
“GENERALIDADES DE ENZIMAS · PDF fileCapítulo 1 Naturaleza y clasificación...
34
Capítulo 1 Naturaleza y clasificación de enzimas 1 “GENERALIDADES DE ENZIMAS” Autores: Martha Ubalde (Laboratorio de Enzimas hidrolíticas, Facultad de Ciencias) Carolina Villadóniga (Laboratorio de Enzimas hidrolíticas, Facultad de Ciencias) Cecilia Giacomini (Cátedra de Bioquímica, Facultad de Química) Valeria Grazú (Cátedra de Bioquímica, Facultad de Química) Responsable: Prof. Ana María Cantera 2002
Transcript of “GENERALIDADES DE ENZIMAS · PDF fileCapítulo 1 Naturaleza y clasificación...

Capítulo 1 Naturaleza y clasificación de enzimas
1
“GENERALIDADES DE
ENZIMAS”
Autores: Martha Ubalde (Laboratorio de Enzimas hidrolíticas, Facultad de Ciencias) Carolina Villadóniga (Laboratorio de Enzimas hidrolíticas, Facultad de
Ciencias)
Cecilia Giacomini (Cátedra de Bioquímica, Facultad de Química)
Valeria Grazú (Cátedra de Bioquímica, Facultad de Química)
Responsable: Prof. Ana María Cantera
2002
Capítulo 1 Naturaleza y clasificación de enzimas
2
1- Naturaleza y clasificación de enzimas.
1.1- Generalidades
Excepto por un grupo de moléculas de RNA catalítico, las enzimas son proteínas. Al igual que otros
catalizadores son efectivas en muy baja concentración (del orden de nM), se recuperan sin alterar luego de
la reacción, no cambian la posición del equilibrio de la reacción que catalizan sino que mediante la
formación de un complejo enzima-sustrato, disminuyen la energía de activación aumentando la velocidad a
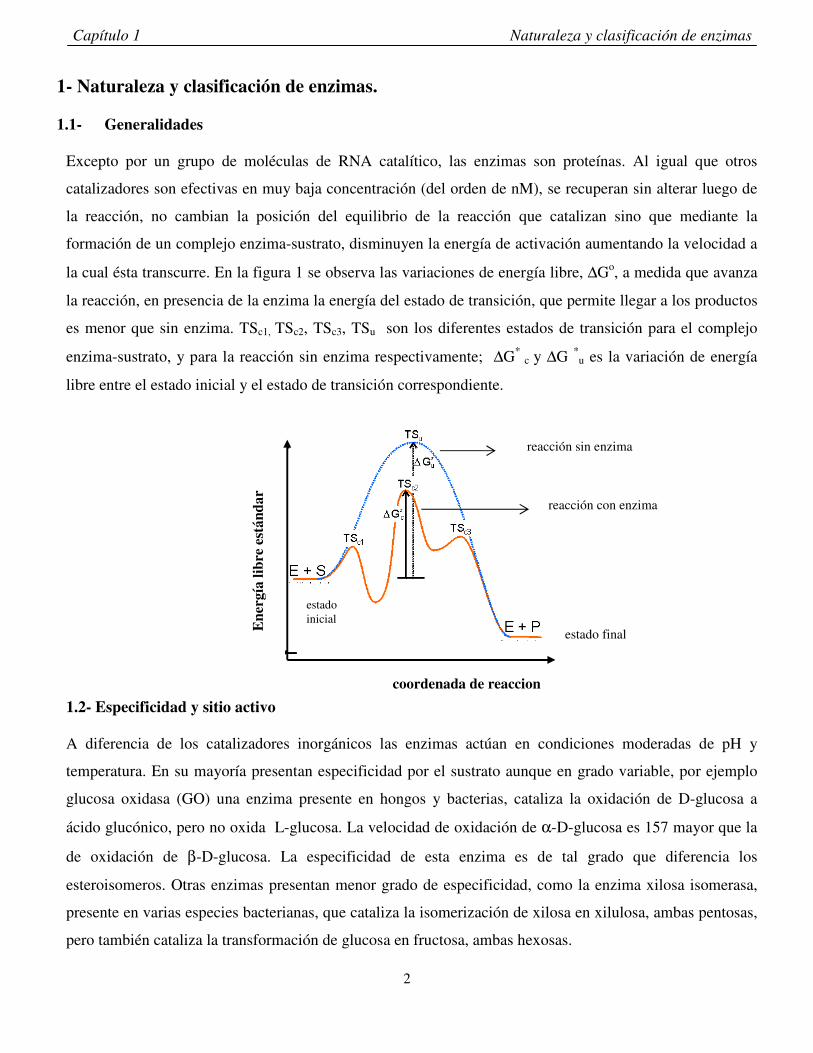
la cual ésta transcurre. En la figura 1 se observa las variaciones de energía libre, ∆Go, a medida que avanza
la reacción, en presencia de la enzima la energía del estado de transición, que permite llegar a los productos
es menor que sin enzima. TSc1, TSc2, TSc3, TSu son los diferentes estados de transición para el complejo
enzima-sustrato, y para la reacción sin enzima respectivamente; ∆G* c y ∆G *
u es la variación de energía
libre entre el estado inicial y el estado de transición correspondiente.
1.2- Especificidad y sitio activo
A diferencia de los catalizadores inorgánicos las enzimas actúan en condiciones moderadas de pH y
temperatura. En su mayoría presentan especificidad por el sustrato aunque en grado variable, por ejemplo
glucosa oxidasa (GO) una enzima presente en hongos y bacterias, cataliza la oxidación de D-glucosa a
ácido glucónico, pero no oxida L-glucosa. La velocidad de oxidación de α-D-glucosa es 157 mayor que la
de oxidación de β-D-glucosa. La especificidad de esta enzima es de tal grado que diferencia los
esteroisomeros. Otras enzimas presentan menor grado de especificidad, como la enzima xilosa isomerasa,
presente en varias especies bacterianas, que cataliza la isomerización de xilosa en xilulosa, ambas pentosas,
pero también cataliza la transformación de glucosa en fructosa, ambas hexosas.
reacción sin enzima
reacción con enzima
estado final
estado inicial
En
erg
ía l
ibre
est
án
da
r
coordenada de reaccion

Capítulo 1 Naturaleza y clasificación de enzimas
3
El sitio activo de las enzimas consiste en un grupo de 3-12 aminoácidos organizados en una estructura
tridimensional, en una zona de la proteína. Este sitio tiene una fuerte afinidad por el sustrato debido a la
naturaleza química de los residuos aminoacídicos que la componen.
Ejemplos de estos grupos reactivos son el grupo tiol de cisteínas, el anillo imidazol de histidina, el grupo
hidroxilo de serina.
1.3- Muchas enzimas necesitan de componentes no enzimáticos Algunas enzimas necesitan de la presencia de componentes adicionales conocidos como cofactores para la
actividad. El complejo enzima-cofactor, catalíticamente activo se denomina holoenzima y la proteína en
ausencia del cofactor se denomina apoenzima.
Los cofactores pueden ser : - iones inorgánicos - coenzimas - grupos prostéticos Iones metálicos inorgánicos
pueden ser parte integral de la estructura de la enzima o pueden asociarse con el sustrato, ayudando a la
unión con la enzima y aumentando la actividad catalítica.
Por ejemplo el Fe +2 se encuentra asociado con el grupo hemo en la peroxidasa y la catalasa encontrándose
fuertemente unido a la enzima, mientras que el Mg +2 se complejea con el ATP 4- y es un componente
esencial en las reacciones que involucran a esta molécula como aquellas catalizadas por fosfotransferasas.
Coenzimas
Son sustancias orgánicas de peso molecular relativamente bajo comparado con la proteína. Muchas
coenzimas contienen moléculas de vitaminas como parte de su estructura. Se encuentran unidas a la proteína
por enlaces débiles y funciona efectivamente como cosustrato de la enzima. Tienen funciones especiales
como la transferencia de hidrógeno ( NAD+ en reacciones de deshidrogenación) o la transferencia de grupos
acilo (coenzimaA en el metabolismo de ácidos grasos). Muchas enzimas que catalizan reacciones diferentes
presentan los mismos coenzimas. Por ejemplo se conocen más de 100 deshidrogenasas que presentan el
NAD+ como coenzima.
Cuando la coenzima se encuentra fuertemente unida a la molécula de enzima y permanece unida a la enzima
luego de finalizado el ciclo catalítico, se denomina grupo prostético. El FAD es otro transportador de átomos
de hidrógeno asociado con enzimas oxidantes como la glucosa oxidasa. El grupo hemo de la catalasa y la
peroxidasa con su anillo de porfirinas también se conoce como grupo prostético.
Capítulo 1 Naturaleza y clasificación de enzimas
4
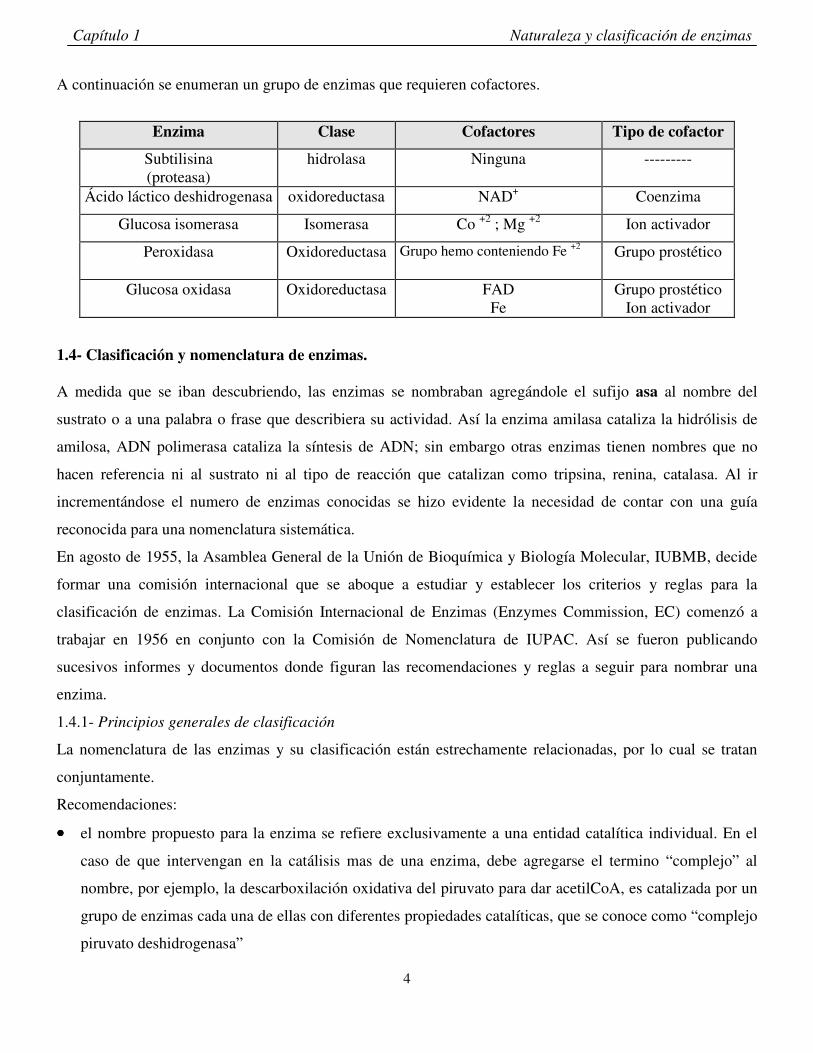
A continuación se enumeran un grupo de enzimas que requieren cofactores.
Enzima Clase Cofactores Tipo de cofactor
Subtilisina (proteasa)
hidrolasa Ninguna ---------
Ácido láctico deshidrogenasa oxidoreductasa NAD+ Coenzima
Glucosa isomerasa Isomerasa Co +2 ; Mg +2 Ion activador
Peroxidasa Oxidoreductasa Grupo hemo conteniendo Fe +2 Grupo prostético
Glucosa oxidasa Oxidoreductasa FAD Fe
Grupo prostético Ion activador
1.4- Clasificación y nomenclatura de enzimas.
A medida que se iban descubriendo, las enzimas se nombraban agregándole el sufijo asa al nombre del
sustrato o a una palabra o frase que describiera su actividad. Así la enzima amilasa cataliza la hidrólisis de
amilosa, ADN polimerasa cataliza la síntesis de ADN; sin embargo otras enzimas tienen nombres que no
hacen referencia ni al sustrato ni al tipo de reacción que catalizan como tripsina, renina, catalasa. Al ir
incrementándose el numero de enzimas conocidas se hizo evidente la necesidad de contar con una guía
reconocida para una nomenclatura sistemática.
En agosto de 1955, la Asamblea General de la Unión de Bioquímica y Biología Molecular, IUBMB, decide
formar una comisión internacional que se aboque a estudiar y establecer los criterios y reglas para la
clasificación de enzimas. La Comisión Internacional de Enzimas (Enzymes Commission, EC) comenzó a
trabajar en 1956 en conjunto con la Comisión de Nomenclatura de IUPAC. Así se fueron publicando
sucesivos informes y documentos donde figuran las recomendaciones y reglas a seguir para nombrar una
enzima.
1.4.1- Principios generales de clasificación
La nomenclatura de las enzimas y su clasificación están estrechamente relacionadas, por lo cual se tratan
conjuntamente.
Recomendaciones:
•••• el nombre propuesto para la enzima se refiere exclusivamente a una entidad catalítica individual. En el
caso de que intervengan en la catálisis mas de una enzima, debe agregarse el termino “complejo” al
nombre, por ejemplo, la descarboxilación oxidativa del piruvato para dar acetilCoA, es catalizada por un
grupo de enzimas cada una de ellas con diferentes propiedades catalíticas, que se conoce como “complejo
piruvato deshidrogenasa”

