
ÍNDICE DE MATERIAS -...
52
2 ÍNDICE DE MATERIAS 1. INTRODUCCIÓN ................................................................................. 2 1.1. Objetivo general ................................................................................. 4 1.2. Objetivos específicos ......................................................................... 4 2. REVISIÓN BIBLIOGRÁFICA ................................................................ 5 2.1. Descripción botánica.......................................................................... 5 2.2. Distribución geográfica ...................................................................... 6 2.3. Citogenética del género Leucocoryne................................................ 7 2.4. Poliploidía en la mejora vegetal ......................................................... 8 2.4.1. Características de la colchicina...................................................... 11 2.4.2. Métodos para determinar ploidia.................................................... 14 2.4.2.1. Determinación por caracteres citológicos ................................... 14 2.4.2.2. Determinación por su morfología y desarrollo............................. 14 2.4.2.3. Cariología.................................................................................... 14 2.4.2.4. Citometría de flujo ....................................................................... 15 3. MATERIALES Y MÉTODOS ................................................................ 16 3.1. Material ............................................................................................. 16 3.1.1. Ubicación del ensayo ..................................................................... 16 3.1.2. Material vegetal .............................................................................. 16 3.1.3. Equipos de invernadero ................................................................. 17 3.1.4. Equipos de laboratorio ................................................................... 17 3.2. Método .............................................................................................. 18 3.2.1. Aplicación de colchicina ................................................................. 18 3.2.2. Polinización .................................................................................... 19 3.2.3. Germinación de semillas ................................................................ 20 3.2.4. Determinación de ploidia................................................................ 21 4. PRESENTACIÓN Y DISCUSIÓN DE RESULTADOS ......................... 24 4.1. Resultados de polinización dirigida ................................................... 24 4.2. Resultados de polinización natural ................................................... 25 4.3. Efecto de la colchicina sobre la geminación de semillas .................. 26 4.4. Análisis de ploidia ............................................................................. 32 5. CONCLUSIONES ................................................................................ 40 6. RESUMEN ........................................................................................... 41 7. ABSTRACT .......................................................................................... 42 8. LITERATURA CITADA ........................................................................ 43
Transcript of ÍNDICE DE MATERIAS -...

2
ÍNDICE DE MATERIAS 1. INTRODUCCIÓN ................................................................................. 2 1.1. Objetivo general................................................................................. 4 1.2. Objetivos específicos ......................................................................... 4 2. REVISIÓN BIBLIOGRÁFICA ................................................................ 5 2.1. Descripción botánica.......................................................................... 5 2.2. Distribución geográfica ...................................................................... 6 2.3. Citogenética del género Leucocoryne................................................ 7 2.4. Poliploidía en la mejora vegetal ......................................................... 8 2.4.1. Características de la colchicina...................................................... 11 2.4.2. Métodos para determinar ploidia.................................................... 14 2.4.2.1. Determinación por caracteres citológicos ................................... 14 2.4.2.2. Determinación por su morfología y desarrollo............................. 14 2.4.2.3. Cariología.................................................................................... 14 2.4.2.4. Citometría de flujo....................................................................... 15 3. MATERIALES Y MÉTODOS................................................................ 16 3.1. Material ............................................................................................. 16 3.1.1. Ubicación del ensayo..................................................................... 16 3.1.2. Material vegetal.............................................................................. 16 3.1.3. Equipos de invernadero ................................................................. 17 3.1.4. Equipos de laboratorio ................................................................... 17 3.2. Método.............................................................................................. 18 3.2.1. Aplicación de colchicina ................................................................. 18 3.2.2. Polinización.................................................................................... 19 3.2.3. Germinación de semillas................................................................ 20 3.2.4. Determinación de ploidia................................................................ 21 4. PRESENTACIÓN Y DISCUSIÓN DE RESULTADOS ......................... 24 4.1. Resultados de polinización dirigida................................................... 24 4.2. Resultados de polinización natural ................................................... 25 4.3. Efecto de la colchicina sobre la geminación de semillas .................. 26 4.4. Análisis de ploidia ............................................................................. 32 5. CONCLUSIONES ................................................................................ 40 6. RESUMEN........................................................................................... 41 7. ABSTRACT.......................................................................................... 42 8. LITERATURA CITADA ........................................................................ 43

3
1. INTRODUCCIÓN
Dentro de las etapas del mejoramiento genético encontramos la variación, la
que se refiere a las distintas combinaciones genéticas que se hallan en los
organismos de una población o al conjunto genético de variabilidad
encontrado en ella. La variación es una etapa importante porque es la base
sobre la cual se hará una selección de aquellos genotipos que tengan la
mejor expresión en caracteres medibles. En ausencia de variación el
mejorador debe aumentarlas mediante la mutación o buscarla en los lugares
de origen de las especies que se van a mejorar.
En el mejoramiento genético de plantas, se hace necesario el uso de
diversas técnicas genéticas, por lo que muchos autores se han dedicado con
interés a la investigación de la citogenética, basados en el uso de poliploidia
que presentan las especies cultivadas y/o sus ancestros o especies
silvestres. La poliploidia∗ tiene significación especial en el fitomejoramiento,
ya que influye en el incremento de la diversidad genética en el reino vegetal.
La poliploidia ofrece al genetista la oportunidad de lograr cambios en los
caracteres de una planta al alterar en número cromosómico, dentro de las
células individuales. También ha sido un factor importante en la evolución de
las especies vegetales. El uso de la poliploidia recibió gran impulso por parte
de los fitogenetistas cuando se descubrió que la Colchicina podría ser
utilizada para aumentar o duplicar el número de cromosomas.
La inducción de poliploidia supone la interrupción de la secuencia normal de
acontecimientos en la división nuclear.
∗ Palabra revisada de Diccionario de Botanica (FONT, 1953).

4
Por definición, un organismo poliploide posee más de dos juegos completos
de cromosomas en sus células somáticas, y el tipo de poliploide está
determinado por el número de juegos y por las relaciones genéticas que
existen entre ellos.
Es una serie poliploide presente en la naturaleza se encuentran niveles de
ploidia 2x, 3x, 4x, 5x, etc., presentando generalmente la especie con el
número cromosómico más alto, mayor vigor, por un aumento del tamaño
individual de la célula, aunque este aumento no siempre se extiende a
órganos y tejidos (STEBBINS, 1971).
La duplicación cromosomal se puede obtener por adición de complementos
cromosómicos de dos especies, o por duplicación natural del número de
cromosomas, y de acuerdo a su origen se denominan alotetraploide y
autotetraploide respectivamente.
La inducción de un poliploide puede realizarse de diversas maneras: golpes
de frío o calor, radiaciones o agentes químicos como él oxido nitroso, el etil-
mercurio y la sulfanilamida. Sin embargo la colchicina es el agente más
efectivo (ELLIOT, 1964).
La colchicina inhibe, en la división celular, el mecanismo del huso y propicia
la formación de células con el doble o más del doble del número de
cromosomas. La sustancia bloquea el ensamblaje de los microtúbulos del
huso mitótico, con esto, se impide el ordenamiento y la segregación de los
cromosomas, pero no su ciclo de duplicación y condensación.

5
En consecuencia mediante la técnica de la colchicina se puede inducir
plantas poliploide en forma sencilla, para el caso del género Leucocoryne, se
pretende desarrollar un método para duplicar el número de cromosomas de
Leucocoryne purpurea (2n = 10) y Leucocoryne sp. (2n = 18), con el
propósito de obtener órganos más grandes como flores, bulbos o varas;
aumento del vigor de la planta y hábitos de floración más tardíos, obteniendo
de esta manera plantas con mayor potencial ornamental y económico.
1.1. Objetivo general:
Desarrollar un método de aplicación de colchicina in vivo en dos especies de
plantas de Leucocoryne, 2n = 10 y 2n = 18, para la obtención de individuos
con el número cromosómico duplicado 4n.
1.2. Objetivos específicos:
• Evaluar una técnica de inducción de plantas poliploides durante el
proceso de meiosis.
• Determinar la concentración óptima de colchicina para la obtención de
plantas poliploides.
• Cuantificar el efecto del tratamiento con colchicina en la germinación
de las semillas de Leucocoryne.

6
2. REVISIÓN BIBLIOGRÁFICA
2.1. Descripción botánica:
MANSUR et al. (2002) señala que el género Leucocoryne comprende un
grupo de plantas herbáceas y bulbosas, endémicas de Chile, está formado
por cerca de 14 especies, las cuales se distinguen por su alta variabilidad
genética en cuanto a forma y aroma de sus flores.
El género Leucocoryne pertenece a la familia Alliaceae, subfamilia Aliodeae,
tribu Brodiaceae (DAHLGREN, CLIFFORD y YEO, 1985), aunque ha sido
clasificado como Liliaceae (ENGLER, 1964, citado por ZÖELLNER, 1972) y
Amaryllidaceae (HUTCHINSON, 1934, citado por ZÖELLNER, 1972). La
clasificación taxonómica basada en las características morfológicas, es difícil
ya que existen poblaciones de este género que presentan gran variabilidad
de forma y colorido tanto en los tépalos como en los estaminodios
(ZÖELLNER, 1972).
El género Leucocoryne se caracteriza por poseer gran cantidad de raicillas
filiformes de color blanco, que nacen del disco basal del bulbo. El bulbo es
esférico o piriforme y está cubierto por membranas o fibras de color castaño.
Las hojas son lineares, angostas y estriadas en número de dos a cuatro.
Desarrollan una umbela por escapo floral y poseen un tubo floral formado por
los tépalos creciendo en su parte inferior y por seis lacinias en dos series, las
que son muy semejantes entre sí (ZÖELLNER, 1972).
La fase de reproducción sexual normalmente se produce cuando el
meristema apical del disco desarrolla a expensas de sustancias de reserva

7
acumuladas en un tallo floral, que al rasgarse en su extremo se remata por
una inflorescencia en umbela (MAROTO, 1989).
MANSUR et. al. (2004) determinó que muchas especies del género
Leucocoryne, tiende claramente hacia la polinización cruzada y a la
autoincompatibilidad, lo que podría explicar su alto nivel de variabilidad
genética.
2.2. Distribución geográfica:
El género Leucocoryne es endémico de Chile. Todas las especies se
distribuyen a lo largo de gran parte del territorio nacional, desde Iquique
(Norte del país, I Región) a la provincia del Bío-Bío (VIII Regíon). En la zona
norte se encuentra desde una altura de 500 – 800 m sobre el nivel del mar en
las quebradas, sobreviviendo gracias a las abundantes neblinas, al igual que
en las cercanías de Antofagasta y Tocopilla, a partir del río Elqui hasta el río
Maipo, las poblaciones se extienden desde la cordillera de la Costa hasta la
cordillera de Los Andes.hacia, al sur del país las poblaciones de Leucocoryne
sólo crecen en los cordones montañosos de la costa a una altura de 400 –
800 m. Desde el Valle de Copiapó hasta el río Elqui, debido a las condiciones
climatológicas más favorables, se presenta la mayor abundancia
poblacional., A partir del río Maipo al sur decrece considerablemente la
presencia del género, y en la provincia del Bío-Bío sólo es posible encontrar
la especie L. alliacea, formando parte del sotobosque (ZÖELLNER, 1972).
Leucocoryne purpurea Gay. (ANEXO Nº1), habita en la zona costera de la IV
Región, abunda en el sector comprendido entre la Bahía de Tongoy y el
Parque Nacional Fray Jorge. Las flores son de color púrpura, que se
intensifica hacia la base (ZÖELLNER, 1972).

