
HYMENOPTERA PARASITOIDES ASOCIADOS A LA POLILLA...
73
HYMENOPTERA PARASITOIDES EN LA RESERVA NATURAL EL HATICO, VALLE DEL CAUCA, COLOMBIA LORENA LÓPEZ QUIÑONEZ UNIVERSIDAD DEL VALLE FACULTAD DE CIENCIAS NATURALES Y EXACTAS PROGRAMA ACADÉMICO DE BIOLOGÍA SANTIAGO DE CALI 2011
Transcript of HYMENOPTERA PARASITOIDES ASOCIADOS A LA POLILLA...

HYMENOPTERA PARASITOIDES EN LA RESERVA NATURAL EL HATICO,
VALLE DEL CAUCA, COLOMBIA
LORENA LÓPEZ QUIÑONEZ
UNIVERSIDAD DEL VALLE
FACULTAD DE CIENCIAS NATURALES Y EXACTAS
PROGRAMA ACADÉMICO DE BIOLOGÍA
SANTIAGO DE CALI
2011

ii
HYMENOPTERA PARASITOIDES EN LA RESERVA NATURAL EL HATICO,
VALLE DEL CAUCA, COLOMBIA
LORENA LÓPEZ QUIÑONEZ
UNIVERSIDAD DEL VALLE
FACULTAD DE CIENCIAS NATURALES Y EXACTAS
PROGRAMA ACADÉMICO DE BIOLOGÍA
SANTIAGO DE CALI
2011

iii
HYMENOPTERA PARASITOIDES EN LA RESERVA NATURAL EL HATICO,
VALLE DEL CAUCA, COLOMBIA
LORENA LÓPEZ QUIÑONEZ
Informe final de Trabajo de grado presentado como requisito parcial para optar al
título de Bióloga con énfasis en Entomología.
Directora
Inge Armbrecht
Bióloga, Ph.D
Codirector
James Montoya Lerma
Biólogo, Ph.D
UNIVERSIDAD DEL VALLE
FACULTAD DE CIENCIAS NATURALES Y EXACTAS
PROGRAMA ACADÉMICO DE BIOLOGÍA
SANTIAGO DE CALI
2011

iv
UNIVERSIDAD DEL VALLE
FACULTAD DE CIENCIAS NATURALES Y EXACTAS
PROGRAMA ACADÉMICO DE BIOLOGÍA
LORENA LÓPEZ QUIÑONEZ (1989)
HYMENOPTERA PARASITOIDES EN LA RESERVA NATURAL EL HATICO,
VALLE DEL CAUCA, COLOMBIA
Temas:
Himenópteros parasitoides
Agroecología
Sistemas silvopastoriles
Herbivoría

v

vi
Podrán cortar todas las flores,
pero no podrán detener la primavera.
Pablo Neruda

vii
AGRADECIMIENTOS
Agradezco a la Universidad del Valle y a la sección de Entomología por la
excelente formación académica brindada y por ofrecerme el espacio para
desarrollarme como profesional.
A mi directora Inge Armbrecht y mi codirector James Montoya por convertirse en
mis guías y un apoyo constante durante el último año; por acogerme, tan
cálidamente, en el Grupo Ecología de Agroecosistemas y Hábitats Naturales
(GEAHNA) y por enseñarme la importancia del trabajo en equipo.
A mis padres Gloria y Salomón, que velaron por mi seguridad, salud y progreso en
mis estudios, por su constante apoyo, ánimo y compañía.
A las excelentes personas que encontré durante los cinco años de carrera y que
se convirtieron en mis grandes amigos: Ana Paola, Jorge Horacio y Ximena, que
no solo me acompañaron durante la jornada sino que además fueron partícipes de
este proyecto.
A mis amigas Angélica, Luisa y Johana que me acompañaron y se divirtieron en
campo durante la realización de este trabajo.
A María del Pilar Hernández y Diego Campos, quienes me brindaron su apoyo y
me ayudaron en la identificación de los parasitoides.

viii
A los demás profesores que se convirtieron en parte importante en mi rumbo
profesional Wilmar, Oscar, Alan, Nancy.
A Karen y Carolina, cuyo trabajo abrió las puertas a este proyecto, por su
colaboración y su entusiasmo al darme a conocer la Reserva y por las facilidades
para idear este proyecto.
Un gran agradecimiento a los hermanos Molina, Carlos Hernando y Enrique José,
quienes abrieron las puertas del proyecto de vida familiar “Reserva Natural El
Hatico”, me ofrecieron apoyo y todas las facilidades para llevar a cabo el trabajo y
a CIPAV por su apoyo incondicional.
Y, por último, aunque no menos importante, agradecimientos a la Sociedad
Colombiana de Entomología, pues este estudio fue financiado por esa entidad al
otorgarme el primer puesto de la “Beca de incentivo a la investigación
entomológica en las universidades colombianas (Beca SOCOLEN)” versión 2010.

ix
TABLA DE CONTENIDO
Página
1. Resumen…………………………………………………………………………..…….1
2. Introducción………………………………………………………………………..……3
3. Antecedentes…………………………………………………………………….……..6
3.1 Sistemas silvopastoriles y sistemas naturales..…………………………………...6
3.2 El algarrobo y la potencial plaga……………………………………………..……..7
3.3 Identificación de la diversidad parasítica…………………………………………..9
4. Objetivos…………………………………………………………………………….…11
4.1 Objetivo general……………………………………………………………………..11
4.2 Objetivos específicos………………………………………………………………..11
4.3 Hipótesis y predicción biológica 1………………………...……………………….11
4.4 Hipótesis y predicción biológica 2…………………………………………………12
5. Materiales y métodos…………………………………………………………………13
5.1 Área de estudio…………………………………………………………………..….13
5.2 Fase de campo…………………………………………………………………...….14
5.3 Determinación taxonómica…………………………………………………………17
5.4 Análisis de datos…………………………………………………………………….18
6. Resultados y discusión…………………………………………………………….…20
6.1 Composición general de parasitoides……………………………………………..20
6.2 Comparación de parasitoides entre sistemas……………………………………24
6.3 Composición vegetal y especies de parasitoides………………………………..30
6.4 Cobertura vegetal y especies de parasitoides……………………………………34
6.5 Niveles de herbivoría y riqueza de parasitoides…………………………………35
6.6 Propiedades físico-químicas del sistema silvopastoril…..………………………38
7. Conclusiones…………………………………………………………………………..41
8. Recomendaciones…………………………………………………………………….43
9. Literatura citada……………………………………………………………………….45
10. Anexos………………………………………………………………………………..50

x
LISTA DE FIGURAS
Página
Figura 1. Esquema de la distribución de los lotes muestreados dentro del
sistema silvopastoril, la ubicación del bosque y la ubicación aproximada del
transecto de muestreo establecido en el mismo (40m hace referencia a la
longitud del transecto sin escala alguna). Modificado del plano original de la
Reserva Natural El Hatico (2003) …………………………………………....……15
Figura 2. Ramas y hoja del algarrobo compuesta por cuatro o seis subraquis.
Fotografías por Lorena López Quiñonez………………………………………….17
Figura 3. Frecuencias de morfoespecies por superfamilia de Hymenoptera
parasítica, colectados en ambas épocas climáticas, incluyendo el sistema
silvopastoril y el sistema natural de la Reserva Natural El Hatico………..……20
Figura 4. Análisis de agrupamiento, basado en el índice de similitud de
Jaccard, para los lotes presentes en el sistema silvopastoril y para el sistema
natural (bosque) en la Reserva Natural El Hatico…………..…………….......…27
Figura 5. Análisis de agrupamiento, basado en el índice de similitud de Bray-
Curtis, para los lotes presentes en el sistema silvopastoril y para el sistema
natural (bosque) en la Reserva Natural El Hatico..…………………………...…28
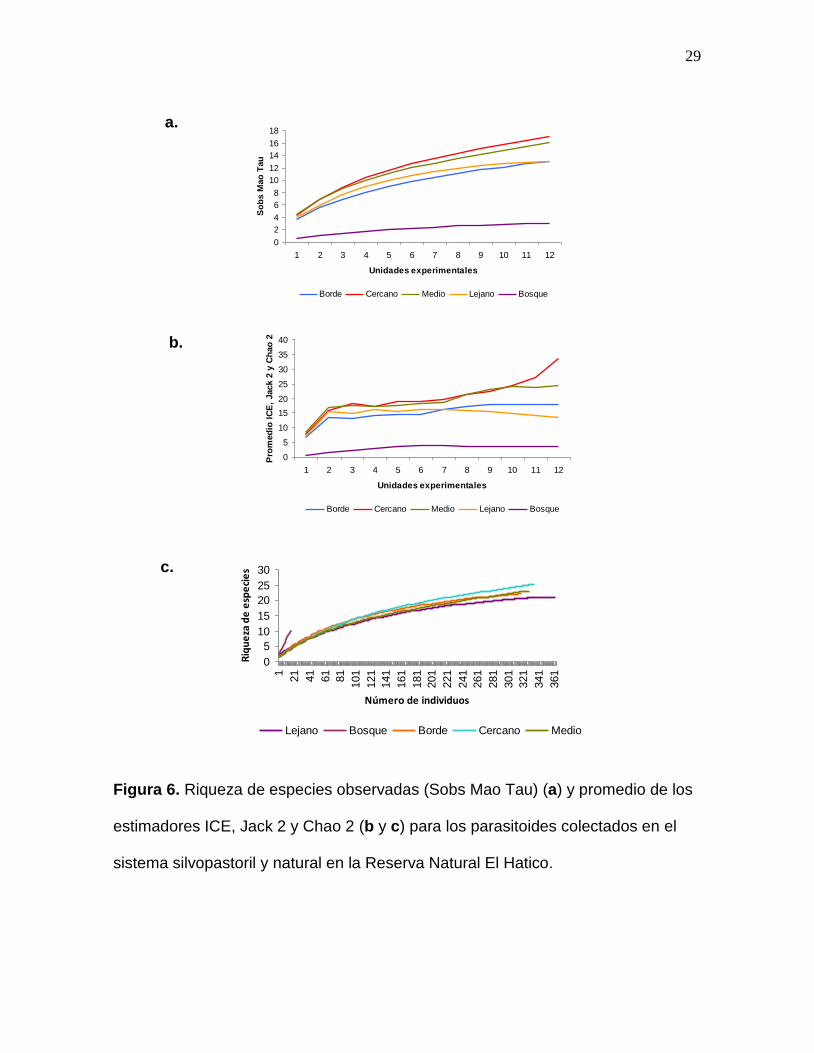
Figura 6. Riqueza de especies observadas (Sobs Mao Tau) (a) y promedio de
los estimadores ICE, Jack 2 y Chao 2 (b y c) para los parasitoides colectados
en el sistema silvopastoril y natural en la Reserva Natural El Hatico………….29

xi
Figura 7. Comparación de la riqueza de plantas en cada lote del sistema
silvopastoril en la Reserva Natural El Hatico. Se incluyen las especies de
plantas encontradas en ambas épocas climáticas…….....................................33
Figura 8. Relación entre la abundancia de parasitoides y la riqueza de plantas
en cada lote del sistema silvopastoril en la Reserva Natural El Hatico…….....33
Figura 9. Relación entre porcentaje de cobertura de los algarrobos y la
abundancia de parasitoides en cada uno de los lotes del sistema silvopastoril
en la Reserva Natural El Hatico.………………………………....………………..35
Figura 10. Diferencias estacionales entre los porcentajes de herbivoría de
(épocas seca y lluviosa) en la Reserva Natural El Hatico.…………..………….36
Figura 11. Porcentajes de herbivoría promedio de los algarrobos de cuatro
lotes del sistema silvopastoril en la Reserva Natural El Hatico, en dos épocas
climáticas diferentes y abundancia promedio de parasitoides en ambas
épocas………………………………………….……………………………………..36
Figura 12. Porcentajes de herbivoría de tres estratos diferentes del follaje de
los algarrobos (alto, medio y bajo), en cuatro lotes del sistema silvopastoril y
dos épocas climáticas: a. seca (febrero de 2010); b. lluviosa (mayo de
2010)…………………………………………………………………………………..38
Figura 13. Temperatura promedio de los cuatro lotes en el sistema silvopastoril
de la Reserva Natural El Hatico y abundancia de parasitoides colectados en los
mismos………………………………………………................................………..39
Figura 14. Temperatura y precipitación promedio en los últimos cuatro años
(2007 - 2010) en la Reserva Natural El Hatico…………………………………...39

xii
LISTA DE TABLAS
Página
Tabla 1. Superfamilias, familias, subfamilias, géneros y/o especies de
Hymenoptera parasítica encontradas en el sistema silvopastoril y el bosque de
la Reserva Natural El Hatico.……………………………………………………….22
Tabla 2. Riqueza de especies, abundancia e índice de Shannon (H’) del
sistema natural (bosque) y cada lote del sistema silvopastoril en la RNH,
teniendo en cuenta ambas épocas climáticas (seca: febrero 2010 y lluviosa:
mayo 2010) …..............................................................................................….25
Tabla 3. Comparación entre los índices de Shannon (H’) del bosque y cada
lote del sistema silvopastoril, mediante prueba t-student. Los valores de p en
rojo indican diferencias significativas entre la diversidad de los lotes…………26
Tabla 4. Familias, subfamilias, géneros y especies de plantas encontradas
dentro del sistema silvopastoril en la Reserva Natural El Hatico…….. …….....32
ANEXO A. Superfamilia, familia, morfoespecie (msp), género y/o especie, y
abundancia de cada una, según el lote (en sistema silvopastoril) incluyendo el
bosque y la época climática en la que fueron colectadas. Los lotes se denotan
según su nombre: Bor, Borde; Cer, Cercano; Med, Medio; Lej, Lejano; y Bos,
Bosque……………………………………………………………………………..…50
ANEXO B. Morfoespecies capturadas en los lotes presentes en el sistema
silvopastoril y en el bosque con su respectiva abundancia………………..…...51
ANEXO C. Morfoespecies capturadas y su abundancia en los cuatros lotes del
sistema silvopastoril, en el bosque y discriminados por época climática...……52

xiii
ANEXO D. Porcentajes de herbivoría (%H) medidos en época seca de los 24
árboles muestreados y de cada uno de sus tres estratos (alto, medio y
bajo)…………………………………………………………………………………...54
ANEXO E. Porcentajes de herbivoría obtenidos en ambas épocas climáticas y
discriminados por lote y estrato…………………………………………………….55
ANEXO F. Porcentajes de herbivoría (%H) medidos en época lluviosa de los
24 árboles muestreados y de cada uno de sus tres estratos (alto, medio y
bajo)………………………………………………………………………………...…56
ANEXO G. Porcentaje de cobertura, fecha y hora a la cual se tomó la
temperatura de los 24 algarrobos muestreados en época seca……………..…58
ANEXO H. Porcentaje de cobertura, fecha y hora a la cual se tomó la
temperatura de los 24 algarrobos muestreados en época lluviosa…………….58
ANEXO I. Altura promedio de los algarrobos muestreados.…………………....59


