
DETECCION Y CARACTERIZACION DEL VIRUS DEL …bdigital.unal.edu.co/63302/1/TESIS.pdf · A Dios, a...
203
Detección y caracterización del virus del Síndrome Reproductivo y Respiratorio Porcino en tres granjas de producción intensiva para el establecimiento del control local de la enfermedad Eliana Mendoza Niño Universidad Nacional de Colombia Facultad de Medicina Veterinaria y de Zootecnia Maestría Salud y Producción Animal Bogotá, Colombia 2015
-
Upload
phungnguyet -
Category
Documents
-
view
215 -
download
0
Transcript of DETECCION Y CARACTERIZACION DEL VIRUS DEL …bdigital.unal.edu.co/63302/1/TESIS.pdf · A Dios, a...

Detección y caracterización del virus
del Síndrome Reproductivo y
Respiratorio Porcino en tres granjas
de producción intensiva para el
establecimiento del control local de la
enfermedad
Eliana Mendoza Niño
Universidad Nacional de Colombia Facultad de Medicina Veterinaria y de Zootecnia
Maestría Salud y Producción Animal Bogotá, Colombia
2015

Detección y caracterización del virus
del Síndrome Reproductivo y
Respiratorio Porcino en tres granjas
de producción intensiva para el
establecimiento del control local de la
enfermedad
Eliana Mendoza Niño
Tesis presentada como requisito parcial para optar el título de:
Magister en Salud y Producción Animal
Director:
Jairo Jaime Correa M.V., M.Sc., Ph.D.
Línea de Investigación
Microbiología y Epidemiología
Grupo de Investigación
Microbiología y Epidemiología
Universidad Nacional de Colombia
Facultad de Medicina Veterinaria y de Zootecnia
Maestría Salud y Producción Animal
Bogotá, Colombia
2015

Dedicatoria
Esta es la suma de
muchos esfuerzos, en los que
he tenido a mi lado seres que
me han inspirado y otros que
me han retado. Gracias a todos
por que sin ellos no tendría la
satisfacción de ver cumplida
esta meta.
Va dedicado a los seres de luz
que me acompañan siempre: a
mi Mamá, a Lola, a mi Tío.
A mi Papá por forjar el ser que
soy.
A mis amigos y a todo aquel
que tuvo que ver en alguna parte de
esta historia.
Y por supuesto a los animales….

Agradecimientos
A Dios, a mis padres, a mi abuela y tío, por hacer de mí quien soy hoy.
A mis amigos por su constante aliento y buen sentido del humor.
Al Doctor Jairo Jaime Correa, por su dirección y orientación
Al Doctor José Dario Mogollón, por su acompañamiento y aportes técnicos.
A los profesores, compañeros y personal administrativo del Posgrado de Medicina
Veterinaria y de Zootecnia de la Universidad Nacional.
A la Asociación Colombiana de Porcicultores – Fondo Nacional de la Porcicultura, por su
apoyo financiero.
A las compañeras del Laboratorio Nacional de Diagnóstico Veterinario (Laboratorio de
Medicina Porcina y Biología Molecular) del Instituto Colombiano Agropecuario, por su
apoyo técnico.
A todo aquel que vivió conmigo, parte de esta experiencia.

Resumen
El presente estudio se realizó con el fin de evaluar el comportamiento del virus del PRRS
y su evolución en tres granjas de tipo intensivo del departamento del Valle del Cauca,
con el fin de orientar un programa de control específico para la condición en cada una de
dichas granjas. Para tal objetivo se evaluaron tres factores: tres granjas, siete grupos
etáreos (madres, reproductores, reemplazos, lactantes, destetos, preceba y ceba)
durante nueve meses, a partir de septiembre de 2012 a mayo de 2013.
En cada granja se tomaron muestras de suero de cada grupo etareo, muestras de fluido
oral de la preceba y ceba, tejidos y lavados bronquio alveolares (LBA) a partir de las
mortalidades que presentaron sintomatología compatible con el PRRS. Para determinar
la respuesta serológica se utilizó la técnica de ELISA (PRRS X 3 de IDEXX), igualmente
se determinó la presencia del agente viral mediante el uso de la qRT-PCR y aislamientos
en células MA-104. Los productos amplificados fueron secuenciados para determinar la
cepa circulante en cada una de las granjas.
Los resultados indicaron un incremento en la circulación del PRRSV en explotaciones
intensivas del departamento del Valle del Cauca, específicamente dentro de los sistemas
de producción de la preceba y ceba. Se observó que el uso de pruebas serológicas junto
con técnicas moleculares para el diagnóstico de PRRSV, permite conocer la dinámica del
virus dentro de cada granja y grupo etáreo. Mediante la secuenciación, se determinó que
las cepas circulantes correspondían al genotipo americano, confirmando los hallazgos
obtenidos en los primeros estudios filogenéticos para PRRSV realizados en el país.
Todos estos resultados permitieron inferir que el comportamiento en cada granja
respecto a PRRSV fue diferente, por tal razón, las recomendaciones igualmente debieron
ser aplicadas en forma individual con el fin de lograr el control adecuado de la
enfermedad.
Palabras clave: Control; Diagnóstico; PCR; PRRS.

Abstract
This study was conducted to determine the PRRS virus activity and its evolution in three
pig intensive production farms from an Colombian region: Valle del Cauca province; in
order to establish a specific monitoring program for each production system. To this aim,
three factors were evaluated: (i) farms (three), (ii) groups of age (mothers, boars,
replacements, piglest, weaned, pre-fattening and fattening) (iii) time ( nine months, from
September 2012 to May 2013).
In each farm serum samples were collect from each group of age, oral fluid samples were
taken from the pre-fattening and fattening groups, tissues and broncho alveolar lavage
(BAL) was recovered from necropsied pigs with PRRS compatible symptoms. To
determine the serological response an antibodies detection ELISA kit (IDEXX PRRS X3)
was employed, also the presence of viral genome was determined by real time RT-PCR
(qRT-PCR) and PRRSV isolation was performed on MA-104 cells. Aditionally, amplified
products from ORF-5 were sequenced to verify circulating strains into each farm.
Results indicated an increase of PRRSV circulation on Valle del Cauca region,
specifically within pre-fattening and fattening groups. The employment of both serological
and tests with molecular tests for PRRSV diagnostic provide understanding about viral
dynamic within each farm and age group. By sequencing it was found that strains
obtained corresponded to the American genotype, confirming findings from the first
PRRSV phylogenetic studies, conducted in the country. These results allowed to infer that
the PRRSV behavior in each farm was different, for this reason, the recommendations
should also be applied individually in order to achieve an adequate control of the disease.
Keywords: Control; Diagnosis; PCR; PRRS.

Contenido
Resumen y Abstract
Lista de tablas
Lista de figuras
Anexos
Abreviaturas y siglas
Introducción
Objetivos
1. Síndrome Reproductivo y Respiratorio Porcino - PPRS
1.1Historia
1.2 Situación mundial
1.3 Situación Colombia
1.4 Impacto económico
1.5 Agente etiológico
1.5.1 Proteínas no estructurales (NSPs)
1.5.2 Proteínas estructurales
1.6 Patogénesis
1.7 Respuesta Inmune
1.7.1 Respuesta inmune innata
1.7.2 Inmunidad adaptativa
1.7.3 Persistencia
1.8 Diagnóstico
1.8.1 Identificación de los signos clínicos
1.8.2 Lesiones microscópicas
1.8.2.1 Histopatología
1.8.3 Detección del virus
1.8.3.1 Aislamiento viral
1.8.3.2 Reacción en Cadena de la Polimerasa - PCR
1.8.4 Secuenciación
1.8.5 Pruebas serológicas
1.8.5.1 ELISA
1.8.5.2 Inmunofluorescencia indirecta – IFA

1.8.5.3 Seroneutralización
1.9 Control del PRRS
Bibliografía
2. Evaluación de la dinámica de anticuerpos frente al virus del Síndrome
Reproductivo y Respiratorio Porcino (PRRS) en tres granjas del departamento del
Valle del Cauca
Resumen
Abstract
Introducción
2.1 Materiales y Métodos
2.1.1 Ubicación geográfica y características de las granjas
2.1.2 Muestras
2.1.3 Evaluación Serológica
2.1.4 Análisis Estadístico
2.2 Resultados
2.2.1 Granja 1
2.2.2 Granja 2
2.2.3 Granja 3
2.3 Discusión
2.4 Conclusiones
2.5 Recomendaciones
Bibliografía
3. Determinación mediante la técnica de RT-PCR en tiempo real (qRT-PCR) de la
presencia del virus del Síndrome Reproductivo y Respiratorio Porcino (PRRSV) en
tres granjas del Valle del Cauca
Resumen
Abstract
Introducción
3.1 Materiales y Métodos
3.1.1 Ubicación geográfica y características de las granjas
3.1.2 Muestras
3.1.2.1 Sueros
3.1.2.2 Fluidos Orales (FO)

3.1.2.3 Lavado Bronquio alveolar (LBA)
3.1.2.4 Tejidos
3.1.3 Reacción en Cadena de la Polimerasa en Tiempo real (RT-qPCR)
3.1.3.1 Virus
3.1.3.2 Extracción ARN viral
3.1.3.3 Síntesis de ADNc
3.1.3.4 Curva Estándar
3.1.4 Análisis estadístico
3.2 Resultados
3.2.1 qRT-PCR en muestras de sueros
3.2.1.1 Granja 1
3.2.1.2 Granja 2
3.2.1.3 Granja 3
3.2.1.4 Comparación resultados por RT-PCR tiempo real y ELISA
3.2.2 qRT-PCR en muestras de Fluido oral (FO)
3.2.2.1 Granja 1
3.2.2.2 Granja 2
3.2.2.3 Granja 3
3.2.3 qRT-PCR en muestras de pulmón y ganglio linfático
3.2.3.1 Granja 1
3.2.3.2 Granja 2
3.2.3.3 Granja 3
3.2.4 qRT-PCR en lavado bronquioalveolar (LBA)
3.2.4.1 Granja 1
3.2.4.2 Granja 2
3.2.4.3 Granja 3
3.3 Discusión
3.4 Conclusiones
3.5 Recomendaciones
Bibliografía

4. Detección y determinación genotípica del virus del Síndrome Reproductivo y
Respiratorio Porcino (PRRSV) mediante aislamiento en células MA-104 y
secuenciación de muestras provenientes de tres granjas del valle del Cauca
Resumen
Abstract
Introducción
4.1 Materiales y Métodos
4.1.1 Ubicación geográfica y características de las granjas
4.1.2 Muestras
4.1.2.1 Lavado bronquioalveolar (LBA)
4.1.2.2 Tejidos
4.1.3 Virus
4.1.4 Aislamiento viral
4.1.4.1 Células
4.1.5 Secuenciación
4.2 Resultados
4.2.1 Aislamiento
4.2.2 Secuenciación
4.3 Discusión
4.4 Conclusiones
4.5 Recomendaciones
Bibliografía
5. Medidas de Control Propuestas
6. Conclusiones y Recomendaciones
6.1 Conclusiones
6.2 Recomendaciones
A. Anexo: Buffer fosfato salino (PBS) pH 7.2 +/- 0.2
B. Anexo: Solución penicilina-estreptomicina (PEST) – 100X
C. Anexo: Buffer fosfato salino (PBS) 10x para cultivo celular
Bibliografía

Lista de tablas
Tabla 2-1 Ubicación geográfica de las tres granjas y de los sitios productivos por granja
Tabla 2-2 Resultados por ELISA en Granja 1
Tabla 2-3 Resultados por ELISA en Granja 2
Tabla 2-4 Resultados por ELISA en Granja 3
Tabla 3-1 Ubicación geográfica de las tres granjas y de los sitios productivos por granja
Tabla 3-2 Sueros procesados por granja durante el tiempo de estudio
Tabla 3-3. FO procesados por granja durante el tiempo de estudio
Tabla 3-4. LBA procesados por granja durante el tiempo de estudio
Tabla 3-5. Tejidos procesados por granja durante el tiempo de estudio
Tabla 3-6. Comparación resultados obtenidos por qRT-PCR y ELISA en sueros en las
tres granjas
Tabla 3-7. Comportamiento serológico de las muestras positivas por qRT-PCR
Tabla 4-1. Descripción de los Primers utilizados en la secuenciación
Tabla 4-2. Descripción de las cepas americanas y europeas utilizadas en la realización
del árbol filogenético de las muestras concenso
Tabla 4-3. Resultados de aislamientos en células MA-104 en tejidos y LBA

Lista de figuras
Figura 1.1. Estructura del virus y del genoma de PRRS
Figura 2-1 Ubicación geográfica de cada granja dentro del Departamento del Valle del
Cauca
Figura 2-2 Comportamiento de los promedios de resultados del grupo ceba dentro de la
Granja 1 durante el tiempo de estudio
Figura 2-3 Comportamiento de los promedios de resultados serológicos del grupo de las
Madres dentro de la Granja 3 durante el tiempo de estudio
Figura 2-4 Comportamiento de los promedios de resultados serológicos del grupo de los
Lactantes dentro de la Granja 3 durante el tiempo de estudio
Figura 2-5. Comportamiento de los promedios de resultados serológicos del grupo de los
Destetos dentro de la Granja 3 durante el tiempo de estudio.
Figura 2-6 Comportamiento de los promedios de resultados serológicos del grupo de los
Reproductores dentro de la granja 3 durante el tiempo de estudio.
Figura 2-7 Comportamiento de los promedios de resultados serológicos del grupo de la
Preceba dentro de la Granja 3 durante el tiempo de estudio.
Figura 2-8 Comportamiento de los promedios de resultados serológicos del grupo de los
Reemplazos dentro de la Granja 3 durante el tiempo de estudio
Figura 2-9 Comportamiento de los promedios de resultados serológicos del grupo de la
Ceba dentro de la Granja 3 durante el tiempo de estudio
Figura 3-1 Ubicación geográfica de cada granja dentro del Departamento del Valle del
Cauca.
Figura 3-2 Curva Estándar por qRT-PCR.
Figura 3-3. Resultados obtenidos por qRT-PCR en sueros dentro de cada granja
evaluada

Figura 3-4. Resultados por qRT-PCR en sueros de cada grupo etáreo en la granja 1.
Figura 3-5. Resultados de qRT-PCR en sueros durante el tiempo en la granja 1.
Figura 3-6. Resultados por qRT-PCR en sueros de cada grupo etareo en la granja 2
Figura 3-7. Resultados de qRT-PCR en sueros durante el tiempo en la granja 2
Figura 3-8. Resultados por qRT-PCR en sueros de cada grupo etareo en la granja 3.
Figura 3-9. Resultados de qRT-PCR en sueros durante el tiempo en la granja 3.
Figura 3-10. Resultados por qRT-PCR en fluídos orales en las tres granjas
Figura 3-11. Resultados por qRT-PCR en FO por grupo etáreo en la granja 1.
Figura 3-12. Resultados por qRT-PCR en FO durante el tiempo en la granja 1.
Figura 3-13. Resultados por qRT-PCR en FO por grupo etáreo en la granja 2
Figura 3-14. Resultados en el tiempo por qRT-PCR en muestras de FO en la granja 2
Figura 3-15. Resultados por qRT-PCR en FO por grupo etáreo en la granja 3
Figura 3-16. Resultados por RT-PCR en muestras de FO en los grupos etáreos
evaluados en la granja 3.
Figura 3-17. Resultados por qRT-PCR en tejidos en las tres granjas evaluadas
Figura 3-18. Resultados por qRT-PCR en tejidos por grupo etareo en la granja 1.
Figura 3-19. Resultados por qRT-PCR en tejidos a través del tiempo en (A) preceba y (B)
ceba en la granja 1.
Figura 3-20. Resultados por qRT-PCR en tejidos por grupo etáreo en la granja 2.
Figura 3-21. Resultados por qRT-PCR en tejidos a través del tiempo en (A) preceba y (B)
ceba en la granja 1.
Figura 3-22. Resultados por qRT-PCR en tejidos por grupo etáreo en la granja 3.

Figura 3-23. Resultados por qRT-PCR en tejidos a través del tiempo en (A) preceba y (B)
ceba de la granja 3.
Figura 3-24. Resultados por qRT-PCR en LBA para las tres granjas evaluadas
Figura 3-25. Resultados por RT-PCR en LBA por grupo etáreo en la granja 1.
Figura 3-26. Resultados por qRT-PCR en LBA a través del tiempo en la granja 1.
Figura 3-27. Resultados por qRT-PCR en LBA por grupo etáreo en la granja 2
Figura 3-28. Resultados por qRT-PCR en LBA por grupo etáreo en la granja 3.
Figura 3-29. Resultados por qRT-PCR de LBA a través del tiempo en la granja 3.
Figura 4-1. Gel de electroforesis a partir de muestras positivas a qRT-PCR
Figura 4-2. Árbol filogenético realizado en Mega 5, mediante Neighborg-Joining
(Bootstrap 1000).

Anexos
Anexo 1. Buffer fosfato salino (PBS) pH 7.2 +/- 0.2
Anexo 2. Solucion penicilina-estreptomicina (PEST) – 100X
Anexo 3. Buffer fosfato salino (PBS) 10x para cultivo celular

Abreviaturas y Siglas
ADE Antibody dependent enhancement – Aumento dependiente de Anticuerpos
ADNc Acido Desoxirribonucleico complementario
ARNd ARN de doble cadena
EC Efecto citopático
ELISA Enzyme-Linked ImmunoSorbent Assay: ensayo por inmunoabsorción
ligado a enzimas
FO Fluido Oral
HP-PRRSV Highly pathogenic porcine reproductive and respiratory syndrome virus
IFA Inmunofluorescencia indirecta
IFN Interferón
IRF3 Interferon regulator factor 3 - Factor 3 regulador de Interferón
ISG Interferon stimulated gene – Gen estimulador de Interferón
IkBα nuclear factor of kappa light polypeptide gene enhancer in B-cells inhibitor,
alpha
kDa Kilodaltons
LBA Lavado Bronquioalveolar
LNDV Laboratorio Nacional de Diagnóstico Veterinario
LV Lelystad virus
MAPs Macrófagos alveolares porcinos
NF-kB Nuclear factor kappa B
MARC 145 African green monkey kidney derived cells
NSP Non-structural protein - Proteína no structural
ORF Open Reading Frame – marco de lectura abierta
OIE Organisation for Animal Health - Organización mundial para la salud
animal
pb Pares de bases
PI Pos infección
PRRSV Porcine reproductive and respiratory syndrome virus – Virus del síndrome
reproductivo y respiratorio porcino
PCR Reacción en cadena de la polimerasa
qPCR Quantitative PCR

RT-PCR Reverse transcription-PCR
VSN Seroneutralización viral

Introducción
El Síndrome Reproductivo y Respiratorio Porcino (PRRS) es la enfermedad infecciosa de
los porcinos que tiene gran importancia a nivel mundial por su alto impacto a nivel
económico (Holtkman et al., 2013). Esta enfermedad genera altos costos en la
producción porcícola, atribuibles especialmente a pérdidas en la productividad, gastos
veterinarios e implementación de mayores medidas de bioseguridad y cambios en las
características de manejo de los animales. En Estados Unidos, se estimó el impacto
económico del PRRSV entre los años 2005 y 2010, observando que la enfermedad le
costaba a los productores americanos US$ 663,91 millones al año, aproximadamente
US$1.8 millones por día.
En Colombia, los primeros reportes de esta enfermedad, datan de 1996, donde mediante
evidencia serológica se determinó la circulación del PRRSV (Mogollón et al., 2003).
Estudios realizados en 1997, reportaron una prevalencia de la enfermedad del 6.5% en
animales de traspatio y un 22.7% en animales confinados. Estos estudios determinaron
que el virus se encontraba ampliamente distribuido en el país de forma no uniforme,
predominando en los sistemas de producción intensiva (Mogollón et al., 2003). De igual
manera el virus fue aislado de tejidos y sueros en diferentes regiones del país y en los
dos sistemas de producción. Las cepas encontradas en el país se ubicaron dentro del
genotipo americano del virus. Para el año 2006 se observó que la prevalencia de la
enfermedad en explotaciones extensivas se redujo al 4.3%, sin embargo se encontró un
mayor número de departamentos afectados con respecto a estudios previos (Cruz et al.,
2006). En Colombia, actualmente la enfermedad es endémica, constituyendo un factor
limitante en el desarrollo de la industria porcina.
El agente etiológico del PRRS, por su parte, es de difícil control debido a que presenta
una gran variación genómica y la capacidad de producir infecciones persistentes. La alta
variabilidad, se traduce en cambios en los patrones de infectividad y en generación de
una respuesta inmune poco efectiva en el huésped. Los dos genotipos del virus, el tipo I
o Europeo, representado por la cepa Lelystad y el tipo II o Americano, representado por
la cepa VR2332, presentan una alta variabilidad tanto entre estos aislados como entre
genotipos.

Teniendo en cuenta, que en los últimos años no se obtuvo información sobre esta
enfermedad en el país y contando con las experiencias a nivel internacional, se realizó el
presente estudio con el fin de verificar el estatus del PRRSV dentro de tres granjas de
tipo intensivo ubicadas en una de las regiones porcícolas más importantes del país, el
departamento del Valle del Cauca. Se tomaron diferentes tipos de muestras que
incluyeron sueros, tejidos, fluidos orales y lavados bronquio alveolares, en los diferentes
grupos etáreos (madres, reproductores, reemplazos, lactantes, destetos, preceba y
ceba), durante un lapso de nueve meses. De igual manera se emplearon varios métodos
diagnósticos para determinar tanto la dinámica del virus, como las cepas circulantes, con
el fin de conocer el estado actual de la enfermedad en estas granjas y por ende en la
zona y así establecer medidas de control para la condición propia de cada una de ellas.
Los resultados obtenidos confirmaron lo reportado en otros países, donde se observó un
comportamiento diferente del PRRSV dentro de cada granja evaluada, dentro de una
zona o región, por lo tanto las medidas debieron ser aplicadas igualmente de forma
particular. Se priorizó el uso de adecuadas medidas de bioseguridad, un apropiado
manejo de los reemplazos y de los lechones, como la verificación del origen del semen
utilizado en la granja.
El conocimiento del estatus sanitario dentro de una granja, como dentro de un país, con
respecto a enfermedades como el PRRS, permite detectar los factores asociados con su
presentación, con el fin de aplicar las medidas adecuadas para el control, favoreciendo
así un aumento en la productividad y en la admisibilidad de los productos de origen
porcino en diversos mercados internacionales.

OBJETIVOS
OBJETIVO GENERAL
Estudiar el comportamiento del virus del PRRS y su evolución en tres granjas de tipo
intensivo de la zona del Valle del Cauca, con el fin de establecer el control para la
condición particular en cada una de ellas.
OBJETIVOS ESPECIFICOS
Determinar la dinámica de anticuerpos frente al virus del PRRS en tres granjas de tipo
intensivo y ciclo completo durante un periodo de nueve meses.
Aislar las cepas circulantes en las granjas, en células permisibles a la replicación del
PRRSV, a partir de muestras de tejidos y lavados bronquio alveolares.
Detectar mediante técnicas de qRT-PCR, la presencia del ARN del virus del PRRS en
muestras de suero, tejidos, fluido oral y lavado bronco alveolar, de porcinos provenientes
de tres granjas intensivas del Departamento del Valle del Cauca.
Secuenciar y caracterizar los aislados obtenidos, mediante estudios filogenéticos con el
fin de confirmar su correspondencia con cepas ya reconocidas o establecer si éstas
corresponden a nuevas cepas circulantes en el país.
Establecer medidas de control de acuerdo con los resultados obtenidos.

1. Síndrome Reproductivo y Respiratorio Porcino El Síndrome Reproductivo y Respiratorio Porcino (PRRS) una enfermedad infecciosa que
afecta a la especie porcina, se caracteriza por ocasionar alteraciones respiratorias
especialmente en lechones y fallas reproductivas en hembras gestantes (Benfield et al.,
1999). El virus de PRRS (PRRSV) evolutivamente ha incrementado su virulencia,
causando una generalizada mortalidad en hembras al final de la gestación y en lechones
en crecimiento de todas las edades; es la enfermedad más importante de los porcinos
alrededor del mundo (Neumann et al., 2005; Zhou and Yang, 2010).
1.1 Historia
A finales de los años 80, en los Estados Unidos surgió una enfermedad no antes
reconocida en porcinos, ocasionando pérdidas reproductivas hacia el final de la
gestación, incremento del número de lechones nacidos débiles, neumonía en neonatos y
lechones, reducción en el crecimiento e incremento de la mortalidad (Hill, 1990; Keffaber,
1989; Loula, 1991). Un síndrome similar fue descrito inicialmente en Alemania en 1990
propagándose a otros países de Europa. Como el agente causal de la enfermedad era
desconocido, recibió en los Estados Unidos el nombre de la “enfermedad misteriosa del
cerdo” y en Europa “síndrome respiratorio y aborto epidémico porcino” o “enfermedad de
las orejas azules del cerdo” (Wensvoort et al., 1991). Una vez, el agente etiológico de la
enfermedad fue descubierto se le denominó como el síndrome respiratorio y reproductivo
porcino (SIRS) en Norteamérica y en Europa como el síndrome respiratorio y
reproductivo porcino (PRRS). La enfermedad actualmente se encuentra en muchos
países afectando la industria porcícola y la seguridad alimenticia alrededor del mundo
(Albina, 1997; Blaha, 2000)
1.2 Situación mundial
Actualmente, el PRRSV es considerado endémico y ha sido reportado en la mayoría de
zonas productoras de porcinos a nivel mundial (Zimmerman, 2003). De acuerdo con la
UN-FAO, únicamente Australia, Nueva Zelanda y Suiza han sido declaradas libres del
virus del PRRS (Dietze et al., 2011). Los brotes más recientes se reportaron en Suecia

(2008), Sudáfrica (2004 y 2007), Rusia, Vietnam y China (2006) (Osorio, 2010). En 2006,
una cepa altamente patógena de PRRS (HP-PRRS), también conocida como la
enfermedad de la fiebre alta del cerdo, fue inicialmente reportada en China y desde
entonces se ha diseminado rápidamente ocasionando mortalidades principalmente en
cerdos jóvenes. Esta cepa también ha sido reportada en Vietnam, Laos, Cambodia,
Myanmar, Filipinas y Rusia (Hu et al., 2009; Li et al., 2011; Tian et al., 2007; Zhou et al.,
2008).
La evolución del PRRSV se ha caracterizado por la aparición de cepas con una virulencia
incrementada, como lo ha demostrado la cepa 1-18-2, la cual surgió en 2007 afectando la
zona norte y centro de Estados Unidos (Li et al., 2009, 2011; Zhou et al., 2008)
En Suramérica, Argentina y Brasil son libres de la enfermedad. Chile en 2013 efectuó
una auto declaración oficial como país libre ante la OIE, sin embargo, en octubre del
mismo año, se detectó un brote de PRRS con una nueva cepa, que afectó a planteles
industriales y no industriales (traspatio) (http://www.sag.cl/ambitos-de-accion/sindrome-
reproductivo-y-respiratorio-porcino-prrs).
1.3 Situación en Colombia
En Colombia, la primera evidencia serológica del PRRSV fue encontrada en 1996
(Mogollón et al., 2003). En 1997 se confirmó ésta reactividad (6.5%) en 17
departamentos con alta producción porcina. Entre 1997 y 1999 se realizaron aislamientos
virales a partir de sueros y muestras de tejido pulmonar recolectados en predios con
historial clínico de PRRS y serología positiva. Este estudio permitió recuperar virus en
diferentes regiones del país y en diferentes sistemas de producción porcina. Paralelo a
los estudios citados, en el año 2000, se inició la evaluación molecular de los aislamientos
obtenidos y se concluyó que estos se encontraban dentro del genotipo americano (Tipo
II), sin embargo, no se descartó la presencia del genotipo Europeo en el país, teniendo
en cuenta que en este estudio solo se emplearon primers específicos para detectar
cepas americanas (Mogollón et al., 2003). Para el año 2006 se observó que la
prevalencia de la enfermedad en explotaciones extensivas se redujo al 4.3%, sin

embargo se encontró un mayor número de departamentos afectados con respecto a
hallazgos previos (Cruz et al., 2006).
1.4 Impacto económico
Se han realizado dos estudios estimando el impacto económico del PRRS en Estados
Unidos. De acuerdo con Neumann et al (2005), la pérdida anual debida a PRRSV se
estima que excede los 560 millones de dólares, donde el 11.9% corresponde a
gestación/maternidad; 35.9% a recría/destete y el 52.2% a engorde. En un segundo
estudio realizado por Holtkamp et al (2013), el total de costos anuales debidos a pérdida
de la productividad en la cría y crecimiento en cerdos, durante el periodo 2005 a 2010,
excedió en aproximadamente 104 millones de dólares a los 560 millones de costo anual
estimados en 2005. Adicional a estos costos, se encontró que el costo anual debido a
gastos veterinarios, que no fueron contemplados en el estudio de 2005, oscilaba en
477.79 millones de dólares anuales.
1.5 Agente etiológico
En 1991, Wensvoort et al en el Instituto Veterinario Central de Lelystad (Holanda)
aislaron en cultivos primarios de macrófagos alveolares porcinos (MAPs) un nuevo virus
al que se denominó virus Lelystad (LV). Posteriormente, en Estados Unidos se aisló un
virus estructural y patogénicamente relacionado con el LV, el cual recibió la
denominación de VR-2332, considerado como la cepa americana de referencia (Benfield
et al., 1992; Collins et al., 1992). Al comparar las secuencias del virus Lelystad,
posteriormente denominado Tipo I y el Norteamericano o Tipo II, se observó que
compartían propiedades similares a la de los virus del orden Nidovirales, familia
Arteriviridae y género Arterivirus, que incluyen a su vez, el Virus de la Arteritis Equina
(VAE), el Virus Elevador de la Lactato-Deshidrogenasa del Ratón (VLD) y el Virus de la
Fiebre Hemorrágica de los Simios (VFHS) (Benfield et al., 1992; Cavanagh, 1997, Dea et
al., 2000). Los virus de la familia Arteriviridae comparten algunas características como
tropismo por los macrófagos, una elevada variabilidad genética ocasionada por
mutaciones o recombinaciones y la capacidad de causar infecciones persistentes
(Cavanagh, 1997).

Se ha demostrado que la cepa Lelystad y la cepa VR-2332 difieren aproximadamente en
un 40% a nivel de la secuencia del genoma y son antigénicamente distintas (Nelson et
al., 1999). Los dos genotipos poseen solo el 70% de identidad a nivel de nucleótidos y las
cepas dentro de cada genotipo varían hasta en un 20% en sus secuencias (Key et al.,
2001; Meng, 2000). La proteína estructural más variable es la GP5 con una homología de
aminoácidos del 50 a 55% entre aislados Norteamericanos y Europeos (Mardassi et al.,
1996; Meng et al., 1995a).
El PRRSV es esférico, presenta una envoltura lipídica y posee un diámetro promedio de
62 nm (Figura 1.1). Su nucleocápside presenta simetría helicoidal y dentro de ella se
encuentra el genoma constituido por un ARN de cadena sencilla y polaridad positiva con
una longitud promedio de 15 kb. Este a su vez consta de nueve marcos de lectura abierta
(Open Reading Frame - ORFs) solapantes que se transcriben en la célula infectada como
ARNm subgenómicos (Snijder and Meulenberg, 1998).
La longitud completa del ARN (ARNm1) es usada para la traducción de dos ORFs:
ORF1a y ORF1b. La traducción del ORF1a produce la poli proteína pp1a, mientras que el
ORF1b es expresado por medio del desplazamiento del fragmento ribosomal (ribosomal
frame shift) llevando a la formación de la poli proteína pp1ab (Meulenberg et al., 1993).
Las poli proteínas pp1a y pp1ab son procesadas por proteasas virales liberando 14
proteínas no estructurales las que incluyen las proteasas (NSP1α, NSP1β, NSP2 y
NSP4), una RNA polimerasa dependiente de RNA (NSP9), una helicasa (NSP10) y una
endonucleasa (NSP11) (Snijder and Meulenberg., 1998).
Los ORFs 2 a 5, codifican para proteínas glicosiladas de la envoltura: la GP2 a GP5; el
ORF6 codifica para una proteína no glicosilada de membrana (M) y el ORF7 la proteína
de la nucleocápside (N) (Dea et al., 2000; Snijder and Meulenberg, 1998). El ORF2b está
incluido dentro de ORF2 y codifica una pequeña proteína no glicosilada E o 2b (Wu et al.,
2001).

Figura 1.1. Estructura del virus y del genoma de PRRS. En el extremo 5´del genoma
se localizan los ORFs que dan lugar a las proteínas no estructurales (NSPs) y hacia el
extremo 3´ aquellos que dan origen a las proteínas estructurales. En la superficie del
virus se observa la distribución de las proteínas estructurales incluida la nueva proteína
obtenida a partir del ORF5. Imagen que hace parte de la presentación del Doctor Iván
Díaz Luque, Investigador del CRESA. Symposium internacional de porcinocultura II.
SEPOR. 2012.
1.5.1 Proteínas no estructurales (NSPs)
En la terminación N de la poli proteína pp1a se encuentran las proteínas NSP1α y NSP1β
que contienen un dominio proteasa cisteína (Chen et al., 2010a). NSP1α contiene tres
dominios, un dominio dedo de zinc N-terminal, un dominio proteasa cisteína como
papaína (PCPα) con un ion zinc ligado al sitio activo, requerido para la actividad
proteolítica y una extensión C-terminal unida al sitio de unión del sustrato del dominio

PCPα (Sun et al., 2009). NSP1β tiene 4 dominios, un domino nucleasa dependiente de
metal N-terminal, un dominio enlazador, un dominio proteasa cisteína como papaína
(PCPβ) y una extensión C-terminal unida al sitio de unión del sustrato del dominio PCPβ
(Xue, et al., 2010). La inactivación de PCPα en NSP1α, bloquea la síntesis de ARNm
subgenómicos sin alterar la replicación, mientras que cuando la actividad de PCPβ es
eliminada de Nsp1β, no se observa síntesis de ARN viral (Kroese et al., 2008).
NSP2 es una proteína grande que varía en tamaño de 1168-1196 residuos entre
diferentes cepas de PRRSV, contiene además un cluster de epítopes lineales de células
B, que son indispensables para la replicación del virus (Oleksiewicz et al., 2001) y a su
vez modulan la respuesta inmune del huésped (Chen et al., 2010b). Esta es la proteína
no estructural más variable con solo 32% de identidad entre subtipos (Allende et al.,
1999). Contiene un dominio proteasa cisteína N terminal, seguido por una región
hipervariable rica en prolina, una región hidrofóbica que contiene cuatro hélices TM
(transmembrana) y un dominio C terminal conservado (Han et al., 2009). La deleción de
alguno de sus epítopes puede atenuar la replicación y la virulencia del PRRSV (Darwich
et al, 2011). NSP3 contiene cuatro hélices TM, que participan en la formación de los
complejos de replicación y en el anclaje de otras proteínas no estructurales en las
membranas de dicho complejo (Han et al., 2009). NSP4 es activada por la proteasa
NSP2 después del clivaje del complejo NSP2-NSP3. Posee una proteasa 3C (3CLP) que
consta de pliegues de quimo tripsina que pueden estar involucrados en la regulación del
procesamiento de la poli proteína viral (Van Aken, et al., 2006).
Los productos del clivaje de pp1a, como son NSP5, NSP6, NSP8 y NSP12 no han sido
totalmente descritos, desconociéndose su estructura y funciones (Doklan, 2010; Yun and
Lee, 2013).
NSP7 contiene un sitio de clivaje interno que da lugar a dos proteínas NSP7α y NSP7β,
cuyas funciones específicas aún se desconocen (Doklan, 2010; Yun and Lee, 2013).
NSP9 a NSP11 son las proteínas más conservadas dentro de los nidovirus, la NSP9
corresponde a la polimerasa viral dependiente de ARN (Gorbalenya et al., 2006).
NSP10 presenta tres dominios, un dominio de unión al zinc N terminal, un

dominio de unión y un dominio helicasa. El dominio de unión (S2429P) ha sido
implicado en la síntesis de ARNs subgenómicos (Van Dinten et al., 1997). NSP11
contiene el dominio endoribonucleasa (NendoU) específico para uridilato, el cual es el
mayor marcador genético único para nidovirus (Fang and Snijder, 2010; Gorbalenya et
al., 2006). Se ha demostrado que la inmunidad innata del huésped mediada por el
interferón (IFN) es modulada por un conjunto de NSPs (NSP1α, NSP1β, NSP2, NSP4 y
NSP11) del PRRSV (Beura et al., 2010).
1.5.2 Proteínas estructurales
Los ORFs 2a, 2b, 3 y 4 localizados en el extremo 3’ del genoma viral, codifican para las
proteínas asociadas al virión (estructurales) denominadas GP2a, 2b, GP3 y GP4.
(Meulenberg et al. 1995, 1997; Meulenberg y Besten, 1996; Wu et al., 2001, 2005).
La GP2 contiene una secuencia señal N-terminal entre los residuos 1-40, seguida por un
ectodominio de 168 residuos con una hélice TM (transmembrana) y un endodominio de
20 residuos. La GP2 presenta dos sitios de glicosilación conservados (Wissink et al.,
2004) y posee una masa molecular de 29-30 kDa. Una vez es sintetizada se localiza a
nivel perinuclear y citoplasmático en la célula hospedadora y se incorpora a la envoltura
del virión en forma de monómeros. Su función es participar en la replicación viral y se
considera un pobre inmunógeno ya que induce niveles bajos de anticuerpos
neutralizantes (Rogan et al., 2000).
La proteína E o 2b, es expresada a partir del ORF2b que se encuentra totalmente
incrustada dentro del ORF2a que codifica para la proteína GP2 (Wu et al., 2001). La
proteína E es de tipo estructural, no glicosilada y tiene una masa molecular de 10 kDa,
está conformada por 73 aminoácidos y es conservada entre aislados europeos y
americanos (Yoon, 2003); forma un canal iónico que incrustado en la membrana viral,
permite la adhesión del virión y la liberación del genoma en el citoplasma (Lee and Yoo,
2006).
GP3 corresponde a una glicoproteína menor de la envoltura con 27-29 kDa de masa
molecular (Mardassi et al., 1995; Murtaugh et al, 1995). Es la proteína del PRRSV más

altamente glicosilada, con seis puntos de glicosilación localizados en el ectodominio
putativo. La homología de secuencias codificantes para esta proteína entre los genotipos
Europeos y Americanos es del 58% y la más alta divergencia se presenta en la región C-
terminal, donde el genotipo Europeo tiene una extensión adicional de 11 aminoácidos; el
ectodominio es idéntico en un 70% entre los dos genotipos (Gonin et al., 1998; Meng et
al., 1995a). Esta ha sido una de las proteínas más controversiales respecto a su papel
como constituyente de la envoltura de la partícula viral, lo cual ha sido comprobado
debido a su habilidad para producir anticuerpos neutralizantes sola o junto a GP4 y GP5
(De Lima et al., 2009; Jiang et al., 2008). La propiedad de GP3 para producir
inmunogenicidad se explica por su habilidad para ser secretada desde las células
infectadas en forma de homodímero (Mardassi et al., 1995).
La proteína GP4 corresponde a una proteína estructural de la envoltura, con una masa
molecular de 19-20 kDa. Contiene un péptido señal clivado desde el residuo 1 a 21 y una
hélice TM del residuo 156 a 177. Presenta cuatro sitios potenciales de glicosilación que
son altamente conservados entre aislados europeos y americanos (Meulenberg et al.,
1997; Murtaugh et al., 1995; Zhang et al., 1998). Parece estar asociada al proceso de
replicación viral ya que su deleción en clones infecciosos de ADNc hace imposible
obtener una nueva progenie vírica (Welch et al., 2004; Wissink et al., 2005). Se ha
demostrado también que GP4 interactúa con otras glicoproteínas resultando en la
formación de complejos de multiproteínas. GP2, GP3 y GP4 son incorporados como
complejos multiméricos en la cubierta de ambos genotipos (Das et al., 2010).
El ORF 5 codifica la proteína GP5 que es la proteína mayoritaria de la envoltura, tiene un
tamaño de 24-25 kDa (Meulenberg et al., 2000) y posee una secuencia señal N-terminal,
seguida por un ectodominio glicosilado (Meulenberg et al., 1995). La GP5 presenta una
región hidrofóbica que incluye entre uno a tres hélices transmembrana (TM) (Dea et al.,
2000; Mardassi et al., 1996) y un largo endodominio C-terminal, lo cual es característico
de los nidovirus. El corto ectodominio de 30 aminoácidos presenta posiblemente los sitios
de N-glicosilación que varían en número dependiendo de la cepa; los aislados europeos
contienen dos N-glicanos mientras que los americanos tres (Mardassi et al., 1995;
Meulenberg, 2000). Los sitios conservados de glicosilación de la proteína GP5 como el
N46 en aislados Europeos y N44 en Americanos, parecen ser críticos en el proceso de

infección por PRRSV (Ansari et al., 2006; Wissink et al., 2004, citados por Van Breedman
et al., 2010).
GP5 está involucrada en la interacción inicial con la célula huésped posiblemente en la
fusión con las membranas celulares (Wissink et al., 2005). La más efectiva respuesta de
anticuerpos neutralizantes es dirigida contra ésta proteína lo que indica que puede ser el
determinante de inmunidad protectiva y puede estar involucrada en la unión a receptores
(Gonin et al., 1999; Ostrowski et al., 2002). De igual manera, es la proteína más variable
de PRRSV, con solo 51-55% de homología de secuencia entre el genotipo Europeo y
Americano (Meulenberg et al., 2000; Murtaugh et al., 1995). La hipervariabilidad en GP5
es probablemente la responsable de la ausencia de una reacción inmune cruzada entre
virus (Meng, 2000a). Esta proteína también forma heterodímeros con la proteína
codificada por ORF6, los que podrían estar relacionados con los procesos de ensamblaje
viral y su unión a la célula hospedadora (Delputte et al, 2002).
La proteína de membrana (M) no es glicosilada y tiene 18-19 kDa de masa molecular
(Mardasii et al., 1995; Meulenberg et al., 2000). Presenta un corto ectodomino N-terminal,
tres segmentos transmembrana (TM) y un endodominio C-terminal. Es la proteína
estructural más conservada de PRRSV (Meng et al., 1995b). La proteína M de los
aislados Americanos contiene 174 aminoácidos, mientras que en los aislados Europeos
constan de 173 aminoácidos (Meulenberg et al., 2000). Se ha demostrado que la
proteína M es altamente antigénica y produce una respuesta de anticuerpos a los 10 días
siguientes a la infección (Loemba et al 1996; Yoon et al., 1995, citado por Yoon, 2003).
En macrófagos alveolares pulmonares porcinos (MAPs) la adhesión inicial del virión es
mediada por interacción de la proteína M viral con el proteoglicano heparan sulfato
(Delputte et al., 2002). Forma heterodímeros con la proteína GP5 lo que podría estar
relacionado con la infectividad del virus, ya que este complejo se une a los macrófagos
por medio de uniones dependientes de ácido siálico a la sialodhesina (Van Breedman et
al., 2010).
El ORF7 codifica para la proteína N de la cápside del virus. Esta no es glicosilada y tiene
una masa molecular de 14-15 kDa. Corresponde al 20-40% del contenido proteíco del
virión, siendo la más abundante (Dea et al., 2000, citado por Yoon, 2003). La mayoría de

anticuerpos producidos en la infección son específicos para esta proteína ya que cuenta
con varios epítopes, algunos específicos para aislados europeos o norteamericanos,
mientras que otros son conservados en ambos grupos. Además, existe un antígeno
inmunodominante en la región central de esta proteína entre los aminoácidos 50-66 y 80-
90 (Meulenberg et al., 2000), siendo adecuada para el diagnóstico de la enfermedad. Se
ha sugerido que esta proteína juega un papel en la citopatogénesis del virus (Rowland et
al., 1999, citado por Yoon, 2003) y de igual manera se ha relacionado su presencia en el
núcleo y nucléolo de la célula con la capacidad replicativa del virus y con la ausencia de
inducción de anticuerpos neutralizantes (Pei et al., 2008). Cepas con mutaciones en la
señal de localización nuclear de la proteína N producen una viremia con bajos títulos
virales, de corta duración pero a su vez produce altos niveles de anticuerpos
neutralizantes y reducida persistencia viral en tonsilas (Lee and Yoo, 2006; Pei et al.,
2008).
Recientemente, Murtaugh et al (2011a), empleando espectrometría de masas
descubrieron un nuevo polipéptido de 51 aminoácidos, que es codificado por un
fragmento de lectura alternativo de mRNA5 subgenómico (sgmRNA5), que está presente
en el PRRSV y en otros arterivirus y se presume está relacionado con la replicación del
virus (Murtaugh et al., 2011b).
1.6 Patogénesis viral
El PRRSV es transmitido por vía horizontal mediante el contacto entre cerdos o por vía
vertical por medio de infección transplacental durante el último tercio de la gestación
(Christianson et al., 1992). La transmisión horizontal del PRRSV se produce
frecuentemente por contacto directo de animales susceptibles con animales infectados a
través de la vía oro-nasal, por heridas en la piel o de las superficies mucosas
(Zimmerman et al., 2006) y mediante el uso de semen contaminado o por medio de la
monta directa (Christopher-Hennings et al., 1996; Reiner et al., 2009). La transmisión
indirecta involucra el contagio a través de fómites como botas y overoles; el empleo de
equipo contaminado como agujas; por personal de la granja y visitantes; vehículos de
transporte contaminados; agua y alimentos; moscas, mosquitos y aerosoles (Cho & Dee,
2006; Dee et al., 2002; Desrosiers, 2011).

En la transmisión vertical, el PRRSV puede atravesar la placenta e infectar el feto hacia
los 90 días de la gestación, siendo el timo el sitio preferencial de replicación, reportes de
campo sugieren que luego de una falla reproductiva inducida por PRRSV, las hembras
desarrollan una inmunidad protectiva, esto se basa en que hembras afectadas tienen
camadas normales a pesar de la circulación del virus dentro de la granja (Stevenson et
al., 1993, citado por Karniychuk and Nauwynck, 2013). Las hembras seropositivas
pueden transmitir anticuerpos maternos a sus camadas vía calostro, sin embargo la
inmunidad pasiva declina y puede llevar a infección luego del destete cuando los
animales son mezclados (OIE, 2008; Rowland, 2010).
El PRRSV se replica principalmente en los macrófagos alveolares porcinos (MAPs),
también en células de la línea germinal monocítica, incluyendo macrófagos
intravasculares, subconjuntos de macrófagos en nódulos linfoides o bazo y macrófagos
intravasculares de la placenta y del cordón umbilical (Lawson et al., 1997).
El primer paso en la infección de los macrófagos, es una débil unión del virus vía heparan
sulfato, este último actúa como un factor de unión de baja afinidad que concentra las
partículas virales sobre la superficie celular (Delputte et al., 2002, 2005, 2007).
Posteriormente el complejo proteína M y GP5 se une al extremo N-terminal de la
sialodesina (CD169), esta interacción es dirigida por el dominio de unión al ácido siálico
en el extremo N-terminal de CD169 y el ácido siálico sobre la superficie del virión (Van
Breedam et al., 2010). Esta unión es seguida por un proceso de endocitosis mediada por
clatrina (Nauwynck et al., 1999). Otros factores del huésped, como la vimentina, una
proteína filamento intermediaria (intermediate-filament (IF) protein) (Kim et al., 2006) y
CD151 un miembro de la superfamilia tetraspanina (Shanmukhappa et al., 2007) han sido
identificados como receptores en células MARC-145, la línea celular susceptible a
PRRSV (Kreutz, 1998).
Una vez internalizadas las partículas virales, estas son transportadas a los endosomas
tempranos donde el genoma viral es liberado en el citoplasma en una reacción que
requiere acidificación endosomal y el receptor scavenger CD163 (Calvert et al., 2007;

Van Gorp et al., 2008; Welch and Calvert, 2010). El papel de CD163 es mediado a través
del dominio 5 rico en cisteína y por la interacción con GP2 y GP4 (Das et al., 2010).
Una vez el genoma entra al citoplasma, ORF1a y ORF1b son traducidos produciendo dos
largas poli proteínas pp1a y pp1ab, las cuales a su vez dan lugar a las NSPs, proceso
que es mediado por proteasa virales (NSP1α, NSP1β, NSP2 y NSP4) (Snijder et al.,
1996). El ensamble de las NSPs se produce bajo un complejo de replicación y
transcripción (RTC) el cual a su vez dirige la amplificación del genoma y la síntesis de
ARNm sub genómicos (Fang and Snijder, 2010). A través de seis de los ARNm sub
genómicos, son expresados los ORFs superpuestos y localizados en la región proximal
3´, los cuales a su vez codifican las proteínas estructurales (Doklan, 2010). En el último
estado de la replicación viral, múltiples copias de proteína N se unen al ARN genómico
sintetizado, formando la nucleocápside (Tijms et al., 2002) la cual surge en el lumen del
retículo endoplasmático y/o complejo de Golgi para luego adquirir las demás proteínas
estructurales (Wissink et al., 2005). Finalmente la progenie de viriones acumulados en los
compartimentos de la membrana intracelular, son liberados en el espacio extracelular
mediante exocitosis (Dea et al., 1995).
En adición, la exitosa infección por el PRRSV depende de la presencia de proteasas
localizadas en el endosoma como son la catepsina E y la serina proteasa (Misinzo et al.,
2008). El rol de las proteasas es escindir y liberar el péptido de fusión de las proteínas de
la envoltura a fin de iniciar la fusión de las membranas (Harrison, 2008).
Posterior a la exposición con el PRRSV, la replicación se presenta primariamente en los
macrófagos alveolares, a partir de los cuales se disemina rápidamente siendo los
pulmones y nódulos linfáticos los sitios con la más alta carga viral. La viremia
generalmente dura cuatro semanas con un pico en el título de virus entre el día 7 y 14
pos infección (PI), este pico usualmente corresponde a la fase clínica de la enfermedad.
Posterior a la fase de viremia el virus persiste en varios tejidos de origen linfoide como
tonsilas y nódulos linfáticos hasta los 157 días PI (Allende eta al., 2000). El ARN viral ha
sido detectado hasta los 250 días PI (Wills et al., 2003). Horter et al (2002) detectaron
virus infeccioso de PRRS en el 84% de animales entre los 63 y 105 días post inoculación.
Rowland et al. (2003) aislaron el PRRSV de la tonsila y nódulos linfoides de cerdos

infectados in útero a los 132 días pos nacimiento. En muchos casos la infección es
eliminada entre los 3 a 4 meses (Molina et al., 2008)
1.7 Respuesta inmune
De forma general, la exposición al PRRSV involucra un largo periodo de viremia
aproximadamente de un mes, seguido por una fase de persistencia de al menos 3 a 4
meses o más. Los cerdos infectados se recuperan de la infección hacia el final de esta
fase de persistencia, lo cual indica un retraso en el desarrollo de una respuesta inmune
adaptativa. Presentan a su vez, una respuesta inmune protectiva contra la re-exposición
a la misma cepa viral (Murtaugh et al., 2002).
1.7.1 Respuesta inmune innata
La respuesta inmune al PRRSV, comienza con una respuesta antiviral innata en el
citoplasma de un macrófago infectado, donde la presencia de una doble cadena de ARN
(ARNds) induce una cascada de eventos intracelulares que inician con la producción de
interferón tipo I (α y β) (Murtaugh et al., 2003). El PRRSV modifica la respuesta inmune
innata del huésped al afectar la inducción de interferón tipo I, por medio de varios
mecanismos: reduce la expresión de TLR3 (Toll-like-receptor) el cual normalmente
produce la activación del factor 3 regulador de interferón (IRF3), de IRF7 y NF-Kb
(Nuclear factor kB) (factor nuclear potenciador de las cadenas ligeras kappa de las
células B activadas) (Wang and Zhang, 2014). Afecta por lo tanto la producción de
citoquinas proinflamatorias y quimioquinas como la IL-6, IL8, el factor de necrosis tumoral
α (TNF- α), RANTES (regulated on activation, normal T cell expressed and secreted) y el
interferón tipo I (IFN α/β) (Yoo et al., 2010).
El PRRSV inhibe la síntesis de interferón tipo I por medio de las proteínas NSP1, NSP2,
NSP11 y N (Kim et al., 2010; Shi et al., 2011). NSP1 se cliva en las subunidades NSP1α
y NSP1β que se van a ubicar en el núcleo de la célula inhibiendo la expresión de IFN-β,
interfiere con la fosforilación y translocación nuclear de IRF3, de igual manera inhibe la
asociación de IRF3 con la proteína de unión (CBP) a CREB (cAMP response element-
binding) y aumenta la degradación de CBP (Chand et al., 2012; Wang and Zhang, 2014).

De igual forma, la proteína NSP2, bloquea la fosforilación y translocación nuclear de IRF3
(Sun et al., 2012b).
NSP11 es una endonucleasa y antagonista del IFN. Esta proteína suprime la activación
del promotor de IFN-β y la expresión de IRF3-mediated genes, de igual manera inhibe la
fosforilación y translocación nuclear de IFR3 (Nedialkova et al 2009; Wang and Zhang,
2014). La proteína N por su parte interfiere con la fosforilación inducida por ARNds y con
la translocación de IRF3 (Wang and Zhang, 2014).
De otro lado, PRRSV inhibe la señalización de NF-kB, inhibiendo la fosforilación de IkBα
(nuclear factor of kappa light polypeptide gene enhancer in B-cells inhibitor, alpha)
llevando al bloqueo de la translocación nuclear de NF-kB (Lee et al., 2005; Song et al.,
2010; Sun et al., 2012a). Inhibe la señalización JAK-STAT y la expresión de ISG
(interferon stimulated gene), por medio de las proteínas no estructurales NSPA1β, NSP7,
NSP12, GP3 y N. Estas proteínas bloquean la translocación nuclear de STAT 1 que
activa el IFN e inhiben la función antiviral de ISG15 (Patel et al., 2010; Wang et al., 2013).
PRRSV interfiere activamente con la inducción de apoptosis lo cual puede ser debido a
su habilidad para inhibir la producción de IFN tipo I especialmente en los estados
tempranos de infección (Sun et al., 2012a). El IPS-1 (IFN-b promoter stimulator-1) induce
la vía de señalización RIG-1(retinoid-inducible gene 1), lo cual lleva a la expresión de IFN
tipo I y a su vez a la apoptosis. La proteína no estructural NSP11 tiene la capacidad de
degradar al ARNm IPS-1 resultando en la supresión de IFN tipo I y por lo tanto inhibiendo
el mecanismo de apoptosis (Sun et al., 2012a). El PRRSV estimula vías antiapoptóticas
en macrófagos al comienzo de la infección pero por otro lado macrófagos infectados
pueden morir por apoptosis al final de la misma (Costers et al., 2008), interfiriendo con la
presentación de antígenos (Loving et al., 2007; Wang et al., 2007), ya que su replicación
en células dendríticas (DC) derivadas de monocitos (Mo-DC), lleva a la muerte de la
célula vía apoptosis y necrosis, lo cual lleva a su vez a una reducida expresión del
complejo mayor de histocompatibilidad (CMH) clase I y clase II y de CD14 y CD11b/c
(Mendoza et al., 2009; Wang et al., 2007).