8
2.3. Citogenética del género Leucocoryne:
CAVE (1939) realizó el primer estudio cariológico en Leucocoryne ixioides, y
reporto que poseía un cariotipo de 2n = 18.
Las observaciones citológicas realizadas en ejemplares de la mayoría de las
especies conocidas de Leucocoryne, han permito establecer que un grupo de
especies posee un cariotipo 2n = 10, en tanto un segundo grupo posee un
cariotipo 2n = 18 (BAHAMONDES y LABARCA, 1994; GRAU 1992; PALMA-
ROJAS, ZEPEDA y LABARCA, 1991; CROSA 1988; CAVE, 1939), estas
observaciones fueron confirmadas por (ARANEDA, SALAS y MANSUR,
2004), incluyendo otras poblaciones no definidas taxonomicamente, de
distintas zonas geográficas, las que presentaron los mismos números
cromosómicos de 2n = 10 y 2n = 18, distribuyéndose equitativamente y
encontró una especie con número cromosómico de 2n = 14.
CROSA (1988) señala que el cariotipo 2n=18 sería explicable por la
poliploidización a partir de especies 2n=10, sugiriendo la ocurrencia de una
hibridación de especies ancestrales de numero cromosómico 2n = 10
originando individuos tetraploides 2n = 20. En éstos habría ocurrido una
disminución del número cromosómico debido a la fusión céntrica entre
cromosomas acrocéntricos.
MORENO y ARANCIO (2001) plantean que de acuerdo a antecedentes
morfológicos, estos indican la existencia de flujo genético entre L. purpurea y
L. coquimbensis que habría generado F1 semifértiles. Éstos en una
retrocruza con los parentales habrían formados individuos fértiles y
semifértiles de las individuos mezclados. SALAS y MANSUR (2004)
establecieron que se produce un flujo genético entre poblaciones donde

9
coexisten individuos de distintos niveles de ploidia originando híbridos
ínterespecíficos con un número cromosómico intermedio 2n = 10, 2n =14, 2n
=18 y 2n = 22, produciendo una poliploidia natural en estas poblaciones.
2.4. Poliploidia en la mejora vegetal:
En las células somáticas se encuentran dos juegos de cromosomas
homólogos, cada uno de los cuales proviene de sus progenitores masculino y
femenino respectivamente, denominándose esta condición como diploide. En
algunas especies, las células somáticas presentan más de dos juegos de
cromosomas, recibiendo el nombre de poliploides. La poliplodización es un
proceso, no un evento (DE WET, 1981)
Los diploides son 2x, los triploides 3x, los tetraploides 4x, los pentaploides 5x
y así sucesivamente. Los miembros que se encuentran en un nivel superior
a los diploides, son llamados poliploides en conjunto (ELLIOT, 1964).
Los genomas de acuerdo a su dotación cromosómica, pueden ser:
Autopoliploide, en que los n juegos cromosómicos son homólogos (AAAA) y
Alopoliploides, en que los juegos cromosómicos pueden ser total o
parcialmente no homólogos y que se definen como Alopoliploides genómicos
(AABB) y Alopoliploides segmentales (AsAsAtAt), respectivamente
(STEBBINS, 1971; GRANT, 1981).
La poliploidia tiene importancia respecto a campos tan diversos como la
citogenética, la fisiología, la reproducción vegetal, la citotaxonomía y la
biogeografía (GRANT, 1989).

10
La poliplodia se constituye como el rasgo más conspicuo de la evolución
cromosómica en las plantas superiores (STEBBINS, 1971). Una alta
proporción de las especies de las plantas superiores son poliploides. Según
AVERETT (1981), en las primeras aproximaciones señala que el 36% de las
angiospermas son poliploides, pero entre un 70 y 80% pueden presentar
poliploidia en su historia evolucionaria. Posteriormente otros cálculos estiman
que los porcentajes serían, 49% de las Dicotiledóneas, 60 % de las
Monocotiledóneas y el 52% de las Angiospermas (GRANT, 1989).
Él término poliploidia se refiere a una relación aritmética especial entre los
números cromosómicos de organismos emparentados que poseen diferentes
números (GRANT, 1989).
El efecto más inmediato y universal de la poliploidia en las plantas es un
aumento del tamaño y masa celular. Este hecho no necesariamente origina
un aumento en el tamaño total de planta, y usualmente el efecto de
gigantismo debido a poliploidia puede estar restringido a flores y semillas.
Puede cambiar también la forma y textura de hojas y pétalos, que
usualmente son más gruesos y firmes que en los progenitores diploides
(STEBBINS, 1971). Otro efecto descrito es la disminución de la velocidad de
crecimiento, como consecuencia de un aumento en la duración del ciclo
celular, aunque esto no parece ser universal, pues en algunas especies con
distintos niveles de ploidia, los tiempos de ciclo no varían significativamente
(REES, 1972).
Los diversos tipos de plantas cultivadas muestran diferencias de
adaptabilidad a la inducción de poliploides. Las plantas con un número bajo
de cromosomas tienen una mayor probabilidad de responder bien a la
duplicación que aquellas plantas con números altos de cromosomas. En la

11
duplicación de cromosomas es más probable que se tenga éxito con plantas
alógamas que con plantas autógamas; las probabilidades de éxito con
poliploides son mayores en plantas que se cultivan por sus partes
vegetativas, que en aquellas que se cultivan por su semilla (ELLIOT, 1964).
Los poliploides generalmente tienen una fertilidad inferior a la de sus
prototipos diploides. En ciertos cultivos hortícolas o de flores, en que los
costos de la semilla son de poca consideración en relación con la belleza y la
vistosidad de flores más grandes, la reducción en la fertilidad no es un factor
crítico (ELLIOT, 1964).
La aplicación práctica de la técnica de la colchicina propuesta por
(BLAKESLEE et al., 1937, citado por HANCOCK, 1997), abrió el camino para
la producción de poliploides en cantidades virtualmente ilimitadadas.
Hacia 1979 se han duplicado con éxito el número cromosómico alrededor de
150 especies, utilizando colchicina (DEWEY, 1979).
HANCOCK (1997) señala que durante el siglo XX, la colchicina ha sido
utilizada en distintas investigaciones para duplicar el número cromosómico
en numerosas especies cultivadas. Indicando por ejemplo, la inducción
poliploides en manzana, uvas, peras, duraznos, arándano agrio y frutillas, la
duplicación de cromosomas en varias especies de flores, como las petunias,
maravillas, lilium entre otras, en hortalizas como lechugas, ají, papas, etc. y
en cultivos industriales como centeno, tabaco, algodón y remolacha
azucarera.

12
2.4.1. Características de la colchicina.
La Colchicina puede utilizarse para duplicar el número de cromosomas. Éste
producto tiene influencia en la producción de poliploidía, al impedir el
desarrollo de los husos acromáticos y paredes celulares sin afectar la división
de los cromosomas. Producto de esto, el número cromosómico aumenta de
diploide a tetraploide, aún cuando todos los cromosomas permanecen
dentro de una célula individual (POEHLMAN, 1965).
Según POEHLMAN (1965), la Colchicinaes es un alcaloide extraído de las
semillas o de las raíces del Colchicum autumnale, es una sustancia soluble
en agua y por consiguiente fácil de utilizar. En concentraciones
relativamente bajas pueden ser soportadas por muchas plantas sin resultar
tóxica (BRAUER, 1969).
Su fórmula química es la siguiente: C22H25NO6 con un peso molecular de
339,4 Es un polvo amarillo pálido con un punto de fusión de 157ºC. Se
oscurece a la luz, muy soluble en alcohol y cloroformo; soluble en agua, y se
utiliza en genética vegetal (POEHLMAN, 1965).
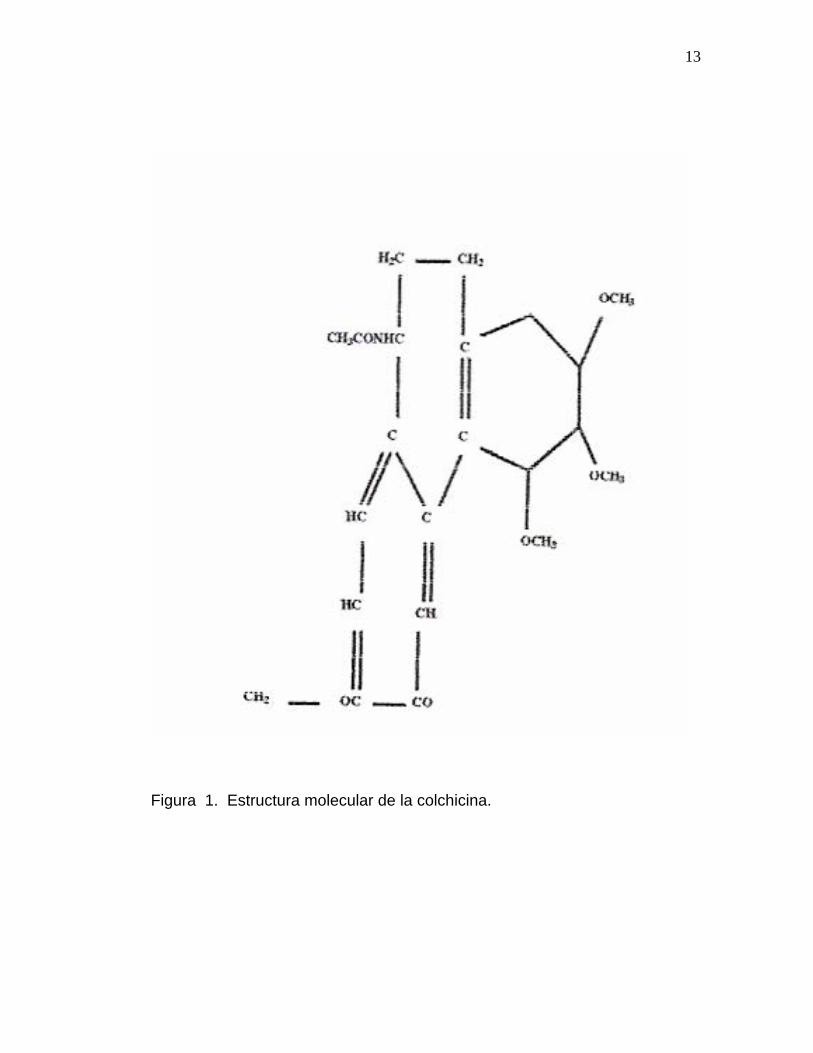
13
Figura 1. Estructura molecular de la colchicina.

