que Codifica para la Fosfolipasa C (PLC) en Raíces ... · PKC: Proteína cinasa C PTK ... En...
74
MAESTRÍA EN CIENCIAS Y BIOTECNOLOGÍA DE PLANTAS Detección de la Presencia del Gen que Codifica para la Fosfolipasa C (PLC) en Raíces Transformadas de Catharanthus roseus (L) G. Don Lucila Aurelia Sánchez Cach Centro de lnvestigación Cientifica de Yucatán, A.C.
Transcript of que Codifica para la Fosfolipasa C (PLC) en Raíces ... · PKC: Proteína cinasa C PTK ... En...

MAESTRÍA EN CIENCIAS YBIOTECNOLOGÍA DE PLANTAS
Detección de la Presencia del Genque Codifica para la Fosfolipasa C(PLC) en Raíces Transformadas de
Catharanthus roseus (L) G. Don
Lucila Aurelia Sánchez CachCentro de lnvestigación Cientifica de Yucatán, A.C.


CENTRO DE INVESTIGACIÓN CIENTÍFICA DE YUCATÁN, A.C.
UNIDAD DE BIOLOGiA EXPERIMENTAL
DETECCIÓN DE LA PRESENCIA DEL GEN QUE CODIFICAPARA LA FOSFOLIPASA C (PLC) EN RAÍCES
TRANSFORMADAS DE CatAaranfAus nose«s (L) G. Don
TESIS QUE PRESENTA:
Q. F. 8. LUCILA AURELIA SÁNCHEZ CACH
PARA OBTENER EL GRADO DEMAESTRÍA EN CIENCIAS Y BIOTECNOLOGÍA DE PLANTAS
IVIÉRIDA, VUCATÁN, MÉXICO2001


DEDICATORIAS
A Í)ií)8, por í;oáo 1o qaQ me hJi ó8:do.
A mi eFposo Carlos por todo el Íj_Qm:po qaecí]"parcimos Qn ql posgraáo, por
¢orzo r:aazir,o me ha 8;:y7ióaóo cin ciBf,e
¢:aóüo, por toda sa pac'iQncia y porCodo Ql amor Tac] com:partjmo8.
A mis Áj:jos, Freidóy Jeisíis r a tiCÁiqTuSSa, porgae Sozi en z7]i vzdó
lo máB vaJmBo.
A mis paóre8 7 mi saQgra, c}8peciíiheD4ea poi madrQ r s mi 3ric)gra porTtiq
siam:prQ paodo cont4r con Q11a8.
A mis hermímas, por todo el cariíio Tae7ue órmda]i
A mis amigo8, porTae Qn t,oáo momefff,o
paciáo comtíir con Qrb8.


RECONOCIMIENTOS
El presente trabajo se realizó bajo la dirección de la Dra. Teresa HemándezSotomayor en la Unidad de Biología Experimental del Centro de lnvestigaciónCientífica de Yucatán, A. C.
Este trabajo fue financiado por el Consejo Nacional de Ciencia y Tecnología(4119P-N9608). Lucila Aurelia Sánchez Cach fue becaria del Consejo Nacional deCiencia y Tecnología (116920).
AGFUDEcllvllENTOS
A la Dra. S. M. Teresa Hemández Sotomayor, a la Dra. Anaid Antaramian,al Dr. Gregorio Godoy Hemández y al Dr. Felipe.Vázquez Flota, por fomar pahedel comité tutoral y por sus sugerencias en la realización de este trabajo.
A la Dra. S. M. Teresa Hemández Sotomayor, a la Dra. Anaid Antaramian,al Dr. Felipe Vázquez Flota, al Dr. Diógenes lnfante Herrera y al Dr Victor M.Suárez Solís, por §us sugerencias y revisión crítica con que enriquecieron estemanuscrito.
De manera muy especial al Dr. Felipe Vázquez, por sus comentarios y todala asesoría otorgada a mi formación y al presente trabaio.
A las autoridades del Centro de lnvestigación Científica de Yucatán A. C.por permitir realizar este trabajo en sus instalaciones, especialmente a la unidadde Biología Experimental.


ABREVIATURAS
ADN: Ac.ido desoxirribonucleicoADNc: Acido desoxirribonucleico complementarioARN: Acido ribonucleicoBLAST: Basic Hnear Alignment Search Tool.bp: pares de basescpm.. cuentas por minutoCTAB: Cetiltrimetil bromuro de amonioDAG: 1,2-diacilglicerolDEPC; Dietilpirocarbonatod NTP: Desoxi rri bonucleótidos trifosfatatadosDTT: DitiotreitolEDTA: Acido etilendiaminotetraacéticoGDP: difosfato de guaninaGTP: Trifosfato de guaninah: horalp3: lnositol 1,4,5-trifosfatokb: KilobaseskDa: K`ilodaltonesmin: minutolvIOPS: 3-(N-Moriolino) propano ácido sulfónicoPCR: Reacción en cadena de la polimerasaPH: Homología a pleckstrinaPl: FosfatidilinositolPIP. Fosfatidilinositol fosfatoPIP2: Fosfatidilinositol 4,5-bifosfatoPl-PLC: Fosfolipasa C específica para fosfatidilinositolPLC: Fosfolipasa CPLD: Fosfolipasa DPKC: Proteína cinasa CPTK: Proteína cinasa de tirosinaÍpm: revoluciones por minutoRT-PCR: Transcripción Reversa-Reacción en Cadena de la PolimerasaSDS: Dodecil sulfato de sodioSH2: Homología a Src 2SH3: Homología a Src 3Src:SSC: Cloruro-citrato de sodioTE: Tris-EDTATBE: Tri s-Bórax-E DTATDC: Triptofano descarboxilasaQ.32PdcTP.. Trifosfato de desoxicitidina


CONTENIDO
Resumen
CAPITULO 1
CAPITULO 2
CAPITULO 3
CAPITULO 4
CAPITULO 5
Abstract
lntroducción
AntecedentesReceptores y transducción de señales
Fosfolipasa C en mamíferos
Regiones estructurales
Dominios X y Y
Dominios SH2 y SH3
Dominios PH
Dominios EF-hand
Dominio C2
Mecanismos de activación y regulación de las PLCs
PLCP
PLcy
PLC6
PLC nuclear
Fosfolipasa C en plantas
Estudios moleculares de PLC en diferentes sistemas
Estudios moleculares de PLC en plantas
Fosfolipasa C de C, roseus
Modelo de estudio Cafharanfhus roseus (L) G. Don
Detección por Southern-blot del gen que codifica para lafosfolipasa C y su expresión en Northern-blot en raícestrans{ormadas de Catharanthus roseus
Análisis de los ADNcs que codifican para la fosfolipasa Cde raíces transformadas de Caíharar}fhus roseusobtenidos mediante estrategias de RT-PCR
Discusión general
Conclusiones y perspectivas
3
5
77
8
10
10
11
11
11
12
12
12
13
14
14
14
15
18
19
20
33


RESUMEN
La fosfolipasa C (PLC) específica de fosfoinosítidos tiene una función crucial enla transducción de la señal que inicia en la superficie celular con la generación de lossegundos mensajeros diacilglicerol e inositol trifosfato a partir de fósfátidilinositolbifosfato. En mamíferos la PLC se clasifiü en tres grupos de acuerdo a su secuenciade aminoácidos, P, y y Ó, y se encuentra asociada a la membrana plasmática,distribuidas en el citosol o en el núcleo celular. En plantas, hace algunos años sedetectó la presencia de esta enzima en forma semejante a la de los mamíferos, esdecir, asociada a la membrana y en el citosol.
Hasta la fecha se han clonado los ADNcs que codifican para esta enzíma de\Tes modelos de es"tiio., Arabidopsis thaliana, Glycine max y Solanum tuberosum`Todos estos ADNcs codifican proteínas que se clasifican dentro del grupo de las PLCtipo Ó de mamíferos.
En raíces transformadas de Caíhafflnínus mseus se ha detectado la actividadde la PLC asociada a la membrana y en el citosol. El objetivo de este trabajo fue laclonación del gen o el ADNc de la PLC. lnicialmente se presenta el análisis de 10clonas parciales aisladas a partir de ARN total de raíces transformadas de C. mseusmediante la técnica de RT-PCR (Transcrjpción Reversa y Reacción en Cadena de laPolimerasa) utilizando un oligonucleótido sentido diseñado contra las secuencias Xconservadas de la PLC y el oligo dT en antisentído. Posteriormente se realizaronexperimentos de Southem y Northem-blot con una sonda de ADNc de PLC de A.Íha//.ana y se determinaron las condiciones para detectar la presencia de la PLC a nivelgenómico, no siendo posible detectar los niveles de expresión a lo largo de un ciclo decultivo.
Para validar las técnicas, se realizaron estos mismos experimentos utilizandouna sonda homóloga a C. n)seus como es la triptofano descarboxilasa (TDC) y uno delos fragmentos obtenidos de las clonas. Estos experimentos fueron positivoslográndose detectar este gen.


ABSTRACT
Phosphoinositide-specific phospholipase C plays a crucial role in initiatingsurfaa=-mediated signal transduction by generating the second messenger moleculesdiacylglycerol and inositol phosphates from phosphatidylinositol bisphosphate. lnmammalian cells, PLC is clasified in P, y and 6 forms depending on their aminoacidsequence. PLC can be membrane-attached or localized in cytosol or in nucleus.
A few years ago, PLC activity was detected in plants, both in plasma membraneand in cytosol, as it happens in mammalian cells. Purification of the enzyme andcDNAs cloning have been reported in Arab/.dops/'s Íha/i.ana, G/i.cyne max ahd So/anumfubemstím. These cDNAs encode PLCÓ-like proteíns.
PLC activity has been also detected in Cafharanínus msetts hairy roots both inplasma-membrane and in cytosol. The main goal of this work was to clone the gene orcDNA related to PLC. We stafted Wth the cloning of PLC gene or cDNA, firsth \^®analized 10 putative PLC fragments obtained from Reverse Transcriptase-PolimeraseChain Reaction (RT-PCR) of C, roseus transformed roots. Southem-blots andNonhern-blots analysis were peíformed using a PLC cDNA clone from A, Íha/Í.ana as aprobe, and conditions to detect PLC presence in genomic DNA were determined bothfor gene level expression along the culture cycle.
To validate these technics the same experiments \^ere done using a C. mseustryptophan decarboxylase (TDC) cDNA clone as a homolog probe. Finally the sameanalysis vvas carried out vrith a putative PLC fragment as a probe.


lNTRODUccloN
En los organismos, el mantenimiento de la homeostasis celular requiere de uncontinuo flujo de información entre las células que lo integran. Este flujo es necesariopara coordinar y regular el desarrollo y la organización dentro de lo§ tejidos, paracontrolar el crecimiento y la división celular y para responder a señales químicas delmedio ambiente. En organismos complejos la comunicación celular se realiza a travésde diversas moléculas transductoras que actuan como señales extraoelulares (Lodishet al.,1995).
En la membrana plasmática existen diversos receptores que. han sidoclasificados en diferentes familias con base a su estructura o de acuerdo a la funciónque desempeñan, como por ejemplo, aquellos que regulan el paso de iones a travésde la membrana (canales iónicos), los que presentan una actividad enzimáticaintrínseca de cinasa o de fosfatasa y los acoplados a proteínas G heterotriméricas(Drayer y Van Haastert,1994).
En plantas y animales las señales extracelulares controlan el crecimiento demuchos tejidos, regulan la síntesis y la secreción de proteínas así como la oomposiciónde los flLiidos del organismo. Las células del organismo detectan lo que sucede en elambiente extracelular, manteniendo su homeostasis y traduciéndolo en respuestafisiológica. Algunas señales inducen una modificación en la actividad de una o másenzimas presentes en la célula. Los mecanismos de transducción de señales en lascélulas animales han sido ampliamente estudiados. Estos inician en receptores de lamembrana plasmática que pueden estar acoplados a proteínas G heterotriméricas, seha observado la participación de diferentes enzimas capaces de producir moléculasdenominadas segundos mensajeros, estos segundos mensajeros se encargan decontinuar el flujo de información al interior de la célula, jnduciendo cambios a nivelintracelular (Drayer y Van Haastert,1994; Lodish et al.,1995).
Un ejemplo clásioo de la generación de segundos mensajeros es aquel queutiliza la hidrólisis de fosfoinosítidos por acción de la fosfolipasa C para producir dossegundos mensajeros intracelulares: el 1,2diacilglicerol de naturaleza hidrofóbica y elinositol 1,4,5-trifosfáto de naturaleza hidrofílica (Shulka,1982; Berridge,1987). EI DAGpemanece en la membrana y activa a la proteína cinasa C (Dekker et al.,1995) y elIP3 se intemaliza en el citosol para inducir la liberación de Ca2. del retículoendoplásmico (Nishizuka,1992; Berridge,1993).
En mamíferos se han identificado 10 isoenzimas de PLC, que de acuerdo a susecuencia de aminoácidos, han sido clasificadas en tres tipos: P, y y 8 (Rh® y Choi,1992; Cockcroft y Thoma8, 1992; Benridge, 1993; Noh et al., 1995) y que seencuentran distribuidas en el citosol (Ryu et al., 1987a y b; Bennet y Crooke] 1987;Rebecchi y Rosen,1987; Fukui et al.,1988; Homma et al.,1988; Meldrum et al.,1989;Waldo et al.,1994), en la membrana plasmática (Banno et al.,1988; Banno y Nozawa,1987; Lee et al., 1987; Baldassare et al., 1989) y en el núcleo celular (Martelli et al„

1992; Kuricki et al„ 1992; Diveclia et al.,1993; Mazzoni. et al.,1992). Todas consistende una sola cadena polipeptídica (Rhee y Bae, 1997) y tienen en común variasregiones estructurales como son: los dominios PH de homología a la pleckstrina,(Tyers et al.,1988; Mayer et al.,1993; Haslam et al.,1993; Musacchio et al.,1993;Shaw,19930), las regiones EF-hand (Bairoch y Cox,1990; lkura,1996), los dominiosX y Y, (Emori et al.,1989; Rhee,1991) los dominios C2 y los dominios adicionales enPLCP (una región de aproximadamente 500 aminoácidos) y en la PLcy los dominiosparecidos a Src (SH2 y SH3) (Emori et al.,1989; Shu et al.,1988; Rhee y Bae,1997).
En plantas se ha detectado la actividad de esta proteína tanto en el citosol comoasociada a membrana en varios modelos de estudio (MCMurray e lrvine,1988; Kamaday Muto,1991; Yotsushima et al.,1992; De Los Santos-Briones et al.,1997); y se hanpurificado y recientemente se han clonado ADNcs que codifican para esta proteína entres modelos de estudio; A Íha/i.ana, S. Íubenosum y soya (Yamamoto et al., 1995;Hirayama et al.,1995; 1997; Shi et al.,1995; Kopka et al.,1998).
En este trabajo se presenta un avance en el proceso de clonación del ADNc quecodifica para la fosfolipasa C de raíces transformadas de Caíharaníbus n]seus.

Capítulo 1
ANTECEDENTES
RECEPTORES Y TIUNSDUCCION DE SEÑALES
La capacidad para responder al ambiente circundante y controlar la entrada ysalida de moléculas a través de la membrana plasmática son características crucialesde una célula. Estos procesos son atrlbuidos a proteínas que residen en la membranaplasmática. La membrana plasmática es impermeable a moléculas hidrosolubles,incluyendo iones, pequeñas moléculas inorgánicas y polipéptidos o prot9Ínas. Paraentrar o influenciar el interior de la célula, una molécula hidrofílica debe reaccionar conuna proteína residente en la membrana plasmátiffi. Esta molécula extracelular esllamada un agonista. La proteína blanco a la que el agonista se une es llamada unreceptor (fig 1,1). Estas interacciones no son necesarias para una moléculaliposoluble, tales como las hormonas esteroidales, las cuales pueden difundir a travésde la membrana plasmática al citoplasma donde se unen a su proteína blanco (Lewin,1994).
Figura 1.1 : Esquema del modelo de transducción de señales, Oomado de Chin-Vera,1995).
La señal es transmitida por medio de un cambio en la conformación de unaproteína de membrana. Como resultado de esta modificación, hay un cambio
7

subsecuente en la actividad del dominio citosólico de la proteína de membrana o conalguna otra proteína con la cual ésta se asocia, fig 1.1, (Lewin,1994),
Cuando un receptor permanece en la membrana y responde a la unión de unagonista, activa un mecanismo de respuesta en el citosol, este proceso es llamadotransducción de señal, (fig 1.1) ya que, en efecto una señal ha sido transmitida através de la membrana (Levm,1994)
La transducción de una señal proporciona un mecanismo para la amplificaciónde la señal original. Cuando un agonista extracelular interactúa con un receptor demembrana, éste se convierte de su forma inactiva a su forma activa. En su forma activael receptor estimula una actividad catalítica que genera una señal citosólica muchomayor que la señal extracelular original Algunas veces la señal citosólica incrementala cantidad de una pequeña molécula dentro de la célula. Una molécula producida enrespuesta a la transducción de una señal extracelular es llamada segundo mensajero(Lewn,1994)
Existen dos mecanismos principales de transducción de señales; uno en el queel receptor puede ser una proteína transmembranal con dominios tanto extracelularescomo citosólicos en el que la unión del agonista en el lado extracelular influyen en laactividad del dominio citosólico, Una reacx=ión típica es la unión del agonista paraactivar a una proteína tirosina cinasa transmembranal, la cual fosforila su propiodominio citciplásmico que capacita a este receptor para asociarse y activar a unaproteína blanco la cual a su vez actúa sobre un nuevo substrato en la célula. En el otromecanismo el receptor interactúa con una proteína G que está asociada a lamembrana. Las proteínas G son llamadas así por su capacidad para unir nucléotidosde guanina. La forma inactiva de la proteína G es un trímero unido a una molécula dedifosfato de guanina. Cuando se une el agonista a un receptor, éste actúa sobre unaproteína G causando la disociación del nucleótido difosfatado que es reemplazado porun nucléotido trifosfatado. Como resultado la proteína G se disocia en una subunidadsimple que lleva el nucléotido (subunidad G) y en un dímero (subunidades Py), estosúltimos activan a otras proteínas blanco que en ocasiones están asociadas a lamembrana y estas proteínas a su vez reaccionan con una molécula blanco en elcitosol. Estos eventos estimulan la producción de segundos mensajeros como es elcaso del inositol l ,4,5-trifosfato y el l ,2-diacilglicerol producidos por la fosfolipasa C(Levm,1994).
FOSFOLIPASA C EN MAIVIÍFEROS
La hidrólisis de fosfolípidos de membrana, entre ellos el fosfatidilinositol 4,5-bifosfato por una fosfolipasa C específica es uno de los primeros eventos en laregulación de varias funciones extracelulares realizadas por más de 100 moléculas deseñalización extracelular (Rhee y Choi, 1992; Cockcroft y Thomas, 1993; Berridge,1993, Noh et al.,1995). Esta reacción producx3, al hidrolizar el PIP2, dos segundosmensaieros intracelulares el 1,2 diacilglicerol de naturaleza hidrofóbica y el inositoll,4,5-trifosfato de naturaleza hidrofílica (Shulka, 1982; Bemdge, 1987). EI DAG
8

ppe3T::T:,=m:ne'naeT:Tobsr:,n:a?aa,:ávuac,:,:a"pbr::::r.:nc::acsta29d(epe.:tí:ru,eotáláo'p:::Lí®e:fig 1.2, (Nishizuka,1992; Berridge,1993)
0 -o-b-o_C,-,-Oo
© i(Po
O=C C=O
_H2C-?-CHI"
00
>
_O-r=O J-
® Ol\P:-o-
Activación de PKC
Liberación de Ca2+
Figum 1.2: Esquema de la reacción que cataliza la PLC. PIP2, Fosfatidilinosnol 4,5-bifosfato;PLC, Fosfolipasa C., DAG, Diacilglicerol; lp3, lnositol 1,4,5-trifostato; PKC, Proteína cinasa C.
A la fecha en mamíferos se han identificado 10 isoenzimas de PLC, todas ellasson cadenas polipeptidícas sencillas que de acuerdo a su secuencia de aminoácidos,han sido clasificadas en tres tipos: P, y y 8. De éstas, cuatro son de tipo PLCP, dos detipo PLcy y cuatro de tipo PLC8, fig 1.3, (Rhee y Choi,1992; Cockcroft y Thomas,1992, Berridge,1993: Noh et al ,1995), cada subgrupo, es nombrado por la adición deun número arábigo después de la letra griega, por ejemplo; PLCSi, PLCP2 (Rhee yChoi,1992).
Se ha observado que las isoenzimas de PLC se loffilizan tanto en el citosol(Ryu et al.,1987a y b., Bennet y Crooke,1987: Rebecchi y Rosen,1987, Fukui et al.,1988; Homma et al ,1988; Meldrum et al ,1989, Waldo et al.,1994), como asociada amembrana plasmática (Bamo et al„ 1988: Banno y Nozawa,1987; Lee et al ,1987.,Baldassare et al , 1989) Por otro lado también existen evidencias de que la PLC selocaliza en el núcleo celular (Martem et al., 1992, Kuricki et al , 1992; Divecha et al.,1993, Mazzoni et al.,1992).
9

