PROPAGACIÓN in vitro DE LOS...
83
PROPAGACIÓN in vitro DE LOS PORTAINJERTOS DE CEREZO (Prunus avium L.) GISELA 5 Y Prunus cerasus.
Transcript of PROPAGACIÓN in vitro DE LOS...

PROPAGACIÓN in vitro DE LOS PORTAINJERTOS DE CEREZO (Prunus avium L.) GISELA 5 Y Prunus cerasus.

ÍNDICE
1. INTRODUCCIÓN 1 2. REVISIÓN BIBLIOGRÁFICA 4 2.1. Características de los patrones de cerezo 4 2.1.1. Gisela 5 4 2.1.2. Prunus cerasus 6 2.2. Propagación del cerezo 6 2.3. Selección del explante 7 2.3.1. Explantes utilizados en Prunus 7 2.3.2. Cultivo de segmentos nodales 8 2.4. Tratamiento de desinfección 8
2.5. Pardeamiento y antioxidantes 9 2.6. Medio de cultivo 11 2.7. Organogénesis 13 2.7.1. Rizogénesis in vitro 13 2.7.2. Neoformación de yemas 14 2.8. Reguladores de crecimiento 14 2.8.1. Auxinas 15 2.8.2. Citoquininas 15 2.9. Proliferación 16 2.10. Enraizamiento 17 2.11. Aclimatación 18
3. MATERIALES Y MÉTODOS 22 3.1. Ubicación de los experimentos 22 3.2. Material vegetal 22 3.3. Medio de cultivo 22 3.4. Manipulación del material vegetal 24 3.5. Mediciones de los parámetros a evaluar 26 3.6. Ensayos 27 4. PRESENTACIÓN Y DISCUSIÓN DE RESULTADOS 44 5. CONCLUSIONES 66 6. RESUMEN 67 7. ABSTRACT 68 8. LITERATURA CITADA 69 ANEXOS

1. INTRODUCCIÓN
Durante los últimos seis años, el cultivo del cerezo ha logrado un importante
crecimiento. Esto se ve reflejado en las cifras que entrega ODEPA (2003) de la
temporada 2001/2002 en cuanto a volumen de producción 32.000 ton, exportación
12.700 ton y superficie 6.200 ha.
La situación anteriormente descrita se ha generado como respuesta a las favorables
condiciones del mercado externo, en conjunto con la aparición e introducción de
nuevas variedades y portainjertos, como así también de un manejo que incorpora
riego presurizado y sistemas de conducción bien definidos, asociados a un aumento
en la densidad de la plantación y a nuevas técnicas orientadas a reducir el
crecimiento vegetativo e inducir una rápida entrada en producción (FUNDACIÓN
CHILE, 2000).
Dado lo anterior, en los últimos años se han desarrollado diversos patrones de
cerezo, respondiendo a una necesidad de renovación de los que se utilizaban
tradicionalmente (HORMAZA y GELLA, 1996). Algunos de estos nuevos
portainjertos de cerezo que se están utilizando comercialmente son el portainjerto
enanizante Gisela 5 y Prunus cerasus, portainjerto propagado vegetativamente, por
yemas o estacas, debido a su estructura genética heterocigótica.
Para responder a la alta demanda de los nuevos portainjertos ha sido necesario
desarrollar otros sistemas de propagación. Uno de estos sistemas es la propagación
in vitro, práctica que se ha establecido en la industria comercial. Esta técnica tiene la
ventaja de lograr un gran número de plantas idénticas, pues se evita la heterocigosis
del árbol, y se obtienen plantas libres de enfermedades; especialmente de virosis.
Además, este sistema de propagación se ha desarrollado como una nueva
alternativa para aquellos portainjertos difíciles de propagar a través de otros
métodos, como son el enraizamiento por estacas o la propagación por semillas.

A través de este taller se realizó la propagación in vitro del portainjerto para cerezo
Gisela 5 y Prunus cerasus, con los que se ensayaron distintos métodos de
desinfección y diferentes medios de cultivo en tres de las cuatro etapas que forman
parte de la propagación in vitro: establecimiento, multiplicación y pretransplante
(enraizamiento y aclimatación) (HARTMANN y KESTER, 1995).
Esta investigación se desarrolló como continuación del taller realizado por VARGAS,
2003.
En el presente taller de investigación de proponen cuatro hipótesis:
• El uso de hipoclorito de sodio y fungicidas en la etapa de establecimiento,
tienen efecto antiséptico sobre los explantes de los portainjertos Gisela 5 y
Prunus cerasus.
• Utilizando en el medio de cultivo citoquininas y auxinas en una relación
favorable a las citoquininas, la multiplicación de los brotes de los
portainjertos Gisela 5 y Prunus cerasus es más efectiva.
• Al reducir la concentración de los macro y microelementos del medio de
cultivo MURASHIGE y SKOOG (1962), y aplicando a este medio ácido
indolbutírico (AIB) se obtiene un óptimo enraizamiento de los brotes del
portainjerto Gisela 5.
• El porcentaje de plantas sobrevivientes en la etapa de aclimatación aumenta
al someter las plantas propagadas in vitro a una preaclimatación en la
cámara de crecimiento.
El objetivo general de esta investigación es establecer un protocolo de propagación
in vitro para portainjertos de cerezo.

Los objetivos específicos son los siguientes:
• Evaluar el efecto de la desinfección de los explantes a distintas
concentraciones de hipoclorito de sodio para iniciar un cultivo aséptico in
vitro.
• Protocolizar un medio de cultivo de los explantes para la proliferación.
• Protocolizar un medio de cultivo de los explantes para el enraizamiento.
• Evaluar el efecto del uso de sellos de polipropileno sobre la aclimatación de
plantas de Gisela 5 propagadas in vitro.

2. REVISIÓN BIBLIOGRÁFICA 2.1. Características de los patrones de cerezo:
2.1.1. Gisela 5
Es un híbrido seleccionado en la Universidad Justus Liebig de Giessen (Alemania)
(LORETI y GIL, 1993).
Su origen es el cruzamiento de Prunus cerasus “Stockton morello” x Prunus
canescens. Se propaga por estaca semileñosa y también in vitro. Induce un vigor
débil, (menos de la mitad del F12-1) (Figura 1), una fructificación precoz, es
resistente a los fríos invernales, no produce sierpes y permite que las variedades
injertadas tengan ángulos de inserción de estructura abierta (PINOCHET y
TORRENTS, 1998).
Otorga una productividad muy buena y muy rápida a la variedad injertada sobre él,
pero puede disminuir el calibre de la fruta (CLAVERIE, 2002).
Pruebas en Alemania indican que no puede ser situado en condiciones anaeróbicas
en suelos arcillosos, donde Phythophtora puede causar daño. Muestra alguna
tolerancia a infecciones por PNRSV (Prunus Necrotic Ringspot Virus – Virus del
anillado necrótico de los Prunus) y PDV (Prune Dwarf Virus – Virus del enanismo de
los Prunus) (WEBSTER y LOONY, 1996).
En vivero, la variedad injertada sobre este patrón se injerta utilizando el injerto en
escudete. Principalmente se realiza a fines de enero. En relación con la afinidad de
injerto aparentemente ha resultado poco satisfactoria en Gisela 1 y Gisela 5, pero
buena en Gisela 10 (LORETI y GIL, 1993).
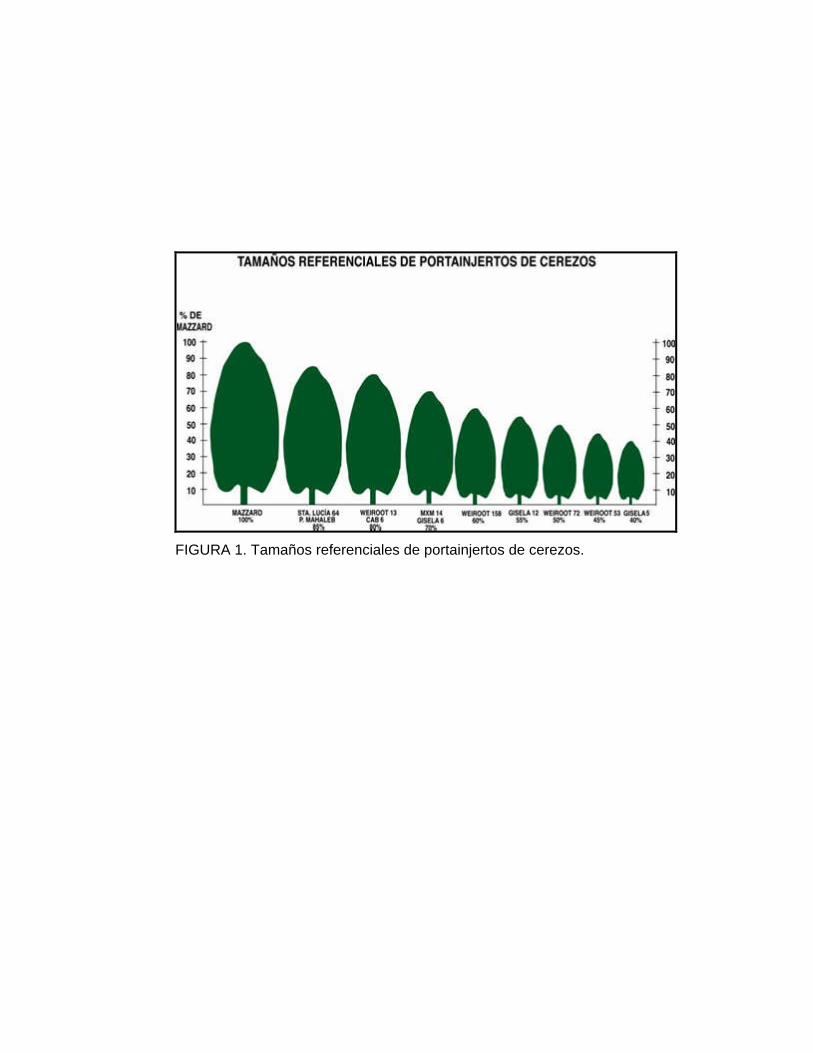
FIGURA 1. Tamaños referenciales de portainjertos de cerezos.

2.1.2. Prunus cerasus
Se desconoce su origen. Se propaga fácilmente por hijuelos o sierpes que nacen de
las raíces de los portainjertos de guindo agrio en huertos de cerezo ya establecidos
o de árboles aislados de la propia especie. Esta facilidad de emisión de rebrotes
representa a la vez una característica negativa desde el punto de vista productivo
del cerezo (JOUBLAN, 2002).
En Chile se usaba en suelos pesados y húmedos donde no prosperan otros
patrones como Mericier o Prunus mahaleb. Este patrón induce una mayor
precocidad en la producción y reduce el desarrollo, lo que permite una mayor
densidad de plantación. Como características negativas muestra un cierto grado de
incompatibilidad que se manifiesta por un mayor diámetro de la variedad que del
patrón. Su arraigamiento es muy superficial y débil por lo que puede presentar un
anclaje insatisfactorio que hace al árbol crecer inclinado. Desde el punto de vista
sanitario es afectado por cáncer bacterial, aunque aparentemente presentaría una
mayor tolerancia que el cerezo. También presenta problemas con algunas virosis
(JOUBLAN, 2002).
2.2 Propagación del cerezo:
Tradicionalmente la propagación se realizaba mediante injerto sobre patrón obtenido
a partir de semilla, con la ventaja de que la semilla evitaba la transmisión de
enfermedades, pero con el inconveniente de que se obtenían patrones vigorosos y
heterogéneos (INFOAGRO, 2004). En el caso de estos portainjertos, las plantas se
cultivan muy juntas en un semillero durante un año, luego se trasladan al vivero,
plantándola a unos 10 cm de distancia y se les deja crecer un segundo año antes de
injertarlos de yema. Si las condiciones de crecimiento son buenas, las semillas se
pueden plantar en forma directa en el vivero, temprano en primavera, y las plántulas

estarán suficientemente grandes para injertarse a fines del verano o principios de
otoño (HARTAMANN y KESTER, 1995).
Actualmente se tiende a hacer el injerto sobre patrones clonales de guindo, Santa
Lucía y cerezo. Tras un año de cría del patrón se realiza el injerto y se deja crecer
un año más antes de llevar a cabo el transplante (INFOAGRO, 2004).
El tipo de injerto utilizado es el de yema en T o injerto de astilla (HARTMANN y
KESTER, 1995).
En los últimos años se ha desarrollado la técnica in vitro para la propagación de
portainjertos y variedades comerciales de cerezo, técnica que se ha desarrollado de
manera comercial en viveros extranjeros y nacionales como es el caso de Vivero
Sur.
2.3. Selección del explante:
2.3.1. Explantes utilizados en Prunus
Existen numerosos informes de ensayos anteriores donde son utilizados diversos
tipos de explantes para la micropropagación de cerezos, obteniendo diversos
resultados.
MUNA et al. (1999), DAL ZOTTO y DOCAMPO (1997), MILLER et al. (1982) y SNIR
(1982) utilizaron ápices de yemas obteniendo muy buenos resultados en el
establecimiento y las fases sucesivas de la propagación in vitro de cerezos.
Por otro lado DRADI, VITO y STANDARDI (1996) usaron segmentos nodales que se
comportaron satisfactoriamente. OZZAMBAK y HEPAKSOY (1997) usaron
segmentos nodales de guinda ácida del cultivar “Heimanns Rubinweichsel”, donde

obtuvieron 1500 brotes a partir de un explante después de cinco meses de periodo
de subcultivo.
2.3.2. Cultivo de segmentos nodales
Con este nombre se conoce el aislamiento de una yema, junto con una porción de
tallo, para obtener un brote a partir de la yema. Cada una de las yemas que se
encuentran en las axilas de las hojas pueden ser aisladas sobre un medio nutritivo,
intentándose así su desarrollo in vitro. La velocidad de propagación depende del
número de yemas disponibles, si este número es pequeño la velocidad de
multiplicación resultará lenta. Cuantas más hojas se inicien, habrá más número de
yemas disponibles y por lo tanto la propagación será más rápida (PIERIK, 1990).
El método de segmentos nodales ha producido un éxito apreciable en muchas
plantas diferentes: papas, Vitis rupestris, peral, rosal, Hedera helix, Salix babilonica,
etc. (PIERIK, 1990).
2.4. Tratamiento de desinfección:
La desinfección del explante es uno de los factores más importantes que afectan el
éxito del establecimiento del explante estéril en un medio de cultivo.
Para la desinfección de explantes de cerezo se han propuesto diversos
tratamientos, entre los cuales se encuentran:
MILLER et al. (1982) remojaron los explantes en una solución de 0.5% de hipoclorito
de sodio con 0.1% de Tween 20 por diez minutos, seguido de dos enjuagues de
cinco minutos cada uno con agua destilada desionizada estéril. Posteriormente,
remojaron los explantes en etanol al 70% por cinco minutos y enjuagaron dos veces,
cada enjuague de cinco minutos, con agua destilada desionizada estéril.

JONES y HOPGOOD (1979), iniciaron la desinfección de explantes provenientes de
Prunus insistia y Prunus avium, lavándolos durante 50 minutos en agua corriente y
luego fueron inmersos en una solución al 10% de Domestos (preparación comercial
de hipoclorito de sodio) durante 25 minutos, seguido por cuatro enjuagues en agua
destilada estéril.
ÖZZAMBACK y HEPAKSOY (1997), obtuvieron buenos resultados en la
desinfección de segmentos nodales de Prunus cerasus al sumergir los explantes
durante 20 minutos en una solución al 2% de hipoclorito de sodio, agregando una
gotas de Tween 20. Posteriormente, realizaron tres enjuagues con agua destilada
estéril por 5 minutos.
VARGAS (2003), determinó que el mejor protocolo de desinfección para segmentos
uninodales de Gisela 5, Santa Lucía 64 y Colt, los tres portainjertos de cerezo,
consiste en lavar los explantes en agua corriente, posteriormente sumergirlos en
etanol al 70% durante 5 segundos, enjuagar con agua destilada estéril con ácido
ascórbico más ácido cítrico y luego en una solución de hipoclorito de sodio al 2,5%
por 15 minutos. Utilizando este protocolo obtuvo porcentajes de sobrevivencia
mayores al 90% durante la siembra realizada en primavera.
2.5. Pardeamiento y antioxidantes:
La iniciación del cultivo es el primer paso crítico en el éxito de la micropropagación
de alguna planta. Una de las causas de pérdida de explantes, además de la
contaminación, es la oxidación de compuestos fenólicos causado por la ruptura
celular debido a la herida. Las quinonas resultantes aparecen como una mancha en
el medio de cultivo (MARKS y SIMPSON, 1990). La polimerización de éstas con
proteínas en los tejidos vegetales causa la inhibición del crecimiento o muerte de los
explantes. La reacción es catalizada por la polifenoloxidasa (PPO), enzima
responsable de la oxidación de los o-difenoles a o-quinonas (MARKS y SIMPSON,

1990). La biosíntesis de estos compuestos fenólicos se ve incrementada por la
acción de la luz (YU y MEREDITH, 1986), es por ello que una de las estrategias
para reducir la difusión de los exudados cafés en el medio de cultivo es mantener
los explantes en oscuridad (ZIV y HALEVY, 1983).
El cultivo in vitro de especies leñosas, frecuentemente se ve afectado por el
necrosamiento y oxidación de sus tejidos, principalmente al comienzo de éste (YU y
MEREDITH, 1986). Para detener este pardeamiento existen diversos métodos:
• Adición de carbón activo al medio.
• Adición de PVP (Polyvinylpyrrolidone) al medio, polímero que absorbe las
sustancias de tipo fenólico.
• Adición de antioxidantes como el ácido cítrico, ácido ascórbico, tiourea o L-
cistina.
• Adición de dietil-ditiocarbamato (Dieca) en los enjuagues, después de la
esterilización.
• Adición de tres aminoácidos (glutamina, arginina y asparagina).
• El repicado frecuente a un medio fresco, a veces frena lentamente la
formación de pigmentos.
• Mantener las bases de los vástagos en la oscuridad durante el cultivo puede
disminuir la oxidación causado por foto-activación.
• Mojar los explantes en agua antes de situarlos en el cultivo también reduce
la oxidación.
• La reducción de sales en el medio de cultivo puede reducir la exudación.
• La eliminación de reguladores de crecimiento puede disminuir la oxidación
(PIERIK, 1990).
Se ha determinado que el pardeamiento puede ser reducido con un pretratamiento a
los explantes con antioxidantes, incorporación de antioxidantes al medio,
incubación de los cultivos en la oscuridad y realizar frecuentes subcultivos a un
medio fresco (YU y MEREDITH 1986) .

