Producción Fitasa Por Aspergillus ficuum Y Otros En SSF
7
Production of phytate hydrolysing enzyme by some fungi S J Howson and R P Davis AR C Fo od R ese arch Institut e, Colney Lane, Norwich NR 4 7UA, UK (Received 9 February 1983) Eighty-four fungi from twenty five species have been examined for the production of extracellular enzymes capable of hydrolysing phytate 3-phytase, myo-inositol hexakisphosph ate 3-phospho- hydrol ase , EC 3.1.3.8, and 6-ph ytase, myo-inositol hexakisphosphate 6-phosphohydrolase, EC 3.1.3.26) when grown in: 1) rapeseed meal RSM); 2) a semisy nthet ie medium containing phyt ate as the sole phosphorus source PSM); 3) potato dextrose broth PDB). Altho ugh 58 active strains showed sub- stant ial acti vity, resul ts in either of the media were of no value in indic ating activ ity in RSM. There was no relationship betw een the ability o f a fungus to hydrolyse phytate and its tax onomi c position. Aspergillus ficuum NRRL 3135 had the greatest activity in the synthetic medium, and was relatively act ive in RSM. The extra cel lul ar enz yme had maximum activity after 10 days growth in PSM and had a temperature opt imum of 55°C. Two pH optima were noted at pH 2.0 and 5. 5. Inorgani c phosphate inhibited enzyme production; ammonia ions were a better nitrogen source than nitrate or urea. Keywords: Rapeseed meal; phytate; p hytate hydrolysing enzyme;Aspergillu s ficuum Introduction Salts of phytic acid myo-inositol hexakisdihydrogen- phosphate) are found in many plant tissues and related food products. 1,2 Phytate constitutes approximately 1-2 by weight of cereals and oilseeds a and phytate phosphorus accounts for between 60 and 90 of the total phosphorus present in these seeds; 4 it is considered to act either as a reserve of phosphorus or myo4nositol or in maintaining a phosphorus balance, s'6 The insoluble ph ytates are closely associated with proteins, especially in oilseeds7 and reduce the bioavailability 4'8 of basic amino acid groups in the proteins. Phytic acid is also a strong chelating agent and readily binds covalent metal ions, rendering them insoluble and thus unavailable for absorption. 3'9 The nutritional con- sequences of phytic acid interactions with protein and minerals are well documented 3'4'9 and methods for its removal from foods of plant origin are desirable. Most research has concentrated on the removal of phytate from soyabean and related products, because of their importance as sources of edible oil and as a protein source for human and animal consumption. 3A° Recently, increased interest has been given to rapeseed, the m ost suitable oil seed for temperate climates and consequently a source of protein, n-14 However, rapeseed meal and protein isolates possess even higher phytic acid levels than ma ny plant foods a and animal feeding trials have highlighted the problems caused by phytate, s-17 Methods have been developed for the removal of phytates from soyabean meal 4 although none has yet achieved economic feasibility. One possibility is the use of phytase enzymes (3-phytase, rnyo-inositol hexakisphosphate 3-phosphohydrolase, EC 3.1.3.8, and 6-phytase, myo-inositol hexakisphosphate 6-phosphohydrolase, EC 3.1.3.26) for the hydrolysis of the phytic acid into phosphoric acid and myo-inositol. Several workers have studied the properties of phytase enzymes, including those isolated from micro- organisms. 18-2° Indeed, phy tate reduction is a characteristic of fermentations such as those carried out by bakers' yeast 21-23 and in the production of soyabean tempeh. 24 The w ork in this laboratory was primarily concerned with the removal of phytate from rapeseed meal by using phytase. This paper reports a survey of microorganisms for the ability to produce a suitable extracellular phytase, some characteristic properties of a partially purified mould phytase and a comparison of this enzyme with wheat 6-phytase. Materials and methods Organisms Cultures were maintained on Difco-Bacto yeast malt agar (YM) slopes, except for strains of Botrytis cinerea and Cladosporium spp., which were kept on Difco-Bacto potato dextrose agar (PDA). To increase spore production, species of Aspergillus were grown on Czapek-Dox agar. Screening programme The optimum moisture content for the production of phytase by growing cultures in rapeseed meal was deter- mined with Rhizopus oligosporus, a mould used in producing temp eh in Indonesia 2s and already shown to reduce the level of phytate during fermentation.24 Spores ofR . oligosporus were harvested from YM slopes, previously incubated for 7 days at 25°C, using 9 ml 0.1 Tween 80. The suspension was filtered through two layers of sterile muslin, centrifuged at 0141--0229/83/050377--06 03.00 © 1983 Butterworth & Co. (Pu bl ish ers) Ltd Enzyme Micr ob. Tech nol. , 1983, Vol . 5, Septe mber 377
-
Upload
marcelo-torres -
Category
Documents
-
view
223 -
download
0
Transcript of Producción Fitasa Por Aspergillus ficuum Y Otros En SSF

8/13/2019 Producción Fitasa Por Aspergillus ficuum Y Otros En SSF
http://slidepdf.com/reader/full/produccion-fitasa-por-aspergillus-ficuum-y-otros-en-ssf 1/6
Pro du ct ion of phytate hydrolys inge n z y m e b y s o m e fu n g i
S J H o w s o n a n d R P D a v i s
AR C Food R esearch Institute, Colney Lane, Norwich NR 4 7UA, UK
( R e c e i v e d 9 F e b r u a r y 1 9 8 3 )
Eighty-four fungi from twenty five species have been examined for the production of extracellular
enzymes capable of hydrolysing phytate 3-phytase, myo-inositol hexakisphosphate 3-phospho-
hydrolase, EC 3.1.3.8, and 6-phytase, myo-inositol hexakisphosphate 6-phosphohydrolase, EC 3.1.3.26)
when grown in: 1) rapeseed meal RSM); 2) a semisynthet ie medium containing phytate as the sole
phosphorus source PSM); 3) potato dextrose broth PDB). Although 58 active strains showed sub-
stantial activity, results in either of the media were of no value in indicating activ ity in RSM. There
was no relationship between the ability o f a fungus to hydrolyse phyt ate and its taxonomic position.
Asper g i l lus f i cuum NRRL 3135 had the greatest activity in the synthetic medium, and was relatively
active in RSM. The extracellular enz yme had max imum activity aft er 10 days growth in PSM and had
a temperature opt imum of 55°C. Two pH optima were noted at pH 2.0 and 5.5. Inorganic phosphate
inhibited enzyme production; ammonia ions were a better nitrogen source than nitrate or urea.