Regiones estructu rales
Las isoenzimas de las PLCs, consisten de una sola cadena polipeptídica conpesos moleculares de 150-154 kDa para la PLCP,145-148 kDa para la PLcy y 85i38kDa para la PLCó (Rhee y Bae,1997) y tienen en común varias regiones estructuralescomo son: los dominios PH de homología a la pleckstrina, las regiones EF-hand, losdominios X y Y, los dominios C2 y los dominios adicionales en PLCP (una región deaproximadamente 500 aminoácidos) y en PLcy los dominios con homología a Src (SH2y SH3), fig 1 3.
Ii!Ñi!;,:__
H2
Figura 1.3: Repi.esemación esquemática de la clasificación y los dominios esú.ucturales
g:;°:tg¡rnuaps°qsu8et¡::ecriFaHéarpg:íga%ed:°Tn::°8:a2+:£';C§#¡nFOFs-h£Tg,',t:g::nsff2n;U3i¡§rstT::ant:::homólogas a Src; C2, regjones homólogas en la familia de proteína cinasas C dependientes de Ca2+ y laGp, extención carboxilo teminal de interacción con la proteína G. (Adaptado cle lívine,1996)
Dominios X y Y
El análisis de la secuencia de aminoácidos de los tres grupos de PLCs revela laexistencia de dos regiones altamente conservadas denominadas dominios X y Y deaproxmadamente 170 y 260 aminoácidos respectivamente La homologia que hayentre los dominios X es de un 60% y el de los dominios Y de 40% en las diferentesespecies. Estos dos dominios son necesarios y suficientes para la actividad catalíticade la enzima (Emori et al., 1989; Rhee, 1991) como ha sido confirmado por laestructura cristalográfica de la PLCói (Essen et al ,1996) Estas regiones, separadasen su secuencia primaria por secuencias de tamaños variables, convergen en unaestructura tridimencional para fomar dos mitades de un "barril TIM" (triosa fosfatoisomerasa) dtstorcionado y cerrado.
10

Dominjos SH2 y SH3
Las isoenzjmas PLCP y las PLCs contienen una corta secuencía de 50-70aminoácidos que separa las regiones X y Y. Las isoenzimas PLcy tienen una largasecuencia de aproximadamente 400 aminoácidos donde están contenidas las regionesadicionales con secuencia homóloga a la proteína Src, dos regiones SH2 y una SH3(Emori et al.,1989; Shu et al.,1988; Rhee y Bae,1997).
Los dominios SH2 y SH3 son pequeños modulos de estructura proteíca quecomprenden aproximadamente 100 y 50 aminoácidos respectivamente que regulaninteracciones proteína-proteína. Estos dos dominios en PLcyi no son necesarios parala actividad enzimática ya que la proteína retiene la actividad cuando estas regionesson elímínadas (Bristol et al.,1988; Emori et al.,1989). A pesar de esto, los dominiosSH2 de la PLcy, están involucrados con la asociación de esta proteína con sitiosespecíficos fosforilados en tirosina presentes en otras proteinas (Anderson et al.,1990; Margolis et al.,1990a y b; Mohammadi et al.,1991; Rotin et al.,1992; Soler etal.,1993).
Se sugiere que esta interacción es un prerrequisito para la fosforilación dePLcyi en residuos de tirosina por tirosina cjnasas activadas. El dominio SH3aparentemente funciona facmtando la asociación de la PLcyi con proteínas decitoesqueleto, particularmente con proteínas ricas en residuos de prolina (Bar-Sagi etal.,1993; Gout et al.,1993).
Dominios PH
El dominio homológo a pleckstrina (proteína sustrato de la proteina cinasa C enplaquetas), denominado dominio PH, es una secuencia regulatoria de alrededor de100 aminoácidos encontrado en varias proteínas involucradas en la transducción deseñales (Tyers et al., 1988; Mayer et al., 1993; Haslam et al., 1993; Musacchio et al.,1993; Shaw,1993),
Todas las isoenzimas de PLC de mamíferos contienen un dominio PH en laregión amino terminal que precede a las regiones EF-hand (Parker et al.,1994; Essenet al., 1996). Las PLcy contjenen otra región PH entre las regiones X y Y la cual esinterrumpida por las secuencias SH2 y SH3. Se sugiere que el dominio PH aminoterminal de la PLC6i es necesario para unirse al PIP2 en vesículas biffipa (Rebecchi etal., 1992). También se ha considerado que el dominio PH ayuda a la translocación dela PLC a la membrana plasmática uniéndose a ésta a través de la interacción con elsubstrato PIP2 con lo cual se ve estimulada la actividad catalítica de la PLC (Cifuenteset al.,1993; Parker et al„ 1994; Harlan et al.,1994).
Dominio EFhand
El motivo estructural "EF-hand" fue descubierto por primera vez en la estructuracristalina de la parvalbúmina por Kretsinger y Nockolds en 1973. Dicha estructura
11

f=rnmsástuend:,t?gsd:-hu:','o=spda',sapuceas2t.asEPs::P:nod,:::'aersT,eunctt:,ya,:e,eu,::vaas:::::r::#:,q::sido identificado en otras numerosas proteínas. muchas de las cuales funcionan comosensoras de Ca2+ en la célula y entre las que se pueden mencionar a la calmodulina yla troponina. Las PLCs poseen una región "EF-hand" localizada en la región amino
;er:T:?na:,ssLgeu,t:enndeon:J£:::tá:dpdHe,u::rtacar2?i,Bó:,r:,cehn:n:náoírT:,áon,,keugr:;a|t:gg)en
Dominio C2Además las PLCs poseen en su dominio carboxilo terminal una secuencia
homóloga a C2 que se puede considerar como una extensión del dominio Y. Los
:,on::n::sccd2e;ueenr:,:J?eesntáf:ücdag:Psmo:rr::,:enr::a:opma:áoiaa:o:an,Lae;aeTá,:n::apdr:tsían2?por las enzimas (Kaibuchi et al.,1989)
Las lsoformas P de la PLC contienen regiones adicionales que confierencaracterísticas regulatorias únicas a estas moléculas; éstas son extensiones carboxiloterminal (de cerca de 500 aminoácidos) que se asocian a las proteínas G y de estemodo son reguladas (Pawson, 1995).
MECANISIVIOS DE ACTIVACION Y REGULACION DE LAS PLCS
PLCP: La función de la proteína G en la regulación de la hidrólisis del PIP2 seconoce a través de la acción de muchas hormonas que utilizan receptores acoplados aproteínas G, mediante la acción de los nucleótidos de guanina en preparaciones libresde células, y en algunos casos, por medio de la atenuación con la toxina pertusis. Lassubunidades ci de todas las proteínas G heterotriméricas activan a las isoenzimas dePLCP pero no a las PLcy o PLCÓ (Rhee y Choi,1992; Cockcroft y Thomas, 1992;Berridge, 1993; Noh et al., 1995, Stemweis y Smrcka, 1992). Los receptores queactivan el meffinismo (Ga-PLcf}) incluyen aquellos para tromboxanos A2, bradiquinina,angiotensina 11, histamina, vasopresina, acetilcolina y agonistas G1-adrenérgicos (Nohet al.,1995; Kuang et al.,1996).
Las subunidades Ga activadas por GTpys (un análogo no hidrolizable del GTP)estimulan la actividad de las isoenzimas de PLCP en el siguiente orden, PLCP, 2PLCÍ}3 > PLCP2 (Noh et al ,1995; Lee y Rhee 1995). La PLCP4 también es activada porlas subunidades ci de las proteínas G pero su actividad basal es inhibida porribonucléotidos entre los que se incluyen GTpys, por lo que se dificulta una estimaciónreal de un aumento en su activ`dad enzimática (Lee et al., 1994).
Las subunidades f)y de las proteínas G han demostrado ser reguladoresefectivos de varios efectores de proteínas G (Sternweis, 1994). La regulación de laactividad de la PLCP por las subunidades Py de las proteínas G se reportó primero enpreparaciones crudas de células HL60 (Camps et al ,1992) y de hígado (Blank et al.,
12

1992) La sensibilidad de las isoenzimas de PLCP por las subunidades By difieren delde las GG y disminuyen en el siguiente orden PLCP3 > PLCP2 > PLCpi (Noh et al.,1995; Lee y Rhee, 1995, Smrcka y Sternweis, 1993; Park et al., 1993) La PLCP4parece no ser sensible a la activación por las subunidades Py (Lee et al.,1994, Jianget al.,1994). La capacidad de las subunidades Py de las proteínas G para activar a laPLCP2 es propiciada en respuesta a una unión con el receptor de la homonaluteinizante, el receptor de vasopresina V2, receptores Pl y P2 adrenérgicos,receptores muscarimcos m2 de acetilcolina y los receptores de interleucina s Todoesto ha sido demostrado usando un sistema de cotransfección en células COS (Noh etal.,1995; Zhu y Birnbaumer,1996., Jiang et al„ 1996).
La región de la PLCB que interactúa con la subunidad a de la proteína G difierede las que responden a las subunidades Py, mientías que la región ffirboxilo terminalque sigue al dominio Y es esencial para la activación de PLCpi y PLCP2 porsubunidades a (Nho et al.,1995; Lee y Rhee 1995), el sitio de interacción de la PLCP2con las subunidades Py está localizado entre los res.iduos Glu435 y Vai6" (Kuang et ai.,1996a).
PLC7: La activación de la PLC t.ipo y ocurre a través de su fosforilación por unaproteína tirosina cinasa o por receptores con actividad de tiros.ina cinasa (Nishibe etal ,1990; Koch et al.,1991) La activación de la PLcy por cinasas se puede dar através de factores de crecimiento, tales como el factor de crecimiento derivado deplaquetas, factor de crecimiento epidérmico, factor de crecimiento de fibroblastos,factor de crecimiento de nervios y factor de crecimiento de hepatocitos. La unión deestos factores de crecimiento a sus receptores induce la activación de una proteínat.irosina cinasa intrinseca del receptor y la consecuente fosforilación en residuos detiro§ina de numerosas proteínas incluyendo al receptor per se y a la PLcy (Rhee yChoi, 1992, Cockcroft y Thomas, 1992., Berridge, 1993; Noh et al., 1995). Laautofosforilación del receptor crea sitios de unión de alta afinidad para varias proteínasque contienen dominios SH2, como la PLcy (Noh et al„ 1995). La fosforilación de laPLcyi por los receptores de factores de crecimiento ocurre en los mismos sitios y son_ ., ._ _.,_,__:__ ..^ ,^ T„.783 hi^^iiaa la
y 1254. La sustitución por fenilalanina de la Tyr783 b|oquea iapor factores de crecimiento derivados de plaquetas en células_ __ ,_ __l:_.__:j.-aiiivauiu.I `IC> .L ...- `~ r -..----.-----
NIH3T3, Io que indica que la fosforilación de la Tyr783 es importante para la activaciónde la PLcy,. (Rhee y Cho.i,1992; Cockcroft y Thomas,1992., Berridge,1993; Noh elal., 1995, Lee et al., 1995). Esta fosforilación en tirosina de la PLcy[ parece promoversu asociación con componentes del citoesqueleto ayudado por el domin.io SH3.
Las PTKs que no son receptores también fosforilan y activan a las isoenzimasde PLcy en respuesta a la unión de ciehos ligandos a receptores de la superficiecelular, tales receptores .incluyen los receptores antigénicos de las células T,inmunoglobulinas de membrana, integrinas y varios receptores de citosinas (Noh et al. ,1995; Keely y Parise, 1996, Gómez-Guerrero et al., 1996, Misra et al., 1995). Lasregiones honmlógas a Src parecen ser responsable de la fosforilación en tirosina de laPLcy en el músculo liso de células vasculares y en plaquetas, ya que laelectroporación de anticuerpos contra Src inhibió la fosforilación de la PLC7 promovida
13
las tirosinas 771, 783activación de la PLC

por angiotensina 11 o factores activadores de plaquetas, en estas células, corrobcirandode esta manera la participación de las regiones homologas a Src en este proceso(Sadoshima e lzumo,1996).
Las isoenzimas de PLcy pueden ser activadas directamente por segundosmensajeros derivados de lípidos en la ausencia de la fosforilación de tirosina El ácidofosfatídico producido por acción de la fosfolipasa D activa a PLcy purifi.cada a travésde un modificador alostérico (Jones y Carpenter,1993) Las isoenzimas de PLcy sontambién estimuladas por ácido araquidónico en presencia de la proteína tau en célulasneuronales o de proteínas semejantes a tau en células no neuronales (Hwang et al„1996) El efecto de tau y el acido araquidónico es específico para isoenzimas de PLcyy es marcadamente inhibida por fosfatidilcolina
PLCS Aún no se conoce el mecanismo de regulación de la PLC8, §in embargoexisten dos mecanismos propuestos, el primero es a través de una proteína que uneGTP llamada Gh (Feng et al,1996) Esta proteína Gh está formada por dossubunidades, una G y una P (lm y Graham,1990) La subunidad GhQ es una proteínamultifuncional que tienen actividad de transglutaminasa y activa formando un complejocon la PLCói purificada (Nakaoka et al,1994, Das et al,1993) El segundomecanismo propuesto involucra a un polipéptido con homología limitada a protei'na§con actividad de Rho-GAP que pueden estimular la actividad de PLCSL pero no PLC0io PLcyi (Homma y Emory,1995)
PLC riucleam La cascada de transducción de §eñales de la PLC también pareceocurm en el núcleo (Divecha e lrvine, 1995) La PLCpi es la i§oforma de PLC másabundante en el núcleo de varios tipos de células La proteína nuclear de PLCpiparece ser activada independientemente de su contraparte en la membrana plasmáticapor un mecanismo desconocido, más activo durante el crecimiento celular que durantela diferenciación (Martem et aL 1992, York y Majerus, 1994, Marmiroh et al , 1994,Divecha et al.,1995) Los cambios en la cantidad nuclear de PLCpi se correlacionancon cámbios en la cantidad de PIP2 hidrolizado en el núcleo Estudios en las células
que carecen de PLcf}i como consecuencia de la mutación del gen correspondiente,revelan que esta proteína es esencial para iniciar la síntesis de ADN en respuesta afactores de crecimiento semejantes a insulina (Manzoh et al , 1997)
FOSFOLIPASA C EN PLANTAS
En células vegetales también se han identificado enzimas con actividad de PLC,tanto citosólicas como membranales (MCMurray e lrvine, 1988; Melin et al , 1987;Pfaffman et al ,1987, Tate et al ,1989, Kamada and Muto,1991, Yotsushima et al ,1992, De Los Santos-Briones et al., 1997) Las enzimas citosólicas hidrolizan
preferencialmente al Pl y PIP y por último al PIP2 en tanto que las membranales tienen
#f;:í;:espá:cp:2P.,ybrep;Pa2ra::Sma:x|::aasct,vc,,á:sdo(,MfficsMu?reacye:,tí:,.:o,ngcsesn,tri?,,:nnee:
:'ém'b::Zá,Y:;sui:::Ta®entcea|t,ra`c:ogn2és`:,,:reo;:,:;ésldge8oc,:2,m,:g;:aiM::::::,:e|9t:P,?
14

Yotsushima et al„ 1992; lrvine et al.,1980; Arz and Grambow,1994; Dr®back,1992;Melin et al.,1992).
En contraste con estos resultados, la PLC de C. /oseus, de tanto la fraccioncitosólica como la membranal, se activan a concentraciones micromolares de Ca2+libre La enzima citosólica tiene preferencia primero por PIP, posteriormente por Pl yPIP2 y la membranal por PIP2 primero y después por PIP y Pl (De Los Santos-Brioneset al.,1997)
ESTUDIOS MOLECULARES DE PLC EN DIFERENTES SISTEMAS
Entre las secuencias que se han aislado para la fosfolipasa C en mamíferos serpeLP:_h,a48unEát:Npcr.::ín¥rseebré:rdaec,::::n:oqruseucdoed;f:=depnacr,aaá:acpar2?t;í:::;|:iTándag:
PLC específica para fosfatidilinositol y presenta algunas regiones que tienen similitudcon el dominio no catalítico de proteínas cinasas que no son receptores (Mark et al.,1988).
Pann-Ghill y colaboradores en 1988a, reportaron el aislamiento de clonas deADNc de cerebro de rata mediante el uso de anticuerpos monoclonales contra PLCsde cerebro de bovino. Estas clonas corresponden a la PLC-l y la PLC-lll, (actualmentePLC 0 y 8), las cuales codifican para dos polipéptidos con pesos moleculares de 138.2y 85.8 kDa respectivamente; la comparación de secuencias de estas dos enzimas conotras isoenzimas de PLC-ll (de peso molecular 148.3 kDa) reveló un bajo nivel dehomología; sin embargo, también se encontró una similitud entre las tres isoformas endos regiones de aproximadamente 150 y 120 aminoácidos. Estos dominiosconservados son separados por una región variable en PLC-ll que tienen homologíacon las regiones no catalíticas de tirosina cinasas que no son receptores. Las PLC-l y111 son cortas y aparentemente no están relacionadas con estas tirosina cinasas.
En c]tro escrutinio de una biblioteca construida a partir de ARN de cerebro derata, utilizando anticuerpos contra la PLC de cerebro de bovino, se encontró un ADNcde 5106 bp, que codifica para una proteína con 1289 aminoácidos con peso molecularcalculado en condiciones desnaturalizantes de 148`4 kDa. Dos regiones en estaproteína exhiben una similitud significativa en su secuencia de aminoácidos con elproducto de varios oncogenes relacionados con tirosina cinasas (yes, src, ®r, ab/, rps,Íes y Íck) (Pann-Hill et al , 1988b). En un escrutinio realizado en una biblioteca deADNc de retina de bovino, utilizando como sondas fragmentos conservados del ADNcdel gen norpA que codifica para la PLC en D/osoph/./a me/anogaster, se encontraroncuatro formas alternativas de PLC del tipo P que son altamente homólogas a laproteína codificada por norpA El grado de conservación de secuencias entre estasproteínas y aquella codificada por norpA es mayor que la que existe con otras PLCscx)nocidas, como es el caso del motivo de GTpasa encontrado en las proteínas de PLCen la retina de bovino y la de norpA, pero no en las otras PLCs (Ferreira et al.,1993)
15

