
GENÉTICA DE LAS DISLIPIDEMIAS EN...
18
125 GENÉTICA DE LAS DISLIPIDEMIAS EN MÉXICO GENETICS OF LIPID DISORDERS IN MEXICO Carlos A. Aguilar Salinas 1 , Marco A. Melgarejo Hernández 1 , Donaji V. Gómez Velasco 1 , Liliana Muñoz Hernández 1 , Luz E. Guillén Pineda 1 , Carmen Moreno Villatoro 1 , Hortensia Moreno Macías 2 , Alicia Huerta Chagoya 2 , Rita A. Gómez Díaz 3 , Roopa Mehta 1 , Olimpia Arellano Campos 1 , Ivette Cruz Bautista 1 , Maria Teresa Tusie Luna 2,4 Departamento de Endocrinología y Metabolismo. Instituto Nacional de Ciencias Médicas y Nutrición Unidad de Biología Molecular y Medicina Genómica. Instituto Nacional de Ciencias Médicas y Nutrición Unidad de Investigación en Epidemiologia Clínica, UMAE Hospital de Especialidades, CMN Siglo XXI, IMSS Instituto de Investigaciones Biomédicas. Universidad Nacional Autónoma de México. Tel: 52-55-54870900 ext 4524. email: [email protected] Resumen La prevalencia de la hipertrigliceridemia y la hipoalfalipoproteinemia es mayor en México que en poblaciones de origen caucásico. Este fenómeno resulta de un estilo de vida no saludable y la frecuencia de variantes genéticas que aumentan la susceptibilidad para tener dichas anormalidades. El objetivo de la revisión es describir el conocimiento vigente sobre la genética de las dislipidemias en mestizos mexicanos. La frecuencia de los alelos de susceptibilidad en los mexicanos es mayor que la encontrada en los europeos para varios de los genes Cárdenas Monroy C, González Andrade M, Guevara Flores A, Lara Lemus R, Matuz Mares D, Molina Jijón E, Torres Durán PV. Mensaje Bioquímico, Vol. XL, 125-142, Depto. de Bioquímica, Facultad de Medicina, Universidad Nacional Autónoma de México. Cd. Universitaria, CDMX.,MÉXICO.,(2016). (http://bioq9c1.fmedic.unam.mx/TAB) (ISSN-0188-137X)
Transcript of GENÉTICA DE LAS DISLIPIDEMIAS EN...

125
GENÉTICA DE LAS DISLIPIDEMIAS EN MÉXICO
GENETICS OF LIPID DISORDERS IN MEXICO
Carlos A. Aguilar Salinas1, Marco A. Melgarejo Hernández1, Donaji V. Gómez Velasco1, Liliana Muñoz Hernández1, Luz E. Guillén Pineda1, Carmen Moreno Villatoro1, Hortensia Moreno Macías2, Alicia Huerta Chagoya2, Rita A. Gómez Díaz3, Roopa Mehta1, Olimpia Arellano Campos1, Ivette Cruz Bautista1, Maria
Teresa Tusie Luna 2,4
Departamento de Endocrinología y Metabolismo. Instituto Nacional de Ciencias Médicas y Nutrición
Unidad de Biología Molecular y Medicina Genómica. Instituto Nacional de Ciencias Médicas y Nutrición
Unidad de Investigación en Epidemiologia Clínica, UMAE Hospital de Especialidades, CMN Siglo XXI, IMSS
Instituto de Investigaciones Biomédicas. Universidad Nacional Autónoma de México.
Tel: 52-55-54870900 ext 4524. email: [email protected]
Resumen
La prevalencia de la hipertrigliceridemia y la hipoalfalipoproteinemia es
mayor en México que en poblaciones de origen caucásico. Este fenómeno resulta
de un estilo de vida no saludable y la frecuencia de variantes genéticas que
aumentan la susceptibilidad para tener dichas anormalidades. El objetivo de la
revisión es describir el conocimiento vigente sobre la genética de las dislipidemias
en mestizos mexicanos. La frecuencia de los alelos de susceptibilidad en los
mexicanos es mayor que la encontrada en los europeos para varios de los genes
Cárdenas Monroy C, González Andrade M, Guevara Flores A, Lara Lemus R, Matuz Mares D, Molina Jijón E, Torres Durán PV. Mensaje Bioquímico, Vol. XL, 125-142, Depto. de Bioquímica, Facultad de Medicina, Universidad Nacional Autónoma de México. Cd. Universitaria, CDMX.,MÉXICO.,(2016). (http://bioq9c1.fmedic.unam.mx/TAB)
(ISSN-0188-137X)

MENSAJE BIOQUÍMICO, VOL. XL (2016)
126
que tienen el mayor efecto sobre los niveles de triglicéridos o de colesterol-HDL.
Tal es el caso de las variantes de APOA5 y MLXIPL para la hipertrigliceridemia y
de ABCA1 y CETP para la hipoalfalipoproteinemia. Asimismo, existen variantes
propias de la población amerindia que tienen efectos mayores sobre la
concentración de los lípidos plasmáticos (Ej. las variantes R230C del transportador
ABCA1, un haplotipo de riesgo en SIK3 y el rs1349411 en APOBEC1). Por otra
parte, el estudio de la hipertrigliceridemia en mexicanos ha dado lugar a varias
contribuciones significativas. Se identificaron asociaciones entre la hiperlipidemia
familiar combinada y USF1, TCF7L2 y HNF4A. En resumen, postulamos que el
estudio de los determinantes genéticos de las dislipidemias y de otras
enfermedades metabólicas en amerindios es un recurso para identificar nuevas
variantes genéticas y vías asociadas con dichos padecimientos.
Palabras clave: triglicéridos, colesterol-HDL, colesterol.
Abstract
The prevalence of hypertriglyceridemia and hypoalphalipoproteinemia is
higher in Mexico than in Caucasian populations. This phenomenon results from an
unhealthy lifestyle and the high frequency of genetic variants that increase
susceptibility to have these abnormalities. The aim of the review is to describe the
current knowledge on the genetics of dyslipidemias in Mexican mestizos. The
frequency of susceptibility alleles in Mexicans is greater than that found in Europe
for several of the genes that have the greatest effect on triglycerides or HDL-
cholesterol. Such is the case for the variants of APOA5, MLXIPL for
hypertriglyceridemia and ABCA1, CETP for hypoalphalipoproteinemia. There are
also own variants of the Amerindian population that have major effects on the
concentration of plasma lipids (e.g. The R230C variant of the transporter ABCA1, a
risk haplotype in SIK3 and rs1349411 in the APOBEC1). Moreover, the study of
hypertriglyceridemia in Mexico has resulted in several significant contributions.
Associations between familial combined hyperlipidemia and USF1, HNF4A and
TCF7L2 were identified. In short, we postulate that the study of the genetic
determinants of dyslipidemia and other metabolic diseases in Amerindians is a
resource to identify new genetic pathways and phenotypes associated with
increased risk for having metabolic disorders.
Keywords: triglycerides HDL-cholesterol, cholesterol.

