
Profesional - UABCSbiblio.uabcs.mx/tesis/TE079.pdf1 i Fue mi propðsito al realizar este trabajo...
128
UNIVERSIDAD AUTONOMA DE BAJA CALIFORNIA SUR ANALlSIS SISTEMATlCO BIOQUlMICO y EVOlUCION ENZIMATICA FUNCIONAL EN CUATRO ESPECIES DE CABRILLAS DEL MAR DE CORTES GENERO Ep inephelus SERRANIDAE EP INEPHELlNAE Tesis Profesional Que poro obtener ti 1iIuIe d BIOlOGO MARINO Presento LUIS GERARDO LOPEZ LEMUS Lo Paz B C s 1985
Transcript of Profesional - UABCSbiblio.uabcs.mx/tesis/TE079.pdf1 i Fue mi propðsito al realizar este trabajo...

UNIVERSIDAD AUTONOMA DE BAJA CALIFORNIA SUR
ANALlSIS SISTEMATlCO BIOQUlMICO y EVOlUCION ENZIMATICA
FUNCIONAL EN CUATRO ESPECIES DE CABRILLAS DEL MAR DE
CORTES GENERO Ep inephelus SERRANIDAE EP INEPHELlNAE
Tesis ProfesionalQue poro obtener ti 1iIuIe d
BIOlOGO MARINO
Presento
LUIS GERARDO LOPEZ LEMUS
Lo Paz B C s 1985

7
r t II I
E
3n

A Ml MADRE
A QUIEN DEBO TODO LO QUE SOY Y POR QUIEN ANHELO SER TODO LO QUE DEBO

LOS ESTUDIOS QUE DAN LUGAR A ESTE TRABAJO FUERO 1 REALIZADOS EN EL
CENTRO DE INVESTIGACIONES BIOLOGICAS DE BAJA CALIFORNIA SUR A C EN
LOS DEPARTAMENTOS DE BIOLOGIA MARINA BIOLOGIA EXPERIMENTAL y
APLICADA Y BIOTECNOLOGIA DURANTE EL PERIODO COMPRENDIDO ENTRE OCTUBRE
DE 1983 Y DICIEMBRE DE 1984

CONTENIDO
l PREFAC 10
11 111ICE DE FIGURAS v
I I I 11ICE DE TABLAS vii
IV LISTA DE ABREVIATURAS viii
V RESUMEN
VI
VII
A
C
D
VI I l
IX
INTRODUCC ION
ANTECEDENTES Y GENERALIDADES
Estudios genØtico bioquímicos en peces
B Especiación y evolución en peces estudiadas mediante el
anÆlisis elect ofo Øtico de p oteinas 15
Inter acc icr es genoma med io amb ierte
Adapt c ión b ioqu imica tempera tu a
MlTER 1ALES y METODOS
2
3
Especies analizadas Y sitios de colecta
Preparación de los extractos tisulares
Electroforesis y tinción de los geles
4 CÆlculo de f ecuencias gØnicas distancias genØticas y
construcc ión de 1 dendrogr ama 36
5 Purificación del isozima lactato deshidrogenasa del
mœsculo esquelØtico M4 LDH 37
6
7
Estudios en la cinØtica de los isozimas M4 LDH
React ivos sus1 arlc ias
RESUL TADOS
1 Datos rroOrfomØtr icos mer íst icos
3
8
8
18
21
27
27
28
30
38
40
56
58

2
3
4
X
1
2
3
4
XI
1
2
3
4
Anålisis electroforetico y distancias genØticas 56
PUl ificlIción del isozima M4LOH
57
Estudios en la cinØtica de los isozimas M4 LDH 57
01 SCUS ION Y CONCLUS 1 OIES 74
l1ed ic iones py oporc ionales 74
Calacteres mer sticos 75
El estudio electroforØtico las distancias genØticas y la
s istemÆt icll de Ep inephe 1 us 76
Estud ios en 1 a c ine1 ica de los isoz imas M4 LDH 82
APENO ICES 8S
Isozimas homomØl icos y he1 elomºricos 8S
El cÆlculo de las frecuencias gØnicas y genot picas S4
Distancia genØtica en1 re poblaciones naturales S6
CinØt ica de sa1 uración enz imå1 ica 100
4a Eficiencia catalltica 101
XII LITERATURA elTfDA 104

PREFACIO
jl erencia de la manera de pen ar tradicional en la biología
predarwinista que abordaba la investigación de la diversidad orgÆnica
de manera descriptiva basÆndose en el estudio de caracteres tomados
como rasgos estables en unos poco individuos el objetivo del biólogo
evolucionista moderno es el estudio causal de esta diver idad Dentro
de se enfoqlJe las idea s centr ales son la de población como unidad de
an li s 1 as de la ar iac ion y la adapt ac ión como fenómenos
aracterlstlcos de las poblaciones
studio de l a IJar iac ión es patr imon io de la genŒtica de
0tlac onea y el estudio de la adaptación se incluye tradicionalmente
p el lrplio terreno de la ecolog a las exigencia del anål is is han
determlnado que ambos estudios hayan tenido un tratamiento
re1 at ivamente independ iente nasta hace pocos aæo Pero e tamos en
pre encia de un momento en el de arrollo de la teorla biológica en el
que se ha ad Jert ido l a nece idad de e tudiar ambos fenómenos como
di 5 t ird c s as p ect os de ur pr oceso un ltario que se man if ies ta en el seno
de las poblaciones naturales De ahí el auge creciente de una biologta
poblacional en la que concluyen la genØtica y la ecolog a de
poblaciones en la bœsqueda de expl icaciones cada vez mÆ profundas de
las causas y de los resultados de las interacciones que tienen lugar en
el seno de las poblaciones y entre estas y el medio ambiente

1 i
Fue mi propðsito al realizar este trabajo tratar un aspecto del
fenómeno de la variacio elect oforØtica y cinØtica de enzlrr s en peces
ma P inos y Sf coyre1 ac iór e01 rl 1 a adap1ac iór Qu i e 11 amar la atencIón
s obr e 1 as POI lb11 idıodes que S lHgen cUıo ndo s e tratan estos fenómer os
dentro del mar cü de confllJerlcia genØti o ecológica eæalada i desta1 ar
11 misrr tiempo algunos problemas de la signIficación de 1 a
adaptaciones bioqy micas y el anålisi electrofóretlco en fiSiología
compa r ada y taxonomía respect ilJamente que todav ˝a no t an rec îb Ido urja
explicaclðn adecuada y que quiz4 puedan ser esclarecidos a travØs de
un in estl3aclón biológico poblacional integrada
Agradezco al Centro de Investigaciones BIOlógicas de Baja CalIfornia
Sur A C CIB el apoyð acad mico econ6mico y admlr istrat ivo
recibid s dlrante la elaboraclón de este trabajo En la misma medida
agradezco al Consejo Nacional de Ciencia y Tecnología CONACyT el
soporte financiero recibido a trav s de una beca tesIs y un convenIo de
apoyo CIB CONACyT No Reg 44182 y PCECCNA el2elS63 respectivamente
Quiero agradecer a los doctores George N Somero Oonald W WIIKIe y
Jeffrey B Graham de la Institución Scrlpps de Ocearograf fa
Uroiversidad de Cal ifornla Sa Olego p su ayuda en la
Ordeterminación definitiva de los objetivos de este trabaja El
Somero facilitó en calidad de prØstamo la col urnr a cromatogråfica
utilizada en este estudio
Agradezco al Dr F lix Córdoba Alva el permitirme utilizar espaCIO

equipo y reactivos de sus laboratorios aslmismo por brindarme
iii
la
Aloportunidad integrarme a su equipo de trabajo en el aæo de 1980
Dr JosØ Luis Ochoa por sus consejos y observaciones Al Dr Francisco
X Arredondo Vega por la asesorla t4cnica recibida durante
inicial del desarrollo de esta investigación
la etapa
A las siguientes personas que me permitieron realizar la parte
correspondiente al trabajo de campo durante sus salidas de colecta
quiero dejar asentado mi sincero agradecimiento
Biólogos Benito 8ermœdez Guadalupe Garc a JosØ de La Cruz Oscar
Tala era Hornero ROdrlguez Garza Jesœs ROdr guez Rorner o Octavio
Maravilla M en C Enrique Gonzålez y al equipo de lancheros de la
U A B C S
Por su paciente e invaluable ayuda en la mecanograf a del manuscrito
expreso mi gratitud a la Srita Cenobia Castillo y en la preparac ión
de las ilustraciones al Sr JosØ A Martinez y a los biólogos Teodoro
Reynoso Carlos Sepœlveda y Carlos Sånchez
Agradezco de una manera muy espeCial al B ió l Felipe Ascencio su
e tusla ta colaboración en la realización de los proced imientos
cromatogråficos aquí descritos Su ayuda jugó un papel
en el desarrollo y finalización de este estudio
muy i ortante
Quier agr adecfor al Biól Jon Elorduy el haberse tomado la molestia

i 1
de r e isar correg ir d la Mejor forma la pr imer a er Ion de este
1 r b 50
Finalmente hago un reconocimiento espec al para el Dr Daniel LI uct
Belda quiØn grðdezco su orien acì n apoyo e InterŁs en mi
f ormac i Ó n y deser lperJo n i i f ï cos al B I Ó l Francisco de Lachlca
Bor i lla por l as val iosas d isc I i es y punto s de v ista durante la
integraci6n final del presente estudio y al Bi l Joaqurn Arvizu por
sus obs vaciones y sugerencia
A la Srita Ana L z Alvar ez y te do aquello amigos q Je me apoyaron
Øl cord inIa r ce n este tr abajo t asta el f iral les estare s iempre muy
agradeCido

y
INDICE DE FIGURAS
1 Efecto de lŁ variación en la afinidad enzima sustrato 26
º Epinephelus Cephalopholis panamensis 41
3 Epinephelus Epinephelus acanthistius 41
4 Epinephelus Epinephelus labriformis 44
Epinephelus Epinephelus analogus 44
E Local izac ión geogråf ica de los s it ios de colecta48
7 Localización geogrÆfica de los sitios decolecta 4S
8 Pez con espinas y radios ilustrando las partes Y todos
de eonteo Y me d i c ion 50
S Aparato de electroforesis de fabricación domØstica 51
Sistema de tinción para MPl
SÜtema de 1 inción para ME
Sistema de tinción para AOH
Sistema de tinción para GPI
Sistema de tinción para LOH
Sistema de tinción para MOH
Sist@ma de tinción para PGt4
Sistema de tinción para 500
Sistema de tinci n par a GOH
IS Fenograma de identidad genØtica entre 4 especies de
cabr illas del Mar de Cort s 68
20 Analogfa estrJctural entre el piruvato y el oxamato 6S
21 SeparaCión de los isozimas M4 y H4 del tejido muscular 70

ˇ
22 Efecto de la temper atur a de ersayo sobre el Km con
respecto al piruvato para los M4 LDH de E analogus
E paramers iE ac anih i sti us 72
23 Der dr gr mas comparati os q Jo rf estran las relaciones
filogen ticas de cuatro especies de cabrillas del Mar de
Cort 88
24 Patrones electrofor ticos que se observarfan en c so de
mono di tr i tetrå meroE 93
25 Velocidad de reac iln graficada contra la co cen raci n
del sUEtrato para lIna reaccl n qlle obedece la cin tlce de
Michae 1 is t ente o de s turac ión 103

vii
INDICE DE TABLAS
l Sistemas enzimÆticos resueltos con Øxito en cuatro
especies de Epinephelus 2
2 Caracteres genØricos y sUbgenØricos de Epinephelus 9
3 Caracteres importantes de las especies de Epinephelus 60
4 Medidas proporcionales de 10 espec menes de E panamensis 61
5 t1ed id s proporcionales de 10 de E acanthistius 62espeClmenes
Med ida proporcionales de 10 espec menes de E 1 abr iformis 63
Medidas proporcionales de 10 espec menes de E analogus 64
Frecuencias alØl icas de los 1 oc i poi imÓrf icos en y entre
6
7
8
cuatro especies de cabrillas del Mar de CortØs 65
9 Oistanci s genØticas y error standard entre cuatro especies
de cabrillas del Mar de CortØs 67
10 Valores de Km con respecto al piruvato para los M4 LOH en
E ac anth i st i us 71
11 Valores de Km con respecto al piruvato para los M4 LOH en
E analogus y E labriformis 71
12 parÆmetr os cin˝tiC lS para los M4 LOH purHicados de tres
especies de cabrillas del Mar de CortØs a 25 C y a las
temperaturas media de los habitats de colecta 73
13 Estructura de las subunidades de isozimas homo ricos y
heter1mØ r ieos en he teroc i 9 ot os 92

LISTA DE ABREVIATURAS
vii i
A
AOH
cm
o
O glucosa
ONA ADtP
E C C E
EBT
EOTA
ft
GOH
GPI
9
g
G l S Dif GOP
GSP
GSPO
HCl
in
Kcat
Km
K fosfato
lOH
Aleta anal
Alcohol deshid ogenasa
centímetros
Aleta dorsal
Forma dextrorrotatoria del monosac rido glucosa
Acido desoxi ribonuclØico
Consejo Enzimtico Enzyme Council
EDTA Bor atos Tr is
Acido etilen dinitrilo tetra ac tico
Pies
Glucosa deshidrogenasa
Glucosa fosfato isome asa
g amos
Aceleración de la gravedad
Glucosa l S difosfato
Glucosa S fosfato
Glucosa S fosfato deshidrogenasa
Acido clorhídrico fórmula del
Pulgadas
Constante de la tasa de catålisis enzimÆtica
Constante aparente de Michaelis
K2HP04 Fosfato diPotåsico
lactato deshidrogenasa

Ix
M Mol ar
t—H Malato deshidrogenasa
I—Hm Forma mitocondrial de I—H
t—Hs Forma somÆtica o citosòlica de I—H
IE Enz ima mÆl ica
MgCl2 Cloruro de magneSio fórmula del
tfI Manosa fosfato isomerasa
MTT Metil tiazolil tetrazolio
m metros
mg mi 1 igramos
lNI mlllamperes
mi mil il itros
mm milimetros
mM mi 1 imo 1 ar
Al microl itros
JM micromoles
NAO Nicotinamida aden n dinucleótido forma oxidada
NAOH NAO forma reducida
NlOP NAO fosfato forma oxidada
NAOPH NAOP forma reducida
N2EOTA EOTA sal disódica
N fosfato Na2HP04 Fosfato disódiCO
nm nanometros
02 Ox geno fórmula de la molØcula de
PC Fosfato c itrato
PGM Fosfoglucomutasa

x
SPG
PMS
Ql0
6 fosfogluconato
Fenazina metosulfato
Se explica en el texto
rpm revoluciones por minuto
Solución amortiguadora para la muestraSB
500
SP
Te
TEM1
sal ie buffer
Superóxido dismutasa
vmax
Sodio fosfato
Tris citrato
Tris EOTA MgCI2 Anhtdrido malØico
Tris hidroximetil aminometano BASE
Unweighted paired group method using arithmetic
averages MØtodo de agrupación pareada no
sesgado que ut il iza med ias ar itmØt icas
Unidades por mililitro
Voltios
velocidad
Velocidad mÆxima
Tr 15
UPGMI
U mi
v
v

RESUMEN
Cuatro especies de cabrillas y r ros gØnero Epinephelus con areas
de distribución solapadas en las regiones media y baja del Mar de
CortØs fueron estudiadas en tØrminos de caracteres
genØtico bioquímicos y funcionales de enzimas El anÆlisis
electroforØtico de estos cuatro congØneres reveló que la especie
Epinephelus Cephalophol is panamensis encuentra justificada su
pOSición dentro de Cephalopholis en base a las diferencias observadas
en los caracteres anatómicos con respecto al resto de las especies sin
embargo sostiene un estrecho parentesco genØtico con labriformis y
el resto del tronco Epinephelus lo cual sugiere una divergencia
reciente de tal tronco Se propone un esquema alternativo en la
filogenia del grupo en base a estos estudios Estas dos espeecies
cercanamente relacionadas se encuentran igualmente emparentadas con E
analogusJ asímismo ellas tres se encuentran igualmente distantes de la
garropa del bajo Golfo acanthistius Las distancias genØticas que
separan a las tres especies templadas y subtropical cercanamente
relacionadas sugieren la ausencia de contacto genØtico entre ellas
durante los pasados 3 5 millones de aæos aproximadamente
Los resultados de los ensayos enzimÆticos con lactato deshidrogenasas
electroforØticamente diferentes del tejido muscular M4 LDH de las
cabrillas tropicales subtropicales y templadas mostraron diferencias
termocompensatorias en las propiedades cinØticas Km aparente con

2
respecto al piruvato la constante de la tasa catalCtica Kcat Estas
adaptaciones cinØticas se reflejan en una fuerte conservaClon de las
caracterfsticas funcionales del enzima a las temperaturas medias de los
habitats de las especies que difieren por 5 S e en promedio Estos
resultados Indican que tan sólo diferencias menores en la temperatura
del habltat cuerpo son suficientes para favorecer la evolución de
adaptaciones funcionales en los enzlmas El uso de especies
congenericas cercanamente relacionadas habitantes de entornos
tØrmlcamente diferentes en estudios de los procesos de adaptación
molecular a escala fina constitu en una valiosa aportaclôn al estudio
de la evolución molecular

3
INTRODUCCION
El en40que principal de lo estudios evolutivos recientes se ha
r ient ado hac h el pape 1 que desempeæan 1 as var iantes enz i t Icas
hoz lmas y aloz lmas en 1 a adaptac 16n med io amb lental y la evo 1 uc lôn
de las espec ies Lewont in 1974 Ayala 1976 Hedr iCK 1 l 19761
So ro 1978 La b Squeda de respuestas a esta Inquietud se ha
abordado desde dos niveles experimentales di4erentes en tØrminos de
cistancias evolutivas entre las especies enzlmas que se estØn
comparando Por una parte se han estudiado los enzimas de organismos
ampliamente distanciados por ejemplO mamf4eros y especies ectotØrmicas
l ow 1973 Borgrnann y Moon 1975 Borgmann II l 19751
Johns ton y Wa 11 esby 1977 Somero 1978 Somero y S iebenall er 1979
Por otra parte muchos intentos se han hecho para determinar cual es el
alor selectivo de las distintas variantes alØlicas de los enzlmas
alozimas en poblaciones de especies œnicas expuestas a di4erentes
cond ic iones med io amb ientales por ejemplo tØrmicas Kohen 1969
Merritt 1972 Place y Powers 1979 Y asI mientras ambos tipos de
estudios han permitido la realización de hallazgos en relación a los
patrones bÆsicos de evolución roolecular el papel de las variantes
enzi ticas en I adaptación medio ambiental a escala 4ina persiste
aun en controversia
Existe una tercer a 40r ma complementar ia para abordar el problema
Esta involucra el examen de enzimas de especies congenØricas

4
cercanamente relacionadas con ecolog as similares pero ocupando
habitats que difieran en temperatura presión Siebenaller y Somero
1978 1979 Somero y Siebenaller 1979 o alguna otra variable capaz
de afectar la función enzimÆtica Las especies congenØricas proveen de
la suficiente divergencia evolutiva para permitir la visualización del
papel que realizan diferencias nzimÆticas funcionales mlnimas pero
significativamente selectivas con un relieve mås claro Y definido Es
as como a diferencia de una situación que involucra a diferentes
poblaciones de una sola especie en la que las d iferenc ias alØl icas
fijadas en las diferentes subpoblaciones no pueden establecerse por
razones de tiempo evolutivo o el flujo genŁtico entre los grupos los
congØneres que habitan medios o ambientes distintos Y que poseen
diferentes formas alØlicas fijadas de una clase particular de enzirM
proveen de un excelente sistema de estudio para explicar cuÆl es la
importanCia de las variantes enzimåticas
ambiental a una escala relativamente fina
en la adaptación medio
De los factores frsicos que frecuentemente varían entre los entornos
de especies cercanamente relacionadas la temperatura es uno de los mås
importantes fisiológica Y bioqufmicamente Existen numerosos casos de
reemplazo de especies a lo largo de gradientes t rmicos en el med io
ambiente marino espeCialmente en organismos ectotŁrmicos Hubbs
1948 El rnan 1953 Como 1 a temperatur a es uno de los pocos par`metros
medio ambientales que afecta directamente a los organismos a nivel
molecular adem s de ser una fuerza selectiva importante en la
evolucion de las proternas segGn se ha demostrado Somero 1978

Hc c I chla y Sorner 1973 1984 podr fa se r que especies congenØricas
c rcaname t elacionadas habitantes de entœrnos con ligeras
diferencias t rmicas proveyeran de una excelente oportunidad para
i uest igar los efectos de peqt eias d iferenc ias med io amb ientales sobre
l Ł c ll ió mol2 1 1 1 funcic nil Y sobre ciertos eventos de evolución y
especiaci6n de las especies involucradas
Los mers y cabrillas del g nero Epinephelus habitantes del ar de
C rt s v de tros res tropicales y subtropicales del mundo proveen
de tal lTodelo pat el estudio de este tipo de pl blerras Las cuatro
especies de Epinephelus consideradas en este trabaja son similares
e 11 óg il merlt c lrllpar ten un numerCt cClns ider able de caracteres
morf 169icos Todas ellas son sol itarias o bien gregarias en algunas
reas ð cier as hor s del día o bien en ciertas pocas del ario
Habitan los arrecifes y sor especies predadoras Smith 1971 Thomson
ø 1979 Srnith 1371 a s bdividido al gran genero Epinephelus
en cinco IJbg e r ero Ep lrephe us Pr ortl i er Ci P S Ce phalopho1is
Dermltolepis y Alphe tes de los cuales Epinephelus contiene a tres de
lðs especies s diadas Y Cephalopholis a la otra Sin embargo debe
Yloi arse ql JE 1 cl sific ci r genår ic de la famil u roo estÆ
e t bil i d y la dis cu ión respecto œn per iste Gosllne 1966
rr i h 1371 Jo dirìs Cln 1983 Randall y Ben TLlvia 1983 Johrson y
ftt r er 1 84 L s Æ ea de di tribyción de estas especies se
r I It r rJ Ol F j 11 p a de esto l abundar e ia de las especies
ir de J rea ð otras TlcIMsor 1979 de tal forma fue
F it le detect r q Je las terrp r tlJr as promedio tu e estos organismos

6
exper imentan d H i@ren El inephe I U El inephe I us analogus y f
Iº tr iformis 18 C E Cephalphol is panamensis 230 C y E E
acanth ist i Js 24 26 C U S Oepar tment of Cornmer ce and Geodet ic
Survey 1952 López Lemus 1984 dato no publ icado
En este trabajo se reportan los resultados de una invest igac ión
electroforØtica de estas cuatro especies de cabrillas el cual ind ica
que han estado separadas por lo menos 3 4 millones de aæos y que el
gØnero Cephalopholis sostiene un estrecho parentezco genØtico con
Epinephelus de acuerdo a las similitudes observadas en las frecuencias
de sus loci enzim ticos con respecto a los del resto de las especies
estudiadas por lo cual se propone un esquema alternativo en la
filogenia y sistematica del grupo Se realizaron estudios relacionados
con el papel de la adaptaci n enzim tica a las variaciones t rmicas a
un nivel de escala fina para uno de los loci genicos en el cual se
detectaron diferentes formas alØl icas fijadas y que codifica para el
isozima del rr sculo esquelØtico M4 o A4 lactato deshidrogenasa LOH
E C 11127 NAO lactato oxidorreductasa Los patrones adaptativos
en la constante aparente de 1ichael is Km con respecto al p iruvato y
la constante con respecto a la tasa cataltica Kcat lnd lcan que las
diferencias en la temperatura de tan s610 algunos grados cent grados
sor suf ic ientes para selecc ionar d ifet encias termocompensator ias er las
propiedades cineticas de los enzimas Estas adaptaciones rr leculares de
las cabrillas se discuten a la luz de las afinidades morfológicas y
genØticas de las distintas especies en cuestión que son I as mas
cOmÚnes dentro del gØnero en el Mar de CortØs Thomsoni 1979 y

