
Rhizobius ventralis -...
61
1. INTRODUCCIÓN Uno de los factores que influyen en el desarrollo de una población es la ausencia o ineficiencia de enemigos naturales que la controlen, por lo tanto, en aquellos países en donde los chanchitos blancos son considerados como nativos, no constituyen un problema fitosanitario, lo que supone la existencia de cierta forma de control natural (YUDELEVICH, 1950). En Chile, las principales plagas entomológicas que afectan a la agricultura son de origen foráneo, las que luego de ingresadas, al encontrar condiciones favorables y huéspedes adecuados, adquieren mayor intensidad que en su país de origen, causando en algunos casos, pérdidas económicas importantes (GONZALEZ y ROJAS, 1966). CAPDEVILLE (1945) señala que los chanchitos blancos son una plaga especialmente susceptible a ser atacada por enemigos naturales de varias especies. Sus cuerpos suaves, la falta de protección, sus movimientos lentos y más particularmente sus hábitos de establecer colonias o grupos, los hace accesibles a los parásitos y predadores. Además, su gran capacidad reproductiva hace posible soportar un alto número de enemigos naturales sin peligro de extinción. Los intentos por controlar diversas plagas mediante el uso de enemigos naturales, se inician en Chile en el año 1903 con la importación desde California de dos coccinélidos, Rhizobius ventralis y Hippodamia convergens,
Transcript of Rhizobius ventralis -...

1. INTRODUCCIÓN
Uno de los factores que influyen en el desarrollo de una población es la
ausencia o ineficiencia de enemigos naturales que la controlen, por lo tanto,
en aquellos países en donde los chanchitos blancos son considerados como
nativos, no constituyen un problema fitosanitario, lo que supone la existencia
de cierta forma de control natural (YUDELEVICH, 1950).
En Chile, las principales plagas entomológicas que afectan a la agricultura
son de origen foráneo, las que luego de ingresadas, al encontrar condiciones
favorables y huéspedes adecuados, adquieren mayor intensidad que en su
país de origen, causando en algunos casos, pérdidas económicas
importantes (GONZALEZ y ROJAS, 1966).
CAPDEVILLE (1945) señala que los chanchitos blancos son una plaga
especialmente susceptible a ser atacada por enemigos naturales de varias
especies. Sus cuerpos suaves, la falta de protección, sus movimientos lentos
y más particularmente sus hábitos de establecer colonias o grupos, los hace
accesibles a los parásitos y predadores. Además, su gran capacidad
reproductiva hace posible soportar un alto número de enemigos naturales sin
peligro de extinción.
Los intentos por controlar diversas plagas mediante el uso de enemigos
naturales, se inician en Chile en el año 1903 con la importación desde
California de dos coccinélidos, Rhizobius ventralis y Hippodamia convergens,

2
predadores de conchuelas (Coccidae) y pulgones (Aphididae)
respectivamente. A partir de 1915 el Ministerio de Agricultura, comenzó
oficialmente la introducción de entomófagos, acción emprendida a través del
Departamento de Sanidad Vegetal para controlar al pulgón lanígero del
manzano (Eriosoma lanigerum). Entre 1929 y 1931, se constató la
presencia de una grave plaga conocida como Conchuela acanalada de los
cítricos (Icerya purchasi Maskell), e intensos ataques de “chanchitos blancos”
(Pseudococcus sp.) en el valle de Quillota. Así nace la necesidad de crear un
organismo dedicado a la introducción, crianza y distribución de insectos
benéficos. Para tal efecto, en 1937, se crea la actual Subestación
Experimental La Cruz, dependiente del Instituto de Investigaciones
Agropecuarias (INIA). Éste, bajo el nombre de Insectario La Cruz, ha
introducido una enorme diversidad de insectos benéficos para el control de
plagas que afectan al país (GONZÁLEZ Y ROJAS, 1966).
ZUÑIGA (1984) señala a lo menos diez especies de enemigos naturales
introducidos para el control de chanchito blanco entre los años 1931 y 1983.
Por otra parte, desde 1980, la Facultad de Agronomía de la Pontificia
Universidad Católica de Valparaíso, ha desarrollado un programa de
multiplicación comercial y liberación masiva de insectos benéficos de las
especies Cryptolaemus montrouzieri, Leptomastidea abnormis, Coccophagus
gurneyi y Sympherobius maculipennis.
LA V Región de Chile, en especial el valle de Quillota, por su importancia
agrícola y su cercanía con el Insectario La Cruz, ha sido la cuna de los
constantes esfuerzos en busca del establecimiento de especies benéficas
que han sido introducidas.

3
La contraparte está conformada por los enemigos naturales nativos, según
YUDELEVICH (1950), de origen desconocido, dejando en claro que podían
haber sido introducidos en forma accidental. Estos, según el mismo autor,
son de escaso interés económico, y aunque su acción no es decisiva, no
deja de ser considerable en el control de la plaga. Pese a las deficiencias
que presentan, desempeñan un rol que no debe desestimarse en el control
biológico general de la plaga. Las desventajas de biología, hiperparásitos y
especificidad escasa son compensadas parcialmente, por su abundante
número y su libre reproducción y distribución. Sin embargo, el rol de estos
agentes, descrito por YUDELEVICH en 1950, no puede ser asemejado a la
realidad actual, ya que en ese entonces, el control biológico en Chile estaba
iniciándose en forma eficiente y la introducción de más enemigos naturales
sería llevada a cabo con posterioridad.
El control biológico, tiende sus bases en un estudio ecológico de las
interacciones entre individuos, de dos o más especies, que conforman un
nicho ecológico. Las relaciones más importantes son el parasitismo y la
predación, que en conjunto reducen los niveles de chanchito blanco. Éstos al
ser limitados generan una nueva relación, la de competencia entre enemigos
naturales.
La competencia puede traducirse en ajustes de equilibrio por parte de dos
especies, podría conducir a que una especie substituya a otra, y la obligue a
ocupar otro espacio o a servirse de otro alimento (ODUM, 1969).

4
En una competencia desigual, se puede producir una severa sobre
explotación de los recursos. En tales circunstancias, algunas especies
tendrían acceso a suficientes recursos para crecer, sobrevivir y reproducirse.
En contraste, la reproducción de las especies menos aptas puede declinar,
las migraciones serán favorecidas o una población puede ser destruida
(SPEIGHT, HUNTER y WATT, 1999).
1.1 Hipótesis de trabajo:
El efecto de la introducción de especies exóticas, la multiplicación y
liberación masiva de alguna de éstas, pueden haber afectado a las
poblaciones de las especies nativas, lo cual no ha sido evaluado. Es posible
suponer, por lo tanto, que la introducción y liberación masiva de insectos
benéficos, ha desplazado a las especies nativas que controlan al chanchito
blanco.
1.2 Objetivo general:
Evaluar los efectos de la introducción y liberación masiva de especies
exóticas, para el control de chanchito blanco sobre las especies de enemigos
naturales nativos asociados a esta plaga.

5
1.3 Objetivos específicos:
- Identificar los enemigos naturales asociados a las especies
Pseudococcus longispinus, Pseudococcus calceolariae y Planococcus
citri en la comuna de Quillota.
- Evaluar la importancia relativa de cada enemigo natural asociado a las
especies Pseudococcus longispinus, Pseudococcus calceolariae y
Planococcus citri en la comuna de Quillota y su comportamiento
estacional.

6
22.. RREEVVIISSIIÓÓNN BBIIBBLLIIOOGGRRÁÁFFIICCAA
2.1 Antecedentes generales:
CAPDEVILLE, (1945) señala que, en cultivos de palto (Persea americana),
P. longispinus causa daño directamente al succionar savia en la parte aérea
de la planta e indirectamente por la secreción de grandes cantidades de
sustancia azucarada, sobre la cual se desarrolla la fumagína (Capnodium
sp.). Este hongo, produce una cubierta negra que interfiere con los procesos
fisiológicos normales de la planta y disminuye la calidad del fruto. Grandes
infestaciones en el pedúnculo y fruto retardan el desarrollo de éste, además
de provocar una disminución del tamaño y caída de frutos.
El Control biológico, consiste en la regulación o supresión del potencial
reproductor de organismos a través de la acción de parásitos, predadores y
patógenos. En el campo de la entomología económica, la aplicación del
control biológico, se orienta hacia la mantención de un organismo perjudicial
bajo el nivel de daño económico, mediante el uso de agentes denominados
colectivamente entomófagos (parásitos y predadores), y patógenos
(GONZALEZ y ROJAS, 1966).
De todas las formas de control de plagas, el control biológico, es la práctica
más eficaz y económica que existe. Comparado con el control químico, el
control biológico posee ventajas de las que éste carece: es permanente y de
costo prácticamente nulo. Además, está libre de todas las desventajas

7
inherentes al control químico, como la contaminación ambiental con daños
para la vida silvestre y riesgos para la salud humana, perturbación del
equilibrio natural con sus resultantes de resurgencias de plagas, provocación
de plagas secundarias, desarrollo de poblaciones resistentes a plaguicidas y
un alto costo económico (BEINGOLEA, 1977).
Hay varias definiciones de competencias que son útiles para comprender la
lucha por recursos limitados. La competencia intraespecífica describe a la
competencia entre individuos de una misma especie por un recurso limitado;
en cambio, la competencia interespecífica refiere a la competencia de
recursos limitados entre individuos de diferentes especies. En el primer caso,
los individuos utilizan los recursos de igual manera, mientras que la
interespecífica implica una competencia desigual por cuanto los recursos
limitados son utilizados en forma desigual, correspondiendo a las
características biológicas de cada especie, como por ejemplo voracidad,
eficiencia, capacidad reproductiva y capacidad de búsqueda (SPEIGHT,
HUNTER y WATT, 1999).
2.2 La plaga:
En nuestro país, se encuentran varias especies de chanchitos blancos, entre
las cuales se puede mencionar a: Pseudococcus affinis Maskell (sin.
Pseudococcus obscurus, Pseudococcus viburni), Pseudococcus maritimus
Ehrhorn, Pseudococcus longispinus Targ. Tozz. (sin. Pseudococcus adonium
(L.)), Pseudococcus calceolarie Maskell (sin. Pseudococcus fragilis,
Pseudococcus gahani) y Planococcus citri Risso. Éstos se distribuyen

