
Linfocitos T - Facultad de Medicina · de la cØlula T. PrÆcticamente cualquier clase de molØcula...
17
Linfocitos T 0 Cátedra de Bioquímica Facultad de Medicina - UNNE Linfocitos T Brandan, Nora Profesora Titular. Cátedra de Bioquímica. Facultad de Medicina. UNNE. Luponio, Alberto Jefe de Trabajos Prácticos. Cátedra de Bioquímica. Facultad de Medicina. UNNE. González, Juan José Ayudante Alumno por Concurso. Cátedra de Bioquímica. Facultad de Medicina. UNNE. González Roibón, Nilda Ayudante Alumna por Concurso. Cátedra de Bioquímica. Facultad de Medicina. UNNE. Klinzuk, Sofía Ayudante Alumna por Concurso. Cátedra de Bioquímica. Facultad de Medicina. UNNE. Introducción El hombre, como todos los seres vivientes, está amenazado constantemente por agentes patógenos (infecciosos o no) que pueden favorecer el desarrollo de una enfermedad. Todos estos organismos poseen mecanismos defensivos protectores que les aseguran cierta integridad, y solamente por falla de los mismos puede desencadenarse un estado patológico. Conceptos: Inmunidad (del latín, “estar libre de carga”): reacción a las sustancias extrañas, incluyendo microorganismos, así como macromoléculas, tales como proteínas y polisacáridos, sin que dicha reacción tenga una repercusión fisiológica o patológica. Sistema Inmunitario: constituido por las células y moléculas responsables de la inmunidad; es extremadamente complejo y posee una diversidad de actividades para mantener la homeostasis de la salud. Respuesta Inmunitaria: es la respuesta global y coordinada tras la introducción de sustancia extrañas. Antígeno: cualquier sustancia capaz de unirse específicamente a un anticuerpo o a un receptor de la célula T. Prácticamente cualquier clase de molécula biológica (metabolitos intermediarios, azúcares, lípidos, autacoides y hormonas, así como también macromoléculas, hidratos de carbono complejos, fosfolípidos como ácidos nucleicos y proteínas) puede actuar como antígeno. Sin embargo, sólo las macromoléculas pueden iniciar la activación linfocitaria necesaria para la activación de anticuerpos; estas moléculas se denominan inmunógenos. Para protegerse contra estas agresiones, el organismo ha desarrollado diferentes mecanismos de defensa: Universidad Nacional del Nordeste Facultad de Medicina Cátedra de Bioquímica
Transcript of Linfocitos T - Facultad de Medicina · de la cØlula T. PrÆcticamente cualquier clase de molØcula...

Linfocitos T 0
Cátedra de Bioquímica Facultad de Medicina - UNNE
Linfocitos TBrandan, NoraProfesora Titular. Cátedra de Bioquímica. Facultad de Medicina. UNNE.
Luponio, AlbertoJefe de Trabajos Prácticos. Cátedra de Bioquímica. Facultad de Medicina. UNNE.
González, Juan JoséAyudante Alumno por Concurso. Cátedra de Bioquímica. Facultad de Medicina. UNNE.
González Roibón, NildaAyudante Alumna por Concurso. Cátedra de Bioquímica. Facultad de Medicina. UNNE.
Klinzuk, SofíaAyudante Alumna por Concurso. Cátedra de Bioquímica. Facultad de Medicina. UNNE.
IntroducciónEl hombre, como todos los seres vivientes, está amenazado constantemente por agentespatógenos (infecciosos o no) que pueden favorecer el desarrollo de una enfermedad.Todos estos organismos poseen mecanismos defensivos protectores que les aseguran ciertaintegridad, y solamente por falla de los mismos puede desencadenarse un estado patológico.Conceptos:Inmunidad (del latín, “estar libre de carga”): reacción a las sustancias extrañas, incluyendomicroorganismos, así como macromoléculas, tales como proteínas y polisacáridos, sin quedicha reacción tenga una repercusión fisiológica o patológica.Sistema Inmunitario: constituido por las células y moléculas responsables de la inmunidad; esextremadamente complejo y posee una diversidad de actividades para mantener lahomeostasis de la salud.Respuesta Inmunitaria: es la respuesta global y coordinada tras la introducción de sustanciaextrañas.Antígeno: cualquier sustancia capaz de unirse específicamente a un anticuerpo o a un receptorde la célula T. Prácticamente cualquier clase de molécula biológica (metabolitosintermediarios, azúcares, lípidos, autacoides y hormonas, así como también macromoléculas,hidratos de carbono complejos, fosfolípidos como ácidos nucleicos y proteínas) puede actuarcomo antígeno. Sin embargo, sólo las macromoléculas pueden iniciar la activación linfocitarianecesaria para la activación de anticuerpos; estas moléculas se denominan inmunógenos.Para protegerse contra estas agresiones, el organismo ha desarrollado diferentes mecanismosde defensa:
Universidad Nacional del NordesteFacultad de MedicinaCátedra de Bioquímica

Linfocitos T 1
Cátedra de Bioquímica Facultad de Medicina - UNNE
Comenzando con la Inmunidad Natural o Innata, inespecífica, que abarca desde lasdenominadas “barreras físicas” (piel, secreciones de las mucosas, enzimas proteolíticas,pH del estómago, etc.) hasta la activación de factores humorales, como el Sistema deComplemento (por la vía alterna), o celulares, como la fagocitosis por macrófagos o laactivación de las células NK.
Si, a pesar de la activación de todos estos mecanismos de la Inmunidad Natural lainfección sigue progresando, el sistema inmune pondrá en marcha el mecanismo de laInmunidad Adquirida, también con la activación de mecanismos humorales (activacióndel Complemento por la vía clásica y anticuerpos producidos por linfocitos B o Ly B) ycelulares (desarrollo de citotoxicidad dependiente de anticuerpos y por linfocitos Tcitotóxicos o Ly Tc).
