![[bovino] Leucosis Enzootica](https://static.fdocuments.es/doc/165x107/55cf8c7a5503462b138cd197/bovino-leucosis-enzootica.jpg)
Las enfermedades animales por retrovirus: leucosis bovina ...
43
Rev. sci. tech. Off. int. Epiz., 1990, 9 (4), 1077-1119 Las enfermedades animales por retrovirus: leucosis bovina enzoótica, anemia infecciosa de los équidos, artritis/encefalitis caprina B. TOMA *, M. ELOIT * y M. SAVEY ** Resumen: Este artículo presenta las características esenciales de tres retrovirosis animales: la leucosis bovina enzoótica, la anemia infecciosa de los équidos y la artritis/encefalitis caprina. La originalidad de estas enfermedades consiste en un estado de infección persistente, que se prolonga durante toda la vida del organismo hospedador y que corresponde a la presencia de un provirus integrado en las células del hospedador. Los animales infectados constituyen por lo tanto fuentes potenciales prolongadas del agente patógeno. Los conocimientos disponibles actualmente sobre la epidemiología y la detección de estas enfermedades son suficientes para la puesta en marcha de métodos de lucha sanitaria eficaces. PALABRAS CLAVE: Anemia infecciosa de los équidos - Artritis/encefalitis caprina - Diagnóstico - Epidemiología - Leucosis bovina enzoótica - Retrovirus. INTRODUCCIÓN El número de enfermedades animales debidas a los retrovirus es elevado y continúa aumentando conforme lo hace la identificación de nuevos agentes en el seno de este grupo, por ejemplo: el virus de la inmunodeficiencia felina en 1987 (60) (véase Cuadro I). En el seno de este conjunto de enfermedades, han sido elegidas tres de las más importantes por sus repercusiones económicas y serán las únicas presentadas en este trabajo. En un número limitado de páginas, es imposible el tratar de manera detallada los diferentes aspectos de una enfermedad, y con mayor razón hacerlo de tres enfermedades. Por este motivo, ha sido necesario seleccionar en cuanto a las nociones presentadas en las líneas que siguen. Han sido preferidos los capítulos sobre los que los conocimientos evolucionan y aquellos que tienen una importancia mayor para la comprensión de las medidas de lucha, es decir: características esenciales del agente patógeno, epidemiología, diagnóstico y profilaxis. Por lo contrario, los síntomas y lesiones no se han desarrollado. * Ecole Nationale Vétérinaire d'Alfort, 7 avenue du Général de Gaulle, 94704 Maisons-Alfort, Francia. ** CNEVA, Laboratoire de pathologie bovine, 31 avenue Tony Garnier, B.P. 7033, 69342 Lyon Cedex 07, Francia.
Transcript of Las enfermedades animales por retrovirus: leucosis bovina ...

Rev. sci. tech. Off. int. Epiz., 1990, 9 (4), 1077-1119
Las enfermedades animales por retrovirus: leucosis bovina enzoótica, anemia infecciosa de
los équidos, artritis/encefalitis caprina B. TOMA *, M. ELOIT * y M. SAVEY **
Resumen: Este artículo presenta las características esenciales de tres retrovirosis animales: la leucosis bovina enzoótica, la anemia infecciosa de los équidos y la artritis/encefalitis caprina.
La originalidad de estas enfermedades consiste en un estado de infección persistente, que se prolonga durante toda la vida del organismo hospedador y que corresponde a la presencia de un provirus integrado en las células del hospedador. Los animales infectados constituyen por lo tanto fuentes potenciales prolongadas del agente patógeno.
Los conocimientos disponibles actualmente sobre la epidemiología y la detección de estas enfermedades son suficientes para la puesta en marcha de métodos de lucha sanitaria eficaces.
PALABRAS CLAVE: Anemia infecciosa de los équidos - Artritis/encefalitis caprina - Diagnóstico - Epidemiología - Leucosis bovina enzoótica - Retrovirus.
I N T R O D U C C I Ó N
El número de enfermedades animales debidas a los retrovirus es elevado y continúa aumentando conforme lo hace la identificación de nuevos agentes en el seno de este grupo, por ejemplo: el virus de la inmunodeficiencia felina en 1987 (60) (véase Cuadro I).
En el seno de este conjunto de enfermedades, han sido elegidas tres de las más importantes por sus repercusiones económicas y serán las únicas presentadas en este trabajo. En un número limitado de páginas, es imposible el tratar de manera detallada los diferentes aspectos de una enfermedad, y con mayor razón hacerlo de tres enfermedades. Por este motivo, ha sido necesario seleccionar en cuanto a las nociones presentadas en las líneas que siguen. H a n sido preferidos los capítulos sobre los que los conocimientos evolucionan y aquellos que tienen una importancia mayor para la comprensión de las medidas de lucha, es decir: características esenciales del agente patógeno, epidemiología, diagnóstico y profilaxis. Por lo contrario, los síntomas y lesiones no se han desarrollado.
* Ecole Nationale Vétérinaire d'Alfort, 7 avenue du Général de Gaulle, 94704 Maisons-Alfort, Francia. ** CNEVA, Laboratoire de pathologie bovine, 31 avenue Tony Garnier, B.P. 7033, 69342 Lyon
Cedex 07, Francia.

1078
Los retrovirus poseen todos una transcriptasa reverse (reverse transcriptasa, de donde el término de retrovirus) responsable de la síntesis de una copia de A D N a partir del A R N viral.
El A D N así formado (provirus), puede conservarse en el núcleo de ciertas células del hospedador de estos virus y esta propiedad original es la causa de las características particulares de las diferentes infecciones debidas a los retrovirus. Se trata de infecciones persistentes, que se prolongan durante toda la vida del organismo hospedador y corresponden a la presencia de la «información» de origen viral integrada en las células del hospedador. Bajo esta forma integrada, el agente patógeno está al abrigo de las defensas inmunitarias de su hospedador. Salvo casos particulares, los retrovirus se presentan en los organismos infectados bajo la forma proviral mucho más que bajo la forma de viriones completos, circulantes en los líquidos del organismo. Po r esta razón, la transmisión se produce habitualmente por la transferencia de sangre o de secreciones que contengan las células infectadas y se ve favorecida por todas las actividades que permitan la inoculación de cantidades, incluso mínimas, de sangre o de otras suspensiones de células: tomas de sangre o inyecciones en serie, cirugía menor, picaduras de ar t rópodos, absorción de calostro, etc.
C U A D R O I
Retrovirus agentes de enfermedades de los animales domésticos (18)
Subfamilia Hospedador Virus
Oncovirinae Pájaros Virus de la leucosis aviar (Avian leukosis virus) Virus del sarcoma aviar (Avian sarcoma virus) Virus de la reticuloendoteliosis (Reticuloendotheliosis virus)
Bovinos Virus de la leucosis bovina enzoótica (Enzootic bovine leukosis virus)
Cerdos Virus del sarcoma del cerdo (Porcine sarcoma virus)
Gatos Virus de la leucosis felina (Feline leukosis virus) Virus del sarcoma felino (Feline sarcoma virus)
Lentivirinae Ovinos Virus maedi-visna (Maedi-visna virus)
Cabras Virus de artritis/encefalitis caprina (Caprine arthritis/encephalitis virus)
Caballos Virus de la anemia infecciosa de los équidos (Equine infectious anemia virus)
Bovinos Virus de la inmunodeficiencia bovina (Bovine immunodeficiency virus)
Gatos Virus de la inmunodeficiencia felina (Feline immunodeficiency virus)
Spumavirinae Bovinos Virus espumoso bovino (Bovine foamy virus)
Gatos Virus espumoso felino (Feline foamy virus)

1079
Los casos particulares corresponden, especialmente, a los mecanismos que permiten a los viriones maduros escapar a las defensas del organismo, como la desviación antigénica registrada, en particular, con el virus de la anemia infecciosa de los équidos.
Este carácter de infección persistente confiere a los retrovirus (lo mismo que a los herpesvirus) una seguridad de estabilidad y de perennidad de la infección de los rebaños. La ausencia de curación espontánea de estas infecciones y la persistencia del peligro de transmisión a los organismos sanos, imponen unas estrategias de lucha adaptadas , cuyos principios son actualmente bien conocidos, pero cuya puesta en práctica se muestra, en ciertos casos, de un costo prohibitivo.
El objetivo de las líneas que siguen es el de presentar lo esencial de los conocimientos actuales sobre estas tres retrovirosis, desde una óptica de lucha contra estas enfermedades. Las nociones que se presentan provienen de la bibliografía y se han beneficiado de la fuente de informaciones que han proporcionado a la OIE los países cuya lista figura en el Cuadro II .
C U A D R O I I
Lista de los países que han dado informaciones a la OIE
Africa Argelia, Botswana, Congo, Egipto, Etiopía, Lesoto, Madagascar, Senegal, Sudáfrica, Zambia
América
Canadá, Chile, Estados Unidos, Haití
Asia
Indonesia, Japón, Jordania, Myanmar, Oman, Sri Lanka, Taiwan R.O.C.
Europa Alemania (Rep. Fed. de), Checoslovaquia, Chipre, Dinamarca, Francia, Irlanda, Italia, Luxemburgo, Noruega, Países Bajos, Reino Unido, Suecia, Suiza, Turquía, Yugoslavia
Oceania Australia
LA LEUCOSIS BOVINA ENZOÓTICA
G E N E R A L I D A D E S
La leucosis bovina enzoótica (LBE) es una enfermedad infecciosa y contagiosa, propia de los bovinos, debida a un virus de la familia de los Retroviridae: el virus leucemógeno bovino (VLB).

1080
Reinando en forma enzoótica en los rebaños bovinos, se desarrolla:
- b i e n sea bajo la forma de una infección inaparente, a veces acompañada de una modificación del hemograma (linfocitosis persistente),
- o bien como una forma tumoral, encontrada principalmente entre los bovinos adultos (de media de 5 a 8 años) y que se define en este caso como una afección neoplásica maligna de la línea linfoide, evolucionando en la mayoría de los casos bajo la forma de un linfosarcoma multicéntrico.
La LBE debe distinguirse de otras dos afecciones leucósicas que afectan severamente de forma esporádica los bovinos y cuya etiología es desconocida:
- la leucosis juvenil (bien sea en la forma tímica, o bien en la forma multicéntrica),
- la leucosis cutánea del adulto.
La distinción entre estos diferentes casos se reagrupa en el Cuadro III .
C U A D R O I I I
Las diferentes leucosis bovinas (18)
La LBE es una enfermedad expandida universalmente.
Aunque no se tenga ninguna certidumbre, parece que la LBE no es transmisible al hombre (13). Ni la investigación del virus entre los profesionales expuestos (veterinarios, criadores, personal de mataderos o de lecherías), ni la investigación de una tal etiología entre los casos de cánceres humanos (leucemia y otros) han sido positivos (7). A pesar de ello, el virus puede multiplicarse en las células de monos y en las células humanas (27) y algunos autores han observado la aparición de anticuerpos específicos en los chimpancés inoculados (90).
Desde el punto de vista económico, la LBE no ha constituido jamás una enfermedad de los rebaños bovinos responsable de pérdidas económicas considerables. Sus repercusiones varían según los países. Solamente los casos tumorales, inexorablemente mortales, constituyen una pérdida económica directa. Su frecuencia depende de las tasas de infección de los animales y varía por lo tanto mucho en función de los países, desde algunas unidades a algunas centenas por cada 100 000 canales.
Formas lesionales Epidemiología Virus de la LBE
Multicéntrica -
Juvenil
Tímica Esporádica -
Cutánea —
Adulta
Multicéntrica Enzo ótica +

1081
Algunos países han logrado una tasa de infección prácticamente nula en sus ganaderías, mientras que otros, por lo contrario, mantienen una tasa de infección de sus ganados superior al 5 0 % .
Las exigencias sanitarias establecidas para la importación de los bovinos por los países que habían dedicado importantes esfuerzos a la erradicación de la LBE, han llevado a todos los países implicados por los cambios internacionales de bovinos a interesarse por la situación epidemiológica de LBE y a adoptar una política de lucha teniendo en cuenta sus objetivos y los apremios económicos.
V I R O L O G Í A
El retrovirus de la LBE es cercano de los virus H T L V I y HTLV II del hombre (39). Sus principales características han sido objeto de una reciente revisión (6).
La totalidad de su secuencia nucleotídica es conocida actualmente y esto ha permitido, jun to a los caracteres generales de los retrovirus, identificar algunas características particulares (65, 66, 67, 79). Así, existe un gene codificado por una proteasa, que imbrica los genes G A G y P O L pero que se lee en un cuadro de lectura diferente (Figura 1). Dos cuadritos abiertos de lectura (SOR y LOR) se sitúan después del gene ENV. El producto de los genes LOR y SOR activa la transcripción de los ARN mensajeros virales (11, 30, 75, 76) y puede ser de algunos A R N mensajeros celulares, lo que aportaría una hipótesis para explicar el poder oncógeno de este virus (6).
FIG. 1
Estructura del genoma del provirus de la leucosis bovina enzoótica (6)
Las proteínas estructurales del virus comprenden las proteínas internas (p l5 , p24, p l 2 , p l4 ) y las glucoproteínas de envoltura (gp30 y la glucoproteína mayor gp51). Diferentes epitopos de la gp51 se han identificado con la aplicación práctica de desarrollar las pruebas de competición ELISA (62), que permiten revelar la presencia de anticuerpos anti gp51 en los bovinos infectados.
Inicialmente, la replicación del virus se ha obtenido mediante el cultivo en linfocitos de bovinos infectados espontáneamente y en estado de linfocitosis persistente (50).