14
CUBERO (1999), señala que el sistema consagrado desde hace mucho
tiempo es la utilización de colchicina, sustancia que produce una mitosis
anormal (llamada c-mitosis) al anular la formación del acromático, esencial
en la separación de los cromatidios hermanos en anafase, y así mismo como
consecuencia la del tabique celular, que debería dividir, en dos hijas la célula
inicial. Ello hace que en vez de emigrar dos 2n cromátidas a cada polo para
formar posteriormente, por la replicación del ADN, sendas células hijas con
2n cromosomas cada una, quedan 4n cromatidios en el centro de la célula
original. Dichas cromatidias completan luego su proceso normal de
reduplicación, convirtiéndose en 4n cromosomas. La célula es pues,
tetraploide, y lo será también el individuo que eventualmente llegue a formar.
Según ALLARD (1967), la colchicina es el agente más efectivo para la
duplicación cromosomal. Ésta al ser aplicada a las plantas en una pasta de
lanolina, embebida en un algodón que se mantiene en contacto con el tejido
meristemático o mediante cualquier otro procedimiento, no se generan
filamentos del huso en muchas células, los cromosomas no se alinean en el
plano ecuatorial, si no, que se dividen sin moverse hacia los polos.
Posteriormente los cromosomas pasan por una telofase normal, formando
una membrana alrededor del núcleo con doble número de cromosomas.
Mientras se mantenga en la célula una concentración crítica de colchicina, la
duplicación se repite varias veces hasta que después de tres o cuatro días
pueden existir varios centenares de cromosomas. Por el contrario, si la
colchicina se aplica por poco tiempo vuelve a formarse el huso y se detiene
la duplicación. Generalmente, sólo las células con número tetraploides, o en
algunos casos octaploides, son capaces de reproducirse y dar lugar a
sectores de tejidos a partir de los cuales se puede perpetuar la raza
poliploide, en programas de mejoramiento genético.

15
2.4.2. Métodos para determinar ploidia
2.4.2.1. Determinación por caracteres citológicos
Es posible identificar tejidos u órganos poliploides a través de sus células las
que suelen ser mayores o de diferente forma, particularmente los estomas y
los granos de polen (CUBERO, 1999).
FRANDSEN (1967) indica la relación entre ploidia y número de cloroplastos
por célula estomática, calculando la densidad promedio tanto para plantas
diploides coma para plantas tetraploides. Las plantas diploides tienen
estomas más densos y de diámetro más pequeño que las plantas
tetraploides.
Las plantas tetraploides generalmente tienen granos de polen más grandes
que las plantas diploides y mayor cantidad de poros (CRAMER, 1999).
2.4.2.2. Determinación por su morfología y desarrollo
El poliploide suele presentar gigantismo, aunque no siempre; existe un nivel
de ploidia por encima del cual ya no hay mas aumento de tamaño. Suelen
ser de menor velocidad de desarrollo que los diploides (CUBERO, 1999).
2.4.2.3. Cariología
El material elemental de la cariología es el conjunto de cromosomas de una
célula en división. Un cariotipo proporciona información sobre el número de
cromosomas o número diploide (2n) (SPOTORNO, 1985).

16
CRAMER (1999) plantea que el estudio cariológico se puede realizar en
células madres del polen, así como también en puntas de raíces en activo
crecimiento.
CHONG y OZIAS-AKINS (1992) consideran que el conteo de cromosomas
en células meristemáticas es el método inequívoco para la determinación del
número de cromosomas y nivel de ploidia de una especie.
2.4.2.4. Citometría de flujo.
La citometría de flujo permite medir el contenido de DNA de un aislado del
núcleo. Si los núcleos se tiñen con un marcador específico para DNA se
puede determinar el tamaño relativo del genoma de los individuos en estudio.
Cuando los niveles de ploidia del testigo son conocidos y se miden de esta
manera, se pude deducir el nivel de ploidia del individuo en estudio. La
ventaja de esta técnica es la rapidez y el procesamiento de datos mediante
un software (OLLITRAULT-SAMMARCELLI et al., 1993).
Un citómetro de flujo típico analiza dos parámetros ópticos básicos de las
partículas en suspensión, o sea, emisión de dispersión frontal y lateral de la
fluorescencia de diferentes longitudes de ondas (DOLEZEL, LUCRETTIN y
MACAS, 1995).
La Citometría de flujo del ADN (COSTICH et al., 1993), es un método rápido
y directo de conocer la ploidia de una planta a partir de la cantidad de ADN
existente en sus células, sin embargo, se necesita de un equipamiento
especial.

17
3. MATERIALES Y MÉTODOS
3.1. Material:
3.1.1. Ubicación del ensayo
El ensayo se realizó en el Laboratorio de Fitogenética y el invernadero frío de
la Facultad de Agronomía de la Pontificia Universidad Católica de Valparaíso,
ubicada en la provincia de Quillota, V región, cuyas coordenadas geográficas
son 32°50’’ Latitud Sur y 71°13’’ Longitud Oeste.
3.1.2. Material vegetal
El material vegetal empleado en este estudio, se obtuvo de plantas que se
encuentran en la colección de germoplasma de la Facultad de Agronomía de
la Pontificia Universidad Católica de Valparaíso y que corresponde a las
especies Leucocoryne purpurea y Leucocoryne spp. (Alcones), genotipo que
aún no es clasificado taxonómicamente y se denomina según su lugar de
colecta (ARANEDA et al., 2004).
Leucocoryne sp. es un ecotipo recolectado en la localidad de Alcones
ubicada en km 370, ruta cinco norte, Comuna de Ovalle, Provincia de Limarí
(38º 46’ S 71º 35’ O). Presenta un color azulado en las lacinias y en el ápice
de los estaminodios, donde el color es más intenso (ANEXO Nº1).
Las plantas se identificaron y fueron distribuidas al azar en un invernadero
frío.

18
CUADRO 1. Listado de material vegetal y plantas tratadas del género Leucocoryne.
Material Vegetal Número
cromosómico
Número total
plantas
Número de plantas
tratadas
L. sp. (tipo Alcones)1 2n = 18 306 154
L. purpurea 2n = 10 230 67
1 Para las presentaciones de cuadro, el ecotipo L. sp. recolectado en la localidad Alcones se designa Tipo Alcones.
3.1.3. Equipos de invernadero
• Sustrato 50% arena y 50% tierra de hoja esterilizada con vapor.
• Maceteros de plásticos de 15 cm. de diámetro.
• Contenedores de polietileno de 1 l.
• Herramientas de jardinería.
• Harneros, etiquetas de identificación.
3.1.4. Equipos de laboratorio.
• Solución de colchicina al 0,05% y 0,1%.
• Agua destilada.
• Pinzas y bisturí.
• Placas Petri.
• Papel filtro y algodón.
• Balanza analítica.
• Pipeta.
• Espátula.
• Guantes quirúrgicos.
• Mascarilla.

19
3.2. Método:
3.2.1. Aplicación de colchicina.
Se prepararon dos soluciones de colchicina con diferentes concentraciones:
0,05% y 0,1%.
Se utilizaron plantas de Leucocoryne que se encontraban en estado de
prefloración, específicamente aquellos escapos o botones florales en estado
temprano de desarrollo cuya longitud aproximada era 3 mm.
CAVE (1939) señala que aparentemente la reducción de las células del polen
ocurre cuando el tallo floral aún no emerge del bulbo. Pero las observaciones
cariológicas desarrolladas en Laboratorio de Fitogenética de la Facultad de
Agronomía de la Pontificia Universidad Católica de Valparaíso, indican que
fenológicamente la emergencia a ras de suelo del escapo floral coincide
fisiológicamente a la ocurrencia de meiosis.
La tierra alrededor del escapo fue removida para realizar pequeñas
incisiones superficiales con un bisturí y favorecer de esta forma la posterior
penetración de la solución de colchicina.
Se utilizó papel filtro cortado en forma rectangular con dimensiones de 5 x 4
cm para formar un cilindro que, posteriormente fue colocado envolviendo el
escapo. Sobre la parte superior del cilindro, se colocó una mota de algodón.
Se aplicó 1 ml de solución de colchicina sobre la parte superior del cilindro y
se marcó el cilindro con una cinta de color indicativo de la dosis aplicada. El
escapo se mantuvo con colchicina durante 12 hr y luego se retiró el cilindro.