Capítulo 1 Naturaleza y clasificación de enzimas
5
•••• la propiedad especifica que diferencia una enzima de otra es la reacción química que cataliza, por lo tanto
es lógico que esto sea la base para su nomenclatura y clasificación. Sin embargo la aplicación de este
criterio tiene algunas consecuencias, no puede asignársele nombre a una enzima hasta no conocer la
reacción que cataliza; también se da el caso de que enzimas que catalizan la misma reacción pero son de
diferente origen (bacterianas, vegetales, animales) figuran como una sola en la clasificación, lo mismo
ocurre con las isoenzimas, que no están diferenciadas.
•••• en base al tipo de reacción que catalizan las enzimas se clasifican en clases, cada enzima se individualiza
según el sustrato sobre el que actúa. Esto exige elegir una dirección para la reacción que se describe, se
toma como criterio escribir la reacción en la dirección que se presume de importancia fisiológica.
1.4.2.- Esquema de clasificación.
A cada enzima se le asigna un número clasificatorio de cuatro dígitos separados por un punto y precedido por
la sigla EC (Enzyme Commission). Se clasifican en 6 clases indicada por el primer dígito, cada clase se
divide en subclases (segundo dígito) que a su vez se divide en sub-subclases (tercer dígito) y dentro de ellas
cada enzima se individualiza con el número ordinal que le corresponde dentro de la sub-subclase (cuarto
dígito).
clase tipo de reacción que catalizan
1. Oxidoreductasas oxido-reducciones
2. Transferasas transferencia de grupos de un compuesto a otro
3. Hidrolasas hidrólisis
4. Liasas adición de un grupo a doble enlace
5. Isomerasas rearreglos intramoleculares
6. Ligasas formación de enlaces utilizando la energía de la hidrólisis de ATP

Capítulo 1 Naturaleza y clasificación de enzimas
6
1.5- Ribozimas Aunque la gran mayoría de las enzimas son proteínas, hoy se sabe que ciertas moléculas de ARN tienen
actividad catalítica; estas se conocen como ribozimas.
En las células eucariotas, el ADN se transcribe en una molécula de ARN que contiene secuencias no
codificantes (intrones) y secuencias codificadoras (exones). Durante el proceso de maduración un intrón
cataliza su propia eliminación y la unión de los exones adyacentes, sin la participación de proteínas. La
estrategia es similar a la vista en las enzimas clásicas, el plegamiento del ARN con una molécula de GTP
genera una estructura tridimensional que facilita la ruptura del enlace fosfodiester entre el intrón y el exón y
la subsecuente unión de los exones.
Otra ribozima bien caracterizada, la Ribonucleasa P de E. coli, tiene un componente proteico de 17500 Da y
un componente ARN de 377 nucleótidos (M1 RNA). Se ha comprobado que la porción M1 RNA es
suficiente para la catálisis cortando precursores de t-RNA en la posición correcta, aparentemente la proteína
solo se necesita para estabilizar el ARN o facilitar su función en condiciones particulares de las células.
El descubrimiento de ARNs catalíticos proporciono nuevos conocimientos y cuestionamientos sobre la
función catalítica en general, así como importantes implicancias sobre el origen de la vida y su evolución.
1.6- Referencias
IUBMB nomenclature Homepage, http://www.chem.qmul.ac.uk/iubmb/ Lehninger, A., Nelson, D., Cox, M. En: Principios de Bioquímica 2 da Ed.; Ediciones Omega, Barcelona (1993) Stryer, L., En: Bioquímica 4ta Ed.; Editorial Reverté, Barcelona, (1995). Teal, A. R., y Wymer, P., En: Enzymes and their role in biotechnology;
http://www.biochemistry.org/education/basc03.htm
Capítulo 2 Caracterización de enzimas
7
2- Caracterización de enzimas La medida de la cantidad de enzima presente en una solución por lo general no es expresada en términos
clásicos tales como mg/ml, ya que la enzima puede representar una pequeña fracción de la muestra. Es así
que las enzimas son cuantificadas en términos de su actividad biológica. La actividad enzimática puede
determinarse midiendo la velocidad de desaparición de sustrato o la velocidad de aparición de producto. En
general es mejor medirla por aparición de producto ya que es más exacto determinar aparición de pequeñas
cantidades de producto que la desaparición de pequeñas cantidades de sustrato. A menudo se utilizan
sustratos artificiales que liberan cromóforos que pueden ser determinados colorimétricamente.
Una forma común y útil de determinar la actividad enzimática es en término de unidades. Las unidades de
enzima en general se definen como:
“ La cantidad de enzima necesaria para producir determinada cantidad de producto (milimoles, µmoles etc.)
por unidad de tiempo (minutos, segundos etc.) bajo determinadas condiciones de temperatura, pH y
concentración de sustrato”
Existen muchas definiciones de unidades enzimáticas, esta depende de la enzima estudiada y de como la
defina el investigador.
La concentración de una enzima en una solución se expresa por lo general como unidades por mililitro.
La actividad específica (AE) de una enzima se calcula como el cociente entre la actividad y la concentración
de proteínas, y se expresa en general como unidades de enzima por milígramo de proteína (U/mg proteína).
Al ser las enzimas moléculas biológicas, su actividad se ve afectada por las condiciones de su entorno tales
como, pH, temperatura, concentración de sustrato, fuerza iónica, presencia de cofactores etc. Por lo tanto
para trabajar con ellas hay que caracterizarlas.
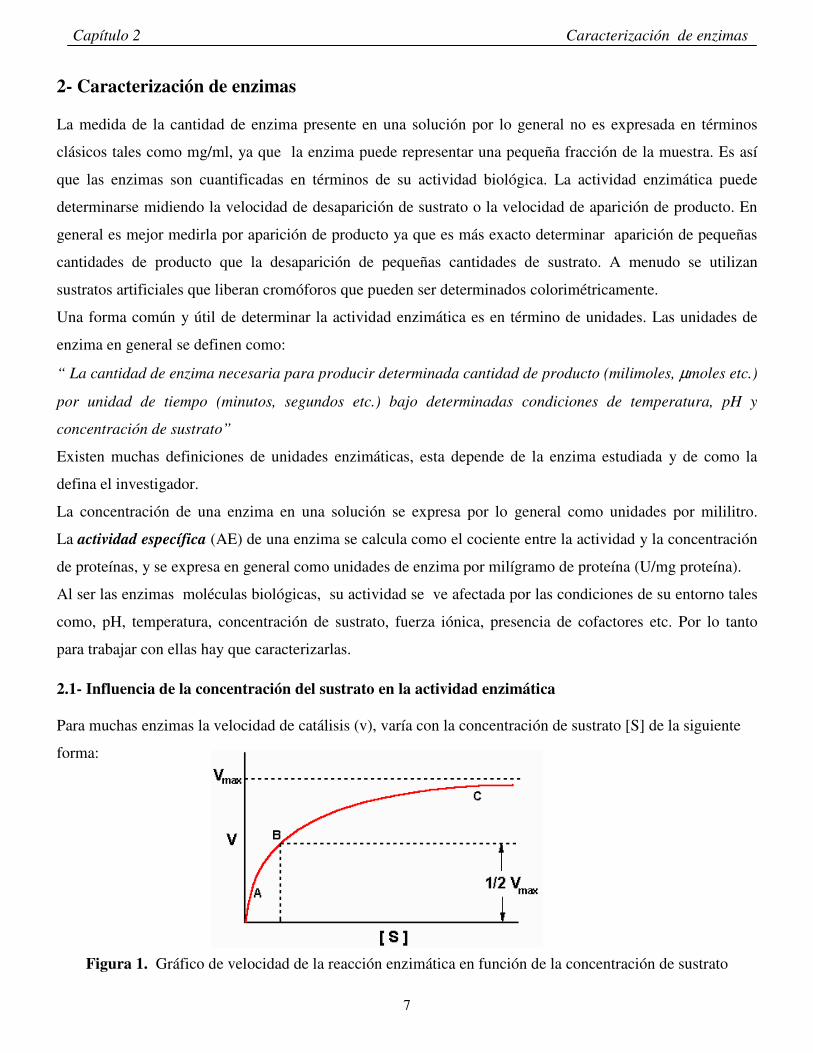
2.1- Influencia de la concentración del sustrato en la actividad enzimática Para muchas enzimas la velocidad de catálisis (v), varía con la concentración de sustrato [S] de la siguiente
forma:
Figura 1. Gráfico de velocidad de la reacción enzimática en función de la concentración de sustrato