7
c n el pape 1 de las f I uctuac iones en Ia te eratura de I Plloceno y
Pleistoceno que pudieron haber influido en la formaciÓn y distribuciön
d estas especies

s
ANTECEDENTES y GEt ERAL1DADES
Ningœn otro grupo de vertebrado experimenta las amplias variaciones
arrbientales que se observan para los peces En el caso de la
ter eratura chos peces de altas latitudes sobreviven cont nuamente a
temperaturas cercanas al punto de congelación del agua de mar
aprox imadamente 18 C Los pecesillos del desierto genero
Cpr inodon del suroeste nortearner icano hab itan aguas con temperaturas
de 4el 43 C Brown Feldmeth 1971 Los peces de las grandes
profund idades tienen temperatur as corporales t p icas cercanas a los
2 4 e ademÆs tales especIes pueden vivir soportando presIones de
varios cientos de atmósferas Los peces son especiales entre los
ver tebrados por su ampl ia gama de es trateg ias osmor r egulator ias
d gase en el contenido osmótico total los tipos de osmal itos
ut lizados en la regulación os tica revisión de Yancel 1982
Los teleósteo dulceacurcolas tienen un contenido osrr tico total
aproximado a un tercio de aquello marino mientras que los
elasmabranquios marinos tienen los flu dos corporale 1 iger amente mas
concentrado que el agua de mar Los peces difieren adem s en el estilo
de vida por ejemplo en sus estr ategias al imenticias capacidades
para poderosos rompimiento locomotores Los atunes por ejerr lo
tienen capacidades locomotoras tremendamente desarrolladas y deben
nadar contfnuamente para satisfacer sus requerimientos de ventllacl n
forzada En e 1 otro ex tremo lCho s peces ab I s ales son perezos os s e
posan e peran o flotanesperan mIentras se alimentan teniendo

8
Icapacida des limitadas para la locomocion Sullivan y Somero 1880
E tas diferencias en los hÆbitos alimenticios Y de locomoción se
reflejan en una arr 1 ia gama de capacidades distintas para la glucóliSis
Y Asc Alar y en necesidade var iantes para asimilar productos finales del
metabolismo Castellini y Somero 1981
Dil do la extr ema diversidad en 105 atr ibutos biológicos Y ambientales
de los peces este grupo de vertebrados ha constituido una colección
ITllI part icul or de e r garl is me s d ispon ibles para el estud io b ioqu mico y
fisiológico de la adaptación De hecho cada una de las caracter sticas
1 istadas arriba terr eratura presión hidroståtica concentración
osmØ t i a y campos ic ión de los osmol itos y hab illdades locomotr ices se
espera Sean manifestadas en el diseæo de los sistemas bioqu micos de
los peces especi lrnente en 1 as propiedades de los sistemas enzimÆticos
contr ctiles El enfoque del presente trabajo es hacia el pr imer
sistema poni ndose especial nfasis en la naturaleza de las principales
estrategias adaptativas que facilitan la conservaclon de 1 as
propiedades m s crIticas en las proternas de acuerdo a las condiciones
tØrmicas rooientales de las espeCies aquí tratadas conSiderando de
Igual forma sus finidades morfológicas y genØticas con el tiempo
voluti Jo
A Estudios genØtico bioqulmicos en peces
Una r ue Ia d lmens ión er el entend imiento de lavariac ión pr oteica tuvo
su base en dos ever tos de le s aæos Seis El modelo de la estructura de

10
la molØcula del DNA propuesto por Watson y CricK 1953 a y b llevó a
la comprensión de la relación directa entre genes y proteínas La
electroforesis en matrices de gel de almidón Smithies 1955 apoyada y
meJorada por la aplicaci6n de metodos de tinción histoquímica Hunter y
MarKert 1957 sumaron simpleza y sensibilidad al estudio de esta
variaci6n Estos eventos condujeron a una explosión de información al
respecto de la variación en las proteínas y su significado que empezó
en los S0s y ha continuado hasta nuestros días El estudio de la
variación genØtica a nivel de proteínas ha hecho contribuciones
importantes a una diversa gama de disciplinas orientadas hacia la
biología Muchos peces se han estudiado electroforØticamente revisado
por de Ligny 1971
Los problemas actuales de investigaci n en esta Ærea tienen sus
raíces en la mitad de los aæos 50s cuando George Ridway del U S
Sureau of Commercial Fisheries ahora el National Marine Fisheries
Service propuso que el salm6n del Pacífico Oncorhynchus que se
capturaba en alta mar pod ía ser identif icado de acuerdo a su or igen
continental por d iferenc ias detectables serológicamente que
reflejarían supuestamente diferencias genØticas El problema fue
abordado a travØs de estudios en las proteínas del suero y antígenos en
erUroc Uos
Se detectaron variantes en las prote nas sØricas en el salmón O
utilizando inmunodifusión e inmunoelectroforesis con antisuer os
obtenidos en conejos principalmente Una variante llamada A o ant geno

11
51 f encor trada en hembras madur as de todas las espec ies de
Col rnón ido examínºo das y se pr obó su ut il idad para identificar madurez
ma IIe para estudie r aciales Un par de antlgenos sØr ícos llamados
I y 11 que pa r ec leron Jar iar rnarcadamente en su d istr ibuc ión entre los
pece de arigen s i it ice amer icano const ituyeron una gran esperanza
para poder aSignar el origen continental de estos peces capturados en
el ocØar o ab ierto Sin embar go las dificultades para producir
cantidades adecuadas de antisueros potentes y espec ficos junto con
fuerte indi aciones de que estas variaciones reflejaban artefactos de
p eser c ión d iferen ial mÆs que d iferencias gen4t icas vÆl idas
ondujeron a descontinuar el estudio de estos anttgenos
Lo ant genos en er itroc itos se examinaron junto con una gran 1 ista
de sust ncias hemaglutinantes incluyendo sueros normales de una gran
la t ied d de animales fitohemaglutininas sueros heteroinmunes
xenogenØic s e iso inmunes alogenØicos s lo con los suerOS
iso inmunes se obtuvo una cierta promesa real para unaf
identificacion
confiable de diferencias genØticas en las especies del salmón del
PacIfico Estos reactivo producidos en la trucha arco iris
galrdneri se util izaron en la identificación de ciertas diferencias
entr e las poblaciones de distintos salmones para ind icar el
entrecr Jzarnier to en una poblaCión de la trucha S clarKi Sin embargo
estos antisueros eran difrciles de producir y purificar en las
cantidades decu das sus patrones de reaccion eran muy complejos y
hab a una p rdida de potencia en lo mismos despuØs de su
lrr cenamiento en congelaCión de un aæo al Siguiente

12
En una reuisi6n detallada de los estudios descritos Hodgins 1972
cot1lUYó
an hecho avances Significativos en conceptos al respecto de la
na uraleza de las poblaciones de peces como resultado de estudios
serológicoS e inmunoquímicos La magnitud de dos grandes problemas
milita en contra de futuros estudios e tensivos al respecto en la
actual idC d 1 problemas tØcnicos tales como la dificultad de
producir cantidades suficientes de antisuero de alta actividad y la
naturaleza crítica de la preser vación de las muestras y 2 problemas
teóricos tales como relacionar los grupos sanguíneos al genotipo Sin
embargo estos procedimientos pueden utilizarse en la diferenciación de
stocKs bajo ciertas condiciones
DespuØs de una dØcada de Øxito limitado y frustraciÓn considerable a
trav s del uso de mØtodos serológicos e intentando la identificación de
diferencias gen ticas en las poblaCiones de peces se hizo necesaria
una nueva forma para abordar el problema As se volvi6 toda la
atención hacia la electroforesis en matr z de gel de almidón
Pronto se hizo aparente que este era el mØtodo a escoger sobre
cualquier otro mØtodo previo que se hubiera utilizado para la
identificación de variaciones genØticas intra especie en
que se estudiaban
las especies
La ventaja principal de los datos genØtico bioqu micos que se

13
obtienen a travØs de la electroforesis en gel en comparaci n con los
datos inmunológicos es que se pueden hacer interpretaciones validas
directamente de los datos sin procesar crudos La expresión
codominante de la mayorla de los alelos variantes se observa en los
geles de almid n posteriormente al corrimiento electroforØtico es
decir en un individuo que contenga diferentes alelos de un locus dado
cada uno de los alelos se expresarÆ como una sola y distinta prote na
Tal situación permite co nmente la designación de los genotipos en
muestras individuales basadas en los patrones de tinción sobre los
geles La frecuencia con la que un gen dado ocurre en una poblaci n de
individuos puede determinarse directamente Y la distribución de los
fenotipos probarse contra las desviaciones de valores esperados
basados en modelos genØticos simples siendo uno de ellos el modelo de
Hardy Weinberg Este establece que en una población de individuos que
se aparean al azar en la ausencia de una cierta variedad de fuerzas que
lo alteren p ej selección mutación etc la distribución esperada
de los genotipos estarÆ determinada por la combinación al azar de sus
alelos
En el caso de dos alelos A B la frecuencia genot pica es asl
q 2 AA I 2q l q AB I 1 q 2 BB
Donde q es la proporción de los alelos A en la poblaciÓn Pueden
hacerse inferencias a partir de estos datos crudos al respecto de la
estructura de la protelna en t rminos de las subunidades que la

14
componen basandose en el numero de bandas observadas para un sistema
particular en individuos heterocigotos Esto se dicute en detalle en el
apØndice No 1 Otras dos ventajas pråcticas del mØtodo de gel de
almidón son que no se requieren antisueros y que la preparación de la
muestra asi como su preservación es mucho mÆs Simple Las ventajas
generales de esta metodologla sobre otras resultaron en un enorme
diferencial de información genØtica al respecto de la variacion
intraespecIfica por unidad de esfuerzo obtenible comparando los mØtodos
anteriores Y el descrito aquí Como un ejemplo hipotØtico pero basado
en la experiencia algœn investigador que se encuentre examinando
especies jamÆs estudiadas previamente pOdría obtener mayor cantidad de
información gen tica vålida en un dIa utilizando mØtodos
electroforØticos en gel de almidón que lo que todo un equipo de
investigadores pOdría obtener utilizando mØtodos de grupos sanguíneos
serol gicos o inmunoquímicos
En 1968 la principal porción de los esfuerzos que se dedicaban hacia
estudios genØticos se volcaron hacia los mØtodos electroforØticos y de
tinciÓn histoquímica Los primeros estudios en esta fase resultaron
sensacionales pues a n no se acostumbraba encontrar las variaciones
genØticas con la facilidad hecha posible gracias al cambio de
metodolog as A partir de aquí cantidades sustanciales de variación en
las proteínas entre las pOblaciones y en las poblaciones se
demostraron con tØcnicas electroforØticas revisado por Powell 1975
Y la existencia de variantes alØlicas que no pueden distinguirse por
todos electroforeticos standard tambiØn han sido reportadas en

IS
comparaciones de poblaciones y especies Johnson 1975 Siebenaller y
Somero 1978 Bernstein 1l 1973 El papel de estas yariantes
alØlicas detectables y crtpticas se ha transformado en el principal
objeto de estudio en el desarrollo del
contemporÆneo Lewontin 1974
conocimiento eyolutiYo
B Especiación y evolución en peces estudiadas mediante el
electroforØtico de protelnas
Para aquellos organismos gonocortsticos y con reproducción sexual
anål is 1s
tales como los peces el concepto de especie estÆ basado en el
aislamiento reproductivo de los grupos de poblaciones que se
entrecruzan yerdaderamente con respecto a otros En la prÆctica las
especies casi siempre de distinguen y describen sobre la base de
diferencias anatómicas Es razonable esperar que casi todas las
especies reconocidas actualmente sean morfológicamente distintas unas
de otras dada esta prÆctica Sin embargo la diferenciación anatómica
no constituye una base ni necesaria ni suficiente para el
reconocimiento de especies separadas La literatura esta llena de
ejemplos de especies que exhiben polimorfismos anatomicos dram1ticos
que son aœn conespeclficos y ejemp 1 os de comp 1 ejos de especies
morfol giCamente crlpticos que son de hecho unidades gen ticas
independientes Grassle y Grassle 1976 Borden l 1977 Gould et
l 1975 Salmon 1979
Un criterio alternatiYo para el reconocimiento de especies distintas

16
aquel de aislamiento reproductivo parece obvio dada la anterior
de inición de especie Sin embargo este criterio es debilitado por
numerosos ejemplos de entrecruzamiento ocasional entre especies bien
reconocidas De hecho tal hibridación interespeci ica bajo condiciones
naturales o de laboratorio est muy bien documentada para los peces
Schwartz 1981 y para otros grupos de animales a tØcnica de
electro oresis en gel para prote nas provee de un examen poderoso
aunque indirecto de la validØz de las especies como tales Dado que
esta t cnica permite la medición de la relaci n genØtica entre los
individuos debido a la expresión codominante de la mayor a de los
alelos en los loci analizados puede servir como un medio para
determinar la exclusividad gen tica de cualquier conjunto de organismOs
por ejemplo la identi icación de distintas especies Este modo de
abordar el problema es particularmente uerte en casos de simpatrià
verdadera en espacio Y tiempo En tales casos las espeCies
genØticamente diferenciadas son åcilmente reconocibles cuando se
detectan di erencias al licas fijadas Aquellas poblaciones simpåtricas
caracterizadas por diferencias al licas fijadas habrÆn evolucionado
tales poblaciones deben considerarse como
reproductivo Asi
especies biológicas
clara y e ectivamente por medio de aislamiento
verdaderas Por otra parte las observaciones de uniformidad genØtica
ya sea en tØrminos de distribuciones de recuencias alŁlicas similares
entre las muestras o espeCialmente en tØrminos de los loci
invariablemente idØnticos en todos los individuos son consistentes
con pero no establecen e ectivamente la naturaleza conespec ica de
tales poblaCiones Graves y Rosenblatt 1980 Sage y Selander 1975

17
Turner y Grosse 198e1 Manooch et 1976
En aquellos casos de alopatrta en espacio y tiempo las
distinciones descritas se vuelven confusas debido a efectos
cofundadores potenciales de diferencias geogrÆficas o temporales en la
composición alØlica de los organismos Geogråficamente esto podrta
tomar la forma de clinales aparentes en la frecuencia al lica o dado
un muestreo discontInuo en tiempo y espacio pOdrían aparecer aœn como
diferencias alØlicas aparentemente fijadas entre las muestras
Aspinwall 1974 Powers Y Place 1978 Se debe tener por ello
extremo cuidado al interpretar tales datos de muestras alopåtricas pues
el aislamiento reproductor resultante de alopatrIa espacial o temporal
puede no tener base biológica alguna Es mejor dicho muy dif1cil
determinar si las poblaciones alopåtricas pueden o no entrecruzarse si
se estableciera el contacto bajo condiciones naturales
Ademas de proveer una medida lo suficientemente fuerte de las
relaciones reproductivas de pOblaciones simpåtricas la aportación
electrofor tica brinda otros beneficios Uno es la identificación
segura de h bridos interespectficos Fl entre dos especies con mÚltiples
diferencias al licas fijadas Esto es resultado directo de la expresión
codominante de los alelos característicos de los loci y es una mejora
clara sobre le USO del criterio morfolÓgico que es generalmente menos
poderoso debido a un tipo de herencia mezclada cuantitativa en la
mayoría de las caracterlsticas anatómicas Un segundo beneficio del
anålisis bioqulmico de las espeCies se deriva de la hipótesis del reloj

18
molecular Nei 19711 Kimura y Ohta 1971 Maxson y Wilson 1974
Wilson l 1977 Carlson 1978 que asume que las proteínas
evolucionan a un ritmo relativamente constante AsI con la propia
calibración es posible estimar el tiempo aproximado de divergencia en
cualquiera de las especies que se analicen Gorman y Renzi 1979 Wyles
y Gorman 1980 Vawter i al 1980 basado en los valores de
distancia gen tlca derivados de los estudios electroforØticos Nei
1972
C Interacciones genoma medio ambiente
En al actualidad existe una cierta incertidumbre al respecto del
papel de la selección natural en el mantenimiento de polimorfismos
bioqu micos en las poblaciones La gran cantidad de polimorfismo
observado en los organismos se ha explicado en base a la selección
PraKash l 1979 y otros factores debidos al azar King y JuKes
1969 La utilidad potencial del polimorfismo genØtico bioqu mico
pOdría extenderse considerablemente con el mejor entendimiento del
grado en el cual las formas alØlicas de distintas proteínas interactœan
con los componentes del medio ambiente Las variantes pOdrían tener
entonces mayor valor al examinar los patrones de dispersi n y
cuestiones de sistemàtica taxonomla
En lo peces como en otros grupos de vertebrados que se han
estudiado poco se sabe al respecto de la magnitud en que las
interacciones genoma medio ambiente y las variantes

19
genØtico bio u micas aportan a la cantidad de variación ue se ha
descrito Kohen y sus asociados han relacionado las formas alØlicas de
esterasas en ciertos tele steos dulceaculcolas con la temperatura
sobre la base de la distribución geogrÆfica y actividad bio urmica
I ohen 1969 Kohen 1970 M S Johnson 1971 1977 relacionó en
base a sus observaciones y experimentos las formas alelicas de LDH en
Anoplarchus purpurescens con diferencias tØrmicas del ambiente
En otros estud ios Johnson et l 1972 1973 se observó un exceso
significativo de individuos heteroci90tos para los loci pOlimórficos
fosfoglucomutasa PGM y glucosa fosfato deshidrogenasa G6PD en
colectas a grandes profundidades en el ocØano Pacifico de espec menes
de Sebastes que no se observaron en colectas a profundidad
menores TambiØn se reportaron asociaciones significativas no aZarosa
entre los genotipos de estos dos sistemas pOlimÓrficos a mayores
PI ofund idades que no se observaron en colectas de aguas someras Estas
observaciones sugirieron que las fuerzas selectivas actuaban sobre
estos dos loci a mayores profundidades y seæalaron la necesidad de
incluir otros productos en el individuo as como para las influencias
externas cuando se consideran los componentes de la selección natural
en un locus particular Esto es un ejemplo en el que las fuerzas
selectivas pudieran actuar de un modo diferencial en segmentos
distintos de la misma poblaci n vease Williams 1966
Los estud ios real izados por Utteri 1974 incluyeron un examen
detallado de la cinØtica de las reacciones enzimÆticas en las formas

20
alØlicas de OH en Q mencionado con anterioridad os datos
indicaron primeramente que las diferencias en las constantes
aparentes de Michaelis Km existen en fenotipos diferentes de OH
hepÆtica de este salmÓn a temperaturas fisiológicas y en segundo
lugar que las formas del enzima que parecen idØnticas
electrofor ticamente tienen diferentes Km Estos resultados sugieren
que los alelos silenciosos Boyer 1972 Johnson 1976 discusión de
este trabajo se reflejan en sustituciones de aminoåcidos de carga
similar presentes y que alteran las propiedades funcionales de los
enzimas sin cambiar sus movilidades electrofor ticas
El obtener una mayor y mejor comprension de las relaciones entre la
variación genØtica bioqu mica y los factores ambientales es la
principal aunque difIcil dirección de las investigaciones que se han
realizado durante los pasados 15 aæos La gran cantidad de datos
descriptivos recogidos durante los 60s y 70s que ilustran la
considerable variación genØtica bioquImica en diversos organismos ha
dado la suficiente base empírica para un cambio en el pensamiento en
relación con la cantidad de variación genØtica que puede tolerarse en
las poblaciones naturales y el papel que desempeæa esta variaciôn en el
proceso evolutivo a meta a n mÆs compleja de demostrar el significado
biológico de esta variacion es un campo abierto a futuros
investigadores
Un fen meno similar se ha descrito Merritt
alØlicos de OH en Pimephales promelas y
norte sur observado para esta especie
1972 para los variantes
su relación al clinal

21
o Adaptacion bioqu mica temperatura
Durante una muy buena porci n de este siglo la bioqu mica pas por un
importante período de investigación descriptiva y en suI
mayor la
empírica Una reversión importante de este enfoque comenzo cuando se
puso en claro la maquinaria bÆsica de la organizaciÓn metabólica y los
t ioquímicos se encararon con el problema de la interpretación desde un
punto de vista funcional de una gran cantidad de conocimiento
emplrico Con este fIn el pensamiento de la bíoqu mica se avocó de una
manera total hacia las teorlas de la evolución y adaptación
fisiológica consecuentemente aquellos caracteres cuya ventaja
selectiva y significación fisiológica que fueron largamente ignorados
se transformaron en los temas centrales de muchas reas de la
investigación en bioqu mica
El enfoque funcional de la bioqutmica se encuentra tal vez mejor
ejemplificado por las actuales teorías de regulación enzimÆtica estos
conceptos estin basados en ideas de adaptación fiSiOlógica y las
contribuciones hechas por diferentes reacciones enzimÆticas son
consideradas en el contexto de necesidades y demandas celulares
AtKinson 196 1966 1968 1977 Stadtman 1968 Cuando el
metabolismo es enfocado desde una perspectiva funcional la importancia
de una reacción enzimÆtica nica no es vista como cat`lisis
sino en tØrminos del papel que esta reacción juega como parte de un
complicado trabajO coordinado de transformaciones metabólicaS
Las teorías actuales de regulación metabólica enzimÆtica pueden

22
resumirse como siguel la contribución que una yía metabólica dada hace
al metabolismo total depende principalmente de las necesidades de la
cŁlula por los productos de tal vra a actividad de una yfa est
estrechamente controlada a travØs de interruptores encendido apagado
en uno o mÆs de los enzimas que funcionen al inic lo de tal v fa En
muchos casos el enzima interruptor es el primer enzima que es
exclusivo de la vía este sitio de control permite obYlamente una
regulación altamente eficiente de la yía metab lica como un todo El
parÆmetro que ha sido electo de una manera general para medir el
papel del interruptor es la afinidad enzima sustrato E S que
normalmente es proporcional al recíproco de la constante aparente de
Michaelis Km con respecto al sustrato a mayoría de los moduladores
positivos conocidos activadores incrementan la afinidad E S los
moduladores negativos tienen el efecto opuesto Estas relaciones se
ilustran en la figura l Una implicaci n importante de la modulación de
la afinidad E S como mecanismo controlador de la actividadI
metabol ica
es que los niveles de sustrato en la cØlula deben estar normalmente
bajo las concentraciones de saturación vmax pues los cambios de Km
podr n afectar las tasas de reacción solamente si la concentración de
sustrato es baja Fig 1 Cuando los niveles de sustrato son bajos es
aparente que pequeæos cambios de Km podr n conducir a cambios grandes
en la velOCidad de reacción Por ello la modulaCión de Km es vista
como un mecanismo altamente eficiente en la regulación de la actividad
metab 1 ica
os problemas bÆsicos de control metaból ico en organ ismos
r1 ì J

