
FUNCIÓN BIOLÓGICA DE LAS PROTEÍNAS · segundo, se conoce como el número de recambio. Tabla1 ....
33
1 FUNCIÓN BIOLÓGICA DE LAS PROTEÍNAS 1. ENZIMAS 1.1 ASPECTOS GENERALES Los enzimas son catalizadores muy potentes y eficaces, químicamente son proteínas Como catalizadores, los enzimas actúan en pequeña cantidad y se recuperan indefinidamente. No llevan a cabo reacciones que sean energéticamente desfavorables, no modifican el sentido de los equilibrios químicos, sino que aceleran su consecución ( Figura 1). Figura 1. Reacción catalizada por una enzima 1.2 CARACTERÍSTICAS DE LA ACCIÓN ENZIMÁTICA La característica más sobresaliente de los enzimas es su elevada especificidad. Esta es doble y explica que no se formen subproductos: 1. Especificidad de sustrato. El sustrato (S) es la molécula sobre la que el enzima ejerce su acción catalítica. El enzima sacarasa actúa sobre la sacarosa, la amilasa sobre el almidón.
Transcript of FUNCIÓN BIOLÓGICA DE LAS PROTEÍNAS · segundo, se conoce como el número de recambio. Tabla1 ....

1
FUNCIOacuteN BIOLOacuteGICA DE LAS PROTEIacuteNAS
1 ENZIMAS
11 ASPECTOS GENERALES
Los enzimas son catalizadores muy potentes y eficaces quiacutemicamente son proteiacutenas
Como catalizadores los enzimas actuacutean en pequentildea cantidad y se recuperan
indefinidamente No llevan a cabo reacciones que sean energeacuteticamente
desfavorables no modifican el sentido de los equilibrios quiacutemicos sino que aceleran
su consecucioacuten ( Figura 1)
Figura 1 Reaccioacuten catalizada por una enzima
12 CARACTERIacuteSTICAS DE LA ACCIOacuteN ENZIMAacuteTICA
La caracteriacutestica maacutes sobresaliente de los enzimas es su elevada especificidad Esta es
doble y explica que no se formen subproductos
1 Especificidad de sustrato El sustrato (S) es la moleacutecula sobre la que el enzima
ejerce su accioacuten cataliacutetica El enzima sacarasa actuacutea sobre la sacarosa la
amilasa sobre el almidoacuten
2
2 Especificidad de accioacuten Cada reaccioacuten estaacute catalizada por un enzima
especiacutefico Por ejemplo el enzima sacarasa es muy especifico rompe el enlace
glucosidico de la sacarosa
Sin embargo hay distintos grados de especificidad La sacarasa es muy especiacutefico
de la sacarosa (sustrato natural) mientras que la maltosa y la isomaltosa son
sustratos anaacutelogos El enzima actuacutea con maacutexima eficacia sobre el sustrato natural
y con menos eficacia sobre los sustratos anaacutelogos
La accioacuten enzimaacutetica se caracteriza por la formacioacuten de un complejo que
representa el estado de transicioacuten
E + S
ES
E + P
El sustrato se une al enzima a traveacutes de numerosas interacciones deacutebiles como son
puentes de hidroacutegeno electrostaacuteticos hidroacutefobos etc en un lugar especiacutefico el
centro activo o sitio activo Este centro es una pequentildea porcioacuten del enzima
constituido por una serie de aminoaacutecidos que interaccionan con el sustrato
13 SITIO ACTIVO
Las moleacuteculas de enzimas contienen hendiduras o cavidades denominadas
sitio activo El sitio activo estaacute formado por las cadenas laterales de residuos
especiacuteficos lo que ocasiona que tenga un arreglo tridimensional particular diferente
al resto de la proteiacutena Este sitio es afiacuten por la estructura tridimensional del sustrato
(Figura 2)
enzima
+ sustrato
sitio activo
sitio activo vaciacuteo ocupado
3
Figura 2 Representacioacuten de la formacioacuten del complejo enzima-sustrato
Es decir el sitio activo esta formado por ciertos aminoaacutecidos que forman un
microambiente caracteriacutestico dentro de la propia cadena y que llevan a cabo la
reaccioacuten generalmente existe soacutelo uno por moleacutecula de enzima
Las enzimas adquieren su poder cataliacutetico cuando presentan una estructura
secundaria y terciaria muy especiacutefica de tal manera que los aminoaacutecidos
componentes del sitio activo se encuentran vecinales formando dicho
microambiente Asiacute por ejemplo la quimotripsina crea su centro activo con los
aminoaacutecidos histidina y serina localizados en las posiciones 57 y 195 respectivamente
se puede decir que forzosamente esta moleacutecula debe adquirir una estructura
tridimensional de tal manera que dichos aminoaacutecidos sean adyacentes
14 EFICIENCIA CATALITICA
La mayoriacutea de las reacciones catalizadas por enzimas son muy eficientes y
transcurren desde 106 hasta 1014 veces maacutes raacutepido que la misma reaccioacuten no
catalizada (Tabla 1) Tiacutepicamente cada moleacutecula de enzima es capaz de transformar
cada segundo de 100 a 1000 moleacuteculas de substrato en producto El nuacutemero de
estas moleacuteculas transformadas a producto por moleacutecula de enzima en cada
segundo se conoce como el nuacutemero de recambio
Tabla1 Eficiencia de las reacciones catalizadas por algunas enzimas
Enzima Velocidad en
ausencia de enzima
Velocidad de
reaccioacuten catalizada
Rendimiento
Anhidrasa carboacutenica Corismato mutasa Triosafosfato isomerasa Carboxipeptidasa A AMP nucleosidasa
Nucleasa estafilococal
13 X 10 ndash1
26 X 10 ndash5
43 X 10 ndash6
30 X 10 ndash9
10 X 10 ndash11
17 X 10 -13
10 X 106
50 4300 578 60
95
77 X 106
19 X 106
10 X 109
19X 1011
60 X 1012
56 X 1014
4
128 NOMENCLATURA DE LAS ENZIMAS
A cada enzima se le asignan dos nombres El primero es corto y es como se
conoce a la proteiacutena de manera coloquial es el nombre sugerido El segundo es maacutes
completo e infiere propiedades sobre la reaccioacuten que la enzima desarrolla se le
conoce como nombre sistemaacutetico este nombre permite reconocer a la enzima sin
ambiguumledad y localizarla en el metabolismo
a) Nombre sugerido
Muchas de las enzimas poseen en su nombre el sufijo ldquo-asardquo unido al nombre
del substrato de la reaccioacuten que cataliza por ejemplo
Ureasa proteiacutena cuyo sustrato es la urea Lactasa proteiacutena cuyo sustrato es la lactosa
Tambieacuten suele utilizarse este sufijo a la descripcioacuten de la reaccioacuten que la enzima cataliza
Lactato deshidrogenada deshidrogena (le quita Hidroacutegenos) al lactato
Adenilato ciclasa hace un ciclo en la adenina
Algunas enzimas poseen nombres que no representan su actividad o sustrato
Lisozima
Tripsina
b) Nombre sistemaacutetico
La Unioacuten Internacional de Bioquiacutemica y Biologiacutea Molecular (IUBMB) desarrolloacute
un sistema de nomenclatura en el cual las enzimas se dividen en seis clases
principales cada una con numerosos subgrupos
127 CLASIFICACIOacuteN DE LAS ENZIMAS
5
En funcioacuten de su accioacuten cataliacutetica especiacutefica los enzimas se clasifican en 6 grandes grupos o clases
1 Oxidorreductasas Reacciones de transferencia de electrones
2 Transferasas Transferencia de grupos funcionales Ej UDP-glucosa-fructosa-
glucotransferasa
3 Hidrolasas Reacciones de hidroacutelisis Ej lipasa proteasa celulasa
4 Liasas Adicioacuten a dobles enlaces Ej carboxilasa fenilalanina amonioliasa
5 Isomerasas Reacciones de isomerizacioacutenEj fosfoglucosa isomerasa
6 Ligasas Se conociacutean como sintetasas Participan en la formacioacuten de enlaces
con hidroacutelisis de ATP
Para todas las clasificaciones se muestran ejemplos representativos
1- OXIDOREDUCTASAS
Catalizan reacciones de oxidacioacuten-reduccioacuten
2e-
2- TRANSFERASAS
Catalizan la transferencia de grupos que contienen C N o P
THF tetrahidrofolato
6
3- HIDROLASAS
Catalizan la ruptura de enlaces por la adicioacuten de una moleacutecula de agua
4- LIASAS
Catalizan la ruptura de enlaces C-C C-S y algunos enlaces C-N
5- ISOMERASAS
Catalizan la racemizacioacuten de isoacutemeros oacutepticos o geomeacutetricos
7
6- LIGASAS
Catalizan la formacioacuten de enlaces entre C y O S N Esta reaccioacuten soacutelo es posible
mediante la energiacutea derivada de fosfatos ricos en energiacutea como el del fosfato de la
moleacutecula de ATP
15 COFACTORES y COENZIMAS Algunas enzimas se asocian con moleacuteculas de caraacutecter no proteiacuteco que son
necesarias para el funcionamiento de la enzima estas moleacuteculas se denominan
cofactores (Figura 3) Comuacutenmente los factores encontrados en las enzimas incluyen
iones metaacutelicos como el Zn2+ o el Fe2+ ( Tabla 2) tambieacuten pueden ser moleacuteculas
orgaacutenicas que se denomina coenzimas como el NAD+ FAD la coenzima A
Generalmente las coenzimas son derivados de las vitaminas A la enzima en ausencia
de su cofactor (cuando lo tiene) se le denomina apoenzima en presencia de su
cofactor (cuando lo tiene) se le denomina holoenzima La apoenzima generalmente
carece de actividad bioloacutegica La diferencia entre un cofactor y un grupo prosteacutetico
como el grupo hemo es que este uacuteltimo estaacute unido de manera covalente a la
enzima mientras que el cofactor puede ser removido de la misma con relativa
facilidad
Se pueden sentildealar algunas caracteriacutesticas de las coenzimas como generalmente son
estables al calor y a otros agentes desnaturalizantes de las proteiacutenas pueden ser
recicladas y algunas provienen de las vitaminas
8
Figura 3 Cofactores y Coenzima
Tabla 2 Algunos enzimas que contienen iones metaacutelicos o los necesitan como
cofactores
Zn+2 Cu+2 (Cu+)
Alcohol- deshidrogenada Tirosinasa Anhidrasa-carboacutenica Citocromo-oxidasa Carboxipeptidasa
Mg+2 K+
Fosfohidrolasa Piruvato- fosfoquinasa Fosfotransferasas
Mn+2 Arginasa Na+ Fosfotransferasas ATPasa de la membrana plasmaacutetica
Fe +2 o Fe+3
9
Citocromos
Peroxidasa Catalasa Ferredoxina
Texto ------- (Tabla 3)
Tabla 3 Relacioacuten entre algunas coenzimas y las vitaminas a partir de las
cuales se producen Enfermedades generadas por la deficiencia
Coenzima Reaccioacuten Fuente vitamiacutenica Enfermedad
Humana por deficiencia
Biocitina Carboxilacioacuten Biotina
Coenzima A Transferencia de acilos Pantotenato
Coenzimas de Cobalamima Alquilacioacuten Cobalamina (B12) anemia perniciosa
Coenzimas de flavina Oxidacioacuten-reduccioacuten Riboflavina (B2)
Acido lipoacuteico Transferencia de acilos -
Coenzimas de nicotinamida Oxidacioacuten-reduccioacuten Nicotinamida (niacina) Pelagra
Fosfato de piridoxal Transferencia de grupo amino Piridoxina (B6)
Tetrahidrofolato Transferencia de un grupo de 1 Carbono Acido foacutelico anemia
megaloblaacutestica
Tiamina pirofosfato Transferencia de aldehiacutedo Tiamina (B1) beriberi
no tiene un nombre especiacutefico su aparicioacuten es muy rara
Otras coenzimas
NADP
Coenzima Q
Biotina
NAD y NADP
10
Coenzima Q (CoQ)
Esta coenzima es sintetizada a partir de farnesil pirofosfato Al igual que el
FMN la CoQ es capaz de aceptar y donar uno o dos electrones porque su forma
semiquinona es estable A su forma oxidada se le conoce como ubiquinona (de
ubicuo lat ubique en todas partes) o quinona A su forma reducida se le conoce
como QH2 ubiquinol o hidroquinona (Figura 2) A la coenzima solo parcialmente
reducida se le conoce como ubisemiquinona semiquinona o QH (Figura 4)
Figura 4 Representacioacuten de la moleacutecula de CoQ
La siacutentesis de la CoQ se lleva a cabo a traveacutes de unidades isoprenoides (Figura 5)
11
Figura 5 La CoQ se produce a partir de isoprenoides
Biotina
La biotina consiste de un anillo imidazol que estaacute unido en forma cis a la
cadena lateral tetrahidrotiofeno del valerato (Figura 6)
12
Figura 6 Estructura de la biotina
La enzima holocarboxilasa sintetasa (HCS) cataliza la activacioacuten mediante
biotinilacioacuten de cinco carboxilasas en ceacutelulas humanas La deficiencia de HCS
produce el siacutendrome deficiencia muacuteltiple de carboxilasas (DMC) Esta enfermedad es
potencialmente mortal y se caracteriza por la deficiencia de la actividad de todas las
carboxilasas La deficiencia de biotina no solo disminuye la actividad de estas enzimas
sino que parece modificar la expresioacuten geneacutetica
126 PROPIEDADES DE LAS ENZIMAS
Las propiedades de las enzimas derivan del hecho de ser proteiacutenas y de actuar como
catalizadores Como proteiacutenas poseen una conformacioacuten natural maacutes estable que las
demaacutes conformaciones posibles Asiacute cambios en la conformacioacuten suelen ir asociados
en cambios en la actividad cataliacutetica Los factores que influyen de manera maacutes directa
sobre la actividad de un enzima son
pH
temperatura
cofactores
A) EFECTO DEL pH
Al comprobar experimentalmente la influencia del pH en la velocidad de las
reacciones enzimaacuteticas se obtienen curvas que indican que los enzimas
presentan un pH oacuteptimo de actividad El pH puede afectar de varias maneras
13
o El centro activo puede contener aminoaacutecidos con grupos ionizados
que pueden variar con el pH
o La ionizacioacuten de aminoaacutecidos que no estaacuten en el centro activo puede
provocar modificaciones en la conformacioacuten de la enzima
o El sustrato puede verse afectado por las variaciones del pH
Algunos enzimas presentan variaciones peculiares La pepsina del estoacutemago
presenta un oacuteptimo a pH=2 la ureasa lo tiene a pH 7 y la fosfatasa alcalina del
intestino un pH= 12
B) EFECTO DE LA TEMPERATURA
Influye en la actividad El punto oacuteptimo representa el maacuteximo de actividad A
temperaturas bajas los enzimas se hallan ldquomuy riacutegidosrdquo y cuando se supera un
valor considerable (mayor de 50) la actividad cae bruscamente porque como
proteiacutena el enzima se desnaturaliza
128 EFECTO DE LOS COFACTORES SOBRE LA ACTIVIDAD
ENZIMATICA
A veces un enzima requiere para su funcioacuten la presencia de sustancias no proteicas
que colaboran en la cataacutelisis los cofactores Casi un tercio de los enzimas conocidos
requieren cofactores Cuando el cofactor es una moleacutecula orgaacutenica se llama
coenzima En la figura inferior podemos observar una moleacutecula de hemoglobina
(proteiacutena que transporta oxiacutegeno) y su coenzima (el grupo hemo) Cuando los
cofactores y las coenzimas se encuentran unidos covalentemente al enzima se llaman
14
grupos prosteacuteticos La forma cataliacuteticamente activa del enzima es decir el enzima
unido a su grupo prosteacutetico se llama holoenzima La parte proteica de un
holoenzima (inactiva) se llama apoenzima de forma que
apoenzima + grupo prosteacutetico= holoenzima
129 MODO DE ACCIOacuteN DE LAS ENZIMAS
En las reacciones espontaacuteneas los productos finales tienen menos energiacutea libre de
Gibbs (∆G) que los reactantes (figura inferior izquierda) Por lo tanto las reacciones
espontaacuteneas liberan energiacutea de Gibbs ((∆G lt 0) Sin embargo el comienzo de la
reaccioacuten requiere un aporte inicial de energiacutea Esta energiacutea inicial que hay que
suministrar a los reactantes para que la reaccioacuten transcurra se llama energiacutea de
activacioacuten (Ea) Cuanto menor es la Ea maacutes faacutecilmente transcurre la reaccioacuten
Perfil energeacutetico de una reaccioacuten espontaacutenea Perfil energeacutetico de una reaccioacuten catalizada
15
La accioacuten de los catalizadores consiste precisamente en disminuir la Ea (Figura superior derecha) Los enzimas
son catalizadores especialmente eficaces ya que disminuyen la Ea auacuten maacutes que los catalizadores inorgaacutenicos
Por ejemplo la descomposicioacuten del agua oxigenada (H2O2) para dar H2O y O2 puede ocurrir sin catalizador
con un catalizador inorgaacutenico (platino) o con un enzima especiacutefico (catalasa) Las respectivas Ea para cada
proceso son 18 12 y 6 Kcalmol Asiacute se puede calcular que el platino acelera la reaccioacuten 20000 veces mientras
que la catalasa la acelera 370000 veces
Para que una reaccioacuten quiacutemica tenga lugar las moleacuteculas de los reactantes deben chocar con una energiacutea y
una orientacioacuten adecuadas La actuacioacuten del enzima (1) permite que los rea ctantes (sustratos) se unan a su
centro activo con una orientacioacuten oacuteptima para que la reaccioacuten se produzca y (2) modifica las propiedades
quiacutemicas del sustrato unido a su centro activo debilitando los enlaces existentes y facilitando la formacioacuten de
otros nuevos (Figuras inferiores)
1210 REGULACIOacuteN DE SU ACTIVIDAD ENZIMATICA
16
Una moleacutecula de enzima no tiene por que actuar siempre a la misma velocidad Su
actividad puede estar modulada por
Cambios en el pH
Cambios en la temperatura
Presencia de cofactores
Las concentraciones del sustrato y de los productos finales
Presencia de inhibidores
Modulacioacuten alosteacuterica
Modificacioacuten covalente
Activacioacuten por proteolisis
Isoenzimas
EFECTO DE LAS CONCENTRACIONES SOBRE LA ACTIVIDAD ENZIMAacuteTICA
La velocidad de una reaccioacuten enzimaacutetica depende de la concentracioacuten de sustrato La Figura de la derecha muestra la velocidad de una reaccioacuten enzimaacutetica a 6 concentraciones distintas de sustrato
Ademaacutes la presencia de los productos finales puede hacer que la reaccioacuten sea maacutes lenta o incluso invertir su sentido (Figura inferior)
EFECTO DE LOS INHIBIDORES SOBRE LA ACTIVIDAD ENZIMAacuteTICA
Ciertas moleacuteculas pueden inhibir la accioacuten cataliacutetica de un enzima son los inhibidores Estos inhibidores bien pueden ocupar temporalmente el centro activo por semejanza estructural con el sustrato original ( inhibidor competitivo) o bien alteran la conformacioacuten espacial del enzima impidiendo su unioacuten al sustrato ( inhibidor no competitivo) (Figuras inferiores)
Inhibidor competitivo Inhibidor no competitivo
17
MODULACIOacuteN ALOSTEacuteRICA DE LA ACTIVIDAD ENZIMAacuteTICA
Hay enzimas que pueden adoptar 2 conformaciones interconvertibles llamadas R (relajada) y T (tensa) R es la forma maacutes activa porque se une al sustrato con maacutes afinidad Las formas R y T se encuentran en equilibrio R lt==gt T (Figura inferior)
Ciertas sustancias tienden a estabilizar la forma R Son los llamados moduladores positivos El propio sustrato es a
menudo un modulador positivo Las moleacuteculas que favorecen la forma R pero que actuacutean sobre una regioacuten del enzima distinta del centro activo son los activadores alosteacutericos (Figura inferior izquierda)
Activador alosteacuterico favorece la unioacuten del sustrato
Inhibidor alosteacuterico impide la unioacuten del sustrato
Las sustancias que favorecen la forma T y disminuyen la actividad enzimaacutetica son los moduladores negativos Si
estos moduladores actuacutean en lugares distintos del centro activo del enzima se llaman inhibidores alosteacutericos
18
(Figura superior derecha
EFECTO DE LA MODIFICACIOacuteN COVALENTE SOBRE LA ACTIVIDAD ENZIMAacuteTICA
Otros enzimas pasan de una forma menos activa a otra maacutes activa unieacutendose covalentemente a un grupo quiacutemico de pequentildeo tamantildeo como el Pi o el AMP Tambieacuten se da el caso inverso en el que un enzima muy activo se desactiva al liberar alguacuten grupo quiacutemico En las enzimas de las viacuteas degradativas del metabolismo la
forma fosforilada es maacutes activa que la no fosforilada mientras que en las viacuteas biosinteacuteticas ocurre lo contrario En las figuras inferiores se ilustra la activacioacuten de una proteiacutena por fosforilacioacuten
Elementos de la reaccioacuten El enzima no fosforilado es inactivo El enzima fosforilado es activo
ACTIVACIOacuteN PROTEOLIacuteTICA DE LA ACTIVIDAD ENZIMAacuteTICA
Algunos enzimas no se sintetizan como tales sino como proteiacutenas precursoras sin actividad enzimaacutetica Estas proteiacutenas se llaman
proenzimas o zimoacutegenos Para activarse los zimoacutegenos sufren un ataque hidroliacutetico que origina la liberacioacuten de uno o varios peacuteptidos El resto de la moleacutecula proteica adopta la conformacioacuten y las propiedades del enzima activo Muchos enzimas digestivos se secretan en forma de zimoacutegenos y en el tubo digestivo
se convierten en la forma activa Es el caso de la -quimotripsina que se sintetiza en forma de quimotripsinoacutegeno (Figura superior) Si estos enzimas se sintetizasen directamente en forma activa destruiriacutean la propia ceacutelula que las
produce Asiacute la tripsina pancreaacutetica (una proteasa) se sintetiza como tripsinoacutegeno (inactivo) Si por alguna razoacuten se activa en el propio paacutencreas la glaacutendula sufre un proceso de autodestruccioacuten (pancreatitis aguda) a
menudo mortal
19
REGULACIOacuteN DE LA ACTIVIDAD ENZIMAacuteTICA POR MEDIO DE ISOENZIMAS
Algunos enzimas tienen distinta estructura molecular aunque su funcioacuten bioloacutegica es similar Se llaman isozimas o isoenzimas Estas diferencias de estructura se traducen en ligeros cambios en sus propiedades de forma que cada isozima se adapta perfectamente a la funcioacuten que debe realizar Asiacute podemos observar la existencia de isoenzimas en funcioacuten de
el tipo de tejido Por ejemplo la lactato deshidrogenasa presenta isozimas distintos en muacutesculo y corazoacuten el compartimento celular donde actuacutea Por ejemplo la malato deshidrogenasa del citoplasma es distinta
de la de la mitocondria el momento concreto del desarrollo del individuo Por ejemplo algunos enzimas de la glicolisis del feto
son diferentes de los mismos enzimas en el adulto
CINEacuteTICA ENZIMAacuteTICA
Los principios generales de las reacciones quiacutemicas se aplican tambieacuten a las reacciones enzimaacuteticas Por este motivo antes de empezar con la cineacutetica quiacutemica se van a repasar algunos conceptos baacutesicos de cineacutetica quiacutemica
A continuacioacuten se describiraacuten los siguientes conceptos
Cineacutetica enzimaacutetica Modelo cineacutetico de Michaelis-Menten Caacutelculo de la KM y la Vmax de un enzima Actividad enzimaacutetica
20
NOMENCLATURA DE LOS ENZIMAS
CATALISIS MOLECULAR
Un catalizador modifica la velocidad de una reaccioacuten quiacutemica sin ser utilizado o aparecer como uno de los productos de la reaccioacuten Una reaccioacuten quiacutemica en la que un substrato(S) se
transforma en un producto(P) S P ocurre por que cierta fraccioacuten de moleacuteculas de S posee mucho maacutes energiacutea que el resto de ellas lo que es suficiente para que alcancen un estado activado en el que pueda formarse o romperse un enlace quiacutemico y se forme el producto(P)
La energiacutea de activacioacuten es la cantidad de energiacutea expresada en caloriacuteas necesaria para que todas las moleacuteculas de un mol a una temperatura dada alcancen el estado reactivo Mientras que el estado de transicioacuten es el estado rico en energiacutea de las moleacuteculas que
interaccionan en la cima de la barrera de activacioacuten La velocidad de una reaccioacuten quiacutemica es proporcional a la concentracioacuten del complejo en el estado de transicioacuten
Una reaccioacuten quiacutemica se puede acelerar de la siguiente forma 1) Al aumentar la temperatura se incrementa la energiacutea cineacutetica por lo que es mayor el nuacutemero de moleacuteculas que alcanzan el
estado de transicioacuten Generalmente el Q10 =2 lo que indica que la velocidad de una reaccioacuten quiacutemica se duplica al aumentar la temperatura en 10
0C 2) Antildeadiendo un catalizador que
disminuye la energiacutea de activacioacuten y aumenta la velocidad de reaccioacuten
La enzima (E) se combina con el substrato (S) formando el complejo de transicioacuten enzima-
substrato (E-S) mediante una reaccioacuten reversible cuya energiacutea de activacioacuten es menor que la de la reaccioacuten no catalizada Cuando se forma el producto de la reaccioacuten (P) se regenera de nuevo la enzima (E) de forma libre la que puede combinarse de nuevo con otra moleacutecula de substrato (S)
Una enzima reduce maacutes eficientemente la energiacutea de activacioacuten(Ea) de una reaccioacuten que un catalizador inorgaacutenico lo que permite que una reaccioacuten se realice a menor temperatura
El siguiente ejemplo ilustra mejor lo que hemos discutido
Reaccioacuten Energiacutea de activacioacuten (Kcalmol-1
)
21
a) el peroacutexido de hidroacutegeno se descompone en
H2 O2 H2 O + O2
18
b) el hierro cataliacutetico (Fe) realiza la reaccioacuten
H2 O2 H2 O + O2
13
c) el platino cataliacutetico (Pt) realiza la reaccioacuten
H2 O2 H2 O + O2
12
d) la catalasa una enzima hepaacutetica la realiza
H2O2 H2O + O2
5
Una enzima no modifica la energiacutea libre ni la constante de equilibrio sino que disminuye la energiacutea de activacioacuten de la reaccioacuten
Ir al principio
EFECTO DE LA CONCENTRACIOacuteN DEL SUBSTRATO SOBRE LA
CATALISIS ENZIMATICA
La actividad de una enzima se puede estudiar in vitro bajo condiciones controladas antildeadieacutendole una enzima a un substrato Siacute se trabaja con la condicioacuten de que la concentracioacuten del substrato sea saturante variando entonces la concentracioacuten de enzima se observa que aumenta el producto de la reaccioacuten a pH y temperatura constantes
22
Si se mantiene la concentracioacuten de la enzima constante y variando la concentracioacuten de substrato se obtiene una curva hiperboacutelica como la de la figura Al principio un aumento de la
concentracioacuten de substrato produce un aumento raacutepi do de la velocidad de reaccioacuten pero si se sigue aumentando la concentracioacuten de substrato la velocidad de reaccioacuten comienza a disminuir y a muy altas concentraciones de substrato se observa que no cambia la velocidad de
reaccioacuten se dice que los centros activos de la enzima se encuentran saturados La velocidad de reaccioacuten que se obtiene a esa alta concentracioacuten de substrato se define como la velocidad maacutexima (V) de la reaccioacuten enzimaacutetica bajo las condiciones especificadas La concentracioacuten de
substrato (S) a la semivelocidad maacutexima de reaccioacuten (V2) se puede determinar de la figura y representa la constante de Michaelis o Km la cual es una caracteriacutestica para cada enzima La inversa de Km o 1Km mide aproximadamente la afinidad de la enzima por el subs trato
Mientras maacutes pequentildeo sea el valor de Km mayor seraacute la afinidad de la enzima por el substrato Si varias enzimas compiten en el metabolismo por el mismo substrato eacuteste seraacute transformado preferentemente por la enzima con mayor afinidad
La ecuacioacuten de Michaelis describe la relacioacuten cuantitativa entre la velocidad de reaccioacuten y la concentracioacuten de substrato [S] si se conoce Vmax o Km
v=
En donde v= es la velocidad de reaccioacuten observada a una concentracioacuten de substrato determinada [S]
Km= constante de Michaelis en molesliacutet ro
23
Vmax= velocidad maacutexima a concentracioacuten saturante de substrato
Si v= Vmax2 entonces
=
Km+[S] = 2[S]
Km =[S]
De tal forma que podemos concluir que Km es igual a la concentracioacuten de substrato a la semivelocidad maacutexima de reaccioacuten
1 Concentraciones relativas de E y S la concentracioacuten de substrato S es
mucho mayor que la de E de tal manera que la cantidad de substrato
unido a la enzima en cualquier momento es muy pequentildea
2 Se asume el estado estacionario ES no cambia con el tiempo esto
quiere decir que la velocidad de formacioacuten de ES es igual a aquella para
su desintegracioacuten En general un intermediario en una serie de reacciones estaacute en estado estacionario cuando su velocidad de siacutentesis
es igual a su velocidad de degradacioacuten 3 Velocidad inicial para el anaacutelisis de reacciones enzimaacuteticas soacutelo se
utiliza la velocidad inicial de la reaccioacuten que es la velocidad ejercida por
la enzima inmediatamente despueacutes de que se ha puesto en contacto con el substrato y hasta antes de que se haya consumido el 10 de la concentracioacuten inicial del mismo La razoacuten de lo anterior es que en ese
momento la concentracioacuten del producto de la reaccioacuten que se ha acumulado en muy pequentildea y por tanto la reaccioacuten en el sentido
inverso es decir la transformacioacuten del producto en el substrato original puede ser ignorada
24
Caracteriacutesticas de Km la constante de Michaelis-Menten es
caracteriacutestica de una enzima y particular para un substrato Refleja la afinidad de la enzima por ese substrato K m es numeacutericamente igual a la concentracioacuten
de substrato a la cual la velocidad de reaccioacuten es la mitad de la Vmax (Km= Vmax2) Este paraacutemetro Km no variacutea con la concentracioacuten de enzima
Significado de una Km pequentildea un valor numeacuterico pequentildeo de Km
refleja una alta afinidad de la enzima por su substrato porque a una baja
concentracioacuten del mismo la enzima a desarrollado ya la mitad de la velocidad maacutexima
Significado de una Km grande el valor numeacuterico grande de Km refleja
una baja afinidad de la enzima por su substrato porque a una concentracioacuten
elevada del mismo la enzima desarrolla la mitad de la velocidad maacutexima
Relacioacuten de la velocidad con la concentracioacuten de enzima la
velocidad de la reaccioacuten es directamente proporcional a la concentracioacuten de la
enzima a cualquier concentracioacuten de substrato Por ejemplo si la concentracioacuten de enzima es disminuida a la mitad la velocidad inicial de la reaccioacuten (v0) es
reducida tambieacuten a la mitad de la original
Orden de la reaccioacuten cuando S es mas pequentildea que la Km la
velocidad de la reaccioacuten es aproximadamente igual a la concentracioacuten de
substrato
100
50
0
Substrato
Km Figura Localizacioacuten de la constante de Michaelis (Km)
25
La velocidad de reaccioacuten se dice en estas condiciones es de primer
orden con respecto al substrato Cuando S es mas grande que la Km la
velocidad es constante e igual a la Vmax La velocidad de la reaccioacuten es independiente de la concentracioacuten de substrato y se dice que es de orden cero
con respecto a la concentracioacuten de substrato
Las enzimas pueden ser aisladas de las ceacutelulas u organismos para estudiar sus
propiedades in vitro es decir en un tubo de ensayo Las diferentes enzimas muestran diferentes respuestas a los cambios en la concentracioacuten de substrato temperatura y pH
Concentracioacuten de substrato
La forma hiperboacutelica de la curva de velocidad vs Substrato
Efecto de la temperatura en la actividad enzimaacutetica
Efecto de la variacioacuten del pH en la actividad enzimaacutetica
Incremento de la velocidad con la temperatura En general la velocidad de la reaccioacuten se incrementa con la temperatura hasta que un punto maacuteximo es alcanzado
Este incremento se debe a que aumenta el nuacutemero de moleacuteculas ricas en energiacutea que pueden pasar la barrera energeacutetica de estado de transicioacuten para formar a los productos
Decremento de la velocidad con la temperatura la elevacioacuten excesiva de la
temperatura del medio que contiene a las enzimas resulta en un decremento de la velocidad como resultado de la desnaturalizacioacuten es decir la peacuterdida de la estructura tridimensional de las enzimas
26
zona de reactivacioacuten maacuteximo de actividad
Zona de
inactivacioacuten
0 Temperatura
Figura dependencia de la actividad enzimaacutetica con la temperatura
Velocidad maacutexima la velocidad de una reaccioacuten (v) es el nuacutemero de
moleacuteculas de substrato convertidas en producto por unidad de tiempo y
usualmente se expresa en micromoles de producto formadas por minuto La velocidad de una reaccioacuten catalizada por una enzima aumenta conforme se incrementa la concentracioacuten de substrato hasta que se llega a una velocidad maacutexima (Vmax) a partir de la cual la velocidad de la reaccioacuten es
independiente de la cantidad de substrato El que la velocidad no pueda
seguirse incrementando refleja la saturacioacuten del sitio activo con el substrato
Vmax
0 Substrato
Figura representacioacuten de una cineacutetica tiacutepicamente michaeliana
Vmax = velocidad de saturacioacuten o maacutexima
27
Cuando se grafica la velocidad de la reaccioacuten v0 contra la concentracioacuten de substrato S no siempre es posible determinar la condicioacuten
en que se ha llegado a la velocidad maacutexima Vmax debido al incremento de la pendiente en la hipeacuterbola a concentraciones de substrato elevadas
Figura experimento cineacutetico para la actividad enzimaacutetica
Se presentan los valores experimentales y los resultados del ajuste al modelo de Michaelis
Auacuten asiacute si se grafica una doble reciacuteproca es decir el inverso de la
velocidad (1v0) contra el inverso de la concentracioacuten de substrato (1 S ) se
obtiene una liacutenea recta El regraacutefico de Lineweaver-Burke tambieacuten se
denomina de ldquodobles reciacuteprocasrdquo y se utiliza para calcular la Km y la Vmax asiacute como para determinar el mecanismo de accioacuten de los diversos tipos de
inhibidores
La ecuacioacuten que describe a la graacutefca de Lineweaver-Burke es
Vmax
1
SVmax
Km
v
1
0 (3)
que describe una recta en donde el intercepto con el eje de las abscisas es igual a ndash1Km y el intercepto con el eje de las ordenadas es igual a 1Vmax
28
Figura experimento cineacutetico para la actividad enzimaacutetica Regraacutefico de
Lineweaver-Burke
Se presentan los valores experimentales y los resultados del ajuste al modelo
REPRESENTACIOacuteN GRAacuteFICA DE LINEWEAVER Y BURK
Si se toma el inverso en ambos miembros de la ecuacioacuten de Michaelis tenemos
=
+
29
que es la ecuacioacuten de una liacutenea recta cuya pendiente es que intercepta al eje de las
ordenadas en y a la abcisa en el punto Lo que es evidente al hacer
ya que se convierte en
Si se hace una graacutefica con los dobles inversos colocando los valores de en el eje de las
ordenadas y en el eje de las abcisas se obtiene una liacutenea recta a partir de la cual se puede calcular faacutecilmente Km
EFECTO DEL pH SOBRE LA ACTIVIDAD ENZIMATICA
Sabiendo que las enzimas son proteinas cualquier cambio brusco de pH puede alterar el caraacutecter ioacutenico de los grupos amino y carboxilo en la superficie proteica afectando asiacute las propiedades cataliacuteticas de una enzima A pH alto o bajo se puede producir la desnaturalizacioacuten
de la enzima y en consecuencia su inactivacioacuten La fosfatasa aacutecida es maacutes activa a pH 50 mientras que la fosfatasa alcalina lo es a pH 90 Muchas enzimas tienen maacutexima actividad cerca de la neutralidad en un rango de pH de 6 a 8
El pH per se no afecta la actividad enzimaacutetica sino la concentracioacuten de protones Los protones
ademaacutes de alterar la estructura de la enzima y el substrato pueden participar tambieacuten en la reaccioacuten como substrato o producto En esos casos la concentracioacuten de protones afecta directamente la velocidad de la reaccioacuten
ENZIMA pH OPTIMO
30
Pepsina 15
Tripsina 77
Catalasa 76
Arginasa 97
Fumarasa 78
Ribonucleasa 78
EFECTO DE ACTIVADORES
Un aumento en la temperatura provoca un aumento de la velocidad de reaccioacuten hasta cierta temperatura oacuteptima ya que despueacutes de aproximadamente 45
0 C se comienza a producir la
desnaturalizacioacuten teacutermica Las enzimas de muchos mamiacuteferos tienen una temperatura oacuteptima
de 370 C por encima de esa temperatura comienzan a inactivarse y se destruyen Sin embargo
existen especies de bacterias y algas que habitan en fuentes de aguas termales y en el otro extremo ciertas bacterias aacuterticas tienen temperaturas oacuteptimas cercanas a 0
0 C
31
ESPECIFICIDAD ABSOLUTA
Cierta enzimas se encuentran como aproenzimas o zimogenos que son inactivos
El tripsinoacutegeno es convertido en tripsina por la misma accioacuten de la tripsina que remueve un hexapeacuteptido
tripsina Tripsinoacutegeno Tripsina + Val (Asp)4 lisina
Otra forma de activacioacuten consiste en mantener los grupos SH reducidos Estos grupos pueden
formar parte de centros activos de ciertas enzimas Si se oxidan estos grupos a -S-S- la enzima se inactiva Asiacute tenemos que la enzima papaina se inactiva despueacutes de exponerla al ox iacutegeno pero cuando se le antildeade un reductor apropiado la enzima se reactiva Los grupos ndashS-S- se convierten en ndashSH activaacutendose la enzima
Otra forma de activacioacuten requiere la presencia de otra substancia que se enlaza al sitio alosteacuterico el compuesto que se enlaza se denomina un efector alosteacuterico El centro alosteacuterico es bien especiacutefico para el efector o modulador alosteacuterico
ESPECIFICIDAD RELATIVA
Oxiacutegeno L-aminoaacutecido a -cetoaacutecido + NH3 + H 2 O 2 L-aminoaacutecido oxidasa Oxiacutegeno D-aminoaacutecido a -cetoaacutecido + NH3 + H 2 O 2 D-aminoaacutecido oxidasa
32
En este ejemplo se muestra la estereo-especificidad Una enzima puede tener una especificidad oacuteptica para isomeros D o L
Hay enzimas que muestran especificidad absoluta como la aspartasa que cataliza la adicioacuten reversible de amoniacuteaco al doble enlace del aacutecido fumaacuterico pero no al de otros aacutecidos insaturados
HOOC ndash CH = CH ndash COO- + NH4
+ HOOC ndash CH2 - CHNH2 - COO
- + H
+
INHIBICIOacuteN ENZIMATICA
Mediante el uso de inhibidores enzimaacuteticos se ha obtenido informacioacuten muy valiosa sobre la conformacioacuten del centro activo de algunas enzimas
a Inhibicioacuten irreversible Este tipo de inhibidores forman un enlace covalente con las enzimas cerca del centro activo Un ejemplo son los gases nerviosos como el
fluorofosfato de diisopropilo (DFP) que forma un complejo con la enzima acetilcolinesterasa Los animales envenenados con este gas quedan paralizados debido a la imposibilidad de transmitir adecuadamente los impulsos nerviosos
Ester di-isopropil fos foacuterico de la enzima (inactivo)
b) Inhibicioacuten competitiva
Es cuando el inhibidor compite con el substrato por la unioacuten con el centro activo de la
enzima Este tipo de inhibicioacuten puede reducirse si se aumenta la concentracioacuten de substrato
-OOC - CH2 - CH2 - COO
- -OOC - CH = CH - COO
- + 2H
+ Succinato Succinato Fumarato
33
Deshidrogenasa COO
- I CH2 Malonato (inhibidor competitivo) I COO
- En la inhibicioacuten competitiva la enzima se combina con el inhibidor para formar un complejo enzima-inhibidor E + I E I en competencia con la reaccioacuten normal E +S ES
La sulfanilamida (bactericida) interfiere con la siacutentesis del aacutecido foacutelico compitiendo con el aacutecido p-aminobenzoico de una forma competitiva
HOOC-sect- NH2 NH2 ndashSO2 - sect- NH2 aacutecido p- amino benzoico sulfanilamida
sect = fenil
a Inhibicioacuten no competitiva
Esta inhibicioacuten se caracteriza por que no se puede revertir el efecto del inhibidor aumentando la concentracioacuten del substrato
Por ejemplo la inhibicioacuten de la deshidrogenasa del gliceraldehido -3-fosfato por la yodo-acetamida
Enzima-SH + I-CH2 CONH2 Enzima-SCH2 CONH2 + IH
La deshidrogenasa del gliceraldehido-3 fosfato puede ser inhibida tambieacuten por el aacutecido iodo-aceacutetico
Enzima-SH + ICH2 COOH Enzima-SCH2 COOH + IH
c) Inhibicioacuten por metales
Ciertos metales como el plomo mercurio y arseacutenico inhiben enzimas que tienen en su centro activo grupos -SH libres

2
2 Especificidad de accioacuten Cada reaccioacuten estaacute catalizada por un enzima
especiacutefico Por ejemplo el enzima sacarasa es muy especifico rompe el enlace
glucosidico de la sacarosa
Sin embargo hay distintos grados de especificidad La sacarasa es muy especiacutefico
de la sacarosa (sustrato natural) mientras que la maltosa y la isomaltosa son
sustratos anaacutelogos El enzima actuacutea con maacutexima eficacia sobre el sustrato natural
y con menos eficacia sobre los sustratos anaacutelogos
La accioacuten enzimaacutetica se caracteriza por la formacioacuten de un complejo que
representa el estado de transicioacuten
E + S
ES
E + P
El sustrato se une al enzima a traveacutes de numerosas interacciones deacutebiles como son
puentes de hidroacutegeno electrostaacuteticos hidroacutefobos etc en un lugar especiacutefico el
centro activo o sitio activo Este centro es una pequentildea porcioacuten del enzima
constituido por una serie de aminoaacutecidos que interaccionan con el sustrato
13 SITIO ACTIVO
Las moleacuteculas de enzimas contienen hendiduras o cavidades denominadas
sitio activo El sitio activo estaacute formado por las cadenas laterales de residuos
especiacuteficos lo que ocasiona que tenga un arreglo tridimensional particular diferente
al resto de la proteiacutena Este sitio es afiacuten por la estructura tridimensional del sustrato
(Figura 2)
enzima
+ sustrato
sitio activo
sitio activo vaciacuteo ocupado
3
Figura 2 Representacioacuten de la formacioacuten del complejo enzima-sustrato
Es decir el sitio activo esta formado por ciertos aminoaacutecidos que forman un
microambiente caracteriacutestico dentro de la propia cadena y que llevan a cabo la
reaccioacuten generalmente existe soacutelo uno por moleacutecula de enzima
Las enzimas adquieren su poder cataliacutetico cuando presentan una estructura
secundaria y terciaria muy especiacutefica de tal manera que los aminoaacutecidos
componentes del sitio activo se encuentran vecinales formando dicho
microambiente Asiacute por ejemplo la quimotripsina crea su centro activo con los
aminoaacutecidos histidina y serina localizados en las posiciones 57 y 195 respectivamente
se puede decir que forzosamente esta moleacutecula debe adquirir una estructura
tridimensional de tal manera que dichos aminoaacutecidos sean adyacentes
14 EFICIENCIA CATALITICA
La mayoriacutea de las reacciones catalizadas por enzimas son muy eficientes y
transcurren desde 106 hasta 1014 veces maacutes raacutepido que la misma reaccioacuten no
catalizada (Tabla 1) Tiacutepicamente cada moleacutecula de enzima es capaz de transformar
cada segundo de 100 a 1000 moleacuteculas de substrato en producto El nuacutemero de
estas moleacuteculas transformadas a producto por moleacutecula de enzima en cada
segundo se conoce como el nuacutemero de recambio
Tabla1 Eficiencia de las reacciones catalizadas por algunas enzimas
Enzima Velocidad en
ausencia de enzima
Velocidad de
reaccioacuten catalizada
Rendimiento
Anhidrasa carboacutenica Corismato mutasa Triosafosfato isomerasa Carboxipeptidasa A AMP nucleosidasa
Nucleasa estafilococal
13 X 10 ndash1
26 X 10 ndash5
43 X 10 ndash6
30 X 10 ndash9
10 X 10 ndash11
17 X 10 -13
10 X 106
50 4300 578 60
95
77 X 106
19 X 106
10 X 109
19X 1011
60 X 1012
56 X 1014
4
128 NOMENCLATURA DE LAS ENZIMAS
A cada enzima se le asignan dos nombres El primero es corto y es como se
conoce a la proteiacutena de manera coloquial es el nombre sugerido El segundo es maacutes
completo e infiere propiedades sobre la reaccioacuten que la enzima desarrolla se le
conoce como nombre sistemaacutetico este nombre permite reconocer a la enzima sin
ambiguumledad y localizarla en el metabolismo
a) Nombre sugerido
Muchas de las enzimas poseen en su nombre el sufijo ldquo-asardquo unido al nombre
del substrato de la reaccioacuten que cataliza por ejemplo
Ureasa proteiacutena cuyo sustrato es la urea Lactasa proteiacutena cuyo sustrato es la lactosa
Tambieacuten suele utilizarse este sufijo a la descripcioacuten de la reaccioacuten que la enzima cataliza
Lactato deshidrogenada deshidrogena (le quita Hidroacutegenos) al lactato
Adenilato ciclasa hace un ciclo en la adenina
Algunas enzimas poseen nombres que no representan su actividad o sustrato
Lisozima
Tripsina
b) Nombre sistemaacutetico
La Unioacuten Internacional de Bioquiacutemica y Biologiacutea Molecular (IUBMB) desarrolloacute
un sistema de nomenclatura en el cual las enzimas se dividen en seis clases
principales cada una con numerosos subgrupos
127 CLASIFICACIOacuteN DE LAS ENZIMAS
5
En funcioacuten de su accioacuten cataliacutetica especiacutefica los enzimas se clasifican en 6 grandes grupos o clases
1 Oxidorreductasas Reacciones de transferencia de electrones
2 Transferasas Transferencia de grupos funcionales Ej UDP-glucosa-fructosa-
glucotransferasa
3 Hidrolasas Reacciones de hidroacutelisis Ej lipasa proteasa celulasa
4 Liasas Adicioacuten a dobles enlaces Ej carboxilasa fenilalanina amonioliasa
5 Isomerasas Reacciones de isomerizacioacutenEj fosfoglucosa isomerasa
6 Ligasas Se conociacutean como sintetasas Participan en la formacioacuten de enlaces
con hidroacutelisis de ATP
Para todas las clasificaciones se muestran ejemplos representativos
1- OXIDOREDUCTASAS
Catalizan reacciones de oxidacioacuten-reduccioacuten
2e-
2- TRANSFERASAS
Catalizan la transferencia de grupos que contienen C N o P
THF tetrahidrofolato
6
3- HIDROLASAS
Catalizan la ruptura de enlaces por la adicioacuten de una moleacutecula de agua
4- LIASAS
Catalizan la ruptura de enlaces C-C C-S y algunos enlaces C-N
5- ISOMERASAS
Catalizan la racemizacioacuten de isoacutemeros oacutepticos o geomeacutetricos
7
6- LIGASAS
Catalizan la formacioacuten de enlaces entre C y O S N Esta reaccioacuten soacutelo es posible
mediante la energiacutea derivada de fosfatos ricos en energiacutea como el del fosfato de la
moleacutecula de ATP
15 COFACTORES y COENZIMAS Algunas enzimas se asocian con moleacuteculas de caraacutecter no proteiacuteco que son
necesarias para el funcionamiento de la enzima estas moleacuteculas se denominan
cofactores (Figura 3) Comuacutenmente los factores encontrados en las enzimas incluyen
iones metaacutelicos como el Zn2+ o el Fe2+ ( Tabla 2) tambieacuten pueden ser moleacuteculas
orgaacutenicas que se denomina coenzimas como el NAD+ FAD la coenzima A
Generalmente las coenzimas son derivados de las vitaminas A la enzima en ausencia
de su cofactor (cuando lo tiene) se le denomina apoenzima en presencia de su
cofactor (cuando lo tiene) se le denomina holoenzima La apoenzima generalmente
carece de actividad bioloacutegica La diferencia entre un cofactor y un grupo prosteacutetico
como el grupo hemo es que este uacuteltimo estaacute unido de manera covalente a la
enzima mientras que el cofactor puede ser removido de la misma con relativa
facilidad
Se pueden sentildealar algunas caracteriacutesticas de las coenzimas como generalmente son
estables al calor y a otros agentes desnaturalizantes de las proteiacutenas pueden ser
recicladas y algunas provienen de las vitaminas
8
Figura 3 Cofactores y Coenzima
Tabla 2 Algunos enzimas que contienen iones metaacutelicos o los necesitan como
cofactores
Zn+2 Cu+2 (Cu+)
Alcohol- deshidrogenada Tirosinasa Anhidrasa-carboacutenica Citocromo-oxidasa Carboxipeptidasa
Mg+2 K+
Fosfohidrolasa Piruvato- fosfoquinasa Fosfotransferasas
Mn+2 Arginasa Na+ Fosfotransferasas ATPasa de la membrana plasmaacutetica
Fe +2 o Fe+3
9
Citocromos
Peroxidasa Catalasa Ferredoxina
Texto ------- (Tabla 3)
Tabla 3 Relacioacuten entre algunas coenzimas y las vitaminas a partir de las
cuales se producen Enfermedades generadas por la deficiencia
Coenzima Reaccioacuten Fuente vitamiacutenica Enfermedad
Humana por deficiencia
Biocitina Carboxilacioacuten Biotina
Coenzima A Transferencia de acilos Pantotenato
Coenzimas de Cobalamima Alquilacioacuten Cobalamina (B12) anemia perniciosa
Coenzimas de flavina Oxidacioacuten-reduccioacuten Riboflavina (B2)
Acido lipoacuteico Transferencia de acilos -
Coenzimas de nicotinamida Oxidacioacuten-reduccioacuten Nicotinamida (niacina) Pelagra
Fosfato de piridoxal Transferencia de grupo amino Piridoxina (B6)
Tetrahidrofolato Transferencia de un grupo de 1 Carbono Acido foacutelico anemia
megaloblaacutestica
Tiamina pirofosfato Transferencia de aldehiacutedo Tiamina (B1) beriberi
no tiene un nombre especiacutefico su aparicioacuten es muy rara
Otras coenzimas
NADP
Coenzima Q
Biotina
NAD y NADP
10
Coenzima Q (CoQ)
Esta coenzima es sintetizada a partir de farnesil pirofosfato Al igual que el
FMN la CoQ es capaz de aceptar y donar uno o dos electrones porque su forma
semiquinona es estable A su forma oxidada se le conoce como ubiquinona (de
ubicuo lat ubique en todas partes) o quinona A su forma reducida se le conoce
como QH2 ubiquinol o hidroquinona (Figura 2) A la coenzima solo parcialmente
reducida se le conoce como ubisemiquinona semiquinona o QH (Figura 4)
Figura 4 Representacioacuten de la moleacutecula de CoQ
La siacutentesis de la CoQ se lleva a cabo a traveacutes de unidades isoprenoides (Figura 5)
11
Figura 5 La CoQ se produce a partir de isoprenoides
Biotina
La biotina consiste de un anillo imidazol que estaacute unido en forma cis a la
cadena lateral tetrahidrotiofeno del valerato (Figura 6)
12
Figura 6 Estructura de la biotina
La enzima holocarboxilasa sintetasa (HCS) cataliza la activacioacuten mediante
biotinilacioacuten de cinco carboxilasas en ceacutelulas humanas La deficiencia de HCS
produce el siacutendrome deficiencia muacuteltiple de carboxilasas (DMC) Esta enfermedad es
potencialmente mortal y se caracteriza por la deficiencia de la actividad de todas las
carboxilasas La deficiencia de biotina no solo disminuye la actividad de estas enzimas
sino que parece modificar la expresioacuten geneacutetica
126 PROPIEDADES DE LAS ENZIMAS
Las propiedades de las enzimas derivan del hecho de ser proteiacutenas y de actuar como
catalizadores Como proteiacutenas poseen una conformacioacuten natural maacutes estable que las
demaacutes conformaciones posibles Asiacute cambios en la conformacioacuten suelen ir asociados
en cambios en la actividad cataliacutetica Los factores que influyen de manera maacutes directa
sobre la actividad de un enzima son
pH
temperatura
cofactores
A) EFECTO DEL pH
Al comprobar experimentalmente la influencia del pH en la velocidad de las
reacciones enzimaacuteticas se obtienen curvas que indican que los enzimas
presentan un pH oacuteptimo de actividad El pH puede afectar de varias maneras
13
o El centro activo puede contener aminoaacutecidos con grupos ionizados
que pueden variar con el pH
o La ionizacioacuten de aminoaacutecidos que no estaacuten en el centro activo puede
provocar modificaciones en la conformacioacuten de la enzima
o El sustrato puede verse afectado por las variaciones del pH
Algunos enzimas presentan variaciones peculiares La pepsina del estoacutemago
presenta un oacuteptimo a pH=2 la ureasa lo tiene a pH 7 y la fosfatasa alcalina del
intestino un pH= 12
B) EFECTO DE LA TEMPERATURA
Influye en la actividad El punto oacuteptimo representa el maacuteximo de actividad A
temperaturas bajas los enzimas se hallan ldquomuy riacutegidosrdquo y cuando se supera un
valor considerable (mayor de 50) la actividad cae bruscamente porque como
proteiacutena el enzima se desnaturaliza
128 EFECTO DE LOS COFACTORES SOBRE LA ACTIVIDAD
ENZIMATICA
A veces un enzima requiere para su funcioacuten la presencia de sustancias no proteicas
que colaboran en la cataacutelisis los cofactores Casi un tercio de los enzimas conocidos
requieren cofactores Cuando el cofactor es una moleacutecula orgaacutenica se llama
coenzima En la figura inferior podemos observar una moleacutecula de hemoglobina
(proteiacutena que transporta oxiacutegeno) y su coenzima (el grupo hemo) Cuando los
cofactores y las coenzimas se encuentran unidos covalentemente al enzima se llaman
14
grupos prosteacuteticos La forma cataliacuteticamente activa del enzima es decir el enzima
unido a su grupo prosteacutetico se llama holoenzima La parte proteica de un
holoenzima (inactiva) se llama apoenzima de forma que
apoenzima + grupo prosteacutetico= holoenzima
129 MODO DE ACCIOacuteN DE LAS ENZIMAS
En las reacciones espontaacuteneas los productos finales tienen menos energiacutea libre de
Gibbs (∆G) que los reactantes (figura inferior izquierda) Por lo tanto las reacciones
espontaacuteneas liberan energiacutea de Gibbs ((∆G lt 0) Sin embargo el comienzo de la
reaccioacuten requiere un aporte inicial de energiacutea Esta energiacutea inicial que hay que
suministrar a los reactantes para que la reaccioacuten transcurra se llama energiacutea de
activacioacuten (Ea) Cuanto menor es la Ea maacutes faacutecilmente transcurre la reaccioacuten
Perfil energeacutetico de una reaccioacuten espontaacutenea Perfil energeacutetico de una reaccioacuten catalizada
15
La accioacuten de los catalizadores consiste precisamente en disminuir la Ea (Figura superior derecha) Los enzimas
son catalizadores especialmente eficaces ya que disminuyen la Ea auacuten maacutes que los catalizadores inorgaacutenicos
Por ejemplo la descomposicioacuten del agua oxigenada (H2O2) para dar H2O y O2 puede ocurrir sin catalizador
con un catalizador inorgaacutenico (platino) o con un enzima especiacutefico (catalasa) Las respectivas Ea para cada
proceso son 18 12 y 6 Kcalmol Asiacute se puede calcular que el platino acelera la reaccioacuten 20000 veces mientras
que la catalasa la acelera 370000 veces
Para que una reaccioacuten quiacutemica tenga lugar las moleacuteculas de los reactantes deben chocar con una energiacutea y
una orientacioacuten adecuadas La actuacioacuten del enzima (1) permite que los rea ctantes (sustratos) se unan a su
centro activo con una orientacioacuten oacuteptima para que la reaccioacuten se produzca y (2) modifica las propiedades
quiacutemicas del sustrato unido a su centro activo debilitando los enlaces existentes y facilitando la formacioacuten de
otros nuevos (Figuras inferiores)
1210 REGULACIOacuteN DE SU ACTIVIDAD ENZIMATICA
16
Una moleacutecula de enzima no tiene por que actuar siempre a la misma velocidad Su
actividad puede estar modulada por
Cambios en el pH
Cambios en la temperatura
Presencia de cofactores
Las concentraciones del sustrato y de los productos finales
Presencia de inhibidores
Modulacioacuten alosteacuterica
Modificacioacuten covalente
Activacioacuten por proteolisis
Isoenzimas
EFECTO DE LAS CONCENTRACIONES SOBRE LA ACTIVIDAD ENZIMAacuteTICA
La velocidad de una reaccioacuten enzimaacutetica depende de la concentracioacuten de sustrato La Figura de la derecha muestra la velocidad de una reaccioacuten enzimaacutetica a 6 concentraciones distintas de sustrato
Ademaacutes la presencia de los productos finales puede hacer que la reaccioacuten sea maacutes lenta o incluso invertir su sentido (Figura inferior)
EFECTO DE LOS INHIBIDORES SOBRE LA ACTIVIDAD ENZIMAacuteTICA
Ciertas moleacuteculas pueden inhibir la accioacuten cataliacutetica de un enzima son los inhibidores Estos inhibidores bien pueden ocupar temporalmente el centro activo por semejanza estructural con el sustrato original ( inhibidor competitivo) o bien alteran la conformacioacuten espacial del enzima impidiendo su unioacuten al sustrato ( inhibidor no competitivo) (Figuras inferiores)
Inhibidor competitivo Inhibidor no competitivo
17
MODULACIOacuteN ALOSTEacuteRICA DE LA ACTIVIDAD ENZIMAacuteTICA
Hay enzimas que pueden adoptar 2 conformaciones interconvertibles llamadas R (relajada) y T (tensa) R es la forma maacutes activa porque se une al sustrato con maacutes afinidad Las formas R y T se encuentran en equilibrio R lt==gt T (Figura inferior)
Ciertas sustancias tienden a estabilizar la forma R Son los llamados moduladores positivos El propio sustrato es a
menudo un modulador positivo Las moleacuteculas que favorecen la forma R pero que actuacutean sobre una regioacuten del enzima distinta del centro activo son los activadores alosteacutericos (Figura inferior izquierda)
Activador alosteacuterico favorece la unioacuten del sustrato
Inhibidor alosteacuterico impide la unioacuten del sustrato
Las sustancias que favorecen la forma T y disminuyen la actividad enzimaacutetica son los moduladores negativos Si
estos moduladores actuacutean en lugares distintos del centro activo del enzima se llaman inhibidores alosteacutericos
18
(Figura superior derecha
EFECTO DE LA MODIFICACIOacuteN COVALENTE SOBRE LA ACTIVIDAD ENZIMAacuteTICA
Otros enzimas pasan de una forma menos activa a otra maacutes activa unieacutendose covalentemente a un grupo quiacutemico de pequentildeo tamantildeo como el Pi o el AMP Tambieacuten se da el caso inverso en el que un enzima muy activo se desactiva al liberar alguacuten grupo quiacutemico En las enzimas de las viacuteas degradativas del metabolismo la
forma fosforilada es maacutes activa que la no fosforilada mientras que en las viacuteas biosinteacuteticas ocurre lo contrario En las figuras inferiores se ilustra la activacioacuten de una proteiacutena por fosforilacioacuten
Elementos de la reaccioacuten El enzima no fosforilado es inactivo El enzima fosforilado es activo
ACTIVACIOacuteN PROTEOLIacuteTICA DE LA ACTIVIDAD ENZIMAacuteTICA
Algunos enzimas no se sintetizan como tales sino como proteiacutenas precursoras sin actividad enzimaacutetica Estas proteiacutenas se llaman
proenzimas o zimoacutegenos Para activarse los zimoacutegenos sufren un ataque hidroliacutetico que origina la liberacioacuten de uno o varios peacuteptidos El resto de la moleacutecula proteica adopta la conformacioacuten y las propiedades del enzima activo Muchos enzimas digestivos se secretan en forma de zimoacutegenos y en el tubo digestivo
se convierten en la forma activa Es el caso de la -quimotripsina que se sintetiza en forma de quimotripsinoacutegeno (Figura superior) Si estos enzimas se sintetizasen directamente en forma activa destruiriacutean la propia ceacutelula que las
produce Asiacute la tripsina pancreaacutetica (una proteasa) se sintetiza como tripsinoacutegeno (inactivo) Si por alguna razoacuten se activa en el propio paacutencreas la glaacutendula sufre un proceso de autodestruccioacuten (pancreatitis aguda) a
menudo mortal
19
REGULACIOacuteN DE LA ACTIVIDAD ENZIMAacuteTICA POR MEDIO DE ISOENZIMAS
Algunos enzimas tienen distinta estructura molecular aunque su funcioacuten bioloacutegica es similar Se llaman isozimas o isoenzimas Estas diferencias de estructura se traducen en ligeros cambios en sus propiedades de forma que cada isozima se adapta perfectamente a la funcioacuten que debe realizar Asiacute podemos observar la existencia de isoenzimas en funcioacuten de
el tipo de tejido Por ejemplo la lactato deshidrogenasa presenta isozimas distintos en muacutesculo y corazoacuten el compartimento celular donde actuacutea Por ejemplo la malato deshidrogenasa del citoplasma es distinta
de la de la mitocondria el momento concreto del desarrollo del individuo Por ejemplo algunos enzimas de la glicolisis del feto
son diferentes de los mismos enzimas en el adulto
CINEacuteTICA ENZIMAacuteTICA
Los principios generales de las reacciones quiacutemicas se aplican tambieacuten a las reacciones enzimaacuteticas Por este motivo antes de empezar con la cineacutetica quiacutemica se van a repasar algunos conceptos baacutesicos de cineacutetica quiacutemica
A continuacioacuten se describiraacuten los siguientes conceptos
Cineacutetica enzimaacutetica Modelo cineacutetico de Michaelis-Menten Caacutelculo de la KM y la Vmax de un enzima Actividad enzimaacutetica
20
NOMENCLATURA DE LOS ENZIMAS
CATALISIS MOLECULAR
Un catalizador modifica la velocidad de una reaccioacuten quiacutemica sin ser utilizado o aparecer como uno de los productos de la reaccioacuten Una reaccioacuten quiacutemica en la que un substrato(S) se
transforma en un producto(P) S P ocurre por que cierta fraccioacuten de moleacuteculas de S posee mucho maacutes energiacutea que el resto de ellas lo que es suficiente para que alcancen un estado activado en el que pueda formarse o romperse un enlace quiacutemico y se forme el producto(P)
La energiacutea de activacioacuten es la cantidad de energiacutea expresada en caloriacuteas necesaria para que todas las moleacuteculas de un mol a una temperatura dada alcancen el estado reactivo Mientras que el estado de transicioacuten es el estado rico en energiacutea de las moleacuteculas que
interaccionan en la cima de la barrera de activacioacuten La velocidad de una reaccioacuten quiacutemica es proporcional a la concentracioacuten del complejo en el estado de transicioacuten
Una reaccioacuten quiacutemica se puede acelerar de la siguiente forma 1) Al aumentar la temperatura se incrementa la energiacutea cineacutetica por lo que es mayor el nuacutemero de moleacuteculas que alcanzan el
estado de transicioacuten Generalmente el Q10 =2 lo que indica que la velocidad de una reaccioacuten quiacutemica se duplica al aumentar la temperatura en 10
0C 2) Antildeadiendo un catalizador que
disminuye la energiacutea de activacioacuten y aumenta la velocidad de reaccioacuten
La enzima (E) se combina con el substrato (S) formando el complejo de transicioacuten enzima-
substrato (E-S) mediante una reaccioacuten reversible cuya energiacutea de activacioacuten es menor que la de la reaccioacuten no catalizada Cuando se forma el producto de la reaccioacuten (P) se regenera de nuevo la enzima (E) de forma libre la que puede combinarse de nuevo con otra moleacutecula de substrato (S)
Una enzima reduce maacutes eficientemente la energiacutea de activacioacuten(Ea) de una reaccioacuten que un catalizador inorgaacutenico lo que permite que una reaccioacuten se realice a menor temperatura
El siguiente ejemplo ilustra mejor lo que hemos discutido
Reaccioacuten Energiacutea de activacioacuten (Kcalmol-1
)
21
a) el peroacutexido de hidroacutegeno se descompone en
H2 O2 H2 O + O2
18
b) el hierro cataliacutetico (Fe) realiza la reaccioacuten
H2 O2 H2 O + O2
13
c) el platino cataliacutetico (Pt) realiza la reaccioacuten
H2 O2 H2 O + O2
12
d) la catalasa una enzima hepaacutetica la realiza
H2O2 H2O + O2
5
Una enzima no modifica la energiacutea libre ni la constante de equilibrio sino que disminuye la energiacutea de activacioacuten de la reaccioacuten
Ir al principio
EFECTO DE LA CONCENTRACIOacuteN DEL SUBSTRATO SOBRE LA
CATALISIS ENZIMATICA
La actividad de una enzima se puede estudiar in vitro bajo condiciones controladas antildeadieacutendole una enzima a un substrato Siacute se trabaja con la condicioacuten de que la concentracioacuten del substrato sea saturante variando entonces la concentracioacuten de enzima se observa que aumenta el producto de la reaccioacuten a pH y temperatura constantes
22
Si se mantiene la concentracioacuten de la enzima constante y variando la concentracioacuten de substrato se obtiene una curva hiperboacutelica como la de la figura Al principio un aumento de la
concentracioacuten de substrato produce un aumento raacutepi do de la velocidad de reaccioacuten pero si se sigue aumentando la concentracioacuten de substrato la velocidad de reaccioacuten comienza a disminuir y a muy altas concentraciones de substrato se observa que no cambia la velocidad de
reaccioacuten se dice que los centros activos de la enzima se encuentran saturados La velocidad de reaccioacuten que se obtiene a esa alta concentracioacuten de substrato se define como la velocidad maacutexima (V) de la reaccioacuten enzimaacutetica bajo las condiciones especificadas La concentracioacuten de
substrato (S) a la semivelocidad maacutexima de reaccioacuten (V2) se puede determinar de la figura y representa la constante de Michaelis o Km la cual es una caracteriacutestica para cada enzima La inversa de Km o 1Km mide aproximadamente la afinidad de la enzima por el subs trato
Mientras maacutes pequentildeo sea el valor de Km mayor seraacute la afinidad de la enzima por el substrato Si varias enzimas compiten en el metabolismo por el mismo substrato eacuteste seraacute transformado preferentemente por la enzima con mayor afinidad
La ecuacioacuten de Michaelis describe la relacioacuten cuantitativa entre la velocidad de reaccioacuten y la concentracioacuten de substrato [S] si se conoce Vmax o Km
v=
En donde v= es la velocidad de reaccioacuten observada a una concentracioacuten de substrato determinada [S]
Km= constante de Michaelis en molesliacutet ro
23
Vmax= velocidad maacutexima a concentracioacuten saturante de substrato
Si v= Vmax2 entonces
=
Km+[S] = 2[S]
Km =[S]
De tal forma que podemos concluir que Km es igual a la concentracioacuten de substrato a la semivelocidad maacutexima de reaccioacuten
1 Concentraciones relativas de E y S la concentracioacuten de substrato S es
mucho mayor que la de E de tal manera que la cantidad de substrato
unido a la enzima en cualquier momento es muy pequentildea
2 Se asume el estado estacionario ES no cambia con el tiempo esto
quiere decir que la velocidad de formacioacuten de ES es igual a aquella para
su desintegracioacuten En general un intermediario en una serie de reacciones estaacute en estado estacionario cuando su velocidad de siacutentesis
es igual a su velocidad de degradacioacuten 3 Velocidad inicial para el anaacutelisis de reacciones enzimaacuteticas soacutelo se
utiliza la velocidad inicial de la reaccioacuten que es la velocidad ejercida por
la enzima inmediatamente despueacutes de que se ha puesto en contacto con el substrato y hasta antes de que se haya consumido el 10 de la concentracioacuten inicial del mismo La razoacuten de lo anterior es que en ese
momento la concentracioacuten del producto de la reaccioacuten que se ha acumulado en muy pequentildea y por tanto la reaccioacuten en el sentido
inverso es decir la transformacioacuten del producto en el substrato original puede ser ignorada
24
Caracteriacutesticas de Km la constante de Michaelis-Menten es
caracteriacutestica de una enzima y particular para un substrato Refleja la afinidad de la enzima por ese substrato K m es numeacutericamente igual a la concentracioacuten
de substrato a la cual la velocidad de reaccioacuten es la mitad de la Vmax (Km= Vmax2) Este paraacutemetro Km no variacutea con la concentracioacuten de enzima
Significado de una Km pequentildea un valor numeacuterico pequentildeo de Km
refleja una alta afinidad de la enzima por su substrato porque a una baja
concentracioacuten del mismo la enzima a desarrollado ya la mitad de la velocidad maacutexima
Significado de una Km grande el valor numeacuterico grande de Km refleja
una baja afinidad de la enzima por su substrato porque a una concentracioacuten
elevada del mismo la enzima desarrolla la mitad de la velocidad maacutexima
Relacioacuten de la velocidad con la concentracioacuten de enzima la
velocidad de la reaccioacuten es directamente proporcional a la concentracioacuten de la
enzima a cualquier concentracioacuten de substrato Por ejemplo si la concentracioacuten de enzima es disminuida a la mitad la velocidad inicial de la reaccioacuten (v0) es
reducida tambieacuten a la mitad de la original
Orden de la reaccioacuten cuando S es mas pequentildea que la Km la
velocidad de la reaccioacuten es aproximadamente igual a la concentracioacuten de
substrato
100
50
0
Substrato
Km Figura Localizacioacuten de la constante de Michaelis (Km)
25
La velocidad de reaccioacuten se dice en estas condiciones es de primer
orden con respecto al substrato Cuando S es mas grande que la Km la
velocidad es constante e igual a la Vmax La velocidad de la reaccioacuten es independiente de la concentracioacuten de substrato y se dice que es de orden cero
con respecto a la concentracioacuten de substrato
Las enzimas pueden ser aisladas de las ceacutelulas u organismos para estudiar sus
propiedades in vitro es decir en un tubo de ensayo Las diferentes enzimas muestran diferentes respuestas a los cambios en la concentracioacuten de substrato temperatura y pH
Concentracioacuten de substrato
La forma hiperboacutelica de la curva de velocidad vs Substrato
Efecto de la temperatura en la actividad enzimaacutetica
Efecto de la variacioacuten del pH en la actividad enzimaacutetica
Incremento de la velocidad con la temperatura En general la velocidad de la reaccioacuten se incrementa con la temperatura hasta que un punto maacuteximo es alcanzado
Este incremento se debe a que aumenta el nuacutemero de moleacuteculas ricas en energiacutea que pueden pasar la barrera energeacutetica de estado de transicioacuten para formar a los productos
Decremento de la velocidad con la temperatura la elevacioacuten excesiva de la
temperatura del medio que contiene a las enzimas resulta en un decremento de la velocidad como resultado de la desnaturalizacioacuten es decir la peacuterdida de la estructura tridimensional de las enzimas
26
zona de reactivacioacuten maacuteximo de actividad
Zona de
inactivacioacuten
0 Temperatura
Figura dependencia de la actividad enzimaacutetica con la temperatura
Velocidad maacutexima la velocidad de una reaccioacuten (v) es el nuacutemero de
moleacuteculas de substrato convertidas en producto por unidad de tiempo y
usualmente se expresa en micromoles de producto formadas por minuto La velocidad de una reaccioacuten catalizada por una enzima aumenta conforme se incrementa la concentracioacuten de substrato hasta que se llega a una velocidad maacutexima (Vmax) a partir de la cual la velocidad de la reaccioacuten es
independiente de la cantidad de substrato El que la velocidad no pueda
seguirse incrementando refleja la saturacioacuten del sitio activo con el substrato
Vmax
0 Substrato
Figura representacioacuten de una cineacutetica tiacutepicamente michaeliana
Vmax = velocidad de saturacioacuten o maacutexima
27
Cuando se grafica la velocidad de la reaccioacuten v0 contra la concentracioacuten de substrato S no siempre es posible determinar la condicioacuten
en que se ha llegado a la velocidad maacutexima Vmax debido al incremento de la pendiente en la hipeacuterbola a concentraciones de substrato elevadas
Figura experimento cineacutetico para la actividad enzimaacutetica
Se presentan los valores experimentales y los resultados del ajuste al modelo de Michaelis
Auacuten asiacute si se grafica una doble reciacuteproca es decir el inverso de la
velocidad (1v0) contra el inverso de la concentracioacuten de substrato (1 S ) se
obtiene una liacutenea recta El regraacutefico de Lineweaver-Burke tambieacuten se
denomina de ldquodobles reciacuteprocasrdquo y se utiliza para calcular la Km y la Vmax asiacute como para determinar el mecanismo de accioacuten de los diversos tipos de
inhibidores
La ecuacioacuten que describe a la graacutefca de Lineweaver-Burke es
Vmax
1
SVmax
Km
v
1
0 (3)
que describe una recta en donde el intercepto con el eje de las abscisas es igual a ndash1Km y el intercepto con el eje de las ordenadas es igual a 1Vmax
28
Figura experimento cineacutetico para la actividad enzimaacutetica Regraacutefico de
Lineweaver-Burke
Se presentan los valores experimentales y los resultados del ajuste al modelo
REPRESENTACIOacuteN GRAacuteFICA DE LINEWEAVER Y BURK
Si se toma el inverso en ambos miembros de la ecuacioacuten de Michaelis tenemos
=
+
29
que es la ecuacioacuten de una liacutenea recta cuya pendiente es que intercepta al eje de las
ordenadas en y a la abcisa en el punto Lo que es evidente al hacer
ya que se convierte en
Si se hace una graacutefica con los dobles inversos colocando los valores de en el eje de las
ordenadas y en el eje de las abcisas se obtiene una liacutenea recta a partir de la cual se puede calcular faacutecilmente Km
EFECTO DEL pH SOBRE LA ACTIVIDAD ENZIMATICA
Sabiendo que las enzimas son proteinas cualquier cambio brusco de pH puede alterar el caraacutecter ioacutenico de los grupos amino y carboxilo en la superficie proteica afectando asiacute las propiedades cataliacuteticas de una enzima A pH alto o bajo se puede producir la desnaturalizacioacuten
de la enzima y en consecuencia su inactivacioacuten La fosfatasa aacutecida es maacutes activa a pH 50 mientras que la fosfatasa alcalina lo es a pH 90 Muchas enzimas tienen maacutexima actividad cerca de la neutralidad en un rango de pH de 6 a 8
El pH per se no afecta la actividad enzimaacutetica sino la concentracioacuten de protones Los protones
ademaacutes de alterar la estructura de la enzima y el substrato pueden participar tambieacuten en la reaccioacuten como substrato o producto En esos casos la concentracioacuten de protones afecta directamente la velocidad de la reaccioacuten
ENZIMA pH OPTIMO
30
Pepsina 15
Tripsina 77
Catalasa 76
Arginasa 97
Fumarasa 78
Ribonucleasa 78
EFECTO DE ACTIVADORES
Un aumento en la temperatura provoca un aumento de la velocidad de reaccioacuten hasta cierta temperatura oacuteptima ya que despueacutes de aproximadamente 45
0 C se comienza a producir la
desnaturalizacioacuten teacutermica Las enzimas de muchos mamiacuteferos tienen una temperatura oacuteptima
de 370 C por encima de esa temperatura comienzan a inactivarse y se destruyen Sin embargo
existen especies de bacterias y algas que habitan en fuentes de aguas termales y en el otro extremo ciertas bacterias aacuterticas tienen temperaturas oacuteptimas cercanas a 0
0 C
31
ESPECIFICIDAD ABSOLUTA
Cierta enzimas se encuentran como aproenzimas o zimogenos que son inactivos
El tripsinoacutegeno es convertido en tripsina por la misma accioacuten de la tripsina que remueve un hexapeacuteptido
tripsina Tripsinoacutegeno Tripsina + Val (Asp)4 lisina
Otra forma de activacioacuten consiste en mantener los grupos SH reducidos Estos grupos pueden
formar parte de centros activos de ciertas enzimas Si se oxidan estos grupos a -S-S- la enzima se inactiva Asiacute tenemos que la enzima papaina se inactiva despueacutes de exponerla al ox iacutegeno pero cuando se le antildeade un reductor apropiado la enzima se reactiva Los grupos ndashS-S- se convierten en ndashSH activaacutendose la enzima
Otra forma de activacioacuten requiere la presencia de otra substancia que se enlaza al sitio alosteacuterico el compuesto que se enlaza se denomina un efector alosteacuterico El centro alosteacuterico es bien especiacutefico para el efector o modulador alosteacuterico
ESPECIFICIDAD RELATIVA
Oxiacutegeno L-aminoaacutecido a -cetoaacutecido + NH3 + H 2 O 2 L-aminoaacutecido oxidasa Oxiacutegeno D-aminoaacutecido a -cetoaacutecido + NH3 + H 2 O 2 D-aminoaacutecido oxidasa
32
En este ejemplo se muestra la estereo-especificidad Una enzima puede tener una especificidad oacuteptica para isomeros D o L
Hay enzimas que muestran especificidad absoluta como la aspartasa que cataliza la adicioacuten reversible de amoniacuteaco al doble enlace del aacutecido fumaacuterico pero no al de otros aacutecidos insaturados
HOOC ndash CH = CH ndash COO- + NH4
+ HOOC ndash CH2 - CHNH2 - COO
- + H
+
INHIBICIOacuteN ENZIMATICA
Mediante el uso de inhibidores enzimaacuteticos se ha obtenido informacioacuten muy valiosa sobre la conformacioacuten del centro activo de algunas enzimas
a Inhibicioacuten irreversible Este tipo de inhibidores forman un enlace covalente con las enzimas cerca del centro activo Un ejemplo son los gases nerviosos como el
fluorofosfato de diisopropilo (DFP) que forma un complejo con la enzima acetilcolinesterasa Los animales envenenados con este gas quedan paralizados debido a la imposibilidad de transmitir adecuadamente los impulsos nerviosos
Ester di-isopropil fos foacuterico de la enzima (inactivo)
b) Inhibicioacuten competitiva
Es cuando el inhibidor compite con el substrato por la unioacuten con el centro activo de la
enzima Este tipo de inhibicioacuten puede reducirse si se aumenta la concentracioacuten de substrato
-OOC - CH2 - CH2 - COO
- -OOC - CH = CH - COO
- + 2H
+ Succinato Succinato Fumarato
33
Deshidrogenasa COO
- I CH2 Malonato (inhibidor competitivo) I COO
- En la inhibicioacuten competitiva la enzima se combina con el inhibidor para formar un complejo enzima-inhibidor E + I E I en competencia con la reaccioacuten normal E +S ES
La sulfanilamida (bactericida) interfiere con la siacutentesis del aacutecido foacutelico compitiendo con el aacutecido p-aminobenzoico de una forma competitiva
HOOC-sect- NH2 NH2 ndashSO2 - sect- NH2 aacutecido p- amino benzoico sulfanilamida
sect = fenil
a Inhibicioacuten no competitiva
Esta inhibicioacuten se caracteriza por que no se puede revertir el efecto del inhibidor aumentando la concentracioacuten del substrato
Por ejemplo la inhibicioacuten de la deshidrogenasa del gliceraldehido -3-fosfato por la yodo-acetamida
Enzima-SH + I-CH2 CONH2 Enzima-SCH2 CONH2 + IH
La deshidrogenasa del gliceraldehido-3 fosfato puede ser inhibida tambieacuten por el aacutecido iodo-aceacutetico
Enzima-SH + ICH2 COOH Enzima-SCH2 COOH + IH
c) Inhibicioacuten por metales
Ciertos metales como el plomo mercurio y arseacutenico inhiben enzimas que tienen en su centro activo grupos -SH libres

