
SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna · del ganadero, costos y facilidad de...
98
SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna Editores: Graciela Quintans José Ignacio Velazco Gonzalo Roig
Transcript of SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna · del ganadero, costos y facilidad de...

1
INIA TREINTA Y TRES SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna
SEMINARIO DE ACTUALIZACIÓNTÉCNICA: Cría Vacuna
Editores: Graciela QuintansJosé Ignacio VelazcoGonzalo Roig

2
SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna INIA TREINTA Y TRES
Título: SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna
Editores: Graciela QuintansJosé Ignacio VelazcoGonzalo Roig
Serie Técnica Nº 174
© 2008, INIA
ISBN: 978-9974-38-251-0
Editado por la Unidad de Comunicacion y Transferencia de Tecnologia del INIA.Andes 1365, Piso 12. Montevideo - UruguayPágina Web: http://www.inia.org.uy
Quedan reservados todos los derechos de la presente edición. Este libro no se podráreproducir total o parcialmente sin expreso consentimiento del INIA.

Instituto Nacional de Investigación Agropecuaria
Integración de la Junta Directiva
Ing. Agr., Dr. Dan Piestun - Presidente
Ing. Agr., Dr. Mario García - Vicepresidente
Ing. Ind. Aparicio Hirschy
Ing. Agr. José Bonica
Ing. Agr. Rodolfo M. Irigoyen
Ing. Agr. Mario Costa

4
SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna INIA TREINTA Y TRES

5
INIA TREINTA Y TRES SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna
INTRODUCCIÓN GENERAL
El sector agropecuario es fundamental para el crecimiento de la economía y el desarrollosocial del País. Si se consideran las exportaciones como uno de los indicadores parademostrar la trascendencia del sector sobre la economía nacional, “los números son con-tundentes”, donde aproximadamente el 70% del total de las exportaciones proviene de loque genera la agroindustria nacional. Por lo tanto, el desarrollo futuro de este sector hacea la prosperidad y bienestar de la población. En este contexto, y si se tienen en cuenta lasexportaciones por sub-sector, el conjunto de las divisas provenientes de la sumatoria decarne bovina y ovina, lana y cueros, constituye el 36% del total exportado, demostrando asíla importancia de “la vaca” en el destino de cada uno de los integrantes de esta Nación.
A nivel primario los números son contundentes en cuanto a la importancia social de laganadería, donde la mayor parte de las explotaciones especializadas en ganadería de car-ne y lana (78.8%) son productores del tipo familiar y que a su vez poseen superficies deexplotación menores a 500 ha, lo cual explica que esta mayoría ocupen sólo el 21.9% delárea y posean el 24.5% del total de las cabezas equivalentes vacunas. De otro punto devista, considerando la orientación familiar de la producción, la mayor cantidad de estableci-mientos familiares y medios se encuentran dedicados a la ganadería de carne y lana,representando un 65.2 y 65.5% respectivamente. Para las explotaciones familiares el 65%de su principal ingreso, lo constituye la ganadería de carne y/o lana, resaltando así laimportancia económica y social del rubro; seguida de la horticultura (12%), lechería (11%),suinocultura (2%) y avicultura (2%).
Estamos en un momento histórico de la agropecuaria nacional, donde se están operandocambios fenomenales, y la ganadería no escapa a esta situación inédita. Los mercados demayor valor están abiertos para las carnes uruguayas, y en otros de gran potencial existenbuenas oportunidades de apertura en el corto ó mediano plazo. La industria nacional se haposicionado frente a esta realidad incrementado sustancialmente la capacidad de faena yde frío y el grado de procesamiento y por ende de generación de mayor valor agregado.
Por otra parte la capitalización y consolidación de este momento es sujeta al aumento delvolumen, eficiencia y consistencia de la elaboración de productos cárnicos diferenciadosorientados a satisfacer las demandas incrementales de los mercados, contemplando as-pectos de bienestar animal, inocuidad alimentaria, palatabilidad y valor nutritivo del pro-ducto y su influencia sobre la salud humana y las preferencias de los consumidores, en uncontexto de respeto del medioambiente y de equidad y responsabilidad social y ética.
Como nunca están dadas las condiciones para favorecer y consolidar “las estrategias dealianzas tempranas” entre productores y la industria. La amplia disponibilidad de mercadosy la falta de producto, están cimentando una planificación de la producción de carne sinprecedentes, donde la industria frigorífica no sólo esta dispuesta a establecer contratoscon los invernadores sino con los propios criadores, en un contexto de elaboración de unproducto definido para abastecer un determinado nicho de mercado. La concreción deestas alianzas, los precios alentadores de la reposición en los dos últimos años, sustentanel pensamiento de que estamos frente a un momento histórico para la ganadería. La incor-poración de “paquetes tecnológicos” encuentra así un marco fértil para un plan de desarro-llo genuino de la ganadería. Este momento genera un marco favorable para dirigir los es-fuerzos y recursos públicos y privados para la promoción del incremento de la producciónde carne de calidad.

6
SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna INIA TREINTA Y TRES
Desde el punto de vista de los horizontes tecnológicos que puede explorar la cría, esposible alcanzar niveles crecientes de tasas de destete, donde existen opciones claras demejora para superar los niveles históricos de 60 y 65%. La combinación de las tecnologíasdisponibles y el orden del uso de las mismas que varia de acuerdo a una serie de factoresque determinan su adopción (entre otros, la escala y orientación productiva del negociodel ganadero, costos y facilidad de implementación de propuestas tecnológicas), infraes-tructura disponible, nivel de educación y edad del productor, capacitación del personal acargo, aversión al riesgo, motivaciones y percepción de negocio en el corto y medianoplazo por parte del ganadero, etc.), permiten aspirar a niveles cercanos al 90%. Todosquienes trabajamos con los ganaderos, en particular los criadores, y teniendo en cuentasu percepción sobre el futuro del negocio de la cría y el rol que juega la incorporación de latecnología, rápidamente comprendemos que el grado de adopción tecnológica es una ecua-ción compleja y con muchos componentes involucrados, y de difícil resolución debido a lanaturaleza de este proceso productivo, donde el factor humano y sobretodo la heterogenei-dad del mismo juega un rol preponderante en el proceso de comprensión de la problemáti-ca. Sin embargo, dentro de esta heterogeneidad, también se puede comprobar que muchosproductores de referencia están alcanzando niveles consistentemente del 85% de destetea lo largo de muchos años. De cualquier manera, es importante destacar que el “paradigmade la cría” ya no es sólo preñar las vacas, sino que otros elementos hacen a la lógicaproductiva, económica, comportamental y social de los sistemas criadores.
En el mencionado escenario, el sector criador, tiene una serie de desafíos que hacen a lacompetitividad del mismo, donde la investigación nacional esta trabajando para encarar yresolver al menos parte de los mismos, particularmente en un contexto de aumento delprecio y de la renta de la tierra, así como del crecimiento competitivo de la agricultura y laforestación. Todo ello, presiona a la necesidad de un aumento de la productividad de la críabovina y a repensar la misma.
En esta publicación se documenta un importante volumen de información científica y expe-riencias generadas en la recría y cría bovina por diferentes investigadores de diferentesdisciplinas pertenecientes a diferentes instituciones. La propia conformación de los equi-pos de trabajo que han llevado adelante los artículos que se presentan en esta publicación,demuestra el enfoque moderno, multidisciplinario, y multinstitucional, generando sinergias,eficiencia y complementariedad entre especialistas e instituciones para proponer solucio-nes tecnológicas a este eslabón tan fundamental para la Cadena Cárnica del Uruguay.
Ing. Agr. PhD. Fabio MontossiDirector Programa Nacional de Carne y Lana

7
INIA TREINTA Y TRES SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna
TABLA DE CONTENIDO
1. GENÉTICA Y REPRODUCCIÓN ......................................................................................91.1. Genética y Fertilidad: transitando un camino común..............................................9
Graciela Quintans1.2. Selección y Reproducción en bovinos de carne .................................................... 11
Jorge I. Urioste1.3. Recursos Genéticos y ambientes de producción en la cría vacuna ..................... 25
Ana Carolina Espasandin, Marcelo Ciria Ruggiero1.4. Reporte total del rodeo: Una nueva etapa de las evaluaciones
genéticas de bovinos para carne ........................................................................... 40Mario Lema, Olga Ravagnolo
1.5. Nuevas oportunidades para el área de cría:Biotecnología molecular aplicada a la producción animal .................................... 47Mariana Carriquiry , Ana Meikle
2. ALGUNAS ESTRATEGIAS PARA DISMINUIR LA EDADAL PRIMER SERVICIO EN VAQUILLONAS ..................................................................... 52
2.1. Recría vacuna: antecedentes y nuevos enfoques ................................................. 53Graciela Quintans
2.2. Principales factores que afectan la aparición de la pubertaden vaquillonas de razas carniceras ........................................................................ 56Graciela Quintans, Gonzalo Roig
2.3. Efecto del manejo nutricional en el primer invierno sobre la apariciónde la pubertad en terneras de raza carnicera (primer año de evaluación) ............ 59Juan Miguel Straumann, Walter Ayala, Ana Inés Vázquez, Graciela Quintans
2.4. Efecto del manejo nutricional en el primer invierno y el biotipo sobrela aparición de la pubertad en terneras de raza carnicera(segundo año de evaluación) .................................................................................. 64Santiago Barreto, Diego Negrín, Walter Ayala, Graciela Quintans
2.5. Efecto de tres ganancias invernales sobre la aparición de la pubertaden terneras de raza carnicera (tercer año de evaluaciÓn) .................................... 70Álvaro Costa, Roberto Moreira, Antonia Scarsi, Walter Ayala, Graciela Quintans
2.6. Manejo nutricional en el primer invierno y manejos diferencialesposteriores para alcanzar similar peso vivo en otoño.Efecto en la aparición de la pubertad (cuarto año de evaluación) ......................... 77Graciela Quintans, Antonia Scarsi, Carlos López, Fernando Pereyra
2.7. La bioestimulación permite disminuir la edad a la pubertaden vaquillonas de carne .......................................................................................... 82Carolina Fiol, Graciela Quintans, Rodolfo Ungerfeld
2.8. Servicio de vaquillonas en otoño a los 20 meses de edad(Resultados preliminares) ...................................................................................... 90Graciela Quintans, José Ignacio Velazco, Gonzalo Roig
3. ALTERNATIVAS DE MANEJO NUTRICIONAL, CONTROL DEL AMAMANTAMIENTOY TRATAMIENTOS HORMONALES DURANTE EL POSPARTO ...................................... 99

8
SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna INIA TREINTA Y TRES
3.1. La alternativa para incrementar la tasa de procreo:disminución del anestro posparto ......................................................................... 99Graciela Quintans
3.2. Herramientas para mejorar la utilización del forraje del campo natural,el ingreso económico de la cría y atenuar los efectos de la variabilidadclimática en sistemas de cría vacuna del Uruguay ............................................. 110
a Impacto de cambios en la estrategia de asignación de forraje sobre laproductividad de la cría con diversos grupos genéticos bajo pastoreode campo natural .................................................................................................. 110Pablo Soca, Fernando Olmos, Ana Espasandín, Dorrel Bentancur,Fernando Pereyra, Victoria Cal, Martín Sosa, Martín Do Carmo
b Empleo del flushing y destete temporario de forma táctica para mejorarindicadores reproductivos y concentración de preñez de vacas primíparas .......120Pablo Soca, Mariana Carriquiry, Graciela Quintans, Carlos Lopez, Ana Espasandín,Ana Inés Trujillo, Maria de Jesús Marichal, Ana Laura Astessiano, Raquel Pérez-Clariget
3.3. Rol de los minerales en la producción de vacunos de carne de Uruguay ..........135Oscar Pittaluga
3.4. Impacto de la mejora nutricional posparto junto a un destete temporariosobre la tasa de preñez en vacas Braford primíparas ........................................147Carolina Jiménez de Aréchaga, Oscar Pittaluga, Graciela Quintans
3.5. Evaluación del destete a corral por 14 días sobreEl desempeño Reproductivo en vacas de carne primíparas y multíparasy el crecimiento de sus terneros .........................................................................153Graciela Quintans, Carolina Jiménez de Aréchaga,José Ignacio Velazco, Ana Inés Vázquez
3.6. Efecto del destete temporario en función de la actividad ovárica sobre elporcentaje de preñez en vacas multíparas y primíparas ....................................165Graciela Quintans, Carolina Jiménez de Aréchaga, José Ignacio Velazco,Gonzalo Roig, Daniel Briano, Juan Andrés López, Santiago Viana,Fernando Pereyra, Carlos López
3.7. Efecto de la condición corporal y la restricción del amamantamientocon y sin presencia del ternero sobre la producción de leche, anestroposparto y crecimiento de los terneros ...............................................................172Graciela Quintans, Georgget Banchero, Mariana Carriquiry,Carlos López, Fernando Baldi
3.8. Un tratamiento para la inducción de la ovulación en vacas con críaal pie asociado a inseminación artificial a tiempo fijo .........................................182Guillermo T. de Nava Silva
3.9. Tratamientos hormonales en vacas para carne (Bos taurus) en anestrocon cría al pie para mejorar su comportamiento productivo y reproductivo ........189Juan Bolivar Rodríguez Blanquet
Nota: las contribuciones realizadas son responsabilidad de sus autores.

9
INIA TREINTA Y TRES SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna
1.1. GENÉTICA Y FERTILIDAD: TRANSITANDO UN CAMINO COMÚN
Graciela Quintans1
1 Ing. Agr., PhD. - Programa Nacional de Producción de Carne y Lana - INIA Treinta y Tres.
centrado en la evaluación y selección porcaracterísticas de crecimiento. En la actua-lidad, se dispone de valores de cría parapesos en diferentes edades (nacimiento,destete, 15 y 18 meses) y para característi-cas indicadoras de calidad de res (área delojo del bife y espesor de grasa), así comotambién de circunferencia escrotal que seha asociado a precocidad sexual en las hem-bras.
Las características asociadas a la performan-ce reproductiva son las económicamentemás importantes en muchos sistemas deproducción, especialmente los netamentepastoriles (Church 1976, citado por Rovira1996; Newman et al., 1992; Urioste et al.,1998). Lamentablemente existe cierta difi-cultad de determinar característ icasreproductivas fácilmente cuantificables quesean genéticamente relacionadas con la fer-tilidad en hembras (Johnston y Bunter, 1996).
Existe cierta controversia a nivel académicoen relación al impacto de la selección deanimales para mayor tamaño y la performan-ce reproductiva y productiva del rodeo. Exis-ten limitantes sobre la interpretación de larelación genética entre las característicasde crecimiento y las característ icasreproductivas. Por ejemplo, en sistemaspastoriles es claro que existe una relacióndesfavorable entre tamaño de la vaca y superformance reproductiva mientras que ensistemas más intensivos esta relación no estan clara.
En la actualidad y en otras partes del mun-do gracias al avance de metodologías y sis-
1. GENÉTICA Y REPRODUCCIÓN
Siempre estuvimos convencidos que lagenética es una herramienta excelente paraidentificar y seleccionar animales que seadapten mejor y expresen de mejor manerasu potencial reproductivo. También, de al-guna forma expresamos nuestra preocupa-ción de la marcada tendencia genética en elincremento de los pesos de nuestros gana-dos jóvenes lo que puede repercutir sin dudaen un mayor tamaño adulto. Sobretodo cuan-do la mayoría de estos animales se desa-rrollan en ambientes restrictivos de produc-ción, llámese campo natural. Esto no quie-re decir que el incremento del tamaño delos animales esté afectando la fertilidad delos mismos, no lo sabemos, simplementeque si el 70% de lo que consume un animalse destina a mantenimiento, un animal conun tamaño adulto mayor, requerirá mayor ali-mentación para cumplir con sus necesida-des básicas.
Parecería que esta inquietud no es nuevapara nuestro medio y en este marco no po-demos dejar de citar al Profesor Jaime Roviraquien ya en 1974 declaraba “…animalesgrandes con mayores exigencias alimenti-cias en condiciones normales de producción(ambientes más pobres) ven resentida la pro-ducción” y se preguntaba por ese entonces“…¿hacia donde conduce esta carrera alo-cada en que están empeñados los criado-res de todas las razas en el sentido de au-mentar el tamaño de todas ellas? ¿No per-deremos en el camino muchas característi-cas muy deseables? ”
Los Programas de Evaluación Genética denuestro país para ganado de carne se han

10
SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna INIA TREINTA Y TRES
temas de registro en diversos programas deevaluación genética, se están utilizando di-ferentes características reproductivas regis-tradas en las hembras como criterios deselección para mejorar la fertilidad. Algunosejemplos son días al parto, largo de gesta-ción, edad al primer parto, intervalo entre elprimer y segundo parto, intervalo entre otrospartos, probabilidad de la vaca de permane-cer en el rodeo y probabilidad de preñez pre-coz.
Por otra parte se nos abren nuevas oportu-nidades de avances en esta área y comple-mentaria a la descrita anteriormente: labiotecnología molecular. La seleccióngenética asist ida por marcadoresmoleculares es una herramienta de apoyo alos programas tradicionales de selecciónsiendo un valioso instrumento en variablesde baja heredabilidad como la fertilidad.
Sabemos que la selección bovina por carac-terísticas de fertilidad es una opción de me-joramiento compleja, larga y desafiante. Peroestamos en un momento óptimo de conoci-miento de nuestra ganadería pastoril que nos
permite plantearnos ese desafío. Los técni-cos y productores estamos todos juntos dis-puestos a transitar ese camino.
Referencias bibliográficas
Johnston, DJ., Bunter, KL. 1996. Days tocalving in Angus catt le: Genetic andenvironmental effects, and covariances withother traits. Livestock Production Science,v.45, n.1, p.13-22.Rovira, J. 1974. Aspectos básicos de la cría,Revista FUCREA, No. 20: 42-49Rovira, J. 1996. Manejo nutritivo de los ro-deos de cría en pastoreo. Montevideo, He-misferio Sur. 288 p.Newman, S., Morris, CA., Baker, RL. andNicoll, GB. 1992. Genetic improvement ofbeef cattle in New Zealand: breedingobjectives. Livestock Production Science 32,111-130.Urioste, JI., Ponzoni, RW., Aguirrezabala,MA., Rovere, G. and Saavedra, D. 1998.Breeding object ives for pasture-fedUruguayan beef cattle. Journal of AnimalBreeding and Genetics 115, 357-373.

11
INIA TREINTA Y TRES SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna
Resumen
Los caracteres reproductivos en vacunos paracarne son económicamente importantes,pero el uso de información reproductiva comoherramienta de selección presenta difi-cultades, especialmente bajo condicionespastoriles extensivas. Se revisa la literaturainternacional sobre características defertilidad posibles de ser incorporadas aprocedimientos estándar de selección. Anivel nacional, siete modelos genéticosfueron desarrollados para interpretar lavariabilidad presente en las característicasdía de parto (DP) y/o Éxito al parto (EP).Registros de parto de 2032 o 3442 vacasAberdeen Angus fueron usados para estimarparámetros genéticos. Las heredabilidadesvariaron entre 0.19 y 0.31 para DP y 0.27 a0.43 para EP, dependiendo de los modelos.Las correlaciones genéticas para DP fueronaltamente posit ivas (0.82-0.88) entredi ferentes partos. Las correlacionesgenéticas entre EP medidos en diferentespartos, pueden ser asumidas comopositivas, de medias a altas (80.56-0.80), apesar de algunas inconsistencias en losanálisis. Correlaciones genéticas negativas(genéticamente favorables) y de medias aaltas (-0.54 a -0.91) fueron encontradas entreDP y EP, sugiriendo que DP podría ser usadocomo característica indicativa del caráctereconómicamente importante EP. Un modelounivariado de repetibilidad para DP, con unapenalización para vacas no paridas essugerido como un modelo útil para laimplementación práctica de evaluacionesgenéticas. El registro de los datos debemejorar en calidad para futuras aplicacionesde evaluaciones genéticas para caracte-rísticas de fertilidad. Los trabajos desarro-llados demuestran la factibilidad de encararprocesos selectivos para la mejora genéticade rasgos reproductivos en bovinos de carne.
Introducción
La eficiencia reproductiva del ganado vacu-no de cría es baja en Uruguay: cifras aporta-das por el Ministerio de Ganadería, Agricul-tura y Pesca muestran una gran variaciónen tasas de destete según año de parto, enun rango que va desde el 50 al 75% en elperiodo 1981-2002 (MGAP-DIEA, 2003). Lasvariables reproductivas están fuertementeinfluenciadas por el manejo y la nutrición,pero también existe una base genética.
En las últimas décadas, el mejoramientogenético de las razas de vacunos de carneha estado centrado en características decrecimiento. Sin embargo, altas tasasreproductivas en un rodeo vacuno están di-rectamente relacionadas con la rentabilidadde la producción de carne. Los caracteresreproduct ivos han mostrado ser lascaracterísticas económicamente másimportantes en muchos sistemas deproducción (Newman et al., 1992; MacNeilet al., 1994; Urioste et al., 1998; Phocas etal., 1998).
¿Se puede seleccionar por fertilidad? Enprincipio, para que una característica sea útilen un esquema de evaluación genética na-cional, debe ser heredable y poco costosade medir y registrar. Normalmente se creeque las características reproductivas tienenbaja heredabilidad, y por esa razón son difí-ciles de mejorar a través de la seleccióndentro de raza (recordemos: la heredabilidadexpresa cuánto de la superioridad o inferio-ridad observada en los padres en una deter-minada característica es dable ser expresa-da en la siguiente generación). Sin embar-go, estudios nacionales y extranjeros sugie-ren que la heredabilidad puede ser mayor delo que se creía para algunas características
1Prof., Ing. Agr., PhD. Catedrático de Mejoramiento Genético Animal. Facultad de Agronomía Universidad de la República.
1.2. SELECCIÓN Y REPRODUCCIÓN EN BOVINOS DE CARNE
Jorge I. Urioste1

12
SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna INIA TREINTA Y TRES
reproductivas. Por lo tanto, rasgos de fertili-dad en vacunos de carne que fueran fácil-mente medibles en la mayoría de las situa-ciones serían muy apropiados para ser in-cluidas en los sistemas nacionales de eva-luación de vacunos de carne.
En la última década, los genetistas austra-lianos (Johnston y Bunter, 1996; Graser etal., 2005) han aplicado con éxito modelosrelativamente simples para la característica“Días hasta el parto”. Esta característicaestá definida como el número de días entrela entrada del toro al servicio y la fecha departo. Esto quiere decir que a mayor núme-ro de días, peor es la habilidad reproductivade la hembra, pues demora en quedar pre-ñada. Los criadores y técnicos australianoshan desarrollado un sistema de registro ade-cuado a su medición, y en principio sólo seusa en casos de monta natural. Cuando unavaca queda vacía se la penaliza con un nú-mero extra de días (21 días más allá de lafecha de parto de la última vaca que pariódentro de la época de parición). Como lasvacas tienen varios partos durante su vida,el seguimiento de la historia reproductiva dela hembra ayuda a tener mayor precisión enla estimación de su capacidad genética parareproducirse.
En Uruguay, el Programa de EvaluaciónGenética de la raza Aberdeen Angus estápermanentemente en desarrollo. Medidas demérito genético (Desvíos Esperados en laProgenie) para características de crecimientoy carcasa son calculados de modo rutina-rio, pero todavía no existen medidas simila-res para la habilidad reproductiva de las hem-bras. En comparación con la situación aus-traliana, la base de datos de Aberdeen Anguses un poco más problemática, pues fue crea-da inicialmente para hacer el seguimientode los productos, pero no de sus madres.Por el momento, la información reproductivade las vacas solo puede ser obtenida a tra-vés de los registros de nacimiento de sushijos, pues la información sobre su compor-tamiento reproductivo no se registraespecíficamente. La situación es similar en
otros países de ganadería semi-extensiva,tales como Sudáfrica o Argentina.
La principal debilidad detectada es que lavaca vacía no queda registrada en el siste-ma. Sin embargo, podemos hacer un segui-miento indirecto (si bien de menor calidad),definiendo la permanencia de la vaca comoel tiempo entre el primero y el último partoregistrado en la base de datos, y asumien-do que quedó vacía cuando un nacimientodentro de ese periodo no es reportado. Enese sentido, el enfoque tradicional de consi-derar la fecha de nacimiento del ternero (el“producto”) se transforma en la “fecha departo” de la vaca (ahora el dato está miradodesde el lado de la hembra).
La obtención de mejoras en la informaciónes particularmente importante en paísescomo el nuestro donde los datos son esca-sos y costosos de obtener. La baja a mediaintensividad de los sistemas de manejo lo-cales impone una restricción sobre el nú-mero de características que se pueden me-dir, así como la frecuencia con la cual pue-den ser medidos. Características que sonfacilmente medibles en la mayoría de las si-tuaciones y a bajo costo serían apropiadaspara su uso en condiciones nacionales.Adicionalmente, en situaciones donde labase de datos es pequeña, la recuperaciónde información perdida a través de supues-tos razonables es vital para una evaluacióngenética.
Existe entonces la necesidad de una mayorcaracterización de caracteres reproductivosque cumplan con estos requisitos. El objeti-vo de este trabajo es a) presentar un marcoconceptual general sobre las posibilidadesde selección en rasgos reproductivos enbovinos de carne; b) Análisis, en la pobla-ción Aberdeen Angus bajo control, de día departo (DP) y éxito al parto (EP) como carac-terísticas biológicas posibles de ser inclui-das en programas de mejoramiento genéticode ganado de carne bajo sistemas extensi-vos o semi-extensivos como los existentesen Uruguay.

13
INIA TREINTA Y TRES SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna
Oportunidades de selección en rasgosreproductivos
El comportamiento reproductivo en vacunosde carne es un carácter complejo, conmuchos subcomponentes, los cualespueden ser separados en diferentes gruposrelativamente fáci les de medir y conheredabilidades más altas que una medidageneral de eficiencia reproductiva. Varios deestos componentes de habilidad reproductivahan sido estudiados por diversos inves-tigadores (Bourdon y Brinks, 1983; Azzamy Nielsen, 1987; Buddenberg et al., 1990;López de Torre y Brinks, 1990; Meyer et al.,1990; Ponzoni, 1992; MacGregor, 1995;MacGregor y Casey, 1999; Gutiérrez et al.,2002), pero todavía no ha aparecido unamedida totalmente satisfactoria para mejorargenéticamente la habilidad repro-ductiva dela vaca.
Rust y Groeneveld (2001) han resumido ydiscutido ventajas y desventajas de diferen-tes componentes, tales como tiempo al pri-mer celo, número de servicios por concep-ción, tasa de preñez, largo de gestación,edad al primer parto, días al parto, fecha departo e intervalo interparto. Algunos ejem-plos de dificultades con las diferentes varia-bles se enumeran a continuación.
Días al primer celo: es difícil de medir encondiciones de campo, implica la observa-ción detallada de todo el rodeo de una ma-nera regular.Número de servicios por concepción: requie-re el registro de cada servicio, raramenteposible en condiciones de monta natural.Tasa de preñez: es un carácter binario, yrequiere detección de preñez en todo el ro-deo.Largo de gestación: requiere observación yregistro del servicio y el parto, tiene pocavariación genética.Edad al primer parto: refleja mayoritariamentedecisiones de manejo, que pueden confun-dirse con dist into mérito genét ico.Adicionalmente, solo representa un compo-
nente inicial en la vida reproductiva de lavaca.Intervalo interparto: sólo existente para va-cas con 2º parto y más; es sólo útil en esta-ciones de parto no restringidas.
Si el apareamiento se produce en el marcode un sistema de registro limitado, a menu-do la única información disponible es si lavaca ha producido un ternero, y su fecha departo. Si la estación de servicio es restringi-da, la fecha de parto también es indicativade la habilidad para una temprana concep-ción en el periodo de servicio (Meyer et al.,1990; Rege y Famula, 1993).
El intervalo interparto ha sido una medidapredominante de comportamientoreproductivo, particularmente en vacunospara leche. El ganado para carne, sin em-bargo, no es apareado todo el año sino enestaciones restringidas de servicio. Se es-pera que aquellas vacas con los intervalosinterparto más cortos sean más fértiles yreproductivamente más eficientes. Sin em-bargo, Bourdon y Brinks (1983) han argu-mentado que esta característica ofrece unamedida sesgada de habilidad reproductivacuando las vacas son servidas durante untiempo restringido. Esto es debido a su aso-ciación negativa con la fecha previa de par-to: vacas con un parto temprano tienen losintervalos interparto siguientes más largos.Selección a favor de intervalos interpartomás cortos podría resultar en una selecciónindirecta para edad más tardía a la puber-tad, pues vacas con los intervalos interpartomás cortos son a menudo aquellas cuyosterneros nacieron más tarde en la estaciónde cría.
Las características reproductivas en gana-do de carne son difíciles de medir, reportare interpretar, y los procedimientos para es-timar el mérito genético de estas caracte-rísticas no son sencillos. La expresión delmérito genético en reproducción está a me-nudo constreñida por el manejo y la nutri-ción, así como un desarrollo inadecuado delos sistemas de registros, todo lo cual hace

14
SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna INIA TREINTA Y TRES
que la estimación de valores genético-aditi-vos sea una tarea difícil. Esto es particular-mente cierto en situaciones pastoriles, don-de la información sobre las vacas de cría esextremadamente limitada. La mayor parte delos animales se reproducirán cuando las con-diciones se encuentren en condiciones próxi-mas a las óptimas, pero en condicionesmenos favorables, solo aquellos con altomérito para su adaptación reproductiva po-drán perpetuarse (Morris, 1980).
Las posibilidades de usar informaciónreproductiva como herramienta de selecciónen programas de mejoramiento genéticopara vacunos de carne son entonces limita-das. Por un lado, es aceptado que lasheredabi l idades para caracteresreproductivos son bajas. A pesar de ello, elresumen de Koots et al., (1994) sugiere quepodrían ser mayores que en ganado leche-ro. Además, los sistemas de registroreproductivo pueden ser afectados por laestructura de edades y el manejo del rodeo.
Información sobre algunas de las caracte-rísticas indicadoras de fertilidad, tales comointervalo interparto o edad al primer parto esobtenida en el marco de sistemas de regis-tro en producción de carne. Sin embargo,un número importante de investigadores(Bourdon y Brinks, 1983; Azzam y Nielsen,1987; Meacham y Notter, 1987; Buddenberget al., 1990; López de Torre y Brinks, 1990;García Paloma et al., 1992) han sugerido eluso de fecha de parto como una medidareproductiva más adecuada, especialmentecuando se practica el apareamiento restrin-gido. Bourdon y Brinks (1983) definieron “fe-cha de parto” como el número de días (a partirde una fecha arbitraria) en el cual la vacatuvo el parto dentro de la estación de partocorrespondiente.
La fecha de parto es fácilmente registrable,y es económicamente importante porque ter-neros más pesados al destete están normal-mente asociados con fechas tempranas departo. Adic ionalmente, puede sersistemáticamente observada en las vacas
paridas y luego reportada cuando los terne-ros son registrados. Conceptualmente, esequivalente a la característica “días al par-to” (Meyer et al., 1990; Johnston y Bunter,1996) o al término “día de parto” utilizadopor Ponzoni (1992).
Día de parto obvia la necesidad de registrarel primer día de servicio, pues se numera apartir del primer parto ocurrido, dentro delmismo rodeo y año. Esto es especialmenteútil en sistemas extensivos o semi-extensi-vos como los existentes en Uruguay, ma-yormente basados en pasturas. Johnston yBunter (1996) demostraron que días al partofue una medida adecuada de habilidadreproductiva para el caso de trabajar congrandes bases de datos de campo, comoes el caso de la situación australiana.
Un aspecto importante en el análisis de Díade Parto es cómo tratar animales que fallanen el parto (registros censurados). La infor-mación sobre vacas falladas debe ser inclui-da en las evaluaciones, para hacer el mejoruso de los datos disponibles para habilidadreproductiva. Una aproximación simple fueusada por Johnston y Bunter (1996). En esteestudio, a las vacas con registros censura-dos (vacas falladas) les fue asignado un va-lor proyectado dentro del mismo grupo con-temporáneo (dichas vacas recibieron todasel mismo valor). Un enfoque alternativo fueaplicado por Donoghue et al., (2004a, b),quienes asumieron una distribución normaltruncada para los registros censurados, ex-trajeron valores al azar de dicha distribucióny obtuvieron un registro para las hembrascensuradas, condicionalmente a la informa-ción disponible. Esto le permite a los datosdeterminar el valor de la característica parahembras censuradas. Forni y Albuquerque(2005), sin embargo, no consideraron a lashembras falladas en su análisis, pues un in-tento de incluirlas, usando una penalización,no mejoró la identificación de diferenciasgenéticas entre animales.
Adic ionalmente, algunas medidasreproductivas (por ejemplo, éxito al parto,

15
INIA TREINTA Y TRES SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna
preñez en vaquillonas, preñez al 2º parto)son típicamente características binarias queno presentan una expresión fenotípica con-tinua, y son registrados como éxito (1) o fra-caso (0). Numerosos trabajos han sido re-portados sobre análisis de datos discretosen el área del mejoramiento genético (e.g.Wright, 1934; Gianola, 1982; Gianola yFoulley, 1983). Según Wright (1934), paraque una característica discontinua se expre-sara, un valor umbral en una escala conti-nua subyacente (escala de susceptibilidad,“liability” en inglés) debía ser cruzado. Estemodelo asume que la variable subyacentees influenciada por efectos ambientales y pormuchos genes que tienen pequeños efec-tos aditivos, razón por la cual se postula unadistribución normal de las “observaciones”.
Estudios realizados en Uruguay
Este capítulo reporta resultados de los si-guientes estudios científicos, los cualesserán referidos por sus numerales RomanosI-IV.
I. Urioste, JI., Chang, YM., Naya, H. andGianola, D. 2007. Genetic variability incalving success in Aberdeen Angus cowsunder extensive recording. Animal 1:1081-1088.
II. Urioste, JI., Misztal, I. and Bertrand, JK.2007a. Fertility traits in spring-calvingAberdeen Angus catt le. 1. Modeldevelopment and genetic parameters.Journal of Animal Science. 85:2854-2860.
III. Urioste, JI., Misztal, I. and Bertrand, JK.2007b. Fertility traits in spring-calvingAberdeen Angus catt le. 2. Modelcomparison. Journal of Animal Science.85:2861-2865.
IV. Urioste, JI., Misztal, I. and Bertrand, JK.2008. Alternative repeatability models forgenetic evaluation of beef fertility data. Aser enviado a la revista científica LivestockScience.
Materiales
El trabajo se realizó en parte en Uruguay(Facultad de Agronomía, Universidad de laRepública) y en parte en Estados Unidos,en las Universidades de Wisconsin yGeorgia, utilizando información del Serviciode Evaluación de Reproductores (SER) deAberdeen Angus. Para cuidar especialmen-te la calidad del análisis, se hizo una selec-ción muy fuerte de datos, donde las vacasdebían tener fecha de nacimiento, el padreidentificado, un primer parto a los 2 o 3 años,no haber sido usadas como donadoras o re-cipientes en transferencia de embriones, ymantener un intervalo interparto razonable(siempre mayor a 280 días), siempre en pri-mavera (solo el 3% de los partos de la basede datos son declarados como de otoño).Solo se tomaron las 3 primeras oportunida-des de parto. El conjunto de datos inicialestaba compuesto por aproximadamente33.000 registros de partos, provenientes de14.000 vacas en 56 rodeos.
En el artículo I, los datos finales consistie-ron en 2.032 registros de vacas teniendo unasegunda oportunidad de parto, nacidas en-tre 1975 y 2000, en 24 rodeos; 1.080 y 947vacas tuvieron una tercera y cuarta oportu-nidad de parto, respectivamente. Un total de444 ancestros machos, incluyendo 409 to-ros con registros de hijas y sus padres yabuelos maternos, fueron incluidos en el ar-chivo de genealogías. En los artículos II, III,y IV, 6.763 registros de 3.442 vacas naci-das de 455 padres, nacidas entre 1975 y2000 en 19 rodeos estuvieron disponiblespara el análisis. El archivo de genealogíastenía 7.748 animales relacionados a la infor-mación recolectada.
Éxito al Parto (EP) fue definido en los estu-dios como una característica binaria: hem-bras que producían un ternero fueron codifi-cadas como uno (éxito), y vacas sin un par-to registrado en un año determinado, peroapareciendo en años posteriores, les fueasignado el valor cero (fracaso) en los valo-res de los años específicos entre dos par-tos identificados.

16
SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna INIA TREINTA Y TRES
Día de Parto (DP) fue definido (artículos II-IV) como el número de días desde el co-mienzo de la estación de parto en un año yrodeo específico hasta la fecha de parto dela vaca. Es una variable continua, fácilmen-te registrable, que asume largo de gestaciónconstante para poder detectar la variaciónen la habilidad reproductiva de las vacas. LaFigura 1 esquematiza la situación de 3 va-cas típicas en un rodeo cualquiera.
Procedimientos estadísticos
En los diferentes estudios se usaron 7 mo-delos estadísticos con diferente grado de so-fisticación. Modelos umbrales (e.g., Gianolay Foulley, 1983) fueron usados en los artí-culos I-IV para el análisis de éxito al partocomo respuesta binaria. El modelo umbralpostula una variable aleatoria, continua ysubyacente, l lamada susceptibi l idad(“liability”), que genera respuestas binariasobservadas (en este caso éxito o fracaso alparto) cuando trasciende un valor umbraldeterminado.
En el artículo I, la variable subyacente éxitoal parto fue expresada como una función li-neal de los efectos del año de nacimientode la vaca, el rodeo y el padre de la vaca.
Figura 1. Ilustración de la variable Día de Parto.
En los artículos II-IV, los tres modelos de-sarrollados incluyeron el efecto de grupocontemporáneo (rodeo x año x grupo demanejo de servicio) en cada una de las tresoportunidades de parto, efectos de edad alparto (3 niveles dentro de cada oportunidadde parto: animales dentro de ± 1 SD poredad, menos de -1 o más de 1 SD), y esta-do fisiológico al servicio (vaca lactando o nolactando). Otras variables explicatorias, ta-les como sexo del ternero gestado y criadofueron inicialmente exploradas, pero luegodescartadas debido a la falta de significancia.Un efecto genético aditivo para cada animal(“modelo animal”) fue ajustado en todos losmodelos, a diferencia del modelo “padre”usado en el Artículo I.
Modelos más simples, como un modelo derepetibilidad para días al parto, usando unapenalización para vacas falladas, ha sidoimplementado con éxito en las evaluacionesgenéticas para ganado de carne en Austra-lia (Graser et al., 2005). En el artículo IV,tres modelos de repetibilidad fueron compa-rados. Todos los modelos incluyeron losefectos de grupo contemporáneo, edad alparto, estado fisiológico al servicio, genéticoaditivo animal, y ambiente permanente.