En otro reporte publicado en 1995 por Cheng y sus colaboradores se describeuna búsqueda en una biblioteca de ADNc de aorta de corazón humano con unfragmento de un ADNc de la PLCói de células de glioma de rata encontrando un ADNcde 2.3 kb el cual codifica una PLC tipo Ól de 756 aminoácidos con un peso molecularde 87 kDa estimado en electroforesjs desnaturalizante de poliacrilamida. EI ADNc dela PLC81 de humano tiene un 95% de homología con el ADNc de la PLCói de rat.a y laproteína es semejante a las PLCs tipo 8, ya que carece de una extención carboxiloterminal después del dominío Y (Cheng et al„ 1995).
Por otro lado, Lee y Rhee reportaron un ADNc que codmca para una PLCÓ4 queno había sido reportada previamente. Este ADNc se clonó a partir de una biblioteca deADNc de cerebro de rata realizando amplificaciones por Reacción en Cadena de laPolimerasa con oligonucleotidos diseñados a partir de las PLCs tipo s conocidas. EsteADNc codifica para una proteína de 772 aminoácidos con un peso molecular calculadoen electroforesis desnaturalizante de poliacrilamida de 88.9 kDa. En su estructuraprimaría es similar a las isoenzimas de PLC tipo 6 con una identidad de 45% para la
::pceóááe7n2coág:rac:2a,:eLfaüpyLC46Z%esp:ir:„::apíacsó3áeL:aap:i::ia;pe.s,p::í,fiJ#.sy::inmunodetección utilizando anticuerpos específicos contra la PLC84, se detectó en stejidos de la rata una proteína de 90 kDa con baja intensidad comparada con la deotras isoenzimas de PLC y otra de 93 kDa fue detectada en testículo pero no en losotros siete tejidos y los autores concluyen que la proteína de 93 kDa es derivada de unacomodo diferencial del ARNm que codifica para la PLC84 de 90 kDa que contiene 32aminoácidos adicionales entre los dominios catalíticos X y Y (Lee y Rhee, 1996).
En una búsqueda en una biblioteü de ADNc de hígado regenerado de rata seencontró una nueva PLC64 que codifica para una proteína de 770 aminoácidos consimilitudes estructurales a las isoenzimas de PLC reportadas. EI ARNm de la PLCó4 seexpresa más en hígado regenerado y en células tumorales que en hígado normal.E§tudios de inmimodetección e inmunocitoquímica demuestran que la PLC64predomina en núcleo e incrementa en la transcición de la fase Gi a la fase S del ciclocelular y continúa así hasta la fase M, desapareciendo cuando las células reinician lafase G,. Esto sugiere que la PLCÓ4 es expresada en el núcleo en respuesta a laestimulación mitogénica y que tiene una importante función en el crecimíemo celularcomo uno de los genes que se expresan tempranamente durante la transcición de lafase Gi a la fase S en el ciclo celular (Liu et al., 1996).
Además de los ADNcs de PLC clonados en mamíferos, también se hanreportado AIDNcs en Saccharomyces cerevisiae (PLC1)` Dictyostelium discoideum(Ddpl_C), Drosophila melanogaster (norpA), Bacillus cereus (PIPLC), Listeriamonocytoger"s (plcA) y Slaphyloooccus aureus Wc).
En S. ce/ev/-s/'ae, el gen PLC7 participa en el crecimiento ülular ya que unainterrupción del mismo da como resultado un lento crecimiento o la muerte de lalevadura. El gen f'LC7 codifica para un polipeptido de 869 aminoácídos con un pesomolecular ffilcülado de 101 kDa. Este polipeptido tiene las regiones X y Y conservadas
16

de todas las fosfolipasas de mamíferos y su estructura es similar a la PICs demamíferos. La expresión de un ADNc de PLCsi de rata suprime el defecto delcrecimientocausadoporlainterrupcióndelgen.(Yoko-Oetal.,1993).
En D. d/sco/.deum, DdpLC tiene fuerte homologia con la PLCs de mamíferos, nose ha descrito una función conocida, sin embargo, en mutaciones (pk9 las célulascrecen y se desarrollan normalmente aunque la activ.idad de PLC no es detectada.Previas observaciones en D. djsco/.deum sugieren que la regulación de la PLC esimportante para la quimotaxis y la diferenciación (Drayer y Van Haasteft 1992,Bominaar et al„ 1991, Bominaar y Van Haastert,1994) sin embargo, en las mutantesyamencionadasseobservaunaquimiotaxisydesarrollonormalylosnivelesdelp3son ligeramente reducidos
En 0 me/anogaster norpA codifica para la PLC. La .identificación del productode norpA como PLC emergió del hallazgo de que mutaciones de no.pA reducenseveramente la actividad de PLC en el ojo (Yoshioka et al ,1985) y que la proteina essimilar en estructura y secuencia de aminoácidos a la PLC de mamíferos (Bloomqu.istet al , 1988). Esta proteína es esenc.ial para la fototransducción y además de serampliamente encontrada en retina, Ios productos del gen son expresados en unavariedad de tejidos que no contienen maquinaria para la fototransducción implicandoquenorpAestáinvolucradotamb.iénenotrosmecanismosdetransducción(Zhuetal,1993). Las mutaciones en este gen a su vez disminuyen el potencial fotoreceptor,reduciendo la visión de la mosca (Hotta and Benzer, 1970). norpA tiene fuertehomología con otros tres genes de PLC tipo P (Bahk et al , 1994; Shortridge et al.,1991, Ferreira et al ,1993)
En 8. ce®tA ha sido comprobado que Pl-PLC, a pesar de tener un pesomolecular de 35 kDa como ha sido determinado por electroforesis en poliacrilamida,está fuertemente relacionada con las otras PLCs de eucar.iotes Se sabe también queestaproteínaessecretadaatravésdelamembranaplasmáticaalmediodecultivoyreconocenlaestructuradelinos.itolphosphatopresenteenelPl,peronohidrolizanlosderivados más fosforilados como P14 fosfato y PIP2 (Kuppe et aL 1989). Se hareportadoquelasPLCsbacterianasmuestranefectossemejantesalainsulinacuandoson incubadas con preparac.iones de membranas sensibles a .insulina o células talescomo membranas de hepatocitos (Saltiel y Cuatrecasas,1986) y hepatomas ( Mato etal.,1987).
En L, Monocyfogenes, p/cA cod.ifica para una proteína de 317 aminoácidos conunpe¥mL¿,gcoun,g:Eg?#'.cS_?:'|.en§J_#on`#ñ_g_r;{Í^pÉLm6úd#e£he==eu£=Btnt#nni?aggeonstsey\PLCs eucarioticas. La actividad de PLC en L. monocyfogenes se ha encontrado en elmedio de cultivo de la bacteria detectándose la mayor actividad en especiespatógenas, por lo que se cree que p/cA esta relacionada con las propiedades devirulencia del bacilo en comb.inación con la toxina listenolisina 0 (Mengaud et al ,1 991 ) .
En S. auyeu6 el gen p/c tiene una secuencia de 984 pb y codifica para unaproteína de un peso molecular aproximado de 341 kDa La cornparación de la
17

secuenciadeaminoácidosniuestraunasimilitudquevadel51al56°/oconlasPLCsde8 c`ereus 8 """ y L Monocyfogenes En S aurieus la actMdad de la PLCes asociada a cepas de estafilococos coagulasa positiva, sin embargo, esta actividadnoserelacionaconlapatogenicidaddelabacteria(DaughertyyLow,1993)
ESTUDIOS MOLECULARES DE PLC EN PLANTAS
Enlosúlti'moscmcDañosaparecieronreportesdelaclonacióndegenesdePLCde plantas superiores entre los que se encuentran tres de Araóíc/opsÍ.s "m (Atha''ana„node§oya(G|yc,nem`*xTit-r¿:`á:-i;£'(:o%nuuGm:%%:oosPuS#),t,%''FZ
AtpLCIS N
EF-hand
At pLCIF N
At pLC2 N
PI-PLcl soya N
St PLCISt pLC2 NSt PLC3
C (Yamamotoet. al„1995)
C (Hirayama et al.1995)
C (Hirayama et al 1997)
C (Shiet al .1995\
C (Kopkaetal„1998)
Figura Lti Represenúcjón esquemáijca de las PLCs de planüs y los dominios
io:ng:5¡Cé:Ii,'exs.yEyF-!::|h,roesginta,í:,nmfsuná,::eLeg::astoerha,::a:rpat:,giscgu,eA!:&eand:ad=E:,:,;aa:ad:,uaT
El pr mer gen de PLC clonado y caracterizado en A fha//am (AfflLCIF) fue de'msohkobyy,?:Egeusraa.uanuaepsr=nteÉ#bacronnná:i?-Ér=5:_ií:_`s_;Í.rFii|dt±F','aaníl:|!!:Tk:,i::f#eerode€,moho y levadura qiie sm embargo, no se expresa en plántulas jóvenes. Este gen seclonó durante la inducción floral y codifica para un polipeptti de 533 aminoácidos(Yamamob et al ,1995) El segundo gen clonado y secuenciado (AíPLCIS), codificapara la PLC específica para PIP2, su expresión se induce por la deshidratación y elestrésporsalinidad.Estaproteínatieneun3442%yun19-33%dehomologíaconlasregiones X y Y conservadas de la PLC6 de mami.feros respectivamente, posee unaregión"EF-hand"enelextremoaminoterminal,tieneunamasamolecularde643kDaylaproteimesdependientedeionesCa2+parasuactividad(Hirayamaetal,1995)EI
18

tercer gen (AfpLC2) codifica para una PLC específica de fosfoinosítidos y esconstitutivamente expresado en tejidos vegetativos y florales, es similar a la PLC deplantas y las de mamífero tipo 6, la identidad de esta PLC con las PLCs de plantas esde 52.0% (AtpLCIS), 50.5% (AtpLCIF) y 70.8% (Pl-PLC soya): a diferencia deAtpLCIS, esta última no posee la región EF-hand en la región amino terminal y comose expresan de manera diferencial, se sugiere especificidad en la función de cada una(Hirayama et al.,1997)
El gen clonado en soya (Pl-PLC), tiene una longitud de 1.8 Kb que codifica parauna proteina de 600 aminoácidos con un peso molecular de 688 kDa y estáfuenemente relacionado con la PLCs de mamíferos, a d.iferencia con el segundo genclonado en A, Íha/i.ana, el gen de soya codifica una proteina que posee un dominio EF-hand entre los dominios X y Y y dada la localización de esta región, los autoresproponen que se trata de una nueva familia de isoenzimas de PLC. La actividad de laproteína codificada por este gen es dependiente de Ca" y su expresión en S.cerev/sí.ae complementa la letalidad que causa la deficiencia del gen en la levadura, yaque tiene un 24.8% de identidad de aminoácidos y 48.3% de sim" con la enzimacodificada en S. cenev/.s/.ae (Shi et al.,1995)
Recientemente se reportó la clonación de tres ADNcs (sÍp/cl, sÍp/c2 y stp/c3) apartir de una biblioteca de ADNc de fragmentos epidérmicos de hojas de papatotalmente expandidas, mediante el uso de una sonda de ADNc de PLC de A. fha//.anaEI ADNc de sfp/c7 es una secuencia de 1981 bp que codifica para una proteína de 596aminoácidos, el ADNc de sfp/c2 tiene 1940 bp con un marco de lectura abierta querepresenta un polipéptido de 565 aminoácidos y el ADNc síp/c3 codifica um proteínade 585 aminoácidos con una secuencia de 2009 bp. La estructura primar.ia de éstasPLCs son similares a sus homológas reportadas en plantas. Las homologías entre lasproteínas de las PLCs de papa indican que StpLC2 y StpLC3 están mágestrechamente relacionadas entre si que con sus homólogas en plantas y que StpLC1
:st,:sd,s;aLnéesmes:tner:,Sapcé:Tf:=as®pna:astafsoáf,á,,T::itíddo:,,asdoenotraacst,::3::,e:odrepé:g+tasaconcentraciones nanomolares y no poseen el motivo EF-hand; m experimentos deNorthern-blotdemostraronqueseexpresandiferencialnienteenlostejidosdelaplantay son expresados en respuesta a estreses ambientales tales como heridas ydeshidratación (Kopka et al ,1998).
FOSFOLIPASA C DE C. mseus
En el laboratorio de la Dra Hernández en la Unidad de Biología Experimentaldel Centro de lnvestigación Científica de Yucatán (CICY), utHizando raicestransformadas de C, noseu§ línea J1, ha sido posible medü y ffiracterizar la activ.idadde la PLC utilizando como subs{rato al PIP2 tanto en la fracción citosólica como en lafracción membranal (De Los Santos-Bnones et al„ 1997). Se han de`eminado losefectos que tienen diferentes detergentes, la dependencia del pH y se han encontradolas condiciones óptimas para la solubilización de la enzima membranal para tener unapreparación semipurificada (De Los Santos-Briones, 1998).
19

Entre los efectos producidos por los detergentes, el {ritón X-100 y el deoxicolatode sodio, se ha observado que el triton X-100 inhibe dramáticamente la actividad enlas dos fracciones Por otra parte, el deoxicolato de sodio estimula la actjvidad de laPLC citosólica a bajas concentraciones y la inhibe a altas concentraciones, en tantoque en la fracción membranal tiene un efecto inhibitorio de manera dosi.s dependienteEl octilglucósido tiene poco efecto en la actividad membranal y un efecto estimulatorioen la fracción citosólica (Hernández-Sotomayor et al. ,1997).
La mejor condición para la solubilización de la actividad membranal se encontrócon 2 M de KCI, por lo que posiblemente esta isoforma está unida a la membrana porinteracción electrostática (Hernández-Sotomayor et al„ 1997).
Otro aspecto en la caracterización de la enzi.ma es la medición del efecto de laneomicina y de varios cationes dívalentes tanto en la fracción soluble como en lafracción membranal En este sistema, la neomicina inhibió a la PLC de rnanera dosisdependiente. El efecto de diferentes cationes divalentes tales como Ni2+, Cu2+ y Zn2+sobre la actividad de la enzima, y la presencia o ausencia de Ca2+, también se estudióy se observó que en la presencia de Ca2+ los tres cationes divalentes inhibieron laactividad de la PLC en ambas fracciones de forma dosis dependiente, encontrándoseque Cu2+ es el inhibidor más potente para la forma membranal En ausencia de Ca2+ serequirieron mayores concentraciones de Cu2+ y Zn2+ para detectar su efecto inhibitorio(Piña-Chablé et al ,1998)
MODELO DE ESTUDIO Calnamnfbus mseus (L) G. Don.
C. roseüs es una angiosperma di.cotiledónea, perteneciente a la familiaApocynaceae, es originaria de Madagascar y se dispersó por cultivo a tift el mundo.Es .un arbusto pequeño y erguido de 30 a 60 cm de altura, perenne y común en lasregiones tropicales (Taylor y Famsworlh, 1975). Es una de las plantas medicinalesmás ampliamente estudiadas (Moreno et al , 1995), estos estudios se han enfocado enel ai.slamiento y semisíntesis de sus alcaloides Se han aislado más de 200 alcaloi.desde diferentes partes de la planta, muchos de ellos con actividad farmacéuticaimportante (Cordell, 1980; Mersey y Cutler, 1986). De estos alcaloides los másimportan{es son la vinblasti.na y la vincristina con propiedades antileucémicas, laajmalicina como antihipertensivo y la serpentina con efectos sedantes Los bajosrendimientos de estos alcaloides en la planta, además de su alü costo comercial, hapromovido la investigación hacia nuevos métodos alternativos para la producción deestos alcaloides tales como la síntesi.s o semisíntesis (Kutney et al ,1988) y el cultivode células y tejidos (Miura e Hirata, 1986)
Ac{ualmente la Unidad de Biología Experimental del Centro de lnvestjgaciónniÉ}nti.fira Ha V„^-+Á. ^,.^_._ ___ _ __Cienti.fica de Yucatán cuenta con varios modelos de estudio paraalcaloides a pariir de cultivos de C. rioseus,. uno de los cualestransformadas li'nea J1, la cual ya ha sido caracterizada en cuanto a
Ia obtención deson las raíces
crecjmiento yde hojas con
_ , `_ ___ .,....,. UU `,qlauICH¿aud tjri cuanto a suproducción de alffiloides Esta línea se ob!uvo medjante la infecciónAgnobacíen.um mí.zogenes, fig 15, (Ciau-Uitz et al.,1994)
20

Figura 1.5: Modelo de estudio: El modelo cle estudio utilizado en este trabajo son las raicestransformadas de C, n]seus. A) aspecto de las flores 8) aspecto de las raíces transfomadas vistasdesde abajo del matráz.
En C mseus se han caracterizado varias enzimas `involucradas en la biosíntesisde los alcaloides indólicos (Moreno et al ,1995). La elucidación de los mecanismos detransducción de señales que pudieran estar involucrados o relacionados con laactivación o expresión de las enzimas de la biosíntesis de alcaloides, permitirá lamanipulación de los diferentes factores que puedan conducir al incremento de laproducción de alcaloides en cultivos de C rioseus.
21

OBJETIVO
lniciar con los procesos de clonación del gen que codifica para la fosfolipasa Cen raíces transformadas de C. noseus línea J1.
OBJETIVOS PARTICULARES
1.-Detectar la presencia del gen que codifica para la fosfolipasa C en raícestransformadas de C. noseus, línea J1.
2 -Evaluar los niveles de expresión de la fosfolipasa C a 1o largo de un ciclo decultivo de raíces transformadas de C, roseus, línea J1.
3.- Realizar la clonación y secuenciación de fragmentos obtenidos medianteestrategias de RT-PCR en C. /oseus con oligonucleótidos diseñados a partir de lasregiones conservadas para la fosfolipasa C.
22

REFERENCIAS BIBLIOGRAFICAS
Anderson D., Koch C. A., Grey L., Ellis C., Moran M. F. y Pawson T. (1990). Bindingof SH2 domairis of phospholipase Cyi, GAP, and Src to activated growth factorreceptors. Science, 250: 979-982.
Arz M. C. y Grambow H. J. (1994). Polyphosphoinositide phospholipase C andevidence for inositol-phosphate-hydrolizyng activities in the plasma-membranefraction from light-grown \^heat ( mí/.cum aesfívum L. ) leaves. Planta,195: 57€2.
Bahk Y. Y., Lee Y. H., Lee T. G., Seo J., Ryu S. H. y Suh P. G. (1994). Two forms ofphospholipase C-beta 1 generated by alternative splicing J. Biol. Chem., 269:8240-8245.
Bairoch A y Cox J. A. (1990) EF-hand motifs in inositol phospholipid-specificphospholipase C. FEBS Lett„ 269i 454456.
Banno Y., Yada Y. y Nozawa Y. (1988). Purification and characterization of nuclearphospholipase C specific for phosphoinositides. J. Biol. Chem., 263: 11459-11465.
Banno Y. y Nozawa Y. (1987). Charactenzation of partially purified phospholipase Cfrom human platelet membranes. Biochem. J., 248: 396401.
Balda§sare J. J., Hender§on P. A y Fisher G. J. (1989). lsolation andcharacterization of one soluble and t\^o membrane associated forms ofphosphoinositide-specific phospholipase C from human platelets. Biochemistry,28: 6010€016.
Bar€agi D., Rotin D., Batzer A., Ma,ndiyan V. y Schlessinger J. (1993). SH3domains direct cellular localization of signaling molecules. Cell, 74: 83-91.
Bennett C. F. y Crooke S. T. (1987). Purification and characterization of aphosphoinositide-specific phospholipase C from guinea pig uterus. J. Biol.Chem., 262: 13789-13797.
Berridge M. J. (1987). lnositol trisphospbate and diacylglycerol: tMo interacting secx)ndmessengers. Annu. Rev. Biochem., 56: 159-193.
Berridge M. J. (1993). lnositol trisphosphate and calcium signalling. Nature, 361: 315-325.
Blank J. L., Brattain K. A. y Exton J. H. (1992) Activation of cytosolicphosphoinositide phospholipase C by G-protein beta gamma subunits. J. Biol.Chem„ 267: 23069-23075.
Bloomquist 8. T., Shortridge R. D., Schneuwly S., Perdew M., Montoll C., St®llerH., Rubin G. y Pak W. L. (1988). lsolation of a putative phospholipase C gene ofDrosophí./a, norpA and its role in phototransduction Cell, 54. 723-733.
Bominaar A. A., Kesbeke F., SnaarJagaiska 8. E., Peters D. J. M., Schaap P. yVan Haastert P. J. M. (1991). Aberrant chemotaxis and differentiation in
23