Con respecto a la inmunidad innata de tipo celular, esta se caracteriza por ser débil, la
respuesta de IFNα, la producción de citoquinas inflamatorias y la activación de células
natural killer (NK) es mínima. A su vez la reducida activación de las NK, reduce la
producción de IFNγ, del factor de necrosis tumoral (TNF) y la actividad citotóxica antiviral
(Murtaugh et a., 2003).
1.7.2 Inmunidad adaptativa
Los anticuerpos anti-PRRS pueden ser detectados en la primera semana PI (Mulupuri et
al., 2008) mientras que los anticuerpos neutralizantes no se detectan antes de 4
semanas PI. Los títulos máximos se alcanzan entre la 10 a 18 semana PI (Yoo et al.,
2010).
Las primeras inmunoglobulinas en aparecer son tipo M detectables desde el día 5-7 con
un pico el día 14 y declinando hasta el día 42 PI (Park et al., 1995). Los niveles de la IgG
presentan un pico entre el día 21 a 35 PI, persistiendo posteriormente en bajas
concentraciones (Mulpuri et al., 2008). Algunos reportes indican que la viremia y la
replicación viral pueden persistir incluso en presencia de anticuerpos neutralizantes
(López et al., 2007), igualmente, la viremia puede ser resuelta aun antes de que los
anticuerpos neutralizantes sean detectados (Mateu and Díaz., 2008; Vezina et al., 1996).
Los anticuerpos específicos son dirigidos específicamente contra las proteínas N y M del
virus, estos aparecen hacia el día 7 PI., seguidos por anticuerpos contra la proteína
GP5. De igual manera, se presenta una respuesta temprana de animales infectados
frente a epitopes de la proteína nsp2 probable proteína inmunodominante del PRRSV
(De Lima et al., 2006; Loemba et al., 1996; Nelson et al., 1994). Muchas pruebas
diagnósticas detectan anticuerpos solamente contra la proteína N. Estos aparecen en la
primera semana PI y permanecen por varios meses (Mateu et al., 2008).
El desarrollo temprano de anticuerpos no neutralizantes tiene un efecto significante en el
desarrollo de la enfermedad. Se ha observado que este tipo de anticuerpos favorecen la
replicación viral en los MAPs a través de un mecanismo denominado “incremento
dependiente de anticuerpos” (antibody-dependent enhancement - ADE) (Yoon et al.,

1996, 1997) donde los anticuerpos no neutralizantes aumentan la replicación viral en los
MAPs. Los blancos de esos anticuerpos no neutralizantes son las proteínas GP5 y N
(Cancel-Tirado et al., 2004; Yoon et al., 1996). De igual manera, este tipo de respuesta
humoral puede actuar como un mecanismo de evasión del sistema inmune mediante la
estrategia del caballo de Troya, ya que los virus son opsonizados e internalizados en los
macrófagos vía endocitosis y las partículas virales son liberadas antes de sufrir el
proceso de degradación a nivel del fagolisosoma, continuando su ciclo replicativo (Mateu
and Díaz, 2008).
Los anticuerpos neutralizantes, son detectados hacia el día 28 PI y están dirigidos
principalmente contra la proteína GP5 la cual posee el epítope mayor de neutralización o
epítope B (Gonin et al., 1999; Nelson et al., 1994; Plagemann et al., 2002). El papel que
juegan los anticuerpos neutralizantes en el control de la infección por PRRSV, ha sido
materia de debate debido a la presencia de viremia en el huésped junto con una actividad
neutralizante. Se han postulado diversas hipótesis para explicar el retraso en el
desarrollo de anticuerpos neutralizantes, dentro de estas se encuentran: la variación en
la expresión de moléculas relacionadas con la presentación de antígeno, lo cual conlleva
a una demora en la movilización de células del sistema inmune resultando en una baja
respuesta de anticuerpos neutralizantes (Chang et al., 2008; Flores-Mendoza et al.,
2008). Una de las hipótesis más aceptadas es la presencia de un “epítope trampa”
(Ostrowski et al., 2002), el cual se localiza cerca al epitope neutralizante de GP5
produciendo una respuesta inmune no protectiva. De otro lado se encuentra el número
de glicosilaciones las cuales actúan como un blindaje ocultando al epítope B, evadiendo
así una respuesta inmune neutralizante; la pérdida de glicosilaciones en el ectodominio
de GP5 puede aumentar la inmunogenicidad y la susceptibilidad a la neutralización
(Jiang et al., 2007).
La respuesta proliferativa de células T se caracteriza principalmente por la expresión de
IFN-γ e IL-2 (López Fuertes et al., 1999). Las células que secretan IFN-γ son
principalmente CD4+ CD8+ y células T citotóxicas CD4-/CD8αβ (Meier et al., 2003).

1.7.3 Persistencia
El mecanismo de persistencia depende de una combinación de factores como son la
presencia de una gruesa superficie glicosilada en la estructura del virión, la redirección
de la respuesta humoral hacia proteínas no superficiales, el drift genético y antigénico y
la subversión del gen que induce interferón (Chand et al., 2012).
El mecanismo por el cual se presenta una débil respuesta contra la proteína GP5 está
vinculado a la presencia de varios sitos de N-glicosilación. El ectodominio de la proteína
posee dos sitos conservados de N-glicosilación, localizados en la posición N44 y N51 en
los virus tipo 2 y en N46 y N53 en los virus tipo I (Plagemann et al., 2002; Wissink et al.,
2004). Adicionalmente, el dominio rico en serina-asparagina entre los aminoácidos 30 y
38 posee una pequeña región que contiene un número variable de potenciales sitios de
N-glicosilación. La infección de cerdos con el PRRSV que presentaban ausencia de N34
y/o N51 resultó en un incremento en la producción de anticuerpos neutralizantes. El
mecanismo básico del blindaje glicano en la resistencia a los anticuerpos se debe a la
protección del conservado epítope B localizado entre los residuos 37-45 (Ostrowski et
al., 2002).
La diversidad genética del PRRSV involucra un drift genético por medio de puntos de
mutación y un shift genético por medio de recombinaciones. La tasa de sustitución de
nucleótidos para PRRSV se encuentra en un rango de 4.7 a 9.8 x 10-2/sitio/año, que es la
más alta encontrada para cualquier virus ARN (Hanada et al., 2005; Jenkins et al., 2002).
Las más altas variaciones en las secuencias de péptidos se encuentran en el
ectodominio de la GP5, flanqueando dos sitios conservados de N-glicosilación. La
hipervariabilidad de GP5 está vinculada a la presencia de un epítope trampa localizado
entre amino ácidos 27 y 30. Otra región que presenta variación en la secuencia peptídica
cercana a un epítope neutralizante se encuentra en la proteína GP4 (Chand et al., 2012).
Mutaciones en NSP1 y NSP2 pueden contribuir a la persistencia, alterando el nivel
general de replicación viral dentro de la célula (Chand et al., 2012).

1.8 Diagnóstico
Con el fin de conocer la actividad del PRRSV en granja, el estatus sanitario por animal y
lograr el control de la enfermedad, se han implementado varios tipos de técnicas
diagnosticas. Inicialmente, las pruebas de rutina se limitaban al aislamiento viral y a las
técnicas serológicas como seroneutralizacion viral (VSN) y la fluorescencia indirecta de
anticuerpos (IFA), pero en los últimos años los avances tecnológicos han permitido
mejorar la detección del virus, estos incluyen el enzimoinmunoanálisis (ELISA), la
reacción en cadena de la polimerasa (PCR), el polimorfismo de longitud de fragmentos
de restricción (RFLP) y el análisis de secuencias nucleotídicas y de aminoácidos
(Christopher-Henings et al., 2002).
Cuando la historia clínica y los hallazgos patológicos sugieren una infección por PRRSV
se requiere la detección del antígeno, del genoma viral o el aislamiento del mismo, a
partir de las muestras de los animales infectados, apoyado a su vez por los niveles de
anticuerpos en el momento del episodio clínico (Yoon et al., 2003)
1.8.1 Identificación de los signos clínicos
La enfermedad se caracteriza inicialmente por un episodio agudo con letargia, apetito
reducido y propagación rápida en la piara entre 7 a 10 días. Los signos clínicos
reproductivos incluyen infertilidad, agalactia, reducción en la tasa de partos, marcado
incremento de abortos en el último tercio de la gestación, lechones nacidos muertos,
momificaciones o lechones nacidos vivos débiles; signos respiratorios también pueden
estar presentes y las cerdas pueden transmitir vía transplacentaria el PRRSV a los
lechones. En lechones que superan la preñez y las fases neonatales, el PRRS se
manifiesta como una enfermedad respiratoria y se acompaña de infecciones secundarias,
con Pasteurella multocida, Circovirus tipo 2 (PCV2), Mycoplasma hyopneumoniae,
Streptococcus suis, Salmonella cholerasuis, Haemophilus parasuis y virus de la influenza
porcina. En lechones pos-destete y cerdos en crecimiento, los signos clínicos incluyen
disnea, anorexia, letargia, hiperemia cutánea y disminución en la ganancia de peso. Las
infecciones secundarias son comunes. Los cerdos adultos presentan un menor impacto a
nivel del sistema respiratorio. La infección subclínica ocurre a menudo en cerdas de

descarte, en la ceba, reproductores, cerdas jóvenes y cerdas adultas; en algunas granjas
la infección es asintomática. En machos infectados, el PRRSV puede aparecer en el
semen, afectando la morfología y función de los espermatozoides (Dietze et al., 2011;
OIE 2008).
1.8.2 Lesiones microscópicas
1.8.2.1 Histopatología
Aunque el PRRSV produce infección multisistémica, las lesiones más características son
usualmente observadas en piel, tejido respiratorio y tejido linfoide, dependiendo de la
cepa viral involucrada, factores iatrogénicos y la presencia de infecciones secundarias
(Dietze et al., 2011). La neumonía intersticial multifocal es la lesión histológica más
característica de infección por el PRRSV. Las lesiones pulmonares se caracterizan por
tres cambios: aumento de grosor de la pared alveolar, hipertrofia e hiperplasia de
neumocitos tipo II con acumulación de material necrótico y células inflamatorias en los
espacios alveolares (Collins et al., 1996; Dietze et al., 2011). Los ganglios linfáticos se
muestran hemorrágicos, con hiperplasia folicular, focos de necrosis folicular y residuos en
los folículos (OIE, 2008, Dietze et al., 2011).
La condición corporal de fetos abortados en el último tercio de gestación varía de frescos
a autolizados, la hemorragia umbilical se reporta como una lesión característica de
infección por PRRS (Dietze et al., 2011).
1.8.3 Detección del virus
La identificación del PRRSV puede obtenerse mediante el aislamiento del virus, la
detección del genoma viral y la detección de proteínas víricas (OIE 2012). Tras la
infección, los cerdos desarrollan una viremia e infección pulmonar que pueden persistir
durante semanas en los animales más jóvenes y días en los adultos, lo cual hace que las
muestras de suero y los lavados pulmonares bronco alveolares sean ideales para
detectar este agente (OIE, 2012).

1.8.3.1 Aislamiento viral
Esta es una prueba confirmatoria en casos sospechosos de PRRS, permitiendo detectar
el virus en muestras de tejidos y sueros. El virus se replica tanto en MAPs que
constituyen el sistema de cultivo más susceptible para los virus de ambos tipos
antigénicos, como en las células MARC-145, un clon de la línea celular MA-104 (ICA,
2014; OIE 2012).
En casos agudos, las muestras de elección son el fluido broquioalveolar, pulmón y suero.
En animales persistentemente infectados, la tonsila, secreción orofaringea y fluido
bronquioalveolar. En caso de aborto, las muestras deben colectarse de las momias,
cerdos abortados y nacimientos débiles (ICA, 2014).
Los cambios típicos (efecto citopático) que presentan las células MARC-145 debido a la
infección con PRRSV, son redondeamiento celular, agrupamiento y lisis, estos cambios
ocurren entre los 2 a 5 días PI. La positividad de la prueba se determina al observar el
efecto citopático del virus en las células. (ICA, 2014).
1.8.3.2 Reacción en cadena de la polimerasa - PCR
Permite la amplificación de diferentes regiones del genoma del PRRSV (ORF5, ORF6 y/o
ORF7) a partir de diferentes tipos de muestras que incluyen tejidos, suero, semen, lavado
bronquio alveolar, fluidos orales e hisopos oro faríngeos (Christopher-Henings., et al.,
2002).
La detección del ácido nucleico del PRRSV puede llevarse a cabo por medio de la
reacción en cadena de la polimerasa con transcripción inversa (RT-PCR), la RT-PCR
anidada y la RT-PCR cuantitativa en tiempo real (qRT-PCR) (Drew, 1995; Kleiboeker et
al., 2005; Larochelle & Magar, 1997; Mardassi et al., 1994; Wasilk et al., 2004, citados
por OIE 2012). Estas pruebas se usan frecuentemente para detectar el ácido nucleico
viral en tejidos, suero o semen (Christopher-Hennings et al., 1997 citado por Manual OIE
2012). Igualmente, se ha diseñado una técnica de PCR múltiple para diferenciar las
cepas tipo I de las de tipo II (Gilbert et al., 1997 citado por OIE 2012).

La qRT-PCR se caracteriza por completar la amplificación y el análisis en un sistema
cerrado lo cual disminuye la contaminación, igualmente permite medir cuantitativamente
el ARN, utiliza menos tiempo y cuenta con alta sensibilidad (Chung et al., 2005; Tian et
al., 2010; Wasilk et al., 2004). La qRT-PCR basado en SYBR Green utiliza un fluoróforo
que se une al ADN bicatenario que se va generando, sin tener en cuenta el origen de la
secuencia de ADN (Martínez et al., 2008).
1.8.4 Secuenciación
La secuenciación del PRRSV comenzó en los años 90, esta técnica permite obtener la
secuencia exacta de una serie de nucleótidos del ORF bajo análisis (Christopher-Henings
et al., 2002). Kapur et al (1996) demostraron la utilidad del análisis de secuencias en
diferenciar aislados del PRRSV. A partir de sus hallazgos se determinó que el gen ORF5
es el gen mas variable y ORF6 el más conservado, esta es la razón por la cual el ORF5
es el gen blanco dentro de esta técnica.
La información de las secuencias de los aislados de PRRS permite diferenciar cepas de
campo de cepas vacunales y monitorear cepas dentro de un sistema de producción ya
que permite determinar la introducción de una variante diferente a las que se encuentran
circulando en granja (Yoon et al.,2003; Murtaug, 2012).
1.8.5 Pruebas serológicas
La serología se lleva a cabo generalmente mediante técnicas como el IPMA, la
inmunofluorescencia, o el enzimoinmunoanálisis- ELISA, sobre el cual se han descrito
varias modificaciones (Albina et al. 1992; Cho et al., 1997; Denac et al., 1997, citados por
OIE 2012).
1.8.5.1 ELISA
La ELISA comercial para detección de anticuerpos contra el virus de PRRS ELISA
(HerdChek: IDEXX Laboratories Inc.Westbrook. Maine) usa un formato indirecto para
detectar anticuerpos en suero contra ciertos antígenos del virus de PRRS. De acuerdo
con las instrucciones del fabricante una muestra se considera positiva cuando el cociente
M/P (muestra/positivo) es igual o superior a 0.40 lo cual indicaría presencia de

anticuerpos (Christopher-Henings et al., 2002). Esta técnica presenta alta sensibilidad y
especificidad, los valores de sensibilidad y especificidad corresponden al 98.8% y 99.9%
respectivamente (HerdChek PRRS (package insert) Westbrook, Maine. IDEXX
Laboratories, 2011).
Los anticuerpos detectados por la técnica de ELISA aparecen entre los 9 a 13 días PI,
alcanzando un máximo entre los 30 a 50 días para luego descender. Se estima que los
anticuerpos pueden ser aún detectados por esta técnica hasta los 4 a 10 meses post
infección (Collins et al., 1996).
En la actualidad existen varios enzimoinmunoanálisis (ELISA) comerciales: ELISA
indirecta, ELISA de bloqueo y ELISA doble, que permiten distinguir entre las reacciones
serológicas a los virus europeo y americano (OIE, 2012).
1.8.5.2 Inmunofluorescencia indirecta - IFA
Esta técnica altamente específica (99.5%) detecta anticuerpos a partir de los 7 a 15 días
hasta los 2 a 3 meses PI. (Yoon et al., 1995) y de igual manera permite determinar el
título del suero. La IFA se utiliza como prueba confirmatoria de muestras que hayan sido
positivas por ELISA (ICA, 2014) en microplacas de 96 pozos utilizando la línea celular
MARC-145, donde las células infectadas con el virus de PRRS se fijan y son usadas
como sustrato celular. Se consideran positivos animales con títulos ≥1:16 y negativos
con títulos ≤ 1:16 (ICA, 2014; OIE, 2012).
1.8.5.3 Seroneutralización
Se basa en la detección de anticuerpos neutralizantes en el suero de animales
sospechosos utilizando cultivos celulares y un título conocido de virus, el cual es
incubado con varias diluciones de las muestras de suero. Después de la incubación, el
suero es adicionado sobre una línea celular sensible para determinar el titulo de
anticuerpos que es capaz de neutralizar el PRRSV. Su sensibilidad es menor que la de
IFA o ELISA debido a que los anticuerpos neutralizantes contra el PRRSV se desarrollan
lentamente posterior a la infección (> 21 días) (Christopher-Henings et al., 2002; OIE,
2012).

1.9 Control del PRRS
El control y prevención del virus se debe basar en (i) minimizar la circulación del PRRSV
dentro de una granja, (ii) limitar los efectos de la circulación viral y co-infecciones
bacterianas secundarias y (iii) maximizar y estabilizar la inmunidad de la piara (Dee,
2003). De igual manera se busca evitar la introducción del virus a una granja negativa o
limitar la introducción de nuevas cepas en granjas ya infectadas, por lo tanto la
bioseguridad debe ser revisada interna y externamente (OIE, 2008).
Las rutas directas de transmisión dentro y entre poblaciones incluyen animales infectados
y semen contaminado, por esta razón todos los machos y hembras primerizas deben
provenir de granjas negativas a PRRSV y ser aislados en cuarentena por lo menos 30
días durante los cuales se practican pruebas serológicas antes de ser introducidos en la
granja, a su vez el semen debe proceder de machos negativos a PRRSV (Papatsiros,
2012).
La introducción de hembras de reemplazo consta de tres periodos: el periodo de
aislamiento, la aclimatación y la recuperación (Pesente et al., 2006; Vashisht et al.,
2008). La duración de cada periodo puede variar entre 30 a 60 días dependiendo de la
edad de la hembra de reemplazo, el estatus dentro como entre granjas, el tipo y tamaño
de la granja. El periodo de aislamiento incluye la realización de pruebas serológicas con
el fin de determinar el estatus de infección por PRRSV de los animales entrantes. La
aclimatación comienza 30 días después de la introducción de las hembras de reemplazo,
con el fin de exponerlas a la cepa circulante en la granja y para tal fin se han reportado
varios métodos que incluyen, la exposición a tejidos de lechones nacidos débiles y de
mortinatos, uso de vacunas, exposición por contacto entre las cerdas jóvenes y cerdos
de destete, que se utilizan como fuentes de virus y la inoculación de los animales de
reemplazo negativos con el suero de cerdos virémicos de la misma granja (Dee, 2003;
Vashisht et al., 2008). Esta última práctica se ha incrementado debido al aumento de la
heterogeneidad genética entre cepas de PPRSV y a que las vacunas no inducen
suficiente protección contra las nuevas cepas identificadas (Fano et al., 2005). De otro
lado la inoculación de una cepa homóloga provee una alto nivel de protección frente a la
exposición con la misma cepa viral (Batista et al., 2002). El desarrollo de un período de

aclimatación de reemplazos lleva a la estabilización de los signos clínicos, a la
producción de lechones negativos a PRRSV al destete, a la prevención de brotes
asociados con cepas endémicas, al desarrollo de una inmunidad específica a las cepas
homólogas de la granja y a la mejora de los parámetros de producción (Pesente et al.,
2006; Vashisht et al., 2008).
Otra estrategia de control es la denominada piara cerrada, que es utilizada para eliminar
la circulación viral reduciendo la diseminación viral y los portadores, en este caso se
finaliza la introducción de reemplazos a la granja durante 4 a 8 meses, esto dependerá
del estatus sanitario de la misma y el flujo de animales. Este sistema puede ocasionar
efectos como una inadecuada distribución de los partos, sin embargo esto se puede
minimizar aplicando proyectos de cría de cerdas jóvenes de reemplazo en instalaciones
apartadas (Torremorel and Christianson, 2002; Cho y Dee, 2006; Zimmerman et al.,
2006).
De otro lado el flujo todo dentro y todo fuera (TD/TF), es efectivo para controlar una
variedad de patógenos respiratorios en cerdos destetos. Consiste en dividir las
instalaciones en áreas individuales lo que permite una limpieza profunda y desinfección
de las instalaciones entre cada grupo de cerdos. Este método reduce la propagación
horizontal del PRRSV desde los cerdos más adultos a aquellos recientemente ingresados
al área (Dee, 2003).
Las medidas McREBEL (Management Changes to Reduce Exposure to Bacteria to
Eliminate Losses) propuestas por McCaw en 1995, constan de una serie de pasos para
reducir la diseminación del PRRSV entre lechones, estos incluyen:
1. Evitar el intercambio de lechones entre camadas
2. Realizar atetes o adopciones sólo en las primeras 24 horas de vida
3. Solo mover los lechones o las cerdas entre las mismas salas.
4. Evitar el uso de nodrizas para salvar a los lechones enfermos o retrasados.
5. Evitar el manejo innecesario de medicamentos en los lechones
6. Evitar el movimiento de los animales retrasados a otras habitaciones con animales
más jóvenes o nodrizas.

7. Aplicar el sistema de todo dentro-todo fuera (TD/TF) en los precebos, dejando 2 o 3
días de vacío sanitario entre lotes.
8. Hacer adopciones de lechones sólo cuando sea absolutamente necesario y en los
casos en que exista la seguridad de que han recibido suficiente calostro.
9. Sacrificar a los lechones enfermos o que tengan un peso significativamente inferior al
normal en el destete.
10. No practicar el "feed-back" (retroalimentación con heces).
11. Las cerdas deben ser lavadas antes de entrar en la sala de partos.
12. El personal debe lavarse las manos o llevar guantes desechables después de
manipular cada camada.
13. Las agujas hipodérmicas y las hojas de los bisturís deben cambiarse después de
cada camada.
14. En camadas grandes, se puede hacer una supervisión de las lactancias, separando
lechones para asegurarse de que todos los lechones reciben calostro de su madre.
Estas medidas facilitan y optimizan el crecimiento de los lechones lactantes, reducen la
mortalidad antes del destete y favorecen la ganancia de peso al destete (McCaw, 2003).
La despoblación parcial por su parte, se basa en el principio que la circulación del
PRRSV se presenta en un grupo específico de producción. Permite interrumpir la
transmisión horizontal y es una buena medida para controlar la enfermedad pos destete o
eliminar el virus desde la población destetada (Dee, 2003).
Los programas regionales de control y eliminación han ganado popularidad a nivel
mundial. En Suecia y Loire en Francia la eliminación de la enfermedad fue posible
minimizando el movimiento de los animales, de vehículos y practicando la despoblación-
repoblación de las piaras infectadas (Le Potier et al., 1997, Carlsson et al., 2009). En
Ontario (Canadá), se han presentado varias iniciativas de control regional (Waddell,
2010). En México-Sonora se realizó un control regional del PRRS que incluía la
participación activa y capacitación constante de los técnicos (Batista et al., 2010). A nivel
de América latina, se han presentado tentativas de control y eliminación regional pero los
resultados aún son preliminares (Corzo et al., 2010).

Bibliografía Albina,E., Leforban, Y., Baron, T., Plana-Duran, J. and Vannier, P. 1992. An enzyme
linked immunosorbent assay (ELISA) for the detection of antibodies to the porcine
reproductive and respiratory syndrome (SDRP) virus. Ann. Rech. Vet. 23, 167–176.
Albina, E., 1997. Epidemiology of porcine respiratory and reproductive syndome (PRRS):
an overview. Vet. Microbiol. 55: 309-316.
Allende, R., Lewis, T.L., Lu, Z., Rock, D. L., Kutish, G. F., Ali, A., Doster, A. R., Osorio,
F.A. 1999. North American and European porcine reproductive and respiratory syndrome
viruses differ in non-structural protein coding regions. J Gen Virol. 80: 307-15.
Allende, R., W. W. Laegreid, G. F. Kutish, J. A. Galeota, R. W. Wills, and F.
A. Osorio. 2000. Porcine reproductive and respiratory syndrome virus: Description of
persistence in individual pigs upon experimental infection. J Virol. 74:10834-7.
Ansari, I.H., Know, B., Osorio, F.A., Pasttnaik, A. K. 2006. Influence of N-linked
glycosylation of porcine reproductive and respiratory syndrome virus GP5 on virus
infectivity, antigenicity and ability to induce neutralizing antibodies. J Virol. 80:3994-4004.
Batista, L., Pijoan, C., Torremorell, M. 2002. Experimental injection of gilts with porcine
reproductive and respiratory syndrome virus (PRRSV) during acclimatization. J Swine
Health Prod., 10: 147–50.
Batista, L., Fernández, M., Miramontes, J., Pliego, R., Mondaca, M. 2010. Advancing
PRRS area regional control in the state of Sonora, Mexico. In: Proc. Int. Pig Vet. Soc.
Congr., Vancouver, Canada, 274.
Bautista, E.M., Molitor, T.W. 1997. Cell-mediated immunity to porcine reproductive and
respiratory syndrome virus in swine. Viral Immunol. 10:83-94.

Benfield, D.A., Nelson, E., Collins, J.E., Harris, L., Goyal, S.M., Robinson, D.,
Christianson, W.T., Morrison, R.B., Gorcyca, D., Chladek, D. 1992. Characterization of
swine infertility and respiratory syndrome (SIRS) virus (isolate ATCC VR-2332). J. Vet.
Diagn. Invest. 4, 127-133.
Benfield, D.A., Collins, J.E., Dee, S.A., Hallbur, P.G., Joo, H.S., Lager, K.M., Mengeling,
W.L., Murtauhgh, M.P., Rossow, K.D., Stevenson, G.W. & Zimmerman J.J. 1999. Porcine
reproductive and respiratory syndrome. In: Diseases of Swine, Eight Edition, Straw B.E.,
D’Allaire S., Mengeling W.L. & Taylor D.J., eds. Iowa State University Press. Ames, Iowa,
USA, 201–232.
Beura, L.K, Sarkar, S.N, Kwon, B, Subramaniam, S, Jones, C, Pattnaik, A.K., Osorio, F.A.
2010. Porcine reproductive and respiratory syndrome virus nonstructural protein 1beta
modulates host innate immune response by antagonizing IRF3 activation. J Virol. 84,
1574-1584.
Blaha, T., 2000. The “colorful” epidemiology of PRRS. Vet. Res. 31: 77-83
Calvert, J., Slade, D.E., Shields, SL., Jolie, R., Mannan, RM., Ankenbauer, RG., Welch,
S.K. 2007. CD163 expression confers susceptibility to porcine reproductive and
respiratory syndrome viruses. J Virol. 81, 7371-7279.
Cancel-Tirado, S.M., Evans, R.B., Yoon, K. 2004. Monoclonal antibody analysis of
porcine reproductive and respiratory syndrome virus epitopes associated with antibody-
dependent enhancement and neutralization of virus infection. Vet. Immunol.
Immunopathol. 102, 249-262.
Carlsson, U., Wallgren, P., Renstrom, L.H., Lindberg, A., Eriksson, H., Thoren, P.,
Eliasson-Selling, L., Lundeheim, N., Norregard, E., Thorn, C., Elvander, M. 2009.
Emergence of porcine reproductive and respiratory syndrome in Sweden: detection,
response and eradication. Transbound Emerg. Dis. 56, 121-131.

Cavanagh, D. 1997. Nidovirales: a new order comprising Coronaviridae and Arteriviridae.
Arch. Virol. 142, 629-633.
Chand, R.R., Trible, B.R., Rowland, R.R.R., 2012. Pathogenesis of porcine reproductive
and respiratory syndrome virus. Current Opinion in Virology, 2:256-263.
Chang, H.C., Peng, Y.T., Chang, H.L., Chaung, H.C., Chung, W.B. 2008. Phenotypic and
functional modulation of bone marrow-derived dendritic cells by porcine reproductive and
respiratory syndromr virus. Vet. Microbiol. 129:281-293.
Chen, J.Z., Lawson, S., Sun, Z., et al. 2010a. “Identification of two autocleavage products
of nonstructural protein 1 (nsp1) in porcine reproductive and respiratory syndrome virus
infected cells: nsp1 function as interferon antagonist,” Virology, 398: 87–97.
Chen, Z., Zhou, X., Lunney, J.K., Lawson, S., Sun, Z., Brown, E., Christopher-Hennings,
J., Knudsen, D., Nelson, E., Fang, Y. 2010b. Inmunodominant epitopes in nsp2 of porcine
reproductive and respiratory syndrome virus are dispensable for replication but play an
important role in modulation of host immune response. J. Gen. Virol. 91, 1047-1057.
Cho, J.G. and Dee, S.A. 2006. Porcine reproductive and respiratory syndrome virus.
Theriogenology, 66; 655–662.
Christianson, W.T., Collins, J.E., Benfield, D.A., Harris, L., Gorcyca, D.E., Chladek, D.W.,
Morrison, R.B., Joo, H.S. 1992. Experimental reproduction of swine infertility and
respiratory syndrome in pregnant sows. Am. J. Vet. Res, 53:485-488.
Christopher-Hennings, J., Nelson, E.A., Benfield, D.A. 1996. Detecting PRRSV in boar
semen. Swine Health and Production, 4: 37-39.
Christopher-Hennings, J., Nelson, E.A., Nelson, J. K. and Benfield, D. A. 1997. Effects of
a modified-live virus vaccine against porcine reproductive and respiratory syndrome in
boars. Am. J. Vet. Res. 58, 40–45.

Christopher-Hennings, J., Faaberg, K.S., Murtaugh, M.P., Nelson, E.A., Roog, M.B.,
Vaughn, E.M., Yoon, K.J., Zimmerman, J.J. 2002. Porcine reproducive and respiratory
syndrome (PRRS) diagnostics: interpretación and limitations, Journal of Swine Health and
Production, 10: 213-218.
Cho, H. J., McNab, B., Dubuc, C., Jordan, L., Afshar, A., Magar, R., Prins, S., and
Eernisse, K. 1997. Comparative study of serological methods for the detection of
antibodies to porcine reproductive and respiratory syndrome virus. Can. J. Vet. Res. 61,
161–166.
Chung, W.B., Chan, W.H., Chaung, H.C., Lien, Y., Wu, C.C., Huang, Y.L. 2005. Real-time
PCR for quantitation of porcine reproductive and respiratory syndrome virus and porcine
circovirus type2 in naturally-infected and challenged pigs. Journal of Virological Methods,
124:11-19.
Collins, J.E., Benfield, D.A., Chritianson, W.T., Harris L., Hennings, J.C., Shaw, D.P.,
Goyal, S.M., NcCullough, S., Morrison, R.B., Joo, H.S. 1992. Islotation of swine infertility
and respiratory syndrome virus (isolate ARCC VR-2332) in North America and
experimental reproduction of the disease in gnotobiotic pigs. J Vet Diagn Invest. Apr
4(2):117-126.
Collins, J., Dee, S., Halbur, P., Keffaber, K., Lautner, B., McCaw, M., Rodibaugh, M.,
Sanford, E., Yesye, P., 1996. Laboratory diagnosis of porcine reproductive and
respiratory syndrome virus (PRRS) infection, Swine health and production, 4: 33-35.
Corzo, C.A., Mondaca, E., Wayne, S., Torremorell, M., Dee, S., Davies, P., Morrison,
R.B., 2010. Control and elimination of porcine reproductive and respiratory syndrome
virus. Virus Res. 154, 185-192.
Costers, S., Lefebvre, D.J., Delputte, P.L., Nauwynck, H.J. 2008. Porcine reproductive
and respiratory syndrome virus modulates apoptosis during replication in alveolar
macrophages. Arch Virol. 2008;153(8):1453-65

Cruz, M.C., Mogollón, J.D., Rincón, M.A., Peña, N.B., Ruiz, S., Lora, A.M. 2006.
Prevalencia serológica del Síndrome Reproductivo y Respiratorio Porcino (PRRS) en
cerdos de explotaciones extensivas de Colombia. Rev. Med. Vet. Zoot. 53:33-41
Darwich, L., Gimeno, M., Sibila, M., Díaz, I., de La Torre, E., Dotti, S., Kuzemtseva, L.,
Martin, M., Pujols, J., Mateu, E. 2011. Genetic and inmunobiological diversities of porcine
reproductive and respiratory síndrome genotype I strains. Vet Mic. 150:49-62
Das, P.B., Dinh, P.X., Ansari, L.H., de Lima, M., Osorio, F.A., Pattnaik, A.K., 2010. The
minor envelope glycoproteins GP2 and GP4 of porcine reproductive and respiratory
syndrome virus interact with the receptor CD163. J.Virol.84: 1731-1740.
Dea, S., Sawyer, N., Alain, R., Athanassious, R. 1995. Ultrastructural characteristics and
morphogenesis of porcine reproductive and respiratory syndrome virus propagated in the
highly permissive MARC-145 cell clone. Adv. Exp. Med. Biol. 380: 95-98.
Dea, S., Gagnon, C.A., Mardassi, H., et al. 2000. Current knowledge on the structural
proteins of porcine reproductive and respiratory syndrome (PRRS) virus: comparison of
the North American and European isolates. Arch Virol. 145: 659-688.
Dee, S. A. 2003. Approaches to prevention, control and eradication. Compendium PRRS:
Second edition, 119-128.
Dee, S., Deen, J., Rossow, K., Weise, C., Eliason, R., Otake, S., Joo, H.S., Pijoan, C.,
2002. Mechanical transmission of porcine reproductive and respiratory syndrome virus
throughout a coordinated sequence of events during warm weather. Can. J. Vet. Res.
66:232-239.
De Lima, M., Pattnaik, A.K., Flores, E.F., Osorio, F.A. 2006. Serologic marker candidates
identified among B-cell liner epitopes of nsp2 and structural proteins of a North American
strain of porcine reproductive and respiratory syndrome. Virology, 353; 410-421.

De Lima, M., Ansari, I.H., Das, P, B., Ku, B.J., Martínez-Lobo, F.J., Osorio, F.A. 2009.
GP3 is a structural componente of the PRRSV type II (US) virion. Virology, 390: 31.36.
Delputte, P.L., Vanderheijden, N., Nauwynck, H.J., Pensaert, MB. 2002. Involvement of
the matrix protein in attachment of porcine reproductive and respiratory syndrome virus to
a heparinlike receptor on porcine alveolar macrophages. J. Virol. 76, 4312-20.
Delputte, P.L., Costers, S and Nauwynck, H.J. 2005. Analisys of porcine reproductive and
respiratory syndrome virus attachment and internalization: distinctive roles for heparan
sulphate and sialodhesin. J Gen Virol. 86:1441-1445.
Delputte, P. L., Van Breedman, W. Delrue, I, Oetke, C, Crocker, P.R and Nauwynck, H.J.
2007. Porcine arterivirus attachment to the macrophage-specific receptor sialodhesin is
dependent on the sialic acid-binding activity of the N-terminal immunoglobulin domain of
sialodhesin.J Virol. 81:9546-9550.
Denac, H., Moser, C., Tratschin J., D., and Hofmann, M., A. 1997. An indirect ELISA
for the detection of antibodies against porcine reproductive and respiratory
syndrome virus using recombinant nucleocapsid protein as antigen. J. Virol.
Methods, 65, 169–181.
Desrosier, R. 2011. Transmission of swine pathogens: different means, different needs.
Animal Health Research Reviews, 12; 1-13.
Drew, T., W. 1995b. Comparative serology of porcine reproductive and respiratory
syndrome in pigs. J. Vet. Med. Sci. 58, 941–946.
Dietze, K., Pinto, J., Wainwright, S., Christopher, H., Khomenko, S. 2011. Porcine
reproductive and respiratory syndrome (PRRS) virulence jumps and persistent circulation
in Southeast Asia disponible en. www.fao.org/docrep/013/al849e/al849e00.pdf
Dokland, T. 2010. The structural biology of PRRS. Virus Research, 154: 86-97.

Fang, Y., and Snijder, E.J. 2010. The PRRSV replicase: exploring the multifunctionality of
an intriguing set of nonstructural proteins. Virus Res. 154:61-76.
Fano, E., Olea, L., Pijoan, C. 2005. Eradication of porcine reproductive and respiratory
syndrome virus by serum inoculation of naïve gilts. Canad J Vet Res. 69: 71-4.
Flores-Mendoza, L., Silva-Campa, E., Resendiz, M., Osorio, F.A., Hernández, J. 2008.
Porcine Reproductive and Respiratory Syndrome Virus infects mature dendritic cells and
up-regulates interleukin-10 production. Clin. Vaccine Immunol. 15: 720-725.
Gilbert, S.A., Larochelle, R., Magar, R., Cho, H. J., Deregt, D. 1997.Typirng of Porcine
Reproductive and Respiratory Syndrome Viruses by multiplex PCR Assay. Journal of
Clinical Microbiology, 35; 264-267.
Gonin, P., Mardassi, H., Gagnon, C.A., Massie, B., Dea, S. 1998. A nonstructural and
antigenic glycoprotein is encoded by ORF3 of the IAF-Klop strain of porcine reproductive
and respiratory syndrome virus. Arch Virol. 143, 1927-40.
Gonin, P., Pirzadeh, B., Gagnon, C.A., Dea S. 1999. Seroneutralization of porcine
reproductive and respiratory syndrome virus correlates with antibody response to the GP5
major envelope glycoprotein. J. Vet. Diagn. Invest. 11, 20-26.
Gorbalenya, A.E., Enjuanes, L., Ziebuhr, J., Snijder, E.J., 2006. Nidovirales: envolving the
largest RNA virus genome. Virus Res. 117:17-37.
Han, J., Rutherford, M.S., Faaberg, K.S. 2009. The porcine reproductive and respiratory
syndrome virus nsp2 cysteine protease domain possesses both trans and cis cleavage
activities. J. Virol. 83, 9449-9463.
Hanada, K., Suzuki, Y., Nakane, T., Hirose, O., Gojobori, T. 2005. The origin and
evolution of porcine reproductive and respiratory syndrome viruses. Mol. Biol. Evol. 22:
1024-1031.

Harrison, S.C., 2008. Viral membrane fusión. Nat. Struct. Mol. Biol. 15:690-698.
HerdChek PRRS (package insert) Westbrook, Maine. IDEXX Laboratories, 2011.
Horter, D.C., Pogranichniy, R.M., Chang, C.C., Evans, R.B., Yoon, K.J., Zimmerman, J.J.
2002. Characterization of the carrier state in porcine reproductive and respiratory
syndrome virus infection. Vet. Microbiol. 86:213-228.
Hill, H., 1990. Overview and history of Mystery Swine Disease (swine infertility/respiratory
syndrome). In: Proceedings of the Mystery Swine Disease Committee Meeting. Livestock
Conversation Institute, Denver, CO, pp. 29-31.
Holthkamp, D.J., Kliebenstein, J.B., Neumann, E.J. 2013. Assessment of the economic
impact of porcine reproductive and respiratory síndrome virus on United States pork
producers. Journal of Swine Health and Production, 21: 72-84.
Hu, H., Li, X., Zhang, Z., Shuai, J., Chen, N., Liu, G., Fang, W. 2009. Porcine reproductive
and respiratory syndrome viruses predominant in southeastern China from 2004 to 2007
were from a common source and underwent further divergence. Arch Virol. 154, 391-398.
ICA. 2014. Técnica de aislamiento del virus de PRRS. Disponible en:
http://portal.ica.gov.co/DocManagerSwift/User/ElementViewer.aspx?LV=1&E=4640&rnd=
0.7680949024628146&HideHeader=true&Tab=w_0.2779094502393862
Jenkins, G.M., Rambaut, A., Pybus, O.G., Holmes, E.C., 2002. Rates of molecular
evolution in RNA viruses: a quantitative phylogenetic analysis. J. Mol. Evol. 54:156-165.
Jiang, W., Jiang, P., Wang, X., Li, Y., Du, Y. 2007. Influence of Porcine reproductive and
respiratory syndrome virus GP5 glycoprotein N-linked glycans on immune responses in
mice. Virus Genes, 35: 663-671.

Jiang, W., Jiang, P., Wang, X., Li, Y and Du, Y. 2008. Enhanced immune responses of
mice inoculated recombinant adenoviruses expressing GP5 by fusion with GP3 and/or
GP4 of PRRS virus. Virus Res. 136(1-2):50-57.
Kleiboeker, S.B., Schommer, S.K., Lee, S.M., Watkins, S., Chittick, W., Polson, D. 2005.
Simultaneous detection of North American and European porcine reproductive and
respiratory syndrome virus using real-time quantitative reverse transcriptase-PCR. J. Vet.
Diagn. Invest. 17,165–170.
Keffaber, K.K., 1989.Reproductive failure of unknown etiology. AASP 1, 2-10.
Key, K.F., Haqshenas, G., Guenette, D.K., et al. 2001. Genetic variation and phylogenetic
analyses of the ORF 5 gene of acute porcine reproductive and respiratory syndrome virus
isolates. Vet Microbiol. 83:249-263.
Kim, J, K., Fahad, A.M., Shanmukhappa, K and Kapil, S. 2006. Defining the cellular
targets of porcine reproductive and respiratory syndrome virus blocking monoclonal
antibody 7G10. J Virol. 80:689-696.
Kim, O., Sun, Y., Lai, F.W., Song, C., and Yoo, D., 2010. “Modulation of type I interferon
induction by porcine reproductive and respiratory syndrome virus and degradation of
CREB-binding protein by non-structural protein 1 in MARC-145 and HeLa cells,” Virology,
402:315–326.
Kreutz, L.C. 1998. Cellular membrane factors are the major determinants of porcine
reproductive and respiratory syndrome virus tropism. Virus Res., 53:121-128.
Kroesse, M.V., Zevenhoven-Dobbe, J.C., Bos de Ruijter, J.N., Peeters, B.P., Meulenberg,
J.J., Conelissen, L.A., Snijder, E.J. 2008. The nsp1 alpha and nsp1 papain-like
autoproteinases are essential for porcine reproductive and respiratory syndrome virus
RNA synthesis. J Gen Virol. 89:494-499.

Larochelle, R., and Magar, R. 1997. Evaluation of the presence of porcine reproductive
and respiratory syndrome virus by polymerase chain reaction. J. Virol. Methods. 47, 273–
278.
Lawson, S.R., Rossow, K.D., Collins, J.E., Benfield, D.A., Rowland, R.R. 1997. Porcine
Reproductive and respiratory syndrome virus infection of gnotobiotic pigs: sites of virus
replication and co-localization with MAC-387 staining. Virus Res, 51, 105-113.
Le Potier, M.F., Blanquefort, P., Morvan, E., Albina, E., 1997. Results of a control
programme for the porcine reproductive and respiratory syndrome in the French 'Pays de
la Loire' region. Vet. Microbiol. 55, 355-360.
Lee, S.M., Kleiboeker, S.B. 2005. Porcine arterivirus activates the NF-kappaB pathway
through IkappaB degradation. Virology, 342, 47–59.
Lee, C., and Yoo, D. 2006. The small envelope protein of porcine reproductive and
respiratory syndrome virus possesses ion channel protein-like properties. Virology 355,
30-43.
Li, B., Xiao, S., Wang, Y., Xu, S., Jiang, Y., Chen, H., Fang, L. 2009. Inmunogenicity of
the highly pathogenic porcine reproductive ans respiratory syndrome virus GP5 protein
encoded by a synthetic ORF5 gene. Vaccine, 27:1957-1963.
Li, B., Fang, L., Guo, X., Gao, J., Song, T., Bi, J., He, K., Chen, H., Xiao, S. 2011.
Epidemiology and evolutionary characteristics of the porcine reproductive and respiratory
syndrome virus in China between 2006 and 2010. J Clin Microbiol, 49:3175-3183.
Loemba, H.D., Mounir, S., Mardassi, H. 1996. Kinetics of humoral immune response to
the major structural proteins of the porcine reproductive and respiratory syndrome virus.
Arch. Virol.141:751-761.

López Fuertes, L., Domenech, N., Alvarez, B., Ezguerra, A., Domínguez, J., Castro, J.M.,
Alonso F. 1999. Analysis of cellular immune response in pigs recovered of porcine
reproductive and respiratory syndrome infection. Virus Research 64: 33-42.
López, O.J., Oliveira, M, F., Álvarez-García, E., Kwon, B.J., Doster, A., Osorio, F, A.
2007. Protection against porcine reproductive and respiratory syndrome virus (PRRSV)
infection through passive transfer of PRRSV-neutralizing antibodies is dose dependent.
Clin Vaccine Immunol. 14:269–75.
Loula, T., 1991. Mystery pig disease. Agri-practice 12, 23-34.
Loving, C.L., Brockmeier, SL., Sacco, R.E. 2007. Differential type I interferon activation
and susceptibility of dendritic cell populations to porcine arterivirus. Inmunology, 120: 217-
229.
McCaw, M. 2003. McREBEL Maanagement. Compendium PRRS: Second edition, 131-
136.
Manual terrestre de la OIE. 2012. Síndrome disgenésico y respiratorio porcino, capítulo
2.8.7. Disponible en:
http://www.oie.int/fileadmin/Home/esp/Health_standards/tahm/2.08.07_PRRS.pdf
Mardassi, H., Wilson, L., Mounir, S., Dea, S. 1994. Detection of porcine reproductive and
respiratory syndrome and efficient differentiation between Canadian and European strains
by reverse transcription and PCR amplification. J Clin Microbiol. 32:2197-2203.
Mardassi, H., Mounir, S., Dea, S. 1995. Molecular analysis of the ORFs3 to 7 of porcine
reproductive and respiratory syndrome virus, Québec reference strain. Arch. Virol. 140:
1405-1418.
Mardassi, H., Massie, B., Dea, S., 1996. Intracellular synthesis,m processing, and
transport of proteins encoded by ORFs 5 to 7 of porcine reproductive and respiratory
syndrome virus. Virology, 221: 98-112.

Mateu, E., Diaz, I. 2008. The challenge of PRRS inmunology. The veterinary journal, 177:
345-351.
Martínez, E., Riera, P., Sitja, M., Fang, Y., Oliveira, S., Maldonado, J. 2008. Simultaneous
detection and genotyping of porcine reproductive and respiratory síndrome virus (PRRSV)
by real-time RT-PCR and amplicon melting curve analysis usin SYBR Green. Research in
Veterinary Science, 85:184-193.
Meier, W.A., Galeota, J., Osorio, F, A., Husmann, R.J., Schnitzlein, W.M., Zuckermann,
F.A. 2003. Gradual development of the interferon-γ response of swine to porcine
reproductive and respiratory syndrome virus infection or vaccination. Virology, 309: 18-31.
Mendoza, L.F., Campa, E.S., Resendiz, M., Mata-Haro, V., Osorio, F.A., Hernandez, J.
2009. Effect of porcine reproductive and respiratory syndrome (PRRS) virus on porcine
monocite-derived dendritic cells. Vet Mex. 40.
Meng, X.J., Paul, P.S., Halburg, P.G., Morozov, I. 1995a. Sequence comparison of open
reading frames 2 to 5 of low and high virulence United States isolates of porcine
reproductive and respiratory syndrome virus. J Gen Virol. 76: 3181-3188.
Meng, X.J., Paul, P.S., Halbur, P.G., Lum, M.A. 1995b. Phylogenetic analyses of the
putative M (ORF6) and N (ORF7) genes of porcine reproductive and respiratory
syndrome virus (PRRSV): implication for the existence of two genotypes of PRRSV in the
U.S.A. and Europe. Arch. Virol. 140:745-755.
Meng, W, J., 2000. Heterogeneity of porcine reproductive and respiratory syndrome virus:
implications for current vaccine efficacy and future vaccine development. Vet. Microbiol.
74, 309-329.
Meulenberg, J.J., Julst, M.M., de Meijer, E.J., Moonen, P.I., den Besten, A., de Khuyver,
E.P., Wensvoort, R., Moorman, R.J., 1993. Lelystad virus, the causative agent of porcine
epidemic abortion and respiratory syndrome (PEARS), is related to LDV and EAV.
Virology, 192: 62-72.

Meulenberg, J.J., Petersen-den Besten, A., De Kluyver, E.P., Moormann, R.J., Schaaper,
W.M., Wensvoort, G. 1995. Characterization of proteins encoded by ORFs 2 to 7 of
Lelystad virus. Virology, 206: 155-163.
Meulenberg, J.J.M., and Besten, A.P. 1996. Identification and characterization of a sixth
structural protein of lelystad virus: the glicoprotein GP2 encoded by ORF2 is incorporated
in virus particles. Virology 225. 44-51.
Meulenberg, J.J.M., Van Nieuwstadt, A.P., Van Essen-Zandbergen, A., Langeveld, J.P.M.
1997. Posttranslational processing and identification of a neutralization domain of the
GP4 protein encoded by ORF4 of Lelystad virus. J. Virol. 71, 6061-67.
Meulenberg, J.M. 2000. PRRS, The virus. Vet. Res. 31:11-21.
Misinzo, G.M., Delputte, P.L., Nauwynck, H.J., 2008. Involvement of proteasas in porcine
reproductive and respiratory syndrome virus uncoating upon internalization in primary
macrophagues. Vet. Res. 39-55.
Mogollón, J.D., Rincón, M.A., Arbeláez, G., Ruiz, S., Rodríguez, V. 2003. PRRS virus in
Colombia. 2003 PRRS Compendium, second edition. 231-233.
Molina, R.M., Cha, S.H., Chittick, W., Lawson, S., Murtaugh, M.P., Nelson, E.A.,
Christopher-Hennings, J., Yonn, K.J., Evans, R., Rowland, R.R.R., Zimmerman, J.J.,
2008. Inmune response against porcine reproductive and respiratory syndrome virus
durin acute and chronic infection. Veterinary immunology and immunopathology, 126:283-
292.
Mulupuri, P., Zimmerman, J.J., Hermann, J., Johnson, C,R., Cano, J.P., Yu, W., Dee,
S.A., Murtaugh, M,P. 2008. Antigen-specific B cell responses to porcine reproductive and
respiratory syndrome virus infection. J Virol. 82:358–70.
Murtaugh, M.P., Elam, M.R., Kakach, L.T. 1995. Comparison of the structural protein
coding sequences of the VR-2332 and Lelystad virus strains of the PRRS virus. Arch.
Virol. 140: 1451-60.

Murtaugh, M.P., Xiao, Z., Zuckermann, F. 2002. Immunological responses of swine to
porcine reproductive and respiratory syndrome virus infection. Viral Immunology, 15: 533-
547.
Murtaugh, M.P, Xiao, Z., Rutherford, M.S and Zuckermann, F. 2003. Inmunology. 2003
PRRS Compendium:second edition, p 193-206.
Murtaugh, M., Johnson, C.R., Griggs, T.F., Gnanandaraja, J. 2011a. Novel structural
protein in porcine reproductive and respiratory syndrome virus encoded by an alternative
ORF5 present in all arteriviruses. J Gen Virol. 92:1107-1116.
Murtaugh, M., Johnson, C., Griggs, T., Robinson, S., Gnanandarajah, J. 2011b. New
protein identification in PRRSV. Leman Swine Conference, 51.
Murtaugh, M.P. 2012. Use and interpretation of sequencing in PRRSV control programs.
Disease Diagnostics. Allen D, Leman Swine Conference.49-56.
Nauwynck, H.J., Duan, X., Favoreel, H.W., Van Oostveldt, P., Pensaert, M.B., 1999. Entry
of porcine reproductive and respiratory syndrome virus into porcine alveolar macrophages
via receptor mediated endocitosis. J. Gen. Virol. 80:297-305.
Nedialkova, D.D., Ulferts, R., Van den Born, E., Lauber, C., Gorbalenya, A.E., Ziebuhr, J.,
Snijder, E.J., 2009. Biochemical characterization of arterivirus nonstructural protein 11
reveals the nidovirus-wide conservation of a replicative endoribonuclease. J. Virol. 83:
5671-5682.
Nelson, E.A., Christopher-Hennings, J., Benfield, D.A. 1994. Serum immune response to
the proteins of porcine reproductive and respiratory syndrome (PRRS) virus. J Vet Diagn
Invest. 6:410-415.
Nelsen, C.J., Murtaugh, M.P., Faaberg, KS. 1999 Porcine reproductive and respiratory
syndrome virus comparison: divergent evolution on two continents. J Virol. 73, 270-80.