Mc COMB (1978) y WALKEY (1972) adicionaron polyvinylpyrrolidone (PVP) al
medio para minimizar la oxidación de explantes en manzano.
ZIV y HALEVY (1983) utilizaron antioxidantes, ácido ascórbico y ácido cítrico en
explantes de Strelitzia inmediatamente después de la excisión, lo cual permitió
reducir el pardeamiento del medio de cultivo.
OYANEDEL (1995), al comparar el efecto antioxidante de PVP, CA, DIECA, ácido
ascórbico y ácido cítrico sobre yemas axilares de palto cvs. Velvick y Lula, en este
último observó un efecto positivo del ácido ascórbico, favoreciendo la
organogénesis. En el caso del cv. Velvick obtuvo solo un 5% de brotación en los
tratamientos con DIECA y ácido ascórbico más ácido cítrico.
SAN MARTÍN (1996), utilizando yemas axilares de palto cv Lula y Velvick evaluó
dos métodos para reducir el pardeamiento y favorecer la brotación. Uno de ellos fue
la transferencia sucesiva de los explantes en el medio WPM (Woody Plant Medium)
incluyendo en éste PVP-360 y PVPP 6755 como antioxidantes y la citoquinina
sintética CPPU (Forclorofenuron) en concentraciones de 0,1 mg/L para el cv. Velvick
y 0,5 mg/L para el cv. Lula. El trasvasije de los explantes no ejerció efecto sobre el
pardeamiento. El otro método utilizado por la autora fue el pretratamiento de
etiolación, con el cual obtuvo un efecto positivo en el cv. Lula, asociado a menores
contenidos de fenoles totales del explante.
2.6. Medio de cultivo:
La elección del medio de cultivo es uno de los principales problemas a solucionar
previo a la siembra de una determinada especie (MARGARA, 1988). Los
ingredientes del medio varían según el tipo de planta y de la etapa de propagación
en que se esté trabajando. Estos ingredientes se pueden agrupar en: sales

inorgánicas, compuestos orgánicos, ingredientes naturales complejos, sostenes
inertes, hormonas y reguladores de crecimiento (HARTMANN y KESTER, 1995).
Para el cultivo in vitro de cerezo se utilizan principalmente dos tipos de medio. El
medio MURASHIGE y SKOOG (1962), usado ampliamente en un buen número de
especies y tipos de cultivo, en particular para plantas herbáceas y para cultivo de
tejidos en general. Este medio fue utilizado por ÖZZAMBAK y HEPAKSOY (1997),
DRADI, VITO y STANDRADI (1996), PEDROTI y CORNU (1991) y BARALDI et al.
(1988), obteniendo muy buenos resultados.
El otro medio alternativo es el Medio para Plantas Leñosas (WPM), (HARTMANN y
KESTER, 1995). PEVALEK-KOZLINA y JELASKA (1987), utilizando como medio
base WPM para cada una de las etapas de la propagación in vitro de Prunus avium,
lograron obtener plantas con crecimiento normal. De la misma forma, HAMMATT y
GRANT (1998), en su trabajo con Prunus avium cvs. F12/1 and Charger y genotipo
1908, y cinco genotipos de Prunus serotina y dos híbridos de P. avium x P.sargentii
obtuvieron desarrollo de brotes utilizando como medio base WPM (Woody Plant
Medium).
TANG et al. (2002), durante las etapas de establecimiento y proliferación utilizaron
el medio WPM para el cultivo de guindo dulce (Prunus avium L.) cv. Burlat,
Hedelfinger, Napoleón, Schneiders, y de guindo ácido (Prunus cerasus L.) cv.
Beutal, Spacher, Rexelle y Morellenfeuer, obteniendo brotes en cada uno de los
cultivares nombrados. Es más, el medio WPM estimuló de mejor manera la
caulogénesis de los brotes que el medio MS.

2.7. Organogénesis:
La organogénesis es la base fundamental de la multiplicación vegetativa, que se
apoya siempre en la formación de nuevos meristemos (MARGARA, 1988).
2.7.1. Rizogénesis in vitro:
Rizogénesis es el fenómeno de organogénesis mas generalmente implicado en la
multiplicación vegetativa (MARGARA, 1988).
La neoformación de los meristemos de raíz, como también la de los meristemos de
tallo resulta siempre de una desdiferenciación celular provocada, que conduce a la
producción de las células meristemáticas primarias y a la organización de un esbozo
meristemático (MARGARA, 1988).
La desdiferenciación procede por etapas sucesivas. Pero, según la naturaleza del
tejido original, existen diversas modalidades. En el caso de las raíces neoformadas
a partir de un callo bajo las condiciones in vitro, provienen de la desdiferenciación
local de ciertos tejidos del explante. Por otra parte, los tejidos en el origen de la
desdiferenciación pueden ser diversos, por ejemplo, parénquima o tejido cambial
(MARGARA, 1988).
En la rizogénesis in vitro pueden presentarse dos situaciones diferentes; el explante
inicial puede ser apto para la rizogénesis, o el explante puede ser inicialmente no
apto para la rizogénesis. En este ejemplo, un cierto número de factores actúan en
interacción para desencadenar la rizogénesis: aportación de auxinas, de un azúcar,
presencia de sales minerales, aportación eventual de ácido giberélico, luz y
temperatura (MARGARA, 1988).

2.7.2. Neoformación de yemas:
La brotación en su sentido más amplio, designa al mismo tiempo la iniciación y el
desarrollo de las yemas terminales, axilares, adventicias o neoformadas sobre un
callo (MARGARA, 1988).
El desencadenamiento de la brotación en cultivo in vitro, frecuentemente
observado, resulta del empleo de citoquininas, eventualmente asociadas a las
auxinas. Como consecuencia de las observaciones sobre la médula del tabaco
(SKOOG, 1971; SKOOG Y MILLER, 1956), frecuentemente confirmadas en otras
especies, se admite generalmente que la neoformación de yemas a partir de un
callo está bajo el control de interacciones entre citoquininas y auxinas. La relación
citoquinina/auxina debe ser elevada para que exista neoformación de yemas
(MARGARA, 1988).
Generalizando, la caulogénesis dependería de una relación óptima entre las
concentraciones totales de citoquininas y de auxinas exógenas y endógenas
(MARGARA, 1988).
2.8. Reguladores de crecimiento:
Los principios de la técnica del cultivo in vitro, reposan en parte en el conocimiento y
utilización de las auxinas. La expansión actual de las investigaciones sobre la
organogénesis y las aplicaciones a la multiplicación vegetativa está ampliamente
ligada a la utilización conjunta o secuencial de auxinas y citoquininas (MARGARA,
1988).

2.8.1. Auxinas:
Es muy importante la elección de la auxina en función de objetivo del cultivo. El
ácido β-indolpropiónico (AIP), el ácido β-indolacético (AIA), y el ácido β-indolbutírico
(AIB) son auxinas relativamente débiles. En los trabajos dirigidos a la multiplicación
vegetativa se reemplaza, con frecuencia y favorablemente, por AIB y AIP
(MARGARA, 1988).
2.8.2. Citoquininas:
Las citoquininas también influyen en la morfogénesis en un cultivo de tejidos. La
conservación del crecimiento de callos indiferenciados es generalmente lograda con
concentraciones aproximadamente similares de citoquinina y auxina. Una alta
relación molar de citoquinina sobre auxina tiende a inducir desarrollo de yemas,
mientras que una alta relación de auxina sobre citoquinina induce el desarrollo de
raíces. (GEORGE y SHERRINGTON, 1984).
A débil concentración (10--7 a 10-6 M), las citoquininas se utilizan frecuentemente
para estimular la proliferación de los tejidos en cultivo. A concentración más elevada
(10--6 a 10 –5 M) se emplean ampliamente para desencadenar la neoformación de
yemas sobre callos. Finalmente, pueden utilizarse a muy fuertes concentraciones,
(10 -5 a 10 –4 M) para favorecer la proliferación in vitro de meristemos axilares en
cultivo de ápices y para permitir la obtención de macollas de yemas, ulteriormente
fragmentadas (MARGARA, 1988).
Debido a que ejercen generalmente un efecto inhibidor sobre la rizogénesis, las
citoquininas se evitan o se emplean en dosis muy débiles en los medios de
enraizamiento (MARGARA, 1988).

Las citoquininas que se emplean con frecuencia son: kinetina (6 furfurilaminopurina),
benciladenina (6 bencilaminopurina), 2 isopentiladenina (2 iP) y zeatina (MARGARA,
1988).
La benciladenina se emplea con frecuencia debido a su gran actividad y su pequeño
costo (MARGARA, 1988).
2.9. Proliferación:
La función de la etapa de proliferación es incrementar el número de propágulos para
su enraizamiento posterior. Los explantes expandidos en la etapa de
establecimiento se cortan y los propágulos se vuelven a cultivar en un nuevo medio.
La multiplicación se repite con intervalos regulares. Si se difiere la trasferencia a
menudo se presenta deterioro y recuperación lenta. Es posible que sea necesario
hacer transferencias, con intervalos de tres a cuatro semanas y de manera ideal
deben hacerse cuando los brotes empiecen a aumentar de longitud (HARTMANN y
KESTER, 1995).
ÖZZAMBAK y HEPAKSOY (1997), lograron establecer que el medio ideal para la
proliferación de Prunus cerasus es el medio MS, conteniendo 1.0 – 1.5 mg/L de
BAP. Utilizando estas concentraciones ÖZAMBAK y HEPAKSOY (1997) obtuvieron
de 4,45 brotes por explante.
BARALDI et al. (1988) obtuvieron la más alta proliferación de los cultivares de
Prunus cerasus; Meteor, Monymorency y Northstar adicionando al medio MS 1 mg/L
de BA.
MUNA et al. (1999) sugieren para propagar in vitro el portainjerto Maxma 14, el
medio MURASHIGE y SKOOG (1962). Para la proliferación de brotes aconsejan el
medio base MS más 4.44 uM de BA con 0.49 uM de AIB.

VARGAS (2003), obtuvo la mayor proliferación de explantes de Gisela 5 utilizando
BAP a la concentración de 1,5 mg/L, logrando un promedio de 5.23 brotes por
explante en un plazo de 60 días.
2.10. Enraizamiento:
El enraizamiento es una de las etapas fundamentales para preparar a la planta
para su plantación y establecimiento fuera del medio de cultivo artificial
(HARTMANN y KESTER, 1995).
Uno de los cambios que se realiza en esta etapa es la reducción completa de la
concentración de citoquininas; utilizados en la etapa de proliferación para aumentar
la provisión de auxina. En este medio el propágulo puede pasar dos a cuatro
semanas para permitir que se desarrollen raíces (HARTMANN y KESTER, 1995).
Otra condición que favorece la iniciación de raíces, particularmente en las plantas
que enraizan con dificultad es la reducción de sales inorgánicas. (HARTMANN y
KESTER, 1995). Lo anterior de debe a que la composición mineral de muchos
medios, en exceso de algunos macroelementos induce un crecimiento anormal,
vitrificación y otros efectos indeseables (RUZIC, et al., 2000).
Los resultados obtenidos por DIMASSI-THERIOU (1995), en sus estudios de
enraizamiento realizados con el portainjerto GF-677, arrojaron que al reducir a la
mitad la concentración mineral del medio MS y agregando 1 mg/L de AIB aumenta
el porcentaje de enraizamiento y estimula la elongación radicular, aunque el número
de raíces y el peso fresco de ellas se ve reducido.
TRAVERS, STARBUCK y NATARELLA (1985) al evaluar el efecto del medio de
enraizamiento sobre el portainjerto de manzano Antonovka 313, encontraron que
reduciendo las concentraciones de los macronutrientes y micronutrientes a un

cuarto y a la mitad de su concentración, respectivamente, y adicionando al medio
0,25 µM de AIB más 1 g/L de carbón activado, se lograron buenos porcentajes de
enraizamiento. De igual manera, DREW (1987), al reducir la concentración mineral
del medio de enraizamiento para Carica papaya L. logró aumentar el número de
raíces y la velocidad de iniciación de éstas.
BARALDI et al. (1988), obtuvieron muy buenos resultados en el enraizamiento de
Prunus cerasus utilizando el medio MURASHIGE Y SKOOG (1962) modificado con
0.5 y 1 mg/L de AIB. Concentraciones mayores a éstas (2 – 4 mg/L) siempre
causaron producción de callo.
JONES, HOPGOOD y O’FARRELL (1977), utilizando el medio MURASHIGE Y
SKOOG (1962), complementado con 0.1 mg/L de GA3, 1 mg/L de AIB y 162 mg/L de
floroglucinol para el enraizamiento de 500 brotes del portainjerto de manzano M26,
obtuvieron un 97% de enraizamiento.
JONES y HOPGOOD, (1979), para enraizar el portainjerto Pixy (Prunus insistia) y F
12/1 (Prunus avium) utilizaron el medio MURASHIGE y SKOOG (1962)
complementado con AIB a 3 ppm obteniendo un buen porcentaje de enraizamiento
en ambos casos.
2.11. Aclimatación:
La etapa de aclimatación es esencial para que la planta sea capaz de sobrevivir el
cambio que significa pasar de un medio aséptico de cultivo al ambiente de vida
natural en el invernadero y luego en su sitio final, es decir, la planta se debe volver
autótrofa (HARTMANN y KESTER, 1995).
Durante este período son esenciales varias condiciones:

• Mantenimiento elevado de la humedad relativa (50 al 100%) durante 2 a 3
semanas para proteger la planta de la desecación y permitir que inicie
nuevas raíces y brotes.
• Medio de enraice, suelo aireado, bien drenado, que permita que las raíces
se desarrollen con rapidez.
• Protección contra diversos organismos patógenos hasta que haya adquirido
algo de resistencia.
• Control del crecimiento durante el transplante para superar o evitar el letargo
y la falta de crecimiento resultante (HARTMANN y KESTER, 1995).
A pesar de otorgar a las plantas todas las condiciones anteriormente nombradas, un
alto porcentaje de plantas micropropagadas se pierde usualmente por daño durante
el período de aclimatación y esto limita la aplicación de las técnicas de
micropropagación y ha restringido el uso extensivo de la micropropagación. Las
razones de esta dificultad incluye la reducida cantidad de cera epicuticular (SUTTER
y LANGHANS, 1982, GROUT, 1975), el pobre desarrollo de cutícula y el inadecuado
funcionamiento de los estomas (BRAINERD y FUCHIGAMI, 1981, FUCHIGAMI,
CHENG, SCELDNER, 1981), que da como resultado la excesiva pérdida de agua,
una pobre capacidad fotosintética (GROUT y MILLAM, 1985, DONALLY, VIDAVER
y COLBOW, 1984), y anormalidades anatómicas (WETZSTEIN y SOMMER, 1982).
Algunas de las formas de mejorar los porcentajes de sobrevivencia en la etapa de
aclimatación se cuentan el uso de retardantes como el paclobutrazol en el medio,
enriquecimiento con CO2 en las cámaras de crecimiento y reducción en la humedad
relativa en los tubos de cultivo (RAVINDRA y THOMAS, 1995). El uso de estas
técnicas tiene ventajas y desventajas. Específicamente, la reducción en la humedad
relativa en los tubos reduce el crecimiento de las plantas, pero se produce una
mayor cantidad de cera sobre la superficie de las hojas en comparación a plantas
cultivadas bajo condiciones de alta humedad relativa. Además, se produce la
reducción de la apertura estomática lo que reduce la pérdida de agua y por
consiguiente puede aumentar la tasa de sobrevivencia en el transplante. En el

trabajo realizado por WARDLE, DOBBS y SHORT (1983), con coliflor demostraron
que la reducción en la humedad relativa durante el cultivo in vitro significativamente
incrementa los niveles de cera en la superficie en hojas de coliflor, causando una
consiguiente disminución en la tasa de pérdida de agua.
En la investigación realizada por BRAINERD y FUCHIGAMI (1981) sobre la
aclimatación de plantas de manzano cv. Mac 9 concluyeron que éstas pueden ser
aclimatadas exponiéndose a una baja humedad relativa (30 – 40%) dentro de cuatro
a cinco días, y que esta baja humedad de aclimatación implica el desarrollo de una
acelerada respuesta estomática.
Lo anterior también se ve apoyado a través de lo realizado por FUCHIGAMI,
CHENG y SOELDER (1981), quienes al comparar grupos de plantas del portainjerto
de ciruelo Pixy sometidas a 100 y 50% de humedad relativa, encontraron que las
plantas sometidas al mayor porcentaje de humedad relativa, sufrieron una mayor
pérdida de agua, en comparación al grupo de plantas sometidas a una humedad del
50%, debido a la carencia de cera epicuticular y a la menor respuesta estomática.
RITCHIE, SHORT y DAVEY (1991), sometieron explantes de crisantemo y
remolacha a 100%, 81% y 58% de humedad relativa durante ocho semanas. En
plantas cultivadas de remolacha y crisantemo, las hojas que se iniciaron y se
desarrollaron con niveles de humedad relativa debajo del 100% fueron más
eficientes en limitar la pérdida de agua, que las que fueron cultivadas a 100% de
humedad relativa, ya que lograron un incremento en la producción de cera
epicuticular, densidad de tricomas y la mejora en el cierre estomático.
JONES y HOPGOOD (1979), después de mantener 3 semanas las plantas de Pixy y
F 12/ 1 en medio de enraizamiento, más del 80% de las plantas enraizadas de
ambos cultivares se establecieron cuando se transfirieron a macetas. Una vez que
las raíces estaban bien desarrolladas, los tubos con las pequeñas plantas fueron
transferidas desde la cámara de crecimiento a un invernadero sombreado por 10

días. Luego las plantas fueron sacadas de los tubos y colocadas en macetas de 8
cm con compost en cajas de propagación húmeda en el sombreadero, y después de
14 días fueron removidos desde las cajas y se desarrollaron normalmente en el
invernadero.
MILLER et al. (1982), luego de enraizar las plantas durante 5 semanas, las
transfirieron a un medio de turba más perlita, posteriormente colocadas en un
invernadero bajo mist intermitente. Como resultado, obtuvieron 73 plantas
sobrevivientes de 120 plantas transferidas.