17
INIA TREINTA Y TRES SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna
Evidencias de variabilidad genética enreproducción
Los principales resultados de los estudiosrealizados se pueden resumir de la siguien-te manera:
• El porcentaje de parición (EP en la po-blación analizada) fue 86.3%, 65.9%, y67.3% para el primer, segundo y tercerparto, respectivamente. Los promediosobtenidos para DP fueron de 36, 59 y 52días para el primero, segundo y tercerparto.
• Las distintas estimaciones de hereda-bilidad para DP variaron entre 0.19 y 0.31,obteniéndose los valores más altos en laprimera oportunidad de parto.
• Las heredabilidades para EP fueron algomás altas, entre 0.27 y 0.43.
• Las correlaciones genéticas entre DPmedidos en los tres primeros partos fuepositiva y de media a alta (alrededor de0.6), indicando que vacas con buena ha-bilidad reproductiva seguirán siendo bue-nas en etapas posteriores de su vida.
• Las correlaciones genéticas entre éxitosal parto en los tres primeros partos fuetambién alta, en el entorno de 0.80.
• Las correlaciones genéticas entre DP y
SP fueron fuertemente negativas (entre -0.54 y -0.91), es decir, favorables: unaselección por menores días de parto con-duciría a un aumento en el suceso al par-to. Este dato es muy importante: quieredecir que si seleccionáramos por meno-res días de parto, indirectamente esta-ríamos mejorando el porcentaje deparición.
Adicionalmente, en el artículo IV se descri-be el cambio genético obtenido a lo largo delos años analizados. El promedio de valoresde cría de DP por año de nacimiento aumen-tó desde 1976 a 1990, reflejando una ten-dencia negativa en la habilidad reproductiva,con un marcado cambio positivo en la se-gunda mitad del periodo considerado (Figu-ra 2).
Discusión general
El desarrollo de esquemas de evaluación ysu aplicación en el mejoramiento genéticoinvolucra varios pasos, tales como la defini-ción de objetivos de selección, elección yaplicación de medidas adecuadas (criteriosde selección), estimación de parámetrosgenéticos, elección de modelos y métodosde análisis, y diseño de procedimientos
Figura 2. Cambio en el valor de cría (VC) promedio para Día de Parto según el año denacimiento de los animales.
VC

18
SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna INIA TREINTA Y TRES
prácticos para la estimación de valores decría y de tendencias genéticas. Finalmente,la selección y uso de animales potencial-mente superiores debe ser llevada a cabo.Algunos de estos pasos han sido tratadosen estos trabajos, en referencia a las carac-terísticas reproductivas; otros, han sido sim-plemente mencionados. Se retoman aquí,para establecer un marco general apropiadode las investigaciones realizadas.
Objetivos y criterios de selección
La definición de objetivos de selección es elpaso inicial en el desarrollo de programasnacionales de mejoramiento genético(Ponzoni y Newman, 1989). En ganado decarne, esta área ha recibido atención en losúltimos años (Newman et al., 1992; Phocaset al., 1998; Urioste et al., 1998; Amer etal., 2001; Kluyts et al., 2003; Fernández-Perea y Alenda Jiménez, 2004; Wolfová yNitter, 2004; Wolfová et al., 2005a, b).
El desarrollo de objetivos de selección su-pone la toma de decisiones de naturalezaeconómica, con poca atención a las conse-cuencias genéticas. En este paso importaidentificar rasgos biológicos que afecten elbenefic io económico de la empresaagropecuaria (ingresos, costos, o ambos).Las consideraciones genéticas se vuelvenrelevantes al momento de la evaluación delos animales como progenitores de la si-guiente generación. Algunos rasgos biológi-cos en el objetivo de selección pueden serdifíciles o costosos de medir, en tanto pue-de haber características que estén altamen-te correlacionadas con los rasgos en el ob-jetivo, pero sin estar necesariamente inclui-dos en él. Este podría ser el caso, si inclu-yéramos DP (criterio de selección) paramejorar EP (objetivo de selección).
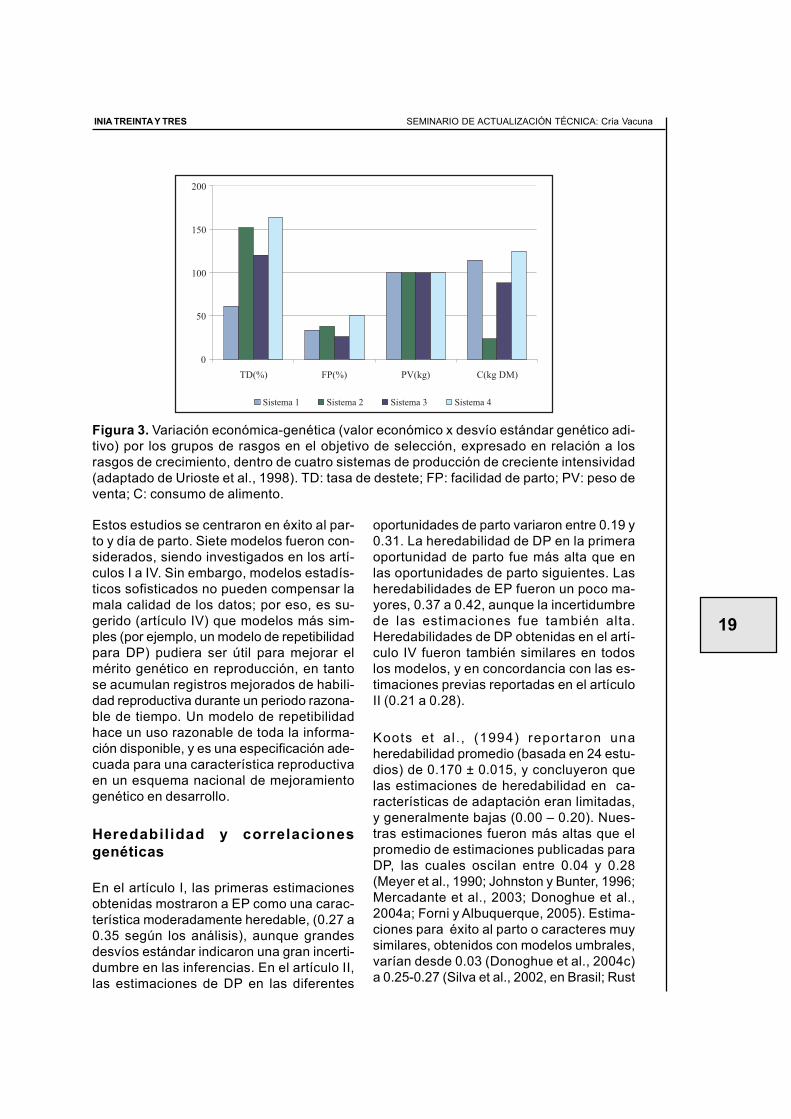
Los rasgos biológicos de interés para la me-jora genética pueden ser groseramente agru-pados en aquellos relacionados a la repro-ducción, el crecimiento, consumo de alimen-to y atributos de carcasa. En Uruguay,Urioste et al., (1998) incluyeron tasa de des-
tete (TD) en los objetivos de selección, ba-sado en su asociación con mayores núme-ros de hijos y vacas de descarte, y tambiéncon mayores costos de alimentación, ma-nejo y comercialización. Los altos valoreseconómicos encontrados para TD (Figura 3)confirmaron que se debe poner un énfasissustancial en la selección para mejorar losrasgos reproductivos en los sistemas uru-guayos de producción. Un examen de losvalores económicos relativos de reproduccióny crecimiento en una serie de estudios endiversos países (Ponzoni y Newman 1989;Newman et al., 1992; Macneil et al., 1994;Phocas et al., 1998; Fernández-Perea yAlenda Jiménez, 2004) también indican quelas tasas reproductivas son al menos tan im-portantes como las características de cre-cimiento desde un punto de vista económico.
Urioste et al., (1998) señalaron la ventajaeconómica de seleccionar por un objetivocompleto (inclusión de todos aquellos ras-gos de importancia económica), comparadocon la selección exclusivamente por pesovivo. Adicionalmente, ellos demostraron elvalor del uso de medidas reproductivas comocriterios de selección. Un índice de selec-ción combinando características de creci-miento y reproducción (circunferenciaescrotal y día de parto) produjo una respues-ta más balanceada en los diferentes gruposde características, con un notable beneficioeconómico. Graser et al., (1994) tambiénconcluyeron que las medidas de fertilidadson valiosos criterios adicionales de selec-ción. Dada la simplicidad relativa de regis-trar variables como circunferencia escrotaly día de parto, su incorporación al sistemade registros ganaderos de Uruguay estáampliamente justificada. Los resultados pre-sentados en el artículo II confirman que laselección por DP más cortos conducirá aaumentos correlacionados en EP.
Modelos estadísticos
Encontrar un modelo apropiado para las ca-racterísticas reproductivas es un componenterelevante de cualquier evaluación genética.
19
INIA TREINTA Y TRES SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna
Figura 3. Variación económica-genética (valor económico x desvío estándar genético adi-tivo) por los grupos de rasgos en el objetivo de selección, expresado en relación a losrasgos de crecimiento, dentro de cuatro sistemas de producción de creciente intensividad(adaptado de Urioste et al., 1998). TD: tasa de destete; FP: facilidad de parto; PV: peso deventa; C: consumo de alimento.
Estos estudios se centraron en éxito al par-to y día de parto. Siete modelos fueron con-siderados, siendo investigados en los artí-culos I a IV. Sin embargo, modelos estadís-ticos sofisticados no pueden compensar lamala calidad de los datos; por eso, es su-gerido (artículo IV) que modelos más sim-ples (por ejemplo, un modelo de repetibilidadpara DP) pudiera ser útil para mejorar elmérito genético en reproducción, en tantose acumulan registros mejorados de habili-dad reproductiva durante un periodo razona-ble de tiempo. Un modelo de repetibilidadhace un uso razonable de toda la informa-ción disponible, y es una especificación ade-cuada para una característica reproductivaen un esquema nacional de mejoramientogenético en desarrollo.
Heredabilidad y correlacionesgenéticas
En el artículo I, las primeras estimacionesobtenidas mostraron a EP como una carac-terística moderadamente heredable, (0.27 a0.35 según los análisis), aunque grandesdesvíos estándar indicaron una gran incerti-dumbre en las inferencias. En el artículo II,las estimaciones de DP en las diferentes
oportunidades de parto variaron entre 0.19 y0.31. La heredabilidad de DP en la primeraoportunidad de parto fue más alta que enlas oportunidades de parto siguientes. Lasheredabilidades de EP fueron un poco ma-yores, 0.37 a 0.42, aunque la incertidumbrede las estimaciones fue también alta.Heredabilidades de DP obtenidas en el artí-culo IV fueron también similares en todoslos modelos, y en concordancia con las es-timaciones previas reportadas en el artículoII (0.21 a 0.28).
Koots et al. , (1994) reportaron unaheredabilidad promedio (basada en 24 estu-dios) de 0.170 ± 0.015, y concluyeron quelas estimaciones de heredabilidad en ca-racterísticas de adaptación eran limitadas,y generalmente bajas (0.00 – 0.20). Nues-tras estimaciones fueron más altas que elpromedio de estimaciones publicadas paraDP, las cuales oscilan entre 0.04 y 0.28(Meyer et al., 1990; Johnston y Bunter, 1996;Mercadante et al., 2003; Donoghue et al.,2004a; Forni y Albuquerque, 2005). Estima-ciones para éxito al parto o caracteres muysimilares, obtenidos con modelos umbrales,varían desde 0.03 (Donoghue et al., 2004c)a 0.25-0.27 (Silva et al., 2002, en Brasil; Rust

20
SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna INIA TREINTA Y TRES
y Groeneveld, 2002, en Sudáfrica). Phocaset al., (2002), usando un modelo padre, re-portaron valores entre 0.07 y 0.16, con gran-des errores estándar. Evans et al., (1999)definieron preñez de vaquillonas como elevento en el cual una vaquillona concibe ypermanece preñada hasta la palpación, dadoque ella haya sido expuesta al servicio. Usan-do datos de 986 vaquillonas Hereford, obtu-vieron una estimación de heredabilidad de0.138 con un error estándar de 0.089. Conmucho mayor número de datos, Eler et al.,(2004) obtuvieron una estimación deheredabilidad de 0.61 (0.10) y 0.68 (0.09)para preñez en vaquillonas.
En el artículo II, se encontraron correlacio-nes genéticas positivas y altas (0.82 a 0.88)entre medidas de DP. Las correlacionesgenéticas entre medidas de EP también fue-ron positivas y de medias a altas (0.56 a0.80). Correlaciones genéticas medias a al-tas y negativas (favorables) entre DP y EP (-0.54 a -0.91) fueron estimadas en este aná-lisis, sugir iendo que el mejoramientogenético por DP (día de parto más tempranodentro de la correspondiente estación departo) conduciría a una mejora en el éxito alparto de las vacas. La asociación negativa(genéticamente favorable) entre DP y EP esun descubrimiento relevante. Esta relaciónpodría ser usada para la implementación delmejoramiento genético en caracteres de fer-tilidad, donde DP podría actuar como carac-terística indicadora de EP. Esta última ca-racterística tiene una expresión fenotípicabinaria, en tanto DP es lineal y continua, ypodría ser fácilmente incorporada dentro delos procedimientos estándar ya desarrolla-dos en el sistema de evaluación genéticade la raza Aberdeen Angus. Es también muyprobable que DP logre un alto grado de com-prensión y aceptación entre los criadores dedicha raza.
Tendencias genéticas
La descripción genética de las poblacionesincluye su cambio a lo largo de periodos detiempo definidos (e.g., MacNeil y Newman,
1994; Mercadante et al., 2002). Para todoslos modelos usados en el artículo IV, el pro-medio de valores de cría de DP por año denacimiento aumentó hasta 1990 (tendencianegativa en la habilidad reproductiva), conun marcado cambio positivo a partir de allí(Figura 2).
No es fácil interpretar estas tendencias. Sepodría argumentar que los criadores no es-taban considerando la habilidad reproductivadentro de sus criterios de selección, y con-secuentemente, los cambios en el tiempode los valores de cría estimados deben serconsecuencia de respuestas correlacionadas.Esos cambios podrían ser reflejo del aumen-to de las importaciones de semen de gana-do de carne desde América del Norte (Esta-dos Unidos y Canadá) en los años 80, épo-ca en la cual se priorizan animales con mé-rito genético sobresaliente en característi-cas de crecimiento.
Adicionalmente, la aparición del primer sis-tema nacional de evaluación genética usan-do técnicas de modelo animal BLUP en losaños 90 puede haber jugado un cierto papelen la conciencia, por parte de los criadores,del tipo de animal necesario para una co-rrecta adaptación a los sistemas locales deproducción. Estas han sido, sin duda, mar-cas decisivas en el camino del cambiogenético de la raza Aberdeen Angus en elUruguay. Este aumento en la capacidad dedefinir el tipo de animal más adaptado a sis-temas pastoriles seguramente pondrá ma-yor énfasis en características biológicas vin-culadas a la reproducción.
¿Es la selección aplicable para rasgosreproductivos?:
Conclusiones
Varias conclusiones pueden ser sacadas apartir de las investigaciones desarrolladas.Además de ser relativamente heredables, DPy EP son baratas y simples de medir. Lainclusión de alguna de ellas en un sistema

21
INIA TREINTA Y TRES SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna
de registros es entonces altamente reco-mendable. La inclusión de DP o EP comocaracteres de fertilidad en una evaluacióngenética, dependiendo de razonesgenéticas, económicas, o cualquiera otra,ha sido recomendada por Ponzoni (1992).Él concluyó que, desde un punto de vistagenético, la diferencia entre usar cualquierade ellos sería pequeña, comparada con elefecto de ignorar completamente la repro-ducción. Una ventaja de usar un modelo paraDP podría ser en el contexto de mayor pro-ducción de por vida (Morris, 1980; GarciaPaloma et al., 1992). Una desventaja del usode EP en evaluaciones genéticas es que lasvacas que paren tardíamente pueden ser in-advertidamente seleccionadas.
En los estudios realizados se usaron mode-los estadísticos con distintos niveles de com-plejidad. El más sencillo de ellos es equiva-lente al actualmente usado en la evaluacióngenética australiana para Días al parto y po-dría implementarse con éxito en el sistemade evaluación del SER. Debería ser usadocon cierto cuidado por un tiempo, en tantose mejora la calidad de los datos y la esti-mación de parámetros genéticos. La elec-ción final de la característica a incluir en laevaluación genética de reproductores debe-ría de todos modos ser influida por conside-raciones de naturaleza no genética, talescomo su facilidad de incorporación al siste-ma de registros de producción o facilidad decomprensión y aceptación por parte de loscriadores. Paralelamente, el sistema de cap-tura de datos tiene que ser pensado desdela óptica de la vaca, llevando un registro com-pleto de su comportamiento reproductivo, in-corporando el registro sistemático de fechasde inseminación o comienzo de servicio,diagnóstico de preñez en vaquillonas y va-cas, tipos de manejos usados en el servicio(inseminación artificial o monta natural), eincluso códigos de refugo de las vacas.
En resumen, la característica económica-mente importante EP parece no ser una op-ción inmediata para evaluación genética porfertilidad, bajo las circunstancias actuales
de calidad de información. Esto podría cam-biar si los registros fueran más precisos ylos modelos genéticos mejorados. La cali-dad de los datos podría ser mejorada, a tra-vés del registro de las fechas de servicio,toros y tipo de servicio usado (inseminaciónartificial o monta natural), estado de preñezen vaquillonas y vacas, fechas de entrada ysalida, resultado final de toda la estación deservicio (incluso si las preñeces finalmenteno se confirman), y códigos de causas deabandono del rodeo de cría. La captura adi-cional de variación genética usando una ca-racterística continua como DP, y a su vezgenéticamente correlacionada con EP, esdeseable. Si estos cambios son adoptadosen la práctica, los criadores tendrán herra-mientas para realizar una selección directasobre los siempre económicamente impor-tantes rasgos de fertilidad. En el futuro, es-timaciones de correlaciones genéticas en-tre DP o EP con datos de crecimiento, ul-trasonido o circunferencia escrotal, actual-mente registrados en el sistema de regis-tros Aberdeen Angus, podría brindar una se-rie de contribuciones decisivas hacia un fu-turo índice de Mérito Total en el ganadoAberdeen Angus de Uruguay.
Agradecimientos
A los dedicados criadores de AberdeenAngus, por permitirme usar sus datos en prode la búsqueda permanente de nueva tecno-logía genética. A mis colegas, ProfesoresDaniel Gianola, Yu-Mei Chang, KeithBertrand e Ignacy Misztal, por recibirmegentilmente y poner a mi disposición todosu conocimiento y capacidad técnica. A losorganizadores de este Taller, por permitirmeaportar elementos para una discusión crucialsobre la mejora de los rodeos de cría a tra-vés de todas las herramientas disponibles.
Referencias bibliográficas
Amer, PR., Simm, G., Keane, MG., Diskin,MG. and Wickham, BW. 2001. Breedingobjectives for beef cattle in Ireland. LivestockProduction Science, 67, 223-239.

22
SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna INIA TREINTA Y TRES
Azzam, SM. and Nielsen, MK. 1987. Geneticparameters for gestation length, birth dateand first breeding date in beef cattle. Journalof Animal Science 64, 348-356.Bourdon, RM. and Brinks, JS. 1983. Calvingdate versus calving interval as a reproductivemeasure in beef cattle. Journal of AnimalScience 57, 1412-1417.Buddenberg, BJ., Brown, CJ. and Brown, AH.1990. Heritability estimates of calving datein Hereford cattle maintained on range undernatural mating. Journal of Animal Science68, 70-74.Donoghue, KA., Rekaya, R. and Bertrand,JK. 2004a. Comparison of methods for han-dling censored records in beef fertility data:simulation study. Journal of Animal Science82, 351-356.Donoghue, KA., Rekaya, R. and Bertrand,JK. 2004b. Comparison of methods for han-dling censored records in beef fertility data:field data. Journal of Animal Science 82,357-361.Donoghue, KA., Rekaya, R., Bertrand, JK.and Misztal, I. 2004c. Threshold-linearanalysis of measures of fertility in artificialinsemination data and days to calving in beefcattle. Journal of Animal Science 82, 987-993.Eler, JP., Silva, JA II V., Evans, JL., Ferraz,JBS., Dias, F. and Golden, BL. 2004. Additivegenetic relat ionships between heiferpregnancy and scrotal circumference inNellore cattle. Journal of Animal Science 82,2519-2527.Evans, JL., Golden, BL., Bourdon, RM. andLong, KL. 1999. Addit ive geneticrelationships between heifer pregnancy andscrotal circumference in Hereford cattle.Journal of Animal Science 77, 2621-2628.Fernández-Perea, MT. and Alenda Jiménez,R. Economic weights for a selection indexin Avileña purebred beef cattle. LivestockProduction Science 89, 223-233.Forni, S. and Albuquerque, LG. 2005.Estimates of genetic correlations betweendays to calving and reproductive and weighttraits in Nelore cattle. Journal of AnimalScience 83, 1511-1515.
Garcia Paloma, JA., Alberio, R. Miquel, MC.,Grondona, MO., Carr i l lo, J. andSchiersmann; G. 1992. Effect of calving dateon lifetime productivity of cows in a wintercalving Aberdeen Angus herd. AnimalProduction 55, 177-184.Gianola, D. and Foulley, JL. 1983. Sireevaluation for ordered categorical data witha threshold model. Genetique, Selection,Evolution 15, 201-223.Gianola, D. 1982. Theory and analysis ofthreshold characters. Journal of AnimalScience 54, 1079-1096.Graser, H-U., Tier, B., Johnston, DJ. andBarwick, SA. 2005. Genetic evaluation forthe beef industry in Australia. AustralianJournal of Experimental Agriculture 45, 913-921.Graser, H-U., Nitter, G. and Barwick, SA.1994. Evaluation of advanced industrybreeding schemes for Australian beef cattle.II. Selection on combinations of growth,reproduction and carcass criteria. AustralianJournal of Agricultural Research 45, 1657-1669.Gutiérrez, JP., Alvarez, I., Fernández, I.,Royo, LJ., Díez, J., and Goyache, F. 2002.Genetic relationships between calving date,calving interval, age at first calving and typetraits in beef cattle. Livestock ProductionScience 78, 215-222.Johnston, DJ. and Bunter, KL. 1996. Daysto calving in Angus cattle: genetic andenvironmental effects, and covariances withother traits. Livestock Production Science45, 13-22.Kluyts, JF., Neser, FWC. and Bradfields, MJ.2003. Development of breeding objectives forbeef cattle breeding: derivation of economicvalues. South African Journal of AnimalScience, 33, 142-158.Koots, KR., Gibson, JP., Smith, C. andWilton, JW. 1994. Analyses of publishedgenetic parameter estimates for beefproduction traits. 1. Heritability. AnimalBreeding Abstracts 62, 309-338.Lopez de Torre, G. and Brinks, JS. 1990.Some alternatives to calving date and intervalas measures of fertility in beef cattle. Journal

23
INIA TREINTA Y TRES SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna
of Animal Science 68, 2650-2657.MacGregor, RG. and Casey, NH. 1999.Evaluation of calving interval and calving dateas measures of reproductive performance ina beef herd. Livestock Production Science57, 181-191.MacGregor, RG. 1995. Evaluation of calvingdate and calving interval as measures ofreproductive efficiency in beef cows. Journalof the South African Veterinary Association66, 235-238.MacNeil , MD. and Newman, S. 1994.Genetic analysis of calving date in Miles CityLine 1 Hereford cattle. Journal of AnimalScience 72, 3073-3079.MacNeil, MD., Newman, S., and Steward-Smith, J. 1994. Relative economic values forCanadian beef production using specializedsire and dam lines. Canadian Journal ofAnimal Science 74, 411-417.Meacham, NS. and Notter, DR., 1987.Heritability estimates for calving date inSimmental cattle. Journal of Animal Science64, 701-705.Mercadante MEZ., Packer IU., Razook, AG.,Cyrillo, JNSG. and Figueiredo, LA. 2003.Direct and correlated responses to selectingfor yearling weight on reproductive performan-ce of Nelore cows. Journal of AnimalScience 81, 376-384.Mercadante, MEZ., Packer IU., Razook,AG., Melo, CMR., Cyri l lo, JNSG. andFigueiredo, LA. 2002. Dias ao parto deFêmeas Nelore de um Experimento deSeleção para crescimento. II – Modelo deregressão aleatória. Revista Brasileira deZootecnia 31, 1726-1733.Meyer, K., Hammond, K., Parnell, PF.,Mackinnon, MJ. and Sivarajasingam, S.1990. Est imates of her i tabi l i ty andrepeatabil ity for reproductive traits inAustralian beef cattle. Livestock ProductionScience 25, 15-30.MGAP-DIEA. 2003. La ganadería en Uru-guay. Contribución a su conocimiento. http:// www.mgap.gub.uy/Diea/Rubros/Ganaderia/Ganaderia_Junio2003.pdf. Acceso: 3 de No-viembre de 2007.Morris, CA. 1980. A review of relationships
between aspects of reproduction in beefheifers and their lifetime production. 2.Associations with relative calving date andwith dystocia. Animal Breeding Abstracts 48,753-767.Newman, S., Morris, CA., Baker, RL. andNicoll, GB. 1992. Genetic improvement ofbeef cattle in New Zealand: breedingobjectives. Livestock Production Science 32,111-130.Phocas, F., and Laloe, D. 2003. Evaluationmodels and genetic parameters for calvingdifficulty in beef cattle. Journal of AnimalScience 81, 933-938.Phocas, F., Vinet, A. and Renand, G. 2002.Genetic variability of reproductive traits inCharolais cows. Proceedings of the 7th WorldCongress on Genetics Applied to LivestockProduction, Montpellier, August 19-23, 2002,29, 513-516.Phocas, F., Bloch, C., Chapelle, P. ,Bécherel, F., Renand, G. and Ménissier, F.1998. Developing a breeding objective for aFrench purebred beef cattle selectionprogramme. Livestock Production Science57, 49-65.Ponzoni, RW. 1992. Which trait for geneticimprovement of beef cattle reproduction:calving rate or calving day? Journal of Ani-mal Breeding and Genetics 109, 119-128.Ponzoni, RW. and Newman, S. 1989.Developing breeding objectives for Australianbeef cattle production. Animal Production 49,35-47.Rege, JEO., and Famula, TR. 1993. Factorsaffecting calving date and its relationship withproduction traits of Hereford dams. AnimalProduction 57, 385-395.Rust, T. and Groeneveld, E. 2002. Variancecomponent estimation of female fertility traitsin two indigenous and two European beefcattle breeds of South Africa. South AfricanJournal of Animal Science 32, 23-29.Rust, T. and Groeneveld, E. 2001. Variancecomponent estimation on female fertilitytraits in beef cattle. South African Journal ofAnimal Science 31, 131-141.Silva, JAII de V., Van Melis, MH., Eler, JP.,Ferraz, JBS. and Oliveira, HN. 2002.

24
SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna INIA TREINTA Y TRES
Heritability for subsequent rebreeding inNelore cows estimated with Bayesianinference. Proceedings of the 7th WorldCongress on Genetics Applied to LivestockProduction, Montpellier, August 19-23, 2002,29, 665-668.Urioste, JI., Ponzoni, RW., Aguirrezabala,MA., Rovere, G. and Saavedra, D. 1998.Breeding object ives for pasture-fedUruguayan beef cattle. Journal of AnimalBreeding and Genetics 115, 357-373.Wolfová, M., Wolf, J., Pribyl, J. Zahrádková,R. and Kica, J. 2005 Breeding objectives forbeef cattle used in different productionsystems: 1. Model development. LivestockProduction Science 95, 201-215.
Wolfová, M., Wolf, J., Zahrádková, R., Pribyl,J., Dano, J., Krupa, E. and Kica, J. 2005,Breeding objectives for beef cattle used indifferent production systems. 2. Modelapplication to production systems with theCharolais breed. Livestock ProductionScience 95, 217-230.Wolfová , M. and Nitter, G. 2004. Relativeeconomic weights of maternal versus directtraits in breeding schemes. LivestockProduction Science 88, 117-127.Wright, S. 1934. An analysis of variability innumber of digits in an inbred strain of guineapigs. Genetics 19, 506-536.

25
INIA TREINTA Y TRES SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna
Introducción
En la mejora genética animal existen dosherramientas a ser utilizadas para lograrcambios perdurables a largo plazo: selec-ción entre y dentro de razas y cruzamientosinterraciales. En Uruguay, tradicionalmentela cría se ha realizado utilizando a la razaHereford y mejorándola por medio de selec-ción. Esta raza, de origen británico, se cla-sifica dentro de las llamadas líneas mater-nas, dados sus menores requerimientos demantenimiento, así como su habilidad endestetar terneros moderadamente pesados.No obstante, los resultados obtenidos en lacría no son satisfactorios y su estancamientoen un 63% de destete revela ineficienciasen el proceso de cría. Las opciones frecuen-temente ofrecidas para paliar esta situaciónse centran en la mejora en la oferta y cali-dad del forraje (con pasturas sembradas omejoradas) o en su utilización, testada me-diante el estado corporal de las vacas y laaltura de la pastura (Soca et al., 1992). Otrasalternativas como destetes temporarios oprecoces (Simeone, 2000; Quintans, 2007),requieren medidas de manejo adicionalespor lo que muchas veces son resistidas porel productor. En contraste con la lentitud enlos cambios que se logran usando la tradi-cional selección de las razas puras, el usode la diversidad genética combinada permi-te la aparición de cambios rápidamente, es-pecialmente en características de bajaheredabilidad como son las inherentes a lareproducción.
En este contexto, los cruzamientos surgencomo una alternativa capaz de mejorar loscaracteres que menos responden a los pro-cesos de selección artificial, explotando el
vigor híbrido y la complementariedad gene-rados por la interacción de los genes prove-nientes de razas de distintos orígenes. Exis-te un gran número de razas bovinas,biológicamente diferentes, que resultan endesempeños variables en las posibles situa-ciones ambientales de nuestro país. Jenkinsy Ferrel (1999) demostraron que dentro deuna misma raza dependiendo del ambienteal cual son expuestos los animales, la efi-ciencia de su producción puede variar, des-tinando mayor o menor proporción de la ener-gía consumida a funciones de mantenimientoo de producción.
Asimismo, tanto razas como cruzas son sen-sibles a las variaciones ambientales, o di-cho de otro modo son afectadas por lainteracción genotipo x ambiente y en espe-cial en caracteres reproductivos debido a labaja influencia de los efectos aditivos parasu expresión. En función de estos antece-dentes, en este trabajo se presentan resul-tados obtenidos a nivel nacional y extranje-ro en variables vinculadas a la reproducciónen diferentes recursos genéticos y escena-rios productivos.
Eficiencia reproductiva en razas puras
La gran mayoría de los trabajos realizadospara evaluar eficiencia reproductiva de razasbovinas carniceras y cruzamientos, se de-sarrollaron en las décadas del ’70 y ’80 enEstados Unidos. En el USDA-MARC (MeatAnimal Research Center) en Nebraska (Es-tados Unidos) se evaluaron en su forma pura,9 razas que diferían en su potencial genéticopara peso a la madurez (cuando alcanza25% de grasa corporal), producción de le-che observada al momento del pico, ganancia
1 Ing. Agr., DSc. Mejoramiento Genético Animal - Depto. Prod. Animal y Pasturas - EEMAC, Facultad de Agronomía.Universidad de la República.
2 Bach. Estudiante en Tesis - Facultad de Agronomía.
1.3. RECURSOS GENÉTICOS Y AMBIENTES DE PRODUCCIÓN EN LACRÍA VACUNA
Ana Carolina Espasandin1 , Marcelo Ciria Ruggiero2
26
SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna INIA TREINTA Y TRES
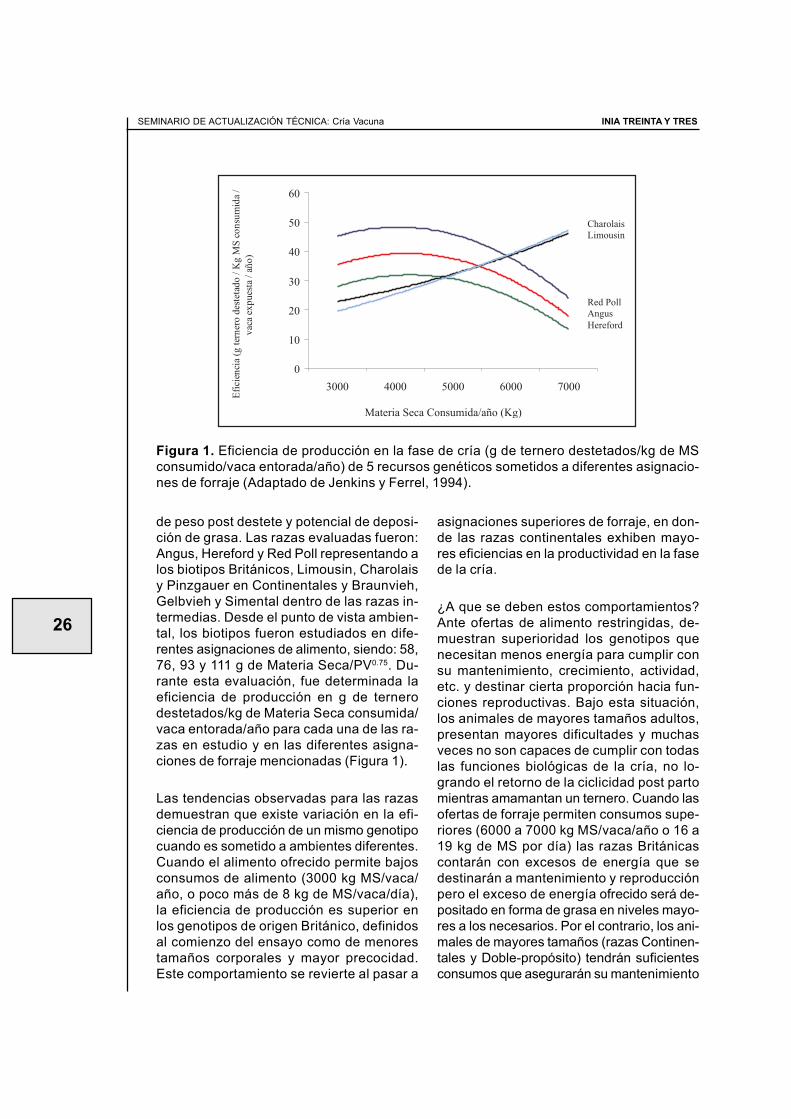
de peso post destete y potencial de deposi-ción de grasa. Las razas evaluadas fueron:Angus, Hereford y Red Poll representando alos biotipos Británicos, Limousin, Charolaisy Pinzgauer en Continentales y Braunvieh,Gelbvieh y Simental dentro de las razas in-termedias. Desde el punto de vista ambien-tal, los biotipos fueron estudiados en dife-rentes asignaciones de alimento, siendo: 58,76, 93 y 111 g de Materia Seca/PV0.75. Du-rante esta evaluación, fue determinada laeficiencia de producción en g de ternerodestetados/kg de Materia Seca consumida/vaca entorada/año para cada una de las ra-zas en estudio y en las diferentes asigna-ciones de forraje mencionadas (Figura 1).
Las tendencias observadas para las razasdemuestran que existe variación en la efi-ciencia de producción de un mismo genotipocuando es sometido a ambientes diferentes.Cuando el alimento ofrecido permite bajosconsumos de alimento (3000 kg MS/vaca/año, o poco más de 8 kg de MS/vaca/día),la eficiencia de producción es superior enlos genotipos de origen Británico, definidosal comienzo del ensayo como de menorestamaños corporales y mayor precocidad.Este comportamiento se revierte al pasar a
Figura 1. Eficiencia de producción en la fase de cría (g de ternero destetados/kg de MSconsumido/vaca entorada/año) de 5 recursos genéticos sometidos a diferentes asignacio-nes de forraje (Adaptado de Jenkins y Ferrel, 1994).
asignaciones superiores de forraje, en don-de las razas continentales exhiben mayo-res eficiencias en la productividad en la fasede la cría.
¿A que se deben estos comportamientos?Ante ofertas de alimento restringidas, de-muestran superioridad los genotipos quenecesitan menos energía para cumplir consu mantenimiento, crecimiento, actividad,etc. y destinar cierta proporción hacia fun-ciones reproductivas. Bajo esta situación,los animales de mayores tamaños adultos,presentan mayores dificultades y muchasveces no son capaces de cumplir con todaslas funciones biológicas de la cría, no lo-grando el retorno de la ciclicidad post partomientras amamantan un ternero. Cuando lasofertas de forraje permiten consumos supe-riores (6000 a 7000 kg MS/vaca/año o 16 a19 kg de MS por día) las razas Británicascontarán con excesos de energía que sedestinarán a mantenimiento y reproducciónpero el exceso de energía ofrecido será de-positado en forma de grasa en niveles mayo-res a los necesarios. Por el contrario, los ani-males de mayores tamaños (razas Continen-tales y Doble-propósito) tendrán suficientesconsumos que asegurarán su mantenimiento

27
INIA TREINTA Y TRES SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna
y reproducción, logrando destetar ternerosmucho más pesados que las razas peque-ñas.
Las variaciones diferenciales en la perfor-mance de las distintas razas dejan en evi-dencia el concepto de interacción genotipox ambiente en las situaciones evaluadas.Trabajos delineados experimentalmente paraestudiar la existencia de interaccionesgenotipo x ambiente fueron dirigidos porKoger et al., (1962; 1975) y Peackock et al.,(1971). En la Estación Experimental delUSDA en Miles City, Montana y en la Uni-versidad de Florida fueron desarrolladas lí-neas de la raza Hereford seleccionadas paraaltas tasas de crecimiento en ambos am-bientes. Con el fin de conocer los límitesbiológicos en la reproducción ante variacio-nes ambientales, se intercambiaron mues-tras aleatorias de machos y hembras de cadalocalidad y se midieron sus desempeños.Las hembras de ambos grupos generarondescendencia evaluándose su posterior vidareproductiva. Los resultados se presentan enel Cuadro 1.
Las diferencias entre las líneas (Local - In-troducida) resultaron significativas (6.7 y6.1% para tasas de preñez y de destete,respectivamente). Las mismas variables fue-ron estudiadas en las hembras nacidas delas vacas transferidas con el fin de removerposibles efectos residuales del ambienteanterior. Nuevamente, las diferencias entrelas líneas locales y las introducidas revela-ron significancia estadística y aún de mayormagnitud que en el caso anterior (11.6 y12.5%, respectivamente). Con estos resul-tados los autores concluyen que existeinteracción genotipo x ambiente dentro de
Cuadro 1. Performance reproductiva de líneas Hereford seleccionadas en Montana y Florida(Koger et al.,1979)
Línea y Ambiente Tasa de Preñez % Tasa de Destete %Montana en Montana 81.6 ± 1.3 73.7 ± 1.3Florida en Montana 82.8 ± 1.6 76.1 ± 1.7Montana en Florida 72.0 ± 1.3 65.0 ± 1.4Florida en Florida 86.5 ± 1.7 79.7 ± 1.8
una misma raza cuando ésta es seleccio-nada en un ambiente específico y que lamisma continúa en las sucesivas generacio-nes.
Utilizando mellizos idénticos y fraternos enla raza Hereford, en la Universidad deWisconsin (Madison) durante el periodo1953-1987 se estudió la performancereproductiva y la producción del leche, ade-más del consumo y la eficiencia de uso delalimento. Kress et al., (1971) reportaron queen ambientes de alta y baja energía en ladieta (aproximadamente 40 NDT de separa-ción entre ambas) no se observaroninteracciones genotipo x ambiente para loscaracteres de crecimiento y eficiencia deconversión. Algunas medidas reproductivascomo edad a primer estro, edad a primeraconcepción y primer parto, mostraron ten-dencias a la interacción, con valores de 14.3a 11% de la variación total y se observaronevidentes interacciones en la producción deleche. No obstante, los autores señalan queel bajo número de pares de mellizos con quecontaron, cuestiona los resultados obteni-dos. Los resultados de estos trabajos nospermiten visualizar que existen recursos masy menos eficientes para cada ambiente deproducción, y que este concepto no es sóloválido cuando se trata de comparacionesentre razas diferentes sino dentro de razas,en las diversas líneas generadas, tantascomo ambientes posibles haya.
En base en estos conceptos, es lógico pen-sar que la mejor vaca debería ser aquellacapaz de proveernos la mayor productividadcon las menores exigencias, y con los me-nores costos atribuidos a sus requerimien-

28
SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna INIA TREINTA Y TRES
tos de mantenimiento. Por tanto parece ob-vio que seleccionar por razas o líneas máseficientes (para cada ambiente) nos llevaríaa resultados exitosos en las empresascriadoras. Afortunadamente, los trabajosrealizados en esta línea indican que losrequerimientos de mantenimiento en las va-cas de cría presentan var iabi l idadesgenéticas medias a altas, sugiriendo así lapertinencia de la selección por mayoreseficiencias biológicas. Sin embargo, losmétodos hasta el momento probados paradeterminar los requerimientos individuales demantenimiento, no sólo son dificultosos sinoque demandan altos costos en su instrumen-tación.
Líneas nuevas de investigación visualizan al“residual feed intake” o consumo residual dealimento como un posible indicador nuevoen eficiencia. El consumo residual de ali-mento (definido como la diferencia entre elconsumo observado menos el consumo es-timado en base a las exigencias de mante-nimiento y crecimiento en un individuo) pa-recería ser un buen estimador de la eficien-cia animal (Archer et al., 2002). La selec-ción por CRA identificaría a animales de me-nores consumos y menores exigencias demantenimiento, sin alterar el peso adulto nila ganancia de peso (Koch et al., 1963,Basarab et al., 2003). Tanto el consumo to-tal como el consumo residual han sido re-portados con heredabilidades medias a al-tas (0.16 a 0.39) en las razas Hereford yAngus, respect ivamente, así comogenéticamente relacionados con la eficien-cia de conversión con una correlacióngenética de 0.31 (Herd y Bishop, 2000; Arthuret al., 2001).
Por otro lado, evidencias recientes (Mooreet al., 2003, Nkrumah et al., 2007) han de-mostrado que las concentraciones en san-gre del factor de crecimiento similar a lainsulina tipo -I (IGF-I) y de leptina estaríanasociadas con características productivascomo crecimiento, características y calidadde carcasa y consumo residual. Asimismo,se han establecido asociaciones entre dife-
rentes polimorfismos en el gen de leptina yen su promotor (Nkrumah et al., 2005;Schenkel et al., 2004) con característicasde carcasa, calidad de carne y consumoresidual.
Si bien no deberíamos abandonar la búsque-da de métodos más sencillos y practicablespara el estudio de la eficiencia de uso de laenergía, estos hallazgos nos explican porque la selección tradicional no es por elmomento una práctica disponible para estacaracterística. Surge entonces la alternati-va de cruzamientos entre razas diferentespara la búsqueda de una mayor eficienciaen la producción en la fase de cría.
Eficiencia reproductiva en animalescruza
Como se mencionara anteriormente, sabe-mos que los caracteres reproductivos pre-sentan heredabilidades de bajas a muy ba-jas. Recordando que la heredabilidad es laproporción de la varianza total de una carac-terística (Aditiva + Dominancia + Epistasis+ Ambiental) en una población debida ex-clusivamente a la variación genética aditiva(óa
2 / óa2 + ód
2 + ó i2 + óe
2). Por lo tanto, unaheredabilidad baja significa que el carácterserá en gran medida dependiente de efec-tos genéticos no aditivos así como ambien-tales.
En una revisión bibliográfica realizada porKoots et al., (1994) enriquecida por unmetanálisis integrando resultados experi-mentales en heredabilidades y correlacionesgenéticas provenientes de muchos trabajos,se observaron magnitudes de heredabilidadde 0.01 a 0.17 para todos los caracteresreproductivos estudiados. A excepción de laedad a la pubertad que presentó altaheredabilidad (0.48), las tasas de concep-ción, intervalos entre partos o fechas de par-to, presentaron heredabilidades de 0.10, 0.06y 0.08, respectivamente.