3
Figura 2 Representacioacuten de la formacioacuten del complejo enzima-sustrato
Es decir el sitio activo esta formado por ciertos aminoaacutecidos que forman un
microambiente caracteriacutestico dentro de la propia cadena y que llevan a cabo la
reaccioacuten generalmente existe soacutelo uno por moleacutecula de enzima
Las enzimas adquieren su poder cataliacutetico cuando presentan una estructura
secundaria y terciaria muy especiacutefica de tal manera que los aminoaacutecidos
componentes del sitio activo se encuentran vecinales formando dicho
microambiente Asiacute por ejemplo la quimotripsina crea su centro activo con los
aminoaacutecidos histidina y serina localizados en las posiciones 57 y 195 respectivamente
se puede decir que forzosamente esta moleacutecula debe adquirir una estructura
tridimensional de tal manera que dichos aminoaacutecidos sean adyacentes
14 EFICIENCIA CATALITICA
La mayoriacutea de las reacciones catalizadas por enzimas son muy eficientes y
transcurren desde 106 hasta 1014 veces maacutes raacutepido que la misma reaccioacuten no
catalizada (Tabla 1) Tiacutepicamente cada moleacutecula de enzima es capaz de transformar
cada segundo de 100 a 1000 moleacuteculas de substrato en producto El nuacutemero de
estas moleacuteculas transformadas a producto por moleacutecula de enzima en cada
segundo se conoce como el nuacutemero de recambio
Tabla1 Eficiencia de las reacciones catalizadas por algunas enzimas
Enzima Velocidad en
ausencia de enzima
Velocidad de
reaccioacuten catalizada
Rendimiento
Anhidrasa carboacutenica Corismato mutasa Triosafosfato isomerasa Carboxipeptidasa A AMP nucleosidasa
Nucleasa estafilococal
13 X 10 ndash1
26 X 10 ndash5
43 X 10 ndash6
30 X 10 ndash9
10 X 10 ndash11
17 X 10 -13
10 X 106
50 4300 578 60
95
77 X 106
19 X 106
10 X 109
19X 1011
60 X 1012
56 X 1014
4
128 NOMENCLATURA DE LAS ENZIMAS
A cada enzima se le asignan dos nombres El primero es corto y es como se
conoce a la proteiacutena de manera coloquial es el nombre sugerido El segundo es maacutes
completo e infiere propiedades sobre la reaccioacuten que la enzima desarrolla se le
conoce como nombre sistemaacutetico este nombre permite reconocer a la enzima sin
ambiguumledad y localizarla en el metabolismo
a) Nombre sugerido
Muchas de las enzimas poseen en su nombre el sufijo ldquo-asardquo unido al nombre
del substrato de la reaccioacuten que cataliza por ejemplo
Ureasa proteiacutena cuyo sustrato es la urea Lactasa proteiacutena cuyo sustrato es la lactosa
Tambieacuten suele utilizarse este sufijo a la descripcioacuten de la reaccioacuten que la enzima cataliza
Lactato deshidrogenada deshidrogena (le quita Hidroacutegenos) al lactato
Adenilato ciclasa hace un ciclo en la adenina
Algunas enzimas poseen nombres que no representan su actividad o sustrato
Lisozima
Tripsina
b) Nombre sistemaacutetico
La Unioacuten Internacional de Bioquiacutemica y Biologiacutea Molecular (IUBMB) desarrolloacute
un sistema de nomenclatura en el cual las enzimas se dividen en seis clases
principales cada una con numerosos subgrupos
127 CLASIFICACIOacuteN DE LAS ENZIMAS
5
En funcioacuten de su accioacuten cataliacutetica especiacutefica los enzimas se clasifican en 6 grandes grupos o clases
1 Oxidorreductasas Reacciones de transferencia de electrones
2 Transferasas Transferencia de grupos funcionales Ej UDP-glucosa-fructosa-
glucotransferasa
3 Hidrolasas Reacciones de hidroacutelisis Ej lipasa proteasa celulasa
4 Liasas Adicioacuten a dobles enlaces Ej carboxilasa fenilalanina amonioliasa
5 Isomerasas Reacciones de isomerizacioacutenEj fosfoglucosa isomerasa
6 Ligasas Se conociacutean como sintetasas Participan en la formacioacuten de enlaces
con hidroacutelisis de ATP
Para todas las clasificaciones se muestran ejemplos representativos
1- OXIDOREDUCTASAS
Catalizan reacciones de oxidacioacuten-reduccioacuten
2e-
2- TRANSFERASAS
Catalizan la transferencia de grupos que contienen C N o P
THF tetrahidrofolato
6
3- HIDROLASAS
Catalizan la ruptura de enlaces por la adicioacuten de una moleacutecula de agua
4- LIASAS
Catalizan la ruptura de enlaces C-C C-S y algunos enlaces C-N
5- ISOMERASAS
Catalizan la racemizacioacuten de isoacutemeros oacutepticos o geomeacutetricos
7
6- LIGASAS
Catalizan la formacioacuten de enlaces entre C y O S N Esta reaccioacuten soacutelo es posible
mediante la energiacutea derivada de fosfatos ricos en energiacutea como el del fosfato de la
moleacutecula de ATP
15 COFACTORES y COENZIMAS Algunas enzimas se asocian con moleacuteculas de caraacutecter no proteiacuteco que son
necesarias para el funcionamiento de la enzima estas moleacuteculas se denominan
cofactores (Figura 3) Comuacutenmente los factores encontrados en las enzimas incluyen
iones metaacutelicos como el Zn2+ o el Fe2+ ( Tabla 2) tambieacuten pueden ser moleacuteculas
orgaacutenicas que se denomina coenzimas como el NAD+ FAD la coenzima A
Generalmente las coenzimas son derivados de las vitaminas A la enzima en ausencia
de su cofactor (cuando lo tiene) se le denomina apoenzima en presencia de su
cofactor (cuando lo tiene) se le denomina holoenzima La apoenzima generalmente
carece de actividad bioloacutegica La diferencia entre un cofactor y un grupo prosteacutetico
como el grupo hemo es que este uacuteltimo estaacute unido de manera covalente a la
enzima mientras que el cofactor puede ser removido de la misma con relativa
facilidad
Se pueden sentildealar algunas caracteriacutesticas de las coenzimas como generalmente son
estables al calor y a otros agentes desnaturalizantes de las proteiacutenas pueden ser
recicladas y algunas provienen de las vitaminas
8
Figura 3 Cofactores y Coenzima
Tabla 2 Algunos enzimas que contienen iones metaacutelicos o los necesitan como
cofactores
Zn+2 Cu+2 (Cu+)
Alcohol- deshidrogenada Tirosinasa Anhidrasa-carboacutenica Citocromo-oxidasa Carboxipeptidasa
Mg+2 K+
Fosfohidrolasa Piruvato- fosfoquinasa Fosfotransferasas
Mn+2 Arginasa Na+ Fosfotransferasas ATPasa de la membrana plasmaacutetica
Fe +2 o Fe+3
9
Citocromos
Peroxidasa Catalasa Ferredoxina
Texto ------- (Tabla 3)
Tabla 3 Relacioacuten entre algunas coenzimas y las vitaminas a partir de las
cuales se producen Enfermedades generadas por la deficiencia
Coenzima Reaccioacuten Fuente vitamiacutenica Enfermedad
Humana por deficiencia
Biocitina Carboxilacioacuten Biotina
Coenzima A Transferencia de acilos Pantotenato
Coenzimas de Cobalamima Alquilacioacuten Cobalamina (B12) anemia perniciosa
Coenzimas de flavina Oxidacioacuten-reduccioacuten Riboflavina (B2)
Acido lipoacuteico Transferencia de acilos -
Coenzimas de nicotinamida Oxidacioacuten-reduccioacuten Nicotinamida (niacina) Pelagra
Fosfato de piridoxal Transferencia de grupo amino Piridoxina (B6)
Tetrahidrofolato Transferencia de un grupo de 1 Carbono Acido foacutelico anemia
megaloblaacutestica
Tiamina pirofosfato Transferencia de aldehiacutedo Tiamina (B1) beriberi
no tiene un nombre especiacutefico su aparicioacuten es muy rara
Otras coenzimas
NADP
Coenzima Q
Biotina
NAD y NADP
10
Coenzima Q (CoQ)
Esta coenzima es sintetizada a partir de farnesil pirofosfato Al igual que el
FMN la CoQ es capaz de aceptar y donar uno o dos electrones porque su forma
semiquinona es estable A su forma oxidada se le conoce como ubiquinona (de
ubicuo lat ubique en todas partes) o quinona A su forma reducida se le conoce
como QH2 ubiquinol o hidroquinona (Figura 2) A la coenzima solo parcialmente
reducida se le conoce como ubisemiquinona semiquinona o QH (Figura 4)
Figura 4 Representacioacuten de la moleacutecula de CoQ
La siacutentesis de la CoQ se lleva a cabo a traveacutes de unidades isoprenoides (Figura 5)
11
Figura 5 La CoQ se produce a partir de isoprenoides
Biotina
La biotina consiste de un anillo imidazol que estaacute unido en forma cis a la
cadena lateral tetrahidrotiofeno del valerato (Figura 6)
12
Figura 6 Estructura de la biotina
La enzima holocarboxilasa sintetasa (HCS) cataliza la activacioacuten mediante
biotinilacioacuten de cinco carboxilasas en ceacutelulas humanas La deficiencia de HCS
produce el siacutendrome deficiencia muacuteltiple de carboxilasas (DMC) Esta enfermedad es
potencialmente mortal y se caracteriza por la deficiencia de la actividad de todas las
carboxilasas La deficiencia de biotina no solo disminuye la actividad de estas enzimas
sino que parece modificar la expresioacuten geneacutetica
126 PROPIEDADES DE LAS ENZIMAS
Las propiedades de las enzimas derivan del hecho de ser proteiacutenas y de actuar como
catalizadores Como proteiacutenas poseen una conformacioacuten natural maacutes estable que las
demaacutes conformaciones posibles Asiacute cambios en la conformacioacuten suelen ir asociados
en cambios en la actividad cataliacutetica Los factores que influyen de manera maacutes directa
sobre la actividad de un enzima son
pH
temperatura
cofactores
A) EFECTO DEL pH
Al comprobar experimentalmente la influencia del pH en la velocidad de las
reacciones enzimaacuteticas se obtienen curvas que indican que los enzimas
presentan un pH oacuteptimo de actividad El pH puede afectar de varias maneras
13
o El centro activo puede contener aminoaacutecidos con grupos ionizados
que pueden variar con el pH
o La ionizacioacuten de aminoaacutecidos que no estaacuten en el centro activo puede
provocar modificaciones en la conformacioacuten de la enzima
o El sustrato puede verse afectado por las variaciones del pH
Algunos enzimas presentan variaciones peculiares La pepsina del estoacutemago
presenta un oacuteptimo a pH=2 la ureasa lo tiene a pH 7 y la fosfatasa alcalina del
intestino un pH= 12
B) EFECTO DE LA TEMPERATURA
Influye en la actividad El punto oacuteptimo representa el maacuteximo de actividad A
temperaturas bajas los enzimas se hallan ldquomuy riacutegidosrdquo y cuando se supera un
valor considerable (mayor de 50) la actividad cae bruscamente porque como
proteiacutena el enzima se desnaturaliza
128 EFECTO DE LOS COFACTORES SOBRE LA ACTIVIDAD
ENZIMATICA
A veces un enzima requiere para su funcioacuten la presencia de sustancias no proteicas
que colaboran en la cataacutelisis los cofactores Casi un tercio de los enzimas conocidos
requieren cofactores Cuando el cofactor es una moleacutecula orgaacutenica se llama
coenzima En la figura inferior podemos observar una moleacutecula de hemoglobina
(proteiacutena que transporta oxiacutegeno) y su coenzima (el grupo hemo) Cuando los
cofactores y las coenzimas se encuentran unidos covalentemente al enzima se llaman
14
grupos prosteacuteticos La forma cataliacuteticamente activa del enzima es decir el enzima
unido a su grupo prosteacutetico se llama holoenzima La parte proteica de un
holoenzima (inactiva) se llama apoenzima de forma que
apoenzima + grupo prosteacutetico= holoenzima
129 MODO DE ACCIOacuteN DE LAS ENZIMAS
En las reacciones espontaacuteneas los productos finales tienen menos energiacutea libre de
Gibbs (∆G) que los reactantes (figura inferior izquierda) Por lo tanto las reacciones
espontaacuteneas liberan energiacutea de Gibbs ((∆G lt 0) Sin embargo el comienzo de la
reaccioacuten requiere un aporte inicial de energiacutea Esta energiacutea inicial que hay que
suministrar a los reactantes para que la reaccioacuten transcurra se llama energiacutea de
activacioacuten (Ea) Cuanto menor es la Ea maacutes faacutecilmente transcurre la reaccioacuten
Perfil energeacutetico de una reaccioacuten espontaacutenea Perfil energeacutetico de una reaccioacuten catalizada
15
La accioacuten de los catalizadores consiste precisamente en disminuir la Ea (Figura superior derecha) Los enzimas
son catalizadores especialmente eficaces ya que disminuyen la Ea auacuten maacutes que los catalizadores inorgaacutenicos
Por ejemplo la descomposicioacuten del agua oxigenada (H2O2) para dar H2O y O2 puede ocurrir sin catalizador
con un catalizador inorgaacutenico (platino) o con un enzima especiacutefico (catalasa) Las respectivas Ea para cada
proceso son 18 12 y 6 Kcalmol Asiacute se puede calcular que el platino acelera la reaccioacuten 20000 veces mientras
que la catalasa la acelera 370000 veces
Para que una reaccioacuten quiacutemica tenga lugar las moleacuteculas de los reactantes deben chocar con una energiacutea y
una orientacioacuten adecuadas La actuacioacuten del enzima (1) permite que los rea ctantes (sustratos) se unan a su
centro activo con una orientacioacuten oacuteptima para que la reaccioacuten se produzca y (2) modifica las propiedades
quiacutemicas del sustrato unido a su centro activo debilitando los enlaces existentes y facilitando la formacioacuten de
otros nuevos (Figuras inferiores)
1210 REGULACIOacuteN DE SU ACTIVIDAD ENZIMATICA
16
Una moleacutecula de enzima no tiene por que actuar siempre a la misma velocidad Su
actividad puede estar modulada por
Cambios en el pH
Cambios en la temperatura
Presencia de cofactores
Las concentraciones del sustrato y de los productos finales
Presencia de inhibidores
Modulacioacuten alosteacuterica
Modificacioacuten covalente
Activacioacuten por proteolisis
Isoenzimas
EFECTO DE LAS CONCENTRACIONES SOBRE LA ACTIVIDAD ENZIMAacuteTICA
La velocidad de una reaccioacuten enzimaacutetica depende de la concentracioacuten de sustrato La Figura de la derecha muestra la velocidad de una reaccioacuten enzimaacutetica a 6 concentraciones distintas de sustrato
Ademaacutes la presencia de los productos finales puede hacer que la reaccioacuten sea maacutes lenta o incluso invertir su sentido (Figura inferior)
EFECTO DE LOS INHIBIDORES SOBRE LA ACTIVIDAD ENZIMAacuteTICA
Ciertas moleacuteculas pueden inhibir la accioacuten cataliacutetica de un enzima son los inhibidores Estos inhibidores bien pueden ocupar temporalmente el centro activo por semejanza estructural con el sustrato original ( inhibidor competitivo) o bien alteran la conformacioacuten espacial del enzima impidiendo su unioacuten al sustrato ( inhibidor no competitivo) (Figuras inferiores)
Inhibidor competitivo Inhibidor no competitivo
17
MODULACIOacuteN ALOSTEacuteRICA DE LA ACTIVIDAD ENZIMAacuteTICA
Hay enzimas que pueden adoptar 2 conformaciones interconvertibles llamadas R (relajada) y T (tensa) R es la forma maacutes activa porque se une al sustrato con maacutes afinidad Las formas R y T se encuentran en equilibrio R lt==gt T (Figura inferior)
Ciertas sustancias tienden a estabilizar la forma R Son los llamados moduladores positivos El propio sustrato es a
menudo un modulador positivo Las moleacuteculas que favorecen la forma R pero que actuacutean sobre una regioacuten del enzima distinta del centro activo son los activadores alosteacutericos (Figura inferior izquierda)
Activador alosteacuterico favorece la unioacuten del sustrato
Inhibidor alosteacuterico impide la unioacuten del sustrato
Las sustancias que favorecen la forma T y disminuyen la actividad enzimaacutetica son los moduladores negativos Si
estos moduladores actuacutean en lugares distintos del centro activo del enzima se llaman inhibidores alosteacutericos
18
(Figura superior derecha
EFECTO DE LA MODIFICACIOacuteN COVALENTE SOBRE LA ACTIVIDAD ENZIMAacuteTICA
Otros enzimas pasan de una forma menos activa a otra maacutes activa unieacutendose covalentemente a un grupo quiacutemico de pequentildeo tamantildeo como el Pi o el AMP Tambieacuten se da el caso inverso en el que un enzima muy activo se desactiva al liberar alguacuten grupo quiacutemico En las enzimas de las viacuteas degradativas del metabolismo la
forma fosforilada es maacutes activa que la no fosforilada mientras que en las viacuteas biosinteacuteticas ocurre lo contrario En las figuras inferiores se ilustra la activacioacuten de una proteiacutena por fosforilacioacuten
Elementos de la reaccioacuten El enzima no fosforilado es inactivo El enzima fosforilado es activo
ACTIVACIOacuteN PROTEOLIacuteTICA DE LA ACTIVIDAD ENZIMAacuteTICA
Algunos enzimas no se sintetizan como tales sino como proteiacutenas precursoras sin actividad enzimaacutetica Estas proteiacutenas se llaman
proenzimas o zimoacutegenos Para activarse los zimoacutegenos sufren un ataque hidroliacutetico que origina la liberacioacuten de uno o varios peacuteptidos El resto de la moleacutecula proteica adopta la conformacioacuten y las propiedades del enzima activo Muchos enzimas digestivos se secretan en forma de zimoacutegenos y en el tubo digestivo
se convierten en la forma activa Es el caso de la -quimotripsina que se sintetiza en forma de quimotripsinoacutegeno (Figura superior) Si estos enzimas se sintetizasen directamente en forma activa destruiriacutean la propia ceacutelula que las
produce Asiacute la tripsina pancreaacutetica (una proteasa) se sintetiza como tripsinoacutegeno (inactivo) Si por alguna razoacuten se activa en el propio paacutencreas la glaacutendula sufre un proceso de autodestruccioacuten (pancreatitis aguda) a
menudo mortal
19
REGULACIOacuteN DE LA ACTIVIDAD ENZIMAacuteTICA POR MEDIO DE ISOENZIMAS
Algunos enzimas tienen distinta estructura molecular aunque su funcioacuten bioloacutegica es similar Se llaman isozimas o isoenzimas Estas diferencias de estructura se traducen en ligeros cambios en sus propiedades de forma que cada isozima se adapta perfectamente a la funcioacuten que debe realizar Asiacute podemos observar la existencia de isoenzimas en funcioacuten de
el tipo de tejido Por ejemplo la lactato deshidrogenasa presenta isozimas distintos en muacutesculo y corazoacuten el compartimento celular donde actuacutea Por ejemplo la malato deshidrogenasa del citoplasma es distinta
de la de la mitocondria el momento concreto del desarrollo del individuo Por ejemplo algunos enzimas de la glicolisis del feto
son diferentes de los mismos enzimas en el adulto
CINEacuteTICA ENZIMAacuteTICA
Los principios generales de las reacciones quiacutemicas se aplican tambieacuten a las reacciones enzimaacuteticas Por este motivo antes de empezar con la cineacutetica quiacutemica se van a repasar algunos conceptos baacutesicos de cineacutetica quiacutemica
A continuacioacuten se describiraacuten los siguientes conceptos
Cineacutetica enzimaacutetica Modelo cineacutetico de Michaelis-Menten Caacutelculo de la KM y la Vmax de un enzima Actividad enzimaacutetica
20
NOMENCLATURA DE LOS ENZIMAS
CATALISIS MOLECULAR
Un catalizador modifica la velocidad de una reaccioacuten quiacutemica sin ser utilizado o aparecer como uno de los productos de la reaccioacuten Una reaccioacuten quiacutemica en la que un substrato(S) se
transforma en un producto(P) S P ocurre por que cierta fraccioacuten de moleacuteculas de S posee mucho maacutes energiacutea que el resto de ellas lo que es suficiente para que alcancen un estado activado en el que pueda formarse o romperse un enlace quiacutemico y se forme el producto(P)
La energiacutea de activacioacuten es la cantidad de energiacutea expresada en caloriacuteas necesaria para que todas las moleacuteculas de un mol a una temperatura dada alcancen el estado reactivo Mientras que el estado de transicioacuten es el estado rico en energiacutea de las moleacuteculas que
interaccionan en la cima de la barrera de activacioacuten La velocidad de una reaccioacuten quiacutemica es proporcional a la concentracioacuten del complejo en el estado de transicioacuten
Una reaccioacuten quiacutemica se puede acelerar de la siguiente forma 1) Al aumentar la temperatura se incrementa la energiacutea cineacutetica por lo que es mayor el nuacutemero de moleacuteculas que alcanzan el
estado de transicioacuten Generalmente el Q10 =2 lo que indica que la velocidad de una reaccioacuten quiacutemica se duplica al aumentar la temperatura en 10
0C 2) Antildeadiendo un catalizador que
disminuye la energiacutea de activacioacuten y aumenta la velocidad de reaccioacuten
La enzima (E) se combina con el substrato (S) formando el complejo de transicioacuten enzima-
substrato (E-S) mediante una reaccioacuten reversible cuya energiacutea de activacioacuten es menor que la de la reaccioacuten no catalizada Cuando se forma el producto de la reaccioacuten (P) se regenera de nuevo la enzima (E) de forma libre la que puede combinarse de nuevo con otra moleacutecula de substrato (S)
Una enzima reduce maacutes eficientemente la energiacutea de activacioacuten(Ea) de una reaccioacuten que un catalizador inorgaacutenico lo que permite que una reaccioacuten se realice a menor temperatura
El siguiente ejemplo ilustra mejor lo que hemos discutido
Reaccioacuten Energiacutea de activacioacuten (Kcalmol-1
)
21
a) el peroacutexido de hidroacutegeno se descompone en
H2 O2 H2 O + O2
18
b) el hierro cataliacutetico (Fe) realiza la reaccioacuten
H2 O2 H2 O + O2
13
c) el platino cataliacutetico (Pt) realiza la reaccioacuten
H2 O2 H2 O + O2
12
d) la catalasa una enzima hepaacutetica la realiza
H2O2 H2O + O2
5
Una enzima no modifica la energiacutea libre ni la constante de equilibrio sino que disminuye la energiacutea de activacioacuten de la reaccioacuten
Ir al principio
EFECTO DE LA CONCENTRACIOacuteN DEL SUBSTRATO SOBRE LA
CATALISIS ENZIMATICA
La actividad de una enzima se puede estudiar in vitro bajo condiciones controladas antildeadieacutendole una enzima a un substrato Siacute se trabaja con la condicioacuten de que la concentracioacuten del substrato sea saturante variando entonces la concentracioacuten de enzima se observa que aumenta el producto de la reaccioacuten a pH y temperatura constantes
22
Si se mantiene la concentracioacuten de la enzima constante y variando la concentracioacuten de substrato se obtiene una curva hiperboacutelica como la de la figura Al principio un aumento de la
concentracioacuten de substrato produce un aumento raacutepi do de la velocidad de reaccioacuten pero si se sigue aumentando la concentracioacuten de substrato la velocidad de reaccioacuten comienza a disminuir y a muy altas concentraciones de substrato se observa que no cambia la velocidad de
reaccioacuten se dice que los centros activos de la enzima se encuentran saturados La velocidad de reaccioacuten que se obtiene a esa alta concentracioacuten de substrato se define como la velocidad maacutexima (V) de la reaccioacuten enzimaacutetica bajo las condiciones especificadas La concentracioacuten de
substrato (S) a la semivelocidad maacutexima de reaccioacuten (V2) se puede determinar de la figura y representa la constante de Michaelis o Km la cual es una caracteriacutestica para cada enzima La inversa de Km o 1Km mide aproximadamente la afinidad de la enzima por el subs trato
Mientras maacutes pequentildeo sea el valor de Km mayor seraacute la afinidad de la enzima por el substrato Si varias enzimas compiten en el metabolismo por el mismo substrato eacuteste seraacute transformado preferentemente por la enzima con mayor afinidad
La ecuacioacuten de Michaelis describe la relacioacuten cuantitativa entre la velocidad de reaccioacuten y la concentracioacuten de substrato [S] si se conoce Vmax o Km
v=
En donde v= es la velocidad de reaccioacuten observada a una concentracioacuten de substrato determinada [S]
Km= constante de Michaelis en molesliacutet ro
23
Vmax= velocidad maacutexima a concentracioacuten saturante de substrato
Si v= Vmax2 entonces
=
Km+[S] = 2[S]
Km =[S]
De tal forma que podemos concluir que Km es igual a la concentracioacuten de substrato a la semivelocidad maacutexima de reaccioacuten
1 Concentraciones relativas de E y S la concentracioacuten de substrato S es
mucho mayor que la de E de tal manera que la cantidad de substrato
unido a la enzima en cualquier momento es muy pequentildea
2 Se asume el estado estacionario ES no cambia con el tiempo esto
quiere decir que la velocidad de formacioacuten de ES es igual a aquella para
su desintegracioacuten En general un intermediario en una serie de reacciones estaacute en estado estacionario cuando su velocidad de siacutentesis
es igual a su velocidad de degradacioacuten 3 Velocidad inicial para el anaacutelisis de reacciones enzimaacuteticas soacutelo se
utiliza la velocidad inicial de la reaccioacuten que es la velocidad ejercida por
la enzima inmediatamente despueacutes de que se ha puesto en contacto con el substrato y hasta antes de que se haya consumido el 10 de la concentracioacuten inicial del mismo La razoacuten de lo anterior es que en ese
momento la concentracioacuten del producto de la reaccioacuten que se ha acumulado en muy pequentildea y por tanto la reaccioacuten en el sentido
inverso es decir la transformacioacuten del producto en el substrato original puede ser ignorada
24
Caracteriacutesticas de Km la constante de Michaelis-Menten es
caracteriacutestica de una enzima y particular para un substrato Refleja la afinidad de la enzima por ese substrato K m es numeacutericamente igual a la concentracioacuten
de substrato a la cual la velocidad de reaccioacuten es la mitad de la Vmax (Km= Vmax2) Este paraacutemetro Km no variacutea con la concentracioacuten de enzima
Significado de una Km pequentildea un valor numeacuterico pequentildeo de Km
refleja una alta afinidad de la enzima por su substrato porque a una baja
concentracioacuten del mismo la enzima a desarrollado ya la mitad de la velocidad maacutexima
Significado de una Km grande el valor numeacuterico grande de Km refleja
una baja afinidad de la enzima por su substrato porque a una concentracioacuten
elevada del mismo la enzima desarrolla la mitad de la velocidad maacutexima
Relacioacuten de la velocidad con la concentracioacuten de enzima la
velocidad de la reaccioacuten es directamente proporcional a la concentracioacuten de la
enzima a cualquier concentracioacuten de substrato Por ejemplo si la concentracioacuten de enzima es disminuida a la mitad la velocidad inicial de la reaccioacuten (v0) es
reducida tambieacuten a la mitad de la original
Orden de la reaccioacuten cuando S es mas pequentildea que la Km la
velocidad de la reaccioacuten es aproximadamente igual a la concentracioacuten de
substrato
100
50
0
Substrato
Km Figura Localizacioacuten de la constante de Michaelis (Km)
25
La velocidad de reaccioacuten se dice en estas condiciones es de primer
orden con respecto al substrato Cuando S es mas grande que la Km la
velocidad es constante e igual a la Vmax La velocidad de la reaccioacuten es independiente de la concentracioacuten de substrato y se dice que es de orden cero
con respecto a la concentracioacuten de substrato
Las enzimas pueden ser aisladas de las ceacutelulas u organismos para estudiar sus
propiedades in vitro es decir en un tubo de ensayo Las diferentes enzimas muestran diferentes respuestas a los cambios en la concentracioacuten de substrato temperatura y pH
Concentracioacuten de substrato
La forma hiperboacutelica de la curva de velocidad vs Substrato
Efecto de la temperatura en la actividad enzimaacutetica
Efecto de la variacioacuten del pH en la actividad enzimaacutetica
Incremento de la velocidad con la temperatura En general la velocidad de la reaccioacuten se incrementa con la temperatura hasta que un punto maacuteximo es alcanzado
Este incremento se debe a que aumenta el nuacutemero de moleacuteculas ricas en energiacutea que pueden pasar la barrera energeacutetica de estado de transicioacuten para formar a los productos
Decremento de la velocidad con la temperatura la elevacioacuten excesiva de la
temperatura del medio que contiene a las enzimas resulta en un decremento de la velocidad como resultado de la desnaturalizacioacuten es decir la peacuterdida de la estructura tridimensional de las enzimas
26
zona de reactivacioacuten maacuteximo de actividad
Zona de
inactivacioacuten
0 Temperatura
Figura dependencia de la actividad enzimaacutetica con la temperatura
Velocidad maacutexima la velocidad de una reaccioacuten (v) es el nuacutemero de
moleacuteculas de substrato convertidas en producto por unidad de tiempo y
usualmente se expresa en micromoles de producto formadas por minuto La velocidad de una reaccioacuten catalizada por una enzima aumenta conforme se incrementa la concentracioacuten de substrato hasta que se llega a una velocidad maacutexima (Vmax) a partir de la cual la velocidad de la reaccioacuten es
independiente de la cantidad de substrato El que la velocidad no pueda
seguirse incrementando refleja la saturacioacuten del sitio activo con el substrato
Vmax
0 Substrato
Figura representacioacuten de una cineacutetica tiacutepicamente michaeliana
Vmax = velocidad de saturacioacuten o maacutexima
27
Cuando se grafica la velocidad de la reaccioacuten v0 contra la concentracioacuten de substrato S no siempre es posible determinar la condicioacuten
en que se ha llegado a la velocidad maacutexima Vmax debido al incremento de la pendiente en la hipeacuterbola a concentraciones de substrato elevadas
Figura experimento cineacutetico para la actividad enzimaacutetica
Se presentan los valores experimentales y los resultados del ajuste al modelo de Michaelis
Auacuten asiacute si se grafica una doble reciacuteproca es decir el inverso de la
velocidad (1v0) contra el inverso de la concentracioacuten de substrato (1 S ) se
obtiene una liacutenea recta El regraacutefico de Lineweaver-Burke tambieacuten se
denomina de ldquodobles reciacuteprocasrdquo y se utiliza para calcular la Km y la Vmax asiacute como para determinar el mecanismo de accioacuten de los diversos tipos de
inhibidores
La ecuacioacuten que describe a la graacutefca de Lineweaver-Burke es
Vmax
1
SVmax
Km
v
1
0 (3)
que describe una recta en donde el intercepto con el eje de las abscisas es igual a ndash1Km y el intercepto con el eje de las ordenadas es igual a 1Vmax
28
Figura experimento cineacutetico para la actividad enzimaacutetica Regraacutefico de
Lineweaver-Burke
Se presentan los valores experimentales y los resultados del ajuste al modelo
REPRESENTACIOacuteN GRAacuteFICA DE LINEWEAVER Y BURK
Si se toma el inverso en ambos miembros de la ecuacioacuten de Michaelis tenemos
=
+
29
que es la ecuacioacuten de una liacutenea recta cuya pendiente es que intercepta al eje de las
ordenadas en y a la abcisa en el punto Lo que es evidente al hacer
ya que se convierte en
Si se hace una graacutefica con los dobles inversos colocando los valores de en el eje de las
ordenadas y en el eje de las abcisas se obtiene una liacutenea recta a partir de la cual se puede calcular faacutecilmente Km
EFECTO DEL pH SOBRE LA ACTIVIDAD ENZIMATICA
Sabiendo que las enzimas son proteinas cualquier cambio brusco de pH puede alterar el caraacutecter ioacutenico de los grupos amino y carboxilo en la superficie proteica afectando asiacute las propiedades cataliacuteticas de una enzima A pH alto o bajo se puede producir la desnaturalizacioacuten
de la enzima y en consecuencia su inactivacioacuten La fosfatasa aacutecida es maacutes activa a pH 50 mientras que la fosfatasa alcalina lo es a pH 90 Muchas enzimas tienen maacutexima actividad cerca de la neutralidad en un rango de pH de 6 a 8
El pH per se no afecta la actividad enzimaacutetica sino la concentracioacuten de protones Los protones
ademaacutes de alterar la estructura de la enzima y el substrato pueden participar tambieacuten en la reaccioacuten como substrato o producto En esos casos la concentracioacuten de protones afecta directamente la velocidad de la reaccioacuten
ENZIMA pH OPTIMO
30
Pepsina 15
Tripsina 77
Catalasa 76
Arginasa 97
Fumarasa 78
Ribonucleasa 78
EFECTO DE ACTIVADORES
Un aumento en la temperatura provoca un aumento de la velocidad de reaccioacuten hasta cierta temperatura oacuteptima ya que despueacutes de aproximadamente 45
0 C se comienza a producir la
desnaturalizacioacuten teacutermica Las enzimas de muchos mamiacuteferos tienen una temperatura oacuteptima
de 370 C por encima de esa temperatura comienzan a inactivarse y se destruyen Sin embargo
existen especies de bacterias y algas que habitan en fuentes de aguas termales y en el otro extremo ciertas bacterias aacuterticas tienen temperaturas oacuteptimas cercanas a 0
0 C
31
ESPECIFICIDAD ABSOLUTA
Cierta enzimas se encuentran como aproenzimas o zimogenos que son inactivos
El tripsinoacutegeno es convertido en tripsina por la misma accioacuten de la tripsina que remueve un hexapeacuteptido
tripsina Tripsinoacutegeno Tripsina + Val (Asp)4 lisina
Otra forma de activacioacuten consiste en mantener los grupos SH reducidos Estos grupos pueden
formar parte de centros activos de ciertas enzimas Si se oxidan estos grupos a -S-S- la enzima se inactiva Asiacute tenemos que la enzima papaina se inactiva despueacutes de exponerla al ox iacutegeno pero cuando se le antildeade un reductor apropiado la enzima se reactiva Los grupos ndashS-S- se convierten en ndashSH activaacutendose la enzima
Otra forma de activacioacuten requiere la presencia de otra substancia que se enlaza al sitio alosteacuterico el compuesto que se enlaza se denomina un efector alosteacuterico El centro alosteacuterico es bien especiacutefico para el efector o modulador alosteacuterico
ESPECIFICIDAD RELATIVA
Oxiacutegeno L-aminoaacutecido a -cetoaacutecido + NH3 + H 2 O 2 L-aminoaacutecido oxidasa Oxiacutegeno D-aminoaacutecido a -cetoaacutecido + NH3 + H 2 O 2 D-aminoaacutecido oxidasa
32
En este ejemplo se muestra la estereo-especificidad Una enzima puede tener una especificidad oacuteptica para isomeros D o L
Hay enzimas que muestran especificidad absoluta como la aspartasa que cataliza la adicioacuten reversible de amoniacuteaco al doble enlace del aacutecido fumaacuterico pero no al de otros aacutecidos insaturados
HOOC ndash CH = CH ndash COO- + NH4
+ HOOC ndash CH2 - CHNH2 - COO
- + H
+
INHIBICIOacuteN ENZIMATICA
Mediante el uso de inhibidores enzimaacuteticos se ha obtenido informacioacuten muy valiosa sobre la conformacioacuten del centro activo de algunas enzimas
a Inhibicioacuten irreversible Este tipo de inhibidores forman un enlace covalente con las enzimas cerca del centro activo Un ejemplo son los gases nerviosos como el
fluorofosfato de diisopropilo (DFP) que forma un complejo con la enzima acetilcolinesterasa Los animales envenenados con este gas quedan paralizados debido a la imposibilidad de transmitir adecuadamente los impulsos nerviosos
Ester di-isopropil fos foacuterico de la enzima (inactivo)
b) Inhibicioacuten competitiva
Es cuando el inhibidor compite con el substrato por la unioacuten con el centro activo de la
enzima Este tipo de inhibicioacuten puede reducirse si se aumenta la concentracioacuten de substrato
-OOC - CH2 - CH2 - COO
- -OOC - CH = CH - COO
- + 2H
+ Succinato Succinato Fumarato
33
Deshidrogenasa COO
- I CH2 Malonato (inhibidor competitivo) I COO
- En la inhibicioacuten competitiva la enzima se combina con el inhibidor para formar un complejo enzima-inhibidor E + I E I en competencia con la reaccioacuten normal E +S ES
La sulfanilamida (bactericida) interfiere con la siacutentesis del aacutecido foacutelico compitiendo con el aacutecido p-aminobenzoico de una forma competitiva
HOOC-sect- NH2 NH2 ndashSO2 - sect- NH2 aacutecido p- amino benzoico sulfanilamida
sect = fenil
a Inhibicioacuten no competitiva
Esta inhibicioacuten se caracteriza por que no se puede revertir el efecto del inhibidor aumentando la concentracioacuten del substrato
Por ejemplo la inhibicioacuten de la deshidrogenasa del gliceraldehido -3-fosfato por la yodo-acetamida
Enzima-SH + I-CH2 CONH2 Enzima-SCH2 CONH2 + IH
La deshidrogenasa del gliceraldehido-3 fosfato puede ser inhibida tambieacuten por el aacutecido iodo-aceacutetico
Enzima-SH + ICH2 COOH Enzima-SCH2 COOH + IH
c) Inhibicioacuten por metales
Ciertos metales como el plomo mercurio y arseacutenico inhiben enzimas que tienen en su centro activo grupos -SH libres

4
128 NOMENCLATURA DE LAS ENZIMAS
A cada enzima se le asignan dos nombres El primero es corto y es como se
conoce a la proteiacutena de manera coloquial es el nombre sugerido El segundo es maacutes
completo e infiere propiedades sobre la reaccioacuten que la enzima desarrolla se le
conoce como nombre sistemaacutetico este nombre permite reconocer a la enzima sin
ambiguumledad y localizarla en el metabolismo
a) Nombre sugerido
Muchas de las enzimas poseen en su nombre el sufijo ldquo-asardquo unido al nombre
del substrato de la reaccioacuten que cataliza por ejemplo
Ureasa proteiacutena cuyo sustrato es la urea Lactasa proteiacutena cuyo sustrato es la lactosa
Tambieacuten suele utilizarse este sufijo a la descripcioacuten de la reaccioacuten que la enzima cataliza
Lactato deshidrogenada deshidrogena (le quita Hidroacutegenos) al lactato
Adenilato ciclasa hace un ciclo en la adenina
Algunas enzimas poseen nombres que no representan su actividad o sustrato
Lisozima
Tripsina
b) Nombre sistemaacutetico
La Unioacuten Internacional de Bioquiacutemica y Biologiacutea Molecular (IUBMB) desarrolloacute
un sistema de nomenclatura en el cual las enzimas se dividen en seis clases
principales cada una con numerosos subgrupos
127 CLASIFICACIOacuteN DE LAS ENZIMAS
5
En funcioacuten de su accioacuten cataliacutetica especiacutefica los enzimas se clasifican en 6 grandes grupos o clases
1 Oxidorreductasas Reacciones de transferencia de electrones
2 Transferasas Transferencia de grupos funcionales Ej UDP-glucosa-fructosa-
glucotransferasa
3 Hidrolasas Reacciones de hidroacutelisis Ej lipasa proteasa celulasa
4 Liasas Adicioacuten a dobles enlaces Ej carboxilasa fenilalanina amonioliasa
5 Isomerasas Reacciones de isomerizacioacutenEj fosfoglucosa isomerasa
6 Ligasas Se conociacutean como sintetasas Participan en la formacioacuten de enlaces
con hidroacutelisis de ATP
Para todas las clasificaciones se muestran ejemplos representativos
1- OXIDOREDUCTASAS
Catalizan reacciones de oxidacioacuten-reduccioacuten
2e-
2- TRANSFERASAS
Catalizan la transferencia de grupos que contienen C N o P
THF tetrahidrofolato
6
3- HIDROLASAS
Catalizan la ruptura de enlaces por la adicioacuten de una moleacutecula de agua
4- LIASAS
Catalizan la ruptura de enlaces C-C C-S y algunos enlaces C-N
5- ISOMERASAS
Catalizan la racemizacioacuten de isoacutemeros oacutepticos o geomeacutetricos
7
6- LIGASAS
Catalizan la formacioacuten de enlaces entre C y O S N Esta reaccioacuten soacutelo es posible
mediante la energiacutea derivada de fosfatos ricos en energiacutea como el del fosfato de la
moleacutecula de ATP
15 COFACTORES y COENZIMAS Algunas enzimas se asocian con moleacuteculas de caraacutecter no proteiacuteco que son
necesarias para el funcionamiento de la enzima estas moleacuteculas se denominan
cofactores (Figura 3) Comuacutenmente los factores encontrados en las enzimas incluyen
iones metaacutelicos como el Zn2+ o el Fe2+ ( Tabla 2) tambieacuten pueden ser moleacuteculas
orgaacutenicas que se denomina coenzimas como el NAD+ FAD la coenzima A
Generalmente las coenzimas son derivados de las vitaminas A la enzima en ausencia
de su cofactor (cuando lo tiene) se le denomina apoenzima en presencia de su
cofactor (cuando lo tiene) se le denomina holoenzima La apoenzima generalmente
carece de actividad bioloacutegica La diferencia entre un cofactor y un grupo prosteacutetico
como el grupo hemo es que este uacuteltimo estaacute unido de manera covalente a la
enzima mientras que el cofactor puede ser removido de la misma con relativa
facilidad
Se pueden sentildealar algunas caracteriacutesticas de las coenzimas como generalmente son
estables al calor y a otros agentes desnaturalizantes de las proteiacutenas pueden ser
recicladas y algunas provienen de las vitaminas
8
Figura 3 Cofactores y Coenzima
Tabla 2 Algunos enzimas que contienen iones metaacutelicos o los necesitan como
cofactores
Zn+2 Cu+2 (Cu+)
Alcohol- deshidrogenada Tirosinasa Anhidrasa-carboacutenica Citocromo-oxidasa Carboxipeptidasa
Mg+2 K+
Fosfohidrolasa Piruvato- fosfoquinasa Fosfotransferasas
Mn+2 Arginasa Na+ Fosfotransferasas ATPasa de la membrana plasmaacutetica
Fe +2 o Fe+3
9
Citocromos
Peroxidasa Catalasa Ferredoxina
Texto ------- (Tabla 3)
Tabla 3 Relacioacuten entre algunas coenzimas y las vitaminas a partir de las
cuales se producen Enfermedades generadas por la deficiencia
Coenzima Reaccioacuten Fuente vitamiacutenica Enfermedad
Humana por deficiencia
Biocitina Carboxilacioacuten Biotina
Coenzima A Transferencia de acilos Pantotenato
Coenzimas de Cobalamima Alquilacioacuten Cobalamina (B12) anemia perniciosa
Coenzimas de flavina Oxidacioacuten-reduccioacuten Riboflavina (B2)
Acido lipoacuteico Transferencia de acilos -
Coenzimas de nicotinamida Oxidacioacuten-reduccioacuten Nicotinamida (niacina) Pelagra
Fosfato de piridoxal Transferencia de grupo amino Piridoxina (B6)
Tetrahidrofolato Transferencia de un grupo de 1 Carbono Acido foacutelico anemia
megaloblaacutestica
Tiamina pirofosfato Transferencia de aldehiacutedo Tiamina (B1) beriberi
no tiene un nombre especiacutefico su aparicioacuten es muy rara
Otras coenzimas
NADP
Coenzima Q
Biotina
NAD y NADP
10
Coenzima Q (CoQ)
Esta coenzima es sintetizada a partir de farnesil pirofosfato Al igual que el
FMN la CoQ es capaz de aceptar y donar uno o dos electrones porque su forma
semiquinona es estable A su forma oxidada se le conoce como ubiquinona (de
ubicuo lat ubique en todas partes) o quinona A su forma reducida se le conoce
como QH2 ubiquinol o hidroquinona (Figura 2) A la coenzima solo parcialmente
reducida se le conoce como ubisemiquinona semiquinona o QH (Figura 4)
Figura 4 Representacioacuten de la moleacutecula de CoQ
La siacutentesis de la CoQ se lleva a cabo a traveacutes de unidades isoprenoides (Figura 5)
11
Figura 5 La CoQ se produce a partir de isoprenoides
Biotina
La biotina consiste de un anillo imidazol que estaacute unido en forma cis a la
cadena lateral tetrahidrotiofeno del valerato (Figura 6)
12
Figura 6 Estructura de la biotina
La enzima holocarboxilasa sintetasa (HCS) cataliza la activacioacuten mediante
biotinilacioacuten de cinco carboxilasas en ceacutelulas humanas La deficiencia de HCS
produce el siacutendrome deficiencia muacuteltiple de carboxilasas (DMC) Esta enfermedad es
potencialmente mortal y se caracteriza por la deficiencia de la actividad de todas las
carboxilasas La deficiencia de biotina no solo disminuye la actividad de estas enzimas
sino que parece modificar la expresioacuten geneacutetica
126 PROPIEDADES DE LAS ENZIMAS
Las propiedades de las enzimas derivan del hecho de ser proteiacutenas y de actuar como
catalizadores Como proteiacutenas poseen una conformacioacuten natural maacutes estable que las
demaacutes conformaciones posibles Asiacute cambios en la conformacioacuten suelen ir asociados
en cambios en la actividad cataliacutetica Los factores que influyen de manera maacutes directa
sobre la actividad de un enzima son
pH
temperatura
cofactores
A) EFECTO DEL pH
Al comprobar experimentalmente la influencia del pH en la velocidad de las
reacciones enzimaacuteticas se obtienen curvas que indican que los enzimas
presentan un pH oacuteptimo de actividad El pH puede afectar de varias maneras
13
o El centro activo puede contener aminoaacutecidos con grupos ionizados
que pueden variar con el pH
o La ionizacioacuten de aminoaacutecidos que no estaacuten en el centro activo puede
provocar modificaciones en la conformacioacuten de la enzima
o El sustrato puede verse afectado por las variaciones del pH
Algunos enzimas presentan variaciones peculiares La pepsina del estoacutemago
presenta un oacuteptimo a pH=2 la ureasa lo tiene a pH 7 y la fosfatasa alcalina del
intestino un pH= 12
B) EFECTO DE LA TEMPERATURA
Influye en la actividad El punto oacuteptimo representa el maacuteximo de actividad A
temperaturas bajas los enzimas se hallan ldquomuy riacutegidosrdquo y cuando se supera un
valor considerable (mayor de 50) la actividad cae bruscamente porque como
proteiacutena el enzima se desnaturaliza
128 EFECTO DE LOS COFACTORES SOBRE LA ACTIVIDAD
ENZIMATICA
A veces un enzima requiere para su funcioacuten la presencia de sustancias no proteicas
que colaboran en la cataacutelisis los cofactores Casi un tercio de los enzimas conocidos
requieren cofactores Cuando el cofactor es una moleacutecula orgaacutenica se llama
coenzima En la figura inferior podemos observar una moleacutecula de hemoglobina
(proteiacutena que transporta oxiacutegeno) y su coenzima (el grupo hemo) Cuando los
cofactores y las coenzimas se encuentran unidos covalentemente al enzima se llaman
14
grupos prosteacuteticos La forma cataliacuteticamente activa del enzima es decir el enzima
unido a su grupo prosteacutetico se llama holoenzima La parte proteica de un
holoenzima (inactiva) se llama apoenzima de forma que
apoenzima + grupo prosteacutetico= holoenzima
129 MODO DE ACCIOacuteN DE LAS ENZIMAS
En las reacciones espontaacuteneas los productos finales tienen menos energiacutea libre de
Gibbs (∆G) que los reactantes (figura inferior izquierda) Por lo tanto las reacciones
espontaacuteneas liberan energiacutea de Gibbs ((∆G lt 0) Sin embargo el comienzo de la
reaccioacuten requiere un aporte inicial de energiacutea Esta energiacutea inicial que hay que
suministrar a los reactantes para que la reaccioacuten transcurra se llama energiacutea de
activacioacuten (Ea) Cuanto menor es la Ea maacutes faacutecilmente transcurre la reaccioacuten
Perfil energeacutetico de una reaccioacuten espontaacutenea Perfil energeacutetico de una reaccioacuten catalizada
15
La accioacuten de los catalizadores consiste precisamente en disminuir la Ea (Figura superior derecha) Los enzimas
son catalizadores especialmente eficaces ya que disminuyen la Ea auacuten maacutes que los catalizadores inorgaacutenicos
Por ejemplo la descomposicioacuten del agua oxigenada (H2O2) para dar H2O y O2 puede ocurrir sin catalizador
con un catalizador inorgaacutenico (platino) o con un enzima especiacutefico (catalasa) Las respectivas Ea para cada
proceso son 18 12 y 6 Kcalmol Asiacute se puede calcular que el platino acelera la reaccioacuten 20000 veces mientras
que la catalasa la acelera 370000 veces
Para que una reaccioacuten quiacutemica tenga lugar las moleacuteculas de los reactantes deben chocar con una energiacutea y
una orientacioacuten adecuadas La actuacioacuten del enzima (1) permite que los rea ctantes (sustratos) se unan a su
centro activo con una orientacioacuten oacuteptima para que la reaccioacuten se produzca y (2) modifica las propiedades
quiacutemicas del sustrato unido a su centro activo debilitando los enlaces existentes y facilitando la formacioacuten de
otros nuevos (Figuras inferiores)
1210 REGULACIOacuteN DE SU ACTIVIDAD ENZIMATICA
16
Una moleacutecula de enzima no tiene por que actuar siempre a la misma velocidad Su
actividad puede estar modulada por
Cambios en el pH
Cambios en la temperatura
Presencia de cofactores
Las concentraciones del sustrato y de los productos finales
Presencia de inhibidores
Modulacioacuten alosteacuterica
Modificacioacuten covalente
Activacioacuten por proteolisis
Isoenzimas
EFECTO DE LAS CONCENTRACIONES SOBRE LA ACTIVIDAD ENZIMAacuteTICA
La velocidad de una reaccioacuten enzimaacutetica depende de la concentracioacuten de sustrato La Figura de la derecha muestra la velocidad de una reaccioacuten enzimaacutetica a 6 concentraciones distintas de sustrato
Ademaacutes la presencia de los productos finales puede hacer que la reaccioacuten sea maacutes lenta o incluso invertir su sentido (Figura inferior)
EFECTO DE LOS INHIBIDORES SOBRE LA ACTIVIDAD ENZIMAacuteTICA
Ciertas moleacuteculas pueden inhibir la accioacuten cataliacutetica de un enzima son los inhibidores Estos inhibidores bien pueden ocupar temporalmente el centro activo por semejanza estructural con el sustrato original ( inhibidor competitivo) o bien alteran la conformacioacuten espacial del enzima impidiendo su unioacuten al sustrato ( inhibidor no competitivo) (Figuras inferiores)
Inhibidor competitivo Inhibidor no competitivo
17
MODULACIOacuteN ALOSTEacuteRICA DE LA ACTIVIDAD ENZIMAacuteTICA
Hay enzimas que pueden adoptar 2 conformaciones interconvertibles llamadas R (relajada) y T (tensa) R es la forma maacutes activa porque se une al sustrato con maacutes afinidad Las formas R y T se encuentran en equilibrio R lt==gt T (Figura inferior)
Ciertas sustancias tienden a estabilizar la forma R Son los llamados moduladores positivos El propio sustrato es a
menudo un modulador positivo Las moleacuteculas que favorecen la forma R pero que actuacutean sobre una regioacuten del enzima distinta del centro activo son los activadores alosteacutericos (Figura inferior izquierda)
Activador alosteacuterico favorece la unioacuten del sustrato
Inhibidor alosteacuterico impide la unioacuten del sustrato
Las sustancias que favorecen la forma T y disminuyen la actividad enzimaacutetica son los moduladores negativos Si
estos moduladores actuacutean en lugares distintos del centro activo del enzima se llaman inhibidores alosteacutericos
18
(Figura superior derecha
EFECTO DE LA MODIFICACIOacuteN COVALENTE SOBRE LA ACTIVIDAD ENZIMAacuteTICA
Otros enzimas pasan de una forma menos activa a otra maacutes activa unieacutendose covalentemente a un grupo quiacutemico de pequentildeo tamantildeo como el Pi o el AMP Tambieacuten se da el caso inverso en el que un enzima muy activo se desactiva al liberar alguacuten grupo quiacutemico En las enzimas de las viacuteas degradativas del metabolismo la
forma fosforilada es maacutes activa que la no fosforilada mientras que en las viacuteas biosinteacuteticas ocurre lo contrario En las figuras inferiores se ilustra la activacioacuten de una proteiacutena por fosforilacioacuten
Elementos de la reaccioacuten El enzima no fosforilado es inactivo El enzima fosforilado es activo
ACTIVACIOacuteN PROTEOLIacuteTICA DE LA ACTIVIDAD ENZIMAacuteTICA
Algunos enzimas no se sintetizan como tales sino como proteiacutenas precursoras sin actividad enzimaacutetica Estas proteiacutenas se llaman
proenzimas o zimoacutegenos Para activarse los zimoacutegenos sufren un ataque hidroliacutetico que origina la liberacioacuten de uno o varios peacuteptidos El resto de la moleacutecula proteica adopta la conformacioacuten y las propiedades del enzima activo Muchos enzimas digestivos se secretan en forma de zimoacutegenos y en el tubo digestivo
se convierten en la forma activa Es el caso de la -quimotripsina que se sintetiza en forma de quimotripsinoacutegeno (Figura superior) Si estos enzimas se sintetizasen directamente en forma activa destruiriacutean la propia ceacutelula que las
produce Asiacute la tripsina pancreaacutetica (una proteasa) se sintetiza como tripsinoacutegeno (inactivo) Si por alguna razoacuten se activa en el propio paacutencreas la glaacutendula sufre un proceso de autodestruccioacuten (pancreatitis aguda) a
menudo mortal
19
REGULACIOacuteN DE LA ACTIVIDAD ENZIMAacuteTICA POR MEDIO DE ISOENZIMAS
Algunos enzimas tienen distinta estructura molecular aunque su funcioacuten bioloacutegica es similar Se llaman isozimas o isoenzimas Estas diferencias de estructura se traducen en ligeros cambios en sus propiedades de forma que cada isozima se adapta perfectamente a la funcioacuten que debe realizar Asiacute podemos observar la existencia de isoenzimas en funcioacuten de
el tipo de tejido Por ejemplo la lactato deshidrogenasa presenta isozimas distintos en muacutesculo y corazoacuten el compartimento celular donde actuacutea Por ejemplo la malato deshidrogenasa del citoplasma es distinta
de la de la mitocondria el momento concreto del desarrollo del individuo Por ejemplo algunos enzimas de la glicolisis del feto
son diferentes de los mismos enzimas en el adulto
CINEacuteTICA ENZIMAacuteTICA
Los principios generales de las reacciones quiacutemicas se aplican tambieacuten a las reacciones enzimaacuteticas Por este motivo antes de empezar con la cineacutetica quiacutemica se van a repasar algunos conceptos baacutesicos de cineacutetica quiacutemica
A continuacioacuten se describiraacuten los siguientes conceptos
Cineacutetica enzimaacutetica Modelo cineacutetico de Michaelis-Menten Caacutelculo de la KM y la Vmax de un enzima Actividad enzimaacutetica
20
NOMENCLATURA DE LOS ENZIMAS
CATALISIS MOLECULAR
Un catalizador modifica la velocidad de una reaccioacuten quiacutemica sin ser utilizado o aparecer como uno de los productos de la reaccioacuten Una reaccioacuten quiacutemica en la que un substrato(S) se
transforma en un producto(P) S P ocurre por que cierta fraccioacuten de moleacuteculas de S posee mucho maacutes energiacutea que el resto de ellas lo que es suficiente para que alcancen un estado activado en el que pueda formarse o romperse un enlace quiacutemico y se forme el producto(P)
La energiacutea de activacioacuten es la cantidad de energiacutea expresada en caloriacuteas necesaria para que todas las moleacuteculas de un mol a una temperatura dada alcancen el estado reactivo Mientras que el estado de transicioacuten es el estado rico en energiacutea de las moleacuteculas que
interaccionan en la cima de la barrera de activacioacuten La velocidad de una reaccioacuten quiacutemica es proporcional a la concentracioacuten del complejo en el estado de transicioacuten
Una reaccioacuten quiacutemica se puede acelerar de la siguiente forma 1) Al aumentar la temperatura se incrementa la energiacutea cineacutetica por lo que es mayor el nuacutemero de moleacuteculas que alcanzan el
estado de transicioacuten Generalmente el Q10 =2 lo que indica que la velocidad de una reaccioacuten quiacutemica se duplica al aumentar la temperatura en 10
0C 2) Antildeadiendo un catalizador que
disminuye la energiacutea de activacioacuten y aumenta la velocidad de reaccioacuten
La enzima (E) se combina con el substrato (S) formando el complejo de transicioacuten enzima-
substrato (E-S) mediante una reaccioacuten reversible cuya energiacutea de activacioacuten es menor que la de la reaccioacuten no catalizada Cuando se forma el producto de la reaccioacuten (P) se regenera de nuevo la enzima (E) de forma libre la que puede combinarse de nuevo con otra moleacutecula de substrato (S)
Una enzima reduce maacutes eficientemente la energiacutea de activacioacuten(Ea) de una reaccioacuten que un catalizador inorgaacutenico lo que permite que una reaccioacuten se realice a menor temperatura
El siguiente ejemplo ilustra mejor lo que hemos discutido
Reaccioacuten Energiacutea de activacioacuten (Kcalmol-1
)
21
a) el peroacutexido de hidroacutegeno se descompone en
H2 O2 H2 O + O2
18
b) el hierro cataliacutetico (Fe) realiza la reaccioacuten
H2 O2 H2 O + O2
13
c) el platino cataliacutetico (Pt) realiza la reaccioacuten
H2 O2 H2 O + O2
12
d) la catalasa una enzima hepaacutetica la realiza
H2O2 H2O + O2
5
Una enzima no modifica la energiacutea libre ni la constante de equilibrio sino que disminuye la energiacutea de activacioacuten de la reaccioacuten
Ir al principio
EFECTO DE LA CONCENTRACIOacuteN DEL SUBSTRATO SOBRE LA
CATALISIS ENZIMATICA
La actividad de una enzima se puede estudiar in vitro bajo condiciones controladas antildeadieacutendole una enzima a un substrato Siacute se trabaja con la condicioacuten de que la concentracioacuten del substrato sea saturante variando entonces la concentracioacuten de enzima se observa que aumenta el producto de la reaccioacuten a pH y temperatura constantes
22
Si se mantiene la concentracioacuten de la enzima constante y variando la concentracioacuten de substrato se obtiene una curva hiperboacutelica como la de la figura Al principio un aumento de la
concentracioacuten de substrato produce un aumento raacutepi do de la velocidad de reaccioacuten pero si se sigue aumentando la concentracioacuten de substrato la velocidad de reaccioacuten comienza a disminuir y a muy altas concentraciones de substrato se observa que no cambia la velocidad de
reaccioacuten se dice que los centros activos de la enzima se encuentran saturados La velocidad de reaccioacuten que se obtiene a esa alta concentracioacuten de substrato se define como la velocidad maacutexima (V) de la reaccioacuten enzimaacutetica bajo las condiciones especificadas La concentracioacuten de
substrato (S) a la semivelocidad maacutexima de reaccioacuten (V2) se puede determinar de la figura y representa la constante de Michaelis o Km la cual es una caracteriacutestica para cada enzima La inversa de Km o 1Km mide aproximadamente la afinidad de la enzima por el subs trato
Mientras maacutes pequentildeo sea el valor de Km mayor seraacute la afinidad de la enzima por el substrato Si varias enzimas compiten en el metabolismo por el mismo substrato eacuteste seraacute transformado preferentemente por la enzima con mayor afinidad
La ecuacioacuten de Michaelis describe la relacioacuten cuantitativa entre la velocidad de reaccioacuten y la concentracioacuten de substrato [S] si se conoce Vmax o Km
v=
En donde v= es la velocidad de reaccioacuten observada a una concentracioacuten de substrato determinada [S]
Km= constante de Michaelis en molesliacutet ro
23
Vmax= velocidad maacutexima a concentracioacuten saturante de substrato
Si v= Vmax2 entonces
=
Km+[S] = 2[S]
Km =[S]
De tal forma que podemos concluir que Km es igual a la concentracioacuten de substrato a la semivelocidad maacutexima de reaccioacuten
1 Concentraciones relativas de E y S la concentracioacuten de substrato S es
mucho mayor que la de E de tal manera que la cantidad de substrato
unido a la enzima en cualquier momento es muy pequentildea
2 Se asume el estado estacionario ES no cambia con el tiempo esto
quiere decir que la velocidad de formacioacuten de ES es igual a aquella para
su desintegracioacuten En general un intermediario en una serie de reacciones estaacute en estado estacionario cuando su velocidad de siacutentesis
es igual a su velocidad de degradacioacuten 3 Velocidad inicial para el anaacutelisis de reacciones enzimaacuteticas soacutelo se
utiliza la velocidad inicial de la reaccioacuten que es la velocidad ejercida por
la enzima inmediatamente despueacutes de que se ha puesto en contacto con el substrato y hasta antes de que se haya consumido el 10 de la concentracioacuten inicial del mismo La razoacuten de lo anterior es que en ese
momento la concentracioacuten del producto de la reaccioacuten que se ha acumulado en muy pequentildea y por tanto la reaccioacuten en el sentido
inverso es decir la transformacioacuten del producto en el substrato original puede ser ignorada
24
Caracteriacutesticas de Km la constante de Michaelis-Menten es
caracteriacutestica de una enzima y particular para un substrato Refleja la afinidad de la enzima por ese substrato K m es numeacutericamente igual a la concentracioacuten
de substrato a la cual la velocidad de reaccioacuten es la mitad de la Vmax (Km= Vmax2) Este paraacutemetro Km no variacutea con la concentracioacuten de enzima
Significado de una Km pequentildea un valor numeacuterico pequentildeo de Km
refleja una alta afinidad de la enzima por su substrato porque a una baja
concentracioacuten del mismo la enzima a desarrollado ya la mitad de la velocidad maacutexima
Significado de una Km grande el valor numeacuterico grande de Km refleja
una baja afinidad de la enzima por su substrato porque a una concentracioacuten
elevada del mismo la enzima desarrolla la mitad de la velocidad maacutexima
Relacioacuten de la velocidad con la concentracioacuten de enzima la
velocidad de la reaccioacuten es directamente proporcional a la concentracioacuten de la
enzima a cualquier concentracioacuten de substrato Por ejemplo si la concentracioacuten de enzima es disminuida a la mitad la velocidad inicial de la reaccioacuten (v0) es
reducida tambieacuten a la mitad de la original
Orden de la reaccioacuten cuando S es mas pequentildea que la Km la
velocidad de la reaccioacuten es aproximadamente igual a la concentracioacuten de
substrato
100
50
0
Substrato
Km Figura Localizacioacuten de la constante de Michaelis (Km)
25
La velocidad de reaccioacuten se dice en estas condiciones es de primer
orden con respecto al substrato Cuando S es mas grande que la Km la
velocidad es constante e igual a la Vmax La velocidad de la reaccioacuten es independiente de la concentracioacuten de substrato y se dice que es de orden cero
con respecto a la concentracioacuten de substrato
Las enzimas pueden ser aisladas de las ceacutelulas u organismos para estudiar sus
propiedades in vitro es decir en un tubo de ensayo Las diferentes enzimas muestran diferentes respuestas a los cambios en la concentracioacuten de substrato temperatura y pH
Concentracioacuten de substrato
La forma hiperboacutelica de la curva de velocidad vs Substrato
Efecto de la temperatura en la actividad enzimaacutetica
Efecto de la variacioacuten del pH en la actividad enzimaacutetica
Incremento de la velocidad con la temperatura En general la velocidad de la reaccioacuten se incrementa con la temperatura hasta que un punto maacuteximo es alcanzado
Este incremento se debe a que aumenta el nuacutemero de moleacuteculas ricas en energiacutea que pueden pasar la barrera energeacutetica de estado de transicioacuten para formar a los productos
Decremento de la velocidad con la temperatura la elevacioacuten excesiva de la
temperatura del medio que contiene a las enzimas resulta en un decremento de la velocidad como resultado de la desnaturalizacioacuten es decir la peacuterdida de la estructura tridimensional de las enzimas
26
zona de reactivacioacuten maacuteximo de actividad
Zona de
inactivacioacuten
0 Temperatura
Figura dependencia de la actividad enzimaacutetica con la temperatura
Velocidad maacutexima la velocidad de una reaccioacuten (v) es el nuacutemero de
moleacuteculas de substrato convertidas en producto por unidad de tiempo y
usualmente se expresa en micromoles de producto formadas por minuto La velocidad de una reaccioacuten catalizada por una enzima aumenta conforme se incrementa la concentracioacuten de substrato hasta que se llega a una velocidad maacutexima (Vmax) a partir de la cual la velocidad de la reaccioacuten es
independiente de la cantidad de substrato El que la velocidad no pueda
seguirse incrementando refleja la saturacioacuten del sitio activo con el substrato
Vmax
0 Substrato
Figura representacioacuten de una cineacutetica tiacutepicamente michaeliana
Vmax = velocidad de saturacioacuten o maacutexima
27
Cuando se grafica la velocidad de la reaccioacuten v0 contra la concentracioacuten de substrato S no siempre es posible determinar la condicioacuten
en que se ha llegado a la velocidad maacutexima Vmax debido al incremento de la pendiente en la hipeacuterbola a concentraciones de substrato elevadas
Figura experimento cineacutetico para la actividad enzimaacutetica
Se presentan los valores experimentales y los resultados del ajuste al modelo de Michaelis
Auacuten asiacute si se grafica una doble reciacuteproca es decir el inverso de la
velocidad (1v0) contra el inverso de la concentracioacuten de substrato (1 S ) se
obtiene una liacutenea recta El regraacutefico de Lineweaver-Burke tambieacuten se
denomina de ldquodobles reciacuteprocasrdquo y se utiliza para calcular la Km y la Vmax asiacute como para determinar el mecanismo de accioacuten de los diversos tipos de
inhibidores
La ecuacioacuten que describe a la graacutefca de Lineweaver-Burke es
Vmax
1
SVmax
Km
v
1
0 (3)
que describe una recta en donde el intercepto con el eje de las abscisas es igual a ndash1Km y el intercepto con el eje de las ordenadas es igual a 1Vmax
28
Figura experimento cineacutetico para la actividad enzimaacutetica Regraacutefico de
Lineweaver-Burke
Se presentan los valores experimentales y los resultados del ajuste al modelo
REPRESENTACIOacuteN GRAacuteFICA DE LINEWEAVER Y BURK
Si se toma el inverso en ambos miembros de la ecuacioacuten de Michaelis tenemos
=
+
29
que es la ecuacioacuten de una liacutenea recta cuya pendiente es que intercepta al eje de las
ordenadas en y a la abcisa en el punto Lo que es evidente al hacer
ya que se convierte en
Si se hace una graacutefica con los dobles inversos colocando los valores de en el eje de las
ordenadas y en el eje de las abcisas se obtiene una liacutenea recta a partir de la cual se puede calcular faacutecilmente Km
EFECTO DEL pH SOBRE LA ACTIVIDAD ENZIMATICA
Sabiendo que las enzimas son proteinas cualquier cambio brusco de pH puede alterar el caraacutecter ioacutenico de los grupos amino y carboxilo en la superficie proteica afectando asiacute las propiedades cataliacuteticas de una enzima A pH alto o bajo se puede producir la desnaturalizacioacuten
de la enzima y en consecuencia su inactivacioacuten La fosfatasa aacutecida es maacutes activa a pH 50 mientras que la fosfatasa alcalina lo es a pH 90 Muchas enzimas tienen maacutexima actividad cerca de la neutralidad en un rango de pH de 6 a 8
El pH per se no afecta la actividad enzimaacutetica sino la concentracioacuten de protones Los protones
ademaacutes de alterar la estructura de la enzima y el substrato pueden participar tambieacuten en la reaccioacuten como substrato o producto En esos casos la concentracioacuten de protones afecta directamente la velocidad de la reaccioacuten
ENZIMA pH OPTIMO
30
Pepsina 15
Tripsina 77
Catalasa 76
Arginasa 97
Fumarasa 78
Ribonucleasa 78
EFECTO DE ACTIVADORES
Un aumento en la temperatura provoca un aumento de la velocidad de reaccioacuten hasta cierta temperatura oacuteptima ya que despueacutes de aproximadamente 45
0 C se comienza a producir la
desnaturalizacioacuten teacutermica Las enzimas de muchos mamiacuteferos tienen una temperatura oacuteptima
de 370 C por encima de esa temperatura comienzan a inactivarse y se destruyen Sin embargo
existen especies de bacterias y algas que habitan en fuentes de aguas termales y en el otro extremo ciertas bacterias aacuterticas tienen temperaturas oacuteptimas cercanas a 0
0 C
31
ESPECIFICIDAD ABSOLUTA
Cierta enzimas se encuentran como aproenzimas o zimogenos que son inactivos
El tripsinoacutegeno es convertido en tripsina por la misma accioacuten de la tripsina que remueve un hexapeacuteptido
tripsina Tripsinoacutegeno Tripsina + Val (Asp)4 lisina
Otra forma de activacioacuten consiste en mantener los grupos SH reducidos Estos grupos pueden
formar parte de centros activos de ciertas enzimas Si se oxidan estos grupos a -S-S- la enzima se inactiva Asiacute tenemos que la enzima papaina se inactiva despueacutes de exponerla al ox iacutegeno pero cuando se le antildeade un reductor apropiado la enzima se reactiva Los grupos ndashS-S- se convierten en ndashSH activaacutendose la enzima
Otra forma de activacioacuten requiere la presencia de otra substancia que se enlaza al sitio alosteacuterico el compuesto que se enlaza se denomina un efector alosteacuterico El centro alosteacuterico es bien especiacutefico para el efector o modulador alosteacuterico
ESPECIFICIDAD RELATIVA
Oxiacutegeno L-aminoaacutecido a -cetoaacutecido + NH3 + H 2 O 2 L-aminoaacutecido oxidasa Oxiacutegeno D-aminoaacutecido a -cetoaacutecido + NH3 + H 2 O 2 D-aminoaacutecido oxidasa
32
En este ejemplo se muestra la estereo-especificidad Una enzima puede tener una especificidad oacuteptica para isomeros D o L
Hay enzimas que muestran especificidad absoluta como la aspartasa que cataliza la adicioacuten reversible de amoniacuteaco al doble enlace del aacutecido fumaacuterico pero no al de otros aacutecidos insaturados
HOOC ndash CH = CH ndash COO- + NH4
+ HOOC ndash CH2 - CHNH2 - COO
- + H
+
INHIBICIOacuteN ENZIMATICA
Mediante el uso de inhibidores enzimaacuteticos se ha obtenido informacioacuten muy valiosa sobre la conformacioacuten del centro activo de algunas enzimas
a Inhibicioacuten irreversible Este tipo de inhibidores forman un enlace covalente con las enzimas cerca del centro activo Un ejemplo son los gases nerviosos como el
fluorofosfato de diisopropilo (DFP) que forma un complejo con la enzima acetilcolinesterasa Los animales envenenados con este gas quedan paralizados debido a la imposibilidad de transmitir adecuadamente los impulsos nerviosos
Ester di-isopropil fos foacuterico de la enzima (inactivo)
b) Inhibicioacuten competitiva
Es cuando el inhibidor compite con el substrato por la unioacuten con el centro activo de la
enzima Este tipo de inhibicioacuten puede reducirse si se aumenta la concentracioacuten de substrato
-OOC - CH2 - CH2 - COO
- -OOC - CH = CH - COO
- + 2H
+ Succinato Succinato Fumarato
33
Deshidrogenasa COO
- I CH2 Malonato (inhibidor competitivo) I COO
- En la inhibicioacuten competitiva la enzima se combina con el inhibidor para formar un complejo enzima-inhibidor E + I E I en competencia con la reaccioacuten normal E +S ES
La sulfanilamida (bactericida) interfiere con la siacutentesis del aacutecido foacutelico compitiendo con el aacutecido p-aminobenzoico de una forma competitiva
HOOC-sect- NH2 NH2 ndashSO2 - sect- NH2 aacutecido p- amino benzoico sulfanilamida
sect = fenil
a Inhibicioacuten no competitiva
Esta inhibicioacuten se caracteriza por que no se puede revertir el efecto del inhibidor aumentando la concentracioacuten del substrato
Por ejemplo la inhibicioacuten de la deshidrogenasa del gliceraldehido -3-fosfato por la yodo-acetamida
Enzima-SH + I-CH2 CONH2 Enzima-SCH2 CONH2 + IH
La deshidrogenasa del gliceraldehido-3 fosfato puede ser inhibida tambieacuten por el aacutecido iodo-aceacutetico
Enzima-SH + ICH2 COOH Enzima-SCH2 COOH + IH
c) Inhibicioacuten por metales
Ciertos metales como el plomo mercurio y arseacutenico inhiben enzimas que tienen en su centro activo grupos -SH libres
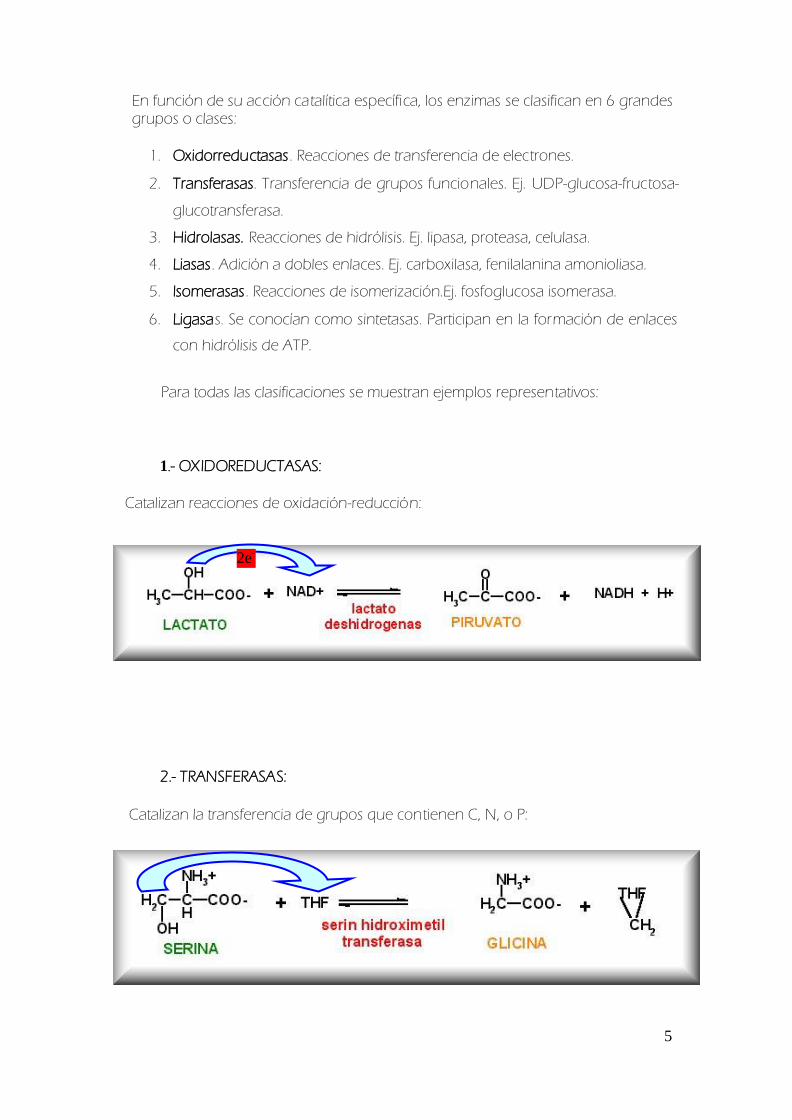
5
En funcioacuten de su accioacuten cataliacutetica especiacutefica los enzimas se clasifican en 6 grandes grupos o clases
1 Oxidorreductasas Reacciones de transferencia de electrones
2 Transferasas Transferencia de grupos funcionales Ej UDP-glucosa-fructosa-
glucotransferasa
3 Hidrolasas Reacciones de hidroacutelisis Ej lipasa proteasa celulasa
4 Liasas Adicioacuten a dobles enlaces Ej carboxilasa fenilalanina amonioliasa
5 Isomerasas Reacciones de isomerizacioacutenEj fosfoglucosa isomerasa
6 Ligasas Se conociacutean como sintetasas Participan en la formacioacuten de enlaces
con hidroacutelisis de ATP
Para todas las clasificaciones se muestran ejemplos representativos
1- OXIDOREDUCTASAS
Catalizan reacciones de oxidacioacuten-reduccioacuten
2e-
2- TRANSFERASAS
Catalizan la transferencia de grupos que contienen C N o P
THF tetrahidrofolato
6
3- HIDROLASAS
Catalizan la ruptura de enlaces por la adicioacuten de una moleacutecula de agua
4- LIASAS
Catalizan la ruptura de enlaces C-C C-S y algunos enlaces C-N
5- ISOMERASAS
Catalizan la racemizacioacuten de isoacutemeros oacutepticos o geomeacutetricos
7
6- LIGASAS
Catalizan la formacioacuten de enlaces entre C y O S N Esta reaccioacuten soacutelo es posible
mediante la energiacutea derivada de fosfatos ricos en energiacutea como el del fosfato de la
moleacutecula de ATP
15 COFACTORES y COENZIMAS Algunas enzimas se asocian con moleacuteculas de caraacutecter no proteiacuteco que son
necesarias para el funcionamiento de la enzima estas moleacuteculas se denominan
cofactores (Figura 3) Comuacutenmente los factores encontrados en las enzimas incluyen
iones metaacutelicos como el Zn2+ o el Fe2+ ( Tabla 2) tambieacuten pueden ser moleacuteculas
orgaacutenicas que se denomina coenzimas como el NAD+ FAD la coenzima A
Generalmente las coenzimas son derivados de las vitaminas A la enzima en ausencia
de su cofactor (cuando lo tiene) se le denomina apoenzima en presencia de su
cofactor (cuando lo tiene) se le denomina holoenzima La apoenzima generalmente
carece de actividad bioloacutegica La diferencia entre un cofactor y un grupo prosteacutetico
como el grupo hemo es que este uacuteltimo estaacute unido de manera covalente a la
enzima mientras que el cofactor puede ser removido de la misma con relativa
facilidad
Se pueden sentildealar algunas caracteriacutesticas de las coenzimas como generalmente son
estables al calor y a otros agentes desnaturalizantes de las proteiacutenas pueden ser
recicladas y algunas provienen de las vitaminas
8
Figura 3 Cofactores y Coenzima
Tabla 2 Algunos enzimas que contienen iones metaacutelicos o los necesitan como
cofactores
Zn+2 Cu+2 (Cu+)
Alcohol- deshidrogenada Tirosinasa Anhidrasa-carboacutenica Citocromo-oxidasa Carboxipeptidasa
Mg+2 K+
Fosfohidrolasa Piruvato- fosfoquinasa Fosfotransferasas
Mn+2 Arginasa Na+ Fosfotransferasas ATPasa de la membrana plasmaacutetica
Fe +2 o Fe+3
9
Citocromos
Peroxidasa Catalasa Ferredoxina
Texto ------- (Tabla 3)
Tabla 3 Relacioacuten entre algunas coenzimas y las vitaminas a partir de las
cuales se producen Enfermedades generadas por la deficiencia
Coenzima Reaccioacuten Fuente vitamiacutenica Enfermedad
Humana por deficiencia
Biocitina Carboxilacioacuten Biotina
Coenzima A Transferencia de acilos Pantotenato
Coenzimas de Cobalamima Alquilacioacuten Cobalamina (B12) anemia perniciosa
Coenzimas de flavina Oxidacioacuten-reduccioacuten Riboflavina (B2)
Acido lipoacuteico Transferencia de acilos -
Coenzimas de nicotinamida Oxidacioacuten-reduccioacuten Nicotinamida (niacina) Pelagra
Fosfato de piridoxal Transferencia de grupo amino Piridoxina (B6)
Tetrahidrofolato Transferencia de un grupo de 1 Carbono Acido foacutelico anemia
megaloblaacutestica
Tiamina pirofosfato Transferencia de aldehiacutedo Tiamina (B1) beriberi
no tiene un nombre especiacutefico su aparicioacuten es muy rara
Otras coenzimas
NADP
Coenzima Q
Biotina
NAD y NADP
10
Coenzima Q (CoQ)
Esta coenzima es sintetizada a partir de farnesil pirofosfato Al igual que el
FMN la CoQ es capaz de aceptar y donar uno o dos electrones porque su forma
semiquinona es estable A su forma oxidada se le conoce como ubiquinona (de
ubicuo lat ubique en todas partes) o quinona A su forma reducida se le conoce
como QH2 ubiquinol o hidroquinona (Figura 2) A la coenzima solo parcialmente
reducida se le conoce como ubisemiquinona semiquinona o QH (Figura 4)
Figura 4 Representacioacuten de la moleacutecula de CoQ
La siacutentesis de la CoQ se lleva a cabo a traveacutes de unidades isoprenoides (Figura 5)
11
Figura 5 La CoQ se produce a partir de isoprenoides
Biotina
La biotina consiste de un anillo imidazol que estaacute unido en forma cis a la
cadena lateral tetrahidrotiofeno del valerato (Figura 6)
12
Figura 6 Estructura de la biotina
La enzima holocarboxilasa sintetasa (HCS) cataliza la activacioacuten mediante
biotinilacioacuten de cinco carboxilasas en ceacutelulas humanas La deficiencia de HCS
produce el siacutendrome deficiencia muacuteltiple de carboxilasas (DMC) Esta enfermedad es
potencialmente mortal y se caracteriza por la deficiencia de la actividad de todas las
carboxilasas La deficiencia de biotina no solo disminuye la actividad de estas enzimas
sino que parece modificar la expresioacuten geneacutetica
126 PROPIEDADES DE LAS ENZIMAS
Las propiedades de las enzimas derivan del hecho de ser proteiacutenas y de actuar como
catalizadores Como proteiacutenas poseen una conformacioacuten natural maacutes estable que las
demaacutes conformaciones posibles Asiacute cambios en la conformacioacuten suelen ir asociados
en cambios en la actividad cataliacutetica Los factores que influyen de manera maacutes directa
sobre la actividad de un enzima son
pH
temperatura
cofactores
A) EFECTO DEL pH
Al comprobar experimentalmente la influencia del pH en la velocidad de las
reacciones enzimaacuteticas se obtienen curvas que indican que los enzimas
presentan un pH oacuteptimo de actividad El pH puede afectar de varias maneras
13
o El centro activo puede contener aminoaacutecidos con grupos ionizados
que pueden variar con el pH
o La ionizacioacuten de aminoaacutecidos que no estaacuten en el centro activo puede
provocar modificaciones en la conformacioacuten de la enzima
o El sustrato puede verse afectado por las variaciones del pH
Algunos enzimas presentan variaciones peculiares La pepsina del estoacutemago
presenta un oacuteptimo a pH=2 la ureasa lo tiene a pH 7 y la fosfatasa alcalina del
intestino un pH= 12
B) EFECTO DE LA TEMPERATURA
Influye en la actividad El punto oacuteptimo representa el maacuteximo de actividad A
temperaturas bajas los enzimas se hallan ldquomuy riacutegidosrdquo y cuando se supera un
valor considerable (mayor de 50) la actividad cae bruscamente porque como
proteiacutena el enzima se desnaturaliza
128 EFECTO DE LOS COFACTORES SOBRE LA ACTIVIDAD
ENZIMATICA
A veces un enzima requiere para su funcioacuten la presencia de sustancias no proteicas
que colaboran en la cataacutelisis los cofactores Casi un tercio de los enzimas conocidos
requieren cofactores Cuando el cofactor es una moleacutecula orgaacutenica se llama
coenzima En la figura inferior podemos observar una moleacutecula de hemoglobina
(proteiacutena que transporta oxiacutegeno) y su coenzima (el grupo hemo) Cuando los
cofactores y las coenzimas se encuentran unidos covalentemente al enzima se llaman
14
grupos prosteacuteticos La forma cataliacuteticamente activa del enzima es decir el enzima
unido a su grupo prosteacutetico se llama holoenzima La parte proteica de un
holoenzima (inactiva) se llama apoenzima de forma que
apoenzima + grupo prosteacutetico= holoenzima
129 MODO DE ACCIOacuteN DE LAS ENZIMAS
En las reacciones espontaacuteneas los productos finales tienen menos energiacutea libre de
Gibbs (∆G) que los reactantes (figura inferior izquierda) Por lo tanto las reacciones
espontaacuteneas liberan energiacutea de Gibbs ((∆G lt 0) Sin embargo el comienzo de la
reaccioacuten requiere un aporte inicial de energiacutea Esta energiacutea inicial que hay que
suministrar a los reactantes para que la reaccioacuten transcurra se llama energiacutea de
activacioacuten (Ea) Cuanto menor es la Ea maacutes faacutecilmente transcurre la reaccioacuten
Perfil energeacutetico de una reaccioacuten espontaacutenea Perfil energeacutetico de una reaccioacuten catalizada
15
La accioacuten de los catalizadores consiste precisamente en disminuir la Ea (Figura superior derecha) Los enzimas
son catalizadores especialmente eficaces ya que disminuyen la Ea auacuten maacutes que los catalizadores inorgaacutenicos
Por ejemplo la descomposicioacuten del agua oxigenada (H2O2) para dar H2O y O2 puede ocurrir sin catalizador
con un catalizador inorgaacutenico (platino) o con un enzima especiacutefico (catalasa) Las respectivas Ea para cada
proceso son 18 12 y 6 Kcalmol Asiacute se puede calcular que el platino acelera la reaccioacuten 20000 veces mientras
que la catalasa la acelera 370000 veces
Para que una reaccioacuten quiacutemica tenga lugar las moleacuteculas de los reactantes deben chocar con una energiacutea y
una orientacioacuten adecuadas La actuacioacuten del enzima (1) permite que los rea ctantes (sustratos) se unan a su
centro activo con una orientacioacuten oacuteptima para que la reaccioacuten se produzca y (2) modifica las propiedades
quiacutemicas del sustrato unido a su centro activo debilitando los enlaces existentes y facilitando la formacioacuten de
otros nuevos (Figuras inferiores)
1210 REGULACIOacuteN DE SU ACTIVIDAD ENZIMATICA
16
Una moleacutecula de enzima no tiene por que actuar siempre a la misma velocidad Su
actividad puede estar modulada por
Cambios en el pH
Cambios en la temperatura
Presencia de cofactores
Las concentraciones del sustrato y de los productos finales
Presencia de inhibidores
Modulacioacuten alosteacuterica
Modificacioacuten covalente
Activacioacuten por proteolisis
Isoenzimas
EFECTO DE LAS CONCENTRACIONES SOBRE LA ACTIVIDAD ENZIMAacuteTICA
La velocidad de una reaccioacuten enzimaacutetica depende de la concentracioacuten de sustrato La Figura de la derecha muestra la velocidad de una reaccioacuten enzimaacutetica a 6 concentraciones distintas de sustrato
Ademaacutes la presencia de los productos finales puede hacer que la reaccioacuten sea maacutes lenta o incluso invertir su sentido (Figura inferior)
EFECTO DE LOS INHIBIDORES SOBRE LA ACTIVIDAD ENZIMAacuteTICA
Ciertas moleacuteculas pueden inhibir la accioacuten cataliacutetica de un enzima son los inhibidores Estos inhibidores bien pueden ocupar temporalmente el centro activo por semejanza estructural con el sustrato original ( inhibidor competitivo) o bien alteran la conformacioacuten espacial del enzima impidiendo su unioacuten al sustrato ( inhibidor no competitivo) (Figuras inferiores)
Inhibidor competitivo Inhibidor no competitivo
17
MODULACIOacuteN ALOSTEacuteRICA DE LA ACTIVIDAD ENZIMAacuteTICA
Hay enzimas que pueden adoptar 2 conformaciones interconvertibles llamadas R (relajada) y T (tensa) R es la forma maacutes activa porque se une al sustrato con maacutes afinidad Las formas R y T se encuentran en equilibrio R lt==gt T (Figura inferior)
Ciertas sustancias tienden a estabilizar la forma R Son los llamados moduladores positivos El propio sustrato es a
menudo un modulador positivo Las moleacuteculas que favorecen la forma R pero que actuacutean sobre una regioacuten del enzima distinta del centro activo son los activadores alosteacutericos (Figura inferior izquierda)
Activador alosteacuterico favorece la unioacuten del sustrato
Inhibidor alosteacuterico impide la unioacuten del sustrato
Las sustancias que favorecen la forma T y disminuyen la actividad enzimaacutetica son los moduladores negativos Si
estos moduladores actuacutean en lugares distintos del centro activo del enzima se llaman inhibidores alosteacutericos
18
(Figura superior derecha
EFECTO DE LA MODIFICACIOacuteN COVALENTE SOBRE LA ACTIVIDAD ENZIMAacuteTICA
Otros enzimas pasan de una forma menos activa a otra maacutes activa unieacutendose covalentemente a un grupo quiacutemico de pequentildeo tamantildeo como el Pi o el AMP Tambieacuten se da el caso inverso en el que un enzima muy activo se desactiva al liberar alguacuten grupo quiacutemico En las enzimas de las viacuteas degradativas del metabolismo la
forma fosforilada es maacutes activa que la no fosforilada mientras que en las viacuteas biosinteacuteticas ocurre lo contrario En las figuras inferiores se ilustra la activacioacuten de una proteiacutena por fosforilacioacuten
Elementos de la reaccioacuten El enzima no fosforilado es inactivo El enzima fosforilado es activo
ACTIVACIOacuteN PROTEOLIacuteTICA DE LA ACTIVIDAD ENZIMAacuteTICA
Algunos enzimas no se sintetizan como tales sino como proteiacutenas precursoras sin actividad enzimaacutetica Estas proteiacutenas se llaman
proenzimas o zimoacutegenos Para activarse los zimoacutegenos sufren un ataque hidroliacutetico que origina la liberacioacuten de uno o varios peacuteptidos El resto de la moleacutecula proteica adopta la conformacioacuten y las propiedades del enzima activo Muchos enzimas digestivos se secretan en forma de zimoacutegenos y en el tubo digestivo
se convierten en la forma activa Es el caso de la -quimotripsina que se sintetiza en forma de quimotripsinoacutegeno (Figura superior) Si estos enzimas se sintetizasen directamente en forma activa destruiriacutean la propia ceacutelula que las
produce Asiacute la tripsina pancreaacutetica (una proteasa) se sintetiza como tripsinoacutegeno (inactivo) Si por alguna razoacuten se activa en el propio paacutencreas la glaacutendula sufre un proceso de autodestruccioacuten (pancreatitis aguda) a
menudo mortal
19
REGULACIOacuteN DE LA ACTIVIDAD ENZIMAacuteTICA POR MEDIO DE ISOENZIMAS
Algunos enzimas tienen distinta estructura molecular aunque su funcioacuten bioloacutegica es similar Se llaman isozimas o isoenzimas Estas diferencias de estructura se traducen en ligeros cambios en sus propiedades de forma que cada isozima se adapta perfectamente a la funcioacuten que debe realizar Asiacute podemos observar la existencia de isoenzimas en funcioacuten de
el tipo de tejido Por ejemplo la lactato deshidrogenasa presenta isozimas distintos en muacutesculo y corazoacuten el compartimento celular donde actuacutea Por ejemplo la malato deshidrogenasa del citoplasma es distinta
de la de la mitocondria el momento concreto del desarrollo del individuo Por ejemplo algunos enzimas de la glicolisis del feto
son diferentes de los mismos enzimas en el adulto
CINEacuteTICA ENZIMAacuteTICA
Los principios generales de las reacciones quiacutemicas se aplican tambieacuten a las reacciones enzimaacuteticas Por este motivo antes de empezar con la cineacutetica quiacutemica se van a repasar algunos conceptos baacutesicos de cineacutetica quiacutemica
A continuacioacuten se describiraacuten los siguientes conceptos
Cineacutetica enzimaacutetica Modelo cineacutetico de Michaelis-Menten Caacutelculo de la KM y la Vmax de un enzima Actividad enzimaacutetica
20
NOMENCLATURA DE LOS ENZIMAS
CATALISIS MOLECULAR
Un catalizador modifica la velocidad de una reaccioacuten quiacutemica sin ser utilizado o aparecer como uno de los productos de la reaccioacuten Una reaccioacuten quiacutemica en la que un substrato(S) se
transforma en un producto(P) S P ocurre por que cierta fraccioacuten de moleacuteculas de S posee mucho maacutes energiacutea que el resto de ellas lo que es suficiente para que alcancen un estado activado en el que pueda formarse o romperse un enlace quiacutemico y se forme el producto(P)
La energiacutea de activacioacuten es la cantidad de energiacutea expresada en caloriacuteas necesaria para que todas las moleacuteculas de un mol a una temperatura dada alcancen el estado reactivo Mientras que el estado de transicioacuten es el estado rico en energiacutea de las moleacuteculas que
interaccionan en la cima de la barrera de activacioacuten La velocidad de una reaccioacuten quiacutemica es proporcional a la concentracioacuten del complejo en el estado de transicioacuten
Una reaccioacuten quiacutemica se puede acelerar de la siguiente forma 1) Al aumentar la temperatura se incrementa la energiacutea cineacutetica por lo que es mayor el nuacutemero de moleacuteculas que alcanzan el
estado de transicioacuten Generalmente el Q10 =2 lo que indica que la velocidad de una reaccioacuten quiacutemica se duplica al aumentar la temperatura en 10
0C 2) Antildeadiendo un catalizador que
disminuye la energiacutea de activacioacuten y aumenta la velocidad de reaccioacuten
La enzima (E) se combina con el substrato (S) formando el complejo de transicioacuten enzima-
substrato (E-S) mediante una reaccioacuten reversible cuya energiacutea de activacioacuten es menor que la de la reaccioacuten no catalizada Cuando se forma el producto de la reaccioacuten (P) se regenera de nuevo la enzima (E) de forma libre la que puede combinarse de nuevo con otra moleacutecula de substrato (S)
Una enzima reduce maacutes eficientemente la energiacutea de activacioacuten(Ea) de una reaccioacuten que un catalizador inorgaacutenico lo que permite que una reaccioacuten se realice a menor temperatura
El siguiente ejemplo ilustra mejor lo que hemos discutido
Reaccioacuten Energiacutea de activacioacuten (Kcalmol-1
)
21
a) el peroacutexido de hidroacutegeno se descompone en
H2 O2 H2 O + O2
18
b) el hierro cataliacutetico (Fe) realiza la reaccioacuten
H2 O2 H2 O + O2
13
c) el platino cataliacutetico (Pt) realiza la reaccioacuten
H2 O2 H2 O + O2
12
d) la catalasa una enzima hepaacutetica la realiza
H2O2 H2O + O2
5
Una enzima no modifica la energiacutea libre ni la constante de equilibrio sino que disminuye la energiacutea de activacioacuten de la reaccioacuten
Ir al principio
EFECTO DE LA CONCENTRACIOacuteN DEL SUBSTRATO SOBRE LA
CATALISIS ENZIMATICA
La actividad de una enzima se puede estudiar in vitro bajo condiciones controladas antildeadieacutendole una enzima a un substrato Siacute se trabaja con la condicioacuten de que la concentracioacuten del substrato sea saturante variando entonces la concentracioacuten de enzima se observa que aumenta el producto de la reaccioacuten a pH y temperatura constantes
22
Si se mantiene la concentracioacuten de la enzima constante y variando la concentracioacuten de substrato se obtiene una curva hiperboacutelica como la de la figura Al principio un aumento de la
concentracioacuten de substrato produce un aumento raacutepi do de la velocidad de reaccioacuten pero si se sigue aumentando la concentracioacuten de substrato la velocidad de reaccioacuten comienza a disminuir y a muy altas concentraciones de substrato se observa que no cambia la velocidad de
reaccioacuten se dice que los centros activos de la enzima se encuentran saturados La velocidad de reaccioacuten que se obtiene a esa alta concentracioacuten de substrato se define como la velocidad maacutexima (V) de la reaccioacuten enzimaacutetica bajo las condiciones especificadas La concentracioacuten de
substrato (S) a la semivelocidad maacutexima de reaccioacuten (V2) se puede determinar de la figura y representa la constante de Michaelis o Km la cual es una caracteriacutestica para cada enzima La inversa de Km o 1Km mide aproximadamente la afinidad de la enzima por el subs trato
Mientras maacutes pequentildeo sea el valor de Km mayor seraacute la afinidad de la enzima por el substrato Si varias enzimas compiten en el metabolismo por el mismo substrato eacuteste seraacute transformado preferentemente por la enzima con mayor afinidad
La ecuacioacuten de Michaelis describe la relacioacuten cuantitativa entre la velocidad de reaccioacuten y la concentracioacuten de substrato [S] si se conoce Vmax o Km
v=
En donde v= es la velocidad de reaccioacuten observada a una concentracioacuten de substrato determinada [S]
Km= constante de Michaelis en molesliacutet ro
23
Vmax= velocidad maacutexima a concentracioacuten saturante de substrato
Si v= Vmax2 entonces
=
Km+[S] = 2[S]
Km =[S]
De tal forma que podemos concluir que Km es igual a la concentracioacuten de substrato a la semivelocidad maacutexima de reaccioacuten
1 Concentraciones relativas de E y S la concentracioacuten de substrato S es
mucho mayor que la de E de tal manera que la cantidad de substrato
unido a la enzima en cualquier momento es muy pequentildea
2 Se asume el estado estacionario ES no cambia con el tiempo esto
quiere decir que la velocidad de formacioacuten de ES es igual a aquella para
su desintegracioacuten En general un intermediario en una serie de reacciones estaacute en estado estacionario cuando su velocidad de siacutentesis
es igual a su velocidad de degradacioacuten 3 Velocidad inicial para el anaacutelisis de reacciones enzimaacuteticas soacutelo se
utiliza la velocidad inicial de la reaccioacuten que es la velocidad ejercida por
la enzima inmediatamente despueacutes de que se ha puesto en contacto con el substrato y hasta antes de que se haya consumido el 10 de la concentracioacuten inicial del mismo La razoacuten de lo anterior es que en ese
momento la concentracioacuten del producto de la reaccioacuten que se ha acumulado en muy pequentildea y por tanto la reaccioacuten en el sentido
inverso es decir la transformacioacuten del producto en el substrato original puede ser ignorada
24
Caracteriacutesticas de Km la constante de Michaelis-Menten es
caracteriacutestica de una enzima y particular para un substrato Refleja la afinidad de la enzima por ese substrato K m es numeacutericamente igual a la concentracioacuten
de substrato a la cual la velocidad de reaccioacuten es la mitad de la Vmax (Km= Vmax2) Este paraacutemetro Km no variacutea con la concentracioacuten de enzima
Significado de una Km pequentildea un valor numeacuterico pequentildeo de Km
refleja una alta afinidad de la enzima por su substrato porque a una baja
concentracioacuten del mismo la enzima a desarrollado ya la mitad de la velocidad maacutexima
Significado de una Km grande el valor numeacuterico grande de Km refleja
una baja afinidad de la enzima por su substrato porque a una concentracioacuten
elevada del mismo la enzima desarrolla la mitad de la velocidad maacutexima
Relacioacuten de la velocidad con la concentracioacuten de enzima la
velocidad de la reaccioacuten es directamente proporcional a la concentracioacuten de la
enzima a cualquier concentracioacuten de substrato Por ejemplo si la concentracioacuten de enzima es disminuida a la mitad la velocidad inicial de la reaccioacuten (v0) es
reducida tambieacuten a la mitad de la original
Orden de la reaccioacuten cuando S es mas pequentildea que la Km la
velocidad de la reaccioacuten es aproximadamente igual a la concentracioacuten de
substrato
100
50
0
Substrato
Km Figura Localizacioacuten de la constante de Michaelis (Km)
25
La velocidad de reaccioacuten se dice en estas condiciones es de primer
orden con respecto al substrato Cuando S es mas grande que la Km la
velocidad es constante e igual a la Vmax La velocidad de la reaccioacuten es independiente de la concentracioacuten de substrato y se dice que es de orden cero
con respecto a la concentracioacuten de substrato
Las enzimas pueden ser aisladas de las ceacutelulas u organismos para estudiar sus
propiedades in vitro es decir en un tubo de ensayo Las diferentes enzimas muestran diferentes respuestas a los cambios en la concentracioacuten de substrato temperatura y pH
Concentracioacuten de substrato
La forma hiperboacutelica de la curva de velocidad vs Substrato
Efecto de la temperatura en la actividad enzimaacutetica
Efecto de la variacioacuten del pH en la actividad enzimaacutetica
Incremento de la velocidad con la temperatura En general la velocidad de la reaccioacuten se incrementa con la temperatura hasta que un punto maacuteximo es alcanzado
Este incremento se debe a que aumenta el nuacutemero de moleacuteculas ricas en energiacutea que pueden pasar la barrera energeacutetica de estado de transicioacuten para formar a los productos
Decremento de la velocidad con la temperatura la elevacioacuten excesiva de la
temperatura del medio que contiene a las enzimas resulta en un decremento de la velocidad como resultado de la desnaturalizacioacuten es decir la peacuterdida de la estructura tridimensional de las enzimas
26
zona de reactivacioacuten maacuteximo de actividad
Zona de
inactivacioacuten
0 Temperatura
Figura dependencia de la actividad enzimaacutetica con la temperatura
Velocidad maacutexima la velocidad de una reaccioacuten (v) es el nuacutemero de
moleacuteculas de substrato convertidas en producto por unidad de tiempo y
usualmente se expresa en micromoles de producto formadas por minuto La velocidad de una reaccioacuten catalizada por una enzima aumenta conforme se incrementa la concentracioacuten de substrato hasta que se llega a una velocidad maacutexima (Vmax) a partir de la cual la velocidad de la reaccioacuten es
independiente de la cantidad de substrato El que la velocidad no pueda
seguirse incrementando refleja la saturacioacuten del sitio activo con el substrato
Vmax
0 Substrato
Figura representacioacuten de una cineacutetica tiacutepicamente michaeliana
Vmax = velocidad de saturacioacuten o maacutexima
27
Cuando se grafica la velocidad de la reaccioacuten v0 contra la concentracioacuten de substrato S no siempre es posible determinar la condicioacuten
en que se ha llegado a la velocidad maacutexima Vmax debido al incremento de la pendiente en la hipeacuterbola a concentraciones de substrato elevadas
Figura experimento cineacutetico para la actividad enzimaacutetica
Se presentan los valores experimentales y los resultados del ajuste al modelo de Michaelis
Auacuten asiacute si se grafica una doble reciacuteproca es decir el inverso de la
velocidad (1v0) contra el inverso de la concentracioacuten de substrato (1 S ) se
obtiene una liacutenea recta El regraacutefico de Lineweaver-Burke tambieacuten se
denomina de ldquodobles reciacuteprocasrdquo y se utiliza para calcular la Km y la Vmax asiacute como para determinar el mecanismo de accioacuten de los diversos tipos de
inhibidores
La ecuacioacuten que describe a la graacutefca de Lineweaver-Burke es
Vmax
1
SVmax
Km
v
1
0 (3)
que describe una recta en donde el intercepto con el eje de las abscisas es igual a ndash1Km y el intercepto con el eje de las ordenadas es igual a 1Vmax
28
Figura experimento cineacutetico para la actividad enzimaacutetica Regraacutefico de
Lineweaver-Burke
Se presentan los valores experimentales y los resultados del ajuste al modelo
REPRESENTACIOacuteN GRAacuteFICA DE LINEWEAVER Y BURK
Si se toma el inverso en ambos miembros de la ecuacioacuten de Michaelis tenemos
=
+
29
que es la ecuacioacuten de una liacutenea recta cuya pendiente es que intercepta al eje de las
ordenadas en y a la abcisa en el punto Lo que es evidente al hacer
ya que se convierte en
Si se hace una graacutefica con los dobles inversos colocando los valores de en el eje de las
ordenadas y en el eje de las abcisas se obtiene una liacutenea recta a partir de la cual se puede calcular faacutecilmente Km
EFECTO DEL pH SOBRE LA ACTIVIDAD ENZIMATICA
Sabiendo que las enzimas son proteinas cualquier cambio brusco de pH puede alterar el caraacutecter ioacutenico de los grupos amino y carboxilo en la superficie proteica afectando asiacute las propiedades cataliacuteticas de una enzima A pH alto o bajo se puede producir la desnaturalizacioacuten
de la enzima y en consecuencia su inactivacioacuten La fosfatasa aacutecida es maacutes activa a pH 50 mientras que la fosfatasa alcalina lo es a pH 90 Muchas enzimas tienen maacutexima actividad cerca de la neutralidad en un rango de pH de 6 a 8
El pH per se no afecta la actividad enzimaacutetica sino la concentracioacuten de protones Los protones
ademaacutes de alterar la estructura de la enzima y el substrato pueden participar tambieacuten en la reaccioacuten como substrato o producto En esos casos la concentracioacuten de protones afecta directamente la velocidad de la reaccioacuten
ENZIMA pH OPTIMO
30
Pepsina 15
Tripsina 77
Catalasa 76
Arginasa 97
Fumarasa 78
Ribonucleasa 78
EFECTO DE ACTIVADORES
Un aumento en la temperatura provoca un aumento de la velocidad de reaccioacuten hasta cierta temperatura oacuteptima ya que despueacutes de aproximadamente 45
0 C se comienza a producir la
desnaturalizacioacuten teacutermica Las enzimas de muchos mamiacuteferos tienen una temperatura oacuteptima
de 370 C por encima de esa temperatura comienzan a inactivarse y se destruyen Sin embargo
existen especies de bacterias y algas que habitan en fuentes de aguas termales y en el otro extremo ciertas bacterias aacuterticas tienen temperaturas oacuteptimas cercanas a 0
0 C
31
ESPECIFICIDAD ABSOLUTA
Cierta enzimas se encuentran como aproenzimas o zimogenos que son inactivos
El tripsinoacutegeno es convertido en tripsina por la misma accioacuten de la tripsina que remueve un hexapeacuteptido
tripsina Tripsinoacutegeno Tripsina + Val (Asp)4 lisina
Otra forma de activacioacuten consiste en mantener los grupos SH reducidos Estos grupos pueden
formar parte de centros activos de ciertas enzimas Si se oxidan estos grupos a -S-S- la enzima se inactiva Asiacute tenemos que la enzima papaina se inactiva despueacutes de exponerla al ox iacutegeno pero cuando se le antildeade un reductor apropiado la enzima se reactiva Los grupos ndashS-S- se convierten en ndashSH activaacutendose la enzima
Otra forma de activacioacuten requiere la presencia de otra substancia que se enlaza al sitio alosteacuterico el compuesto que se enlaza se denomina un efector alosteacuterico El centro alosteacuterico es bien especiacutefico para el efector o modulador alosteacuterico
ESPECIFICIDAD RELATIVA
Oxiacutegeno L-aminoaacutecido a -cetoaacutecido + NH3 + H 2 O 2 L-aminoaacutecido oxidasa Oxiacutegeno D-aminoaacutecido a -cetoaacutecido + NH3 + H 2 O 2 D-aminoaacutecido oxidasa
32
En este ejemplo se muestra la estereo-especificidad Una enzima puede tener una especificidad oacuteptica para isomeros D o L
Hay enzimas que muestran especificidad absoluta como la aspartasa que cataliza la adicioacuten reversible de amoniacuteaco al doble enlace del aacutecido fumaacuterico pero no al de otros aacutecidos insaturados
HOOC ndash CH = CH ndash COO- + NH4
+ HOOC ndash CH2 - CHNH2 - COO
- + H
+
INHIBICIOacuteN ENZIMATICA
Mediante el uso de inhibidores enzimaacuteticos se ha obtenido informacioacuten muy valiosa sobre la conformacioacuten del centro activo de algunas enzimas
a Inhibicioacuten irreversible Este tipo de inhibidores forman un enlace covalente con las enzimas cerca del centro activo Un ejemplo son los gases nerviosos como el
fluorofosfato de diisopropilo (DFP) que forma un complejo con la enzima acetilcolinesterasa Los animales envenenados con este gas quedan paralizados debido a la imposibilidad de transmitir adecuadamente los impulsos nerviosos
Ester di-isopropil fos foacuterico de la enzima (inactivo)
b) Inhibicioacuten competitiva
Es cuando el inhibidor compite con el substrato por la unioacuten con el centro activo de la
enzima Este tipo de inhibicioacuten puede reducirse si se aumenta la concentracioacuten de substrato
-OOC - CH2 - CH2 - COO
- -OOC - CH = CH - COO
- + 2H
+ Succinato Succinato Fumarato
33
Deshidrogenasa COO
- I CH2 Malonato (inhibidor competitivo) I COO
- En la inhibicioacuten competitiva la enzima se combina con el inhibidor para formar un complejo enzima-inhibidor E + I E I en competencia con la reaccioacuten normal E +S ES
La sulfanilamida (bactericida) interfiere con la siacutentesis del aacutecido foacutelico compitiendo con el aacutecido p-aminobenzoico de una forma competitiva
HOOC-sect- NH2 NH2 ndashSO2 - sect- NH2 aacutecido p- amino benzoico sulfanilamida
sect = fenil
a Inhibicioacuten no competitiva
Esta inhibicioacuten se caracteriza por que no se puede revertir el efecto del inhibidor aumentando la concentracioacuten del substrato
Por ejemplo la inhibicioacuten de la deshidrogenasa del gliceraldehido -3-fosfato por la yodo-acetamida
Enzima-SH + I-CH2 CONH2 Enzima-SCH2 CONH2 + IH
La deshidrogenasa del gliceraldehido-3 fosfato puede ser inhibida tambieacuten por el aacutecido iodo-aceacutetico
Enzima-SH + ICH2 COOH Enzima-SCH2 COOH + IH
c) Inhibicioacuten por metales
Ciertos metales como el plomo mercurio y arseacutenico inhiben enzimas que tienen en su centro activo grupos -SH libres
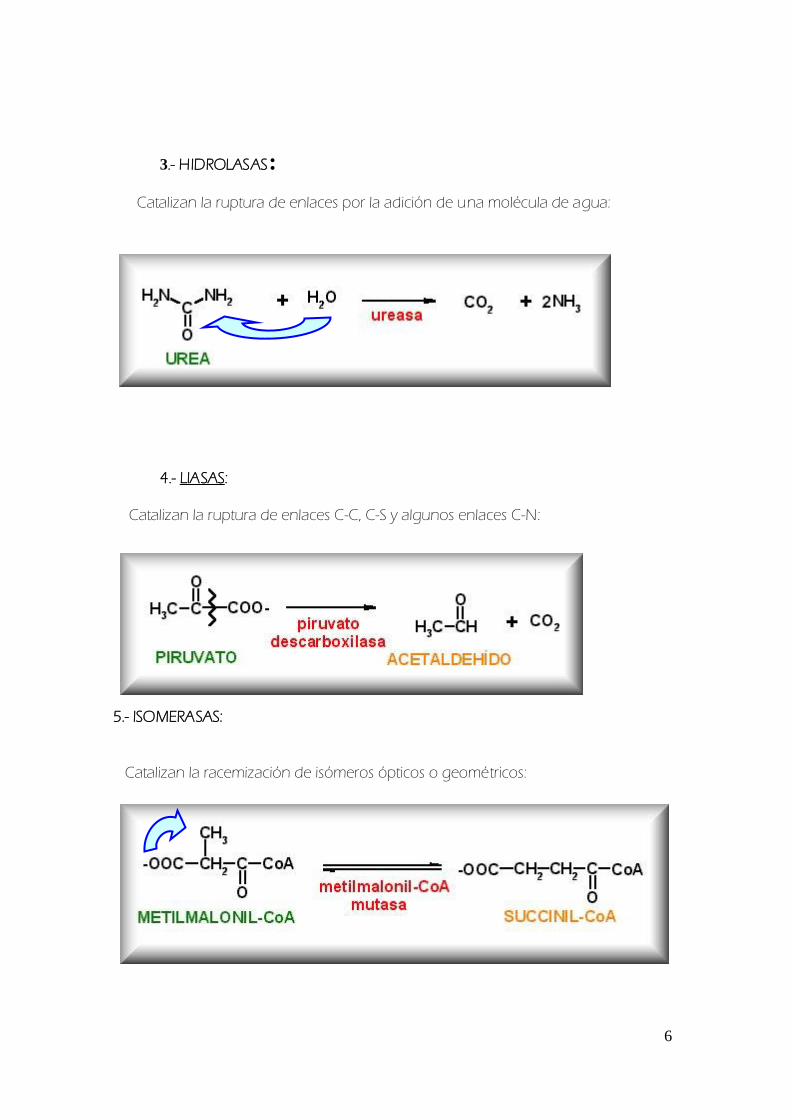
6
3- HIDROLASAS
Catalizan la ruptura de enlaces por la adicioacuten de una moleacutecula de agua
4- LIASAS
Catalizan la ruptura de enlaces C-C C-S y algunos enlaces C-N
5- ISOMERASAS
Catalizan la racemizacioacuten de isoacutemeros oacutepticos o geomeacutetricos
7
6- LIGASAS
Catalizan la formacioacuten de enlaces entre C y O S N Esta reaccioacuten soacutelo es posible
mediante la energiacutea derivada de fosfatos ricos en energiacutea como el del fosfato de la
moleacutecula de ATP
15 COFACTORES y COENZIMAS Algunas enzimas se asocian con moleacuteculas de caraacutecter no proteiacuteco que son
necesarias para el funcionamiento de la enzima estas moleacuteculas se denominan
cofactores (Figura 3) Comuacutenmente los factores encontrados en las enzimas incluyen
iones metaacutelicos como el Zn2+ o el Fe2+ ( Tabla 2) tambieacuten pueden ser moleacuteculas
orgaacutenicas que se denomina coenzimas como el NAD+ FAD la coenzima A
Generalmente las coenzimas son derivados de las vitaminas A la enzima en ausencia
de su cofactor (cuando lo tiene) se le denomina apoenzima en presencia de su
cofactor (cuando lo tiene) se le denomina holoenzima La apoenzima generalmente
carece de actividad bioloacutegica La diferencia entre un cofactor y un grupo prosteacutetico
como el grupo hemo es que este uacuteltimo estaacute unido de manera covalente a la
enzima mientras que el cofactor puede ser removido de la misma con relativa
facilidad
Se pueden sentildealar algunas caracteriacutesticas de las coenzimas como generalmente son
estables al calor y a otros agentes desnaturalizantes de las proteiacutenas pueden ser
recicladas y algunas provienen de las vitaminas
8
Figura 3 Cofactores y Coenzima
Tabla 2 Algunos enzimas que contienen iones metaacutelicos o los necesitan como
cofactores
Zn+2 Cu+2 (Cu+)
Alcohol- deshidrogenada Tirosinasa Anhidrasa-carboacutenica Citocromo-oxidasa Carboxipeptidasa
Mg+2 K+
Fosfohidrolasa Piruvato- fosfoquinasa Fosfotransferasas
Mn+2 Arginasa Na+ Fosfotransferasas ATPasa de la membrana plasmaacutetica
Fe +2 o Fe+3
9
Citocromos
Peroxidasa Catalasa Ferredoxina
Texto ------- (Tabla 3)
Tabla 3 Relacioacuten entre algunas coenzimas y las vitaminas a partir de las
cuales se producen Enfermedades generadas por la deficiencia
Coenzima Reaccioacuten Fuente vitamiacutenica Enfermedad
Humana por deficiencia
Biocitina Carboxilacioacuten Biotina
Coenzima A Transferencia de acilos Pantotenato
Coenzimas de Cobalamima Alquilacioacuten Cobalamina (B12) anemia perniciosa
Coenzimas de flavina Oxidacioacuten-reduccioacuten Riboflavina (B2)
Acido lipoacuteico Transferencia de acilos -
Coenzimas de nicotinamida Oxidacioacuten-reduccioacuten Nicotinamida (niacina) Pelagra
Fosfato de piridoxal Transferencia de grupo amino Piridoxina (B6)
Tetrahidrofolato Transferencia de un grupo de 1 Carbono Acido foacutelico anemia
megaloblaacutestica
Tiamina pirofosfato Transferencia de aldehiacutedo Tiamina (B1) beriberi
no tiene un nombre especiacutefico su aparicioacuten es muy rara
Otras coenzimas
NADP
Coenzima Q
Biotina
NAD y NADP
10
Coenzima Q (CoQ)
Esta coenzima es sintetizada a partir de farnesil pirofosfato Al igual que el
FMN la CoQ es capaz de aceptar y donar uno o dos electrones porque su forma
semiquinona es estable A su forma oxidada se le conoce como ubiquinona (de
ubicuo lat ubique en todas partes) o quinona A su forma reducida se le conoce
como QH2 ubiquinol o hidroquinona (Figura 2) A la coenzima solo parcialmente
reducida se le conoce como ubisemiquinona semiquinona o QH (Figura 4)
Figura 4 Representacioacuten de la moleacutecula de CoQ
La siacutentesis de la CoQ se lleva a cabo a traveacutes de unidades isoprenoides (Figura 5)
11
Figura 5 La CoQ se produce a partir de isoprenoides
Biotina
La biotina consiste de un anillo imidazol que estaacute unido en forma cis a la
cadena lateral tetrahidrotiofeno del valerato (Figura 6)
12
Figura 6 Estructura de la biotina
La enzima holocarboxilasa sintetasa (HCS) cataliza la activacioacuten mediante
biotinilacioacuten de cinco carboxilasas en ceacutelulas humanas La deficiencia de HCS
produce el siacutendrome deficiencia muacuteltiple de carboxilasas (DMC) Esta enfermedad es
potencialmente mortal y se caracteriza por la deficiencia de la actividad de todas las
carboxilasas La deficiencia de biotina no solo disminuye la actividad de estas enzimas
sino que parece modificar la expresioacuten geneacutetica
126 PROPIEDADES DE LAS ENZIMAS
Las propiedades de las enzimas derivan del hecho de ser proteiacutenas y de actuar como
catalizadores Como proteiacutenas poseen una conformacioacuten natural maacutes estable que las
demaacutes conformaciones posibles Asiacute cambios en la conformacioacuten suelen ir asociados
en cambios en la actividad cataliacutetica Los factores que influyen de manera maacutes directa
sobre la actividad de un enzima son
pH
temperatura
cofactores
A) EFECTO DEL pH
Al comprobar experimentalmente la influencia del pH en la velocidad de las
reacciones enzimaacuteticas se obtienen curvas que indican que los enzimas
presentan un pH oacuteptimo de actividad El pH puede afectar de varias maneras
13
o El centro activo puede contener aminoaacutecidos con grupos ionizados
que pueden variar con el pH
o La ionizacioacuten de aminoaacutecidos que no estaacuten en el centro activo puede
provocar modificaciones en la conformacioacuten de la enzima
o El sustrato puede verse afectado por las variaciones del pH
Algunos enzimas presentan variaciones peculiares La pepsina del estoacutemago
presenta un oacuteptimo a pH=2 la ureasa lo tiene a pH 7 y la fosfatasa alcalina del
intestino un pH= 12
B) EFECTO DE LA TEMPERATURA
Influye en la actividad El punto oacuteptimo representa el maacuteximo de actividad A
temperaturas bajas los enzimas se hallan ldquomuy riacutegidosrdquo y cuando se supera un
valor considerable (mayor de 50) la actividad cae bruscamente porque como
proteiacutena el enzima se desnaturaliza
128 EFECTO DE LOS COFACTORES SOBRE LA ACTIVIDAD
ENZIMATICA
A veces un enzima requiere para su funcioacuten la presencia de sustancias no proteicas
que colaboran en la cataacutelisis los cofactores Casi un tercio de los enzimas conocidos
requieren cofactores Cuando el cofactor es una moleacutecula orgaacutenica se llama
coenzima En la figura inferior podemos observar una moleacutecula de hemoglobina
(proteiacutena que transporta oxiacutegeno) y su coenzima (el grupo hemo) Cuando los
cofactores y las coenzimas se encuentran unidos covalentemente al enzima se llaman
14
grupos prosteacuteticos La forma cataliacuteticamente activa del enzima es decir el enzima
unido a su grupo prosteacutetico se llama holoenzima La parte proteica de un
holoenzima (inactiva) se llama apoenzima de forma que
apoenzima + grupo prosteacutetico= holoenzima
129 MODO DE ACCIOacuteN DE LAS ENZIMAS
En las reacciones espontaacuteneas los productos finales tienen menos energiacutea libre de
Gibbs (∆G) que los reactantes (figura inferior izquierda) Por lo tanto las reacciones
espontaacuteneas liberan energiacutea de Gibbs ((∆G lt 0) Sin embargo el comienzo de la
reaccioacuten requiere un aporte inicial de energiacutea Esta energiacutea inicial que hay que
suministrar a los reactantes para que la reaccioacuten transcurra se llama energiacutea de
activacioacuten (Ea) Cuanto menor es la Ea maacutes faacutecilmente transcurre la reaccioacuten
Perfil energeacutetico de una reaccioacuten espontaacutenea Perfil energeacutetico de una reaccioacuten catalizada
15
La accioacuten de los catalizadores consiste precisamente en disminuir la Ea (Figura superior derecha) Los enzimas
son catalizadores especialmente eficaces ya que disminuyen la Ea auacuten maacutes que los catalizadores inorgaacutenicos
Por ejemplo la descomposicioacuten del agua oxigenada (H2O2) para dar H2O y O2 puede ocurrir sin catalizador
con un catalizador inorgaacutenico (platino) o con un enzima especiacutefico (catalasa) Las respectivas Ea para cada
proceso son 18 12 y 6 Kcalmol Asiacute se puede calcular que el platino acelera la reaccioacuten 20000 veces mientras
que la catalasa la acelera 370000 veces
Para que una reaccioacuten quiacutemica tenga lugar las moleacuteculas de los reactantes deben chocar con una energiacutea y
una orientacioacuten adecuadas La actuacioacuten del enzima (1) permite que los rea ctantes (sustratos) se unan a su
centro activo con una orientacioacuten oacuteptima para que la reaccioacuten se produzca y (2) modifica las propiedades
quiacutemicas del sustrato unido a su centro activo debilitando los enlaces existentes y facilitando la formacioacuten de
otros nuevos (Figuras inferiores)
1210 REGULACIOacuteN DE SU ACTIVIDAD ENZIMATICA
16
Una moleacutecula de enzima no tiene por que actuar siempre a la misma velocidad Su
actividad puede estar modulada por
Cambios en el pH
Cambios en la temperatura
Presencia de cofactores
Las concentraciones del sustrato y de los productos finales
Presencia de inhibidores
Modulacioacuten alosteacuterica
Modificacioacuten covalente
Activacioacuten por proteolisis
Isoenzimas
EFECTO DE LAS CONCENTRACIONES SOBRE LA ACTIVIDAD ENZIMAacuteTICA
La velocidad de una reaccioacuten enzimaacutetica depende de la concentracioacuten de sustrato La Figura de la derecha muestra la velocidad de una reaccioacuten enzimaacutetica a 6 concentraciones distintas de sustrato
Ademaacutes la presencia de los productos finales puede hacer que la reaccioacuten sea maacutes lenta o incluso invertir su sentido (Figura inferior)
EFECTO DE LOS INHIBIDORES SOBRE LA ACTIVIDAD ENZIMAacuteTICA
Ciertas moleacuteculas pueden inhibir la accioacuten cataliacutetica de un enzima son los inhibidores Estos inhibidores bien pueden ocupar temporalmente el centro activo por semejanza estructural con el sustrato original ( inhibidor competitivo) o bien alteran la conformacioacuten espacial del enzima impidiendo su unioacuten al sustrato ( inhibidor no competitivo) (Figuras inferiores)
Inhibidor competitivo Inhibidor no competitivo
17
MODULACIOacuteN ALOSTEacuteRICA DE LA ACTIVIDAD ENZIMAacuteTICA
Hay enzimas que pueden adoptar 2 conformaciones interconvertibles llamadas R (relajada) y T (tensa) R es la forma maacutes activa porque se une al sustrato con maacutes afinidad Las formas R y T se encuentran en equilibrio R lt==gt T (Figura inferior)
Ciertas sustancias tienden a estabilizar la forma R Son los llamados moduladores positivos El propio sustrato es a
menudo un modulador positivo Las moleacuteculas que favorecen la forma R pero que actuacutean sobre una regioacuten del enzima distinta del centro activo son los activadores alosteacutericos (Figura inferior izquierda)
Activador alosteacuterico favorece la unioacuten del sustrato
Inhibidor alosteacuterico impide la unioacuten del sustrato
Las sustancias que favorecen la forma T y disminuyen la actividad enzimaacutetica son los moduladores negativos Si
estos moduladores actuacutean en lugares distintos del centro activo del enzima se llaman inhibidores alosteacutericos
18
(Figura superior derecha
EFECTO DE LA MODIFICACIOacuteN COVALENTE SOBRE LA ACTIVIDAD ENZIMAacuteTICA
Otros enzimas pasan de una forma menos activa a otra maacutes activa unieacutendose covalentemente a un grupo quiacutemico de pequentildeo tamantildeo como el Pi o el AMP Tambieacuten se da el caso inverso en el que un enzima muy activo se desactiva al liberar alguacuten grupo quiacutemico En las enzimas de las viacuteas degradativas del metabolismo la
forma fosforilada es maacutes activa que la no fosforilada mientras que en las viacuteas biosinteacuteticas ocurre lo contrario En las figuras inferiores se ilustra la activacioacuten de una proteiacutena por fosforilacioacuten
Elementos de la reaccioacuten El enzima no fosforilado es inactivo El enzima fosforilado es activo
ACTIVACIOacuteN PROTEOLIacuteTICA DE LA ACTIVIDAD ENZIMAacuteTICA
Algunos enzimas no se sintetizan como tales sino como proteiacutenas precursoras sin actividad enzimaacutetica Estas proteiacutenas se llaman
proenzimas o zimoacutegenos Para activarse los zimoacutegenos sufren un ataque hidroliacutetico que origina la liberacioacuten de uno o varios peacuteptidos El resto de la moleacutecula proteica adopta la conformacioacuten y las propiedades del enzima activo Muchos enzimas digestivos se secretan en forma de zimoacutegenos y en el tubo digestivo
se convierten en la forma activa Es el caso de la -quimotripsina que se sintetiza en forma de quimotripsinoacutegeno (Figura superior) Si estos enzimas se sintetizasen directamente en forma activa destruiriacutean la propia ceacutelula que las
produce Asiacute la tripsina pancreaacutetica (una proteasa) se sintetiza como tripsinoacutegeno (inactivo) Si por alguna razoacuten se activa en el propio paacutencreas la glaacutendula sufre un proceso de autodestruccioacuten (pancreatitis aguda) a
menudo mortal
19
REGULACIOacuteN DE LA ACTIVIDAD ENZIMAacuteTICA POR MEDIO DE ISOENZIMAS
Algunos enzimas tienen distinta estructura molecular aunque su funcioacuten bioloacutegica es similar Se llaman isozimas o isoenzimas Estas diferencias de estructura se traducen en ligeros cambios en sus propiedades de forma que cada isozima se adapta perfectamente a la funcioacuten que debe realizar Asiacute podemos observar la existencia de isoenzimas en funcioacuten de
el tipo de tejido Por ejemplo la lactato deshidrogenasa presenta isozimas distintos en muacutesculo y corazoacuten el compartimento celular donde actuacutea Por ejemplo la malato deshidrogenasa del citoplasma es distinta
de la de la mitocondria el momento concreto del desarrollo del individuo Por ejemplo algunos enzimas de la glicolisis del feto
son diferentes de los mismos enzimas en el adulto
CINEacuteTICA ENZIMAacuteTICA
Los principios generales de las reacciones quiacutemicas se aplican tambieacuten a las reacciones enzimaacuteticas Por este motivo antes de empezar con la cineacutetica quiacutemica se van a repasar algunos conceptos baacutesicos de cineacutetica quiacutemica
A continuacioacuten se describiraacuten los siguientes conceptos
Cineacutetica enzimaacutetica Modelo cineacutetico de Michaelis-Menten Caacutelculo de la KM y la Vmax de un enzima Actividad enzimaacutetica
20
NOMENCLATURA DE LOS ENZIMAS
CATALISIS MOLECULAR
Un catalizador modifica la velocidad de una reaccioacuten quiacutemica sin ser utilizado o aparecer como uno de los productos de la reaccioacuten Una reaccioacuten quiacutemica en la que un substrato(S) se
transforma en un producto(P) S P ocurre por que cierta fraccioacuten de moleacuteculas de S posee mucho maacutes energiacutea que el resto de ellas lo que es suficiente para que alcancen un estado activado en el que pueda formarse o romperse un enlace quiacutemico y se forme el producto(P)
La energiacutea de activacioacuten es la cantidad de energiacutea expresada en caloriacuteas necesaria para que todas las moleacuteculas de un mol a una temperatura dada alcancen el estado reactivo Mientras que el estado de transicioacuten es el estado rico en energiacutea de las moleacuteculas que
interaccionan en la cima de la barrera de activacioacuten La velocidad de una reaccioacuten quiacutemica es proporcional a la concentracioacuten del complejo en el estado de transicioacuten
Una reaccioacuten quiacutemica se puede acelerar de la siguiente forma 1) Al aumentar la temperatura se incrementa la energiacutea cineacutetica por lo que es mayor el nuacutemero de moleacuteculas que alcanzan el
estado de transicioacuten Generalmente el Q10 =2 lo que indica que la velocidad de una reaccioacuten quiacutemica se duplica al aumentar la temperatura en 10
0C 2) Antildeadiendo un catalizador que
disminuye la energiacutea de activacioacuten y aumenta la velocidad de reaccioacuten
La enzima (E) se combina con el substrato (S) formando el complejo de transicioacuten enzima-
substrato (E-S) mediante una reaccioacuten reversible cuya energiacutea de activacioacuten es menor que la de la reaccioacuten no catalizada Cuando se forma el producto de la reaccioacuten (P) se regenera de nuevo la enzima (E) de forma libre la que puede combinarse de nuevo con otra moleacutecula de substrato (S)
Una enzima reduce maacutes eficientemente la energiacutea de activacioacuten(Ea) de una reaccioacuten que un catalizador inorgaacutenico lo que permite que una reaccioacuten se realice a menor temperatura
El siguiente ejemplo ilustra mejor lo que hemos discutido
Reaccioacuten Energiacutea de activacioacuten (Kcalmol-1
)
21
a) el peroacutexido de hidroacutegeno se descompone en
H2 O2 H2 O + O2
18
b) el hierro cataliacutetico (Fe) realiza la reaccioacuten
H2 O2 H2 O + O2
13
c) el platino cataliacutetico (Pt) realiza la reaccioacuten
H2 O2 H2 O + O2
12
d) la catalasa una enzima hepaacutetica la realiza
H2O2 H2O + O2
5
Una enzima no modifica la energiacutea libre ni la constante de equilibrio sino que disminuye la energiacutea de activacioacuten de la reaccioacuten
Ir al principio
EFECTO DE LA CONCENTRACIOacuteN DEL SUBSTRATO SOBRE LA
CATALISIS ENZIMATICA
La actividad de una enzima se puede estudiar in vitro bajo condiciones controladas antildeadieacutendole una enzima a un substrato Siacute se trabaja con la condicioacuten de que la concentracioacuten del substrato sea saturante variando entonces la concentracioacuten de enzima se observa que aumenta el producto de la reaccioacuten a pH y temperatura constantes
22
Si se mantiene la concentracioacuten de la enzima constante y variando la concentracioacuten de substrato se obtiene una curva hiperboacutelica como la de la figura Al principio un aumento de la
concentracioacuten de substrato produce un aumento raacutepi do de la velocidad de reaccioacuten pero si se sigue aumentando la concentracioacuten de substrato la velocidad de reaccioacuten comienza a disminuir y a muy altas concentraciones de substrato se observa que no cambia la velocidad de
reaccioacuten se dice que los centros activos de la enzima se encuentran saturados La velocidad de reaccioacuten que se obtiene a esa alta concentracioacuten de substrato se define como la velocidad maacutexima (V) de la reaccioacuten enzimaacutetica bajo las condiciones especificadas La concentracioacuten de
substrato (S) a la semivelocidad maacutexima de reaccioacuten (V2) se puede determinar de la figura y representa la constante de Michaelis o Km la cual es una caracteriacutestica para cada enzima La inversa de Km o 1Km mide aproximadamente la afinidad de la enzima por el subs trato
Mientras maacutes pequentildeo sea el valor de Km mayor seraacute la afinidad de la enzima por el substrato Si varias enzimas compiten en el metabolismo por el mismo substrato eacuteste seraacute transformado preferentemente por la enzima con mayor afinidad
La ecuacioacuten de Michaelis describe la relacioacuten cuantitativa entre la velocidad de reaccioacuten y la concentracioacuten de substrato [S] si se conoce Vmax o Km
v=
En donde v= es la velocidad de reaccioacuten observada a una concentracioacuten de substrato determinada [S]
Km= constante de Michaelis en molesliacutet ro
23
Vmax= velocidad maacutexima a concentracioacuten saturante de substrato
Si v= Vmax2 entonces
=
Km+[S] = 2[S]
Km =[S]
De tal forma que podemos concluir que Km es igual a la concentracioacuten de substrato a la semivelocidad maacutexima de reaccioacuten
1 Concentraciones relativas de E y S la concentracioacuten de substrato S es
mucho mayor que la de E de tal manera que la cantidad de substrato
unido a la enzima en cualquier momento es muy pequentildea
2 Se asume el estado estacionario ES no cambia con el tiempo esto
quiere decir que la velocidad de formacioacuten de ES es igual a aquella para
su desintegracioacuten En general un intermediario en una serie de reacciones estaacute en estado estacionario cuando su velocidad de siacutentesis
es igual a su velocidad de degradacioacuten 3 Velocidad inicial para el anaacutelisis de reacciones enzimaacuteticas soacutelo se
utiliza la velocidad inicial de la reaccioacuten que es la velocidad ejercida por
la enzima inmediatamente despueacutes de que se ha puesto en contacto con el substrato y hasta antes de que se haya consumido el 10 de la concentracioacuten inicial del mismo La razoacuten de lo anterior es que en ese
momento la concentracioacuten del producto de la reaccioacuten que se ha acumulado en muy pequentildea y por tanto la reaccioacuten en el sentido
inverso es decir la transformacioacuten del producto en el substrato original puede ser ignorada
24
Caracteriacutesticas de Km la constante de Michaelis-Menten es
caracteriacutestica de una enzima y particular para un substrato Refleja la afinidad de la enzima por ese substrato K m es numeacutericamente igual a la concentracioacuten
de substrato a la cual la velocidad de reaccioacuten es la mitad de la Vmax (Km= Vmax2) Este paraacutemetro Km no variacutea con la concentracioacuten de enzima
Significado de una Km pequentildea un valor numeacuterico pequentildeo de Km
refleja una alta afinidad de la enzima por su substrato porque a una baja
concentracioacuten del mismo la enzima a desarrollado ya la mitad de la velocidad maacutexima
Significado de una Km grande el valor numeacuterico grande de Km refleja
una baja afinidad de la enzima por su substrato porque a una concentracioacuten
elevada del mismo la enzima desarrolla la mitad de la velocidad maacutexima
Relacioacuten de la velocidad con la concentracioacuten de enzima la
velocidad de la reaccioacuten es directamente proporcional a la concentracioacuten de la
enzima a cualquier concentracioacuten de substrato Por ejemplo si la concentracioacuten de enzima es disminuida a la mitad la velocidad inicial de la reaccioacuten (v0) es
reducida tambieacuten a la mitad de la original
Orden de la reaccioacuten cuando S es mas pequentildea que la Km la
velocidad de la reaccioacuten es aproximadamente igual a la concentracioacuten de
substrato
100
50
0
Substrato
Km Figura Localizacioacuten de la constante de Michaelis (Km)
25
La velocidad de reaccioacuten se dice en estas condiciones es de primer
orden con respecto al substrato Cuando S es mas grande que la Km la
velocidad es constante e igual a la Vmax La velocidad de la reaccioacuten es independiente de la concentracioacuten de substrato y se dice que es de orden cero
con respecto a la concentracioacuten de substrato
Las enzimas pueden ser aisladas de las ceacutelulas u organismos para estudiar sus
propiedades in vitro es decir en un tubo de ensayo Las diferentes enzimas muestran diferentes respuestas a los cambios en la concentracioacuten de substrato temperatura y pH
Concentracioacuten de substrato
La forma hiperboacutelica de la curva de velocidad vs Substrato
Efecto de la temperatura en la actividad enzimaacutetica
Efecto de la variacioacuten del pH en la actividad enzimaacutetica
Incremento de la velocidad con la temperatura En general la velocidad de la reaccioacuten se incrementa con la temperatura hasta que un punto maacuteximo es alcanzado
Este incremento se debe a que aumenta el nuacutemero de moleacuteculas ricas en energiacutea que pueden pasar la barrera energeacutetica de estado de transicioacuten para formar a los productos
Decremento de la velocidad con la temperatura la elevacioacuten excesiva de la
temperatura del medio que contiene a las enzimas resulta en un decremento de la velocidad como resultado de la desnaturalizacioacuten es decir la peacuterdida de la estructura tridimensional de las enzimas
26
zona de reactivacioacuten maacuteximo de actividad
Zona de
inactivacioacuten
0 Temperatura
Figura dependencia de la actividad enzimaacutetica con la temperatura
Velocidad maacutexima la velocidad de una reaccioacuten (v) es el nuacutemero de
moleacuteculas de substrato convertidas en producto por unidad de tiempo y
usualmente se expresa en micromoles de producto formadas por minuto La velocidad de una reaccioacuten catalizada por una enzima aumenta conforme se incrementa la concentracioacuten de substrato hasta que se llega a una velocidad maacutexima (Vmax) a partir de la cual la velocidad de la reaccioacuten es
independiente de la cantidad de substrato El que la velocidad no pueda
seguirse incrementando refleja la saturacioacuten del sitio activo con el substrato
Vmax
0 Substrato
Figura representacioacuten de una cineacutetica tiacutepicamente michaeliana
Vmax = velocidad de saturacioacuten o maacutexima
27
Cuando se grafica la velocidad de la reaccioacuten v0 contra la concentracioacuten de substrato S no siempre es posible determinar la condicioacuten
en que se ha llegado a la velocidad maacutexima Vmax debido al incremento de la pendiente en la hipeacuterbola a concentraciones de substrato elevadas
Figura experimento cineacutetico para la actividad enzimaacutetica
Se presentan los valores experimentales y los resultados del ajuste al modelo de Michaelis
Auacuten asiacute si se grafica una doble reciacuteproca es decir el inverso de la
velocidad (1v0) contra el inverso de la concentracioacuten de substrato (1 S ) se
obtiene una liacutenea recta El regraacutefico de Lineweaver-Burke tambieacuten se
denomina de ldquodobles reciacuteprocasrdquo y se utiliza para calcular la Km y la Vmax asiacute como para determinar el mecanismo de accioacuten de los diversos tipos de
inhibidores
La ecuacioacuten que describe a la graacutefca de Lineweaver-Burke es
Vmax
1
SVmax
Km
v
1
0 (3)
que describe una recta en donde el intercepto con el eje de las abscisas es igual a ndash1Km y el intercepto con el eje de las ordenadas es igual a 1Vmax
28
Figura experimento cineacutetico para la actividad enzimaacutetica Regraacutefico de
Lineweaver-Burke
Se presentan los valores experimentales y los resultados del ajuste al modelo
REPRESENTACIOacuteN GRAacuteFICA DE LINEWEAVER Y BURK
Si se toma el inverso en ambos miembros de la ecuacioacuten de Michaelis tenemos
=
+
29
que es la ecuacioacuten de una liacutenea recta cuya pendiente es que intercepta al eje de las
ordenadas en y a la abcisa en el punto Lo que es evidente al hacer
ya que se convierte en
Si se hace una graacutefica con los dobles inversos colocando los valores de en el eje de las
ordenadas y en el eje de las abcisas se obtiene una liacutenea recta a partir de la cual se puede calcular faacutecilmente Km
EFECTO DEL pH SOBRE LA ACTIVIDAD ENZIMATICA
Sabiendo que las enzimas son proteinas cualquier cambio brusco de pH puede alterar el caraacutecter ioacutenico de los grupos amino y carboxilo en la superficie proteica afectando asiacute las propiedades cataliacuteticas de una enzima A pH alto o bajo se puede producir la desnaturalizacioacuten
de la enzima y en consecuencia su inactivacioacuten La fosfatasa aacutecida es maacutes activa a pH 50 mientras que la fosfatasa alcalina lo es a pH 90 Muchas enzimas tienen maacutexima actividad cerca de la neutralidad en un rango de pH de 6 a 8
El pH per se no afecta la actividad enzimaacutetica sino la concentracioacuten de protones Los protones
ademaacutes de alterar la estructura de la enzima y el substrato pueden participar tambieacuten en la reaccioacuten como substrato o producto En esos casos la concentracioacuten de protones afecta directamente la velocidad de la reaccioacuten
ENZIMA pH OPTIMO
30
Pepsina 15
Tripsina 77
Catalasa 76
Arginasa 97
Fumarasa 78
Ribonucleasa 78
EFECTO DE ACTIVADORES
Un aumento en la temperatura provoca un aumento de la velocidad de reaccioacuten hasta cierta temperatura oacuteptima ya que despueacutes de aproximadamente 45
0 C se comienza a producir la
desnaturalizacioacuten teacutermica Las enzimas de muchos mamiacuteferos tienen una temperatura oacuteptima
de 370 C por encima de esa temperatura comienzan a inactivarse y se destruyen Sin embargo
existen especies de bacterias y algas que habitan en fuentes de aguas termales y en el otro extremo ciertas bacterias aacuterticas tienen temperaturas oacuteptimas cercanas a 0
0 C
31
ESPECIFICIDAD ABSOLUTA
Cierta enzimas se encuentran como aproenzimas o zimogenos que son inactivos
El tripsinoacutegeno es convertido en tripsina por la misma accioacuten de la tripsina que remueve un hexapeacuteptido
tripsina Tripsinoacutegeno Tripsina + Val (Asp)4 lisina
Otra forma de activacioacuten consiste en mantener los grupos SH reducidos Estos grupos pueden
formar parte de centros activos de ciertas enzimas Si se oxidan estos grupos a -S-S- la enzima se inactiva Asiacute tenemos que la enzima papaina se inactiva despueacutes de exponerla al ox iacutegeno pero cuando se le antildeade un reductor apropiado la enzima se reactiva Los grupos ndashS-S- se convierten en ndashSH activaacutendose la enzima
Otra forma de activacioacuten requiere la presencia de otra substancia que se enlaza al sitio alosteacuterico el compuesto que se enlaza se denomina un efector alosteacuterico El centro alosteacuterico es bien especiacutefico para el efector o modulador alosteacuterico
ESPECIFICIDAD RELATIVA
Oxiacutegeno L-aminoaacutecido a -cetoaacutecido + NH3 + H 2 O 2 L-aminoaacutecido oxidasa Oxiacutegeno D-aminoaacutecido a -cetoaacutecido + NH3 + H 2 O 2 D-aminoaacutecido oxidasa
32
En este ejemplo se muestra la estereo-especificidad Una enzima puede tener una especificidad oacuteptica para isomeros D o L
Hay enzimas que muestran especificidad absoluta como la aspartasa que cataliza la adicioacuten reversible de amoniacuteaco al doble enlace del aacutecido fumaacuterico pero no al de otros aacutecidos insaturados
HOOC ndash CH = CH ndash COO- + NH4
+ HOOC ndash CH2 - CHNH2 - COO
- + H
+
INHIBICIOacuteN ENZIMATICA
Mediante el uso de inhibidores enzimaacuteticos se ha obtenido informacioacuten muy valiosa sobre la conformacioacuten del centro activo de algunas enzimas
a Inhibicioacuten irreversible Este tipo de inhibidores forman un enlace covalente con las enzimas cerca del centro activo Un ejemplo son los gases nerviosos como el
fluorofosfato de diisopropilo (DFP) que forma un complejo con la enzima acetilcolinesterasa Los animales envenenados con este gas quedan paralizados debido a la imposibilidad de transmitir adecuadamente los impulsos nerviosos
Ester di-isopropil fos foacuterico de la enzima (inactivo)
b) Inhibicioacuten competitiva
Es cuando el inhibidor compite con el substrato por la unioacuten con el centro activo de la
enzima Este tipo de inhibicioacuten puede reducirse si se aumenta la concentracioacuten de substrato
-OOC - CH2 - CH2 - COO
- -OOC - CH = CH - COO
- + 2H
+ Succinato Succinato Fumarato
33
Deshidrogenasa COO
- I CH2 Malonato (inhibidor competitivo) I COO
- En la inhibicioacuten competitiva la enzima se combina con el inhibidor para formar un complejo enzima-inhibidor E + I E I en competencia con la reaccioacuten normal E +S ES
La sulfanilamida (bactericida) interfiere con la siacutentesis del aacutecido foacutelico compitiendo con el aacutecido p-aminobenzoico de una forma competitiva
HOOC-sect- NH2 NH2 ndashSO2 - sect- NH2 aacutecido p- amino benzoico sulfanilamida
sect = fenil
a Inhibicioacuten no competitiva
Esta inhibicioacuten se caracteriza por que no se puede revertir el efecto del inhibidor aumentando la concentracioacuten del substrato
Por ejemplo la inhibicioacuten de la deshidrogenasa del gliceraldehido -3-fosfato por la yodo-acetamida
Enzima-SH + I-CH2 CONH2 Enzima-SCH2 CONH2 + IH
La deshidrogenasa del gliceraldehido-3 fosfato puede ser inhibida tambieacuten por el aacutecido iodo-aceacutetico
Enzima-SH + ICH2 COOH Enzima-SCH2 COOH + IH
c) Inhibicioacuten por metales
Ciertos metales como el plomo mercurio y arseacutenico inhiben enzimas que tienen en su centro activo grupos -SH libres