Keywords: Rapeseed meal; phytate; p hytate hydrolysing enzyme;Aspergillus ficuum
I n t r o d u c t i o n
S a l t s o f phy t ic ac id myo-inositol h e x a k i s d i h y d r o g e n -
p h o s p h a t e ) a r e f o u n d i n m a n y p l a n t t i s s u es a n d r e l a te d f o o d
p r o d u c t s . 1,2 P h y t a t e c o n s t i t u t e s a p p r o x i m a t e l y 1 - 2 b y
w e i g h t o f c e re a l s a n d o i l se e d s a a n d p h y t a t e p h o s p h o r u s
a c c o u n t s fo r b e t w e e n 6 0 a n d 9 0 o f t h e t o t a l p h o s p h o r u s
pr esen t in these s eeds ;4 i t is cons ide r ed to ac t e i the r a s ar e se r v e o f p h o s p h o r u s o r myo4nositol o r i n m a i n t a i n i n g
a p h o s p h o r u s b a l a n c e , s ' 6 T h e i n s o l u b l e p h y t a t e s a r e c l o s e ly
as soc ia ted wi th p r o te ins , e spec ia l ly in o i l s eeds7 and r educe
the b ioava i lab i l i ty4 ' 8 o f bas ic am ino ac id g r oups in the
pr o te ins . P hyt ic ac id i s a l so a s t r ong che la t ing agen t and
r e a d i ly b i n d s c o v a l e n t m e t a l i o n s , re n d e r i n g t h e m i n s o lu b l e
a n d t h u s u n a v a i la b l e f o r a b s o r p t i o n . 3 '9 T h e n u t r i t i o n a l c o n -
s e q u e n c e s o f p h y t i c a c i d i n t e ra c t i o n s w i t h p r o t e i n a n d
m i n e r a ls a r e w e l l d o c u m e n t e d 3 '4 ' 9 a n d m e t h o d s f o r i ts
r e m o v a l f r o m f o o d s o f p l a n t o r i g i n a r e d e s ir a b le . M o s t
r e s e a rc h h a s c o n c e n t r a t e d o n t h e r e m o v a l o f p h y t a t e f r o m
s o y a b e a n a n d r e l a t e d p r o d u c t s , b e c a u s e o f t h e i r i m p o r t a n c e
a s so u r c e s o f e d i b l e o i l a n d a s a p r o t e i n s o u r c e f o r h u m a na n d a n i m a l c o n s u m p t i o n . 3 A °
R e c e n t l y , i n c r e a s ed i n t e r e s t h a s b e e n g i v e n t o r a p e s e e d ,
t h e m o s t s u i t a b l e o i l s e e d f o r t e m p e r a t e c l i m a t e s a n d
c o n s e q u e n t l y a s o u r c e o f p r o t e i n ,n - 1 4 H o w e v e r , r a p e s e e d
m ea l and pr o te in i so la te s pos ses s even h ighe r phy t ic ac id
l e v el s t h a n m a n y p l a n t f o o d s a a n d a n i m a l f e e d i n g t r ia l s
h a v e h i g h li g h t e d t h e p r o b l e m s c a u s e d b y p h y t a t e , s - 1 7
M e t h o d s h a v e b e e n d e v e l o p e d f o r th e r e m o v a l o f p h y t a t e s
f r o m s o y a b e a n m e a l 4 a lt h o u g h n o n e h a s y e t a c h i e v e d
e c o n o m i c f e a s i b i l it y . O n e p o s s i b i li t y is t h e u s e o f p h y t a s e
e n z y m e s ( 3 - p h y t as e , rnyo-inositol h e x a k i s p h o s p h a t e
3 - p h o s p h o h y d r o l a s e , E C 3 . 1 . 3 .8 , a n d 6 - p h y t a s e , myo-inositol
h e x a k i s p h o s p h a t e 6 - p h o s p h o h y d r o l a s e , E C 3 . 1 . 3 . 2 6 ) f o r t h e
h y d r o l y s i s o f th e p h y t i c a c i d i n t o p h o s p h o r i c a c i d a n d
myo-inositol. S e v e ra l w o r k e r s h a v e s t u d i ed t h e p r o p e r t i e s
o f p h y t a s e e n z y m e s , i n c lu d i n g t h o s e i s o l a t e d f r o m m i c r o -
o r g a n i s m s . 1 8 - 2 ° I n d e e d , p h y t a t e r e d u c t i o n i s a c h a r a c t er i s ti c
o f f e r m e n t a t i o n s s u c h a s th o s e c a r r i ed o u t b y b a k e r s 'y e a s t 2 1 -2 3 a n d i n t h e p r o d u c t i o n o f s o y a b e a n t e m p e h . 24
T h e w o r k i n t h is l a b o r a t o r y w a s p r i m a r i l y c o n c e r n e d
w i t h t h e r e m o v a l o f p h y t a t e f r o m r a p e s e e d m e a l b y u s in g
p h y t a s e . T h i s p a p e r r e p o r t s a s u r v e y o f m i c r o o r g a n i s m s
f o r t h e a b i l i t y t o p r o d u c e a s u i ta b l e e x t r a c e ll u l a r p h y t a s e ,
s o m e c h a r a c t e r i s ti c p r o p e r t i e s o f a p a r t i a ll y p u r i f i e d m o u l d
p h y t a s e a n d a c o m p a r i so n o f t h i s e n z y m e w i t h w h e a t
6 - p h y t a s e .
M a t er ia l s a nd met ho ds
Organisms
C u l t u r es w e r e m a i n t a i n e d o n D i f c o - B a c t o y e a s t m a l t
a g a r ( Y M ) s l o p e s, e x c e p t f o r s t r a in s o f Botrytis cinerea
a n d Cladosporium s p p . , w h i c h w e r e k e p t o n D i f c o - B a c t op o t a t o d e x t r o s e a g a r ( P D A ) . T o i n c r e as e s p o r e p r o d u c t i o n ,
species of Aspergillus w e r e g r o w n o n C z a p e k - D o x a g a r.
Screen ing program me
T h e o p t i m u m m o i s t u r e c o n t e n t f o r t h e p r o d u c t i o n o f