1082
Una producción viral, sin efecto citopatógeno, puede obtenerse bien sea en línea continua de linfocitos, o bien por co-cultivo de linfocitos infectados con las células efectrices (células de murciélago, células esplénicas de embrión de bovino, células renales de fetos de cordero o FLK: Fetal Lamb Kidney).
La infección por el virus de la LBE entraña la aparición de anticuerpos neutralizantes; estos no tienen, no obstante, ningún efecto protector contra el desarrollo de un linfosarcoma. Los anticuerpos anti gp51 inyectados a un cordero permiten protegerle frente a una prueba virulenta (44).
Has ta el momento presente, los estudios destinados a la puesta a punto de una vacuna contra la LBE no han sido decisivos (concentración de antígeno gp51 y adsorción sobre adyuvante, recombinación con el virus de la vacuna, introducción en los complejos inmunoestimulantes ISCOM, etc.) (55, 68) pero parece que pueden tenerse esperanzas (6).
P A T O G E N I A
La patogenia de la LBE es compleja y permanece oscura en numerosos puntos .
La infección por el virus leucemógeno bovino se traduce por tres estados sucesivos y acumulativos: la infección inaparente, la linfocitosis persistente y el linfosarcoma.
La infección inaparente
El animal no presenta ningún signo clínico ni hematológico, únicamente su respuesta serológica es positiva.
La infección puede adquirirse antes del nacimiento (pequeño porcentaje de infección in útero); la tasa de infección en los rebaños leucósicos aumenta con la edad.
Después de la infección, el plazo de seroconversión varía de 2 a 8 semanas y depende sin duda, en parte, de la carga viral del inoculo. Po r ejemplo, este plazo es de 3 a 4 semanas después de la inoculación de 5.10 6 linfocitos (o sea el equivalente de 1 mililitro de sangre) por vía intradérmica, intratraqueal o subcutánea. El plazo es del mismo orden después de la inyección de 50 microlitros de sangre. En condiciones naturales, no obstante, la respuesta serológica de algunos bovinos puede no hacerse positiva en inmunodifusión más que después de tres meses de la infección.
La linfocitosis persistente
La fórmula sanguínea de un bovino afectado está per turbada por un aumento persistente de los linfocitos. La linfocitosis persistente aparece raramente antes de la edad de los 2 años. Según los rebaños, alcanza del 10 al 90% de los animales infectados.
Lo más frecuentemente persiste varios años, hasta la muerte del animal. A veces, esta linfocitosis precede a la aparición de los tumores, siendo entonces la duración de la evolución variable, entre algunas semanas a algunos años. Puede también desaparecer antes de la aparición de los tumores.

1083
La linfocitosis persistente corresponde a una proliferación policlonal de linfocitos B, caracterizada por una presencia simultánea de numerosos clones linfocitarios distinguibles por las zonas diferentes de integración de los provirus en los cromosomas. No se t ra ta por tan to de células tumorales, ya que su capacidad de ser multiplicadas in vitro es diferente de la de las células t ransformadas (6). De manera inversa, las células tumorales derivan lo más corrientemente de un solo clon celular, aunque, en función de los tumores, las zonas de integración cromosómica del provirus aparezcan diferentes (34). Finalmente, las células tumorales, aunque integren el A D N proviral en sus cromosomas, no sintetizan (o lo hacen muy poco) las proteínas virales (6).
El incremento de los linfocitos afecta también a los linfocitos T (96). E n los casos de linfocitosis persistente, la tasa de anticuerpos aumenta al mismo tiempo que el número de leucocitos.
El linfosarcoma
Es ésta la única forma clínicamente visible y se caracteriza por la aparición de tumores, asociada a una linfocitosis persistente y a una respuesta serológica positiva.
El linfosarcoma aparece en general en los animales entre S y 8 años. N o se desarrolla más que sobre un escaso porcentaje de los bovinos infectados, o sea, cada año, el 0,5 al 1 % de los animales infectados. La evolución se hace rápidamente hacia la muerte .
La respuesta inmunitaria frente al virus leucemógeno bovino no ejerce ningún efecto protector frente al desarrollo tumoral. En un animal infectado, las tasas de anticuerpos son generalmente más elevadas cuando se desarrolla un linfosarcoma que en el caso de solamente una leucositosis persistente.
Los animales afectados de linfosarcoma no presentan una inmunosupresión. Así mismo, tampoco se ha observado inmunosupresión en los animales infectados en el curso de su vida fetal (81).
E P I D E M I O L O G Í A
Epidemiología descriptiva
L a infección por el virus de la LBE ha sido señalada en la mayor parte de los países que lo han investigado y donde ella se presenta de forma enzoótica en ciertos rebaños o regiones.
La situación actual es muy variable en función de los países (Cuadro IV). En el seno de un mismo país, las tasas de infección de los rebaños pueden ser muy diferentes de una región a otra. En los diferentes países, la difusión de la infección está en relación con la importación de reproductores.
En las ganaderías infectadas, las tasas de infección de los animales son muy variables, desde algunos individuos, al 30-50%, o incluso más.
E n los países templados, las conversiones serológicas de los animales son más numerosas al final del verano (47). Los casos tumorales aparecen en cualquier momento del año .

1084
C U A D R O I V
Principales informaciones relativas a la epidemiología y a las medidas de lucha oficiales dadas por los países
que han enviado un informe a la OIE sobre la leucosis bovina enzoótica
País Epidemiología Medidas de lucha oficiales
Africa Argelia Casos esporádicos en el Enfermedad de declaración Dirección de los este del país, esencialmente obligatoria. Servicios veterinarios sobre ganado importado Control serológico a la
importación Botswana Enfermedad no identificada Departamento de los por aislamiento de virus Servicos Veterinarios (M.G. Mosienyane) Congo Enfermedad no señalada Dirección de Producción Animal (J. Bansimba Maringa) Egipto No señalada Organización General de los Servicios Veterinarios (A.A. Moussa) Etiopía No señalada Departamento de Servicios Veterinarios (Z. Dagnachew) Lesoto Ausencia Departamento de Salud animal (O.L. Letuka) Madagascar Enfermedad no señalada Dirección de Cría (V.R. Ranatvoson) Sudafrica Enfermedad enzoótica. Tasa Directorate of Animal de infección de los animales Health pudiendo alcanzar el 90% en (D.W. Verwoerd) los rebaños lecheros; incidencia
más baja en las razas de carne Zambia Ausencia Departamento de Servicios Veterinarios y de Control Tsetse (K.L. Samui)
América Canadá Situación sin duda parecida No es enfermedad de declaración Dirección General, a la señalada por Keller a obligatoria. Se considera que Producción e inspección la OIE en 1981: infección para la exportación una de los alimentos del 41% de los rebaños serologia negativa individual (N.G. Willis) lecheros (9,3% de infección es una garantía suficiente sin
de los animales) y del 10% que sea necesario que el de los rebaños de carne rebaño de origen esté indemne (0,5% de infección de los de LBE animales)

C U A D R O I V (cont.)
1085
País Epidemiología Medidas de lucha oficiales
Chile Puesta en marcha de un sistema División Protección de certificación de los rebaños pecuaria indemnes de LBE en las regiones (J. Benavides Muñoz) 9 y 10
Asia Indonesia Tasa de infección de los Director General de animales variable según Servicios de Cría las razas de 0,5% a 3% (Soehadji) Myanmar Presencia Cría de ganado y departamento veterinario (Than Tint) Oman Enfermedad no señalada Departamento de Salud Animal (Sultán Ahmed Al Sultán) Sri Lanka Enfermedad no señalada Oficina del Ministerio de Estado para el desarrollo ganadero y producción lechera (S.B. Dhanapala) Taiwàn R.O.C. Aumento de la prevalencia. Consejo de Agricultura Introducción del virus por (R.C.T. Lee) los animales importados
Europa Alemania (Rep. Fed. de) Linfocitosis persistente en Enfermedad de declaración Escuela Veterinaria el 50% de los animales obligatoria. Eliminación de Hannóver infectados. Tumor en de los animales con (U. Truyen, L. Haas y 0,1 al 10% de los animales serologia positiva. O.R. Kaaden) infectados. La RFA está casi indemne de
En 1979: 13 900 focos LBE En 1989: 63 focos
Checoslovaquia Tasa de infección media de Programa de lucha intensiva Administración Estatal los animales: 1,13% en trance de aplicación Veterinaria
los animales: 1,13% en trance de aplicación
(J. Krecek) Dinamarca Desde enero de 1988 a (P. Have y octubre de 1989, se han R. Hoff-J0rgensen) identificado tres rebaños
infectados, mediante la prueba ELISA sobre sueros individuales (719 490 sueros)
Irlanda Introducción de la Departamento de infección por un lote de Agricultura y 80 bovinos importados de Alimentación Canadá en 1974. Detección (R.G. Cullen) de todos los animales infectados
en los rebaños correspondientes y sacrificio. Desde 1979, ausencia de respuesta positiva

1086
C U A D R O I V (cont.)
Epidemiología analítica
Fuentes de virus
La fuente de virus casi exclusiva está representada por los bovinos infectados. El virus leucemógeno bovino está presente en los linfocitos de los animales infectados; de aquí que toda materia extraída de un bovino infectado y que contenga los linfocitos, puede ser virulenta.
a) La sangre
La fracción celular de la sangre y sólamente ella alberga al virus. No obstante, en el caso de almacenamiento prolongado de las muestras (2 semanas a + 4°C), el virus puede aislarse también del plasma sanguíneo, debido sin duda al hecho de una lisis celular (69). El nivel de la «viremia» puede apreciarse investigando la cantidad mínima de sangre de bovino infectado necesaria para transmitir la enfermedad: 1/100° de gota puede ser suficiente (57). Roberts y col. (74) han demostrado que la sangre de un bovino puede ser infecciosa unos quince días antes de la aparición de los anticuerpos séricos. La transmisión de la infección se asegura de forma más eficaz con la ayuda de los linfocitos infectados que con una suspensión de partículas virales (35).
País Epidemiología Medidas de lucha oficiales
Luxemburgo No parece existir Enfermedad de declaración Administración de los obligatoria Servicios Veterinarios (J. Kremer)
Suecia Tasa de infección de los Programa de lucha a Instituto Nacional animales variable según base del voluntariado Veterinario las regiones, máximum en (A. Eng valí y el sudeste M. Wierup)
Suiza Escasa prevalencia Laboratorio del Servicio serológica: 3 respuestas Cantonal Veterinario positivas sobre 11 398 (A.F. Gachet-Piguet) ELISA en leche de mezcla
Turquía Casos esporádicos Director General de Producción y control (E. Istanbulluoglu)
Oceania Australia Tasas de infección de los Enfermedad de declaración Departamento de rebaños lecheros en obligatoria en todos Agricultura Queensland: 70,9% (tasa los Estados. Programa oficial (T.M. Ellis) de prevalencia de la de lucha solamente en el
infección entre los Estado de Queensland animales: 13,7%). Tasa de infección de los animales de razas de carne: 0,4%

1087
b) El calostro y la leche
El virus ha sido puesto en evidencia en la leche y en el calostro de vacas infectadas (33, 54). Fal tan datos sobre el carácter perenne o no de esta excreción durante la lactación así como sobre su nivel. El estudio cinético de la aparición del virus ha mostrado que se efectúa de forma concomitante en la leche y en la sangre, es decir, en los quince días que siguen la inoculación (89).
c) El esperma
La investigación del virus en el esperma ha sido objeto de numerosos estudios en razón del temor de una posible diseminación del virus a partir de toros infectados de los centros de inseminación (3, 89, 41 , 40). Parece que el esperma no es virulento en las condiciones normales. N o obstante, las lesiones traumáticas o inflamatorias podrían permitir, en ciertos casos, su contaminación por intermedio de los linfocitos.
d) Otras secreciones y excreciones
- Orina y heces
La investigación del virus por la inoculación al cordero se ha mostrado siempre negativa (52, 64).
- Saliva
Su virulencia se ha demostrado en 5 bovinos de 17 infectados (61).
- Secreciones nasales y brónquicas
El virus ha sido aislado de la fracción celular del líquido de lavado brónquico a partir de 6 bovinos infectados de entre 9 (72). E n dos casos de seis, las secreciones nasales se han revelado virulentas (fracción celular únicamente). Estos resultados no son sorprendentes a priori por razón del flujo linfocitario permanente que existe entre la circulación general y el pu lmón profundo. Queda no obstante posible que una contaminación accidental por los elementos sanguíneos haya podido falsear la experiencia.
En resumen, la presencia de los linfocitos en una secreción o excreción condiciona su virulencia. Una extravasación sanguínea o una lesión inflamatoria local pueden engendrar o aumentar la virulencia de una materia normalmente poco o nada virulenta.
Esta explicación se deduce de ciertos resultados contradictorios a propósito del poder infeccioso del esperma, de la saliva o de las orinas de bovinos infectados. Por otra parte , parece lógico pensar que las mamitis pueden contribuir a aumentar la carga viral de la leche de las vacas infectadas.
Es necesario recordar que, en las condiciones habituales, la sangre sobre todo y la leche, siguen siendo las materias virulentas más importantes.
Receptividad
Se debe distinguir la receptividad a la infección y la de la expresión de una linfocitosis persistente o de la forma tumoral .
- L a receptividad intrínseca de los animales a la infección es sin duda muy parecida. Diferentes factores pueden jugar un papel:

1088
La presencia de anticuerpos calostrales juega un papel protector en los terneros procedentes de vacas infectadas (93, 36).
Las condiciones de cría pueden tener un papel determinante al facilitar o no la transmisión en función de las precauciones tomadas en ciertas circunstancias: descornado, cirugía menor, tomas de sangre en serie, etc. Las ocasiones de contacto estrecho entre animales intervienen sin duda igualmente para favorecer la transmisión (Lassauzet, comunicación personal, 1989). Este factor podría ser en principio la causa de la diferencia sistemáticamente señalada entre la tasa de infección de los rebaños lecheros claramente superior a la de la infección de los rebaños lactantes (en el momento actual, especialmente en Australia, en Canadá , los Estados Unidos, en Sudáfrica, etc.) (véase los informes de estos países).
Las condiciones climáticas pueden favorecer la transmisión cuando son favorables a la multiplicación de los insectos.
- Los casos familiares de linfocitosis persistente son actualmente bien conocidos. En ciertos rebaños, la infección, tanto como la enfermedad clínicamente expresada, tiene una incidencia particularmente importante .
Es verosímil que la capacidad de desarrollar una linfocitosis persistente, incluso un linfosarcoma, depende de factores genéticos. E n esta hipótesis, su naturaleza y sus mecanismos de acción no son conocidos (38).
Transmisión
a) Transmisión directa
- Vía oral
La transmisión por vía oral ha sido estudiada sobre todo en el marco de una infección del ternero por el calostro o la leche. H a sido demostrada en condiciones experimentales (92, 93).
Pero el papel de la leche y del calostro, a pesar de su carácter potencialmente virulento, parece limitado en condiciones naturales (vía oral) en relación a los factores de contacto (20).
Dos hipótesis han sido evocadas para explicarlo: la primera hace jugar un papel protector a los anticuerpos de origen calostral absorbidos por el ternero; la segunda contempla la impermeabilidad de la mucosa intestinal a los linfocitos infectados por el VLB después de las 24 a 36 horas primeras de la vida del ternero. Estos dos factores pueden evidentemente jugar de forma concomitante. La primera hipótesis ha sido confirmada, especialmente por Van der Maaten y col. en 1981 (93) y Lassauzet y col. en 1989 (36).
- Via respiratoria
La instilación de un aerosol virulento por vía intranasal permite reproducir la infección (91). Por otra parte, la inoculación intratraqueal de 5.10 6 linfocitos infectados ha provocado en cuatro casos sobre cuatro la infección de los bovinos (71).
Siendo las materias de expectoración de los bovinos infectados potencialmente virulentas, parece que esta vía de transmisión pueda jugar un papel.

1089
- V í a venérea
Los resultados de las experiencias son contradictorios según que el i n c u l o depositado en el tractus genital de las vacas fuese: los linfocitos aislados de un bovino infectado (4 vacas de cada 6 se infectan) (51), o una mezcla de esperma bovino y de linfocitos infectados (1 vaca de cada 4 se infecta) (71).
Parece pues existir un agente espermático inactivante del virus que explicaría las dificultades de aislamiento del virus en el esperma de los bovinos infectados.
El papel de la vía venérea parece pues menor. Por otra parte, ninguna publicación manifiesta actualmente casos probados de transmisión venérea de la LBE en condiciones naturales.
- T r a n s m i s i ó n in útero
La transmisión del virus de la madre al feto no plantea actualmente ninguna duda. Sólo las tasas de infección in útero en el seno de una ganadería infectada varían según los autores. La metodología de la encuesta consiste en efectuar una serologia sobre los terneros recién nacidos antes de la t oma del calostro (31). La investigación del virus en los linfocitos del recién nacido es un método más seguro.
Las tasas de transmisión in útero recogidas en la bibliografía varían de un máximo de un 14 a 2 5 % de las hembras infectadas en un rebaño conocido por su gran receptividad al virus, hasta 3 a 6% de las hembras infectadas en los rebaños representativos de la media del censo bovino (8). Esta transmisión sobrevendría por la vía transplacentaria durante los 6 últimos meses de la vida intrauterina.
Este modo de transmisión, sin ser despreciable, no representa pues más que un aspecto sin duda menor, salvo casos excepcionales, de la difusión del virus en el seno de un rebaño.
La transmisibilidad de la infección por los gametos del huevo, es decir in ovo, ha sido regularmente invalidada. En un estudio de transferencia de 21 embriones de 6 a 7 días, obtenidos de 8 vacas infectadas, ningún ternero estaba infectado (58). La observación de casos familiares debe estar pues mucho más relacionada con la influencia de los factores genéticos y el contagio precoz.
b) Transmisión indirecta
Se basa en la virulencia de la sangre de los animales infectados.
- T r a n s m i s i ó n por los ar t rópodos picadores
Consideraciones de tipo epidemiológico habían llevado a pensar que los artrópodos pueden jugar un papel en la transmisión de la LBE: en los Estados Unidos y en el Japón, la incidencia de la infección aparecía maximalmente durante la estación cálida en la que los ar t rópodos son más numerosos (8). Estas observaciones han sido confirmadas en Francia (47).
Los mosquitos no jugarían más que un papel limitado en la transmisión de la LBE (10), contrariamente a los tabánidos, por dos razones: por una parte , su escaso tamaño y, por otra , sus hábitos alimentarios que les hacen generalmente comenzar y terminar una comida sanguínea sobre el mismo hospedador . Se puede comparar estas constataciones con las, idénticas, señaladas para la anemia infecciosa de los équidos.

1090
Foil y col. (23) han transmitido la infección a los corderos y a las cabras a partir de una vaca afectada de linfocitosis persistente por interrupción de la comida sanguínea de Tabanus fuscicostatus; la picadura de 50 a 100 tábanos ha transmitido la infección mientras que la de 10 o 25 tábanos no lo ha permitido. Oshima y col. (56) han obtenido resultados semejantes.
Más recientemente (24), estos mismos autores han obtenido la transmisión después de la picadura de 10 a 20 tábanos . Un volumen de 0,1 ml de sangre de una vaca en estado de linfocitosis permanente fue suficiente para transmitir la infección. Una cantidad del orden de 1 500 linfocitos ha permitido la transmisión.
Finalmente, más que por simples transportadores mecánicos, las garrapatas pueden jugar el papel de vectores en la transmisión de la LBE gracias a una transmisión transtadial (32).
En resumen, el papel vectorial de los artrópodos en la transmisión de la LBE y especialmente el de los tabánidos se manifiesta cada vez más claramente (47).
- Transmisión iatrógena
La posibilidad de transmisión de linfocitos infectados de bovino a bovino en ocasión de las tomas de muestras o de inyecciones múltiples con una misma aguja ha sido sospechada desde hace t iempo (95). De esta manera, una cantidad de sangre residual en la luz de la aguja es suficiente para reproducir la infección (29). Ciertas prácticas veterinarias con intenciones profilácticas, el empleo de jeringas, de agujas (70), e incluso a veces de instrumentos quirúrgicos, de un animal infectado a otro , pueden ser culpadas, incluso aunque algunos ensayos no hayan permitido demostrar la transmisión del virus (94).
El papel del tatuaje ha sido claramente puesto en evidencia (59, 42, 37).
En resumen, se puede concluir, como indicaban Burridge y Thurmond en 1981 (8) que:
- A l r e d e d o r del 5 % de terneros nacidos de madre infectada, se infectan in útero.
- La transmisión del virus por ingestión de calostro o de leche parece limitada.
- La gran mayoría de los bovinos se infectan por contacto con los animales ya infectados, según modalidades diferentes: promiscuidad, insectos picadores, intervenciones del veterinario sin precauciones, etc.
D I A G N Ó S T I C O Y D E T E C C I Ó N
Pasaremos en silencio los diferentes exámenes de laboratorio distintos de los serológicos ya que ellos son muy clásicos (examen histopatológico), o son poco utilizados (examen hematológico) desde la puesta a punto de técnicas serológicas (investigación del virus) (informe de Italia a la OIE, 1989).
La detección de la LBE es exclusivamente serológica y se encamina a las tomas de muestras de sangre o de leche, individuales o en forma de mezclas.

1091
Bases
Después de la infección, el plazo de seroconversión varía en una media de 2 a 8 semanas (53) pero puede a veces sobrepasar los 3 meses (20). Los anticuerpos séricos persisten durante toda la vida económica de los animales, pero su título sufre fluctuaciones en particular al final de la gestación o al comienzo de la lactación o bien puede bajar de forma importante hasta pasar por debajo del nivel mínimo de detección (49, 5).
Po r otra parte, de manera excepcional, parece que algunos bovinos nacidos de madres infectadas, mantenidos en estricto aislamiento, pueden no dar una respuesta positiva más que después de varios meses, e incluso 3 años después de la desaparición de los anticuerpos de origen materno (83). Estos hechos, que son rarísimos, no deben pues hacer dudar de la noción de base que consiste en considerar que el plazo máximo de aparición de una tasa detectable de anticuerpos es del orden de 3 meses.
E n el ternero nacido de madre infectada, la desaparición de los anticuerpos de origen materno, no es efectiva más que después de 3 a 7 meses (2, 9, 21 , 84, 58).
Técnicas
Actualmente las dos técnicas mayores de diagnóstico y de la detección serológica de la LBE son la inmunodifusión en gelosa IDG y la prueba ELISA.
La inmunodifusión en gelosa
Es ampliamente usada. La técnica ha sido codificada en algunos países (por ejemplo en la CEE) y los estuches están disponibles en el comercio. Las modalidades de control de los reactivos han sido precisadas (anexo G de la Directiva 88 /406 /CEE del 14 de junio 1988).
Esta técnica es utilizable exclusivamente para los sueros (individuales o en forma de pequeñas mezclas). Se t rata de una prueba simple de realizar, específica y sensible. N o obstante debe ser leída por un técnico entrenado y se presta mal al t ratamiento y a la lectura de un gran número de sueros.
La prueba ELISA
Es cada vez más utilizada para el descubrimiento serológico de la LBE (1 , 4, 19, 26, 48, 63, 85, 86, 45, 88, 28, 25, 62). Los pocilios o estuches están disponibles en el comercio y llevan diversas modalidades técnicas (ELISA directo o por competición) recurriendo o no a los anticuerpos monoclonales (22).
La prueba ELISA puede aplicarse a los sueros o a las leches (individuales o en forma de mezclas). Dentro de las diversidades de las técnicas comercializadas, cada unidad es acompañada por los protocolos de control de los estuches que permiten garantizar niveles satisfactorios de sensibilidad y de especifidad (17). Así, en el seno de la CEE, el nivel mínimo exigido como nivel de sensibilidad o detectabilidad de los anticuerpos es análogo al de la inmunodifusión en gelosa para los sueros y se establece en función de un suero de referencia (suero E4 al 1/10°).
Pa ra la leche, un nivel mínimo análogo se exige, destinado a permitir obtener al menos las mismas prestaciones que las resultantes del análisis por IDG del suero del animal del que se ha estudiado la leche en ELISA.

1092
Esta exigencia está basada en un razonamiento que admite que en términos medios el título en anticuerpos de la LBE es 25 veces más débil en la leche que en el suero de la misma vaca [10 veces más bajo para Florent y col. (22); 27 veces más bajo para Mammerickx y col. (46); 26 veces más débil, informe de Dinamarca a la OIE, 1989]. Se t rata por supuesto de una media ya que los diferentes estudios (87, 14) han demostrado perfectamente las fluctuaciones de los títulos de anticuerpos de la LBE durante la lactación.
Pa ra una leche individual, la prueba ELISA debe pues revelar al mínimo la cantidad de anticuerpos contenidos en el suero E4 diluido al 1/250 (dilución 25 veces más fuerte que el 1/10).
Pa ra las leches de mezcla, el nivel de detectabilidad exigible es el mismo, lo que implica tener en cuenta el número de vacas de las que proviene la mezcla de leches o recurrir a una técnica de concentración del lactosuero. Pa ra Forschner y col. (25), un pocilio ELISA comercializado permite desde ahora obtener una respuesta positiva sobre una leche de mezcla de 100 vacas, de las que sólo una está infectada.
Las ventajas de la prueba ELISA residen en su rapidez de ejecución referida a un número elevado de muestras que se pueden estudiar en una sesión, su lectura objetiva y sus posibilidades de automatización. Un control sistemático de los lotes de cada productor permite obtener una sensibilidad y una especificidad satisfactorias (17) y, en estas condiciones, la prueba ELISA se muestra como un sistema económico y eficaz para la detección sistemática de la infección de los rebaños lecheros, así como para el control de rutina del estado satisfactorio de los rebaños sanos o saneados (15, 16).
La interpretación
La interpretación de los resultados serológicos obtenidos sobre el suero de un animal debe tener en cuenta la edad del animal, la fecha del último contacto con un animal infectado y el estatus serológico de la madre en el caso de los terneros jóvenes. Se resume en el Cuadro V.
A nivel individual se tendrá en cuenta que todo bovino de más de 7 meses con una serología positiva, debe considerársele como infectado y por lo tanto como una fuente de contaminación potencial. No se puede dar fiabilidad a una respuesta negativa obtenida al final de la gestación o en el comienzo de la lactación en una vaca que pertenezca a una ganadería infectada.
A nivel de una ganadería, se puede recordar la siguiente regla: un rebaño no se le puede considerar como indemne de LBE desde el punto de vista médico, más que si dos exáménes serológicos del conjunto de los animales de edad superior a los 7 meses, practicados con más de tres meses de intervalo, se han mostrado negativos, en ausencia de posibilidad de contaminación durante este periodo.
P R O F I L A X I S S A N I T A R I A
Siendo la LBE una enfermedad infecciosa provocada por un virus que se transmite, de manera esencial, por contacto entre un animal infectado y un animal sano, es posible combatir la extensión de esta enfermedad mediante medidas estrictamente sanitarias.