7
6- LIGASAS
Catalizan la formacioacuten de enlaces entre C y O S N Esta reaccioacuten soacutelo es posible
mediante la energiacutea derivada de fosfatos ricos en energiacutea como el del fosfato de la
moleacutecula de ATP
15 COFACTORES y COENZIMAS Algunas enzimas se asocian con moleacuteculas de caraacutecter no proteiacuteco que son
necesarias para el funcionamiento de la enzima estas moleacuteculas se denominan
cofactores (Figura 3) Comuacutenmente los factores encontrados en las enzimas incluyen
iones metaacutelicos como el Zn2+ o el Fe2+ ( Tabla 2) tambieacuten pueden ser moleacuteculas
orgaacutenicas que se denomina coenzimas como el NAD+ FAD la coenzima A
Generalmente las coenzimas son derivados de las vitaminas A la enzima en ausencia
de su cofactor (cuando lo tiene) se le denomina apoenzima en presencia de su
cofactor (cuando lo tiene) se le denomina holoenzima La apoenzima generalmente
carece de actividad bioloacutegica La diferencia entre un cofactor y un grupo prosteacutetico
como el grupo hemo es que este uacuteltimo estaacute unido de manera covalente a la
enzima mientras que el cofactor puede ser removido de la misma con relativa
facilidad
Se pueden sentildealar algunas caracteriacutesticas de las coenzimas como generalmente son
estables al calor y a otros agentes desnaturalizantes de las proteiacutenas pueden ser
recicladas y algunas provienen de las vitaminas
8
Figura 3 Cofactores y Coenzima
Tabla 2 Algunos enzimas que contienen iones metaacutelicos o los necesitan como
cofactores
Zn+2 Cu+2 (Cu+)
Alcohol- deshidrogenada Tirosinasa Anhidrasa-carboacutenica Citocromo-oxidasa Carboxipeptidasa
Mg+2 K+
Fosfohidrolasa Piruvato- fosfoquinasa Fosfotransferasas
Mn+2 Arginasa Na+ Fosfotransferasas ATPasa de la membrana plasmaacutetica
Fe +2 o Fe+3
9
Citocromos
Peroxidasa Catalasa Ferredoxina
Texto ------- (Tabla 3)
Tabla 3 Relacioacuten entre algunas coenzimas y las vitaminas a partir de las
cuales se producen Enfermedades generadas por la deficiencia
Coenzima Reaccioacuten Fuente vitamiacutenica Enfermedad
Humana por deficiencia
Biocitina Carboxilacioacuten Biotina
Coenzima A Transferencia de acilos Pantotenato
Coenzimas de Cobalamima Alquilacioacuten Cobalamina (B12) anemia perniciosa
Coenzimas de flavina Oxidacioacuten-reduccioacuten Riboflavina (B2)
Acido lipoacuteico Transferencia de acilos -
Coenzimas de nicotinamida Oxidacioacuten-reduccioacuten Nicotinamida (niacina) Pelagra
Fosfato de piridoxal Transferencia de grupo amino Piridoxina (B6)
Tetrahidrofolato Transferencia de un grupo de 1 Carbono Acido foacutelico anemia
megaloblaacutestica
Tiamina pirofosfato Transferencia de aldehiacutedo Tiamina (B1) beriberi
no tiene un nombre especiacutefico su aparicioacuten es muy rara
Otras coenzimas
NADP
Coenzima Q
Biotina
NAD y NADP
10
Coenzima Q (CoQ)
Esta coenzima es sintetizada a partir de farnesil pirofosfato Al igual que el
FMN la CoQ es capaz de aceptar y donar uno o dos electrones porque su forma
semiquinona es estable A su forma oxidada se le conoce como ubiquinona (de
ubicuo lat ubique en todas partes) o quinona A su forma reducida se le conoce
como QH2 ubiquinol o hidroquinona (Figura 2) A la coenzima solo parcialmente
reducida se le conoce como ubisemiquinona semiquinona o QH (Figura 4)
Figura 4 Representacioacuten de la moleacutecula de CoQ
La siacutentesis de la CoQ se lleva a cabo a traveacutes de unidades isoprenoides (Figura 5)
11
Figura 5 La CoQ se produce a partir de isoprenoides
Biotina
La biotina consiste de un anillo imidazol que estaacute unido en forma cis a la
cadena lateral tetrahidrotiofeno del valerato (Figura 6)
12
Figura 6 Estructura de la biotina
La enzima holocarboxilasa sintetasa (HCS) cataliza la activacioacuten mediante
biotinilacioacuten de cinco carboxilasas en ceacutelulas humanas La deficiencia de HCS
produce el siacutendrome deficiencia muacuteltiple de carboxilasas (DMC) Esta enfermedad es
potencialmente mortal y se caracteriza por la deficiencia de la actividad de todas las
carboxilasas La deficiencia de biotina no solo disminuye la actividad de estas enzimas
sino que parece modificar la expresioacuten geneacutetica
126 PROPIEDADES DE LAS ENZIMAS
Las propiedades de las enzimas derivan del hecho de ser proteiacutenas y de actuar como
catalizadores Como proteiacutenas poseen una conformacioacuten natural maacutes estable que las
demaacutes conformaciones posibles Asiacute cambios en la conformacioacuten suelen ir asociados
en cambios en la actividad cataliacutetica Los factores que influyen de manera maacutes directa
sobre la actividad de un enzima son
pH
temperatura
cofactores
A) EFECTO DEL pH
Al comprobar experimentalmente la influencia del pH en la velocidad de las
reacciones enzimaacuteticas se obtienen curvas que indican que los enzimas
presentan un pH oacuteptimo de actividad El pH puede afectar de varias maneras
13
o El centro activo puede contener aminoaacutecidos con grupos ionizados
que pueden variar con el pH
o La ionizacioacuten de aminoaacutecidos que no estaacuten en el centro activo puede
provocar modificaciones en la conformacioacuten de la enzima
o El sustrato puede verse afectado por las variaciones del pH
Algunos enzimas presentan variaciones peculiares La pepsina del estoacutemago
presenta un oacuteptimo a pH=2 la ureasa lo tiene a pH 7 y la fosfatasa alcalina del
intestino un pH= 12
B) EFECTO DE LA TEMPERATURA
Influye en la actividad El punto oacuteptimo representa el maacuteximo de actividad A
temperaturas bajas los enzimas se hallan ldquomuy riacutegidosrdquo y cuando se supera un
valor considerable (mayor de 50) la actividad cae bruscamente porque como
proteiacutena el enzima se desnaturaliza
128 EFECTO DE LOS COFACTORES SOBRE LA ACTIVIDAD
ENZIMATICA
A veces un enzima requiere para su funcioacuten la presencia de sustancias no proteicas
que colaboran en la cataacutelisis los cofactores Casi un tercio de los enzimas conocidos
requieren cofactores Cuando el cofactor es una moleacutecula orgaacutenica se llama
coenzima En la figura inferior podemos observar una moleacutecula de hemoglobina
(proteiacutena que transporta oxiacutegeno) y su coenzima (el grupo hemo) Cuando los
cofactores y las coenzimas se encuentran unidos covalentemente al enzima se llaman
14
grupos prosteacuteticos La forma cataliacuteticamente activa del enzima es decir el enzima
unido a su grupo prosteacutetico se llama holoenzima La parte proteica de un
holoenzima (inactiva) se llama apoenzima de forma que
apoenzima + grupo prosteacutetico= holoenzima
129 MODO DE ACCIOacuteN DE LAS ENZIMAS
En las reacciones espontaacuteneas los productos finales tienen menos energiacutea libre de
Gibbs (∆G) que los reactantes (figura inferior izquierda) Por lo tanto las reacciones
espontaacuteneas liberan energiacutea de Gibbs ((∆G lt 0) Sin embargo el comienzo de la
reaccioacuten requiere un aporte inicial de energiacutea Esta energiacutea inicial que hay que
suministrar a los reactantes para que la reaccioacuten transcurra se llama energiacutea de
activacioacuten (Ea) Cuanto menor es la Ea maacutes faacutecilmente transcurre la reaccioacuten
Perfil energeacutetico de una reaccioacuten espontaacutenea Perfil energeacutetico de una reaccioacuten catalizada
15
La accioacuten de los catalizadores consiste precisamente en disminuir la Ea (Figura superior derecha) Los enzimas
son catalizadores especialmente eficaces ya que disminuyen la Ea auacuten maacutes que los catalizadores inorgaacutenicos
Por ejemplo la descomposicioacuten del agua oxigenada (H2O2) para dar H2O y O2 puede ocurrir sin catalizador
con un catalizador inorgaacutenico (platino) o con un enzima especiacutefico (catalasa) Las respectivas Ea para cada
proceso son 18 12 y 6 Kcalmol Asiacute se puede calcular que el platino acelera la reaccioacuten 20000 veces mientras
que la catalasa la acelera 370000 veces
Para que una reaccioacuten quiacutemica tenga lugar las moleacuteculas de los reactantes deben chocar con una energiacutea y
una orientacioacuten adecuadas La actuacioacuten del enzima (1) permite que los rea ctantes (sustratos) se unan a su
centro activo con una orientacioacuten oacuteptima para que la reaccioacuten se produzca y (2) modifica las propiedades
quiacutemicas del sustrato unido a su centro activo debilitando los enlaces existentes y facilitando la formacioacuten de
otros nuevos (Figuras inferiores)
1210 REGULACIOacuteN DE SU ACTIVIDAD ENZIMATICA
16
Una moleacutecula de enzima no tiene por que actuar siempre a la misma velocidad Su
actividad puede estar modulada por
Cambios en el pH
Cambios en la temperatura
Presencia de cofactores
Las concentraciones del sustrato y de los productos finales
Presencia de inhibidores
Modulacioacuten alosteacuterica
Modificacioacuten covalente
Activacioacuten por proteolisis
Isoenzimas
EFECTO DE LAS CONCENTRACIONES SOBRE LA ACTIVIDAD ENZIMAacuteTICA
La velocidad de una reaccioacuten enzimaacutetica depende de la concentracioacuten de sustrato La Figura de la derecha muestra la velocidad de una reaccioacuten enzimaacutetica a 6 concentraciones distintas de sustrato
Ademaacutes la presencia de los productos finales puede hacer que la reaccioacuten sea maacutes lenta o incluso invertir su sentido (Figura inferior)
EFECTO DE LOS INHIBIDORES SOBRE LA ACTIVIDAD ENZIMAacuteTICA
Ciertas moleacuteculas pueden inhibir la accioacuten cataliacutetica de un enzima son los inhibidores Estos inhibidores bien pueden ocupar temporalmente el centro activo por semejanza estructural con el sustrato original ( inhibidor competitivo) o bien alteran la conformacioacuten espacial del enzima impidiendo su unioacuten al sustrato ( inhibidor no competitivo) (Figuras inferiores)
Inhibidor competitivo Inhibidor no competitivo
17
MODULACIOacuteN ALOSTEacuteRICA DE LA ACTIVIDAD ENZIMAacuteTICA
Hay enzimas que pueden adoptar 2 conformaciones interconvertibles llamadas R (relajada) y T (tensa) R es la forma maacutes activa porque se une al sustrato con maacutes afinidad Las formas R y T se encuentran en equilibrio R lt==gt T (Figura inferior)
Ciertas sustancias tienden a estabilizar la forma R Son los llamados moduladores positivos El propio sustrato es a
menudo un modulador positivo Las moleacuteculas que favorecen la forma R pero que actuacutean sobre una regioacuten del enzima distinta del centro activo son los activadores alosteacutericos (Figura inferior izquierda)
Activador alosteacuterico favorece la unioacuten del sustrato
Inhibidor alosteacuterico impide la unioacuten del sustrato
Las sustancias que favorecen la forma T y disminuyen la actividad enzimaacutetica son los moduladores negativos Si
estos moduladores actuacutean en lugares distintos del centro activo del enzima se llaman inhibidores alosteacutericos
18
(Figura superior derecha
EFECTO DE LA MODIFICACIOacuteN COVALENTE SOBRE LA ACTIVIDAD ENZIMAacuteTICA
Otros enzimas pasan de una forma menos activa a otra maacutes activa unieacutendose covalentemente a un grupo quiacutemico de pequentildeo tamantildeo como el Pi o el AMP Tambieacuten se da el caso inverso en el que un enzima muy activo se desactiva al liberar alguacuten grupo quiacutemico En las enzimas de las viacuteas degradativas del metabolismo la
forma fosforilada es maacutes activa que la no fosforilada mientras que en las viacuteas biosinteacuteticas ocurre lo contrario En las figuras inferiores se ilustra la activacioacuten de una proteiacutena por fosforilacioacuten
Elementos de la reaccioacuten El enzima no fosforilado es inactivo El enzima fosforilado es activo
ACTIVACIOacuteN PROTEOLIacuteTICA DE LA ACTIVIDAD ENZIMAacuteTICA
Algunos enzimas no se sintetizan como tales sino como proteiacutenas precursoras sin actividad enzimaacutetica Estas proteiacutenas se llaman
proenzimas o zimoacutegenos Para activarse los zimoacutegenos sufren un ataque hidroliacutetico que origina la liberacioacuten de uno o varios peacuteptidos El resto de la moleacutecula proteica adopta la conformacioacuten y las propiedades del enzima activo Muchos enzimas digestivos se secretan en forma de zimoacutegenos y en el tubo digestivo
se convierten en la forma activa Es el caso de la -quimotripsina que se sintetiza en forma de quimotripsinoacutegeno (Figura superior) Si estos enzimas se sintetizasen directamente en forma activa destruiriacutean la propia ceacutelula que las
produce Asiacute la tripsina pancreaacutetica (una proteasa) se sintetiza como tripsinoacutegeno (inactivo) Si por alguna razoacuten se activa en el propio paacutencreas la glaacutendula sufre un proceso de autodestruccioacuten (pancreatitis aguda) a
menudo mortal
19
REGULACIOacuteN DE LA ACTIVIDAD ENZIMAacuteTICA POR MEDIO DE ISOENZIMAS
Algunos enzimas tienen distinta estructura molecular aunque su funcioacuten bioloacutegica es similar Se llaman isozimas o isoenzimas Estas diferencias de estructura se traducen en ligeros cambios en sus propiedades de forma que cada isozima se adapta perfectamente a la funcioacuten que debe realizar Asiacute podemos observar la existencia de isoenzimas en funcioacuten de
el tipo de tejido Por ejemplo la lactato deshidrogenasa presenta isozimas distintos en muacutesculo y corazoacuten el compartimento celular donde actuacutea Por ejemplo la malato deshidrogenasa del citoplasma es distinta
de la de la mitocondria el momento concreto del desarrollo del individuo Por ejemplo algunos enzimas de la glicolisis del feto
son diferentes de los mismos enzimas en el adulto
CINEacuteTICA ENZIMAacuteTICA
Los principios generales de las reacciones quiacutemicas se aplican tambieacuten a las reacciones enzimaacuteticas Por este motivo antes de empezar con la cineacutetica quiacutemica se van a repasar algunos conceptos baacutesicos de cineacutetica quiacutemica
A continuacioacuten se describiraacuten los siguientes conceptos
Cineacutetica enzimaacutetica Modelo cineacutetico de Michaelis-Menten Caacutelculo de la KM y la Vmax de un enzima Actividad enzimaacutetica
20
NOMENCLATURA DE LOS ENZIMAS
CATALISIS MOLECULAR
Un catalizador modifica la velocidad de una reaccioacuten quiacutemica sin ser utilizado o aparecer como uno de los productos de la reaccioacuten Una reaccioacuten quiacutemica en la que un substrato(S) se
transforma en un producto(P) S P ocurre por que cierta fraccioacuten de moleacuteculas de S posee mucho maacutes energiacutea que el resto de ellas lo que es suficiente para que alcancen un estado activado en el que pueda formarse o romperse un enlace quiacutemico y se forme el producto(P)
La energiacutea de activacioacuten es la cantidad de energiacutea expresada en caloriacuteas necesaria para que todas las moleacuteculas de un mol a una temperatura dada alcancen el estado reactivo Mientras que el estado de transicioacuten es el estado rico en energiacutea de las moleacuteculas que
interaccionan en la cima de la barrera de activacioacuten La velocidad de una reaccioacuten quiacutemica es proporcional a la concentracioacuten del complejo en el estado de transicioacuten
Una reaccioacuten quiacutemica se puede acelerar de la siguiente forma 1) Al aumentar la temperatura se incrementa la energiacutea cineacutetica por lo que es mayor el nuacutemero de moleacuteculas que alcanzan el
estado de transicioacuten Generalmente el Q10 =2 lo que indica que la velocidad de una reaccioacuten quiacutemica se duplica al aumentar la temperatura en 10
0C 2) Antildeadiendo un catalizador que
disminuye la energiacutea de activacioacuten y aumenta la velocidad de reaccioacuten
La enzima (E) se combina con el substrato (S) formando el complejo de transicioacuten enzima-
substrato (E-S) mediante una reaccioacuten reversible cuya energiacutea de activacioacuten es menor que la de la reaccioacuten no catalizada Cuando se forma el producto de la reaccioacuten (P) se regenera de nuevo la enzima (E) de forma libre la que puede combinarse de nuevo con otra moleacutecula de substrato (S)
Una enzima reduce maacutes eficientemente la energiacutea de activacioacuten(Ea) de una reaccioacuten que un catalizador inorgaacutenico lo que permite que una reaccioacuten se realice a menor temperatura
El siguiente ejemplo ilustra mejor lo que hemos discutido
Reaccioacuten Energiacutea de activacioacuten (Kcalmol-1
)
21
a) el peroacutexido de hidroacutegeno se descompone en
H2 O2 H2 O + O2
18
b) el hierro cataliacutetico (Fe) realiza la reaccioacuten
H2 O2 H2 O + O2
13
c) el platino cataliacutetico (Pt) realiza la reaccioacuten
H2 O2 H2 O + O2
12
d) la catalasa una enzima hepaacutetica la realiza
H2O2 H2O + O2
5
Una enzima no modifica la energiacutea libre ni la constante de equilibrio sino que disminuye la energiacutea de activacioacuten de la reaccioacuten
Ir al principio
EFECTO DE LA CONCENTRACIOacuteN DEL SUBSTRATO SOBRE LA
CATALISIS ENZIMATICA
La actividad de una enzima se puede estudiar in vitro bajo condiciones controladas antildeadieacutendole una enzima a un substrato Siacute se trabaja con la condicioacuten de que la concentracioacuten del substrato sea saturante variando entonces la concentracioacuten de enzima se observa que aumenta el producto de la reaccioacuten a pH y temperatura constantes
22
Si se mantiene la concentracioacuten de la enzima constante y variando la concentracioacuten de substrato se obtiene una curva hiperboacutelica como la de la figura Al principio un aumento de la
concentracioacuten de substrato produce un aumento raacutepi do de la velocidad de reaccioacuten pero si se sigue aumentando la concentracioacuten de substrato la velocidad de reaccioacuten comienza a disminuir y a muy altas concentraciones de substrato se observa que no cambia la velocidad de
reaccioacuten se dice que los centros activos de la enzima se encuentran saturados La velocidad de reaccioacuten que se obtiene a esa alta concentracioacuten de substrato se define como la velocidad maacutexima (V) de la reaccioacuten enzimaacutetica bajo las condiciones especificadas La concentracioacuten de
substrato (S) a la semivelocidad maacutexima de reaccioacuten (V2) se puede determinar de la figura y representa la constante de Michaelis o Km la cual es una caracteriacutestica para cada enzima La inversa de Km o 1Km mide aproximadamente la afinidad de la enzima por el subs trato
Mientras maacutes pequentildeo sea el valor de Km mayor seraacute la afinidad de la enzima por el substrato Si varias enzimas compiten en el metabolismo por el mismo substrato eacuteste seraacute transformado preferentemente por la enzima con mayor afinidad
La ecuacioacuten de Michaelis describe la relacioacuten cuantitativa entre la velocidad de reaccioacuten y la concentracioacuten de substrato [S] si se conoce Vmax o Km
v=
En donde v= es la velocidad de reaccioacuten observada a una concentracioacuten de substrato determinada [S]
Km= constante de Michaelis en molesliacutet ro
23
Vmax= velocidad maacutexima a concentracioacuten saturante de substrato
Si v= Vmax2 entonces
=
Km+[S] = 2[S]
Km =[S]
De tal forma que podemos concluir que Km es igual a la concentracioacuten de substrato a la semivelocidad maacutexima de reaccioacuten
1 Concentraciones relativas de E y S la concentracioacuten de substrato S es
mucho mayor que la de E de tal manera que la cantidad de substrato
unido a la enzima en cualquier momento es muy pequentildea
2 Se asume el estado estacionario ES no cambia con el tiempo esto
quiere decir que la velocidad de formacioacuten de ES es igual a aquella para
su desintegracioacuten En general un intermediario en una serie de reacciones estaacute en estado estacionario cuando su velocidad de siacutentesis
es igual a su velocidad de degradacioacuten 3 Velocidad inicial para el anaacutelisis de reacciones enzimaacuteticas soacutelo se
utiliza la velocidad inicial de la reaccioacuten que es la velocidad ejercida por
la enzima inmediatamente despueacutes de que se ha puesto en contacto con el substrato y hasta antes de que se haya consumido el 10 de la concentracioacuten inicial del mismo La razoacuten de lo anterior es que en ese
momento la concentracioacuten del producto de la reaccioacuten que se ha acumulado en muy pequentildea y por tanto la reaccioacuten en el sentido
inverso es decir la transformacioacuten del producto en el substrato original puede ser ignorada
24
Caracteriacutesticas de Km la constante de Michaelis-Menten es
caracteriacutestica de una enzima y particular para un substrato Refleja la afinidad de la enzima por ese substrato K m es numeacutericamente igual a la concentracioacuten
de substrato a la cual la velocidad de reaccioacuten es la mitad de la Vmax (Km= Vmax2) Este paraacutemetro Km no variacutea con la concentracioacuten de enzima
Significado de una Km pequentildea un valor numeacuterico pequentildeo de Km
refleja una alta afinidad de la enzima por su substrato porque a una baja
concentracioacuten del mismo la enzima a desarrollado ya la mitad de la velocidad maacutexima
Significado de una Km grande el valor numeacuterico grande de Km refleja
una baja afinidad de la enzima por su substrato porque a una concentracioacuten
elevada del mismo la enzima desarrolla la mitad de la velocidad maacutexima
Relacioacuten de la velocidad con la concentracioacuten de enzima la
velocidad de la reaccioacuten es directamente proporcional a la concentracioacuten de la
enzima a cualquier concentracioacuten de substrato Por ejemplo si la concentracioacuten de enzima es disminuida a la mitad la velocidad inicial de la reaccioacuten (v0) es
reducida tambieacuten a la mitad de la original
Orden de la reaccioacuten cuando S es mas pequentildea que la Km la
velocidad de la reaccioacuten es aproximadamente igual a la concentracioacuten de
substrato
100
50
0
Substrato
Km Figura Localizacioacuten de la constante de Michaelis (Km)
25
La velocidad de reaccioacuten se dice en estas condiciones es de primer
orden con respecto al substrato Cuando S es mas grande que la Km la
velocidad es constante e igual a la Vmax La velocidad de la reaccioacuten es independiente de la concentracioacuten de substrato y se dice que es de orden cero
con respecto a la concentracioacuten de substrato
Las enzimas pueden ser aisladas de las ceacutelulas u organismos para estudiar sus
propiedades in vitro es decir en un tubo de ensayo Las diferentes enzimas muestran diferentes respuestas a los cambios en la concentracioacuten de substrato temperatura y pH
Concentracioacuten de substrato
La forma hiperboacutelica de la curva de velocidad vs Substrato
Efecto de la temperatura en la actividad enzimaacutetica
Efecto de la variacioacuten del pH en la actividad enzimaacutetica
Incremento de la velocidad con la temperatura En general la velocidad de la reaccioacuten se incrementa con la temperatura hasta que un punto maacuteximo es alcanzado
Este incremento se debe a que aumenta el nuacutemero de moleacuteculas ricas en energiacutea que pueden pasar la barrera energeacutetica de estado de transicioacuten para formar a los productos
Decremento de la velocidad con la temperatura la elevacioacuten excesiva de la
temperatura del medio que contiene a las enzimas resulta en un decremento de la velocidad como resultado de la desnaturalizacioacuten es decir la peacuterdida de la estructura tridimensional de las enzimas
26
zona de reactivacioacuten maacuteximo de actividad
Zona de
inactivacioacuten
0 Temperatura
Figura dependencia de la actividad enzimaacutetica con la temperatura
Velocidad maacutexima la velocidad de una reaccioacuten (v) es el nuacutemero de
moleacuteculas de substrato convertidas en producto por unidad de tiempo y
usualmente se expresa en micromoles de producto formadas por minuto La velocidad de una reaccioacuten catalizada por una enzima aumenta conforme se incrementa la concentracioacuten de substrato hasta que se llega a una velocidad maacutexima (Vmax) a partir de la cual la velocidad de la reaccioacuten es
independiente de la cantidad de substrato El que la velocidad no pueda
seguirse incrementando refleja la saturacioacuten del sitio activo con el substrato
Vmax
0 Substrato
Figura representacioacuten de una cineacutetica tiacutepicamente michaeliana
Vmax = velocidad de saturacioacuten o maacutexima
27
Cuando se grafica la velocidad de la reaccioacuten v0 contra la concentracioacuten de substrato S no siempre es posible determinar la condicioacuten
en que se ha llegado a la velocidad maacutexima Vmax debido al incremento de la pendiente en la hipeacuterbola a concentraciones de substrato elevadas
Figura experimento cineacutetico para la actividad enzimaacutetica
Se presentan los valores experimentales y los resultados del ajuste al modelo de Michaelis
Auacuten asiacute si se grafica una doble reciacuteproca es decir el inverso de la
velocidad (1v0) contra el inverso de la concentracioacuten de substrato (1 S ) se
obtiene una liacutenea recta El regraacutefico de Lineweaver-Burke tambieacuten se
denomina de ldquodobles reciacuteprocasrdquo y se utiliza para calcular la Km y la Vmax asiacute como para determinar el mecanismo de accioacuten de los diversos tipos de
inhibidores
La ecuacioacuten que describe a la graacutefca de Lineweaver-Burke es
Vmax
1
SVmax
Km
v
1
0 (3)
que describe una recta en donde el intercepto con el eje de las abscisas es igual a ndash1Km y el intercepto con el eje de las ordenadas es igual a 1Vmax
28
Figura experimento cineacutetico para la actividad enzimaacutetica Regraacutefico de
Lineweaver-Burke
Se presentan los valores experimentales y los resultados del ajuste al modelo
REPRESENTACIOacuteN GRAacuteFICA DE LINEWEAVER Y BURK
Si se toma el inverso en ambos miembros de la ecuacioacuten de Michaelis tenemos
=
+
29
que es la ecuacioacuten de una liacutenea recta cuya pendiente es que intercepta al eje de las
ordenadas en y a la abcisa en el punto Lo que es evidente al hacer
ya que se convierte en
Si se hace una graacutefica con los dobles inversos colocando los valores de en el eje de las
ordenadas y en el eje de las abcisas se obtiene una liacutenea recta a partir de la cual se puede calcular faacutecilmente Km
EFECTO DEL pH SOBRE LA ACTIVIDAD ENZIMATICA
Sabiendo que las enzimas son proteinas cualquier cambio brusco de pH puede alterar el caraacutecter ioacutenico de los grupos amino y carboxilo en la superficie proteica afectando asiacute las propiedades cataliacuteticas de una enzima A pH alto o bajo se puede producir la desnaturalizacioacuten
de la enzima y en consecuencia su inactivacioacuten La fosfatasa aacutecida es maacutes activa a pH 50 mientras que la fosfatasa alcalina lo es a pH 90 Muchas enzimas tienen maacutexima actividad cerca de la neutralidad en un rango de pH de 6 a 8
El pH per se no afecta la actividad enzimaacutetica sino la concentracioacuten de protones Los protones
ademaacutes de alterar la estructura de la enzima y el substrato pueden participar tambieacuten en la reaccioacuten como substrato o producto En esos casos la concentracioacuten de protones afecta directamente la velocidad de la reaccioacuten
ENZIMA pH OPTIMO
30
Pepsina 15
Tripsina 77
Catalasa 76
Arginasa 97
Fumarasa 78
Ribonucleasa 78
EFECTO DE ACTIVADORES
Un aumento en la temperatura provoca un aumento de la velocidad de reaccioacuten hasta cierta temperatura oacuteptima ya que despueacutes de aproximadamente 45
0 C se comienza a producir la
desnaturalizacioacuten teacutermica Las enzimas de muchos mamiacuteferos tienen una temperatura oacuteptima
de 370 C por encima de esa temperatura comienzan a inactivarse y se destruyen Sin embargo
existen especies de bacterias y algas que habitan en fuentes de aguas termales y en el otro extremo ciertas bacterias aacuterticas tienen temperaturas oacuteptimas cercanas a 0
0 C
31
ESPECIFICIDAD ABSOLUTA
Cierta enzimas se encuentran como aproenzimas o zimogenos que son inactivos
El tripsinoacutegeno es convertido en tripsina por la misma accioacuten de la tripsina que remueve un hexapeacuteptido
tripsina Tripsinoacutegeno Tripsina + Val (Asp)4 lisina
Otra forma de activacioacuten consiste en mantener los grupos SH reducidos Estos grupos pueden
formar parte de centros activos de ciertas enzimas Si se oxidan estos grupos a -S-S- la enzima se inactiva Asiacute tenemos que la enzima papaina se inactiva despueacutes de exponerla al ox iacutegeno pero cuando se le antildeade un reductor apropiado la enzima se reactiva Los grupos ndashS-S- se convierten en ndashSH activaacutendose la enzima
Otra forma de activacioacuten requiere la presencia de otra substancia que se enlaza al sitio alosteacuterico el compuesto que se enlaza se denomina un efector alosteacuterico El centro alosteacuterico es bien especiacutefico para el efector o modulador alosteacuterico
ESPECIFICIDAD RELATIVA
Oxiacutegeno L-aminoaacutecido a -cetoaacutecido + NH3 + H 2 O 2 L-aminoaacutecido oxidasa Oxiacutegeno D-aminoaacutecido a -cetoaacutecido + NH3 + H 2 O 2 D-aminoaacutecido oxidasa
32
En este ejemplo se muestra la estereo-especificidad Una enzima puede tener una especificidad oacuteptica para isomeros D o L
Hay enzimas que muestran especificidad absoluta como la aspartasa que cataliza la adicioacuten reversible de amoniacuteaco al doble enlace del aacutecido fumaacuterico pero no al de otros aacutecidos insaturados
HOOC ndash CH = CH ndash COO- + NH4
+ HOOC ndash CH2 - CHNH2 - COO
- + H
+
INHIBICIOacuteN ENZIMATICA
Mediante el uso de inhibidores enzimaacuteticos se ha obtenido informacioacuten muy valiosa sobre la conformacioacuten del centro activo de algunas enzimas
a Inhibicioacuten irreversible Este tipo de inhibidores forman un enlace covalente con las enzimas cerca del centro activo Un ejemplo son los gases nerviosos como el
fluorofosfato de diisopropilo (DFP) que forma un complejo con la enzima acetilcolinesterasa Los animales envenenados con este gas quedan paralizados debido a la imposibilidad de transmitir adecuadamente los impulsos nerviosos
Ester di-isopropil fos foacuterico de la enzima (inactivo)
b) Inhibicioacuten competitiva
Es cuando el inhibidor compite con el substrato por la unioacuten con el centro activo de la
enzima Este tipo de inhibicioacuten puede reducirse si se aumenta la concentracioacuten de substrato
-OOC - CH2 - CH2 - COO
- -OOC - CH = CH - COO
- + 2H
+ Succinato Succinato Fumarato
33
Deshidrogenasa COO
- I CH2 Malonato (inhibidor competitivo) I COO
- En la inhibicioacuten competitiva la enzima se combina con el inhibidor para formar un complejo enzima-inhibidor E + I E I en competencia con la reaccioacuten normal E +S ES
La sulfanilamida (bactericida) interfiere con la siacutentesis del aacutecido foacutelico compitiendo con el aacutecido p-aminobenzoico de una forma competitiva
HOOC-sect- NH2 NH2 ndashSO2 - sect- NH2 aacutecido p- amino benzoico sulfanilamida
sect = fenil
a Inhibicioacuten no competitiva
Esta inhibicioacuten se caracteriza por que no se puede revertir el efecto del inhibidor aumentando la concentracioacuten del substrato
Por ejemplo la inhibicioacuten de la deshidrogenasa del gliceraldehido -3-fosfato por la yodo-acetamida
Enzima-SH + I-CH2 CONH2 Enzima-SCH2 CONH2 + IH
La deshidrogenasa del gliceraldehido-3 fosfato puede ser inhibida tambieacuten por el aacutecido iodo-aceacutetico
Enzima-SH + ICH2 COOH Enzima-SCH2 COOH + IH
c) Inhibicioacuten por metales
Ciertos metales como el plomo mercurio y arseacutenico inhiben enzimas que tienen en su centro activo grupos -SH libres

