishuangbanna, southwest China. Applied Soil Ecology, 22, pp.241–
253.
Oliveira, R.S. et al., 2010. Genetic, phenotypic and functional variation within a Glomus geosporum
isolate cultivated with or without the stress of a highly alkaline anthropogenic sediment.
Applied Soil Ecology, 45(1), pp.39–48. Disponible en:
http://linkinghub.elsevier.com/retrieve/pii/S0929139310000132 [Consultado Febrero 27,
2013].
Ospina, B. & Ceballos, H., 2002. La Yuca en el Tercer Milenio: Sistemas modernos de producción,
procesamiento, utilización y comercialización, Cali: Publicaciones CIAT. Disponible en:
www.clayuca.org/.../27-la-yuca-en-el-tercer-milenio-sistemas-modernos.
Oyetunji, O.J. & Osonubi, O., 2007. Assessment of Influence of Alley Cropping System and
Arbuscular Mycorrhizal ( AM ) Fungi on Cassava Productivity in Derived Savanna Zone of
Nigeria. World, 3(4), pp.489–495.
Prammanee, S. et al., 2010. Growth and starch content evaluation on newly released cassava
cultivars, Rayong 9, Rayong 7 and Rayong 80 at different harvest times. Kasetsart Journal -
Natural Science, 44(4), pp.558–563.
Quiñones-Aguilar, E.E., López-Pérez, L. & Rincón-Enríquez, G., 2014. Growth Dynamics of
Papaya Due To Mycorrhizal Inoculation and Phosphorous Fertilization. Revista Chapingo
Serie Horticultura, XX(2), pp.223–237. Disponible en:
http://www.chapingo.mx/revistas/horticultura/contenido.php?id_articulo=1759&id_revistas=1.

168 Funcionalidad de la simbiósis entre variedades de yuca y genotipos de
Rhizophagus irregularis en la Orinoquía Colombiana
Raju, P.S. et al., 1990. Benefit and cost analysis and phosphorus efficiency of VA mycorrhizal
fungi colonizations with sorghum (Sorghum bicolor) genotypes grown at varied phosphorus
levels. Plant and Soil, 124(2), pp.199–204. Disponible en:
http://link.springer.com/10.1007/BF00009260.
Remy, W. et al., 1994. Four hundred-million-year-old vesicular arbuscular mycorrhizae. Plant
biology (Stuttgart, Germany) Biology, 91(December), pp.11841–11843.
Requena, N. et al., 2007. Plant signals and fungal perception during arbuscular mycorrhiza
establishment. Phytochemistry, 68(1), pp.33–40. Disponible en:
http://www.ncbi.nlm.nih.gov/pubmed/17095025 [Consultado March 16, 2012].
Rivas, L. et al., 2004. Manejo y usos de suelos de la Altillanura Colombiana. Proyecto de
evaluación de impacto convenio M.-C. Convenio de evaluación de impacto, ed.,
Publicaciones CIAT.
Rodriguez, A. & Sanders, I.R., 2016. Ciencia Y Tecnología Colombo-Suiza Ayuda a Alimentar El
Planeta: De La Revolución Verde a La Revolución Microbiana. Acta Biológica Colombiana,
21(1Supl), pp.297–303. Disponible en:
http://www.revistas.unal.edu.co/index.php/actabiol/article/view/50856.
Sadeghian, S., 2009. Impacto de la ganadería sobre el suelos. Alternativas sostenible de manejo.
Publicaciones de Cenibanano, p.15 p.
Sanchez, P.A. & Salinas, J.G., 1981. Low-input technology for managing Oxisols and Ultisols in
tropical America. Advances in agronomy, 34, pp.279–406.
Sánchez, T. & Alonso, L., 2002. Conservación y acondicionamiento de las raíces frescas. In CIAT,
ed. La yuca en el tercer milenio. Sistemas modernos de producción, procesamiento,
utilización y comercialización. Cali, Colombia: CIAT, pp. 503–526.
Sanders, I.R., 2010. “Designer” mycorrhizas?: Using natural genetic variation in AM fungi to
increase plant growth. The ISME journal, 4(9), pp.1081–3. Disponible en:
http://www.ncbi.nlm.nih.gov/pubmed/20613789 [Consultado Noviembre 20, 2014].

Bibliografía 169
Sanders, I.R. & Croll, D., 2010. Arbuscular mycorrhiza: the challenge to understand the genetics of
the fungal partner. Annual review of genetics, 44, pp.271–92. Disponible en:
http://www.ncbi.nlm.nih.gov/pubmed/20822441 [Consultado Marzo 13, 2012].
Sanders, I.R. & Rodriguez, A., 2016. Aligning molecular studies of mycorrhizal fungal diversity with
ecologically important levels of diversity in ecosystems. pp.1–7. Disponible en:
http://dx.doi.org/10.1038/ismej.2016.73.
Santisopasri, V., 2001. Impact of water stress on yield and quality of cassava starch. Industrial
crops. pp.115–129. Disponible en:
http://www.sciencedirect.com/science/article/pii/S0926669000000583.
Schüβler, A., Schwarzott, D. & Walker, C., 2001. A new fungal phylum, the Glomeromycota :
phylogeny and evolution. 105 (December), pp.1413–1421.
SM. H. Shamshiri, K. Usha, and Bhupinder Singh, “Growth and Nutrient Uptake Responses of
Kinnow to Vesicular Arbuscular Mycorrhizae,” ISRN Agronomy, vol. 2012, Article ID 535846,
7 pages, 2012. doi:10.5402/2012/535846
Sieverding, E., 1991. Vesicular–arbuscular mycorrhiza management in tropical agrosystems. In
Eschborn: Deutsche Gesellschaft für Technische Zusammenarbeit (GTZ). p. 371.
Sipsa, MinAgricultura & DANE, 2014. Comportamiento de los precios. Informe de contexto,
Bogotá. Disponible en:
http://www.dane.gov.co/files/investigaciones/agropecuario/sipsa/Semana_29dic_04ene_2013
.pdf.
Smith, S.E. et al., 2004. Functional diversity in arbuscular mycorrhizal ( AM ) symbioses : the
contribution of the mycorrhizal P uptake pathway is not correlated with mycorrhizal responses
in growth or total P uptake. , pp.511–524.
Smith, S.E. & Read, D.J., 2008. Mycorrhizal Symbiosis Third edit. Elsevier, ed., New York: Elsevier
B.V.
Smith, S.E. & Smith, F.A., 2011. Roles of arbuscular mycorrhizas in plant nutrition and growth: new
paradigms from cellular to ecosystem scales. Annual review of plant biology, 62, pp.227–50.