1093
C U A D R O V
Interpretación de los resultados serológicos de diagnóstico individual de la LBE
Se pueden distinguir las medidas a establecer a nivel de un rebaño, a proteger o sanear, y en plan regional o nacional.
Lucha a nivel de un rebaño
Protección de un rebaño sano
a) Introducción de animales en el rebaño
La medida más evidente y la más eficaz es sin duda el evitar la introducción de un animal infectado en el seno del rebaño.
Durante la puesta en cuarentena de un animal comprado, una petición de diagnóstico serológico es por lo tanto necesaria: para los animales comprados en ganaderías indemnes, es suficiente un solo examen serológico. Po r lo contrario, para los animales procedentes de rebaños infectados o de situación sanitaria desconocida, será imperativo el observar una cuarentena de 3 meses practicando un control serológico al comienzo y al fin de la misma. Una duración de cuarentena tan larga suele ser difícilmente realizable en la práctica, por lo que es deseable el no introducir en los rebaños indemnes más que animales procedentes de ganaderías sanas.
Bovino Resultado serológico
Ultimo contacto con un animal
infectado Interpretación
< 7 meses
+
• Si nace de madre infectada: distinción con anticuerpos de origen calostral imposible; recomenzar el examen después de los 7 meses de edad
• Si nace de madre indemne o ausencia de toma de calostro: animal infectado
-
< 3 meses
> 3 meses
• Repetir un examen más de 3 meses después del último contacto con un animal infectado
• Animal sano
+ Animal infectado
> 7 meses -
< 3 meses
> 3 meses
• Repetir un examen más de 3 meses después del último contacto con un animal infectado
• Animal sano

1094
b) Material de los veterinarios
Debe llamarse la atención de los veterinarios sobre la necesidad absoluta de cambiar sistemáticamente de aguja en su material de toma de sangre o en la intravenosa entre dos explotaciones.
c) Pastoreo de los animales
La cohabitación de ganados sanos e infectados en los pastos comunales puede considerarse como un factor de riesgo en las regiones en las que los tábanos o, en menor importancia, los mosquitos, son numerosos. Por analogía con la anemia infecciosa de los équidos, una distancia de unos 50 metros entre el rebaño infectado y el rebaño sano limitará fuertemente los riesgos de transmisión por los ar t rópodos.
La periódica verificación de la ausencia de infección del rebaño, será deseable. E n los rebaños lecheros ésta puede asegurarse simplemente por la prueba ELISA aplicada regularmente a la leche de mezcla de la explotación.
E n los rebaños lactantes, la detección puede hacerse anualmente sobre mezclas de sueros tomados para otras pruebas profilácticas, como por ejemplo, de la brucelosis bovina.
Saneamiento de rebaños infectados
Las medidas a tomar se relacionan con dos categorías:
- Por una parte, la utilización de algunas reglas profilácticas simples que permitan limitar la diseminación del virus en el seno del rebaño.
- Por otra, la puesta en marcha de un plan de lucha con vistas a la erradicación de la infección.
a) Las reglas profilácticas
- A g u j a s e instrumentos de los veterinarios
Las reglas profilácticas t ra tan sobre todo de evitar toda clase de transporte de sangre de un animal infectado a otro: es por lo tanto indispensable, cuando se realize una toma de sangre o una inyección intravenosa, el utilizar agujas de uso único. Los datos experimentales son suficientemente sólidos para que la responsabilidad del veterinario clínico pueda ser comprometida si una tal precaución no se ha respetado.
Por lo mismo, la desinfección de todos los instrumentos quirúrgicos utilizados «en serie» es necesaria (descornado, castración) y el cambio entre cada animal de los guantes de plástico utilizados en las exploraciones rectales, es una buena precaución. Di Giacomo y col. (12) han demostrado la utilidad del respeto de precauciones durante el descornado y han obtenido una disminución de la transmisión del virus. Ruppaner y col. (78) han resumido las precauciones a tener en cuenta para evitar la transmisión de sangre de un animal a ot ro:

1095
Precauciones a respetar para evitar la transmisión de sangre de un animal a otro
- Ar t rópodos
Una atención particular debe ponerse en la lucha contra los ar t rópodos picadores. Existen, en particular, unas plaquitas auriculares de efecto repelente que son eficaces durante varios meses y cuyo empleo se puede aconsejar.
- Calostros
La toma calostral, en los terneros nacidos de madre infectada, no presenta riesgos mayores frente al beneficio para el animal ya que representa para éste la adquisición de una buena inmunidad pasiva. No obstante, cuando ello sea posible, es deseable la pasteurización del calostro ya que preserva la actividad de las inmunoglobulinas, suprimiendo en cambio la infecciosidad viral (73, 77). Se puede también utilizar el calostro de vacas indemnes.
b) Plan de lucha
Hay dos tipos de planes de lucha que pueden permitir la erradicación de la infección en un rebaño: la eliminación por matanza de los bovinos infectados, o la puesta en marcha de un plan de separación total de los bovinos infectados en establos independientes.
- Eliminación de los bovinos infectados
Podr ía proponerse el siguiente modelo: control serológico de todos los bovinos de más de 7 meses, cada 3 a 6 meses; aislamiento y sacrificio de los animales con serología positiva; el rebaño se considera como saneado cuando todos los animales que hayan eliminado los anticuerpos calostrales propios, han dado una respuesta serológica negativa 2 veces con 6 meses de intervalo.
Acciones Precauciones
Intervención con la Los instrumentos deben ser limpiados y lavados con agua ayuda de instrumentos caliente, sumergidos en lejía y después en agua de javel
Tatuaje y colocación Los instrumentos deben ser desinfectados entre cada de marcas en la oreja ternero
Descornado Sólo 2 a 3 animales deben descornarse al mismo tiempo, y después deben mantenerse separados para evitar que se froten uno contra otro. La hemostasia debe asegurarse y la herida tratada con un desinfectante y un repelente para los artrópodos
Ablación del pezón Es necesario emplear una hoja de bisturí diferente para cada animal. La hemostasia debe ser correcta
Vacunación y Es preciso cambiar de aguja para cada animal detección global para las tomas de sangre, las vacunaciones y las
tuberculinizaciones
Exploración rectal Es necesario cambiar de guantes de plástico para cada vaca

1096
Este método da excelentes resultados. Su eficacia es menor cuando un número de fallos determinado se produce en el protocolo: falta de identificación de los animales, sustitución de animales, introducción de animales infectados, tomas de sangre en serie con la misma aguja, etc. Por esto es necesaria una perfecta coordinación entre los 4 niveles implicados en el proceso de lucha: la explotación ganadera, el veterinario clínico, los laboratorios de diagnóstico y el Servicio Veterinario oficial (43).
Por otra parte, el éxito de las medidas de lucha está disminuido también cuando la tasa incial de animales infectados y / o el porcentaje de animales jóvenes infectados son elevados.
La mayor dificultad de saneamiento de los rebaños fuertemente infectados, como se conoce bien para la brucelosis y la tuberculosis bovinas, justifica plenamente el sacrificio de entrada de la totalidad de los animales del rebaño.
Este tipo de profilaxia, aunque tiene la ventaja de ser eficaz (80), sencillo de poner en marcha, y aplicable a todos los tipos de explotaciones, sigue siendo por lo menos de un coste prohibitivo para el ganadero si no hay una política de subvenciones para los sacrificios. Conduce efectivamente a eliminar los animales cuyo porvenir económico no habría sido más que raramente comprometido por la aparición de una forma tumoral .
Por consiguiente, este modelo de profilaxis puede difícilmente ser aplicado por un criador a título individual (al menos en los casos de rebaños muy infectados), pero encuentra plenamente su indicación en el cuadro de una acción colectiva.
- Separación de los animales infectados
La creación de un rebaño sano a partir de unos efectivos infectados ha sido estudiada y ha tenido éxito (82) aislando los terneros indemnes del resto de rebaño.
Este tipo de profilaxis, con el inconveniente importante de poder disponer de locales adecuados y de extremar el rigor en el manejo del rebaño, puede ya permitir a un ganadero motivado, el eliminar la infección leucósica de su ganadería. Es preciso, en este caso, disponer de técnicas serológicas muy sensibles (o de repetir los exámenes), de forma que se evite la introducción en la ganadería sana de un ternero considerado como indemne mientras él se encuentre al comienzo de la seroconversión.
Lucha a escala regional o nacional
De manera general, el saneamiento de un país frente a una enfermedad de incubación larga como la LBE (o la tuberculosis) no se puede hacer más que teniendo en cuenta el conjunto de animales infectados, potencialmente fuentes de contaminación durante un periodo largo, sin limitarse a ver sólo la parte emergente del iceberg que representan los focos donde se manifiesta la enfermedad. Esto es tanto más cierto en el caso de la LBE donde muchos animales infectados pueden llegar al final de su vida económica sin exteriorizar los síntomas de LBE (o sin que sean reconocidos como infectados).
Cada país debe de definir su política de lucha en función de la situación epidemiológica y de los objetivos a alcanzar.
Pa ra algunos países, el objetivo puede limitarse a la exportación de reproductores, sin contemplar por lo tanto la erradicación de la infección, particularmente por razón del elevado costo de un tal programa.

1097
Pa ra otros países, el objetivo de erradicación puede ser contemplado, con un calendario de lucha y con etapas variables en función de la situación epidemiológica y de los recursos de todo orden a dedicar a la lucha contra esta enfermedad. N o existe ningún programa valedero para todo . Cada país debe definir su programa, teniendo en cuenta los criterios señalados anteriormente y a la luz de las informaciones suministradas por los países que ya han llegado a la situación de erradicación.
La puesta a punto de pruebas ELISA de gran sensibilidad (pero conservando una especificidad satisfactoria) aplicables a las mezclas de muestras (leches o sueros) ha sido un decisivo progreso para una realización económica de las primeras etapas de la investigación de la infección en un país y de la ulterior vigilancia de los rebaños sanos. El saneamiento de los rebaños infectados pasa obligadamente por un diagnóstico individual.
La experiencia de los países europeos que han comenzado la lucha hace varios años, muestra que la aplicación de las medidas sanitarias clásicas permite hacer desaparecer la enfermedad. El plazo para obtener la erradicación es tan to más largo cuanto que la tasa de infección inicial de los rebaños sea elevada y que las medidas aplicadas sean incompletas.
Cuando las medidas son empleadas correctamente y la tasa de infección de los animales no es muy elevada, el saneamiento de los rebaños infectados se obtiene rápidamente.
El número de países que han consagrado sus esfuerzos y sumas importantes a la lucha contra la LBE aumentará progresivamente. Estos países pedirán un respeto en la aplicación de las normas que permitan la garantía de la ausencia de riesgo de recontaminación de sus ganados a partir de animales importados, conforme al Código Zoosanitario Internacional de la OIE.
LA ANEMIA INFECCIOSA DE LOS ÉQUIDOS
La anemia infecciosa de los équidos (AIE) es una virosis propia de los équidos, de evolución lo más corrientemente crónica con episodios agudos, que se traduce por fiebre, anemia, abatimiento y edemas.
Esta enfermedad se caracteriza por la perennidad de la viremia durante años, la formación de lesiones de origen inmunológico y la plasticidad antigénica aparente de sus virus.
Señalada por primera vez en Francia, en 1843, por Lignée, fue estudiada en el curso de la segunda mitad del siglo XIX y los trabajos de Vallée y Carré (143) llegaron a la posibilidad del aislamiento del virus y al conocimiento de las modalidades del contagio. Posteriormente, la AIE ha sido identificada sucesivamente en la mayor parte de los países y se la encuentra actualmente con una frecuencia y una gravedad variables en numerosos países de Africa, América, Asia, Australia y en Europa .