p h y t a s e b y g r o w i n g c u l t u r e s i n r a p e s e e d m e a l w a s d e t e r -
m i n e d w i t h Rhizopus oligosporus, a m o u l d u s e d i np r o d u c i n g t e m p e h i n I n d o n e s i a 2 s a n d a l r e a d y s h o w n t o
r e d u c e t h e l e v e l o f p h y t a t e d u r i n g f e r m e n t a t i o n . 2 4
S p o r e s o f R . oligosporus w e r e h a r v e s t e d f r o m Y M
slopes , p r ev iou s ly incuba te d f o r 7 days at 25°C , us ing
9 m l 0 .1 T w e e n 8 0 . T h e s u s p e n s i o n w a s f i lt e r e d
t h r o u g h t w o l a y e r s o f s t e ri l e m u s l in , c e n t r i fu g e d a t
0141 --0229/83/050377--06 03.00© 1983 Butterworth & Co. (Publishers) Ltd Enzyme Microb. Technol. , 1983, Vol . 5, September 377
8/13/2019 Producción Fitasa Por Aspergillus ficuum Y Otros En SSF
http://slidepdf.com/reader/full/produccion-fitasa-por-aspergillus-ficuum-y-otros-en-ssf 2/6
P a p e r s
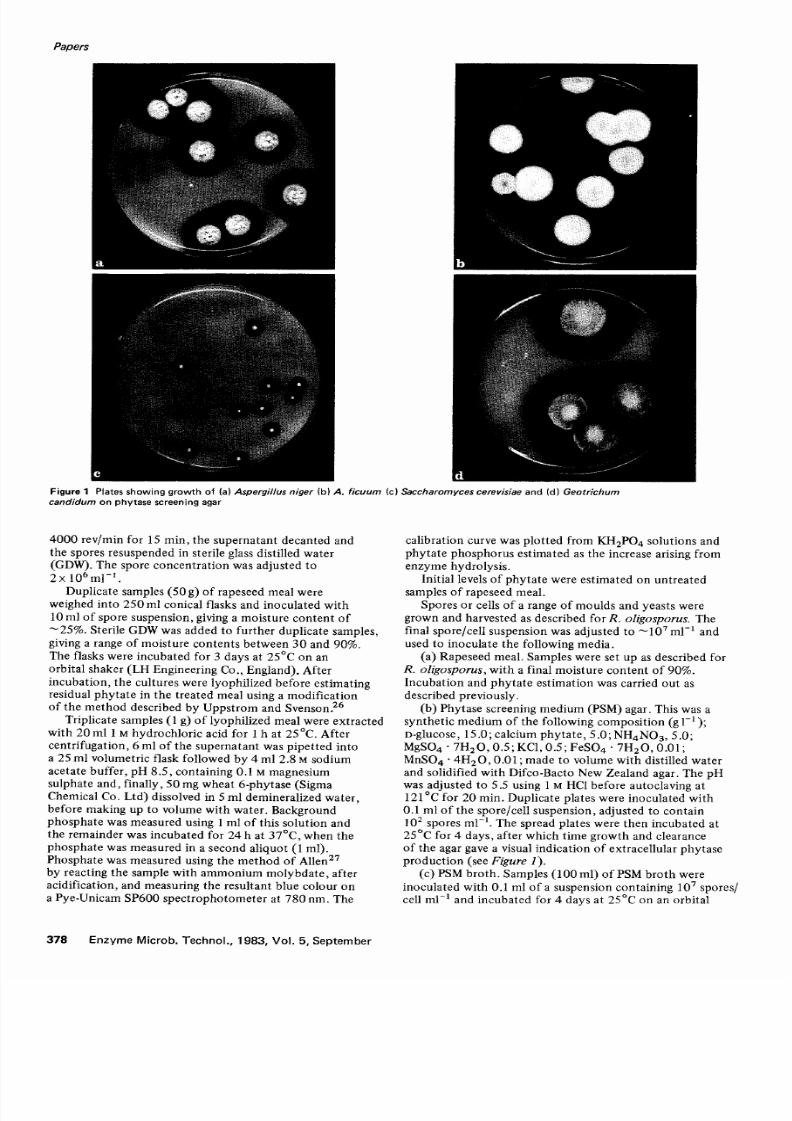
Figure 1 P la tes show ing grow th o f a) Aspergillus niger b) A f i cu u m c) Saccharomyces cerevis iae and d) Geot r ichum
c n d i d u m on phyt se screening g r
4 0 0 0 r e v / m i n f o r 1 5 m i n , t h e s u p e r n a t a n t d e c a n t e d a n d
the spores re suspe nded in s t e r il e glas s d i s ti l led w a te r
( G D W ) . T h e s p o r e c o n c e n t r a t i o n w a s a d ju s t e d t o
2 × 1 0 6 m l - ~ .
D u p l i c a t e s a m p l e s ( 5 0 g ) o f r a p e s e e d m e a l w e r e
w e i g h e d i n t o 2 5 0 m l c o n i c a l f l as k s an d i n o c u l a t e d w i t h
1 0 m l o f s p o r e s u s p e n s i o n , gi vi ng a m o i s t u r e c o n t e n t o f
~ 2 5 . S t e ri l e G D W w a s a d d e d t o f u r t h e r d u p l i c a t e sa m p l e s ,
g iv in g a r a n g e o f m o i s t u r e c o n t e n t s b e t w e e n 3 0 a n d 9 0 .
T h e f l a sk s w e r e i n c u b a t e d f o r 3 d a y s a t 2 5 ° C o n a n
o r b i t a l s h a k e r ( L H E n g i n e e ri n g C o . , E n g l a n d ) . A f t e ri n c u b a t i o n , t h e c u l t u r e s w e r e ly o p h i l i z e d b e f o r e e s t i m a t i n gr e s id u a l p h y t a t e i n t h e t r e a t e d m e a l u s i ng a m o d i f i c a t i o n
o f t h e m e t h o d d e sc r ib e d b y U p p s t r o m a n d S v e n s o n.26
T r i p l ic a t e s a m p l e s ( 1 g ) o f l y o p h i l i z e d m e a l w e r e e x t r a c t e d
w i t h 2 0 m l 1 M h y d r o c h l o r i c a c i d f o r 1 h a t 2 5 ° C . A f t e r
c e n t r i f u g a t i o n , 6 m l o f t h e s u p e r n a t a n t w a s p i p e t t e d i n t o
a 2 5 m l v o l u m e t r i c f l a s k f o l lo w e d b y 4 m l 2 . 8 M s o d i u m
a c e t a t e b u f f e r , p H 8 . 5 , c o n t a i n i n g 0 .1 M m a g n e s i u m
s u l p h a te a n d , f i n a l l y , 5 0 m g w h e a t 6 - p h y t a s e ( S i g m a
C h e m i c a l C o . L t d ) d i s so l v e d i n 5 m l d e m i n e r a l i z e d w a t e r ,
b e f o r e m a k i n g u p t o v o l u m e w i t h w a t e r . B a c k g r o u n d
p h o s p h a t e w a s m e a s u r e d u s i n g 1 m l o f t h i s s o l u t i o n a n d
t h e r e m a i n d e r w a s i n c u b a t e d f o r 2 4 h a t 3 7 ° C , w h e n t h e
p h o s p h a t e w a s m e a s u r e d i n a s e c o n d a l i q u o t ( 1 m l ) .
P h o s p h a t e w a s m e a s u r e d u s in g t h e m e t h o d o f A l l e n 2 7
b y r e a c ti n g t h e s a m p l e w i t h a m m o n i u m m o l y b d a t e , a f t e r
a c i d i f ic a t i o n , an d m e a s u r i n g t h e r e s u l t a n t b l u e c o l o u r o n
a P y e- U n i c a m S P6 0 0 s p e c t r o p h o t o m e t e r a t 7 8 0 n m . T h e
c a l i b r a ti o n c u r v e w a s p l o t t e d f r o m KH PO 4 so lu t ions and
p h y t a t e p h o s p h o r u s e s t i m a t e d a s th e i n c r e a s e a ri s in g f r o m
e n z y m e h y d r o l y s i s .
I n i t ia l le v e ls o f p h y t a t e w e r e e s t i m a t e d o n u n t r e a t e d
s a m p l e s o f r a p e s e e d m e a l .
S p o r e s o r c e l ls o f a r a n g e o f m o u l d s a n d y e a s t s w e r e
g r o w n a n d h a r v e s t e d a s d e sc r i b e d f o r R. oligosporus. T h e
f i n a l sp o r e / c e l l s u sp e n s i o n w a s a d j u s te d t o ~ 1 0 7 m 1 - 1 a n d
u s e d t o i n o c u l a t e t h e f o l lo w i n g m e d i a .
( a ) R a p e s e e d m e a l . S a m p l e s w e r e s e t u p a s d e s c r i b e d f o r
R. oligosporus, w i t h a f i n a l m o i s t u r e c o n t e n t o f 9 0 .I n c u b a t i o n a n d p h y t a t e e s t i m a t i o n w a s c a r r ie d o u t a s
desc r ibed prev ious ly .