29
INIA TREINTA Y TRES SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna
Debemos agregar a esta situación la esca-sez de caracteres reproductivos evaluadosgenéticamente en el mundo. Es conocido elpor qué de esto, radicando en la dificultadde medición de los mismos, asociado a lostiempos que se requieren para el registro dealgunas variables (ej. vida productiva,“Stayability”, día de parto, entre otras). Con-forme mencionábamos en secciones ante-riores, las herramientas genéticas capacesde promover los efectos genéticos no aditi-vos son los cruzamientos interraciales. Lasinterferencias producidas entre y dentro dealelos provenientes de razas diferentes seexpresan fenotípicamente en el conocidoefecto de Heterosis. En su definición, laheterosis es la diferencia existente entre laperformance de los individuos cruza F1 y laperformance de individuos de las razas pu-ras paternas del cruzamiento, criadoscontemporáneamente y en condiciones am-bientales semejantes (F1-Media de razaspuras). Su valor puede expresarse en térmi-nos absolutos (en la unidad en que se ex-presa la característica), o en términos rela-tivos (como porcentaje de las razas puras).Los estudios realizados en el MARC tam-bién incluyeron diversos cruzamientos en-tre razas productoras de carne. Vale aclararque las respuestas obtenidas permiten co-nocer el desempeño de las diferentes com-binaciones, no así determinar valores deheterosis debido a los delineamientos ex-perimentales definidos. Vacas de las razasHereford y Angus fueron apareadas con to-ros de razas Británicas (Hereford, Angus yRed Pol entre otras), Continentales
Cuadro 2. Desempeño durante la fase de cría en hijas de madres Angus y Hereford contoros de razas Británicas, Continentales y Cebuinas (Koch et al., 1994)
Genotipo Largo de Peso al Partos Peso al Peso a la Edad a ladel padre gestación nacer distócicos destete pubertad pubertad
(días) (kg) % (kg) (kg)* (días)*Británico 284.5 36 3.5 194 273 347
Continental 288 40 13 205 315 384Cebuino 293 39 8 201 307 422
Madres AA 285 37 7 206 288 351Madres HH 288 39 10 192 291 379
*De las hembras cruza
(Limousin, Charolais, Chianina), Cebuinos(Brahman, Sahiwal), así como de doble pro-pósito. El largo de gestación, peso al nacer,la incidencia de partos distócicos, el pesoal destete, y la edad y peso con que alcan-zaron la pubertad estas combinaciones ra-ciales, se presentan en la Cuadro 2. Asimis-mo, se agregan los desempeños de madresHereford y Angus por separado (independien-temente de la raza del padre utilizada).
En el estudio de Koch et al., (1994), los ani-males de todas las razas fueron sometidosa condiciones ambientales semejantes, condietas cuya oferta energética fue moderada.Las hembras hijas de toros con mayorestamaños adultos (Continentales y Cebuinos)alcanzaron la pubertad a edades y pesosmayores a las hijas de toros Hereford yAngus (en promedio necesitaron 38 kg másde peso y 56 días (casi la duración de 3 ci-clos) más para alcanzar la pubertad en com-paración con las cruzas derivadas dentro delas razas Británicas. Las cruzas con razasíndicas, en este caso Brahman y Sahiwal,fueron las menos precoces en alcanzar lapubertad, excediendo su edad en 38 díasrespecto a las Continentales. Koch et al.,(1994) adjudican estos resultados a presio-nes de selección diferenciales a las que hanestado sujetas las ramas Bos Taurus y BosIndicus. Sin embargo, la raza índica Nelore,luego de su introducción y selección (aun-que mucho más reciente) en Brasil, actual-mente se destaca por su precocidad sexual,siendo capaz de alcanzar la pubertad a los14 meses de edad (Silva et al., 2003).
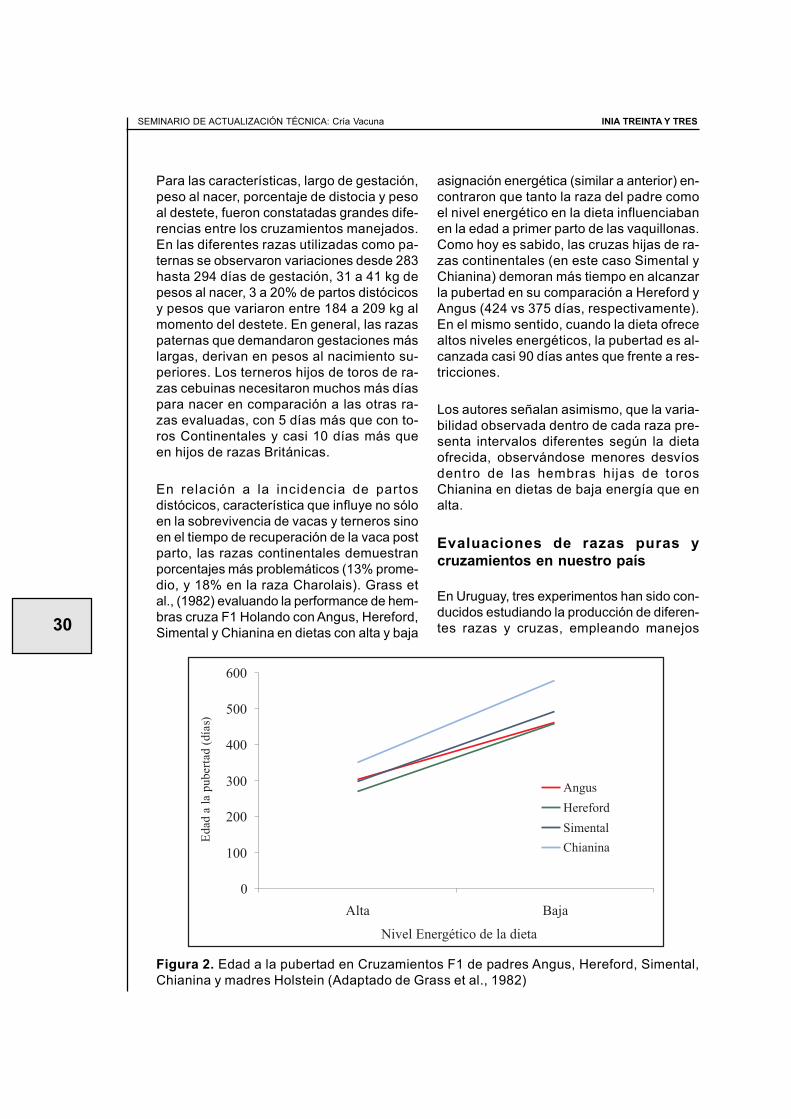
30
SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna INIA TREINTA Y TRES
Para las características, largo de gestación,peso al nacer, porcentaje de distocia y pesoal destete, fueron constatadas grandes dife-rencias entre los cruzamientos manejados.En las diferentes razas utilizadas como pa-ternas se observaron variaciones desde 283hasta 294 días de gestación, 31 a 41 kg depesos al nacer, 3 a 20% de partos distócicosy pesos que variaron entre 184 a 209 kg almomento del destete. En general, las razaspaternas que demandaron gestaciones máslargas, derivan en pesos al nacimiento su-periores. Los terneros hijos de toros de ra-zas cebuinas necesitaron muchos más díaspara nacer en comparación a las otras ra-zas evaluadas, con 5 días más que con to-ros Continentales y casi 10 días más queen hijos de razas Británicas.
En relación a la incidencia de partosdistócicos, característica que influye no sóloen la sobrevivencia de vacas y terneros sinoen el tiempo de recuperación de la vaca postparto, las razas continentales demuestranporcentajes más problemáticos (13% prome-dio, y 18% en la raza Charolais). Grass etal., (1982) evaluando la performance de hem-bras cruza F1 Holando con Angus, Hereford,Simental y Chianina en dietas con alta y baja
Figura 2. Edad a la pubertad en Cruzamientos F1 de padres Angus, Hereford, Simental,Chianina y madres Holstein (Adaptado de Grass et al., 1982)
asignación energética (similar a anterior) en-contraron que tanto la raza del padre comoel nivel energético en la dieta influenciabanen la edad a primer parto de las vaquillonas.Como hoy es sabido, las cruzas hijas de ra-zas continentales (en este caso Simental yChianina) demoran más tiempo en alcanzarla pubertad en su comparación a Hereford yAngus (424 vs 375 días, respectivamente).En el mismo sentido, cuando la dieta ofrecealtos niveles energéticos, la pubertad es al-canzada casi 90 días antes que frente a res-tricciones.
Los autores señalan asimismo, que la varia-bilidad observada dentro de cada raza pre-senta intervalos diferentes según la dietaofrecida, observándose menores desvíosdentro de las hembras hijas de torosChianina en dietas de baja energía que enalta.
Evaluaciones de razas puras ycruzamientos en nuestro país
En Uruguay, tres experimentos han sido con-ducidos estudiando la producción de diferen-tes razas y cruzas, empleando manejos
31
INIA TREINTA Y TRES SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna
extensivos bajo pastoreo de campo naturaly eventualmente el uso de mejoramientos encobertura. En orden cronológico, los prime-ros estudios comenzaron en la Estación Ex-perimental del Norte perteneciente al INIA-Tacuarembó y fueron conducidos por el Ing.Agr. Oscar Pittaluga a partir de 1979 mo-mento en el que se permitió el ingreso demateriales genéticos de origen cebuino a Uru-guay, hasta entonces prohibido. La inclusiónde estas razas en sistemas del noroeste delpaís (suelos arenosos de Rivera yTacuarembó) se fundamentó en el logro demayores respuestas productivas basadas enla mayor adaptación de los genotiposcebuinos a las condiciones climáticas (ca-lor) y pastoriles (forrajes más toscos) domi-nantes en esta región del país. Los resulta-dos de estos trabajos durante la fase de críase resumieron en varias publicaciones(Pittaluga et al., 1996, 2005).
En la década del ’90 la Facultad de Agrono-mía en convenio con la Caja Notarial de Ju-bilaciones y Pensiones instaló dos sistemasde cruzamientos diseñados por el Ing. Agr.Diego Gimeno y publicados por Gimeno etal., (2002) y Espasandin et al., (2006). Elprimero de ellos, de 10 años de duracióncomenzó cruzando vientres de la raza
Hereford con las razas Angus, Salers yNelore y las progenies generadas se estu-diaron en su vida productiva. Así, fueron es-tudiados machos y hembras Hereford puros,sus cruzas F1, F2 y retrocruzas a lo largode todo el ciclo de producción (desde el na-cimiento hasta la faena). Estos trabajos sereal izaron en los departamentos dePaysandú en las fases de cría y recría y enSoriano durante la fase de engorde de losanimales.Paralelamente, en el año ’94, en la Esta-ción Experimental Bernardo Rosengurtt dela Facultad de Agronomía (ex Bañado deMedina) en el departamento de Cerro Largo,comenzó un experimento de cruzamientosentre las razas Hereford y Angus, bajo undiseño dialélico también de 10 años de du-ración y evaluando todo el ciclo de produc-ción de carne vacuna. Durante la fase de críafueron evaluadas las variables largo de ges-tación, pesos al nacer y destete, porcenta-jes de parición en vacas y vaquillonas de 1er
y 2do entore, facilidad de ocurrencia del pri-mer parto, así como indicadores globales dela cría vacuna. Los resultados para la varia-ble largo de gestación, en las diferentes cru-zas generadas en ambos experimentos (CajaNotarial y Bañado de Medina) son presenta-dos en la Figura 3.
Figura 3. Largo de Gestación en diferentes razas y cruzas en Uruguay (Adaptado de Gimenoet al., 2002 y Espasandin et al., 2006)
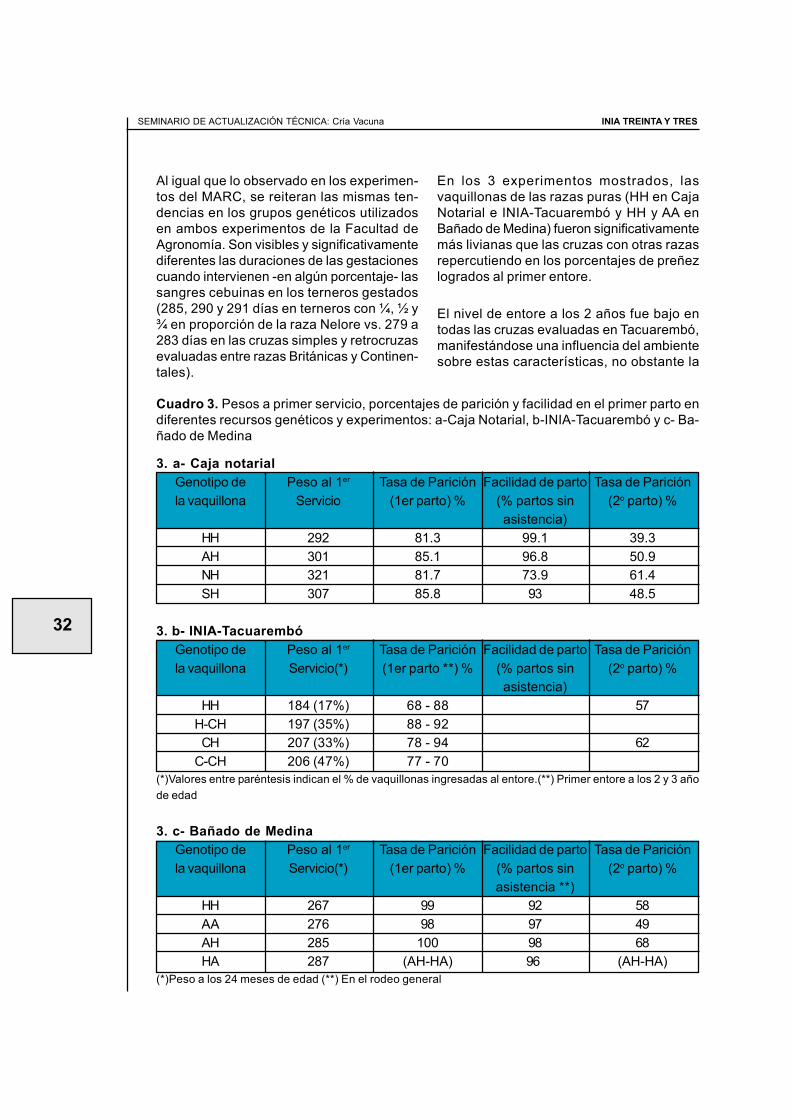
32
SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna INIA TREINTA Y TRES
Al igual que lo observado en los experimen-tos del MARC, se reiteran las mismas ten-dencias en los grupos genéticos utilizadosen ambos experimentos de la Facultad deAgronomía. Son visibles y significativamentediferentes las duraciones de las gestacionescuando intervienen -en algún porcentaje- lassangres cebuinas en los terneros gestados(285, 290 y 291 días en terneros con ¼, ½ y¾ en proporción de la raza Nelore vs. 279 a283 días en las cruzas simples y retrocruzasevaluadas entre razas Británicas y Continen-tales).
Cuadro 3. Pesos a primer servicio, porcentajes de parición y facilidad en el primer parto endiferentes recursos genéticos y experimentos: a-Caja Notarial, b-INIA-Tacuarembó y c- Ba-ñado de Medina
En los 3 experimentos mostrados, lasvaquillonas de las razas puras (HH en CajaNotarial e INIA-Tacuarembó y HH y AA enBañado de Medina) fueron significativamentemás livianas que las cruzas con otras razasrepercutiendo en los porcentajes de preñezlogrados al primer entore.
El nivel de entore a los 2 años fue bajo entodas las cruzas evaluadas en Tacuarembó,manifestándose una influencia del ambientesobre estas características, no obstante la
3. a- Caja notarialGenotipo de Peso al 1er Tasa de Parición Facilidad de parto Tasa de Pariciónla vaquillona Servicio (1er parto) % (% partos sin (2o parto) %
asistencia)HH 292 81.3 99.1 39.3AH 301 85.1 96.8 50.9NH 321 81.7 73.9 61.4SH 307 85.8 93 48.5
3. b- INIA-TacuarembóGenotipo de Peso al 1er Tasa de Parición Facilidad de parto Tasa de Pariciónla vaquillona Servicio(*) (1er parto **) % (% partos sin (2o parto) %
asistencia)HH 184 (17%) 68 - 88 57
H-CH 197 (35%) 88 - 92CH 207 (33%) 78 - 94 62
C-CH 206 (47%) 77 - 70(*)Valores entre paréntesis indican el % de vaquillonas ingresadas al entore.(**) Primer entore a los 2 y 3 añode edad
3. c- Bañado de MedinaGenotipo de Peso al 1er Tasa de Parición Facilidad de parto Tasa de Pariciónla vaquillona Servicio(*) (1er parto) % (% partos sin (2o parto) %
asistencia **)HH 267 99 92 58AA 276 98 97 49AH 285 100 98 68HA 287 (AH-HA) 96 (AH-HA)
(*)Peso a los 24 meses de edad (**) En el rodeo general

33
INIA TREINTA Y TRES SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna
proporción ingresada al entore incrementacon el grado de sangre cebuina llegandohasta 47% en la proporción ¾ Cebú - ¼Hereford. Desde el punto de vista ambien-tal, este problema no se verificó en los tra-bajos de Bañado de Medina y Caja Notarial,en donde los porcentajes de parición regis-tradas fueron superiores a 81%.
La superioridad en peso vivo inherente a loscebuinos también se observa en el experi-mento de Caja Notarial, con diferencias sig-nificativas en el peso al primer servicio enlas vaquillonas F1 Nelore-Hereford respectoa las cruzas contemporáneas (321 vs. 304kg para NH vs AH-SH, respectivamente) laque se incrementa en forma aún más mar-cada en la comparación con la raza Herefordpura, con casi 30 kg menos de peso vivo.Siguiendo en la misma línea, las hembrasde cruzas recíprocas entre las razas Britá-nicas (Hereford y Angus) alcanzaron los 2años de edad, con 15 kg más en compara-ción con la media de las razas puras. Noobstante, estas diferencias en peso no re-percutieron en los indicadores de preñez yparición en ambos experimentos. Cabe men-cionar que el peso adulto que alcanzaron lasvacas en Bañado de Medina, fue 422, 437 y436 kg en las razas Angus, Hereford y F1,respectivamente.
Las di f icultades al parto también seincrementan significativamente en los cru-zamientos de hembras Hereford con padresNelore. Gimeno et al., (2002) reportan
heterosis de 18% en la dificultad al parto enlas condiciones evaluadas en nuestro país.En el Cuadro 4 se presentan los porcenta-jes medios de parición observados en vacasprimíparas pertenecientes a los mismos ex-perimentos descriptos.
Son conocidas las razones fisiológicas quecausan bajas performances en esta catego-ría. Mientras que en vacas adultas la ener-gía consumida se destina al mantenimien-to, lactación y con menor prioridad a repro-ducción, en las vaquillonas de 2do entoreademás de estas prioridades se debe desti-nar una porción de energía para el crecimien-to animal. En Bañado de Medina, las prue-bas de chi-cuadrado revelaron diferenciassignificativas entre los grupos genéticos delas vacas primíparas (P<0.01), con clara su-perioridad en las F1 (AH y HA), con 67.5%de parición, en relación a las razas purascon 53%. Estas diferencias resultan enheterosis porcentuales de 27% para el por-centaje de preñez en esta categoría.
Similares tendencias se observaron entre lascruzas y Hereford en los ensayos de CajaNotarial, con performances reproductivasmuy destacadas en las vacas Nelore-Hereford (con 61% de preñez vs. 49 en lasotras cruzas y 39% en las Hereford puras).Una vez más es posible comprobar los efec-tos que el ambiente pastoril impuso en losexperimentos de INIA-Tacuarembó, en don-de las tasas de parición logradas en el se-gundo parto no superaron el 40%, tanto en
Cuadro 4. Porcentajes de parición en vacas primíparas (2do. entore) para las razas Hereford,Angus y sus cruzas F1
Grupo Genético Bañado de Medina Caja Notarial INIA Tacuarembó% parición % parición % parición
HH 58 39.3 39.8AH 68 50.9HA 67AA 49SH 48.5NH 61.4
Cebú-H 37
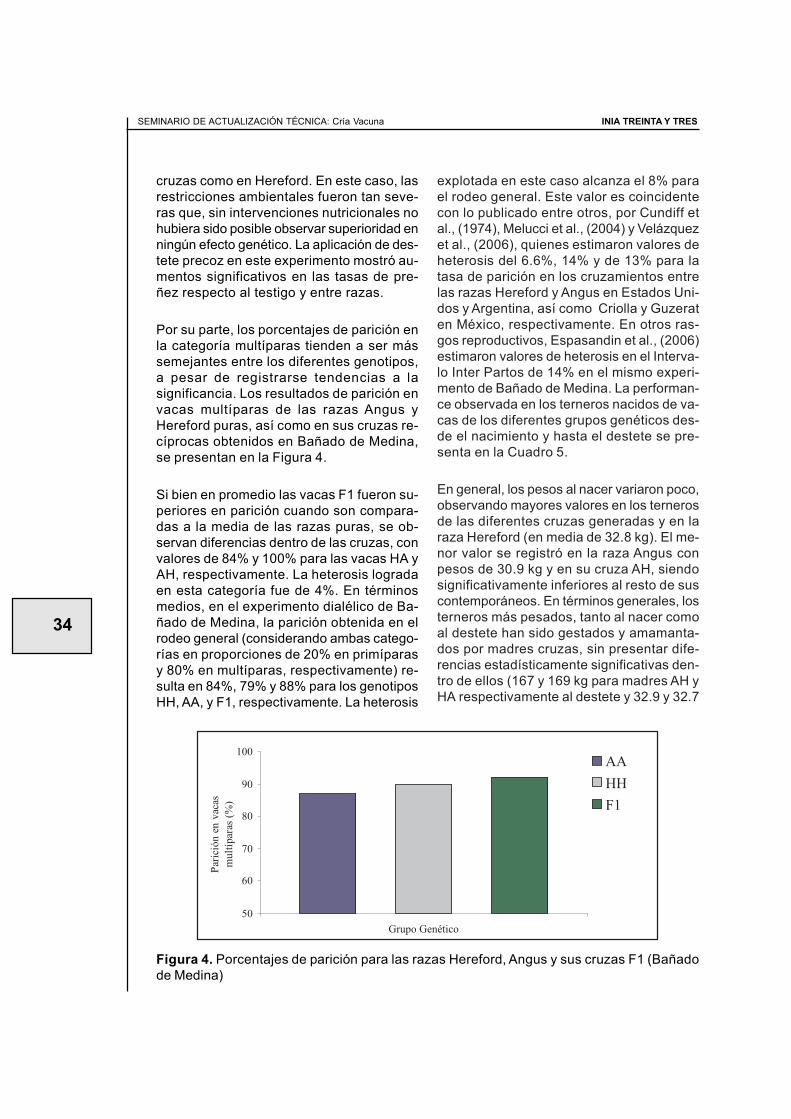
34
SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna INIA TREINTA Y TRES
cruzas como en Hereford. En este caso, lasrestricciones ambientales fueron tan seve-ras que, sin intervenciones nutricionales nohubiera sido posible observar superioridad enningún efecto genético. La aplicación de des-tete precoz en este experimento mostró au-mentos significativos en las tasas de pre-ñez respecto al testigo y entre razas.
Por su parte, los porcentajes de parición enla categoría multíparas tienden a ser mássemejantes entre los diferentes genotipos,a pesar de registrarse tendencias a lasignificancia. Los resultados de parición envacas multíparas de las razas Angus yHereford puras, así como en sus cruzas re-cíprocas obtenidos en Bañado de Medina,se presentan en la Figura 4.
Si bien en promedio las vacas F1 fueron su-periores en parición cuando son compara-das a la media de las razas puras, se ob-servan diferencias dentro de las cruzas, convalores de 84% y 100% para las vacas HA yAH, respectivamente. La heterosis logradaen esta categoría fue de 4%. En términosmedios, en el experimento dialélico de Ba-ñado de Medina, la parición obtenida en elrodeo general (considerando ambas catego-rías en proporciones de 20% en primíparasy 80% en multíparas, respectivamente) re-sulta en 84%, 79% y 88% para los genotiposHH, AA, y F1, respectivamente. La heterosis
Figura 4. Porcentajes de parición para las razas Hereford, Angus y sus cruzas F1 (Bañadode Medina)
explotada en este caso alcanza el 8% parael rodeo general. Este valor es coincidentecon lo publicado entre otros, por Cundiff etal., (1974), Melucci et al., (2004) y Velázquezet al., (2006), quienes estimaron valores deheterosis del 6.6%, 14% y de 13% para latasa de parición en los cruzamientos entrelas razas Hereford y Angus en Estados Uni-dos y Argentina, así como Criolla y Guzeraten México, respectivamente. En otros ras-gos reproductivos, Espasandin et al., (2006)estimaron valores de heterosis en el Interva-lo Inter Partos de 14% en el mismo experi-mento de Bañado de Medina. La performan-ce observada en los terneros nacidos de va-cas de los diferentes grupos genéticos des-de el nacimiento y hasta el destete se pre-senta en la Cuadro 5.
En general, los pesos al nacer variaron poco,observando mayores valores en los ternerosde las diferentes cruzas generadas y en laraza Hereford (en media de 32.8 kg). El me-nor valor se registró en la raza Angus conpesos de 30.9 kg y en su cruza AH, siendosignificativamente inferiores al resto de suscontemporáneos. En términos generales, losterneros más pesados, tanto al nacer comoal destete han sido gestados y amamanta-dos por madres cruzas, sin presentar dife-rencias estadísticamente significativas den-tro de ellos (167 y 169 kg para madres AH yHA respectivamente al destete y 32.9 y 32.7

35
INIA TREINTA Y TRES SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna
kg al nacer, para madres AH y HA, respecti-vamente). La inclusión de genética cebuina,tanto en Tacuarembó como en Caja Notarialgenera animales con mayores potencialesde crecimiento (heterosis individual) asícomo madres capaces de ofrecer mejorescondiciones ambientales a sus hijos duran-te la lactancia (heterosis materna).
El uso de madres cruza resulta en mayoreshabilidades maternas expresadas en suproducción de leche y su comportamientomaternal (mayor vínculo madre-hijo). Trabajosposteriores sobre el mismo sistema dialélicoobservaron producciones de leche de 3.1,4.2 y 4.5 litros por día para vacas primíparasHereford, Angus y F1 (Gioia y Licha, s/p).En este caso, la producción de las vacascruza y Angus puras no difirió y sí sedistanciaron significativamente de las vacasHereford. Sumado a esto, el ternero cruzaposee por causa de su heterosis, una mayorhabilidad para crecer y para aprovechar laleche producida por su madre a través de sudiferente comportamiento de amaman-tamiento. Es de aclarar que en estas mediasparticipan también los efectos de la razapaterna (HH o AA), en donde se visualiza unmayor aporte en efectos individuales de HHque de AA. Indepen-dientemente de la madre
utilizada, los hijos de toros HH promediaron163 kg, mientras que los hijos de toros AAalcanzaron 161.5 kg al destete, pero estadiferencia no fue significativa (P<0.05). Esimportante destacar, que los mayores pesosal nacer ofrecidos por las madres cruza noregistraron dificultades al parto, en donde lasfrecuencias de partos sin dificultad fueronde 99%, 98.5%, 97% y 100% en las madresAA, HH, AH y HA, respectivamente.
La comparación entre los animales puros(hijos de toros y vacas AA o HH) resultó sig-nificativa (P<0.05), siendo superior el pesoalcanzado por los terneros de la raza Angus(156 vs. 151 para Angus y Hereford respec-tivamente), dado principalmente por la des-tacada habilidad materna de la raza Angus.En la cruza simple entre las razas Angus yHereford (F1), hubieron diferencias significa-tivas (P<0.05) en los pesos al destete se-gún el padre o madre utilizados. La combi-nación padre Hereford con madre Angus ob-tuvo 10 kg más al destete que su recíproca,con 164 kg y 154 kg, respectivamente. Eneste caso, también se atribuyen estas dife-rencias a la producción de leche de las ma-dres Angus. En síntesis, los pesos al des-tete superiores correspondieron a los terne-ros hijos de madres cruza (sin diferencias
Cuadro 5. Pesos al nacer y al destete en terneros hijos de diferentes razas y cruzas(Bañado de Medina y Caja Notarial) (Adaptado de Pittaluga et al., 1996; Gimeno et al.,2002 y Espasandin et al., 2006).
Peso al nacer (kg) Peso al destete (kg)Grupo genéticodel ternero
HHAH y A-AH
H-AHHA y H-HA
A-HAAA
SH y S-SHH-SH
NH y N-NHH-NH
Cebú-H
33.1 32.5 132 151 12532.5-32.1 30.3-32.4 154-167 134-147
33.7 32.1 169 14632.8-32.9 164-169
32.4 16730.9 156
32.3-33.5 136-15733.4 154
34.9-32.7 149-16931.3 176
171
Bañado de Caja Bañado de Caja INIAMedina notarial Medina notarial Tacuarembó

36
SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna INIA TREINTA Y TRES
entre AH o HA), seguidos por las cruzas F1(los hijos de madres Angus presentaronmayores pesos al destete) y por último, en-tre los terneros de razas puras, aparece unasuperioridad en la raza Angus.
El resultado global del proceso de cría eneste experimento se resume en el Cuadro6, presentando los kg de ternero destetadospor cada vaca entorada en cada año. Estevalor surge de la conjunción de las variablesPorcentaje de parición con los kilogramosal destete en cada cruza realizada.
Las tendencias son en términos generales,similares a las presentadas anteriormentepara los pesos al destete. Dados los por-centajes de parición registrados, así comolos pesos al destete, en las retrocruzas (AA/HH x F1) observamos los mayores índicesen la variable kg de ternero destetado/vaca/año, con 148 kg promedio, para la cruza depadres Hereford y Angus con madres cruza(AH y HA). Para el trabajo de Bañado deMedina y considerando el rodeo general (va-cas primíparas y multíparas) la superioridadalcanzada en esta variable fue de 3.6% parala cruza simple y de 18% para lasretrocruzas. Si bien este valor representa ala heterosis individual en la cruza simple (AAx HH o viceversa), en las retrocruzas com-prende no sólo la heterosis individual, sinola materna, además de los efectos racialesde Hereford y Angus (los que estarán en pro-porciones de ¾ y ¼, según la combinaciónrealizada).
Cuadro 6. Kg de ternero destetados/vaca/año y Eficiencia global de lacría en diferentes sistemas de cruzamientos del Uruguay
Grupo Genético Grupo Genético Bañado de INIAdel Padre de la madre Medina Tacuarembó (#)
HH HH 127 1AA AA 123HH AA 130AA HH 129
AA/HH F1 148- Cebú - HH 1.4
(#) Eficiencia = (Tasa de Destete x Peso al Destete)/ Peso de la madre al destete0.75
La superioridad de las madres cruza tam-bién fue observada en los experimentos deTacuarembó, en donde estimaron laseficiencias de producción en la cría con va-lores de 1 y 1.4 para madres Hereford purasy Cebú-Hereford, respectivamente. Estosvalores observados en nuestros ambientesde producción son semejantes a los obser-vados por Melucci et al., (2004), trabajandocon las mismas razas en Argentina, así comoCundiff et al., (1974) quienes reportan valo-res de 14% para las cruzas entre estas ra-zas. Arthur, et al., (1999) observaronheterosis de 26% para esta característicapero en cruzamientos entre las razasBrahman y Hereford, valor superior explica-do en parte a la mayor distancia genéticaexistente entre estas razas. Vale destacarque estos autores enfatizan en la sensibili-dad que este parámetro presenta ante lasinteracciones genotipo x ambiente, atribu-yendo a ésta las diferencias encontradas enla literatura. Sin duda, y tal como la teoría loapunta, los niveles de heterosis logrados enlos diferentes cruzamientos y ambientes enlos caracteres reproductivos son muy supe-riores a los obtenibles en variables de creci-miento. A su vez, dentro de las cruzas eva-luadas, es también comprobado que la ma-yor heterosis se alcanza cruzando razas demayor distancia genética, en este caso tau-rinas y cebuinas.
Es importante conocer la heterosis que unacaracterística genera cuando se cruzan dos

37
INIA TREINTA Y TRES SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna
razas para saber su importancia real e inde-pendiente de los efectos raciales. Al com-parar animales cruza respecto a puros, nopodemos olvidar que además de la heterosiscon que cuentan los primeros, también seincluyen los efectos de cada una de las ra-zas participantes tanto en sus efectos aditi-vos raciales como en la complementariedadque se puede generar en las distintas com-binaciones. Los efectos raciales soncuantificables (los conocidos gi) y repercuti-rán en forma positiva o negativa, según elcarácter y la raza considerados. No obstan-te, otros -y tal vez- los más importantes noson posibles de contabilizar ya que se refie-ren a la complementación de características.
Consideraciones finales
Existen en el mundo y en nuestro país, re-cursos genéticos de diferentes orígenes quehoy cuentan con evaluaciones de sus des-empeños en diversos ambientes de produc-ción. Una visión conjunta de los trabajosrealizados expone la variación en la produc-tividad de razas y cruzas frente a distintosambientes. Las características reproductivasson más sujetas a estas variaciones ambien-tales respecto a caracteres de crecimientoy terminación, dada su baja heredabilidad.Por esa razón es posible obtener mejorasrápidas mediante el uso de cruzamientos,en donde los mayores niveles de heterosishan sido comprobados en la fase de cría.En nuestro país, si bien no comparables, losresultados de los diferentes sistemas decruzamientos realizados permiten orientar-nos en relación al alcance potencial de al-gunos recursos genéticos para distintas si-tuaciones de producción. Por último es im-portante conocer además de la heterosis,las contribuciones de cada raza en un sis-tema de cruzamientos, algunas de ellas po-sitivas y otras negativas.
Referencias bibliográficas
Archer, JA., Reverter, A., Herd, RM.,Johnston, DJ. and Arthur, PF. 2002. Geneticvariation in feed intake and efficiency ofmature beef cows and relationships withpost-weaning measurements. Proceedings ofthe 7th World Congress on Genetics Appliedto Livestock Production. Montpellier, France,Comm. 10-7.Arthur, PF., Archer, JA., Johnston., DJ.,Herd, RM., Richardson, EC. and Parnell, PF.2001. Genetic and phenotypic variance andcovariance components for feed intake, feedefficiency, and other postweaning traits inAngus cattle. Journal of Animal Science, v.79, p. 2805-2811.Arthur PF., Hearnshaw H. and StephensonPD. 1999. Direct and maternal additive andheterosis effects from crossing Bos indicusand Bos taurus cattle: cow and calf perfor-mance in two environments. LivestockProduction Science, v. 57, n. 3, p. 231-241.Basarab, JA., Price, MA., Aalhus, JA., Okine,EK., Snelling, WM. and Lyle, KL. 2003. Re-sidual feed intake and body composition inyoung growing cattle. Canadian Journal ofAnimal Science, v. 83, p. 189-204.Cundiff, LV., Gregory, KE. and Koch, RM.1974. Effects of Heterosis on Reproductionin Hereford, Angus and Shorthorn Cattle.Journal of Animal Science, 38:711-727.Espasandin, AC., Franco, J., Olivera, G.,Bentancur, O., Gimeno, D., Pereyra, F. yRogberg, M. 2006. Impacto productivo y eco-nómico del uso del cruzamiento entre lasrazas Hereford y Angus en el Uruguay. XXXIVJornadas Uruguayas de Buiatría. Paysandú.p. 41-51.Gimeno, D., Aguilar, I., Franco, J. y Feed,O. 2002. Rasgos productivos y reproductivosde hembras cruza. Cruzamientos en bovinospara carne. Seminario de Actualización Téc-nica: Serie de Actividades de Difusión, 295INIA.

38
SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna INIA TREINTA Y TRES
Grass, JA., Hansen, PJ., Rutledje, JJ. andHauser, ER. 1982. Genotype x Environmentinteractions on reproductive traits in bovinefemales. I. Age at puberty as influenced bybreed of sire, dietary regimen and season.Journal of Animal Science, v. 55, p. 1441.Herd, RM. and Bishop, SC. 2000. Geneticvariation in residual feed intake and itsassociation with other production traits inBritish Hereford cattle. Livestock ProductionScience, v. 63, p.111-119.Jenkins, TG. and Ferrel, CL. 1999. Beef Cowefficiency - Revisted. Beef ImprovementFederation. Proceedings, 1999. Disponblelen: http://beefimprovement.org. Consultadoen: 10/03/2006.Jenkins, TG. and Ferre l, CL. 1994.Productivity through weaning of nine breedsof cattle under varying feed availabilities: I.Inic ial Evaluat ion. Journal of AnimalScience, v. 72, n. 11, p. 2787-2797.Koch, RM., Cundiff, LV. and Gregory, KE.1994. Heterosis and Breed Effects onReproduction. Factors Affecting Calf Crop.Ed. M.J. Fields y R. S. Sand. CRC Press,Florida, USA, p. 223-242.Koch, RM., Swiger, LA., Chambers, D. andGregory, KE. 1963. Efficiency of feed use inbeef cattle. Journal of Animal Science, v. 22,p.486-494.Koger, M., Reynolds, WL., Kirk, WG.,Peacock FM. and Warnick, AC. 1979.Reproductive performance of crossbreed andstraight bred cattle in different pastureprograms in Florida. Journal of AnimalScience, 21, n. 1, p.14-19.Koger, M., Peacock, FM., Kirk, WG. andCrockett, JR. 1975. Heterosis effects onweaning performance of Brahman andShorthorn calves. Journal of AnimalScience, v. 40 p. 826Koger, M., Burns, WT., Panish, OF. andButts, WTG. 1962. E interact ions inHererford cattle. I. Reproductive traits.Journal of Animal Science, v. 49, p. 396.Koots, KR., Gibson, JP., Smith, C. andWilton, JW. 1994. Analyses of publishedgenetic parameter estimates for beefproduction traits: 1. Heritability. Animal
Breeding Abstracts, v. 62, n. 5, p. 309-38.Kress, DD., England, B., Hauser, ER. andChapman, AB. 1971. Genetic environmentalinteractions in identical and fraternal twinbeef cattle. II. Feed Efficiency, reproductiveperformance, conformation score and fatthickness. Journal of Animal Science, v. 33,p. 1186.Melucci, LM., Fernandez Miganne, J.,Mezzadra, CA. y Vil lareal, EL. 2004.Heterosis y efectos raciales para caracte-res reproductivos en el cruzamiento entreAngus y Hereford. XXV Reunión Argentina deFisiología Vegetal, Santa Rosa, La Pampa,22-24 de Septiembre de 2004.Moore, KL., Johnston, DF., Herd, RM. andGraser, HU. 2003. Genetic and non-geneticeffects on plasma insulin-like growth factor-1 (IGF-1) concentration and production traitsin Angus cattle. Proceedings of thew 15thConference of the Assoc. Adv. Anim Breedingand Genet. Melbourne, Australia p. 222-226.Nkrumah, JD., Keisler, DH. Crews, JR.,Basarab, DH., Wang, Z., Li, C., Price, MA.,Okine, EK. and Moore, SS. 2007. Geneticand phenotypic relationships of serum leptinconcentration with performance, efficiency ofgain, and carcass merit of feedlot cattle.Journal of Animal Science, v. 85, n. 9, p.2147–2155.Nkrumah, JD., Li, C., Yu, J., Hansen, C.,Keis ler, DH. and Moore, SS. 2005.Polymorphisms in the bovine leptin promoterassociated with serum leptin concentration,growth, feed intake, feeding behavior, andmeasures of carcass merit. Journal of Ani-mal Science, v. 83, n. 1, p. 20 – 28.Pittaluga, O. 2005. Inclusión de genéticaCebuina para la mejora de la productividaden la fase de cría. Revista INIA Nº 4, p. 6-9.Pittaluga, O. y De Mattos, D. 1996. Cruza-mientos Cebú-Hereford en Rodeos de Críadel Norte del País. Serie Técnica Nº 79 INIA-Tacuarembó. Ed. INIA, Montevideo.Quintans, G. 2007. Efecto de distintas téc-nicas de control del amamantamiento nsobreparámetros reproductivos y productivos.XXXV Jornadas Uruguayas de Buiatría.Paysandú, 7-9 de Junio de 2007. p. 158-166.