23
poiquilote mos p oYienen del hecho de que la mayo ia de las funciones
reguladoras descritas arriba son afectadas directamente por cambios en
los parÆmet os tales como la temperatura por ejemplo la afinidad E S
puede variar de muy diversas y complejas maneras con la temperatura
Baldwin 1971 Somero 1969 a y b Baldwin y HochachKa 1970
1969HochachKa y Somero 1968 1973 1984 Somero y HochachKa 1968
Para algunos enzimas las interacciones enzima modulador son altamente
importantes problemas para el
los cambios de este tipo presentan
control del metabol ismo Behr isch y
sensibles a la tempe atura
HochachKa 1969 a y b En otros casos las funciones reguladoras
parecen se independientes de la temperatura Somero 1969a Somero y
Hochachla 1968 HochachKa y Somero 1973 1984El flujo de carbono a
travØs de las diversas ramas metabólicas es particularmente sensible a
cambios tØrmicos y puede mostrar características an6malas debidas a la
temperatura HochachKa 1968a Dean 1969 Las concentraciones iAnicas
supuestamente su compartamentalizaci6n pueden cambiar en respuesta
a las variaciones en la temperatura HeinicKe y Houston 1965
Hochachla y Somero 1971 HicKman et l 1964 Estos hallazgos
indican que los complejOS mecanismos que permiten a un organismo
regular estrechamente su actividad enzimÆtica tambiØn someten al
organismo a una alta vulnerabilidad y a los efectos deletØreos de
cambios repentinos en las condiciones ambientales Sin embargo el
hecho de que estas funciones enzimÆticas sean directamente afectadas
por la temperatura deja una posibil idad si los cambios inducidos por
el ambiente temperatura sobre las propiedades enzimÆticas ocurren en
Ulla d irecc ión adaptat iva y a un ritmo adaptat ivo entonces es pos ible

24
que el organismo sea capaz de hacer uso de una manera positiva de los
cambios ambientales que pueden parecer de et4reos en terrenos a priori
La elaboración de mecanismos de control que se han discutido hasta
aqu pOdr a servir de materia prima para que la selección diseæeN
mecanismos homeost4ticos que permitan al organismo adaptarse a los
cambios del ambiente En la discusión de este trabajo se encontraran
puntos de vista al respecto de la manera en que los poiquilotermos
pueden abordar estos procesos
serÆ importante tener en mente a lo largo del presente estudio que
las estrategias adaptativas empleadas por 10 pOiquilotermos var lan
considerablemente entre los diferentes organismos por ejemplo si se
encuentran disponibles comportamientos alternativos de escape de las
c ir cunstanc ias amb ientales daæ inas entonces la neces idad de adaptac ión
bicqutmica extensiva se ve disminu da ade s el rango de capacidades
adaptativas mostradas por un organismo pOdr4n ser dependientes de la
complejidad de su propio entorno Los eces que viven a temperaturas
casi constantes en los ocØanos antÆrticos muestran una habilidad menor
para aclimatar su metabolismo que los peces euritØrmicos como la trucha
o el dorado Somero 1968 Esta complejidad en las respuestas
adaptativas de los organismos podrÆ depender del ritmo y extension en
que los parÆmetros ambientales tales como la temper atura gases
disueltos y salinidad fluct en
Consideraciones de este tipo han llevado a los fiSiÓ1ogos a
considerar un cierto nœmero de diferentes direcciones en el tiempo con

25
respecto a la adaptación ambiental En un extremo estÆ la adaptaci n
evolutiva un proceso que requiere de muchas generaciones para su
terminación Un ejemplo de esto es el ajuste latitudinal de las tasas
n tab licas de los peces Wohlschlang 1964 Somero l 1968
Hemmingsen 1969
Respuestas fisiológicas similares a la temperatura se observan
comœnmente sobre una base estacional Roberts 1964 1967 Este
proceso de adaptac ión que ocurre en e1 curso de d as semanas o meses
se llama aclimatación o aclimatizacion
Finalmente los biólogos han reconocido recientemente que para por
10 menos algunos organismos y procesos fisiolÓgicos la adaptación al
entorno es inmediata por ejemplo Q10 aproximados a la unidad se han
teportado para muchos organ ismos de 1 a zona de entremareas Newe 11
1966 1967 Baldwin 1968
Hasta hace pocos aæos los bioqu micos comparativos han iniciado una
investigacion intensiva de los camb ios b ioqu micos que estÆn
involucrados en cada uno de estos procesos de adaptaci n
Las adaptaciones de este tipo que ocurren en el haÞitat natural del
organismo donde una variedad de factores ambientales ademAs de aquel
de interØs pudiese variar se llaman aclimatizaciones Las
adaptaciones inducidas en el laboratorio en respuesta a la variación de
un paråmetro ambiental se llaman aclimataciones Prosser 1973 Wilson
1979 HochachKa y Somero 1984
Definido como la razön entre las velocidades de reacción con una
di rencia tØrmica de 10 C

26
Vma
OeO
uo VmaeL
2
2 34 5 10 15 20
SubS1fa10 J 104 M
FIG l El efecto de la variaclon de la afinidad enzima sustrato defini
do como el recíproco de la constante de Michaelis Km sobre la
actividad enzimÆtica Los enzimas A y B catalizan la misma reacción
y exhiben la misma actividad a la concentración de saturación del
sustrato v Los enzimas difieren en el doble de su Km A con
centracione afisiológicas del sustrato indicadas por las líneas
punteadas el enzima A es mucho mÆs activa que B En tØrminos de
la teoría de la regulación enzimÆtica B podría representar un es
tado desactivado del enzima y A representar un estado activado
B podría ser convertido en A mediante el acoplamiento de un modula
dor positivo por el contrario A podría convertirse en B bajo la
influencia de un modulador negativo Hochachka y Somero 1971

27
MATERIALES y METODOS
I Especies analizadas y sitios de colecta
Las especies utilizadas en este estudio se ilustran en las figuras 2
3 4 y 5 se incluyen aspectos biológicos generales para cada
especie Los espec menes fueron colectados mediante buceo libre y
arpón anzuelo o redes en arrecifes rocosos de fÆcil acceso a lo largo
de la zona occidental del bajo Golfo de California incluyØndose la
porcion mas sUre a de la parte media del mismo Los sitios de colecta
fueron los siguie tes
Punta Mechudo Is1 a San JosØ e Islote San Francis uito Complejo
Insular Esp ritu Santo La Partida Las Cruces San Juan de los Planes y
Cueva de Le n La Rivera y Cabo Pulmo Los Barriles y la región de los
Cabos todos los sitios en el estado de Saja California Sur Fig 6 y
7 En algunas ocasiones se obtuvieron organismos mediante compra
directa a los pescadores en las distintas localidades
Una vez colectados los organismos se tomaron los datos mertsticos y
morfomØtr icos de uso co n en Ict 1010g a Sm1th 19711 Thomsonl l
1979 listados a continuación
Mertsticos Fig S
Filas transversas de escamas
Escamas sobre la línea lateral

28
Escarr s bajo la línea lateral
Escamas alrededor del pedœnculo caudal
Espinas y radios en las aletas
Radios branqui6stegos
Arcos branquiales
MorfomØtricos Fig 8
Profundidad del pedœnculo caudal
Del origen de la aleta dorsal a
la punta del hocico
Longitud de la base de la aleta
dorsal
Longitud patrón
Longitud cefålica
Amplitud cef lica
Longitud del hocico
Amplitud suborbital
Longitud cef lica postorbital
Oi metro orbital
Longitud de las mandtbulas
Profundidad del cuerpo
Amplitud del cuerpo
Longitud de la aleta caudal
Longitud de la base de la aleta
anal
Longitud del ped nculo caudal
Longitud de la aleta pectoral
Longitudes de espinas dorsales
1 III y la œltima
Longitud de las espinas anales
Se tomaron de igual forma los datos de campo pertinentes temperatura
del agua profundidad de la colecta tipo de sustrato actiyidad del
organismo y hora del dia
Se examinó un total de S0 espectmenes 15 de E acanthistius y I de
E analogus 25 de E labriformis y 25 de E Cephalopholis panamensis

29
2 Preparación de los extractos tisulares
Tomados estos datos los especímenes enteros o bien muestra de ojo
corazón hígado y mœsculo esquelØtico de los individuos colectados se
n9lðron inmediatamente en hieler as de campo comunes para evitar la
descomposición de los tejidos y consecuente desnaturalizaci6n de los
enzimas contenidos en ellos Una vez congeladas se transportaron al
laboratorio donde se preservaron a 20 C hasta su procesamiento Se
procuró que las muestras fueran analizadas dentro de las 3 semanas
posteriores a su colecta pues pasado tal período se obtenían resultados
poco fiables
Los tejidos deseados mósculo esquelØtico mÚsculo card aco h gado
y ojo fueron cortados en trozos pequeæos a una cantidad aproximada de
g tal cantidad de tejido fue homogenizada mec nicamente en 2
volœmenes de solución amortiguadora para las muestras S81 0 1M
Tris HCI pH 7 0 utilizando un disruptor de tejidos Polytron 8rinKman
Instruments y manteniendo baja la temperatura con un baæo de hielo
durante el proceso
Los homogenados así obtenidos se centrifugaron a 3800 rpm por 30
minutos para retirar fibras y restos de los tejidos que no fueron
hornogen izados Poster iormente 1 5 mI del sobrenadante obtenido se
transfirió a microtubos y se centrifugó a 15600 g durante 5 minutos
Los sobrenadantes as obtenidos se utilizaron para electroforesis o
bien se mantuvieron a temperatura de congelación hasta su anÆlisis Se
consideró el mismo periodo de confiabilidad antes mencionado

30
Los proced imientos descr i tos corresponden a Gorman 1 ll 1975
ligeramente modificados de Selander et al 1971 mismos que se
utilixaron en los estudios electroforØticos protocolos de
electroforesis e interpretación de las bandas en el gel con algunas
modificaciones de acuerdo a la disponibilidad de tiempo equipo y
reactivos Estos procedimientos se describen en la siguiente sección
3 Electroforesis y tinción de los geles
Las electroforesis se llevaron a cabo en sistemas de gel
horixontales similares a aquellOS empleadOS en otros laboratorios que
utilixan electroforesis en gel de almidôn para un gran nœmero de
organismos Fig 9 El equipo utilixado fue de fabricaci6n domØstica
LoS geles contuvieron almidôn hidrolixado para electroforesis
Connaught Labs al 12 en solución amortiguadora correspondiente de
acuerdo al sistema de electroforesis que se utilice para el an lisis
del marcador gen4t leo enxima deseado Tab 1
Los geles se prepararon de acuerdo a las instrucciones del fabricante
Connaught Labs Smlthies 1955 dej ndoseles enfriar a temperatura
ambiente por espacio de media hora entonces se colocaron dentro del
refrigerador 3 4 C durante media hora mas Se hixo una inclslon
transversal al gel a 5 cm de un extremo las dimensiones de los geles
fueron siempre 30 cm x 15 cm x 0 5 cm donde se colocaron de 12 a 14
muestras pequeæos cuadrados de 0 5 cm de lado de papel filtro Whatman

31
3 impregnados con el sobrenadante resultante del proceso descrito en
la secci6n anterior
Los geles con la muestra de tejido se sometieron al paso de una
cantidad dada de corriente elØctrica electroforesis usualmente entre
los 125 y 140 V Y 30 20 mA por espacio de 18 20 horas Cada protelna
de la muestra migra a travØs del gel en una dirección y a una velocidad
que dependen de su carga el ctrica neta y su tamaæo molecular
Todos los ensayos electroforeticos se llevaron a cabo dentro de un
cuarto frio 5 10 C y con refrigeraciòn directa sobre el gel en
corrimiento utilizando un sistema de l quido frIo circulante
Fr igomix
Una vez cumplido el tiempo de electroforesis los geles se recortaron
en dos rebanadas longitudinales Cada una de ellas se trató con una
solución qu mica que contiene un substrato especifico para el enzima
deseado y una sal Los geles se incubaron con esta solución en la
obscuridad a una temperatura de 35 37 C durante 20 minutos En este
periodo de incubaclón el enzima catallza la conversion del sustrato
hacia el producto que entonces se acopla con la sal para dar bandas
coloreadas en los puntos hasta donde ha mlgrado el enzima Dado que los
enzimas que son codificados por diferentes alelos pueden tener
diferente estructura molecular y distinta carga y por ello diferente
movilidad en el campo elØctrico la composición genØtica en el locus
genico que codifica para un enzima dado pudo establecerse para cada

32
Individuo a partir del nœmero y posición de las bandas electroforØticas
ver apØndice NO 1
Cada pez fue analizado para 9 enzlmas deflnl ndose un total de 16
loci por marcador por pez Tales loci fueron los siguientes
E r s Locr
Ldh aLdh bLdh c
h sMdh a sMdh b
Me
j
Pgm a Pgm b
Gpi a Gpi b
Adh
Sod a Sod b
Gdh
Lactato deshidrogenasa
Malato deshidrogenasa
Enzima malica
Manosa fosfato isomerasa
Fosfoglucomutasa
Glucosa fosfato isomerasa
Alcohol deshidrogenasa
Superoxido dismutasa
Glucosa deshidrogenasa
Se utilizaron cuatro sistemas de electroforesis para el an`lisis de
tales marcadores genØtícos enzimas que se describen en las pÆginas
Siguientes

l Sistema para corrimiento electroforØtico Tel
Gell 12X almidón peso volumen enl
0 0175 M Tris
0 0058 M CUrato
pH 7 0
Amortiguador para electrodosl
8 075 M TI is
0 1325 M C trato
pH 7 0
Electroforesis 18 horas 125 V 35 mAl 4 C
11 Sistema para corrimiento electroforØtico EBTI
Gel 12X almidbn peso volumen
8 042 M TI is
0 023 M Borato
0 00089 M EOTA
pH 8 6
Amortiguador para
En el citodo0 18 M TI Is
0 1 M Borato
6 0036 M EOTA
enl
electrodosl
En el ånodo0 129 M TI is
8 671 M Bora to
6 0026 M EOTA
pH 8 6
Electrofores is 18 horas 150 V 18 mAl 4 C
111 Sistema para corrimiento electroforØtico TEMM
Gel 12X almidón peso volumen enl
Soluci n amortiguadora para electrodos dilulda 1110Amortiguador para electrodosl
El 1 M TI h
0 1 M Maleato
0 01 M Na2 EOTA
0 01 M MgCl2
pH 7 4
Electroforesisl 18 horas 125 V35 mAl 4 e
IV Sistema para corrimiento electroforØtico pel
Gell 12X almidón peso volumen enl
6 87 mM K fosfato
1 21 mM Cltrato
pH 7 6
33
e

34
Amortiguador para electrodos
0 214 M K fosfato
0 027 M Cltrato
pH 6 7
Electroforesis 17 18 horas 12 V 3 mAl 4 e
En algunas ocasiones se util izó un quinto sistema que a continuaciónse describel
V Sistema para corrimiento electroforØtico SP
Gel 12 almidón p eso volumen en
Soluc ión amort iguador a para electrodos d ilu da 1 10
Amortiguador para electrodos0 01 M Na fosfato
pH 6
Electrofor esis 18 hor as 150 V 30 32 4 e
Como se ha mencionado al finalizar el corrimiento electroforØtico
los geles se trataror con una solucibn qutmica o mezcla reactar te
apropiada para el enzima que se deseaba visual izar la composicion de
ta es mezclas se describe a continuación
1 Glucosa fosfato Isomerasa GP 1 2 Lactato deshidrogenasa LOH
0 03 M Tr is pH 8 0 20 mi 0 05 M Tris pH 8 0 25 mi
Fr uctosa 6 fosfato 20 mg NO 10 mg
Glucosa 6 fosfato Icido l`ctico 0 5 mI
deshidrogenasa 100 U mi 10 1 MTT S mg ml 0 5 mI
NOP 5 mg PMS S mg ml 0 2 mI
MTT 5 mg ml 0 5 mi Agar 2 30 mI
PMS 5 mg ml 0 2 mI
Agar 2 30 mI
Incubados en la obscur idad a 37 e durante 5 10 min
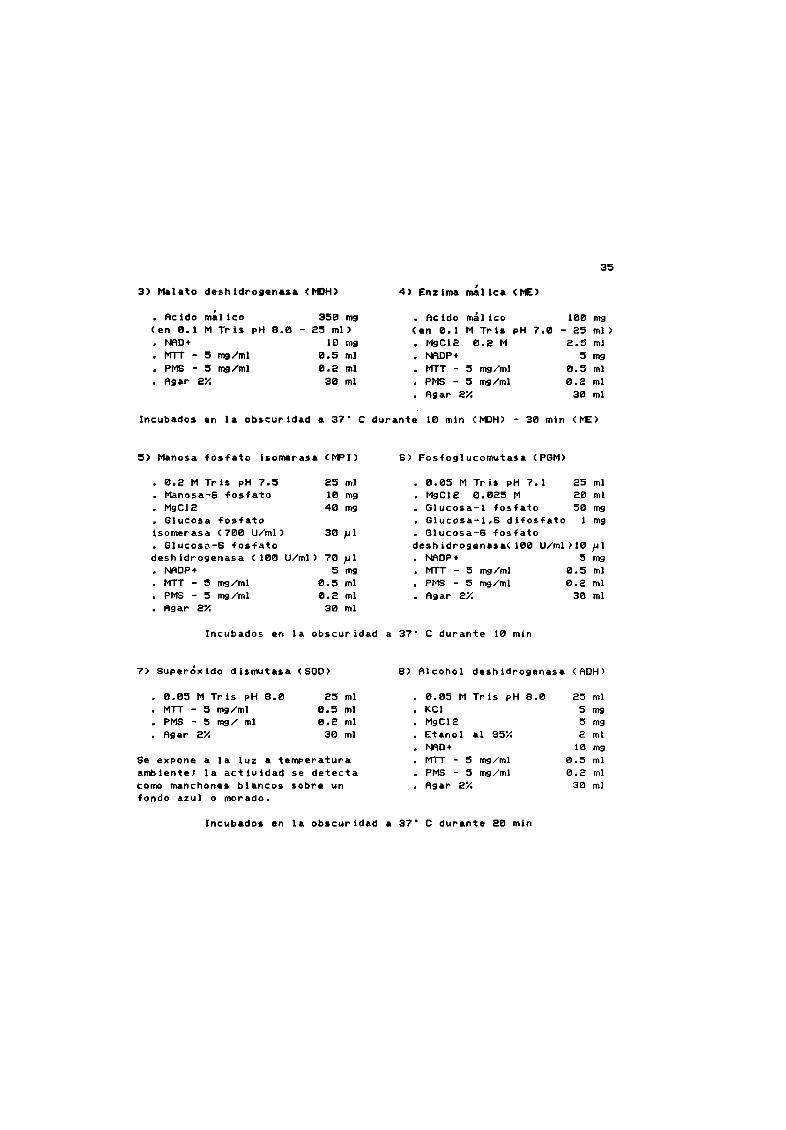
35
3 Malato deshidrogenasa tIlH 4 Enzima fIEmalica
Acido millco 356 mg Acido mÆlico 100 mg
en el 1 M Tr is pH e 0 25 mi en 0 1 M Tris pH 7 0 25 mi
NAO 10 mg MgCl2 0 2 M 2 5 mi
MTT 5 mg ml 0 5 mi NAOP 5 mgPMS 5 mg ml 0 2 mi MTT 5 mg ml 0 5 mi
Agar 2Y 30 mi PMS 5 rng ml 0 2 mi
Agar 2Y 30 mi
Incubados en la obscuridad a 37 C durante 10 mln tIlH 30 mln ME
5 Manosa fosfato lsomerasa MPI 6 Fosfoglucomutasa PGM
0 2 M Tris pH 7 5
Manosa S fosfato
MgCl2
Glucosa fosfato
isomerasa 700 U mi
Glucosß 6 fosfato
deshldrogenasa 100 U mi
NAOP
MTT 5 rng ml
PMS 5 rng ml
Agar 2Y
25 mi
10 mg40 mg
0 05 M Tris pH 7 1 25
MgCl2 0 025 M 20
Glucosa l fosfato 50
Glucosa I 6 difosfato 1
Glucosa S fosfato
deshidrogenasa 100 U ml 10
NAOP 5
MTT 5 rng ml 0 5
PMS 5 rng ml 0 2
Agar 2Y 30
mi
mi
mg
mg
30 J 1
70 J1
5mg
0 5 mi
0 2 mi
30 mi
Jlrng
mi
mi
mi
Incubados en la obscuridad a 37 C durante 10 min
7 Super6xido dismutasa SOO e Alcohol deshidrogenasa AOH
0 05 M Tris pH e 0
MTT 5 mg ml
PMS 5 mg mi
Agar 2Y
25 mi
0 5 mi
0 2 mi
30 mi
0 05 M Tris pH 8 0
KCI
MgCl2
Etanol al 95X
NAO
MTT 5 mg ml
PMS 5 rng ml
Agar 2Y
25 mi
5mg
5 mg
2 mi
10 mg
0 5 mi
0 2 mi
30 mi
Se expone a la luz a temperatura
ambiente la actividad se detecta
como manchones blancos sobre un
fondo azul o morado
Incubados en la obscuridad a 37 C durante 20 min

36
9 Glucosa deshidrogenasa GOH
0 0 M K fosfato pH 7 5
O glucosa 1 0 M
NAO
MTT 5 rng ml
PMS 5 mg ml
Agar 2
25 mi
9 9
20 mg
0 5 mi
0 2 mi
30 mi
Incubado en la obscuridad a 37 e durante 3 5 horas
Las mezclas reactantes se prepararon justamente antes de usarse dada
su inestabilidad química
Los sistemas de tinción para los enzimas descritos se ilustran en las
figuras 10 18
4 CÆlculos de frecuencias gØnicas distancias genØticas y construcción
del dendrograma
A partir de las bandas coloreadas contenidas en el gel se anotaron
los genotipos de los individuos y se calcularon las frecuencias
alØlicas de acuerdo al mØtodo de Stansfield 1969 la distancia
genØtica entre las poblaciones Nei 1971 1972 1978 su correlacion
con el tiempo Carlson 1978 Y finalmente la construcción del
dendrograma con el metodo UPG SoKal y Sneath 1963
Los procedimientos para tales cÆlculos se describen en los apendices
2 y 3

37
5 PUl if icac ión del isozima lactato deshidrogenasa del MÚsculo
M4 LDHesqueletico
El homopol mero del isoz ima lactato deshidrogenasa del mœsculo
esquel tico M4 LDH se purificð por cromatografla de af in idad
utilizando una columna de Sepharosa 4B oxamato Pharmacia Co de
acuerdo a los procedimientos de Yancel y Somero 1978 1 igeramente
modificados de aquellos de O Carra et L 1974 Estos procedimientos
se describen a continuación
Muestras de MÚsculo esquelØtico se homogenizaron en solución
aroortiguadora 50 mM fosfato de potasio pH 6 8 en una proporción de 3
g 17 mi Se mantuvo baja la te eratura durante la homogenización
utilizando una baæo de hielo El homogenizado se centrifugó a 10000 g
durante 30 min 4 C
El sobrenadante se llevo a 0 5 M KCl respecto al volumen total y a
0 4 mM respecto a NAOH aplic ndose posteriormente a una columna de
Sepharosa oxamato 10 mi prev iamente equilibrada con solución
amortiguadora de fosfato de potasio pH 6 8 conteniendo 0 2 mM NAOH
La columna fue lavada con este amortiguador de equilibrio hasta que la
absorbancia del eluido calese hasta 0 a 280 nm
El isozima M4 LDH se eluyó de la columna con solución amortiguadora
de fosfato de potasio 50 mM pH 6 8 conteniendo 0 25 M KCl y 2 5 mM
NAO