Respuesta adquirida. Mecanismos celulares mediados por Ly T
Dentro de este sistema celular existen dos tipos de linfocitos T, que se diferencian por laexpresión en membrana de dos moléculas de superficie diferentes (CD4+ o CD8+) que van adeterminar sus funciones efectoras:
Ly T colaboradores o helper, CD4+ (Ly Th), cuya principal función es la secreción decitoquinas, por medio de las cuales estimula la producción de anticuerpos por los Ly B ylas actividades microbicidas de los macrófagos, que constituyen los dos mecanismosprincipales para la eliminación de microorganismos extracelulares o que viven envesículas fagocíticas.
Ly T citolíticos o citotóxicos, CD8+, que lisan las células que presentan antígenosextraños en su membrana; es por ello que son útiles en la eliminación de virus y
Linfocitos T 2
Cátedra de Bioquímica Facultad de Medicina - UNNE
bacterias que infectan y viven en el interior de distintos tipos celulares, así comotambién de células neoplásicas.
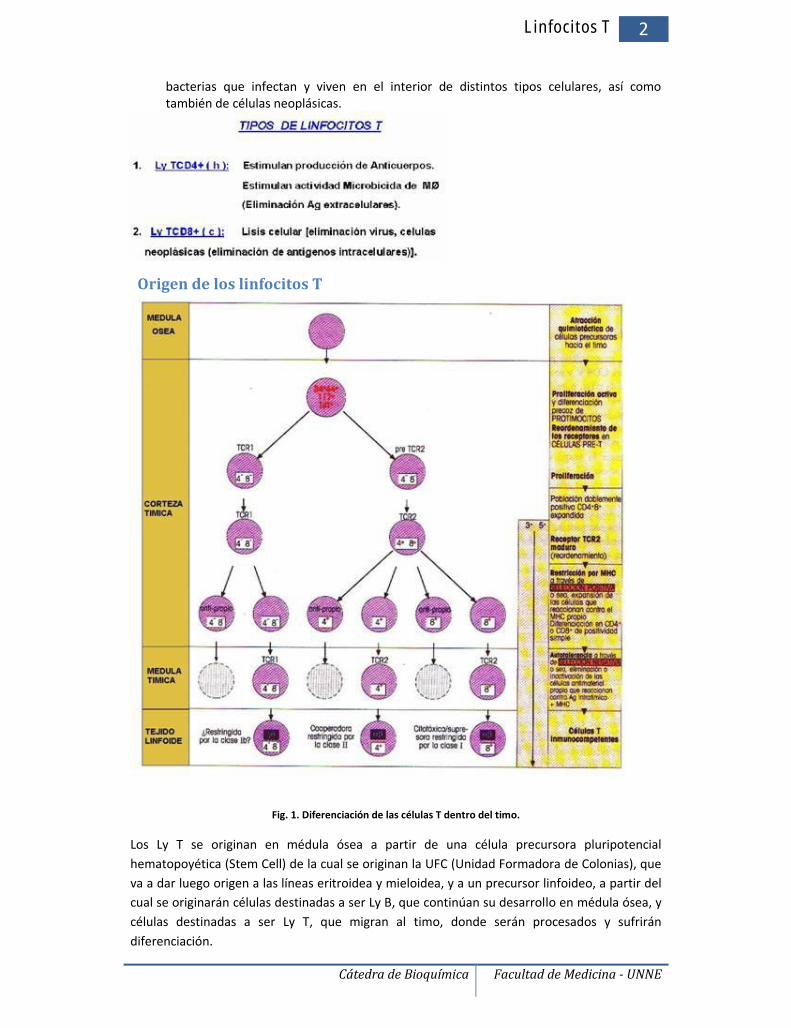
Origen de los linfocitos T
Fig. 1. Diferenciación de las células T dentro del timo.
Los Ly T se originan en médula ósea a partir de una célula precursora pluripotencialhematopoyética (Stem Cell) de la cual se originan la UFC (Unidad Formadora de Colonias), queva a dar luego origen a las líneas eritroidea y mieloidea, y a un precursor linfoideo, a partir delcual se originarán células destinadas a ser Ly B, que continúan su desarrollo en médula ósea, ycélulas destinadas a ser Ly T, que migran al timo, donde serán procesados y sufrirándiferenciación.

Linfocitos T 3
Cátedra de Bioquímica Facultad de Medicina - UNNE
Sólo el 5% de los linfocitos presentes en el timo permanecen viables para una adecuadarespuesta inmune y pasan a la circulación (el 95% restante muere por mecanismos deapoptosis celular). Los Ly T que dejan el timo y alcanzan el torrente sanguíneo, van adiseminarse por todo el organismo para alojarse en el tejido linfoideo corporal, másextensamente en los ganglios linfáticos, vigilante ante la entrada de algún agente agresor.
Fases de la Respuesta InmuneCuando una noxa ingresa, la respuesta inmune mediada por estas células se desarrollará entres fases:
Reconocimiento: unión de antígenos extraños a receptores específicos de linfocitosmaduros.
Activación: proliferación, expansión clonal y diferenciación hacia células efectoras(acontecimientos como consecuencia del reconocimiento). Requiere de la participaciónde dos señales, el antígeno y células colaboradoras, con participación de moléculascoestimuladoras.
Fase efectora: se desarrollan las funciones que llevan a la eliminación del antígeno.
ReconocimientoLos Ly T responden únicamente ante antígenos proteicos unidos a proteínas de superficie deotras células que “muestran” el antígeno a este linfocito (es decir, no responden anteproteínas solubles o circulantes).
Estos antígenos proteicos deben ser procesados, ya que estos linfocitos sólo pueden reconocerlos fragmentos peptídicos derivados de éstos, y, específicamente, la secuencia de aminoácidos.Estos fragmentos peptídicos se unen únicamente a proteínas de membrana codificadas porgenes CMH (Complejo Mayor de Histocompatibilidad). Se dice además, que este proceso está“restringido por el CMH propio” (los linfocitos sólo responden ante aquéllos antígenos unidosa moléculas que puedan ser reconocidas como propias).
Existen dos clases de moléculas CMH de membrana: Clase I, presente en las células nucleadasy Clase II, presente en células del Sistema Inmune (Ly, macrófagos, células dendríticas, etc.)