39
INIA TREINTA Y TRES SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna
Schenkel, FS., Miller, SP. and Wilton, JW.2004. Genetic parameters and breeddifferences for feed efficiency, growth, andbody composition traits of young beef bulls.Canadian Journal of Animal Science, v. 84,p. 177-185.Silva, JA., Melis, MHV., Eler, JP. y Ferraz,JBS . 2003. Estimação de parâmetrosgenéticos para probabilidade de prenhez aos14 meses e altura da garupa em bovinos daraça Nelore. Revista Brasileira de Zootec-nia, v.32, n.5, p.1141-1146.Simeone, A. 2000. Destete Temporario, des-tete precoz y comportamiento reproductivoen vacas de Ciria en Uruguay. En: Estrate-gia para acortar el anestro post-parto en va-cas de carne.Velazquez, GM., Montaño-Bermudez, M. yFranquez, JAP. 2006. Efectos genéticos di-rectos, maternos y heterosis individual paratasas de estro, gestación, parición y deste-te de vacas Criollo, Guzerat y sus cruzasF1. Tec. Pecu. Méx. 42(2):143-154.

40
SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna INIA TREINTA Y TRES
1.4. REPORTE TOTAL DEL RODEO: UNA NUEVA ETAPA DE LASEVALUACIONES GENÉTICAS DE BOVINOS PARA CARNE
Mario Lema1 , Olga Ravagnolo2
1 Ing. Agr., MSc.- Programa Nacional de Producción de Carne y Lana - INIA La Estanzuela2 Ing. Agr., PhD.- Programa Nacional de Producción de Carne y Lana - INIA Las Brujas
luación, no siendo válidas las comparacio-nes entre animales de diferentes razas.
Las evaluaciones genéticas nacionales parabovinos de carne son una herramienta am-pliamente difundida y utilizada en la selec-ción de reproductores en nuestro país. Paralas principales razas de carne, Hereford yAberdeen Angus, las evaluaciones tuvieronsus comienzos a final de la década del 80con trabajos de preparación y levantamien-to de registros, siendo publicados los pri-meros catálogos nacionales dereproductores a comienzos de la década del90.
En el cuadro 1 se presenta la evolución dealgunos indicadores de las evaluacionesgenéticas nacionales para las razasHereford, Aberdeen Angus y Braford.
En los años transcurridos desde la primeraevaluación genética hasta el momento nosolo se ha incrementado el número de ca-bañas y animales participantes de las tresrazas, sino que se ha realizado una acumu-lación importante de datos de genealogía,de comprensión por parte de los usuariosde los resultados y de utilización de estosresultados en la selección de losreproductores. Los años de experiencia acu-mulados han solidificado el proceso de lasevaluaciones genéticas permitiendo su ma-yor difusión y utilización por parte de losproductores.
De forma general, salvando particularidadesentre razas, las evaluaciones proporcionanEPDs para:
Introducción
Los programas de mejoramiento genético deuna raza tienen como objetivo identificar ypromocionar los animales que mejor se adap-ten a las condiciones de producción exis-tentes y que al mismo tiempo mejoren el be-neficio económico de las explotaciones.Para esto es necesario valerse de informa-ción objet iva y precisa sobre losreproductores, que permita tomar decisionesde selección y hacer un uso diferencial delos mismos (Aguilar et al., 2005).
La materia prima de las evaluacionesgenéticas son los registros, entendiéndosepor éstos la información de identificación in-dividual, genealógica y productiva. En estaetapa se define la calidad potencial de lasevaluaciones genéticas, siendo necesariocontar con información completa y precisapara obtener resultados confiables. Esta in-formación junto a las circunstancias de pro-ducción (rodeo, grupo contemporáneo, edad,manejo alimenticio, edad de la madre, etc.)son combinadas para el cálculo de los valo-res genéticos, que son expresados en tér-minos de diferencia esperada en la progenie(EPD “Expected Progeny Differences” o DEP“Diferencias Esperadas en la Progenie”).Estos valores genéticos proveen de una pre-dicción del potencial genético futuro de laprogenie de un individuo, para una determi-nada característica.
Los EPDs proporcionan la mejor descripcióngenética de un animal y permite compararlocon animales de otros rodeos, de otras eda-des e incluso con antepasados. Las com-paraciones a través de EPDs son válidas en-tre animales pertenecientes a la misma eva-

41
INIA TREINTA Y TRES SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna
Hereford (2)
Pesos alnacer/año
Nº cabañasPesos al
nacer /cabTotal
registrosRegistros/cab/añoA.AngusPesos alnacer/año
Nº cabañasPesos al
nacer/cabaña
Totalregistros
Registros/cab/añoBraford
Pesos alnacer/año
Nº cabañas
1) Características de crecimiento (peso alnacer, al destete, a los 15 y 18 meses).
2) Habilidad lechera, característica asocia-da al crecimiento de los ternerosdestetados por las hijas de un determi-nado toro.
3) Características de calidad de carne ta-les como área de ojo del bife y espesorde gordura subcutánea.
4) Circunferencia escrotal, característicaasociada a fertilidad de machos y edad ala pubertad de las hembras.
2000 2001 2002 2003 2004 2005 2006(1) 2007(1)
6.249 6.676 6.188 6.557 6.940 6.491 7.034 7.524
111 111 112 109 117 126 127 138
56 60 55 60 59 52 55 55
26.771 30.006 29.631 32.932 35.299 34.367 37.866 45.178
241 270 265 302 302 273 298 327
1.805 2.573 2.943 3.367 3.796 3.745 3.405
21 28 43 45 46 57
86 92 68 75 83 66
5.713 8.228 14.229 15.189 17.516 17.528
272 294 331 338 381 308
93 381 396 476 383 552 418 587
7 7 8 7 8 8 8 8
Años
(1) Al momento de la realización de este artículo no se encuentra disponible información de nacimientos2007 y de pesadas de 15 y/o 18 meses para la raza Aberdeen Angus y Braford. (2) Incluye información deHereford puro registrado
Cuadro 1. Evolución del número de cabañas participantes en las evaluaciones, númerototal de animales y registros evaluados, y número promedio de registros por cabaña paralas razas Hereford, Aberdeen Angus y Braford (2000/2007).
Las primeras evaluaciones genéticas nacio-nales proporcionaron inicialmente EPDs parapesos al destete, 15 y 18 meses y habilidadlechera. Posteriormente fueron incorporadasen etapas sucesivas peso al nacimiento, cir-cunferencia escrotal, área de ojo del bife yespesor de grasa subcutánea. En la actua-lidad se llevan registros además de facilidadal parto para la raza Hereford, de grasaintramuscular a los 18 meses para las ra-zas Hereford y Aberdeen Angus y registrode condición corporal y peso de las vacas al

42
SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna INIA TREINTA Y TRES
momento de destetar los terneros. Esta in-formación permitirá en el mediano plazo po-der incorporar más características con EPDs.
Las características y sus heredabilidades
Si bien se han incorporado varias caracte-rísticas a las evaluaciones genéticas, de for-ma general, éstas han sido asociadas al cre-cimiento de los animales. Este tipo de ca-racterísticas presentan heredabilidades me-dias a altas (Figura 1).
En la parte inferior del gráfico (color verde)se presentan las heredabilidades para ca-racterísticas de crecimiento y calidad decanal. Se observa que para las característi-cas evaluadas en nuestro país lasheredabilidades se sitúan en un intervalo de0.26 a 0.45. Este factor asociado a la facili-dad de medición que tienen estas caracte-rísticas (por ejemplo peso al destete), de-terminan que el progreso logrado por selec-
Figura 1. Heredabilidades directas promedio para características productivasFuente: AAABG (2008) en base a varios trabajos.
ción a través de las mismas sea más rápidoy visible que cuando es realizado por carac-terísticas de baja heredabilidad.
Las características reproductivas (partesuperior del gráfico - color gris) son de difícilmedición, siendo afectadas en formaimportante por el ambiente (nutrición,manejo, sanidad, etc.). Presentan bajasheredabilidades por lo que es esperableobtener menores progresos genéticos alseleccionar por ellas. El complejo decaracterísticas reproductivas es de difícildefinición y problemático para obtener EPDsdesde el punto de vista estadístico (conobservaciones categóricas como la preñez)y desde el punto de vista práctico (conobservaciones disponibles tarde en la vidade los candidatos seleccionados para serpadres) (Garrick, 2005). Según este autor,ese sería el principal motivo por el que losEPDs fueron focalizados inicialmente encaracterísticas de fácil medición como lospesos a diferentes edades.

43
INIA TREINTA Y TRES SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna
El desempeño reproductivo es un componen-te importante de los beneficios de los siste-mas de producción. Garrick (2005) sostieneque la tasa de preñez de las vaquillonas, lalongevidad y los requerimientos nutricionalesde la vaca adulta son determinantes del in-greso económico de los productores. Estastres características han sido incorporadas alas evaluaciones genéticas de algunas ra-zas en Estados Unidos.
Garrick (2006) utilizando modelos de simu-lación concluye que solo realizar énfasis enlos componentes de crecimiento puede de-terminar una sobreestimación de su impac-to en el ingreso de los sistemas cuando escomparado con visiones más integrales quecontemplen crecimiento y reproducción.
Realizar evaluaciones genéticas para carac-terísticas asociadas a la reproducción per-mitiría conocer las relaciones genéticas en-tre éstas y las demás características eva-luadas. El conocimiento de estas relacionesgenéticas para nuestras condiciones produc-tivas es relevante al momento de estudiar ydefinir objetivos de selección y al momentode elaborar índices de selección para lasevaluaciones genéticas nacionales de bovi-nos de carne.
Características reproductivas
Existe una variedad de características quepueden ser asociadas a la eficienciareproductiva. Sin embargo existen diferen-cias entre ellas en relación al tipo de infor-mación necesaria para su obtención, dificul-tad y costo de medición, grado de asocia-ción con fertilidad, variabilidad genética, etc.
Las posibles características a ser incluidasen la evaluación genética son las siguientes:• Edad al primer y segundo parto• Periodo de gestación• Intervalo entre partos• Número de días para el parto• Probabilidad de permanecer en el rodeo
- Habilidad de permanencia en el rodeo(stayability)
• Probabilidad de preñez de la vaquillona auna edad determinada
Algunas de éstas características se expre-san luego de varios años en el rodeo, ejem-plo típico, la probabilidad de permanecer enel rodeo. Esto hace que sea indispensablecontar con información de muchos años paraevaluar esta característica.
Reporte Total de Rodeo
En países donde han sido incorporadas ca-racterísticas reproductivas a las evaluacio-nes genéticas (por ejemplo Hereford en NuevaZelanda y Angus en Australia) la forma derecoger la información se conoce como Re-porte Total de Rodeo del inglés Whole HerdReporting.
El objetivo del Reporte Total del Rodeo esacumular información reproductiva y de des-empeño individual a lo largo de toda la vidaproductiva del animal. Para esto es necesa-rio que todos los animales del rodeo poseanregistros. En este tipo de reporte, el eje dela información es la vaca, desde su naci-miento, pasando por todas las instancias dereproducción hasta el momento de su refugo.
El sistema de evaluación genética actualpara bovinos de carne recoge información decaracterísticas que se expresan en un de-terminado momento de la vida del animal,tiene como eje del mismo el momento deldestete del animal. Tomando como ejemploel peso al destete, este es medido una solavez en la vida del animal. Los terneros queson refugados al destete no son pesados,por lo que no queda constancia de su peso,de los motivos de refugo ni del hecho de quesu madre logró destetar un ternero.
Esta forma de colectar la información deter-mina que no queden registros de las causaspor los cuales algunos individuos no fueron

44
SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna INIA TREINTA Y TRES
medidos (muerte, refugo, venta) y como con-secuencia esta información no es disponi-ble para las madres de estos animales. Estepunto constituye una clara limitante para laevaluación de características reproductivas,donde todos los eventos relacionados a lavida productiva de una vaca y sus hijos sonimportantes. Es también relevante disponerde información de fallas al entore y de otrosmovimientos del rodeo como muertes y ven-ta de vacas y/o terneros.
Para poder realizar comparaciones justas deldesempeño reproductivo de las vacas duran-te su permanencia en el rodeo de cría, secrea la necesidad de incorporar informaciónadicional. La obtención de estos datos im-plica modificaciones al sistema actual de re-gistros, al manejo de las bases de datos yal sistema de ingreso de registros. Si bienel volumen de información necesario es ma-yor, esto no determina un cambio radical enel sistema de recolección de información queactualmente se lleva a cabo en nuestro país.
Recolección de información para elReporte Total del Rodeo
El Reporte Total de Rodeo funciona en baseal mantenimiento de inventarios y registrode los eventos importantes que ocurren enla vida de los animales. Para lograr esto serealizan formularios con listados de los ani-males que deben ser completados en mo-mentos determinados.
En la práctica para el productor el ReporteTotal de Rodeo implica el registro de ciertainformación adicional. A continuación se pre-senta en forma resumida la información aser completada por los productores. Mayo-res detalles pueden ser encontrados enBREEDPLAN, BIO, 2006, BIF, 2002 oNBCEC, 2008.
1) Al entore
Previo al inicio del entore son enviadas alproductor planillas con un listado de todas
las vacas del rodeo y de las vaquillonas quepotencialmente puedan ser entoradas oinseminadas. En las mismas se deberá in-dicar si la vaca entrará o no en el entore (oservicio) y en caso de no hacerlo se deberáindicar el motivo (serán proporcionados có-digos de refugo).
En las planillas se deberá indicar además:• Fecha de comienzo y fin del entore• Fecha de inseminación (si corresponde)• HBU del toro o semen• Fecha de repasos• Fecha de refugo• Código del motivo del refugo• Código de manejo / Lote• Reiterar información tantas veces sea
necesario
2) Parición y destete
En la parición se debe completar una plani-lla elaborada a partir de la información pro-veniente del entore o de la inseminación.
En la planilla de parición se deberá indicar:• Identificación de la vaca (preimpresa, ela-
borada a partir de las planillas de entoreo inseminación)
• Fecha de nacimiento de vaca (día, mes yaño) (preimpreso)
• Fecha de parto• Identificación del ternero• Sexo del ternero• Peso del ternero al nacer• Facilidad al parto (evaluada en 5 catego-
rías: sin dificultad, dificultad menor (al-guna asistencia), dificultad mayor (nor-malmente asistencia mecánica), cesáreay presentación anómala
• Motivo por el que no presenta ternero(aborto, muerta, venta, falla al entore,etc.)
• Código de refugo del ternero (nacimien-to)
• Código de manejo / lote

45
INIA TREINTA Y TRES SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna
Al momento del destete se deberá comple-tar la planilla con las vacas indicadas regis-trando la siguiente información:• Peso de la vaca al destete• Condición corporal de la vaca al destete• Fecha del destete (día, mes y año)• Peso al destete del ternero• Código de refugo del ternero (destete)• Código de manejo / lote
Adicionalmente deben ser identificados losanimales que dejan el rodeo y los motivos(refugo, venta, patas, ubre) y los animalesque se incorporan al rodeo (compras). En elcaso de los establecimientos que realicendiagnóstico de preñez este podrá ser incor-porado a la información de cada vaca.
Esta en estudio la conveniencia o no de ela-borar planillas específicas de parición paraevitar acumular tanta información en las pla-nillas de destete. Los grupos de manejo tan-to en la parición, entore y previo al destetedeben ser especificados para cada vaca.
Para no ser desafectada de los inventarios,en cada periodo de 12 meses, todas las va-cas que pertenecen al rebaño deben contarcon alguno de los siguientes datos:
• registro de parto• razón por la cual la vaca no produjo un
ternero (ejemplos: programa de transfe-rencia de embriones, movimiento parapróxima época de entore)
Para desafectar vacas del inventario se en-vía a los productores un listado de las vacassin progenie para que sea verificado y de esaforma la vaca pueda ser eliminada del ro-deo.
3) Planillas de pesadas a los 15 y 18 meses
Son enviadas planillas al productor con ellistado de animales a ser pesados a estasedades para ser completadas de la mismaforma en que lo son actualmente.
Beneficios del Reporte Total deRodeo
El principal objetivo del Reporte Total delRodeo es permitir la obtención de EPDs paracaracterísticas reproductivas. Ejemplos deestos son presentados a continuación:
EPD de preñez de vaquillonas: estima la di-ferencia entre individuos en la probabilidadde que sus hijas queden preñadas a los dosaños de edad.
EPD para habilidad de permanencia en elrodeo: cuantifica las diferencias esperadasentre los individuos de que sus hijas perma-nezcan el rodeo por lo menos hasta los 6años de edad. La razón principal de que unahembra sea refugada antes de los 6 años esque falle al entore. Consecuentemente la ha-bilidad de permanencia en el rodeo es enprimer lugar una estimación de la fertilidadsostenida de una hembra.
EPD para facilidad al parto: expresa la dife-rencia entre individuos en la proporción es-perada de partos no asistidos. Puede serdividido en EPD directo que estima la dife-rencia entre individuos en la facilidad al par-to de su progenie y EPD maternal que refle-ja la diferencia entre individuos en la facili-dad que tendrán sus hijas en parir.Adicionalmente a la obtención de EPDs elReporte Total del Rodeo permitirá identificarproblemas de los toros o del rodeo en gene-ral que servirán para que los productorestomen medidas para mejorar el desempeñoreproductivo.
A partir de la información recolectada sepodrán elaborar indicadores reproductivos losque permitirán realizar un seguimiento de losmismos detectando problemas y dificultadesen cada establecimiento. Ejemplos de es-tos indicadores son número de vacas ex-puestas a reproducción, porcentaje de pre-ñez, porcentaje de terneros nacidos vivos,% de destete, kg de ternero destetado porvaca expuesta a reproducción.

46
SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna INIA TREINTA Y TRES
Consideraciones finales
Luego de acumulados algunos años de in-formación obtenida a través del Reporte Totaldel Rodeo se podrán obtener los primerosEPDs nacionales para característicasreproductivas en bovinos de carne.
Colectar información a través del ReporteTotal del Rodeo permitirá a los productoresobtener informaciones útiles para el manejode sus rodeos. En tanto, el ordenamientode los registros reproductivos de los rodeospor parte de los productores que lleven ade-lante este sistema, permitirá visualizar susventajas como herramienta de gestión y enel mediano plazo ayudar a su difusión haciarodeos comerciales.
Agradecimientos
Agradecemos las instancias de discusiónrealizadas con el equipo técnico del Progra-ma de Producción de Carne y Lana así comoal Ing. Agr. Jorge Urioste por sus aportestanto en discusión como por sus trabajosrealizados en la materia. Las mismas apor-taron y seguirán aportando en la toma dedecisiones necesarias para llevar a caboestas modificaciones al sistema de regis-tros en bovinos para carne.
Referencias bibliográficas
AAABG. 2008. Associat ion for theadvancement of animal breeding and genetics.Disponible en http://www.gparm.csiro.au/Aguilar, I., Ravagnolo, O., Pravia, MI. yCiappesoni, G. 2005. Mejoramiento genéticode ganado de carne. Revista Aru, Nº 68.pag.50-53.BIF. 2002. Guidelines for uniform BeefImprovement Programs.161p. disponible enhttp://www.beefimprovement.org/library.htmlBIO. 2006. Herd recording. 60pBREEDPLAN. User Manual.110 p.Garrick, DJ. 2006. Genetic improvement ofbeef cattle – assessing the ramifications of
genetic change. In: 8th world congress ongenetic applied to livestock production. BeloHorizonte, Brasil. pp.13-18.Garrick, DJ. 2005. Trends and developmentsin genetic evaluation of beef cattle in UnitedStates. National beef cattle evaluation - Co-lorado State University, Cornel University,University of Georgia. Consultado 3 de may.2008. Disponib le en http:/ /www.ansci.cornell.edu/nbcec/waf1.pdf.NBCEC. 2008. Beef Sire Selection Manual.National Beef Cattle Evaluation Consortium. 78p. Disponible en http:// www.ansci.cornell.edu/nbcec/ sire_selection_manual.html

47
INIA TREINTA Y TRES SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna
Introducción
La competitividad del sector ganadero en elUruguay es cada vez mayor y esto deman-da la adopción de nuevas tecnologías de bajocosto que mejoren la productividad y la ren-tabilidad de la empresa ganadera. Hoy éstatiene su mayor amenaza en la competenciapor el uso del recurso suelo frente a otrasactividades más rentables por unidad de su-perficie como la agricultura, forestación y le-chería fundamentalmente, lo cual la obliga aaumentar mucho su eficiencia en el uso deeste recurso, o sea es obligatorio transfor-mar más eficientemente el pasto en produc-to animal.
La selección de reproductores por crecimien-to acelerado y mayores pesos vivos ha ge-nerado un incremento en el peso vivo a ma-durez y por lo tanto un incremento del tama-ño adulto promedio de las diferentes razasy de sus requerimientos de mantenimiento,determinando mayores costos de produc-ción, una menor eficiencia en uso de los re-cursos alimenticios, y un aumento potencialen problemas ambientales (heces, emisiónde gases). Por otro lado, este avancegenético en caracteres de crecimiento haincidido en forma negativa en los indicadoresreproduct ivos de los rodeos de cría,traduciéndose en el estancado porcentaje dedestete, en promedio de 63% (Pereira ySoca, 2003).
Luego de años de investigación y desarro-llo, el uso de biotecnología molecular paraproducción animal es una realidad. En nues-tro país, existen compañías comerciales queofrecen productos únicos a través de lagenotipificación que permiten clasificar ani-males de acuerdo a su potencial productivo
(características fenotípicas de interés pro-ductivo) según los resultados del ADN. Sinembargo, no es de extrañar que se genereconfusión en cuanto a la utilidad e interpre-tación de estos resultados, ya que técnicosy productores estamos aprendiendo en con-junto el uso e interpretación de ésta infor-mación. El objetivo de este artículo es clari-ficar algunos conceptos relacionados con elgenotipificado de ADN de los animales y susutilidades.
¿Qué es la biotecnología molecular?
La biotecnología molecular, es la ciencia demanipulación del ADN (ácido desoxirribo-nucleico, que contiene las instruccionesgenéticas para el desarrollo y funcionamientode los organismos vivos) a través del cualmejoradores genéticos y demás científicospueden:
– entender mejor y buscar soluciones a laslimitantes de producción (ej. eficienciaalimenticia, reproducción).
– mejorar características nutricionales devarios productos agropecuarios que de locontrario es difícil hacerlo mediante me-joramiento convencional.
– controlar, prevenir y diagnosticar enferme-dades emergentes y actuales.
– caracter izar, conservar y manejarsustentablemente así como utilizar es-tratégicamente la diversidad genética.
¿Qué son los polimorfismos de una solabase (SNPs) o marcadores moleculares?
El ADN esta constituido por cuatro bases(nucleótidos) adenina, guanina, citosina ytimina que se recombinan de una manera
1 Ing. Agr., PhD. Departamento de Producción Animal y Pasturas. Facultad de Agronomía. Universidad de la República.2DMV., PhD. Facultad de Veterinaria. Universidad de la República.
1.5. NUEVAS OPORTUNIDADES PARA EL ÁREA DE CRÍA:BIOTECNOLOGÍA MOLECULAR APLICADA A LA PRODUCCIÓN ANIMAL
Mariana Carriquiry1 , Ana Meikle2

48
SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna INIA TREINTA Y TRES
única en cada organismo viviente. Lospolimorfismos de una sola base (SNPs) sonuna variación en la secuencia de ADN queafecta a un solo nucleótido del genoma, yexplica la variación entre los individuos, pu-diendo afectar la respuesta de los mismosa factores de nutrición, manejo, enfermeda-des, fármacos, etc.
Un marcador molecular es una variación es-pecífica en el ADN entre individuos que hasido asociado con alguna característica deinterés (ej. mayor terneza de la carne). Lasvariantes genéticas son conocidas comoalelos y el genotipificado (determinación deun marcador de ADN/genético) significa es-tablecer que alelo posee un individuo deter-minado para un marcador molecular especí-fico. El SNP puede estar localizado en elgen de interés o en regiones cercanas a él;para que sea un marcador molecular debemodificar una acción biológica que resulteen beneficio económico. La seleccióngenética asist ida por marcadoresmoleculares es una herramienta para asistiry no para sustituir los programas de selec-ción tradicionales (Van Eenennaam, 2007).
Un gran número de genes asociados con ca-racterísticas de crecimiento y productividadhan sido identificados a través del mapeodel genoma, lo cual constituye una exce-lente oportunidad para la selección asistidapor marcadores (Zhang, 2005). Sin embar-go, en nuestro país, si bien el uso depolimorfismos de genes específicos se hautilizado de forma aislada e individual paraincrementar el valor de los reproductores anivel comercial, su uso como complementode los métodos de selección tradicionalesno se ha realizado. El uso de marcadoresmoleculares es bastante directo cuando setrata de seleccionar por caracteres queinvolucran un número reducido de genes (ca-rácter simple) como por ejemplo, color depelaje, presencia/ausencia de cuernos, pre-sencia/ausencia de enfermedades o carac-terísticas detrimentales hereditarias (BLAD,DUMPS), pero no es sencillo cuando losmarcadores moleculares están relacionados
con características complejas que involucranmuchos genes. Marcadores moleculares decaracteres complejos como por ejemplo, fer-tilidad, eficiencia alimenticia, marmoleado,etc, están relacionados con solo uno de losmuchos genes que contribuyen con esa ca-racterística y por lo tanto la presencia o au-sencia de numerosos genes “no marcados”y el medio ambiente/manejo de producciónafectará la expresión o no del fenotipo de-seado (ej. menor consumo residual de ali-mento, mayor preñez, mayor marmoleado).Por esta razón, es importante que para latoma de decisiones, la información sobremarcadores moleculares se combine y eva-lúe conjuntamente con los valores de críaya que estos últimos estiman la contribu-ción de los genes “no marcados” para elcarácter deseado (Van Eenennaam, 2007).
Asimismo es esencial que una vez que laasociación entre alelos de un marcadormolecular y fenotipos de una característicade interés es identificada, estudios de vali-dación en poblaciones independientes seandesarrollados de manera de comprobar queesa asociación es real. La mayoría de losestudios de asociación y validación son rea-lizados en condiciones de estabulación enlos cuales los animales reciben dietas total-mente mezcladas ad libitum. Es así que,antes de conocer la utilidad y/o eficienciadel uso de determinado marcador moleculary/o sistema de clasificación basado en lagenotipificación de animales, es necesarioestudiar si existe o no asociación entre mar-cadores moleculares publicados a nivel in-ternacional y/o usados a nivel comercial conlas respuestas productivas y/o reproductivasde ganado en nuestras condiciones de pro-ducción.
¿Cuál es el beneficio del uso de losmarcadores moleculares?
Los beneficios potenciales de una selecciónasistida por marcadores serán en aquelloscaracteres que tengan baja heredabilidad(fertilidad), difícil o muy caros de medir (re-sistencia a enfermedades), que no pueden

49
INIA TREINTA Y TRES SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna
medirse hasta que el animal haya contribui-do a la siguiente generación (característicasde carcasa), no son seleccionadas en el pre-sente porque no son medidas rutinarias (ter-neza), están genéticamente correlacionadoscon alguna característica que no se quiereincrementar (Parret, 2005).
Considerando la dificultad relativa en la re-colección de los datos de performance, lamagnitud relativa de la heredabilidad y de lavariación observada entre los fenotipos delos caracteres, la cantidad de informacióndisponible en el presente, y el momento deobtención de la información en el ciclo vitalde un animal, las siguientes característicasserán las que obtendrán de mayor a menorgrado el beneficio de la selección asistidapor marcadores moleculares (Parret, 2005):1) Resistencia a enfermedades, 2) Calidadde carcasa y atributos de palatabilidad, 3)Fertilidad y eficiencia reproductiva, 4) Canti-dad y rendimiento de carcasa, 5)Producciónde leche y habilidad materna, 6) Caracterís-ticas de crecimiento
En este sentido, para el área de cría, técni-cos de las Facultades de Agronomía, Vete-rinaria e INIA hemos propuesto trabajar enla validación de marcadores molecularespara:
1) Fertilidad
El uso de SNPs de genes específicos aso-ciados a mejores indicadores reproductivoses incipiente a nivel mundial y los resulta-dos hasta ahora publicados se han obtenidoen sistemas de producción intensivos conestabulación completa. Las limitantes dellogro reproductivo son la longitud del anestroposparto y la mortalidad embrionaria tem-prana. El correcto funcionamiento del ejehipotálamo-hipofisiario-gonadal, regulador dela reproducción, requiere la integración deseñales periféricas que indican el estatusfisiológico y nutricional de la vaca e identifi-ca a la misma como pronta para concebir yllevar adelante una gestación. Señales cla-ves incluyen los componentes del eje
somatotrófico (hormona de crecimiento, GH;insul ina; factor de crecimiento t ipoinsulinémico, IGF-I; sus 6 proteínas deunión, IGFBPs, y proteasas de BPs), loscuales se ven afectados por alteraciones enel metabolismo energético. El sistema IGF-I y la insulina afectan tanto el crecimiento,desarrollo y sobrevivencia de los folículos(Lucy, 2000; Webb et al., 2004) y varios es-tudios apoyan la hipótesis de que los nive-les circulantes de IGF-I en el periparto sonbuenos indicadores de la capacidad de re-torno a la ciclicidad ovárica o sea del largode anestro postparto (Roberts et al., 1997;Beam y Butler,1998; Meikle et al., 2004). Lamayor parte de las mortal idadesembrionarias tienen lugar en el primer mesde gestación debido a defectos en el meca-nismo de reconocimiento materno de la ges-tación. El IGF-I - cuya biodisponiblidad estaregulada por la GH, las IGFBPs y susproteasas- es un potente estimulador delcrecimiento embrionario (Thatcher et al.,2003). Debido al predominante rol que jue-gan los componentes del eje somatróficosobre el crecimiento folicular y embrionario,constituyen genes candidatos para identifi-car y validar marcadores moleculares aso-ciados a fertilidad en sistemas de produc-ción pastoriles, generando estrategiasinnovadoras en los programas de mejoranacionales.
2) Consumo residual de alimentos
El consumo residual de alimentos (CRA)(Koch et al., 1963) se calcula como la dife-rencia entre el consumo real y la cantidadde alimento que un animal debería comer enbase a su peso vivo y su velocidad de ga-nancia de peso (según las tablas de reque-rimientos). Los animales más eficientes tie-nen un CRA negativo (o sea el consumo ob-servado es menor que el predicho) y losmenos eficientes tienen un CRA positivo (osea el consumo observado es mayor que elpredicho). Evidencias recientes (Moore et al.,2003, Nkrumah et al., 2007) han demostra-do que las concentraciones en sangre deIGF-I y de leptina estarían asociadas con

50
SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna INIA TREINTA Y TRES
características productivas como crecimien-to, características y calidad de carcasa,consumo de materia seca y consumo resi-dual de alimento. Es así, que conjuntamen-te con la GH (reguladora de la síntesis IGF-I), son potencialmente buenos genes candi-datos para el estudio de marcadoresgenéticos. Se han establecido asociacionesentre diferentes polimorfismos en el gen deleptina y en su promotor (Nkrumah et al.,2005; Schenkel et al., 2005) con caracterís-ticas de carcasa, calidad de carne y consu-mo residual de alimento. Mas recientemen-te un SNP en el gen ghrelin tendió a mostrarefectos en el CRA, conversión alimenticia yeficiencia de crecimiento parcial (Shermanet al., 2008). Todos estos trabajos han sidogenerados en Norteamérica, Europa o Aus-tralia, no hemos encontrado reportes al res-pecto en la región.
Consideraciones finales
La selección asistida por marcadores tieneel potencial de ser un instrumento valiosopara complementar la evaluación genética,principalmente en aquellos caracteres eva-luados que tienen baja heredabilidad o sondifíciles de medir durante la vida del animal,como podrían ser la fertilidad, el consumoresidual de alimentos y calidad de carne. Esimprescindible que las asociaciones entrelos SNPs y las respuestas productivas y/oreproductivas del ganado sean validadas ennuestras condiciones de producción. El avan-ce biotecnológico, el estudio acabado delpotencial del mismo conjuntamente con avan-ces en trazabilidad y sanidad animal, nospermitiría en un futuro cercano mejorar laeficiencia productiva y posicionarnos a nivelregional e internacional con mejores produc-tos.
Referencias bibliográficas
Beam, SW. y Butler, WR. 1998. Energy Ba-lance, Metabolic Hormones, and EarlyPostpartum Follicular Development in DairyCows Fed Prilled Lipid. Journal of DairyScience, 91: 121-131.
Koch, RM., Swiger, LA., Chambers, D. andGregory, KE. 1963. Efficiency of feed use inbeef cattle. Journal of Animal Science,22:486-494.Lucy, M. 2000. Regulation of ovarian folliculargrowth by somatotropin and insulin-likegrowthfactors in cattle. Journal of DairyScience, 83:1635-1647.Meikle, A., Kulcsar, M., Chilliard, Y., Febel,H., Delavaud, C., Cavestany, D. andChilibroste, P. 2004. Effects of Parity andBody Condit ion Score at Calving onEndocrine and Reproductive Parameters ofthe Dairy Cow under Grazing Conditions.Reproduction 127, 727-737.Moore, KL., Johnston, DF., Herd, RM. andGraser, HU. 2003. Genetic and non-geneticeffects on plasma insulin-like growth factor-1 (IGF-1) concentration and production traitsin Angus cattle. Proceedings of thew 15thConference of the Assoc. Adv. AnimBreeding and Genet. Melbourne, Australia p.222-226Nkrumah, JD., Li, C., Yu, J., Hansen, C.,Keis ler, DH. and Moore, SS. 2005.Polymorphisms in the bovine leptin promoterassociated with serum leptin concentration,growth, feed intake, feeding behavior, andmeasures of carcass merit. Journal of Ani-mal Science, 83(1): 20 – 28.Nkrumah, JD., Keisler, D.H. Crews, JR.,Basarab, JA., Wang, Z., Li, C., Price,MA.,Okine, EK. and Moore, SS. 2007.Genetic and phenotypic relationships ofserum leptin concentration with performan-ce, efficiency of gain, and carcass merit offeedlot cattle. Journal of Animal Science,85(9): 2147 – 2155.Parret, D. 2005. Profit Pointers-Marker-Assisted Selection in Beef Cattle http: //www.livestocktrail. uiuc .edu /beefnet/paperDisplay.cfm? ContentID = 6992Pereira, G., Soca, P. 2003. Aspectos rele-vantes de la cría vacuna en el Uruguay. Fa-cultad de Agronomía/Universidad de la Re-pública. http://www.fagro.edu.uyRoberts, AJ., Nugent, RA., Klindt, III J. andJenkins, TG. 1997. Circulating insulin-likegrowth factor I, insulin-like growth factor

51
INIA TREINTA Y TRES SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna
binding proteins, growth hormone, andresumption of estrus in postpartum cowssubjected to dietary energy restriction.Journal of Animal Science, 75:1909–1917.Schenkel, FS., Miller, SP. and Wilton, JW.2004. Genetic parameters and breeddifferences for feed efficiency, growth, andbody composition traits of young beef bulls.Canadian Journal of Animal Science, 84:177-185, 2004.Thatcher,WW., Guzeloglu, A., Meikle, A.,Kamimura, S., Bilby, T. and Kowalski, AA .2003. Regulation of embryo survival in cattle.Reproduction 61: 253-260Van Eenennaam, A. 2007. DNA Markers:Explanation of Validation and Utilization http:// animalscience .ucdavis.edu/animalbiotech/Outreach/Marker Assisted Selection in BeefCattle.pdf.Webb, R., Garnsworthy, PC., Gong, JG andArmstrong. DG. 2004. Control of folliculargrowth: Local interactions and nutritionalinfluences. Journal of Animal Science, 82(E.Suppl.):E63–E74Zhang, G. 2005. Molecular Biology: Animal.Encyclopedia of Animal Science. 653-656.