20
3.2.2. Polinización
Los cruzamientos fueron dirigidos en forma intraespecífica y entre plantas
tratadas con las mismas dosis de colchicina. Del total de plantas tratadas se
polinizó el 20%. Este procedimiento, se realizó para asegurar una mejor
polinización en las plantas tratadas, ya que existe una tendencia en el genero
Leucocoryne a la auto incompatibilidad.
El resto de las plantas se dejo actuar la polinización natural de la especie
esperando que se cumpla la tendencia natural a formar poliploides. Cabe
destacar que en el invernadero donde florecieron las plantas tratadas, existen
una gran cantidad de plantas del genero Leucocoryne de distintas especies
asegurando una gran diversidad de polen, existente en el medio.
El protocolo de polinización manual ha sido definido como parte del proyecto
de mejoramiento genético para plantas del género Leucocoryne en la
Facultad de Agronomía de la Pontificia Universidad Católica de Valparaíso
que se resume en las siguientes etapas:
Emasculación de la flor escogida como parental femenino, la cual consiste en
hacer una incisión usando unas pinzas desde el ápice hasta un nivel medio
del tubo floral, abriendo el corte para proceder a remover los estambres,
tomándolas con las mismas pinzas. Proceso que se ejecuta en estado de
botón floral con tépalos coloreados (según la especie o ecotipo) con el ápice
de los mismos levemente abiertos, en esta etapa los estambres se
encuentran viables sin polen maduro en las anteras.
Remoción de anteras maduras desde la flor escogida como parental
masculino. Utilizando el mismo procedimiento anterior de corte del tubo

21
floral, para extraer las anteras o bien introduciendo las pinzas por la boca del
tubo para tomar las anteras y jalando hacia arriba suavemente para
extraerlas, procedimiento que se realiza usando estados avanzados del
desarrollo floral para asegurar madurez de las anteras y la dehiscencia.
Posteriormente se deposita el polen sobre el estigma de la flor parental
femenina. Con la antera extraída de la flor parental masculina, tomada con
pinzas, se procede a tomar el estigma de la flor parental femenina hasta
dejarlo de una coloración amarilla, que revela la presencia de abundante
polen. Luego se pliegan los tépalos hacia el centro y se coloca una cinta de
enmascarar para evitar la entrada de insectos polinizantes que distorsionen
el tratamiento.
3.2.3. Germinación de semillas
Todas las semillas se obtuvieron de frutos maduros (secos). Las semillas
fueron seleccionas por uniformidad de tamaño, se contaron y guardaron en
sobres de papel.
Las semillas se pusieron a germinar sobre placas Petri cubiertas con papel
absorbente empapados con agua destilada. Las placas se almacenaron en
una cámara sin luz y con temperatura constante de 15ºC, las perdidas por
evaporación fueron repuestas cada dos días, una semilla se considera
germinada cuando emerge la radícula, los conteos de semillas germinadas
se realizaron cada dos días, y la germinación de toda la población se asumió
cuando ninguna de las semillas germinó luego de seis días consecutivos.
• Test de germinación.

22
Se utilizó la ecuación ocupada por (DE LA CUADRA, 2002):
p = A [1 – exp{-k(t –t0)}]
Donde:
p: porcentaje de germinación en el tiempo t.
A: porcentaje final de germinación.
t0: tiempo estimado de germinación de la primera semilla.
k: medida de extensión de tiempo de germinación.
El transplante se realizó a mediada que las semillas alcanzaron una longitud
de radícula de 2 mm. Se colocaron en maceteros y se llevaron a invernadero
para su desarrollo. Las plantas fueron regadas una vez por semana hasta
que alcanzaron una altura aproximada de 10 cm, coincidente con 1 cm2 de
área foliar para ser analizadas.
3.2.4. Determinación de ploidia
El tejido utilizado para el análisis de citometría de flujo, corresponde a una
pequeña porción de hoja de tamaño mínimo de 1cm2, las hojas se obtuvieron
en dos estados de las plantas, ya que el análisis también se realizó en dos
etapas:
El análisis de ploidia mediante citrometría de flujo, se basa en la iluminación
de la muestra con una lámpara de mercurio, la que excita las moléculas
teñidas con DAPI, las que emiten fluorescencia, que es captada por un foto
multiplicador que convierte la fluorescencia en pulsos electrónicos, los cuales
son digitalizados y convertidos en números binarios y almacenados en
histogramas unidimensionales.

23
En la etapa preliminar, éste se realizó sobre hojas jóvenes, de plántulas
obtenidas de semillas. Este primer análisis incluyó ambas especies, pero no
todos los tratamientos ya que en dos de ellos (0.05 colchicina+polinización
dirigida y 0.1colchicina+polinización dirigida) las plántulas no sobrevivieron
después del transplante o las hojas no alcanzaron el tamaño mínimo
requerido para ser analizadas.
Posteriormente se hizo crecer los bulbillos obtenidos de la primera etapa,
estos se cosecharon y se mantuvieron en cámara según las
recomendaciones de manejo propuestas por OHKAWA et. al. (1996).
Concluido el receso se plantaron durante una temporada de crecimiento con
el objeto que los bulbos acumulen reservas y aumenten su tamaño Una vez
completada la temporada de crecimiento fueron nuevamente llevadas a una
cámara de receso vegetativo.
En la segunda etapa, en el análisis se utilizó hojas obtenidas de plantas post-
juvenil obtenidas de bulbos plantados en maceteros individuales crecidas en
invernadero. Este análisis se realizó solo para la especie Leucocoryne
purpurea, ya que se obtuvieron mayor número de bulbos y los resultados de
duplicación fueron mejores.
Las plantas se analizaron mediante Citometría de Flujo en la Empresa
PLANT CYTOMETRY SERVICES domiciliada en Europalaan 74 - 5481 JG
Schijndel, Holanda. El protocolo de análisis de citometría de flujo utilizado por
la empresa Plant Cytometry Services. Esta explicado en el (ANEXO Nº2)
Las plántulas fueron cosechadas vaciando los maceteros sobre una serie de
harneros de diferente tamiz, para separar los bulbos de la parte aérea. Los
bulbos fueron almacenados en sobres de papel y llevados a cámara a 20ºC.

24
por cuatro meses. Posteriormente fueron plantados en maceteros
independientes con el objeto de aumentar su tamaño y asegurar su viabilidad
en ensayos posteriores.
Las hojas primarias fueron colocadas en un sobre de papel absorbente
humedecido con agua destilada, cada sobre fue etiquetado e introducido en
bolsas de plásticos herméticas, en grupos de diez plantas por bolsa. Éstas
bolsas fueron enviadas en sobres con cojinetes de aire para evitar ser
aplastadas, el transporte fue por medio un servicio de Courier internacional.

25
4. PRESENTACIÓN Y DISCUSIÓN DE RESULTADOS.
4.1. Resultado polinización dirigida:
Los resultados de la polinización intraespecífica y de plantas tratadas con la
misma dosis de colchicina se indican en el Cuadro 2 y 3.
CUADRO 2. Porcentaje de cruzamientos exitosos, para los genotipos y distintas dosis colchicina.
Genotipo Dosis
Colchicina (%)
Total Cruzamientos
Cruzamientos exitosos
Porcentaje de éxito
L. spp. (Alcones) 0.05 19 12 63% L. spp. (Alcones) 0,1 18 4 22% L. spp. (Alcones) Testigo 20 12 60% L. purpurea 0,05 7 6 86% L. purpurea 0,1 11 10 91% L. purpurea Testigo 20 17 85%
Los porcentajes de éxito varían entre 22% a 91%, encontrándose en la
especie Leucocoryne purpurea, los mayores valores.
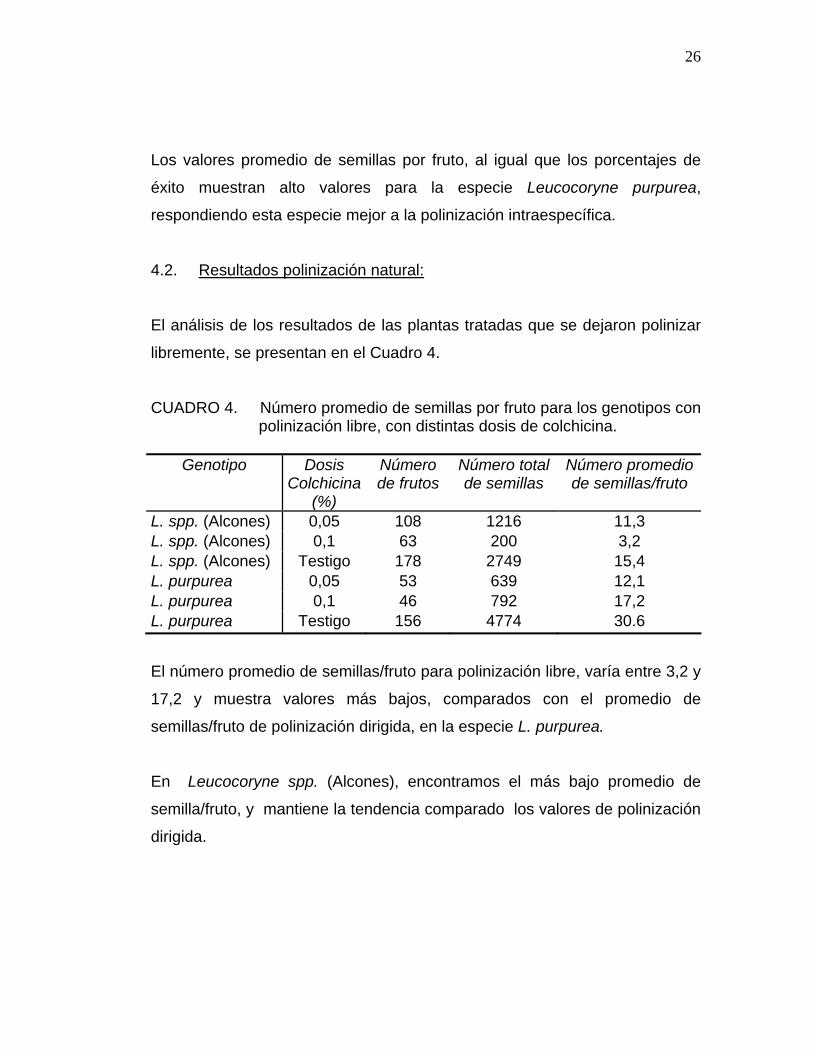
CUADRO 3. Número promedio de semillas por fruto para los genotipos con
polinización dirigida, con distintas dosis de colchicina.
Genotipo Dosis Colchicina
(%)
Número de frutos
Número total de semillas
Número promedio de semillas/fruto
L. spp. (Alcones) 0,05 12 83 6,9 L. spp. (Alcones) 0,1 4 29 7,3 L. spp. (Alcones) Testigo 12 211 17,6 L. purpurea 0,05 6 345 57,5 L. purpurea 0,1 10 554 55,4 L. purpurea Testigo 17 966 56,8
26
Los valores promedio de semillas por fruto, al igual que los porcentajes de
éxito muestran alto valores para la especie Leucocoryne purpurea,
respondiendo esta especie mejor a la polinización intraespecífica.
4.2. Resultados polinización natural:
El análisis de los resultados de las plantas tratadas que se dejaron polinizar
libremente, se presentan en el Cuadro 4.
CUADRO 4. Número promedio de semillas por fruto para los genotipos con polinización libre, con distintas dosis de colchicina.
Genotipo Dosis
Colchicina (%)
Número de frutos
Número total de semillas
Número promedio de semillas/fruto
L. spp. (Alcones) 0,05 108 1216 11,3 L. spp. (Alcones) 0,1 63 200 3,2 L. spp. (Alcones) Testigo 178 2749 15,4 L. purpurea 0,05 53 639 12,1 L. purpurea 0,1 46 792 17,2 L. purpurea Testigo 156 4774 30.6
El número promedio de semillas/fruto para polinización libre, varía entre 3,2 y
17,2 y muestra valores más bajos, comparados con el promedio de
semillas/fruto de polinización dirigida, en la especie L. purpurea.
En Leucocoryne spp. (Alcones), encontramos el más bajo promedio de
semilla/fruto, y mantiene la tendencia comparado los valores de polinización
dirigida.