1
1. RESUMEN
El término “silvopastoril”, define un sistema de producción que involucra diferentes
estratos arbóreos que sirven de alimento al ganado. En el sistema silvopastoril de
la Reserva Natural El Hatico, El Cerrito, Valle, los frutos del árbol de algarrobo
(Prosopis juliflora, Fabaceae) son utilizados comúnmente como alimento para
bovinos. Sin embargo, la estabilidad del sistema, desde 2006, está amenazada
por la polilla Eccopsis galapagana Razowski & Landry (Lepidoptera: Tortricidae)
que afecta su follaje y la producción legumbres. Además, existe un fragmento de
bosque primario junto al sistema silvopastoril que constituye el ecosistema natural
de la Reserva. Considerando la vocación agroecológica de la misma y motivados
por el brote poblacional de la polilla del algarrobo, se propuso este estudio cuyo
objetivo fue identificar las especies de parasitoides, su distribución, su densidad y
relacionarlas con su entorno, con miras a detectar características promisorias en el
diseño de estrategias de control biológico. Se realizaron dos salidas de campo,
espaciadas tres meses, de cuatro días cada una. Se llevó un registro individual de
24 árboles de algarrobo escogidos al azar, en cuatro lotes dentro del sistema
silvopastoril. Para la captura de himenópteros se utilizaron platos amarillos
impregnados con aceite de ricino ubicados en cada árbol muestreado y, al interior
del bosque primario (sistema natural). Se midió el porcentaje promedio de
herbivoría, ocasionado específicamente por la polilla del algarrobo; la cobertura
del dosel de cada árbol muestreado, la temperatura y se determinó la composición
vegetal de cada lote. Se colectó un total de 1376 parasitoides, pertenecientes a
siete superfamilias, 18 familias y 42 morfoespecies, de las cuales el 71% se
identificó hasta género. La superfamilia más representativa fue Chalcidoidea con

2
el mayor número de morfoespecies y la familia más abundante fue Encyrtidae con
el 94% de la abundancia total. En el sistema silvopastoril, el lote más lejano con
respecto al bosque (900m), presentó el mayor valor de diversidad de parasitoides,
superado por el bosque (sistema natural). Finalmente se encontró relación positiva
entre composición y cobertura vegetal y abundancia de parasitoides (r=0.8 y r=0.6,
respectivamente), pero no hubo relación alguna entre porcentaje de herbivoría y
abundancia de parasitoides. Se concluye que la presencia de diversidad vegetal
contribuye al sostenimiento de una diversidad alta de parasitoides y que ésta
variedad, en asocio con sus altas poblaciones, probablemente contribuyen al
control natural y resiliencia del sistema orgánico, libre de pesticidas que se maneja
en la Reserva Natural El Hatico.

3
2. INTRODUCCIÓN
En agroecología, el planteamiento de programas fitosanitarios eficientes se puede
alcanzar mediante el estudio de las relaciones entre la diversificación de la
vegetación, la dinámica poblacional de herbívoros y sus enemigos naturales
asociados en agroecosistemas particulares (Haddad et al. 2001). Estas iniciativas
representan la mejor esperanza de conservar la biodiversidad en áreas del mundo
donde la mayor parte del hábitat natural se ha transformado en sistemas agrícolas
o pecuarios (Altieri 1999). Los sistemas silvopastoriles son medios productivos en
los cuales se utilizan especies leñosas (árboles, arbustos, palmas) en asociación
con cultivos agrícolas o con animales en el mismo terreno (Mahecha 1998). La
presencia de animales pastando-ramoneando entre o bajo los árboles, que
pueden ser de diferente clase, corresponde a este tipo específico de
agroecosistema el cual garantiza la estabilidad de la productividad del suelo, logra
una alta producción, variado rendimiento y aumenta la provisión de forraje nutritivo
a la ganadería (Mahecha 1998). Este manejo se practica en la actualidad dentro
de la Reserva Natural El Hatico, El Cerrito (Valle del Cauca), donde se exhiben
cultivos orgánicos de caña de azúcar, intercalados con un amplio sistema
silvopastoril con árboles forrajeros integrados al componente pecuario y adyacente
a un guadual. Esta zona cuanta con alta diversidad por la presencia de plantas
ornamentales, un amplio jardín de varias décadas y con un fragmento de bosque
primario de aproximadamente 15 ha.
El algarrobo, Prosopis juliflora (Sw.) DC. (Fabaceae: Mimosoideae) juega un papel
importante dentro del sistema silvopastoril de la Reserva El Hatico, porque

4
representa un tercer estrato de tipo arbóreo en el perfil vegetal. Esta leguminosa
produce frutos apetecibles que, en épocas de sequía, se constituyen en alimento
invaluable para el ganado, con producciones promedio desde 18.4 (Santos et al.
1996) hasta 35 k/árbol, (Ramírez 1997) medidos en Cundinamarca y en la
Reserva Natural El Hatico, respectivamente. En este último sitio, se ha reportado
el brote de un lepidóptero Tortricidae, muy recientemente determinado por Dr. J.
Brown, Laboratorio de taxonomía de Missisipi, como Eccopsis galapagana
Razowski & Landry, descrita en 2008 y se creía endémica de las islas Galápagos,
Ecuador (Razowski et al. 2008). En la Reserva Natural El Hatico, sus larvas se
alimentan sobre las hojas de P. juliflora y aunque no cuantificado, su ataque ha
generado una disminución aproximada del 80% en la producción de legumbres en
los últimos tres años (C.H. Molina, com. pers. 2010). Por constituirse en una plaga
potencial, se requiere también identificar sus potenciales enemigos naturales,
específicamente himenópteros parasitoides que permitan plantear estrategias de
control consecuentes con el manejo ecológico que se lleva a cabo dentro del
sistema.
El presente estudio representa una primera exploración conducente a evaluar la
diversidad de himenópteros parasíticos y a la búsqueda de un manejo ecológico
de la polilla del algarrobo en la Reserva Natural El Hatico. Dado que los fitófagos y
parasitoides comprenden una alta proporción de la diversidad de insectos (Gibson
et al.1997), entonces, las interacciones planta-herbívoro-parasitoide constituyen el
mayor ensamblaje alimenticio terrestre (Sugiura 2007). Para entender o estudiar
dichas interacciones es necesario identificar los elementos inherentes a ellas. Se

5
han registrado en la actualidad varias especies de himenópteros parasitoides
asociados a la polilla del algarrobo (Reyes 2010). Además, eventualmente, se han
observado niveles de daño menor en algunos algarrobos adyacentes al fragmento
de bosque primario presente dentro del sistema (C.H. Molina, com. pers. 2010).
Todo lo anterior, conlleva a preguntarse si las especies de himenópteros
parasitoides se encuentran asociadas a los sistemas manejados dentro de la
reserva, a la polilla del algarrobo y si la presencia y abundancia de los mismos se
explica por la existencia de un bosque primario adyacente a los sistemas o por las
características inherentes del mismo.

6
3. ANTECEDENTES
3.1 Sistemas silvopastoriles y sistemas naturales
En la Reserva Natural El Hatico, se tienen sistemas silvopastoriles en los que
además de gramíneas como el pasto estrella, Cynodon plectostachyus (K. Schum)
Pilg. (Mahecha 1998), pasto guinea Panicum maximum (Jacq) B.K. Simon &
S.W.L. Jacobs, pasto argentina Cynodon dactylon (L.) Pers, se integran varios
estratos arbóreos tipo Leucaena leucocephala (Lamk) de Wit. In: 't Mannetje, L.;
algarrobo (P. juliflora), chiminango (Pithecellobium dulce Benth. (Dakhani babul,
Vilati imli), aromo (Acacia sp.), matarratón (Glicirida sepium (Jacq.) Kunth ex.
Griseb) entre otros, para ramoneo. Las arvenses en caña de azúcar son
manejadas por la integración de animales pequeños como ovinos. La zona más
alejada del bosque (900m) de la Reserva, es considerada como un sistema
manejado de forma distinta pues no es intensamente utilizada para el ramoneo e
integra otras plantas, árboles como samanes (Albizia saman (Jacq.) F. Muell) y
palmas (Syagrus sancona H. Karst., Roystonea regia (Kunth) O.F. Cook) que han
hecho durante varias décadas que esta zona se diversifique en comunidades vivas
(C.H. Molina, com. pers. 2010). En el caso del relicto de bosque, considerado
como el sistema natural de la Reserva, es uno de los fragmentos de bosque seco
más importantes del Valle del Cauca junto con los bosques de El Vínculo y el
bosque del Medio (Armbrecht & Chacón de Ulloa 1999), y presenta una amplia
zona de platanillos, helechos, árboles de gran altura y edad como caracolíes
(Anacardium excelsum L.), ceibas (Ceiba pentandra (L.) Gaertn.) y ficus (Ficus
sp.).

7
Los agroecosistemas son ambientes que corresponden a los estados iniciales de
la sucesión ecológica y, por lo tanto, son poco estables (Gliessman 2002). Según
Price y Waldbauer (1975), el número de especies de artrópodos en una
comunidad depende de factores como la diversidad de especies y estructural de
las plantas y distancia entre la comunidad y las fuentes de colonización. Por lo
tanto, es importante conocer los factores intrínsecos del agroecosistema, sus
límites y su matriz. Lo anterior conduce al escenario de la reserva El Hatico, en
donde se combinan una serie de elementos que involucran los factores
mencionados. Dado que los himenópteros parasitoides son artrópodos existe la
posibilidad que a medida que aumenta la complejidad taxonómica y estructural de
la vegetación se encuentre más variedad y abundancia de estos insectos.
3.2 El algarrobo y la potencial plaga
El adulto de E. galapagana es pequeño (5 -15mm), de color marrón claro con un
patrón de bandas transversales café (Reyes 2010). Sus larvas son típicas
enrolladoras de hojas. Son de color oscuro en los primeros ínstares y pardo claro
en los últimos. Cuando son perturbadas se dejan caer al suelo, hacen rápidos
movimientos laterales o caminan rápidamente para esconderse. Las pupas de
esta familia (Tortricidae) generalmente se localizan en los foliolos enrollados del
árbol formando una cámara pupal que tapizan interiormente con seda (Madrigal-
Cardeño 2003). Los adultos son de hábitos nocturnos, revolotean frecuentemente
cerca del suelo en el cual se posan camuflándose eficientemente (op. cit.).

8
Para obtener resultados favorables tales como regulación de plagas se requiere,
primero, identificar ensamblajes de biodiversidad, identificar las especies fitófagas,
especies plagas y plagas potenciales, depredadores y parasitoides. Cada sistema
tiene particularidades que se deben identificar en una primera etapa. Además,
identificar ensamblajes faunísticos, determinar la riqueza de especies y
composición vegetal, es igualmente importante para establecer medidas de
control, ya sea preventivo o de respuesta, pues dichos controles permiten el
manejo ecológico y tienen efectos significativos sobre las comunidades de
insectos (Haddad et al. 2001). Estos son algunos de los mayores desafíos para los
agroecólogos; aún así es indispensable identificar los parámetros que describen la
afinidad planta-hospedero-parasitoide para la evaluación de potenciales agentes
de control como especies de insectos o de plantas para la regulación de plagas
(Altieri 1999, Rincón & López-Ávila 1999).
Para mantener la estabilidad de los paisajes fragmentados es indispensable la
migración y el flujo de especies. Para la conformación de metapoblaciones viables
se requiere un flujo mínimo de individuos reproductivos a través de los parches de
hábitat que sostienen las subpoblaciones. Estos factores se pueden cumplir
cuando se mantienen las condiciones de la matriz que circunda áreas con
importantes niveles de biodiversidad (e.g. relictos de bosque, bosques en
regeneración) y se asegura su funcionamiento y permanencia en el tiempo
(Elzinga et al. 2007, Armbrecht 2008). Esta migración y movimiento se ha logrado,
en el caso de especies de insectos, gracias a la estrategia de “push – pull” para el
manejo integrado de plagas (MIP). Esta técnica usa un estímulo modificador del

9
comportamiento para manipular la distribución y abundancia de plagas y/o
insectos benéficos para su control pues consiste en la siembra de plantas que
atraen, alejan, interceptan y/o retienen insectos blanco (especies plaga o
parasitoides) para reducir el daño en los cultivos (Cook et al. 2007, Shelton &
Badenes-Perez 2006). La plaga se repele fuera del recurso (push) y
simultáneamente los enemigos naturales son atraídos (pull) hacia otros recursos
(e.g. plantas nectaríferas, sitios de nidación). La mayoría del trabajo utilizando esta
estrategia se ha enfocado en evaluar el comportamiento y el control de especies
plaga (Cook et al. 2007) pero también se puede evaluar si las especies de plantas
ya establecidas dentro de zonas con baja intensificación agrícola como
agroforestales, cumplen o no el papel de amortiguadores.
3.3 Identificación de la diversidad parasítica
Haddad y colegas (2001) evaluaron la relación entre los diferentes gremios de
Insecta y la riqueza y composición vegetal; encontrando efectos positivos y
significativos entre la abundancia y riqueza de un amplio número de órdenes de
insectos y la diversidad vegetal. Sus resultados se apoyan en estudios previos que
igualmente han demostrado que una mayor densidad y diversidad de plantas
presenta altas concentraciones de recursos, un gran potencial de atracción y una
gran diversidad de insectos, entre ellos presas, depredadores y parasitoides (op.
cit.). En Ecuador Tylianakis et al. (2005) al evaluar la diversidad de himenópteros
en cinco tipos de hábitat diferentes, desde monocultivos hasta fragmentos de
bosque secundario con niveles medios de perturbación, encontraron que incluso
hábitats con intenso manejo, como sistemas agroforestales, presentan una

10
diversidad de himenópteros que hace un aporte importante de especies al
mosaico de los paisajes fragmentados. Estos autores no solo se enfocaron en la
distribución espacial de las especies, sino también en la fluctuación temporal de
sus poblaciones, encontrando que los fragmentos de bosque cumplen una
importante función en el recambio de especies durante las diferentes épocas del
año. Esto se debe a la oferta temporal de recursos tanto en los sistemas
perturbados como en los mismos fragmentos de bosque. Cronin (2003), al evaluar
cómo la matriz afecta el movimiento, la oviposición y la distribución espacial de
Anagrus columbi (Chalcidoidea: Mymaridae), parasitoide de huevos de una
especie de saltahojas, en un paisaje fragmentado, encontró mayor porcentaje de
parasitismo dentro de los fragmentos de bosque que en el borde de dichos
fragmentos. Comprobó, además, que la calidad y la composición vegetal de la
matriz que los rodea es igualmente importante para mantener grandes
poblaciones de parasitoides y sobretodo en el flujo entre fragmentos. Como
conclusión, este autor afirma que la especificidad de hábitat de las especies de
himenópteros disminuye con el grado de intervención del hábitat.
Los tres estudios mencionados, evaluaron la importancia del flujo de especies en
paisajes fragmentados y cómo se afecta la diversidad de especies de
himenópteros con el grado de intervención del ecosistema. Este abordaje asume
que la intensificación agrícola (monocultivo) o grado de intervención lleva implícito
un cambio (disminución) en la diversidad vegetal y que, por tanto, la identidad y
diversidad de especies de parasitoides se relacionan con diferentes especies
plagas asociadas a diferentes manejos o agroecosistemas.