D/cíyosíe/Í.um mutant fgdc wth a defective regulation of receptor-stimulatedphosphoinositidase C J Cell Sci.,100: 825-831.
Bominaar A. A. y Van Haastert P. J. M. (1994). Phospholipase C in D/.síyoste/Í.umc//.sc\o/deum, ldentification of stimulatory and inhibitory surfacx3 receptors and G-proteins Biochem. J , 297' 189-193
Bristol A., Ha S. Wl., Kriz R. W., Stahl lvl. L., Fan Y. S., Byer M. G., Eddy R. L.,Shows T. 8. y Knopf J. L. (1988). Phospholipase C-148: chromosomal locationand deletion mapping of functional domains Cold Spring Harbor Symp QuantBiol., 53. 915-920
Camps Ivl., Hou C., Sidiropoulos D., Stock J. 8., Jakobs K. H. y Gierschik P.(1992) Stimulation of phospholipase C by guanine-nucleotide-binding protein Pysubunits Eur. J Biochem., 206: 821-831
Ciau-Uitz R., Miranda-Ham M. L., Coello{oello J., Chi 8., Pacheco L. M. y Loyola-Vargas V. lvl. (1994). lndole alkaloid production by transformed and non-transformed root cultures of Caíha/aníhus roseus. ln Vitro Cell Dev. Biol., 30:84-88.
Cifuentes M. E., Honkanen L. y Rebecchi M. J. (1993) Proteolytic fragments of
phospholipase-§pecific Cól. J, Biol. Chem., 26811586-11593.
Cheng H. F., Jiang " J., Chen C. L., Liu S. M., Wong L. P., Lomasney J. W. y KingK. (1995). Cloning and identifícation of amíno acid residues of humanPhospholipase C81 essential for catalysis J Biol Chem., 270 5495-5505.
Chin-Vera J. (1995). Función del inositol 1, 4, 5-trifosfato (lp3) cx)mo segundomensajero en diferentes cultivos de Caíharaníhus roseus. Tesis de Licenciatura.Universidad Autónoma de Yucatán, Mérida Yuc , México.
Cockcroft S. G. y Thomas G. M. H. (1992). lnositol-lipid-specific phospholipase Cisoenzymes and their diffential regulation by receptors. Biochem J., 288 1-14.
Cordell G. A. (1980). The botanical, chemical, biosynthetic and pharmacx)logicalaspects of Caíharaníhus /oseus (L) G. Don. (Apocynaceae). En RecentAdvances in Natural Product Research. Editado por Woo, W S and Han, 8 H ,pp 65-72-
Das T., Baek K. J., Gray C. e lm M. J. (1993) Evidence that the Gh protein is a signalmedjator from G1-adrenoreceptor to a phospholipase C. 11. Purification andcharacterization of a Ghcoupled 64-kDa phospholipase C and reconstitution ofcil-adrenoreceptor, Gh family, and phospholipase C. J Biol, Chem , 268: 27398-27405
Daugheriy S. y Low Ivl. G. (1993) Cloning, expression, and mutagenesis ofphosphatidylinositol-specific phospholipase C from Síaphy/ococcus auneus: apotential staphylococcal virulence factor. lnfection and lmmunity, 61 : 5078-5089.
Dekker L. V., Palmer R. H. y Parker P. J. (1995). The protein kinase C and proteinkinase C related gene families. Curr. Opin. Struct Biol., 5: 396402.
24

De Los Santos-Briones C., lvluñozsánchez J. A„ Cliin-Vera J., Loyola-Vargas V.M. y Hemándezsotomayor S. M. T. (1997) Phosphatidylinositol 4, 5-biphosphate-phospholipase C activity during the growng phase of Caíharaníhusroseus transformed roots. J. Plant Physiol„ 34: 175-180.
De Los Santos-Briones C. (1998) Estudio de la enzima fosfolipasa C en raícestransformadas de Caíharaníhus noseus, Tesis de Doctorado. Centro delnvestigación Científica de Yucatán Mérida, Yuc , México
Divecha N,, Banfic H. e lrvine R. F, (1993). lnositides and the nucleus and inositidesin the nucleus. Cell, 74. 405407
Divecha N. e lrvine R. F, (1995) Phospholipid signalling Cell, 80. 269-278.
Divecha N., Letcher A. J., Banfic H. H., Rhee S. G. e lrvine R. F. (1995) Changes inthe components of a nuclear inositide cycle during differentiation in murineerythroleukaemia cells Biochem. J , 312 63€7
Drayer A. L. y Van Haasterl P. J. M. (1992) Molecular cloning and expression of aphosphoinositide-specific phospholipase C of Dicfyosíe/Í.um c}oscoÍ.c/eum. J BiolChem., 267: 18387-18392.
Drayer A. L. y Van Haastert P. J. M. (1994) Transmembrane signalling in eukaryotes.phosphoinositide-specific phospholipase C of D/cíyosfe//um di`sco/.deum J. Biol.Chem , 267: 18387-18392.
DÍ©bak 8. K. (1992) The plant phosphoinositide system. Biochem. J , 288 697-712.
Emori Y., Homma Y., Sorimachi H., Kawasaki H., Nakanishi D., Suzuki K. yTakenawa T. (1989). A second type of rat phosphoinositide-specificphospholipase C containing a src-related sequence not essential forphosphoinositide-hydrolizyng activity. J Biol Chem , 264: 21885-21890.
Essen L. 0., Perisic 0., Cheung R., Katan M. y Williams R. L. (1996). Crystalstructure of a mammalian phosphoinositide-specific phospholipase C6 Nature,380: 595€02
Feng J. F„ Rhee S. G. e lm MU. (1996) Evidence that phospholipase Ól is theeffector in the Gh (transglutaminase ll)-mediated signalling. J Biol. Chem , 27116451 -16454.
Ferreira P. A., Shortridge R. D. y Pak W. L. (1993) Distinctive subtypes of bovinephospholipase C that have preferential expression in the retine and highhomology to the norpA gene product of Drosopní/a Proc Natl Acad Sci. USA„90. 6042J5046
Fukui T., Lutz R. J. y Lowenstein J. M. (1988). Purification of a phospholipase C fromrat liver cyto§ol that acts on pho§phatidylinositol 4,5-bisphosphate andphosphatidylinositol 4-phosphate J Biol. Chem , 263 17730-17737.
GómezJ3uerrero C., Duque N. y Egido J. (1996) Stimulation of Fc (alpha) receptorsinduces tyrosine phosphorylation of phospholipase C-gamma(1),
25

phosphatidylinositol phosphate hydrolysis, and Ca2+ mobilization in rat andhuman mesangial cells. J. lmmunol.,156: 4369.4376.
Gout 1., Dhand R„ Hiles 1. D., Fry M. J., Panayotou G., Das P., Troung 0., Tofty N.F., Hsuan J„ Booker G. W., Campbell 1. D. y Wateitield M. D. (1993) TheGTpase dynamin binds to and is activated by a subset of SH3 domains. Cell, 75:25-36.
Harlan J. E., Hadjuk P. J., Yoon H. S. y Fesik S. W. (1994) Pleckstrin homologydomains bind to phosphstidylinositol4,5-biphosphate. Nature, 371..168-170.
Haslam R. J., Koide H. 8. Y Hemmings 8. A. (1993). Pleckstrin domain homology.Nature, 363: 309-310.
Hemández€otomayor S. lvl. T., De Los Santos-Briones C., Muñoz€ánchez J. A.,PiñaJ3hable M. L. y Loyola-Vargas V. M. (1997). Phospholipase C inCaíharaníhus roseus transformed roots En Radical Biology, editado por FloresH. E., Lynch J. P. y Eissenstat D., pp. 422425.
Hii.ayama T., Otho Ch., Ivlizoguchi T. y Shinozaki K. (1995) A gene encoding aphosphaditylinositol-specific phospholipase C is induced by dehydration and saltstress in Arabí.dops/.s Íha/Í.ana. Proc. Natl Acad. Sci. USA., 92: 3903-3907.
Hirayama T„ Mitsukawa N., Shibata D. y Shínozaki K. (1997) AfpLC2, a geneencoding phosphoinositide-specific phospholipase C, is constitutively expressedin vegetative and floral tissues in Arab/dopsí's Íha//ana. Plant Molecular Biology,34.175-180.
Homma Y. y Emori Y. (1995). A dual functicmal signal mediator showing Rho-GAP and
phospholipase Ci} stimulating activities. EMBO J.,14: 286-291.
Homma Y., lmaki J., Nakanishi 0. y Takenawa T. (1988) lsolation andcharacterization of a y-type phosphoinositide-specific phospholipase C (PLct2)J. Biol. Chem., 263. 6592€598
Hotta Y. y Benzer S. (1970). Genetic dissection of the Orosoph//a nervous system bymeans of mosaics. Proc. Natl. Acad. Sci. USA , 67: 1156-1163.
Hwang S. C., Jhon D. Y., Bae Y. S., Kim J. H. y Rhee S. G. (1996). Activation of
phospholipase Ct by the concer[ed action of tau proteins and arachidonic acid.J. Biol. Chem., 271: 18342-18349.
lkura lvl. (1996). Calcium binding and conformational response in EF-hand proteins.Trends Bioch Sci., 21.14-17.
Im M. J. y Graham R. M. (1990) A novel guanine nucleotide-binding protein coupled tothe cil-adrenergic receptor. J. Biol. Chem., 265: 18944-18955.
lrvine R. F., Letcher A. J. y Dawson R. M. C. (1980). Phosphatidylinositol
phosphodiesterase in higher plants. Biochem. J.,192: 279-283.
lrvine R. (1996)` Takjng stock of Pl-PLC. Nature, 380 581-583.
26

Jiang H., Kuang Y„ Wu Y., Smi.cka A., Simon M.1 y Wu D. (1996) Pertusis toxin-sensitive activation of phospholipase C by the C5ci and fMet-Leu-Phe receptorsJ. Biol. Chem , 271 : 13430-13434.
Jiang H. P., Wu D. Q. y Simon M.1. (1994) Activation of phospholipase C beta 4 byheterotrimeric GTP-binding proteins. J. Biol Chem., 269: 7593-7596.
Jones G. A., y Carpenter G. (1993) The regulation of phospholipase C-gamma 1 byphosphatidic acid Assessment of kinetic parameters. J Biol Chem , 26820845-20850.
Kaibuchi K., Fukumoto Y., Oku N., Takai Y., Arai K. L. y Muramatsu Wl. A. (1989)Molecular genetic analysis of the regulatory and catalytic domains of prciteinkinase C J Biol. Chem., 264.13489-13496.
Kamada Y. y Ivluto S. (1991) Ca2+ regulation of phosphatidylinositol turnover in the
plasma membrane of tobacco suspension culture cells. Biochim. Biophys. Acta,1093: 72-79.
Keely P. J. y Parise L. V. (1996). The ci2Pi integrm is a necessary co-recx=ptor forcollagen-induced activation of syk and the subsequent phosphorylation ofphospholipase Cy2 in platelets. J. Bjol. Chem., 271 : 26668-26676
Koch A. C., Anderson D. A., Moran M. F., EIlis C. y Pawson T. (1991 ) SH2 and SH3domains: elements that control interactions of cytoplasmic signalling molecules.Science, 252: 668€74.
Kopka J., Pical C., Gray J. E y Müller-Róber 8. (1998). Molecular and enzymaticcharacterization of three phosphoinisite-specific phospholipase C isoforms frompotato. Plant Physiol.,116: 239-250
Kretsinger R H y Nockolds C E. (1973) Carp Muscle Calcium-binding Protein J. BiolChem., 248. 3313-3326.
Kuang Y., Wu Y., Jiang H. y Wu D. (1996) Selective G protein coupling by C-Cchemokjne receptors. J. Biol. Chem , 271. 3975-3978`
Kuang Y., Wu Y., Smrcka A., Jiang H. y Wu D. (1996a). ldentification of a
phospholipase CP2 region that interacts wth Gpy Proc. Natl Acad. Sci USA ,93: 2964-2968.
Kuppe A., Evans L. M., MCMillen D. A. y Griffith H. (1989). Phosphatidylinositol-specific phospholipase C of Bac/.//us cereus. Cloning, sequencing, andrelationship to other phosphoplipases. Journal of Bacteriology, 171. 6077€083
Kuricki H., Tamiya-Koizumi K., Asano lvl., Yoshida S., Kojima K. y Nimura Y.(1992) Existence of phosphoinositide-specific phospholipase C in rat liver nucleiand lts change during liver regeneration. J Biochem.,111 : 283-286.
Kutney J. P., Choi L. S. L., Nakano J., Tsukamoto H., Mchugh lvl. y Boulet C. A.(1988), A highly efficient and commercially important synthesis of the antitumorCaíharaníhus alkaloids vinblastine and leurosidine from catharanthine andvindoline. Heterocycles, 27: 1845-1853.
27

Lee C. W. Lee K. H., Lee S. 8., Park D. y Rhee S. G. (1994). Regulation ofphospholipase C-beta 4 by ribonucleotides and the alpha subunit of Gq. J. Biol.Chem„ 269: 25335-25338.
Lee K. Y., Ryu S. H., Suh P. G., Choi W. C. y Rhee S. G. (1987). Phospholipase Cassociated with particulate fractions of bovine brain. Proc. Natl. Acad. Sci. USA.,84: 5540-5544.
Lee S. 8. y Rhee S. G. (1995). Significance of PIP2 hydrolisis and regulation of
phospholipase C isoenzymes. Curr. Opin Cell Biol., 7: 183-189.
Lee S. 8. y Rhee S. G. (1996) Molecular cloning, splice variants, expression, and
purification of phospholipase C-84. J. Biol Chem , 271 : 25-31
Lewin 8. (1994). Genes V„ Editado por Oxford University., pp 319-336
Liu N., Fukami K., Yu H. y Takenawa T. (1996). A new phosphollpase C 64 ls inducedat s-phase of the cell cycle and appears in the nucleus. J Biol Chem., 271 : 335-360
Lodish H., Baaltimore D., Berk A., Zipurski S. L., Matsudaria P y Damell. J. (1995)Molecular Cell Biology. Scientific Amencan Books p` 873-924.
Manzoli L., Billi A. M., Rubbini S., Bavelloni A., Gilmour R. S., Rhee S. G., y CoccoL. (1997). Essential role for nuclear phospholipase C betal in insulin-like growthfactor l-induced mitogenesis Cancer Res., 57. 2137-2139.
Margolis 8., Li N., Koch A., Mohammadi M., Hurwitz D., Ullrich A., Zilberstein A.,Pawson T. y Schlessinger J. (1990a). The tyrosine phosphorilated carboxyterminus of the EGF receptor is a binding site for GAP and PLcy. EMBO J , 94375-4380.
Margolis 8., Bellot A. M., Haneyer A. M., Ullrich A., Schlessinger J. y Silberstein A.(1990b). Tyrosine kinase activity is essential for the association c>fphospholipase C-y with the epidermal growth factor receptor Mol Cell. Biol.,10435-441 .
Marmiroli S., Ognibene A., Bavelloni A., Cinti C., Cocco L. y Maraldi N. M. (1994).lnterleukin 1 alpha stimulates nuclear phospholipase C m human osteosarcomaSaos-2 cells. J Biol Chem., 269 13-16.
Martelli A. M., Gilmour R. S., Bertagnolo V„ Neri L. lvl., Manzoli L. y Cocco L.
(1992) Nuclear localization and signalling activity of phospholipase CP in Swiss3T3 cells. Nature, 358 242-245.
Mato J, M,, Kelly A., Abler A. y Jarrett L. (1987). ldentification of a novel insuline-sensitive glycophospholipid from H35 hepatoma cells. J Biol. Chem., 262. 2131-2137
Mayer 8. J., Ren R„ Clark K. L. y Baltimore D. (1993). A putative modular domainpresent in diverse signalling proteins. Cell, 73: 629€30.
Mazzoni M., Bertagnolo V., Neri L. Wl. Carini C., Marchisio M., Milani D., Manzoli F.A. y Capitani S. (1992). Discrete subcellular localization of phosphoinisitadase
28

C P, y and Ó in PC12 rat pheochromocytoma cells BÍochem. Biophys. ResCommun.,187: 114-120.
WlcMurray W. C., e lrvine R. F. (1988). Phosphatídylinosi.tol 4,5+bisphosphate
phosphodiesterase in higher plants. Biochem. J„ 249: 877-881.
Meldrum E., Katan M., y Parker P. (1989) A novel inositol-phospholipid-specific
phospholipase C Rapid purification and characterization Eur J Biochem ,182ÍS]3ÍS]7 .
Melin P. M.. Sommarin M.. Sandelius A. S„ Jei.gil 8. (1987) ldentification of Ca2+stimulated polyphosphoinositide pho§pholipase C in isolated plant plasmamembranes. FEBS Lett., 223: 87-91.
lvlelin P. M., Pical C., Jergil 8. y Sommarin M. (1992) Polyphosphoinositide
phospholipase C in Wieat root plasma membranes Panial purification andcharacterization. Biochim Biophys. Acta,1123163-169
Mengaud J., Bi.aun-Brelon y Cassart P. (1991) Identification of phosphatydili.nositol-specific phospholipase C activity in L/stena monocytogenes: a novel type ofvirulence factor? Molecular Microbiology, 5. 367-372.
Mersey 8. G. y Cutler A. J. (1986). Differential distribution of specific indole alkaloidsin leaves of Caíhanenfhus noseus Can J. Bot., 64: 1039-1045.
Misra U. K., Gawdi G. y Pizzo S. V. (1995). Ligation of the alpha 2-macroglobulinsignalling receptor on macrophages induces protein phosphorylation and anincrease in cytosolic pH Biochem J , 309.151-158
Miui.a Y. e Hii.ata K. (1986) An organ culture of Caíharanf^us ,oseus capable ofproducing substantial amount of indole alkaloids Eur. Pat. Appl , EP 0200225A2
Mohammadi M., Honneger A. M., Rotin D., Fishcher R., Bellot F., Li W., Dionne C.A., Jaye M., Rubinstein M. y Schlessinger J. (1991). A tyrosine-phosphorylated carboxi-terminal peptide of the fibroblast growth factor receptor(Flg) is a binding site for the SH2 domain of phospholipase Ctl Mol Cell Biol ,1 1 : 5068-5078.
Morono P. R. H., Van der Heii.den R. y Verpoorte R. (1995). Cell and tissue culturesof Caínaranffius rioseus: a literature survey. Plant Cell Tiss Org. Cult„ 42: 1-25.
Musacchio A., Gibson T., Rice P., Thompson J. y Saraste M. (1993). The PHdomaín: a common piece in the structural patchwork of signalli.ng proteins.Trends Biochem. Sci.,18 343-348.
Nakaoka H., Pérez D. M., Baek K. J., Das T., Husain A., Misono K., lm M. J. yGraham R. M. (1994) Gh A GTP-Binding Proteín wth transglutaminase activityand receptor signalling function. Science, 264.1593-1596.
Nishibe s., Wahl M. l., Hernández-Sotomayors. M. T., Tonks N. K., Rhee s.G. yCarpenter G. (1990). Increase of the catalytic activity of phospholipase C+1 bytyrosine phosphorylation Sciencx3, 250 1253-1256
29