1098
V I R O L O G Í A
El virus de la AIE ha sido mal conocido durante mucho t iempo. Este relativo escaso conocimiento fue debido a la ausencia prolongada de un sistema práctico de cultivo in vivo o en cultivo celular; por otra parte , las primeras células que han permitido la multiplicación del virus en cultivo, los leucocitos de caballo, comportan muchas dificultades técnicas de cultivo (como la necesidad de una fuerte concentración de suero en el medio de cultivo, 50% o más , así como la frecuente contaminación por los citomegalovirus) y el cultivo sobre un sustrato celular cómodo, la línea Equina Dermis, no se ha puesto a punto más que muy tardíamente (121).
Los caracteres generales del virus de la AIE han sido objeto de una revisión reciente (112). Posee los caracteres estructurales de los lentivirus y brota a partir de las membranas citoplásmicas. Su transcriptasa reverse es M g + + dependiente.
Su composición proteica está bien determinada actualmente. El complejo ribonucleoproteína comprende la proteína básica p l 1. Las proteínas internas son p9, p l 5 (proteína fosforilada) y p26. La envoltura está compuesta de glucoproteínas gp90 y gp45 (107, 123, 124) (Figura 2).
FlG. 2
Representación esquemática del virus de la AIE (107, 123, 124)
La organización genética del virus de la AIE es comparable a la de otros retrovirus (Figura 3). El A R N viral es copiado en A D N por la transcriptasa reverse. Está flanqueado por dos regiones terminales repetidas (Long Terminal Repeats) conteniendo las señales de regulación de la expresión de los genes. El gene G A G

1099
codificado por las proteínas internas y comprendiendo 1 458 bases. Las proteínas derivan de un precursor único que es una poliproteína ordenada de la siguiente manera: pl5-p26-pentapéptido-pl l -p9.
FiG. 3
Organización genética del virus de la AIE (113)
El gene P O L es codificado por la transcriptasa reverse. Se lee en un cuadro de lectura diferente del gene G A G y le recubre sobre 251 bases. Por otra parte , el gene ENV (2 577 bases) es codificado por las glucoproteínas de envoltura. N o recubre el gene P O L (114, 139, 131, 102, 127). El genoma del virus posee también tres cuadros cortos de lectura S1, S2, y S3. SI y S2 están situados en la región intergénica POL-ENV y S3 en el gene ENV (113, 139). El producto de expresión de S2 activa la expresión del genoma del virus actuando sobre una secuencia blanco situada entre las posiciones -31 y + 2 2 en relación con el lugar de iniciación de la transcripción de los A R N mensajeros virales (103).
El virus se cultiva por siembra en cultivo de macrófagos de caballos infectados o infectando los macrófagos de caballos sanos. En estas células, el virus de la AIE posee un efecto citopatógeno que se manifiesta por la destrucción de las células. Este procedimiento sigue siendo difícil de emplear ya que es necesario poseer una fuente de macrófagos y de suero exentos de citomegalovirus. Las células de la línea continua Equine Dermis aseguran igualmente una replicación del virus de la AIE sin presentar lesiones y pueden quedar infectadas crónicamente en el curso de pasadas sucesivas (121). La línea de t imo de perro Cf2Th presentaría las mismas características (97).
Las propiedades antigénicas del virus dependen de numerosos grupos de antígenos con propiedades diferentes. Las proteínas internas p26 y p l 5 inducen la formación de anticuerpos revelables por las técnicas de fijación de complemento, de inmunofluorescencia y sobre todo de inmunodifusión en gelosa. Estas proteínas son estables de una cepa viral a otra y constituyen por lo tan to los antígenos utilizados en el diagnóstico. Por otra parte, p26 posee una comunidad antigénica con la proteína homóloga del virus HIV y de los lentivirus ovinos y caprinos. La proteína p l 5 posee igualmente una homología antigénica con la proteína p l 7 del virus HIV (122, 145). Las glucoproteínas de envoltura (gp90 y gp45) intervienen en la reacción de neutralización. Kono y col. (119) habían mostrado que en el curso de la infección en un caballo, los antígenos periféricos del virión sufrían una modificación antigénica que suponía la aparición de variantes antigénicas a las que el organismo se adapta produciendo, con cierto desfase, anticuerpos que neutralizan la nueva especificidad. Este fenómeno permite el comprender la coexistencia casi constante, en la sangre de

1100
un caballo infectado, de virus y de anticuerpos anti virus AIE . Los mecanismos de este desfase antigénico son cada vez más estudiados y los principales resultados serán presentados en las líneas siguientes.
La infección experimental de un caballo lleva a la aparición de crisis cíclicas hipertermizantes. La viremia es máxima al momento de las crisis, y en general indetectable por inoculación en cultivos celulares entre las crisis. Existe un plazo variable entre el pico de la viremia correspondiente y la síntesis de anticuerpos que neutralicen una variante. Po r regla general, no existe ninguna neutralización cruzada entre las diferentes variantes. Las gp90 y / o gp45 de cada variante presentan modificaciones estructurales, detectadas por una movilidad diferente en electroforesis. Por lo contrario, la proteína p26 no se modifica. El análisis de los péptidos y de los glucopéptidos trípsicos de cada variante muestra varios hechos. Las modificaciones no se suman para las variantes de aparición sucesiva en un mismo caballo; las cartas glucopeptídicas de gp90 y gp45 muestran que existe para cada glucoproteína dos tipos de glucosilación diferentes y que algunas variantes presentan pues la misma carta; las cartas peptídicas de p26, p l 5 y p9 son idénticas en todas las variantes (132).
El análisis de los epitopos de gp90 y de gp45 de varios aislados consecutivos derivados de una misma cepa inicial, con la ayuda de anticuerpos monoclonales, ha sido realizado (106). Dos epitopos se conservan en todas las cepas y se les reconoce por los anticuerpos monoclonales no neutralizantes. Los otros epitopos son de expresión variable (en particular todos aquellos reconocidos por los anticuerpos monoclonales neutralizantes). Prácticamente todos los aislados son diferentes desde el criterio del perfil de epitopos expresados. Los aislados idénticos sobre este criterio difieren sobre la base del examen de sus péptidos y glucopéptidos trípsicos o de sus cartas oligonucleotídicas. Algunos epitopos neutralizantes en la cepa inicial sufren alteraciones ligeras entre variantes que, aunque no impiden la fijación del anticuerpo monoclonal correspondiente, no le permiten realizar la neutralización de la variante.
Recientemente, se han apor tado informaciones sobre las modificaciones del gene ENV y sobre el estudio de sus correlaciones con las modificaciones de estructura antigénica (128). Los subfragmentos del gene ENV se han expresado bajo la forma de proteína de fusión en Escherichia coli. El estudio en Western Blot de estos polipéptidos con los anticuerpos monoclonales descritos precedentemente ha permitido localizar aproximadamente las secuencias que codifican estos epitopos. Algunos de estos epitopos que corresponden a los anticuerpos no neutralizantes son codificados por regiones constantes, y se revelan como conservados entre las diferentes cepas. Los epitopos reconocidos por los anticuerpos neutralizantes se conservan poco y son codificados por las regiones variables del gene. Se han obtenido resultados superponibles con los sueros de caballos experimental o naturalmente infectados. Un epitopo en la parte COOH-terminal de la gp45, es reconocido fuertemente por todos los sueros, mientras que los epitopos codificados por la parte variable de la proteína suministran respuestas diversas según los sueros. Un epitopo en la parte NH 2 - terminal de la gp90 es reconocido débilmente. En resumen, los epitopos conservados (no neutralizantes) se sitúan en las partes NH2 y COOH-terminales de la proteína codificada por el gene ENV. Los epitopos variables (neutralizantes) son codificados por la parte variable del gene ENV. Resultados comparables se han descrito para el virus HIV.

1101
L A E N F E R M E D A D
La duración de la incubación puede variar de 5 a 7 dias hasta más de tres meses. La forma más clásica de la enfermedad corresponde a una sucesión de crisis hipertermizantes durante las cuales la viremia es máxima. En el curso de estas crisis, el animal presenta una fiebre intensa, una anorexia, una anemia que pueden estar acompañadas de signos inconstantes (síndrome hepatorenal, síndrome gastrointestinal, m i o c a r d i t i s , men ing i t i s ) . Las crisis d e s a p a r e c e n g e n e r a l m e n t e al c a b o aproximadamente de un año, quedando el animal portador asintomático de virus (99). Según los casos, la evolución clínica puede ser aguda, subaguda o crónica. Por último, existen formas inaparentes, bien sea de golpe, bien después de una forma clínica expresada. Las lesiones macroscópicas son variables según la forma clínica: hepatomegalia y esplenomegalia, miocarditis, hemorragias (petequias, sufusiones) sobre las serosas y las mucosas. Las lesiones microscópicas consisten en una proliferación de células linfoides y infiltración de los diferentes órganos (bazo, hígado...), así como en una acumulación de sideroleucocitos (macrófagos conteniendo hemosiderina, catabolito de la hemoglobina).
P A T O G E N I A
La patogenia de la AIE es cada vez mejor conocida.
- L a anemia es consecuencia de dos mecanismos: por una parte , una hemólisis de naturaleza verosímilmente inmunológica, intra y extravascular, y por otra parte , una disminución del funcionamiento de la médula ósea.
Sentsui y Kono (133, 134) han demostrado que la hemólisis resulta verosímilmente de la adherencia de la hemaglutinina viral a los glóbulos rojos: estos son entonces rápidamente fagocitados (sideroleucocitos) e igualmente hemolizados bajo la acción del complemento. Esta hemólisis se acelera en presencia de anticuerpos dirigidos contra el virus de la A I E .
- E n la glomerulitis de los caballos afectados de una forma aguda de la AIE , se pueden poner en evidencia las IgG anti AIE y del C 3 . Esto sugeriría que esta lesión es debida al depósito en el r iñón de los complejos circulantes virus-anticuerpos. McGuire y col. (120) han señalado la presencia de estos complejos circulantes y han mostrado que el 99% del virus infeccioso presente en el suero se encuentra bajo la forma de tales complejos. Paralelamente a la fijación de C3 sobre los hematíes y los glomérulos se nota una disminución de la tasa de C 3 circulante en los caballos enfermos.
Hemos indicado en el capítulo de virología que las modificaciones del gene ENV eran identificables en las diferentes variantes antigénicas de aparición sucesiva en un caballo infectado.
Varias hipótesis pueden justificar las modificaciones de la secuencia del gene ENV. La más probable corresponde a la aparición de mutaciones puntuales durante la replicación por una falta de fidelidad de la transcriptasa reverse. La selección de nuevas variantes necesita que la respuesta inmunitaria del hospedador sea funcional (129)

1102
y ha sido corrientemente explicada por la presencia de anticuerpos que neutralizan las variantes preexistentes. Se ha podido, no obstante, mostrar que muchos ciclos de fiebre y de viremia, asociados al aislamiento de variantes diferentes, existen incluso antes de la aparición de anticuerpos neutralizantes. Los sueros precoces correspondientes pueden distinguir estos aislados en inmunofluorescencia de membrana frente a las células infectadas. Estos resultados sugieren que la variación antigénica resulta igualmente del reconocimiento y de la destrucción de las células infectadas por ciertas variantes y no únicamente de la presión de selección correspondiente a la existencia de anticuerpos neutralizantes (118, 119, 98).
E P I D E M I O L O G Í A
Epidemiología descriptiva
En muchos países, la incidencia anual de los casos clínicamente manifiestos así como las serologías positivas, está en regresión. El Cuadro VI presenta las informaciones que han sido aportadas por los diferentes países. Existen generalmente claras fluctuaciones estacionarias de la incidencia, correspondientes a la estación de actividad máxima de los ar t rópodos. La distribución en el espacio de la enfermedad varía según los países. Normalmente la AIE es más frecuente en las regiones cálidas y húmedas.
La enfermedad no tiene tendencia a una difusión rápida, pero los casos pueden observarse a gran distancia de un foco inicial.
Epidemiología analítica
Las fuentes del virus están representadas esencialmente por los équidos infectados. Los animales enfermos, durante las crisis hipertermizantes, representan un peligro importante ya que su viremia es máxima y puede estimarse aproximadamente en unos 10 6 /ml (110, 132). Entre las crisis, el título viral es habitualmente muy bajo pero puede permanecer elevado en algunos animales. En los caballos infectados de manera inaparente, sin historia de síntomas declarados, el título viral es habitualmente muy bajo, indetectable incluso por inoculación de 300 mi de sangre a un caballo sano (113). Algunas secreciones y excreciones (leche, calostro, moco. . . ) son también virulentas en el momento de las crisis y pueden jugar un papel en las modalidades de transmisión directa (119).
El modo habitual de transmisión del virus de la AIE a partir de la fuente virulenta esencial, la sangre, está representado por la picadura de artrópodos hematófagos o de las agujas utilizadas en las inyecciones. Los conocimientos actuales referentes a la transmisión del virus por los artrópodos han sido revisados recientemente por Issel y col. (113).
Los ar t rópodos responsables pertenecen a los géneros Tabanus, Stomoxys, Chrysops e Hybomitra. La transmisión es exclusivamente mecánica, ya que el virus de la AIE no se multiplica en el ar t rópodo (137). En la eficacia de la transmisión intervienen diferentes variables. En particular, es preponderante el estado del caballo donante . En un caballo en fase aguda de la enfermedad, ha podido ser lograda la transmisión a otro caballo por intermedio de un solo artrópodo (105). Por lo contrario,

1103
C U A D R O V I
Situación epidemiológica de la AIE y medidas de lucha en los diferentes países que han informado a la OIE
País Epidemiología Medidas de lucha oficiales
Africa Argelia Indemne Enfermedad de declaración
Control a la importación obligatoria
Lesoto Indemne Departamento de Cría, Maseru (O.L. Letuka)
Sudafrica Indemne Departamento de Agricultura, Economía y Mercado, Servicios Veterinarios, Pretoria
Zambia Indemne Departamento de Servicios Veterinarios y de Control Tsetse, Lusaka (K.L. Samui)
América Canadá Enzoótica en el norte de Enfermedad de declaración Producción de alimentos Canadá, esporádica en las obligatoria. e inspección, otras regiones. Tasa de Programas voluntarios de Ottawa prevalencia anual de los erradicación. Compensación (N.G. Willis) caballos con serologia financiera por el sacrificio
positiva = 0,2% de los caballos infectados
Chile En 1981, 23 ganaderías A partir de 1981: Servicio Agrícola y infectadas de las 570 • Controles serológicos a la Ganadero controladas; 144 caballos venta, antes del transporte División Protección infectados entre 19 904 y antes de la introducción Pecuaria, controlados. en un efectivo Santiago A partir de 1986: casos • Controles serológicos cada
esporádicos 6 meses en los rebaños indemnes
• Saneamiento de los rebaños infectados por controles serológicos mensuales y sacrificio de los caballos con serologia positiva
• Importación de caballos después de dos controles serológicos con 1 mes de intervalo
Desde 1986: aligeramiento de las medidas y sólo control de los caballos de introducción en un rebaño