27
4.3. Efecto de la colchicina sobre la germinación de semillas:
Las curvas de germinación se ajustaron por regresión lineal simple a la
ecuación exponencial p = A [1-exp{k(t-t0)}] y los valores de los coeficientes de
determinación (R2) se encuentran en un rango que varia entre 0,93 a 1,0
indicando que las curvas se comportan de forma similar (Cuadro 5)
CUADRO 5. Valores de coeficiente de determinación (R2) obtenidos de
curvas de germinación ajustadas por regresión lineal simple.
Genotipo Tratamiento R2 L. spp. (Alcones) 0,05% colchicina + polinización dirigida 0,96 L. spp. (Alcones) 0,05% colchicina + polinización libre 0,99 L. spp. (Alcones) 0,1% colchicina + polinización dirigida 0,96 L. spp. (Alcones) 0,1% colchicina + polinización libre 0,98 L. spp. (Alcones) Testigo 0,97 L. purpurea 0,05% colchicina + polinización dirigida 0,93 L. purpurea 0,05% colchicina + polinización libre 0,99 L. purpurea 0,1% colchicina + polinización dirigida 1 L. purpurea 0,1% colchicina + polinización libre 0,99 L. purpurea Testigo 0,93
El porcentaje final de germinación (A), tiempo en dias, estimado de
germinación de la primera semilla (to), extensión en la población del tiempo
de germinación (k), y velocidad de germinación de la mitad del total de
semillas que germinan (1/tA/2), se presentan en el Cuadro 6.

28
CUADRO 6. Efecto de la colchicina y tipo de polinización sobre los parámetros que describen la curva de germinación.
Genotipo Tratamiento Total de semillas
A(%)1 to 2 K 2 1/tA/2 2
L. spp. (Alcones)
0,05% colchicina + polinización dirigida 83
22 b 5,29 a 0,20 ab 0,11 a
L. spp. (Alcones)
0,05% colchicina + polinización libre 1216
24 b 6,51 a 0,21 a 0,10 a
L. spp. (Alcones)
0,1% colchicina + polinización dirigida 29
30 ab 7,07 a 0,20 ab 0,10 a
L. spp. (Alcones)
0,1% colchicina + polinización libre 346
30 ab 6,01 a 0,16 b 0,10 a
L. spp. (Alcones)
Testigo 200
62 a 7,64 a 0,26 a 0,10 a
L. purpurea 0,05% colchicina + polinización dirigida 345
28 c 5,91 a 0,25 ab 0,11 a
L. purpurea 0,05% colchicina + polinización libre 639
33 bc 4,13 a 0,28 a 0,15 a
L. purpurea 0,1% colchicina + polinización dirigida 554
36 bc 4,90 a 0,26 a 0,13 a
L. purpurea 0,1% colchicina + polinización libre 792
38 b 4,84 a 0,17 b 0,11 a
L. purpurea Testigo 200 88 a 5,23 a 0,30 a 0,13 a 1 Valores seguidos con letras iguales no presentan diferencias significativas con P<0.05 según Test de diferencia entre dos proporciones. 2 Valores seguidos con letras iguales no presentas diferencias significativas con P<0.05 según Intervalo de confianza.
Los porcentajes de germinación para Leucocoryne spp. (Alcones) muestran
que existen diferencias significativas entre el testigo y dos de los
tratamientos, pero no existe diferencia entre ellos.
Para Leucocoryne purpurea, los porcentajes de germinación del testigo
presentan diferencias significativas en todos los tratamientos, y también
existen diferencias entre los tratamientos.
Respecto a los tratamientos con colchicina existe un efecto sobre el
porcentaje final de germinación, siendo más efectivo en Leucocoryne
purpurea donde todos los tratamientos disminuyen en forma significativa

29
respecto al testigo, mientras que para Leucocoryne spp. (Alcones) el
porcentaje final del testigo es más bajo respecto a Leucocoryne purpurea y
sólo se diferencia en dos de los tratamientos, donde la dosis de colchicina
corresponde a 0,05%, para polinización libre y polinización dirigida.
Trabajos anteriores señalan que uno de los efectos recurrentes en el uso de
la colchicina es el comportamiento errático, con disminución de la fertilidad
de las plantas y la viabilidad de la semilla (BREMER-REINDERS y BREMER,
1952).
En tetraploides inducidos por colchicina en Eustoma grandiflorum
encontraron que su única característica negativa era su baja fertilidad de
semilla de un 2%, pero indican que esta característica se puede revertir con
cruzamientos dirigidos y selección (GRIESBACH y BHAT, 1990)
Los valores del tiempo estimado de germinación de la primera semilla (to) o
de latencia, estadísticamente, no muestran diferencias significativas entre los
tratamientos y el testigo, para ambas especies estudiadas, siendo menor los
valores que presenta Leucocoryne purpurea, sin embargo, las dos especies
presentan comportamientos homogéneos en el tiempo de germinación de la
primera semilla.
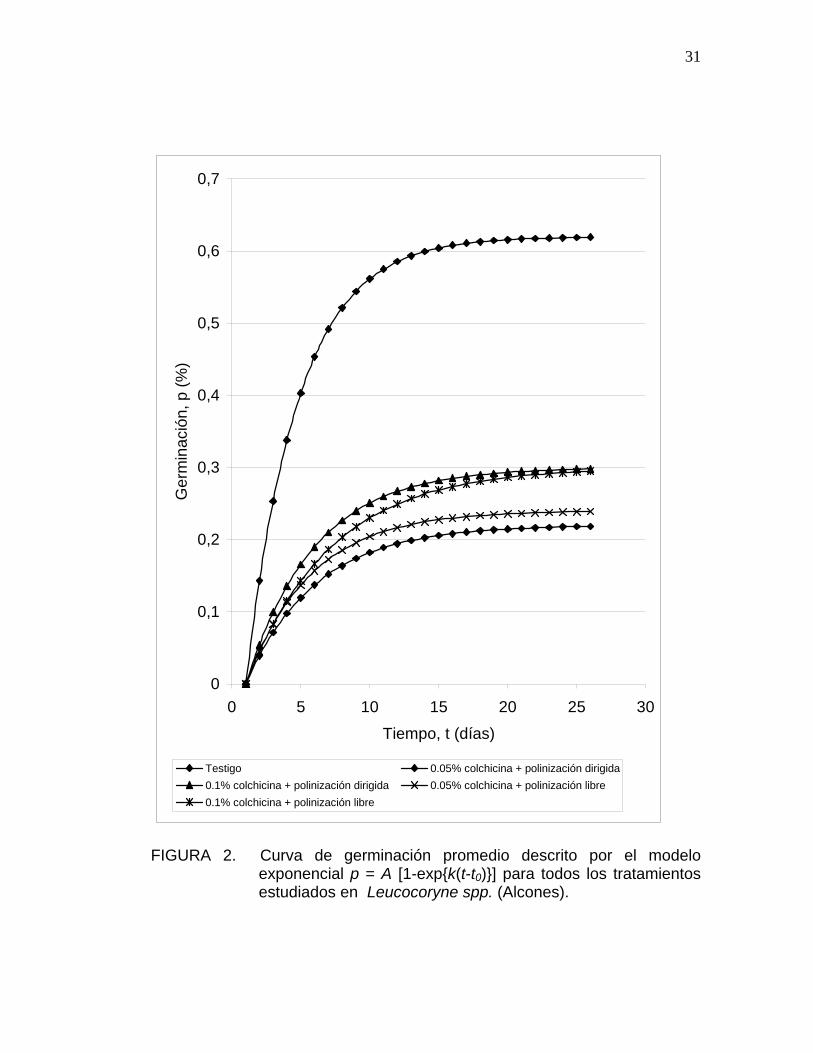
El parámetro k muestra valores similares, sólo presentan diferencias al
comparar el testigo con respecto al tratamiento 0,1% de colchicina y
polinización libre, tanto para Leucocoryne purpurea como para Leucocoryne
spp. (Alcones), los valores de k indican la duración del tiempo entre la
germinación de la primera y la última semilla, por lo tanto, los valores de
estos tratamientos se traducen en mayor desuniformidad en la emergencia

30
de las plántulas. Una mejor forma de visualizar este parámetro es mediante
las curvas de germinación (Figura 2 y 3).
Los efectos sobre la tasa de desarrollo han sido descritos por (HANCOCK,
1997) y señala que existe una disminución de la velocidad de crecimiento
como consecuencia de un aumento en la duración del ciclo celular. Esto no
parece ser universal, pues en otras especies con distintos niveles de ploidia,
los tiempos de ciclo no varían significativamente (REES, 1972).
31
0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0 5 10 15 20 25 30
Tiempo, t (días)
Ger
min
ació
n, p
(%)
Testigo 0.05% colchicina + polinización dirigida0.1% colchicina + polinización dirigida 0.05% colchicina + polinización libre0.1% colchicina + polinización libre
FIGURA 2. Curva de germinación promedio descrito por el modelo
exponencial p = A [1-exp{k(t-t0)}] para todos los tratamientos estudiados en Leucocoryne spp. (Alcones).