Nishizuka Y. (1992) lntracellular signalling by hydrolisis of phospholipids andactivation of protein kinase C. Science, 258: 607€14.
Noh D. Y., Shin, S. H. y Rhee S. G. (1995) Phosphoinositide-specific phospholipase Cand mitogenic signalling Biochm Biophys Acta,1242: 99-114
Pann€hill S., Ryu S. H., Moon K. H., Shu H. W. y Rhee S. G. (1988a) Cloning andsequence of multiple forms of phospholipase C Cell, 54.161-169
Pann€hill S., Ryu S. H., Moon K. H., Shu H. W. y Rhee S. G. (1988b) lnositolphospholipid-specific phospholipase C complete cDNA and protein sequencesand sequence homology to tyrosine kinase-related oncogene products ProcNatl Acad. Sci. USA., 85: 5419-5423
Park D., Jhon D.-Y., Lee C. W., Lee K. H. y Rhee S. G. (1993) Activation ofphospholipase C isoenzymes by G protein beta gamma subunits J. Biol Chem ,268: 45734576.
Parker P. J., Hemmings 8. A., Gierschik P. (1994) PH domains and phospholipases-a meaningful relationship? Trends Biochem Sci.,19 54-55
Pawson T. (1995) Protein modules and signalling networks Nature, 373: 573-580
Pfaffmann H., Hartmann E., Brightman A. 0., y Morre D. J. (1987)Phosphatidylinositol specific Phospholipase C of plant stems: membraneassociated activity concentrated in plasma membranes. Plant Physiol , 85 1151 -1155_
Piña€hable M. L., De Los Santos-Briones C., Muñoz€ánchez J. A. y Hemández-Sotomayor S. M. T. (1998) Effect of different inhibitors on phospholipase Cactivity in CaíharanínLÍs roseus transformed roots Prostaglandins & Other LiptdsMediators, 56: 19-31.
Rebecchi M. J. y Rosen 0. lvl. (1987) Purification of a prio§phoinositide-specificphospholipase C from bovme brain. J. Biol Chem , 262.12526-12532
Rebecchi M., Peterson A. y MCLaughlin S. (1992). Phosphoinositide-specificphospholipase Ci}1 binds wth high affinity to phospholipid vesicles containingphosphatidylinositol 4,5-bisphosphate Biochemistry, 31.12742-12747.
Rhee S. G. y Crioi K. D. (1992) Regulation of inositol phospholipid-specific
phospholipase C isozymes. J Biol. Chem , 267: 12393-12396.
Rhee S. G (1991 ). lnositol phospholipid-specific phospholipase C: lnteraction of the yiisoform with tyrosine kinase. Trends Bioch. Sci„ 16: 297-301.
Rhee S. G. y Bae Y. S. (1997) Regulation of phosphoinositide-specific phospholipaseC isozymes. J Biol. Chem., 272.15045-15048.
Rotin D., Margolis 8., Ivlohammadi M., Daly R. J., Daum G., Li N., Fishcher E. H.,Burgess W. H., Ullrich A. y Schlessinger J. (1992). SH2 domains preventtyrosine dephosphorylation of the EGF receptor: identification ot tyr 992 as thehigh affinity binding sites for the SH2 domains of phospholipase CJy EMB0 J.,11 : 559-567`
30

Ryu S. H., Cho K. S. Lee K. Y., Suh P. G. y Rhee S. G. (1987a) Purification andCharactertzation of two inmunologically distinct phosphoinositide-specificphospholipase C from bovine brain. J Biol. Chem., 262.12511-12518
Ryu S. H., Suh P. G., Cho, K. S., Lee K. Y. y Rhee S. G. (1987b). Bovine brain cytosolcontains three inmunologically distinct forms of inositolphospholipid-specific
phospholipase C Proc Natl Acad. Sci USA , 84. 6649€653
Sadoshima J. e lzumo S. (1996). The heterotrimeric Gq protein-coupled angiotensin 11receptor activates p21 'as via the tyrosine kinase-shc-Grb2-sos pat"y in cardiacmyocytes EMBO J ,15. 775-787
Saltiel A. R. y Cuatrecasas P. (1986) lnsuline stimulates the generation from hepaticplasma membranes of modulators derived from an inositol glycolipid. Proc NatlAcad Sci USA., 83. 5793-5797.
Shaw G. (1993) ldentification of novel pleckstrin homology (PH) domains provides ahypothesis for PH domain function Biochem Biophys Res. Commun., 195.1145-1151.
Shi J„ González R. A. y Bhattacharrya M. K. (1995) Characterization of a plasmamembrane-associated phosphoinositide-specific phospholipase C from soybeanThe Plant Journal, 8: 381-390.
Shortridge R. D,, Yoon J. Y., Rlending C., Bloomqui§t 8. T., Perdew M. y Pak W.L.(1991) A Drosophila Phospholipase C gene that is expressed in the centralnervous system J Biol. Chem , 266.12474-12480
Shulka S. D. (1982). Phosphatidylinositol specific phosphol.ipase C. Life Sci., 30: 1323-1326.
Smrcka A. V. y Stemweis P. C. (1993) Regulation of purified subtypes ofphosphatidylinositol-specific phospholipase C beta by G protein alpha and betagamma subunits J. Biol Chem , 268 9667-9674.
Soler C., Beguinot L., Sokin A. y Carpenter G. (1993). lndividual epidermal growthfactor receptor autophosphorylation sites do not stringently define associationmotifs for several SH€ontaining proteins. J. Biol. Chem„ 268 22010-22019.
Stahl M. L., Ferenz C. R., Kelleher L. K., Kriz R. W. y Knopf J. L. (1988) Sequencesimilarity of phospholipase C wth the non-catalytic region src. Nature, 332: 269-272_
Stemweis P. C. (1994) The active role of Py in signal transduction Curr Opin Cell.Biol., 6.198-203.
Sternweis P. C. y Smrcka A. V. (1992) Regulation of phospholipase C by G proteins.Trends. Biochem. Sci„ 17: 502-506
Suh P. G., Ryu S. H„ Moon K. H., Sur[ H. W. y Rhee S. G. (1988) Clomng andsequence of multiple forms of phospholipase C. Cell, 54: 161-169.
31

Tate 8. F., Schalier G. E., Sussman lvl. R., Crain R. C. (1989). Characterization of apolyphosphoinositide phospholipase C from the plasma membrane of Avenasaf/`va. Plant Physiol„ 91 -1275-1279.
Taylor W.1. y Famsworlh N. R. (1975) The Catharanthus alkaloids Marcell Dekkerlnc NewYork
Tyers M., Rachubinski R. A., Steward M. 1, Varricho A. lvl. Shorr R. G. L. Haslam R.J. y Harley C. 8. (1988) Molecular cloning and expression of the major proteinkinase C substrate of plateles. Nature, 333: 470473
Waldo G. L., Morris A. J. y Harden T. K. (1994) Purification of G-protein-regulatedphospholipase C from Turkey erythrocytes Methods in Enzymology, 238. 195-207
Yamamoto Y. T., Conkling M. A., Sussex 1. M. e lrish V. F. (1995) An ArabidopsiscDNA related to animal Phosphoinosotide-specific phospholipase C genes PlantPhysiol„ 107: 1029-1030.
Yokoú T., Matsui Y., Yagisawa H., Nojima H., Uno 1. y Tohe A. (1993) The putativephosphoinositide-specific phospholipase C gene, PLC7, of the yeastSacchanomyces cemví.siae is important for cxgll growth Proc. Natl. Acad SciUSA , 90. 1804- 1808.
Yotsushima K., Nakamura K., Mitsui T., lgaue 1. (1992). Purification andcharacterization of phosphatidylinositol-specific phospholipase C in suspension-cultured cells on rice (On.za safí.va L.). Biosci Biotech. Biochem , 56. 1247-1251 _
Yoshioka T., lnoue H. y Hotta Y. (1985) Absence of phosphatidylinositolphosphodiesterase in the head of a drosophila visual mutant, norpA (no receptorpotential A).J. Biochem., (Tokio) 97: 1251-1254.
York J. D. y Majerus P. W. (1994) Nuclear phosphatydilinositols decrease during s-phase of the cell cycle in HeLa cells J Biol. Chem , 269: 7847-7850.
Zhu L., Mckay R. R. y Shortridge R. D. (1993) Tissue-specific expression ofphospholipase C encoded by the norpA gene of Dnosoph//a me/anogasíer J.Biol. Chem , 268: 15994-16001.
Zhu X. y Bimbaumer L. (1996) G protein subunits and stimulation of phospholipase Cby Gs- and Gi-coupled receptors Lack of receptor selectivity of Gais andevidence for a synergic interaction between Gpy and the a subunit of a receptor-activated G protein. Proc. Natl. Acad. Sci. USA„ 93: 2827-2831.
32

Capítulo 2
Detección por Southern-blot del gen que codifica para la fosfolipasa C yde su expresión por Northern-blot en raíce§ transformadas de CafhafflnlAusroseus.
lNTRODUCCIÓN
Las técnicas de análisis por transferencia (blot) son un método para detectarbiomoléculas específicas en muestras de composición compleja Esta técnica puedeser aplicada a biomoléculas que se adhieren establemente a un soporte de matenalsólido tales como nitrocelulosa, nylon o papel y son capaces de unirse a su ligandoanálogo. Usualmente las biomoléculas de interés son separadas primero de acuerdo asu tamaño o carga antes de transferirse a la membrana (Memelink et al , 1994)
Este método fue originalmente implementado para ADN por Edwn MSouthem (Southern,1975) quien demostró que los fragmentos de restricción del ADNque habían sido electroforétiffimente frac€ionados en geles de agarosa podían sertransferidos a un soporte sólido y detectados como bandas discretas de§pués de lahibndación cm una sonda de ADN complementario Cuando el método de Southern seaplicó al ARN, este fue llamado Northern-blot, mientras que el análisis de Western-blotse refiere a la transferencia de proteínas a un soporte sólido y a su detección medianteel uso de anticuerpos Muchas variantes, incluyendo el Southwestern (Detección deproteínas inmovilizadas con sondas de ADN) y el far Western (detección de proteínasinmovilizadas con sondas de proteínas que no son anticuerpos) han sido descritosAdemás de la transferencia a partir de geles, las muestras pueden ser directamenteaplicadas sobre la membrana sin una separación, ya sea en forma de círculos o rayas(dot y slot-blot respectivamente) (Memelink et al„ 1994).
Las técnicas de transferencia no han cambiado mucho desde su primeradescripción, excepto que la disponibmdad de membranas sofisticadas se haincrementado y que han aparecido equipos para marcaje de sondas así como aparatospara electroforesis y transferencias Aunque las membranas originalmente usadaspara la transferencia de ADN y ARN fueron hechas de nitrocelulosa, hoy en día lasmembranas de nylon son mejores debido a su alta capacidad de unión de ácidosnucleicos a su resistencia y que se pueden reutilizar (Memelink et al ,1994)
En el presente trabajo con el fin de detectar la presencia y la expresión del genque codtfica para la fosfolipasa C en las raíces transformadas de C. roseus, seutilizaron las técnicas Southern-blot y Northern-blot, seguida de una hibridación conuna sonda de PLC de Arab/.dopsí.s Íha//ana y con el fragmento obtenido por RT-PCR deC. roseus (Cap 3, fig 2). Posteriormente, para validar las técnicas empleadas serealizaron hibridaciones con una sonda homóloga de C, mseus, y de la triptofanodescarboxilasa.
33

MATERIALES Y MÉTODOS
Material vegetal: Como material biológico se usaron raíces transformadas deC roseus línea J1, transformadas con Agnoóacíenum rh/.zogenes, las cuales sonsubcultivadas en medio 85 de Gamborg (Gamborg et al ,1968), a la mitad de su fuerzaiónica adicionado con 3% de sacarosa y a pH 5 7 antes de autoclavear Las raícesfueron resembradas cada 14 días con un inóculo inicial de 0 5 g. de tejido en 100 ml demedio estéril, las raíces se deiaron crecer a 25 °C +/- 2 °C en la obscuridad y enagitación continua de 100 rpm`
Sonda de ADNc do PLC: La sonda de PLC de A fha/Í.ana utilizada para estetrabaio se generó de la digestión con la enzíma Wco 1 del plásmido pNH495, el cual tieneel ADNc de AtpLCI S (fosfolipasa C de A Íha//.ana inducible por estrés) en el sitio Eco Rldel pBSSKH, el fragmento generado es de 1 2 kb Este, se utilizó como sonda y semarcó radiactivamente utilizando el equipo de Random primer de GIBCO siguiendo los
pasos indicados en el manual y alcanzó una actividad específica de 3.5 x 107cpm/LigADN. Este plásmido fue donado por el Dr Hirayama Takashi del Laboratorio de BiologíaMolecular de Plantas de Toyota Japón (Hirayama et al ,1995)
Sonda de ADNc de TDC: La sonda de TDC de C. noseus utilizada para validarlas técnicas se generó de la digestión con la enzima Eco Rl del plásmido pTDC5, elcual contenía la longitud completa del ADNc de TDC, en el sitio Eco Rl del pBSSKllEste plásmido fue enviado por el Dr Vincenzo De Luca (De Luffi et al., 1989) dellnstituto de lnvestigación en Biología Vegetal del departamento de Ciencias Biológicasde la Universidad de Montreal El marcaje de la sonda se realizó utilizando el equipo deRandom primer de GIBCO siguiendo los pasos indicados en el manual
Extracción de ADN: La extracción de ADN genómico de células vegetales sellevó a cabo utilizando la técnica de Cetil Trimetil Bromuro de Amonio Se pesaron 2 gde raíces y se congelaron con nitrógeno líquido, se pulverizaron en un mortero y elpulverizado se transfirió a un tubo de centrífuga, se le añadió amortiguador A deextracción (2% CTAB,100 mM Tris-Hcl pH 8 0, 20 mM EDTA pH 8.0,14 M Nacl) y seincubó a 65 °C. Se extrajo con cloroformo. alcohol isoamílico (24.1), se adicionóamortiguador 8 de extracción (10% CTAB, 0.7M Nacl), se extrajo con la mezcla desolventes y se precipitó con isopropanol Los ácidos nucleicos se resuspendieron enamortiguador TE (10 mM Tris-Hcl pH 8 0,1 mM EDTA pH 8 0) y se precipitaron conacetato de amonio 5 M y etanol 100%, se lavó la pastilla con etanol al 70%, se dejósecar y se resuspendió en H20 estéril (Rogers y Bendich, 1994)
Extracción cle ARN: La extracción de ARN se realizó de acuerdo a lametodología descrita por Schuler y Zielinski en 1989. Se pesaron 2 g. de materialvegetal, se congelaron con nitrógeno líquido, se molieron hasta formar un polvo fino yse transfirió a tubos de centrífuga conteniendo amortiguador de extracción (50 mM Tris-Hcl pH 8.5,10 mM EDTA pH SO, 200 mM Nacl), se realizó una extracción confenol.cloroformo:alcohol isoamílico (5050:1) y una segunda extracción concloroformo.alcohol isoamílico (50.1 ), Ia fase acuosa se precipitó cx)n isopropanol, y sedejó secar la pastilla, durante 5 min La pastilla se resuspendió en amortiguador TE (10mM Tris+Hcl pH 7 5, 1 mM EDTA pH 8.0) y se precipitó con Nacl 5 M y etanol al 100%.La pastilla se resuspendió en TE y se precipitó con s M de urea y 10 M de Licl durante
34

toda la noche Se centrifugó y se dejó secar la pastilla la cual se resuspendió en 2 mlde TE para repetm la precipitación etanólica Finalmente, el ARN se resuspendió enagua, se cuantificó por espectrofotometría y se fraccionó en una electroforesis enagarosan-BE (89 mM tris base, 89 mM ácido bórico, 2 mM EDTA pH 8.0) para observarsu integridad Las muestras integras §e mantienen almacx=nadas a -80 °C Todas lassoluciones utilizadas en la extracción fueron preparadas con agua tratada con (DEPC)
Hibridación ADN-ADN (Southem-blot): Para hacer el Southern-blot (Southern1975) de ADN genómico de C. roseus, se tomaron 50 LLg de ADN y se digirieron con lasenzimas de restricción Eco RI, Eco RV, H/.nc} 111, Bam Hl, Sac 1, durante 48 h Losfragmentos obtenidos se separaron electroforéticamente en geles de agarosa al1 %n-BE (89 mM Tris base, 89 mM ácido bórico, 2 mM EDTA pH 8 0), por un espaciode 14 h a 10 volts y se transfirieron a membranas de nylon, se fijaron con luz UV en uncrosslinking stratagene (1 200 KJ/cm2) y las membranas se guardaron para su posteriorhibridación La prehibridación de las membranas conteniendo ADN genómico digeridose realizó en amortiguador de fosfatos [250 mM de amohiguador de fosfatos pH 7 0,7% SDS, 1 mM EDTA pH 8 0, 1 % albúmina sérica bovina], (Church y Gilbert 1984) a 42°C durante 4 h y la hibridación se realizó en la misma solución añadiendo una alícuota
de la sonda marcada con ci-32P-dcTP. La hibridación se realizó a 65 °C para sondashomólogas y 42 °C para la sonda heteróloga y los lavados a alta astringencia con 2XSSC, 01% SDS durante 20 min a 55 °C durante 30 min. y un lavado a 55 °C durante15 min y lo§ lavados de baia astringencia con 2X SSC, 0 1 % SDS durante 20 min a 37°C 2 veces Se detectó la hibridación exponiendo en una película de autorradiografía.
Hibridación ARNADN (Northem-blot): Para el Northern-blot se utilizaronmuestras de 20 LLg de ARN total correspondientes a los días de 0 a 10,14 y 21 del ciclode cultivo de raíces transformadas de C. roseus, Ias cuales, se separaron en un gel deelectroforesis desnaturalizante en [1X de amortiguador MOPS (0.1 M 3-(N-Morfolino)propano ácido sulfónico (MOPS) pH 7.0, 40 mM acetato de Na, 5 mM EDTA pH 8.0),6 6% formaldehído, 0 8% agarosa], el ARN total, se desnaturalizó con 2.2 vol deformamida, 0.44 vol de MOPS amortiguador lox y 0.77 vol formaldehído al 37.5%, semezcló y adicionó 1 til de bromuro de etidio de 1 mg/ml y 0.2 vol de amort.iguador deseparación para ARN. Esta mezcla se incubó durante 5 min a 65 °C, se enfrió en hieloy las muestras se separaron en una electroforesis a 40 volts, al terminar laelectroforesis se transfirió el gel por capilaridad a una membrana de nylon con 10XSSC, al terminar la transferencia se tomó la membrana y se fijó con luz UV en uncrosslinking stratagene a 1 200 KJ/cm2 (Sambrock et al 1989).
La prehibridación y la hibridación con la sonda se llevaron a cabo a las mismasconcentraciones de las soluciones utilizadas para el Southern-blot , la temperatura quese utilizó fue de 50 °C y los lavados de las membranas fueron hechos gomo sedescribió para el Southern-blot, posteriormente las membranas se expusieron enpelículas de autorradiografía (Church y Gilbeh, 1984).
35
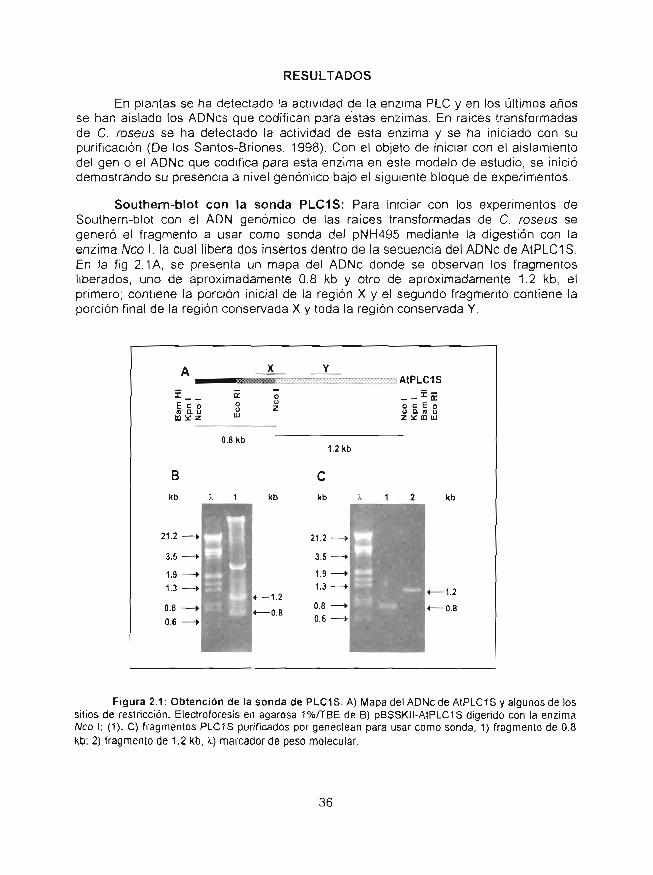
RESULTADOS
En plantas se ha detectado la actividad de la enzima PLC y en los últimos añosse han aislado los ADNcs que codifican para estas enzimas En raíces transformadasde C. n)seus se ha detectado la actividad de esta enzima y se ha iniciado con supurificación (De los Santos-Briones, 1998) Con el obieto de iniciar con el aislamientodel gen o el ADNc que codifica para esta enzima en este modelo de estudio, se iniciódemostrando su presencia a nivel genómico bajo el siguiente bloque de experimentos
Southem-blot con la sonda PLCIS: Para iniciar con los experimentos deSouthern-blot con el ADN genómico de las raíces transformadas de C. roseL6 segeneró el fragmento a usar como sonda del pNH495 mediante la dige§tión con laenzima Wco 1, la cual libera dos insertos dentro de la secuencia del ADNc de AtpLCI S.En la fig 21A, se presenta un mapa del ADNc donde se observan los fragmentosliberados, uno de aproximadamente Os kb y otro de aproximadamente 12 kb, elprimero, contiene la porción inicial de la región X y el segundo fragmento contiene laporción final de la región conservada X y toda la región conservada Y
Figura 2.1 : Obtención de la sonda de PLCIS. A) Mapa del ADNc de AtpLCI S y algunos de lossilios de restricción. Electroforesis en agarosa 1 %ffBE de 8) pBSSKll-AtpLCIS digerido con la enzimaWco 1, (1) C) fragmentos PLCIS punficados por geneclean para usar como sorida,1) fragmento de 0 skb, 2) fragmento de 1.2 kb, ),) marcadcm de peso molecular.
36

