La corteza entorrinal (formación del hipocampo) en el ...
9
La corteza entorrinal (formación del hipocampo) en el envejecimiento y en la enfermedad de Alzheimer. Interpretación neuroanatómica T. Sobre vi el a, A. lnsau st i, A. Salinas, L .M. Gon zalo y R. ln sau st i Departamento de Anatomía. Facultad de Medicina. Universidad de Navarra RESUM EN. En este trabajo se re aliza un análi sis de los cambios a nivel neuronal que presenta la corteza entorrinal a lo largo del envejecimiento (30 casos) y en la enfermedad de Alzheimer (17 casos). Los cam- bios en ambos casos indi can una pérdida neuronal (medida por un índice de la atrofia cortical) de la cor- teza entorrinal. Aq uell as porciones que se relacionan más intensamente con la corteza cerebral de asocia- ción polisensorial apenas muestran variación con el envejecimiento, mientras que sufren una pérdida neuronal y una atrofia muy intensa en los casos con enfermedad de Al zheimer. Esta situación es interpre- tada a la luz de los conocimientos actuales de las co- nexiones neuroanatómicas de la corteza entorrinal y del resto de la formación del hipocampo y de su pa- pel en las funciones de memoria. SUMMARY. This paper deals with the neuronal changes shown by the entorhinal cortex in aging (30 cases) and in Alzheimer's disease (17 cases). In both instances, changes show a neuronal loss (measured as an index of cortical atrophy). The en- torhinal cortex more closely related to polymodal association cortex hardly shows any variation with age, while Alzheimer cases present very intense neuronal loss. This pattern is interpretad tal<ing into account the present knowledge about the connecti- vity of the entorhinal cortex and the remainder of the hippocampal formation and their role in memory processing. (Hev Med Univ Navar"' 19')7; 41: 19·27). Pa la bras clave Coteza entorrinal, Lóbulo te mporal, senescencia, en- f erme dad ele Al zheimer, atrofia cortica l. 19 Key wor ds Entorhinal cortex; human; tempora ll obe; ag ing; Alz- heimer's disease; co rtical atroph y. Cor respondencia Dr. H. Tn saus ti Departamento de An atomía Universidad de Navar ra Apdo. 273, 31080 Pamplo na I ntroducción Desde que a comi enzos el e siglo Bechterew (1900) desc ribiera camb i os en la memoria asociad os a lesio- n es neuropatológicas demostrabl es en la p arte medial del lób ulo te mporal, esta zona cerebral se ha asociado a dicha función. D esde entonces son numerosos los estudios que han constatado y extendido esta simple asociación entre la memo ri a y la formac ió n de l h ip o- campo. E spec ialmente no ta ble fu e la descripción de un paciente (paciente H.M.) al que , pa ra aliviar de una epile psia intratable, se resecó la parte anterior y medial el e a mbo s lóbulos te mporales , es tudio publica do en 1957 por Scoville y Milne r. La resección incluía el po- lo temporal, la a mí gdala y la mayor pa rte ele la forma- ci ón del hipocamp o'. Este paciente, el cu al aú n vive, presentó d esde el postope ra torio y hasta la actualidad, una incapaci da d pa ra formar nuevos recuerdos. La re- la ción entre la formación del hi pocampo y la me moria anterógrada fue p os teriormente r eforzada en un caso d escrito por Zola-Mor ga n y co la bora d ores en 1986, el 1 Incluimos d término rorm:tción del hipoc:un¡K, una serie hx::a1iz01das la cara medi:tl del lóbulo en gr.m cone-ctadas esto cs. no prcsent:m rt"dproc:.s :-.inu a través dt! un01 serie tlt! p:.· S('·' inlt!rmcd ios. Dt!stle las zon a:-. mf ts ;t l:t corteza ('t-rebí.l.l tempoml l as m:í s alejada.-.. cstru<l uras son 1:- l t:oneza cntorrin:al, el para.,uhku lo. r suhínt- lo, los <· ampos CA 1, CA2, y Ud hipocampo propi<uncntt! dk· ho y la cirum- \·olución dentada REVISTA DE MEDICINA DE LA UNIVERSI DAD DE NAVARRA ENERO.MARZO 1997 19
Transcript of La corteza entorrinal (formación del hipocampo) en el ...

La corteza entorrinal (formación del hipocampo) en el envejecimiento y en la enfermedad de Alzheimer.
Interpretación neuroanatómica
T. Sobrevie la, A. lnsausti, A. Salinas, L .M. Gonzalo y R. l nsausti
Departamento de Anatomía. Facultad de Medicina. Universidad de Navarra
RESUMEN. En este trabajo se realiza un análisis de los cambios a nivel neuronal que presenta la corteza entorrinal a lo largo del envejecimiento (30 casos) y en la enfermedad de Alzheimer (17 casos). Los cambios en ambos casos indican una pérdida neuronal (medida por un índice de la atrofia cortical) de la corteza entorrinal. Aquellas porciones que se relacionan más intensamente con la corteza cerebral de asociación polisensorial apenas muestran variación con el envejecimiento, mientras que sufren una pérdida neuronal y una atrofia muy intensa en los casos con enfermedad de Alzheimer. Esta situación es interpretada a la luz de los conocimientos actuales de las conexiones neuroanatómicas de la corteza entorrinal y del resto de la formación del hipocampo y de su papel en las funciones de memoria. SUMMARY. This paper deals with the neuronal changes shown by the entorhinal cortex in aging (30 cases) and in Alzheimer's disease (17 cases). In both instances, changes show a neuronal loss (measured as an index of cortical atrophy). The entorhinal cortex more closely related to polymodal association cortex hardly shows any variation with age, while Alzheimer cases present very intense neuronal loss. This pattern is interpretad tal<ing into account the present knowledge about the connectivity of the entorhinal cortex and the remainder of the hippocampal formation and their role in memory processing.