3. MATERIALES Y MÉTODOS
3.1. Ubicación de los experimentos:
Los experimentos se realizaron en el Laboratorio de Propagación Profesor Gregorio
Rosenberg de la Facultad de Agronomía de la Pontificia Universidad Católica de
Valparaíso, ubicada en La Palma, Quillota, Quinta Región, entre enero del 2003 y
enero del 2004.
3.2. Material vegetal:
El material vegetal a utilizar proviene del huerto de colección de variedades y
portainjertos de cerezos, ubicada en La Estación Experimental La Palma, Quillota.
Los patrones utilizados como plantas donadoras de explantes fueron Gisela 5 y
Prunus cerasus. Ambos portainjertos fueron plantados el año 1997.
Gisela 5 se encuentra conducido en copa y Prunus cerasus es utilizado como
portainjerto de variedades de guindo dulce.
3.3. Medio de cultivo:
El medio de cultivo base a utilizar en cada uno de los ensayos relacionados con las
etapas previas a la aclimatación de las plantitas fue el descrito por MURASHIGE Y
SKOOG (1962) más 7,5 gr/L de agar Algas Marinas. (Cuadro 1).
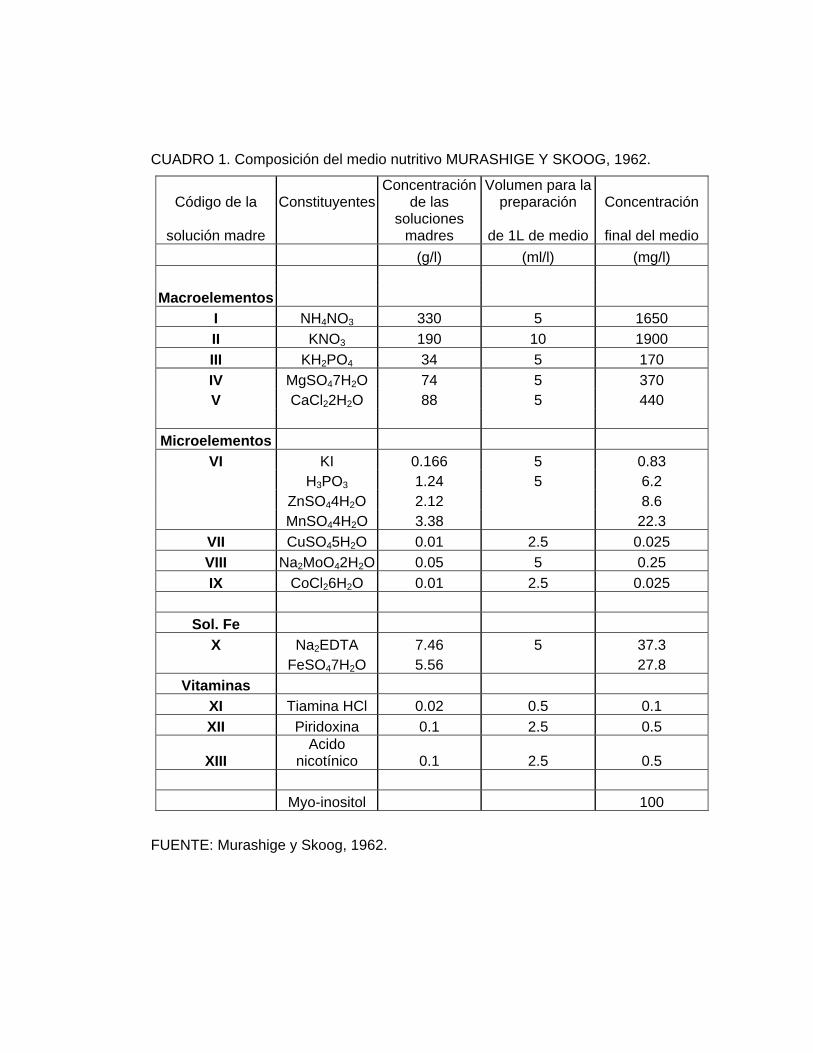
CUADRO 1. Composición del medio nutritivo MURASHIGE Y SKOOG, 1962.
Código de la ConstituyentesConcentración
de las Volumen para la
preparación Concentración
solución madre soluciones
madres de 1L de medio final del medio (g/l) (ml/l) (mg/l)
Macroelementos I NH4NO3 330 5 1650 II KNO3 190 10 1900 III KH2PO4 34 5 170 IV MgSO47H2O 74 5 370 V CaCl22H2O 88 5 440
Microelementos VI KI 0.166 5 0.83 H3PO3 1.24 5 6.2 ZnSO44H2O 2.12 8.6 MnSO44H2O 3.38 22.3
VII CuSO45H2O 0.01 2.5 0.025 VIII Na2MoO42H2O 0.05 5 0.25 IX CoCl26H2O 0.01 2.5 0.025
Sol. Fe X Na2EDTA 7.46 5 37.3 FeSO47H2O 5.56 27.8
Vitaminas XI Tiamina HCl 0.02 0.5 0.1 XII Piridoxina 0.1 2.5 0.5
XIII Acido
nicotínico 0.1 2.5 0.5 Myo-inositol 100
FUENTE: Murashige y Skoog, 1962.

Las sales minerales y compuestos orgánicos se pesaron en una balanza digital. En
la medición de líquidos se utilizaron pipetas graduadas, matraces de aforo y
matraces de Erlenmeyer.
El pH de los medios se ajustó a 5.7, con alícuotas de ácido clorhídrico 0.1 N ó
hidróxido de potasio 0.1 N.
Se emplearon tubos de vidrio de 40 ml, disponiendo de 10 ml de medio por unidad,
cubriéndolos con un cuadrado de aluminio laminado de aproximadamente 25 cm2 y
frascos de 150 ml llenados con 40 ml de medio tapados con su respectiva tapa.
Los tubos y frascos fueron esterilizados en un autoclave de control manual, a 121ºC
durante 15 minutos.
3.4. Manipulación del material vegetal:
La manipulación del material vegetal se realizó bajo una cámara de flujo laminar
(Figura 2), con aire filtrado de 0.2 micras, disponiendo de un explante por tubo o
frasco según sea el caso.
El corte de los explantes se realizó con un bisturí y una pinza previamente
flameados en alcohol.
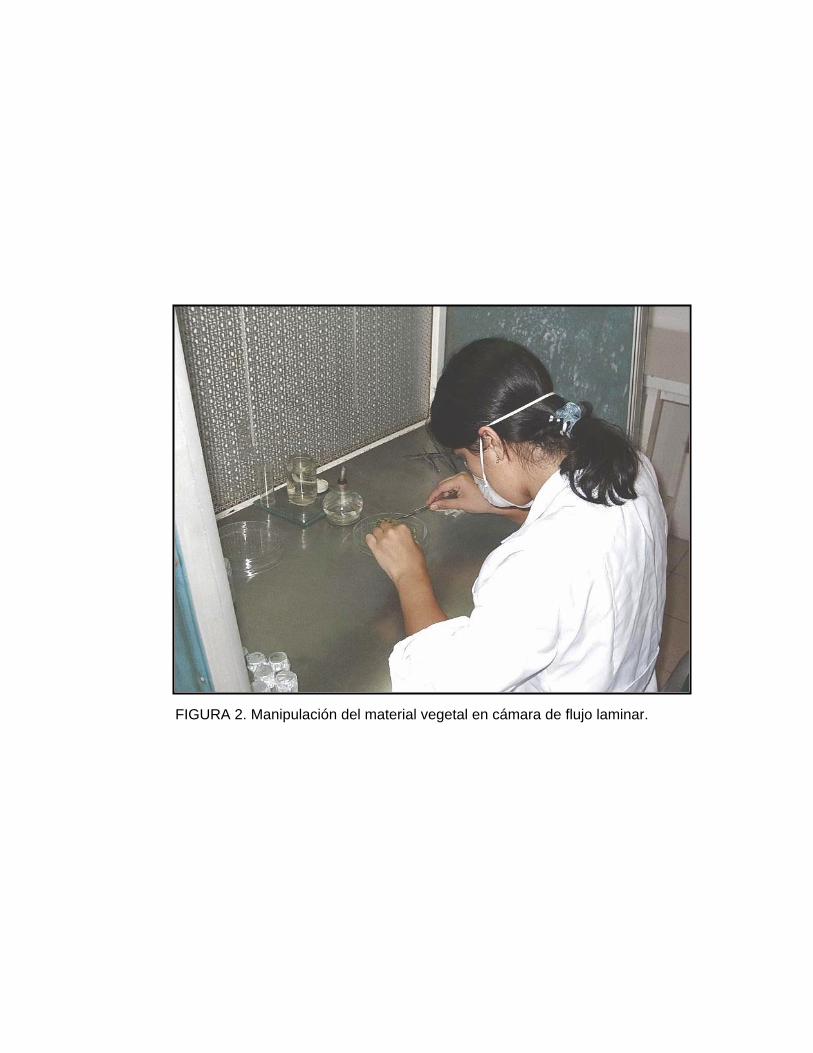
FIGURA 2. Manipulación del material vegetal en cámara de flujo laminar.

3.5. Mediciones de los parámetros a evaluar:
Las mediciones de longitud fueron realizadas con un pie de metro digital marca
“Mitutoyo”, Digimatic Caliper, código 500-144.
Para evaluar el grado de oxidación se utilizó la Escala Relativa de Pardeamiento
(ERP), citada por OYANEDEL (1995), asignando una calificación a cada estado del
explante:
1: Tejido con pardeamiento en la zona de corte.
2: Tejido con pardeamiento en la zona del corte y necrosis apical.
3: Necrosis apical del explante.
El color fue medido de acuerdo a la tabla Munsell (MUNSELL, 1998) asignando un
número a cada color del explante:
1: 5 GY 4/8 verde oscuro
2: 5 GY 5/6 verde de la especie
3: 5 GY 6/8 verde claro
4: 5 GY 7/8 verde amarillo

3.6. Ensayos:
Ensayo 1: Evaluación del efecto de la desinfección en segmentos uninodales de dos patrones de cerezo recolectados en marzo.
El material vegetal del portainjerto Gisela 5 consistió en ramillas de crecimientos
vegetativos del año, de 15-30 cm de largo, de vigor equivalente, obtenidas de los
cuatro puntos cardinales del árbol donador del material. Para el caso del portainjerto
Prunus cerasus se utilizaron sierpes originadas en el verano, de 15-30 cm de largo,
de vigor equivalente.
Tanto las ramillas como las sierpes fueron seleccionadas al azar, cortadas con una
tijera de podar previamente desinfectada con una solución de hipoclorito de sodio al
1 %.
Para ambos casos se emplearon segmentos uninodales de 1 cm de longitud (Figura
3 y 4).
La desinfección de los explantes se realizó de la siguiente manera: utilizando una
tijera de podar se eliminaron las hojas de las ramillas y sierpes, luego fueron
cortadas en segmentos nodales para ser cada uno cepillado bajo el agua corriente.
Una vez cepillados los explantes, fueron inmersos en etanol al 70 % durante cinco
segundos y enjuagados con una solución de agua estéril más antioxidantes (ácido
ascórbico y ácido cítrico en una concentración de 400 mg/L cada uno). Enseguida,
los explantes fueron sumergidos en hipoclorito de sodio a diferentes
concentraciones según el tratamiento, disuelto en agua estéril con antioxidantes
(ácido ascórbico y ácido cítrico en una concentración de 400 mg/L cada uno), más
una gota de Tween 20 agitándolos durante 15 minutos. La solución de hipoclorito de
sodio estaba contenida en un matraz de Erlenmeyer de 250 ml. Posteriormente los
explantes se enjuagaron tres veces con agua destilada estéril más ácido cítrico y

FIGURA 3. Segmento uninodal de 1 cm de longitud, proveniente de hijuelos de
Prunus cerasus.

FIGURA 4. Segmentos uninodales de 1 cm de longitud provenientes de ramillas del
portainjerto Gisela 5.

ácido ascórbico a una concentración de 400mg/L cada uno. Estos enjuagues se
realizaron bajo la cámara de flujo laminar.
Los tratamientos fueron los siguientes:
T1: Desinfección con hipoclorito de sodio al 1.5 %
T2: Desinfección con hipoclorito de sodio al 2 %
T3: Desinfección con hipoclorito de sodio al 2.5 %
Una vez desinfectados los explantes, fueron sembrados en tubos individuales que
contenían 10 ml del medio de cultivo MURASHIGE y SKOOG (1962),
complementado con 30 gr/L de sacarosa y más 7,5 gr/L de agar algas marinas.
Sembrados los explantes, los tubos se sellaron como se indicó en el punto 3.3.
La siembra se realizó en una cámara de flujo laminar, inoculando un explante por
tubo. El corte de los explantes se realizó a medio cm sobre y bajo la yema con
bisturí y pinzas previamente flameadas. Una vez sembrados, los explantes se
llevaron a cámara de crecimiento. Durante los primeros siete días de incubación los
tubos se mantuvieron en oscuridad total. Pasado este período los tubos se llevaron
paulatinamente a la luz para finalmente ser sometidos a un fotoperíodo de 16 horas
dentro de la cámara de crecimiento.
Como diseño estadístico se utilizó un Diseño Completamente al Azar (DCA)
unifactorial. Como se señaló anteriormente, los tratamientos aplicados fueron tres,
cada uno con 33 repeticiones.
Los parámetros a medir fueron el grado de oxidación evaluados a los 10 días de
realizada la siembra y el porcentaje de sobrevivencia evaluado a los 10 y 20 días de
sembrado cada explante.

El grado de oxidación se analizó mediante el test no paramétrico de Kruskal-Wallis y
el porcentaje de sobrevivencia a través del test de comparación de proporciones.
Para ambos test se aplicó un nivel de significancia del 5%.

Ensayo 2: Efecto de tres concentraciones de Acido Indol Butírico (AIB) en el medio de enraizamiento sobre el comportamiento in vitro de Gisela 5.
El material vegetal utilizado para el desarrollo de este ensayo corresponde a plantas
del patrón Gisela 5, obtenidas por VARGAS (2003). De estas plantas se tomaron
100 al azar y se colocaron a enraizar. Para la elección de estas plantas se
consideraron aquellas que tuvieran un crecimiento mínimo de 2 cm y 5 hojas
verdaderas.
Como medio de enraizamiento se utilizó el medio MURASHIGE y SKOOG (1962)
(MS) con su composición mineral a la concentración normal o reducida a la mitad
según el tratamiento, complementado con AIB en distintas concentraciones
dependiendo del tratamiento:
T1: MS completo + 0.5 mg/L de AIB
T2: MS completo + 1 mg/L de AIB
T3: MS reducido a la mitad + 0.5 mg/L de AIB
T4: MS reducido a la mitad + 1 mg/L de AIB
El medio de enraizamiento fue depositado en frascos de 150 ml y tapados. Cada
frasco se llenó con 40 ml de medio.
El manejo de las plantas se realizó bajo la cámara de flujo laminar, realizando los
cortes bajo agua estéril más antioxidantes (400 mg/L de ácido ascórbico y 400 mg/L
de ácido cítrico). Una vez realizado este proceso, los frascos se llevaron a la cámara
de crecimiento donde se mantuvieron 42 días en enraizamiento.
Se utilizó un Diseño Completamente al Azar (DCA) multifactorial, puesto que se
evaluaron dos factores. Cada tratamiento se constituyó de 25 repeticiones.

Los parámetros a medir fueron el tiempo que demoraron en aparecer las primeras
raíces, para lo cual se realizaron observaciones diarias, a partir de la segunda
semana después de realizada la siembra, número y longitud de las raíces evaluados
al mes de realizada la siembra. Estos tres parámetros fueron sometidos a un
análisis de varianza con un 5% de significancia. Dado el resultado de este análisis,
los tres parámetros fueron contrastados con el Test de Tukey con un nivel de
significancia del 5%.

Ensayo 3: Efecto de tres tipos de sellos sobre la aclimatación del patrón de cerezo Gisela 5.
Como material vegetal se utilizaron las plantas de Gisela 5 que enraizaron en el
ensayo 2.
Antes de llevar las plantas al sombreadero para su aclimatación, se realizó la
preaclimatación de las plantas dentro de la cámara de crecimiento. Para ello, una
vez que las plantas del patrón Gisela 5 enraizaron se procedió a cambiar la cubierta
de los frascos (tapas de aluminio) por sellos de polipropileno. El material se obtuvo
de las mascarillas que utilizan los médicos, las cuales están constituidas de tres
capas de distinta porosidad (Figura 5), las mismas que utilizó FIGUEROA (2003).
El cambio de sellos se realizó dentro de la cámara de flujo laminar.
Los tratamientos fueron los siguientes:
T0: Frasco con tapa de aluminio
T1: Frasco con capa 1 + capa 3 (Figura 6)
T2: Frasco con capa 1 + capa 2
El grado de porosidad de las capas es el siguiente:
Capa 1: Más porosa
Capa 2: Menos porosa
Capa 3: Porosidad intermedia
Después de 20 días de preaclimatación, las plantas fueron llevadas al invernadero
donde cada planta se transplantó a un vaso de 200 cc utilizando como sustrato
turba más perlita en la relación 2:1. Una vez realizado el transplate, los vasos con
plantas fueron envueltos en plástico y sellados. A partir de la fecha de transplante

FIGURA 5. Representación de la porosidad de las capas de polipropileno utilizadas
en la preaclimatación.
CAPA 1
CAPA 2
CAPA 3
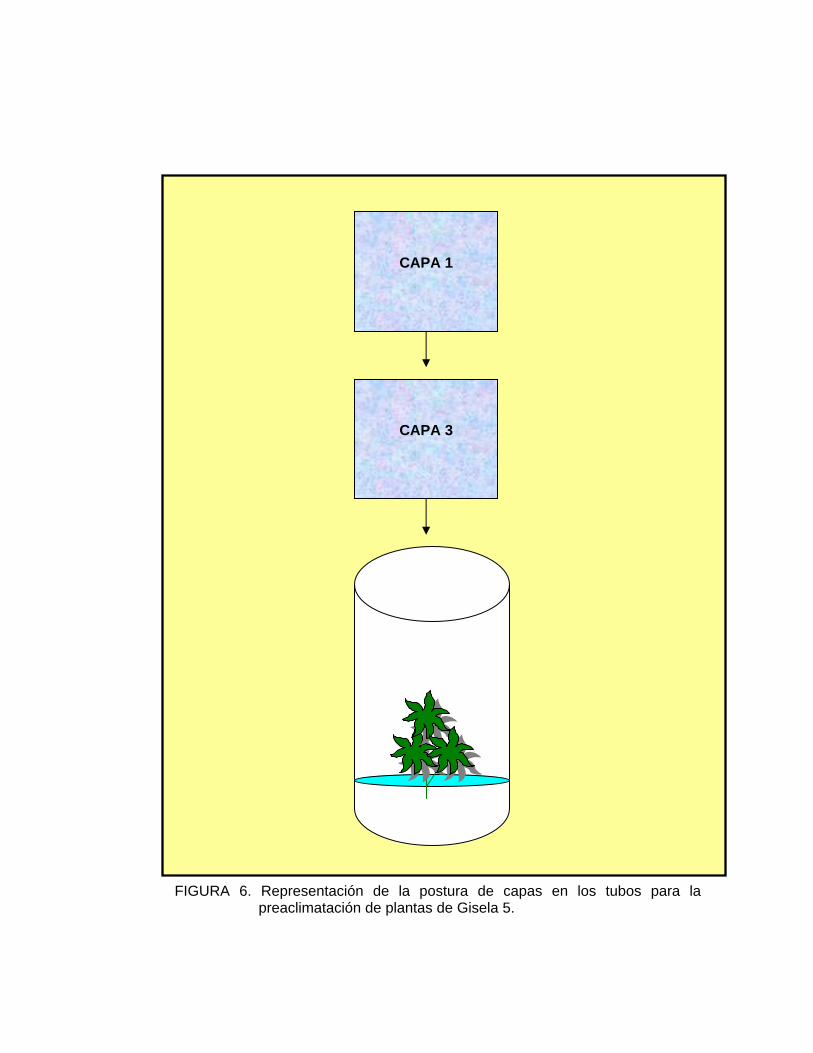
FIGURA 6. Representación de la postura de capas en los tubos para la preaclimatación de plantas de Gisela 5.
CAPA 3
CAPA 1

se cortó una punta de la bolsa plástica cada semana, y finalmente en la tercera
semana se abrió completamente la bolsa.
Se utilizó un Diseño Completamente al Azar. Cada tratamiento estuvo constituido
por 14 repeticiones, siendo la unidad muestral cada plantita.
Los parámetros a evaluar fueron la proporción de plantas sobrevivientes, el
crecimiento de las plantas (cm), para lo cual se utilizó una regla, y el número de
hojas nuevas. La evaluación se realizó un mes después de transferir las plantas de
la cámara de crecimiento al invernadero.
La proporción de plantas sobrevivientes se analizó mediante el test de comparación
de proporciones. El crecimiento de las plantas y el número de hojas nuevas se
analizaron a través de un análisis de varianza y después se sometieron al Test de
Tukey. Ambos test se realizaron con un nivel de significancia de 5%.