8
Figura 3 Cofactores y Coenzima
Tabla 2 Algunos enzimas que contienen iones metaacutelicos o los necesitan como
cofactores
Zn+2 Cu+2 (Cu+)
Alcohol- deshidrogenada Tirosinasa Anhidrasa-carboacutenica Citocromo-oxidasa Carboxipeptidasa
Mg+2 K+
Fosfohidrolasa Piruvato- fosfoquinasa Fosfotransferasas
Mn+2 Arginasa Na+ Fosfotransferasas ATPasa de la membrana plasmaacutetica
Fe +2 o Fe+3
9
Citocromos
Peroxidasa Catalasa Ferredoxina
Texto ------- (Tabla 3)
Tabla 3 Relacioacuten entre algunas coenzimas y las vitaminas a partir de las
cuales se producen Enfermedades generadas por la deficiencia
Coenzima Reaccioacuten Fuente vitamiacutenica Enfermedad
Humana por deficiencia
Biocitina Carboxilacioacuten Biotina
Coenzima A Transferencia de acilos Pantotenato
Coenzimas de Cobalamima Alquilacioacuten Cobalamina (B12) anemia perniciosa
Coenzimas de flavina Oxidacioacuten-reduccioacuten Riboflavina (B2)
Acido lipoacuteico Transferencia de acilos -
Coenzimas de nicotinamida Oxidacioacuten-reduccioacuten Nicotinamida (niacina) Pelagra
Fosfato de piridoxal Transferencia de grupo amino Piridoxina (B6)
Tetrahidrofolato Transferencia de un grupo de 1 Carbono Acido foacutelico anemia
megaloblaacutestica
Tiamina pirofosfato Transferencia de aldehiacutedo Tiamina (B1) beriberi
no tiene un nombre especiacutefico su aparicioacuten es muy rara
Otras coenzimas
NADP
Coenzima Q
Biotina
NAD y NADP
10
Coenzima Q (CoQ)
Esta coenzima es sintetizada a partir de farnesil pirofosfato Al igual que el
FMN la CoQ es capaz de aceptar y donar uno o dos electrones porque su forma
semiquinona es estable A su forma oxidada se le conoce como ubiquinona (de
ubicuo lat ubique en todas partes) o quinona A su forma reducida se le conoce
como QH2 ubiquinol o hidroquinona (Figura 2) A la coenzima solo parcialmente
reducida se le conoce como ubisemiquinona semiquinona o QH (Figura 4)
Figura 4 Representacioacuten de la moleacutecula de CoQ
La siacutentesis de la CoQ se lleva a cabo a traveacutes de unidades isoprenoides (Figura 5)
11
Figura 5 La CoQ se produce a partir de isoprenoides
Biotina
La biotina consiste de un anillo imidazol que estaacute unido en forma cis a la
cadena lateral tetrahidrotiofeno del valerato (Figura 6)
12
Figura 6 Estructura de la biotina
La enzima holocarboxilasa sintetasa (HCS) cataliza la activacioacuten mediante
biotinilacioacuten de cinco carboxilasas en ceacutelulas humanas La deficiencia de HCS
produce el siacutendrome deficiencia muacuteltiple de carboxilasas (DMC) Esta enfermedad es
potencialmente mortal y se caracteriza por la deficiencia de la actividad de todas las
carboxilasas La deficiencia de biotina no solo disminuye la actividad de estas enzimas
sino que parece modificar la expresioacuten geneacutetica
126 PROPIEDADES DE LAS ENZIMAS
Las propiedades de las enzimas derivan del hecho de ser proteiacutenas y de actuar como
catalizadores Como proteiacutenas poseen una conformacioacuten natural maacutes estable que las
demaacutes conformaciones posibles Asiacute cambios en la conformacioacuten suelen ir asociados
en cambios en la actividad cataliacutetica Los factores que influyen de manera maacutes directa
sobre la actividad de un enzima son
pH
temperatura
cofactores
A) EFECTO DEL pH
Al comprobar experimentalmente la influencia del pH en la velocidad de las
reacciones enzimaacuteticas se obtienen curvas que indican que los enzimas
presentan un pH oacuteptimo de actividad El pH puede afectar de varias maneras
13
o El centro activo puede contener aminoaacutecidos con grupos ionizados
que pueden variar con el pH
o La ionizacioacuten de aminoaacutecidos que no estaacuten en el centro activo puede
provocar modificaciones en la conformacioacuten de la enzima
o El sustrato puede verse afectado por las variaciones del pH
Algunos enzimas presentan variaciones peculiares La pepsina del estoacutemago
presenta un oacuteptimo a pH=2 la ureasa lo tiene a pH 7 y la fosfatasa alcalina del
intestino un pH= 12
B) EFECTO DE LA TEMPERATURA
Influye en la actividad El punto oacuteptimo representa el maacuteximo de actividad A
temperaturas bajas los enzimas se hallan ldquomuy riacutegidosrdquo y cuando se supera un
valor considerable (mayor de 50) la actividad cae bruscamente porque como
proteiacutena el enzima se desnaturaliza
128 EFECTO DE LOS COFACTORES SOBRE LA ACTIVIDAD
ENZIMATICA
A veces un enzima requiere para su funcioacuten la presencia de sustancias no proteicas
que colaboran en la cataacutelisis los cofactores Casi un tercio de los enzimas conocidos
requieren cofactores Cuando el cofactor es una moleacutecula orgaacutenica se llama
coenzima En la figura inferior podemos observar una moleacutecula de hemoglobina
(proteiacutena que transporta oxiacutegeno) y su coenzima (el grupo hemo) Cuando los
cofactores y las coenzimas se encuentran unidos covalentemente al enzima se llaman
14
grupos prosteacuteticos La forma cataliacuteticamente activa del enzima es decir el enzima
unido a su grupo prosteacutetico se llama holoenzima La parte proteica de un
holoenzima (inactiva) se llama apoenzima de forma que
apoenzima + grupo prosteacutetico= holoenzima
129 MODO DE ACCIOacuteN DE LAS ENZIMAS
En las reacciones espontaacuteneas los productos finales tienen menos energiacutea libre de
Gibbs (∆G) que los reactantes (figura inferior izquierda) Por lo tanto las reacciones
espontaacuteneas liberan energiacutea de Gibbs ((∆G lt 0) Sin embargo el comienzo de la
reaccioacuten requiere un aporte inicial de energiacutea Esta energiacutea inicial que hay que
suministrar a los reactantes para que la reaccioacuten transcurra se llama energiacutea de
activacioacuten (Ea) Cuanto menor es la Ea maacutes faacutecilmente transcurre la reaccioacuten
Perfil energeacutetico de una reaccioacuten espontaacutenea Perfil energeacutetico de una reaccioacuten catalizada
15
La accioacuten de los catalizadores consiste precisamente en disminuir la Ea (Figura superior derecha) Los enzimas
son catalizadores especialmente eficaces ya que disminuyen la Ea auacuten maacutes que los catalizadores inorgaacutenicos
Por ejemplo la descomposicioacuten del agua oxigenada (H2O2) para dar H2O y O2 puede ocurrir sin catalizador
con un catalizador inorgaacutenico (platino) o con un enzima especiacutefico (catalasa) Las respectivas Ea para cada
proceso son 18 12 y 6 Kcalmol Asiacute se puede calcular que el platino acelera la reaccioacuten 20000 veces mientras
que la catalasa la acelera 370000 veces
Para que una reaccioacuten quiacutemica tenga lugar las moleacuteculas de los reactantes deben chocar con una energiacutea y
una orientacioacuten adecuadas La actuacioacuten del enzima (1) permite que los rea ctantes (sustratos) se unan a su
centro activo con una orientacioacuten oacuteptima para que la reaccioacuten se produzca y (2) modifica las propiedades
quiacutemicas del sustrato unido a su centro activo debilitando los enlaces existentes y facilitando la formacioacuten de
otros nuevos (Figuras inferiores)
1210 REGULACIOacuteN DE SU ACTIVIDAD ENZIMATICA
16
Una moleacutecula de enzima no tiene por que actuar siempre a la misma velocidad Su
actividad puede estar modulada por
Cambios en el pH
Cambios en la temperatura
Presencia de cofactores
Las concentraciones del sustrato y de los productos finales
Presencia de inhibidores
Modulacioacuten alosteacuterica
Modificacioacuten covalente
Activacioacuten por proteolisis
Isoenzimas
EFECTO DE LAS CONCENTRACIONES SOBRE LA ACTIVIDAD ENZIMAacuteTICA
La velocidad de una reaccioacuten enzimaacutetica depende de la concentracioacuten de sustrato La Figura de la derecha muestra la velocidad de una reaccioacuten enzimaacutetica a 6 concentraciones distintas de sustrato
Ademaacutes la presencia de los productos finales puede hacer que la reaccioacuten sea maacutes lenta o incluso invertir su sentido (Figura inferior)
EFECTO DE LOS INHIBIDORES SOBRE LA ACTIVIDAD ENZIMAacuteTICA
Ciertas moleacuteculas pueden inhibir la accioacuten cataliacutetica de un enzima son los inhibidores Estos inhibidores bien pueden ocupar temporalmente el centro activo por semejanza estructural con el sustrato original ( inhibidor competitivo) o bien alteran la conformacioacuten espacial del enzima impidiendo su unioacuten al sustrato ( inhibidor no competitivo) (Figuras inferiores)
Inhibidor competitivo Inhibidor no competitivo
17
MODULACIOacuteN ALOSTEacuteRICA DE LA ACTIVIDAD ENZIMAacuteTICA
Hay enzimas que pueden adoptar 2 conformaciones interconvertibles llamadas R (relajada) y T (tensa) R es la forma maacutes activa porque se une al sustrato con maacutes afinidad Las formas R y T se encuentran en equilibrio R lt==gt T (Figura inferior)
Ciertas sustancias tienden a estabilizar la forma R Son los llamados moduladores positivos El propio sustrato es a
menudo un modulador positivo Las moleacuteculas que favorecen la forma R pero que actuacutean sobre una regioacuten del enzima distinta del centro activo son los activadores alosteacutericos (Figura inferior izquierda)
Activador alosteacuterico favorece la unioacuten del sustrato
Inhibidor alosteacuterico impide la unioacuten del sustrato
Las sustancias que favorecen la forma T y disminuyen la actividad enzimaacutetica son los moduladores negativos Si
estos moduladores actuacutean en lugares distintos del centro activo del enzima se llaman inhibidores alosteacutericos
18
(Figura superior derecha
EFECTO DE LA MODIFICACIOacuteN COVALENTE SOBRE LA ACTIVIDAD ENZIMAacuteTICA
Otros enzimas pasan de una forma menos activa a otra maacutes activa unieacutendose covalentemente a un grupo quiacutemico de pequentildeo tamantildeo como el Pi o el AMP Tambieacuten se da el caso inverso en el que un enzima muy activo se desactiva al liberar alguacuten grupo quiacutemico En las enzimas de las viacuteas degradativas del metabolismo la
forma fosforilada es maacutes activa que la no fosforilada mientras que en las viacuteas biosinteacuteticas ocurre lo contrario En las figuras inferiores se ilustra la activacioacuten de una proteiacutena por fosforilacioacuten
Elementos de la reaccioacuten El enzima no fosforilado es inactivo El enzima fosforilado es activo
ACTIVACIOacuteN PROTEOLIacuteTICA DE LA ACTIVIDAD ENZIMAacuteTICA
Algunos enzimas no se sintetizan como tales sino como proteiacutenas precursoras sin actividad enzimaacutetica Estas proteiacutenas se llaman
proenzimas o zimoacutegenos Para activarse los zimoacutegenos sufren un ataque hidroliacutetico que origina la liberacioacuten de uno o varios peacuteptidos El resto de la moleacutecula proteica adopta la conformacioacuten y las propiedades del enzima activo Muchos enzimas digestivos se secretan en forma de zimoacutegenos y en el tubo digestivo
se convierten en la forma activa Es el caso de la -quimotripsina que se sintetiza en forma de quimotripsinoacutegeno (Figura superior) Si estos enzimas se sintetizasen directamente en forma activa destruiriacutean la propia ceacutelula que las
produce Asiacute la tripsina pancreaacutetica (una proteasa) se sintetiza como tripsinoacutegeno (inactivo) Si por alguna razoacuten se activa en el propio paacutencreas la glaacutendula sufre un proceso de autodestruccioacuten (pancreatitis aguda) a
menudo mortal
19
REGULACIOacuteN DE LA ACTIVIDAD ENZIMAacuteTICA POR MEDIO DE ISOENZIMAS
Algunos enzimas tienen distinta estructura molecular aunque su funcioacuten bioloacutegica es similar Se llaman isozimas o isoenzimas Estas diferencias de estructura se traducen en ligeros cambios en sus propiedades de forma que cada isozima se adapta perfectamente a la funcioacuten que debe realizar Asiacute podemos observar la existencia de isoenzimas en funcioacuten de
el tipo de tejido Por ejemplo la lactato deshidrogenasa presenta isozimas distintos en muacutesculo y corazoacuten el compartimento celular donde actuacutea Por ejemplo la malato deshidrogenasa del citoplasma es distinta
de la de la mitocondria el momento concreto del desarrollo del individuo Por ejemplo algunos enzimas de la glicolisis del feto
son diferentes de los mismos enzimas en el adulto
CINEacuteTICA ENZIMAacuteTICA
Los principios generales de las reacciones quiacutemicas se aplican tambieacuten a las reacciones enzimaacuteticas Por este motivo antes de empezar con la cineacutetica quiacutemica se van a repasar algunos conceptos baacutesicos de cineacutetica quiacutemica
A continuacioacuten se describiraacuten los siguientes conceptos
Cineacutetica enzimaacutetica Modelo cineacutetico de Michaelis-Menten Caacutelculo de la KM y la Vmax de un enzima Actividad enzimaacutetica
20
NOMENCLATURA DE LOS ENZIMAS
CATALISIS MOLECULAR
Un catalizador modifica la velocidad de una reaccioacuten quiacutemica sin ser utilizado o aparecer como uno de los productos de la reaccioacuten Una reaccioacuten quiacutemica en la que un substrato(S) se
transforma en un producto(P) S P ocurre por que cierta fraccioacuten de moleacuteculas de S posee mucho maacutes energiacutea que el resto de ellas lo que es suficiente para que alcancen un estado activado en el que pueda formarse o romperse un enlace quiacutemico y se forme el producto(P)
La energiacutea de activacioacuten es la cantidad de energiacutea expresada en caloriacuteas necesaria para que todas las moleacuteculas de un mol a una temperatura dada alcancen el estado reactivo Mientras que el estado de transicioacuten es el estado rico en energiacutea de las moleacuteculas que
interaccionan en la cima de la barrera de activacioacuten La velocidad de una reaccioacuten quiacutemica es proporcional a la concentracioacuten del complejo en el estado de transicioacuten
Una reaccioacuten quiacutemica se puede acelerar de la siguiente forma 1) Al aumentar la temperatura se incrementa la energiacutea cineacutetica por lo que es mayor el nuacutemero de moleacuteculas que alcanzan el
estado de transicioacuten Generalmente el Q10 =2 lo que indica que la velocidad de una reaccioacuten quiacutemica se duplica al aumentar la temperatura en 10
0C 2) Antildeadiendo un catalizador que
disminuye la energiacutea de activacioacuten y aumenta la velocidad de reaccioacuten
La enzima (E) se combina con el substrato (S) formando el complejo de transicioacuten enzima-
substrato (E-S) mediante una reaccioacuten reversible cuya energiacutea de activacioacuten es menor que la de la reaccioacuten no catalizada Cuando se forma el producto de la reaccioacuten (P) se regenera de nuevo la enzima (E) de forma libre la que puede combinarse de nuevo con otra moleacutecula de substrato (S)
Una enzima reduce maacutes eficientemente la energiacutea de activacioacuten(Ea) de una reaccioacuten que un catalizador inorgaacutenico lo que permite que una reaccioacuten se realice a menor temperatura
El siguiente ejemplo ilustra mejor lo que hemos discutido
Reaccioacuten Energiacutea de activacioacuten (Kcalmol-1
)
21
a) el peroacutexido de hidroacutegeno se descompone en
H2 O2 H2 O + O2
18
b) el hierro cataliacutetico (Fe) realiza la reaccioacuten
H2 O2 H2 O + O2
13
c) el platino cataliacutetico (Pt) realiza la reaccioacuten
H2 O2 H2 O + O2
12
d) la catalasa una enzima hepaacutetica la realiza
H2O2 H2O + O2
5
Una enzima no modifica la energiacutea libre ni la constante de equilibrio sino que disminuye la energiacutea de activacioacuten de la reaccioacuten
Ir al principio
EFECTO DE LA CONCENTRACIOacuteN DEL SUBSTRATO SOBRE LA
CATALISIS ENZIMATICA
La actividad de una enzima se puede estudiar in vitro bajo condiciones controladas antildeadieacutendole una enzima a un substrato Siacute se trabaja con la condicioacuten de que la concentracioacuten del substrato sea saturante variando entonces la concentracioacuten de enzima se observa que aumenta el producto de la reaccioacuten a pH y temperatura constantes
22
Si se mantiene la concentracioacuten de la enzima constante y variando la concentracioacuten de substrato se obtiene una curva hiperboacutelica como la de la figura Al principio un aumento de la
concentracioacuten de substrato produce un aumento raacutepi do de la velocidad de reaccioacuten pero si se sigue aumentando la concentracioacuten de substrato la velocidad de reaccioacuten comienza a disminuir y a muy altas concentraciones de substrato se observa que no cambia la velocidad de
reaccioacuten se dice que los centros activos de la enzima se encuentran saturados La velocidad de reaccioacuten que se obtiene a esa alta concentracioacuten de substrato se define como la velocidad maacutexima (V) de la reaccioacuten enzimaacutetica bajo las condiciones especificadas La concentracioacuten de
substrato (S) a la semivelocidad maacutexima de reaccioacuten (V2) se puede determinar de la figura y representa la constante de Michaelis o Km la cual es una caracteriacutestica para cada enzima La inversa de Km o 1Km mide aproximadamente la afinidad de la enzima por el subs trato
Mientras maacutes pequentildeo sea el valor de Km mayor seraacute la afinidad de la enzima por el substrato Si varias enzimas compiten en el metabolismo por el mismo substrato eacuteste seraacute transformado preferentemente por la enzima con mayor afinidad
La ecuacioacuten de Michaelis describe la relacioacuten cuantitativa entre la velocidad de reaccioacuten y la concentracioacuten de substrato [S] si se conoce Vmax o Km
v=
En donde v= es la velocidad de reaccioacuten observada a una concentracioacuten de substrato determinada [S]
Km= constante de Michaelis en molesliacutet ro
23
Vmax= velocidad maacutexima a concentracioacuten saturante de substrato
Si v= Vmax2 entonces
=
Km+[S] = 2[S]
Km =[S]
De tal forma que podemos concluir que Km es igual a la concentracioacuten de substrato a la semivelocidad maacutexima de reaccioacuten
1 Concentraciones relativas de E y S la concentracioacuten de substrato S es
mucho mayor que la de E de tal manera que la cantidad de substrato
unido a la enzima en cualquier momento es muy pequentildea
2 Se asume el estado estacionario ES no cambia con el tiempo esto
quiere decir que la velocidad de formacioacuten de ES es igual a aquella para
su desintegracioacuten En general un intermediario en una serie de reacciones estaacute en estado estacionario cuando su velocidad de siacutentesis
es igual a su velocidad de degradacioacuten 3 Velocidad inicial para el anaacutelisis de reacciones enzimaacuteticas soacutelo se
utiliza la velocidad inicial de la reaccioacuten que es la velocidad ejercida por
la enzima inmediatamente despueacutes de que se ha puesto en contacto con el substrato y hasta antes de que se haya consumido el 10 de la concentracioacuten inicial del mismo La razoacuten de lo anterior es que en ese
momento la concentracioacuten del producto de la reaccioacuten que se ha acumulado en muy pequentildea y por tanto la reaccioacuten en el sentido
inverso es decir la transformacioacuten del producto en el substrato original puede ser ignorada
24
Caracteriacutesticas de Km la constante de Michaelis-Menten es
caracteriacutestica de una enzima y particular para un substrato Refleja la afinidad de la enzima por ese substrato K m es numeacutericamente igual a la concentracioacuten
de substrato a la cual la velocidad de reaccioacuten es la mitad de la Vmax (Km= Vmax2) Este paraacutemetro Km no variacutea con la concentracioacuten de enzima
Significado de una Km pequentildea un valor numeacuterico pequentildeo de Km
refleja una alta afinidad de la enzima por su substrato porque a una baja
concentracioacuten del mismo la enzima a desarrollado ya la mitad de la velocidad maacutexima
Significado de una Km grande el valor numeacuterico grande de Km refleja
una baja afinidad de la enzima por su substrato porque a una concentracioacuten
elevada del mismo la enzima desarrolla la mitad de la velocidad maacutexima
Relacioacuten de la velocidad con la concentracioacuten de enzima la
velocidad de la reaccioacuten es directamente proporcional a la concentracioacuten de la
enzima a cualquier concentracioacuten de substrato Por ejemplo si la concentracioacuten de enzima es disminuida a la mitad la velocidad inicial de la reaccioacuten (v0) es
reducida tambieacuten a la mitad de la original
Orden de la reaccioacuten cuando S es mas pequentildea que la Km la
velocidad de la reaccioacuten es aproximadamente igual a la concentracioacuten de
substrato
100
50
0
Substrato
Km Figura Localizacioacuten de la constante de Michaelis (Km)
25
La velocidad de reaccioacuten se dice en estas condiciones es de primer
orden con respecto al substrato Cuando S es mas grande que la Km la
velocidad es constante e igual a la Vmax La velocidad de la reaccioacuten es independiente de la concentracioacuten de substrato y se dice que es de orden cero
con respecto a la concentracioacuten de substrato
Las enzimas pueden ser aisladas de las ceacutelulas u organismos para estudiar sus
propiedades in vitro es decir en un tubo de ensayo Las diferentes enzimas muestran diferentes respuestas a los cambios en la concentracioacuten de substrato temperatura y pH
Concentracioacuten de substrato
La forma hiperboacutelica de la curva de velocidad vs Substrato
Efecto de la temperatura en la actividad enzimaacutetica
Efecto de la variacioacuten del pH en la actividad enzimaacutetica
Incremento de la velocidad con la temperatura En general la velocidad de la reaccioacuten se incrementa con la temperatura hasta que un punto maacuteximo es alcanzado
Este incremento se debe a que aumenta el nuacutemero de moleacuteculas ricas en energiacutea que pueden pasar la barrera energeacutetica de estado de transicioacuten para formar a los productos
Decremento de la velocidad con la temperatura la elevacioacuten excesiva de la
temperatura del medio que contiene a las enzimas resulta en un decremento de la velocidad como resultado de la desnaturalizacioacuten es decir la peacuterdida de la estructura tridimensional de las enzimas
26
zona de reactivacioacuten maacuteximo de actividad
Zona de
inactivacioacuten
0 Temperatura
Figura dependencia de la actividad enzimaacutetica con la temperatura
Velocidad maacutexima la velocidad de una reaccioacuten (v) es el nuacutemero de
moleacuteculas de substrato convertidas en producto por unidad de tiempo y
usualmente se expresa en micromoles de producto formadas por minuto La velocidad de una reaccioacuten catalizada por una enzima aumenta conforme se incrementa la concentracioacuten de substrato hasta que se llega a una velocidad maacutexima (Vmax) a partir de la cual la velocidad de la reaccioacuten es
independiente de la cantidad de substrato El que la velocidad no pueda
seguirse incrementando refleja la saturacioacuten del sitio activo con el substrato
Vmax
0 Substrato
Figura representacioacuten de una cineacutetica tiacutepicamente michaeliana
Vmax = velocidad de saturacioacuten o maacutexima
27
Cuando se grafica la velocidad de la reaccioacuten v0 contra la concentracioacuten de substrato S no siempre es posible determinar la condicioacuten
en que se ha llegado a la velocidad maacutexima Vmax debido al incremento de la pendiente en la hipeacuterbola a concentraciones de substrato elevadas
Figura experimento cineacutetico para la actividad enzimaacutetica
Se presentan los valores experimentales y los resultados del ajuste al modelo de Michaelis
Auacuten asiacute si se grafica una doble reciacuteproca es decir el inverso de la
velocidad (1v0) contra el inverso de la concentracioacuten de substrato (1 S ) se
obtiene una liacutenea recta El regraacutefico de Lineweaver-Burke tambieacuten se
denomina de ldquodobles reciacuteprocasrdquo y se utiliza para calcular la Km y la Vmax asiacute como para determinar el mecanismo de accioacuten de los diversos tipos de
inhibidores
La ecuacioacuten que describe a la graacutefca de Lineweaver-Burke es
Vmax
1
SVmax
Km
v
1
0 (3)
que describe una recta en donde el intercepto con el eje de las abscisas es igual a ndash1Km y el intercepto con el eje de las ordenadas es igual a 1Vmax
28
Figura experimento cineacutetico para la actividad enzimaacutetica Regraacutefico de
Lineweaver-Burke
Se presentan los valores experimentales y los resultados del ajuste al modelo
REPRESENTACIOacuteN GRAacuteFICA DE LINEWEAVER Y BURK
Si se toma el inverso en ambos miembros de la ecuacioacuten de Michaelis tenemos
=
+
29
que es la ecuacioacuten de una liacutenea recta cuya pendiente es que intercepta al eje de las
ordenadas en y a la abcisa en el punto Lo que es evidente al hacer
ya que se convierte en
Si se hace una graacutefica con los dobles inversos colocando los valores de en el eje de las
ordenadas y en el eje de las abcisas se obtiene una liacutenea recta a partir de la cual se puede calcular faacutecilmente Km
EFECTO DEL pH SOBRE LA ACTIVIDAD ENZIMATICA
Sabiendo que las enzimas son proteinas cualquier cambio brusco de pH puede alterar el caraacutecter ioacutenico de los grupos amino y carboxilo en la superficie proteica afectando asiacute las propiedades cataliacuteticas de una enzima A pH alto o bajo se puede producir la desnaturalizacioacuten
de la enzima y en consecuencia su inactivacioacuten La fosfatasa aacutecida es maacutes activa a pH 50 mientras que la fosfatasa alcalina lo es a pH 90 Muchas enzimas tienen maacutexima actividad cerca de la neutralidad en un rango de pH de 6 a 8
El pH per se no afecta la actividad enzimaacutetica sino la concentracioacuten de protones Los protones
ademaacutes de alterar la estructura de la enzima y el substrato pueden participar tambieacuten en la reaccioacuten como substrato o producto En esos casos la concentracioacuten de protones afecta directamente la velocidad de la reaccioacuten
ENZIMA pH OPTIMO
30
Pepsina 15
Tripsina 77
Catalasa 76
Arginasa 97
Fumarasa 78
Ribonucleasa 78
EFECTO DE ACTIVADORES
Un aumento en la temperatura provoca un aumento de la velocidad de reaccioacuten hasta cierta temperatura oacuteptima ya que despueacutes de aproximadamente 45
0 C se comienza a producir la
desnaturalizacioacuten teacutermica Las enzimas de muchos mamiacuteferos tienen una temperatura oacuteptima
de 370 C por encima de esa temperatura comienzan a inactivarse y se destruyen Sin embargo
existen especies de bacterias y algas que habitan en fuentes de aguas termales y en el otro extremo ciertas bacterias aacuterticas tienen temperaturas oacuteptimas cercanas a 0
0 C
31
ESPECIFICIDAD ABSOLUTA
Cierta enzimas se encuentran como aproenzimas o zimogenos que son inactivos
El tripsinoacutegeno es convertido en tripsina por la misma accioacuten de la tripsina que remueve un hexapeacuteptido
tripsina Tripsinoacutegeno Tripsina + Val (Asp)4 lisina
Otra forma de activacioacuten consiste en mantener los grupos SH reducidos Estos grupos pueden
formar parte de centros activos de ciertas enzimas Si se oxidan estos grupos a -S-S- la enzima se inactiva Asiacute tenemos que la enzima papaina se inactiva despueacutes de exponerla al ox iacutegeno pero cuando se le antildeade un reductor apropiado la enzima se reactiva Los grupos ndashS-S- se convierten en ndashSH activaacutendose la enzima
Otra forma de activacioacuten requiere la presencia de otra substancia que se enlaza al sitio alosteacuterico el compuesto que se enlaza se denomina un efector alosteacuterico El centro alosteacuterico es bien especiacutefico para el efector o modulador alosteacuterico
ESPECIFICIDAD RELATIVA
Oxiacutegeno L-aminoaacutecido a -cetoaacutecido + NH3 + H 2 O 2 L-aminoaacutecido oxidasa Oxiacutegeno D-aminoaacutecido a -cetoaacutecido + NH3 + H 2 O 2 D-aminoaacutecido oxidasa
32
En este ejemplo se muestra la estereo-especificidad Una enzima puede tener una especificidad oacuteptica para isomeros D o L
Hay enzimas que muestran especificidad absoluta como la aspartasa que cataliza la adicioacuten reversible de amoniacuteaco al doble enlace del aacutecido fumaacuterico pero no al de otros aacutecidos insaturados
HOOC ndash CH = CH ndash COO- + NH4
+ HOOC ndash CH2 - CHNH2 - COO
- + H
+
INHIBICIOacuteN ENZIMATICA
Mediante el uso de inhibidores enzimaacuteticos se ha obtenido informacioacuten muy valiosa sobre la conformacioacuten del centro activo de algunas enzimas
a Inhibicioacuten irreversible Este tipo de inhibidores forman un enlace covalente con las enzimas cerca del centro activo Un ejemplo son los gases nerviosos como el
fluorofosfato de diisopropilo (DFP) que forma un complejo con la enzima acetilcolinesterasa Los animales envenenados con este gas quedan paralizados debido a la imposibilidad de transmitir adecuadamente los impulsos nerviosos
Ester di-isopropil fos foacuterico de la enzima (inactivo)
b) Inhibicioacuten competitiva
Es cuando el inhibidor compite con el substrato por la unioacuten con el centro activo de la
enzima Este tipo de inhibicioacuten puede reducirse si se aumenta la concentracioacuten de substrato
-OOC - CH2 - CH2 - COO
- -OOC - CH = CH - COO
- + 2H
+ Succinato Succinato Fumarato
33
Deshidrogenasa COO
- I CH2 Malonato (inhibidor competitivo) I COO
- En la inhibicioacuten competitiva la enzima se combina con el inhibidor para formar un complejo enzima-inhibidor E + I E I en competencia con la reaccioacuten normal E +S ES
La sulfanilamida (bactericida) interfiere con la siacutentesis del aacutecido foacutelico compitiendo con el aacutecido p-aminobenzoico de una forma competitiva
HOOC-sect- NH2 NH2 ndashSO2 - sect- NH2 aacutecido p- amino benzoico sulfanilamida
sect = fenil
a Inhibicioacuten no competitiva
Esta inhibicioacuten se caracteriza por que no se puede revertir el efecto del inhibidor aumentando la concentracioacuten del substrato
Por ejemplo la inhibicioacuten de la deshidrogenasa del gliceraldehido -3-fosfato por la yodo-acetamida
Enzima-SH + I-CH2 CONH2 Enzima-SCH2 CONH2 + IH
La deshidrogenasa del gliceraldehido-3 fosfato puede ser inhibida tambieacuten por el aacutecido iodo-aceacutetico
Enzima-SH + ICH2 COOH Enzima-SCH2 COOH + IH
c) Inhibicioacuten por metales
Ciertos metales como el plomo mercurio y arseacutenico inhiben enzimas que tienen en su centro activo grupos -SH libres

9
Citocromos
Peroxidasa Catalasa Ferredoxina
Texto ------- (Tabla 3)
Tabla 3 Relacioacuten entre algunas coenzimas y las vitaminas a partir de las
cuales se producen Enfermedades generadas por la deficiencia
Coenzima Reaccioacuten Fuente vitamiacutenica Enfermedad
Humana por deficiencia
Biocitina Carboxilacioacuten Biotina
Coenzima A Transferencia de acilos Pantotenato
Coenzimas de Cobalamima Alquilacioacuten Cobalamina (B12) anemia perniciosa
Coenzimas de flavina Oxidacioacuten-reduccioacuten Riboflavina (B2)
Acido lipoacuteico Transferencia de acilos -
Coenzimas de nicotinamida Oxidacioacuten-reduccioacuten Nicotinamida (niacina) Pelagra
Fosfato de piridoxal Transferencia de grupo amino Piridoxina (B6)
Tetrahidrofolato Transferencia de un grupo de 1 Carbono Acido foacutelico anemia
megaloblaacutestica
Tiamina pirofosfato Transferencia de aldehiacutedo Tiamina (B1) beriberi
no tiene un nombre especiacutefico su aparicioacuten es muy rara
Otras coenzimas
NADP
Coenzima Q
Biotina
NAD y NADP
10
Coenzima Q (CoQ)
Esta coenzima es sintetizada a partir de farnesil pirofosfato Al igual que el
FMN la CoQ es capaz de aceptar y donar uno o dos electrones porque su forma
semiquinona es estable A su forma oxidada se le conoce como ubiquinona (de
ubicuo lat ubique en todas partes) o quinona A su forma reducida se le conoce
como QH2 ubiquinol o hidroquinona (Figura 2) A la coenzima solo parcialmente
reducida se le conoce como ubisemiquinona semiquinona o QH (Figura 4)
Figura 4 Representacioacuten de la moleacutecula de CoQ
La siacutentesis de la CoQ se lleva a cabo a traveacutes de unidades isoprenoides (Figura 5)
11
Figura 5 La CoQ se produce a partir de isoprenoides
Biotina
La biotina consiste de un anillo imidazol que estaacute unido en forma cis a la
cadena lateral tetrahidrotiofeno del valerato (Figura 6)
12
Figura 6 Estructura de la biotina
La enzima holocarboxilasa sintetasa (HCS) cataliza la activacioacuten mediante
biotinilacioacuten de cinco carboxilasas en ceacutelulas humanas La deficiencia de HCS
produce el siacutendrome deficiencia muacuteltiple de carboxilasas (DMC) Esta enfermedad es
potencialmente mortal y se caracteriza por la deficiencia de la actividad de todas las
carboxilasas La deficiencia de biotina no solo disminuye la actividad de estas enzimas
sino que parece modificar la expresioacuten geneacutetica
126 PROPIEDADES DE LAS ENZIMAS
Las propiedades de las enzimas derivan del hecho de ser proteiacutenas y de actuar como
catalizadores Como proteiacutenas poseen una conformacioacuten natural maacutes estable que las
demaacutes conformaciones posibles Asiacute cambios en la conformacioacuten suelen ir asociados
en cambios en la actividad cataliacutetica Los factores que influyen de manera maacutes directa
sobre la actividad de un enzima son
pH
temperatura
cofactores
A) EFECTO DEL pH
Al comprobar experimentalmente la influencia del pH en la velocidad de las
reacciones enzimaacuteticas se obtienen curvas que indican que los enzimas
presentan un pH oacuteptimo de actividad El pH puede afectar de varias maneras
13
o El centro activo puede contener aminoaacutecidos con grupos ionizados
que pueden variar con el pH
o La ionizacioacuten de aminoaacutecidos que no estaacuten en el centro activo puede
provocar modificaciones en la conformacioacuten de la enzima
o El sustrato puede verse afectado por las variaciones del pH
Algunos enzimas presentan variaciones peculiares La pepsina del estoacutemago
presenta un oacuteptimo a pH=2 la ureasa lo tiene a pH 7 y la fosfatasa alcalina del
intestino un pH= 12
B) EFECTO DE LA TEMPERATURA
Influye en la actividad El punto oacuteptimo representa el maacuteximo de actividad A
temperaturas bajas los enzimas se hallan ldquomuy riacutegidosrdquo y cuando se supera un
valor considerable (mayor de 50) la actividad cae bruscamente porque como
proteiacutena el enzima se desnaturaliza
128 EFECTO DE LOS COFACTORES SOBRE LA ACTIVIDAD
ENZIMATICA
A veces un enzima requiere para su funcioacuten la presencia de sustancias no proteicas
que colaboran en la cataacutelisis los cofactores Casi un tercio de los enzimas conocidos
requieren cofactores Cuando el cofactor es una moleacutecula orgaacutenica se llama
coenzima En la figura inferior podemos observar una moleacutecula de hemoglobina
(proteiacutena que transporta oxiacutegeno) y su coenzima (el grupo hemo) Cuando los
cofactores y las coenzimas se encuentran unidos covalentemente al enzima se llaman
14
grupos prosteacuteticos La forma cataliacuteticamente activa del enzima es decir el enzima
unido a su grupo prosteacutetico se llama holoenzima La parte proteica de un
holoenzima (inactiva) se llama apoenzima de forma que
apoenzima + grupo prosteacutetico= holoenzima
129 MODO DE ACCIOacuteN DE LAS ENZIMAS
En las reacciones espontaacuteneas los productos finales tienen menos energiacutea libre de
Gibbs (∆G) que los reactantes (figura inferior izquierda) Por lo tanto las reacciones
espontaacuteneas liberan energiacutea de Gibbs ((∆G lt 0) Sin embargo el comienzo de la
reaccioacuten requiere un aporte inicial de energiacutea Esta energiacutea inicial que hay que
suministrar a los reactantes para que la reaccioacuten transcurra se llama energiacutea de
activacioacuten (Ea) Cuanto menor es la Ea maacutes faacutecilmente transcurre la reaccioacuten
Perfil energeacutetico de una reaccioacuten espontaacutenea Perfil energeacutetico de una reaccioacuten catalizada
15
La accioacuten de los catalizadores consiste precisamente en disminuir la Ea (Figura superior derecha) Los enzimas
son catalizadores especialmente eficaces ya que disminuyen la Ea auacuten maacutes que los catalizadores inorgaacutenicos
Por ejemplo la descomposicioacuten del agua oxigenada (H2O2) para dar H2O y O2 puede ocurrir sin catalizador
con un catalizador inorgaacutenico (platino) o con un enzima especiacutefico (catalasa) Las respectivas Ea para cada
proceso son 18 12 y 6 Kcalmol Asiacute se puede calcular que el platino acelera la reaccioacuten 20000 veces mientras
que la catalasa la acelera 370000 veces
Para que una reaccioacuten quiacutemica tenga lugar las moleacuteculas de los reactantes deben chocar con una energiacutea y
una orientacioacuten adecuadas La actuacioacuten del enzima (1) permite que los rea ctantes (sustratos) se unan a su
centro activo con una orientacioacuten oacuteptima para que la reaccioacuten se produzca y (2) modifica las propiedades
quiacutemicas del sustrato unido a su centro activo debilitando los enlaces existentes y facilitando la formacioacuten de
otros nuevos (Figuras inferiores)
1210 REGULACIOacuteN DE SU ACTIVIDAD ENZIMATICA
16
Una moleacutecula de enzima no tiene por que actuar siempre a la misma velocidad Su
actividad puede estar modulada por
Cambios en el pH
Cambios en la temperatura
Presencia de cofactores
Las concentraciones del sustrato y de los productos finales
Presencia de inhibidores
Modulacioacuten alosteacuterica
Modificacioacuten covalente
Activacioacuten por proteolisis
Isoenzimas
EFECTO DE LAS CONCENTRACIONES SOBRE LA ACTIVIDAD ENZIMAacuteTICA
La velocidad de una reaccioacuten enzimaacutetica depende de la concentracioacuten de sustrato La Figura de la derecha muestra la velocidad de una reaccioacuten enzimaacutetica a 6 concentraciones distintas de sustrato
Ademaacutes la presencia de los productos finales puede hacer que la reaccioacuten sea maacutes lenta o incluso invertir su sentido (Figura inferior)
EFECTO DE LOS INHIBIDORES SOBRE LA ACTIVIDAD ENZIMAacuteTICA
Ciertas moleacuteculas pueden inhibir la accioacuten cataliacutetica de un enzima son los inhibidores Estos inhibidores bien pueden ocupar temporalmente el centro activo por semejanza estructural con el sustrato original ( inhibidor competitivo) o bien alteran la conformacioacuten espacial del enzima impidiendo su unioacuten al sustrato ( inhibidor no competitivo) (Figuras inferiores)
Inhibidor competitivo Inhibidor no competitivo
17
MODULACIOacuteN ALOSTEacuteRICA DE LA ACTIVIDAD ENZIMAacuteTICA
Hay enzimas que pueden adoptar 2 conformaciones interconvertibles llamadas R (relajada) y T (tensa) R es la forma maacutes activa porque se une al sustrato con maacutes afinidad Las formas R y T se encuentran en equilibrio R lt==gt T (Figura inferior)
Ciertas sustancias tienden a estabilizar la forma R Son los llamados moduladores positivos El propio sustrato es a
menudo un modulador positivo Las moleacuteculas que favorecen la forma R pero que actuacutean sobre una regioacuten del enzima distinta del centro activo son los activadores alosteacutericos (Figura inferior izquierda)
Activador alosteacuterico favorece la unioacuten del sustrato
Inhibidor alosteacuterico impide la unioacuten del sustrato
Las sustancias que favorecen la forma T y disminuyen la actividad enzimaacutetica son los moduladores negativos Si
estos moduladores actuacutean en lugares distintos del centro activo del enzima se llaman inhibidores alosteacutericos
18
(Figura superior derecha
EFECTO DE LA MODIFICACIOacuteN COVALENTE SOBRE LA ACTIVIDAD ENZIMAacuteTICA
Otros enzimas pasan de una forma menos activa a otra maacutes activa unieacutendose covalentemente a un grupo quiacutemico de pequentildeo tamantildeo como el Pi o el AMP Tambieacuten se da el caso inverso en el que un enzima muy activo se desactiva al liberar alguacuten grupo quiacutemico En las enzimas de las viacuteas degradativas del metabolismo la
forma fosforilada es maacutes activa que la no fosforilada mientras que en las viacuteas biosinteacuteticas ocurre lo contrario En las figuras inferiores se ilustra la activacioacuten de una proteiacutena por fosforilacioacuten
Elementos de la reaccioacuten El enzima no fosforilado es inactivo El enzima fosforilado es activo
ACTIVACIOacuteN PROTEOLIacuteTICA DE LA ACTIVIDAD ENZIMAacuteTICA
Algunos enzimas no se sintetizan como tales sino como proteiacutenas precursoras sin actividad enzimaacutetica Estas proteiacutenas se llaman
proenzimas o zimoacutegenos Para activarse los zimoacutegenos sufren un ataque hidroliacutetico que origina la liberacioacuten de uno o varios peacuteptidos El resto de la moleacutecula proteica adopta la conformacioacuten y las propiedades del enzima activo Muchos enzimas digestivos se secretan en forma de zimoacutegenos y en el tubo digestivo
se convierten en la forma activa Es el caso de la -quimotripsina que se sintetiza en forma de quimotripsinoacutegeno (Figura superior) Si estos enzimas se sintetizasen directamente en forma activa destruiriacutean la propia ceacutelula que las
produce Asiacute la tripsina pancreaacutetica (una proteasa) se sintetiza como tripsinoacutegeno (inactivo) Si por alguna razoacuten se activa en el propio paacutencreas la glaacutendula sufre un proceso de autodestruccioacuten (pancreatitis aguda) a
menudo mortal
19
REGULACIOacuteN DE LA ACTIVIDAD ENZIMAacuteTICA POR MEDIO DE ISOENZIMAS
Algunos enzimas tienen distinta estructura molecular aunque su funcioacuten bioloacutegica es similar Se llaman isozimas o isoenzimas Estas diferencias de estructura se traducen en ligeros cambios en sus propiedades de forma que cada isozima se adapta perfectamente a la funcioacuten que debe realizar Asiacute podemos observar la existencia de isoenzimas en funcioacuten de
el tipo de tejido Por ejemplo la lactato deshidrogenasa presenta isozimas distintos en muacutesculo y corazoacuten el compartimento celular donde actuacutea Por ejemplo la malato deshidrogenasa del citoplasma es distinta
de la de la mitocondria el momento concreto del desarrollo del individuo Por ejemplo algunos enzimas de la glicolisis del feto
son diferentes de los mismos enzimas en el adulto
CINEacuteTICA ENZIMAacuteTICA
Los principios generales de las reacciones quiacutemicas se aplican tambieacuten a las reacciones enzimaacuteticas Por este motivo antes de empezar con la cineacutetica quiacutemica se van a repasar algunos conceptos baacutesicos de cineacutetica quiacutemica
A continuacioacuten se describiraacuten los siguientes conceptos
Cineacutetica enzimaacutetica Modelo cineacutetico de Michaelis-Menten Caacutelculo de la KM y la Vmax de un enzima Actividad enzimaacutetica
20
NOMENCLATURA DE LOS ENZIMAS
CATALISIS MOLECULAR
Un catalizador modifica la velocidad de una reaccioacuten quiacutemica sin ser utilizado o aparecer como uno de los productos de la reaccioacuten Una reaccioacuten quiacutemica en la que un substrato(S) se
transforma en un producto(P) S P ocurre por que cierta fraccioacuten de moleacuteculas de S posee mucho maacutes energiacutea que el resto de ellas lo que es suficiente para que alcancen un estado activado en el que pueda formarse o romperse un enlace quiacutemico y se forme el producto(P)
La energiacutea de activacioacuten es la cantidad de energiacutea expresada en caloriacuteas necesaria para que todas las moleacuteculas de un mol a una temperatura dada alcancen el estado reactivo Mientras que el estado de transicioacuten es el estado rico en energiacutea de las moleacuteculas que
interaccionan en la cima de la barrera de activacioacuten La velocidad de una reaccioacuten quiacutemica es proporcional a la concentracioacuten del complejo en el estado de transicioacuten
Una reaccioacuten quiacutemica se puede acelerar de la siguiente forma 1) Al aumentar la temperatura se incrementa la energiacutea cineacutetica por lo que es mayor el nuacutemero de moleacuteculas que alcanzan el
estado de transicioacuten Generalmente el Q10 =2 lo que indica que la velocidad de una reaccioacuten quiacutemica se duplica al aumentar la temperatura en 10
0C 2) Antildeadiendo un catalizador que
disminuye la energiacutea de activacioacuten y aumenta la velocidad de reaccioacuten
La enzima (E) se combina con el substrato (S) formando el complejo de transicioacuten enzima-
substrato (E-S) mediante una reaccioacuten reversible cuya energiacutea de activacioacuten es menor que la de la reaccioacuten no catalizada Cuando se forma el producto de la reaccioacuten (P) se regenera de nuevo la enzima (E) de forma libre la que puede combinarse de nuevo con otra moleacutecula de substrato (S)
Una enzima reduce maacutes eficientemente la energiacutea de activacioacuten(Ea) de una reaccioacuten que un catalizador inorgaacutenico lo que permite que una reaccioacuten se realice a menor temperatura
El siguiente ejemplo ilustra mejor lo que hemos discutido
Reaccioacuten Energiacutea de activacioacuten (Kcalmol-1
)
21
a) el peroacutexido de hidroacutegeno se descompone en
H2 O2 H2 O + O2
18
b) el hierro cataliacutetico (Fe) realiza la reaccioacuten
H2 O2 H2 O + O2
13
c) el platino cataliacutetico (Pt) realiza la reaccioacuten
H2 O2 H2 O + O2
12
d) la catalasa una enzima hepaacutetica la realiza
H2O2 H2O + O2
5
Una enzima no modifica la energiacutea libre ni la constante de equilibrio sino que disminuye la energiacutea de activacioacuten de la reaccioacuten
Ir al principio
EFECTO DE LA CONCENTRACIOacuteN DEL SUBSTRATO SOBRE LA
CATALISIS ENZIMATICA
La actividad de una enzima se puede estudiar in vitro bajo condiciones controladas antildeadieacutendole una enzima a un substrato Siacute se trabaja con la condicioacuten de que la concentracioacuten del substrato sea saturante variando entonces la concentracioacuten de enzima se observa que aumenta el producto de la reaccioacuten a pH y temperatura constantes
22
Si se mantiene la concentracioacuten de la enzima constante y variando la concentracioacuten de substrato se obtiene una curva hiperboacutelica como la de la figura Al principio un aumento de la
concentracioacuten de substrato produce un aumento raacutepi do de la velocidad de reaccioacuten pero si se sigue aumentando la concentracioacuten de substrato la velocidad de reaccioacuten comienza a disminuir y a muy altas concentraciones de substrato se observa que no cambia la velocidad de
reaccioacuten se dice que los centros activos de la enzima se encuentran saturados La velocidad de reaccioacuten que se obtiene a esa alta concentracioacuten de substrato se define como la velocidad maacutexima (V) de la reaccioacuten enzimaacutetica bajo las condiciones especificadas La concentracioacuten de
substrato (S) a la semivelocidad maacutexima de reaccioacuten (V2) se puede determinar de la figura y representa la constante de Michaelis o Km la cual es una caracteriacutestica para cada enzima La inversa de Km o 1Km mide aproximadamente la afinidad de la enzima por el subs trato
Mientras maacutes pequentildeo sea el valor de Km mayor seraacute la afinidad de la enzima por el substrato Si varias enzimas compiten en el metabolismo por el mismo substrato eacuteste seraacute transformado preferentemente por la enzima con mayor afinidad
La ecuacioacuten de Michaelis describe la relacioacuten cuantitativa entre la velocidad de reaccioacuten y la concentracioacuten de substrato [S] si se conoce Vmax o Km
v=
En donde v= es la velocidad de reaccioacuten observada a una concentracioacuten de substrato determinada [S]
Km= constante de Michaelis en molesliacutet ro
23
Vmax= velocidad maacutexima a concentracioacuten saturante de substrato
Si v= Vmax2 entonces
=
Km+[S] = 2[S]
Km =[S]
De tal forma que podemos concluir que Km es igual a la concentracioacuten de substrato a la semivelocidad maacutexima de reaccioacuten
1 Concentraciones relativas de E y S la concentracioacuten de substrato S es
mucho mayor que la de E de tal manera que la cantidad de substrato
unido a la enzima en cualquier momento es muy pequentildea
2 Se asume el estado estacionario ES no cambia con el tiempo esto
quiere decir que la velocidad de formacioacuten de ES es igual a aquella para
su desintegracioacuten En general un intermediario en una serie de reacciones estaacute en estado estacionario cuando su velocidad de siacutentesis
es igual a su velocidad de degradacioacuten 3 Velocidad inicial para el anaacutelisis de reacciones enzimaacuteticas soacutelo se
utiliza la velocidad inicial de la reaccioacuten que es la velocidad ejercida por
la enzima inmediatamente despueacutes de que se ha puesto en contacto con el substrato y hasta antes de que se haya consumido el 10 de la concentracioacuten inicial del mismo La razoacuten de lo anterior es que en ese
momento la concentracioacuten del producto de la reaccioacuten que se ha acumulado en muy pequentildea y por tanto la reaccioacuten en el sentido
inverso es decir la transformacioacuten del producto en el substrato original puede ser ignorada
24
Caracteriacutesticas de Km la constante de Michaelis-Menten es
caracteriacutestica de una enzima y particular para un substrato Refleja la afinidad de la enzima por ese substrato K m es numeacutericamente igual a la concentracioacuten
de substrato a la cual la velocidad de reaccioacuten es la mitad de la Vmax (Km= Vmax2) Este paraacutemetro Km no variacutea con la concentracioacuten de enzima
Significado de una Km pequentildea un valor numeacuterico pequentildeo de Km
refleja una alta afinidad de la enzima por su substrato porque a una baja
concentracioacuten del mismo la enzima a desarrollado ya la mitad de la velocidad maacutexima
Significado de una Km grande el valor numeacuterico grande de Km refleja
una baja afinidad de la enzima por su substrato porque a una concentracioacuten
elevada del mismo la enzima desarrolla la mitad de la velocidad maacutexima
Relacioacuten de la velocidad con la concentracioacuten de enzima la
velocidad de la reaccioacuten es directamente proporcional a la concentracioacuten de la
enzima a cualquier concentracioacuten de substrato Por ejemplo si la concentracioacuten de enzima es disminuida a la mitad la velocidad inicial de la reaccioacuten (v0) es
reducida tambieacuten a la mitad de la original
Orden de la reaccioacuten cuando S es mas pequentildea que la Km la
velocidad de la reaccioacuten es aproximadamente igual a la concentracioacuten de
substrato
100
50
0
Substrato
Km Figura Localizacioacuten de la constante de Michaelis (Km)
25
La velocidad de reaccioacuten se dice en estas condiciones es de primer
orden con respecto al substrato Cuando S es mas grande que la Km la
velocidad es constante e igual a la Vmax La velocidad de la reaccioacuten es independiente de la concentracioacuten de substrato y se dice que es de orden cero
con respecto a la concentracioacuten de substrato
Las enzimas pueden ser aisladas de las ceacutelulas u organismos para estudiar sus
propiedades in vitro es decir en un tubo de ensayo Las diferentes enzimas muestran diferentes respuestas a los cambios en la concentracioacuten de substrato temperatura y pH
Concentracioacuten de substrato
La forma hiperboacutelica de la curva de velocidad vs Substrato
Efecto de la temperatura en la actividad enzimaacutetica
Efecto de la variacioacuten del pH en la actividad enzimaacutetica
Incremento de la velocidad con la temperatura En general la velocidad de la reaccioacuten se incrementa con la temperatura hasta que un punto maacuteximo es alcanzado
Este incremento se debe a que aumenta el nuacutemero de moleacuteculas ricas en energiacutea que pueden pasar la barrera energeacutetica de estado de transicioacuten para formar a los productos
Decremento de la velocidad con la temperatura la elevacioacuten excesiva de la
temperatura del medio que contiene a las enzimas resulta en un decremento de la velocidad como resultado de la desnaturalizacioacuten es decir la peacuterdida de la estructura tridimensional de las enzimas
26
zona de reactivacioacuten maacuteximo de actividad
Zona de
inactivacioacuten
0 Temperatura
Figura dependencia de la actividad enzimaacutetica con la temperatura
Velocidad maacutexima la velocidad de una reaccioacuten (v) es el nuacutemero de
moleacuteculas de substrato convertidas en producto por unidad de tiempo y
usualmente se expresa en micromoles de producto formadas por minuto La velocidad de una reaccioacuten catalizada por una enzima aumenta conforme se incrementa la concentracioacuten de substrato hasta que se llega a una velocidad maacutexima (Vmax) a partir de la cual la velocidad de la reaccioacuten es
independiente de la cantidad de substrato El que la velocidad no pueda
seguirse incrementando refleja la saturacioacuten del sitio activo con el substrato
Vmax
0 Substrato
Figura representacioacuten de una cineacutetica tiacutepicamente michaeliana
Vmax = velocidad de saturacioacuten o maacutexima
27
Cuando se grafica la velocidad de la reaccioacuten v0 contra la concentracioacuten de substrato S no siempre es posible determinar la condicioacuten
en que se ha llegado a la velocidad maacutexima Vmax debido al incremento de la pendiente en la hipeacuterbola a concentraciones de substrato elevadas
Figura experimento cineacutetico para la actividad enzimaacutetica
Se presentan los valores experimentales y los resultados del ajuste al modelo de Michaelis
Auacuten asiacute si se grafica una doble reciacuteproca es decir el inverso de la
velocidad (1v0) contra el inverso de la concentracioacuten de substrato (1 S ) se
obtiene una liacutenea recta El regraacutefico de Lineweaver-Burke tambieacuten se
denomina de ldquodobles reciacuteprocasrdquo y se utiliza para calcular la Km y la Vmax asiacute como para determinar el mecanismo de accioacuten de los diversos tipos de
inhibidores
La ecuacioacuten que describe a la graacutefca de Lineweaver-Burke es
Vmax
1
SVmax
Km
v
1
0 (3)
que describe una recta en donde el intercepto con el eje de las abscisas es igual a ndash1Km y el intercepto con el eje de las ordenadas es igual a 1Vmax
28
Figura experimento cineacutetico para la actividad enzimaacutetica Regraacutefico de
Lineweaver-Burke
Se presentan los valores experimentales y los resultados del ajuste al modelo
REPRESENTACIOacuteN GRAacuteFICA DE LINEWEAVER Y BURK
Si se toma el inverso en ambos miembros de la ecuacioacuten de Michaelis tenemos
=
+
29
que es la ecuacioacuten de una liacutenea recta cuya pendiente es que intercepta al eje de las
ordenadas en y a la abcisa en el punto Lo que es evidente al hacer
ya que se convierte en
Si se hace una graacutefica con los dobles inversos colocando los valores de en el eje de las
ordenadas y en el eje de las abcisas se obtiene una liacutenea recta a partir de la cual se puede calcular faacutecilmente Km
EFECTO DEL pH SOBRE LA ACTIVIDAD ENZIMATICA
Sabiendo que las enzimas son proteinas cualquier cambio brusco de pH puede alterar el caraacutecter ioacutenico de los grupos amino y carboxilo en la superficie proteica afectando asiacute las propiedades cataliacuteticas de una enzima A pH alto o bajo se puede producir la desnaturalizacioacuten
de la enzima y en consecuencia su inactivacioacuten La fosfatasa aacutecida es maacutes activa a pH 50 mientras que la fosfatasa alcalina lo es a pH 90 Muchas enzimas tienen maacutexima actividad cerca de la neutralidad en un rango de pH de 6 a 8
El pH per se no afecta la actividad enzimaacutetica sino la concentracioacuten de protones Los protones
ademaacutes de alterar la estructura de la enzima y el substrato pueden participar tambieacuten en la reaccioacuten como substrato o producto En esos casos la concentracioacuten de protones afecta directamente la velocidad de la reaccioacuten
ENZIMA pH OPTIMO
30
Pepsina 15
Tripsina 77
Catalasa 76
Arginasa 97
Fumarasa 78
Ribonucleasa 78
EFECTO DE ACTIVADORES
Un aumento en la temperatura provoca un aumento de la velocidad de reaccioacuten hasta cierta temperatura oacuteptima ya que despueacutes de aproximadamente 45
0 C se comienza a producir la
desnaturalizacioacuten teacutermica Las enzimas de muchos mamiacuteferos tienen una temperatura oacuteptima
de 370 C por encima de esa temperatura comienzan a inactivarse y se destruyen Sin embargo
existen especies de bacterias y algas que habitan en fuentes de aguas termales y en el otro extremo ciertas bacterias aacuterticas tienen temperaturas oacuteptimas cercanas a 0
0 C
31
ESPECIFICIDAD ABSOLUTA
Cierta enzimas se encuentran como aproenzimas o zimogenos que son inactivos
El tripsinoacutegeno es convertido en tripsina por la misma accioacuten de la tripsina que remueve un hexapeacuteptido
tripsina Tripsinoacutegeno Tripsina + Val (Asp)4 lisina
Otra forma de activacioacuten consiste en mantener los grupos SH reducidos Estos grupos pueden
formar parte de centros activos de ciertas enzimas Si se oxidan estos grupos a -S-S- la enzima se inactiva Asiacute tenemos que la enzima papaina se inactiva despueacutes de exponerla al ox iacutegeno pero cuando se le antildeade un reductor apropiado la enzima se reactiva Los grupos ndashS-S- se convierten en ndashSH activaacutendose la enzima
Otra forma de activacioacuten requiere la presencia de otra substancia que se enlaza al sitio alosteacuterico el compuesto que se enlaza se denomina un efector alosteacuterico El centro alosteacuterico es bien especiacutefico para el efector o modulador alosteacuterico
ESPECIFICIDAD RELATIVA
Oxiacutegeno L-aminoaacutecido a -cetoaacutecido + NH3 + H 2 O 2 L-aminoaacutecido oxidasa Oxiacutegeno D-aminoaacutecido a -cetoaacutecido + NH3 + H 2 O 2 D-aminoaacutecido oxidasa
32
En este ejemplo se muestra la estereo-especificidad Una enzima puede tener una especificidad oacuteptica para isomeros D o L
Hay enzimas que muestran especificidad absoluta como la aspartasa que cataliza la adicioacuten reversible de amoniacuteaco al doble enlace del aacutecido fumaacuterico pero no al de otros aacutecidos insaturados
HOOC ndash CH = CH ndash COO- + NH4
+ HOOC ndash CH2 - CHNH2 - COO
- + H
+
INHIBICIOacuteN ENZIMATICA
Mediante el uso de inhibidores enzimaacuteticos se ha obtenido informacioacuten muy valiosa sobre la conformacioacuten del centro activo de algunas enzimas
a Inhibicioacuten irreversible Este tipo de inhibidores forman un enlace covalente con las enzimas cerca del centro activo Un ejemplo son los gases nerviosos como el
fluorofosfato de diisopropilo (DFP) que forma un complejo con la enzima acetilcolinesterasa Los animales envenenados con este gas quedan paralizados debido a la imposibilidad de transmitir adecuadamente los impulsos nerviosos
Ester di-isopropil fos foacuterico de la enzima (inactivo)
b) Inhibicioacuten competitiva
Es cuando el inhibidor compite con el substrato por la unioacuten con el centro activo de la
enzima Este tipo de inhibicioacuten puede reducirse si se aumenta la concentracioacuten de substrato
-OOC - CH2 - CH2 - COO
- -OOC - CH = CH - COO
- + 2H
+ Succinato Succinato Fumarato
33
Deshidrogenasa COO
- I CH2 Malonato (inhibidor competitivo) I COO
- En la inhibicioacuten competitiva la enzima se combina con el inhibidor para formar un complejo enzima-inhibidor E + I E I en competencia con la reaccioacuten normal E +S ES
La sulfanilamida (bactericida) interfiere con la siacutentesis del aacutecido foacutelico compitiendo con el aacutecido p-aminobenzoico de una forma competitiva
HOOC-sect- NH2 NH2 ndashSO2 - sect- NH2 aacutecido p- amino benzoico sulfanilamida
sect = fenil
a Inhibicioacuten no competitiva
Esta inhibicioacuten se caracteriza por que no se puede revertir el efecto del inhibidor aumentando la concentracioacuten del substrato
Por ejemplo la inhibicioacuten de la deshidrogenasa del gliceraldehido -3-fosfato por la yodo-acetamida
Enzima-SH + I-CH2 CONH2 Enzima-SCH2 CONH2 + IH
La deshidrogenasa del gliceraldehido-3 fosfato puede ser inhibida tambieacuten por el aacutecido iodo-aceacutetico
Enzima-SH + ICH2 COOH Enzima-SCH2 COOH + IH
c) Inhibicioacuten por metales
Ciertos metales como el plomo mercurio y arseacutenico inhiben enzimas que tienen en su centro activo grupos -SH libres
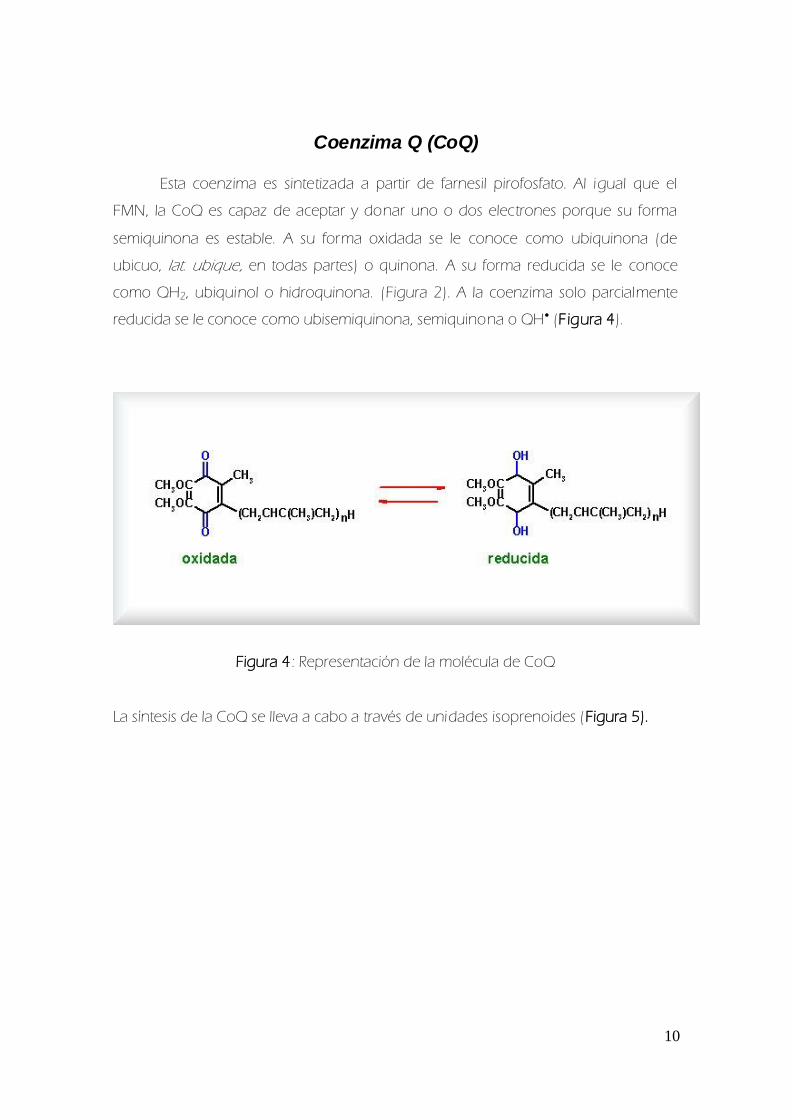
10
Coenzima Q (CoQ)
Esta coenzima es sintetizada a partir de farnesil pirofosfato Al igual que el
FMN la CoQ es capaz de aceptar y donar uno o dos electrones porque su forma
semiquinona es estable A su forma oxidada se le conoce como ubiquinona (de
ubicuo lat ubique en todas partes) o quinona A su forma reducida se le conoce
como QH2 ubiquinol o hidroquinona (Figura 2) A la coenzima solo parcialmente
reducida se le conoce como ubisemiquinona semiquinona o QH (Figura 4)
Figura 4 Representacioacuten de la moleacutecula de CoQ
La siacutentesis de la CoQ se lleva a cabo a traveacutes de unidades isoprenoides (Figura 5)
11
Figura 5 La CoQ se produce a partir de isoprenoides
Biotina
La biotina consiste de un anillo imidazol que estaacute unido en forma cis a la
cadena lateral tetrahidrotiofeno del valerato (Figura 6)
12
Figura 6 Estructura de la biotina
La enzima holocarboxilasa sintetasa (HCS) cataliza la activacioacuten mediante
biotinilacioacuten de cinco carboxilasas en ceacutelulas humanas La deficiencia de HCS
produce el siacutendrome deficiencia muacuteltiple de carboxilasas (DMC) Esta enfermedad es
potencialmente mortal y se caracteriza por la deficiencia de la actividad de todas las
carboxilasas La deficiencia de biotina no solo disminuye la actividad de estas enzimas
sino que parece modificar la expresioacuten geneacutetica
126 PROPIEDADES DE LAS ENZIMAS
Las propiedades de las enzimas derivan del hecho de ser proteiacutenas y de actuar como
catalizadores Como proteiacutenas poseen una conformacioacuten natural maacutes estable que las
demaacutes conformaciones posibles Asiacute cambios en la conformacioacuten suelen ir asociados
en cambios en la actividad cataliacutetica Los factores que influyen de manera maacutes directa
sobre la actividad de un enzima son
pH
temperatura
cofactores
A) EFECTO DEL pH
Al comprobar experimentalmente la influencia del pH en la velocidad de las
reacciones enzimaacuteticas se obtienen curvas que indican que los enzimas
presentan un pH oacuteptimo de actividad El pH puede afectar de varias maneras
13
o El centro activo puede contener aminoaacutecidos con grupos ionizados
que pueden variar con el pH
o La ionizacioacuten de aminoaacutecidos que no estaacuten en el centro activo puede
provocar modificaciones en la conformacioacuten de la enzima
o El sustrato puede verse afectado por las variaciones del pH
Algunos enzimas presentan variaciones peculiares La pepsina del estoacutemago
presenta un oacuteptimo a pH=2 la ureasa lo tiene a pH 7 y la fosfatasa alcalina del
intestino un pH= 12
B) EFECTO DE LA TEMPERATURA
Influye en la actividad El punto oacuteptimo representa el maacuteximo de actividad A
temperaturas bajas los enzimas se hallan ldquomuy riacutegidosrdquo y cuando se supera un
valor considerable (mayor de 50) la actividad cae bruscamente porque como
proteiacutena el enzima se desnaturaliza
128 EFECTO DE LOS COFACTORES SOBRE LA ACTIVIDAD
ENZIMATICA
A veces un enzima requiere para su funcioacuten la presencia de sustancias no proteicas
que colaboran en la cataacutelisis los cofactores Casi un tercio de los enzimas conocidos
requieren cofactores Cuando el cofactor es una moleacutecula orgaacutenica se llama
coenzima En la figura inferior podemos observar una moleacutecula de hemoglobina
(proteiacutena que transporta oxiacutegeno) y su coenzima (el grupo hemo) Cuando los
cofactores y las coenzimas se encuentran unidos covalentemente al enzima se llaman
14
grupos prosteacuteticos La forma cataliacuteticamente activa del enzima es decir el enzima
unido a su grupo prosteacutetico se llama holoenzima La parte proteica de un
holoenzima (inactiva) se llama apoenzima de forma que
apoenzima + grupo prosteacutetico= holoenzima
129 MODO DE ACCIOacuteN DE LAS ENZIMAS
En las reacciones espontaacuteneas los productos finales tienen menos energiacutea libre de
Gibbs (∆G) que los reactantes (figura inferior izquierda) Por lo tanto las reacciones
espontaacuteneas liberan energiacutea de Gibbs ((∆G lt 0) Sin embargo el comienzo de la
reaccioacuten requiere un aporte inicial de energiacutea Esta energiacutea inicial que hay que
suministrar a los reactantes para que la reaccioacuten transcurra se llama energiacutea de
activacioacuten (Ea) Cuanto menor es la Ea maacutes faacutecilmente transcurre la reaccioacuten
Perfil energeacutetico de una reaccioacuten espontaacutenea Perfil energeacutetico de una reaccioacuten catalizada
15
La accioacuten de los catalizadores consiste precisamente en disminuir la Ea (Figura superior derecha) Los enzimas
son catalizadores especialmente eficaces ya que disminuyen la Ea auacuten maacutes que los catalizadores inorgaacutenicos
Por ejemplo la descomposicioacuten del agua oxigenada (H2O2) para dar H2O y O2 puede ocurrir sin catalizador
con un catalizador inorgaacutenico (platino) o con un enzima especiacutefico (catalasa) Las respectivas Ea para cada
proceso son 18 12 y 6 Kcalmol Asiacute se puede calcular que el platino acelera la reaccioacuten 20000 veces mientras
que la catalasa la acelera 370000 veces
Para que una reaccioacuten quiacutemica tenga lugar las moleacuteculas de los reactantes deben chocar con una energiacutea y
una orientacioacuten adecuadas La actuacioacuten del enzima (1) permite que los rea ctantes (sustratos) se unan a su
centro activo con una orientacioacuten oacuteptima para que la reaccioacuten se produzca y (2) modifica las propiedades
quiacutemicas del sustrato unido a su centro activo debilitando los enlaces existentes y facilitando la formacioacuten de
otros nuevos (Figuras inferiores)
1210 REGULACIOacuteN DE SU ACTIVIDAD ENZIMATICA
16
Una moleacutecula de enzima no tiene por que actuar siempre a la misma velocidad Su
actividad puede estar modulada por
Cambios en el pH
Cambios en la temperatura
Presencia de cofactores
Las concentraciones del sustrato y de los productos finales
Presencia de inhibidores
Modulacioacuten alosteacuterica
Modificacioacuten covalente
Activacioacuten por proteolisis
Isoenzimas
EFECTO DE LAS CONCENTRACIONES SOBRE LA ACTIVIDAD ENZIMAacuteTICA
La velocidad de una reaccioacuten enzimaacutetica depende de la concentracioacuten de sustrato La Figura de la derecha muestra la velocidad de una reaccioacuten enzimaacutetica a 6 concentraciones distintas de sustrato
Ademaacutes la presencia de los productos finales puede hacer que la reaccioacuten sea maacutes lenta o incluso invertir su sentido (Figura inferior)
EFECTO DE LOS INHIBIDORES SOBRE LA ACTIVIDAD ENZIMAacuteTICA
Ciertas moleacuteculas pueden inhibir la accioacuten cataliacutetica de un enzima son los inhibidores Estos inhibidores bien pueden ocupar temporalmente el centro activo por semejanza estructural con el sustrato original ( inhibidor competitivo) o bien alteran la conformacioacuten espacial del enzima impidiendo su unioacuten al sustrato ( inhibidor no competitivo) (Figuras inferiores)
Inhibidor competitivo Inhibidor no competitivo
17
MODULACIOacuteN ALOSTEacuteRICA DE LA ACTIVIDAD ENZIMAacuteTICA
Hay enzimas que pueden adoptar 2 conformaciones interconvertibles llamadas R (relajada) y T (tensa) R es la forma maacutes activa porque se une al sustrato con maacutes afinidad Las formas R y T se encuentran en equilibrio R lt==gt T (Figura inferior)
Ciertas sustancias tienden a estabilizar la forma R Son los llamados moduladores positivos El propio sustrato es a
menudo un modulador positivo Las moleacuteculas que favorecen la forma R pero que actuacutean sobre una regioacuten del enzima distinta del centro activo son los activadores alosteacutericos (Figura inferior izquierda)
Activador alosteacuterico favorece la unioacuten del sustrato
Inhibidor alosteacuterico impide la unioacuten del sustrato
Las sustancias que favorecen la forma T y disminuyen la actividad enzimaacutetica son los moduladores negativos Si
estos moduladores actuacutean en lugares distintos del centro activo del enzima se llaman inhibidores alosteacutericos
18
(Figura superior derecha
EFECTO DE LA MODIFICACIOacuteN COVALENTE SOBRE LA ACTIVIDAD ENZIMAacuteTICA
Otros enzimas pasan de una forma menos activa a otra maacutes activa unieacutendose covalentemente a un grupo quiacutemico de pequentildeo tamantildeo como el Pi o el AMP Tambieacuten se da el caso inverso en el que un enzima muy activo se desactiva al liberar alguacuten grupo quiacutemico En las enzimas de las viacuteas degradativas del metabolismo la
forma fosforilada es maacutes activa que la no fosforilada mientras que en las viacuteas biosinteacuteticas ocurre lo contrario En las figuras inferiores se ilustra la activacioacuten de una proteiacutena por fosforilacioacuten
Elementos de la reaccioacuten El enzima no fosforilado es inactivo El enzima fosforilado es activo
ACTIVACIOacuteN PROTEOLIacuteTICA DE LA ACTIVIDAD ENZIMAacuteTICA
Algunos enzimas no se sintetizan como tales sino como proteiacutenas precursoras sin actividad enzimaacutetica Estas proteiacutenas se llaman
proenzimas o zimoacutegenos Para activarse los zimoacutegenos sufren un ataque hidroliacutetico que origina la liberacioacuten de uno o varios peacuteptidos El resto de la moleacutecula proteica adopta la conformacioacuten y las propiedades del enzima activo Muchos enzimas digestivos se secretan en forma de zimoacutegenos y en el tubo digestivo
se convierten en la forma activa Es el caso de la -quimotripsina que se sintetiza en forma de quimotripsinoacutegeno (Figura superior) Si estos enzimas se sintetizasen directamente en forma activa destruiriacutean la propia ceacutelula que las
produce Asiacute la tripsina pancreaacutetica (una proteasa) se sintetiza como tripsinoacutegeno (inactivo) Si por alguna razoacuten se activa en el propio paacutencreas la glaacutendula sufre un proceso de autodestruccioacuten (pancreatitis aguda) a
menudo mortal
19
REGULACIOacuteN DE LA ACTIVIDAD ENZIMAacuteTICA POR MEDIO DE ISOENZIMAS
Algunos enzimas tienen distinta estructura molecular aunque su funcioacuten bioloacutegica es similar Se llaman isozimas o isoenzimas Estas diferencias de estructura se traducen en ligeros cambios en sus propiedades de forma que cada isozima se adapta perfectamente a la funcioacuten que debe realizar Asiacute podemos observar la existencia de isoenzimas en funcioacuten de
el tipo de tejido Por ejemplo la lactato deshidrogenasa presenta isozimas distintos en muacutesculo y corazoacuten el compartimento celular donde actuacutea Por ejemplo la malato deshidrogenasa del citoplasma es distinta
de la de la mitocondria el momento concreto del desarrollo del individuo Por ejemplo algunos enzimas de la glicolisis del feto
son diferentes de los mismos enzimas en el adulto
CINEacuteTICA ENZIMAacuteTICA
Los principios generales de las reacciones quiacutemicas se aplican tambieacuten a las reacciones enzimaacuteticas Por este motivo antes de empezar con la cineacutetica quiacutemica se van a repasar algunos conceptos baacutesicos de cineacutetica quiacutemica
A continuacioacuten se describiraacuten los siguientes conceptos
Cineacutetica enzimaacutetica Modelo cineacutetico de Michaelis-Menten Caacutelculo de la KM y la Vmax de un enzima Actividad enzimaacutetica
20
NOMENCLATURA DE LOS ENZIMAS
CATALISIS MOLECULAR
Un catalizador modifica la velocidad de una reaccioacuten quiacutemica sin ser utilizado o aparecer como uno de los productos de la reaccioacuten Una reaccioacuten quiacutemica en la que un substrato(S) se
transforma en un producto(P) S P ocurre por que cierta fraccioacuten de moleacuteculas de S posee mucho maacutes energiacutea que el resto de ellas lo que es suficiente para que alcancen un estado activado en el que pueda formarse o romperse un enlace quiacutemico y se forme el producto(P)
La energiacutea de activacioacuten es la cantidad de energiacutea expresada en caloriacuteas necesaria para que todas las moleacuteculas de un mol a una temperatura dada alcancen el estado reactivo Mientras que el estado de transicioacuten es el estado rico en energiacutea de las moleacuteculas que
interaccionan en la cima de la barrera de activacioacuten La velocidad de una reaccioacuten quiacutemica es proporcional a la concentracioacuten del complejo en el estado de transicioacuten
Una reaccioacuten quiacutemica se puede acelerar de la siguiente forma 1) Al aumentar la temperatura se incrementa la energiacutea cineacutetica por lo que es mayor el nuacutemero de moleacuteculas que alcanzan el
estado de transicioacuten Generalmente el Q10 =2 lo que indica que la velocidad de una reaccioacuten quiacutemica se duplica al aumentar la temperatura en 10
0C 2) Antildeadiendo un catalizador que
disminuye la energiacutea de activacioacuten y aumenta la velocidad de reaccioacuten
La enzima (E) se combina con el substrato (S) formando el complejo de transicioacuten enzima-
substrato (E-S) mediante una reaccioacuten reversible cuya energiacutea de activacioacuten es menor que la de la reaccioacuten no catalizada Cuando se forma el producto de la reaccioacuten (P) se regenera de nuevo la enzima (E) de forma libre la que puede combinarse de nuevo con otra moleacutecula de substrato (S)
Una enzima reduce maacutes eficientemente la energiacutea de activacioacuten(Ea) de una reaccioacuten que un catalizador inorgaacutenico lo que permite que una reaccioacuten se realice a menor temperatura
El siguiente ejemplo ilustra mejor lo que hemos discutido
Reaccioacuten Energiacutea de activacioacuten (Kcalmol-1
)
21
a) el peroacutexido de hidroacutegeno se descompone en
H2 O2 H2 O + O2
18
b) el hierro cataliacutetico (Fe) realiza la reaccioacuten
H2 O2 H2 O + O2
13
c) el platino cataliacutetico (Pt) realiza la reaccioacuten
H2 O2 H2 O + O2
12
d) la catalasa una enzima hepaacutetica la realiza
H2O2 H2O + O2
5
Una enzima no modifica la energiacutea libre ni la constante de equilibrio sino que disminuye la energiacutea de activacioacuten de la reaccioacuten
Ir al principio
EFECTO DE LA CONCENTRACIOacuteN DEL SUBSTRATO SOBRE LA
CATALISIS ENZIMATICA
La actividad de una enzima se puede estudiar in vitro bajo condiciones controladas antildeadieacutendole una enzima a un substrato Siacute se trabaja con la condicioacuten de que la concentracioacuten del substrato sea saturante variando entonces la concentracioacuten de enzima se observa que aumenta el producto de la reaccioacuten a pH y temperatura constantes
22
Si se mantiene la concentracioacuten de la enzima constante y variando la concentracioacuten de substrato se obtiene una curva hiperboacutelica como la de la figura Al principio un aumento de la
concentracioacuten de substrato produce un aumento raacutepi do de la velocidad de reaccioacuten pero si se sigue aumentando la concentracioacuten de substrato la velocidad de reaccioacuten comienza a disminuir y a muy altas concentraciones de substrato se observa que no cambia la velocidad de
reaccioacuten se dice que los centros activos de la enzima se encuentran saturados La velocidad de reaccioacuten que se obtiene a esa alta concentracioacuten de substrato se define como la velocidad maacutexima (V) de la reaccioacuten enzimaacutetica bajo las condiciones especificadas La concentracioacuten de
substrato (S) a la semivelocidad maacutexima de reaccioacuten (V2) se puede determinar de la figura y representa la constante de Michaelis o Km la cual es una caracteriacutestica para cada enzima La inversa de Km o 1Km mide aproximadamente la afinidad de la enzima por el subs trato
Mientras maacutes pequentildeo sea el valor de Km mayor seraacute la afinidad de la enzima por el substrato Si varias enzimas compiten en el metabolismo por el mismo substrato eacuteste seraacute transformado preferentemente por la enzima con mayor afinidad
La ecuacioacuten de Michaelis describe la relacioacuten cuantitativa entre la velocidad de reaccioacuten y la concentracioacuten de substrato [S] si se conoce Vmax o Km
v=
En donde v= es la velocidad de reaccioacuten observada a una concentracioacuten de substrato determinada [S]
Km= constante de Michaelis en molesliacutet ro
23
Vmax= velocidad maacutexima a concentracioacuten saturante de substrato
Si v= Vmax2 entonces
=
Km+[S] = 2[S]
Km =[S]
De tal forma que podemos concluir que Km es igual a la concentracioacuten de substrato a la semivelocidad maacutexima de reaccioacuten
1 Concentraciones relativas de E y S la concentracioacuten de substrato S es
mucho mayor que la de E de tal manera que la cantidad de substrato
unido a la enzima en cualquier momento es muy pequentildea
2 Se asume el estado estacionario ES no cambia con el tiempo esto
quiere decir que la velocidad de formacioacuten de ES es igual a aquella para
su desintegracioacuten En general un intermediario en una serie de reacciones estaacute en estado estacionario cuando su velocidad de siacutentesis
es igual a su velocidad de degradacioacuten 3 Velocidad inicial para el anaacutelisis de reacciones enzimaacuteticas soacutelo se
utiliza la velocidad inicial de la reaccioacuten que es la velocidad ejercida por
la enzima inmediatamente despueacutes de que se ha puesto en contacto con el substrato y hasta antes de que se haya consumido el 10 de la concentracioacuten inicial del mismo La razoacuten de lo anterior es que en ese
momento la concentracioacuten del producto de la reaccioacuten que se ha acumulado en muy pequentildea y por tanto la reaccioacuten en el sentido
inverso es decir la transformacioacuten del producto en el substrato original puede ser ignorada
24
Caracteriacutesticas de Km la constante de Michaelis-Menten es
caracteriacutestica de una enzima y particular para un substrato Refleja la afinidad de la enzima por ese substrato K m es numeacutericamente igual a la concentracioacuten
de substrato a la cual la velocidad de reaccioacuten es la mitad de la Vmax (Km= Vmax2) Este paraacutemetro Km no variacutea con la concentracioacuten de enzima
Significado de una Km pequentildea un valor numeacuterico pequentildeo de Km
refleja una alta afinidad de la enzima por su substrato porque a una baja
concentracioacuten del mismo la enzima a desarrollado ya la mitad de la velocidad maacutexima
Significado de una Km grande el valor numeacuterico grande de Km refleja
una baja afinidad de la enzima por su substrato porque a una concentracioacuten
elevada del mismo la enzima desarrolla la mitad de la velocidad maacutexima
Relacioacuten de la velocidad con la concentracioacuten de enzima la
velocidad de la reaccioacuten es directamente proporcional a la concentracioacuten de la
enzima a cualquier concentracioacuten de substrato Por ejemplo si la concentracioacuten de enzima es disminuida a la mitad la velocidad inicial de la reaccioacuten (v0) es
reducida tambieacuten a la mitad de la original
Orden de la reaccioacuten cuando S es mas pequentildea que la Km la
velocidad de la reaccioacuten es aproximadamente igual a la concentracioacuten de
substrato
100
50
0
Substrato
Km Figura Localizacioacuten de la constante de Michaelis (Km)
25
La velocidad de reaccioacuten se dice en estas condiciones es de primer
orden con respecto al substrato Cuando S es mas grande que la Km la
velocidad es constante e igual a la Vmax La velocidad de la reaccioacuten es independiente de la concentracioacuten de substrato y se dice que es de orden cero
con respecto a la concentracioacuten de substrato
Las enzimas pueden ser aisladas de las ceacutelulas u organismos para estudiar sus
propiedades in vitro es decir en un tubo de ensayo Las diferentes enzimas muestran diferentes respuestas a los cambios en la concentracioacuten de substrato temperatura y pH
Concentracioacuten de substrato
La forma hiperboacutelica de la curva de velocidad vs Substrato
Efecto de la temperatura en la actividad enzimaacutetica
Efecto de la variacioacuten del pH en la actividad enzimaacutetica
Incremento de la velocidad con la temperatura En general la velocidad de la reaccioacuten se incrementa con la temperatura hasta que un punto maacuteximo es alcanzado
Este incremento se debe a que aumenta el nuacutemero de moleacuteculas ricas en energiacutea que pueden pasar la barrera energeacutetica de estado de transicioacuten para formar a los productos
Decremento de la velocidad con la temperatura la elevacioacuten excesiva de la
temperatura del medio que contiene a las enzimas resulta en un decremento de la velocidad como resultado de la desnaturalizacioacuten es decir la peacuterdida de la estructura tridimensional de las enzimas
26
zona de reactivacioacuten maacuteximo de actividad
Zona de
inactivacioacuten
0 Temperatura
Figura dependencia de la actividad enzimaacutetica con la temperatura
Velocidad maacutexima la velocidad de una reaccioacuten (v) es el nuacutemero de
moleacuteculas de substrato convertidas en producto por unidad de tiempo y
usualmente se expresa en micromoles de producto formadas por minuto La velocidad de una reaccioacuten catalizada por una enzima aumenta conforme se incrementa la concentracioacuten de substrato hasta que se llega a una velocidad maacutexima (Vmax) a partir de la cual la velocidad de la reaccioacuten es
independiente de la cantidad de substrato El que la velocidad no pueda
seguirse incrementando refleja la saturacioacuten del sitio activo con el substrato
Vmax
0 Substrato
Figura representacioacuten de una cineacutetica tiacutepicamente michaeliana
Vmax = velocidad de saturacioacuten o maacutexima
27
Cuando se grafica la velocidad de la reaccioacuten v0 contra la concentracioacuten de substrato S no siempre es posible determinar la condicioacuten
en que se ha llegado a la velocidad maacutexima Vmax debido al incremento de la pendiente en la hipeacuterbola a concentraciones de substrato elevadas
Figura experimento cineacutetico para la actividad enzimaacutetica
Se presentan los valores experimentales y los resultados del ajuste al modelo de Michaelis
Auacuten asiacute si se grafica una doble reciacuteproca es decir el inverso de la
velocidad (1v0) contra el inverso de la concentracioacuten de substrato (1 S ) se
obtiene una liacutenea recta El regraacutefico de Lineweaver-Burke tambieacuten se
denomina de ldquodobles reciacuteprocasrdquo y se utiliza para calcular la Km y la Vmax asiacute como para determinar el mecanismo de accioacuten de los diversos tipos de
inhibidores
La ecuacioacuten que describe a la graacutefca de Lineweaver-Burke es
Vmax
1
SVmax
Km
v
1
0 (3)
que describe una recta en donde el intercepto con el eje de las abscisas es igual a ndash1Km y el intercepto con el eje de las ordenadas es igual a 1Vmax
28
Figura experimento cineacutetico para la actividad enzimaacutetica Regraacutefico de
Lineweaver-Burke
Se presentan los valores experimentales y los resultados del ajuste al modelo
REPRESENTACIOacuteN GRAacuteFICA DE LINEWEAVER Y BURK
Si se toma el inverso en ambos miembros de la ecuacioacuten de Michaelis tenemos
=
+
29
que es la ecuacioacuten de una liacutenea recta cuya pendiente es que intercepta al eje de las
ordenadas en y a la abcisa en el punto Lo que es evidente al hacer
ya que se convierte en
Si se hace una graacutefica con los dobles inversos colocando los valores de en el eje de las
ordenadas y en el eje de las abcisas se obtiene una liacutenea recta a partir de la cual se puede calcular faacutecilmente Km
EFECTO DEL pH SOBRE LA ACTIVIDAD ENZIMATICA
Sabiendo que las enzimas son proteinas cualquier cambio brusco de pH puede alterar el caraacutecter ioacutenico de los grupos amino y carboxilo en la superficie proteica afectando asiacute las propiedades cataliacuteticas de una enzima A pH alto o bajo se puede producir la desnaturalizacioacuten
de la enzima y en consecuencia su inactivacioacuten La fosfatasa aacutecida es maacutes activa a pH 50 mientras que la fosfatasa alcalina lo es a pH 90 Muchas enzimas tienen maacutexima actividad cerca de la neutralidad en un rango de pH de 6 a 8
El pH per se no afecta la actividad enzimaacutetica sino la concentracioacuten de protones Los protones
ademaacutes de alterar la estructura de la enzima y el substrato pueden participar tambieacuten en la reaccioacuten como substrato o producto En esos casos la concentracioacuten de protones afecta directamente la velocidad de la reaccioacuten
ENZIMA pH OPTIMO
30
Pepsina 15
Tripsina 77
Catalasa 76
Arginasa 97
Fumarasa 78
Ribonucleasa 78
EFECTO DE ACTIVADORES
Un aumento en la temperatura provoca un aumento de la velocidad de reaccioacuten hasta cierta temperatura oacuteptima ya que despueacutes de aproximadamente 45
0 C se comienza a producir la
desnaturalizacioacuten teacutermica Las enzimas de muchos mamiacuteferos tienen una temperatura oacuteptima
de 370 C por encima de esa temperatura comienzan a inactivarse y se destruyen Sin embargo
existen especies de bacterias y algas que habitan en fuentes de aguas termales y en el otro extremo ciertas bacterias aacuterticas tienen temperaturas oacuteptimas cercanas a 0
0 C
31
ESPECIFICIDAD ABSOLUTA
Cierta enzimas se encuentran como aproenzimas o zimogenos que son inactivos
El tripsinoacutegeno es convertido en tripsina por la misma accioacuten de la tripsina que remueve un hexapeacuteptido
tripsina Tripsinoacutegeno Tripsina + Val (Asp)4 lisina
Otra forma de activacioacuten consiste en mantener los grupos SH reducidos Estos grupos pueden
formar parte de centros activos de ciertas enzimas Si se oxidan estos grupos a -S-S- la enzima se inactiva Asiacute tenemos que la enzima papaina se inactiva despueacutes de exponerla al ox iacutegeno pero cuando se le antildeade un reductor apropiado la enzima se reactiva Los grupos ndashS-S- se convierten en ndashSH activaacutendose la enzima
Otra forma de activacioacuten requiere la presencia de otra substancia que se enlaza al sitio alosteacuterico el compuesto que se enlaza se denomina un efector alosteacuterico El centro alosteacuterico es bien especiacutefico para el efector o modulador alosteacuterico
ESPECIFICIDAD RELATIVA
Oxiacutegeno L-aminoaacutecido a -cetoaacutecido + NH3 + H 2 O 2 L-aminoaacutecido oxidasa Oxiacutegeno D-aminoaacutecido a -cetoaacutecido + NH3 + H 2 O 2 D-aminoaacutecido oxidasa
32
En este ejemplo se muestra la estereo-especificidad Una enzima puede tener una especificidad oacuteptica para isomeros D o L
Hay enzimas que muestran especificidad absoluta como la aspartasa que cataliza la adicioacuten reversible de amoniacuteaco al doble enlace del aacutecido fumaacuterico pero no al de otros aacutecidos insaturados
HOOC ndash CH = CH ndash COO- + NH4
+ HOOC ndash CH2 - CHNH2 - COO
- + H
+
INHIBICIOacuteN ENZIMATICA
Mediante el uso de inhibidores enzimaacuteticos se ha obtenido informacioacuten muy valiosa sobre la conformacioacuten del centro activo de algunas enzimas
a Inhibicioacuten irreversible Este tipo de inhibidores forman un enlace covalente con las enzimas cerca del centro activo Un ejemplo son los gases nerviosos como el
fluorofosfato de diisopropilo (DFP) que forma un complejo con la enzima acetilcolinesterasa Los animales envenenados con este gas quedan paralizados debido a la imposibilidad de transmitir adecuadamente los impulsos nerviosos
Ester di-isopropil fos foacuterico de la enzima (inactivo)
b) Inhibicioacuten competitiva
Es cuando el inhibidor compite con el substrato por la unioacuten con el centro activo de la
enzima Este tipo de inhibicioacuten puede reducirse si se aumenta la concentracioacuten de substrato
-OOC - CH2 - CH2 - COO
- -OOC - CH = CH - COO
- + 2H
+ Succinato Succinato Fumarato
33
Deshidrogenasa COO
- I CH2 Malonato (inhibidor competitivo) I COO
- En la inhibicioacuten competitiva la enzima se combina con el inhibidor para formar un complejo enzima-inhibidor E + I E I en competencia con la reaccioacuten normal E +S ES
La sulfanilamida (bactericida) interfiere con la siacutentesis del aacutecido foacutelico compitiendo con el aacutecido p-aminobenzoico de una forma competitiva
HOOC-sect- NH2 NH2 ndashSO2 - sect- NH2 aacutecido p- amino benzoico sulfanilamida
sect = fenil
a Inhibicioacuten no competitiva
Esta inhibicioacuten se caracteriza por que no se puede revertir el efecto del inhibidor aumentando la concentracioacuten del substrato
Por ejemplo la inhibicioacuten de la deshidrogenasa del gliceraldehido -3-fosfato por la yodo-acetamida
Enzima-SH + I-CH2 CONH2 Enzima-SCH2 CONH2 + IH
La deshidrogenasa del gliceraldehido-3 fosfato puede ser inhibida tambieacuten por el aacutecido iodo-aceacutetico
Enzima-SH + ICH2 COOH Enzima-SCH2 COOH + IH
c) Inhibicioacuten por metales
Ciertos metales como el plomo mercurio y arseacutenico inhiben enzimas que tienen en su centro activo grupos -SH libres
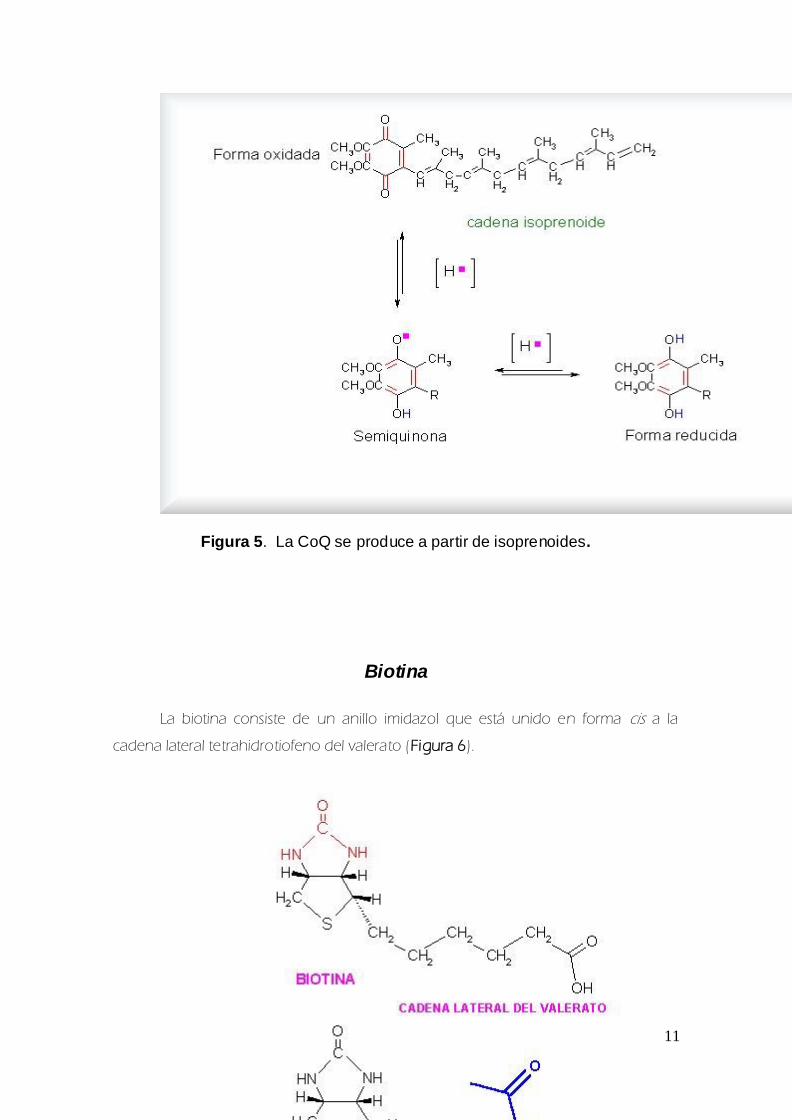
11
Figura 5 La CoQ se produce a partir de isoprenoides
Biotina
La biotina consiste de un anillo imidazol que estaacute unido en forma cis a la
cadena lateral tetrahidrotiofeno del valerato (Figura 6)
12
Figura 6 Estructura de la biotina
La enzima holocarboxilasa sintetasa (HCS) cataliza la activacioacuten mediante
biotinilacioacuten de cinco carboxilasas en ceacutelulas humanas La deficiencia de HCS
produce el siacutendrome deficiencia muacuteltiple de carboxilasas (DMC) Esta enfermedad es
potencialmente mortal y se caracteriza por la deficiencia de la actividad de todas las
carboxilasas La deficiencia de biotina no solo disminuye la actividad de estas enzimas
sino que parece modificar la expresioacuten geneacutetica
126 PROPIEDADES DE LAS ENZIMAS
Las propiedades de las enzimas derivan del hecho de ser proteiacutenas y de actuar como
catalizadores Como proteiacutenas poseen una conformacioacuten natural maacutes estable que las
demaacutes conformaciones posibles Asiacute cambios en la conformacioacuten suelen ir asociados
en cambios en la actividad cataliacutetica Los factores que influyen de manera maacutes directa
sobre la actividad de un enzima son
pH
temperatura
cofactores
A) EFECTO DEL pH
Al comprobar experimentalmente la influencia del pH en la velocidad de las
reacciones enzimaacuteticas se obtienen curvas que indican que los enzimas
presentan un pH oacuteptimo de actividad El pH puede afectar de varias maneras
13
o El centro activo puede contener aminoaacutecidos con grupos ionizados
que pueden variar con el pH
o La ionizacioacuten de aminoaacutecidos que no estaacuten en el centro activo puede
provocar modificaciones en la conformacioacuten de la enzima
o El sustrato puede verse afectado por las variaciones del pH
Algunos enzimas presentan variaciones peculiares La pepsina del estoacutemago
presenta un oacuteptimo a pH=2 la ureasa lo tiene a pH 7 y la fosfatasa alcalina del
intestino un pH= 12
B) EFECTO DE LA TEMPERATURA
Influye en la actividad El punto oacuteptimo representa el maacuteximo de actividad A
temperaturas bajas los enzimas se hallan ldquomuy riacutegidosrdquo y cuando se supera un
valor considerable (mayor de 50) la actividad cae bruscamente porque como
proteiacutena el enzima se desnaturaliza
128 EFECTO DE LOS COFACTORES SOBRE LA ACTIVIDAD
ENZIMATICA
A veces un enzima requiere para su funcioacuten la presencia de sustancias no proteicas
que colaboran en la cataacutelisis los cofactores Casi un tercio de los enzimas conocidos
requieren cofactores Cuando el cofactor es una moleacutecula orgaacutenica se llama
coenzima En la figura inferior podemos observar una moleacutecula de hemoglobina
(proteiacutena que transporta oxiacutegeno) y su coenzima (el grupo hemo) Cuando los
cofactores y las coenzimas se encuentran unidos covalentemente al enzima se llaman
14
grupos prosteacuteticos La forma cataliacuteticamente activa del enzima es decir el enzima
unido a su grupo prosteacutetico se llama holoenzima La parte proteica de un
holoenzima (inactiva) se llama apoenzima de forma que
apoenzima + grupo prosteacutetico= holoenzima
129 MODO DE ACCIOacuteN DE LAS ENZIMAS
En las reacciones espontaacuteneas los productos finales tienen menos energiacutea libre de
Gibbs (∆G) que los reactantes (figura inferior izquierda) Por lo tanto las reacciones
espontaacuteneas liberan energiacutea de Gibbs ((∆G lt 0) Sin embargo el comienzo de la
reaccioacuten requiere un aporte inicial de energiacutea Esta energiacutea inicial que hay que
suministrar a los reactantes para que la reaccioacuten transcurra se llama energiacutea de
activacioacuten (Ea) Cuanto menor es la Ea maacutes faacutecilmente transcurre la reaccioacuten
Perfil energeacutetico de una reaccioacuten espontaacutenea Perfil energeacutetico de una reaccioacuten catalizada
15
La accioacuten de los catalizadores consiste precisamente en disminuir la Ea (Figura superior derecha) Los enzimas
son catalizadores especialmente eficaces ya que disminuyen la Ea auacuten maacutes que los catalizadores inorgaacutenicos
Por ejemplo la descomposicioacuten del agua oxigenada (H2O2) para dar H2O y O2 puede ocurrir sin catalizador
con un catalizador inorgaacutenico (platino) o con un enzima especiacutefico (catalasa) Las respectivas Ea para cada
proceso son 18 12 y 6 Kcalmol Asiacute se puede calcular que el platino acelera la reaccioacuten 20000 veces mientras
que la catalasa la acelera 370000 veces
Para que una reaccioacuten quiacutemica tenga lugar las moleacuteculas de los reactantes deben chocar con una energiacutea y
una orientacioacuten adecuadas La actuacioacuten del enzima (1) permite que los rea ctantes (sustratos) se unan a su
centro activo con una orientacioacuten oacuteptima para que la reaccioacuten se produzca y (2) modifica las propiedades
quiacutemicas del sustrato unido a su centro activo debilitando los enlaces existentes y facilitando la formacioacuten de
otros nuevos (Figuras inferiores)
1210 REGULACIOacuteN DE SU ACTIVIDAD ENZIMATICA
16
Una moleacutecula de enzima no tiene por que actuar siempre a la misma velocidad Su
actividad puede estar modulada por
Cambios en el pH
Cambios en la temperatura
Presencia de cofactores
Las concentraciones del sustrato y de los productos finales
Presencia de inhibidores
Modulacioacuten alosteacuterica
Modificacioacuten covalente
Activacioacuten por proteolisis
Isoenzimas
EFECTO DE LAS CONCENTRACIONES SOBRE LA ACTIVIDAD ENZIMAacuteTICA
La velocidad de una reaccioacuten enzimaacutetica depende de la concentracioacuten de sustrato La Figura de la derecha muestra la velocidad de una reaccioacuten enzimaacutetica a 6 concentraciones distintas de sustrato
Ademaacutes la presencia de los productos finales puede hacer que la reaccioacuten sea maacutes lenta o incluso invertir su sentido (Figura inferior)
EFECTO DE LOS INHIBIDORES SOBRE LA ACTIVIDAD ENZIMAacuteTICA
Ciertas moleacuteculas pueden inhibir la accioacuten cataliacutetica de un enzima son los inhibidores Estos inhibidores bien pueden ocupar temporalmente el centro activo por semejanza estructural con el sustrato original ( inhibidor competitivo) o bien alteran la conformacioacuten espacial del enzima impidiendo su unioacuten al sustrato ( inhibidor no competitivo) (Figuras inferiores)
Inhibidor competitivo Inhibidor no competitivo
17
MODULACIOacuteN ALOSTEacuteRICA DE LA ACTIVIDAD ENZIMAacuteTICA
Hay enzimas que pueden adoptar 2 conformaciones interconvertibles llamadas R (relajada) y T (tensa) R es la forma maacutes activa porque se une al sustrato con maacutes afinidad Las formas R y T se encuentran en equilibrio R lt==gt T (Figura inferior)
Ciertas sustancias tienden a estabilizar la forma R Son los llamados moduladores positivos El propio sustrato es a
menudo un modulador positivo Las moleacuteculas que favorecen la forma R pero que actuacutean sobre una regioacuten del enzima distinta del centro activo son los activadores alosteacutericos (Figura inferior izquierda)
Activador alosteacuterico favorece la unioacuten del sustrato
Inhibidor alosteacuterico impide la unioacuten del sustrato
Las sustancias que favorecen la forma T y disminuyen la actividad enzimaacutetica son los moduladores negativos Si
estos moduladores actuacutean en lugares distintos del centro activo del enzima se llaman inhibidores alosteacutericos
18
(Figura superior derecha
EFECTO DE LA MODIFICACIOacuteN COVALENTE SOBRE LA ACTIVIDAD ENZIMAacuteTICA
Otros enzimas pasan de una forma menos activa a otra maacutes activa unieacutendose covalentemente a un grupo quiacutemico de pequentildeo tamantildeo como el Pi o el AMP Tambieacuten se da el caso inverso en el que un enzima muy activo se desactiva al liberar alguacuten grupo quiacutemico En las enzimas de las viacuteas degradativas del metabolismo la
forma fosforilada es maacutes activa que la no fosforilada mientras que en las viacuteas biosinteacuteticas ocurre lo contrario En las figuras inferiores se ilustra la activacioacuten de una proteiacutena por fosforilacioacuten
Elementos de la reaccioacuten El enzima no fosforilado es inactivo El enzima fosforilado es activo
ACTIVACIOacuteN PROTEOLIacuteTICA DE LA ACTIVIDAD ENZIMAacuteTICA
Algunos enzimas no se sintetizan como tales sino como proteiacutenas precursoras sin actividad enzimaacutetica Estas proteiacutenas se llaman
proenzimas o zimoacutegenos Para activarse los zimoacutegenos sufren un ataque hidroliacutetico que origina la liberacioacuten de uno o varios peacuteptidos El resto de la moleacutecula proteica adopta la conformacioacuten y las propiedades del enzima activo Muchos enzimas digestivos se secretan en forma de zimoacutegenos y en el tubo digestivo
se convierten en la forma activa Es el caso de la -quimotripsina que se sintetiza en forma de quimotripsinoacutegeno (Figura superior) Si estos enzimas se sintetizasen directamente en forma activa destruiriacutean la propia ceacutelula que las
produce Asiacute la tripsina pancreaacutetica (una proteasa) se sintetiza como tripsinoacutegeno (inactivo) Si por alguna razoacuten se activa en el propio paacutencreas la glaacutendula sufre un proceso de autodestruccioacuten (pancreatitis aguda) a
menudo mortal
19
REGULACIOacuteN DE LA ACTIVIDAD ENZIMAacuteTICA POR MEDIO DE ISOENZIMAS
Algunos enzimas tienen distinta estructura molecular aunque su funcioacuten bioloacutegica es similar Se llaman isozimas o isoenzimas Estas diferencias de estructura se traducen en ligeros cambios en sus propiedades de forma que cada isozima se adapta perfectamente a la funcioacuten que debe realizar Asiacute podemos observar la existencia de isoenzimas en funcioacuten de
el tipo de tejido Por ejemplo la lactato deshidrogenasa presenta isozimas distintos en muacutesculo y corazoacuten el compartimento celular donde actuacutea Por ejemplo la malato deshidrogenasa del citoplasma es distinta
de la de la mitocondria el momento concreto del desarrollo del individuo Por ejemplo algunos enzimas de la glicolisis del feto
son diferentes de los mismos enzimas en el adulto
CINEacuteTICA ENZIMAacuteTICA
Los principios generales de las reacciones quiacutemicas se aplican tambieacuten a las reacciones enzimaacuteticas Por este motivo antes de empezar con la cineacutetica quiacutemica se van a repasar algunos conceptos baacutesicos de cineacutetica quiacutemica
A continuacioacuten se describiraacuten los siguientes conceptos
Cineacutetica enzimaacutetica Modelo cineacutetico de Michaelis-Menten Caacutelculo de la KM y la Vmax de un enzima Actividad enzimaacutetica
20
NOMENCLATURA DE LOS ENZIMAS
CATALISIS MOLECULAR
Un catalizador modifica la velocidad de una reaccioacuten quiacutemica sin ser utilizado o aparecer como uno de los productos de la reaccioacuten Una reaccioacuten quiacutemica en la que un substrato(S) se
transforma en un producto(P) S P ocurre por que cierta fraccioacuten de moleacuteculas de S posee mucho maacutes energiacutea que el resto de ellas lo que es suficiente para que alcancen un estado activado en el que pueda formarse o romperse un enlace quiacutemico y se forme el producto(P)
La energiacutea de activacioacuten es la cantidad de energiacutea expresada en caloriacuteas necesaria para que todas las moleacuteculas de un mol a una temperatura dada alcancen el estado reactivo Mientras que el estado de transicioacuten es el estado rico en energiacutea de las moleacuteculas que
interaccionan en la cima de la barrera de activacioacuten La velocidad de una reaccioacuten quiacutemica es proporcional a la concentracioacuten del complejo en el estado de transicioacuten
Una reaccioacuten quiacutemica se puede acelerar de la siguiente forma 1) Al aumentar la temperatura se incrementa la energiacutea cineacutetica por lo que es mayor el nuacutemero de moleacuteculas que alcanzan el
estado de transicioacuten Generalmente el Q10 =2 lo que indica que la velocidad de una reaccioacuten quiacutemica se duplica al aumentar la temperatura en 10
0C 2) Antildeadiendo un catalizador que
disminuye la energiacutea de activacioacuten y aumenta la velocidad de reaccioacuten
La enzima (E) se combina con el substrato (S) formando el complejo de transicioacuten enzima-
substrato (E-S) mediante una reaccioacuten reversible cuya energiacutea de activacioacuten es menor que la de la reaccioacuten no catalizada Cuando se forma el producto de la reaccioacuten (P) se regenera de nuevo la enzima (E) de forma libre la que puede combinarse de nuevo con otra moleacutecula de substrato (S)
Una enzima reduce maacutes eficientemente la energiacutea de activacioacuten(Ea) de una reaccioacuten que un catalizador inorgaacutenico lo que permite que una reaccioacuten se realice a menor temperatura
El siguiente ejemplo ilustra mejor lo que hemos discutido
Reaccioacuten Energiacutea de activacioacuten (Kcalmol-1
)
21
a) el peroacutexido de hidroacutegeno se descompone en
H2 O2 H2 O + O2
18
b) el hierro cataliacutetico (Fe) realiza la reaccioacuten
H2 O2 H2 O + O2
13
c) el platino cataliacutetico (Pt) realiza la reaccioacuten
H2 O2 H2 O + O2
12
d) la catalasa una enzima hepaacutetica la realiza
H2O2 H2O + O2
5
Una enzima no modifica la energiacutea libre ni la constante de equilibrio sino que disminuye la energiacutea de activacioacuten de la reaccioacuten
Ir al principio
EFECTO DE LA CONCENTRACIOacuteN DEL SUBSTRATO SOBRE LA
CATALISIS ENZIMATICA
La actividad de una enzima se puede estudiar in vitro bajo condiciones controladas antildeadieacutendole una enzima a un substrato Siacute se trabaja con la condicioacuten de que la concentracioacuten del substrato sea saturante variando entonces la concentracioacuten de enzima se observa que aumenta el producto de la reaccioacuten a pH y temperatura constantes
22
Si se mantiene la concentracioacuten de la enzima constante y variando la concentracioacuten de substrato se obtiene una curva hiperboacutelica como la de la figura Al principio un aumento de la
concentracioacuten de substrato produce un aumento raacutepi do de la velocidad de reaccioacuten pero si se sigue aumentando la concentracioacuten de substrato la velocidad de reaccioacuten comienza a disminuir y a muy altas concentraciones de substrato se observa que no cambia la velocidad de
reaccioacuten se dice que los centros activos de la enzima se encuentran saturados La velocidad de reaccioacuten que se obtiene a esa alta concentracioacuten de substrato se define como la velocidad maacutexima (V) de la reaccioacuten enzimaacutetica bajo las condiciones especificadas La concentracioacuten de
substrato (S) a la semivelocidad maacutexima de reaccioacuten (V2) se puede determinar de la figura y representa la constante de Michaelis o Km la cual es una caracteriacutestica para cada enzima La inversa de Km o 1Km mide aproximadamente la afinidad de la enzima por el subs trato
Mientras maacutes pequentildeo sea el valor de Km mayor seraacute la afinidad de la enzima por el substrato Si varias enzimas compiten en el metabolismo por el mismo substrato eacuteste seraacute transformado preferentemente por la enzima con mayor afinidad
La ecuacioacuten de Michaelis describe la relacioacuten cuantitativa entre la velocidad de reaccioacuten y la concentracioacuten de substrato [S] si se conoce Vmax o Km
v=
En donde v= es la velocidad de reaccioacuten observada a una concentracioacuten de substrato determinada [S]
Km= constante de Michaelis en molesliacutet ro
23
Vmax= velocidad maacutexima a concentracioacuten saturante de substrato
Si v= Vmax2 entonces
=
Km+[S] = 2[S]
Km =[S]
De tal forma que podemos concluir que Km es igual a la concentracioacuten de substrato a la semivelocidad maacutexima de reaccioacuten
1 Concentraciones relativas de E y S la concentracioacuten de substrato S es
mucho mayor que la de E de tal manera que la cantidad de substrato
unido a la enzima en cualquier momento es muy pequentildea
2 Se asume el estado estacionario ES no cambia con el tiempo esto
quiere decir que la velocidad de formacioacuten de ES es igual a aquella para
su desintegracioacuten En general un intermediario en una serie de reacciones estaacute en estado estacionario cuando su velocidad de siacutentesis
es igual a su velocidad de degradacioacuten 3 Velocidad inicial para el anaacutelisis de reacciones enzimaacuteticas soacutelo se
utiliza la velocidad inicial de la reaccioacuten que es la velocidad ejercida por
la enzima inmediatamente despueacutes de que se ha puesto en contacto con el substrato y hasta antes de que se haya consumido el 10 de la concentracioacuten inicial del mismo La razoacuten de lo anterior es que en ese
momento la concentracioacuten del producto de la reaccioacuten que se ha acumulado en muy pequentildea y por tanto la reaccioacuten en el sentido
inverso es decir la transformacioacuten del producto en el substrato original puede ser ignorada
24
Caracteriacutesticas de Km la constante de Michaelis-Menten es
caracteriacutestica de una enzima y particular para un substrato Refleja la afinidad de la enzima por ese substrato K m es numeacutericamente igual a la concentracioacuten
de substrato a la cual la velocidad de reaccioacuten es la mitad de la Vmax (Km= Vmax2) Este paraacutemetro Km no variacutea con la concentracioacuten de enzima
Significado de una Km pequentildea un valor numeacuterico pequentildeo de Km
refleja una alta afinidad de la enzima por su substrato porque a una baja
concentracioacuten del mismo la enzima a desarrollado ya la mitad de la velocidad maacutexima
Significado de una Km grande el valor numeacuterico grande de Km refleja
una baja afinidad de la enzima por su substrato porque a una concentracioacuten
elevada del mismo la enzima desarrolla la mitad de la velocidad maacutexima
Relacioacuten de la velocidad con la concentracioacuten de enzima la
velocidad de la reaccioacuten es directamente proporcional a la concentracioacuten de la
enzima a cualquier concentracioacuten de substrato Por ejemplo si la concentracioacuten de enzima es disminuida a la mitad la velocidad inicial de la reaccioacuten (v0) es
reducida tambieacuten a la mitad de la original
Orden de la reaccioacuten cuando S es mas pequentildea que la Km la
velocidad de la reaccioacuten es aproximadamente igual a la concentracioacuten de
substrato
100
50
0
Substrato
Km Figura Localizacioacuten de la constante de Michaelis (Km)
25
La velocidad de reaccioacuten se dice en estas condiciones es de primer
orden con respecto al substrato Cuando S es mas grande que la Km la
velocidad es constante e igual a la Vmax La velocidad de la reaccioacuten es independiente de la concentracioacuten de substrato y se dice que es de orden cero
con respecto a la concentracioacuten de substrato
Las enzimas pueden ser aisladas de las ceacutelulas u organismos para estudiar sus
propiedades in vitro es decir en un tubo de ensayo Las diferentes enzimas muestran diferentes respuestas a los cambios en la concentracioacuten de substrato temperatura y pH
Concentracioacuten de substrato
La forma hiperboacutelica de la curva de velocidad vs Substrato
Efecto de la temperatura en la actividad enzimaacutetica
Efecto de la variacioacuten del pH en la actividad enzimaacutetica
Incremento de la velocidad con la temperatura En general la velocidad de la reaccioacuten se incrementa con la temperatura hasta que un punto maacuteximo es alcanzado
Este incremento se debe a que aumenta el nuacutemero de moleacuteculas ricas en energiacutea que pueden pasar la barrera energeacutetica de estado de transicioacuten para formar a los productos
Decremento de la velocidad con la temperatura la elevacioacuten excesiva de la
temperatura del medio que contiene a las enzimas resulta en un decremento de la velocidad como resultado de la desnaturalizacioacuten es decir la peacuterdida de la estructura tridimensional de las enzimas
26
zona de reactivacioacuten maacuteximo de actividad
Zona de
inactivacioacuten
0 Temperatura
Figura dependencia de la actividad enzimaacutetica con la temperatura
Velocidad maacutexima la velocidad de una reaccioacuten (v) es el nuacutemero de
moleacuteculas de substrato convertidas en producto por unidad de tiempo y
usualmente se expresa en micromoles de producto formadas por minuto La velocidad de una reaccioacuten catalizada por una enzima aumenta conforme se incrementa la concentracioacuten de substrato hasta que se llega a una velocidad maacutexima (Vmax) a partir de la cual la velocidad de la reaccioacuten es
independiente de la cantidad de substrato El que la velocidad no pueda
seguirse incrementando refleja la saturacioacuten del sitio activo con el substrato
Vmax
0 Substrato
Figura representacioacuten de una cineacutetica tiacutepicamente michaeliana
Vmax = velocidad de saturacioacuten o maacutexima
27
Cuando se grafica la velocidad de la reaccioacuten v0 contra la concentracioacuten de substrato S no siempre es posible determinar la condicioacuten
en que se ha llegado a la velocidad maacutexima Vmax debido al incremento de la pendiente en la hipeacuterbola a concentraciones de substrato elevadas
Figura experimento cineacutetico para la actividad enzimaacutetica
Se presentan los valores experimentales y los resultados del ajuste al modelo de Michaelis
Auacuten asiacute si se grafica una doble reciacuteproca es decir el inverso de la
velocidad (1v0) contra el inverso de la concentracioacuten de substrato (1 S ) se
obtiene una liacutenea recta El regraacutefico de Lineweaver-Burke tambieacuten se
denomina de ldquodobles reciacuteprocasrdquo y se utiliza para calcular la Km y la Vmax asiacute como para determinar el mecanismo de accioacuten de los diversos tipos de
inhibidores
La ecuacioacuten que describe a la graacutefca de Lineweaver-Burke es
Vmax
1
SVmax
Km
v
1
0 (3)
que describe una recta en donde el intercepto con el eje de las abscisas es igual a ndash1Km y el intercepto con el eje de las ordenadas es igual a 1Vmax
28
Figura experimento cineacutetico para la actividad enzimaacutetica Regraacutefico de
Lineweaver-Burke
Se presentan los valores experimentales y los resultados del ajuste al modelo
REPRESENTACIOacuteN GRAacuteFICA DE LINEWEAVER Y BURK
Si se toma el inverso en ambos miembros de la ecuacioacuten de Michaelis tenemos
=
+
29
que es la ecuacioacuten de una liacutenea recta cuya pendiente es que intercepta al eje de las
ordenadas en y a la abcisa en el punto Lo que es evidente al hacer
ya que se convierte en
Si se hace una graacutefica con los dobles inversos colocando los valores de en el eje de las
ordenadas y en el eje de las abcisas se obtiene una liacutenea recta a partir de la cual se puede calcular faacutecilmente Km
EFECTO DEL pH SOBRE LA ACTIVIDAD ENZIMATICA
Sabiendo que las enzimas son proteinas cualquier cambio brusco de pH puede alterar el caraacutecter ioacutenico de los grupos amino y carboxilo en la superficie proteica afectando asiacute las propiedades cataliacuteticas de una enzima A pH alto o bajo se puede producir la desnaturalizacioacuten
de la enzima y en consecuencia su inactivacioacuten La fosfatasa aacutecida es maacutes activa a pH 50 mientras que la fosfatasa alcalina lo es a pH 90 Muchas enzimas tienen maacutexima actividad cerca de la neutralidad en un rango de pH de 6 a 8
El pH per se no afecta la actividad enzimaacutetica sino la concentracioacuten de protones Los protones
ademaacutes de alterar la estructura de la enzima y el substrato pueden participar tambieacuten en la reaccioacuten como substrato o producto En esos casos la concentracioacuten de protones afecta directamente la velocidad de la reaccioacuten
ENZIMA pH OPTIMO
30
Pepsina 15
Tripsina 77
Catalasa 76
Arginasa 97
Fumarasa 78
Ribonucleasa 78
EFECTO DE ACTIVADORES
Un aumento en la temperatura provoca un aumento de la velocidad de reaccioacuten hasta cierta temperatura oacuteptima ya que despueacutes de aproximadamente 45
0 C se comienza a producir la
desnaturalizacioacuten teacutermica Las enzimas de muchos mamiacuteferos tienen una temperatura oacuteptima
de 370 C por encima de esa temperatura comienzan a inactivarse y se destruyen Sin embargo
existen especies de bacterias y algas que habitan en fuentes de aguas termales y en el otro extremo ciertas bacterias aacuterticas tienen temperaturas oacuteptimas cercanas a 0
0 C
31
ESPECIFICIDAD ABSOLUTA
Cierta enzimas se encuentran como aproenzimas o zimogenos que son inactivos
El tripsinoacutegeno es convertido en tripsina por la misma accioacuten de la tripsina que remueve un hexapeacuteptido
tripsina Tripsinoacutegeno Tripsina + Val (Asp)4 lisina
Otra forma de activacioacuten consiste en mantener los grupos SH reducidos Estos grupos pueden
formar parte de centros activos de ciertas enzimas Si se oxidan estos grupos a -S-S- la enzima se inactiva Asiacute tenemos que la enzima papaina se inactiva despueacutes de exponerla al ox iacutegeno pero cuando se le antildeade un reductor apropiado la enzima se reactiva Los grupos ndashS-S- se convierten en ndashSH activaacutendose la enzima
Otra forma de activacioacuten requiere la presencia de otra substancia que se enlaza al sitio alosteacuterico el compuesto que se enlaza se denomina un efector alosteacuterico El centro alosteacuterico es bien especiacutefico para el efector o modulador alosteacuterico
ESPECIFICIDAD RELATIVA
Oxiacutegeno L-aminoaacutecido a -cetoaacutecido + NH3 + H 2 O 2 L-aminoaacutecido oxidasa Oxiacutegeno D-aminoaacutecido a -cetoaacutecido + NH3 + H 2 O 2 D-aminoaacutecido oxidasa
32
En este ejemplo se muestra la estereo-especificidad Una enzima puede tener una especificidad oacuteptica para isomeros D o L
Hay enzimas que muestran especificidad absoluta como la aspartasa que cataliza la adicioacuten reversible de amoniacuteaco al doble enlace del aacutecido fumaacuterico pero no al de otros aacutecidos insaturados
HOOC ndash CH = CH ndash COO- + NH4
+ HOOC ndash CH2 - CHNH2 - COO
- + H
+
INHIBICIOacuteN ENZIMATICA
Mediante el uso de inhibidores enzimaacuteticos se ha obtenido informacioacuten muy valiosa sobre la conformacioacuten del centro activo de algunas enzimas
a Inhibicioacuten irreversible Este tipo de inhibidores forman un enlace covalente con las enzimas cerca del centro activo Un ejemplo son los gases nerviosos como el
fluorofosfato de diisopropilo (DFP) que forma un complejo con la enzima acetilcolinesterasa Los animales envenenados con este gas quedan paralizados debido a la imposibilidad de transmitir adecuadamente los impulsos nerviosos
Ester di-isopropil fos foacuterico de la enzima (inactivo)
b) Inhibicioacuten competitiva
Es cuando el inhibidor compite con el substrato por la unioacuten con el centro activo de la
enzima Este tipo de inhibicioacuten puede reducirse si se aumenta la concentracioacuten de substrato
-OOC - CH2 - CH2 - COO
- -OOC - CH = CH - COO
- + 2H
+ Succinato Succinato Fumarato
33
Deshidrogenasa COO
- I CH2 Malonato (inhibidor competitivo) I COO
- En la inhibicioacuten competitiva la enzima se combina con el inhibidor para formar un complejo enzima-inhibidor E + I E I en competencia con la reaccioacuten normal E +S ES
La sulfanilamida (bactericida) interfiere con la siacutentesis del aacutecido foacutelico compitiendo con el aacutecido p-aminobenzoico de una forma competitiva
HOOC-sect- NH2 NH2 ndashSO2 - sect- NH2 aacutecido p- amino benzoico sulfanilamida
sect = fenil
a Inhibicioacuten no competitiva
Esta inhibicioacuten se caracteriza por que no se puede revertir el efecto del inhibidor aumentando la concentracioacuten del substrato
Por ejemplo la inhibicioacuten de la deshidrogenasa del gliceraldehido -3-fosfato por la yodo-acetamida
Enzima-SH + I-CH2 CONH2 Enzima-SCH2 CONH2 + IH
La deshidrogenasa del gliceraldehido-3 fosfato puede ser inhibida tambieacuten por el aacutecido iodo-aceacutetico
Enzima-SH + ICH2 COOH Enzima-SCH2 COOH + IH
c) Inhibicioacuten por metales
Ciertos metales como el plomo mercurio y arseacutenico inhiben enzimas que tienen en su centro activo grupos -SH libres