8
ampliamente en Chile, siendo importantes en diversos frutales, entre los que
se destaca a cítricos (Rutaceae), chirimoyos (Annona cherimola Mill), paltos
(Persea americana) y la vid (Vitis vinifera) (GONZÁLEZ, 1991).
Los chanchitos blancos deben su nombre común a la sustancia blanca,
cérea, a menudo en forma de polvo, filamentos, proyecciones o láminas que
cubren el cuerpo de la hembra a partir del tercer estadío larvario
(CAPDEVILLE, 1945).
YUDELEVICH (1950), describe a los chanchitos blancos como un gran
género de insectos perjudiciales. Alargados u ovalados, con segmentación
bien diferenciada. Generalmente cubiertos por una secreción harinosa o
algodonosa, que corrientemente sobresalen de los bordes del cuerpo en
forma de filamentos y prolongaciones más o menos regulares, de forma y
tamaño relativamente variables para cada especie. Las hembras
caracterizadas por sus extremidades bien desarrolladas y una libre
movilización en todo su ciclo evolutivo. Los machos son pequeños, frágiles y
alados, provistos de dos o más filamentos caudales de secreción cerosa y
longitud específicamente variable. Los estados larvarios antes de la primera
muda carecen de partes bucales y cerarios.
La duración del ciclo biológico de los chanchitos blancos, según
YUDELEVICH (1950), es de 61 días en verano, para la zona de Santiago, sin
especificar la especie. Además, señala este autor, que los ciclos biológicos y,
por lo tanto, el número de generaciones anuales varían apreciablemente
dependiendo de la época y estación del año. El mismo autor presume cuatro

9
generaciones al año para el caso de Planococcus citri en el área de Quillota.
A partir de diciembre, hay una alta superposición de generaciones.
2.3 Enemigos naturales de los chanchitos blancos:
YUDELEVICH (1950) en ese entonces, clasifica a los enemigos naturales de
los chanchitos blancos en dos grupos. El primero conformado por aquellos
que considera de escasa trascendencia económica en Chile, en el cual, cita
a Xenoleucopis olalquiagay Blanchard, Sympherobius pallidum Gay y
Baccha sp. En el segundo, los de importancia económica en Chile, se
encuentra a Cryptolaemus montrouzieri Muls., Coccophagus gurneyi Comp.,
Leptomastidea abnormis Gir., Tetracnemus pretiosus Timberlake y
Pseudaphycus sp.
El mismo autor recuperó enemigos naturales de chanchitos blancos, del cual,
se expondrán los datos obtenidos en la localidad de Quillota, La Cruz y
Limache. YUDELEVICH (1950) tomó en conjunto a todos los chanchitos
blancos existentes: P. citri, P. calceolarie, P longispinus y P. maritimus, este
último solo representó el 1.7 % de los chanchitos blancos y no tuvo
parasitismo, por lo que se hace irrelevante para el análisis posterior. Los
enemigos naturales encontrados en aquella época y su ponderación
correspondieron a: Xenoleucopis (Leucopis sp.) (35,5%), Sympherobius sp.
(26,7%), Coccophagus gurneyi (8,8%), Cryptolaemus montrouzieri (18,4%) y
Leptomastidea abnormis (10,3%).

10
HARRISON (1993) recupera en muestras de Pseudococcus longispinus,
recolectadas entre los meses de octubre y abril de un huerto de palto en el
área de Quillota, los siguientes enemigos naturales: Tetracnemoidea
brevicornis, Coccophagus gurneyi, Scymnus sp. y Cryptolaemus
montrouzieri. Éste autor señala que, en cuanto a proporción en que se
encontraron las cuatro especies mencionadas, la mayor importancia la tiene
Coccophagus gurneyi y Tetracnemoidea brevicornis. Éstas presentan dos
aumentos poblacionales, el mayor está entre los meses de diciembre y
enero, siendo mas pronunciado para C. gurneyi; el segundo, para T.
brevicornis, ocurre a mediados de enero, y para C. gurneyi, a fines de enero.
A fines de febrero ocurre una notable disminución de ambos parasitoides.
QUIROS (1998) recupera parasitoides de P. calceolarie y P. longispinus en
palto (Persea americana). En el primer caso, menciona a C. gurneyi y T.
brevicornis, además, un individuo de Aenasius sp. encontrado el 28 enero.
Se conoce en Chile a A. punctatus Compere (Neodiscoides), insecto nativo,
muy escaso que parasita a P. longispinus. No se descarta la posibilidad de
otros Aenasius que parasiten a P. calceolarie. Su detección sobre P.
calceolarie es la primera citada para Chile. C. gurneyi presentó una alta
predominancia durante todo el período muestreado, con dos aumentos
poblacionales, en octubre y enero; la última fue más importante con un 48 %
del parasitismo. A su vez, T. brevicornis tuvo dos aumentos poblacionales,
en octubre y en la segunda quincena de noviembre y primera quincena de
diciembre. El último más importante con un 34,29 % del parasitismo y
coincidiendo con una disminución de C. gurneyi. Para P. longispinus,
QUIROS (1998) señala a C. gurneyi y T. brevicornis.

11
DUOTT (1981), destaca que la capacidad de los parasitoides para descubrir
a sus huéspedes, aún cuando exista una baja densidad de parasitoides, es la
cualidad más importante que determina su efectividad como un agente de
control.
DUOTT (1981), señala que esta habilidad de búsqueda de los parasitoides
está ligada a varios factores, de los cuales menciona: su poder de
movimiento, percepción, sobrevivencia, agresividad y persistencia. Además,
la alta capacidad reproductiva de los parasitoides, que puede determinar
cuán rápido estos son capaces de controlar a sus huéspedes.
El mismo autor señala, que existen muchos factores que determinan la
existencia y el mantenimiento de una relación particular entre el huésped y el
parasitoide. Estos factores son los que se encuentran entre los problemas de
investigación más absorbentes en la biología de los parasitoides. Para
obtener una relación huésped-parasitoide, las dos especies deben cubrir los
requisitos iniciales: ser temporal, geográfica y ecológicamente coincidentes.
Pero, aún cuando estos requisitos se cubran, la relación parasítica puede no
establecerse si existen barreras físicas, fisiológicas, psicológicas o
nutricionales.
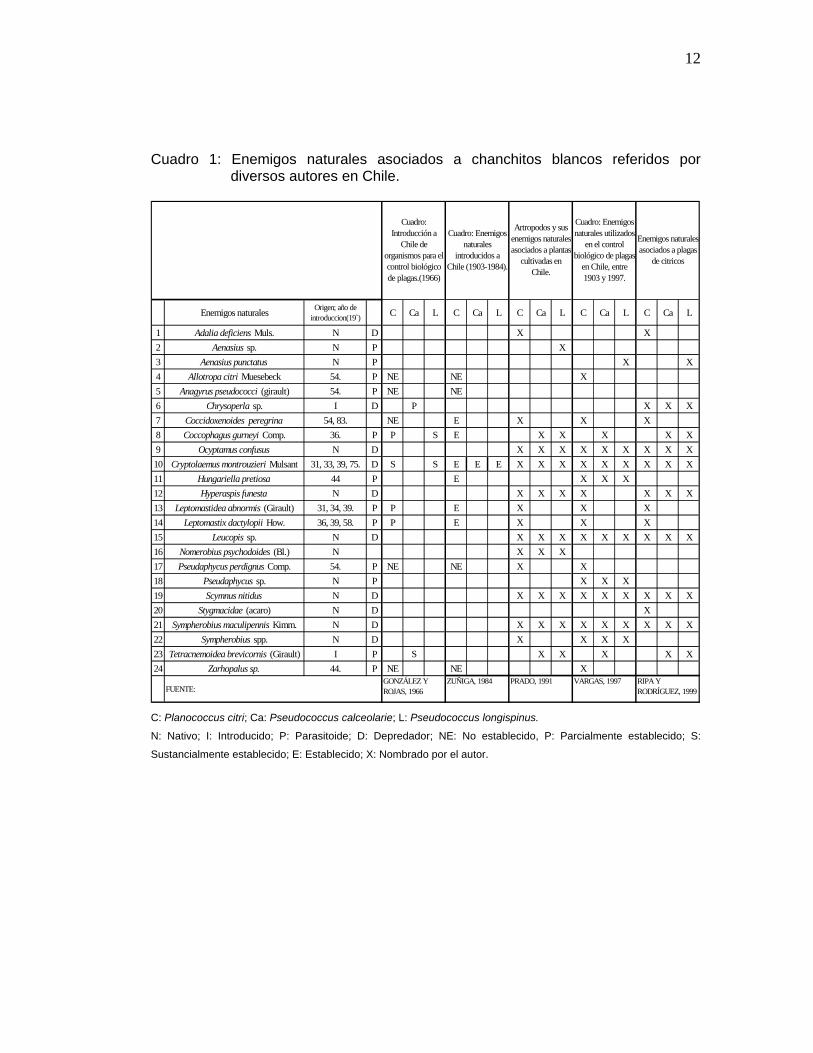
En el Cuadro 1 se muestran los esfuerzos de diferentes autores en el
reconocimiento de las diferentes especies de enemigos naturales, que se
han encontrado asociados a los chanchitos blancos en estudio.
12
Cuadro 1: Enemigos naturales asociados a chanchitos blancos referidos por diversos autores en Chile.
Enemigos naturales Origen; año de introduccion(19`) C Ca L C Ca L C Ca L C Ca L C Ca L
1 Adalia deficiens Muls. N D X X2 Aenasius sp. N P X3 Aenasius punctatus N P X X4 Allotropa citri Muesebeck 54. P NE NE X5 Anagyrus pseudococci (girault) 54. P NE NE6 Chrysoperla sp. I D P X X X7 Coccidoxenoides peregrina 54, 83. NE E X X X8 Coccophagus gurneyi Comp. 36. P P S E X X X X X9 Ocyptamus confusus N D X X X X X X X X X10 Cryptolaemus montrouzieri Mulsant 31, 33, 39, 75. D S S E E E X X X X X X X X X11 Hungariella pretiosa 44 P E X X X12 Hyperaspis funesta N D X X X X X X X13 Leptomastidea abnormis (Girault) 31, 34, 39. P P E X X X14 Leptomastix dactylopii How. 36, 39, 58. P P E X X X15 Leucopis sp. N D X X X X X X X X X16 Nomerobius psychodoides (Bl.) N X X X17 Pseudaphycus perdignus Comp. 54. P NE NE X X18 Pseudaphycus sp. N P X X X19 Scymnus nitidus N D X X X X X X X X X20 Stygmacidae (acaro) N D X21 Sympherobius maculipennis Kimm. N D X X X X X X X X X22 Sympherobius spp. N D X X X X23 Tetracnemoidea brevicornis (Girault) I P S X X X X X24 Zarhopalus sp. 44. P NE NE X
FUENTE:
Enemigos naturales asociados a plagas
de citricos
GONZÁLEZ Y ROJAS, 1966
ZUÑIGA, 1984 PRADO, 1991 VARGAS, 1997 RIPA Y RODRÍGUEZ, 1999
Cuadro: Introducción a
Chile de organismos para el control biológico de plagas.(1966)
Cuadro: Enemigos naturales
introducidos a Chile (1903-1984).
Artropodos y sus enemigos naturales asociados a plantas
cultivadas en Chile.
Cuadro: Enemigos naturales utilizados
en el control biológico de plagas
en Chile, entre 1903 y 1997.
C: Planococcus citri; Ca: Pseudococcus calceolarie; L: Pseudococcus longispinus.
N: Nativo; I: Introducido; P: Parasitoide; D: Depredador; NE: No establecido, P: Parcialmente establecido; S:
Sustancialmente establecido; E: Establecido; X: Nombrado por el autor.