(Hev Med Univ Navar"' 19')7; 41: 19·27).
Palabras c la ve Coteza entorrina l, Lóbulo temporal, senescencia, en
fermedad ele Alzheimer, atrofia cortical.
19
Key w ords Entorhinal cortex; human; temporallobe; aging; Alz
heimer's d isease; cortical atrophy.
Corresponden c ia Dr. H. Tnsausti Departamento de Anatomía Universidad de Navarra Apdo. 273, 31080 Pamplona
Introducción Desde que a comienzos ele siglo Bechterew (1900)
describiera cambios e n la memoria asociados a lesiones neuropatológicas demostrables en la parte medial de l lóbulo temporal, esta zona cerebral se ha asociado a dicha función. Desde entonces son numerosos los estud ios que han constatado y extendido esta s imple asociación entre la memoria y la formación del h ipocampo. Especialmente notable fue la descripción de un paciente (paciente H.M.) al que, para aliviar de una epilepsia in tratable, se resecó la parte anterior y medial ele ambos lóbulos te mporales, estudio publicado en 1957 por Scoville y Milner. La resección incluía el polo temporal , la amígdala y la mayor parte ele la formación del hipocampo'. Este paciente, el cual aún vive, presentó desde el postoperatorio y hasta la actualidad , una incapacidad para formar nuevos recuerdos. La relación entre la formación del hipocampo y la memoria anterógrada fue posteriormente reforzada en un caso descrito por Zola-Mo rgan y colaboradores en 1986, el
1 Incluimos ha~, d término rorm:tción del hipoc:un¡K, una serie d~ ~.st ructur:ls hx::a1iz01das ~n la cara medi:tl del lóbulo t~mporal qu~ ~.,t;ín en gr.m m~dida cone-ctadas unidir~don:-1 -
m~ntt-, esto cs. qu~ no prcsent:m concxion~s rt"dproc:.s :-.inu a través dt! un01 serie tlt! p:.· S('·' inlt!rmcdios. Dt!stle las zon a:-. mfts pró.xim:t~ ;t l:t corteza ('t-rebí.l.l tempoml h:~st a las m:ís alejada.-.. c~stas cs tru<luras son 1:-l t:oneza cntorrin:al, el para.,uhkulo. pr~.,l&hkulo r suhíntlo, los <·ampos CA 1, CA2, y CA.~ Ud hipocampo propi<uncntt! dk·ho y final m~ntc la cirum\·olución dentada
REVISTA DE MEDICINA DE LA UNIVERSIDAD DE NAVARRA ENERO.MARZO 1997 19

cual presentaba un déficit de memoria anterógracla tras un fallo cardíaco que lesionó selectivamente el sector CAl del hipocampo. No se encontraron otras lesiones estructurales capaces ele explicar el déficit de memoria en este caso. Por consiguiente, la alteración de l flujo normal ele los impulsos nerviosos a través del hipocampo mediante la lesión selectiva ele uno de sus campos fue suficiente para impedir que las experiencias nuevas se consolidaran como recuerdos. Muy recientemente el mismo grupo de investigadores (RempelCiower y cols., 1996) ha publicado tres casos más en los que se hace una exhaustiva caracterización neuropsicológica, neuroradiológica y neuroanatómica de estos pacientes.
De un modo independiente Hyman y cols., (22) publicaron un estudio en el que mostraban cómo e n la enfermedad de Alzheimer, la lesión de otro componente ele la formación del hipocampo, en este caso la corteza entorrinal, mostraba una clara correspondencia con los datos neuroanatómicos experimentales obtenidos en primates. Aunque la neuropatología de la corteza entorrinal en la enfermedad de Alzheimer ya e ra conocida ante riormente, este trabajo puso de manifiesto que la destrucción predominante de las capas 11 y V de la corteza entorrinal aislaba el hipocampo del resto de la corteza cerebral. La base neuroanatómica última ya e ra conocida desde los estudios de Caja!. La corteza ento rrinal, por medio de la vía perforante es la principal fuente de aferencias del hipocampo. El estudio concluía que esta desconexió n a islaba el hipocampo del resto de la corteza cerebral, atribuyéndose a este hecho los transtornos de memoria tan característicos de la enfermedad de Alzheimer.
Estos dos e jemplos no hacen sino reforzar la noción de que la formación de l hipocampo contribuye ele un modo decisivo en la memoria denominada declarativa2. Esta elaboración, cualquiera que sean sus mecanismos celulares y moleculares últimos, requiere la integridad de los circuitos neuroanatómicos de la formación del hipocampo, y cuando por efecto de una noxa se lesiona alguno de los eslabones ele esta cadena surge un dete rioro ele la memoria susceptible de ser detectado por pruebas neuropsicológicas.
La investigación experimental, especialmente en primates, ha puesto de manifiesto de modo incontrover-
1 Aunqut: l:a :tCCI>Ción no es uní\'cx.-a y admite divisiones o~ at:uerdo coo dif<.>rcntcs inte rpret:aeion~s psicológic~s. :tq\IÍ se ha seguido b clnbor.u.:ion eJe Squire (1987) el cu:ll distingue dos tipos de mcmori;a rund:1rn~ntales: memoria d~darativ::t y no dcclnrativa. T:m solo la menlori;;l dcd1rativ:t, t:mto ~n el conocimiento de h~t:h()S com o en el de cxperiendas, ~s sensible :1 In lc.si(m d~ la formación <k·\ hipo<.-:ttnpo.