(b ) P hytase s c reen ing m edium (P S M) aga r . Th i s was a
s y n t h e t i c m e d i u m o f t h e f o l l o w i n g c o m p o s i t i o n ( g 1 -1 ) ;
D - g l uc o s e, 1 5 . 0 ; c a l c i u m p h y t a t e , 5 . 0 ; N H 4 N O a , 5 . 0 ;
M g S O 4 • 7 H 2 0 , 0 . 5 ; K C 1, 0 . 5 ; F e S O 4 • 7 H 2 0 , 0 . 0 1 ;
M n S O 4 • 4 H 2 0 , 0 . 01 ; m a d e t o v o l u m e w i t h d i st i ll e d w a t e r
a n d s o l i d if i e d w i t h D i f c o - B a c t o N e w Z e a l a n d a g a r . T h e p H
was ad jus ted to 5 .5 us ing 1 M HCI be fo re au toc lav ing a t
1 2 1 ° C f o r 2 0 m i n . D u p l i c a t e p l a t es w e r e i n o c u l a t e d w i t h
0 .1 m l o f t h e s p o r e / c e ll s u s p e n s io n , a d j u s t e d t o c o n t a i n
l 2 spores m 1-1 . The spread p la te s were then incuba ted a t
2 5 ° C f o r 4 d a y s , a f te r w h i c h t i m e g r o w t h a n d c l e a r a n c eof the aga r gave a v i sua l ind ica t ion o f ex t race l lu la r phy tase
p r o d u c t i o n ( s e e Figure 1 .( c ) P SM b r o t h . S a m p l e s ( 1 0 0 m l ) o f PS M b r o t h w e r e
inocu la ted w i th 0 .1 m l o f a suspens ion con ta in ing 107 spores /
c e ll m 1- 1 a n d i n c u b a t e d f o r 4 d a y s a t 2 5 ° C o n a n o r b i t a l
3 7 8 E n z y m e M i c r o b . T e c h n o l . , 1 9 8 3 , V o l . 5 , S e p t e m b e r
8/13/2019 Producción Fitasa Por Aspergillus ficuum Y Otros En SSF
http://slidepdf.com/reader/full/produccion-fitasa-por-aspergillus-ficuum-y-otros-en-ssf 3/6
s h a k e r . T h e c u l t u r e w a s t h e n f i l t e r ed u s i n g W h a t m a n N o . 1
f i lt e r p a p e r a n d t h e f i l t r a t e c e n t r i f u g e d a t 1 2 0 0 0 r e v / m i n a t5 ° C f o r 3 0 m i n . T h e s u p e r n a t a n t s w e r e t h e n s t o r e d a t - - 2 0 ° C
p r i o r t o a s s ay i n g p h y t a s e a c t i v i t y .
I n t o t e s t t u b e s w e r e p i p e t t e d 0 . 5 m l o f 0 .2 M s o d i u m
a c e t a t e b u f f e r ( p H 4 . 4 ) , 0 . 5 m l s o d i u m p h y t a t e i n 0 . 2 M
s o d i u m a c e t a t e b u f f e r f o l l o w e d b y 1 m l o f c u l t u r e f i l t r a t e ,
g iv ing a f ina l pH o f 5 . 5 . Dupl ica te s am ples were incub a ted
i n a w a t e r b a t h a t 4 0 0 C f o r 0 a n d 4 5 m i n , b e f o r e t h e
r e a c t i o n w a s s t o p p e d b y a d d i n g 2 m l 1 5 T C A . O r t h o -
p h o s p h a t e i n t h e r e a c t i o n m i x t u r e w a s e s t i m a t e d a c c o r d i n g
t o A l l en 2 7 a n d p h y t a s e a c t i v i t y m e a s u r e d a s t h e a m o u n t o f
p h o s p h a t e r e l e a s ed d u r i n g t h e i n c u b a t i o n t i m e . O n e u n i t
o f p h y t a s e a c t i v i t y w a s d e f i n e d as t h e a m o u n t o f e n z y m e
c a p a b l e o f r e l e as i n g o n e m o l e o f i n o r g a n ic p h o s p h o r u s i n
o n e m i n u t e u n d e r t h e d e f i n e d r e a c t i o n c o n d i t i o n s .
( d ) D i f c o - B a c t o P o t a t o d e x t r o s e b r o t h ( P D B ) . I n o c u l a t i o n ,
i n c u b a t i o n , r e c o v e r y a n d p h y t a s e a s s a y o f c u l t u r e f i l t r a t e s
in P DB was ca r r i ed ou t a s fo r P S M bro th .
Ch a r a c te r iz a ti o n o f t h e p h y ta s e o fA s p e r g i l l u s f i c u u m
F l a s k s co n t a i n i n g 1 0 0 m l P S M b r o t h w e r e i n o c u l a t e d
w i t h 1 0 6 s p o r e s o f A . ficuum N R R L 3 1 3 5 a s d e s cr i be d
a b o v e , a n d i n c u b a t e d o n a n o r b i t a l s h a k e r a t 2 5 ° C . D u p l i-
c a t e c u l t u r e s w e r e h a r v e s te d d a i l y u p t o 1 6 d a y s , t h e
f i l t ra t e p u r i f i e d a n d a s s a y e d f o r p h y t a s e a c t i v i ty a s d e s c r ib e d .
T h e t e m p e r a t u r e p r o f i l e o f p h y t a s e a c t i v i t y i n P SM
c u l t u r e f i l t r a te s o f A . fieuum w a s d e t e r m i n e d b y a s s a y s o f
t r ip l i ca te s am ples a s desc r ibed p rev iou s ly ove r a range of
2 0 ° C t o 7 0 ° C . B l a n k s a n d te s t s w e r e b o t h p e r f o r m e d a t
e a c h t e m p e r a t u r e a f t e r e q u i l i b r a t i o n o f a l l c o m p o n e n t s
p r i o r t o t h e r e a c t i o n .
T h e p H p r o f i l e o f p h y t a s e a c t i v i t y w a s d e t e r m i n e d a t
4 0 ° C . T o a c h i e v e a w i d e p H r a n g e , t h e s o d i u m p h y t a t e
c o n c e n t r a t i o n w a s r e d u c e d t o 1 0 m g m 1 - 1 o f r e l e v a n t
b u f f e r . B u f f e r s u se d w e r e : S o r e n s e n s g l y c in e I ( p H 2 . 1 5 -
3 . 0 5 ); K o l th o f f s b o r a x - s u c c i n a t e ( p H 3 . 8 7 - 7 . 1 4 ) a n d
S o r e n s e n s c i t r a te I I ( p H 7 . 2 5 - 8 . 0 ) . 2 8 T h e a s s a y w a s
c o n d u c t e d u s in g d u p l i c a t e s a m p l e s a s d e s c r i b e d , a n d t h e
p H o f t h e f i n a l r e a c t i o n m i x t u r e w a s re c o r d e d .
T h e e f f e c t o f i n o r g a n i c p h o s p h a t e o n t h e d e v e l o p m e n t
o f p h y t a s e a c t i v i t y d u r i n g g r o w t h w a s t e st e d b y i n c u b a t i n g
d u p l i c a t e s a m p l e s o f A . fieuum a t 25°C for 4 days in P S M
b r o t h c o n t a i n i n g d i f f e r e n t c o n c e n t r a t i o n s o f K H 2 P O 4 a s
t h e o n l y s o u r c e o f i n o r g a n ic p h o s p h a t e Table 4).T h e e f f e c t o f v a r i a t io n o f t h e n i t r o g e n s o u r c e o n e n z y m e
p r o d u c t i o n w a s t e s t e d b y a l t e r i n g t h e n i t r o g e n o u s c o m -
p o u n d s p r e s e n t i n t h e b a s i c P SM b r o t h Table 5). The f ina l
l e ve l o f n i t r o g e n i n e a c h m e d i u m w a s t h e s a m e a s in t h e
bas ic P S M bro th a s desc r ibed ea r l i e r . The phytase a s say
w a s c a r r ie d o u t a s a l r e a d y d e s c r ib e d a f t e r a d j u s t m e n t o f t h e
p H o f t h e c u l t u r e f i l t r a te s t o 5 .5 u s i ng 0 .1 M N a O H o r