13
2.3.1 Depredadores nativos
Adalia deficiens Mulsant.
Coccinélido, principalmente es un depredador de diferentes especies de
áfidos presentes en cítricos (Aphis spiraecola, Toxoptera aurantii, Aphis
gossypii, Macrosiphum euphorbiae), ocasionalmente cuando el alimento es
escaso, se alimenta de chanchitos blancos y arañitas. El adulto, es una
chinita de cabeza oscura y antenas cortas, los élitros pueden ser
completamente rojos, o bien presentar manchas negras. En las especies de
coccinélidos, es normal que exista una gran variabilidad en el diseño de
colores, ésta se debe a factores genéticos y ambientales. Los huevos con
forma de huso, de color amarillo o anaranjado y un poco más grandes que en
otras especies de coccinélidos, son depositados en pequeños grupos de 6 a
13 (INIA, 2001).
Ocyptamus confusus Goot (=Baccha valdiviana Philippi) Sírfido, depredador de Planococcus citri,
Pseudococcus calceolariae, P. longispinus y P. viburni. La larva da origen a
una pupa periforme, blanquecina con manchas circulares de color verde
pizarra y de tamaño cercano a 3 mm (INIA, 2001).

14
YUDELEVICH (1950), le atribuye un rol secundario, ya que no se encuentra
en grandes cantidades, actúa también sobre otros cóccidos y es parasitado
por varias especies de himenópteros.
Hyperaspis funesta Germain
Coccinélido, depredador de toda las especies de chanchito blanco. Adultos
pequeños de tamaño no mayor a 2 mm, con alas negras con coloraciones
rojizas más o menos redondeadas. Cuando las larvas alcanzan un tamaño
medio de 4 a 5 mm se asemejan a las larvas de Cryptolaemus, aunque de
menor tamaño y mayor abundancia de penachos de cera blanca, de
preferencia se alimentan de huevos de chanchito blanco. La poca
abundancia que se observa de larvas de Hyperaspis, se debe probablemente
a la existencia de un hiperparasitoide que causa la mortalidad de las pupas.
Cuando el alimento es escaso, los coccinélidos en general pueden
alimentarse de huevos y larvas pequeñas de lepidópteros, arañitas, trips e
incluso presentar canibalismo, alimentándose de huevos y larvas pequeñas
de su misma especie (INIA, 2001).
Leucopis sp.
Chamaemyiidae, depredador de Planococcus citri, Pseudococcus
calceolariae, P. longispinus y P. viburni. Los adultos son de un típico color
gris y tamaño aproximado de 2 mm. Las hembras colocan pequeños huevos
de un color blanco puro entre las colonias de chanchitos. Las larvas son

15
ciegas, carecen de extremidades y son de un color blanquecino y un tamaño
no superior a 1,5 mm. Se mueven activamente entre los individuos de la
colonia de chanchitos alimentándose de huevos y ninfas. Cuando alcanzan
su máximo desarrollo, se transforman en pupas de una característica
coloración parda a castaño y tamaño cercano a 2,5 mm, éstas permanecen
adheridas a la superficie y son algo difíciles de observar, pues
frecuentemente se encuentran cubiertas por restos de la colonia de
chanchitos blancos (INIA, 2001).
CAPDEVILLE (1945) describe a Leucopis sp., como uno de los enemigos
más importantes de Pseudococcus en el sur de California. El díptero
predador llega a ser muy abundante cuando el ataque de la plaga es grande
y de larga duración, pero su acción no es lo suficientemente rápida como
para impedir que un serio ataque de chanchitos blancos cause daño.
Las larvas aparecen en gran cantidad entre las masas algodonosas y no se
ha visto atacando a otras especies que no sean los chanchitos blancos
(YUDELEVICH, 1950).
Scymnus nitidus Philippi
Coccinélido depredador de numerosas especies de chanchito blanco. El
cuerpo del adulto presenta un tamaño de 1,8 a 2 mm de largo por 1mm de
ancho, es de color café claro, con antenas y extremidades cortas. Cada élitro
presenta dos manchas más claras de forma redondeada. Las larvas son de

16
color blanquecino y presentan el cuerpo completamente cubierto por
vellosidades (INIA, 2001).
En Chile, se ha observado un número muy pequeño del coccinélido
depredador Scymnus nitidus (Philippi), sus larvas poseen la capacidad de
encontrar Pseudococcus que se encuentran bien protegidos (INIA, 2001).
Sympherobius maculipennis Kimm
Hemeróbido depredador de todas las especies de chanchito blanco. Los
adultos poseen grandes alas membranosas con una venación muy marcada,
ojos sobresalientes, antenas muy largas y filamentosas. El cuerpo tiene una
coloración parda. Las hembras ponen muchos huevos. Las larvas son
alargadas con un engrosamiento en la parte central del cuerpo. Poseen un
aparato bucal adaptado para perforar y succionar el contenido de sus presas
(INIA, 2001). Este depredador es hiperparasitado por varias especies de
microhimenópteros, como Solenofigitis lautus De Santi. El tejido del capullo
de Sympherobius permite observar el hiperparasitoide (YUDELEVICH, 1950).
Entre los años 1980 y 1985, fue criado y liberado masivamente en huertos de
la Quinta Región, a través de un programa de Control Biológico comercial

17
desarrollado por la Facultad de Agronomía de la Pontificia Universidad
Católica de Valparaíso.
2.3.2. Parasitoides nativos
Aenasius punctatus (Compere)
Encírtido. Tiene una reducida acción controladora sobre Pseudococcus
longispinus. Tanto las hembras como los machos son de un color negruzco,
el cuerpo del adulto es de tamaño algo mayor a 1 mm. Las hembras
parasitan ninfas de tercer estadio. Las ninfas parasitadas son de un color
amarillento y fácilmente distinguibles por su tamaño, dan lugar a una momia
de mayor tamaño que la originada por Tetracnemoidea y Coccophagus (INIA,
2001).
Pseudaphycus sp. Nr. Angelicus
Encírtido parasitoide de Pseudococcus longispinus, endémico de Chile. Su
presencia es escasa al igual que su acción de control sobre chanchito blanco
(INIA, 2001).
Éste posee una notable capacidad de ubicar a los Pseudococcus y
parasitarlos. Introduce en sus cuerpos un número variable de huevos, según

18
sea el tamaño del chanchito blanco. Una vez que completa el desarrollo en el
interior, emergen de uno hasta 60 individuos por chanchito blanco
parasitado. Parasita desde los individuos más pequeños hasta las hembras
que han comenzado su oviposición (INIA, 2001).
2.3.3 Depredadores introducidos
Chrysoperla sp.
Crisópido, depredador generalista que muestra mayor actividad en áfidos
(Aphis spiraecola, Toxoptera aurantii y Aphis gossyppii) y chanchito blanco
(Planococcus citri, Pseudococcus longispinus. Pseudococcus calceolariae y
Pseudococcus viburni). Los individuos de esta especie presentan un cuerpo
muy delicado de color verde pálido, los adultos poseen grandes alas
membranosas con una venación muy marcada, ojos sobresalientes y
antenas muy largas y filamentosas. El cuerpo del adulto puede alcanzar un
tamaño de 20 mm. La hembra, usualmente, pone los huevos aislados y
ocasionalmente en grupos, hileras o al azar sobre las hojas. Cada huevo
permanece alejado de la superficie foliar, sostenido por un delgado pedicelo.
Los huevos son ovales, de tamaño menor a 1 mm, inicialmente de un color
verde pálido que se torna blanco grisáceo con el tiempo. La larva es alargada
con un engrosamiento en la parte central del cuerpo, puede desarrollarse de
1 mm hasta 6 a 8 mm, de color gris o pardo, presenta extremidades bien
desarrolladas. Al nacer, es inmediatamente capaz de alimentarse de su
presa; posee un aparato bucal adaptado con mandíbulas muy desarrolladas
de forma curvada para perforar, inyectar un veneno paralizante y succionar

19
los fluidos corporales de sus presas. Todos los estados larvales de esta
especie son activos depredadores, llegando a consumirse entre ellos si no
hay recursos disponibles. El último estadío larval teje un capullo esférico y
compacto de seda blanquecina en lugares protegidos de la planta y pupa. La
emergencia del adulto la efectúa a través de un orificio circular. Los adultos
son activos voladores, sobre todo en la tarde y en la noche. Existe una
dificultad por parte de este depredador de encontrar sus presas en cultivos
con presencia de pilosidades y sustancias pegajosas (INIA, 2001).
Cryptolaemus montrouzieri Mulsant
Coccinélido importado desde Estados Unidos entre el año 1931 y 1946. En
1995, se importó desde California una muestra colectada desde una zona
templada en Australia, y en 1996 una muestra que se alimentaba casi
exclusivamente de Saissetia oleae. Es una de las primeras especies de
coccinélidos introducidos al país que mostró una notable efectividad en el
control de chanchitos blancos, ocasionalmente se encuentra depredando
ninfas de la Saissetia oleae, sin embargo, su reproducción es mejor sobre
chanchitos blancos. Al provenir de regiones más cálidas, tiene dificultades de
aclimatación durante el invierno, desde la Región Metropolitana al sur, por lo
que debe ser permanentemente liberado. Las internaciones más recientes de
individuos provenientes de zonas más templadas, han mostrado una mejor
adaptación a las condiciones invernales que se presentan en el área citrícola
de la Zona Central. El adulto de Cryptolaemus mide entre 2,5 y 3 mm, tiene
los élitros de un color azul oscuro a negro con una franja anaranjada en el
borde posterior, cabeza y protórax rojizo. La larva, se encuentra cubierta de
penachos de cera blanca, de forma similar al chanchito blanco, aunque de