Los antígenos derivados de proteínas citosólicas, son presentados en unión con el CMHI y reconocidos por el linfocito T CD8+. Los antígenos extracelulares, son procesados y presentados unidos al CMH II yreconocidos por los CD4+.

Linfocitos T 4
Cátedra de Bioquímica Facultad de Medicina - UNNE
El reconocimiento del antígeno en ambos casos (CD4+ o CD8+) se hace mediante el TCR(Receptor de Células T), que no distingue entre antígenos extra e intracelulares, por lo que laespecificidad depende de su unión al CMH correspondiente.
Estructura del TCREl receptor para los complejos péptido-CMH en la mayoría de las células T es un heterodímero
formado por dos cadenas polipeptídicas transmembrana llamadas y, unidascovalentemente entre sí mediante puentes disulfuro.
Ambas cadenas presentan porciones extracelulares que contienen un dominio variable (V), N-terminal, y una región constante (C), proximal a la membrana. Los dominios V tienen unaestructura terciaria similar a los dominios V de las inmunoglobulinas, y el dominio C de la
cadena es similar a los dominios C de éstas. Esta homología que contienen los dominios lesda la particularidad de ser miembros de una familia de moléculas que constituyen la“superfamilia de inmunoglobulinas”
Las regiones V de las cadenas y del TCR poseen secuencias que son altamente variablesentre los diferentes clones de células T, lo que refleja su papel en el reconocimiento delantígeno.
Además, las cadenas y contienen una región “bisagra” corta, adyacente a la cara exteriorde la membrana plasmática. Un residuo de cisteína en cada región bisagra contribuye a unenlace disulfuro que une las dos cadenas.
En ambas cadenas la porción transmembrana presenta residuos aminoacídicos hidrófobos.Una característica poco habitual de estas porciones es la presencia de residuos aminoacídicos
cargados positivamente, entre ellos un residuo de lisina (cadena ), o un residuo de lisina y
arginina (cadena ), que interaccionan con residuos cargados negativamente de la porcióntransmembrana de los polipéptidos CD3 (ver más adelante).
Las cadenas y poseen colas citoplasmáticas C-terminales de 5 a 12 aminoácidos delongitud, las cuales son demasiado pequeñas como para mostrar una actividad enzimáticaintrínseca, por eso requieren otras moléculas asociadas físicamente al TCR para que funcionencomo traductores de señales. (esquema).
A diferencia de las inmunoglobulinas, el TCR no es secretado y de ahí que no realice funcionesefectoras por sí mismo, sino que, una vez que se ha unido a los complejos péptido-CMH, poneen marcha señales que activan funciones efectoras de la célula T.
Tanto las cadenas como las del TCR contribuyen a la especificidad en el reconocimiento delcomplejo antígeno peptídico-CMH.
Las regiones altamente variables de estas cadenas se denominan “regiones de alta diversidad”(existen al menos tres) y corresponden a las regiones determinantes de la complementariedad(CDR) de las inmunoglobulinas. Ambas cadenas poseen CDR1 y CDR2 dentro de los dominios V,mientras que el CDR3 no se encuentra en su totalidad dentro de este dominio.

Linfocitos T 5
Cátedra de Bioquímica Facultad de Medicina - UNNE
Por lo tanto, la superficie de unión al antígeno de un TCR consta de tres bucles horquilla
hipervariables que aporta la cadena y tres, la cadena , dando un total de seis CDR.
La molécula del TCR confiere a la célula T la capacidad de reconocer antígenos peptídicosunidos a moléculas del CMH, pero tanto su expresión como su función en la activación deestas células depende de otras cinco proteínas transmembrana que se asocian no
covalentemente al heterodímero , formando el llamado “Complejo TCR Funcional”. Tresproteínas de este complejo se denominan moléculas CD3 e incluyen a miembros de la
superfamilia de las inmunoglobulinas de alta homología, llamados y. Además, en un 80 a
un 90 %, contiene un homodímero unido por puentes disulfuro, cadena , que no pertenece ala superfamilia.
Fig. 2. Componentes del complejo receptor de la célula T (TCR).
Estructura y función de las proteínas CD3, y Las cadenas y son entre sí muy homólogas, presentan regiones extracelulares N-terminales que contienen un dominio variable único, semejante al de las inmunoglobulinas.
Los segmentos transmembrana de todas las cadenas CD3 contienen un residuo de ácidoaspártico, que tiene carga negativa, característica importante para su asociación física con las
cadenas y del TCR.
Los dominios citoplasmáticos tienen una longitud que varía de 44 a 81 residuos aminoacídicos,tamaño suficiente para traducir señales al interior de la célula. Cada una de las colascitoplasmáticas posee una copia de una secuencia motivo, denominada “motivo de activacióndel inmunorreceptor vía tirosina” (ITAM) o “motivo de activación para el reconocimiento delantígeno” (ARAM), importante para las funciones de señalización. Este está compuesto por 26residuos aminoacídicos, en los que la secuencia tir-x-x-leucina aparece dos veces con unaseparación de seis a ocho residuos (x= aminoácidos sin especificar).
Linfocitos T 6
Cátedra de Bioquímica Facultad de Medicina - UNNE
Los residuos de tirosina son fosforilados en respuesta al reconocimiento del antígeno por elTCR; esto permite la unión de las proteínas que contienen dominios SH2 a los ITAM y lainiciación de una cascada de señales intracelulares.
Los dominios extracelulares de las cadenas y son cortos, de 9 residuos; los residuostransmembrana contienen ácido aspártico y los dominios citoplasmáticos son largos, de 113 a
115 residuos. Las cadenas y difieren entre sí por sus colas citoplasmáticas. Además, la
cadena posee tres ITAM.
Funciones de las proteínas CD3 y .Cuando un antígeno se une al TCR, las cadenas asociadas, CD3 y traducen señales alcitoplasma de la célula que conducen a su activación funcional. Además de estas proteínasasociadas al complejo TCR, se requieren de otras moléculas, coestimuladores adicionales, queinteractúan con moléculas de superficie de la célula T distintas del complejo TCR.