11
4. OBJETIVOS
4.1 Objetivo general
Establecer si la abundancia y diversidad de himenópteros parasitoides asociados
al sistema silvopastoril y al sistema natural de la Reserva Natural El Hatico se
puede explicar por su composición y cobertura vegetal.
4.2 Objetivos específicos
Identificar y cuantificar la riqueza y abundancia de parasitoides en el sistema
silvopastoril de la Reserva.
Comparar la riqueza y abundancia de parasitoides del sistema silvopastoril con
un bosque primario (sistema natural y control positivo) presente dentro del
sistema.
Relacionar la abundancia de parasitoides con cobertura y composición vegetal.
Relacionar la abundancia de los parasitoides con los niveles de ataque de la
polilla del algarrobo (Lepidoptera:Tortricidae) sobre los árboles de algarrobo.
4.3 Hipótesis biológica 1: La diversidad y abundancia de especies de
parasitoides se relacionan con la cobertura y composición vegetal del sistema.
Predicción 1: Las diferencias en diversidad y abundancia de parasitoides dentro
del sistema silvopastoril están correlacionadas a los cambios en cobertura y
composición vegetal en distintos puntos del mismo.

12
4.4 Hipótesis biológica 2: La riqueza y abundancia de las especies de avispas
parasitoides que habitan dentro del sistema silvopastoril son afectados
favorablemente, por el fragmento de bosque primario.
Predicción 2: A menor distancia del fragmento de bosque (en el mismo hábitat),
mayor será la riqueza y abundancia de especies de avispas asociadas al sistema
silvopastoril de la Reserva.

13
5. MATERIALES Y MÉTODOS
5.1 Área de estudio
El trabajo de campo se llevó a cabo en la Reserva Natural El Hatico, reserva de la
Sociedad Civil, de 288 ha, localizada en el municipio de El Cerrito, departamento
del Valle del Cauca, Colombia (3º 27`N y 76º 32`O), a 1000msnm; con
precipitación promedio anual de 750mm, distribuida bimodalmente (abril - junio y
octubre - noviembre); temperatura promedio de 24ºC y humedad relativa de 75%
(Mahecha 1998). La Reserva se sitúa en una zona ecológica, según Holdridge
(1978), de Bosque Seco Tropical (bs- T).
Dentro de la reserva existe un fragmento de bosque primario de 15 hectáreas, el
cual comparte límites con los sistemas manejados, específicamente cultivo
orgánico de caña y sistema silvopastoril. Este último consiste en una amplia red de
lotes y la zona más alejada del bosque (900m), además de ser utilizada para
pastoreo, tiene una variedad de plantas ornamentales como palmas sanconas
(Syagrus sancona H. Karst.), palmas reales (Roystonea regia (Kunth) O.F. Cook),
samanes (Albizia saman (Jacq.) F. Muell), pomarrosos (Syzygium malaccense
(L.) Merr. & L. M. Perry.), entre otras. Esta zona es muy diversa y comprende todo
el jardín y la vivienda que ha sido usada durante nueve generaciones, por esto
este punto se considera un “sitio especial”. Los lotes del sistema silvopastoril
contienen pasto estrella (Cynodon plectostachyus (K. Schum) Pilg.) y pasto guinea
(Panicum maximum) (Jacq) B.K. Simon & S.W.L. Jacobs, intercalados con más de
18 especies de árboles y arbustos incluyendo leucaena (Leucaena leucocephala
(Lam.) De Wit.) para pastoreo y algarrobo (Prosopis juliflora (Sw.) DC.); sin

14
embargo, para este estudio no se tuvo en cuenta lotes que incluyeran leucaena
como complemento del pasto estrella y el pasto guinea.
5.2 Fase de campo
Se llevó a cabo una salida piloto (de tres días) en la cual se realizaron actividades
de organización y evaluación de las condiciones del sistema silvopastoril y el
sistema natural (bosque). Con base en el plano de manejo de la reserva (año
2003) se escogieron los lotes a muestrear y con un GPS (marca Garmin ) se
midieron las coordenadas geográficas y se realizó un conteo de árboles de P.
juliflora, en cada uno de ellos. Se escogieron, marcaron y georeferenciaron
aquellos algarrobos sobre los cuales se realizaron los muestreos subsiguientes.
Con el fin de obtener datos de ambos periodos climáticos y, por tanto, una mayor
representatividad de la fauna de Hymenoptera parasítica, se realizaron dos salidas
de campo, cada una de cuatro días, en las últimas semanas de febrero (periodo
seco) y mayo (periodo de lluvias) de 2010. Además del bosque, en cada salida se
muestrearon cuatro lotes de, aproximadamente, 8000m2 c/u, a diferentes
distancias del fragmento de bosque presente en la reserva (Figura 1) uno al borde
del bosque (10m), los siguientes dos lotes a una distancia cercana (225m) y media
(450m) y, el último, con la distancia máxima posible con relación al bosque
(900m). Se midió la distancia entre el punto medio del bosque y el punto medio de
cada uno de los lotes de muestreo mediante el uso de GPS.

15
Figura 1. Esquema de la distribución de los lotes muestreados dentro del sistema
silvopastoril, la ubicación del bosque y la ubicación aproximada del transecto de
muestreo establecido en el mismo (40m hace referencia a la longitud del transecto
sin escala alguna). Modificado del plano original de la Reserva Natural El Hatico
(2003).
10m
225m
450m
900m
40m

16
En cada lote se escogieron, de forma aleatoria, seis árboles (para un total de 24
en los cuatro lotes) y se marcaron con placas metálicas de 3 x 4cm para llevar un
registro individual. Se tomaron nueve muestras de trozos de ramas de
aproximadamente 20cm cortadas con desjarretadora (unida a una pértiga de 13m)
a partir de tres estratos verticales, de 33% cada uno, del área de follaje del árbol,
así: el estrato bajo o área inferior del árbol, el estrato alto o área del dosel y el
estrato medio en el cual se encuentra el área media del follaje del árbol. De cada
muestra se obtuvo el porcentaje de herbivoría, definido como hojas afectadas por
la polilla del algarrobo sobre el total de hojas en cada trozo de rama.
Además, con el objeto de capturar himenópteros parasitoides, en cada árbol
marcado, se sujetaron con tachuelas, a dos metros de altura, platos plásticos
esféricos amarillos (pan traps) de 20cm de radio, impregnados con aceite de ricino
(Briceño et al. 2006, García 2003). Otros seis platos se distribuyeron en árboles,
escogidos al azar, a lo largo de un transecto de 40m, a 20m en el interior del
bosque, como muestreo testigo. De este modo, se pudo estimar el número de
especies al interior de los sistemas, incluyendo las potencialmente asociadas a la
polilla del algarrobo.
Dado el pequeño tamaño de los folíolos de P. Juliflora, pues sus hojas son
compuestas y bipinnadas (Figura 2) el porcentaje de herbivoría se calculó
mediante el conteo de cada hoja (conjunto de foliolos, óvalo marcado en la Figura
2) que presentó alguna evidencia de daño por masticador (específicamente la

17
polilla del algarrobo) y se dividió sobre el número total de hojas en el trozo de 20
cm de la rama colectada (n=3 por cada estrato del árbol).
Por último, se tomaron medidas de porcentaje de cobertura de dosel debajo de
cada árbol de algarrobo con un densitómetro esférico de vegetación y se registró
en cada lote la temperatura a, aproximadamente, a un metro de distancia con
respecto al tronco del árbol y a la misma distancia del suelo.
Figura 2. Ramas y hoja del algarrobo compuesta por cuatro o seis subraquis.
Fotografías por Lorena López Quiñonez.
5.3 Determinación taxonómica
Se colectó e identificó hasta el nivel de género y/o especie las arvenses, arbustos
y árboles dentro del lote, diferentes al algarrobo. Se identificaron con la ayuda del
personal encargado del herbario de la Universidad del Valle y confirmado por
comparaciones con las plantas presentes en el mismo. Las plantas colectadas
reposan en el Herbario de la Universidad del Valle (CUVC).

18
Los himenópteros colectados, se montaron en seco y se identificaron hasta el nivel
taxonómico más bajo posible mediante las claves de Gibson et al. (1997) y
Fernández & Sharkey (2006). La colección en seco resultante, debidamente
rotulada, se depositó en el Museo de Entomología de la Universidad del Valle
(MUSENUV).
5.4 Análisis de datos
Para los análisis estadísticos se tuvo en cuenta dos factores: época climática y
sistemas de manejo (silvopastoril y bosque). Por otro lado, dentro del sistema
silvopastoril se consideraron cuatro condiciones de acuerdo a la distancia respecto
al bosque: Borde, Cercano, Medio y Lejano.
Para poner a prueba la primera hipótesis biológica se calculó el índice de
diversidad de Shannon (H’) para los himenópteros parasitoides colectados en los
cuatro lotes del sistema silvopastoril y los capturados en el sistema natural
(bosque primario); se realizó también una prueba t-student con los cinco índices
obtenidos comparándolos en pares. Se llevaron a cabo dos análisis de
agrupamiento basados en los índices de similitud de Jaccard y Bray Curtis,
altamente sensibles a la riqueza de especies y abundancia de especies,
respectivamente. Para los análisis se utilizó el programa estadístico PAST 1.73b
(Hammer et al. 2001).
Para conocer el nivel de efectividad del muestreo en los diferentes sistemas, se
realizaron curvas de rarefacción para cada uno (sistema silvopastoril que incluye

19
los lotes Borde, Cercano, Medio y Lejano, y bosque), con los valores observados
de abundancia (Sobs Mao Tau), promediando los estimadores ICE, Chao 2 y
Jacknife 2 (Estimates 800, Colwell 1997) y, por último, con base en la riqueza de
especies y el número de especímenes colectados (PAST 1.73b, Hammer et al.
2001).
Los porcentajes de herbivoría obtenidos en las diferentes épocas y sistemas
fueron analizados mediante una ANOVA factorial con el objeto de examinar si
existía interacción entre los dos factores evaluados (clima y sistema de manejo) o
si se presentaban diferencias estadísticamente significativas para cada factor por
separado. Esto se hizo empleando el paquete Statistica 7.0. Además, se calculó
el índice de correlación de Pearson (r) para corroborar si existe algún tipo de
relación entre cobertura vegetal, riqueza de plantas y abundancia de parasitoides.
Por último, mediante Statistica 7.0, se realizó una prueba t-student para buscar
diferencias entre la cobertura vegetal medida en las dos épocas climáticas y una
ANOVA de una vía para identificar diferencias entre la cobertura vegetal de los
diferentes lotes y sistemas.

20
6. RESULTADOS Y DISCUSIÓN
6.1 Composición general de parasitoides
Se capturaron 1376 especímenes de Hymenoptera, más específicamente del
grupo conocido como “Hymenoptera parasítica”, agrupadas en siete superfamilias,
18 familias y 42 morfoespecies (Tabla 1), de las cuales 30 (71%) fueron
determinadas hasta género y dos (5%) hasta especie. Chalcidoidea fue la
superfamilia más representativa con nueve familias, 20 morfoespecies
identificadas y 1295 especímenes colectados, correspondiente al 48% de las
morfoespecies (Figura 3) y al 94.11% del número total de especímenes
capturados.
Platygastroidea
24%
Chalcidoidea
49%Chrysidoidea
8%
Mymmaromatoidea
3%
Ichneumonoidea
3%
Diaprioidea
3%
Ceraphronoidea
10%
Figura 3. Frecuencias de morfoespecies por superfamilia de Hymenoptera
parasítica, colectados en ambas épocas climáticas, incluyendo el sistema
silvopastoril y el sistema natural de la Reserva Natural El Hatico.

21
Encyrtidae presentó el mayor número de morfoespecies (5) dentro de
Chalcidoidea (Tabla 1) y la mayor abundancia con 1138 especímenes capturados
que equivalen al 83% de la abundancia total. A su vez, a los géneros Acerophagus
cf. y Pentelicus, les correspondió, respectivamente, el 69 y 11% de la abundancia
total (Anexo A), siendo los géneros más abundantes en el sistema, seguidos por
Alaptus, de Mymaridae, con el 3.42% (Anexo A). Vale la pena resaltar que
Acerophagus cf. y Pentelicus fueron, respectivamente, los más abundantes en
época seca (febrero 2010) y lluviosa (mayo 2010).
En su mayoría, la fauna parasítica identificada dentro de la Reserva corresponde a
familias y géneros que han sido comúnmente utilizados como agentes de control
biológico en Colombia (Fernández & Sharkey 2006). La familia Encyrtidae ocupa
el segundo puesto en términos de éxito en programas de control biológico clásico
(Madrigal-Cardeño 2001) y de Acerophagus, que presentó mayor abundancia
dentro de la reserva en época seca, se han reportado, desde 1989, varias
especies parasitando huevos de diferentes hospederos en el Valle del Cauca
(Fernández & Sharkey 2006). Mymaridae constituye una familia en la cual todas
las especies conocidas son parasitoides de huevos (Madrigal-Cardeño 2001) y al
igual la mayoría de las familias de parasitoides encontradas dentro de la Reserva
son endoparasitoides de huevos.

22
Tabla 1. Superfamilias, familias, subfamilias, géneros y/o especies de
Hymenoptera parasítica encontradas en el sistema silvopastoril y el bosque de la
Reserva Natural El Hatico.
Superfamilia Familia Subfamilia Género y/o especie Msp*
Ceraphronoidea Ceraphronidae Ceraphron Cer 1 Ceraphronoidea Ceraphronidae Aphanogmus Cer 2 Ceraphronoidea Ceraphronidae Ceraphron Cer 3 Ceraphronoidea Ceraphronidae Aphanogmus Cer 4 Ceraphronoidea Megaspilidae Dendrocerus Meg 1
Chalcidoidea Aphelinidae Coccophaginae Encarsia Aph 1 Chalcidoidea Aphelinidae Coccophaginae Encarsia pergandiella
Chalcidoidea Chalcididae Chalcidinae Brachymeria Cha 1 Chalcidoidea Chalcididae Chalcidinae Chalcis Cha 2 Chalcidoidea Chalcididae Chalcidinae Trigonura Cha 3
Chalcidoidea Encyrtidae Encyrtinae Acerophagus cf. Enc 1 Chalcidoidea Encyrtidae Encyrtinae Cerchysiella cf. Enc 2 Chalcidoidea Encyrtidae Encyrtinae Encyrtus Enc 3 Chalcidoidea Encyrtidae Encyrtinae Plagiomerus cf. Enc 4 Chalcidoidea Encyrtidae Encyrtinae Pentelicus Enc 5
Chalcidoidea Eulophidae Eul 1 Chalcidoidea Eulophidae Eul 2
Chalcidoidea Eurytomidae Eur 1
Chalcidoidea Mymaridae Gonatocerinae Alaptus Mym 1 Chalcidoidea Mymaridae Gonatocerinae Camptoptera Mym 2 Chalcidoidea Mymaridae Gonatocerinae Gonatocerus Mym 3
Chalcidoidea Pteromalidae Pte 1
Chalcidoidea Signiphoridae Signiphora Sig 1
Chalcidoidea Trichogrammatidae Tri 1 Chalcidoidea Trichogrammatidae Tri 2
Chrysidoidea Bethylidae Bet 1 Chrysidoidea Bethylidae Bet 2
Chrysidoidea Chrysididae Chr 1
Chrysidoidea Dryinidae Dry 1
Diaprioidea** Diapriidae Diapriinae Doliopria Dia 1
Ichneumonoidea Braconidae Microgastrinae Bra 1
Mymarommatoidea Mymarommatidae Mma 1
Platygastroidea** Platygastridae Platygastrinae Isostasius Pla 1 Platygastroidea Platygastridae Platygastrinae Synopeas Pla 2 Platygastroidea Platygastridae Scelioninae Idris Pla 3 Platygastroidea Platygastridae Telenominae Trissolcus Pla 4 Platygastroidea Platygastridae Telenominae Telenomus Pla 5 Platygastroidea Platygastridae Telenominae Telenomus longicornis Platygastroidea Platygastridae Telenominae Telenomus Pla 7 Platygastroidea Platygastridae Scelioninae Calotelea Pla 8 Platygastroidea Platygastridae Telenominae Trissolcus Pla 9 Platygastroidea Platygastridae Telenominae Telenomus Pla 10
*A las morfoespecies que no se determinaron hasta el nivel de especie, se les asignó un nombre de acuerdo a su familia y número de morfoespecie. **sensu Sharkey (2007).