Neumann, E.J.,Kliebenstein, J.B., Johnson, C.D., Mabry, J.W., Bush, E.J., Seitzinger,
A.H., Green, A.L. & Zimmerman, J.J. 2005. Assessment of the economic impact of
porcine reproductive and respiratory syndrome on swine production in the United
States.Journal of the American Veterinary Medical Association. 227(3): 385-392.
Oleksiewicz, M.B., Botner, A., Toft, P., Normann, P., Storgaard, T., 2001. Epitope
mapping porcine reproductive and respiratory syndrome virus by phage display: the nsp2
fragment of the replicase polyprotein contains a cluster of B-cells epitopes. J. Virol. 75:
3277-3290.
Osorio, F. 2010. PRRSV infections: a world update. Acta Scientiae Veterinariae. 38 (supl
1): s269-s27.
Ostrowski, M., Galeota, JA., Jar, AM., Platt, KB., Osorio, FA., Lopez, O. 2002.
Identification of neutralizing and nonneutralizing epitopes in the porcine reproductive and
respiratory syndrome virus GP5 ectodomain. J. Virol. 76: 4241-50.
Park, B.K., Joo, H.S., Dee, S.S., Pijoan, C. 1995. Evaluation of an indirect fluorescent IgM
antibody test for the detection of pigs with recent infection of porcine reproductive and
respiratory syndrome virus. J. Vet. Diagn. Invest. 7: 544-546.
Papatsiros, V.G. 2012. Porcine Herd Health Management Practices for the Control of
PRRSV Infection. Clinic of Medicine, Faculty of Veterinary Medicine, University of
Thessaly,Greece.
Patel, D., Nan, Y., Shen, M., Ritthipichai, K., Zhu, X., and Zhang, Y. 2010. “Porcine
reproductive and respiratory syndrome virus inhibits type I interferon signaling by blocking
STAT1/STAT2 nuclear translocation,” Journal of Virology, vol. 84, 21; 11045– 11055.
Pei, Y., Hodgins, D.C., Lee, C., Calvert, J.G., Welch, S.K., Jolie, R., Keith, M., Yoo, D.
2008. Functional mapping of the porcine reproductive and repiratory syndrome virus

capsid protein nuclear localization signal and its pathogenic association. Virus Res. 135:
107-114.
Plagemann, P.G.W., Rowland, R.R.R., Faaberg, K.S., 2002. The primary neutralization
epitope of porcine reproductive and respiratory syndrome virus strain VR-2332 is located
in the middle of the GP5 ectodomain. Arch. Virol., 147: 2327-2347.
Pesente, P., Rebonato, V., Sandri, G., Giovanardi, D., SperatiRuffoni, L. and Torriani, S.
2006. Phylogenetic analysis of ORF5 and ORF7 sequences of porcine reproductive and
respiratory syndrome virus (PRRSV) from PRRS-positive Italian farms: A showcase for
PRRSV epidemiology and its consequences on farm management. Veterinary
Microbiology, 114: 214-224.
PRRS: the disease, its diagnosis, prevention and control. Appendices IV and V of the
report of the OIE ad hoc group on porcine reproductive respiratory syndrome Paris, 9 - 11
June 2008.
Reiner, G., Fresen, C., Bronnert, S., Willems, H. 2009. Porcine reproductive and
respiratory syndrome virus (PRRSV infection in wild boars. Veterinary Microbiology,
136:250-258.
Rogan, D., Levere, J., Yoo, D. 2000. Immune response for the open reading frame 2
(ORF2) membrane glycoprotein of porcine reproductive and respiratory syndrome virus
(PRRSV) delivered in pigs by recombinant vaccinia virus. 16th International Pig
Veterinary Society Congress, Melbourne, Autstralia, p. 643.
Rowland, R.R., Kervin, R., Kuckleburg, C., et al., 1999. The localization of porcine
reproductive and respiratory syndrome virus nucleocapsid protein to the nucleolus of
infected cells and identification of a potential nucleolar localization signal sequence. Virus
Res. 64:1-12.

Rowland, R.R.R., Lawson, S., Rossow, K., Benfield, D.A., 2003. Lymphoid tissue tropism
of porcine reproductive and respiratory syndrome virus replication during persistent
infection of pigs originally exposed to virus in utero. Vet. Microbiol. 96:219-235.
Rowland, R.R. 2010. The interaction between PRRSV and the late gestation pig fetus.
Virus Res, 154(1-2):114-22.
Shanmukhappa, K., Kim, J.K., Kapil, S. 2007. Role of CD151, A tetraspanin, in porcine
reproductive and respiratory syndrome virus infection. Virol. J. 4: 62-73.
Shi, W., Wang, L., Lietal, X., 2011. “Endoribonuclease activities of porcine reproductive
and respiratory syndrome virus nsp11 was essential for nsp11 to inhibit IFN-β induction,”
Molecular Immunology, 48:1568–1572.
Síndrome disgenésico y respiratorio porcino. 2012. Manual Terrestre de la OIE,
capitulo2.8.7.Disponible en:
http://www.oie.int/fileadmin/Home/esp/Health_standards/tahm/2.08.07_PRRS.pdf
Síndrome reproductivo y respiratorio porcino (PRRS). Servicio Agrícola y Ganadero
(SAG).Disponible en: (http://www.sag.cl/ambitos-de-accion/sindrome-reproductivo-y-
respiratorio-porcino-prrs).
Snijder, E.J., Wassenaar, A.L., Van Dinten, L.C., Spaan, W.J., Gorbalenya, A.E. 1996.
The arterivirus nsp4 protease is the prototype of a novel group of chymotrypsin-like
enzimes, the 3C-like serine proteases. J. Biol. Chem. 271: 4864:4871.
Snijder, E.J., Meulenberg, J.J. 1998. The molecular biology of arteriviruses. J Gen Virol.
69: 971-979.
Snijder and Meulenberg. 2001. Arteriviruses in Fields. Virology, 1:1205-1220.
Song, C., Krell, P., Yoo, D. 2010. Nonstructural protein 1alpha subunit-based inhibition of
NF-kappaB activation and suppression of interferon-beta production by porcine
reproductive and respiratory syndrome virus. Virology, 407: 268–280.

Stevenson, G.W., Van Alstine, W.G., Kanitz C.L., Keffaber, K.K. 1993. Endemic porcine
reproductive and respiratory syndrome virus infection of nursery pigs in two swine herds
without current reproductive failure. J Vet Diagn Invest. 5:432–434.
Sun, Y., Xue, F., Guo, Y., Ma, M., Hao, N., Zhang, X.C., Lou, Z., Li, X., and Rao, Z., 2009.
Crystal structure of porcine reproductive and respiratory syndrome virus leader protease
Nsp1 alpha. J. Virol. 83: 10931-10940.
Sun, Y., Han, M., Kim, CH., Calvert, J, G., Y, D., 2012a. Interplay between Interferon-
Mediated Innate Immunity and Porcine Reproductive and Respiratory Syndrome Virus.
Viruses, 4: 424-446.
Sun, Z., Ransburgh, R., Snijder, E.J., and Fang, Y. 2012b. “Nonstructural Protein 2 of
porcine reproductive and respiratory syndrome virus inhibits the antiviral function of
interferon-stimulated gene 15,” Journal of Virology, vol. 86(7): 3839–3850.
Tian, H., Wu, J.Y., Shang, Y.J., Cheng, Y., Liu, W.T. 2010. The developmente of a rapid
SYBR one step real-time RT-PCR for detection of porcine reproductive and respiratory
syndrome virus. Virology Journal, 7: 1-7.
Tijms, M.A., Van Der Meer, Y., Snijder, E.J. 2002. Nuclear localization of non-structural
protein 1 and nucleocapsid protein of equine arteitis virus. J.Gen. Virol. 83:795-800.
Torremorell, M., and Christianson, W, T. 2002. PRRS eradication by herd closure. Adv
Pork Prod., 13: 169–76.
Van Aken, D., Snijder, E.J., and Gorbalenya, A.E., 2006. Mutagenesis analysis of the
nsp4 main proteinase reveals determinants of arterivirus replicase polyprotein
autoprocessing. J. Virol. 80: 3428-3437.
Van Breedman, W., Van Gorp, H., Zhang, J.Q., Crocker, P.R., Delputte, P.L., Nauwynck,
H.J. 2010. The M/GP5 Glycoprotein complex of porcine reproductive and respiratory

syndrome virus binds the Sialodhesin receptor in a Sialic Acid-dependent manner. Plos
Pathogens. 6: 1-11.
Van Dinten, L.C., deen Boon, J.A., Wassennar, A.L., Spaan, W.J., Snijder, E.J., 1997. An
infectious aerterivirus cDNA clone: identification of a replicase point mutation that
abolishes discontinuos mRNA transcription. Proc Natl Acad Sci USA, 64: 991-996.
Van Gorp, H., Van Breedam, W., Delputte, P.L., Nauwynck, H.J. 2008. Sialodhesin and
CD163 join forces during entry of the porcine reproductive and respiratory syndrome
virus. J. Gen. Virol. 89: 2943-2953.
Van Reeth, K., Nauwynck, H. 2000. Proinflammatory cytokines and viral respiratory
disease in pigs. Vet. Res. 31: 187-213.
Vashisht, K., Erlandson, K.R., Firkins, L.D., Zuckermann, F.A. and Goldberg, T.L. 2008.
Evaluation of contact exposure as a method for acclimatizing growing pigs to porcine
reproductive and respiratory syndrome virus. Journal of the American Veterinary Medical
Association, 232:1530-1535.
Vezina, S.A., Loemba, H., Foutnier, M., Dea, S., Archibault, D. 1996. Antibody production
and blastogenesis response in pigs experimentally infected with porcine reproductive and
respiratory syndrome virus. Can J Vet Res. 60:94–99.
Waddell, J., 2010. PRRS area regional control and elimination: Why is Nebraska a good
place to start? In: Proc. George A Young Swine Health Management Conf., South Sioux,
Nebraska, 54-59.
Wang, X., Eaton, M., Mayer, M., Li, H., He, D., Nelson, E., et al. 2007. Porcine
reproductive and respiratory syndrome virus productively infects monocyte–derived
dendritic cells and compromises their antigen–presenting ability. Arch Virol. 152:289–
303.

Wang, R., Nan, Y., Yu, Y., Yang, Z., and Zhang, Y. 2013. “Variable interference with
interferon signal transduction by different strains of porcine reproductive and respiratory
syndrome virus,” Veterinary Microbiology, vol. 166 (3-4): 493–503.
Wang, R and Zhang, Y.J. 2014. Antagonizing Interferon-mediated immune response by
porcine reproductive and respiratory syndrome virus. BioMed Research International,
315470: 1-9.
Wasilk, A., Callahan, J.D., Christopher-Hennings, J., Gay, T.A., Fang, Y., Dammen, M.,
Reos, M.E., Torremorell, M., Polson, D., Mellencamp, M., Nelson E., and Nelson, W.M.
2004. Detection of U.S., Lelystad, and European-like porcine reproductive and respiratory
syndrome viruses and relative quantitation in boar semen and serum samples by real-
time PCR. J. Clin. Microbiol., 42, 4453–4461.
Welch, S.K., Calvert, J.G., 2010. A brief review of CD163 and its role in PRRSV infection.
Virus Research, 154(1-2):98-103.
Welch, S.W., Jolie, R., Pearce, D.S., Koertje, W.D., Fuog, E., Shields, S.L., Yoo, D.,
Calvert, J.G. 2004. Construction and evaluation of genetically engineered replication-
defective porcine reproductive and respiratory syndrome virus vaccine candidates. Vet.
Immunol. Immunopathol. 102: 277-290.
Wensvoort, G., Terpstra, D., Pol, J.M.A., Ter Laak, E.A., Bloemraad, M., de Kluyver, E.P.,
Kragten, D., Van Buiten, L., Den Besten, A., Wagenaar, F., Broekhuijsen, J.M., Moonen,
P.L.J.M., Zetstra, T., De Boer, E.A., Tibben, H.J., de Jong, M.F., Van´t Veld, P.,
Groenland, G.J.R., Van Gennep, J.A., Voets, M.Th., Verheijden, J.H.M., Braamskamp, J.
1991. Mystery swine disease in the Netherlands: the isolation of Lelystad virus. Vet. Q.
13: 121-130.
Wensvoort, G. 1993. Lelystad virus and the porcine epidemic abortion and respiratory
syndrome. Vet Res. 24: 117-124.

Wills, R.W., Doster, A.R., Galeota, J.A., Sur, J.H., Osorio, F.A. 2003. Duration of infection
and proportion of pigs persistently infected with porcine reproductive and respiratory
syndrome virus. J Clin Microbiol. 41:58–62.
Wissink, E.H.J., Kroese, M.V., Maneschijn-Bonsing, J.G., Meulenberg, J.J.M., van Rijn,
P.A., Rijsewijk, F.A.M., Rottier, P.J.M., 2004. Significance of the oligosaccharides of the
porcine reproductive and respiratory syndrome virus glycoproteins GP2a y GP5 for
infectious virus production. J. Gen. Virol. 85: 3715-3723.
Wisssink, E.H.J., Kroese, M.V., Van Wijk, H.A.R., Rijsewijk, F.A.M., Meulenberg, J.J.M.,
Rottier, P.J.M. 2005. Envelope protein requirements for the assembly of infectious virions
of porcine reproductive and respiratory syndrome virus. J. Virol. 79: 13495-12506.
Wu, W., Fang, Y., Farwell, R., Steffen-Bien, M., Rowland, R.R., Christopher-Hennings, J.,
Nelson, E.A. 2001. A 10-kDa structural protein of porcine reproductive and respiratory
syndrome virus encoded by ORF2b. Virology 287: 183-191.
Wu, W., Fang, Y., Rowland, R.R., Lawson, SR., Christopher-Hennings, J., Yoon, K.,
Nelson, E.A. 2005. The 2b protein as a minor structural component of PRRSV. Virus Res.
114, 177-181.
Xue, F., Sun, Y., Yan, L., Zhao, C., Chen, J., Bartlam, M., Li, X., Lou, Z., and Rao, Z.,
2010. The crystal structure of of porcine reproductive and respiratory syndrome virus
nonstructural protein Nsp1beta reveals a novel metal-dependent nuclease. J. Virol.
84:6461-6471.
Yoo, D., Song, C., Sun, Y., Du, Y., Kim, O., Liu, H.C. 2010. Modulation of host cell
responses and evasion strategies for porcine reproductive and respiratory syndrome
virus. Virus Res. 154:48–60.
Yoon, K.J., Zimmerman, J.J., McGinley, M.J. 1995. Failure to consider the antigenic
diversity of porcine reproductive and respiratory syndrome PRRS virus isolates may lead
to misdiagnosis. J Vet Diagn. Invest. 7:386-387.

Yoon, K.J., Zimmerman, J.J., Wu, L.L., Platt, K.B. 1996. Antibody-dependent
enhancement (ADE) of porcine reproductive and respiratory syndrome virus (PRRS)
infection in pigs. Virol Inmunol. 9:51-63.
Yoon, K., Wu, L., Zimmerman, J.J., Platt, K.B. 1997. Field isolates of porcine reproductive
and respiratory syndrome virus (PRRSV) vary in their susceptibility to antibody dependent
enhancement (ADE) of infection. Vet. Microb. 55: 277-287.
Yoon, K.J. 2003. Virology. PRRS Compendium. Second edition, p163-189.
Yoon, K.J, .Christopher-Hennings, J., Nelson, E.A. 2003. Diagnosis. PRRS Compendium.
Second edition, p59-74.
Yun, S.I., and Lee, Y.M., 2013. Overview: Replication of porcine reproductive and
respiratory syndrome virus. Journal of Microbiology, 51:711-723.
Zhang, Y., Sharma, R.D., Paul, P.S. 1998. Monoclonal antibodies against
conformationally dependent epitopes on porcine reproductive and respiratory syndrome
virus. Vet. Microb. 63: 125-136.
Zhou, L., and Yang, H. 2010. Porcine reproductive and respiratory syndrome virus in
China. Virus Res. 154: 31-37.
Zhou Y.J., Hao, X.F., Tian, Z.J., Tong, G.Z., Yoo, D., An, T.Q., Zhou, G.X., Li, G.X., Qiu,
H.J., Wei, T.C., and Yuan, X.F. 2008. Highly virulent porcine reproductive and respiratory
syndrome virusemerged in China. Transbound Emerg Dis. 55:152-164.
Zhou, L., Chen, S., Zhang, J., Zeng, J., Guo, X., Ge, X., Zhang, D., Yang, H. 2009.
Molecular variation analysis of porcine reproductive and respiratory syndrome virus in
China. Virus Research, 145:97-105.

Zimmerman, J.J., Yonn, K.J., Wills, R.W., Swenson, S.L., 1997. General overview of
PRRS: a perspective from the United States. Vet. Microbiol. 55:187-196.
Zimmerman J. 2003. Epidemiology and Ecology. 2003 PRRS Compendium. Second
edition. P. 27-50.
Zimmerman, J., Benfield, D.A., Murtaugh, M.P., Osorio, F., Stenson, G.W., Torremorell,
M., 2006. Porcine reproductive and respiratory syndrome virus (porcine arterivirus).
Diseases of swine, 387-418. Blackwell publishing, Ames, Iowa.

Evaluación de la dinámica de anticuerpos
frente al virus del Síndrome Reproductivo y
Respiratorio Porcino (PRRSV) en tres granjas
del departamento del Valle del Cauca
Resumen
El Síndrome Reproductivo y Respiratorio Porcino (PRRS), es una enfermedad endémica
en Colombia. A partir del año 1997 se comenzaron a realizar estudios serológicos para
determinar la actividad del virus en los sistemas de producción y los diferentes grupos
etáreos. Estos estudios demostraron la presencia de actividad del PRRSV en el país,
especialmente en sistemas intensivos. El presente estudio fue realizado entre 2012 y
2013, en una zona de producción intensiva como el Valle del Cauca, utilizando diferentes
granjas en cuanto a manejo y medidas de bioseguridad, diferentes grupos etáreos y
muestreos durante el tiempo, utilizando como prueba serológica la técnica de ELISA de
IDEXX, PRRS x 3. Los resultados obtenidos confirmaron las diferencias en cuanto a
actividad viral dentro de cada granja, identificando el grupo de la ceba como el de mayor
seropositividad. De igual manera, se evidencio la necesidad de utilizar conjuntamente
con las técnicas serológicas, técnicas moleculares para determinar la dinámica de la
infección y por ende tomar las medidas adecuadas de control.
Palabras clave: Diagnóstico, ELISA, PRRS, Serología
Abstract
Porcine reproductive and respiratory syndrome (PRRS), is an endemic disease in
Colombia. Since 1997 serological studies began to determine virus activity in the
production systems and on the different groups of age. These studies showed the
presence and different activity levels of PRRSV on the country, especially in production
intensive systems. The present study was carried out between 2012 and 2013, on the
Valle del Cauca province, one of the most important pig production region in Colombia.

Three farms were selected based on different practices of management and measures of
biosafety. It was colleted serum samples during nine months and on differents groups of
age. An antibodies detecting ELISA kit (IDEXX, PRRS ELISA test x 3) was employed to
evaluate humoral immune response. Results confirmed differences in terms of viral
activity within each farm, identifying the fattening group as this that presented higher
seropositivity.
As in Colombia there is not vaccination against PRRS, any serological response is proof
of viral activity; in this study all farms had PRRSV antibodies response confirming viral
presence and disease manifestation. We recommended to employ both serological and
molecular techniques to determine infection dynamics and thus to take appropriates
control measures.
Key words: Diagnostic, ELISA, PRRS, Serology.
Introducción
El Síndrome Reproductivo y Respiratorio Porcino (PRRS) es una enfermedad infecciosa
que ocasiona importantes pérdidas económicas a la industria porcina alrededor del
mundo (Holtkamp et al., 2013). Se caracteriza, por ocasionar abortos en el último tercio
de la gestación y producir neumonía intersticial en lechones (Collins et al., 1992;
Wensvoort et al., 1991). El virus de PRRS (PRRSV) se divide en dos genotipos, Europeo
o Tipo I y Americano o Tipo II, representados por las cepas Lelystad (LV) y VR-2332,
respectivamente (Allende et al., 1999; Nelsen et al., 1999).
El PRRSV es un Arterivirus, que posee un genoma ARN de cadena sencilla y polaridad
positiva de 15 kb de longitud, presenta 10 marcos de lectura abierta (ORFs). Los ORF1a
y ORF1b abarcan el 80% del genoma y codifican para enzimas involucradas en la
replicación viral; los polipéptidos de estos ORFs dan origen a 14 proteínas no
estructurales (NSPs) que incluyen NSP1α, NSP1β, NSP2, NSP2TF y NSP3 a NSP12.
Los ORF2a, ORF2b, ORF3 a ORF7 y ORF5a, codifican para 8 proteínas estructurales,
GP2, proteína E, GP3, GP4, GP5, proteína de la membrana (M), proteína de la

nucleocápside (N) y proteína ORF5a (Faaberg et al., 2011; Snijder and Meulenberg,
1998; Snijder et al., 2013).
La exposición inicial con el PRRSV se caracteriza en muchos casos por la presencia de
síntomas clínicos a nivel respiratorio y una abundante replicación en macrófagos
alveolares y tisulares. Esta fase aguda se caracteriza por una alta carga viral en tejidos
como tonsila, pulmón, órganos linfoides y presencia de viremia. Posterior a la fase aguda,
sobreviene una fase de infección persistente que involucra un bajo nivel de replicación
viral a nivel del tejido linfoide (Zimmerman et al., 1992). El virus eventualmente es
eliminado del organismo; el proceso de infección por PRRSV se estima que tiene una
duración de al menos 150 días (Allende et al., 2003).
Las principales células blanco para la infección por el PRRSV son los macrófagos
alveolares pulmonares porcinos (MAPs) los cuales juegan un papel importante en la
inmunidad del huésped, sin embargo una vez estos son infectados, el PRRSV desarrolla
mecanismos para interferir con la respuesta inmune celular (Sun et al., 2012a). Uno de
los mayores componentes de la inmunidad innata antiviral es el interferón (INF) tipo I (α
y β) que cumple diversas funciones, entre ellas, la generación de proteínas antivirales
(PAVs) y la estimulación de la actividad citotóxica de las células T-CD8 (Pestka, 2007); el
PRRSV puede obstaculizar la producción del IFN tipo I de varias formas entre ellas,
reduciendo la expresión de TLR3 (Toll-like receptor 3) (Sang et al., 2008), antagonizando
la inducción de IFN por medio de proteínas no estructurales como las NSP1, NSP2,
NSP11 y N (Kim et al., 2010; Shi et al., 2011), interfiriendo con las vías de señalización
JAK/STAT (Patel et al., 2010), inhibiendo la señalización NF-kB (nuclear factor-kappa-
beta), suprimiendo la respuesta del IFN tipo I o activando esta señalización para
optimizar la replicación viral, la apoptosis y la modulación de citoquinas (Lee et al., 2005).
Igualmente, inhibe la función antiviral de ISG15 (IFN- stimulated gene 15) (Sun et al.,
2012 b) y modifica la expresión de moléculas involucradas en la presentación de
antígeno (Mateu and Díaz, 2008).
En el desarrollo de la inmunidad adaptativa contra PRRSV, los anticuerpos circulantes
son detectables entre los días 5 a 7 post infección (PI), con seroconversión a los 14 días
PI (Yoon et al., 1995 citado por Mateu and Díaz, 2008). Las primeras inmunoglobulinas

en surgir son tipo M (IgM) detectables entre el día 5 a 7 con un pico el día 14 declinando
hacia el día 42 PI. La inmunoglobulina tipo G aparece hacia la segunda semana PI, con
un pico entre los días 21 y 42 (Vezina et al., 1996 citado por Mateu and Díaz, 2008).
Para el caso de PRRS, los anticuerpos de aparición temprana son dirigidos
específicamente contra las proteínas N y M, estos aparecen hacia el día 7 PI, seguidos
por anticuerpos contra la proteína GP5. De igual manera, en animales infectados, se
presenta una respuesta temprana frente a epítopes de la proteína nsp2, probablemente
la proteína inmunodominante del virus (De Lima et al., 2006; Oleksiewicz et al., 2001).
Distintas pruebas diagnósticas detectan anticuerpos principalmente contra la proteína N,
estos persisten por varios meses pero no están relacionados con protección
inmunológica (Mateu and Díaz, 2008).
De otro lado, los anticuerpos no neutralizantes son responsables del aumento de
replicación viral dependiente de anticuerpos (ADE) (Yoon et al., 1996, 1997, citado por
Darwich et al., 2010), este fenómeno se describe como un incremento en la
internalización de partículas virales en los macrófagos como resultado de la
opsonización. Los epítopes responsables de inducir anticuerpos relacionados con ADE
se ubican en las proteínas N y GP5 (Cancel-Tirado et al., 2004; Gonin et al., 1999;
Nelson et al., 1994; Ostrowski et al., 2002). El efecto de esta respuesta inmune no
neutralizante actúa como mecanismo de evasión del sistema inmune mediante la
estrategia de caballo de Troya, ya que permite el aumento de la internalización de las
partículas virales en los macrófagos (Mateu and Díaz, 2008). Estos anticuerpos son
detectables hacia el día 28 post infección (PI) (Diaz et al., 2005; Meier et al., 2003; Yoon
et al., 1994).
Se han postulado varias hipótesis sobre el retraso y el bajo título de anticuerpos
neutralizantes, entre ellas se propone la presencia de un epítope denominado “trampa”
(epítope A) localizado cerca al epítope neutralizante (epitope B) de la glicoproteína GP5,
los anticuerpos generados contra el epitíope A no inducen una respuesta inmune
protectiva (Ostrowski et al., 2002). De otro, lado la glicosilación puede afectar la
antigenicidad de GP5, esto se conoce como "blindaje glicano", el cual posiblemente
reduce la capacidad de los anticuerpos para neutralizar PRRSV. Se ha observado, que

la eliminación de los sitios de glicosilación N34, N51y N34+51 produce una mayor
sensibilidad a la neutralización por anticuerpos (Ansari et al., 2006).
El PRRSV induce una inmunidad protectiva contra la re-exposición a la misma cepa viral,
de igual manera ocasiona una prolongada viremia e infección persistente durante la cual
el huésped puede continuar eliminando el virus (Murtaugh et al., 2002).
Las técnicas serológicas para el diagnóstico del PRRSV, permiten evaluar el estatus y la
dinámica de la infección en las explotaciones porcícolas donde no se evidencian signos
clínicos de la enfermedad y en este caso técnicas como ELISA permiten monitorear la
presencia del virus en granja (Mattheu et al., 2006; Wellenberg, 2006).
En Colombia en 1996, se evidenció por primera vez la circulación del PRRSV
empleando la técnica de ELISA, demostrando la presencia de anticuerpos contra este
virus. Posteriormente, el virus fue aislado a partir de muestras de suero y de tejido
pulmonar. Estudios de caracterización molecular situaron los aislados virales dentro del
genotipo Americano (Mogollón et al., 2003).
Teniendo en cuenta el tiempo transcurrido desde la realización del último estudio
serológico en diferentes zonas del país, el presente proyecto se desarrolló con el fin de
establecer la dinámica de anticuerpos frente al PRRSV en tres granjas localizadas en
una zona de alta producción porcícola del país, determinando así el comportamiento del
virus dentro de cada granja, entre granjas y en diferentes grupos etáreos durante un
periodo de nueve meses.
2.1 Materiales y Métodos
2.1.1 Ubicación geográfica y características de las granjas
El estudio se realizó en tres granjas de tipo intensivo y ciclo completo, ubicadas en el
Departamento del Valle del Cauca, localizado en el sur occidente del país a una altitud de
1400-1600 msnm, una temperatura promedio de 23-24°C y una humedad relativa del
65% a 75%. Esta zona se caracteriza por una producción porcícola de tipo intensivo, que
presentó en los meses previos al estudio, reportes de animales con sintomatología
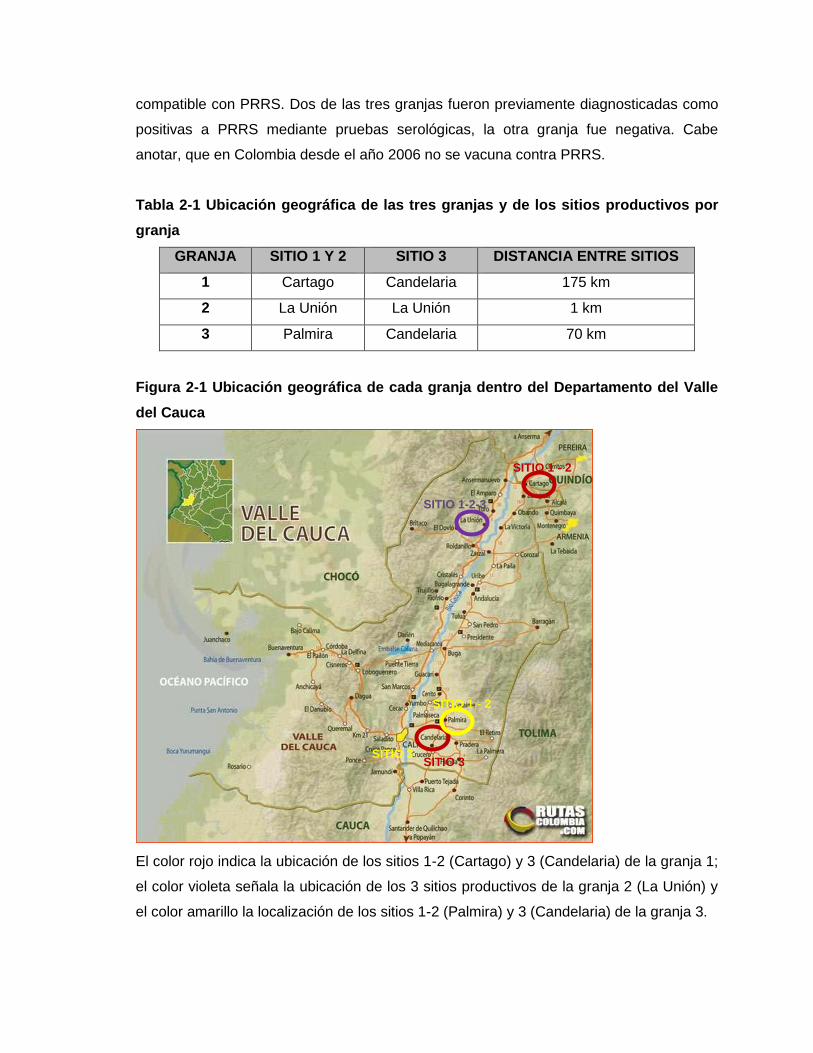
compatible con PRRS. Dos de las tres granjas fueron previamente diagnosticadas como
positivas a PRRS mediante pruebas serológicas, la otra granja fue negativa. Cabe
anotar, que en Colombia desde el año 2006 no se vacuna contra PRRS.
Tabla 2-1 Ubicación geográfica de las tres granjas y de los sitios productivos por
granja
GRANJA SITIO 1 Y 2 SITIO 3 DISTANCIA ENTRE SITIOS
1 Cartago Candelaria 175 km
2 La Unión La Unión 1 km
3 Palmira Candelaria 70 km
Figura 2-1 Ubicación geográfica de cada granja dentro del Departamento del Valle
del Cauca
El color rojo indica la ubicación de los sitios 1-2 (Cartago) y 3 (Candelaria) de la granja 1;
el color violeta señala la ubicación de los 3 sitios productivos de la granja 2 (La Unión) y
el color amarillo la localización de los sitios 1-2 (Palmira) y 3 (Candelaria) de la granja 3.
SITIO 1 - 2
SITIO 3
SITIO 1-2-3
SITIO 3
SITIO 1 - 2

2.1.2 Muestras
Se tomaron muestras de sangre mediante venopunción (vena cava), en tubos
individuales (BD Vacutainer), éstas fueron enviadas el mismo día al Laboratorio de
Virología de la Facultad de Medicina Veterinaria y de Zootecnia de la Universidad
Nacional. Fueron centrifugadas a 1000xg durante 10 minutos, el suero obtenido se
almacenó a -20°C, hasta su procesamiento. Los sueros provenían de los diferentes
grupos etáreos presentes en las granjas: madres, reproductores, reemplazos, lactantes,
lechones destetos (20-21 días), preceba (7-9 semanas) y ceba (12-14 semanas y 18-22
semanas). Se obtuvieron 5 muestras de suero por cada grupo etáreo, por granja y por
cada mes, durante los 9 meses en que se desarrolló el muestreo, desde septiembre de
2012 a mayo de 2013.
2.1.3 Evaluación serológica
Con el fin de evaluar la presencia y comportamiento de los anticuerpos frente al PRRSV,
se utilizó el kit comercial de ELISA HerdCheck* IDEXX PRRSx3, realizándose el
procedimiento especificado en el inserto por el laboratorio productor.
Se emplearon placas revestidas con antígenos recombinantes del virus del PRRS donde
fueron adicionados 100 µl de una predilución 1:40 de cada muestra y posteriormente 100
µl de los controles negativo y positivo, se incubaron y se realizaron tres lavados,
posteriormente se adicionaron 100 µl de anticuerpo anti porcino conjugado con
peroxidasa (HRPO) y se incubó durante 30 minutos. Luego de tres lavados se
adicionaron 100 µl de substrato TMB durante quince minutos y se detuvo la reacción con
la solución de frenado. La lectura se realizó en un lector de micro placas BioTekPower
Wave HT, a una densidad óptica (DO) de 650 nm. La presencia o ausencia de
anticuerpos contra el virus del PRRS se determinó calculando el cociente
muestra/positivo (M/P). Una muestra se consideró negativa si el cociente M/P fue inferior
a 0.40 y positiva si el cociente M/P fue igual o superior a 0.40.

2.1.4 Análisis estadístico
Para la variable M/P se realizó un análisis de varianza utilizando el programa SAS,
teniendo como fuentes de variación, granja (1, 2 y 3), grupo etáreo (madres,
reproductores, reemplazos, lactantes, destetos, preceba y ceba) y meses (septiembre de
2012 a mayo de 2013). Al presentarse efectos significativos, se empleó una prueba de
comparación múltiple de Tukey.
2.2 Resultados
Al evaluar el nivel de significancia de las fuentes de variación, se observó una interacción
altamente significativa entre los tres factores analizados en el estudio: granja, grupo
etáreo y tiempo. Se observaron diferencias altamente significativas entre grupo y granja
durante los meses del estudio específicamente en las granjas 1 y 3; la granja 2 no
presentó interacciones significativas entre estos factores. En total, se analizaron 910
muestras de suero; se evaluaron 45 muestras por grupo etáreo. En la granja 2 no se
analizaron muestras del mes de Septiembre. Los datos fueron analizados por grupo
etáreo, por granja, durante un lapso de nueve meses.
2.2.1 Granja 1
Se analizaron 315 muestras y siete grupos etáreos. De acuerdo con los promedios de
resultados obtenidos, la ceba presentó el mayor número de muestras positivas (Tabla 2-
2). Con relación a la interacción grupos etareos-granjas-meses, se observó una
diferencia altamente significativa (<.0001) dentro de la granja 1 con respecto al grupo de
la ceba durante los meses del estudio.
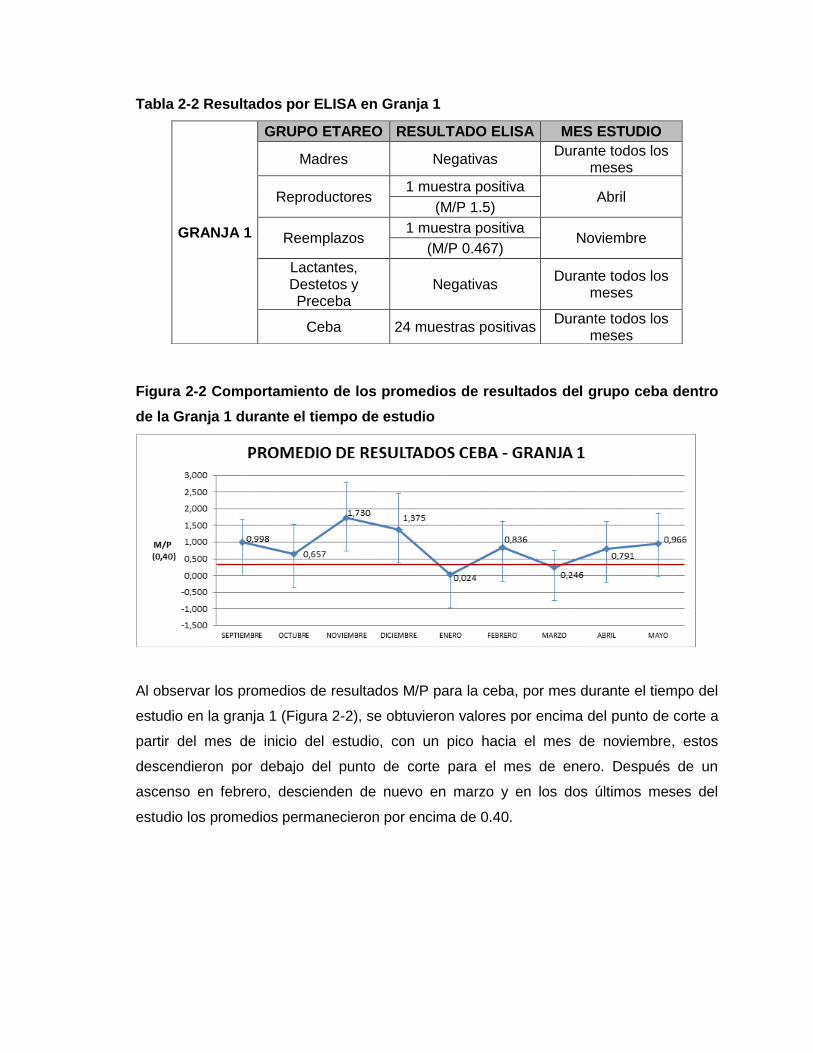
Tabla 2-2 Resultados por ELISA en Granja 1
Figura 2-2 Comportamiento de los promedios de resultados del grupo ceba dentro
de la Granja 1 durante el tiempo de estudio
Al observar los promedios de resultados M/P para la ceba, por mes durante el tiempo del
estudio en la granja 1 (Figura 2-2), se obtuvieron valores por encima del punto de corte a
partir del mes de inicio del estudio, con un pico hacia el mes de noviembre, estos
descendieron por debajo del punto de corte para el mes de enero. Después de un
ascenso en febrero, descienden de nuevo en marzo y en los dos últimos meses del
estudio los promedios permanecieron por encima de 0.40.
GRANJA 1
GRUPO ETAREO RESULTADO ELISA MES ESTUDIO
Madres Negativas Durante todos los
meses
Reproductores 1 muestra positiva
Abril (M/P 1.5)
Reemplazos 1 muestra positiva
Noviembre (M/P 0.467)
Lactantes, Destetos y Preceba
Negativas Durante todos los
meses
Ceba 24 muestras positivas Durante todos los
meses
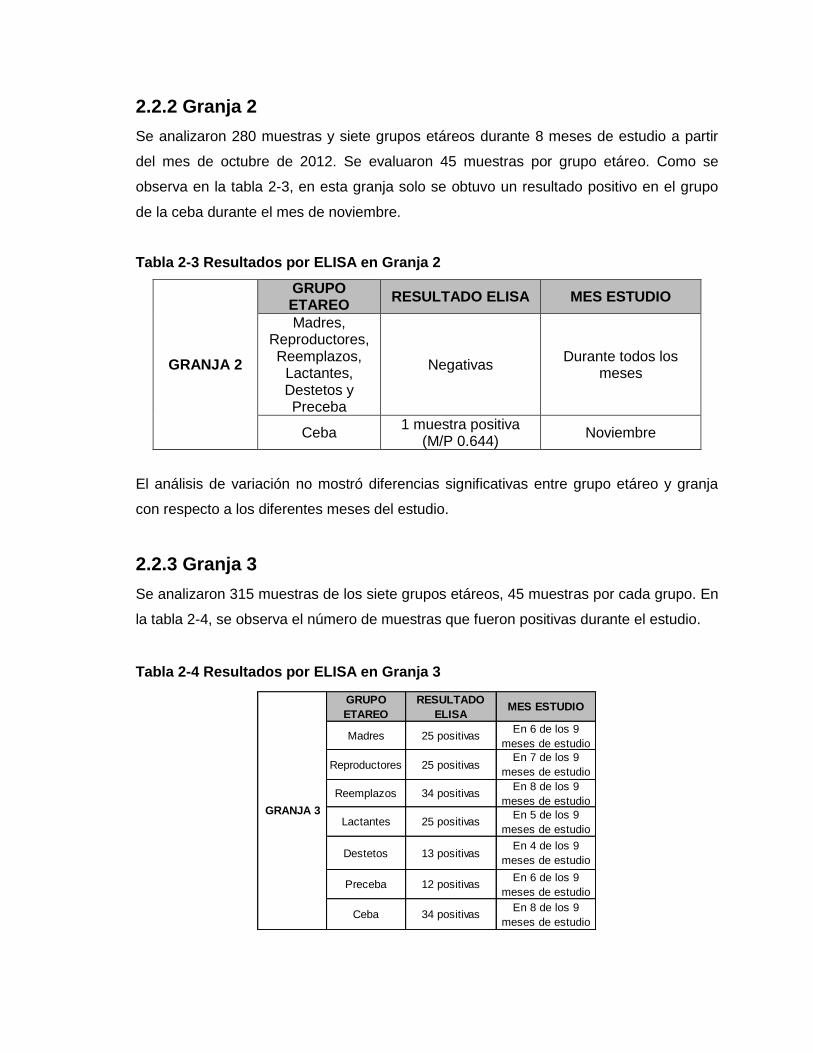
2.2.2 Granja 2
Se analizaron 280 muestras y siete grupos etáreos durante 8 meses de estudio a partir
del mes de octubre de 2012. Se evaluaron 45 muestras por grupo etáreo. Como se
observa en la tabla 2-3, en esta granja solo se obtuvo un resultado positivo en el grupo
de la ceba durante el mes de noviembre.
Tabla 2-3 Resultados por ELISA en Granja 2
GRANJA 2
GRUPO ETAREO
RESULTADO ELISA MES ESTUDIO
Madres, Reproductores, Reemplazos,
Lactantes, Destetos y Preceba
Negativas Durante todos los
meses
Ceba 1 muestra positiva
(M/P 0.644) Noviembre
El análisis de variación no mostró diferencias significativas entre grupo etáreo y granja
con respecto a los diferentes meses del estudio.
2.2.3 Granja 3
Se analizaron 315 muestras de los siete grupos etáreos, 45 muestras por cada grupo. En
la tabla 2-4, se observa el número de muestras que fueron positivas durante el estudio.
Tabla 2-4 Resultados por ELISA en Granja 3
GRUPO
ETAREO
RESULTADO
ELISAMES ESTUDIO
Madres 25 positivasEn 6 de los 9
meses de estudio
Reproductores 25 positivasEn 7 de los 9
meses de estudio
Reemplazos 34 positivasEn 8 de los 9
meses de estudio
Lactantes 25 positivasEn 5 de los 9
meses de estudio
Destetos 13 positivasEn 4 de los 9
meses de estudio
Preceba 12 positivasEn 6 de los 9
meses de estudio
Ceba 34 positivasEn 8 de los 9
meses de estudio
GRANJA 3

De acuerdo con la tabla 2-4, se observó que todos los grupos etáreos de la granja 3
presentaron resultados positivos, en mayor número dentro del grupo de los reemplazos y
la ceba. El análisis de variación mostró diferencias altamente significativas (<.0001) entre
grupo etáreo y granja con respecto a los diferentes meses del estudio.
En las figuras 2-3, 2-4, 2-5, correspondientes a los grupos de las madres, lactantes y
destetos, respectivamente, se observa un promedio de resultados de M/P superior a 0.40
a partir del mes de inicio del estudio, presentando un pico entre octubre y noviembre, con
un descenso hacia diciembre y enero con valores cercanos o por debajo del punto de
corte, posteriormente vuelven a incrementarse, con un descenso hacia el final del
estudio.
Figura 2-3 Comportamiento de los promedios de resultados serológicos del grupo
de las madres dentro de la granja 3 durante el tiempo de estudio
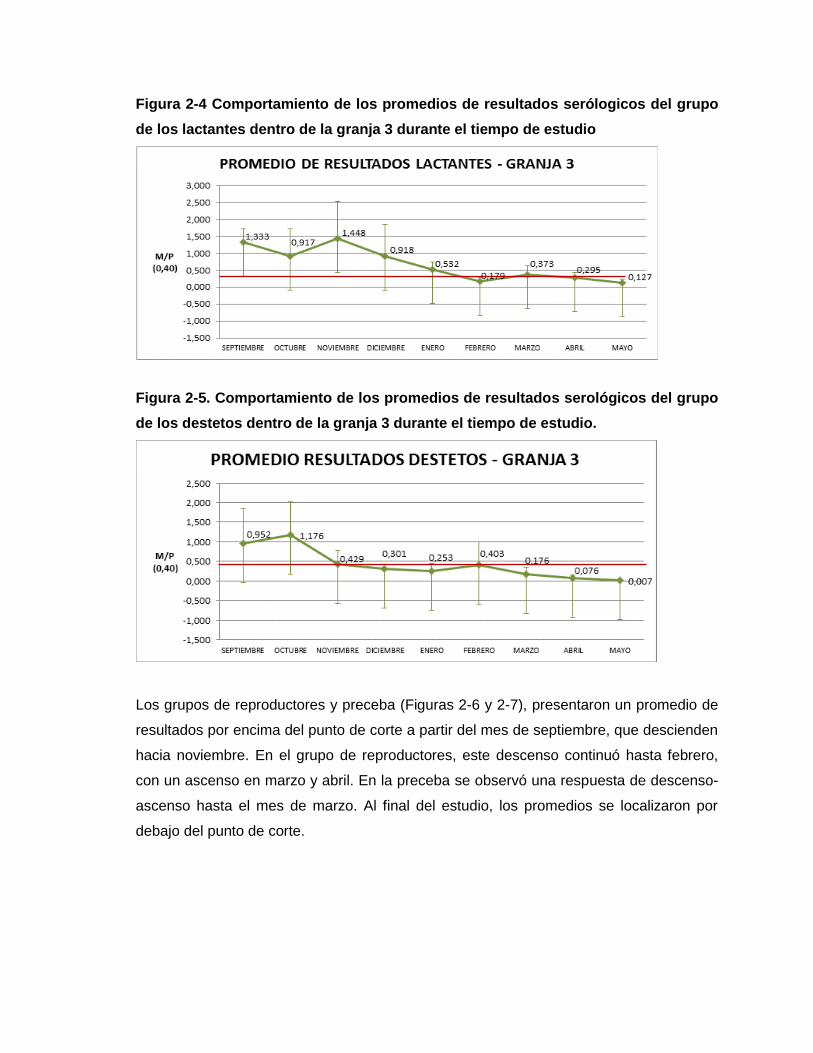
Figura 2-4 Comportamiento de los promedios de resultados serólogicos del grupo
de los lactantes dentro de la granja 3 durante el tiempo de estudio
Figura 2-5. Comportamiento de los promedios de resultados serológicos del grupo
de los destetos dentro de la granja 3 durante el tiempo de estudio.
Los grupos de reproductores y preceba (Figuras 2-6 y 2-7), presentaron un promedio de
resultados por encima del punto de corte a partir del mes de septiembre, que descienden
hacia noviembre. En el grupo de reproductores, este descenso continuó hasta febrero,
con un ascenso en marzo y abril. En la preceba se observó una respuesta de descenso-
ascenso hasta el mes de marzo. Al final del estudio, los promedios se localizaron por
debajo del punto de corte.

Figura 2-6 Comportamiento de los promedios de resultados serológicos del grupo
de los reproductores dentro de la granja 3 durante el tiempo de estudio.
Figura 2-7 Comportamiento de los promedios de resultados serológicos del grupo
de la preceba dentro de la granja 3 durante el tiempo de estudio.
En las gráficas 2-8 y 2-9, se observa que los grupos de los reemplazos y ceba,
conservaron los promedios de resultados durante cada mes por encima del punto de
corte, excepto en la ceba que presentó para el mes de Mayo un descenso por debajo de
0.40.
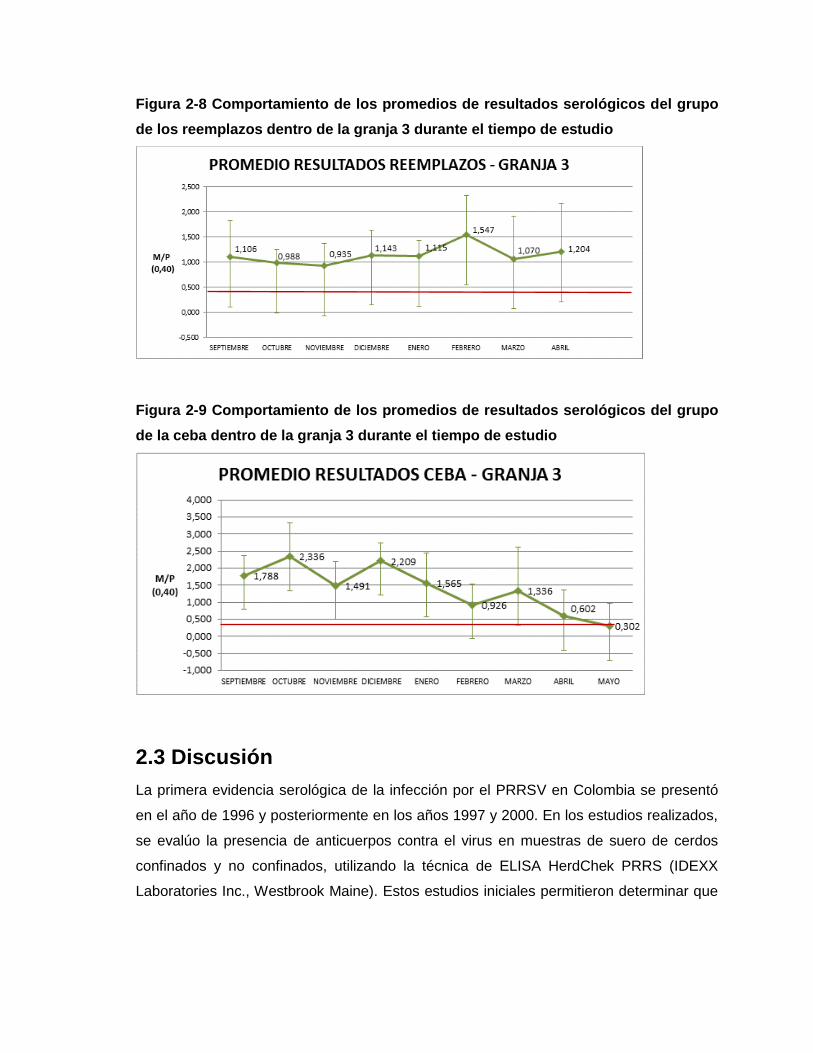
Figura 2-8 Comportamiento de los promedios de resultados serológicos del grupo
de los reemplazos dentro de la granja 3 durante el tiempo de estudio
Figura 2-9 Comportamiento de los promedios de resultados serológicos del grupo
de la ceba dentro de la granja 3 durante el tiempo de estudio
2.3 Discusión
La primera evidencia serológica de la infección por el PRRSV en Colombia se presentó
en el año de 1996 y posteriormente en los años 1997 y 2000. En los estudios realizados,
se evalúo la presencia de anticuerpos contra el virus en muestras de suero de cerdos
confinados y no confinados, utilizando la técnica de ELISA HerdChek PRRS (IDEXX
Laboratories Inc., Westbrook Maine). Estos estudios iniciales permitieron determinar que