1 M HC1 as neces sa ry .
Co m p a r i s o n o f a c t i v i ty o f wh e a t 6 - p h y ta s e a n dA . f i c u u m p h y ta s e o n R S M
S i x l it r es o f P SM w a s i n o c u l a t e d w i t h 5 m l o f a s p o r e
s u s p e n s i o n o f A . fieuum conta in ing 107 spores m l -x . The
c u l t u r e w a s i n c u b a t e d a t 2 5 ° C o n a n o r b i t a l s h a k e r( 2 5 0 r e v ] m i n ) f o r 1 0 d a y s . I t w a s th e n f i l te r e d t h r o u g h
W h a t m a n N o . 1 fi l te r p a p e r a n d t h e s u p e r n a t a n t c e n t r i -
f u g ed a t 1 2 0 0 0 r e v / m i n fo r 3 0 m i n a t 4 ° C . T h e 4 9 0 0 m l
o f c u l t u r e f i l t r a te o b t a i n e d w a s ly o p h i l i z e d ( E d w a r d S h e l f
M o d e l E G 6 ( S ) ) a n d t h e r e s u l t a n t p r e p a r a t i o n w a s s u s p e n d e d
S t u d ie s o f m i c r o b i a l p h y t a s e : S J H o w s o n a n d R P D a v is
i n 4 0 m l o f g l a s s -d i s ti l le d w a t e r . T h i s w a s a s sa y ed a n d f o u n d
t o h a v e a n a c t i v i t y o f 0 . 6 3 4 U m 1 - 1.
T h is p h y t a s e w a s th e n c o m p a r e d w i t h a c o m m e r c i a l
p r e p a r a t i o n o f w h e a t 6 - p h y t a s e ( S ig m a C h e m i c a l C o . ) .
T h e w h e a t 6 - p h y t a s e h a d a n a c t i v i t y o f 0 . 0 1 8 U m g - 1 so l i d.
T o 5 g o f m e a l w a s a d d ed 5 0 0 m g o f o n e o f t h e p h y t a s e
p r e p a r a t i o n s a n d 4 0 m l o f g l a ss - di s ti ll e d w a t e r ( e q u i v a l e n t
m o i s t u r e c o n t e n t 9 0 ) . D u p l i c a t e sa m p l e s w e r e in c u b a t e d
in 100 m l con ica l f l a sks a t 40°C in a shak ing incu ba to r( N e w B r u n s w i c k L t d ) . T h e c o n d i t i o n s o f i n c u b a t i o n v a r i e d
a s f o l lo w s : ( a ) 2 4 h ; ( b ) 4 8 h ; ( c ) a f t e r 2 4 h a n o t h e r 5 0 0 m g
o f e n z y m e w a s a d d e d a n d i n c u b a t e d f o r a f u r t h e r 2 4 h ;
( d ) a s f o r (c ) e x c e p t 5 g m e a l w a s a d d e d r a t h e r t h a n t h e
e n z y m e .
A f t e r i n c u b a t i o n t h e s e s a m p l e s w e r e t h e n e x t r a c t e d a n d
as sayed a s desc r ibed prev ious ly .
R e s u l t s a n d d i s c u s s i o n
T h e o p t i m u m m o i s t u r e c o n t e n t f o r p h y t a s e a c t i v it y , a s
s h o w n b y t h e r e d u c t i o n o f p h y t a t e , i n r a p e s ee d m e a l in c u -
b a t e d f o r 3 d a y s a t 2 5 ° C a f t e r i n o c u l a t i o n w i t h s p o r e s o fR. oligosporusw a s 8 0 Table 1). Reinh old 29 a l so
r e p o r t e d i n c r e a s e d r e m o v a l o f p h y t a t e f r o m b r e a d d o u g h s
i n c u b a t e d w i t h Saccharom yces cerevisiaea t h i g h m o i s t u r e
c o n t e n t s . G l a ss a n d G e d d e s a ° f o u n d t h a t p h y t a s e a c t i v i ty
w a s n o t d e t e c t a b l e i n d r y d o r m a n t w h e a t g r a in s w h i ls t
ac t iv i ty inc reased wi th r i s ing m ois tu re con ten t dur ing
s to rage .
T h e r e d u c t i o n o f p h y t a t e i n r a p e s e e d m e a l a c h i e v e d b y
a r a n g e o f f u n g i a n d y e a s t s , a f t e r i n c u b a t i o n u n d e r s i m il a r
cond i t ions , va r ied be tw een 10 and 80 . A s im i la r degree
o f v a r i a t io n i n t h e r e d u c t i o n o f p h y t a t e w a s o b s e r v e d f o r
d i f fe ren t s t ra ins o f a g iven spec ies ; da ta fo r Aspergillus nigera re shown in Table 2. E x t r a c e l h i la r p h y t a s e a c t i v i ty w a sde tec ted in 33 ou t o f 84 s t ra ins t e s ted in P S M and in 37
s t ra ins in P DB Table 3). A l l s t ra i n s w h i c h s h o w e d e n z y m e
able 1 R e d u c t i o n o f p h y t a t e i n r a pe s e e d m e a l i n o c u l a t e d w i t h
c o n i d i a o f hizopus oligosporusa n d i n c u b a t e d f o r 3 d a y s at 3 0 ° C
P h y t a t e c o n t e n t
M o i s t u r e c o n t e n t o f m e a l( ) ( m g g - l ) r e d u c t i o n
2 5 3 6 . 0 1 8
3 0 3 7 . 2 1 5
4 0 3 8 .1 1 4
5 O 3 6 .8 1 66 0 3 5 . 0 2 0
7 0 3 1 . 2 2 9
8 0 1 6 . 0 6 4
9 0 2 0 . 5 5 4
ab le 2 Effec t of cul ture of various stra ins of A niger on thephyta te c on ten t o f rapeseed mea l
Res idua l phy ta tein meal Phytate reduc t ion
St ra in (mg g- l ) ( w/w )
L25 7.7 37.9CS180 8.2 34.3
L9 8.4 32.3CSl 4 4 .2 66 .0UD31 4 9 .7 21 .8LIO 10.9 12.2L26 10.9 12.1CS14 3 .8 69 .4
E n z y m e M i c r o b . T e c h n o l . , 1 9 8 3, V o l . 5 , S e p t e m b e r 3 7 9
8/13/2019 Producción Fitasa Por Aspergillus ficuum Y Otros En SSF
http://slidepdf.com/reader/full/produccion-fitasa-por-aspergillus-ficuum-y-otros-en-ssf 4/6
P a p e r s
Table 3 S u r v e y o f m i c r o o r g a n i s m s f o r p h y t a t e h y d r o l y s i n g a c t i v i ty
i n c u l t u r e f i l t r a t e s
S t r a in s e x h i b i t i n g S t r a i n s e x h i b i t i n g
a c t i v i t y i n P S M a c t i v i t y i n P D B
P o s i t i ve Ne g a t i ve P o s i t i ve Ne g a t i ve
Asp e rg i l l u s n ig e r 0
A f i c u m 1
A p e t r a k i i 0
A c a n d i d u s 3
A f l a vu s 0
A w e n t i i 0
A v e r s i c o l o r 2
A s y d o w i 3
A re p e n s 2
A ch e va l ie r i 0
A a m s t e l o d a m i 3
S a c c h a r o m y c e s
cerevisiae 4
S u v a r u m 0
R h i z o p u s o r y z a e 2
R o l i g o s p o r u s 1
R s t o l o n i f e r 2
R a r r h i z u s 0
M u c o r r a c e m o s us 3
M p i r i f o r m i s 4
G e o t r i c h u m c a n d i d u m 1
B o t r y t i s c i n e r e a 2
C l a d os p o r i u m
c l a d o s p o r o i d e s 0
R h o d o t o r u l l a
g r a m i n i s 0
A o r y z a e 0
7 3 4
0 1 0
2 0 2
2 0 5
6 4 2
1 1 0
3 1 4
0 3 0
2 3 1
2 1 1
2 3 2
8 6 6
1 0 1
3 2 3
0 0 1
2 2 2
1 0 I
2 4 1
0 2 2
0 0 1
3 1 4
1 0 1
1 0 1
2 0 2
0 03
-~ 0 .0 2-g
>
~- o . o
T h e r e w a s l i t t l e c o r r e l a t i o n b e t w e e n p h y t a t e h y d r o l y s i n g
a c t i v i t y i n t h e t w o m e d i a F i gure 2 ) . M o r e o v e r , t h e r e w a s
l i t t l e c o r r e l a t i o n b e t w e e n t h e a b i l i t y to r e d u c e p h y t a t e i n
R S M a n d t o p r o d u c e a n e x t r a c e l l u l a r e n z y m e i n P S M o r
P D B F i gure s 3 a n d 4 .