23
Muy pocos ejemplares fueron colectados de las restantes superfamilias (Tabla 1,
Anexo A).
Entre los géneros encontrados se destaca Gonatocerus (Mymaridae) y Telenomus
(Platigastridae) por haber sido utilizadas exitosamente para el control biológico de
varias especies plaga, desde los años sesenta (Fernández & Sharkey 2006,
Madrigal-Cardeño 2001). La presencia de cuatro morfoespecies de Telenomus,
dentro de la Reserva, sugiere el establecimiento de poblaciones naturales de este
parasitoide en la región o que, posiblemente, sea resultado de continuas
liberaciones que aún se llevan a cabo.
Por otro lado, se conoce que más de cinco especies de Encarsia, incluyendo a E.
pergandiella Howard, que atacan a la mosca blanca (Hemiptera: Aleyrodidae) han
sido liberadas regularmente en el Valle, en las últimas tres décadas (Myartseva et
al. 2007). Por lo tanto, el hallazgo de E. pergandiella, al igual que una especie
desconocida del mismo género, dentro de la Reserva sea explicada por la
presencia de mosca blanca en algunos algarrobos cercanos al relicto de bosque.
No obstante, se conoce que estos parasitoides tienen un amplio rango de
hospederos.
El género Signiphora (Signiphoridae) contiene especies parásitas y hiperparásitas
de cóccidos y algunas especies de homópteros de la familia Aleyrodidae y de
algunos de sus parasitoides (Myartseva et al. 2007, Fernández & Sharkey 2006).

24
Se identificaron endoparasitoides de larvas y pupas como bracónidos
(Microgastrinae) y calcídidos de los géneros Trigonura, Chalcis y Brachymeria.
Además, se pudo observar enjambres de estos últimos rodeando las larvas y
pupas de la polilla del algarrobo, en época seca, especialmente, cuando la larva
del algarrobo se encontraba en los últimos ínstares de su ciclo larval. Aunque
Reyes (2010) reportó la emergencia de Conura (Chalcididae) a partir de pupas de
la larva del algarrobo, no se capturó ningún individuo de este género.
Sólo se tiene el reporte de que cuatro de las morfoespecies de parasitoides
identificadas (i.e. Brachymeria, Pteromalidae: Pte 1, Trichogrammatidae: Tri 1 e
Isostasius) atacan directamente a la polilla (Reyes 2010). Sin embargo, la baja
captura de estas cuatro especies no permitió relacionar claramente el vínculo
parasitoide-polilla-herbivoría.
6.2 Comparación de parasitoides entre sistemas
En los cuatro lotes muestreados dentro del sistema silvopastoril se colectaron 41
morfoespecies y un total de 1358 himenópteros parasitoides que corresponden al
97% de la abundancia total. La abundancia promedio de los tres lotes más
cercanos al bosque: Borde (10m), Cercano (225m) y Medio (450m) (sistema
silvopastoril) fue 331 (Tabla 2) y 39 (93%) especies se colectaron en estos lotes
(Anexo A). Los índices de diversidad (Tabla 2) son comparables con varios
registros de himenópteros en sistemas agroforestales que muestran índices de
diversidad (H’) entre 1 y 2 (Cronin 2003, Haddad et al. 2001). No obstante, no se
encontraron diferencias significativas entre los índices calculados (Tabla 3). El lote

25
ubicado a 900 metros de distancia del bosque y considerado como una zona de
alta diversidad floral y artropodiana, presentó el mayor número de individuos
colectados (n=364, Tabla 2), que representa el 26% de la abundancia total. Este
lote comparte la riqueza de especies con los demás lotes, con excepción de la
morfoespecie de Braconidae (subfamilia Microgastrinae, Anexo A), capturada
exclusivamente en este lote y en el sistema natural. Vale la pena resaltar que se
encontró una morfoespecie de Pteromalidae con un único individuo capturado en
el bosque primario (Figura 1, Anexo A).
Tabla 2. Riqueza de especies, abundancia e índice de Shannon (H’) del sistema
natural (bosque) y cada lote del sistema silvopastoril en la RNH, teniendo en
cuenta ambas épocas climáticas (seca: febrero 2010 y lluviosa: mayo 2010).
Bosque Borde (10m) Cercano (225m) Medio (450m) Lejano (900m) Total
Riqueza 11 23 26 24 22 42*
Abundancia 18 319 345 330 364 1376
H’ 2.139 1.22 1.13 1.053 1.486
* Número total de morfoespecies capturadas, NO es una cifra acumulada según los valores de la
tabla.
El índice de Shannon del lote Lejano fue el mayor valor del sistema silvopastoril
(H’= 1.486), inferior al registrado en el bosque (sistema natural), con el cual no
presentó diferencias significativas (Tabla 3). Sin embargo este fue
significativamente diferente al obtenido en los otros lotes muestreados (Tabla 3).

26
Tabla 3. Comparación entre los índices de Shannon (H’) del bosque y cada lote
del sistema silvopastoril, mediante prueba t-student. Los valores de p en negrilla
indican diferencias significativas entre la diversidad de los lotes.
Borde Cercano Medio Lejano Bosque
Borde 1
Cercano 0.49825 1
Medio 0.21384 0.57533 1
Lejano 0.025057 0.002731 0.00025807 1
Bosque 0.0099225 0.0039852 0.0018159 0.098324 1
El bosque presentó la menor riqueza de especies (26%), la menor abundancia
(1.3%, Tabla 2) pero el índice de diversidad más alto (H’= 2.139). Esto indica que
el fragmento de bosque muestreado tiene un alto nivel de heterogeneidad, es
decir, no hay dominancia de unas pocas especies, por el contrario se encuentran
equitativamente repartidos entre ellas (Zar 1984). Lo anterior explica por qué la
abundancia (18) es, aproximadamente, igual al número de especies (11). Este
índice de diversidad es congruente con estudios realizados en paisajes
fragmentados que evalúan el recambio y diversidad de especies de himenópteros
en un mosaico paisajístico. Generalmente, los índices de diversidad (H’) de dichos
fragmentos no son menores a 2 (Cronin 2003, Haddad et al. 2001). Aún así, no
hubo diferencias con el índice del lote Lejano pero sí fue significativamente
diferente con el índice de diversidad de los demás lotes del sistema silvopastoril.
El análisis de agrupamiento basado en el índice de similitud de Jaccard (Figura 4)
el cual es sensible a la riqueza de especies de cada área, reveló que el lote Lejano

27
comparte, el 60% de las especies con el lote Cercano, mientras que el bosque,
sólo comparte el 20% de las especies con los lotes del sistema silvopastoril
(Figura 4).
Figura 4. Análisis de agrupamiento, basado en el índice de similitud de Jaccard,
para los lotes presentes en el sistema silvopastoril y para el sistema natural
(bosque) en la Reserva Natural El Hatico.
De igual manera, el análisis de agrupamiento, basado en el índice de similitud de
Bray-Curtis (Figura 5), el cual es sensible a la abundancia de cada una de las
especies colectadas, mostró que el lote Lejano comparte con los demás lotes del
sistema silvopatoril (Borde, Cercano y Medio) el 60% de la abundancia de
especímenes colectados, mientras que el bosque comparte con los mismos
menos del 10% de la abundancia total (Figura 5).
00
.61.2
1.82
.43
3.6
4.2
4.8
5.4
6
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9 1
Similarity
Bosque
Cercano
Lejano
Medio
Borde

28
Con respecto a las épocas climáticas, se colectó el 45.42% (624) del total de
especímenes en la época seca (febrero 2010), siendo mayor el número de
himenópteros parasitoides colectados en la época lluviosa con un 54.57% (752);
no obstante el número de morfoespecies colectadas en ambas épocas fue el
mismo (33).
Figura 5. Análisis de agrupamiento, basado en el índice de similitud de Bray-
Curtis, para los lotes presentes en el sistema silvopastoril y para el sistema
natural (bosque) en la Reserva Natural El Hatico.
Con respecto a la riqueza observada en cada área (Figura 6 a), se encontró que el
bosque mantuvo el menor nivel a medida que aumentan las unidades de muestreo
a diferencia de los lotes presentes en el sistema silvopastoril los cuales se
mantuvieron en un corto rango de 12 a 16 especies de himenópteros.
00
.61.2
1.82
.43
3.6
4.2
4.8
5.4
6
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9 1
Similarity
Bosque
Lejano
Cercano
Medio
Borde
29
a.
b.
c.
Figura 6. Riqueza de especies observadas (Sobs Mao Tau) (a) y promedio de los
estimadores ICE, Jack 2 y Chao 2 (b y c) para los parasitoides colectados en el
sistema silvopastoril y natural en la Reserva Natural El Hatico.
0
2
4
6
8
10
12
14
16
18
1 2 3 4 5 6 7 8 9 10 11 12
Unidades experimentales
So
bs M
ao
Tau
Borde Cercano Medio Lejano Bosque
0
5
10
15
20
25
30
35
40
1 2 3 4 5 6 7 8 9 10 11 12
Unidades experimentales
Pro
med
io I
CE
, Jack 2
y C
hao
2
Borde Cercano Medio Lejano Bosque
0
5
10
15
20
25
30
1
21
41
61
81
101
121
141
161
181
201
221
241
261
281
301
321
341
361
Número de individuos
Riq
ue
za d
e e
spe
cie
s
Lejano Bosque Borde Cercano Medio

30
La riqueza de especies estimada (Figura 6 b), mostró que tanto la curva del lote
Lejano como las de los lotes Borde y Medio, tenían tendencia a ser constantes,
mostrando que el muestreo en estos lotes fue el que más se acercó al valor
estimado. Por el contrario, la curva del lote Cercano se observó ascendente e
irregular (Figura 6 a), lo cual indica que, probablemente, se debe hacer un mayor
esfuerzo de muestreo en dicha zona para alcanzar los valores de riqueza de
especies estimados.
La curva del bosque obtenida con el promedio de los estimadores (Figura 6 b), se
mantuvo baja y con tendencia a ser constante pero al compararse con aquella
obtenida con base en el número de especímenes colectados (Figura 6 c), indica
que aunque la curva de estimadores parezca constante, se debe a que el esfuerzo
de muestreo debe ser aún mayor en el sistema natural que el realizado en el
sistema silvopastoril pues los individuos colectados en el primero (18
especímenes) fueron considerablemente bajos en comparación con los colectados
en los últimos (340 especímenes en promedio). Por su parte, los lotes Lejano y
Cercano mostraron las curvas con mayor número de especies y mayor número de
individuos (Figura 6 c).
6.3 Composición vegetal y especies de parasitoides
Se registró un total de 21 familias y 34 morfoespecies de plantas presentes dentro
del sistema silvopastoril (los cuatro lotes muestreados) y el 91% se identificó hasta
especie (Tabla 4). De éstas el 53% fueron arvenses (plantas herbáceas de 1m de
altura o menos) y el 47% restante árboles y arbustos (plantas de más de 1m de

31
altura). Las arvenses dominantes fueron Lantana camara L., Parthenium
hysterophorus L. y Sida acuta Brum. F., encontradas en la mayoría de los lotes y
en altas densidades. La diversidad de arvenses es de gran importancia ya que
cumplen el papel de atrayentes de enemigos naturales al ofrecer recursos de
microhábitat y alimento que, en el caso específico de los parasitoides, puede ser
determinante en la riqueza de especies, en la longevidad y el nivel de parasitismo
de las especies asociadas a dichas plantas (Araj et al. 2008).
Los árboles que se encontraron con mayor abundancia, especialmente por la
presencia de corredores en los bordes de varios de los lotes de muestreo, fueron
aromo (Acacia farnesiana (L.) Willd), mataratón (Gliricidia sepium (Jacq.) Kunth ex
Walp), chiminango (Pithecellobium dulce (Roxb.) Benth) y flor amarillo (Senna
spectabilis (DC.) Irwin & Barneby).
Los mayores valores de riqueza de plantas se encontraron en los lotes Cercano y
Lejano, con 18 especies cada uno, por lo cual no se encontraron diferencias
significativas entre la riqueza de estos dos lotes, contrario a lo obtenido al
compararlos con la riqueza encontrada en los dos lotes restantes (p= 0.041123,
gl=3) (Figura 7).