La fig 218 ilustra el resultado de la digestión con la enzima Nco 1, estos dosfragmentos se purificaron como se describió en la metodología y se almacenaron parausar como sonda, la fig 2.1C ilustra una electroforesis de los dos fragmentospurificados.
Después de generar las sondas se decidió utilizar el fragmento de 1 2 kb para lahibridación de la membrana preparada con ADN genómico de C. roseus digerido conlas enzimas de restricción Eco Rl, H/nd 111, Bam Hl y Sac 1 (fig 2 2A), la membrana sehibridó con la sonda mencionada marcada con ct-32P-dcTP La temperatura dehibndación utilizada en esta ocasión fue de 42 °C por tratarse de una sonda heterólogaLa fig 2.28 muestra los resultados obtenidos al hibridar las membranas a 42 °C y hacerlos lavados a 37 °C. A temperaturas mayores (entre 45 y 65 °C) no se observaronbandas de hibridación entre el ADN genómicx) y la sonda marcada (datos nomostrados). Los lavados para esta membrana también fueron de baja astringenciasimilar a lo que reporta Hirayama et al ,1997, es decir 2X de SSC, 0.1% SDS a 37 °Cdos veces 20 min , es así cx)mo en la fig 2.28 se observa la hibridación de la sondaheteróloga de A Ína//'ana con el ADN genómico de C roseus
Figura 2.2: Southern-blot de C. rosei/s con la sonda de PLCIS A) Electroforesis en agarosa0 8%n-BE de ADN genómico de C mseus digerido con las enzimas 1 -4) Eco Rl, Hind 111, Bam Hl y Sac 1,5) Fragmento obtenido por RT-PCR de C roseüs, 6) fragmento de PLC de 0.8 kb y 7) fragmento de PLCde 1.2 Kb 8) Autorradiografía de la membrana A hibndada con la sorida PLCIS de 1.2 kb marcada conct.32P-dcTP, » marcador de peso molecular.
37

Como se observa, existe por lo menos un fragmento que hibridó con la sondaheteróloga en cada digestión, excepto con la enzima Bam HI Con Eco Rl y Sac 1obtuvo un fragmento ep cada caso correspondiendo a tamaños de 4.3 y 52respectivamente y con Ht'nd 111, dos fragmentos de 5 9 y s 0 kb como se observa en laautorradiografía de la fig 2 28, de este modo puede decirse, que a baja astringencia setiene una evidencia de la posible presencia del gen que codifica para la PLC en rai.cestransformadas de 0. rioseus Anteriormente en este traba/o se habían realizadoexperimentos de Sou(hern-blot con temperaturas de entre 45 y 60 °C y los lavados aalta astringencia desde 2X de SSC, 01% de SDS hasta 0 lx de SSC, 01% SDS a lamisma temperatura de hibridación y no se había logrado observar la hibridación dealgún fragmento del ADN genómico de las raícxgs transformadas de C. rioseus En estaocasión no se realizaron los lavados a alta astringencia ya que se trata de una sondaheteróloga e incluso Hirayama et al , 1997, reporta condiciones de baja astringencia a25 y 37 °C, por experiencias anteriores con este tipo de análisis, como se mencionóanteriormente, se sabe que si los lavados en la membrana con el ADN de C roseus, sehacen más astringentes, i.ncluso a 38 °C, se despegan los fragmentos que hibridaronLos resultados de los lavados a 38 °C o más no §e muestran, puesto que sonnegativos.
Validación de la lécnica con la sonda de TDC: lnicialmente, como semencionó, los experimentos de hibridación con la sonda PLCIS se realizaban encondiciones de mediana astringencia, es decir, entre 45 a 60 °C y los lavados de 40 a55 °C en 2X y 0 2X SSC, 0.1% SDS como reporta Hirayama et al 1997. Como se hamencionado no se observó hibridación alguna en el modelo de estudio por lo quesurgió la idea de validar la técnica empleando una sonda homóloga a C roseus. Lasonda selecci`onada fue el ADNc de la TDC Se sabe que este gen está presente en elmodelo de estudio ya que su producto, la enzima triptofano descarboxilasa estainvolucrada en el metabolismo de los alcaloides. En la fig 2 3A se observa la fo{ografíade la electroforesis del ADN genómico de C nosew digerido con las enzimas derestricción Eco RI, Ht'nd 111, Bam HI, Eco RV y Sac 1 como se señala. Posteriormente serealizó la hibridación de acuerdo a como reporta Goddijn et al`,1994
El resultado de la hibridación se observa en la fig 38 donde se puede observarque los fragmentos hibri`dados concuerdan con los reportados anteriormente (Goddijnet al , 1994) Io que indica que el protocolo empleado en la metodología es biénmanejado y si anteriormente no se ha observado hibridación con la sonda de PLC esporque se trata de una sonda heteróloga y que es necesario obtener una sondahomóloga para poder tener la certeza de cuales son los fragmentos de PLC en elgenóma y poder estimar el número de copias de este gen en el modelo de estudio.
38

Figura 2.3: Southern-blot de C. /ose«s con sonda de TDC. A) Electroforesis en agarosaO.8%nBE de ADN genómico de C. /osetís digerido con las enzimas 1-5) Eco Rl, H/.nd 111, Bam Hl, Eco
:ev|y7Sf:La6)c:rágmcoennt:-3d2ep-TdEip?'^)A:':=ái:rgáaefi;ed£l:oTeecT,barr:naAhibridadaconlasondadeTDC
Northem-blot de ARN de C. mseus. Como se mencionó en la metodología seextrajo ARN total de las raíces transformadas de C. noseus de los días 0 a 10,14 y 21del ciclo de cultivo, ya que en los primeros días de acuerdo a antecedentes dellaboratorio se encuentra la máxima actividad de la enzima de las fracx=iones citosólicay membranal (De los Santos et al.,1997). En la fig 2.4 se observa una fotografía delARN extraído y desnaturalizado que se transfirió a la membrana para las hibridaciones.Se observa que el ARN es integro y sin embargo, no se logró la hibridación con lasonda PLCI S utilizada en este trabajo. La astringencia fue de 50 °C y aún así sólo selogró hibridación con las subunidades ribosomales (no se incluyen los resultadosnegativos). No se disminuyó la temperatura de hibridación como se realjzó con elSouthern-blot ya que en este caso, como se ha mencionado, se tiene el inconvenientede las subunidades ribosomales y aunque la hibridación se hiciera a baja temperaturahabría que lavar la membrana a alta temperatura para eliminar la inespecififfidad de lassubunidades ribosomales
39

Figura 2.4: Electroforesis de ARN total de C. roseLis. Electroforesis en agarosa 0.8% del ARNtotal de C. mseus correspondientes a los días del ciclo de cultivo indicados en el texto, las muestras sefraccionaron eii condiciones desnaturalizantes con fomaldehído.
DISCUSIÓN
En células vegetales se han aislado ADNcs que codifican para la PLC, todasellas con características similares a las PLC6 de otros modelos de estudio.
El presente trabajo es un esfuerzo por demostrar la presencia de este gen en C.rioseL/s. Se realizaron experimentos de Southern-blot y Northern-blot utilizando la sondaPLCI S (Hirayama et al.,1995)
En los Southern-blot, por tratarse de una sonda heteróloga se inició con lashibridaciones a 55 °C y los lavados se hacían a la misma temperatura, sin embargo, alobservar que no se lograba obtener la hibridación se fue bajando la temperatura de lahibrídación y los lavados, hasta Hegar a 42 °C de hibridación y 40 °C de lavados.Después de varios intentos fallidos, se decidió validar la técnica utilizando una sondahomóloga, la del ADNc que codifica para la TDC, cuyo producto convierte el triptofancia triptamina en C. /oseus y otras plantas que producen alcaloides indólicos (Goddijn etal., 1994). La hibridación con la sonda de TDC realizada como se encontró en laliteratura resultó positiva, descartando de este modo un mal manejo de la técnica ysatisfaciendo la necesidad de demostrar que si se era capaz de lograr la hibridación deun gen que se sabía estaba presente en las raíces transformadas de C. roseus.
40

Posteriormente, al revisar la literatura se realizó la hibridación con la sonda dePLCIS a 42 °C y los lavados a 37 °C de forma similar a 1o que reporta Hirayama ycolaboradores en 1997. Estos autores reportan la obtención de un ADNc (AtpLC2) quese expresa constitutivamente en tejidos vegetativos y florales y que en un análisis deSouthern-blot con las sondas AtpLCIS y AtpLC2 a alta y baja astringencia,encx)ntraron fragmentos que no corresponden entre sÍ bajo las dos condiciones, esdecir que la primera sonda no cruza con la segunda y viceversa en condiciones de altay baja astringencia por lo que es de esperarse que sea muy difícil lograr la hibridacióncon una sonda de un ADNc que es inducible bajo ciertas condiciones como es el casode PLCIS que se expresa en cx)ndiciones de deshidratación y estrés salino, en otromodelo de estudio. El resultado obtenido hasta este momento con el Southern-blot deC noseus, que es a muy baia astringencia, no es totalmente satisfactorio pero si es unavance en el proceso de hibridación con la sonda heteróloga, ya que se sabe que ni amediana ni a alta astringencia hibrida con la sonda PLCIS y esto sólo se logróutilizando una muy baia astringencia que ya ha sido reportada y quizá se puedameiorar hasta el momento de tener una sonda homóloga al modelo de estudio. Esposible también que la secuencia en los nucleótidos de la sonda sea muy diferente a ladel modelo de estudio y por lo tanto la homología en los nucleótidos sea muy pobre
Con respecto al Northern-blot, no se logró la hibridación con la sonda PLCISaún utilizando una mediana astringencia ya que en éstas condiciones, la sonda PLCIShibrida inespecíficamente a los ARN ribosomales y no se observa ninguna otra bandaque indique ser el mensajero de la PLC de C. roseus a no ser que el tamaño de estosmensajeros coincidan con la de las subunidades ribosomales Aunque aquí también sepresenta la misma problemática que se tiene con el Southern-blot y, sobre todo con lasonda PLCIS, o quizá es un poco más grave ya que la detección en el ADN genómicoimplica sólo presencia en el genoma en tanto que en el Northern-blot se requiere quelos mensaieros tengan la homología suficiente con el ADNc que se está usando comosonda o que en un momento dado éstos estén presentes en el momento de laextracción y si éstos, como se sabe, son muy específicos a una respuesta y la sondautilizada es inducible por estrés y el ARN estudiado está en condiciones basales, e§ unpoco difícil lograr la hibridación cruzada En este trabajo no se logró más éxito que elobtenido con el Southern-blot, quizá si las raíces se hubieran sometido adeshidratación o estrés salino durante los diferentes días de cultivo o durante diferentestiempos se hubiese podido identificar algún nivel de expresión de la PLC de C. roseuscon la sonda que se utilizó. Otra estrategia para lograr la hibridación sería lapurificación de los mensaieros y su fraccionamiento en agarosa para su posteriorhibridación.
41

REFERENCIAS BIBLIOGRAFICAS
Church G. Y Gilbeit W. (1984). Genomic sequencing. Proc Natl. Affid. Sci. USA , 81 :1991
De los Santos-Briones C. (1998) Estudio de la enzima fosfolipasa C en raícestransformadas de Caíharaníhus roseus. Tesis de Doctorado. Centro delnvestigación Científica de Yucatán Mérida, Yuc. , México.
De los Santos-Briones C., Muñoz€ánchez J. A., Chin-Vera J.. Loyola-Vargas V. M.y Hemández€otomayor S. M. T. (1997). Phosphatidylinositol 4, 5-bjphosphate-phospholipase C activity during the growng phase of Caíharaníhus noseustransformed roots. J. Plant Physiol., 34: 175-180
De Luca V., Marineau C. y Brisson N. (1989). Molecular cloning and analisis of cDNAa plant tryptophan decarboxylase. Comparison with animal dopa decarboxilases.Proc. Natl. Acad. Sci. USA., 86: 2582-2586.
Gamborg 0. L., lvljller R. A, y Ojima K. (1968) Nutrient requeriments of suspensionculture of soybean root cells. Exp. Cell Res., 50: 151-158.
Goddijn 0. J. M., Lohman F. P., Kam de R. J., Schilperoort R. A. y Hoge J. H. C.(1994). Nucleótide sequence of the tryptophan decarboxilase gene ofCaíhananíhus noseus and expresssion of tdc-gusA gene fusíons in M.coír.anaÍabacum. Mol. Gen genet., 242: 217-225.
Hirayama T.. Otr`o Ch., Mizoguchi T. y Shinozakj K. (1995). A gene encoding aphosphaditylinositol-specific phospholipase C is induced by dehydration and saltstress in Arabi.dops/.s Íha/Í'ana. Proc. Natl. Acad. Sci. USA., 92: 3903-3907.
Hirayama T., Mitsukawa N., Shibata D. y Shínozaki K. (1997). AÍPLC2, a geneencoding phosphoinositide-specific phospholipase C, is constitutively expressedin vegetative and floral tissues in Afflb/.dopsí.s fha//.ana. Plant Molecular Biology. ,34: 175-180.
Memelink J., Swords K. M. M., Staehelin 1. A. y Hoge J. H C. (1994). Southern,Northern and Western blot analisís. En Plant Molecular Biology Manual editadopor GeMn S. 8 y Schilperoort R. A. F1: 1-23.
Rogers S. 0 y Bendich A. J. (1994). Extraction of total celular DNA from plants, algaeand fungi En Plant Molecular Biology Manual editado por Gelvin S. 8 ySchilperoort R. A. D1: 1-8.
Sambrock J, Fritsh El, Maniatis T. (1989). Molecular Cloning Laboratory Manual. 2aed., Cold Spring Harbor Laboratory Press. U.S.A
Schuler M. A. y Zielinski R. E. (1989). A plant DNA minipreparation. Version 11. EnMethods in Plant Molecular Biology. pp. 89-96
Southem E. M. (1975) Detectjon of specific sequences among DNA fragmentsseparated by gel electrophoresis J. Biol Chem., 98.. 503-517.
42

Capítulo 3
Análisis de los ADNcs que codifican para la fosfolipasa C de raícestransformadas de CafAamntAus mseus obtenidos mediante estrategias de RT-PCR.
lNTRODUCCIÓN
Los métodos sensibles para la detección y análisis de moléculas de ARN son unimportante aspecto de muchos de los estudios en biología molecular. Entre losmétodos comunmente usados se encuentran; la hibridación /'n sffu, Northern-blot, dot oslot-blot, ensayos de nucleasa Sl y estudios de protección de ARNasa A (Davis et al„1986; Ausubel et al ,1987; Berger y Kimmel,1987).
La adaptación de la tecnología del PCR ha proporcionado un importanteprogreso en el área. Seeburg y sus colaboradores fueron los primeros en describir eluso de la amplificación de secuencias de ARN mensajero (P. Seeburg et al., 1986,simposio en UCLA, no publicado).
Una de las estrategias utilizadas para la clonación de ADNcs o fragmentos deADNc que codifican para genes específicos es la RT-PCR. Esta reacción consiste dedos pasos. El primer paso, la Transcripción Reversa (RT), se inicia con el ARN total delmodelo estudiado donde se realizará la amplificación de todos los ARN mensajerospresentes en la población utilizando como oligonucleótido iniciador a un oligo dT. EIsegundo paso, la Reacción en Cadena de la Polimerasa (PCR), se llevará a cabo parala amplificación del fragmento de ADNc de interés ya que en esta reacción se utilizanoligonucleótidos especificos para el ADN que se desea obtener.
En el presente trabajo, con el fin de amplififfir un fragmento de ADNc de C.mseus que tuviera similitud con las fosfolipasas C (PLCs) reportadas, en la reacción dePCR se empleó un juego de oligonucleotidos degenerados en sentido y antisentido delas regiones conservadas X y Y de las PLCs de plantas y levaduras. Losoligonucleótidos sentido son PLC1 (5' GGNGTNMGNGTNATHGARYTNGAYATHTG 3')y PLCA (5' CGGATCCTYCCAICAITCYAAYTCLAIRCAICKRCAICC 3') y los antisentidoPLC2 (5' YTGCATRTTRAANGCNACCATYTGNGCNCC 3') y PLCB ( 5' GGAAITCTCICp:WAA:wNÍ;rrrpm¡m3;).
MATERIALES Y MÉTODOS
Material vegetal: Como material biológioo se utilizaron raíces transformadas deC, mset/s, linea Jl transformadas con Agmbacíerium rh/2ogenes, las cuales sonsubcultivadas en medio 85 de Gamborg (Gamborg et al 1968), a la mitad de su fuerzaiónica adicionado con 3% de sacarosa y a pH 5.7 antes de autoclavear. Las raícesfueron resembradas cada 14 días con un inóculo inicial de 0.5 g. de tejido en 100 ml demedio estéril, las raíces se dejaron crecer a 25 °C +/- 2 °C en la obscuridad y enagitación continua de 100 rpm.
43