PRRSV estaba ampliamente distribuido en el país de forma no uniforme, predominando
en los sistemas de producción intensiva. Para el caso particular del Valle del Cauca, en
1997 se evaluaron 544 muestras procedentes de 7 granjas, donde se obtuvieron 93
muestras positivas (17.1%) (Mogollón et al., 2003). Para el año 2006 se realizó un nuevo
estudio serológico con el fin de actualizar la situación de PRRSV en el país, se incluyeron
para este caso los departamentos donde se manejaban producciones extensivas o de
traspatio, tomando muestras en mataderos (cerdos de descarte y de ceba). Las muestras
se analizaron con el kit HerdChek ELISA PPRS x 2 (Laboratorios IDEXX), obteniéndose
un total de 71 reactores positivos de 1.658 sueros analizados y una prevalencia del 4.3%.
Específicamente, en el departamento del Valle se encontraron 6 reactores en 170
muestras evaluadas y una prevalencia de la enfermedad del 3.5% (Cruz et al., 2006).
En el presente estudio realizado en el departamento del Valle del Cauca, se analizaron
tres factores dentro de una misma zona: granja (3), grupo etáreo (7) y tiempo (9 meses),
los que presentaron una interacción estadística altamente significativa, particularmente
en las granjas 1 y 3. Por su parte la granja 2, no presentó esta interacción
permaneciendo seronegativa durante el tiempo de estudio. Es de notar el
comportamiento diferente entre granjas dentro de una misma zona geográfica. En
general, se obtuvo un porcentaje de positividad del 21.43% de un total de 910 muestras
evaluadas en las tres granjas y un 44.26% de positividad de 130 muestras evaluadas en
la ceba, lo cual, con respecto a los resultados obtenidos en los años 2000 y 2006 (Cruz
et al., 2006; Mogollón et al., 2003), evidencia un aumento en los porcentajes de
positividad, especialmente dentro de este grupo etáreo. Lo anterior indica inicialmente
que hay una mayor actividad del PRRSV en la zona, posiblemente asociado al
crecimiento de la industria porcina intensiva como extensiva
(http://asoporcicultores.co/porcicultores/), que conllevó a la presencia de un mayor
número de granjas colindantes, lo que incrementa las posibilidades de trasmisión del
virus. Igualmente, debe tenerse en cuenta que no existen protocolos de bioseguridad
unificados entre granjas, como también que hay una mayor movilización de animales.
Las diferencias encontradas entre granjas podrían estar relacionadas con varios factores,
entre ellos los tipos de manejo productivo en cada una de ellas; en Colombia la
producción se hace mayoritariamente en flujo continuo y no bajo el concepto de todo

dentro todo fuera. Así, las granjas 1 y 3, que presentaron la mayor positividad tienen el
sitio 1 (madres, lactantes, destetos, reemplazos y reproductores) y el sitio 2 (preceba) en
un solo lugar, mientras que el sitio 3 (ceba) se encuentra en otra ubicación geográfica, en
este caso a 175 Km y 70 Km para las granjas 1 y 3 respectivamente (Figura 2-1). Por el
contrario, la granja 2 presenta los tres sitios en un solo lugar y fue negativa durante todo
el estudio. Esto puede llevar a plantear que la movilización de los animales entre sitios
puede ser un factor que posibilita la transmisión indirecta del PRRSV. Este tipo de
transmisión ha sido controversial, sin embargo datos recolectados durante un brote en
Inglaterra evidenciaron que el virus se puede diseminar por medio de aerosoles en un
radio de hasta 3 Km. (Edwards et al., 1992). Información obtenida de un estudio
epidemiológico a gran escala en Dinamarca, sugirió a los aerosoles como una potencial
ruta de transmisión indirecta dentro de regiones productoras de porcinos (Mortensen et
al., 2002). De otro lado, también se plantea que las movilizaciones por largos trayectos a
temperaturas muy elevadas, como es el caso de estas granjas, pueden estar
relacionadas con cuadros de estrés, lo que puede generar una inmunosupresión que
favorecería la infección por diferentes patógenos, entre ellos PRRSV.
El proceso de transporte, per se, ha sido considerado como un factor de riesgo
importante para la entrada de patógenos en granjas porcícolas. Esto debido al probable
contacto de porcinos susceptibles con virus como el de PRRS, al interior de camiones
contaminados (Dee et al., 2004; 2005). De acuerdo con Poumian (1995), todos los
camiones, trailers y otros vehículos usados para transportar animales, productos
animales, productos alimenticios o equipos contaminados, son un riesgo potencial en la
diseminación de enfermedades.
En lo referente a factores intrínsecos en una granja seropositiva, es posible encontrar
otras fuentes de transmisión, como fómites, insectos y otras especies animales (Cho and
Dee, 2006). Igualmente, se ha demostrado que el uso de agujas o instrumental
contaminado con fluidos orgánicos puede transmitir el PRRSV (Otake et al., 2002a,
2002b). Con respecto a los insectos, en algunos estudios se ha evaluado el papel del
mosquito Aedes vexans en la transmisión del virus, para esto, mosquitos capturados a
partir de planteles de cerdos infectados y puestos en contacto con cerdos centinelas,
demostraron dicha transmisión (Otake et al., 2002c). Por otra parte, Shurrer et al. (2004)

revelaron que las moscas pueden contaminarse con el virus y transportarlo a una
distancia de 1.7 km. Se puede inferir por lo tanto, que los insectos pueden convertirse en
posibles agentes diseminadores del virus entre planteles porcinos bajo condiciones de
campo. Con respecto a otras especies animales (perros, ratones, gatos) no se ha
demostrado su capacidad como vectores mecánicos o biológicos del virus de PRRS
(Wills et al, 2000). En Colombia, no se han realizado estudios sobre estos factores que
pueden ser determinantes para la diseminación y mantenimiento del PRRSV,
particularmente por la condición tropical del país donde la diversidad de insectos y
roedores es muy amplia.
Con respecto al comportamiento de los anticuerpos en la granja 1, se observó
seropositividad en el sitio 3, mientras que el grupo de la preceba fue seronegativo
durante todo el estudio. Se podría inferir, que los animales del sitio 2 se infectaron
durante el transporte o directamente en el sitio 3. Sin embargo, al correlacionar los
resultados de serología con los de qRT-PCR (datos presentados en el siguiente capítulo),
se pudo establecer la presencia del genoma del PRRSV en el sitio 2 más no en el sitio 3;
esto indica que los animales ya se encontraban infectados y seroconvirtieron en el sitio 3.
De acuerdo con lo anterior, la serología permitió establecer la circulación viral pero no fue
eficiente para determinar el momento de la infección, lo que nos lleva a proponer que el
diagnóstico de PRRSV debe soportarse en la serología como también en la detección del
agente viral, esto con el fin de hacer un análisis más preciso de la dinámica del virus y de
la respuesta inmune, conducente a tomar las medidas de control adecuadas.
Como se observa en la Figura 2-2, el grupo de la ceba en la granja 1, presentó
seropositividad en los primeros cuatro meses (septiembre a diciembre), descendiendo a
zona de seronegatividad a partir del mes de enero y marzo, ascendiendo hacia el final del
estudio. Aquí es importante tener en cuenta que en Colombia hacia el final de año se
incrementa significativamente el consumo y comercialización de carne de cerdo, lo que
se traduce en una gran salida de animales de la granja, esto puede explicar el hecho de
la reducción en el número de seropositivos en el mes de enero. Una vez se reactivó la
producción en la ceba, se volvió a presentar un M/P mayor a 0.4 en abril y mayo, lo que
lleva a pensar que la infección continuo a partir del sitio 2 y se reactivó en el sitio 3 al ser
incrementada la población. Este nuevo episodio de seropositividad igualmente pudo estar

relacionado con la infección con una cepa heteróloga posiblemente por ingreso de
animales. Esto último puede sustentarse en el hecho de que este virus induce una
inmunidad protectiva efectiva contra la re exposición a la misma cepa viral (Murtaugh et
al., 2002).
De otro lado, teniendo en cuenta el kit de ELISA empleado en este estudio sobre el cual
se ha reportado que detecta anticuerpos principalmente contra la proteína N y de
acuerdo con Nelson et al (1994) quienes estudiaron la cinética de la respuesta humoral
del cerdo frente al genotipo americano del PRRSV, donde se determinó que los
anticuerpos tempranos fueron dirigidos contra la proteína N (15 KDa), se puede sugerir
que los anticuerpos detectados en este estudio, fueron indicativo del comienzo de una
infección por PRRSV en la granja. Adicionalmente, en un estudio llevado a cabo en la
Universidad de Hannover, donde se evalúo la respuesta de anticuerpos seguida a la
vacunación, se compararon nueve kits para diagnóstico de PRRS de los cuales
HerdChek PRRS 2xR y PRRSx3, detectaron la exposición temprana al virus (Strutzberg-
Minder et al. 2012).
De igual manera, el descenso en la seropositividad en la ceba de la granja 1 durante el
tiempo, pudo obedecer a que los anticuerpos circulantes en este momento no fueron
detectados por la técnica de ELISA empleada, ya que no correspondían a una respuesta
a una infección temprana por el PRRSV. De otro lado, esta seropositividad pudo
igualmente corresponder a una infección pasada si se tiene en cuenta el tiempo de
persistencia de los anticuerpos antiPRRSV. Estos son detectables por ELISA entre los 9
a 13 días PI (Yoon et al., 1995, citado por Evans et al., 2008) y decaen en el tiempo
pudiendo persistir hasta por 28 meses (Desrosiers and Boutin, 2002). También se sabe
que muchos cerdos liberan el virus durante 3 a 4 meses pos exposición (Wills et al.,
2003) así, los animales seropositivos al PRRSV, pueden ser negativos a la presencia del
virus y en consecuencia la seropositividad es indicativa de una infección pasada (Evans
et al., 2008).
Con relación a los resultados serológicos en la granja 2 (Tabla 2-3), donde se observó
seropositividad únicamente en una muestra en la ceba durante los 9 meses de estudio,
se puede especular en primer lugar que dicho resultado (0.64) correspondió a un falso

positivo ya que ocurrió al inicio del estudio, mientras que durante el resto del muestreo, la
granja y este grupo etáreo en particular fue negativo. De otro lado, las prácticas de
manejo en esta granja, donde no se realizó movilización de animales de un sitio a otro; la
aplicación de medidas de bioseguridad con respecto al control de insectos y roedores; el
manejo de las hembras de reemplazo, las cuales fueron monitoreadas y comprobadas
como seronegativas al entrar a la granja y durante el periodo de cuarentena, llevaron a
identificar a esta granja como negativa dentro de este estudio.
Con respecto a la granja 3, se encontró seropositividad en todos los grupos etáreos
(Tabla 2-4). La presencia de anticuerpos en el grupo de animales jóvenes de la granja,
reveló la presencia del PRRSV y una transmisión activa del virus. En el grupo de las
madres, los reproductores, los reemplazos y ceba, la seropositividad pudo estar
relacionada, ya sea con la introducción viral, generalmente por el ingreso de animales de
reemplazo a la granja o a la presencia de animales persistentemente infectados dentro
de la misma (Evans et al., 2008).
La introducción viral pudo originarse por la entrada de hembras de reemplazo que
presentaron previa exposición al virus e ingresaron a la granja transmitiendo el agente a
los animales susceptibles; de otro lado, que siendo seronegativas se infectaron con la
cepa viral circulante en la granja ya sea por contacto directo con los porcinos de la granja
o por medio de la exposición con muestras (placentas) procedentes de animales
positivos como parte de las prácticas de infección que se realizan para el control de la
enfermedad.
Es posible que en la granja 3 se encontraran animales persistentemente infectados,
entendiéndose la persistencia como la presencia del virus infectivo durante periodos
prolongados en determinados órganos como tonsilas y nódulos linfáticos a partir de los
cuales continua siendo eliminado en niveles bajos, presentando una posterior
disminución durante el transcurso del tiempo (Allende et al., 2000). Se ha detectado virus
en tonsilas y nódulos linfoides a los 132 días después del nacimiento en cerdos
infectados in útero (Rowland et al., 2003) y desde los 105 a 157 días PI en cerdos
jóvenes (Horter et al., 2002). A su vez, Evans et al (2008), concluyeron que la
persistencia está asociada con granjas de gran tamaño, localizadas en regiones con alta

densidad en cerdos y una continua introducción de lotes de animales infectados. La
granja 3 es considerada una granja intensiva con más de 7000 animales, ubicada en una
zona de alta producción porcícola del país, además de acuerdo con los resultados por
qRT-PCR (datos presentados en el siguiente capítulo) se obtuvieron muestras positivas
en ganglios linfáticos, órgano donde se produce la persistencia del virus, lo cual podría
relacionarse de igual manera con la presencia de animales con esta condición,
ocasionada por el PRRSV.
Con relación al grupo de las madres, reportes de campo sugieren que luego de una falla
reproductiva inducida por PRRSV, las hembras desarrollan una inmunidad protectiva,
esto se basa en que hembras afectadas tienen camadas normales a pesar de la
circulación del virus dentro de la granja (Stevenson et al., 1993, citado por Karniychuk
and Nauwynck, 2013). Las hembras seropositivas pueden transmitir anticuerpos
maternos a sus camadas vía calostro, sin embargo la inmunidad pasiva declina y puede
llevar a infección luego del destete cuando los animales son mezclados (OIE, 2008), Esto
puede explicar los resultados positivos obtenidos dentro del grupo de los destetos en el
presente estudio.
De igual manera, no se descarta la posibilidad de transmisión del PRRSV por medio de
semen contaminado o por medio de la monta directa con machos de las granjas, que
igualmente fueron seropositivos (Christopher-Hennings et al., 1996).
En general, en el grupo de las madres, lactantes y destetos (Figuras 2-3, 2-4, 2-5) en la
granja 3, se observó un comportamiento de anticuerpos similar durante los meses de
estudio. Básicamente en el mes de enero comenzaron a descender los valores de M/P
manteniéndose por debajo del punto de corte 0.40 hasta el mes de mayo. Esta reducción
de los niveles de anticuerpos detectables por ELISA, puede indicar que la infección
avanzo a la fase crónica de la enfermedad caracterizada por disminución en los niveles
de anticuerpos; de otro lado, el estudio de Evans et al. (2008) indico que cerdas de varios
partos, que habían sido seropositivas debido a una pasada transmisión viral, presentaron
posteriormente una disminución gradual del virus y por lo tanto de dicha seropositividad.

En el grupo de los reproductores y preceba (Figura 2-6, 2-7), se observó de igual manera
un descenso en el nivel de anticuerpos para los meses de Febrero y Enero
respectivamente, sin embargo se vuelve a presentar seropositividad en los meses
siguientes, lo cual pudo estar asociado a reinfección con una cepa viral diferente o con la
misma cepa circulante en granja si se tienen en cuenta los valores de M/P cercanos al
punto de corte.
Para el caso de los reemplazos y ceba (Figura 2-8, 2-9), se presentó seropositividad
durante ocho de los nueve meses del estudio. La presencia de anticuerpos en hembras
de cría se relaciona con previa exposición al virus e introducción en una granja; a la
exposición con la cepa circulante en la granja y a su vez al contacto con muestras de
animales positivos a PRRSV como parte de las técnicas de manejo de reemplazos. Con
respecto a los resultados en la ceba, los cerdos del sitio 2 pudieron infectarse en el sitio 3
o estar previamente infectados y durante el traslado o la posible presentación de un
cuadro de estrés, seroconvertir en el sitio 3, presentando estos resultados seropositivos.
A pesar de la seropositividad observada durante la mayor parte de tiempo del estudio,
todos los grupos presentaron seronegatividad para el mes de Mayo cuando culmino el
muestreo, lo cual pudo obedecer a la aplicación de medidas de bioseguridad y manejo
adecuadas.
En general, aun cuando las granjas se encontraban localizadas en una misma zona
productiva y geográfica, mediante el presente estudio se observó un comportamiento
serológico diferente dentro de cada una, lo cual podría estar relacionado con las
condiciones de bioseguridad y/o manejo particulares. De igual manera, cada grupo
etáreo exhibió una respuesta específica, donde el grupo de la ceba presentó los valores
de positividad más significativos, debido posiblemente a las diferencias en manejo y
movilidad de los animales hacia los sitios correspondientes. Los hallazgos de este
estudio igualmente, concordaron con aquellos encontrados por Salinas et al., (2008)
quienes evaluaron cinco granjas (Nuevo León-México) en diferentes etapas de
producción (destete, iniciación, finalización, gestantes, lactantes y monta-servicio), donde
encontraron diferencias en cuanto a la seroprevalencia entre granjas (6.7% a 45%), lo
cual se relacionó básicamente con factores de manejo y bioseguridad. La mayor

prevalencia de seropositivos al PRRSV se encontró en el área de producción en
comparación con el área de reproducción. La etapa con más alta seroprevalencia
correspondió a la finalización con 56%, seguida del inicio con 36%.
Lo anterior, confirma las diferencias existentes en cuanto a actividad viral, no solo dentro
de las granjas sino entre las mismas y dentro de grupos etáreos, por lo anterior y
teniendo en cuenta la variabilidad genética del virus, es conveniente revisar el
comportamiento del PRRSV en forma particular para cada granja.
2.4 Conclusiones
Se confirman la circulación del PRRSV en explotaciones porcícolas de tipo intensivo en
el país. Se observó un incremento en la actividad del virus tanto en la zona del Valle del
Cauca, como en los diferentes grupos etáreos evaluados, principalmente en la ceba.
2.5 Recomendaciones
El estudio demostró que el comportamiento del virus de PRRS en Colombia es particular
para cada granja, por tal razón, es importante revisar las medidas de bioseguridad, el
impacto de las movilizaciones a grandes distancias entre los sitios productivos de los
animales y realizar junto con estudios serológico pruebas moleculares con el fin de
determinar la dinámica el PRRSV dentro de cada granja.

Bibliografía
Allende, R., Lewis, T. L., Lu, Z., Rock, D.L., Kutish, G.F., Ali, A., Doster, AR., Osorio FA.
1999. North American and European porcine reproductive and respiratory syndrome
viruses differ in non-structural protein coding regions. J Gen Virol. 80, 307-15.
Allende, R., Laegreid, W.W., Kutish, G.F., Galeota, J. A.,Wills, R.W and Osorio, F.A.
2003. Porcine reproductive and respiratory syndrome virus: description of persistence in
individual pigs upon experimental infection. J Virol; 72: 10834-7.
Ansari, I.H., Kwon, B., Osorio, F.A., Pattnaik, A.K., 2006. Influence of N-linked
glycosylation of porcine reproductive and respiratory syndrome virus GP5 on virus
infectivity, antigenicity and ability to induce neutralizing antibodies. J Virol. 80: 3994-4004.
Cancel-Tirado, S.M., Evans, R.B., Yoon, K. 2004. Monoclonal antibody analysis of
porcine reproductive and respiratory syndrome virus epitopes associated with antibody-
dependent enhancement and neutralization of virus infection. Vet. Immunol.
Immunopathol. 102: 249-262.
Collins, J.E., Benfield, D.A., Chritianson, W.T., Harris, L., Hennings, J.C., Shaw, D.P.,
Goyal, S.M., NcCullough, S., Morrison, R.B, Joo, H.S. 1992. Islotation of swine infertility
and respiratory syndrome virus (isolate ARCC VR-2332) in North America and
experimental reproduction of the disease in gnotobiotic pigs. J Vet Diagn Invest. Apr
4(2):117-126.
Christopher-Hennings, J., Nelson, E.A., Benfield, D.A. 1996. Detecting PRRSV in boar
semen. Swine Health and Production, 4: 37-39.
Cho, J.G., Dee, S.A. 2006. Porcine reproductive and respiratory syndrome virus.
Theriogenology, 66: 655-662.

Cruz, M.C., Mogollón, J.D., Rincón, M.A., Peña, N.B., Ruiz, S., Lora, A.M., 2006.
Prevalencia serológica del síndrome reproductivo y respiratorio porcino (PRRS) en
cerdos de explotaciones extensivas de Colombia. Rev. Med. Vet. Zoot. 53: 33-41.
Darwich, L., Diaz, I., Mateu, E. 2010. Certainties, doubts and hypotheses in porcine
reproductive and respiratory virus immunobiology. Virus Research, 154: 123-132.
De Lima, M., Pattnaik, A.K., Flores, E.F., Osorio, F.A., 2006. Serologic marker candidates
identified among B-cell linear epitopes of Nsp2 and structural proteins of a North
American strain of porcine reproductive and respiratory syndrome virus. Virology, 30:410-
421.
Dee, S. A., Deen, J., Otake, S., Pijoan, C. 2004.An assessment of transport vehicles as a
source of porcine reproductive and respiratory syndrome virus transmission to susceptible
pigs. Can J Vet Res. 68:124-133.
Dee, S.A., Deen, J., Burns, D., Douthiht, G., Pijoan, C. 2005. An evaluation of
desinfectants of the sanitation of porcine reproductive and respiratory virus contaminated
transport vehicles at cold temtperatures. Can. J. Vet. Res. 69: 64-71-0.
Desrosiers, R., Boutin, M. 2002. An attempt to eradicate porcine reproductive and
respiratory syndrome virus (PRRSV) after an outbreak in a breeding herd: Eradication
strategy and persistence of antobody titers in sows. Journal of SwineHealth and
Production, 10 : 23-25.
Diaz, I., Darwich, L., Papaterra, G., Pujols, J., Mateu, E., 2005. Inmune responses of pigs
after experimental infection with European strain of porcine reproductive and respiratory
syndrome virus. Journal of General Virology, 86:1943-1951.
Edwards, S., Robertson, I.B., Wilesmith, J. W. 1992. PRRS blue-eared pig disease in
Great Britain. Am. Assoc. Swine. Pract. News, 4:32-6.

Evans, Ch. M., Medley, G, F., Green, L.E. 2008. Porcine reproductive and respiratory
syndrome virus (PRRSV) in GB pig herds: farm characteristics associated with
heterogeneity in seroprevalence. BMC Veterinary Research, 4: 48.
Faaberg, K.S., Balasuriya, U.B.R., Brinton, M.A., et al., 2011. “Family Arteriviridae”, in
Virus Taxonomy, ninth report of the International Committee on Taxonomy of Viruses,
A.M.Q King, Ed., Elsevier Academic Press, London, UK.
Gonin, P., Pirzadeh, B., Gagnon, C.A., Dea, S., 1999. Seroneutralization of porcine
reproductive and respiratory syndrome virus correlates with antibody response to the GP5
major envelope glycoprotein. Journal of Veterinary Diagnostic Investigation, 11: 20-26.
Holtkamp, D.J., Kliebenstein, J.B., Neumann, E.J., 2013. Assessment of the economic
impact of porcine reproductive and respiratory syndrome virus on United States pork
producers. Journal of Swine Health and Production, 21: 72-84.
Karniychuk, U.U and Nauwynck, H.J. 2013. Pathogenesis and prevention of placental and
transplacental porcine Reproductive and respiratory syndrome virus infection.
VeterinaryResearch, 44:95.
Kim, O., Sun, Y., Lai, F.W., Song, C. and Yoo, D. 2010. “Modulationof type I interferon
induction by porcine reproductive andrespiratory syndrome virus and degradation of
CREB-bindingprotein by non-structural protein 1 in MARC-145 and HeLacells,” Virology,
402: 315–326.
Lee, S.M., Kleiboeker, S.B. 2005. Porcine arterivirus activates the NF-kappaB pathway
through IkappaB degradation. Virology, 342: 47–59.
Mattheu, E., Tello, M., Coll, A., Casal, J., Martin, M., 2006. Comparison of three ELISAs
for the diagnosis of porcine reproductive and respiratory syndrome. Vet Rec. 159: 717-
718.

Mateu, E., Diaz, I., 2008. The challenge of PRRS inmunology. The veterinary Journal,
117: 345-351.
Meier, W.A., Galeota, A., J., Osorio, F.A., Husmann, R.J., Schnitzlein, W.M and
Zuckerman, F.A. 2003. Gradual development of the interferon-gamma response of swine
to porcine reproductive and respiratory syndrome virus infection or vacunation.Virology,
309: 18-31.
Mogollón, J.D., Rincón, M.A., Arbeláez, G., Ruiz, S., Rodríguez, V. 2003. PRRS in
Colombia.PRSS Compendium Producer Edition.National Pork Board.Des Moines, Iowa,
USA, section 6.6.
Mortensen, S., Sogaard, R., Boklund, A., Stark, K., D., Christensen, J, et al. 2002. Risk
factors for infection of sow herds with porcine reproductive and respiratory syndrome
(PRRS) virus. Prev. Vet. Med. 53: 83-101.
Murtaugh, M.P., Xiao, Z and Zuckermann, F. 2002. Inmunological responses of swine to
porcine reproductive and respiratory syndrome virus infection. Viral Inmunol. 15: 533-547.
Nelsen, C. J., Murtaugh, M.P., Faaberg, KS. 1999 Porcine reproductive and respiratory
syndrome virus comparison: divergent evolution on two continents. J Virol. 73: 270-80.
Nelson, E.A., Christopher-Hennings, J., Benfield, D.A., 1994. Serum immune responses
to the proteins of porcine reproductive and respiratory syndrome (PRRS) virus. Journal of
Veterinary Diagnostic Investigation, 6:410-415.
Oleksiewicz, M.B., Botner, A., Toft, P., Normann, P.,Storgaard, t., 2001. Epitope mapping
porcine reproductive and respiratory syndrome virus by phage display: the nsp2 fragment
of the replicase polyprotein contains a cluster of B-cells epitopes. J. Virol. 75, 3277-3290.
Ostrowski, M.J., Galeota, J.A., Jar, A.M., Platt, K.B., Osorio, F.A and Lopez, O, J. 2002.
Identification of neutralizing and nonnutralizing epitopes in the porcine reproductive and
respiratory syndrome virus GP5 ectodomain. J. Virol. 76: 4241-4250.

Otake, S., Dee, S. A., Rossow, K. D., Deen, J., Joo, H. S., Molitor, T. W. 2002a.
Transmission of porcine reproductive and respiratory syndrome virus by fomites (bootas
and coveralls). Swine Health Prod. 10:59-65.
Otake, S., Dee, S.A.,Rossow, K.D. 2002b. Transmission of porcine reproductive and
respiratory syndrome virus by needles. Vet. Rec., 150: 114-115.
Otake, S., Dee, S.A., Rossow, K.D, Moon, R.D., Pijoan, C. 2002c. Mechanical
transmission of porcine reproductive and respiratory syndrome virus by mosquitoes
Aedesvexans. Can. J. Vet. Res. 66: 191-195.
Patel, D., Nan, Y., Shen, M., Ritthipichai, K., Zhu, X., Zhang, Y.J. 2010. Porcine
reproductive andrespiratory syndrome virus inhibits type I interferon signaling by blocking
STAT1/STAT2 nuclear translocation. J. Virol. 84; 11045–11055.
Pestka, S. 2007. The interferons: 50 years after their Discovery, there is much more to
learn, The Journal of Biological Chemistry, 282: 20047-20051.
Poumian, M. 1995. Disinfection of trucks and trailers. Revue Scientifique et Teccnique de
I’OIE, 19:165-170 et 171-176.
PRRS: the disease, its diagnosis, prevention and control. Appendices IV and V of the
report of the OIE ad hoc group on porcine reproductive respiratory syndrome Paris, 9 - 11
June 2008.
Revista Porcicultura Colombiana. Asociación Colombiana de Porcicultores. En línea.
Edición 156 de 2011, Disponible en: (http://asoporcicultores.co/porcicultores/)
Salinas, J.A., Lara, J., Flores, H., Avalos, R., Zarate, J.J., Riojas, V., Segura, J.C.2008.
Presencia de Animales seropositivos al Síndrome Reproductivo y Respiratorio Porcino en
Nuevo León. Vet. Mex. 39: 215-221.

Sang, Y., Ross, C.R., Rowland, R.R., Blecha, F. 2008. Toll-like receptor 3 activation
decreases porcine arterivirus infection. Viral. Immunol. 21:303–313.
Shi, X., Wang, L., Li, X., et al. 2011. “Endoribonuclease activities of porcine reproductive
and respiratory syndrome virus nsp11was essential for nsp11 to inhibit IFN-β induction,”
Molecular Immunology, 48: 1568–1572.
Snijder, E.J, Meulenberg, J.J. 1998. The molecular biology of arteriviruses. J Gen Virol.
69: 971-979.
Snijder, E.J., Kikkert, M., Fang, Y. 2013. “Arterivirus molecular biology and pathogenesis”
Journal of General Virology, 94: 2141-2163.
Stevenson, G.W., Van Alstine, W.G., Kanitz, C.L., Keffaber, K.K. 1993. Endemic porcine
reproductive and respiratory syndrome virus infection of nursery pigs in two swine herds
without current reproductive failure. J Vet Diagn Invest. 5:432–434.
Sun, Y., Han, M., Kim, Ch., Calvert, J. G., Yoo, D. 2012 a. Interplay between Interferon-
Mediated Innate Immunity and Porcine Reproductive and Respiratory Syndrome Virus.
Viruses, 4, 424-446.
Sun, Z., Ransburgh, R., Snijder, E.J., and Fang, Y. 2012 b. “Nonstructuralprotein 2 of
porcine reproductive and respiratory syndromevirus inhibits the antiviral function of
interferon-stimulatedgene 15,” Journal of Virology, 86; 3839–3850.
Yoon, I.J., Joo, H.S., Goyal, S.M., Molitor, T.W., 1994. A modified serum neutralization
test for the detection of antibody to porcine reproductive and respiratory syndrome virus in
swine sera. Journal of Veterinary Diagnostic Investigating, 6:289-292.
Yoon, K,J., Zimmerman, J.J., Swenson, S.L., McGinley, M.J., Eeernisse, K.A., Brekiv, A.,
Rhinehart, L.L., Frey, M.L., Hill, H.T., Platt, K.B., 1995. Characterization of the humoral
response to porcine reproductive and respiratory syndrome (PRRS) virus infection.

Journal of Veterinary diagnostic investigation: official publication of the American
Association of Veterinary Laboratory Diagnosticians, Inc. 7:305-312.
Yoon KJ, Zimmerman JJ, Wu LL, Platt KB. 1996. Antibody-dependent enhancement
(ADE) of porcine reproductive and respiratory syndrome virus (PRRS) infection in pigs.
VirolInmunol 9:51-63.
Yoon, K., Wu, L., Zimmerman, JJ. Platt, KB. 1997. Field isolates of porcine reproductive
and respiratory syndrome virus (PRRSV) vary in their susceptibility to antibody dependent
enhancement (ADE) of infection. Vet. Microb. 55, 277-287.
Vezina, S.A., Loemba, H., Fournier, M., Dea, S., Archambault, D. 1996. Antibody
production and blastogenic response in pigs experimentally infected with porcine
reproductive and respiratory syndrome virus. Canadian Journal of Veterinary Research,
60: 94-99.
Wellenberg, G.J., 2006. Review: diagnostic methods for the detection of porcine
reproductive and respiratory syndrome virus (PRRSV) infections. Tijdschr. Diergeneeskd,
131 (16): 566-572.
Wensvoort, G., Terpstra, D., Pol, J.M.A., TerLaak, E.A., Bloemraad, M., De Kluyver, E.P.,
Kragten, D., Van Buiten, L., Den Besten, A., Wagenaar, F., Broekhuijsen, J.M., Moonen,
P.L.J.M., Zetstra, T., De Boer, E.A., Tibben, H.J., De Jong, M.F., Van´t Veld, P.,
Groenland, G.J.R., Van Gennep, J.A., Voets, M.T., Verheijden, J.H.M., Braamskamp, J.
1991. Mystery swine disease in the Netherlands: the isolation of Lelystad virus. Vet. Q.
13: 121-130.
Wills, R., Osorio, F.A., Doster, A. 2000. Susceptibility of selected non-swine species to
infection with porcine reproductive and respiratory syndrome virus. In: Proceedings of the
31st Ann Meet Am Assoc Swine Pract. 411-413.

Wills, R.W., Doster, A.R., Galeota, J.A., Sur, J.H., Osorio, F.A., 2003. Duration of infection
and proportion of pigs persistently infected with porcine reproductive and respiratory
syndrome virus. J Clin Microbiology, 41: 58-62.
Zimmerman, J., Sanderson, T., Eernisse, K.,Hill, H., Frey, M. 1992. Transmission of SIRS
virus in convalescent animals to commingled penmates under experimental conditions.
Am. Assoc. Swine Pract.Newslett. 4, 25.

Determinación mediante la técnica de RT-
PCR en tiempo real (qRT-PCR) de la
presencia del virus del Síndrome
Reproductivo y Respiratorio Porcino (PRRSV)
en tres granjas del Valle del Cauca
Resumen El Síndrome Reproductivo y Respiratorio Porcino (PRRS), ocasiona un alto impacto
económico en las explotaciones porcícolas a nivel mundial, para su control se requiere de
la implementación de técnicas diagnósticas que permitan la detección temprana del
agente viral dentro de una explotación. El objetivo de este estudio fue detectar mediante
la técnica de PCR tiempo real (qRT-PCR) la presencia del ARN del PRRSV en muestras
de suero, tejidos, fluido oral y lavado bronco alveolar de porcinos proveniente de tres
granjas intensivas del Departamento del Valle. Los resultados permitieron determinar la
presencia de actividad viral en las tres granjas, con mayor carga viral en aquellas que
realizaban desplazamientos entre sitios productivos y dentro de los grupos de la preceba
y ceba. Los diferentes tipos de muestra evaluadas y el uso de la técnica de qRT-PCR,
evidenciaron diferentes estados de la infección por el PRRSV en cada una de las
granjas, lo que indica que aún en la misma región o zona, el comportamiento de la
enfermedad es particular para cada una, por lo tanto, el control y medidas de manejo
deben ser aplicados de igual forma.
Palabras clave: ARN, Diagnóstico, PCR, PRRS.

Abstract
Porcine reproductive and respiratory syndrome (PRRS), causes a high economic losses
on pig farms around the world, its control requires the implementation of diagnostic
techniques that allow early viral detection within farms. The objective of this study was to
detect PRRSV genome through the PCR technique using real-time (qRT-PCR,). Viral
RNA was measured in samples of serum, tissues, oral fluids and broncho alveolar lavage
(BAL) coming from three intensive productión farms from the Valle del Cauca province.
The results allowed the presence of viral genome whitin the three farms with higher viral
loads, especially on those who travel between production sites and within the pre-
fattening and fattening groups. The qRT-PCR technique on different kind of samples,
showed different stages of infection by PRRSV in each one of the farms, which indicates
that even on the same region or zone, the behavior of the disease is particular to each
farm and therefore control and management measures must be applied in the same way.
Keywords: Diagnostic, PCR, PRRS, RNA.

Introducción El Síndrome Reproductivo y Respiratorio Porcino (PRRS) es actualmente una de las
enfermedades con mayor impacto económico que afecta a la industria porcina alrededor
del mundo (Neumann et al., 2005). Esta enfermedad que emergió casi simultáneamente
entre los años 1980 y 1990 en Norteamérica y Europa, se caracteriza por ocasionar
enfermedad respiratoria en lechones y fallas reproductivas en cerdas, estos últimos
síntomas incluyen abortos, muerte fetal y nacimiento de lechones débiles (Benfield et al.,
1992; Wensvoort et al., 1991).
El PRRSV, se divide principalmente en genotipo I o tipo Europeo y genotipo II o tipo
Norteamericano. Estos comparten un 60% de identidad entre nucleótidos (Allende et al.,
1999; Nelsen et al., 1999). Las cepas representativas de cada uno de los genotipos,
Lelystad (LV) y VR-2332, respectivamente (Benfield et al., 1992; Wensvoort et al., 1991)
difieren aproximadamente en 40% a nivel genómico (Meulenberg et al., 1993; Nelson et
al., 1993; Nelsen et al., 1999). De igual manera, los aislados dentro de cada genotipo
pueden variar aproximadamente en el 20% a nivel de nucleótidos (Meng, 2000; Nelsen et
al., 1999).
El genoma del PRRSV es ARN de cadena sencilla y polaridad positiva, con una longitud
promedio de 15 kb. Pertenece a la familia Arteriviridae, género Arterivirus. Dentro de esta
familia también se encuentran el virus de la Arteritis Equina (EAV), el virus Elevador de la
Lactato deshidrogenasa del ratón (LDV) y el virus de la Fiebre Hemorrágica del Simio
(SHFV) (Cavanagh, 1997). El genoma viral consta de una región UTR- 5´ no traducida,
10 marcos de lectura abierta (ORFs) y una región UTR- 3´ (Britton and Cavanagh, 2008;
Johnson et al., 2011). Los ORFs 1a y 1b constituyen aproximadamente el 75% del
genoma y codifican para dos poli proteínas, las replicasas pp1a y pp1b. Estas a su vez,
son clivadas en 14 proteínas no estructurales (NSPs) que están involucradas en la
replicación y en la transcripción del ARNm subgenómico viral, además presentan
actividad replicasa, proteasa y polimerasa. Las GP2a, GP2b, GP3 y GP4, son proteínas
menores de la envoltura, son N-glicosiladas y forman heterotrímeros mediante uniones
disulfuro (Wissink et al., 2005). Las proteínas GP5 y M, proteínas mayores de la cubierta,
forman heterodímeros mediante uniones disulfuro a nivel del retículo endoplasmático

(ER). La proteína de la nucleocápside (N) es la más abundante, corresponde al
componente inmunogénico del virión y existe como dímero. Recientemente fue
descubierto un polipéptido de 51 aminoácidos el cual es codificado por un ORF
alternativo del ARNm5, el cual precede al sitio de iniciación de la proteína mayor de la
cubierta, GP5 (Johnson et al., 2011). El ORF5a, está presente en una amplia variedad de
aislados del genotipo I y II del PRRSV y su función está relacionada con la replicación del
virus. De otro lado, se ha evidenciado la expresión de bajos niveles de anticuerpos anti-
proteína ORF5a posterior a la infección de cerdos con PRRSV y no se descarta que ésta
induzca una respuesta inmune mediada predominantemente por células (Jeong et al.,
2010).
El ORF6 es altamente conservado entre los aislados norteamericanos con un 100% de
identidad entre aminoácidos y a su vez es el más conservado entre aislados americanos
y europeos con 70-81% de identidad (Allende et al., 1999; Murtaugh et al., 1995). El
ORF7 es igualmente conservado entre aislados Norteamericanos con 95-100% de
homología y el 57-59% de identidad de secuencia de aminoácidos con el virus Lelystad
(Allende et al., 1999; Meng et al., 1995a, 1995b; Murtaugh et al., 1995). Por lo anterior, el
ORF7 es un blanco robusto para diagnosticar PRRSV a través de la amplificación por
RT-PCR y para diferenciar entre los genotipos americano y europeo (Le Gall et al., 1998;
Suárez et al., 1996, citados por Martínez et al., 2008).
La alta tasa de mutación, rápida evolución y variabilidad genética de las cepas de
PRRSV, han dificultado el desarrollo de técnicas diagnósticas seguras a largo plazo y
como consecuencia de esto, resultados falsos negativos han sido reportados mediante
las pruebas disponibles comercialmente (Harmon et al., 2012; Wernike et al., 2012). Por
tal razón, desde 1995 varios métodos de RT-PCR convencional han sido desarrollados
para la detección rápida y específica de PRRSV en cerdos, no obstante, estos requieren
manipulación post PCR ya que el análisis y visualización de los productos se verifican
posteriormente mediante la utilización de electroforesis en gel de agarosa, lo que puede
aumentar el riesgo de contaminación.
En contraste, el RT-PCR tiempo real (qRT-PCR), completa la amplificación y el análisis
en un sistema cerrado permitiendo detectar y cuantificar las secuencias específicas de

ácidos nucleicos mediante el uso de reporteros fluorescentes en la reacción. La qRT-
PCR ha sido ampliamente empleada para el diagnóstico temprano de enfermedades
animales de origen viral (Shaw et al., 2007) ya que los datos de la curva melting permiten
verificar los productos de amplificación, disminuyendo así mismo el potencial riesgo de
contaminación y por lo tanto la presentación de resultados falsos positivos (García et al.,
2007; Mackay et al., 2002; Tian et al., 2010).
Un método no específico para la detección de los productos amplificados por qRT-PCR,
incluye el uso de moléculas intercalantes que tienen afinidad por ADN de doble cadena y
que al ser oxidados generan una señal fluorescente; esta fluorescencia emitida es
capturada en la etapa de extensión de cada ciclo y es proporcional al número de copias
de ADN de doble cadena obtenidas en la PCR. El reportero más usado para este fin es el
SYBR Green (Zipper et al., 2004), molécula cargada positivamente que al unirse al surco
menor del ADN incrementa su fluorescencia (Martínez et al., 2008). Este método a su vez
se ha relacionado con la detección de virus ARN que presentan alta variabilidad
genómica y ha permitido diferenciar cepas altamente patógenas de PRRSV (HP-PRRSV)
de cepas clásicas del genotipo II del virus (Chai et al., 2013).
La curva de disociación de la técnica, permite distinguir entre positivos, negativos y
dímeros, corrigiendo el error que comete el SYBR al unirse a todo ADN bicatenario
(Rubio et al., 2007). El SYBR Green qRT-PCR, puede detectar pequeñas variaciones en
la secuencia blanco y esas variaciones pueden indicar la presencia de una nueva cepa
de PRRSV, esto genera una ventaja sobre el PCR convencional basado en gel, con el
cual no se detectan estas diferencias; de igual manera ha demostrado ser un sistema de
detección temprana de PRRSV, ya que ha permitido detectar y tipificar el virus en suero a
los dos días post infección (Martínez et al., 2008).
Las muestras más comunes usadas para monitorear PRRSV incluyen suero, hisopos
sanguíneos, fluidos orales y semen. Estudios han mostrado que el ARN de PRRSV
puede ser detectado en suero, fluido oral e hisopos sanguíneos entre 24 y 48 horas PI y
en muestras de semen entre 48 a 120 horas PI (Kittawornrat et al., 2010).

Colecciones de sueros, son usados regularmente para monitorear el estatus de PRRSV
por RT-PCR, sin embargo, un estudio relacionado demostró un decremento en la
sensibilidad cuando fueron utilizados pooles de tres a cinco muestras de suero y sangre,
especialmente durante los primeros días de infección (Rovira et al., 2007).
Recientemente, el uso de las muestras de fluido oral para vigilancia y diagnóstico se ha
incrementado debido a la facilidad en el método de colección (Ramirez et al., 2012). La
presencia del PRRSV en fluido oral fue reportado por primera vez en 1997, a partir de
hisopados bucales colectados de cerdos jóvenes infectados experimentalmente (Wills et
al., 1997). Otros estudios han revelado tasas de detección del ARN de PRRSV similares,
tanto en suero como fluido oral de animales infectados experimentalmente (Kittawornrat
et al., 2010). Se ha reportado que el fluido oral contiene factores endógenos antivirales y
antibacteriales que pueden teóricamente afectar la estabilidad de la PCR (Shugars,
1999). Sin embargo, de acuerdo con Prickett et al (2010), el ARN del PRRSV es
relativamente resistente a la degradación en fluidos orales, no se requiere el uso de
antimicrobianos para preservar la muestra, es suficiente un adecuado manejo de
almacenamiento y transporte a temperaturas de congelación o refrigeración a 4°C para
mantener la integridad del ARN en la muestra de fluido oral.
En general, las pruebas basadas en PCR son útiles para detectar circulación inmediata
de patógenos, mientras que las pruebas basadas en anticuerpos, brindan información
con respecto a la inmunidad de la granja y a la historia de la infección en una población
(Olsen et al., 2013). Es por lo anterior y teniendo en cuenta las ventajas diagnósticas del
qRT-PCR en la detección de infecciones por el PRRSV, que se realizó el presente
estudio, con el fin determinar la actividad viral en tres granjas intensivas del Valle del
Cauca, utilizando muestras de suero, tejidos (pulmón y ganglios), lavados bronco
alveolares y fluido oral durante un periodo de nueve meses.

3.1 Materiales y Métodos
3.1.1 Ubicación geográfica y características de las granjas
El estudio se realizó en tres granjas de tipo intensivo y ciclo completo, ubicadas en el
departamento del Valle del Cauca, localizado en el sur occidente de Colombia, con una
altura de 1400-1600 msnm, una temperatura promedio de 23-24°C y una humedad
relativa del 65% a 75%. Esta zona se caracteriza por una producción porcícola de tipo
intensivo, que presentó en los meses previos al estudio reportes de animales con
sintomatología compatible con PRRS. En la Tabla 3-1 y Figura 3-1, se indican cómo se
encuentran distribuidas cada una de las granjas, su localización y distancia dentro del
departamento. Cabe aclarar, que en Colombia desde el año 2006 no se vacuna contra
PRRS.
Tabla 3-1 Ubicación geográfica de las tres granjas y de los sitios productivos por
granja
GRANJA SITIO 1 Y 2 SITIO 3 DISTANCIA ENTRE SITIOS
1 Cartago Candelaria 175 km
2 La Unión La Unión 1 km
3 Palmira Candelaria 70 km

Figura 3-1 Ubicación geográfica de cada granja dentro del Departamento del Valle
del Cauca
El color rojo indica la ubicación de los sitios 1-2 (Cartago) y 3 (Candelaria) de la granja 1;
el color azul señala la ubicación de los 3 sitios productivos de la granja 2 (La Unión) y el
color amarillo la localización de los sitios 1-2 (Palmira) y 3 (Candelaria) de la granja 3.
3.1.2 Muestras Se tomaron muestras de sueros, fluidos orales (FO), lavados bronquio alveolares (LBA) y
tejidos (pulmón y ganglios). Los sueros empleados se seleccionaron a partir de aquellos
que por ELISA mostraron los resultados positivos más altos (mayor a 0.4) y negativos
más bajos (menor a 0.4), en cada grupo etareo y en cada granja. Las muestras se
seleccionaron de los grupos de madres, reproductores, reemplazos, lactantes, destetos
(20-21 días), preceba (7-9 semanas) y ceba (12-14 semanas y 18-22 semanas). Los FO
se tomaron de la preceba (semana 4-8) y ceba (semana 10- 14, 18 y 22), en cada una de
las tres granjas. Los LBA y los tejidos se obtuvieron a partir de las mortalidades que se
SITIO 1 - 2
SITIO 1 - 2
SITIO 1 – 2 - 3
SITIO 3
SITIO 3

presentaron en los días de muestreo o a través del sacrificio voluntario en cada granja
(proceso aprobado por el Comité de Bioética de la Facultad de Medicina Veterinaria y de
Zootecnia de la Universidad Nacional de Colombia), lo que correspondió a 2 ó 3 animales
durante cada mes. Las muestras se recolectaron a partir de Septiembre de 2012 hasta
Mayo de 2013.
3.1.2.1 Sueros
La muestra de sangre fue colectada mediante venopunción (vena cava), en tubos
individuales (BD Vacutainer). Las muestras fueron centrifugadas a 1000 x g durante 10
minutos y almacenadas a -70°C hasta su posterior procesamiento. Se evaluaron un total
de 355 muestras, en la tabla 3-2, se discrima el número de sueros evaluados dentro de
cada granja.
Tabla 3-2. Sueros procesados por granja durante el tiempo de estudio
Granja Número sueros
1 112
2 81
3 162
Total 355
3.1.2.2 Fluidos Orales (FO)
Estas muestras se obtuvieron mediante el uso de un lazo de algodón suspendido durante
30 minutos en corrales que alojaban entre 20 a 25 animales. Se recolectó la muestra
presionando el lazo dentro de una bolsa y posteriormente el contenido se trasladó a un
tubo de 15 ml, este se mantuvo en refrigeración hasta su ingreso al laboratorio. Las
muestras de fluido fueron centrifugadas a 1000 x g durante 10 minutos, se alicuotaron y
almacenaron a -70°C hasta su procesamiento. Se evaluaron en total 305 muestras, en la
tabla 3-3, se especifica el número de FO procesados por cada granja.
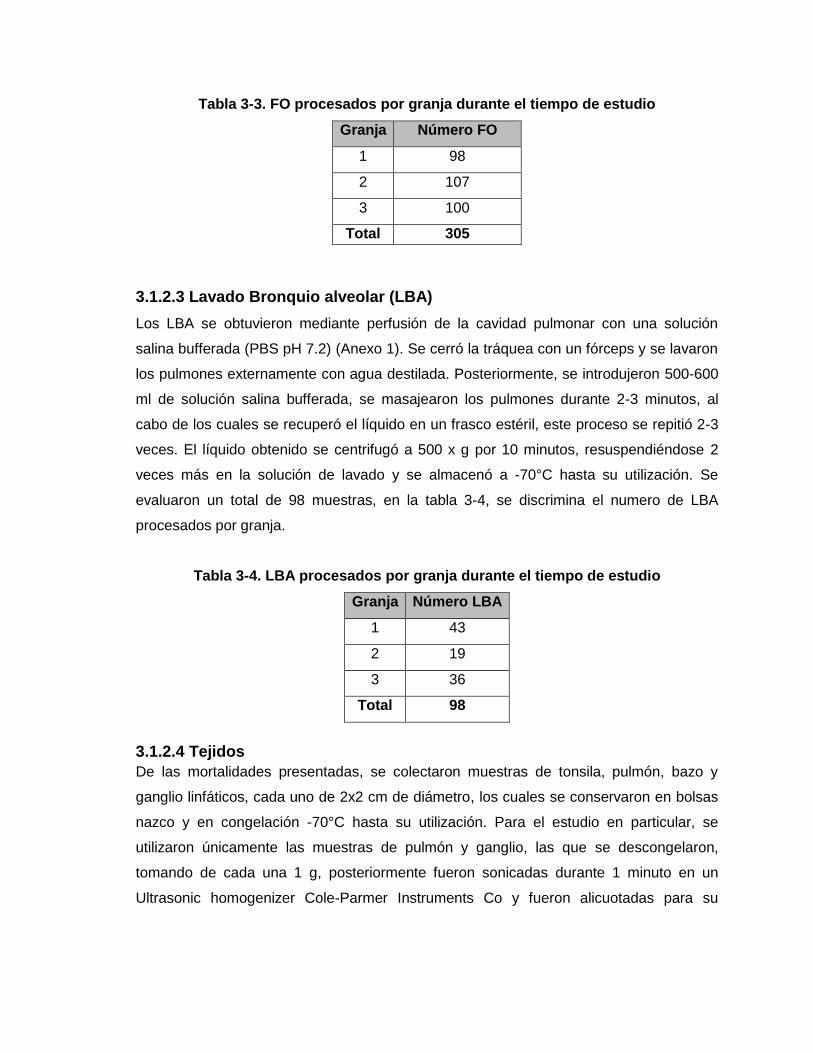
Tabla 3-3. FO procesados por granja durante el tiempo de estudio
Granja Número FO
1 98
2 107
3 100
Total 305
3.1.2.3 Lavado Bronquio alveolar (LBA)
Los LBA se obtuvieron mediante perfusión de la cavidad pulmonar con una solución
salina bufferada (PBS pH 7.2) (Anexo 1). Se cerró la tráquea con un fórceps y se lavaron
los pulmones externamente con agua destilada. Posteriormente, se introdujeron 500-600
ml de solución salina bufferada, se masajearon los pulmones durante 2-3 minutos, al
cabo de los cuales se recuperó el líquido en un frasco estéril, este proceso se repitió 2-3
veces. El líquido obtenido se centrifugó a 500 x g por 10 minutos, resuspendiéndose 2
veces más en la solución de lavado y se almacenó a -70°C hasta su utilización. Se
evaluaron un total de 98 muestras, en la tabla 3-4, se discrimina el numero de LBA
procesados por granja.
Tabla 3-4. LBA procesados por granja durante el tiempo de estudio
Granja Número LBA
1 43
2 19
3 36
Total 98
3.1.2.4 Tejidos
De las mortalidades presentadas, se colectaron muestras de tonsila, pulmón, bazo y
ganglio linfáticos, cada uno de 2x2 cm de diámetro, los cuales se conservaron en bolsas
nazco y en congelación -70°C hasta su utilización. Para el estudio en particular, se
utilizaron únicamente las muestras de pulmón y ganglio, las que se descongelaron,
tomando de cada una 1 g, posteriormente fueron sonicadas durante 1 minuto en un
Ultrasonic homogenizer Cole-Parmer Instruments Co y fueron alicuotadas para su

empleo en las pruebas de RT-qPCR y aislamiento viral. Se evaluaron 205 muestras (103
pulmones y 102 ganglios linfáticos). En la tabla 3-5, se especifica el número de tejidos
evaluados dentro de cada granja.
Tabla 3-5. Tejidos procesados por granja durante el tiempo de estudio
Granja Tejido Número
1
Pulmón 43
Ganglio 45
2
Pulmón 23
Ganglio 17
3
Pulmón 37
Ganglio 40
Total 205
3.1.3 Reacción en Cadena de la Polimerasa en Tiempo real (qRT-
PCR)
3.1.3.1 Virus
Se utilizó la cepa viral PRRS 3P (genotipo americano) Referencia PDV0401, Lote 1,
proveniente del National Veterinary Service Laboratories (NVSL) APHIS-USDA.
3.1.3.2 Extracción ARN viral
La extracción del ARN se realizó con el QIAamp Viral RNA Mini kit (QIAGEN®, Cat. No.
52906). Para la preparación de los reactivos y purificación del ARN viral se siguieron las
instrucciones del productor contenidas en el inserto.
3.1.3.3 Síntesis de ADNc
Se preparó la mezcla maestra utilizando el High Capacity cDNA Reverse Transcription Kit
(Cat. 4368813). Para un volumen total de 20 µl se utilizaron 2.0 µl de 10X RT buffer, 0.8
µl de 25XdNTP Mix (100Mm), 2.0 µl de 10X RT Random Primers, 1.0 µl de la
transcriptasa reversa Multiscribe RNase Inhibitor, 4.2 µl de agua y 10 µl de ARN. Las
muestras fueron procesadas en un termociclador BIO-RAD, DNA Engine, Peltier Thermal

Cycler. Se emplearon las siguientes condiciones: un ciclo a 25°C por 10 minutos, un ciclo
a 37°C por 120 minutos y un ciclo a 85°C por 5 minutos.
3.1.3.4 Curva Estándar
Para la curva patrón para la qRT-PCR, se empleó la cepa viral VR-2332. Inicialmente, se
realizó una RT-PCR convencional para amplificar una porción del genoma de 197 pb,
ubicada entre el ORF6 y el ORF7. Se empleo el primer forward (AAC CAC GCA TTT
GTC GTC) y reverse (TGG CAC AGC TGA TTG ACT GG), sugeridos por el Doctor
Michael Murtaugh (ICA, 2012). Para un volumen final de 25µl se utilizaron 14.25 µl agua,
2,5 µl de 10X Buffer, 1,25 µl de MgCl, 1,25 µl dNTPS, 1.25 µl de cada primer, 0.25 µl de
taq-pol y 3 µl cDNA. Las muestras se sometieron a las siguientes condiciones : 1 ciclo a
94°C por 5 minutos, 30 ciclos a 94°C por 1 minuto; 57.8°C por 1 minuto; 72°C por 1
minuto; 1 ciclo 72°C por 1 minuto y 4°C por 1 minuto. Una vez se obtuvo un producto de
197 pb, se realizó su purificación a partir de electroforesis en gel de agarosa (1.5%) y
posteriormente, mediante cuantificación (Nanodrop NYXTechnik Iluminyx) se obtuvieron
20.25 ng/µL lo cual correspondió según el programa (www.scienceprimer.com/copy-
number-calculator-for-realtime-pcr) a 9.38 x 1010 número de copias de la secuencia.
Posteriormente, a partir de este producto se realizaron diluciones desde 10-1 hasta 10-10.
Se preparó la mezcla maestra para qPCR. Para un volumen de 20 µl: 7.2 µl agua, 0.4 µl
de cada primer, 10 µl de la SYBR® Green PCR Master Mix (SYBR® Green I Dye,
AmpliTaq Gold® DNA Polymerase, dNTPs con dUTP, Passive Reference y optimized
buffer) y 2 µl del ADNc. La qRT-PCR basada en SYBR Green, fue realizada en un
termociclador Lightcycler 480 II Roche, usando las siguientes condiciones: 1 ciclo de
95°C por 1 minuto, 45 ciclos de 95°C por 3 segundos, 60°C por 30 segundos, 72°C por
30 segundos, 1 ciclo 95°C por 30 segundos y 70°C por 1 minuto, 1 ciclo de 37°C por 10
segundos. Las diluciones se prepararon por duplicado y fueron detectables hasta 10-8. De
acuerdo con la curva estándar (Figura 3.2), los valores de Ct (cycle threshold)
considerados positivos se encontraron entre 15 y 32 (La estandarización de la curva
patrón fue realizada por Mancipe, L.F., datos no publicados).

Figura 3-2 Curva Estándar por qRT-PCR
(Mancipe, 2013)
3.1.4 Análisis estadístico Se realizó un análisis de varianza utilizando el programa SAS, teniendo como fuentes de
variación, granja (1, 2 y 3), grupo etáreo (madres, reproductores, reemplazos, lactantes,
destetos, preceba y ceba) y meses (Septiembre de 2012 a Mayo de 2013). Al
presentarse efectos significativos, se empleó una prueba de comparación múltiple de
Tukey.
3.2 Resultados Con respecto a los resultados estadísticos, en las muestras de suero y FO se obtuvieron
resultados significativos del mes para cada granja y grupo (p<0.05 y p<.0001
respectivamente). Con respecto a los tejidos, en los pulmones, no se presentó efecto
significativo ni interacción (p>0.05). En los resultados de las muestras de ganglios, el
único efecto significativo fue mes (p<.0001) y no se presentaron significancias en las
interacciones (p>0.05). En los LBA, se presento efecto significativo del mes en la granja
1 en los dos grupos etareos (p=0.0026 y p=0.0157) y en la granja 3 grupo 7 (p<.0001).