32
Tabla 4. Familias, subfamilias, géneros y especies de plantas encontradas dentro
del sistema silvopastoril en la Reserva Natural El Hatico.
Familia Subfamilia Género Especie Nombre Común
Arv
ense
s
Amarantaceae Achyrantes Achyrantes aspera Bledo
Asclepiadaceae Asclepia Asclepia curassavica Algodoncillo
Asteraceae Eirmocephala Eirmocephala brachiata Tabaquillo
Asteraceae Emilia Emilia fosbergii Pincelito
Asteraceae Parthenium Parthenium hysterophorus Marihuana macho
Campanulaceae Hypobroma Hypobroma longiflora
Ciperaceae Cyperus Cyperus hermafhroditus Coquito
Euphorbiaceae Acalypha Acalypha setosa
Fabaceae Cesalpinioidae Senna Senna hirsuta
Fabaceae Faboidae Crotalaria Crotalaria palida
Fabaceae Mimosoidae Mimosa Mimosa pudica Dormidera
Loasaceae Nasa
Malvaceae Sida Sida acuta Escoba
Phytollacaceae Trichostigma Trichostigma octandrum
Phytollacaeae Rivina Rivina humilis
Solanaceae Capsicum Ají
Solanaceae Solanum Lulo de perro
Verbenaceae Lantana Lantana camara Lantana
Achatocarpaceae Achatocarpus Achatocarpus nigricans Totocal
Bombacaceae Ceiba Ceiba pentandra Ceiba
Árb
ole
s y
arb
usto
s
Fabaceae Cesalpinioidae Senna Senna spectabilis Flor amarillo
Fabaceae Faboidae Erythrina Erythrina poeppigiana Písamo Fabaceae Faboidae Gliricida Gliricidia sepium Matarratón Fabaceae Mimosoidae Acacia Acacia farnesiana Aromo Fabaceae Mimosoidae Leucaena Leucaena leucocephala Leucaena Fabaceae Mimosoidae Pithecellobium Pithecellobium dulce Chiminango Fabaceae Mimosoidae Saman Saman samanea Saman Meliaceae Guarea Guarea guidonia Guarea o ceibo Moraceae Maclura Maclura tinctoria Dinde Myrtaceae Sizigium Sizigium malaccense Pomarrosa Palmae Siagrus Siagrus sancona Palma sancona Rutaceae Zanthoxylum Zanthoxylum roifolium Tachuelo Sterculiaceae Guazuma Guazuma ulmifolia Guázimo Urticaceae Urera Urera baccifera Urera

33
Borde Cercano Medio Lejano
Lote
6
8
10
12
14
16
18
20
22
Riq
ueza
Figura 7. Comparación de la riqueza de plantas en cada lote del sistema
silvopastoril en la Reserva Natural El Hatico. Se incluyen las especies de plantas
encontradas en ambas épocas climáticas.
Se observó una ligera tendencia a la correlación entre la abundancia de
parasitoides dentro del sistema silvopastoril y la vegetación (Figura 8), ya que los
lotes con mayor riqueza de plantas (Cercano y Lejano) mostraron también las
mayores abundancias de parasitoides.
290
300
310
320
330
340
350
360
370
Borde Cercano Medio Lejano
0
2
4
6
8
10
12
14
16
18
20
Abundancia de parasitoides Riqueza de plantas
Figura 8. Relación entre la abundancia de parasitoides y la riqueza de plantas en
cada lote del sistema silvopastoril en la Reserva Natural El Hatico.

34
El coeficiente de correlación de Pearson obtenido (r = 0.8) indica que sí existe una
relación positiva y significativa entre la abundancia de parasitoides y la riqueza de
especies de plantas de cada lote. Lo anterior es consecuente con la enorme
importancia que tiene la diversidad floral sobre la diversificación de insectos en
agroforestales. Se ha puesto a prueba en diversos estudios diferentes modelos de
policultivos, de cultivos artesanales, entre otros y, en su mayoría, se ha
comprobado que la composición y diversidad de plantas dentro o alrededor de los
sistemas productivos aumenta significativamente el número de enemigos
naturales, incluyendo parasitoides (Letourneau et al. 2010, Araj et al. 2008). Del
mismo modo, la presencia algunas especies de plantas florales y/o ornamentales
pueden hacer el papel de amortiguadores influenciando el movimiento de
herbívoros evitando que lleguen hasta el cultivo o atrayéndolos fuera del mismo
(Letourneau et al. 2010).
6.4 Cobertura vegetal y especies de parasitoides
Al medir el porcentaje de cobertura vegetal en épocas seca (febrero de 2010) y
lluviosa (mayo de 2010) (Figura 9), de cada algarrobo presente en los cuatro lotes
muestreados dentro del sistema silvopastoril no se encontró diferencias
significativas entre ellas (p= 0.057039, gl= 46).

35
0
20
40
60
80
100
Borde Cercano Medio Lejano
Lote%
Co
bert
ura
290
300
310
320
330
340
350
360
370
Ab
un
dan
cia
Época seca Época de lluvias Abundancia
Figura 9. Relación entre porcentaje de cobertura de los algarrobos y la
abundancia de parasitoides en cada uno de los lotes del sistema silvopastoril en la
Reserva Natural El Hatico.
Al examinar si existe relación entre el porcentaje de cobertura de los algarrobos de
cada lote y el número de parasitoides capturados en los mismos (Figura 9)
mediante un análisis de varianza (ANOVA) de una vía, no se encontraron
diferencias estadísticamente significativas entre los porcentajes de cobertura de
los algarrobos de los cuatro lotes (p= 0.360852, gl=3). A su vez, el coeficiente de
correlación (r= 0.6), confirmó la relación positiva y significativa entre las dos
variables (Figura 9).
6.5 Niveles de herbivoría y riqueza de parasitoides
Se encontraron diferencias significativas entre mediciones de herbivoría de la
polilla Tortricidae durante las dos épocas climáticas del año (seca-febrero 2010 y
lluviosa-mayo 2010) (p= 0.024365, gl=1) (Figura 10), lo cual puede ser claramente
observado en campo ya que al llegar la época de lluvias el follaje de los algarrobos

36
es reestablecido, debido a una menor presencia de esta plaga por las condiciones
climáticas que la afectan.
Seca Lluviosa
Época
70
75
80
85
90
95
100
% H
erb
ivo
ría
Figura 10. Diferencias estacionales entre los porcentajes de herbivoría de (épocas
seca y lluviosa) en la Reserva Natural El Hatico.
0
20
40
60
80
100
Borde Cercano Medio Lejano
Lote
% H
erb
ivo
ría
280
300
320
340
360
380
Ab
un
dan
cia
Época seca Época de lluvias Abundancia de parasitoides
Figura 11. Porcentajes de herbivoría promedio de los algarrobos de cuatro lotes
del sistema silvopastoril en la Reserva Natural El Hatico, en dos épocas climáticas
diferentes y abundancia promedio de parasitoides en ambas épocas.

37
No se observó ningún tipo de relación entre las variables porcentaje de herbivoría
obtenido en los algarrobos con la riqueza y la abundancia de parasitoides en cada
lote del sistema silvopastoril (Figura 11).
A pesar de la disminución en la herbivoría en época lluviosa vale la pena recordar
que no se observó el mismo patrón con la riqueza ni abundancia de parasitoides.
Se conoce que los ensamblajes y el tamaño en las poblaciones de insectos
cambian según las condiciones climáticas (Letourneau et al. 2009) y, aunque las
poblaciones de la polilla del algarrobo disminuyeron en época de lluvias, fue
notorio un recambio entre las especies de la familia Encyrtidae, específicamente
entre aquellas de Acerophagus y Pentelicus, ya que las poblaciones de ésta última
aumentaron considerablemente en la época húmeda y las de Acerophagus
bajaron un poco no abrupta, permitiendo mantener, de ambas especies,
poblaciones con un alto número de individuos (Anexo A).
Las mediciones de herbivoría en tres estratos verticales para el follaje de los
algarrobos del sistema (33% del mismo) no revelaron diferencias significativas
entre ninguno de los estratos ni los lotes (p= 0.614324, gl=2; p=0.237154, gl=3,
estratos y lotes de la época seca y p= 0.132240, gl=2; p=0.071513, gl=3, estratos
y lotes de la época lluviosa, ANOVA de dos vías) (Figura 12).

38
a.
b.
Figura 12. Porcentajes de herbivoría de tres estratos diferentes del follaje de los
algarrobos (alto, medio y bajo), en cuatro lotes del sistema silvopastoril y dos
épocas climáticas: a. seca (febrero de 2010); b. lluviosa (mayo de 2010).
6.6 Propiedades físico-químicas del sistema silvopastoril
Al evaluar otras propiedades implícitas en el sistema silvopastoril se encontró que,
el lote Medio es el que presentó mayores niveles de temperatura tanto en época
seca (30 ºC) como lluviosa (28 ºC) y vale la pena resaltar que este fue también el
lote en el cual se capturó el menor número de parasitoides en ambas épocas
(Figura 13).
0
20
40
60
80
100
Borde Cercano Medio Lejano
Lote
% H
erb
ivo
ría
Alto Medio Bajo
0
20
40
60
80
100
Borde Cercano Medio Lejano
Lote
% H
erb
ivo
ría
Alto Medio Bajo

39
24
25
26
27
28
29
30
31
Borde Cercano Lejano Casa
Lote
Tem
pera
tura
(ºC
)290
300
310
320
330
340
350
360
370
Ab
un
dan
cia
Época seca Época lluviosa Abundancia
Figura 13. Temperatura promedio de los cuatro lotes en el sistema silvopastoril de
la Reserva Natural El Hatico, y abundancia de parasitoides colectados en los
mismos.
En contraste, el lote Lejano presentó los más bajos niveles de temperatura (entre
29 – 30 ºC en época seca y 26 – 27 ºC en época lluviosa) (Figura 13) y, a su vez,
el mayor número de parasitoides.
Figura 14. Temperatura y precipitación promedio en los últimos cuatro años (2007
- 2010) en la Reserva Natural El Hatico.
0
50
100
150
200
250
E F M A M J J A S O N D E F M A M J J A S O N D E F M A M J J A S O N D E F M A M J J A21
21,5
22
22,5
23
23,5
24
24,5
25
25,5
Precipitación Temperatura Media 2007-2010
2007 2007
2008 2009 2010
Pre
cip
itac
ión
mm
/me
s
Tem
pera
tura
(ºC)

40
Los muestreos se realizaron durante dos épocas climáticas muy marcadas. En la
época seca (febrero de 2010), la Reserva se encontraba bajo el fenómeno del
niño; se registraron los mayores niveles de temperatura y los menores niveles de
precipitación en los últimos cuatro años (Figura 14). Por el contrario, en la época
de lluvias (mayo de 2010), estaba iniciando el fenómeno de la niña, unos de los
periodos más húmedos en la historia de la Reserva, registrando los mayores
niveles de precipitación en los últimos cuatros años (C.H. Molina, com. pers. 2010)
(Figura 14).

41
7. CONCLUSIONES
En la Reserva Natural El Hatico, se presenta alta abundancia, riqueza y diversidad
de especies de himenópteros parasitoides. Posiblemente como producto de más
de tres décadas de implementar un manejo ecológico, existen poblaciones de
enemigos naturales, específicamente parasitoides, con gran potencial para
implementar estrategias de control biológico contra E. galapagana. Aún así, los
resultados obtenidos solo apoyan una de las hipótesis biológicas enunciadas al
comienzo de este estudio. El cambio en la composición vegetal entre épocas
climáticas (seca y lluviosa) y su variación entre los lotes muestreados, está
relacionado con la riqueza y con la densidad de especies de parasitoides en la
Reserva Natural El Hatico. Por el contrario, la distancia con respecto al bosque no
influye significativamente en su abundancia ni en su riqueza.
La totalidad de las especies de parasitoides identificadas en la Reserva, no está
relacionada con la densidad poblacional de la polilla del algarrobo, por lo cual se
necesita cuantificar el número de especies que se alimentan de E. galapagana en
la Reserva Natural El Hatico y sus dinámicas poblacionales.
La regulación de herbívoros, como la polilla del algarrobo, no parece estar
influenciada únicamente por factores climáticos, es aparentemente resultado de la
sinergia entre factores bióticos tales como enemigos naturales, estructura y
diversidad floral, entre otros, y factores abióticos como la precipitación.

42
No se observa evidencia de que la disminución en los porcentajes de herbivoría en
época lluviosa, esté relacionada con cambios en la abundancia o número de
especies de parasitoides. Esta parece estar más relacionada con la cantidad de
precipitación que se presenta en el área de manejo silvopastoril.

43
8. RECOMENDACIONES
Es necesario un mayor esfuerzo de muestreo dentro del relicto de bosque de la
Reserva para cubrir un área de muestreo, mucho más extensa del sistema natural.
La cría y obtención de los parasitoides que atacan la polilla del algarrobo sería un
aporte muy significativo para implementar estrategias de control dentro de la
Reserva.
Se deben realizar estudios más detallados con la polilla, conocer más sobre su
comportamiento, su biología y sus plantas nutricias.
Llevar a cabo ensayos controlados de liberación de parasitoides comerciales, tipo
Trichogramma o Telenomus, midiendo los porcentajes de parasitismo como primer
paso para la disminución de la plaga dentro del sistema silvopastoril.
Es importante darle continuidad a los muestreos en el tiempo para poder
determinar el comportamiento de una plaga que ha encontrado su máxima
expresión en las variaciones climáticas que se han venido acentuando
principalmente en períodos del fenómeno del niño. Adicionalmente a esto, el
presente trabajo de campo coincidió con la época final este fenómeno (el año más
seco de toda la historia en la RNH) y el inicio de la época del fenómeno de la niña
(el año más lluvioso de la historia).

44
A pesar que este trabajo le da continuidad a otro proyecto de investigación en la
Reserva Natural El Hatico, aún quedan muchas inquietudes por despejar, frente a
una plaga que ha hecho su primera aparición desde el año 2006, causando
problemas de estabilidad en el sistema de producción.

45
9. LITERATURA CITADA
ALTIERI, M. A. 1999. Agroecología. Bases científicas para una agricultura
sustentable. Editorial Nordan–Comunidad. pp. 15 – 19.
ARAJ, S. E., S. WRATTEN, A. LISTER & H. BUCKLEY. 2008. Flora diversity,
parasitoids and hyperparasitoids – A laboratory approach. Basic and Applied
Ecology 9: 588 – 597.
ARMBRECHT, I. & P. ULLOA-CHACÓN. 1999. Rareza y diversidad de hormigas
en fragmentos de bosque seco colombianos y sus matrices. Biotropica 31: 646 –
653.
ARMBRECHT, I. 2008. El papel de la matriz rural como conector entre reservas.
El cafetal tradicional como hábitat hospitalario y conector en el paisaje. pp. 117 -
127. En: Gustavo Kattan & L. G. Naranjo (eds). Regiones Biodiversas.
Herramientas para la planificación de Sistemas regionales de áreas Protegidas.
Fundación EcoAndina, WCS y WWF. Cali, Colombia.
BRICEÑO, R. A.; D. TORRES & L. ROMERO. 2006. Biodiversidad y distribución
de la subfamilia Alysiinae (Hymenoptera: Braconidae) en tres ecosistemas
naturales de Venezuela. Entomotropica 21 (2): 73 – 82.
COLWELL, R. K. 1997. User´s guide to Estimates 800. Statistical estimation of
species richness and shared species from samples.
http://viceroy.eeb.uconn.edu/estimates.
COOK, S. M., Z. R. KHAN & J. A. PICKETT. 2007. The use of push-pull strategies
in integrated pest management. Annual Review of Entomology 52: 375 – 400.
CRONIN, J. T. 2003. Matrix heterogeneity and host-parasitoid interactions in
space. Ecology 84 (6): 1506 – 1516.

