
Mutaciones Homeoticas
31
Pregunta: ¿Qué son las mutaciones homeóticas y como es que su estudio contribuye al entendimiento del control genético del desarrollo floral? ¿Cómo se distinguen los genes homeóticos vegetales de los genes homeóticos de la Drosophila? Presenta: Noé Vaquera Enero 2009
-
Upload
guestc9c1f6 -
Category
Education
-
view
11.237 -
download
0
Transcript of Mutaciones Homeoticas

Pregunta:¿Qué son las mutaciones homeóticas y como es que su estudio contribuye al entendimiento del
control genético del desarrollo floral? ¿Cómo se distinguen los genes homeóticos vegetales de los
genes homeóticos de la Drosophila?
Presenta: Noé Vaquera
Enero 2009

Genes homeóticos
Identidad de los órganos o partes del cuerpo.
Investigación en genética en más de 100 años.
William Bateston: Tratado de variación genética (Bateston 1894).
“Homeosis”. “Modificación de la antena de un insecto a pata, o un ojo de un crustáceo en antena, o un pétalo a estambre, y aquello similar a esto”.
Flores dobles, en número de pétalos, descritas por los naturalistas por siglos.

Mutaciones homeóticas
William Bateston
Carlos Linneo
J. W. von Goethe

Mutaciones homeóticas
Linnaeus y Goethe, s. XVIII, casi los primeros en considerar que las flores monstruos o anormales podrían proveer información importante acerca de las reglas que gobiernan la heredabilidad genética (revisado de Meyerowitz et al., 1989).
Goethe (1790): “metamorfosis anormal” podía permitirnos “el revelar los secretos que la metamorfosis normal nos oculta, y para ver de manera distinta que, del curso regular de desarrollo, nosotros podemos solo inferir”. (Arber, 1946) Declaración profética, 200 años despues.

Genes homeóticos
Mutaciones en Drosophila, Identificación de un set de genes homeóticosCodifican para los factores de transcripción que determinan
los sitios donde deben desarrollarse las estructuras específicas.
Switches importantes de desarrollo que activan el programa genético completo para una estructura particular.
La expresión de genes homeóticos entonces, da a los órganos su identidad.

Mutaciones homeóticas

Genes homeóticos
• Los genes que determinan la identidad de los órganos florales fueron descubiertos gracias las mutantes homeóticas florales.
• Las flores dicotiledóneas constan de sucesivos verticilos (whorls) de órganos que se forman como resultado de la actividad de los meristemos florales: sépalos, pétalos, estambres, y carpelos.

Verticilos Florales
1. Sépalos
4. Carpelos
2. Pétalos
3. Estambres
Organizado en 4 verticilos (‘whorls’)

Identidad de los órganos florales en mutantes homeóticas
• Una función principal de los genes de identidad del meristemo floral es el activar un pequeño set de genes que especifiquen la identidad de los órganos florales.
• La identidad de los órganos florales fue originalmente identificada en A. thaliana y en A. majus en la base de sus fenotipos mutantes, los cuales están caracterizados en las transformaciones homeótica de un tipo de órgano floral en otro.

Mutaciones homeóticas

Mutantes homeóticas• La mutante A. thaliana apetala 2 (ap2) contiene carpelos
en las posiciones que están normalmente ocupadas por sépalos, y estambres en las posiciones que están normalmente ocupadas por pétalos (fig. 1b).
• apetala 3 (ap3) y las flores pistillata (pi) muestran transformaciones homeóticas de pétalos a sépalos, y estambres en carpelos (fig. 1c), mientras que las flores agamous (ag) forman pétalos en lugar de estambres y sépalos en lugar de carpelos (fig.1d).
• Además, el meristemo floral de las flores ag falla al terminar la producción de los órganos del cuarto verticilo. En su lugar, las flores ag están indeterminadas y continúan produciendo nuevas células que son incorporadas dentro de muchos extra verticilos de sépalos y pétalos.