Aguilar Salinas CA, Melgarejo Hernández MA, et al.
127
Introducción
Las enfermedades cardiovasculares que resultan de la aterosclerosis
son un contribuyente mayor de la mortalidad y la discapacidad en México [1].
La aterosclerosis es un proceso multifactorial originado por la presencia de
diversos factores de riesgo. Estudios en modelos animales, en cohortes con
seguimiento a largo plazo y con el empleo de terapias hipolipemiantes
demuestran que las concentraciones anormales de colesterol, triglicéridos,
colesterol-LDL y colesterol-No HDL son un determinante mayor del desarrollo
de cardiopatía isquémica [2]. Los lípidos sanguíneos tienen una alta
heredabilidad; más de 60 genes determinan su concentración plasmática. Las
anormalidades de los valores de los lípidos plasmáticos se integran en el
término “dislipidemias”. Pueden ser resultado de patologías primarias del
metabolismo de las lipoproteínas (como la hipercolesterolemia familiar o la
hiperlipidemia familiar combinada) o secundarias a otras enfermedades que
alteren la producción o eliminación de las lipoproteínas (como la diabetes y el
hipotiroidismo). Su prevalencia varía entre las poblaciones y los grupos
étnicos. En México, las dislipidemias son el factor de riesgo cardiovascular
más frecuente [3].
El objetivo del manuscrito es describir el estado del conocimiento de la
genética de las dislipidemias en México, ya que el componente amerindio de
la población mexicana contribuye a la patogénesis de las dislipidemias con
alelos de susceptibilidad inexistentes en otras poblaciones.
Epidemiología de las dislipidemias en México.
Nuestro país cuenta con tres encuestas nacionales (1994, 2000 y 2006)
en que se han medido la concentración de los lípidos sanguíneos en los
adultos mayores de 20 años [3-6]. La hipoalfalipoproteinemia (colesterol-HDL <
40 mg/dL) es la dislipidemia más común (población total 60.5%, hombres
68.1%, mujeres 53.9%). Esta anormalidad es más frecuente en el sur del
país, en individuos con sobrepeso o diabetes. La prevalencia encontrada en
México es una de las más altas de las informadas en encuestas con cobertura
nacional; la tasa no se ha modificado en el periodo comprendido por las
encuestas. La hipercolesterolemia es la segunda anormalidad más frecuente
(población total 43.6%, hombres 39.3%, mujeres 47.2%). El número de
casos ha aumentado, en especial, entre las mujeres mayores de 50 años. Su
prevalencia es mayor en el norte y el centro del país [6]. La prevalencia de
hipercolesterolemia es menor en México comparado con la informada en
Estados Unidos, sin embargo, la diferencia desaparece al ajustar por edad. La

MENSAJE BIOQUÍMICO, VOL. XL (2016)
128
prevalencia de la hipertrigliceridemia es 31.5%, y es más común en los
hombres, en las personas con obesidad o diabetes. La prevalencia de
hipertrigliceridemia es proporcional a la edad y es mayor en los hombres (36.9%
vs 26.9%). El tabaquismo, el sedentarismo, la diabetes y un índice de masa
corporal superior a 25 kg/m2 son condiciones asociadas a la hipertrigliceridemia.
La prevalencia es mayor en México que la observada en naciones con
etnicidad Caucásica. En los Estados Unidos de América, los hispanos es el
grupo étnico con la prevalencia más alta.
Dislipidemias poligénicas en México
Para postular posibles explicaciones para la mayor prevalencia de la
hipoalfalipoproteinemia y la hipertrigliceridemia observada en México y
poblaciones hispanas residentes en los Estados Unidos, se necesita un enfoque
integrador. La interacción entre variantes genéticas comunes (con efecto
pequeño), variantes raras (con gran efecto) y factores ambientales determinan
su ocurrencia [7]. Una parte de la explicación se encuentra en los patrones de
alimentación de la población, la adopción de estilos de vida poco saludables y la
prevalencia creciente de la diabetes y la obesidad. Sin embargo, la respuesta a los
factores ambientales es modulada por la prevalencia de alelos de susceptibilidad,
la cual difiere entre los grupos étnicos. Las poblaciones de origen amerindio no
han sido incluidas en los consorcios internacionales que han identificado los alelos
de susceptibilidad y las mutaciones causales de las dislipidemias. Por ello, su
estudio en mexicanos es una oportunidad para generar conocimiento nuevo.
Se han publicado varios meta-análisis de los estudios de escrutinio
completo del genoma con múltiples marcadores (Genome Wide Association Study,
GWAS por sus siglas en inglés) [8,9]. En la versión más reciente, se incluyeron
muestras de 188,577 europeos y 7,898 individuos no caucásicos de África y Asia
[28]. Las concentraciones de los lípidos plasmáticos se asociaron con variaciones
de 157 loci. Los niveles de triglicéridos se asocian con variantes localizadas en 37
genes; dieciséis de ellas no están asociadas con ningún otro rasgo lipídico. Pese a
lo anterior, un gran porcentaje de los determinantes genéticos queda por ser
descrito ya que las asociaciones actualmente conocidos explican menos del 10%
de la varianza de la concentración de los triglicéridos. Los genes con las
asociaciones más fuertes con las concentraciones de triglicéridos fueron: TRIB1,
ANGPTL3, LPL, GCKR, FADS1-2-3 y MLXIPL. Estos resultados no pueden
extenderse directamente a cada población, debido a las diferencias interétnicas en
la arquitectura genética. Por ejemplo, SORT1 y LDLR tienen variantes específicas
en africanos; el mismo fenómeno se observó para APOA5 y CETP en asiáticos del