20 REVISTA DE MEDICINA DE LA UNIVERSIDAD DE NAVARRA ENERO-MARZO 1997
tibie esta afirmación. Numerosos trabajos, especialmente los de Van Hoesen y cols. ( 43, 44) han sentado las bases del conocimiento neuroanatómico del papel de la corteza entorrinal en los circuitos ele la formación del hipocampo. Posteriormente, estudios neuroanatómicos realizados con técnicas más sensibles (1 , 25) pone de manifiesto dos hechos importantes: 1) el hipocampo propiamente dicho (circunvolución dentada, campos CA3, CA2 y CAl) no recibe aferencias directas desde e l neocórtex; 2) la información procedente de l neocórtex termina en la corteza entorrinal desde donele es transmitida hacia el hipocampo propiamente dicho; asimismo, es interesante resaltar que las proyecciones corticales que terminan en la corteza entorrina l se originan únicamente en áreas corticales ele asociación multimodal (esto es, que responden a estímulos polisensoriales), pero no ele las unimoclales. Esto sitúa a la corteza entorrinal como un centro en donde converge la información cortical que ha ele alcanzar el hipocampo propiamente dicho. Por consiguiente el flujo de la actividad neuronal ante un estímulo comienza en las áreas sensitivas primarias y se extiende a las áreas sensitivas de asociación unimoclal y posteriormente a las multimoclales (27), destacando la corteza perirrinal (áreas 35 y 36 ele Brodmann, (12)) y parahipocámpica posterior (á reas TF y TH de Von Economo (46)) para confluir finalmente en la corteza entorrinal, dentro de un eje neocortico-entorrino-bipocampal necesario para la funció n de memoria.
Partiendo de este planteamiento, hemos examinado la corteza entorrinal humana, así como las limítrofes (corteza perirrinal y parahipocámpica posterior) en una serie amplia de casos con el fin de determinar la homología entre el primate y e l hombre, y anal izar los cambios asociados al envejecimiento en casos control (sin evidenCia ele alteraciones neurológicas o psiquiát:ricas) y los provocados por la enfermedad de Alzheimer. La evidencia de que tienen lugar cambios e n la corteza entorrinal asociados al envejecimiento y a la enfe rmedad de Alzheimer es conocido en la literatura (4, 5, 7-9, 11 , 13, 15-19, 20-22, 29, 31, 34-36,40,42,45, 49). A modo de resumen se puede afirmar que las dos lesiones más características ele la enfermedad de Alzheime r, las placas neuríticas y la degeneración neurofibrilar se encuentran de modo constante en la corteza entorrinal, aunque no en las mismas capas. Además de las lesiones ante riores, se asocia una gran pérdida neuronal (16) en particular ele las capas TI (reducción de un 60% ) y ele la capa V (reducción de un 40%) que son las que presentan mayor degeneración neurofibri-
20

lar, así como gliosis acompananclo a una profunda atrofia ele la corteza entorrinal.
A diferencia de los estudios anteriores, en nuestro planteamiento hemos querido precisar más la distribució n regional ele la pé rdida neuronal dentro ele la corteza entorrinal, aunque sin plantearnos una dete rminació n cuantitativa ele la misma. Sin embargo, sí que hemos aplicado un paráme tro global que objetiva la inrensidacl ele la atrofia de la corteza ento rrinal. Estudios anteriores en el primate (3), posteriormente elaborados en la corteza entorrinal humana (26) muestran una heterogene idad de la corteza entorrinal desde el punto de vista estructural que guarda bastante paralelismo en las dos especies (7 campos en la corteza entorrinal del primate, y 8 en el hombre). Cada uno de los campos ciroarquitectónicos de la corteza entorrinal recibe conexiones de áreas corticales distintas (24, 25) y a su vez proyecta selectivamente sobre la circunvolución dentada (48) y los campos CA3, CAZ y CAl del hipocampo (47) . Por consiguiente, nuestro planteamiento desde el punto ele vista ne uroanatómico ele la pérdida neuronal y atrofia consiguiente ele la corteza cntorrinal se centra en aplicar los conocimientos existentes acerca ele la conectividad de la corteza entorrinal en el primate a los campos citoarquilectónicos homólogos en el hombre, y ele este modo profundizar en la comprensión de l dé ficit de memoria que presentan los pacientes co n enfermedad ele Alzhe imer.
Materia l y Métodos Hemos estudiado la corteza entorrinal en 30 casos
control (rango 12-110 ai1os) , así como en 17 casos (rango 69-98 años) con clemencia y diagnóstico postmOttem ele enfermedad de Alzheimer acorde con los criterios de Khachaturian (28) y CEl{AJ) (30) . El estadio evolutivo de la enfermedad fue dete rminado mediante la ap licación de las escalas de los test de Folstein y cols. (14), y el test de comportamiento de Crichton y Robinson (1965). Los cerebros fueron fijados por inmersión en formol al lOo/o o bien perfundidos por vía carotídea con paraformaldehído al 4o/o (26). Tras ser fotogral1aclos los cerebros se seccionaron seriadamente en el plano coronal a 50 ¡.~.m
y teñidos con tionina para la evaluación ele los campos citoarquitectónicos de la co1teza entorrinal, además de las muestras destinadas para el examen neuropatológico. Las secciones conteniendo las estructuras de interés fueron dibujados a cámara clara con un cstereomicroscopio, eldimitándose los campos de la corteza entoninal , así como los adyacentes ele la corteza perirrinal y parahipo-
21
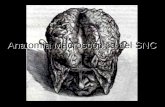
Fotografía de la cara ventromedial del lóbulo temporal en un ce· rebro humano. El tronco del encéfalo ha sido seccionado y elimi· nado con el fin de mostrar la totalidad de la circunvolución pa· ra hipocámpica. Abreviaturas: EC, campo caudal; ECL, campo caudallimitante; Ele, campo lateral caudal; Elr, campo lateral ros· tral; El, campo intermedio; EMI, campo medial intermedio; EO, campo olfatorio; ER, campo rostral (todos ellos de la corteza en· torrinal); ga, gyrus ambiens; M, mesencephalon; PT, polo tempo· rol; se, sulcus collateralis; ssa, sulcus semiannularis.