Modelo ABCE

Modelo ABC• Los análisis de estas mutantes homeóticas de A.
thaliana y sus contrapartes en A. majus condujeron en 1991 a la formulación de modelo clásico ABC para la especificación de la identidad de los órganos florales.
• El modelo ABC postula que las tres funciones reguladoras -A, B y C- trabajan de una manera combinatoria para conferir una identidad a los órganos en cada verticilo (fig 1).
• La función A, conferida por la los genes homeóticos clase A AP1 y AP2 en A. thaliana y los genes redundantes tipo AP2, LIPLESS 1 y 2 (LIP1 y LIP2) en A. majus, que especifica la identidad de sépalos en el verticilo 1. Una función combinada con B, conferida por los genes clase B, AP3 y PI en A. thaliana y DEFICIENS (DEF) y GLOBOSA (GLO) en A. majus, especifica la identidad de pétalo en el verticilo 2.


Modelo ABC• La función B combinada con C, conferida por el gen
clase C, AG en Arabidopsis y PLENA (PLE) y FARINELLI (FAR) en A. majus, especifica la identidad de estambre en el verticilo 3: la función C sola especifica la identidad de carpelos en el verticilo 4 y también confiere la determinación floral.
• Una segunda faceta clave en el modelo ABC es que la función A y la función C son mutualmente antagonistas tal que la actividad clase C se expande en las flores mutantes y viceversa. Aunque los detalles difieren, el programa básico de desarrollo de los patrones de los órganos florales que es encapsulado por el modelo ABC parece ser ampliamente conservado entre las especies de plantas que han sido estudiadas extensamente, incluyendo el tulipán, la petunia, la primavera, y aun plantas con plantas menos vistosas como son arroz y maíz.

Extendiendo el modelo: Genes clase E
• Extendiendo el modelo: Genes clase E. los estudios de la redundancia genética han llevado a la identificaron de los genes homeóticos florales clase E. Los genes SEP clase E de A. thaliana codifican proteínas relacionadas que son redundantes y requeridas para la especificidad de pétalos, estambres y carpelos, como las flores triple mutante sep1 sep2 sep3 que contiene solo sépalos. SEP4 es requerida redundantemente con los otras tres genes SEP para conferir la identidad de sépalos (fig.1e), y contribuye al desarrollo de los otros tres tipos de órganos.

Genes Clase E
• Las mutantes cuádruples sep muestran la conversión de todos los cuatro tipos florales a estructura tipo-hoja con algo de carácter carpeloide, y recuerda a las triples mutantes ap2 ap3 ag que carecen de la función ABC, Las ortólogas de SEP se han identificado en muchas especies de plantas divergentes. En petunia, las ortólogas de SEP FBP2 PBP5 (FLORAL BINDING PROTEÍNA 2 y 5) especifica la identidad pétalo, estambre y carpelo en una forma redundante, con FBP2 proveyendo la correspondiente función E a SEP3 en A. thaliana.

Modelo ABCE

Genes homeóticos• Muchos genes homeóticos se han clonado en plantas y
animales, y la mayor parte de ellos se ha encontrado que codifican para factores de transcripción. En animales, estas proteínas también albergan un sitio de unión específica al DNA llamado el homeodominio y esta codificado por los genes llamados homeobox. Las plantas también contienen genes homeobox, como aquellos que codifican las proteínas de homeodominio KNOTTED y KNOX que regulan el destino celular y los patrones en las hojas.
• Sin embargo, la mayor parte de los genes que han sido identificados son genes MADS box, los cuales codifican para una clase diferente de factores de transcripción que contiene un sitio de unión al DNA llamado dominio MADS DNA unión/dimerización (Reichmann y Meyerowitz, 1997; Ng y Yanofsky, 2001).