52
SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna INIA TREINTA Y TRES
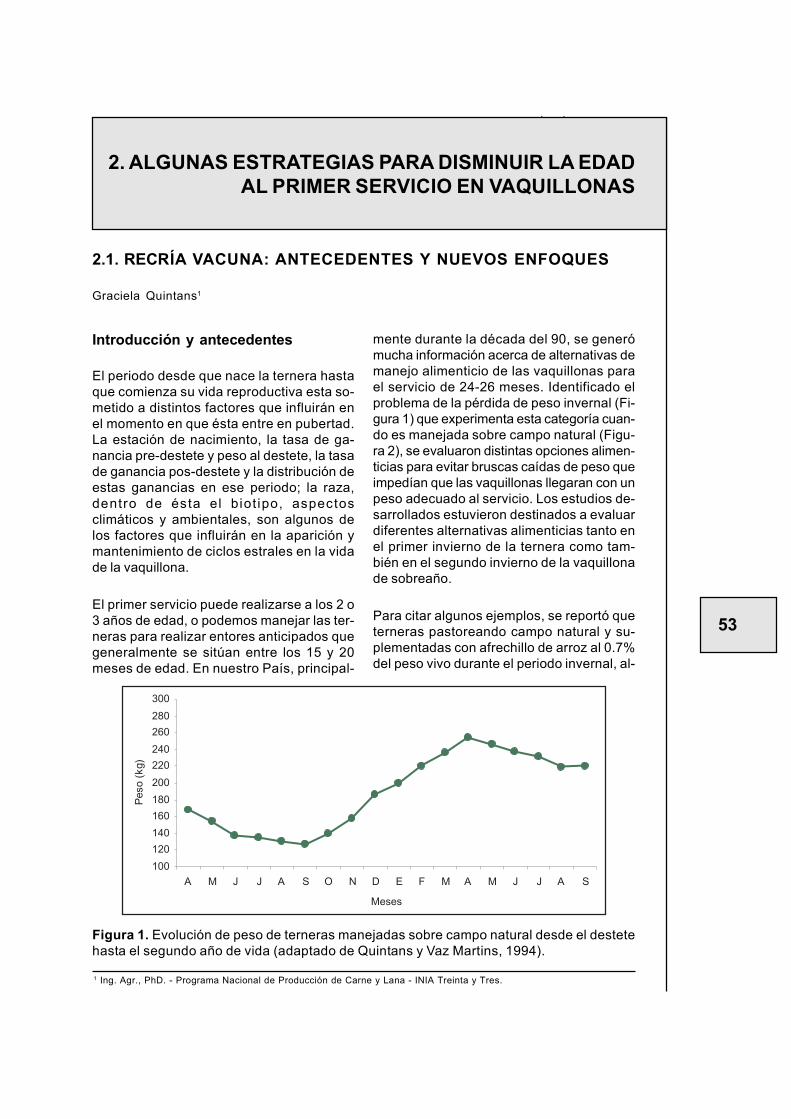
53
INIA TREINTA Y TRES SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna
2.1. RECRÍA VACUNA: ANTECEDENTES Y NUEVOS ENFOQUES
Graciela Quintans1
mente durante la década del 90, se generómucha información acerca de alternativas demanejo alimenticio de las vaquillonas parael servicio de 24-26 meses. Identificado elproblema de la pérdida de peso invernal (Fi-gura 1) que experimenta esta categoría cuan-do es manejada sobre campo natural (Figu-ra 2), se evaluaron distintas opciones alimen-ticias para evitar bruscas caídas de peso queimpedían que las vaquillonas llegaran con unpeso adecuado al servicio. Los estudios de-sarrollados estuvieron destinados a evaluardiferentes alternativas alimenticias tanto enel primer invierno de la ternera como tam-bién en el segundo invierno de la vaquillonade sobreaño.
Para citar algunos ejemplos, se reportó queterneras pastoreando campo natural y su-plementadas con afrechillo de arroz al 0.7%del peso vivo durante el periodo invernal, al-
1 Ing. Agr., PhD. - Programa Nacional de Producción de Carne y Lana - INIA Treinta y Tres.
Figura 1. Evolución de peso de terneras manejadas sobre campo natural desde el destetehasta el segundo año de vida (adaptado de Quintans y Vaz Martins, 1994).
2. ALGUNAS ESTRATEGIAS PARA DISMINUIR LA EDADAL PRIMER SERVICIO EN VAQUILLONAS
Introducción y antecedentes
El periodo desde que nace la ternera hastaque comienza su vida reproductiva esta so-metido a distintos factores que influirán enel momento en que ésta entre en pubertad.La estación de nacimiento, la tasa de ga-nancia pre-destete y peso al destete, la tasade ganancia pos-destete y la distribución deestas ganancias en ese periodo; la raza,dentro de ésta el biot ipo, aspectosclimáticos y ambientales, son algunos delos factores que influirán en la aparición ymantenimiento de ciclos estrales en la vidade la vaquillona.
El primer servicio puede realizarse a los 2 o3 años de edad, o podemos manejar las ter-neras para realizar entores anticipados quegeneralmente se sitúan entre los 15 y 20meses de edad. En nuestro País, principal-
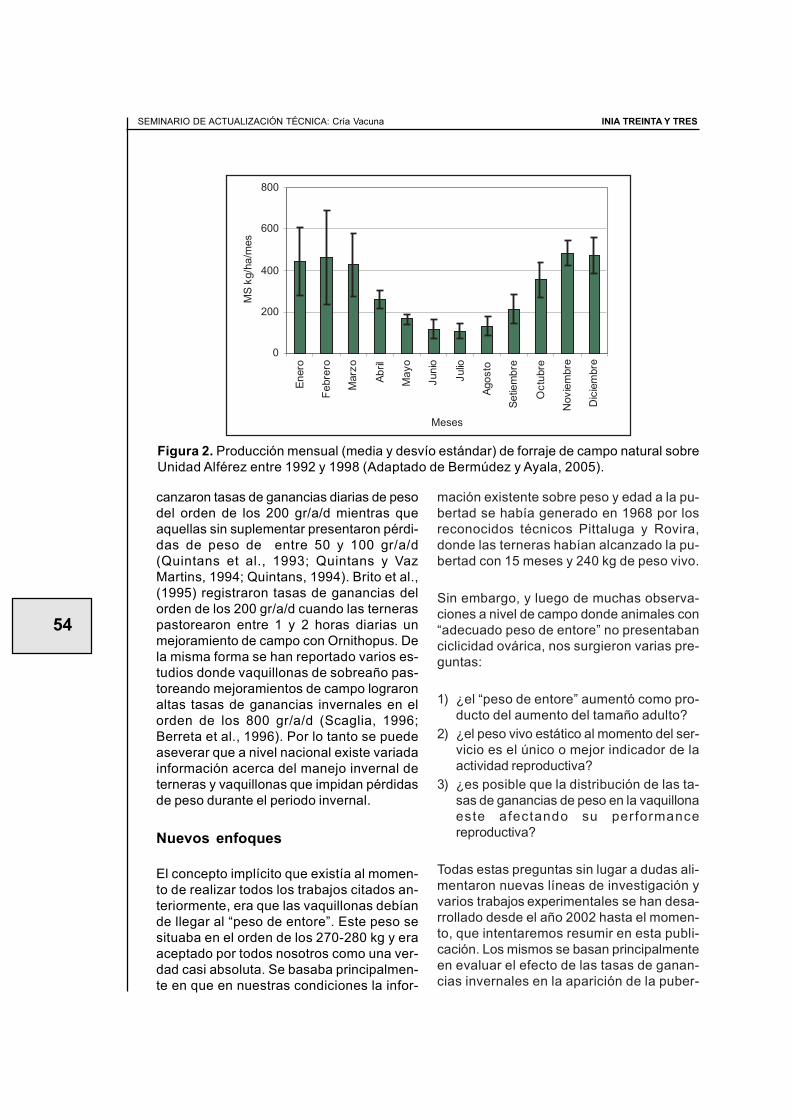
54
SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna INIA TREINTA Y TRES
canzaron tasas de ganancias diarias de pesodel orden de los 200 gr/a/d mientras queaquellas sin suplementar presentaron pérdi-das de peso de entre 50 y 100 gr/a/d(Quintans et al., 1993; Quintans y VazMartins, 1994; Quintans, 1994). Brito et al.,(1995) registraron tasas de ganancias delorden de los 200 gr/a/d cuando las terneraspastorearon entre 1 y 2 horas diarias unmejoramiento de campo con Ornithopus. Dela misma forma se han reportado varios es-tudios donde vaquillonas de sobreaño pas-toreando mejoramientos de campo lograronaltas tasas de ganancias invernales en elorden de los 800 gr/a/d (Scaglia, 1996;Berreta et al., 1996). Por lo tanto se puedeaseverar que a nivel nacional existe variadainformación acerca del manejo invernal deterneras y vaquillonas que impidan pérdidasde peso durante el periodo invernal.
Nuevos enfoques
El concepto implícito que existía al momen-to de realizar todos los trabajos citados an-teriormente, era que las vaquillonas debíande llegar al “peso de entore”. Este peso sesituaba en el orden de los 270-280 kg y eraaceptado por todos nosotros como una ver-dad casi absoluta. Se basaba principalmen-te en que en nuestras condiciones la infor-
Figura 2. Producción mensual (media y desvío estándar) de forraje de campo natural sobreUnidad Alférez entre 1992 y 1998 (Adaptado de Bermúdez y Ayala, 2005).
mación existente sobre peso y edad a la pu-bertad se había generado en 1968 por losreconocidos técnicos Pittaluga y Rovira,donde las terneras habían alcanzado la pu-bertad con 15 meses y 240 kg de peso vivo.
Sin embargo, y luego de muchas observa-ciones a nivel de campo donde animales con“adecuado peso de entore” no presentabanciclicidad ovárica, nos surgieron varias pre-guntas:
1) ¿el “peso de entore” aumentó como pro-ducto del aumento del tamaño adulto?
2) ¿el peso vivo estático al momento del ser-vicio es el único o mejor indicador de laactividad reproductiva?
3) ¿es posible que la distribución de las ta-sas de ganancias de peso en la vaquillonaeste afectando su performancereproductiva?
Todas estas preguntas sin lugar a dudas ali-mentaron nuevas líneas de investigación yvarios trabajos experimentales se han desa-rrollado desde el año 2002 hasta el momen-to, que intentaremos resumir en esta publi-cación. Los mismos se basan principalmenteen evaluar el efecto de las tasas de ganan-cias invernales en la aparición de la puber-

55
INIA TREINTA Y TRES SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna
tad (primer evento reproductivo en la vida dela vaquillona), y frente a distintas alternati-vas nutricionales, cuales son las probabili-dades de realizar entores a edad temprana.También se presenta un trabajo acerca delmanejo de la bioestimulación como otra al-ternativa más para adelantar la pubertad. He-mos comenzado a evaluar servicios de oto-ño a los 18-20 meses de edad y los resulta-dos preliminares también se presentan acontinuación.
Existe otra información generada en estostrabajos y es la determinación y actualiza-ción de los pesos y edades a los que lasterneras alcanzan la pubertad frente a nue-vos biotipos tanto puros como cruzas. Caberecordar que, entre otros factores, la terne-ra debería alcanzar entre el 65 y 70% de supeso adulto para comenzar a ciclar. No de-bemos olvidar que en los últimos 30 añosse ha incorporado genética importada debiotipos desarrollados en condiciones dife-rentes a nuestros sistemas de producciónpor lo tanto recobra vital importancia tenerlas variables reproductivas caracterizadas yevaluadas en nuestros propios animales ysistemas productivos.
Referencias bibliográficas
Bermúdez, R. y Ayala, W. 2005. Producciónde forraje de un campo natural de la zona delomadas del este. En: Seminario de actuali-zación técnica en manejo de campo natu-ral. Serie Técnica Nº 151. INIA Treinta y TresBerreta, E., Pittaluga, O., Brito, G, Pigurina,G. y Risso, D. 1996. Efecto de la adminis-
tración de forraje sobre el crecimiento deterneras y vaquillonas en campo naturalmejorado de Basalto. INIA. Actividades deDifusión nº. 108. pp. IX-5-IX-8.Brito, G., Risso, D., Olmos, F. y Zarza, A.1995. Registro de pastoreo de Ornithopuscompressus cv. «Encantada». INIA. Activi-dades de Difusión nº. 65. pp. 24-26Pittaluga, O. y Rovira, J. 1968. Influencia delnivel nutricional predestete sobre el creci-miento y pubertad de terneras Hereford. Bo-letín Técnico, 5 (2): 68-78. Facultad de Agro-nomía, Estación Experimental de Paysandú.Quintans, G. y Vaz Martins, D. 1994. Efectode diferentes fuentes de suplemento sobreel comportamiento de terneras. En: Bovinospara Carne: Avances en la Suplementaciónde la Recría e Invernada Intensiva. SerieActividades de Difusión Nº 34. INIA Treinta yTres.Quintans, G. 1994. Suplementación de ter-neras y vaquillonas con afrechillo de arrozdesgrasado. En: Bovinos para Carne: Avan-ces en Suplementación de la Recría eInvernada Intensiva. Serie Actividades deDifusión 34. INIA Treinta y Tres.Quintans, G., Vaz Martins, D. y Carriquiry,E. 1993. Efecto de la suplementación inver-nal sobre el comportamiento de terneras. En:Campo Natural: Estrategia Invernal Manejoy Suplementación. Resultados Experimen-tales. INIA Treinta y Tres. p 35-53.Scaglia, G. 1996. Alternativas para la alimen-tación de la vaca de cría durante el periodoinvernal. En: Producción Animal. UnidadExperimental Palo a Pique. Serie Activida-des de Difusión Nº 110. INIA Treinta y Tres
Figura 3. Vaquillonas pastoreando campo natural.

56
SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna INIA TREINTA Y TRES
2.2. PRINCIPALES FACTORES QUE AFECTAN LA APARICIÓN DE LAPUBERTAD EN VAQUILLONAS DE RAZAS CARNICERAS
Graciela Quintans1 , Gonzalo Roig2
gonadotropinas (particularmente LH) y laausencia de estimulación ovárica (Imakawaet al., 1986, Wiltbank et al., 2002). Por lotanto el estatus metabólico de la ternera seconectará con el eje reproductivo en un com-plejo diálogo en el cual varios interlocutoresjuegan roles preponderantes, complejos ymucha veces interaccionando entre sí.
Para citar sólo algunos ejemplos, las res-tricciones de la dieta y balances energéti-cos negativos disminuyen las concentracio-nes de insulina en la circulación (Diskin etal., 2003) y ésta, entre otras funciones, esuna señal metabólica que influye en la se-creción de LH desde la hipófisis y regula lasensibilidad del ovario a las gonadotropinas.Por otra parte, concentraciones bajas de IGF-I están asociadas con el atraso de la puber-tad (Granger et al. citado por Diskin et al.,2003). Bossis et al., (2000) reportaron unadisminución lineal en las concentraciones enplasma de IGF-I a partir del comienzo de larestricción alimenticia hasta el anestro. Tam-bién la leptina estaría involucrada en esteproceso (hormona sintetizada y secretadapor el tejido adiposo) y su concentración espredictor del status nutricional (reserva deenergía en el tejido adiposo) aunque su rolparecería ser mas permisivo (Blache et al.,2006).
En nuestras condiciones pastoriles es co-mún observar animales que entran en celo(pubertad) en el otoño temprano y luego caenen anestro en invierno, principalmente porbajos niveles nutricionales. Existe informa-ción que vaquillonas perdiendo entre un 4 y23% del peso vivo inicial, pueden caer enanestro (Imakawa et al., 1986; Vizcarra etal., 1991; Vizcarra y Wetteman, 1993), y enel estudio de Vizcarra y Wetteman (1993) el
1 Ing. Agr., PhD. - Programa Nacional de Producción de Carne y Lana - INIA Treinta y Tres.2 DMV. - Programa Nacional de Producción de Carne y Lana. - INIA Treinta y Tres.
Según Robinson (1977) la pubertad se defi-ne como el proceso por el cual los animalesse vuelven capaces de reproducirse y Rovira(1996) la define como el momento en que semanifiesta el primer estro o estro acompa-ñado de la ovulación correspondiente. Laedad a la pubertad y al primer parto son ele-mentos que afectan la producción de unavaca en todo su ciclo de vida (Lesmeister etal., 1973) ya que hembras ciclando más tem-prano son más eficientes en subsecuentesentores y en su posterior vida productiva(Rege y Famula, 1993).
En las hembras vacunas, muchos son losfactores que pueden influir en la edad y pesoal cual se alcanza la pubertad. Entre ellosse destacan el plano nutricional (expresadoen el peso al destete y manejo pos-deste-te), la genética (variando el peso y la edad ala pubertad entre razas y entre líneas de unamisma raza), e l fotoper iodo, labioestimulación, los tratamientos hormona-les y la sanidad.
Bajo circunstancias normales, una hembrano entrará en pubertad hasta que no alcan-ce un estado de desarrollo que le asegure lacapacidad de tener y cuidar una cría. Poresta razón la pubertad está más pobremen-te relacionada con la edad cronológica y másfuertemente con el estado de desarrollo, pesovivo y presencia de adecuadas reservas cor-porales (Lindsay et al., citados por Quintans,2002).
La restricción nutricional potencia el meca-nismo inhibitorio de los estrógenos (fee-dback negativo prepuberal), generando unadisminución de la liberación de GnRH des-de el hipotálamo con la consecuente dismi-nución de la frecuencia de secreción de

57
INIA TREINTA Y TRES SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna
66% de las vaquillonas recobraron laciclicidad 4.5 semanas después de ser re-alimentadas pero tuvieron que alcanzar unpeso 12% superior al inicial. Esta es una delas posibles causas de que vaquillonas quetienen pesos similares al momento del ser-vicio puedan presentar estatus ovárico dife-rente, reafirmando el concepto de que la evo-lución de peso previo al momento del servi-cio puede afectar la actividad ovárica. Estose ve más acentuado en condicionespastoriles y ambientales como las de nues-tra región.
Con respecto al biotipo, se ha observado quelas vaquillonas Bos taurus son más preco-ces en llegar a la pubertad que las vaquillonasBos indicus (Gregory et al., 1979; Freetly yCundiff, 1997; Thallman et al., 1999;Rodrígues et al., 2002). Gregory et al.,(1979) encontraron que vaquillonas Brahmanx Hereford alcanzaron la pubertad 76 díasmás tarde y con 36 kg mas de peso que elpromedio de las cruzas Hereford x AberdeenAngus. Las vaquillonas Hereford x Angus pre-sentaron 90% de celo a los 340 kg mientrasque las Brahman x Hereford presentaron un86% de celo a los 370 kg. El cruzamientode razas de aptitud carnicera suele lograr lapubertad a edad más temprana dado por unefecto propio de la heterosis (Gregory et al.,1978). La expresión más temprana de la pu-bertad recobra mayor impacto cuando se rea-lizan entores anticipados (15 o 18 meses).También es importante destacar que dentrode una misma raza pueden existir variacio-
Figura 1. Terneras de destete durante su primer invierno de vida pastoreando mejoramien-tos de campo
nes de peso a la pubertad, debidas princi-palmente al manejo nutricional previo (comose citó anteriormente) o a diferencias en ta-maño adulto que implican requerimientos di-ferentes.
La bioestimulación ha sido identificada comoun factor que podría adelantar el comienzode la actividad reproductiva cuando ternerasprepúberes son expuestas a machos o hem-bras androgenizadas. Para citar algunosejemplos, Kunkle y Sand (1993) reportaronque vaquillonas prepúberes expuestas a to-ros adelantaron la pubertad 40 días y 50%de ellas se preñaron durante los primeros21 días de servicio, comparado con sólo un16.5% de preñez en hembras no expuestasal efecto macho. Por otra parte Oliveira etal., (2004) encontraron que 62.8% de lasvaquillonas Nelore expuestas al efecto ma-cho alcanzaron la pubertad mientras que lascompañeras no expuestas lo hicieron en un45.7%.
Respecto al fotoperiodo, es interesante des-tacar que la época del año ejerce una in-fluencia en la entrada en pubertad y Hafez,(1996) indica que las condiciones invernalesdurante el periodo prepuberal retrasan lapubertad. En ese mismo sentido, el mes denacimiento de la ternera influiría en el mo-mento que pueda alcanzar su primer celo yexiste alguna evidencia que indica que ter-neras nacidas en otoño tienden a alcanzarla pubertad antes que las nacidas en prima-vera (Senger, 2004).

58
SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna INIA TREINTA Y TRES
Referencias bibliográficas
Blache, D., Chagas, LM. and Martin, GB.2006. Nutritional inputs into reproductiveneuroendocrine control system-amultidimensional perspective. In: Reproductionin Domestic Ruminants VI. NottinghamUniversity Press. Ed. Juengel, JL., Murray, JF.and Smith, MF. pp. 122-139.Bossis, I., Wettemann, RP., Welty, SD.,Vizcarra, JA., Spicer, LJ. and Diskin, MG.1999. Nutritionally induced anovulation in beefheifers: ovarian and endocrine functionpreceding cessation of ovulation. Journal ofAnimal Science, 77: 1536-1546.Diskin, MG., Mackey, DR., Roche, JF. andSreenan, JM. 2003. Effects of nutrition andmetabolic status on circulating hormones andovarian follicle development in cattle. AnimalReproduction Science, 78: 345-370.Freetly, HC. y Cundiff, LV. 1997. Postweaninggrowth and reproduction characteristics ofheifers sired by bulls of seven breeds andraised on different levels of nutrition. Journalof Animal Science, 75: 2841-2851.Gregory, KE., Laster, DB., Cundiff, LV., Smith,GM. and Koch, RM. 1979. Characterization ofbiological types of cattle-cycles III: II. Growthrate and puberty in females. Journal of AnimalScience, 49:461-471.Gregory, KE., Laster, DB., Cundiff, LV., Koch,RM. and Smith, GM. 1978. Heterosis and breedmaternal and transmitted effects in beef cattle.II. Growth and puberty in females. Journal ofAnimal Science. 47: 1042-1053.Hafez, ESE. 1996. Reproducción e insemina-ción artificial en animales. 6ª. ed. México,Interamericana - Mc Graw-Hill. 542 p.Imakawa, K., Day, ML. and Zaleky, DD. 1987.Effect of estradiol and diets varying in energyon secretion of luteinizing hormone in beefheifers. Journal of Animal Science, 64: 805-815.Kunkle, WE. y Sand, RS. 1993. Nutrition andmanagement for the replacement heifer. En:Northwest Beef Production Conference (1993,Quincy). Proceedings. Florida, University of Flo-rida. pp.49-57. (NFREC Extension Report 93-2)Lesmeister, JL., Burfening, PJ. and Blackwell,RL. 1973. Date of first calving in beef cowsand subsequent calf production. Journal of
Animal Science, 36: 1-6.Oliveira, CMG., Oliveira Filho, BD., Gambarini,ML. and Viu, MA. 2004. Influence ofbiostimulation on puberty age and pregnancyrate in Nelore heifers. En: InternationalCongress on Animal Reproduction (15º, 2004,Porto Seguro). Abstracts. Porto Seguro, Brazil,CBRA, vol: 1, p. 20Quintans, G. 2002. Manejo de la recría vacu-na en sistemas ganaderos. En: Seminario deActualización Técnica (2002, Treinta y Tres).Cría y recría ovina y vacuna. Treinta y Tres,INIA. pp. 47-56 (Actividades de Difusión Nº. 288).Rege, JEO., y Famula, TR. 1993. Factorsaffecting calving date and its relationship withproduction traits of Hereford dams. AnimalProduction, 57; 385-395.Robinson, TJ. 1977. Reproduction in cattle. En:Reproduction in domestic animals. HH. Cole;PT. Cupps eds. 3rd. ed. New York, AcademicPress. pp. 433-441.Rodrigues, HD., Kinder, JE., and Fitzpatrick,LA. 2002. Estradiol regulation of luteinizinghormone secretion in heifers of two breed typesthat reach puberty at different ages. Biology ofReproduction, 66: 603-609.Rovira, J. 1996. Manejo nutritivo de los rodeosde cría en pastoreo. Montevideo, HemisferioSur. 288 p.Senger, PL. 2003. Pathways to pregnancy andparturition. 2n. ed. New York, Cadmus. 384 p.Thallman, RM., Cundiff, LV., Gregory, KE. andKoch, RM. 1999. Germplasm evaluation in beefcattle-Cycle IV: postweaning growth andpuberty of heifers. Journal of Animal Science,77: 2651-2659.Vizcarra, JA. and Wetteman, RP. 1993.Relationship between body weight changes inpostpuberal heifers and cessation of lutealactivity. Okla. Agr. Exp. Sta. Res. Rep.Vizcarra, JA. 1991. Relationship between theoccurrence of puberty in heifers and thecessation of luteal activity after nutritionalrestriction. Okla. Agr. Exp. Sta. Res. Rep. MP-134: 44.Wiltbank, MC.; Gumen, A.; Sartori, R. 2002.Clasificación fisiológica de condicionesanovulatorias en Bovinos. Theriogenology, 57(1): 21-53.

59
INIA TREINTA Y TRES SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna
2.3. EFECTO DEL MANEJO NUTRICIONAL EN EL PRIMER INVIERNOSOBRE LA APARICIÓN DE LA PUBERTAD EN TERNERAS DE RAZACARNICERA (PRIMER AÑO DE EVALUACIÓN)
Juan Miguel Straumann1 , Walter Ayala2 , Ana Inés Vázquez3 , Graciela Quintans4
te 162 días sobre un mejoramiento de cam-po hasta el final del experimento (Febrero),con el objetivo que realizaran altas ganan-cias de peso vivo. Las terneras fueron pesa-das cada 14 días durante todo el periodoexperimental. A partir del manejo conjunto(Setiembre) los animales fueron sangradossemanalmente para determinar el inicio dela actividad ovárica en base a la concentra-ción de progesterona en sangre. Se tomócomo criterio dos muestras sucesivas conniveles de progesterona superiores a 1 ng/ml como inicio de la actividad ovárica. En lapastura se determinó: disponibilidad y altu-ra de forraje, tasa de crecimiento del camponatural utilizando jaulas de exclusión y com-posición botánica. Se realizaron análisis dedigestibilidad In Vitro, proteína cruda, fibradetergente ácido y cenizas. Para el análisisde las variables continuas (ganancias depeso, edad, y peso) se utilizó un modelo li-neal general mediante el procedimiento GLMdel paquete estadístico SAS (1999). Paraanalizar las variables discretas (porcentajede vaquillonas que entraron en pubertad) seutilizó el procedimiento GENMOD del mis-mo paquete estadístico. Los valores se pre-sentan como promedio ± error de la media(em) y el nivel de significancia utilizado fueP <0.05.
Resultados y discusión
Durante el manejo invernal, las terneras quepastoreaban campo natural (T) perdieron casi19 kg en los 92 días de invierno, a razón de-0.220 kg/a/d (Cuadro 1). Estas pérdidas fue-ron superiores a las encontradas por
1 Ing. Agr. Realizó tesis de grado en INIA Treinta y Tres.2 Ing Agr., PhD. -Director de Programa Nacional de Pasturas y Forrajes - INIA Treinta y Tres.3 Ing. Agr. University of Wisconsin-Madison. 1675 Observatory Dr. Madison, WI 53706. USA.4 Ing. Agr., PhD. - Programa Nacional de Producción de Carne y Lana.- INIA Treinta y Tres.
Introducción
El momento en que las terneras entran enpubertad determina su capacidad para re-producirse y la posibilidad de que entren alrodeo como hembra productiva. En serviciosa edades más tempranas que las tradicio-nales (2 o 3 años de edad) este eventoreproductivo es de suma importancia. Elobjetivo del presente trabajo fue evaluar di-ferentes tasas de ganancias invernales so-bre el peso y edad a la pubertad en ternerasde razas carniceras (Tesis de Facultad deAgronomía - Straumann 2006).
Materiales y métodos
El experimento fue realizado entre Junio del2002 y Febrero del 2003, en la Unidad Ex-perimental Palo a Pique (UEPP), pertene-ciente a la Estación Experimental del Este,INIA Treinta y Tres. Treinta y seis ternerasde destete (8 meses de edad; 155 ± 3.4 kg),cruza Aberdeen Angus x Hereford, fueronasignadas a tres tratamientos de alimenta-ción invernal durante 92 días (4/6 al 3/9). Lostratamientos alimenticios fueron designadospara lograr tres tasas de ganancias diferen-tes: Grupo T (Testigo, perdiendo peso; n=12)pastoreando campo natural; Grupo M (Man-teniendo peso; n=12) y Grupo G (Ganandopeso; n=12). El manejo invernal en los gru-pos G y M se realizó sobre mejoramientosde campo con una mezcla de Trifoliumrepens, Lotus corniculatus y Loliummultiflorum con distintas asignaciones deforraje. Finalizada esta etapa las ternerasfueron manejadas en forma conjunta duran-

60
SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna INIA TREINTA Y TRES
Quintans et al., (1993) de -0.100 kg/a/d, parala misma categoría y similar base forrajera.Cabe destacar que la altura de la pasturalimitó de forma importante la cosecha delforraje (Cuadro 2). Las terneras del grupo Mlograron leves tasas de ganancias de pesodiarias (GMD) culminando dicho periodo concasi 10 kg más (GMD de 0.116 kg/a/d) cum-pliéndose el objetivo de obtener leves ganan-cias para este periodo. Las terneras del gru-po G ganaron 55 kg en el mismo periodo(GMD de 0.650 kg/a/d). Estas últimas GMDse asemejan a las encontradas por Short yBellows, (1971) de 0.630 kg/a/d, trabajandocon terneras de destete cruza Hereford-Angus, manejadas con un plano alto de ali-mentación post-destete. A nivel nacionalSoca et al., (2001) encontraron tasas deganancia de 0.670 kg/a/d recriando ternerassobre mejoramientos de Lotus Maku en igualestación del año con una asignación de fo-rraje de 16% del peso vivo.
A partir del manejo conjunto primaveral (3/9), las terneras presentaron tasas de ganan-cia de peso elevadas independientemente decual había sido su manejo nutricional ante-rior. Es remarcable la alta tasa de gananciade peso que realizaron en este periodo losanimales del grupo T (0.947 kg/día), ganan-cias superiores a las encontradas por
Quintans et al., (1993) para igual estacióndel año e igual categoría, pero sobre camponatural (0.696 kg/día). Estas tasas de ga-nancia determinaron que las terneras aumen-taran en este periodo 82 kg de peso vivo.Llama la atención la menor ganancia diariapara el periodo del grupo M, ya que Lawrencey Pearce, (1971), citado por Frick y Borges,(2003) invernando terneros a un nivel mediode restricción, obtuvieron ganancias diariasde peso en la realimentación de 0.980 kg/día. A campo, se pudo observar en este pe-riodo de reagrupamiento de los lotes un efec-to de dominancia de las terneras pertene-cientes al grupo G sobre las terneras delgrupo M. Sin embargo, es solo una observa-ción que no permite extraer una conclusión.El grupo G presentó tasas de gananciaestadísticamente iguales al grupo T.
Durante el periodo estival se observó unacaída en las ganancias diarias contrastantescon las ganancias citadas por otros autorespara igual periodo en vaquillonas (0.588 kg/d; Frick y Borges 2003) y (0.548 kg/d;Quintans et al., 1993). Esta caída en la tasade ganancia diaria (Cuadro 4) fue atribuida auna disminución en la calidad del forraje conun bajo porcentaje de proteína cruda y au-mentos en la fracción de fibra detergenteácido (Cuadro 5).
Cuadro 1. Peso y edad inicial, peso final y ganancia media diaria (+em) para manejo invernalPeso inicial Edad inicial Peso final Ganancia diariainvernal (kg) invernal (Días) invernal (kg) invernal (kg/día)
T 155.5 ± 3.4a 245.5 ± 3.2a 136.8 ± 4.7a -0.220 ± 0.04aM 155.5 ± 3.4a 244.0 ± 2.8a 165.3 ± 4.7b 0.116 ± 0.03bG 154.5 ± 3.4a 249.3 ±2.6a 209.2 ± 4.7c 0.650 ± 0.03c
T: grupo perdiendo peso. M: grupo manteniendo peso. G: grupo ganando peso. Letras diferentes en la mismacolumna expresan diferencias estadísticamente significativas con P< 0.05
Cuadro 2. Principales características del forraje ofrecido durante el periodo invernalDisponible* Altura inicial DMO* FDA* PC* AF*(kg MS/ha) forraje* (cm) (%) (%) (%) (%)
T 1575 4.5 38.7 46.2 8.5 5M 3662 16.2 48.2 45.1 13.7 3G 3317 18.5 51.9 44.2 13.3 15
T: grupo perdiendo peso. M: grupo manteniendo peso. G: grupo ganando peso. DMO: digestibilidad de la materiaorgánica. FDA: fibra detergente ácido. PC: proteína cruda. AF: asignación de forraje. *datos promedio
Tratamiento
Tratamiento
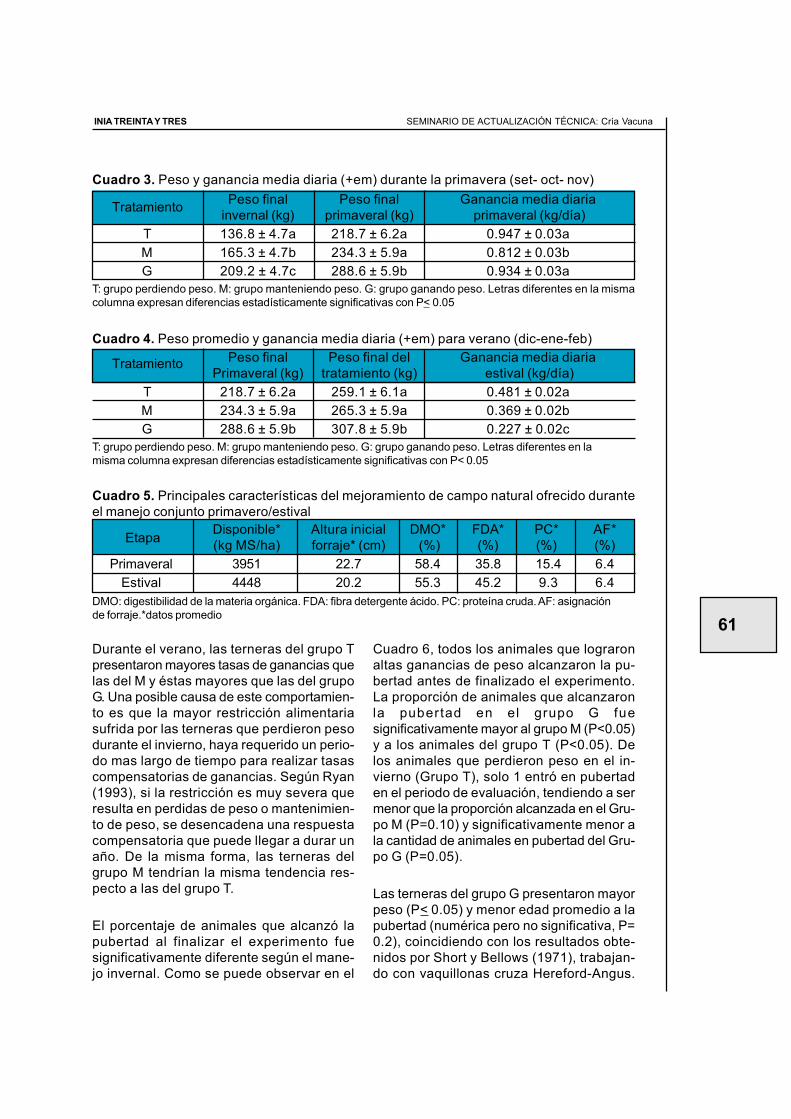
61
INIA TREINTA Y TRES SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna
Durante el verano, las terneras del grupo Tpresentaron mayores tasas de ganancias quelas del M y éstas mayores que las del grupoG. Una posible causa de este comportamien-to es que la mayor restricción alimentariasufrida por las terneras que perdieron pesodurante el invierno, haya requerido un perio-do mas largo de tiempo para realizar tasascompensatorias de ganancias. Según Ryan(1993), si la restricción es muy severa queresulta en perdidas de peso o mantenimien-to de peso, se desencadena una respuestacompensatoria que puede llegar a durar unaño. De la misma forma, las terneras delgrupo M tendrían la misma tendencia res-pecto a las del grupo T.
El porcentaje de animales que alcanzó lapubertad al finalizar el experimento fuesignificativamente diferente según el mane-jo invernal. Como se puede observar en el
Cuadro 6, todos los animales que lograronaltas ganancias de peso alcanzaron la pu-bertad antes de finalizado el experimento.La proporción de animales que alcanzaronla pubertad en el grupo G fuesignificativamente mayor al grupo M (P<0.05)y a los animales del grupo T (P<0.05). Delos animales que perdieron peso en el in-vierno (Grupo T), solo 1 entró en pubertaden el periodo de evaluación, tendiendo a sermenor que la proporción alcanzada en el Gru-po M (P=0.10) y significativamente menor ala cantidad de animales en pubertad del Gru-po G (P=0.05).
Las terneras del grupo G presentaron mayorpeso (P< 0.05) y menor edad promedio a lapubertad (numérica pero no significativa, P=0.2), coincidiendo con los resultados obte-nidos por Short y Bellows (1971), trabajan-do con vaquillonas cruza Hereford-Angus.
Cuadro 5. Principales características del mejoramiento de campo natural ofrecido duranteel manejo conjunto primavero/estival
Disponible* Altura inicial DMO* FDA* PC* AF*(kg MS/ha) forraje* (cm) (%) (%) (%) (%)
Primaveral 3951 22.7 58.4 35.8 15.4 6.4Estival 4448 20.2 55.3 45.2 9.3 6.4
DMO: digestibilidad de la materia orgánica. FDA: fibra detergente ácido. PC: proteína cruda. AF: asignaciónde forraje.*datos promedio
Etapa
Cuadro 3. Peso y ganancia media diaria (+em) durante la primavera (set- oct- nov)Peso final Peso final Ganancia media diaria
invernal (kg) primaveral (kg) primaveral (kg/día)T 136.8 ± 4.7a 218.7 ± 6.2a 0.947 ± 0.03aM 165.3 ± 4.7b 234.3 ± 5.9a 0.812 ± 0.03bG 209.2 ± 4.7c 288.6 ± 5.9b 0.934 ± 0.03a
T: grupo perdiendo peso. M: grupo manteniendo peso. G: grupo ganando peso. Letras diferentes en la mismacolumna expresan diferencias estadísticamente significativas con P< 0.05
Tratamiento
Cuadro 4. Peso promedio y ganancia media diaria (+em) para verano (dic-ene-feb)Peso final Peso final del Ganancia media diaria
Primaveral (kg) tratamiento (kg) estival (kg/día)T 218.7 ± 6.2a 259.1 ± 6.1a 0.481 ± 0.02aM 234.3 ± 5.9a 265.3 ± 5.9a 0.369 ± 0.02bG 288.6 ± 5.9b 307.8 ± 5.9b 0.227 ± 0.02c
T: grupo perdiendo peso. M: grupo manteniendo peso. G: grupo ganando peso. Letras diferentes en lamisma columna expresan diferencias estadísticamente significativas con P< 0.05
Tratamiento

62
SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna INIA TREINTA Y TRES
Figura 1. Evolución de peso según tratamiento durante el periodo experimental
Figura 2. Terneras en el mes de agosto sobre campo natural y sobre pasturas mejoradas
Cuadro 6. Porcentaje, peso, y edad en días (media ± em), de las hembras que lograron lapubertad según manejo invernal
No de animales que Peso pubertad Edad pubertadalcanzaron la pubertad (kg) (Días)
T 9.1 (1/11)b* 246 ± 17.5a 486 ± 21.2aM 41.7 (5/12)b 278 ± 7.8ab 466 ± 9.5aG 100 (12/12)a 294 ± 5.0b 452 ± 6.1a
T: grupo perdiendo peso. M: grupo manteniendo peso. G: grupo ganando peso. Letras diferentes en la mismacolumna expresan diferencias estadísticamente significativas con P< 0.05. Nota: Para la variable % animalespúberes T vs. M: P = 0.10. * murió una ternera durante el manejo conjunto
Tratamiento

63
INIA TREINTA Y TRES SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna
Estos autores encontraron que a medida queaumentaba la ganancia diaria, las vaquillonasalcanzaban la pubertad con menor edad y amayor peso vivo. A pesar de registrarse pe-sos similares al final del periodo experimen-tal en los grupos T y M (atribuibles al creci-miento compensatorio) la act iv idadreproduct iva fue mayor en aquel lasvaquillonas que no tuvieron restriccionesnutricionales severas durante el invierno.
Consideraciones finales
La alimentación durante el primer inviernopos-destete afectó la proporción, peso y edada la que los animales entraron en pubertad.Las altas tasas de ganancia durante la pri-mavera y verano no fueron suficientes pararevertir el efecto que tuvo el primer inviernosobre aspectos reproductivos, dentro del pe-riodo evaluado. Similares resultados fueronobtenidos por Frick y Borges, (2003), quie-nes encontraron que la ganancia diaria in-vernal (a diferencia del peso al entore) es unbuen indicador de la futura fertilidad envaquillonas con servicio a los 18 meses loque también debería ser tenido en cuentapara entores mas tardíos. De todas formascabe destacar que deben repetirse este tipode experimentos teniendo en cuenta la va-riabilidad que existe en los resultados cuan-do se realizan trabajos en condicionespastoriles.
Referencias bibliográficas
Frick, F., y Borges, M. 2003. Factores queafectan la fertilidad de vaquillonas Herefordy Brahman x Hereford entoradas a los 18meses de edad. Tesis Ing. Agr. Montevideo,Uruguay. Facultad de Agronomía. 140 p.Quintans, G., Vaz Martins, D. y Carriquiry,E. 1993. Efecto de la suplementación inver-nal sobre el comportamiento de terneras.INIA- Campo Natural. Estrategia invernal -Manejo y Suplementación. Resultados Ex-perimentales.Ryan, WJ., Williams, IH. and Moir, RJ. 1993.Compensatory grown in sheep and cattle. 1-
Growth pattern and feed intake. AustralianJournal of Agricultural Research, 44 1609-1621.Short, RE. and Bel lows, RA. 1971.Relationships among weigth gains, age atpuberty and reproductive performance inheifers. Journal of Animal Science. 32: 127-131.Soca, P., Bermúdez, R., Ayala, W.,Mancuello, C., Arrarte, D., Pereira, G., Leiva,G., Fernández, M. y Hernández, P. 2001.Utilización de mejoramientos de campo conlotus El Rincón y lotus Maku para la recríavacuna en la zona este del país. En: Jorna-da de Difusión de Resultados. (2001, Ro-cha). Utilización de mejoramientos de cam-po con lotus El Rincón y lotus Maku para larecría vacuna en la zona este del país Mon-tevideo, INIA. pp. 7-23.Straumann, JM. 2006. Efecto del manejonutricional en el primer invierno sobre la apa-rición de la pubertad en terneras de razacarnicera. Primer año de evaluación. Tesisde grado de la Facultad de Agronomía, Uni-versidad de la República, p 99.