Capítulo 2 Caracterización de enzimas
8
Para una determinada concentración de enzima la velocidad (v) es casi proporcional a la concentración de
sustrato [S] , cuando [S] es pequeña. Cuando [S] es elevada , la velocidad de catálisis es prácticamente
independiente de [S]. En este caso decimos que estamos en condiciones de saturación. Michaelis y Menten
propusieron un modelo sencillo que explica estas características cinéticas.
k1 k3 E + S ES E + P k2
La transformación de sustrato en producto implica la formación de un complejo enzima-sustrato (ES) , que
ocurre a través del sitio activo de la enzima. Este sitio presenta una estructura tal que permite una unión
específica y de alta afinidad con el sustrato, además de proveer de un ambiente que favorece los eventos
catalíticos.
El complejo ES tiene dos destinos posibles: i) puede disociarse hasta E y S con una constante de disociación
k2, o ii) puede continuar hasta formar un producto P con una constante de velocidad k3. Se supone que nada
del producto revierte al sustrato inicial, una condición que se cumple en el estado inicial de la reacción, antes
de que la concentración de producto sea apreciable.
La formación catalítica del producto con regeneración de la enzima, es una reacción simple de primer orden
y su velocidad está determinada por la concentración del complejo enzima-sustrato y el valor de k3: v = k3 [ES]
Suponiendo que el complejo ES se encuentra en condiciones de estado estacionario, es posible llegar a la
siguiente ecuación de velocidad, más conocida como ecuación de Michaelis-Menten:
v = Vmax [S] / Km + [S] donde Km = k2 +k3 / k1 y Vmax = K3 [ET] El desarrollo matemático de esta deducción se puede encontrar en cualquier texto de Bioquímica. En términos experimentales Km es numéricamente igual a la concentración de sustrato a la cual la velocidad
de la reacción es la mitad de la velocidad máxima (Vmax ) Cuando [S]= Km entonces v = Vmax / 2 Km es una característica útil y fundamental de cada enzima y un sustrato en particular. También puede verse
como un índice de la afinidad de la enzima por su sustrato bajo condiciones de temperatura, pH y fuerza
iónica determinadas. Cuanto menor es el valor de Km mayor es la afinidad de la enzima por el sustrato. Si el
sitio activo de la enzima es capaz de unirse y reaccionar con varias moléculas de estructura similar, entonces

Capítulo 2 Caracterización de enzimas
9
hay un número de sustratos potenciales para la enzima, y para cada sustrato, la enzima puede presentar un
valor de Km diferente.
Por ejemplo la enzima β-galactosidasa presenta diferentes sustratos naturales y artificiales y distintos valores
de Km para cada uno de ellos
Sustrato Km
Lactosa 1 x 10 –3M
p-nitrofenil-β-galactósido 2 x 10 –4M
p-aminofenil-β-galactósido 4 x 10 –3 M
4 – metil umbelliferil β-galactósido 2 x 10 –4 M
Tabla 1. Valores de Km de la enzima β-galactosidasa para distintos sustratos
Los valores de Km de los enzimas varían ampliamente. Para la mayoría de los enzimas varían entre 10-1y
10-7 M.
Para determinar la actividad enzimática, el sustrato debe estar presente en exceso, de forma de asegurarnos
que la enzima se encuentra saturada y que por lo tanto la velocidad enzimática es independiente de la
concentración de sustrato (punto C de la Figura 1). Se considera que estamos en condiciones de saturación
cuando la concentración de sustrato es mayor o igual a 10 Km.
Concentración de sustrato relativa a Km Velocidad observada relativa a velocidad
1000 Km 1.0 Vmax
100 Km 0.99 Vmax
10 Km 0.91 Vmax
3 Km 0.75 Vmax
1 Km 0.50 Vmax
0.3 Km 0.25 Vmax
0.1 Km 0.091 Vmax
0.01 Km 0.01 Vmax
0.001 Km 0.001 Vmax
Tabla 2. Relación entre concentración de sustrato y velocidad de la reacción enzimática
La Vmax , sin embargo no es una característica fundamental para una enzima y su valor va a depender de la
cantidad de enzima presente. Si esto se estandariza a un mol de enzima, el valor teórico de Vmax obtenido es
el número de recambio o actividad molar. Esta es una medida útil del poder catalítico de una enzima. Su

Capítulo 2 Caracterización de enzimas
10
valor cuantifica el número de moléculas de sustrato transformadas por molécula de enzima por minuto. La
anhidrasa carbónica tiene la mayor actividad molar de las enzimas conocidas. El valor del número de
recambio de varias enzimas conocidas de se muestra a continuación.
Enzima Nº de Recambio
(min-1
)
anhidrasa Carbónica 36 x 10 6
catalasa 5.6 x 10 6
β-galactosidasa 12 x 10 3
quimiotripsina 6 x 10 3
lisozima 60
Tabla 3. Números de recambio de algunas enzimas
2.1.1- Determinación de parámetros cinéticos. La gráfica de velocidad en función de la concentración de sustrato (Figura 1) es una hipérbola rectangular.
No es útil en la práctica trabajar con esta curva dado que es difícil dibujar una hipérbola rectangular en forma
exacta, colocar las asíntotas correctamente y a su vez detectar desviaciones de la curva esperada.
Para determinar los parámetros cinéticos de una enzima conviene entonces reordenar la ecuación de
Michaelis-Menten de manera que proporcione una gráfica lineal. Hay varios tipos de linealizaciones de esta
ecuación pero la que más se utiliza es una representación doble inversa, también llamada representación de
Lineweaver-Burk.
1 = 1 + Km x 1
v Vmax Vmax [S]
Figura 2. Representación de Lineweaver-Burk de la ecuación de Michaelis-Menten
Capítulo 2 Caracterización de enzimas
11
2.2- Medida de velocidad
Al reordenar la ecuación de Michaelis-Menten de la siguiente forma: v = k3 x [ET] 1 + Km
[S] queda claro que la velocidad de la reacción es directamente proporcional a la concentración de la enzima,
siempre y cuando se trabaje en condiciones iniciales; dado que sólo en ese caso se puede hacer la suposición
de que la [S] es constante.
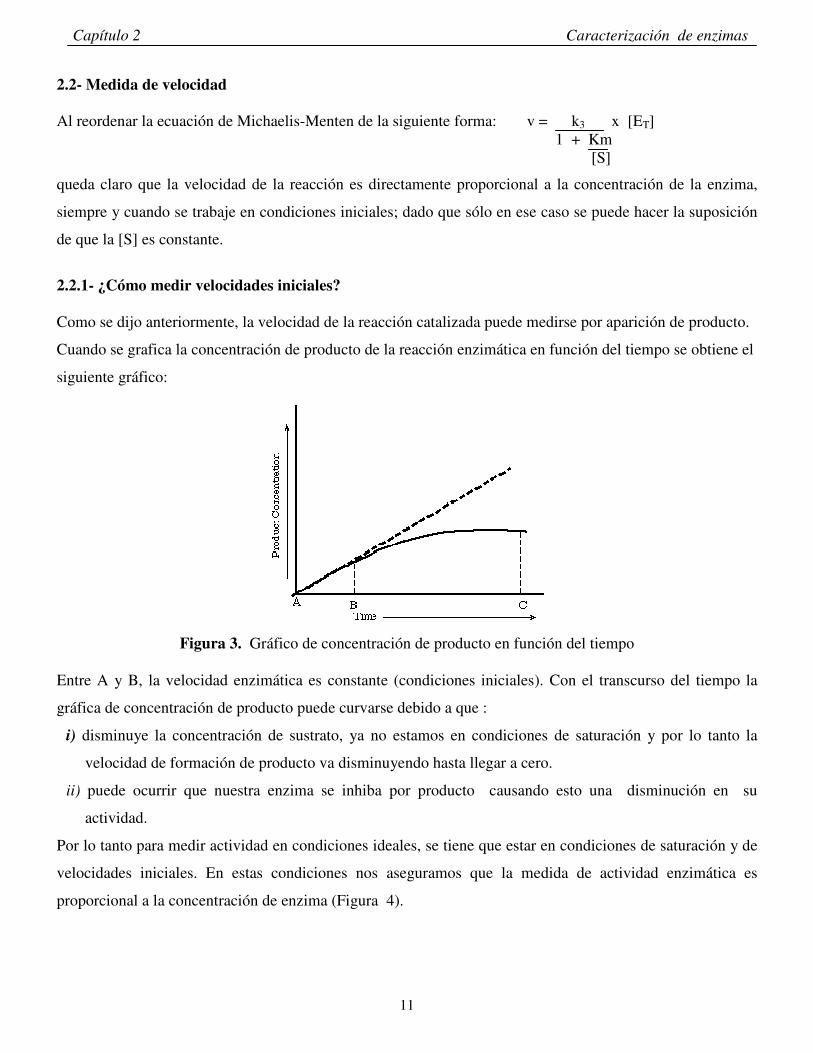
2.2.1- ¿Cómo medir velocidades iniciales? Como se dijo anteriormente, la velocidad de la reacción catalizada puede medirse por aparición de producto.
Cuando se grafica la concentración de producto de la reacción enzimática en función del tiempo se obtiene el
siguiente gráfico:
Figura 3. Gráfico de concentración de producto en función del tiempo
Entre A y B, la velocidad enzimática es constante (condiciones iniciales). Con el transcurso del tiempo la
gráfica de concentración de producto puede curvarse debido a que :
i) disminuye la concentración de sustrato, ya no estamos en condiciones de saturación y por lo tanto la
velocidad de formación de producto va disminuyendo hasta llegar a cero.
ii) puede ocurrir que nuestra enzima se inhiba por producto causando esto una disminución en su
actividad.
Por lo tanto para medir actividad en condiciones ideales, se tiene que estar en condiciones de saturación y de
velocidades iniciales. En estas condiciones nos aseguramos que la medida de actividad enzimática es
proporcional a la concentración de enzima (Figura 4).

Capítulo 2 Caracterización de enzimas
12
Figura 4. Efecto de la concentración de la enzima en la velocidad de reacción.
2.3- Influencia del pH y la temperatura
Las enzimas son moléculas muy sensibles a cambios en el medio circundante. Es así que un cambio en el pH
o la temperatura, pueden afectar profundamente la actividad y estabilidad de una enzima. pH o temperatura óptima: es aquel pH o temperatura a la cual la enzima presenta su máxima actividad. Estabilidad térmica y con el pH: es el rango de pH o temperatura en el cual la enzima es capaz de retener su
actividad catalítica.
2.3.1- Efecto de la temperatura
2.3.1.1- Actividad enzimática en función de la temperatura
La influencia de la temperatura en la actividad de las enzimas se representa generalmente en las llamadas
curvas de temperatura optima (Figura 5), en las que se grafica la actividad en función de la temperatura.
Figura 5. Gráfico de velocidad en función de la temperatura.