cámpica posterior. A patt ir ele este material se realizaron reconstrucciones bidimensionales de estas áreas corticales, las cuales fueron cuantificadas planjmétricamentc. Series adicionales teñidas con distintos métodos para mielina, acetilcoli nesterasa, placas neuríticas, así como frente a distintos anticuerpos (proteína Tau, (-amiloide , ncurofilamcntos no fosforilaclos) fueron analizadas en algunos casos.
Resultados
Localización y cat·acterísticas de la cot·teza entoninal.
La corteza entorrinal se sitúa en la cara ventromecl ial del lóbulo temporal (Fig. 1). Sus límites no están demasiado definidos macroscópicamente, pero aproximadamente sobrepasa e l uncus en unos mm. desde su límite anterior al posterior. El límite medial se sitCta con la corteza periamigclalina por medio del surco semianular, para continuarse posteriormente con la circunvolución ambiens y la fisura del hipocampo. El límite lateral lo forma e l surco colateral, habitualmente en la palte media ele su labio medial. Limita con la corteza pe rirrinal por la parte anterior y lateral , la cual se continúa con la corteza pa-
REVISTA DE MEDICINA DE LA UNIVERSIDAD DE NAVARRA ENERO-MARZO 1997 21

. . r
rahipocámpica posterior. Por la pa1te medial limita con la corteza periamigclalina y el complejo subicular (26).
l listológicamente se definen una serie ele capas que han recibido dife re ntes nombres segú n los au tores. Nosotros (26) distinguimos seis capas: 1) capa molecular; IT) capa de células estre lladas; 111) capa piramidal externa; IV) lam ina dissecans; V) capa piramidal interna; VI) capa polimorfa. La presencia, modificación o ausencia de d ist intas capas justi fica la divis ión citoarquitectónica e n S campos entorrinales. Estos campos son ele rostral a caudal y de medial a lateral: 1) campo olfatorio (EO); 2) campo rostral (El{); 3) campo lateral rostral (ELR); 4) campo mecli ~tl inte rmedio (El\111); 5) campo intermedio (El); 6) campo lateral caudal (Ele); 7) campo caudal (EC); 8) campo cauda l limitante (ECL). Esta d ivisión ele la corteza e nto rrinal humana guarda un paralelismo a la anteriormendc descrita e n el primate (3), facilitando ele este modo la extrapolación ele los elatos experimentales.
Variación de la corteza entorrinal en el envejecimiento y enfet·medad de Alzheimer
El envejecimie nto normal no supone alteraciones cualitativas importantes en la corteza entorrinal. El aspecto
Figura 2
1' 1 1·
histológico es comparable a Jo largo de las décadas, y es Cmicamente en las dos últimas décadas examinadas (91-100 y 101-110 ai'los) cuando se aprecia una d isminución del número ele células aparente por la reducción e n grosor de la corteza entorrinal , aunque todas las capas están presentes (37). En resumen, el envejecimiento no conlleva una clesestructuración ele la corteza entorrinal , sino que se mantienen, independientemente ele que el mímero absoluto ele neuronas disminuya.
En la serie ele casos con enfermedad ele Alzheimer la corteza entorrinal pudo ser delimitada en general con bastante precisión. En algunos casos sin embargo, la desaparición de las capas corticales, que también afectaba a las las áreas co1ticales perirrinal y parahipocámpica posterior, impedía demarcarla de estas otras áreas vecinas. En general, el mayor clespoblamianto neuronal correspondía a los casos en los que la e!Úermedad tenía una mayor duración, aunque también había excepciones.
Estado de los campos citoarquitectónicos de la corteza entorrinal en la enfermedad de Alzheimer.
La afectación de los distintos campos de la corteza entorrinal no era homogénea ya que las porciones ros-
' . 11
Elr
Serie de microfotografías de la corteza entorrina l comparativas de la senescencia y de la enfermedad de Alzheimer. La identificación de cada campo y su laminación queda indicada en cada microfotografía, así como su pertenencia al grupo control (C) o a l grupo con enfer· medad de Alzheimer (A) (foto superior, foto superior e inferior página 6).
22 REVISTA DE MEDICINA DE LA UNIVERSIDAD DE NAVARRA ENERO-MARZO 1997 22

1 ., •. • E~l iu ·. ·: : ~, . t· !~ l , .