Otra función de estas proteínas es facilitar la expresión en superficie del complejo TCR en sutotalidad.
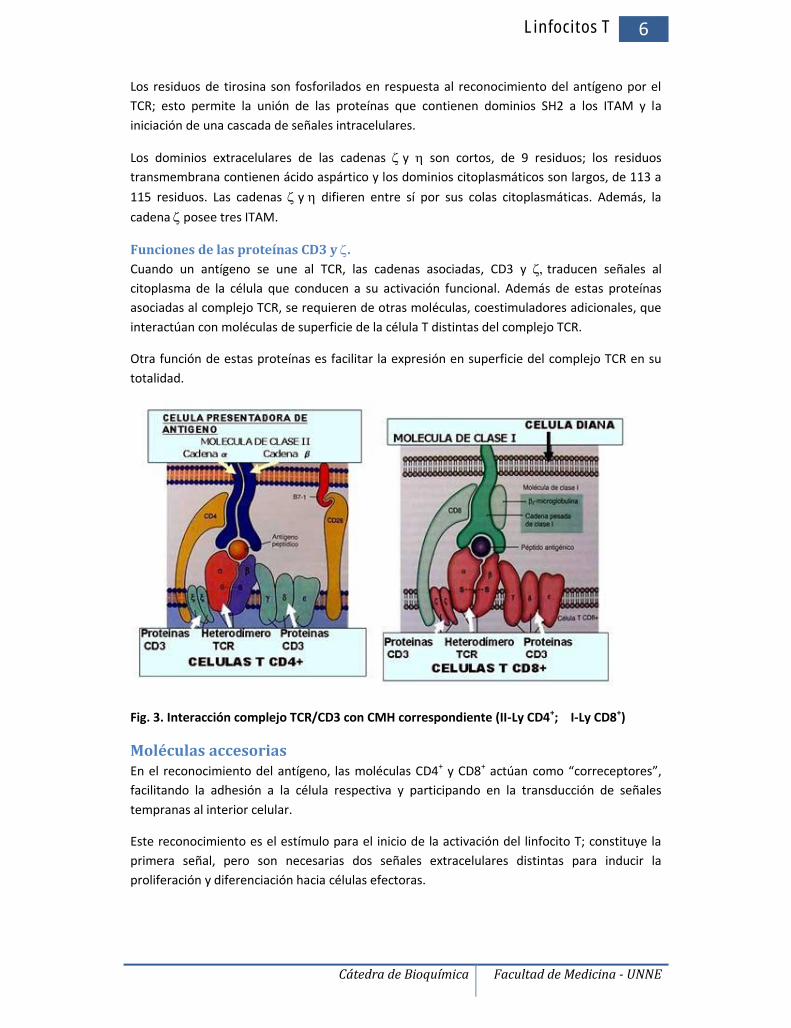
Fig. 3. Interacción complejo TCR/CD3 con CMH correspondiente (II-Ly CD4+; I-Ly CD8+)
Moléculas accesoriasEn el reconocimiento del antígeno, las moléculas CD4+ y CD8+ actúan como “correceptores”,facilitando la adhesión a la célula respectiva y participando en la transducción de señalestempranas al interior celular.
Este reconocimiento es el estímulo para el inicio de la activación del linfocito T; constituye laprimera señal, pero son necesarias dos señales extracelulares distintas para inducir laproliferación y diferenciación hacia células efectoras.

Linfocitos T 7
Cátedra de Bioquímica Facultad de Medicina - UNNE
Esta segunda señal va a estar dada por las “moléculas coestimuladoras”, que son moléculas desuperficie presentes en células presentadoras de antígeno (APC) y/o diana, y que interactúancon receptores específicos presentes en el linfocito T.
Estructura y funciónCD4+: es una glucoproteína transmembrana, miembro de la superfamilia de inmunoglobulinas,que se unen al CMH-II con funciones de adhesión y señalización, y se expresa en el 65 % de losLy T circulantes. Es un monómero que además puede expresarse en monocitos y macrófagos;tiene cuatro dominios globulares extracelulares, incluyendo un dominio N-terminal, tipo V ytres que no son ni de tipo V ni C. Hay además una región transmembrana hidrofóbica y unacola citoplasmática de 38 residuos aminoácidos básicos principalmente.
Fig. 4. Superfamilia de las IG.
CD8+: es una glucoproteína transmembrana que se une a moléculas CMH-I, y es expresada enun 35 % de los Ly T circulantes; sus funciones son similares a las de CD4+. Estructuralmente es
un heterodímero unidos por puentes disulfuro compuestos por CD8 y CD8, cada una poseeun dominio globular extracelular, una región hidrofóbica transmembrana y una colacitoplasmática o intracelular de 25 residuos aminoácidos básicos principalmente.
En cuanto a las funciones, tanto CD4+ como CD8+ facilitan la adhesión entre células T y las APC
o células diana. El CD4+ se une al dominio 2 del CMH-II, CD8+ se une al dominio 3 del CMH-I;además cumplen funciones de traducción de señales a través de una proteína llamada ICK, con

Linfocitos T 8
Cátedra de Bioquímica Facultad de Medicina - UNNE
actividad de tir-quinasa, que se asocia en forma no covalente a las colas citoplasmáticas de
CD4+ y CD8+. ICK fosforila residuos de tirosina en los ITAM de CD3 y .
Si bien los Ly T reaccionan con antígenos unidos al CMH, para una completa estimulaciónrequieren moléculas coestimuladoras, junto a las señales inducidas por el antígeno. Una faltade coestimulación puede inducir a un estado de falta de respuesta denominado “anergia”.
La expresión de estos coestimuladores se halla regulada por proteínas proinflamatorias y esogarantiza que los linfocitos sean activados en el momento y lugar adecuados.
En condiciones fisiológicas también contribuyen a los fenómenos de tolerancia inmunológica.
CD28: es una de las moléculas de superficie más importantes en la coestimulación para laactivación del linfocito T. Se une a CD80 o B7-1 y CD86 o B7-2, expresadas en la APC; es unhomodímero de la superfamilia de las inmunoglobulinas que se expresa en el 80 % de lascélulas T CD4+ y en el 50 % de las CD8+.