20
mayor tamaño y desplazamiento más rápido. Tanto las larvas como los
adultos son depredadores de huevos, ninfas migratorias, ninfas y adultos de
diferentes especies de Pseudococcus, en especial de las especies ovíparas.
Las hembras adultas oviponen en las masas algodonosas de huevos de
chanchito blanco, en especial, en hojarascas secas y ramas del tronco en
árboles frutales. Los huevos son de color blanquecino, de un tamaño entre
0,3 y 0,4 mm. Se considera en el país, el depredador más importante de
chanchito blanco. Muestra una elevada capacidad de depredación (INIA,
2001).
Se ha observado que tanto larvas, como adultos permanecen mayor tiempo
buscando chanchitos blancos en hojas con mielecilla. Una vez que la plaga
desaparece, deben existir hospederos alternativos, de los cuales no se
tienen antecedentes, y que permiten que este depredador no muera (INIA
2001).
Cryptolaemus montrouzieri Mulsant, es considerado el predador más
importante de los chanchitos blancos. Su acción es similar sobre todas las
especies del género, salvo el caso de P. longispinus, cuyo control no produce
el resultado esperado (CAPDEVILLE, 1945; YUDELEVICH, 1950).
C. montrouzieri se considera muy efectivo en el control de ataques fuertes de
chanchitos blancos, pero su acción en focos pequeños no es suficiente para
evitar la reinfestación. Hace descender un ataque hasta cierto límite en el
cual la plaga continúa siendo un problema serio, por lo tanto, este

21
controlador no es capaz de erradicar localmente la plaga (YUDELEVICH,
1950).
Desde 1980 ha sido criado y liberado masivamente cada año en huertos de
chirimoyos, cítricos y paltos, especialmente entre Ovalle y Peumo a través de
un programa llevado a cabo por la Facultad de Agronomía de la Pontificia
Universidad Católica de Valparaíso (1980 – 1995) y luego por Xilema S.A.
(1995 a la fecha).
2.3.4 Parasitoides introducidos
Coccophagus gurneyi Compere
Afelínido importado desde Estados Unidos en 1936. Destacado parasitoide
de Pseudococcus calceolariae y, en menor grado, de P. longispinus. La
hembra es de color negro con una banda amarillenta transversal al comienzo
del abdomen, el cuerpo en el estadio adulto mide alrededor de 1mm de largo.
El macho es completamente negro y algo más pequeño que la hembra. Las
hembras oviponen en ninfas de segundo estadio. El chanchito blanco
parasitado se transforma en una momia de forma aguzada con un leve color
oscuro o verde petróleo. Se encuentra muy bien establecido a lo largo de
todo el país (INIA, 2001).
En el año 1927, fue observada la oviposición de C. gurneyi sobre

22
Pseudococcus gahani. Posteriormente, al comprobarse la efectividad del
parasitoide sobre la plaga, C. gurneyi fue enviado a California, efectuándose
las primeras liberaciones en junio del año 1928. Después de este año, no fue
necesaria ninguna liberación posterior, ya que se consideraba a la especie
definitivamente establecida (YUDELEVICH, 1950).
La misma situación ocurrió en Chile, después de ser traído al país en 1936,
por el Departamento de Sanidad Vegetal, no siendo necesario liberaciones
posteriores del parasitoide. Esto da un índice de su completa y total
aclimatación (YUDELEVICH, 1950).
Sin embargo, y a pesar de estar establecido, entre los años 1980 y 1995 fue
criado en la Facultad de Agronomía de la Pontificia Universidad Católica de
Valparaíso y liberado masivamente en huertos de la Quinta Región
Actualmente, es considerado parasitoide primario de P. gahani, pese a que el
primer ejemplar fue obtenido de P. longispinus. También, se le ha observado
oviponiendo libremente sobre P. citri y P. maritimus, pero en ambos casos,
C. gurneyi no ha terminado normalmente su desarrollo, lo que hace suponer
que los huevos del parasitoide son destruidos por un proceso de fagocitosis
(YUDELEVICH, 1950).
La hembra es fácilmente identificable por su coloración característica, su
cuerpo es negro con una franja amarilla en la base del abdomen. El macho
es completamente negro.

23
YUDELEVICH, (1950) señala que C. gurneyi ovipone sobre chanchitos
blancos de los dos sexos y en todos los estadios evolutivos, evitando al
parecer, los primeros estadios larvales. Normalmente, un solo huevo del
parasitoide debería ser depositado en cada huésped, pero generalmente las
hembras colocan dos o más, al parecer, C. gurneyi no distingue entre un
huésped parasitado y otro que no lo está, por lo tanto, un mismo huésped
tiene la posibilidad de ser atacado por más de una hembra, aún se produce
comúnmente el caso de una misma hembra parasitando dos o más veces un
mismo chanchito blanco, por consiguiente, el superparasitismo es muy
frecuente.
La duración del ciclo biológico, en condiciones de campo, no es uniforme ya
que varía por una serie de circunstancias, entre las cuales se encuentra la
época, temperatura, edad del huésped, abundancia de alimento y presencia
de otros parasitoides (YUDELEVICH, 1950).
Según CAPDEVILLE (1945), el ciclo de vida de C. gurneyi, desde huevo a
adulto, demora aproximadamente 27 días. YUDELEVICH, (1950) observa
que bajo condiciones de laboratorio, la emergencia de los parasitoides
adultos de C. gurneyi es entre los 25 y 31 días siguientes a la oviposición.
La duración del ciclo biológico va en función de la temperatura a la cual se
desarrolle el insecto. En experimentos realizados en laboratorio a una
temperatura constante de 23 ºC, se obtiene una duración del ciclo biológico
de 15 a 18 días.

24
Leptomastidea abnormis (Girault)
Encírtido importado desde Estados Unidos entre los años 1931 y 1939, se ha
establecido muy bien dentro del país. Endoparasitoide solitario de
Planococcus citri. Los individuos de esta especie son pequeñas avispas de
color pardo, el cuerpo del adulto mide alrededor de 0,75 mm, con antenas
largas. Las delicadas alas presentan bandas transversales de color
negruzco, los individuos en reposo colocan las alas en forma perpendicular al
cuerpo. Las hembras adultas oviponen en los estadios primarios de los
chanchitos blancos. Los huevos fertilizados, dan origen a hembras y los no
fertilizados, a machos. Una vez que nace la larva, comienza a alimentarse de
la hemolinfa del chanchito blanco y en sus últimos estadios termina por
consumir totalmente el contenido del cuerpo, ocasionando su muerte. El
cuerpo del chanchito blanco parasitado sufre ciertos cambios y pasa a
denominarse momia. La momia de Leptomastidea es de color anaranjado, de
tamaño similar a la de Coccidoxenoides peregrina, y se mantiene entre los
individuos de la colonia, o bien, separada de éstos (INIA, 2001).
Su importación a Chile ha sido considerada como altamente beneficiosa y su
propagación y reproducción no constituyen un problema. YUDELEVICH
(1950), lo consideró casi desaparecido.
En la decada de los 80, fue criado masivamente y liberado en huertos de
chirimoyos y cítricos en la Quinta Región en el único programa de Control
Biológico comercial desarrollado por la Facultad de Agronomía de la
Pontificia Universidad Católica de Valparaíso.

25
Leptomastix dactylopii Howard
Encírtido nativo de Brasil, importado desde Estados Unidos, entre los años
1936 y 1958. Endoparasitoide solitario específico de Planococcus citri. No se
reproduce bien en otras especies. Pequeña avispa de color pardo
amarillento, el cuerpo del adulto mide aproximadamente 3 mm de largo. Las
hembras son más grandes que los machos y tienen antenas largas, rectas y
sin pilosidades, mientras que en los machos son peludas y ligeramente
torcidas. Las hembras adultas son atraídas por el olor de plantas hospederas
infestadas y de chanchos blancos no parasitados. Pueden discriminar entre
chanchitos blancos no parasitados y parasitados, evitando oviponer en los
últimos. El hospedero parasitazo, es rechazado por simple contacto de
antenas, por comportamientos defensivos del hospedero o por la detección
de un conducto que se prolonga desde el huevo colocado por el parasitoide
anterior que emerge por la superficie del hospedero. Si no lo rechaza de
inmediato, lo hace después de insertar el ovipositor. Las hembras oviponen
de preferencia en ninfas de tercer estadio y hembras preadultas, pero no en
adultos poniendo huevos. La larva en desarrollo, transforma al chanchito
blanco parasitado en una momia de color ámbar a amarillo verdoso, de un
tamaño que varía entre 1,5 y 2 mm. El adulto abandona la momia abriendo
un orificio en uno de sus extremos. Una vez que el adulto ha emergido se
alimenta de la mielecilla producida por los chanchitos blancos, la que actúa,
como fuente de carbohidratos suplementaria. Sin embargo, no se alimenta
de la hemolinfa de su hospedero como lo hacen otros parasitoides. Los
adultos de este parasitoide presentan una excelente capacidad de búsqueda,
pudiendo detectar los individuos aislados (INIA, 2001).

26
Pese a las repetidas liberaciones efectuadas del parasitoide en la V y VI
Regiones del país, no ha logrado establecerse debido a bajas temperaturas
invernales que no permiten su sobrevivencia, pudiendo encontrarse sólo en
la Zona Norte del país e Isla de Pascua. Esta especie, se produce
comercialmente en otros países para controlar Planococcus citri (INIA, 2001).
Coccidoxenoides peregrina Timberlake
(=Pauridea peregrina Timberiake). Encírtido nativo del sur de China,
importada desde Estados Unidos en 1954, y desde Israel, en 1986.
Parasitoide solitario de Planococcus citri. El adulto, es una pequeña avispita
negruzca, de un tamaño que varía entre 0,5 y 0,7 mm, con antenas cortas.
Normalmente, es partenogenético y raramente se ven los machos. Las
hembras adultas, rápidamente después de su emergencia, oviponen en los
primeros estadios ninfales del chanchito blanco. La momia, es de forma
ovoidal, de color blanquecino con una mancha oscura en uno de sus
extremos y de tamaño inferior a 1 mm, ésta se mantiene entre los individuos
de la colonia de chanchito blanco. Esta especie, se produce comercialmente
en otros países para el control de P. citri. Gracias a su reducido tamaño, es
capaz de ubicar y alcanzar aún aquellos chanchitos blancos que se
encuentran en lugares muy protegidos (INIA, 2001).