Capítulo 2 Caracterización de enzimas
13
Los efectos de la temperatura en la actividad de una enzima son complicados y pueden ser considerados
como dos fuerzas que actúan en forma simultanea pero en sentidos opuestos. A medida que la temperatura
aumenta, la velocidad de reacción aumenta pero a su vez ocurre una inactivación progresiva
(desnaturalización) de la proteína. Este efecto es más pronunciado a medida que aumenta la temperatura.
La aceleración de la reacción por aumento de la temperatura es causada porque a mayor temperatura una
mayor fracción de moléculas tienen la energía suficiente para proveer la energía de activación de la reacción.
Por lo general cada 10º C de incremento en al temperatura, la velocidad de la reacción se duplica. El
fenómeno de inactivación es debido a que a altas temperaturas las moléculas de enzima vibran y se tuercen
tan rápido que algunos de los enlaces no covalentes se rompe. Cuando la temperatura destruye la estructura
terciaria, las moléculas de enzima se inactivan o desnaturalizan. Algunas enzimas se desnaturalizan a
temperaturas sólo un poco mayor que la temperatura del cuerpo humano, pero algunas pocas son estables
incluso a la temperatura de ebullición del agua.
Por lo tanto lo que se observa es una “Temperatura óptima aparente”.
La desnaturalización térmica de una enzima es dependiente del tiempo, por lo que el término “Temperatura
óptima” tiene muy poco significado a menos que se tome el cuenta el tiempo de exposición. Cuanto más
corto sea el tiempo de exposición la temperatura óptima de la reacción enzimática puede aumentar.
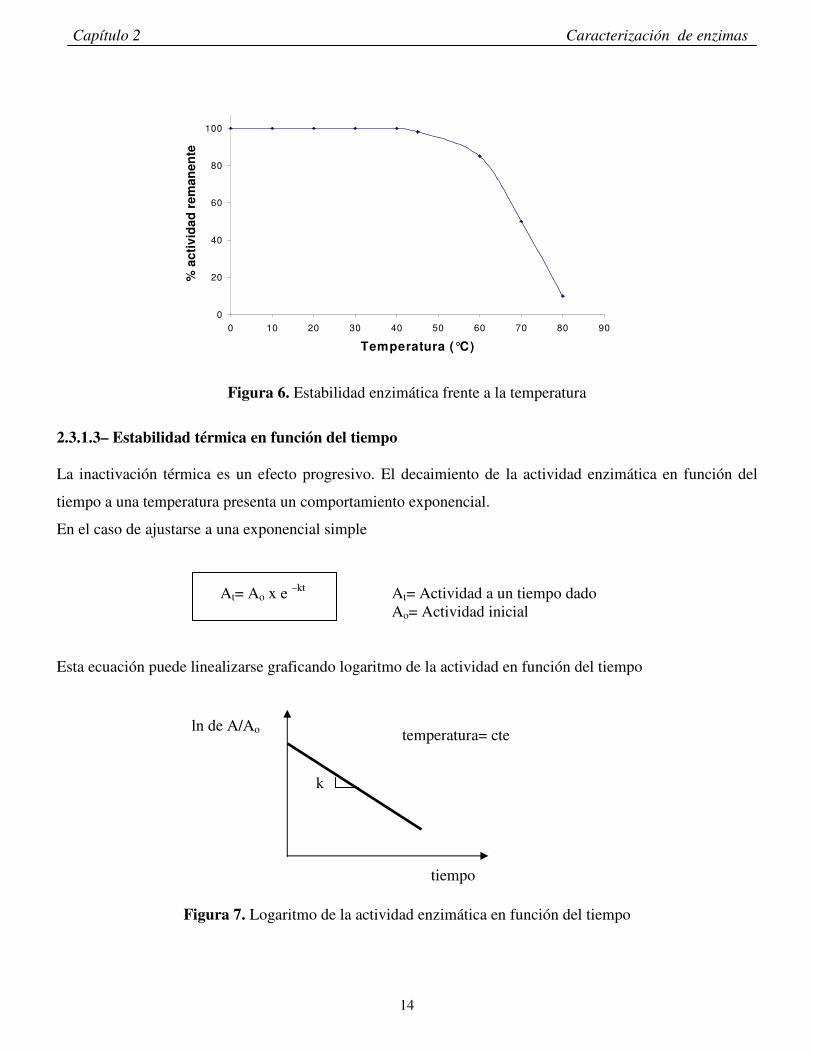
2.3.1.2- Estabilidad de la enzima en función de la temperatura El efecto de la temperatura en la estabilidad de una enzima puede ser determinado exponiendo a la enzima
en ausencia de sustrato a varias temperaturas por un tiempo determinado. Una vez transcurrido dicho tiempo
se mide la actividad remanente de la misma a una temperatura prefijada y se la compara con la actividad
inicial a dicha temperatura (Figura 6).
Capítulo 2 Caracterización de enzimas
14
Figura 6. Estabilidad enzimática frente a la temperatura
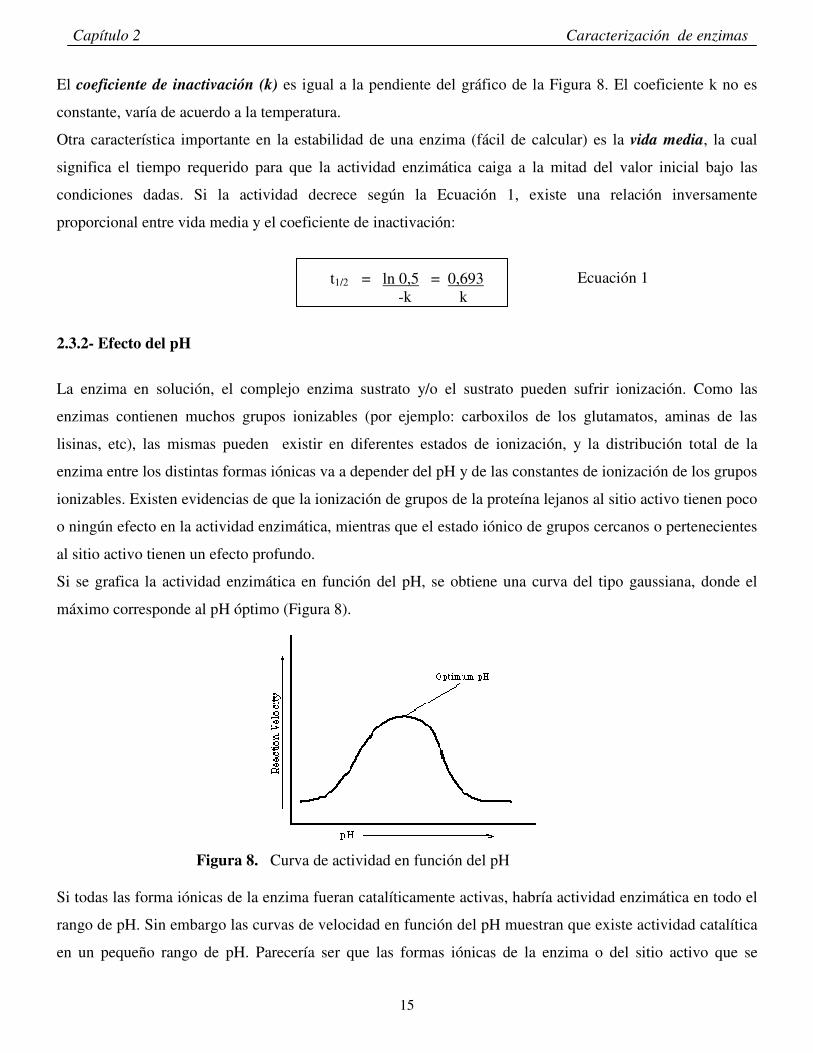
2.3.1.3– Estabilidad térmica en función del tiempo La inactivación térmica es un efecto progresivo. El decaimiento de la actividad enzimática en función del
tiempo a una temperatura presenta un comportamiento exponencial.
En el caso de ajustarse a una exponencial simple
At= Ao x e –kt At= Actividad a un tiempo dado
Ao= Actividad inicial Esta ecuación puede linealizarse graficando logaritmo de la actividad en función del tiempo
Figura 7. Logaritmo de la actividad enzimática en función del tiempo
tiempo
ln de A/Ao temperatura= cte
k
0
20
40
60
80
100
120
0 10 20 30 40 50 60 70 80 90
Temperatura (°C)
% a
cti
vid
ad
re
ma
ne
nte
Capítulo 2 Caracterización de enzimas
15
El coeficiente de inactivación (k) es igual a la pendiente del gráfico de la Figura 8. El coeficiente k no es
constante, varía de acuerdo a la temperatura.
Otra característica importante en la estabilidad de una enzima (fácil de calcular) es la vida media, la cual
significa el tiempo requerido para que la actividad enzimática caiga a la mitad del valor inicial bajo las
condiciones dadas. Si la actividad decrece según la Ecuación 1, existe una relación inversamente
proporcional entre vida media y el coeficiente de inactivación:
t1/2 = ln 0,5 = 0,693
-k k
2.3.2- Efecto del pH
La enzima en solución, el complejo enzima sustrato y/o el sustrato pueden sufrir ionización. Como las
enzimas contienen muchos grupos ionizables (por ejemplo: carboxilos de los glutamatos, aminas de las
lisinas, etc), las mismas pueden existir en diferentes estados de ionización, y la distribución total de la
enzima entre los distintas formas iónicas va a depender del pH y de las constantes de ionización de los grupos
ionizables. Existen evidencias de que la ionización de grupos de la proteína lejanos al sitio activo tienen poco
o ningún efecto en la actividad enzimática, mientras que el estado iónico de grupos cercanos o pertenecientes
al sitio activo tienen un efecto profundo.
Si se grafica la actividad enzimática en función del pH, se obtiene una curva del tipo gaussiana, donde el
máximo corresponde al pH óptimo (Figura 8).
Figura 8. Curva de actividad en función del pH
Si todas las forma iónicas de la enzima fueran catalíticamente activas, habría actividad enzimática en todo el
rango de pH. Sin embargo las curvas de velocidad en función del pH muestran que existe actividad catalítica
en un pequeño rango de pH. Parecería ser que las formas iónicas de la enzima o del sitio activo que se
Ecuación 1

Capítulo 2 Caracterización de enzimas
16
encuentran en forma mayoritaria a ese pH son catalíticamente activas.
La causa de que la actividad decaiga a ambos lados del pH óptimo puede ser debido a:
i) el hecho de la enzima, el sustrato y/o el complejo enzima-sustrato se encuentren mayoritariamente en
una forma iónica no activa
ii) que el pH afecte la estabilidad de la enzima, ocurriendo desnaturalización irreversible.
iii) combinación de ambos efectos.
Es necesario por lo tanto comparar estas curvas con las de estabilidad de la enzima en función del pH para
eliminar “pHs óptimos falsos”. El efecto del pH en la estabilidad de la enzima es posible determinarlo
experimentalmente. Para esto se incuba la enzima en ausencia de sustrato a distintos pH por un período
determinado de tiempo. Al cabo del mismo, se procede a medir la actividad de la enzima luego de retornar el
pH a un valor de referencia.
Si se observa que en los rangos de pH donde la enzima es estable, la misma muestra una disminución de su
actividad catalítica, esta disminución sólo puede ser atribuida a formas iónicas no activas. Puede llegar a
ocurrir que el pH óptimo para la actividad enzimática no coincida con el rango de pH en que la enzima es
más estable.
El valor de pH óptimo varía considerablemente de una enzima a otra, y a su vez depende del origen de la
enzima (Tabla 4).
Enzima pH óptimo
lipasa (páncreas) 8.0 lipasa (estómago) 4.0-5.0
pepsina 1.5-1.6 tripsina 7.8-8.7 ureasa 7.0
invertasa 4.5 maltasa 6.1-7.0
amilasa (páncreas) 6.7-7.0 amilasa (malta) 4.6-5.2
catalasa 7.0
Tabla 4. pH óptimos de actividad de distintas enzimas
La evolución a relacionado a las enzimas con sus ambientes naturales. Por ejemplo, la enzima pepsina que
participa en la digestión de proteínas y se encuentra sólo en el estómago, es más activa a los valores bajos de
pH que prevalecen en el estómago luego de una comida. Por el contrario, la amilasa que se encuentra en la
saliva trabaja mejor a pH neutros, que es el pH característico de la boca.