Ensayo 4: Evaluación del efecto de la desinfección en segmentos uninodales de dos patrones de cerezo recolectados en septiembre.
El material vegetal del portainjerto Gisela 5 consistió en ramillas de crecimientos
vegetativos del año, de 15-30 cm de largo, de vigor equivalente, obtenidas de los
cuatro puntos cardinales del árbol donador del material. Para el caso del portainjerto
Prunus cerasus se utilizaron sierpes generadas en la primavera 2003, de 15-30 cm
de largo, de vigor equivalente.
Tanto las ramillas como las sierpes fueron seleccionadas al azar, cortadas con una
tijera de podar previamente desinfectada con una solución de hipoclorito de sodio al
1 %.
Para ambos casos se emplearon segmentos uninodales de 1 cm de longitud.
La desinfección de los explantes se realizó de la siguiente manera: utilizando una
tijera de podar se eliminaron las hojas de las ramillas y sierpes, luego fueron
cortadas en segmentos nodales para ser cada uno cepillado bajo el agua corriente.
Una vez cepillados los explantes fueron inmersos durante 15 min en una solución de
Benlate más Captan a la concentración de 1,2 gr/L cada uno, luego el proceso
continuó igual que en el ensayo 1.
Los tratamientos fueron los siguientes:
T1: Benlate/Captan + hipoclorito de sodio al 1.5 %
T2: Benlate/Captan + hipoclorito de sodio al 2 %
T3: Benlate/Captan + hipoclorito de sodio al 2.5 %
Una vez desinfectados los explantes, fueron sembrados en tubos individuales que
contenían 10 ml del medio de cultivo MURASHIGE y SKOOG (1962),

complementado con 30 gr/L de sacarosa y más 7,5 gr/L de agar algas marinas.
Sembrados los explantes, los tubos se sellaron como se indicó en el punto 3.3.
La siembra se realizó en una cámara de flujo laminar, inoculando un explante por
tubo. El corte de los explantes se realizó a medio cm sobre y bajo la yema con
bisturí y pinzas previamente flameadas. Una vez sembrados, los explantes se
llevaron a cámara de crecimiento. Durante los primeros siete días de incubación los
tubos se mantuvieron en oscuridad total. Pasado este período los tubos se llevaron
paulatinamente a la luz para finalmente ser sometidos a un fotoperíodo de 16 horas
dentro de la cámara de crecimiento.
Como diseño estadístico se utilizó un Diseño Completamente al Azar (DCA)
unifactorial. Como se señaló anteriormente, los tratamientos aplicados fueron tres,
cada uno con 30 repeticiones.
Los parámetros a medir fueron, el grado de oxidación evaluados a los 10 días de
realizada la siembra y el porcentaje de sobrevivencia evaluado a los 10 y 20 días de
sembrado cada explante.
El grado de oxidación se analizó mediante el test no paramétrico de Kruskal-Wallis y
el porcentaje de sobrevivencia a través del test de comparación de proporciones.
Para ambos test se utilizó un nivel de significancia del 5%.

Ensayo 5: Evaluación del efecto de la desinfección en segmentos uninodales del patrón de cerezo Gisela 5 recolectados en octubre.
La recolección del material vegetal del portainjerto Gisela 5 se realizó de la misma
manera que en el ensayo 1.
Como explantes se utilizaron segmentos uninodales de 1 cm de longitud.
Los tratamientos fueron los siguientes:
T1: Benlate (1,2 gr/L)/Captan (1,2 gr/L) + Hipoclorito de sodio al 2,5 % + Phyton
(0,75 cc/250 ml)
T2: Benlate (1,2 gr/L)/Captan (1,2 gr/L) + hipoclorito de sodio al 2,5 %
T3: Hipoclorito de sodio al 2.5 % + Phyton (0,75 cc/250 ml)
La desinfección de los explantes se realizó de la siguiente manera: utilizando una
tijera de podar se eliminaron las hojas de las ramillas, luego fueron cortadas en
segmentos nodales para ser cada uno cepillado bajo el agua corriente.
En los tres tratamientos los explantes fueron sumergidos en etanol al 70% durante 5
seg para luego ser enjuagados con agua destilada estéril más antioxidantes (ácido
ascórbico y ácido cítrico en una concentración de 400 mg/L cada uno). En los
tratamientos uno y dos el paso por el etanol fue después de colocar los explantes en
la solución de Benlate más Captan, donde se mantuvieron 15 min. En el tratamiento
tres el etanol se aplicó antes de colocar los explantes en la solución de Hipoclorito
de Sodio. El paso de los explantes por el hipoclorito de sodio en todos los
tratamientos fue de 15 min, luego de este tiempo los explantes se enjuagaron tres
veces con agua destilada estéril más antioxidantes (ácido ascórbico y ácido cítrico
en una concentración de 400 mg/L cada uno).

Los explantes que fueron colocados en una solución de Phyton (funguicida-
bactericida), se mantuvieron en ésta durante 15 min. La preparación de esta
solución se realizó en la cámara de flujo laminar, utilizando como disolvente agua
destilada estéril.
La desinfección con Hipoclorito de sodio y Phyton se realizó dentro de la cámara de
flujo laminar, utilizando como material de vidrio matraces de Erlenmeyer de 250 ml.
El medio utilizado y el método de siembra es el mismo que el utilizado en el ensayo
1.
Como diseño estadístico se utilizó un Diseño Completamente al Azar (DCA)
unifactorial. Como se señaló anteriormente, los tratamientos aplicados fueron tres,
cada uno con 30 repeticiones.
Los parámetros a medir fueron el porcentaje y grado de oxidación evaluados a los
10 días de realizada la siembra y el porcentaje de sobrevivencia evaluado a los 10 y
20 días de sembrado cada explante.
Los parámetros a medir fueron el grado de oxidación evaluados a los 10 días de
realizada la siembra y el porcentaje de sobrevivencia evaluado a los 10 y 20 días de
sembrado cada explante.
El grado de oxidación se analizó mediante el test no paramétrico de Kruskal-Wallis y
el porcentaje de sobrevivencia a través del test de comparación de proporciones.
Para ambos test se utilizó un nivel de significancia del 5%.

Ensayo 6: Efecto de tres concentraciones de Bencil-aminopurina (BAP) en el medio de proliferación sobre el comportamiento in vitro de Gisela 5 y Prunus cerasus.
El material vegetal utilizado fueron explantes de Prunus cerasus y Gisela 5
provenientes de la etapa de establecimiento, es decir, segmentos nodales de 1 cm
de longitud.
Como medio de cultivo se usó el medio MURASHIGE y SKOOG (1962) tanto en su
parte mineral como orgánica (Cuadro 1), suplementado con 0.5 mg/L de AIB, 30 g/L
de sacarosa y tres concentraciones de BAP, dependiendo del tratamiento.
Los tratamientos fueron los siguientes:
T1: Medio de establecimiento + 0.5 mg/L AIB + 1.0 mg/L BAP
T2: Medio de establecimiento + 0.5 mg/L AIB + 1.5 mg/L BAP
T3: Medio de establecimiento + 0.5 mg/L AIB + 2.0 mg/L BAP
Se realizaron repiques cada 20 días, realizando los cortes bajo agua estéril más
antioxidantes (ácido ascórbico y ácido cítrico en una concentración de 400 mg/L
cada uno).
Se utilizó un Diseño Completamente al Azar. Cada tratamiento estaba compuesto
de 30 repeticiones.
Los parámetros evaluados fueron el crecimiento del brote (mm), número de hojas,
color de hojas, el grado y porcentaje de oxidación, medidos al final de la etapa de
proliferación (40 días).
El número de brotes por explante, el crecimiento del brote y número de hojas se
analizaron mediante un análisis de varianza (ANDEVA). Dado el resultado de este

análisis, los tres parámetros fueron contrastados con el Test de Tukey con un nivel
de significancia del 5%. Mientras que el color de las hojas se analizó mediante el
test no paramétrico de Kruskal-Wallis con un nivel de significancia del 5%.
4. PRESENTACIÓN Y DISCUSIÓN DE RESULTADOS Ensayo 1. Evaluación del efecto de la desinfección de segmentos uninodales
de dos patrones de cerezo recolectados en marzo sobre la oxidación y porcentaje de sobrevivencia.
Para este ensayo, se utilizaron los portainjertos de cerezo Gisela 5 y Prunus
cerasus.
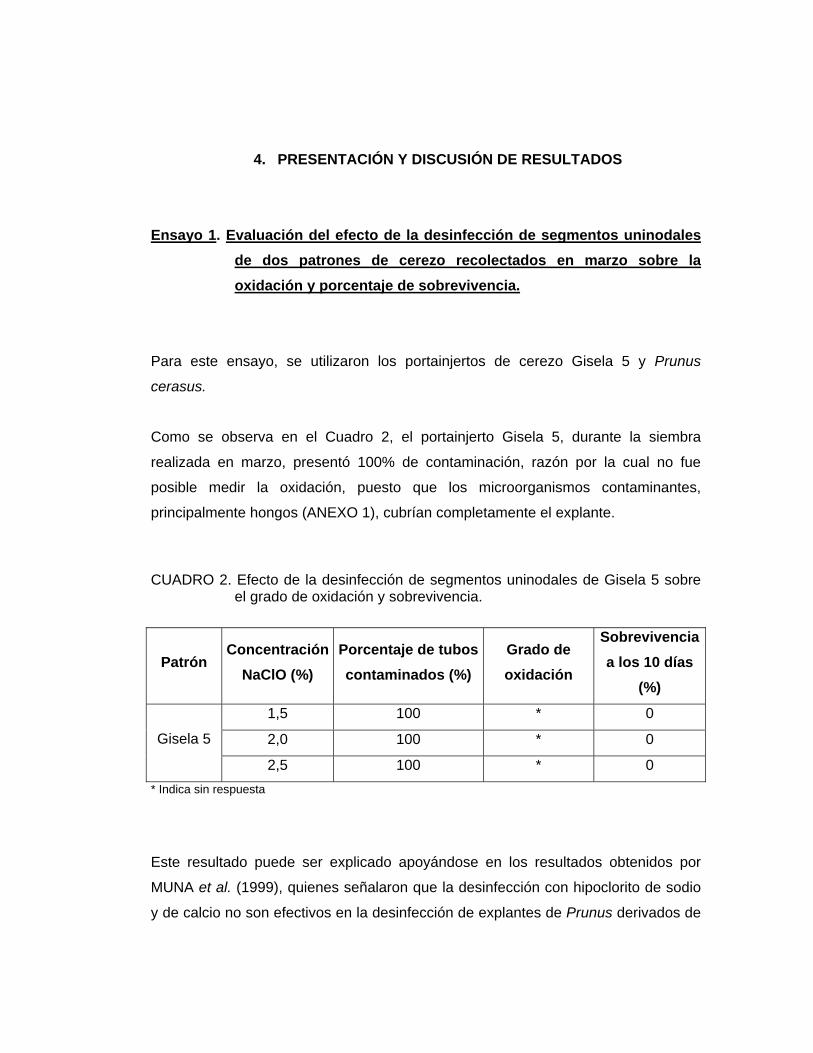
Como se observa en el Cuadro 2, el portainjerto Gisela 5, durante la siembra
realizada en marzo, presentó 100% de contaminación, razón por la cual no fue
posible medir la oxidación, puesto que los microorganismos contaminantes,
principalmente hongos (ANEXO 1), cubrían completamente el explante.
CUADRO 2. Efecto de la desinfección de segmentos uninodales de Gisela 5 sobre
el grado de oxidación y sobrevivencia.
* Indica sin respuesta
Este resultado puede ser explicado apoyándose en los resultados obtenidos por
MUNA et al. (1999), quienes señalaron que la desinfección con hipoclorito de sodio
y de calcio no son efectivos en la desinfección de explantes de Prunus derivados de
Patrón Concentración
NaClO (%) Porcentaje de tubos contaminados (%)
Grado de oxidación
Sobrevivencia a los 10 días
(%)
1,5 100 * 0
2,0 100 * 0 Gisela 5
2,5 100 * 0

árboles adultos crecidos bajo condiciones de campo, comparado con plantas
madres que crecen bajo invernadero. Debido a esto, es que estos investigadores
sugieren desinfectar los explantes provenientes de árboles adultos crecidos bajo
condiciones de campo con cloruro mercúrico al 0,01% a pesar de ser tóxico para las
plantas y para el operador, ya que ha sido determinado por la EPA (Environmental
Protection Agency) como un posible carcinógeno (PROBLEMAS DE QUIMICA,
2004). De igual manera, DAL ZOTTO y DOCAMPO (1997), obtuvieron un porcentaje
de contaminación en yemas de Mariana 2624 y Pixy tomadas del exterior mayor al
80%, mientras que el de yemas procedentes de plantas mantenidas en invernadero
no superó el 35%.
La nula sobrevivencia de los segmentos nodales utilizados como explantes puede
deberse al tamaño de éstos, que fue de 1 cm, explantes que ya contaban con una
yema bastante desarrollada, con un alto número de escamas; condición muy
favorable para la permanencia de microorganismos y que hace más difícil la
eliminación de éstos. Esto concuerda con lo señalado por GEORGE y
SHERRINGTON (1984) quienes señalan que explantes grandes son difíciles de
descontaminar de manera efectiva.
En el caso del patrón Prunus cerasus, en el Cuadro 3, se puede observar que no
existe diferencia significativa entre las proporciones de plantas sobrevivientes
comparadas, es decir, el uso de cualquiera de las tres concentraciones de
hipoclorito de sodio da como resultado una proporción similar de plantas
sobrevivientes.
La causa de contaminación de los explantes de este portainjerto fue tanto de
hongos como de bacterias (ANEXO 2).
La sobrevivencia de los explantes de Prunus cerasus, al compararla con los
explantes de Gisela 5 fue notoriamente superior. Esta diferencia se debió a que los
segmentos nodales de Prunus cerasus aún no tenían la yema visiblemente

desarrollada como Gisela 5, condición que favoreció la desinfección de los
explantes, puesto que no habían escamas de yemas que permitieran el alojamiento
de los microorganismos, encontrándose solo en la superficie de los segmentos
nodales. Según GEORGE y SHERRINGTON (1984), los contaminantes están
presente principalmente en la superficie de las partes de las plantas utilizadas como
donadoras de explantes, aunque pueden estar alojados en grietas, entre las
escamas de las yemas o en otras partes, resultando bastante difícil eliminarlos.
CUADRO 3. Efecto de la desinfección sobre la sobrevivencia de segmentos uninodales de Prunus cerasus sembrados en marzo.
* Letras iguales indican que no existe diferencia significativa entre los tratamientos, según Test de Comparación de Proporciones (P= 0,05) Esto indica que el uso de hipoclorito de sodio es efectivo para la desinfección de los
explantes de diversas especies de Prunus, puesto que otros investigadores como
OZZAMBAK y HEPAKSOY (1997), DAL ZOTTO y DOCAMPO (1997), MILLER et al.
(1982) y JONES y HOPOOG (1979) obtuvieron una alta sobrevivencia de explantes
utilizando como agente desinfectante hipoclorito de sodio. Sin embargo, esto se
contrapone a lo expresado por MUNA, AHMAD y ABDUL-ARHMAN (1999), pero
como se señaló anteriormente, la efectividad del agente desinfectante depende en
gran parte de las condiciones en las que se encuentra la planta donadora del
explante (bajo invernadero o en condiciones de campo) y de las características del
explante.
Sobrevivencia (%) Patrón
Concentración NaClO (%) 10 días 20 días
1,5 84.8 a* 60.6 a*
2,0 69.7 a* 57.6 a* Prunus cerasus
2,5 78.8 a* 57.6 a*