3.2.1 qRT-PCR en muestras de sueros Se evaluaron en total 355 muestras de sueros, las cuales para su análisis fueron
divididas en dos grupos, las negativas y positivas por ELISA (valor M/P por debajo y por
encima de 0.40 respectivamente). En la figura 3-3 se comparan los resultados obtenidos
teniendo en cuenta todos los grupos etareos en las tres granjas.
Figura 3-3. Resultados obtenidos por qRT-PCR en sueros dentro de cada granja
evaluada
En la figura 3-3, se observa que el promedio de valores Ct encontrados en las tres
granjas estuvo por encima del punto de corte 15-32; sin embargo, las desviaciones
estándar muestran que se encontaron valores dentro del punto de corte o muy próximos
al mismo, lo que indica presencia de genoma viral tanto en muestras de ELISA positivas
como negativas.
3.2.1.1 Granja 1
Se evaluaron 112 muestras de sueros encontrándose 11 (9.82%) positivos por qRT-PCR.
En la figura 3-4, se observa el comportamiento de resultados por qRT-PCR, de acuerdo
al resultado por ELISA y por grupo etareo.
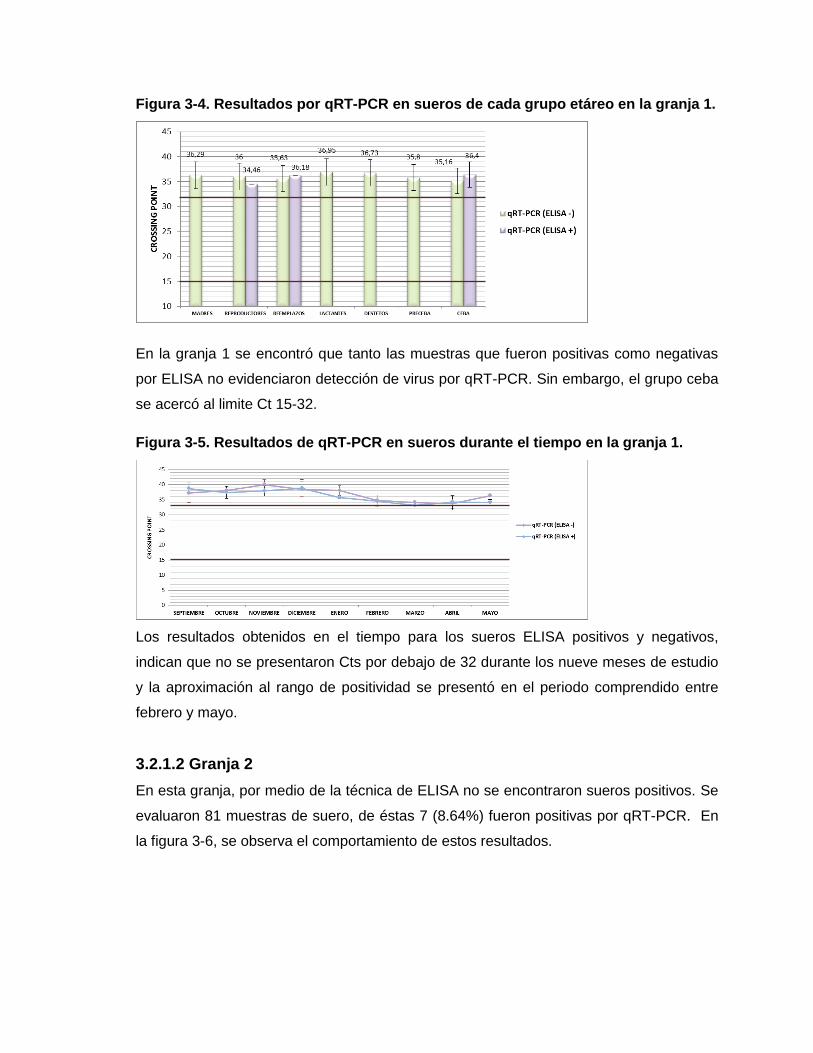
Figura 3-4. Resultados por qRT-PCR en sueros de cada grupo etáreo en la granja 1.
En la granja 1 se encontró que tanto las muestras que fueron positivas como negativas
por ELISA no evidenciaron detección de virus por qRT-PCR. Sin embargo, el grupo ceba
se acercó al limite Ct 15-32.
Figura 3-5. Resultados de qRT-PCR en sueros durante el tiempo en la granja 1.
Los resultados obtenidos en el tiempo para los sueros ELISA positivos y negativos,
indican que no se presentaron Cts por debajo de 32 durante los nueve meses de estudio
y la aproximación al rango de positividad se presentó en el periodo comprendido entre
febrero y mayo.
3.2.1.2 Granja 2
En esta granja, por medio de la técnica de ELISA no se encontraron sueros positivos. Se
evaluaron 81 muestras de suero, de éstas 7 (8.64%) fueron positivas por qRT-PCR. En
la figura 3-6, se observa el comportamiento de estos resultados.
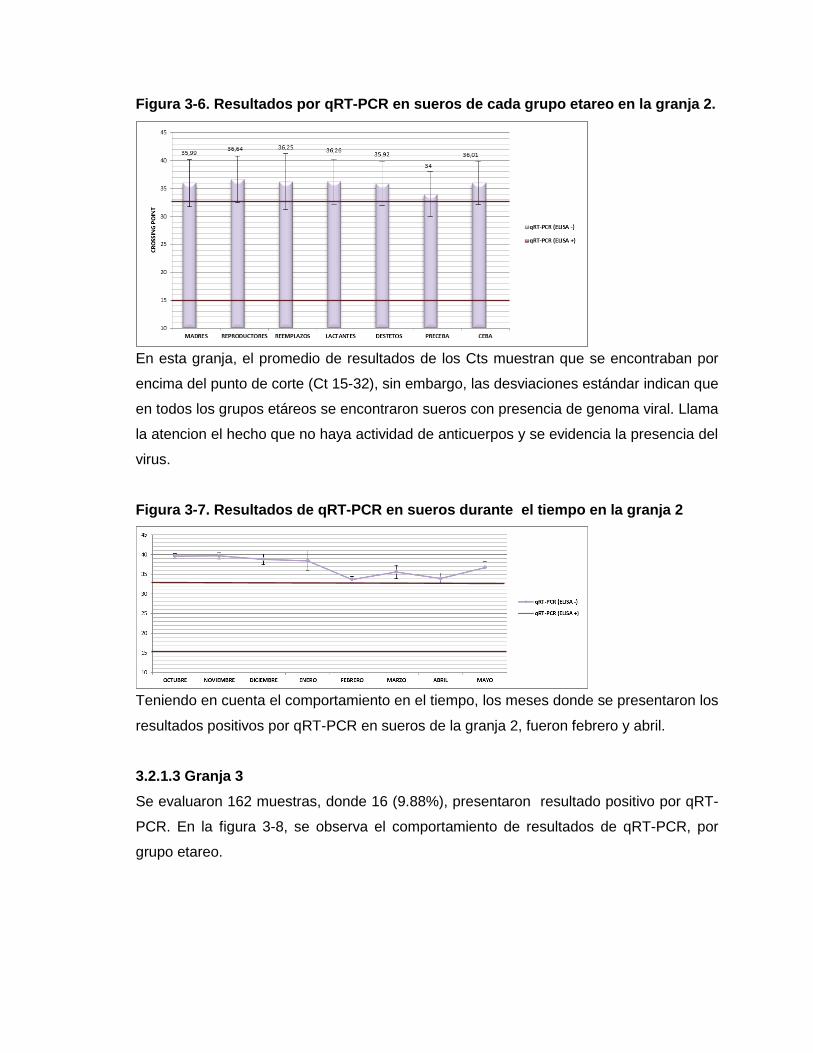
Figura 3-6. Resultados por qRT-PCR en sueros de cada grupo etareo en la granja 2.
En esta granja, el promedio de resultados de los Cts muestran que se encontraban por
encima del punto de corte (Ct 15-32), sin embargo, las desviaciones estándar indican que
en todos los grupos etáreos se encontraron sueros con presencia de genoma viral. Llama
la atencion el hecho que no haya actividad de anticuerpos y se evidencia la presencia del
virus.
Figura 3-7. Resultados de qRT-PCR en sueros durante el tiempo en la granja 2
Teniendo en cuenta el comportamiento en el tiempo, los meses donde se presentaron los
resultados positivos por qRT-PCR en sueros de la granja 2, fueron febrero y abril.
3.2.1.3 Granja 3
Se evaluaron 162 muestras, donde 16 (9.88%), presentaron resultado positivo por qRT-
PCR. En la figura 3-8, se observa el comportamiento de resultados de qRT-PCR, por
grupo etareo.
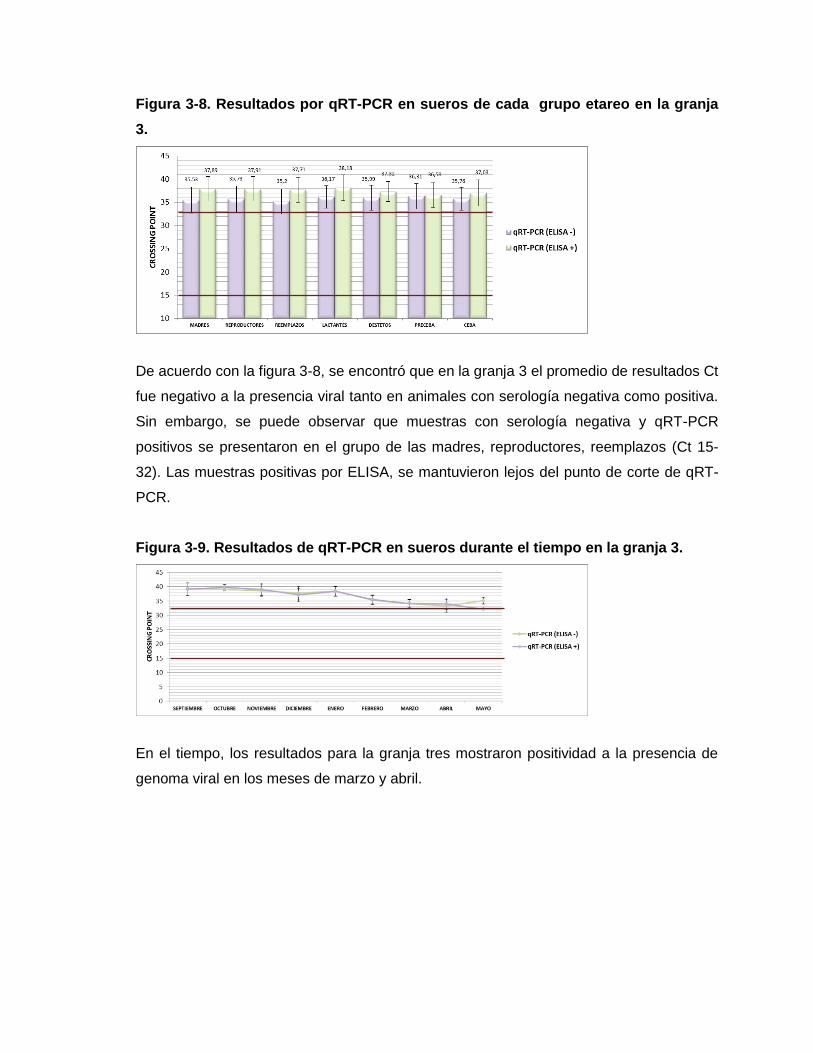
Figura 3-8. Resultados por qRT-PCR en sueros de cada grupo etareo en la granja
3.
De acuerdo con la figura 3-8, se encontró que en la granja 3 el promedio de resultados Ct
fue negativo a la presencia viral tanto en animales con serología negativa como positiva.
Sin embargo, se puede observar que muestras con serología negativa y qRT-PCR
positivos se presentaron en el grupo de las madres, reproductores, reemplazos (Ct 15-
32). Las muestras positivas por ELISA, se mantuvieron lejos del punto de corte de qRT-
PCR.
Figura 3-9. Resultados de qRT-PCR en sueros durante el tiempo en la granja 3.
En el tiempo, los resultados para la granja tres mostraron positividad a la presencia de
genoma viral en los meses de marzo y abril.
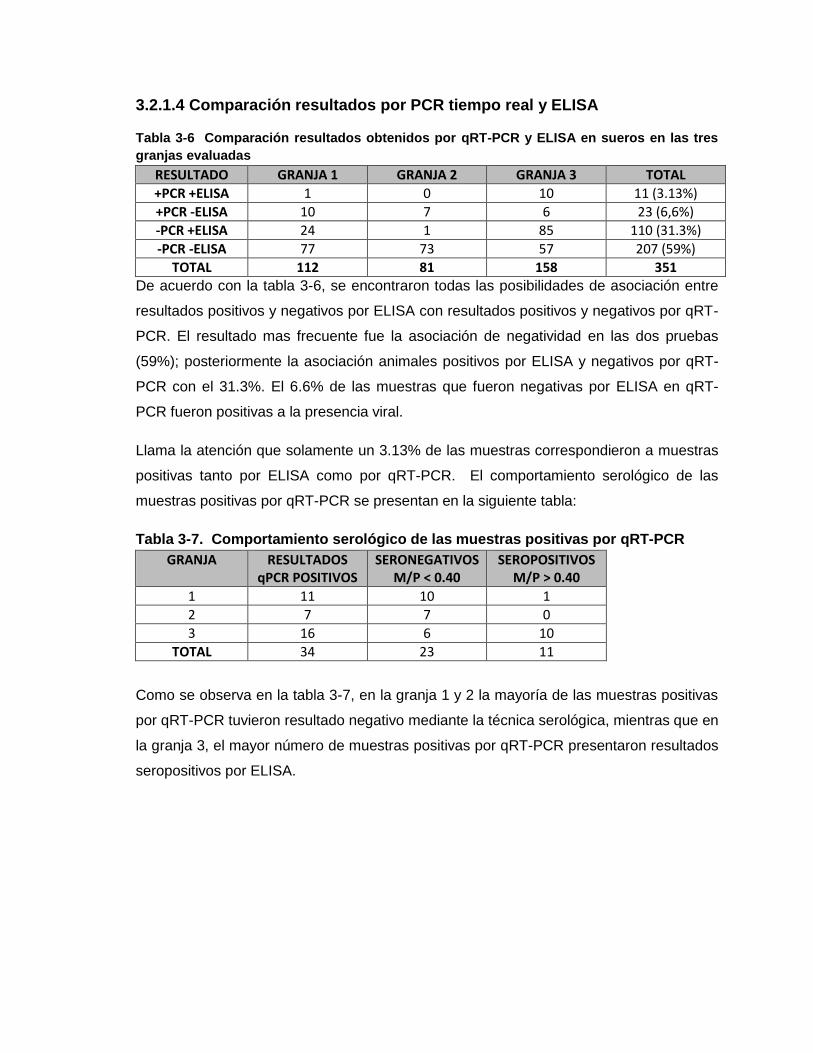
3.2.1.4 Comparación resultados por PCR tiempo real y ELISA
Tabla 3-6 Comparación resultados obtenidos por qRT-PCR y ELISA en sueros en las tres
granjas evaluadas
RESULTADO GRANJA 1 GRANJA 2 GRANJA 3 TOTAL
+PCR +ELISA 1 0 10 11 (3.13%)
+PCR -ELISA 10 7 6 23 (6,6%)
-PCR +ELISA 24 1 85 110 (31.3%)
-PCR -ELISA 77 73 57 207 (59%)
TOTAL 112 81 158 351
De acuerdo con la tabla 3-6, se encontraron todas las posibilidades de asociación entre
resultados positivos y negativos por ELISA con resultados positivos y negativos por qRT-
PCR. El resultado mas frecuente fue la asociación de negatividad en las dos pruebas
(59%); posteriormente la asociación animales positivos por ELISA y negativos por qRT-
PCR con el 31.3%. El 6.6% de las muestras que fueron negativas por ELISA en qRT-
PCR fueron positivas a la presencia viral.
Llama la atención que solamente un 3.13% de las muestras correspondieron a muestras
positivas tanto por ELISA como por qRT-PCR. El comportamiento serológico de las
muestras positivas por qRT-PCR se presentan en la siguiente tabla:
Tabla 3-7. Comportamiento serológico de las muestras positivas por qRT-PCR
GRANJA RESULTADOS qPCR POSITIVOS
SERONEGATIVOS M/P < 0.40
SEROPOSITIVOS M/P > 0.40
1 11 10 1
2 7 7 0
3 16 6 10
TOTAL 34 23 11
Como se observa en la tabla 3-7, en la granja 1 y 2 la mayoría de las muestras positivas
por qRT-PCR tuvieron resultado negativo mediante la técnica serológica, mientras que en
la granja 3, el mayor número de muestras positivas por qRT-PCR presentaron resultados
seropositivos por ELISA.
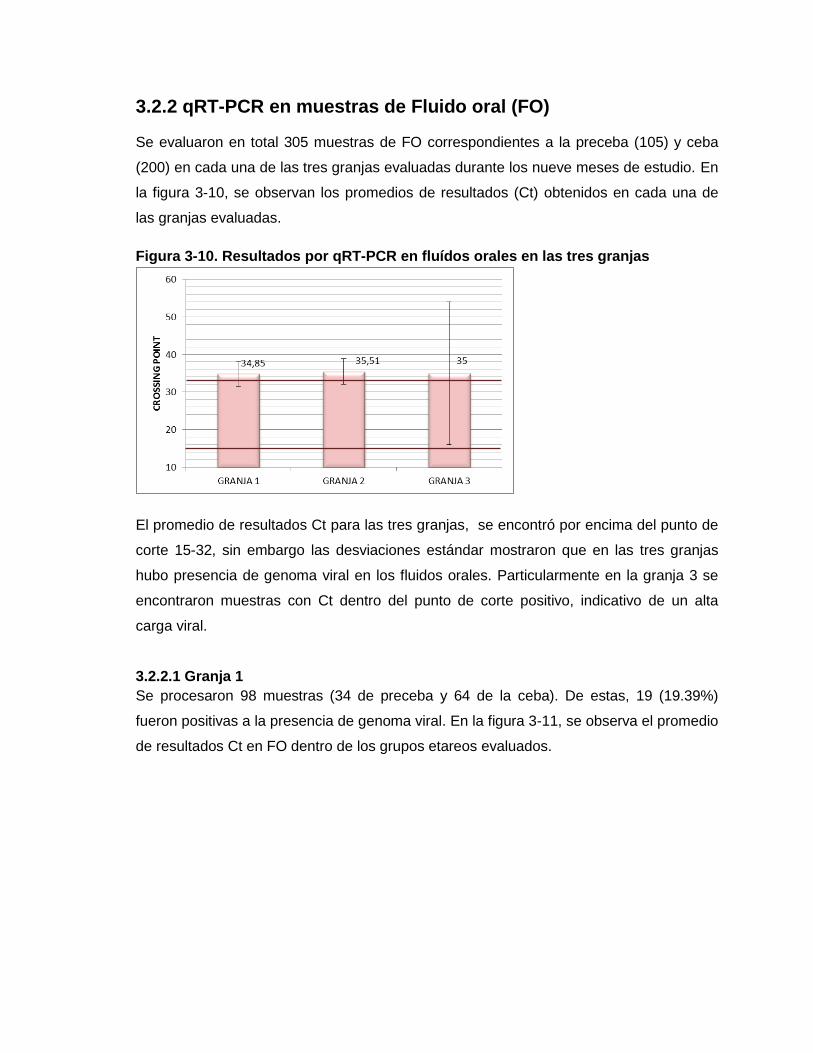
3.2.2 qRT-PCR en muestras de Fluido oral (FO)
Se evaluaron en total 305 muestras de FO correspondientes a la preceba (105) y ceba
(200) en cada una de las tres granjas evaluadas durante los nueve meses de estudio. En
la figura 3-10, se observan los promedios de resultados (Ct) obtenidos en cada una de
las granjas evaluadas.
Figura 3-10. Resultados por qRT-PCR en fluídos orales en las tres granjas
El promedio de resultados Ct para las tres granjas, se encontró por encima del punto de
corte 15-32, sin embargo las desviaciones estándar mostraron que en las tres granjas
hubo presencia de genoma viral en los fluidos orales. Particularmente en la granja 3 se
encontraron muestras con Ct dentro del punto de corte positivo, indicativo de un alta
carga viral.
3.2.2.1 Granja 1
Se procesaron 98 muestras (34 de preceba y 64 de la ceba). De estas, 19 (19.39%)
fueron positivas a la presencia de genoma viral. En la figura 3-11, se observa el promedio
de resultados Ct en FO dentro de los grupos etareos evaluados.
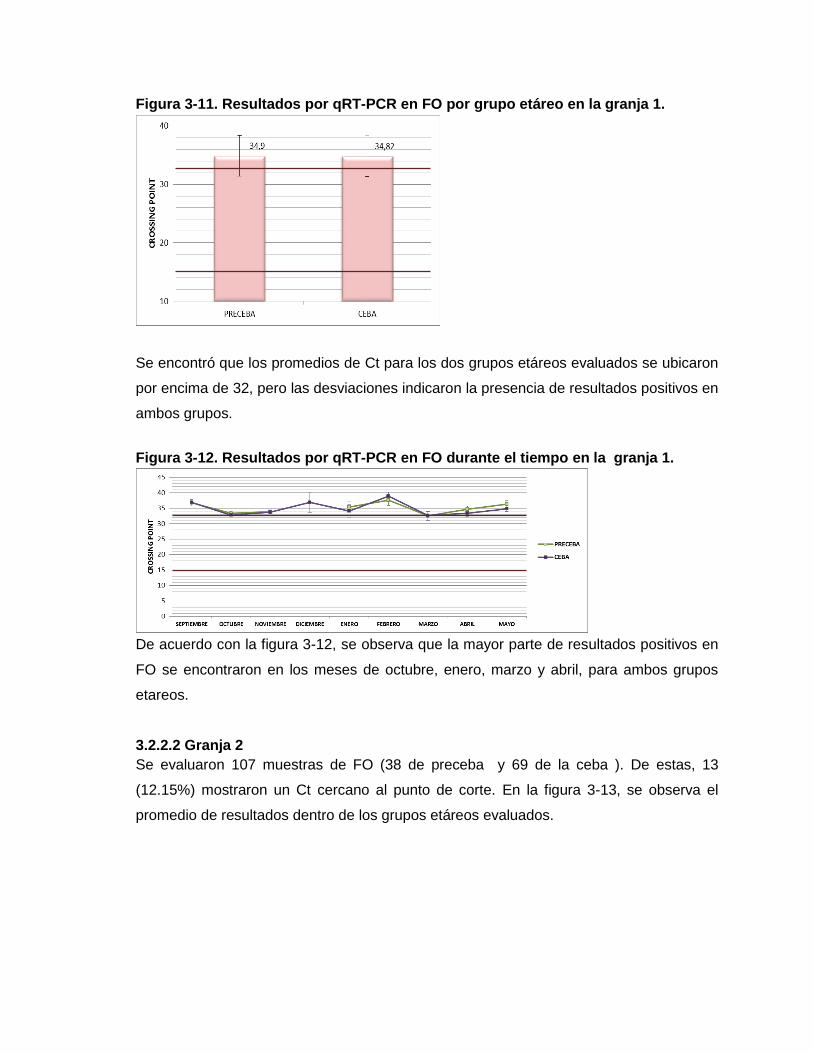
Figura 3-11. Resultados por qRT-PCR en FO por grupo etáreo en la granja 1.
Se encontró que los promedios de Ct para los dos grupos etáreos evaluados se ubicaron
por encima de 32, pero las desviaciones indicaron la presencia de resultados positivos en
ambos grupos.
Figura 3-12. Resultados por qRT-PCR en FO durante el tiempo en la granja 1.
De acuerdo con la figura 3-12, se observa que la mayor parte de resultados positivos en
FO se encontraron en los meses de octubre, enero, marzo y abril, para ambos grupos
etareos.
3.2.2.2 Granja 2
Se evaluaron 107 muestras de FO (38 de preceba y 69 de la ceba ). De estas, 13
(12.15%) mostraron un Ct cercano al punto de corte. En la figura 3-13, se observa el
promedio de resultados dentro de los grupos etáreos evaluados.
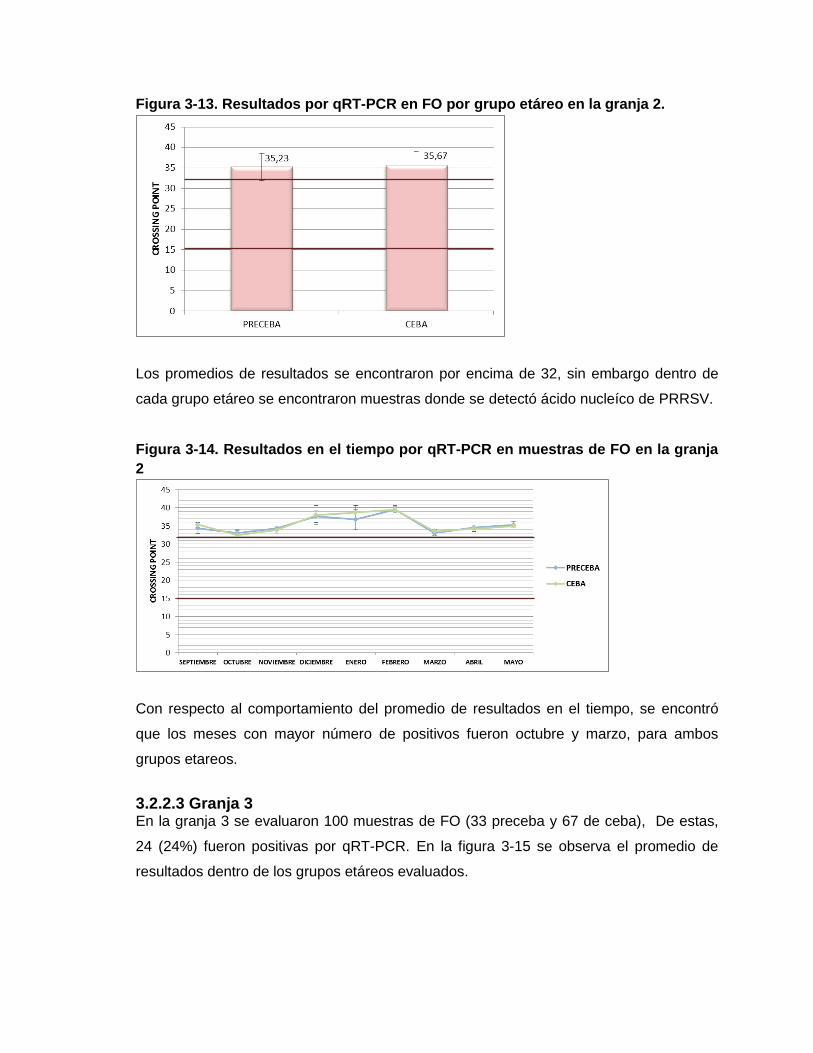
Figura 3-13. Resultados por qRT-PCR en FO por grupo etáreo en la granja 2.
Los promedios de resultados se encontraron por encima de 32, sin embargo dentro de
cada grupo etáreo se encontraron muestras donde se detectó ácido nucleíco de PRRSV.
Figura 3-14. Resultados en el tiempo por qRT-PCR en muestras de FO en la granja
2
Con respecto al comportamiento del promedio de resultados en el tiempo, se encontró
que los meses con mayor número de positivos fueron octubre y marzo, para ambos
grupos etareos.
3.2.2.3 Granja 3 En la granja 3 se evaluaron 100 muestras de FO (33 preceba y 67 de ceba), De estas,
24 (24%) fueron positivas por qRT-PCR. En la figura 3-15 se observa el promedio de
resultados dentro de los grupos etáreos evaluados.
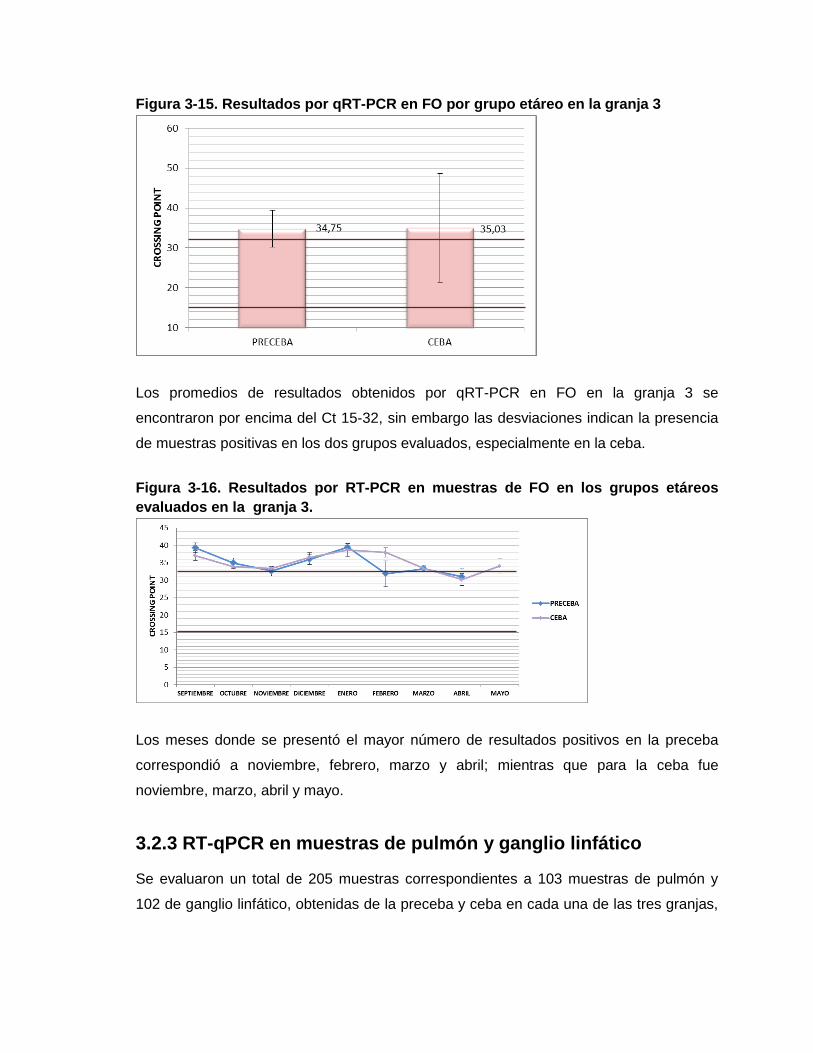
Figura 3-15. Resultados por qRT-PCR en FO por grupo etáreo en la granja 3
Los promedios de resultados obtenidos por qRT-PCR en FO en la granja 3 se
encontraron por encima del Ct 15-32, sin embargo las desviaciones indican la presencia
de muestras positivas en los dos grupos evaluados, especialmente en la ceba.
Figura 3-16. Resultados por RT-PCR en muestras de FO en los grupos etáreos
evaluados en la granja 3.
Los meses donde se presentó el mayor número de resultados positivos en la preceba
correspondió a noviembre, febrero, marzo y abril; mientras que para la ceba fue
noviembre, marzo, abril y mayo.
3.2.3 RT-qPCR en muestras de pulmón y ganglio linfático
Se evaluaron un total de 205 muestras correspondientes a 103 muestras de pulmón y
102 de ganglio linfático, obtenidas de la preceba y ceba en cada una de las tres granjas,
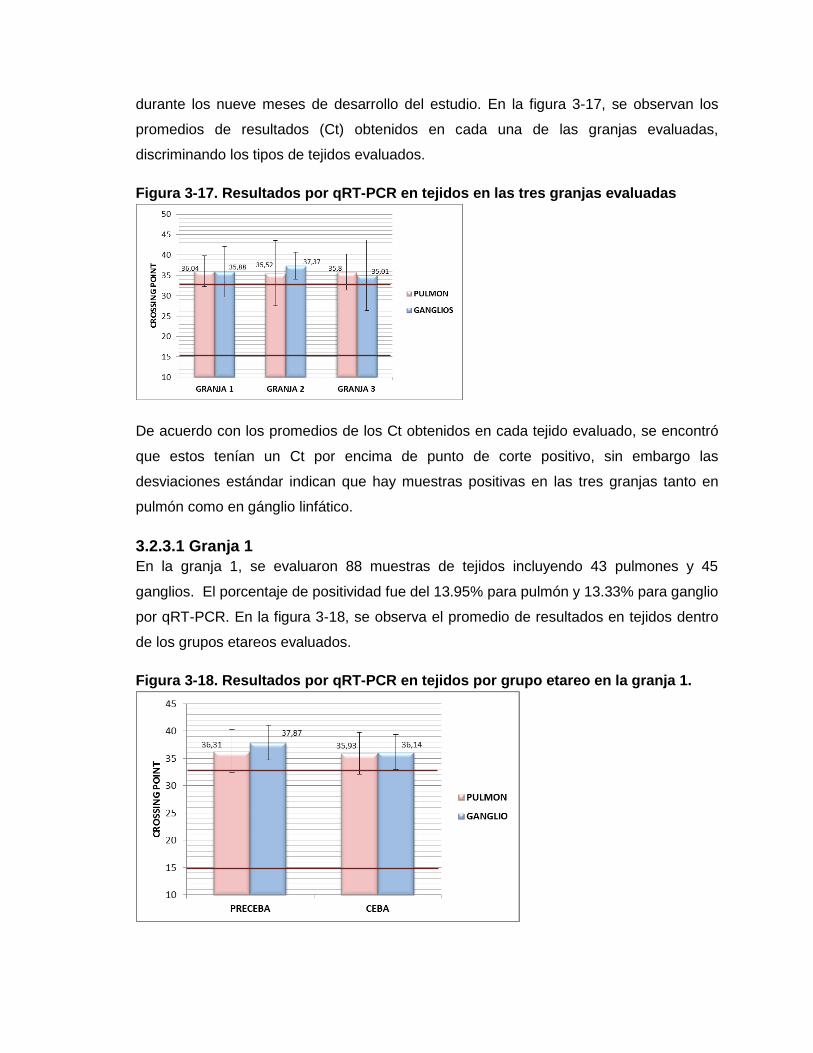
durante los nueve meses de desarrollo del estudio. En la figura 3-17, se observan los
promedios de resultados (Ct) obtenidos en cada una de las granjas evaluadas,
discriminando los tipos de tejidos evaluados.
Figura 3-17. Resultados por qRT-PCR en tejidos en las tres granjas evaluadas
De acuerdo con los promedios de los Ct obtenidos en cada tejido evaluado, se encontró
que estos tenían un Ct por encima de punto de corte positivo, sin embargo las
desviaciones estándar indican que hay muestras positivas en las tres granjas tanto en
pulmón como en gánglio linfático.
3.2.3.1 Granja 1
En la granja 1, se evaluaron 88 muestras de tejidos incluyendo 43 pulmones y 45
ganglios. El porcentaje de positividad fue del 13.95% para pulmón y 13.33% para ganglio
por qRT-PCR. En la figura 3-18, se observa el promedio de resultados en tejidos dentro
de los grupos etareos evaluados.
Figura 3-18. Resultados por qRT-PCR en tejidos por grupo etareo en la granja 1.
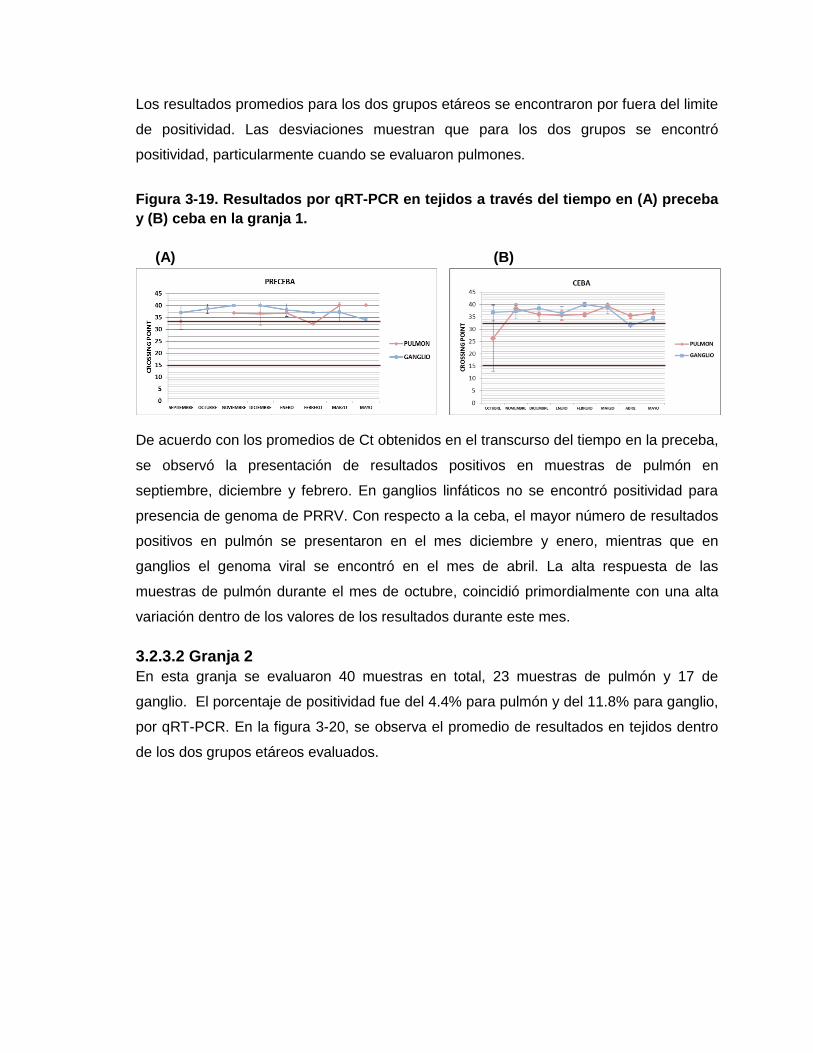
Los resultados promedios para los dos grupos etáreos se encontraron por fuera del limite
de positividad. Las desviaciones muestran que para los dos grupos se encontró
positividad, particularmente cuando se evaluaron pulmones.
Figura 3-19. Resultados por qRT-PCR en tejidos a través del tiempo en (A) preceba
y (B) ceba en la granja 1.
De acuerdo con los promedios de Ct obtenidos en el transcurso del tiempo en la preceba,
se observó la presentación de resultados positivos en muestras de pulmón en
septiembre, diciembre y febrero. En ganglios linfáticos no se encontró positividad para
presencia de genoma de PRRV. Con respecto a la ceba, el mayor número de resultados
positivos en pulmón se presentaron en el mes diciembre y enero, mientras que en
ganglios el genoma viral se encontró en el mes de abril. La alta respuesta de las
muestras de pulmón durante el mes de octubre, coincidió primordialmente con una alta
variación dentro de los valores de los resultados durante este mes.
3.2.3.2 Granja 2
En esta granja se evaluaron 40 muestras en total, 23 muestras de pulmón y 17 de
ganglio. El porcentaje de positividad fue del 4.4% para pulmón y del 11.8% para ganglio,
por qRT-PCR. En la figura 3-20, se observa el promedio de resultados en tejidos dentro
de los dos grupos etáreos evaluados.
(A) (B)
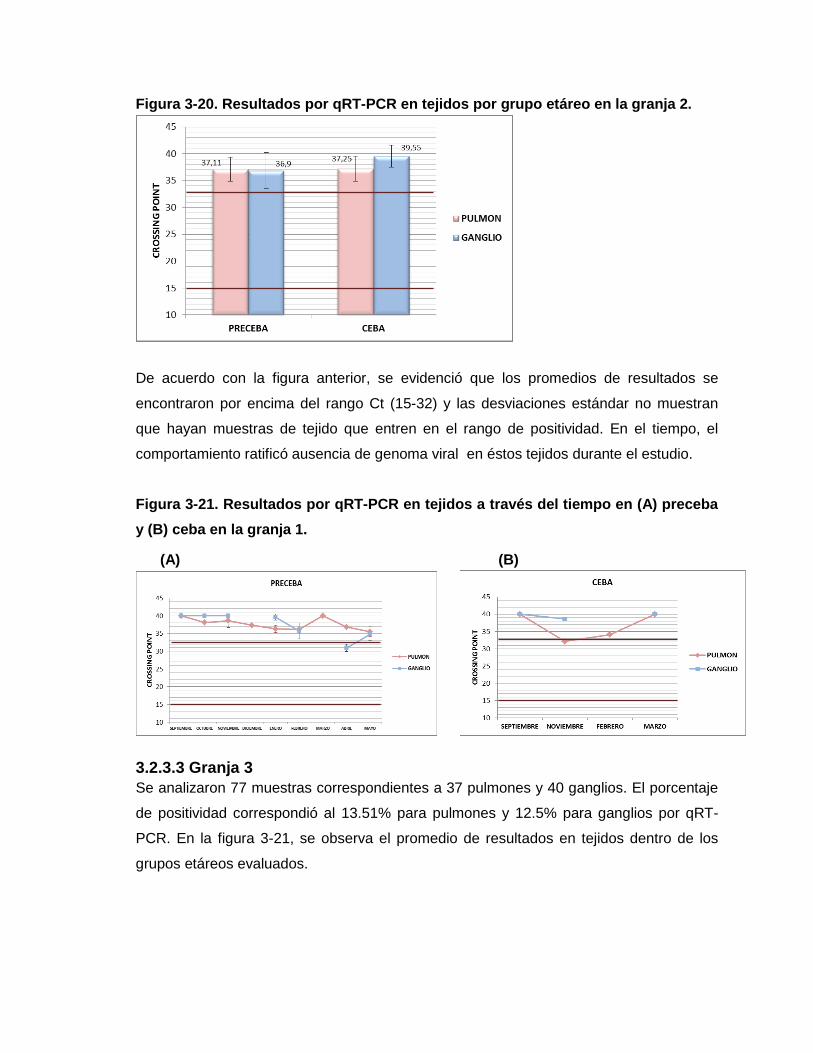
Figura 3-20. Resultados por qRT-PCR en tejidos por grupo etáreo en la granja 2.
De acuerdo con la figura anterior, se evidenció que los promedios de resultados se
encontraron por encima del rango Ct (15-32) y las desviaciones estándar no muestran
que hayan muestras de tejido que entren en el rango de positividad. En el tiempo, el
comportamiento ratificó ausencia de genoma viral en éstos tejidos durante el estudio.
Figura 3-21. Resultados por qRT-PCR en tejidos a través del tiempo en (A) preceba
y (B) ceba en la granja 1.
3.2.3.3 Granja 3
Se analizaron 77 muestras correspondientes a 37 pulmones y 40 ganglios. El porcentaje
de positividad correspondió al 13.51% para pulmones y 12.5% para ganglios por qRT-
PCR. En la figura 3-21, se observa el promedio de resultados en tejidos dentro de los
grupos etáreos evaluados.
(A) (B)
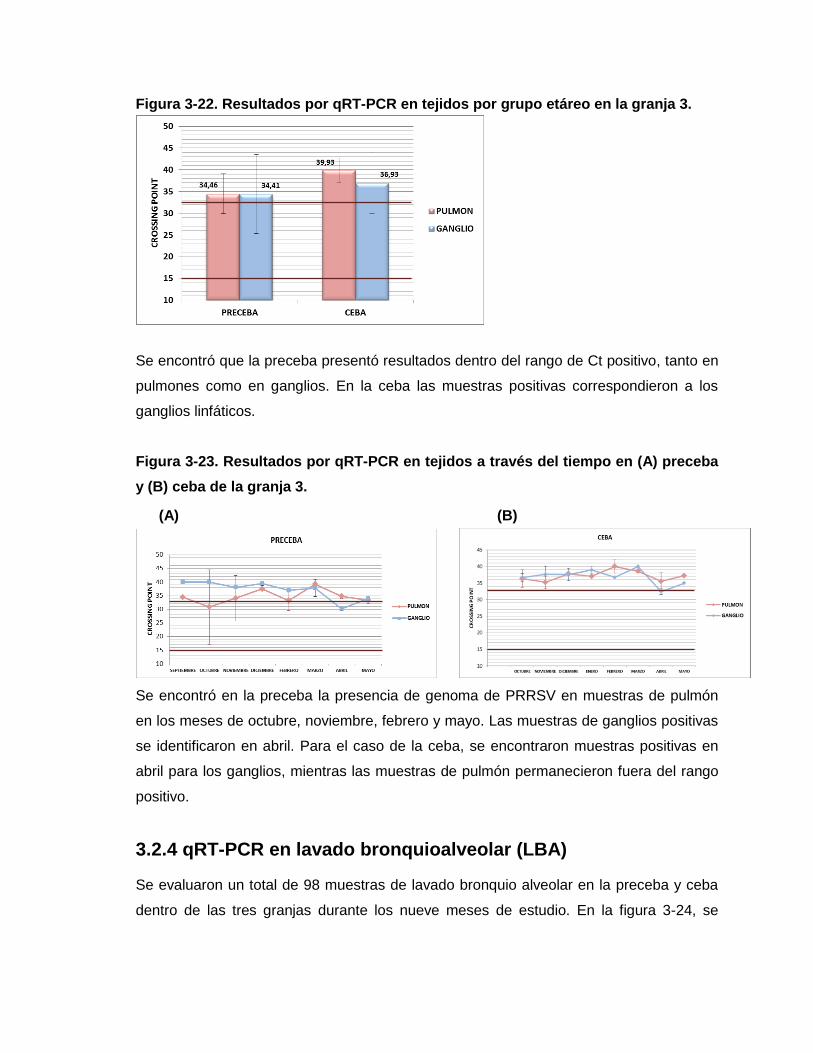
Figura 3-22. Resultados por qRT-PCR en tejidos por grupo etáreo en la granja 3.
Se encontró que la preceba presentó resultados dentro del rango de Ct positivo, tanto en
pulmones como en ganglios. En la ceba las muestras positivas correspondieron a los
ganglios linfáticos.
Figura 3-23. Resultados por qRT-PCR en tejidos a través del tiempo en (A) preceba
y (B) ceba de la granja 3.
Se encontró en la preceba la presencia de genoma de PRRSV en muestras de pulmón
en los meses de octubre, noviembre, febrero y mayo. Las muestras de ganglios positivas
se identificaron en abril. Para el caso de la ceba, se encontraron muestras positivas en
abril para los ganglios, mientras las muestras de pulmón permanecieron fuera del rango
positivo.
3.2.4 qRT-PCR en lavado bronquioalveolar (LBA)
Se evaluaron un total de 98 muestras de lavado bronquio alveolar en la preceba y ceba
dentro de las tres granjas durante los nueve meses de estudio. En la figura 3-24, se
(A) (B)
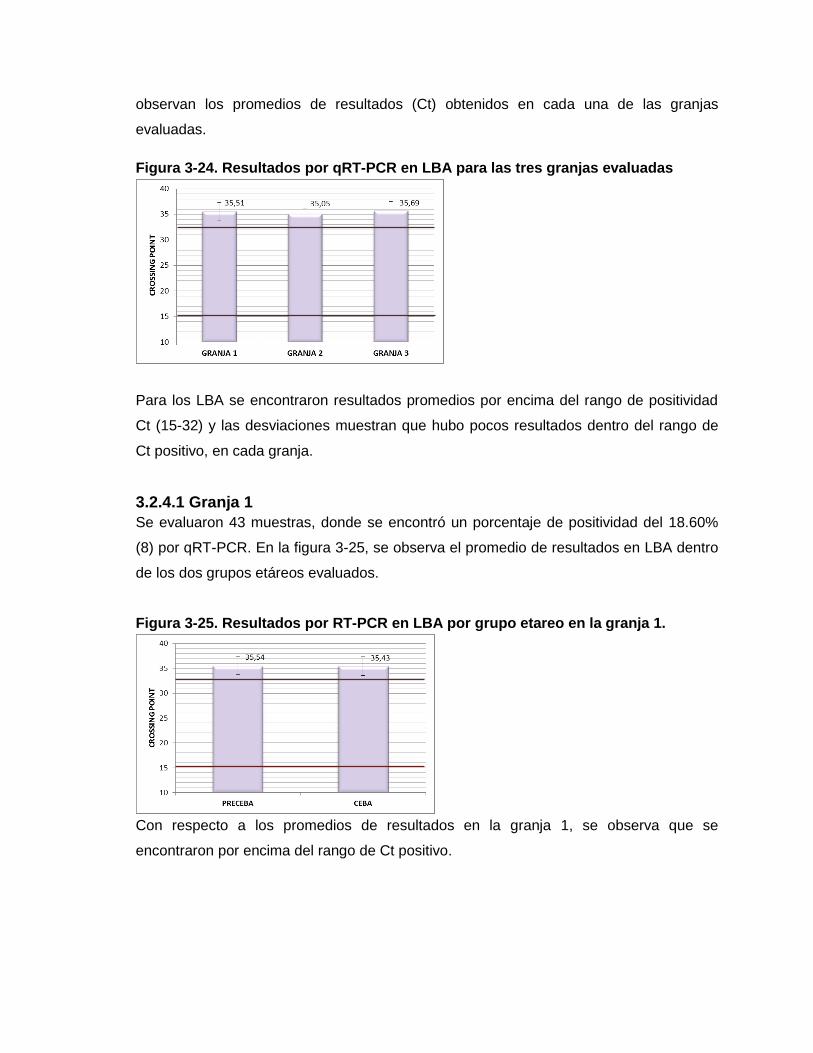
observan los promedios de resultados (Ct) obtenidos en cada una de las granjas
evaluadas.
Figura 3-24. Resultados por qRT-PCR en LBA para las tres granjas evaluadas
Para los LBA se encontraron resultados promedios por encima del rango de positividad
Ct (15-32) y las desviaciones muestran que hubo pocos resultados dentro del rango de
Ct positivo, en cada granja.
3.2.4.1 Granja 1
Se evaluaron 43 muestras, donde se encontró un porcentaje de positividad del 18.60%
(8) por qRT-PCR. En la figura 3-25, se observa el promedio de resultados en LBA dentro
de los dos grupos etáreos evaluados.
Figura 3-25. Resultados por RT-PCR en LBA por grupo etareo en la granja 1.
Con respecto a los promedios de resultados en la granja 1, se observa que se
encontraron por encima del rango de Ct positivo.
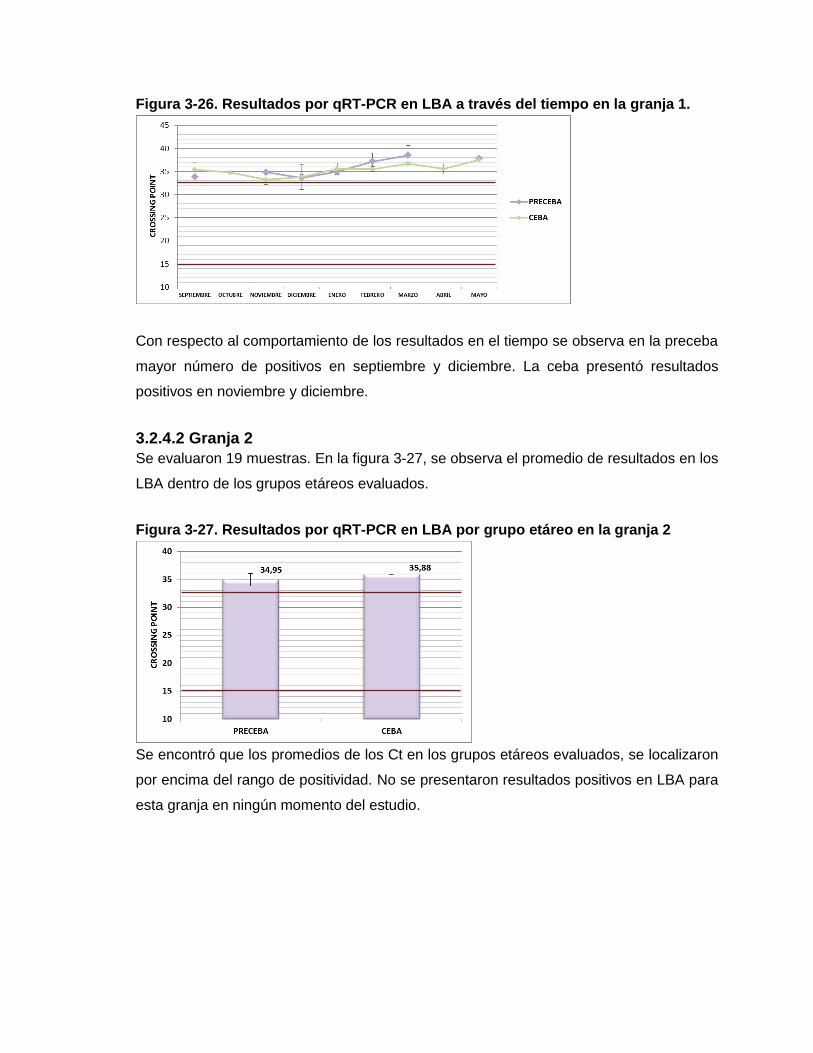
Figura 3-26. Resultados por qRT-PCR en LBA a través del tiempo en la granja 1.
Con respecto al comportamiento de los resultados en el tiempo se observa en la preceba
mayor número de positivos en septiembre y diciembre. La ceba presentó resultados
positivos en noviembre y diciembre.
3.2.4.2 Granja 2
Se evaluaron 19 muestras. En la figura 3-27, se observa el promedio de resultados en los
LBA dentro de los grupos etáreos evaluados.
Figura 3-27. Resultados por qRT-PCR en LBA por grupo etáreo en la granja 2
Se encontró que los promedios de los Ct en los grupos etáreos evaluados, se localizaron
por encima del rango de positividad. No se presentaron resultados positivos en LBA para
esta granja en ningún momento del estudio.
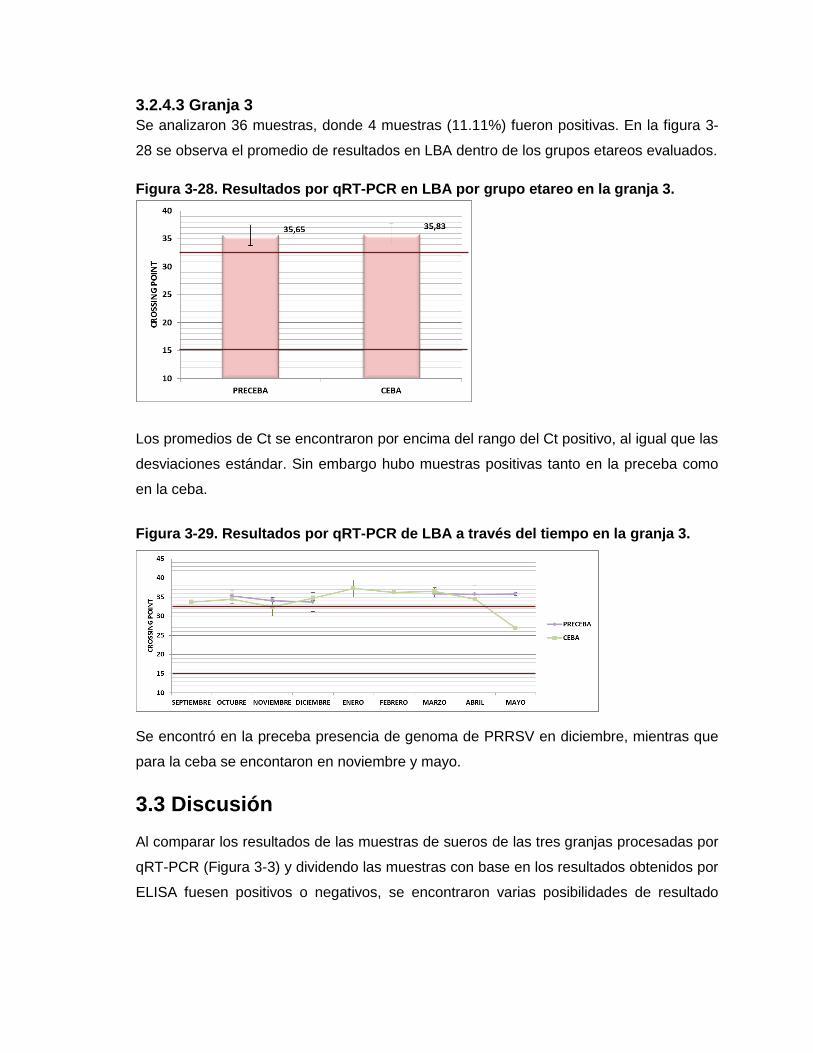
3.2.4.3 Granja 3
Se analizaron 36 muestras, donde 4 muestras (11.11%) fueron positivas. En la figura 3-
28 se observa el promedio de resultados en LBA dentro de los grupos etareos evaluados.
Figura 3-28. Resultados por qRT-PCR en LBA por grupo etareo en la granja 3.
Los promedios de Ct se encontraron por encima del rango del Ct positivo, al igual que las
desviaciones estándar. Sin embargo hubo muestras positivas tanto en la preceba como
en la ceba.
Figura 3-29. Resultados por qRT-PCR de LBA a través del tiempo en la granja 3.
Se encontró en la preceba presencia de genoma de PRRSV en diciembre, mientras que
para la ceba se encontaron en noviembre y mayo.
3.3 Discusión
Al comparar los resultados de las muestras de sueros de las tres granjas procesadas por
qRT-PCR (Figura 3-3) y dividendo las muestras con base en los resultados obtenidos por
ELISA fuesen positivos o negativos, se encontraron varias posibilidades de resultado

(Tabla 3-6). Uno de ellos fue la presencia de anticuerpos sin detección viral, lo cual
puede indicar que el periodo de viremia fue superado, se produjo una respuesta inmune
efectiva (particularmente de tipo humoral) y posiblemente el virus se localizó en tejidos,
tal y como se observa en la Figura 3-17, donde se determinó la presencia de genoma
viral tanto en pulmón como ganglio, en las tres granjas.
De otro lado, una infección típica por PRRSV se caracteriza por un estado inicial de
viremia aguda que permanece aproximadamente 4 semanas, seguida por un estado
donde alcanza niveles indetectables (Islam et al., 2013). La seropositividad en los
animales puede ser producto de una infección antigua si se tiene en cuenta el tiempo de
persistencia de los anticuerpos antiPRRSV en un animal. Estos son detectables por
ELISA entre los 9 a 13 días PI (Yoon et al., 1995, citado por Evans et al., 2008) y decaen
en el tiempo pudiendo persistir hasta por 28 meses (Desrosiers and Boutin, 2002).
También se sabe que muchos cerdos liberan el virus durante 3 a 4 meses pos exposición
(Wills et al., 2003) así, los animales seropositivos al PRRSV, pueden ser negativos a la
presencia del virus y en consecuencia la seropositividad es indicativa de una infección
pasada (Evans et al., 2008). Al no haber evidencia de la presencia de genoma viral en
las muestras que por ELISA fueron positivas se puede afirmar que no hay animales en
estado de viremia y que la respuesta humoral no se puede asociar con presencia activa
de enfermedad.
Otra posibilidad de respuesta encontrada, fue la presencia de virus en sueros que por
ELISA fueron negativos. Mediante PCR, la viremia puede ser detectada desde el día 1
hasta la semana 4 o 5 PI (Wasilk et al., 2004), aunque la presencia de estos resultados
positivos por PCR no indican necesariamente la presencia de virus viable (Wills et al.,
2003).
Al respecto de la viremia en diferentes grupos, Boddicker et al (2012), al desafiar cerdos
con una cepa altamente virulenta, demostraron sustanciales diferencias entre los perfiles
virémicos individuales y totales, clasificando los animales en dos categorías (non-
rebounders y rebounders) de acuerdo con los perfiles virales en suero. Esto lleva a
especular en el presente estudio, que algunos animales al momento de tomar la muestra
no presentaban viremia, pero días más tarde pudieron diseminar virus infeccioso y

presentar posiblemente resultados positivos a la prueba de qRT-PCR. Lo anterior se
explica ya que cerdos expuestos al PRRSV presentan una prolongada viremia donde
luego de una inicial remoción del PRRSV en la sangre, la viremia periódicamente re-
aparece con los tejidos linfoides como el sitio primario de replicación (Chand et al., 2012).
Lo anterior, en el caso del presente estudio, implicaría que las viremias detectadas no
corresponderían únicamente al producto de una infección temprana, si no a un estado de
infeccion persistente.
En un estudio llevado a cabo por Martínez et al (2008), encontraron por RT-qPCR -
SYBR Green, la detección temprana del PRRSV en suero a los 2 días PI. Chai et al
(2013) por su parte, reportaron que la tasa de detección de positivos por qRT-PCR-SYBR
Green fue mucho más alta que la obtenida con la PCR convencional. Teniendo en
cuenta lo anterior y los resultados obtenidos en el presente estudio en sueros, donde se
utilizo qRT-PCR-SYBR Green, se estableció la presencia de viremia y por lo tanto, la
posibilidad de una infección temprana dentro de los grupos etareos en las tres granjas
evaluadas al igual que la posible presencia de virus en bajas cantidades, que podría no
ser infeccioso (Figura 3-4, 3-6 y 3-8).
Al analizar la presencia de virus en los reemplazos y madres, se puede especular que
esta se encuentra relacionada ya sea con la introducción de una nueva cepa viral o con
la posible presencia de animales persistentemente infectados. En el caso de los
reemplazos, estos pudieron ingresar infectados a la granja o fueron expuestos a la cepa
viral circulante mediante las practicas de manejo tradicionales como el uso de placentas
de animales enfermos. Es importante tener en cuenta que dentro de las granjas el virus
se mantiene endémico por medio de un ciclo de transmisión de animales portadores a
susceptibles a través de nacimientos o compra de los mismos (Horter et al., 2002). En
cuanto a las madres, la viremia en éstas, puede llevar a la replicación del PRRSV en el
endometrio con la subsecuente infección fetal a través de la placenta (Karniychuk and
Nauwynck, 2013). A su vez, las madres seropositivas pueden transmitir anticuerpos
maternos a sus camadas vía calostro, sin embargo, cuando la inmunidad pasiva declina
puede llevar a infección luego del destete cuando los animales son mezclados (OIE,
2008) como se observó en este caso en la preceba y posteriormente en la ceba. De otro
lado, las cerdas que se infectan en la fase final de la gestación pueden infectar a sus

crías de manera transplacentaria lo cual propicia el nacimiento de animales
persistentemente infectados; de igual manera el PRRSV, puede ser eliminado a través
del calostro y la leche (Wagstrom et al., 2000), favoreciendo así la diseminación, lo cual
fue evidente en los resultados positivos obtenidos en destetos y lactantes de este
estudio.
Los resultados positivos obtenidos en la preceba y la ceba dentro de las tres granjas,
indican la circulación del virus dentro de las mismas, aun sin la presencia de signos
clínicos, por lo anterior, la técnica de RT-PCR tiene ventajas para determinar la presencia
de dicha circulación viral (Duinhof et al., 2011).
Teniendo en cuenta la presencia de diferentes sitios productivos (Tabla y Figura 3-1) que
implicaba desplazamientos entre diferentes municipios, se puede inferir que algunos
animales previamente infectados o en estado virémico, al ingresar al sitio 3, evidenciaron
la enfermedad, esto como parte del traslado y las condiciones de estrés asociadas con el
mismo. La diseminación del PRRSV entre granjas es causada por la introducción de
animales infectados o por el uso de semen contaminado, mientras que la diseminación
dentro de las granjas es debida principalmente al movimiento entre corrales de animales
portadores o infectados o cuando los cerdos son mezclados durante los diferentes
periodos de producción (Kittawornrat et al., 2011).
Evans et al. (2008) y Zimmerman et al. (2006) caracterizaron la diseminación del PRRSV
dentro de las granjas bajo dos diferentes patrones: el primero donde al menos el 90 % de
los animales destetos son infectados a las 8.5 semanas de edad y el segundo donde,
entre el 20 al 40%, son infectados entre las 10 a 12 semanas de edad. Lo anterior
confirma los hallazgos obtenidos en el presente estudio donde los grupos de los
destetos, preceba y la ceba presentaron resultados positivos . Por otro lado, en un
estudio llevado a cabo en 10 granjas porcícolas en Holanda, se encontró que los grupos
con edades entre 9 a 16 semanas presentaron el más alto Odds Ratio (posibilidad que
una condición o enfermedad se presente en un grupo de población frente al riesgo de
que ocurra en otro), indicando a su vez que estos grupos son los más adecuados para la
detección de la circulación de PRRSV por RT-PCR (Duinhof et al., 2011).