1104
C U A D R O V I (cont.)
País Epidemiología Medidas de lucha oficiales
Asia Myanmar Cría de ganado y Departamento Veterinario (T. Tint)
Se han descrito casos
Europa Alemania (Rep. Fed. de) Instituto de Virología Escuela Veterinaria, Hannóver (0 . Trayen, L. Haas, O.R. Kaaden) Checoslovaquia
Indemne
Indemne
Enfermedad de declaración obligatoria
Controles serológicos antes de la venta, los transportes, etc.
Irlanda Departamento de Agricultura y Alimentos Dublin (R.G. Cullen) Italia Ministerio de Sanidad Dirección General de Servicios Veterinarios División III
Luxemburgo Administración de los Servicios Veterinarios (J. Kremer) Suecia
Yugoslavia
2 500 controles serológicos por año. Ninguna serologia positiva desde 1975
Casos esporádicos (tasa de prevalencia de los caballos con serologia positiva en 1989 = 0,04%)
Indemne
Indemne
469 animales infectados en 1988
Control serológico de los caballos de carreras, de deporte ecuestre y de turismo ecuestre. Existencia de programas facultativos de erradicación en las ganaderías. Aislamiento y sacrificio de los caballos de serologia positiva Enfermedad de declaración obligatoria
Enfermedad de declaración obligatoria Controles serológicos regulares de los efectivos de caballos. Controles individuales en caso de agrupación de caballos. Sacrifico de los animales infectados. Controles a la importación
Oceania Australia Departamento de Agricultura (T.M. Ellis)
Enzoótica en el norte y el oeste de Queensland. Esporádica y no identificada en las otras regiones
Ningún programa de lucha oficial. Enfermedad de declaración obligatoria en ciertos estados

1105
en un caballo afebril, entre dos crisis, los fracasos de la transmisión experimental han sido numerosos (116, 101). Otra variable importante es la distancia que separa un caballo infectado de un caballo sano. En efecto, el a r t rópodo no puede jugar su papel de vector más que en el caso de que su comida sanguínea sobre un caballo infectado se interrumpa y que la continúe sobre un caballo sano, ya que la persistencia de la infectividad viral en el a r t rópodo no parece pasar de las 4 horas (105). Los trabajos de Foil (104) han most rado que el 99% de los tábanos interrumpidos en su comida sanguínea re tornaban sobre el mismo caballo cuando ningún otro caballo estaba presente en un radio de 50 metros. Así, una distancia de seguridad de unos 200 metros entre caballo sano y otros caballos parece ser suficiente para limitar el riesgo de transmisión por los ar t rópodos (113).
La transmisión del virus es igualmente posible por todo material manchado por la sangre de un animal infectado (agujas, material quirúrgico. . .) , el virus puede sobrevivir varios días sobre agujas contaminadas (144). Por últ imo, la transmisión in útero es posible (115) pero parece tener un impacto epidemiológico limitado (111).
D I A G N Ó S T I C O
En ausencia de técnicas de rutina utilizables que permitan el aislamiento del virus, las técnicas de diagnóstico tratan de poner en evidencia los anticuerpos postinfecciosos.
La puesta a punto de un test de inmunodifusión en gelosa, el test de Coggins (100), ha facilitado mucho la identificación de los caballos infectados. El antígeno utilizado ha sido primero producido a partir de extractos de bazo de caballos enfermos en fase aguda (126), y después a partir de virus producido en cultivos celulares de línea Equina Dermis (121). La proteína mayor interna p26 es la constituyente esencial del antígeno. El interés de su utilización ha sido posteriormente confirmado en razón de su estabilidad antigénica entre las distintas cepas virales (132). Inversamente, las glucoproteínas de envoltura son difícilmente utilizables, aunque el suero de los caballos infectados las reconoce fuertemente (125, 128), su variabilidad antigénica es importante. No obstante, la puesta en evidencia de un epitopo de la gp45 fuertemente reconocido por el suero de caballos infectados y conservado entre las diferentes cepas estudiadas (128) podría abrir nuevas perspectivas. La proteína p l 5 es también un constituyente menor de los antígenos utilizados en IDG y puede a veces dar una segunda línea de precipitado en la realización del test de Coggins (142). Las técnicas ELISA que ponen en evidencia los anticuerpos contra la p26 han sido también descritas (135, 138, 140) y algunas están actualmente comercializadas permitiendo obtener un resultado en unos quince minutos . Por últ imo, una técnica de Western Blot ha sido también desarrollada (130).
La prueba o test de Coggins es una técnica sencilla de realizar, sensible y específica. N o obstante, algunos sueros de escaso título en anticuerpos dan resultados de difícil lectura. Algunos caballos infectados suministran una respuesta en anticuerpos de muy bajo nivel (109, 141). La proporción de estos animales en relación con el conjunto de animales infectados es muy baja, inferior al 1% (113). Estas nociones no deben enmascarar que el test de Coggins representa una excelente técnica diagnóstica que permite identificar en destacadas condiciones de especificidad la casi totalidad de los caballos infectados.

1106
La técnica de ejecución de la prueba o test de Coggins está explicada in extenso en el Código Zoosanitario Internacional ( 5 a edición, 368-372) y en el Manual de métodos recomendados para el diagnóstico y los productos biológicos de la OIE (I, 16). Existe un suero internacional de referencia representando el nivel mínimo de detección que debe alcanzar todo laboratorio que pratique la prueba de Coggins, y que ha sido definido después de un estudio de sueros de concentración en anticuerpos variada realizado por diferentes laboratorios a través del mundo .
La interpretación de la prueba de Coggins está basada sobre la cinética de la evolución de los anticuerpos precipitantes que aparecen muy generalmente lo más tarde 2 meses después de la infección. Ellos están habitualmente presentes en el momento de la aparición de los síntomas, pero a veces pueden no aparecer más que en los días siguientes (como máximo 10 dias después del comienzo de los síntomas). Po r otra parte, la detección serológica puede estar dificultada en los potros nacidos de madre con serologia positiva en razón de una persistencia de anticuerpos calostrales que se eliminan a lo más tardar a la edad de seis meses. Sobre la base de estos datos, la interpretación de los resultados de la prueba de Coggins en las diferentes situaciones se resume en el Cuadro VII .
E n un animal enfermo, el recuento de los sideroleucocitos puede tener un buen valor pronóst ico. E n el caballo sano, esta tasa es inferior a 7 por 100 000 leucocitos, mientras que en los días que siguen a una crisis de AIE, la tasa puede ser de algunos cientos a 1 000 sideroleucocitos por 100 000 leucocitos.
P R O F I L A X I S
El Cuadro VI resume las informaciones enviadas por los diferentes países.
La protección de los países pasa por el control de los équidos importados, mediante un test serológico efectuado recientemente, y la procedencia de rebaños donde ningún caso de AIE se haya identificado en los 3 meses precedentes (Código Zoosanitario Internacional de la OIE, 5 a edición, 229-230).
La protección de las explotaciones indemnes necesita un control de los animales introducidos. Es posible introducir en ellos los équidos de serologia negativa procedentes de efectivos donde todos los équidos tengan igualmente una serologia negativa. En ausencia de conocimientos de la situación sanitaria del ganado de origen, será deseable no introducir más que los équidos con serología negativa, puestos en cuarentena en la explotación durante 40 a 65 días (plazo máximo de conversión serológica) y que den una respuesta negativa al final de este periodo.
El saneamiento de ganaderías infectadas necesita un aislamiento de los enfermos hasta su eliminación. Los caballos infectados de manera inaparente deben igualmente ser detectados por un test serológico y ser aislados antes de su eliminación. Este control serológico debe repetirse cada 30 a 45 días, con la aplicación de las mismas medidas en los animales de respuesta positiva. Los controles serológicos pueden detenerse cuando dos controles con 60 días de intervalo hayan dado un resultado negativo sobre la totalidad de los caballos. Los potros nacidos de yeguas infectadas deben ser considerados como potencialmente infectados hasta que un control realizado 2 meses después del destete resulte negativo. La utilización de jeringas y de agujas de uso
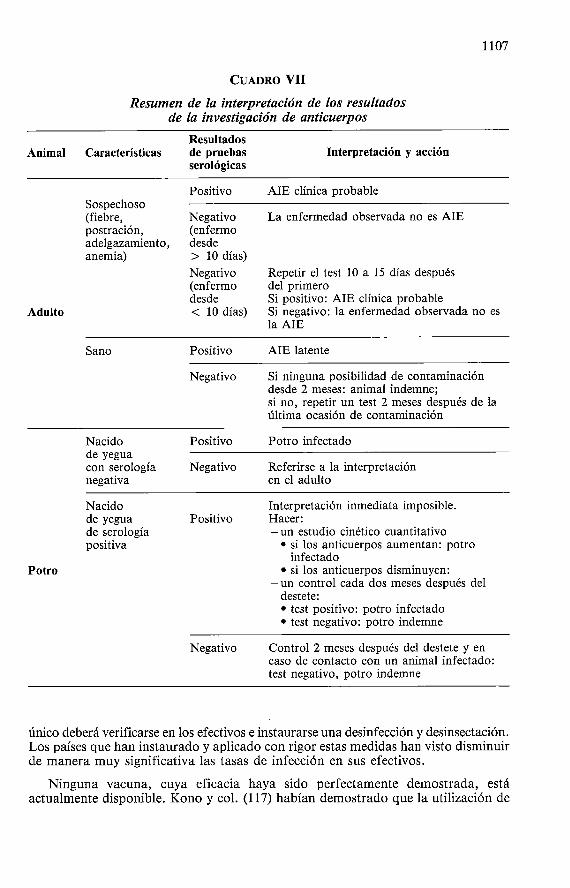
1107
C U A D R O VII
Resumen de la interpretación de los resultados de la investigación de anticuerpos
Animal Características Resultados de pruebas serológicas
Interpretación y acción
Sospechoso (fiebre, postración, adelgazamiento, anemia)
Positivo AIE clínica probable
Adulto
Sospechoso (fiebre, postración, adelgazamiento, anemia)
Negativo (enfermo desde > 10 días) Negativo (enfermo desde < 1 0 días)
La enfermedad observada no es AIE
Repetir el test 10 a 15 días después del primero Si positivo: AIE clínica probable Si negativo: la enfermedad observada no es la AIE
Sano Positivo
Negativo
AIE latente
Si ninguna posibilidad de contaminación desde 2 meses: animal indemne; si no, repetir un test 2 meses después de la última ocasión de contaminación
Nacido de yegua con serologia negativa
Positivo
Negativo
Potro infectado
Referirse a la interpretación en el adulto
Potro
Nacido de yegua de serologia positiva
Positivo
Negativo
Interpretación inmediata imposible. Hacer: - u n estudio cinético cuantitativo
• si los anticuerpos aumentan: potro infectado
• si los anticuerpos disminuyen: - u n control cada dos meses después del
destete: • test positivo: potro infectado • test negativo: potro indemne
Control 2 meses después del destete y en caso de contacto con un animal infectado: test negativo, potro indemne
único deberá verificarse en los efectivos e instaurarse una desinfección y desinsectación. Los países que han instaurado y aplicado con rigor estas medidas han visto disminuir de manera muy significativa las tasas de infección en sus efectivos.
Ninguna vacuna, cuya eficacia haya sido perfectamente demostrada, está actualmente disponible. Kono y col. (117) habían demostrado que la utilización de

1108
una cepa atenuada protegía contra una prueba virulenta por la cepa homóloga, pero no para una cepa heteróloga. Una vacuna con virus vivo se utiliza en China con resultados aparentemente favorables (136).
En conclusión, las medidas de lucha contra la AIE son actualmente bien conocidas. La eficacia depende del rigor con el que ellas se aplican, al menos en los países en los que la tasa de infección permite realizar el sacrificio de los caballos infectados.
LA ARTRITIS/ENCEFALITIS CAPRINA
La artritis/encefalitis caprina (AEC) es una enfermedad que existe desde hace muchos años, pero su estudio científico no había conocido un pleno desarrollo verdadero más que desde hace unos diez años (véase historia). Afección muy cosmopolita debida a un virus próximo al del SIDA del hombre , representa a la vez una dominante patológica de la crianza caprina y un modelo en patología comparada .
Se comprende pues fácilmente la multiplicidad de los trabajos que le han sido dedicados. En esta breve revisión, no es posible nombrarlos y censarlos todos . Pondremos el acento, pues, por una parte, sobre su presentación geográfica y su importancia, gracias a las informaciones enviadas por varios países para la Sesión General de Mayo de 1990 (Cuadro VIII), y por otra parte, sobre ciertos aspectos esenciales para su comprensión y su control (virología, patogenia, epidemiología, métodos de lucha).
D E F I N I C I Ó N - N O S O L O G Í A
Se t ra ta de una afección que se traduce clínicamente:
- E n el adulto, por artritis y periartritis simétricas (lo más corrientemente afectando los carpos o «rodillas», pero también los codos, más raramente los corvejones o otras articulaciones), las inflamaciones de las bolsas serosas (en particular bolsas atlanto-occipital y supra-espinal) y probablemente las esclerosis mamarias difusas provocando una disimetría mamaria . En la cabra primípara, la aparición rápida después del parto de una induración difusa de la mama («mama de madera») es también para ciertos autores una manifestación de la A E C .
- En el jóven entre 2 y 6 meses, por una ataxia posterior evolucionando hacia una cuadriplegía rápidamente mortal (leucoencefalomielitis).
Existen por otra parte afecciones mucho más raras (pulmonar de los jóvenes y de los adultos, nerviosa del adulto) asociadas o no a los precedentes (consúltese diagnóstico).