• ,..,. '·. i;_ ~ • 14 ~ ~ ••• t • • 1
' .. EM I
traJes estaban en genera l mejor preservadas (esto es, la reclucciún neuronal en las capas JI y V no e ra tan acusada). El campo olfatorio (EO) presentó una pé rdida neuronal intensa en tan sólo un caso de la serie, mientras que en el resto se d ife renciaban las características citoarquitcctó nicas, aún con una glios is más acentua-
23
da. El campo rostral (Fig. 2) presentó una franca d isminución neuronal en tres casos, mientras que el resto se mostró con una relativa indemnidad. Los ca mpos lateral rostra l (Elr, Fig. 2) y el lateral cauda l (ELe) resultaron afectados en todos los casos, siendo imposihle delimitarlos en la mitad ele e llos. La reducción del
REVISTA DE MEDICINA DE 111 UNIVERSIDAD DE NAVARRA ENERO.MARZO 1997 23

número ele neuronas fue muy acusada en las capas li y V. Por otra parte, la transición entre estos campos laterales y la corteza perirrinal, inmediatamente adyacente por fuera , resultaba imposible ele delimitar en los casos más marcados. El campo medial intermedio (EMI, Fig. 2) presentó una pérdida neuronal más intensa en la capa V que en la capa II , la cual se reconoció en todos los casos. El campo intermedio (El, Fig. 2) mostró pérdida neuronal muy importante en las capas II y V, fácilmente reconocibles por la laminación precisa que este campo entorrinal presenta en casos normales. Algunos de los casos con enfermedad de Alzhe imer también presentaban pérdida neuronal apreciable en la capa III. Los campos caudal (EC, Fig. 2) y caudal limitante (ECL, Fig.2), cuya transición no siempre es fácil de diferenciar, estaban profundamente afectados en la serie ele Alzheimer, por lo que sólo se pudieron diferenciar en una cuarta parte ele los casos. La capa V presentaba gran pérdida neuronal en toda su extensión mediolateral. La capa II también estaba claramente clisminuícla en cuanto al número ele neuronas, pero en algunos casos se obse1varon grupos celulares ele la capa JI relativamente indemnes en la parte más medial de la corteza entorrina l. Las capas III y VI estaban, al igual que en el resto de los campos, más respetadas.
Análisis cuantitativo de la atrofia de la corteza entorrinal en el envejecimiento y enfermedad de Alzheimer.
El análisis cuantitativo de la atrofia de la corteza entorrinal en el envejecimiento y enfermedad de Alzheimer se realizó por medio ele las reconstrucciones bidimensionales y mensurando su extensión. Este método descrito por Van Essen y Maunssell ( 41) y adaptado por nosotros posteriormente (25) consiste basicamente en desarrollar (en su acepción ele extender lo que está arrollado) sobre líneas a intervalos conocidos (equivalentes al espaciamiento de las secciones) sobre las cuales se marcan la longitud de cada campo citoarquitectónico. Uniendo las líneas se obtiene el desarrollo ele la corteza entorrinal y sus diferentes campos citoarquitectónicos en dos dimensiones.
Los resultados mostraron que e l área calculada (es decir, la extensión de la corteza entorrinal medida planimétricamente tras la reconstrucción bidimensional) de la corteza entorrinal disminuyó un 11.4% al comparar los casos mayores y menores ele 65 años en la serie control. Sin embargo, esta disminución no afectaba a ningún campo específico, sino que todos ellos esta-
24 REVISTA DE MEDICINA DE LA UNIVERSIDAD DE NAVARRA ENERo-MARZO 1997
ban clisminuíclos uniformemente, a excepoon de los campos laterales ELr y Ele, los cuales mostraban una menor reducción, o dicho ele otro modo, persisten inalterados durante el envejecimiento en lo que a extensión se refiere.
En la serie de casos ele Alzheimer la disminución del área calculada ele la corteza entorrinal alcanzó un 20 % comparativamente con el grupo control correspondien te (casos por encima de 65 años). La delimitación por campos citoarquitectónicos ofreció dificultades adicionales al ser imposible en ciertos casos delimitar algunos campos adyacentes. Los resultados mostraron una tendencia opuesta a lo mostrado en la serie de envejecimiento. Los campos olfatorio (EO), rostral (ER), y medial intermedio (EMI) disminuyeron mucho menos (de hecho, su extensión relativa , esto es, con respecto a la totalidad ele la extensión de la corteza entorrinal, se incrementó). En cambio, los campos intermed io (EI), lateral (ELr más ELe) y caudal (EC más ECL) sufrieron una disminución mayor.
La diferencia entre área calculada total a lo largo ele las diferentes décadas y la enfermedad de Alzheimer se puede apreciar globalmente en la Fig. 3. Puede observarse cómo hay una aproximación de los valores a partir de la séptima década, para hacerse similares en la novena década.
Figura 3
CORTEZA ENTORRINAL, AREA CALCULADA
Sr-----------------------------------,
4
3
2
o o 2 3 4 5 6 7 8 9 10 11 12 EDAD (en décadas)
a CASOS CONTROL. 9 CASOS ALZHEIMER
Representación de la va riación con la edad de las áreas calcula· das (ver texto) de la corteza entorrinal, así como con la enferme· dad de Alzheimer. Se puede observar cómo es unicamente a par· tir de la s~ década cuando la atrofia de la corteza entorrinal se hace patente hasta alcanzar valores comparables a los de la en· fermedad de Al:zheimer.