Los meses de mayor presentación de resultados positivos en este estudio fueron febrero,
marzo y abril, lo que pudo coincidir con el movimiento de animales destetos o al traslado
al sitio 3 donde se presentaba la mezcla de animales infectados con animales
susceptibles. De igual manera, hacia el mes de abril se presentan normalmente cambios
climáticos en la zona tropical (incremento de lluvias) que posiblemente podrían estar
relacionados con la diseminación de patógenos en esta zona. Los anteriores hallazgos
coinciden con los obtenidos en el análisis de varianza donde se encontró un efecto
significativo del mes para cada granja y grupo etareo, donde la situación de bioseguridad
y manejo propia de cada granja se ve reflejada en los resultados obtenidos en los
diferentes grupos etareos y a su vez en los meses de mayor presentación de positivos.
Con respecto a las muestras de FO (Figura 3-10), las tres granjas presentaron resultados
dentro del punto de corte (15-32). Estos resultados indican la presencia de uno o más
animales infectados dentro de las diferentes poblaciones muestreadas, preceba y ceba,
lo cual confirma la circulación del PRRSV dentro de las granjas. A este respecto, en un
estudio llevado a cabo por Ramírez et al (2012) quienes realizaron vigilancia
epidemiológica en poblaciones porcinas usando FO, observaron mediante muestreos
consecutivos en el tiempo, que el estado sanitario dentro de los establos era muy
variable incluso entre establos en un mismo sistema productivo, como se observa en el
presente estudio.
En la infección temprana, la cantidad de virus en el FO es menor que en sangre, mientras
que en las etapas posteriores de viremia se encuentra más virus en el FO (Kittawornrat et
al., 2010). Esto puede sugerir que algunos de los animales con resultados de FO
positivos por qRT-PCR, presentaban una infección con PRRSV más avanzada,
pudiéndose encontrar igualmente dentro de las granjas, grupos de animales virémicos y
otros diseminando virus por medio de FO. Los datos obtenidos en un estudio reportado
por Kittawornrat et al (2014), donde evaluaron muestras de FO en el pre destete,
indicaron el estado endémico de la enfermedad, la cual involucra infección subclínica en
lactantes con la posterior transmisión a la población en crecimiento que ocurre cuando
son trasladados o mezclados después del destete. Con respecto a estas observaciones,
es posible especular que los grupos de lactantes y destetos evaluados en el estudio del

Valle, pudieron presentar resultados positivos al qRT-PCR en FO, favoreciendo la
circulación viral y explicando los resultados positivos en la preceba y ceba.
Los fluidos orales en forma inherente contienen factores endógenos antivirales y
antibacteriales que pueden afectar la estabilidad de los blancos de la PCR (Prickett et al.,
2010), sin embargo los virus RNA son relativamente resistentes a la degradación en
fluidos orales y el adecuado manejo de la muestra como el almacenamiento a -70°C
mantiene la integridad del genoma del PRRSV, tal como se realizó en el presente
estudio.
Los meses en los cuales se presentaron los resultados positivos para FO fueron octubre,
noviembre, abril y marzo, correspondiendo al inicio y al final del estudio lo que pudo
coincidir con el traslado de los animales al sitio 3 donde se obtuvo la mayor cantidad de
resultados positivos; a los cambios en el clima asociados con el comienzo de épocas de
lluvia y a la salida de animales para comercialización. Tal como se observó en los
hallazgos del análisis de varizanza, se presentó un efecto significativo del mes para cada
granja y grupo etáreo en estas muestras.
Las muestras de tejidos, como ya se mencionó, se tomaron de las mortalidades y de los
sacrificios de animales enfermos, si se tiene en cuenta que el origen de las muestras de
tejidos proceden de la preceba y ceba, se confirma la circulación del PRRSV dentro de
las granjas. Sin embargo, se observaron diferencias con respecto a los promedios de Ct
por granja, lo cual puede estar asociado a las diferencias de manejo, bioseguridad y al
desplazamiento entre sitios.
La detección del PRRSV en pulmón se presume asociada con la replicación del virus en
los MAPs durante la fase aguda de infección tal como se observa en la figura 3-17,
donde aunque los Ct se encontraron por encima del punto de corte, las desviaciones
indicaron presencia de genoma viral en pulmón en la granja 1, 3 y en mayor proporción
en la preceba. La cantidad de PRRSV en animales infectados, alcanza un máximo en
pulmón y sangre ente los 5 a 9 días PI y luego declina. En el pulmón la infección declina
hacia el día 20 PI, aunque el virus puede ser aislado de fluidos pulmonares por periodos
extendidos de tiempo (Johnson et al., 2004). Posterior a esta fase aguda de infección,

continua un período de persistencia viral en nódulos linfoides en un proceso que
involucra replicación. Los sitios primarios de persistencia incluyen tonsila y nódulos
linfáticos (Horter et al., 2002: Xiao et al., 2004). Se ha reportado PRRSV en tonsilas y
nódulos linfáticos hacia los 132 días después del nacimiento, en cerdos infectados in
útero (Rowland et al., 2003) y desde los 105 a 157 días en cerdos jóvenes PI (Horter et
al., 2002).
De acuerdo con los resultados obtenidos, se comprobó la presencia del virus a nivel
pulmonar lo cual pudo estar relacionado con el posible comienzo de la infección en la
preceba y ceba. A su vez, se presentaron resultados positivos en ganglios lo cual podría
corresponder a la presencia de animales persistentemente infectados en estos sitios
productivos. De acuerdo con lo observado en la figura 3-17, las granjas 1 y 3 presentaron
la presencia de genoma viral en pulmón y ganglio lo cual podría indicar, la presencia de
una posible persistencia y/o infección dentro de las mismas; por su parte la granja 2
evidenció presencia de virus en pulmón, indicando posiblemente la presencia únicamente
de infección. Lo anterior puede deberse a los efectos de los diferentes tipos de manejo
en cada una de las granjas.
Se observaron resultados positivos en las muestras de pulmón en diferentes meses del
estudio lo cual, teniendo en cuenta los resultados del análisis de varianza, indican que no
se presentó un efecto significativo del mes para cada granja y grupo etáreo analizado.
Con respecto a la respuesta de los ganglios, la positividad se evidenció principalmente
hacia abril en las tres granjas y al tener en cuenta el análisis de varianza el único efecto
significativo fue mes sin otras interacciones significativas entre los demas factores, granja
y grupo etáreo. Con respecto a lo anterior, lo que se evidencia en las muestras de tejidos
es que el mes donde se presentaron los resultados no tiene efecto sobre los resultados
obtenidos en cada una de las granjas y las interacciones obtenidas no son tan
significativas como aquellas observadas en los sueros y FO, quizá por que las muestras
de tejidos no fueron representativas en cada una de las granjas y tampoco dentro de
cada uno de los grupos etáreos presentes.
Con relación a los lavados bronquio alveolares, las tres granjas presentaron un promedio
de Ct fuera del punto de corte, donde la granja 2 fue negativa para detección viral en esta

muestra. Las desviaciones estuvieron muy cerca del punto de corte en granja 1 y 3
(figura 3-25 y 3-28). Las muestras positivas se hallaron hacia final del año en noviembre
y diciembre (Figura 3-29), lo cual pudo estar relacionado con el transporte hacia el sitio 3
o ceba para su comercialización durante el fin de año, por lo tanto, los animales podrían
estar comenzando la infección o encontrarse en la fase de persistencia. Estos hallazgos
coincidieron con los obtenidos en el correspondiente análisis estadistico, donde se
presentó un efecto significativo del mes en la granja 1, tanto para preceba como para
ceba y en la granja 3 específicamente en la ceba.
Es evidente, con relación a los diferentes tipos de muestras y técnicas, las diferencias
encontradas con cada una de ellas. A nivel serológico, es posible evidenciar que
resultados negativos por si solos no indican que la granja no presenta circulación viral, a
su vez, resultados positivos pueden estar indicando una antigua infeccion, por tal razón
es necesario usar técnicas conjuntas como ELISA con RT-PCR para conocer la
dinámica de la infección por el PRRSV. Con respecto al FO, esta es una muestra
representativa, fácil de obtener y que permite conocer la situación sanitaria de una
población, que en el caso del presente estudio permitió determinar la presencia del virus
principalmente en la preceba y ceba en las granjas 1 y 3, las cuales a su vez mostraron
condiciones de manejo similares. La presencia de virus en las muestras de tejidos y LBA,
permitió evidenciar diferentes fases de infeción en cada una de las granjas. Por lo
anterior, un programa de control podría establecerse inicialmente con la toma rutinaria de
muestras de FO para monitorear primordialmente poblaciones de destetos, preceba y
ceba por qRT-PCR. Los tejidos a partir de las mortalidades, sacrificios de animales
enfermos o colas, permitirían conocer el estado de infección en que se encuentra la
granja, ya sea en el inicio de la enfermedad o en la fase de persistencia, por tal razón
tanto el pulmón como los ganglios son muestras que permiten orientar el estado de
infección en el cual se encuentra una población dentro de una explotación. Con respecto
a los sueros, se podrían analizar rutinariamente los correspondientes a las madres,
reproductores, reemplazos, lactantes, de igual manera serían útiles para monitorear la
población en general cuando ya se han tomado diferentes medidas de bioseguridad y de
manejo, o cuando se han obtenido resultados positivos en FO y tejidos.

3.4 Conclusiones Es necesario el uso conjunto de técnicas serológicas y moleculares en el diagnóstico de
PRRSV, ya que la evaluación de cada una permite conocer la fase de infección en la cual
se encuentra el virus en una explotación. Por otro lado, las tres granjas evaluadas en
este estudio presentaron resultados positivos por qRT-PCR en diferentes tipos de
muestras lo que demostró la presencia del PRRSV en diferentes estados de la
enfermedad, donde los grupos más afectados fueron la preceba y ceba confirmando la
circulación del virus dentro de cada granja. La detección de ARN en tejidos como pulmón
y ganglios linfáticos indicaron a su vez la presencia tanto de animales virémicos, como
persistentemente infectados dentro de los grupos productivos. Los diferentes tipos de
muestras analizadas pueden ser utilizadas en las diferentes fases de un programa de
control del PRRSV.
3.5 Recomendaciones Utilizar las técnicas moleculares junto con pruebas serológicas que permitan reflejar las
diferentes fases de la infección y permitan conocer la dinámica del virus en una población
con el fin de tomar las acciones de control adecuadas.
Se recomienda hacer uso de los diferentes tipos de muestras como FO para monitoreo
rutinario de los grupos productivos de la preceba y ceba especialmente por qRT-PCR; los
tejidos como pulmón y ganglios de las mortalidades o animales enfermos o colas, que
permiten orientar el estado de infección en el cual se encuentra una población dentro de
una granja. Por otro lado, los sueros permiten monitorear las madres, reemplazos,
reproductores y lactantes, o la población total cuando se han establecido diferentes
medidas de manejo y bioseguridad o en el caso de obtenerse resultados positivos en FO
y/o tejidos.

Bibliografía
Allende, R., Lewis, T.L., Lu, Z., Rock, D.L., Kutish, G.F., Ali, A., Doster, A.R., Osorio, F.A.
1999. North American and European porcine reproductive and respiratory síndrome virus
differ in non-structural protein coding regions. J. Gen. Virol. 80: 307-315.
Benfield, D.A., Nelson, E., Collins, J.E., Harris, L., Goyal, S.M., Robison, D., Christianson,
W.T., Morrison, R.B., Gorcyca, D., Chladek, D., 1992. Characterization of swine infertility
and respiratory syndrome (SIRS) virus (isolate VR-2332). J. Vet. Diagn. Invest. 4, 127–
133.
Boddicker, N., Waide, E., Rowland, R., Lunney, J., Garrick, D., et al. 2012. Evidence for a
major QTL associated with host response to porcine reproductive and respiratory
syndrome virus challenge. Journal of Animal Science 90: 1733–1746.
Britton, P. and Cavanagh, D. 2008. Nidovirus genome organization and expression
mechanisms. In Perlam, S., Gallagher, T., and Snijder, E.J. (eds.), Nidoviruses, pp. 29-
46. American society for Microbiology Press, Washington, District of Columbia.
Cavanagh, D. 1997. Nidovirales: a new order comprising Coronaviridae and
Arteriviridae. Arch. Virol. 142, 629-633.
Chai, Z., Ma, W., Fu, F., Lang, Y., Wang, W., Tong, G., Liu, Q., Cai, X., Li, X. 2013. A
SYBR Green-based real-time RT-PCR assay for simple and rapid detection and
differentiation of highly pathogenic and classical type 2 porcine reproductive and
respiratory syndrome virus circulating in China. Arch Virol. 158(2): 407-15.
Chand, R.R., Trible, B.R., Rowland, R.R.R., 2012. Pathogenesis of porcine reproductive
and respiratory syndrome virus. Current Opinion in Virology, 2:256-263.

Duinhof, T.F., van Schaik, G., van Esch, E.J.B., Wellenberg, G.J., 2011. Detection of
PRRSV circulation in herds without clinical signs of PRRS: comparison of five age groups
to asses to preferred age group and simple size. Vet. Microbiol; 150: 180-184.
Evans, CH. M., Medley, G,F., Green, L.E. 2008. Porcine reproductive and respiratory
syndrome virus (PRRSV) in GB pig herds: farm characteristics associated with
heterogeneity in seroprevalence. BMC Veterinary Research; 4: 48.
García, A.R., Rodríguez, V., Kukielka, D. 2007. Aplicación de la RT-PCR en tiempo real
para el diagnóstico de PRRS. RCCV, 1(2); 646-653.
Harmon, K.M., Abate, S.A., Chriswell, A.J., Strait, W.A.E., 2012. Comparison of two
commercial real-time reverse transcriptase polymerase chain reaction assays for porcine
reproductive and respiratory syndrome virus. Swine Health Prod., 20: 184-188.
Horter, D.C., Pogranichniy, R.M., Chang, C.C., Evans, R.B., Yoon, K.J., Zimmerman, J.J.,
2002. Characterization of the carrier state in porcine reproductive and respiratory
syndrome virus infection. Vet. Microbiol. 86:213-228.
Murtaugh, M. 2012. International molecular and virology diagnostic workshop for PRRSV.
ICA. October 28-November 2.
Islam, Z.U., Bishop, S.C., Savill, N.J., Rowland, R.R., Lunney, J.K., Trible, B., Doeschl-
Wilson, A.B. 2013. Quantitative analysis of porcine reproductive and respiratory syndrome
(PRRS) viremia profiles from experimental infection: a statistical modelling approach.
PLoS One, 8(12): 1-13
Jeong, H. J., Song, Y. J., Lee, S. W., Lee, J. B., Park, S. Y., Song, C. S., Ha, G. W., Oh,
J. S., Oh, Y. K. & Choi, I. S. 2010. Comparative measurement of cell-mediated immune
responses of swine to the M and N proteins of porcine reproductive and respiratory
syndrome virus. Clin Vaccine Immunol. 17: 503–512.

Johnson, W., Roof, M., Vaughn, E., Christopher-Hennings, J., Johnson, C.R., Murtaugh,
M.P., 2004. Pathogenic and humoral responses to porcine reproductive and respiratory
syndrome virus (PRRSV) are related to viral load in acute infection. Vet. Inmunol.
Inmunopathol- 102:233-247.
Johnson, C.R., Griggs, T.F., Gnanandarajah, J., Murtaugh, M.P. 2011. Novel structural
protein in porcine reproductive and respiratory syndrome virus encoded by an alternative
ORF5 present in all arteriviruses. Journal of General Virology, 92: 1107–1116.
Karniychuk, U.U and Nauwynck, H.J. 2013. Pathogenesis and prevention of placental and
transplacental porcine Reproductive and respiratory syndrome virus infection.
VeterinaryResearch, 44:95.
Kittawornrat, A., Prickett, J., Chittick, W., Wang, C., Engle, M., Johnson, J., Patnayak, D.,
Schwartz, T., Whitney, D., Olsen, C., Schwartz, K., Zimmerman, J., 2010. Porcine
reproductive and respiratory syndrome virus (PRRSV) in serum and oral fluid samples
from individual boars: Will oral fluid replace serum for PRRSV surveillance? Virus
Research 154, 170–176.
Kiyyawornrat, A., Panyasing, Y., Irwin, C., Rademacher, c., Hoogland, M., Wang, C.,
Johnson, J., Main, R., Rowland, R., Zimmerman, J. 2011. PRRS virus antibody detection
in oral fluid samples: The 1st generation PRRS oral fluid ELISA is ready for prime time.
International symposium on emerging and re-emerging pig disease. Allen D. Leman
Swine Conference.
Kiyyawornrat, A., Panyasing, Y., Irwin, Goodell, C., Wang, C., Gauger, P., Harmon, K.,
Rauh, R., Desfresne, L., Levis, I., Zimmerman, J. 2014. Porcine reproductive and
repiratory syndrome virus (PRRSV) surveillance usin pre weaning oral fluid samples
detects circulation of wild type PRRSV. Veterinary microbiology, 168: 331-339.

Le Gall, A.O., Legeay, O., Bourhy, H., Arnauld, C., Albina, E., Jestin, A. 1998. Molecular
variation in the nucleoprotein gene (ORF7) of the porcine reproductive and respiratory
syndrome virus (PRRSV). Virus Research, 54: 9-21.
Mackay, I.M., Arden, K.E., Nitsche, A. 2002. Real-time PCR in virology. Nucleic Acids
Res; 30:1292-1305.
Martínez, E., Riera, P., Sitja, M., Fang, Y., Oliveira, S., Maldonado, J., 2008.
Simultaneous detection and genotyping of porcine reproductive and respiratory síndrome
virus (PRRSV) by real-time RT-PCR and amplicon melting curve analysis using SYBR
Green. Research in Veterinary Science; 85: 184-193.
Meng, X. J., Paul, P.S., Halburg, P.G., Morozov, I. 1995a. Sequence comparison of open
reading frames 2 to 5 of low and high virulence United States isolates of porcine
reproductive and respiratory syndrome virus. J Gen Virol. 76: 3181-3188.
Meng, X. J., Paul, P.S., Halbur, P.G., Lum, M.A. 1995b. Phylogenetic analyses of the
putative M (ORF6) and N (ORF7) genes of porcine reproductive and respiratory
syndrome virus (PRRSV): implication for the existence of two genotypes of PRRSV in the
U.S.A. and Europe. Arch. Virol. 140: 745-755.
Meng, W, J., 2000. Heterogeneity of porcine reproductive and respiratory syndrome virus:
implications for current vaccine efficacy and future vaccine development. Vet. Microbiol.
74: 309-329.
Murtaugh, M.P., Elam, M.R., Kakach, L.T. 1995. Comparison of the structural protein
coding sequences of the VR-2332 and Lelystad virus strains of the PRRS virus. Arch.
Virol. 140: 1451-60.
Nelsen, C.J., Murtaugh, M.P., Faaberg, K.S.1999. Porcine reproductive and respiratory
síndrome virus comparison: divergent evolution on two continents. J. Virol. 73: 270-280.

Nelson, E.A., Christopher-Hennings, J., Drew, T., Wensvoort, G., Collins, J.E., Benfield,
D.A., 1993. Differentiation of U.S. and European isolates of porcine reproductive and
respiratory syndrome virus by monoclonal antibodies. J Clin Microbiol; 31: 3184:3189.
Neumann, E.J.,Kliebenstein, J.B., Johnson, C.D., Mabry, J.W., Bush, E.J., Seitzinger,
A.H., Green, A.L. & Zimmerman, J.J. 2005. Assessment of the economic impact of
porcine reproductive and respiratory syndrome on swine production in the United
States.Journal of the American Veterinary Medical Association. 227(3): 385-392.
Olsen, C., Wang, Ch., Christopher-Hennings, J., Doolitle, K., Harmon, K., Abate, S.,
Kittawornrat, A., Lizano, S., Main, R., Nelson, E., Otterson, T., Panyasing, Y.,
Rademacher, C., Rauh, R., Saha, R., Zimmerman, J. 2013. Probability of detecting
porcine reproductive and respiratory syndrome virus infection using pen-based swine oral
fluid specimens as a function of within-pen prevalence. Journal of Veterinary Diagnostic
Investigation, XX(X); 1-8.
Prickett, J. R., Cutler, S., Kinyon, J.M., Naberhaus, N., Stensland, W.R., Yoon, K.J.,
Zimmerman, J.J. 2010. Stability of porcine reproductive and respiratory syndrome virus
and antibody in swine oral fluid. Journal of swine health and production, 18: 187-195.
PRRS: the disease, its diagnosis, prevention and control. Appendices IV and V of the
report of the OIE ad hoc group on porcine reproductive respiratory syndrome Paris, 9 - 11
June 2008.
Ramírez, A., Wang, C., Prickett, J.R., Pogranichniy, R., Yonn, K.J., Main, R., Johnson,
J.K., Rademacher, C., Hoogland, M., Hoffman, P., Kurtz, A., Kurtz, E., Zimmerman, J.,
2012. Efficient surveillance of pig populations using oral fluids. Prev. Vet. Med. 104:292-
300.
Rovira, A., Clement, T., Christopher-Hennings, J., Thompson, B., Engle, M., Reicks, D.,
Muñoz-Zanzi, C. 2007. Evaluation of sensitivity of reverse-transcription polymerase chain
reaction to detect porcine reproductive and respiratory syndrome virus on individual and
pooled samples from boars. J. Vet. Diagn. Invest., 19: 502-509.

Rowland, R.R.R., Lawson, S., Rossow, K., Benfield, D.A., 2003. Lymphoid tissue tropism
of porcine reproductive and respiratory syndrome virus replication during persistent
infection of pigs originally exposed to virus in utero. Vet. Microbiol, 96:219-235.
Rubio, A., Rodriguez, V., Kukielka, D. 2007. Aplicación de la RT-PCR en tiempo real para
el diagnóstico de PRRS. RCCV, 1: 646-653.
Shaw, A.E., Reid, S.M., Ebert, K., Hutchings, G.H., Ferris, N.P., King, D.P. 2007.
Implementation of a one-step-real-time RT-PCR protocol for diagnosis of foot-and-mouth
disease. Journal of Virology Methodsd, 143: 81-85.
Shugars, D.C. 1999. Endogenous mucosal antiviral factors of the oral cavity. J. Infect.
Dis., 179: 431-435.
Suarez, P., Zardoya, R., Martin, M.J., Prieto, C., Dopazo, J., Solana, A., Castro, J.M.
1996. Phylogenetic relationships of European strains of porcine reproductive and
respiratory syndrome virus (PRRSV) inferred from DNA sequences of putative ORF5 and
ORF7 genes. Virus Research, 42: 159-165.
Tian, H., Wu, J.Y., Shang, Y.J.,Cheng, Y., Liu, X.,T.2010. The developmente of rapid
SYBR one step real-time RT-PCR for detection of porcine reproductive and respiratory
syndrome virus. Virology Journal, 7;90.
Wangstrom, E.A., Chang, C.C., Yoon, K.J., and Zimmermann, J. J. 2000. Shedding of
porcine reproductive and respiratory syndrome virus in mammaria gland secretions of
sows. Am. J. Vet. Res., 62: 1876-1880.
Wasilk, A., Callahan, J.D., Christopher-Hennings, J., Gay, T.A., Fang, Y., Dammen, M.,
Reos, M.E., Torremorell, M., Polson, D., Mellencamp, M., Nelson, E., Nelson, W.M.,
2004. Detection of U.S., Lelystad, and European-like porcine reproductive and respiratory
syndrome viruses and relative quantitation in boar semen and serum samples by real-
time PCR. Journal of Clinical Microbiology 42, 4453–4461.

Wensvoort, G., Terpstra, D., Pol, J.M.A., Ter Laak, E.A., Bloemraad, M., de Kluyver, E.P.,
Kragten, D., Van Buiten, L., Den Besten, A., Wagenaar, F., Broekhuijsen, J.M., Moonen,
P.L.J.M., Zetstra, T., De Boer, E.A., Tibben, H.J., de Jong, M.F., Van´t Veld, P.,
Groenland, G.J.R., Van Gennep, J.A., Voets, M.Th., Verheijden, J.H.M., Braamskamp, J.
1991. Mystery swine disease in the Netherlands: the isolation of Lelystad virus. Vet. Q.
13: 121-130.
Wernike K., Bonilauri, P., Dauber, M., Errington, J., LeBlanc, N., Revilla-Fernandez, S.,
Hjulsager, C., Isaksson, M., Stadejek, T., Beer, M., Hoffmann, B. 2012. Porcine
reproductive and respiratory syndrome virus. Interlaboratory ring trial to evaluate real-time
reverse transcription polymerase chain reaction detection method. J. Vet. Diagn. Invest.,
24: 855-866.
Wills, R.W.,Zimmerman, J.J. 1997. Porcine reproductive and respiratory syndrome virus:
a persistent infection. Vet Microbiol, 55:231-240.
Wills, R.W., Doster, A.R., Galeota, J.A., Sur, J.H., Osorio, F.A., 2003. Duration of infection
and proportion of pigs persistently infected with porcine reproductive and respiratory
síndrome virus. J Clin Microbiology; 41: 58-62.
Wissink, E.H.J., Kroese, M.V., van Wijk, H.A.R., Rijsewijk, F.A.M., Meulenberg, J.J.,
Rottier, P.J.M., 2005. Envelope protein requirements for the assembly of infectious virions
or porcine reproductive and respiratory syndrome virus. J. Virol. 79, 12495–12506.
Xiao, Z., Batista, L., Dee, S., Halbur, P., Murtaugh, M.P., 2004. The level of virus-specific
T-cell and macrophage recruitment in porcine reproductive and respiratory syndrome
virus infection in pigs is independent of virus load. J. Virol. 78, 5923–5933.
Zimmerman, J., Benfield, D.A., Murtaugh, M.P., Osorio, F., Stenson, G.W., Torremorell,
M., 2006. Porcine reproductive and respiratory syndrome virus (porcine arterivirus).
Diseases of swine, 9th 387-418. Blackwell publishing, Ames, Iowa.

Zipper,H., Brunner, H., Bernhagen, J., Vitzthum, F. 2004. Investigations on DNA
intercalation and surface binding by SYBR green I, its structure determination and
methodological implications. Nucleic Acids Res. 32: 103.

Detección y determinación genotípica del
virus del Síndrome Reproductivo y
Respiratorio Porcino (PRRSV) mediante
aislamiento en células MA - 104 y
secuenciación de muestras provenientes de
tres granjas del Valle del Cauca.
Resumen En Colombia a finales de los años noventa, se realizó un estudio para determinar el
genotipo al cual correspondían las cepas circulantes en granjas intensivas y extensivas
del país, los resultados indicaron que estas pertenecían al genotipo americano, al mismo
tiempo se obtuvieron aislados en MAPs y células MARC 145, lo cual demostró la
capacidad de recuperación del virus en diferentes regiones y sistemas de producción. El
presente estudio buscó aislar las cepas actualmente circulantes en tres granjas de una
zona de alta produción porcícola en el país y con condiciones de manejo y bioseguridad
diferentes. Para lo anterior se tomaron muestras de sueros, tejidos y lavados
bronquioalveolares. De igual manera se intentó aislar el virus en estas muestras
utilizando la línea celular MA104. Los resultados obtenidos confirmaron que las cepas
aisladas en estas granjas correspondían al genotipo americano, sin embargo estos
estudios deben ser realizados frecuentemente para determinar las variaciones de las
cepas existentes. De igual manera aunque la línea celular utilizada fue susceptible al
PRRSV, es conveniente utilizar adicionalmente otro tipo celular, para aislar
eficientemente cepas de campo.
Palabras clave: Análisis filogenético, PRRSV, Secuenciación.

Abstract
In Colombia at the end of the 1990s, a study was conducted to determine the PRRSV
genotype circulating in intensive and extensive farms of the country. Results indicated
that isolated strains obtained on MAPs and MARC 145 cells belonged to the American
genotype, which showed resilience of the virus in different regions and production
systems. The goal of the present study was to isolate viral strains currently circulating
within three farms with different management and biosecurity conditions in an area of high
pig production in Colombia. Samples of tissues and broncho alveolar lavages were
employed to try viral isolation on MA104 cells. Them Results confirmed that strains
isolated in these farms corresponded to the American genotype, however these kind of
studies must be performed frequently to determine variations on existing strains. In the
same way, although the cell line employed is susceptible to the PRRSV infection it is
convenient to use another cell types as primary culture of pulmonary macrophages to
obtain a more efficient system to isolate PRRSV field strains.
Keywords: Phylogenetic analyses, PRRSV, Sequencing
Introducción
El Síndrome Reproductivo y Respiratorio Porcino (PRRS) es una enfermedad que afecta
a la especie porcina alrededor del mundo ocasionando significativas pérdidas
económicas (Neumann et al., 2005). Las manifestaciones clínicas de la enfermedad
incluyen fallas reproductivas caracterizadas por presencia de abortos, mortinatos y
momificaciones, como alteraciones respiratorias asociadas a neumonía intersticial linfo-
mononuclear (Rossow et al., 1994).
El virus del PRRS pertenece a la familia Arteriviridae, orden Nidovirales que incluye el
virus de la Arteritis Equina (EAV), el virus Elevador de la Deshidrogenasa Láctica del
Ratón (LDV) y el virus de la Fiebre Hemorrágica del Simio (SHFV) (Conzelman et al.,
1993). El PRRSV fue identificado a finales de los años ochenta, simultáneamente en
Europa (Wensvoort et al., 1991) y en los Estados Unidos (Benfield et al., 1992), desde

entonces las diferencias genómicas y antigénicas han llevado a distinguir dos diferentes
genotipos, el Europeo o tipo I (cepa Lelystad) y el Americano o tipo II (cepa VR-2332)
(Meng et al.,1995). Los dos genotipos comparten alrededor del 70% de identidad a nivel
de nucleótidos (Allende et al., 2000) y 50-80% de similaridad entre aminoácidos
(Forsberg, 2005). El genotipo II ha sido rutinariamente aislado en células de simio MA-
104 y su subclon MARC-145 mientras que el genotipo I ha presentado dificultades para
adaptarse a estas células (Murtaugh et al., 2010). Con respecto a la presentación clínica,
el tipo I se ha caracterizado por causar enfermedad reproductiva en el último tercio de la
gestación, mientras que el II causa enfermedad tanto reproductiva como respiratoria. Las
formas más virulentas se han observado en las cepas del genotipo II y la mayor
diversidad genética se presenta en el genotipo I (Stadejek et al., 2006, 2008). La
divergencia no solo se presenta entre los dos genotipos, los aislados individuales de
PRRSV y los subtipos dentro de cada genotipo puede variar aproximadamente en un
20% a nivel de nucleótidos (Han et al.,2006), por ejemplo variantes de PRRSV tipo I
(subtipos 2, 3 y 4) identificadas en el oriente de Europa (Stadejek et al., 2006, 2002) son
más virulentas que el subtipo I, debido probablemente a un aumento en la actividad de
citoquinas en el huésped (Morgan et al., 2013; Weesendorp et al., 2013). Una
manifestación importante de la variabilidad genómica del PRRSV, especialmente dentro
de las poblaciones de cerdos que albergan múltiples genotipos circulantes, es la
diversidad antigénica resultante (Liao et al., 2013).
El genoma viral es lineal, ARN de cadena sencilla y polaridad positiva, con una
estructura CAP en el extremo 5´y un tallo poly (A) en el extremo 3´. Posee al menos diez
fragmentos de lectura abierta (ORFs) (Johnson et al., 2011; Firth et al., 2011). Los
primeros ORF´s (ORF1a y ORF1b) codifican poli-proteínas no estructurales relacionadas
con la transcripción y replicación del genoma (Snijder and Meulenberg., 2001). Las
proteínas estructurales codificadas en ORF2 a ORF7 son expresadas a partir de seis
RNAm que presentan estructuras CAP y son poliadenilados. La mayor proteína de la
cubierta es la glicoproteína GP5 codificada por ORF5 del genoma viral (Mardassi et al.,
1995; Meulenberg et al., 2000).
El PRRSV se caracteriza por su inestabilidad genética en ciertos segmentos del genoma;
las secuencias de nucleótidos cambian frecuentemente debido a puntos de mutación,

deleciones, inserciones o recombinaciones. El ORF5 es usado para diferenciar el
PRRSV, ya que esta porción de la cubierta del virus se expone tanto al sistema inmune
como a la presión del medio y permite así diferenciar aquellas cepas que mantienen su
proceso replicativo de otras cepas o de otros patógenos (Dee et al., 2001; Chang et al.,
2002). De igual manera la diversidad genética que muestra el gen ORF5, es utilizado
como blanco principal en secuenciación del PRRSV permitiendo diferenciar aislados de
campo de aislados vacunales (Murtaugh, 2012).
El PRRSV infecta in vivo células de la línea macrófago-monocito en particular
macrófagos alveolares porcinos (MAPs); in vitro, el virus ha sido aislado en cultivos
primarios de MAPs y líneas celulares inmortalizadas como MARC-145 y CL2621
(subclones de MA-104 línea celular de riñón de mono) (Bautista et al., 1993; Kim et al.,
1993; Mengeling et al., 1996). El PRRSV se une a las células blanco mediante la
interacción entre la glicoproteína viral GP5 y la sialodhesina porcina, mientras que la
internalización y decapsidación requieren la presencia del trímero formado por las
proteínas GP2, GP3 y GP4 que interactúan con el CD163 del huésped (Lu et al., 2014).
En Colombia entre los años de 1997 y 1999 se realizaron aislamientos del PRRSV a
partir de muestras de sueros y pulmones en células MARC-145; se obtuvieron un total de
45 aislamientos, de los cuales 24 correspondieron a animales confinados y 9 a no
confinados; 14 aislamientos se obtuvieron de muestras de sueros provenientes de
animales con edades entre 6-8 semanas. A su vez se realizaron análisis filogenéticos
que demostraron que en ese momento las cepas circulantes en el país correspondían al
genotipo Americano (Mogollón et al., 2003).
Debido al tiempo transcurrido desde la realización de dichos aislamientos y análisis
filogenético, se estableció la realización de este estudio con el fin de aislar las cepas del
PRRSV circulantes en tres granjas ubicadas en el Valle del Cauca, con el fin de
determinar posteriormente mediante secuenciación el genotipo predominante dentro de
cada una de ellas y asÍ confirmar los hallazgos obtenidos previamente.

4.1 Materiales y Métodos
4.1.1 Ubicación geográfica y características de las granjas
El estudio se realizó en tres granjas de tipo intensivo y ciclo completo, ubicadas en el
Departamento del Valle del Cauca, localizado en el sur occidente del país con una altura
de 1400-1600 msnm, una temperatura promedio de 23-24°C y una humedad relativa del
65% a 75% (Figura 2-1). Esta zona se caracteriza por una producción porcícola de tipo
intensivo, que presento en los meses previos al estudio reportes de animales con
sintomatología compatible con PRRS.
4.1.2 Muestras Las muestras utilizadas fueron tejidos (pulmón - ganglios) y lavados bronquioalveolares;
estos se obtuvieron a partir de las mortalidades que se presentaron en los días de
muestreo o a través del sacrificio voluntario en cada granja. Este procedimiento fue
avalado por el Comité de Bioética de la Facultad de Medicina Veterinaria y de Zootecnia
de la Universidad Nacional de Colombia. Se tomaron muestras de 2 o 3 animales en
cada mes durante la duración del estudio. Las muestras se recolectaron a partir de
Septiembre de 2012 hasta Mayo de 2013.
4.1.2.1 Lavados Bronquioalveolares (LBA)
Las muestras de LBA se obtuvieron por perfusión de la cavidad pulmonar mediante
lavado bronco alveolar con un Buffer Fosfato Salino (PBS) pH 7.2+/-0.2 (Anexo 1), más
solución de penicilina-estreptomicina (PEST) (Anexo 2). Una vez expuesta la cavidad
torácica, se cerró la tráquea con un fórceps y los pulmones se lavaron externamente con
agua destilada. Posteriormente, se introdujeron 500-600 ml de solución salina, se
frotaron los pulmones durante 2-3 minutos al cabo de los cuales el líquido del lavado se
recuperó en un frasco estéril, este proceso se repitió 2-3 veces. El líquido obtenido se
centrifugo a 5000 x g por 10 minutos, se elimino el pellet y se repitió dos veces más,
posteriormente se almaceno a -70°C hasta su utilización.

4.1.2.2 Tejidos
De las mortalidades presentadas, se colectaron muestras de tonsila, pulmón, bazo y
ganglio linfáticos, cada uno de 2x2 cm de diámetro que se conservaron en bolsas nazco
y en congelación -70°C hasta su utilización. Para el estudio en particular, se utilizaron
únicamente las muestras de pulmón y ganglio (1 g), las que se sonicaron durante 1
minuto en un Ultrasonic homogenizer Cole-Parmer Instruments Co., adicionando luego 1
ml de medio MEM 199 (Gibco-Ref 31100-035) sin suplementar. En el caso de las
muestras para aislamiento se prepararon previamente, adicionando a 1 ml de muestra, 9
ml de MEM con 1% de PEST (Pen Strep-Gibco-Ref 15140), se centrifugaron a 3000 g x
10 minutos, el sobrenadante se filtro en membrana 0.40, para la posterior inoculación de
las células. Las muestras para qRT-PCR fueron centrifugadas a 5000 g x 3 minutos y el
sobrenadante fue utilizado para el proceso de extracción correspondiente.
4.1.3 Virus
Se utilizó la cepa viral PRRS 3P (genotipo americano), Referencia PDV0401, Lote 1,
proveniente del National Veterinary Service Laboratories (NVSL) APHIS-USDA. Se
realizó una dilución 1:10 del virus con medio MEM sin suplementar para su posterior uso
como control positivo en la prueba de aislamiento.
4.1.4 Aislamiento viral
4.1.4.1 Células
Se utilizaron monocapas de células MA-104 a una concentración inicial de 2 x 106 en
placas de 24 pozos. Estas fueron cultivadas en en medio MEM 199 (Gibco-Ref 31100-
035) y suplementadas con 10% de Suero Fetal Bovino (SFB) (Gibco-Cat 26250-084) y
0.1% de Penicilina-Estreptomicina, (Pen Strep-Gibco-Ref 15140). Luego de 24-48 horas
de incubación, y con un 80% de confluencia, las células fueron inoculadas con las
muestras previamente preparadas (ítem 4.1.2) de pulmón, ganglio, lavado bronquio
alveolar y con el control del PRRSV.
Los inoculos se utilizaron tanto puros como en dilución 1:10, en medio MEM sin
suplementar. Para la inoculación, inicialmente se eliminó el medio de cultivo y se realizó
el lavado de las celulas con 200 µl de Buffer Fosfato Salino (PBS) para cultivo celular
(Anexo 3), posteriormente se adicionaron 200 µl de los inoculos (puros y 1:10). Se

emplearon dos pozos como control positivo y dos como control celular. Luego de una
hora de adsorción a 37°C y 5% de CO2 se adicionó 1 ml por pozo del medio MEM
suplementado con 5% de SFB y 1% de antibióticos (penicilina-estreptomicina) (Anexo 2).
Las muestras se incubaron entre 48-72 horas a 37°C y 5% de CO2, observando
diariamente la monocapa para determinar la presencia de efecto citopático. Se colectó el
sobrenadante por muestra, se conservó a -70°C para la posterior infección de nuevas
monocapas de MA-104. Se realizaron tres pasajes para cada muestra. No se realizo
congelación de células ya que no se contaba con el correspondiente anticuerpo
monoclonal para la detección mediante fluorescencia.
4.1.5 Secuenciación A partir de las muestras de sueros, tejidos, fluidos y lavados bronquio alveolares positivas
por qRT-PCR, se realizó la extracción del ARN viral con el kit RNeasy mini (74106) de
QUIAGEN. Para la realización de la transcripción reversa se utilizó el kit M-MLV Reverse
Transcriptase (28025-013) de Invitrogen, donde para un volumen final de 10 µl se
adicionaron 5 µl de los Hexámeros (7.5 ng/µl) y 5 µl de ARN. Las condiciones dadas
fueron 1 ciclo a 75°C por 3 minutos. Posteriormente para un volumen final de 20 µl se
mezclaron 4 µl de Agua, 4 µl de Buffer 5x, 1 µl de dNTPs 10mM. 0.5 µl de Inhibidor de
RNAsas, 0.5 µl de Enzima MMLV 200U/ µl y 10 µl de ARN más hexámeros. Las
condiciones dadas fueron 1 ciclo a 37°C por 50 minutos y 1 ciclo a 70°C por 15 minutos.
Los primers utilizados en la PCR convencional fueron los siguientes:
Tabla 4-1. Descripción de los Primers utilizados en la secuenciación
PRIMER SECUENCIA (5´- 3´) ORF5 FEd (Ext) ATG TGC GRY TGC TYC ATT TCA TGA CAC C (R= A, G – Y= C, T)
ORF5 REd (Ext) AGG TGC ARA AGC CCY AGC AGT CGG CCG C (R= A, G – Y= C, T)
ORF5 FId (Int) TCA TGA CAC CTG ARR CCA TGA GGT GGG C (R=A,G)
ORF5 RId (Int) GCC AAD AKC ACC TTT TST GGA GSC GTG C (K=G,T) (D=A,G,T) (S=G,C)
Se realizó una PCR anidada con el kit Go Taq Flexi DNA Polymerase (M829) de
Promega. Para la PCR externa se mezclaron 10.66 µl de agua, 5 µl de Buffer Green 5x,
2 µl de Cloruro de Magnesio 25 mM, 0.5 µl de dNTPs 10mM, 0.87 µl de primer FEd
20pmol/ µl, 0.87 µl de primer REd 20pmol/ µl, 0.1 µl de Enzima 5U/ µl y 5 µl de ADNc
para un volumen final de 25 µl. Las condiciones utilizadas fueron: 1 ciclo a 94°C durante
2 minutos, 30 ciclos a 94°C por 30 segundos, 60 °C por 30 segundos, 72°C por 45
segundos, 1 ciclo por 72°C por 10 minutos. Para la PCR interna se mezclaron 30.52 µl de

agua, 11 µl de Buffer Green 5X, 4.4 µl de Cloruro de Magnesio, 1.1 µl de dNTPs, 1.914 µl
de primer FId, 1.914 µl de primer RId, 0.22 µl de Enzima Taq Polimerasa y 4.4 µl del
producto de la PCR externa, para un volumen final de 55 µl. Las condiciones utilizadas
fueron: 1 ciclo a 94°C durante 2 minutos, 30 ciclos a 94°C por 30 segundos, 65 °C por 30
segundos, 72°C por 45 segundos, 1 ciclo por 72°C por 10 minutos.
A partir de la PCR se tomo un volumen de 3 µl, que se utilizó para la realización de la
electroforesis en gel de agarosa al 1.5%, con el fin de determinar la banda de 729 pb
correspondientes al amplicon de la región ORF5. Para la purificación se tomaron 50 µl
de producto de la PCR interna utilizando el QIAquick PCR purification kit (Quiagen, Cat.
281206). La seuenciacion se hizo utilizando el kit Big Dye Terminator v 1.1, v 3.1, 5x
Sequencing Buffer (Applied Biosystems, Ref. 4336697), donde el producto se purifico de
nuevo utilizando el BigDye X Terminator (Applied Biosystems, Cat. 4376486),
posteriormente se sometió a agitación durante una hora a 1200 rpm por 1 minuto, se
centrifugó a 3000 rpm por 3 minutos y a partir de aquí se tomaron 10 µl que se llevaron a
una placa de 96 pozos para secuenciar en una Analizador genético 3130 Applied
Biosystems. Una vez obtenidos los correspondientes concensos de las muestras
analizadas, estos fueron alineados frente a varias cepas tanto americanas como
europeas mediante el programa Clustal W2 y posteriormente se realizó el árbol
filogenético por medio del programa Mega 5 (Figura 4-2). Las muestras fueron
secuenciadas en el Laboratorio Nacional de Diagnóstico Veterinario (LNDV) del ICA.