38
Las fracciones ac ivas se reun ier on lo concen raron por
ultrafil ración con una membrana AMICON PM30 Se dializaron contra un
amortiguador saturado de sulfato de amonio y se preservaron a 4 C como
un precipitado de sulfato de amonio al 80 conteniendo 2 3 mg ml de
prote lna
Estos precipitados se dializaron contra 00 volœmenes de solución
amortiguadora 80 mM Tris HC1 pH 7 a C antes de USArse en los
estudios cineticos que se describiran mas tarde
La pureza de cada preparaCión se probó utilizando electroforesis en
gel de poliacrilamida dodecilsulfato de sodio SDS PAGE de acuerdo a
los procedimientos de Laemmli 1970 y FairbanKs et al IS71
descritos ampl iamente por S nchez 1983 Las preparaciones de los
enzimas M4 LDH de las distintas especies de Epinephelus mostraron una
banda œnica que comigró con M4 LDH de conejo comercial Sigma Chemical
Co
6 Estudios en la cinØtica de los isozimas M4 LDH
La actividad enzimåtica se registró en un espectro fotómetro 8ausch
Lomb Spectronic 2000 Las constantes aparentes de Michaelis Km con
respecto al p i l IJVato se obtuv ieron c inØt icamente determinando 1 a
relación entre la velocidad inicial de reacción y la concentración de
piruvato Esta reacci n provee una aproximación de la afinidad entre
el enzima y el sustrato a las diferentes ter eraturas La solucion para

39
el ensayo enzimÆtico 5 0 mI contuvo 200 M NADH Y concentraciones de
piruvato entre 0 2 y 1 0 mM Se utilizó una solución amortiguadora con
80 mM imidazolHC1 pH S 98 a 2 C para asegurar que el pH en cada
temperatura de medición fuera fisiol giCamente apropiado el pH de los
amortiguadores de imidazol cambia aproximadamente 0 017 unidades de pH
por gradO cent grado aumentado una relación de incremento de
pH incremento de temperatura similar a aquella encontrada en los
flu dos de la mayor a de los ectotermos Reeves 1977 Yancey Y Somero
1978 Graves y Somero 1982 Las soluciones patrón stOCK se
prepararon frescas cada tres horas
Los valores de Km aparentes determinados grÆficamente SE
determinaron gråf icamente a cinco temperaturas 10 15 20 25 Y 30 C
La temperatura en las cubetas de reaccion se mantuvo controlada
utilizando un baæo de agua con una variación de 0 8 C Se hicieron
mediciones por duplicado en cada una de
piruvato utilizadas
las 8 concentraciones de
La constante catalitica Kcat una expresion de la maxima tasa de
recambio por sitio activo del enzima o eficiencia catal tica måxiIT la
veloc idad m x ima de reacc iór que una molØcula de enz ima puede genel ar
fue determinada para todas 1 as espec ies excepto para el isoz iIT t14 LDH
de labriformis Para las mediciones de Kcat una pequeæa al icuota de
las fracciones reunidas de la elución de la columna que tuvier a la
måxima actividad para LDH fue dilu da y probada del modo descrito
dentro de los 30 minutos posteriores a la purificación para asegurar un

40
m nimo en la perdida de ac ividad os valores de Kcat estan basados en
los valores te6ricos de vmax Ambos valores te6ricos de Km y Kcat
fueron determinados utilizando el mØtodo de regresi n de gråficos
Lineweaver BurK de acuerdo a WilKinson 1961
La concentración de prote nas de las fracciones reunidas en el piCO
de actividad fue determinada espectrofoto tricamente midiendo la
absorbancia a 280 nm Y computando la concentración de DH usando un
coeficiente de extinción de 1 29 para una solución de LOH con 1 mg de
proteína por mililitro Stinson y HolbrooK 1973 a solución de
enzima fue exhaustivamente dializada para quitar todo resto de NAO
que tarooien absorbe fuer emente a 280 nm proveniente de los
procedimientos cromatogråficos Se juZg terminada la di11isis cuando
la absorbancia alcanzó 1 8 en una razón de 280 260 nm que es tIpica
para la absorción de protefnas en la ausencia de contribución por
nucleotidos a al absorbancia Graves y Somero 19821 Graves l
1983
7 Reac ivos y sustancias
Todos los enzimas y coenzimas sustratos y colorantes fueron de Sigma
Chemical Co Sto Louis MI U 5 A El resto de sustancias y reactivos
provienen de diversas casas comerciales y fueron simpre de grado
analC ico
Para una explicación detallada al respecto de Km Y Kcat
apØndice 4
vease el

FIGURAS 2 Y 3
Af 41Þ lA I p AIJ n 1NV
411 If
1 trnti i W o c F
t i 1f 0 r I ii i i
u lf0 t ti e i
1tl Lli
pinephe us Cłph pholis ponamensis Steindochner 1876
flA y
1 i ˛ tl A
8 jv
t f Z t i ti 1u
I oO
oKf
Jrv
w
iiS
Epinephe u Epinephelus oconfhislius Gilbert 1892
41

42
Figura 2
ENJAMBRE
ENJAMBRE Panama graysby
Epinephelus Cephalopholis panamensis Steindachner 1876
Especímen ilustrado
Adulto Joven 160 mm 6 3 in por Tor Hansen talla mÆxima que
alcanzan estas espec ies 305 mm 12 in
Características distintivas
Esta pequeæa cabrilla puede distinguirse de otras por su aleta caudal
redondeada de 9 a 10 bandas obscuras que cruzan su cuerpo y numerosas
manchas azules y anaranjadas a los lados de la cabeza Los Juvenile
son similares a los adultos en la coloración excepto que el adulto
tiene una gran mancha negra Justo por detrÆ del ojo E a cabrilla ha
s ido descr i ta como Petrometopon panamens is D IX 14 A II1 a
Distribución
El Ærea de distribuciön del enjambre va desde
Cal ifornia Roca Consang ha ta Colombia Isla
Galåpagos Es muy comœn en arrecifes poco profundos
Golfo de California
el propio Golfo de
del Coco e Isla
en el centro y bajo
Ecolog a
El enjambre es un pez solitario y reservado que e encuentra a poca
profundidad o hasta los 75 m 250 ft Esta cabrilla permanece cerca
del fondo escondida en recovecos saliendo solamente para capturar
pequeæos peces y crustÆceos En caut iver io atacarÆ e intentarÆ comer
peces de su misma talla Se le colecta frecuentemente a lo largo de las
costas rocosas del Golfo de California y puede que sea el serrånido mÆsabundante en ciertos arrec ifes tal es como los de Sah a Concepc lóna c s
Una especie similar a E panamensis ocurre en el AtlÆnticoOccidental El enjambre E cruentatus Lacepede es la cabrilla mÆcomœn en las Sahamas y a pesar de su pequeæo tamaæo 30 cm tiene
cierta importancia allÆ como especie comestible y a diferencia de fpanamensis es poco temerosa de los buzos
Referencia Thomson 1979

43
Fig 3
BAQUETA
BAQUETA Glf coney
Epinephellls Epinephelus acanthistius Gilbert 1892
Especimen i 1 ustado
Adtllto 7lel mm 28 in por Alex Kers1 itch
mayor de 0 m 3 ft
alcanzan na longitud
Calacter s1 icas distintivas
Esta garropa se caracteriza por su gran cabeza su cuerpo robusto y
color rosado achocolatado Dependiendo de la edad y la profundidad
l color var a en intensidad Las espinas dorsales anteriores
especialmente la tercera y la cuarta son alargadas y la merrbrar a
ntre ellas se encuentra hendida Una prominente barra negr a obl icua
bigote corre por encima de la mand bJla sJPerior las aletas son
e bscuras distalmente Hasta hace poco había s ido colocada dentro del
gØr ero Cephalop lolis D I 17 A III 9
Distr ibuc ión
La baqueta se distribuye por todo el Golfo de California hasta
Es frecuente que aparezca como fauna de acompaæamiento en las
camar Ø neras tan al norte como Puerto Peæasco
Perœredes
Ecolog a
La baqueta antes considerada como una espee ie t ara es un pez comœnde los fe r dos a todo lo lar go del Golfo de Cal ifornia Esta cabrilla
hab itante de las profundidades moderadas se encuentra por los
arrecifes aislados y fondos arenosos cercanos a las costas normalmente
a pr ofundidades que exceden los 45 m 150 ft Es muy raro observarla
mie tras se bucea por los arrecifes pero es capturada frecuentemente
por los anzuelos de los pescadores a profndidades de 90 m 300 ft
Dada su preferencia por las profundidades poco se sabe de su biolog a
A veces se vende come pargo colorado en la pescader as de Puerto
Pe1asco
Referencia Thomson l 1979

44FIGURAS 4 Y S
c
f
Epinephe us Epinephe u Jobriformis Jenyns t 843
o
EpinePhe u Epinephe us ana ogus Gill t 1864

45
Figura 4
CABRILLA PIEORERA
CABRILLA PIEDRERA Flag cabrilla
Ep inephe l us Ep inephe 1 1S 1 abr iformis Jenyns lB43
E5pecir n ilustrado
Adulto joven 230 mm 9 in por Alex Kerstitch alcanzan una
longitud de alrededor de 510 mm 20 in
Características distintivas
Epinephelus labriformis es una cabrilla de talla mediana ue puede
distinguirse f cilmente de otras por su patrón de coloraci6n tan
especial el cuerpo va del verde olivo al cafØ rojizo manchas blancas
de varios tamaæos y formas irregulares la cubren y las partes blandas
de las aletas dorsal anal y caudal son rojas en los rgenes dorsales
y ventrales Una mancha obscura a manera de silla de montar aparece
por encima del ped nculo caudal El interior de la boca es rojo Tiene
once espinas dorsales y sus pe ueæas aletas pØlvicas no llegan hasta el
ano D XI 16 17 A 111 8
Oistr ibuc ton
La cabrilla piedrera se distribuye desde Bahía Magdalena en elPacífiCO hasta la parte mÆs septentrional del Golfo de California
Puerto Lobos y hacia el sur hasta Perœ incluyendo las islas alejadas
de las costas Es muy comun en el Golfo medio y bajo en arrecifes poco
profundos
Ecol09ta
La cabrilla piedrera es un predador solitario ue parece alimentarse
día y noche tomando peces pe ueæos en su mayoría durante el d a y
crusticeos al atardecer Un pez reservado E labriformis abandona su
guarida para capturar a su presa pero permanece cercana al fondo
rocoso Los juveniles aparecen en el arrecife de octubre a diciembre
lo cual indica ue el desove ocurre avanzado el verano A pesar de ue
los adultos son los mÆs abundantes a poca profundidad ocurren hasta
los 30 m 100 ft cuando menos
Referencia Thomson j 1979

46
Figura 5
CABRILLA PINTA
CABRILLA PINTA Spotted cabrilla
Epinephelus Epinephe us analogus Gill 864
Especirren ilustrado
Juven il 195 mm 7 7 ir por Tor Hansen se d ice qUe alcanza una
talla de 0 8 m 2 1 2 ft 9 Kg 20 lb
Caracter sticas distintivas
Esta garropa de talla mediana puede distinguirse de otras por las
siguientes caracter sticas diez espinas dorsales aleta caudal
redondeada y numerosas manchas obscuras entre barras p lidas
cubriØndole el cuerpo El color es cafØ rOjizo con grandes manchascafØs que recuerdan el patrón de coloración del mero jew fish E
itajara Su gemelo del Atlåntico Occidental es la cabrilla de roca
adsencionis OsbeK y el nombre analogus similar se refiere a su
fuerte semejanza con esta especie D X 16 16 A 111 8
Di5tribución
La cabrilla pinta se distribuye desde el sur de California y losar ec ifes rocosos mÆs norteæos de 1 Go 1 fo has ta Per y 1 as Is 1 as
Galåpagos La pinta es la garropa mÆs abundante en el Golfo superior
Ecologra
El habitat preferido de la pinta poco frecuente en la cercanía de
las costas parece ser aquel de los arrecifes aislados en la lejanía de
las mismas donde es abundante y frecuentemente capturada por los
anzuelos de los pescadores Por este hecho de agregarse en los
arrecifes alejados de las costas a prOfundidades moderadas su ecolog a
ha sido poco estudiada La cabrilla de roca que antes se mencionaba
por el contrario es una especie de aguas poco profundas
Las cabrillas pintas son voraces depredadores que se alimentan de
crustÆceos y peces en gran parte sobre los fondos rocosos y arenosos
En el inv ierno cuando 1 a te eratura de l agua de 1 Go 1 fo es baj a es tos
peces se acercan a la costa hacia aguas someras donde se allmentan de
Iangostilla el Kr ill langostero o cangrejo rojo peli˝gicoPleuroncodes planipes que es muy abundante en esa Øpoca
A pesar de ser uno de los manjares rr s finos del Golfo la pinta no
ocupa un lugar muy importante entre los pescadores con caæa pues es
f Cil de capturar y cuando es anzueleado no opone una resistencia muy

47
espectacular En lugar de eso nada hacia el fondo y se esconde en
hendiduras y recovecos siendo dificil para el pescador el desenganche
Sin embargo los pescadores locales explotan con Øxito esta especie
Referencia Thomson 1 1979

110030
Islote Los AnImas
250 OO
Son Fronc squ to
òO
Cl
lO1i
VSl
V
V
lV
ISI
Partido
t
1510Es plritu Santo
FIG 6 Localización geogrÆfica de los sitios de colecta
25000
240 30
48
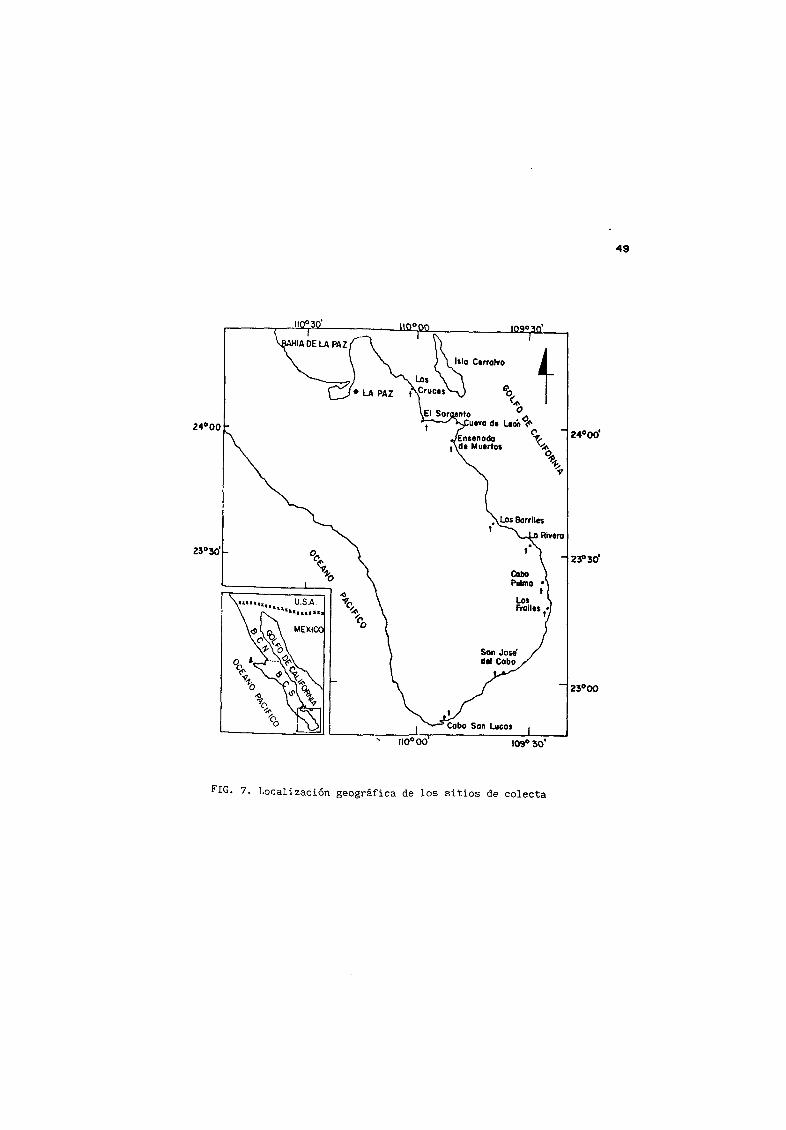
o
l c
tC
onto
ðu vo d le0˝l
24000
O
23030 o
10
2330
o1
10
23000
FIG 7 Localización geogrÆfica de los sitios de colecta
48

Figura 8 Un pez conespinas y radios ilustrando las partes y mØtodos del conteo y medición l interorbital 2 occipital 3 nuca 4 longitud de la cabeza S longitud predorsal6 longitud estÆndar 7 longitud al escote 8 longitud total 9 longitud de la base de la
primera aleta dorsal oaleta espinosa lO una de las espinas de la primera aleta dorsal espina de la segunda aieta dorsal 12 altura de la segunda aleta dorsal 13 longitud del bordedistal externo o libre de la segunda aleta dorsal 14 uno de los radios blandos de lasegundaaleta dorsal 15 longitud del hocico lS longitud del ojo 17 longitud cefÆlica postorbital18 escamas sobre la línea lateral o series laterales que son contadas 19 altura mÆxima del
cuerpo 20 uno de los poros de la línea lateral en una linea lateral completa 21 una de lasescamas laterales que con las dEmÆs forma la serie de la linea lateral 22 longitud de la base dela segunda aleta dorsal o aleta blanda 23 altura mínima odel pedœnculo caudal 24 la aletapectoral 25 uno de los radios blandos de la aleta pectoral 26 región abdominal vientre27 escamas bajo la línea lateral o series lateralesque son contadas 28 longitud de la base dela aleta anal 29 longitud del pedœnculo caudal 3 longitud de la mandíbula superior31 istmo 32 pecho 33 altura de la espina pØlvica 34 altura de la aleta pØlvica 35 uno
de los radios blandos de la aleta pØlvica 36 espinas de la aleta anal 37 radios blandes de laaleta anal 38 radios rudimentarios de la aleta caudal 39 uno de los principales radios blandos sin ramificar de la aleta caudal 40 radio blando ramificado de la aleta caudal 41 la aletacaudal con el nœmero en lo mÆs profundo de escote de la misma
50

1
4
FIG 9 Aparato de electroforØsis de uso convencional
1 Bandas coloreadas que indican la posición de los
enzimas posteriormente al corrimiento electroforØti
co 2 Ranura para el inicio del corrimiento 3
Gel matriz de almidón hidrolizado 4 Electrodo
5 Puente de contacto 6 Cubeta para la solución
amortiguadora o tampón

Tabla l Sistemas EnzimÆt1cos resueltos con Øxito en
cuatro espec1es de Epinephelus
protena Sistema ElectrofNo E C Loci Estructura
Oxidorreductasas
Lactato de8hidrogenasa 11127 1 4 TetrÆmero TC EBT
Ilalato deshidrogenasa 11137 1 3 D mero TC TEMM
Enzima mil1ca 1 1140 1 2 TetrÆmero TEHM
Super6x1do dismutasa 115 11 1 2 D mero TC TEMM EBT
Glucosa desh1drogenasa 11147 1 D mero SP
Alcohol desh1drogenasa 1111 1 2 D mero EBT
Transferasas
Fosfog1ucomutasa 2 7 5 1 1 2 Mon6mero TEHM TC
Isomera8as
Ilanosa fosfato 180m 5 3 18 1 Mon6mero EBT
Glucosa fosfato 1som 5 3 19 1 2 D mero EBT TC
52

ANOSA 6 FOSFATO
f MPI
FRUCTOSA 6 FOSFATO
G6P
6PG
IN AOpl
NAOPH
10 Sistema de tinción poro MPI
IL MALATO
PIRUVATO
Figll
INAOpl
NAOPH
Sistema de tinción poro ME
I ETANOL I
ACETALOEHIOO NAOH
FíO 12 Sistema de tinción para A O H
formazón
1 MTTI
formazan
I MTTI
formazón
r MTT I
3

UCTO A 6 FOSFATOIGPI
G6P
6PG
Fig 13 Sistema
NADP
NAOPH0
de tincian poro GPI
IL LACTATOI
PIRUVATO
I NAOI
NADH
Fil 14 Sistema de tinción poro LDH
MALATOJ
OXALACETATO
INAOI
NAOH
Fig 15 Sistema de tincion paro MOH
farmazón
l MT
formozón
IMTTI
formazån
IMTTI
4

LJCOSA I FOSFATO 1PGM G I 6 oiFl
G6P
6PG
INAOP armazón
tll MTT INAOPH
Fig 16 Sistema de Tinción poro PG M
I MTT I lI LUZ I
I PMSIformazcˇnmorado obscuro
bandosamarillo cloro
01
FíO 17 Sistema de tinacion poro SOO
lGLUCOSA 1
GLUCONATO
INAO p 1 farmazón
IMTT INAO P H
Fig 18 Sistema de tinción poro GDH
55

56
RESULTADOS
l Datos morfomØtr icos y mer sticos
En el presente estud io se ha puesto ha compar at ivamente poco Ønfas is
In los datos mer sticos Las cu ntas que se han utilizadoI
estan
Useæadas para establecer los nor rnales para las especies Y para dar
Ina indicación del r ango de Jar iacion El material disponible no fue
suficiente para j Jstificar un estudio exhaust ivo de variaclon
geogrÆfica en estos tØrminos Las cuentas de los r adios de las aletas
sin embargo son lo bastante jt Iles corno caracteres taxonómicos y se
determinaron siempre Tab 2 Y 3
Los datos morfomØtr icos en las cabr i 11 as fuero n mÆs satisfactorios
que los mer sticos en cierta forma pues la mayor a de los espec rnenes
fueron lo suf ic ientemente grandes para obtener considerable exactitud
en las medidas AdemÆs las especies anal izadas tienen caracteres
dhtintivos expresables a traves de las medidas tomadas Se muestran
los valores obten idos de 10 espec menes por espec ie col ectados en las
distintas local idades Tabs 4 7
2 Anål isis electrofor t ico y d istanc las genØt icas
I
Los resultados de los anal isis electroforeticos se presentan en la
tabla a Los valores de d istanc ia genØt ica Ne i 1972 calculados a
I
partir de los datos electroforet icos estan dados en la tabla 9 Se