27
Tetracnemoidea brevicornis Girault
(=Tetracnemus pretiosus Timberlake). Encírtido importado desde Estados
Unidos, en 1944. Parasitoide frecuente de Pseudococcus longispinus a lo
largo de todo el país y de P. calceolariae. Las hembras, son pequeñas
avispas de color negro azulado, cuyo tamaño bordea 1 mm. Los machos, son
de color negro, ligeramente más grandes que las hembras y con antenas
ramificadas en cuatro prolongaciones. Las hembras parasitan ninfas
pequeñas, dando origen a momias pequeñas, de un color blanquecino con
una mancha negruzca en uno de sus extremos (INIA, 2001).
YUDELEVICH (1950), describe a T. brevicornis como uno de los valiosísimos
parasitoides de Pseudococcus calceolariae, cuya principal y notable
característica consiste en sus antenas ramificadas y plumosas en los
machos. El mismo autor señala que el origen de T. brevicornis, en Chile, es
desconocido. Su existencia puede explicarse por la introducción hecha en
1944 y que se consideró prácticamente perdida, o bien, llegó casualmente en
una importación de otros parasitoides de Pseudococcus. Fue observado por
primera vez por Compere en 1927, parasitando a Pseudococcus gahani. En
California, se demostró ser tan eficaz como Coccophagus gurneyi, con la
ventaja de que las dos especies se complementaban en el control de la plaga
en los diversos niveles o intensidades, en que ésta se presenta. Ambas
especies, son valiosísimos en la destrucción de focos iniciales e individuos
solitarios de P. longispinus, condiciones en que C. montrouzieri es
prácticamente inútil (YUDELEVICH, 1950).

28
T. brevicornis es predominante en Australia, donde los focos de chanchitos
blancos son sumamente pequeños, y C. gurneyi predomina en California,
donde los ataques son más fuertes y abundantes. Por lo tanto, algunos
autores han llegado a la conclusión que la mayor o menor efectividad de
estos parasitoides, va en relación a las diferentes intensidades de los brotes
de la plaga. C. gurneyi, es eficaz en los focos de cierta intensidad, en los que
desplaza a T. brevicornis hasta que la plaga desciende hasta cierto nivel,
desde el cual comienza a su vez a ser desplazado por T. brevicornis. Este
último, a su vez, supera progresivamente a C. gurneyi a medida que los
brotes son más pequeños y en los casos de individuos solitarios.
Por consiguiente, la acción combinada y técnicamente dirigida de los dos
parasitoides traería como consecuencia el control económico ideal:
Coccophagus bajaría el nivel de los chanchitos blancos, dejándolos a la
altura de la óptima actividad de Tetracnemus. En el caso de que la plaga
rebotara quedaría nuevamente a nivel de ataque ideal de Coccophagus,
dejando las liberaciones del depredador Cryptolaemus montrouzieri para
cuando la plaga adquiera mayores densidades (YUDELEVICH, 1950).
Zarhopalus sp.
A pesar de haber sido importado y liberado, prácticamente no debe darse
como existente en el país (YUDELEVICH, 1950).

29
33.. MMAATTEERRIIAALLEESS YY MMÉÉTTOODDOOSS
3.1 Lugar del estudio:
El estudio fue realizado en huertos de paltos, cítricos y chirimoyos infestados
con chanchitos blancos, ubicados en la comuna de Quillota, V Región. La
duración de esta prospección fué de marzo 2003 a marzo 2004, período en
el cual, se realizó, un muestreo mensual por cada especie de chanchito
blanco en estudio.
3.2 Especie plaga estudiada:
Se estudiaron las siguientes especies de Chanchitos blancos: Planococcus
citri (Risso), Pseudococcus calceolariae (Maskell), Pseudococcus longispinus
(Targioni & Tozzetti).
3.3 Material colectado:
Se colectó material vegetal con colonias de chanchito blanco, de una sola
especie y en todos sus estadios. Este provino de huertos con presencia de
chanchito blanco, ya sea en cítricos, paltos o chirimoyos.

30
Las muestras, consistentes en ramas o frutos infectados con chanchito
blanco, fueron llevadas al laboratorio de Control Biológico de la Facultad de
Agronomía de la Pontificia Universidad Católica de Valparaíso.
Se procedió a limpiar cada muestra, separando en primer lugar, a los
enemigos naturales encontrados de carácter predador. Estos se clasificaron
en forma inmediata evaluando el porcentaje de incidencia. Aquellos estados
juveniles difíciles de identificar se criaron a temperatura controlada (15ºC -
25ºC), humedad relativa cercana al 60% y con alimentación hasta obtener
el estado adulto.
Las muestras se colocaron en una batería de crianza completamente sellada
y con ventilación apropiada. Las baterías fueron ubicadas en salas de
crianzas a temperatura controlada (15ºC-25ºC) y humedad relativa cercana
al 60%. Cada batería fue rotulada según: fecha de recolección, especie frutal
afectada, especie de chanchito blanco.
Se supervisaron dos veces por semana las baterías para la recuperación de
enemigos naturales parasitoides que fueron naciendo. La extracción de estos
se realizó mediante un aspirador entomológico. Las muestras fueron
desechadas a los 40 días, para evitar contabilizar segundas generaciones.
Los parasitoides recuperados por muestra fueron conservados en alcohol
(50%) para su posterior clasificación.

31
3.4 Identificación de las especies asociadas a chanchito blanco:
La clasificación de las especies encontradas fue llevada a cabo por
académicos de la Pontificia Universidad Católica de Valparaíso y consultoría
a personal del INIA La Cruz.
Los datos fueron tabulados y confrontados con los antecedentes preliminares
sobre diversidad de especies benéficas asociadas a chanchitos blancos en
Chile.
3.5 Evaluación de la importancia de cada enemigo natural o nivel de
participación:
Se evaluaron en forma separada cada especie de chanchito blanco con sus
controladores asociados.
Para cada fecha de muestreo, la importancia relativa de cada insecto
benéfico encontrado, se evaluó en base al total de insectos encontrados en
la muestra, diferenciando parasitoides de depredadores.

32
3.6 Determinación del comportamiento estacional de cada enemigo natural.
El comportamiento estacional de cada especie fue referido a su importancia
relativa.
Para cada enemigo natural encontrado, se esquematizó su nivel de
participación mensual, durante el año de estudio.
Para determinar el efecto de los enemigos naturales introducidos, se hizo un
análisis según los datos obtenidos en cuanto a cantidad de individuos,
participación y antecedentes bibliográficos.

33
4. PRESENTACIÓN Y DISCUSIÓN DE RESULTADOS
4.1 Identificación de las especies de enemigos naturales encontradas sobre
chanchitos blancos:
Sobre las tres especies de chanchitos blancos estudiadas, se encontró un
total de diez especies de enemigos naturales, de las cuales, correspondieron
a cuatro especies nativas: Ocyptamus confusus, Hyperaspis funesta,
Sympherobius sp., Leucopis sp.; y Seis especies introducidas: Cryptolaemus
montrouzieri, Chrysoperla sp, Leptomastidea abnormis, Coccidoxenoides
peregrina, Coccophagus gurneyi, Tetracnemoidea brevicornis (CUADRO 2),
resultados que difieren en parte, a lo encontrado por YUDELEVICH, en 1950.
CUADRO 2. Enemigos naturales asociados a Pseudococcus en la Comuna de
Quillota.
YUDELEVICH (1950) recuperó sobre chanchitos blancos, un total de cinco
especies, dos nativas: Sympherobius sp., Leucopis sp. y tres introducidas:
Cryptolaemus montrouzieri, Leptomastidea abnormis y Coccophagus gurneyi.
Todas las especies mencionadas fueron encontradas en el presente estudio.
Plaga Planococcus citri Pseudococcus calceolarie Pseudococcus longispinusOcyptamus confusus Cryptolaemus montrouzieri Cryptolaemus montrouzieri
Cryptolaemus montrouzieri Hyperaspis funesta Hyperaspis funestaDepredadores Chrysoperla sp. Leucopis sp.
Hyperaspis funestaSympherobius sp.
Leptomastidea abnormis Coccophagus gurneyi Coccophagus gurneyiCoccidoxenoides peregrina Tetracnemoidea brevicornis Tetracnemoidea brevicornis
Parasitoides

34
De las especies de enemigos naturales que YUDELEVICH (1950) no
encontró en chanchitos blancos y que si fueron identificadas en el presente
estudio, se señala a Tetracnemoidea brevicornis, que siendo considerada
introducida en aquella época, según el autor, existió un cierto desorden e
improvisación en las introducciones efectuadas a la fecha y se carecía de
experiencia en la cría artificial de insectos. Coccidoxenoides peregrina fue
introducida en 1954, no teniendo YUDELEVICH (1950), antecedentes de
este insecto; Baccha sp., hoy llamada Ocyptamus confusus, Sympherobius
sp. y Leucopis sp, especies descritas por él, como poco abundantes pero
que se observan a menudo. En cambio, de Hyperaspis funesta, dicho autor
no tenia antecedentes.
HARRISON (1993), sobre Pseudococcus longispinus, recupera a
Tetracnemoidea brevicornis, Coccophagus gurneyi, Scymnus sp. y
Cryptolaemus montrouzieri. De estas especies, tres fueron recuperadas en el
presente estudio sobre la misma especie de chanchito blanco:
Tetracnemoidea brevicornis, Coccophagus gurneyi y Cryptolaemus
montrouzieri. El mimo autor no menciona a Hyperaspis funesta, encontrado
en la presente prospección sobre las tres especies de chanchito blanco
estudiadas, pero si agrega a Scymnus sp. coccinélido depredador nativo de
muchas especies de chanchitos blancos, que aparece en forma ocasional
(INIA, 2001). Scymnus sp. no fue encontrado en el presente estudio sobre
ninguna especie de chanchito blanco.
Por su parte QUIROS (1998), quien estudió a los parasitoides de chanchitos
blancos en palto, menciona a Coccophagus gurneyi y Tetracnemoidea
brevicornis como los parasitoides que comparten P. longispinus y P.

35
calceolarie. Además, agrega la primera detección de un individuo de
Aenasius sp. sobre P. calceolarie, parasitoide nativo muy escaso y que sólo
aparece citado por PRADO (1991) como controlador de P. longispinus.
De las especies encontradas por QUIROS (1991), en el presente estudio
fueron recuperadas Coccophagus gurneyi y Tetracnemoidea brevicornis, no
siendo encontrada la especie Aenasius sp.
De HARRISON (1993) Y QUIROS (1998), cuyos estudios fueron realizados
en la misma zona, se infiere que la recuperación de enemigos naturales de
chanchitos blancos, no refleja una situación estable y general, ya que ciertos
enemigos naturales especialmente nativos, aparecen en forma ocasional y
su apreciación puede estar determinada por eventos circunstanciales. Esto
se extiende al análisis posterior, en el cual, se evaluará la importancia de los
enemigos naturales encontrados.
Toda las especies mencionadas hasta el momento son citadas por PRADO
(1991), quien analiza a todos los enemigos naturales de chanchito blanco en
el país, el autor además, incluye a Nomerobius pychodoides, Hemerobius
hageni y Leptomastix dactilopi, este último característico de la zona norte.