Tabla 4-2 Descripción de las cepas americanas y europeas utilizadas en la realización del
árbol filogenético de las muestras concenso IDENTIFICACION NCBI CEPAS
EF442773.1|:1716-2318
Porcine respiratory and reproductive syndrome virus strain JA142 glycosylated minor envelope protein (GP2a), non-glycosylated minor envelope protein (GP2b), glycosylated minor envelope protein (GP3), glycosylated minor envelope protein (GP4), major envelope protein (GP5), membrane protein (M), and nucleocapsid protein (N) genes, complete
cds
DQ988080.1|:13790-14392 Porcine respiratory and reproductive syndrome virus isolate Ingelvac ATP, complete genome
AY424271.1|:13790-14392 Porcine reproductive and respiratory syndrome virus strain JA142, complete genome
AF494042.1|:13772-14374 Porcine reproductive and respiratory syndrome virus isolate P129, complete genome
EF112445.1|:13698-14300 Porcine respiratory and reproductive syndrome virus strain JXA1, complete genome
AY032626.1|:13789-14391 Porcine reproductive and respiratory syndrome virus strain CH-1a, complete genome
AF184212.1|:13897-14499 Porcine reproductive and respiratory syndrome virus strain SP, complete genome
DQ176019.1|:13396-13998 Porcine respiratory and reproductive syndrome virus isolate MN184A, complete genome
83588787|gb|DQ306878.1| Porcine respiratory and reproductive syndrome virus isolate 98-3403 glycoprotein 5 gene, complete cds
83588785|gb|DQ306877.1| Porcine respiratory and reproductive syndrome virus isolate 98-3298 glycoprotein 5 gene, complete cds
83588789|gb|DQ306879.1| Porcine respiratory and reproductive syndrome virus isolate 99-3584 glycoprotein 5 gene, complete cds
AF331831.1|:13786-14388 Porcine reproductive and respiratory syndrome virus BJ-4, complete genome
AF066183.4|:13789-14391 Porcine reproductive and respiratory syndrome virus RespPRRS MLV, complete genome
AF176348.2|:13788-14390 Porcine reproductive and respiratory syndrome virus isolate PA8 complete genome
U87392.3|PRU87392:13788-14390 Porcine reproductive and respiratory syndrome virus strain ATCC VR-2332, complete genome
343795953|gb|JN411262.1| Porcine reproductive and respiratory syndrome virus isolate snu100057 major envelope protein gene, complete cds
63109218|gb|DQ009647.1| Porcine respiratory and reproductive syndrome virus strain CRESA-VP21 envelope glycoprotein (ORF5) gene, complete cds
M96262.2|LEYPOLYENV:13494-14099
Lelystad virus, complete genome
58532741|gb|AY743931.1| Porcine reproductive and respiratory syndrome virus isolate Porcilis GP5 gene, complete cds
JF802085.1|:13405-14010 Porcine reproductive and respiratory syndrome virus strain lena, complete genome
GU047345.1|:13464-14069 Porcine reproductive and respiratory syndrome virus isolate NMEU09-1, complete genome
EU076704.1|:13470-14075 Porcine respiratory and reproductive syndrome virus isolate HKEU16, complete genome
AY366525.1|:13443-14048 Porcine reproductive and respiratory syndrome virus strain EuroPRRSV, complete genome
KC862567.1|:13494-14099 Porcine reproductive and respiratory syndrome virus isolate DK-2011-05-11-14, complete genome
AF325691.1|:13766-14368 Porcine reproductive and respiratory syndrome virus isolate NVSL 97-7985 IA 1-4-2, complete genome
KC862577.1|:13788-14390 Porcine reproductive and respiratory syndrome virus isolate DK-2011-030311-1, complete genome
DQ473474.1|:13788-14390 Porcine respiratory and reproductive syndrome virus isolate LMY, complete genome
192806901:13778-14380 Porcine respiratory and reproductive syndrome virus genomic RNA, complete genome, strain: EDRD-1
JN660150.1|:13345-13947 Porcine reproductive and respiratory syndrome virus strain NADC31, complete genome
KF611905.1|:13397-13999 Porcine reproductive and respiratory syndrome virus isolate HENAN-XINX, complete genome
EU807840.1|:13786-14388 Porcine respiratory and reproductive syndrome virus isolate CH-1R, complete genome
EU262603.1|:13584-14186 Porcine respiratory and reproductive syndrome virus strain Em2007, complete genome
KF287140.1|:13764-14366 Porcine reproductive and respiratory syndrome virus isolate HK13, complete genome
JQ715697.1|:13661-14263 Porcine reproductive and respiratory syndrome virus strain NVDC-GD2-2011, complete genome
EU939312.1|:13698-14300 Porcine respiratory and reproductive syndrome virus strain JSyx, complete genome
GQ374442.1|:13698-14300 Porcine reproductive and respiratory syndrome virus strain GDBY1, complete genome
JQ663556.1|:13698-14300 Porcine reproductive and respiratory syndrome virus strain 10-10QN, complete genome
HQ315837.1|:13698-14300 Porcine reproductive and respiratory syndrome virus isolate SY0909, complete genome
EF635006.1|:13698-14300 Porcine respiratory and reproductive syndrome virus strain HUN4, complete genome
FJ889130.1|:13697-14299 Porcine respiratory and reproductive syndrome virus strain CWZ-1-F3, complete genome
JX512910.1|:13698-14300 Porcine reproductive and respiratory syndrome virus strain SRV07, complete genome
JQ663545.1|:13698-14300 Porcine reproductive and respiratory syndrome virus strain 10-10BJ-5, complete genome
GQ857656.1|:13698-14300 Porcine reproductive and respiratory syndrome virus strain SX-1, complete genome
FJ349261.1|:13434-14039 Porcine respiratory and reproductive syndrome virus strain KNU-07, complete genome
AY588319.1|:13494-14099 PRRSV LV4.2.1, complete genome
DQ489311.1|:13443-14048 Porcine reproductive and respiratory syndrome virus strain SD01-08, complete genome
GQ461593.1|:13494-14099 Porcine reproductive and respiratory syndrome virus strain SHE, complete genome
EF484033.1|:13789-14391 Porcine respiratory and reproductive syndrome virus clone pMLV, complete genome
CONCENSO 1 Lavado bronquioalveolar Granja 1 preceba
CONCENSO 2 Lavado bronquioalveolar Granja 1 ceba
CONCENSO 3 Lavado bronquioalveolar Granja 3 Ceba
4.2 Resultados
4.2.1 Aislamiento Se analizaron 205 muestras de tejidos (103 pulmones y 102 ganglios linfáticos) y 98
muestras de LBA. Las muestras donde fue posible el aislamiento del virus se presentan
en la Tabla 4-4.
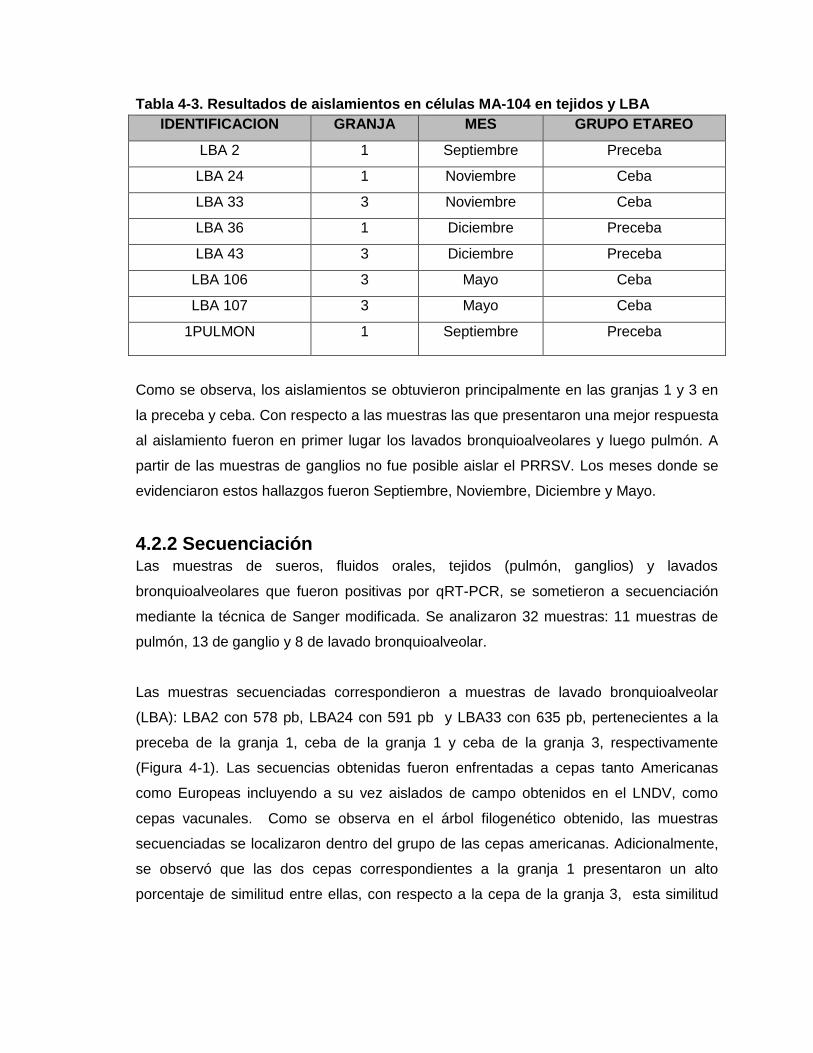
Tabla 4-3. Resultados de aislamientos en células MA-104 en tejidos y LBA
IDENTIFICACION GRANJA MES GRUPO ETAREO
LBA 2 1 Septiembre Preceba
LBA 24 1 Noviembre Ceba
LBA 33 3 Noviembre Ceba
LBA 36 1 Diciembre Preceba
LBA 43 3 Diciembre Preceba
LBA 106 3 Mayo Ceba
LBA 107 3 Mayo Ceba
1PULMON 1 Septiembre Preceba
Como se observa, los aislamientos se obtuvieron principalmente en las granjas 1 y 3 en
la preceba y ceba. Con respecto a las muestras las que presentaron una mejor respuesta
al aislamiento fueron en primer lugar los lavados bronquioalveolares y luego pulmón. A
partir de las muestras de ganglios no fue posible aislar el PRRSV. Los meses donde se
evidenciaron estos hallazgos fueron Septiembre, Noviembre, Diciembre y Mayo.
4.2.2 Secuenciación Las muestras de sueros, fluidos orales, tejidos (pulmón, ganglios) y lavados
bronquioalveolares que fueron positivas por qRT-PCR, se sometieron a secuenciación
mediante la técnica de Sanger modificada. Se analizaron 32 muestras: 11 muestras de
pulmón, 13 de ganglio y 8 de lavado bronquioalveolar.
Las muestras secuenciadas correspondieron a muestras de lavado bronquioalveolar
(LBA): LBA2 con 578 pb, LBA24 con 591 pb y LBA33 con 635 pb, pertenecientes a la
preceba de la granja 1, ceba de la granja 1 y ceba de la granja 3, respectivamente
(Figura 4-1). Las secuencias obtenidas fueron enfrentadas a cepas tanto Americanas
como Europeas incluyendo a su vez aislados de campo obtenidos en el LNDV, como
cepas vacunales. Como se observa en el árbol filogenético obtenido, las muestras
secuenciadas se localizaron dentro del grupo de las cepas americanas. Adicionalmente,
se observó que las dos cepas correspondientes a la granja 1 presentaron un alto
porcentaje de similitud entre ellas, con respecto a la cepa de la granja 3, esta similitud
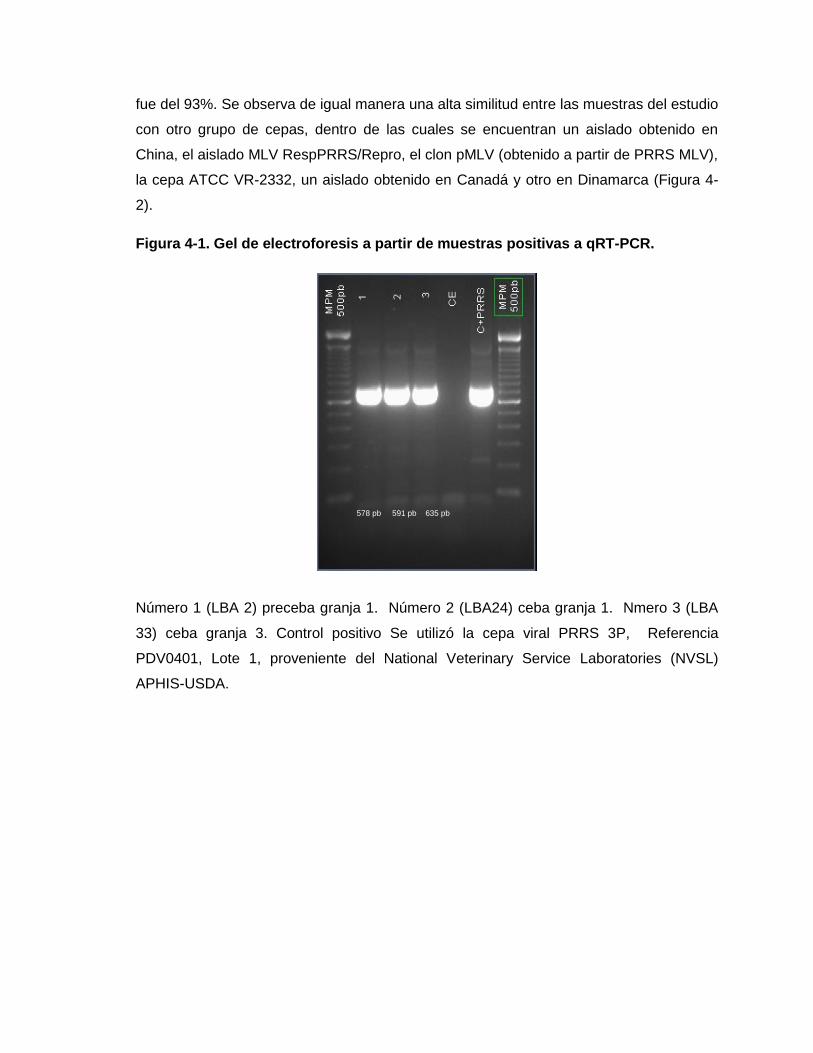
fue del 93%. Se observa de igual manera una alta similitud entre las muestras del estudio
con otro grupo de cepas, dentro de las cuales se encuentran un aislado obtenido en
China, el aislado MLV RespPRRS/Repro, el clon pMLV (obtenido a partir de PRRS MLV),
la cepa ATCC VR-2332, un aislado obtenido en Canadá y otro en Dinamarca (Figura 4-
2).
Figura 4-1. Gel de electroforesis a partir de muestras positivas a qRT-PCR.
Número 1 (LBA 2) preceba granja 1. Número 2 (LBA24) ceba granja 1. Nmero 3 (LBA
33) ceba granja 3. Control positivo Se utilizó la cepa viral PRRS 3P, Referencia
PDV0401, Lote 1, proveniente del National Veterinary Service Laboratories (NVSL)
APHIS-USDA.
578 pb 591 pb 635 pb
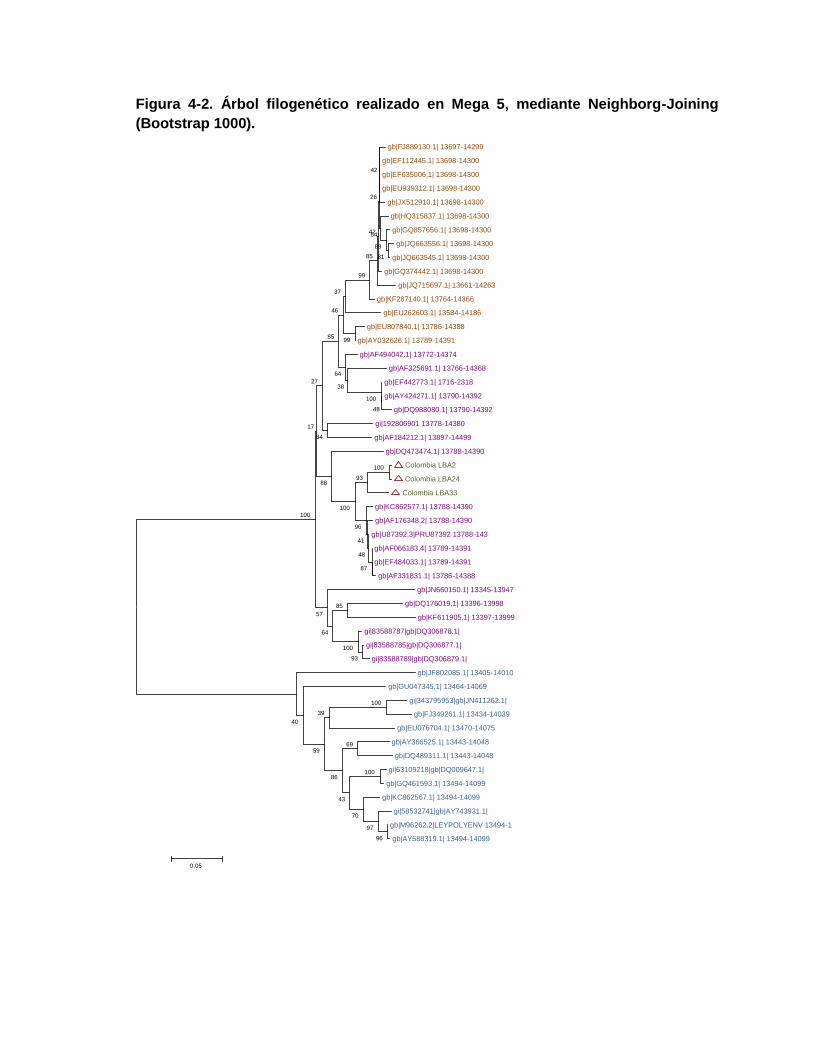
Figura 4-2. Árbol filogenético realizado en Mega 5, mediante Neighborg-Joining
(Bootstrap 1000).
gb|FJ889130.1| 13697-14299 gb|EF112445.1| 13698-14300 gb|EF635006.1| 13698-14300 gb|EU939312.1| 13698-14300
gb|JX512910.1| 13698-14300 gb|HQ315837.1| 13698-14300 gb|GQ857656.1| 13698-14300 gb|JQ663556.1| 13698-14300
gb|JQ663545.1| 13698-14300 gb|GQ374442.1| 13698-14300
gb|JQ715697.1| 13661-14263 gb|KF287140.1| 13764-14366
gb|EU262603.1| 13584-14186 gb|EU807840.1| 13786-14388
gb|AY032626.1| 13789-14391 gb|AF494042.1| 13772-14374
gb|AF325691.1| 13766-14368 gb|EF442773.1| 1716-2318 gb|AY424271.1| 13790-14392
gb|DQ988080.1| 13790-14392 gi|192806901 13778-14380 gb|AF184212.1| 13897-14499
gb|DQ473474.1| 13788-14390 Colombia LBA2 Colombia LBA24
Colombia LBA33 gb|KC862577.1| 13788-14390 gb|AF176348.2| 13788-14390
gb|U87392.3|PRU87392 13788-143 gb|AF066183.4| 13789-14391 gb|EF484033.1| 13789-14391 gb|AF331831.1| 13786-14388
gb|JN660150.1| 13345-13947 gb|DQ176019.1| 13396-13998
gb|KF611905.1| 13397-13999 gi|83588787|gb|DQ306878.1| gi|83588785|gb|DQ306877.1|
gi|83588789|gb|DQ306879.1| gb|JF802085.1| 13405-14010
gb|GU047345.1| 13464-14069 gi|343795953|gb|JN411262.1|
gb|FJ349261.1| 13434-14039 gb|EU076704.1| 13470-14075
gb|AY366525.1| 13443-14048 gb|DQ489311.1| 13443-14048
gi|63109218|gb|DQ009647.1| gb|GQ461593.1| 13494-14099
gb|KC862567.1| 13494-14099 gi|58532741|gb|AY743931.1|
gb|M96262.2|LEYPOLYENV 13494-1 gb|AY588319.1| 13494-14099
100
96
100
97 70
69
43
86
39
59
40
100
93 100
87
93
48 41
96
100
85
48 100
64
88
57
100
17 34
27
99
38 64
85
46
37
99
81 89
85
54 42
26
42
0.05

El color naranja corresponde a cepas aisladas en Asia de origen americano, el color
violeta a las cepas aisladas en América y las azules a las cepas Europeas. En color rojo
y verde se encuentran las muestras aisladas dentro del estudio, denominadas Colombia
LBA2, LBA24 y LBA33.
4.3 Discusión Del total de las muestras empleadas para intentar aislamiento, solo 8 fueron viables para
obtener resultados positivos evidenciados por la presencia de efectos citopáticos típicos
de la infección, caracterizados por pérdida de continuidad de la monocapa,
redondeamiento celular y agrupamiento. Como ya se ha documentado, el genotipo II de
PRRSV es rutinariamente aislado en células MA-104 y su subclon MARC-145, mientras
que el genotipo I no se ha adaptado a estas líneas celulares (Murtaugh et al., 2010) esto
confirma inicialmente que estas cepas pertenecían al genotipo americano, lo cual fue
posteriormente verificado mediante los resultados de la secuenciación.
Con respecto al tipo celular utilizado existen varias referencias al respecto. En un estudio
realizado por Bautista et al (1993) donde se evaluaron las diferencias en el aislamiento
de PRRSV en células MA104 (también denominadas CL2621) y macrófagos alveolares
pulmonares (MAPs), se logró concluir que aunque los MAPs son relativamente más
sensibles para el aislamiento de PRRSV, ambos tipos celulares pueden ser empleados.
En un estudio más reciente (Fuentes de Abin, et al., 2009) reportaron diferencias en la
eficiencia de infección por PRRSV tanto en células MA-104 como en PAMs; al día 6 post
inoculación el 70% de las cepas de campo crecieron en los MAPs mientras solo el 2% en
células MA104. También se ha evidenciado que las células MA-104 no son permisivas al
PRRSV de tipo silvestre, esto ha sido relacionado a la insuficiente presencia de CD163
para producir la infección en las células huésped (Calvert et al., 2007). En un hallazgo
reportado por Fuentes de Abin et al (2009), se aislaron 20 cepas de campo en PAMs a
las 72 horas de incubación y 25 cepas adicionales a los 6 días de incubación, solo una
cepa de campo fue aislada en células MA104 a los 14 días. Con relación a lo anterior y a
los resultados obtenidos en este estudio, se podría asegurar que no todas las cepas
obtenidas a partir de las granjas se pudieron replicar en las células MA-104, esto pudo
deberse a factores como una baja sensibilidad de las células a los virus de campo y/o a
una concentración baja de virus. Se debe tener en cuenta el número de pasajes hechos

(3) para obtener la manifestación del EC, ya que se ha reportado que se puede encontrar
efecto en periodos de incubación de hasta 14 días por pasaje en células MA-104
(Fuentes de Abin et al., 2009).
Los aislados se obtuvieron a partir de lavados bronquioalveolares y pulmón, lo cual
puede indicar que en el momento de la toma de las muestras, los animales se
encontraban en la fase aguda de la enfermedad y el virus se encontraba replicando a
nivel de MAPs, esto a su vez indica circulación del PRRSV en las granjas 1 y 3. El grupo
de la preceba pudo adquirir el virus a partir de reemplazos o de los animales adultos de
los sitios 1 y 2 transportándolo posteriormente al sitio 3 o adquiriendo el virus que allí se
encontraba circulando.
Dentro del presente estudio, al comparar los resultados obtenidos de los aislados en MA-
104 por qRT-PCR, se comprueba la presencia de ARN viral en las muestras. Sin
embargo, resultados qRT-PCR positivos no siempre indican la presencia de virus viable
(Wills et al., 2003). De otro lado la prueba qRT-PCR es más sensible que el aislamiento
viral para identificar cerdos infectados por el PRRSV (Wills et al., 2003).
Los aislamientos se obtuvieron a partir de muestras tomadas al comienzo del estudio,
básicamente hacia el final del año (Septiembre, Noviembre, Diciembre) y al final del
estudio en Mayo. Estas circunstancias pudieron obedecer posiblemente al traslado y
mezcla de animales ya sea durante el destete o cuando fueron trasladados al sitio 3, o a
la comercialización que se presenta durante el fin del año. Lo anterior pudo conllevar al
inicio de la infeccion y por ende a una mayor carga viral a nivel pulmonar o a la presencia
de persistencia a nivel de los líquidos pulmonares.
En cuanto a las secuencias obtenidas en el estudio, al ser enfrentadas con cepas
aisladas en Asia (primordialmente americanas), América y Europa, éstas se ubicaron
dentro del grupo de las cepas de origen americano, lo que corrobora los hallazgos de los
estudios realizados en Colombia entre 1997 y 1999, donde se concluyó que las cepas
circulantes en el país correspondían a este mismo genotipo. Sin embargo, del total de
muestras secuenciadas solo se lograron amplificar tres, lo cual, no corresponde a una
muestra representativa que permita determinar que únicamente se encuentran circulando
cepas americanas en la población evaluada. De igual manera, se requeriría la

caracterización continua del virus en la zona para identificar posibles nuevas variantes
(Torrison et al, 2003), ya que los cambios en el virus de PRRS se presentan a una tasa
de 0.5% a 1% por año en campo (Murtaugh, 2012). En el año 2012, en el Taller
Internacional de Diagnóstico Virológico y Molecular para PRRSV realizado en el
Laboratorio Nacional de Diagnóstico Veterinario del ICA, se secuenciaron una serie de
muestras que incluyeron sueros, fluidos orales y tejidos positivos por RT-PCR, en las
cuales se identificaron cepas de alta virulencia dentro de los clusters, sin embargo el
doctor Murtaugh, aclaró que estos resultados correspondían a datos preliminares y
debían ser interpretados cuidadosamente (Murtaugh, 2012a).
Las diferencias en similitud indican por un lado, que la misma cepa se encuentra
circulando entre la preceba y ceba de la granja 1, pero existe variación con respecto a la
cepa ubicada en la granja 3, aun cuando las granjas se localizan en la misma zona.
De igual manera, se observó similitud con otro grupo de cepas dentro de las cuales se
encuentran la cepa representativa del genotipo americano ATCC-VR 2332 y una cepa
vacunal viva modificada (MLV) Resp/PRRS Repro que fue posiblemente utilizada en las
granjas, ya que actualmente no se vacuna en Colombia contra PRRSV.
4.4 Conclusiones
Se confirmó en las granjas evaluadas, la presencia del genotipo Americano del PRRSV,
confirmando los hallazgos obtenidos en los muestreos realizados a finales de los años
noventa. La movilización, medidas de manejo y bioseguridad en las granjas influyeron en
las diferencias en cuanto a resultados positivos obtenidos en cada una. Por su parte, los
grupos etáreos de la preceba y ceba fueron los grupos donde fueron aisladas las cepas
del PRRSV, confirmando que existe circulación viral en las granjas 1 y 3.
4.5 Recomendaciones
Realizar los aislamientos en células MARC-145 y/o MAPs, las cuales son más sensibles
a las cepas de campo. De igual manera realizar secuenciaciones periódicamente con el
fin de descartar la posibilidad de circulación de cepas de tipo Europeo.

Bibliografía
Allende, R., Kutish, G, F., Laegreid, W., Lu, Z., Lewis, T.L., Rock, D.L., Friesen, J.,
Galeota, J.A., Doster, A.R., Osorio, F.A. 2000. Mutations in the genome of porcine
reproductive and respiratory síndrome virus responsable for the attenuation phenotype,
Arch. Virol, 145; 1149-1161.
Bautista, E.M., Goyal, S.M., Yoon, I.J., Joo, H.S., Collins, J.E. 1993. Comparison of
porcine alveolar macrophages and CL 2621 for detection of porcine reproductive and
respiratory syndrome (PRRS) virus and anti-PRRS antibody. J. Vet. Diagn. Invest. 5: 163-
165.
Benfield, DA., Nelson, E., Collins, JE., Harris, L., Goyal, SM., Robinson, D., Christianson,
WT., Morrison, RB., Gorcyca, D., Chladek, D. 1992. Characterization of swine infertility
and respiratory syndrome (SIRS) virus (isolate ATCC VR-2332). J. Vet. Diagn. Invest. 4,
127-133.
Calvert, J.G., Slade, D.E., Shields, S.L., 2007. CD163 expression confers susceptibility to
porcine reproductive and respiratory porcine syndrome viruses. J. Virol; 81: 7371-7379.
Chang, C.C., Yoon, K.J., Zimmerman, J.J., Harmon, K.M., Dixon, P.M., Dvorak, C.M.T.,
Murtaugh, M.P. 2002 Evolution of porcine reproductive and respiratory syndrome virus
during sequential passages in pigs. J Virol. 76:4750-4763.
Conzelman, K., Visser, N., van Woensel, P., Thiel, H. 1993. Molecular characterization of
porcine reproductive and respiratory syndrome virus, a member of the Arterivirus group.
Virology; 193:329-339.
Dee, S.A., et al. 2001. Identification of genetically diverse sequences (ORF5) of porcine
reproductive and respiratory syndrome virus in a swine herd. Can J Vet Res., 65: 254-
260.

Firth, A.E., Zevebhoven-Dobbe, J.C., Wills, N.M., Go, Y.Y., Balasuriya, U.B., Atkins, J.F.,
Snijder, RE.J., Posthuma, C.C. 2011. Discovery of a small arterivirus gene that overlaps
the GP5 coding sequence and is important for virus production. J. Gen. Virol; 92: 1097-
1106.
Forsberg, R. 2005. Divergence time of porcine reproductive and respiratory syndrome
virus subtypes. Mol. Biol. Evol. 22:2131-2134.
Fuentes de Abin, M., Spronk, G., Wagner, M., Fitzsmmons, M., Abrahante, J., Murtaugh,
M.P. 2009. Comparative infection efficiency of porcine reproductive and respiratory
syndrome (PRRS) virus field isolates on MA-104 cells and porcine alveolar macrophages.
Can J Vet Res; 73(3):200-204.
Han, J., Wang, Y., Faaberg, K.S., 2006. Complete genome analysis of RFLP 184
isolatesof porcine reproductive and respiratory syndrome virus. Virus Res. 122 (1–2),175–
182.
Johnson, C.R., Griggs, T.F., Gnanandarajah, J., Murtaugh, M.P. 2011. Novel structural
protein in porcine reproductive and respiratory síndrome virus encoded by alternative
ORF5 present in all arterivirus. J. Gen. Virol. 92:1107-1116.
Kim, H.S., Kwang, J., Yoon, I.J., Joo, H.S., Frey, M.L.,1993. Enhanced replication of
porcine reproductive and respiratory syndrome (PRRS) virus in a homogeneous
subpopulstion of MA-104 cell line. Arch Virol; 133: 477-483.
Liao, Y.C., Lin, H.H., Lin, C.H., Chung, W.B. 2013. Identification of cytotoxic T lymphocyte
epitopes on swine viruses: multi-epitope design for universal T cell vaccine. PLOS ONE,
8(12):e84443.
Lu, Z, H., Archibald, A,L., Ait-Ali, T. 2014. Beyond the whole genome consensus:
Unravelling of PRRS phylogenomics using next generation sequencing technologies.
Virus Research, http://dx.doi.org/10.1016/j.virusres.2014.10.004

Mardassi, H., Mounir, S., Dea, S. 1995. Molecular analysis of the ORFs3 to 7 of porcine
reproductive and respiratory syndrome virus, Québec reference strain. Arch. Virol. 140:
1405-1418.
Mengeling, W.L., Vorwald, A.C., Larger, K., M. Brockmeier, S.L. 1996. Diagnosis of
porcine reproductive and respiratory syndrome using infected alveolar macrophages
collected from live pigs. Vet Microbiol; 49: 105-115.
Meng, X.J., Paul, P.S., Halburg, P.G., Morozov, I. 1995. Sequence comparison of open
reading frames 2 to 5 of low and high virulence United States isolates of porcine
reproductive and respiratory syndrome virus. J Gen Virol. 76: 3181-3188.
Meulenberg, J.J., 2000. PRSSV, the virus. Vet Res; 31:11-21.
Mogollón, J.D., Rincón, M.A., Arbeláez, G., Ruiz, S., Rodríguez, V. 2003. PRRS in
Colombia.PRSS Compendium Producer Edition.National Pork Board.Des Moines, Iowa,
USA, section 6.6.
Morgan, S.B., Graham, S.P., Salguero, F.J., Sánchez Cordón, P.J., Mokhtar, H.,
Rebel,J.M.J., Weesendorp, E., Bodman-Smith, K.B., Steinbach, F., Frossard, J.P.,
2013.Increased pathogenicity of European porcine reproductive and respiratory syn-
drome virus is associated with enhanced adaptive responses and viral clearance.Vet.
Microbiol. 163 (1–2), 13–22.
Murtaugh, M.P., Stadejek, T., Abrahante, J,E., Lam, T.T.Y., Leung, C.C.F. 2010. The
ever- expanding diversity of porcine reproductive and respiratory syndrome virus. Virus
Research, 154:18-30.
Murtaugh, M.P. 2012. Use and interpretation of sequencing in PRRSV control programs.
Allen D. Leman Swine Conference; 49-55.

Murtaugh, M. 2012a. International molecular and virology diagnostic workshop for
PRRSV. ICA. October 28-November 2.
Neumann, E.J., Kliebestein, J.B., Johnson, C.D., Mabry, J.W., Bush, E.J.,Seitzinger, A.H.,
Green, A.L., Zimmerman, J.J.,2005. Assessment of the economic impact of porcine
reproductive and respiratory síndrome on swine production in the United States. Journal
of American Veternary Medical Association; 227:385-392.
Rossow, K.D., Bautista, E.M., Goyal, S.M., Molitor, T.W., Murtaugh, M.P., Morrison, R.B.,
Benfield, D.A., Collins, J.E. 1994. Experimental porcine reproductive and respiratory
síndrome virus infection in one, four and 10 weeks old pigs. J. Vet. Diagn Invest, 6:3-12.
Snijder and Meulenberg. 2001. Arteriviruses in Fields. Virology 1:1205-1220.
Stadejek, T., Stankevicius, A., Storgaard, T., Oleksiewicz, M.B., Belak, S., Drew,
T.W.,Pejsak, Z., 2002. Identification of radically different variants of porcine repro-ductive
and respiratory syndrome virus in Eastern Europe: towards a commonancestor for
European and American viruses. J. Gen. Virol. 83 (Pt 8), 1861–1873.
Stadejek, T., Oleksiewicz, M.B., Potapchuk, D., Podgorska, K., 2006. Porcine
reproductive and respiratory syndrome virus strains of exceptional diversity in eastern
Europe support the definition of new genetic subtypes. J. Gen. Virol. 87, 1835–1841.
Stadejek, T., Oleksiewicz, M.B., Scherbakov, A.V., Timina, A.M., Krabbe, J.S., Chabros,
K., Potapchuk, D., 2008. Definition of subtypes in the European genotype of porcine
reproductive and respiratory syndrome virus: nucleocapsid characteristics and
geographical distribution in Europe. Arch. Virol. 153 (8), 1479–1488.
Torrison, J., Rossow, K,. Yeske, P. 2003. Use of PRRS virus sequence information within
herds. Allen D. Leman Swine Conference.
Weesendorp, E., Morgan, S., Stockhofe-Zurwieden, N., Popma-De Graaf, D.J., Gra-ham,
S.P., Rebel, J.M., 2013. Comparative analysis of immune responses following

experimental infection of pigs with European porcine reproductive and respi-ratory
syndrome virus strains of differing virulence. Vet. Microbiol. 163 (1–2),1–12.
Wills R.W., Doster, A.R., Galeota, J.A., Sur, J.H., Osorio, F.A., 2003. Duration of infection
and proportion of pigs persistently infected with porcine reproductive and respiratory
syndrome virus. Journal of Clinical Microbiology; 41: 58-62.
5. Medidas de Control Propuestas
El presente estudio, permitió evidenciar la presencia de circulación del PRRSV en las
granjas evaluadas, con particularidades para cada una de ellas. Es significativo que la
granja 2 fue seronegativa y no se evidenció presencia de genoma viral en muestras de
de sueros y tejidos.
Los resultados obtenidos, permiten sugerir medidas de control a ser implementadas en
estas granjas, las que incluyen evitar que haya infección o reinfección en la granjas y/o
disminuir la circulación del virus, evitar la entrada de una nueva cepa viral, así como
evitar la propagación de la enfermedad a otras explotaciones. Para lo anterior, se
recomienda aplicar y/o extremar las medidas de bioseguridad y manejo, las cuales fueron
mencionadas en el numeral correspondiente al control del PRRS (1.9), sin embargo se
hará mención de algunas en particular.
Es aconsejable extremar las medidas de bioseguridad en las tres granjas, limitando la
entrada de visitantes a las mismas e imponer el baño y cambio de ropa al entrar y salir de
estas. Ya que se cuenta con sitios productivos que requieren el transporte continuo de
animales e insumos, se debe instaurar el lavado y desinfección de camiones al entrar y
salir de las instalaciones. Establecer control de roedores e insectos mediante el uso de
rodenticidas, trampas o láminas adherentes, a su vez mantener una adecuada limpieza y
desinfección de las instalaciones. Igualmente, se recomienda evitar el ingreso de
animales domésticos a las instalaciones y asegurar que estos se encuentren en
adecuadas condiciones sanitarias. También emplear machos certificados como negativos
a PRRSV para la monta, o semen procedente de centros negativos a la enfermedad.

De otro lado se deben obtener reemplazos a partir de granjas negativas a PRRSV y
llevar a cabo adecuadas medidas de manejo para dichos reemplazos las cuales fueron
citadas en el numeral 1.9 del presente estudio. Exponer los animales de reemplazo al
virus circulante en la granja mediante el contacto con animales confirmados como
positivos, solicitando asistencia técnica para el uso de otro tipo de prácticas como
exposición a las placentas o inoculación de suero. Incluir el uso de las Medidas McRebel,
ya que como se evidenció, los grupos productivos más jóvenes evidenciaron la presencia
del PRRSV.
De igual manera, se deben establecer flujos de animales como un sistema todo dentro-
todo fuera para evitar la diseminación del virus al momento de la mezcla de los animales
al destete, en la preceba y ceba. De igual manera se debería plantear la posibilidad de
disponer adecuadamente de los animales positivos mediante sacrificio, con el fin de
evitar la diseminación de la enfermedad.
Los resultados de este trabajo, permiten igualmente proponer dentro de las medidas de
control de la enfermedad, la importancia del monitoreo del PRRSV en las granjas,
mediante el uso conjunto de técnicas diagnósticas serológicas y moleculares. Esto para
confirmar con exactitud la presencia del virus en la explotación, ya que se pueden
presentar casos como el de la granja 2, la cual fue negativa por serología pero a través
de qRT-PCR se demostró la presencia del genoma viral en en muestras de FO; de igual
forma, por serología se determinó como positivo el grupo de la ceba en la granja 1, sin
embargo, se evidenció la presencia del virus en la preceba y destetos los cuales fueron
seronegativos. Se recomienda igualmente, realizar un monitoreo de la presencia del virus
a partir de la toma y evaluación de diferentes tipos de tejidos provenientes de las
mortalidades y sacrificios, así es posible establecer la presencia de animales virémicos
como persistentemente infectados en las granjas, como también la fase de infección en
la cual se encuentra la población, lo que permite determinar cuál podría ser el origen de
dicha infección y asi establecer medidas de eliminación de la población persistente para
evitar la continua diseminación viral.

6. Conclusiones y recomendaciones
6.1 Conclusiones
Se confirmó la circulación del PRRSV en explotaciones porcícolas de tipo intensivo en el
país. De igual manera, se encontró un incremento en la actividad del virus tanto en la
zona del Valle del Cauca, como en los diferentes grupos etáreos evaluados,
principalmente en la ceba.
Es necesario el uso conjunto de técnicas serológicas y moleculares en el diagnóstico de
PRRSV, ya que los resultados que arroja cada una, permite conocer la fase de infección
en la cual se encuentra el virus en una explotación.
Las tres granjas evaluadas en este estudio presentaron resultados positivos por qRT-
PCR en diferentes tipos de muestras, lo que demostró la presencia del PRRSV en
diferentes estados de infeccion. Los grupos más afectados fueron la preceba y ceba
confirmando la circulación del virus dentro de cada granja.
La detección de ARN viral en sueros, tejidos como pulmón y ganglios linfáticos indicaron
la posible presencia de animales virémicos como persistentemente infectados dentro de
los grupos productivos. Los diferentes tipos de muestras analizadas pueden ser utilizadas
en las diferentes fases de diagnóstico, dentro de un programa de control del PRRSV.
Se confirmó la presencia, en las granjas evaluadas, del genotipo Americano del PRRSV,
confirmando los hallazgos obtenidos en los muestreos realizados a finales de los años
noventa. La movilización, medidas de manejo y bioseguridad en las granjas influyeron en
las diferencias en cuanto a resultados positivos obtenidos en cada una.
La preceba y ceba fueron los grupos donde fueron aisladas las cepas del PRRSV,
confirmando la circulación viral en las granjas 1 y 3.

6.2 Recomendaciones El estudio demostró que el comportamiento del virus de PRRS en Colombia es particular
para cada granja, por tal razón, es importante revisar las medidas de bioseguridad, el
impacto de las movilizaciones a grandes distancias entre los sitios productivos de los
animales y realizar junto con estudios serológico pruebas moleculares con el fin de
determinar la dinámica el PRRSV dentro de cada granja para aplicar las medidas de
control adecuadas.
Utilizar las técnicas moleculares junto con pruebas serológicas que permitan reflejar las
diferentes fases de la infección y que permitan conocer la dinámica del virus en una
población con el fin de tomar las acciones de control adecuadas.
Se recomienda hacer uso de los diferentes tipos de muestras como FO para monitoreo
rutinario de los grupos productivos de la preceba y ceba especialmente por qRT-PCR.
Los tejidos como pulmón y ganglios de las mortalidades o animales enfermos o colas,
permiten orientar el estado de infección en el cual se encuentra una población dentro de
una granja. Por otro lado, los sueros permiten monitorear las madres, reemplazos,
reproductores y lactantes, o la población total cuando se han establecido diferentes
medidas de manejo y bioseguridad o en el caso de obtenerse resultados positivos en FO
y/o tejidos.
Realizar los aislamientos en células MARC-145 y/o MAPs, las cuales son más sensibles
a las cepas de campo, de igual manera, se deben realizar secuenciaciones periódicas
con el fin de descartar la posibilidad de circulación de cepas de tipo Europeo.
Es de importancia determinar mediante estudios regionales, las cepas que se encuentran
circulando, ya que como se ha evidenciado en este estudio existen diferencias incluso
dentro de una misma granja y esto influye en los mecanismos de control que deben ser
utilizados. De igual manera, debe establecerse un programa de monitoreo periódico para
establecer tanto la dinámica del virus en una zona, como las cepas circulantes y sus
variaciones en el tiempo.


Anexos
Anexo A. Buffer fosfato salino (PBS) pH 7.2 +/- 0.2
Cloruro de sodio (NaCl) 8.00 g
Cloruro de Potasio (KCl) 0.20 g
Fosfato dibásico de sodio anhidro (Na2HPO4) 1.15 g
Agua destilada csp 1 litro
Ajustar pH 7.2 +/- 0-2
Esterilizar
Almacenar a 4°C
Anexo B. Solución penicilina-estreptomicina (PEST) –
100X
Penicilina 1.000.000U 1 frasco
Estreptomicina 1 g
Agua destilada csp 100 ml
Filtrar por millipore 0.22u
Envasar en fraco de vidrio y mantener en congelación (-22°C).
Anexo C. Buffer fosfato salino (PBS) 10x para cultivo
celular
Cloruro de sodio (NaCl) 8.00g
Cloruro de potasio (KCl) 16 g
Fosfato dibásico de sodio anhidro (Na2HPO4) 1.92 g
Fosfato monobásico de potasio (KH2PO4) 2.40 g
Glucosa 80 g

Rojo de fenol 0.80 g
Bicarbonato de sodio (NaHCO3) 12 g
Agua destilada csp 4 litros
Diluir a 1x tomando 1 litro de PBS 10x más 9 litros de agua
Filtrar por millipore 0.22u
Envasar en frasco de vidrio
Realizar control de esterilidad
Mantener en refrigeración

Bibliografía
Albina,E., Leforban, Y., Baron, T., Plana-Duran, J. and Vannier, P. 1992. An enzyme
linked immunosorbent assay (ELISA) for the detection of antibodies to the porcine
reproductive and respiratory syndrome (SDRP) virus. Ann. Rech. Vet. 23, 167–176.
Albina, E., 1997. Epidemiology of porcine respiratory and reproductive syndome (PRRS):
an overview. Vet. Microbiol. 55: 309-316.
Allende, R., Lewis, T.L., Lu, Z., Rock, D. L., Kutish, G. F., Ali, A., Doster, A. R., Osorio,
F.A. 1999. North American and European porcine reproductive and respiratory syndrome
viruses differ in non-structural protein coding regions. J Gen Virol. 80: 307-15.
Allende, R., W. W. Laegreid, G. F. Kutish, J. A. Galeota, R. W. Wills, and F.A. Osorio.
2000. Porcine reproductive and respiratory syndrome virus: Description of persistence in
individual pigs upon experimental infection. J Virol. 74:10834-7.
Allende, R., Kutish, G, F., Laegreid, W., Lu, Z., Lewis, T.L., Rock, D.L., Friesen, J.,
Galeota, J.A., Doster, A.R., Osorio, F.A. 2000. Mutations in the genome of porcine
reproductive and respiratory síndrome virus responsable for the attenuation phenotype,
Arch. Virol, 145; 1149-1161.
Allende, R., Laegreid, W.W., Kutish, G.F., Galeota, J. A.,Wills, R.W and Osorio, F.A.
2003. Porcine reproductive and respiratory syndrome virus: description of persistence in
individual pigs upon experimental infection. J Virol; 72: 10834-7.
Ansari, I.H., Know, B., Osorio, F.A., Pasttnaik, A. K. 2006. Influence of N-linked
glycosylation of porcine reproductive and respiratory syndrome virus GP5 on virus
infectivity, antigenicity and ability to induce neutralizing antibodies. J Virol. 80:3994-4004.

Batista, L., Pijoan, C., Torremorell, M. 2002. Experimental injection of gilts with porcine
reproductive and respiratory syndrome virus (PRRSV) during acclimatization. J Swine
Health Prod., 10: 147–50.
Batista, L., Fernández, M., Miramontes, J., Pliego, R., Mondaca, M. 2010. Advancing
PRRS area regional control in the state of Sonora, Mexico. In: Proc. Int. Pig Vet. Soc.
Congr., Vancouver, Canada, 274.
Bautista, E.M., Molitor, T.W. 1997. Cell-mediated immunity to porcine reproductive and
respiratory syndrome virus in swine. Viral Immunol. 10:83-94.
Bautista, E.M., Goyal, S.M., Yoon, I.J., Joo, H.S., Collins, J.E. Comparison of porcine
alveolar macrophages and CL 2621 for detection of porcine reproductive and respiratory
syndrome (PRRS) virus and anti-PRRS antibody. J. Vet. Diagn. Invest. 5: 163-165.
Benfield, D.A., Nelson, E., Collins, J.E., Harris, L., Goyal, S.M., Robinson, D.,
Christianson, W.T., Morrison, R.B., Gorcyca, D., Chladek, D. 1992. Characterization of
swine infertility and respiratory syndrome (SIRS) virus (isolate ATCC VR-2332). J. Vet.
Diagn. Invest. 4, 127-133.
Benfield, D.A., Christopher-Henings, J., Nelson, E.A., Rowland, R.R., Nelson, J.K., Chase
C.C.I. and Rossow, K.D., 1997. Persistent fetal infection of porcine reproductive and
respiratory syndrome (PRRS) virus, p. 455-458. In Proceedings of the American
Association of Swine Practitioners. American Association of Swine Practitioners, Quebec
City, Qubec, Canada.
Benfield, D.A., Collins, J.E., Dee, S.A., Hallbur, P.G., Joo, H.S., Lager, K.M., Mengeling,
W.L., Murtauhgh, M.P., Rossow, K.D., Stevenson, G.W. & Zimmerman J.J. 1999. Porcine
reproductive and respiratory syndrome. In: Diseases of Swine, Eight Edition, Straw B.E.,
D’Allaire S., Mengeling W.L. & Taylor D.J., eds. Iowa State University Press. Ames, Iowa,
USA, 201–232.

Beura, L.K, Sarkar, S.N, Kwon, B, Subramaniam, S, Jones, C, Pattnaik, A.K., Osorio, F.A.
2010. Porcine reproductive and respiratory syndrome virus nonstructural protein 1beta
modulates host innate immune response by antagonizing IRF3 activation. J Virol. 84,
1574-1584.
Blaha, T., 2000. The “colorful” epidemiology of PRRS. Vet. Res. 31: 77-83
Boddicker, N., Waide, E., Rowland, R., Lunney, J., Garrick, D., et al. 2012. Evidence for a
major QTL associated with host response to porcine reproductive and respiratory
syndrome virus challenge. Journal of Animal Science 90: 1733–1746.
Britton, P. and Cavanagh, D. 2008. Nidovirus genome organization and expression
mechanisms. In Perlam, S., Gallagher, T., and Snijder, E.J. (eds.), Nidoviruses, pp. 29-
46. American society for Microbiology Press, Washington, District of Columbia.
Calvert, J., Slade, D.E., Shields, SL., Jolie, R., Mannan, RM., Ankenbauer, RG., Welch,
S.K. 2007. CD163 expression confers susceptibility to porcine reproductive and
respiratory syndrome viruses. J Virol. 81, 7371-7279.
Cancel-Tirado, S.M., Evans, R.B., Yoon, K. 2004. Monoclonal antibody analysis of
porcine reproductive and respiratory syndrome virus epitopes associated with antibody-
dependent enhancement and neutralization of virus infection. Vet. Immunol.
Immunopathol. 102, 249-262.
Carlsson, U., Wallgren, P., Renstrom, L.H., Lindberg, A., Eriksson, H., Thoren, P.,
Eliasson-Selling, L., Lundeheim, N., Norregard, E., Thorn, C., Elvander, M. 2009.
Emergence of porcine reproductive and respiratory syndrome in Sweden: detection,
response and eradication. Transbound Emerg. Dis. 56, 121-131.
Cavanagh, D. 1997. Nidovirales: a new order comprising Coronaviridae and Arteriviridae.
Arch. Virol. 142, 629-633.

Chai, Z., Ma, W., Fu, F., Lang, Y., Wang, W., Tong, G., Liu, Q., Cai, X., Li, X. 2013. A
SYBR Green-based real-time RT-PCR assay for simple and rapid detection and
differentiation of highly pathogenic and classical type 2 porcine reproductive and
respiratory syndrome virus circulating in China. Arch Virol. 158(2): 407-15.
Chand, R.R., Trible, B.R., Rowland, R.R.R., 2012. Pathogenesis of porcine reproductive
and respiratory syndrome virus. Current Opinion in Virology, 2:256-263.
Chang, C.C., Yoon, K.J., Zimmerman, J.J., Harmon, K.M., Dixon, P.M., Dvorak, C.M.T.,
Murtaugh, M.P. 2002 Evolution of porcine reproductive and respiratory syndrome virus
during sequential passages in pigs. J Virol. 76:4750-4763.
Chang, H.C., Peng, Y.T., Chang, H.L., Chaung, H.C., Chung, W.B. 2008. Phenotypic and
functional modulation of bone marrow-derived dendritic cells by porcine reproductive and
respiratory syndromr virus. Vet. Microbiol. 129:281-293.
Chen, J.Z., Lawson, S., Sun, Z., et al. 2010a. “Identification of two autocleavage products
of nonstructural protein 1 (nsp1) in porcine reproductive and respiratory syndrome virus
infected cells: nsp1 function as interferon antagonist,” Virology, 398: 87–97.
Chen, Z., Zhou, X., Lunney, J.K., Lawson, S., Sun, Z., Brown, E., Christopher-Hennings,
J., Knudsen, D., Nelson, E., Fang, Y. 2010b. Inmunodominant epitopes in nsp2 of porcine
reproductive and respiratory syndrome virus are dispensable for replication but play an
important role in modulation of host immune response. J. Gen. Virol. 91, 1047-1057.
Cho, J.G. and Dee, S.A. 2006. Porcine reproductive and respiratory syndrome virus.
Theriogenology, 66; 655–662.
Christianson, W.T., Collins, J.E., Benfield, D.A., Harris, L., Gorcyca, D.E., Chladek, D.W.,
Morrison, R.B., Joo, H.S. 1992. Experimental reproduction of swine infertility and
respiratory syndrome in pregnant sows. Am. J. Vet. Res, 53:485-488.

Christopher-Hennings, J., Nelson, E.A., Benfield, D.A. 1996. Detecting PRRSV in boar
semen. Swine Health and Production, 4: 37-39.
Christopher-Hennings, J., Nelson, E.A., Nelson, J. K. and Benfield, D. A. 1997. Effects of
a modified-live virus vaccine against porcine reproductive and respiratory syndrome in
boars. Am. J. Vet. Res. 58, 40–45.
Christopher-Hennings, J., Faaberg, K.S., Murtaugh, M.P., Nelson, E.A., Roog, M.B.,
Vaughn, E.M., Yoon, K.J., Zimmerman, J.J. 2002. Porcine reproducive and respiratory
syndrome (PRRS) diagnostics: interpretación and limitations, Journal of Swine Health and
Production, 10: 213-218.
Cho, H. J., McNab, B., Dubuc, C., Jordan, L., Afshar, A., Magar, R., Prins, S., and
Eernisse, K. 1997. Comparative study of serological methods for the detection of
antibodies to porcine reproductive and respiratory syndrome virus. Can. J. Vet. Res. 61,
161–166.
Chung, W.B., Chan, W.H., Chaung, H.C., Lien, Y., Wu, C.C., Huang, Y.L. 2005. Real-time
PCR for quantitation of porcine reproductive and respiratory syndrome virus and porcine
circovirus type2 in naturally-infected and challenged pigs. Journal of Virological Methods,
124:11-19.
Collins, J.E., Benfield, D.A., Chritianson, W.T., Harris L., Hennings, J.C., Shaw, D.P.,
Goyal, S.M., NcCullough, S., Morrison, R.B., Joo, H.S. 1992. Islotation of swine infertility
and respiratory syndrome virus (isolate ARCC VR-2332) in North America and
experimental reproduction of the disease in gnotobiotic pigs. J Vet Diagn Invest. Apr
4(2):117-126.
Collins, J., Dee, S., Halbur, P., Keffaber, K., Lautner, B., McCaw, M., Rodibaugh, M.,
Sanford, E., Yesye, P., 1996. Laboratory diagnosis of porcine reproductive and
respiratory syndrome virus (PRRS) infection, Swine health and production, 4: 33-35.

Conzelman, K., Visser, N., van Woensel, P., Thiel, H. 1993. Molecular characterization of
porcine reproductive and respiratory syndrome virus, a member of the Arterivirus group.
Virology; 193:329-339.
Corzo, C.A., Mondaca, E., Wayne, S., Torremorell, M., Dee, S., Davies, P., Morrison,
R.B., 2010. Control and elimination of porcine reproductive and respiratory syndrome
virus. Virus Res. 154, 185-192.
Costers, S., Lefebvre, D.J., Delputte, P.L., Nauwynck, H.J. 2008. Porcine reproductive
and respiratory syndrome virus modulates apoptosis during replication in alveolar
macrophages. Arch Virol. 2008;153(8):1453-65
Cruz, M.C., Mogollón, J.D., Rincón, M.A., Peña, N.B., Ruiz, S., Lora, A.M. 2006.
Prevalencia serológica del Síndrome Reproductivo y Respiratorio Porcino (PRRS) en
cerdos de explotaciones extensivas de Colombia. Rev. Med. Vet. Zoot. 53:33-41
Darwich, L., Diaz, I., Mateu, E. 2010. Certainties, doubts and hypotheses in porcine
reproductive and respiratory virus immunobiology. Virus Research, 154: 123-132.
Darwich, L., Gimeno, M., Sibila, M., Díaz, I., de La Torre, E., Dotti, S., Kuzemtseva, L.,
Martin, M., Pujols, J., Mateu, E. 2011. Genetic and inmunobiological diversities of porcine
reproductive and respiratory síndrome genotype I strains. Vet Mic. 150:49-62
Das, P.B., Dinh, P.X., Ansari, L.H., de Lima, M., Osorio, F.A., Pattnaik, A.K., 2010. The
minor envelope glycoproteins GP2 and GP4 of porcine reproductive and respiratory
syndrome virus interact with the receptor CD163. J.Virol.84: 1731-1740.
Dea, S., Sawyer, N., Alain, R., Athanassious, R. 1995. Ultrastructural characteristics and
morphogenesis of porcine reproductive and respiratory syndrome virus propagated in the
highly permissive MARC-145 cell clone. Adv. Exp. Med. Biol. 380: 95-98.

Dea, S., Gagnon, C.A., Mardassi, H., et al. 2000. Current knowledge on the structural
proteins of porcine reproductive and respiratory syndrome (PRRS) virus: comparison of
the North American and European isolates. Arch Virol. 145: 659-688.
Dee, S.A., et al. 2001. Identification of genetically diverse sequences (ORF5) of porcine
reproductive and respiratory syndrome virus in a swine herd. Can J Vet Res., 65: 254-
260.
Dee, S. A. 2003. Approaches to prevention, control and eradication. Compendium PRRS:
Second edition, 119-128.
Dee, S., Deen, J., Rossow, K., Weise, C., Eliason, R., Otake, S., Joo, H.S., Pijoan, C.,
2002. Mechanical transmission of porcine reproductive and respiratory syndrome virus
throughout a coordinated sequence of events during warm weather. Can. J. Vet. Res.
66:232-239.
Dee, S. A., Deen, J., Otake, S., Pijoan, C. 2004. An assessment of transport vehicles as
a source of porcine reproductive and respiratory syndrome virus transmission to
susceptible pigs. Can J Vet Res. 68:124-133.
Dee, S.A., Deen, J., Burns, D., Douthiht, G., Pijoan, C. 2005. An evaluation of
desinfectants of the sanitation of porcine reproductive and respiratory virus contaminated
transport vehicles at cold temtperatures. Can. J. Vet. Res. 69: 64-71-0.
De Lima, M., Pattnaik, A.K., Flores, E.F., Osorio, F.A., 2006. Serologic marker candidates
identified among B-cell linear epitopes of Nsp2 and structural proteins of a North
American strain of porcine reproductive and respiratory syndrome virus. Virology, 30:410-
421.
De Lima, M., Ansari, I.H., Das, P, B., Ku, B.J., Martínez-Lobo, F.J., Osorio, F.A. 2009.
GP3 is a structural componente of the PRRSV type II (US) virion. Virology, 390: 31.36.