Las B7-1 y B7-2 son glucoproteínas homólogas de cadena lateral sencilla, cada una con dosdominios extracelulares tipo inmunoglobulina, una región transmembrana y una colacitoplasmática; además también pertenecen a la superfamilia de inmunoglobulinas con igualescaracterísticas y funciones que CD28.
No se conoce por completo cómo CD28 estimula la activación de los Ly T, tampoco cómoestimula la producción de IL-2 y preserva a la célula de la apoptosis.
CD45 o T200: está formado por varias moléculas de diferente tipo (8 en total), codificadas porun solo gen. Son proteínas intrínsecas con un dominio citoplasmático de 705 residuosaminoácidos, con actividad de tir-fosfatasa y que participa en la regulación de la respuesta.
También hay integrinas implicadas en la interacción de adhesión de células T y otras células,como la LFA-1, que se asocia a la molécula de adhesión intercelular 1 o ICAM-1, que es unaglucoproteína transmembrana con cinco dominios extracelulres tipo inmunoglobulina. Otramolécula que media función de adhesión y estimulación es el CD2 o LFA-2, glucoproteínaperteneciente a la superfamilia que se asocia con LFA-3 o CD52, presente en las céluas T.
Fig. 5. Participación de las moléculas de superficie (complejo TCR/CD3, correceptores y moléculasaccesorias).

Linfocitos T 9
Cátedra de Bioquímica Facultad de Medicina - UNNE
Como consecuencia de estas interacciones se desencadenan una serie de señalesintracelulares.
Acontecimientos intracelulares en la activación de la célula TEl reconocimiento del antígeno y las señales coestimuladoras activan a las células T vírgenes,que sufren expansión clonal y diferenciación hacia células T efectoras y de memoria. Laestimulación antigénica de células T efectoras conduce a una expansión clonal posterior y aldesarrollo de funciones efectoras, como secreción de citoquinas y actividades citolíticas.
Cuando el TCR se une al complejo péptido-CMH, se activan proteínas tir-quinasa (PTK)asociadas al TCR; una de ellas, la ICK, sería la responsable de la fosforilación temprana de losITAM en las células T. Esta es una proteína de la familia de ser-treonina quinasa que se asociaestrechamente a las colas citoplasmáticas de las moléculas CD4+ y CD8+ del linfocito T.
Una vez que los ITAM del CD3 y las cadenas son fosforiladas en sus residuos de tirosina por laICK, se convierten en “sitios específicos de acoplamiento”, que se unen a proteínascitoplasmáticas con dominios SH2. Una de ellas, de gran importancia, es la denominada ZAP-70, la cual para activarse debe ser fosforilada en un residuo de tirosina interno, lo cual ocurre
rápidamente tras su unión a la cadena .
Una vez activada la ZAP-70, puede autofosforilarse en otros residuos de tirosina y entoncesfuncionar como una molécula armazón para el reclutamiento de otras moléculas deseñalización con dominios SH2.
Fig. 6. Las señales a través del complejo TCR/CD3/CD4/CD8 inician una cascada de tirosina proteína quinasa (TPKo PTK).
Linfocitos T 10
Cátedra de Bioquímica Facultad de Medicina - UNNE
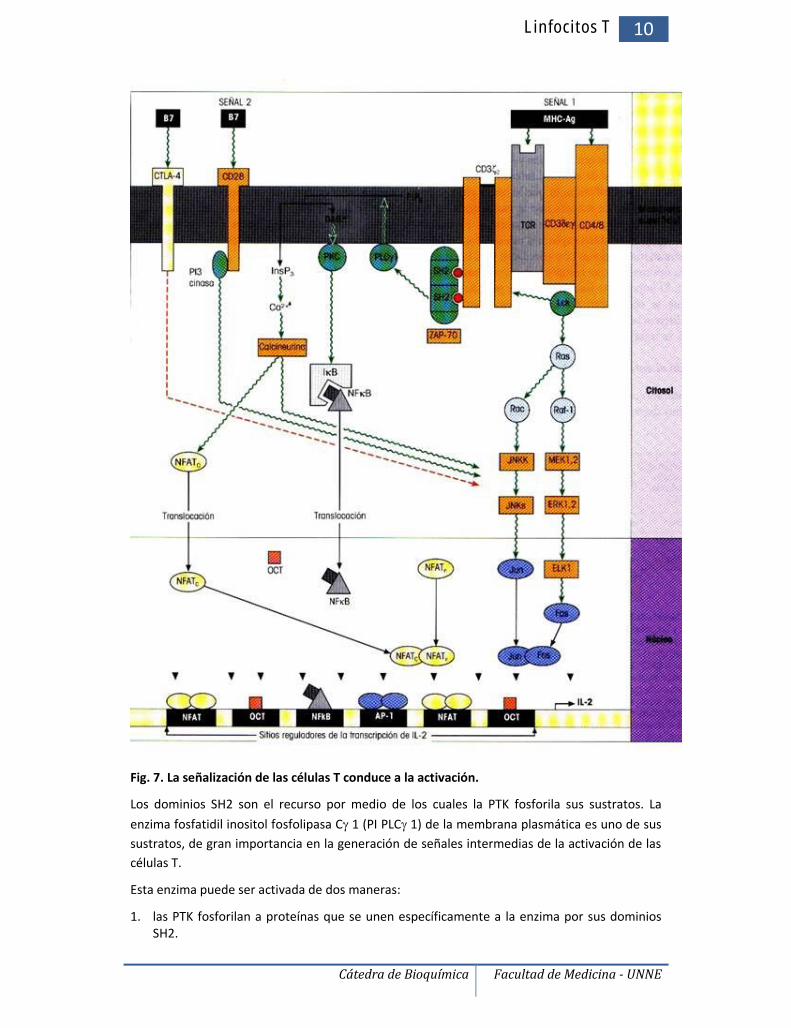
Fig. 7. La señalización de las células T conduce a la activación.