36
4.2 Importancia relativa de cada enemigo natural asociado a las especies
Pseudococcus longispinus, Pseudococcus calceolariae y Planococcus
citri en la comuna de Quillota:
Se recolectó un total de 884 enemigos naturales, tanto en estados larvales,
como pupas y adultos. De este total, correspondieron 493 enemigos
naturales para P. citri, 213 para P. calceolarie y 232 para P. longispinus,
obedeciendo, al tamaño de las muestras recolectadas, para cada especie y a
la disponibilidad de estas, sobre todo en los meses de invierno (ANEXO 1 y
2).
Por lo anterior, la importancia asignada a cada especie de enemigo natural,
se expone en forma distinta según la especie plaga estudiada. La
ponderación asignada para cada especie de enemigos naturales se muestra
en el Cuadro 3.
CUADRO 3. Importancia asignada para cada enemigo natural y su
comportamiento(*), para cada especie plaga. Planococcus citri % Pseudococcus calceolarie % Pseudococcus longispinus %
Ocyptamus confusus 62,8 Cryptolaemus montrouzieri 85 Cryptolaemus montrouzieri 88,9Cryptolaemus montrouzieri 35,2 Hyperaspis funesta 5 Hyperaspis funesta 11,1Chrysoperla sp. 0,7 Leucopis sp. 10Hyperaspis funesta 0,7Sympherobius sp. 0,7
Total depredadores: 33,6 9,5 3,9
Leptomastidea abnormis 77,6 Coccophagus gurneyi 70,7 Coccophagus gurneyi 66,4Coccidoxenoides peregrina 22,4 Tetracnemoidea brevicornis 29,3 Tetracnemoidea brevicornis 33,6
Total parasitoides: 66,4 90,5 96,1 Nota: Las especies consideradas Nativas, se muestran en color rojo.

37
4.2.1 Depredadores nativos
Ocyptamus confusus, encontrado sólo en P. citri, representó al único
depredador nativo que mostró una importancia considerable. O. confusus,
que YUDELEVICH (1950) no lo encontró en sus prospecciones, pero que lo
cita como de escaso interés económico, ponderó un 62.8% de los
depredadores encontrados para esta plaga. Según PRADO (1991), este
díptero se presenta sobre las tres plagas estudiadas. O. confusus fue
encontrado por SAA (2004) en P. calceolarie y en P. citri, representando un
29.4% y un 32.3% de los depredadores encontrados respectivamente.
A pesar de la importancia relativa obtenida para O. confusus, su
característica predatoria restringida sólo a su estado larvario y la presencia
de varias especies hiperparásitas que reducen su acción, hacen de O.
confusus, un controlador limitado (YUDELEVICH, 1950).
En el presente estudio, O. confusus fue recuperado una vez alcanzado el
estado adulto en las baterías de crianza, sin embargo, a pesar de lo descrito,
no se recuperó ningún hiperparasito.
Sympherobius sp., H. funesta y Leucopis sp., se presentan en pequeña
proporción, no superando el 11.1% de los depredadores encontrados en
cada especie plaga. PRADO, 1991 y RIPA Y RODRÍGUEZ, 1999 citan a
estas especies como controladores generalistas de todos los chanchitos
blancos, además de otras plagas, por lo que, a pesar de su poca

38
participación, no es inferible una extinción y su poca presencia puede ser
atribuible a la preferencia por otros huéspedes.
4.2.2 Depredadores introducidos
Chrysoperla sp, es la especie foránea menos relevante, encontradose un
solo ejemplar sobre P. citri, representó el 0.7% de los depredadores
encontrados. Especie generalista de los chanchitos blancos, fue introducida
para el control de pulgones (Aphididae) (RIPA Y RODRÍGUEZ, 1999).
Chrysoperla sp, controla además, ácaros y mosquitas blancas por lo que su
participación en el complejo de enemigos naturales de chanchitos blancos es
baja.
La ponderación obtenida para Cryptolaemus montrouzieri varia según la
especie plaga, de esta forma: 35.2% para P. citri, 85% para P. calceolarie y
88.9% para P. longispinus. El menor porcentaje atribuido a P. citri, obedeció
a la aparición de O. confusus.
A pesar de la ponderación asignada de C. montrouzieri en P. longispinus,
una menor cantidad de individuos fue encontrada con respecto a las demás
plagas en estudio. Esto es corroborado por SAA (2004), y atribuible según
INIA (2001), a la preferencia de C. montrouzieri por masas algodonosas de
chanchitos blancos.

39
4.2.2 Parasitoides introducidos:
Los parasitoides tuvieron un nivel de participación alto, encontrándose en
forma abundante en las tres plagas en estudio. Todas las especies
parasíticas encontradas son foráneas según GONZALEZ y ROJAS (1966). P.
calceolarie y P. longispinus comparten a Coccophagus gurneyi y
Tetracnemoidea brevicornis. En P. citri se encontró a Leptomastidea
abnormis y Coccidoxenoides peregrina.
Coccophagus gurneyi, es el controlador mas abundante para P. calceolarie y
P. longispinus. Al ponderar un poco mas de un 70% de los parasitoides
encontrados en ambos casos, se sitúa a más de treinta puntos de
Tetracnemoidea brevicornis. En P. citri, el parasitoide mas abundante
correspondió a Leptomastidea abnormis, el que se encontró en un 77.6% de
los casos frente a Coccidoxenoides peregrina que apareció en un 22.4%.
De las referencias mas antiguas que se tienen al respecto, YUDELEVICH
(1950), sólo encontró como parasitoides a Coccophagus gurneyi y a
Leptomastidea abnormis, especies que hasta ahora se mantienen como las
más importantes, y que superan ampliamente a toda las restantes especies
de enemigos naturales. Por otra parte, CAPDEVILLE (1945), señala a L.
abnormis como un parasitoide definitivamente establecido en Chile.

40
YUDELEVICH (1950), señala además, la característica hiperparásita de C.
gurneyi sobre T. brevicornis y de parasitoide primario, que justifican su mayor
prevalencia en las muestras.
La mayor importancia de C. gurneyi sobre T. brevicornis es señalada,
además, por HARRISON (1993), QUIROS (1998) y SAA, (2004) sobre P.
calceolarie y P. longispinus. En el caso de P. citri, SAA (2004) sólo
encuentra a L. abnormis.
Del total de enemigos naturales encontrados, y para las tres plagas en
estudio, los parasitoides en su conjunto conforman el grupo más abundante,
ponderando un 66.4% sobre P. citri, 90.5% sobre P. calceolarie y 96.1%
sobre P. longispinus. Dado que YUDELEVICH (1950), ponderó a los
parasitoides en un 19% del total de enemigos naturales encontrados sobre
las tres plagas en estudio, se puede afirmar un incremento significativo de las
especies parasíticas a la fecha, todas ellas introducidas oficialmente. Por
consiguiente, y considerando que las especies depredadoras son
mayoritariamente nativas, se da en manifiesto una drástica reducción en la
participación de las especies nativas que controlan a los chanchitos blancos.
DUOTT (1981) señala las ventajas de los parasitoides, como su capacidad
de búsqueda, alta tasa reproductiva, su agresividad y persistencia, que
determinan el éxito en su establecimiento.

41
4.3 Estacionalidad de los enemigos naturales encontrados.
No se encontraron muestras de las tres especies de chanchito blanco en
estudio en los meses invernales. Según RIPA y RODRÍGUEZ (1999), los
chanchitos blancos reducen su población y suelen invernar bajo las cortezas
apegadas al tronco y hojarascas. Esto correspondió a los meses de junio a
agosto en P. citri, julio a septiembre en P. calceolarie y en P. longispinus, a
los meses de junio y julio.
Debido a pudriciones en las baterías de crianzas, no aparecen datos de las
muestras de P. citri en los meses de mayo y septiembre, de P. calceolarie en
el mes de abril. y de P. longispinus en los meses de abril, mayo, septiembre
y noviembre.
4.3.1 Depredadores
La importancia relativa atribuida a cada especie depredadora, en cada mes,
y para cada plaga en estudio, se muestra en la Figura 1.

42
Planococcus citri
0
20
40
60
80
100
M A M J J A S O N D E F
H. funestaO. confususCrisoperla sp.Sympherobius sp.C. montrouzieri
Pseudococcus calceolarie
0
20
40
60
80
100
M A M J J A S O N D E F
Leucopis sp.H. funestaC. montrouzieri
Pseudococcus longispinus
0
20
40
60
80
100
M A M J J A S O N D E F
H. funesta
C. montrouzieri
FIGURA 1. Importancia relativa de los depredadores encontrados, asignada para
cada mes y según la plaga en estudio.
%
%
%

43
Hyperaspis funesta apareció en el mes de marzo sobre P. citri y P.
longispinus y en el mes de enero sobre P. calceolarie. Leucopis sp. en el
mes de enero sobre P. calceolarie y Sympherobius sp. sobre P. citri, en el
mes de marzo. Estas especies representan a las especies nativas menos
relevantes, la importancia relativa asignada a estas especies varió desde un
3.7% en el caso de H. funesta y Sympherobius sp. sobre P. citri, hasta un
40% en el caso de Leucopis sp. sobre P. longispinus. Sin embargo, la
aparición de las especies mencionadas no sobrepasó a los dos individuos en
ninguno de los meses en que fueron encontrados. Por consiguiente, y
tomando en cuenta que durante el periodo en estudio, se encontraron en
forma dispersa, es deducible su aparición ocasional.
Según SAA (2004), siendo el nivel de participación de Leucopis sp. bajo con
respecto a los demás enemigos naturales, su aparición es mas frecuente. El
autor recupera a esta especie en los meses de octubre, noviembre y
diciembre en P. longispinus. El mismo autor, no menciona a H. funesta y
Sympherobius sp.
Ocyptamus confusus se encontró sólo en P. citri, entre los meses de
diciembre y abril, periodo más corto que el del resto de los enemigos
naturales de importancia en la especie plaga señalada. Su mayor
participación, se observó en el mes de enero, representando el 84.9% de la
totalidad de los depredadores encontrados para ese mes. SAA (2004),
encuentra a Ocyptamus confusus, en los meses de marzo, abril, septiembre
y octubre sobre P. citri. y en los meses de abril y junio sobre P. calceolarie.