T h e Aspergi l lus s p e c ie s a p p e a r e d t o b e t h e b e s t s o u r c e
o f e x t r a c e l l u l a r p h y t a s e e n z y m e s , w h i l s t A spe rg i ll u s f i c uu m
90
8 0
7 0
6 0
50 •
4 o •
3 t
A 4 L • A
t . & •
t I
0 0.01
I I I I I
0.07 O.07
PSM activity (units ml t)
Figure 3 R e d u c t i o n o f p h y t a t e c o n te n t o f R S M c o m p a r e d w i th
e x t r a c e l l u l a r p h y t a s e a c t i v i t y o f m o u l d s a n d y e a s t s w h e n g r o w n
in PSM
9O
8o
70
6 0
= = = •
• • ~ 50
0 0 . O I O . 0 2 O . 0 3 g 4 0
P D B a c t i v i t y ( u n i t s m l - I ) ~ •
Figure 2 E x t r a c e l l u l a r p h y t a s e a c t i v i t y o f v a r i o u s s t r a in s o f m o u l d s
a n d y e a s ts w h e n g r o w n i n P S M a n d P D B ~ 3 0
a c t i v i t y i n P S M b r o t h c u l t u r e s w e r e a l so s e e n to p r o d u c e z o ~ •
z o n e s o f c l e a r a n c e in a d v a n c e o f m y c e l i a l g r o w t h o n P S M
a g a r p l a t e s Figure 1) . T w e l v e s t r a i n s g a v e p h y t a s e a c t i v i t y •
i n b o t h P S M f i l t r a t e s a n d P D B , i n d i c a t i n g t h a t t h e e n z y m e to
w a s c o n s t i t u t i v e , w h i ls t 2 1 s t r a i n s s h o w e d p h y t a s e a c t i v i t y
o n l y i n PS M b r o t h i n d i c a t i n g t h a t t h e e n z y m e i s i n d u c e d b y L, jt h e p r e s e n c e o f t h e s u b s t r a t e . H o w e v e r , 2 5 s t r a in s t e s t e d o
s h o w e d a c t i v i t y o n l y i n P D B f i l t r a t e s a n d n o t i n P SM ,
w h i c h m a y h a v e b e e n a n e f f e c t o f s u b s t ra t e i n h i b i t i o n .
T h e t o t a l n u m b e r o f a c t i v e s t r a i n s w a s 5 8 ; 2 6 s t r a i n s
s h o w e d n o a c t i v i t y i n t h e c u l t u r e f i l t ra t e s .
| | I |
0 .0 4 0 .0 7
PDB activity un its ml l)
F igure 4 R e d u c t i o n o f p h y t a t e c o n t e n t o f R S M b y s t r a in s o f
m o u l d s a n d y e a st s w h e n c o m p a r e d t o t h e i r e x t r a c e l l u l a r p h y ta s e
a c t i v i t y i n P D B
3 8 0 E n z y m e M i c r o b . T e c h n o h , 1 9 8 3 , V o l . 5 , S e p t e m b e r
8/13/2019 Producción Fitasa Por Aspergillus ficuum Y Otros En SSF
http://slidepdf.com/reader/full/produccion-fitasa-por-aspergillus-ficuum-y-otros-en-ssf 5/6
pr ov ide d the m o s t ac t ive cu l tu r e f i l t r a te s . S h ieh and W ar e 18
a l so f o u n d t h a t Aspergillus s p e ci e s c o m m o n l y p o s s e s s e d
p h y t a s e e n z y m e s a n d t h a t t h e m o s t a c t i v e p h y t a s e e x t r a c t s
w e r e p ro d u c e d b y Aspergil lus f icuum. B o t h i n t e r - a n d
in t r a spec ie s va r ia t ion was obse r ved .
P r o p e r t ie s o f e x t r ac e l lu l a r p h y ta s e f r o mA s p e r g i l l u s f i c u u m
The h ighes t ex t r ace l lu la r phy tase ac t iv i ty in P S M cu l tu r e
f i l tr a t e s o f A . f i e u u m w a s r e c o r d e d a f t e r 1 0 d a y s i n c u b a t i o n
a t 2 5 ° C (Figure 5), w h e n t h e a c t i v i t y o f t h e c u l t u r e f i l tr a t e
w a s a p p r o x i m a t e l y 6 t im e s g r e a t e r t h a n a t 4 d a y s .
S k o w r o n s k i 31 f o u n d t h a t p h y t a s e a c t i v i ty i n c u l t u r e f i l tr a t e s
o f t h e c l o s e ly r e l a t ed f u n g u s A. niger r e a c h e d a p e a k a f t e r
11 to 12 days . Th e s i tua t ion va r ie s wi t h spec ie s . S t r a ins o f
Ge o t r ie h u m e a n d i d u m s h o w e d o p t i m u m a c t iv i t y a t 4 d a y s
u n d e r t h e s a m e c o n d i t i o n s ; a f t e r t h is t i m e , a c t i v i t y w a s
r e d u ce d b y > 6 0 ( d a t a n o t s h o w n ) .
T h e i n c r e a s e in a c t i v i t y w i t h t i m e r e c o r d e d i n c u l t u r e
f i l tr a t e s o f A . f i e u u m c o u l d h a v e b e e n a r e su l t o f a u t o l y s i s
r a t h e r t h a n i n d u c t i o n , a s th e e n z y m e a p p e a r s t o b e c o n s t i -
t u t i v e . T h e m y c e l i a l m a t d i s i n te g r a t e d a f t e r 1 1 - 1 2 d a y s ,
i n d i c a ti n g t h a t a u t o l y s i s o f t h e m y c e l i u m o c c u r r e d a n d
c o u l d r e s u l t i n l e a k a g e o f i n t ra c e l lu l a r e n z y m e i n t o t h e
m e d i u m . S k o w r o n s k i 3 1 r e a c h e d a s i m i la r c o n c l u s i o n b y
e x c l u d i n g s u b s t r a t e i n d u c t i o n b y c o m p o n e n t s o f t h e
c u l t u r e m e d i u m .