Los dominios SH2 son el recurso por medio de los cuales la PTK fosforila sus sustratos. La
enzima fosfatidil inositol fosfolipasa C 1 (PI PLC 1) de la membrana plasmática es uno de sussustratos, de gran importancia en la generación de señales intermedias de la activación de lascélulas T.
Esta enzima puede ser activada de dos maneras:
1. las PTK fosforilan a proteínas que se unen específicamente a la enzima por sus dominiosSH2.

Linfocitos T 11
Cátedra de Bioquímica Facultad de Medicina - UNNE
2. las PTK fosforilan directamente a la PI PLC 1. Entonces, esta última activada cataliza lahidrólisis de un fosfolípido de membrana, llamado “fosfatidil inositol 4-5 bifosfato”,formando de este modo, los segundos mensajeros diacilglicerol (DAG) e inositol 1-4-5trifosfato (IP3).
El IP3 estimula la liberación de calcio de los depósitos intracelulares, provocando una rápidaelevación del ión libre, que se une rápidamente a una proteína reguladora dependiente decalcio, denominada “calmodulina”. Los complejos Ca2+-calmodulina pueden activar variasenzimas, entre ellas algunas fosfatasas importantes para la activación de factores detranscripción (FT).
La función del DAG es activar ciertas isoformas de proteínas quinasas, que junto con el Ca2+
estimulan la producción de enzimas para la activación de FT.
La PTK no solamente induce la escisión de fosfolípidos, sino que también activa vías deseñalización a través de la cascada RAS, las cuales sirven para unir receptores de la membranacon quinasas que actúan a favor de corriente, incluyendo las cascadas de quinasas deproteínas activadas por mitógenos (MAP). Al final, estas vías activadas, catalizan lafosforilación y activación directa de FT o de sus componentes.
Por último, estos FT activados conducen a la transcripción de varios genes que codificancitoquinas, receptores para citoquinas, y otras proteínas necesarias para la función efectora delas células T.
Funciones efectorasComo consecuencia de la activación, las célulasproliferan y se diferencian hacia células efectoras,que van a desarrollar sus funciones específicas:
Los Ly T helper secretan citoquinas (IL-2), queactúan sobre las APC estimulando su proliferacióny expansión clonal; y sobre sí mismo dondeestimula proliferación y expansión clonal.Dependiendo del tipo de antígeno (intra oextracelular), también participan otras citoquinas(IL-12 o IL-4) que estimulan la diferenciación del LyT precursor (P), hacia 2 subgrupos celulares (LyTh1 o Th2) y células de memoria (M). El Ly Th 1,
secreta Interferon (INF )Factor de NecrosisTumoral (TNF), IL-2. Estas citoquinas actúan luegosobre macrófagos, estimulando su activación yfagocitosis (Inmunidad mediada por células). El LyTh 2, secreta una amplia gama de citoquinas queactúan sobre Ly B, estimulando su proliferación,expansión clonal y diferenciación haciaplasmocitos productores de anticuerpos y Ly B(Inmunidad humoral).
Fig. 8. Generación de los subgrupos CD4+ Th1 y Th2.
Linfocitos T 12
Cátedra de Bioquímica Facultad de Medicina - UNNE
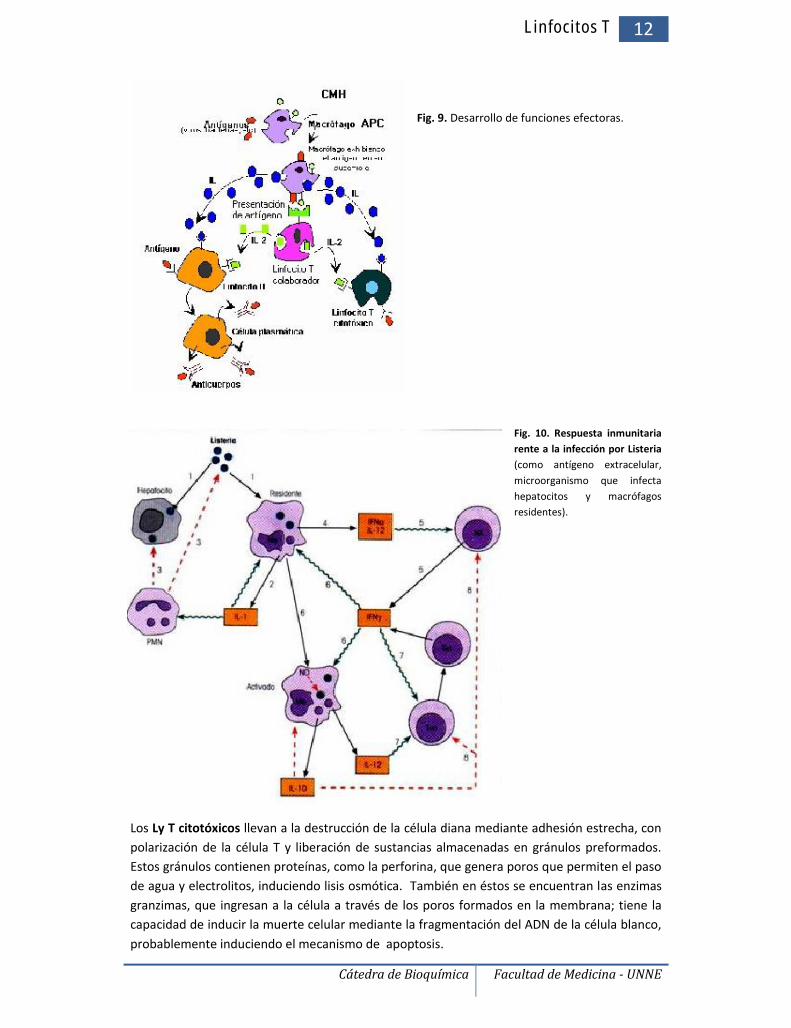
Fig. 9. Desarrollo de funciones efectoras.
Fig. 10. Respuesta inmunitariarente a la infección por Listeria(como antígeno extracelular,microorganismo que infectahepatocitos y macrófagosresidentes).