Aguilar Salinas CA, Melgarejo Hernández MA, et al.
129
sur. Estas observaciones refuerzan la necesidad de incluir las poblaciones
multiétnicas en el estudio de los rasgos complejos.
Dos GWAS han incluido muestras de mexicanos, sin embargo, en uno de
estos informes se utilizaron las muestras de México sólo para la replicación.
Kooner y colaboradores [10] identificaron la contribución de MLXIPL en un GWAS
que incluyó, en la etapa inicial, 1,005 hombres europeos y 1,006 hombres
asiáticos. Cerca de 900 SNPs fueron seleccionados para la segunda fase, en la
que participaron 4,568 personas. De ellos, 2,528 fueron mexicanos. Se
encontraron asociaciones con concentraciones de triglicéridos con 13 SNPs;
cuatro de ellos en MLXIPL. Los SNPs adicionales están en las proximidades del
cluster APOA1/C3/ A4/A5 y en LPL. De los tres grupos étnicos estudiados, los
mexicanos tienen la mayor prevalencia del alelo de riesgo de los cuatro SNPs de
MLXIPL. Por otra parte, Weissglass-Volkov y colaboradores llevaron a cabo un
GWAS de dos etapas en el que se incluyeron exclusivamente muestras de casos y
controles mexicanos (Figura 1) [11]. El estudio tenía por objetivo la replicación de
las asociaciones descritas en caucásicos y la identificación de nuevas
asociaciones específicas a nuestra población. La etapa inicial incluyó 2,240
muestras, 563,599 SNPs analizados y 769,042 SNPs imputados. Los casos se
definieron como individuos mestizos con los triglicéridos plasmáticos > 200 mg /
dL, libres del empleo de hipolipemiantes, diabetes, obesidad mórbida o cualquier
causa secundaria de dislipidemia. Los controles fueron sujetos sanos con
triglicéridos <150 mg / dL. La segunda etapa incluyó 1,235 SNPs y 2,121
participantes. Las principales asociaciones reportadas en caucásicos fueron
replicadas. Las señales con mayor fuerza de asociación fueron identificadas en
APOA5, GCKR y LPL, seguidas por ANGPTL3, TIMD4-HAVCR1, MLXIPL y
CILP2. La prevalencia del alelo de susceptibilidad de APOAV, ANGPTL3 y TIMD4
fue significativamente mayor en los mexicanos que en los caucásicos. Cerca del
50% de las asociaciones reportadas en caucásicos son generalizables a los
mexicanos. Se identificaron algunas variantes específicas para nuestra población.
Esta señal se encuentra en el cromosoma 18q11 (P = 2.43 x 10-8). El alelo de
susceptibilidad es común en México y raro en caucásicos. Mediante el análisis de
la expresión de los genes regulados por este locus en muestras de grasa se
identificó que el gen de Niemann-Pick C1 como el candidato más probable para
explicar la asociación. La asociación fue replicada en una muestra independiente,
además se observó que el patrón de ligamiento de las variantes es distinto en
algunos de los genes asociados a la hipertrigliceridemia. Esta aseveración aplica
para APOA5, MLXIPL y CILP2. Este fenómeno permitió descartar la existencia de
asociación con variantes con significancia limítrofe y delimitar con precisión las
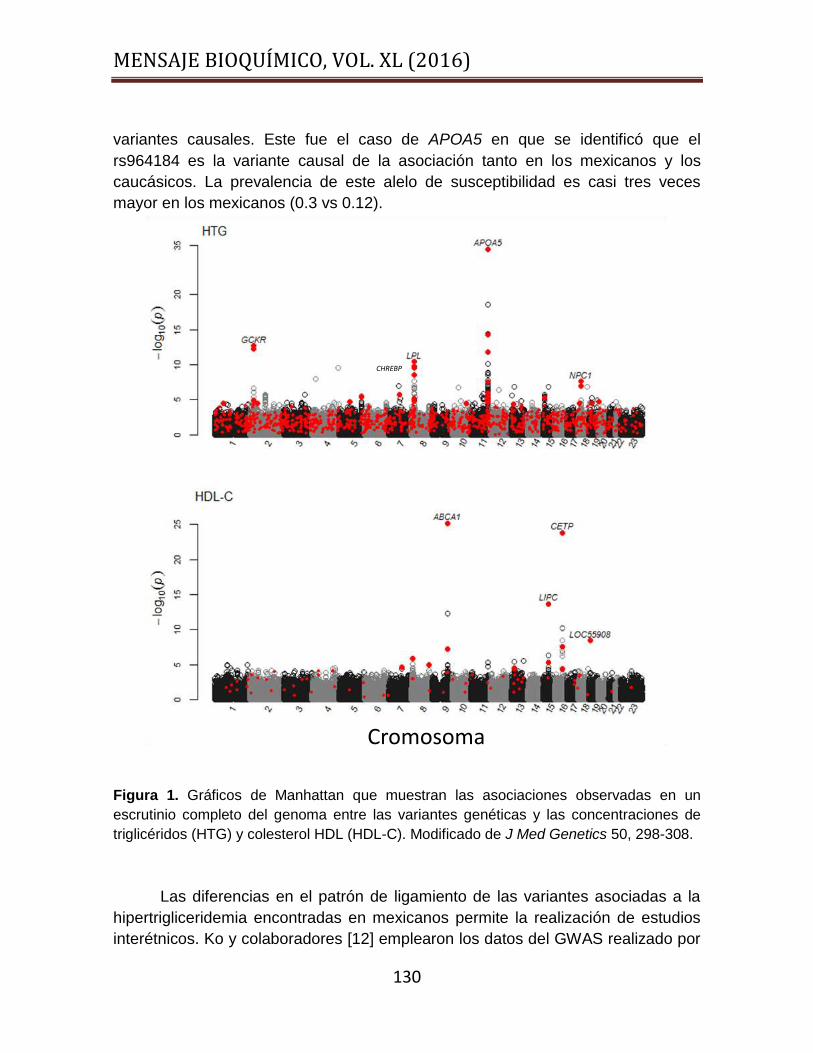
MENSAJE BIOQUÍMICO, VOL. XL (2016)
130
variantes causales. Este fue el caso de APOA5 en que se identificó que el
rs964184 es la variante causal de la asociación tanto en los mexicanos y los
caucásicos. La prevalencia de este alelo de susceptibilidad es casi tres veces
mayor en los mexicanos (0.3 vs 0.12).
Figura 1. Gráficos de Manhattan que muestran las asociaciones observadas en un
escrutinio completo del genoma entre las variantes genéticas y las concentraciones de
triglicéridos (HTG) y colesterol HDL (HDL-C). Modificado de J Med Genetics 50, 298-308.
Las diferencias en el patrón de ligamiento de las variantes asociadas a la
hipertrigliceridemia encontradas en mexicanos permite la realización de estudios
interétnicos. Ko y colaboradores [12] emplearon los datos del GWAS realizado por
CHREBP
Cromosoma