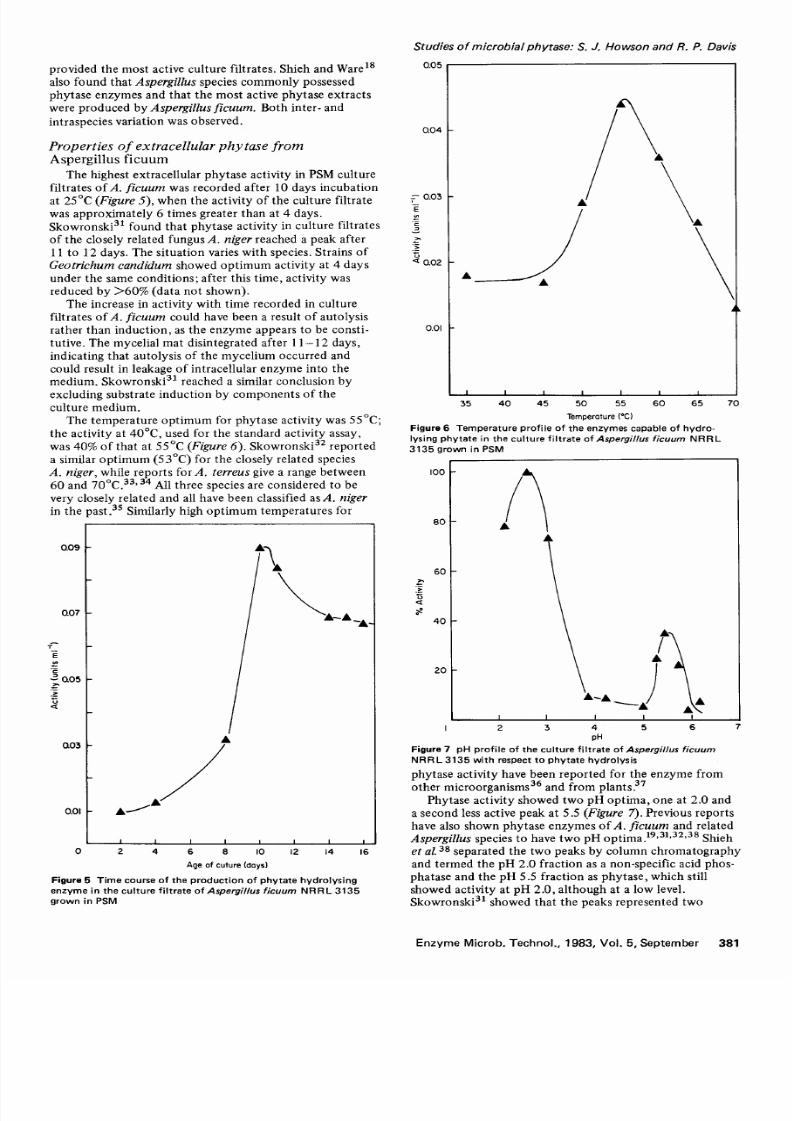
T h e t e m p e r a t u r e o p t i m u m f o r p h y t a s e a c ti v i ty w a s 5 5 ° C ;
the ac t iv i ty a t 40°C, used f o r the s tanda r d ac t iv i ty a s say ,
w a s 4 0 o f t h a t at 5 5 ° C (Figure 6). S k o w r o n s k i 32 r e p o r t e d
a s i m il a r o p t i m u m ( 5 3 ° C ) f o r t h e c l o s e l y r e la t e d s p e c i e s
A. niger, w h i l e r e p o r t s f o r A . terreus give a r ange be tw een
60 and 70°C . 33 ' 34 Al l th r ee spec ie s a r e cons ide r ed to be
ve r y c lose ly r e la ted and a ll have been c la s s if i ed a s A. nigerin th e ~ast.35 S im i l a rl y h ig h o p t i m u m t e m p e r a t u r e s f o r
0 0 9
0 0 7
0 0 5
a o 3
0.OI
O 2 4 6 8 I 0 12 14 16
Age of cuture days)
F i g u r e 5 T i m e c o u r s e o f th e p r o d u c t i o n o f p h y t a t e h y d r o l y s i n g
e n z y m e i n t h e c u l t u r e f i l t r a t e o f s p e r g i ll u s f ic u u m N R R L 3 1 3 5
g r o w n i n P S M
S t u d i es o f m i c r o b i a l p h y t a s e : S J H o w s o n a n d R P D a v is
0.05
0 0 4
O 03
< 0.02
0.01
I I I I I I
35 40 45 50 55 60 65
Temperature *C)
F i g u r e 6 Temperature p r o f i l e o f t h e e n z y m e s c a p a b l e o f hydro-
lysing p h y t a t e in the culture filtrate of sperg i l lus f i cuum N R R L
3135 grown in PSM
70
8O
6O
_>
4O
2
I I = t I
2 3 4 5 6 7pH
F i g u r e 7 pH p r o f i l e o f t h e c u l t u r e f i lt r a t e o f Aspergillus ficuumN R R L 3 1 3 5 w i t h r e sp e c t t o p h y t a t e h y d ro l y s is
p h y t a s e a c t i v it y h av e b e e n r e p o r t e d f o r t h e e n z y m e f r o m
o t h e r m i c r o o r g a n i s m s 3 6 a n d f r o m p l a n t s .37
P h y t a s e a c t i v i t y s h o w e d t w o p H o p t i m a , o n e a t 2 . 0 a n d
a second le s s ac t ive peak a t 5 . 5 (Figure 7). P r e v i o u s r e p o r t s
h a v e a ls o s h o w n p h y t a s e e n z y m e s o f A . f i c u u m and r e la ted
Aspergillus s p e ci e s t o h a v e t w o p H op tim a. 19 31 32 3S S hiehet al. 3 8 s e p a ra t e d t h e t w o p e a k s b y c o l u m n c h r o m a t o g r a p h y
a n d t e r m e d t h e p H 2 . 0 f r a c t i o n a s a n o n - s p e c i fi c a ci d p h o s -
p h a t a s e a n d t h e p H 5 .5 f r a c t i o n a s p h y t a s e , w h i c h s t i ll
s h o w e d a c t i v i t y a t p H 2 . 0 , a l t h o u g h a t a l o w l e v e l.
S k o w r o n s k i 31 s h o w e d t h a t t h e p e a k s r e p r e s e n t e d t w o
E n z y m e M i c r o b . T e c h n o l . 1 9 8 3 V o l . 5 S e p t e m b e r 3 8 1

8/13/2019 Producción Fitasa Por Aspergillus ficuum Y Otros En SSF
http://slidepdf.com/reader/full/produccion-fitasa-por-aspergillus-ficuum-y-otros-en-ssf 6/6
Papers
T a b l e 4 E f f e c t o f v a r i o u s c o n c e n t r a t i o n s o f i n o r g a n i c p h o s p h a t e o n
e x t r a c e l lu l a r p h y ta s e a c t i v i ty o f A ficuum in PSM
In o rg a n ic p h o s p h a t e c o n c e n t r a t i o n A c t i v i t y
(mg d l -~ ) (u n i t s m l -~ )
2 0 .2 4
5 0 .2 0
1 0 0 . 1 2
1 5 0 . 0 5
2 0 0 . 0 2
2 5 0
5 0 0
T a b l e 5 E f f e c t o f v a r i o u s n i t r o g e n so u rc e s o n e x t r a c e l l u l a r p h y t a s e
p r o d u c t i o n i n A . ficuum
A c t i v i t y R e l a t i v e a c t i v i t yN i t ro g e n so u rce (u n i t s m l - l ) ( ) P S M
( N H 4 ) 2 S O 4 0 . 0 2 6 1 7 3 . 3
U r e a 0 . 0 0 4 2 6 . 7
N H 4 H 2 P O 4 0 . 0 2 6 1 7 3 . 3
N a N O 3 0 . 0 1 2 8 0 . 0
K N O 3 0 . 0 1 4 9 3 . 4
N H , N O 3 ( s t a n d a r d P S M ) 0 . 0 1 5 1 0 0
T a b l e 6 C o m p a r i s o n o f r e d u c t i o n o f p h y t a t e i n R S M b y w h e a t
6 - p h y t a s e a n d A ficuum p h y t a s e u n d e r d i f f e r e n t c o n d i t i o n s
( 5 0 0 m g e n z y m e + 4 0 m l G D W + 5 g m e a l i n c u ba t e d a t 4 0 ° C )
C o n d i t i o n s r e d u c t i o n
W h e a t 6 p h y t a s e
2 4 h 7 3 .4
4 8 h 8 0 . 3
2 4 h + 5 0 0 m g e n z y m e 8 7 . 0
2 4 h + 5 0 0 m g m e a l 7 3 . 0
A ficuum p h y t a s e
2 4 h 6 5 . 2
4 8 h 6 5 . 9
2 4 h + 5 0 0 m g e n z y m e 8 4 . 0
2 4 h + 5 0 0 mg me a l 8 0 .2
enzymes as they exhibited markedly different heat stability
propert ies. In con tras t, Irving and Cosgrove 19 fou nd it
impossible to discern whether the two peaks represented
one or two enzymes as they failed to separate the two
active centres either b y gel electrophoresis or the
visualization method .