12
Figura 6 Estructura de la biotina
La enzima holocarboxilasa sintetasa (HCS) cataliza la activacioacuten mediante
biotinilacioacuten de cinco carboxilasas en ceacutelulas humanas La deficiencia de HCS
produce el siacutendrome deficiencia muacuteltiple de carboxilasas (DMC) Esta enfermedad es
potencialmente mortal y se caracteriza por la deficiencia de la actividad de todas las
carboxilasas La deficiencia de biotina no solo disminuye la actividad de estas enzimas
sino que parece modificar la expresioacuten geneacutetica
126 PROPIEDADES DE LAS ENZIMAS
Las propiedades de las enzimas derivan del hecho de ser proteiacutenas y de actuar como
catalizadores Como proteiacutenas poseen una conformacioacuten natural maacutes estable que las
demaacutes conformaciones posibles Asiacute cambios en la conformacioacuten suelen ir asociados
en cambios en la actividad cataliacutetica Los factores que influyen de manera maacutes directa
sobre la actividad de un enzima son
pH
temperatura
cofactores
A) EFECTO DEL pH
Al comprobar experimentalmente la influencia del pH en la velocidad de las
reacciones enzimaacuteticas se obtienen curvas que indican que los enzimas
presentan un pH oacuteptimo de actividad El pH puede afectar de varias maneras
13
o El centro activo puede contener aminoaacutecidos con grupos ionizados
que pueden variar con el pH
o La ionizacioacuten de aminoaacutecidos que no estaacuten en el centro activo puede
provocar modificaciones en la conformacioacuten de la enzima
o El sustrato puede verse afectado por las variaciones del pH
Algunos enzimas presentan variaciones peculiares La pepsina del estoacutemago
presenta un oacuteptimo a pH=2 la ureasa lo tiene a pH 7 y la fosfatasa alcalina del
intestino un pH= 12
B) EFECTO DE LA TEMPERATURA
Influye en la actividad El punto oacuteptimo representa el maacuteximo de actividad A
temperaturas bajas los enzimas se hallan ldquomuy riacutegidosrdquo y cuando se supera un
valor considerable (mayor de 50) la actividad cae bruscamente porque como
proteiacutena el enzima se desnaturaliza
128 EFECTO DE LOS COFACTORES SOBRE LA ACTIVIDAD
ENZIMATICA
A veces un enzima requiere para su funcioacuten la presencia de sustancias no proteicas
que colaboran en la cataacutelisis los cofactores Casi un tercio de los enzimas conocidos
requieren cofactores Cuando el cofactor es una moleacutecula orgaacutenica se llama
coenzima En la figura inferior podemos observar una moleacutecula de hemoglobina
(proteiacutena que transporta oxiacutegeno) y su coenzima (el grupo hemo) Cuando los
cofactores y las coenzimas se encuentran unidos covalentemente al enzima se llaman
14
grupos prosteacuteticos La forma cataliacuteticamente activa del enzima es decir el enzima
unido a su grupo prosteacutetico se llama holoenzima La parte proteica de un
holoenzima (inactiva) se llama apoenzima de forma que
apoenzima + grupo prosteacutetico= holoenzima
129 MODO DE ACCIOacuteN DE LAS ENZIMAS
En las reacciones espontaacuteneas los productos finales tienen menos energiacutea libre de
Gibbs (∆G) que los reactantes (figura inferior izquierda) Por lo tanto las reacciones
espontaacuteneas liberan energiacutea de Gibbs ((∆G lt 0) Sin embargo el comienzo de la
reaccioacuten requiere un aporte inicial de energiacutea Esta energiacutea inicial que hay que
suministrar a los reactantes para que la reaccioacuten transcurra se llama energiacutea de
activacioacuten (Ea) Cuanto menor es la Ea maacutes faacutecilmente transcurre la reaccioacuten
Perfil energeacutetico de una reaccioacuten espontaacutenea Perfil energeacutetico de una reaccioacuten catalizada
15
La accioacuten de los catalizadores consiste precisamente en disminuir la Ea (Figura superior derecha) Los enzimas
son catalizadores especialmente eficaces ya que disminuyen la Ea auacuten maacutes que los catalizadores inorgaacutenicos
Por ejemplo la descomposicioacuten del agua oxigenada (H2O2) para dar H2O y O2 puede ocurrir sin catalizador
con un catalizador inorgaacutenico (platino) o con un enzima especiacutefico (catalasa) Las respectivas Ea para cada
proceso son 18 12 y 6 Kcalmol Asiacute se puede calcular que el platino acelera la reaccioacuten 20000 veces mientras
que la catalasa la acelera 370000 veces
Para que una reaccioacuten quiacutemica tenga lugar las moleacuteculas de los reactantes deben chocar con una energiacutea y
una orientacioacuten adecuadas La actuacioacuten del enzima (1) permite que los rea ctantes (sustratos) se unan a su
centro activo con una orientacioacuten oacuteptima para que la reaccioacuten se produzca y (2) modifica las propiedades
quiacutemicas del sustrato unido a su centro activo debilitando los enlaces existentes y facilitando la formacioacuten de
otros nuevos (Figuras inferiores)
1210 REGULACIOacuteN DE SU ACTIVIDAD ENZIMATICA
16
Una moleacutecula de enzima no tiene por que actuar siempre a la misma velocidad Su
actividad puede estar modulada por
Cambios en el pH
Cambios en la temperatura
Presencia de cofactores
Las concentraciones del sustrato y de los productos finales
Presencia de inhibidores
Modulacioacuten alosteacuterica
Modificacioacuten covalente
Activacioacuten por proteolisis
Isoenzimas
EFECTO DE LAS CONCENTRACIONES SOBRE LA ACTIVIDAD ENZIMAacuteTICA
La velocidad de una reaccioacuten enzimaacutetica depende de la concentracioacuten de sustrato La Figura de la derecha muestra la velocidad de una reaccioacuten enzimaacutetica a 6 concentraciones distintas de sustrato
Ademaacutes la presencia de los productos finales puede hacer que la reaccioacuten sea maacutes lenta o incluso invertir su sentido (Figura inferior)
EFECTO DE LOS INHIBIDORES SOBRE LA ACTIVIDAD ENZIMAacuteTICA
Ciertas moleacuteculas pueden inhibir la accioacuten cataliacutetica de un enzima son los inhibidores Estos inhibidores bien pueden ocupar temporalmente el centro activo por semejanza estructural con el sustrato original ( inhibidor competitivo) o bien alteran la conformacioacuten espacial del enzima impidiendo su unioacuten al sustrato ( inhibidor no competitivo) (Figuras inferiores)
Inhibidor competitivo Inhibidor no competitivo
17
MODULACIOacuteN ALOSTEacuteRICA DE LA ACTIVIDAD ENZIMAacuteTICA
Hay enzimas que pueden adoptar 2 conformaciones interconvertibles llamadas R (relajada) y T (tensa) R es la forma maacutes activa porque se une al sustrato con maacutes afinidad Las formas R y T se encuentran en equilibrio R lt==gt T (Figura inferior)
Ciertas sustancias tienden a estabilizar la forma R Son los llamados moduladores positivos El propio sustrato es a
menudo un modulador positivo Las moleacuteculas que favorecen la forma R pero que actuacutean sobre una regioacuten del enzima distinta del centro activo son los activadores alosteacutericos (Figura inferior izquierda)
Activador alosteacuterico favorece la unioacuten del sustrato
Inhibidor alosteacuterico impide la unioacuten del sustrato
Las sustancias que favorecen la forma T y disminuyen la actividad enzimaacutetica son los moduladores negativos Si
estos moduladores actuacutean en lugares distintos del centro activo del enzima se llaman inhibidores alosteacutericos
18
(Figura superior derecha
EFECTO DE LA MODIFICACIOacuteN COVALENTE SOBRE LA ACTIVIDAD ENZIMAacuteTICA
Otros enzimas pasan de una forma menos activa a otra maacutes activa unieacutendose covalentemente a un grupo quiacutemico de pequentildeo tamantildeo como el Pi o el AMP Tambieacuten se da el caso inverso en el que un enzima muy activo se desactiva al liberar alguacuten grupo quiacutemico En las enzimas de las viacuteas degradativas del metabolismo la
forma fosforilada es maacutes activa que la no fosforilada mientras que en las viacuteas biosinteacuteticas ocurre lo contrario En las figuras inferiores se ilustra la activacioacuten de una proteiacutena por fosforilacioacuten
Elementos de la reaccioacuten El enzima no fosforilado es inactivo El enzima fosforilado es activo
ACTIVACIOacuteN PROTEOLIacuteTICA DE LA ACTIVIDAD ENZIMAacuteTICA
Algunos enzimas no se sintetizan como tales sino como proteiacutenas precursoras sin actividad enzimaacutetica Estas proteiacutenas se llaman
proenzimas o zimoacutegenos Para activarse los zimoacutegenos sufren un ataque hidroliacutetico que origina la liberacioacuten de uno o varios peacuteptidos El resto de la moleacutecula proteica adopta la conformacioacuten y las propiedades del enzima activo Muchos enzimas digestivos se secretan en forma de zimoacutegenos y en el tubo digestivo
se convierten en la forma activa Es el caso de la -quimotripsina que se sintetiza en forma de quimotripsinoacutegeno (Figura superior) Si estos enzimas se sintetizasen directamente en forma activa destruiriacutean la propia ceacutelula que las
produce Asiacute la tripsina pancreaacutetica (una proteasa) se sintetiza como tripsinoacutegeno (inactivo) Si por alguna razoacuten se activa en el propio paacutencreas la glaacutendula sufre un proceso de autodestruccioacuten (pancreatitis aguda) a
menudo mortal
19
REGULACIOacuteN DE LA ACTIVIDAD ENZIMAacuteTICA POR MEDIO DE ISOENZIMAS
Algunos enzimas tienen distinta estructura molecular aunque su funcioacuten bioloacutegica es similar Se llaman isozimas o isoenzimas Estas diferencias de estructura se traducen en ligeros cambios en sus propiedades de forma que cada isozima se adapta perfectamente a la funcioacuten que debe realizar Asiacute podemos observar la existencia de isoenzimas en funcioacuten de
el tipo de tejido Por ejemplo la lactato deshidrogenasa presenta isozimas distintos en muacutesculo y corazoacuten el compartimento celular donde actuacutea Por ejemplo la malato deshidrogenasa del citoplasma es distinta
de la de la mitocondria el momento concreto del desarrollo del individuo Por ejemplo algunos enzimas de la glicolisis del feto
son diferentes de los mismos enzimas en el adulto
CINEacuteTICA ENZIMAacuteTICA
Los principios generales de las reacciones quiacutemicas se aplican tambieacuten a las reacciones enzimaacuteticas Por este motivo antes de empezar con la cineacutetica quiacutemica se van a repasar algunos conceptos baacutesicos de cineacutetica quiacutemica
A continuacioacuten se describiraacuten los siguientes conceptos
Cineacutetica enzimaacutetica Modelo cineacutetico de Michaelis-Menten Caacutelculo de la KM y la Vmax de un enzima Actividad enzimaacutetica
20
NOMENCLATURA DE LOS ENZIMAS
CATALISIS MOLECULAR
Un catalizador modifica la velocidad de una reaccioacuten quiacutemica sin ser utilizado o aparecer como uno de los productos de la reaccioacuten Una reaccioacuten quiacutemica en la que un substrato(S) se
transforma en un producto(P) S P ocurre por que cierta fraccioacuten de moleacuteculas de S posee mucho maacutes energiacutea que el resto de ellas lo que es suficiente para que alcancen un estado activado en el que pueda formarse o romperse un enlace quiacutemico y se forme el producto(P)
La energiacutea de activacioacuten es la cantidad de energiacutea expresada en caloriacuteas necesaria para que todas las moleacuteculas de un mol a una temperatura dada alcancen el estado reactivo Mientras que el estado de transicioacuten es el estado rico en energiacutea de las moleacuteculas que
interaccionan en la cima de la barrera de activacioacuten La velocidad de una reaccioacuten quiacutemica es proporcional a la concentracioacuten del complejo en el estado de transicioacuten
Una reaccioacuten quiacutemica se puede acelerar de la siguiente forma 1) Al aumentar la temperatura se incrementa la energiacutea cineacutetica por lo que es mayor el nuacutemero de moleacuteculas que alcanzan el
estado de transicioacuten Generalmente el Q10 =2 lo que indica que la velocidad de una reaccioacuten quiacutemica se duplica al aumentar la temperatura en 10
0C 2) Antildeadiendo un catalizador que
disminuye la energiacutea de activacioacuten y aumenta la velocidad de reaccioacuten
La enzima (E) se combina con el substrato (S) formando el complejo de transicioacuten enzima-
substrato (E-S) mediante una reaccioacuten reversible cuya energiacutea de activacioacuten es menor que la de la reaccioacuten no catalizada Cuando se forma el producto de la reaccioacuten (P) se regenera de nuevo la enzima (E) de forma libre la que puede combinarse de nuevo con otra moleacutecula de substrato (S)
Una enzima reduce maacutes eficientemente la energiacutea de activacioacuten(Ea) de una reaccioacuten que un catalizador inorgaacutenico lo que permite que una reaccioacuten se realice a menor temperatura
El siguiente ejemplo ilustra mejor lo que hemos discutido
Reaccioacuten Energiacutea de activacioacuten (Kcalmol-1
)
21
a) el peroacutexido de hidroacutegeno se descompone en
H2 O2 H2 O + O2
18
b) el hierro cataliacutetico (Fe) realiza la reaccioacuten
H2 O2 H2 O + O2
13
c) el platino cataliacutetico (Pt) realiza la reaccioacuten
H2 O2 H2 O + O2
12
d) la catalasa una enzima hepaacutetica la realiza
H2O2 H2O + O2
5
Una enzima no modifica la energiacutea libre ni la constante de equilibrio sino que disminuye la energiacutea de activacioacuten de la reaccioacuten
Ir al principio
EFECTO DE LA CONCENTRACIOacuteN DEL SUBSTRATO SOBRE LA
CATALISIS ENZIMATICA
La actividad de una enzima se puede estudiar in vitro bajo condiciones controladas antildeadieacutendole una enzima a un substrato Siacute se trabaja con la condicioacuten de que la concentracioacuten del substrato sea saturante variando entonces la concentracioacuten de enzima se observa que aumenta el producto de la reaccioacuten a pH y temperatura constantes
22
Si se mantiene la concentracioacuten de la enzima constante y variando la concentracioacuten de substrato se obtiene una curva hiperboacutelica como la de la figura Al principio un aumento de la
concentracioacuten de substrato produce un aumento raacutepi do de la velocidad de reaccioacuten pero si se sigue aumentando la concentracioacuten de substrato la velocidad de reaccioacuten comienza a disminuir y a muy altas concentraciones de substrato se observa que no cambia la velocidad de
reaccioacuten se dice que los centros activos de la enzima se encuentran saturados La velocidad de reaccioacuten que se obtiene a esa alta concentracioacuten de substrato se define como la velocidad maacutexima (V) de la reaccioacuten enzimaacutetica bajo las condiciones especificadas La concentracioacuten de
substrato (S) a la semivelocidad maacutexima de reaccioacuten (V2) se puede determinar de la figura y representa la constante de Michaelis o Km la cual es una caracteriacutestica para cada enzima La inversa de Km o 1Km mide aproximadamente la afinidad de la enzima por el subs trato
Mientras maacutes pequentildeo sea el valor de Km mayor seraacute la afinidad de la enzima por el substrato Si varias enzimas compiten en el metabolismo por el mismo substrato eacuteste seraacute transformado preferentemente por la enzima con mayor afinidad
La ecuacioacuten de Michaelis describe la relacioacuten cuantitativa entre la velocidad de reaccioacuten y la concentracioacuten de substrato [S] si se conoce Vmax o Km
v=
En donde v= es la velocidad de reaccioacuten observada a una concentracioacuten de substrato determinada [S]
Km= constante de Michaelis en molesliacutet ro
23
Vmax= velocidad maacutexima a concentracioacuten saturante de substrato
Si v= Vmax2 entonces
=
Km+[S] = 2[S]
Km =[S]
De tal forma que podemos concluir que Km es igual a la concentracioacuten de substrato a la semivelocidad maacutexima de reaccioacuten
1 Concentraciones relativas de E y S la concentracioacuten de substrato S es
mucho mayor que la de E de tal manera que la cantidad de substrato
unido a la enzima en cualquier momento es muy pequentildea
2 Se asume el estado estacionario ES no cambia con el tiempo esto
quiere decir que la velocidad de formacioacuten de ES es igual a aquella para
su desintegracioacuten En general un intermediario en una serie de reacciones estaacute en estado estacionario cuando su velocidad de siacutentesis
es igual a su velocidad de degradacioacuten 3 Velocidad inicial para el anaacutelisis de reacciones enzimaacuteticas soacutelo se
utiliza la velocidad inicial de la reaccioacuten que es la velocidad ejercida por
la enzima inmediatamente despueacutes de que se ha puesto en contacto con el substrato y hasta antes de que se haya consumido el 10 de la concentracioacuten inicial del mismo La razoacuten de lo anterior es que en ese
momento la concentracioacuten del producto de la reaccioacuten que se ha acumulado en muy pequentildea y por tanto la reaccioacuten en el sentido
inverso es decir la transformacioacuten del producto en el substrato original puede ser ignorada
24
Caracteriacutesticas de Km la constante de Michaelis-Menten es
caracteriacutestica de una enzima y particular para un substrato Refleja la afinidad de la enzima por ese substrato K m es numeacutericamente igual a la concentracioacuten
de substrato a la cual la velocidad de reaccioacuten es la mitad de la Vmax (Km= Vmax2) Este paraacutemetro Km no variacutea con la concentracioacuten de enzima
Significado de una Km pequentildea un valor numeacuterico pequentildeo de Km
refleja una alta afinidad de la enzima por su substrato porque a una baja
concentracioacuten del mismo la enzima a desarrollado ya la mitad de la velocidad maacutexima
Significado de una Km grande el valor numeacuterico grande de Km refleja
una baja afinidad de la enzima por su substrato porque a una concentracioacuten
elevada del mismo la enzima desarrolla la mitad de la velocidad maacutexima
Relacioacuten de la velocidad con la concentracioacuten de enzima la
velocidad de la reaccioacuten es directamente proporcional a la concentracioacuten de la
enzima a cualquier concentracioacuten de substrato Por ejemplo si la concentracioacuten de enzima es disminuida a la mitad la velocidad inicial de la reaccioacuten (v0) es
reducida tambieacuten a la mitad de la original
Orden de la reaccioacuten cuando S es mas pequentildea que la Km la
velocidad de la reaccioacuten es aproximadamente igual a la concentracioacuten de
substrato
100
50
0
Substrato
Km Figura Localizacioacuten de la constante de Michaelis (Km)
25
La velocidad de reaccioacuten se dice en estas condiciones es de primer
orden con respecto al substrato Cuando S es mas grande que la Km la
velocidad es constante e igual a la Vmax La velocidad de la reaccioacuten es independiente de la concentracioacuten de substrato y se dice que es de orden cero
con respecto a la concentracioacuten de substrato
Las enzimas pueden ser aisladas de las ceacutelulas u organismos para estudiar sus
propiedades in vitro es decir en un tubo de ensayo Las diferentes enzimas muestran diferentes respuestas a los cambios en la concentracioacuten de substrato temperatura y pH
Concentracioacuten de substrato
La forma hiperboacutelica de la curva de velocidad vs Substrato
Efecto de la temperatura en la actividad enzimaacutetica
Efecto de la variacioacuten del pH en la actividad enzimaacutetica
Incremento de la velocidad con la temperatura En general la velocidad de la reaccioacuten se incrementa con la temperatura hasta que un punto maacuteximo es alcanzado
Este incremento se debe a que aumenta el nuacutemero de moleacuteculas ricas en energiacutea que pueden pasar la barrera energeacutetica de estado de transicioacuten para formar a los productos
Decremento de la velocidad con la temperatura la elevacioacuten excesiva de la
temperatura del medio que contiene a las enzimas resulta en un decremento de la velocidad como resultado de la desnaturalizacioacuten es decir la peacuterdida de la estructura tridimensional de las enzimas
26
zona de reactivacioacuten maacuteximo de actividad
Zona de
inactivacioacuten
0 Temperatura
Figura dependencia de la actividad enzimaacutetica con la temperatura
Velocidad maacutexima la velocidad de una reaccioacuten (v) es el nuacutemero de
moleacuteculas de substrato convertidas en producto por unidad de tiempo y
usualmente se expresa en micromoles de producto formadas por minuto La velocidad de una reaccioacuten catalizada por una enzima aumenta conforme se incrementa la concentracioacuten de substrato hasta que se llega a una velocidad maacutexima (Vmax) a partir de la cual la velocidad de la reaccioacuten es
independiente de la cantidad de substrato El que la velocidad no pueda
seguirse incrementando refleja la saturacioacuten del sitio activo con el substrato
Vmax
0 Substrato
Figura representacioacuten de una cineacutetica tiacutepicamente michaeliana
Vmax = velocidad de saturacioacuten o maacutexima
27
Cuando se grafica la velocidad de la reaccioacuten v0 contra la concentracioacuten de substrato S no siempre es posible determinar la condicioacuten
en que se ha llegado a la velocidad maacutexima Vmax debido al incremento de la pendiente en la hipeacuterbola a concentraciones de substrato elevadas
Figura experimento cineacutetico para la actividad enzimaacutetica
Se presentan los valores experimentales y los resultados del ajuste al modelo de Michaelis
Auacuten asiacute si se grafica una doble reciacuteproca es decir el inverso de la
velocidad (1v0) contra el inverso de la concentracioacuten de substrato (1 S ) se
obtiene una liacutenea recta El regraacutefico de Lineweaver-Burke tambieacuten se
denomina de ldquodobles reciacuteprocasrdquo y se utiliza para calcular la Km y la Vmax asiacute como para determinar el mecanismo de accioacuten de los diversos tipos de
inhibidores
La ecuacioacuten que describe a la graacutefca de Lineweaver-Burke es
Vmax
1
SVmax
Km
v
1
0 (3)
que describe una recta en donde el intercepto con el eje de las abscisas es igual a ndash1Km y el intercepto con el eje de las ordenadas es igual a 1Vmax
28
Figura experimento cineacutetico para la actividad enzimaacutetica Regraacutefico de
Lineweaver-Burke
Se presentan los valores experimentales y los resultados del ajuste al modelo
REPRESENTACIOacuteN GRAacuteFICA DE LINEWEAVER Y BURK
Si se toma el inverso en ambos miembros de la ecuacioacuten de Michaelis tenemos
=
+
29
que es la ecuacioacuten de una liacutenea recta cuya pendiente es que intercepta al eje de las
ordenadas en y a la abcisa en el punto Lo que es evidente al hacer
ya que se convierte en
Si se hace una graacutefica con los dobles inversos colocando los valores de en el eje de las
ordenadas y en el eje de las abcisas se obtiene una liacutenea recta a partir de la cual se puede calcular faacutecilmente Km
EFECTO DEL pH SOBRE LA ACTIVIDAD ENZIMATICA
Sabiendo que las enzimas son proteinas cualquier cambio brusco de pH puede alterar el caraacutecter ioacutenico de los grupos amino y carboxilo en la superficie proteica afectando asiacute las propiedades cataliacuteticas de una enzima A pH alto o bajo se puede producir la desnaturalizacioacuten
de la enzima y en consecuencia su inactivacioacuten La fosfatasa aacutecida es maacutes activa a pH 50 mientras que la fosfatasa alcalina lo es a pH 90 Muchas enzimas tienen maacutexima actividad cerca de la neutralidad en un rango de pH de 6 a 8
El pH per se no afecta la actividad enzimaacutetica sino la concentracioacuten de protones Los protones
ademaacutes de alterar la estructura de la enzima y el substrato pueden participar tambieacuten en la reaccioacuten como substrato o producto En esos casos la concentracioacuten de protones afecta directamente la velocidad de la reaccioacuten
ENZIMA pH OPTIMO
30
Pepsina 15
Tripsina 77
Catalasa 76
Arginasa 97
Fumarasa 78
Ribonucleasa 78
EFECTO DE ACTIVADORES
Un aumento en la temperatura provoca un aumento de la velocidad de reaccioacuten hasta cierta temperatura oacuteptima ya que despueacutes de aproximadamente 45
0 C se comienza a producir la
desnaturalizacioacuten teacutermica Las enzimas de muchos mamiacuteferos tienen una temperatura oacuteptima
de 370 C por encima de esa temperatura comienzan a inactivarse y se destruyen Sin embargo
existen especies de bacterias y algas que habitan en fuentes de aguas termales y en el otro extremo ciertas bacterias aacuterticas tienen temperaturas oacuteptimas cercanas a 0
0 C
31
ESPECIFICIDAD ABSOLUTA
Cierta enzimas se encuentran como aproenzimas o zimogenos que son inactivos
El tripsinoacutegeno es convertido en tripsina por la misma accioacuten de la tripsina que remueve un hexapeacuteptido
tripsina Tripsinoacutegeno Tripsina + Val (Asp)4 lisina
Otra forma de activacioacuten consiste en mantener los grupos SH reducidos Estos grupos pueden
formar parte de centros activos de ciertas enzimas Si se oxidan estos grupos a -S-S- la enzima se inactiva Asiacute tenemos que la enzima papaina se inactiva despueacutes de exponerla al ox iacutegeno pero cuando se le antildeade un reductor apropiado la enzima se reactiva Los grupos ndashS-S- se convierten en ndashSH activaacutendose la enzima
Otra forma de activacioacuten requiere la presencia de otra substancia que se enlaza al sitio alosteacuterico el compuesto que se enlaza se denomina un efector alosteacuterico El centro alosteacuterico es bien especiacutefico para el efector o modulador alosteacuterico
ESPECIFICIDAD RELATIVA
Oxiacutegeno L-aminoaacutecido a -cetoaacutecido + NH3 + H 2 O 2 L-aminoaacutecido oxidasa Oxiacutegeno D-aminoaacutecido a -cetoaacutecido + NH3 + H 2 O 2 D-aminoaacutecido oxidasa
32
En este ejemplo se muestra la estereo-especificidad Una enzima puede tener una especificidad oacuteptica para isomeros D o L
Hay enzimas que muestran especificidad absoluta como la aspartasa que cataliza la adicioacuten reversible de amoniacuteaco al doble enlace del aacutecido fumaacuterico pero no al de otros aacutecidos insaturados
HOOC ndash CH = CH ndash COO- + NH4
+ HOOC ndash CH2 - CHNH2 - COO
- + H
+
INHIBICIOacuteN ENZIMATICA
Mediante el uso de inhibidores enzimaacuteticos se ha obtenido informacioacuten muy valiosa sobre la conformacioacuten del centro activo de algunas enzimas
a Inhibicioacuten irreversible Este tipo de inhibidores forman un enlace covalente con las enzimas cerca del centro activo Un ejemplo son los gases nerviosos como el
fluorofosfato de diisopropilo (DFP) que forma un complejo con la enzima acetilcolinesterasa Los animales envenenados con este gas quedan paralizados debido a la imposibilidad de transmitir adecuadamente los impulsos nerviosos
Ester di-isopropil fos foacuterico de la enzima (inactivo)
b) Inhibicioacuten competitiva
Es cuando el inhibidor compite con el substrato por la unioacuten con el centro activo de la
enzima Este tipo de inhibicioacuten puede reducirse si se aumenta la concentracioacuten de substrato
-OOC - CH2 - CH2 - COO
- -OOC - CH = CH - COO
- + 2H
+ Succinato Succinato Fumarato
33
Deshidrogenasa COO
- I CH2 Malonato (inhibidor competitivo) I COO
- En la inhibicioacuten competitiva la enzima se combina con el inhibidor para formar un complejo enzima-inhibidor E + I E I en competencia con la reaccioacuten normal E +S ES
La sulfanilamida (bactericida) interfiere con la siacutentesis del aacutecido foacutelico compitiendo con el aacutecido p-aminobenzoico de una forma competitiva
HOOC-sect- NH2 NH2 ndashSO2 - sect- NH2 aacutecido p- amino benzoico sulfanilamida
sect = fenil
a Inhibicioacuten no competitiva
Esta inhibicioacuten se caracteriza por que no se puede revertir el efecto del inhibidor aumentando la concentracioacuten del substrato
Por ejemplo la inhibicioacuten de la deshidrogenasa del gliceraldehido -3-fosfato por la yodo-acetamida
Enzima-SH + I-CH2 CONH2 Enzima-SCH2 CONH2 + IH
La deshidrogenasa del gliceraldehido-3 fosfato puede ser inhibida tambieacuten por el aacutecido iodo-aceacutetico
Enzima-SH + ICH2 COOH Enzima-SCH2 COOH + IH
c) Inhibicioacuten por metales
Ciertos metales como el plomo mercurio y arseacutenico inhiben enzimas que tienen en su centro activo grupos -SH libres

13
o El centro activo puede contener aminoaacutecidos con grupos ionizados
que pueden variar con el pH
o La ionizacioacuten de aminoaacutecidos que no estaacuten en el centro activo puede
provocar modificaciones en la conformacioacuten de la enzima
o El sustrato puede verse afectado por las variaciones del pH
Algunos enzimas presentan variaciones peculiares La pepsina del estoacutemago
presenta un oacuteptimo a pH=2 la ureasa lo tiene a pH 7 y la fosfatasa alcalina del
intestino un pH= 12
B) EFECTO DE LA TEMPERATURA
Influye en la actividad El punto oacuteptimo representa el maacuteximo de actividad A
temperaturas bajas los enzimas se hallan ldquomuy riacutegidosrdquo y cuando se supera un
valor considerable (mayor de 50) la actividad cae bruscamente porque como
proteiacutena el enzima se desnaturaliza
128 EFECTO DE LOS COFACTORES SOBRE LA ACTIVIDAD
ENZIMATICA
A veces un enzima requiere para su funcioacuten la presencia de sustancias no proteicas
que colaboran en la cataacutelisis los cofactores Casi un tercio de los enzimas conocidos
requieren cofactores Cuando el cofactor es una moleacutecula orgaacutenica se llama
coenzima En la figura inferior podemos observar una moleacutecula de hemoglobina
(proteiacutena que transporta oxiacutegeno) y su coenzima (el grupo hemo) Cuando los
cofactores y las coenzimas se encuentran unidos covalentemente al enzima se llaman
14
grupos prosteacuteticos La forma cataliacuteticamente activa del enzima es decir el enzima
unido a su grupo prosteacutetico se llama holoenzima La parte proteica de un
holoenzima (inactiva) se llama apoenzima de forma que
apoenzima + grupo prosteacutetico= holoenzima
129 MODO DE ACCIOacuteN DE LAS ENZIMAS
En las reacciones espontaacuteneas los productos finales tienen menos energiacutea libre de
Gibbs (∆G) que los reactantes (figura inferior izquierda) Por lo tanto las reacciones
espontaacuteneas liberan energiacutea de Gibbs ((∆G lt 0) Sin embargo el comienzo de la
reaccioacuten requiere un aporte inicial de energiacutea Esta energiacutea inicial que hay que
suministrar a los reactantes para que la reaccioacuten transcurra se llama energiacutea de
activacioacuten (Ea) Cuanto menor es la Ea maacutes faacutecilmente transcurre la reaccioacuten
Perfil energeacutetico de una reaccioacuten espontaacutenea Perfil energeacutetico de una reaccioacuten catalizada
15
La accioacuten de los catalizadores consiste precisamente en disminuir la Ea (Figura superior derecha) Los enzimas
son catalizadores especialmente eficaces ya que disminuyen la Ea auacuten maacutes que los catalizadores inorgaacutenicos
Por ejemplo la descomposicioacuten del agua oxigenada (H2O2) para dar H2O y O2 puede ocurrir sin catalizador
con un catalizador inorgaacutenico (platino) o con un enzima especiacutefico (catalasa) Las respectivas Ea para cada
proceso son 18 12 y 6 Kcalmol Asiacute se puede calcular que el platino acelera la reaccioacuten 20000 veces mientras
que la catalasa la acelera 370000 veces
Para que una reaccioacuten quiacutemica tenga lugar las moleacuteculas de los reactantes deben chocar con una energiacutea y
una orientacioacuten adecuadas La actuacioacuten del enzima (1) permite que los rea ctantes (sustratos) se unan a su
centro activo con una orientacioacuten oacuteptima para que la reaccioacuten se produzca y (2) modifica las propiedades
quiacutemicas del sustrato unido a su centro activo debilitando los enlaces existentes y facilitando la formacioacuten de
otros nuevos (Figuras inferiores)
1210 REGULACIOacuteN DE SU ACTIVIDAD ENZIMATICA
16
Una moleacutecula de enzima no tiene por que actuar siempre a la misma velocidad Su
actividad puede estar modulada por
Cambios en el pH
Cambios en la temperatura
Presencia de cofactores
Las concentraciones del sustrato y de los productos finales
Presencia de inhibidores
Modulacioacuten alosteacuterica
Modificacioacuten covalente
Activacioacuten por proteolisis
Isoenzimas
EFECTO DE LAS CONCENTRACIONES SOBRE LA ACTIVIDAD ENZIMAacuteTICA
La velocidad de una reaccioacuten enzimaacutetica depende de la concentracioacuten de sustrato La Figura de la derecha muestra la velocidad de una reaccioacuten enzimaacutetica a 6 concentraciones distintas de sustrato
Ademaacutes la presencia de los productos finales puede hacer que la reaccioacuten sea maacutes lenta o incluso invertir su sentido (Figura inferior)
EFECTO DE LOS INHIBIDORES SOBRE LA ACTIVIDAD ENZIMAacuteTICA
Ciertas moleacuteculas pueden inhibir la accioacuten cataliacutetica de un enzima son los inhibidores Estos inhibidores bien pueden ocupar temporalmente el centro activo por semejanza estructural con el sustrato original ( inhibidor competitivo) o bien alteran la conformacioacuten espacial del enzima impidiendo su unioacuten al sustrato ( inhibidor no competitivo) (Figuras inferiores)
Inhibidor competitivo Inhibidor no competitivo
17
MODULACIOacuteN ALOSTEacuteRICA DE LA ACTIVIDAD ENZIMAacuteTICA
Hay enzimas que pueden adoptar 2 conformaciones interconvertibles llamadas R (relajada) y T (tensa) R es la forma maacutes activa porque se une al sustrato con maacutes afinidad Las formas R y T se encuentran en equilibrio R lt==gt T (Figura inferior)
Ciertas sustancias tienden a estabilizar la forma R Son los llamados moduladores positivos El propio sustrato es a
menudo un modulador positivo Las moleacuteculas que favorecen la forma R pero que actuacutean sobre una regioacuten del enzima distinta del centro activo son los activadores alosteacutericos (Figura inferior izquierda)
Activador alosteacuterico favorece la unioacuten del sustrato
Inhibidor alosteacuterico impide la unioacuten del sustrato
Las sustancias que favorecen la forma T y disminuyen la actividad enzimaacutetica son los moduladores negativos Si
estos moduladores actuacutean en lugares distintos del centro activo del enzima se llaman inhibidores alosteacutericos
18
(Figura superior derecha
EFECTO DE LA MODIFICACIOacuteN COVALENTE SOBRE LA ACTIVIDAD ENZIMAacuteTICA
Otros enzimas pasan de una forma menos activa a otra maacutes activa unieacutendose covalentemente a un grupo quiacutemico de pequentildeo tamantildeo como el Pi o el AMP Tambieacuten se da el caso inverso en el que un enzima muy activo se desactiva al liberar alguacuten grupo quiacutemico En las enzimas de las viacuteas degradativas del metabolismo la
forma fosforilada es maacutes activa que la no fosforilada mientras que en las viacuteas biosinteacuteticas ocurre lo contrario En las figuras inferiores se ilustra la activacioacuten de una proteiacutena por fosforilacioacuten
Elementos de la reaccioacuten El enzima no fosforilado es inactivo El enzima fosforilado es activo
ACTIVACIOacuteN PROTEOLIacuteTICA DE LA ACTIVIDAD ENZIMAacuteTICA
Algunos enzimas no se sintetizan como tales sino como proteiacutenas precursoras sin actividad enzimaacutetica Estas proteiacutenas se llaman
proenzimas o zimoacutegenos Para activarse los zimoacutegenos sufren un ataque hidroliacutetico que origina la liberacioacuten de uno o varios peacuteptidos El resto de la moleacutecula proteica adopta la conformacioacuten y las propiedades del enzima activo Muchos enzimas digestivos se secretan en forma de zimoacutegenos y en el tubo digestivo
se convierten en la forma activa Es el caso de la -quimotripsina que se sintetiza en forma de quimotripsinoacutegeno (Figura superior) Si estos enzimas se sintetizasen directamente en forma activa destruiriacutean la propia ceacutelula que las
produce Asiacute la tripsina pancreaacutetica (una proteasa) se sintetiza como tripsinoacutegeno (inactivo) Si por alguna razoacuten se activa en el propio paacutencreas la glaacutendula sufre un proceso de autodestruccioacuten (pancreatitis aguda) a
menudo mortal
19
REGULACIOacuteN DE LA ACTIVIDAD ENZIMAacuteTICA POR MEDIO DE ISOENZIMAS
Algunos enzimas tienen distinta estructura molecular aunque su funcioacuten bioloacutegica es similar Se llaman isozimas o isoenzimas Estas diferencias de estructura se traducen en ligeros cambios en sus propiedades de forma que cada isozima se adapta perfectamente a la funcioacuten que debe realizar Asiacute podemos observar la existencia de isoenzimas en funcioacuten de
el tipo de tejido Por ejemplo la lactato deshidrogenasa presenta isozimas distintos en muacutesculo y corazoacuten el compartimento celular donde actuacutea Por ejemplo la malato deshidrogenasa del citoplasma es distinta
de la de la mitocondria el momento concreto del desarrollo del individuo Por ejemplo algunos enzimas de la glicolisis del feto
son diferentes de los mismos enzimas en el adulto
CINEacuteTICA ENZIMAacuteTICA
Los principios generales de las reacciones quiacutemicas se aplican tambieacuten a las reacciones enzimaacuteticas Por este motivo antes de empezar con la cineacutetica quiacutemica se van a repasar algunos conceptos baacutesicos de cineacutetica quiacutemica
A continuacioacuten se describiraacuten los siguientes conceptos
Cineacutetica enzimaacutetica Modelo cineacutetico de Michaelis-Menten Caacutelculo de la KM y la Vmax de un enzima Actividad enzimaacutetica
20
NOMENCLATURA DE LOS ENZIMAS
CATALISIS MOLECULAR
Un catalizador modifica la velocidad de una reaccioacuten quiacutemica sin ser utilizado o aparecer como uno de los productos de la reaccioacuten Una reaccioacuten quiacutemica en la que un substrato(S) se
transforma en un producto(P) S P ocurre por que cierta fraccioacuten de moleacuteculas de S posee mucho maacutes energiacutea que el resto de ellas lo que es suficiente para que alcancen un estado activado en el que pueda formarse o romperse un enlace quiacutemico y se forme el producto(P)
La energiacutea de activacioacuten es la cantidad de energiacutea expresada en caloriacuteas necesaria para que todas las moleacuteculas de un mol a una temperatura dada alcancen el estado reactivo Mientras que el estado de transicioacuten es el estado rico en energiacutea de las moleacuteculas que
interaccionan en la cima de la barrera de activacioacuten La velocidad de una reaccioacuten quiacutemica es proporcional a la concentracioacuten del complejo en el estado de transicioacuten
Una reaccioacuten quiacutemica se puede acelerar de la siguiente forma 1) Al aumentar la temperatura se incrementa la energiacutea cineacutetica por lo que es mayor el nuacutemero de moleacuteculas que alcanzan el
estado de transicioacuten Generalmente el Q10 =2 lo que indica que la velocidad de una reaccioacuten quiacutemica se duplica al aumentar la temperatura en 10
0C 2) Antildeadiendo un catalizador que
disminuye la energiacutea de activacioacuten y aumenta la velocidad de reaccioacuten
La enzima (E) se combina con el substrato (S) formando el complejo de transicioacuten enzima-
substrato (E-S) mediante una reaccioacuten reversible cuya energiacutea de activacioacuten es menor que la de la reaccioacuten no catalizada Cuando se forma el producto de la reaccioacuten (P) se regenera de nuevo la enzima (E) de forma libre la que puede combinarse de nuevo con otra moleacutecula de substrato (S)
Una enzima reduce maacutes eficientemente la energiacutea de activacioacuten(Ea) de una reaccioacuten que un catalizador inorgaacutenico lo que permite que una reaccioacuten se realice a menor temperatura
El siguiente ejemplo ilustra mejor lo que hemos discutido
Reaccioacuten Energiacutea de activacioacuten (Kcalmol-1
)
21
a) el peroacutexido de hidroacutegeno se descompone en
H2 O2 H2 O + O2
18
b) el hierro cataliacutetico (Fe) realiza la reaccioacuten
H2 O2 H2 O + O2
13
c) el platino cataliacutetico (Pt) realiza la reaccioacuten
H2 O2 H2 O + O2
12
d) la catalasa una enzima hepaacutetica la realiza
H2O2 H2O + O2
5
Una enzima no modifica la energiacutea libre ni la constante de equilibrio sino que disminuye la energiacutea de activacioacuten de la reaccioacuten
Ir al principio
EFECTO DE LA CONCENTRACIOacuteN DEL SUBSTRATO SOBRE LA
CATALISIS ENZIMATICA
La actividad de una enzima se puede estudiar in vitro bajo condiciones controladas antildeadieacutendole una enzima a un substrato Siacute se trabaja con la condicioacuten de que la concentracioacuten del substrato sea saturante variando entonces la concentracioacuten de enzima se observa que aumenta el producto de la reaccioacuten a pH y temperatura constantes
22
Si se mantiene la concentracioacuten de la enzima constante y variando la concentracioacuten de substrato se obtiene una curva hiperboacutelica como la de la figura Al principio un aumento de la
concentracioacuten de substrato produce un aumento raacutepi do de la velocidad de reaccioacuten pero si se sigue aumentando la concentracioacuten de substrato la velocidad de reaccioacuten comienza a disminuir y a muy altas concentraciones de substrato se observa que no cambia la velocidad de
reaccioacuten se dice que los centros activos de la enzima se encuentran saturados La velocidad de reaccioacuten que se obtiene a esa alta concentracioacuten de substrato se define como la velocidad maacutexima (V) de la reaccioacuten enzimaacutetica bajo las condiciones especificadas La concentracioacuten de
substrato (S) a la semivelocidad maacutexima de reaccioacuten (V2) se puede determinar de la figura y representa la constante de Michaelis o Km la cual es una caracteriacutestica para cada enzima La inversa de Km o 1Km mide aproximadamente la afinidad de la enzima por el subs trato
Mientras maacutes pequentildeo sea el valor de Km mayor seraacute la afinidad de la enzima por el substrato Si varias enzimas compiten en el metabolismo por el mismo substrato eacuteste seraacute transformado preferentemente por la enzima con mayor afinidad
La ecuacioacuten de Michaelis describe la relacioacuten cuantitativa entre la velocidad de reaccioacuten y la concentracioacuten de substrato [S] si se conoce Vmax o Km
v=
En donde v= es la velocidad de reaccioacuten observada a una concentracioacuten de substrato determinada [S]
Km= constante de Michaelis en molesliacutet ro
23
Vmax= velocidad maacutexima a concentracioacuten saturante de substrato
Si v= Vmax2 entonces
=
Km+[S] = 2[S]
Km =[S]
De tal forma que podemos concluir que Km es igual a la concentracioacuten de substrato a la semivelocidad maacutexima de reaccioacuten
1 Concentraciones relativas de E y S la concentracioacuten de substrato S es
mucho mayor que la de E de tal manera que la cantidad de substrato
unido a la enzima en cualquier momento es muy pequentildea
2 Se asume el estado estacionario ES no cambia con el tiempo esto
quiere decir que la velocidad de formacioacuten de ES es igual a aquella para
su desintegracioacuten En general un intermediario en una serie de reacciones estaacute en estado estacionario cuando su velocidad de siacutentesis
es igual a su velocidad de degradacioacuten 3 Velocidad inicial para el anaacutelisis de reacciones enzimaacuteticas soacutelo se
utiliza la velocidad inicial de la reaccioacuten que es la velocidad ejercida por
la enzima inmediatamente despueacutes de que se ha puesto en contacto con el substrato y hasta antes de que se haya consumido el 10 de la concentracioacuten inicial del mismo La razoacuten de lo anterior es que en ese
momento la concentracioacuten del producto de la reaccioacuten que se ha acumulado en muy pequentildea y por tanto la reaccioacuten en el sentido
inverso es decir la transformacioacuten del producto en el substrato original puede ser ignorada
24
Caracteriacutesticas de Km la constante de Michaelis-Menten es
caracteriacutestica de una enzima y particular para un substrato Refleja la afinidad de la enzima por ese substrato K m es numeacutericamente igual a la concentracioacuten
de substrato a la cual la velocidad de reaccioacuten es la mitad de la Vmax (Km= Vmax2) Este paraacutemetro Km no variacutea con la concentracioacuten de enzima
Significado de una Km pequentildea un valor numeacuterico pequentildeo de Km
refleja una alta afinidad de la enzima por su substrato porque a una baja
concentracioacuten del mismo la enzima a desarrollado ya la mitad de la velocidad maacutexima
Significado de una Km grande el valor numeacuterico grande de Km refleja
una baja afinidad de la enzima por su substrato porque a una concentracioacuten
elevada del mismo la enzima desarrolla la mitad de la velocidad maacutexima
Relacioacuten de la velocidad con la concentracioacuten de enzima la
velocidad de la reaccioacuten es directamente proporcional a la concentracioacuten de la
enzima a cualquier concentracioacuten de substrato Por ejemplo si la concentracioacuten de enzima es disminuida a la mitad la velocidad inicial de la reaccioacuten (v0) es
reducida tambieacuten a la mitad de la original
Orden de la reaccioacuten cuando S es mas pequentildea que la Km la
velocidad de la reaccioacuten es aproximadamente igual a la concentracioacuten de
substrato
100
50
0
Substrato
Km Figura Localizacioacuten de la constante de Michaelis (Km)
25
La velocidad de reaccioacuten se dice en estas condiciones es de primer
orden con respecto al substrato Cuando S es mas grande que la Km la
velocidad es constante e igual a la Vmax La velocidad de la reaccioacuten es independiente de la concentracioacuten de substrato y se dice que es de orden cero
con respecto a la concentracioacuten de substrato
Las enzimas pueden ser aisladas de las ceacutelulas u organismos para estudiar sus
propiedades in vitro es decir en un tubo de ensayo Las diferentes enzimas muestran diferentes respuestas a los cambios en la concentracioacuten de substrato temperatura y pH
Concentracioacuten de substrato
La forma hiperboacutelica de la curva de velocidad vs Substrato
Efecto de la temperatura en la actividad enzimaacutetica
Efecto de la variacioacuten del pH en la actividad enzimaacutetica
Incremento de la velocidad con la temperatura En general la velocidad de la reaccioacuten se incrementa con la temperatura hasta que un punto maacuteximo es alcanzado
Este incremento se debe a que aumenta el nuacutemero de moleacuteculas ricas en energiacutea que pueden pasar la barrera energeacutetica de estado de transicioacuten para formar a los productos
Decremento de la velocidad con la temperatura la elevacioacuten excesiva de la
temperatura del medio que contiene a las enzimas resulta en un decremento de la velocidad como resultado de la desnaturalizacioacuten es decir la peacuterdida de la estructura tridimensional de las enzimas
26
zona de reactivacioacuten maacuteximo de actividad
Zona de
inactivacioacuten
0 Temperatura
Figura dependencia de la actividad enzimaacutetica con la temperatura
Velocidad maacutexima la velocidad de una reaccioacuten (v) es el nuacutemero de
moleacuteculas de substrato convertidas en producto por unidad de tiempo y
usualmente se expresa en micromoles de producto formadas por minuto La velocidad de una reaccioacuten catalizada por una enzima aumenta conforme se incrementa la concentracioacuten de substrato hasta que se llega a una velocidad maacutexima (Vmax) a partir de la cual la velocidad de la reaccioacuten es
independiente de la cantidad de substrato El que la velocidad no pueda
seguirse incrementando refleja la saturacioacuten del sitio activo con el substrato
Vmax
0 Substrato
Figura representacioacuten de una cineacutetica tiacutepicamente michaeliana
Vmax = velocidad de saturacioacuten o maacutexima
27
Cuando se grafica la velocidad de la reaccioacuten v0 contra la concentracioacuten de substrato S no siempre es posible determinar la condicioacuten
en que se ha llegado a la velocidad maacutexima Vmax debido al incremento de la pendiente en la hipeacuterbola a concentraciones de substrato elevadas
Figura experimento cineacutetico para la actividad enzimaacutetica
Se presentan los valores experimentales y los resultados del ajuste al modelo de Michaelis
Auacuten asiacute si se grafica una doble reciacuteproca es decir el inverso de la
velocidad (1v0) contra el inverso de la concentracioacuten de substrato (1 S ) se
obtiene una liacutenea recta El regraacutefico de Lineweaver-Burke tambieacuten se
denomina de ldquodobles reciacuteprocasrdquo y se utiliza para calcular la Km y la Vmax asiacute como para determinar el mecanismo de accioacuten de los diversos tipos de
inhibidores
La ecuacioacuten que describe a la graacutefca de Lineweaver-Burke es
Vmax
1
SVmax
Km
v
1
0 (3)
que describe una recta en donde el intercepto con el eje de las abscisas es igual a ndash1Km y el intercepto con el eje de las ordenadas es igual a 1Vmax
28
Figura experimento cineacutetico para la actividad enzimaacutetica Regraacutefico de
Lineweaver-Burke
Se presentan los valores experimentales y los resultados del ajuste al modelo
REPRESENTACIOacuteN GRAacuteFICA DE LINEWEAVER Y BURK
Si se toma el inverso en ambos miembros de la ecuacioacuten de Michaelis tenemos
=
+
29
que es la ecuacioacuten de una liacutenea recta cuya pendiente es que intercepta al eje de las
ordenadas en y a la abcisa en el punto Lo que es evidente al hacer
ya que se convierte en
Si se hace una graacutefica con los dobles inversos colocando los valores de en el eje de las
ordenadas y en el eje de las abcisas se obtiene una liacutenea recta a partir de la cual se puede calcular faacutecilmente Km
EFECTO DEL pH SOBRE LA ACTIVIDAD ENZIMATICA
Sabiendo que las enzimas son proteinas cualquier cambio brusco de pH puede alterar el caraacutecter ioacutenico de los grupos amino y carboxilo en la superficie proteica afectando asiacute las propiedades cataliacuteticas de una enzima A pH alto o bajo se puede producir la desnaturalizacioacuten
de la enzima y en consecuencia su inactivacioacuten La fosfatasa aacutecida es maacutes activa a pH 50 mientras que la fosfatasa alcalina lo es a pH 90 Muchas enzimas tienen maacutexima actividad cerca de la neutralidad en un rango de pH de 6 a 8
El pH per se no afecta la actividad enzimaacutetica sino la concentracioacuten de protones Los protones
ademaacutes de alterar la estructura de la enzima y el substrato pueden participar tambieacuten en la reaccioacuten como substrato o producto En esos casos la concentracioacuten de protones afecta directamente la velocidad de la reaccioacuten
ENZIMA pH OPTIMO
30
Pepsina 15
Tripsina 77
Catalasa 76
Arginasa 97
Fumarasa 78
Ribonucleasa 78
EFECTO DE ACTIVADORES
Un aumento en la temperatura provoca un aumento de la velocidad de reaccioacuten hasta cierta temperatura oacuteptima ya que despueacutes de aproximadamente 45
0 C se comienza a producir la
desnaturalizacioacuten teacutermica Las enzimas de muchos mamiacuteferos tienen una temperatura oacuteptima
de 370 C por encima de esa temperatura comienzan a inactivarse y se destruyen Sin embargo
existen especies de bacterias y algas que habitan en fuentes de aguas termales y en el otro extremo ciertas bacterias aacuterticas tienen temperaturas oacuteptimas cercanas a 0
0 C
31
ESPECIFICIDAD ABSOLUTA
Cierta enzimas se encuentran como aproenzimas o zimogenos que son inactivos
El tripsinoacutegeno es convertido en tripsina por la misma accioacuten de la tripsina que remueve un hexapeacuteptido
tripsina Tripsinoacutegeno Tripsina + Val (Asp)4 lisina
Otra forma de activacioacuten consiste en mantener los grupos SH reducidos Estos grupos pueden
formar parte de centros activos de ciertas enzimas Si se oxidan estos grupos a -S-S- la enzima se inactiva Asiacute tenemos que la enzima papaina se inactiva despueacutes de exponerla al ox iacutegeno pero cuando se le antildeade un reductor apropiado la enzima se reactiva Los grupos ndashS-S- se convierten en ndashSH activaacutendose la enzima
Otra forma de activacioacuten requiere la presencia de otra substancia que se enlaza al sitio alosteacuterico el compuesto que se enlaza se denomina un efector alosteacuterico El centro alosteacuterico es bien especiacutefico para el efector o modulador alosteacuterico
ESPECIFICIDAD RELATIVA
Oxiacutegeno L-aminoaacutecido a -cetoaacutecido + NH3 + H 2 O 2 L-aminoaacutecido oxidasa Oxiacutegeno D-aminoaacutecido a -cetoaacutecido + NH3 + H 2 O 2 D-aminoaacutecido oxidasa
32
En este ejemplo se muestra la estereo-especificidad Una enzima puede tener una especificidad oacuteptica para isomeros D o L
Hay enzimas que muestran especificidad absoluta como la aspartasa que cataliza la adicioacuten reversible de amoniacuteaco al doble enlace del aacutecido fumaacuterico pero no al de otros aacutecidos insaturados
HOOC ndash CH = CH ndash COO- + NH4
+ HOOC ndash CH2 - CHNH2 - COO
- + H
+
INHIBICIOacuteN ENZIMATICA
Mediante el uso de inhibidores enzimaacuteticos se ha obtenido informacioacuten muy valiosa sobre la conformacioacuten del centro activo de algunas enzimas
a Inhibicioacuten irreversible Este tipo de inhibidores forman un enlace covalente con las enzimas cerca del centro activo Un ejemplo son los gases nerviosos como el
fluorofosfato de diisopropilo (DFP) que forma un complejo con la enzima acetilcolinesterasa Los animales envenenados con este gas quedan paralizados debido a la imposibilidad de transmitir adecuadamente los impulsos nerviosos
Ester di-isopropil fos foacuterico de la enzima (inactivo)
b) Inhibicioacuten competitiva
Es cuando el inhibidor compite con el substrato por la unioacuten con el centro activo de la
enzima Este tipo de inhibicioacuten puede reducirse si se aumenta la concentracioacuten de substrato
-OOC - CH2 - CH2 - COO
- -OOC - CH = CH - COO
- + 2H
+ Succinato Succinato Fumarato
33
Deshidrogenasa COO
- I CH2 Malonato (inhibidor competitivo) I COO
- En la inhibicioacuten competitiva la enzima se combina con el inhibidor para formar un complejo enzima-inhibidor E + I E I en competencia con la reaccioacuten normal E +S ES
La sulfanilamida (bactericida) interfiere con la siacutentesis del aacutecido foacutelico compitiendo con el aacutecido p-aminobenzoico de una forma competitiva
HOOC-sect- NH2 NH2 ndashSO2 - sect- NH2 aacutecido p- amino benzoico sulfanilamida
sect = fenil
a Inhibicioacuten no competitiva
Esta inhibicioacuten se caracteriza por que no se puede revertir el efecto del inhibidor aumentando la concentracioacuten del substrato
Por ejemplo la inhibicioacuten de la deshidrogenasa del gliceraldehido -3-fosfato por la yodo-acetamida
Enzima-SH + I-CH2 CONH2 Enzima-SCH2 CONH2 + IH
La deshidrogenasa del gliceraldehido-3 fosfato puede ser inhibida tambieacuten por el aacutecido iodo-aceacutetico
Enzima-SH + ICH2 COOH Enzima-SCH2 COOH + IH
c) Inhibicioacuten por metales
Ciertos metales como el plomo mercurio y arseacutenico inhiben enzimas que tienen en su centro activo grupos -SH libres