32
0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0,8
0,9
1
0 5 10 15 20 25 30Tiempo, t (días)
Ger
min
ació
n, p
(%)
Testigo 0.05% colchicina + polinización dirigida
0.1% colchicina + polinización dirigida 0.05% colchicina + polinización libre
0.1% colchicina + polinización libre
FIGURA 3. Curva de germinación promedio descrito por el modelo exponencial p = A [1-exp{k(t-t0)}] para todos los tratamientos estudiados en Leucocoryne purpurea.

33
4.4. Análisis de ploidia:
Los resultados obtenidos del análisis de citometría de flujo, se realizó sólo
para el grupo de plántulas que lograron sobrevivir posterior al transplante, ya
que se produjo una importante pérdida de individuos en esta etapa.
El Cuadro 7 indica los resultados de citometría de flujo, para plántulas
obtenidas de semillas, donde incluyen las dos especies estudiadas y los
niveles encontrados. La citometría de flujo entrega niveles de ploidia para
individuos diploides (2n), tetraploides (4n) y mixaploides (2n – 4n) y se
presentan a continuación.
Solo se logro duplicación del número de cromosomas en Leucocoryne
purpurea. En Leucocoryne spp. (Alcones) se produjo una duplicación parcial
logrando solo individuos mixaploides. Los individuos mixaploides presentan
niveles intermedios de ploidia entre diploides y tetraploides. Todos los
mixaploides analizados presentan entre 10% y 20% de cromosomas
duplicados.
El número de plantas analizadas para la especie Leucocoryne spp. (Alcones)
es bajo, debido a que se produjo una gran pérdida de plantas en el estudio
de germinación, y un bajo establecimiento post transplante, lo que se puede
deber a que solo se logra duplicación parcial originando semillas inviables o
plántulas de tipo quimeras, estas no logran desarrollar la totalidad de sus
tejidos.

34

35
Es imposible comparar si se cumple lo señalado por (ALLARD, 1967),
respecto a la adaptabilidad de las plantas cultivadas a la inducción de
poliploides, que aquellas plantas con número bajo de cromosomas es más
probable que respondan bien a la duplicación que aquellas plantas con
números altos de cromosomas.
Esto indicaría que Leucocoryne purpurea 2n = 10 tiene mayor probabilidad
de duplicación cromosomal, pero es necesario un estudio más acabado de
Leucocoryne spp. (Alcones) 2n = 18 para afirmar que esta situación se
presenta en este estudio.
Estos propuestos si se cumplen en los estudios realizados en Lilium donde
destacan que la diferencia en sensibilidad al tratamiento con colchicina entre
genotipos puede ser muy grande (VAN TUYL, 1989)
Por otro lado (GRANT, 1989) señala que los poliploides contienen mucha
duplicación de material genético en los núcleos, pueden tolerar, a menudo, la
pérdida de uno o más pares de cromosomas. Esto lleva a series poliploides
modificadas que terminan con lo que se ha denominado como reducción de
la poliploidia, lo que puede explicar que solo se logra encontrar individuos
mixaploides.
Leucocoryne purpurea presenta duplicación parcial y total del número de
cromosomas, pero las plantas tetraploides no sobrevivieron, ya que el
análisis de citometría de flujo fue destructivo y solo sobrevivieron las plantas
no analizadas, las que fueron cosechadas en el momento que la primera hoja
muere.

36
Debido a que Leucocoryne purpurea presento mejores porcentajes de
duplicación, las plantas que sobrevivieron y lograron formar un bulbito fueron
analizadas después de alcanzar un tamaño adecuado, logrado en dos
temporadas de crecimiento, lo que nos aseguraría la obtención de individuos
vivos con su número cromosómico duplicado.
En el Cuadro 8, se presentan los resultados del segundo análisis de
citometría de flujo, realizado sobre hojas obtenidas de bulbos jóvenes, para
la especie L. purpurea.
Los niveles de ploidia para el genotipo Leucocoryne purpurea, van desde un
alto porcentaje de plantas diploides, hasta la duplicación parcial, encontrando
plantas triploides y mixaploides en bajos porcentajes; y obteniendo
individuos duplicados al nivel de tetraploides y hexaploides, pero también en
porcentajes bajos.
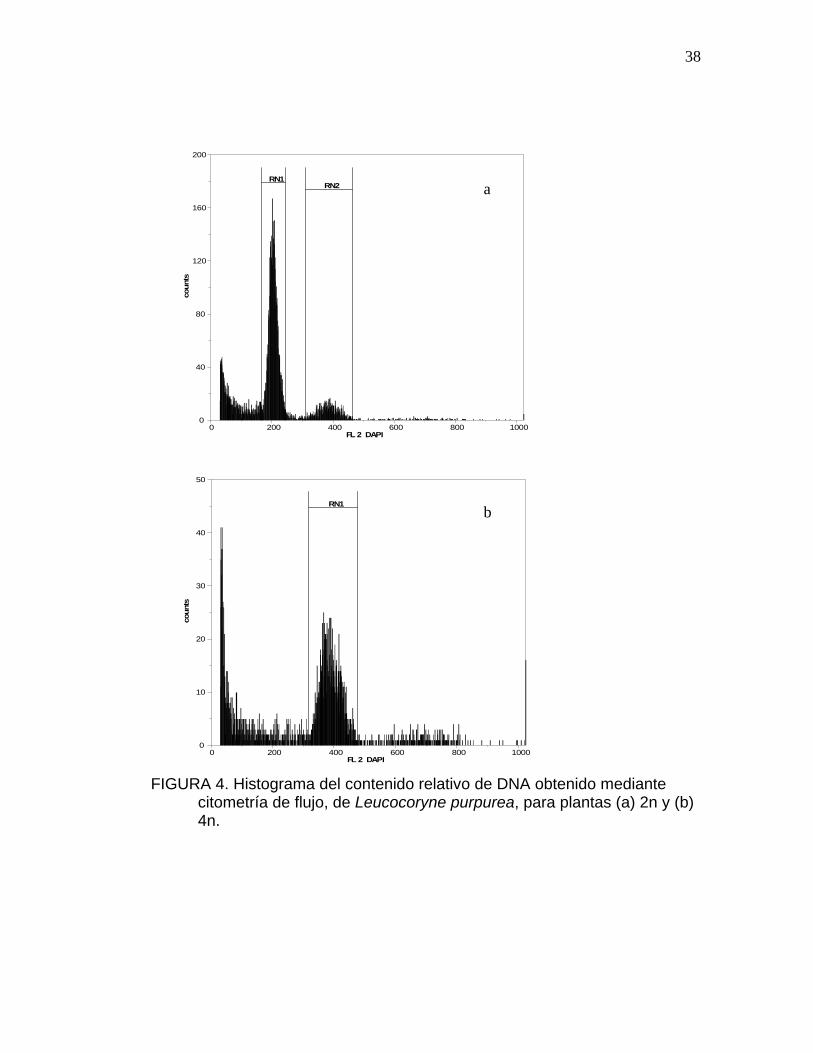
En la Figura 4 se presentan los histogramas de los niveles diploides y
tetraploides. La intensidad de la fluorescencia esta correlacionada
directamente a la cantidad de DNA que es teñido con DAPI.
Ambas concentraciones de colchicina lograron generar duplicación parcial y
total del número de cromosomas, siendo los mejores porcentajes de
individuos mixaploides y de individuos tetraploides de un 5.15%, con una
concentración 0.05%.

37
38
FIGURA 4. Histograma del contenido relativo de DNA obtenido mediante
citometría de flujo, de Leucocoryne purpurea, para plantas (a) 2n y (b) 4n.
0 200 400 600 800 10000
40
80
120
160
200
FL 2 DAPI
coun
ts
0 200 400 600 800 10000
40
80
120
160
200
FL 2 DAPI
coun
tsRN1
RN2 a
0 200 400 600 800 10000
10
20
30
40
50
FL 2 DAPI
coun
ts
0 200 400 600 800 10000
10
20
30
40
50
FL 2 DAPI
coun
ts
RN1b

39
Respecto a otros estudios (HALINAR, 1990), señala que cuando se realiza
duplicación de cromosomas utilizando tratamientos con colchicina, en
general está condicionado a éxito o fracaso, y a menudo resultan quimeras o
mixaploides inestables más que conversiones verdaderos tetraploides
estables; por lo cual no se aconseja depender solo de colchicina , si no que
también se debería poner a prueba el uso de otros agentes antimitóticos
como los herbecidas oryzalin y trifluralin, puesto que son menos toxicos y
tienen mejor afinidad para las células de las plantas (CHAUVIN et al., 2003).
Comparando los tipos de polinización, se obtuvieron mejores porcentajes en
cuando se dejo actuar la polinización natural, ya que en los individuos
obtenidos de cruzamientos solo encontramos duplicación parcial.
Lo que se tendría relación con lo postulado por (GRANT, 1981 y STEBBINS,
1971), que factores tanto internos como externos involucrados en la
generación y posterior establecimiento de poliploide natural. Básicamente, es
fundamental que se de la combinación de tres condiciones. La primera es la
existencia de especies diploides que posean diferentes genomas o
subgenomas. La segunda es que exista hibridación natural entre estas
especies, la tercera es que sean plantas perennes de vida larga, aumentando
la posibilidad de duplicaciones cromosómicas somaticas, o bien que sean
plantas anuales autocompatibles para así aumentar las posibilidades de
unión de gametos no reducidos.
Lo que nos indica que las plantas duplicadas exitosamente, se puede haber
debido a un proceso de autopolinizacion. Sin embargo, esto contrastaría con
la tendencia de la especie a la autoimcompatibilidad, que presenta la
especie. (MANSUR et al., 2004).

40
Estos resultados indican la limitancia de desarrollar un programa de
producción de poliploides, con el método propuesto es este estudio, ya que
solo seria valido en los genotipos donde la autoimcompatibilidad no es
importante, siendo este un grupo menor dentro el género.
Por otro lado resultaría interesante realizar un estudio de la frecuencia de
producción de gametos no reducidos, por una parte, de los que se producen
por aplicación de colchicina u otro agente antimitótico y por otro lado, a las
distintas especies, para poder determinar su real tendencia ha producir
poliploidia. Estos estudios se han desarrollados en programas de
poliploidización de Lilium (HALINAR, 1990).