Shieh e t a l . 38 showed that increased levels of inorganic
phosphorus in the growth medium suppressed synthesis of
both enzy mes although the ratio of the pH 2.0 to the
pH 5.0 peaks was reversed, and the phytase enzymebecame the most active enzy me produced. This was con-
firmed by our experime nts in which differe nt levels of
inorganic phospho rus were added to PSM brot h, which
contains negligible inorganic phosphorus. Inhi biti on of
enzyme pr oduc tion occurre d above 0.2 mg m1-1 of inorganic
phosphorus , as can be seen from the results in T a b l e 4 .
Phytate production is also influenced by nitrogen
source, with ammonium ions being markedly superior to
nitrate and urea ( T a b l e 5 ) . We found that urea supported
little growth. Ammo niu m salts were also foun d to be the
best nitrogen source in the medium for subsequent phytase
production by A . t e r r e u s cultures.34
It was of interest to compare the activity towardsphytate in RSM of phytate degrading enzymes from
A . f i c u u m and from wheat which is commercially available.
Both enzymes decreased the phytic acid content to a sub-
stantial degree, with the p reparat ion from wheat being
somewhat more active.Residual phytate was present in
RSM even after prolonged in cubat ion. This result does not
reflect loss of activity by the en zyme for the additio n to
the i ncubation mixture containing enzyme from either
source of more RSM led to fu rther removal of phytate
( T a b l e 6 ) . It may be that some proportion of the phytate
in RSM is inaccessible to the enzy me, perhaps by its
occlusion within cells or by cellular const ituent s. The
addition of further enzyme to meal in which phytate had
already been substa ntially degraded by earlier enzyme
addition did not result in complete breakdown of phytate
(data not shown).
e f e r e n c e s
1 Webster, . E.J. Agr ic . Re s . 1928, 37,12 3-12 52 Oberleas,D. and Harland, B. F.J. A m . D i e t . A s s o c . 1981,
79,433-4363 Erdman, J. W. J. Am. Oi l . Che m. Soc . 1979, 56,73 6-7414 Cheryan, M. C R C C n t . R e v . F o o d S cL N u t r . 1980, 13,
297-3355 Cosgrove, D. J. R e v . P u r e A p p l . C h e m . 1966, 16,2096 Asada,K., Tanaka, K. and Kasai, Z . P l a n t C e l l P h y s i o l . 1968,
9,1957 Fontaine, T. D., Pens, W. A. and Irving, G. W. J. Biol . Che m.
1946, 16 4,487-5 078 Barre,R. and van Huot, N.Bul l . Soc . Chim . B io l . 1965, 47,
1399-14179 Erdman, J. W. and Forbes, R. M. F o o d P r o d u c t D e v . 1977,
11, 46-4810 Molina, M. R. and Bressani, R. D e v . F o o d S c L 1979, 2, 39-4511 Ohlson, R. and Sepp, R. in F o o d P r o t e i n S o u r c e s (Pirie, N. W.,
ed.) Cambridge University Press, 197512 Ohlson, R. andAnjou, K.J. Am. O i l C h e m . S o c . 1979,56,
431-43713 Rutkowski, A. and Koslowska, M. J. A m . O i l C h e m . S o c . 1979,
56,475-47714 Jones, J.D.J . Am. O i l C h e m . S o c . 1979,56,716-72115 N w o k o l o , E . N . a n d B r a g g , D . B . C a n . J . A n i m . S c i . 1977,57,
47516 D a v ie s , N . T . a n d O l p i n , S . E . B r . J . N u t r . 1979,41,591-60317 Lo, G. S., Settle, S. L., Steinke, F. H. and Hopkins, D. T.
J . Nutr . 1981, 111, 2223-223518 Shieh,T. R. and Ware, J. H.A p p l . M i c r o b i o l. 1968, 16, 1348-
135119 I r vi n g , G . C . J . a n d C o s g r o v e , D . J . A u s t . J . B i o l. Sc L 1974,
27, 361-36820 Bitar, K. and Reinhold, J. B i o c h i m . B i o p h y s . A c t a 1972,
268,442-45221 R e i n h o l d , J . G . J . A m . D i e t . A s s o c . 1975,66,3822 Harland, B. F. and Harland, J. Cereal Chem. 1980, 57, 22723 Knorr, D., Watkins, T. R. and Carlson, B. L.J. Food Sc i .
1981,46, 1866-186924 Sudarmadji, S. and Markakis, P. J. S c L F o o d A g r i c . 1977, 28,
381-383
25 Wang,H. L. and Hesseltine, C. W. i n M i c r o b i a l T e c h n o l o g y(Peppier, H. J. and Perlman, D., eds) Academic Press, NewYork, 1979, vol. 11, ch. 4, pp. 95-1 28
26 Uppstrom, B. and Svensson, R.J. S c L F o o d A g r i c . 1980, 31,651-656
27 A l l en , R . J . L . B i o c h em . J . 1940,34,858-86528 H a n d b o o k o f B i o c h e m i s tr y (Sober, H. A., ed.) CRC Publishing
Co., Cleveland, 196829 Reinhold, J. G. E c o l. F o o d N u t r . 1972, 1, 18730 Glass,R. L. and Geddes, W. F. Ce re al Che m. 1959, 36,18631 S k o w r o n s k i , T . P o l is h J. S o i l S c L 1976,9, 145-15032 Skowronski, T . A c t a M i c r o b i o l . P o l. 1978, 27, 41-4833 Yamada, K., Miroda, Y. and Yamamoto, S. Agric. BioL Che m.
1968,32, 1275-128234 Yamada, K., Minoda, Y., Kobayashi, T., Hidaka, Y., Matuo, H.
and Kobayashi, M.J. F e r m e n t . T e c h n o l . 1968, 46,858-862
35 Raper, K. B. and Fennel, D. I. Theg e n u s A s p e r g i l lu s Williamsand Wilkins, Baltimore, 196536 Greaves,M. P., Anderson, G. and Webley, D. M. B i o c h i m .
B i o p h y s . A c t a 1967, 132,412-41837 Pe e rs , F . G . B ioc he m . J . 1953,53,102-11038 Shieh,T. R., Wodzinski, R. J. and Ware, J. M.J. Bac te r io l .
1969, 100, 1161-1165
3 8 2 E n z y m e M i c r o b . T e c h n o l . , 1 9 8 3 , V o l . 5 , S e p t e m b e r