14
grupos prosteacuteticos La forma cataliacuteticamente activa del enzima es decir el enzima
unido a su grupo prosteacutetico se llama holoenzima La parte proteica de un
holoenzima (inactiva) se llama apoenzima de forma que
apoenzima + grupo prosteacutetico= holoenzima
129 MODO DE ACCIOacuteN DE LAS ENZIMAS
En las reacciones espontaacuteneas los productos finales tienen menos energiacutea libre de
Gibbs (∆G) que los reactantes (figura inferior izquierda) Por lo tanto las reacciones
espontaacuteneas liberan energiacutea de Gibbs ((∆G lt 0) Sin embargo el comienzo de la
reaccioacuten requiere un aporte inicial de energiacutea Esta energiacutea inicial que hay que
suministrar a los reactantes para que la reaccioacuten transcurra se llama energiacutea de
activacioacuten (Ea) Cuanto menor es la Ea maacutes faacutecilmente transcurre la reaccioacuten
Perfil energeacutetico de una reaccioacuten espontaacutenea Perfil energeacutetico de una reaccioacuten catalizada
15
La accioacuten de los catalizadores consiste precisamente en disminuir la Ea (Figura superior derecha) Los enzimas
son catalizadores especialmente eficaces ya que disminuyen la Ea auacuten maacutes que los catalizadores inorgaacutenicos
Por ejemplo la descomposicioacuten del agua oxigenada (H2O2) para dar H2O y O2 puede ocurrir sin catalizador
con un catalizador inorgaacutenico (platino) o con un enzima especiacutefico (catalasa) Las respectivas Ea para cada
proceso son 18 12 y 6 Kcalmol Asiacute se puede calcular que el platino acelera la reaccioacuten 20000 veces mientras
que la catalasa la acelera 370000 veces
Para que una reaccioacuten quiacutemica tenga lugar las moleacuteculas de los reactantes deben chocar con una energiacutea y
una orientacioacuten adecuadas La actuacioacuten del enzima (1) permite que los rea ctantes (sustratos) se unan a su
centro activo con una orientacioacuten oacuteptima para que la reaccioacuten se produzca y (2) modifica las propiedades
quiacutemicas del sustrato unido a su centro activo debilitando los enlaces existentes y facilitando la formacioacuten de
otros nuevos (Figuras inferiores)
1210 REGULACIOacuteN DE SU ACTIVIDAD ENZIMATICA
16
Una moleacutecula de enzima no tiene por que actuar siempre a la misma velocidad Su
actividad puede estar modulada por
Cambios en el pH
Cambios en la temperatura
Presencia de cofactores
Las concentraciones del sustrato y de los productos finales
Presencia de inhibidores
Modulacioacuten alosteacuterica
Modificacioacuten covalente
Activacioacuten por proteolisis
Isoenzimas
EFECTO DE LAS CONCENTRACIONES SOBRE LA ACTIVIDAD ENZIMAacuteTICA
La velocidad de una reaccioacuten enzimaacutetica depende de la concentracioacuten de sustrato La Figura de la derecha muestra la velocidad de una reaccioacuten enzimaacutetica a 6 concentraciones distintas de sustrato
Ademaacutes la presencia de los productos finales puede hacer que la reaccioacuten sea maacutes lenta o incluso invertir su sentido (Figura inferior)
EFECTO DE LOS INHIBIDORES SOBRE LA ACTIVIDAD ENZIMAacuteTICA
Ciertas moleacuteculas pueden inhibir la accioacuten cataliacutetica de un enzima son los inhibidores Estos inhibidores bien pueden ocupar temporalmente el centro activo por semejanza estructural con el sustrato original ( inhibidor competitivo) o bien alteran la conformacioacuten espacial del enzima impidiendo su unioacuten al sustrato ( inhibidor no competitivo) (Figuras inferiores)
Inhibidor competitivo Inhibidor no competitivo
17
MODULACIOacuteN ALOSTEacuteRICA DE LA ACTIVIDAD ENZIMAacuteTICA
Hay enzimas que pueden adoptar 2 conformaciones interconvertibles llamadas R (relajada) y T (tensa) R es la forma maacutes activa porque se une al sustrato con maacutes afinidad Las formas R y T se encuentran en equilibrio R lt==gt T (Figura inferior)
Ciertas sustancias tienden a estabilizar la forma R Son los llamados moduladores positivos El propio sustrato es a
menudo un modulador positivo Las moleacuteculas que favorecen la forma R pero que actuacutean sobre una regioacuten del enzima distinta del centro activo son los activadores alosteacutericos (Figura inferior izquierda)
Activador alosteacuterico favorece la unioacuten del sustrato
Inhibidor alosteacuterico impide la unioacuten del sustrato
Las sustancias que favorecen la forma T y disminuyen la actividad enzimaacutetica son los moduladores negativos Si
estos moduladores actuacutean en lugares distintos del centro activo del enzima se llaman inhibidores alosteacutericos
18
(Figura superior derecha
EFECTO DE LA MODIFICACIOacuteN COVALENTE SOBRE LA ACTIVIDAD ENZIMAacuteTICA
Otros enzimas pasan de una forma menos activa a otra maacutes activa unieacutendose covalentemente a un grupo quiacutemico de pequentildeo tamantildeo como el Pi o el AMP Tambieacuten se da el caso inverso en el que un enzima muy activo se desactiva al liberar alguacuten grupo quiacutemico En las enzimas de las viacuteas degradativas del metabolismo la
forma fosforilada es maacutes activa que la no fosforilada mientras que en las viacuteas biosinteacuteticas ocurre lo contrario En las figuras inferiores se ilustra la activacioacuten de una proteiacutena por fosforilacioacuten
Elementos de la reaccioacuten El enzima no fosforilado es inactivo El enzima fosforilado es activo
ACTIVACIOacuteN PROTEOLIacuteTICA DE LA ACTIVIDAD ENZIMAacuteTICA
Algunos enzimas no se sintetizan como tales sino como proteiacutenas precursoras sin actividad enzimaacutetica Estas proteiacutenas se llaman
proenzimas o zimoacutegenos Para activarse los zimoacutegenos sufren un ataque hidroliacutetico que origina la liberacioacuten de uno o varios peacuteptidos El resto de la moleacutecula proteica adopta la conformacioacuten y las propiedades del enzima activo Muchos enzimas digestivos se secretan en forma de zimoacutegenos y en el tubo digestivo
se convierten en la forma activa Es el caso de la -quimotripsina que se sintetiza en forma de quimotripsinoacutegeno (Figura superior) Si estos enzimas se sintetizasen directamente en forma activa destruiriacutean la propia ceacutelula que las
produce Asiacute la tripsina pancreaacutetica (una proteasa) se sintetiza como tripsinoacutegeno (inactivo) Si por alguna razoacuten se activa en el propio paacutencreas la glaacutendula sufre un proceso de autodestruccioacuten (pancreatitis aguda) a
menudo mortal
19
REGULACIOacuteN DE LA ACTIVIDAD ENZIMAacuteTICA POR MEDIO DE ISOENZIMAS
Algunos enzimas tienen distinta estructura molecular aunque su funcioacuten bioloacutegica es similar Se llaman isozimas o isoenzimas Estas diferencias de estructura se traducen en ligeros cambios en sus propiedades de forma que cada isozima se adapta perfectamente a la funcioacuten que debe realizar Asiacute podemos observar la existencia de isoenzimas en funcioacuten de
el tipo de tejido Por ejemplo la lactato deshidrogenasa presenta isozimas distintos en muacutesculo y corazoacuten el compartimento celular donde actuacutea Por ejemplo la malato deshidrogenasa del citoplasma es distinta
de la de la mitocondria el momento concreto del desarrollo del individuo Por ejemplo algunos enzimas de la glicolisis del feto
son diferentes de los mismos enzimas en el adulto
CINEacuteTICA ENZIMAacuteTICA
Los principios generales de las reacciones quiacutemicas se aplican tambieacuten a las reacciones enzimaacuteticas Por este motivo antes de empezar con la cineacutetica quiacutemica se van a repasar algunos conceptos baacutesicos de cineacutetica quiacutemica
A continuacioacuten se describiraacuten los siguientes conceptos
Cineacutetica enzimaacutetica Modelo cineacutetico de Michaelis-Menten Caacutelculo de la KM y la Vmax de un enzima Actividad enzimaacutetica
20
NOMENCLATURA DE LOS ENZIMAS
CATALISIS MOLECULAR
Un catalizador modifica la velocidad de una reaccioacuten quiacutemica sin ser utilizado o aparecer como uno de los productos de la reaccioacuten Una reaccioacuten quiacutemica en la que un substrato(S) se
transforma en un producto(P) S P ocurre por que cierta fraccioacuten de moleacuteculas de S posee mucho maacutes energiacutea que el resto de ellas lo que es suficiente para que alcancen un estado activado en el que pueda formarse o romperse un enlace quiacutemico y se forme el producto(P)
La energiacutea de activacioacuten es la cantidad de energiacutea expresada en caloriacuteas necesaria para que todas las moleacuteculas de un mol a una temperatura dada alcancen el estado reactivo Mientras que el estado de transicioacuten es el estado rico en energiacutea de las moleacuteculas que
interaccionan en la cima de la barrera de activacioacuten La velocidad de una reaccioacuten quiacutemica es proporcional a la concentracioacuten del complejo en el estado de transicioacuten
Una reaccioacuten quiacutemica se puede acelerar de la siguiente forma 1) Al aumentar la temperatura se incrementa la energiacutea cineacutetica por lo que es mayor el nuacutemero de moleacuteculas que alcanzan el
estado de transicioacuten Generalmente el Q10 =2 lo que indica que la velocidad de una reaccioacuten quiacutemica se duplica al aumentar la temperatura en 10
0C 2) Antildeadiendo un catalizador que
disminuye la energiacutea de activacioacuten y aumenta la velocidad de reaccioacuten
La enzima (E) se combina con el substrato (S) formando el complejo de transicioacuten enzima-
substrato (E-S) mediante una reaccioacuten reversible cuya energiacutea de activacioacuten es menor que la de la reaccioacuten no catalizada Cuando se forma el producto de la reaccioacuten (P) se regenera de nuevo la enzima (E) de forma libre la que puede combinarse de nuevo con otra moleacutecula de substrato (S)
Una enzima reduce maacutes eficientemente la energiacutea de activacioacuten(Ea) de una reaccioacuten que un catalizador inorgaacutenico lo que permite que una reaccioacuten se realice a menor temperatura
El siguiente ejemplo ilustra mejor lo que hemos discutido
Reaccioacuten Energiacutea de activacioacuten (Kcalmol-1
)
21
a) el peroacutexido de hidroacutegeno se descompone en
H2 O2 H2 O + O2
18
b) el hierro cataliacutetico (Fe) realiza la reaccioacuten
H2 O2 H2 O + O2
13
c) el platino cataliacutetico (Pt) realiza la reaccioacuten
H2 O2 H2 O + O2
12
d) la catalasa una enzima hepaacutetica la realiza
H2O2 H2O + O2
5
Una enzima no modifica la energiacutea libre ni la constante de equilibrio sino que disminuye la energiacutea de activacioacuten de la reaccioacuten
Ir al principio
EFECTO DE LA CONCENTRACIOacuteN DEL SUBSTRATO SOBRE LA
CATALISIS ENZIMATICA
La actividad de una enzima se puede estudiar in vitro bajo condiciones controladas antildeadieacutendole una enzima a un substrato Siacute se trabaja con la condicioacuten de que la concentracioacuten del substrato sea saturante variando entonces la concentracioacuten de enzima se observa que aumenta el producto de la reaccioacuten a pH y temperatura constantes
22
Si se mantiene la concentracioacuten de la enzima constante y variando la concentracioacuten de substrato se obtiene una curva hiperboacutelica como la de la figura Al principio un aumento de la
concentracioacuten de substrato produce un aumento raacutepi do de la velocidad de reaccioacuten pero si se sigue aumentando la concentracioacuten de substrato la velocidad de reaccioacuten comienza a disminuir y a muy altas concentraciones de substrato se observa que no cambia la velocidad de
reaccioacuten se dice que los centros activos de la enzima se encuentran saturados La velocidad de reaccioacuten que se obtiene a esa alta concentracioacuten de substrato se define como la velocidad maacutexima (V) de la reaccioacuten enzimaacutetica bajo las condiciones especificadas La concentracioacuten de
substrato (S) a la semivelocidad maacutexima de reaccioacuten (V2) se puede determinar de la figura y representa la constante de Michaelis o Km la cual es una caracteriacutestica para cada enzima La inversa de Km o 1Km mide aproximadamente la afinidad de la enzima por el subs trato
Mientras maacutes pequentildeo sea el valor de Km mayor seraacute la afinidad de la enzima por el substrato Si varias enzimas compiten en el metabolismo por el mismo substrato eacuteste seraacute transformado preferentemente por la enzima con mayor afinidad
La ecuacioacuten de Michaelis describe la relacioacuten cuantitativa entre la velocidad de reaccioacuten y la concentracioacuten de substrato [S] si se conoce Vmax o Km
v=
En donde v= es la velocidad de reaccioacuten observada a una concentracioacuten de substrato determinada [S]
Km= constante de Michaelis en molesliacutet ro
23
Vmax= velocidad maacutexima a concentracioacuten saturante de substrato
Si v= Vmax2 entonces
=
Km+[S] = 2[S]
Km =[S]
De tal forma que podemos concluir que Km es igual a la concentracioacuten de substrato a la semivelocidad maacutexima de reaccioacuten
1 Concentraciones relativas de E y S la concentracioacuten de substrato S es
mucho mayor que la de E de tal manera que la cantidad de substrato
unido a la enzima en cualquier momento es muy pequentildea
2 Se asume el estado estacionario ES no cambia con el tiempo esto
quiere decir que la velocidad de formacioacuten de ES es igual a aquella para
su desintegracioacuten En general un intermediario en una serie de reacciones estaacute en estado estacionario cuando su velocidad de siacutentesis
es igual a su velocidad de degradacioacuten 3 Velocidad inicial para el anaacutelisis de reacciones enzimaacuteticas soacutelo se
utiliza la velocidad inicial de la reaccioacuten que es la velocidad ejercida por
la enzima inmediatamente despueacutes de que se ha puesto en contacto con el substrato y hasta antes de que se haya consumido el 10 de la concentracioacuten inicial del mismo La razoacuten de lo anterior es que en ese
momento la concentracioacuten del producto de la reaccioacuten que se ha acumulado en muy pequentildea y por tanto la reaccioacuten en el sentido
inverso es decir la transformacioacuten del producto en el substrato original puede ser ignorada
24
Caracteriacutesticas de Km la constante de Michaelis-Menten es
caracteriacutestica de una enzima y particular para un substrato Refleja la afinidad de la enzima por ese substrato K m es numeacutericamente igual a la concentracioacuten
de substrato a la cual la velocidad de reaccioacuten es la mitad de la Vmax (Km= Vmax2) Este paraacutemetro Km no variacutea con la concentracioacuten de enzima
Significado de una Km pequentildea un valor numeacuterico pequentildeo de Km
refleja una alta afinidad de la enzima por su substrato porque a una baja
concentracioacuten del mismo la enzima a desarrollado ya la mitad de la velocidad maacutexima
Significado de una Km grande el valor numeacuterico grande de Km refleja
una baja afinidad de la enzima por su substrato porque a una concentracioacuten
elevada del mismo la enzima desarrolla la mitad de la velocidad maacutexima
Relacioacuten de la velocidad con la concentracioacuten de enzima la
velocidad de la reaccioacuten es directamente proporcional a la concentracioacuten de la
enzima a cualquier concentracioacuten de substrato Por ejemplo si la concentracioacuten de enzima es disminuida a la mitad la velocidad inicial de la reaccioacuten (v0) es
reducida tambieacuten a la mitad de la original
Orden de la reaccioacuten cuando S es mas pequentildea que la Km la
velocidad de la reaccioacuten es aproximadamente igual a la concentracioacuten de
substrato
100
50
0
Substrato
Km Figura Localizacioacuten de la constante de Michaelis (Km)
25
La velocidad de reaccioacuten se dice en estas condiciones es de primer
orden con respecto al substrato Cuando S es mas grande que la Km la
velocidad es constante e igual a la Vmax La velocidad de la reaccioacuten es independiente de la concentracioacuten de substrato y se dice que es de orden cero
con respecto a la concentracioacuten de substrato
Las enzimas pueden ser aisladas de las ceacutelulas u organismos para estudiar sus
propiedades in vitro es decir en un tubo de ensayo Las diferentes enzimas muestran diferentes respuestas a los cambios en la concentracioacuten de substrato temperatura y pH
Concentracioacuten de substrato
La forma hiperboacutelica de la curva de velocidad vs Substrato
Efecto de la temperatura en la actividad enzimaacutetica
Efecto de la variacioacuten del pH en la actividad enzimaacutetica
Incremento de la velocidad con la temperatura En general la velocidad de la reaccioacuten se incrementa con la temperatura hasta que un punto maacuteximo es alcanzado
Este incremento se debe a que aumenta el nuacutemero de moleacuteculas ricas en energiacutea que pueden pasar la barrera energeacutetica de estado de transicioacuten para formar a los productos
Decremento de la velocidad con la temperatura la elevacioacuten excesiva de la
temperatura del medio que contiene a las enzimas resulta en un decremento de la velocidad como resultado de la desnaturalizacioacuten es decir la peacuterdida de la estructura tridimensional de las enzimas
26
zona de reactivacioacuten maacuteximo de actividad
Zona de
inactivacioacuten
0 Temperatura
Figura dependencia de la actividad enzimaacutetica con la temperatura
Velocidad maacutexima la velocidad de una reaccioacuten (v) es el nuacutemero de
moleacuteculas de substrato convertidas en producto por unidad de tiempo y
usualmente se expresa en micromoles de producto formadas por minuto La velocidad de una reaccioacuten catalizada por una enzima aumenta conforme se incrementa la concentracioacuten de substrato hasta que se llega a una velocidad maacutexima (Vmax) a partir de la cual la velocidad de la reaccioacuten es
independiente de la cantidad de substrato El que la velocidad no pueda
seguirse incrementando refleja la saturacioacuten del sitio activo con el substrato
Vmax
0 Substrato
Figura representacioacuten de una cineacutetica tiacutepicamente michaeliana
Vmax = velocidad de saturacioacuten o maacutexima
27
Cuando se grafica la velocidad de la reaccioacuten v0 contra la concentracioacuten de substrato S no siempre es posible determinar la condicioacuten
en que se ha llegado a la velocidad maacutexima Vmax debido al incremento de la pendiente en la hipeacuterbola a concentraciones de substrato elevadas
Figura experimento cineacutetico para la actividad enzimaacutetica
Se presentan los valores experimentales y los resultados del ajuste al modelo de Michaelis
Auacuten asiacute si se grafica una doble reciacuteproca es decir el inverso de la
velocidad (1v0) contra el inverso de la concentracioacuten de substrato (1 S ) se
obtiene una liacutenea recta El regraacutefico de Lineweaver-Burke tambieacuten se
denomina de ldquodobles reciacuteprocasrdquo y se utiliza para calcular la Km y la Vmax asiacute como para determinar el mecanismo de accioacuten de los diversos tipos de
inhibidores
La ecuacioacuten que describe a la graacutefca de Lineweaver-Burke es
Vmax
1
SVmax
Km
v
1
0 (3)
que describe una recta en donde el intercepto con el eje de las abscisas es igual a ndash1Km y el intercepto con el eje de las ordenadas es igual a 1Vmax
28
Figura experimento cineacutetico para la actividad enzimaacutetica Regraacutefico de
Lineweaver-Burke
Se presentan los valores experimentales y los resultados del ajuste al modelo
REPRESENTACIOacuteN GRAacuteFICA DE LINEWEAVER Y BURK
Si se toma el inverso en ambos miembros de la ecuacioacuten de Michaelis tenemos
=
+
29
que es la ecuacioacuten de una liacutenea recta cuya pendiente es que intercepta al eje de las
ordenadas en y a la abcisa en el punto Lo que es evidente al hacer
ya que se convierte en
Si se hace una graacutefica con los dobles inversos colocando los valores de en el eje de las
ordenadas y en el eje de las abcisas se obtiene una liacutenea recta a partir de la cual se puede calcular faacutecilmente Km
EFECTO DEL pH SOBRE LA ACTIVIDAD ENZIMATICA
Sabiendo que las enzimas son proteinas cualquier cambio brusco de pH puede alterar el caraacutecter ioacutenico de los grupos amino y carboxilo en la superficie proteica afectando asiacute las propiedades cataliacuteticas de una enzima A pH alto o bajo se puede producir la desnaturalizacioacuten
de la enzima y en consecuencia su inactivacioacuten La fosfatasa aacutecida es maacutes activa a pH 50 mientras que la fosfatasa alcalina lo es a pH 90 Muchas enzimas tienen maacutexima actividad cerca de la neutralidad en un rango de pH de 6 a 8
El pH per se no afecta la actividad enzimaacutetica sino la concentracioacuten de protones Los protones
ademaacutes de alterar la estructura de la enzima y el substrato pueden participar tambieacuten en la reaccioacuten como substrato o producto En esos casos la concentracioacuten de protones afecta directamente la velocidad de la reaccioacuten
ENZIMA pH OPTIMO
30
Pepsina 15
Tripsina 77
Catalasa 76
Arginasa 97
Fumarasa 78
Ribonucleasa 78
EFECTO DE ACTIVADORES
Un aumento en la temperatura provoca un aumento de la velocidad de reaccioacuten hasta cierta temperatura oacuteptima ya que despueacutes de aproximadamente 45
0 C se comienza a producir la
desnaturalizacioacuten teacutermica Las enzimas de muchos mamiacuteferos tienen una temperatura oacuteptima
de 370 C por encima de esa temperatura comienzan a inactivarse y se destruyen Sin embargo
existen especies de bacterias y algas que habitan en fuentes de aguas termales y en el otro extremo ciertas bacterias aacuterticas tienen temperaturas oacuteptimas cercanas a 0
0 C
31
ESPECIFICIDAD ABSOLUTA
Cierta enzimas se encuentran como aproenzimas o zimogenos que son inactivos
El tripsinoacutegeno es convertido en tripsina por la misma accioacuten de la tripsina que remueve un hexapeacuteptido
tripsina Tripsinoacutegeno Tripsina + Val (Asp)4 lisina
Otra forma de activacioacuten consiste en mantener los grupos SH reducidos Estos grupos pueden
formar parte de centros activos de ciertas enzimas Si se oxidan estos grupos a -S-S- la enzima se inactiva Asiacute tenemos que la enzima papaina se inactiva despueacutes de exponerla al ox iacutegeno pero cuando se le antildeade un reductor apropiado la enzima se reactiva Los grupos ndashS-S- se convierten en ndashSH activaacutendose la enzima
Otra forma de activacioacuten requiere la presencia de otra substancia que se enlaza al sitio alosteacuterico el compuesto que se enlaza se denomina un efector alosteacuterico El centro alosteacuterico es bien especiacutefico para el efector o modulador alosteacuterico
ESPECIFICIDAD RELATIVA
Oxiacutegeno L-aminoaacutecido a -cetoaacutecido + NH3 + H 2 O 2 L-aminoaacutecido oxidasa Oxiacutegeno D-aminoaacutecido a -cetoaacutecido + NH3 + H 2 O 2 D-aminoaacutecido oxidasa
32
En este ejemplo se muestra la estereo-especificidad Una enzima puede tener una especificidad oacuteptica para isomeros D o L
Hay enzimas que muestran especificidad absoluta como la aspartasa que cataliza la adicioacuten reversible de amoniacuteaco al doble enlace del aacutecido fumaacuterico pero no al de otros aacutecidos insaturados
HOOC ndash CH = CH ndash COO- + NH4
+ HOOC ndash CH2 - CHNH2 - COO
- + H
+
INHIBICIOacuteN ENZIMATICA
Mediante el uso de inhibidores enzimaacuteticos se ha obtenido informacioacuten muy valiosa sobre la conformacioacuten del centro activo de algunas enzimas
a Inhibicioacuten irreversible Este tipo de inhibidores forman un enlace covalente con las enzimas cerca del centro activo Un ejemplo son los gases nerviosos como el
fluorofosfato de diisopropilo (DFP) que forma un complejo con la enzima acetilcolinesterasa Los animales envenenados con este gas quedan paralizados debido a la imposibilidad de transmitir adecuadamente los impulsos nerviosos
Ester di-isopropil fos foacuterico de la enzima (inactivo)
b) Inhibicioacuten competitiva
Es cuando el inhibidor compite con el substrato por la unioacuten con el centro activo de la
enzima Este tipo de inhibicioacuten puede reducirse si se aumenta la concentracioacuten de substrato
-OOC - CH2 - CH2 - COO
- -OOC - CH = CH - COO
- + 2H
+ Succinato Succinato Fumarato
33
Deshidrogenasa COO
- I CH2 Malonato (inhibidor competitivo) I COO
- En la inhibicioacuten competitiva la enzima se combina con el inhibidor para formar un complejo enzima-inhibidor E + I E I en competencia con la reaccioacuten normal E +S ES
La sulfanilamida (bactericida) interfiere con la siacutentesis del aacutecido foacutelico compitiendo con el aacutecido p-aminobenzoico de una forma competitiva
HOOC-sect- NH2 NH2 ndashSO2 - sect- NH2 aacutecido p- amino benzoico sulfanilamida
sect = fenil
a Inhibicioacuten no competitiva
Esta inhibicioacuten se caracteriza por que no se puede revertir el efecto del inhibidor aumentando la concentracioacuten del substrato
Por ejemplo la inhibicioacuten de la deshidrogenasa del gliceraldehido -3-fosfato por la yodo-acetamida
Enzima-SH + I-CH2 CONH2 Enzima-SCH2 CONH2 + IH
La deshidrogenasa del gliceraldehido-3 fosfato puede ser inhibida tambieacuten por el aacutecido iodo-aceacutetico
Enzima-SH + ICH2 COOH Enzima-SCH2 COOH + IH
c) Inhibicioacuten por metales
Ciertos metales como el plomo mercurio y arseacutenico inhiben enzimas que tienen en su centro activo grupos -SH libres
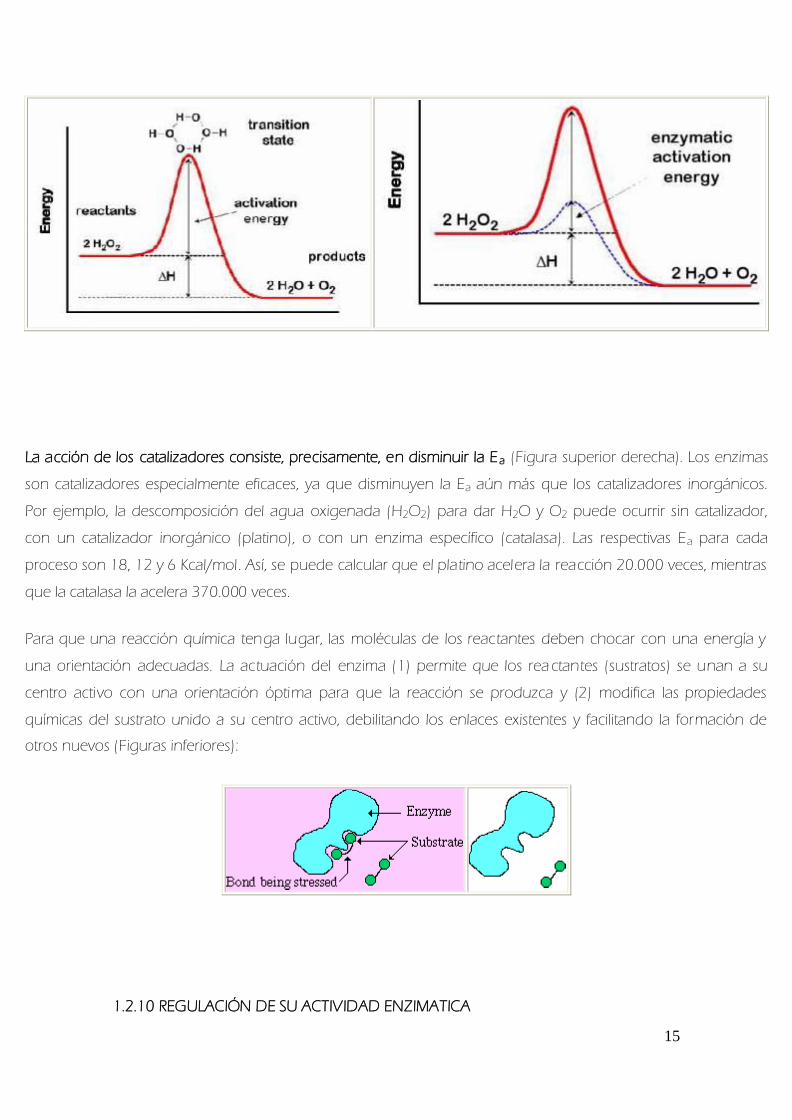
15
La accioacuten de los catalizadores consiste precisamente en disminuir la Ea (Figura superior derecha) Los enzimas
son catalizadores especialmente eficaces ya que disminuyen la Ea auacuten maacutes que los catalizadores inorgaacutenicos
Por ejemplo la descomposicioacuten del agua oxigenada (H2O2) para dar H2O y O2 puede ocurrir sin catalizador
con un catalizador inorgaacutenico (platino) o con un enzima especiacutefico (catalasa) Las respectivas Ea para cada
proceso son 18 12 y 6 Kcalmol Asiacute se puede calcular que el platino acelera la reaccioacuten 20000 veces mientras
que la catalasa la acelera 370000 veces
Para que una reaccioacuten quiacutemica tenga lugar las moleacuteculas de los reactantes deben chocar con una energiacutea y
una orientacioacuten adecuadas La actuacioacuten del enzima (1) permite que los rea ctantes (sustratos) se unan a su
centro activo con una orientacioacuten oacuteptima para que la reaccioacuten se produzca y (2) modifica las propiedades
quiacutemicas del sustrato unido a su centro activo debilitando los enlaces existentes y facilitando la formacioacuten de
otros nuevos (Figuras inferiores)
1210 REGULACIOacuteN DE SU ACTIVIDAD ENZIMATICA
16
Una moleacutecula de enzima no tiene por que actuar siempre a la misma velocidad Su
actividad puede estar modulada por
Cambios en el pH
Cambios en la temperatura
Presencia de cofactores
Las concentraciones del sustrato y de los productos finales
Presencia de inhibidores
Modulacioacuten alosteacuterica
Modificacioacuten covalente
Activacioacuten por proteolisis
Isoenzimas
EFECTO DE LAS CONCENTRACIONES SOBRE LA ACTIVIDAD ENZIMAacuteTICA
La velocidad de una reaccioacuten enzimaacutetica depende de la concentracioacuten de sustrato La Figura de la derecha muestra la velocidad de una reaccioacuten enzimaacutetica a 6 concentraciones distintas de sustrato
Ademaacutes la presencia de los productos finales puede hacer que la reaccioacuten sea maacutes lenta o incluso invertir su sentido (Figura inferior)
EFECTO DE LOS INHIBIDORES SOBRE LA ACTIVIDAD ENZIMAacuteTICA
Ciertas moleacuteculas pueden inhibir la accioacuten cataliacutetica de un enzima son los inhibidores Estos inhibidores bien pueden ocupar temporalmente el centro activo por semejanza estructural con el sustrato original ( inhibidor competitivo) o bien alteran la conformacioacuten espacial del enzima impidiendo su unioacuten al sustrato ( inhibidor no competitivo) (Figuras inferiores)
Inhibidor competitivo Inhibidor no competitivo
17
MODULACIOacuteN ALOSTEacuteRICA DE LA ACTIVIDAD ENZIMAacuteTICA
Hay enzimas que pueden adoptar 2 conformaciones interconvertibles llamadas R (relajada) y T (tensa) R es la forma maacutes activa porque se une al sustrato con maacutes afinidad Las formas R y T se encuentran en equilibrio R lt==gt T (Figura inferior)
Ciertas sustancias tienden a estabilizar la forma R Son los llamados moduladores positivos El propio sustrato es a
menudo un modulador positivo Las moleacuteculas que favorecen la forma R pero que actuacutean sobre una regioacuten del enzima distinta del centro activo son los activadores alosteacutericos (Figura inferior izquierda)
Activador alosteacuterico favorece la unioacuten del sustrato
Inhibidor alosteacuterico impide la unioacuten del sustrato
Las sustancias que favorecen la forma T y disminuyen la actividad enzimaacutetica son los moduladores negativos Si
estos moduladores actuacutean en lugares distintos del centro activo del enzima se llaman inhibidores alosteacutericos
18
(Figura superior derecha
EFECTO DE LA MODIFICACIOacuteN COVALENTE SOBRE LA ACTIVIDAD ENZIMAacuteTICA
Otros enzimas pasan de una forma menos activa a otra maacutes activa unieacutendose covalentemente a un grupo quiacutemico de pequentildeo tamantildeo como el Pi o el AMP Tambieacuten se da el caso inverso en el que un enzima muy activo se desactiva al liberar alguacuten grupo quiacutemico En las enzimas de las viacuteas degradativas del metabolismo la
forma fosforilada es maacutes activa que la no fosforilada mientras que en las viacuteas biosinteacuteticas ocurre lo contrario En las figuras inferiores se ilustra la activacioacuten de una proteiacutena por fosforilacioacuten
Elementos de la reaccioacuten El enzima no fosforilado es inactivo El enzima fosforilado es activo
ACTIVACIOacuteN PROTEOLIacuteTICA DE LA ACTIVIDAD ENZIMAacuteTICA
Algunos enzimas no se sintetizan como tales sino como proteiacutenas precursoras sin actividad enzimaacutetica Estas proteiacutenas se llaman
proenzimas o zimoacutegenos Para activarse los zimoacutegenos sufren un ataque hidroliacutetico que origina la liberacioacuten de uno o varios peacuteptidos El resto de la moleacutecula proteica adopta la conformacioacuten y las propiedades del enzima activo Muchos enzimas digestivos se secretan en forma de zimoacutegenos y en el tubo digestivo
se convierten en la forma activa Es el caso de la -quimotripsina que se sintetiza en forma de quimotripsinoacutegeno (Figura superior) Si estos enzimas se sintetizasen directamente en forma activa destruiriacutean la propia ceacutelula que las
produce Asiacute la tripsina pancreaacutetica (una proteasa) se sintetiza como tripsinoacutegeno (inactivo) Si por alguna razoacuten se activa en el propio paacutencreas la glaacutendula sufre un proceso de autodestruccioacuten (pancreatitis aguda) a
menudo mortal
19
REGULACIOacuteN DE LA ACTIVIDAD ENZIMAacuteTICA POR MEDIO DE ISOENZIMAS
Algunos enzimas tienen distinta estructura molecular aunque su funcioacuten bioloacutegica es similar Se llaman isozimas o isoenzimas Estas diferencias de estructura se traducen en ligeros cambios en sus propiedades de forma que cada isozima se adapta perfectamente a la funcioacuten que debe realizar Asiacute podemos observar la existencia de isoenzimas en funcioacuten de
el tipo de tejido Por ejemplo la lactato deshidrogenasa presenta isozimas distintos en muacutesculo y corazoacuten el compartimento celular donde actuacutea Por ejemplo la malato deshidrogenasa del citoplasma es distinta
de la de la mitocondria el momento concreto del desarrollo del individuo Por ejemplo algunos enzimas de la glicolisis del feto
son diferentes de los mismos enzimas en el adulto
CINEacuteTICA ENZIMAacuteTICA
Los principios generales de las reacciones quiacutemicas se aplican tambieacuten a las reacciones enzimaacuteticas Por este motivo antes de empezar con la cineacutetica quiacutemica se van a repasar algunos conceptos baacutesicos de cineacutetica quiacutemica
A continuacioacuten se describiraacuten los siguientes conceptos
Cineacutetica enzimaacutetica Modelo cineacutetico de Michaelis-Menten Caacutelculo de la KM y la Vmax de un enzima Actividad enzimaacutetica
20
NOMENCLATURA DE LOS ENZIMAS
CATALISIS MOLECULAR
Un catalizador modifica la velocidad de una reaccioacuten quiacutemica sin ser utilizado o aparecer como uno de los productos de la reaccioacuten Una reaccioacuten quiacutemica en la que un substrato(S) se
transforma en un producto(P) S P ocurre por que cierta fraccioacuten de moleacuteculas de S posee mucho maacutes energiacutea que el resto de ellas lo que es suficiente para que alcancen un estado activado en el que pueda formarse o romperse un enlace quiacutemico y se forme el producto(P)
La energiacutea de activacioacuten es la cantidad de energiacutea expresada en caloriacuteas necesaria para que todas las moleacuteculas de un mol a una temperatura dada alcancen el estado reactivo Mientras que el estado de transicioacuten es el estado rico en energiacutea de las moleacuteculas que
interaccionan en la cima de la barrera de activacioacuten La velocidad de una reaccioacuten quiacutemica es proporcional a la concentracioacuten del complejo en el estado de transicioacuten
Una reaccioacuten quiacutemica se puede acelerar de la siguiente forma 1) Al aumentar la temperatura se incrementa la energiacutea cineacutetica por lo que es mayor el nuacutemero de moleacuteculas que alcanzan el
estado de transicioacuten Generalmente el Q10 =2 lo que indica que la velocidad de una reaccioacuten quiacutemica se duplica al aumentar la temperatura en 10
0C 2) Antildeadiendo un catalizador que
disminuye la energiacutea de activacioacuten y aumenta la velocidad de reaccioacuten
La enzima (E) se combina con el substrato (S) formando el complejo de transicioacuten enzima-
substrato (E-S) mediante una reaccioacuten reversible cuya energiacutea de activacioacuten es menor que la de la reaccioacuten no catalizada Cuando se forma el producto de la reaccioacuten (P) se regenera de nuevo la enzima (E) de forma libre la que puede combinarse de nuevo con otra moleacutecula de substrato (S)
Una enzima reduce maacutes eficientemente la energiacutea de activacioacuten(Ea) de una reaccioacuten que un catalizador inorgaacutenico lo que permite que una reaccioacuten se realice a menor temperatura
El siguiente ejemplo ilustra mejor lo que hemos discutido
Reaccioacuten Energiacutea de activacioacuten (Kcalmol-1
)
21
a) el peroacutexido de hidroacutegeno se descompone en
H2 O2 H2 O + O2
18
b) el hierro cataliacutetico (Fe) realiza la reaccioacuten
H2 O2 H2 O + O2
13
c) el platino cataliacutetico (Pt) realiza la reaccioacuten
H2 O2 H2 O + O2
12
d) la catalasa una enzima hepaacutetica la realiza
H2O2 H2O + O2
5
Una enzima no modifica la energiacutea libre ni la constante de equilibrio sino que disminuye la energiacutea de activacioacuten de la reaccioacuten
Ir al principio
EFECTO DE LA CONCENTRACIOacuteN DEL SUBSTRATO SOBRE LA
CATALISIS ENZIMATICA
La actividad de una enzima se puede estudiar in vitro bajo condiciones controladas antildeadieacutendole una enzima a un substrato Siacute se trabaja con la condicioacuten de que la concentracioacuten del substrato sea saturante variando entonces la concentracioacuten de enzima se observa que aumenta el producto de la reaccioacuten a pH y temperatura constantes
22
Si se mantiene la concentracioacuten de la enzima constante y variando la concentracioacuten de substrato se obtiene una curva hiperboacutelica como la de la figura Al principio un aumento de la
concentracioacuten de substrato produce un aumento raacutepi do de la velocidad de reaccioacuten pero si se sigue aumentando la concentracioacuten de substrato la velocidad de reaccioacuten comienza a disminuir y a muy altas concentraciones de substrato se observa que no cambia la velocidad de
reaccioacuten se dice que los centros activos de la enzima se encuentran saturados La velocidad de reaccioacuten que se obtiene a esa alta concentracioacuten de substrato se define como la velocidad maacutexima (V) de la reaccioacuten enzimaacutetica bajo las condiciones especificadas La concentracioacuten de
substrato (S) a la semivelocidad maacutexima de reaccioacuten (V2) se puede determinar de la figura y representa la constante de Michaelis o Km la cual es una caracteriacutestica para cada enzima La inversa de Km o 1Km mide aproximadamente la afinidad de la enzima por el subs trato
Mientras maacutes pequentildeo sea el valor de Km mayor seraacute la afinidad de la enzima por el substrato Si varias enzimas compiten en el metabolismo por el mismo substrato eacuteste seraacute transformado preferentemente por la enzima con mayor afinidad
La ecuacioacuten de Michaelis describe la relacioacuten cuantitativa entre la velocidad de reaccioacuten y la concentracioacuten de substrato [S] si se conoce Vmax o Km
v=
En donde v= es la velocidad de reaccioacuten observada a una concentracioacuten de substrato determinada [S]
Km= constante de Michaelis en molesliacutet ro
23
Vmax= velocidad maacutexima a concentracioacuten saturante de substrato
Si v= Vmax2 entonces
=
Km+[S] = 2[S]
Km =[S]
De tal forma que podemos concluir que Km es igual a la concentracioacuten de substrato a la semivelocidad maacutexima de reaccioacuten
1 Concentraciones relativas de E y S la concentracioacuten de substrato S es
mucho mayor que la de E de tal manera que la cantidad de substrato
unido a la enzima en cualquier momento es muy pequentildea
2 Se asume el estado estacionario ES no cambia con el tiempo esto
quiere decir que la velocidad de formacioacuten de ES es igual a aquella para
su desintegracioacuten En general un intermediario en una serie de reacciones estaacute en estado estacionario cuando su velocidad de siacutentesis
es igual a su velocidad de degradacioacuten 3 Velocidad inicial para el anaacutelisis de reacciones enzimaacuteticas soacutelo se
utiliza la velocidad inicial de la reaccioacuten que es la velocidad ejercida por
la enzima inmediatamente despueacutes de que se ha puesto en contacto con el substrato y hasta antes de que se haya consumido el 10 de la concentracioacuten inicial del mismo La razoacuten de lo anterior es que en ese
momento la concentracioacuten del producto de la reaccioacuten que se ha acumulado en muy pequentildea y por tanto la reaccioacuten en el sentido
inverso es decir la transformacioacuten del producto en el substrato original puede ser ignorada
24
Caracteriacutesticas de Km la constante de Michaelis-Menten es
caracteriacutestica de una enzima y particular para un substrato Refleja la afinidad de la enzima por ese substrato K m es numeacutericamente igual a la concentracioacuten
de substrato a la cual la velocidad de reaccioacuten es la mitad de la Vmax (Km= Vmax2) Este paraacutemetro Km no variacutea con la concentracioacuten de enzima
Significado de una Km pequentildea un valor numeacuterico pequentildeo de Km
refleja una alta afinidad de la enzima por su substrato porque a una baja
concentracioacuten del mismo la enzima a desarrollado ya la mitad de la velocidad maacutexima
Significado de una Km grande el valor numeacuterico grande de Km refleja
una baja afinidad de la enzima por su substrato porque a una concentracioacuten
elevada del mismo la enzima desarrolla la mitad de la velocidad maacutexima
Relacioacuten de la velocidad con la concentracioacuten de enzima la
velocidad de la reaccioacuten es directamente proporcional a la concentracioacuten de la
enzima a cualquier concentracioacuten de substrato Por ejemplo si la concentracioacuten de enzima es disminuida a la mitad la velocidad inicial de la reaccioacuten (v0) es
reducida tambieacuten a la mitad de la original
Orden de la reaccioacuten cuando S es mas pequentildea que la Km la
velocidad de la reaccioacuten es aproximadamente igual a la concentracioacuten de
substrato
100
50
0
Substrato
Km Figura Localizacioacuten de la constante de Michaelis (Km)
25
La velocidad de reaccioacuten se dice en estas condiciones es de primer
orden con respecto al substrato Cuando S es mas grande que la Km la
velocidad es constante e igual a la Vmax La velocidad de la reaccioacuten es independiente de la concentracioacuten de substrato y se dice que es de orden cero
con respecto a la concentracioacuten de substrato
Las enzimas pueden ser aisladas de las ceacutelulas u organismos para estudiar sus
propiedades in vitro es decir en un tubo de ensayo Las diferentes enzimas muestran diferentes respuestas a los cambios en la concentracioacuten de substrato temperatura y pH
Concentracioacuten de substrato
La forma hiperboacutelica de la curva de velocidad vs Substrato
Efecto de la temperatura en la actividad enzimaacutetica
Efecto de la variacioacuten del pH en la actividad enzimaacutetica
Incremento de la velocidad con la temperatura En general la velocidad de la reaccioacuten se incrementa con la temperatura hasta que un punto maacuteximo es alcanzado
Este incremento se debe a que aumenta el nuacutemero de moleacuteculas ricas en energiacutea que pueden pasar la barrera energeacutetica de estado de transicioacuten para formar a los productos
Decremento de la velocidad con la temperatura la elevacioacuten excesiva de la
temperatura del medio que contiene a las enzimas resulta en un decremento de la velocidad como resultado de la desnaturalizacioacuten es decir la peacuterdida de la estructura tridimensional de las enzimas
26
zona de reactivacioacuten maacuteximo de actividad
Zona de
inactivacioacuten
0 Temperatura
Figura dependencia de la actividad enzimaacutetica con la temperatura
Velocidad maacutexima la velocidad de una reaccioacuten (v) es el nuacutemero de
moleacuteculas de substrato convertidas en producto por unidad de tiempo y
usualmente se expresa en micromoles de producto formadas por minuto La velocidad de una reaccioacuten catalizada por una enzima aumenta conforme se incrementa la concentracioacuten de substrato hasta que se llega a una velocidad maacutexima (Vmax) a partir de la cual la velocidad de la reaccioacuten es
independiente de la cantidad de substrato El que la velocidad no pueda
seguirse incrementando refleja la saturacioacuten del sitio activo con el substrato
Vmax
0 Substrato
Figura representacioacuten de una cineacutetica tiacutepicamente michaeliana
Vmax = velocidad de saturacioacuten o maacutexima
27
Cuando se grafica la velocidad de la reaccioacuten v0 contra la concentracioacuten de substrato S no siempre es posible determinar la condicioacuten
en que se ha llegado a la velocidad maacutexima Vmax debido al incremento de la pendiente en la hipeacuterbola a concentraciones de substrato elevadas
Figura experimento cineacutetico para la actividad enzimaacutetica
Se presentan los valores experimentales y los resultados del ajuste al modelo de Michaelis
Auacuten asiacute si se grafica una doble reciacuteproca es decir el inverso de la
velocidad (1v0) contra el inverso de la concentracioacuten de substrato (1 S ) se
obtiene una liacutenea recta El regraacutefico de Lineweaver-Burke tambieacuten se
denomina de ldquodobles reciacuteprocasrdquo y se utiliza para calcular la Km y la Vmax asiacute como para determinar el mecanismo de accioacuten de los diversos tipos de
inhibidores
La ecuacioacuten que describe a la graacutefca de Lineweaver-Burke es
Vmax
1
SVmax
Km
v
1
0 (3)
que describe una recta en donde el intercepto con el eje de las abscisas es igual a ndash1Km y el intercepto con el eje de las ordenadas es igual a 1Vmax
28
Figura experimento cineacutetico para la actividad enzimaacutetica Regraacutefico de
Lineweaver-Burke
Se presentan los valores experimentales y los resultados del ajuste al modelo
REPRESENTACIOacuteN GRAacuteFICA DE LINEWEAVER Y BURK
Si se toma el inverso en ambos miembros de la ecuacioacuten de Michaelis tenemos
=
+
29
que es la ecuacioacuten de una liacutenea recta cuya pendiente es que intercepta al eje de las
ordenadas en y a la abcisa en el punto Lo que es evidente al hacer
ya que se convierte en
Si se hace una graacutefica con los dobles inversos colocando los valores de en el eje de las
ordenadas y en el eje de las abcisas se obtiene una liacutenea recta a partir de la cual se puede calcular faacutecilmente Km
EFECTO DEL pH SOBRE LA ACTIVIDAD ENZIMATICA
Sabiendo que las enzimas son proteinas cualquier cambio brusco de pH puede alterar el caraacutecter ioacutenico de los grupos amino y carboxilo en la superficie proteica afectando asiacute las propiedades cataliacuteticas de una enzima A pH alto o bajo se puede producir la desnaturalizacioacuten
de la enzima y en consecuencia su inactivacioacuten La fosfatasa aacutecida es maacutes activa a pH 50 mientras que la fosfatasa alcalina lo es a pH 90 Muchas enzimas tienen maacutexima actividad cerca de la neutralidad en un rango de pH de 6 a 8
El pH per se no afecta la actividad enzimaacutetica sino la concentracioacuten de protones Los protones
ademaacutes de alterar la estructura de la enzima y el substrato pueden participar tambieacuten en la reaccioacuten como substrato o producto En esos casos la concentracioacuten de protones afecta directamente la velocidad de la reaccioacuten
ENZIMA pH OPTIMO
30
Pepsina 15
Tripsina 77
Catalasa 76
Arginasa 97
Fumarasa 78
Ribonucleasa 78
EFECTO DE ACTIVADORES
Un aumento en la temperatura provoca un aumento de la velocidad de reaccioacuten hasta cierta temperatura oacuteptima ya que despueacutes de aproximadamente 45
0 C se comienza a producir la
desnaturalizacioacuten teacutermica Las enzimas de muchos mamiacuteferos tienen una temperatura oacuteptima
de 370 C por encima de esa temperatura comienzan a inactivarse y se destruyen Sin embargo
existen especies de bacterias y algas que habitan en fuentes de aguas termales y en el otro extremo ciertas bacterias aacuterticas tienen temperaturas oacuteptimas cercanas a 0
0 C
31
ESPECIFICIDAD ABSOLUTA
Cierta enzimas se encuentran como aproenzimas o zimogenos que son inactivos
El tripsinoacutegeno es convertido en tripsina por la misma accioacuten de la tripsina que remueve un hexapeacuteptido
tripsina Tripsinoacutegeno Tripsina + Val (Asp)4 lisina
Otra forma de activacioacuten consiste en mantener los grupos SH reducidos Estos grupos pueden
formar parte de centros activos de ciertas enzimas Si se oxidan estos grupos a -S-S- la enzima se inactiva Asiacute tenemos que la enzima papaina se inactiva despueacutes de exponerla al ox iacutegeno pero cuando se le antildeade un reductor apropiado la enzima se reactiva Los grupos ndashS-S- se convierten en ndashSH activaacutendose la enzima
Otra forma de activacioacuten requiere la presencia de otra substancia que se enlaza al sitio alosteacuterico el compuesto que se enlaza se denomina un efector alosteacuterico El centro alosteacuterico es bien especiacutefico para el efector o modulador alosteacuterico
ESPECIFICIDAD RELATIVA
Oxiacutegeno L-aminoaacutecido a -cetoaacutecido + NH3 + H 2 O 2 L-aminoaacutecido oxidasa Oxiacutegeno D-aminoaacutecido a -cetoaacutecido + NH3 + H 2 O 2 D-aminoaacutecido oxidasa
32
En este ejemplo se muestra la estereo-especificidad Una enzima puede tener una especificidad oacuteptica para isomeros D o L
Hay enzimas que muestran especificidad absoluta como la aspartasa que cataliza la adicioacuten reversible de amoniacuteaco al doble enlace del aacutecido fumaacuterico pero no al de otros aacutecidos insaturados
HOOC ndash CH = CH ndash COO- + NH4
+ HOOC ndash CH2 - CHNH2 - COO
- + H
+
INHIBICIOacuteN ENZIMATICA
Mediante el uso de inhibidores enzimaacuteticos se ha obtenido informacioacuten muy valiosa sobre la conformacioacuten del centro activo de algunas enzimas
a Inhibicioacuten irreversible Este tipo de inhibidores forman un enlace covalente con las enzimas cerca del centro activo Un ejemplo son los gases nerviosos como el
fluorofosfato de diisopropilo (DFP) que forma un complejo con la enzima acetilcolinesterasa Los animales envenenados con este gas quedan paralizados debido a la imposibilidad de transmitir adecuadamente los impulsos nerviosos
Ester di-isopropil fos foacuterico de la enzima (inactivo)
b) Inhibicioacuten competitiva
Es cuando el inhibidor compite con el substrato por la unioacuten con el centro activo de la
enzima Este tipo de inhibicioacuten puede reducirse si se aumenta la concentracioacuten de substrato
-OOC - CH2 - CH2 - COO
- -OOC - CH = CH - COO
- + 2H
+ Succinato Succinato Fumarato
33
Deshidrogenasa COO
- I CH2 Malonato (inhibidor competitivo) I COO
- En la inhibicioacuten competitiva la enzima se combina con el inhibidor para formar un complejo enzima-inhibidor E + I E I en competencia con la reaccioacuten normal E +S ES
La sulfanilamida (bactericida) interfiere con la siacutentesis del aacutecido foacutelico compitiendo con el aacutecido p-aminobenzoico de una forma competitiva
HOOC-sect- NH2 NH2 ndashSO2 - sect- NH2 aacutecido p- amino benzoico sulfanilamida
sect = fenil
a Inhibicioacuten no competitiva
Esta inhibicioacuten se caracteriza por que no se puede revertir el efecto del inhibidor aumentando la concentracioacuten del substrato
Por ejemplo la inhibicioacuten de la deshidrogenasa del gliceraldehido -3-fosfato por la yodo-acetamida
Enzima-SH + I-CH2 CONH2 Enzima-SCH2 CONH2 + IH
La deshidrogenasa del gliceraldehido-3 fosfato puede ser inhibida tambieacuten por el aacutecido iodo-aceacutetico
Enzima-SH + ICH2 COOH Enzima-SCH2 COOH + IH
c) Inhibicioacuten por metales
Ciertos metales como el plomo mercurio y arseacutenico inhiben enzimas que tienen en su centro activo grupos -SH libres

16
Una moleacutecula de enzima no tiene por que actuar siempre a la misma velocidad Su
actividad puede estar modulada por
Cambios en el pH
Cambios en la temperatura
Presencia de cofactores
Las concentraciones del sustrato y de los productos finales
Presencia de inhibidores
Modulacioacuten alosteacuterica
Modificacioacuten covalente
Activacioacuten por proteolisis
Isoenzimas
EFECTO DE LAS CONCENTRACIONES SOBRE LA ACTIVIDAD ENZIMAacuteTICA
La velocidad de una reaccioacuten enzimaacutetica depende de la concentracioacuten de sustrato La Figura de la derecha muestra la velocidad de una reaccioacuten enzimaacutetica a 6 concentraciones distintas de sustrato
Ademaacutes la presencia de los productos finales puede hacer que la reaccioacuten sea maacutes lenta o incluso invertir su sentido (Figura inferior)
EFECTO DE LOS INHIBIDORES SOBRE LA ACTIVIDAD ENZIMAacuteTICA
Ciertas moleacuteculas pueden inhibir la accioacuten cataliacutetica de un enzima son los inhibidores Estos inhibidores bien pueden ocupar temporalmente el centro activo por semejanza estructural con el sustrato original ( inhibidor competitivo) o bien alteran la conformacioacuten espacial del enzima impidiendo su unioacuten al sustrato ( inhibidor no competitivo) (Figuras inferiores)
Inhibidor competitivo Inhibidor no competitivo
17
MODULACIOacuteN ALOSTEacuteRICA DE LA ACTIVIDAD ENZIMAacuteTICA
Hay enzimas que pueden adoptar 2 conformaciones interconvertibles llamadas R (relajada) y T (tensa) R es la forma maacutes activa porque se une al sustrato con maacutes afinidad Las formas R y T se encuentran en equilibrio R lt==gt T (Figura inferior)
Ciertas sustancias tienden a estabilizar la forma R Son los llamados moduladores positivos El propio sustrato es a
menudo un modulador positivo Las moleacuteculas que favorecen la forma R pero que actuacutean sobre una regioacuten del enzima distinta del centro activo son los activadores alosteacutericos (Figura inferior izquierda)
Activador alosteacuterico favorece la unioacuten del sustrato
Inhibidor alosteacuterico impide la unioacuten del sustrato
Las sustancias que favorecen la forma T y disminuyen la actividad enzimaacutetica son los moduladores negativos Si
estos moduladores actuacutean en lugares distintos del centro activo del enzima se llaman inhibidores alosteacutericos
18
(Figura superior derecha
EFECTO DE LA MODIFICACIOacuteN COVALENTE SOBRE LA ACTIVIDAD ENZIMAacuteTICA
Otros enzimas pasan de una forma menos activa a otra maacutes activa unieacutendose covalentemente a un grupo quiacutemico de pequentildeo tamantildeo como el Pi o el AMP Tambieacuten se da el caso inverso en el que un enzima muy activo se desactiva al liberar alguacuten grupo quiacutemico En las enzimas de las viacuteas degradativas del metabolismo la
forma fosforilada es maacutes activa que la no fosforilada mientras que en las viacuteas biosinteacuteticas ocurre lo contrario En las figuras inferiores se ilustra la activacioacuten de una proteiacutena por fosforilacioacuten
Elementos de la reaccioacuten El enzima no fosforilado es inactivo El enzima fosforilado es activo
ACTIVACIOacuteN PROTEOLIacuteTICA DE LA ACTIVIDAD ENZIMAacuteTICA
Algunos enzimas no se sintetizan como tales sino como proteiacutenas precursoras sin actividad enzimaacutetica Estas proteiacutenas se llaman
proenzimas o zimoacutegenos Para activarse los zimoacutegenos sufren un ataque hidroliacutetico que origina la liberacioacuten de uno o varios peacuteptidos El resto de la moleacutecula proteica adopta la conformacioacuten y las propiedades del enzima activo Muchos enzimas digestivos se secretan en forma de zimoacutegenos y en el tubo digestivo
se convierten en la forma activa Es el caso de la -quimotripsina que se sintetiza en forma de quimotripsinoacutegeno (Figura superior) Si estos enzimas se sintetizasen directamente en forma activa destruiriacutean la propia ceacutelula que las
produce Asiacute la tripsina pancreaacutetica (una proteasa) se sintetiza como tripsinoacutegeno (inactivo) Si por alguna razoacuten se activa en el propio paacutencreas la glaacutendula sufre un proceso de autodestruccioacuten (pancreatitis aguda) a
menudo mortal
19
REGULACIOacuteN DE LA ACTIVIDAD ENZIMAacuteTICA POR MEDIO DE ISOENZIMAS
Algunos enzimas tienen distinta estructura molecular aunque su funcioacuten bioloacutegica es similar Se llaman isozimas o isoenzimas Estas diferencias de estructura se traducen en ligeros cambios en sus propiedades de forma que cada isozima se adapta perfectamente a la funcioacuten que debe realizar Asiacute podemos observar la existencia de isoenzimas en funcioacuten de
el tipo de tejido Por ejemplo la lactato deshidrogenasa presenta isozimas distintos en muacutesculo y corazoacuten el compartimento celular donde actuacutea Por ejemplo la malato deshidrogenasa del citoplasma es distinta
de la de la mitocondria el momento concreto del desarrollo del individuo Por ejemplo algunos enzimas de la glicolisis del feto
son diferentes de los mismos enzimas en el adulto
CINEacuteTICA ENZIMAacuteTICA
Los principios generales de las reacciones quiacutemicas se aplican tambieacuten a las reacciones enzimaacuteticas Por este motivo antes de empezar con la cineacutetica quiacutemica se van a repasar algunos conceptos baacutesicos de cineacutetica quiacutemica
A continuacioacuten se describiraacuten los siguientes conceptos
Cineacutetica enzimaacutetica Modelo cineacutetico de Michaelis-Menten Caacutelculo de la KM y la Vmax de un enzima Actividad enzimaacutetica
20
NOMENCLATURA DE LOS ENZIMAS
CATALISIS MOLECULAR
Un catalizador modifica la velocidad de una reaccioacuten quiacutemica sin ser utilizado o aparecer como uno de los productos de la reaccioacuten Una reaccioacuten quiacutemica en la que un substrato(S) se
transforma en un producto(P) S P ocurre por que cierta fraccioacuten de moleacuteculas de S posee mucho maacutes energiacutea que el resto de ellas lo que es suficiente para que alcancen un estado activado en el que pueda formarse o romperse un enlace quiacutemico y se forme el producto(P)
La energiacutea de activacioacuten es la cantidad de energiacutea expresada en caloriacuteas necesaria para que todas las moleacuteculas de un mol a una temperatura dada alcancen el estado reactivo Mientras que el estado de transicioacuten es el estado rico en energiacutea de las moleacuteculas que
interaccionan en la cima de la barrera de activacioacuten La velocidad de una reaccioacuten quiacutemica es proporcional a la concentracioacuten del complejo en el estado de transicioacuten
Una reaccioacuten quiacutemica se puede acelerar de la siguiente forma 1) Al aumentar la temperatura se incrementa la energiacutea cineacutetica por lo que es mayor el nuacutemero de moleacuteculas que alcanzan el
estado de transicioacuten Generalmente el Q10 =2 lo que indica que la velocidad de una reaccioacuten quiacutemica se duplica al aumentar la temperatura en 10
0C 2) Antildeadiendo un catalizador que
disminuye la energiacutea de activacioacuten y aumenta la velocidad de reaccioacuten
La enzima (E) se combina con el substrato (S) formando el complejo de transicioacuten enzima-
substrato (E-S) mediante una reaccioacuten reversible cuya energiacutea de activacioacuten es menor que la de la reaccioacuten no catalizada Cuando se forma el producto de la reaccioacuten (P) se regenera de nuevo la enzima (E) de forma libre la que puede combinarse de nuevo con otra moleacutecula de substrato (S)
Una enzima reduce maacutes eficientemente la energiacutea de activacioacuten(Ea) de una reaccioacuten que un catalizador inorgaacutenico lo que permite que una reaccioacuten se realice a menor temperatura
El siguiente ejemplo ilustra mejor lo que hemos discutido
Reaccioacuten Energiacutea de activacioacuten (Kcalmol-1
)
21
a) el peroacutexido de hidroacutegeno se descompone en
H2 O2 H2 O + O2
18
b) el hierro cataliacutetico (Fe) realiza la reaccioacuten
H2 O2 H2 O + O2
13
c) el platino cataliacutetico (Pt) realiza la reaccioacuten
H2 O2 H2 O + O2
12
d) la catalasa una enzima hepaacutetica la realiza
H2O2 H2O + O2
5
Una enzima no modifica la energiacutea libre ni la constante de equilibrio sino que disminuye la energiacutea de activacioacuten de la reaccioacuten
Ir al principio
EFECTO DE LA CONCENTRACIOacuteN DEL SUBSTRATO SOBRE LA
CATALISIS ENZIMATICA
La actividad de una enzima se puede estudiar in vitro bajo condiciones controladas antildeadieacutendole una enzima a un substrato Siacute se trabaja con la condicioacuten de que la concentracioacuten del substrato sea saturante variando entonces la concentracioacuten de enzima se observa que aumenta el producto de la reaccioacuten a pH y temperatura constantes
22
Si se mantiene la concentracioacuten de la enzima constante y variando la concentracioacuten de substrato se obtiene una curva hiperboacutelica como la de la figura Al principio un aumento de la
concentracioacuten de substrato produce un aumento raacutepi do de la velocidad de reaccioacuten pero si se sigue aumentando la concentracioacuten de substrato la velocidad de reaccioacuten comienza a disminuir y a muy altas concentraciones de substrato se observa que no cambia la velocidad de
reaccioacuten se dice que los centros activos de la enzima se encuentran saturados La velocidad de reaccioacuten que se obtiene a esa alta concentracioacuten de substrato se define como la velocidad maacutexima (V) de la reaccioacuten enzimaacutetica bajo las condiciones especificadas La concentracioacuten de
substrato (S) a la semivelocidad maacutexima de reaccioacuten (V2) se puede determinar de la figura y representa la constante de Michaelis o Km la cual es una caracteriacutestica para cada enzima La inversa de Km o 1Km mide aproximadamente la afinidad de la enzima por el subs trato
Mientras maacutes pequentildeo sea el valor de Km mayor seraacute la afinidad de la enzima por el substrato Si varias enzimas compiten en el metabolismo por el mismo substrato eacuteste seraacute transformado preferentemente por la enzima con mayor afinidad
La ecuacioacuten de Michaelis describe la relacioacuten cuantitativa entre la velocidad de reaccioacuten y la concentracioacuten de substrato [S] si se conoce Vmax o Km
v=
En donde v= es la velocidad de reaccioacuten observada a una concentracioacuten de substrato determinada [S]
Km= constante de Michaelis en molesliacutet ro
23
Vmax= velocidad maacutexima a concentracioacuten saturante de substrato
Si v= Vmax2 entonces
=
Km+[S] = 2[S]
Km =[S]
De tal forma que podemos concluir que Km es igual a la concentracioacuten de substrato a la semivelocidad maacutexima de reaccioacuten
1 Concentraciones relativas de E y S la concentracioacuten de substrato S es
mucho mayor que la de E de tal manera que la cantidad de substrato
unido a la enzima en cualquier momento es muy pequentildea
2 Se asume el estado estacionario ES no cambia con el tiempo esto
quiere decir que la velocidad de formacioacuten de ES es igual a aquella para
su desintegracioacuten En general un intermediario en una serie de reacciones estaacute en estado estacionario cuando su velocidad de siacutentesis
es igual a su velocidad de degradacioacuten 3 Velocidad inicial para el anaacutelisis de reacciones enzimaacuteticas soacutelo se
utiliza la velocidad inicial de la reaccioacuten que es la velocidad ejercida por
la enzima inmediatamente despueacutes de que se ha puesto en contacto con el substrato y hasta antes de que se haya consumido el 10 de la concentracioacuten inicial del mismo La razoacuten de lo anterior es que en ese
momento la concentracioacuten del producto de la reaccioacuten que se ha acumulado en muy pequentildea y por tanto la reaccioacuten en el sentido
inverso es decir la transformacioacuten del producto en el substrato original puede ser ignorada
24
Caracteriacutesticas de Km la constante de Michaelis-Menten es
caracteriacutestica de una enzima y particular para un substrato Refleja la afinidad de la enzima por ese substrato K m es numeacutericamente igual a la concentracioacuten
de substrato a la cual la velocidad de reaccioacuten es la mitad de la Vmax (Km= Vmax2) Este paraacutemetro Km no variacutea con la concentracioacuten de enzima
Significado de una Km pequentildea un valor numeacuterico pequentildeo de Km
refleja una alta afinidad de la enzima por su substrato porque a una baja
concentracioacuten del mismo la enzima a desarrollado ya la mitad de la velocidad maacutexima
Significado de una Km grande el valor numeacuterico grande de Km refleja
una baja afinidad de la enzima por su substrato porque a una concentracioacuten
elevada del mismo la enzima desarrolla la mitad de la velocidad maacutexima
Relacioacuten de la velocidad con la concentracioacuten de enzima la
velocidad de la reaccioacuten es directamente proporcional a la concentracioacuten de la
enzima a cualquier concentracioacuten de substrato Por ejemplo si la concentracioacuten de enzima es disminuida a la mitad la velocidad inicial de la reaccioacuten (v0) es
reducida tambieacuten a la mitad de la original
Orden de la reaccioacuten cuando S es mas pequentildea que la Km la
velocidad de la reaccioacuten es aproximadamente igual a la concentracioacuten de
substrato
100
50
0
Substrato
Km Figura Localizacioacuten de la constante de Michaelis (Km)
25
La velocidad de reaccioacuten se dice en estas condiciones es de primer
orden con respecto al substrato Cuando S es mas grande que la Km la
velocidad es constante e igual a la Vmax La velocidad de la reaccioacuten es independiente de la concentracioacuten de substrato y se dice que es de orden cero
con respecto a la concentracioacuten de substrato
Las enzimas pueden ser aisladas de las ceacutelulas u organismos para estudiar sus
propiedades in vitro es decir en un tubo de ensayo Las diferentes enzimas muestran diferentes respuestas a los cambios en la concentracioacuten de substrato temperatura y pH
Concentracioacuten de substrato
La forma hiperboacutelica de la curva de velocidad vs Substrato
Efecto de la temperatura en la actividad enzimaacutetica
Efecto de la variacioacuten del pH en la actividad enzimaacutetica
Incremento de la velocidad con la temperatura En general la velocidad de la reaccioacuten se incrementa con la temperatura hasta que un punto maacuteximo es alcanzado
Este incremento se debe a que aumenta el nuacutemero de moleacuteculas ricas en energiacutea que pueden pasar la barrera energeacutetica de estado de transicioacuten para formar a los productos
Decremento de la velocidad con la temperatura la elevacioacuten excesiva de la
temperatura del medio que contiene a las enzimas resulta en un decremento de la velocidad como resultado de la desnaturalizacioacuten es decir la peacuterdida de la estructura tridimensional de las enzimas
26
zona de reactivacioacuten maacuteximo de actividad
Zona de
inactivacioacuten
0 Temperatura
Figura dependencia de la actividad enzimaacutetica con la temperatura
Velocidad maacutexima la velocidad de una reaccioacuten (v) es el nuacutemero de
moleacuteculas de substrato convertidas en producto por unidad de tiempo y
usualmente se expresa en micromoles de producto formadas por minuto La velocidad de una reaccioacuten catalizada por una enzima aumenta conforme se incrementa la concentracioacuten de substrato hasta que se llega a una velocidad maacutexima (Vmax) a partir de la cual la velocidad de la reaccioacuten es
independiente de la cantidad de substrato El que la velocidad no pueda
seguirse incrementando refleja la saturacioacuten del sitio activo con el substrato
Vmax
0 Substrato
Figura representacioacuten de una cineacutetica tiacutepicamente michaeliana
Vmax = velocidad de saturacioacuten o maacutexima
27
Cuando se grafica la velocidad de la reaccioacuten v0 contra la concentracioacuten de substrato S no siempre es posible determinar la condicioacuten
en que se ha llegado a la velocidad maacutexima Vmax debido al incremento de la pendiente en la hipeacuterbola a concentraciones de substrato elevadas
Figura experimento cineacutetico para la actividad enzimaacutetica
Se presentan los valores experimentales y los resultados del ajuste al modelo de Michaelis
Auacuten asiacute si se grafica una doble reciacuteproca es decir el inverso de la
velocidad (1v0) contra el inverso de la concentracioacuten de substrato (1 S ) se
obtiene una liacutenea recta El regraacutefico de Lineweaver-Burke tambieacuten se
denomina de ldquodobles reciacuteprocasrdquo y se utiliza para calcular la Km y la Vmax asiacute como para determinar el mecanismo de accioacuten de los diversos tipos de
inhibidores
La ecuacioacuten que describe a la graacutefca de Lineweaver-Burke es
Vmax
1
SVmax
Km
v
1
0 (3)
que describe una recta en donde el intercepto con el eje de las abscisas es igual a ndash1Km y el intercepto con el eje de las ordenadas es igual a 1Vmax
28
Figura experimento cineacutetico para la actividad enzimaacutetica Regraacutefico de
Lineweaver-Burke
Se presentan los valores experimentales y los resultados del ajuste al modelo
REPRESENTACIOacuteN GRAacuteFICA DE LINEWEAVER Y BURK
Si se toma el inverso en ambos miembros de la ecuacioacuten de Michaelis tenemos
=
+
29
que es la ecuacioacuten de una liacutenea recta cuya pendiente es que intercepta al eje de las
ordenadas en y a la abcisa en el punto Lo que es evidente al hacer
ya que se convierte en
Si se hace una graacutefica con los dobles inversos colocando los valores de en el eje de las
ordenadas y en el eje de las abcisas se obtiene una liacutenea recta a partir de la cual se puede calcular faacutecilmente Km
EFECTO DEL pH SOBRE LA ACTIVIDAD ENZIMATICA
Sabiendo que las enzimas son proteinas cualquier cambio brusco de pH puede alterar el caraacutecter ioacutenico de los grupos amino y carboxilo en la superficie proteica afectando asiacute las propiedades cataliacuteticas de una enzima A pH alto o bajo se puede producir la desnaturalizacioacuten
de la enzima y en consecuencia su inactivacioacuten La fosfatasa aacutecida es maacutes activa a pH 50 mientras que la fosfatasa alcalina lo es a pH 90 Muchas enzimas tienen maacutexima actividad cerca de la neutralidad en un rango de pH de 6 a 8
El pH per se no afecta la actividad enzimaacutetica sino la concentracioacuten de protones Los protones
ademaacutes de alterar la estructura de la enzima y el substrato pueden participar tambieacuten en la reaccioacuten como substrato o producto En esos casos la concentracioacuten de protones afecta directamente la velocidad de la reaccioacuten
ENZIMA pH OPTIMO
30
Pepsina 15
Tripsina 77
Catalasa 76
Arginasa 97
Fumarasa 78
Ribonucleasa 78
EFECTO DE ACTIVADORES
Un aumento en la temperatura provoca un aumento de la velocidad de reaccioacuten hasta cierta temperatura oacuteptima ya que despueacutes de aproximadamente 45
0 C se comienza a producir la
desnaturalizacioacuten teacutermica Las enzimas de muchos mamiacuteferos tienen una temperatura oacuteptima
de 370 C por encima de esa temperatura comienzan a inactivarse y se destruyen Sin embargo
existen especies de bacterias y algas que habitan en fuentes de aguas termales y en el otro extremo ciertas bacterias aacuterticas tienen temperaturas oacuteptimas cercanas a 0
0 C
31
ESPECIFICIDAD ABSOLUTA
Cierta enzimas se encuentran como aproenzimas o zimogenos que son inactivos
El tripsinoacutegeno es convertido en tripsina por la misma accioacuten de la tripsina que remueve un hexapeacuteptido
tripsina Tripsinoacutegeno Tripsina + Val (Asp)4 lisina
Otra forma de activacioacuten consiste en mantener los grupos SH reducidos Estos grupos pueden
formar parte de centros activos de ciertas enzimas Si se oxidan estos grupos a -S-S- la enzima se inactiva Asiacute tenemos que la enzima papaina se inactiva despueacutes de exponerla al ox iacutegeno pero cuando se le antildeade un reductor apropiado la enzima se reactiva Los grupos ndashS-S- se convierten en ndashSH activaacutendose la enzima
Otra forma de activacioacuten requiere la presencia de otra substancia que se enlaza al sitio alosteacuterico el compuesto que se enlaza se denomina un efector alosteacuterico El centro alosteacuterico es bien especiacutefico para el efector o modulador alosteacuterico
ESPECIFICIDAD RELATIVA
Oxiacutegeno L-aminoaacutecido a -cetoaacutecido + NH3 + H 2 O 2 L-aminoaacutecido oxidasa Oxiacutegeno D-aminoaacutecido a -cetoaacutecido + NH3 + H 2 O 2 D-aminoaacutecido oxidasa
32
En este ejemplo se muestra la estereo-especificidad Una enzima puede tener una especificidad oacuteptica para isomeros D o L
Hay enzimas que muestran especificidad absoluta como la aspartasa que cataliza la adicioacuten reversible de amoniacuteaco al doble enlace del aacutecido fumaacuterico pero no al de otros aacutecidos insaturados
HOOC ndash CH = CH ndash COO- + NH4
+ HOOC ndash CH2 - CHNH2 - COO
- + H
+
INHIBICIOacuteN ENZIMATICA
Mediante el uso de inhibidores enzimaacuteticos se ha obtenido informacioacuten muy valiosa sobre la conformacioacuten del centro activo de algunas enzimas
a Inhibicioacuten irreversible Este tipo de inhibidores forman un enlace covalente con las enzimas cerca del centro activo Un ejemplo son los gases nerviosos como el
fluorofosfato de diisopropilo (DFP) que forma un complejo con la enzima acetilcolinesterasa Los animales envenenados con este gas quedan paralizados debido a la imposibilidad de transmitir adecuadamente los impulsos nerviosos
Ester di-isopropil fos foacuterico de la enzima (inactivo)
b) Inhibicioacuten competitiva
Es cuando el inhibidor compite con el substrato por la unioacuten con el centro activo de la
enzima Este tipo de inhibicioacuten puede reducirse si se aumenta la concentracioacuten de substrato
-OOC - CH2 - CH2 - COO
- -OOC - CH = CH - COO
- + 2H
+ Succinato Succinato Fumarato
33
Deshidrogenasa COO
- I CH2 Malonato (inhibidor competitivo) I COO
- En la inhibicioacuten competitiva la enzima se combina con el inhibidor para formar un complejo enzima-inhibidor E + I E I en competencia con la reaccioacuten normal E +S ES
La sulfanilamida (bactericida) interfiere con la siacutentesis del aacutecido foacutelico compitiendo con el aacutecido p-aminobenzoico de una forma competitiva
HOOC-sect- NH2 NH2 ndashSO2 - sect- NH2 aacutecido p- amino benzoico sulfanilamida
sect = fenil
a Inhibicioacuten no competitiva
Esta inhibicioacuten se caracteriza por que no se puede revertir el efecto del inhibidor aumentando la concentracioacuten del substrato
Por ejemplo la inhibicioacuten de la deshidrogenasa del gliceraldehido -3-fosfato por la yodo-acetamida
Enzima-SH + I-CH2 CONH2 Enzima-SCH2 CONH2 + IH
La deshidrogenasa del gliceraldehido-3 fosfato puede ser inhibida tambieacuten por el aacutecido iodo-aceacutetico
Enzima-SH + ICH2 COOH Enzima-SCH2 COOH + IH
c) Inhibicioacuten por metales
Ciertos metales como el plomo mercurio y arseacutenico inhiben enzimas que tienen en su centro activo grupos -SH libres

17
MODULACIOacuteN ALOSTEacuteRICA DE LA ACTIVIDAD ENZIMAacuteTICA
Hay enzimas que pueden adoptar 2 conformaciones interconvertibles llamadas R (relajada) y T (tensa) R es la forma maacutes activa porque se une al sustrato con maacutes afinidad Las formas R y T se encuentran en equilibrio R lt==gt T (Figura inferior)
Ciertas sustancias tienden a estabilizar la forma R Son los llamados moduladores positivos El propio sustrato es a
menudo un modulador positivo Las moleacuteculas que favorecen la forma R pero que actuacutean sobre una regioacuten del enzima distinta del centro activo son los activadores alosteacutericos (Figura inferior izquierda)
Activador alosteacuterico favorece la unioacuten del sustrato
Inhibidor alosteacuterico impide la unioacuten del sustrato
Las sustancias que favorecen la forma T y disminuyen la actividad enzimaacutetica son los moduladores negativos Si
estos moduladores actuacutean en lugares distintos del centro activo del enzima se llaman inhibidores alosteacutericos
18
(Figura superior derecha
EFECTO DE LA MODIFICACIOacuteN COVALENTE SOBRE LA ACTIVIDAD ENZIMAacuteTICA
Otros enzimas pasan de una forma menos activa a otra maacutes activa unieacutendose covalentemente a un grupo quiacutemico de pequentildeo tamantildeo como el Pi o el AMP Tambieacuten se da el caso inverso en el que un enzima muy activo se desactiva al liberar alguacuten grupo quiacutemico En las enzimas de las viacuteas degradativas del metabolismo la
forma fosforilada es maacutes activa que la no fosforilada mientras que en las viacuteas biosinteacuteticas ocurre lo contrario En las figuras inferiores se ilustra la activacioacuten de una proteiacutena por fosforilacioacuten
Elementos de la reaccioacuten El enzima no fosforilado es inactivo El enzima fosforilado es activo
ACTIVACIOacuteN PROTEOLIacuteTICA DE LA ACTIVIDAD ENZIMAacuteTICA
Algunos enzimas no se sintetizan como tales sino como proteiacutenas precursoras sin actividad enzimaacutetica Estas proteiacutenas se llaman
proenzimas o zimoacutegenos Para activarse los zimoacutegenos sufren un ataque hidroliacutetico que origina la liberacioacuten de uno o varios peacuteptidos El resto de la moleacutecula proteica adopta la conformacioacuten y las propiedades del enzima activo Muchos enzimas digestivos se secretan en forma de zimoacutegenos y en el tubo digestivo
se convierten en la forma activa Es el caso de la -quimotripsina que se sintetiza en forma de quimotripsinoacutegeno (Figura superior) Si estos enzimas se sintetizasen directamente en forma activa destruiriacutean la propia ceacutelula que las
produce Asiacute la tripsina pancreaacutetica (una proteasa) se sintetiza como tripsinoacutegeno (inactivo) Si por alguna razoacuten se activa en el propio paacutencreas la glaacutendula sufre un proceso de autodestruccioacuten (pancreatitis aguda) a
menudo mortal
19
REGULACIOacuteN DE LA ACTIVIDAD ENZIMAacuteTICA POR MEDIO DE ISOENZIMAS
Algunos enzimas tienen distinta estructura molecular aunque su funcioacuten bioloacutegica es similar Se llaman isozimas o isoenzimas Estas diferencias de estructura se traducen en ligeros cambios en sus propiedades de forma que cada isozima se adapta perfectamente a la funcioacuten que debe realizar Asiacute podemos observar la existencia de isoenzimas en funcioacuten de
el tipo de tejido Por ejemplo la lactato deshidrogenasa presenta isozimas distintos en muacutesculo y corazoacuten el compartimento celular donde actuacutea Por ejemplo la malato deshidrogenasa del citoplasma es distinta
de la de la mitocondria el momento concreto del desarrollo del individuo Por ejemplo algunos enzimas de la glicolisis del feto
son diferentes de los mismos enzimas en el adulto
CINEacuteTICA ENZIMAacuteTICA
Los principios generales de las reacciones quiacutemicas se aplican tambieacuten a las reacciones enzimaacuteticas Por este motivo antes de empezar con la cineacutetica quiacutemica se van a repasar algunos conceptos baacutesicos de cineacutetica quiacutemica
A continuacioacuten se describiraacuten los siguientes conceptos
Cineacutetica enzimaacutetica Modelo cineacutetico de Michaelis-Menten Caacutelculo de la KM y la Vmax de un enzima Actividad enzimaacutetica
20
NOMENCLATURA DE LOS ENZIMAS
CATALISIS MOLECULAR
Un catalizador modifica la velocidad de una reaccioacuten quiacutemica sin ser utilizado o aparecer como uno de los productos de la reaccioacuten Una reaccioacuten quiacutemica en la que un substrato(S) se
transforma en un producto(P) S P ocurre por que cierta fraccioacuten de moleacuteculas de S posee mucho maacutes energiacutea que el resto de ellas lo que es suficiente para que alcancen un estado activado en el que pueda formarse o romperse un enlace quiacutemico y se forme el producto(P)
La energiacutea de activacioacuten es la cantidad de energiacutea expresada en caloriacuteas necesaria para que todas las moleacuteculas de un mol a una temperatura dada alcancen el estado reactivo Mientras que el estado de transicioacuten es el estado rico en energiacutea de las moleacuteculas que
interaccionan en la cima de la barrera de activacioacuten La velocidad de una reaccioacuten quiacutemica es proporcional a la concentracioacuten del complejo en el estado de transicioacuten
Una reaccioacuten quiacutemica se puede acelerar de la siguiente forma 1) Al aumentar la temperatura se incrementa la energiacutea cineacutetica por lo que es mayor el nuacutemero de moleacuteculas que alcanzan el
estado de transicioacuten Generalmente el Q10 =2 lo que indica que la velocidad de una reaccioacuten quiacutemica se duplica al aumentar la temperatura en 10
0C 2) Antildeadiendo un catalizador que
disminuye la energiacutea de activacioacuten y aumenta la velocidad de reaccioacuten
La enzima (E) se combina con el substrato (S) formando el complejo de transicioacuten enzima-
substrato (E-S) mediante una reaccioacuten reversible cuya energiacutea de activacioacuten es menor que la de la reaccioacuten no catalizada Cuando se forma el producto de la reaccioacuten (P) se regenera de nuevo la enzima (E) de forma libre la que puede combinarse de nuevo con otra moleacutecula de substrato (S)
Una enzima reduce maacutes eficientemente la energiacutea de activacioacuten(Ea) de una reaccioacuten que un catalizador inorgaacutenico lo que permite que una reaccioacuten se realice a menor temperatura
El siguiente ejemplo ilustra mejor lo que hemos discutido
Reaccioacuten Energiacutea de activacioacuten (Kcalmol-1
)
21
a) el peroacutexido de hidroacutegeno se descompone en
H2 O2 H2 O + O2
18
b) el hierro cataliacutetico (Fe) realiza la reaccioacuten
H2 O2 H2 O + O2
13
c) el platino cataliacutetico (Pt) realiza la reaccioacuten
H2 O2 H2 O + O2
12
d) la catalasa una enzima hepaacutetica la realiza
H2O2 H2O + O2
5
Una enzima no modifica la energiacutea libre ni la constante de equilibrio sino que disminuye la energiacutea de activacioacuten de la reaccioacuten
Ir al principio
EFECTO DE LA CONCENTRACIOacuteN DEL SUBSTRATO SOBRE LA
CATALISIS ENZIMATICA
La actividad de una enzima se puede estudiar in vitro bajo condiciones controladas antildeadieacutendole una enzima a un substrato Siacute se trabaja con la condicioacuten de que la concentracioacuten del substrato sea saturante variando entonces la concentracioacuten de enzima se observa que aumenta el producto de la reaccioacuten a pH y temperatura constantes
22
Si se mantiene la concentracioacuten de la enzima constante y variando la concentracioacuten de substrato se obtiene una curva hiperboacutelica como la de la figura Al principio un aumento de la
concentracioacuten de substrato produce un aumento raacutepi do de la velocidad de reaccioacuten pero si se sigue aumentando la concentracioacuten de substrato la velocidad de reaccioacuten comienza a disminuir y a muy altas concentraciones de substrato se observa que no cambia la velocidad de
reaccioacuten se dice que los centros activos de la enzima se encuentran saturados La velocidad de reaccioacuten que se obtiene a esa alta concentracioacuten de substrato se define como la velocidad maacutexima (V) de la reaccioacuten enzimaacutetica bajo las condiciones especificadas La concentracioacuten de
substrato (S) a la semivelocidad maacutexima de reaccioacuten (V2) se puede determinar de la figura y representa la constante de Michaelis o Km la cual es una caracteriacutestica para cada enzima La inversa de Km o 1Km mide aproximadamente la afinidad de la enzima por el subs trato
Mientras maacutes pequentildeo sea el valor de Km mayor seraacute la afinidad de la enzima por el substrato Si varias enzimas compiten en el metabolismo por el mismo substrato eacuteste seraacute transformado preferentemente por la enzima con mayor afinidad
La ecuacioacuten de Michaelis describe la relacioacuten cuantitativa entre la velocidad de reaccioacuten y la concentracioacuten de substrato [S] si se conoce Vmax o Km
v=
En donde v= es la velocidad de reaccioacuten observada a una concentracioacuten de substrato determinada [S]
Km= constante de Michaelis en molesliacutet ro
23
Vmax= velocidad maacutexima a concentracioacuten saturante de substrato
Si v= Vmax2 entonces
=
Km+[S] = 2[S]
Km =[S]
De tal forma que podemos concluir que Km es igual a la concentracioacuten de substrato a la semivelocidad maacutexima de reaccioacuten
1 Concentraciones relativas de E y S la concentracioacuten de substrato S es
mucho mayor que la de E de tal manera que la cantidad de substrato
unido a la enzima en cualquier momento es muy pequentildea
2 Se asume el estado estacionario ES no cambia con el tiempo esto
quiere decir que la velocidad de formacioacuten de ES es igual a aquella para
su desintegracioacuten En general un intermediario en una serie de reacciones estaacute en estado estacionario cuando su velocidad de siacutentesis
es igual a su velocidad de degradacioacuten 3 Velocidad inicial para el anaacutelisis de reacciones enzimaacuteticas soacutelo se
utiliza la velocidad inicial de la reaccioacuten que es la velocidad ejercida por
la enzima inmediatamente despueacutes de que se ha puesto en contacto con el substrato y hasta antes de que se haya consumido el 10 de la concentracioacuten inicial del mismo La razoacuten de lo anterior es que en ese
momento la concentracioacuten del producto de la reaccioacuten que se ha acumulado en muy pequentildea y por tanto la reaccioacuten en el sentido
inverso es decir la transformacioacuten del producto en el substrato original puede ser ignorada
24
Caracteriacutesticas de Km la constante de Michaelis-Menten es
caracteriacutestica de una enzima y particular para un substrato Refleja la afinidad de la enzima por ese substrato K m es numeacutericamente igual a la concentracioacuten
de substrato a la cual la velocidad de reaccioacuten es la mitad de la Vmax (Km= Vmax2) Este paraacutemetro Km no variacutea con la concentracioacuten de enzima
Significado de una Km pequentildea un valor numeacuterico pequentildeo de Km
refleja una alta afinidad de la enzima por su substrato porque a una baja
concentracioacuten del mismo la enzima a desarrollado ya la mitad de la velocidad maacutexima
Significado de una Km grande el valor numeacuterico grande de Km refleja
una baja afinidad de la enzima por su substrato porque a una concentracioacuten
elevada del mismo la enzima desarrolla la mitad de la velocidad maacutexima
Relacioacuten de la velocidad con la concentracioacuten de enzima la
velocidad de la reaccioacuten es directamente proporcional a la concentracioacuten de la
enzima a cualquier concentracioacuten de substrato Por ejemplo si la concentracioacuten de enzima es disminuida a la mitad la velocidad inicial de la reaccioacuten (v0) es
reducida tambieacuten a la mitad de la original
Orden de la reaccioacuten cuando S es mas pequentildea que la Km la
velocidad de la reaccioacuten es aproximadamente igual a la concentracioacuten de
substrato
100
50
0
Substrato
Km Figura Localizacioacuten de la constante de Michaelis (Km)
25
La velocidad de reaccioacuten se dice en estas condiciones es de primer
orden con respecto al substrato Cuando S es mas grande que la Km la
velocidad es constante e igual a la Vmax La velocidad de la reaccioacuten es independiente de la concentracioacuten de substrato y se dice que es de orden cero
con respecto a la concentracioacuten de substrato
Las enzimas pueden ser aisladas de las ceacutelulas u organismos para estudiar sus
propiedades in vitro es decir en un tubo de ensayo Las diferentes enzimas muestran diferentes respuestas a los cambios en la concentracioacuten de substrato temperatura y pH
Concentracioacuten de substrato
La forma hiperboacutelica de la curva de velocidad vs Substrato
Efecto de la temperatura en la actividad enzimaacutetica
Efecto de la variacioacuten del pH en la actividad enzimaacutetica
Incremento de la velocidad con la temperatura En general la velocidad de la reaccioacuten se incrementa con la temperatura hasta que un punto maacuteximo es alcanzado
Este incremento se debe a que aumenta el nuacutemero de moleacuteculas ricas en energiacutea que pueden pasar la barrera energeacutetica de estado de transicioacuten para formar a los productos
Decremento de la velocidad con la temperatura la elevacioacuten excesiva de la
temperatura del medio que contiene a las enzimas resulta en un decremento de la velocidad como resultado de la desnaturalizacioacuten es decir la peacuterdida de la estructura tridimensional de las enzimas
26
zona de reactivacioacuten maacuteximo de actividad
Zona de
inactivacioacuten
0 Temperatura
Figura dependencia de la actividad enzimaacutetica con la temperatura
Velocidad maacutexima la velocidad de una reaccioacuten (v) es el nuacutemero de
moleacuteculas de substrato convertidas en producto por unidad de tiempo y
usualmente se expresa en micromoles de producto formadas por minuto La velocidad de una reaccioacuten catalizada por una enzima aumenta conforme se incrementa la concentracioacuten de substrato hasta que se llega a una velocidad maacutexima (Vmax) a partir de la cual la velocidad de la reaccioacuten es
independiente de la cantidad de substrato El que la velocidad no pueda
seguirse incrementando refleja la saturacioacuten del sitio activo con el substrato
Vmax
0 Substrato
Figura representacioacuten de una cineacutetica tiacutepicamente michaeliana
Vmax = velocidad de saturacioacuten o maacutexima
27
Cuando se grafica la velocidad de la reaccioacuten v0 contra la concentracioacuten de substrato S no siempre es posible determinar la condicioacuten
en que se ha llegado a la velocidad maacutexima Vmax debido al incremento de la pendiente en la hipeacuterbola a concentraciones de substrato elevadas
Figura experimento cineacutetico para la actividad enzimaacutetica
Se presentan los valores experimentales y los resultados del ajuste al modelo de Michaelis
Auacuten asiacute si se grafica una doble reciacuteproca es decir el inverso de la
velocidad (1v0) contra el inverso de la concentracioacuten de substrato (1 S ) se
obtiene una liacutenea recta El regraacutefico de Lineweaver-Burke tambieacuten se
denomina de ldquodobles reciacuteprocasrdquo y se utiliza para calcular la Km y la Vmax asiacute como para determinar el mecanismo de accioacuten de los diversos tipos de
inhibidores
La ecuacioacuten que describe a la graacutefca de Lineweaver-Burke es
Vmax
1
SVmax
Km
v
1
0 (3)
que describe una recta en donde el intercepto con el eje de las abscisas es igual a ndash1Km y el intercepto con el eje de las ordenadas es igual a 1Vmax
28
Figura experimento cineacutetico para la actividad enzimaacutetica Regraacutefico de
Lineweaver-Burke
Se presentan los valores experimentales y los resultados del ajuste al modelo
REPRESENTACIOacuteN GRAacuteFICA DE LINEWEAVER Y BURK
Si se toma el inverso en ambos miembros de la ecuacioacuten de Michaelis tenemos
=
+
29
que es la ecuacioacuten de una liacutenea recta cuya pendiente es que intercepta al eje de las
ordenadas en y a la abcisa en el punto Lo que es evidente al hacer
ya que se convierte en
Si se hace una graacutefica con los dobles inversos colocando los valores de en el eje de las
ordenadas y en el eje de las abcisas se obtiene una liacutenea recta a partir de la cual se puede calcular faacutecilmente Km
EFECTO DEL pH SOBRE LA ACTIVIDAD ENZIMATICA
Sabiendo que las enzimas son proteinas cualquier cambio brusco de pH puede alterar el caraacutecter ioacutenico de los grupos amino y carboxilo en la superficie proteica afectando asiacute las propiedades cataliacuteticas de una enzima A pH alto o bajo se puede producir la desnaturalizacioacuten
de la enzima y en consecuencia su inactivacioacuten La fosfatasa aacutecida es maacutes activa a pH 50 mientras que la fosfatasa alcalina lo es a pH 90 Muchas enzimas tienen maacutexima actividad cerca de la neutralidad en un rango de pH de 6 a 8
El pH per se no afecta la actividad enzimaacutetica sino la concentracioacuten de protones Los protones
ademaacutes de alterar la estructura de la enzima y el substrato pueden participar tambieacuten en la reaccioacuten como substrato o producto En esos casos la concentracioacuten de protones afecta directamente la velocidad de la reaccioacuten
ENZIMA pH OPTIMO
30
Pepsina 15
Tripsina 77
Catalasa 76
Arginasa 97
Fumarasa 78
Ribonucleasa 78
EFECTO DE ACTIVADORES
Un aumento en la temperatura provoca un aumento de la velocidad de reaccioacuten hasta cierta temperatura oacuteptima ya que despueacutes de aproximadamente 45
0 C se comienza a producir la
desnaturalizacioacuten teacutermica Las enzimas de muchos mamiacuteferos tienen una temperatura oacuteptima
de 370 C por encima de esa temperatura comienzan a inactivarse y se destruyen Sin embargo
existen especies de bacterias y algas que habitan en fuentes de aguas termales y en el otro extremo ciertas bacterias aacuterticas tienen temperaturas oacuteptimas cercanas a 0
0 C
31
ESPECIFICIDAD ABSOLUTA
Cierta enzimas se encuentran como aproenzimas o zimogenos que son inactivos
El tripsinoacutegeno es convertido en tripsina por la misma accioacuten de la tripsina que remueve un hexapeacuteptido
tripsina Tripsinoacutegeno Tripsina + Val (Asp)4 lisina
Otra forma de activacioacuten consiste en mantener los grupos SH reducidos Estos grupos pueden
formar parte de centros activos de ciertas enzimas Si se oxidan estos grupos a -S-S- la enzima se inactiva Asiacute tenemos que la enzima papaina se inactiva despueacutes de exponerla al ox iacutegeno pero cuando se le antildeade un reductor apropiado la enzima se reactiva Los grupos ndashS-S- se convierten en ndashSH activaacutendose la enzima
Otra forma de activacioacuten requiere la presencia de otra substancia que se enlaza al sitio alosteacuterico el compuesto que se enlaza se denomina un efector alosteacuterico El centro alosteacuterico es bien especiacutefico para el efector o modulador alosteacuterico
ESPECIFICIDAD RELATIVA
Oxiacutegeno L-aminoaacutecido a -cetoaacutecido + NH3 + H 2 O 2 L-aminoaacutecido oxidasa Oxiacutegeno D-aminoaacutecido a -cetoaacutecido + NH3 + H 2 O 2 D-aminoaacutecido oxidasa
32
En este ejemplo se muestra la estereo-especificidad Una enzima puede tener una especificidad oacuteptica para isomeros D o L
Hay enzimas que muestran especificidad absoluta como la aspartasa que cataliza la adicioacuten reversible de amoniacuteaco al doble enlace del aacutecido fumaacuterico pero no al de otros aacutecidos insaturados
HOOC ndash CH = CH ndash COO- + NH4
+ HOOC ndash CH2 - CHNH2 - COO
- + H
+
INHIBICIOacuteN ENZIMATICA
Mediante el uso de inhibidores enzimaacuteticos se ha obtenido informacioacuten muy valiosa sobre la conformacioacuten del centro activo de algunas enzimas
a Inhibicioacuten irreversible Este tipo de inhibidores forman un enlace covalente con las enzimas cerca del centro activo Un ejemplo son los gases nerviosos como el
fluorofosfato de diisopropilo (DFP) que forma un complejo con la enzima acetilcolinesterasa Los animales envenenados con este gas quedan paralizados debido a la imposibilidad de transmitir adecuadamente los impulsos nerviosos
Ester di-isopropil fos foacuterico de la enzima (inactivo)
b) Inhibicioacuten competitiva
Es cuando el inhibidor compite con el substrato por la unioacuten con el centro activo de la
enzima Este tipo de inhibicioacuten puede reducirse si se aumenta la concentracioacuten de substrato
-OOC - CH2 - CH2 - COO
- -OOC - CH = CH - COO
- + 2H
+ Succinato Succinato Fumarato
33
Deshidrogenasa COO
- I CH2 Malonato (inhibidor competitivo) I COO
- En la inhibicioacuten competitiva la enzima se combina con el inhibidor para formar un complejo enzima-inhibidor E + I E I en competencia con la reaccioacuten normal E +S ES
La sulfanilamida (bactericida) interfiere con la siacutentesis del aacutecido foacutelico compitiendo con el aacutecido p-aminobenzoico de una forma competitiva
HOOC-sect- NH2 NH2 ndashSO2 - sect- NH2 aacutecido p- amino benzoico sulfanilamida
sect = fenil
a Inhibicioacuten no competitiva
Esta inhibicioacuten se caracteriza por que no se puede revertir el efecto del inhibidor aumentando la concentracioacuten del substrato
Por ejemplo la inhibicioacuten de la deshidrogenasa del gliceraldehido -3-fosfato por la yodo-acetamida
Enzima-SH + I-CH2 CONH2 Enzima-SCH2 CONH2 + IH
La deshidrogenasa del gliceraldehido-3 fosfato puede ser inhibida tambieacuten por el aacutecido iodo-aceacutetico
Enzima-SH + ICH2 COOH Enzima-SCH2 COOH + IH
c) Inhibicioacuten por metales
Ciertos metales como el plomo mercurio y arseacutenico inhiben enzimas que tienen en su centro activo grupos -SH libres