MADS-box• Los genes MADS-box codifican para factores de
transcripción y se han encontrado plantas, animales y fungi. En plantas, los genes MADS-box incluyen genes regulatorios del desarrollo comparables a genes homeobox en animales. La región de la proteína codificada por la región altamente conservada MADS-box es llamada el dominio MADS y es parte del dominio de unión al DNA. Esta compuesta por 55 aminoácidos. Se ha propuesto que hay por lo menos 2 linajes (tipo I y tipo II) de genes MADS-box en plantas, animales y hongos (fig1; Alvarez-Buylla et al. 2000b). la mayor parte de los genes bien estudiados en plantas son del tipo II y tienen tres dominios mas que el tipo I: dominio de intervención (I) con cerca de 30 codones, el dominio coiled-coil tipo keratina (K) de 70 codones aproximadamente, y el C-terminal de longitud variable. Estos genes son llamados el tipo MICK y son específicos de plantas. Estos a su vez se dividen en 9 clases.

MADS-box

Antigüedad y evolución de los genes MADS-box• En un estudio reciente (Nam et al.) que tuvo como objeto
tener una visión de los roles de estos genes en la evolución morfólogica, se investigo el origen y la diversificación de los genes MADS-box florales mediante análisis de genética molecular evolutiva. Sus resultados sugieren que el ancestro común mas reciente de los genes MADS-box de hoy en día evolucionaron hace 650 m.a., mucho antes de la explosión Cámbrica [La explosión cámbrica hace referencia a la aparición repentina de organismos macroscópicos multicelulares complejos hace 542/530 m.a. Este período marca una brusca transición en el registro fósil con la aparición de los miembros más primitivos de muchos phylum de metazoos (animales multicelulares)].
• Este estudio apoya la idea de que existen dos linajes evolutivos (tipo I y II) de MADS-box en plantas y animales

Homeobox• Un homeobox es una secuencia de DNA que forma
parte de genes implicados en la regulación del desarrollo (morfogénesis) de los animales.
• Forman la familia de genes homeóticos HOM/Hox. Los genes homeobox codifican proteínas que le indican a las células de distintos segmentos del embrión en desarrollo "qué clase de estructuras tienen que hacer" (por ejemplo, antenas para la cabeza y patas para los tres segmentos torácicos). Genes homeobox casi idénticos también fueron encontrados en los genomas de ratones y de seres humanos.
• Los homeobox fueron descubiertos de forma independiente en 1983 por Walter Gebring, de la Universidad de Basilea, y por Matthew Scott y Amy Weiner, de la U. de Indiana.

Homebox
• Homeodominio. Un homeobox tiene una extensión de unos 180 pares de bases y codifica una secuencia de aminoácidos o dominio proteíco que puede adherirse a segmentos del ADN.
• Los genes homeobox codifican factores de transcripción que inician cascadas de otros genes, como todos los necesarios para la construcción de una extremidad.
• Subfamilias: La familia de genes homeóticos en vertebrados puede dividirse en dos sub-familias:
4. Acomplejados, llamados genes HOX o clase I
5. No-acomplejados, o genes homeóticos divergentes