44
De Chrysoperla sp., sólo se encontró un ejemplar en el mes de diciembre.
SAA (2004) no encuentra ningún ejemplar de Chrysoperla sp. sobre P. citri,
P. calceolarie o P. longispinus, lo que corrobora su carácter ocasional.
Cryptolaemus montrouzieri apareció en los meses de marzo y abril y
posteriormente entre los meses de octubre y febrero sobre P. citri; en los
meses de marzo, mayo, enero y febrero sobre P. calceolarie y en los meses
de marzo, enero y febrero sobre P. longispinus. Por consiguiente, su
distribución en el tiempo fue mas amplia en P. citri, en donde se encontró en
toda las muestras recolectadas mientras que sobre P. longispinus, su
participación se redujo a tres meses.
En una muestra de P. calceolarie, en el mes de diciembre, fueron
recolectados desde las baterías de crianza cerca de 100 larvas de C.
montrouzieri, cantidad que supera con creses a lo recolectado en todo el año
de estudio y que representó el 100% de los enemigos naturales encontrados
en ese mes. Sin embargo, la muestra fue rechazada ya que no permitió la
recuperación de parasitoides, pero queda en manifiesto, la capacidad
depredadora de C. montrouzieri sobre colonias de chanchito blanco,
precedido por una postura de una hembra sobre masas de huevos. Algo
similar señala SAA (2004), quien recolecta en una muestra de P. citri ciento
nueve individuos de C. montrouzieri (marzo 2002), correspondiendo al 56%
de los enemigos naturales en esa fecha.

45
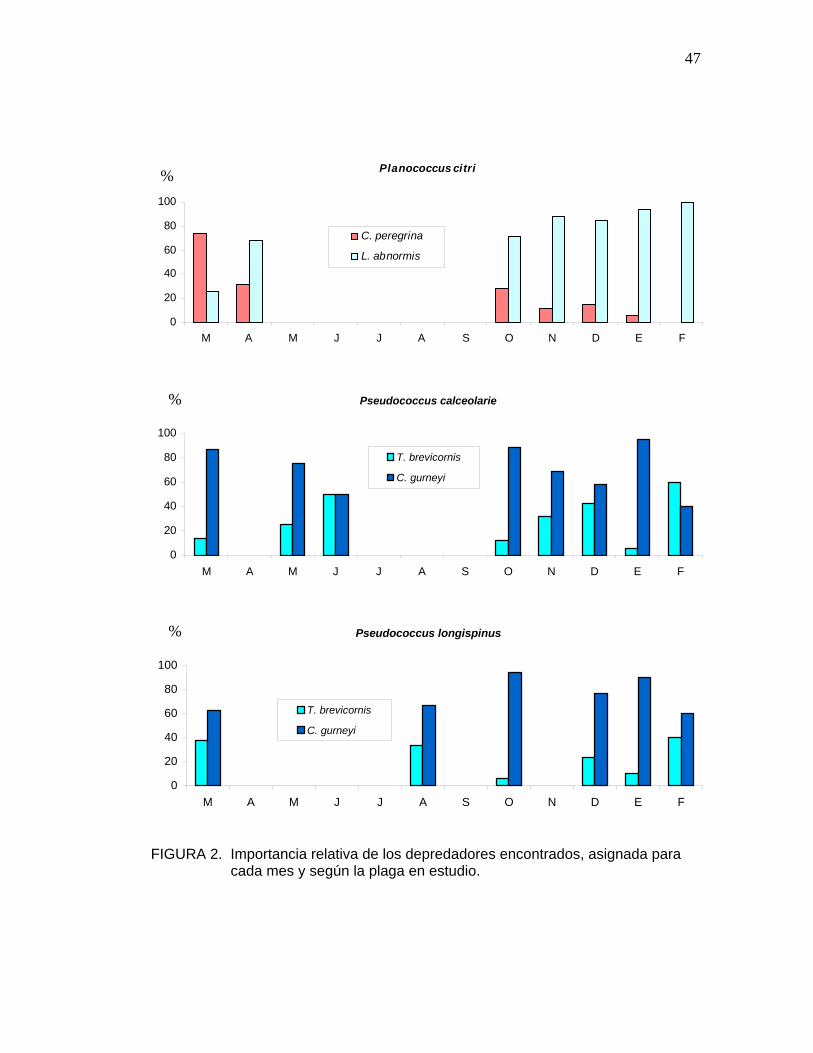
4.3.2 Parasitoides
La importancia relativa atribuida a cada especie parasitoide, en cada mes, y
para cada plaga en estudio, se muestra en la Figura 2.
Los enemigos naturales parasitoides fueron encontrados en toda las
muestras de las tres especies de chanchito blanco en estudio, por lo que
presentan una distribución en el tiempo mas amplia que aquellos de carácter
predador, o igual, en el caso de C. montrouzieri sobre P. citri.
En P. citri, las especies Leptomastidea abnormis y Coccidoxenoides
peregrina fueron encontradas en los meses de marzo y abril y de octubre a
enero, predominando la primera especie en importancia en la mayoría de los
meses, a excepción del mes de marzo. En el mes de febrero, sólo apareció
L. abnormis.
En el caso de P. calceolarie y P. longispinus, en todo los meses en que se
recuperaron parasitoides, aparecieron las especies C. gurneyi y T.
brevicornis, actuando en conjunto y mostrándose una clara dominancia de C.
gurneyi en las dos plagas mencionadas, a excepción, del mes de febrero en
P. calceolarie. Los parasitoides mencionados, fueron encontrados en los
meses de marzo, mayo y junio y, desde octubre a febrero en P. calceolarie.
En P. longispinus se encontraron en los meses de marzo, agosto, octubre y,
en los meses diciembre enero y febrero. QUIROS (1998), recupera a C.
gurneyi entre los meses de septiembre y enero, sobre P. calceolarie y P.

46
longispinus, mientras que a T. brevicornis, entre los meses de septiembre y
diciembre.
Un análisis más detenido se muestra en el Cuadro 4, el cual, se refiere a los
antecedentes reunidos para cada especie en particular y lo concluido por el
autor.
47
Planococcus citri
0
20
40
60
80
100
M A M J J A S O N D E F
C. peregrina
L. abnormis
Pseudococcus calceolarie
0
20
40
60
80
100
M A M J J A S O N D E F
T. brevicornis
C. gurneyi
Pseudococcus longispinus
0
20
40
60
80
100
M A M J J A S O N D E F
T. brevicornis
C. gurneyi
FIGURA 2. Importancia relativa de los depredadores encontrados, asignada para cada mes y según la plaga en estudio.
%
%
%

48
CUADRO 4. Análisis individual de cada enemigo natural, sus referencias y resultados.
ESPECIE REFERENCIAS RESULTADOS
1 Allotropa citri I P GONZÁLEZ Y ROJAS, 1966 y ZUÑIGA, 1984 como no establecido para P. citri.
No encontrado.
2Anagyrus
pseudococci I PGONZÁLEZ Y ROJAS, 1966 y ZUÑIGA, 1984 como no establecido para P. citri.
No encontrado.
3Coccophagus
gurneyi I P
GONZÁLEZ Y ROJAS, 1966 como parcialmente establecido para P. citri y sustancialmente establecido para longispinus. PRADO, 1991y RIPA Y RODRÍGUEZ para P. longispinus.
Encontrado en casi todo los meses representando el parasitoide mas importante de P. calceolarie y P. longispinus.
4 Chrysoperla sp. I D
GONZÁLEZ Y ROJAS, 1966 como parcialmente establecido para P. calceolarie; RIPA Y RODRÍGUEZ para Pseudococcus .
Solo un ejemplar encontrado en P. citri.
5Cryptolaemus montrouzieri I D
GONZÁLEZ Y ROJAS, 1966 como sustancialmente establecido para P. citri y P. longispinus. ZUÑIGA, 1984;PRADO, 1991; VARGAS, 1997 y RIPA Y RODRÍGUEZ para Pseudococcus .
35,2% de participación sobre P. citri , 85% en P. calceolarie y 88,9% en P. longispinus, con respecto al resto de depredadores encontrados.
6Hungariella
pretiosa I PZUÑIGA, 1984 como establecido para P. citri .VARGAS, 1997 para Pseudococcus.
No encontrado.
7Leptomastidea
abnormis I P
GONZÁLEZ Y ROJAS, 1966 como parcialmente establecido para P. citri . ZUÑIGA, 1984 como parcialmente establecido para P. citri . PRADO, 1991; VARGAS, 1997 y RIPA Y RODRÍGUEZ para P. citri
Aparece como el parasitoide mas importante sobre P. citri.
8Leptomastix dactylopii I P
GONZÁLEZ Y ROJAS, 1966 como parcialmente establecido para P. citri . ZUÑIGA, 1984 como parcialmente establecido para P. citri .PRADO, 1991; VARGAS, 1997 y RIPA Y RODRÍGUEZ para P. citri
No encontrado.
9Coccidocenoides
peregrina I P
GONZÁLEZ Y ROJAS, 1966 como no establecido para P. citri y P. longispinus. ZUÑIGA, 1984 como establecido para P. citri. PRADO, 1991; VARGAS, 1997 y RIPA Y RODRIGUES para P. citri
Encontrado en P. citri, de menor importancia que L. abnormis
10Pseudaphycus
perdignus I P
GONZALEZ Y ROJAS, 1966 y ZUÑIGA, 1984 como no establecido para P. citri. PRADO, 1991 y VARGAS, 1997 para P. citri.
No encontrado.
11Tetracnemoidea
brevicorni I P
GONZÁLEZ Y ROJAS, 1966 como sustancialmente establecido para P. calceolarie. PRADO, 1991 para P. calceolarie y P longispinus. VARGAS, 1997 para P. calceolarie y RIPA Y RODRIGUES para P. calceolarie y P longispinus.
Desde casi desconocido en los años 50, hoy representa al segundo parasitoide de P. calceolarie y P. longispinus.
12 Zarhopalus sp. I PGONZÁLEZ Y ROJAS, 1966 y ZUÑIGA, 1984 como no establecido para P. citri. VARGAS, 1997 para P. citri.
No encontrado.

49
CUADRO 4. (continuación) ESPECIE REFERENCIAS RESULTADOS
1 Adalia deficiens N D PRADO, 1991 para P. citri No encontrado
2Aenasius punctatus N P
PRADO, 1991 (Aenasius sp); VARGAS, 1997 y RIPA Y RODRÍGUEZ para P. longispinus.
No encontrado
3Hyperaspis
funesta N DPRADO, 1991 y RIPA Y RODRÍGUEZ para Pseudococcus.
Aparece en forma casual con una importancia muy baja en las tres especies de chanchito blanco.
4 Leucopis sp. N D PRADO, 1991; VARGAS, 1997 y RIPA Y RODRÍGUEZ para Pseudococcus .
Encontrado en poca cantidad, su importancia se ha reducido notablemente desde los años 50.
5Ocyptamus confusus N D
PRADO, 1991; VARGAS, 1997 y RIPA Y RODRÍGUEZ para Pseudococcus .
Encontrado sólo en P. citri con una participación del 62,8% respecto al resto de los depredadores encontrados.
6 Pseudaphycus sp. N PVARGAS, 1997 para Pseudococcus . No encontrado
7 Scymnus nitidus N D PRADO, 1991; VARGAS, 1997 y RIPA Y RODRÍGUEZ para Pseudococcus .
No encontrado
8 Stygmacida N D RIPA Y RODRÍGUEZ para P. citri. No encontrado.
9Sympherobius maculipennis N D
PRADO, 1991; VARGAS, 1997 y RIPA Y RODRÍGUEZ para Pseudococcus .
Clasficado como Sympherobius sp.
10 Sympherobius sp. N DPRADO, 1991 para P citri. VARGAS, 1997 para Pseudococcus .
Encontrado en P. citri, con baja importancia.
I: introducido; N: nativo; P: parasitoide; D: depredador.