18
(Figura superior derecha
EFECTO DE LA MODIFICACIOacuteN COVALENTE SOBRE LA ACTIVIDAD ENZIMAacuteTICA
Otros enzimas pasan de una forma menos activa a otra maacutes activa unieacutendose covalentemente a un grupo quiacutemico de pequentildeo tamantildeo como el Pi o el AMP Tambieacuten se da el caso inverso en el que un enzima muy activo se desactiva al liberar alguacuten grupo quiacutemico En las enzimas de las viacuteas degradativas del metabolismo la
forma fosforilada es maacutes activa que la no fosforilada mientras que en las viacuteas biosinteacuteticas ocurre lo contrario En las figuras inferiores se ilustra la activacioacuten de una proteiacutena por fosforilacioacuten
Elementos de la reaccioacuten El enzima no fosforilado es inactivo El enzima fosforilado es activo
ACTIVACIOacuteN PROTEOLIacuteTICA DE LA ACTIVIDAD ENZIMAacuteTICA
Algunos enzimas no se sintetizan como tales sino como proteiacutenas precursoras sin actividad enzimaacutetica Estas proteiacutenas se llaman
proenzimas o zimoacutegenos Para activarse los zimoacutegenos sufren un ataque hidroliacutetico que origina la liberacioacuten de uno o varios peacuteptidos El resto de la moleacutecula proteica adopta la conformacioacuten y las propiedades del enzima activo Muchos enzimas digestivos se secretan en forma de zimoacutegenos y en el tubo digestivo
se convierten en la forma activa Es el caso de la -quimotripsina que se sintetiza en forma de quimotripsinoacutegeno (Figura superior) Si estos enzimas se sintetizasen directamente en forma activa destruiriacutean la propia ceacutelula que las
produce Asiacute la tripsina pancreaacutetica (una proteasa) se sintetiza como tripsinoacutegeno (inactivo) Si por alguna razoacuten se activa en el propio paacutencreas la glaacutendula sufre un proceso de autodestruccioacuten (pancreatitis aguda) a
menudo mortal
19
REGULACIOacuteN DE LA ACTIVIDAD ENZIMAacuteTICA POR MEDIO DE ISOENZIMAS
Algunos enzimas tienen distinta estructura molecular aunque su funcioacuten bioloacutegica es similar Se llaman isozimas o isoenzimas Estas diferencias de estructura se traducen en ligeros cambios en sus propiedades de forma que cada isozima se adapta perfectamente a la funcioacuten que debe realizar Asiacute podemos observar la existencia de isoenzimas en funcioacuten de
el tipo de tejido Por ejemplo la lactato deshidrogenasa presenta isozimas distintos en muacutesculo y corazoacuten el compartimento celular donde actuacutea Por ejemplo la malato deshidrogenasa del citoplasma es distinta
de la de la mitocondria el momento concreto del desarrollo del individuo Por ejemplo algunos enzimas de la glicolisis del feto
son diferentes de los mismos enzimas en el adulto
CINEacuteTICA ENZIMAacuteTICA
Los principios generales de las reacciones quiacutemicas se aplican tambieacuten a las reacciones enzimaacuteticas Por este motivo antes de empezar con la cineacutetica quiacutemica se van a repasar algunos conceptos baacutesicos de cineacutetica quiacutemica
A continuacioacuten se describiraacuten los siguientes conceptos
Cineacutetica enzimaacutetica Modelo cineacutetico de Michaelis-Menten Caacutelculo de la KM y la Vmax de un enzima Actividad enzimaacutetica
20
NOMENCLATURA DE LOS ENZIMAS
CATALISIS MOLECULAR
Un catalizador modifica la velocidad de una reaccioacuten quiacutemica sin ser utilizado o aparecer como uno de los productos de la reaccioacuten Una reaccioacuten quiacutemica en la que un substrato(S) se
transforma en un producto(P) S P ocurre por que cierta fraccioacuten de moleacuteculas de S posee mucho maacutes energiacutea que el resto de ellas lo que es suficiente para que alcancen un estado activado en el que pueda formarse o romperse un enlace quiacutemico y se forme el producto(P)
La energiacutea de activacioacuten es la cantidad de energiacutea expresada en caloriacuteas necesaria para que todas las moleacuteculas de un mol a una temperatura dada alcancen el estado reactivo Mientras que el estado de transicioacuten es el estado rico en energiacutea de las moleacuteculas que
interaccionan en la cima de la barrera de activacioacuten La velocidad de una reaccioacuten quiacutemica es proporcional a la concentracioacuten del complejo en el estado de transicioacuten
Una reaccioacuten quiacutemica se puede acelerar de la siguiente forma 1) Al aumentar la temperatura se incrementa la energiacutea cineacutetica por lo que es mayor el nuacutemero de moleacuteculas que alcanzan el
estado de transicioacuten Generalmente el Q10 =2 lo que indica que la velocidad de una reaccioacuten quiacutemica se duplica al aumentar la temperatura en 10
0C 2) Antildeadiendo un catalizador que
disminuye la energiacutea de activacioacuten y aumenta la velocidad de reaccioacuten
La enzima (E) se combina con el substrato (S) formando el complejo de transicioacuten enzima-
substrato (E-S) mediante una reaccioacuten reversible cuya energiacutea de activacioacuten es menor que la de la reaccioacuten no catalizada Cuando se forma el producto de la reaccioacuten (P) se regenera de nuevo la enzima (E) de forma libre la que puede combinarse de nuevo con otra moleacutecula de substrato (S)
Una enzima reduce maacutes eficientemente la energiacutea de activacioacuten(Ea) de una reaccioacuten que un catalizador inorgaacutenico lo que permite que una reaccioacuten se realice a menor temperatura
El siguiente ejemplo ilustra mejor lo que hemos discutido
Reaccioacuten Energiacutea de activacioacuten (Kcalmol-1
)
21
a) el peroacutexido de hidroacutegeno se descompone en
H2 O2 H2 O + O2
18
b) el hierro cataliacutetico (Fe) realiza la reaccioacuten
H2 O2 H2 O + O2
13
c) el platino cataliacutetico (Pt) realiza la reaccioacuten
H2 O2 H2 O + O2
12
d) la catalasa una enzima hepaacutetica la realiza
H2O2 H2O + O2
5
Una enzima no modifica la energiacutea libre ni la constante de equilibrio sino que disminuye la energiacutea de activacioacuten de la reaccioacuten
Ir al principio
EFECTO DE LA CONCENTRACIOacuteN DEL SUBSTRATO SOBRE LA
CATALISIS ENZIMATICA
La actividad de una enzima se puede estudiar in vitro bajo condiciones controladas antildeadieacutendole una enzima a un substrato Siacute se trabaja con la condicioacuten de que la concentracioacuten del substrato sea saturante variando entonces la concentracioacuten de enzima se observa que aumenta el producto de la reaccioacuten a pH y temperatura constantes
22
Si se mantiene la concentracioacuten de la enzima constante y variando la concentracioacuten de substrato se obtiene una curva hiperboacutelica como la de la figura Al principio un aumento de la
concentracioacuten de substrato produce un aumento raacutepi do de la velocidad de reaccioacuten pero si se sigue aumentando la concentracioacuten de substrato la velocidad de reaccioacuten comienza a disminuir y a muy altas concentraciones de substrato se observa que no cambia la velocidad de
reaccioacuten se dice que los centros activos de la enzima se encuentran saturados La velocidad de reaccioacuten que se obtiene a esa alta concentracioacuten de substrato se define como la velocidad maacutexima (V) de la reaccioacuten enzimaacutetica bajo las condiciones especificadas La concentracioacuten de
substrato (S) a la semivelocidad maacutexima de reaccioacuten (V2) se puede determinar de la figura y representa la constante de Michaelis o Km la cual es una caracteriacutestica para cada enzima La inversa de Km o 1Km mide aproximadamente la afinidad de la enzima por el subs trato
Mientras maacutes pequentildeo sea el valor de Km mayor seraacute la afinidad de la enzima por el substrato Si varias enzimas compiten en el metabolismo por el mismo substrato eacuteste seraacute transformado preferentemente por la enzima con mayor afinidad
La ecuacioacuten de Michaelis describe la relacioacuten cuantitativa entre la velocidad de reaccioacuten y la concentracioacuten de substrato [S] si se conoce Vmax o Km
v=
En donde v= es la velocidad de reaccioacuten observada a una concentracioacuten de substrato determinada [S]
Km= constante de Michaelis en molesliacutet ro
23
Vmax= velocidad maacutexima a concentracioacuten saturante de substrato
Si v= Vmax2 entonces
=
Km+[S] = 2[S]
Km =[S]
De tal forma que podemos concluir que Km es igual a la concentracioacuten de substrato a la semivelocidad maacutexima de reaccioacuten
1 Concentraciones relativas de E y S la concentracioacuten de substrato S es
mucho mayor que la de E de tal manera que la cantidad de substrato
unido a la enzima en cualquier momento es muy pequentildea
2 Se asume el estado estacionario ES no cambia con el tiempo esto
quiere decir que la velocidad de formacioacuten de ES es igual a aquella para
su desintegracioacuten En general un intermediario en una serie de reacciones estaacute en estado estacionario cuando su velocidad de siacutentesis
es igual a su velocidad de degradacioacuten 3 Velocidad inicial para el anaacutelisis de reacciones enzimaacuteticas soacutelo se
utiliza la velocidad inicial de la reaccioacuten que es la velocidad ejercida por
la enzima inmediatamente despueacutes de que se ha puesto en contacto con el substrato y hasta antes de que se haya consumido el 10 de la concentracioacuten inicial del mismo La razoacuten de lo anterior es que en ese
momento la concentracioacuten del producto de la reaccioacuten que se ha acumulado en muy pequentildea y por tanto la reaccioacuten en el sentido
inverso es decir la transformacioacuten del producto en el substrato original puede ser ignorada
24
Caracteriacutesticas de Km la constante de Michaelis-Menten es
caracteriacutestica de una enzima y particular para un substrato Refleja la afinidad de la enzima por ese substrato K m es numeacutericamente igual a la concentracioacuten
de substrato a la cual la velocidad de reaccioacuten es la mitad de la Vmax (Km= Vmax2) Este paraacutemetro Km no variacutea con la concentracioacuten de enzima
Significado de una Km pequentildea un valor numeacuterico pequentildeo de Km
refleja una alta afinidad de la enzima por su substrato porque a una baja
concentracioacuten del mismo la enzima a desarrollado ya la mitad de la velocidad maacutexima
Significado de una Km grande el valor numeacuterico grande de Km refleja
una baja afinidad de la enzima por su substrato porque a una concentracioacuten
elevada del mismo la enzima desarrolla la mitad de la velocidad maacutexima
Relacioacuten de la velocidad con la concentracioacuten de enzima la
velocidad de la reaccioacuten es directamente proporcional a la concentracioacuten de la
enzima a cualquier concentracioacuten de substrato Por ejemplo si la concentracioacuten de enzima es disminuida a la mitad la velocidad inicial de la reaccioacuten (v0) es
reducida tambieacuten a la mitad de la original
Orden de la reaccioacuten cuando S es mas pequentildea que la Km la
velocidad de la reaccioacuten es aproximadamente igual a la concentracioacuten de
substrato
100
50
0
Substrato
Km Figura Localizacioacuten de la constante de Michaelis (Km)
25
La velocidad de reaccioacuten se dice en estas condiciones es de primer
orden con respecto al substrato Cuando S es mas grande que la Km la
velocidad es constante e igual a la Vmax La velocidad de la reaccioacuten es independiente de la concentracioacuten de substrato y se dice que es de orden cero
con respecto a la concentracioacuten de substrato
Las enzimas pueden ser aisladas de las ceacutelulas u organismos para estudiar sus
propiedades in vitro es decir en un tubo de ensayo Las diferentes enzimas muestran diferentes respuestas a los cambios en la concentracioacuten de substrato temperatura y pH
Concentracioacuten de substrato
La forma hiperboacutelica de la curva de velocidad vs Substrato
Efecto de la temperatura en la actividad enzimaacutetica
Efecto de la variacioacuten del pH en la actividad enzimaacutetica
Incremento de la velocidad con la temperatura En general la velocidad de la reaccioacuten se incrementa con la temperatura hasta que un punto maacuteximo es alcanzado
Este incremento se debe a que aumenta el nuacutemero de moleacuteculas ricas en energiacutea que pueden pasar la barrera energeacutetica de estado de transicioacuten para formar a los productos
Decremento de la velocidad con la temperatura la elevacioacuten excesiva de la
temperatura del medio que contiene a las enzimas resulta en un decremento de la velocidad como resultado de la desnaturalizacioacuten es decir la peacuterdida de la estructura tridimensional de las enzimas
26
zona de reactivacioacuten maacuteximo de actividad
Zona de
inactivacioacuten
0 Temperatura
Figura dependencia de la actividad enzimaacutetica con la temperatura
Velocidad maacutexima la velocidad de una reaccioacuten (v) es el nuacutemero de
moleacuteculas de substrato convertidas en producto por unidad de tiempo y
usualmente se expresa en micromoles de producto formadas por minuto La velocidad de una reaccioacuten catalizada por una enzima aumenta conforme se incrementa la concentracioacuten de substrato hasta que se llega a una velocidad maacutexima (Vmax) a partir de la cual la velocidad de la reaccioacuten es
independiente de la cantidad de substrato El que la velocidad no pueda
seguirse incrementando refleja la saturacioacuten del sitio activo con el substrato
Vmax
0 Substrato
Figura representacioacuten de una cineacutetica tiacutepicamente michaeliana
Vmax = velocidad de saturacioacuten o maacutexima
27
Cuando se grafica la velocidad de la reaccioacuten v0 contra la concentracioacuten de substrato S no siempre es posible determinar la condicioacuten
en que se ha llegado a la velocidad maacutexima Vmax debido al incremento de la pendiente en la hipeacuterbola a concentraciones de substrato elevadas
Figura experimento cineacutetico para la actividad enzimaacutetica
Se presentan los valores experimentales y los resultados del ajuste al modelo de Michaelis
Auacuten asiacute si se grafica una doble reciacuteproca es decir el inverso de la
velocidad (1v0) contra el inverso de la concentracioacuten de substrato (1 S ) se
obtiene una liacutenea recta El regraacutefico de Lineweaver-Burke tambieacuten se
denomina de ldquodobles reciacuteprocasrdquo y se utiliza para calcular la Km y la Vmax asiacute como para determinar el mecanismo de accioacuten de los diversos tipos de
inhibidores
La ecuacioacuten que describe a la graacutefca de Lineweaver-Burke es
Vmax
1
SVmax
Km
v
1
0 (3)
que describe una recta en donde el intercepto con el eje de las abscisas es igual a ndash1Km y el intercepto con el eje de las ordenadas es igual a 1Vmax
28
Figura experimento cineacutetico para la actividad enzimaacutetica Regraacutefico de
Lineweaver-Burke
Se presentan los valores experimentales y los resultados del ajuste al modelo
REPRESENTACIOacuteN GRAacuteFICA DE LINEWEAVER Y BURK
Si se toma el inverso en ambos miembros de la ecuacioacuten de Michaelis tenemos
=
+
29
que es la ecuacioacuten de una liacutenea recta cuya pendiente es que intercepta al eje de las
ordenadas en y a la abcisa en el punto Lo que es evidente al hacer
ya que se convierte en
Si se hace una graacutefica con los dobles inversos colocando los valores de en el eje de las
ordenadas y en el eje de las abcisas se obtiene una liacutenea recta a partir de la cual se puede calcular faacutecilmente Km
EFECTO DEL pH SOBRE LA ACTIVIDAD ENZIMATICA
Sabiendo que las enzimas son proteinas cualquier cambio brusco de pH puede alterar el caraacutecter ioacutenico de los grupos amino y carboxilo en la superficie proteica afectando asiacute las propiedades cataliacuteticas de una enzima A pH alto o bajo se puede producir la desnaturalizacioacuten
de la enzima y en consecuencia su inactivacioacuten La fosfatasa aacutecida es maacutes activa a pH 50 mientras que la fosfatasa alcalina lo es a pH 90 Muchas enzimas tienen maacutexima actividad cerca de la neutralidad en un rango de pH de 6 a 8
El pH per se no afecta la actividad enzimaacutetica sino la concentracioacuten de protones Los protones
ademaacutes de alterar la estructura de la enzima y el substrato pueden participar tambieacuten en la reaccioacuten como substrato o producto En esos casos la concentracioacuten de protones afecta directamente la velocidad de la reaccioacuten
ENZIMA pH OPTIMO
30
Pepsina 15
Tripsina 77
Catalasa 76
Arginasa 97
Fumarasa 78
Ribonucleasa 78
EFECTO DE ACTIVADORES
Un aumento en la temperatura provoca un aumento de la velocidad de reaccioacuten hasta cierta temperatura oacuteptima ya que despueacutes de aproximadamente 45
0 C se comienza a producir la
desnaturalizacioacuten teacutermica Las enzimas de muchos mamiacuteferos tienen una temperatura oacuteptima
de 370 C por encima de esa temperatura comienzan a inactivarse y se destruyen Sin embargo
existen especies de bacterias y algas que habitan en fuentes de aguas termales y en el otro extremo ciertas bacterias aacuterticas tienen temperaturas oacuteptimas cercanas a 0
0 C
31
ESPECIFICIDAD ABSOLUTA
Cierta enzimas se encuentran como aproenzimas o zimogenos que son inactivos
El tripsinoacutegeno es convertido en tripsina por la misma accioacuten de la tripsina que remueve un hexapeacuteptido
tripsina Tripsinoacutegeno Tripsina + Val (Asp)4 lisina
Otra forma de activacioacuten consiste en mantener los grupos SH reducidos Estos grupos pueden
formar parte de centros activos de ciertas enzimas Si se oxidan estos grupos a -S-S- la enzima se inactiva Asiacute tenemos que la enzima papaina se inactiva despueacutes de exponerla al ox iacutegeno pero cuando se le antildeade un reductor apropiado la enzima se reactiva Los grupos ndashS-S- se convierten en ndashSH activaacutendose la enzima
Otra forma de activacioacuten requiere la presencia de otra substancia que se enlaza al sitio alosteacuterico el compuesto que se enlaza se denomina un efector alosteacuterico El centro alosteacuterico es bien especiacutefico para el efector o modulador alosteacuterico
ESPECIFICIDAD RELATIVA
Oxiacutegeno L-aminoaacutecido a -cetoaacutecido + NH3 + H 2 O 2 L-aminoaacutecido oxidasa Oxiacutegeno D-aminoaacutecido a -cetoaacutecido + NH3 + H 2 O 2 D-aminoaacutecido oxidasa
32
En este ejemplo se muestra la estereo-especificidad Una enzima puede tener una especificidad oacuteptica para isomeros D o L
Hay enzimas que muestran especificidad absoluta como la aspartasa que cataliza la adicioacuten reversible de amoniacuteaco al doble enlace del aacutecido fumaacuterico pero no al de otros aacutecidos insaturados
HOOC ndash CH = CH ndash COO- + NH4
+ HOOC ndash CH2 - CHNH2 - COO
- + H
+
INHIBICIOacuteN ENZIMATICA
Mediante el uso de inhibidores enzimaacuteticos se ha obtenido informacioacuten muy valiosa sobre la conformacioacuten del centro activo de algunas enzimas
a Inhibicioacuten irreversible Este tipo de inhibidores forman un enlace covalente con las enzimas cerca del centro activo Un ejemplo son los gases nerviosos como el
fluorofosfato de diisopropilo (DFP) que forma un complejo con la enzima acetilcolinesterasa Los animales envenenados con este gas quedan paralizados debido a la imposibilidad de transmitir adecuadamente los impulsos nerviosos
Ester di-isopropil fos foacuterico de la enzima (inactivo)
b) Inhibicioacuten competitiva
Es cuando el inhibidor compite con el substrato por la unioacuten con el centro activo de la
enzima Este tipo de inhibicioacuten puede reducirse si se aumenta la concentracioacuten de substrato
-OOC - CH2 - CH2 - COO
- -OOC - CH = CH - COO
- + 2H
+ Succinato Succinato Fumarato
33
Deshidrogenasa COO
- I CH2 Malonato (inhibidor competitivo) I COO
- En la inhibicioacuten competitiva la enzima se combina con el inhibidor para formar un complejo enzima-inhibidor E + I E I en competencia con la reaccioacuten normal E +S ES
La sulfanilamida (bactericida) interfiere con la siacutentesis del aacutecido foacutelico compitiendo con el aacutecido p-aminobenzoico de una forma competitiva
HOOC-sect- NH2 NH2 ndashSO2 - sect- NH2 aacutecido p- amino benzoico sulfanilamida
sect = fenil
a Inhibicioacuten no competitiva
Esta inhibicioacuten se caracteriza por que no se puede revertir el efecto del inhibidor aumentando la concentracioacuten del substrato
Por ejemplo la inhibicioacuten de la deshidrogenasa del gliceraldehido -3-fosfato por la yodo-acetamida
Enzima-SH + I-CH2 CONH2 Enzima-SCH2 CONH2 + IH
La deshidrogenasa del gliceraldehido-3 fosfato puede ser inhibida tambieacuten por el aacutecido iodo-aceacutetico
Enzima-SH + ICH2 COOH Enzima-SCH2 COOH + IH
c) Inhibicioacuten por metales
Ciertos metales como el plomo mercurio y arseacutenico inhiben enzimas que tienen en su centro activo grupos -SH libres

19
REGULACIOacuteN DE LA ACTIVIDAD ENZIMAacuteTICA POR MEDIO DE ISOENZIMAS
Algunos enzimas tienen distinta estructura molecular aunque su funcioacuten bioloacutegica es similar Se llaman isozimas o isoenzimas Estas diferencias de estructura se traducen en ligeros cambios en sus propiedades de forma que cada isozima se adapta perfectamente a la funcioacuten que debe realizar Asiacute podemos observar la existencia de isoenzimas en funcioacuten de
el tipo de tejido Por ejemplo la lactato deshidrogenasa presenta isozimas distintos en muacutesculo y corazoacuten el compartimento celular donde actuacutea Por ejemplo la malato deshidrogenasa del citoplasma es distinta
de la de la mitocondria el momento concreto del desarrollo del individuo Por ejemplo algunos enzimas de la glicolisis del feto
son diferentes de los mismos enzimas en el adulto
CINEacuteTICA ENZIMAacuteTICA
Los principios generales de las reacciones quiacutemicas se aplican tambieacuten a las reacciones enzimaacuteticas Por este motivo antes de empezar con la cineacutetica quiacutemica se van a repasar algunos conceptos baacutesicos de cineacutetica quiacutemica
A continuacioacuten se describiraacuten los siguientes conceptos
Cineacutetica enzimaacutetica Modelo cineacutetico de Michaelis-Menten Caacutelculo de la KM y la Vmax de un enzima Actividad enzimaacutetica
20
NOMENCLATURA DE LOS ENZIMAS
CATALISIS MOLECULAR
Un catalizador modifica la velocidad de una reaccioacuten quiacutemica sin ser utilizado o aparecer como uno de los productos de la reaccioacuten Una reaccioacuten quiacutemica en la que un substrato(S) se
transforma en un producto(P) S P ocurre por que cierta fraccioacuten de moleacuteculas de S posee mucho maacutes energiacutea que el resto de ellas lo que es suficiente para que alcancen un estado activado en el que pueda formarse o romperse un enlace quiacutemico y se forme el producto(P)
La energiacutea de activacioacuten es la cantidad de energiacutea expresada en caloriacuteas necesaria para que todas las moleacuteculas de un mol a una temperatura dada alcancen el estado reactivo Mientras que el estado de transicioacuten es el estado rico en energiacutea de las moleacuteculas que
interaccionan en la cima de la barrera de activacioacuten La velocidad de una reaccioacuten quiacutemica es proporcional a la concentracioacuten del complejo en el estado de transicioacuten
Una reaccioacuten quiacutemica se puede acelerar de la siguiente forma 1) Al aumentar la temperatura se incrementa la energiacutea cineacutetica por lo que es mayor el nuacutemero de moleacuteculas que alcanzan el
estado de transicioacuten Generalmente el Q10 =2 lo que indica que la velocidad de una reaccioacuten quiacutemica se duplica al aumentar la temperatura en 10
0C 2) Antildeadiendo un catalizador que
disminuye la energiacutea de activacioacuten y aumenta la velocidad de reaccioacuten
La enzima (E) se combina con el substrato (S) formando el complejo de transicioacuten enzima-
substrato (E-S) mediante una reaccioacuten reversible cuya energiacutea de activacioacuten es menor que la de la reaccioacuten no catalizada Cuando se forma el producto de la reaccioacuten (P) se regenera de nuevo la enzima (E) de forma libre la que puede combinarse de nuevo con otra moleacutecula de substrato (S)
Una enzima reduce maacutes eficientemente la energiacutea de activacioacuten(Ea) de una reaccioacuten que un catalizador inorgaacutenico lo que permite que una reaccioacuten se realice a menor temperatura
El siguiente ejemplo ilustra mejor lo que hemos discutido
Reaccioacuten Energiacutea de activacioacuten (Kcalmol-1
)
21
a) el peroacutexido de hidroacutegeno se descompone en
H2 O2 H2 O + O2
18
b) el hierro cataliacutetico (Fe) realiza la reaccioacuten
H2 O2 H2 O + O2
13
c) el platino cataliacutetico (Pt) realiza la reaccioacuten
H2 O2 H2 O + O2
12
d) la catalasa una enzima hepaacutetica la realiza
H2O2 H2O + O2
5
Una enzima no modifica la energiacutea libre ni la constante de equilibrio sino que disminuye la energiacutea de activacioacuten de la reaccioacuten
Ir al principio
EFECTO DE LA CONCENTRACIOacuteN DEL SUBSTRATO SOBRE LA
CATALISIS ENZIMATICA
La actividad de una enzima se puede estudiar in vitro bajo condiciones controladas antildeadieacutendole una enzima a un substrato Siacute se trabaja con la condicioacuten de que la concentracioacuten del substrato sea saturante variando entonces la concentracioacuten de enzima se observa que aumenta el producto de la reaccioacuten a pH y temperatura constantes
22
Si se mantiene la concentracioacuten de la enzima constante y variando la concentracioacuten de substrato se obtiene una curva hiperboacutelica como la de la figura Al principio un aumento de la
concentracioacuten de substrato produce un aumento raacutepi do de la velocidad de reaccioacuten pero si se sigue aumentando la concentracioacuten de substrato la velocidad de reaccioacuten comienza a disminuir y a muy altas concentraciones de substrato se observa que no cambia la velocidad de
reaccioacuten se dice que los centros activos de la enzima se encuentran saturados La velocidad de reaccioacuten que se obtiene a esa alta concentracioacuten de substrato se define como la velocidad maacutexima (V) de la reaccioacuten enzimaacutetica bajo las condiciones especificadas La concentracioacuten de
substrato (S) a la semivelocidad maacutexima de reaccioacuten (V2) se puede determinar de la figura y representa la constante de Michaelis o Km la cual es una caracteriacutestica para cada enzima La inversa de Km o 1Km mide aproximadamente la afinidad de la enzima por el subs trato
Mientras maacutes pequentildeo sea el valor de Km mayor seraacute la afinidad de la enzima por el substrato Si varias enzimas compiten en el metabolismo por el mismo substrato eacuteste seraacute transformado preferentemente por la enzima con mayor afinidad
La ecuacioacuten de Michaelis describe la relacioacuten cuantitativa entre la velocidad de reaccioacuten y la concentracioacuten de substrato [S] si se conoce Vmax o Km
v=
En donde v= es la velocidad de reaccioacuten observada a una concentracioacuten de substrato determinada [S]
Km= constante de Michaelis en molesliacutet ro
23
Vmax= velocidad maacutexima a concentracioacuten saturante de substrato
Si v= Vmax2 entonces
=
Km+[S] = 2[S]
Km =[S]
De tal forma que podemos concluir que Km es igual a la concentracioacuten de substrato a la semivelocidad maacutexima de reaccioacuten
1 Concentraciones relativas de E y S la concentracioacuten de substrato S es
mucho mayor que la de E de tal manera que la cantidad de substrato
unido a la enzima en cualquier momento es muy pequentildea
2 Se asume el estado estacionario ES no cambia con el tiempo esto
quiere decir que la velocidad de formacioacuten de ES es igual a aquella para
su desintegracioacuten En general un intermediario en una serie de reacciones estaacute en estado estacionario cuando su velocidad de siacutentesis
es igual a su velocidad de degradacioacuten 3 Velocidad inicial para el anaacutelisis de reacciones enzimaacuteticas soacutelo se
utiliza la velocidad inicial de la reaccioacuten que es la velocidad ejercida por
la enzima inmediatamente despueacutes de que se ha puesto en contacto con el substrato y hasta antes de que se haya consumido el 10 de la concentracioacuten inicial del mismo La razoacuten de lo anterior es que en ese
momento la concentracioacuten del producto de la reaccioacuten que se ha acumulado en muy pequentildea y por tanto la reaccioacuten en el sentido
inverso es decir la transformacioacuten del producto en el substrato original puede ser ignorada
24
Caracteriacutesticas de Km la constante de Michaelis-Menten es
caracteriacutestica de una enzima y particular para un substrato Refleja la afinidad de la enzima por ese substrato K m es numeacutericamente igual a la concentracioacuten
de substrato a la cual la velocidad de reaccioacuten es la mitad de la Vmax (Km= Vmax2) Este paraacutemetro Km no variacutea con la concentracioacuten de enzima
Significado de una Km pequentildea un valor numeacuterico pequentildeo de Km
refleja una alta afinidad de la enzima por su substrato porque a una baja
concentracioacuten del mismo la enzima a desarrollado ya la mitad de la velocidad maacutexima
Significado de una Km grande el valor numeacuterico grande de Km refleja
una baja afinidad de la enzima por su substrato porque a una concentracioacuten
elevada del mismo la enzima desarrolla la mitad de la velocidad maacutexima
Relacioacuten de la velocidad con la concentracioacuten de enzima la
velocidad de la reaccioacuten es directamente proporcional a la concentracioacuten de la
enzima a cualquier concentracioacuten de substrato Por ejemplo si la concentracioacuten de enzima es disminuida a la mitad la velocidad inicial de la reaccioacuten (v0) es
reducida tambieacuten a la mitad de la original
Orden de la reaccioacuten cuando S es mas pequentildea que la Km la
velocidad de la reaccioacuten es aproximadamente igual a la concentracioacuten de
substrato
100
50
0
Substrato
Km Figura Localizacioacuten de la constante de Michaelis (Km)
25
La velocidad de reaccioacuten se dice en estas condiciones es de primer
orden con respecto al substrato Cuando S es mas grande que la Km la
velocidad es constante e igual a la Vmax La velocidad de la reaccioacuten es independiente de la concentracioacuten de substrato y se dice que es de orden cero
con respecto a la concentracioacuten de substrato
Las enzimas pueden ser aisladas de las ceacutelulas u organismos para estudiar sus
propiedades in vitro es decir en un tubo de ensayo Las diferentes enzimas muestran diferentes respuestas a los cambios en la concentracioacuten de substrato temperatura y pH
Concentracioacuten de substrato
La forma hiperboacutelica de la curva de velocidad vs Substrato
Efecto de la temperatura en la actividad enzimaacutetica
Efecto de la variacioacuten del pH en la actividad enzimaacutetica
Incremento de la velocidad con la temperatura En general la velocidad de la reaccioacuten se incrementa con la temperatura hasta que un punto maacuteximo es alcanzado
Este incremento se debe a que aumenta el nuacutemero de moleacuteculas ricas en energiacutea que pueden pasar la barrera energeacutetica de estado de transicioacuten para formar a los productos
Decremento de la velocidad con la temperatura la elevacioacuten excesiva de la
temperatura del medio que contiene a las enzimas resulta en un decremento de la velocidad como resultado de la desnaturalizacioacuten es decir la peacuterdida de la estructura tridimensional de las enzimas
26
zona de reactivacioacuten maacuteximo de actividad
Zona de
inactivacioacuten
0 Temperatura
Figura dependencia de la actividad enzimaacutetica con la temperatura
Velocidad maacutexima la velocidad de una reaccioacuten (v) es el nuacutemero de
moleacuteculas de substrato convertidas en producto por unidad de tiempo y
usualmente se expresa en micromoles de producto formadas por minuto La velocidad de una reaccioacuten catalizada por una enzima aumenta conforme se incrementa la concentracioacuten de substrato hasta que se llega a una velocidad maacutexima (Vmax) a partir de la cual la velocidad de la reaccioacuten es
independiente de la cantidad de substrato El que la velocidad no pueda
seguirse incrementando refleja la saturacioacuten del sitio activo con el substrato
Vmax
0 Substrato
Figura representacioacuten de una cineacutetica tiacutepicamente michaeliana
Vmax = velocidad de saturacioacuten o maacutexima
27
Cuando se grafica la velocidad de la reaccioacuten v0 contra la concentracioacuten de substrato S no siempre es posible determinar la condicioacuten
en que se ha llegado a la velocidad maacutexima Vmax debido al incremento de la pendiente en la hipeacuterbola a concentraciones de substrato elevadas
Figura experimento cineacutetico para la actividad enzimaacutetica
Se presentan los valores experimentales y los resultados del ajuste al modelo de Michaelis
Auacuten asiacute si se grafica una doble reciacuteproca es decir el inverso de la
velocidad (1v0) contra el inverso de la concentracioacuten de substrato (1 S ) se
obtiene una liacutenea recta El regraacutefico de Lineweaver-Burke tambieacuten se
denomina de ldquodobles reciacuteprocasrdquo y se utiliza para calcular la Km y la Vmax asiacute como para determinar el mecanismo de accioacuten de los diversos tipos de
inhibidores
La ecuacioacuten que describe a la graacutefca de Lineweaver-Burke es
Vmax
1
SVmax
Km
v
1
0 (3)
que describe una recta en donde el intercepto con el eje de las abscisas es igual a ndash1Km y el intercepto con el eje de las ordenadas es igual a 1Vmax
28
Figura experimento cineacutetico para la actividad enzimaacutetica Regraacutefico de
Lineweaver-Burke
Se presentan los valores experimentales y los resultados del ajuste al modelo
REPRESENTACIOacuteN GRAacuteFICA DE LINEWEAVER Y BURK
Si se toma el inverso en ambos miembros de la ecuacioacuten de Michaelis tenemos
=
+
29
que es la ecuacioacuten de una liacutenea recta cuya pendiente es que intercepta al eje de las
ordenadas en y a la abcisa en el punto Lo que es evidente al hacer
ya que se convierte en
Si se hace una graacutefica con los dobles inversos colocando los valores de en el eje de las
ordenadas y en el eje de las abcisas se obtiene una liacutenea recta a partir de la cual se puede calcular faacutecilmente Km
EFECTO DEL pH SOBRE LA ACTIVIDAD ENZIMATICA
Sabiendo que las enzimas son proteinas cualquier cambio brusco de pH puede alterar el caraacutecter ioacutenico de los grupos amino y carboxilo en la superficie proteica afectando asiacute las propiedades cataliacuteticas de una enzima A pH alto o bajo se puede producir la desnaturalizacioacuten
de la enzima y en consecuencia su inactivacioacuten La fosfatasa aacutecida es maacutes activa a pH 50 mientras que la fosfatasa alcalina lo es a pH 90 Muchas enzimas tienen maacutexima actividad cerca de la neutralidad en un rango de pH de 6 a 8
El pH per se no afecta la actividad enzimaacutetica sino la concentracioacuten de protones Los protones
ademaacutes de alterar la estructura de la enzima y el substrato pueden participar tambieacuten en la reaccioacuten como substrato o producto En esos casos la concentracioacuten de protones afecta directamente la velocidad de la reaccioacuten
ENZIMA pH OPTIMO
30
Pepsina 15
Tripsina 77
Catalasa 76
Arginasa 97
Fumarasa 78
Ribonucleasa 78
EFECTO DE ACTIVADORES
Un aumento en la temperatura provoca un aumento de la velocidad de reaccioacuten hasta cierta temperatura oacuteptima ya que despueacutes de aproximadamente 45
0 C se comienza a producir la
desnaturalizacioacuten teacutermica Las enzimas de muchos mamiacuteferos tienen una temperatura oacuteptima
de 370 C por encima de esa temperatura comienzan a inactivarse y se destruyen Sin embargo
existen especies de bacterias y algas que habitan en fuentes de aguas termales y en el otro extremo ciertas bacterias aacuterticas tienen temperaturas oacuteptimas cercanas a 0
0 C
31
ESPECIFICIDAD ABSOLUTA
Cierta enzimas se encuentran como aproenzimas o zimogenos que son inactivos
El tripsinoacutegeno es convertido en tripsina por la misma accioacuten de la tripsina que remueve un hexapeacuteptido
tripsina Tripsinoacutegeno Tripsina + Val (Asp)4 lisina
Otra forma de activacioacuten consiste en mantener los grupos SH reducidos Estos grupos pueden
formar parte de centros activos de ciertas enzimas Si se oxidan estos grupos a -S-S- la enzima se inactiva Asiacute tenemos que la enzima papaina se inactiva despueacutes de exponerla al ox iacutegeno pero cuando se le antildeade un reductor apropiado la enzima se reactiva Los grupos ndashS-S- se convierten en ndashSH activaacutendose la enzima
Otra forma de activacioacuten requiere la presencia de otra substancia que se enlaza al sitio alosteacuterico el compuesto que se enlaza se denomina un efector alosteacuterico El centro alosteacuterico es bien especiacutefico para el efector o modulador alosteacuterico
ESPECIFICIDAD RELATIVA
Oxiacutegeno L-aminoaacutecido a -cetoaacutecido + NH3 + H 2 O 2 L-aminoaacutecido oxidasa Oxiacutegeno D-aminoaacutecido a -cetoaacutecido + NH3 + H 2 O 2 D-aminoaacutecido oxidasa
32
En este ejemplo se muestra la estereo-especificidad Una enzima puede tener una especificidad oacuteptica para isomeros D o L
Hay enzimas que muestran especificidad absoluta como la aspartasa que cataliza la adicioacuten reversible de amoniacuteaco al doble enlace del aacutecido fumaacuterico pero no al de otros aacutecidos insaturados
HOOC ndash CH = CH ndash COO- + NH4
+ HOOC ndash CH2 - CHNH2 - COO
- + H
+
INHIBICIOacuteN ENZIMATICA
Mediante el uso de inhibidores enzimaacuteticos se ha obtenido informacioacuten muy valiosa sobre la conformacioacuten del centro activo de algunas enzimas
a Inhibicioacuten irreversible Este tipo de inhibidores forman un enlace covalente con las enzimas cerca del centro activo Un ejemplo son los gases nerviosos como el
fluorofosfato de diisopropilo (DFP) que forma un complejo con la enzima acetilcolinesterasa Los animales envenenados con este gas quedan paralizados debido a la imposibilidad de transmitir adecuadamente los impulsos nerviosos
Ester di-isopropil fos foacuterico de la enzima (inactivo)
b) Inhibicioacuten competitiva
Es cuando el inhibidor compite con el substrato por la unioacuten con el centro activo de la
enzima Este tipo de inhibicioacuten puede reducirse si se aumenta la concentracioacuten de substrato
-OOC - CH2 - CH2 - COO
- -OOC - CH = CH - COO
- + 2H
+ Succinato Succinato Fumarato
33
Deshidrogenasa COO
- I CH2 Malonato (inhibidor competitivo) I COO
- En la inhibicioacuten competitiva la enzima se combina con el inhibidor para formar un complejo enzima-inhibidor E + I E I en competencia con la reaccioacuten normal E +S ES
La sulfanilamida (bactericida) interfiere con la siacutentesis del aacutecido foacutelico compitiendo con el aacutecido p-aminobenzoico de una forma competitiva
HOOC-sect- NH2 NH2 ndashSO2 - sect- NH2 aacutecido p- amino benzoico sulfanilamida
sect = fenil
a Inhibicioacuten no competitiva
Esta inhibicioacuten se caracteriza por que no se puede revertir el efecto del inhibidor aumentando la concentracioacuten del substrato
Por ejemplo la inhibicioacuten de la deshidrogenasa del gliceraldehido -3-fosfato por la yodo-acetamida
Enzima-SH + I-CH2 CONH2 Enzima-SCH2 CONH2 + IH
La deshidrogenasa del gliceraldehido-3 fosfato puede ser inhibida tambieacuten por el aacutecido iodo-aceacutetico
Enzima-SH + ICH2 COOH Enzima-SCH2 COOH + IH
c) Inhibicioacuten por metales
Ciertos metales como el plomo mercurio y arseacutenico inhiben enzimas que tienen en su centro activo grupos -SH libres

20
NOMENCLATURA DE LOS ENZIMAS
CATALISIS MOLECULAR
Un catalizador modifica la velocidad de una reaccioacuten quiacutemica sin ser utilizado o aparecer como uno de los productos de la reaccioacuten Una reaccioacuten quiacutemica en la que un substrato(S) se
transforma en un producto(P) S P ocurre por que cierta fraccioacuten de moleacuteculas de S posee mucho maacutes energiacutea que el resto de ellas lo que es suficiente para que alcancen un estado activado en el que pueda formarse o romperse un enlace quiacutemico y se forme el producto(P)
La energiacutea de activacioacuten es la cantidad de energiacutea expresada en caloriacuteas necesaria para que todas las moleacuteculas de un mol a una temperatura dada alcancen el estado reactivo Mientras que el estado de transicioacuten es el estado rico en energiacutea de las moleacuteculas que
interaccionan en la cima de la barrera de activacioacuten La velocidad de una reaccioacuten quiacutemica es proporcional a la concentracioacuten del complejo en el estado de transicioacuten
Una reaccioacuten quiacutemica se puede acelerar de la siguiente forma 1) Al aumentar la temperatura se incrementa la energiacutea cineacutetica por lo que es mayor el nuacutemero de moleacuteculas que alcanzan el
estado de transicioacuten Generalmente el Q10 =2 lo que indica que la velocidad de una reaccioacuten quiacutemica se duplica al aumentar la temperatura en 10
0C 2) Antildeadiendo un catalizador que
disminuye la energiacutea de activacioacuten y aumenta la velocidad de reaccioacuten
La enzima (E) se combina con el substrato (S) formando el complejo de transicioacuten enzima-
substrato (E-S) mediante una reaccioacuten reversible cuya energiacutea de activacioacuten es menor que la de la reaccioacuten no catalizada Cuando se forma el producto de la reaccioacuten (P) se regenera de nuevo la enzima (E) de forma libre la que puede combinarse de nuevo con otra moleacutecula de substrato (S)
Una enzima reduce maacutes eficientemente la energiacutea de activacioacuten(Ea) de una reaccioacuten que un catalizador inorgaacutenico lo que permite que una reaccioacuten se realice a menor temperatura
El siguiente ejemplo ilustra mejor lo que hemos discutido
Reaccioacuten Energiacutea de activacioacuten (Kcalmol-1
)
21
a) el peroacutexido de hidroacutegeno se descompone en
H2 O2 H2 O + O2
18
b) el hierro cataliacutetico (Fe) realiza la reaccioacuten
H2 O2 H2 O + O2
13
c) el platino cataliacutetico (Pt) realiza la reaccioacuten
H2 O2 H2 O + O2
12
d) la catalasa una enzima hepaacutetica la realiza
H2O2 H2O + O2
5
Una enzima no modifica la energiacutea libre ni la constante de equilibrio sino que disminuye la energiacutea de activacioacuten de la reaccioacuten
Ir al principio
EFECTO DE LA CONCENTRACIOacuteN DEL SUBSTRATO SOBRE LA
CATALISIS ENZIMATICA
La actividad de una enzima se puede estudiar in vitro bajo condiciones controladas antildeadieacutendole una enzima a un substrato Siacute se trabaja con la condicioacuten de que la concentracioacuten del substrato sea saturante variando entonces la concentracioacuten de enzima se observa que aumenta el producto de la reaccioacuten a pH y temperatura constantes
22
Si se mantiene la concentracioacuten de la enzima constante y variando la concentracioacuten de substrato se obtiene una curva hiperboacutelica como la de la figura Al principio un aumento de la
concentracioacuten de substrato produce un aumento raacutepi do de la velocidad de reaccioacuten pero si se sigue aumentando la concentracioacuten de substrato la velocidad de reaccioacuten comienza a disminuir y a muy altas concentraciones de substrato se observa que no cambia la velocidad de
reaccioacuten se dice que los centros activos de la enzima se encuentran saturados La velocidad de reaccioacuten que se obtiene a esa alta concentracioacuten de substrato se define como la velocidad maacutexima (V) de la reaccioacuten enzimaacutetica bajo las condiciones especificadas La concentracioacuten de
substrato (S) a la semivelocidad maacutexima de reaccioacuten (V2) se puede determinar de la figura y representa la constante de Michaelis o Km la cual es una caracteriacutestica para cada enzima La inversa de Km o 1Km mide aproximadamente la afinidad de la enzima por el subs trato
Mientras maacutes pequentildeo sea el valor de Km mayor seraacute la afinidad de la enzima por el substrato Si varias enzimas compiten en el metabolismo por el mismo substrato eacuteste seraacute transformado preferentemente por la enzima con mayor afinidad
La ecuacioacuten de Michaelis describe la relacioacuten cuantitativa entre la velocidad de reaccioacuten y la concentracioacuten de substrato [S] si se conoce Vmax o Km
v=
En donde v= es la velocidad de reaccioacuten observada a una concentracioacuten de substrato determinada [S]
Km= constante de Michaelis en molesliacutet ro
23
Vmax= velocidad maacutexima a concentracioacuten saturante de substrato
Si v= Vmax2 entonces
=
Km+[S] = 2[S]
Km =[S]
De tal forma que podemos concluir que Km es igual a la concentracioacuten de substrato a la semivelocidad maacutexima de reaccioacuten
1 Concentraciones relativas de E y S la concentracioacuten de substrato S es
mucho mayor que la de E de tal manera que la cantidad de substrato
unido a la enzima en cualquier momento es muy pequentildea
2 Se asume el estado estacionario ES no cambia con el tiempo esto
quiere decir que la velocidad de formacioacuten de ES es igual a aquella para
su desintegracioacuten En general un intermediario en una serie de reacciones estaacute en estado estacionario cuando su velocidad de siacutentesis
es igual a su velocidad de degradacioacuten 3 Velocidad inicial para el anaacutelisis de reacciones enzimaacuteticas soacutelo se
utiliza la velocidad inicial de la reaccioacuten que es la velocidad ejercida por
la enzima inmediatamente despueacutes de que se ha puesto en contacto con el substrato y hasta antes de que se haya consumido el 10 de la concentracioacuten inicial del mismo La razoacuten de lo anterior es que en ese
momento la concentracioacuten del producto de la reaccioacuten que se ha acumulado en muy pequentildea y por tanto la reaccioacuten en el sentido
inverso es decir la transformacioacuten del producto en el substrato original puede ser ignorada
24
Caracteriacutesticas de Km la constante de Michaelis-Menten es
caracteriacutestica de una enzima y particular para un substrato Refleja la afinidad de la enzima por ese substrato K m es numeacutericamente igual a la concentracioacuten
de substrato a la cual la velocidad de reaccioacuten es la mitad de la Vmax (Km= Vmax2) Este paraacutemetro Km no variacutea con la concentracioacuten de enzima
Significado de una Km pequentildea un valor numeacuterico pequentildeo de Km
refleja una alta afinidad de la enzima por su substrato porque a una baja
concentracioacuten del mismo la enzima a desarrollado ya la mitad de la velocidad maacutexima
Significado de una Km grande el valor numeacuterico grande de Km refleja
una baja afinidad de la enzima por su substrato porque a una concentracioacuten
elevada del mismo la enzima desarrolla la mitad de la velocidad maacutexima
Relacioacuten de la velocidad con la concentracioacuten de enzima la
velocidad de la reaccioacuten es directamente proporcional a la concentracioacuten de la
enzima a cualquier concentracioacuten de substrato Por ejemplo si la concentracioacuten de enzima es disminuida a la mitad la velocidad inicial de la reaccioacuten (v0) es
reducida tambieacuten a la mitad de la original
Orden de la reaccioacuten cuando S es mas pequentildea que la Km la
velocidad de la reaccioacuten es aproximadamente igual a la concentracioacuten de
substrato
100
50
0
Substrato
Km Figura Localizacioacuten de la constante de Michaelis (Km)
25
La velocidad de reaccioacuten se dice en estas condiciones es de primer
orden con respecto al substrato Cuando S es mas grande que la Km la
velocidad es constante e igual a la Vmax La velocidad de la reaccioacuten es independiente de la concentracioacuten de substrato y se dice que es de orden cero
con respecto a la concentracioacuten de substrato
Las enzimas pueden ser aisladas de las ceacutelulas u organismos para estudiar sus
propiedades in vitro es decir en un tubo de ensayo Las diferentes enzimas muestran diferentes respuestas a los cambios en la concentracioacuten de substrato temperatura y pH
Concentracioacuten de substrato
La forma hiperboacutelica de la curva de velocidad vs Substrato
Efecto de la temperatura en la actividad enzimaacutetica
Efecto de la variacioacuten del pH en la actividad enzimaacutetica
Incremento de la velocidad con la temperatura En general la velocidad de la reaccioacuten se incrementa con la temperatura hasta que un punto maacuteximo es alcanzado
Este incremento se debe a que aumenta el nuacutemero de moleacuteculas ricas en energiacutea que pueden pasar la barrera energeacutetica de estado de transicioacuten para formar a los productos
Decremento de la velocidad con la temperatura la elevacioacuten excesiva de la
temperatura del medio que contiene a las enzimas resulta en un decremento de la velocidad como resultado de la desnaturalizacioacuten es decir la peacuterdida de la estructura tridimensional de las enzimas
26
zona de reactivacioacuten maacuteximo de actividad
Zona de
inactivacioacuten
0 Temperatura
Figura dependencia de la actividad enzimaacutetica con la temperatura
Velocidad maacutexima la velocidad de una reaccioacuten (v) es el nuacutemero de
moleacuteculas de substrato convertidas en producto por unidad de tiempo y
usualmente se expresa en micromoles de producto formadas por minuto La velocidad de una reaccioacuten catalizada por una enzima aumenta conforme se incrementa la concentracioacuten de substrato hasta que se llega a una velocidad maacutexima (Vmax) a partir de la cual la velocidad de la reaccioacuten es
independiente de la cantidad de substrato El que la velocidad no pueda
seguirse incrementando refleja la saturacioacuten del sitio activo con el substrato
Vmax
0 Substrato
Figura representacioacuten de una cineacutetica tiacutepicamente michaeliana
Vmax = velocidad de saturacioacuten o maacutexima
27
Cuando se grafica la velocidad de la reaccioacuten v0 contra la concentracioacuten de substrato S no siempre es posible determinar la condicioacuten
en que se ha llegado a la velocidad maacutexima Vmax debido al incremento de la pendiente en la hipeacuterbola a concentraciones de substrato elevadas
Figura experimento cineacutetico para la actividad enzimaacutetica
Se presentan los valores experimentales y los resultados del ajuste al modelo de Michaelis
Auacuten asiacute si se grafica una doble reciacuteproca es decir el inverso de la
velocidad (1v0) contra el inverso de la concentracioacuten de substrato (1 S ) se
obtiene una liacutenea recta El regraacutefico de Lineweaver-Burke tambieacuten se
denomina de ldquodobles reciacuteprocasrdquo y se utiliza para calcular la Km y la Vmax asiacute como para determinar el mecanismo de accioacuten de los diversos tipos de
inhibidores
La ecuacioacuten que describe a la graacutefca de Lineweaver-Burke es
Vmax
1
SVmax
Km
v
1
0 (3)
que describe una recta en donde el intercepto con el eje de las abscisas es igual a ndash1Km y el intercepto con el eje de las ordenadas es igual a 1Vmax
28
Figura experimento cineacutetico para la actividad enzimaacutetica Regraacutefico de
Lineweaver-Burke
Se presentan los valores experimentales y los resultados del ajuste al modelo
REPRESENTACIOacuteN GRAacuteFICA DE LINEWEAVER Y BURK
Si se toma el inverso en ambos miembros de la ecuacioacuten de Michaelis tenemos
=
+
29
que es la ecuacioacuten de una liacutenea recta cuya pendiente es que intercepta al eje de las
ordenadas en y a la abcisa en el punto Lo que es evidente al hacer
ya que se convierte en
Si se hace una graacutefica con los dobles inversos colocando los valores de en el eje de las
ordenadas y en el eje de las abcisas se obtiene una liacutenea recta a partir de la cual se puede calcular faacutecilmente Km
EFECTO DEL pH SOBRE LA ACTIVIDAD ENZIMATICA
Sabiendo que las enzimas son proteinas cualquier cambio brusco de pH puede alterar el caraacutecter ioacutenico de los grupos amino y carboxilo en la superficie proteica afectando asiacute las propiedades cataliacuteticas de una enzima A pH alto o bajo se puede producir la desnaturalizacioacuten
de la enzima y en consecuencia su inactivacioacuten La fosfatasa aacutecida es maacutes activa a pH 50 mientras que la fosfatasa alcalina lo es a pH 90 Muchas enzimas tienen maacutexima actividad cerca de la neutralidad en un rango de pH de 6 a 8
El pH per se no afecta la actividad enzimaacutetica sino la concentracioacuten de protones Los protones
ademaacutes de alterar la estructura de la enzima y el substrato pueden participar tambieacuten en la reaccioacuten como substrato o producto En esos casos la concentracioacuten de protones afecta directamente la velocidad de la reaccioacuten
ENZIMA pH OPTIMO
30
Pepsina 15
Tripsina 77
Catalasa 76
Arginasa 97
Fumarasa 78
Ribonucleasa 78
EFECTO DE ACTIVADORES
Un aumento en la temperatura provoca un aumento de la velocidad de reaccioacuten hasta cierta temperatura oacuteptima ya que despueacutes de aproximadamente 45
0 C se comienza a producir la
desnaturalizacioacuten teacutermica Las enzimas de muchos mamiacuteferos tienen una temperatura oacuteptima
de 370 C por encima de esa temperatura comienzan a inactivarse y se destruyen Sin embargo
existen especies de bacterias y algas que habitan en fuentes de aguas termales y en el otro extremo ciertas bacterias aacuterticas tienen temperaturas oacuteptimas cercanas a 0
0 C
31
ESPECIFICIDAD ABSOLUTA
Cierta enzimas se encuentran como aproenzimas o zimogenos que son inactivos
El tripsinoacutegeno es convertido en tripsina por la misma accioacuten de la tripsina que remueve un hexapeacuteptido
tripsina Tripsinoacutegeno Tripsina + Val (Asp)4 lisina
Otra forma de activacioacuten consiste en mantener los grupos SH reducidos Estos grupos pueden
formar parte de centros activos de ciertas enzimas Si se oxidan estos grupos a -S-S- la enzima se inactiva Asiacute tenemos que la enzima papaina se inactiva despueacutes de exponerla al ox iacutegeno pero cuando se le antildeade un reductor apropiado la enzima se reactiva Los grupos ndashS-S- se convierten en ndashSH activaacutendose la enzima
Otra forma de activacioacuten requiere la presencia de otra substancia que se enlaza al sitio alosteacuterico el compuesto que se enlaza se denomina un efector alosteacuterico El centro alosteacuterico es bien especiacutefico para el efector o modulador alosteacuterico
ESPECIFICIDAD RELATIVA
Oxiacutegeno L-aminoaacutecido a -cetoaacutecido + NH3 + H 2 O 2 L-aminoaacutecido oxidasa Oxiacutegeno D-aminoaacutecido a -cetoaacutecido + NH3 + H 2 O 2 D-aminoaacutecido oxidasa
32
En este ejemplo se muestra la estereo-especificidad Una enzima puede tener una especificidad oacuteptica para isomeros D o L
Hay enzimas que muestran especificidad absoluta como la aspartasa que cataliza la adicioacuten reversible de amoniacuteaco al doble enlace del aacutecido fumaacuterico pero no al de otros aacutecidos insaturados
HOOC ndash CH = CH ndash COO- + NH4
+ HOOC ndash CH2 - CHNH2 - COO
- + H
+
INHIBICIOacuteN ENZIMATICA
Mediante el uso de inhibidores enzimaacuteticos se ha obtenido informacioacuten muy valiosa sobre la conformacioacuten del centro activo de algunas enzimas
a Inhibicioacuten irreversible Este tipo de inhibidores forman un enlace covalente con las enzimas cerca del centro activo Un ejemplo son los gases nerviosos como el
fluorofosfato de diisopropilo (DFP) que forma un complejo con la enzima acetilcolinesterasa Los animales envenenados con este gas quedan paralizados debido a la imposibilidad de transmitir adecuadamente los impulsos nerviosos
Ester di-isopropil fos foacuterico de la enzima (inactivo)
b) Inhibicioacuten competitiva
Es cuando el inhibidor compite con el substrato por la unioacuten con el centro activo de la
enzima Este tipo de inhibicioacuten puede reducirse si se aumenta la concentracioacuten de substrato
-OOC - CH2 - CH2 - COO
- -OOC - CH = CH - COO
- + 2H
+ Succinato Succinato Fumarato
33
Deshidrogenasa COO
- I CH2 Malonato (inhibidor competitivo) I COO
- En la inhibicioacuten competitiva la enzima se combina con el inhibidor para formar un complejo enzima-inhibidor E + I E I en competencia con la reaccioacuten normal E +S ES
La sulfanilamida (bactericida) interfiere con la siacutentesis del aacutecido foacutelico compitiendo con el aacutecido p-aminobenzoico de una forma competitiva
HOOC-sect- NH2 NH2 ndashSO2 - sect- NH2 aacutecido p- amino benzoico sulfanilamida
sect = fenil
a Inhibicioacuten no competitiva
Esta inhibicioacuten se caracteriza por que no se puede revertir el efecto del inhibidor aumentando la concentracioacuten del substrato
Por ejemplo la inhibicioacuten de la deshidrogenasa del gliceraldehido -3-fosfato por la yodo-acetamida
Enzima-SH + I-CH2 CONH2 Enzima-SCH2 CONH2 + IH
La deshidrogenasa del gliceraldehido-3 fosfato puede ser inhibida tambieacuten por el aacutecido iodo-aceacutetico
Enzima-SH + ICH2 COOH Enzima-SCH2 COOH + IH
c) Inhibicioacuten por metales
Ciertos metales como el plomo mercurio y arseacutenico inhiben enzimas que tienen en su centro activo grupos -SH libres

21
a) el peroacutexido de hidroacutegeno se descompone en
H2 O2 H2 O + O2
18
b) el hierro cataliacutetico (Fe) realiza la reaccioacuten
H2 O2 H2 O + O2
13
c) el platino cataliacutetico (Pt) realiza la reaccioacuten
H2 O2 H2 O + O2
12
d) la catalasa una enzima hepaacutetica la realiza
H2O2 H2O + O2
5
Una enzima no modifica la energiacutea libre ni la constante de equilibrio sino que disminuye la energiacutea de activacioacuten de la reaccioacuten
Ir al principio
EFECTO DE LA CONCENTRACIOacuteN DEL SUBSTRATO SOBRE LA
CATALISIS ENZIMATICA
La actividad de una enzima se puede estudiar in vitro bajo condiciones controladas antildeadieacutendole una enzima a un substrato Siacute se trabaja con la condicioacuten de que la concentracioacuten del substrato sea saturante variando entonces la concentracioacuten de enzima se observa que aumenta el producto de la reaccioacuten a pH y temperatura constantes
22
Si se mantiene la concentracioacuten de la enzima constante y variando la concentracioacuten de substrato se obtiene una curva hiperboacutelica como la de la figura Al principio un aumento de la
concentracioacuten de substrato produce un aumento raacutepi do de la velocidad de reaccioacuten pero si se sigue aumentando la concentracioacuten de substrato la velocidad de reaccioacuten comienza a disminuir y a muy altas concentraciones de substrato se observa que no cambia la velocidad de
reaccioacuten se dice que los centros activos de la enzima se encuentran saturados La velocidad de reaccioacuten que se obtiene a esa alta concentracioacuten de substrato se define como la velocidad maacutexima (V) de la reaccioacuten enzimaacutetica bajo las condiciones especificadas La concentracioacuten de
substrato (S) a la semivelocidad maacutexima de reaccioacuten (V2) se puede determinar de la figura y representa la constante de Michaelis o Km la cual es una caracteriacutestica para cada enzima La inversa de Km o 1Km mide aproximadamente la afinidad de la enzima por el subs trato
Mientras maacutes pequentildeo sea el valor de Km mayor seraacute la afinidad de la enzima por el substrato Si varias enzimas compiten en el metabolismo por el mismo substrato eacuteste seraacute transformado preferentemente por la enzima con mayor afinidad
La ecuacioacuten de Michaelis describe la relacioacuten cuantitativa entre la velocidad de reaccioacuten y la concentracioacuten de substrato [S] si se conoce Vmax o Km
v=
En donde v= es la velocidad de reaccioacuten observada a una concentracioacuten de substrato determinada [S]
Km= constante de Michaelis en molesliacutet ro
23
Vmax= velocidad maacutexima a concentracioacuten saturante de substrato
Si v= Vmax2 entonces
=
Km+[S] = 2[S]
Km =[S]
De tal forma que podemos concluir que Km es igual a la concentracioacuten de substrato a la semivelocidad maacutexima de reaccioacuten
1 Concentraciones relativas de E y S la concentracioacuten de substrato S es
mucho mayor que la de E de tal manera que la cantidad de substrato
unido a la enzima en cualquier momento es muy pequentildea
2 Se asume el estado estacionario ES no cambia con el tiempo esto
quiere decir que la velocidad de formacioacuten de ES es igual a aquella para
su desintegracioacuten En general un intermediario en una serie de reacciones estaacute en estado estacionario cuando su velocidad de siacutentesis
es igual a su velocidad de degradacioacuten 3 Velocidad inicial para el anaacutelisis de reacciones enzimaacuteticas soacutelo se
utiliza la velocidad inicial de la reaccioacuten que es la velocidad ejercida por
la enzima inmediatamente despueacutes de que se ha puesto en contacto con el substrato y hasta antes de que se haya consumido el 10 de la concentracioacuten inicial del mismo La razoacuten de lo anterior es que en ese
momento la concentracioacuten del producto de la reaccioacuten que se ha acumulado en muy pequentildea y por tanto la reaccioacuten en el sentido
inverso es decir la transformacioacuten del producto en el substrato original puede ser ignorada
24
Caracteriacutesticas de Km la constante de Michaelis-Menten es
caracteriacutestica de una enzima y particular para un substrato Refleja la afinidad de la enzima por ese substrato K m es numeacutericamente igual a la concentracioacuten
de substrato a la cual la velocidad de reaccioacuten es la mitad de la Vmax (Km= Vmax2) Este paraacutemetro Km no variacutea con la concentracioacuten de enzima
Significado de una Km pequentildea un valor numeacuterico pequentildeo de Km
refleja una alta afinidad de la enzima por su substrato porque a una baja
concentracioacuten del mismo la enzima a desarrollado ya la mitad de la velocidad maacutexima
Significado de una Km grande el valor numeacuterico grande de Km refleja
una baja afinidad de la enzima por su substrato porque a una concentracioacuten
elevada del mismo la enzima desarrolla la mitad de la velocidad maacutexima
Relacioacuten de la velocidad con la concentracioacuten de enzima la
velocidad de la reaccioacuten es directamente proporcional a la concentracioacuten de la
enzima a cualquier concentracioacuten de substrato Por ejemplo si la concentracioacuten de enzima es disminuida a la mitad la velocidad inicial de la reaccioacuten (v0) es
reducida tambieacuten a la mitad de la original
Orden de la reaccioacuten cuando S es mas pequentildea que la Km la
velocidad de la reaccioacuten es aproximadamente igual a la concentracioacuten de
substrato
100
50
0
Substrato
Km Figura Localizacioacuten de la constante de Michaelis (Km)
25
La velocidad de reaccioacuten se dice en estas condiciones es de primer
orden con respecto al substrato Cuando S es mas grande que la Km la
velocidad es constante e igual a la Vmax La velocidad de la reaccioacuten es independiente de la concentracioacuten de substrato y se dice que es de orden cero
con respecto a la concentracioacuten de substrato
Las enzimas pueden ser aisladas de las ceacutelulas u organismos para estudiar sus
propiedades in vitro es decir en un tubo de ensayo Las diferentes enzimas muestran diferentes respuestas a los cambios en la concentracioacuten de substrato temperatura y pH
Concentracioacuten de substrato
La forma hiperboacutelica de la curva de velocidad vs Substrato
Efecto de la temperatura en la actividad enzimaacutetica
Efecto de la variacioacuten del pH en la actividad enzimaacutetica
Incremento de la velocidad con la temperatura En general la velocidad de la reaccioacuten se incrementa con la temperatura hasta que un punto maacuteximo es alcanzado
Este incremento se debe a que aumenta el nuacutemero de moleacuteculas ricas en energiacutea que pueden pasar la barrera energeacutetica de estado de transicioacuten para formar a los productos
Decremento de la velocidad con la temperatura la elevacioacuten excesiva de la
temperatura del medio que contiene a las enzimas resulta en un decremento de la velocidad como resultado de la desnaturalizacioacuten es decir la peacuterdida de la estructura tridimensional de las enzimas
26
zona de reactivacioacuten maacuteximo de actividad
Zona de
inactivacioacuten
0 Temperatura
Figura dependencia de la actividad enzimaacutetica con la temperatura
Velocidad maacutexima la velocidad de una reaccioacuten (v) es el nuacutemero de
moleacuteculas de substrato convertidas en producto por unidad de tiempo y
usualmente se expresa en micromoles de producto formadas por minuto La velocidad de una reaccioacuten catalizada por una enzima aumenta conforme se incrementa la concentracioacuten de substrato hasta que se llega a una velocidad maacutexima (Vmax) a partir de la cual la velocidad de la reaccioacuten es
independiente de la cantidad de substrato El que la velocidad no pueda
seguirse incrementando refleja la saturacioacuten del sitio activo con el substrato
Vmax
0 Substrato
Figura representacioacuten de una cineacutetica tiacutepicamente michaeliana
Vmax = velocidad de saturacioacuten o maacutexima
27
Cuando se grafica la velocidad de la reaccioacuten v0 contra la concentracioacuten de substrato S no siempre es posible determinar la condicioacuten
en que se ha llegado a la velocidad maacutexima Vmax debido al incremento de la pendiente en la hipeacuterbola a concentraciones de substrato elevadas
Figura experimento cineacutetico para la actividad enzimaacutetica
Se presentan los valores experimentales y los resultados del ajuste al modelo de Michaelis
Auacuten asiacute si se grafica una doble reciacuteproca es decir el inverso de la
velocidad (1v0) contra el inverso de la concentracioacuten de substrato (1 S ) se
obtiene una liacutenea recta El regraacutefico de Lineweaver-Burke tambieacuten se
denomina de ldquodobles reciacuteprocasrdquo y se utiliza para calcular la Km y la Vmax asiacute como para determinar el mecanismo de accioacuten de los diversos tipos de
inhibidores
La ecuacioacuten que describe a la graacutefca de Lineweaver-Burke es
Vmax
1
SVmax
Km
v
1
0 (3)
que describe una recta en donde el intercepto con el eje de las abscisas es igual a ndash1Km y el intercepto con el eje de las ordenadas es igual a 1Vmax
28
Figura experimento cineacutetico para la actividad enzimaacutetica Regraacutefico de
Lineweaver-Burke
Se presentan los valores experimentales y los resultados del ajuste al modelo
REPRESENTACIOacuteN GRAacuteFICA DE LINEWEAVER Y BURK
Si se toma el inverso en ambos miembros de la ecuacioacuten de Michaelis tenemos
=
+
29
que es la ecuacioacuten de una liacutenea recta cuya pendiente es que intercepta al eje de las
ordenadas en y a la abcisa en el punto Lo que es evidente al hacer
ya que se convierte en
Si se hace una graacutefica con los dobles inversos colocando los valores de en el eje de las
ordenadas y en el eje de las abcisas se obtiene una liacutenea recta a partir de la cual se puede calcular faacutecilmente Km
EFECTO DEL pH SOBRE LA ACTIVIDAD ENZIMATICA
Sabiendo que las enzimas son proteinas cualquier cambio brusco de pH puede alterar el caraacutecter ioacutenico de los grupos amino y carboxilo en la superficie proteica afectando asiacute las propiedades cataliacuteticas de una enzima A pH alto o bajo se puede producir la desnaturalizacioacuten
de la enzima y en consecuencia su inactivacioacuten La fosfatasa aacutecida es maacutes activa a pH 50 mientras que la fosfatasa alcalina lo es a pH 90 Muchas enzimas tienen maacutexima actividad cerca de la neutralidad en un rango de pH de 6 a 8
El pH per se no afecta la actividad enzimaacutetica sino la concentracioacuten de protones Los protones
ademaacutes de alterar la estructura de la enzima y el substrato pueden participar tambieacuten en la reaccioacuten como substrato o producto En esos casos la concentracioacuten de protones afecta directamente la velocidad de la reaccioacuten
ENZIMA pH OPTIMO
30
Pepsina 15
Tripsina 77
Catalasa 76
Arginasa 97
Fumarasa 78
Ribonucleasa 78
EFECTO DE ACTIVADORES
Un aumento en la temperatura provoca un aumento de la velocidad de reaccioacuten hasta cierta temperatura oacuteptima ya que despueacutes de aproximadamente 45
0 C se comienza a producir la
desnaturalizacioacuten teacutermica Las enzimas de muchos mamiacuteferos tienen una temperatura oacuteptima
de 370 C por encima de esa temperatura comienzan a inactivarse y se destruyen Sin embargo
existen especies de bacterias y algas que habitan en fuentes de aguas termales y en el otro extremo ciertas bacterias aacuterticas tienen temperaturas oacuteptimas cercanas a 0
0 C
31
ESPECIFICIDAD ABSOLUTA
Cierta enzimas se encuentran como aproenzimas o zimogenos que son inactivos
El tripsinoacutegeno es convertido en tripsina por la misma accioacuten de la tripsina que remueve un hexapeacuteptido
tripsina Tripsinoacutegeno Tripsina + Val (Asp)4 lisina
Otra forma de activacioacuten consiste en mantener los grupos SH reducidos Estos grupos pueden
formar parte de centros activos de ciertas enzimas Si se oxidan estos grupos a -S-S- la enzima se inactiva Asiacute tenemos que la enzima papaina se inactiva despueacutes de exponerla al ox iacutegeno pero cuando se le antildeade un reductor apropiado la enzima se reactiva Los grupos ndashS-S- se convierten en ndashSH activaacutendose la enzima
Otra forma de activacioacuten requiere la presencia de otra substancia que se enlaza al sitio alosteacuterico el compuesto que se enlaza se denomina un efector alosteacuterico El centro alosteacuterico es bien especiacutefico para el efector o modulador alosteacuterico
ESPECIFICIDAD RELATIVA
Oxiacutegeno L-aminoaacutecido a -cetoaacutecido + NH3 + H 2 O 2 L-aminoaacutecido oxidasa Oxiacutegeno D-aminoaacutecido a -cetoaacutecido + NH3 + H 2 O 2 D-aminoaacutecido oxidasa
32
En este ejemplo se muestra la estereo-especificidad Una enzima puede tener una especificidad oacuteptica para isomeros D o L
Hay enzimas que muestran especificidad absoluta como la aspartasa que cataliza la adicioacuten reversible de amoniacuteaco al doble enlace del aacutecido fumaacuterico pero no al de otros aacutecidos insaturados
HOOC ndash CH = CH ndash COO- + NH4
+ HOOC ndash CH2 - CHNH2 - COO
- + H
+
INHIBICIOacuteN ENZIMATICA
Mediante el uso de inhibidores enzimaacuteticos se ha obtenido informacioacuten muy valiosa sobre la conformacioacuten del centro activo de algunas enzimas
a Inhibicioacuten irreversible Este tipo de inhibidores forman un enlace covalente con las enzimas cerca del centro activo Un ejemplo son los gases nerviosos como el
fluorofosfato de diisopropilo (DFP) que forma un complejo con la enzima acetilcolinesterasa Los animales envenenados con este gas quedan paralizados debido a la imposibilidad de transmitir adecuadamente los impulsos nerviosos
Ester di-isopropil fos foacuterico de la enzima (inactivo)
b) Inhibicioacuten competitiva
Es cuando el inhibidor compite con el substrato por la unioacuten con el centro activo de la
enzima Este tipo de inhibicioacuten puede reducirse si se aumenta la concentracioacuten de substrato
-OOC - CH2 - CH2 - COO
- -OOC - CH = CH - COO
- + 2H
+ Succinato Succinato Fumarato
33
Deshidrogenasa COO
- I CH2 Malonato (inhibidor competitivo) I COO
- En la inhibicioacuten competitiva la enzima se combina con el inhibidor para formar un complejo enzima-inhibidor E + I E I en competencia con la reaccioacuten normal E +S ES
La sulfanilamida (bactericida) interfiere con la siacutentesis del aacutecido foacutelico compitiendo con el aacutecido p-aminobenzoico de una forma competitiva
HOOC-sect- NH2 NH2 ndashSO2 - sect- NH2 aacutecido p- amino benzoico sulfanilamida
sect = fenil
a Inhibicioacuten no competitiva
Esta inhibicioacuten se caracteriza por que no se puede revertir el efecto del inhibidor aumentando la concentracioacuten del substrato
Por ejemplo la inhibicioacuten de la deshidrogenasa del gliceraldehido -3-fosfato por la yodo-acetamida
Enzima-SH + I-CH2 CONH2 Enzima-SCH2 CONH2 + IH
La deshidrogenasa del gliceraldehido-3 fosfato puede ser inhibida tambieacuten por el aacutecido iodo-aceacutetico
Enzima-SH + ICH2 COOH Enzima-SCH2 COOH + IH
c) Inhibicioacuten por metales
Ciertos metales como el plomo mercurio y arseacutenico inhiben enzimas que tienen en su centro activo grupos -SH libres
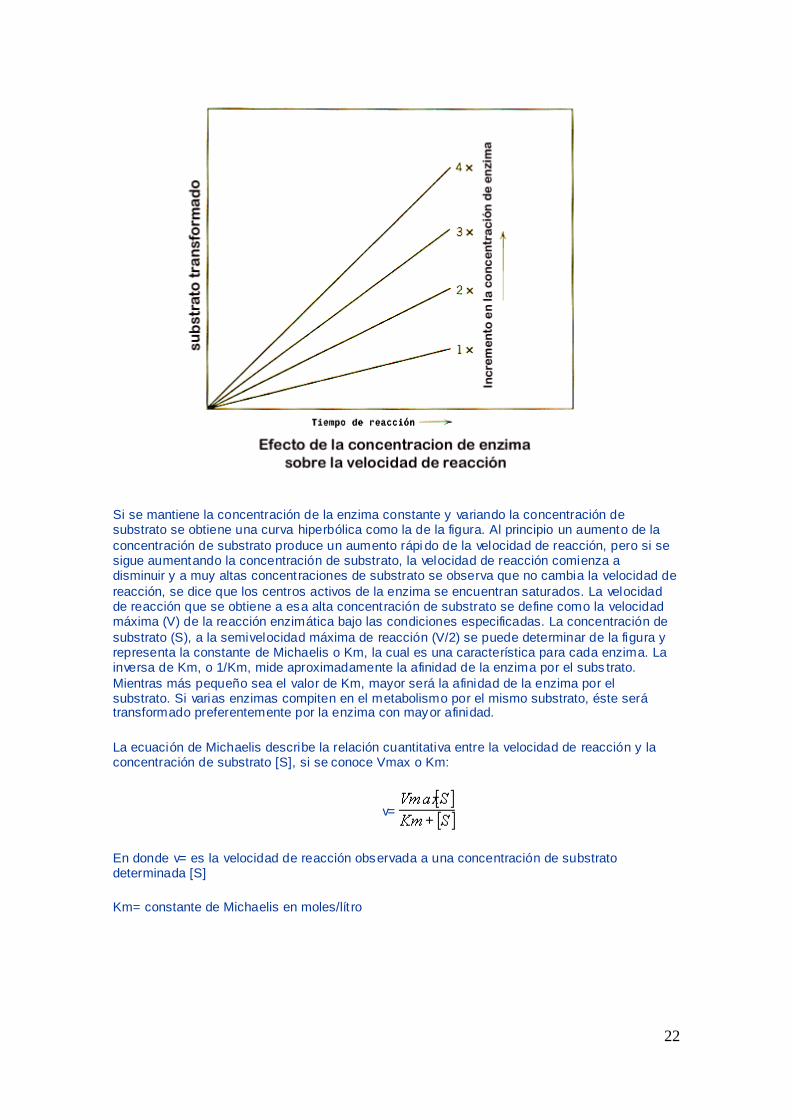
22
Si se mantiene la concentracioacuten de la enzima constante y variando la concentracioacuten de substrato se obtiene una curva hiperboacutelica como la de la figura Al principio un aumento de la
concentracioacuten de substrato produce un aumento raacutepi do de la velocidad de reaccioacuten pero si se sigue aumentando la concentracioacuten de substrato la velocidad de reaccioacuten comienza a disminuir y a muy altas concentraciones de substrato se observa que no cambia la velocidad de
reaccioacuten se dice que los centros activos de la enzima se encuentran saturados La velocidad de reaccioacuten que se obtiene a esa alta concentracioacuten de substrato se define como la velocidad maacutexima (V) de la reaccioacuten enzimaacutetica bajo las condiciones especificadas La concentracioacuten de
substrato (S) a la semivelocidad maacutexima de reaccioacuten (V2) se puede determinar de la figura y representa la constante de Michaelis o Km la cual es una caracteriacutestica para cada enzima La inversa de Km o 1Km mide aproximadamente la afinidad de la enzima por el subs trato
Mientras maacutes pequentildeo sea el valor de Km mayor seraacute la afinidad de la enzima por el substrato Si varias enzimas compiten en el metabolismo por el mismo substrato eacuteste seraacute transformado preferentemente por la enzima con mayor afinidad
La ecuacioacuten de Michaelis describe la relacioacuten cuantitativa entre la velocidad de reaccioacuten y la concentracioacuten de substrato [S] si se conoce Vmax o Km
v=
En donde v= es la velocidad de reaccioacuten observada a una concentracioacuten de substrato determinada [S]
Km= constante de Michaelis en molesliacutet ro
23
Vmax= velocidad maacutexima a concentracioacuten saturante de substrato
Si v= Vmax2 entonces
=
Km+[S] = 2[S]
Km =[S]
De tal forma que podemos concluir que Km es igual a la concentracioacuten de substrato a la semivelocidad maacutexima de reaccioacuten
1 Concentraciones relativas de E y S la concentracioacuten de substrato S es
mucho mayor que la de E de tal manera que la cantidad de substrato
unido a la enzima en cualquier momento es muy pequentildea
2 Se asume el estado estacionario ES no cambia con el tiempo esto
quiere decir que la velocidad de formacioacuten de ES es igual a aquella para
su desintegracioacuten En general un intermediario en una serie de reacciones estaacute en estado estacionario cuando su velocidad de siacutentesis
es igual a su velocidad de degradacioacuten 3 Velocidad inicial para el anaacutelisis de reacciones enzimaacuteticas soacutelo se
utiliza la velocidad inicial de la reaccioacuten que es la velocidad ejercida por
la enzima inmediatamente despueacutes de que se ha puesto en contacto con el substrato y hasta antes de que se haya consumido el 10 de la concentracioacuten inicial del mismo La razoacuten de lo anterior es que en ese
momento la concentracioacuten del producto de la reaccioacuten que se ha acumulado en muy pequentildea y por tanto la reaccioacuten en el sentido
inverso es decir la transformacioacuten del producto en el substrato original puede ser ignorada
24
Caracteriacutesticas de Km la constante de Michaelis-Menten es
caracteriacutestica de una enzima y particular para un substrato Refleja la afinidad de la enzima por ese substrato K m es numeacutericamente igual a la concentracioacuten
de substrato a la cual la velocidad de reaccioacuten es la mitad de la Vmax (Km= Vmax2) Este paraacutemetro Km no variacutea con la concentracioacuten de enzima
Significado de una Km pequentildea un valor numeacuterico pequentildeo de Km
refleja una alta afinidad de la enzima por su substrato porque a una baja
concentracioacuten del mismo la enzima a desarrollado ya la mitad de la velocidad maacutexima
Significado de una Km grande el valor numeacuterico grande de Km refleja
una baja afinidad de la enzima por su substrato porque a una concentracioacuten
elevada del mismo la enzima desarrolla la mitad de la velocidad maacutexima
Relacioacuten de la velocidad con la concentracioacuten de enzima la
velocidad de la reaccioacuten es directamente proporcional a la concentracioacuten de la
enzima a cualquier concentracioacuten de substrato Por ejemplo si la concentracioacuten de enzima es disminuida a la mitad la velocidad inicial de la reaccioacuten (v0) es
reducida tambieacuten a la mitad de la original
Orden de la reaccioacuten cuando S es mas pequentildea que la Km la
velocidad de la reaccioacuten es aproximadamente igual a la concentracioacuten de
substrato
100
50
0
Substrato
Km Figura Localizacioacuten de la constante de Michaelis (Km)
25
La velocidad de reaccioacuten se dice en estas condiciones es de primer
orden con respecto al substrato Cuando S es mas grande que la Km la
velocidad es constante e igual a la Vmax La velocidad de la reaccioacuten es independiente de la concentracioacuten de substrato y se dice que es de orden cero
con respecto a la concentracioacuten de substrato
Las enzimas pueden ser aisladas de las ceacutelulas u organismos para estudiar sus
propiedades in vitro es decir en un tubo de ensayo Las diferentes enzimas muestran diferentes respuestas a los cambios en la concentracioacuten de substrato temperatura y pH
Concentracioacuten de substrato
La forma hiperboacutelica de la curva de velocidad vs Substrato
Efecto de la temperatura en la actividad enzimaacutetica
Efecto de la variacioacuten del pH en la actividad enzimaacutetica
Incremento de la velocidad con la temperatura En general la velocidad de la reaccioacuten se incrementa con la temperatura hasta que un punto maacuteximo es alcanzado
Este incremento se debe a que aumenta el nuacutemero de moleacuteculas ricas en energiacutea que pueden pasar la barrera energeacutetica de estado de transicioacuten para formar a los productos
Decremento de la velocidad con la temperatura la elevacioacuten excesiva de la
temperatura del medio que contiene a las enzimas resulta en un decremento de la velocidad como resultado de la desnaturalizacioacuten es decir la peacuterdida de la estructura tridimensional de las enzimas
26
zona de reactivacioacuten maacuteximo de actividad
Zona de
inactivacioacuten
0 Temperatura
Figura dependencia de la actividad enzimaacutetica con la temperatura
Velocidad maacutexima la velocidad de una reaccioacuten (v) es el nuacutemero de
moleacuteculas de substrato convertidas en producto por unidad de tiempo y
usualmente se expresa en micromoles de producto formadas por minuto La velocidad de una reaccioacuten catalizada por una enzima aumenta conforme se incrementa la concentracioacuten de substrato hasta que se llega a una velocidad maacutexima (Vmax) a partir de la cual la velocidad de la reaccioacuten es
independiente de la cantidad de substrato El que la velocidad no pueda
seguirse incrementando refleja la saturacioacuten del sitio activo con el substrato
Vmax
0 Substrato
Figura representacioacuten de una cineacutetica tiacutepicamente michaeliana
Vmax = velocidad de saturacioacuten o maacutexima
27
Cuando se grafica la velocidad de la reaccioacuten v0 contra la concentracioacuten de substrato S no siempre es posible determinar la condicioacuten
en que se ha llegado a la velocidad maacutexima Vmax debido al incremento de la pendiente en la hipeacuterbola a concentraciones de substrato elevadas
Figura experimento cineacutetico para la actividad enzimaacutetica
Se presentan los valores experimentales y los resultados del ajuste al modelo de Michaelis
Auacuten asiacute si se grafica una doble reciacuteproca es decir el inverso de la
velocidad (1v0) contra el inverso de la concentracioacuten de substrato (1 S ) se
obtiene una liacutenea recta El regraacutefico de Lineweaver-Burke tambieacuten se
denomina de ldquodobles reciacuteprocasrdquo y se utiliza para calcular la Km y la Vmax asiacute como para determinar el mecanismo de accioacuten de los diversos tipos de
inhibidores
La ecuacioacuten que describe a la graacutefca de Lineweaver-Burke es
Vmax
1
SVmax
Km
v
1
0 (3)
que describe una recta en donde el intercepto con el eje de las abscisas es igual a ndash1Km y el intercepto con el eje de las ordenadas es igual a 1Vmax
28
Figura experimento cineacutetico para la actividad enzimaacutetica Regraacutefico de
Lineweaver-Burke
Se presentan los valores experimentales y los resultados del ajuste al modelo
REPRESENTACIOacuteN GRAacuteFICA DE LINEWEAVER Y BURK
Si se toma el inverso en ambos miembros de la ecuacioacuten de Michaelis tenemos
=
+
29
que es la ecuacioacuten de una liacutenea recta cuya pendiente es que intercepta al eje de las
ordenadas en y a la abcisa en el punto Lo que es evidente al hacer
ya que se convierte en
Si se hace una graacutefica con los dobles inversos colocando los valores de en el eje de las
ordenadas y en el eje de las abcisas se obtiene una liacutenea recta a partir de la cual se puede calcular faacutecilmente Km
EFECTO DEL pH SOBRE LA ACTIVIDAD ENZIMATICA
Sabiendo que las enzimas son proteinas cualquier cambio brusco de pH puede alterar el caraacutecter ioacutenico de los grupos amino y carboxilo en la superficie proteica afectando asiacute las propiedades cataliacuteticas de una enzima A pH alto o bajo se puede producir la desnaturalizacioacuten
de la enzima y en consecuencia su inactivacioacuten La fosfatasa aacutecida es maacutes activa a pH 50 mientras que la fosfatasa alcalina lo es a pH 90 Muchas enzimas tienen maacutexima actividad cerca de la neutralidad en un rango de pH de 6 a 8
El pH per se no afecta la actividad enzimaacutetica sino la concentracioacuten de protones Los protones
ademaacutes de alterar la estructura de la enzima y el substrato pueden participar tambieacuten en la reaccioacuten como substrato o producto En esos casos la concentracioacuten de protones afecta directamente la velocidad de la reaccioacuten
ENZIMA pH OPTIMO
30
Pepsina 15
Tripsina 77
Catalasa 76
Arginasa 97
Fumarasa 78
Ribonucleasa 78
EFECTO DE ACTIVADORES
Un aumento en la temperatura provoca un aumento de la velocidad de reaccioacuten hasta cierta temperatura oacuteptima ya que despueacutes de aproximadamente 45
0 C se comienza a producir la
desnaturalizacioacuten teacutermica Las enzimas de muchos mamiacuteferos tienen una temperatura oacuteptima
de 370 C por encima de esa temperatura comienzan a inactivarse y se destruyen Sin embargo
existen especies de bacterias y algas que habitan en fuentes de aguas termales y en el otro extremo ciertas bacterias aacuterticas tienen temperaturas oacuteptimas cercanas a 0
0 C
31
ESPECIFICIDAD ABSOLUTA
Cierta enzimas se encuentran como aproenzimas o zimogenos que son inactivos
El tripsinoacutegeno es convertido en tripsina por la misma accioacuten de la tripsina que remueve un hexapeacuteptido
tripsina Tripsinoacutegeno Tripsina + Val (Asp)4 lisina
Otra forma de activacioacuten consiste en mantener los grupos SH reducidos Estos grupos pueden
formar parte de centros activos de ciertas enzimas Si se oxidan estos grupos a -S-S- la enzima se inactiva Asiacute tenemos que la enzima papaina se inactiva despueacutes de exponerla al ox iacutegeno pero cuando se le antildeade un reductor apropiado la enzima se reactiva Los grupos ndashS-S- se convierten en ndashSH activaacutendose la enzima
Otra forma de activacioacuten requiere la presencia de otra substancia que se enlaza al sitio alosteacuterico el compuesto que se enlaza se denomina un efector alosteacuterico El centro alosteacuterico es bien especiacutefico para el efector o modulador alosteacuterico
ESPECIFICIDAD RELATIVA
Oxiacutegeno L-aminoaacutecido a -cetoaacutecido + NH3 + H 2 O 2 L-aminoaacutecido oxidasa Oxiacutegeno D-aminoaacutecido a -cetoaacutecido + NH3 + H 2 O 2 D-aminoaacutecido oxidasa
32
En este ejemplo se muestra la estereo-especificidad Una enzima puede tener una especificidad oacuteptica para isomeros D o L
Hay enzimas que muestran especificidad absoluta como la aspartasa que cataliza la adicioacuten reversible de amoniacuteaco al doble enlace del aacutecido fumaacuterico pero no al de otros aacutecidos insaturados
HOOC ndash CH = CH ndash COO- + NH4
+ HOOC ndash CH2 - CHNH2 - COO
- + H
+
INHIBICIOacuteN ENZIMATICA
Mediante el uso de inhibidores enzimaacuteticos se ha obtenido informacioacuten muy valiosa sobre la conformacioacuten del centro activo de algunas enzimas
a Inhibicioacuten irreversible Este tipo de inhibidores forman un enlace covalente con las enzimas cerca del centro activo Un ejemplo son los gases nerviosos como el
fluorofosfato de diisopropilo (DFP) que forma un complejo con la enzima acetilcolinesterasa Los animales envenenados con este gas quedan paralizados debido a la imposibilidad de transmitir adecuadamente los impulsos nerviosos
Ester di-isopropil fos foacuterico de la enzima (inactivo)
b) Inhibicioacuten competitiva
Es cuando el inhibidor compite con el substrato por la unioacuten con el centro activo de la
enzima Este tipo de inhibicioacuten puede reducirse si se aumenta la concentracioacuten de substrato
-OOC - CH2 - CH2 - COO
- -OOC - CH = CH - COO
- + 2H
+ Succinato Succinato Fumarato
33
Deshidrogenasa COO
- I CH2 Malonato (inhibidor competitivo) I COO
- En la inhibicioacuten competitiva la enzima se combina con el inhibidor para formar un complejo enzima-inhibidor E + I E I en competencia con la reaccioacuten normal E +S ES
La sulfanilamida (bactericida) interfiere con la siacutentesis del aacutecido foacutelico compitiendo con el aacutecido p-aminobenzoico de una forma competitiva
HOOC-sect- NH2 NH2 ndashSO2 - sect- NH2 aacutecido p- amino benzoico sulfanilamida
sect = fenil
a Inhibicioacuten no competitiva
Esta inhibicioacuten se caracteriza por que no se puede revertir el efecto del inhibidor aumentando la concentracioacuten del substrato
Por ejemplo la inhibicioacuten de la deshidrogenasa del gliceraldehido -3-fosfato por la yodo-acetamida
Enzima-SH + I-CH2 CONH2 Enzima-SCH2 CONH2 + IH
La deshidrogenasa del gliceraldehido-3 fosfato puede ser inhibida tambieacuten por el aacutecido iodo-aceacutetico
Enzima-SH + ICH2 COOH Enzima-SCH2 COOH + IH
c) Inhibicioacuten por metales
Ciertos metales como el plomo mercurio y arseacutenico inhiben enzimas que tienen en su centro activo grupos -SH libres