64
SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna INIA TREINTA Y TRES
2.4. EFECTO DEL MANEJO NUTRICIONAL EN EL PRIMER INVIERNO YEL BIOTIPO SOBRE LA APARICIÓN DE LA PUBERTAD EN TERNERASDE RAZA CARNICERA (SEGUNDO AÑO DE EVALUACIÓN)
Santiago Barreto1 , Diego Negrín1, Walter Ayala2 , Graciela Quintans3
corniculatus y Lolium multiflorum que teníaal momento del experimento más de diezaños de instalados y un 20% de legumino-sas (entre 10 y 14% de proteína cruda). Fi-nalizada esta etapa, las terneras fueronmanejadas en forma conjunta durante 196días sobre el mismo mejoramiento de cam-po hasta el final del experimento (Marzo),con el objetivo que realizaran altas ganan-cias de peso vivo. Los animales fueron pe-sados cada 14 días y fueron sangrados se-manalmente desde el comienzo del manejoconjunto (11/9/03) (11.2 meses de edad pro-medio) hasta el final del experimento (26/3/04) (17.9 meses de edad promedio) paradeterminación de progesterona en sangre.El criterio empleado para determinar el ini-cio de la pubertad fue la primera muestra dedos consecutivas con concentraciones deprogesterona por encima de 1 ng/ml. Duran-te los últimos 24 días del experimento serealizó detección de celos dos veces al díadurante una hora y los días 16 y 26 de mar-zo se realizaron ecografías para determinaractividad ovárica. En la pastura se determi-nó: disponibilidad y altura de forraje, rela-ción verde-seco y composición botánica. Serealizaron análisis de digestibilidad In Vitro,proteína cruda, fibra detergente ácido y ce-nizas. Las variables continuas se analiza-ron mediante el análisis de un modelo linealgeneral, utilizando el procedimiento GLM delSistema de Análisis Estadístico SAS (1999).Para las variables discretas se utilizó el pro-grama GENMOD de SAS (1999) con distri-bución binomial. Los valores se presentancomo promedio ± error de la media (em) y elnivel de significancia utilizado fue P <0.01.
1 Ing. Agr. Realizó tesis de grado en INIA Treinta y Tres.2 Ing Agr., PhD. - Director de Programa Nacional de Pasturas y Forrajes - INIA Treinta y Tres.3 Ing. Agr., PhD. - Programa Nacional de Producción de Carne y Lana - INIA Treinta y Tres.
Introducción
En el trabajo anterior (primer año de evalua-ción dentro de la línea de investigación depubertad), se evaluó el efecto de diferentestasas de ganancias invernales sobre la edad,peso y proporción de animales que entranen pubertad. El objetivo del presente trabajofue evaluar el efecto de dos tasas de ganan-cias invernales en dos biotipos carniceros(Hereford puro y cruzas Aberdeen Angus xHereford) sobre la aparición de la pubertad(Tesis de Facultad de Agronomía - Barreto yNegrín 2005). Las condiciones del experi-mento fueron muy similares al trabajo delprimer año.
Materiales y métodos
El experimento fue realizado entre Junio del2003 y Marzo del 2004, en la Unidad Expe-rimental Palo a Pique (UEPP), pertenecien-te a la Estación Experimental del Este, INIATreinta y Tres. Treinta y seis terneras dedestete Hereford (H, n=18) y cruzasAberdeen Angus con Hereford (AH, n=18)(edad promedio: 250 ± 4.4 días, peso pro-medio: 145.5 ± 14.5 kg) fueron asignadas ados tratamientos de alimentación invernaldurante 97 días (6/6 al 11/9). El diseño ex-perimental fue un factorial 2x2, siendo losfactores el biotipo (B) y la tasa de gananciainvernal (TG). Los tratamientos alimenticiosfueron asignados para lograr dos tasas deganancias contrastantes: pérdidas de pesoinvernal (PPI, n=18) y ganancias de pesoinvernal (GPI, n=18). El manejo invernal serealizó sobre mejoramientos de campo conuna mezcla de Trifolium repens, Lotus

65
INIA TREINTA Y TRES SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna
Resultados y discusión
El forraje verde invernal disponible mínimofue de 1250 kg MS/ha en julio y el máximo afinales de agosto fue de 1950 kg MS/ha. Acausa de un mayor grado de degradacióndel mejoramiento (restos secos, gramilla),se decidió aumentar la asignación de forrajeinvernal (kg MS/100 kg PV/día) en ambostratamientos en comparación con el experi-mento del año anterior (2002). Las asigna-ciones de forraje manejadas para los trata-mientos de pérdida (PPI) y ganancia (GPI)de peso fueron en promedio durante los 97días del periodo invernal de 6 y 18% respec-tivamente. Este manejo fue exitoso en lo-grar diferencias significativas en las tasasde ganancias durante el invierno (P<0.01).Las terneras que pastorearon la menor asig-nación (PPI) perdieron 8.3 kg en promedioen los 97 días de invierno (0.116 ± 0.02 kg/a/d) mientras que las terneras con una ma-
yor asignación (GPI) ganaron casi 40.6 kgen el mismo periodo (0.398 ± 0.02 kg/a/d).Respecto al biotipo, las terneras HH tendie-ron (P=0.07) a presentar mayores tasas deganancia invernal que las AH (0.165 ± 0.02vs. 0.116 ± 0.02 kg/a/d) (Cuadro 2).
El periodo de 196 días de manejo conjuntosobre campo mejorado, fue divido para suanálisis en los primeros 98 días (primavera)y últimos 98 días (verano). La disponibilidadpromedio de las pasturas manejadas duran-te el periodo primaveral fue próxima a los5000 kg MS/ha con un porcentaje de legu-minosas promedio de 21% y con una dismi-nución en la proporción de restos secos enrelación al invierno. La tasa de gananciamedia diaria de las terneras en este periodofue de 1.162 ± 0.03 y 1.102 ± 0.03 kg/a/dpara PPI y GPI respectivamente (P= 0.12).Al comparar las tasas de ganancias diariasentre los biotipos, estas fueron similares,
Disponible(kg MS/ha)
% de forraje verdedel total disponibleLeguminosas (%)
Altura (cm)% Digestibilidad
% PC% FDA% FDN
% Cenizas
Característica Invierno Primavera Verano
3080.5 4943.3 3536.7
49.8 78.7 64.0
11.7 21 3.711.3 15.5 13.333.3 52.1 48.110.1 12.1 8.0847.3 42.4 47.673.0 61.2 73.110.3 11.1 8.5
PC: Proteína cruda. FDA: fibra detergente acida. FDN: fibra detergente neutra.
Cuadro 1. Principales características del forraje ofrecido durante el periodo experimental
Etapa TG (kg/día) Biotipo (B) PPPI GPI H AH TG B
Invernal1 0.116± 0.02 0.398±0.02 0.165±0.02 0.116± 0.02 <0.001 0.07Primaveral2 1.162 ±0.03 1.103±0.03 1.113±0.03 1.152± 0.03 0.12 0.30
Estival3 0.540 ±0.02 0.419±0.02 0.460±0.02 0.499± 0.02 <0.001 0.16TG: tasa de ganancia diaria. 1 periodo de 97 dias (6/6-11/9) 2 periodo de 98 dias(11/9-18/12) 3 periodo de 98dias (18/12 -25/3)
Cuadro 2. Tasas de ganancia (TG) de peso vivo (kg/a/d) en los tres periodos evaluados(media ± em)

66
SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna INIA TREINTA Y TRES
siendo las mismas de 1.113 ± 0.03 y 1.152± 0.03 kg/a/d para las terneras H y AH res-pectivamente (P = 0.30).
Para el verano se observó una caída en lasganancias diarias en ambos grupos con res-pecto a la primavera, concordantes con lasganancias citadas por otros autores paraigual periodo en vaquillonas (Frick y Borges,2003; Quintans et al., 1993), esta disminu-ción en las ganancias generales, se asociaa una caída de la calidad de las pasturas enverano, producto de la floración del trébolblanco, raigras y las gramíneas anuales, conaumento en la proporción de restos secos.Durante este periodo también se observó unadiferencia en la tasa de ganancia debida altratamiento invernal donde los animales delgrupo PPI ganaron 0.540 ± 0.02 kg/a/d au-mentando 52 kg su peso vivo en promedio,mientras que las terneras del grupo GPI ga-naron 0.419 ± 0.02 kg/a/d (P<0.01)incrementando su peso vivo en 41 kg pro-medio.
Los animales que perdieron peso en invier-no comenzaron a realizar una tasa de ga-nancia compensatoria hacia el verano, sinembargo dicho periodo no fue suficiente pararevertir las diferencias generadas en el in-vierno, finalizando el periodo experimental(25/3/04) con terneras significativamentemas pesadas en el grupo GPI que en el PPI(Cuadro 3).
Según Frick y Borges, (2003) se puedeconsiderar una ganancia diaria de 0.250 kg/día como un valor de ganancia invernal
“umbral” por debajo de la cual existecrecimiento compensatorio. Por otro lado,Ryan (1990) demostró que la duración de lacompensación es directamente proporcionala la severidad de la restricción. La respuestacompensatoria de los animales restringidosuna vez comenzada la realimentación estáen función tanto del grado en que su tasa decrecimiento aumenta por encima de los norestringidos, como del tiempo en quepersiste esa mayor tasa de crecimiento. Alaumentar la severidad de la restricción esprobable que aumente el largo del periodocompensator io, más que la tasa decrecimiento durante la compensación (Ryan,1990). Esto podría estar explicando porquéen primavera las terneras del grupo PPI nolograron mayor tasa de crecimiento, aunqueexistió una tendencia (P= 0.12), la cual setransformó en mayor tasa de ganancia en elverano (P<0.01).
El porcentaje de animales que alcanzó lapubertad al finalizar el experimento fuesignificativamente diferente según el mane-jo invernal (Cuadro 4). Tan solo 7/18 terne-ras con PPI alcanzaron la pubertad al finali-zar el periodo experimental mientras que 15/18 terneras con GPI lograron llegar a la pu-bertad al final del ensayo (39% vs. 83% paraPPI y GPI respectivamente) (P<0.01). A suvez, las terneras con GPI, lo hicieron 34 díasantes que las terneras del grupo PPI en elmismo periodo. Por otra parte, se registróuna interacción entre tasa de ganancia ybiotipo para el porcentaje de animales púberes(Cuadro 5). El porcentaje de animales que al-canzaron la pubertad en el tratamiento con
Cuadro 3. Evolución de peso (kg) para ambos tratamientos durante el transcurso del expe-rimento
Inicio Trat 0 Fin Invierno 1 Fin Primavera 2 Fin Verano 3
PPI 145.5 ± 15.5a 137.2 ± 17.0a 245.1 ± 24.8a 299.3 ± 26.7a
GPI 145.4 ± 13.4a 186.0 ± 19.3b 289.2 ± 22.0b 332.1 ± 23.3bDif. Peso* 0.1 48.8 44.1 32.8
PPI: perdida de peso invernal. GPI: ganancia de peso invernal. 0 Inicio Tratamiento: 6/6/03. 1 Fin Invierno:11/9/03. 2 Fin Primavera: 18/12/03. 3 Fin Verano: 25/3/04. * Diferencia de peso en kg (GPI – PPI). Diferentesletras en la misma columna presentan diferencias estadísticamente significativas al 1%.
Tratamientoinvernal
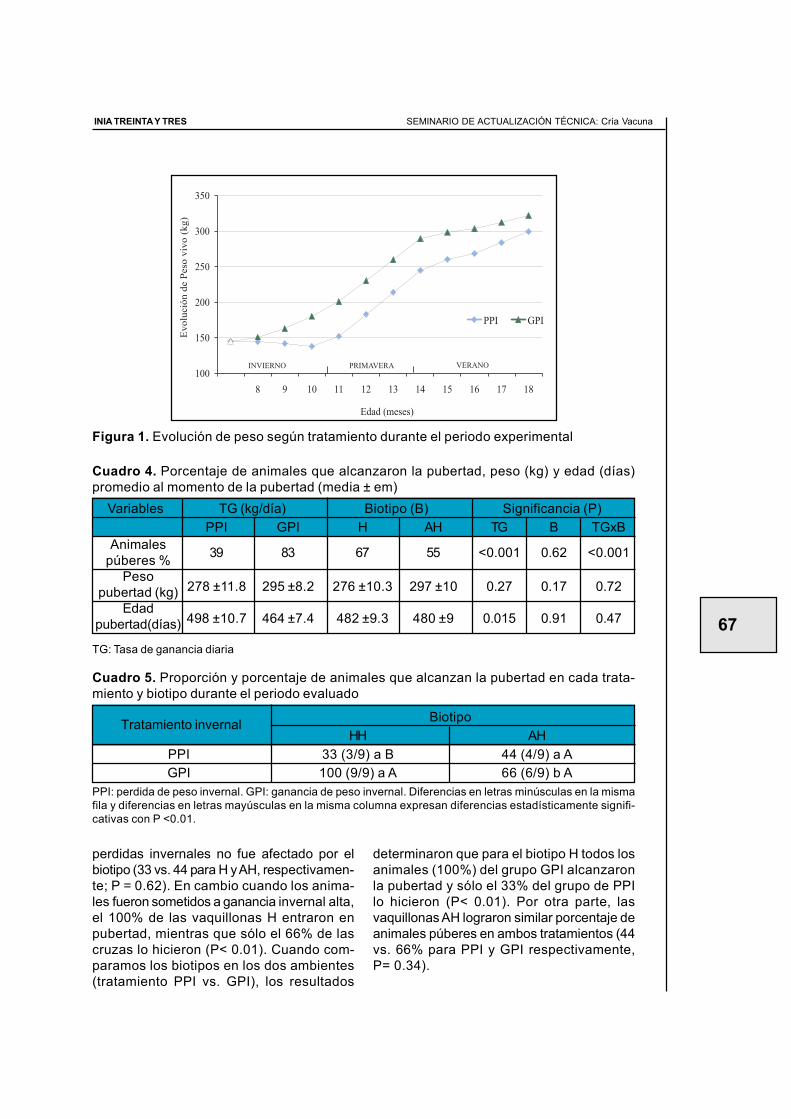
67
INIA TREINTA Y TRES SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna
perdidas invernales no fue afectado por elbiotipo (33 vs. 44 para H y AH, respectivamen-te; P = 0.62). En cambio cuando los anima-les fueron sometidos a ganancia invernal alta,el 100% de las vaquillonas H entraron enpubertad, mientras que sólo el 66% de lascruzas lo hicieron (P< 0.01). Cuando com-paramos los biotipos en los dos ambientes(tratamiento PPI vs. GPI), los resultados
Figura 1. Evolución de peso según tratamiento durante el periodo experimental
Animalespúberes %
Pesopubertad (kg)
Edadpubertad(días)
Cuadro 4. Porcentaje de animales que alcanzaron la pubertad, peso (kg) y edad (días)promedio al momento de la pubertad (media ± em)
Variables TG (kg/día) Biotipo (B) Significancia (P)PPI GPI H AH TG B TGxB
39 83 67 55 <0.001 0.62 <0.001
278 ±11.8 295 ±8.2 276 ±10.3 297 ±10 0.27 0.17 0.72
498 ±10.7 464 ±7.4 482 ±9.3 480 ±9 0.015 0.91 0.47
TG: Tasa de ganancia diaria
determinaron que para el biotipo H todos losanimales (100%) del grupo GPI alcanzaronla pubertad y sólo el 33% del grupo de PPIlo hicieron (P< 0.01). Por otra parte, lasvaquillonas AH lograron similar porcentaje deanimales púberes en ambos tratamientos (44vs. 66% para PPI y GPI respectivamente,P= 0.34).
Cuadro 5. Proporción y porcentaje de animales que alcanzan la pubertad en cada trata-miento y biotipo durante el periodo evaluado
BiotipoHH AH
PPI 33 (3/9) a B 44 (4/9) a AGPI 100 (9/9) a A 66 (6/9) b A
PPI: perdida de peso invernal. GPI: ganancia de peso invernal. Diferencias en letras minúsculas en la mismafila y diferencias en letras mayúsculas en la misma columna expresan diferencias estadísticamente signifi-cativas con P <0.01.
Tratamiento invernal

68
SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna INIA TREINTA Y TRES
Los animales del grupo PPI (7/18) alcanza-ron la pubertad con un peso promedio de 278kg. Sin embargo, al finalizar el periodo ex-perimental, los animales de dicho grupo queno habían alcanzado la pubertad (11/18)promediaron un peso de 295 kg, similar alpeso con el que los animales del grupo GPI(15/18) lograron su primera actividad luteal.Esto indicaría que la tasa de ganancia inver-nal estaría influyendo en la posibilidad dealcanzar la pubertad y que el peso vivo porsí mismo no sería un buen indicador de laactividad reproductiva en esta categoría deanimales.
Dos animales que entraron en pubertad nomanifestaron celo durante el mes de marzo(Cuadro 6). Esto también fue confirmado porlos bajos niveles de progesterona en sangreque presentaban al final del periodo de eva-luación, así como las ecografías ováricasque manifestaban ausencia de cuerpo lúteo.Es común observar que los animales luegode entrar en pubertad caen en anestro yasea por causas nutricionales o climáticas,entre otras. En el caso de este experimen-to, a fines del verano y principio de otoño seregistró una sequía que podría haber afecta-do el comportamiento de estos animales.
Consideraciones finales
La tasa de ganancia invernal fue el factormás importante en determinar el porcentajede animales púberes. Las terneras con ga-nancias de peso invernal lograron entrar an-tes en pubertad y en mayor proporción queaquellas que perdieron peso durante dichoperiodo. Se manifestó una interacción entretasa de ganancia invernal y biotipo en el
porcentaje de animales que alcanzaron lapubertad en el periodo evaluado. Las terne-ras cruzas AH no presentaron diferencias enel porcentaje de pubertad según tasas deganancia invernal. Sin embargo las ternerasHereford puras lograron un mayor porcenta-je de pubertad cuando las tasas de ganan-cias invernales fueron al tas ysignificativamente menor porcentaje de lasmismas alcanzó la pubertad cuando perdie-ron peso en invierno. Si bien las terneras queperdieron peso en invierno presentaron unamayor tasa de ganancia en el verano res-pecto a las que ganaron peso, ésta no fuesuficiente para revertir las diferencias gene-radas durante el invierno. Por otra parte, sibien ambos grupos de terneras finalizaronel experimento con pesos significativamentediferentes (299 vs. 332 kg para PPI y GPIrespectivamente) el peso logrado al final delexperimento por las terneras del grupo PPIque no habían entrado en pubertad (295 kg,n=11/18, 61%) fue el mismo que el alcan-zado por las terneras del grupo GPI a la pu-bertad (295 kg, n= 15/18, 83%). Esto esta-ría indicando que el peso por sí mismo en elotoño no es el mejor indicador de la ciclicidadovárica en esta categoría de animales jóve-nes. Esta información es coincidente conresultados obtenidos en el experimento an-terior (2002-2003). A su vez, Frick y Borges,(2003) también concluyeron que es másimportante el peso al final del invierno resul-tante de la tasa de ganancia invernal y delpeso al destete, que el peso al servicio alos 18 meses para predecir el comportamien-to reproductivo, ya que el peso al serviciono considera la distribución estacional de laganancia. Por lo tanto, para entores antici-pados la distribución de las tasas de ganan-cias deben ser consideradas y dentro de
Cuadro 6. Proporción y porcentaje de terneras que alcanzaron la pubertad y que fuerondetectadas en celo durante el último mes de evaluación
Tratamiento Proporción de Proporción de animalesanimales púberes % en celo (marzo) %
PPI 7/18 39b 6/18 33bGPI 15/18 83a 14/18 77a
Diferentes letras en la misma columna presentan diferencias estadísticamente significativas al 1%.

69
INIA TREINTA Y TRES SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna
este marco, la tasa de ganancia invernalparecería ser un factor determinante paraalcanzar la performance reproductiva adecua-da.
Referencias bibliográficas
Barreto, S. y Negrín, D. 2005. Efecto delmanejo nutricional en el primer invierno so-bre la aparición de la pubertad en ternerasde raza carnicera. Segundo año de evalua-ción. Tesis de grado de la Facultad de Agro-nomía, Universidad de la Republica, p107.Frick, F. y Borges, M. 2003. Factores queafectan la fertilidad de vaquillonas Herefordy Brahman x Hereford entoradas a los 18meses de edad. Tesis Ing. Agr. Montevideo,Uruguay. Facultad de Agronomía. 140 p.Quintans, G., Vaz Martins, D. y Carriquiry,E. 1993. Efecto de la suplementación inver-nal sobre el comportamiento de terneras.INIA- Campo Natural. Estrategia invernal-Manejo y Suplementación. Resultados Ex-perimentales.Ryan, WJ. 1990. Compensatory Growth insheep and cattle. PhD. Thesis, School ofAgriculture. The University of Western Aus-tralia.Straumann, JM. 2006. Efecto del manejonutricional en el primer invierno sobre la apa-rición de la pubertad en terneras de razacarnicera. Primer año de evaluación. Tesisde grado de la Facultad de Agronomía, Uni-versidad de la República, p 99.

70
SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna INIA TREINTA Y TRES
2.5. EFECTO DE TRES GANANCIAS INVERNALES SOBRE LAAPARICIÓN DE LA PUBERTAD EN TERNERAS DE RAZA CARNICERA(TERCER AÑO DE EVALUACIÓN)
Álvaro Costa1 , Roberto Moreira1, Antonia Scarsi1, Walter Ayala2 , Graciela Quintans3
rimento (Marzo), con el objetivo que realiza-ran altas ganancias de peso vivo. Los ani-males fueron pesados cada 14 días y fueronsangrados semanalmente a partir de 7/10hasta el final del experimento (31/3/04) paradeterminación de progesterona en sangre.El criterio empleado para determinar el ini-cio de la pubertad fue la primera muestra dedos consecutivas con concentraciones deprogesterona por encima de 1 ng/ml. Desdenoviembre hasta la finalización del experi-mento (31/3/05) se realizó detección de ce-los dos veces al día durante 45 minutos ylos días 21 y 31 de marzo se realizaronecografías para determinar actividad ovárica.Al final del ensayo (31/3/05) se efectuó unaecografía midiendo espesor de grasa dorsal,para determinar la relación entre el manejoalimenticio, el espesor de grasa dorsal y laaparición de la pubertad. En la pastura sedeterminó: disponibilidad, rechazo y alturade forraje, relación verde-seco y composi-ción botánica. Se realizaron análisis dedigestibilidad In Vitro, proteína cruda, fibradetergente ácido y cenizas. Las variablescontinuas (peso vivo, ganancia de peso yespesor de grasa dorsal) se analizaron me-diante un modelo lineal general, utilizandoel procedimiento GLM del sistema de análi-sis estadístico SAS versión 9.03 (2005).Para las variables discretas se utilizó el pro-grama GENMOD de SAS con distribuciónbinomial para pubertad y ciclicidad, y distri-bución de Poisson para número de celos.Los valores se presentan como promedio ±error de la media (em) y el nivel designificancia utilizado fue P<0.05.
1 Ing. Agr. Realizó tesis de grado en INIA Treinta y Tres.2 Ing Agr., PhD. - Director de Programa Nacional de Pasturas y Forrajes - INIA Treinta y Tres.3 Ing. Agr., PhD. - Programa Nacional de Producción de Carne y Lana - INIA Treinta y Tres.
Introducción
Este trabajo se enmarca dentro de la mis-ma línea de investigación que se viene pre-sentando, donde los efectos de diferentesmanejos pastoriles sobre la performance pro-ductiva y reproductiva de terneras son eva-luados. En este estudio en particular se in-corporaron nuevas variables como fue la de-tección de celo por un periodo de 6 meses yuna medición de espesor de grasa dorsal alfinalizar el trabajo (Tesis Facultad de Agro-nomía – Costa, Moreira y Scarsi 2007).
Materiales y métodos
El experimento, fue realizado entre Junio del2004 y Marzo del 2005, en la Unidad Expe-rimental Palo a Pique (UEPP), pertenecien-te a la Estación Experimental del Este, INIATreinta y Tres. Treinta y seis terneras dedestete Aberdeen Angus x Hereford (AH) (8meses de edad; 138.7 ± 3.1 kg) fueron asig-nadas a tres tratamientos de alimentacióninvernal durante 111 días (4/6 al 23/9). Lostratamientos alimenticios fueron designadospara lograr tres tasas de ganancias diferen-tes: Grupo B (Baja, perdiendo peso; n=12)pastoreando campo natural; Grupo M (Me-dia, manteniendo peso; n=12) y Grupo A(Alta, ganando peso; n=12). El manejo in-vernal en los grupos A y M se realizó sobremejoramientos de campo con una mezclade Trifolium repens, Lotus corniculatus yLolium multiflorum, con distintas asignacio-nes de forraje. Finalizada esta etapa (23/9),las terneras fueron manejadas en forma con-junta durante 189 días sobre el mismo me-joramiento de campo hasta el final del expe-

71
INIA TREINTA Y TRES SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna
Resultados y discusión
El manejo nutricional invernal fue exitoso enlograr diferencias significativas en las tasasde ganancias durante el invierno (P<0.05).Si bien es esperable una perdida de peso,las terneras que pastoreaban campo natural(B) ganaron 11 kg durante los 111 días delperiodo invernal, a razón de 0.134 kg/a/d conuna asignación de forraje promedio de 5.6%.Cabe destacar la mayor tasa de gananciadiaria de éstas terneras en comparación conel periodo invernal del primer año de experi-mento (2002-2003, Straumann, 2006), en elcual terneras sobre campo natural con asig-nación de forraje similar (5%) registraronperdidas de 0.220 kg/a/d. Estas diferenciasestán asociadas a las bajas temperaturas,al mayor porcentaje de heladas y a proble-mas por anegamiento de potreros durante elinvierno del primer año.
Las terneras del grupo M lograron gananciasde peso de 0.385 kg/a/d con una asignaciónde forraje del 8%, culminando dicho periodocon 37.3 kg más. Estas ganancias fueronsimilares a las obtenidas en el periodo in-vernal del segundo año de experimento(0.398 kg/a/d) (2003-2004 Barreto y Negrín,2005) en terneras sobre mejoramientos decampo pero con una asignación del 18%.Esa menor tasa de ganancia diaria en rela-ción a la asignación de forraje se explica poruna mayor proporción de forraje seco ygramilla en comparación con el presenteexperimento.
Las terneras del grupo A ganaron 56.2 kg enel mismo periodo (0.535 kg/a/d), manejadasen el mismo mejoramiento pero con unaasignación de forraje del 16%. Durante elprimer año (2002-2003) se registraron ma-yores ganancias invernales (0.650 kg/a/d)sobre mejoramiento de campo con una asig-nación del 15%, a causa de una mejor cali-dad del mejoramiento con mayor porcentajede leguminosas y mayor nivel de proteínacruda en comparación con el presente tra-bajo.
El periodo de 189 días de manejo conjuntosobre campo mejorado, fue divido para suanálisis en los primeros 88 días (primavera)y últimos 101 días (verano). Debido a lascondiciones estivales que se presentaron enel año 2005 para un mejor análisis se dividióel verano en dos: verano antes de seca (Va/s) correspondiendo a la información recaba-da hasta el 15 de enero del 2005 (26 días) yverano después de seca (Vd/s) que corres-ponde al periodo restante hasta el otoño (68días).
Durante la primavera disminuyó la proporciónde restos secos en el mejoramiento, aumen-tando el nivel de proteína cruda asociado aun mayor porcentaje de leguminosas. Lostres grupos registraron aumentos en las ta-sas de ganancia media diaria primaveral sinobservarse diferencias estadísticas entregrupos de terneras (0.808; 0.807 y 0.804 kg/a/d para baja, media y alta ganancia inver-nal respectivamente).
Figura 1. Diferentes determinaciones en la pastura (altura, disponibilidad, composiciónbotánica)
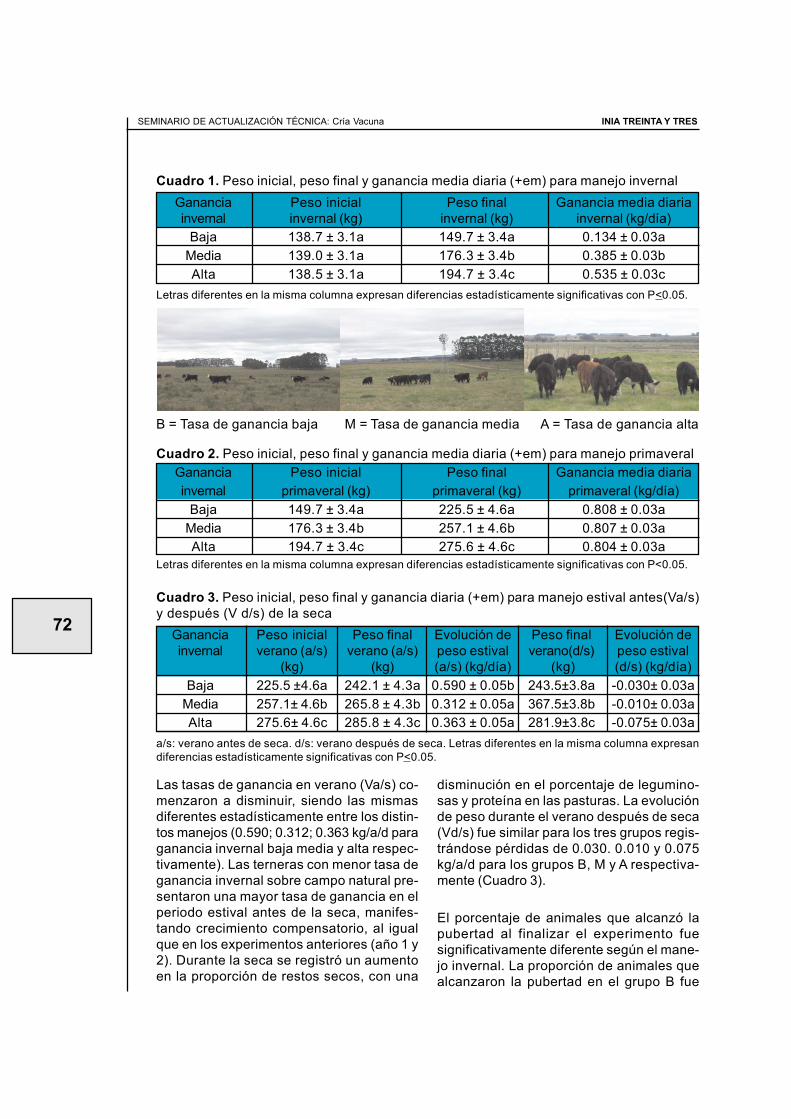
72
SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna INIA TREINTA Y TRES
B = Tasa de ganancia baja M = Tasa de ganancia media A = Tasa de ganancia alta
Cuadro 1. Peso inicial, peso final y ganancia media diaria (+em) para manejo invernalGanancia Peso inicial Peso final Ganancia media diariainvernal invernal (kg) invernal (kg) invernal (kg/día)
Baja 138.7 ± 3.1a 149.7 ± 3.4a 0.134 ± 0.03aMedia 139.0 ± 3.1a 176.3 ± 3.4b 0.385 ± 0.03bAlta 138.5 ± 3.1a 194.7 ± 3.4c 0.535 ± 0.03c
Letras diferentes en la misma columna expresan diferencias estadísticamente significativas con P<0.05.
Las tasas de ganancia en verano (Va/s) co-menzaron a disminuir, siendo las mismasdiferentes estadísticamente entre los distin-tos manejos (0.590; 0.312; 0.363 kg/a/d paraganancia invernal baja media y alta respec-tivamente). Las terneras con menor tasa deganancia invernal sobre campo natural pre-sentaron una mayor tasa de ganancia en elperiodo estival antes de la seca, manifes-tando crecimiento compensatorio, al igualque en los experimentos anteriores (año 1 y2). Durante la seca se registró un aumentoen la proporción de restos secos, con una
Cuadro 3. Peso inicial, peso final y ganancia diaria (+em) para manejo estival antes(Va/s)y después (V d/s) de la seca
Ganancia Peso inicial Peso final Evolución de Peso final Evolución deinvernal verano (a/s) verano (a/s) peso estival verano(d/s) peso estival
(kg) (kg) (a/s) (kg/día) (kg) (d/s) (kg/día)Baja 225.5 ±4.6a 242.1 ± 4.3a 0.590 ± 0.05b 243.5±3.8a -0.030± 0.03a
Media 257.1± 4.6b 265.8 ± 4.3b 0.312 ± 0.05a 367.5±3.8b -0.010± 0.03aAlta 275.6± 4.6c 285.8 ± 4.3c 0.363 ± 0.05a 281.9±3.8c -0.075± 0.03a
a/s: verano antes de seca. d/s: verano después de seca. Letras diferentes en la misma columna expresandiferencias estadísticamente significativas con P<0.05.
disminución en el porcentaje de legumino-sas y proteína en las pasturas. La evoluciónde peso durante el verano después de seca(Vd/s) fue similar para los tres grupos regis-trándose pérdidas de 0.030. 0.010 y 0.075kg/a/d para los grupos B, M y A respectiva-mente (Cuadro 3).
El porcentaje de animales que alcanzó lapubertad al finalizar el experimento fuesignificativamente diferente según el mane-jo invernal. La proporción de animales quealcanzaron la pubertad en el grupo B fue
Cuadro 2. Peso inicial, peso final y ganancia media diaria (+em) para manejo primaveralGanancia Peso inicial Peso final Ganancia media diariainvernal primaveral (kg) primaveral (kg) primaveral (kg/día)
Baja 149.7 ± 3.4a 225.5 ± 4.6a 0.808 ± 0.03aMedia 176.3 ± 3.4b 257.1 ± 4.6b 0.807 ± 0.03aAlta 194.7 ± 3.4c 275.6 ± 4.6c 0.804 ± 0.03a
Letras diferentes en la misma columna expresan diferencias estadísticamente significativas con P<0.05.
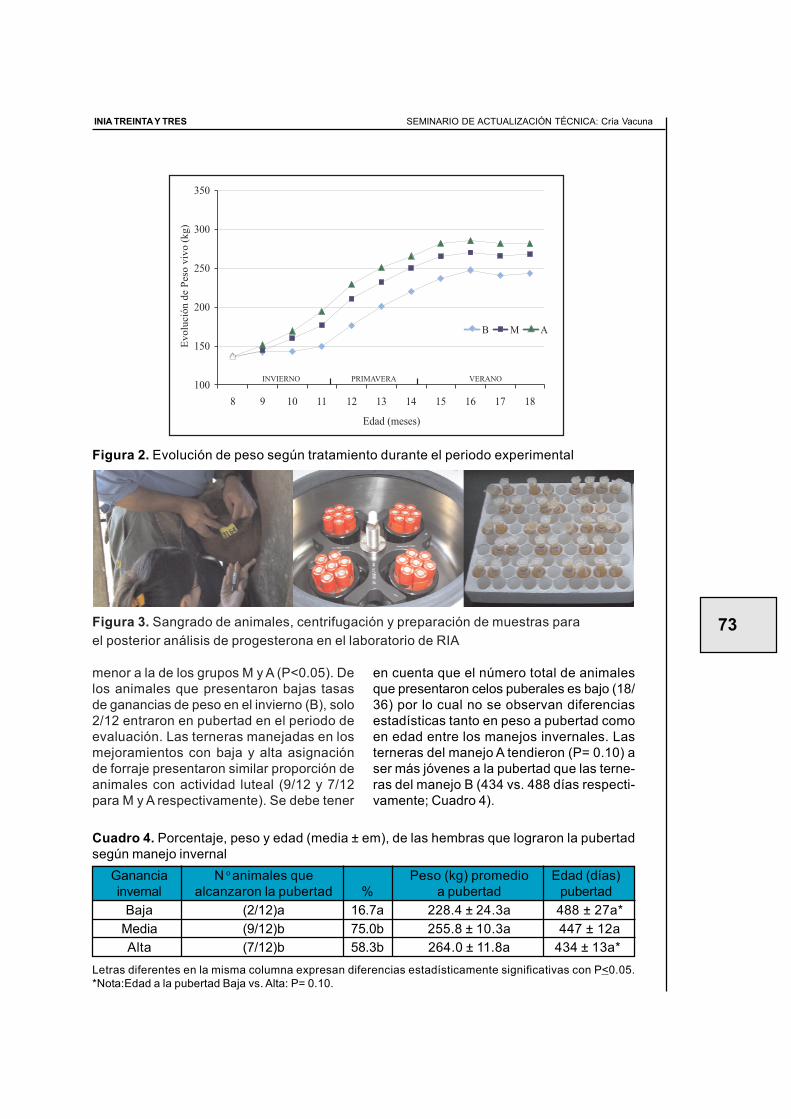
73
INIA TREINTA Y TRES SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna
Figura 2. Evolución de peso según tratamiento durante el periodo experimental
menor a la de los grupos M y A (P<0.05). Delos animales que presentaron bajas tasasde ganancias de peso en el invierno (B), solo2/12 entraron en pubertad en el periodo deevaluación. Las terneras manejadas en losmejoramientos con baja y alta asignaciónde forraje presentaron similar proporción deanimales con actividad luteal (9/12 y 7/12para M y A respectivamente). Se debe tener
Figura 3. Sangrado de animales, centrifugación y preparación de muestras parael posterior análisis de progesterona en el laboratorio de RIA
Cuadro 4. Porcentaje, peso y edad (media ± em), de las hembras que lograron la pubertadsegún manejo invernal
Ganancia N o animales que Peso (kg) promedio Edad (días)invernal alcanzaron la pubertad % a pubertad pubertad
Baja (2/12)a 16.7a 228.4 ± 24.3a 488 ± 27a*Media (9/12)b 75.0b 255.8 ± 10.3a 447 ± 12aAlta (7/12)b 58.3b 264.0 ± 11.8a 434 ± 13a*
Letras diferentes en la misma columna expresan diferencias estadísticamente significativas con P<0.05.*Nota:Edad a la pubertad Baja vs. Alta: P= 0.10.
en cuenta que el número total de animalesque presentaron celos puberales es bajo (18/36) por lo cual no se observan diferenciasestadísticas tanto en peso a pubertad comoen edad entre los manejos invernales. Lasterneras del manejo A tendieron (P= 0.10) aser más jóvenes a la pubertad que las terne-ras del manejo B (434 vs. 488 días respecti-vamente; Cuadro 4).

74
SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna INIA TREINTA Y TRES
Cuando analizamos las terneras agrupándo-las en púberes y no púberes (Cuadro 5) seobserva que las terneras que alcanzaron lapubertad presentaron tasas de ganancias depeso vivo superiores (aproximadamente 40%superior) durante el periodo invernal en com-paración con aquellas que permanecieron sinmanifestación puberal.
Los animales que integran el grupo depúberes esta compuesto por un 88% de ani-males que pasaron su primer invierno enmejoramientos logrando tasas de gananciaen promedio de 0.414 kg/a/d, mientras queel grupo de no púberes esta compuesto enun 55% de animales manejados en su pri-mer invierno sobre campo natural con tasasde ganancia de 0.289 kg/a/d.
Durante la sequía el 42% de los animalespúberes cayeron en anestro, por lo que seevaluó el efecto del momento de entrar apubertad (temprana vs. tardía) sobre la pos-terior caída en anestro. Se observó que soloun 39% (7/18) de los animales presentaronpubertad en forma temprana; y frente alestrés ambiental (Vd/s) tan sólo el 28.5%(2/7) cayeron en anestro. En las vaquillonasque entraron a pubertad de forma tardía(61%; 11/18) un 45.5% (5/11) cayeron en
Cuadro 5. Ganancia promedio (kg/a/d) por estación según comportamiento reproductivoComportamiento reproductivo Invierno Primavera Verano a/s
Púberes 0.414 0.800 0.377No púberes 0.289 0.813 0.467Diferencias* 0.125 -0.013 -0.090
* Diferencias en ganancias (Púberes - no púberes) (kg/a/día)
Figura 4. Detección de celo durante el experimento
anestro frente a un estrés ambiental. Seobservó que fueron necesarios más días deocurrencia de condiciones adversas para quelas hembras con pubertades más tempra-nas caigan en anestro. En la misma líneade razonamiento, Vizcarra y Wettemann,(1995) reportaron que las vaquillonas quehabían entrado antes en pubertad tardaron185 días en caer en anestro con restricciónalimenticia mientras que aquellas que ha-bían entrado de forma tardía a la pubertadnecesitaron 106 días de restricción alimen-ticia para caer en anestro, marcando unamisma tendencia que la encontrada en elpresente trabajo.
Respecto al espesor de grasa dorsal (EGD)medido al final del periodo experimental(Cuadro 6), se apreció una diferencia de 0.67mm de EGD entre los manejos M y A. Enacuerdo con los resultados de Hopper et al.,(1993), animales con mayor cantidad de fo-rraje ofrecido en el invierno presentaron unamayor deposición de grasa. Las ternerasmanejadas sobre campo natural durante elinvierno (B) presentaron un valor de EGD in-termedio entre M y A. Una posible explica-ción es lo expresado por Hall et al., (1995)quien atribuye las diferencias al aumento dela relación músculo/hueso en la carcasa.

75
INIA TREINTA Y TRES SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna
Los animales púberes tendieron a presen-tar mayor espesor de grasa dorsal (P=0.09)(Cuadro 7). Esta pequeña diferencia podríaestar indicando que los animales que pre-sentaron pubertad exhibieron un mayor ba-lance energético, lo que les permitió unamayor deposición de grasa influyendo deforma positiva sobre el inicio de la pubertad.De todas formas son datos preliminares eneste tipo de variable que no permite extraerconclusiones.
A pesar de haberse observado una tenden-cia a un crecimiento compensatorio del gru-po de vaquillonas no púberes durante el ve-rano (Va/s), las menores tasas de gananciainvernales pueden haber afectado el compor-tamiento reproductivo al igual que en los ex-perimentos anteriores (años 1 y 2). En acuer-do con Wiltbank et al., (2002) las diferen-
Ganancia invernal EGD (mm)Baja 2.30 ± 0.22ab
Media 1.89 ± 0.19aAlta 2.56 ± 0.16b
Cuadro 6. Espesor de grasa dorsal (EGD) en milímetros (media ± em) según manejo invernal
Letras diferentes en la misma columna expresan diferencias estadísticamente significativas con P<0.05.
Celo Puberal EGD (mm)Si 2.44 ± 0.16a*No 2.05 ± 0.15a*
Cuadro 7. EGD en milímetros (media ± em) según presencia o ausencia de celo puberal
Letras diferentes en la misma columna expresan diferencias estadísticamente significativas con P<0.05.;Nota * P=0.09
Figura 5. Determinaciones de espesor de grasa dorsal (EGD)
cias entre los aportes de energía y proteínametabolizable según tratamiento invernal,actuarían de formas distintas sobre elhipotálamo (síntesis y secreción de GnRH)en la adenohipófisis (síntesis y secreción deFSH y LH), a nivel ovárico regulando el cre-cimiento fol icular y en la síntesis deesteroides. Por lo tanto se podría inferir queel balance nutricional estaría influenciandode alguna forma el crecimiento y desarrollode la ternera a través de los procesos ante-riormente mencionados y en consecuenciaen su performance reproductiva. En base alos resultados obtenidos durante estos tresaños, el peso estático tiene importancia endeterminar la probabilidad de que una terne-ra comience a ciclar, así como también ladistribución estacional de las ganancias depeso jugaría un rol determinante.