170 Funcionalidad de la simbiósis entre variedades de yuca y genotipos de
Rhizophagus irregularis en la Orinoquía Colombiana
Disponible en: http://www.ncbi.nlm.nih.gov/pubmed/21391813 [Consultado Septiembre 27,
2014].
Smith, S.E., Smith, F.A. & Jakobsen, I., 2003. Mycorrhizal fungi can dominate phosphate supply to
plants irrespective of growth responses. Plant physiology, 133(1), pp.16–20.
Solaiman, M.Z. & Hirata, H., 1997. Effect of arbuscular mycorrhizal fungi inoculation of rice
seedlings at the nursery stage upon performance in the paddy field and greenhouse. Plant
and Soil, 191(1), pp.1–12.
Stockinger, H., Walker, C. & Schüssler, A., 2009. “Glomus intraradices DAOM197198”, a model
fungus in arbuscular mycorrhiza research, is not Glomus intraradices. The New phytologist,
183(4), pp.1176–87. Disponible en: http://www.ncbi.nlm.nih.gov/pubmed/19496945
[Consultado Julio 18, 2012].
Tamasloukht, M.B. et al., 2003. Root Factors Induce Mitochondrial-Related Gene Expression and
Fungal Respiration during the Developmental Switch from Asymbiosis to Presymbiosis in the
Arbuscular Mycorrhizal Fungus Gigaspora rosea. 131(March), pp.1468–1478.
Thrall, P. et al., 2007. Coevolution of symbiotic mutualists and parasites in a community context.
Trends in Ecology & Evolution, 22, pp.120–126.
Tisserant, E. et al., 2013. Genome of an arbuscular mycorrhizal fungus provides insight into the
oldest plant symbiosis. Proceedings of the National Academy of Sciences of the United
States of America, 110(50), pp.20117–22. Disponible en:
http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=3864322&tool=pmcentrez&rendert
ype=abstract.
Toro, J.C. & Cañas, A., 1983. Determinación del contenido de material seca y almidón en yuca por
el sistema de gravedad específica En:Domíngu., Cali, Colombia: PNUD-CIAT.
Vallejo, F.. & Estrada, E.I., 2002. Mejoramiento Genético de Plantas, Palmira: Impresora Feriva
S.A. p. 85 p.
Vierheilig, H. et al., 1998. Ink and vinegar, a simple staining technique for arbuscular-mycorrhizal

Bibliografía 171
fungi. Applied and environmental microbiology, 64(12), pp.5004–7. Disponible en:
http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=90956&tool=pmcentrez&rendertyp
e=abstract.
Vierheilig, H. et al., 1998. Ink and vinegar, a simple staining technique for arbuscular-mycorrhizal
fungi. Applied and environmental microbiology, 64(12), pp.5004–7. Disponible en:
http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=90956&tool=pmcentrez&rendertyp
e=abstract.
Vogelsang, K.M. et al., 2013. Mycorrhizal fungal identity and diversity productivity of a tallgrass the
prairie system. , 172(3), pp.554–562.
Wang, B. & Qiu, Y.L., 2006. Phylogenetic distribution and evolution of mycorrhizas in land plants.
Mycorrhiza, 16(5), pp.299–363.
Wiseman, P.E. & Wells, C.E., 2009. Arbuscular Mycorrhizal Inoculation Affects Root Development
of Acer and Magnolia Species. J. Environ. Hort, 27(2), pp.70–79.
Wu, Y., Liu, T. & He, X., 2009. Mycorrhizal and dark septate endophytic fungi under the canopies
of desert plants in Mu Us Sandy Land of China. Frontiers of Agriculture in China, 3(2),
pp.164–170.
Wyss, T. et al., 2016. Population genomics reveals that within-fungus polymorphism is common
and maintained in populations of the mycorrhizal fungus Rhizophagus irregularis. The ISME
journal, pp.1–13.
Yocum, D.., 1981. Quantifications of costs and benefits accrued by onion plants from vesicular-
arbuscular mycorrhizal associations. In Fifth North Am. Conf. Mycorrhizae. Quebec, Canada:
University Laval, p. 21.
Yost, R. & Fox, R., 1979. Contribution of Mycorrhizae to P nutrition of crops growing on an Oxisol.
Agronomy Journal, 71, pp.903–908.


