Sonda de ADNc de PLC: La sonda de PLC de A. fha/Í.ana utilizada para estetrabajo se generó de la digestión con la enzima Eco Rl del plásmido pNH495, el cualtiene el ADNc de AtpLCI S (fosfolipasa C de A. íha/Íana inducible por estrés) en el sitioEco Rl del pBSSKll, el fragmento generado es de 1.6 kb. Este plásmido fue donado porel Dr Hirayama Takashi del Laboratorio de Biología Molecular de Plantas de ToyotaJapón (Hirayama et al ,1995). El marcaje de la sonda (PLCIS) se realizó utilizando elequipo de Random primer (iniciador al azar) de GIBCO siguiendo los pasos indicadosen el manual, se obtuvo una actividad específica de 9.1 X 106 cpm/Lig
oseusss°en:batu::':read:amn::|::,::esT,¡oqn°®P:|aReTn-z::::EEifiigd::3::s#dmoó;°8:T8:,:,fragmento utilizado fue de aproximadamente 600 bp correspondiente a un fragmentodesconocido.
Extracción de ARN: La extracción de ARN se realizó de acuerdo a lametodología descrita por Schuler y Zielinski en 1989 Brevemente se pesaron 2 g dematerial vegetal se congelaron con nitrógeno líquido, se molieron hasta formar un polvofino y se transfirió a tubos de centrífuga conteniendo amortiguador de extracción (50mM Tris-Hcl pH 8.5,10 mM EDTA pH S 0, 200 mM NacI), se realizó una extraccióncon fenol cloroformo:alcohol isoamílico (50:50:1) y una segunda extracción concloroformo:alcohol isoamílico (50:1 ), la fase acuosa se precipitó con isopropanol, y sedejó secar la pastilla, durante 5 min. La pastilla se resuspendió en amortiguador TE (10mM Tris-Hcl pH 7.5, 1 mM EDTA pH 8.0) y se precipitó con Nacl 5 M y etanol al 100%La pastilla se resuspendió en TE y se precipitó con s M de urea y 10 M de Licl durantetoda la noche. Se centrifugó y dejó secar la pastjlla la cual se resuspendió en 2 ml deTE para repetir la precipitación etanólica Finalmente el ARN se resuspendió en aguaesténl, se cuantificó por espectrofotometría y se fraccionó en una electroforesis enagarosa/TBE (89 mM Tris base, 89 mM ácido bórico, 2 mM EDTA pH 80) paraobservar su integridad, Las muestras integras se mantienen almacenadas a Ú0 °C.Todas las muestras utilizadas en la extracción fueron preparadas con agua tratada conDEPC.
Extracción de ADN: La extracción de ADN genómico de raíces transformadasde C, noseus, se llevó a ffibo utilizando la técnica de Cetil Trimetil Bromuro de Amonio(CTAB) Se pesaron 2 g. de raíces y se congelaron con nitrógeno líquido, sepulverizaron en un mortero y el pulverizado se transfirió a un tubo de centrífuga, se leañadió amortiguador A de extracción (2% CTAB, 100 mM Tris-Hcl pH S 0, 20 mMEDTA pH 8.0, 1.4 M NacI) y se incubó a 65 °C. Se extrajo con cloroformo. alcoholisoamílico (24'1), se adicionó amortiguador 8 de extracción (10% CTAB, 0.7M Nacl),se extrajo con la mezcla de solventes y se precipitó con isopropanol. Los ácidosnucleícos se resuspendieron en amortiguaclor TE (10 mM Tris-Hcl pH 8.0, 1 mM EDTApH 8 0) y se precipitaron con acetato de amonio 5 M y etanol 100%, se lavó la pastillacon etanol 70%, se dejó secar y se resuspendió en H20 estéril (Rogers y Bendich,1994).
RT-PCR: Con el ARN total extraído de raíces transfomadas de C. mseus del día2 del ciclo de cultivo, se realizó un RT-PCR La primera parte de la reacción latranscripción reversa se llevó a cabo utiljzando 2 LLg de ARN total,10 ug/ml de oligo dT,
44

0.5 mM dNTPs, 10 mM DTT, 100 U de enzima transcriptasa reversa y lx deamortiguador de reacción en un volumen de 20 Ltl, la reacción de transcripción se dejó1 h a 37 °C. Posteriormente, de este producto se tomó una alícuota de 10 Lil y seadicionó 20 LiM de dNTPs, 5 tig de cada oligonucleótido en las siguientescombinaciones (PLC1-PLC2; PLCA-PLCB, PLC1-PLCB, PLCA-PLC2; PLC1-oligo dT yPLCA-oligo dT),1.5 mM Mgc12, 2.5 U de Taq polimerasa,1X de amortiguador reaccióny dio inició la PCR bajo las siguientes condiciones 94 °C, 5min, 94 °C, 1 min; 45 °C, 1 5min, 72 °C,15 min. durante 40 ciclos y 72 °C 10 min Los fragmentos amplificados se
i'ngv:trroongednee:c:,e::ostoa,;acgTeMt;d:'::í:ngfeosrci'á:o:nbae:teT,::uda:Ee'coT,rp:r':n;:ogpak:ta,d;conservar el plásmido con inserto.
Análisis de los productos ligados: El análisis de las transformantes de laposible PLC de C mseus, se realizó como sigue, se extraio el plásmido, de acuerdo alas indicaciones del manual del equipo (Qiagen plasmid midi kit) y posteriormente sedigirió con la enzima Eco Rl; posteriormente se realizó un Southern-blot (Southern,1975), con la sonda PLCI S
Secuenciación de fragmentos: El proceso preparativo de las muestras para lasecuenciación se llevó a cabo de acuerdo a las indicaciones del equipo Taqsequencing de Boehringer.
Hibridación ADN-ADN (Southem-blot): El experimento de hibridación(Southern,1975) que se realizó fue el del fragmento de ADNc obtenido por RT-PCRcon la sonda PLCIS marcada con ci-32P-clcTP y con el ADN genómico de raícestransformadas de C. roseus. En el primer caso, 25 LLg de plásmido correspondientes alas transformantes digeridos con Eco Rl, un control negativo (pHSV106 digerido conEco Rl) y 3 ng pBSSKll-AtpLCIS digerido con Eco Rl, se separaron en geles de
:g:áoasap:'c'|o€e:aTr=Eáaseár:n:f:£:r_odnc:gn(aa::vTd:rdandaed?Ty'£n|yosech;#:;o;ceonnL:segundo caso, ADN genómico digerido con las enzimas Eco Rl, H/.nc/ 111, Bam Hl, EcoRV y Sac 1 se separó en una electroforesis, se transfirió a una membrana de nylon y sehibridó con el fragmento obtenido por RT-PCR marcado con a-32P-dcTP (actividad de17 X 108 cpm/ug). La hibridación se llevó a cabo en el amortiguador de fosfatos (250mM de amortiguador de fosfatos pH 7 0, 7% SDS,1 mM EDTA pH 8.0,1% albúminasérica bovina), (Church y Gilbert,1984) a 50 y 65 °C respectivamente durante 24 h,terminado el tiempo de hibridación se lavaron las membranas con 2X SSC, 01% SDSdurante 20 mtn a 50 y 55 °C, otra vez durante 30 min y una vez más a 60 °C durante30 min se detectó la hibridación exponiendo en una película de autorradiografía.
RESULTADOS
RT-PCR: La estrategia seguida en este trabajo se basó en la técnica de RT-PCR mediante cual se obtuvieron fragmentos de aproximadamente 600 pares de bases(bp). Se pahió del ARN total del día dos del ciclo de cultivo de raíces transformadas deC. noseus, después de la reacción de RT se llevó a cabo la PCR utilizando diferentes
45
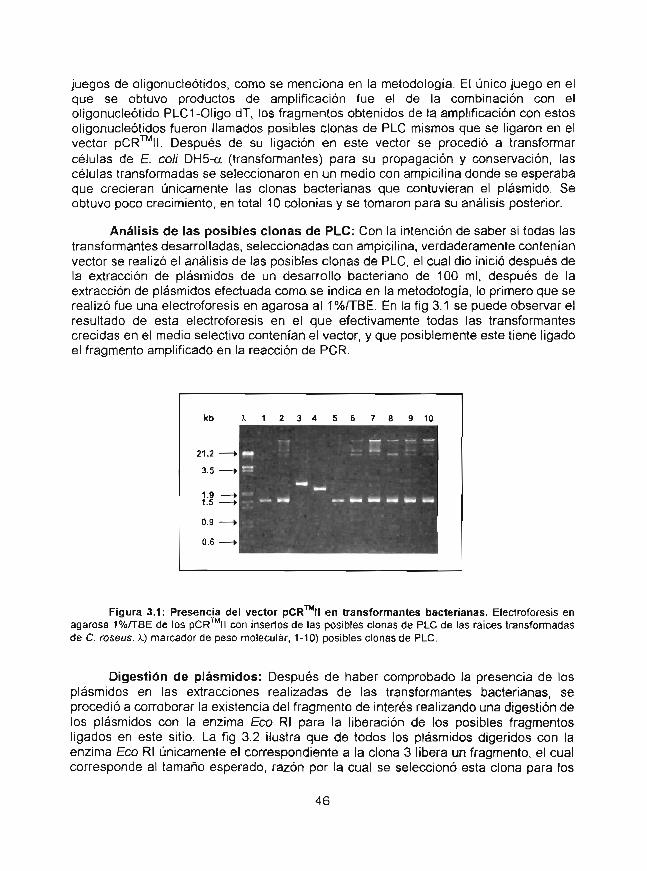
iuegos de oligonucleótidos, como se menciona en la metodología. El único juego en elque se obtuvo productos de amplificación fue el de la combinación con eloligonucleótido PLC1 -Oligo dT, los fragmentos obtenidos de la amplificación con estosoligonucleótidos fueron llamados posibles clonas de PLC mismos que se ljgaron en elvector pCRTMii. Después de su ligación en este vector se procedió a transformarcélulas de E. co/Í. DH5u (transformantes) para su propagación y conservación, lascélulas transformadas se seleccionaron en un medio con ampicilina donde se esperabaque crecieran únicamente las clonas bacterianas que contuvieran el plásmido. Seobtuvo poco crecimiento, en total 10 colonias y se tomaron para su análisis posterior.
Análisis de las posibles clonas de PLC: Con la intención de saber si todas lastransformantes desarrolladas, seleccionadas con ampicilina, verdaderamente conteníanvector se realizó el análisis de las posibles clonas de PLC, el cual dio inició después dela extraa=ión de plásmidos de un desarrollo bacteriano de 100 ml, después de laextracción de plásmidos efectuada comcL se indica en la metodología, lo primero que serealizó fue una electroforesis en agarosa al 1 %nBE. En la fig 3.1 se puede observar elresultado de esta electroforesis en el que efectivamente todas las transfomiantescrecidas en el medio selectivo contenían el vector, y que posiblemente este tiene ligadoel fragmento amplificado en la reacción de PCR.
Figura 3.1 : Pre3encia del vector pCR"ll en tmnsformantes bacterianas. Eleclroforesis enagarosa 1 %HBE de los pCR"ll con irsertos de las posibles clonas de PLC de las raíces transformadasde C. roseus. ^) marcador de peso molecular.1-10) posibles c]onas de PLC.
Digestión de plásmidos: Después de haber comprobado la presencia de losplásmidos en las extracciones realizadas de las transformantes bacterianas, seprocedió a corroborar la existencia del fragmento de interés realizando una digestión delos plásmidos con la enzima Eco Rl para la liberación de los posibles fragmentosligados en este sitio La fig 3 2 ilustra que de todos los plásmidos digeridos con laenzima Eco Rl únicamente el correspondiente a la clona 3 libera un fragmento, el cualcorresponde al tamaño esperado, razón por la cual se seleccionó esta clona para los
46

experimentos posteriores También se observa que no todos los plásmidos digeridosestán a la misma altura o al mismo peso molecular, lo cual indica una digestiónincompleta, en este caso se tendría que hacer una dilución del plásmido y digerirlo denuevo, y si se observa en el peso del vector quiere decir que no se tiene fragmentoligado en el mismo
Figura 3.2: Digestión de los vectores pCR"ll con la enzima Eco Rl. Electroforesis enagarosa 1%nBE de los plásmidos digeridos con la enzima Eoo Rl ^) marcadorde peso molecular,1-10)posibles clonas de PLC.
Southem-blot de la posible clona de PLC: Debido a que la clona 3 es la únicaque liberó el inserto clonado se decidió continuar trabaiando con ella,1o primero que serealizó fue un Southern-blot, se requirió de la sonda PLCI S derivada de la digestióncon la enzima Eco Rl del plásmido pNH495 Este plásmido contenía la longitudcompleta del ADNc de AtpLCI S (fosfolipasa C de A, fha//ana inducible por estrés) en elsitio Eco Rl del pBSSKII. La digestión con la enzima liberó un fragmento deaproximadamente 1.6 kb como puede observarse en la fig 3.3, posteriormente estefragmento liberado se purif.icó como se describió en la metodología y se marcóradiactivamente para u§ar como sonda. La fig 3.3A es un esquema de mapa del ADNcde AtpLCIS con algunos de sus sitios de restricción La fig 338 ilustra unaelectroforesis del fragmento liberado por la digestión con la enzima Eco Rl y finalmentela fig 3 38 muestra una imagen del gel donde se observa el fragmento purificado a usarcomo sonda.
EI Southern-blot con la transformante 3 se realizó después de digerirla con EcoRl. La transformante 8, que no poseía inserto también fue digerida con Eco Rl, uncontrol negativo (pHSV106 digerido con Eco Rl) Como control positivo pBSSKll-AtpLCI S digerido con Eco RI Se procedió como se indica en materiales y métodos y lamembrana se hibridó con la sonda PLCIS marcada con a-32P-dcTP En la fig 3 4A sepuede observar una fotografía de la electroforesis de los plásmidos digeridos yfraccionados que se transfirieron a la membrana de nylon, y en la fig 3.48 la
47

autorradiografía de la hibridación en la que se aprecia que el fragmento de latransfomante 3 hibndó con la sonda PLCIS con lo cual se tendría posiblemente unÍragmento de C, mset/s que sea homológo a PLC.
AXY _AtpLCIS
¡E3 E £ 3EEi1.6kb
Bc
kb 1 1 kb kb ^ 1 kb
21.2 -
¥_-
212-
-1.63.5-1.3=0.9-0.6- 3.5--3.21.§_
-" :..:=
Figura 3.3: Obtención de la sonda de PLCIS. Electroforesis en agarosa 1%nBE de A)pBSSKll-AtpLCIS digendo con la enzima Eco Rl, 8) fragmento PLCIS purificado por geneclean parausar como soncla. ^) marcador de peso molecular.
Sin embargo para tener la certeza de que este fragmento que hibridó con la PLCde A. Íha/Í.ana realmente correspondía a una PLC fue necesario secuenciarlo Sesecuenció como se describió en la metodología, se observa también en estaautorradiografía la hibridación con el fragmento del control positivo y con los vectoresde la transformante s y del control positivo, esto es debido a que al generar la sonda ladigestión se realizó con la misma enzima (Eco Rl) lo cual incluía un fragmento delpolilinker del vector que se marcó e hibridó con estos vectores y que da la hibridacióninespecífica del plásmido, para eliminar esta inespecificidad se tendría que lavar acondiciones más astringentes la membrana, quizá no tan solo con temperatura sinotambién disminuir el contenido de sales del SSC o generar una sonda más pequeñaque excluyera los segmentos del polilinker, como es el caso de la digestión con laenzima Wco 1.
48

Figura 3.4: Soutl`ern-blot de la transformante 3. A) electroforesis en agarosa 1 %nBE de 1)transfomante 3 digerida con Eco Rl, 2) transformante s digenda con Eco Rl, 3) pHSV106 digerido con
:oo:d:,pyL4c,,:B::#-dAatp=:1a:#g-:ádTopf#ae=:mo:dEec:eR::)oteü==fiogmfíademibridadón®nla
Se realizó el análisis visual de la secuencia obtenida, localizando los sitíos decorte de la enzima Eco Rl. Como se observa en la fig 3.5, está presente un sitio decorte para esta enzima correspondiente al vector en que se encuentra ligado, este es elsitio de inicio de la secuencia clonada y en este caso solo se tiene uno de estos sitiosya que se secuenció hasta donde termina la secuencia clonada y al final de la mismase puede observar una señal de poliadenilación que corresponde a la complementariaal oligo dT utilizado en la amplificación.
6AATTC H AACTGGATCCACTGCTTCAAACCCGGGCATCACGCTAAAAACCGTTACCTGTTGGTCGGGTTCCCGGAACGAACGCAAGAGCATCAGTGGcccGAccGGAGCTccTATGCTGnGTCGCTAGAGGACACAGAGGACAAGCAAGAAGCAACAGTAACCACACCAGAGGTCCCGACGGTAGAGGTTCGTCTATCGCACTGGACAccrrAccAAGTATGAATTGCCCAGCcAcCGCAGTACCGAGCCGCTACTACCATCTTCCCCGATAAAACCTACCTGAACAGAACATCCATCCTCAGCCAGTAAAGTGGTGATTGACAACTGGTCACATTCGGTCCC 1 1 1 1 1 1 ATCITCGGAGACTCCTCTGACceACCACGCATGGTCGTCTTCITCGGCCAATCACGCATGGCGCCAACAGGAGAGACTGACTCATTTGATACAACAGACATGGAGACTCCTGCCCCAGGAAACAGACGTGACAGCAATGGCACAGACCCGACCACGAGGTACTGCH-GCAGTGCCAGCGGCCTCCATCGTCAGAATCTGGGCTCTCGCCGGAACAGTGGAACATACTTCTAAAllTGITGGGCAGCAAAAAA
Figura 3.5: Secuencia do la clona 3. Análisis de la secuencia de nucleótidos de latransformante 3 que se sometió a la base de datos utilizando el programa BLAST, subrayado: secuenciade cohe de la enzima Eoo Rl oorrespondiente al vector; enmarcado: secuencia correspondiente al vector.
49