46
ELZINGA, J. A., S. V. NOUHUYS, D. J. V. LEEUWEN & A. BIERE. 2007.
Distribution and colonisation ability of three parasitoids and their herbivorous host
in a fragmented Landscape. Basic and Applied Ecology 8: 75 - 88.
FERNÁNDEZ, F. & M. J. SHARKEY (eds.). 2006. Introducción a los Hymenoptera
de la región neotropical. Sociedad Colombiana de Entomología y Universidad
Nacional de Colombia, Bogota. Editorial Guadalupe Ltda, Bogotá D.C.
GARCÍA, J. L. 2003. Comparación de la captura de Hymenoptera (Insecta)
mediante cuatro métodos de muestreo, en los cerros Yaví y Yutajé del Pantepui
venezolano. Entomotropica 8(1): 27-35.
GIBSON G. A. P.; J. T. HUBER & J. B. WOOLLEY (eds.). 1997. Annotated keys to
the genera of Nearctic Chalcidoidea (Hymenoptera). NRC Research Press.
Ottawa, Ontario, Canadá. pp. 1 – 4.
GLIESSMAN, S. R. 2002. Agroecología: procesos ecológicos en agricultura
sostenible. LUTICAT, Turrialba, Costa Rica. pp. 26.
HADDAD, N. M.; D. TILMAN; J. HAARSTAD; M. RITCHIE & J. M. H. KNOPS.
2001. Contrasting effects of plant richness and composition on insect communities:
a field experiment. The American Naturalist 158 (1): 17 – 35.
HAMMER, O., D. HARPER & P. D. RYAN. 2001. PAST: Paleontological statistics
software package for education and data analysis. Paleontología Electrónica 4: 1 –
9.
HOLDRIDGE, L. R. 1978. Ecología basada en zonas de vida. Instituto
Interamericano de Cooperación para la Agricultura (IICA): Serie de libros y
materiales educativos No. 34.

47
LETOURNEAU, D. K., J. A. JEDLICKA, S. G. BOTHWELL & C. R. MORENO.
2009. Effects of natural enemy biodiversity on the supression of arthropod
herbivores in terrestrial ecosystems. Annual Review of Ecology 40: 573 – 592.
LETOURNEAU, D. K., I. ARMBRECHT, B. SALGUERO-RIVERA, J. MONTOYA-
LERMA, E. JIMÉNEZ-CARMONA, M. C. DAZA, S. ESCOBAR, V. GALINDO, C.
GUITIÉRREZ, S. DUQUE-LÓPEZ, J. LÓPEZ-MEJÍA, A. M. ACOSTA-RANGEL, L.
RIVERA, C. A. SAAVEDRA, A. M. TORRES & A. REYES-TRUJILLO. 2010. Does
plant diversity Benedit agroecosystems? A synthetic review. Ecologycal
Applications doi: 10.1890/09-2026.
MADRIGAL-CARDEÑO, A. 2001. Fundamentos del control biológico de plagas. 1ª
ed. Medellín. Centro de publicaciones Universidad Nacional de Colombia, sede
Medellín. pp. 91 - 120.
MADRIGAL-CARDEÑO, A. 2003. Insectos forestales en Colombia. Biología,
hábitos, ecología y manejo. 1ª ed. Medellín, Editorial Marín Vieco Ltda. pp. 361.
MAHECHA, L. 1998. Análisis de la relación planta-animal desde el punto de vista
nutrición en un sistema silvopastoril de pasto estrella africana Cynodon
plectostachyus, leucaena Leucaena leucocephala y algarrobo Prosopis juliflora en
el Valle del Cauca. Tesis de Magíster. Universidad del Valle, Facultad de Ciencias.
pp. 4 - 9, 25.
MYARTSEVA, S. N., E. RUÍZ-CANCINO & M. CORONADO-BLANCO. 2007. A
review of parasitoids (Hymenoptera: Chalcidoidea) of Trialeurodes floridensis
(Hemiptera: Aleyrodidae) with description of a new species from Mexico. Florida
Entomologist 90(4): 635 – 640.

48
PRINCE, P. W. & G. P. WALDBAUER. 1975. Ecologycal aspects of pest
management. Pp. 587 En: Metcalf, R.L. & W.H. Luckman (eds.). Introduction to
insect pests management.. Nueva York, Ed. John Willey.
RAMÍREZ, H. 1997. Evaluación de dos sistemas silvopastoriles integrados por
Cynodon plectostachyus, Leucaena leucocephala y Prosopis juliflora. Seminario
Internacional de sistemas sostenibles de producción agropecuaria. Cali. Colombia.
CIPAV.
RAZOWSKI, J., B. LANDRY & L. ROQUE-ALBELO. 2008. The Tortricidae
(Lepidoptera) of the galapagos islands, Ecuador. Revue Suisse de Zoologie 115
(1): 185 – 220.
REYES, L. K., C. GIRALDO, J. MONTOYA-LERMA, E. J. MOLINA, C. H. MOLINA
& Z. CALLE. 2010. Ciclo biológico de la polilla defoliadora de Prosopis juliflora
(algarrobo) en un sistema silvopastoril del Valle del Cauca, Colombia. Memorias VI
Congreso Latinoamericano de agroforestería para la producción pecuaria
sostenible. 28 al 30 de septiembre 2010, Panamá, Hotel El Panamá.
RINCÓN, C. & A. LÓPEZ-ÁVILA. 1999. Estudios biológicos del parasitoide
Trichogrammatidae lopezandinensis (Hymenoptera: Trichogrammatidae)
orientados al control de la polilla guatemalteca de la papa Tecia solanivora
(Lepidoptera: Gelechiidae). Revista Colombiana de Entomología 25 (1-2): 68.
SANTOS, L. D., C. A. BERNAL & J. H. DUARTE. 1996. Introducción a la
evaluación de la producción de algarrobo (Prosopis juliflora) en áreas de bosque
seco tropical, Alto Magdalena, Cundinamarca, Colombia. Seminario Internacional
de sistemas sostenibles de producción agropecuaria. Cali. Colombia. CIPAV.
SHARKEY, M. J. 2007. Phylogeny and classification of Hymenoptera. Zootaxa
1668: 521 – 548.

49
SHELTON, A. M. & F. R. BADENES-PEREZ. 2006. Concepts and application of
trap cropping in the pest management. Annual Review of Entomology 51: 285 –
308.
SUGIURA, S. 2007. Structure of a herbivore–parasitoid community: are parasitoids
shared by different herbivore guilds? Basic and Applied Ecology 8: 544 – 551.
TRIAPITSYN, S. V., D. B. VICKERMAN, J. M. HERATY & G. A. LOGARZO. 2006.
Anew species of Gonatocerus (Hymenoptera: Mymaridae) parasitic on proconiine
sharpshooters (Hemiptera: Cicadellidae) in the New World. Zootaxa 1158: 55 – 67.
TYLIANAKIS, J. M., A. M. KLEIN & T. TSCHARNTKE. 2005. Spatiotemporal
variation in the diversity of Hymenoptera across a tropical habitat gradient. Ecology
86 (12): 3296 – 3302.
ZAR, J. H. 1984. Biostatistical analysis. 2ª ed. Nueva Jersey, Prentice-Hall. pp. 32
– 34.
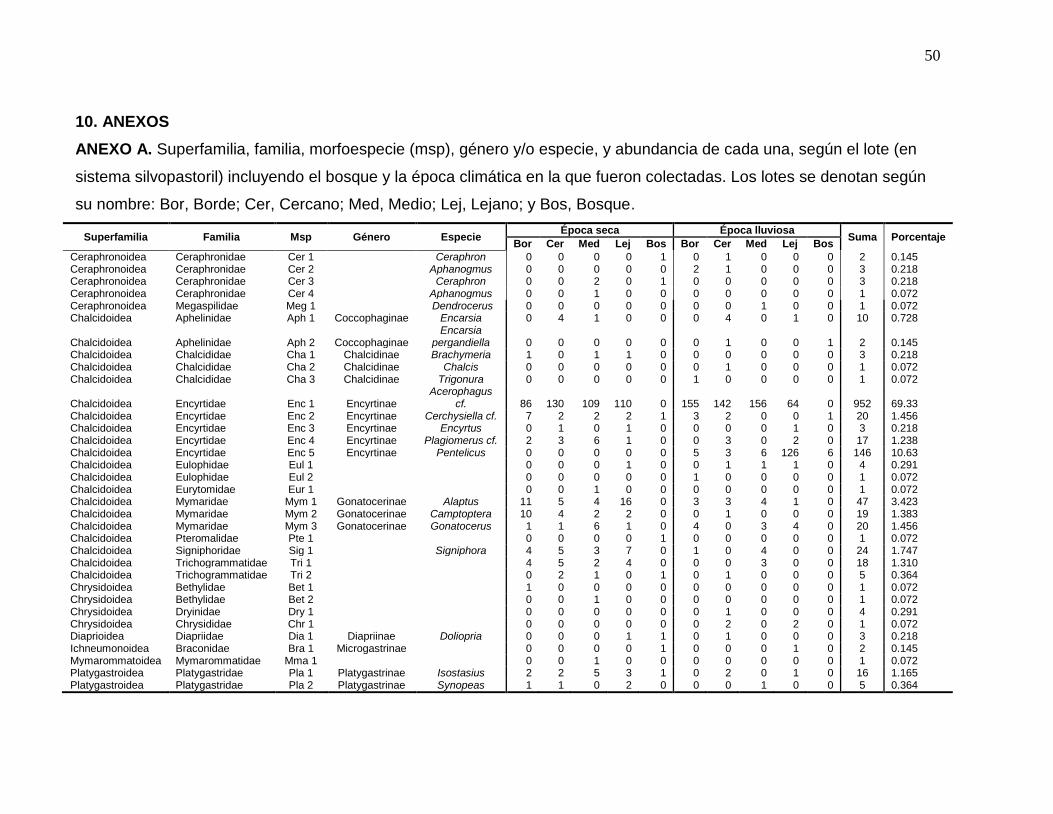
50
10. ANEXOS
ANEXO A. Superfamilia, familia, morfoespecie (msp), género y/o especie, y abundancia de cada una, según el lote (en
sistema silvopastoril) incluyendo el bosque y la época climática en la que fueron colectadas. Los lotes se denotan según
su nombre: Bor, Borde; Cer, Cercano; Med, Medio; Lej, Lejano; y Bos, Bosque.
Superfamilia Familia Msp Género Especie Época seca Época lluviosa
Suma Porcentaje Bor Cer Med Lej Bos Bor Cer Med Lej Bos
Ceraphronoidea Ceraphronidae Cer 1 Ceraphron 0 0 0 0 1 0 1 0 0 0 2 0.145 Ceraphronoidea Ceraphronidae Cer 2 Aphanogmus 0 0 0 0 0 2 1 0 0 0 3 0.218 Ceraphronoidea Ceraphronidae Cer 3 Ceraphron 0 0 2 0 1 0 0 0 0 0 3 0.218 Ceraphronoidea Ceraphronidae Cer 4 Aphanogmus 0 0 1 0 0 0 0 0 0 0 1 0.072 Ceraphronoidea Megaspilidae Meg 1 Dendrocerus 0 0 0 0 0 0 0 1 0 0 1 0.072 Chalcidoidea Aphelinidae Aph 1 Coccophaginae Encarsia 0 4 1 0 0 0 4 0 1 0 10 0.728
Chalcidoidea Aphelinidae Aph 2 Coccophaginae Encarsia
pergandiella 0 0 0 0 0 0 1 0 0 1 2 0.145 Chalcidoidea Chalcididae Cha 1 Chalcidinae Brachymeria 1 0 1 1 0 0 0 0 0 0 3 0.218 Chalcidoidea Chalcididae Cha 2 Chalcidinae Chalcis 0 0 0 0 0 0 1 0 0 0 1 0.072 Chalcidoidea Chalcididae Cha 3 Chalcidinae Trigonura 0 0 0 0 0 1 0 0 0 0 1 0.072
Chalcidoidea Encyrtidae Enc 1 Encyrtinae Acerophagus
cf. 86 130 109 110 0 155 142 156 64 0 952 69.33 Chalcidoidea Encyrtidae Enc 2 Encyrtinae Cerchysiella cf. 7 2 2 2 1 3 2 0 0 1 20 1.456 Chalcidoidea Encyrtidae Enc 3 Encyrtinae Encyrtus 0 1 0 1 0 0 0 0 1 0 3 0.218 Chalcidoidea Encyrtidae Enc 4 Encyrtinae Plagiomerus cf. 2 3 6 1 0 0 3 0 2 0 17 1.238 Chalcidoidea Encyrtidae Enc 5 Encyrtinae Pentelicus 0 0 0 0 0 5 3 6 126 6 146 10.63 Chalcidoidea Eulophidae Eul 1 0 0 0 1 0 0 1 1 1 0 4 0.291 Chalcidoidea Eulophidae Eul 2 0 0 0 0 0 1 0 0 0 0 1 0.072 Chalcidoidea Eurytomidae Eur 1 0 0 1 0 0 0 0 0 0 0 1 0.072 Chalcidoidea Mymaridae Mym 1 Gonatocerinae Alaptus 11 5 4 16 0 3 3 4 1 0 47 3.423 Chalcidoidea Mymaridae Mym 2 Gonatocerinae Camptoptera 10 4 2 2 0 0 1 0 0 0 19 1.383 Chalcidoidea Mymaridae Mym 3 Gonatocerinae Gonatocerus 1 1 6 1 0 4 0 3 4 0 20 1.456 Chalcidoidea Pteromalidae Pte 1 0 0 0 0 1 0 0 0 0 0 1 0.072 Chalcidoidea Signiphoridae Sig 1 Signiphora 4 5 3 7 0 1 0 4 0 0 24 1.747 Chalcidoidea Trichogrammatidae Tri 1 4 5 2 4 0 0 0 3 0 0 18 1.310 Chalcidoidea Trichogrammatidae Tri 2 0 2 1 0 1 0 1 0 0 0 5 0.364 Chrysidoidea Bethylidae Bet 1 1 0 0 0 0 0 0 0 0 0 1 0.072 Chrysidoidea Bethylidae Bet 2 0 0 1 0 0 0 0 0 0 0 1 0.072 Chrysidoidea Dryinidae Dry 1 0 0 0 0 0 0 1 0 0 0 4 0.291 Chrysidoidea Chrysididae Chr 1 0 0 0 0 0 0 2 0 2 0 1 0.072 Diaprioidea Diapriidae Dia 1 Diapriinae Doliopria 0 0 0 1 1 0 1 0 0 0 3 0.218 Ichneumonoidea Braconidae Bra 1 Microgastrinae 0 0 0 0 1 0 0 0 1 0 2 0.145 Mymarommatoidea Mymarommatidae Mma 1 0 0 1 0 0 0 0 0 0 0 1 0.072 Platygastroidea Platygastridae Pla 1 Platygastrinae Isostasius 2 2 5 3 1 0 2 0 1 0 16 1.165 Platygastroidea Platygastridae Pla 2 Platygastrinae Synopeas 1 1 0 2 0 0 0 1 0 0 5 0.364
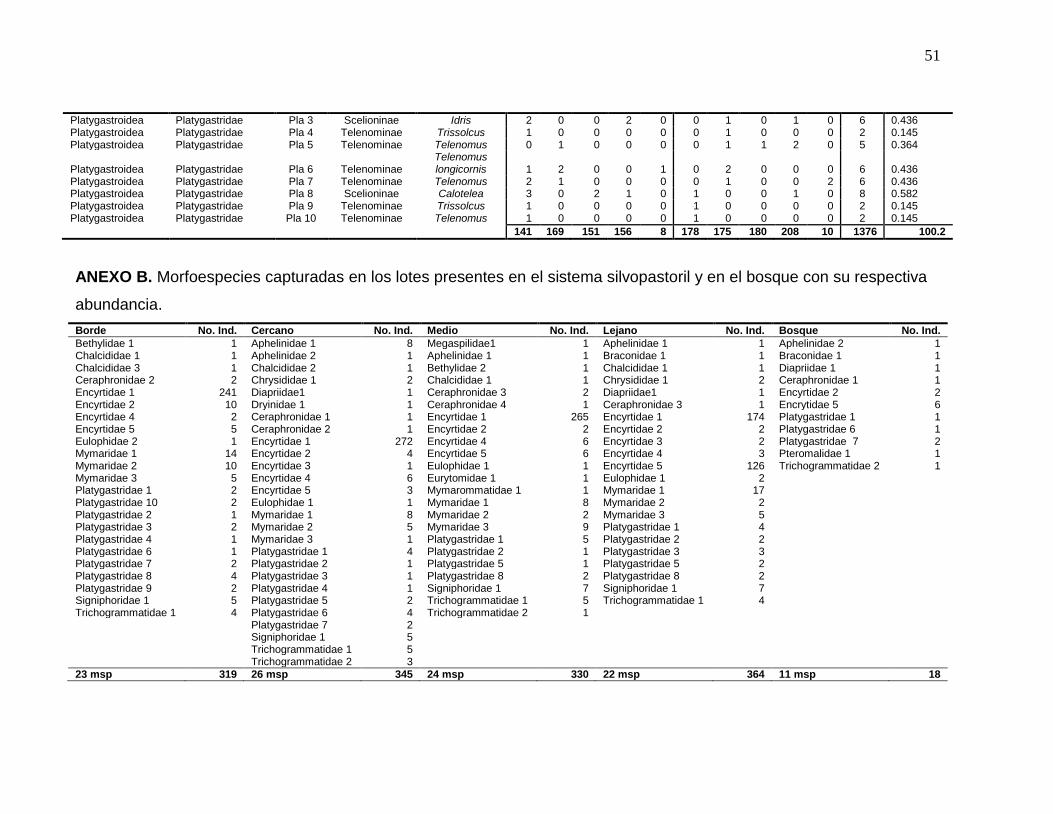
51
Platygastroidea Platygastridae Pla 3 Scelioninae Idris 2 0 0 2 0 0 1 0 1 0 6 0.436 Platygastroidea Platygastridae Pla 4 Telenominae Trissolcus 1 0 0 0 0 0 1 0 0 0 2 0.145 Platygastroidea Platygastridae Pla 5 Telenominae Telenomus 0 1 0 0 0 0 1 1 2 0 5 0.364
Platygastroidea Platygastridae Pla 6 Telenominae Telenomus longicornis 1 2 0 0 1 0 2 0 0 0 6 0.436
Platygastroidea Platygastridae Pla 7 Telenominae Telenomus 2 1 0 0 0 0 1 0 0 2 6 0.436 Platygastroidea Platygastridae Pla 8 Scelioninae Calotelea 3 0 2 1 0 1 0 0 1 0 8 0.582 Platygastroidea Platygastridae Pla 9 Telenominae Trissolcus 1 0 0 0 0 1 0 0 0 0 2 0.145 Platygastroidea Platygastridae Pla 10 Telenominae Telenomus 1 0 0 0 0 1 0 0 0 0 2 0.145
141 169 151 156 8 178 175 180 208 10 1376 100.2
ANEXO B. Morfoespecies capturadas en los lotes presentes en el sistema silvopastoril y en el bosque con su respectiva
abundancia.
Borde No. Ind. Cercano No. Ind. Medio No. Ind. Lejano No. Ind. Bosque No. Ind.
Bethylidae 1 1 Aphelinidae 1 8 Megaspilidae1 1 Aphelinidae 1 1 Aphelinidae 2 1 Chalcididae 1 1 Aphelinidae 2 1 Aphelinidae 1 1 Braconidae 1 1 Braconidae 1 1 Chalcididae 3 1 Chalcididae 2 1 Bethylidae 2 1 Chalcididae 1 1 Diapriidae 1 1 Ceraphronidae 2 2 Chrysididae 1 2 Chalcididae 1 1 Chrysididae 1 2 Ceraphronidae 1 1 Encyrtidae 1 241 Diapriidae1 1 Ceraphronidae 3 2 Diapriidae1 1 Encyrtidae 2 2 Encyrtidae 2 10 Dryinidae 1 1 Ceraphronidae 4 1 Ceraphronidae 3 1 Encrytidae 5 6 Encyrtidae 4 2 Ceraphronidae 1 1 Encyrtidae 1 265 Encyrtidae 1 174 Platygastridae 1 1 Encyrtidae 5 5 Ceraphronidae 2 1 Encyrtidae 2 2 Encyrtidae 2 2 Platygastridae 6 1 Eulophidae 2 1 Encyrtidae 1 272 Encyrtidae 4 6 Encyrtidae 3 2 Platygastridae 7 2 Mymaridae 1 14 Encyrtidae 2 4 Encyrtidae 5 6 Encyrtidae 4 3 Pteromalidae 1 1 Mymaridae 2 10 Encyrtidae 3 1 Eulophidae 1 1 Encyrtidae 5 126 Trichogrammatidae 2 1 Mymaridae 3 5 Encyrtidae 4 6 Eurytomidae 1 1 Eulophidae 1 2 Platygastridae 1 2 Encyrtidae 5 3 Mymarommatidae 1 1 Mymaridae 1 17 Platygastridae 10 2 Eulophidae 1 1 Mymaridae 1 8 Mymaridae 2 2 Platygastridae 2 1 Mymaridae 1 8 Mymaridae 2 2 Mymaridae 3 5 Platygastridae 3 2 Mymaridae 2 5 Mymaridae 3 9 Platygastridae 1 4 Platygastridae 4 1 Mymaridae 3 1 Platygastridae 1 5 Platygastridae 2 2 Platygastridae 6 1 Platygastridae 1 4 Platygastridae 2 1 Platygastridae 3 3 Platygastridae 7 2 Platygastridae 2 1 Platygastridae 5 1 Platygastridae 5 2 Platygastridae 8 4 Platygastridae 3 1 Platygastridae 8 2 Platygastridae 8 2 Platygastridae 9 2 Platygastridae 4 1 Signiphoridae 1 7 Signiphoridae 1 7 Signiphoridae 1 5 Platygastridae 5 2 Trichogrammatidae 1 5 Trichogrammatidae 1 4 Trichogrammatidae 1 4 Platygastridae 6 4 Trichogrammatidae 2 1 Platygastridae 7 2 Signiphoridae 1 5 Trichogrammatidae 1 5 Trichogrammatidae 2 3
23 msp 319 26 msp 345 24 msp 330 22 msp 364 11 msp 18