57
Jestr el e ograma de identidad genØtica entre las cuatro especies de
oÞrillas est Jdiadas en la figura 19
3 Plrj fi ción del isozima M4 LDIl
El isozima M4 LDH del mœsculo esquelØtico de las especies estudiadas
H copla fuer temente a l oxamato inmov i 1 izado corl la resina
cromatogråfica Sept arose 48 a qui n reconoce como un analogo del
pirtlvato en pr esenci de NAOH y se eluye fÆcilmente a bajas
oncentr aciones sal iras Kel 6 25 t1 NADt 2 5 mM cuando el NADH se
s JSpende del irrigard e I igs 20 y 21
4 Estudios de la cinØtica de los isozimŁl s M4 LOH
Par a deter minar si tIa ocur r idc evolución molecular funcional
si9nificativa en respuesta a los diferentes entornos t rmicos de estas
especies se investigó la respuesta funcional de un locus con distintos
alelos en ellas Iuios sÜtemas enzimà1 icos mostrar on diferencias
llel icas errtre de s o ma s de las cabrillas Tab 8 Se decidió
restrigir los an lis is al enzima LDH del tipc muscular M4 o A4 por su
9 ð cti idad en el mo sculo bla nco del pez su estabilidad en el
tejido congelado y porque ex iste una gran cant idad de irformac lón de
apoyo en la l iteratLlra Yarlce y Se rrero 1878 a b SClmerc
Slebealler 1879 Go a es y Somero 1882 Graves l 1983 Esto
vltirO l no es el c so de clJ3 1quietotro enzima que mostr j E difererlcias
aiØl cas fijadas ertre las especies de Epineptelus Tres formas

58
electrofor ticamerlte distintas del enzima M4 IDH se resolvieron er las
cuatro especies Tab 8 Cada especie fue rilonorrórfica para s u alelo
particular el tamaæo de las muestras se lista en la tabla 8 f
analogus y E 1 abr Hormis las dos especies de aguas templadas
compart ieron un aleI o con mov i 1 id d anhd ica lenta mientras que la
subtrop ical E pð namens is la tropical E acanthistius se
encontraron ij adas para los ale1 os de mov i I idad intermed ia l råp ida
respect i lamente
Los valores de Km con respecto al p iruvato de valios ind iv iduos de la
misma especie se muestran en la tabla le Se hicieron rØplicas de los
ensayos enz imÆt icos ut il izando I a mismaI
pr eparac lon del enz ima o
preparaciones ltiples del mismo individuo resultando en una
reproducibilidad del SŒ
Los ísozimH M4 LDH electrofor ticarnente id nticos de E analogus l
E abriformis tambien mostraron los mismos valores de Km con respecto
al piruvato Tab 11
Sin embargo se observaron diferencias notables enI
la cinetica de los
tres alozirnas electroforØticamente distintos Fig 22 que fueron
resueltas en el anÆl is is prev iamente descr ito Tab 8 Mås aun los
yalores de Kcat de los tres M4 LDH examinados fueron notablemente
diferentes a 25 C Tab 12

59
rabll2 CARACTERES GENERICOS y SUBGENERICOS DE Epinephelus
Epinephelus
Epinephelus Cephalopholis
Longitud cefll ica gr ande grande
Aleh caudal redonda o cuadrada redonda o cuadrada
Hueso supramx i l al presente presente
Crestas fronto
parietales craneanas
convergente s
anter iormente
convergentes
anteriormente
Pared supra etmo idea presente o ausente ausente
AITlPI itud suborb ital grande grande
Proceso postocular pl esente presente
Espina antrorsa 1 ausente ausente
Radios dorsales IX XI 14 18 IX 13 16
Radios anales III 7 10 111 8 9
Radios pectorales 32 40 32 38
L neas transversas de
escamas
80 128 65 84
Branquispinas totales 21 28 16 17
Se describen nicamente el gØnero y subg neros que contienen a las
espec ies cons ider adas en este estud io
Con respecto al preopØrculo

alal al al al alØ
Ø Ø Ø Ø ØIl al ll ll ll alel Q Q Q QI 60Ø g Ø Ø Øal e g c
o o oQ Ø C C el C
QI Q QI QIo
co
Ø11 11 11 11ł
en I I I ICI CI CI CI
ł1Il alØ Cal
Ø o al alc al C C
11 Q Cc C C
Q C C c cS al c QI QI
i Q c cO c
I oØ
jl IIIal Q o ł Q Q QIel Ic lo
e c cQ Q QI
C en C 1Il mQ Q c Q l
ł III loQ al llIt
I I m I I oo c al al l lo QI
Nll 1Il o Q llfIl e o o al 1Il Q c 1Ilil O e 0 co lI o m l el III
al o Q o c u e kti el k lI k lI QlI mfIl al al k al lI
oc c lll al al IIIU o o C o OC C c kfIl k C 1Il m c C CIl CIl III CO o III11 o o m ll Q m Q ll al alkQ mti u k o k C k mcocofIlO
ti C
j lo O lO 0 III1Il
8 I Q11 CI O ON N N III
ti
alfIl0 o lO mł CI CI
í alk I I I QJ CCI CI 11 0 C alCI CI CI lOO C III C
l III lal lO O
el c IIIk 0 0 lO lO Qti No lo
rJ lDal ml QI llO QIfIl U k
3 c QI al al U
í lO l C QIC k l lo
CO 4 CIl c III m III m
QJ lo1 el Qm1Il lO III lO m
m CIl lOIII QI o al Q III Q
ll m c Cm1Il ll O c QI III Ql 1Il O C 1Il loQ om ClO lolD alClIl CIQ III e c C CCIl m 1Il m III III l O III llQI
o l c o O O C c C alc llO c Oc ooO C C CalC SaQ c lO m E cQ lO o lO c1Il
Izl el III c lOlO lO lo NCI 4 11 0 lO

TABLA 4 rFOIDAS PROPORCIONALES DE 10 ESÞECIMENES DE Epinephelus panamensis
Medición 1 2 3 4 5 6 7 8 9 10 media
Long i tud patrón mm 124 160 210 220 250 254 262 278 296 310
Longitud cefålica 389 413 416 400 391 413 407 400 401 411 4v4 1
AlII litud cefåUca 158 171 168 168 145 154 160 173 174 177 164 8Profundidad cefålica 253 252 272 269 265 275 279 275 272 276 268 8
Longitud del hocico 86 94 96 83 84 99 104 102 102 103 95 3
Amplitud subolbital 24 32 46 37 36 39 44 38 42 51 38 6
AllIp litud intel orbital 56 53 47 53 47 54 48 59 56 60 53 3
Longitud orbital 86 66 83 76 76 80 68 76 66 72 74 9Postorbital cefÆlica 231 252 245 236 200 250 251 243 230 251 238 9Mui lar 180 192 192 186 200 213 202 204 210 214 1993
t1 nd bula inhrior 191 201 20 1 200 201 220 210 212 211 228 207 8Hocico al preopØrc Jlo a7 290 293 284 287 290 293 296 287 300 289 7
Long i tud sup rama xi lar 60 81 70 63 67 60 64 60 60 61 646
Amplitud madlar 38 45 43 46 47 52 42 46 45 46 45 0
Amplitud del cuerpo 150 148 143 163 130 132 166 160 170 186 154 8Profundidad del cuerpo 334 365 360 365 357 345 367 362 363 363 358 1Profundidad pedo caud 136 138 144 148 141 101 136 140 141 145 137 O
Hocico origen dorsal 381 400 401 392 395 412 410 387 410 399 O
Hocico base pectoral 380 373 372 380 373 380 380 382 381 400 380 1Hotlco base p˛lvica 400 395 402 450 387 405 392 408 440 442 412 1
Long i tud base dorsal 515 552 519 555 540 530 545 545 505 488 529 9Oorul depr imida 620 680 f33 690 665 640 665 640 615 605 645 8
Base ana1 159 173 191 180 175 170 170 175 170 172 173 2
Anal deprin ida 31 335 340 344 320 304 318 305 298 310 318 6Fin dorsal inic caudal 150 146 146 135 150 135 138 135 140 140 141 5
Longitud pedo caudal 190 180 201 190 182 182 180 185 191 170 185 4
A1eh pector l 250 260 277 268 256 240 251 243 243 235 252 8
Aleta pØlvica 190 192 202 185 191 170 185 175 170 173 183 3
Espina dorsal 1 55 60 60 65 60 52 55 58 59 57 58 1
Espina dorsal III 120 132 120 130 130 120 113 125 113 120 122 3
Espina dorsal IX 110 125 132 120 120 100 120 110 113 100 1150
Espina anal 1 55 88 77 63 64 70 63 71 70 66 68 7
Espina anll II 135 140 175 156 140 132 120 140 125 120 138 3
Espina anal 111 135 140 152 130 140 125 122 126 115 110 129 5B caudll radios supo 220 240 245 236 225 224 215 214 205 200 222 4
B caudal radios med 255 265 282 268 235 240 230 225 220 225 244 5
8 clIudal radios ini 22 245 230 225 223 230 185 230 200 205 229 5
Todas las proporciones se expresan en milØsimos de la longitud patr n Los 15
pec menes provienen de Columna 1 Los Frailes Columnas 2 4 Cabo Pulmo 5 6
e Los llarril E Santo El Mechudo 7 10 Las Cruces
61

TARLA 5 nIDAS PROPQRCIONAL S DE 10 ESPECIMENES DE Epinephelus acanthistius
4
Mediciôn 2 3 4 5 6 7 8 9 10 media
Long i tud patrón mm 408 532 567 570 573 580 604 637 642 720
Long i tud cef lica 416 420 425 426 423 428 333 438 434 435 417 8
Amplitud cef4˛lica 190 183 170 164 180 170 183 199 196 195 183 OProfundidad cefålica 320 310 305 323 320 320 312 328 315 315 316 8
Longitud del hocico 95 97 97 103 94 94 187 100 100 105 97 2
Amplitud suborbital 32 26 26 22 30 30 30 36 40 40 31 2
Amplitud interQrbital 64 70 70 65 72 60 61 75 80 77 69 4
Long i tud orbital 100 100 95 103 87 92 105 83 88 802 93 3Postorbital cefílica 228 238 235 235 248 235 250 200 260 262 239 1
Ma xi lar 200 207 207 207 210 195 210 190 215 210 205 1Mandíbula inferior 182 210 200 205 200 186 210 200 217 217 202
Hocico al preopØrculo 0 290 300 303 306 255 300 290 307 308 295 9
Longitud supNlmaxi lar 70 86 81 83 72 70 68 73 79 72 75 4
Amp 1 i tud ma x ilar 50 52 50 52 53 54 55 50 48 48 51 2
Amplitud del cuerpo 156 174 154 166 155 163 167 160 172 172 163 9
Profund idad del cuerpo 410 406 400 407 397 385 383 390 466 420 406 4Profundidad pedo caud 125 120 125 126 126 126 130 132 130 140 128 0Hocico origen dorsal 330 393 390 400 400 380 407 394 400 405 393 9
Hocico base pectoral 394 425 426 405 390 390 390 373 400 405 399 8Hocico base p lvica 425 436 450 416 425 420 400 405 434 437 424 8
Longitud base dorsal 553 540 536 567 540 540 540 553 530 526 544 4
Dorsal deprimida 695 687 677 717 673 700 702 696 664 644 675 5
Base ana 1 2CO 196 205 201 197 195 186 196 190 200 196 6Anal deprimida 370 362 385 380 344 370 372 370 336 296 358 5
Fin dorsal inic caudal 122 121 130 120 120 118 130 127 120 125 123 3
Long i tud ped caudal 175 183 180 190 190 180 182 186 182 200 184 8
Aleta pedoral 310 310 330 320 300 300 313 312 295 300 309 O
Aleta pØlvica 275 280 276 300 256 282 276 282 246 244 271
Espina do al 1 60 62 70 57 62 60 64 64 61 52 61 Ü
Espina dorsal II 162 153 154 150 169 142 180 150 160 160 158
Espina dOI sal IX 163 170 170 163 165 163 173 170 164 143 164 1
Espina anal 1 70 58 69 57 62 58 66 64 60 60 62 1
Espina anal II 142 126 181 186 140 125 127 127 118 103 127 2
Espina anal III 152 146 152 150 147 140 166 150 142 121 146 6
B caudal radios s up 263 310 283 272 246 258 282 296 263 266 273 9
B cauda 1 rad ios med 287 267 320 312 292 293 293 309 290 284 294 7
B caudal radios inf 266 273 260 266 256 260 293 282 270 280 270 6
4
Todas las proporc lones se expresan en milØsimos de la longitud patrôn Los es
pecím nes provi ene de Columnas 1 2 Y 3 El Mechudo Los Planes La R1VEH dl
4 10 Cabo Pulmo Los Frailes
óz

ABLA 6 I IOAS PROPORCIONALES 1 1 ESPES It1ENES DE Epin phelus labriiormis40
r1edlC LCO 2 3 4 5 6 7 8 9 10 media
o liA patrón mmi 15S 226 242 280 298 319 344 367 378 383
Long i ud cefÆlica 427 446 C 428 410 430 420 400 437 427 425 7
mpli ud cefÆlica 177 193 2C 207 186 10 120 170 210 187 192 OProtlJnd t jad cefÆlica 242 265 2CJ 2ò7 242 265 252 242 270 260 257 2
LO i i tJ j del h 00 i c o 92 110 1 103 100 110 11 196 106 100 102 7
Arti i t d uborbltal 26 39 44 44 42 48 46 50 47 42 4
wp 1 i t Jd i nterorb i ta 1 50 88 6 9 2 O J 60 60 ó8 66 64 2Long 1 tud orbital 94 90 87 7 70 66 67 69 68 75 8Postorbltal ceTllitd 242 266 260 62 251 260 255 255 266 266 258 3t JJ ì lar 186 193 194 lQl 186 190 190 183 184 200 189 711and bula inferior 190 202 162 202 192 196 200 186 193 210 193 3
H JClçO al precpØrculo 286 290 223 292 288 2C12 286 286 285 293 288 6
Long tud suprama i la 57 57 59 63 60 57 56 48 56 63 57 5
r 11 t Jd i axllar 47 47 48 43 44 43 43 45 41 47 44 8
mp 11 tud del uerpo 1 6 166 223 207 156 192 146 1 T5 206 166 177 3
P f Jnd idad dlfl cuerpo 320 338 2E 27l 342 346 328 333 368 344 343 5
ProtlJna ilj d pedo caLd 117 103 l 120 110 11 1Ü6 112 110 103 1108
hOCIC J origen d 01 sa 1 El 406 313 4 393 387 390 385 384 403 389 0H t t base peçtoT al 400 382 40 36 360 377 360 366 340 360 370 9HOC1C1 base p lvica 453 428 S 413 4lJJ 426 4 07 415 407 414 423 4
onglttld ase dorsal 112 572 570 612 590 590 576 588 593 588 588 1CO Hl dØprímida 68 632 610 677 ö65 ó55 700 644 660 700 665 93a e illal 171 164 178 171 171 UO 158 170 167 163 167 2
AnJl aepr l lda 320 275 290 301 273 270 277 285 270 264 282 4
In d Jrsal iI11C caLdal 198 102 lO 100 1 02 103 136 120 960 103 106 4Jn 1 t A ped caUdil 186 185 18 leC 190 178 20C1 190 180 200 186 9AI til pectoral 241 217 217 22u 206 213 207 210 312 190 213 3Aleta p@lvica 193 172 17 80 166 156 161 155 157 157 166 7
Espha dorsal 1 70 52 60 70 55 50 50 58 48 40 55 3
Esp 1 1orsal III 148 133 1J 1 126 132 1 1 32 110 126 131 9
Espi a dorsal XI 135 102 115 1 6 101 104 194 193 191 135 107 6
Espi 13 anal 1 80 60 67 64 57 57 52 57 53 58 60 5
Espi il anal II 143 113 125 113 126 103 192 106 191 193 110 5
Espha anal III 144 122 12 11 6 112 107 107 107 105 110 115 3
e cau 1ill radios supo 221 210 05 d43 215 210 215 220 210 210 216 O
2 çauda1 radios ed 236 210 212 228 218 205 211 220 200 203 214 3
S c udil1 radios i ni 218 214 217 23 216 215 223 227 204 209 217 5
Tod J las proporciones se expresan e nilØsi lloS de la longitud patrón Los esI
pT VierH f de ee1 U ala 1 Los F all s ea 1 umr aS 4 6 8 10 1 EP Cllll es Ij
Santo Las Cruces Cueva de Leon 7 Cabo Pu 1 0 9 El Mechuôü
bj
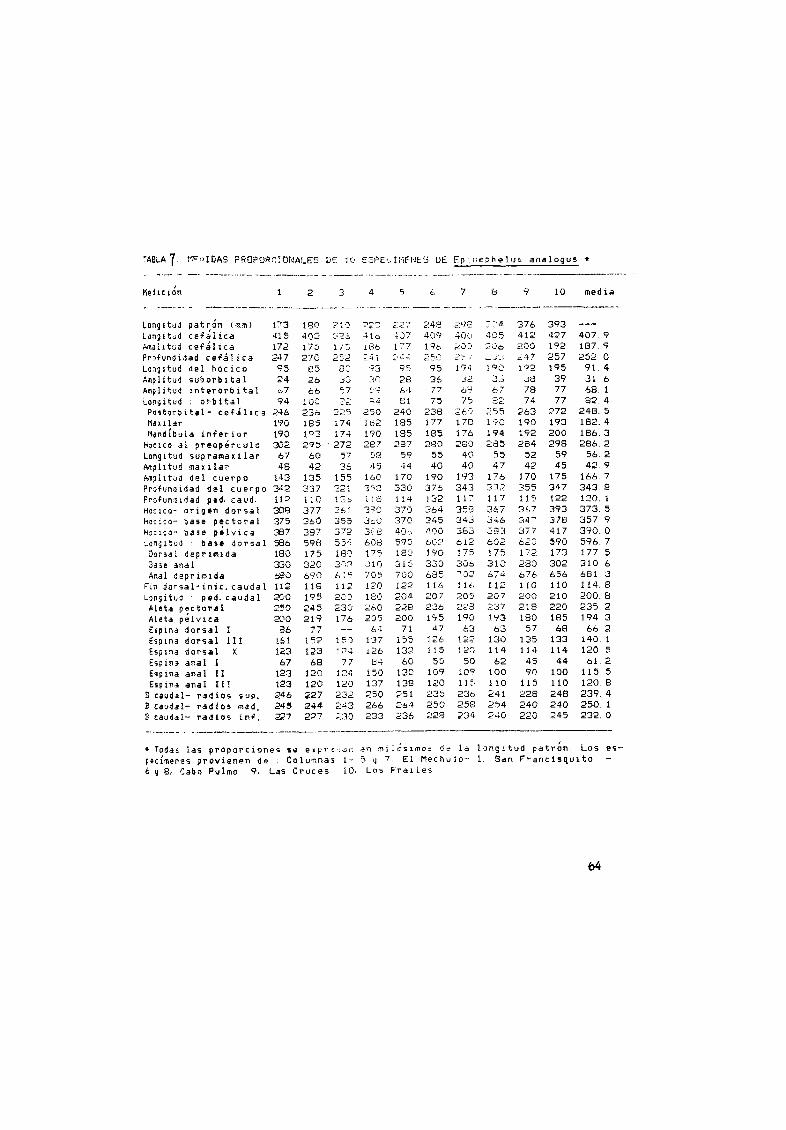
ABIA 7 DIOAS PROPORCCNA ES DE le ESf E 1 tr FNES DE Ep lnepnellJs analogus
liedlcíón 2 3 4 5 6 7 8 9 10 media
M
Lon9 í tu patr ón TI 173 180 o 23 Z 27 481
j 2 376 393
Lon í tA cef líca 415 402 116 107 409 40iJ 405 412 427 407 9
11 01 i tlJd cefº˝llca 172 175 175 186 177 1 1 d0 20 200 192 187 9
Prot JMí jad ce Ælica 247 270 2 d 1 Lrt J 247 257 252 O
Ll í tud del hocíco 9S e5 O i3 95 95 194 190 192 195 91 4
A p 1 í tJJ su olbítal 24 26 j iC 28 36 32 38 39 31 6
Amplítud Interorbítal 7 66 7 64 n ó9 i 7 78 77 68 1
Long ltud orbítal 94 10 c 84 81 75 I 82 74 77 82 4
Po torbítal cef˝l1ca 246 236 225 250 240 238 2t 255 263 272 248 5
tia ular 190 185 174 182 185 177 178 190 190 193 182 4
tiand bula ínferíor 190 193 17t 190 185 185 176 194 192 200 186 3
HOCl c o al preopculo 32 29 272 287 287 280 280 285 284 298 286 2
Longítud supl ama 11ar 67 60 57 08 59 55 40 55 52 59 56 2
AIp 11 tud ma i 141 48 42 35 45 44 40 40 47 42 45 4 9
A ill í tu j del cuerpo 143 135 155 160 170 190 193 176 170 175 166 7
ratuM ídad del cuerpo 342 337 321 3 8 330 376 343 332 355 347 343 8
PrO JM Idad pedo caud lP llC 13 118 114 132 117 117 115 122 120 1
HOCl e0 origen dar sa 1 338 377 2f 390 370 864 358 367 3b7 393 373 5
Mace c a base p ctoral 375 360 355 360 370 345 343 346 347 378 357 9
MacIc base pt lvíca 387 387 372 3C 8 40 41 0 383 33 377 417 390 0
on ttuj ba1t dorsal 586 598 5 i 608 595 tJ 2 612 602 620 590 596 7
Dorsa 1 deprlmída 180 175 18 175 180 190 175 175 172 173 177 5
Sa e 3nal 330 320 3 31 315 330 306 310 280 302 310 6
Anal d pr irnda 080 690 6 5 705 700 685 2 674 676 656 681 3
FlO dor al ínic caudal 112 118 112 120 1 116 116 112 110 110 114 8
Laogituj pedo caudal 20 195 2QQ 180 204 207 205 207 200 210 200 8
Aleta p ctoral 250 245 230 260 228 236 2 8 237 218 220 235 2
Aleta pelvlca Ó0 219 176 205 200 115 190 193 180 185 194 3
Espín dorsal 1 36 77 64 73 47 63 63 57 68 66 2
E p In dorsll II 161 1 15 137 155 126 122 130 135 133 140 1
EspIna dorsal X 123 123 1 4 126 132 115 12t 114 114 114 120 5
E p ín 3na 1 1 67 68 77 84 60 55 50 62 45 44 61 2
E p1na anal 11 123 120 124 150 130 109 109 100 90 100 115 5
Esp í f13 anal II 123 120 12 137 138 120 11 110 115 110 120 8
S caudal radios s p 246 227 232 250 251 235 23b 241 228 248 239 4
B cauda 1 radios mæd 245 244 213 266 264 250 258 254 240 240 250 1
a ca Jdal radíos i nf 227 2 7 J J 233 236 228 234 240 220 245 232 O
ToddS las ploporcíones se ei pf at 1 milØ51mOS d la longItud patr n Los es
pecím@r s províenen de Columnas 1 5 J El Mec hu j o San Francisquitob a Cabo Pulmo 9 Las Cruces lO Los Fralles
64

jibia ð FT l culncias alØlicas dt los loci polimól ticos en y entT P cuatro espechs dI cabT illiH del 1 1 de COT t Aquf llos loci neo 1 istadoaquíse I ncontT aTon fijados paTa el mismo alelo en las cuat o esperitosLos valorls pTóximos a los nombreeper Ficos indican el tamaæo de
las muestras
4
Lar Ale E analog 15 panamlnsi s 5 E acanthistius 15 b labl il oTffil ı 2
ldh a a 1 00b 1 00
100 1 00
Mdh a 1 00b 1 00 1 00
100
slldh a 1 00b 100 1 00 100
Me a 100 100 100b 1 00
Mpi a a 100 1 00 100b 1 00
gm b a 1 00 00 1 00b 1 00
Gp i a a 0 20b 0 90 1 00 O 80 0 84
O 10 0 16
Gpi b a 1 00b 100 100
100
Adh a 100 100b 1 00 1 00
Sod a a 100 100 1 00b 100
SOd b a 1 00b 100 1 00 100
Gdh a 100 100 100b 100
NOlllbrl s abrlviaturas de los loci investigadolactato deshidrogr nasa
ILdh a Ldh b Ldh c malato deshidrogenasa m Mdh sMdh a s Id h b en
65

lima mÆ1icil Me manosa fosfilto ìsofnl rasa Mpi —J fosfog1ucomuta iJ Pgm a
pgm b glucosa fosfato ìsomel asa lpi a Gpì bJ alcohol deshidr09fnilSa Adh
superondo dismutasa Sod a Sod b 91l1 05 d hidroYlnaSΠGdhl
b