41
5. CONCLUSIONES
El método de aplicación de colchicina utilizado en plantas del género
Leucocoryne logra una duplicación total o parcial del número de
cromosomas, influyendo en cierto grado en el proceso de meiosis, sin
embargo no se puede determinar el momento exacto de aplicación, sin un
estudio citológico de la meiosis.
Ambas concentraciones utilizadas, 0.05 y 0.1 de colchicina, producen
modificación del número cromosomal y no resultan tóxicas para la planta.
Pero los porcentajes obtenidos de individuos tetraploides son muy bajos.
La colchicina afecta el porcentaje final de germinación (A) y la extensión del
tiempo de germinación (k), es decir, afecta la viabilidad de las semillas y la
tasa de desarrollo de éstas.
La polinización natural o libre favorece la formación de individuos
tetraploides, sin embargo no seria interesante aplicar el método en un
programa extensivo de poliploidización, ya que es contrario a la tendencia del
género por la autoimcompatibilidad.

42
6. RESUMEN La duplicación de cromosomas mediante la utilización de colchicina, sigue siendo una técnica atractiva y viable en el mejoramiento de plantas ornamentales. El género Leucocoryne endémico de Chile presenta gran potencial ornamental como flor de corte y planta de maceta, por lo tanto se estudió un método de aplicación de colchicina in-vivo para obtener plantas tetraploides. En este estudio, se utilizaron dos dosis de colchicina 0,1% y 0,05% y se aplicaron en prefloración, sobre escapos florales en estados tempranos de desarrollo durante 12 horas. Una proporción de estas plantas se polinizaron mediante cruzamientos dirigidos y el resto fueron dejadas para que actuara la polinización natural de la especie. A las semillas obtenidas se les practicó un test de germinación utilizando el modelo de regresión lineal simple descrito por la ecuación exponencial p = A [1-exp{k(t-t0)}] y se evaluó el nivel de ploidia mediante citometría de flujo. Se concluyó que el método de aplicación y las dosis de colchicina utilizadas lograron duplicar total o parcialmente en número de cromosomas y que existe un efecto detrimental en la germinación de semillas.

43
7. ABSTRACT
Induction of polyplody in plants of the Leucocoryne genus. Chromosome duplication using colchicine continues to be an attractive and viable technique in the breeding of ornamental crops. The Leucocoryne genus, endemic to Chile, has great potential for ornamental use as both cut flowers and pot plants. Therefore a method for the application of colchicine in-vivo to obtain tetraploid plants was studied. In this study two colchicine doses, 0.1% and 0.05%, were applied before flowering to Leucocoryne scapes in early stages of development, for 12 hours. Some of these plants were pollinated manually under supervised crosses and the rest were left to pollinate naturally. The seeds were then tested for germination using a simple linear regression model described by the exponential equation p = A [1-exp{k(t-t0)}], and the ploidy level was evaluated using flow cytometry. It was concluded that the application method and the colchicine doses utilized were able to partially or completely duplicate the chromosome number and that there was a detrimental effect on seed germination.

44
8. LITERATURA CITADA ALLARD, A. 1967. Principios de la mejora genética de las plantas.
Barcelona, Omega. 498p. ARANEDA, L.; SALAS, P.; MANSUR L. 2004. Chromosome numbers in the
Chilean endemic genus Leucocoryne (Huilli). Journal Am. Soc. Hort. Science 129(1): 77-80.
ARUMAGANATHAN, K. 1991. Estimation of nuclear DNA content of plants
by flow cytometry. Plant Molecular Biology Reporter 9 (3): 229-233. AVERETT, J. 1981. Polyploidy in plant taxa. Summary. In: Lewis Walter H.
ed.. Polyploidy: Biological relevance. New York. Plenanm Press. BAHAMONDES, N. y LABARCA, C. 1994. Citogenética y probables
mecanismos de evolución en especies de los géneros Leucocoryne y Pabellonia (Alliaceae). Tesis de licenciatura. Departamento de Biología. Facultad de Ciencias. Universidad de La Serena.
BRAUER, C. 1969. Fitogenética aplicada: los conocimientos de la herencia
vegetal al servicio de la humanidad. México D. F., Limusa. 598p. BREMER-REINDERS, D. E. and BREMER, G. 1952. Methods used for
producing polyploid agricultural plants. Euphytica 1: 87-94. CAVE, M. 1939. Macrosporogenesis in Leucocoryne ixioides Lindl.
Cytology 8: 407-411. CHAUVIN, J.; SOUCHET, C.; DTEC, J. AND ELLISSECHE, D. 2003.
Chromosome doubling of 2x Solanum species by oryzalin: method development and comparison with spontaneous chromosome doubling in vitro. Plant, Cell, Tissue and Organ Culture 73:65-73.
CHONG, S; OZIAS-AKINS, P. 1992. Rapid estimation of ploidy levels in vitro
regenerated interespecific Arachis hybrids and fertile triploids. Euphytica 64: 183-188
COSTICH, D.E.; ORTIZ, R.; MEAGLER, T.R.; BREDERTE, L.P. 1993.
Determination of ploidy level and nuclear DNA content in blueberry by flow cytometry. Theoretical and Applied Genetics 86: 1001-1006

45
CRAMER, S. 1999. Laboratory techniques for determining ploidy in plants. HortTechnology 9(4): 594-596.
CROSA, O. 1988. Los cromosomas de nueve especies del género
Leucocoryne Lindley, (Alliae – Alliacea). Boletín insvestigaciones. 17: 1– 2.
CUBERO, J. 1999. Introducción a la mejora genética vegetal. Córdova.
Mundi-Prensa. 365p. DAHLGREN, R. CLIFFORD, H.; Yeo, P. 1985. The families of the
monocotyledons. Berlin. Springer-Verlag. 520p. DE LA CUADRA C.; MANSUR, L.; VERDUGO, G. AND ARRAIGADA, L.
2002. Deterioro de las semillas de Leucocoryne spp. en función del tiempo de almacenaje. Agricultura Técnica 62.(1):46-55.
DE WET, J. 1980. Origins of polyploids. In: Polyploidy, Biological Relevance.
Levis W. (ed.). New York. Plenam Press. pp 3-16. DEWEY, D.R. 1979, Some applications and misapplications of poliploidy, In:
Polyploidy.Lewis W. (ed).. New York. Plenam Press. pp 445-470. DOLEZEL, J., LUCRETTI, S. and MACAS J. 1995. Flow cytometric analysis
and sorting of plant chromosome. In: Brandham P. E. and Bennett M. D. eds. Kew chromosome conference IV, p185-200. Royal Botanic Gardens, Kew.
FRANDSEN, N.O. 1967. Haploidproducktion aus einem
kartoffelzuchtmaterial mit intensiver wildareingrenzung. Max-Planck-Instituf für züchtungsforshchung. Köl-Vogelsang. Berlin. Springer-Verlag. Genetica Breeding Research 37(3): 120-134.
FONT QUER, P. 1953. Diccionario de Botanica. Barcelona, ed. Labor S.A.: 1244p ELLIOT, F. 1964. Citogenética y mejoramiento de plantas. México,
Continental. 474p. GRANT, V. 1989. Especiación vegetal. México, D.F., Limusa. 587p. ______. 1981. Plant speciation. Colombia, N.Y. University Press. 563p.

46
GRAU, J. 1992. Eine neue Art der Gattung Leucocoryne (Alliaceae) aus Mittelchile. Mitt. Bot. Staatssamml 30: 431 – 437.
GRIESBACH, R. J.; BHAT, R. N. 1990. Colchicine-induced polyploidy in
Eustoma grandiflorum. HortScience 25(10): 1284-1286. HALINAR, J. C. 1990. Polyploidy and unreduced gametes. The Daylily
Journal 45(4): 339-346. HANCOCK, J. F. 1997. The colchicine story. HortScience 32(6): 1011-1012. MANSUR L.; GONZALEZ M.; ROJAS I.; SALAS P. 2004. Self Incompatibility
in the Chilean Endemic Genus Leucocoryne (Huilli). Journal Am. Soc. Hort. Sience 129(6):836:838.
MANSUR, L.; ZOELLNER, O.; RIELDEMENN, P.; VERDUGO, G. y
HARRISON, C. 2002. Leucocoryne un género nativo chileno y su uso como planta de jardín. Viña del Mar, ed Reimco. 49 p. Serie Manuales de Innovación Tecnológica para la Agricultura. Manual n°1. Universidad Católica de Valparaíso.
MAROTO, J.V. 1989. Horticultura herbácea especial. Madrid, Mundi-Prensa.
590p. MORENO, R. y ARANCIO, G. 2001. Posibles híbridos naturales entre dos
especies del género Leucocoryne. Gayana 58(1): 86-87. OHKAWA, K.; KIM, H.H.; NITTA, E. and FUKOZAWA, Y. 1996. Storage
temperature and duration effect flower bud development, shoot emergence and flowering of Leucocoryne coquimbensis F. Phil. J. Amer. Soc. Hort. Sci. 123 (4): 586-591
OLLITRAULT-SAMMARCELLI, F., LEGAVE, J. M., MICHAUX-FERRIERE,
N., HIRSH, A. M. 1993. Use of flow cytometry for rapid determination of ploidy level in the genus Actinidia. Sciencia Horticulturae 57(1994): 303-313.
PALMA-ROJAS, C., ZEPEDA, S., y LABARCA, M. 1991. Citogenética
comparada en Leucocoryne (Liliaceae). Universidad de Santiago. VIII Reunión Nacional de Botánica. Santiago, del 16 al 19 de octubre de 1991. Chile. pp 85.

47
POEHLMANN, J.M. 1965. Mejoramiento genético de las cosechas. México D. F., Limusa. 453p.
REES, H. 1972. DNA in higher plants. Brookhaven symposia in Biology. 23:
394-418. SALAS P.; and MANSUR L. 2004. Gene flow between parents with different
ploidy levels in a natural population of Leucocoryne. Journal Am. Soc. Hort. Science 129(6):833-835.
SPOTORNO, A. 1985. Conceptos en cariología descriptiva y comparada.
In: Fernández-Donoso, R., ed. El núcleo, los cromosomas y la evolución, Santiago, Unesco. pp 135-165.
STEBBINS, G.L. 1971. Chromosomal evolution in higher plants. London, ed.
Edward Arnold Ltda. 216p VAN TUYL, J. M. 1989. Research on mitotic and meiotic polyploidization in
Lily breeding. Herbertia 45(1-2):97-103. ZÖELLNER, O. 1972. El género Leucocoryne. Anales Museo de Historia
Natural 5: 9-83.