52
ANEXO C. Morfoespecies capturadas y su abundancia en los cuatros lotes del sistema silvopastoril, en el bosque y
discriminados por época climática.
ÉPOCA SECA
Borde No. Ind. Cercano No. Ind. Medio No. Ind. Lejano No. Ind. Bosque No. Ind.
Bethylidae 1 1 Aphelinidae 1 4 Aphelinidae 1 1 Chalcididae 1 1 Braconidae 1 1 Chalcididae 1 1 Encyrtidae 1 130 Bethylidae 2 1 Diapriidae 1 1 Diapriidae 1 1 Encyrtidae 1 86 Encyrtidae 2 2 Chalcididae 1 1 Ceraphronidae 3 1 Ceraphronidae 1 1 Encyrtidae 2 7 Encyrtidae 3 1 Ceraphronidae 3 2 Encyrtidae 1 110 Encyrtidae 2 1 Encyrtidae 4 2 Encyrtidae 4 3 Ceraphronidae 4 1 Encyrtidae 2 2 Platygastridae 1 1 Mymaridae 1 11 Mymaridae 1 5 Encyrtidae 1 109 Encyrtidae 3 1 Platygastridae 6 1 Mymaridae 2 10 Mymaridae 2 4 Encyrtidae 2 2 Encyrtidae 4 1 Pteromalidae 1 1 Mymaridae 3 1 Mymaridae 3 1 Encyrtidae 4 6 Eulophidae 1 1 Trichogrammatidae 2 1 Platygastridae 1 2 Platygastridae 1 2 Eurytomidae 1 1 Mymaridae 1 16 Platygastridae 2 1 Platygastridae 2 1 Mymarommatidae 1 1 Mymaridae 2 2 Platygastridae 3 2 Platygastridae 5 1 Mymaridae 1 4 Mymaridae 3 1 Platygastridae 4 1 Platygastridae 6 2 Mymaridae 2 2 Platygastridae 1 3 Platygastridae 6 1 Platygastridae 7 1 Mymaridae 3 6 Platygastridae 2 2 Platygastridae 7 2 Signiphoridae 1 5 Platygastridae 1 5 Platygastridae 3 2 Platygastridae 8 3 Trichogrammatidae 1 5 Platygastridae 8 2 Platygastridae 8 1 Platygastridae 9 1 Trichogrammatidae 2 2 Signiphoridae 1 3 Signiphoridae 1 7 Platygastridae 10 1 Trichogrammatidae 1 2 Trichogrammatidae 1 4 Signiphoridae 1 4 Trichogrammatidae 2 1 Trichogrammatidae 1 4
19 msp 141 16 msp 169 19 msp 150 17 msp 156 8 msp 8
Total abundancia época seca 624
ÉPOCA LLUVIOSA
Borde No. Ind. Cercano No. Ind. Medio No. Ind. Lejano No. Ind. Bosque No. Ind.
Chalcididae 3 1 Aphelinidae 1 4 Megaspilidae 1 1 Aphelinidae 1 1 Aphelinidae 2 1 Ceraphronidae 2 2 Aphelinidae 2 1 Encyrtidae 1 156 Braconidae 1 1 Encyrtidae 2 1 Encyrtidae 1 155 Chalcididae 2 1 Encyrtidae 5 6 Chrysididae 1 2 Encyrtidae 5 6 Encyrtidae 2 3 Chrysididae 1 2 Eulophidae 1 1 Encyrtidae 1 64 Platygastridae 7 2 Encyrtidae 5 5 Diapriidae 1 1 Mymaridae 1 4 Encyrtidae 3 1 Eulophidae 2 1 Dryinidae 1 1 Mymaridae 3 3 Encyrtidae 4 2 Mymaridae 1 3 Ceraphronidae 2 1 Platygastridae 2 1 Encyrtidae 5 126 Mymaridae 3 4 Encyrtidae 1 142 Platygastridae 5 1 Eulophidae 1 1 Platygastridae 8 1 Encyrtidae 2 2 Signiphoridae 1 4 Mymaridae 1 1 Platygastridae 9 1 Encyrtidae 4 3 Trichogrammatidae 1 3 Mymaridae 3 4 Platygastridae 10 1 Encyrtidae 5 3 Platygastridae 1 1 Signiphoridae 1 1 Eulophidae 1 1 Platygastridae 3 1 Mymaridae 1 3 Platygastridae 5 2 Mymaridae 2 1 Platygastridae 8 1 Platygastridae 1 2 Platygastridae 3 1 Platygastridae 4 1
53
Platygastridae 5 1 Platygastridae 6 2 Platygastridae 7 1 Trichogrammatidae 2 1
12 msp 178 22 msp 176 10 msp 180 14 msp 208 4 msp 10
Total abundancia época lluviosa 752
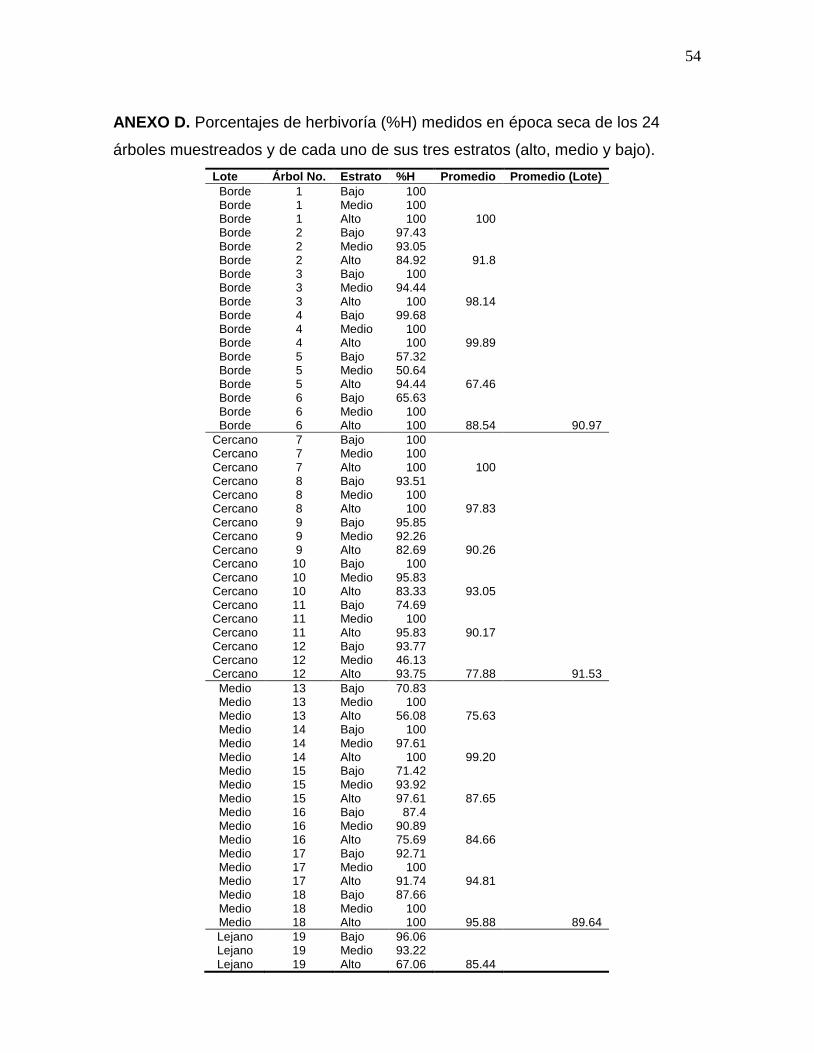
54
ANEXO D. Porcentajes de herbivoría (%H) medidos en época seca de los 24
árboles muestreados y de cada uno de sus tres estratos (alto, medio y bajo).
Lote Árbol No. Estrato %H Promedio Promedio (Lote)
Borde 1 Bajo 100 Borde 1 Medio 100 Borde 1 Alto 100 100 Borde 2 Bajo 97.43 Borde 2 Medio 93.05 Borde 2 Alto 84.92 91.8 Borde 3 Bajo 100 Borde 3 Medio 94.44 Borde 3 Alto 100 98.14 Borde 4 Bajo 99.68 Borde 4 Medio 100 Borde 4 Alto 100 99.89 Borde 5 Bajo 57.32 Borde 5 Medio 50.64 Borde 5 Alto 94.44 67.46 Borde 6 Bajo 65.63 Borde 6 Medio 100 Borde 6 Alto 100 88.54 90.97
Cercano 7 Bajo 100 Cercano 7 Medio 100 Cercano 7 Alto 100 100 Cercano 8 Bajo 93.51 Cercano 8 Medio 100 Cercano 8 Alto 100 97.83 Cercano 9 Bajo 95.85 Cercano 9 Medio 92.26 Cercano 9 Alto 82.69 90.26 Cercano 10 Bajo 100 Cercano 10 Medio 95.83 Cercano 10 Alto 83.33 93.05 Cercano 11 Bajo 74.69 Cercano 11 Medio 100 Cercano 11 Alto 95.83 90.17 Cercano 12 Bajo 93.77 Cercano 12 Medio 46.13 Cercano 12 Alto 93.75 77.88 91.53
Medio 13 Bajo 70.83 Medio 13 Medio 100 Medio 13 Alto 56.08 75.63 Medio 14 Bajo 100 Medio 14 Medio 97.61 Medio 14 Alto 100 99.20 Medio 15 Bajo 71.42 Medio 15 Medio 93.92 Medio 15 Alto 97.61 87.65 Medio 16 Bajo 87.4 Medio 16 Medio 90.89 Medio 16 Alto 75.69 84.66 Medio 17 Bajo 92.71 Medio 17 Medio 100 Medio 17 Alto 91.74 94.81 Medio 18 Bajo 87.66 Medio 18 Medio 100 Medio 18 Alto 100 95.88 89.64
Lejano 19 Bajo 96.06 Lejano 19 Medio 93.22 Lejano 19 Alto 67.06 85.44