Homeobox
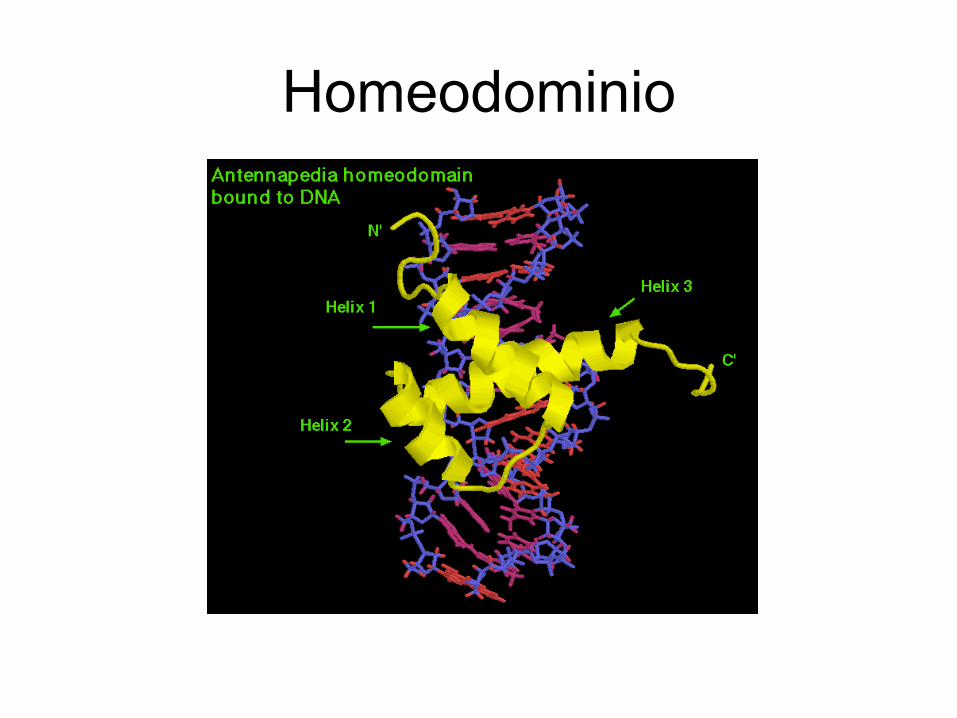
Homeodominio


Drosophila• Especificación del destino de órgano en Drosophila:
como Antennapedia distingue entre destino antena y pata.
• Una de las más impresionantes mutaciones en Drosophila es la conversión de antenas en patas por la mutación de ganancia de función dominante, Antennapedia. Se ha mostrado que las mutaciones recesivas que carecen de Antennapedia tienen un fenotipo con una antena apareciendo de los discos de las patas (Struhl, 1981). Los estudios moleculares mostraron que la mutación dominante ocasionó que Antennapedia sea mal expresada en los discos imaginales de las antenas, y produciendo patas en su lugar (Schneuwley et al., 1987).

Drosophila
• Entonces parece que Antennapedia fue el gen cuyo producto causo la especificación de patas.
• Un nuevo estudio de Casares y Mann(1998) reta esta interpretación y dice que la proteína normal de Antennapedia no esta involucrada en especificar patas. Mas probablemente, parece ser responsable de inhibir la producción de antenas.

Distinct features of Development• Animals• Homeo box gene for
pattern formation, but has MADS genes
• Dorsal-ventral specification: similar logic but use distinct genes
• Both Tyr and Ser/Thr membrane receptor kinases present
• Cytosol or nuclear receptors for steroid hormone
• Plants• MADS genes for pattern
formation, but has homeo box genes for other patterns
• Dorsal-ventral specification: similar logic but use distinct genes
• >400 membrane receptor Ser/Thr kinases, but no classic Tyr kinases
• Membrane receptor Ser/Thr kinases for steroid hormone

• Referencias• Krizek, B. and Fletcher,C. Molecular mechanisms of flower development: an
armchair guide. Nature Reviews Genetics 6, 688-698 (September 2005) | doi:10.1038/nrg1675 .
• Eckardt, N. MADS Monsters: Controlling Floral Organ Identity. The Plant Cell, Vol. 15, 803, April 2003, www.plantcell.org © 2003 American Society of Plant Biologists.
• Lincoln Taiz and Eduardo Zeiger. Plant Physiology, 3rd. Ed.; Sinauer Associates (Aug 30 2002).
• Nam et al. Antiquity and Evolution of the MADS-Box Gene Family Controlling Flower Development in Plants. Mol. Biol. Evol. 20(9):1435-1447. 2003
• Lemons, D. and McGinnis, W. Genomic Evolution of Hox Gene Clusters. Science 313, 1918 (2006); DOI: 10.1126/science.1132040
• http://en.wikipedia.org/wiki/Hox_genes. • http://zygote.swarthmore.edu/droso4.html. Specification of organ fate in
drosophila: how antennapedia distinguishes between antennal and leg fates• http://4e.plantphys.net/categories.php?t=t. Plant Physiology, Fourth Edition
by Lincoln Taiz and Eduardo Zeiger. Created by Sinauer Associates Inc.• http://en.wikipedia.org/wiki/Homeobox• Ainsworth, C. Flowering and its Manipulation; Annual Plant Reviews. 2006.


