48
ANEXOS

49
ANEXO 1. (a) Leucocoryne spp. (Tipo Alcones). (b) Leucocoryne purpurea.
a
b

50
ANEXO 2. Protocolo de análisis de citrometría de flujo, utilizado por Plant Cytometry Services. ISOLATION OF NUCLEI. Starting with a certain amount of fresh plant material (preferably young leaves), the first step is to isolate the nuclei from the plant cells and keep them intact in a solution.To reach this, leaf material (a few cm
2 /20-50 mgr)) is
"chopped" with a sharp razor blade in an icecold buffer, in a plastic petri disc. DNA buffer (stored at 4 ºC) a little modified after.(ARUMUGANATHAN AND EARLE 1991) 5 mM Hepes 10 mM Magnesium sulphate heptahydrate 50 mM Potasium chloride 0,2 % Triton X-100 2% DTT (Dithiothreitol) 4 mgr/liter DAPI pH 8 DAPI is a fluorescent dye which selectively complexes with double-stranded DNA to give a product that fluoresce at 465 nm. DAPI has specific DNA-binding properties with preference for adenine-thymine (AT)-rich sequences. After chopping, the buffer (ca. 2 ml.), containing cell constituents and large tissue remnants, is passed through a nylon filter of 40 micrometer mesh size. With this method it is possible to get thousands of nuclei from a leaf of a few cm2. FLOW CYTOMETRY. The solution with stained nuclei is send through the flowcytometer. The fluorescence of the stained nuclei, passing through the focus of a lightbeam from a high-pressure mercurylamp, is measured by a photomultiplier and converted into voltagepulses. These voltagepulses are electronically processed to yield integral and peak signals and can be processed by a computer. When the samples are run with the appropriate filter-settings for excitation and emission, DNA histograms can be produced. MATERIAL. Flowcytometer: PAS II (Partec GmbH, Otto Hahnstrasse 32, D-4400 Münster, Germany) with a high pressure mercury lamp, OSRAM HBO 100 long life Filtercombination with DAPI:

51
Heat protection filter KG-1 Excitation-filters: UG-1and BG-38. Dichroic mirrors: TK 420 and TK 560. Emission-filter: GG 435 DNA histograms are performed on a normal PC. Software: Flows 2.00 (development and distribution by Plant Cytometry Services)
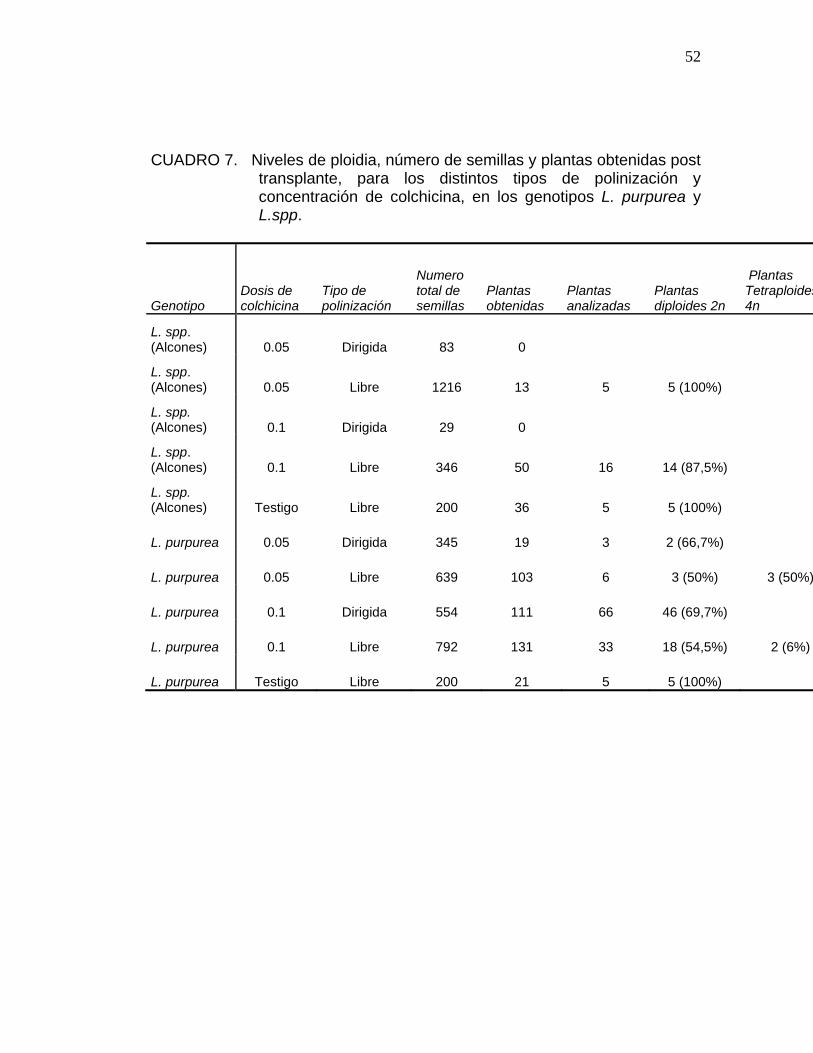
52
CUADRO 7. Niveles de ploidia, número de semillas y plantas obtenidas post transplante, para los distintos tipos de polinización y concentración de colchicina, en los genotipos L. purpurea y L.spp.
Genotipo Dosis de colchicina
Tipo de polinización
Numero total de semillas
Plantas obtenidas
Plantas analizadas
Plantas diploides 2n
Plantas Tetraploides4n
L. spp. (Alcones) 0.05 Dirigida 83 0
L. spp. (Alcones) 0.05 Libre 1216 13 5 5 (100%)
L. spp. (Alcones) 0.1 Dirigida 29 0
L. spp. (Alcones) 0.1 Libre 346 50 16 14 (87,5%)
L. spp. (Alcones) Testigo Libre 200 36 5 5 (100%)
L. purpurea 0.05 Dirigida 345 19 3 2 (66,7%)
L. purpurea 0.05 Libre 639 103 6 3 (50%) 3 (50%)
L. purpurea 0.1 Dirigida 554 111 66 46 (69,7%)
L. purpurea 0.1 Libre 792 131 33 18 (54,5%) 2 (6%)
L. purpurea Testigo Libre 200 21 5 5 (100%)
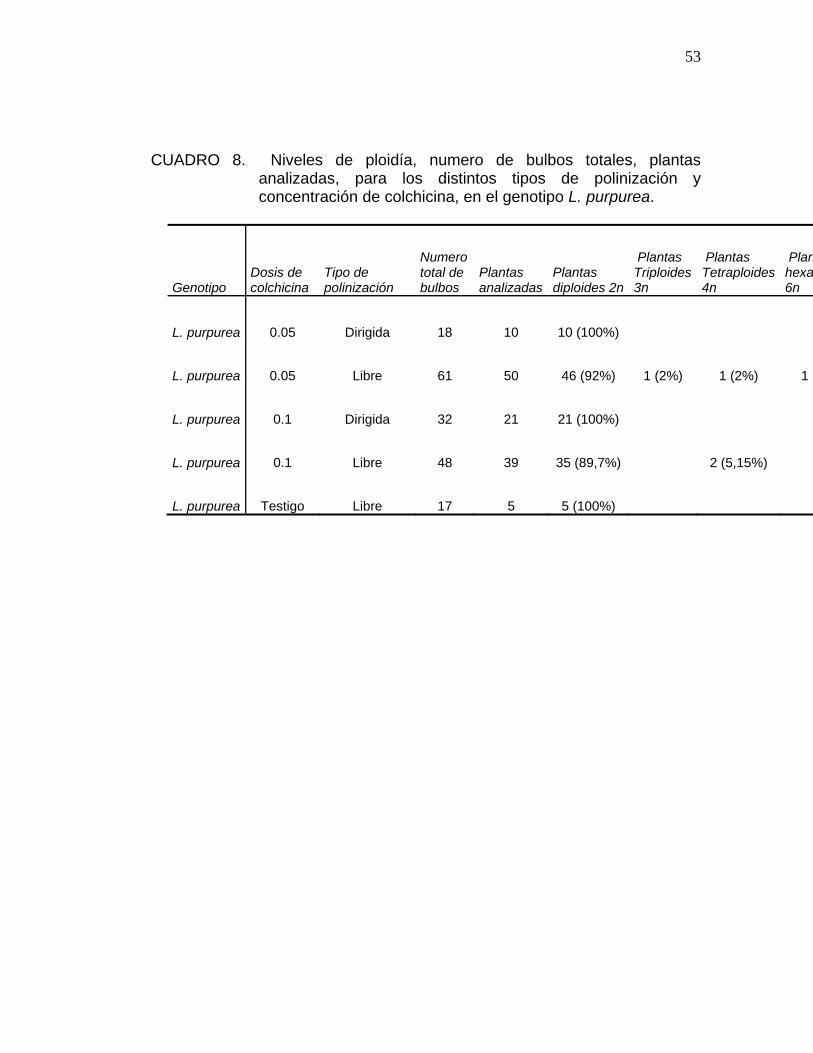
53
CUADRO 8. Niveles de ploidía, numero de bulbos totales, plantas analizadas, para los distintos tipos de polinización y concentración de colchicina, en el genotipo L. purpurea.
Genotipo Dosis de colchicina
Tipo de polinización
Numero total de bulbos
Plantas analizadas
Plantas diploides 2n
Plantas Triploides 3n
Plantas Tetraploides 4n
Planhexap6n
L. purpurea 0.05 Dirigida 18 10 10 (100%)
L. purpurea 0.05 Libre 61 50 46 (92%) 1 (2%) 1 (2%) 1 (
L. purpurea 0.1 Dirigida 32 21 21 (100%)
L. purpurea 0.1 Libre 48 39 35 (89,7%) 2 (5,15%)
L. purpurea Testigo Libre 17 5 5 (100%)


