Delputte, P.L., Vanderheijden, N., Nauwynck, H.J., Pensaert, MB. 2002. Involvement of
the matrix protein in attachment of porcine reproductive and respiratory syndrome virus to
a heparinlike receptor on porcine alveolar macrophages. J. Virol. 76, 4312-20.
Delputte, P.L., Costers, S and Nauwynck, H.J. 2005. Analisys of porcine reproductive and
respiratory syndrome virus attachment and internalization: distinctive roles for heparan
sulphate and sialodhesin. J Gen Virol. 86:1441-1445.
Delputte, P. L., Van Breedman, W. Delrue, I, Oetke, C, Crocker, P.R and Nauwynck, H.J.
2007. Porcine arterivirus attachment to the macrophage-specific receptor sialodhesin is
dependent on the sialic acid-binding activity of the N-terminal immunoglobulin domain of
sialodhesin.J Virol. 81:9546-9550.
Denac, H., Moser, C., Tratschin J., D., and Hofmann, M., A. 1997. An indirect ELISA
for the detection of antibodies against porcine reproductive and respiratory syndrome
virus using recombinant nucleocapsid protein as antigen. J. Virol. Methods, 65, 169–181.
Desrosiers, R., Boutin, M. 2002. An attempt to eradicate porcine reproductive and
respiratory syndrome virus (PRRSV) after an outbreak in a breeding herd: Eradication
strategy and persistence of antobody titers in sows. Journal of SwineHealth and
Production, 10: 23-25.
Desrosier, R. 2011. Transmission of swine pathogens: different means, different needs.
Animal Health Research Reviews, 12; 1-13.
Drew, T., W. 1995b. Comparative serology of porcine reproductive and respiratory
syndrome in pigs. J. Vet. Med. Sci. 58, 941–946.
Diaz, I., Darwich, L., Papaterra, G., Pujols, J., Mateu, E., 2005. Inmune responses of pigs
after experimental infection with European strain of porcine reproductive and respiratory
syndrome virus. Journal of General Virology, 86:1943-1951.

Dietze, K., Pinto, J., Wainwright, S., Christopher, H., Khomenko, S. 2011. Porcine
reproductive and respiratory syndrome (PRRS) virulence jumps and persistent circulation
in Southeast Asia disponible en. www.fao.org/docrep/013/al849e/al849e00.pdf
Dokland, T. 2010. The structural biology of PRRS. Virus Research, 154: 86-97.
Duinhof, T.F., van Schaik, G., van Esch, E.J.B., Wellenberg, G.J., 2011. Detection of
PRRSV circulation in herds without clinical signs of PRRS: comparison of five age groups
to asses to preferred age group and simple size. Vet. Microbiol; 150: 180-184.
Edwards, S., Robertson, I.B., Wilesmith, J. W. 1992. PRRS blue-eared pig disease in
Great Britain. Am. Assoc. Swine. Pract. News, 4:32-6.
Evans, Ch. M., Medley, G, F., Green, L.E. 2008. Porcine reproductive and respiratory
syndrome virus (PRRSV) in GB pig herds: farm characteristics associated with
heterogeneity in seroprevalence. BMC Veterinary Research, 4: 48.
Faaberg, K.S., Balasuriya, U.B.R., Brinton, M.A., et al., 2011. “Family Arteriviridae”, in
Virus Taxonomy, ninth report of the International Committee on Taxonomy of Viruses,
A.M.Q King, Ed., Elsevier Academic Press, London, UK.
Fang, Y., and Snijder, E.J. 2010. The PRRSV replicase: exploring the multifunctionality of
an intriguing set of nonstructural proteins. Virus Res. 154:61-76.
Fano, E., Olea, L., Pijoan, C. 2005. Eradication of porcine reproductive and respiratory
syndrome virus by serum inoculation of naïve gilts. Canad J Vet Res. 69: 71-4.
Firth, A.E., Zevebhoven-Dobbe, J.C., Wills, N.M., Go, Y.Y., Balasuriya, U.B., Atkins, J.F.,
Snijder, RE.J., Posthuma, C.C. 2011. Discovery of a small arterivirus gene that overlaps
the GP5 coding sequence and is important for virus production. J. Gen. Virol; 92: 1097-
1106.

Flores-Mendoza, L., Silva-Campa, E., Resendiz, M., Osorio, F.A., Hernández, J. 2008.
Porcine Reproductive and Respiratory Syndrome Virus infects mature dendritic cells and
up-regulates interleukin-10 production. Clin. Vaccine Immunol. 15: 720-725.
Forsberg, R. 2005. Divergence time of porcine reproductive and respiratory syndrome
virus subtypes. Mol. Biol. Evol. 22:2131-2134.
Fuentes de Abin, M., Spronk, G., Wagner, M., Fitzsmmons, M., Abrahante, J., Murtaugh,
M.P. 2009. Comparative infection efficiency of porcine reproductive and respiratory
syndrome (PRRS) virus field isolates on MA-104 cells and porcine alveolar macrophages.
Can J Vet Res; 73(3):200-204.
García, A.R., Rodríguez, V., Kukielka, D. 2007. Aplicación de la RT-PCR en tiempo real
para el diagnóstico de PRRS. RCCV, 1(2); 646-653.
Gilbert, S.A., Larochelle, R., Magar, R., Cho, H. J., Deregt, D. 1997.Typirng of Porcine
Reproductive and Respiratory Syndrome Viruses by multiplex PCR Assay. Journal of
Clinical Microbiology, 35; 264-267.
Gonin, P., Mardassi, H., Gagnon, C.A., Massie, B., Dea, S. 1998. A nonstructural and
antigenic glycoprotein is encoded by ORF3 of the IAF-Klop strain of porcine reproductive
and respiratory syndrome virus. Arch Virol. 143, 1927-40.
Gonin, P., Pirzadeh, B., Gagnon, C.A., Dea S. 1999. Seroneutralization of porcine
reproductive and respiratory syndrome virus correlates with antibody response to the GP5
major envelope glycoprotein. J. Vet. Diagn. Invest. 11, 20-26.
Gorbalenya, A.E., Enjuanes, L., Ziebuhr, J., Snijder, E.J., 2006. Nidovirales: envolving the
largest RNA virus genome. Virus Res. 117:17-37.
Han, J., Wang, Y., Faaberg, K.S., 2006. Complete genome analysis of RFLP 184
isolatesof porcine reproductive and respiratory syndrome virus. Virus Res. 122 (1–2),175–
182.

Han, J., Rutherford, M.S., Faaberg, K.S. 2009. The porcine reproductive and respiratory
syndrome virus nsp2 cysteine protease domain possesses both trans and cis cleavage
activities. J. Virol. 83, 9449-9463.
Hanada, K., Suzuki, Y., Nakane, T., Hirose, O., Gojobori, T. 2005. The origin and
evolution of porcine reproductive and respiratory syndrome viruses. Mol. Biol. Evol. 22:
1024-1031.
Harmon, K.M., Abate, S.A., Chriswell, A.J., Strait, W.A.E., 2012. Comparison of two
commercial real-time reverse transcriptase polymerase chain reaction assays for porcine
reproductive and respiratory syndrome virus. Swine Health Prod., 20: 184-188.
Harrison, S.C., 2008. Viral membrane fusión. Nat. Struct. Mol. Biol. 15:690-698.
HerdChek PRRS (package insert) Westbrook, Maine. IDEXX Laboratories, 2011).
Holtkamp, D.J., Kliebenstein, J.B., Neumann, E.J., 2013. Assessment of the economic
impact of porcine reproductive and respiratory syndrome virus on United States pork
producers. Journal of Swine Health and Production, 21: 72-84.
Horter, D.C., Pogranichniy, R.M., Chang, C.C., Evans, R.B., Yoon, K.J., Zimmerman, J.J.
2002. Characterization of the carrier state in porcine reproductive and respiratory
syndrome virus infection. Vet. Microbiol. 86:213-228.
Hill, H., 1990. Overview and history of Mystery Swine Disease (swine infertility/respiratory
syndrome). In: Proceedings of the Mystery Swine Disease Committee Meeting. Livestock
Conversation Institute, Denver, CO, pp. 29-31.
Holthkamp, D.J., Kliebenstein, J.B., Neumann, E.J. 2013. Assessment of the economic
impact of porcine reproductive and respiratory síndrome virus on United States pork
producers. Journal of Swine Health and Production, 21: 72-84.

Hu, H., Li, X., Zhang, Z., Shuai, J., Chen, N., Liu, G., Fang, W. 2009. Porcine reproductive
and respiratory syndrome viruses predominant in southeastern China from 2004 to 2007
were from a common source and underwent further divergence. Arch Virol. 154, 391-398.
ICA. 2014. Técnica de aislamiento del virus de PRRS. Disponible en:
http://portal.ica.gov.co/DocManagerSwift/User/ElementViewer.aspx?LV=1&E=4640&rnd=
0.7680949024628146&HideHeader=true&Tab=w_0.2779094502393862
Islam, Z.U., Bishop, S.C., Savill, N.J., Rowland, R.R., Lunney, J.K., Trible, B., Doeschl-
Wilson, A.B. 2013. Quantitative analysis of porcine reproductive and respiratory syndrome
(PRRS) viremia profiles from experimental infection: a statistical modelling approach.
PLoS One, 8(12): 1-13
Jenkins, G.M., Rambaut, A., Pybus, O.G., Holmes, E.C., 2002. Rates of molecular
evolution in RNA viruses: a quantitative phylogenetic analysis. J. Mol. Evol. 54:156-165.
Jeong, H. J., Song, Y. J., Lee, S. W., Lee, J. B., Park, S. Y., Song, C. S., Ha, G. W., Oh,
J. S., Oh, Y. K. & Choi, I. S. 2010. Comparative measurement of cell-mediated immune
responses of swine to the M and N proteins of porcine reproductive and respiratory
syndrome virus. Clin Vaccine Immunol. 17: 503–512.
Jiang, W., Jiang, P., Wang, X., Li, Y., Du, Y. 2007. Influence of Porcine reproductive and
respiratory syndrome virus GP5 glycoprotein N-linked glycans on immune responses in
mice. Virus Genes, 35: 663-671.
Jiang, W., Jiang, P., Wang, X., Li, Y and Du, Y. 2008. Enhanced immune responses of
mice inoculated recombinant adenoviruses expressing GP5 by fusion with GP3 and/or
GP4 of PRRS virus. Virus Res. 136(1-2):50-57.
Johnson, W., Roof, M., Vaughn, E., Christopher-Hennings, J., Johnson, C.R., Murtaugh,
M.P., 2004. Pathogenic and humoral responses to porcine reproductive and respiratory
syndrome virus (PRRSV) are related to viral load in acute infection. Vet. Inmunol.
Inmunopathol- 102:233-247.

Johnson, C.R., Griggs, T.F., Gnanandarajah, J., Murtaugh, M.P. 2011. Novel structural
protein in porcine reproductive and respiratory syndrome virus encoded by an alternative
ORF5 present in all arteriviruses. Journal of General Virology, 92: 1107–1116.
Karniychuk, U.U and Nauwynck, H.J. 2013. Pathogenesis and prevention of placental and
transplacental porcine Reproductive and respiratory syndrome virus infection.
VeterinaryResearch, 44:95.
Kleiboeker, S.B., Schommer, S.K., Lee, S.M., Watkins, S., Chittick, W., Polson, D. 2005.
Simultaneous detection of North American and European porcine reproductive and
respiratory syndrome virus using real-time quantitative reverse transcriptase-PCR. J. Vet.
Diagn. Invest. 17,165–170.
Keffaber, K.K., 1989.Reproductive failure of unknown etiology. AASP 1, 2-10.
Key, K.F., Haqshenas, G., Guenette, D.K., et al. 2001. Genetic variation and phylogenetic
analyses of the ORF 5 gene of acute porcine reproductive and respiratory syndrome virus
isolates. Vet Microbiol. 83:249-263.
Kim, H.S., Kwang, J., Yoon, I.J., Joo, H.S., Frey, M.L.,1993. Enhanced replication of
porcine reproductive and respiratory syndrome (PRRS) virus in a homogeneous
subpopulstion of MA-104 cell line. Arch Virol; 133: 477-483.
Kim, J, K., Fahad, A.M., Shanmukhappa, K and Kapil, S. 2006. Defining the cellular
targets of porcine reproductive and respiratory syndrome virus blocking monoclonal
antibody 7G10. J Virol. 80:689-696.
Kim, O., Sun, Y., Lai, F.W., Song, C., and Yoo, D., 2010. “Modulation of type I interferon
induction by porcine reproductive and respiratory syndrome virus and degradation of
CREB-binding protein by non-structural protein 1 in MARC-145 and HeLa cells,” Virology,
402:315–326.

Kittawornrat, A., Prickett, J., Chittick, W., Wang, C., Engle, M., Johnson, J., Patnayak, D.,
Schwartz, T., Whitney, D., Olsen, C., Schwartz, K., Zimmerman, J., 2010. Porcine
reproductive and respiratory syndrome virus (PRRSV) in serum and oral fluid samples
from individual boars: Will oral fluid replace serum for PRRSV surveillance? Virus
Research 154, 170–176.
Kiyyawornrat, A., Panyasing, Y., Irwin, C., Rademacher, c., Hoogland, M., Wang, C.,
Johnson, J., Main, R., Rowland, R., Zimmerman, J. 2011. PRRS virus antibody detection
in oral fluid samples: The 1st generation PRRS oral fluid ELISA is ready for prime time.
International symposium on emerging and re-emerging pig disease. Allen D. Leman
Swine Conference.
Kiyyawornrat, A., Panyasing, Y., Irwin, Goodell, C., Wang, C., Gauger, P., Harmon, K.,
Rauh, R., Desfresne, L., Levis, I., Zimmerman, J. 2014. Porcine reproductive and
repiratory syndrome virus (PRRSV) surveillance usin pre weaning oral fluid samples
detects circulation of wild type PRRSV. Veterinary microbiology, 168: 331-339.
Kreutz, L.C. 1998. Cellular membrane factors are the major determinants of porcine
reproductive and respiratory syndrome virus tropism. Virus Res., 53:121-128.
Kroesse, M.V., Zevenhoven-Dobbe, J.C., Bos de Ruijter, J.N., Peeters, B.P., Meulenberg,
J.J., Conelissen, L.A., Snijder, E.J. 2008. The nsp1 alpha and nsp1 papain-like
autoproteinases are essential for porcine reproductive and respiratory syndrome virus
RNA synthesis. J Gen Virol. 89:494-499.
Larochelle, R., and Magar, R. 1997. Evaluation of the presence of porcine reproductive
and respiratory syndrome virus by polymerase chain reaction. J. Virol. Methods. 47, 273–
278.
Lawson, S.R., Rossow, K.D., Collins, J.E., Benfield, D.A., Rowland, R.R. 1997. Porcine
Reproductive and respiratory syndrome virus infection of gnotobiotic pigs: sites of virus
replication and co-localization with MAC-387 staining. Virus Res, 51, 105-113.

Le Gall, A.O., Legeay, O., Bourhy, H., Arnauld, C., Albina, E., Jestin, A. 1998. Molecular
variation in the nucleoprotein gene (ORF7) of the porcine reproductive and respiratory
syndrome virus (PRRSV). Virus Research, 54: 9-21.
Le Potier, M.F., Blanquefort, P., Morvan, E., Albina, E., 1997. Results of a control
programme for the porcine reproductive and respiratory syndrome in the French 'Pays de
la Loire' region. Vet. Microbiol. 55, 355-360.
Lee, S.M., Kleiboeker, S.B. 2005. Porcine arterivirus activates the NF-kappaB pathway
through IkappaB degradation. Virology, 342, 47–59.
Lee, C., and Yoo, D. 2006. The small envelope protein of porcine reproductive and
respiratory syndrome virus possesses ion channel protein-like properties. Virology 355,
30-43.
Li, B., Xiao, S., Wang, Y., Xu, S., Jiang, Y., Chen, H., Fang, L. 2009. Inmunogenicity of
the highly pathogenic porcine reproductive ans respiratory syndrome virus GP5 protein
encoded by a synthetic ORF5 gene. Vaccine, 27:1957-1963.
Li, B., Fang, L., Guo, X., Gao, J., Song, T., Bi, J., He, K., Chen, H., Xiao, S. 2011.
Epidemiology and evolutionary characteristics of the porcine reproductive and respiratory
syndrome virus in China between 2006 and 2010. J Clin Microbiol, 49:3175-3183.
Liao, Y.C., Lin, H.H., Lin, C.H., Chung, W.B. 2013. Identification of cytotoxic T lymphocyte
epitopes on swine viruses: multi-epitope design for universal T cell vaccine. PLOS ONE,
8(12):e84443
Loemba, H.D., Mounir, S., Mardassi, H. 1996. Kinetics of humoral immune response to
the major structural proteins of the porcine reproductive and respiratory syndrome virus.
Arch. Virol.141:751-761.

López Fuertes, L., Domenech, N., Alvarez, B., Ezguerra, A., Domínguez, J., Castro, J.M.,
Alonso F. 1999. Analysis of cellular immune response in pigs recovered of porcine
reproductive and respiratory syndrome infection. Virus Research 64: 33-42.
López, O.J., Oliveira, M, F., Álvarez-García, E., Kwon, B.J., Doster, A., Osorio, F, A.
2007. Protection against porcine reproductive and respiratory syndrome virus (PRRSV)
infection through passive transfer of PRRSV-neutralizing antibodies is dose dependent.
Clin Vaccine Immunol. 14:269–75.
Loula, T., 1991. Mystery pig disease. Agri-practice 12, 23-34.
Loving, C.L., Brockmeier, SL., Sacco, R.E. 2007. Differential type I interferon activation
and susceptibility of dendritic cell populations to porcine arterivirus. Inmunology, 120: 217-
229.
Lu, Z, H., Archibald, A,L., Ait-Ali, T. 2014. Beyond the whole genome consensus:
Unravelling of PRRS phylogenomics using next generation sequencing technologies.
Virus Research, http://dx.doi.org/10.1016/j.virusres.2014.10.004
Mackay, I.M., Arden, K.E., Nitsche, A. 2002. Real-time PCR in virology. Nucleic Acids
Res; 30:1292-1305.
McCaw, M. 2003. McREBEL Maanagement. Compendium PRRS: Second edition, 131-
136.
Manual terrestre de la OIE. 2012. Síndrome disgenésico y respiratorio porcino, capítulo
2.8.7. Disponible en:
http://www.oie.int/fileadmin/Home/esp/Health_standards/tahm/2.08.07_PRRS.pdf
Mardassi, H., Wilson, L., Mounir, S., Dea, S. 1994. Detection of porcine reproductive and
respiratory syndrome and efficient differentiation between Canadian and European strains
by reverse transcription and PCR amplification. J Clin Microbiol. 32:2197-2203.

Mardassi, H., Mounir, S., Dea, S. 1995. Molecular analysis of the ORFs3 to 7 of porcine
reproductive and respiratory syndrome virus, Québec reference strain. Arch. Virol. 140:
1405-1418.
Mardassi, H., Massie, B., Dea, S., 1996. Intracellular synthesis,m processing, and
transport of proteins encoded by ORFs 5 to 7 of porcine reproductive and respiratory
syndrome virus. Virology, 221: 98-112.
Mattheu, E., Tello, M., Coll, A., Casal, J., Martin, M., 2006. Comparison of three ELISAs
for the diagnosis of porcine reproductive and respiratory syndrome. Vet Rec. 159: 717-
718.
Mateu, E., Diaz, I. 2008. The challenge of PRRS inmunology. The veterinary journal, 177:
345-351.
Martínez, E., Riera, P., Sitja, M., Fang, Y., Oliveira, S., Maldonado, J. 2008. Simultaneous
detection and genotyping of porcine reproductive and respiratory síndrome virus (PRRSV)
by real-time RT-PCR and amplicon melting curve analysis usin SYBR Green. Research in
Veterinary Science, 85:184-193.
Meier, W.A., Galeota, J., Osorio, F, A., Husmann, R.J., Schnitzlein, W.M., Zuckermann,
F.A. 2003. Gradual development of the interferon-γ response of swine to porcine
reproductive and respiratory syndrome virus infection or vaccination. Virology, 309: 18-31.
Men, W.J., Paul, P.S.,Halbur, P.G., Lum, M.A., 1995. Phylogenetic analyses of the
putative M (ORF 6) and N (ORF 7) genes of porcine reproductive and respiratory
syndrome virus (PRRSV): implication for the existence of two genotypes of PRRSV in the
USA and Europe. Archives of Virology; 140: 745-755.
Mendoza, L.F., Campa, E.S., Resendiz, M., Mata-Haro, V., Osorio, F.A., Hernandez, J.
2009. Effect of porcine reproductive and respiratory syndrome (PRRS) virus on porcine
monocite-derived dendritic cells. Vet Mex. 40.

Mengeling, W.L., Vorwald, A.C., Larger, K., M. Brockmeier, S.L. 1996. Diagnosis of
porcine reproductive and respiratory syndrome using infected alveolar macrophages
collected from live pigs. Vet Microbiol; 49: 105-115.
Meng, X.J., Paul, P.S., Halburg, P.G., Morozov, I. 1995a. Sequence comparison of open
reading frames 2 to 5 of low and high virulence United States isolates of porcine
reproductive and respiratory syndrome virus. J Gen Virol. 76: 3181-3188.
Meng, X.J., Paul, P.S., Halbur, P.G., Lum, M.A. 1995b. Phylogenetic analyses of the
putative M (ORF6) and N (ORF7) genes of porcine reproductive and respiratory
syndrome virus (PRRSV): implication for the existence of two genotypes of PRRSV in the
U.S.A. and Europe. Arch. Virol. 140:745-755.
Meng, W, J., 2000. Heterogeneity of porcine reproductive and respiratory syndrome virus:
implications for current vaccine efficacy and future vaccine development. Vet. Microbiol.
74, 309-329.
Meulenberg, J.J., Julst, M.M., de Meijer, E.J., Moonen, P.I., den Besten, A., de Khuyver,
E.P., Wensvoort, R., Moorman, R.J., 1993. Lelystad virus, the causative agent of porcine
epidemic abortion and respiratory syndrome (PEARS), is related to LDV and EAV.
Virology, 192: 62-72.
Meulenberg, J.J., Petersen-den Besten, A., De Kluyver, E.P., Moormann, R.J., Schaaper,
W.M., Wensvoort, G. 1995. Characterization of proteins encoded by ORFs 2 to 7 of
Lelystad virus. Virology, 206: 155-163.
Meulenberg, J.J.M., and Besten, A.P. 1996. Identification and characterization of a sixth
structural protein of lelystad virus: the glicoprotein GP2 encoded by ORF2 is incorporated
in virus particles. Virology 225. 44-51.
Meulenberg, J.J.M., Van Nieuwstadt, A.P., Van Essen-Zandbergen, A., Langeveld, J.P.M.
1997. Posttranslational processing and identification of a neutralization domain of the
GP4 protein encoded by ORF4 of Lelystad virus. J. Virol. 71, 6061-67.

Meulenberg, J.M. 2000. PRRS, The virus. Vet. Res. 31:11-21.
Misinzo, G.M., Delputte, P.L., Nauwynck, H.J., 2008. Involvement of proteasas in porcine
reproductive and respiratory syndrome virus uncoating upon internalization in primary
macrophagues. Vet. Res. 39-55.
Mogollón, J.D., Rincón, M.A., Arbeláez, G., Ruiz, S., Rodríguez, V. 2003. PRRS virus in
Colombia. 2003 PRRS Compendium, second edition. 231-233.
Mogollón, J.D., Rincón, M.A., Arbeláez, G., Ruiz, S., Rodríguez, V. 2003. PRRS in
Colombia.PRSS Compendium Producer Edition.National Pork Board.Des Moines, Iowa,
USA, section 6.6.
Molina, R.M., Cha, S.H., Chittick, W., Lawson, S., Murtaugh, M.P., Nelson, E.A.,
Christopher-Hennings, J., Yonn, K.J., Evans, R., Rowland, R.R.R., Zimmerman, J.J.,
2008. Inmune response against porcine reproductive and respiratory syndrome virus
durin acute and chronic infection. Veterinary immunology and immunopathology, 126:283-
292.
Morgan, S.B., Graham, S.P., Salguero, F.J., Sánchez Cordón, P.J., Mokhtar, H.,
Rebel,J.M.J., Weesendorp, E., Bodman-Smith, K.B., Steinbach, F., Frossard, J.P.,
2013.Increased pathogenicity of European porcine reproductive and respiratory syn-
drome virus is associated with enhanced adaptive responses and viral clearance.Vet.
Microbiol. 163 (1–2), 13–22.
Mortensen, S., Sogaard, R., Boklund, A., Stark, K., D., Christensen, J, et al. 2002. Risk
factors for infection of sow herds with porcine reproductive and respiratory syndrome
(PRRS) virus. Prev. Vet. Med. 53: 83-101.
Mulupuri, P., Zimmerman, J.J., Hermann, J., Johnson, C,R., Cano, J.P., Yu, W., Dee,
S.A., Murtaugh, M,P. 2008. Antigen-specific B cell responses to porcine reproductive and
respiratory syndrome virus infection. J Virol. 82:358–70.

Murtaugh, M.P., Elam, M.R., Kakach, L.T. 1995. Comparison of the structural protein
coding sequences of the VR-2332 and Lelystad virus strains of the PRRS virus. Arch.
Virol. 140: 1451-60.
Murtaugh, M.P., Xiao, Z., Zuckermann, F. 2002. Immunological responses of swine to
porcine reproductive and respiratory syndrome virus infection. Viral Immunology, 15: 533-
547.
Murtaugh, M.P, Xiao, Z., Rutherford, M.S and Zuckermann, F. 2003. Inmunology. 2003
PRRS Compendium:second edition, p 193-206.
Murtaugh, M.P., Stadejek, T., Abrahante, J,E., Lam, T.T.Y., Leung, C.C.F. 2010. The
ever- expanding diversity of porcine reproductive and respiratory syndrome virus. Virus
Research, 154:18-30.
Murtaugh, M., Johnson, C.R., Griggs, T.F., Gnanandaraja, J. 2011a. Novel structural
protein in porcine reproductive and respiratory syndrome virus encoded by an alternative
ORF5 present in all arteriviruses. J Gen Virol. 92:1107-1116.
Murtaugh, M., Johnson, C., Griggs, T., Robinson, S., Gnanandarajah, J. 2011b. New
protein identification in PRRSV. Leman Swine Conference, 51.
Murtaugh, M.P. 2012. Use and interpretation of sequencing in PRRSV control programs.
Disease Diagnostics. Allen D, Leman Swine Conference.49-56.
Murtaugh, M. 2012. International molecular and virology diagnostic workshop for PRRSV.
ICA. October 28-November 2.
Nauwynck, H.J., Duan, X., Favoreel, H.W., Van Oostveldt, P., Pensaert, M.B., 1999. Entry
of porcine reproductive and respiratory syndrome virus into porcine alveolar macrophages
via receptor mediated endocitosis. J. Gen. Virol. 80:297-305.

Nedialkova, D.D., Ulferts, R., Van den Born, E., Lauber, C., Gorbalenya, A.E., Ziebuhr, J.,
Snijder, E.J., 2009. Biochemical characterization of arterivirus nonstructural protein 11
reveals the nidovirus-wide conservation of a replicative endoribonuclease. J. Virol. 83:
5671-5682.
Nelsen, C.J., Murtaugh, M.P., Faaberg, KS. 1999 Porcine reproductive and respiratory
syndrome virus comparison: divergent evolution on two continents. J Virol. 73, 270-80.
Nelson, E.A., Christopher-Hennings, J., Drew, T., Wensvoort, G., Collins, J.E., Benfield,
D.A., 1993. Differentiation of U.S. and European isolates of porcine reproductive and
respiratory syndrome virus by monoclonal antibodies. J Clin Microbiol; 31: 3184:3189.
Nelson, E.A., Christopher-Hennings, J., Benfield, D.A. 1994. Serum immune response to
the proteins of porcine reproductive and respiratory syndrome (PRRS) virus. J Vet Diagn
Invest. 6:410-415.
Neumann, E.J.,Kliebenstein, J.B., Johnson, C.D., Mabry, J.W., Bush, E.J., Seitzinger,
A.H., Green, A.L. & Zimmerman, J.J. 2005. Assessment of the economic impact of
porcine reproductive and respiratory syndrome on swine production in the United
States.Journal of the American Veterinary Medical Association. 227(3): 385-392.
Olsen, C., Wang, Ch., Christopher-Hennings, J., Doolitle, K., Harmon, K., Abate, S.,
Kittawornrat, A., Lizano, S., Main, R., Nelson, E., Otterson, T., Panyasing, Y.,
Rademacher, C., Rauh, R., Saha, R., Zimmerman, J. 2013. Probability of detecting
porcine reproductive and respiratory syndrome virus infection using pen-based swine oral
fluid specimens as a function of within-pen prevalence. Journal of Veterinary Diagnostic
Investigation, XX(X); 1-8.
Oleksiewicz, M.B., Botner, A., Toft, P., Normann, P., Storgaard, T., 2001. Epitope
mapping porcine reproductive and respiratory syndrome virus by phage display: the nsp2
fragment of the replicase polyprotein contains a cluster of B-cells epitopes. J. Virol. 75:
3277-3290.

Osorio, F. 2010. PRRSV infections: a world update. Acta Scientiae Veterinariae. 38 (supl
1): s269-s27.
Ostrowski, M., Galeota, JA., Jar, AM., Platt, KB., Osorio, FA., Lopez, O. 2002.
Identification of neutralizing and nonneutralizing epitopes in the porcine reproductive and
respiratory syndrome virus GP5 ectodomain. J. Virol. 76: 4241-50.
Otake, S., Dee, S. A., Rossow, K. D., Deen, J., Joo, H. S., Molitor, T. W. 2002a.
Transmission of porcine reproductive and respiratory syndrome virus by fomites (bootas
and coveralls). Swine Health Prod. 10:59-65.
Otake, S., Dee, S.A.,Rossow, K.D. 2002b. Transmission of porcine reproductive and
respiratory syndrome virus by needles. Vet. Rec., 150: 114-115.
Otake, S., Dee, S.A., Rossow, K.D, Moon, R.D., Pijoan, C. 2002c. Mechanical
transmission of porcine reproductive and respiratory syndrome virus by mosquitoes
Aedesvexans. Can. J. Vet. Res. 66: 191-195.
Park, B.K., Joo, H.S., Dee, S.S., Pijoan, C. 1995. Evaluation of an indirect fluorescent IgM
antibody test for the detection of pigs with recent infection of porcine reproductive and
respiratory syndrome virus. J. Vet. Diagn. Invest. 7: 544-546.
Papatsiros, V.G. 2012. Porcine Herd Health Management Practices for the Control of
PRRSV Infection. Clinic of Medicine, Faculty of Veterinary Medicine, University of
Thessaly,Greece.
Patel, D., Nan, Y., Shen, M., Ritthipichai, K., Zhu, X., and Zhang, Y. 2010. “Porcine
reproductive and respiratory syndrome virus inhibits type I interferon signaling by blocking
STAT1/STAT2 nuclear translocation,” Journal of Virology, vol. 84, 21; 11045– 11055.
Pei, Y., Hodgins, D.C., Lee, C., Calvert, J.G., Welch, S.K., Jolie, R., Keith, M., Yoo, D.
2008. Functional mapping of the porcine reproductive and repiratory syndrome virus

capsid protein nuclear localization signal and its pathogenic association. Virus Res. 135:
107-114.
Plagemann, P.G.W., Rowland, R.R.R., Faaberg, K.S., 2002. The primary neutralization
epitope of porcine reproductive and respiratory syndrome virus strain VR-2332 is located
in the middle of the GP5 ectodomain. Arch. Virol., 147: 2327-2347.
Pesente, P., Rebonato, V., Sandri, G., Giovanardi, D., SperatiRuffoni, L. and Torriani, S.
2006. Phylogenetic analysis of ORF5 and ORF7 sequences of porcine reproductive and
respiratory syndrome virus (PRRSV) from PRRS-positive Italian farms: A showcase for
PRRSV epidemiology and its consequences on farm management. Veterinary
Microbiology, 114: 214-224.
Pestka, S. 2007. The interferons: 50 years after their Discovery, there is much more to
learn, The Journal of Biological Chemistry, 282: 20047-20051.
PRRS: the disease, its diagnosis, prevention and control. Appendices IV and V of the
report of the OIE ad hoc group on porcine reproductive respiratory syndrome Paris, 9 - 11
June 2008.
Poumian, M. 1995. Disinfection of trucks and trailers. Revue Scientifique et Teccnique de
I’OIE, 19:165-170 et 171-176.
Prickett, J. R., Cutler, S., Kinyon, J.M., Naberhaus, N., Stensland, W.R., Yoon, K.J.,
Zimmerman, J.J. 2010. Stability of porcine reproductive and respiratory syndrome virus
and antibody in swine oral fluid. Journal of swine health and production, 18: 187-195.
Ramírez, A., Wang, C., Prickett, J.R., Pogranichniy, R., Yonn, K.J., Main, R., Johnson,
J.K., Rademacher, C., Hoogland, M., Hoffman, P., Kurtz, A., Kurtz, E., Zimmerman, J.,
2012. Efficient surveillance of pig populations using oral fluids. Prev. Vet. Med. 104:292-
300.

Reiner, G., Fresen, C., Bronnert, S., Willems, H. 2009. Porcine reproductive and
respiratory syndrome virus (PRRSV infection in wild boars. Veterinary Microbiology,
136:250-258.
Revista Porcicultura Colombiana. Asociación Colombiana de Porcicultores. En línea.
Edición 156 de 2011, Disponible en: (http://asoporcicultores.co/porcicultores/)
Rogan, D., Levere, J., Yoo, D. 2000. Immune response for the open reading frame 2
(ORF2) membrane glycoprotein of porcine reproductive and respiratory syndrome virus
(PRRSV) delivered in pigs by recombinant vaccinia virus. 16th International Pig
Veterinary Society Congress, Melbourne, Autstralia, p. 643.
Rossow, K.D., Bautista, E.M., Goyal, S.M., Molitor, T.W., Murtaugh, M.P., Morrison, R.B.,
Benfield, D.A., Collins, J.E. 1994. Experimental porcine reproductive and respiratory
síndrome virus infection in one, four and 10 weeks old pigs. J. Vet. Diagn Invest, 6:3-12.
Rovira, A., Clement, T., Christopher-Hennings, J., Thompson, B., Engle, M., Reicks, D.,
Muñoz-Zanzi, C. 2007. Evaluation of sensitivity of reverse-transcription polymerase chain
reaction to detect porcine reproductive and respiratory syndrome virus on individual and
pooled samples from boars. J. Vet. Diagn. Invest., 19: 502-509.
Rowland, R.R., Kervin, R., Kuckleburg, C., et al., 1999. The localization of porcine
reproductive and respiratory syndrome virus nucleocapsid protein to the nucleolus of
infected cells and identification of a potential nucleolar localization signal sequence. Virus
Res. 64:1-12.
Rowland, R.R.R., Lawson, S., Rossow, K., Benfield, D.A., 2003. Lymphoid tissue tropism
of porcine reproductive and respiratory syndrome virus replication during persistent
infection of pigs originally exposed to virus in utero. Vet. Microbiol. 96:219-235.
Rowland, R.R. 2010. The interaction between PRRSV and the late gestation pig fetus.
Virus Res, 154(1-2):114-22.

Rubio, A., Rodriguez, V., Kukielka, D. 2007. Aplicación de la RT-PCR en tiempo real para
el diagnóstico de PRRS. RCCV, 1: 646-653.
Salinas, J.A., Lara, J., Flores, H., Avalos, R., Zarate, J.J., Riojas, V., Segura, J.C.2008.
Presencia de Animales seropositivos al Síndrome Reproductivo y Respiratorio Porcino en
Nuevo León. Vet. Mex. 39: 215-221.
Sang, Y., Ross, C.R., Rowland, R.R., Blecha, F. 2008. Toll-like receptor 3 activation
decreases porcine arterivirus infection. Viral. Immunol. 21:303–313.
Shanmukhappa, K., Kim, J.K., Kapil, S. 2007. Role of CD151, A tetraspanin, in porcine
reproductive and respiratory syndrome virus infection. Virol. J. 4: 62-73.
Shaw, A.E., Reid, S.M., Ebert, K., Hutchings, G.H., Ferris, N.P., King, D.P. 2007.
Implementation of a one-step-real-time RT-PCR protocol for diagnosis of foot-and-mouth
disease. Journal of Virology Methodsd, 143: 81-85.
Shi, W., Wang, L., Lietal, X., 2011. “Endoribonuclease activities of porcine reproductive
and respiratory syndrome virus nsp11 was essential for nsp11 to inhibit IFN-β induction,”
Molecular Immunology, 48:1568–1572.
Shugars, D.C. 1999. Endogenous mucosal antiviral factors of the oral cavity. J. Infect.
Dis., 179: 431-435.
Síndrome disgenésico y respiratorio porcino. 2012. Manual Terrestre de la OIE,
capitulo2.8.7.Disponible en:
http://www.oie.int/fileadmin/Home/esp/Health_standards/tahm/2.08.07_PRRS.pdf
Síndrome reproductivo y respiratorio porcino (PRRS). Servicio Agrícola y Ganadero
(SAG).Disponible en: (http://www.sag.cl/ambitos-de-accion/sindrome-reproductivo-y-
respiratorio-porcino-prrs).

Snijder, E.J., Wassenaar, A.L., Van Dinten, L.C., Spaan, W.J., Gorbalenya, A.E. 1996.
The arterivirus nsp4 protease is the prototype of a novel group of chymotrypsin-like
enzimes, the 3C-like serine proteases. J. Biol. Chem. 271: 4864:4871.
Snijder, E.J., Meulenberg, J.J. 1998. The molecular biology of arteriviruses. J Gen Virol.
69: 971-979.
Snijder and Meulenberg. 2001. Arteriviruses in Fields. Virology, 1:1205-1220.
Snijder, E.J., Kikkert, M., Fang, Y. 2013. “Arterivirus molecular biology and pathogenesis”
Journal of General Virology, 94: 2141-2163.
Song, C., Krell, P., Yoo, D. 2010. Nonstructural protein 1alpha subunit-based inhibition of
NF-kappaB activation and suppression of interferon-beta production by porcine
reproductive and respiratory syndrome virus. Virology, 407: 268–280.
Stadejek, T., Stankevicius, A., Storgaard, T., Oleksiewicz, M.B., Belak, S., Drew,
T.W.,Pejsak, Z., 2002. Identification of radically different variants of porcine repro-ductive
and respiratory syndrome virus in Eastern Europe: towards a commonancestor for
European and American viruses. J. Gen. Virol. 83 (Pt 8), 1861–1873.
Stadejek, T., Oleksiewicz, M.B., Potapchuk, D., Podgorska, K., 2006b. Porcine
reproductive and respiratory syndrome virus strains of exceptional diversity in eastern
Europe support the definition of new genetic subtypes. J. Gen. Virol. 87, 1835–1841.
Stadejek, T., Oleksiewicz, M.B., Scherbakov, A.V., Timina, A.M., Krabbe, J.S., Chabros,
K., Potapchuk, D., 2008. Definition of subtypes in the European genotype of porcine
reproductive and respiratory syndrome virus: nucleocapsid characteristics and
geographical distribution in Europe. Arch. Virol. 153 (8), 1479–1488.

Stevenson, G.W., Van Alstine, W.G., Kanitz C.L., Keffaber, K.K. 1993. Endemic porcine
reproductive and respiratory syndrome virus infection of nursery pigs in two swine herds
without current reproductive failure. J Vet Diagn Invest. 5:432–434
Suárez, P., Zardoya, R., Martin, M.J., Prieto, C., Dopazo, J., Solana, A., Castro, J.M.
1996. Phylogenetic relationships of European strains of porcine reproductive and
respiratory syndrome virus (PRRSV) inferred from DNA sequences of putative ORF5 and
ORF7 genes. Virus Research, 42: 159-165.
Sun, Y., Xue, F., Guo, Y., Ma, M., Hao, N., Zhang, X.C., Lou, Z., Li, X., and Rao, Z., 2009.
Crystal structure of porcine reproductive and respiratory syndrome virus leader protease
Nsp1 alpha. J. Virol. 83: 10931-10940.
Sun, Y., Han, M., Kim, CH., Calvert, J, G., Y, D., 2012a. Interplay between Interferon-
Mediated Innate Immunity and Porcine Reproductive and Respiratory Syndrome Virus.
Viruses, 4: 424-446.
Sun, Z., Ransburgh, R., Snijder, E.J., and Fang, Y. 2012b. “Nonstructural Protein 2 of
porcine reproductive and respiratory syndrome virus inhibits the antiviral function of
interferon-stimulated gene 15,” Journal of Virology, vol. 86(7): 3839–3850.
Tian, H., Wu, J.Y., Shang, Y.J., Cheng, Y., Liu, W.T. 2010. The developmente of a rapid
SYBR one step real-time RT-PCR for detection of porcine reproductive and respiratory
syndrome virus. Virology Journal, 7: 1-7.
Tijms, M.A., Van Der Meer, Y., Snijder, E.J. 2002. Nuclear localization of non-structural
protein 1 and nucleocapsid protein of equine arteitis virus. J.Gen. Virol. 83:795-800.
Torremorell, M., and Christianson, W, T. 2002. PRRS eradication by herd closure. Adv
Pork Prod., 13: 169–76.
Torrison, J., Rossow, K,. Yeske, P. 2003. Use of PRRS virus sequence information within
herds. Allen D. Leman Swine Conference.

Van Aken, D., Snijder, E.J., and Gorbalenya, A.E., 2006. Mutagenesis analysis of the
nsp4 main proteinase reveals determinants of arterivirus replicase polyprotein
autoprocessing. J. Virol. 80: 3428-3437.
Van Breedman, W., Van Gorp, H., Zhang, J.Q., Crocker, P.R., Delputte, P.L., Nauwynck,
H.J. 2010. The M/GP5 Glycoprotein complex of porcine reproductive and respiratory
syndrome virus binds the Sialodhesin receptor in a Sialic Acid-dependent manner. Plos
Pathogens. 6: 1-11.
Van Dinten, L.C., deen Boon, J.A., Wassennar, A.L., Spaan, W.J., Snijder, E.J., 1997. An
infectious aerterivirus cDNA clone: identification of a replicase point mutation that
abolishes discontinuos mRNA transcription. Proc Natl Acad Sci USA, 64: 991-996.
Van Gorp, H., Van Breedam, W., Delputte, P.L., Nauwynck, H.J. 2008. Sialodhesin and
CD163 join forces during entry of the porcine reproductive and respiratory syndrome
virus. J. Gen. Virol. 89: 2943-2953.
Van Reeth, K., Nauwynck, H. 2000. Proinflammatory cytokines and viral respiratory
disease in pigs. Vet. Res. 31: 187-213.
Vashisht, K., Erlandson, K.R., Firkins, L.D., Zuckermann, F.A. and Goldberg, T.L. 2008.
Evaluation of contact exposure as a method for acclimatizing growing pigs to porcine
reproductive and respiratory syndrome virus. Journal of the American Veterinary Medical
Association, 232:1530-1535.
Vezina, S.A., Loemba, H., Foutnier, M., Dea, S., Archibault, D. 1996. Antibody production
and blastogenesis response in pigs experimentally infected with porcine reproductive and
respiratory syndrome virus. Can J Vet Res. 60:94–99.
Waddell, J., 2010. PRRS area regional control and elimination: Why is Nebraska a good
place to start? In: Proc. George A Young Swine Health Management Conf., South Sioux,
Nebraska, 54-59.

Wang, X., Eaton, M., Mayer, M., Li, H., He, D., Nelson, E., et al. 2007. Porcine
reproductive and respiratory syndrome virus productively infects monocyte–derived
dendritic cells and compromises their antigen–presenting ability. Arch Virol. 152:289–303.
Wang, R., Nan, Y., Yu, Y., Yang, Z., and Zhang, Y. 2013. “Variable interference with
interferon signal transduction by different strains of porcine reproductive and respiratory
syndrome virus,” Veterinary Microbiology, vol. 166 (3-4): 493–503.
Wang, R and Zhang, Y.J. 2014. Antagonizing Interferon-mediated immune response by
porcine reproductive and respiratory syndrome virus. BioMed Research International,
315470: 1-9.
Wangstrom, E.A., Chang, C.C., Yoon, K.J., and Zimmermann, J. J. 2000. Shedding of
porcine reproductive and respiratory syndrome virus in mammaria gland secretions of
sows. Am. J. Vet. Res., 62: 1876-1880.
Wasilk, A., Callahan, J.D., Christopher-Hennings, J., Gay, T.A., Fang, Y., Dammen, M.,
Reos, M.E., Torremorell, M., Polson, D., Mellencamp, M., Nelson E., and Nelson, W.M.
2004. Detection of U.S., Lelystad, and European-like porcine reproductive and respiratory
syndrome viruses and relative quantitation in boar semen and serum samples by real-
time PCR. J. Clin. Microbiol., 42, 4453–4461.
Welch, S.K., Calvert, J.G., 2010. A brief review of CD163 and its role in PRRSV infection.
Virus Research, 154(1-2):98-103.
Welch, S.W., Jolie, R., Pearce, D.S., Koertje, W.D., Fuog, E., Shields, S.L., Yoo, D.,
Calvert, J.G. 2004. Construction and evaluation of genetically engineered replication-
defective porcine reproductive and respiratory syndrome virus vaccine candidates. Vet.
Immunol. Immunopathol. 102: 277-290.
Weesendorp, E., Morgan, S., Stockhofe-Zurwieden, N., Popma-De Graaf, D.J., Gra-ham,
S.P., Rebel, J.M., 2013. Comparative analysis of immune responses

followingexperimental infection of pigs with European porcine reproductive and respi-
ratory syndrome virus strains of differing virulence. Vet. Microbiol. 163 (1–2),1–12.
Wellenberg, G.J., 2006. Review: diagnostic methods for the detection of porcine
reproductive and respiratory syndrome virus (PRRSV) infections. Tijdschr. Diergeneeskd,
131 (16): 566-572.
Wensvoort, G., Terpstra, D., Pol, J.M.A., Ter Laak, E.A., Bloemraad, M., de Kluyver, E.P.,
Kragten, D., Van Buiten, L., Den Besten, A., Wagenaar, F., Broekhuijsen, J.M., Moonen,
P.L.J.M., Zetstra, T., De Boer, E.A., Tibben, H.J., de Jong, M.F., Van´t Veld, P.,
Groenland, G.J.R., Van Gennep, J.A., Voets, M.Th., Verheijden, J.H.M., Braamskamp, J.
1991. Mystery swine disease in the Netherlands: the isolation of Lelystad virus. Vet. Q.
13: 121-130.
Wensvoort, G. 1993. Lelystad virus and the porcine epidemic abortion and respiratory
syndrome. Vet Res. 24: 117-124.
Wernike K., Bonilauri, P., Dauber, M., Errington, J., LeBlanc, N., Revilla-Fernandez, S.,
Hjulsager, C., Isaksson, M., Stadejek, T., Beer, M., Hoffmann, B. 2012. Porcine
reproductive and respiratory syndrome virus. Interlaboratory ring trial to evaluate real-time
reverse transcription polymerase chain reaction detection method. J. Vet. Diagn. Invest.,
24: 855-866.
Wills, R.W.,Zimmerman, J.J. 1997. Porcine reproductive and respiratory syndrome virus:
a persistent infection. Vet Microbiol, 55:231-240.
Wills, R., Osorio, F.A., Doster, A. 2000. Susceptibility of selected non-swine species to
infection with porcine reproductive and respiratory syndrome virus. In: Proceedings of the
31st Ann Meet Am Assoc Swine Pract. 411-413.
Wills, R.W., Doster, A.R., Galeota, J.A., Sur, J.H., Osorio, F.A. 2003. Duration of infection
and proportion of pigs persistently infected with porcine reproductive and respiratory
syndrome virus. J Clin Microbiol. 41:58–62.

Wissink, E.H.J., Kroese, M.V., Maneschijn-Bonsing, J.G., Meulenberg, J.J.M., van Rijn,
P.A., Rijsewijk, F.A.M., Rottier, P.J.M., 2004. Significance of the oligosaccharides of the
porcine reproductive and respiratory syndrome virus glycoproteins GP2a y GP5 for
infectious virus production. J. Gen. Virol. 85: 3715-3723.
Wisssink, E.H.J., Kroese, M.V., Van Wijk, H.A.R., Rijsewijk, F.A.M., Meulenberg, J.J.M.,
Rottier, P.J.M. 2005. Envelope protein requirements for the assembly of infectious virions
of porcine reproductive and respiratory syndrome virus. J. Virol. 79: 13495-12506.
Wu, W., Fang, Y., Farwell, R., Steffen-Bien, M., Rowland, R.R., Christopher-Hennings, J.,
Nelson, E.A. 2001. A 10-kDa structural protein of porcine reproductive and respiratory
syndrome virus encoded by ORF2b. Virology 287: 183-191.
Wu, W., Fang, Y., Rowland, R.R., Lawson, SR., Christopher-Hennings, J., Yoon, K.,
Nelson, E.A. 2005. The 2b protein as a minor structural component of PRRSV. Virus Res.
114, 177-181.
Xiao, Z., Batista, L., Dee, S., Halbur, P., Murtaugh, M.P., 2004. The level of virus-specific
T-cell and macrophage recruitment in porcine reproductive and respiratory syndrome
virus infection in pigs is independent of virus load. J. Virol. 78, 5923–5933.
Xue, F., Sun, Y., Yan, L., Zhao, C., Chen, J., Bartlam, M., Li, X., Lou, Z., and Rao, Z.,
2010. The crystal structure of of porcine reproductive and respiratory syndrome virus
nonstructural protein Nsp1beta reveals a novel metal-dependent nuclease. J. Virol.
84:6461-6471.
Yoo, D., Song, C., Sun, Y., Du, Y., Kim, O., Liu, H.C. 2010. Modulation of host cell
responses and evasion strategies for porcine reproductive and respiratory syndrome
virus. Virus Res. 154:48–60.

Yoon, I.J., Joo, H.S., Goyal, S.M., Molitor, T.W., 1994. A modified serum neutralization
test for the detection of antibody to porcine reproductive and respiratory syndrome virus in
swine sera. Journal of Veterinary Diagnostic Investigating, 6:289-292.
Yoon, K.J., Zimmerman, J.J., McGinley, M.J. 1995. Failure to consider the antigenic
diversity of porcine reproductive and respiratory syndrome PRRS virus isolates may lead
to misdiagnosis. J Vet Diagn. Invest. 7:386-387.
Yoon, K,J., Zimmerman, J.J., Swenson, S.L., McGinley, M.J., Eeernisse, K.A., Brekiv, A.,
Rhinehart, L.L., Frey, M.L., Hill, H.T., Platt, K.B., 1995. Characterization of the humoral
response to porcine reproductive and respiratory syndrome (PRRS) virus infection.
Journal of Veterinary diagnostic investigation: official publication of the American
Zimmerman, J.J., Association of Veterinary Laboratory Diagnosticians, Inc. 7:305-312.
Yoon, K.J., Wu, L.L., Platt, K.B. 1996. Antibody-dependent enhancement (ADE) of
porcine reproductive and respiratory syndrome virus (PRRS) infection in pigs. Virol
Inmunol. 9:51-63.
Yoon, K., Wu, L., Zimmerman, J.J., Platt, K.B. 1997. Field isolates of porcine reproductive
and respiratory syndrome virus (PRRSV) vary in their susceptibility to antibody dependent
enhancement (ADE) of infection. Vet. Microb. 55: 277-287.
Yoon, K.J. 2003. Virology. PRRS Compendium. Second edition, p163-189.
Yoon, K.J, .Christopher-Hennings, J., Nelson, E.A. 2003. Diagnosis. PRRS Compendium.
Second edition, p59-74.
Yun, S.I., and Lee, Y.M., 2013. Overview: Replication of porcine reproductive and
respiratory syndrome virus. Journal of Microbiology, 51:711-723.
Zhang, Y., Sharma, R.D., Paul, P.S. 1998. Monoclonal antibodies against
conformationally dependent epitopes on porcine reproductive and respiratory syndrome
virus. Vet. Microb. 63: 125-136.

Zhou, L., and Yang, H. 2010. Porcine reproductive and respiratory syndrome virus in
China. Virus Res. 154: 31-37.
Zhou Y.J., Hao, X.F., Tian, Z.J., Tong, G.Z., Yoo, D., An, T.Q., Zhou, G.X., Li, G.X., Qiu,
H.J., Wei, T.C., and Yuan, X.F. 2008. Highly virulent porcine reproductive and respiratory
syndrome virusemerged in China. Transbound Emerg Dis. 55:152-164.
Zhou, L., Chen, S., Zhang, J., Zeng, J., Guo, X., Ge, X., Zhang, D., Yang, H. 2009.
Molecular variation analysis of porcine reproductive and respiratory syndrome virus in
China. Virus Research, 145:97-105.
Zimmerman, J., Sanderson, T., Eernisse, K.,Hill, H., Frey, M. 1992. Transmission of SIRS
virus in convalescent animals to commingled penmates under experimental conditions.
Am. Assoc. Swine Pract.Newslett. 4, 25.
Zimmerman, J.J., Yonn, K.J., Wills, R.W., Swenson, S.L., 1997. General overview of
PRRS: a perspective from the United States. Vet. Microbiol. 55:187-196.
Zimmerman J. 2003. Epidemiology and Ecology. 2003 PRRS Compendium. Second
edition. P. 27-50.
Zimmerman, J., Benfield, D.A., Murtaugh, M.P., Osorio, F., Stenson, G.W., Torremorell,
M., 2006. Porcine reproductive and respiratory syndrome virus (porcine arterivirus).
Diseases of swine, 387-418. Blackwell publishing, Ames, Iowa.
Zipper, H., Brunner, H., Bernhagen, J., Vitzthum, F. 2004. Investigations on DNA
intercalation and surface binding by SYBR green I, its structure determination and
methodological implications. Nucleic Acids Res. 32: 103.



