CUADRO 4. Efecto de la desinfección sobre la el grado de oxidación de segmentos uninodales de Prunus cerasus sembrados en marzo a los 10 días de sembrados.
Concentración de NaClO (%) Promedio grado de oxidación
(ERP)**
1.5% 1.1 a*
2% 1.2 a
2.5% 1.2 a
* Letras iguales indican que no existe diferencia significativa entre los tratamientos, según Test de Kruskal Wallis (P= 0,05) **ERP: Escala Relativa de Pardeamiento
Con respecto a la oxidación, en el Cuadro 4 se puede observar que no existe
diferencia significativa entre los tratamientos. Además, se observa que el promedio
del grado de oxidación fue muy cercano a uno, es decir, según la Escala Relativa de
Pardeamiento (ERP), los explantes sólo presentaban pardeamiento en la zona de
corte.
En este caso se utilizaron como antioxidantes ácido cítrico y ácido ascórbico. Estos
mismos fueron utilizados por ZIV y HALEVY (1983) sobre explantes de Strelitzia, y
por DRADI, VITO y STANDARDI (1996) quienes adicionaron el ácido ascórbico y
ácido cítrico al medio de establecimiento para 11 ecotipos de P. mahaleb,
obteniendo muy bajos porcentajes de oxidación.
Ensayo 2: Efecto del medio base y de dos concentraciones de Ácido Indol Butírico (AIB) en el enraizamiento in vitro de Gisela 5.
Durante el proceso de enraizamiento del portainjerto Gisela 5, llevado a cabo en
invierno, se observaron diferencias entre los tratamientos según las variables
medidas.
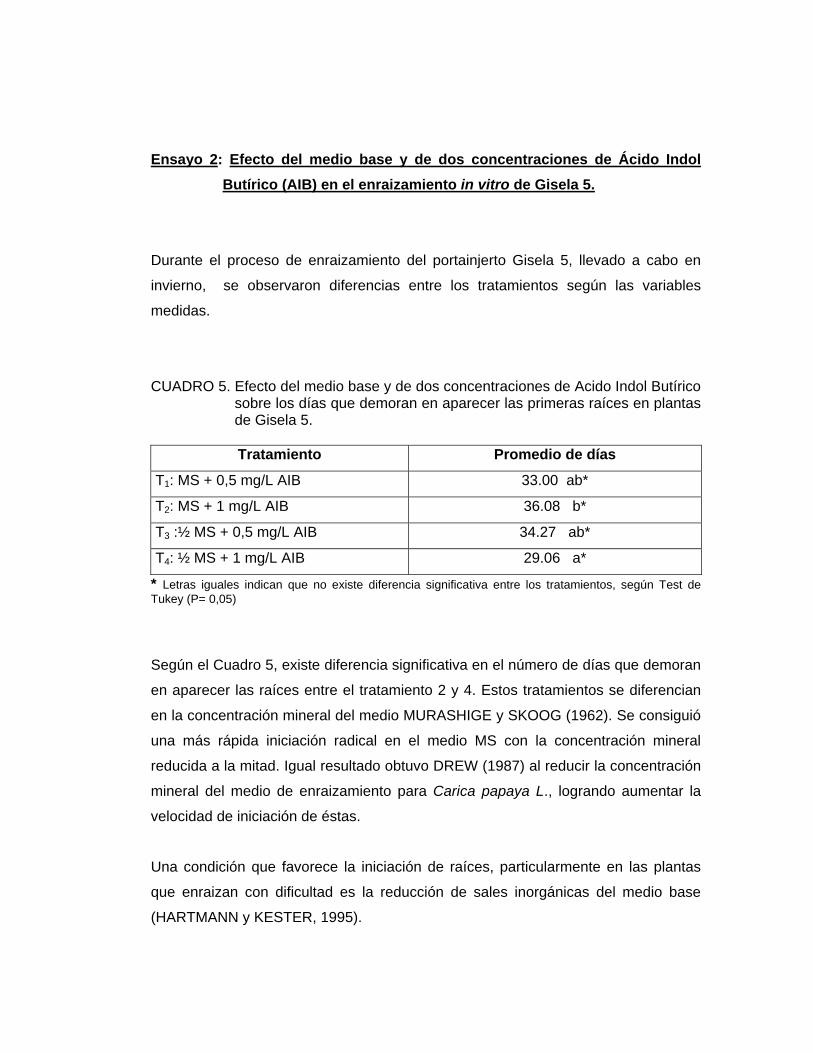
CUADRO 5. Efecto del medio base y de dos concentraciones de Acido Indol Butírico sobre los días que demoran en aparecer las primeras raíces en plantas de Gisela 5.
Tratamiento Promedio de días
T1: MS + 0,5 mg/L AIB 33.00 ab*
T2: MS + 1 mg/L AIB 36.08 b*
T3 :½ MS + 0,5 mg/L AIB 34.27 ab*
T4: ½ MS + 1 mg/L AIB 29.06 a*
* Letras iguales indican que no existe diferencia significativa entre los tratamientos, según Test de Tukey (P= 0,05)
Según el Cuadro 5, existe diferencia significativa en el número de días que demoran
en aparecer las raíces entre el tratamiento 2 y 4. Estos tratamientos se diferencian
en la concentración mineral del medio MURASHIGE y SKOOG (1962). Se consiguió
una más rápida iniciación radical en el medio MS con la concentración mineral
reducida a la mitad. Igual resultado obtuvo DREW (1987) al reducir la concentración
mineral del medio de enraizamiento para Carica papaya L., logrando aumentar la
velocidad de iniciación de éstas.
Una condición que favorece la iniciación de raíces, particularmente en las plantas
que enraizan con dificultad es la reducción de sales inorgánicas del medio base
(HARTMANN y KESTER, 1995).
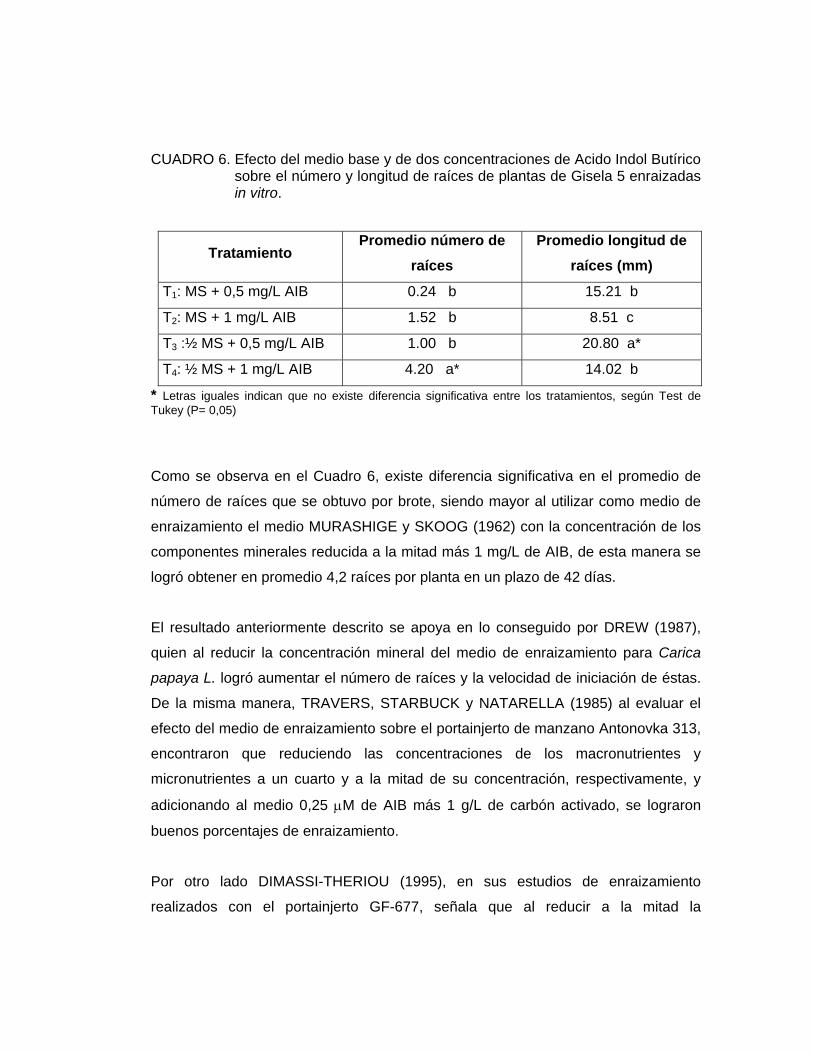
CUADRO 6. Efecto del medio base y de dos concentraciones de Acido Indol Butírico sobre el número y longitud de raíces de plantas de Gisela 5 enraizadas in vitro.
* Letras iguales indican que no existe diferencia significativa entre los tratamientos, según Test de Tukey (P= 0,05)
Como se observa en el Cuadro 6, existe diferencia significativa en el promedio de
número de raíces que se obtuvo por brote, siendo mayor al utilizar como medio de
enraizamiento el medio MURASHIGE y SKOOG (1962) con la concentración de los
componentes minerales reducida a la mitad más 1 mg/L de AIB, de esta manera se
logró obtener en promedio 4,2 raíces por planta en un plazo de 42 días.
El resultado anteriormente descrito se apoya en lo conseguido por DREW (1987),
quien al reducir la concentración mineral del medio de enraizamiento para Carica
papaya L. logró aumentar el número de raíces y la velocidad de iniciación de éstas.
De la misma manera, TRAVERS, STARBUCK y NATARELLA (1985) al evaluar el
efecto del medio de enraizamiento sobre el portainjerto de manzano Antonovka 313,
encontraron que reduciendo las concentraciones de los macronutrientes y
micronutrientes a un cuarto y a la mitad de su concentración, respectivamente, y
adicionando al medio 0,25 µM de AIB más 1 g/L de carbón activado, se lograron
buenos porcentajes de enraizamiento.
Por otro lado DIMASSI-THERIOU (1995), en sus estudios de enraizamiento
realizados con el portainjerto GF-677, señala que al reducir a la mitad la
Tratamiento Promedio número de
raíces Promedio longitud de
raíces (mm)
T1: MS + 0,5 mg/L AIB 0.24 b 15.21 b
T2: MS + 1 mg/L AIB 1.52 b 8.51 c
T3 :½ MS + 0,5 mg/L AIB 1.00 b 20.80 a*
T4: ½ MS + 1 mg/L AIB 4.20 a* 14.02 b

concentración mineral del medio MS y agregando 1 mg/L de AIB el número de
raíces y el peso fresco de ellas se vio reducido.
Como se dijo anteriormente, el mayor número de raíces se obtuvo con el medio MS
con su concentración mineral reducida a la mitad más 1 mg/L de AIB. Mientras que
el portainjerto Mariana 2624, en el medio MS a concentración mineral completa más
1 mg/L de AIB, logró desarrollar un promedio de 4 raíces por planta (DAL ZOTTO y
DOCAMPO, 1997). Esto pone de manifiesto que la respuesta a un determinado
medio depende en gran medida de la especie y de la variedad, ya que se obtuvo la
misma respuesta, pero con un medio distinto para cada caso.
Otro factor que influye en el enraizamiento, según NORTON y NORTON (1988), es
la estación en la cual se lleva a cabo el proceso. En su experiencia con especies del
género Prunus y Spiraea, obtuvieron un mayor número de raíces por brote
enraizando en verano (6.42 raíces/planta) que en invierno (4.17 raíces/planta).
Como se observa en el Cuadro 6, el número de raíces por planta, obtenido en el
ensayo es bajo, aun en el tratamiento 4, si lo comparamos con los resultados
obtenidos por NORTON y NORTON (1988) en verano. Por lo que en este caso la
estación pudo tener un efecto en el enraizamiento, pues ésta fue realizada en
invierno.
En cuanto a la longitud de las raíces, en el Cuadro 6, se observa que existe
diferencia significativa según los tratamientos, es decir, existe diferencia en usar uno
u otro tratamiento en cuanto a la longitud promedio de las raíces. Es decir, al reducir
la concentración mineral del medio MS las raíces desarrollan una mayor longitud.
Esto coincide con lo obtenido por DIMASSI-THERIOU (1995), quien al reducir la
concentración del medio MS obtuvo un aumento significativo en la longitud promedio
de las raíces.
En este ensayo, se aprecia que la menor longitud de raíces fue de 8.51 mm,
utilizando como medio de enraizamiento el medio MS a concentración completa más

1 mg/L AIB, en cambio DAL ZOTTO y DOCAMPO (1997) en plantas del portainjerto
Mariana 2624, enraizadas en el mismo medio obtuvieron raíces con una longitud
promedio de 60 mm. Esto deja claro que las especies vegetales no responden de
igual manera a las mismas condiciones, aunque pertenezcan al mismo género.
Otro fenómeno que se observó durante el enraizamiento fue que en aquellos
tratamientos con el medio MURASHIGE y SKOOG (1962) con sus macro y
micronutrientes a concentración normal, muchas de las plantas se vitrificaron. Esto
pone en evidencia que la composición mineral de muchos medios, en especial a lo
referido a los macronutrientes, causa efectos indeseables, siendo uno de ellos la
vitrificación, además de otros efectos indeseables, como el crecimiento anormal de
las plantas, observados por RUZIC et al. (2000) en su trabajo con el portainjerto
Gisela 5.
De esta manera se pone de manifiesto que la reducción de la concentración mineral
del medio base (MS) para el enraizamiento del portainjerto Gisela 5 es muy
importante para conseguir un rápido y adecuado enraizamiento.
El uso de AIB en el medio de enraizamiento a la concentración de 1 mg/L, es
favorable para el desarrollo de las raíces in vitro del portainjerto Gisela 5, así como
para otros portainjertos del género Prunus. Ejemplo de ello se observa en los
trabajos realizados por DAL ZOTTO y DOCAMPO (1997), DIMASSI-THERIOU
(1995), BARALDI et al. (1988) y JONES, HOPGOOD y O’FARRELL (1977).
En el caso de la propagación in vitro, es preferible tener plantas con mayor número
de raíces que plantas con raíces de gran longitud, ya que al tener un mayor número
de raíces la planta tiene una mayor superficie de absorción y al momento de
transplantar la planta a un recipiente para su aclimatación, las raíces le otorgarán
una mayor sujeción al sustrato.

Ensayo 3: Efecto de tres tipos de sellos sobre la aclimatación del patrón de cerezo Gisela 5.
En este ensayo se sometió a un período de preaclimatación y posterior aclimatación
plantas del portainjerto Gisela 5.
En el Cuadro 7, según test de comparación de proporciones, se observa que no
existe diferencia significativa entre las proporciones de plantas sobrevivientes.
CUADRO 7. Efecto de la preaclimatación in vitro sobre la sobrevivencia de plantas
de Gisela 5.
Tratamientos Sobrevivencia
(%)
Frasco con tapa de aluminio 28.57 a*
Frasco con capa 1 + capa 3
25.63 a
Frasco con capa 1 + capa 2
21.43 a
* Letras iguales indican que no existe diferencia significativa entre los tratamientos, según Test de Comparación de Proporciones (P= 0,05)
La idea de utilizar cubiertas de polipropileno tiene por objetivo aumentar la
ventilación al interior de los tubos de cultivo, con el fin de que un mayor número de
plantas sobrevivan a la aclimatación. Sin embargo, los resultados que se obtienen
depende en gran medida de la especie. Un ejemplo de ello lo constituye el trabajo
realizado por MURPHY et al. (1998), en el cual, utilizando plantas de Hosta y
Delphinium, registraron resultados distintos entre ambas especies al modificar el
ambiente al interior de los tubos de cultivo. Con Hosta, no había efecto de las

pequeñas aberturas en los sellos de los tubos sobre la sobrevivencia, en cambio las
plantas de Delphinium bajo las mismas condiciones que la especie anterior
mostraron aumento de la sobrevivencia.
El resultado de este ensayo es contrario al resultado obtenido por SAFFIE (2002),
quien al preaclimatar plantas de vid propagadas in vitro, logró una tasa de
sobrevivencia mayor que no preaclimatando. De la misma manera, FIGUEROA
(2003), utilizando cubiertas de polipropileno, obtuvo una mayor sobrevivencia de
plantas que utilizando cubiertas de papel aluminio.
Además, ZOBAYED, ARMSTRONG y ARMSTRONG (2001) observaron que las
plantas de tubos con un sistema de ventilación, tienen tasas de sobrevivencia más
altas que las plantas de frascos sin ventilación, al ser llevadas a las condiciones ex
vitro.
Sin embargo, WARDLE, DOBBS y SHORT (1983) señalan que, en las plantas de
crisantemo cultivadas en tubos con una menor humedad relativa, se observó un alto
porcentaje de mortalidad y aquellas que sobrevivían eran más pequeñas y con
pocas raíces.
Según lo observado en el Cuadro 8, no existe diferencia significativa en la longitud
de las plantas de los tratamientos 1, 2 y 3 sobrevivientes al proceso de aclimatación.
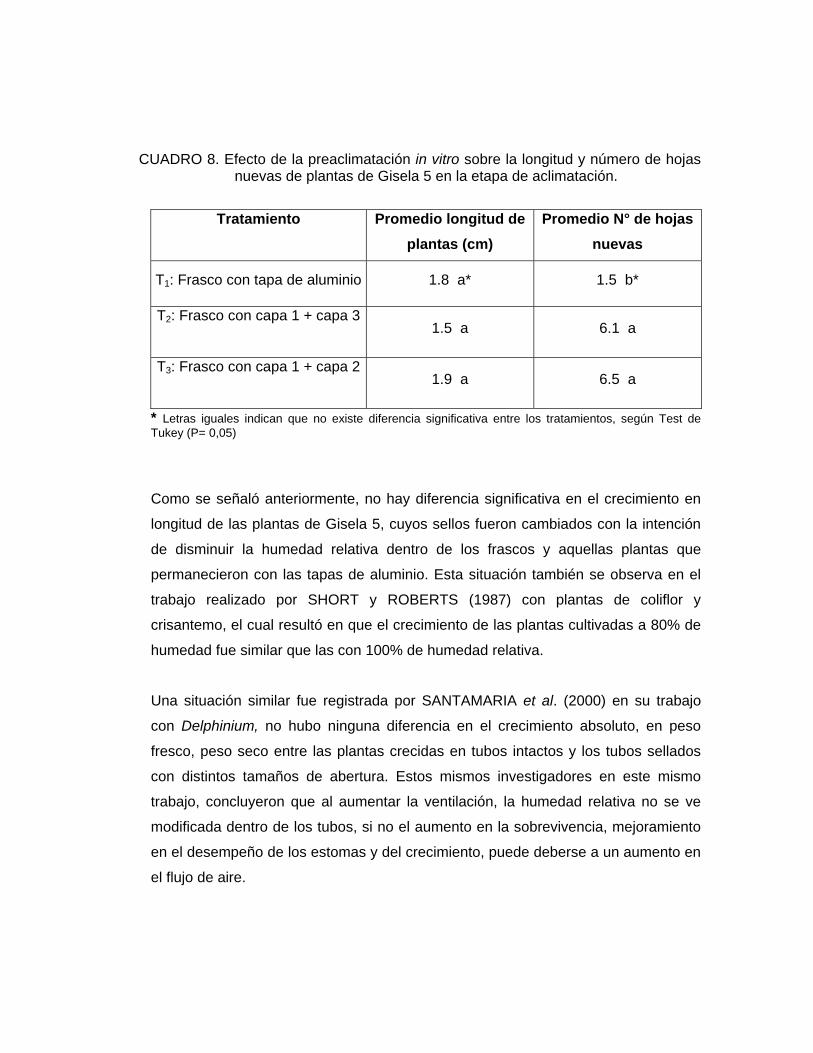
CUADRO 8. Efecto de la preaclimatación in vitro sobre la longitud y número de hojas nuevas de plantas de Gisela 5 en la etapa de aclimatación.
Tratamiento Promedio longitud de plantas (cm)
Promedio N° de hojas nuevas
T1: Frasco con tapa de aluminio 1.8 a* 1.5 b*
T2: Frasco con capa 1 + capa 3
1.5 a 6.1 a
T3: Frasco con capa 1 + capa 2
1.9 a 6.5 a
* Letras iguales indican que no existe diferencia significativa entre los tratamientos, según Test de Tukey (P= 0,05) Como se señaló anteriormente, no hay diferencia significativa en el crecimiento en
longitud de las plantas de Gisela 5, cuyos sellos fueron cambiados con la intención
de disminuir la humedad relativa dentro de los frascos y aquellas plantas que
permanecieron con las tapas de aluminio. Esta situación también se observa en el
trabajo realizado por SHORT y ROBERTS (1987) con plantas de coliflor y
crisantemo, el cual resultó en que el crecimiento de las plantas cultivadas a 80% de
humedad fue similar que las con 100% de humedad relativa.
Una situación similar fue registrada por SANTAMARIA et al. (2000) en su trabajo
con Delphinium, no hubo ninguna diferencia en el crecimiento absoluto, en peso
fresco, peso seco entre las plantas crecidas en tubos intactos y los tubos sellados
con distintos tamaños de abertura. Estos mismos investigadores en este mismo
trabajo, concluyeron que al aumentar la ventilación, la humedad relativa no se ve
modificada dentro de los tubos, si no el aumento en la sobrevivencia, mejoramiento
en el desempeño de los estomas y del crecimiento, puede deberse a un aumento en
el flujo de aire.