Aguilar Salinas CA, Melgarejo Hernández MA, et al.
131
Weissglass-Volkov y los analizó en conjunto con un GWAS implementado en
finlandeses (en total, 19,273 participantes). Se incluyeron en el análisis a las
variantes que tenían una frecuencia alélica al comparar los dos grupos étnicos; a
este abordaje se le denominó CPAS (cross-population allele screen). Con tal
estrategia se logró la identificación en población mexicana de nuevas asociaciones
con las concentraciones de triglicéridos. Entre éstas, se identificó una variante en
SIK 3 (rs139961185). SIK3 (Salt-inducible kinase 3) es una cinasa relacionada a la
cinasa de proteínas activada por AMP. Los modelos murinos en que se elimina la
expresión de este gen tienen un fenotipo de lipodistrofia, hipolipidemia,
hipoglucemia y una mayor sensibilidad a la insulina, acompañada por colestasis y
colelitiasis [13]. Estos hallazgos sugieren que SIK3 es un nuevo regulador de
energía que modula el metabolismo del colesterol y ácidos biliares. Un análisis
secundario en un subgrupo de 37 sujetos, encontró una asociación del alelo de
riesgo de la variante rs139961185 con las concentraciones de triglicéridos
después de la exposición a una dieta alta en grasas. Los sujetos con el alelo de
riesgo (alelo A/A o A/G) mostraron una concentración de triglicéridos mayor
durante toda la curva (8 horas), en contraste con los que no presentaban dicho
alelo (G/G) (P=0.03 para el área bajo la curva de triglicéridos), y que además fue
independiente de la concentración basal de triglicéridos, sugiriendo que este
cambio en SIK3 puede retardar el aclaramiento de triglicéridos después de una
dieta alta en grasa.
Los determinantes genéticos de la concentración del colesterol-HDL
también fueron motivo de estudio en los GWAS arriba mencionados (Figura 1).
Los genes que codifican la apolipoproteína AI, lecitina: colesterol aciltransferasa,
la proteína de transferencia de ésteres de colesterol, la lipasa hepática y la
proteína de unión de cassette ATP A1 (ABCA1) son actores clave en el
metabolismo del colesterol-HDL. Pese a ello, las variantes identificadas a la fecha
explican solo el 8.05% de la variación interindividual en el colesterol HDL. En
mexicanos, se identificó la contribución mayor que tiene la variante R230C del
transportador ABC-A1 en la concentración del colesterol HDL en los GWAS
publicados por Kooner y Weissglass-Volkov. Dicha variante había sido identificada
como determinante de la concentración del colesterol-HDL en varios estudios de
casos y controles. Los genotipos R230C y C230C fueron más frecuentes en
personas con colesterol-HDL bajo (≤10 percentil de la población mexicana)
comparado contra un grupo con colesterol-HDL alto (≥90 percentil) (45% vs. 2.9%,
P= 0.00006, P= 0.0005 después de ajustar por la etnicidad) [14,15]. La variante
R230C disminuye en 30% la actividad del transportador [16], el cual juega un
papel central en la génesis de las HDL y en el transporte en reversa del colesterol.