Capítulo 2 Caracterización de enzimas
17
2.4- Efecto de los inhibidores sobre las enzimas.
Los inhibidores son sustancias que reducen la actividad de una enzima. Los inhibidores pueden ser
componentes propios de la célula o extraños a la misma. En el primer caso pueden ser un elemento
importante para la regulación del metabolismo celular.
Los inhibidores pueden clasificarse en:
2.4.1- Reversibles
Cuando reaccionan en forma reversible con la enzima estableciendo un equilibrio entre la enzima libre y el
complejo enzima-inhibidor (EI y/o ESI). La constante de equilibrio para la disociación del complejo enzima-
inhibidor se conoce como KI. KI= [E][I]
[EI]
Este tipo de inhibición siempre puede ser revertida mediante la eliminación del inhibidor (por ej: por
diálisis). A su vez, al tratarse de un equilibrio, que usualmente se alcanza muy rápidamente, el grado de
inhibición es aparentemente independiente del tiempo.
Existen varios tipos de inhibición reversible; todos ellos implican la unión no covalente de un inhibidor a la
enzima pero difieren en los mecanismos por medio de los cuales reducen la actividad enzimática y en la
forma en que afectan la cinética de la reacción.
i) inhibidor competitivo
Es un inhibidor estructuralmente similar al sustrato por lo que puede unirse en forma reversible al sitio
activo. Durante la fracción de tiempo en que la molécula de inhibidor está ocupando el sitio activo, la enzima
no está disponible para la catálisis. El efecto global es como si la enzima no pudiera unirse al sustrato tan
bien cuando está presente el inhibidor. Así pues, cabe prever que la enzima actúe como si su Km se
incrementara por la presencia del inhibidor. Por lo que la forma de revertir este tipo de inhibición es
desplazando el inhibidor aumentando la concentración de sustrato.
Por lo general, los productos de una reacción enzimática son los inhibidores competitivos más comunes. Esto
es importarte tenerlo en cuenta en el caso de desarrollar un proceso industrial, debido a que si el producto es
un inhibidor competitivo, para poder alcanzar la máxima eficiencia es necesario remover los productos por
ejemplo por ultrafiltración.

Capítulo 2 Caracterización de enzimas
18
ii) no competitivo
Este tipo de inhibidor reversible se une a la enzima en un sitio distinto del sitio activo. La unión del inhibidor
no interfiere con la unión del sustrato pero previene su procesamiento, por lo tanto se observa una
disminución de la Vmax.
Estos inhibidores no tienen porque estar relacionados estructuralmente con el sustrato, por lo que la
inhibición no puede ser revertida por incremento de la concentración del mismo.
iii) acompetitivo
Este tipo de inhibidor se une reversiblemente al complejo ES en sitios diferentes al del sitio activo, dando un
complejo ESI inactivo. El inhibidor no se une a la enzima libre, debido a que no tiene un sitio
complementario de unión. Es la unión del sustrato con la enzima libre lo que provoca un cambio
conformacional que desenmascara o forma el sitio de unión del inhibidor.
Este tipo de inhibición es poco común en sistemas con un único sustrato, pero si es un tipo común de
inhibición por producto en sistemas con más de un producto y sustrato.
En la Figura 9 podemos encontrar las linealizaciones de Lineweaver-Burk correspondientes a los distintos
tipos de inhibición reversible.
Figura 9. Linealizaciones de Lineveawer-Burk de los distintos tipos de inhibición reversible. Panel A: enzima sin inhibidor; Panel B: inhibición competitiva; Panel C: inhibición no competitiva; Panel C: inhibición acompetitiva.

Capítulo 2 Caracterización de enzimas
19
La Tabla 5 es un resumen de las principales características de los distintos tipos de inhibición reversible.
Tipo de inhibidor Esquema de reacción Efecto cinético
inhibidor competitivo k1 k3 E + S ES E + P + k2
I k4 k5 KI = k5 / k4 EI v= Vmax x S / KMapp + S
KMapp= KM (1 +I / KI)
- Km se ve aumentada - Vmax no es afectada
inhibidor no competitivo k1 k3 E + S ES E + P + k2 + I I k4 k5 k4 k5 k1 EI ESI k2 KI = k5 / k4 v= Vmax app x S / KM + S
Vapp app= Vmax / (1 +I / KI)
- Km no es alterada - Vmax disminuye en forma proporcional a la concentra-ción de inhibidor.
inhibidor acompetitivo k1 k3 E + S ES E + P k2 +
I k4 k5
ESI
KI = k4 / k5 v= Vmax app x S / KM app + S Vmax app= Vmax / (1+ I/KI) KM app= KM / (1 +I / KI)
- Km disminuye - Vmax disminuye
Tabla 5. Principales características de los distintos tipos de inhibición irreversible
2.4.2- Irreversibles
En el caso de la inhibición irreversible el inhibidor no se encuentra en equilibrio con el complejo enzima-
inhibidor, por lo tanto no se reactiva la enzima removiendo el inhibidor mediante diálisis y otros métodos, a
diferencia de lo que sucede con los inhibidores reversibles. El efecto cinético de un inhibidor irreversible es
disminuir la concentración de enzima activa, lo cual lleva a una disminución de la velocidad de la reacción.
La inhibición irreversible se caracteriza por un aumento progresivo en el tiempo, llegando en última instancia

Capítulo 2 Caracterización de enzimas
20
a la inhibición completa aún con el inhibidor muy diluído, siempre que esté en exceso respecto a la
concentración de la enzima presente. La efectividad del inhibidor no se expresa como una constante de
equilibrio, sino como una constante de velocidad, que determina la fracción de la enzima inhibida en un
período determinado de tiempo para una cierta concentración de inhibidor.
Los inhibidores de este tipo generalmente causan inactivación por modificación covalente de la estructura de
la enzima. En la mayoría de los casos estas sustancias reaccionan con algún grupo funcional del sitio activo
para bloquear el lugar de unión del sustrato o para dejarlo catalíticamente inactivo. El cianuro es un clásico
ejemplo; el mismo se une covalentemente a la citocromo oxidasa mitocondrial, inhibiendo de esta forma
todas las reacciones asociadas con el transporte de electrones.
Referencias Cornish-Bowden, A., En: Fundamentals of enzyme kinetics; Portland Press, Ltd., London, (1995).
Dixon, M., Webb, E. C., En: Enzymes 3era Ed.; Dixon, M., Webb, . C., (Eds), Academic Press, New York, (1979). Hartmeier, W., En: Immobilized biocatalysists; Springer-Verlag, Berlin, (1988). Mathews, C., y Van Holde, K., En: Bioquímica 2da Ed., McGraw-Hill, Interamericana, Madrid, (1998). Messing, R., En: Immobilized enzymes for industrial reactors; Academic Press, Inc, London, (1975).
Purves, W., Orians, G., Craig, H., y Sadava, D., En: Life: the science of biology, W. H. Freeman & Company, (Eds), (1998). Stryer, L., En: Bioquímica 4ta Ed.; Editorial Reverté, Barcelona, (1995). Teal, A. R., y Wymer, P., En: Enzymes and their role in biotechnology;
http://www.biochemistry.org/education/basc03.htm

Capítulo 3 Extracción y purificación de enzimas
21
3- Extracción y purificación de enzimas
3.1- Producción de enzimas
Las enzimas son producidas a partir de células, donde cumplen su función metabólica. Más de 2500 enzimas
diferentes han sido aisladas y caracterizadas hasta el momento, pero esto sólo refleja un 10 % del potencial
enzimático existente en la naturaleza.
Las células constituyen hasta hoy el único recurso para la obtención de biocatalizadores, aunque no puede
descartarse en un futuro su fabricación por síntesis química. Las enzimas pueden proceder de tejidos de
organismos diferenciados (células animales y vegetales) y microorganismos (procariotes y eucariotes).
Un esquema general de producción de enzimas consiste en cuatro etapas:
1) Generación o producción Fermentación en el caso de enzimas microbianas;
producción agropecuaria o cultivo en el caso de enzimas
de tejidos
2) Recuperación Separación, concentración, extracción
3) Purificación varía según el tipo de catalizador enzimático que se use;
puede incluir varias etapas o no existir
4) Formulación acabado y normalización del producto enzimático
Los procesos productivos, de acuerdo al origen de la enzima, difieren principalmente en la etapa de
generación. Las etapas restantes, son en cambio esencialmente independientes al mismo, siendo en general
más simple la extracción de enzimas de tejidos que la de enzimas intracelulares microbianas, debido a la
mayor dificultad de disrupción de la célula.
3.1.1- Generación
Es la primer etapa del proceso de producción y es donde hay mayores diferencias dependiendo del origen de
la enzima. Las enzimas de origen vegetal se obtienen a partir de subproductos de la actividad agrícola,
mientras que las de origen animal de subproductos de matadero. A su vez las enzimas microbianas se
obtienen como productos metabólicos en un proceso de fermentación. Por lo general las enzimas microbianas
de interés comercial son productos del metabolismo aeróbico y cumplen con una función catabólica, estando
su producción asociada al crecimiento.
Si bien existe hoy en día un gran auge de las enzimas microbianas, aún se produce para la industria un
número importante de enzimas de tejidos animales y vegetales, ya que los esfuerzos por reemplazar estas
enzimas por contrapartes de origen microbiano han sido infructuosos (por ejemplo las amilasas para la
preparación de mostos de cerveza, etc).
Capítulo 3 Extracción y purificación de enzimas
22
3.1.2- Recuperación
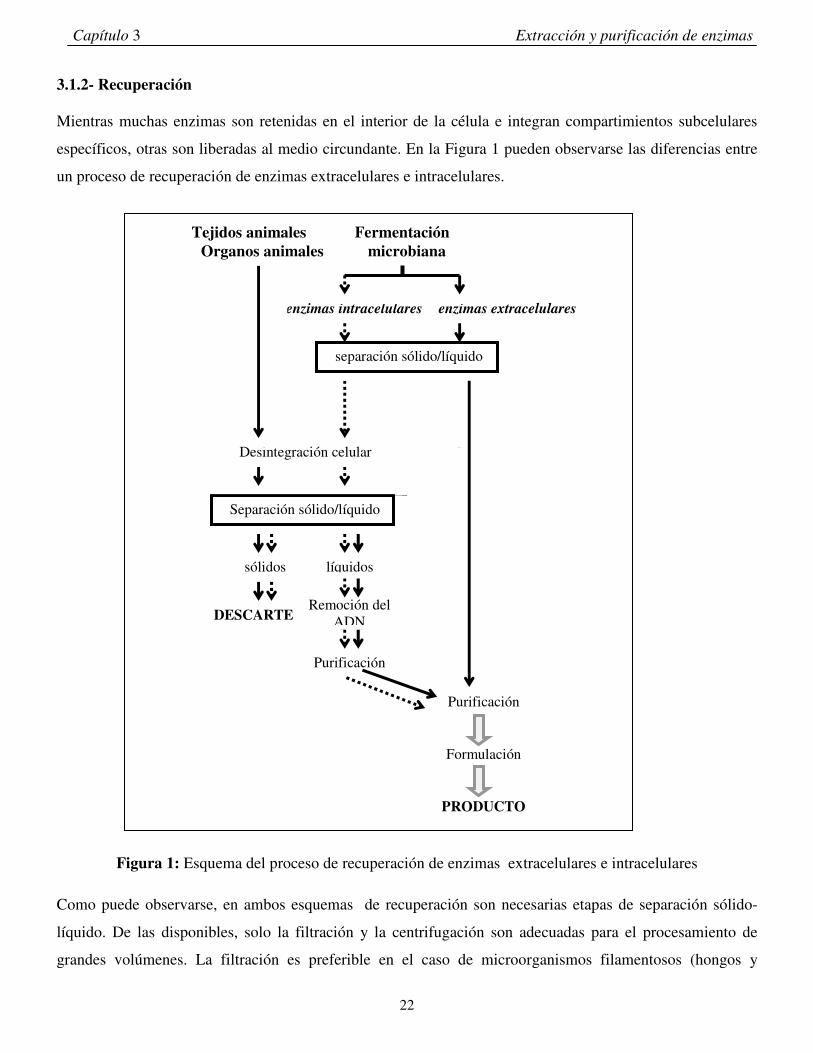
Mientras muchas enzimas son retenidas en el interior de la célula e integran compartimientos subcelulares
específicos, otras son liberadas al medio circundante. En la Figura 1 pueden observarse las diferencias entre
un proceso de recuperación de enzimas extracelulares e intracelulares.
Figura 1: Esquema del proceso de recuperación de enzimas extracelulares e intracelulares
Como puede observarse, en ambos esquemas de recuperación son necesarias etapas de separación sólido-
líquido. De las disponibles, solo la filtración y la centrifugación son adecuadas para el procesamiento de
grandes volúmenes. La filtración es preferible en el caso de microorganismos filamentosos (hongos y
Tejidos animales
Organos animales
Fermentación
microbiana
enzimas intracelulares enzimas extracelulares
separación sólido/líquido
Separación sólido/líquido
Desintegración celular
sólidos líquidos
DESCARTE Remoción del
ADN
Purificación
Purificación
Formulación
PRODUCTO