El Cuadro 8 indica que el mayor número de hojas nuevas al finalizar el período de
aclimatación se observó en las plantas provenientes del tratamiento 1 y 2, es decir,
en aquellas plantas que fueron preaclimatadas utilizando cubiertas de polipropileno,
y el menor valor en el tratamiento testigo. Esto indica que en el tratamiento 1 y 2
hubo un importante aumento en el área foliar de las plantas.
Esto coincide con el resultado obtenido por FIGUEROA (2003), ya que obtuvo un
mayor número total de hojas en plantas que fueron preaclimatadas utilizando
cubiertas de polipropileno.
Un fenómeno que se observó durante el proceso de preaclimatación en la cámara
de crecimiento, fue que las plantas con la cubierta de aluminio mostraron un mayor
desarrollo in vitro que las provenientes de los tratamientos con cubiertas de
polipropileno. Esto concuerda con lo señalado por ZOBAYED, ARMSTRONG y
ARMSTRONG (2001), quienes observaron que el tamaño de las plantas in vitro
estaba inversamente relacionado con el grado de ventilación del tubo de cultivo.
Además, SHORT y ROBERTS (1987) señalaron que al disminuir la humedad
relativa al interior del tubo de cultivo, se reduce el crecimiento de las plántulas.
Lo descrito anteriormente, coincide con lo señalado por SANTAMARÍA et al. (2000)
quienes observaron una mayor área foliar en las plantas de Delphinium
provenientes de tubos con una menor tasa de intercambio gaseoso.
Otro fenómeno que se observó durante el proceso de preaclimatación de las plantas
de Gisela 5, ocurrido dentro de la cámara de crecimiento, fue la notable disminución
del volumen del medio de cultivo contenido en los frascos con cubierta de
polipropileno, mientras que en los frascos con cubierta de aluminio, esta disminución
fue casi nula. Similares resultados obtuvieron MURPHY et al. (1998) en plantas de
Delphinium y Hosta, quienes observaron que la pérdida de peso del medio se debía
a una ganancia de peso de la planta y a la evapotranspiración, pero en tubos con

mayor ventilación, la ganancia de peso era reducida y la pérdida de peso del medio
era casi cinco veces mayor que en los tubos con papel aluminio.
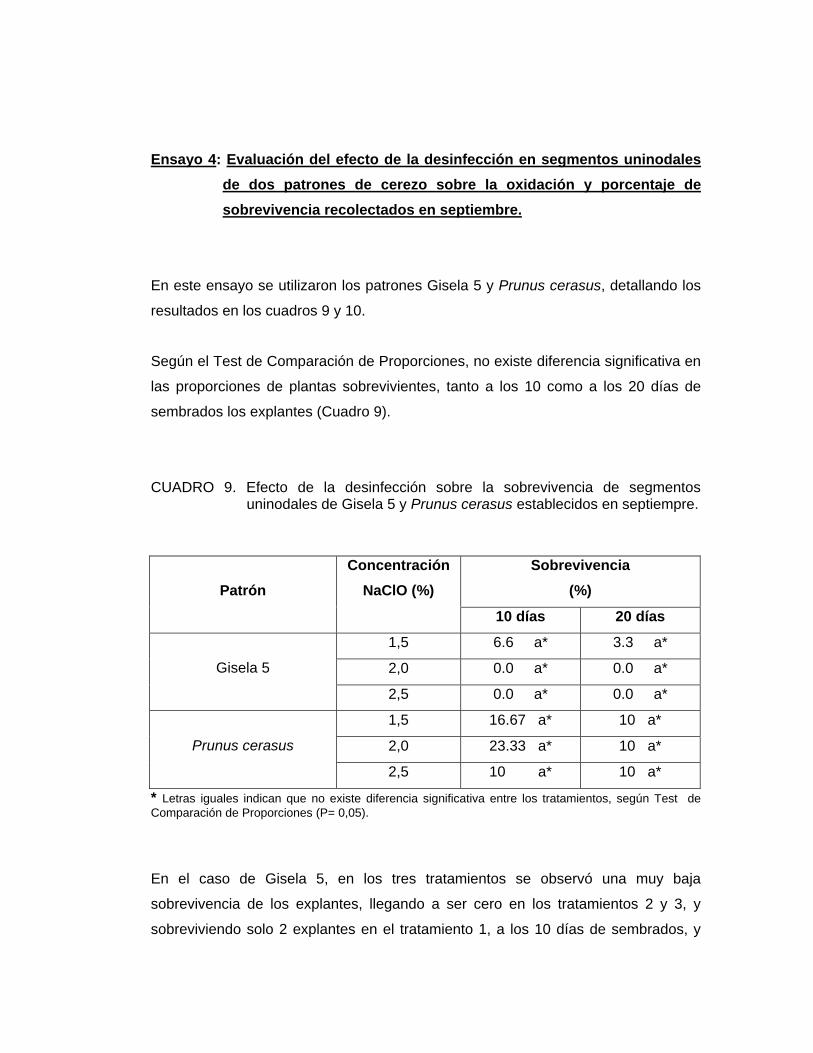
Ensayo 4: Evaluación del efecto de la desinfección en segmentos uninodales de dos patrones de cerezo sobre la oxidación y porcentaje de sobrevivencia recolectados en septiembre.
En este ensayo se utilizaron los patrones Gisela 5 y Prunus cerasus, detallando los
resultados en los cuadros 9 y 10.
Según el Test de Comparación de Proporciones, no existe diferencia significativa en
las proporciones de plantas sobrevivientes, tanto a los 10 como a los 20 días de
sembrados los explantes (Cuadro 9).
CUADRO 9. Efecto de la desinfección sobre la sobrevivencia de segmentos uninodales de Gisela 5 y Prunus cerasus establecidos en septiempre.
* Letras iguales indican que no existe diferencia significativa entre los tratamientos, según Test de Comparación de Proporciones (P= 0,05).
En el caso de Gisela 5, en los tres tratamientos se observó una muy baja
sobrevivencia de los explantes, llegando a ser cero en los tratamientos 2 y 3, y
sobreviviendo solo 2 explantes en el tratamiento 1, a los 10 días de sembrados, y
Sobrevivencia (%) Patrón
Concentración NaClO (%)
10 días 20 días
1,5 6.6 a* 3.3 a*
2,0 0.0 a* 0.0 a* Gisela 5
2,5 0.0 a* 0.0 a*
1,5 16.67 a* 10 a*
2,0 23.33 a* 10 a* Prunus cerasus
2,5 10 a* 10 a*

disminuyendo a 1 explante sobreviviente a los 20 días de sembrados para el mismo
tratamiento (ANEXO 3). Debido a la baja sobrevivencia de los explantes, no fue
posible medir la oxidación de éstos.
La principal causa de contaminación fueron agentes bacterianos, disminuyendo
notoriamente la contaminación por hongos con respecto al ensayo 1 (ANEXOS 3 y
4). Esto demuestra la efectividad del uso de la combinación de fungicidas Benlate
más Captan en la desinfección de los explantes.
Esta baja sobrevivencia de explantes, hace valer el hecho que es más difícil
desinfectar explantes provenientes de plantas crecidas bajo condiciones de campo,
que de aquellas desarrolladas bajo invernadero. Por esto mismo, MUNA et al.
(1999) sugieren desinfectar las yemas axilares y ápices de brotes que provenían de
árboles adultos de siete a ocho años de edad del patrón de cerezo Maxma 14 que
crecían bajo condiciones de campo, con un desinfectante más poderoso como el
HgCl2 (cloruro de mercurio), pues el hipoclorito de sodio y de calcio para desinfectar
superficialmente los explantes no fueron efectivos, mientras que el HgCl2 fue un
buen desinfectante, a pesar de ser tóxico para las plantas y el operador como se
indicó anteriormente.
Para el patrón Prunus cerasus, la situación fue muy similar al caso de Gisela 5. Es
así como se puede observar en el Cuadro 9 que no existe diferencia significativa
entre las proporciones de plantas sobrevivientes.
La contaminación que se generó en este caso fue causada principalmente por
bacterias internas, pues la aplicación de Benlate más Captan en cada uno de los
tratamientos disminuyó a casi cero la población de hongos contaminantes, al igual
que ocurrió con Gisela 5. Esta situación provocó la no brotación de los explantes, a
pesar de estar en primavera, época considerada óptima para la explantación tanto
por OZZAMBAK y HEPAKSOY (1997) y NORTON y NORTON (1988). Esto se debe

a que ciertos contaminantes bacterianos presentes en el interior de la planta pueden
inhibir de gran manera el potencial de crecimiento (HARTMANN y KESTER, 1995).
A pesar que la proporción de plantas sobrevivientes fue baja a los 10 días de
sembrados los explantes, fue posible la medición de la oxidación en todos ellos, ya
que los microorganismos permitieron la visualización de ésta.
En el Cuadro 10, se puede observar que no existe diferencia significativa entre los
tratamientos con respecto al grado de oxidación de los explantes.
CUADRO 10. Efecto de la desinfección sobre la el grado de oxidación de segmentos
uninodales de Prunus cerasus sembrados en septiembre medidos a los 10 días de sembrados.
* Letras iguales indican que no existe diferencia significativa entre los tratamientos, según Test de Kruskal Wallis (P= 0,05).
Como se aprecia en el Cuadro 10, el grado de oxidación de los explantes fue muy
cercano a 2, es decir, que éstos presentaban pardeamiento en la zona de corte y
necrosis en la zona apical.
Similar grado de oxidación obtuvo VARGAS (2003) al desinfectar con hipoclorito de
sodio al 2,5% explantes de Gisela 5, cuyo valor promedio fue de 1,5. En cambio, los
explantes de Colt desarrollaron un grado de oxidación mayor a pesar de haber sido
establecidos en primavera al igual que Gisela 5.
Concentración NaClO (%) Promedio grado de oxidación (ERP)
1.5% 1.6 a*
2% 1.6 a
2.5% 1.7 a

Ensayo 5: Evaluación del efecto de la desinfección de segmentos uninodales del patrón de cerezo Gisela 5 sobre la oxidación y porcentaje de sobrevivencia recolectados en octubre.
En este ensayo se sometió a desinfección con Benlate/Captan, Phyton e hipoclorito
de sodio el portainjerto Gisela 5.
En el Cuadro 11 se puede observar que existe diferencia significativa entre el
tratamiento tres y los tratamientos uno y dos, es decir, el tratamiento 3 entrega una
proporción de plantas sobrevivientes significativamente mayor al resto de los
tratamientos de desinfección.
CUADRO 11. Efecto de la desinfección sobre la sobrevivencia de segmentos
uninodales de Gisela 5 establecidos en octubre.
* Letras iguales indican que no existe diferencia significativa entre los tratamientos, según Test de Comparación de Proporciones (P= 0,05).
La principal causa de contaminación de los explantes y del medio fueron las
bacterias (ANEXO 5).
Con la aplicación del tratamiento 3, se logró obtener un alto número de explantes
sobrevivientes. En cuanto a los otros tratamientos, la sobrevivencia fue muy baja, lo
que fue causado principalmente por agentes bacterianos internos, ya que la
Sobrevivencia (%) Patrón Tratamientos
10 días 20 días
T1: Benlate/Captan + NaClO + Phyton 6.67 b* 3.3 b*
T2: Benlate/Captan + NaClO 10.0 b* 6.67 b* Gisela 5
T3: NaClO + Phyton 86.66 a* 83.33 a*

contaminación se observó hacia el interior del medio en forma de manchas que se
generaban desde el explante. Esto pone en evidencia que eliminar agentes
contaminantes internos a la planta requiere de otros métodos, como es el cultivo de
meristemas, y no sólo el uso de desinfectantes superficiales (HARTMANN y
KESTER, 1995).
En el tratamiento 3 se utilizó un bactericida – fungicida sistémico bajo el nombre
comercial de Pitón, cuyo ingrediente activo es sulfato de cobre pentahidratado. Con
este producto se realizó el enjuague de los explantes dentro de la cámara de flujo
laminar. El uso de este producto provocó la diferencia entre los tres tratamientos, ya
que el ser sistémico, tiene la capacidad que al ser absorbido por el explante pueda
eliminar microorganismos dentro de éste.
RAMÍREZ y SALAZAR (1997), lograron establecer segmentos uninodales de plantas
adultas, mediante el uso de antibiótico, fungicida y antioxidantes en el medio de
cultivo y/o mediante un lavado de los explantes inmediatamente antes de realizada
la siembra.
Lo anterior deja en claro que el estado sanitario de la planta madre donadora de
explantes es muy importante para establecer con éxito el cultivo (HARTMANN y
KESTER, 1995).
Los explantes que sobrevivieron brotaron, confirmando que la primavera es la mejor
época de explantación para las especies de Prunus, según lo señalado por
OZZAMBAK y HEPAKSOY (1997), SIMONETTI (1995) y NORTON y NORTON
(1988).
En el Cuadro 12 se puede observar que no existe diferencia significativa entre los
tratamientos con respecto al grado de oxidación de los explantes. Como se puede
ver también, el grado de pardeamiento después de 10 días de realizada la siembra

es muy cercana a 1, es decir, los explantes sólo desarrollaron pardeamiento en la
zona de corte.
CUADRO 12. Efecto de la desinfección sobre la el grado de oxidación de segmentos uninodales de Gisela 5 establecidos en octubre medidos a los 10 días de sembrados.
* Letras iguales indican que no existe diferencia significativa entre los tratamientos, según Test de Kruskal Wallis (P= 0,05).
El grado de oxidación obtenido por Gisela 5 en este ensayo es muy similar al
obtenido por el mismo portainjerto en el ensayo realizado por VARGAS (2003),
quien también realizó la siembra de los explantes en octubre y utilizó hipoclorito de
sodio al 2,5% para la desinfección de ellos. En cambio, al utilizar una concentración
de hipoclorito de sodio a menor concentración, obtuvo un grado de oxidación mayor
(2,5), pero con la diferencia que el establecimiento lo realizó en otoño. Esto deja en
claro que la estación en la cual se realiza el establecimiento es un factor importante
a considerar.
Concentración de NaClO (%) Promedio grado de oxidación (ERP)
Benlate/Captan + NaClO + Phyton 1.3 a*
Benlate/Captan + NaClO 1.3 a
NaClO + Phyton 1.3 a
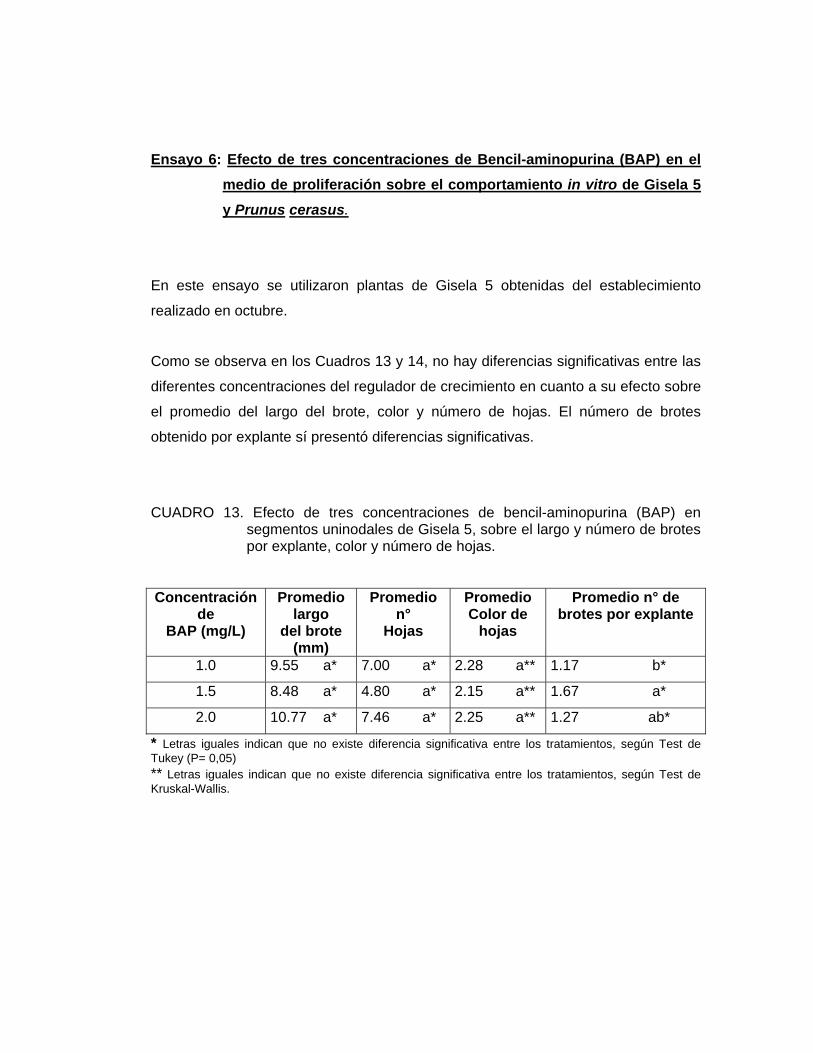
Ensayo 6: Efecto de tres concentraciones de Bencil-aminopurina (BAP) en el medio de proliferación sobre el comportamiento in vitro de Gisela 5 y Prunus cerasus.
En este ensayo se utilizaron plantas de Gisela 5 obtenidas del establecimiento
realizado en octubre.
Como se observa en los Cuadros 13 y 14, no hay diferencias significativas entre las
diferentes concentraciones del regulador de crecimiento en cuanto a su efecto sobre
el promedio del largo del brote, color y número de hojas. El número de brotes
obtenido por explante sí presentó diferencias significativas.
CUADRO 13. Efecto de tres concentraciones de bencil-aminopurina (BAP) en
segmentos uninodales de Gisela 5, sobre el largo y número de brotes por explante, color y número de hojas.
Concentración
de BAP (mg/L)
Promedio largo
del brote (mm)
Promedio n°
Hojas
Promedio Color de
hojas
Promedio n° de brotes por explante
1.0 9.55 a* 7.00 a* 2.28 a** 1.17 b*
1.5 8.48 a* 4.80 a* 2.15 a** 1.67 a*
2.0 10.77 a* 7.46 a* 2.25 a** 1.27 ab*
* Letras iguales indican que no existe diferencia significativa entre los tratamientos, según Test de Tukey (P= 0,05) ** Letras iguales indican que no existe diferencia significativa entre los tratamientos, según Test de Kruskal-Wallis.
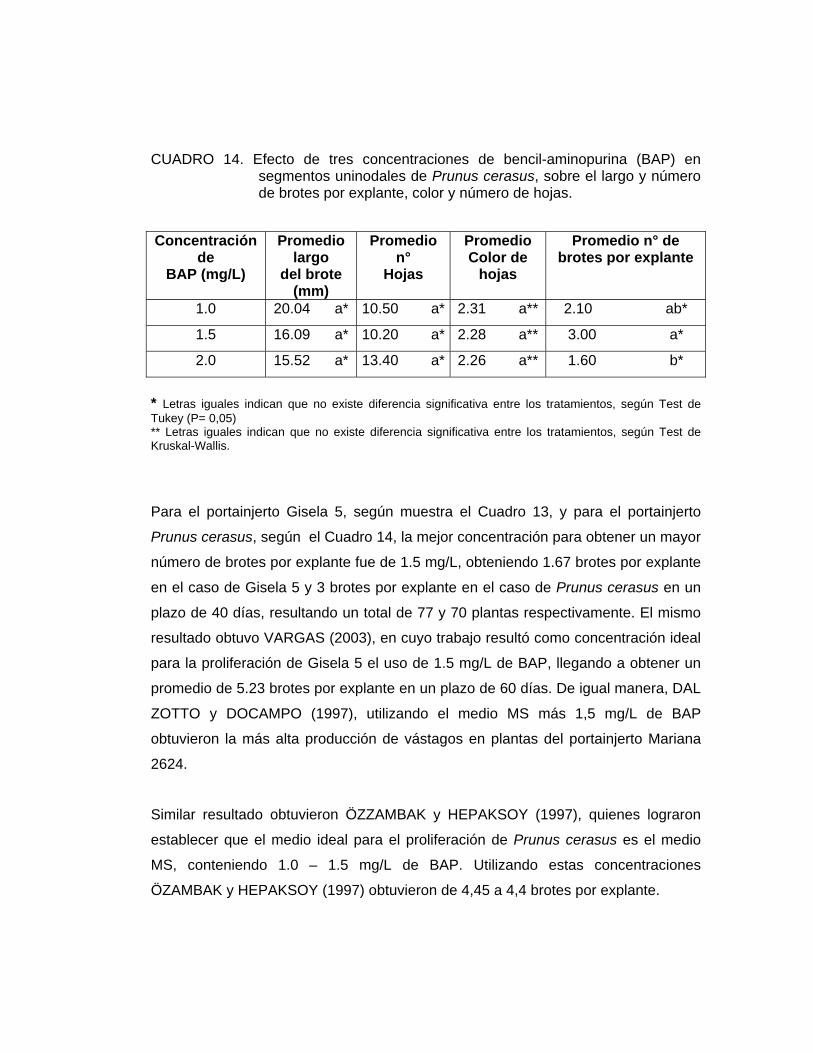
CUADRO 14. Efecto de tres concentraciones de bencil-aminopurina (BAP) en segmentos uninodales de Prunus cerasus, sobre el largo y número de brotes por explante, color y número de hojas.
* Letras iguales indican que no existe diferencia significativa entre los tratamientos, según Test de Tukey (P= 0,05) ** Letras iguales indican que no existe diferencia significativa entre los tratamientos, según Test de Kruskal-Wallis.
Para el portainjerto Gisela 5, según muestra el Cuadro 13, y para el portainjerto
Prunus cerasus, según el Cuadro 14, la mejor concentración para obtener un mayor
número de brotes por explante fue de 1.5 mg/L, obteniendo 1.67 brotes por explante
en el caso de Gisela 5 y 3 brotes por explante en el caso de Prunus cerasus en un
plazo de 40 días, resultando un total de 77 y 70 plantas respectivamente. El mismo
resultado obtuvo VARGAS (2003), en cuyo trabajo resultó como concentración ideal
para la proliferación de Gisela 5 el uso de 1.5 mg/L de BAP, llegando a obtener un
promedio de 5.23 brotes por explante en un plazo de 60 días. De igual manera, DAL
ZOTTO y DOCAMPO (1997), utilizando el medio MS más 1,5 mg/L de BAP
obtuvieron la más alta producción de vástagos en plantas del portainjerto Mariana
2624.
Similar resultado obtuvieron ÖZZAMBAK y HEPAKSOY (1997), quienes lograron
establecer que el medio ideal para el proliferación de Prunus cerasus es el medio
MS, conteniendo 1.0 – 1.5 mg/L de BAP. Utilizando estas concentraciones
ÖZAMBAK y HEPAKSOY (1997) obtuvieron de 4,45 a 4,4 brotes por explante.
Concentración de
BAP (mg/L)
Promedio largo
del brote (mm)
Promedio n°
Hojas
Promedio Color de
hojas
Promedio n° de brotes por explante
1.0 20.04 a* 10.50 a* 2.31 a** 2.10 ab*
1.5 16.09 a* 10.20 a* 2.28 a** 3.00 a*
2.0 15.52 a* 13.40 a* 2.26 a** 1.60 b*