1109
C U A D R O VIII
Artritis/Encefalitis Caprina (AEC): Situación epidemiológica y medidas de lucha
en los diferentes países que han enviado una respuesta a la OIE
• Epidemiología Medidas de lucha oficiales Posibilidad de Posibilidad de plan
País Porcentage de infección calificación de erradicación Porcentage de infección oficialmente oficialmente controlada de controlado en los
Animal Rebaños los rebaños rebaños infectados
Africa Argelia CAEV - reconocimiento
de las cabras importadas
Lesoto AEC - ausencia Ministerio de Agricultura (O.L. Letuka)
Zambia AEC - ausencia Ministerio de Agricultura y cooperativas (K.L. Samui)
América Canadá Enzoótica No Agricultura Canadá 77% * Dirección General Producción e Inspección de alimentos (N.G. Willis)
Estados Unidos de 30% de las cabras de No No América origen europeo U.S.D.A. 81% globalmente * Servicio de Inspección de salud de animales y plantas (A.B. Thiermann)
Haití Reconocimiento Ministerio de de la AEC después Agricultura de recursos de la importación naturales y de desarrollo rural Servicio de salud animal (J.H. Jolivet Toussaint)
Asia Jordania Ministerio de Agricultura
0% 0%

1110
C U A D R O VIII (cont.)
Epidemiología Medidas de lucha oficiales Posibilidad de Posibilidad de plan
País Porcentage de infección calificación de erradicación Porcentage de infección oficialmente oficialmente controlada de controlado en los
Animal Rebaños los rebaños rebaños infectados
Europa Alemania (Rep. Fed. de) No No Instituto de Virología, 6% a 30% Hannóver (U. Truyen, L. Haas, O.R. Kaaden)
Francia Sí * Probable en el CNEVA/ Estación 65% Voluntariado de porvenir * Regional de patología los ganaderos Voluntariado de caprina los ganaderos (G. Perrin) CNEVA/ Laboratorio 77% * de Patología de Pequeños Rumiantes y de Abejas (P. Russo y C. Vitu)
Gran Bretaña Sí No * Ministerio de 4% 10% 1. Esquema de Agricultura, Pesca y 9,5% salud para Alimentación ovejas y Jefe del cabras (MAFF) Servicio Veterinario 2. British Goat (K.C. Meldrum) Society *
Irlanda Sí Tests a la Departamento de 0% 0% Voluntariado de importación y a Agricultura y los ganaderos la exportación Alimentos obligatorios (R.Q. Cullen)
Italia No No Facultad de Medicina Variable según las Veterinaria de Turin regiones y las razas. (E. Maglione, Las razas importadas P. Neblia, S. Rosati) parecen más
infectadas que las Facultad de Medicina razas locales Veterinaria de Bari (F. Marsilio, M. Tempesta, P. G. Tes car)
Luxemburgo Administración de los Servicios Veterinarios (J.P. Kremer)
CAEV serologia

1111
C U A D R O VIII (cont.)
Estas diferentes formas clínicas pertenecen a un mismo conjunto y pueden reproducirse (al menos para las formas articulares y nerviosas) por inoculación a los animales sensibles de un retrovirus designado con el nombre de virus de la Artritis/Encefalitis Caprina o Caprine/Arthri t is Encephalitis Virus (CAEV). Los órganos blancos son la sede de una intensa infiltración por las células mononucleadas (linfocitos, monocitos y macrófagos).
La A E C pertenece al grupo de las «enfermedades a virus lento» (Slow virus diseases) (207), con otras infecciones provocadas por los lentivirus (161, 197), como la maedi-visna de los ovinos.
La infección de un caprino por el CAEV no provoca sistemáticamente una enfermedad con síntomas y lesiones, aunque esta enfermedad persiste a lo largo de su vida. Se realiza lo más corrientemente durante los primeros meses de la vida y provoca una seroconversión frente al CAEV en algunas semanas. Un cierto número de animales infectados (del 5 % al 7 5 % , según los rebaños) presentarán después de un largo periodo de incubación (1 año a 6 o 7 años) una afección clínica que se traduce por la aparición de los signos precedentemente descritos en el adulto. Estos signos persistirán toda la vida del animal, conduciéndole normalmente a una degradación de su estado general, asociada con una disminución de la producción, que justifica su eliminación. La forma nerviosa observada en los jóvenes es mucho menos frecuente que la que afecta a los adultos. Es generalmente identificada en los rebaños muy infectados (más del 90% de adultos seropositives).
Epidemiología Medidas de lucha oficiales Posibilidad de Posibilidad de plan
País Porcentage de infección calificación de erradicación Porcentage de infección oficialmente oficialmente controlada de controlado en los
Animal Rebaños los rebaños rebaños infectados
Suecia No No Instituto Veterinario Nacional - Uppsala (A. Engvall, M. Wierup) Suiza Sí Sí Oficina Veterinaria 75% Desde 1983 Desde 1984 Federal Voluntariado de Voluntariado de (D. Riggenbach) los ganaderos los ganaderos
Oceania Australia Sí No Departamento de Según los Estados Según los Poco probable Agricultura del 6% a 44% 46% a 82% Estados en el porvenir Oeste Australiano de los de los rebaños Voluntariado (T.M. Ellis) animales lecheros de los
lecheros 0% a 7% de ganaderos 0% a 2% de los rebaños los Angora Angora
* Informaciones complementarias obtenidas de la bibliografía

1112
La fuerte prevalencia de la infección en los numerosos países así como sus consecuencias sobre la productividad de los rebaños, en particular, en la producción lechera, así como el obstáculo que representa para el comercio de los reproductores han generado numerosos estudios para su conocimiento y su control. Este está organizado en algunos Estados por las autoridades oficiales y las organizaciones representativas de los ganaderos.
H I S T O R I A
La presencia de «rodillas gruesas» asociadas o no a las cojeras y a una degradación del estado general en las cabras adultas ha sido desde hace tiempo señalado por los veterinarios y los criadores. Los estudios suizos (210) y japoneses (189) han descrito las afecciones totalmente comparables a la A E C en su forma articular en el adulto.
Linda Cork, al describir en los Estados Unidos una leucoencefalomielitis infecciosa de los cabritos (LEIC) (159), a priori sin relación con las observaciones anteriores, iba a relanzar el interés de la comunidad científica ya que la LEIC fue rápidamente identificada en otros países [para una revisión véase Knight y Jokinen (180); Al-Ani y Vestwer (151)]. El mismo equipo (160) identificó el virus responsable de la LEIC probando que era también responsable de una poliartritis crónica de los caprinos adultos enteramente comparable a las descripciones suiza y japonesa. Numerosos trabajos [para una revisión véase Robinson y Ellis (204)] iban a continuación a probar la existencia de la infección por el CAEV y la presencia de la A E C en numerosos países.
V I R O L O G Í A
El CAEV es un virus que pertenece a la familia de los retroviridae y a la subfamilia de los lentivirinae. Posee un cierto número de propiedades comunes a todos los lentivirinae, que comparte en particular con los virus maedi-visna (VVM) (192).
- S u genoma está constituido de A R N poliadenilo de polaridad positiva conteniendo de 9 000 a 10 000 pares de bases para un peso molecular de 5,5.10 6
daltons.
- 3 genes estructurales GAG, P O L y ENV codifican respectivamente para las proteínas internas de bajo peso molecular (p28, p l 9 , p l 6 y pl4) , la transcriptasa reverse y cuatro glucoproteínas de envoltura como la gp l35 .
- Existe una variabilidad antigénica de las glucoproteínas de envoltura de las cepas aisladas del terreno, revelada por el estudio de los anticuerpos dirigidos contra estas glucoproteínas en la seroneutralización (191). La aparición de variantes espontáneas en el curso del tiempo en una cabra infectada, ha sido también descrito (168).
Po r lo contrario los dos virus (CAEV y VVM) llevan en su genoma las secuencias nucleotídicas que permiten diferenciarlos (202, 201). La homología de las secuencias es del orden del 15 al 70% entre el CAEV y el VVM (171) cuando las condiciones de hibridación son muy específicas, ésta es más elevada (201) cuando lo son menos.

1113
- E l CAEV replica débilmente en las células de origen ovino, sin efecto citopatógeno marcado y posee un tropismo para los tejidos sinoviales. A la inversa, el VVM produce una infección lítica de las células de origen ovino y caprino (192).
- No ha sido posible poner en evidencia la diferencia entre las cepas aisladas de las cabras afectadas de artritis o de neumopat ía crónica por los diferentes criterios estudiados (análisis de las proteínas estructurales, de las propiedades antigénicas y de cultivo in vitro, etc.) (169).
E n medios extracelulares las partículas completas tienen una talla de 80 a 100 nm con un núcleo denso a los electrones y formando una banda homogénea en la ultracentrifugación isopícnica en sacarosa a 1,15 g m / c m 2 (156).
El virus se aisla generalmente a partir de explantes de la membrana sinovial de una articulación afectada (160, 193, 205), pero también a partir de otros tejidos (plexo coroideo, pulmón, mama, leucocitos) (163, 157) de animales enfermos. Se ha detectado un efecto citopatógeno al comienzo de uno a cinco pases y se traduce por la formación de sincitios en los cuales se distingue en el microcopio electrónico las partículas de núcleos densos y los fenómenos de protrusión de los viriones en la superficie de la membrana citoplásmica. El virus puede ser pasado en serie sobre células primarias (membrana sinovial, plexo coroideo, riñón, timo) obtenidas de cabritos recién nacidos antes de que hayan bebido el calostro (190).
Se puede señalar su relativa fragilidad en el medio exterior y el hecho de que queda completamente inactivado en los líquidos biológicos (leche y calostro) por un calentamiento a 56°C durante una hora (147).
P A T O G E N I A
El hecho esencial es la persistencia de una infección s u b c l í c a durante largos periodos (varios años en los adultos, algunos meses en los cabritos).
Lo más frecuentemente, el virus entra en el organismo por la vía digestiva y después alcanza a las células blanco que son de la línea monocito-macrófaga (161, 192).
Su replicación en la mayoría de las células no va más allá del estadio de provirus que se integra o es estrechamente asociado al A D N celular. La restricción concierne a la etapa de la transcripción de los A R N mensajeros.
El virus, adoptando una estrategia tipo caballo de Troya, se disemina por el organismo a través de los monocitos (174).
Los anticuerpos neutralizantes (no producidos o a un título muy bajo) no participan en absoluto en el control de la replicación viral.
El CAEV es reencontrado en el sistema nervioso central, las sinoviales, y también en el t imo, los nódulos linfáticos, la mama y los pulmones. Las lesiones tisulares son una consecuencia de la replicación viral, ella misma relacionada con la transformación de los monocitos en macrófagos. Una mediación inmunológica al menos parcial interviene en el desarrollo de las lesiones ya que las cabras inmunizadas con un CAEV inactivado y después infectadas por un CAEV salvaje, desarrollan más rápidamente

1114
lesiones más severas que los testigos no inmunizados (184); de la misma manera, De Martini y col. (164) han demostrado que las cabras infectadas por el CAEV tienen respuestas a los mitogenos de los linfocitos T más importantes que las de los animales no infectados.
Se demostrará seguramente en el futuro, como para los otros lentivirus (161) que los factores genéticos propios de la especie caprina vienen a modular el desarrollo de estas lesiones. Ya se ha demostrado la acción de ciertos factores externos del ambiente (187) o de las enfermedades intercurrentes [Ellis, en Smith y col., 1985 (209)].
El CAEV se distingue de otros lentivirinae porque no induce la formación de anticuerpos neutralizantes. Esta situación estaría relacionada con el modo de glucosilación de la glucoproteína de envoltura que podría disminuir la avidez de los anticuerpos o disimular los determinantes antigénicos. Po r lo contrario, induce la formación de anticuerpos frente a las proteínas internas, en particular la p28.
Las manifestaciones clínicas precoces observadas entre los cabritos podrían ser la manifestación de una infección in útero, asociada a una exposición postnatal a partir del calostro y de la leche (192).
E P I D E M I O L O G Í A
Epidemiología descriptiva
Extensión de la infección
Parece estar muy ampliamente extendida en los países desarrollados (149) (Cuadro VIII) , en particular en América del Norte (EEUU y Canadá) y en Europa : Francia (187), Noruega, Suiza, Gran Bretaña que se distingue por su baja tasa de infección (162). La infección parece extendida en Italia y en ciertas regiones españolas (172). Por lo contrario, la República de Ir landa y la Ir landa del Norte (146) parecen indemnes. En Africa, la infección parece ausente de Sudáfrica, de Somalia, de Sudán; en Kenia la prevalencia es muy baja (149) y esencialmente relacionada con la importación de cabras y machos cabríos en los programas de mejora genética (148). En Nigeria, la seroprevalencia está comprendida entre 0 y el 18% de los animales, según las regiones (153).
En Australia, la infección está ampliamente extendida en los rebaños lecheros y en cambio mucho menos en las cabras de Angora , Cachemira (155, 173); y es nula en las cabras vueltas al estado salvage (211, 166).
En Nueva Zelanda, la seroprevalencia parece muy baja (16% de las cabras y el 1,5% de los rebaños) (182) y esencialmente relacionada con la importación; lo mismo ocurre en las Islas Fidji (149).
En América Latina (Perú y México) (149), la seroprevalencia es baja y la infección está relacionada con las importaciones lo mismo que en Haití . Globalmente, los ganados lecheros de América del Norte, de Europa , y de Australia y aquellos en los que se han introducido reproductores mejoradores procedentes de estos países parecen ser los más infectados. Las cabras explotadas por su vellón y las de los continentes latinoámericano y africano parecen poco infectadas.