MENSAJE BIOQUÍMICO, VOL. XL (2016)
132
Un hallazgo mayor es que la variante R230C es propia de las poblaciones de
origen amerindio. El alelo C230 tiene una frecuencia de 0.109 en mestizos
residentes en el centro de México. La prevalencia es mayor en los mayas (0.28),
purépechas (0.214), yaquis (0.203) y en los teenek (0.179). En contraste, el alelo
C230 no se ha encontrado en África, Europa, China, las poblaciones del sur de
Asia o en Inuits [17]. Su presencia tiene un efecto negativo significativo en los
niveles de colesterol-HDL (-4.2%). El tamaño del efecto es mayor en hombres. El
riesgo atribuible a la variante a nivel poblacional fue medido en la Encuesta
Nacional de Salud 2000; el 12.2% de la variabilidad del colesterol HDL en adultos
mexicanos es atribuible a la presencia de la variante R230C [18]. Finalmente,
usando la estrategia CPAS se identificaron variantes del gen RORA como
determinante de la concentración del colesterol-HDL. Las variantes identificadas
no están presentes en otras poblaciones.
En suma, la evidencia derivada de estudios de asociación genética
demuestra que existen variantes específicas de la población mestiza mexicana
asociadas con la hipertrigliceridemia y la hipoalfalipoproteinemia. El mismo
fenómeno fue informado en el proyecto SIGMA para la diabetes tipo 2 [19]. Estas
observaciones son acordes con el intenso proceso de selección que ha sufrido la
población amerindia, por lo que postulamos que el fenómeno puede ser resultado
de un efecto de supervivencia. Las variantes se preservan en estas poblaciones
porque proporcionaron alguna ventaja en el pasado (es decir, mantener el
colesterol en el interior de las células durante tiempos de hambre). La exposición a
un estilo de vida afluente y un aporte excesivo de nutrimentos resulta en el efecto
opuesto al que dio origen a la conservación de la variante.
Dislipidemias oligogénicas o monogénicas en México.
Hiperlipidemia familiar combinada (HLFC): Es la hiperlipidemia primaria más
común. Se caracteriza por la presencia de diferentes fenotipos de lípidos dentro de
una familia; los casos deben tener una concentración de la apolipoproteina B por
arriba de la percentila 90 de la población (mayor de 108 mg/dL en hombres y 98
mg/dL en mujeres mexicanas; mayor de 120 mg/dL en caucásicos) [20]. Su
expresión más común es la hiperlipidemia mixta (es decir colesterol > 200 mg/dL
más triglicéridos >150 mg/dL). Se asocia frecuentemente al síndrome metabólico.
El estudio de la genética de la hiperlipidemia familiar combinada ha atraído la
atención de varios grupos [21]. A pesar de ello, el conocimiento de la fisiopatología
de esta condición es incompleto [22]. Una proporción significativa de los estudios
han sido realizada en poblaciones caucásicas; algunas de las asociaciones han
sido originalmente identificadas o replicadas en mexicanos. La HLFC ha sido

Aguilar Salinas CA, Melgarejo Hernández MA, et al.
133
consistentemente relacionada con un locus en el cromosoma (1q21-23) en los
caucásicos, asiáticos y mexicanos [23]. Existen varios candidatos en esta región
para explicar la asociación (TXNIP, RXRG, CRABP2, ATF6 y USF1). Los
resultados obtenidos en finlandeses sugieren que las variantes en USF1 son
responsables de la asociación [24]. El mismo haplotipo de riesgo fue identificado
en familias mexicanas, con la asociación más significativa observada con el rasgo
“triglicéridos” (P= 0.0009). El producto del gen USF1 es un factor de transcripción
que regula cerca de 40 genes implicados en el metabolismo de lípidos (APOA5,
APOC3, APOA2, APOE, HSL y PNPLA2). USF1 forma un complejo con USF2
para iniciar la transcripción de los genes diana [25]. Bajo la estimulación de
insulina, USF1 es fosforilada, bloqueando su capacidad para promover la
transcripción de genes. Sin embargo, ninguno de los SNPs de USF1 asociados
con la HLFC (es decir, rs3737787) en finlandeses o mexicanos cambian la
secuencia del factor de transcripción. Por lo tanto, se necesitan estudios
adicionales para establecer relaciones causales. El genotipo rs3737787 se asocia
con diferencias en la expresión de los genes diana de USF1 en grasa o
linfoblastos [26]. Las expresiones de dos módulos de genes asocian la HLFC y la
concentración de triglicéridos. Las variantes de riesgo del rs3737787 se asocian
con cambios en la expresión de los mismos módulos a los que se relaciona la
HLFC. Uno de los módulos, llamado URFA (USF1-regulada, asociada a HLFC)
contiene 504 genes. Las variaciones en la expresión del módulo URFA explica una
parte significativa de las variables de la HLFC (fenotipo HLFC 10%, 6% colesterol,
triglicéridos 17% y apolipoproteína B 9%). Trece genes del módulo estaban
asociados tanto a la HLFC como a la concentración de triglicéridos. Entre ellos se
encuentran FADS3, el cual se identificó previamente en un GWAS como
determinante de la concentración de triglicéridos. FADS3 está involucrada en la
síntesis de ácidos grasos omega-3 y omega-6. También ABCC6, AKT2, GCLM y
HSD11B1 son parte del módulo URFA; todos ellos regulan el metabolismo de
lipoproteínas o de los carbohidratos.
Otras asociaciones entre variantes genéticas y HLFC han sido identificadas
en mexicanos (Figura 2). El producto del gen TCF7L2 es un factor de transcripción
que juega un papel importante en la vía de señalización de Wnt. Las proteínas
Wnt regulan la proliferación y diferenciación celular mediante la activación o
represión de múltiples genes; algunos de ellos están involucrados en el
metabolismo de lípidos y carbohidratos. Las variantes rs7903146 y rs12255372 de
TCF7L2 se asocian con un riesgo mayor de diabetes tipo 2. Debido a la
coexistencia de la HLFC y la diabetes, Huertas-Vázquez y cols [27] exploraron si
las variantes rs7903146 y rs12255372 se asocian con algunos de los fenotipos de

MENSAJE BIOQUÍMICO, VOL. XL (2016)
134
la HLFC en las familias mexicanas y finlandesas. Estas variantes se asociaron con
la concentración de triglicéridos en la población mexicana; el hallazgo se repitió en
los finlandeses. Se observaron diferencias en la expresión de TCF7L2 en el tejido
adiposo subcutáneo asociadas con el alelo de susceptibilidad. La misma
conclusión se planteó cuando se compararon los casos con o sin HLFC. Otra
asociación descrita en mexicanos es la observada entre variantes de HNF4A y
HLFC. HNF4 alfa es uno de los genes causales de la diabetes tipo MODY
(Maturity onset diabetes of the young, por sus siglas en inglés), y regula la
expresión de USF1 y APOB (principal componente de las lipoproteínas
aterogénicas) [28]. Los haplotipos asociados con HLFC fueron los mismos en
ambas mexicanos y finlandeses. HNF4alfa es un regulador transcripcional de
USF1.
Figura 2: Tres factores transcripcionales participan en la fisiopatología de la Hiperlipidemia familiar combinada (HLFC).
Los hallazgos de los estudios anteriormente descritos fueron
complementados con un GWAS en familias mexicanas con HLFC (n = 52,567
sujetos) [29]. Como muestra de replicación, se estudiaron 1,446 individuos no
relacionados con o sin hipertrigliceridemia. La concentración de la Apo B fue
seleccionada como la variable de resultado principal. Los participantes tenían una
proporción amerindia de 0.5. Niveles de Apo B no se asociaron con la etnicidad.
Se replicaron varios loci anteriormente identificados como asociados con la HLFC