24

Discusión La descripción clásica ele la patología presente en la
corteza entorrinal en el e nvejecimie nto y en la enfermedad ele Alzhcimer ha sido tratada por diversos autores, algunos ele ellos citados en la introducción. F.n su mayor parte adolecen ele la limitacié>n del análisis a una pcquei'la muestra de la cmteza entorrinal. Nuestro enfoque del problema trata de avanzar más allá de la mera consratación de la clisminucié>n ne uronal y su preponderante localizaci6n en las capas TI y V ele la corteza e ntorrina l, pa ra hacer un estudio que abarcara la totalidad ele la m isma. Nuestros resulwclos nos permiten concluir que la corteza entorrinal en la serie de envejecimiento y en la serie de Alzheimer difieren en que en el primer caso la atrofia cortical (especialmente acusada en la novena y décim a décadas) es armónica en e l conjunto ele la corteza entorrinal, mientras que en la enfermedad de Alzheime r afecta de un modo más intenso a las regiones laterales y caudales. Si examinamos la cmteza entorrinal del primate y las conexiones corticales que presenta podemos observar que la mayoría ele las aferencias corticales terminan en los campos laterales y caudales (24, 25, 39), homé>logas a las descritas en la corteza entmrinal humana . Haciendo una extrapolación se puede afirmar que la mayor afectación de estos campos deja al neocórtex virtualmente desconectado ele la formación del hipocampo, como ya apuntaran Hyman y cols. en 1984 (22). La topografía de la p royección entorrino-hipocam pal explmada e n el primate por Willer y cols. , (46) permite hacer la predicción de que es la parte del cue rpo y la cola del hipocampo las regiones que más desconectadas quedarían. La distribucié>n la minar de la patología y desaparición neuronal, centrada fundamentalmente en la capa 11 y V nos permiten analiza r aún más en profundidad el problema. La capa II es el origen principal de la vía perfmante (47, 48), terminando en la capa molecular de la circunvolución dentada, la cual sufriría las m a y ores consecuencias de la cleafe rentización. La capa V es e l origen principal de la proyección de retorno desde la corteza enrorrinal hacia el neocé>rtex (Mui1oz e Tnsausti, en preparación), y a su vez recibe proyecciones desde el hipocampo, a partir del campo CA 1 y subículo (32). Estas relaciones ele la formación del hipocampo pueden apreciarse en la Figura 4. Por consiguiente, el neocé>rtex queda desconectado ele la formación del h ipocampo doblemente porque al desaparecer las neuronas ele la capa V ele la corteza entorrinal las eferencias ele CAl y subículo no alcanzan su objetivo, y por la misma razón el neocóitex deja ele recibir los el Dujo normal de los impulsos ner-
25
Figura 4
AMI(;DALA CL Al!STlU\1 NUCLEOS nrtAU:S SUCU:O I ASAL (l>fU'"E:Jin t'UCU:OS SUPRAt.IA\ULARES lAL.0,\10 \ILDIAL ,UU A lf:C.~fl: ... TAl- \'l:t<o"Tl::Al NUCUU DEL JtAF'E
\ ' ~ ~ ~ ~
·. ', .. .. .. ~ . ',. J:EGION[5 Ol..F'AlOIUAS Cl.<\l'STitUM A\IIGDALA "'Unt:oS SEPT ALU N!;CU:O ACC\ll>tll"\tf, CAUDADOIPUT A).ll; 11. BI,.OT,U.,VIO 'IUCU:OS MA\nLA1US
....... ,'! 1 1
' ' ' . ' ' . ' '
. ' ' 1
CORTI'L\ 1)_RIAJ.I\AL,Uu ~~ • ) 6 GlltO t'"ll.\ilii'OCA!I.IP ICO Urrn Tf • 111 COJ;;T EZ " CI\Gl:L-\R CORHZ.o, PIJUfOR)l[ COI.li:ZA I .. Sl!LAJ: CORTr.l4. O RBITOf RO\IAt
Representación esquemática de las conexiones de la corteza en· torrinal en el primate, en los que se han indicado (doble raya in· terrumpiendo a lgunas de las flechas) los puntos de las vías ner· viosas en los que la enfermedad de Alzheimer provoca una destrucción particula r. Modificado de Amara! e lnsausti (2).
viosos proceclenres ele la corteza entorrinal. Los trabajos de Hraak y cols. (11) indican que la secuencia ele lesiones aparece en primer lugar en la corteza entorrinal y perirrinal, con lo cual no sorprende, a la luz de lo expuesto, que entre los primeros y más prominentes síntomas ele la enfenn edacl ele Alzheimer destaque la pérdida ele memoria. Esto no implica que e l progresivo deterioro de las capacidades intelectuales no afecte a otras regiones ele la corteza cerebral, como hemos pod ido comprobar (23, 33) sino que esta profunda afectaci6n ele la formación del hipocampo es el componente principal y precoz de una cascada ele sucesos neurodegene rativos que finalmente conducen a la catastrófica sit\tación de un e nfermo ele Alzheimer en situaci6 n te rminal.
Agradecimientos Q ue remos expresar nuestra gratitud en primer lugar
a las famil ias ele los enfermos que clonaron los cerebros para el estudio. Nuestro agradecimiento a C. García Gortari , P. Botín, M.A. Erdozáin y J. García por la asistencia técnica. Sufragado en parte por subvenciones del De partamento ele Salud, Gobierno de Navarra a R. T. y FIS proyectos 86/ 1129 y 88/ 1682.
REVISTA DE MEDICINA DE LA UNIVERSIDAD DE NAVARRA ENERO.MARZO 1997 25

l. Ammal, D.G. :~ncl Cowan. W.lvl. Cl9HO): Subcortical atfcrcnrs to the hippocampal formation in monke}'· .J. Comp. Neurol. 189: 57j.
2. Amara!, D.G. ;1ncl lnsausti, It (1992): Ilippocampal ronna tion. G. 1':~
xinos Editor, "The Human nervous System··, Academic Pres~. San Diego, 711.
j. Amara!, D.G. , In~austi, It ancl Cowan, W.lvl. 0987): The enlorhinal conex of thc monkey l. Cytoarchitectonic orga
nizalion . .J. Comp. Ncurol. 22tí: 307.
4. Armstrong, R.A., lvlyers, D., Smilh, C.U.H (1992): The clistribution of senile plaques, neurofibril l¡11y tangles ancl be
la/A4 dcposits in thc hippocampu~ in Alzheimer's disease. Neurosci. Hes. Commun. 10: 87.
5. Ball, M.]. 0977): Neuronal loss, neurofibrilla1y tangle~ ami granulovacuoloar clegeneration in the hippocampus with aging and demcntia. Acta Neuropal hol. 36: 111.