Los Ly T citotóxicos llevan a la destrucción de la célula diana mediante adhesión estrecha, conpolarización de la célula T y liberación de sustancias almacenadas en gránulos preformados.Estos gránulos contienen proteínas, como la perforina, que genera poros que permiten el pasode agua y electrolitos, induciendo lisis osmótica. También en éstos se encuentran las enzimasgranzimas, que ingresan a la célula a través de los poros formados en la membrana; tiene lacapacidad de inducir la muerte celular mediante la fragmentación del ADN de la célula blanco,probablemente induciendo el mecanismo de apoptosis.
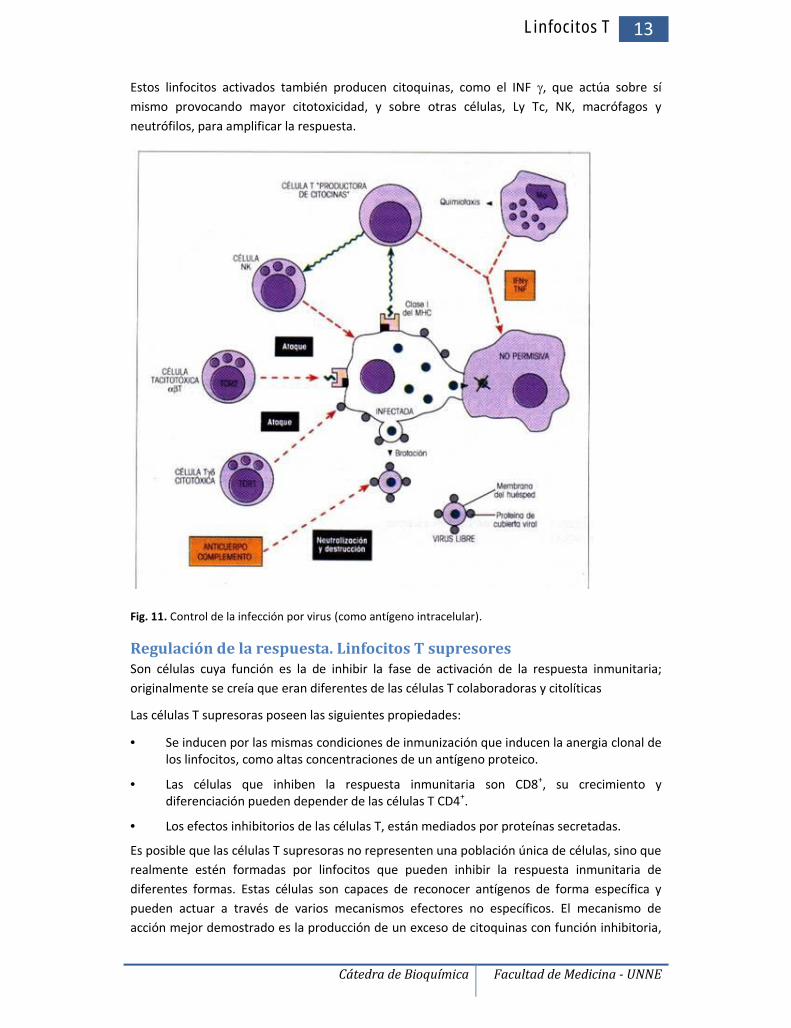
Linfocitos T 13
Cátedra de Bioquímica Facultad de Medicina - UNNE
Estos linfocitos activados también producen citoquinas, como el INF , que actúa sobre símismo provocando mayor citotoxicidad, y sobre otras células, Ly Tc, NK, macrófagos yneutrófilos, para amplificar la respuesta.
Fig. 11. Control de la infección por virus (como antígeno intracelular).
Regulación de la respuesta. Linfocitos T supresoresSon células cuya función es la de inhibir la fase de activación de la respuesta inmunitaria;originalmente se creía que eran diferentes de las células T colaboradoras y citolíticas
Las células T supresoras poseen las siguientes propiedades:
Se inducen por las mismas condiciones de inmunización que inducen la anergia clonal delos linfocitos, como altas concentraciones de un antígeno proteico.
Las células que inhiben la respuesta inmunitaria son CD8+, su crecimiento ydiferenciación pueden depender de las células T CD4+.
Los efectos inhibitorios de las células T, están mediados por proteínas secretadas.
Es posible que las células T supresoras no representen una población única de células, sino querealmente estén formadas por linfocitos que pueden inhibir la respuesta inmunitaria dediferentes formas. Estas células son capaces de reconocer antígenos de forma específica ypueden actuar a través de varios mecanismos efectores no específicos. El mecanismo deacción mejor demostrado es la producción de un exceso de citoquinas con función inhibitoria,

Linfocitos T 14
Cátedra de Bioquímica Facultad de Medicina - UNNE
por ejemplo el Factor Transformador de Crecimiento (TGF), potente inhibidor de laproliferación de células B y T.
En otras situaciones, diferentes citoquinas pueden inhibir distintos tipos de respuestainmunitaria, por ejemplo la IL-4, IL-10 y IL-13 producidas por el Ly Th 2, inhiben la activación demacrófagos; por lo tanto, el Ly Th2 puede inhibir la inmunidad mediada por células y lahipersensibilidad de tipo retardada.
Las células Th 1 productoras de INF pueden actuar como supresoras en la producción de Ig E.Además pueden llevar a cabo sus efectos destruyendo otras células implicadas en la respuestainmunitaria.
La conclusión importante, es que varias poblaciones de células T son capaces de suprimirdiferentes respuestas inmunitarias y puede no existir una única población de célulassupresoras.
Sin esta supresión, el Sistema Inmunológico seguiría trabajando después de la infección.
Fig. 12. Regulación de la respuesta inmune.
Aplicaciones clínicasSe han descripto diversas enfermedades causadas o asociadas a deficiencias que afectan enforma selectiva a ciertas moléculas del complejo TCR-CD3 (defectos estructurales) o bien a latransmisión de señales de activación al interior celular (defectos funcionales).