Aguilar Salinas CA, Melgarejo Hernández MA, et al.
135
(2p.25, p21.31-14.2, 4p16.2-16.1 entre otros) o con otros rasgos de lípidos (por
triglicéridos, 4p15-16, 4q34, 13q22.33- 31.1, 14q11.2-12 y 20q13.11). Se confirmó
la asociación HNF4A y con otros genes (FABP2, FOLH1, MADD, NR1H3,
plantados, CETP, LCAT, APOE y PLTP). Además, se encontraron dos
asociaciones nuevas (rs1424032 ubicados en 16q21 y rs1349411 en 12p13.31). El
locus 16q21 es una región no codificante altamente conservada, y el locus
12p13.31 incluye el gen APOBEC1, un gen implicado en la edición del RNAm de
APOB en el intestino delgado.
En resumen, los estudios interétnicos han sido útiles para identificar y / o
replicar asociaciones entre variantes genéticas y la HLFC. Las variaciones de la
estructura de desequilibrio de ligamiento genómico de las poblaciones con un
origen étnico diferente podrían ser una barrera o un recurso para generar nuevos
conocimientos. Una asociación del mismo haplotipo en la misma dirección en dos
grupos con diferente origen étnico es una evidencia confirmatoria sólida.
Disbetalipoproteinemia: Se caracteriza por la acumulación plasmática de
remanentes de quilomicrones e IDL. Como resultado, tanto el colesterol y las
concentraciones de triglicéridos son anormales (por lo general alrededor de 300
mg / dL). Es causada, en la mayoría de los casos, por el estado homocigótico para
apoE2, una isoforma de la apolipoproteína E que tiene una afinidad disminuida por
sus receptores. Este trastorno es poco común en los hispanos debido a que la
frecuencia de los alelos de apoE2 es baja en los amerindios [30]. Aunque es
controversial, la apoE2 es un marcador de la etnicidad. En las poblaciones
mestizas, la prevalencia de la variante E2 es baja (3%) en comparación con la que
se encuentra en los caucásicos (7-13%).
Hipercolesterolemia familiar: La hipercolesterolemia familiar (HF) es la
dislipidemia que se asocia a un mayor riesgo de aterosclerosis [31]. Existen dos
variantes: la forma autosómica dominante (resultante de mutaciones en los genes
del receptor de LDL, de PCSK9 y de la apolipoproteina B) y la forma autosómica
recesiva (resultante de mutaciones en el gen ARH, también llamado LDLRAP). La
mayoría de los casos corresponden a las formas heterocigotas de la variante
autosómica dominante. La prevalencia de HF es variable en distintas poblaciones.
En la mayoría de los grupos étnicos, es de 1/500 habitantes para los heterocigotos
y para los homocigotos 1/1,000,000. En nuestro país no existen datos
epidemiológicos al respecto, sin embargo, datos derivados de la Encuesta
Nacional de Enfermedades Crónicas sugieren que perfiles de lípidos compatibles
con este padecimiento se observan en menos del 1% de la población adulta

MENSAJE BIOQUÍMICO, VOL. XL (2016)
136
urbana [3]. En algunos grupos étnicos como los Franco-Canadienses de Quebec,
la prevalencia es mayor (1:270); en ellos, la prevalencia se debe a un efecto
"fundador", que resulta del hecho que los europeos que colonizaron esta región
tenían la enfermedad y la transmitieron a las siguientes generaciones. En México,
la mayoría de los casos provienen de comunidades rurales de los estados de
Puebla, Oaxaca y Veracruz.
Más de 1,740 mutaciones se han descrito en el receptor de LDL
(cromosoma 19p) (http://www.ucl.ac.uk/fh), 163 en PCSK9, 319 en LDLRAP y tres
en el gen de la apoproteina B. Es probable que existan otros genes causales de
HF. En ningún estudio ha sido posible identificar todas las mutaciones causales
del fenotipo. El porcentaje es aún mayor en poblaciones no caucásicas (varía del
30-60% de los casos). Aunque errores metodológicos (en las técnicas de biología
molecular o en el diagnóstico) pueden explicar un porcentaje, es poco probable
que esto sea el motivo principal por lo que no se encuentran defectos en los genes
relacionados a la HF. En México se han publicado dos estudios genéticos en
pacientes con hipercolesterolemia familiar. Robles Osorio y colaboradores
describieron las mutaciones del receptor de LDL encontrados en 46 probandos
mexicanos no relacionados, provenientes de tres centros hospitalarios localizados
en la Cd. de México [32]. Se estudiaron a todos los familiares disponibles, lo cual
fue posible en 32 probandos. Entre los familiares se detectaron 68 casos nuevos.
Todos los casos incluidos tenían la presentación autosómica dominante. Se
identificaron 17 casos con mutaciones en el receptor de LDL. Su existencia fue
confirmada al encontrar la mutación en otros miembros de la familia. Sólo dos
mutaciones fueron encontradas en más de un probando; la más común
(encontrada en cuatro sujetos) es una inserción en el exón 4. De las mutaciones
previamente informadas, todas habían sido previamente publicadas en sujetos
españoles. Sin embargo, casi la mitad de los defectos informados eran mutaciones
nuevas. Tres de ellas resultan en proteínas truncadas (Fs92ter108, Q718X y
Fs736ter743). La primera de estas mutaciones fue encontrada en cuatro
probandos distintos que fueron detectados en diferentes centros. Las mutaciones
puntuales fueron la anormalidad más común (n=9), seguida de una inserción
pequeña (n=1) y una deleción pequeña (n=1). Los exones 4 y 6 fueron los que
tuvieron mayor número de mutaciones (5 y 4 respectivamente). Se encontró un
caso con la mutación R3500Q en el gen de la apoB. No se encontraron
mutaciones en el gen PCSK9 pese a que en tres familias se encontró ligamiento
con el locus donde está localizado dicho gen. Por lo tanto, se descartó la
existencia de defectos en el gen del receptor LDL, apoB y PCSK9 en 25
probandos. Los casos sin mutación identificada tuvieron concentraciones