Capítulo 3 Extracción y purificación de enzimas
23
actinomicetes), mientras que la centrifugación es utilizada preferentemente con microorganismos
unicelulares (levaduras y bacterias).
También puede observarse en la Figura 1, la mayor complejidad del proceso de recuperación de las enzimas
intracelulares. Esto es debido a que es necesario la disrupción de las células para poder extraer la enzima de
interés. Es importante hacer notar que si bien el esquema de recuperación de enzimas extracelulares es más
sencillo, tiene el inconveniente de la necesidad obligatoria de una etapa de concentración, la cual puede
lograrse por evaporación al vacío o por ultrafiltración.
Extracción de enzimas intracelulares
Cuando la proteína de interés se encuentra en el medio intracelular, ya sea de una célula animal, bacteriana o
vegetal, el primer paso es extraerla en forma soluble en un buffer de pH y fuerza iónica similar al medio
donde se encontraba originalmente. Para lograr esto hay descriptos un variado número de protocolos básicos
que permiten una extracción sin dañar la proteína buscada, aunque en muchos casos es necesario introducir
algunas modificaciones para aumentar el rendimiento de la extracción y mantener la actividad funcional de la
proteína.
Como primer paso, se debe eliminar todo aquello que contamina la muestra: i) si se va a trabajar con tejidos
animales o vegetales estos se lavan con buffer, ii) en el caso de células bacteriana estas deben separarse del
medio de cultivo centrifugando, luego se resuspende en buffer y se centrifuga nuevamente.
Luego es necesaria la lísis celular para así poder recuperar la enzima. Esta operación es relativamente simple
para el caso de tejidos, pues las células animales son extremadamente frágiles por ausencia de pared celular,
y aunque las células vegetales tienen gruesas paredes celulósicas, están son rígidas y por lo tanto muy
sensibles a fuerzas cortantes. En cambio, la disrupción de células de microorganismos es más compleja
debido a la estructura de su pared celular. La disrupción de células de microorganismos es una etapa crítica
dentro del proceso, no porque no haya sistemas eficientes de disrupción, sino porque éstos deben
compatibilizar una alta eficiencia de ruptura celular con la preservación de la actividad enzimática de interés.
Los distintos métodos de disrupción de las paredes celulares microbianas se encuentran divididos entre: a)
aquellos que provocan la fragmentación del envoltorio celular por aplicación de fuerzas cortantes y b)
aquellos que producen degradación del mismo.
A continuación se da una lista de algunos de los posibles métodos de lísis celular.

Capítulo 3 Extracción y purificación de enzimas
24
Sistema Principio de ruptura Materia prima adecuada Aplicación a gran escala
sonicación Cavitación suspensiones celulares nula
molienda Compresión y esfuerzo cortante
bacterias, tejidos vegetales reducida
homogeinizador
mecánico
Esfuerzo de corte, cavitación
mayoría de tejidos animales y vegetales
más amplia
homogeinizador manual Esfuerzo de corte, cavitación
tejidos vegetales --------
french press Compresión, esfuerzo de corte
bacterias, levaduras, células vegetales
poco probable
digestión alcalina Hidrólisis pared celular bacterias, levaduras muy reducida
digestión enzimática hidrólisis pared celular y ruptura osmótica
bacterias, levaduras más amplia
Tabla 1. Distintos métodos de lísis celular
El sonicador crea vibraciones que rompen las células, estas deben estar en solución en un medio con baja
viscosidad, el tiempo de lísis va de 5 a 10 minutos.
En la molienda se utiliza junto con un tampón que ya es el medio para la extracción, un abrasivo como
alúmina o arena, es muy apropiado para romper paredes de células vegetales, 15 minutos suele ser suficiente
para la extracción del contenido celular.
En el caso de la French press o French Pressure Cell la muestra se somete a alta presión e inmediatamente a
presión atmosférica, este cambio rápido en las condiciones causa que la células estallen y se obtiene así el
extracto con el que se continua trabajando.
El extracto obtenido luego de la lísis, homogenato, se centrifuga para eliminar los restos de pared celular o
los coadyuvantes para la lísis que se utilizan en algunos casos. Dependiendo del material de partida las
condiciones de centrifugación van desde 10 minutos a 15000xg hasta 1 hora a 100000xg, en lo posible se
utilizan centrifugas refrigeradas que permiten mantener el extracto a 4 ºC. El sobrenadante es el extracto
crudo que se separa del resto del material que no interesa, pellet. Muchas veces el extracto crudo se filtra por
algodón o lana de vidrio para eliminar partículas en suspensión.
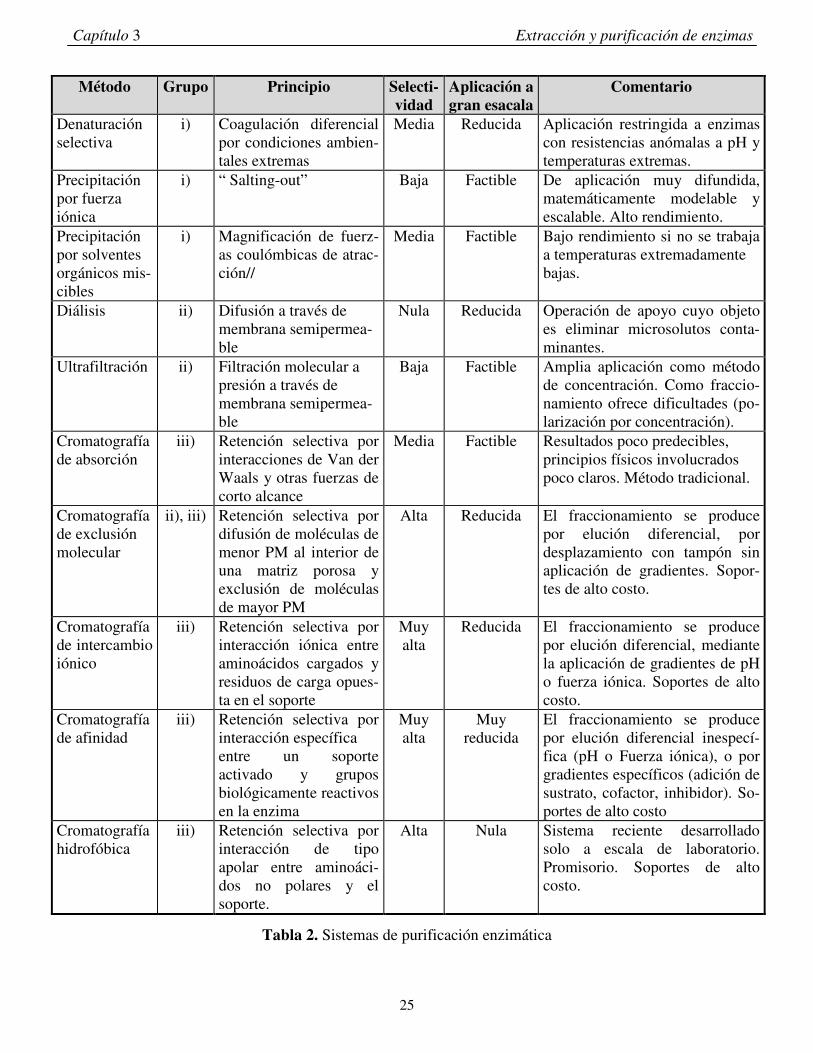
3.1.3- Purificación. Las enzimas se encuentran generalmente en asociación con otras proteínas, ácidos nucleicos, polisacáridos y
lípidos. Existe una gran variedad de métodos usados para remover material contaminante, de esta manera la
enzima es purificada y su actividad específica aumenta (Tabla 2). Los distintos métodos de purificación
pueden agruparse en tres categorías: i) separación en base a solubilidad, ii) separación en base a tamaño, y
iii) separación por retención selectiva (cromatografía).
Capítulo 3 Extracción y purificación de enzimas
25
Método Grupo Principio Selecti-
vidad Aplicación a
gran esacala Comentario
Denaturación selectiva
i) Coagulación diferencial por condiciones ambien-tales extremas
Media Reducida Aplicación restringida a enzimas con resistencias anómalas a pH y temperaturas extremas.
Precipitación por fuerza iónica
i) “ Salting-out” Baja Factible De aplicación muy difundida, matemáticamente modelable y escalable. Alto rendimiento.
Precipitación por solventes orgánicos mis- cibles
i) Magnificación de fuerz-as coulómbicas de atrac-ción//
Media Factible Bajo rendimiento si no se trabaja a temperaturas extremadamente bajas.
Diálisis ii) Difusión a través de membrana semipermea- ble
Nula Reducida Operación de apoyo cuyo objeto es eliminar microsolutos conta-minantes.
Ultrafiltración ii) Filtración molecular a presión a través de membrana semipermea- ble
Baja Factible Amplia aplicación como método de concentración. Como fraccio-namiento ofrece dificultades (po-larización por concentración).
Cromatografía de absorción
iii) Retención selectiva por interacciones de Van der Waals y otras fuerzas de corto alcance
Media Factible Resultados poco predecibles, principios físicos involucrados poco claros. Método tradicional.
Cromatografía de exclusión molecular
ii), iii) Retención selectiva por difusión de moléculas de menor PM al interior de una matriz porosa y exclusión de moléculas de mayor PM
Alta Reducida El fraccionamiento se produce por elución diferencial, por desplazamiento con tampón sin aplicación de gradientes. Sopor-tes de alto costo.
Cromatografía de intercambio iónico
iii) Retención selectiva por interacción iónica entre aminoácidos cargados y residuos de carga opues-ta en el soporte
Muy alta
Reducida El fraccionamiento se produce por elución diferencial, mediante la aplicación de gradientes de pH o fuerza iónica. Soportes de alto costo.
Cromatografía de afinidad
iii) Retención selectiva por interacción específica entre un soporte activado y grupos biológicamente reactivos en la enzima
Muy alta
Muy reducida
El fraccionamiento se produce por elución diferencial inespecí-fica (pH o Fuerza iónica), o por gradientes específicos (adición de sustrato, cofactor, inhibidor). So-portes de alto costo
Cromatografía hidrofóbica
iii) Retención selectiva por interacción de tipo apolar entre aminoáci-dos no polares y el soporte.
Alta Nula Sistema reciente desarrollado solo a escala de laboratorio. Promisorio. Soportes de alto costo.
Tabla 2. Sistemas de purificación enzimática