55
Lejano 20 Bajo 96.96 Lejano 20 Medio 91.35 Lejano 20 Alto 95.06 94.45 Lejano 21 Bajo 88.16 Lejano 21 Medio 100 Lejano 21 Alto 100 96.05 Lejano 22 Bajo 95.13 Lejano 22 Medio 97.91 Lejano 22 Alto 78.7 90.58 Lejano 23 Bajo 24.04 Lejano 23 Medio 51.95 Lejano 23 Alto 38.09 38.02 Lejano 24 Bajo 100 Lejano 24 Medio 85.4 Lejano 24 Alto 96.55 93.98 83.09
Porcentaje promedio en época seca 88.81
ANEXO E. Porcentajes de herbivoría obtenidos en ambas épocas climáticas y
discriminados por lote y estrato.
ÉPOCA SECA ÉPOCA LLUVIOSA
Lote Estrato %H Promedio %H Lote Estrato %H Promedio estrato
Borde Alto 100 Borde Alto 71.94 Borde Alto 84.92 Borde Alto 86.4 Borde Alto 100 Borde Alto 100 Borde Alto 100 Borde Alto 100 Borde Alto 94.44 Borde Alto 100 Borde Alto 100 96.56 Borde Alto 96.29 30.81
Borde Medio 100 Borde Medio 60.42 Borde Medio 93.05 Borde Medio 87.98 Borde Medio 94.44 Borde Medio 90.34 Borde Medio 100 Borde Medio 86.14 Borde Medio 50.64 Borde Medio 100 Borde Medio 100 89.68 Borde Medio 100 29.15
Borde Bajo 100 Borde Bajo 45.44 Borde Bajo 97.43 Borde Bajo 97.1 Borde Bajo 100 Borde Bajo 73.21 Borde Bajo 99.68 Borde Bajo 77.94 Borde Bajo 57.32 Borde Bajo 93.64 Borde Bajo 65.63 86.67 Borde Bajo 70.23 25.41
Promedio lote 90.97 Promedio lote 85.39
Lote Estrato %H Lote Estrato %H
Cercano Alto 100 Cercano Alto 52.31 Cercano Alto 100 Cercano Alto 86.95 Cercano Alto 82.69 Cercano Alto 98.76 Cercano Alto 83.33 Cercano Alto 22.52 Cercano Alto 95.83 Cercano Alto 96.96 Cercano Alto 93.75 92.6 Cercano Alto 100 25.41
Cercano Medio 100 Cercano Medio 57.52 Cercano Medio 100 Cercano Medio 85.18 Cercano Medio 92.26 Cercano Medio 96.66 Cercano Medio 95.83 Cercano Medio 77.75 Cercano Medio 100 Cercano Medio 79.76 Cercano Medio 46.13 89.03 Cercano Medio 90.47 27.07
Cercano Bajo 100 Cercano Bajo 56.37 Cercano Bajo 93.51 Cercano Bajo 90.24 Cercano Bajo 95.85 Cercano Bajo 89.22 Cercano Bajo 100 Cercano Bajo 100 Cercano Bajo 74.69 Cercano Bajo 37.16

56
Cercano Bajo 93.77 92.97 Cercano Bajo 58.24 23.95
Promedio lote 91.53 Promedio lote 76.44
Lote Estrato %H Lote Estrato %H Promedio estrato
Medio Alto 56.08 Medio Alto 58.45 Medio Alto 100 Medio Alto 86.24 Medio Alto 97.61 Medio Alto 76.17 Medio Alto 75.69 Medio Alto 94.44 Medio Alto 91.74 Medio Alto 69.35 Medio Alto 100 86.85 Medio Alto 32.84 23.19
Medio Medio 100 Medio Medio 87.48 Medio Medio 97.61 Medio Medio 95.55 Medio Medio 93.92 Medio Medio 58.46 Medio Medio 90.89 Medio Medio 95 Medio Medio 100 Medio Medio 96.96 Medio Medio 100 97.07 Medio Medio 25.46 25.49
Medio Bajo 70.83 Medio Bajo 80.06 Medio Bajo 100 Medio Bajo 56.19 Medio Bajo 71.42 Medio Bajo 72.75 Medio Bajo 87.4 Medio Bajo 78.97 Medio Bajo 92.71 Medio Bajo 46.36 Medio Bajo 87.66 85.00 Medio Bajo 95.83 23.89
Promedio lote 89.64 Promedio lote 72.58
Lote Estrato %H Lote Estrato %H Promedio estrato
Lejano Alto 67.06 Lejano Alto 95.23 Lejano Alto 95.06 Lejano Alto 90.15 Lejano Alto 100 Lejano Alto 90.47 Lejano Alto 78.7 Lejano Alto 97.91 Lejano Alto 38.09 Lejano Alto 52 Lejano Alto 96.55 79.24 Lejano Alto 92.78 28.80
Lejano Medio 93.22 Lejano Medio 96.32 Lejano Medio 91.35 Lejano Medio 25.43 Lejano Medio 100 Lejano Medio 91.15 Lejano Medio 97.91 Lejano Medio 100 Lejano Medio 51.95 Lejano Medio 75.07 Lejano Medio 85.4 86.63 Lejano Medio 80 25.99
Lejano Bajo 96.06 Lejano Bajo 92.14 Lejano Bajo 96.96 Lejano Bajo 12.94 Lejano Bajo 88.16 Lejano Bajo 94.16 Lejano Bajo 95.13 Lejano Bajo 94.36 Lejano Bajo 24.04 Lejano Bajo 70 Lejano Bajo 100 83.39 Lejano Bajo 93.33 25.38
Promedio lote 83.09 Promedio lote 80.19
ANEXO F. Porcentajes de herbivoría (%H) medidos en época lluviosa de los 24
árboles muestreados y de cada uno de sus tres estratos (alto, medio y bajo).
Lote Muestra No. Estrato %H Promedio (árbol) Promedio (Lote)
Borde 1 Bajo 45.44 Borde 1 Medio 60.42 Borde 1 Alto 71.94 59.26 Borde 2 Bajo 97.1 Borde 2 Medio 87.98 Borde 2 Alto 86.4 90.49 Borde 3 Bajo 73.21 Borde 3 Medio 90.34 Borde 3 Alto 100 87.85 Borde 4 Bajo 77.94 Borde 4 Medio 86.14 Borde 4 Alto 100 88.02

57
Borde 5 Bajo 93.64 Borde 5 Medio 100 Borde 5 Alto 100 97.88 Borde 6 Bajo 70.23 Borde 6 Medio 100 Borde 6 Alto 96.29 88.84 85.39
Cercano 7 Bajo 56.37 Cercano 7 Medio 57.52 Cercano 7 Alto 52.31 55.4 Cercano 8 Bajo 90.24 Cercano 8 Medio 85.18 Cercano 8 Alto 86.95 87.45 Cercano 9 Bajo 89.22 Cercano 9 Medio 96.66 Cercano 9 Alto 98.76 94.88 Cercano 10 Bajo 100 Cercano 10 Medio 77.75 Cercano 10 Alto 22.52 66.75 Cercano 11 Bajo 37.16 Cercano 11 Medio 79.76 Cercano 11 Alto 96.96 71.29 Cercano 12 Bajo 58.24 Cercano 12 Medio 90.47 Cercano 12 Alto 100 82.90 76.44
Medio 13 Bajo 80.06 Medio 13 Medio 87.48 Medio 13 Alto 58.45 75.33 Medio 14 Bajo 56.19 Medio 14 Medio 95.55 Medio 14 Alto 86.24 79.32 Medio 15 Bajo 72.75 Medio 15 Medio 58.46 Medio 15 Alto 76.17 69.12 Medio 16 Bajo 78.97 Medio 16 Medio 95 Medio 16 Alto 94.44 89.47 Medio 17 Bajo 46.36 Medio 17 Medio 96.96 Medio 17 Alto 69.35 70.89 Medio 18 Bajo 95.83 Medio 18 Medio 25.46 Medio 18 Alto 32.84 51.37 72.58
Lejano 19 Bajo 92.14 Lejano 19 Medio 96.32 Lejano 19 Alto 95.23 94.56 Lejano 20 Bajo 12.94 Lejano 20 Medio 25.43 Lejano 20 Alto 90.15 42.84 Lejano 21 Bajo 94.16 Lejano 21 Medio 91.15 Lejano 21 Alto 90.47 91.92 Lejano 22 Bajo 94.36 Lejano 22 Medio 100 Lejano 22 Alto 97.91 97.42 Lejano 23 Bajo 70 Lejano 23 Medio 75.07 Lejano 23 Alto 52 65.69 Lejano 24 Bajo 93.33 Lejano 24 Medio 80 Lejano 24 Alto 92.78 88.70 80.19
Porcentaje promedio en época lluviosa 78.65

58
ANEXO G. Porcentaje de cobertura, fecha y hora a la cual se tomó la temperatura
de los 24 algarrobos muestreados en época seca.
Lote Árbol No. Fecha Hora Temp. (ºC) Promedio %C Promedio
Borde 1 20 - Feb - 2010 2:30 PM 29.7 75 Borde 2 21 - Feb - 2010 9:29 AM 27.7 30 Borde 3 21 - Feb - 2010 3:15 PM 33.5 70 Borde 4 22 - Feb - 2010 10:35 AM 29.5 85 Borde 5 22 - Feb - 2010 4:00 PM 31.8 50 Borde 6 23 - Feb - 2010 11:50 AM 25.5 29.61 90 66.66
Cercano 7 20 - Feb - 2010 3:30 PM 30.1 75 Cercano 8 21 - Feb - 2010 10:10 AM 30 75 Cercano 9 21 - Feb - 2010 2:48 PM 32.4 70 Cercano 10 22 - Feb - 2010 4:26 PM 31.8 75 Cercano 11 22 - Feb - 2010 10:05 AM 28.7 85 Cercano 12 23 - Feb - 2010 10:30 AM 27.7 30.11 95 79.16
Medio 13 20 - Feb - 2010 4:45 PM 30.7 50 Medio 14 21 - Feb - 2010 10:38 AM 30.7 75 Medio 15 21 - Feb - 2010 2:28 PM 35.5 60 Medio 16 22 - Feb - 2010 9:16 AM 28.2 85 Medio 17 22 - Feb - 2010 4:50 PM 31.4 65 Medio 18 23 - Feb - 2010 9:45 AM 26.1 30.43 65 66.66
Lejano 19 20 - Feb - 2010 6:00 PM 29.9 70 Lejano 20 21 - Feb - 2010 11:10 AM 31.5 85 Lejano 21 21 - Feb - 2010 2:05 PM 36 85 Lejano 22 22 - Feb - 2010 8:45 AM 27.7 90 Lejano 23 22 - Feb - 2010 5:30 PM 30.5 90 Lejano 24 23 - Feb - 2010 8:50 AM 24.4 30 40 76.66
Promedio 30.04 72.29
ANEXO H. Porcentaje de cobertura, fecha y hora a la cual se tomó la temperatura
de los 24 algarrobos muestreados en época lluviosa.
Lote Árbol No. Fecha Hora Temp. (ºC) Promedio %C Promedio
Borde 1 22 - May - 2010 5:50 PM 24.8 90 Borde 2 23 - May - 2010 11:45 AM 25.3 85 Borde 3 23 - May - 2010 9:00 AM 25.1 95 Borde 4 24 - May - 2010 3:40 PM 27.2 85 Borde 5 24 - May - 2010 2:35 PM 30.1 35 Borde 6 24 - May - 2010 3:10 PM 29.2 26.95 90 80
Cercano 7 22 - May - 2010 4:40 PM 25.9 85 Cercano 8 23 - May - 2010 11:45 AM 26.3 90 Cercano 9 23 - May - 2010 4:10 PM 26 85 Cercano 10 24 - May - 2010 10:00 AM 26 85 Cercano 11 24 - May - 2010 4:08 PM 29.6 95 Cercano 12 24 - May - 2010 3:45 PM 29.8 27.26 95 89.16
Medio 13 22 - May - 2010 4:32 PM 27.3 50 Medio 14 23 - May - 2010 9:40 AM 24.7 80 Medio 15 23 - May - 2010 4:30 PM 25.9 65 Medio 16 24 - May - 2010 10:40 AM 30 75 Medio 17 24 - May - 2010 5:00 PM 29.6 95 Medio 18 24 - May - 2010 4:30 PM 31.3 28.13 90 75.83
Lejano 19 22 - May - 2010 2:30 PM 27 75 Lejano 20 23 - May - 2010 8:30 AM 23.3 90 Lejano 21 23 - May - 2010 5:10 PM 25.4 95 Lejano 22 24 - May - 2010 11:20 AM 29.5 75 Lejano 23 24 - May - 2010 5:50 PM 26.4 95 Lejano 24 24 - May - 2010 5:25 PM 27.5 26.51 55 80.83
Promedio 27.21 81.45

59
ANEXO I. Altura promedio de los algarrobos muestreados.
Lote Altura (m) Promedio
Borde 25.53 Borde 15.03 Borde 22.17 Borde 25.53 Borde 22.17 Borde 19.4 21.63
Cercano 10.19 Cercano 11.64 Cercano 12.03 Cercano 22.17 Cercano 20.72 Cercano 27.15 17.31
Medio 18.18 Medio 22.17 Medio 15.03 Medio 16.53 Medio 17.06 Medio 12.03 16.83
Lejano 16.53 Lejano 16.53 Lejano 16.01 Lejano 18.18 Lejano 14.11 Lejano 17.06 16.40
Promedio 18.04


