Aguilar Salinas CA, Melgarejo Hernández MA, et al.
137
significativamente menores de colesterol (325±64 vs 390± 48mg/dL, P0.001) y
colesterol-LDL (258±65 vs 315±46 mg/dL, P0.001) comparado con los casos con
mutaciones en el receptor de LDL pese a que eran de mayor edad (44.8 ± 16 vs
36.3 ± 19 años, P<0.05). En el segundo reporte [32], se estudiaron 62 probandos.
Se identificaron 25 mutaciones (24 en el gen del receptor de LDL, 1 en PCSK9 y 1
en el gen de la apolipoproteina B). Cuatro mutaciones del receptor de LDL no
habían sido reportadas con anterioridad. Se encontraron tres mutaciones que
representaron 19 de 38 probandos.
La ausencia de un grupo de mutaciones dominantes en pacientes
mexicanos con hipercolesterolemia impide el uso de herramientas de diagnóstico
genético diseñadas en otros países. Por ende, es necesaria la secuenciación de
los genes candidatos para el diagnóstico molecular en pacientes mexicanos.
Perspectivas y conclusiones
La prevalencia de las enfermedades crónicas, incluyendo las dislipidemias,
ha aumentado en las décadas recientes. Las razones de su incremento se
encuentran en las modificaciones del estilo de vida de la población. Sin embargo,
los cambios sociales han ocurrido por igual en la mayoría de los países del mundo
occidental sin que se alcancen las frecuencias informadas en México. Esta
observación sugiere que existen factores genéticos que aumentan la
susceptibilidad de nuestra población para tenerlas. Los antepasados de las
poblaciones de origen amerindio eran sobrevivientes de múltiples infecciones,
hambre y guerras. La exposición a un estilo de vida afluente es un nuevo reto que
afrontar. Este fenómeno socioeconómico es una oportunidad para generar nuevos
conocimientos sobre la patogenia de la dislipidemias, la obesidad y la diabetes tipo
2. En esta revisión, hemos descrito el conocimiento actual de la genética de las
dislipidemias en mexicanos. La información disponible muestra que es un área
prometedora para la investigación.
El estudio de la genética de las dislipidemias en poblaciones mestizas
enfrenta varios desafíos. El uso de paneles validados de marcadores de etnicidad
y de estrategias adecuadas de análisis son requisitos obligados en todo estudio
que incluya pacientes de origen amerindio. Existe el riesgo de postular
asociaciones espurias de no tomar tales precauciones. Las diferencias de nuestra
población en comparación con otros grupos étnicos sobre su estructura genómica,
en el desequilibrio de ligamiento, en la prevalencia de los alelos de riesgo son
oportunidades para generar conocimiento nuevo. El estudio de la
hipertrigliceridemia en los mexicanos ha dado lugar a varias contribuciones

MENSAJE BIOQUÍMICO, VOL. XL (2016)
138
significativas. Se identificaron asociaciones entre la HLFC y los genes USF1,
TCF7L2 y HNF4A entre otros. Aún más, existen variantes propias de nuestra
población. Tal es el caso de las variantes R230C del transportador ABCA1, un
haplotipo de riesgo en el gen SIK3 y el rs1349411 en el gen APOBEC1. De
particular importancia es la variante del transportador ABCA1 la cual explica un
porcentaje significativo de la variabilidad del colesterol HDL a nivel poblacional. El
mismo fenómeno ha sido informado con variantes de SLC16A11 y HNF1A en la
diabetes tipo 2. La identificación de variantes propias de la población amerindia
asociadas a enfermedades metabólicas abre la oportunidad de proponer
herramientas de diagnóstico y nuevas vertientes de la fisiopatología.
Se necesitan estudios adicionales para identificar las razones para el
aumento de la susceptibilidad para tener la hipertrigliceridemia y la
hipoalfalipoproteinemia de la población hispana [33]. Sin embargo, algunas pistas
se podrían encontrar en los estudios revisados aquí. La frecuencia de los alelos de
susceptibilidad en los mexicanos es mayor que la encontrada en los europeos
para varios de los genes que tienen el mayor efecto sobre los niveles de
triglicéridos o de colesterol-HDL. Ejemplo de ello son las variantes informadas en
los genes APOA5, MLXIPL para la hipertrigliceridemia y en ABCA1 y CETP para la
hipoalfalipoproteinemia. Algunos de los genes antes descritos pueden participar en
el fenotipo ahorrador.
En resumen, postulamos que el estudio de los determinantes genéticos de
las dislipidemias y otras enfermedades metabólicas en amerindios es un recurso
para identificar nuevas variantes genéticas y vías asociadas con los fenotipos en
estudio. A pesar de ello, los amerindios han sido insuficientemente representados
en los consorcios genéticos internacionales. El reconocimiento de este fenómeno
puede resultar en un mayor número de colaboraciones internacionales, estudios
interétnicos y el intercambio de conocimientos e infraestructura entre las
instituciones.
Referencias 1. Córdova-Villalobos JA, Barriguete-Meléndez JA, Lara-Esqueda A, et al
(2008). Salud Publica Mex. 50,419-427. 2. Lusis AJ (2012) Trends Genet 28, 267-275 3. Aguilar-Salinas CA, Gómez Pérez FJ, Rull JA, et al (2010) Salud Pública
Méx. 52 (supl1), S44-S53. 4. Aguilar-Salinas Carlos A, Olaiz G,Valles V, Ríos JM, et al (2001). J Lipid
Res 42, 1298-1307.