50
4. CONCLUSIONES
De las veinticuatro especies de enemigos naturales, citados en Chile para el
control del chanchito blanco, sólo se recuperaron diez, en la Comuna de
Quillota. Estas corresponden a seis especies introducidas: Leptomastidea
abnormis, Coccidoxenoides peregrina, Coccophagus gurneyi,
Tetracnemoidea brevicornis, Cryptolaemus montrouzieri y Chrysoperla sp. y
seis nativas: Ocyptamus confusus, Hyperaspis funesta, Sympherobius sp. y
Leucopis sp.
De las especies nativas o de origen desconocido, Ocyptamus confusus
aparece en forma importante mientras que Hyperaspis funesta, Leucopis sp.
y Sympherobius sp. en forma escasa.
Ocyptamus confusus siendo la especie nativa más importante, aparece por
un tiempo no muy prolongado. En cambio, Hyperaspis funesta, Leucopis sp.
y Sympherobius sp. sólo aparecieron en forma ocasional.
La importancia de los depredadores en general, ha disminuido notablemente,
lo que indica que los parasitoides, todos ellos introducidos y siendo
específicos, han logrado un aumento importante. De las especies nativas, se
tienen los antecedentes de estas son mayoritariamente depredadores
generalistas, por lo que, a pesar de presentarse en escasa cantidad, no es
deducible la extinción de alguna de estas causada por la supresión de las
especies exóticas, que luego de introducidas, fueron criadas y liberadas en
forma masiva.

51
La introducción de especies depredadoras y parasitoides de Pseudococcus
a Chile, no ha suprimido a las especies nativas asociadas a esta plaga.

52
6. RESUMEN
Existen cerca de veinticinco especies de insectos encontradas en la literatura, que actúan como controladores del chanchito blanco en Chile. Un grupo importante pertenece a las especies introducidas oficialmente a través de programas de control biológico clásico. El otro grupo, esta conformado por todas aquellas especies de origen desconocido o consideradas nativas. Se realizó una prospección de los enemigos naturales de chanchitos blancos, para lo cual, fueron recolectadas muestras mensuales, por un periodo de un año, de P. citri, P calceolarie y P. longispinus. De estas, se recuperaron parasitoides y depredadores y se determinó el nivel de participación de estos, a modo de inferir el estado en que se encuentran las especies que no han sido introducidas oficialmente, en comparación, con las foráneas que has sido criadas y liberadas en forma masiva con la colaboración de agentes públicos y privados. Se encontraron cuatro especies nativas de carácter depredador: Ocyptamus confusus, Hyperaspis funesta, Sympherobius sp. y Leucopis sp, Éstas, mostraron una baja participación y una aparición ocasional, a excepción de Ocyptamus confusus, que apareciendo por un periodo de cinco meses desde diciembre, tuvo una participación de un 62,8% en P. citri, con respecto al resto de los depredadores encontrados sobre esta plaga. En ningún caso, puede afirmarse que la introducción y liberación de especies exóticas han logrado eliminar a las especies nativas que controlan al chanchito blanco. El resto de las especies depredadoras encontradas son: Cryptolaemus montrouzieri y Chrysoperla sp, mientras que C. montrouzieri representó el 35,2% de los depredadores encontrados sobre P. citri; 85% en P. calceolarie y 88,9% en P. longispinus. Chrysoperla sp., se encontró solo en P. citri ponderando un 0,7% de los depredadores encontrados en esta especie de chanchito blanco. Los parasitoides Leptomastidea abnormis y Coccidoxenoides peregrina fueron encontrados sobre P. citri, siendo el primero el más importante con un 77,6% respecto a un 22,4%. A su vez, Coccophagus gurneyi y Tetracnemoidea brevicornis se recuperaron de P. calceolarie y P. longispinus. En ambos casos, C. gurneyi representa cerca de un 63% mientras que Tetracnemoidea brevicornis no supera el 32% de los enemigos naturales encontrados.

53
7. ABSTRACT There are nearly twenty five species found in literature, that act as Pseudococus controllers in Chile. One important group belongs to species inserted officially through classic biological control programs. The other one it’s formed by those species which don’t have a known origin or called natives. A research was made about the Pseudococus’s natural enemies. To achieve this, monthly samples were recolected, during a year. Among this samples were P. citri, P calceolarie y P. longispinus. From these, predators and parasites were recovered and it set the participation level of these species, to decide the current condition of the native species that haven’t been officially inserted, in comparison with the foreign ones that have been raised and released plentifully with private and public agents colaboration. Four predator native species were found: Ocyptamus confusus, Hyperaspis funesta, Sympherobius sp. and Leucopis sp. These showed a low participation level and one occasional appearance, except for Ocyptamus confusus, that had a 62,8 % participation from december to april in P. Citri, regarding the rest of the predators found about this plague. It can’t be stated in any way that the introduction and release of exotic species have been able to eliminate native species that control Pseudococus. The rest of the predator species found are: Cryptolaemus montruozieri and Chrysoperla sp. While the first one represented 35,2% of predators found in P. citri, 85% in P. calceolarie and 88,9% in P. longispinus. Chrysoperla sp. was found only in P. citri, pondering 0,7% of predators found in this Pseudococus specie. Parasites Leptomastidea abnormis and Coccidoxenoides peregrina were found in P. Citri. The most important was the first one, with 77,6% against 22,4%. In another case, Coccophagus gurneyi and Tetracnemoidea brevicornis were recovered from P. calceolarie and P. longispinus. In both cases, C. gurneyi represents about 63% while Tetracmoidea brevicornis doesn’t exceed 32% of natural enemies found.

54
8. LITERATURA CITADA
BEINGOLEA, O. 1977 Consideraciones sobre el control biológico y
predación. Revista Peruana de Entomología 20(1): 33-48.
CAPDEVILLE, C. 1945. Plagas de la agricultura en Chile. Quillota, Imprenta
Pacífico. 358 p.
DUOTT, R. y DE BACH, P. 1981. Algunos conceptos y preguntas sobre el
control biológico. In: De BACH, P. Ed. Control biológico de las
plagas de insectos y malas hierbas. Méjico, D.F. C.E.C.S.A. pp 151-
172.
DUOTT, R. 1981. Características biológicas de los adultos entomófagos. In:
DeBACH, P. Ed. Control biológico de las plagas de insectos y malas
hierbas. Méjico, D.F. C.E.C.S.A. pp 179-202.
GONZÁLEZ, R.H. y ROJAS, S. 1966. Estudio analítico del control biológico
de plagas agrícolas en Chile. Agricultura Técnica 26(4): 133-147.
GONZÁLEZ, R. 1991. Chanchitos blancos (Homóptera: Pseudococcidae),
una nueva plaga de ciruelos en Chile. Rev. Frutícola 12(1): 3-7.
HARRISON, M. C. 1993. Control biológico de Pseudococcus longispinus.
Tesis Ing. Agr. Quillota, Universidad Católica de Valparaíso.
Facultad de Agronomía. 74 p.

55
INIA 2001. Manejo Integrado de Plagas en Cítricos, (on line),
www.mipcitricos.cl.
ODUM E. 1969. Ecology. México. D.F. Editorial Interamericana. 412p.
PRADO, E. 1991. Artrópodos y sus enemigos naturales asociados a plantas
cultivadas en Chile. Santiago, Instituto de Investigaciones
Agropecuarias. 207p.
QUIROS, S. 1998. Comportamiento estacional del chanchito blanco,
Hemíptera: Pseudococcidae y sus parasitoides en palto. Tesis Ing.
Agr. Quillota, Universidad Católica de Valparaíso. Facultad de
Agronomía. 178p.
RIPA R. y RODRÍGUEZ 1999. Plagas de cítricos, sus enemigos naturales
y manejo. Santiago, Instituto de Investigaciones Agropecuarias.
151p.
SAA, M. 2004. Determinación de especies benéficas asociadas a
Pseudococcus en la comuna de Quillota. Tesis Ing. Agr. Quillota,
Universidad Católica de Valparaíso. Facultad de Agronomía. 51 p.
SPEIGHT M., HUNTER M. y WATT A. 1999. Ecology of insects, concepts
and applications. Oxford. Editorial Blackwell Science. 350p.
VARGAS N. 1997. Enemigos naturales utilizados en el control biológico de
plagas en Chile. Agricultura Técnica 54(3): 85-89.

56
YUDELEVICH, M, 1950. Control biológico de los Pseudococcus en Chile.
Tesis Ing. Agr. Santiago, Universidad de Chile, Facultad de
Agronomía. 126p.
ZUÑIGA, E. 1987. El control biológico: concepto y algunas características.
IPA La Platina 44: 8-17.
ZUÑIGA, E. 1984. Ochenta años de control biológico en Chile. Revisión
histórica y evaluación de los proyectos desarrollados (1903-1083).
Agricultura Técnica 45(3): 175-183.

57
ANEXOS
58
ANEXO 1. Resultados obtenidos en el presente estudio.
Fecha L. abnormis P. peregrina C. montrouzieri B. valdiviana Crisoperla sp. H. funesta Sympherobius sp.M 10 29 10 15 0 1 1A 32 15 8 8 0 0 0MJJASO 15 6 1 0 0 0 0N 23 3 4 0 0 0 0D 34 6 7 3 1 0 0E 82 5 11 62 0 0 0F 26 0 10 3 0 0 0
Pseudococcus calceolarieFecha C. gurneyi T. brevicornis C. montrouzieri H. funesta Leucopis sp.
M 13 2 5 0 0AM 18 6 8 0 0J 16 16 0 0 0JASO 30 4 0 0 0N 22 10 0 0 0D 15 11 0 0 0E 17 1 2 1 2F 4 6 2 0 0
Fecha C. gurneyi T. brevicornis C. montrouzieri H. funestaM 100 61 2 1AMJJA 2 1 0 0SO 15 1 0 0ND 10 3 0 0E 9 1 2 0F 12 8 4 0
Planococcus citri
Pseudococcus longispinus

59
ANEXO 2. Enemigos naturales encontrados en el presente estudio.

60
ANEXO 2. (continuación)
Fuente: INIA 2001.

61


