Al someter esta secuencia en formato FASTA al banco de datos con el programaBLAST no se encontró similitud con alguna PLC reportada, ni con alguna otrasecuencia de importancia que se encuentre registrada en el banco, dados estosresultados el siguiente paso sería caracterizar a este fragmento.
lniciando con la caracterización y con el objeto de encontrar alguna relevanciade este fragmento en el genoma de las raíces transformadas de C. mseus se realizó unSouthern-blot, utilizando este fragmento como sonda Lo primero que se realizó fue unadigestión de este fragmento con Eoo Rl y que liberó el fragmento clonado deaproximadamente 600 bp como se observa en la fig 3.6A, postertormente estefragmento se purificó cx)mo se describió en la metodología y una alícuota del fragmentopurificado se ilustra en una electroforesis en la fig 3.68.
Figura 3.6: Obtención de la sonda del ffagmento obtenido por RTJ]CR. Electroforesis enagarosa 1%nBE de A) PCR"ll de la transíomante 3 cligerido con la enzima Eoo Rl, 8) fragmentoliberado de la digestión purificado por geneclean para usar como sonda. 1) marcador de peso molecular.
Obtenida la sonda del fragmento obtenido por RT-PCR, se procedió a realizar elSouthern-blot con el ADN genómico de raíces transformadas de C roseus digerido conlas enzimas de restricción Eco Rl, H/nd 111, Bam Hl, Eco RV y Sac 1, como se observaen la fig 3.7A. La sonda marcada con ci-32P-dcTP alcanzó una actividad de 17 X 108cpm/ug La hibridación se realizó a 65 °C por tratarse de una sonda homóloga y elresultado de esta hibridación se ilustra en la autorradiografía de la fig 3.78. Los lavadosrealizado§ en este Southern fueron como sigue 2 veces en 2X SSC, 0 1 % SDS a 50 °Cpor 20 y 30 min. y una vez a 60 °C durante 30 min. y como se puede observar en la fig3.78 tiene varias regiones homologas en el genoma de las raíces transformada§ de C.noseus lnteresante sería saber de que gen o secuencia se trata este fragmento ya quea pesar de haberse secuenciado (fig 3 5) no reporta similitud con los bancos de datos
50
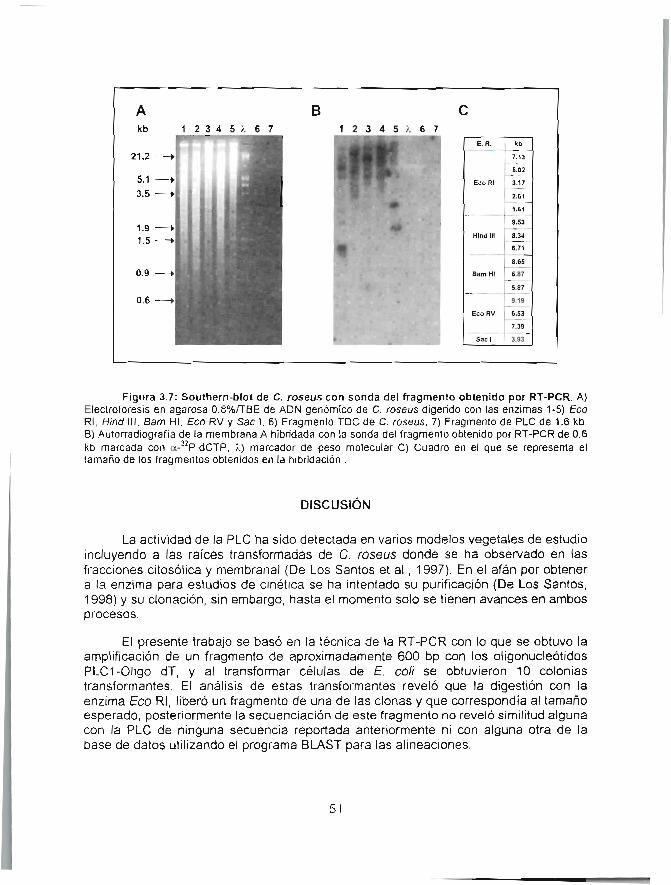
Figura 3.7: Sou.hern-blot de C. /oseus con sonda del fragmento ol)ünido por RT-PCR A)Electroforesis en agarosa 0 8%nBE de ADN genómico de C roseus digerido con las enzimas 1-5) EcoRl. Hmd 111, Bam Hl, Eco RV y Sac 1, 6) Fragmento TDC de C mseus, 7) Fragmenm de PLC de 1.6 kD.
:;Amu::£addáog¿anfi:-g?p,-adg;Fb:;n::t:3r::addeage::ams:,nedcau,::,:r,agcT::;:oeTeen,,dqou:o:eRT;:rce:edn:aoe6,tamaño de los fragmentos obtenidos en la hibndación
DISCUSIÓN
La actividad de la PLC ha sido detectada en varios modelos vegetales de estudioincluyendo a las raíces transformadas de C roseus donde se ha observado en lasfracciones citosólica y membranal (De Los Santos et al„ 1997) En el afán por obtenera la enzima para estudios de cinética se ha intentado su purificac.ión (De Los Santos,1998) y su clonación, sin embargo, hasta el momento solo se tienen avances en ambosprocx3sos.
El presente trabaio se basó en la técnica de la RT-PCR con lo que se obtuvo laamplificación de un fragmento de aproximadamente 600 bp con los ol.igonucleótidosPLC1-Oligo dT, y al transformar células de E coM se obtuvieron 10 coloniastransformantes El análisis de estas transformantes reveló que la digestión con laenzima Eco Rl, liberó un fragmento de una de las clonas y que correspondía al tamañoesperado,posteriormentelasecuenciacióndeestefragmentonorevelósimilitudalgunacon la PLC de ninguna secuencia reportada anteriormente n.i con alguna otra de labase de datos utilizando el programa BLAST para las alineaciones.
51

El análisis de Southem-blot realjzado de este fragmento con la sonda PLCI S, apesar de haber sido de una mediana as{ringencia reveló una hibridación, lo que llevaríaa especular que este fragmen{o tiene similitud con el fragmento utilizado como sonda ycomo consecuencia que tiene similitud con la PLC de A. fha//.ana, sin embargo, al hacer
rnnes::a:'í'f:££áns,opsr°pfránsdm°,d::dees,teveedx:re;'8ÉEffi'ysaeq::::Tfgeqsi%neá:Sti:shát:'sd;íJ3:u{ilizados se reali.zó con Eco Rl al igual que la sonda y que é§ta contenía un fragmentodel sitio de clonación, es decir del polilinker, que también se marcó y que a su vezhibridó con el polilinker del con{rol negativo y con el del vector del control positivo
En el Southern-blot realizado con el ADN genómico de las raíces transformadasde C. rioseus, utilizando como sonda al fragmento obtenido por RT-PCR se observóque varios fragmentos de las digestiones del genoma dieron señal positiva con estasonda a alta astring`encia lo cual indica que este fragmento esta presente en el genomay que es recomendable utilizarlo como sonda en una biblioteca para aislar el ADNccompleto o el gen y pos(eriormente teminar de caracterizarlo, podría encontrarse talvez que se trata de alguna enzima que es importante para alguna función en C. /oseusSin embargo, por el momento el objetivo principal es trabajar con la PLC de C. roseusde modo que este fragmento de aparente interés se mantendrá reservado para trabaiosfuturos.
52

REFERENCIAS BIBLIOGRAFICAS
Ausubel F. M., Brent R., Kingston R., Moore D. E., Seidman J. G., SiTiith J. G., SlruiK. (1987). Current Protocols in Molecular Biology Greene Publishing Associatesand Wiley-lnterscience, New York.
Berger S. L. y Kimmel A. R. (1987). Guide to molecular cloning techniques. MethodsEnzymol.,152: 215-304.
Church G. Y Gilbert W. (1984) Genomic sequencing. Proc Natl Acad. Sci. USA., 81 '1991.
Davis L. G., Dibner lvl. D. y Battey J. F. (1986). Basic methods in molecular biology.Elsevier Science Publishing Co , lnc„ New York.
De Los Santos-Briones (1998). Estudio de la enzima fosfolipasa C en raícestransformadas de Caíharaníhus roseus, Tesis de Doctorado Centro delnvestigación Científica de Yucatán Mérida, Yuc , México
De Los San.os-Briones C., Muñozsánchez J. A., Chin-Vera J., Loyola-Vargas V.M. y Hemández-Sotomayor S. M. T. (1997). Phosphatidylinositol 4, 5-biphosphate-phospholipase C activity during the gro ng phase of Caíharaníhusroseu§ transformed roots. J. Plant Physiol , 34`.175-180.
Gamborg 0. L., Miller R. A. y Ojima K. (1968). Nutrient requirements of suspensionculture of soybean root cells Exp. Cell Res. 50 151-158
Hirayama T., Otho Cli., Mizoguclii T. y Shinozaki K. (1995) A gene encoding aphosphaditylinositol-specific phospholipase C is induced by dehydration and saltstress in Araoi.c/opsi.s íha//.ana. Proc. Natl Acad. Sci. USA„ 92: 3903-3907.
Rogers S. 0 y Bendich A. J. (1994) Extraction of total celular DNA from plants, algaeand fungi. En Plant Molecular Biology Manual editado por GeMn S. 8 ySchilperooft R A. D1: 1-8
Schuler M. A. y Zielinski R. E. (1989). A plant DNA minipreparation Version n EnMethods in Plant Molecular Biology. pp 89-96
Southem E. M. (1975). Detection of specific sequences among DNA fragmentsseparated by gel electrophoresis J. Biol. Chem , 98 503-517
53


Capítulo 4
DISCUSION GENEFUL
Los organismos vivos reaccionan a señales de su medio ambiente para podersobrevivir y adaptarse a sus condiones de vida. Los organismos multicelularesnecesitan comunicación intercelular a través de un continúo flujo de información entrelas células que lo integran (Evans 1998). Las señales extracelulares son transmitidas através de la membrana plasmática por la acción de segundos mensajeros, es decir, serequiere que una molécula agonista se una a receptores específicos en la membranaplasmática para iniciar la producción de segundos mensajeros, los cuales Ínicianreacciones intracelulares que regulan una serie de procesos dentro de la célula.
La hidrólisis de fosfolípidos de membrana, entre ellos el fosfatidilinositol 4,5-bifosfato (PIP2) por una fosfolipasa C (PLC) específica, es uno de los primeros eventosen la regulación de varias funciones extracelulares por más de 100 moléculas deseñalización extracelular (Rhee y Choi 1992). Esta reacción produce al hidrolizar elPIP2t dos segundos mensajeros intracelulares: el 1,2 diacilglicerol (DAG) de naturalezahidrofóbica y el inositol 1,4,5-trifosfato (lp3) de naturaleza hidrofílica (Berridge 1987). EI
P9A9G5)P;rga,npe3=eern{:rTae::r3r%,Yo:#'Vpaaraa':n3L°ct,:íraa,Íg::::,osn(:eK%)a£Pedke|erre:ítí:óendoplásmico (Nishizuka 1992).
El metabolismo de fosfoinositidos es un fenómeno que ria quedado demostradoen plantas por la presencia de enzimas que pahicipan en el como son las fosfolipasasA, C y D (PLA, PLC y PLD respectivamente) (Chapman 1998). La clonación molecularde ADNcs que codifican para múltiples isoenzimas de PLC en plantas han sidoreportadas en los últimos cinoo años y comprenden los de A. Íha//.ana (Hirayama et al.;1995, 1997, Yamamoto et al ; 1995); G/i.cy.ne max (Shi et al.; 1995) y So/anumÍuberosum (Kopka et al.; 1998). Comparadas con las PLCs de otros sistemas las PLCsde plantas aisladas hasta ahora parecen formar parte del grupo de las PLCs ya queexmben una alta homología con éstas. Tienen homología en los dominios X y Y, loscuales constituyen el sitio catalítico, dos de ellas poseen la región EF-hand, la cual esun sitio de unión para Ca2+ y las últimas pertenecientes a S. Íubemsum que poseen el
foosT:fi,L:d:s2coe: p::teríenga,í:eg:ernbdo,:,l:e:eá:,::!+que Son dom,n,oS de ,nteracx=,ón de
En el presente trabajo con el fin de detectar a nivel genómico la presencia y laexpresión del gen que codifica para la fosfolipasa C en las raíces transfomadas de C.noseus, se utilizaron las técnicas Southern-blot y Northem-blot, seguida de unahibridación con una sonda heteróloga de PLC de A. Íha/Í.ana y con el fragmentoobtenido por RT-PCR de C. roseus.
Después de muchos intentos y hasta que se reduio la astringencia y latemperatura de los lavados se pudo observar hibridación positiva que indican lapresencia de este gen, la temperatura usada para los lavados como se reporta en elcapitulo 2 fue de 37 °C, tal como lo repoha Hirayama et al.; 1997, en el cual al utilizar
55

dos ADNcs que se obtuvieron en diferentes condiciones, Ia basal y la inducible porestrés (sonda PLCIS) observaron a alta y baja astringencía señales positivas que nocorrespondían entre sÍ cuando usaban las sondas. Esto coincide con [o señalado porKopka et al.;1998 que al obtener tres ADNcs para la PLC de papa, las cuales seexpresan diferencialmente en tejidos a señales ambientales no tienen híbridizacióncruzada entre sí, como lo demuestran en un experimento de dot-blot. Esto es una pautapara pensar que quízá las PLCs de plantas son muy específicas y es necesario utjljzaruna sonda homóloga para poder detectar una señal positiva a alta o medianaastringencia con el ADN de las raíces transformadas de C. mseus.
En el Northern-blot tampooo se obtuvo la hibridación de mensajeros a lo largodel ciclo de cultivo con la sonda utilizada, esto puede deberse a que la sonda utilizadaes de un ADNc que se aisló bajo condiciones de estrés y el ARN que se utilizó se aislóen condicíones básales, es importante hacer esta aclaración pues Hirayama et. al.;1998, no obtuvo riibridización con sus sondas aún tratándose de sondas provenientesde la misma especie de planta, es decir, una sonda homóloga. Esto indica queposiblemente en plantas sea necesario utilizar sondas homólogas para la detección deotros genes de PLC que se expresan diferencialmente en la misma planta y, aún así,es necesario utilizar bajas temperaturas de lavado como reportaron Hirayama et al.;1997 utilizando dos sondas homólogas.
En la RT-PCR. se obtuvo un fragmento amplificado de aproximadamente 600 bp,que finalmente no tuvo similitud con las PLCs e incluso con ningún otro gen reportadoen la base de datos, posiblemente se trate de un fragmento de un gen que tengaalguna función en las células de las raíces de C. mseus, no se obtuvo la amplificaciónde un fragmento similar al de la PLC quizá por el grado de degeneración deloligonucleótido utilizado, una altemativa para solucionar este problema seria el diseñode nuevos oligonucleótidos que sean diseñados a partir de secuencias conservadas delas PLCs de plantas, es decir a)nsiderando la secuencia de las PLCs de las 3 especiesconocidas hasta ahora y no incluir las secuencias de levadura o mamíferos.
Existen otras fomas en que pudiera lograrse la amplificación de fragmentosutilizando únicamente los oligonucleótidos específicos para la PLC, uno de estos es laPCR anjdada, un procedimiento de dos pasos, en el que los productos de un primerPCR usando iniciadores UexterncN3" son reamplificados usando otro juego de iniciadores"intemos". Otra técnica podría ser el de utilizar dos temperaturas de elongación en el
proceso de PRC.
56

REFERENCIAS BIBLIOGRAFICAS
Berridge lvl, J. (1987). lnositol trisphosphate and diacylglycerol: t\m i.nteracting secondmessengers. Annu. Rev Biochem., 56: 159-193.
Berridge M. J. (1993) lnositol Triphosphate and Calcium Signalling. Nature. 361: 315-325.
Chapman D. K. (1998). Phospholipase activity during plant growth and developmentand m response to environmental stress. Trends in plant science, 3: 419.426.
Dekker L. V., Palmer R. H. y Parker P. J. (1995). The protein kinase C and proteinkinase C related gene famnjes. Curr. Opin. Struct Biol., 5: 396402.
Evans R. M. (1998) The steroid and thryroid hormone receptor superfamily. Science„240: 889-895.
Hirayama T., Otho Ch., Mizoguchi T. y Shinozaki K. (1995). A gene encoding aphosphaditylinositol-specific phospholipase C is índuced by dehydration and saltstress in Arab/.dops/'s Íha//.ana. Proc. Natl. Acad. Sci. USA., 92: 3903-3907.
Hirayama T., lvlitsukawa N., Shibata D. y Shínozaki K. (1997). AíPLC2, a geneencoding phosphoinositide-specific phospholipase C, is constitutively expressedin vegetative and floral tissues in Arabí.dops/.s Íha//.ana Plant Molecular Biology.,34: 175-180.
Kopka J., Pical C., Gray J. E. y Müller-Róber 8. (1998). Molecular and enzymaticcharacterization of three phosphoinositide-specific phospholipase C isoformsfrom potato. Plant Physiol.,116: 239-250.
Njshizuka Y. (1992). lntracellular signalling by hydrolisis of phospholipids and activationof protein kinase C. Science., 258: 607J514.
Rhee S, G. y Choi K. D. (1992). Regulation of inositol phospholipid-specific
phospholipase C isozymes. J. Btol. Chem., 267: 12393-12396.
Shi J., González R. A. y Bhattacliarrya M. K. (1995). Characterization of a plasmamembrane-associated phosphoinositíde-specific phospholipase C from soybean.The Plant Journal„ 8:381-390.
Yamamoto Y. T., Conkling M. A., Sussex 1. M. e lrish V. F. (1995). An ArabidopsiscDNA related to animal Phosphoinositide-specific phospholipase C genes. PlantPhysiol„ 107: 1029.1030.
57


Capítulo 5
CONCLUSIONES y PERSPECTníAS
Conclusjones generales
En el presente trabajo se planteó la clonación del gen o el ADNc que codificapara la PLC de raíces transformadas de Caínafflníhus noseus. En base a los resultadosobtenidos se concluye que..
• En el Southern-blot con el ADN genómico de raíces transfomadas de C.mseus empleando la sonda heteróloga PLCI S de A, Íha/i.ana, sólo se logró,hibridacióna baja astringencia.
• La validación de la técnica de Southem-blot empleando una sonda homólogade C. mseus, (TDC) y el producto obtenido por RT-PCR, demostró que el manejo de lamjsma era adecuado.
• En el Nonhem-blot con el ARN total de raíces transfomadas de C. ®seus, nose logró la hibridacjón con la sonda heteróloga.
• En los experimentos de RT-PCR, se logró ampljficar un fragmento de 0.6 kbutilizando los oligonucleótidos, PLC1-Oligo dT, que se clonó en el vector pCR"ll, alrealizar la digestión de este vector se liberó el inserto.
• La secuenciación del fragmento clonado no reportó similitud con la PLC ni conalgún otro fragmento reportado en la ba§e de datos a pesar de tener mbridación con lasonda heteróloga PLCI S de A, Íha/7.ana.
• EI Southem-blot con el ADN genómico de raíces transfomadas de C. /oseusutilizando como sonda el fragmenlo obtenido por RT-PCR, reveló varios fragmentosque tuvieron hibrjdación cruzada a alta astringencia.
Perspectivas del trabajo
Trabajos previos reportan haber detectado la activjdad de PLC en raícestransformadas de C. mseus, sin embargo, demostrar su presencia a nivel genómico hasido un proceso difícjl de lograr.
Definitivamente la estrategia para poder mejorar las condiciones de hibridacióny así poder asegurar que los fragmentos hibridados efectivamente son de una PLC,sería la obtención de una sonda homóloga que podría consegujrse con las siguientesindicaciones:
59

• El diseño de oligonucleótidos específicos para PLC de plantas, basandoseúnicamente en las secuencias más conservadas reponadas para las mismas yamplificar a partir de ADN genómico.
• Realizar el PCR de la RT, solamente con oligonucleótidos específicos y noemplear el oligo dT y para mejorar sería conveniente realizar el RT-PCR conmensajeros purificados y no con el ARN total.
• Seguir utilizando la sonda heteróloga y repetir varias veces que coincidan lostamaños de hibridación con el ADN genómico digerido con las mismas enzimas derestricción y eluir los fragmentos de los tamaños que concuerden de un gel para haceruna biblioteca subgenómica.
• Hacer una biblioteca de ADNc de uno de los primeros días de ciclo de cultivo yempacar en bacteriofagos que también expresen proteínas, esto con la intención deque si se tiene la proteína pura se pueden sintetizar anticuerpos y realizar uninmunoescrutinio en la misma para poder obtener la clona de ADNc de la PLC.
• Por otra parte al tener a la proteína pura, se secuenciaría y con esta secuenciase podrían diseñar oligonucleótidos específicos para la PLC de raíces transtormadasde C. nDseu6 y hacer PCR para la obtención de una sonda homóloga de ADN y serealizaria el escrutinio en la misma biblioteca.
60


=-=CICY
CENTRO DE INVESTIGACIÓN CIENTÍFICA DE YUCATÁN, A.C.