Estas deficiencias producen alteraciones en el sistema inmunitario, como disminución delnúmero de linfocitos, inhibición de la proliferación celular y reducción de la síntesis decitoquinas, entre otras. Las consecuencias clínicas que se derivan de ésto son variadas ybastante graves (infecciones recurrentes, enfermedades autoinmunes, etc.).
Es necesario aclarar que las inmunodeficiencias se pueden clasificar en:

Linfocitos T 15
Cátedra de Bioquímica Facultad de Medicina - UNNE
Inmunodeficiencia primaria: pueden ser congénitas o adquiridas; en la actualidad seclasifican según el modo de transmisión y en función de si el defecto afecta a las célulasT, B o ambas.
Inmunodeficiencia secundaria: que no se deben a anomalías intrínsecas del desarrollo ode la función de células B o T, sino que pueden ser consecuencia de otras enfermedadesque indirectamente afectan el sistema inmunitario.
Inmunodeficiencias de células T
Síndrome de DiGeorge: se debe a una alteración del desarrollo de los elementosepiteliales del timo, derivados de la tercera y cuarta bolsas faríngeas. El defecto genéticose ha localizado en el cromosoma 22q11. Se caracteriza por la presencia de cardiopatíascongénitas, malformaciones faciales, etc. Los niveles de células T generalmente sonbajos, en un número que puede o no ser suficiente para ofrecer una defensa adecuada.
Deficiencia del receptor de células T: cualquier alteración en la expresión y la función delos componentes del TCR (CD3, y ) puede alterar el desarrollo y función decélulas T; por ejemplo mutaciones hereditarias de CD3, que dan lugar a un déficitselectivo de células T CD8+; o mutaciones de CD3, que provoca una reducciónpreferente de células T CD4+ lo que implica diferencia en la función de traducción de laseñal de cada uno de los componentes CD3.
Deficiencias de CMH-II: Las APC de los pacientes con ésta rara alteración no expresanlas moléculas de clase II en la superficie, por lo tanto las células T CD4+ colaboradoras noencuentran el antígeno en la periferia. Los pacientes sufren infeccionesbroncopulmonares a repetición, diarreas crónicas e infecciones virales mortales antes delos cuatro años de edad.
Deficiencia de tir-quinasa ZAP-70: Es un componente esencial en la cascada detraducción de señal TCR-CD3, que puede mutar en forma hereditaria dando lugar adeficiencias de células T CD8+ y alteraciones funcionales de las células T CD4+.
Síndrome de Inmunodeficiencia Adquirida (SIDA): Es una infección viral causada por unvirus RNA perteneciente a la familia retroviridae que gracias a la codificación de unatranscriptasa inversa es capaz de integrarse al genoma de la célula huésped estimulandola transcripción y la formación de los nuevos viriones. Posee un marcado efecto citolíticoe infecta preferentemente a la subpoblación de células T CD4+ tras unirse a los propiosreceptores de CD4+. Tiene menor afinidad por macrófagos y células gliales del sistemanervioso central (SNC). Se caracteriza por deterioro progresivo de la respuestainmunitaria por disminución del número de Ly T CD4+, con afectación de casi todos losórganos de la economía; se evidencian síntomas gastrointestinales variados, infeccionespor parásitos oportunistas, como Pneumocistis carinni, sarcoma de Kapossi, neoplasiasasociadas y en los últimos estadíos de evolución, síntomas de demencia, por afectacióndel SNC.
Las 10 señales de Deficiencias Inmunológicas Primarias:
1. Ocho o más infecciones de oído distintas en un año.2. Dos o más infecciones serias de sinusitis en un año.3. Dos o más meses con antibióticos y con poco efecto.4. Dos o más neumonías en un año.5. Un niño que no sube de peso o que no crezca adecuadamente.6. Abscesos recurrentes profundos en la piel o en otros órganos.7. Infecciones por hongo en la boca o en la piel después del año de edad.8. Que necesite antibióticos intravenosos para eliminar la infección.

Linfocitos T 16
Cátedra de Bioquímica Facultad de Medicina - UNNE
9. Dos o más infecciones graves como meningitis, osteomielitis, o sepsis.10. Historia en la familia de deficiencias inmunológicas primarias.
Rechazo de transplantes
El rechazo de un injerto se produce por el reconocimiento que el huésped hace del tejidoinjertado como extraño. En el hombre, los antígenos responsables del rechazo son los delSistema Principal de Histocompatibilidad (HLA o CMH). Es un proceso complejo en el queintervienen tanto la inmunidad de tipo celular como los anticuerpos circulantes.
El papel de las células T en el rechazo de los transplantes fue comprobado tanto en el hombrecomo en los animales de experimentación. La destrucción del injerto ocurre con laparticipación de células CD8+ y CD4+. No se conocen por completo las bases celulares ymoleculares del reconocimiento por parte de las células T del huésped
La inmunidad adquirida es producto del sistema linfocitario del organismo. Las personas quetienen una falla genética de linfocitos o cuyos linfocitos han sido destruidos por radiaciones desustancias químicas no pueden desarrollarla; días después mueren por infecciones bacterianasfulminantes. Por tanto, está claro que estas células son esenciales para la supervivencia del serhumano.
Bibliografía1. Patología Estructural y Funcional. Robbins et al. Sexta edición. Editorial Interamericana. 2000.2. Inmunología Celular y Molecular. Abul K. Abbas. Tercera edición.3. Tratado de Fisiología Médica. Guyton-Hall. Novena edición. Editorial Interamericana. 1998.4. Medicina Interna. Farreras-Rozman. Decimocuarta edición. Vol. II. Editorial Harcourt. 2002.5. Principios de Medicina Interna. Harrison y col. Decimocuarta edición. Vol. II. Editorial McGraw-Hill
Interamericana.6. La Respuesta Inmune. Ricardo A. Margni.. Ciencia Hoy, vol. 6 Nº 36. 1997.7. Inmunología. Fundamentos. Ivan Roitt. Novena edición. Editorial Panamericana.8. Fundamentos de Inmunología e Inmunoquímica. Margni. Quinta edición. Editorial Panamericana.
1996.


