6. Bechterew, W.W. (1900): Demonstralion cines Gehirns mil Zerstorung der vorcleren und inneren Theile der ll irnrindc beicler Schlafenlapen. Neurol. Zentralbl. 19: 990.
7. Benzing, W.C., Hracly, D.R., Muf
son, E.J. and Armstrong, O.M. 0993): Eviclence thal lransmitter-conlaining clystror hic neurites precede those conla ining p<1ired helical fi lamellls within seni
le plaques in the entorhinal cortex of nonclemented clclerly and Alzheimer's discase patients. Brain Res. 619: 55.
8. Bouras, C., Hof, I'.R., Giannakopoulos, P., Michcl, J.P. , Morrison, J.! l. (1994): Hegional cli~lribution of neurofibrillary tangles ancl senile plaques in the cerebral cortex of clclerly palienls - i\
quantitativc cvaluation of a one-year autopsy population from a gerialric hosp ital. Cerebral. Cortex. 4 (2, l'vlar-Apr), 138-150.
9. Uraak , H.E. ancl f3raak, E. 0985): On arcas of transition bcrwcen entorhinal alloconex ancl temporal isocortcx in thc human brain. Normal morphology ancl lamina specific neuropathology in
~LTOGL0\FI.A 1
Alzheimcr's disease. Acta Neuropathol.
68: 325.
10. Uraak, 1-l .E. ancl Uraak, E. 0990): Neurofibrillary changes confincd to the entorhinal rcgion ancl an abunclance of cortical amyloicl in cases of presenile and senile demcnria. Acta Ncuropathol. 80:
479.
11. Rraak. H.E. and Rraak, E. 0992): Thc human entorhinal cortex. Normal
morpho log}' and lamina-specific parhology. Neurosci. Res. 15: 6.
12. 13rodmann, K. (1909): Vergleichende Lokalisationslhere der Groshirnrinde. 13arth, Lcip7.ig.
13. Dclaere, P., He, Y., Fayet, G., Duyckaerts, C.. llauw, .J.J. ( 1993): llelaAtí deposils are constanl in thc brain uf the olclest old - An immunocytochemical stucly of 20 frcnch centen¡¡rians. Neurohiology uf Aging. I-1: 191.
·14. Folstcin, M.F., Folstein, S.E. and Me 1 lugh, P.R. (]975): Mini mental state: a practica ! method for grading thc cognilivc state of palicnts for the clinician . .J Psichiat. Hes. 12: 189.
15. Gentelman, S.lVI.. Williams, B., Royston, i\-I.C. , .Jagoc, R. Clinton, .J. , Peny, IU 1. , l nce, P.G., i\llsop, D. , Polak, j.M., Robcrts, G.\X'. 0992): Quantification of beta-Atí protein depositon in the medial temporal lobe - A comparison of Alzheimer's discase and senile dcmentia of the Lewy bocly rype. reurosci. Lctt. HZ: 9.
16. Gómez-Isla, T. , l'rice,j.L., McKccl, D.W. , l'vlorris, J.C., Growclon, j.H. and Hyman, R.T. (1996): Profouncl loss of !ayer II cntorhinal conex ncurons occurs in very mild i\lzheimer's discase. J. Neurosci. 16: 4491.
17. Hanscn, L A., Mllsliah, E., Quijaclafawcell, S., Hexin, D. 0991): Entorhinal neurofibrillmy tangles in i\lzheimer clisease with Lc'i\')' boclies. Neurosci. Lett. 129: 269.
18. Heinsen, 11. , 1 lenn, R., Eiscnmenger, W/., Golz, M., Hohl, ]., Behke, B., Lockemann, U., l'uschel, K. (1994):
26 REVISTA DE MEDICINA DE LA UNIVERSIDAD DE NAVARRA ENERO-MARZO 1997
Quantilativc investigations on the hu
man entorhinal area: Lcft-right asymmetry and age-relatcd changes. Anal. Embryol. 190: 181.
19. Hirano, A. and Zimmerman, li.M. (1962): Alzhcimer's neurofibrillary changes: a lopographic stucly. i\rch. Neurol.
7: 227.
20. llof, P.R., Bierer, L.M., Perl, D.P., Delacourte, A., 13uee, L. , Bouras, C. , Morrison, .J.ll. (1992): Eviclence for early vulncrabiliry of the medial and inferior aspccts of the temporal lobe in an 82 -year-old patienl with preclincial signs of demcntia - Regional ancl laminar distribulion of neurofibrillary tangles and scnile plaques. Arch. Neurol. 1¡9: 946.
21. 1-looper, M. W. and Vogel, S. (1976): The limbic system in Alzheimcr's discase. A m.]. Pathol. 85: l.
22. Hyman, B.T., Van Hoesen, G.W. Damasio, A.R., and Barnes, C.L. (1984) Alzhcimer's disease: ccll-specific palhology isolates the hippocampal formation . Science 225: 11 68.
23. lnsausti, A. 0992): E.~tuclio cito y mieloarquitccrónico de la circunvolución
parahipocámpica posterior del hombre y sus cambios en la senescencia normal y en la enfermedad ele Alzheimer. Tesis doctoral. Universidad ele Navarra.
21. lnsausti, R. and Ammal, D.G. (1988): Distribution of cortical projcctions to the monkey entorhinal cortex: an autoradiographic study. Soc. Neuros
ci. Abstr. 14: 858.
25. Insausti, R., Amara!, D.G. and Cowan, W.M. 0987): The entorhinal cortex of the monkcy. 11. Cortical affcrcnts. ]. Comp. Neurol. 264: 356.
26. lnsausti , R., Tuñón, T., Sobrcviela, T., lnsausl i, A.M. ancl Gonzalo, L.M. (1995): The human cntorhinal conex: a cytoarchitectonic analysis . .J. Comp. Neurol. 355: 17"1.