76
SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna INIA TREINTA Y TRES
Consideraciones finales
Los resultados obtenidos son consistentescon los encontrados hasta el momento enlos experimentos anteriores. Sin embargo altrabajar a cielo abierto en condicionespastoriles, los animales están sujetos a en-frentarse a distintas tasas de gananciasestacionales que no solo dependen de labase forrajera por sí misma, sino tambiéndel efecto directo que ocasiona el clima enlos propios animales. Una sequía importan-te registrada en este trabajo no solo implicóuna disminución en la calidad de la pasturasino que también incorporó un estrés térmi-co a los animales. Durante ese corto perio-do de tiempo, con falta de lluvia y alta tem-peratura, más del 40% de los animales de-jaron de presentar ciclos estrales o celo,confirmando que es una categoría muy sen-sible a los efectos ambientales y la gran re-percusión de estos efectos sobre aspectosproductivos.
Referencias bibliográficas
Barreto, S. y Negrín, D. 2005. Efecto delmanejo nutricional en el primer invierno so-bre la aparición de la pubertad en ternerasde raza carnicera. Segundo año de evalua-ción. Tesis de grado de la Facultad de Agro-nomía, Universidad de la Republica, p107.Costa. AJ., Moreira, RB. y Scarsi, MA. 2007.Efecto del manejo nutricional en el primerinvierno sobre la aparición de la pubertad enterneras de raza carnicera. Tercer año deevaluación. Tesis de grado de la Facultadde Agronomía, Universidad de la Republica106.p.Hall, B., Staigmiller, RB., Bellows, RA.,Short, RE., Moseley, WM. and Bellows, SE.1995. Body composition and metabolicprofiles associated with puberty in beefheifers. Journal of Animal Science, 73: 3409-3420.Hopper, HW., Williams, SE., Berkley, DJ.,Ahmed, PO., Rollosson, MM. and Kises TE.1993. Effect of prepuberal body weight gainand breed on carcass composition at pubertyin beef heifers. Journal of Animal Science,
71: 1104-1111.Straumann, JM. 2006. Efecto del manejonutricional en el primer invierno sobre la apa-rición de la pubertad en terneras de razacarnicera. Primer año de evaluación. Tesisde grado de la Facultad de Agronomía, Uni-versidad de la República, p 99.Vizcarra, JA. and Wettemann, RP.1993.Relationship between body weight changesin postpuberal heifers and cessation of lutealactivity. Oklahoma Agricultural ExperimentStation. Internal Report. 134: 44.Wiltbank, MC., Gumen, A. y Sartori, R. 2002.Clasificación fisiológica de condicionesanovulatorias en Bovinos. Theriogenology.

77
INIA TREINTA Y TRES SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna
Introducción
El objetivo del presente trabajo fue evaluaren condiciones de pastoreo el efecto de di-ferentes manejos nutricionales durante el in-vierno en terneras cruzas Aberdeen Angus xHereford (AH), las recíprocas Hereford xAberdeen Angus-Hereford (HAH) y su efectoen el porcentaje de animales en pubertad alfinal del verano, así como su peso y edad.Se incorpora un elemento nuevo en este di-seño: los animales una vez finalizado el ma-nejo diferencial en invierno, se manejan deforma diferente para lograr similares pesosen el otoño. Por lo tanto no dejamos queexpresaran su potencial de crecimiento de-finido por las tasas de ganancia previas(como los tres trabajos anteriores), sino quelos animales con menor peso al finalizar elinvierno se manejaron en pasturas mejoresque aquellos que finalizaron el invierno conpesos superiores. De esta forma todos losanimales lograron similares pesos en otoño.
Materiales y métodos
El trabajo de campo, se realizó en la UnidadExperimental Palo a Pique de INIA Treinta yTres. El periodo de evaluación fue desde el13 de junio del 2005 al 29 de mayo de 2006(350 días). Se seleccionaron 42 terneras desimilar peso y edad (148.9 ± 2.2 kg y 246.3± 4.2 días). El experimento se subdividió en3 etapas, denominadas manejo invernal,manejo diferencial y manejo conjunto. Parael manejo invernal, de 84 días de duración,las 42 terneras se sortearon por peso y edad
en tres tratamientos o niveles nutricionalesinvernales: Grupo B (Bajo, n= 14; 149.9 ±2.2 kg; 242.7 ± 4.4 días) pastoreando cam-po natural, Grupo M (Medio, n= 14; 146.0 ±2.3 kg; 252.4 ± 4.4 días) pastoreando cam-po natural mejorado, y Grupo A (Alto, n= 14;150.9 ± 2.2; kg, 243.8 ± 4.1 días) pastorean-do campo natural mejorado con una mayorasignación que el grupo anterior. Finalizadaesta etapa se manejaron en potreros conpraderas en forma diferencial para que lasterneras de los distintos grupos llegaran apesos similares al final del verano; esta eta-pa se extendió 175 días. A partir del 6 demarzo del 2006 hasta el final del experimen-to (84 días) las terneras fueron manejadasde forma conjunta en una pradera de 6 ha.Las terneras fueron pesadas cada 14 días ya partir del 12 de Setiembre fueron sangra-das semanalmente hasta el final del experi-mento (37 sangrados). Las muestras de san-gre fueron tomadas para evaluar los nivelesde progesterona en sangre. El criterio utili-zado para inicio de actividad ovárica (puber-tad) fue una concentración de progesteronamayor a 1 ng/ml en dos muestreos conse-cutivos y con actividad cíclica posterior (endos tratamientos se analizaron 13 animalesde 14 por no tener los resultados del labora-torio). En las pasturas se determinó dispo-nibilidad, altura y composición botánica(gramíneas, leguminosas, malezas, restossecos). El 27 de abril del 2006 se realizóecografía para medir espesor de grasa dor-sal entre la 12da y 13er costilla (EGD) y en elpunto P8 (inserción entre músculo glúteo me-dio y bíceps femoral). Las variables continuasse analizaron utilizando modelos mixtos (Proc
1 Ing. Agr., PhD. - Programa Nacional de Producción de Carne y Lana - INIA Treinta y Tres.2 Ing. Agr. Depto de Prod. Animal y Pasturas. Facultad de Agronomía, Universidad de la República.3 DMV. Depto de Prod. Animal y Pasturas, EEBR. Facultad de Agronomía, Universidad de la República.4 Ing. Agr. Depto de Prod. Animal y Pasturas, EEBR. Facultad de Agronomía, Universidad de la República.
2.6. MANEJO NUTRICIONAL EN EL PRIMER INVIERNO Y MANEJOSDIFERENCIALES POSTERIORES PARA ALCANZAR SIMILAR PESOVIVO EN OTOÑO. EFECTO EN LA APARICIÓN DE LA PUBERTAD(CUARTO AÑO DE EVALUACIÓN)
Graciela Quintans1 , Antonia Scarsi2 , Carlos López3 , Fernando Pereyra4

78
SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna INIA TREINTA Y TRES
Mixed) del paquete estadística SAS (versión8.1). Para el análisis de las variables dis-cretas se utilizaron modelos lineales gene-ralizados (Proc GLM) del mismo paqueteestadístico.
Resultados y discusión
A través del manejo invernal se lograron ta-sas de ganancia de peso vivo positivas y di-ferentes entre las terneras que pastorearoncampo natural y las que lo hicieron en cam-po mejorado. Las terneras que pastorearoncampo natural ganaron 25 kg en los 84 díasdel periodo invernal a razón de 0.300 kg/a/d.Esta ganancia fue superior a las encontra-das por Straumann, (2006) de -0.222 kg/a/día y Costa et al., (2007) de 0.134 kg/a/día,para la misma categoría, base forrajera yperiodo. Cabe resaltar que dicho campo na-tural presentaba al inicio del periodo inver-nal una disponibilidad de 1474 kg MS/ha yuna altura de 4.7 cm. La composición botá-nica fue de 48% gramíneas, 10% malezas y42% restos secos. La disponibilidad al finalde esta etapa fue de 1085 kg MS/ha conuna altura promedio de 2.6 cm (Cuadro 2).La tasa de crecimiento de estas pasturasnaturales según la bibliografía es aproxima-damente de 5 kg MS/ha/día (Bermúdez etal., 2003). Las terneras de los grupos M y A
lograron ganancias de peso elevadas paraeste periodo con 47 kg más (GMD de 0.850kg/a/d). Estas tasas de ganancia tambiénfueron superiores a las halladas por Barretoy Negrín, (2005), Straumann, (2006), Costaet al., (2007) (Cuadro 3). Estos animales semanejaron en potreros de 2 ha con una dis-ponibilidad de 1550 y 1675 kg MS/ha pro-medio en esta estación. Los potreros pre-sentaron, en promedio, 4% de leguminosas(Lotus corniculatus, Trifolium repens) y 35%de raigras (Lolium multiflorum).
El 5 de Setiembre comenzó la segunda eta-pa del experimento. Como se puede apre-ciar en la Figura 1 las terneras del grupo Bfinalizaron el invierno con pesos inferiores alos grupos M y A, sin embargo no se obser-varon diferencias de peso entre estos últi-mos (173.4 ± 3.3; 219.7 ± 3.4 y 221.1 ± 3.3kg PV para B, M y A respectivamente). Lapequeña diferencia en asignación de forrajedurante el invierno entre los grupos M y A nopermitió que los animales expresaran nin-guna diferencia en performance productiva,como por ejemplo tasas de ganancia o pesofinal. Las terneras del grupo B pastorearonpraderas con una asignación de forraje del7% del PV mientras que los otros dos gru-pos (M y A) lo hicieron a una asignación deforraje del 2.7% del PV.
Cuadro 2. Características de la base forrajera en el periodo invernal según manejo nutricionalManejo Disponibilidad Altura Rechazo Altura AFinvernal (kg MS/ha) disponible(cm) (kg MS/ha) rechazo (cm) (%PV)
Baja 1474 4.7 1085 2.6 10Media 1548 4.7 1317 4.3 10Alta 1673 7.5 1416 4.4 13
AF: asignación de forraje.
Cuadro 1. Peso inicial, final y ganancia media diaria (media ± em) para periodo invernalManejo Peso inicial Peso final Ganancia media diariainvernal (kg) (kg) invernal (kg/día)
Baja 149.9 ± 2.2a 173.4 ± 3.3a 0.297 ± 0.02aMedia 146.0 ± 2.3a 219.7 ± 3.4b 0.877 ± 0.02bAlta 150.9 ± 2.2a 221.1 ± 3.3b 0.830 ± 0.02b
Letras diferentes en la misma columna expresan diferencias estadísticamente significativas con P = 0.05.
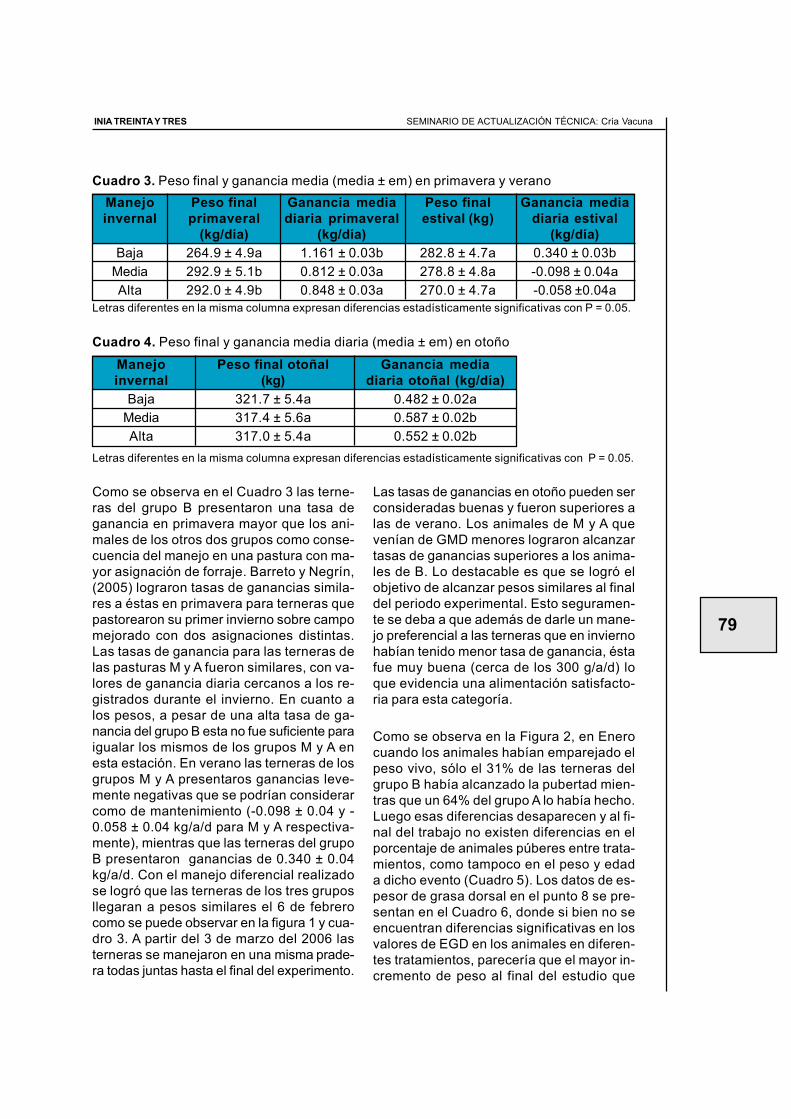
79
INIA TREINTA Y TRES SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna
Cuadro 3. Peso final y ganancia media (media ± em) en primavera y veranoManejo Peso final Ganancia media Peso final Ganancia mediainvernal primaveral diaria primaveral estival (kg) diaria estival
(kg/día) (kg/día) (kg/día)Baja 264.9 ± 4.9a 1.161 ± 0.03b 282.8 ± 4.7a 0.340 ± 0.03b
Media 292.9 ± 5.1b 0.812 ± 0.03a 278.8 ± 4.8a -0.098 ± 0.04aAlta 292.0 ± 4.9b 0.848 ± 0.03a 270.0 ± 4.7a -0.058 ±0.04a
Letras diferentes en la misma columna expresan diferencias estadísticamente significativas con P = 0.05.
Como se observa en el Cuadro 3 las terne-ras del grupo B presentaron una tasa deganancia en primavera mayor que los ani-males de los otros dos grupos como conse-cuencia del manejo en una pastura con ma-yor asignación de forraje. Barreto y Negrín,(2005) lograron tasas de ganancias simila-res a éstas en primavera para terneras quepastorearon su primer invierno sobre campomejorado con dos asignaciones distintas.Las tasas de ganancia para las terneras delas pasturas M y A fueron similares, con va-lores de ganancia diaria cercanos a los re-gistrados durante el invierno. En cuanto alos pesos, a pesar de una alta tasa de ga-nancia del grupo B esta no fue suficiente paraigualar los mismos de los grupos M y A enesta estación. En verano las terneras de losgrupos M y A presentaros ganancias leve-mente negativas que se podrían considerarcomo de mantenimiento (-0.098 ± 0.04 y -0.058 ± 0.04 kg/a/d para M y A respectiva-mente), mientras que las terneras del grupoB presentaron ganancias de 0.340 ± 0.04kg/a/d. Con el manejo diferencial realizadose logró que las terneras de los tres gruposllegaran a pesos similares el 6 de febrerocomo se puede observar en la figura 1 y cua-dro 3. A partir del 3 de marzo del 2006 lasterneras se manejaron en una misma prade-ra todas juntas hasta el final del experimento.
Las tasas de ganancias en otoño pueden serconsideradas buenas y fueron superiores alas de verano. Los animales de M y A quevenían de GMD menores lograron alcanzartasas de ganancias superiores a los anima-les de B. Lo destacable es que se logró elobjetivo de alcanzar pesos similares al finaldel periodo experimental. Esto seguramen-te se deba a que además de darle un mane-jo preferencial a las terneras que en inviernohabían tenido menor tasa de ganancia, éstafue muy buena (cerca de los 300 g/a/d) loque evidencia una alimentación satisfacto-ria para esta categoría.
Como se observa en la Figura 2, en Enerocuando los animales habían emparejado elpeso vivo, sólo el 31% de las terneras delgrupo B había alcanzado la pubertad mien-tras que un 64% del grupo A lo había hecho.Luego esas diferencias desaparecen y al fi-nal del trabajo no existen diferencias en elporcentaje de animales púberes entre trata-mientos, como tampoco en el peso y edada dicho evento (Cuadro 5). Los datos de es-pesor de grasa dorsal en el punto 8 se pre-sentan en el Cuadro 6, donde si bien no seencuentran diferencias significativas en losvalores de EGD en los animales en diferen-tes tratamientos, parecería que el mayor in-cremento de peso al final del estudio que
Manejo Peso final otoñal Ganancia mediainvernal (kg) diaria otoñal (kg/día)
Baja 321.7 ± 5.4a 0.482 ± 0.02aMedia 317.4 ± 5.6a 0.587 ± 0.02bAlta 317.0 ± 5.4a 0.552 ± 0.02b
Cuadro 4. Peso final y ganancia media diaria (media ± em) en otoño
Letras diferentes en la misma columna expresan diferencias estadísticamente significativas con P = 0.05.
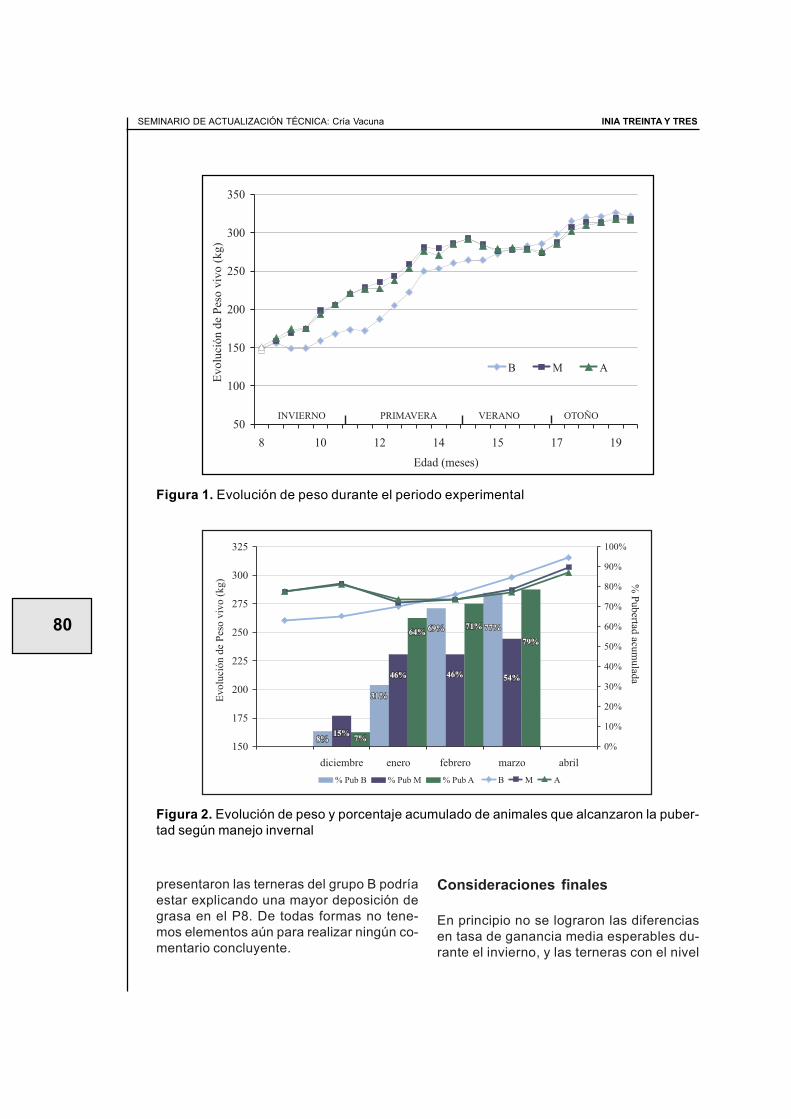
80
SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna INIA TREINTA Y TRES
Figura 1. Evolución de peso durante el periodo experimental
Figura 2. Evolución de peso y porcentaje acumulado de animales que alcanzaron la puber-tad según manejo invernal
presentaron las terneras del grupo B podríaestar explicando una mayor deposición degrasa en el P8. De todas formas no tene-mos elementos aún para realizar ningún co-mentario concluyente.
Consideraciones finales
En principio no se lograron las diferenciasen tasa de ganancia media esperables du-rante el invierno, y las terneras con el nivel
81
INIA TREINTA Y TRES SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna
Gananciainvernal
Cuadro 5. Porcentaje, peso y edad de animales que alcanzaron la pubertad
% Pubertad Peso (kg) Edad (días)
Baja 77 (10/13)a 281.3 ± 4.1a 483.5 ± 7.6aMedia 54 (7/13)a 292.2 ± 5.4a* 476.3 ± 9.6aAlta 79 (11/14)a 277.7 ± 4.0a* 473.9 ± 7.3a
Letras diferentes en la misma columna expresan diferencias estadísticamente significativascon P = 0.05. *Nota: Media y Alta tienden a ser diferentes con un P= 0.06
Cuadro 6. Espesor de grasa dorsal (EGD) y espesor de grasa de cadera(EGC P8) al finalizar el experimento
EGD (mm) EGC P8 (mm)
Baja 4.38 ± 0.16a 4.31 ± 0.20bMedia 4.38 ± 0.16a 3.85 ± 0.20abAlta 4.21 ± 0.16a 3.71 ± 0.19a
Letras diferentes en la misma columna expresan diferencias estadísticamente significativas con P = 0.05.
Gananciainvernal
alimenticio mas bajo presentaron buenastasas de ganancia (0.297 kg/a/d). No seencontraron diferencias en el porcentaje deanimales púberes al final del experimento,lo que reafirma la idea que cuando las terne-ras no son sometidas a pérdidas de peso enel invierno y logran alcanzar moderadas ta-sas de ganancias (en el orden de los 0.200 -0.300 kg/a/d), éstas alcanzan la pubertaden el otoño siguiente, a edades que oscilanentre los 15 y 16 meses. Similares resulta-dos fueron encontrados por Barreto y Negrín,(2005) y Costa et al., (2007). Los valores depeso vivo al que las terneras alcanzaron lapubertad vuelven a situarse en el orden delos 280-290 kg reafirmando que para elbiotipo en estudio, este sería un rango depeso vivo adecuado para alcanzar la puber-tad. Por otra parte el nivel de engrasamientono mostró diferencias entre tratamientos; sinembargo la grasa evaluada en el punto P8fue mayor en los animales B que en los A,lo que podría estar determinando un gradode deposición diferente ya que la distribu-ción de las ganancias de peso vivo en eltiempo fue diferente. Se requiere mayor in-formación sobre este punto y su interaccióncon variables productivas y reproductivas.
Referencias bibliográficas
Barreto, S. y Negrín, D. 2005. Efecto delmanejo nutricional en el primer invierno so-bre la aparición de la pubertad en ternerasde raza carnicera Segundo año de evalua-ción. Tesis de grado de la Facultad de Agro-nomía, Universidad de la República, p107.Bermúdez, R., Ayala, W., Ferrés, S. yQueheille, P. 2003. Opciones forrajeras parala región Este. En: Producción de carne va-cuna y ovina de calidad. Actividades de Di-fusión 317.INIA Treinta y Tres. pp. 1-10.Costa. AJ., Moreira, RB. y Scarsi, MA. 2007.Efecto del manejo nutricional en el primerinvierno sobre la aparición de la pubertad enterneras de raza carnicera. Tercer año deevaluación. Tesis de grado de la Facultadde Agronomía, Universidad de la República106.p.Straumann, JM. 2006 Efecto del manejonutricional en el primer invierno sobre la apa-rición de la pubertad en terneras de razacarnicera. Primer año de evaluación. Tesisde grado de la Facultad de Agronomía, Uni-versidad de la República, p 99.

82
SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna INIA TREINTA Y TRES
2.7. LA BIOESTIMULACIÓN PERMITE DISMINUIR LA EDAD A LAPUBERTAD EN VAQUILLONAS DE CARNE
Carolina Fiol1 , Graciela Quintans2 , Rodolfo Ungerfeld3
(0.10 ± 0.09) y las vaquillonas de menoresrangos de pesos (BEPM: 0.06 ± 0.03, P=0.01; BEPB: 0.05 ± 0.03. P= 0.03). El IA delas vaquillonas que alcanzaron la pubertadfue mayor que el de aquellas que no lo hi-cieron en BEPM (0.09 ± 0.05 vs. 0.05 ± 0.02,P= 0.01), y tendió a ser mayor en las BEPA(0.12 ± 0.10 vs. 0.06 ± 0.02, P= 0.09). Laexposición de vaquillonas prepúberes de 12meses de edad a novillos androgenizadospor 35 días fue efectiva en lograr un adelan-to en el inicio de la actividad cíclica. La res-puesta se relacionó positivamente con elmayor peso corporal inicial de lasvaquillonas, las que a su vez tuvieron unmayor vínculo físico con los machos.
Introducción
En Uruguay la elevada edad al primer entorede las vaquillonas (50 % a los 3 años) esuno de los causantes de la baja eficienciareproductiva de nuestros rodeos (Orcasberro,1990; Pereira, 2003). La posibilidad de ade-lantar la edad en que las vaquillonas inicianla pubertad, y por lo tanto la edad al primerentore, es de gran importancia económicaya que implica disminuir las categorías im-productivas del predio y aumentar el núme-ro de terneros producidos en la vida del ani-mal. Aplicando las tecnologías basadas entratamientos hormonales es posible lograruna menor edad al primer entore, pero losefectos negativos derivados de su uso, tan-to a nivel medio ambiental (disruptores am-bientales) como del comercio de productos(posibles dificultades para el acceso a cier-tos mercados) han ido en aumento. Por esto,en la actualidad la bioestimulación surgecomo una alternativa de creciente interés.
1 DMV. Estudiante de Postgrado. Facultad de Veterinaria. Universidad de la República.2 Ing. Agr., PhD. - Programa Nacional de Producción de Carne y Lana - INIA Treinta y Tres.3 Lic., MSc., PhD. Facultad de Veterinaria. Universidad de la República.
Resumen
El objetivo del presente trabajo fue determi-nar s i la exposición de vaquil lonasprepúberes a novillos androgenizados por 35días lograba un adelanto en el inicio de lapubertad, y si la respuesta se relacionabacon el peso corporal de las hembras al ini-cio de la exposición y con el vínculo físicoentre machos y hembras. Se utilizaron 131vaquillonas de 12 meses de edad y 226.0 ±0.7 kg de peso vivo, que fueron divididas en2 grupos homogéneos en edad y peso, 1)expuestas a novillos androgenizados por 35días (BE, n= 66) y 2) aisladas de todo con-tacto con toros o novillos (distancia mínima=800 m) (NE, n= 65). Las vaquillonas fuerondivididas en 3 categorías de acuerdo al pesoinicial: bajo (= 211 kg, BEPB: n= 22; NEPB:n= 22), medio (211–236 kg, BEPM: n= 24;NEPM: n= 22) y alto (> 236 kg, BEPA: n=20;NEPA: n= 21). El inicio de la actividad cícli-ca se determinó mediante la detección decelos diaria (am/pm) y ecografías ováricassemanales para la visualización de cuerposlúteos. En el grupo BE se determinó un índi-ce de asociación (IA) de cada hembra conlos novillos hasta el comienzo de la puber-tad, registrándose la proximidad física decada novillo con las vaquillonas 2 veces pordía (2 h en cada ocasión), 3 veces por se-mana. Las vaquillonas BE presentaron unamayor proporción de ciclicidad final (20/66vs. 9/65; P= 0.02) que las NE y mayor fre-cuencia acumulada de ciclicidad a partir delDía 21 del ensayo (15.2% vs 3.1%, BE yNE, respectivamente, P= 0.02). Al conside-rar los rangos de pesos iniciales, las dife-rencias únicamente se encontraron entreBEPA y NEPA (11/20 vs. 2/21; P= 0.002). ElIA únicamente resultó diferente entre BEPA

83
INIA TREINTA Y TRES SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna
Su utilización es de alta potencialidad enresultados, fácil aplicación, bajo costo parael productor y ausencia de efectos adversos.
La bioestimulación, o efecto macho o toro, sedefine como el estímulo provocado por la pre-sencia de los machos, induciendo el estro y laovulación mediante estímulos genitales,feromonas u otras señales químicas(Chenoweth, 1983). La efectividad de la utiliza-ción del efecto macho para disminuir el anestroposparto en vacas de cría ha sido bien docu-mentada a nivel internacional (Zalesky et al.,1984; Alberio et al., 1987; Berardinelli y Joshi,2005) y nacional (Miller y Ungerfeld, 2008). Sinembargo, los antecedentes de su uso paraadelantar la pubertad en vaquillonas de carneson muy escasos.
Varios factores parecen influir en la respuestade las vaquillonas a la bioestimulación: algu-nos propios de las hembras (edad y peso almomento del estímulo), y otros vinculados a laintensidad del estímulo (edad de los machos,proporción machos-hembras, duración de laexposición, etc). Se han encontrado resulta-dos positivos con periodos cortos y largos deexposición, tanto en Bos taurus (Izard yVandenbergh, 1982; Roberson et al., 1991)como en Bos indicus (Rekwot et al., 2000;Goncalvez-Oliveira, 2003). La exposición amachos antes del inicio de un tratamiento desincronización de celos también logró mejo-rar el porcentaje de ciclicidad y las tasas depreñez finales (Ferreira de Cuadros y Piva Lo-bato, 2004). En contraposición a lo expuesto,otros no encontraron efecto de labioestimulación sobre la edad de inicio de lapubertad, tanto con periodos cortos (Berardinelliet al., 1978; Macmillan et al., 1979) como lar-gos de exposición (Roberson et al., 1987). Anivel nacional, se realizó un ensayo convaquillonas de 18 - 22 meses, las que fueronclasificadas de acuerdo al peso en 3 rangos yexpuestas o no a los toros por un periodo de 2semanas previo al comienzo de un entore deotoño tardío. El porcentaje de preñez seincrementó en el grupo preestimulado con losmachos, pero únicamente dentro del rango depesos mayores a 280 kg (Ungerfeld, datos nopublicados).
El objetivo del presente trabajo fue determinarsi la exposición a novillos androgenizados du-rante 35 días adelanta la pubertad envaquillonas prepúberes de carne. Dos objeti-vos complementarios fueron determinar si exis-te un rango de peso corporal necesario paralograr una respuesta positiva, y si la proximi-dad física entre machos y hembras -la que serelacionaría con la intensidad del estímulo- in-fluye en los resultados reproductivos encon-trados.
Materiales y métodos
Animales
El ensayo se realizó en la Unidad Experimen-tal de Palo a Pique, INIA Treinta y Tres, entrelos meses de octubre y diciembre de 2007. Seutilizaron 131 vaquillonas prepúberes Herefordy Hereford x Aberdeen Angus de 12 meses deedad y 226.0 ± 0.7 kg (media ± em) y 8 novi-llos androgenizados. Al inicio del ensayo seobservaron los ovarios 2 veces con 15 días dediferencia mediante ecografía ovárica, determi-nándose que ninguna hembra presentaba cuer-pos lúteos. El Día 0 el grupo de vaquillonas seseparó en 2 grupos homogéneos en edad ypeso, 1) expuestas a novillos androgenizadospor 35 días (BE, n= 66) y 2) aisladas de todocontacto con toros o novillos (distancia míni-ma= 800 m) (NE, n= 65). Ambos grupos semantuvieron en potreros separados 1000 m,evitando el contacto visual, táctil u olfativo en-tre los mismos.
Los novillos fueron androgenizados mediantela inyección de testosterona intramuscular (800mg, Ciclopentilpropionato de Testosterona,Dispert, Uruguay) desde una semana previa ala introducción y a intervalos semanales du-rante toda la exposición. Los 4 primeros novi-llos utilizados fueron cambiados por los otros4 el Día 14, manteniendo la misma relaciónmachos-hembras.
Alimentación y pesos corporales
La disponibilidad promedio de materia secaa lo largo del ensayo fue de 2012 kg MS/ha

84
SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna INIA TREINTA Y TRES
y 1173 kg MS/ha en los potreros de los gru-pos BE y NE respectivamente, ajustándosela carga para que la disponibilidad de mate-ria seca por animal fuera similar. Lasvaquillonas fueron divididas en 3 categoríasde acuerdo al peso inicial: bajo (168-211 kg,BEPB: n= 22; NEPB: n= 22), medio (212-236 kg, BEPM: n= 24; NEPM: n= 22) y alto(237-302 kg, BEPA: n= 20; NEPA: n= 21).La evolución del peso vivo se registró en formasemanal en todas las vaquillonas. Las ganan-cias de las vaquillonas a lo largo de los 35 díasdel periodo de exposición fueron de 45.9 ± 7.8kg y 43.0 ± 8.6 kg, en BE y NE, respectiva-mente. Los pesos al finalizar el ensayo (Día42) fueron de 271.4 kg para BE y 269.4 kgpara NE.
Determinación de ciclicidad
La detección de celos se realizó diariamentedurante 40 min en ambos grupos, 2 veces pordía (am/pm) hasta el Día 42. Se realizaronecografías ováricas para determinar presenciade cuerpos lúteos en forma semanal en todaslas vaquillonas de ambos grupos. Se conside-ró que una vaquillona estaba ciclando cuandose visualizaba un cuerpo lúteo en 2 ecografíassucesivas ó al detectarse celo y posterior vi-sualización del cuerpo lúteo.
Índice de Asociación
En el grupo BE se determinó el índice de aso-ciación (IA; Martin y Bateson, 1993) de cadahembra con los novillos, registrándose la dis-tancia machos-hembras cada 10 minutos por4 horas/día, con una frecuencia de 3 veces/semana, 2 h en la mañana (9:00 a 11:00) y 2 hen la tarde (16:00 a 18:00). Se consideró un IAde 1, 0.33 ó 0 para cada vaquillona según seencontrara a menos de 1, entre 1 y 3 ó a másde 3 cuerpos de distancia de los novillos, res-pectivamente. En cada día de registro se de-terminó un IA promedio para cada vaquillona, apartir de los cuales se calculó un IA promediototal. Para ello se incluyó solamente la infor-mación obtenida previamente al día que seconsideró como inicio de la ciclicidad.
Análisis Estadístico
Las frecuencias de vaquillonas ciclando secompararon con el test de chi2, y los IA fue-ron comparados con ANOVA previa normali-zación de los datos por la transformaciónde Bliss.
Resultados
La frecuencia acumulada de vaquillonas cícli-cas fue mayor en el grupo expuesto a los novi-llos androgenizados en comparación a lasvaquillonas no expuestas a partir del Día 21del ensayo (15.2% vs 3.1%, BE y NE, respec-tivamente, P= 0.02), manteniéndose éstas di-ferencias hasta 1 semana luego de finalizadoel periodo de exposición (30.3% vs 13.8%, BEy NE, respectivamente, P=0.02) (Figura 1).
Al comparar los animales de acuerdo al rangode pesos, las vaquillonas BEPA alcanzaron lapubertad durante el periodo de exposición alos novillos (D0 a D35) en mayor proporciónque las NEPA (11/20 vs 2/21; P= 0.002), noexistiendo diferencias entre los grupos de pe-sos medios y bajos. A su vez, también la fre-cuencia acumulada de ciclicidad mostró dife-rencias significativas entre BEPA y NEPA enlos días 21, 28, 35 y 42 del ensayo (Figura 2).
El IA de las vaquillonas de mayor peso(BEPA: 0.10 ± 0.09) fue significativamentemayor que las de peso medio y bajo (BEPM:0.06 ± 0.03, P= 0.01, y BEPB: 0.05 ± 0.03,P= 0.03). El IA de las vaquillonas BEPM yBEPB fue similar (P= 0.28). A su vez, el IAde las vaquillonas que alcanzaron la puber-tad fue mayor que el de aquellas que no lohicieron en BEPM (0.09 ± 0.05 vs. 0.05 ±0.02, P= 0.01), y tendió a ser mayor en lasBEPA (0.12 ± 0.10 vs. 0.06 ± 0.02; P= 0.09).No hubo diferencias en el IA entre los distin-tos rangos de pesos en las vaquillonas quesí iniciaron la actividad cíclica.
Discusión
La exposición a los machos por un periodode 35 días logró adelantar el inicio de la ac-

85
INIA TREINTA Y TRES SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna
tividad cíclica en vaquillonas prepúberes de12 meses de edad. Esto reafirma resultadosanteriores obtenidos en otras condiciones demanejo y a otras edades y rangos de pesos(Izard y Vandenbergh, 1982; Roberson et al.,1991; Ferreira de Cuadros y Piva Lobato,2004). Las diferencias en el inicio deciclicidad entre ambos grupos de vaquillonascomenzaron a manifestarse a las 3 sema-nas de exposición a los novi l losandrogenizados. Como ya se observó enotros trabajos (Goncalvez-Oliveira, 2003), larespuesta a la bioestimulación en bovinospresentaría un patrón de dispersión mayoren el tiempo que el observado en ovinos ycaprinos (Walkden-Brown et al., 1999). Sinembargo, en este trabajo se observaron res-puestas positivas a la bioestimulación lue-go de periodos de exposición más cortosque los reportados por otros autores(Roberson et al., 1991; Goncalvez-Oliveira,2003). También en vacas posparto el efectopositivo en el reinicio de la ciclicidad se ob-servó entre 40 y 90 días luego del comienzode la exposición (Zalesky et al., 1984; Custeret al., 1990). Por otra parte, las diferenciasobservadas en la semana posterior al retiro
Figura 1. Porcentaje de vaquillonas cíclicas a lo largo del ensayo. Barras verdes: vaquillonasen contacto con novillos androgenizados; barras blancas: vaquillonas que permanecieronaisladas de los machos. Día 0: entrada de novillos androgenizados. Nota: * P<0.05; **P<0.01.
de los machos podrían deberse tanto al efec-to previo que ejercieron los novillos sobre lasBE, que habría sido registrado en forma pos-terior por las variables utilizadas, y/ó a unposible efecto de estimulación hembra cícli-ca-hembra prepúber. Este último se genera-ría por el manejo conjunto de ambos gruposde vaquil lonas, a part i r del que lasvaquillonas cíclicas ejercerían un estímulopositivo sobre la ciclicidad de aquellas hem-bras que aún no habían comenzado la pu-bertad (Izard y Vandenbergh, 1982; Wrightet al., 1994).
En el presente estudio se estableció por pri-mera vez la efectividad del uso de novillosandrogenizados para el estímulo de laciclicidad. Si bien existía información sobreel uso de vacas androgenizadas (Burns ySpitzer, 1992), y de machos castradosandrogenizados para estimular la ovulaciónen ovinos (Fulkerson et al., 1981) y caprinos(Mellado y Hernández, 1996), en los bovi-nos no había información publicada al res-pecto. Esto confirma que las vías deestimulación de los machos hacia las hembras
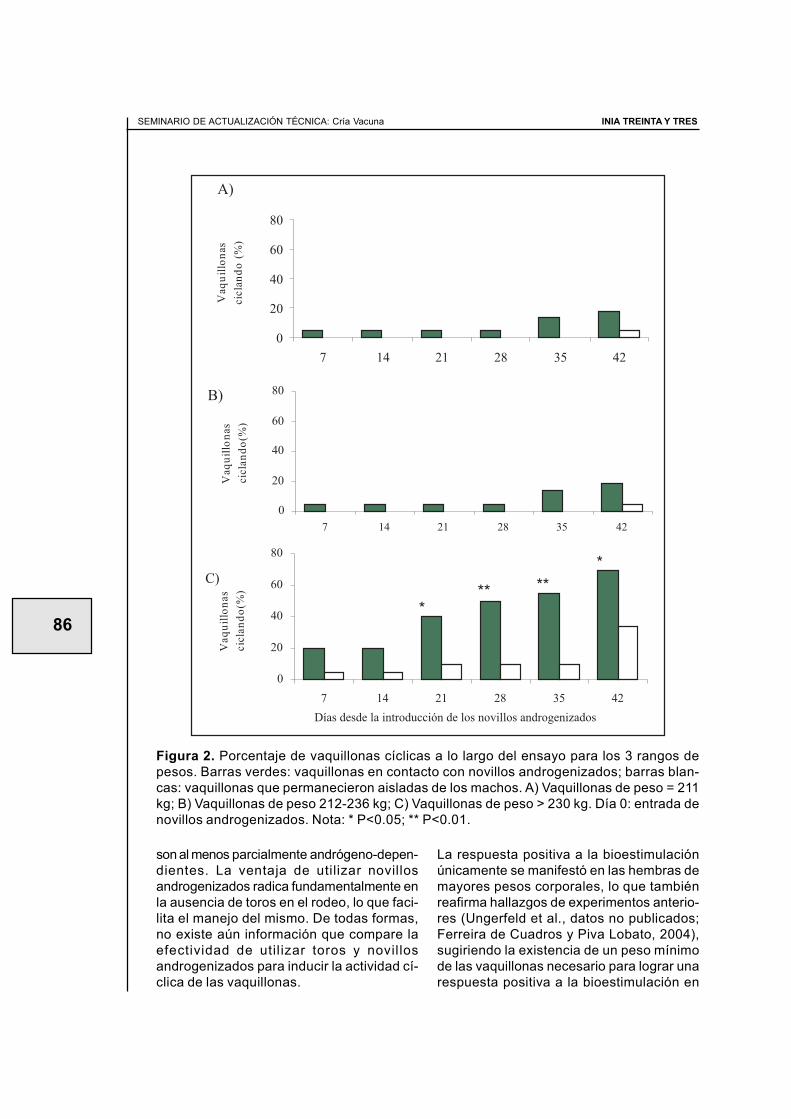
86
SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna INIA TREINTA Y TRES
Figura 2. Porcentaje de vaquillonas cíclicas a lo largo del ensayo para los 3 rangos depesos. Barras verdes: vaquillonas en contacto con novillos androgenizados; barras blan-cas: vaquillonas que permanecieron aisladas de los machos. A) Vaquillonas de peso = 211kg; B) Vaquillonas de peso 212-236 kg; C) Vaquillonas de peso > 230 kg. Día 0: entrada denovillos androgenizados. Nota: * P<0.05; ** P<0.01.
son al menos parcialmente andrógeno-depen-dientes. La ventaja de utilizar novillosandrogenizados radica fundamentalmente enla ausencia de toros en el rodeo, lo que faci-lita el manejo del mismo. De todas formas,no existe aún información que compare laefectividad de util izar toros y novil losandrogenizados para inducir la actividad cí-clica de las vaquillonas.
La respuesta positiva a la bioestimulaciónúnicamente se manifestó en las hembras demayores pesos corporales, lo que tambiénreafirma hallazgos de experimentos anterio-res (Ungerfeld et al., datos no publicados;Ferreira de Cuadros y Piva Lobato, 2004),sugiriendo la existencia de un peso mínimode las vaquillonas necesario para lograr unarespuesta positiva a la bioestimulación en