Capítulo 3 Extracción y purificación de enzimas
26
Un único paso de purificación rara vez es adecuado para purificar una enzima completamente. Por lo
general, si se quiere alcanzar un alto grado de pureza, es necesario recurrir a varios métodos diferentes que se
basen en diferentes propiedades de la enzima en cuestión. Desafortunadamente, el establecimiento de un
protocolo de purificación es un problema de prueba y error: hay que probar procedimientos y luego
seleccionar. Una aproximación es partir de protocolos ya probados aunque las condiciones experimentales
deben ajustarse para cada muestra en particular.
Aparentemente los distintos métodos pueden usarse en cualquier orden y el mejor orden se determina
experimentalmente. Sin embargo, existen ciertas restricciones que hacen que haya un orden lógico. En
general se empieza por métodos que tienen una alta capacidad, que son rápidos y fáciles de desarrollar, y se
sigue con métodos de baja capacidad y alta resolución. La capacidad se refiere a la cantidad de muestra que
puede procesarse y la resolución se refiere a la habilidad de un método para separa la proteína de interés de
otras presentes en el extracto. Normalmente a medida que aumenta la resolución de la técnica el tiempo que
insume también aumenta y la capacidad disminuye (Tabla 3).
Método Generalmente
Decrece capacidad aumenta resolución aumenta tiempo y esfuerzo
• diferencia de solubilidad
• intercambio iónico • adsorción • hidrofobicidad • electroforesis • HPLC Excepciones • gel filtración (baja capacidad, baja resolución)
• cromatografía de afinidad (depende del ligando) Tabla 3. Cuadro comparativo de la capacidad y resolución de distintas técnicas de fraccionamiento proteico
Generalmente los métodos de precipitación (salina, por solventes orgánicos e isoeléctrica), son utilizados en
las etapas iniciales del proceso de purificación; debido a que permiten concentrar la muestra y por lo tanto
disminuir su volumen. En cambio, los procedimientos cromatográficos (intercambio iónico, cromatografía de
exclusión molecular, cromatografía de adsorción, etc), son utilizados luego que la enzima a sido parcialmente
purificada por una técnica de precipitación.

Capítulo 3 Extracción y purificación de enzimas
27
Control de la purificación
Para evaluar que tan efectivo fue un proceso de purificación en su conjunto o en cada una de sus etapas, se
definen dos parámetros que cuantifican la eficacia del proceso, porcentaje de recuperación o
rendimiento (%R) y factor de purificación (FP).
El porcentaje de recuperación indica cuanta proteína de interés había en el extracto original y cuanto hay al
final del proceso o en cada una de las etapas, esto es bastante fácil en la purificación de enzimas ya que estos
cálculos se realizan respecto a la enzima activa.
% R = actividad total de la fracción x 100
actividad total del extracto
El factor de purificación es la relación entre la cantidad de proteína de interés frente a la cantidad de
proteínas totales en el extracto original y lo obtenido al final del proceso o en cada una de las etapas, el caso
de enzimas es la relación entre la actividad específica en el crudo y lo obtenido al final; indica cuanta
proteína contaminante se pudo eliminar.
FP = actividad específica (AE) de la fracción
actividad específica (AE) del extracto
Estos resultados suelen mostrarse en forma de tabla o cuadro de purificación (Tabla 4). Los aspectos
cualitativos de un proceso de purificación se analizan en una elecroforesis, donde se “ve” efectivamente si
hay o no otras proteínas.
Etapa de la
purificación
vol.
Proteínas
Actividad % R FP
mg/mL mg totales U/mL U totales A E
Tabla 4. Cuadro de purificación
Por lo general un proceso de purificación consiste en una secuencia de operaciones o etapas en las cuales los
contaminantes (especialmente proteicos) van siendo subsecuentemente removidos aumentando la actividad

Capítulo 3 Extracción y purificación de enzimas
28
específica de la enzima. Cada etapa de purificación significa, sin embargo, una pérdida de actividad
enzimática. Por lo tanto se ha de llegar a un compromiso entre el factor de purificación y el rendimiento
deseado. A nivel industrial, la tendencia es sacrificar pureza en beneficio de rendimiento; en usos analíticos
o médicos , a la inversa, el rendimiento tendrá una importancia relativa menor.
Criterios de homogeneidad
Cuando una purificación enzimática ha alcanzado la etapa donde posteriores purificaciones o pasos no
producen un incremento en la actividad específica, pueden investigarse con métodos analíticos la
homogeneidad y pureza de la preparación obtenida. Pueden utilizarse métodos cromatográficos,
electroforesis, ultracentrifugación, focalización isoeléctrica, espectrometría de masas, identificación del
residuo amino terminal. La obtención de un único pico proteico es indicativo de homogeneidad, si esto ha
sido comprobado por varios de los métodos mencionados.
Protección de la actividad enzimática durante el proceso de purificación Es importante que se retenga la máxima actividad durante el proceso de purificación de la enzima de interés.
Dado que las enzimas son sustancias relativamente frágiles es esencial evitar condiciones en las cuales son
inestables. Es así que las purificaciones deben llevarse a cabo sin pérdidas inútiles de tiempo, y lo más
conveniente, en general, es guardar las preparaciones en heladera. Si el fin de la purificación es estudiar las
propiedades funcionales y estructurales de la enzima, es necesario obtenerla y mantenerla en su forma nativa.
Si en cambio la enzima purificada va a ser utilizada para estudiar su secuencia aminoacídica, la
desnaturalización tiene una mínima influencia.
Con el fin de mantener a la enzima en su forma activa, deben evitarse:
- altas temperaturas y acidez o alcalinidad extrema: durante el aislamiento y la purificación conviene, en
general, trabajar entre 0 y 4ºC, y evitar pH mayor que 9 o menor que 5.
- formación de espuma: esto es un indicador de desnaturalización proteica. Es así que cuando se transfiere
una solución de enzima de un recipiente a otro debe ser vertida por las paredes del recipiente.
- proteólisis: esto es más probable que ocurra en las primeras etapas de extracción y purificación cuando las
proteasas endógenas responsables del recambio proteico en las células vivientes aún se encuentran
presentes. La mejor forma de evitar la proteólisis es remover las proteasas contaminantes rápidamente o
inhibir su actividad proteolítica. Mientras esto no se hace, es importante mantener las preparaciones
enzimáticas a baja temperatura.
- contaminación: manteniendo la limpieza del material utilizado, dado que las proteínas en soluciones
acuosas son excelentes sistemas nutritivos para los microorganismos

Capítulo 3 Extracción y purificación de enzimas
29
3.1.4- Formulación
Esta etapa es necesaria en el caso de un proceso de producción enzimática a nivel industrial. Consiste en el
acabado y normalización del producto enzimático.
El acabado tiene por objeto dar forma definitiva al producto enzimático y preservar su actividad durante su
período de almacenamiento y comercialización. El mismo incluye las operaciones de desalinización,
esterilización, concentración y/o secado, estabilización por adición de preservantes, y recubrimiento. Dentro
de los preservantes habituales se encuentran: sales inorgánicas, proteínas inertes, polialcoholes, azúcares y
glicoles. Existen además preservantes específicos como sustratos, cofactores e inhibidores disociables,
agentes quelantes, inhibidores de proteasas, reactivos sulfhidrilo para el caso de enzimas que tengan en su
sitio activo grupos oxidables (como es el caso de la ß-galactosidasa); y agentes antimicrobianos.
La normalización consiste en especificar la estabilidad de almacenamiento y la actividad específica del
producto enzimático, utilizando la unidad internacional (UI) como medida de la actividad enzimática. A su
vez la estabilidad deberá especificarse en función del tiempo de vida media a determinada temperatura.
Referencias.
Bollag, D., Rozycki, M., Edelstein, S., En: Protein Methods, 2 da edición, Wiley- Liss Inc, (1996). Grazú, V., En: “β-galactosidasas: distribución, propiedades y aplicaciones”, Trabajo Especial I de la Licenciatura en Bioquímica, Facultad de Ciencias, Universidad de la República Oriental de Uruguay, (1997).
Illanes, A., En: Biotecnología de Enzimas, Monografía Nº 35 de la OEA, Ediciones Universitarias de Valparaíso de la Universidad Católica de Valparaíso, (1994). Jakoby, W., En: Enzyme purification and related techniques. Methods in Enzimology Vol XX Secc VIII: Large Scale Methods, Academic Press, New York, (1981).
Teal, A. R., y Wymer, P., En: Enzymes and their role in biotechnology;
http://www.biochemistry.org/education/basc03.htm