1 Þla9 Distancias genØticas N i 1972 error standard entre cuatro especies de cabrillas del Mar d Cort s La estimaciollse basan en 16
loc i comunes aparentemente a las CUijt o esp ec ies Ap arer en entre
parØntesis bajo los valores de distancia genØtica los tiempos de
divergencia en millones de aeos basadt s 01 la corH lación de SarlCh
1977 entre los valores de distancia genØtica y los tiempos de di
vergencia para vertebrados modiFicado por Carlson 1 21 1 178
S labrilormi s b acantllistius panampnsi s
malogus O 302 1 0 075 0 b05 1 O 110
5 75 1 1 40 11 32 t Oó
1 panamen s i s 0 190 t 0 055 O 780 1 0 1P
3 55 100 14 75 1 2 51
1 acanthistiu5 O b35 t O 1bO
12 00 1 2 15
O 365 t O oao6 85 1 50
b7
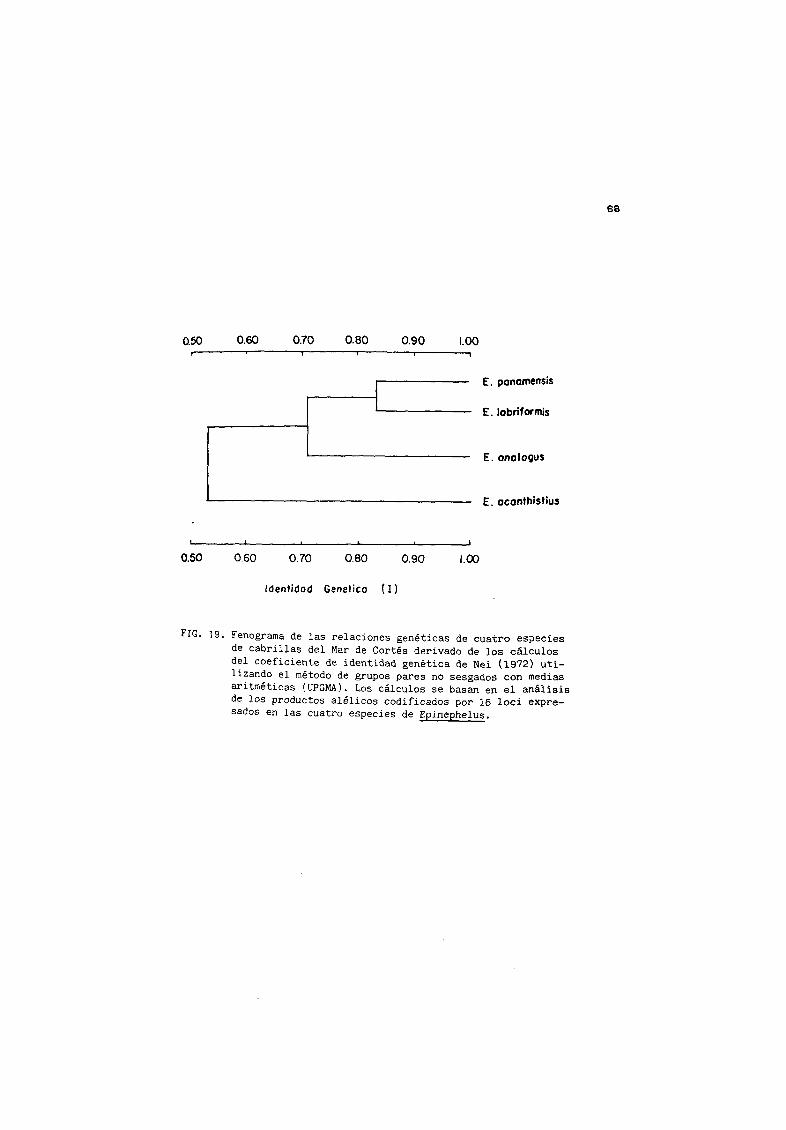
0 50 0 60 0 70 0 80 0 90
11
0 50 0 900 60 0 80
100
E ponomensis
E lobrlformis
E onoJoous
E oconthistlu
100
FIG 19 Fenograma de las relaciones genØticas de cuatro especiesde cabrillas del Mar de CortØs derivado de los cÆlculosdel coeficiente de identidad genØtica de Nei 1972 utilizando el mØtodo de grupos pares no sesgados con mediasaritmØticas UPGMA Los cÆlculos se basan en el anÆlisisde los productos alØlicos codificados por 16 locí expresados en las cuatro especies de Epinephelus
0 70
Identidad Genetica 1
ss

CH2C
C2 C
CH2NH ICH2
NH
IC O
Ic
e O
ti
o
Clsaal
en
CH3o
C
IC
el 0
NH20
Cr
IC
e 0
Oxomolo Oxomotoinmovilizodo
FIG 20 Analogía estructural entre el piruvato el oxamatoy el derivado del oxamato acoplado a la matríz desoporte Sepharose O Carra y Barry 1972
Piruvoto
6S

roJ
sin irrigonte
GIg
goO
16 mM NAO
u
t
16 mM NAO
o 4 8 20
Volumen de Elusión u ˛ colurmo
FIG 21 Separación de los isozimas M4 punteada y H4 continua del
tejido muscular de las distintas especies de Epinephelusmediante cromatografía de afinidad en columna de Sepharoseoxamato
70
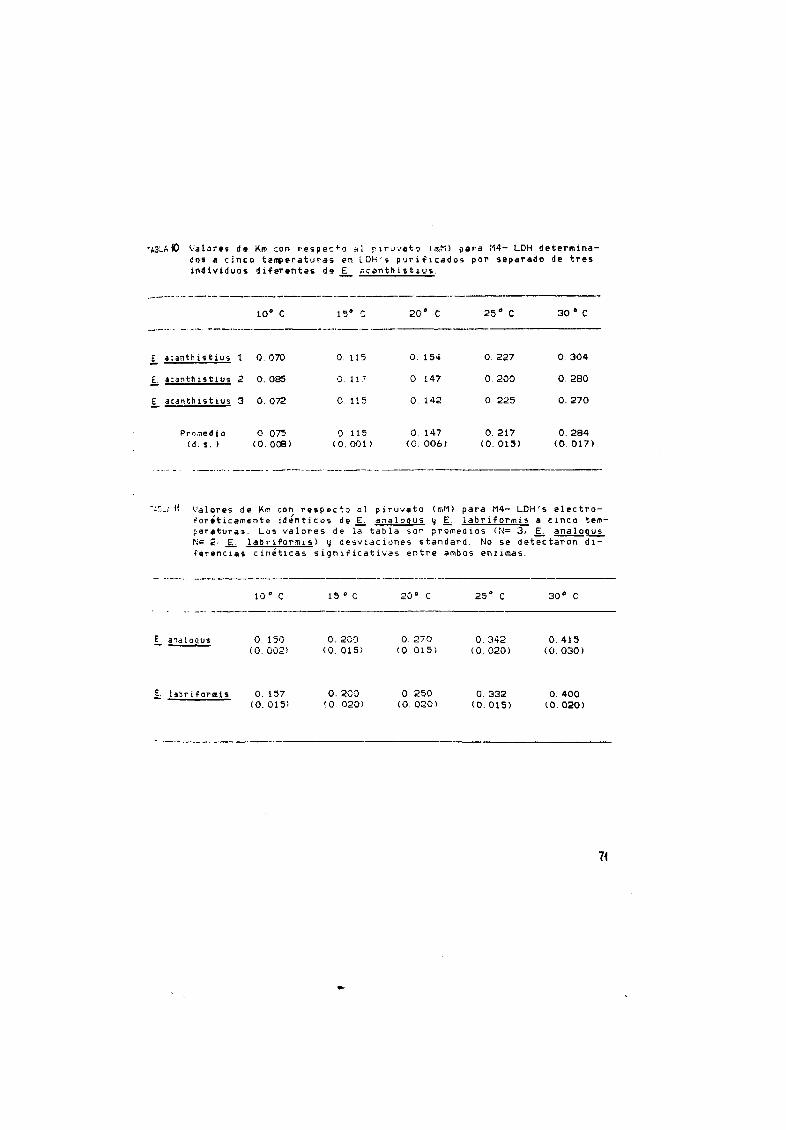
ASLAIO Valores de Km con Tespec l prJ ato cr M para 1 4 LOH determinados a cinco temperatur as en LDH s pCJriflcados por separado de tresindividuos diferentes de canthistlus
10 e 15 S 20 e 25 e 30 e
J a anthistius O 070 0 115 0 154 0 227 O 304
J a anthistius 2 0 085 0 117 O 147 o 200 0 280
L acanthistius 3 0 072 o 115 o 142 o 225 0 270
Prc 1ed io o 075 0 115 o 147 0 217 0284d s 0 oca o 001 0 006 0 015 0 017
3 i ti aloTes de Km con Tespecto al piruvato r M para M4 LOH s electrofOTØtica1 nte idØnticos de a aloQus y g labriformis a cinco tem
peTaturas Los valores de la tabla son promeaios N 3 E analogusN 2 L labrifor llis y desviaciones standard No se det t dTferenclas cin ticas signlficatlvas entre ambos enzimas
leł
e 15 e 200 e 25 e 30 e
E a dlo IUS O 1500 002
0 2000 015
0 2700 015
0 342
O 0200 4150 030
1 labriformiS 0 1570 015
0 200O 020
0 2500 020
0 3320 015
0 4000 020
1i
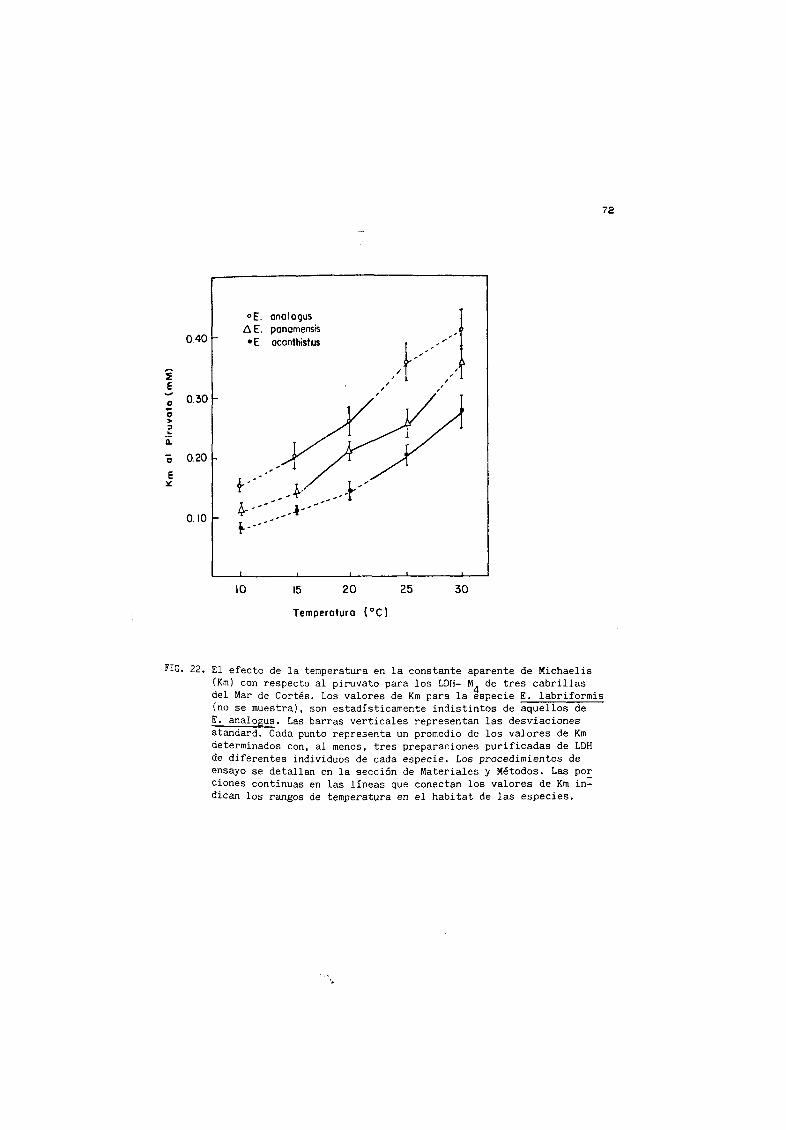
72
iE
0 30
j040
oE analogusð E panamensis
E acanthistils
o
Õ
ä
º 0 20
EC t X L
Xr
f
t0 10
10 15 20 25 30
Temperatura Oe
FIG 22 El efecto de la temperatura en la constante aparente de MichaelisKm con respecto al piruvato para los LDH M4 de tres cabrillas
del Mar de CortØs Los valores de Km para la especie E labriformis
no se muestra son estadísticamente indistintos de aquellos deE analogus Las barras verticales representan las desviaciones
standard Cada punto representa un promedio de los valores de Km
determinados con al menos tres preparaciones purificadas de LDH
de diferentes individuos de cada especie Los procedimientos de
ensayo se detallan en la sección de Materiales y MØtodos Las porciones contínuas en las líneas que conectan los valores de Km in
dican los rangos de temperatura en el habitat de las especies

laÞlalZ ParÆmetl os cinØticos dI los t14 LDH Plll ificado de trt s cabl illas del
Mar de cortØs a S C J las temperatuTas media de ltJs habitats decolecta TI1 paT a cada espec ie LtJs valul E pilNl LDH de L 1 01 1 FtJT ll
no se muestran no di fiertn significativmente de aouello de
analogus Kcat no se determlnó par a el lDH de E labri fOl mis Se daneViaciones standard para los paråmetT os 7inØtilos a 2S C Las
estimaciones de Km est6n basads en las mediciones hechas con cincopreparaciones separadas de LDH de cintu indivlduos de cada especlelas estimaciones de Kcat estÆn basadas en tres purlflcacione de LDH
para cada especie LtJs valores de Kcat y Km a las temperaturas TM
son interpolaciones de grÆfiCOS de Arrhenius J grÆficos de temperatura respectivamente
analog pallampnf i 5 ßac
mthistius K1ll piruvato 2S C O 35 t0 02 roM O 25 t O 0 mM 020 t 0 02
mM Kcat I25 C 895 t 54 seg 1730 t 37 g l 660 t 19 se9
1 Temp lIled TM lS C 23 C 26
e Km piruvato TM 0 25 roM 025lÑ 1 O 24
mM Keat I TM 670 seg l 685 eg 1700 seg
17

74
OISCUSION y CONCLUSXONES
t Med lc lones proporc ional es
Si se encontraban disponibles diez espec lmenes de cada especie
fJeron medidos para establecey sus proporciones corporales Las
mediciones se transformaron a milØsiroos de la longitud patrón y vienen
dadas en las tablas 4 7 Corr el propósito del presente estudio no fue
la investigación de la variación geogrÆfica los especimenes fueron
selecc ionados para su med ic ión s in importar su procedencia Las 37
medidas tomadas se hicieron con el fín de enr iquecer el conjunto de
datos qtle habr ía de contener 1 as py inc ipales caracter sticas anatómicas
de las espec ies Para fa cilita r la comparac ión ce y otros espec imenes
las mediciones estan dadas para los individuos junto con sus promedios
Algunas medidas fueron practicamente constantes para todas las
especies otras revelan tendencias dentro del gtnero o muestran
resemblanzas dentro de los grupos especie que 1 as incluye Smith
IS7D Las espec ies valias de Ep inephe 1 us y los grupos especie del
mismo gØnero mostraron divergencias notables en algunos caracteres
Tabs 4 7 Con r especto a la long itud del max i 1 al E acanthistius
difiere de una manera pronunciada en relaciÓn con las otras especies
De iglJal manera la longitud de la aleta pØlvica es un caracter bien
Grupo especie l Epinephelus Cephalopolis cruentatus f Epanamensis 1 gr Jpo especie 11 E Epinephelus niveatus L Laeanthistius grupo especie IlI E E guaza E E labriformis
grupo especie IV adsensiıis fE anïogu

ns
deflniclo Pl I esta espec le y para el grupo al ue pertenece tal
longitucl es negao tilamente alomØtr ica para ellos
E panº mer s is mostro la tercera esp ina dor sal corta y por el
contrar io 1 a pr imer a esp inao anal larga con respecte a las otras
espec ies
La significaci n sistemÆtica de estos caracteres se discutira en
conjunto con otros rM S tarde
e Caracteres me l sticos
Algunos caracteres contables de es t as ca br ill as fuer en lo
suficientemente constantes para ser de util id ad el la distinción de los
gØreros y espec ies aunque pa r a este tr abaj o impe r taba mÆs cual es elan
edos caracteres mer sticos pr opios I tiles P ara identificar sus
relac iones interespec f icas S in embargo como todos los caracteres
taxonomicos los datos mer sticos lar tan independientemente de tales
relaciones Por ello la pØrdida de espinas dorsales en aralogus y
acanthistius s lo podrÆ evaluarse dentro de toda una estructura que
incluya a todos los caracteres para estas especies y aquellas
relacionadas Tabs 2 y 3

76
S El eshldio electroforØtico las distancias genØticas y la
sistemÆtica de Epinephelus
I
Los resultados del estudio electroforetico presentados en la tabla 8
las d istanc ias genØt icas Ne i 1972 calculadas de los datos
electroforØticos mostradas en la tabla 9 y el fenograma de ident idad
genØ1ica de I as cuatro cabr i 11 as pr esentado en 1 a figura 19 hacen
evidente que las dos cabrillas que se presentan en aguas templadas
López Lemus op cit se encuentran mas cercanamente relac ionadas
entre sI que con la especie tropical acanthistius Las distancias
senØticas entre las corooinaciones pares del grupo L anal ogus f
F anarnensis labrHormis son de alrededor de un medio con respecto a
las distancias entre cualquier miembro de ese grupo y acanthistius
Las distancias geneticas calculadas a partir del estudio
eleetr ofe rØt ico de estas cuatro espec ies de cabr illas del I1ar de
Cortes se prestan a discusión con aquellas afinidades basadas en
anÆlisis mOrfológicos L acanthistius difiere de las otras especies
por su gran long i tud max il ar 1 argos rad ios en la aleta pØlvica y
euerpo muy profundo alJnque no compr imido d ir ase muy bien que es
robusto Las cuentas de los radios de la aleta dorsal son altos en
relaci n a las de su grupo especie tiene 17 pero nicamente 9 espinas
SI la pØrdida de espinas ocurrio en el borde posterior de la parte
up inosa de la dorsa l entonces esto pudo ser acompaæado por la
ganancia en la misma I enea de dos r ad i os suaves La reduccicSn de
esp Inas en la dorsal dentro de esta misma 1 nea es un hecho que
tambiØn se observa en E anal09lJS qlJe tiene una espina dorsal menos en

77
relación al res o de sus congØneres del grupo especie al que per enece
acan h is ius fue r efer ido al gener c Cephal ophol is duran e rwcho
tiempo MeeK y Hildebrand 1925 Smi h 1971 por ener 9 espinas
dorsales sir embargo por el res o de sus carac eres ha sido
claramen e ð S igrado al gr 1Jpo espec ie b n ivea us del genero
Epinephelus por compart Ir la gos rad ios pØlvicos cuerpo profundo
coloración roj iza ac ocola ada y aletas pelvicas obscuras CSmi h
1971
Existe una gran can idad de rabajos que discu en Łt un 1 as afinidades
diferencias entre las cabrillas Las especies del Pacífico Oriental
inclu do el Mar de Cor Øs han s ido tra adas en ex ensas revisiones Y
diversas publ icaciones CGosl ine 1966 Johnson 1983 Snli h 1961
191 51 1971 Wal dman 19841 Wal ford 1937 Johnson y Keener 1984
Randall y Ben TJvia 1983 Robins y StarcK 1961 Rosenbla y
Zahur anec 19671 Smith y Young 19661 Thomson ll 1979 En todas
ellas se encuentran discrepancias en cuan o a los niveles taxonómlcos
de espec ies c gØneros ya que mien rð s unos reconocen generos o ros
reconocen a es os como subgeneros Tal es el caso de Cephalopholis
Sm1th 1971 Rar dall Ben Tuvia 1983
En el presen e rabaJo se es udio una especie de es e genero o
sUbgØnero de Epinephelus segun S ith 1971 Cephºdopholis difiere del
r esto de los EpinephellJs por tener 9 espinas dor sales œnicamente y
pequeæas modificaciones en el cr neo Este g nero cor rende a una sola

78
upec i en el PilC f ico Or iental fpanamens is que muestra una
cercana r elación con L k labriformis de acuerdo con los resultados
aquí presentados
Es pertinente hacer una obsevación al respecto de las evidencias
discutidas hasta aquí es evidente que si la clasificación de E
acant istius fuera correcta dentro de Cephalopholis MeeK y Hildebrand
1925 cl br la esperar In parentesco genØt ico mÆs cercano con E
panamens is y s in embargo se observa que no es as Fig 19 mÆ s aur
las i Hn idades genØt icas de panamens is son mayores hac ia la espec ie
L IabrHormis y L analogus de aguas templadas que al mismo E
acanth ist i Js
Con el objeto de esclarecer los puntos de discusión considØrese la
figura 23
Para que sean Ýtiles en la determinación de I neas filØticas los
caracteres empleados deberÆn ser consistentes en y entre los grupos
lnvolucrados y deber n ajustarse a un m n imo de var iac ion indiidual
En las cabrillas los car acteres mer sticos son œtiles y los rasgos
o teológ icos son mw importantes 111 gunas de 1 as med ic iones
proporcionales sirvieron para separar grupos principales e ind icar
dem s las tender c ias de estos grupos mej or que con otros caracteres
La coloración provee del mÆs œtil de los caracteres en el campo pero es
de valor 1 imitado en la indicaci n de filogenias El patr n juvenil en
pa rticulðr es œt il en el al ineamiento de ciertas espec ies