27. jones, E.G . ancl Powell, T.P.S. (1970): An anatomical stucly of conver
ging scns01y pathways within the cerebral cortcx of the monkey. Brain 93: 793.
26

28. Khachaturian, 7..S. (1985): Diagnosis of Al7.heimer's disease. Arch. Neurol. 42: 1097.
29. l.ippa, C.F. , liamos, ].E., Pulaskisalo, D. , Oegennaro, L.J., Drachman, D.A. (1992): Al7.heimer's disease and aging - Effecrs on perforant pathway perikarya ami syanpses. Neurobiol. Aging 13: 405.
30. Mirra, S.S., Heyman, A. , McKeel , D., Sumi, S.M., Crain, J. , llrownlee. L.M. , Vogel, F.S., Hughes, J.l'., van 13elle, G. ancl 13erg, L. (1991): The consortium to establish a registry for Alzheime r's clisease (CERAD). Part 11 . Sranclarization of the neuropathologic assessment of Alzheimer's disease. Neurology 41: 479.
31. !'rice, J. L. , Da vis, P.R. , Morris, J.C., White, D.L. (1991): The Distribution of Tangles, Plaques and Rclated Immunohistochemical Markers in Healthy Aging and Al7.heimer's Disease. Neurobio l. Aging 12: 295.
32. Rosene, D.L. ami Van Iloesen, G. W. (1987): The hippocampal formation of the primate bra in. A review of some comparative aspects of cytoarchitectu re ancl conncctions. En "Cerebral Cortex" vol. 6, Eds., Jones EG and Peters A. l'lenum Press. New York. p 345.
33. Salinas, A. 0995): EstJu ctura ele la cortc7.a pe rirrinal humana. Modificaciones con e l envejecimiento y la enfermedad de Al7.heimer. Tesis doctoral. Universidad de Navarra.
34. Samuel, \Y/., Masliah, E., llill , I .. R., Buttcrs, N., Teny, R. (1991t): Hippocampal connectivily and Al7.heimer's demcntia: Effects of synapse loss and tangle fre-
quency in a two-compone nt model. Neurology. 44 (11 , Nov.), 2081-208R.
35. Scheff, S.W., Sparks, L., Price, D.A. 0993): Quantitative assessment of S}'
naptic density in the entorhin:~l cortex in Alzheimer's disease. Ann. Neurol. 34: 356.
36. Senut, !vl.C. , Roudier, l\1!., Davous, P., Falletbianco, C., Lamour, Y. 0991): senilc dementia of the Alzheimer type -ls there a corre lation bet ween cntorhinal cortcx and dentate gyrus lesions. Acta Neuropathol. 82:306.
37. Sobrevicla. T. 0991): Esntdio de los cambios citoarquitect6nicos e inmunohistoquímicos de la corteza entorrinal en la enfermedad de Al7.heimer. Tesis Doctoral. Universidad de Nava rra.
3R. Squire, L.R. (1987): Memory and thc brain. Oxford Univesity Press, Oxforcl.
39. Suzuki, \Y/.A. ancl Amaral, D.G. 0994): Topographic organization of the reciproca! connections between thc monkey entorhinal cortex and the perirhinal and parahippocampal cortices. J. Neurosci. 14: 1R56.
40. Trillo, L. and Gon7.alo, L.M. (1992): Ageing of thc human entorhinal cottex and subicular complex. llistol. Histopatho l. 7: 17.
lt'l . Van Essen, D.C. and Maunsell , J .H.R. (19RO): Two dimensional maps of the cerebral cortex. J. Comp. Neurol. 191: 255.
42. Van lloesen, G.W., llyman, B.T. and Damasio, A.R. 0991): Entorhinal cortex pathology in Alzheimer's disease. Hippocampus 1:1
..f3. Van Hoesen, G.\'íl ., Pandya, D.N.
and Burrers, N. C1975a): Some connec
tions of the entorhinal (area 28) and pe
rirhinal (arca 35) cortices of thc rhesus
rnonkey. l. Temporal lobc afferems.
lira in Res. 95: l.
44 . Van Hoesen, G.W., Pandya, D.N.
and Rutters, N. 0975b): Some connec
tions of the of thc entorhinal (arca 28)
and perirhinal (arca 35) cortices of rhe
rhesus monkey. 11. rronta l lobe affcrents.
Rrain Res. 95: 25.
45. Vennersch, P., Frigard , B., David, ].P., Fallctbianco, C., Dclacourre, A.
0992): l'resence of abnormal phosphorylatccl Tau proteins in the entorhinal
cortex of aged non-demcnted subjects. Neurosci. Lctt. 144: 143.
46. Von Economo (1929): The Cytoar
chitectonics of the Human Cerebral Cortex. Oxford University Press. l.ondon and
New York.
47. Witter, M.P. and Arnaral, D.G. (1991): Entorhinal cortex of the monkey.
V. Projections to the dentare gyrus, hippocampus, and subicular complex. ].
Comp. Neurol. 307: 437.
48. Willer, M.P., Van Hocsen, G.W.
and Amara!, D.G. (1989): Topographical
organi7.ation of the entorhinal projection
to the dentare gyrus of the rnonkey. J. Neurosci. 9: 216.
49. Wakabayashi, K., Iloner, W.G. ,
Masliah, E. 0994): Synapse alterations in the hippocampal-entorhinal formation in
Alzheimer's disease with and w ithout
l.ewy body discase. Brain Res. 667: 2.-f.
REVISTA DE MEDICINA DE LA UNIVERSIDAD DE NAVARRA ENERO-MARZO 1997 'll