87
INIA TREINTA Y TRES SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna
el adelanto de la pubertad. Durante elposparto de vacas de carne también se ob-servó una respuesta di ferencial a labioestimulación de acuerdo a la condicióncorporal de las hembras (Stumpf et al., 1992).A partir de los resultados obtenidos en elpresente estudio podría esperarse que laexposic ión a machos en hembrasprepúberes no fuera efectiva en hembras depesos muy bajos respecto al peso mínimonecesario para comenzar la pubertad. A suvez, si bien no fue posible verificarlo en nues-tro trabajo, es probable que no hubiera efec-tos positivos en vaquillonas de muy altospesos corporales que ya estén por iniciarsu actividad cíclica.
Las ganancias de peso en ambos grupos devaquillonas resultaron elevadas, consideran-do que los animales se encontraban en con-diciones extensivas a campo natural. Sinembargo, las vaquillonas que inicialmentepresentaron menores pesos corporales norespondieron a la bioestimulación pese a lasaltas tasas de ganancias de peso. Por tan-to, el peso inicial que presentan lasvaquillonas sería una limitante para obtenerbuenos porcentajes de respuesta que nopuede ser sobrepuesta por altas tasas deganancia de peso.
El IA de las hembras que finalmente ciclaronfue mayor que el de las que no ciclaron. Enbovinos no están claramente determinadaslas vías de estimulación. Considerando queno existe información anterior acerca de larelación existente entre la intensidad delestímulo que ejercen los machos y la res-puesta reproductiva a la bioestimulación, lamayor proximidad física macho-hembra po-dría estar reflejando una mayor interacción,y por tanto un mayor estímulo hacia esasvaquillonas, determinando un inicio más tem-prano de la ciclicidad. Similarmente, en ca-bras se ha reportado que el tiempo de res-puesta al efecto macho se vincula con laproximidad física entre machos y hembras(Alvarez et al., 2003). El mayor IA registra-do en BEPA con respecto a BEPM y BEPBimplicaría que las vaquillonas de pesos su-
periores recibieron un mayor estímulo porparte de los novillos androgenizados que lasde pesos corporales menores. De todas for-mas, el diseño experimental no permite dis-criminar si esta mayor proximidad fue con-secuencia de una preferencia por parte delos novillos hacia hembras de mayor peso,o una mayor proceptividad de éstas hacialos novillos. El hecho de que no existierandiferencias en los IA entre las vaquillonasque efectivamente ciclaron en los distintosrangos de pesos reafirma que la proximidadmacho-hembra es un buen indicador de po-sible respuesta a la bioestimulación.
Consideraciones finales
Los resultados del presente estudio nos per-miten afirmar que la bioestimulación resultauna herramienta efectiva para lograr el ade-lanto del inicio de la pubertad en vaquillonasprepúberes de 12 meses de edad, en com-paración a las hembras que se mantienenaisladas. La respuesta estaría relacionadapositivamente con el mayor peso corporalde las hembras al inicio de la exposición, ya su vez, estaría relacionada con un mayorvínculo físico con los machos en lasvaquillonas de mayores pesos iniciales.
Referencias bibliográficas
Alberio, RH., Schiersmann, G., Carou, N. andMestre, J. 1987. Effect of teaser bull onovarian and behavioural activity of sucklingbeef cows. Animal Reproduction Science, 14;263.Álvarez, L., Martin, GB., Galindo, F. andZarco, LA. 2003. Social dominance offemales goats affect their response to themale effect. Applied Animal BehaviourScience, 84; 119.Berardinelli, JG., Fogwell, RI. and Inskeep,EK. 1978. Effect of electrical stimulation orpresence of a bull on puberty in beef heifers.Theriogenology 9, nº 2; 133.Berardinelli, JG. and Joshi, PS. 2005.Introduction of bulls at different dayspostpartum on resumption of ovarian cycling

88
SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna INIA TREINTA Y TRES
activity in primiparous beef cows. 83; 2106.Burns, PD. and Spitzer, C. 1992. Influenceof biost imulat ion on reproduct ion inpostpartum beef cows., 70; 358.Chenoweth, PJ. 1983. Reproduct ivemanagement procedures in control ofbreeding. Animal Production in Australia, 15;28.Custer, EE., Berardinelli, JG., Short, RE.,Wehrman, M. and Adair, R. 1990.Postpartum interval to estrus and patternsof LH and progesterone in first calf-suckledbeef cows exposed to mature bulls. Journalof Animal Science, 68; 1370.Ferreira de Quadros, SA. y Piva Lobato, JF.2004. Bioestimulcao e comportamentoreprodutivo de novilhas de corte. RevistaBrasileira de Zootecnia 33, nº3; 679.Fulkerson, WJ., Adams, NR. and Gherardi,PB. 1981. Ability of castrate male sheeptreated with oestrogen or testosterone to in-duce and detect oestrus in ewes. AppliedAnimal Ethology, 7; 57.Goncalvez Oliveira, CM. 2003. Tesis deMaestría. Influencia da bioestimulacao e dasuplementacao alimentar sobre a idade apuberdade e taxa de gestacao em novilhasNelore. Universidade Federal de Goias,Escola de Veterinaria.Izard, MK. and Vandenbergh, JG. 1982. Theeffects of bull urine on puberty and calvingdate in crossbred beef heifers. Journal ofAnimal Science, 55, nº 5; 1160.Izard, MK. and Vandenbergh, JG. 1982.Priming pheromones from oestrous cowsincrease synchronization of oestrus in dairyheifers after PGF-2á injection. Journal ofReproduction and Fertility, 66; 189.Macmillan, KL., Allison, AJ. and Struthers,GA. 1979. Some effects of running bulls withsuckling cows or heifers during the prematingperiod. New Zealand Journal of Experimen-tal Agriculture, 7; 121.Mar t in , GB. and Bateson, P. 1993 .Measuring behaviour. An introductoryguide. 2da Edición, Cambridge UniversityPress.
Mellado, M. and Hernández, JR. 1996. Abilityof androgenized goat wethers and does toinduce estrus in goats under extensiveconditions during anestrus and breedingseasons. Small Ruminants Research,. 23;37.Miller, V. and Ungerfeld, R. 2008. Weeklybull exchange shortens postpartum anestrusin suckled beef cows. Theriogenology, acep-tado para su publicación.Orcasberro, R. 1990. Manejo para mejorarla eficiencia reproductiva de los rodeos decría. Revista Plan Agropecuario 70.Pereira, G. 2003. La ganadería en Uruguay.Contribución a su conocimiento. DIEA,MGAP; 80.Rekwot PI., Ogwy, D., Oyedipe, E. andSekoni, V. 2000. Effects of bull exposure andbody growth on onset of puberty in Bunajiiand Friesian x Bunajii heifers. ReproductionNutrition Development, 40; 359.Roberson, MS., Wolfe, MW., Stumpf, TT.,Werth, LA., Cupp, AS., Kojima, N., WolfePL.and Kittok, PJ. 1991. Influence of growthrate and exposure to bulls on age at pubertyin beef heifers. Journal of Animal Science,69; 2092.Roberson, MS., Ansotegui, JG., Berardinelli,JG., Whitman, RW., McInerney, and MJ.1987. Influence of biostimulation by maturebulls on occurrence of puberty in beef heifers.Journal of Animal Science, 64; 1601.Stumpf, TT., Wolfe, MW., Wolfe, PL., Day,ML., Kittok, RJ. and Kinder, JE. 1992. Weightchanges prepartum and presence of bullspostpartum interact to affect duration ofpostpartum anestrus in cows. Journal ofAnimal Science, 70; 3133.Walkden-Brown, SW., Martin, GB. andRestall, BJ. 1999. Role of male-femaleinteractions in regulating reproduction insheep and goats. Journal of Reproductionand Fertility. (Suppl.)52; 243.Wright, IA., Rhind, SM., Smith, AJ. andWhyte, TK. 1994. Female-female influencesin the duration of the post-partum anoestrousperiod in beef heifers. Animal Production, 59;49.

89
INIA TREINTA Y TRES SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna
Zalesky, DD., Day, ML., Garcia Winder, M.,Imakawa, K., Kittok, RJ., D´Occhio, MJ.andKinder JE. 1984. Influence of exposure tobulls on resumption of estrous cyclesfollowing parturition in beef cows. Journal ofAnimal Science, 59, nº5; 1135.

90
SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna INIA TREINTA Y TRES
2.8. SERVICIO DE VAQUILLONAS EN OTOÑO A LOS 20 MESES DEEDAD (RESULTADOS PRELIMINARES)
Graciela Quintans1 , José Ignacio Velazco2 , Gonzalo Roig3
les y su posterior vida productiva serán eva-luados. En esta ocasión, se presentaránsólo los resultados preliminares desde eldestete de las terneras hasta su primer ser-vicio como vaquillonas de 20 meses.
Materiales y métodos
El manejo fue realizado entre Junio del 2006y Febrero del 2008 en la Unidad Experimen-tal Palo a Pique (UEPP), perteneciente a laEstación Experimental del Este, INIA Trein-ta y Tres. Se utilizaron 107 terneras cruzaAberdeen Angus x Hereford destetadas a los5.7 ± 0.07 meses y con un peso de 172.0 ±2.2 kg. Las terneras fueron asignadas a dosniveles distintos de alimentación inverno-pri-maveral durante 141 días (12/6 al 31/10). Losgrupos fueron designados teniendo en cuentapesos diferentes al destete y distintas ta-sas de ganancia diaria invernal. Este proce-dimiento se seleccionó para que las terne-ras llegaran al servicio con pesos homogé-neos. El grupo de mayor peso al destete fueasignado al tratamiento de tasa de ganan-cia invernal MEDIA (M; n= 49; 190.3 ± 2.0kg y 177.3 ± 3.6 días de edad al destete).Se manejó sobre mejoramientos de campocon la finalidad de obtener ganancias mode-radas durante el invierno, en un área totalde 24 ha constituido por 4 potreros de 6 hacada uno. Las terneras ocupaban dichospotreros por un periodo de 7 días permitien-do así un descanso a la pastura de 21 días.Las terneras más livianas al destete fueronasignadas al tratamiento de tasa de ganan-cia ALTA (A; n= 58; 156.4 ± 1.9 kg y 172 ±2.3 días de edad al destete) y rotaron enpraderas de achicoria, raigras y trébol rojo(3 ha) y raigras puro, con el fin de obteneraltas ganancias diarias invernales. La asig-
1 Ing. Agr., PhD. - Programa Nacional de Producción de Carne y Lana - INIA Treinta y Tres.2 Ing. Agr. - Programa Nacional de Producción de Carne y Lana - INIA Treinta y Tres.3 DMV. - Programa Nacional de Producción de Carne y Lana - INIA Treinta y Tres.
Introducción
Como se pudo observar en los trabajos an-teriormente presentados, una adecuada tasade ganancia invernal, que podría situarse enel orden de los 0.300 kg/a/d con un poste-rior crecimiento durante la primavera-veranoy parte del otoño, permitiría alcanzar un altoporcentaje de vaquillonas púberes con 18 a20 meses de edad. La pregunta obligada essi a esos animales los servimos en el otoñoo dejamos transcurrir seis o siete meses máspara realizar el servicio de primavera. Si bieneste es un sistema muy utilizado en Argen-tina y Brasil, poca es la información genera-da en nuestras condiciones y la que logra-mos encontrar es informal y producto deexperiencias personales y aisladas de va-rios productores.
Este sistema a priori tendría una desventajaa destacar: las vacas parirán a fines de ve-rano, principio de otoño y transcurrirán el in-vierno lactando. Por otra parte este sistemano propone volver a servir a la vaquillona enel otoño siguiente sino por el contrariodestetarla en la primavera temprano y ser-virla en el servicio de primavera (ej. noviem-bre) junto con el resto de las categorías.Esto traería asociado un servicio devaquillonas de primera cría pero sin cría alpie, detalle no menor teniendo en cuenta labaja tasa de procreo que en general se pre-senta en esta categoría.
El objetivo del presente trabajo fue evaluaren nuestras condiciones un servicio antici-pado de vaquillonas en el otoño temprano,cuando éstas alcanzan los 20 meses deedad. Las ventajas y desventajas de estetipo de servicio, el desarrollo de los anima-

91
INIA TREINTA Y TRES SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna
nación de forraje varió desde 4.5% hasta 10%dependiendo de la pradera y tiempo de ocu-pación. Una vez finalizado el periodo inverno-primaveral, los animales se manejaron deforma conjunta sobre campo natural hastael otoño del 2007, donde fueron servidas. Lasterneras fueron pesadas periódicamente du-rante el transcurso del trabajo.
Al promediar los 18 meses de edad (20/3/07), se realizó una ecografía ovárica paradeterminar la proporción de animalesciclando. A fines de mayo, promediando los20 meses de edad, las terneras fueronsincronizadas con el protocolo de 6 días deinseminación artificial (IA) y una dosis deprostaglandina. Una vez finalizada la etapade servicio, se hizo una segunda ecografíaen los animales que no fueron detectadosen celo durante la IA y se realizó un repasocon toros por un periodo de 21 días, tam-bién con detección de celo diario. Dos me-ses luego de finalizado el servicio (14/8/07),se realizó el diagnóstico de gestación me-diante ecografía. Todas las vaquillonas va-cías fueron servidas a los 26 meses de edadbajo idéntico protocolo descrito anteriormen-te (IA + 21 días de repaso con toros).
A partir de la inseminación (20 meses) y has-ta el final del trabajo, se evaluó condicióncorporal y dentición en todas las vaquillonas.Desde el comienzo del tratamiento invernal(12/6/06) hasta la inseminación a los 26 me-ses, se registró altura de anca en cm. A los12, 17 y 20 meses se midió área pélvica encm2 utilizando pelvímetro, y se realizaronecografías para determinar espesor de gra-sa dorsal en mm (EGD) entre la 12ª y 13ªcostilla y espesor de grasa de cadera (EGCP8) en mm tomando la imagen desde la pun-ta de la cadera hacia la región caudal en elpunto de unión de los músculos bícepsfemoral y glúteo medio.
Las variables continuas (peso, edad, condi-ción corporal, área pélvica, altura de anca,espesor de grasa dorsal y espesor de grasade cadera) se analizaron mediante un mo-delo lineal general, utilizando el procedimien-
to GLM. La variable continua (ganancia depeso) se analizó por el modelo Proc Mixedcon una estructura de covar ianzaautoregresiva de orden 1. Los valores se pre-sentan como promedio ± error de la media(em) y el nivel de significancia utilizado fueP <0.05. Para las variables discretas (por-centaje de ciclicidad, animales inseminados,preñez y repaso) se utilizó el programaGENMOD con distribución binomial. Lascorrelaciones entre variables se analizaronpor el método de coeficiente de correlacio-nes de Pearson. Todos los procedimientosse realizaron con el sistema de análisis es-tadístico SAS versión 9.03 (2005).
Resultados y discusión
Evolución de peso y tasa de gananciaestacional
El manejo nutricional invernal generó dife-rencias significativas en las tasas de ganan-cias entre tratamientos para este periodo(P<0.05). Las terneras con alto peso al des-tete y que pastoreaban un mejoramiento decampo natural (tratamiento ganancia Media)ganaron 15.8 kg durante los 84 días del pe-riodo invernal, a razón de 0.176 ± 0.04 kg/a/d. Las terneras con moderado peso al des-tete y que pastoreaban praderas (tratamien-to ganancia ALTA), promediaron gananciasde 0.760 ± 0.03 kg/a/d finalizando el invier-no con 58.1 kg de ganancia total (Cuadro1).
Si bien ambos grupos presentaron pesossignificativamente distintos al inicio del pe-riodo primaveral (P<0.05), luego de transcu-rrida la primavera (115 días), los pesos fue-ron similares entre grupos (Cuadro 2)(P>0.05). Cabe destacar que a mediados deeste periodo (31/10/06) se comenzó con elmanejo conjunto de las terneras sobre cam-po natural. Los animales con ganancia dia-ria invernal Media, presentaron mayores ga-nancias en el periodo primaveral (1.303 ±0.03 kg/a/d) en comparación con las terne-ras con Alta ganancia diaria invernal (0.998± 0.03 kg/a/d) (P<0.05). Una ganancia diaria

92
SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna INIA TREINTA Y TRES
de 0.250 kg/d se puede considerar como unvalor de ganancia invernal “umbral” por de-bajo de la cual existe crecimiento compen-satorio (Frick y Borges 2003). Por lo tanto,las terneras con ganancias invernales pordebajo del umbral habrían estado expresan-do crecimiento compensatorio durante laprimavera siguiente.
Durante el periodo estival, ambos lotes deterneras presentaron ganancias de peso si-milares (Alta= 0.717 ± 0.05 kg/a/d; Media=0.792 ± 0.06 kg/a/d) (P>0.05), culminandoel verano con pesos similares entre trata-mientos (Cuadro 3) (P>0.05). La disminuciónen la calidad del forraje natural durante elverano generó una disminución en las ga-nancias diarias en ambos grupos en com-paración con las registradas durante la pri-mavera. Las ganancias diarias registradasen el otoño fueron diferentesestadísticamente entre grupos. Los anima-les con alta ganancia invernal presentaronganancias de 0.148 ± 0.03 kg/a/d, mientrasque los de media ganancia invernal registra-ron ganancias significativamente superiores
Cuadro 1. Peso inicial, peso final y ganancia media diaria (+em) para manejo invernalGanancia Peso inicial Peso final Ganancia diariainvernal invernal (kg) invernal (kg) invernal (kg/día)
Alta 151.9 ± 1.4a 210 ± 2.1a 0.760 ± 0.03aMedia 180.5 ± 1.5b 196.3 ± 3.5b 0.176 ± 0.04b
Letras diferentes en la misma columna expresan diferencias estadísticamente significativas con P<0.05.
Cuadro 2. Peso inicial, peso final y ganancia media diaria (+em) para periodo primaveralGanancia Peso inicial Peso final Ganancia diariaprimaveral primaveral (kg) primaveral (kg) primaveral (kg/día)
Alta 210.0 ± 2.1a 287.4 ± 2.9a 0.998 ± 0.03aMedia 196.3 ± 3.5b 290.4 ± 3.5a 1.303 ± 0.03b
Letras diferentes en la misma columna expresan diferencias estadísticamente significativas con P<0.05.
Cuadro 3. Peso final y ganancia media diaria (+em) para periodo estival y otoñalGanancia Peso final Ganancia diaria Peso final Ganancia diariainvernal estival (kg) estival (kg/día) otoñal (kg) otoñal (kg/día)
Alta 308.8 ± 2.7a 0.717 ± 0.05a 322.3 ± 3.2a 0.148 ± 0.03aMedia 314.6 ± 3.5a 0.792 ± 0.06a 337.7 ± 4.3b 0.313 ± 0.03b
Letras diferentes en la misma columna expresan diferencias estadísticamente significativas con P<0.05.
(0.313 ± 0.03 kg/a/d) (P< 0.05). Ryan (1990)demostró que la duración de la compensa-ción es directamente proporcional a la seve-ridad de la restr icción. La respuestacompensatoria de los animales restringidosuna vez comenzada la realimentación estáen función del grado en que su tasa de cre-cimiento aumenta por encima de los no res-tringidos y del tiempo en que persiste esamayor tasa de crecimiento. Se podría espe-cular que durante el otoño las terneras conmoderada ganancia media invernal podríanhaber seguido expresando algún grado decrecimiento compensatorio y terminaron elperiodo con 15.4 kg más que las ternerasde alta ganancia invernal (P<0.05). Sin em-bargo durante el verano, las tasas de ganan-cias fueron aceptables y similares.
Inseminación a los 20 meses de edad
Ambos grupos de animales presentaron pe-sos similares cuando se realizó la ecografíaovárica en otoño (20/03/07) con 17.6 mesesde edad promedio. Las hembras con altaganancia diaria invernal pesaron 308.8 ± 2.7

93
INIA TREINTA Y TRES SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna
Figura 1. Evolución de peso vivo según tratamiento invernal
Nota: * corresponde a diferencias estadísticamente significativas P<0.05.
kg, mientras que las de moderada gananciainvernal pesaron 314.6 ± 3.5 kg (P>0.05).La proporción de animales ciclando no fuediferente entre tratamientos (P>0.05). El86.2%, de las terneras de alta ganancia in-vernal ciclaban al momento de la ecografíamientras que el 73.5% de las terneras demoderada ganancia diaria invernal estabanciclando al mismo momento.
Al momento de la inseminación (a partir del24/05/07), los animales del lote de alta ga-nancia invernal presentaban un pesosignificativamente menor a los del grupo demedia ganancia (Alta= 321.5 ± 3.2 kg; Me-dia= 338.4 ± 4.0 kg; P<0.05). Si tomamosen cuenta únicamente la proporción de ani-males inseminados, las diferencias a favordel grupo de ganancia moderada invernal fue-ron mayores aún (Alta= 325.9 ± 3.8 kg; Me-dia= 347.3 ± 4.5 kg; P<0.05). La proporciónde animales inseminados no dif i r ió
significativamente entre tratamientos, un72.4% de las terneras de alta ganancia dia-ria invernal fueron inseminadas, mientras queun 69.4% de las terneras de moderada ga-nancia diaria invernal también se inseminaron(P>0.05). El porcentaje de repetición de celofue similar entre tratamientos, con un 21%de animales para repaso en el lote de altaganancia invernal y 32% en el lote de ga-nancia invernal media. El porcentaje de pre-ñez al diagnóstico de gestación (14/08/2007)fue diferente para ambos tratamientos. Lasterneras con alta ganancia diaria invernalalcanzaron un 77.6% de preñez mientras quelas terneras con media ganancia diaria in-vernal registraron un 59.2% de preñez(P<0.05). A modo descriptivo, al momentodel diagnóstico de gestación, tan solo el 8.8%(3/34) de las terneras falladas habían corta-do o estaban cortando los 2 dientes, mien-tras que de los animales preñados al diag-nóstico, el 30.1% (22/73) ya había cortado2 dientes.
Cuadro 4. Peso, porcentaje de ciclicidad, animales inseminados, repetición de celo ypreñez al final del repaso según tratamiento invernal
Ganancia % Animales % Animales Peso % Rep. de Preñezinvernal ciclando Ins. (1erserv) IA Celo general %
Alta (n= 58) 86.2(50/58)a 72.4 (42/58)a 321.5 ±3.2a 21 (9/42)a 77.6(45/58)aMedia (n= 49) 73.5(36/49)a 69.4 (34/49)a 338.4 ±4.0b 32(11/34)a 59.2(29/49)b
Letras diferentes en la misma columna expresan diferencias estadísticamente significativas con P<0.05.

94
SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna INIA TREINTA Y TRES
Espesor de grasa dorsal (EGD) y espesorde grasa de cadera (EGC) en el punto P8
Las terneras con alta ganancia diaria inver-nal tuvieron mayor espesor de grasa dorsaly de cadera en la primavera siguiente (12meses) en comparación con las terneras conganancia invernal moderada (Cuadro 5), sien-do estas diferencias de tipo significativo(P<0.05). En los últimos 2 registros (17 y20 meses de edad) las diferencias en el EGDy el P8 no fueron significativas entre trata-mientos (P>0.05).
Las altas tasas de ganancia invernal expli-carían el mayor espesor de grasa dorsal yde cadera para el lote de Alta ganancia. Amedida que pasaron los meses, las terne-ras con menor ganancia diaria invernal reali-zaron ganancias diarias superiores a las dellote de alta (crecimiento compensatorio), loque llevo a que se igualaran los valores deespesor de grasa. Cabes destacar que losniveles de espesor de grasa se mantuvierondentro del rango para animales en desarro-llo, en los que las altas tasas de gananciadiaria no se reflejan tanto en la deposiciónde grasa sino en la formación de tejido mus-cular. Di Marco et al., (2007) reportaron quese requieren niveles mínimos de gordura enel punto P8 de entre 1.8 - 2.00 mm para quelas vaquillonas puedan expresar pubertad.Los valores de nuestro trabajo fueron supe-riores a los reportados por estos autores.
Altura de anca
Según Baker et al., (1988) la altura de ancaes una de las formas mas convenientes dedescribir el tamaño esquelético de los bovi-
Cuadro 5. EGD y EGC (P8) (+em) en mm a los 12, 17 y 20 meses de edadGanancia 12 17 20invernal EGD mm P8 mm EGD mm P8 mm EGD mm P8 mm
Alta 3.326±0.1a 3.758±0.1a 3.526±0.1a 4.282±0.2a 2.843±0.1a 3.732±0.1aMedia 2.690±0.1b 2.846±0.1b 3.771±0.2a 4.371±0.2a 3.133±0.1a 3.888±0.2a
EGD: espesor de grasa dorsal. P8: espesor de grasa de cadera punto 8. Letras diferentes en la mismacolumna expresan diferencias estadísticamente significativas con P<0.05.
nos de carne. Es una medida que ajustadapor la edad del animal nos permite determi-nar su “frame” o tamaño corporal adulto. Laaltura de anca obviamente esta muyinfluenciada por la nutrición. Es importantedestacar que los animales con mayor es-tructura corporal, más altos y de mayor ta-maño adulto pueden tener impactos negati-vos en las variables reproductivas, como porejemplo edad a la pubertad (Di Marco et al.,2007).
Las terneras con ganancia moderada duran-te el invierno (y mayor peso al destete) tu-vieron una altura de anca superior en com-paración con las terneras con alta gananciainvernal (y menor peso al destete). Al iniciolas diferencias entre grupos fueron mayores(Alta= 102.1 ± 0.3; Media= 106.3 ± 0.4;P<0.05), como reflejo de que los animalesen Alta eran más pesados respecto a losanimales en Media. A medida que pasaronlos meses los valores se fueron emparejan-do (Alta= 122.0 ± 0.4; Media= 123.2 ± 0.5,P>0.05) (en el último registro), de la mismaforma que también los pesos vivos de am-bos grupos fueron homogeneizándose.
Área Pélvica
La medición del área pélvica (cm2) es utili-zada comúnmente durante el preserviciocomo herramienta para evitar partosdistócicos y como uno de los criterios deselección de terneras de reposición. Algu-nos autores han demostrado una moderadacorrelación entre las ganancias de pesoposdestete y el área pélvica (Short yBellows, 1971; Fleck et al., 1980; Buskirket al., 1995). Sin embargo estos autores

95
INIA TREINTA Y TRES SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna
Nota: * corresponde a diferencias estadísticamente significativas con P<0.05
Figura 2. Evolución de altura de anca según tratamiento invernal
reportan que en comparación con el pesovivo o la condición corporal, el efecto de lasganancias diarias sobre el desarrollo esque-lético no seria tan marcado.
Durante el presente estudio, en ninguno delos tres registros (12, 17, 20 meses) exis-tieron diferencias significativas entre trata-mientos (P>0.05). Las terneras con alta ga-nancia invernal tuvieron valores de 103.2 ±1.5; 108.3 ± 1.3; 109.9 ± 1.0, mientras quelas terneras con media ganancia invernal mi-dieron 106.3 ± 1.4; 108.8 ± 1.2; 111.1 ± 1.3cm2, para los 12, 17 y 20 meses respectiva-mente.
Inseminación a los 26 meses de edad (lasque no se preñaron a los 20 meses)
Las vaquillonas que no se preñaron a los 20meses, fueron inseminadas a los 26 meses(a partir del 21/11/07). Los animales del gru-po Alta fueron mas livianos a la insemina-ción (393.3 ± 6.4 kg) en comparación conlas vaquillonas con ganancia invernal Media
(413.4 ± 5.9 kg) siendo estas diferenciasestadísticamente significativas (P<0.05). Laedad a la inseminación fue similar para am-bos grupos (Alta= 773.5 ± 5.5 días; Media=779.9 ± 4.9 días; P>0.05).
De las 14 vaquillonas del tratamiento de altaganancia diaria invernal se inseminaron 13(92.9%) quedando preñadas únicamente 6(42.9%). En el tratamiento de media ganan-cia diaria invernal los resultados fueron si-milares, de 21 vaquillonas se inseminaron20 (95.2%) y quedaron preñadas 10 (47.6%).Las vaquillonas falladas a la inseminaciónrecibieron un repaso con toros, en el cualse preño casi el total de los animales (Alta=92.9%; Media= 90.5%; P>0.05).
Vaquillonas ciclando vs. vaquillonasen anestro
Otra forma de análisis posible de la informa-ción es agrupar los animales en función deaquellos que estaban ciclando y aquellos quepermanecieron en anestro a los 18 meses

96
SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna INIA TREINTA Y TRES
Cuadro 6. Porcentaje de animales inseminados, preñez a la inseminación y preñezgeneral a los 26 meses de edad según tratamiento invernal
Ganancia % Animales Ins. Preñez IA Preñez Generalinvernal (1erserv) % %
Alta (n= 14) 92.9 (13/14)a 42.9 (6/14)a 92.9 (13/14)aMedia (n=21) 95.2 (20/21)a 47.6 (10/21)a 90.5 (19/21)a
Letras diferentes en la misma columna expresan diferencias estadísticamente significativas con P<0.05
de edad, independientemente de los trata-mientos a los que fueron asignados. Cabedestacar las limitaciones que existen en estetipo de análisis al contar con gruposdesbalanceados.
Es así que las vaquillonas se agruparon enfunción de su estado reproductivo a los 18meses, en terneras ciclando a la ecografía(n= 86); y terneras en anestro a la ecografía(n= 21). Este reagrupamiento se realizó bus-cando una mayor asociación entre las varia-bles registradas durante el trabajo y el esta-do reproductivo al año y medio. Las terne-ras que ciclaban a la ecografía registraronun peso al nacer significativamente menor(37.5 ± 0.6 kg) al de las terneras en anestro(42.1 ± 1.6 kg) (P<0.05). Al momento deldestete ambos grupos presentaron pesossimilares (Ciclando= 170.9 ± 2.3; Anestro=176.5 ± 5.4; P>0.05), pero las terneras en
Ciclando(n=86)
Anestro(n= 21)
Cuadro 7. Peso inicial (+em) para periodo invernal, primaveral, estival y otoñal, y peso a lainseminación artificial (IA), según ciclicidad
Actividad Peso inicial Peso inicial Peso inicial Peso inicial Peso a laovárica invernal (kg) primaveral (kg) verano (kg) otoño (kg) I.A (kg)
164.5 ± 1.9a 206.6 ± 2.2b 291.6 ± 2.4b 313.7 ± 2.4b 332.4 ± 3.0b
167.0 ± 3.9a 191.9 ± 4.6a 277.3 ± 4.8a 302.8 ± 4.5a 316.1 ± 4.7a
Letras diferentes en la misma columna expresan diferencias estadísticamente significativas con P<0.05.
Cuadro 8. Ganancia media diaria (+em) para manejo invernal, primaveral, estival y otoñal,según actividad ovárica
Actividad Ganancia diaria Ganancia diaria Ganancia diaria Ganancia diariaovárica invernal (kg/día) primaveral (kg/día) estival (kg/día) otoñal (kg/día)
Ciclando 0.554 ± 0.03a 1.13 ± 0.02a 0.742 ± 0.04a 0.239 ± 0.02aAnestro 0.377 ± 0.06b 1.18 ± 0.05a 0.787 ± 0.09a 0.161 ± 0.05a
Letras diferentes en la misma columna expresan diferencias estadísticamente significativas con P<0.05.
anestro fueron mas jóvenes al destete encomparación con las que cic laban(Ciclando= 177.0 ± 2.2 días; Anestro= 164.3± 4.9; P<0.05).
Ambos grupos de terneras partieron de pe-sos similares al inicio del invierno (Cuadro7). Durante el mismo las tasas de gananciadiaria fueron diferentes entre grupos(Ciclando= 0.554 ± 0.03; Anestro= 0.377 ±0.06; P<0.05). La diferencia en las tasas deganancia invernal generaron pesos distintosal inicio de la primavera (P<0.05). Durantela primavera y el verano, las tasas de ga-nancia diaria fueron similares entre grupos(P>0.05), por lo que al inicio del periodo oto-ñal las diferencias de peso vivo se mantuvie-ron (P<0.05). Al momento de la ecografía,las terneras que ciclaban pesaron 313.7 ±2.4 kg mientras que las que se encontraronen anestro pesaban 302.8 ± 4.5 kg (P<0.05).

97
INIA TREINTA Y TRES SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna
La tasa de ganancia diaria otoñal fue mayor(aunque no significativa) para los animalesciclando (Ciclando= 0.239 ± 0.02; Anestro=0.161 ± 0.05; P>0.05), lo que generó dife-rencias significativas entre grupos para elpeso a la inseminación (Ciclando= 332.4 ±2.9; Anestro= 316.1 ± 4.7; P<0.05). Las ter-neras en anestro partieron de pesos simila-res al primer invierno y tuvieron gananciasmenores pero buenas (0.377 kg/a/d) duran-te esta etapa. Durante el resto del manejonunca lograron igualar (compensar) el pesode las terneras que ciclaron, llegando conmenor peso a la ecografía y a la insemina-ción. Seguramente otros factores distintosal peso y tasas de ganancias también impe-raron para lograr las diferencias en el esta-do reproductivo.
Por otra parte a lo largo de todos estos tra-bajos hemos observado que animales conlas mismas características de peso y edad,presentan diferencias individuales a la horade transformar el consumo de alimento enproducto (éste puede ser peso vivo, ciclicidadovárica, preñez, etc.) lo que denota clara-mente la diferencia en “eficiencia” que exis-te entre animales. Este es un tema muy com-plejo ya que el grado de eficiencia puede
establecerse en diferentes etapas de la ca-dena productiva (cosecha de forraje, trans-formación del alimento, diferencias en envíode señales metabólicas al eje reproductivo,etc.)
Espesor de grasa dorsal (EGD) y espesorde grasa de cadera en el punto P8 enanimales que ciclaron y en animales enanestro
Los valores de espesor de grasa dorsal yespesor de grasa de cadera (P8) tuvierondiferencias significativas a lo largo de todoslos registros (P<0.05). Los animales queciclaron registraron un mayor espesor de gra-sa dorsal y espesor de grasa de cadera encomparación con los animales en anestro.Cabe destacar que las terneras que ciclaron,tuvieron similar peso al destete y mayoresganancias diarias invernales lo que las llevoa ser mas pesadas a la ecografía y a la in-seminación. Buckley et al., (1990) demos-traron que las terneras tienen un incremen-to significativo en la deposición de grasaentre los 8 y 14 meses de edad, momentoen el cual la mayoría de ellas en condicio-nes óptimas de alimentación, presentan pu-bertad. Por ejemplo Lammoglia et al., (2000)
Nota: Las diferencias de EGD y P8 (ECG) entre vacas ciclando y en anestro fue estadísticamente significa-tiva a los 12, 17 y 20 meses con P<0.05
Figura 3. Espesor de grasa dorsal (EGD) y Espesor de grasa de cadera P8 (EGC) a los12, 17 y 20 meses de edad según actividad ovárica a la ecografía de 20 meses

98
SEMINARIO DE ACTUALIZACIÓN TÉCNICA: Cría Vacuna INIA TREINTA Y TRES
encontraron que el 75% de las vaquillonasHereford evaluadas alcanzaron la pubertadcon 13 meses de edad y entre 357 y 360 kgde peso vivo con un espesor de grasa de 4.5mm.
Con respecto a la evolución de la altura deanca (cm) y del área pélvica (cm2), no seregistraron diferencias significativas entre gru-pos (anestro vs. ciclando) en ninguno de losregistros.
Consideraciones finales
Este es un trabajo preliminar que permiteexplorar las variables que pueden estarinvolucradas en la determinación del éxitoen la tasa de preñez en vaquillonas jóvenesen servicios a edades más tempranas. Serequiere mayor información, especialmentede parámetros metabólicos y hormonales,que nos ofrezcan más elementos para po-der realizar un análisis más exhaustivo delas causas que conllevan al éxito en estetipo de manejos.
Referencias bibliográficas
Baker, JF., Stewart, TS., Long, CR. andCartwright. TC. 1988. Multiple regression andprincipal components analysis of puberty andgrowth in cattle. Journal of Animal Science,66:2147-2158.Buckley, BA., Baker, JF., Dickerson, GE. andJenkins, TG. 1990. Body composition andtissue distribution from birth to 14 monthsfor three biological types of beef heifersJournal of Animal Science, 68: 3109-3123.Buskirk, DD., Faulkner, DB. and Ireland, FA.1995. Increased postweaning gain of beefheifers enhances fert i l i ty and mi lkproduction. Journal of Animal Science, 73:937-946.Di Marco, ON., Barcelos, JOJ. y Da Costa,EC. 2007. Crescimento de bovinos de corte.Porto Alegre, Universidade Federal Do RioGrande Do Sul. 276 p.Fleck, AT., Schalles, RR. and Kiracofe, GH.
1980. Effect of growth rate through 30months on reproductive performance of beefheifers. Journal of Animal Science, 51:816.Frick, F. y Borges, M. 2003. Factores queafectan la fertilidad de vaquillonas Herefordy Brahman x Hereford entoradas a los 18meses de edad. Tesis Ing. Agr. Montevideo,Uruguay. Facultad de Agronomía. 140 p.Lammoglia, MA., Bellows, RA., Grings, EE.,Bergman, JW., Bellows, SE., Hallford, DM.and Randel, RD. 2000. Effects of dietary fatand sire breed on puberty, weight, andreproductive traits of F1 beef heifers. Journalof Animal Science, 78: 2244-2252.Ryan, WJ. 1990. Compensatory Growth insheep and cattle. PhD Thesis, School ofAgriculture. The University of Western Aus-tralia.Short, RE. and Bel lows, RA. 1971.Relationship among weight gains, age atpuberty and reproductive performance inheifers. Journal of Animal Science, 32: 127-131.


