7S
En ocas ione se le da demas iada importanc ia a la apar ienc ia general
de los organismos pero tale impresiones subjetivas requier er de apoyo
por parte de otros r asgo que puedan responder mejor a una
cuantificación mÆs precisa
El esquema evolutivo del tronco Epinephelus en sI mismo dista mucho
de ser completo para las especies anal izadas en base a caracteres
morfo tricos mer sticos ShaKleeœ 1982 Smith 1971 Fig
23A identifica dos linajes distintos para e tas especies una l nea
rrJs e pec ial izada de Ep inephe lllS a Cephal opho 1 is cc mc sllbgØner os
las observaciones aqll realizadas concuerdan con aquellas de Smith en
cuanto a caracteres anat micos Dentro de ellosflabr iformis Cll a
afinidades son difíciles de establece es colocada pOt Smith 1871
CHcana al grllpo especie E triatlls por Sll semejanza estrllctllral
aurque tamb i4n sug ier e qtle sea ur forma inmigrante de la region del
lndo Pac f ico s in par ientes cercanos en Ar rica Finalmente propone
una soluci r mÆs viable a esta incógnita que E labriformis es llna
ramificaci n reciente del propio tronco Epinephellls hecho qlle
concuerda de una manera parcial con el esqllema qlle aqui se propone
Fig 238 en base a la información obtenida a partir de los estudios
descritos
El grupo especie al qlle pertenece analoglls tampoco posee parientes
cercanos en Am rica y tambiØn evolucion6 probabler nte de la r eg lon
Indo Pac f iCil Pc el contrar io aquel gr llpo e pec ie que ont iene a E
acanth ist ius a otras espec ies evolllC ionb en ag Jas amer icanas

80
p obablemente De aqu su ant iguedad de acuerdo al esquema aqu
propuesto con respecto al resto de las especies estudiadas y de origen
no ðmer icano seg n las propos ic iones de Smi th 1971
De lo anteriormente seæalado se desprende que i analogus y i
acanthtstius evolucionaron como dos 1 neas diferentes e independiente
tal afirmaciJn va de acuerdo con el esq lema 238 hasta cier to punto el
dendrograma que se propone presenta a E acanthistius como miembro de
una I nea que seI
segrego independientemente del tronco comun
Ep inephe IIs hace un s 15 mi 11 ones de aæos F ig 18 pero E analogus
proviene de una ancestro cor caracteres de panamens is labr iformis
y los prop ios desde ese mismo t ie o d iverg iendo hac ia la forma actual
de L anal ogls poster iormente
acanthistius de distingue como ya se ha dicho por sus 1 ar9 os
rldios pØ I J icos Y cuen1as dorsa les carac1er sticas IX 11
Otra 1 nea ident if icada por una r educc i n secundar ia del rll meo de
esp Inas dorsal es a 9 dio origen a laI
apar lC lon del sub gØnero
Cephalopholis mismo que incluye a la especie L panamensis tratada
aquí cuyas grandes afinidades genØticas con el resto de las especies
de cabr i 1I as sug leren que Cephal opho 1 is es una I enea rec iente der ivada
del mismo sub9 nero Epinephelus desde hace unos 3 5 millones de aæos
LaI
invest igac ion electroforØtica de es1 as especies revele la
t istencia de suficientes diferencias al licas entre ellas hecho que

81
reconf Irma su sttus de espec ies separadas s in embargo el parentesco
de E panamens is con E I abr ifc rrl1is s ugier e ura reciente
divers if irae itn del 1 ron o Epinephelus El trabajo de los
sistematieistas taxonomos de tratarla dentro de un grupo diferente en
base a sus caracteres anatamicos o como una especie dentro del propio
9 nero Epinephelus nicamente tomando como bases sus afinidades
ger ticas const ituye l JrJ impor tante punto de discusion en el cual
deber tomarse rr y en cuenta que la reciente definición evolutiva del
gØnero al n no muestra 1 a suf íc ierlte divergencia genetica a nivel
1OO1ecular en cc mpar acitn cor la notable divergencia de car acteres
anatomicos De tal forma ser vÆlido llamar Ia Ep inephe 1 IS y o
Cephalophol is panamens is hasta que desPŁlØs de tomar en cuenta toda la
Información que se encuentre y discutirla de la manera mas amplia se
llegue al acuerdo sistem tico que habr de definir las categor as de
los gØneros v especies del tronco Epinephelus
Las dos especies de aguas templadas analogus y labriformis y
aquella subtropical E panamensis muestran afinidades genØticas aunque
no anat micas a lo largo de la zona costera del bajo Golfo
Sudcal iforn iano Hubbs 1952 seæal que tales distribuciones de
afinidad son comunes en los peces del Pacífico Drier tal y son
resultado probablemente de cruzamientos ecuatoriales en los perfodos
de compresiJn tropical durante el Pleistcceno Sin embargo al aplicar
la correlaci n distancia genØtica tiempo de divergencia de Carlson
L 1979 modH icada de aq Jell a de Sar ich 1977 obtenida para otros
vertebrados a las distancias gen ticas que separan a las cabrlllas se

8a
obtiene un tien o de divergencia de 3 5 millones de aæos para el par
s cercanamente relacionado panamensis y E labriformis La
distancia genØtica 0 190 que separa a estas dos especies es ademas
cercana a la distancia media entre especies pares de peces separados
por la emers i r de 1 Istmo de Panarnt Vawter II 1980 evento que
ocurr i hace unos 3 mi 11 ones de aæos Ke igw in 1978 As r a pesar de
Iue las cabrillas terl4 ladas Y subtropicales pueden mostrar ciertas
afinidades y o diferenciaos gen ticas y o morfológicas los tierl4os de
divergencia obtenidos de las distancias gen ticas indican que la
separac ion ocurr io durante el Plioceno que la separacion de E
analogus OCllrr iÓ en c iertaI
epoca anterior del mismo Plioceno 5 7
millones de aæos y que E acanthistius se separ de todas ellas
durante el Mioceno Pl ioceno 11 15 mi 11 OMes de a1l 05
4 Estudios el la cir Øtica de los ise zimas M4 LOH
Al comienzo de las comparaciones cinØticas de estos alozimas t14 LOH
diferentes se determin la variabilidad de los patrones cin ticos de
los isozimas purificados de un numero dado de individuos de la misma
espeCie Las replicas de los Km con respecto la piruvato utilizando la
misma preparaci5n o preparaCiones ltiples del mismo ind iv iduo
demostraron una reproducibilidad del 92X Como se muestra en la tabla
10 los valores de Km de val ios ind iv iduos de la misma especie caen
tambi n dentro de este estrecho rango AsI con todo y las limitaciones
de 1 a t cn ica y los tamaæos de muestra no parece existir valiac ion
electrofor tica funcional de caracter crlptico en los M4 LOH de cada

S
UM je 1 c la tr œ espec ies al examinadas
Los M4 LDH electroforØticamente idØ nt icos de E anal09us y f
lat r iformis mo tr æ r orl los mi mC1S valclre para Km con respecto al
piruvato Tab 11 A todas las temperatura s de experimentación no se
e cont aron dif re cias otables entre las dos especies A cada
temperatura los valot es de fm co inc idieran con el 92 de
reprc duibilid ad jo a mer cionado As es plausible decir que existe
ident idad electroforØt ica y c inØt ica entre lc s t44 LDH de das congØner es
1ve habitan arrbientes templados en simpatr la Figs 6 jo 7 19 p ro
C n 5 millores de ar c de divergencia e olutia pe r lo menos
Sin errbargo entre los alozirr s electroforØticamente distintos se
encontrarQn diferenias cinØticamente nQtables Fig 22 A las cinco
temperatur as de ens a o los valQres de Km Con respecto al p iruato de
lo tres alc Ł imas fueron aprec iablemente d ist intos Sola a 30 e lQS
errores standard de E ana 1 ogus E panamens is se solapan
mar9 inalmen te AÝr mŒ s lCI s va le r es de Kcat de lQS tres M4 LDH al ut
examinadQs fueron diferentes a 25 C Tab 12 Con lo anterior los
tres alelos electroforØticamente distintos mostraron d iferenc ias
f lrcion les nQtat les en tØrminos de eficiencia catalftica K t en
Km con r espectQ al pirvvato
Estas diferencias funcionalei entre los alozimas 114 LDH parecen se
compensa cior es a la temperatur e irterpretables en tØrrr inos de los
diferp ntes r es mnes tØr mlCOS exper imentado por las espeCles de

84
cab 111as Pa a los valores Kcat el enzima de la especie templada E
analogus mostr un valor mayor a 25 e que el enzima de la especie
subt opical panamensis Esta 61tima en carrbio tiene un valor mayor
de Kcat que el enz ima de la espec ie màs trop ical E acanth ist ius Tab
12 Se observa as l que 1 a eficiencia catal h ica de los LOH esta
inversamente relacionada con la temperatura promedio del habitat
temperatu a corporal relaciJn que tiende a compensar los efectos de
la tellferatura sobre las reacc iones metabch icas N6tese que a las
temperaturas corporales promediO de las especies los valores de Kcat
de los tres aloz imas LOH son muy paree idos Tab 12 Los datos
refe entes a Kcat de 1 a tab1 a proveen de 1 a ev idencia de que
diferencias menores 4 7 C en las temperaturas corporales promedio
son de suficiente importanCia selectiva en el favorecimiento de
diferencias termocompensatorias en la eficiencia catal tica
A n mÆs pronunc iada es la conservac ión observada para los valores de
la constante aparente de Michael is Km con respecto al piruvato a
tellferaturas fisiológicas para los LDH electrofor ticamente distintos
de estas cabrillas Fig 22 A pesar de que el Km con respecto al
pi uvato fue depend iente de 1 a temperatura en todos los casos los
valores de Km observados a lo largo de los rangos de temperatura en los
habitats de las cuatro especies fueron extremadamente parecidos De
hecho el Km con respecto al p iruvato en el punto med io de cada rango de
temperatura para cada espec ie fue v irtualmente e 1 mismo er todas e11 as
Tab 12 Fig 22 Los valores de Km coro respecto al piruvato
encont ados en los LOH de estas cabrillas cayeron dentro del rango de

85
valores observados para otros M4 LDH de vertebrados probados a
te eraturas fisiológicas y pH mismos que aqu se realizaron 0 15 0 30
ntIde p irUlato Yancey y Somero 1978 a y b S in errbargo en este caso
la conservac ion no es una tendenc ia ampl iamente d istr ibu ida entre
orsar ismos filogen ticamente distantes sino un efecto a escala fina
e tre congener es que v i en erl amb iertes 1 igerarnente d iferentes Como en
el caso de las adaptac iones de Kcat 1 as d iferenc ias t r micas mln lmas
ya mencionadas son de suficiente importancia selectiva para favorecer
cambios evolutivos en los valores de Km
En conclusi n se descubrieron diferencias alØlicas funcionales entre
los M4 LDH de las especies analizadas Los datos de la cinØtica de los
enzimas Tab 12 muestran la existencia de diferencias en ambos
par metros determinados Km con respecto al piruvato y Kcat entre las
e pecies que abundan en los habitats templados y aquellas en habitats
mas tropicales en todos los casos los enzimas de las primeras
especies parecen estar perfectamente adaptadas a aguas mas fr as en
relaciÓn con los enzimas de las œltimas Hecho que tarrbi n explica las
diferencias en abundancia de ciertas especies en ciertas localidades
donde las aguas o bien son templadas o bien tibias As de acuerdo
con estud ios real izados en otros ver tebrados mar cadamer te diferentes
peces reptiles mam feros y que tienen temperaturas corporales
extremas entre 2 C pecs antÆrtico y 46 47 C reptiles del
desierto los LDH de las especies que abundan en aguas frías tinen
inherentemente valores mayores de Km con respecto al piruvato a
cualquier temperatura dada que aquellOS LDH de congeneres adaptados

86
hac ia aguas rMS t ibias Estas d iferenc ias re1 ac ionadas con la
temperatura y los Km favorecer una IM rcada estabil ización del mismo
temperaturas fisiolÓgicas coroo puede observarse en la figura 22 Este
al o grado de conservación de Km en las diferentes especies a sus
temperaturð S corporales prop iadas se t a in t rpretado como una
adaptac ión que aseg lra a los enimas la retención de una cierta
capac idad de reserva para in rementar la tasa de catÆl is is al elevarse
la concentració del sustr ato dtlrante activaciones metaból icas por
eJemplo dJrante el ejerc ic io v i90roso As estab i 1 izando los valores
de I lY en un rango que es cer cano a o de algœr modo mayor que las
concentraciones fisiológicas del sustrato el enz ima sera capaz de
obtener una fracción significativa de su velocidad potencial vmax
bajo cordiciones fisiológicas constntes ademas incrementar su
actividad en tanto los niveles de sustrato se eleven Fersht 1977
Gra es omero 1982 Gra ves 1983 Cabe observar aqui que
s a fuerte conservación de Km con respecto al piruvato se percibe sólo
C uando el medio de ensayovitro tiene el pH apropiado para la
Fledic ión de temperatura Yancey y Somero 19813 Somer o 1981 De esta
forrna la detección de diferencias funcionales significativas entre
enzi as variantes dependerÆ del uso de las condiciones de ensayo que se
asemejen en lo pos it le a la rea l idad espec ialmente en pH y tempel atura
Place y Powens 1979
Las diferencias en la constante catal tica Kcat entre las distintas
especies son tambi n termocompensatorias Las especies abundantes en
agua templadas poseen la variante de LDH con las ma ores tasas de

87
cat41is is
all 90S
potenciales Estos valores de Kcat concuerdan con aquellos
realizados durante el estudio de especies diversas de
vertebrados los cuales mostraron que los valores de Kcat para unð
I
amplia variedad de reacciones enzimaticas incluyendo aquella para LDH
varían inversamente con la adaptac iÓn tØrmica Somero i978 Borgmann
Moon 19751 Graves y Somero 1982 Johnston Wal1 esb 1977 Low
Somero 1976 Somero y S iebenall er I 1979 Graves i 1983 Low II
l 1973
As L 1 as d iferenc ias entre los M4 LDH de los congeneresi
aqu 1
estudiados sugieren quel pequeæas diferencias en la temperatura del
cuerpo son sufucientes para favorecer la selecciÓn de diferencias
adaptativas hacia la temperatura en las propiedades cineticas de los
enz imas un hecho cons istente con 1 as comparac iones hechas en
congŁneres de barracudas con d iferenc ias de 5 8 e promed io en sus
temperaturas corporales GravesSomero 1982
Para final izarI
la conservacion en los valores de Km a 1 as
temperaturas fisiol gicaS observadas en este estudio as 1 como en
otros indican la importancia de este tipo de adaptaciones cinØticas de
los enzimas en evoluci n molecular
El examen de loci enzimas adicionales en estos cuatro congener es y
en otros ectotermos de distribución antitropical podr n sacar a la luz
profundos conocimientos importantes de la adaptación molecular a escala
fina y la evoluci n de las especies

r CephalophOlis E panamensis 1
1
1 EpinepheluS 1TI ID líl rE acaothistius TI
L EpinepheIUS LElobriformis mI
E onalogus nł
CephoIOPhOIiS E ponomensis
1m
ID E lobriformis
IillN
E onalogus
II E oconthistius
FIG 23 Dendrogramas comparativos que muestran las posibles relaciones
filogenØticas de cuatro especies de cabrillas del Mar de CortØsA en base a las característcas anatómicas Smith 1971 8
en base a los caracteres genØticos analizados en el presenteestudio 1 11 111 Y IV son los grupos especie Los subgØneros
aparecen entre parØntesis
ae

88
APENO ICE No 1
hoz lmas homomØr icos y heterom r icos Harr is 1880
Muchas prote nas tienen una estructura multimJrica esto es se han
formado por dos o mas unidades de cadenas po 1 ipept d icas Estas
unidades pueden ser id nticas en cuanto a la secuencia de sus
mino c idos hoz imas homomØr icos o dicha secuenc ia puede d iferir
isozimas hetero r icos El patón isoenzimÆtico ue se observa
espu s del corrimiento electroforJtico depende en general del numero
de subunidades ue ocurr en ero las moltculas de isozimas y del n mero de
cadenas pollpept dicas estructuralrnente distintas ue hayan sido
intet izadas
Si se considera un solo locus en el cual puedan ocurrir dos o mas
alelos y ue cada alelo codifi ue para una version estructural mente
distinta de la cadena polipept dica en cuestitn entonces tendremos ue
un individuo homoci90to en el ue los dos alelos presentes son los
mismos solamente sintetiza una forma de cadena polipept dica pero si
el indi idJO fuese heteroc i90to tendr dos o mÆs alelos diferentes
presentes y por 10 tanto sintetiza dos cadenas polipept dicas
distintas
En el caso de los enzimas monomØricos el patron de bandas isoz imas
que se observan en el heteroci9oto representa una mezcla sencilla de

90
las dos formas que ocurren por sl mismas en cada uno de los homocigotos
correspondientes Sin embargo en el caso de enzimas multi ricos se
forman i oz imas heteromØr icos h lbr idos en el individuo heterocigoto
esto es asumiendo que una vez sintetizadas las cadenas polipept dicas
diferentes se p Jeden cornb inar para formar mal cu 1 as de isoz imas
multimericos y esto por lo general es regla
La estructura de las subunidades de isozimas primarios que pueden
originarse en tales heterocigotos varía de acuerdo al nœmero de
ubunidades que se sucedan en las molØculas de isozimas
En el caso de los dímeros existen dos formas homomer icas y una
heteromer ica er los trlmeros hay dos formas homor ricas Y dos
hetero rica y en los tetràmeros hay dos formas honJ meras y tres
h ertmeras
En general si el numero de subunidades identicas sintetizadas en el
soz ima pr imar io de un indiv iduo homoc igoto es Un entonces en los
eterocigotos se esperara n l de isozimas primarios siendo dos
t omomer Icos Y rl I heterorner icos
Si los polipeptidos determinados por dos distintos alelos difieren
en la carga neta entonces los isozimas primarios forrr dos en
heterocigotos muestran patrones característicos como los de la figura
24 y patrones estructurales tales corr los de la tabla 13

91
Con lost
rT tonornecls hay dos componentes con los drmeros tres con los
tI meros cuatro etc
En cada caso las bandas mas externas corresponden a las formas
thomomer icas I las bardas inter med ias a 1 as heter orner icas
Los patrones s imetr icos de isoz imas como los de la figura 24 se
obsevan nicamente si los pOlipØptidos determinados por los dos alelos
contr ibuyen de igual forma a 1 a ct iv idad erz irroÆt ica y si las
J
subun idades e comb inar al azar y s in r estr icc ion alguna
Si esto sucede la proporci n de actividad de los distintos isoz imas
formados en heterocigotos ser
1 1 para monomeros
para d meros
para tI meros
para tetrÆmeros
12 1
1 3 3 1
1 4 6 4 1
y as suces ivamente
De tal modo un 5ØX de la actividad total esta dada por formas
homo ricas en los d meros en los tr1meros sólo un 25X y en los
Jtetrarneros solo un 12 5

T U 13 Estructura de las subunidades de isozimas homomØricos y heteromØricosen heterocigotos
Monómero Oí mero Trímero letrame ro
Homómero 81 8181 818181 81818181
Helero mero 818281 81 82
828281
81 8 81 828181828282 82828
Homo mero 82 8282 828282 82828282
2

Monomero
HomociQoto
HelerociQoto
Homocigoto
Dimero Trímero
93
Tetrómero
FIG 24 Patrones electroforØticos que se observarían en casos de monómeros
dímero trímeros y tetrÆmeros en individuos heterocigotos

94
APEf JD ICE No 2
t 1 cÆlC ll r de frecuencias 9 nlc 5 genot picð S Roughgarden 1979
Notae i6n
ConsidØrese una poblacion que contiene variación genØtica en un locus
con dos alelos AO y a Entonces los genotipos posibles son AA o
Aa y aa I Introdœzcase la notación como sigue los n6meros
genot fp icos denotados como NAA Nla y Naa son e 1 nœmero de organ ismos
de cada genot ipo respect ivamente Sea el tamaæo total de la pOblac ión
NI entonces
N NlA Nla Naa
Aœn es necesaria una notaci6n mÆs precisa Se definen las frecuencias
genot ip ieas o fracc iones como 1
o NAA N
H NAa N
R Naa N
La suma de O H y R debe ser igual a la unidad esta notación se
utiliza aunque A no sea dominante Finalmente se definen las
fr ecuenc ias gØicas como pag ina s igu iente I

95
p 2NAA NAa 2N
q 2N a NAa 2N
De acuerdo a estas formulas p es la fraccion de los alelos A en la
poza gen ica y q e la fracc i rl de los alelos a Si existen N
individuos entonces existe un total de 2N genes en la poza genica para
el locu A Ahora cada homoc ijloto AA tiene dos alelos A y cada
heteroc igoto tiene un alelo A as p la fracc i n de alelos AI
debera
ser
p 2NAA NAa 2N
La Slma de p y q es la unid Õld Es importante entender la distinci n
entre frecuencias genotfpicas y frecuencias gØnicas Estas ltimas s lo
se refieren a la variaci n gen tica dentro de toda la poza genetica
mientras que 1 as pr imer as ð demas al modo como se encuentra organ izada
esa variacion en los genotipos

96
APENDICE No 3
Distancia genetica entre poblaciones naturales Nei 1972
Resumen
Se formula una medida de distancia genetica O basada en la
identidad de ge es entre poblaciones Esta medida se define como
D In 1 donde I es la identidad normalizada entre los genes de las
poblaciones dos La distancia genetica mide las diferencias alel icas
acu Jladas por locus Si la tasa de sustitucion de genes es constante
la medida se correlaciona linealmente con el tiempo de divergencia
entre las poblaciones bajo aislamiento reproductivo Tamb ien se
orrelaciona de manera lineal con la distancia geografica o el area en
I
ciertos modelos de migracion Ya que D es una medida del numel o
acumulado de diferencias codon por locus tambien puede estimarse a
partir de datos de secuencias de amino cidos en prote nas de especies
lejanamente relacionadas Asf si se dispone de suficientes datos la
distancia genetica entre cualquier par de organismos puede medirse en
t rminos de D Esta medida es aplicable a cualquier tipo de organismos
sin importar su n mero crorr s mico Ploid a o estrategia reproductora
La identidad normalizada de genes entre poblaciones es equivalente
a la identidad prote nica
El numero acumulado de sustituciones de genes o diferencias por
locus es distancia genØtica

97
Proced imiento
Sean X y Y dos poblaciones que se entrecruzan al azar diploide y
con alelos mœltiples que se segregan en cada locus
x i e y i frecuencia s del i esimo alelo n X e Y
j SUI1 x i 2 probabilid d de identidad de dos genes
eseog idos al azar en la pOblac ión ì
j y SUI y i 2 pr bab i 1 ida j de ident idad de dos genes
escogidos al azar en la poblaci6n Y
j XY SUIx i i pr c babilidad de ident idad de un gen en la
poblacion X y un gen en la población Y
La ident ida d nOI m izada de genes entr X e Y con respecto a este
OCUS 9en 5 i no e iste seleccicn J cada alelo se der iva de un
mu1ac ión irflpl en una gener ac i6n ð Flce1 t al es ta def in ida por
1 j j XY SOR j Xjy
Esta cantidad es l unidad cuando las dos poblaciones tIenen los
mismos alelos en frecuencias idØnticas y es cero cuando no tienen
a lelos coml nes
La identidad normalizada de los genes entre X e Y con respecto a
todos 1 o I oe i se def ine corno
JXY SQR JXJY
Donde JX JY JXY son las medias de jX jY y jXY respectivamente
sobre todos los 1 oc i incl ti dos aquellos monomórf icos

98
a distancia gen tica ent e X e Y se define entonces como
o In 1
Esta def in ic itn es lo suficientemente ap op i ada si la tasa de
sustituciones de genes po locus es la misma pa a todos los 1 oc i En
este caso O mide el numero acumulado de sustituciones de genes po
locus
Cuando la tasa de sustituciones de genes varra con el locus Y los
lj s son muchos una medida mÆs apropiada de distancia gen tica viene
dada por
O 1ni
Donde l J XY SGR J X J Y en los que J X J Y y J XY son las
Iiedias geom tr icas de jX jY y jXY respectivamente La f rmula tambi n
puede e presa se como sigue
O SUM dj n
Oond dj In Ij en el j esimo locus y n es el nume o de loci la
SUMatoria va desde j hasta n