En tanto, la concentración de BAP menos eficiente para la proliferación de brotes en
los explantes de Gisela 5 y Prunus cerasus fue de 1 mg/L y 2 mg/L,
respectivamente. El caso de Prunus cerasus se opone al resultado obtenido por
PEÑALOZA (2002), cuyo mejor resultado de proliferación para Prunus persicae lo
logró utilizando 2 mg/L de BAP, dejando en claro la diferencia que existe entre las
especies al momento de preparar un medio de cultivo. En tanto, para JONARD,
LUKMAN y VILLEMUR (1990) la adición de BAP a una concentración de 1 mg/L
indujo una mayor proliferación de brotes en todos los cultivares de Prunus cerasus
probados.
Así mismo, BARALDI et al. (1988) obtuvieron la más alta proliferación de los
cultivares de Prunus cerasus: Meteor, Monymorency y Northstar adicionando al
medio MS 1 mg/L de BA, resultado que no coincide con lo sucedido en esta
investigación, debido a que el regulador de crecimiento utilizado es distinto al usado
por estos investigadores. Ambas son citoquininas, pero su composición química es
distinta, lo que las lleva a ejercer el mismo efecto, pero a concentraciones distintas.
Los resultados obtenidos confirman que el desencadenamiento de la brotación en
cultivo in vitro, resulta del empleo de citoquininas, eventualmente asociadas a las
auxinas, pero la relación citoquinina/auxina debe ser elevada para que exista
neoformación de yemas (MARGARA, 1988). Así mismo, GEORGE y
SHERRINGTON (1984) indican que una alta relación molar de citoquinina sobre
auxina tiende a inducir desarrollo de yemas.
Aunque no se realizó una comparación entre ambos portainjertos, según los
resultados, se puede ver que la respuesta es muy similar ante las distintas
concentraciones de BAP. Esto deja entre ver que el origen de Gisela 5; patrón
resultante del cruzamiento de Prunus cerasus “Stockton morello” x Prunus
canescens, juega un rol importante, pues ambos patrones comparten igual material
genético, por tener Gisela 5, dado su origen, material genético de Prunus cerasus,
lo que los lleva a responder antes ciertos factores de manera similar.

5. CONCLUSIONES
• El mejor protocolo de desinfección para explantes de cerezo var. Gisela 5 en
la época primaveral, se logró utilizando etanol al 70% durante 5 segundos,
luego hipoclorito de sodio al 2,5% por 15 minutos, más la inmersión de los
explantes en Phyton (0,75 cc/250 ml) por otros 15 minutos.
• Para el portainjerto Prunus cerasus, ninguno de los protocolos de
desinfección utilizados para el establecimiento en la época primavera,
mostró ser más eficiente que el otro.
• El mejor medio de cultivo para la proliferación de explantes de los
portainjertos Gisela 5 y Prunus cerasus está constituido por el medio base
MURASHIGE y SKOOG (1962) suplementado con AIB (0,5 mg/L) más BAP
(1,5 mg/L).
• En la etapa de enraizamiento para Gisela 5, el mejor medio está constituido
por el medio MURASHIGE y SKOOG (1962) con la concentración mineral
reducida a la mitad complementado con 1 mg/L de AIB.
• El uso de sellos de polipropileno, no tuvo efecto sobre la sobrevivencia y
crecimiento de las plantas aclimatadas. El efecto se observó en el número de
hojas nuevas originadas durante el proceso de aclimatación.

6. RESUMEN En el Laboratorio de Propagación Profesor Gregorio Rosenberg de la Facultad de Agronomía de la Pontificia Universidad Católica de Valparaíso, ubicada en la localidad de La Palma, Quillota, V región, se llevó a cabo la propagación in vitro del portainjerto Gisela 5, abarcando las cuatro etapas de esta técnica (establecimiento, proliferación, enraizamiento y aclimatación), y del portainjerto Prunus cerasus, con el cual se trabajó en las dos primeras etapas de la propagación in vitro. Se utilizaron segmentos uninodales como explantes iniciales, los cuales se establecieron en otoño y en primavera, siendo esta última la mejor época de explantación. Los dos patrones utilizados brotaron en el medio de establecimiento MURASHIGE y SKOOG (1962). Se probaron diferentes protocolos de desinfección para los explantes, debido a la alta contaminación esperada, resultando ser mejor para el caso de Gisela 5, aquél donde se utilizó etanol al 70%, hipoclorito de sodio al 2,5% y Phyton (0,75 cc/250 ml). En el caso del portainjerto Prunus cerasus, ninguno de los protocolos utilizados mostró ser más eficiente que el otro, razón por la cual, para lograr establecer explantes de este patrón y posteriormente ser traspasados a la etapa de proliferación se utilizó el protocolo señalado como exitoso para Gisela 5. Para la proliferación de los portainjertos Gisela 5 y Prunus cerasus se evaluó la acción de tres concentraciones de BAP (1,0 mg/L, 1,5 mg/L, 2 mg/L) contenidas en un medio constituido por el medio base MS y 0,5 mg/L de AIB, resultando como mejor alternativa el uso de BAP a la concentración de 1,5 mg/L; obteniendo un mayor número de brotes por explante. Los medios de enraizamiento para las plantas de Gisela 5 se obtuvieron combinando los siguientes factores: El medio MURASHIGE y SKOOG (1962) a concentración normal (MS) y a concentración mineral reducida a la mitad (1/2 MS), y dos concentraciones de AIB (0.5 mg/L, 1 mg/L). Con el medio ½ MS más 1 mg/L de AIB se consiguió el mayor número de raíces por explante y las raíces demoraron menos tiempo en aparecer. Mientras que utilizando ½ MS más 0,5 mg/L de AIB se obtuvo la mayor longitud de raíces. En el ensayo de preaclimatación, se comparó el efecto del uso de cubiertas de polipropileno con el uso de la tapa de aluminio. No se observó diferencia en cuanto a la sobrevivencia y crecimiento de las plantas durante la aclimatación, entre los tratamientos. Se obtuvo un mayor número de hojas nuevas en las plantas con cubiertas de polipropileno.

7. ABSTRACT In vitro propagation of the rootstocks Gisela 5 and Prunus cerasus was done in the Professor Gregorio Rosenberg Propagation Laboratory located in the Agronomy Faculty of the Pontificia Universidad Católica de Valparaíso, La Palma, Quillota. V Region. In the case of Gisela 5, four stages of this technique were considered (establishment, proliferation, rooting and acclimatization) while only the first two stages were considered for Prunus cerasus. Single-node stem segments were used as initial explants, which were established in autumn and spring as these are considered the best seasons for explanting. Both rootstocks sprouted in the establishment medium used by MURASHIGE and SKOOG (1962). Different disinfection protocols for explants were tested, due to the expected high risk of contamination. The method that sowed the best results for Gisela 5 was the treatment using 70% Ethanol, 2.5 % sodium hypochloride and Phyton (0.75 cc/250 milliliter). In the case Prunus cerasus, neither protocol showed a higher efficiency, and for this reason, the method that was more successful for Gisela 5 was also used to establish this rootstock, and later used for transfer to the proliferation stage.
Three concentrations of BAP (1,0 mg/L, 1,5 mg/L, 2 mg/L) contained in a medium which had MS and 0.5 mg/L of IBA as base medium, were evaluated for the proliferation of the rootstocks Gisela 5 and Prunus cerasus. The best results were obtained with the use of 1.5 mg/L BAP, which resulted in a higher number of shoots per explant.
The rooting mediums for Gisela 5 rootstocks were obtained by combining the following facts: the MURASHIGE and SKOOG (1962) medium at full-strength (MS) and half-strength (1/2 MS), and two different concentrations of IBA (0.5 mg/L, 1 mg/L). A higher number of roots per explants in a shorter time, was obtained using half-strength MS with 1 mg/L of IBA, while longer roots were obtained using half-strength MS with 0.5 mg/L of IBA.
During the preacclimatization stage, the effect of polypropylene covers was compared with the use of aluminum covers. There was no apparent difference regarding plant survival and growth during the acclimation stage of the treatments. There were a higher number of leaves in plants with polypropylene covers.

8. LITERATURA CITADA
BARALDI, R.; MARINO, G.; ROSATI, P.; and GOLINELLI, A. 1988. Sour cherry
(Prunus cerasus) micropropagation. Acta Horticulturae. (227):405-407.
BRAINERD, K. and FUCHIGAMI, L. 1981. Acclimatization of aseptically cultured
apple plants to low relative humidity. J. Amer. Soc. Hort. Sci. 106 (4): 515-518.
CLAVERIE, J. 2002. Portainjertos de cerezos desarrollados en Francia.
Universidad Católica de Valparaíso. Cultivo del cerezo en la zona centro-norte de Chile. Quillota, 27-28 de noviembre, 2002. s.p.
DAL ZOTTO, A. y DOCAMPO, D. 1997. Micropropagación de los portainjertos de
ciruelo Mariana 2624 (Prunus cerasifera x Prunus musionana) y Pixy (Prunus insistia L.) de sanidad controlada. Phyton. (60):127-135.
DIMASSI-THERIOU, K. 1995. In vitro rooting of rootstock GF-677 (Prunus
amygdalus x Prunus persica) as influenced by mineral concentration of the nutrient medium and type of culture-tube sealing material. Journal of Horticultural Science 70(1)105-108.
DONALLY, D.; VIDAVER, W. and COLBOW, K. Fixation of CO2
14 in tissue cultured red raspberry prier to and after tranfer to soil. Plant Cell Tissue Organ. Cult. 3:313-317.
DRADI, G.; VITO, G.; y STANDRADI, A. 1996. In Vitro Mass propagation of eleven
Prunus mahaleb Ecotypes. Acta Horticultutare. (410): 477-483. DREW, R.A. 1987. The effects of medium composition and cultural conditions on in
vitro root initiation and growth of papaya (Carica papaya L.). Journal of Horticultural Science 62(4):551-556.
FIGUEROA, N. 2003. Preaclimatación in vitro de plantas de violeta africana
(Saintpaulia ionantha Wendl.) y su efecto sobre la aclimatación. Taller

de Licenciatura Ing. Agr. Quillota. Universidad Católica de Valparaíso. Facultad de Agronomía. 64p
FUCHIGAMI, L.; CHENG, T. and SOELDER, A. 1981. Abaxial transpiration and
water loss in aseptically cultured plum. J. Amer. Soc. Hort. Sci. 106 (4): 519-522.
FUNDACION CHILE. 2000. Cerezas: Crecen en volúmenes de exportación.
Agroeconómico (59): 20-26. GEORGE, E and SHERRINGTON, P. 1984. Plant propagation by tissue culture.
1a. ed. Basingstoke, Exegetics Limited. 709p. GROUT, B. 1975. Wax development on leaf surfaces on Brassica oleracea var.
Currawong regenerated from meristem culture. Plant Sci. Lett 5:401-405.
GROUT, B. and MILLAM, S. 1985. Photosynthetic development of
micropropagated strawberry plantlets following transplanting. Ann. Bot. 55:129-131.
HAMMAT, N and GRANT, N. 1998. Shoot regeneration from leaves of Prunus
serotina Ehrh. (black cherry) and P. avium L. (wild cherry). Plant Cell Reports. 17 n°6/7:525-530.
HARTMANN, H. y KESTER, E. 1995. Propagación de plantas. Principios y
Prácticas. 4a. ed. México, C.E.C.S.A. 760p. HORMAZA, J. y GELLA, R. 1996. Situación Actual de los patrones de Cerezos.
Fruticultura Profesional. 88: 5-15. INFORMACIÓN AGROPECUARIA. 2004. El cultivo de la cereza. 2004,. (on line).
www.infoagro.com/frutas/frutas_tradicionales/cereza.htm

JONARD, R.; LUKMAN, D. and VILLEMUR, P. 1990. Early testing of graft incompatibilities in apricot and lemon trees using in vitro techniques. Scienta Horticulturae. 43:117-128.
JONES, O.; HOPGOOD, M. and O’FARRELL. 1977. Propagation in vitro of M.26 apples rootstock. J. Hort. Sci. 52: 235-238.
JONES, O. and HOPGOOD, M. 1979. The successful propagation in vitro of two rootstock of Prunus: the plum rootstock Pixy (P. insistia) and the cherry rootstock F 12/1 (P. avium). J. Hort. Sci. 54: 63-66.
JOUBLAN, J.P. 2002. Avances en cerezo en el sur de Chile. Universidad Católica de Valparaíso. Cultivo del cerezo en la zona centro-norte de Chile. Quillota. 27-28 de noviembre, 2002. s.p.
LORETI, F y GIL, G. 1993. Los portainjertos del ciruelo y del cerezo: Presente y
futuro. Revista Frutícola. 14: 103-111.
MARGARA, J. 1988. Multiplicación Vegetativa y Cultivo in vitro. Madrid, Mundi-Prensa. 236p.
MARKS, T.R. and SIMPSON, S.E. 1990. Reduced phenolic oxidation at culture
initiation in vitro following the exposure of field-grow stockplants to darkness or levels of irradiance. J. Hort. Sci. 65(2): 103-111.
Mc COMB, J. 1978. Clonal propagation of woody plants using tissue culture, with special reference to apples. Combined Proceedings of the International Plant Propagators Society. 28: 413-426.
MILLER, G.; COSTON, C.; DENNY, E. and ROMEO, M. 1982. In vitro Propagation of “Nemaguard” Peach Rootstock. HortScience 17(2): 194.
MUNA, A.; AHMAD, A. y ABDUL-ARHMAN, K. 1999. In vitro Propagation of a Semi-Dwarfing Cherry Rootstock. Plant Cell, Tissue and Organ Culture. 59 (3). 203-208.