23
Vmax= velocidad maacutexima a concentracioacuten saturante de substrato
Si v= Vmax2 entonces
=
Km+[S] = 2[S]
Km =[S]
De tal forma que podemos concluir que Km es igual a la concentracioacuten de substrato a la semivelocidad maacutexima de reaccioacuten
1 Concentraciones relativas de E y S la concentracioacuten de substrato S es
mucho mayor que la de E de tal manera que la cantidad de substrato
unido a la enzima en cualquier momento es muy pequentildea
2 Se asume el estado estacionario ES no cambia con el tiempo esto
quiere decir que la velocidad de formacioacuten de ES es igual a aquella para
su desintegracioacuten En general un intermediario en una serie de reacciones estaacute en estado estacionario cuando su velocidad de siacutentesis
es igual a su velocidad de degradacioacuten 3 Velocidad inicial para el anaacutelisis de reacciones enzimaacuteticas soacutelo se
utiliza la velocidad inicial de la reaccioacuten que es la velocidad ejercida por
la enzima inmediatamente despueacutes de que se ha puesto en contacto con el substrato y hasta antes de que se haya consumido el 10 de la concentracioacuten inicial del mismo La razoacuten de lo anterior es que en ese
momento la concentracioacuten del producto de la reaccioacuten que se ha acumulado en muy pequentildea y por tanto la reaccioacuten en el sentido
inverso es decir la transformacioacuten del producto en el substrato original puede ser ignorada
24
Caracteriacutesticas de Km la constante de Michaelis-Menten es
caracteriacutestica de una enzima y particular para un substrato Refleja la afinidad de la enzima por ese substrato K m es numeacutericamente igual a la concentracioacuten
de substrato a la cual la velocidad de reaccioacuten es la mitad de la Vmax (Km= Vmax2) Este paraacutemetro Km no variacutea con la concentracioacuten de enzima
Significado de una Km pequentildea un valor numeacuterico pequentildeo de Km
refleja una alta afinidad de la enzima por su substrato porque a una baja
concentracioacuten del mismo la enzima a desarrollado ya la mitad de la velocidad maacutexima
Significado de una Km grande el valor numeacuterico grande de Km refleja
una baja afinidad de la enzima por su substrato porque a una concentracioacuten
elevada del mismo la enzima desarrolla la mitad de la velocidad maacutexima
Relacioacuten de la velocidad con la concentracioacuten de enzima la
velocidad de la reaccioacuten es directamente proporcional a la concentracioacuten de la
enzima a cualquier concentracioacuten de substrato Por ejemplo si la concentracioacuten de enzima es disminuida a la mitad la velocidad inicial de la reaccioacuten (v0) es
reducida tambieacuten a la mitad de la original
Orden de la reaccioacuten cuando S es mas pequentildea que la Km la
velocidad de la reaccioacuten es aproximadamente igual a la concentracioacuten de
substrato
100
50
0
Substrato
Km Figura Localizacioacuten de la constante de Michaelis (Km)
25
La velocidad de reaccioacuten se dice en estas condiciones es de primer
orden con respecto al substrato Cuando S es mas grande que la Km la
velocidad es constante e igual a la Vmax La velocidad de la reaccioacuten es independiente de la concentracioacuten de substrato y se dice que es de orden cero
con respecto a la concentracioacuten de substrato
Las enzimas pueden ser aisladas de las ceacutelulas u organismos para estudiar sus
propiedades in vitro es decir en un tubo de ensayo Las diferentes enzimas muestran diferentes respuestas a los cambios en la concentracioacuten de substrato temperatura y pH
Concentracioacuten de substrato
La forma hiperboacutelica de la curva de velocidad vs Substrato
Efecto de la temperatura en la actividad enzimaacutetica
Efecto de la variacioacuten del pH en la actividad enzimaacutetica
Incremento de la velocidad con la temperatura En general la velocidad de la reaccioacuten se incrementa con la temperatura hasta que un punto maacuteximo es alcanzado
Este incremento se debe a que aumenta el nuacutemero de moleacuteculas ricas en energiacutea que pueden pasar la barrera energeacutetica de estado de transicioacuten para formar a los productos
Decremento de la velocidad con la temperatura la elevacioacuten excesiva de la
temperatura del medio que contiene a las enzimas resulta en un decremento de la velocidad como resultado de la desnaturalizacioacuten es decir la peacuterdida de la estructura tridimensional de las enzimas
26
zona de reactivacioacuten maacuteximo de actividad
Zona de
inactivacioacuten
0 Temperatura
Figura dependencia de la actividad enzimaacutetica con la temperatura
Velocidad maacutexima la velocidad de una reaccioacuten (v) es el nuacutemero de
moleacuteculas de substrato convertidas en producto por unidad de tiempo y
usualmente se expresa en micromoles de producto formadas por minuto La velocidad de una reaccioacuten catalizada por una enzima aumenta conforme se incrementa la concentracioacuten de substrato hasta que se llega a una velocidad maacutexima (Vmax) a partir de la cual la velocidad de la reaccioacuten es
independiente de la cantidad de substrato El que la velocidad no pueda
seguirse incrementando refleja la saturacioacuten del sitio activo con el substrato
Vmax
0 Substrato
Figura representacioacuten de una cineacutetica tiacutepicamente michaeliana
Vmax = velocidad de saturacioacuten o maacutexima
27
Cuando se grafica la velocidad de la reaccioacuten v0 contra la concentracioacuten de substrato S no siempre es posible determinar la condicioacuten
en que se ha llegado a la velocidad maacutexima Vmax debido al incremento de la pendiente en la hipeacuterbola a concentraciones de substrato elevadas
Figura experimento cineacutetico para la actividad enzimaacutetica
Se presentan los valores experimentales y los resultados del ajuste al modelo de Michaelis
Auacuten asiacute si se grafica una doble reciacuteproca es decir el inverso de la
velocidad (1v0) contra el inverso de la concentracioacuten de substrato (1 S ) se
obtiene una liacutenea recta El regraacutefico de Lineweaver-Burke tambieacuten se
denomina de ldquodobles reciacuteprocasrdquo y se utiliza para calcular la Km y la Vmax asiacute como para determinar el mecanismo de accioacuten de los diversos tipos de
inhibidores
La ecuacioacuten que describe a la graacutefca de Lineweaver-Burke es
Vmax
1
SVmax
Km
v
1
0 (3)
que describe una recta en donde el intercepto con el eje de las abscisas es igual a ndash1Km y el intercepto con el eje de las ordenadas es igual a 1Vmax
28
Figura experimento cineacutetico para la actividad enzimaacutetica Regraacutefico de
Lineweaver-Burke
Se presentan los valores experimentales y los resultados del ajuste al modelo
REPRESENTACIOacuteN GRAacuteFICA DE LINEWEAVER Y BURK
Si se toma el inverso en ambos miembros de la ecuacioacuten de Michaelis tenemos
=
+
29
que es la ecuacioacuten de una liacutenea recta cuya pendiente es que intercepta al eje de las
ordenadas en y a la abcisa en el punto Lo que es evidente al hacer
ya que se convierte en
Si se hace una graacutefica con los dobles inversos colocando los valores de en el eje de las
ordenadas y en el eje de las abcisas se obtiene una liacutenea recta a partir de la cual se puede calcular faacutecilmente Km
EFECTO DEL pH SOBRE LA ACTIVIDAD ENZIMATICA
Sabiendo que las enzimas son proteinas cualquier cambio brusco de pH puede alterar el caraacutecter ioacutenico de los grupos amino y carboxilo en la superficie proteica afectando asiacute las propiedades cataliacuteticas de una enzima A pH alto o bajo se puede producir la desnaturalizacioacuten
de la enzima y en consecuencia su inactivacioacuten La fosfatasa aacutecida es maacutes activa a pH 50 mientras que la fosfatasa alcalina lo es a pH 90 Muchas enzimas tienen maacutexima actividad cerca de la neutralidad en un rango de pH de 6 a 8
El pH per se no afecta la actividad enzimaacutetica sino la concentracioacuten de protones Los protones
ademaacutes de alterar la estructura de la enzima y el substrato pueden participar tambieacuten en la reaccioacuten como substrato o producto En esos casos la concentracioacuten de protones afecta directamente la velocidad de la reaccioacuten
ENZIMA pH OPTIMO
30
Pepsina 15
Tripsina 77
Catalasa 76
Arginasa 97
Fumarasa 78
Ribonucleasa 78
EFECTO DE ACTIVADORES
Un aumento en la temperatura provoca un aumento de la velocidad de reaccioacuten hasta cierta temperatura oacuteptima ya que despueacutes de aproximadamente 45
0 C se comienza a producir la
desnaturalizacioacuten teacutermica Las enzimas de muchos mamiacuteferos tienen una temperatura oacuteptima
de 370 C por encima de esa temperatura comienzan a inactivarse y se destruyen Sin embargo
existen especies de bacterias y algas que habitan en fuentes de aguas termales y en el otro extremo ciertas bacterias aacuterticas tienen temperaturas oacuteptimas cercanas a 0
0 C
31
ESPECIFICIDAD ABSOLUTA
Cierta enzimas se encuentran como aproenzimas o zimogenos que son inactivos
El tripsinoacutegeno es convertido en tripsina por la misma accioacuten de la tripsina que remueve un hexapeacuteptido
tripsina Tripsinoacutegeno Tripsina + Val (Asp)4 lisina
Otra forma de activacioacuten consiste en mantener los grupos SH reducidos Estos grupos pueden
formar parte de centros activos de ciertas enzimas Si se oxidan estos grupos a -S-S- la enzima se inactiva Asiacute tenemos que la enzima papaina se inactiva despueacutes de exponerla al ox iacutegeno pero cuando se le antildeade un reductor apropiado la enzima se reactiva Los grupos ndashS-S- se convierten en ndashSH activaacutendose la enzima
Otra forma de activacioacuten requiere la presencia de otra substancia que se enlaza al sitio alosteacuterico el compuesto que se enlaza se denomina un efector alosteacuterico El centro alosteacuterico es bien especiacutefico para el efector o modulador alosteacuterico
ESPECIFICIDAD RELATIVA
Oxiacutegeno L-aminoaacutecido a -cetoaacutecido + NH3 + H 2 O 2 L-aminoaacutecido oxidasa Oxiacutegeno D-aminoaacutecido a -cetoaacutecido + NH3 + H 2 O 2 D-aminoaacutecido oxidasa
32
En este ejemplo se muestra la estereo-especificidad Una enzima puede tener una especificidad oacuteptica para isomeros D o L
Hay enzimas que muestran especificidad absoluta como la aspartasa que cataliza la adicioacuten reversible de amoniacuteaco al doble enlace del aacutecido fumaacuterico pero no al de otros aacutecidos insaturados
HOOC ndash CH = CH ndash COO- + NH4
+ HOOC ndash CH2 - CHNH2 - COO
- + H
+
INHIBICIOacuteN ENZIMATICA
Mediante el uso de inhibidores enzimaacuteticos se ha obtenido informacioacuten muy valiosa sobre la conformacioacuten del centro activo de algunas enzimas
a Inhibicioacuten irreversible Este tipo de inhibidores forman un enlace covalente con las enzimas cerca del centro activo Un ejemplo son los gases nerviosos como el
fluorofosfato de diisopropilo (DFP) que forma un complejo con la enzima acetilcolinesterasa Los animales envenenados con este gas quedan paralizados debido a la imposibilidad de transmitir adecuadamente los impulsos nerviosos
Ester di-isopropil fos foacuterico de la enzima (inactivo)
b) Inhibicioacuten competitiva
Es cuando el inhibidor compite con el substrato por la unioacuten con el centro activo de la
enzima Este tipo de inhibicioacuten puede reducirse si se aumenta la concentracioacuten de substrato
-OOC - CH2 - CH2 - COO
- -OOC - CH = CH - COO
- + 2H
+ Succinato Succinato Fumarato
33
Deshidrogenasa COO
- I CH2 Malonato (inhibidor competitivo) I COO
- En la inhibicioacuten competitiva la enzima se combina con el inhibidor para formar un complejo enzima-inhibidor E + I E I en competencia con la reaccioacuten normal E +S ES
La sulfanilamida (bactericida) interfiere con la siacutentesis del aacutecido foacutelico compitiendo con el aacutecido p-aminobenzoico de una forma competitiva
HOOC-sect- NH2 NH2 ndashSO2 - sect- NH2 aacutecido p- amino benzoico sulfanilamida
sect = fenil
a Inhibicioacuten no competitiva
Esta inhibicioacuten se caracteriza por que no se puede revertir el efecto del inhibidor aumentando la concentracioacuten del substrato
Por ejemplo la inhibicioacuten de la deshidrogenasa del gliceraldehido -3-fosfato por la yodo-acetamida
Enzima-SH + I-CH2 CONH2 Enzima-SCH2 CONH2 + IH
La deshidrogenasa del gliceraldehido-3 fosfato puede ser inhibida tambieacuten por el aacutecido iodo-aceacutetico
Enzima-SH + ICH2 COOH Enzima-SCH2 COOH + IH
c) Inhibicioacuten por metales
Ciertos metales como el plomo mercurio y arseacutenico inhiben enzimas que tienen en su centro activo grupos -SH libres
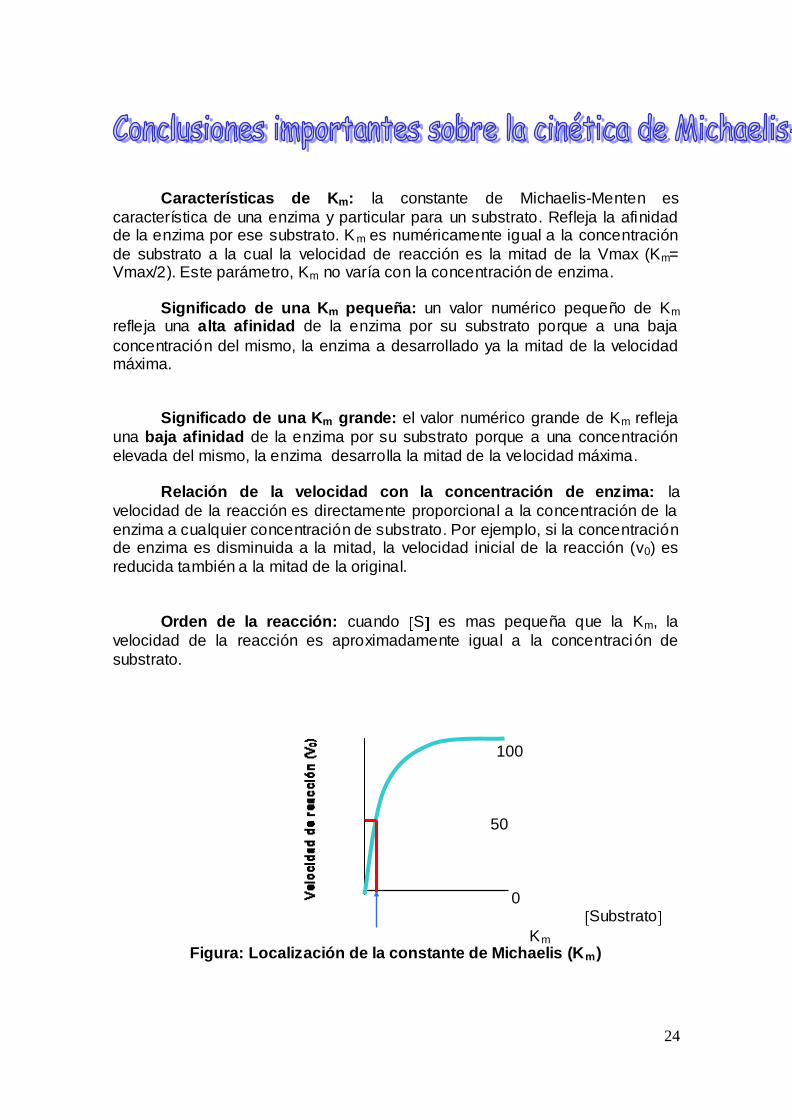
24
Caracteriacutesticas de Km la constante de Michaelis-Menten es
caracteriacutestica de una enzima y particular para un substrato Refleja la afinidad de la enzima por ese substrato K m es numeacutericamente igual a la concentracioacuten
de substrato a la cual la velocidad de reaccioacuten es la mitad de la Vmax (Km= Vmax2) Este paraacutemetro Km no variacutea con la concentracioacuten de enzima
Significado de una Km pequentildea un valor numeacuterico pequentildeo de Km
refleja una alta afinidad de la enzima por su substrato porque a una baja
concentracioacuten del mismo la enzima a desarrollado ya la mitad de la velocidad maacutexima
Significado de una Km grande el valor numeacuterico grande de Km refleja
una baja afinidad de la enzima por su substrato porque a una concentracioacuten
elevada del mismo la enzima desarrolla la mitad de la velocidad maacutexima
Relacioacuten de la velocidad con la concentracioacuten de enzima la
velocidad de la reaccioacuten es directamente proporcional a la concentracioacuten de la
enzima a cualquier concentracioacuten de substrato Por ejemplo si la concentracioacuten de enzima es disminuida a la mitad la velocidad inicial de la reaccioacuten (v0) es
reducida tambieacuten a la mitad de la original
Orden de la reaccioacuten cuando S es mas pequentildea que la Km la
velocidad de la reaccioacuten es aproximadamente igual a la concentracioacuten de
substrato
100
50
0
Substrato
Km Figura Localizacioacuten de la constante de Michaelis (Km)
25
La velocidad de reaccioacuten se dice en estas condiciones es de primer
orden con respecto al substrato Cuando S es mas grande que la Km la
velocidad es constante e igual a la Vmax La velocidad de la reaccioacuten es independiente de la concentracioacuten de substrato y se dice que es de orden cero
con respecto a la concentracioacuten de substrato
Las enzimas pueden ser aisladas de las ceacutelulas u organismos para estudiar sus
propiedades in vitro es decir en un tubo de ensayo Las diferentes enzimas muestran diferentes respuestas a los cambios en la concentracioacuten de substrato temperatura y pH
Concentracioacuten de substrato
La forma hiperboacutelica de la curva de velocidad vs Substrato
Efecto de la temperatura en la actividad enzimaacutetica
Efecto de la variacioacuten del pH en la actividad enzimaacutetica
Incremento de la velocidad con la temperatura En general la velocidad de la reaccioacuten se incrementa con la temperatura hasta que un punto maacuteximo es alcanzado
Este incremento se debe a que aumenta el nuacutemero de moleacuteculas ricas en energiacutea que pueden pasar la barrera energeacutetica de estado de transicioacuten para formar a los productos
Decremento de la velocidad con la temperatura la elevacioacuten excesiva de la
temperatura del medio que contiene a las enzimas resulta en un decremento de la velocidad como resultado de la desnaturalizacioacuten es decir la peacuterdida de la estructura tridimensional de las enzimas
26
zona de reactivacioacuten maacuteximo de actividad
Zona de
inactivacioacuten
0 Temperatura
Figura dependencia de la actividad enzimaacutetica con la temperatura
Velocidad maacutexima la velocidad de una reaccioacuten (v) es el nuacutemero de
moleacuteculas de substrato convertidas en producto por unidad de tiempo y
usualmente se expresa en micromoles de producto formadas por minuto La velocidad de una reaccioacuten catalizada por una enzima aumenta conforme se incrementa la concentracioacuten de substrato hasta que se llega a una velocidad maacutexima (Vmax) a partir de la cual la velocidad de la reaccioacuten es
independiente de la cantidad de substrato El que la velocidad no pueda
seguirse incrementando refleja la saturacioacuten del sitio activo con el substrato
Vmax
0 Substrato
Figura representacioacuten de una cineacutetica tiacutepicamente michaeliana
Vmax = velocidad de saturacioacuten o maacutexima
27
Cuando se grafica la velocidad de la reaccioacuten v0 contra la concentracioacuten de substrato S no siempre es posible determinar la condicioacuten
en que se ha llegado a la velocidad maacutexima Vmax debido al incremento de la pendiente en la hipeacuterbola a concentraciones de substrato elevadas
Figura experimento cineacutetico para la actividad enzimaacutetica
Se presentan los valores experimentales y los resultados del ajuste al modelo de Michaelis
Auacuten asiacute si se grafica una doble reciacuteproca es decir el inverso de la
velocidad (1v0) contra el inverso de la concentracioacuten de substrato (1 S ) se
obtiene una liacutenea recta El regraacutefico de Lineweaver-Burke tambieacuten se
denomina de ldquodobles reciacuteprocasrdquo y se utiliza para calcular la Km y la Vmax asiacute como para determinar el mecanismo de accioacuten de los diversos tipos de
inhibidores
La ecuacioacuten que describe a la graacutefca de Lineweaver-Burke es
Vmax
1
SVmax
Km
v
1
0 (3)
que describe una recta en donde el intercepto con el eje de las abscisas es igual a ndash1Km y el intercepto con el eje de las ordenadas es igual a 1Vmax
28
Figura experimento cineacutetico para la actividad enzimaacutetica Regraacutefico de
Lineweaver-Burke
Se presentan los valores experimentales y los resultados del ajuste al modelo
REPRESENTACIOacuteN GRAacuteFICA DE LINEWEAVER Y BURK
Si se toma el inverso en ambos miembros de la ecuacioacuten de Michaelis tenemos
=
+
29
que es la ecuacioacuten de una liacutenea recta cuya pendiente es que intercepta al eje de las
ordenadas en y a la abcisa en el punto Lo que es evidente al hacer
ya que se convierte en
Si se hace una graacutefica con los dobles inversos colocando los valores de en el eje de las
ordenadas y en el eje de las abcisas se obtiene una liacutenea recta a partir de la cual se puede calcular faacutecilmente Km
EFECTO DEL pH SOBRE LA ACTIVIDAD ENZIMATICA
Sabiendo que las enzimas son proteinas cualquier cambio brusco de pH puede alterar el caraacutecter ioacutenico de los grupos amino y carboxilo en la superficie proteica afectando asiacute las propiedades cataliacuteticas de una enzima A pH alto o bajo se puede producir la desnaturalizacioacuten
de la enzima y en consecuencia su inactivacioacuten La fosfatasa aacutecida es maacutes activa a pH 50 mientras que la fosfatasa alcalina lo es a pH 90 Muchas enzimas tienen maacutexima actividad cerca de la neutralidad en un rango de pH de 6 a 8
El pH per se no afecta la actividad enzimaacutetica sino la concentracioacuten de protones Los protones
ademaacutes de alterar la estructura de la enzima y el substrato pueden participar tambieacuten en la reaccioacuten como substrato o producto En esos casos la concentracioacuten de protones afecta directamente la velocidad de la reaccioacuten
ENZIMA pH OPTIMO
30
Pepsina 15
Tripsina 77
Catalasa 76
Arginasa 97
Fumarasa 78
Ribonucleasa 78
EFECTO DE ACTIVADORES
Un aumento en la temperatura provoca un aumento de la velocidad de reaccioacuten hasta cierta temperatura oacuteptima ya que despueacutes de aproximadamente 45
0 C se comienza a producir la
desnaturalizacioacuten teacutermica Las enzimas de muchos mamiacuteferos tienen una temperatura oacuteptima
de 370 C por encima de esa temperatura comienzan a inactivarse y se destruyen Sin embargo
existen especies de bacterias y algas que habitan en fuentes de aguas termales y en el otro extremo ciertas bacterias aacuterticas tienen temperaturas oacuteptimas cercanas a 0
0 C
31
ESPECIFICIDAD ABSOLUTA
Cierta enzimas se encuentran como aproenzimas o zimogenos que son inactivos
El tripsinoacutegeno es convertido en tripsina por la misma accioacuten de la tripsina que remueve un hexapeacuteptido
tripsina Tripsinoacutegeno Tripsina + Val (Asp)4 lisina
Otra forma de activacioacuten consiste en mantener los grupos SH reducidos Estos grupos pueden
formar parte de centros activos de ciertas enzimas Si se oxidan estos grupos a -S-S- la enzima se inactiva Asiacute tenemos que la enzima papaina se inactiva despueacutes de exponerla al ox iacutegeno pero cuando se le antildeade un reductor apropiado la enzima se reactiva Los grupos ndashS-S- se convierten en ndashSH activaacutendose la enzima
Otra forma de activacioacuten requiere la presencia de otra substancia que se enlaza al sitio alosteacuterico el compuesto que se enlaza se denomina un efector alosteacuterico El centro alosteacuterico es bien especiacutefico para el efector o modulador alosteacuterico
ESPECIFICIDAD RELATIVA
Oxiacutegeno L-aminoaacutecido a -cetoaacutecido + NH3 + H 2 O 2 L-aminoaacutecido oxidasa Oxiacutegeno D-aminoaacutecido a -cetoaacutecido + NH3 + H 2 O 2 D-aminoaacutecido oxidasa
32
En este ejemplo se muestra la estereo-especificidad Una enzima puede tener una especificidad oacuteptica para isomeros D o L
Hay enzimas que muestran especificidad absoluta como la aspartasa que cataliza la adicioacuten reversible de amoniacuteaco al doble enlace del aacutecido fumaacuterico pero no al de otros aacutecidos insaturados
HOOC ndash CH = CH ndash COO- + NH4
+ HOOC ndash CH2 - CHNH2 - COO
- + H
+
INHIBICIOacuteN ENZIMATICA
Mediante el uso de inhibidores enzimaacuteticos se ha obtenido informacioacuten muy valiosa sobre la conformacioacuten del centro activo de algunas enzimas
a Inhibicioacuten irreversible Este tipo de inhibidores forman un enlace covalente con las enzimas cerca del centro activo Un ejemplo son los gases nerviosos como el
fluorofosfato de diisopropilo (DFP) que forma un complejo con la enzima acetilcolinesterasa Los animales envenenados con este gas quedan paralizados debido a la imposibilidad de transmitir adecuadamente los impulsos nerviosos
Ester di-isopropil fos foacuterico de la enzima (inactivo)
b) Inhibicioacuten competitiva
Es cuando el inhibidor compite con el substrato por la unioacuten con el centro activo de la
enzima Este tipo de inhibicioacuten puede reducirse si se aumenta la concentracioacuten de substrato
-OOC - CH2 - CH2 - COO
- -OOC - CH = CH - COO
- + 2H
+ Succinato Succinato Fumarato
33
Deshidrogenasa COO
- I CH2 Malonato (inhibidor competitivo) I COO
- En la inhibicioacuten competitiva la enzima se combina con el inhibidor para formar un complejo enzima-inhibidor E + I E I en competencia con la reaccioacuten normal E +S ES
La sulfanilamida (bactericida) interfiere con la siacutentesis del aacutecido foacutelico compitiendo con el aacutecido p-aminobenzoico de una forma competitiva
HOOC-sect- NH2 NH2 ndashSO2 - sect- NH2 aacutecido p- amino benzoico sulfanilamida
sect = fenil
a Inhibicioacuten no competitiva
Esta inhibicioacuten se caracteriza por que no se puede revertir el efecto del inhibidor aumentando la concentracioacuten del substrato
Por ejemplo la inhibicioacuten de la deshidrogenasa del gliceraldehido -3-fosfato por la yodo-acetamida
Enzima-SH + I-CH2 CONH2 Enzima-SCH2 CONH2 + IH
La deshidrogenasa del gliceraldehido-3 fosfato puede ser inhibida tambieacuten por el aacutecido iodo-aceacutetico
Enzima-SH + ICH2 COOH Enzima-SCH2 COOH + IH
c) Inhibicioacuten por metales
Ciertos metales como el plomo mercurio y arseacutenico inhiben enzimas que tienen en su centro activo grupos -SH libres

25
La velocidad de reaccioacuten se dice en estas condiciones es de primer
orden con respecto al substrato Cuando S es mas grande que la Km la
velocidad es constante e igual a la Vmax La velocidad de la reaccioacuten es independiente de la concentracioacuten de substrato y se dice que es de orden cero
con respecto a la concentracioacuten de substrato
Las enzimas pueden ser aisladas de las ceacutelulas u organismos para estudiar sus
propiedades in vitro es decir en un tubo de ensayo Las diferentes enzimas muestran diferentes respuestas a los cambios en la concentracioacuten de substrato temperatura y pH
Concentracioacuten de substrato
La forma hiperboacutelica de la curva de velocidad vs Substrato
Efecto de la temperatura en la actividad enzimaacutetica
Efecto de la variacioacuten del pH en la actividad enzimaacutetica
Incremento de la velocidad con la temperatura En general la velocidad de la reaccioacuten se incrementa con la temperatura hasta que un punto maacuteximo es alcanzado
Este incremento se debe a que aumenta el nuacutemero de moleacuteculas ricas en energiacutea que pueden pasar la barrera energeacutetica de estado de transicioacuten para formar a los productos
Decremento de la velocidad con la temperatura la elevacioacuten excesiva de la
temperatura del medio que contiene a las enzimas resulta en un decremento de la velocidad como resultado de la desnaturalizacioacuten es decir la peacuterdida de la estructura tridimensional de las enzimas
26
zona de reactivacioacuten maacuteximo de actividad
Zona de
inactivacioacuten
0 Temperatura
Figura dependencia de la actividad enzimaacutetica con la temperatura
Velocidad maacutexima la velocidad de una reaccioacuten (v) es el nuacutemero de
moleacuteculas de substrato convertidas en producto por unidad de tiempo y
usualmente se expresa en micromoles de producto formadas por minuto La velocidad de una reaccioacuten catalizada por una enzima aumenta conforme se incrementa la concentracioacuten de substrato hasta que se llega a una velocidad maacutexima (Vmax) a partir de la cual la velocidad de la reaccioacuten es
independiente de la cantidad de substrato El que la velocidad no pueda
seguirse incrementando refleja la saturacioacuten del sitio activo con el substrato
Vmax
0 Substrato
Figura representacioacuten de una cineacutetica tiacutepicamente michaeliana
Vmax = velocidad de saturacioacuten o maacutexima
27
Cuando se grafica la velocidad de la reaccioacuten v0 contra la concentracioacuten de substrato S no siempre es posible determinar la condicioacuten
en que se ha llegado a la velocidad maacutexima Vmax debido al incremento de la pendiente en la hipeacuterbola a concentraciones de substrato elevadas
Figura experimento cineacutetico para la actividad enzimaacutetica
Se presentan los valores experimentales y los resultados del ajuste al modelo de Michaelis
Auacuten asiacute si se grafica una doble reciacuteproca es decir el inverso de la
velocidad (1v0) contra el inverso de la concentracioacuten de substrato (1 S ) se
obtiene una liacutenea recta El regraacutefico de Lineweaver-Burke tambieacuten se
denomina de ldquodobles reciacuteprocasrdquo y se utiliza para calcular la Km y la Vmax asiacute como para determinar el mecanismo de accioacuten de los diversos tipos de
inhibidores
La ecuacioacuten que describe a la graacutefca de Lineweaver-Burke es
Vmax
1
SVmax
Km
v
1
0 (3)
que describe una recta en donde el intercepto con el eje de las abscisas es igual a ndash1Km y el intercepto con el eje de las ordenadas es igual a 1Vmax
28
Figura experimento cineacutetico para la actividad enzimaacutetica Regraacutefico de
Lineweaver-Burke
Se presentan los valores experimentales y los resultados del ajuste al modelo
REPRESENTACIOacuteN GRAacuteFICA DE LINEWEAVER Y BURK
Si se toma el inverso en ambos miembros de la ecuacioacuten de Michaelis tenemos
=
+
29
que es la ecuacioacuten de una liacutenea recta cuya pendiente es que intercepta al eje de las
ordenadas en y a la abcisa en el punto Lo que es evidente al hacer
ya que se convierte en
Si se hace una graacutefica con los dobles inversos colocando los valores de en el eje de las
ordenadas y en el eje de las abcisas se obtiene una liacutenea recta a partir de la cual se puede calcular faacutecilmente Km
EFECTO DEL pH SOBRE LA ACTIVIDAD ENZIMATICA
Sabiendo que las enzimas son proteinas cualquier cambio brusco de pH puede alterar el caraacutecter ioacutenico de los grupos amino y carboxilo en la superficie proteica afectando asiacute las propiedades cataliacuteticas de una enzima A pH alto o bajo se puede producir la desnaturalizacioacuten
de la enzima y en consecuencia su inactivacioacuten La fosfatasa aacutecida es maacutes activa a pH 50 mientras que la fosfatasa alcalina lo es a pH 90 Muchas enzimas tienen maacutexima actividad cerca de la neutralidad en un rango de pH de 6 a 8
El pH per se no afecta la actividad enzimaacutetica sino la concentracioacuten de protones Los protones
ademaacutes de alterar la estructura de la enzima y el substrato pueden participar tambieacuten en la reaccioacuten como substrato o producto En esos casos la concentracioacuten de protones afecta directamente la velocidad de la reaccioacuten
ENZIMA pH OPTIMO
30
Pepsina 15
Tripsina 77
Catalasa 76
Arginasa 97
Fumarasa 78
Ribonucleasa 78
EFECTO DE ACTIVADORES
Un aumento en la temperatura provoca un aumento de la velocidad de reaccioacuten hasta cierta temperatura oacuteptima ya que despueacutes de aproximadamente 45
0 C se comienza a producir la
desnaturalizacioacuten teacutermica Las enzimas de muchos mamiacuteferos tienen una temperatura oacuteptima
de 370 C por encima de esa temperatura comienzan a inactivarse y se destruyen Sin embargo
existen especies de bacterias y algas que habitan en fuentes de aguas termales y en el otro extremo ciertas bacterias aacuterticas tienen temperaturas oacuteptimas cercanas a 0
0 C
31
ESPECIFICIDAD ABSOLUTA
Cierta enzimas se encuentran como aproenzimas o zimogenos que son inactivos
El tripsinoacutegeno es convertido en tripsina por la misma accioacuten de la tripsina que remueve un hexapeacuteptido
tripsina Tripsinoacutegeno Tripsina + Val (Asp)4 lisina
Otra forma de activacioacuten consiste en mantener los grupos SH reducidos Estos grupos pueden
formar parte de centros activos de ciertas enzimas Si se oxidan estos grupos a -S-S- la enzima se inactiva Asiacute tenemos que la enzima papaina se inactiva despueacutes de exponerla al ox iacutegeno pero cuando se le antildeade un reductor apropiado la enzima se reactiva Los grupos ndashS-S- se convierten en ndashSH activaacutendose la enzima
Otra forma de activacioacuten requiere la presencia de otra substancia que se enlaza al sitio alosteacuterico el compuesto que se enlaza se denomina un efector alosteacuterico El centro alosteacuterico es bien especiacutefico para el efector o modulador alosteacuterico
ESPECIFICIDAD RELATIVA
Oxiacutegeno L-aminoaacutecido a -cetoaacutecido + NH3 + H 2 O 2 L-aminoaacutecido oxidasa Oxiacutegeno D-aminoaacutecido a -cetoaacutecido + NH3 + H 2 O 2 D-aminoaacutecido oxidasa
32
En este ejemplo se muestra la estereo-especificidad Una enzima puede tener una especificidad oacuteptica para isomeros D o L
Hay enzimas que muestran especificidad absoluta como la aspartasa que cataliza la adicioacuten reversible de amoniacuteaco al doble enlace del aacutecido fumaacuterico pero no al de otros aacutecidos insaturados
HOOC ndash CH = CH ndash COO- + NH4
+ HOOC ndash CH2 - CHNH2 - COO
- + H
+
INHIBICIOacuteN ENZIMATICA
Mediante el uso de inhibidores enzimaacuteticos se ha obtenido informacioacuten muy valiosa sobre la conformacioacuten del centro activo de algunas enzimas
a Inhibicioacuten irreversible Este tipo de inhibidores forman un enlace covalente con las enzimas cerca del centro activo Un ejemplo son los gases nerviosos como el
fluorofosfato de diisopropilo (DFP) que forma un complejo con la enzima acetilcolinesterasa Los animales envenenados con este gas quedan paralizados debido a la imposibilidad de transmitir adecuadamente los impulsos nerviosos
Ester di-isopropil fos foacuterico de la enzima (inactivo)
b) Inhibicioacuten competitiva
Es cuando el inhibidor compite con el substrato por la unioacuten con el centro activo de la
enzima Este tipo de inhibicioacuten puede reducirse si se aumenta la concentracioacuten de substrato
-OOC - CH2 - CH2 - COO
- -OOC - CH = CH - COO
- + 2H
+ Succinato Succinato Fumarato
33
Deshidrogenasa COO
- I CH2 Malonato (inhibidor competitivo) I COO
- En la inhibicioacuten competitiva la enzima se combina con el inhibidor para formar un complejo enzima-inhibidor E + I E I en competencia con la reaccioacuten normal E +S ES
La sulfanilamida (bactericida) interfiere con la siacutentesis del aacutecido foacutelico compitiendo con el aacutecido p-aminobenzoico de una forma competitiva
HOOC-sect- NH2 NH2 ndashSO2 - sect- NH2 aacutecido p- amino benzoico sulfanilamida
sect = fenil
a Inhibicioacuten no competitiva
Esta inhibicioacuten se caracteriza por que no se puede revertir el efecto del inhibidor aumentando la concentracioacuten del substrato
Por ejemplo la inhibicioacuten de la deshidrogenasa del gliceraldehido -3-fosfato por la yodo-acetamida
Enzima-SH + I-CH2 CONH2 Enzima-SCH2 CONH2 + IH
La deshidrogenasa del gliceraldehido-3 fosfato puede ser inhibida tambieacuten por el aacutecido iodo-aceacutetico
Enzima-SH + ICH2 COOH Enzima-SCH2 COOH + IH
c) Inhibicioacuten por metales
Ciertos metales como el plomo mercurio y arseacutenico inhiben enzimas que tienen en su centro activo grupos -SH libres
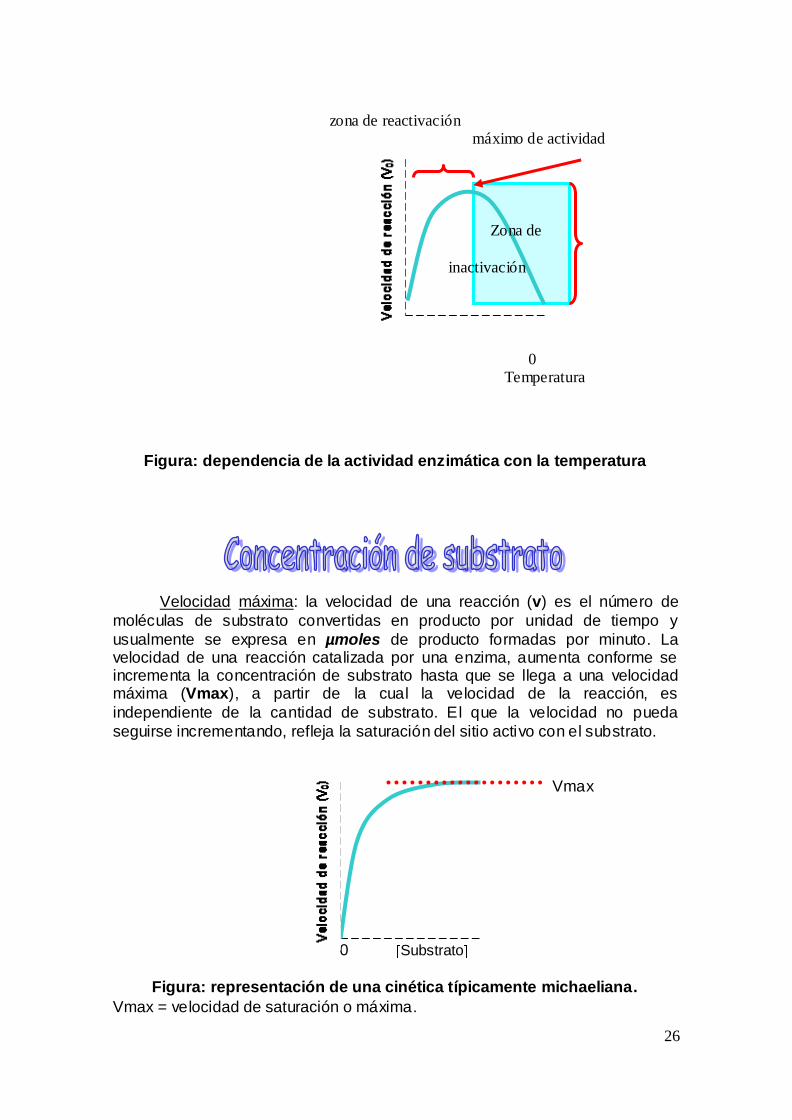
26
zona de reactivacioacuten maacuteximo de actividad
Zona de
inactivacioacuten
0 Temperatura
Figura dependencia de la actividad enzimaacutetica con la temperatura
Velocidad maacutexima la velocidad de una reaccioacuten (v) es el nuacutemero de
moleacuteculas de substrato convertidas en producto por unidad de tiempo y
usualmente se expresa en micromoles de producto formadas por minuto La velocidad de una reaccioacuten catalizada por una enzima aumenta conforme se incrementa la concentracioacuten de substrato hasta que se llega a una velocidad maacutexima (Vmax) a partir de la cual la velocidad de la reaccioacuten es
independiente de la cantidad de substrato El que la velocidad no pueda
seguirse incrementando refleja la saturacioacuten del sitio activo con el substrato
Vmax
0 Substrato
Figura representacioacuten de una cineacutetica tiacutepicamente michaeliana
Vmax = velocidad de saturacioacuten o maacutexima
27
Cuando se grafica la velocidad de la reaccioacuten v0 contra la concentracioacuten de substrato S no siempre es posible determinar la condicioacuten
en que se ha llegado a la velocidad maacutexima Vmax debido al incremento de la pendiente en la hipeacuterbola a concentraciones de substrato elevadas
Figura experimento cineacutetico para la actividad enzimaacutetica
Se presentan los valores experimentales y los resultados del ajuste al modelo de Michaelis
Auacuten asiacute si se grafica una doble reciacuteproca es decir el inverso de la
velocidad (1v0) contra el inverso de la concentracioacuten de substrato (1 S ) se
obtiene una liacutenea recta El regraacutefico de Lineweaver-Burke tambieacuten se
denomina de ldquodobles reciacuteprocasrdquo y se utiliza para calcular la Km y la Vmax asiacute como para determinar el mecanismo de accioacuten de los diversos tipos de
inhibidores
La ecuacioacuten que describe a la graacutefca de Lineweaver-Burke es
Vmax
1
SVmax
Km
v
1
0 (3)
que describe una recta en donde el intercepto con el eje de las abscisas es igual a ndash1Km y el intercepto con el eje de las ordenadas es igual a 1Vmax
28
Figura experimento cineacutetico para la actividad enzimaacutetica Regraacutefico de
Lineweaver-Burke
Se presentan los valores experimentales y los resultados del ajuste al modelo
REPRESENTACIOacuteN GRAacuteFICA DE LINEWEAVER Y BURK
Si se toma el inverso en ambos miembros de la ecuacioacuten de Michaelis tenemos
=
+
29
que es la ecuacioacuten de una liacutenea recta cuya pendiente es que intercepta al eje de las
ordenadas en y a la abcisa en el punto Lo que es evidente al hacer
ya que se convierte en
Si se hace una graacutefica con los dobles inversos colocando los valores de en el eje de las
ordenadas y en el eje de las abcisas se obtiene una liacutenea recta a partir de la cual se puede calcular faacutecilmente Km
EFECTO DEL pH SOBRE LA ACTIVIDAD ENZIMATICA
Sabiendo que las enzimas son proteinas cualquier cambio brusco de pH puede alterar el caraacutecter ioacutenico de los grupos amino y carboxilo en la superficie proteica afectando asiacute las propiedades cataliacuteticas de una enzima A pH alto o bajo se puede producir la desnaturalizacioacuten
de la enzima y en consecuencia su inactivacioacuten La fosfatasa aacutecida es maacutes activa a pH 50 mientras que la fosfatasa alcalina lo es a pH 90 Muchas enzimas tienen maacutexima actividad cerca de la neutralidad en un rango de pH de 6 a 8
El pH per se no afecta la actividad enzimaacutetica sino la concentracioacuten de protones Los protones
ademaacutes de alterar la estructura de la enzima y el substrato pueden participar tambieacuten en la reaccioacuten como substrato o producto En esos casos la concentracioacuten de protones afecta directamente la velocidad de la reaccioacuten
ENZIMA pH OPTIMO
30
Pepsina 15
Tripsina 77
Catalasa 76
Arginasa 97
Fumarasa 78
Ribonucleasa 78
EFECTO DE ACTIVADORES
Un aumento en la temperatura provoca un aumento de la velocidad de reaccioacuten hasta cierta temperatura oacuteptima ya que despueacutes de aproximadamente 45
0 C se comienza a producir la
desnaturalizacioacuten teacutermica Las enzimas de muchos mamiacuteferos tienen una temperatura oacuteptima
de 370 C por encima de esa temperatura comienzan a inactivarse y se destruyen Sin embargo
existen especies de bacterias y algas que habitan en fuentes de aguas termales y en el otro extremo ciertas bacterias aacuterticas tienen temperaturas oacuteptimas cercanas a 0
0 C
31
ESPECIFICIDAD ABSOLUTA
Cierta enzimas se encuentran como aproenzimas o zimogenos que son inactivos
El tripsinoacutegeno es convertido en tripsina por la misma accioacuten de la tripsina que remueve un hexapeacuteptido
tripsina Tripsinoacutegeno Tripsina + Val (Asp)4 lisina
Otra forma de activacioacuten consiste en mantener los grupos SH reducidos Estos grupos pueden
formar parte de centros activos de ciertas enzimas Si se oxidan estos grupos a -S-S- la enzima se inactiva Asiacute tenemos que la enzima papaina se inactiva despueacutes de exponerla al ox iacutegeno pero cuando se le antildeade un reductor apropiado la enzima se reactiva Los grupos ndashS-S- se convierten en ndashSH activaacutendose la enzima
Otra forma de activacioacuten requiere la presencia de otra substancia que se enlaza al sitio alosteacuterico el compuesto que se enlaza se denomina un efector alosteacuterico El centro alosteacuterico es bien especiacutefico para el efector o modulador alosteacuterico
ESPECIFICIDAD RELATIVA
Oxiacutegeno L-aminoaacutecido a -cetoaacutecido + NH3 + H 2 O 2 L-aminoaacutecido oxidasa Oxiacutegeno D-aminoaacutecido a -cetoaacutecido + NH3 + H 2 O 2 D-aminoaacutecido oxidasa
32
En este ejemplo se muestra la estereo-especificidad Una enzima puede tener una especificidad oacuteptica para isomeros D o L
Hay enzimas que muestran especificidad absoluta como la aspartasa que cataliza la adicioacuten reversible de amoniacuteaco al doble enlace del aacutecido fumaacuterico pero no al de otros aacutecidos insaturados
HOOC ndash CH = CH ndash COO- + NH4
+ HOOC ndash CH2 - CHNH2 - COO
- + H
+
INHIBICIOacuteN ENZIMATICA
Mediante el uso de inhibidores enzimaacuteticos se ha obtenido informacioacuten muy valiosa sobre la conformacioacuten del centro activo de algunas enzimas
a Inhibicioacuten irreversible Este tipo de inhibidores forman un enlace covalente con las enzimas cerca del centro activo Un ejemplo son los gases nerviosos como el
fluorofosfato de diisopropilo (DFP) que forma un complejo con la enzima acetilcolinesterasa Los animales envenenados con este gas quedan paralizados debido a la imposibilidad de transmitir adecuadamente los impulsos nerviosos
Ester di-isopropil fos foacuterico de la enzima (inactivo)
b) Inhibicioacuten competitiva
Es cuando el inhibidor compite con el substrato por la unioacuten con el centro activo de la
enzima Este tipo de inhibicioacuten puede reducirse si se aumenta la concentracioacuten de substrato
-OOC - CH2 - CH2 - COO
- -OOC - CH = CH - COO
- + 2H
+ Succinato Succinato Fumarato
33
Deshidrogenasa COO
- I CH2 Malonato (inhibidor competitivo) I COO
- En la inhibicioacuten competitiva la enzima se combina con el inhibidor para formar un complejo enzima-inhibidor E + I E I en competencia con la reaccioacuten normal E +S ES
La sulfanilamida (bactericida) interfiere con la siacutentesis del aacutecido foacutelico compitiendo con el aacutecido p-aminobenzoico de una forma competitiva
HOOC-sect- NH2 NH2 ndashSO2 - sect- NH2 aacutecido p- amino benzoico sulfanilamida
sect = fenil
a Inhibicioacuten no competitiva
Esta inhibicioacuten se caracteriza por que no se puede revertir el efecto del inhibidor aumentando la concentracioacuten del substrato
Por ejemplo la inhibicioacuten de la deshidrogenasa del gliceraldehido -3-fosfato por la yodo-acetamida
Enzima-SH + I-CH2 CONH2 Enzima-SCH2 CONH2 + IH
La deshidrogenasa del gliceraldehido-3 fosfato puede ser inhibida tambieacuten por el aacutecido iodo-aceacutetico
Enzima-SH + ICH2 COOH Enzima-SCH2 COOH + IH
c) Inhibicioacuten por metales
Ciertos metales como el plomo mercurio y arseacutenico inhiben enzimas que tienen en su centro activo grupos -SH libres

27
Cuando se grafica la velocidad de la reaccioacuten v0 contra la concentracioacuten de substrato S no siempre es posible determinar la condicioacuten
en que se ha llegado a la velocidad maacutexima Vmax debido al incremento de la pendiente en la hipeacuterbola a concentraciones de substrato elevadas
Figura experimento cineacutetico para la actividad enzimaacutetica
Se presentan los valores experimentales y los resultados del ajuste al modelo de Michaelis
Auacuten asiacute si se grafica una doble reciacuteproca es decir el inverso de la
velocidad (1v0) contra el inverso de la concentracioacuten de substrato (1 S ) se
obtiene una liacutenea recta El regraacutefico de Lineweaver-Burke tambieacuten se
denomina de ldquodobles reciacuteprocasrdquo y se utiliza para calcular la Km y la Vmax asiacute como para determinar el mecanismo de accioacuten de los diversos tipos de
inhibidores
La ecuacioacuten que describe a la graacutefca de Lineweaver-Burke es
Vmax
1
SVmax
Km
v
1
0 (3)
que describe una recta en donde el intercepto con el eje de las abscisas es igual a ndash1Km y el intercepto con el eje de las ordenadas es igual a 1Vmax
28
Figura experimento cineacutetico para la actividad enzimaacutetica Regraacutefico de
Lineweaver-Burke
Se presentan los valores experimentales y los resultados del ajuste al modelo
REPRESENTACIOacuteN GRAacuteFICA DE LINEWEAVER Y BURK
Si se toma el inverso en ambos miembros de la ecuacioacuten de Michaelis tenemos
=
+
29
que es la ecuacioacuten de una liacutenea recta cuya pendiente es que intercepta al eje de las
ordenadas en y a la abcisa en el punto Lo que es evidente al hacer
ya que se convierte en
Si se hace una graacutefica con los dobles inversos colocando los valores de en el eje de las
ordenadas y en el eje de las abcisas se obtiene una liacutenea recta a partir de la cual se puede calcular faacutecilmente Km
EFECTO DEL pH SOBRE LA ACTIVIDAD ENZIMATICA
Sabiendo que las enzimas son proteinas cualquier cambio brusco de pH puede alterar el caraacutecter ioacutenico de los grupos amino y carboxilo en la superficie proteica afectando asiacute las propiedades cataliacuteticas de una enzima A pH alto o bajo se puede producir la desnaturalizacioacuten
de la enzima y en consecuencia su inactivacioacuten La fosfatasa aacutecida es maacutes activa a pH 50 mientras que la fosfatasa alcalina lo es a pH 90 Muchas enzimas tienen maacutexima actividad cerca de la neutralidad en un rango de pH de 6 a 8
El pH per se no afecta la actividad enzimaacutetica sino la concentracioacuten de protones Los protones
ademaacutes de alterar la estructura de la enzima y el substrato pueden participar tambieacuten en la reaccioacuten como substrato o producto En esos casos la concentracioacuten de protones afecta directamente la velocidad de la reaccioacuten
ENZIMA pH OPTIMO
30
Pepsina 15
Tripsina 77
Catalasa 76
Arginasa 97
Fumarasa 78
Ribonucleasa 78
EFECTO DE ACTIVADORES
Un aumento en la temperatura provoca un aumento de la velocidad de reaccioacuten hasta cierta temperatura oacuteptima ya que despueacutes de aproximadamente 45
0 C se comienza a producir la
desnaturalizacioacuten teacutermica Las enzimas de muchos mamiacuteferos tienen una temperatura oacuteptima
de 370 C por encima de esa temperatura comienzan a inactivarse y se destruyen Sin embargo
existen especies de bacterias y algas que habitan en fuentes de aguas termales y en el otro extremo ciertas bacterias aacuterticas tienen temperaturas oacuteptimas cercanas a 0
0 C
31
ESPECIFICIDAD ABSOLUTA
Cierta enzimas se encuentran como aproenzimas o zimogenos que son inactivos
El tripsinoacutegeno es convertido en tripsina por la misma accioacuten de la tripsina que remueve un hexapeacuteptido
tripsina Tripsinoacutegeno Tripsina + Val (Asp)4 lisina
Otra forma de activacioacuten consiste en mantener los grupos SH reducidos Estos grupos pueden
formar parte de centros activos de ciertas enzimas Si se oxidan estos grupos a -S-S- la enzima se inactiva Asiacute tenemos que la enzima papaina se inactiva despueacutes de exponerla al ox iacutegeno pero cuando se le antildeade un reductor apropiado la enzima se reactiva Los grupos ndashS-S- se convierten en ndashSH activaacutendose la enzima
Otra forma de activacioacuten requiere la presencia de otra substancia que se enlaza al sitio alosteacuterico el compuesto que se enlaza se denomina un efector alosteacuterico El centro alosteacuterico es bien especiacutefico para el efector o modulador alosteacuterico
ESPECIFICIDAD RELATIVA
Oxiacutegeno L-aminoaacutecido a -cetoaacutecido + NH3 + H 2 O 2 L-aminoaacutecido oxidasa Oxiacutegeno D-aminoaacutecido a -cetoaacutecido + NH3 + H 2 O 2 D-aminoaacutecido oxidasa
32
En este ejemplo se muestra la estereo-especificidad Una enzima puede tener una especificidad oacuteptica para isomeros D o L
Hay enzimas que muestran especificidad absoluta como la aspartasa que cataliza la adicioacuten reversible de amoniacuteaco al doble enlace del aacutecido fumaacuterico pero no al de otros aacutecidos insaturados
HOOC ndash CH = CH ndash COO- + NH4
+ HOOC ndash CH2 - CHNH2 - COO
- + H
+
INHIBICIOacuteN ENZIMATICA
Mediante el uso de inhibidores enzimaacuteticos se ha obtenido informacioacuten muy valiosa sobre la conformacioacuten del centro activo de algunas enzimas
a Inhibicioacuten irreversible Este tipo de inhibidores forman un enlace covalente con las enzimas cerca del centro activo Un ejemplo son los gases nerviosos como el
fluorofosfato de diisopropilo (DFP) que forma un complejo con la enzima acetilcolinesterasa Los animales envenenados con este gas quedan paralizados debido a la imposibilidad de transmitir adecuadamente los impulsos nerviosos
Ester di-isopropil fos foacuterico de la enzima (inactivo)
b) Inhibicioacuten competitiva
Es cuando el inhibidor compite con el substrato por la unioacuten con el centro activo de la
enzima Este tipo de inhibicioacuten puede reducirse si se aumenta la concentracioacuten de substrato
-OOC - CH2 - CH2 - COO
- -OOC - CH = CH - COO
- + 2H
+ Succinato Succinato Fumarato
33
Deshidrogenasa COO
- I CH2 Malonato (inhibidor competitivo) I COO
- En la inhibicioacuten competitiva la enzima se combina con el inhibidor para formar un complejo enzima-inhibidor E + I E I en competencia con la reaccioacuten normal E +S ES
La sulfanilamida (bactericida) interfiere con la siacutentesis del aacutecido foacutelico compitiendo con el aacutecido p-aminobenzoico de una forma competitiva
HOOC-sect- NH2 NH2 ndashSO2 - sect- NH2 aacutecido p- amino benzoico sulfanilamida
sect = fenil
a Inhibicioacuten no competitiva
Esta inhibicioacuten se caracteriza por que no se puede revertir el efecto del inhibidor aumentando la concentracioacuten del substrato
Por ejemplo la inhibicioacuten de la deshidrogenasa del gliceraldehido -3-fosfato por la yodo-acetamida
Enzima-SH + I-CH2 CONH2 Enzima-SCH2 CONH2 + IH
La deshidrogenasa del gliceraldehido-3 fosfato puede ser inhibida tambieacuten por el aacutecido iodo-aceacutetico
Enzima-SH + ICH2 COOH Enzima-SCH2 COOH + IH
c) Inhibicioacuten por metales
Ciertos metales como el plomo mercurio y arseacutenico inhiben enzimas que tienen en su centro activo grupos -SH libres
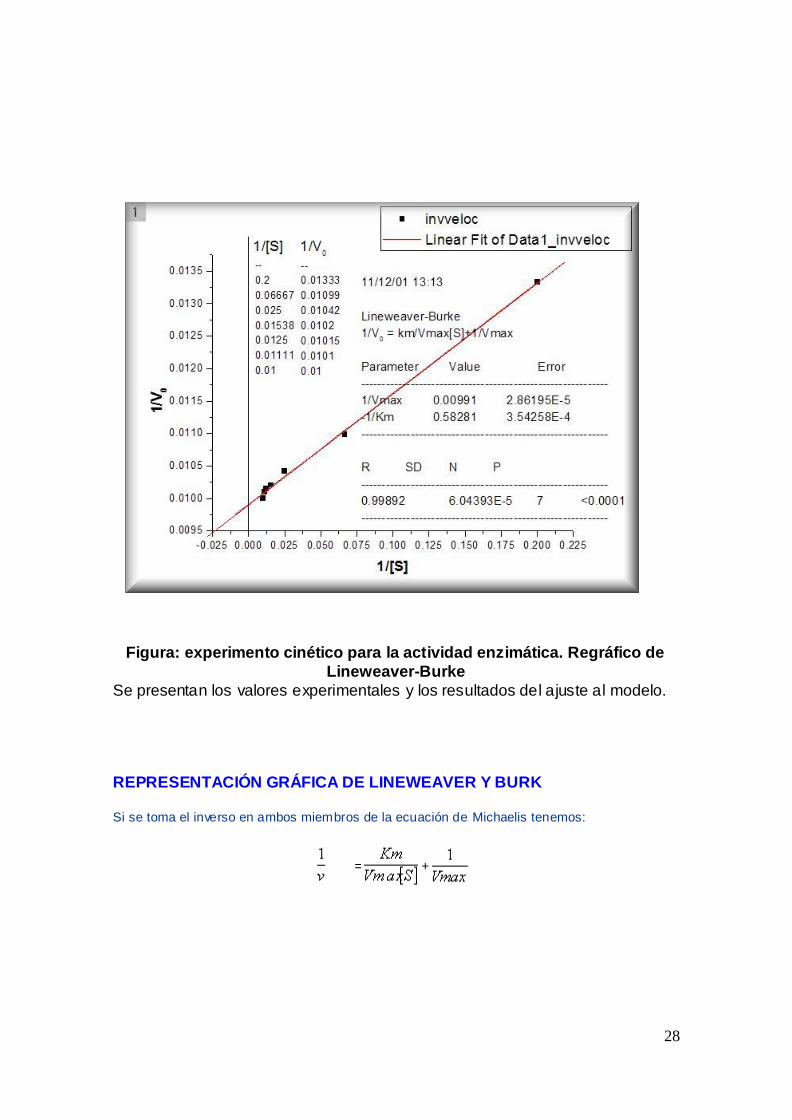
28
Figura experimento cineacutetico para la actividad enzimaacutetica Regraacutefico de
Lineweaver-Burke
Se presentan los valores experimentales y los resultados del ajuste al modelo
REPRESENTACIOacuteN GRAacuteFICA DE LINEWEAVER Y BURK
Si se toma el inverso en ambos miembros de la ecuacioacuten de Michaelis tenemos
=
+
29
que es la ecuacioacuten de una liacutenea recta cuya pendiente es que intercepta al eje de las
ordenadas en y a la abcisa en el punto Lo que es evidente al hacer
ya que se convierte en
Si se hace una graacutefica con los dobles inversos colocando los valores de en el eje de las
ordenadas y en el eje de las abcisas se obtiene una liacutenea recta a partir de la cual se puede calcular faacutecilmente Km
EFECTO DEL pH SOBRE LA ACTIVIDAD ENZIMATICA
Sabiendo que las enzimas son proteinas cualquier cambio brusco de pH puede alterar el caraacutecter ioacutenico de los grupos amino y carboxilo en la superficie proteica afectando asiacute las propiedades cataliacuteticas de una enzima A pH alto o bajo se puede producir la desnaturalizacioacuten
de la enzima y en consecuencia su inactivacioacuten La fosfatasa aacutecida es maacutes activa a pH 50 mientras que la fosfatasa alcalina lo es a pH 90 Muchas enzimas tienen maacutexima actividad cerca de la neutralidad en un rango de pH de 6 a 8
El pH per se no afecta la actividad enzimaacutetica sino la concentracioacuten de protones Los protones
ademaacutes de alterar la estructura de la enzima y el substrato pueden participar tambieacuten en la reaccioacuten como substrato o producto En esos casos la concentracioacuten de protones afecta directamente la velocidad de la reaccioacuten
ENZIMA pH OPTIMO
30
Pepsina 15
Tripsina 77
Catalasa 76
Arginasa 97
Fumarasa 78
Ribonucleasa 78
EFECTO DE ACTIVADORES
Un aumento en la temperatura provoca un aumento de la velocidad de reaccioacuten hasta cierta temperatura oacuteptima ya que despueacutes de aproximadamente 45
0 C se comienza a producir la
desnaturalizacioacuten teacutermica Las enzimas de muchos mamiacuteferos tienen una temperatura oacuteptima
de 370 C por encima de esa temperatura comienzan a inactivarse y se destruyen Sin embargo
existen especies de bacterias y algas que habitan en fuentes de aguas termales y en el otro extremo ciertas bacterias aacuterticas tienen temperaturas oacuteptimas cercanas a 0
0 C
31
ESPECIFICIDAD ABSOLUTA
Cierta enzimas se encuentran como aproenzimas o zimogenos que son inactivos
El tripsinoacutegeno es convertido en tripsina por la misma accioacuten de la tripsina que remueve un hexapeacuteptido
tripsina Tripsinoacutegeno Tripsina + Val (Asp)4 lisina
Otra forma de activacioacuten consiste en mantener los grupos SH reducidos Estos grupos pueden
formar parte de centros activos de ciertas enzimas Si se oxidan estos grupos a -S-S- la enzima se inactiva Asiacute tenemos que la enzima papaina se inactiva despueacutes de exponerla al ox iacutegeno pero cuando se le antildeade un reductor apropiado la enzima se reactiva Los grupos ndashS-S- se convierten en ndashSH activaacutendose la enzima
Otra forma de activacioacuten requiere la presencia de otra substancia que se enlaza al sitio alosteacuterico el compuesto que se enlaza se denomina un efector alosteacuterico El centro alosteacuterico es bien especiacutefico para el efector o modulador alosteacuterico
ESPECIFICIDAD RELATIVA
Oxiacutegeno L-aminoaacutecido a -cetoaacutecido + NH3 + H 2 O 2 L-aminoaacutecido oxidasa Oxiacutegeno D-aminoaacutecido a -cetoaacutecido + NH3 + H 2 O 2 D-aminoaacutecido oxidasa
32
En este ejemplo se muestra la estereo-especificidad Una enzima puede tener una especificidad oacuteptica para isomeros D o L
Hay enzimas que muestran especificidad absoluta como la aspartasa que cataliza la adicioacuten reversible de amoniacuteaco al doble enlace del aacutecido fumaacuterico pero no al de otros aacutecidos insaturados
HOOC ndash CH = CH ndash COO- + NH4
+ HOOC ndash CH2 - CHNH2 - COO
- + H
+
INHIBICIOacuteN ENZIMATICA
Mediante el uso de inhibidores enzimaacuteticos se ha obtenido informacioacuten muy valiosa sobre la conformacioacuten del centro activo de algunas enzimas
a Inhibicioacuten irreversible Este tipo de inhibidores forman un enlace covalente con las enzimas cerca del centro activo Un ejemplo son los gases nerviosos como el
fluorofosfato de diisopropilo (DFP) que forma un complejo con la enzima acetilcolinesterasa Los animales envenenados con este gas quedan paralizados debido a la imposibilidad de transmitir adecuadamente los impulsos nerviosos
Ester di-isopropil fos foacuterico de la enzima (inactivo)
b) Inhibicioacuten competitiva
Es cuando el inhibidor compite con el substrato por la unioacuten con el centro activo de la
enzima Este tipo de inhibicioacuten puede reducirse si se aumenta la concentracioacuten de substrato
-OOC - CH2 - CH2 - COO
- -OOC - CH = CH - COO
- + 2H
+ Succinato Succinato Fumarato
33
Deshidrogenasa COO
- I CH2 Malonato (inhibidor competitivo) I COO
- En la inhibicioacuten competitiva la enzima se combina con el inhibidor para formar un complejo enzima-inhibidor E + I E I en competencia con la reaccioacuten normal E +S ES
La sulfanilamida (bactericida) interfiere con la siacutentesis del aacutecido foacutelico compitiendo con el aacutecido p-aminobenzoico de una forma competitiva
HOOC-sect- NH2 NH2 ndashSO2 - sect- NH2 aacutecido p- amino benzoico sulfanilamida
sect = fenil
a Inhibicioacuten no competitiva
Esta inhibicioacuten se caracteriza por que no se puede revertir el efecto del inhibidor aumentando la concentracioacuten del substrato
Por ejemplo la inhibicioacuten de la deshidrogenasa del gliceraldehido -3-fosfato por la yodo-acetamida
Enzima-SH + I-CH2 CONH2 Enzima-SCH2 CONH2 + IH
La deshidrogenasa del gliceraldehido-3 fosfato puede ser inhibida tambieacuten por el aacutecido iodo-aceacutetico
Enzima-SH + ICH2 COOH Enzima-SCH2 COOH + IH
c) Inhibicioacuten por metales
Ciertos metales como el plomo mercurio y arseacutenico inhiben enzimas que tienen en su centro activo grupos -SH libres
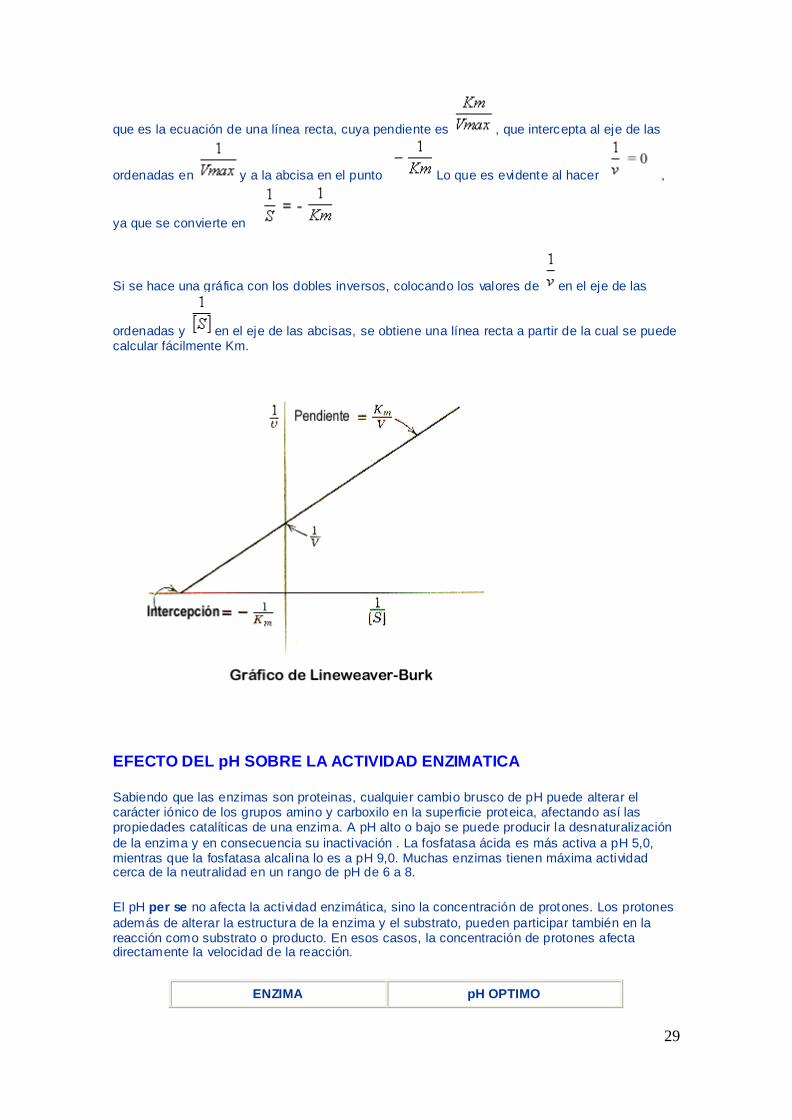
29
que es la ecuacioacuten de una liacutenea recta cuya pendiente es que intercepta al eje de las
ordenadas en y a la abcisa en el punto Lo que es evidente al hacer
ya que se convierte en
Si se hace una graacutefica con los dobles inversos colocando los valores de en el eje de las
ordenadas y en el eje de las abcisas se obtiene una liacutenea recta a partir de la cual se puede calcular faacutecilmente Km
EFECTO DEL pH SOBRE LA ACTIVIDAD ENZIMATICA
Sabiendo que las enzimas son proteinas cualquier cambio brusco de pH puede alterar el caraacutecter ioacutenico de los grupos amino y carboxilo en la superficie proteica afectando asiacute las propiedades cataliacuteticas de una enzima A pH alto o bajo se puede producir la desnaturalizacioacuten
de la enzima y en consecuencia su inactivacioacuten La fosfatasa aacutecida es maacutes activa a pH 50 mientras que la fosfatasa alcalina lo es a pH 90 Muchas enzimas tienen maacutexima actividad cerca de la neutralidad en un rango de pH de 6 a 8
El pH per se no afecta la actividad enzimaacutetica sino la concentracioacuten de protones Los protones
ademaacutes de alterar la estructura de la enzima y el substrato pueden participar tambieacuten en la reaccioacuten como substrato o producto En esos casos la concentracioacuten de protones afecta directamente la velocidad de la reaccioacuten
ENZIMA pH OPTIMO
30
Pepsina 15
Tripsina 77
Catalasa 76
Arginasa 97
Fumarasa 78
Ribonucleasa 78
EFECTO DE ACTIVADORES
Un aumento en la temperatura provoca un aumento de la velocidad de reaccioacuten hasta cierta temperatura oacuteptima ya que despueacutes de aproximadamente 45
0 C se comienza a producir la
desnaturalizacioacuten teacutermica Las enzimas de muchos mamiacuteferos tienen una temperatura oacuteptima
de 370 C por encima de esa temperatura comienzan a inactivarse y se destruyen Sin embargo
existen especies de bacterias y algas que habitan en fuentes de aguas termales y en el otro extremo ciertas bacterias aacuterticas tienen temperaturas oacuteptimas cercanas a 0
0 C
31
ESPECIFICIDAD ABSOLUTA
Cierta enzimas se encuentran como aproenzimas o zimogenos que son inactivos
El tripsinoacutegeno es convertido en tripsina por la misma accioacuten de la tripsina que remueve un hexapeacuteptido
tripsina Tripsinoacutegeno Tripsina + Val (Asp)4 lisina
Otra forma de activacioacuten consiste en mantener los grupos SH reducidos Estos grupos pueden
formar parte de centros activos de ciertas enzimas Si se oxidan estos grupos a -S-S- la enzima se inactiva Asiacute tenemos que la enzima papaina se inactiva despueacutes de exponerla al ox iacutegeno pero cuando se le antildeade un reductor apropiado la enzima se reactiva Los grupos ndashS-S- se convierten en ndashSH activaacutendose la enzima
Otra forma de activacioacuten requiere la presencia de otra substancia que se enlaza al sitio alosteacuterico el compuesto que se enlaza se denomina un efector alosteacuterico El centro alosteacuterico es bien especiacutefico para el efector o modulador alosteacuterico
ESPECIFICIDAD RELATIVA
Oxiacutegeno L-aminoaacutecido a -cetoaacutecido + NH3 + H 2 O 2 L-aminoaacutecido oxidasa Oxiacutegeno D-aminoaacutecido a -cetoaacutecido + NH3 + H 2 O 2 D-aminoaacutecido oxidasa
32
En este ejemplo se muestra la estereo-especificidad Una enzima puede tener una especificidad oacuteptica para isomeros D o L
Hay enzimas que muestran especificidad absoluta como la aspartasa que cataliza la adicioacuten reversible de amoniacuteaco al doble enlace del aacutecido fumaacuterico pero no al de otros aacutecidos insaturados
HOOC ndash CH = CH ndash COO- + NH4
+ HOOC ndash CH2 - CHNH2 - COO
- + H
+
INHIBICIOacuteN ENZIMATICA
Mediante el uso de inhibidores enzimaacuteticos se ha obtenido informacioacuten muy valiosa sobre la conformacioacuten del centro activo de algunas enzimas
a Inhibicioacuten irreversible Este tipo de inhibidores forman un enlace covalente con las enzimas cerca del centro activo Un ejemplo son los gases nerviosos como el
fluorofosfato de diisopropilo (DFP) que forma un complejo con la enzima acetilcolinesterasa Los animales envenenados con este gas quedan paralizados debido a la imposibilidad de transmitir adecuadamente los impulsos nerviosos
Ester di-isopropil fos foacuterico de la enzima (inactivo)
b) Inhibicioacuten competitiva
Es cuando el inhibidor compite con el substrato por la unioacuten con el centro activo de la
enzima Este tipo de inhibicioacuten puede reducirse si se aumenta la concentracioacuten de substrato
-OOC - CH2 - CH2 - COO
- -OOC - CH = CH - COO
- + 2H
+ Succinato Succinato Fumarato
33
Deshidrogenasa COO
- I CH2 Malonato (inhibidor competitivo) I COO
- En la inhibicioacuten competitiva la enzima se combina con el inhibidor para formar un complejo enzima-inhibidor E + I E I en competencia con la reaccioacuten normal E +S ES
La sulfanilamida (bactericida) interfiere con la siacutentesis del aacutecido foacutelico compitiendo con el aacutecido p-aminobenzoico de una forma competitiva
HOOC-sect- NH2 NH2 ndashSO2 - sect- NH2 aacutecido p- amino benzoico sulfanilamida
sect = fenil
a Inhibicioacuten no competitiva
Esta inhibicioacuten se caracteriza por que no se puede revertir el efecto del inhibidor aumentando la concentracioacuten del substrato
Por ejemplo la inhibicioacuten de la deshidrogenasa del gliceraldehido -3-fosfato por la yodo-acetamida
Enzima-SH + I-CH2 CONH2 Enzima-SCH2 CONH2 + IH
La deshidrogenasa del gliceraldehido-3 fosfato puede ser inhibida tambieacuten por el aacutecido iodo-aceacutetico
Enzima-SH + ICH2 COOH Enzima-SCH2 COOH + IH
c) Inhibicioacuten por metales
Ciertos metales como el plomo mercurio y arseacutenico inhiben enzimas que tienen en su centro activo grupos -SH libres
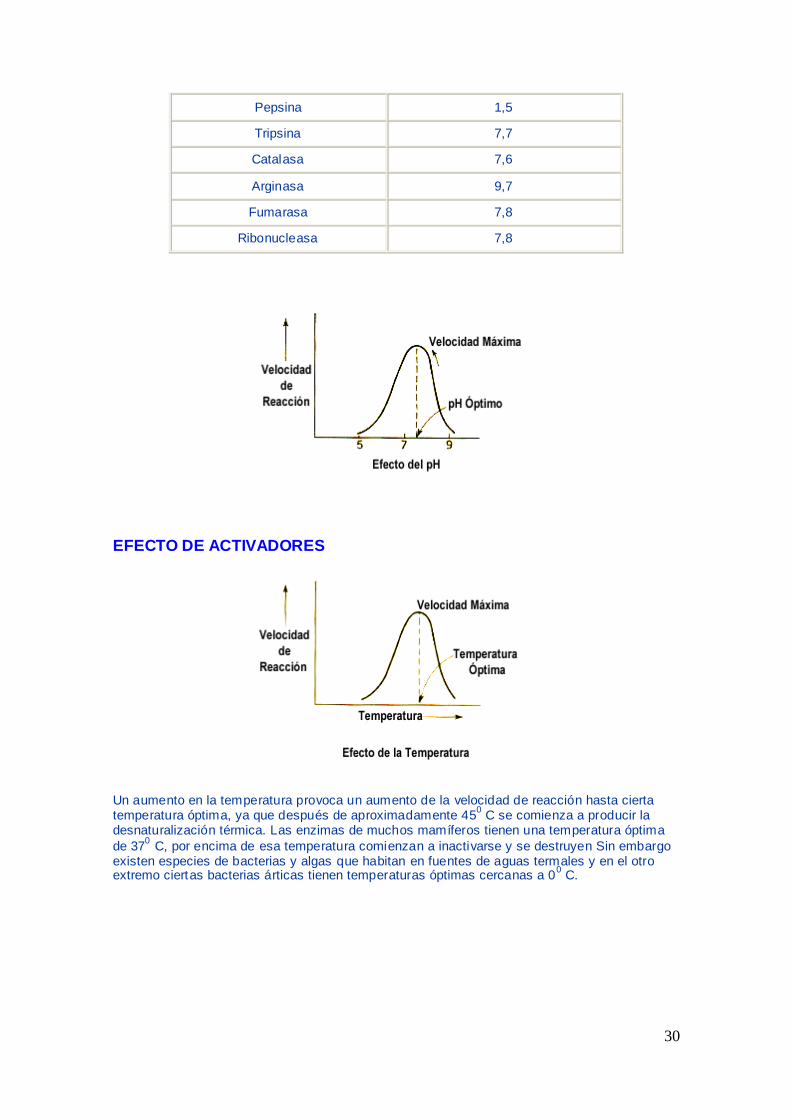
30
Pepsina 15
Tripsina 77
Catalasa 76
Arginasa 97
Fumarasa 78
Ribonucleasa 78
EFECTO DE ACTIVADORES
Un aumento en la temperatura provoca un aumento de la velocidad de reaccioacuten hasta cierta temperatura oacuteptima ya que despueacutes de aproximadamente 45
0 C se comienza a producir la
desnaturalizacioacuten teacutermica Las enzimas de muchos mamiacuteferos tienen una temperatura oacuteptima
de 370 C por encima de esa temperatura comienzan a inactivarse y se destruyen Sin embargo
existen especies de bacterias y algas que habitan en fuentes de aguas termales y en el otro extremo ciertas bacterias aacuterticas tienen temperaturas oacuteptimas cercanas a 0
0 C
31
ESPECIFICIDAD ABSOLUTA
Cierta enzimas se encuentran como aproenzimas o zimogenos que son inactivos
El tripsinoacutegeno es convertido en tripsina por la misma accioacuten de la tripsina que remueve un hexapeacuteptido
tripsina Tripsinoacutegeno Tripsina + Val (Asp)4 lisina
Otra forma de activacioacuten consiste en mantener los grupos SH reducidos Estos grupos pueden
formar parte de centros activos de ciertas enzimas Si se oxidan estos grupos a -S-S- la enzima se inactiva Asiacute tenemos que la enzima papaina se inactiva despueacutes de exponerla al ox iacutegeno pero cuando se le antildeade un reductor apropiado la enzima se reactiva Los grupos ndashS-S- se convierten en ndashSH activaacutendose la enzima
Otra forma de activacioacuten requiere la presencia de otra substancia que se enlaza al sitio alosteacuterico el compuesto que se enlaza se denomina un efector alosteacuterico El centro alosteacuterico es bien especiacutefico para el efector o modulador alosteacuterico
ESPECIFICIDAD RELATIVA
Oxiacutegeno L-aminoaacutecido a -cetoaacutecido + NH3 + H 2 O 2 L-aminoaacutecido oxidasa Oxiacutegeno D-aminoaacutecido a -cetoaacutecido + NH3 + H 2 O 2 D-aminoaacutecido oxidasa
32
En este ejemplo se muestra la estereo-especificidad Una enzima puede tener una especificidad oacuteptica para isomeros D o L
Hay enzimas que muestran especificidad absoluta como la aspartasa que cataliza la adicioacuten reversible de amoniacuteaco al doble enlace del aacutecido fumaacuterico pero no al de otros aacutecidos insaturados
HOOC ndash CH = CH ndash COO- + NH4
+ HOOC ndash CH2 - CHNH2 - COO
- + H
+
INHIBICIOacuteN ENZIMATICA
Mediante el uso de inhibidores enzimaacuteticos se ha obtenido informacioacuten muy valiosa sobre la conformacioacuten del centro activo de algunas enzimas
a Inhibicioacuten irreversible Este tipo de inhibidores forman un enlace covalente con las enzimas cerca del centro activo Un ejemplo son los gases nerviosos como el
fluorofosfato de diisopropilo (DFP) que forma un complejo con la enzima acetilcolinesterasa Los animales envenenados con este gas quedan paralizados debido a la imposibilidad de transmitir adecuadamente los impulsos nerviosos
Ester di-isopropil fos foacuterico de la enzima (inactivo)
b) Inhibicioacuten competitiva
Es cuando el inhibidor compite con el substrato por la unioacuten con el centro activo de la
enzima Este tipo de inhibicioacuten puede reducirse si se aumenta la concentracioacuten de substrato
-OOC - CH2 - CH2 - COO
- -OOC - CH = CH - COO
- + 2H
+ Succinato Succinato Fumarato
33
Deshidrogenasa COO
- I CH2 Malonato (inhibidor competitivo) I COO
- En la inhibicioacuten competitiva la enzima se combina con el inhibidor para formar un complejo enzima-inhibidor E + I E I en competencia con la reaccioacuten normal E +S ES
La sulfanilamida (bactericida) interfiere con la siacutentesis del aacutecido foacutelico compitiendo con el aacutecido p-aminobenzoico de una forma competitiva
HOOC-sect- NH2 NH2 ndashSO2 - sect- NH2 aacutecido p- amino benzoico sulfanilamida
sect = fenil
a Inhibicioacuten no competitiva
Esta inhibicioacuten se caracteriza por que no se puede revertir el efecto del inhibidor aumentando la concentracioacuten del substrato
Por ejemplo la inhibicioacuten de la deshidrogenasa del gliceraldehido -3-fosfato por la yodo-acetamida
Enzima-SH + I-CH2 CONH2 Enzima-SCH2 CONH2 + IH
La deshidrogenasa del gliceraldehido-3 fosfato puede ser inhibida tambieacuten por el aacutecido iodo-aceacutetico
Enzima-SH + ICH2 COOH Enzima-SCH2 COOH + IH
c) Inhibicioacuten por metales
Ciertos metales como el plomo mercurio y arseacutenico inhiben enzimas que tienen en su centro activo grupos -SH libres

31
ESPECIFICIDAD ABSOLUTA
Cierta enzimas se encuentran como aproenzimas o zimogenos que son inactivos
El tripsinoacutegeno es convertido en tripsina por la misma accioacuten de la tripsina que remueve un hexapeacuteptido
tripsina Tripsinoacutegeno Tripsina + Val (Asp)4 lisina
Otra forma de activacioacuten consiste en mantener los grupos SH reducidos Estos grupos pueden
formar parte de centros activos de ciertas enzimas Si se oxidan estos grupos a -S-S- la enzima se inactiva Asiacute tenemos que la enzima papaina se inactiva despueacutes de exponerla al ox iacutegeno pero cuando se le antildeade un reductor apropiado la enzima se reactiva Los grupos ndashS-S- se convierten en ndashSH activaacutendose la enzima
Otra forma de activacioacuten requiere la presencia de otra substancia que se enlaza al sitio alosteacuterico el compuesto que se enlaza se denomina un efector alosteacuterico El centro alosteacuterico es bien especiacutefico para el efector o modulador alosteacuterico
ESPECIFICIDAD RELATIVA
Oxiacutegeno L-aminoaacutecido a -cetoaacutecido + NH3 + H 2 O 2 L-aminoaacutecido oxidasa Oxiacutegeno D-aminoaacutecido a -cetoaacutecido + NH3 + H 2 O 2 D-aminoaacutecido oxidasa
32
En este ejemplo se muestra la estereo-especificidad Una enzima puede tener una especificidad oacuteptica para isomeros D o L
Hay enzimas que muestran especificidad absoluta como la aspartasa que cataliza la adicioacuten reversible de amoniacuteaco al doble enlace del aacutecido fumaacuterico pero no al de otros aacutecidos insaturados
HOOC ndash CH = CH ndash COO- + NH4
+ HOOC ndash CH2 - CHNH2 - COO
- + H
+
INHIBICIOacuteN ENZIMATICA
Mediante el uso de inhibidores enzimaacuteticos se ha obtenido informacioacuten muy valiosa sobre la conformacioacuten del centro activo de algunas enzimas
a Inhibicioacuten irreversible Este tipo de inhibidores forman un enlace covalente con las enzimas cerca del centro activo Un ejemplo son los gases nerviosos como el
fluorofosfato de diisopropilo (DFP) que forma un complejo con la enzima acetilcolinesterasa Los animales envenenados con este gas quedan paralizados debido a la imposibilidad de transmitir adecuadamente los impulsos nerviosos
Ester di-isopropil fos foacuterico de la enzima (inactivo)
b) Inhibicioacuten competitiva
Es cuando el inhibidor compite con el substrato por la unioacuten con el centro activo de la
enzima Este tipo de inhibicioacuten puede reducirse si se aumenta la concentracioacuten de substrato
-OOC - CH2 - CH2 - COO
- -OOC - CH = CH - COO
- + 2H
+ Succinato Succinato Fumarato
33
Deshidrogenasa COO
- I CH2 Malonato (inhibidor competitivo) I COO
- En la inhibicioacuten competitiva la enzima se combina con el inhibidor para formar un complejo enzima-inhibidor E + I E I en competencia con la reaccioacuten normal E +S ES
La sulfanilamida (bactericida) interfiere con la siacutentesis del aacutecido foacutelico compitiendo con el aacutecido p-aminobenzoico de una forma competitiva
HOOC-sect- NH2 NH2 ndashSO2 - sect- NH2 aacutecido p- amino benzoico sulfanilamida
sect = fenil
a Inhibicioacuten no competitiva
Esta inhibicioacuten se caracteriza por que no se puede revertir el efecto del inhibidor aumentando la concentracioacuten del substrato
Por ejemplo la inhibicioacuten de la deshidrogenasa del gliceraldehido -3-fosfato por la yodo-acetamida
Enzima-SH + I-CH2 CONH2 Enzima-SCH2 CONH2 + IH
La deshidrogenasa del gliceraldehido-3 fosfato puede ser inhibida tambieacuten por el aacutecido iodo-aceacutetico
Enzima-SH + ICH2 COOH Enzima-SCH2 COOH + IH
c) Inhibicioacuten por metales
Ciertos metales como el plomo mercurio y arseacutenico inhiben enzimas que tienen en su centro activo grupos -SH libres

32
En este ejemplo se muestra la estereo-especificidad Una enzima puede tener una especificidad oacuteptica para isomeros D o L
Hay enzimas que muestran especificidad absoluta como la aspartasa que cataliza la adicioacuten reversible de amoniacuteaco al doble enlace del aacutecido fumaacuterico pero no al de otros aacutecidos insaturados
HOOC ndash CH = CH ndash COO- + NH4
+ HOOC ndash CH2 - CHNH2 - COO
- + H
+
INHIBICIOacuteN ENZIMATICA
Mediante el uso de inhibidores enzimaacuteticos se ha obtenido informacioacuten muy valiosa sobre la conformacioacuten del centro activo de algunas enzimas
a Inhibicioacuten irreversible Este tipo de inhibidores forman un enlace covalente con las enzimas cerca del centro activo Un ejemplo son los gases nerviosos como el
fluorofosfato de diisopropilo (DFP) que forma un complejo con la enzima acetilcolinesterasa Los animales envenenados con este gas quedan paralizados debido a la imposibilidad de transmitir adecuadamente los impulsos nerviosos
Ester di-isopropil fos foacuterico de la enzima (inactivo)
b) Inhibicioacuten competitiva
Es cuando el inhibidor compite con el substrato por la unioacuten con el centro activo de la
enzima Este tipo de inhibicioacuten puede reducirse si se aumenta la concentracioacuten de substrato
-OOC - CH2 - CH2 - COO
- -OOC - CH = CH - COO
- + 2H
+ Succinato Succinato Fumarato
33
Deshidrogenasa COO
- I CH2 Malonato (inhibidor competitivo) I COO
- En la inhibicioacuten competitiva la enzima se combina con el inhibidor para formar un complejo enzima-inhibidor E + I E I en competencia con la reaccioacuten normal E +S ES
La sulfanilamida (bactericida) interfiere con la siacutentesis del aacutecido foacutelico compitiendo con el aacutecido p-aminobenzoico de una forma competitiva
HOOC-sect- NH2 NH2 ndashSO2 - sect- NH2 aacutecido p- amino benzoico sulfanilamida
sect = fenil
a Inhibicioacuten no competitiva
Esta inhibicioacuten se caracteriza por que no se puede revertir el efecto del inhibidor aumentando la concentracioacuten del substrato
Por ejemplo la inhibicioacuten de la deshidrogenasa del gliceraldehido -3-fosfato por la yodo-acetamida
Enzima-SH + I-CH2 CONH2 Enzima-SCH2 CONH2 + IH
La deshidrogenasa del gliceraldehido-3 fosfato puede ser inhibida tambieacuten por el aacutecido iodo-aceacutetico
Enzima-SH + ICH2 COOH Enzima-SCH2 COOH + IH
c) Inhibicioacuten por metales
Ciertos metales como el plomo mercurio y arseacutenico inhiben enzimas que tienen en su centro activo grupos -SH libres

33
Deshidrogenasa COO
- I CH2 Malonato (inhibidor competitivo) I COO
- En la inhibicioacuten competitiva la enzima se combina con el inhibidor para formar un complejo enzima-inhibidor E + I E I en competencia con la reaccioacuten normal E +S ES
La sulfanilamida (bactericida) interfiere con la siacutentesis del aacutecido foacutelico compitiendo con el aacutecido p-aminobenzoico de una forma competitiva
HOOC-sect- NH2 NH2 ndashSO2 - sect- NH2 aacutecido p- amino benzoico sulfanilamida
sect = fenil
a Inhibicioacuten no competitiva
Esta inhibicioacuten se caracteriza por que no se puede revertir el efecto del inhibidor aumentando la concentracioacuten del substrato
Por ejemplo la inhibicioacuten de la deshidrogenasa del gliceraldehido -3-fosfato por la yodo-acetamida
Enzima-SH + I-CH2 CONH2 Enzima-SCH2 CONH2 + IH
La deshidrogenasa del gliceraldehido-3 fosfato puede ser inhibida tambieacuten por el aacutecido iodo-aceacutetico
Enzima-SH + ICH2 COOH Enzima-SCH2 COOH + IH
c) Inhibicioacuten por metales
Ciertos metales como el plomo mercurio y arseacutenico inhiben enzimas que tienen en su centro activo grupos -SH libres


