99
Not
Existen dos estimaciones diferentes del valor correspondiente de la
distancia gen tica con aquel del tiempo evolutivo Un valor de
distancia genØtica de 1 es equivaler te a 5 millones de anos Nei
1975 mientras que otros investigadores lo aproximan a 18 millones de
aæos Gor man et l 197E Esta l tima pa rece acercar se mð s a los
eventos geolóic conocidos Y que han sido descritos en otros estudios
Gor man et al 19 6 Vaþlte et al 1886 Es esta misma medida la que
aquí se ilizado de acuerd Ias modificaciones ya mencionadas en
el texto

100
APENDXCE No 4
Cinetica de saturacion enzimatica HochachKa y Somero 1984
Las reacciones químicas catalizadas por enzimas pueden ser descritas
d acuerdo a lo siguiente
E SeS ES EP E P
donde S es el sustrato E es el enz ima ES es el complejo
enzima sustrato ES es el complejo activado EP es el comp 1 ejo
enzima producto y P es el producto La velocidad de tales reacciones
catal izadas por enzimas muestra una dependenc ia cr t ica sobre la
concentración del enzima y de iglJal forma el sustrato Cuando estas
se varían experimentalmente se encuentra que la velocidad inicial de
I
reaccion v es directamente proporcional a la concentracion del enzima
EO esto significa que las tasas mÆs altas de reaccion
metab6lica ocurren cuando la concentraci n del enzima se eleva Sin
embargo v sigue una cin tica de saturaci6n con respecto a la
concentrac i n de 1 sustrato S F ig 25 a baja S v
1 inealmente con S en tanto que si S J se incrementa
comienza a quebrantarse y v se incrementa menos rapido
se ir crementa
esta relacion
que S Jcue al saturarse S J v alcanza un valor límite llamado vmax
hasta

101
Estas relaciones entre S l v se describen por la ecuaclon de
Michaelis Ment en
v vmax s Km S
El t rmino Km constante de Michaelis Ment en es la concentración de
sustrato en la que v 1 2 vmax
Eficiencia catal t ica HochachKa y Somero 1984
El t rmino eficiencia catalftica se ha definido de muchas formas
En todas ellas el criterio com n es que tan r Pidamente el enzima puede
convertir el sustrato en el producto expresando resto sobre la base la
cantidad convertida por mol cula de enzima Una de las expresiones se
basa en el numero de recambio medido frecuentemente como los moles de
sus trato convertidos en producto por mol de enzima por unidad de
tiempo generalmente minuto Una medida alternativa es la cons tante
catal tica Kcat que expresa el recambio por sitio activo del enzima
Ambas medidas conllevan la funci n del enzima a su velocidad max ima
posible vmax Una tercera forma para estimar el poder catal tico de
un enzima involucra la razon de Kcat con Km esta constante de primer
orden provee de una estimaci n m s real de la actividad catal tica bajo
concentraciones de sustrato que se encuentran tfpicamente por
debajo de la saturacion Sin embargo para estimar la eficiencia
inherente de un enzima bajo condiciones optimas vmax las primeras

102
dos definiciones son preferibles ya que corresponde con la energla
libre de actiYaci n de la reacci n la cual es la medida termodtnåmica
de cómo puede un enzima reducir la barrera de energ a libre a su
nivel de reacci n

v
v Vmox Kcot Eov sJKm
Vmox2
Km S
FIG 25 La velocidad de reaCC10n graficada contra la concentración del
sustrato para una reacción que obedece la cinØtica de MichaelisMenten o de saturación
eficiencia catalítica
103

184
LITERATURA CXTADA
Aspinwall N 1974 Genetic analysis of North Ar rican populations of
the pin salmon Oncorhynchus gorbus ha possible evidence for the
neutral rrJtat ie n dr 1ft h pothes is Evolut ion 28 295 305
AtKinson D E 1965 Biologieal feedbacK control at the molecular
level Science 150 851 857
1966 Regulation of enzyme activity Ann Rev
Biochem 35 85 124
1968 Citrate and the citrate cycle in the regulation
o enelgymetabolism T W Goodwin Ed Metabolic Roles of
Citrate pp 23 40 Academic Press N Y
1977 Cellular Energy Metabolism and its Regulation
Aeademic Press N Y
Ayala F J 1976 Molecular Evolution Sinauer Associates IncSunderland
Baldwin S 1968 Manometrie measurements of respiratory aetivity in
Aemaea digitalis and Aemaea scabra Veliger 11 79 82
Bald J in J 1971 Adaptat ie n of enzymes to
Acetyleholinesterases in the central nervous system of
Biochem Physiol 4el 181 187
temper ature
f ishes Comp
Baldwin J y P W HoehachKa 1970 Funetional signifieanee of
isoenzymes in thermal aeelimatization Acetylcholinesterase from trout
brain Biochem l 116 883 887
Behr iseh H W y P W HoehachKa 1969a Temperature and the
regulation of enzyme activity in poiKilotherms Properties of rainbow
trout fructose diphosphatase Biocnem J 111 287 295
1969b Temperature and the regulation of enzyme activity
in poiKilotherms Properties of lungfish fructose diphosphatase
Bioehem J 112 601 607
Ber nstein S C L H Throcmor ton J L Hubby 1973
genetie ariabilit in r at Jr al populations Pr oc Nat Acad
7a 3928 3931
Still
Sc 1
mot e
USA
or mann ti 1 J Laidler y T J Moon 1975
t ermodynamics of 1 aetate deh drogenases from beef heart
and fl u der musele Can J Bioct em 53 1196 1206
Kinetics and
beef rnusc 1 e

105
Borgmann IJ y T W t oon 1975 A cOfT1 arison ofd hydrogenases from an ectothermic and an endothermic animal
Biochem 53 898 1004
1 actate
Can J
r i y l E T t i 11 er G Wh i tt y l L
Electrophoretic analysis of evolutionary relationships in
Evolution 31 81 102
Nanney 1977
Tetrahymena
Boyer S H 1972 Silent alleles
453 454
Form and function Nature 239
Brown J H y C R
fluetuating environments
Cyprinodon Evolution 25
Feldmeth 1871 Evolution
Thermal toleranees of
390 388
in constant and
dfsert pupf ish
Car1sorJ S S A C Wilson y R D Maxson 1978 00 alhtAn eloeKs
run in time A r epl y Se ienee 200 1183 1185
Castell ini M A y G N Somero 1981 Buffering capacity of
vertebrate musele Correlations with potentials for anaerobic
funetion J Comp Physiol 143 1 191 198
Dean J M 1969 The metabolism of tissues of thermal1y aeelimated
trout gairdneri Comp Bioehem Physlol 29 185 196
EKman S 1853 Zoogeography of the Sea Sed jcK a d JacKson London
FairbanKs G T L SteeK y D F H Wallaeh
analysis of the major polypeptides of the human
810chemistry 10 2608 2617
1971 Eiee rophoretic
eryt rDcyte membrane
Fersht A 1977 Enzyme StrucÌure and Meehanism W H Freman San
Fr ane isco
Gorman G C M Soule S Y Yang Y E NevQ
genet ies of insular Adr iat ies 1 izards EvolutIon 29
1975 Evolutionary
52 7
Gorman GC Y Kim y J Rubinoff 1976 Genetic
three speeies of Bathygobius from the Atlantic and
Panama Copeia 1976 361 364
relat l nshlps
Par 1 f C s ideô
of
of
GorrM n G C y J Renzi Jr 1879 Gene tlr istalce and
heterozygosity stimates in electrophor etie studies I E et Of safT1 le
size Copeia 1878 242 248
Gosline W A 1966 The limits of the fish family Ser anidae with
notes on other lower pereoids Proe Ca1if Acad Sel 33 9 1 2

106
Id S J O S Woodruff y
r phom tr les of e r Ion a1 Pongo
U ls Y1lgtic l r d nall Sy t
J P Mart in 1973 Ger et ies
er pet A r ew systema t ie pproaeh
Zool 23 518 535
and
to
Grassle J L y J F Grassle 1976 Sibling speeus in
pf lution Ir dieator ea pitell Polychaeta Seienee 192
the marlne
367 369
3 J E R H Pos rlblatt G N Somero 1983 Kinetic and
ectrophor tie diff r ntiation of lactte dehydrogenases of teleost
srecies pair fro the Atlantic and Paeifie eoasts of Panarr Evol tion
31 30 37
GrB es J E y R H Rosenblatt 1980 Genel ie relatlonshlPs of the
color morphs of the serr nid fist Hypoplectrus unicolor Evolution 3
240 245
Gr Jes J E y G Somer o 1982 Eleetrophoretic
enzymle JolutloYl In four species of eastern Pacific
dlffer nt thermal enJlronments Evolution 36 97 106
arld funct ional
bar racudas from
Har rjH 1980 The Pr inciples e f Human Biochemic1 Genetics
Else l˝er North Holland Biomedical Press Tt e Netherlands
Hedr I K P
p ll r lrph ism in
1 32
w M E Ginel ð rl v E
he er0geneous environments
P Ewlng 1976 Genetic
Ann Rev Ecol Syst 7
Yeiniee E A A H Houston 1965 Effcts of thermal acellmationd lublethal heat shocK upon lonic goldflsh earassius auratus L J
F sh Res Board Can 22 1455 1476
Yemr IYlg er E A E L OO Jglas y G C Grl9g 1969 Oxygen
ronsllmpt lor in Antart ie haemogl ob in free f Ish Pagetops is maeropterus
and In three sp cies of Notothenia Comp Bioehem Physiol 29 467
490
HlrKman C P Jr R A t Nabb J S Nelson E O Van Sreeman y O
Comforl 1964 Effects of cold accl Imatlon on electrolyte distributionIr rainbo tr ul gairdrdHi Can J Zool 42 557 597
Hoct B t KB P t J 1968a Acion of ternperature on branch
gl Jcose and acette metat oli m Comf Biet em Physiol 25
po ints in
107 118
1968b Lacl ate dehydrogenase function in Electrophorus
sLJim bl dder and ir the lur gflst I ng Comp Biochem Phsiol 27
613 61
HochachKa P 1 y G N Somero 1968 The adapta tion of enzymes to
temperatre Cornp 8iochem Physiol 27 859 668

107
HoehaehKa P W y G N Somero 1971 B ioehemieal adaptat ion to the
environment k I Hoar W S y D J Randall Eds Fish Physiology
Vol 6 pp 99 158 Aeademie Press N Y
1973 Strategies of Bioehemieal Adaptation Saunders
P A
1984 Bloehemieal Adaptation Prineeton University
Press Prineeton
Hodgins H O 1972 The nature of flsh populations Simon R
C y P A LarKin Eds The StoeK Coneept in Paeifie Salrnon PP 199
208 H R MaeMillan Leetures in Fisherles University of British
Columbia Vancouver B C
Hubbs C L 1948 Changes in fish fauna of western North Ameriea as
eorre 1 ated w i th ehanges in oeean temperature J Mar Res Sears
Foundation 7 459 482
1952 Antitropieal distribution of
organisms Proe 7 Paeifie Sei Congo 3 324 329
fishes and other
Johnson A G F M Utter y H O Hodglns 1972 Eleetrophoretie
investigation of the family Seorpaenidae US Natl Mar Fish Serv
Flsh Bull 70 I 403 413
1973 Stimates of genetle polymorphlsm and heterozygoslty
in three speeies of roeKflsh genus Sebastes Comp Biochem Physiol
44B 397 406
Johnson G B 1976 Hidden alleles at the alf glycerol phosphate
dehydrogenase locus in Collas butterflles Geneti 83 149 167
Johnson G O 1983 Niphon splnosus A prlmitlvr pinephel ine
serranid wlth comments on the monophyly and lnterrelatlon hlps of the
Serranldae Copela 1983 777 787
Johnson G O y P Keener 1984 Ald
American grouper larvae Bull Mar Scl 34
to the identif lcat ion
106 134
of
Johnston I A Y J WallesbY 1977 Molecular mechanisms of
temperature adaptation in fish myofibrillar protelns adenosine
trlphophatases J Co Physiol 119 195 206
Keigwin J O Jr 1978 Plioeene closing of the
based on biostratigraphic evidence from nearby
Caribbean Sea cores Geol09Y 6 630 654
1 s thmus
Pac if ic
of Panama
Oeean and
Kimura M y T Ohta 1971 Prote in polymorph ism as a phase of
molecular evolution Nature 229 467 469

les
1 llg J T JuKes 1969 Non darwlnian evolutlon Seienee 164
778 798
Kohen R K 1969 Esterase heterogeneity
polymorphiSffi Seienee 163 943 944
dynamics of a
1970 Funetional and evolutionary dynamies of
po 1 ymorph le sterases in eatostomid f ishes Trans Am F ish Soe 99
e19 ee6
LaellT l 1 U K 197e CI eavage of structural prote ins dur ing the
assembly f the head of baeteriophage Nature e27 680 685
Lewontil R C 1974
Columbia N Y
The Genetic Sasis of Evolutionary Change
L igny W de Ed 1971 Spec ial Meet ing on the
Serological Identification of Fish StocKs Rapp P
Perm Int Explor Mer 161 1 179
Bioehemieal and
V Reun Cons
Low P S J L Bada y G N Somero 1973 Temperature adaptation
of enzymes Roles of the free energy the enthalpy and enthropy of
activation Proc Nat Aead Sei USA 70 430 432
Low P S y G N Somer o 1976 Adaptation of muscle pyruvate Kinaseto en irc nmertal temperatures and pressures J Exp Zool 198 1 12
Manooch C S III G R Huntsman B Sullivan y J Elliot 1976
Conspecific status of the sparid fishes Pagrus sedecim Glnsburg and
Linnaeus Copela 1976 678 684
Maxsor L R y A C Wllson 1974
evolutlon detected by studylng proteins of
speeles group Selenee 185 66 68
Con Jergent
tr ee frogs
morphological
in the ti
MeeK S E y S F Hildebrand 1ge5 The marlne flshes of Panama
Publ Flelds Mus Nat Hist ZoolSer
No e49 PP xv xix 331
707 e5 71 pis
Merritt R B 197e Geographic distributin and enzymatlc
of lactate dehydrogenase allozymes in the fathead minnow
promelas Amer Nat 196 173 184
properties
P imephales
Nel M 1971 Interspeclflc gere dlffer ences and evolutlonary
stlmated from electrophoretlc data 01 proteln ldentlty Amer Nat
385 397
time
105
197e Genetlc dlstance between populations Amer Nat
06 e83 e92

IElS
Nei M 1878 Est imat ion Di average heter ozY9osit
distance from a small nvrrber of individuals Genetics 8S
arld genet ic
593 5SEl
Newell R C 1966 The effect of ternper at vre on
pioKilotherms Nature 212 427 428
the metabo1 ism oi
1867 Oxidat ive activity of pOlKilotherm mitochondria
3 S a function of tempel atvre J Zoal London 151 28S 311
o erra P y S Sarr y 1972 Lactate dehydrogenase
Specific ligand approach FESS Lett 21 281 29Ø
pvrification
O Car ra P S Sar r y y E Cor coran 1974 Affinit
d ifferent iat ion of 1 actate deh drogenase isoenzyrnes on
abort ive complex fc rmat ion FESS Lett 43 163 168
chromatography
the differential
Place A R Y D E Powers 1979 Genetic variation and
catal ytic eff ie ier c ies Laetate dehydrogenase S allozyrnes In
heteroclit s Ploc Nat Aead Sei USA 76 2354 2358
relative
Fundulvs
POIJell J R 1975 Prote in var lat ion
anima1s Evol Siol 3 79 118
in natur al popvlations of
Powers D A y A R Place 1878 Siochemieal genetics of Fvndulus
het eroel itlJS L l Temporal and spat ial var iat l n in gene frequenc ies
of Ld I B IYIdh A Gpi S and Pgm A Sioehem Genet 16 593 607
PraKash S R C Lewontin
polymorphism on the basis of natural
J L Hubby 1969 Biochemicalselect ion Geneties 61 841 658
prosser C L 1973 Comparative Animal P lysiology W 8 Saunder s
P A
Randall 1 E yA Sen Tuvia 1983 A r eview of groupers Pisces
Serranidae Epinephelinae of the Red Sea with description of a new
species of Cephalopholis Bul1 Mar Sci 33 373 426
Reeves R B 1977 The interaction of bod ter erature and acid base
balance in ectothermic vertebrates Ann Rev Physiol 3S 55S 586
Roberts J L 1964 Metabol ie
seasonal photoperiods and
Meeresvntersue 14 451 465
respcFses If
ternperatvres
fresh water sunflsh to
Helgolander Wiss
1987 Metabol ie compen at ions
sunfis h I2 Molecular Mechanisms of Temper ature
No 84 PP 245 262 AAAS Meeting Washington D
for terr perature in
Adaptations Publ
C
Rob in C 1 y W A StareK 11 1961 t1ater ia1 for a re ision of
Serranus related fish genera Proc Nat Aed Sel Philadelphia 113
259 314

o
118
Ro enblatt R H y B J Zahuranec 1967 The ealOtern Pa ifi
groupers of the genulO Mycteroperca including new species Calif FilOh
Game 53 1 228 245
Roughgarden J 1979 Theory of Population GeneticlO and EvolutionalY
Ecology An Introduction MacMillan Publishing Co lnc London
Sage R D y R K Selander 1975 Trophic radiation
pc lymorphism in cichlid fishes Proc Nat Acad Sci USA 72
4673
through4669
Sa 1 mo n M S D Fe r 1 i s D Joh nson G Hy att y G S Wh i tt 1979
Sehavioral and biochemical evidence for species distinctiveneslO in the
f lddler crabs spec iosa and SP in icarpa Evolut ion 33 182 191
Sanchez R 1983 Estudio sobre las caracteristicas y propiedades dela bacteria halofilica CI8 2 Tesis Profesional Inst Tec La Paz
IEXICO
Sarich V M 1977 Rates sample sizes and the neutrality hypothesis
for electrophoresis in evolutionary studies Nature 265 1 24 28
SChwartz F J 198 World literature to fish hybrids with an
analysis by family species and hybrid Gulf Coast Res Lab Publ 3 1
1 328
Selander R K M H Smith S Y Yang W E Johnsor
Gentry 1971 B iochemical po 1 ymor ph ism and systemat ics in
Peromyscus 1 Variation in the old field mouse
polionotus Univ Texas Publ 7103 1 49 90
y J 8
the genus
Peromyscus
ShaKlee J S C S Tamaru y R S Waples 1982 Speciation and
eJolution of marine fishes studied by the electrophoretic analysis of
proteins Paco Sci 36 141 157
Siebenaller J F 1984 Pressure adaptative differences in
NAD dependent dehydrogenases of congeneric marine fishes living at
different depths J Comp Physiol S 154 1 443 448
Siebenaller J F y G N Somero 1979 Pressure adapta tive
differences in the binding and catalytlc properties of muscle type M4
lactate dehydrogenases of shallow and depth living marine fishelO J
Comp Physiol 129 295 300
1978 Pressure adaptative differences in lactate
dehydrogenases of congeneric fishes living at different depths Science
201 1 255 257
Smith C L 1961 Synopsis
Epinephelus and allied genera of
Fish Bio1 Synopsis No 23 61 pp
of
the
biological datawestern North
on groupers
Atlantic FAO

112
Stadtmn E R 1968 The role of INJUiple enzymes in the
of branched metabolic pathways Ann N Y Aead Sei 1 1
regulation
16 530
Stansfield W D 1969 Geneties McGraw Hil1 BOOKeo
N Y
Stinson R A y J J HolbrooK 1973 Equilibrium blnding of
n ieot inamide nue 1 eotides to 1 aetate dehydrogenases B iochem J 1311
719 728
Sul1ivan K M y G N Somero 1980 Enzyme aet ivities of
sKeletal INJscle and brain as influenced by depth of occurrenee
habits of feeding and locomotion Mar Biol 60 91 99
fishand
Thomson D A L T Findley y A N Kerstitch 1979 Reef Fishes of
the Sea of Cortez The RocKy shore Fishes of the Gulf of California
John Wyley Sons N Y
Turner B J y D J Grosse 1980 Troph ic
l yodon a genU of strea m dwelling goodeid fishes
ecological polymorphism Evolution 34 1 259 270
differentiation of
Speciation versus
US Department of Commerce 1952 Surface water temperatures
stations Pacific Coast Special Publ No 280 Coast and
Survey Washington D C
at t ide
Geodetic
Vawter A Tu R H Rosenblatt y G C Gorman 1980 Genetic
diversity among fishes of the eastern Pacific and the Caribbean
Support for the molecular ClocK Evolution 34 1 705 711
Utter F M
genetic studies
Malins D C y
Perspect ives in
Y
H O Hodg ins Y F W All endorf 1974 8iochemieal
of fishes Potentialities and 1 imitations En
J R Sargent Eds Biochemical and Biophys leal
Marine Biology Vol 1 pp 213 238 Aeademic Press N
Walford L A 1937 Mar ine game f ishes of the Pae if ic
AlasKa to Equator University Calif Press BerKeley xxix
69 pIs
coast
20
from
pp
Waldman J R 1984 Stripped bass in Gregory s fish sKulls
actually a grouper Copeia 1984 793 794
Watson J D y F H C cricK 19 3a Molecular structure of nueleie
acids Nature 171 737 738
19 3b The strueture of DHA Cold Spring Harbor Symp
QJ ant B i 01 18 I 123 131
WilKinson G N 1961 Statistical stimations
Bioehem J S0 324 332
in enzyme Kineties

113
William G C 1966 Adaptation and Natural Seleetion Prineeton
Univer ity Pre Prineeton
Wì1 on A C S S Carl of y T J Whi1 e
evolution Ann Rev Bioehem 46 573 639
1977 Bioehemiea1
Wi1 on J Al Jblhhing Co
1979 Principies
lne N Y
of An i ma 1 Physiology MaeMi 11 an
Woh1sehlag D E 1964 Respir a1ory metabolism and eeologieal
eharae1 er is1 ies of some f is es of McMurdo Zone Antart iea M OLee Ed 8i010gy of 1he Antar tie Seas Vol 1 pp 38 62 Am Geophys
Union Washing1 on O C
Wyles J S y G C Gorman 198Ø The albumin immunolog ieal and Ne i
elee1 rophoretie distanee eorrelation A ealibra1 ion for 1he saurian
genus Iguanidae Copeia 1980 66 71
Yaneey P H M E ClarK S C Hand R D Bowlus y G N
1982 Living wl1h wa1 er s1 ress The evolu1 ion of osmolyte
Seienee 217 1214 1222
Somero
systems
Yaneey P H y G N Somero 1978a Temperature dependenee of
intraeellular pH Its role in the eonservation of pyruva1e apparent Km
values of vertebra1 es lae1ate dehydrogenases J Comp Physiol 125
129 134
1978b Urea requ iring 1 aetate dehydt ogenases of mar ine
elasmobrar eh fishes J Comp Physiol 125 135 141
1980 Methy 1 amire osmor egu1 at ory so 1 utes of
elasmobraneh fishes coun1eraet urea inhibition of enzymes J Expl
2001 212 205 213


