MUNSELL. 1998. Color charts for plant tissues. New York, Munsell Color. s.p. MURASHIGE, T and SKOOG. 1962. A revised medium for rapid growth and
bioassays with tobacco tissue cultures. Phys. Plant. 15: 473-497. MURPHY, K.; SANTAMARÍA, J.; DAVIES, W. and LUMSDEN, P. 1998. Ventilation
of cultured vessels: I. Increased growth in vitro and survival ex vitro Delphinium. Journal of Horticultural Science and Biotechnology 73 (6): 725-729.
NORTON, M. and NORTON, C. 1988. The role of season in determining explant
suitability for in vitro culture of Prunus and Spiraea. Acta Horticulturae N°227:450-452.
OFICINA DE ESTUDIOS Y POLÍTICAS AGRARIAS. 2003. Base de datos
agronómicos. www.odepa.cl (on line).
OYANEDEL, E. 1995. Propagación in vitro de portainjertos de palto (Persea americana Mill.) resistentes a la salinidad, cv. Velvick y Lula. Taller de Licenciatura Ing. Agr. Quillota, Universidad Católica de Valparaíso. Facultad de Agronomía. 82p.
ÖZZAMBAK, E. and HEPAKSOY, S. 1997. Investigations on In Vitro Proliferation of Sour Cherry cv. Heimanns Rubinweichsel. Acta Horticulturae. 447:155-156.
PEDROTI, E and CORNU, D. 1991. Buds Formation From Roots of
Micropropagated Prunus avium L. Plantlets. Acta Horticulturae. 289:141-142.
PEÑALOZA, P. 2002. Obtención de plantas libres de virus y propagación in vitro de
durazno (Prunus persicae, (L) Batch), (on line). www.siza.gemtel.com.mx/libros/delaros
PEVALEK, B. and JELASKA, S. 1987. Microclonal propagation of Prunus avium L.
Acta Hort. 212:599-602

PIERIK, R. 1990. Cultivo in vitro de las plantas superiores. Madrid. Mundi-Prensa. 326p.
PINOCHET, J y TORRENTS, J. 1998. Portainjertos de ciruelo, cerezo y
albaricoquero desde la perspectiva de la replantación y patógenos del suelo. Fruticultura Profesional. (96):6-10.
RAVINDRA, M. and THOMAS, P. 1995. Sachet technique an efficient method for
the acclimatization of micropropagated grapes (Vitis vinifera L.). Current Science 68 (5): 546-548.
RAMIREZ, M. y SALAZAR, E. 1997. Establecimiento in vitro de segmentos nodales
de guayabo (Psidium guajaba L.). Rev. Fac. Agrom. 14:497-506. RITCHIE, G.; SHORT, K. and DAVEY, M. 1991. In vitro acclimatization of
chrysanthemum and sugar beet plantlets by treatment with paclobutrazol and exposure to reduced humidity. Journal of experimental botany 42 (245): 1557-1563.
RUZIC, D.; SARIC, M.; CEROVIC, R. and CULAFIC, L. 2000. Relationship between
the concentration of macroelements, their uptake and multiplication of cherry rootstock Gisela 5 in vitro. Plant Cell, tissue and Organ Culture. 63:9-14.
SAFFIE, D. 2002. Aclimatación de plantas de vid (Vitis vinifera) propagadas in vitro.
Taller de Licenciatura Ing. Agr. Quillota, Universidad Católica de Valparaíso. Facultad de Agronomía. 47p.
SAN MARTÍN, S. 1996. Propagación in vitro de portainjertos de palto (Persea
americana Mill.) cvs. Lula y Velvick, resistentes a la salinidad: enraizamiento y aclimatación. Taller de Licenciatura Ing. Agr. Quillota, Universidad Católica de Valparaíso. Facultad de Agronomía. 111p.
SANTAMARÍA, J.; MURPHY, K.; LEIFERT, C. and LUMSDEN, P. 2000. Ventilation
of cultured vessels: I. Increased water movement rather than reduced concentrations of ethylene and CO2 is responsible for improved

growth and development of Delphinium in vitro. Journal of Horticultural Science and Biotechnology. 75 (3): 320-327.
SHORT, K. and ROBERTS, A. 1987. In vitro hardening of cultured cauliflower and
chrysanthemum plantlets to humidity. Acta Horticulturae. 212: 329-334.
SIMONETTI, M. 1995. Efecto de la época de establecimiento, tipo de tejido y
medio nutritivo en la propagación in vitro del portainjerto GF 677 (Prunus persica x Prunus amigdalus). Taller de Licenciatura Ing. Agr. Quillota, Universidad Católica de Valparaíso. Facultad de Agronomía. 88p
SKOOG, F and MILLER, C. 1956. Chemical regulation of growth and organ
formation in plant tissues cultivated in vitro. Symp. Soc. Exp. Biol. 9:118-131.
SKOOG, M. 1971. Aspects of growth factors interactions in morphogenesis of
tobacco tissue cultures. In: Les cultures de tissues de plantes. Collog. Int. CNRS. 193:115-136.
SNIR, I. 1982. In vitro Propagation of Sweet Cherry Cultivars. HortScience. 12:
545-547. SRIKANDARAJAH, S and MULLINS, G. 1981. Micropropagation on Granny Smith
apple: factors affecting root formation in vitro. Journal of Horticultural science. 56 (1): 71-76.
SUTTER, E. and LANGHANS, R. 1982. Formation of epicuticular wax and its effect
on water loss in cabbage plants regenerated from shoot-tip culture. Can. J. Bot. 60:2896-2092.
TANG, H.; REN, Z.; REUSTLE, G. and KRCZAL, G. 2002. Plant regeneration from leaves of sweet and sour cherry cultivars. Scienta Horticulturae. 93:235-244

TRAVERS, J.N.; STARBUCK, C.J. and NATARELLA, N.J. 1985. Effects of Culture Medium on In Vitro Rooting of Antonovka 313 Apple. HortScience. 20 (6):1051-1052.
VARGAS, G. 2003. Propagación in vitro de Portainjertos de Cerezo. Taller de
Licenciatura Ing. Agr. Universidad Católica de Valparaíso. Facultad de Agronomía. 57p.
WALKEY, D. 1972. Production of apple plantlets from axillary bud meristems.
Canadian Journal of Plant Science. 53: 1085-1087.
WARDLE, K.; DOBBS, E. and SHORT, K. 1983. In vitro acclimatization of aseptically cultured plantlets to humidity. J. Amer. Soc. Hort. Sci. 108 (3): 386 – 389.
WEBSTER, A. and LOONY, N. 1996. Cherries: Crops Physiology, Production and
Uses. 1a. ed. Wallingford, Cab International. 513p. WETZSTEIN, H. and SOMMER, H. 1982. Leaf anatomy of tissue-cultured
Liquidambar styracifera (Hamamalidaceae) during acclimatization. Am. J. Bot. 69:1579-1586.
WARDLE, K.; DOBBS, E. and SHORT, K. 1983. In vitro acclimatization of
aseptically cultured plantlets to humidity. J. Amer. Soc. Hort. Sci. 108 (3): 386 – 389.
YU, D. and MEREDITH, C. 1996. The influence of explant origin on tissue browning
and shoot producction in shoot tip cultures of grapevine. J. Amer. Soc. Hort. Sci. 111(6):972-975.
ZIV, M and HAVELY, A.H. 1983. Control of oxidate browning and in vitro
propagation of Strelitzia reginae. HortScience 18(4):434-436. ZOBAYED, S.; ARMSTRONG, J. and ARMSTRONG, W. 2001. Leaf anatomy of in
vitro tobacco and cauliflower plantlets as affected by different types of ventilation. Plant Science 161 (3): 537-548.
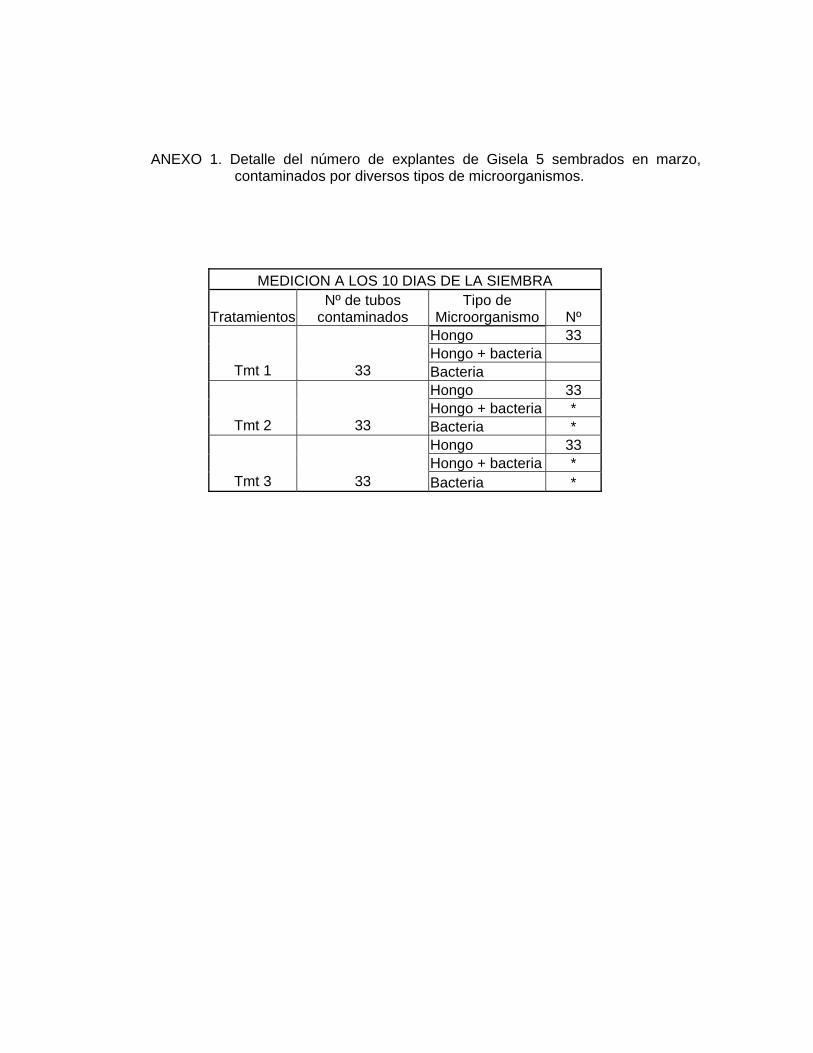
ANEXO 1. Detalle del número de explantes de Gisela 5 sembrados en marzo, contaminados por diversos tipos de microorganismos.
MEDICION A LOS 10 DIAS DE LA SIEMBRA
Tratamientos Nº de tubos
contaminados Tipo de
Microorganismo Nº Hongo 33 Hongo + bacteria
Tmt 1 33 Bacteria Hongo 33 Hongo + bacteria *
Tmt 2 33 Bacteria * Hongo 33 Hongo + bacteria *
Tmt 3 33 Bacteria *
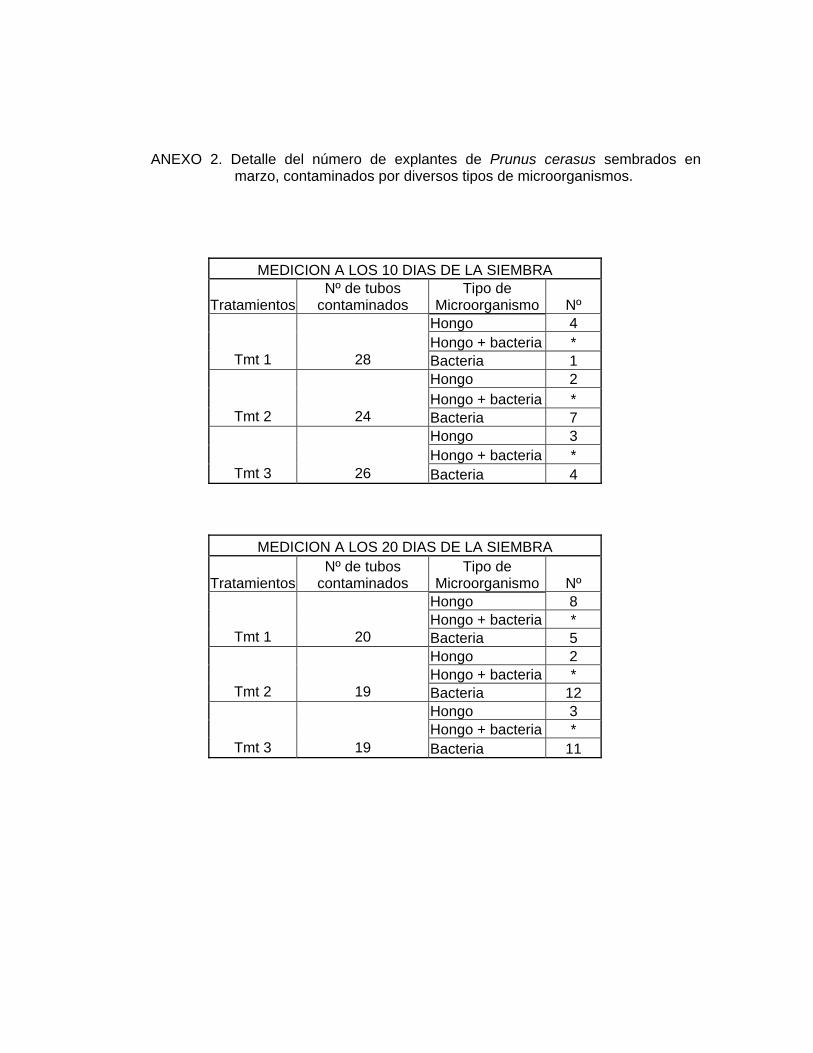
ANEXO 2. Detalle del número de explantes de Prunus cerasus sembrados en marzo, contaminados por diversos tipos de microorganismos.
MEDICION A LOS 10 DIAS DE LA SIEMBRA
Tratamientos Nº de tubos
contaminados Tipo de
Microorganismo Nº Hongo 4 Hongo + bacteria *
Tmt 1 28 Bacteria 1 Hongo 2 Hongo + bacteria *
Tmt 2 24 Bacteria 7 Hongo 3 Hongo + bacteria *
Tmt 3 26 Bacteria 4
MEDICION A LOS 20 DIAS DE LA SIEMBRA
Tratamientos Nº de tubos
contaminados Tipo de
Microorganismo Nº Hongo 8 Hongo + bacteria *
Tmt 1 20 Bacteria 5 Hongo 2 Hongo + bacteria *
Tmt 2 19 Bacteria 12 Hongo 3 Hongo + bacteria *
Tmt 3 19 Bacteria 11
ANEXO 3. Detalle del número de explantes de Gisela 5 sembrados en septiembre, contaminados por diversos tipos de microorganismos.
MEDICION A LOS 10 DIAS DE LA SIEMBRA
Tratamientos Nº de tubos
contaminados Tipo de
Microorganismo Nº Hongo 9 Hongo + bacteria 4
Tmt 1 28 Bacteria 15 Hongo 16 Hongo + bacteria 6
Tmt 2 30 Bacteria 8 Hongo 6 Hongo + bacteria 8
Tmt 3 30 Bacteria 16
MEDICION A LOS 20 DIAS DE LA SIEMBRA
Tratamientos Nº de tubos
contaminados Tipo de
Microorganismo Nº Hongo 10 Hongo + bacteria 4
Tmt 1 29 Bacteria 15 Hongo 16 Hongo + bacteria 6
Tmt 2 30 Bacteria 8 Hongo 6 Hongo + bacteria 8
Tmt 3 30 Bacteria 16
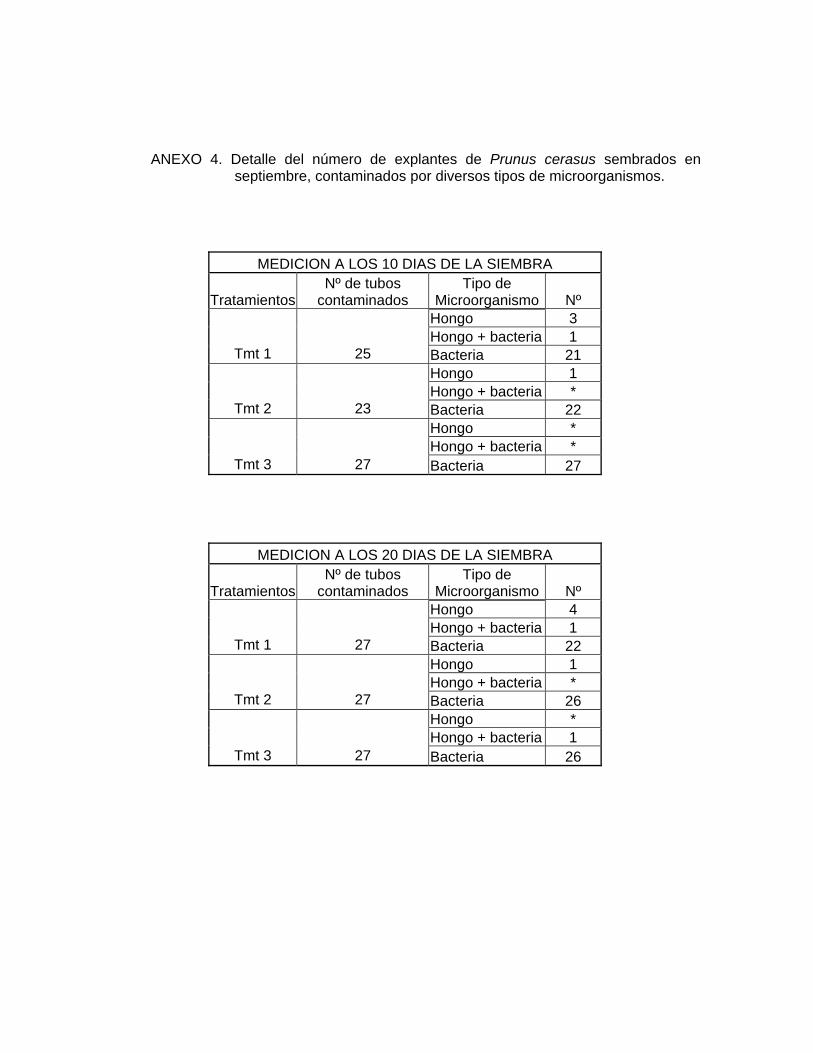
ANEXO 4. Detalle del número de explantes de Prunus cerasus sembrados en septiembre, contaminados por diversos tipos de microorganismos.
MEDICION A LOS 10 DIAS DE LA SIEMBRA
Tratamientos Nº de tubos
contaminados Tipo de
Microorganismo Nº Hongo 3 Hongo + bacteria 1
Tmt 1 25 Bacteria 21 Hongo 1 Hongo + bacteria *
Tmt 2 23 Bacteria 22 Hongo * Hongo + bacteria *
Tmt 3 27 Bacteria 27
MEDICION A LOS 20 DIAS DE LA SIEMBRA
Tratamientos Nº de tubos
contaminados Tipo de
Microorganismo Nº Hongo 4 Hongo + bacteria 1
Tmt 1 27 Bacteria 22 Hongo 1 Hongo + bacteria *
Tmt 2 27 Bacteria 26 Hongo * Hongo + bacteria 1
Tmt 3 27 Bacteria 26
ANEXO 5. Detalle del número de explantes de Gisela 5 sembrados en octubre, contaminados por diversos tipos de microorganismos.
MEDICION A LOS 10 DIAS DE LA SIEMBRA
Tratamientos Nº de tubos
contaminados Tipo de
Microorganismo Nº Hongo * Hongo + bacteria *
Tmt 1 28 Bacteria 28 Hongo * Hongo + bacteria *
Tmt 2 27 Bacteria 27 Hongo * Hongo + bacteria *
Tmt 3 4 Bacteria 4
MEDICION A LOS 20 DIAS DE LA SIEMBRA
Tratamientos Nº de tubos
contaminados Tipo de
Microorganismo Nº Hongo * Hongo + bacteria *
Tmt 1 30 Bacteria 30 Hongo * Hongo + bacteria *
Tmt 2 28 Bacteria 28 Hongo * Hongo + bacteria *
Tmt 3 5 Bacteria 5