Aguilar Salinas CA, Melgarejo Hernández MA, et al.
139
5. Barquera S, Flores M, Olaiz-Fernández G, et al (2007). Salud Publica Mex 49 supl 3, S338-S347.
6. Gómez-Pérez FJ, Rojas R, Villalpando S, et al (2010). Salud Pública Méx. 52 (supl1) S54-S63
7. Brunzell J. (2007) N Engl J Med 357,1009-1017. 8. Global Lipids Genetics Consortium, Willer CJ, Schmidt EM et al. (2013) Nat
Genet. 45, 1274-1283. 9. Weissglas-Volkov D, Aguilar-Salinas CA, Nikkola E,et al. (2013) J Med
Genetics 50, 298-308. 10. Kooner J, Chambers J, Aguilar-Salinas CA, et al (2008). Nature Genetics
40,149-151 11. Weissglas-Volkov D, Aguilar-Salinas CA, Nikkola E,et al. (2013) J Med
Genetics 50, 298-308. 12. Ko A, Cantor RM, Weissglas-Volkov D, et al (2014) Nat Commun. 5:3983-
3991. 13. Uebi T, Itoh Y, Hatano O, et al (2012) PLoS One.7:e37803 14. Villarreal-Molina MT, Aguilar-Salinas CA, Rodríguez-Cruz M, et al. (2007)
Diabetes 56, 1881-1887 15. Villarreal-Molina MT, Flores-Dorantes MT, Arellano-Campos O, et al. (2008)
Diabetes 57, 509-513 16. Flores-Dorantes T, Arellano-Campos O, Posadas-Sánchez R, et al (2010)
Clin Chim Acta. 411, 1214-1217 17. Acuña-Alonzo V, Flores-Dorantes T, Kruit JK, et al (2010) Hum Mol Genet.
19, 2877-2885. 18. Aguilar-Salinas CA, Canizales-Quinteros S, Rojas-Martínez R, et al (2011).
Atherosclerosis 216, 146-150. 19. The SIGMA (Slim Initiative in Genomic Medicine for the Americas) Type 2
Diabetes Consortium et al (2014). Nature 506, 97-101 20. Aguilar Salinas CA, Zamora M, Gómez- Díaz RA, et al (2004). Sem Vasc
Med 4, 203-209 21. Pajukanta P, Nuotio I, Terwilliger JD et al (1998). Nat Genet.18,369-73. 22. Brouwers M, van Greevenbroek M, Stehouwer C, et al (2012). Nat Rev
Endocrinol. 8, 352-362 23. Huertas-Vázquez A, del Rincón-Jarero JP, Canizales-Quinteros S,et al
(2004). Ann Human Genet 68, 419-427. 24. Pajukanta P, Lilja HE, Sinsheimer JS, et al.(2004) Nat Genet 36,371-6. 25. Huertas-Vazquez A, Aguilar-Salinas C, Lusis AJ, et al (2005). Arterioscler
Thromb Vasc Biol.25, 1985-1991 26. Lee JC, Lusis AJ, Pajukanta P (2006). Curr Opin Lipidol. 17,101-109. 27. Plaisier CL, Horvath S, Huertas-Vazquez A, et al (2009). PLoS Genet. 5,
e1000642. 28. Huertas-Vazquez A, Plaisier C, Weissglas-Volkov D, et al (2008).
Diabetologia 51, 62-69 29. Weissglas D, Huertas Vazquez A, Canizalez S, et al (2006). Diabetes 55,
1970-1977.

MENSAJE BIOQUÍMICO, VOL. XL (2016)
140
30. Aguilar CA, Botet JP, Ordovas JM, et al (1999) Atherosclerosis 142, 409-414.
31. Vallejo-Vaz AJ, Kondapally Seshasai SR, Cole D, et al (2015). Atherosclerosis 243, 257-259
32. Robles L, Huerta A, Ordoñez ML, et al (2006). Arch Med Res 37, 102-108 33. Vaca G, Vàzquez A,Magaña MT, et al (2011) Atherosclerosis 218, 391-396. 34. Kuller LH (2004) Cur Opin Lipidol 15, 109-113.

Aguilar Salinas CA, Melgarejo Hernández MA, et al.
141
Semblanza del Dr. Carlos Alberto Aguilar Salinas
Médico especialista en Medicina
Interna y Endocrinología, egresado del
Instituto Nacional de Ciencias Médicas y
Nutrición. Doctorado en Investigación
Médica. Realizó una estancia de
investigación (1992-1994) en Washington
University, School of Medicine (St Louis MO,
EUA) sobre el estudio del metabolismo de las
lipoproteinas, bajo la tutoría del Dr Gustav
Schonfeld. Fue profesor invitado en la misma
institución en el año 2000. Investigador en
Ciencias Médicas F del Departamento de
Endocrinología y Metabolismo del Instituto Nacional de Ciencias Médicas y
Nutrición. Su línea de investigación principal es la epidemiología, fisiopatología y el
tratamiento de las dislipidemias y de los componentes del síndrome metabólico.
Ha publicado 250 artículos en revistas arbitradas y/o indexadas. El índice H de las
publicaciones es 46. Ha sido citado en 7065 ocasiones. Es investigador nacional
por el SNI (nivel III) y por los Institutos Nacionales de Salud (nivel F). Es coeditor
de la Revista de la Asociación Latinoamericana de Diabetes. Bajo su gestión se
logró la indexación de la revista en Latindex. Además, es miembro del comité
editorial de las revistas Metabolism, Current Diabetes Reports, Current Diabetes
Reviews y la Gaceta Médica de México. Es miembro de la Academia Nacional de
Medicina. Fue director de exámenes del Consejo Mexicano de Endocrinología.
Ocupó la presidencia de la Sociedad Mexicana de Nutrición y Endocrinología
(SMNE) durante el 2009.

MENSAJE BIOQUÍMICO, VOL. XL (2016)
142


