1115
Reparto de la enfermedad
La leucoencefalomielitis de los jóvenes no ha sido descrita más que en un pequeño número de países ya que su frecuencia de aparición es débil en los rebaños infectados. H a sido observada en Australia, en Canadá y en los Estados Unidos [para una revisión véase Robinson y Ellis (204)] así como en Suiza (170) y en Francia (154, 198) en donde su incidencia aparece muy limitada (alrededor del 5 % de la clase de edad); se la encuentra en general en los rebaños muy infectados.
La forma poliartrítica del adulto ha sido reencontrada en todos los países donde existe la forma leucoencefalomielítica. H a sido también encontrada en los países o en los rebaños en los que las tasas de seroprevalencia son moderadas (163, 154, 186) y donde no existe la forma juvenil.
Esta constituye la forma más banal de la expresión clínica de la A E C .
La incidencia de las otras formas clínicas (nerviosa, pulmonar del adulto, mamaria de la primípara) es más difícil de apreciar (véase diagnóstico).
Epidemiología analítica
E n condiciones naturales, la transmisión de la infección se hace esencialmente de un modo horizontal de la madre a los cabritos por el calostro y la leche (147, 167). Este modo de transmisión se ve ciertamente favorecido por la administración del calostro de mezcla en las granjas de caprino lechero intensivo, lo que podría explicar que en las explotaciones en que esta práctica no se utiliza (crianza tradicional, cría de Angora) , la prevalencia de la infección es menos elevada (204).
Los anticuerpos presentes en el calostro y que son absorbidos en los tres primeros días de la vida no protegen a los cabritos y no interfieren ni con la producción activa de anticuerpos, ni con el desarrollo de la infección por el CAEV.
Los cabritos nacidos de madres infectadas y que no hayan bebido el calostro, pueden ser también contaminados (en una proporción mucho menos importante) si han sido mantenidos algunas horas en contacto con las madres (contaminación por las secreciones genitales, digestivas o respiratorias) (véase Cuadro IX) .
C U A D R O I X
Eficacia de la contaminación al nacimiento de los cabritos nacidos de madres infectadas, según las modalidades de crianza (147)
Número de cabritos en el grupo
Modo de nacimiento Fuente de calostro y de leche
Contacto postnatal
con cabras infectadas
% de cabritos positivos/CAEV
18 Natural Mezcla Sí 100 9 Natural Madre Sí 78
17 Natural Lamido por la madre
Vaca No 17
10 Natural Separación inmediata de la madre
Vaca No 10

1116
La transmisión horizontal entre adultos es posible pero requiere un contacto prolongado y estrecho; para una pequeña cantidad de animales, puede existir al final de dos meses (204), pero necesita lo más corrientemente de 11 a 14 meses para el 50% de los animales (147, 204).
La transmisión parece ser más rápida en rebaños lecheros ya que en una experiencia (147) el 60% de los animales en riesgo se han infectado en 10 meses; estos resultados sugieren una responsabilidad del ordeño en la transmisión que parece confirmada por una encuesta epidemiológica realizada en una explotación lechera (187).
El contacto entre un macho enfermo y las cabras no infectadas, en el curso de la monta , no parece ser eficaz. Tampoco parece existir transmisión por aerosol (147). A diferencia de otras retrovirosis (leucosis bovina enzoótica, SIDA humano), la sangre no parece ser una fuente eficaz para la transmisión.
La contaminación vertical (en un sentido estricto) en el útero materno parece ser muy rara pero no imposible (147) temendo en cuenta que lesiones idénticas a las observadas en otros tejidos blanco, han sido identificadas en los úteros de animales que padecían la AEC (152) y que este tipo de contaminaciones existe también, raramente, entre los ovinos afectados de maedi . Esto podría explicar la aparición del síndrome leucoencefalomielítico en los rebaños fuertemente infectados y conteniendo numerosos animales clínicamente afectados. Su impacto parecería, no obstante, muy limitado en el desarrollo de la infección y de la enfermedad en los rebaños de producción.
La cuestión de la eventual contaminación de los ovinos por las cabras infectadas y / o enfermas es importante, para los países como Nueva Zelanda o Australia donde los ovinos están indemnes de maedi (lo que es también el caso de la República de Irlanda e Ir landa del Norte) y para todos los países donde la cohabitación de las dos especies puede existir. Las inoculaciones experimentales de CAEV a los corderos han dado resultados negativos después de 16 semanas de observación (vía intratoràcica) (208) o positivos (infección sin enfermedad después de la inoculación a la vez por vía intravenosa, intracerebral e intraarticular) (194). Los corderos alimentados durante tres meses con leche de cabras infectadas por el CAEV han presentado (en cuatro casos sobre seis) una seroconversión frente al CAEV sin signos clínicos después de 13 meses de observación (195). Un contacto estrecho y prolongado (diez y ocho meses) de cabras infectadas con las ovejas no ha producido la infección de estas últimas (195).
Smith y col. (209) han demostrado que los corderos de raza merina en contacto, durante más de 24 meses, con las cabras infectadas, no han presentado ni seroconversión ni lesiones. Por lo contrario, de entre 12 corderos merinos que habían recibido una inoculación de CAEV por vía intratoràcica, 9 hicieron una seroconversión entre la 14 y la 30 a semana post inyección; observados durante 67 a 142 semanas (más de 31 meses), no presentaron ningún signo clínico ni ninguna lesión, aunque 7 de entre ellos teman una viremia detectable. Dos de estos últimos, observados durante un nuevo periodo de 140 semanas, no presentaron ni signos clínicos, ni lesiones macroscópicas o microscópicas (166).
Se puede por lo tanto concluir que el CAEV no es patógeno para las líneas de corderos utilizados en las experiencias australianas, que la contaminación de las ovejas por las cabras por cohabitación estrecha es imposible en las condiciones habituales y que la contaminación queda como poco probable en los corderos salvo que estén alimentándose durante tres meses con leche de cabra infectada, lo que no es una práctica corriente.

1117
Estas conclusiones adquiridas experimentalmente in vivo son coherentes con las constataciones hechas in vitro (véase más arriba la virología).
D I A G N Ó S T I C O
Para este capítulo nos limitaremos a un análisis sucinto, remitiendo al lector especializado a las referencias indicadas.
De la infección
Lo más corrientemente se t ra ta de la puesta en evidencia por la inmunodifusión en gelosa (IDG) de los anticuerpos específicos contra la glucoproteína 135 o la proteína 28. También son disponibles las pruebas ELISA (204, 212, 150, 206).
En Europa, el antígeno utilizado para la IDG es lo más corrientemente preparado a partir del VVM en razón de las comunidades antigénicas estrechas existentes entre los dos virus (163, 199, 172). U n a prueba ELISA con un antígeno CAEV ha sido también propuesta [Houwers en Dawson y col., 1983 (163); 212].
En Australia, en los Estados Unidos y en Nueva Zelanda, los antígenos se preparan a partir de CAEV cualquiera que sea el método utilizado.
L a prueba ELISA parece bastante específica pero más sensible que la IDG (206).
De la enfermedad
El diagnóstico está bien codificado (162), incluso en materia de diagnóstico diferencial, para la leucoencefalomielitis de los jóvenes (158) y la poliartritis de los adultos (180, 204, 172, 200). índices clínicos (179, 164, 188) han sido propuestos y sometidos a pruebas para objetivar la afección articular.
Las formas nerviosas del adulto de la misma forma que las pulmonares, muy raras, son de identificación más delicada (204) sobre todo cuando no están asociadas con lesiones articulares (203, 196).
La m a m a aparece como un órgano blanco para la replicación del CAEV (213, 178, 181) sin que se pueda señalar con certeza al A E C como responsable de trastornos como el desequilibrio de la mama y el pezón de madera («Hard udder») (204). En efecto, si este tipo de patología está corrientemente asociado con la infección por el CAEV, o con la evolución de las otras formas clínicas del A E C , la asociación se ha constatado (200, 187) sin poder ser verdaderamente explicada ni reproducida experimentalmente. Los estudios realizados entre los ovinos con el virus maedi-visna deberían aportar elementos de apreciación (165, 177). Futuros estudios serán necesarios para precisar este aspecto de la A E C cuyo impacto económico podría ser particularmente importante .
Desde ahora se dispone de resultados que relacionan la gravedad de la A E C , enfermedad valorada por el índice clínico, con las pérdidas de producción lechera (187). Sería interesante distinguir en estas pérdidas, las que son relacionadas a la afección puramente locomotriz de las relacionadas con la afección mamaria .

1118
En caso de sospecha en un rebaño, se apreciará clínicamente la incidencia de las deformaciones articulares y sinoviales sobre los animales adultos (de edad mayor de un año) y se buscará la existencia eventual de otras manifestaciones, en particular de las formas nerviosas entre los jóvenes. La confirmación de la sospecha debe tener en cuenta la eventualidad de otras causas de poliartritis entre los adultos (micoplasmas, clamidias), los resultados obtenidos por el estudio de las lesiones (macroscópicas y microscópicas) y la tasa de infección del rebaño. En efecto, globalmente, cuanto más elevada es la tasa, más frecuentes son las formas clínicamente presentes (186, 187, 199). E n la mayoría de los rebaños en los que la enfermedad existe de forma significativa (más del 5 % de los animales afectados) las tasas de infección están comprendidas entre el 50 y el 100% (187). Es preciso señalar que, como en el maedi, un porcentaje no despreciable [25 a 3 5 % para Vitu y col. (212); 5 8 % para Grewal y col. (173)] de animales clínicamente afectados son seronegativos. Es necesario precisar que algunos autores describen en el seno de un mismo rebaño infectado, un aumento de la tasa de infección con la edad (187), mientras que otros (173) no la encuentran. Por último, ciertos autores (212) describen las caídas de anticuerpos séricos en el pico de la lactación en el momento en que aparecen las lesiones articulares.
P R O F I L A X I S
Tan sólo trataremos aquí de los principios del control de la infección; el lector interesado encontrará en la bibliografía y en el Cuadro VIII las medidas propias a los diferentes esquemas de control adoptados según los países.
La única forma de obtener animales no infectados es principalmente evitando toda contaminación perinatal del cabrito y suprimiendo todo contacto con la madre y proveyéndole de un calostro y una leche sanos sin CAEV y después criarlos sin contacto con los animales infectados.
Los trabajos de Adams y col. en 1983 (147) y de McKenzie y col. en 1987 (183) han suministrado las bases científicas y técnicas de los métodos actualmente disponibles. Todos ellos se inspiran en los que se utilizaron para el control de la infección por el VVM en los ovinos [para una revisión, véase Houwers y col. (176) y Hameury (175)].
El calostro o la leche de cabra se esterilizan frente al CAEV por calentamiento a 56°C durante una hora (147) o a 72°C durante 15 segundos o a 57°C durante 10 minutos y manteniéndolo después en botellas termostáticas durante una hora (183). Lo más corrientemente, en la práctica, se administra al cabrito recién nacido el calostro calentado a 56°C durante una hora y después, a partir del tercer día, leche de yaca reconstituida a partir de leche en polvo. Los resultados son excelentes (147, 183) si los cabritos sometidos a este tratamiento se separan inmediatamente de las madres después del par to para evitar toda lamida o toda tetada, y si se les mantienen sin ningún contacto directo con los animales infectados. Si esta última precaución no se toma, los resultados son menos satisfactorios (199). Los anticuerpos anti CAEV adquiridos por el cabrito se eliminan en dos a tres meses. Es preciso señalar que el calostro y la leche de una cabra seronegativa frente al CAEV pueden, no obstante, contener el CAEV y transmitir la infección (147).

1119
Apoyándose sobre estos hechos, muchos países han puesto en marcha los esquemas de calificación de los rebaños y de los métodos de saneamiento (véase el Cuadro VIII) (182, 162, 185) lo más corrientemente basados sobre la voluntariedad de los ganaderos, con la ayuda de un control oficial.
Cada uno de estos esquemas tiene previstas las modalidades de acreditación de los rebaños, sin organizar oficialmente el saneamiento de los rebaños infectados. En cada esquema, se distinguen tres etapas:
- U n a primera etapa que tiende a caracterizar la situación del rebaño frente a la infección.
- Una segunda etapa se asegura mediante pruebas repetidas de la no presencia de infección en el rebaño, y esto lleva a una calificación del rebaño como oficialmente indemne de infección por el CAEV.
- U n a tercera etapa que define las modalidades del mantenimiento de esta calificación.
La primera etapa está basada en un sondeo serológico sobre una parte significativa de los caprinos del rebaño (en Francia de edad mayor de los 12 meses, en el Reino Unido de más de 9 meses), o sobre todos los animales destetados (Nueva Zelanda). Un segundo test que alcanza a la totalidad de los animales (en Francia y en Gran Bretaña) se requiere antes de abordar la segunda etapa, todos los animales del rebaño deben ser entonces seronegatives.
La segunda etapa consiste en una nueva prueba o test serológico del conjunto del ganado efectuado dos veces con 4 a 6 meses de intervalo (Gran Bretaña) o una sola vez (Nueva Zelanda, Francia) . Si los resultados siguen siendo enteramente negativos, el rebaño obtiene la calificación de «oficialmente indemne de infección por el CAEV».
El mantenimiento de la calificación se asegura por el control serológico anual de, al menos, el 2 5 % de las hembras del rebaño (Francia) o de todos los animales cada año (Francia, Nueva Zelanda) o con un intervalo de 18 a 24 meses.
Los resultados parecen ser satisfactorios y permiten contemplar el porvenir con un optimismo razonable, sin forzar a adoptar las medidas de erradicación global insoportables económicamente.
* * *
A consecuencia de la presentación de este informe y después de una discusión, se aprobó la Resolución N° XV durante la 58 a Sesión General de la OIE en mayo de 1990.
*
BIBLIOGRAFÍA (véase págs. 1025-1037)


