Capítulo 4 Aplicaciones industriales de las enzimas
30
4- Aplicaciones de las enzimas.
La producción y aplicación de enzimas es una de los principales intereses de la biotecnología moderna. El
uso de enzimas en procesos industriales es cada vez más importante.
Las aplicaciones comerciales e industriales de las enzimas se realizan desde hace siglos. Actualmente,
debido a la expansión de su uso y a los avances biotecnológicos, se cuenta con un mayor numero de enzimas
disponibles a un costo relativamente más bajo y un mayor conocimiento de sus cualidades y aplicaciones
potenciales.
Un ejemplo de esto, es el caso de las proteasas (enzimas que hidrolizan proteínas) de origen pancreático que
fueron patentadas para ese uso en el lavado de ropa en 1913 pero presentaban baja estabilidad en la
preparación. En 1953-64, las Industrias Novo de Dinamarca han optimizado la producción industrial de la
proteasa alcalina, subtilisina, secretada en grandes cantidades por Bacillus subtilis, para su uso en
detergentes bioactivos. Sus propiedades de alta estabilidad a pH alcalinos en la presencia de fosfato y otros
ingredientes de los detergentes y mantener su actividad proteolítica hasta la temperatura de 60ºC la hacen
particularmente adecuada para este fin.
Las aplicaciones más importantes de las enzimas son en las siguientes áreas:
• Industria alimentaria y agrícola
• Industria farmacéutica
• Industria química
• Métodos analíticos
• Investigación médica.
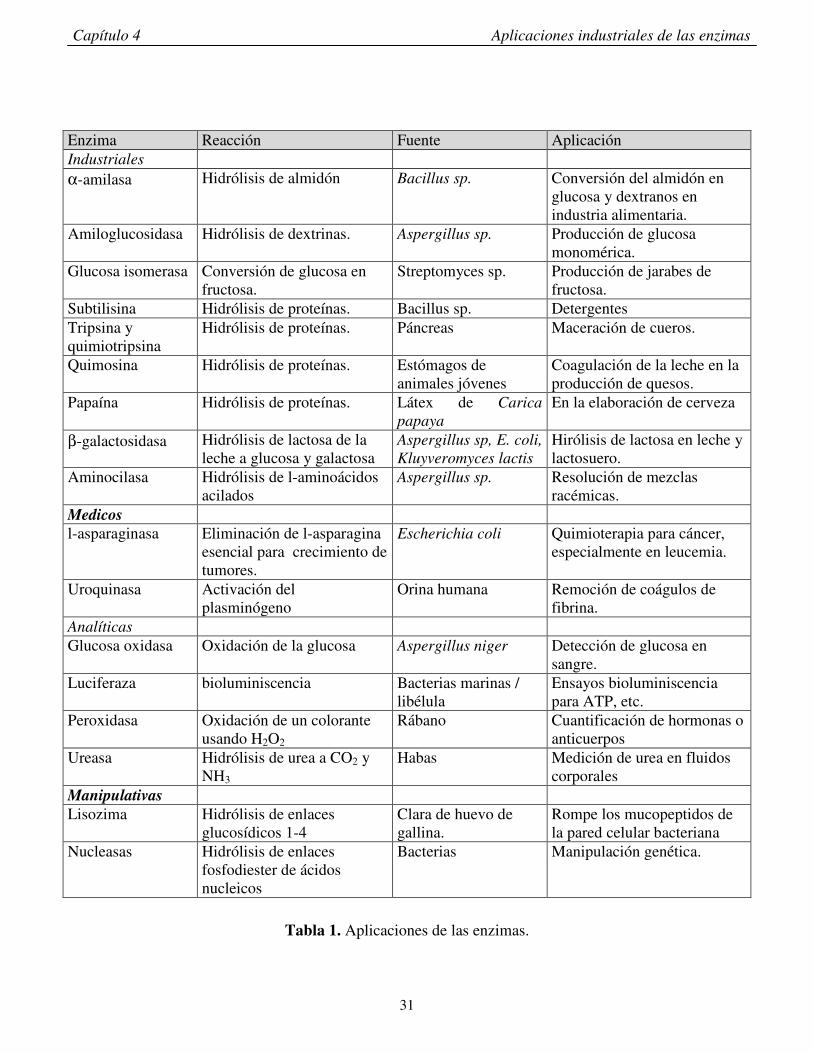
Los principales usos de las enzimas en éstas áreas se resumen en la tabla I.
Capítulo 4 Aplicaciones industriales de las enzimas
31
Enzima Reacción Fuente Aplicación Industriales α-amilasa Hidrólisis de almidón Bacillus sp. Conversión del almidón en
glucosa y dextranos en industria alimentaria.
Amiloglucosidasa Hidrólisis de dextrinas. Aspergillus sp. Producción de glucosa monomérica.
Glucosa isomerasa Conversión de glucosa en fructosa.
Streptomyces sp. Producción de jarabes de fructosa.
Subtilisina Hidrólisis de proteínas. Bacillus sp. Detergentes Tripsina y quimiotripsina
Hidrólisis de proteínas. Páncreas Maceración de cueros.
Quimosina Hidrólisis de proteínas. Estómagos de animales jóvenes
Coagulación de la leche en la producción de quesos.
Papaína Hidrólisis de proteínas. Látex de Carica
papaya
En la elaboración de cerveza
β-galactosidasa Hidrólisis de lactosa de la leche a glucosa y galactosa
Aspergillus sp, E. coli,
Kluyveromyces lactis
Hirólisis de lactosa en leche y lactosuero.
Aminocilasa Hidrólisis de l-aminoácidos acilados
Aspergillus sp. Resolución de mezclas racémicas.
Medicos l-asparaginasa Eliminación de l-asparagina
esencial para crecimiento de tumores.
Escherichia coli Quimioterapia para cáncer, especialmente en leucemia.
Uroquinasa Activación del plasminógeno
Orina humana Remoción de coágulos de fibrina.
Analíticas Glucosa oxidasa Oxidación de la glucosa Aspergillus niger Detección de glucosa en
sangre. Luciferaza bioluminiscencia Bacterias marinas /
libélula Ensayos bioluminiscencia para ATP, etc.
Peroxidasa Oxidación de un colorante usando H2O2
Rábano Cuantificación de hormonas o anticuerpos
Ureasa Hidrólisis de urea a CO2 y NH3
Habas Medición de urea en fluidos corporales
Manipulativas
Lisozima Hidrólisis de enlaces glucosídicos 1-4
Clara de huevo de gallina.
Rompe los mucopeptidos de la pared celular bacteriana
Nucleasas Hidrólisis de enlaces fosfodiester de ácidos nucleicos
Bacterias Manipulación genética.
Tabla 1. Aplicaciones de las enzimas.

Capítulo 4 Aplicaciones industriales de las enzimas
32
De los cientos de enzimas utilizadas industrialmente, más del 50% provienen de hongos y levaduras, por
encima del 33% son bacterianas y el resto se divide entre las de origen animal (8%) y vegetal (4%).
4.1- Enzimas microbianas.
Las enzimas provenientes de hongos, levaduras y bacterias pertenecen a un numero limitado de éstos
microorganismos, predominando Aspergillus sp., Bacillus sp. y Kluyveromyces sp. Las causas de esta
preferencia son varias:
• Las enzimas animales y vegetales son más difíciles de recolectar y almacenar. El crecimiento de
microorganismos y la producción de enzimas es fácilmente controlado. Es posible manejar la
capacidad de los fermentadores de acuerdo a las fluctuaciones en la demanda del producto.
• En general resulta más económico producir enzimas microbianas que animales o vegetales.
• Los microorganismos pueden utilizar para crecer un amplio rango de sustratos. Se pueden incorporar
moléculas inductoras a los fermentadores para inducir la síntesis de enzima. La inhibición de la
síntesis de enzima por retroalimentación puede evitarse al tener la posibilidad de limitar la
acumulación del producto y de reemplazar los nutrientes que se consumen más rápidamente.
• Se pude realizar mutaciones para producir cambios menores en la estructura de la enzima que resulten
en una característica alterada de la enzima como por ejemplo insensibilidad a un inhibidor natural.
Esto permite la producción potenciada de enzima en grandes cantidades.
Un ejemplo de enzimas microbianas aplicadas en procesos industriales es la β-galactosidasa, la enzima que
hidroliza la lactosa. Esta enzima se produce comercialmente para su uso en la industria láctea. La lactosa,
o azúcar de la leche, es relativamente insoluble resultando un problema en muchos productos lácteos. En la
producción de helado, la hidrólisis de lactosa, mejora las propiedades de textura, dulzor y tendencia a
cristalizar. Algunas personas carecen de β-galactosidasa, la enzima que hidroliza la lactosa en el intestino
delgado, por lo tanto no pueden digerir la lactosa de la leche o de los productos lácteos que contengan
lactosa. El pre-tratamiento de la leche genera un producto libre de lactosa adecuada para el consumo por
parte de la población intolerante a éste carbohidrato.
4.2- Enzimas animales.
Las enzimas animales son poco aplicadas en procesos industriales. Ellas son utilizadas frecuentemente en la
producción de agentes terapéuticos o reactivos analíticos.
Como ejemplo se puede citar los activadores del plasminógeno. Es un grupo de enzimas proteolíticas que
actúan sobre una molécula precursora inactiva presente en la sangre llamada plasminógeno que al ser

Capítulo 4 Aplicaciones industriales de las enzimas
33
removida parte de su secuencia se libera la enzima activa plasmina. La plasmina destruye la red de fibrina de
un coágulo sanguíneo. La uroquinasa es el activador del plasminógeno humano más conocido pero su
producción es muy cara. Para realizar un tratamiento sería necesario extraer uroquinasa de 500 L de orina.
Los activadores de plasminógeno producidos por tejidos son más específicos en su acción y pueden ser
producidos en cultivo de células humanas. Aunque esto representa una alternativa, para la producción de una
droga más económica y eficiente, los cultivos de células animales presentan mayores problemas que los de
microorganismos. La estreptoquinasa, una enzima bacteriana, también puede ser utilizada como activador del
plasminógeno, pero es relativamente inespecífica en su acción y puede producir reacciones inmunológicas
severas en el paciente.
4.3- Enzimas recombinantes. Los recientes desarrollos de la genética microbiana han creado un nuevo potencial para la producción de
enzimas. Pueden usarse técnicas de ingeniería genética para manipular el DNA de forma que pueden generarse
copias múltiples de un gen particular que codifique para una enzima de interés comercial. Esto se logra
insertando el gen que interesa a una pequeña molécula de DNA bacteriana (plásmido) que puede replicarse
independientemente de los cromosomas. El gen es integrado dentro del plásmido formando una molécula de
DNA recombinante que puede ser insertada en un huésped adecuado. Los plásmidos pueden replicarse a una
velocidad superior que el DNA cromosomal produciendo varios cientos de copias por célula. Pueden
producirse grandes cantidades de la proteína codificada por el gen clonado. Los genes procarióticos pueden ser
expresados en células bacterianas si se encuentran presentes las secuencias reguladoras adecuadas antes de la
secuencia codificante.
Los genes eucariotas también pueden ser clonados en células bacterianas pero no siempre ocurre la producción
de proteínas activas debido a que muchas de éstas enzimas para estar activas deben ser modificadas luego de la
traducción. Los procesamientos post-traduccionales pueden involucrar glucosidaciones o hidrólisis de
secuencias peptídicas que las células procariotas no pueden realizar. Existe la alternativa del uso de células de
levaduras y hongos como huéspedes para el clonado que sí pueden realizar este tipo de procesamiento debido a
que son células eucariotas. Actualmente, son producidas en células de levadura enzimas recombinantes de
mamíferos biológicamente activas entre las que se destacan: lisozima, quimiosina y el activador del
plasminógeno.
El clonado de genes también permite re-diseñar enzimas en un forma racional (ingeniería genética). Se altera
un aminoácido preciso de la enzima nativa para mejorar sus características. Existen métodos que permiten
cambiar una base específica en una hebra de DNA (mutagénesis dirigida) y luego clonar el gen y expresarlo
en un huésped adecuado para producir la enzima re-diseñada.

Capítulo 4 Aplicaciones industriales de las enzimas
34
Estos desarrollos han sido aplicados a enzimas de interés industrial.
La quimosina es una de las proteasas más importantes en la industria alimentaria. Esta enzima produce la
coagulación de la leche en la manufactura del queso mediante proteólisis limitada. Tradicionalmente, la
quimosina era obtenida del estómago de terneros jóvenes, pero se encuentra en baja concentración. Esto ha
llevado al uso de sustitutos, generalmente de origen microbiano. Actualmente se produce de forma
recombinante.
La α-amilasa de B.subtilis se desnaturaliza por calentamiento. Esta enzima cataliza la conversión de
almidón en glucosa que se ve acelerada a altas temperaturas. Se ha obtenido una α-amilasa recombinante
termoestable mediante clonado en B. subtilis del gen de la α-amilasa de un organismo termoestable.
La subtilisina, utilizada actualmente en detergentes bioactivos, se ha mejorado genéticamente obteniéndose
una enzima con mayor resistencia a la oxidación.
Bibliografía.
Enzimas-campos de aplicación. En: Folleto Técnico de Novo-Nordisk a/s (1989). Dinamarca.
Teal, A. R., y Wymer, P., En: Enzymes and their role in biotechnology;
http://www.biochemistry.org/education/basc03.htm


















